Embed Size (px)
Citation preview

Aus dem Institut für Tierernährung der Tierärztlichen Hochschule Hannover
___________________________________________________________________
Untersuchungen zur Entwicklung eines Screening-Tests zur Beurteilung substituierter Lipasen
im Modell pankreasgangligiertes Miniaturschwein
INAUGURAL-DISSERTATION
Zur Erlangung des Grades einer
Doktorin der Veterinärmedizin
(Dr. med. vet.)
durch die Tierärztliche Hochschule Hannover
Vorgelegt von
Sonka Tieneke Zantz aus Leer
Hannover 2006

Wissenschaftliche Betreuung: Univ.-Prof. Dr. J. Kamphues
1. Gutachter: Univ.-Prof. Dr. J. Kamphues
2. Gutachter: Prof. Dr. B. Schröder
Tag der mündlichen Prüfung: 21. November 2006

Meiner Familie


INHALTSVERZEICHNIS
------------------------------------------------------------------------------------------------------------------------
Inhaltsverzeichnis 1 EINLEITUNG 1 2 SCHRIFTTUM 3 2.1 Vorbemerkung 3 2.1.1 Beteiligung von Verdauungsenzymen verschiedenen
Ursprungs an der Nährstoffspaltung 3 2.2 Die exokrine Pankreasinsuffizienz 4 2.2.1 Vorkommen und Entstehung 4 2.2.2 Folgen des Mangels pankreatischer Verdauungsenzyme 6 2.2.3 Auswirkungen der EPI auf die Verdaulichkeit der Fette 7 2.3 Therapie der EPI 9 2.3.1 Enzymatische Substitutionstherapie in der EPI-Therapie 9 2.3.2 Indikationen für den Einsatz von Enzymen bei Vorliegen einer EPI 10 2.3.3 Diätetik 10 2.3.4 Einsatz von Multi- und Monoenzympräparaten in der Therapie der
EPI 11 2.3.4.1 Multienzympräparate 11 2.3.4.2 Monoenzympräparate 14 2.4 Entwicklung neuer Enzymprodukte 17 2.4.1 Wirkstoffneuentwicklung in der pharmazeutischen Industrie 17 2.4.2 Ablauf bzw. Phasen einer Wirkstoffneuentwicklung 18 2.4.2.1 Anfangsphase 18 2.4.2.2 Die vorklinische Entwicklungsphase 19 2.4.3 In-vivo-Studien am pankreasgangligierten Miniaturschwein als
Modell für den an EPI erkrankten Menschen 20 2.4.3.1 Methoden der Verdaulichkeitsbestimmung zur Bewertung der
Wirksamkeit von Enzympräparaten 21 2.4.3.2 Vor- und Nachteile der Überprüfung der Wirksamkeit substituierter
lipolytischer Enzyme anhand der Bestimmung der Fettverdaulichkeit mittels herkömmlicher Methoden im pankreasgangligierten Miniaturschwein 24
2.4.3.3 Nachteile herkömmlicher Verdaulichkeitsstudien (Bestimmung von praecaecaler und totaler Verdaulichkeit) in der Produktentwicklung von Enzymen zur Substitutionstherapie 25

INHALTSVERZEICHNIS
------------------------------------------------------------------------------------------------------------------------
2.4.4 Die klinische Entwicklungsphase von Enzymen zur
Substitutionstherapie 25 2.4.5 Zulassung und Markteinführung einer pharmazeutischen
Neuentwicklung 27 2.5 Vorteile durch den Einsatz von Screening-Verfahren 28 2.5.1 Möglichkeiten und Grenzen eines Screening-Verfahrens für
Proteasen und Amylasen als ergänzende Methode 29 2.5.2 Überlegungen zur Entwicklung eines Lipase-Screenings 30 3 EIGENE UNTERSUCHUNGEN 32 3.1 Material und Methoden 32 3.1.1 Versuchsziel 32 3.1.2 Versuchstiere 33 3.1.3 Aufstallung und Haltung 35 3.1.4 Statistische Methoden 35 3.2 Versuchsdurchführung 36 3.2.1 Übersicht über die durchgeführten Versuche 36 3.2.2 Methodik der Chymuskollektion 38 3.2.2.1 Exkurs: Begriffsbestimmung „Verschwindensrate“ 39 3.2.3 Enzymprodukte und -substitution 40 3.2.4 Versuchsfutter 41 3.2.5 Versuchsablauf 42 3.2.6 Probengewinnung und -entnahme 45 3.2.7 Analytik und angewandte Untersuchungsparameter 46 3.2.8 Verwendete Berechnungsansätze zur Einschätzung der
praecaecalen Fettverdaulichkeit 54 3.2.9 Entwicklung eines In-vivo-Screening-Tests zur Einschätzung der
Wirksamkeit von Lipasen anhand eines bekannten Multienzymproduktes 57
3.2.10 Anforderungen an eine Testmahlzeit für ein Lipase-Screening 58 3.2.11 Entwicklung einer geeigneten Testmahlzeit 61 3.2.11.1 1. Versuch zur Optimierung einer flüssigen Testmahlzeit 62 3.2.11.2 2. Versuch zur Optimierung einer flüssigen Testmahlzeit 65 3.2.11.3 3. Versuch zur Optimierung einer flüssigen Testmahlzeit 68 3.2.11.4 4. Versuch zur Optimierung einer flüssigen Testmahlzeit 72 3.2.11.5 5. Versuch zur Optimierung einer flüssigen Testmahlzeit 77 3.2.11.6 6. Versuch zur Optimierung einer flüssigen Testmahlzeit 80 3.2.11.7 7. Versuch zur Optimierung einer flüssigen Testmahlzeit 84 3.2 Ergebnisse 91 3.2.1 Screening-Test für Lipasen 91 3.2.2 Allgemeine Beobachtungen 92 3.2.3 Untersuchte Parameter im Rahmen des Screening-Tests 92

INHALTSVERZEICHNIS
------------------------------------------------------------------------------------------------------------------------
3.2.3.1 Absolute Anflutung von Chymus (Trockensubstanz) 92 3.2.3.2 Rohfettkonzentration im Chymus 97 3.2.3.3 Absolute Anflutung von Rohfett 101 3.2.3.4 Chromoxid-Konzentration im Chymus 105 3.2.3.5 Chromoxid-Wiederfindung 106 3.2.3.6 Verhältnis der Konzentrationen von Rohfett und Chromoxid 108 3.2.3.7 Trockensubstanz-Verschwindensrate (TS-VR) 112 3.2.3.8 Rohfett-Verschwindensrate (Rfe-VR) 116 3.2.3.9 Relative Anflutung von Trockensubstanz 120 3.2.3.10 Relative Anflutung von Rohfett 121 3.2.3.11 Relative Trockensubstanz-Verschwindensrate 123 3.2.3.12 Relative Rohfett-Verschwindensrate 126 4 DISKUSSION 128 4.1 Kritik der Methoden 128 4.1.1 Anzahl der Tiere 128 4.1.2 Validierung des Schnelltests 130 4.1.3 Eignung der erstmalig angewandten Methode zur Fettbestimmung
(AnkomHCl Hydrolysis System®) 130 4.2 Erörterung der eigenen Ergebnisse 132 4.2.1 Wie muss eine Testmahlzeit beschaffen sein, um In-vivo die
Wirksamkeit von Lipasen in einem Schnelltest überprüfen zu können? 132
4.2.1.1 Modifikation des Trockensubstanz- und Rohfasergehaltes 133 4.2.1.2 Verwendung von Chromoxid als Marker 134 4.2.1.3 Einsatz eines flüssigen Markers am Vorabend des Screening-Tests 135 4.2.1.4 Stabilisierung der Testmahlzeit (Homogenisierung) 136 4.2.2 Ist es möglich, neu entwickelte Lipasen mit einer optimierten
Testmahlzeit vergleichend zu prüfen und hinsichtlich ihrer Effektivität zu rangieren? 137
4.2.2.1 Vorteile eines Screening-Verfahrens 137 4.2.2.2 Bedeutung der relativen Flussrate für die Berechnung der
„Verschwindensrate“ 138 4.2.2.3 Beurteilung der Parameter 139 4.2.3 Korrelieren die Ergebnisse aus dem Schnelltest mit denen aus
etablierten Verdaulichkeitsversuchen? 142 4.2.3.1 Vergleich der Ergebnisse des Screening-Tests mit denen
herkömmlicher Gesamtverdaulichkeitsstudien 143 4.2.3.2 Vergleich der im Screening-Verfahren aufgestellten Rangierung mit
In-vitro-Ergebnissen 147 4.2.3.3 Kombinationsversuch 149 4.3 Ausblick 151

INHALTSVERZEICHNIS
------------------------------------------------------------------------------------------------------------------------
4.4 Schlussfolgerungen 152 5 ZUSAMMENFASSUNG 153 6 SUMMARY 156 7 LITERATURVERZEICHNIS 159 8 TABELLENANHANG 176

ABKÜRZUNGSVERZEICHNIS
------------------------------------------------------------------------------------------------------------------------
Abkürzungsverzeichnis ® eingetragenes Warenzeichen % Prozent °C Grad Celsius BfArM Bundesinstitut für Arzneimittel und Medizinprodukte BSE Bovine Spongiforme Enzephalopathie ca crude ash (= Rohasche) cfa crude fat (= Rohfett) CF cystische Fibrose cfi crude fibre (= Rohfaser) cp crude protein (= Rohprotein) Cr2O3 Chromoxid d Tag D1 niedrigste Dosierung = 500 IE D2 mittlere Dosierung = 5 000 IE D3 höchste Dosierung = 75 000 IE E1 Testlipase Nr.1 E2 Testlipase Nr.2 E3 Testlipase Nr.3 E4 Testlipase Nr.4 E5 Testlipase Nr.5 EMEA European Medicine Evaluation Agency EPI exokrine Pankreasinsuffizienz FDA Federation of Drug Administration F&E Forschung und Entwicklung FS Fettsäure g Gramm Ges. Gesamt GIT Gastrointestinaltrakt h Stunde HF Haltungsfutter IE Internationale Einheit (hier: pancreatic FIP Pharm. Eur. Unit) IND Investigational New Drug Application kcal Kilokalorien kg Kilogramm KH Kohlenhydrat KJ Kilojoule KT Kontrolltier LCFA long chain fatty acids LFS langkettige Fettsäuren

ABKÜRZUNGSVERZEICHNIS
------------------------------------------------------------------------------------------------------------------------
LPS Lipopolysaccharide min Minuten MJ Megajoule ml Milliliter ML Mikrolipid MW arithmetischer Mittelwert n. F. nach Farbwechsel NfE Stickstoff-freie Extraktstoffe OL Olivenöl oS organische Substanz p Irrtumswahrscheinlichkeit PEI pancreatic exocrine insufficiency (= Exokrine Pankreasinsuffizienz) DM dry matter (= Trockensubstanz) DR disappearence rate (= Verschwindensrate) H2O water (= Wasser) IU international unit (= Internationale Einheit) mM Millimolar pH potentia hydrogenii PK Pankreon® PL-Tier pankreasgangligiertes Tier ppr. postprandial Ra Rohasche Rfa Rohfaser Rfe Rohfett Rp Rohprotein SCFA short chain fatty acids STABW Standardabweichung sV scheinbare Verdaulichkeit TS Trockensubstanz uS ursprüngliche Substanz VDLUFA Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten VFA Verband Forschender Arzneimittelhersteller e. V. VR Verschwindensrate vs versus

EINLEITUNG ------------------------------------------------------------------------------------------------------------------------
1
1. Einleitung
Ein Mangel an pankreatischen Verdauungsenzymen führt -zumeist infolge chronischer
Pankreatitis- zum Bild der exokrinen Pankreasinsuffizienz (EPI). Folgen sind eine
Malabsorption und Maldigestion der Kohlenhydrate, Proteine und besonders der Fette,
wodurch das klinische Leitsymptom der EPI, die Steatorrhoe, ausgelöst wird (DIMAGNO
et al. 1977). Die aktuellen Behandlungsempfehlungen basieren auf der oralen Substitution von
Verdauungsenzymen zumeist porcinen Ursprungs. Bestrebungen, Produkte mikrobiellen
(fungalen bzw. bakteriellen) und biotechnologischen Ursprungs zur Marktreife
weiterzuentwickeln (LAYER u. KELLER 2003), erhöhen die Vielzahl zu untersuchender
Substanzen. Als Vorteile mikrobieller Präparate werden eine höhere Effizienz, ein
verbesserter Verbraucherschutz sowie die Möglichkeit einer gezielten Weiterentwicklung
erachtet. Mikrobiell bzw. biotechnologisch gewonnene Substanzen können im Gegensatz
zu Produkten tierischen Ursprungs durch genetische Veränderung den Anforderungen
entsprechend modifiziert und optimiert werden.
Zur In-vivo-Prüfung der Wirksamkeit substituierter Verdauungsenzyme wurde in
zahlreichen Studien (TABELING 1998; FASSMANN 2001; HELDT 2001; MANDISCHER
2002; FUENTE-DEGE 2003; KAMMLOTT 2003; KARTHOFF 2004; BECKER 2005) das
„pankreasgangligierte, ileocaecal-fistulierte Miniaturschwein“ als geeignetes Modell
etabliert. Die dazu vorgenommenen Verdaulichkeitsuntersuchungen erfordern neben
einem hohen Arbeits-, Zeit- und Kostenaufwand größere Mengen der zu prüfenden
Enzyme, die allerdings erst zu einem relativ späten Zeitpunkt der Entwicklung zur
Verfügung stehen, woraus bei ungenügender In-vivo-Wirksamkeit erhebliche zeitliche und
finanzielle Verluste resultieren können. Im Gegensatz dazu erfordern In-vitro-Tests zwar
einen relativ geringen Aufwand, die Ergebnisse korrelieren jedoch oftmals nur
unzureichend mit den Ergebnissen aus In-vivo-Untersuchungen. Daher besteht ein Bedarf
an zeit- und kostengünstigen In-vivo-Verfahren zur Überprüfung der Wirksamkeit
substituierter Enzyme unter Verwendung geringer Enzymmengen. BECKER (2005) gelang
die Etablierung eines Schnelltests für Amylasen und Proteasen gemäß diesen
Anforderungen.

EINLEITUNG ------------------------------------------------------------------------------------------------------------------------
2
Im Rahmen der hier vorliegenden Studie sollte ein Screening-Verfahren für Lipasen
entwickelt werden, welches den oben genannten Anforderungen genügt (Einsparung von
Zeit und Kosten sowie insbesondere geringer Verbrauch an dem zu prüfenden Enzym)
und eine erste Einschätzung und Rangierung der In-vivo-Effizienz bisher ungeprüfter
Lipasen erlaubt. Etablierte Verdaulichkeitsstudien, die deutlich aufwändiger sind und einen
sehr viel höheren Enzymverbrauch bedingen, wären dann lediglich für die aufgrund der
Rangierung ausgewählten, vielversprechenden Enzyme zur näheren Quantifizierung der
Wirksamkeit notwendig.
Vor diesem Hintergrund sollten in der vorliegenden Arbeit folgende Fragen geklärt
werden:
• Wie muss eine Testmahlzeit beschaffen sein, um In-vivo die Wirksamkeit von
Lipasen in einem Schnelltest überprüfen zu können?
• Ist es möglich, neu entwickelte Lipasen mit einer optimierten Testmahlzeit
vergleichend zu prüfen und entsprechend ihrer Effektivität zu rangieren?
• Korrelieren die Ergebnisse des Schnelltests für Lipasen mit denen aus etablierten
Verdaulichkeitsuntersuchungen?

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
3
2 Schrifttum
2.1 Vorbemerkung
Die vorliegende Studie entstand in Fortsetzung einer Reihe von Untersuchungen
(TABELING 1998; FASSMANN 2001; HELDT 2001; MANDISCHER 2002; FUENTE-
DEGE 2003; KAMMLOTT 2003; KARTHOFF 2004; BECKER 2005) am
pankreasgangligierten Miniaturschwein, welches als Modell für den an exokriner
Pankreasinsuffizienz erkrankten Menschen dient. Während in früheren Arbeiten dieses
Projektes insbesondere die Effekte der Pankreasgangligatur auf die Verdauung der
Nährstoffe und die Milieubedingungen im Gastrointestinaltrakt untersucht wurden,
fokussierten sich die späteren Arbeiten vorwiegend auf methodische Fragestellungen und
alternative Untersuchungsmethoden zur Beurteilung der Effizienz substituierter Enzyme.
Neben etablierten Verdauungsversuchen kamen beispielsweise Tests unter Verwendung
verschiedener Blutparameter (KARTHOFF 2004) sowie ein neu entwickelter Screening-
Test (BECKER 2005) zur Überprüfung der Wirksamkeit von Proteasen und Amylasen zur
Anwendung.
Im Folgenden wird auf einige Aspekte der Fettverdauung sowie die Substitutionstherapie
des an EPI erkrankten Menschen eingegangen, da diese die Symptome bzw. den
Behandlungserfolg entscheidend mitbestimmen.
2.1.1 Beteiligung von Verdauungsenzymen verschiedenen Ursprungs an der Nährstoffspaltung
An der Digestion und Absorption der Nahrung sind bei Mensch und Tier verschiedene
Organsysteme beteiligt. Neben dem Verdauungskanal selbst tragen auch die
Darmanhangsdrüsen (Pankreas und Leber) entscheidend zu Abbau und Resorption der
Nahrungsbestandteile bei. Für die enzymatische Spaltung der Nährstoffe sezerniert der

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
4
exokrine Anteil des Pankreas in erheblichem Umfang Verdauungsenzyme, die in den
Azinizellen gebildet werden und über den Ausführungsgang der Bauchspeicheldrüse ins
Duodenum gelangen (RINDERKNECHT 1993). Des Weiteren produziert das Pankreas in
seinen Gangzellen eine natriumbikarbonatreiche Flüssigkeit (SCHARRER u. WOLFFRAM
2000), welche die in der Ingesta enthaltene Magensäure neutralisiert und die Viskosität
des Chymus herabsetzt. Die Neutralisation stellt sicher, dass die pankreatischen Enzyme
nicht durch die im Chymus enthaltene Säure gastralen Ursprungs denaturiert werden und
im Dünndarm ein für die Aktivität der sezernierten Enzyme optimales Milieu entsteht. Es
sind jedoch auch Enzyme am Abbau der Nahrung beteiligt, die nicht körpereigener
Herkunft sind, sondern der intestinalen Mikroflora des Dünn- und Dickdarms entstammen
(EHRLEIN 2000a, b). Der Umfang der Fermentationsvorgänge der Intestinalflora ist unter
anderem abhängig von der Lokalisation (SCHARRER u. WOLFFRAM 2000), der
Substratverfügbarkeit sowie der Passagerate des Substrates im Gastrointestinaltrakt
(READ et al. 1984; LAYER et al. 1990; KELLER et al. 1997a; KELLER et al. 1997b;
LAYER et al. 1997). Nicht zuletzt ist auch die Dauer der Adaptation an das jeweilige
Substrat für die Intensität der mikrobiellen Fermentation bedeutsam (SCHÜRCH 1969;
KIRCHGESSNER 2004).
Bei AuSCFAll einzelner Funktionen oder Organe kommt es zu teilweise schwerwiegenden
Einschränkungen in der Verdauungsleistung. Infolge von Erkrankungen des Pankreas
kann der Mangel pankreatischer Verdauungsenzyme zur Ausbildung einer exokrinen
Pankreasinsuffizienz (EPI) führen.
2.2 Die exokrine Pankreasinsuffizienz
2.2.1 Vorkommen und Entstehung Die exokrine Pankreasinsuffizienz (EPI) entsteht beim Menschen zumeist aus einer akuten
Entzündung des Pankreasparenchyms (MAYERLE u. LERCH 2001). Die Ursachen für die
akute Pankreatitis sind nach LAYER und KELLER (2003) dabei vielfältig.

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
5
Mögliche Auslöser der akuten Pankreatitis sind nach DIMAGNO et al. 1977; PEREZ et al.
1983; LAYER et al. 1994; LEIDINGER 1997; DIMAGNO et al. 1999; KELLER et al. 2000;
DEGEN et al. 2001; FRIESS et al. 2001; DIMAGNO 2003; LAYER u. KELLER 2003:
• exzessiver Alkoholkonsum
• Mukoviszidose (Cystische Fibrose)
• infektiöse Genese
• Traumata des Pankreas
• Adenokarzinom des Ausführungsganges der Bauchspeicheldrüse
(Pankreasgangobstruktionen)
• Autoimmune Genese
• Morbus Crohn
• postoperative Zustände nach gastrointestinalen Operationen, z.B.
Gastroektomien und intestinale Ektomien
• duodenale Hyperacidierungen, z.B. beim Zollinger-Ellison-Syndrom
In Deutschland erkranken jährlich 0,01- 0,046% der Bevölkerung an dieser akuten
Entzündung. In ca. 1% der Fälle verläuft die Erkrankung letal, während sich aus 13-27%
der akuten Fälle eine chronische EPI entwickelt, die bei ca. 80% der Patienten durch
Gallensteine oder exzessiven Alkoholkonsum ausgelöst wird (MAYERLE u. LERCH). Des
Weiteren scheint die Prävalenz mit zunehmendem Alter anzusteigen (ROTHENBACHER
et al. 2005). Zudem tritt die EPI bei Männern tendenziell häufiger auf als bei Frauen
(ROTHENBACHER et al. 2005). In vielen Regionen der Welt stellt jedoch übermäßiger
Alkoholkonsum die häufigste Ursache dar (SINGER u. MÜLLER 1995). Bei der EPI infolge
eines Alkohol-Abusus tritt im Vergleich zu der idiopathischen Form (DIMAGNO 2001;
STEINBERG et al. 2003) verstärkt Schmerzhaftigkeit (AMMAN et al. 1987), Kalzifikation
(HAYAKAWA et al. 1989), progressiver FunktionsauSCFAll (LAYER u. DIMAGNO 1999),
Diabetes mellitus (WILLIAMS u. MINNICH 1990; RADUN et al. 1997) sowie eine erhöhte
Todesrate auf (HAYAKAWA et al. 1989).

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
6
Aber auch beim Tier sind Fälle von EPI beschrieben: So kommt beispielsweise beim
Deutschen Schäferhund und beim rauhaarigen Collie eine autoimmune lymphozytäre
Pankreatitis (PAA) vor (RIMAILA-PÄRÄNEN u. WESTERMARCK 1982; LEIDINGER 1997;
WIBERG 2004), die eine Atrophie der enzymproduzierenden Pankreasazini nach sich
zieht. Es existieren auch Fallberichte (HASHOLT 1972; WESTERMARCK 1980; WATSON
et al. 1981, LECHOWSKI et al.1991; RITCHEY et al. 1997; BROWNING 1998) über das
Auftreten einer EPI bei weiteren Tierarten (Katze, Giraffe, Wellensittich und
Gelbnackenamazone).
2.2.2 Folgen des Mangels pankreatischer Verdauungsenzyme
Das Fehlen pankreatischer Verdauungsenzyme infolge einer exokrinen
Pankreasinsuffizienz kann nur zum Teil durch extrapankreatische Enzyme (Speichel,
Magen, Dünndarmmukosa sowie Intestinalflora) kompensiert werden. Daher treten beim
Krankheitsbild der exokrinen Pankreasinsuffizienz (EPI) Maldigestion und Malabsorption
auf (FREUDIGER 1991). Das Ausmaß der bei einer EPI auftretenden Symptome hängt
dabei erheblich von der Nährstoffzusammensetzung der aufgenommenen Nahrung ab.
Während die Verdaulichkeit der Kohlenhydrate und Proteine in wesentlich geringerem
Maße reduziert ist, werden die Folgen der EPI insbesondere in einer dramatisch
eingeschränkten Fettverdaulichkeit deutlich (ABRAMS et al. 1987; MÖSSELER et al.
2006a). Daraus können Steatorrhoe, Abmagerung, Konditionsverlust, Meteorismus
(LÖSER u. FÖLSCH 1995), im Endstadium zusätzlich Beeinträchtigung des
Glukosestoffwechsels (RADUN et al. 1997), Motilitätsstörungen des GIT (MALLINSON
1968; KNOX u. MALLINSON 1971) sowie Mangelerscheinungen an Spurenelementen und
fettlöslichen Vitaminen resultieren (FREUDIGER 1971; DUTTA et al. 1982; ROBERTS
1989; WESTERMARCK et al. 1993; NAKAMURA et al. 1996; LAYER et al. 1997; RUTZ et
al. 2000; LARK et al 2001; LAYER u. KELLER 2003). Die Intensität der Steatorrhoe, dem
Leitsymptom der EPI, ist dabei zum einen abhängig von der Fettquantität (Relation
aufgenommene Fettmenge zu residualer Lipaseaktivität), zum anderen von der
Fettqualität: kurzkettige und mittelkettige Fettsäuren können beispielsweise auch in
Abwesenheit pankreatischer Lipasen aufgrund ihrer hydrophileren Eigenschaften direkt

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
7
von den Enterocyten des Dünndarms resorbiert werden (SHIAU 1987; BEITZ u. ALLEN
1993). Bei Ferkeln erreicht die Fettverdaulichkeit von Sauenmilch post natum
beispielsweise Werte von bis zu 96 %, obgleich die lipolytische Aktivität des Pankreas zu
diesem Zeitpunkt noch nicht vollständig ausgeprägt ist. Die hohe Fettverdaulichkeit ist
nach CRANWELL und MOUGHAN (1989) sowie JENSEN et al. (1997) neben der
gastrischen Lipaseaktivität vor allem durch den hohen Anteil kurzkettiger Fettsäuren im
Milchfett zu erklären.
2.2.3 Auswirkungen der EPI auf die Verdaulichkeit der Fette
Klinische Symptome einer EPI treten erst bei einem AuSCFAll von über 90% des
sekretorisch aktiven Gewebes auf (DIMAGNO et al. 1973; FREUDIGER 1991). Bisher
wurde angenommen, dass die physiologische Sekretionsleistung des Pankreas den
Bedarf an Verdauungsenzymen um ein VieLCFAches übersteigt und daher die
Verringerung der Sekretmenge lange ohne klinische Symptome bleibt (DIMAGNO et al.
1973). Neuerdings wird jedoch diskutiert, ob die hohe Sekretionsleistung eine Folge der
niedrigen spezifischen Aktivität der humanen pankreatische Lipase ist (CARRIERE et al.
2005).
Physiologischerweise sind Spaltung und Resorption der meisten Nährstoffe am Ende des
Dünndarms abgeschlossen (PAYNE et al. 1968; KEYS u. DEBARTHE 1974; BRAUDE et
al. 1976; ZEBROWSKA et al. 1980; LEIBOLZ et al. 1986; KIES et al. 1986). Bei AuSCFAll
des exokrinen Pankreas und einem daraus resultierenden Mangel pankreatischer
Verdauungsenzyme sind die Kompensationsmechanismen für die Verdauung der
einzelnen Nährstofffraktionen unterschiedlich ausgeprägt: Die praecaecal existierenden
extrapankreatischen Enzyme (Speichel-, Magen- und Dünndarmenzyme) können das
Fehlen der pankreatischen Enzyme teilweise kompensieren. Dieses gilt insbesondere
bezüglich der Kohlenhydratfraktion (MÖSSELER et al. 2005), aber in gewissen Umfang
auch für Proteine. Der Umfang der kompensatorischen Fettverdauung durch
extrapankreatische lipolytische Enzyme ist jedoch deutlich geringer (ABRAMS et al. 1987).
Durch die beträchtliche Einschränkung der Fettverdaulichkeit manifestiert sich die EPI
zuerst in der Ausbildung einer Steatorrhoe als Ausdruck der Fettmalabsorption (DIMAGNO

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
8
et al. 1977). Die vergleichsweise frühe Beeinträchtigung der Fettverdauung und das
Einsetzen der Steatorrhoe wird vor allem der Empfindlichkeit der Lipasen, insbesondere
gegenüber reduzierten pH-Werten infolge der EPI (DIMAGNO et al. 1977; REGAN et al.
1977; STERNBY et al. 1992; LAYER et al. 2001) und gegenüber vorzeitiger Zerstörung
durch endogene Proteasen zugeordnet (THIRUVENGADAM u. DIMAGNO 1988; LAYER
et al. 1990). Bei den an EPI erkrankten Menschen ist dennoch eine Fettverdaulichkeit von
bis zu 30% beschrieben, die durch die kompensatorisch gesteigerte Aktivität der
gastrischen Lipase erklärt wird (MOREAU et al. 1988; STERNBY et al. 1992; CARRIERE
et al. 1993; CARRIERE et al. 2005). Dennoch ist durch die extrapankreatischen Lipasen
(weder durch linguale noch gastrische Lipasen) keine vollständige Kompensation des
AuSCFAlls der pankreatischen Lipasen möglich. Nach STERNBY et al. (1992) existiert
zudem kein triglyceridspaltendes Enzymsystem an der Bürstensaummembran. Der
niedrige luminale pH-Wert infolge des Bicarbonatmangels bei Vorliegen einer EPI
begünstigt durch Ausfällung der Gallensalze zusätzlich die Fettmalabsorption (REGAN et
al. 1977; NAKAMURA et al. 1994).
Die bei einer EPI auftretenden Maldigestion und Malabsorption führen zu einem Anstieg
der Nährstoffkonzentrationen im Chymus, die am Ileumende in den Dickdarm übertreten
und damit einen erhöhten Nährstoffeinstrom im Dickdarm nach sich ziehen (TABELING
1998; MÖSSELER et al. 2006a,b). Dieser bedingt eine forcierte mikrobielle Fermentation,
die – verglichen mit dem Abbau durch körpereigene Enzyme - zu energetischen Verlusten
führt (KAMPHUES et al. 2004). Der erhöhte Nährstoffeinstrom bedingt Veränderungen der
Zusammensetzung der mikrobiellen Flora, insbesondere im Dickdarm (LAYER et al. 1986;
LADAS et al. 1993; LÖSER u. FÖLSCH 1995; LAYER u. KELLER 1999). Die
entstehenden Fermentationsprodukte können dabei einerseits von Nutzen (FFS),
andererseits (z.B. Ammoniak) aber auch eine Belastung für den Stoffwechsel des Tieres
darstellen (TABELING 1998).
Oben genannte Veränderungen konnten wiederholt auch an Schweinen mit experimentell
ausgelöster EPI bestätigt werden (TABELING 1998; HELDT 2001; FUENTE-DEGE 2003).

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
9
2.3 Therapie der EPI
2.3.1 Enzymatische Substitutionstherapie in der EPI-Therapie
Da unabhängig von der Ätiologie bei Vorliegen einer EPI ein absoluter Mangel an
Verdauungsenzymen vorliegt, besteht die Behandlung der Wahl, sowohl in der Human- als
auch in der Veterinärmedizin, seit Jahrzehnten in der oralen Substitution von
Verdauungsenzymen. Der Therapieansatz verfolgt vor allem den Ersatz bzw. die
Ergänzung der fehlenden Enzyme, insbesondere die Erzielung einer ausreichenden
Lipaseaktivität im Dünndarm (LÖSER u. FÖLSCH 1995). Besonders die deutliche
Einschränkung der Fettverdaulichkeit kann in den meisten Fällen durch entsprechenden
Enzymeinsatz korrigiert werden. Folglich bildet sich auch das klinische Bild der
Steatorrhoe zurück (PAP et al. 1990; LAYER u. KELLER 2003). Durch die empfohlene
Verabreichung eines porcinen Multienzympräparates mit 25.000 - 40.000 IE lipolytischer
Aktivität, 1.700-2.700 IE proteolytische Aktivität sowie 25.000 - 40.000 IE amylolytische
Aktivität pro Mahlzeit (LAYER u. HOLTMANN 1994; LAYER et al. 2001) wird eine
signifikante Verbesserung der Fett-, Protein- und Stärkeverdaulichkeit erreicht (RADUN et
al. 1997; LAYER u. KELLER 2003). Trotz deutlicher Steigerung der Fettverdaulichkeit
kann jedoch nicht von einer Normalisierung der Fettverdaulichkeit gesprochen werden, da
auch nach Enzymsubstitution die Fettverdaulichkeit die bei gesunden Individuen
ermittelten Werte von ca. 95% (KOLBEL et al. 1986) nicht erreicht. Die Supplementierung
beeinflusst zusätzlich die Verdaulichkeit fettlöslicher Vitamine (KARTHOFF 2004) und
zahlreicher Mineralstoffe (KAMMLOTT 2003) durch Verminderung der Seifenbildung sowie
Verbesserung der Micellenbildung positiv. Dadurch wird ein Anstieg der Resorptionsraten
erzielt, die aber meist weiterhin unter dem Niveau gesunder Individuen bleiben
(KARTHOFF 2004).

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
10
2.3.2 Indikationen für den Einsatz von Enzymen bei Vorliegen einer EPI
Ein therapeutisches Eingreifen ist nach LANKISCH (1993), MÖSSNER (1994), LÖSER u.
FÖLSCH (1995) stets dann erforderlich, wenn klinische Symptome einer EPI,
insbesondere Steatorrhoe, auftreten. Auch in der postoperativen Phase nach Eingriffen
am GIT werden Enzyme zur Entlastung supplementiert, da exogen zugeführte
Pankreasenzyme die Sekretion des Pankreas reduzieren (WALKOWIAK et al. 2003). Bei
schmerzhaften Zuständen des Pankreas, die vorwiegend in Zusammenhang mit
Entzündungsreaktionen auftreten, ist eine Enzymsubstitution im Sinne einer
„Schmerztherapie“ angezeigt (SLAFF et al. 1984), da infolge von Feedback-Mechanismen
die endogene Enzymsekretionsrate sinkt, wodurch eine Druckverminderung im Pankreas
entsteht (DOBRILLA 1989; LEBENSTHAL et al. 1994).
2.3.3 Diätetik
Unterstützend zur Substitutionstherapie werden Diäten mit leichtverdaulichen Fetten
(kurzkettige und mittelkettige Fettsäuren; BRUNO et al. 1995; RADUN u.
MALFERTHEINER 1996; SOMMER 1997) und die Aufnahme zahlreicher kleinerer
Mahlzeiten empfohlen (READ et al. 1984). Die Substitution fettlöslicher Vitamine ist
ebenfalls angezeigt (GOODCHILD u. DODGE 1986; DODGE et al. 1990). Zusätzlich wird
empfohlen, die Fettmenge zugunsten des Kohlenhydratanteils in der Diät zu reduzieren
(BOIVIN et al. 1990); ausgenommen davon sind Patienten mit Cystischer Fibrose
(Mukoviszidose), für die bei reduziertem Fettgehalt der Nahrung eine ausreichende
Energieversorgung oftmals nicht sichergestellt werden kann (HEYMANS 1989; KAWCHAK
et al. 1996).

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
11
2.3.4 Einsatz von Multi- und Monoenzympräparaten in der Therapie der EPI
Die Enzympräparate in der Substitutionstherapie zur Behandlung der EPI werden in Multi-
und Monoenzympräparate eingeteilt. Multienzympräparate enthalten mindestens zwei
Enzyme, zumeist jedoch alle drei Gruppen von Verdauungsenzymen (Lipasen, Proteasen
und Amylasen), während Monoenzympräparate nur eine Enzymgruppe (z.B. Lipasen)
beinhalten. Für die Bewertung von Enzympräparaten für die Substitutionstherapie bei EPI
ist vor allem die lipolytische Aktivität maßgebend (SHAW u. BARBEZAT 1982).
Des Weiteren ist eine Differenzierung bezüglich der Herkunft möglich: Es wird
unterschieden zwischen Produkten tierischen (porcinen und bovinen) und mikrobiellen
(fungal und bakteriell) Ursprungs. Es ist davon auszugehen, dass in Zukunft die
Entwicklung und Herstellung von Enzymen unter Verwendung der Gentechnik an
Bedeutung gewinnt, so dass diese zukünftig in der EPI-Behandlung an Bedeutung
gewinnen dürften (LAYER u. KELLER 1999; LANKISCH 2001).
2.3.4.1 Multienzympräparate
In neueren Behandlungsempfehlungen (LAYER u. KELLER 2003) wird eine orale
Supplementationstherapie unter Verwendung eines Multienzymproduktes, einer
Kombination proteolytisch, amylolytisch und lipolytisch wirksamer Substanzen,
befürwortet. In der Therapie der EPI ist der Einsatz pankreatischer Multienzymprodukte
eine etablierte und bewährte Vorgehensweise, wobei bislang fast ausschließlich Produkte
porciner Herkunft eingesetzt werden.
Enzyme porcinen Ursprungs:
Die überwiegende Zahl der Multienzymprodukte zur Substitutionstherapie bei EPI besteht
aus einem aufgereinigtem Extrakt von Pankreasgewebe porcinen Ursprungs. Das porcine
Pankreassekret ist dem des Menschen in seiner Zusammensetzung sehr ähnlich
(MOUGHAN et al. 1994) und eignet sich somit in besonderem Maße. Daher sind auf dem
Markt eine Vielzahl verschiedener Produkte aus porcinem Pankreasextrakt in

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
12
unterschiedlichen galenischen Zubereitungen und Dosierungen erhältlich wie
beispielsweise Cotazym®, Pankrease®, Ultrase®, Viokase®, Panzyat®, Enzym-Lefax®, Ku-
Zymase®, Ilozymase®, Panokase®, Kreon® und Pankreon®. Multienzymprodukte gehören
zu den derzeit am weitesten verbreiteten Enzympräparaten der Substitutionstherapie. Die
Einnahme der Präparate führt zumeist zur Verminderung der Steatorrhoe (GRAHAM 1977;
SCHNEIDER et al. 1985; KOLBEL et al. 1986). Die Enzyme werden nach Denaturierung
durch Verdauungssäfte oder nach bakteriellem Abbau mit den Faeces ausgeschieden
(GRAHAM 1977; LÖSER u. FÖLSCH 1991; LAYER u. KELLER 2003). Die Kombination
porciner Pankreasextrakte mit H2-Blockern oder Protonen-Pumpen-Hemmern verbessert
bei zusätzlich vorliegender Hyperacidierung des Magens den Therapieerfolg aufgrund der
pH-Wert-Erhöhung (DOBRILLA 1989; LAYER u. KELLER 2003; PROESMANS u. DE
BOECK 2003). Zum Schutz der Enzyme vor Denaturierung im Magen wird auch der
Einsatz gecoateter Enzyme empfohlen, die erst bei einem pH-Wert von > 6 freigesetzt
werden (LAYER u. KELLER 2003).
Enzyme bovinen Ursprungs:
In bestimmten Bevölkerungsgruppen ist die Akzeptanz von Produkten porcinen Ursprungs
aus religiösen Gründen (z.B. im Islam und im Judentum) sehr gering. Eine potentielle
Alternative in der Therapie der EPI bietet der Einsatz boviner Pankreasenzyme (LAYER u.
KELLER 2003). Die Zusammensetzung des Pankreassaftes der Wiederkäuer (als
Herbivor) ist jedoch sehr divergent zu der des Menschen. Studien ergaben eine
lipolytische Aktivität boviner pankreatischer Enzymextrakte von nur 25% im Vergleich zu
porcinen (LAYER u. KELLER 2003). Demzufolge müssen wesentlich höhere
Enzymmengen zugeführt werden, um vergleichbare Wirkungen zu erreichen. Daher ist der
therapeutische Nutzen boviner Pankreasprodukte limitiert. Zusätzlich tragen insbesondere
in der westlichen Welt Bedenken über Kontamination mit übertragbaren Pathogenen (z.B.
BSE) zur geringen Verbreitung boviner Multienzympräparate bei (LAYER u. KELLER
2003).

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
13
Nachteile der Substitutionstherapie eingesetzter Multienzymprodukte:
Obgleich porcine Multienzymprodukte heute die Therapie der Wahl darstellen, werden
zukünftig vermutlich vermehrt „alternative Enzymquellen“ zum Einsatz kommen. Denn
trotz der marktbeherrschenden Stellung der porcinen Multienzymprodukte bestehen
Nachteile in der Gewinnung und Zubereitung aus Schlachtkörpern:
Einerseits besteht aufgrund des möglichen Auftretens von Zoonosen bzw. aus
seuchenhygienischer Sicht das Bestreben, alternative Therapieansätze zu Medikamenten
tierischen Ursprungs zu verfolgen. In den letzten Jahren standen wiederholt Tierseuchen,
wie beispielsweise BSE und MKS, im Interesse der Öffentlichkeit, woraufhin die
Verbrauchereinstellung und die öffentliche Meinung gegenüber Produkten tierischer
Herkunft insgesamt kritischer wurden (Zwischenruf: Leibniz Gemeinschaft,
Umweltforschung für politische Praxis, Heft 2/2005). So wird vermutet, dass die
Zurückhaltung der Bevölkerung gegenüber Produkten tierischer Herkunft nicht nur
Lebensmittel, sondern auch Medikamente betrifft. Des Weiteren ist es trotz erheblicher
technologischer Fortschritte weiterhin nicht gelungen, eine standardisierte Produktion, die
eine gleichbleibende Qualität und Aktivität der Enzyme sicherstellt, zu garantieren
(LEBENTHAL et al. 1994). Mängel in der Qualitätssicherung ergeben sich durch die
eingeschränkte Lagerstabilität tierischer Produkte sowie durch große
Chargenunterschiede bezüglich der Enzymaktivitäten (pers. Mitteilung GREGORY 2006).
Vermutlich abhängig von der letzten Fütterung, eventuell auch von der
Futterzusammensetzung und tierindividuellen Unterschieden ist der Gehalt an Enzymen,
der aus dem Pankreas von Schlachttieren gewonnen werden kann, großen
Schwankungen unterworfen (PEKAS et al. 1964; ZABIELSKI et al. 1993, 1997). Um
dennoch möglichst gleichbleibende Enzymaktivitäten sicherzustellen (LEBENTHAL et al.
1994), müssen große Pankreasextraktmengen miteinander vermengt werden (pers.
Mitteilung GREGORY 2006). Dabei ergibt sich die Schwierigkeit, die Kontamination großer
Pankreasmengen durch einzelne verunreinigte Proben bzw. Chargen (z.B. Parvoviren)
auszuschließen. Dieser Ausschluss wird jedoch von den Zulassungsbehörden in
zunehmendem Maße gefordert, sogar für bisher unbekannte Krankheitserreger (z.B.
Tansmissible-spongiforme-Enzephalopathie-Erreger). Bei den sich verschärfenden
Zulassungskriterien könnten sich in der Zukunft als Folge dieser Nachweislücke

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
14
argumentatorische Probleme in Zulassungsverfahren ergeben. Hinzu kommt, dass
Enzyme tierischen Ursprung, im Gegensatz zu gentechnisch veränderbaren mikrobiellen
Produkten, keine bzw. in deutlich geringerem Umfang Modifikationsmöglichkeiten bieten
(z.B. die Veränderung des pH-Wert-Toleranzbereiches) und somit einer erwünschten
Steigerung der Wirksamkeit Grenzen gesetzt sind. Letztlich ist es auch wegen ethischer
Vorbehalte bestimmter religiöser Gruppierungen gegenüber der Einnahme von
Medikamenten porcinen Ursprungs wünschenswert, alternative Enzymquellen zu
erschließen, zu nutzen und auf dem Weltmarkt anzubieten.
2.3.4.2 Monoenzympräparate
Im Folgenden sollen zum Teil neue Ansätze zur Behandlung der EPI vorgestellt werden,
die sich in unterschiedlichem Stadien der Entwicklung bzw. Ausreifung befinden und in der
Zukunft Ergänzungen bzw. Alternativen zu Enzymprodukten tierischer Herkunft darstellen
könnten. Enzyme biotechnologischen Ursprungs weisen aufgrund der Produktion unter
standardisierten Bedingungen vor allem Vorteile in Bezug auf die Homogenität des
Produktes, der standardisierten Aktivität aber auch bezüglich Hygiene und
Verbraucherschutz auf. Außerdem werden durch diese Präparate auch für Gruppen mit
ethischen Ressentiments gegenüber porcinen Produkten (s. o.) therapeutische
Möglichkeiten mit entsprechender Akzeptanz geschaffen. Sie werden bisher als
Monoenzymprodukte verwendet.
An dieser Stelle soll lediglich auf Lipasen näher eingegangen werden. Nach LAYER u.
KELLER (2003) können die Enzyme entsprechend ihrem Ursprung unterteilt werden in:
Enzyme fungalen Ursprungs
Enzyme bakteriellen Ursprungs
Enzyme humanen Ursprungs

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
15
Enzyme fungalen Ursprungs:
Verschiedene Pilze (z.B. Rhizopus arrhizus und Aspergillus niger) produzieren lipolytisch
wirksame Enzyme. Ein wesentlicher Vorteil fungaler Lipasen gegenüber den Enzymen
tierischer Herkunft ist ihre zumeist beachtliche Säurestabilität und hohe Aktivität in einem
weiten pH-Wert-Bereich (z.B. Rhizopus arrhizus: pH-Optimum nach MOREAU et al.
(1988) zwischen 3,5-7,4), die eine Magenpassage ohne wesentlichen Aktivitätsverlust
ermöglicht (POINTER u. FLEGEL 1975), so dass auf ein Coating verzichtet werden könnte
(SUZUKI et al. 1997; LAYER u. KELLER 2003). Des Weiteren ist zur Entfaltung der
lipolytischen Aktivität keine Aktivierung durch Colipasen notwendig (LAYER u. KELLER
2003). Ein Nachteil besteht bisher jedoch in der sehr schnellen Inhibition fungaler Enzyme
durch Gallensalze (MOREAU et al. 1988; RAIMONDO u. DIMAGNO 1994) und Proteasen
(MOREAU et al. 1988; THIRUVENGADAM u. DIMAGNO 1988). Infolgedessen können die
Lipasen ihre Wirksamkeit im Duodenum, wo die Absorption der Lipolyseprodukte maximal
ausgeprägt ist (LAYER u. KELLER 2003), nicht optimal entfalten. Fungale Lipasen wirken
also hauptsächlich im Magen und konnten infolge ihrer geringen In-vivo-Aktivität bisher
nicht als Alternative zu porcinen Präparaten etabliert werden (SCHNEIDER et al. 1985;
MOREAU et al. 1988; ZENTER-MUNRO et al. 1992; FASSMANN 2001; MANDISCHER
2002; FUENTE-DEGE 2003; HELDT 2003). Sollte die Produktion gallensalzstabiler
Mutanten gelingen, könnten fungale Lipasen eine größere Rolle in der EPI-Behandlung
erlangen (pers. Mitteilung GREGORY 2006).
Enzyme bakteriellen Ursprungs:
Bakterien wie Burkholderia plantarii bilden Enzyme mit lipolytischer Aktivität. Diese weisen
eine um 100-fache höhere spezifische Aktivität auf als porcines Pankreasextrakt, so dass
die Menge des verabreichten Enzyms um 99% reduziert werden könnte (SUZUKI et al.
1997, 1999). Um vergleichbare Effekte zu erlangen, müssten Patienten nur noch wenige
Milligramm statt wie bisher mehrere Gramm des Enzymproduktes bzw. -präparates zu sich
nehmen. Dieses würde beispielsweise Mukoviszidosepatienten entlasten, die sich infolge
des generalisierten Leidens einer Vielzahl von Behandlungen gleichzeitig und dauerhaft
unterziehen müssen. Demzufolge ist gerade für CF-Patienten eine geringere Tablettenzahl

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
16
bei gleicher Wirksamkeit (GRIFFIN et al. 1989; MORRISON et al. 1992; SANTINI et al.
2000) von größtem Interesse.
Zudem gelten bakterielle Enzyme sowohl als säurestabil gegenüber einer Denaturierung
im Magen (großer pH-Wert-Toleranzbereich) und durch Proteasen, als auch als stabil
gegenüber Gallensalzen (RAIMONDO u. DIMAGNO 1994). Sowohl in In-vitro-, als auch in
ersten In-vivo-Experimenten mit Hunden erwiesen sie sich den porcinen Enzymen in
Bezug auf ihre lipolytische Wirksamkeit überlegen (SUZUKI et al. 1999). Daraus leitet sich
die Hoffnung ab, in näherer Zukunft potente Alternativen zu den Enzymen tierischer
Herkunft entwickeln zu können (MÖSSNER 1994; LÖSER u. FÖLSCH 1995; LAYER u.
KELLER 2003).
Enzyme humanen Ursprungs:
• Gentransfer:
In der Zukunft könnte eine Expression der pankreatischen Lipase im hepatobiliären
System von EPI-Patienten möglich werden. Die pankreatischen Lipasegene sind
bereits In-vitro in einer menschlichen Gallenblasenzellinie expremiert worden, indem
ein rekombiniertes Adenovirus als Carrier benutzt wurde (KUHEL et al. 2000). Auch ex
vivo gelang die Expression in der Gallenblase eines Schafes, In-vivo erfolgte sie
zudem in den Gallengängen von Ratten (MAEDA et al. 1994, KUHEL et al. 2000). Bei
allen diesen Versuchen gelang eine Produktion von humaner pankreatischer Lipase,
ohne dass sie im Serum nachgewiesen werden konnte, also keine unerwünschte
Sekretion der Enzyme ins Blut stattfand.
• biotechnologisch hergestellte Enzyme:
Nach LÜTHEN u. NIEDERAU (1994) und VILLE et al. (2002) stellt der Einsatz einer
gastrischen Lipase als EPI-Therapeutikum einen interessanten Aspekt dar. Die
ektopische, synthetische Herstellung einer säurestabilen humanen gastrischen Lipase
könnte deren Einsatz in der Substitutionstherapie ermöglichen und damit eine weitere
Alternative zu Enzymprodukten tierischer Herkunft darstellen (LÖSER u. FÖLSCH
1995; LAYER u. KELLER 1999; LANKISCH 2001).

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
17
2.4 Entwicklung neuer Enzymprodukte
2.4.1 Wirkstoffneuentwicklung in der Pharmazeutischen Industrie
Die Entwicklung eines neuen Medikamentes bedeutet für ein pharmazeutisches
Unternehmen ein immer zeit- und kostenintensiver werdendes Unterfangen (GRABOWSKI
2004). Die mittleren Kosten einer Neuentwicklung für den humanmedizinischen Markt
belaufen sich auf ca. 500-800 Millionen US-Dollar für einen neuen Wirkstoff (KUMAR et al.
2001; VFA-Statistics 2005). Dabei entfallen ca. 70% der Gesamtsumme auf abgebrochene
Entwicklungsprojekte (HINZE et al. 2001). Der Entwicklungszeitraum beträgt ab dem
Zeitpunkt der Entdeckung einer Substanz im Mittel 12 Jahre (VFA-Statistics 2005). In
Europa wurden 2002 16 % des Umsatzes der Pharmazeutischen Industrie in Forschung
und Entwicklung (F&E) re-investiert. Die F&E-Ausgaben der Pharmazeutischen Industrie
der USA, Japans und Europas belaufen sich zusammen auf insgesamt 58 Milliarden US
Dollar (VFA-Statistics 2005). Während seit den siebziger Jahren die F&E-Aufwendungen
der OECD von 5 % auf 10 % anstiegen, sanken die Forschungsinvestionen der
Pharmazeutischen Industrie in Deutschland im gleichen Zeitraum von 6,5 % auf unter 5 %
(OECD, ANBERD DATABASE; VFA-Statistics 2005).
Forschung und Entwicklung Nachfolgend soll die Vorgehensweise in der Pharmazeutischen Industrie während einer
Produktneuentwicklung dargestellt werden, um den Nutzen beschleunigter und
vereinfachter Verfahren besonders in der In-vivo-Phase zu verdeutlichen.
Vorbemerkung: Notwendigkeit der Entwicklung eines In-vivo-Verfahrens zur Testung von Lipasen Während sich die Entwicklung von pankreatischen Enzymprodukten in den vergangenen
Jahrzehnten fast ausschließlich auf die Optimierung der galenischen Zubereitung von
porcinen Pankreaspräparationen bzw. Extrakten beschränkte, werden in jüngster

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
18
Vergangenheit neue Ansätze erkennbar, die als Alternativen erforscht und weiter
entwickelt werden (s. o.: fungale und bakterielle sowie gentechnisch oder biotechnologisch
hergestellte humane Lipase). Infolge der VieLCFAlt der Enzyme unterschiedlicher
Herkunft und der Möglichkeit der Entwicklung verschiedener biotechnologischer Varianten
entsteht in Zukunft eine Vielzahl zu testender Enzyme. Daher besteht mehr denn je das
Interesse und der Bedarf nach einem zeit- und materialsparendem In-vivo Verfahren zur
Testung einer Vielzahl von Substanzen, beispielsweise durch die Entwicklung von In-vivo
Screening-Verfahren, wie sie bereits für Amylasen und Proteasen entwickelt wurden
(BECKER 2005). Ein In-vivo Screening-Verfahren zur Testung von Lipasen steht bisher
jedoch noch nicht zur Verfügung.
2.4.2 Ablauf bzw. Phasen einer Wirkstoffneuentwicklung
2.4.2.1 Anfangsphase
Am Anfang eines neuen Entwicklungsprozesses steht im Allgemeinen die Suche nach
einem so genannten „Target“, dem Angriffspunkt im Krankheitsgeschehen, an dem ein
Medikament ansetzen könnte. Im Fall der EPI bedeutet dieses die Suche nach Enzymen,
die möglichst wirkungsvoll die Spaltung der Nährstoffe anstelle der fehlenden
körpereigenen Pankreasenzyme übernehmen.
Eine Vielzahl grundsätzlich bzw. theoretisch in Betracht kommender Substanzen wird
zunächst mittels In-vitro-Testsystemen getestet. In den letzten zehn Jahren wurden für
diese Aufgabe hocheffiziente, z.T. vollautomatische In-vitro-Hochdurchsatz-Verfahren
(„High-throughput Screenings“) entwickelt. Bei den hochtechnologisierten Verfahren
werden Roboter eingesetzt, die bis zu zwei Millionen chemischer Stoffe pro „Target“ testen
(HINZE et al. 2001). Jede Substanz, die einen gewünschten Effekt erzielt, wird daraufhin
als „Hit“ bzw. „Treffer“ bezeichnet (LÖSCHER 2006, Vortrag).
Für einen „Hit“ gibt es auf dem Weg zur Markteinführung jedoch neben der Effektivität sehr
viele weitere Bedingungen, von deren Erfüllung eine Weiterentwicklung abhängt,
beispielsweise möglichst geringe bzw. fehlende Toxizität, Teratogenität und Genotoxität.

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
19
Von den ca. 5 000-10 000 Substanzen mit positiven Effekten werden in zunehmendem
Maße durch molekularbiologische und gentechnische Verfahren verschiedene Varianten
erzeugt, um ihre Eigenschaften gezielt zu verbessern; Computerprogramme werden
unterstützend eingesetzt, um diese Eigenschaften vorherzusagen. Die optimierten
Varianten, auch als Leitstrukturen oder „leads“ bezeichnet (HINZE et al. 2001; LÖSCHER
2006, Vortrag), werden erneut getestet und gegebenenfalls weiter verbessert. Nach
mehreren „Optimierungsrunden“ steht im bestmöglichen Fall eine Substanz zu Verfügung,
die als Wirkstoffkandidat zum Patent angemeldet wird (VFA-Statistics 2005).
2.4.2.2 Die vorklinische Entwicklungsphase
In der nun folgenden, ca. 3-5 Jahre dauernden vorklinischen Entwicklungsphase (JONES
1999) wird die Metabolisierung der Substanz im Gesamtorganismus erforscht. Unter
anderem wird geprüft, ob die Substanz den Krankheitsverlauf im Körper, d.h. In-vivo,
tatsächlich positiv beeinflusst. Daher erfolgen in dieser Phase In-vivo-Untersuchungen zur
Wirksamkeit an Versuchstieren, wobei parallel auch toxikologische Studien, ebenfalls In-
vivo, durchgeführt werden. Bisher werden zur Überprüfung der Wirksamkeit substituierter
Enzyme mittels In-vivo-Verdaulichkeitsstudien im Modelltier Schwein (mit experimentell
ausgelöster EPI) mehrere Kilogramm des zu prüfenden, neuen Enzyms benötigt. Für die
In-vitro-Verfahren werden hingegen lediglich wenige Milligramm bis Gramm pro Test
benötigt. In der Gesamtheit der notwendigen Analysen liegt der In-vitro-Bedarf je nach
Enzym in einem Bereich einiger Kilogramm (POTTHOFF 2006, persönliche Mitteilung).
Allerdings ist die Herstellung entsprechender Mengen in der vorklinischen Phase
schwierig, da zumeist weder industrielle Produktionsverfahren noch großtechnische
Produktionseinheiten zu Verfügung stehen.
Dennoch wird der möglichst frühe Einsatz von Substanzen – in diesem Falle Enzymen – in
In-vivo-Testsystemen aus folgenden Gründen angestrebt:
Zwischen In-vitro- und In-vivo-Ergebnissen bestehen zum Teil erhebliche Unterschiede in
der Bewertung der Wirksamkeit verschiedener „potentieller Wirkstoff-Kandidaten“.
Demzufolge ist die Übertragung der Ergebnisse von In-vitro-Tests auf In-vivo-Tests nicht
unproblematisch. Um also die Wirksamkeit der zu testenden Enzyme frühzeitig

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
20
einschätzen zu können, ist es notwendig, die Korrelation der In-vivo- und In-vitro-
Ergebnisse zu verbessern. Vorteilhaft wäre es, In-vitro-Verfahren zu etablieren, die schon
vor Einsatz des Testenzyms im Tier zu vergleichbaren Bewertungen gelangen. Infolge
mangelhafter Simulationsmöglichkeit der Resorptionsvorgänge im Darm konnten mit
bisher eingesetzten In-vitro-Methoden, wie z.B. TIM®, Rustitec®, Cositec®, Daisy® oder
HFT®, nur teilweise zufrieden stellende Ergebnisse erzielt werden (SCHÜRCH 1969,
DREYER 1989). In jüngster Vergangenheit sind allerdings Fortschritte bei der In-vitro / In-
vivo-Korrelation bezüglich vergleichender Untersuchungen zur Effektivität von
Monoenzymen gelungen (BECKER 2005). Dennoch wird es auch in Zukunft nicht möglich
sein, auf Tierversuche zu verzichten (KIRCHGESSNER 1992; KAMPHUES 2004). Durch
einen möglichst frühzeitigen Einsatz im Tier während des Entwicklungsprozesses können
ungeeignete Präparate von der weiteren Entwicklung ausgeschlossen werden, so dass
eine gezielte und selektive Weiterentwicklung Erfolg versprechender Substanzen
ermöglicht wird.
2.4.3 In-vivo-Studien am pankreasgangligierten Miniaturschwein als Modell für den an EPI erkrankten Menschen
Das Schwein ist nach Einschätzung vieler Autoren das am Besten geeignete Modelltier für
den Menschen zur Durchführung von ernährungsphysiologischen Studien (POND u.
HOUPT 1978; RERAT 1983; FLEMING u. WALEWSKI 1984 und FLORES et al. 1988).
Grund dafür ist die Ähnlichkeit mit dem Menschen im Bezug auf anatomische Verhältnisse
im Gastrointestinaltrakt (GIT), Verdauungsphysiologie sowie Nährstoffbedarf (BOOK u.
BUSTAD 1974; POND u. HOUPT 1978). Hinzu kommen Übereinstimmungen bezüglich
der omnivoren Ernährungsweise, der Nahrungsaufnahmerhythmik und der Körpermasse
(STEVENS 1977; PARRA 1978; ABELLO et al. 1989; DESPORT et al. 1997).
Zur Untersuchung der Auswirkungen von Erkrankungen bzw. FunktionsauSCFAll des
exokrinen Pankreas wurde das In-vivo-Modell „pankreasgangligiertes Miniaturschwein“
etabliert. Dieses Modell wurde bereits in zahlreichen Studien zu Untersuchungen der
Effekte der EPI auf die Verdaulichkeit von Nährstoffen und der Überprüfung der Effekte

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
21
einer Enzymsubstitution eingesetzt (TABELING 1998; FASSMANN 2001; HELDT 2001;
MANDISCHER 2002; FUENTE-DEGE 2003; KAMMLOTT 2003; KARTHOFF 2004;
BECKER 2005). Die Ligatur des Ausführungsganges des Pankreas bedingt eine Atrophie
und bindegewebige Durchsetzung des Parenchyms (RAHKO et al. 1987), in deren Folge
die Produktion und Sekretion von pankreatischen Verdauungsenzymen zum Erliegen
kommt; bemerkenswerter Weise bleibt jedoch die endokrine Funktion (Insulinproduktion)
des Pankreas beim Schwein bestehen (PITÄRANTA et al. 1989). Im Gegensatz zu
anderen Spezies (z.B. Mensch und Hund) wird bei Pankreasgangligatur beim Schwein
kein Diabetes mellitus provoziert (ANDERSON u. ASH 1971; RAHKO et al. 1987;
BERKHOFF et al. 1988; PITÄRANTA et al. 1989), so dass die Versuchsergebnisse nicht
von einer diabetischen Stoffwechsellage beeinflusst werden. Unter Verwendung von
pankreasgangligierten Minischweinen mit ileo-caecaler Umleitungsfistel besteht folglich die
Möglichkeit, den Einfluss der EPI sowohl auf praecaecale als auch auf postileale
Verdauungsprozesse zu quantifizieren (TABELING 1998; FASSMANN 2001; HELDT
2001; MANDISCHER 2002; FUENTE-DEGE 2003; KAMMLOTT 2003; KARTHOFF 2004;
BECKER 2005). Durch Bestimmung der Verdaulichkeit der Nährstoffe bei
supplementierten und unsupplementierten Individuen kann unter Verwendung dieses
Modells die Wirksamkeit verschiedener Enzympräparate bzw. verschiedener galenischer
Zubereitungen vergleichend untersucht werden (TABELING 1998; FASSMANN 2001;
HELDT 2001; MANDISCHER 2002; FUENTE-DEGE 2003; KAMMLOTT 2003;
KARTHOFF 2004; BECKER 2005). Da es sich hierbei zumeist um Studien handelte, in
denen das Schwein als Modelltier für den Menschen eingesetzt wurde, orientierte sich die
verwendete Nahrung (d.h. das Futter) oftmals an der Humanernährung (MANDISCHER
2002; KAMMLOTT 2003; KARTHOFF 2004).
2.4.3.1 Methoden der Verdaulichkeitsbestimmung zur Bewertung der Wirksamkeit von Enzympräparaten
In vorangegangenen Studien des Projektes pankreasgangligiertes Minischwein wurde die
Effektivität verschiedener Enzymprodukte durch Untersuchungen zur Verdaulichkeit von
Nährstoffen im pankreasgangligierten Tier geprüft (TABELING 1998; FASSMANN 2001;

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
22
HELDT 2001; MANDISCHER 2002; FUENTE-DEGE 2003; KAMMLOTT 2003;
KARTHOFF 2004; BECKER 2005). Dabei wurde die Effektivität eines Enzyms danach
bewertet, inwieweit durch dessen Substitution die bei pankreasgangligierten Tieren
reduzierte Verdaulichkeit der Nährstoffe (bzw. des entspr. Nährstoffes) verbessert bzw.
evtl. sogar auf das Niveau von Kontrolltieren angehoben werden konnte.
Grundsätzlich stehen zur Bestimmung der Verdaulichkeit verschiedene Verfahren zur
Verfügung:
Kollektionsmethoden:
quantitativ vollständige Sammlung von Chymus einer gewählten Lokalisation im GIT
/ Faeces über einen definierten Zeitraum
Indikatormethoden:
durch Einsatz eines nicht resorbierbaren Markers kann aufgrund von
Konzentrationsänderungen zwischen resorbierbaren Nährstoffen und nicht
resorbierbarem Marker an gewählter Lokalisation im GIT die bis zur
entsprechenden Lokalisation erreichte Verdaulichkeit kalkuliert werden („Spot-
Sampling“)
Die Verdaulichkeit einer Ration bzw. eines Nährstoffes kann - unabhängig von der
verwendeten Methode - in verschiedenen Abschnitten des GIT bestimmt werden. Dafür
wird, je nach Fragestellung, Probenmaterial aus verschiedenen Lokalisationen des GIT
gewonnen (z.B. über Fisteln).
Die am weitesten verbreiteten Methoden der Verdaulichkeitsbestimmung beim Schwein
(bezogen auf die Lokalisation der Kollektion des Probenmaterials) sind:
1. Bestimmung der Gesamtverdaulichkeit
2. Bestimmung der praecaecalen Verdaulichkeit
Die Bestimmung der Verdaulichkeit eines Nährstoffes über den gesamten
Verdauungstrakt („Gesamtverdaulichkeit“) eignet sich nur eingeschränkt für die

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
23
Untersuchung der Wirksamkeit substituierter pankreatischer Enzyme. Die Verdauung von
Proteinen, Fetten und den meisten Kohlenhydraten durch körpereigene Enzyme ist bei
gesunden Individuen am Ende des Dünndarms nahezu vollständig abgeschlossen
(DROCHNER 1984). Bei ausschließlicher Betrachtung der Gesamtverdaulichkeit kann es
jedoch infolge mikrobieller Fermentationsprozesse im Dickdarm zu Fehleinschätzungen
kommen, besonders im Bezug auf die Kohlenhydrat- und Proteinverdauung (OELFKE
1978; DROCHNER 1987; KAMPHUES 1987), da Kohlenhydrate und Proteine, die infolge
von Maldigestion und –absorption den Dickdarm erreichen, dort einem bakteriellen Abbau
unterliegen. Es ist also unmöglich zwischen Effekten der eingesetzten Enzyme und denen
einer mikrobiellen Umsetzung zu unterscheiden. Durch das gesteigerte Substratangebot
im Dünndarm entsteht bei einem Teil der pankreasgangligierten Schweine und der an EPI
erkrankten Menschen (ca. 30%) ein „bacterial overgrowth“, welcher unter anderem anhand
der im Vergleich zu Kontrolltieren deutlich erhöhten LPS-Werte im Chymus diagnostiziert
werden kann (Anstieg der LPS-Werte bei dem Modelltier „PL-Schwein“ rationsabhängig
um den Faktor 7,6 bis 20 im Vergleich zu Kontrolltieren; MANDISCHER 2002; TABELING
1998). Die Bestimmung des LPS-Gehaltes wird dabei als Indikator für die Masse von
gramnegativen Keimen genutzt (KAMPHUES 1987). Für die grampositive Flora wird
hingegen l-Laktat als Indikator eingesetzt. Bei PL-Tieren wurden im Vergleich zu
Kontrolltieren, z.B. bei einer stärkereichen Diät, um den Faktor 48 erhöhte l-Laktat-Werte
ermittelt (MANDISCHER 2002). Um die Effizienz ergänzter pankreatischer
Verdauungsenzyme unter Ausschluss der Dickdarmfermentation einzuschätzen, ist es
daher zwingend notwendig, die Verdaulichkeit der Nährstoffe vor Eintritt in den mikrobiell
stark besiedelten Dickdarm (also praecaecal) zu bestimmen (KARTHOFF 2004).
Vorstehendes gilt vor allem für die Kohlenhydrat- und Proteinverdauung; Fette hingegen
werden im Dickdarm lediglich in geringem Maße fermentativ umgesetzt (FUENTE-DEGE
2003), so dass praecaecale Verdaulichkeiten und Gesamtverdaulichkeiten oft sehr
ähnlicher Werte ergeben.

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
24
2.4.3.2 Vor- und Nachteile der Überprüfung der Wirksamkeit substituierter lipolytischer Enzyme anhand der Bestimmung der Fettverdaulichkeit mittels herkömmlicher Methoden im pankreasgangligierten Miniaturschwein
In zahlreichen vorangegangenen Studien des Projektes „pankreasgangligiertes
Miniaturschwein“ (TABELING 1998; FASSMANN 2001; HELDT 2001; MANDISCHER
2002; FUENTE-DEGE 2003; KAMMLOTT 2003; KARTHOFF 2004) wurde die
Fettverdaulichkeit sowohl bei gesunden, als auch bei Tieren mit experimentell ausgelöster
EPI vergleichend praecaecal und über den gesamten Verdauungstrakt quantifiziert. Die
Ergebnisse zeigten hohe praecaecale Fettverdaulichkeiten (88,2-97,6 %) bei den
Kontrolltieren und deutlich geringere Verdaulichkeitswerte bei den pankreasgangligierten
Tieren (9,3-43 %) (TABELING 1998; FASSMANN 2001; HELDT 2001; MANDISCHER
2002; FUENTE-DEGE 2003). Da postileal praktisch keine Verdauung bzw. Absorption von
Rohfett stattfindet (FUENTE-DEGE 2003), wird nahezu die gesamte ileal angeflutete
Menge faecal ausgeschieden (MÖSSELER et al. 2006a). Demzufolge ist es zulässig,
anhand der Fettverdaulichkeit über den gesamten Verdauungstrakt
(Gesamtverdaulichkeit) auf die praecaecale Verdaulichkeit zu schließen (FUENTE-DEGE
2003). Einschränkend muss jedoch erwähnt werden, dass bei Bestimmung der
Gesamtverdaulichkeit eine Fehleinschätzung im Sinne einer geringgradigen
Unterschätzung der Fettverdaulichkeit infolge einer mikrobiellen Fettsynthese und
endogener Ausscheidungen der Mukosa (Zellabschilferungen) im Dickdarm möglich ist
(HERTEL et al. 1970; DROCHNER u. MEIER 1991; TABELING 1998; FASSMANN 2001;
HELDT 2001; FUENTE-DEGE 2003; KAMMLOTT 2003). Beide Verfahren sind jedoch für
die Überprüfung der Wirkung lipolytischer Enzyme prinzipiell geeignet und in zahlreichen
Arbeiten (TABELING 1998; HELDT 2001; FUENTE-DEGE 2003; KAMMLOTT 2003;
KARTHOFF 2004) erfolgreich genutzt worden.

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
25
2.4.3.3 Nachteile herkömmlicher Verdaulichkeitsstudien (Bestimmung von praecaecaler und totaler Verdaulichkeit) in der Produktentwicklung von Enzymen zur Substitutionstherapie
Bei herkömmlichen Verdaulichkeitsstudien am pankreasgangligierten Miniaturschwein zur
Überprüfung der Effizienz von in ihrer In-vivo-Wirksamkeit unbekannten Enzymen entsteht
ein immens hoher Zeit- und Enzymbedarf durch die mehrtägige Anfütterungsphase auf die
eine mehrtägige Kot- und Chymussammlung folgt. Auf diese Art und Weise kann innerhalb
von 13 Tagen lediglich ein Enzym bzw. eine Dosierung getestet werden.
Dementsprechend sind im Gegensatz zu In-vitro-Verfahren für die In-vivo-Testung
vergleichsweise große Enzymmengen nötig. In frühen Entwicklungsstadien sind jedoch die
verfügbaren Mengen des zu testenden Enzyms allgemein sehr begrenzt, so dass
üblicherweise nur relativ geringe Enzymmengen (ca. 5-10 g) zur Verfügung stehen
(LÖSCHER 2006, Vortrag). Demzufolge sind In-vivo-Versuche, insbesondere wenn diese
mit Schweinen durchgeführt werden, erst zu einem relativ späten Zeitpunkt der
vorklinischen Phase durchführbar, in der die Enzymproduktion entsprechend
fortgeschritten ist. Da Ergebnisse zur Wirksamkeit aus In-vivo-Verfahren jedoch als
Entscheidungsgrundlage für die Weiterentwicklung eines Enzyms dienen und als
richtungsweisend für die Wirksamkeit im Menschen gewertet werden, entstehen bei
Misserfolg / Entwicklungsabbrüchen kosten- und zeitintensive Verzögerungen.
2.4.4 Die klinische Entwicklungsphase von Enzymen zur Substitutionstherapie
Die Testung der in Frage kommenden Substanzen am Menschen ist in Deutschland bei
der Zulassungsbehörde BfArM (= Bundesinstitut für Arzneimittel und Medizinprodukte)
anzumelden und benötigt zusätzlich die Zustimmung der lokalen Ethikkommission (HINZE
et al. 2001). In den USA ist ein Antrag (IND= Investigational New Drug Application) bei der
FDA (Federation of Drug Administration) zu stellen. Dafür müssen die bisherigen
Ergebnisse aus der vorklinischen Phase offen gelegt und weitreichende

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
26
pharmakodynamische und –kinetische sowie den Herstellungsprozess betreffende
Informationen über die Substanz präsentiert werden (VFA-Statistics 2005). Spätestens zu
diesem Zeitpunkt sollten Möglichkeiten zur Produktion des Enzyms in größeren Mengen
und in hohem Reinheitsgrad bestehen, da in den folgenden, mehrjährigen klinischen
Testphasen bereits große Mengen (mehrere hundert Kilogramm) des Enzyms verbraucht
werden.
Der klinische Teil der Neuentwicklung wird in drei Unterbereiche gegliedert:
Phase I (Dauer ca. 1 Jahr):
Die Substanz wird erstmalig an eine kleine Gruppe (20-100) gesunder Probanden
verabreicht, um mögliche Nebenwirkungen erkennen zu können, den „safe dosage range“,
die Dosierungsrichtlinien, festzulegen und um Metabolisierung und Distribution zu
beobachten. Es wird also überprüft, ob die am Tier gewonnenen Erkenntnisse über den
Wirkstoff auf den gesunden Menschen zu übertragen sind (VFA-Statistics 2005).
Phase II (Dauer ca. 2 Jahre):
In dieser Phase erhält erstmals eine Gruppe von 100-500 Patienten, die an der
entsprechenden Krankheit (z.B. EPI) leiden, die neue Substanz. Ihre Effektivität,
Sicherheit und Nebenwirkungen werden näher untersucht. Weiterhin erfolgt die Auswahl
optimaler Dosierungen (HINZE et al. 2001).
Phase III (Dauer ca. 3 Jahre):
Eine Gruppe von 1000-3000 Patienten erhält unter ärztlicher Begutachtung die Substanz
unter Klinik- und Praxisbedingungen. Hierbei soll die Unbedenklichkeit des Wirkstoffes
auch bei Langzeiteinnahme an einer hinreichend großen Patientenzahl überprüft werden
(HINZE et al. 2001). Der Wirkstoff wird möglichst in Doppeltblind-Studien gegen Placebo
und Konkurrenzprodukte getestet (LÖSCHER 2006, Vortrag).
Die Phase III der klinischen Entwicklung ist mit einem Anteil von über 20% der
Gesamtkosten zusammen mit der vorklinischen Entwicklung (> 30% der Gesamtkosten)
der kostenintensivste Bereich einer Neuentwicklung. Deshalb ist vor jedem Eintritt in eine
neue Entwicklungsphase, besonders aber vor der Phase II sowie III genau abzuwägen, ob
eine Weiterentwicklung aussichtsreich ist (s. Tabelle 1).

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
27
Tabelle 1: Verteilung der Gesamtkosten einer pharmazeutischen Neuentwicklung (%) auf die verschiedenen Phasen der Entwicklung (HINZE et al. 2001)
Vorklinik Phase I Phase II Phase III Zulassung Phase IV Sonstiges
31,9 6,8 11,1 23,3 12,0 10,7 4,2
2.4.5 Zulassung und Markteinführung einer pharmazeutischen Neuentwicklung
Insgesamt werden nur ca. 10 % der Substanzen, die in die klinische Phase kommen,
weiter entwickelt. Diese gelangen dann in das offizielle Zulassungsverfahren (NDA= New
Drug Application) der EMEA (European Medicines Evaluation Agency) in London
(zentrales Verfahren) oder alternativ dezentral zum BfArM bzw. zum Paul-Ehrlich-Institut.
In den USA ist die FDA zuständig. Ein solches Zulassungsverfahren nimmt ca. 1-1,5 Jahre
in Anspruch (TUFTS CENTER FOR THE STUDY OF DRUG DEVELOPMENT 1999).
International arbeitende Pharmazeutische Firmen bemühen sich im Allgemeinen um
Zulassungen für den US-amerikanischen, europäischen und japanischen Markt, da hier
der Großteil des Umsatzes erwirtschaftet wird (s. Tabelle 2).
Tabelle 2: Erwirtschafteter Umsatz aus Medikamenten in % (VFA STATISTICS 2005)
USA Europa Japan Rest der Welt
70 19 4 7
Phase IV:
Nach erfolgreicher Zulassung, die im Mittel ca. 12 Jahren nach Patentierung des
Wirkstoffes erfolgt (s. Abbildung 1), wird das neue Medikament auf dem Markt eingeführt.
Dennoch wird das neue Produkt weiterhin kritisch überwacht, um seltene Nebenwirkungen
und Langzeiteffekte zu überwachen (ALLIANCE PHARMACEUTICAL CORP.- PHASES
OF PRODUCT DEVELOPMENT 2006).

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
28
20 Jahre nach Patentanmeldung verfällt das Patent, bzw. kann maximal um weitere 5
Jahre verlängert werden. Damit verbleiben nach Markteinführung in der Regel nur noch 8
Jahre Patentschutz. Nach Ablauf des Patentschutzes ist Generikern die Nachahmung
erlaubt (VFA-Statistics 2005). Beim Originalhersteller verbleiben nach Ablauf des
Patentschutzes schließlich lediglich ca. 30% des Marktanteils (pers. Mitteilung
IFFLÄNDER 2006).
Abbildung 1: Organigramm zur Entwicklung eines neuen Medikamentes (modifiziert nach LÖSCHER 2006, Vortrag)
2.5 Vorteile durch den Einsatz von Screening-Verfahren
Der Prozess vom Beginn der F&E bis zur Markteinführung eines neuen Produktes ist
durch gesetzliche Vorgaben klar in die oben genannten Phasen eingeteilt. Es existiert nur
wenig Spielraum zur Reduzierung der langen Entwicklungsdauer. Eine der Möglichkeiten
Substanz „Treffer“ LeitstrukturKandidaten
f. klin. Studie
Kandidaten f. vorklin.
Studie
100 000 100 10 3 - 4 1 -2
Erstes Screening
Zweites Screening
Optimierung d. Leitstruktur
Vorklinische Prüfung
Klinische Prüfung
Synthese Isolierung /
Aufreinigung Charakterisier-
ung
In vivoPharmakologie
Toxikologie/ Toxikokinetik
Pharmakokinetik
Phase I: Sicherheit und
Toleranz
Phase II: Dosisfindung
Phase III: Effektivität
In vitro:Pharmakologie Metabolisierung
In vivo:
erste Pharmakokinetik erste Toxikologiestudien
Hoch-Durchsatz-Verfahren:
Rezeptor-bindung
Enzymatische und funktionale
Tests
Funktionelle Suche nach dem Angriffspunkt:
Identifikation Funktionale Analyse
Testentwicklung
1-2 Jahre 3-4 Jahre
5-6 Jahre
Zielsetzung Angriffspunkt
Pharmakologie Toxikologie
Toxikokinetik Pharmakokinetik

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
29
besteht in einer zunehmenden Rationalisierung der Forschungs- und Entwicklungsphase
selbst. Insbesondere im Rahmen von In-vivo-Verfahren ermöglicht der Einsatz von
Screening-Verfahren relevante Zeiteinsparungen, die im IdeaLCFAll eine frühere
Zulassung und Markteinführung erlauben. Dadurch ergibt sich die von der
Pharmazeutischen Industrie angestrebte längere Vermarktungszeit innerhalb der
Patentschutzfrist.
2.5.1 Möglichkeiten und Grenzen eines Screening-Verfahrens für Proteasen und Amylasen als ergänzende Methode
In dem Projekt „pankreasgangligiertes Miniaturschwein“ wurden in vorangegangenen
Studien mittels oben genannter etablierter Methoden der Verdaulichkeitsbestimmung
verlässliche Ergebnisse zur Quantifizierung der Wirksamkeit substituierter
Verdauungsenzyme gewonnen. Um sowohl Zeit- und Arbeitsaufwand als auch
Materialverbrauch zu minimieren, wurde im Rahmen dieses Projektes von BECKER
(2005) ein Screening-Test entwickelt, welcher als ergänzende Methode eine erste
Einschätzung der Wirkung von Amylasen bzw. Proteasen auf die Stärke- bzw.
Proteinverdaulichkeit ermöglicht. Bei diesem Verfahren wird auf eine Anfütterungsphase
(sonst ca. 7-10 Tage) gänzlich verzichtet: die enzymhaltige Testmahlzeit wird nur einmalig
verabreicht. Hierdurch gelingt eine erhebliche Zeit- und Materialersparnis (BECKER 2005).
Bedeutendster Vorteil ist jedoch, dass in diesem Testverfahren aufgrund des Verzichtes
auf eine Anfütterungsphase sowie längere Chymussammlung (3-5 Tage) nur sehr geringe
Mengen des zu testenden Enzyms notwendig sind. Somit kann schon früh mit
Wirksamkeitsstudien am Tier begonnen werden so dass ein Einsatz der Enzyme bereits in
einer sehr frühen Phase der Entwicklung möglich ist, wenn allgemein noch keine großen
Enzymmengen vorhanden sind.
Mit diesem Screening-Verfahren entstand die Option, die In-vivo-Effizienz einer Dosierung
einer Protease oder Amylase innerhalb eines Versuchstages zumindest tendenziell
einschätzen zu können. Nachdem mehrere Testenzyme in üblicherweise jeweils drei
Dosierungen im Screening eingesetzt worden sind, können die Ergebnisse der

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
30
verschiedenen Enzyme (z.B. einer Gruppe von Test-Amylasen) vergleichend bewertet
werden. Über die sich ergebene Beurteilung wird eine Rangierung der zu prüfenden
Enzyme der Testenzyme ermöglicht.
Um zu verdeutlichen, dass die gewonnenen Ergebnisse nicht den allgemeinen
Bedingungen von „Verdaulichkeitswerten“ entsprechen, verwendete BECKER (2005) den
Terminus „Verschwindensrate“.
2.5.2 Überlegungen zur Entwicklung eines Lipase-Screenings
Die Entwicklung eines Screening-Tests für Lipasen ist von besonderem Interesse, da
Lipasen in der Substitutionstherapie eine bedeutende Rolle innehaben und daher einen
wichtigen Baustein in der Entwicklung eines neuen Präparates zur Substitutionstherapie in
der Behandlung der EPI darstellen.
Vor diesem Hintergrund ist zu prüfen, inwieweit das von BECKER (2005) entwickelte
Versuchsdesign (einmalige Fütterung der Testmahlzeit ohne Anfütterungsperiode, Beginn
der Kollektion mit dem erstem Erscheinen von markerhaltigem Chymus an der Ileumfistel,
Kollektionsdauer) für ein Lipase-Screening anwendbar ist. Ergebnisse bereits vorhandener
Versuche (BECKER 2006, persönliche Mitteilung) zeigten, dass beim Einsatz der
Standard-Fett-Diät des Projektes (70 g Rfe / 225 g TS pro Mahlzeit) das Fett dem Chymus
aus der entsprechenden Testmahlzeit zum Teil voraus floss. Dadurch entstand ein
inhomogener Fluss von Marker, Fett und restlichen Bestandteilen der Testmahlzeit. Der
Verzicht auf eine Anfütterungsphase ist jedoch nur bei parallelem Fluss der
Rationsbestandteile sinnvoll bzw. möglich. Für eine optimale Enzymwirkung ist ein
homogener Chymus zur Bereitstellung einer möglichst hohen Substratverfügbarkeit, hier
Fett, erforderlich, um die Aktivität der zu prüfenden Lipasen nicht durch mangelnde
Substratverfügbarkeit zu limitieren und um eine gleichmäßige Verteilung des Enzyms in
der Testmahlzeit zu gewährleisten. Außerdem wird durch einen homogenen Chymusfluss
ausgeschlossen, dass Futter vorheriger Mahlzeiten zusammen mit der Testmahlzeit
anflutet. Die Homogenität des Chymus ist Voraussetzung für eine Kalkulation der Rohfett-
Verschwindensrate anhand des gleichzeitig anflutenden Markers der Testmahlzeit.

SCHRIFTUM ------------------------------------------------------------------------------------------------------------------------
31
Der für die Testung von Lipasen notwendige hohe Fettgehalt in der Testmahlzeit kann
Störungen bzw. Verzögerungen der Magenentleerung bedingen (HUNT u. STUBBS 1975;
READ 1984), die durch Rückkopplungsmechanismen des Dünndarms als Reaktion auf
nicht hydrolysierte Triglyceride (so genannte „Duodenum-, Jejunum-, und Ileumbremse“)
entstehen. Die Entleerung des Magens wird laut BRAUDE et al. (1970), HUNT u. STUBBS
(1975), READ (1984) und DECUYPERE et al. (1985) durch die Verabreichung einer
flüssigen Diät positiv beeinflusst, d.h. beschleunigt. In einer flüssigen Mahlzeit erhöht sich
jedoch die Gefahr für eine Entmischung des Markers von der Testmahlzeit, insbesondere
wenn ein Marker mit relativ hohem spezifischem Gewicht (Chromoxid) eingesetzt wird. Um
der Separation entgegen zu wirken, ist es zu erwägen, die Diät als „stabile Suspension“
mit partikulären Strukturen bzw. Rohfaser anzureichern, obwohl durch einen erhöhten Rfa-
Anteil praecaecal mit einer Retention der Passage zu rechnen ist (DROCHNER 1984).

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
32
3 Eigene Untersuchungen
3.1 Material und Methoden
3.1.1 Versuchsziel
Im Rahmen der vorliegenden Dissertation sollte ein In-vivo-Screening-Test zur
vergleichenden Untersuchung der Effizienz von Lipasen entwickelt werden. Hierzu wurde
die etablierte Methode einer Verdaulichkeitsuntersuchung modifiziert. Dabei konnte auf
Erfahrungen einer vorangegangenen Arbeit (BECKER 2005) dieses Projektes aufgebaut
werden, in der mittels eines „Schnelltests“ eine erste Einschätzung der Wirksamkeit von
Amylasen und Proteasen erfolgte.
Ein entsprechender Schnelltest oder Screening-Test für Lipasen sollte folgende
Anforderungen erfüllen:
• kurzfristige Durchführung (= Zeitersparnis),
• geringster Verbrauch des zu prüfenden Enzyms,
• eine vergleichende Bewertung der zu prüfenden Enzyme ermöglichen.
Gleichzeitig sollte das neu zu entwickelnde Screening-Verfahren eine In-vivo-Simulation
(Magenpassage, Einwirkungen von Restaktivitäten von Enzymen und Mikroflora) im
Dünndarm ermöglichen.
Aufgrund des geringen Enzymaufwandes wäre es möglich, In-vivo-Screening-Tests schon
zu einem vergleichsweise früheren Zeitpunkt der Entwicklung eines zukünftigen
Enzymproduktes durchzuführen. Hierdurch wären erhebliche Einsparungen zu erwarten:
eine Weiterentwicklung fände nur für Prototypen statt, die im In-vivo-Screening-Test als
effektiv beurteilt wurden. Dadurch könnten die zu einem späteren Entwicklungszeitpunkt

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
33
zeit- und materialaufwendigen klassischen Verdaulichkeitsstudien auf die Testung viel
versprechender Enzyme reduziert werden.
Wie bei anderen als Screening eingesetzten Methoden ist auch bei dem hier neu zu
entwickelndem Screening-Test eine realistische „Einschätzung“ in einem wenig
aufwändigem Verfahren oberstes Ziel, während im Anschluss an das Screening für Erfolg
versprechende Produkte eine Absicherung in weitergehenden Untersuchungen mit
Verfahren höherer Genauigkeit erfolgen sollte.
Die Überprüfung der im Screening-Test ermittelten Ergebnisse erfolgt durch Vergleich mit
Ergebnissen aus „klassischen“ Verdaulichkeitsuntersuchungen ausgewählter Lipasen, die
bereits von KARTHOFF (2004) und BECKER (2005) untersucht wurden.
Im Rahmen dieser Arbeit sollte somit ein In-vivo-Screening-Test zur Einschätzung der
Wirksamkeit lipolytischer Enzyme entwickelt werden. Während der Entwicklungsphase
wurde ein etabliertes Multienzympräparat verwendet, das in verschiedenen
vorangegangenen Untersuchungen als hochwirksam befunden wurde. Eine Überprüfung
des neu zu entwickelnden Tests sollte anhand klassischer Verdaulichkeitsversuche
stattfinden, indem zwei ausgewählte Testlipasen mit beiden Verfahren untersucht wurden.
Alle Versuche waren von der Bezirksregierung Hannover genehmigt worden.
3.1.2 Versuchstiere
In den vorliegenden Untersuchungen wurden insgesamt 13 adulte weibliche Göttinger
Miniaturschweine der Zuchtlinie Ellegaard eingesetzt. Allen Tieren wurde operativ eine
ileocaecale Umleitungsfistel implantiert, um die Verdaulichkeit der Nährstoffe bis zum
Ende des Ileums quantifizieren zu können. Bei 10 Schweinen (PL-Tiere) erfolgte
zusätzlich eine Ligatur des Ductus pancreaticus accessorius zur Simulation einer
chronischen exokrinen Pankreasinsuffizienz. Bei 3 Tieren blieb der
Pankreasausführungsgang unversehrt, so dass diese als Kontrollgruppe (KT =
Kontrolltiere) Verwendung fanden.
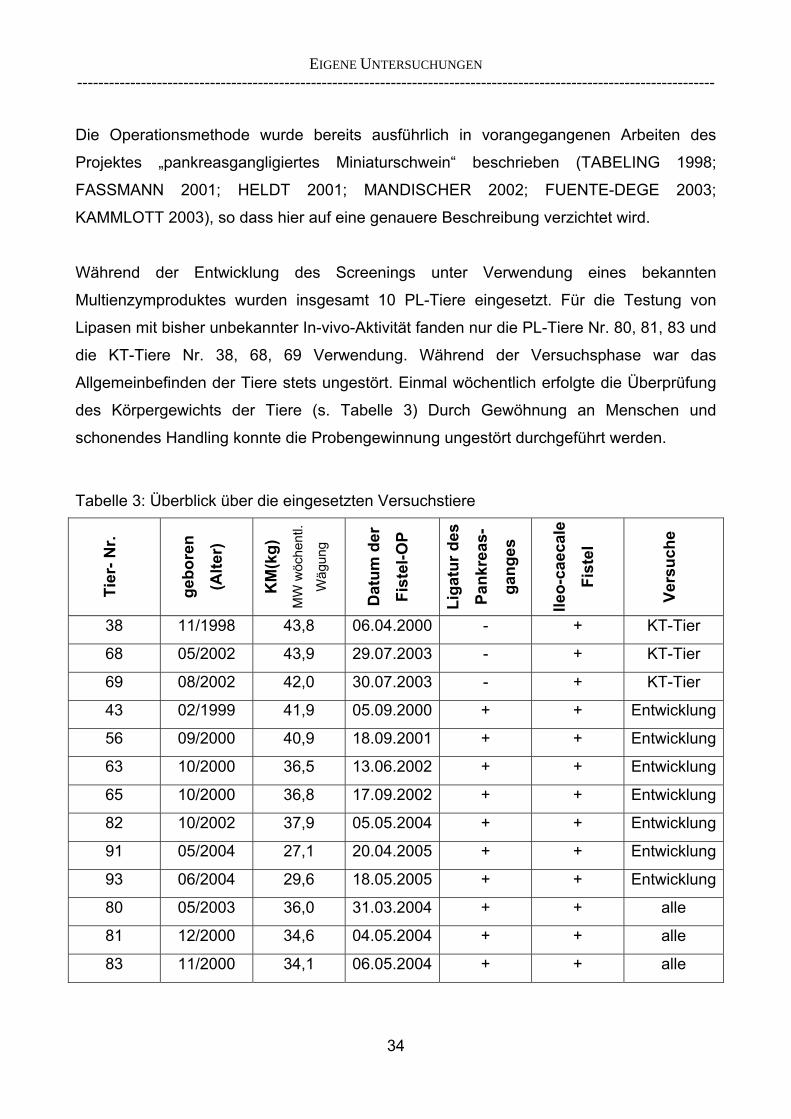
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
34
Die Operationsmethode wurde bereits ausführlich in vorangegangenen Arbeiten des
Projektes „pankreasgangligiertes Miniaturschwein“ beschrieben (TABELING 1998;
FASSMANN 2001; HELDT 2001; MANDISCHER 2002; FUENTE-DEGE 2003;
KAMMLOTT 2003), so dass hier auf eine genauere Beschreibung verzichtet wird.
Während der Entwicklung des Screenings unter Verwendung eines bekannten
Multienzymproduktes wurden insgesamt 10 PL-Tiere eingesetzt. Für die Testung von
Lipasen mit bisher unbekannter In-vivo-Aktivität fanden nur die PL-Tiere Nr. 80, 81, 83 und
die KT-Tiere Nr. 38, 68, 69 Verwendung. Während der Versuchsphase war das
Allgemeinbefinden der Tiere stets ungestört. Einmal wöchentlich erfolgte die Überprüfung
des Körpergewichts der Tiere (s. Tabelle 3) Durch Gewöhnung an Menschen und
schonendes Handling konnte die Probengewinnung ungestört durchgeführt werden.
Tabelle 3: Überblick über die eingesetzten Versuchstiere
Tier
- Nr.
gebo
ren
(Alte
r)
KM
(kg)
M
W w
öche
ntl.
Wäg
ung
Dat
um d
er
Fist
el-O
P
Liga
tur d
es
Pank
reas
-
gang
es
Ileo-
caec
ale
Fist
el
Vers
uche
38 11/1998 43,8 06.04.2000 - + KT-Tier
68 05/2002 43,9 29.07.2003 - + KT-Tier
69 08/2002 42,0 30.07.2003 - + KT-Tier
43 02/1999 41,9 05.09.2000 + + Entwicklung
56 09/2000 40,9 18.09.2001 + + Entwicklung
63 10/2000 36,5 13.06.2002 + + Entwicklung
65 10/2000 36,8 17.09.2002 + + Entwicklung
82 10/2002 37,9 05.05.2004 + + Entwicklung
91 05/2004 27,1 20.04.2005 + + Entwicklung
93 06/2004 29,6 18.05.2005 + + Entwicklung
80 05/2003 36,0 31.03.2004 + + alle
81 12/2000 34,6 04.05.2004 + + alle
83 11/2000 34,1 06.05.2004 + + alle

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
35
3.1.3 Aufstallung und Haltung
Die Aufstallung und Haltung erfolgten in Einzelboxen ohne Einstreu. Auf eine detaillierte
Beschreibung wurde an dieser Stelle verzichtet, da diese denen in den vorangegangenen
Arbeiten des Projektes entsprach (TABELING 1998; FASSMANN 2001; HELDT 2001;
MANDISCHER 2002; FUENTE-DEGE 2003; KAMMLOTT 2003; KARTHOFF 2004;
BECKER 2005).
3.1.4 Statistische Methoden
Zur statistischen Auswertung der Daten wurden folgende Methoden angewandt:
• Berechnung des arithmetischen Mittels
• Berechnung der Standardabweichung
• Varianzanalyse mit repeated measurements nach Tukey Kramer
(Vergleich der verschiedenen Behandlungen)
• t-Test für unverbundene Stichproben
(Vergleich KT-Tiere versus PL-Tiere)
Sowohl die Berechnung der arithmetischen Mittelwerte als auch der
Standardabweichungen erfolgte unter Anwendung des Tabellenkalkulationsprogrammes
Exel’ 2000. Die statistischen Vergleiche wurden mit Hilfe des Statistik Programms SAS
nach Auswahl der statistischen Methoden durch das Institut für Biometrie der Stiftung der
Tierärztlichen Hochschule Hannover durchgeführt.
Signifikante Unterschiede zwischen den Mittelwerten mit einer Irrtumswahrscheinlichkeit
von p ≤ 0,05 wurden durch unterschiedliche Buchstaben bzw. Symbole gekennzeichnet.

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
36
3.2 Versuchsdurchführung
3.2.1 Übersicht über die durchgeführten Versuche
Die Versuche unterteilten sich in folgende Abschnitte:
• Entwicklung (Entwicklung der Testmahlzeit) eines In-vivo-Screening-Tests zur
Einschätzung der Wirksamkeit lipolytischer Enzyme unter Verwendung eines
etablierten Multienzympräparates mit bekannter Wirkung
• Verwendung des neu entwickelten Screening Verfahrens zur Überprüfung der
Effektivität von 5 Lipasen mit noch unbekannter In-vivo-Wirksamkeit
• Rangierung der getesteten Lipasen anhand der ermittelten Ergebnisse
bezüglich:
⇒ Anflutung von Trockensubstanz und Rohfett in einem definiertem
Zeitraum
(→ Maß für Wirksamkeit der Enzyme, Maß für Passage)
⇒ Rohfett-Konzentration im Chymus
(→ Maß für Wirksamkeit der Enzyme)
⇒ Cr2O3-Konzentration im Chymus
(→ Maß für Wirksamkeit der Enzyme, Maß für Passage)
⇒ Rohfett / Cr2O3-Konzentration im Chymus
(→ Maß für Wirksamkeit der Enzyme)
⇒ Cr2O3-Wiederfindung im Chymus
(→ Maß für Passage)
⇒ „Verschwindensrate“ von Trockensubstanz und Rohfett
(→ Maß für Wirksamkeit der Enzyme)
⇒ Relativer Anflutung von Trockensubstanz und Rohfett
(→ Maß für Wirksamkeit der Enzyme)
⇒ Relative Trockensubstanz- und Rohfett-„Verschwindensrate“
(→ Maß für Wirksamkeit der Enzyme)

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
37
Der erste Abschnitt umfasste Versuche, die zur Entwicklung eines Screening-Tests für Lipasen führten. Die im Rahmen des Screenings für Amylasen und Proteasen (BECKER
2005) verwendeten Mischfuttermittel enthielten eine absolute Rohfettmenge von ca. 5g /
Mahlzeit (entsprechend 2 %). Diese Fettmenge pro Mahlzeit erwies sich für die Testung
von Lipasen als zu gering. Demzufolge sollte eine wesentlich fettreichere Ration zum
Einsatz kommen. Als fettreiche Mahlzeit stand die Standard-Fettdiät (30 % Rohfett in der
TS / 70 g Rfe je Mahlzeit), die schon in vorangegangenen Arbeiten des Projektes
verwendet wurde, zur Verfügung. Ihr testweiser Einsatz ergab jedoch, dass
Modifikationen, insbesondere im Bezug auf die Zusammensetzung und Zubereitung der
Testmahlzeit, notwendig sein würden, da die Ergebnisse auf Entmischungen zwischen
Futter und Marker sowie Durchmischungen mit der vorherigen Mahlzeit hindeuteten.
Daraufhin folgte die Entwicklung einer geeigneten Versuchsdiät. Bis zur Festsetzung der
endgültigen Testmahlzeit wurden insgesamt 20 Versuche mit unterschiedlich
zusammengesetzten Mahlzeiten durchgeführt. Davon werden in dem Abschnitt „Versuche
zur Entwicklung einer geeigneten Testmahlzeit“ acht Diäten exemplarisch (zur
Demonstration der Zwischenschritte) dargestellt.
Obgleich dieser Abschnitt sehr umfangreich ist, werden die Untersuchungen zur
„Entwicklung einer geeigneten Testmahlzeit“ und die entsprechenden Ergebnisse im
Teil „Material und Methoden“ dieser Arbeit dargestellt, da die Kenntnis der entwickelten
Vorgehensweisen bzw. Methoden für das Verständnis des eigentlichen Screening-Tests
für Lipasen (Ergebnisse: Ergebnisteil) zwingend notwendig ist.
Im zweiten Abschnitt wurde die Effizienz von fünf mikrobiellen Lipasen sowie des
Multienzymproduktes PK mit dem neu entwickelten Screening-Test untersucht und die
Enzyme entsprechend ihrer In-vivo-Wirksamkeit rangiert.

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
38
3.2.2 Methodik der Chymuskollektion
Die Methodik der Chymuskollektion sowie der Probenaufbereitung folgt grundsätzlich der
von BECKER (2005) beschriebenen Vorgehensweise (s. Abbildung 2):
• Vollständiger Verzicht auf eine Anfütterungsphase
• einmalige Verabreichung einer markerhaltigen Testmahlzeit
• Beginn der Chymuskollektion erst mit beginnender Anflutung der markerhaltigen
Ingesta an der ileocaecalen Umleitungsfistel
• Chymussammlung in zweistündigen Kollektionsintervallen
• Kollektionsdauer: 8 Stunden nach Erscheinen des Markers
• Mindestabstand zwischen zwei Testmahlzeiten von 48 Stunden
* fettfreie, energiereiche Mahlzeit (ProvideXtra®) mit hohem Kohlenhydrat- und Protein-Anteil
Abbildung 2: Schematische Übersicht zum Vorgehen bei der Fütterung und Chymuskollektion im Screening-Test für Lipasen
Versuchsdesign: Chymuskollektion (s. BECKER 2005):
Getrennte & fraktionierte Kollektion
Fütterung der
Testmahlzeit
Morgen des
Versuchstag
Abend des
Versuchstages
Kollektion des „braunen Chymus“
(Kollektionsintervall:
Fütterung bis „Farbumschlag“)
Kollektion des
„grünen“ Chymus
Fütterung
Vorabend* Fütterung von
Haltungsfutter

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
39
Kollektion des „braunen“ Chymus:
Kollektionsintervall: Fütterung bis „Farbwechsel“
Kollektion des „grünen“ (= markerhaltigen) Chymus:
Kollektionsintervall: Beginn: „Farbwechsel“; Dauer: 8 Stunden; Sammlung des Chymus
fraktioniert; ⇒ 4 Proben aus je 2 Stunden je Tier und Tag
Analog zu BECKER (2005) wurde die Effektivität der zu testenden Enzyme sowohl anhand
der absoluten Anflutung des nicht umgesetzten Substrates innerhalb eines definierten
Zeitraums an der ilealen Fistel als auch anhand der kalkulierten „Verschwindensrate“ des
Substrates bewertet.
3.2.2.1 Exkurs: Begriffsbestimmung „ Verschwindensrate“
Aufgrund des Verzichts auf eine Anfütterungsphase und auf
eine mehrtägige Sammlung sind nicht alle Bedingungen im
Sinne eines herkömmlichen Verdaulichkeitsversuchs erfüllt.
Insbesondere befinden sich die Tiere nicht in einem „steady-
state“ bezüglich des Chromoxids. Deshalb sollte bei
Verwendung dieser abgewandelten Versuchsdurchführung im
Screening-Test nicht von „Verdaulichkeiten“ gesprochen
werden, sondern vielmehr von „Verschwindensraten“. Mit der
Nutzung des Begriffes „Verschwindensrate“ (VR) soll zudem
betont werden, dass es sich hierbei um Werte für eine erste
Einschätzung der Wirksamkeit bzw. Effektivität von
Monoenzympräparaten handelt. Verdaulichkeitswerte im
engeren Sinne müssen weiterhin in herkömmlich
durchgeführten Verdaulichkeitsuntersuchungen (inklusive
Anfütterungszeit) und mehrtägiger Kollektion ermittelt werden.
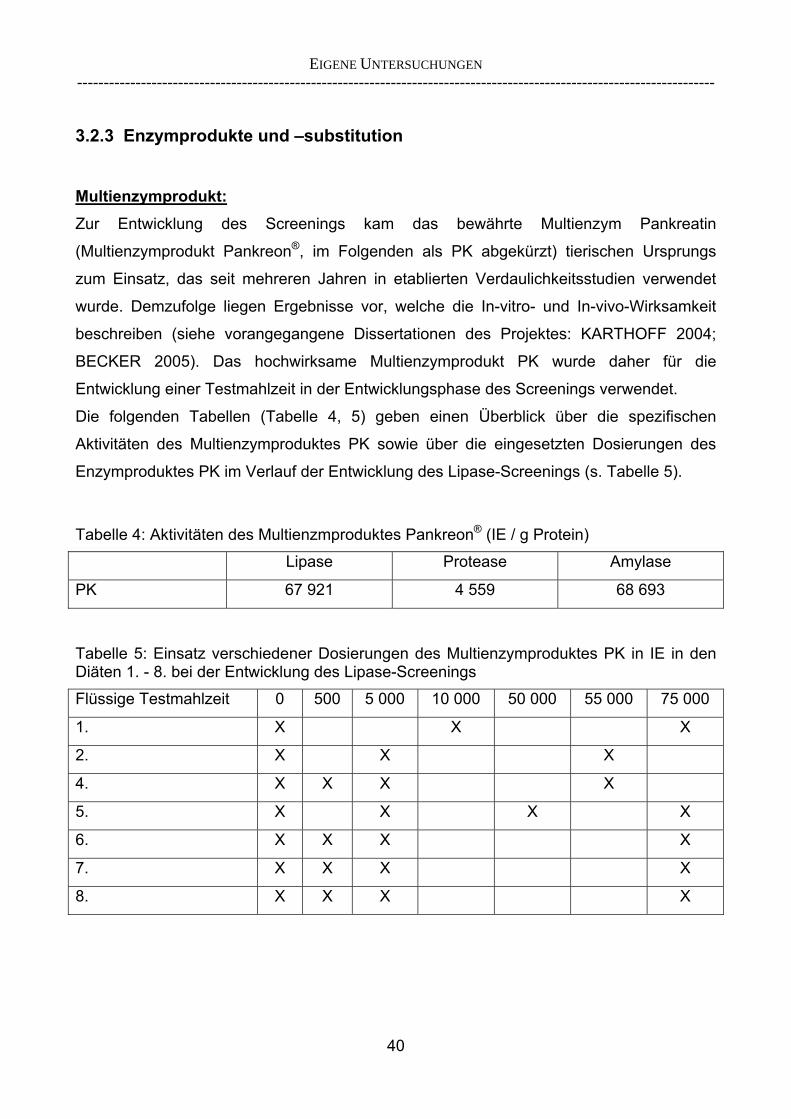
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
40
3.2.3 Enzymprodukte und –substitution
Multienzymprodukt: Zur Entwicklung des Screenings kam das bewährte Multienzym Pankreatin
(Multienzymprodukt Pankreon®, im Folgenden als PK abgekürzt) tierischen Ursprungs
zum Einsatz, das seit mehreren Jahren in etablierten Verdaulichkeitsstudien verwendet
wurde. Demzufolge liegen Ergebnisse vor, welche die In-vitro- und In-vivo-Wirksamkeit
beschreiben (siehe vorangegangene Dissertationen des Projektes: KARTHOFF 2004;
BECKER 2005). Das hochwirksame Multienzymprodukt PK wurde daher für die
Entwicklung einer Testmahlzeit in der Entwicklungsphase des Screenings verwendet.
Die folgenden Tabellen (Tabelle 4, 5) geben einen Überblick über die spezifischen
Aktivitäten des Multienzymproduktes PK sowie über die eingesetzten Dosierungen des
Enzymproduktes PK im Verlauf der Entwicklung des Lipase-Screenings (s. Tabelle 5).
Tabelle 4: Aktivitäten des Multienzmproduktes Pankreon® (IE / g Protein)
Lipase Protease Amylase
PK 67 921 4 559 68 693
Tabelle 5: Einsatz verschiedener Dosierungen des Multienzymproduktes PK in IE in den Diäten 1. - 8. bei der Entwicklung des Lipase-Screenings
Flüssige Testmahlzeit 0 500 5 000 10 000 50 000 55 000 75 000
1. X X X
2. X X X
4. X X X X
5. X X X X
6. X X X X
7. X X X X
8. X X X X

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
41
Monopräparate: Zur Testung innerhalb des Screenings gelangten Lipasen mikrobiellen Ursprungs.
Wenngleich ihre In-vitro-Wirksamkeit zuvor getestet, also bekannt war, stand ihre In-vivo-
Wirksamkeit bis zu diesem Zeitpunkt noch in Frage.
Zu Beginn jeder Versuchsphase wurde die spezifische Aktivität der eingesetzten
Präparate bestimmt. Die eingesetzten Dosierungen richten sich nach den ermittelten
enzymatischen Aktivitäten, die sich auf die lipolytischen Aktivität des bekannten
Pankreon® in IE / g Protein beziehen. Die mikrobiellen lipolytischen Aktivitäten (s. Tabelle
6) wurden anhand eines für Pankreon® optimierten In-vitro-Verfahrens ermittelt. Die
mikrobiellen Lipasen kamen im Screening Test in den Dosierungen 500, 5 000 sowie
75 000 IE lipolytischer Aktivität zum Einsatz (s. Tabelle 7).
Tabelle 6: Lipolytische Aktivität der Testlipasen (IE / g Protein)
E1 E2 E3 E4 E5
3 580 000 3 720 000 3 720 000 4 180 000 220 000
Tabelle 7: Im Lipase-Screening-Test eingesetzte Dosierungen des Multienzymproduktes PK und der Testlipasen E1 - E5
500 IE (D1) 5 000 IE (D2) 75 000 IE (D3)
PK*; E 1; E 2; E 3; E 4; E 5 PK*; E 1; E 2; E 3; E 4; E 5 PK*; E 1; E 2; E 3; E 4; E 5
* zusätzlich 10 00, 50 000 und 55 000 IE
3.2.4 Versuchsfutter
Um die Lipasen im Screening testen zu können, musste eine spezielle Diät eingesetzt
werden. Bei einer TS-Menge von 146,9 g, einer Gesamtfettmenge von 51,6 g und einer
Cr2O3-Menge von 0,368 g / Mahlzeit wurde eine Portion Calshakepulver® (Beutelinhalt =
87g) mit 240 ml Milch (1,5 % Fettanteil, haltbare Vollmilch) und 29,37 g Olivenöl der Firma
Roth angerührt. Durch Verwendung eines Ultra-turrax 900® konnte eine stabile Emulsion
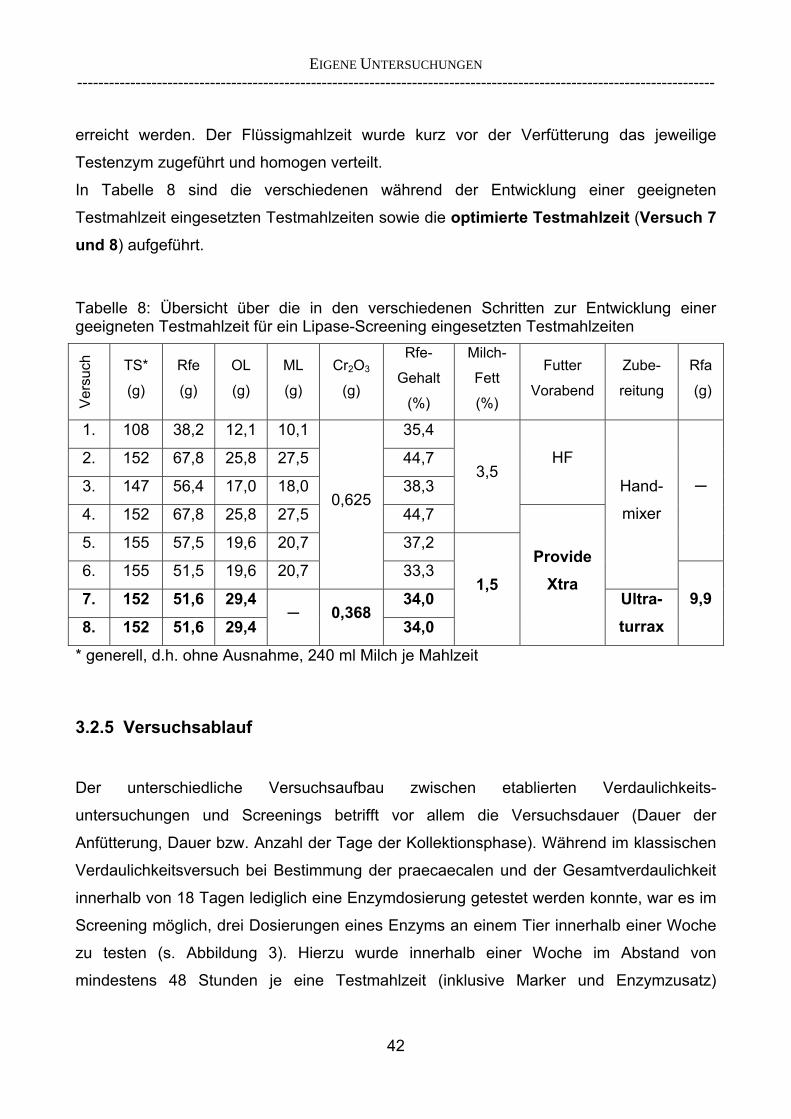
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
42
erreicht werden. Der Flüssigmahlzeit wurde kurz vor der Verfütterung das jeweilige
Testenzym zugeführt und homogen verteilt.
In Tabelle 8 sind die verschiedenen während der Entwicklung einer geeigneten
Testmahlzeit eingesetzten Testmahlzeiten sowie die optimierte Testmahlzeit (Versuch 7 und 8) aufgeführt.
Tabelle 8: Übersicht über die in den verschiedenen Schritten zur Entwicklung einer geeigneten Testmahlzeit für ein Lipase-Screening eingesetzten Testmahlzeiten
Ver
such
TS*
(g)
Rfe
(g)
OL
(g)
ML
(g)
Cr2O3
(g)
Rfe-
Gehalt
(%)
Milch-
Fett
(%)
Futter
Vorabend
Zube-
reitung
Rfa
(g)
1. 108 38,2 12,1 10,1 35,4
2. 152 67,8 25,8 27,5 44,7
3. 147 56,4 17,0 18,0 38,3
HF
4. 152 67,8 25,8 27,5 44,7
3,5
5. 155 57,5 19,6 20,7 37,2
─
6. 155 51,5 19,6 20,7
0,625
33,3
Hand-
mixer
7. 152 51,6 29,4 34,0
8. 152 51,6 29,4 ─ 0,368
34,0
1,5
Provide Xtra
Ultra- turrax
9,9
* generell, d.h. ohne Ausnahme, 240 ml Milch je Mahlzeit
3.2.5 Versuchsablauf
Der unterschiedliche Versuchsaufbau zwischen etablierten Verdaulichkeits-
untersuchungen und Screenings betrifft vor allem die Versuchsdauer (Dauer der
Anfütterung, Dauer bzw. Anzahl der Tage der Kollektionsphase). Während im klassischen
Verdaulichkeitsversuch bei Bestimmung der praecaecalen und der Gesamtverdaulichkeit
innerhalb von 18 Tagen lediglich eine Enzymdosierung getestet werden konnte, war es im
Screening möglich, drei Dosierungen eines Enzyms an einem Tier innerhalb einer Woche
zu testen (s. Abbildung 3). Hierzu wurde innerhalb einer Woche im Abstand von
mindestens 48 Stunden je eine Testmahlzeit (inklusive Marker und Enzymzusatz)

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
43
gefüttert. Aufgrund der technischen und personellen Gegebenheiten in der
Versuchsanlage konnten pro Versuchswoche maximal drei Tiere das Screening
durchlaufen. Somit ließen sich pro Woche 3 x 3 Dosierungen testen.
Bei der Durchführung des Screenings wurden die zu testenden Enzyme in einer Dosierung
von 500 (D1), 5 000 (D2) und 75 000 IE pankreatische Einheiten (D3) eingesetzt (s.
Tabelle 7). Das zu prüfende Enzym wurde der Testmahlzeit lediglich am Morgen des
Screenings zugegeben. Die Prüfung der verschiedenen Dosierungen eines Enzyms
erfolgte im 3 x 3 Lateinischen Quadrat, um den Einfluss tagesabhängiger Schwankungen
zu minimieren. Das Auffangen von Chymus nach Farbwechsel erfolgte in zweistündigen
Intervallen. Nach Gefriertrocknung und Homogenisierung wurde der Rfe-, TS- und Cr2O3-
Gehalt der Proben ermittelt.
Klassischer Verdaulichkeitsversuch: Bestimmung der Bestimmung der
Gesamt- praecaecale
Anfütterungsphase verdaulichkeit: Verdaulichkeit Kollektion 12h: 8-20 Uhr
1.Tag
10. Tag 15.Tag 18.Tag
Screening-Verfahren: 3 Versuchstage je 1 Dos. 3 Versuchstage je 1 Dos. 3 Versuchstage je 1 Dos.
Enzym 1 Enzym 2 Enzym 3
…48 h…
1.d ….. 7.d 1.d …. 7.d 1.d …..
Abbildung 3: Schematische Übersicht über den zeitlichen Ablauf klassischer Verdaulichkeitsversuche und den zeitlichen Ablauf des Screening-Verfahrens

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
44
Fütterung in der Phase „Versuchstage“:
Die Tiere erhielten am Morgen des Versuchstages um 745 Uhr die Testmahlzeit. Direkt vor
Aufnahme der Versuchsration erfolgte die Zumischung des Markers und des jeweils zu
testenden Enzyms. Nach Beendigung der Kollektionsphase erhielten die Tiere 250 g
„Haltungsfutter“, angemischt mit 1000 ml Wasser (ohne Enzym- und Markerzulagen). Allen
Tieren stand jederzeit Tränkwasser über Nippeltränken ad libitum zur Verfügung.
Fütterung in der Phase „Versuchsfreie Tage“:
Wegen des Verzichts auf eine mehrtägige Anfütterung wurden die Versuchstiere zwischen
den einzelnen Versuchstagen mit „Haltungsfutter“ (HF) der Firma Altromin® gefüttert (s.
Tabelle 9). Dieses Alleinfuttermittel für Minischweine basiert auf verschiedenen
Getreideschroten und entsprechenden Nährstoffergänzungen. Die Deckung des Bedarfs
von Rohnährstoffen, Mengen- und Spurenelementen sowie Vitaminen war damit
gewährleistet. Um eine Verstopfung der Fisteln zu vermeiden, wurden die
Ausgangskomponenten doppelt gemahlen. Die Verabreichung von 250 g des Futters,
angemischt mit 1000 ml Wasser, erfolgte in einem Rhythmus von 12 h (745 / 1945). Die PL-
Tiere erhielten zusätzlich zweimal täglich 336 000 IE lipolytischer Aktivität Pankreatin®
(PK), welches jeweils drei Tage vor Versuchsbeginn abgesetzt wurde, um eine
„Verschleppung“ von enzymatischer Restaktivität in der jeweiligen Testphase zu
vermeiden. Das HF enthielt keinen Marker (Cr2O3).
Tabelle 9: Analysenwerte der Rohnährstoffe des „Haltungsfutter“ (g / kg TS)
Ra oS Rp Rfe Rfa Nfe Stärke Zucker
52,7 947 189 49,8 30,3 677 477 62,9
Am Vorabend jedes Versuchstages wurde den Versuchstieren anstelle des
„Haltungsfutters“ 400 ml eines fettfreien, kohlenhydratreichen, eiweißreichen flüssigen
„Energiedrinks“ (ProvideXtra®: 12 % Rp, 88 % KH, 0 % Rfe) in der Geschmacksrichtung
Karotte / Apfel angeboten (s. Tabelle 10).

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
45
Tabelle 10: Zusammensetzung 100 ml des „Energiedrinks“ ProvideXtra® (Geschmackrichtung: Apfel / Karotte)
KJ (kcal) Rp (g) KH(g) Zucker(g) Rfe (g) Ges. FS(g) Rfa (g) Wasser(ml)
525 (125) 3,75 2 7 , 5 5,5 0 0 1 80
3.2.6 Probengewinnung und –aufbereitung
Am Morgen des Versuchstages wurden die ileocaecalen Umleitungsfisteln um 745 Uhr
geöffnet und auf ihre Durchgängigkeit überprüft. An dem ilealen Abschnitt der Fistel wurde
mittels einer Metallklammer ein Behältnis zum Auffangen des Chymus befestigt. Die
caecale Fistel wurde für die Versuchsdauer mit einer Schraubkappe verschlossen.
Gleichzeitig erfolgte die Fütterung, woraufhin die Chymussammlung begann. Der
anflutende Chymus wurde in zweistündigen Intervallen gesammelt und währenddessen
sofort bei -20°C eingefroren.
Auffälliger Weise flutete in den ersten Stunden nach der Aufnahme der markerhaltigen
Diät ausnahmslos brauner Chymus an. Wie BECKER (2005) feststellte, handelte es sich
dabei nahezu ausschließlich um Anflutung von markerlosem Haltungsfutter aus der
Vorabendmahlzeit. Analytisch konnte diese Vermutung bestätigt werden. Zusätzlich wurde
diese These durch eine verringerte TS-Anflutung von „braunem“ Chymus bei Einsatz von
ProvideXtra® gestützt. Der zunächst anflutende braune Chymus wurde gesondert
aufgefangen. Da sich aber der Wechsel zu dem grün -und damit markerhaltigen- Chymus
scharf vollzog, war davon auszugehen, dass die markerhaltige Testmahlzeit zu diesem
Zeitpunkt das Ende des Ileums erreichte. Dieser Farbwechsel im Chymus fand bei den
Tieren zu individuell unterschiedlichen Zeitpunkten, aber im Allgemeinen 4-8 h nach
Fütterung, statt. Mit dem Farbwechsel von „braunem“ zu „grünem“ Chymus begann für das
jeweilige Tier ein neues Sammlungsintervall und die für das eigentliche Screening
relevante Sammlung von Chymus. Der Kollektionszeitraum (siehe BECKER 2005),
welcher in zweistündige Intervalle unterteilt wurde, betrug insgesamt acht Stunden (nach
Farbwechsel). Die Sammlung des Chymus erfolgte in austarierten Plastikschälchen. Ab

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
46
einer Masse von ca. 50 g wurden diese aus verfahrenstechnischen Gründen
(Gefriertrocknung) mit dem dazugehörigen Deckel verschlossen und zur Ermittlung der
ursprünglichen Substanz gewogen. Während des Sammelns und nach dem Verschließen
fand die Aufbewahrung der Proben bei -20°C statt. Am Ende jedes zweistündigen
Intervalls begann auch bei Nichterreichen des maximalen Befüllungszustandes stets die
Verwendung eines neuen Plastikschälchens. Nach Versuchsende wurden die Proben bis
zur Weiterverarbeitung bei -80°C gelagert.
Nach zweitägiger Gefriertrocknung und Ermittlung des Gewichtes wurden die Proben
jedes einzelnen Tieres und jeden Intervalls getrennt unter Verwendung einer Messermühle
(Grindomix GM 200®, Firma Retsch, Haan) gemahlen und homogenisiert.
3.2.7 Analytik und angewandte Untersuchungsparameter
Lediglich die „nach Farbwechsel“ (n. F.) gewonnenen Chymusproben wurden analysiert,
so dass pro Versuchstag und Tier jeweils vier Proben der Analyse zugeführt wurden. Von
den zuvor gefriergetrockneten und gemahlenen Chymusproben wurden der
Trockensubstanz- (TS)-, Rohfett- (Rfe)- und Chromoxid- (Cr2O3)-Gehalt ermittelt. In der
verwendeten Diät wurde zusätzlich der Gehalt an Rohfaser (Rfa), Rohasche (Ra), Zucker
und Stärke bestimmt. Die Ermittlung der N-freien Extraktstoffe (NfE) und organischen
Substanz (oS) erfolgte rechnerisch.
Trockensubstanz (TS):
Die Analyse der Trockensubstanz erfolgte mittels Weender Futtermittelanalyse nach den
Vorschriften der VDLUFA in der Fassung von NAUMANN und BASSELER (2004).
Nach Ermittlung der ursprünglichen Substanz und Zwischenlagerung bei -80°C erfolgte die
Trocknung der Proben in einer Vakuumgefriertrocknungsanlage (alpha 1-4, Firma Christ
und Lyovac GT 2, Firma Finn-Aqua) bis zur Gewichtskonstanz. Nach erneuter
Bestimmung des Gewichtes und Homogenisierung folgte die Entnahme einer Teilprobe,
welche im Trockenschrank bei 103°C mindestens 8 Stunden bis zur Gewichtskonstanz
nachgetrocknet wurde. Über Gewichtsdifferenz wurde der TS-Gehalt ermittelt.

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
47
Rohasche (Ra):
Der Gehalt an Rohasche wurde nach einer zwölfstündigen Veraschung der Probe bei
550°C im Muffelofen ermittelt. Als Rohasche wird dabei der Anteil der Probe bezeichnet,
der nach der Veraschung im Veraschungstiegel verbleibt.
Organische Substanz (oS):
Der Gehalt an organischer Substanz wurde rechnerisch mit Hilfe folgender Formel
bestimmt:
oS = TS – Ra
Rohprotein (Rp):
Die Bestimmung des Rohprotein-Gehaltes erfolgte anhand des Gesamtstickstoff-Gehaltes
mit dem Vario Max® der Firma Elementar (Hanau). Das Gerät arbeitet nach dem Prinzip
der katalytischen Rohrverbrennung der Probe unter Sauerstoffzufuhr und hohen
Temperaturen.
Das hierdurch entstehende N2 wird mit Hilfe eines Wärmeleitfähigkeitsdetektors bestimmt.
Rohfaser (Rfa):
Nach 30-minütiger Vorentfettung wurden die Proben mit jeweils 1,25 %iger Schwefelsäure
und 1,25 % Natronlauge für je eine halbe Stunde in einem Rohfaserbestimmungsgerät
(Fibertec System M1020 Hot Extraktor®, Firma Tecator) gekocht. Nachdem der Rückstand
gewaschen, getrocknet und gewogen worden war, erfolgte eine Veraschung im Muffelofen
bei 550°C. Die dabei auftretenden Massenverluste entsprechen dem Rohfasergehalt der
Probe.
N-freie Extraktstoffe (NfE):
Dieser Parameter wurde rein rechnerisch nach folgender Formel bestimmt:
NfE= TS- (Ra+ Rp+ Rfe+ Rfa)

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
48
Stärke:
Die Ermittlung des Stärkegehaltes basierte auf einer doppelten Bestimmung nach der
amtlichen Methode des VDLUFA (NAUMANN u. BASSLER 2004).
Bestimmung des Hauptwertes: Säurehydrolyse durch Kochen der Probe mit verdünnter
Salzsäure. Nach Abkühlung der Probe folgte Klärung, Filtrierung und polarimetrische
Messung der Lösung. Bestimmung des Blindwertes: Extraktion der Probe mit 40 %igem
Ethanol. Nach Filtration, Säurehydrolyse und anschließende Klärung mit abermaliger
Filtration wurde die optische Drehung der Probenlösung analog zum Hauptwert ermittelt.
Die Multiplikation der Differenz beider Werte mit einem bekannten Faktor für Stärkegehalte
in Mischfuttermitteln von 10,87 ergab den Stärkegehalt der Probe.
Zucker:
Die Methode von LUFF-SCHORL nach Vorschriften der VDLUFA (2004) wurde zur
Bestimmung des Zuckergehaltes verwendet. Die Reduktion des Kupfer(II)-salzes erfolgte
mittels reduzierten Zuckers zu Kupfer(I)-oxid. Durch jodometrische Titration des
unverbrauchten Kupfers konnte auf den Zuckergehalt der Probe geschlossen werden.
Chromoxid:
Die Cr2O3-Gehalte wurden mit Hilfe der Methode nach PETRY und RAPP (1970)
bestimmt. Im Laufe einer Nassveraschung erfolgte die Überführung von Chromoxid
anhand eines Oxidationsgemisches aus Schwefelsäure, Perchlorsäure und
Natriummolybdat in Chromat. Die Messung der Extinktion gegen einen Blindwert wurde im
Photometer bei 365 nm durchgeführt, nachdem die Probe durch Natronlauge alkalisiert
worden war. Der Cr2O3-Gehalt der Probe wurde mit Hilfe eines durch eine Eichreihe
ermittelten Extinktionskoeffizienten errechnet.
Rohfett (Rfe):
Zu Beginn der Entwicklung einer Testmahlzeit im Rahmen dieser Arbeit erfolgte die
Validierung eines neuen Analysengerätes namens AnkomHCl Hydrolysis System® der
Firma Ankom Technology Corporation zur Bestimmung des Rohfettgehaltes.

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
49
Die analytische Genauigkeit der Ergebnisse betrug laut Angaben der Firma ± 0,1 mg. Die
Analyse erfolgte dabei in zwei Schritten:
1. Hydrolyse:
Im ersten Schritt wurden unter Verwendung der Filterbag-Säurehydrolyse-Technik® (Filter
Bag Technique Acid Hydrolysis) die löslichen Nichtfette aus der Probe heraus gelöst. Die
zu lösende Rfe-Fraktion bestand vornehmlich aus Triglyceriden.
Zunächst wurden 0,3 - 0,6 g (je nach vorhandener Probenmenge) Probenmaterial in
säureresistente Filterbags mit einer Porengröße von 1 µm eingewogen, die zwar der
Salzsäurelösung den Durchtritt gewähren, nicht aber das Probenmaterial austreten lassen.
Der Einwaage wurden ca. 0,25 g einer Filtrierhilfe aus Kieselgur (Hyflo Super Cel®
medium der Firma Fluka Chemika) zugegeben. Nach Verschließen der Filterbags mit Hilfe
des so genannten Heatsealers oder Filterbag-Schweißgerätes wurden bis zu 20 Filterbags
in einem Probenständer in der Kammer des Hydrolysegerätes gleichmäßig verteilt.
Grundsätzlich erfolgte eine Doppelbestimmung der Proben. Daraufhin wurden 500 ml 3N
HCl (= 12,7 %ige Salzsäurelösung) in die Kammer gegeben und der Deckel verschlossen.
Der Vorgang wurde auf eine Hydrolysezeit von 40 Minuten und 6 Spülgänge mit
Leitungswasser programmiert. Nach ca. 1,5 Stunden erfolgte die Probenentnahme und die
Filterbags wurden nochmals unter laufendem destilliertem Wasser nachgespült.
Anschließend wurden die Filterbags einzeln auf zuvor ausgewogene Uhrgläser verteilt und
in einen 80°C heißen Trockenschrank überführt. Nach mindestens 6 Stunden folgte ihre
Entnahme und Abkühlung in Exsiccatoren.
2. Extraktion:
Die Extraktion der Rohfette erfolgte in diesem zweiten Analysenschritt. In dem hierfür
benutzten Gerät AnkomXT15Extraktor® der Firma Ankom wurde Petrolether zum Lösen der
fetthaltigen Bestandteile verwendet.
Maximal 15 der nach Punkt 1. bearbeiteten Filterbags wurden in dem Extraktionskessel
platziert. Dieser bestand aus einem verschließbaren und druckstabilen Kammersystem.
Nach 50-minütiger Extrationszeit mit Petrolether wurden die Proben gespült und das
heraus gelöste Extraktionsgemisch automatisch aus dem Kessel abgesaugt.

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
50
Nach Beendigung des Vorganges wurden die Proben wiederum bei 80°C im
Trockenschrank für drei Stunden getrocknet und nach Abkühlung im Exsiccator gewogen.
Durch Differenzbildung zwischen zweitem und dritten Wiegen und Division durch das
eingewogene Anfangsgewicht errechnete sich der prozentuale Rohfett-Anteil der Probe.
Validierung des alternativen Verfahrens zur Rohfettbestimmung (AnkomHCl Hydrolysis Systems®)
Zur Analyse des Rohfettgehaltes in den Chymus- und Futtermittelproben wurde eine
andere Methode als in den vorangegangenen Dissertationen des Projektes (Verwendung
der Soxhlet-Apparatur nach Vorschriften der Weender-Analyse) benutzt. Zur Überprüfung
der Anwendbarkeit bzw. Eignung der neuen Methode sowie zur Bestimmung der
Vergleichbarkeit der Ergebnisse der beiden Verfahren erfolgten zu Beginn der Versuche
zahlreiche parallele Bestimmungen unter Verwendung beider Methoden.
In diesem Abschnitt werden die Ergebnisse der zwei verschiedene Verfahren zur
Bestimmung des Rfe-Gehaltes gegenübergestellt: Die Chymus- und Futterproben wurden
sowohl mit der erstmalig in diesem Projekt verwendeten Ankom-Apparatur, als auch mit
der herkömmlich benutzten Methode der Rohfettbestimmung unter Verwendung der
Soxhlet-Apparatur untersucht und deren Ergebnisse verglichen. Um die
Reproduzierbarkeit der Ergebnisse der neuen Methode nachzuweisen, werden
Mehrfachwiederholungen einzelner Proben ausgewertet.
Anfangs wurden 22 Proben, parallel sowohl mit der etablierten Soxhlet-Methode als auch
mit der neuen Methoden des Hydrolysator- und Extraktionsgerätes von Ankom® (=
% Rfe = 100 x ((Gew. des Beutels nach Hydrolyse – Gew. des Uhrglases) – Gew. des Beutels nach Extraktion)
Einwaagegewicht

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
51
AnkomHCl Hydrolysis System® (Filterbag-Säurehydrolyse-Technik® und AnkomXT15
Extraktor®)) analysiert, wobei stets eine Doppelbestimmung erfolgte.
Es muss hier daraufhin gewiesen werden, dass zum Zeitpunkt dieser Validierung noch
eine geringfügig andere Geräteeinstellung benutzt wurde (Hydrolyse: mit 550 ml HCl für
40 min, 2x spülen mit H2O / Extraktion: 40 min). Während der Versuche wurden hingegen
einige Modifikationen hinzugefügt: es wurden lediglich 500 ml HCl benutzt und die Anzahl
der Spülgänge wurde auf sechs im Gerät und einer weiteren manuellen Spülung mit
destilliertem H2O angehoben. Des Weiteren wurde die Extraktionszeit von 40 auf 50
Minuten erhöht. Diese Veränderungen fanden statt, da sich in den Proben nach
Bearbeitung noch Reste von HCl fanden. Durch diese Modifikationen verbesserte sich die
Übereinstimmung der Doppelwerte deutlich bzw. die Abweichungen bei
Doppelbestimmungen wurden deutlich reduziert.
Vergleich der Methoden: Der Vergleich der Mittelwerte der Doppelwerte zwischen herkömmlicher (Soxhlet) und
neuer Methode (Ankom-Technologie) zeigte, dass die Ergebnisse des Ankom-Gerätes
nicht exakt denen der Soxhlet-Methode entsprechen.
Die mit der Ankom-Apparatur ermittelten Ergebnisse ergaben Werte, die zwischen 1-29 %,
im Mittel aber 10 %, höher lagen als die mit dem Soxhlet-Verfahren gewonnenen
Vergleichswerte. Es ließ sich also eine gerichtete Abweichung der Rfe-Ergebnisse der
„Ankom-Methode“ gegenüber der Werte der „Weender-Methode“ (Soxhlet) ablesen (s. Tabelle 13).
Die Ermittlung der relative Abweichung der Einzelwerte voneinander nach der jeweiligen
Methode betrachtet, ergab vergleichbare Abweichung vom Mittelwert: Bei Bestimmung
des Rohfett-Gehaltes nach der Weender-Methode (Soxhlet) wurden 3,68 % relative
Abweichung vom Mittelwert berechnet, während nach Verwendung der Ankom-Geräte
eine Wert von 3,86 % kalkuliert wurde (s. Tabelle 11).
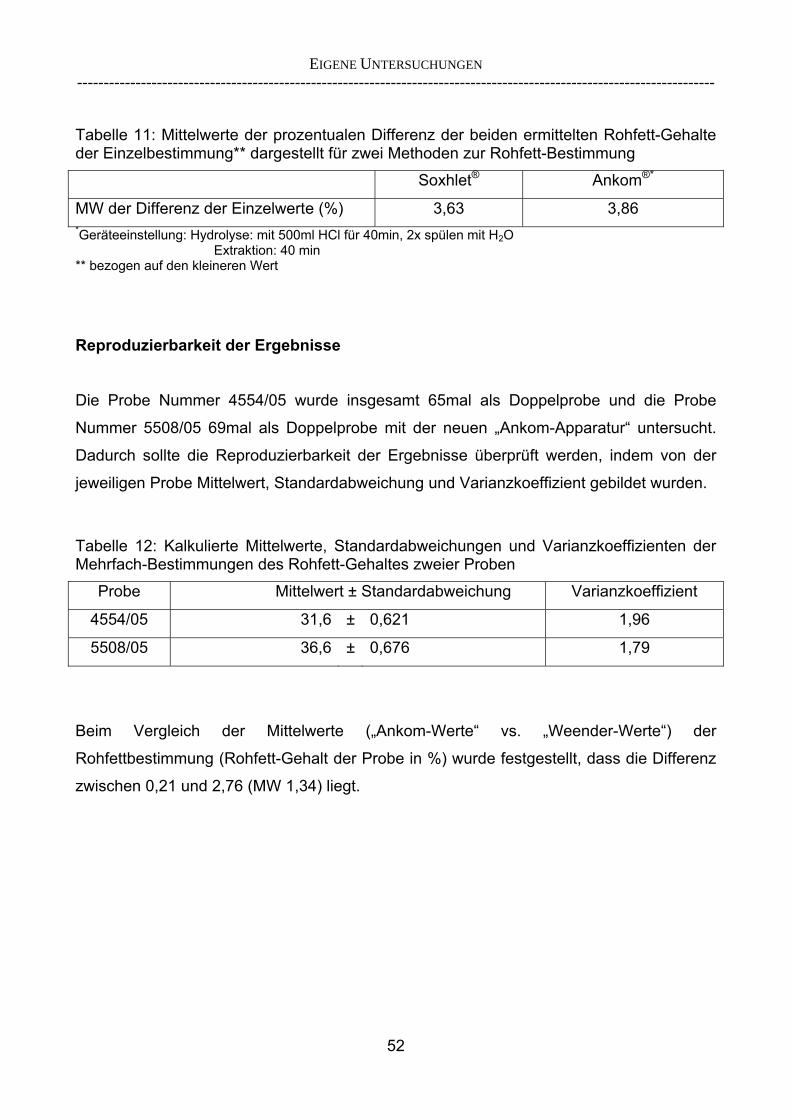
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
52
Tabelle 11: Mittelwerte der prozentualen Differenz der beiden ermittelten Rohfett-Gehalte der Einzelbestimmung** dargestellt für zwei Methoden zur Rohfett-Bestimmung
Soxhlet® Ankom®*
MW der Differenz der Einzelwerte (%) 3,63 3,86 *Geräteeinstellung: Hydrolyse: mit 500ml HCl für 40min, 2x spülen mit H2O Extraktion: 40 min ** bezogen auf den kleineren Wert
Reproduzierbarkeit der Ergebnisse
Die Probe Nummer 4554/05 wurde insgesamt 65mal als Doppelprobe und die Probe
Nummer 5508/05 69mal als Doppelprobe mit der neuen „Ankom-Apparatur“ untersucht.
Dadurch sollte die Reproduzierbarkeit der Ergebnisse überprüft werden, indem von der
jeweiligen Probe Mittelwert, Standardabweichung und Varianzkoeffizient gebildet wurden.
Tabelle 12: Kalkulierte Mittelwerte, Standardabweichungen und Varianzkoeffizienten der Mehrfach-Bestimmungen des Rohfett-Gehaltes zweier Proben
Probe Mittelwert ± Standardabweichung Varianzkoeffizient
4554/05 31,6 ± 0,621 1,96
5508/05 36,6 ± 0,676 1,79
Beim Vergleich der Mittelwerte („Ankom-Werte“ vs. „Weender-Werte“) der
Rohfettbestimmung (Rohfett-Gehalt der Probe in %) wurde festgestellt, dass die Differenz
zwischen 0,21 und 2,76 (MW 1,34) liegt.
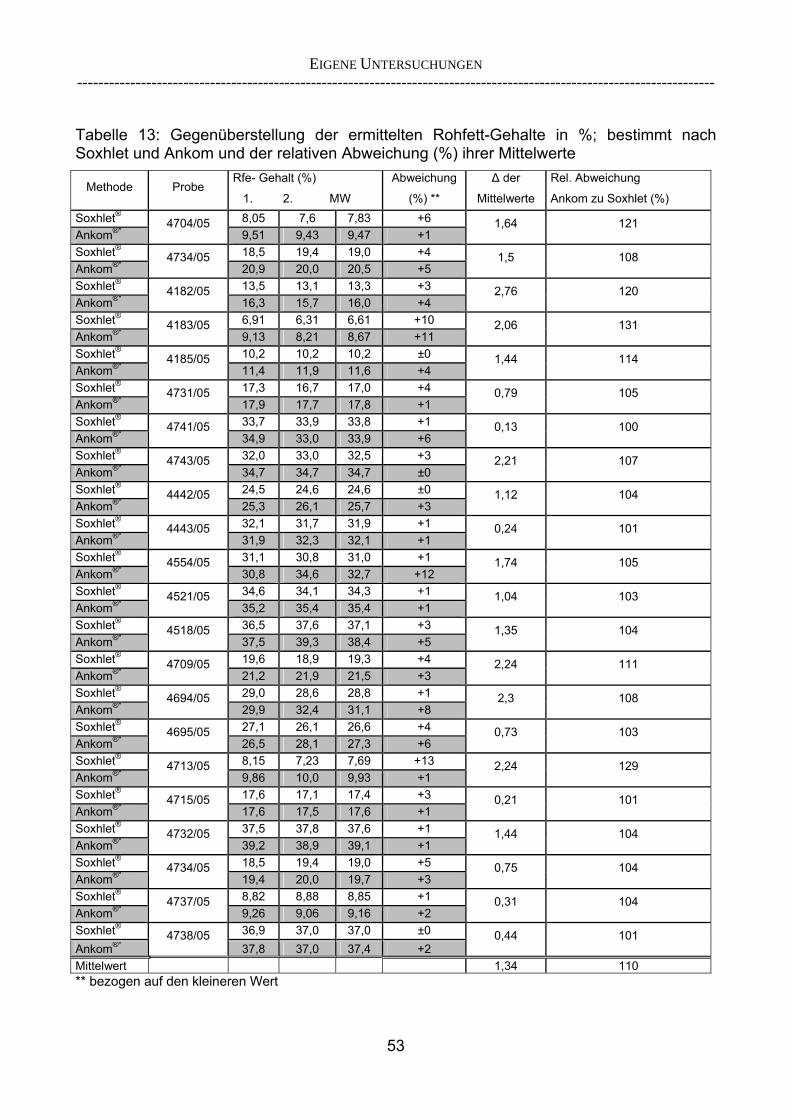
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
53
Tabelle 13: Gegenüberstellung der ermittelten Rohfett-Gehalte in %; bestimmt nach Soxhlet und Ankom und der relativen Abweichung (%) ihrer Mittelwerte
Methode Probe Rfe- Gehalt (%)
1. 2. MW
Abweichung
(%) **
∆ der
Mittelwerte
Rel. Abweichung
Ankom zu Soxhlet (%) Soxhlet® 8,05 7,6 7,83 +6 Ankom®*
4704/05 9,51 9,43 9,47 +1
1,64 121
Soxhlet® 18,5 19,4 19,0 +4 Ankom®*
4734/05 20,9 20,0 20,5 +5
1,5 108
Soxhlet® 13,5 13,1 13,3 +3 Ankom®*
4182/05 16,3 15,7 16,0 +4
2,76 120
Soxhlet® 6,91 6,31 6,61 +10 Ankom®*
4183/05 9,13 8,21 8,67 +11
2,06 131
Soxhlet® 10,2 10,2 10,2 ±0 Ankom®*
4185/05 11,4 11,9 11,6 +4
1,44 114
Soxhlet® 17,3 16,7 17,0 +4 Ankom®*
4731/05 17,9 17,7 17,8 +1
0,79 105
Soxhlet® 33,7 33,9 33,8 +1 Ankom®*
4741/05 34,9 33,0 33,9 +6
0,13 100
Soxhlet® 32,0 33,0 32,5 +3 Ankom®*
4743/05 34,7 34,7 34,7 ±0
2,21 107
Soxhlet® 24,5 24,6 24,6 ±0 Ankom®*
4442/05 25,3 26,1 25,7 +3
1,12 104
Soxhlet® 32,1 31,7 31,9 +1 Ankom®*
4443/05 31,9 32,3 32,1 +1
0,24 101
Soxhlet® 31,1 30,8 31,0 +1 Ankom®*
4554/05 30,8 34,6 32,7 +12
1,74 105
Soxhlet® 34,6 34,1 34,3 +1 Ankom®*
4521/05 35,2 35,4 35,4 +1
1,04 103
Soxhlet® 36,5 37,6 37,1 +3 Ankom®*
4518/05 37,5 39,3 38,4 +5
1,35 104
Soxhlet® 19,6 18,9 19,3 +4 Ankom®*
4709/05 21,2 21,9 21,5 +3
2,24 111
Soxhlet® 29,0 28,6 28,8 +1 Ankom®*
4694/05 29,9 32,4 31,1 +8
2,3 108
Soxhlet® 27,1 26,1 26,6 +4 Ankom®*
4695/05 26,5 28,1 27,3 +6
0,73 103
Soxhlet® 8,15 7,23 7,69 +13 Ankom®*
4713/05 9,86 10,0 9,93 +1
2,24 129
Soxhlet® 17,6 17,1 17,4 +3 Ankom®*
4715/05 17,6 17,5 17,6 +1
0,21 101
Soxhlet® 37,5 37,8 37,6 +1 Ankom®*
4732/05 39,2 38,9 39,1 +1
1,44 104
Soxhlet® 18,5 19,4 19,0 +5 Ankom®*
4734/05 19,4 20,0 19,7 +3
0,75 104
Soxhlet® 8,82 8,88 8,85 +1 Ankom®*
4737/05 9,26 9,06 9,16 +2
0,31 104
Soxhlet® 36,9 37,0 37,0 ±0 Ankom®*
4738/05 37,8 37,0 37,4 +2
0,44 101
Mittelwert 1,34 110 ** bezogen auf den kleineren Wert

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
54
3.2.8 Verwendete Berechnungsansätze zur Einschätzung der praecaecalen Fettverdaulichkeit
Im Folgenden werden drei unterschiedliche Berechnungsansätze zur Ermittlung der
Fettverdaulichkeit sowie ein Ansatz für die Chromoxid-Wiederfindung angegeben.
1. Anflutung:
1a. Trockensubstanz (TS)- und Rohfett (Rfe)-Anflutung absolut (g):
In dem 8-stündigen Kollektionszeitraum wurden in zweistündigen Intervallen sowohl die
angeflutete Trockensubstanz- als auch Rohfettmenge in Gramm ermittelt.
Die Multiplikation von Chymusmenge (TS) und des Rohfettgehaltes (g / kg TS) ergibt die
tatsächlich angeflutete Fettmenge:
Rfe-Anflutung (g) = Rfe im Chymus (g/kg) x Chymusmenge (g)/1000
1b. Flussrate relativ:
Bildung des prozentualen Verhältnisses aus Einzelintervall zu Gesamtintervall des
Kollektionszeitraumes für Rfe.
Die Bestimmung der relativen Flussrate für TS und Cr2O3 erfolgte analog.
Chymusmenge (uS) (g) x TS-Gehalt im Chymus (g/kg)
100TS-Anflutung im Chymus (g) =
Rfe rel. Fluss = Rfe (g) Anflutung im jeweiligen Intervall (2 h)
Rfe (g) Anflutung im Gesamtintervall 0-8 h n.F.
( X 100)

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
55
1c. rel. Anflutung:
Bildung der Relation der absoluten Rohfettanflutung bei PL-0-Tieren mit und ohne
Enzymsubstitution
2. Konzentrationen: Im folgenden Abschnitt werden die zur Berechnung der Konzentrationen von Rohfett und
Chromoxid (Cr2O3) im Chymus genutzten Formeln dargestellt:
2a. – Fettkonzentration
2b. Relation von Rohfett-Konzentration zu Cr2O3-Konzentration
(das aufgenommene Futter enthielt generell eine Rfe- zu Cr2O3-Konzentration Relation
von 140)
Rfe Anflutungsrelation = Rfe Anflutung (g) nach Enzymverabreichung
Rfe Anflutung (g) ohne Enzymverabreichung( ) X 100
Cr2O3- Konzentration (%) = ( Cr2O3 im Chymus (g)
Chymusmenge (g)) X 1000
Relation Rfe-Konz. / Cr2O3-Konz. (%) = Rfe-Konzentration (%)
Cr2O3- Konzentration (%)) X 1000(
Rfe-Konzentration (%) (% der TS)
(Rfe im Chymus (g)
Chymusmenge (g)) X 1000=

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
56
3. Indikatormethode: Diese Berechnung dient der Ermittlung der scheinbaren praecaecalen Verdaulichkeit.
Obwohl diese Methode die Grundlage der Berechnung der Verdaulichkeiten in dieser
Arbeit darstellt, wird im Folgenden von Verschwindensrate gesprochen werden, da die
Bedingungen eines herkömmlichen Verdaulichkeitsversuches nicht erfüllt wurden (s.
vorher).
Chromoxid-Wiederfindung:
1) im Zeitraum 0-8h
sV (%) = 100 - ( % Indikator im Futter
% Indikator im Chymusx
% Nährstoff im Chymus
% Nährstoff im Futter) X 100
Cr2O3-Wiederfindung (%) =Cr2O3- Anflutung im Chymus (g) 1)
Cr2O3 im Futter (g) / 147 g TSX 100

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
57
3.2.9 Entwicklung eines In-vivo-Screening-Tests zur Einschätzung der Wirksamkeit von Lipasen anhand eines bekannten Multienzym-produktes
Fragestellung: Welche Anforderungen werden an eine Diät gestellt, die für einen Lipase-Screening-Test geeignet ist? Der von BECKER (2005) für Proteasen und Amylasen entwickelte Schnelltest sollte, wenn
möglich, auch für die Testung von Lipasen zum Einsatz kommen. Zu diesem Zwecke war
es zum einen notwendig, das Verhältnis von Substrat (Fett) und Enzym (Lipase) in der
Testmahlzeit so auszubalancieren, dass die Substratverfügbarkeit nicht den limitierenden
Faktor darstellte und zum anderen dosisabhängige Effekte erkennbar waren.
Bei Verabreichung fettreicher Mahlzeiten ohne Anfütterungsphase waren die
nachstehenden Probleme zu befürchten:
• Veränderungen bzw. Störungen der Magen-Darm-Motorik und
Passagebeeinträchtigungen durch mangelnde Adaptation infolge des Verzichts
auf eine Anfütterungsphase
• Nicht parallele Anflutung der Nährstoffe und des Markers an der Fistel durch
Entmischungsprozesse (Entmischung des Markers von der Testmahlzeit sowie
Entmischungen der einzelnen Komponenten der Testmahlzeit)
Es stellte sich heraus, dass die Verwendung der von BECKER (2005) eingesetzten
Testmahlzeit, eine Modifikation auf der Basis einer herkömmlichen gebräuchlichen
Mahlzeit (BECKER 2005), wegen der Besonderheiten des Lipase-Screenings nicht zum
Erfolg führte. Es war also notwendig, eine gänzlich neue Testmahlzeit zu entwickeln,
um die vorgenannten Probleme zu minimieren bzw. zu beseitigen.

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
58
3.2.10 Anforderungen an eine Testmahlzeit für ein Lipase-Screening
• paralleler Fluss von Trockensubstanz, Rohfett, Enzym und Marker
(= keine Entmischung)
• physiologische Verhältnisse bezüglich Magenentleerung und Passageraten
(physiologische Verweildauer der Nahrung inklusive der Enzyme in den
verschiedenen Abschnitten des GIT; damit „realistische“ Bedingungen für
Enzym)
• uneingeschränkte Substratverfügbarkeit durch hohen Fettgehalt der
Testmahlzeit
• Fettqualität
• Orientierung an Humanernährung (z.B. Einsatz von Milchfett) als Fettquelle
• Anpassung an In-vitro-Untersuchungen (z.B. Einsatz von Olivenöl) als Fettquelle
(Anforderung ergab sich aus der Verwendung von Olivenöl (in-vitro) und dem
Ziel der Verbesserung der Übereinstimmung der Ergebnisse der In-vivo und In-
vitro-Tests)
• erhöhter Anteil langkettiger Fettsäuren (hoher Olivenölanteil)
Als Basisdiät wurde eine pulverförmige, in Milch zu lösende Mahlzeit mit dem Charakter
eines Milchshakes, der so genannte „Calshake®“ der Firma Fresenius, Bad Homburg,
ausgewählt, der einen hohen Fettgehalt aufwies. Dieser hochkalorische und
leichtverdauliche Milchshake findet als Nahrungsergänzungsmittel Anwendung bei
Personen, die von Malabsorption und Maldigestion (z.B. durch Mukoviszidose, Tumor-
Erkrankungen, Essstörungen etc.) betroffen sind. Bei Zubereitung nach Packungsbeilage
wird das Calshakepulver (g / kg Pulver: Rp 43; Rfe 244; KH 649; Rfa 0; 20 770 kJ / kg)
eines Beutels (= 87 g) mit 240 ml Milch (mit einem Milchfettgehalt von 3,5 %) vermischt. Je
100 ml des auf diese Weise zubereiteten Calshakes enthalten: 3,8 g Rp; 9,6 Rfe g; 21,7 g
KH, 0 Rfa g sowie 788 kJ.
Im Laufe der Entwicklungsphase der Testmahlzeit wurde die Notwendigkeit einiger
Modifikationen bzw. weiterer Additive deutlich, die im Folgenden vorgestellt werden:
Die Hydrolyse und Resorption kurz- und mittelkettiger Fettsäuren erfolgt zum Teil auch
ohne pankreatische Verdauungsenzyme (z.B. Spaltung durch gastrische Lipasen). Um

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
59
insbesondere das lipolytische Vermögen der pankreatischen Lipasen zu beurteilen,
wurden deshalb vermehrt langkettige Fettsäuren eingesetzt, zumal diese einen
bedeutenden Anteil der humanen Fettaufnahme bilden.
Außerdem sollte die Erhöhung des Fettgehaltes der Diät mittels einer in In-vitro-Versuchen
genutzten Fettquelle stattfinden, um die Vergleichbarkeit von Screening-Ergebnissen mit
jenen der In-vitro-Versuche zu verbessern. Diese vorgenannten Gründe führten zu der
Entscheidung, auch Olivenöl als Fettquelle zu nutzen. Durch den Zusatz von Olivenöl
wurde der Fettanteil in der Testmahlzeit auf ca. 35% der TS angehoben, wodurch der
Fettgehalt mit jenem der in früheren Arbeiten des Projektes verwendeten fettreichen
Ration (TABELING 1998) vergleichbar war. Aufgrund der flüssigen Zubereitung bzw. der
Applikation als Emulsion wurde trotz vergleichbarem Fettgehalt eine Verringerung der
oben genannten Probleme (insbesondere der Entmischungsprozesse) erwartet.
Der in der Testmahlzeit eingesetzte Marker Chromoxid (Firma Aldrich®, Steinheim)
erfüllte analog zu dem von BECKER entwickelten Screening-Verfahren zweierlei
Aufgaben:
Optische Kontrolle der Anflutung: Nach BECKER (2005) flutete in den ersten Stunden
nach der morgendlichen Fütterung der Versuchsmahlzeit „brauner“ Chymus aus dem
Haltungsfutter der vorabendlichen Mahlzeit an der Fistel an. Durch die Markierung der
Versuchsmahlzeit mit nichtabsorbierbarem grünem Cr2O3 konnte der aus der Testmahlzeit
stammende Chymus von jenem, der aus dem Futter vom Vorabend bestand, abgegrenzt
werden. Demzufolge war es möglich, die auffällige grüne Farbe des Chymus zu nutzen,
um den Zeitpunkt der Ankunft des Versuchsfutters zu bestimmen und somit den
Chymus des Haltungsfutters von dem des Versuchsfutters zu unterscheiden. Dieser im
Folgenden als „Farbwechsel“ bezeichnete Wechsel der Farbe von „braunem“ zu „grünem“
Chymus wurde also als „Startsignal“ für den Beginn der Chymuskollektion genutzt.
Markerkorrigierte Berechnung der Verschwindensrate (= Indikatormethode) zur Kalkulation
der im jeweiligen Kollektionszeitraum resorbierten Nährstoffmenge mit Hilfe des Markers.
Infolge einer parallel verlaufenden relativen Flussrate von Nährstoffen und Marker kann
am Ileumende die „Verschwindensrate“ von Trockensubstanz und Rohfett berechnet
werden. Die Berechnung der „Verschwindensrate“ bietet den Vorteil, dass anhand der

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
60
markerkorrigierten Kalkulation differenziert werden kann, ob beispielsweise eine geringe
Chymus- bzw. Nährstoffanflutung aus einer hohen Verdaulichkeit oder aber aus einer
geringen Flussrate resultiert.
Im Verlauf der Entwicklung einer geeigneten Testmahlzeit erschien es außerdem
zweckmäßig, eine Ballaststoffquelle (= Methocel®: Methylcellulose (ca. 30 %
Methoxylgruppen und 70 % Cellulose)) hinzu zu fügen. Zum einen führte der Einsatz einer
Rohfaser-Quelle zur Erhöhung der Chymusmenge, da keine beachtenswerte Verdauung
von Rfa im praecaecalen Bereich beim Schwein stattfindet und somit ein Füllungseffekt
des GIT auftrat (und damit indirekt über Dehnungsreize die Passagerate beeinflusst
wurde). Zum anderen erhöhte sich die Chymusanflutung an der ilealen Fistel und damit
die Menge des für Analysen zur Verfügung stehenden Materials. Des Weiteren sollte mit
Zulage des Methocels (= partikuläre Struktur) der Entmischung von Marker, Enzym und
Testmahlzeit entgegen gewirkt werden. Zudem bestand die Annahme, dass die Erhöhung
der TS-Aufnahme zu einem ausgeprägterem Sättigungsgefühl der Tiere betragen würde.
Zusätzlich sollte geprüft werden, ob der Zusatz von Rohfaser („Ballaststoffen“) einen
Einfluss auf die Magenentleerung im Sinne einer Förderung der Magenpassage hat.
Auf die anfangs eingesetzte Fettquelle Mikrolipid (Rohfett-Gehalt: 50 %; emulgierte Fette,
die zur Sondenernährung in der Humanmedizin eingesetzt werden) musste im weiteren
Verlauf der Studie wegen mangelnder Verfügbarkeit verzichtet werden.
Um eine stabile Emulsion zu erlangen wurde im Verlauf der Versuche der Mixer Ultra-turrax 900® anstelle eines Handrührgerätes eingesetzt, da dieser eine feinere Verteilung
der Bestandteile erzeugte. Die verbesserte Homogenität sollte sich stabilisierend auf die
Emulsion der Diät auswirken.
Nach Testung verschiedener Varianten des Calshakes, insbesondere in Bezug auf
Zusammensetzung und Zubereitung, wurde schließlich eine Erfolg versprechende
Rezeptur entwickelt (s. Tabelle 14, 15). Die Testung der Mahlzeit erfolgte ebenso wie die
Auswahl der drei zu testenden Dosierungen, um dosisabhängige Wirkungen für die

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
61
Testung unbekannter Lipasen festzulegen, anhand des gut untersuchten
Multienzympräparates PK (Pankreon®).
Tabelle 14: Menge der Rohnährstoffe und Cr2O3 (g) in der zubereiteten Testmahlzeit
TS Ra Rp Rfe Rfa NfE Cr2O3
147 33,5 11,3 51,6 6,80 73,8 0,368
Tabelle 15: Zubereitungsrezept der Testmahlzeit (modifizierter Calshake®)
Einwaage der Einzelkomponenten und Homogenisierung mittels Ultra-turrax 900® (9500
U / min.): 247,2 g H-Milch (1,5 % Fettanteil) und 0,368 g Chromoxid für 1 Minute
Zugabe von 29,9 g Olivenöl und nochmaligen Homogenisieren mittels Ultra-turrax 900®
für zwei Minuten
Zugabe von einem Beutel Calshake (87 g) und Durchmischung für zwei Minuten mit
Handmixer
Zugabe von 9,88 g Methocel; langsam dazugeben während ca. dreiminütigen Rührens
mit dem Ultra-turrax 900®, bis eine klumpenfreie Emulsion entsteht.
3.2.11 Entwicklung einer geeigneten Testmahlzeit
Im Folgenden wird eine Zusammenstellung der Modifikationen und Bewertungen
maßgeblicher Komponenten der zu entwickelnden Testmahlzeit dargestellt. Ausgehend
von der Idee einer flüssigen Mahlzeit mit hohem Fettanteil (Calshake®) entwickelte sich die
oben genannte Versuchsdiät schrittweise über sieben Optimierungsstufen (flüssige
Testmahlzeit I-VII) zur endgültigen Versuchsmahlzeit:
Parallel zur Entwicklung der Testmahlzeit erfolgten erste Versuche zur Ermittlung
geeigneter Enzymdosierungen, wobei das bereits in zahlreichen In-vivo-Versuchen
(Bestimmung der praecaecalen Verdaulichkeit sowie der Gesamtverdaulichkeit)
eingesetzte Multienzymprodukt Pankreon genutzt wurde.
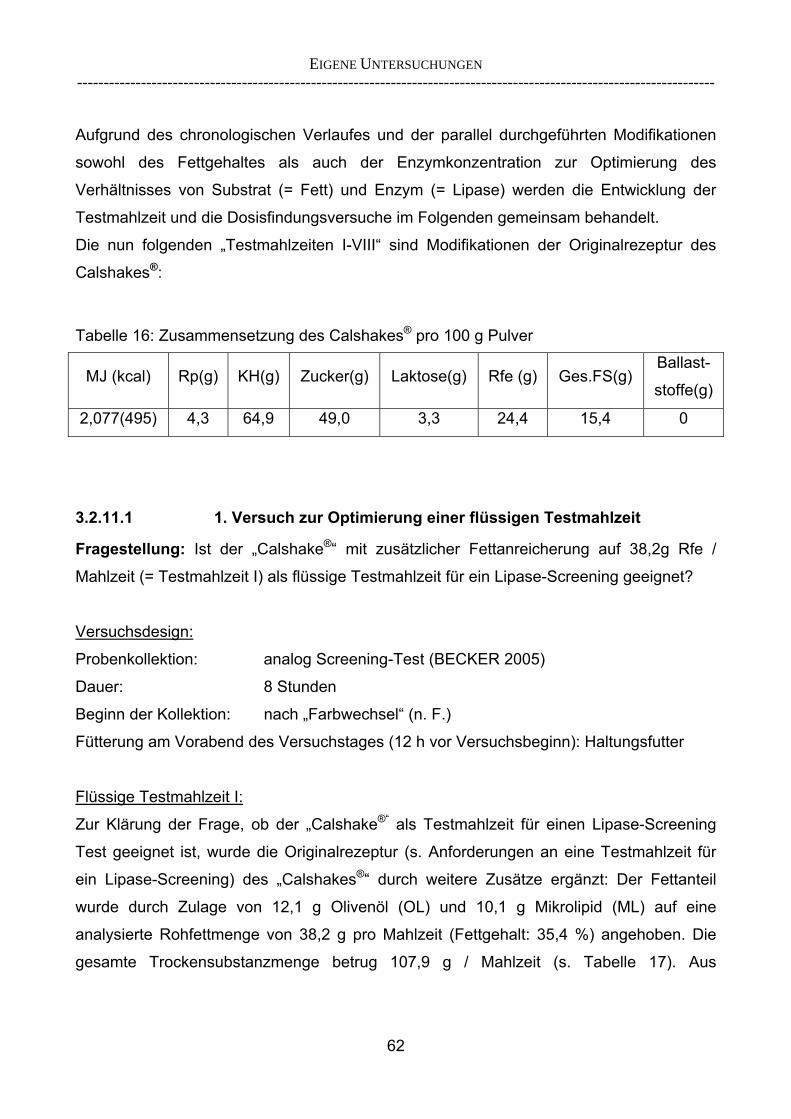
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
62
Aufgrund des chronologischen Verlaufes und der parallel durchgeführten Modifikationen
sowohl des Fettgehaltes als auch der Enzymkonzentration zur Optimierung des
Verhältnisses von Substrat (= Fett) und Enzym (= Lipase) werden die Entwicklung der
Testmahlzeit und die Dosisfindungsversuche im Folgenden gemeinsam behandelt.
Die nun folgenden „Testmahlzeiten I-VIII“ sind Modifikationen der Originalrezeptur des
Calshakes®:
Tabelle 16: Zusammensetzung des Calshakes® pro 100 g Pulver
MJ (kcal) Rp(g) KH(g) Zucker(g) Laktose(g) Rfe (g) Ges.FS(g) Ballast-
stoffe(g)
2,077(495) 4,3 64,9 49,0 3,3 24,4 15,4 0
3.2.11.1 1. Versuch zur Optimierung einer flüssigen Testmahlzeit
Fragestellung: Ist der „Calshake®“ mit zusätzlicher Fettanreicherung auf 38,2g Rfe /
Mahlzeit (= Testmahlzeit I) als flüssige Testmahlzeit für ein Lipase-Screening geeignet?
Versuchsdesign:
Probenkollektion: analog Screening-Test (BECKER 2005)
Dauer: 8 Stunden
Beginn der Kollektion: nach „Farbwechsel“ (n. F.)
Fütterung am Vorabend des Versuchstages (12 h vor Versuchsbeginn): Haltungsfutter
Flüssige Testmahlzeit I:
Zur Klärung der Frage, ob der „Calshake®“ als Testmahlzeit für einen Lipase-Screening
Test geeignet ist, wurde die Originalrezeptur (s. Anforderungen an eine Testmahlzeit für
ein Lipase-Screening) des „Calshakes®“ durch weitere Zusätze ergänzt: Der Fettanteil
wurde durch Zulage von 12,1 g Olivenöl (OL) und 10,1 g Mikrolipid (ML) auf eine
analysierte Rohfettmenge von 38,2 g pro Mahlzeit (Fettgehalt: 35,4 %) angehoben. Die
gesamte Trockensubstanzmenge betrug 107,9 g / Mahlzeit (s. Tabelle 17). Aus
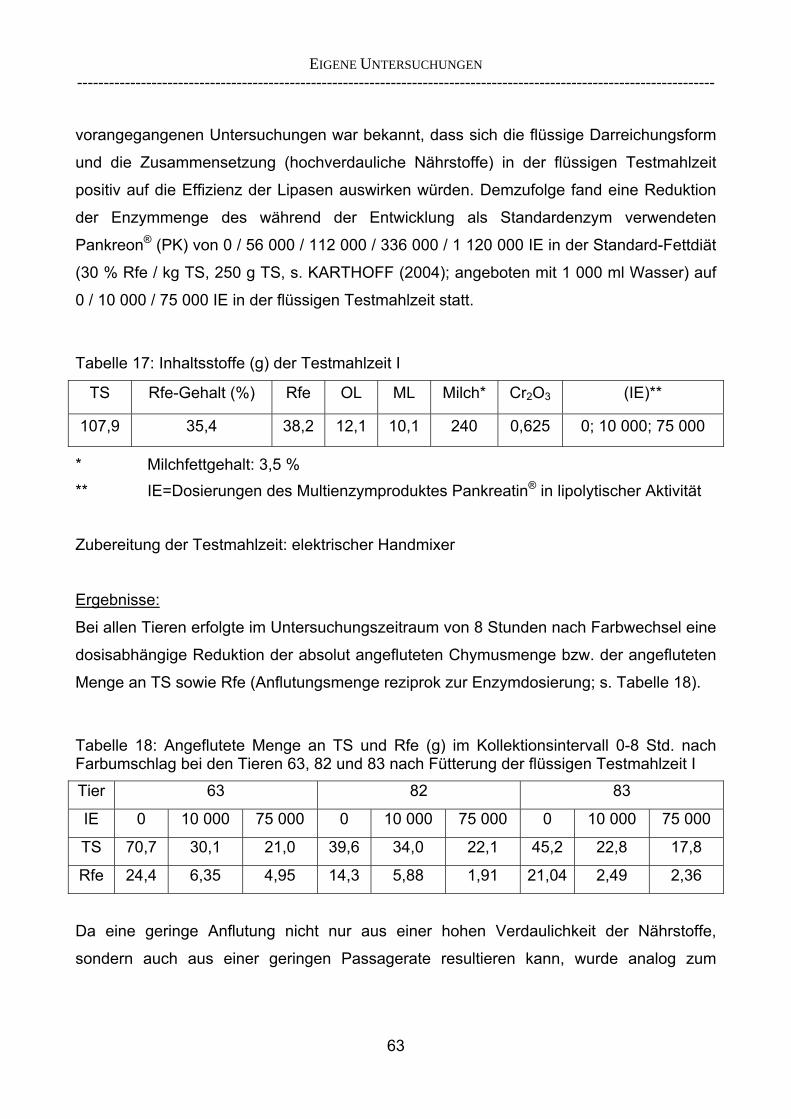
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
63
vorangegangenen Untersuchungen war bekannt, dass sich die flüssige Darreichungsform
und die Zusammensetzung (hochverdauliche Nährstoffe) in der flüssigen Testmahlzeit
positiv auf die Effizienz der Lipasen auswirken würden. Demzufolge fand eine Reduktion
der Enzymmenge des während der Entwicklung als Standardenzym verwendeten
Pankreon® (PK) von 0 / 56 000 / 112 000 / 336 000 / 1 120 000 IE in der Standard-Fettdiät
(30 % Rfe / kg TS, 250 g TS, s. KARTHOFF (2004); angeboten mit 1 000 ml Wasser) auf
0 / 10 000 / 75 000 IE in der flüssigen Testmahlzeit statt.
Tabelle 17: Inhaltsstoffe (g) der Testmahlzeit I
TS Rfe-Gehalt (%) Rfe OL ML Milch* Cr2O3 (IE)**
107,9 35,4 38,2 12,1 10,1 240 0,625 0; 10 000; 75 000
* Milchfettgehalt: 3,5 % ** IE=Dosierungen des Multienzymproduktes Pankreatin® in lipolytischer Aktivität
Zubereitung der Testmahlzeit: elektrischer Handmixer
Ergebnisse:
Bei allen Tieren erfolgte im Untersuchungszeitraum von 8 Stunden nach Farbwechsel eine
dosisabhängige Reduktion der absolut angefluteten Chymusmenge bzw. der angefluteten
Menge an TS sowie Rfe (Anflutungsmenge reziprok zur Enzymdosierung; s. Tabelle 18).
Tabelle 18: Angeflutete Menge an TS und Rfe (g) im Kollektionsintervall 0-8 Std. nach Farbumschlag bei den Tieren 63, 82 und 83 nach Fütterung der flüssigen Testmahlzeit I
Tier 63 82 83
IE 0 10 000 75 000 0 10 000 75 000 0 10 000 75 000
TS 70,7 30,1 21,0 39,6 34,0 22,1 45,2 22,8 17,8
Rfe 24,4 6,35 4,95 14,3 5,88 1,91 21,04 2,49 2,36
Da eine geringe Anflutung nicht nur aus einer hohen Verdaulichkeit der Nährstoffe,
sondern auch aus einer geringen Passagerate resultieren kann, wurde analog zum
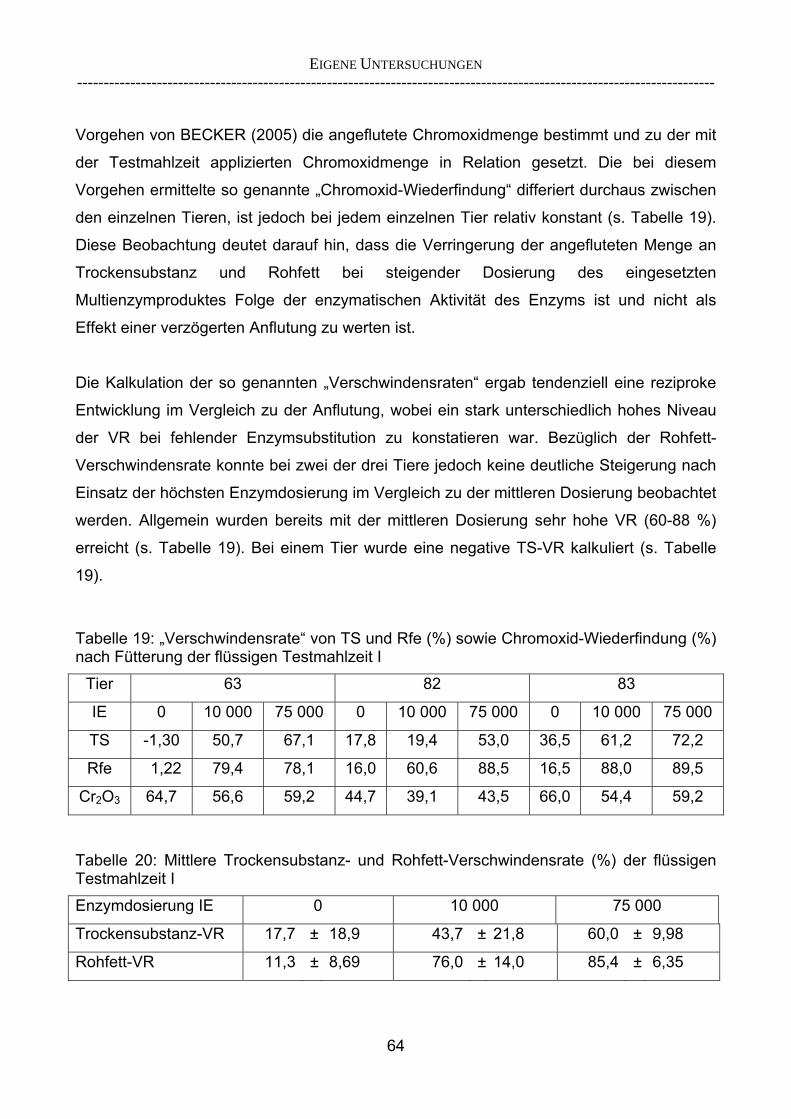
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
64
Vorgehen von BECKER (2005) die angeflutete Chromoxidmenge bestimmt und zu der mit
der Testmahlzeit applizierten Chromoxidmenge in Relation gesetzt. Die bei diesem
Vorgehen ermittelte so genannte „Chromoxid-Wiederfindung“ differiert durchaus zwischen
den einzelnen Tieren, ist jedoch bei jedem einzelnen Tier relativ konstant (s. Tabelle 19).
Diese Beobachtung deutet darauf hin, dass die Verringerung der angefluteten Menge an
Trockensubstanz und Rohfett bei steigender Dosierung des eingesetzten
Multienzymproduktes Folge der enzymatischen Aktivität des Enzyms ist und nicht als
Effekt einer verzögerten Anflutung zu werten ist.
Die Kalkulation der so genannten „Verschwindensraten“ ergab tendenziell eine reziproke
Entwicklung im Vergleich zu der Anflutung, wobei ein stark unterschiedlich hohes Niveau
der VR bei fehlender Enzymsubstitution zu konstatieren war. Bezüglich der Rohfett-
Verschwindensrate konnte bei zwei der drei Tiere jedoch keine deutliche Steigerung nach
Einsatz der höchsten Enzymdosierung im Vergleich zu der mittleren Dosierung beobachtet
werden. Allgemein wurden bereits mit der mittleren Dosierung sehr hohe VR (60-88 %)
erreicht (s. Tabelle 19). Bei einem Tier wurde eine negative TS-VR kalkuliert (s. Tabelle
19).
Tabelle 19: „Verschwindensrate“ von TS und Rfe (%) sowie Chromoxid-Wiederfindung (%) nach Fütterung der flüssigen Testmahlzeit I
Tier 63 82 83
IE 0 10 000 75 000 0 10 000 75 000 0 10 000 75 000
TS -1,30 50,7 67,1 17,8 19,4 53,0 36,5 61,2 72,2
Rfe 1,22 79,4 78,1 16,0 60,6 88,5 16,5 88,0 89,5
Cr2O3 64,7 56,6 59,2 44,7 39,1 43,5 66,0 54,4 59,2
Tabelle 20: Mittlere Trockensubstanz- und Rohfett-Verschwindensrate (%) der flüssigen Testmahlzeit I
Enzymdosierung IE 0 10 000 75 000
Trockensubstanz-VR 17,7 ± 18,9 43,7 ± 21,8 60,0 ± 9,98
Rohfett-VR 11,3 ± 8,69 76,0 ± 14,0 85,4 ± 6,35

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
65
Schlussfolgerungen:
Der erste Einsatz einer flüssigen Testmahlzeit (Testmahlzeit I) als einmalige Versuchsdiät
für die Überprüfung der lipolytischen Aktivität zugesetzter Enzyme war grundsätzlich
positiv zu bewerten (dosisabhängige Effekte bezüglich der absoluten Anflutung von
Trockensubstanz und Rohfett; bis auf wenige Ausnahmen dosisabhängige Steigerung der
kalkulierten Verschwindensraten von Trockensubstanz und Rohfett). Die Chromoxid-
Wiederfindung war mit mittleren Werten von ~60 % leicht niedriger als die von BECKER
(2005) ermittelten Werte. Insgesamt waren bezüglich der Chromoxid-Wiederfindung
weniger Effekte der Enzymzulage bzw. tagesabhängige Schwankungen als vielmehr
tierindividuelle Einflüsse zu vermuten.
Schwerpunkt in der weiteren Optimierung einer Testmahlzeit war die Minderung möglicher
Entmischungsvorgänge (Separation von Marker, Testmahlzeit und zugesetztem Fett). Die
z.T. hohe TS-Anflutung an der ilealen Fistel (TS-Anflutung Tier 63; 0 Enzym: 70,7 g;
negative TS-VR) könnte zudem auf eine Vermischung der flüssigen Testmahlzeit und dem
schrotförmigen Futter des Vorabends hindeuten.
Des Weiteren wurden trotz deutlicher Reduktion der Enzymdosierungen (s. o.) -im
Vergleich zu vorangegangenen Arbeiten des Projektes, in denen fettreiche Mischfutter auf
der Basis von Getreideschrot verwendet wurden- bereits in der geringeren Dosierung (10
000 I.E. / Mahlzeit) sehr hohe Rohfett-Verschwindensraten von im Mittel 76 % erreicht, so
dass im Folgenden eine weitere Reduktion der Enzymdosierung und / oder eine Erhöhung
der Fettmenge pro Mahlzeit notwendig erschien.
3.2.11.2 2. Versuch zur Optimierung einer flüssigen Testmahlzeit
Fragestellung: Können dosisabhängige Effekte der Rohfett-Verschwindensrate durch
Erhöhung des Fettanteils bei gleichzeitiger Reduzierung der Enzymdosierung erreicht
werden?
Versuchsdesign:
Probenkollektion: analog Screening-Test BECKER (2005)
Dauer: 8 Stunden
Beginn der Kollektion: nach „Farbwechsel“
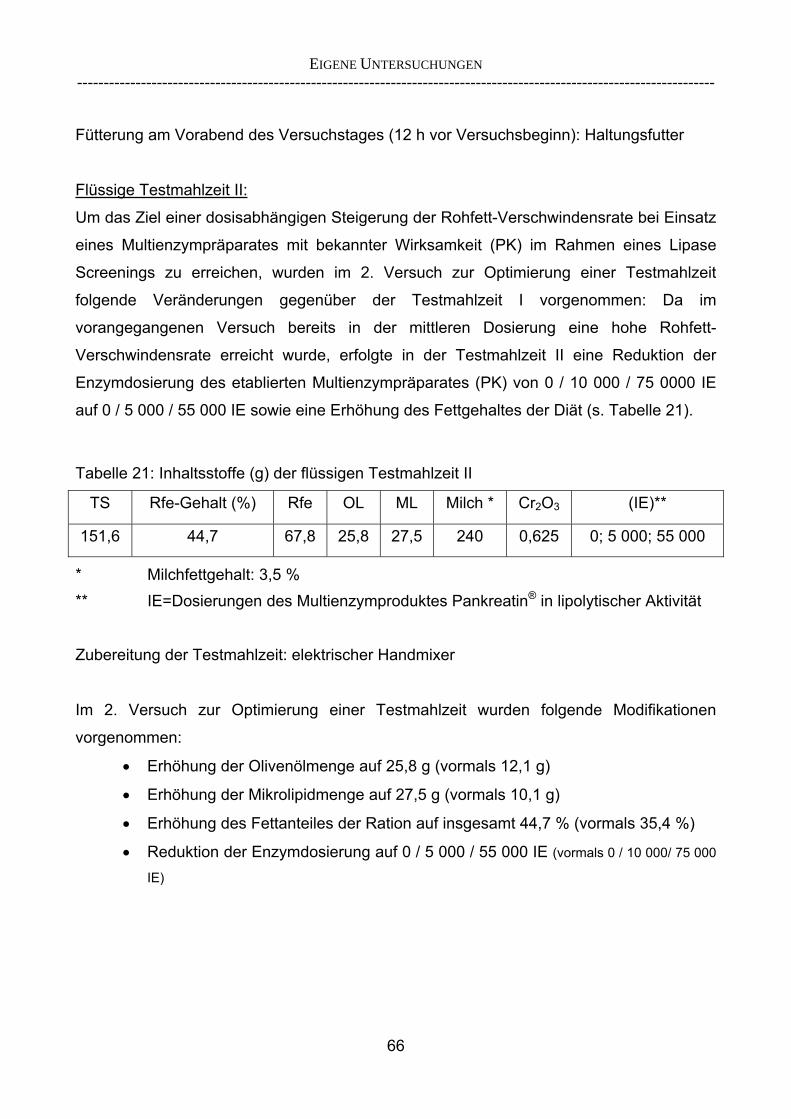
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
66
Fütterung am Vorabend des Versuchstages (12 h vor Versuchsbeginn): Haltungsfutter
Flüssige Testmahlzeit II:
Um das Ziel einer dosisabhängigen Steigerung der Rohfett-Verschwindensrate bei Einsatz
eines Multienzympräparates mit bekannter Wirksamkeit (PK) im Rahmen eines Lipase
Screenings zu erreichen, wurden im 2. Versuch zur Optimierung einer Testmahlzeit
folgende Veränderungen gegenüber der Testmahlzeit I vorgenommen: Da im
vorangegangenen Versuch bereits in der mittleren Dosierung eine hohe Rohfett-
Verschwindensrate erreicht wurde, erfolgte in der Testmahlzeit II eine Reduktion der
Enzymdosierung des etablierten Multienzympräparates (PK) von 0 / 10 000 / 75 0000 IE
auf 0 / 5 000 / 55 000 IE sowie eine Erhöhung des Fettgehaltes der Diät (s. Tabelle 21).
Tabelle 21: Inhaltsstoffe (g) der flüssigen Testmahlzeit II
TS Rfe-Gehalt (%) Rfe OL ML Milch * Cr2O3 (IE)**
151,6 44,7 67,8 25,8 27,5 240 0,625 0; 5 000; 55 000
* Milchfettgehalt: 3,5 % ** IE=Dosierungen des Multienzymproduktes Pankreatin® in lipolytischer Aktivität
Zubereitung der Testmahlzeit: elektrischer Handmixer
Im 2. Versuch zur Optimierung einer Testmahlzeit wurden folgende Modifikationen
vorgenommen:
• Erhöhung der Olivenölmenge auf 25,8 g (vormals 12,1 g)
• Erhöhung der Mikrolipidmenge auf 27,5 g (vormals 10,1 g)
• Erhöhung des Fettanteiles der Ration auf insgesamt 44,7 % (vormals 35,4 %)
• Reduktion der Enzymdosierung auf 0 / 5 000 / 55 000 IE (vormals 0 / 10 000/ 75 000
IE)
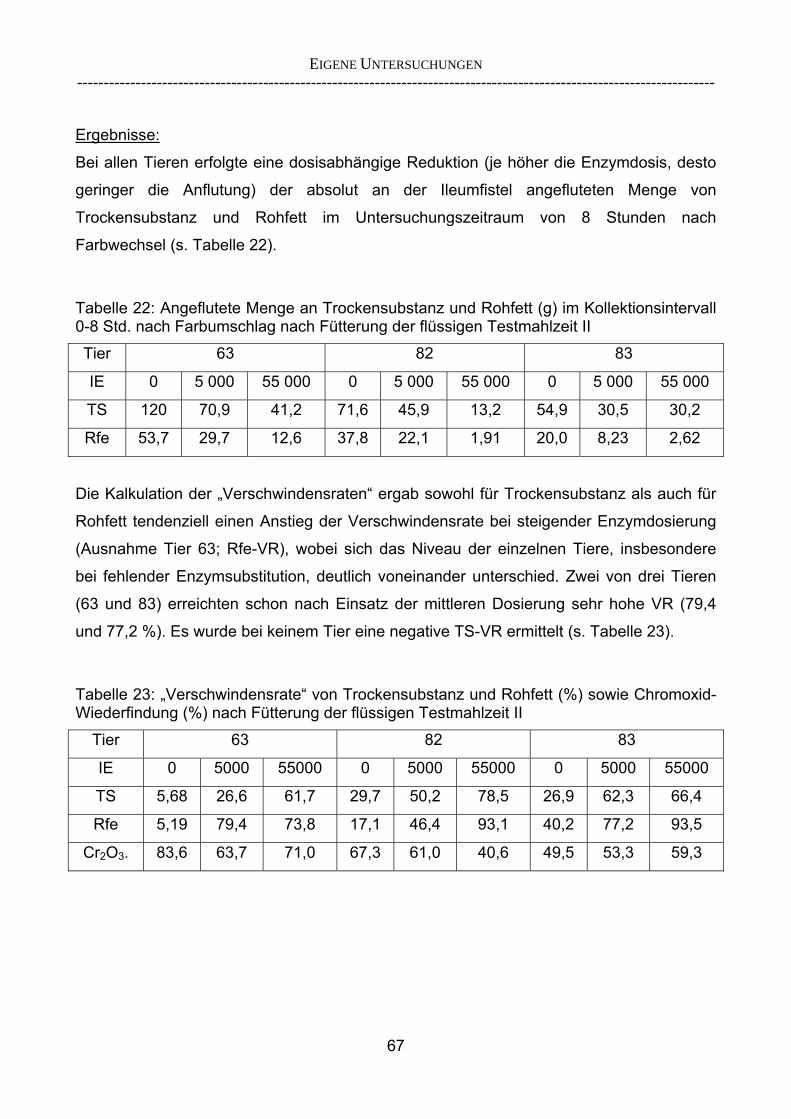
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
67
Ergebnisse:
Bei allen Tieren erfolgte eine dosisabhängige Reduktion (je höher die Enzymdosis, desto
geringer die Anflutung) der absolut an der Ileumfistel angefluteten Menge von
Trockensubstanz und Rohfett im Untersuchungszeitraum von 8 Stunden nach
Farbwechsel (s. Tabelle 22).
Tabelle 22: Angeflutete Menge an Trockensubstanz und Rohfett (g) im Kollektionsintervall 0-8 Std. nach Farbumschlag nach Fütterung der flüssigen Testmahlzeit II
Tier 63 82 83
IE 0 5 000 55 000 0 5 000 55 000 0 5 000 55 000
TS 120 70,9 41,2 71,6 45,9 13,2 54,9 30,5 30,2
Rfe 53,7 29,7 12,6 37,8 22,1 1,91 20,0 8,23 2,62
Die Kalkulation der „Verschwindensraten“ ergab sowohl für Trockensubstanz als auch für
Rohfett tendenziell einen Anstieg der Verschwindensrate bei steigender Enzymdosierung
(Ausnahme Tier 63; Rfe-VR), wobei sich das Niveau der einzelnen Tiere, insbesondere
bei fehlender Enzymsubstitution, deutlich voneinander unterschied. Zwei von drei Tieren
(63 und 83) erreichten schon nach Einsatz der mittleren Dosierung sehr hohe VR (79,4
und 77,2 %). Es wurde bei keinem Tier eine negative TS-VR ermittelt (s. Tabelle 23).
Tabelle 23: „Verschwindensrate“ von Trockensubstanz und Rohfett (%) sowie Chromoxid-Wiederfindung (%) nach Fütterung der flüssigen Testmahlzeit II
Tier 63 82 83
IE 0 5000 55000 0 5000 55000 0 5000 55000
TS 5,68 26,6 61,7 29,7 50,2 78,5 26,9 62,3 66,4
Rfe 5,19 79,4 73,8 17,1 46,4 93,1 40,2 77,2 93,5
Cr2O3. 83,6 63,7 71,0 67,3 61,0 40,6 49,5 53,3 59,3

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
68
Tabelle 24: Mittlere Trockensubstanz- und Rohfett-Verschwindensrate (%) der flüssigen Testmahlzeit II
Enzymdosierung 0 5 000 55 000
Trockensubstanz-VR 20,8 ± 13,1 46,4 ± 18,2 70,1 ± 11,9
Rohfett-VR 20,9 ± 17,8 67,7 ± 18,5 86,8 ± 11,3
Schlussfolgerungen:
Trotz Erhöhung der Fettkonzentration der Mahlzeit bei gleichzeitiger Reduktion der
Enzymdosis ergaben sich nach wie vor sehr hohe Rohfett-Verschwindensraten in der
mittleren (~80 %, außer Tier 82) und hohen Dosierung (~93 %, außer Tier 63). Dies deutet
darauf hin, dass die maximale Enzymwirkung schon fast bei der mittleren Dosierung
erreicht war.
3.2.11.3 3. Versuch zur Optimierung einer flüssigen Testmahlzeit
Fragestellung: Kann durch Verfütterung einer flüssigen Mahlzeit am Vorabend eines
Screening-Versuchs eine „Verschleppung“ von Vorabendfutter in die Kollektionsphase
verhindert werden? Ist es weiterhin möglich, durch Reduktion der mittleren Dosierung eine
deutlichere Abgrenzung der Wirkung der verschiedenen Enzymdosierungen zu erhalten?
Versuchsdesign:
Probenkollektion: analog Screening-Test (BECKER 2005)
Dauer: 8 Stunden
Beginn der Kollektion: nach „Farbwechsel“
Fütterung am Vorabend des Versuchstages (12 h vor Versuchsbeginn):
400 ml ProvideXtra® pro Schwein
Testmahlzeit III:
Die Dosierung des Multienzymproduktes wurde dahingehend modifiziert, dass eine
weitere, sehr niedrige Enzymdosierung (500 IE Pankreatin) hinzugefügt wurde (s. Tabelle
25). Zusätzlich wurde am Vorabend der jeweiligen Versuche statt des schrotförmigen
Haltungsfutters auf Getreidebasis (Breifütterung) 400 ml einer flüssigen Mahlzeit

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
69
(ProvideXtra®) verabreicht. ProvideXtra® ist ein fettfreier Energiedrink (Protein 3,75 g /
Kohlenhydrate 27,5 g, davon Zucker 5,5 g / Fett 0 g / Ballaststoffe 0 g und 80 ml Wasser
je 100 ml fertigem Getränk).
Die vorabendliche Verabreichung von ProvideXtra® wurde erwogen, da bei Gabe einer
flüssigen, fettfreien Mahlzeit mit einer zügigen Magenentleerung gerechnet werden
konnte. Somit sollte eine Anflutung von Rfe aus der Vorabendmahlzeit an der Ileumfistel
im Kollektionsintervall verhindert werden.
Tabelle 25: Inhaltsstoffe (g) der flüssigen Testmahlzeit III
TS Rfe-Gehalt (%) Rfe OL ML Milch* Cr2O3 (IE)**
151,6 44,7 67,8 25,8 27,5 240 0,625 0, 500; 5 000; 55 000
* Milchfettgehalt: 3,5 % ** IE=Dosierungen des Multienzymproduktes Pankreatin® in lipolytischer Aktivität
Zubereitung der Testmalzeit: elektrischer Handmixer
Im 3. Versuch zur Optimierung einer Testmahlzeit wurden folgende Modifikationen
vorgenommen:
• Einsatz einer Flüssignahrung (ProvideXtra®) am Vorabend des Versuches
• Einführung einer neuen Enzymdosierung: 0/ 500/ 5 000/ 55 000 IE (vormals 0 /
5 000 / 55 000 IE )
Ergebnisse:
Aus versuchstechnischen Gründen lagen keine Ergebnisse für Tier 82 und 83 (0 Enzym)
sowie für Tier 43 und 93 in der höchsten Dosierung (55 000 IE) vor. Nicht in allen Fällen
konnte eine dosisabhängige Reduktion der Anflutung verzeichnet werden (s. Tabelle 26).
Da in diesen Fällen jedoch gleichzeitig eine höhere Cr2O3-Wiederfindung vorlag (s. Tabelle
27), ist davon auszugehen, dass diese erhöhte Anflutung aus einer höheren Passagerate
resultierte.
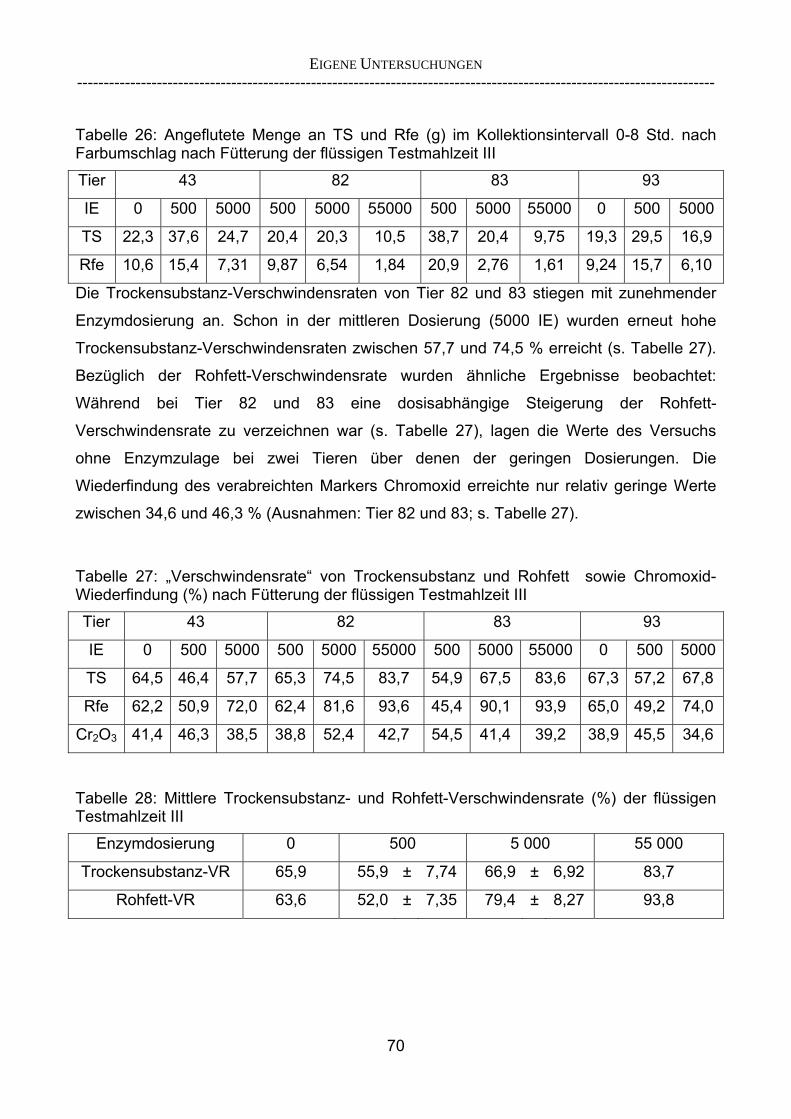
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
70
Tabelle 26: Angeflutete Menge an TS und Rfe (g) im Kollektionsintervall 0-8 Std. nach Farbumschlag nach Fütterung der flüssigen Testmahlzeit III
Tier 43 82 83 93
IE 0 500 5000 500 5000 55000 500 5000 55000 0 500 5000
TS 22,3 37,6 24,7 20,4 20,3 10,5 38,7 20,4 9,75 19,3 29,5 16,9
Rfe 10,6 15,4 7,31 9,87 6,54 1,84 20,9 2,76 1,61 9,24 15,7 6,10
Die Trockensubstanz-Verschwindensraten von Tier 82 und 83 stiegen mit zunehmender
Enzymdosierung an. Schon in der mittleren Dosierung (5000 IE) wurden erneut hohe
Trockensubstanz-Verschwindensraten zwischen 57,7 und 74,5 % erreicht (s. Tabelle 27).
Bezüglich der Rohfett-Verschwindensrate wurden ähnliche Ergebnisse beobachtet:
Während bei Tier 82 und 83 eine dosisabhängige Steigerung der Rohfett-
Verschwindensrate zu verzeichnen war (s. Tabelle 27), lagen die Werte des Versuchs
ohne Enzymzulage bei zwei Tieren über denen der geringen Dosierungen. Die
Wiederfindung des verabreichten Markers Chromoxid erreichte nur relativ geringe Werte
zwischen 34,6 und 46,3 % (Ausnahmen: Tier 82 und 83; s. Tabelle 27).
Tabelle 27: „Verschwindensrate“ von Trockensubstanz und Rohfett sowie Chromoxid-Wiederfindung (%) nach Fütterung der flüssigen Testmahlzeit III
Tier 43 82 83 93
IE 0 500 5000 500 5000 55000 500 5000 55000 0 500 5000
TS 64,5 46,4 57,7 65,3 74,5 83,7 54,9 67,5 83,6 67,3 57,2 67,8
Rfe 62,2 50,9 72,0 62,4 81,6 93,6 45,4 90,1 93,9 65,0 49,2 74,0
Cr2O3 41,4 46,3 38,5 38,8 52,4 42,7 54,5 41,4 39,2 38,9 45,5 34,6
Tabelle 28: Mittlere Trockensubstanz- und Rohfett-Verschwindensrate (%) der flüssigen Testmahlzeit III
Enzymdosierung 0 500 5 000 55 000
Trockensubstanz-VR 65,9 55,9 ± 7,74 66,9 ± 6,92 83,7
Rohfett-VR 63,6 52,0 ± 7,35 79,4 ± 8,27 93,8
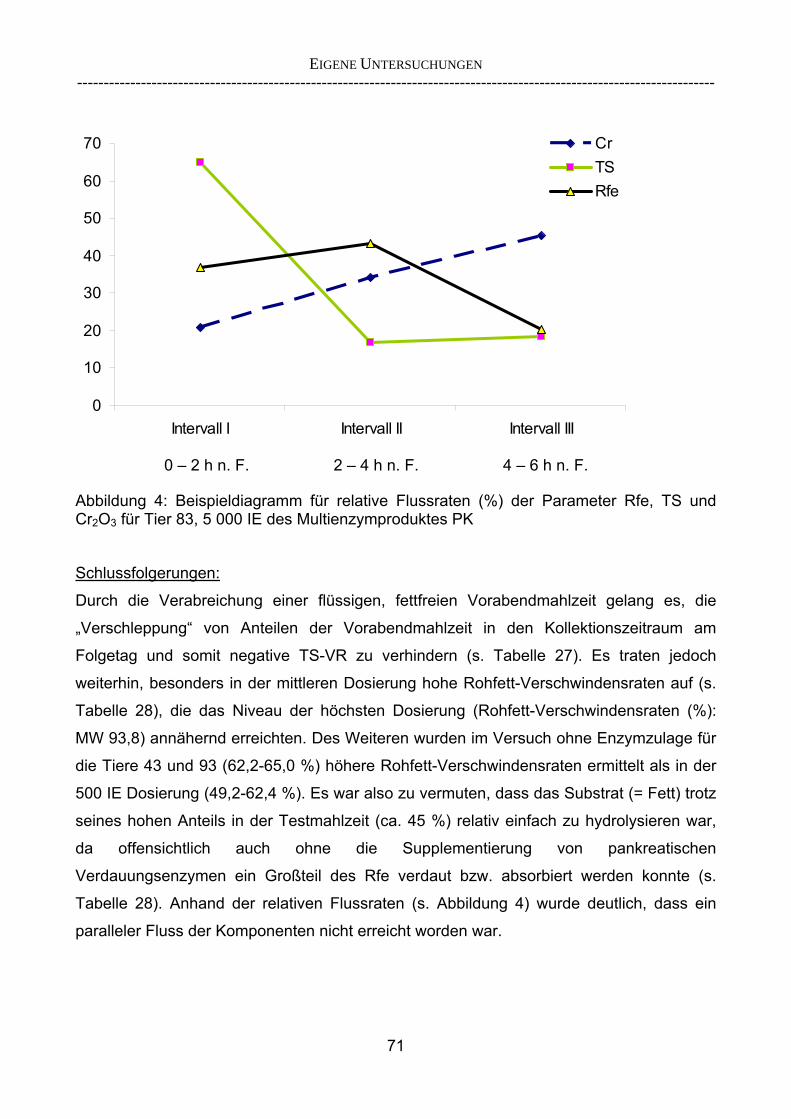
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
71
0
10
20
30
40
50
60
70
Intervall I Intervall II Intervall III
CrTSRfe
0 – 2 h n. F. 2 – 4 h n. F. 4 – 6 h n. F.
Abbildung 4: Beispieldiagramm für relative Flussraten (%) der Parameter Rfe, TS und Cr2O3 für Tier 83, 5 000 IE des Multienzymproduktes PK
Schlussfolgerungen:
Durch die Verabreichung einer flüssigen, fettfreien Vorabendmahlzeit gelang es, die
„Verschleppung“ von Anteilen der Vorabendmahlzeit in den Kollektionszeitraum am
Folgetag und somit negative TS-VR zu verhindern (s. Tabelle 27). Es traten jedoch
weiterhin, besonders in der mittleren Dosierung hohe Rohfett-Verschwindensraten auf (s.
Tabelle 28), die das Niveau der höchsten Dosierung (Rohfett-Verschwindensraten (%):
MW 93,8) annähernd erreichten. Des Weiteren wurden im Versuch ohne Enzymzulage für
die Tiere 43 und 93 (62,2-65,0 %) höhere Rohfett-Verschwindensraten ermittelt als in der
500 IE Dosierung (49,2-62,4 %). Es war also zu vermuten, dass das Substrat (= Fett) trotz
seines hohen Anteils in der Testmahlzeit (ca. 45 %) relativ einfach zu hydrolysieren war,
da offensichtlich auch ohne die Supplementierung von pankreatischen
Verdauungsenzymen ein Großteil des Rfe verdaut bzw. absorbiert werden konnte (s.
Tabelle 28). Anhand der relativen Flussraten (s. Abbildung 4) wurde deutlich, dass ein
paralleler Fluss der Komponenten nicht erreicht worden war.
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
72
3.2.11.4 4. Versuch zur Optimierung einer flüssigen Testmahlzeit
Fragestellung: Können durch eine Veränderung der Fettqualität bzw. –zusammensetzung
deutlichere dosisabhängige Effekte erzeugt werden?
Versuchsdesign:
Probenkollektion: analog Screening-Test BECKER (2005)
Dauer: 8 Stunden
Beginn der Kollektion: nach „Farbwechsel“
Fütterung am Vorabend des Versuchstages (12 h vor Versuchsbeginn):
400 ml ProvideXtra® pro Schwein
Testmahlzeit IV:
Bei der flüssigen Testmahlzeit IV wurde eine Veränderung der Fettzusammensetzung
angestrebt. Es wurde Milch mit einem Milchfettanteil von 1,5 %, anstelle der bisher
gebräuchlichen 3,5 %igen verwendet. Infolge der verringerten Milchfettmenge reduzierte
sich die Gesamtfettmenge in der Ration auf 57,5 g (s. Tabelle 29). Hierdurch erhöhte sich
der prozentuale Anteil des Olivenöls in der Testmahlzeit, wodurch eine erhöhte Zufuhr von
langkettigen Fettsäuren erreicht und der Anteil des leicht zu hydrolysierenden Milchfetts
reduziert wurde.
Tabelle 29: Inhaltsstoffe (g) der flüssigen Testmahlzeit IV
TS Rfe-Gehalt (%) Rfe OL ML Milch* Cr2O3 (IE)**
154,6 37,2 57,5 19,6 20,7 240 0,625 0; 5 00; 5 000, 75 000
* Milchfettgehalt: 1,5 % ** IE=Dosierungen des Multienzymproduktes Pankreatin® in lipolytischer Aktivität Zubereitung der Testmahlzeit: elektrischer Handmixer

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
73
Im 4. Versuch zur Optimierung einer Testmahlzeit wurden folgende Modifikationen
vorgenommen:
• Einsatz von Milch mit einem Milchfettgehalt von 1,5 % (vormals 3,5 %)
• Verringerung der Fettmenge / Mahlzeit auf 57,5 g (vormals 67,8 g)
• Verringerung der Mikrolipidmenge auf 20,7 g (vormals 27,5 g)
• Reduktion des Fettanteiles der Ration auf insgesamt 37,2 % Rfe (vormals 44,7
%)
Ergebnisse:
Auch im 4. Versuch zur Optimierung einer Testmahlzeit waren bezüglich der
Trockensubstanz- und Rohfett-Anflutung nicht bei allen Tieren gerichteten Effekte der
verabreichten Enzymdosierungen zu verzeichnen (s. Tabelle 30). Es ist jedoch in diesem
Zusammenhang darauf hinzuweisen, dass z.T. die beobachtete erhöhte Anflutung von TS
bzw. Rfe anhand der gleichzeitig hohen Chromoxidwiederfindung auf eine erhöhte
Passage zurückzuführen ist (s. Tabelle 30; 31; Tier 43, Tier 93). Bezüglich der
Verschwindensraten von TS und Rfe sind im Allgemeinen hingegen dosisabhängige
Effekte zu verzeichnen (s. Tabelle 31).
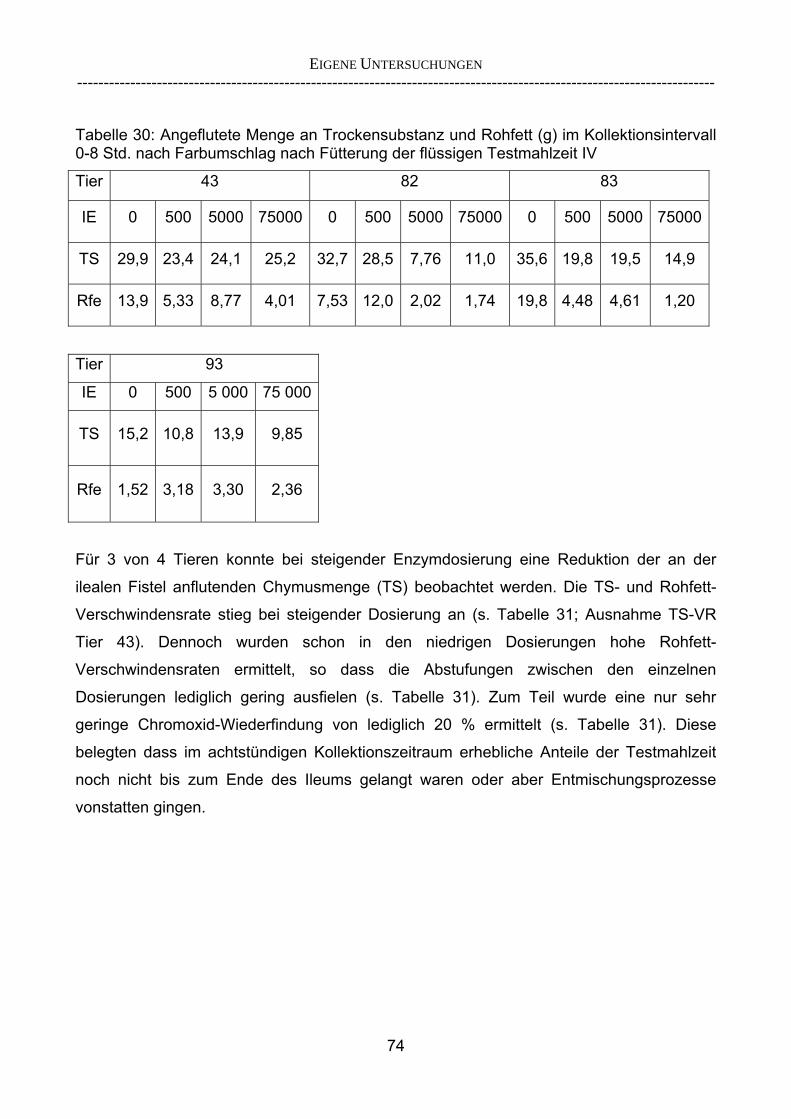
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
74
Tabelle 30: Angeflutete Menge an Trockensubstanz und Rohfett (g) im Kollektionsintervall 0-8 Std. nach Farbumschlag nach Fütterung der flüssigen Testmahlzeit IV
Tier 43 82 83
IE 0 500 5000 75000 0 500 5000 75000 0 500 5000 75000
TS 29,9 23,4 24,1 25,2 32,7 28,5 7,76 11,0 35,6 19,8 19,5 14,9
Rfe 13,9 5,33 8,77 4,01 7,53 12,0 2,02 1,74 19,8 4,48 4,61 1,20
Tier 93
IE 0 500 5 000 75 000
TS 15,2 10,8 13,9 9,85
Rfe 1,52 3,18 3,30 2,36
Für 3 von 4 Tieren konnte bei steigender Enzymdosierung eine Reduktion der an der
ilealen Fistel anflutenden Chymusmenge (TS) beobachtet werden. Die TS- und Rohfett-
Verschwindensrate stieg bei steigender Dosierung an (s. Tabelle 31; Ausnahme TS-VR
Tier 43). Dennoch wurden schon in den niedrigen Dosierungen hohe Rohfett-
Verschwindensraten ermittelt, so dass die Abstufungen zwischen den einzelnen
Dosierungen lediglich gering ausfielen (s. Tabelle 31). Zum Teil wurde eine nur sehr
geringe Chromoxid-Wiederfindung von lediglich 20 % ermittelt (s. Tabelle 31). Diese
belegten dass im achtstündigen Kollektionszeitraum erhebliche Anteile der Testmahlzeit
noch nicht bis zum Ende des Ileums gelangt waren oder aber Entmischungsprozesse
vonstatten gingen.
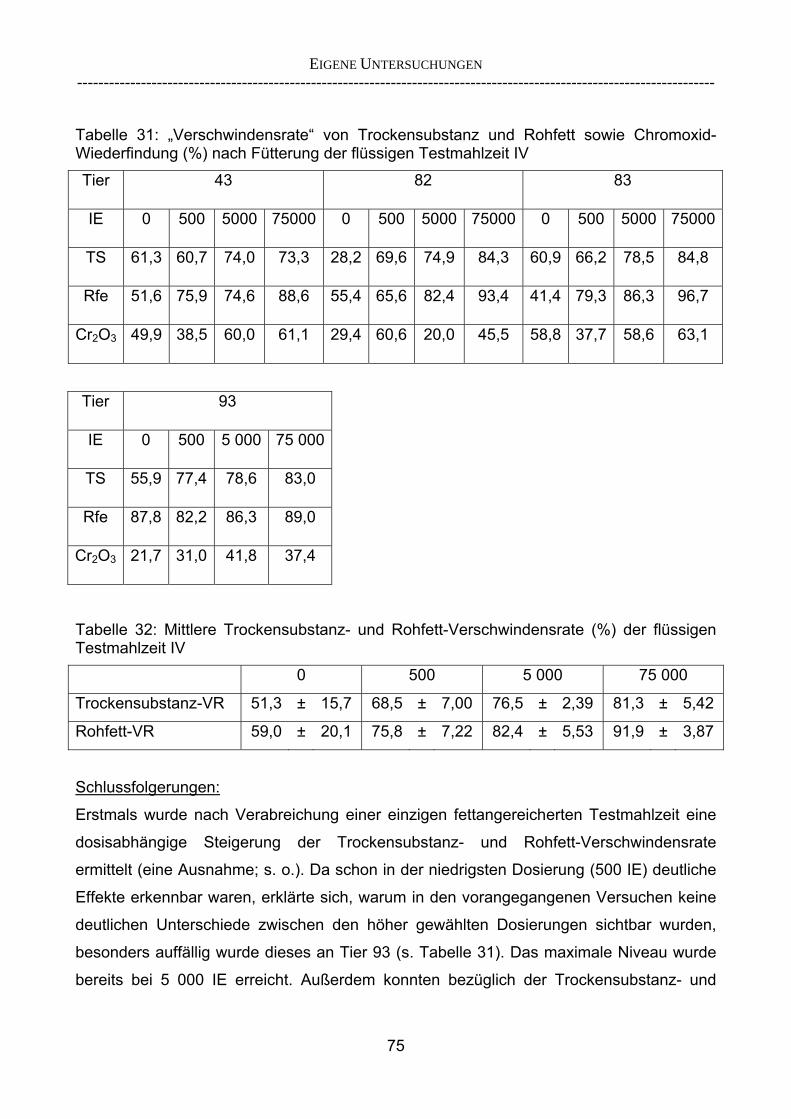
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
75
Tabelle 31: „Verschwindensrate“ von Trockensubstanz und Rohfett sowie Chromoxid-Wiederfindung (%) nach Fütterung der flüssigen Testmahlzeit IV
Tier 43 82 83
IE 0 500 5000 75000 0 500 5000 75000 0 500 5000 75000
TS 61,3 60,7 74,0 73,3 28,2 69,6 74,9 84,3 60,9 66,2 78,5 84,8
Rfe 51,6 75,9 74,6 88,6 55,4 65,6 82,4 93,4 41,4 79,3 86,3 96,7
Cr2O3 49,9 38,5 60,0 61,1 29,4 60,6 20,0 45,5 58,8 37,7 58,6 63,1
Tier 93
IE 0 500 5 000 75 000
TS 55,9 77,4 78,6 83,0
Rfe 87,8 82,2 86,3 89,0
Cr2O3 21,7 31,0 41,8 37,4
Tabelle 32: Mittlere Trockensubstanz- und Rohfett-Verschwindensrate (%) der flüssigen Testmahlzeit IV
0 500 5 000 75 000
Trockensubstanz-VR 51,3 ± 15,7 68,5 ± 7,00 76,5 ± 2,39 81,3 ± 5,42
Rohfett-VR 59,0 ± 20,1 75,8 ± 7,22 82,4 ± 5,53 91,9 ± 3,87
Schlussfolgerungen:
Erstmals wurde nach Verabreichung einer einzigen fettangereicherten Testmahlzeit eine
dosisabhängige Steigerung der Trockensubstanz- und Rohfett-Verschwindensrate
ermittelt (eine Ausnahme; s. o.). Da schon in der niedrigsten Dosierung (500 IE) deutliche
Effekte erkennbar waren, erklärte sich, warum in den vorangegangenen Versuchen keine
deutlichen Unterschiede zwischen den höher gewählten Dosierungen sichtbar wurden,
besonders auffällig wurde dieses an Tier 93 (s. Tabelle 31). Das maximale Niveau wurde
bereits bei 5 000 IE erreicht. Außerdem konnten bezüglich der Trockensubstanz- und
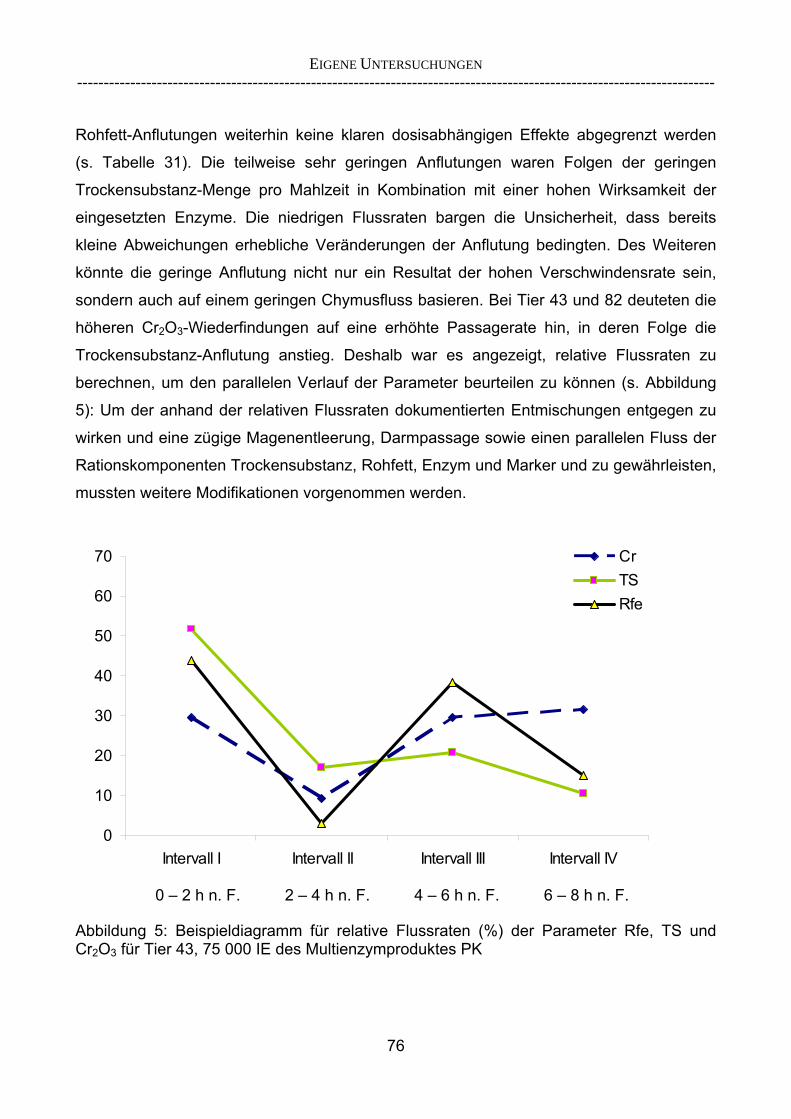
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
76
Rohfett-Anflutungen weiterhin keine klaren dosisabhängigen Effekte abgegrenzt werden
(s. Tabelle 31). Die teilweise sehr geringen Anflutungen waren Folgen der geringen
Trockensubstanz-Menge pro Mahlzeit in Kombination mit einer hohen Wirksamkeit der
eingesetzten Enzyme. Die niedrigen Flussraten bargen die Unsicherheit, dass bereits
kleine Abweichungen erhebliche Veränderungen der Anflutung bedingten. Des Weiteren
könnte die geringe Anflutung nicht nur ein Resultat der hohen Verschwindensrate sein,
sondern auch auf einem geringen Chymusfluss basieren. Bei Tier 43 und 82 deuteten die
höheren Cr2O3-Wiederfindungen auf eine erhöhte Passagerate hin, in deren Folge die
Trockensubstanz-Anflutung anstieg. Deshalb war es angezeigt, relative Flussraten zu
berechnen, um den parallelen Verlauf der Parameter beurteilen zu können (s. Abbildung
5): Um der anhand der relativen Flussraten dokumentierten Entmischungen entgegen zu
wirken und eine zügige Magenentleerung, Darmpassage sowie einen parallelen Fluss der
Rationskomponenten Trockensubstanz, Rohfett, Enzym und Marker und zu gewährleisten,
mussten weitere Modifikationen vorgenommen werden.
0
10
20
30
40
50
60
70
Intervall I Intervall II Intervall III Intervall IV
CrTSRfe
0 – 2 h n. F. 2 – 4 h n. F. 4 – 6 h n. F. 6 – 8 h n. F.
Abbildung 5: Beispieldiagramm für relative Flussraten (%) der Parameter Rfe, TS und Cr2O3 für Tier 43, 75 000 IE des Multienzymproduktes PK
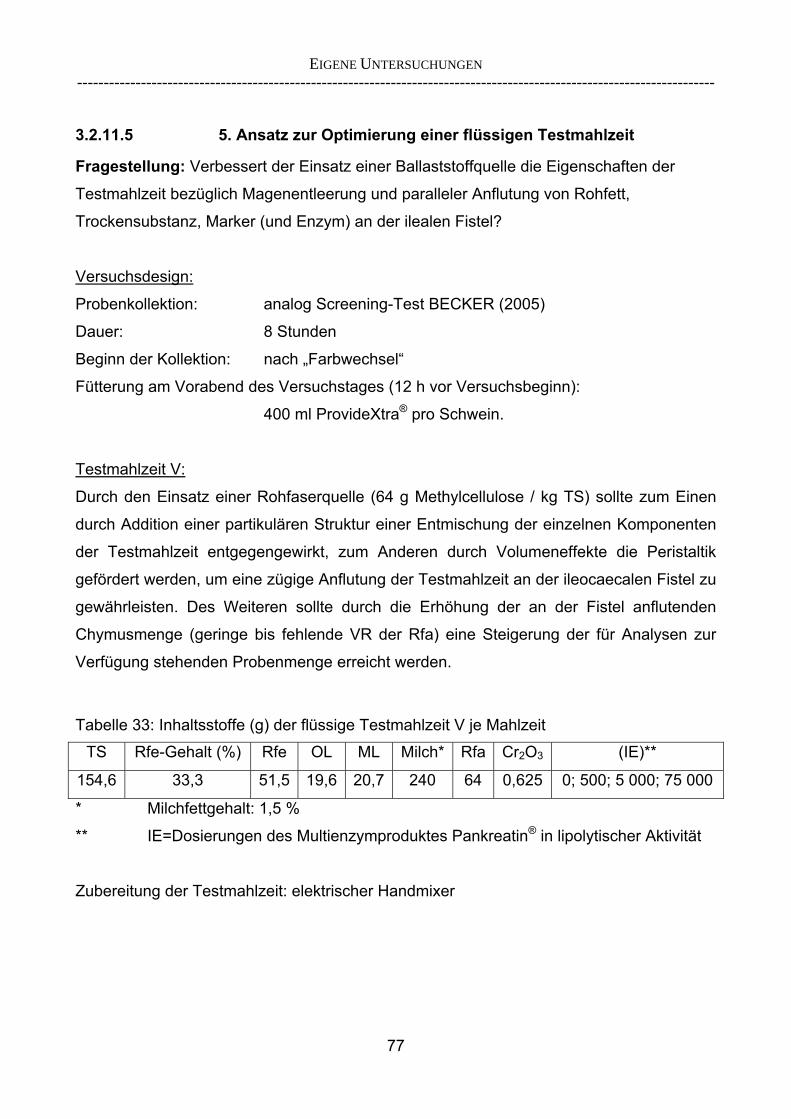
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
77
3.2.11.5 5. Ansatz zur Optimierung einer flüssigen Testmahlzeit
Fragestellung: Verbessert der Einsatz einer Ballaststoffquelle die Eigenschaften der
Testmahlzeit bezüglich Magenentleerung und paralleler Anflutung von Rohfett,
Trockensubstanz, Marker (und Enzym) an der ilealen Fistel?
Versuchsdesign:
Probenkollektion: analog Screening-Test BECKER (2005)
Dauer: 8 Stunden
Beginn der Kollektion: nach „Farbwechsel“
Fütterung am Vorabend des Versuchstages (12 h vor Versuchsbeginn):
400 ml ProvideXtra® pro Schwein.
Testmahlzeit V:
Durch den Einsatz einer Rohfaserquelle (64 g Methylcellulose / kg TS) sollte zum Einen
durch Addition einer partikulären Struktur einer Entmischung der einzelnen Komponenten
der Testmahlzeit entgegengewirkt, zum Anderen durch Volumeneffekte die Peristaltik
gefördert werden, um eine zügige Anflutung der Testmahlzeit an der ileocaecalen Fistel zu
gewährleisten. Des Weiteren sollte durch die Erhöhung der an der Fistel anflutenden
Chymusmenge (geringe bis fehlende VR der Rfa) eine Steigerung der für Analysen zur
Verfügung stehenden Probenmenge erreicht werden.
Tabelle 33: Inhaltsstoffe (g) der flüssige Testmahlzeit V je Mahlzeit
TS Rfe-Gehalt (%) Rfe OL ML Milch* Rfa Cr2O3 (IE)**
154,6 33,3 51,5 19,6 20,7 240 64 0,625 0; 500; 5 000; 75 000
* Milchfettgehalt: 1,5 %
** IE=Dosierungen des Multienzymproduktes Pankreatin® in lipolytischer Aktivität
Zubereitung der Testmahlzeit: elektrischer Handmixer

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
78
Im 5. Versuch zur Optimierung einer Testmahlzeit wurden folgende Modifikationen
vorgenommen:
• Verringerung der Fettmenge / Mahlzeit auf 51,5 g (vormals 57,5 g)
• Reduktion des Fettanteils der Ration auf insgesamt 33,3 % Rfe (vormals 37,2
%)
• Einsatz einer Ballaststoffquelle (Methylcellulose) in einer Dosierung von 9,9 g /
Mahlzeit (64,0 g / kg TS)
Ergebnisse:
Eine dosisabhängige Verringerung sowohl der Trockensubstanz- als auch der Rohfett-
Anflutung konnte lediglich bei zwei der vier Tiere beobachtet werden (s. Tabelle 34).
Hinzuweisen ist jedoch auf die bei allen Tieren sehr deutliche Reduktion der anflutenden
Trockensubstanz- und Rohfett-Mengen bereits nach Einsatz der geringsten Dosierung des
Multienzympräparates PK.
Tabelle 34: Angeflutete Menge an Trockensubstanz und Rohfett (g) im Kollektionsintervall 0-8 Std. nach Farbumschlag nach Fütterung der flüssigen Testmahlzeit V
Tier 43 80 81
IE 0 500 5000 75000 0 500 5000 75000 0 500 5000 75000
TS 57,3 16,2 16,6 22,0 41,5 14,0 10,8 15,7 55,4 32,7 33,0 11,5
Rfe 21,5 2,60 2,88 1,58 19,9 2,49 1,59 1,17 28,4 10,9 10,1 1,27
Tier 83
IE 0 500 5 000 75 000
TS 38,3 23,5 24,7 17,2
Rfe 22,6 5,63 7,45 2,09
Bezüglich der Verschwindensrate von Rohfett und Trockensubstanz wurde bei allen
eingesetzten Tieren eine dosisabhängige Steigerung verzeichnet (s. Tabelle 35). Erstmals
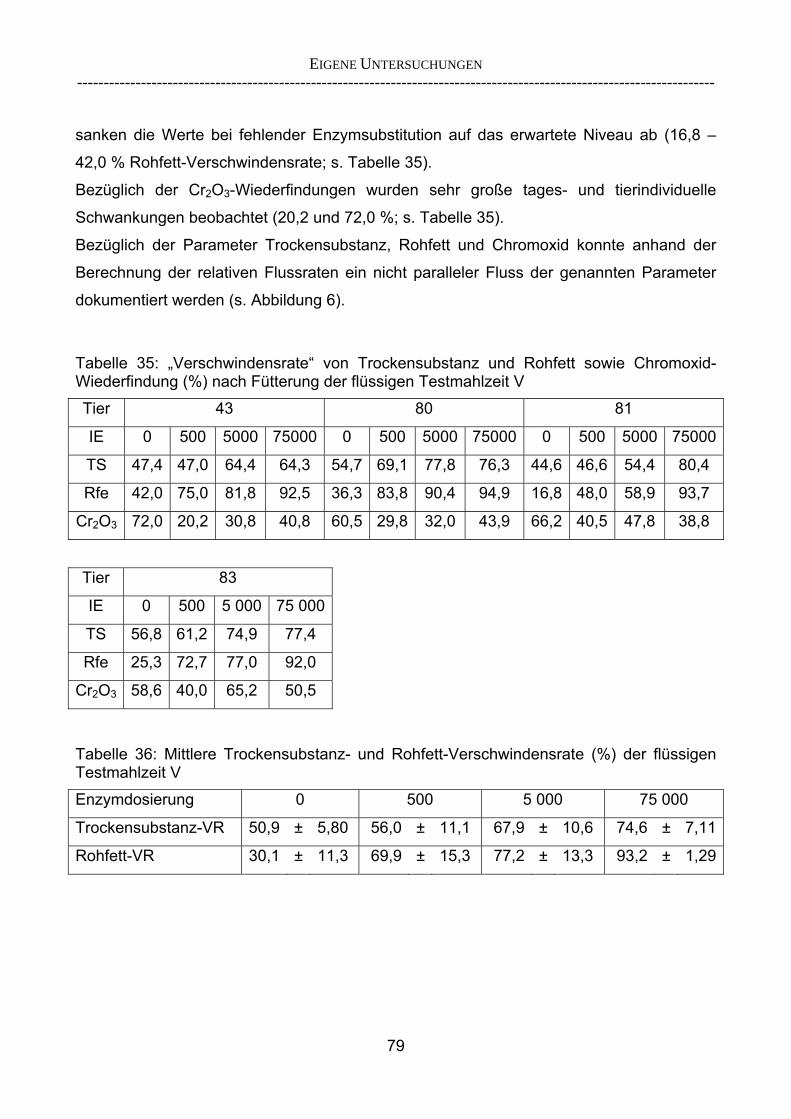
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
79
sanken die Werte bei fehlender Enzymsubstitution auf das erwartete Niveau ab (16,8 –
42,0 % Rohfett-Verschwindensrate; s. Tabelle 35).
Bezüglich der Cr2O3-Wiederfindungen wurden sehr große tages- und tierindividuelle
Schwankungen beobachtet (20,2 und 72,0 %; s. Tabelle 35).
Bezüglich der Parameter Trockensubstanz, Rohfett und Chromoxid konnte anhand der
Berechnung der relativen Flussraten ein nicht paralleler Fluss der genannten Parameter
dokumentiert werden (s. Abbildung 6).
Tabelle 35: „Verschwindensrate“ von Trockensubstanz und Rohfett sowie Chromoxid-Wiederfindung (%) nach Fütterung der flüssigen Testmahlzeit V
Tier 43 80 81
IE 0 500 5000 75000 0 500 5000 75000 0 500 5000 75000
TS 47,4 47,0 64,4 64,3 54,7 69,1 77,8 76,3 44,6 46,6 54,4 80,4
Rfe 42,0 75,0 81,8 92,5 36,3 83,8 90,4 94,9 16,8 48,0 58,9 93,7
Cr2O3 72,0 20,2 30,8 40,8 60,5 29,8 32,0 43,9 66,2 40,5 47,8 38,8
Tier 83
IE 0 500 5 000 75 000
TS 56,8 61,2 74,9 77,4
Rfe 25,3 72,7 77,0 92,0
Cr2O3 58,6 40,0 65,2 50,5
Tabelle 36: Mittlere Trockensubstanz- und Rohfett-Verschwindensrate (%) der flüssigen Testmahlzeit V
Enzymdosierung 0 500 5 000 75 000
Trockensubstanz-VR 50,9 ± 5,80 56,0 ± 11,1 67,9 ± 10,6 74,6 ± 7,11
Rohfett-VR 30,1 ± 11,3 69,9 ± 15,3 77,2 ± 13,3 93,2 ± 1,29
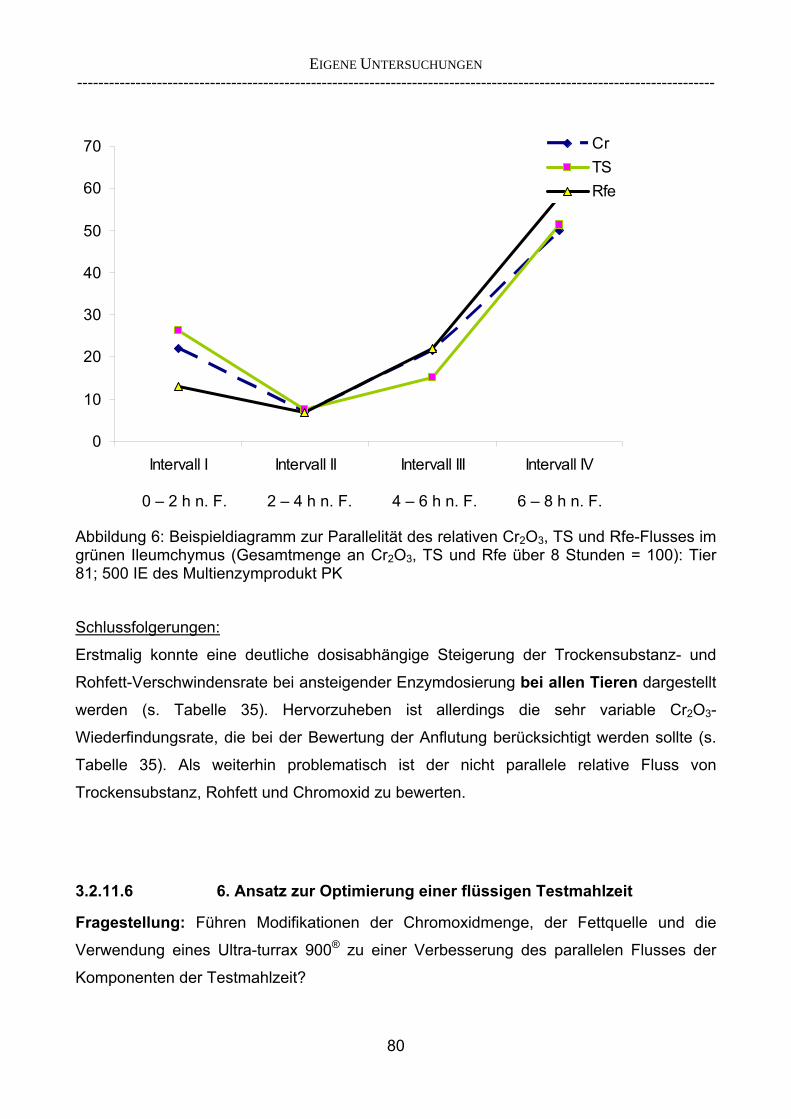
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
80
0
10
20
30
40
50
60
70
Intervall I Intervall II Intervall III Intervall IV
CrTSRfe
0 – 2 h n. F. 2 – 4 h n. F. 4 – 6 h n. F. 6 – 8 h n. F.
Abbildung 6: Beispieldiagramm zur Parallelität des relativen Cr2O3, TS und Rfe-Flusses im grünen Ileumchymus (Gesamtmenge an Cr2O3, TS und Rfe über 8 Stunden = 100): Tier 81; 500 IE des Multienzymprodukt PK
Schlussfolgerungen:
Erstmalig konnte eine deutliche dosisabhängige Steigerung der Trockensubstanz- und
Rohfett-Verschwindensrate bei ansteigender Enzymdosierung bei allen Tieren dargestellt
werden (s. Tabelle 35). Hervorzuheben ist allerdings die sehr variable Cr2O3-
Wiederfindungsrate, die bei der Bewertung der Anflutung berücksichtigt werden sollte (s.
Tabelle 35). Als weiterhin problematisch ist der nicht parallele relative Fluss von
Trockensubstanz, Rohfett und Chromoxid zu bewerten.
3.2.11.6 6. Ansatz zur Optimierung einer flüssigen Testmahlzeit
Fragestellung: Führen Modifikationen der Chromoxidmenge, der Fettquelle und die
Verwendung eines Ultra-turrax 900® zu einer Verbesserung des parallelen Flusses der
Komponenten der Testmahlzeit?

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
81
Versuchsdesign:
Probenkollektion: analog Screening-Test BECKER (2005)
Dauer: 8 Stunden
Beginn der Kollektion: nach „Farbwechsel“
Fütterung am Vorabend des Versuchstages (12 h vor Versuchsbeginn):
400 ml ProvideXtra® pro Schwein.
Testmahlzeit:
Da bisher die Vergleichbarkeit mit voraus gegangenen Versuchen im Vordergrund stand,
wurde bei der flüssigen Testmahlzeit I-V die bisher in diesem Projekt verwendete Menge
Chromoxid (= 0,625g / Mahlzeit) eingesetzt. Aufgrund der im Vergleich zu üblichen
Rationen geringeren Trockensubstanz-Menge je Mahlzeit resultierte aus der Beibehaltung
der Cr2O3-Menge eine erhöhte Chromoxidkonzentration von 4,1 g / kg anstelle von 2,5 g /
kg TS. Da wegen des flüssigen Charakters der Mahlzeit ohnehin die Problematik der
Cr2O3-Entmischung (hohes spezifisches Gewicht) bestand, wurde die Chromoxidmenge
mit dieser Maßnahme an die reduzierte TS-Menge angepasst. Im Folgenden wurden
daher 0,368 g Chromoxid statt 0,625 g pro Versuchsmahlzeit verabreicht (s. Tabelle 37).
Infolge mangelnder Verfügbarkeit (Produktion wurde eingestellt) musste außerdem im
weiteren Verlauf der Versuche auf den Einsatz von Mikrolipid® verzichtet werden. Um die
im Mikrolipid® enthaltenden Fettmengen zu ersetzen, wurde die Olivenöl-Menge von 19,6
g auf nunmehr 29,4 g angehoben (s. Tabelle 37).
Tabelle 37: Inhaltsstoffe (g) der flüssigen Testmahlzeit VI
TS Rfe-Gehalt (%) RfE OL ML Milch* Rfa Cr2O3 (IE)**
151,6 34,6 51,6 29,4 --- 240 64 0,368 0; 500; 5 000; 75 000
* Milchfettgehalt: 1,5 %
** IE=Dosierungen des Multienzymproduktes Pankreatin® in lipolytischer Aktivität
Zubereitung der Testmahlzeit: Ultra-turrax 900®

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
82
Im 6. Versuch zur Optimierung einer Testmahlzeit wurden folgende Modifikationen
vorgenommen:
• Homogenisierung / Zubereitung der Testmahlzeit mittels Ultra-turrax 900®
anstelle eines Handrührgerätes
• Reduzierung der Chromoxidmenge je Mahlzeit (vormals 4,1 g / kg TS) auf 2,5 g
/ kg TS
• Verzicht auf den Einsatz von Mikrolipid® und Ersatz durch Erhöhung der
Olivenöl-Menge, um trotz Verzichts auf Mikrolipid® identische Fettmenge wie in
der flüssigen Testmahlzeit Nr.5 zu erreichen
Ergebnisse:
Bei allen Tieren (Ausnahme Tier 65) wurde bei steigender Enzymdosierung sowohl eine
Reduktion der innerhalb des achtstündigen Kollektionszeitraumes ileal anflutenden
Chymusmenge (TS) als auch der Rohfettmenge beobachtet (s. Tabelle 38).
Tabelle 38: Angeflutete Menge an Trockensubstanz und Rohfett (g) im Kollektionsintervall 0-8 Std. nach Farbumschlag nach Fütterung der flüssigen Testmahlzeit VI
Tier 56 65 82
IE 0 500 5000 75000 0 500 5000 75000 0 500 5000 75000
TS 51,5 42,5 23,7 21,3 61,1 65,8 34,2 26,7 63,0 36,8 29,9 22,5
Rfe 26,1 21,1 6,31 2,67 24,3 18,4 10,3 3,83 34,5 17,8 9,88 2,61
Bei allen eingesetzten Tieren konnte (bis auf wenige Ausnahmen) eine dosisabhängige
Steigerung der Trockensubstanz- und Rohfett-Verschwindensrate verzeichnet werden (s.
Tabelle 39; Ausnahmen Tier 56 und Tier 65). Die Cr2O3-Wiederfindung variierte zwischen
35,8 und 74,1 % (s. Tabelle 39).
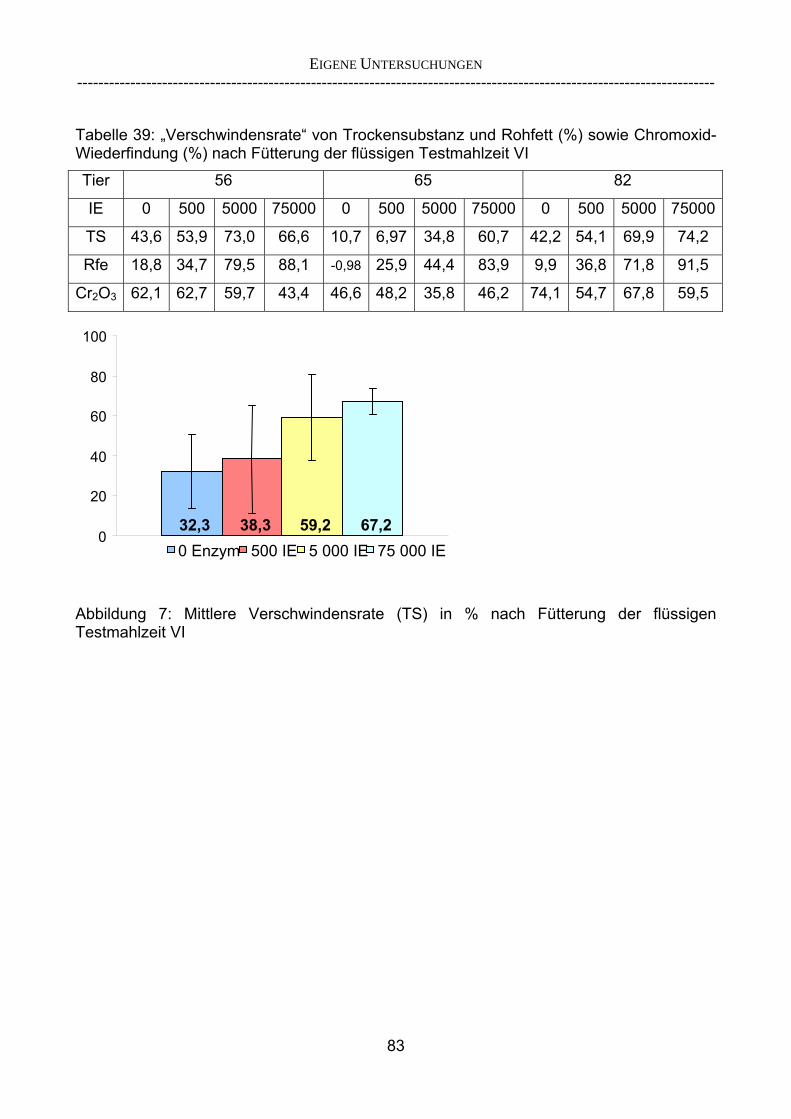
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
83
Tabelle 39: „Verschwindensrate“ von Trockensubstanz und Rohfett (%) sowie Chromoxid-Wiederfindung (%) nach Fütterung der flüssigen Testmahlzeit VI
Tier 56 65 82
IE 0 500 5000 75000 0 500 5000 75000 0 500 5000 75000
TS 43,6 53,9 73,0 66,6 10,7 6,97 34,8 60,7 42,2 54,1 69,9 74,2
Rfe 18,8 34,7 79,5 88,1 -0,98 25,9 44,4 83,9 9,9 36,8 71,8 91,5
Cr2O3 62,1 62,7 59,7 43,4 46,6 48,2 35,8 46,2 74,1 54,7 67,8 59,5
Abbildung 7: Mittlere Verschwindensrate (TS) in % nach Fütterung der flüssigen Testmahlzeit VI
32,3 38,3 59,2 67,20
20
40
60
80
100
0 Enzym 500 IE 5 000 IE 75 000 IE
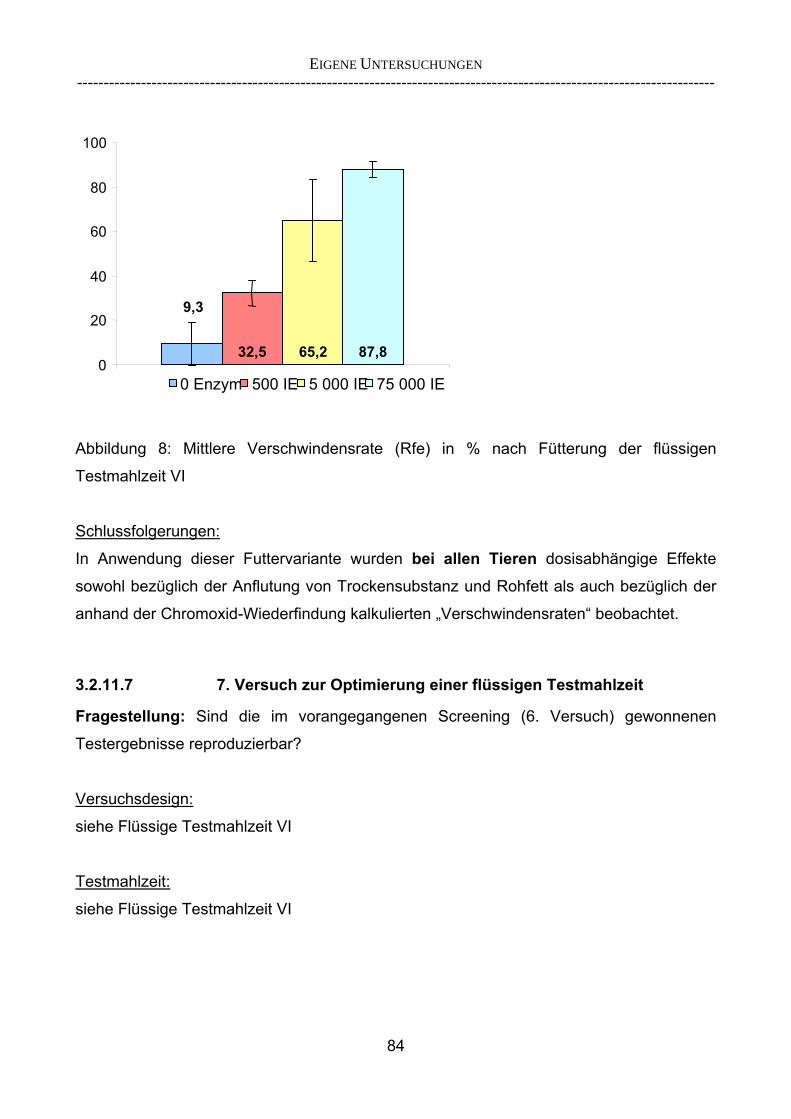
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
84
Abbildung 8: Mittlere Verschwindensrate (Rfe) in % nach Fütterung der flüssigen
Testmahlzeit VI
Schlussfolgerungen:
In Anwendung dieser Futtervariante wurden bei allen Tieren dosisabhängige Effekte
sowohl bezüglich der Anflutung von Trockensubstanz und Rohfett als auch bezüglich der
anhand der Chromoxid-Wiederfindung kalkulierten „Verschwindensraten“ beobachtet.
3.2.11.7 7. Versuch zur Optimierung einer flüssigen Testmahlzeit
Fragestellung: Sind die im vorangegangenen Screening (6. Versuch) gewonnenen
Testergebnisse reproduzierbar?
Versuchsdesign:
siehe Flüssige Testmahlzeit VI
Testmahlzeit:
siehe Flüssige Testmahlzeit VI
9,3
32,5 65,2 87,80
20
40
60
80
100
0 Enzym 500 IE 5 000 IE 75 000 IE

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
85
Um die Wiederholbarkeit der Ergebnisse zu testen wurde die Testmahlzeit aus dem 6.
Versuch zur Entwicklung einer Testmahlzeit in identischer Form bei einer weiteren
Tiergruppe eingesetzt.
Bei allen Tieren, die im 7. Versuch zur Entwicklung einer Testmahlzeit verwendet wurden,
konnte eine dosisabhängige Verringerung der Trockensubstanz- und Rohfett-Anflutung (s.
Tabelle 40) ermittelt werden (Ausnahme Tier 81; vermutlich erhöhte Passagerate, s.
Cr2O3-Wdf. s. Tabelle 41).
Tabelle 40: Angeflutete Menge an Trockensubstanz und Rohfett (g) im Kollektionsintervall 0-8 Std. nach Farbumschlag nach Fütterung der flüssigen Testmahlzeit VI
Tier 43 80 81
IE 0 500 5000 75000 0 500 5000 75000 0 500 5000 75000
TS 74,9 51,8 32,4 20,9 67,2 38,7 32,3 18,5 61,5 39,6 35,4 37,4
Rfe 31,2 23,1 9,14 3,77 29,4 18,8 10,6 3,65 33,1 18,1 9,76 9,23
Tier 83
IE 0 500 5 000 75 000
TS 71,7 30,5 30,4 25,9
Rfe 39,6 12,6 7,88 4,60
Durch den Einsatz der Testmahlzeit VI konnte bei allen Tieren eine deutliche Steigerung
der Trockensubstanz- und Rohfett-Verschwindensrate (%) bewirkt werden (s. Tabelle 41).
Erstmals wurden in allen Versuchen Chromoxid-Wiederfindungen von mehr als 50 %
beobachtet (MW in %: 64,4 ± 9,6; s. Tabelle 41).
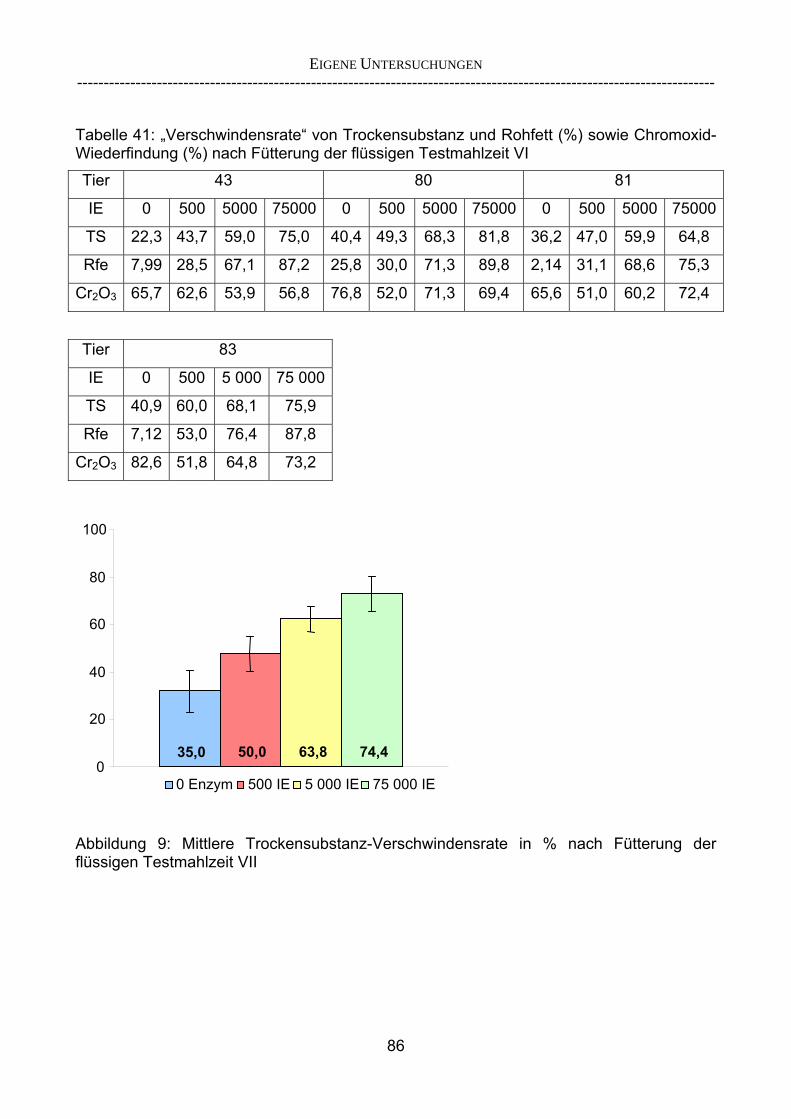
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
86
Tabelle 41: „Verschwindensrate“ von Trockensubstanz und Rohfett (%) sowie Chromoxid-Wiederfindung (%) nach Fütterung der flüssigen Testmahlzeit VI
Tier 43 80 81
IE 0 500 5000 75000 0 500 5000 75000 0 500 5000 75000
TS 22,3 43,7 59,0 75,0 40,4 49,3 68,3 81,8 36,2 47,0 59,9 64,8
Rfe 7,99 28,5 67,1 87,2 25,8 30,0 71,3 89,8 2,14 31,1 68,6 75,3
Cr2O3 65,7 62,6 53,9 56,8 76,8 52,0 71,3 69,4 65,6 51,0 60,2 72,4
Tier 83
IE 0 500 5 000 75 000
TS 40,9 60,0 68,1 75,9
Rfe 7,12 53,0 76,4 87,8
Cr2O3 82,6 51,8 64,8 73,2
Abbildung 9: Mittlere Trockensubstanz-Verschwindensrate in % nach Fütterung der flüssigen Testmahlzeit VII
35,0 50,0 63,8 74,40
20
40
60
80
100
0 Enzym 500 IE 5 000 IE 75 000 IE
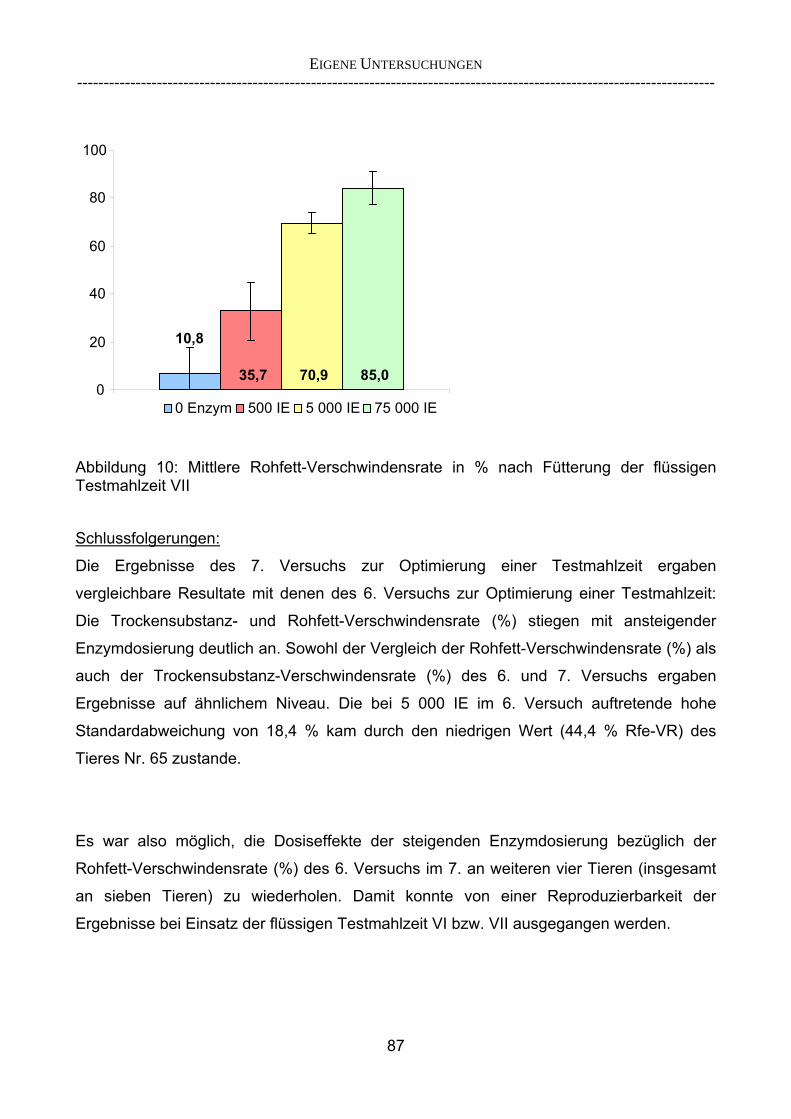
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
87
Abbildung 10: Mittlere Rohfett-Verschwindensrate in % nach Fütterung der flüssigen Testmahlzeit VII Schlussfolgerungen:
Die Ergebnisse des 7. Versuchs zur Optimierung einer Testmahlzeit ergaben
vergleichbare Resultate mit denen des 6. Versuchs zur Optimierung einer Testmahlzeit:
Die Trockensubstanz- und Rohfett-Verschwindensrate (%) stiegen mit ansteigender
Enzymdosierung deutlich an. Sowohl der Vergleich der Rohfett-Verschwindensrate (%) als
auch der Trockensubstanz-Verschwindensrate (%) des 6. und 7. Versuchs ergaben
Ergebnisse auf ähnlichem Niveau. Die bei 5 000 IE im 6. Versuch auftretende hohe
Standardabweichung von 18,4 % kam durch den niedrigen Wert (44,4 % Rfe-VR) des
Tieres Nr. 65 zustande.
Es war also möglich, die Dosiseffekte der steigenden Enzymdosierung bezüglich der
Rohfett-Verschwindensrate (%) des 6. Versuchs im 7. an weiteren vier Tieren (insgesamt
an sieben Tieren) zu wiederholen. Damit konnte von einer Reproduzierbarkeit der
Ergebnisse bei Einsatz der flüssigen Testmahlzeit VI bzw. VII ausgegangen werden.
10,8
35,7 70,9 85,00
20
40
60
80
100
0 Enzym 500 IE 5 000 IE 75 000 IE
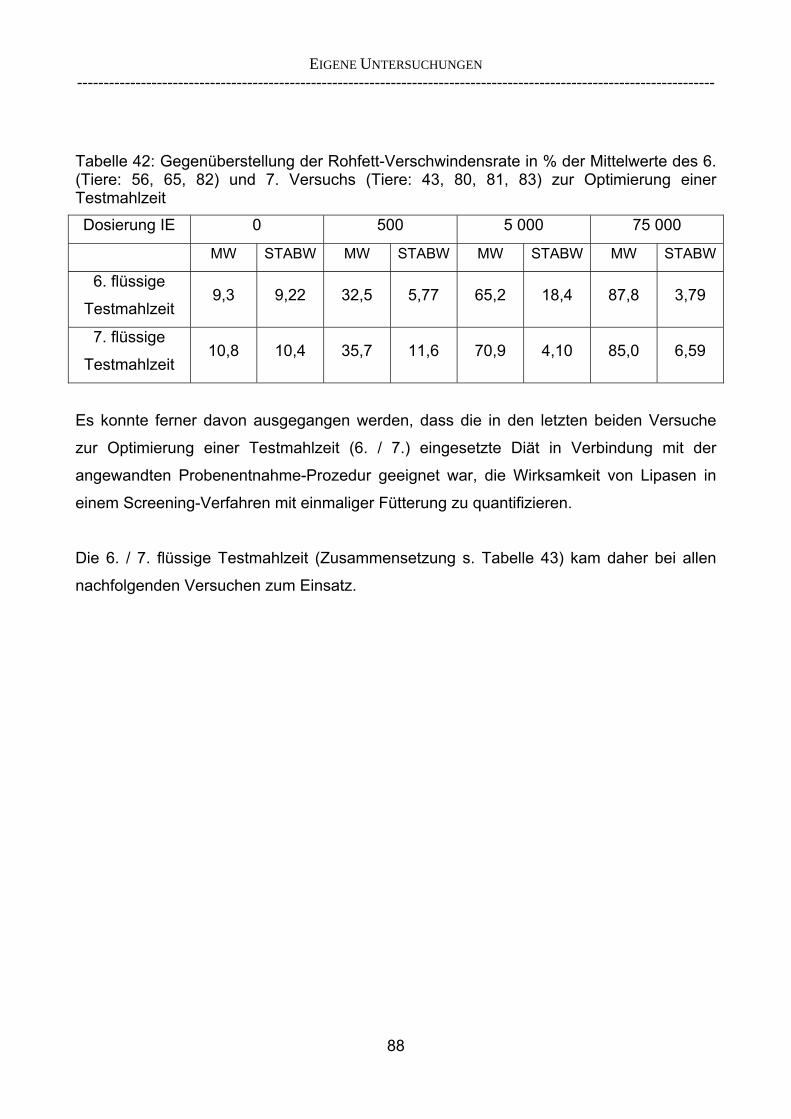
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
88
Tabelle 42: Gegenüberstellung der Rohfett-Verschwindensrate in % der Mittelwerte des 6. (Tiere: 56, 65, 82) und 7. Versuchs (Tiere: 43, 80, 81, 83) zur Optimierung einer Testmahlzeit
Dosierung IE 0 500 5 000 75 000
MW STABW MW STABW MW STABW MW STABW
6. flüssige
Testmahlzeit 9,3 9,22 32,5 5,77 65,2 18,4 87,8 3,79
7. flüssige
Testmahlzeit 10,8 10,4 35,7 11,6 70,9 4,10 85,0 6,59
Es konnte ferner davon ausgegangen werden, dass die in den letzten beiden Versuche
zur Optimierung einer Testmahlzeit (6. / 7.) eingesetzte Diät in Verbindung mit der
angewandten Probenentnahme-Prozedur geeignet war, die Wirksamkeit von Lipasen in
einem Screening-Verfahren mit einmaliger Fütterung zu quantifizieren.
Die 6. / 7. flüssige Testmahlzeit (Zusammensetzung s. Tabelle 43) kam daher bei allen
nachfolgenden Versuchen zum Einsatz.
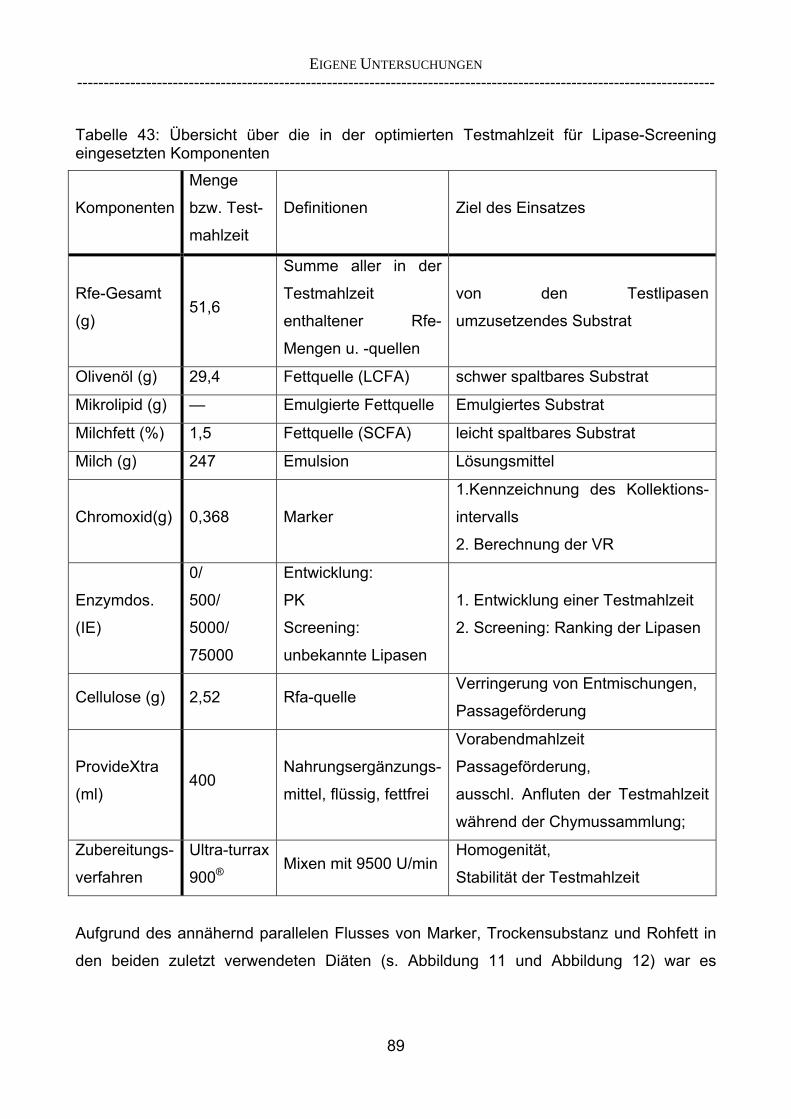
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
89
Tabelle 43: Übersicht über die in der optimierten Testmahlzeit für Lipase-Screening eingesetzten Komponenten
Komponenten
Menge
bzw. Test-
mahlzeit
Definitionen Ziel des Einsatzes
Rfe-Gesamt
(g) 51,6
Summe aller in der
Testmahlzeit
enthaltener Rfe-
Mengen u. -quellen
von den Testlipasen
umzusetzendes Substrat
Olivenöl (g) 29,4 Fettquelle (LCFA) schwer spaltbares Substrat
Mikrolipid (g) — Emulgierte Fettquelle Emulgiertes Substrat
Milchfett (%) 1,5 Fettquelle (SCFA) leicht spaltbares Substrat
Milch (g) 247 Emulsion Lösungsmittel
Chromoxid(g) 0,368 Marker
1.Kennzeichnung des Kollektions-
intervalls
2. Berechnung der VR
Enzymdos.
(IE)
0/
500/
5000/
75000
Entwicklung:
PK
Screening:
unbekannte Lipasen
1. Entwicklung einer Testmahlzeit
2. Screening: Ranking der Lipasen
Cellulose (g) 2,52 Rfa-quelle Verringerung von Entmischungen,
Passageförderung
ProvideXtra
(ml) 400
Nahrungsergänzungs-
mittel, flüssig, fettfrei
Vorabendmahlzeit
Passageförderung,
ausschl. Anfluten der Testmahlzeit
während der Chymussammlung;
Zubereitungs-
verfahren
Ultra-turrax
900® Mixen mit 9500 U/min
Homogenität,
Stabilität der Testmahlzeit
Aufgrund des annähernd parallelen Flusses von Marker, Trockensubstanz und Rohfett in
den beiden zuletzt verwendeten Diäten (s. Abbildung 11 und Abbildung 12) war es
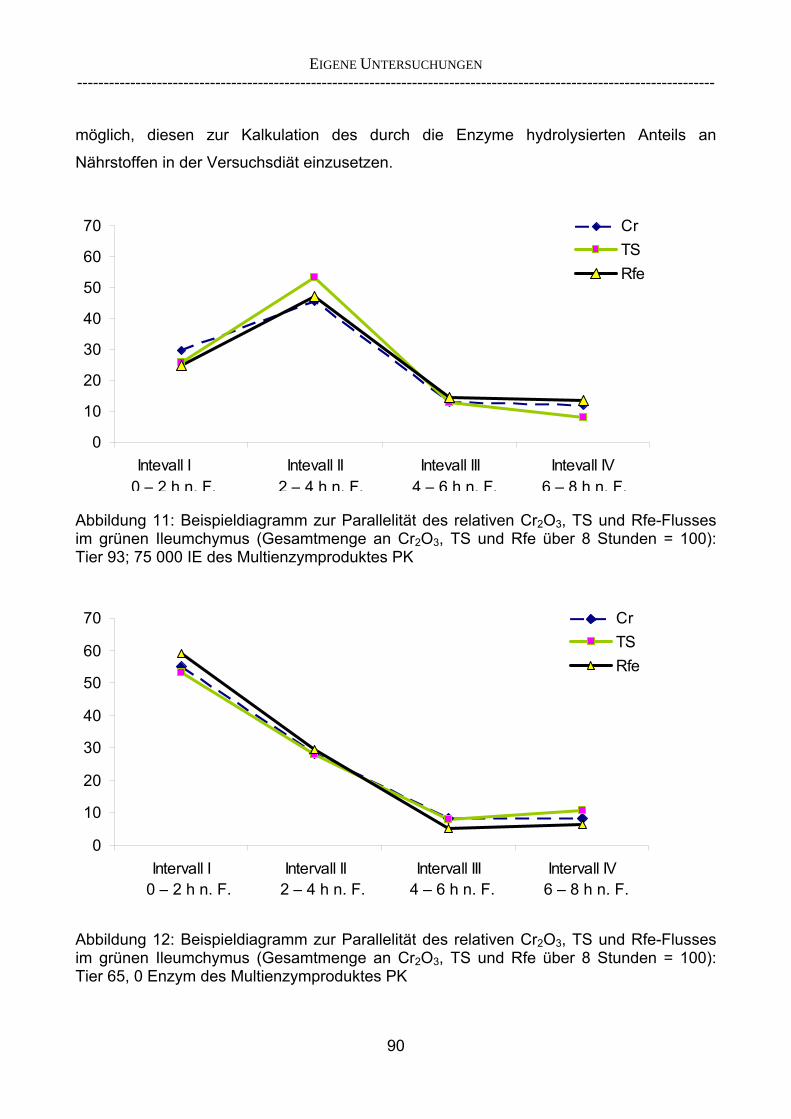
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
90
möglich, diesen zur Kalkulation des durch die Enzyme hydrolysierten Anteils an
Nährstoffen in der Versuchsdiät einzusetzen.
0
10
20
30
40
50
60
70
Intevall I Intevall II Intevall III Intevall IV
CrTSRfe
Abbildung 11: Beispieldiagramm zur Parallelität des relativen Cr2O3, TS und Rfe-Flusses im grünen Ileumchymus (Gesamtmenge an Cr2O3, TS und Rfe über 8 Stunden = 100): Tier 93; 75 000 IE des Multienzymproduktes PK
0
10
20
30
40
50
60
70
Intervall I Intervall II Intervall III Intervall IV
CrTSRfe
Abbildung 12: Beispieldiagramm zur Parallelität des relativen Cr2O3, TS und Rfe-Flusses im grünen Ileumchymus (Gesamtmenge an Cr2O3, TS und Rfe über 8 Stunden = 100): Tier 65, 0 Enzym des Multienzymproduktes PK
0 – 2 h n. F. 2 – 4 h n. F. 4 – 6 h n. F. 6 – 8 h n. F.
0 – 2 h n. F. 2 – 4 h n. F. 4 – 6 h n. F. 6 – 8 h n. F.

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
91
3.3 Ergebnisse
3.3.1 Screening-Test für Lipasen
Nachfolgend wird auf die Versuche eingegangen, die zur Beurteilung der Wirksamkeit von
Monolipasen durchgeführt wurden (Screening-Test für Lipasen). Die Versuche, die im
Rahmen der Entwicklung des Lipase-Screening-Tests stattfanden, sind inklusive ihrer
Ergebnisse im Kapitel „Material und Methoden“ beschrieben.
In den im Kollektionszeitraum (0-8 Stunden n. F.) gewonnenen Chymusproben wurden
folgende Parameter bestimmt:
• Durch Wiegen bestimmte bzw. analysierter Werte:
⇒ Trockensubstanz-Menge (g; uS-Masse x TS-Gehalt)
⇒ Rohfettkonzentration (g / kg) in der TS
⇒ Absolute Anflutung von Rohfett (g; TS-Masse x Rfe-Gehalt)
• Marker-bezogene Werte
⇒ Chromoxid-Konzentration (g / kg TS)
⇒ Chromoxid-Wiederfindung (%)
⇒ Verhältnis der Konzentrationen von Rohfett und Chromoxid
• Sonstige kalkulatorisch abgeleitete Werte
⇒ Trockensubstanz-Verschwindensrate in % (TS-VR)
⇒ Rohfett-Verschwindensrate in % (Rfe-VR)
⇒ Relative Anflutung von TS und Rfe
(Werte im Vergleich zu denen von PL-0-Tieren)
⇒ Relative TS- und Rfe-VR
(Werte im Vergleich zu denen von PL-0-Tieren)
Die Ergebnisse werden im Folgenden abschnittsweise für die verschiedenen Enzyme
einzeln vorgestellt. Innerhalb eines jeden Abschnitts werden zuerst die Kontrolltiere den
pankreasgangligierten Tieren ohne Enzymzulage (PL-0-Tiere) gegenübergestellt.

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
92
Anschließend wird die Wirkung der Enzymzulage auf die verschiedenen Parameter (s. o.)
betrachtet; dabei erfolgt zunächst eine vergleichende Darstellung der Dosis-
Wirkungsbeziehung jedes einzelnen Enzyms (D1, D2, D3 sowie im Vergleich mit KT und
PL-0). Nachfolgend erfolgt eine vergleichende Darstellung der Effekte verschiedener
Lipasen innerhalb einer Dosierung. Abschließend erfolgt eine Rangierung der Enzyme
anhand des jeweiligen Parameters. Als effektivstes Enzym wird dabei jeweils dasjenige
bezeichnet, welches die durch die Pankreasgangligatur bedingten Veränderungen der PL-
Tiere am deutlichsten reduziert bzw. zu einer „Normalisierung“ im Sinne einer Angleichung
an die Werte der Kontrolltiere führt.
Die aufgeführten Werte stellen -soweit nicht anders gekennzeichnet- Mittelwerte der
Ergebnisse von mindestens drei Tieren dar (Darstellung als Mittelwert ±
Standardabweichung). Die Einzelwerte können dem Tabellenanhang entnommen werden.
Signifikante Unterschiede (p ≤ 0,05) zwischen den verschiedenen Enzymen gleicher
Dosierung werden durch Großbuchstaben (z.B. A, B) gekennzeichnet. Signifikante
Unterschiede (p ≤ 0,05) zwischen den verschiedenen Dosierungen eines Enzyms werden
durch unterschiedliche Kleinbuchstaben (z.B. a, b) gekennzeichnet.
3.3.2 Allgemeine Beobachtungen
Das Allgemeinbefinden aller Versuchstiere war während der Versuche ungestört. Das
angebotene Versuchsfutter wurde von den Tieren ausnahmslos vollständig und innerhalb
weniger Minuten aufgenommen.
3.3.3 Untersuchte Parameter
3.3.3.1 Absolute Anflutung von Chymus (Trockensubstanz)
Die aus dem absoluten Chymusfluss (uS) und dem Trockensubstanzgehalt des Chymus
resultierende, über 8 h n. F. am terminalen Ileum anflutende TS-Menge (g) betrug bei den

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
93
Kontrolltieren im Mittel 24,2 (± 1,53) g und war somit signifikant geringer als die ileale TS-
Anflutung der pankreasgangligierten Tiere ohne Enzymzulage (68,8 ± 5,82 g; s. Tabelle
44).
Tabelle 44: Trockensubstanz-Anflutung (g) bei KT und PL-0 –Tieren innerhalb von 8 h n. F. KT 24,2 ± 1,53 aA PL-0 68,8 ± 5,82 bB
Wurden den PL-Tieren Lipasen (E1 - E5) substituiert, kam es zu einer Reduktion der
Trockensubstanz-Anflutung (s. Tabelle 45). Ein dosisabhängiger Effekt konnte bei allen
Enzymen festgestellt, jedoch nicht immer statistisch abgesichert werden (s. Tabelle 45).
Nach Zulage der niedrigsten Dosierung (D1) konnte lediglich durch das Multienzymprodukt
PK eine im Vergleich zu den PL-0-Tieren signifikante Reduktion der anflutenden Chymus
(TS)-Menge beobachtet werden. Durch keines der Enzyme wurde das Niveau der
Kontrolltiere erreicht. In der mittleren Dosierung (D2) bewirkten nur Enzym E1, E4 und PK
eine signifikante Reduktion der Trockensubstanz-Anflutung im Vergleich zu den
pankreasgangligierten Tieren ohne Enzymzulage. Aufgrund hoher Standardabweichungen
konnten E1, E3, E4 und E5 allerdings nicht statistisch von den KT-Tieren abgegrenzt
werden. Obwohl durch das Multienzymprodukt PK die größten Effekte erzielt wurden,
wurde durch dieses, ebenso wie von E2, das Niveau der Kontrolltiere nicht erreicht (s.
Tabelle 45). In der höchsten Dosierung (D3) führten alle verabreichten Enzyme zu einer
signifikanten Verringerung der Trockensubstanz-Anflutung im Vergleich zu PL-0-Tieren.
Durch alle verabreichten Testenzyme mit Ausnahme von E5 wurde das Niveau der KT-
Tiere erreicht. Dosisabhängige Effekte zwischen D1 und D3 waren lediglich für E1, E3 und
E4 zu verzeichnen.
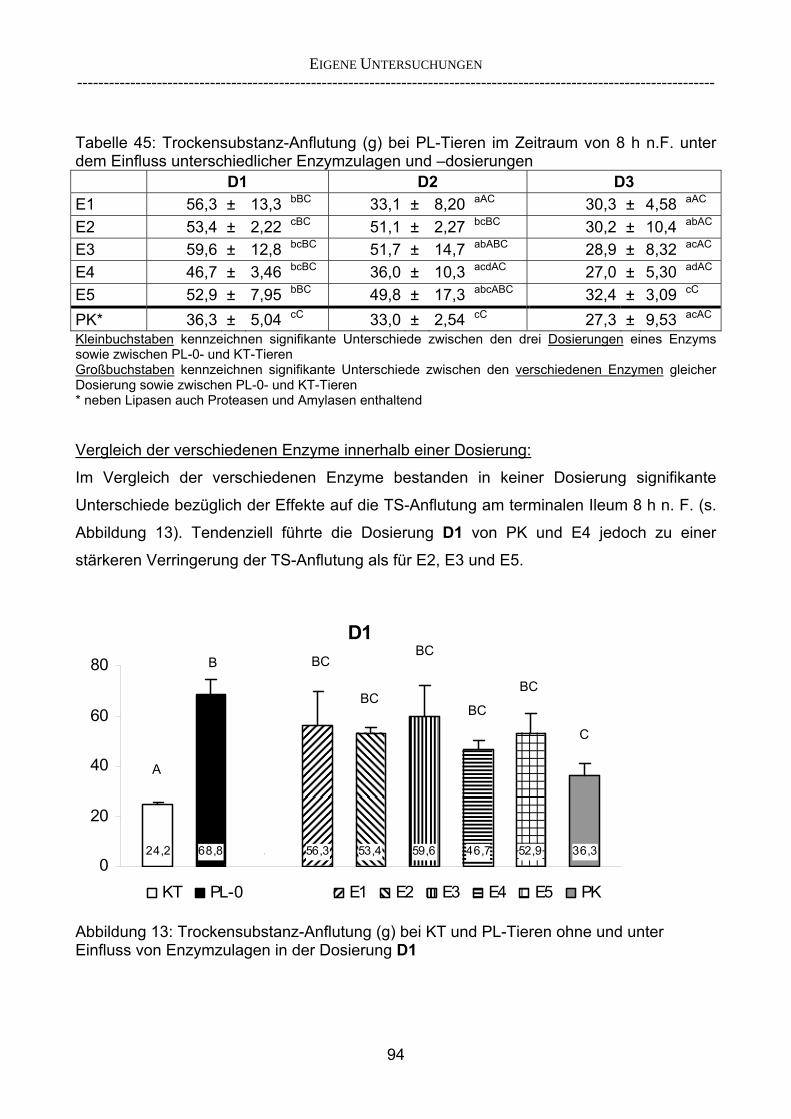
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
94
Tabelle 45: Trockensubstanz-Anflutung (g) bei PL-Tieren im Zeitraum von 8 h n.F. unter dem Einfluss unterschiedlicher Enzymzulagen und –dosierungen D1 D2 D3 E1 56,3 ± 13,3 bBC 33,1 ± 8,20 aAC 30,3 ± 4,58 aAC E2 53,4 ± 2,22 cBC 51,1 ± 2,27 bcBC 30,2 ± 10,4 abAC
E3 59,6 ± 12,8 bcBC 51,7 ± 14,7 abABC 28,9 ± 8,32 acAC E4 46,7 ± 3,46 bcBC 36,0 ± 10,3 acdAC 27,0 ± 5,30 adAC
E5 52,9 ± 7,95 bBC 49,8 ± 17,3 abcABC 32,4 ± 3,09 cC PK* 36,3 ± 5,04 cC 33,0 ± 2,54 cC 27,3 ± 9,53 acAC Kleinbuchstaben kennzeichnen signifikante Unterschiede zwischen den drei Dosierungen eines Enzyms sowie zwischen PL-0- und KT-Tieren Großbuchstaben kennzeichnen signifikante Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung sowie zwischen PL-0- und KT-Tieren * neben Lipasen auch Proteasen und Amylasen enthaltend
Vergleich der verschiedenen Enzyme innerhalb einer Dosierung:
Im Vergleich der verschiedenen Enzyme bestanden in keiner Dosierung signifikante
Unterschiede bezüglich der Effekte auf die TS-Anflutung am terminalen Ileum 8 h n. F. (s.
Abbildung 13). Tendenziell führte die Dosierung D1 von PK und E4 jedoch zu einer
stärkeren Verringerung der TS-Anflutung als für E2, E3 und E5.
D1
24,2 0 53,4 46,768,8 56,3 59,6 52,9 36,30
20
40
60
80
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 13: Trockensubstanz-Anflutung (g) bei KT und PL-Tieren ohne und unter Einfluss von Enzymzulagen in der Dosierung D1
A
B BC
BC
BC
BC
BC
C
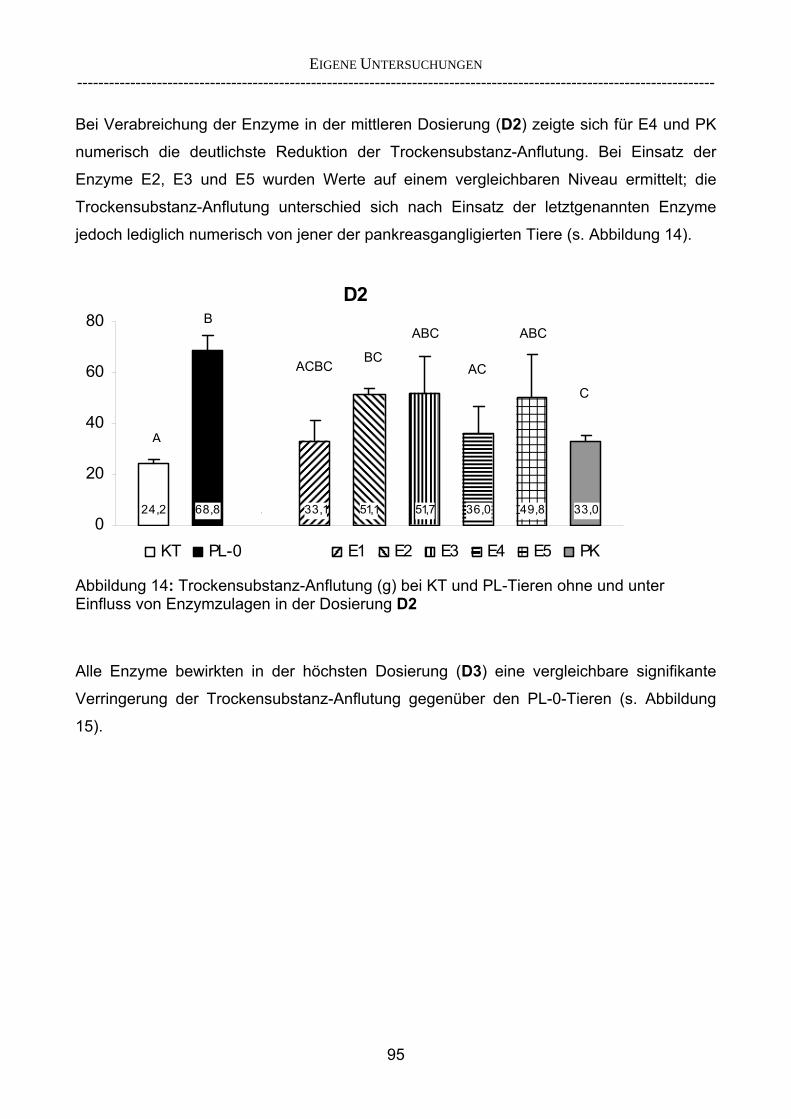
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
95
Bei Verabreichung der Enzyme in der mittleren Dosierung (D2) zeigte sich für E4 und PK
numerisch die deutlichste Reduktion der Trockensubstanz-Anflutung. Bei Einsatz der
Enzyme E2, E3 und E5 wurden Werte auf einem vergleichbaren Niveau ermittelt; die
Trockensubstanz-Anflutung unterschied sich nach Einsatz der letztgenannten Enzyme
jedoch lediglich numerisch von jener der pankreasgangligierten Tiere (s. Abbildung 14).
D2
24,2 68,8 0 33,1 51,1 51,7 36,0 49,8 33,00
20
40
60
80
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 14: Trockensubstanz-Anflutung (g) bei KT und PL-Tieren ohne und unter Einfluss von Enzymzulagen in der Dosierung D2
Alle Enzyme bewirkten in der höchsten Dosierung (D3) eine vergleichbare signifikante
Verringerung der Trockensubstanz-Anflutung gegenüber den PL-0-Tieren (s. Abbildung
15).
A
B
ACBCBC
ABC
AC
ABC
C
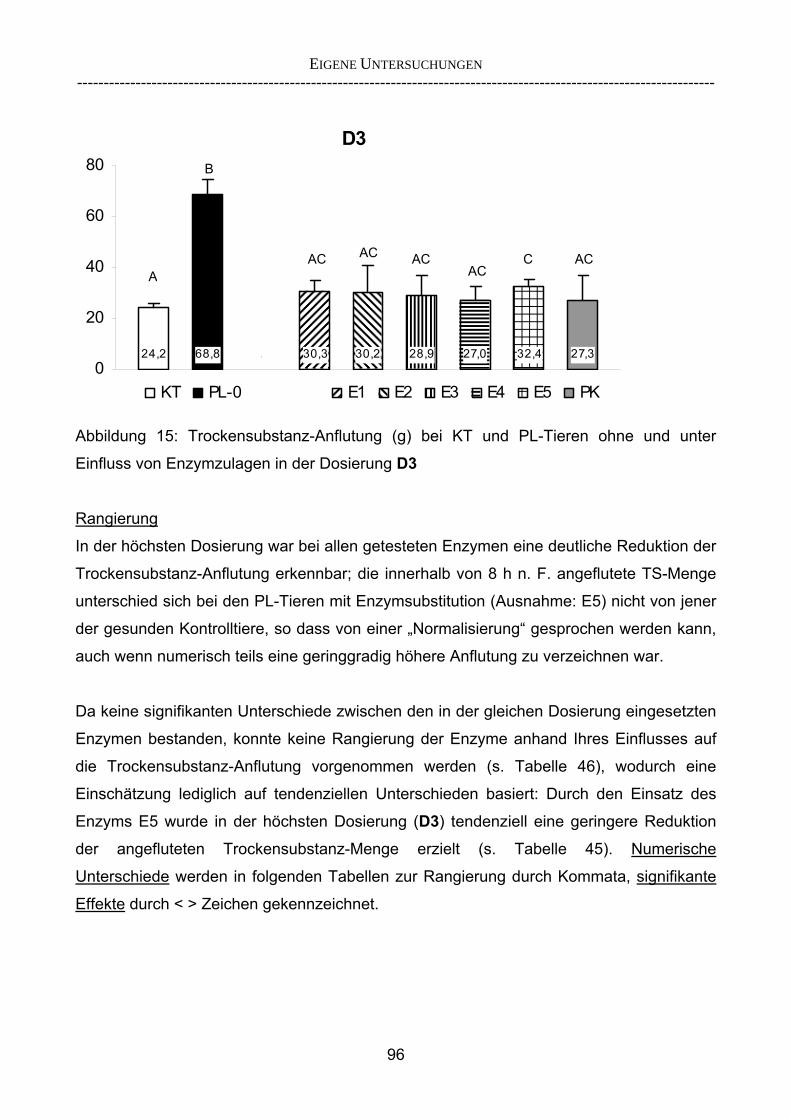
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
96
D3
24,2 68,8 0 30,2 28,9 27,0 27,330,3 32,40
20
40
60
80
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 15: Trockensubstanz-Anflutung (g) bei KT und PL-Tieren ohne und unter
Einfluss von Enzymzulagen in der Dosierung D3
Rangierung
In der höchsten Dosierung war bei allen getesteten Enzymen eine deutliche Reduktion der
Trockensubstanz-Anflutung erkennbar; die innerhalb von 8 h n. F. angeflutete TS-Menge
unterschied sich bei den PL-Tieren mit Enzymsubstitution (Ausnahme: E5) nicht von jener
der gesunden Kontrolltiere, so dass von einer „Normalisierung“ gesprochen werden kann,
auch wenn numerisch teils eine geringgradig höhere Anflutung zu verzeichnen war.
Da keine signifikanten Unterschiede zwischen den in der gleichen Dosierung eingesetzten
Enzymen bestanden, konnte keine Rangierung der Enzyme anhand Ihres Einflusses auf
die Trockensubstanz-Anflutung vorgenommen werden (s. Tabelle 46), wodurch eine
Einschätzung lediglich auf tendenziellen Unterschieden basiert: Durch den Einsatz des
Enzyms E5 wurde in der höchsten Dosierung (D3) tendenziell eine geringere Reduktion
der angefluteten Trockensubstanz-Menge erzielt (s. Tabelle 45). Numerische
Unterschiede werden in folgenden Tabellen zur Rangierung durch Kommata, signifikante
Effekte durch < > Zeichen gekennzeichnet.
A
B
AC AC ACAC
C AC

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
97
Tabelle 46: Rangierung der Enzyme bezüglich der Fähigkeit zur Reduktion der TS-Anflutung (g) am terminalen Ileum Dosierung Wirksamkeit (numerisch) Wirksamkeit (statistische Unterschiede) D1 PK, E4, E5, E2, E1, E3 ─ D2 PK, E1, E4, E5, E2, E3 ─ D3 E4, PK, E3, E2, E1, E5 ─
3.3.3.2 Rohfettkonzentration im Chymus
Im Vergleich zu den Kontrolltieren, die im Ileumchymus innerhalb von 8 Stunden n. F. eine
mittlere Rohfett-Konzentration von 169 (± 35,6) g / kg TS aufwiesen (s. Tabelle 47), war
die Rohfett-Konzentration bei pankreasgangligierten Tieren massiv erhöht (509 ± 62,6 g /
kg TS).
Hinweis: Die verabreichte Testmahlzeit enthielt eine Rohfett-Konzentration von 351 g / kg
TS.
Tabelle 47: Rohfett-Konzentration (g / kg TS) im ilealen Chymus von KT und PL-0 –Tieren (Zeitraum: über 8 h n. F.) KT 169 ± 35,6 aA PL-0 509 ± 62,6 bB Alle zugelegten Enzymprodukte bewirkten eine dosisabhängige Reduktion der Rohfett-
Konzentration des am terminalen Ileum anflutenden Chymus, die jedoch nicht in allen
Fällen statistisch abgesichert werden konnte (s. Tabelle 48): In der niedrigsten Dosierung
(D1) führten alle Enzyme lediglich zu einer tendenziellen Verringerung der Rohfett-
Konzentration im Chymus des terminalen Ileums. Das Niveau der Kontrolltiere wurde bei
dieser Dosierung von keinem der Testenzyme erreicht (s. Tabelle 48). Nach Erhöhung der
Enzymdosierung (D2) ergaben sich lediglich nach Einsatz der Enzyme PK und E1
signifikante Verringerungen der Rohfett-Konzentration des Chymus im Vergleich zu den
PL-Tieren ohne Enzymzulage, wobei auch hier das Niveau der KT-Tiere durch keines der
Enzyme erreicht wurde. In der höchsten Dosierung (D3) führten vier der sechs geprüften
Enzyme zu einer signifikanten Reduktion der Rohfett-Konzentration gegenüber den PL-0-
Tieren (s. Tabelle 48). In dieser Dosierung wurde nach Einsatz der Enzyme PK, E1, E3
und E5 bezüglich der Rohfett-Konzentration im Chymus das Niveau der KT-Tiere erreicht.
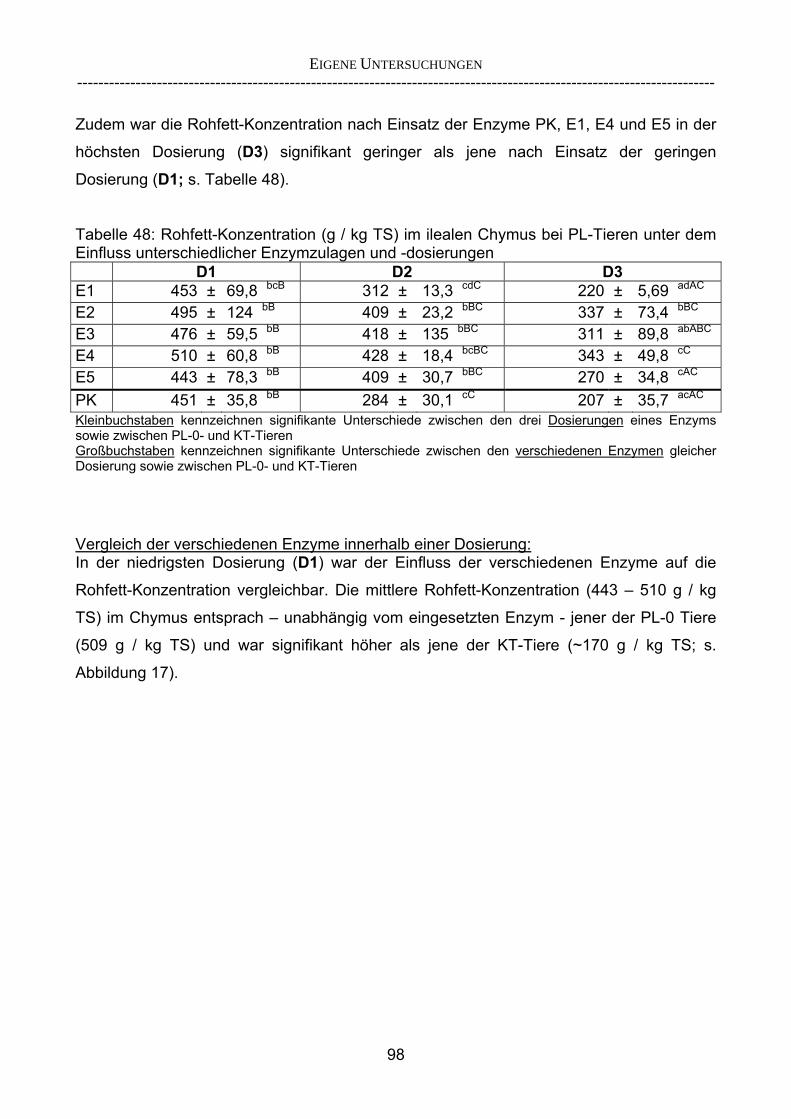
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
98
Zudem war die Rohfett-Konzentration nach Einsatz der Enzyme PK, E1, E4 und E5 in der
höchsten Dosierung (D3) signifikant geringer als jene nach Einsatz der geringen
Dosierung (D1; s. Tabelle 48).
Tabelle 48: Rohfett-Konzentration (g / kg TS) im ilealen Chymus bei PL-Tieren unter dem Einfluss unterschiedlicher Enzymzulagen und -dosierungen D1 D2 D3 E1 453 ± 69,8 bcB 312 ± 13,3 cdC 220 ± 5,69 adAC E2 495 ± 124 bB 409 ± 23,2 bBC 337 ± 73,4 bBC E3 476 ± 59,5 bB 418 ± 135 bBC 311 ± 89,8 abABC E4 510 ± 60,8 bB 428 ± 18,4 bcBC 343 ± 49,8 cC E5 443 ± 78,3 bB 409 ± 30,7 bBC 270 ± 34,8 cAC PK 451 ± 35,8 bB 284 ± 30,1 cC 207 ± 35,7 acAC Kleinbuchstaben kennzeichnen signifikante Unterschiede zwischen den drei Dosierungen eines Enzyms sowie zwischen PL-0- und KT-Tieren Großbuchstaben kennzeichnen signifikante Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung sowie zwischen PL-0- und KT-Tieren Vergleich der verschiedenen Enzyme innerhalb einer Dosierung: In der niedrigsten Dosierung (D1) war der Einfluss der verschiedenen Enzyme auf die
Rohfett-Konzentration vergleichbar. Die mittlere Rohfett-Konzentration (443 – 510 g / kg
TS) im Chymus entsprach – unabhängig vom eingesetzten Enzym - jener der PL-0 Tiere
(509 g / kg TS) und war signifikant höher als jene der KT-Tiere (~170 g / kg TS; s.
Abbildung 17).
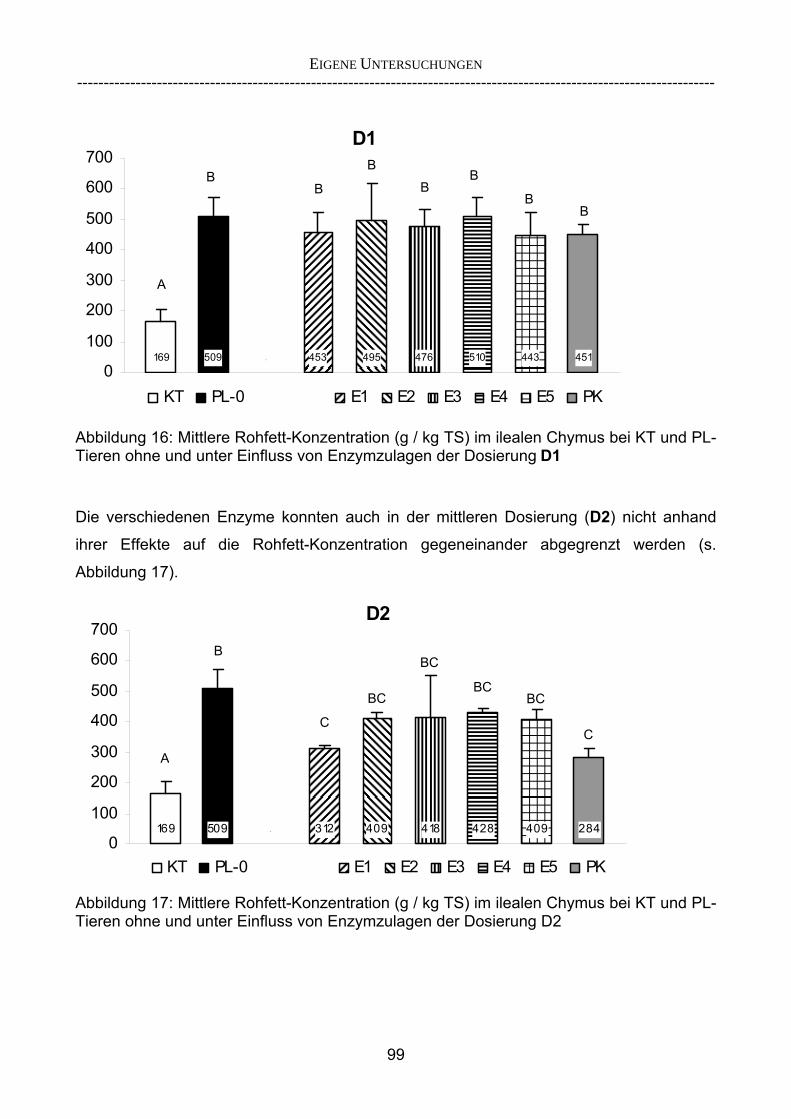
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
99
D1
169 509 0 453 495 476 510 443 4510
100
200
300
400
500
600
700
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 16: Mittlere Rohfett-Konzentration (g / kg TS) im ilealen Chymus bei KT und PL-Tieren ohne und unter Einfluss von Enzymzulagen der Dosierung D1 Die verschiedenen Enzyme konnten auch in der mittleren Dosierung (D2) nicht anhand
ihrer Effekte auf die Rohfett-Konzentration gegeneinander abgegrenzt werden (s.
Abbildung 17).
D2
169 509 0 312 418 428 409 2844090
100
200
300
400
500
600
700
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 17: Mittlere Rohfett-Konzentration (g / kg TS) im ilealen Chymus bei KT und PL-Tieren ohne und unter Einfluss von Enzymzulagen der Dosierung D2
A
B B
B
BB
BB
A
B
C
BC
BC
BCBC
C
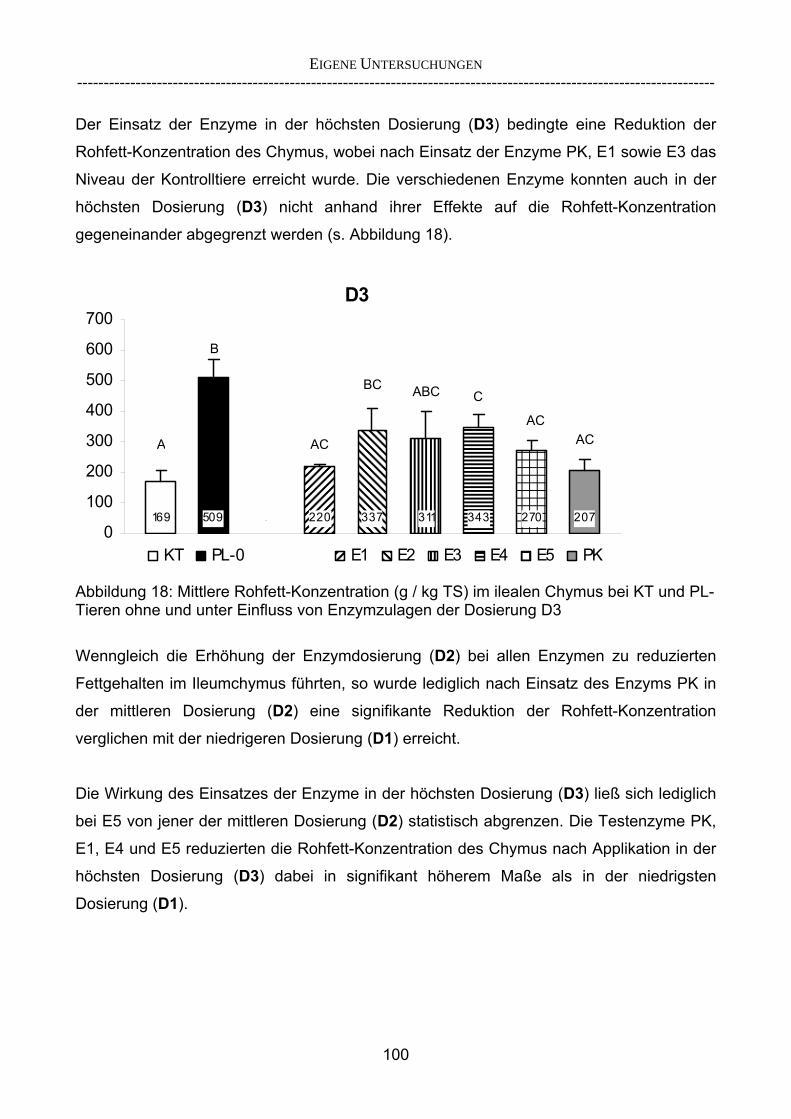
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
100
Der Einsatz der Enzyme in der höchsten Dosierung (D3) bedingte eine Reduktion der
Rohfett-Konzentration des Chymus, wobei nach Einsatz der Enzyme PK, E1 sowie E3 das
Niveau der Kontrolltiere erreicht wurde. Die verschiedenen Enzyme konnten auch in der
höchsten Dosierung (D3) nicht anhand ihrer Effekte auf die Rohfett-Konzentration
gegeneinander abgegrenzt werden (s. Abbildung 18).
D3
169 509 0 220 337 311 343 270 2070
100
200
300
400
500
600
700
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 18: Mittlere Rohfett-Konzentration (g / kg TS) im ilealen Chymus bei KT und PL-Tieren ohne und unter Einfluss von Enzymzulagen der Dosierung D3
Wenngleich die Erhöhung der Enzymdosierung (D2) bei allen Enzymen zu reduzierten
Fettgehalten im Ileumchymus führten, so wurde lediglich nach Einsatz des Enzyms PK in
der mittleren Dosierung (D2) eine signifikante Reduktion der Rohfett-Konzentration
verglichen mit der niedrigeren Dosierung (D1) erreicht.
Die Wirkung des Einsatzes der Enzyme in der höchsten Dosierung (D3) ließ sich lediglich
bei E5 von jener der mittleren Dosierung (D2) statistisch abgrenzen. Die Testenzyme PK,
E1, E4 und E5 reduzierten die Rohfett-Konzentration des Chymus nach Applikation in der
höchsten Dosierung (D3) dabei in signifikant höherem Maße als in der niedrigsten
Dosierung (D1).
A
B
AC
BC ABC C
ACAC

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
101
Rangierung:
Alle Enzyme hatten eine dosisabhängige Reduktion der Rohfett-Konzentration im ileal
anflutendem Chymus zur Folge. Dennoch sank lediglich in der höchsten Dosierung bei
vier Enzymen (E1, E5, E3 und PK) die Rohfett-Konzentration des ilealen Chymus auf das
Niveau der KT-Tiere ab. Aufgrund mangelnder signifikanter Unterschiede zwischen den
Effekten der einzelnen Enzyme innerhalb einer Dosierung konnte eine Rangierung der
Enzyme nur unter Einbeziehung von lediglich tendenziellen Effekten erfolgen (s. Tabelle
49). Das Multienzympräparat PK verringerte die Rohfett-Konzentration am effektivsten.
Von den Monolipasen war E1 in den Dosierungen D1 und D2 am wirkungsvollsten
bezüglich der Reduktion der Rohfett-Konzentration des Chymus. In der höchsten
Dosierung (D3) wurde durch dieses Enzym die Rohfett-Konzentration im Chymus im
Vergleich zu den PL-0-Tieren mehr als halbiert (Reduktion von 509 g / kg TS auf 220 g /
kg TS).
Tabelle 49: Rangierung der Enzyme bezüglich der Reduktion der Rohfett-Konzentration(g / kg TS) im Chymus des terminalen Ileums Dosierung Wirksamkeit (numerisch) Wirksamkeit (statistische Unterschiede) D1 E5, PK, E1, E3, E2, E4 ─ D2 PK, E1, E5, E2, E3, E4 ─ D3 PK, E1, E5, E3, E2, E4 ─
3.3.3.3 Absolute Anflutung von Rohfett
Die am terminalen Ileum innerhalb des Zeitraumes 8 h n. F. angeflutete Rohfett-Menge (g)
betrug bei den Kontrolltieren 4,05 (± 0,70) g. Bei den PL-0-Tieren flutete hingegen ohne
Enzymzulage mit 33,3 (± 4,44) g eine um den Faktor 8,2 signifikant erhöhte Menge an
Rohfett an (s. Tabelle 50).
Tabelle 50: Rohfett-Anflutung (g) am terminalen Ileum bei KT- und PL-0 –Tieren (Zeitraum: über 8 h n. F.) KT 4,05 ± 0,70 aA PL-0 33,3 ± 4,44 bB

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
102
Alle eingesetzten Enzyme führten zu einer dosisabhängigen Verringerung der Rohfett-
Anflutung im Vergleich zu den PL-0-Tieren (s. Tabelle 50). In der geringsten Dosierung
(D1) bewirkte lediglich das Multienzymprodukt PK eine signifikante Reduktion der Rohfett-
Anflutung im Vergleich zu den PL-0-Tieren. Das Niveau der KT-Tiere wurde in der
geringsten Dosierung (D1) durch keines der Testenzyme erreicht. Während es nach
Einsatz von PK, E1, E2 und E4 in der mittleren Dosierung (D2) zu einer signifikanten
Reduktion der Rohfett-Anflutung im Vergleich zu den PL-0-Tieren kam, wurde diese durch
Gabe der Enzyme E3 und E5 nicht signifikant reduziert. Infolge der hohen
Standardabweichung konnte die Rohfett-Anflutung nach Einsatz der Enzyme E4 und E5
nicht von dem Niveau der KT-Tiere abgegrenzt werden.
Die Werte, die nach Einsatz des Multienzymproduktes PK und der Monolipase E3 in der
höchsten Dosierung (D3) ermittelt wurden, waren nicht von den Werten der Kontrolltiere
abzugrenzen.
Zwischen der niedrigsten und der mittleren Dosierung (D1 und D2) bestanden lediglich bei
E1 signifikante Unterschiede in der Rohfett-Anflutung. Ebenso konnte für jedes der
getesteten Enzyme durch Applikation der höchsten Dosierung (D3) eine signifikante
Reduktion der Rohfett-Anflutung gegenüber jener nach Einsatz der niedrigsten Dosierung
(D1) erreicht werden (s. Tabelle 51).
Tabelle 51: Absolute Rohfett-Anflutung (g) bei PL-Tieren unter dem Einfluss unterschiedlicher Enzymzulagen und -dosierungen D1 D2 D3 E1 25,6 ± 6,88 bBC 10,4 ± 2,89 cC 6,66 ± 0,91 cC E2 26,3 ± 6,17 bcBC 21,0 ± 0,20 cdC 9,71 ± 1,41 dC E3 28,9 ± 8,05 bBC 20,9 ± 6,64 bcBC 9,03 ± 3,95 acAC E4 23,9 ± 3,00 bBC 15,5 ± 5,04 acdAC 9,13 ± 0,82 cdC E5 23,2 ± 3,20 bBC 20,6 ± 7,89 abcABC 8,84 ± 1,96 cC PK 16,5 ± 3,39 cC 9,40 ± 1,37 cdC 5,83 ± 2,99 adAC Kleinbuchstaben kennzeichnen signifikante Unterschiede zwischen den drei Dosierungen eines Enzyms sowie zwischen PL-0- und KT-Tieren Großbuchstaben kennzeichnen signifikante Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung sowie zwischen PL-0- und KT-Tieren
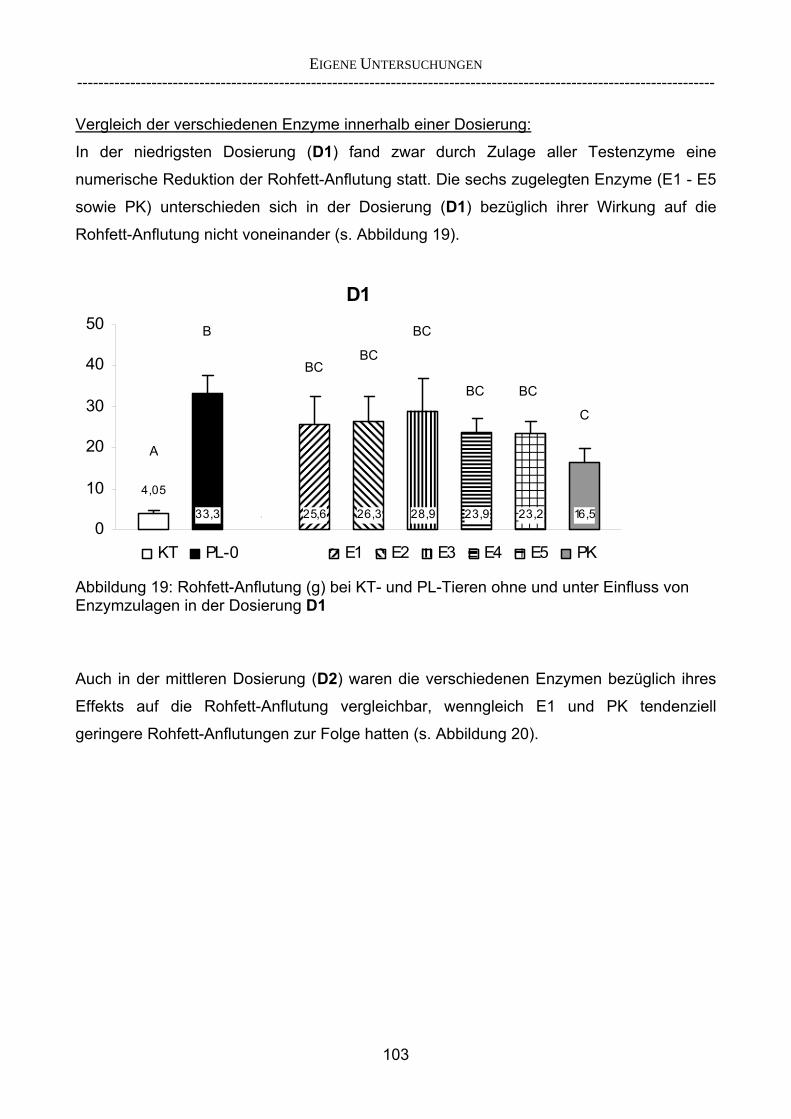
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
103
Vergleich der verschiedenen Enzyme innerhalb einer Dosierung:
In der niedrigsten Dosierung (D1) fand zwar durch Zulage aller Testenzyme eine
numerische Reduktion der Rohfett-Anflutung statt. Die sechs zugelegten Enzyme (E1 - E5
sowie PK) unterschieden sich in der Dosierung (D1) bezüglich ihrer Wirkung auf die
Rohfett-Anflutung nicht voneinander (s. Abbildung 19).
D1
0
4,05
33,3 25,6 26,3 28,9 23,9 23,2 16,50
10
20
30
40
50
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 19: Rohfett-Anflutung (g) bei KT- und PL-Tieren ohne und unter Einfluss von Enzymzulagen in der Dosierung D1
Auch in der mittleren Dosierung (D2) waren die verschiedenen Enzymen bezüglich ihres
Effekts auf die Rohfett-Anflutung vergleichbar, wenngleich E1 und PK tendenziell
geringere Rohfett-Anflutungen zur Folge hatten (s. Abbildung 20).
A
B
BC BC
BC
BC BC
C
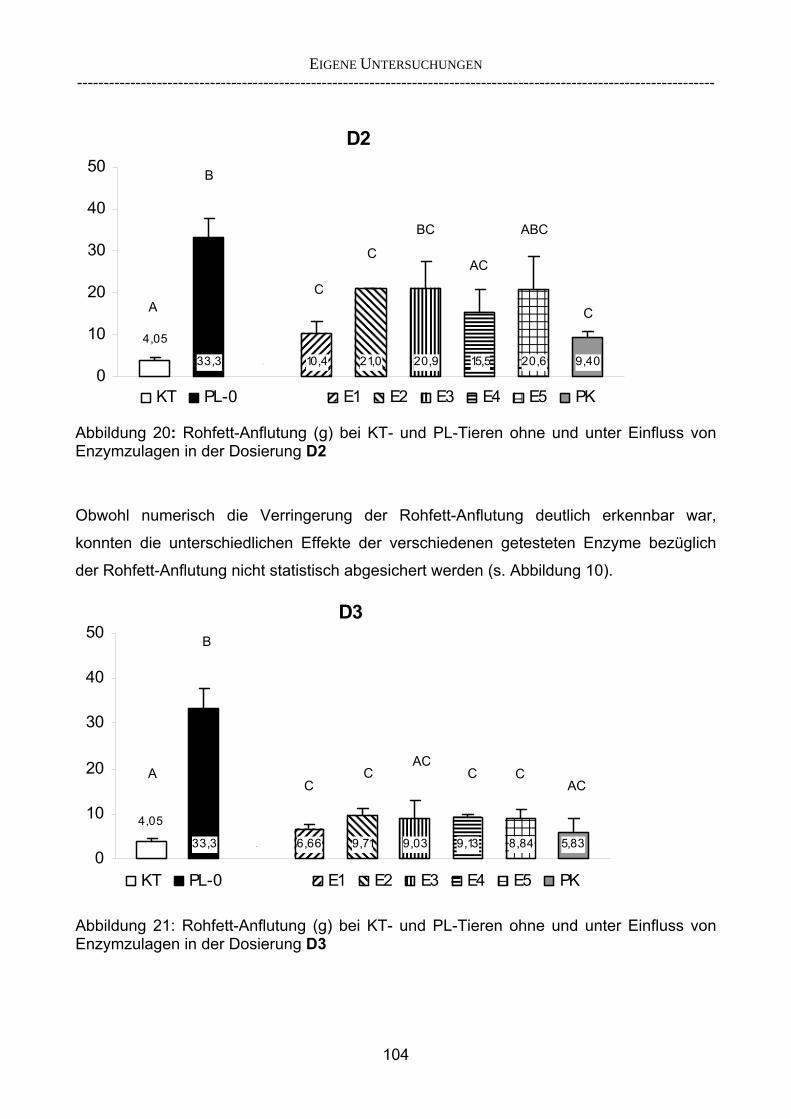
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
104
D2
33,3 0 10,4 21,0 20,9 15,5 20,6 9,40
4,05
0
10
20
30
40
50
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 20: Rohfett-Anflutung (g) bei KT- und PL-Tieren ohne und unter Einfluss von Enzymzulagen in der Dosierung D2
Obwohl numerisch die Verringerung der Rohfett-Anflutung deutlich erkennbar war,
konnten die unterschiedlichen Effekte der verschiedenen getesteten Enzyme bezüglich
der Rohfett-Anflutung nicht statistisch abgesichert werden (s. Abbildung 10).
D3
33,3 6,66 9,71 9,03 9,13 8,84 5,83
4,05
0
0
10
20
30
40
50
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 21: Rohfett-Anflutung (g) bei KT- und PL-Tieren ohne und unter Einfluss von Enzymzulagen in der Dosierung D3
A
B
C
C
BC
AC
ABC
C
A
B
C C
ACC C
AC

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
105
Rangierung:
In keiner Dosierung (D1, D2, D3) bestanden bezüglich der Effekte auf die Rohfett-
Anflutung signifikante Unterschiede zwischen den verschiedenen Enzymen (s. Tabelle
51). In der höchsten Dosierung zeigten alle eingesetzten Enzyme eine weitgehende
Reduktion der angefluteten Rohfett-Mengen, wobei bei Einsatz der Testenzyme PK und
E3 sogar das Niveau der Kontrolltiere erreicht wurde. Eine Rangierung der Enzyme
hinsichtlich der Effektivität der Verringerung der Rohfett-Anflutung konnte infolge
mangelnder statistischer Effekte nicht erfolgen. Numerisch bedingten jedoch in der
niedrigsten Dosierung (D1) PK, E5 und E4 sowie PK und E1 in der mittleren (D2) und
höchsten (D3) Dosierung die vergleichsweise deutlichste Reduktionen der angefluteten
Rohfett-Menge (s. Tabelle 52).
Tabelle 52: Rangierung der Enzyme bezüglich der Reduktion der absoluten Rohfett-Anflutung (g) am terminalen Ileum
Dosierung Wirksamkeit (numerisch) Wirksamkeit (statistische Unterschiede) D1 PK, E5, E4, E1, E2, E3 ─ D2 PK, E1, E4, E5, E3, E2 ─ D3 PK, E1, E5, E3, E4, E2 ─
3.3.3.4 Chromoxid-Konzentration im Chymus
Mit zunehmender Verdauung und Absorption von Nährstoffen ist generell eine steigende
Konzentration des Markers zu erwarten, d.h. der Marker reichert sich bei Erhöhung der
Verdaulichkeit der Nährstoffe zunehmend im Chymus an. Die im Chymus ermittelte Cr2O3-
Konzentration (8 h n. F.) war bei den PL-0-Tieren signifikant geringer (s. Tabelle 53) als
bei den Kontrolltieren.
Tabelle 53: Chromoxid-Konzentrationen (g / kg TS) am terminalen Ileum bei Kontroll- und PL-0 –Tieren (Zeitraum: über 8 h n. F.) KT 8,44 ± 0,54 a PL-0 3,94 ± 0,11 b Tendenziell ergab sich nach Einsatz der verschiedenen substituierten Enzyme eine
dosisabhängige Steigerung der Cr2O3-Konzentration (s. Tabelle 54). In der niedrigsten
Dosierung (D1) wurde nach Einsatz von E4 eine signifikant höhere Cr2O3-Konzentration

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
106
ermittelt als bei E3. Des Weiteren konnte PK in der mittleren Dosierung (D2) anhand der
Cr2O3-Konzentration im Chymus gegenüber E1, E2 sowie E5 statistisch abgegrenzt
werden (s. Tabelle 54). Bei Einsatz der höchsten Dosierung (D3) wurde nach Einsatz von
E4 eine signifikant höhere Cr2O3-Konzentration verzeichnet als nach Substitution von E5.
Lediglich nach Verabreichung der höchsten Dosierung (D3) wurde von PK, E1, E2 und E3
das Niveau der KT-Tiere erreicht.
Tabelle 54: Chromoxid-Konzentrationen (g / kg TS) bei PL-Tieren unter dem Einfluss unterschiedlicher Enzymzulagen und -dosierungen D1 (PL-0 AB) D2 (PL-0 AD) D3 (PL-0 A) E1 4,16 ± 0,675 AB 5,88 ± 0,781 CE 7,14 ± 1,70 ABC E2 3,97 ± 1,04 AB 3,93 ± 0,782 AE 6,50 ± 1,36 ABC E3 4,22 ± 0,571 A 4,77 ± 0,940 ABC 7,42 ± 2,00 ABC E4 4,58 ± 0,518 B 5,13 ± 0,660 BCD 6,47 ± 0,908 B E5 3,52 ± 0,511 AB 4,63 ± 0,939 AC 5,59 ± 0,70 C PK 5,00 ± 0,781 AB 7,05 ± 0,864 B 9,87 ± 3,12 ABC Kleinbuchstaben kennzeichnen signifikante Unterschiede zwischen den drei Dosierungen eines Enzyms sowie zwischen PL-0- und KT-Tieren Großbuchstaben kennzeichnen signifikante Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung sowie zwischen PL-0- und KT-Tieren Tendenziell wurden beim Einsatz von PK, E1, E4 und E3 höhere Cr2O3-Konzentrationen
im Chymus verzeichnet (s. Tabelle 55).
Tabelle 55: Rangierung der Enzyme bezüglich der Cr2O3-Konzentration (g / kg TS) im ilealen Chymus Dosierung Wirksamkeit (numerisch) Wirksamkeit (statistische Unterschiede) D1 PK, E4, E3, E1, E2, E5 E4 > E3 D2 PK, E1, E4, E3, E5, E2 PK > E1/ PK > E2/ PK > E5 D3 PK, E3, E1, E2, E4, E5 E4 > E5
3.3.3.5 Chromoxid-Wiederfindung
Die Bestimmung der Cr2O3–Wiederfindung erfolgte unter Verwendung der in Kapitel
3.1.5.8 beschriebenen Formel. Im Mittel betrug die Chromoxid-Wiederfindung im
Kollektionszeitraum (0-8 h nach Farbumschlag) 56 % (s. Tabelle 56).
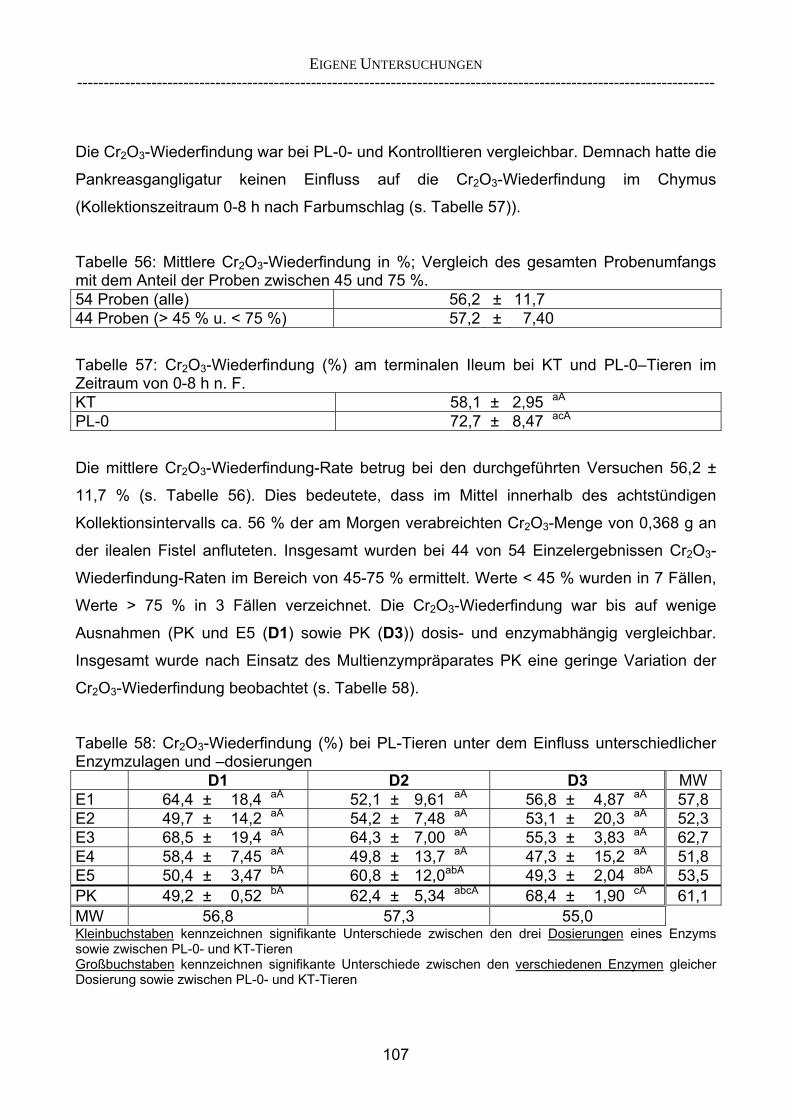
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
107
Die Cr2O3-Wiederfindung war bei PL-0- und Kontrolltieren vergleichbar. Demnach hatte die
Pankreasgangligatur keinen Einfluss auf die Cr2O3-Wiederfindung im Chymus
(Kollektionszeitraum 0-8 h nach Farbumschlag (s. Tabelle 57)).
Tabelle 56: Mittlere Cr2O3-Wiederfindung in %; Vergleich des gesamten Probenumfangs mit dem Anteil der Proben zwischen 45 und 75 %. 54 Proben (alle) 56,2 ± 11,7 44 Proben (> 45 % u. < 75 %) 57,2 ± 7,40
Tabelle 57: Cr2O3-Wiederfindung (%) am terminalen Ileum bei KT und PL-0–Tieren im Zeitraum von 0-8 h n. F. KT 58,1 ± 2,95 aA PL-0 72,7 ± 8,47 acA
Die mittlere Cr2O3-Wiederfindung-Rate betrug bei den durchgeführten Versuchen 56,2 ±
11,7 % (s. Tabelle 56). Dies bedeutete, dass im Mittel innerhalb des achtstündigen
Kollektionsintervalls ca. 56 % der am Morgen verabreichten Cr2O3-Menge von 0,368 g an
der ilealen Fistel anfluteten. Insgesamt wurden bei 44 von 54 Einzelergebnissen Cr2O3-
Wiederfindung-Raten im Bereich von 45-75 % ermittelt. Werte < 45 % wurden in 7 Fällen,
Werte > 75 % in 3 Fällen verzeichnet. Die Cr2O3-Wiederfindung war bis auf wenige
Ausnahmen (PK und E5 (D1) sowie PK (D3)) dosis- und enzymabhängig vergleichbar.
Insgesamt wurde nach Einsatz des Multienzympräparates PK eine geringe Variation der
Cr2O3-Wiederfindung beobachtet (s. Tabelle 58).
Tabelle 58: Cr2O3-Wiederfindung (%) bei PL-Tieren unter dem Einfluss unterschiedlicher Enzymzulagen und –dosierungen D1 D2 D3 MW E1 64,4 ± 18,4 aA 52,1 ± 9,61 aA 56,8 ± 4,87 aA 57,8 E2 49,7 ± 14,2 aA 54,2 ± 7,48 aA 53,1 ± 20,3 aA 52,3 E3 68,5 ± 19,4 aA 64,3 ± 7,00 aA 55,3 ± 3,83 aA 62,7 E4 58,4 ± 7,45 aA 49,8 ± 13,7 aA 47,3 ± 15,2 aA 51,8 E5 50,4 ± 3,47 bA 60,8 ± 12,0abA 49,3 ± 2,04 abA 53,5 PK 49,2 ± 0,52 bA 62,4 ± 5,34 abcA 68,4 ± 1,90 cA 61,1 MW 56,8 57,3 55,0 Kleinbuchstaben kennzeichnen signifikante Unterschiede zwischen den drei Dosierungen eines Enzyms sowie zwischen PL-0- und KT-Tieren Großbuchstaben kennzeichnen signifikante Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung sowie zwischen PL-0- und KT-Tieren

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
108
3.3.3.6 Verhältnis der Konzentrationen von Rohfett und Chromoxid
Bezüglich des Verhältnisses der Rohfett-Konzentration zur Chromoxid-Konzentration im
Chymus wirkte sich die Ligatur des Pankreasganges (129 ± 17,4) im Vergleich zu den
Kontrolltieren (20,1 ± 4,77) signifikant aus. Die Relation Rfe / Cr2O3-Konzentration war bei
den PL-0-Tieren mit nahezu 130 im Vergleich zu den KT-Tieren um den Faktor 6 x erhöht
(s. Tabelle 59).
Tabelle 59: Relation der Rohfett / Cr2O3-Konzentration im ilealen Chymus bei KT- und PL-0 –Tieren (Zeitraum: über 8 h n. F.) KT 20,1 ± 4,77 aA PL-0 129 ± 17,4 bB
Alle Enzyme bedingten eine dosisabhängige Reduktion der Relation von Rohfett-
Konzentration zur Cr2O3-Konzentration im ilealen Chymus, wobei diese Effekte
unterschiedlich stark ausgeprägt waren und nur zum Teil abgesichert werden konnten (s.
Tabelle 60).
In der niedrigsten Dosierung (D1) erzielte keines der eingesetzten Enzyme einen
signifikanten Effekt auf die Rohfett- / Cr2O3-Konzentration verglichen mit den PL-0-Tieren;
ebenfalls wurde durch keines der Testenzyme das Niveau der KT-Tiere erreicht (s. Tabelle
60 und Abbildung 22). Erst in der mittleren Dosierung (D2) konnte nach Einsatz des
Multienzymproduktes PK sowie der Lipase E1 eine signifikante Reduktion der Relation Rfe
/ Cr2O3-Konzentration im Vergleich zu den PL-Tieren ohne Enzymzulage beobachtet
werden. Nach Einsatz des Enzyms E1 wurde bezüglich dieses Parameters sogar das
Niveau der KT-Tiere erreicht. In der höchsten Dosierung (D3) resultierte aus der Zulage
aller Enzyme eine im Vergleich zu den PL-0-Tieren signifikante Reduktion der Relation der
Rfe- / Cr2O3-Konzentration im ileal anflutenden Chymus. Das Niveau der Rfe- / Cr2O3-
Konzentration der KT-Tiere wurde durch Substitution der Enzyme PK, E2 und E3 in der
höchsten Dosierung (D3) erreicht (s. Tabelle 60). Bei PK und E1 war die Reduktion der
Relation in D2 signifikant größer als in D1. Im Vergleich von D3 zu D2 unterschieden sich
E2, E3 und E5 signifikant voneinander. Außerdem wurde durch alle Testenzyme in D3
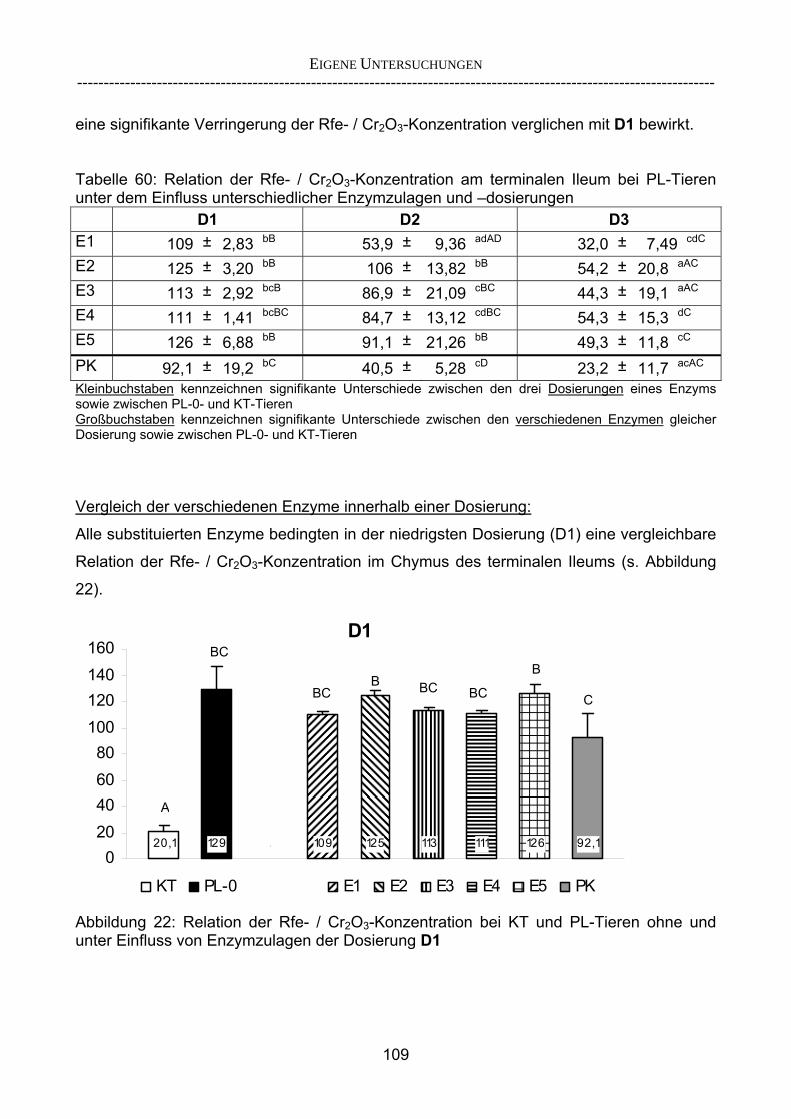
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
109
eine signifikante Verringerung der Rfe- / Cr2O3-Konzentration verglichen mit D1 bewirkt.
Tabelle 60: Relation der Rfe- / Cr2O3-Konzentration am terminalen Ileum bei PL-Tieren unter dem Einfluss unterschiedlicher Enzymzulagen und –dosierungen D1 D2 D3 E1 109 ± 2,83 bB 53,9 ± 9,36 adAD 32,0 ± 7,49 cdC E2 125 ± 3,20 bB 106 ± 13,82 bB 54,2 ± 20,8 aAC E3 113 ± 2,92 bcB 86,9 ± 21,09 cBC 44,3 ± 19,1 aAC E4 111 ± 1,41 bcBC 84,7 ± 13,12 cdBC 54,3 ± 15,3 dC E5 126 ± 6,88 bB 91,1 ± 21,26 bB 49,3 ± 11,8 cC PK 92,1 ± 19,2 bC 40,5 ± 5,28 cD 23,2 ± 11,7 acAC Kleinbuchstaben kennzeichnen signifikante Unterschiede zwischen den drei Dosierungen eines Enzyms sowie zwischen PL-0- und KT-Tieren Großbuchstaben kennzeichnen signifikante Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung sowie zwischen PL-0- und KT-Tieren
Vergleich der verschiedenen Enzyme innerhalb einer Dosierung:
Alle substituierten Enzyme bedingten in der niedrigsten Dosierung (D1) eine vergleichbare
Relation der Rfe- / Cr2O3-Konzentration im Chymus des terminalen Ileums (s. Abbildung
22).
D1
20,1 129 0 109 113 111 126 92,11250
20406080
100120140160
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 22: Relation der Rfe- / Cr2O3-Konzentration bei KT und PL-Tieren ohne und unter Einfluss von Enzymzulagen der Dosierung D1
A
BC
BCB BC BC
B
C
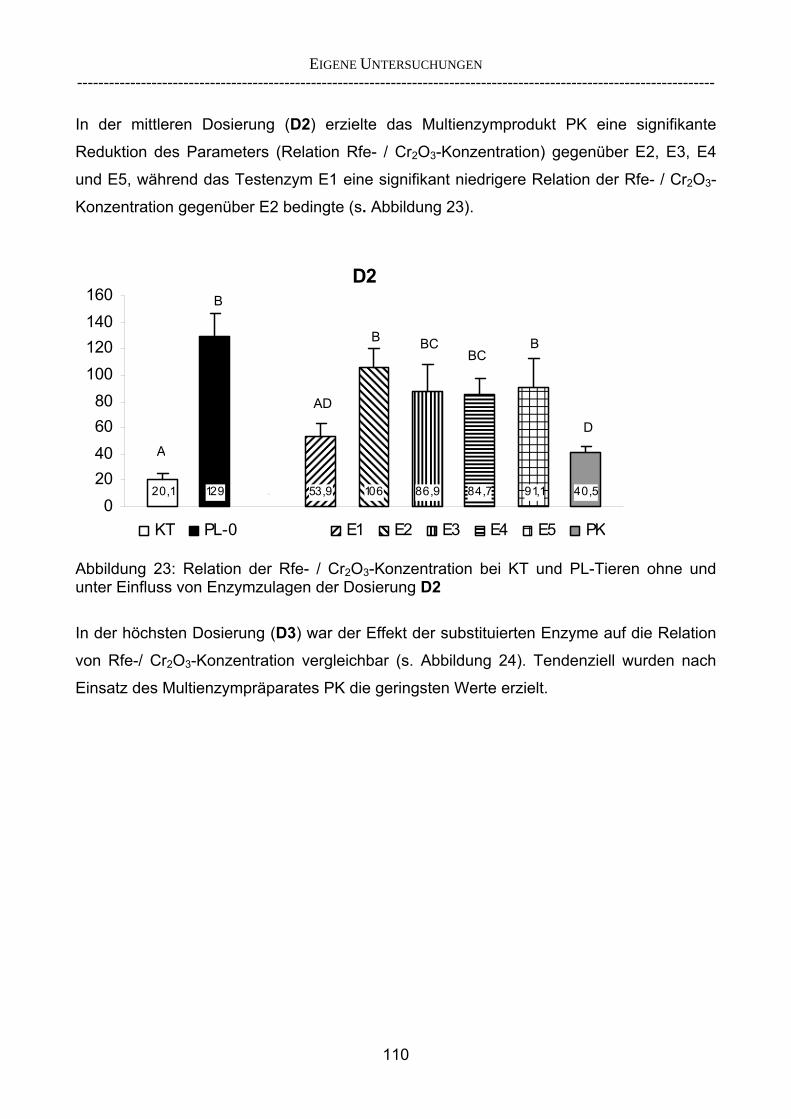
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
110
In der mittleren Dosierung (D2) erzielte das Multienzymprodukt PK eine signifikante
Reduktion des Parameters (Relation Rfe- / Cr2O3-Konzentration) gegenüber E2, E3, E4
und E5, während das Testenzym E1 eine signifikant niedrigere Relation der Rfe- / Cr2O3-
Konzentration gegenüber E2 bedingte (s. Abbildung 23).
D2
20,1 129 0 53,9 106 86,9 84,7 91,1 40,50
20406080
100120140160
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 23: Relation der Rfe- / Cr2O3-Konzentration bei KT und PL-Tieren ohne und unter Einfluss von Enzymzulagen der Dosierung D2
In der höchsten Dosierung (D3) war der Effekt der substituierten Enzyme auf die Relation
von Rfe-/ Cr2O3-Konzentration vergleichbar (s. Abbildung 24). Tendenziell wurden nach
Einsatz des Multienzympräparates PK die geringsten Werte erzielt.
A
B
AD
B BCBC
B
D
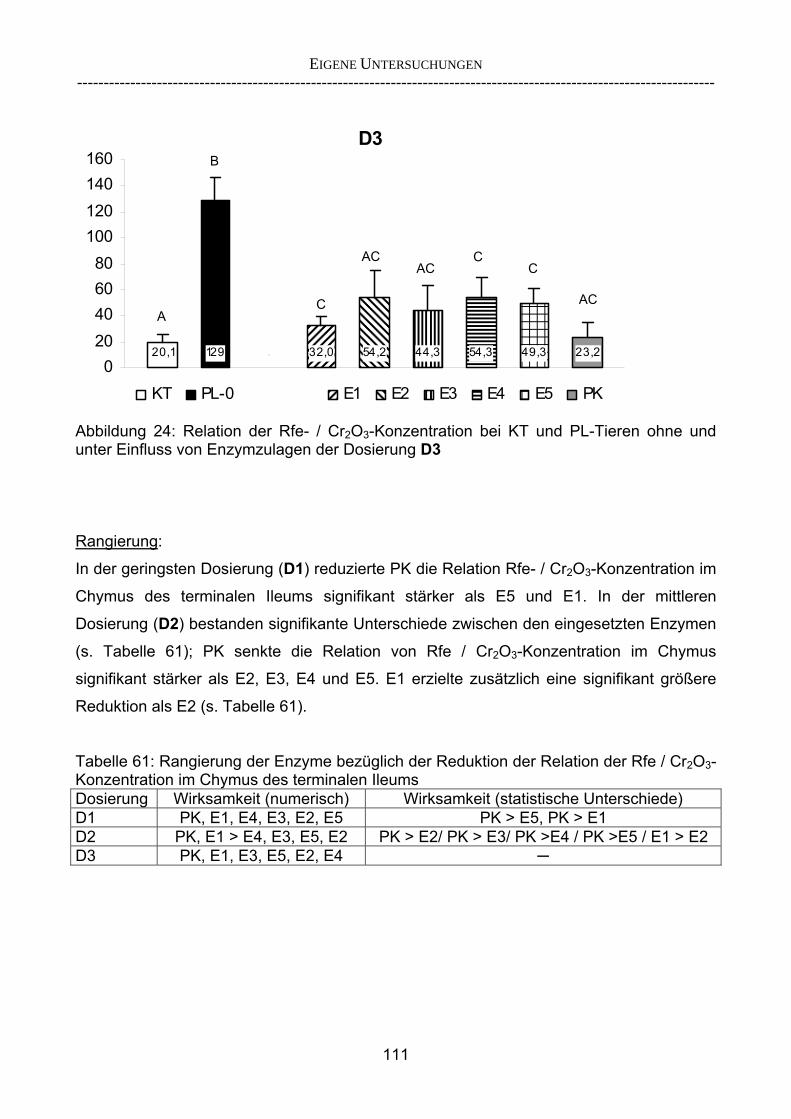
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
111
D3
20,1 129 0 32,0 54,2 44,3 54,3 49,3 23,20
20406080
100120140160
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 24: Relation der Rfe- / Cr2O3-Konzentration bei KT und PL-Tieren ohne und unter Einfluss von Enzymzulagen der Dosierung D3 Rangierung:
In der geringsten Dosierung (D1) reduzierte PK die Relation Rfe- / Cr2O3-Konzentration im
Chymus des terminalen Ileums signifikant stärker als E5 und E1. In der mittleren
Dosierung (D2) bestanden signifikante Unterschiede zwischen den eingesetzten Enzymen
(s. Tabelle 61); PK senkte die Relation von Rfe / Cr2O3-Konzentration im Chymus
signifikant stärker als E2, E3, E4 und E5. E1 erzielte zusätzlich eine signifikant größere
Reduktion als E2 (s. Tabelle 61).
Tabelle 61: Rangierung der Enzyme bezüglich der Reduktion der Relation der Rfe / Cr2O3-Konzentration im Chymus des terminalen Ileums Dosierung Wirksamkeit (numerisch) Wirksamkeit (statistische Unterschiede) D1 PK, E1, E4, E3, E2, E5 PK > E5, PK > E1 D2 PK, E1 > E4, E3, E5, E2 PK > E2/ PK > E3/ PK >E4 / PK >E5 / E1 > E2 D3 PK, E1, E3, E5, E2, E4 ─
A
B
C
ACAC
CC
AC

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
112
3.3.3.7 Trockensubstanz-Verschwindensrate (TS-VR)
Im Vergleich zu den Kontrolltieren, bei denen im Kollektionszeitraum (8 h n. F.) eine
Trockensubstanz-Verschwindensrate von 71,6 (± 1,50) % kalkuliert wurde, war die
Trockensubstanz-Verschwindensrate bei den PL-0-Tieren mit 35,0 (± 8,68) % signifikant
geringer (s. Tabelle 62).
Tabelle 62: Trockensubstanz-Verschwindensrate (%) am terminalen Ileum bei Kontroll- und PL-0 –Tieren KT 71,6 ± 1,50 aA PL-0 35,0 ± 8,68 bB
Durch die Zulage der Enzymprodukte konnte im Vergleich zu den PL-0-Tieren ein
dosisabhängiger Anstieg der TS-Verschwindensrate verzeichnet werden, der jedoch nicht
immer statistisch abzusichern war (s. Tabelle 63). Eine signifikante Erhöhung der
Trockensubstanz-Verschwindensrate gegenüber jener der PL-0-Tieren wurde in der
geringsten Dosierung (D1) lediglich durch PK, in der mittleren Dosierung (D2) durch PK,
E1, E3 und E4 erreicht. In der höchsten Dosierung (D3) wurde bei Einsatz von vier der
sechs Enzyme eine signifikante Steigerung der Trockensubstanz-Verschwindensrate
gegenüber den PL-0-Tieren erreicht. Während in der geringsten Dosierung (D1) lediglich E2 -aufgrund der hohen Standardabweichung- nicht von den Kontrolltiere abzugrenzen
war, wurde das Niveau der Kontrolltiere jedoch von E3 in der mittleren Dosierung (D2)
sowie durch Einsatz der Enzyme E1, E2, E3, und PK in der höchsten Dosierung (D3)
erreicht (s. Tabelle 63).
Es wurde eine signifikante Steigerung der Werte der Trockensubstanz-Verschwindensrate
nach Einsatz der Enzyme PK und E1 in der Dosierung D2 gegenüber der geringsten
Dosierung (D1) nachgewiesen; die höchste Dosierung (D3) bewirkte bei allen Enzymen
(außer E2) eine Steigerung der Trockensubstanz-Verschwindensrate gegenüber der
geringsten Dosierung (D1). Zwischen der mittleren Dosierung (D2) und der höchsten
Dosierung (D3) konnte lediglich für E3 ein signifikanter Effekt der Dosis beobachtet
werden.
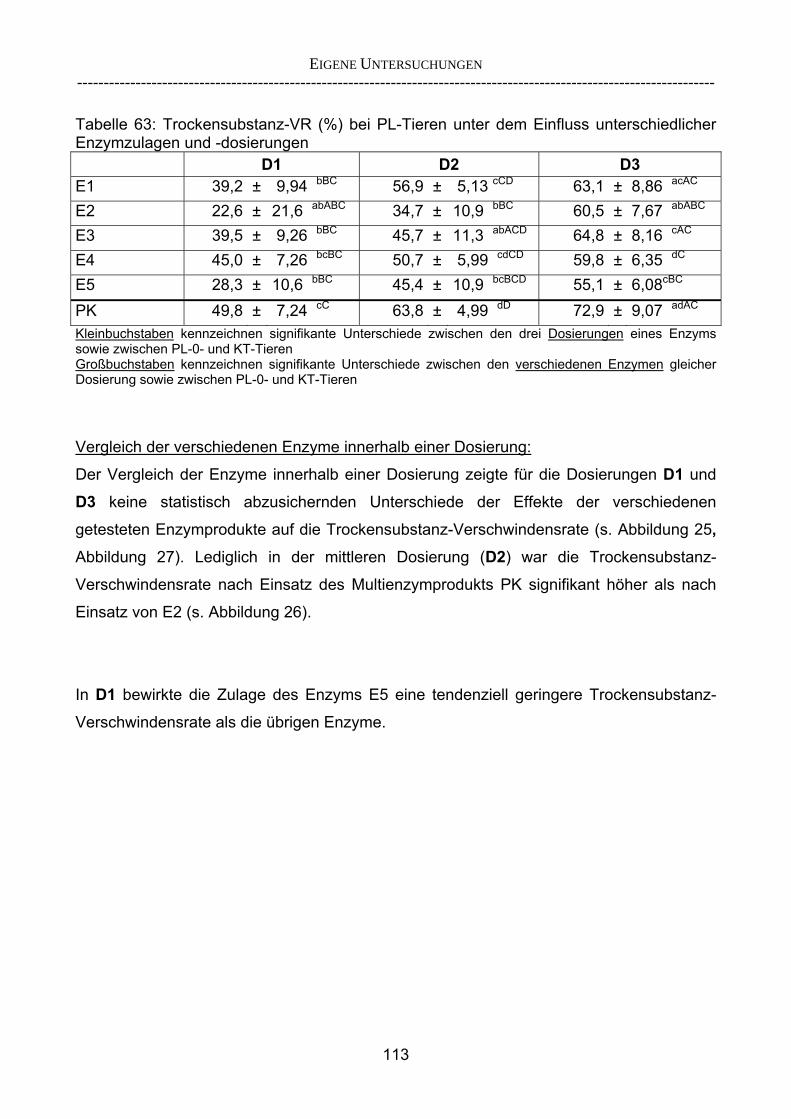
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
113
Tabelle 63: Trockensubstanz-VR (%) bei PL-Tieren unter dem Einfluss unterschiedlicher Enzymzulagen und -dosierungen D1 D2 D3 E1 39,2 ± 9,94 bBC 56,9 ± 5,13 cCD 63,1 ± 8,86 acAC E2 22,6 ± 21,6 abABC 34,7 ± 10,9 bBC 60,5 ± 7,67 abABC E3 39,5 ± 9,26 bBC 45,7 ± 11,3 abACD 64,8 ± 8,16 cAC E4 45,0 ± 7,26 bcBC 50,7 ± 5,99 cdCD 59,8 ± 6,35 dC E5 28,3 ± 10,6 bBC 45,4 ± 10,9 bcBCD 55,1 ± 6,08cBC PK 49,8 ± 7,24 cC 63,8 ± 4,99 dD 72,9 ± 9,07 adAC Kleinbuchstaben kennzeichnen signifikante Unterschiede zwischen den drei Dosierungen eines Enzyms sowie zwischen PL-0- und KT-Tieren Großbuchstaben kennzeichnen signifikante Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung sowie zwischen PL-0- und KT-Tieren
Vergleich der verschiedenen Enzyme innerhalb einer Dosierung:
Der Vergleich der Enzyme innerhalb einer Dosierung zeigte für die Dosierungen D1 und
D3 keine statistisch abzusichernden Unterschiede der Effekte der verschiedenen
getesteten Enzymprodukte auf die Trockensubstanz-Verschwindensrate (s. Abbildung 25, Abbildung 27). Lediglich in der mittleren Dosierung (D2) war die Trockensubstanz-
Verschwindensrate nach Einsatz des Multienzymprodukts PK signifikant höher als nach
Einsatz von E2 (s. Abbildung 26).
In D1 bewirkte die Zulage des Enzyms E5 eine tendenziell geringere Trockensubstanz-
Verschwindensrate als die übrigen Enzyme.
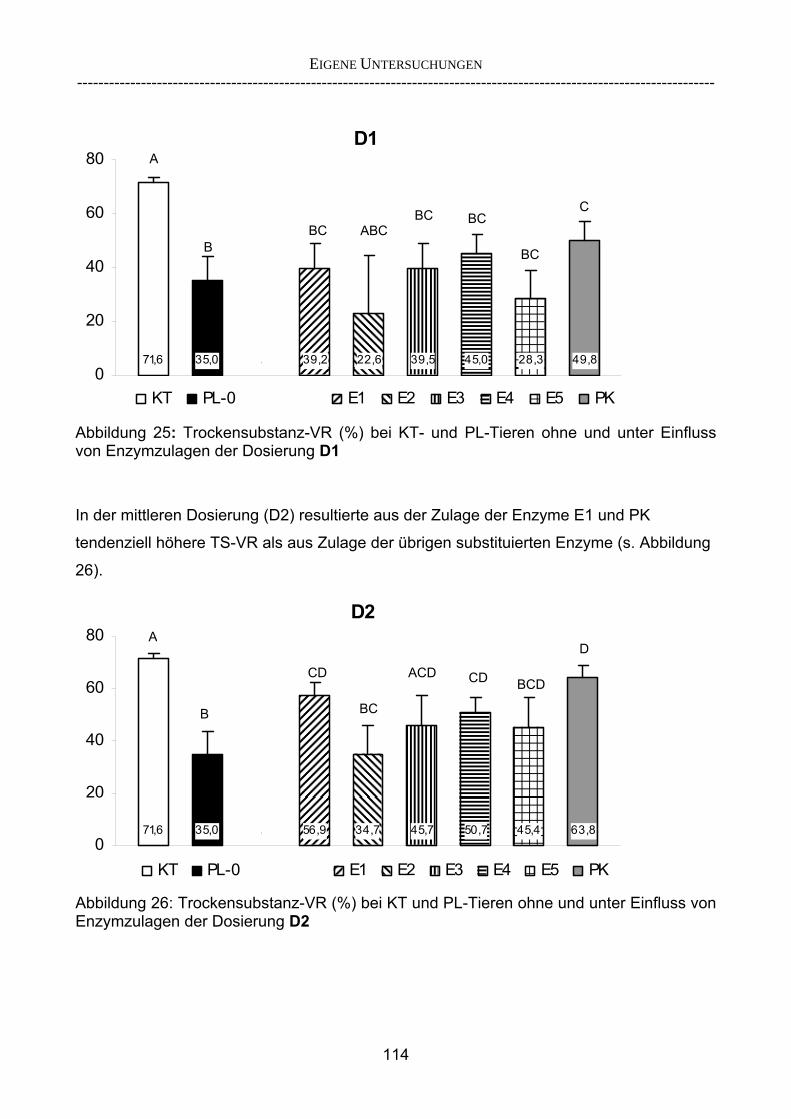
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
114
D1
71,6 35,0 0 39,2 22,6 39,5 45,0 28,3 49,80
20
40
60
80
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 25: Trockensubstanz-VR (%) bei KT- und PL-Tieren ohne und unter Einfluss von Enzymzulagen der Dosierung D1
In der mittleren Dosierung (D2) resultierte aus der Zulage der Enzyme E1 und PK
tendenziell höhere TS-VR als aus Zulage der übrigen substituierten Enzyme (s. Abbildung
26).
D2
71,6 35,0 0 56,9 45,7 50,7 45,4 63,834,70
20
40
60
80
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 26: Trockensubstanz-VR (%) bei KT und PL-Tieren ohne und unter Einfluss von Enzymzulagen der Dosierung D2
A
B BC ABC
BC BC
BC
C
A
B
CD
BC
ACD CD BCD
D
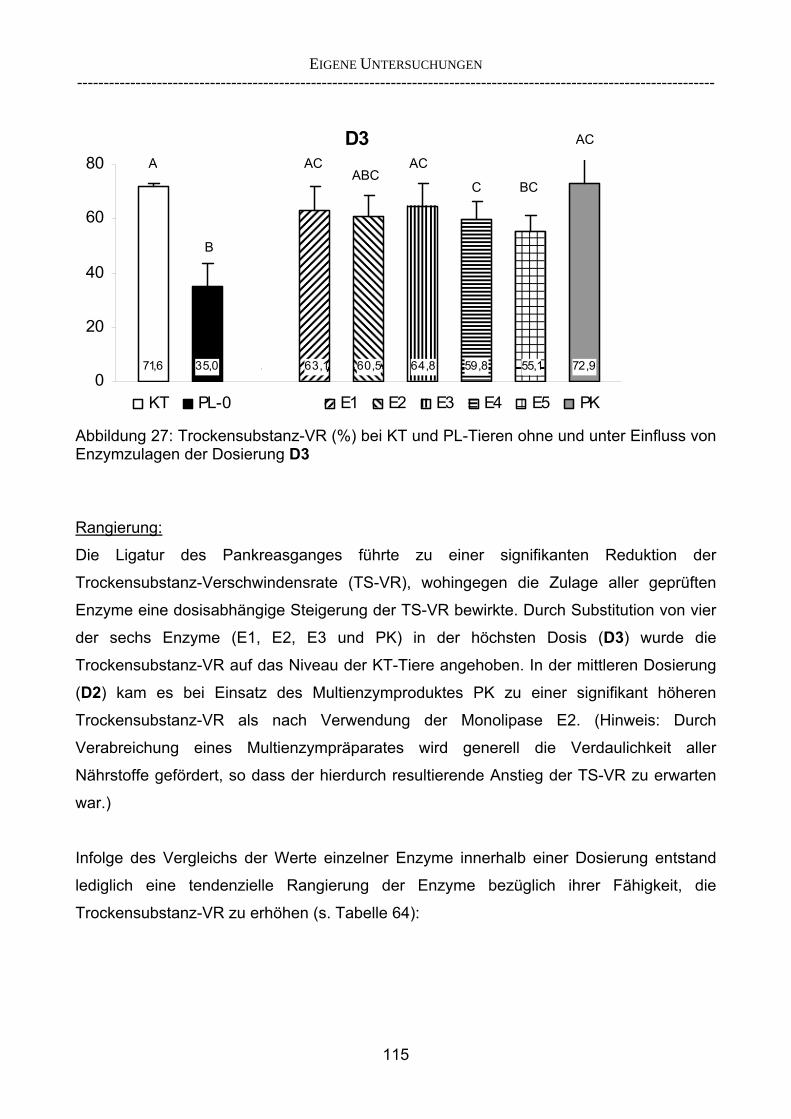
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
115
D3
71,6 35,0 0 63,1 64,8 59,8 55,1 72,960,50
20
40
60
80
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 27: Trockensubstanz-VR (%) bei KT und PL-Tieren ohne und unter Einfluss von Enzymzulagen der Dosierung D3
Rangierung:
Die Ligatur des Pankreasganges führte zu einer signifikanten Reduktion der
Trockensubstanz-Verschwindensrate (TS-VR), wohingegen die Zulage aller geprüften
Enzyme eine dosisabhängige Steigerung der TS-VR bewirkte. Durch Substitution von vier
der sechs Enzyme (E1, E2, E3 und PK) in der höchsten Dosis (D3) wurde die
Trockensubstanz-VR auf das Niveau der KT-Tiere angehoben. In der mittleren Dosierung
(D2) kam es bei Einsatz des Multienzymproduktes PK zu einer signifikant höheren
Trockensubstanz-VR als nach Verwendung der Monolipase E2. (Hinweis: Durch
Verabreichung eines Multienzympräparates wird generell die Verdaulichkeit aller
Nährstoffe gefördert, so dass der hierdurch resultierende Anstieg der TS-VR zu erwarten
war.)
Infolge des Vergleichs der Werte einzelner Enzyme innerhalb einer Dosierung entstand
lediglich eine tendenzielle Rangierung der Enzyme bezüglich ihrer Fähigkeit, die
Trockensubstanz-VR zu erhöhen (s. Tabelle 64):
A
B
AC ABC
AC
C BC
AC

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
116
Tabelle 64: Rangierung der Enzyme bezüglich der Erhöhung der Trockensubstanz-VR (%) Dosierung Wirksamkeit (numerisch) Wirksamkeit (statistische Unterschiede) D1 PK, E4, E3, E1, E5, E2 ─ D2 PK, E1, E4, E3, E5, E2 PK > E2 D3 PK, E3, E1, E2, E4, E5 ─
3.3.3.8 Rohfett-Verschwindensrate (Rfe-VR)
Im Vergleich zu den Kontrolltieren (86,4 ± 3,04 %) wiesen die pankreasgangligierten Tiere
(10,8 ± 10,4 %) eine signifikant geringere Rohfett-Verschwindensrate im
Kollektionsintervall 0-8 h n. F. auf (s. Tabelle 65).
Tabelle 65: Rohfett-Verschwindensrate (%) bei KT- und PL-0–Tieren (Zeitraum: 0-8 h n. F.) KT 86,4 ± 3,04 aA PL-0 10,8 ± 10,4 bBC Alle zugelegten Enzyme (Ausnahme E2; D1) bedingten einen dosisabhängigen Anstieg
der Rohfett-Verschwindensrate verglichen mit den PL-0-Tieren, der jedoch nur zum Teil
statistisch abgesichert werden konnte (s. Tabelle 66). In der geringsten Dosierung (D1) war enzymunabhängig kein signifikanter Effekt auf die Rohfett-Verschwindensrate zu
erkennen (Ausnahme: PK signifikante Steigerung). Vier der sechs geprüften
Enzymprodukte bedingten in der mittleren Dosierung (D2) eine signifikante Erhöhung der
Rohfett-Verschwindensrate im Vergleich zu den PL-0-Tieren. Durch keines der Enzyme
konnte bei Substitution in der mittleren Dosierung das Niveau der Kontrolltiere erreicht
werden. In der höchsten Dosierung (D3) erbrachten alle substituierten Enzyme eine
signifikante Steigerung der Rohfett-Verschwindensrate gegenüber den PL-Tieren ohne
Enzymsubstitution; zudem wurde die Rohfett-Verschwindensrate auf das Niveau der
Kontrolltiere angehoben (Ausnahme: E2 und E4).
Bei Verabreichung von PK und E1 wurde ein signifikanter dosisabhängiger Anstieg
(Vergleich D1 und D2) der Rohfett-Verschwindensrate in verzeichnet (s. Tabelle 66).
Zwischen den Dosierungen D2 und D3 wurde bei Einsatz von E2, E3 und E5 eine
signifikante Steigerung der Rohfett-Verschwindensrate beobachtet. Die Rohfett-
Verschwindensrate war enzymunabhängig nach Einsatz in der höchsten Dosierung (D3)
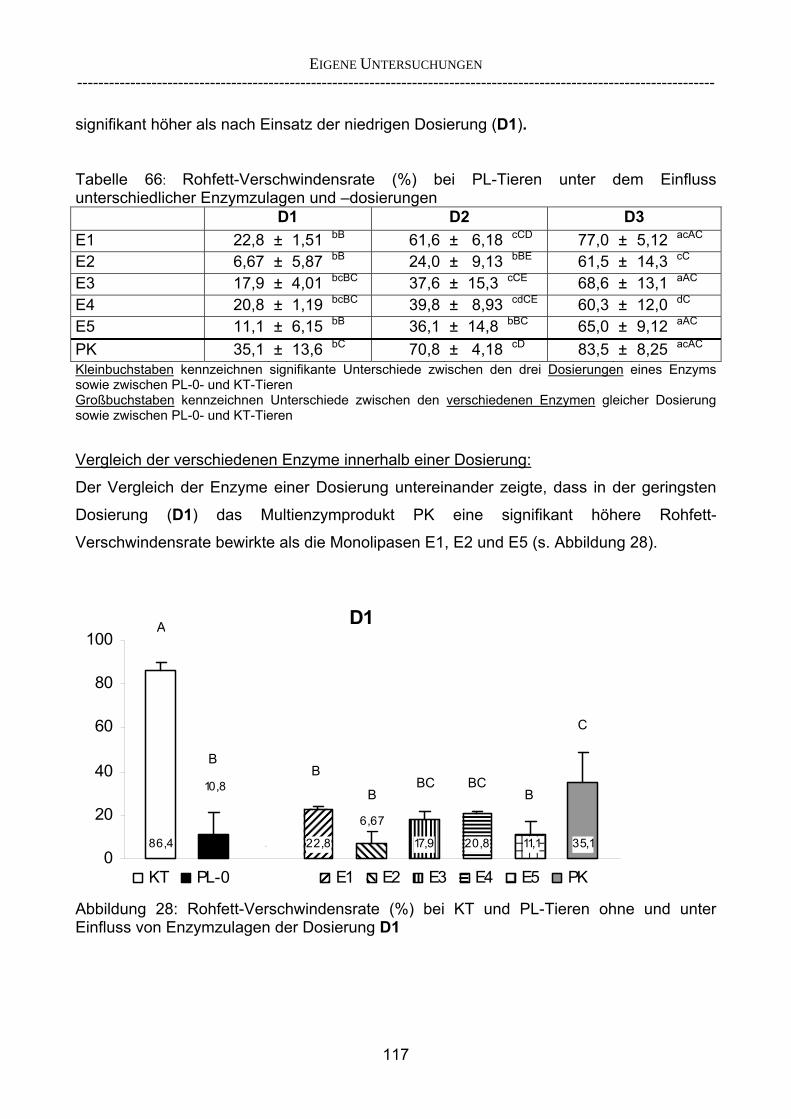
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
117
signifikant höher als nach Einsatz der niedrigen Dosierung (D1).
Tabelle 66: Rohfett-Verschwindensrate (%) bei PL-Tieren unter dem Einfluss unterschiedlicher Enzymzulagen und –dosierungen
D1 D2 D3 E1 22,8 ± 1,51 bB 61,6 ± 6,18 cCD 77,0 ± 5,12 acAC E2 6,67 ± 5,87 bB 24,0 ± 9,13 bBE 61,5 ± 14,3 cC E3 17,9 ± 4,01 bcBC 37,6 ± 15,3 cCE 68,6 ± 13,1 aAC E4 20,8 ± 1,19 bcBC 39,8 ± 8,93 cdCE 60,3 ± 12,0 dC E5 11,1 ± 6,15 bB 36,1 ± 14,8 bBC 65,0 ± 9,12 aAC PK 35,1 ± 13,6 bC 70,8 ± 4,18 cD 83,5 ± 8,25 acAC Kleinbuchstaben kennzeichnen signifikante Unterschiede zwischen den drei Dosierungen eines Enzyms sowie zwischen PL-0- und KT-Tieren Großbuchstaben kennzeichnen Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung sowie zwischen PL-0- und KT-Tieren
Vergleich der verschiedenen Enzyme innerhalb einer Dosierung:
Der Vergleich der Enzyme einer Dosierung untereinander zeigte, dass in der geringsten
Dosierung (D1) das Multienzymprodukt PK eine signifikant höhere Rohfett-
Verschwindensrate bewirkte als die Monolipasen E1, E2 und E5 (s. Abbildung 28).
D1
86,4 0 22,8 17,9 20,8 11,1 35,1
10,8
6,67
0
20
40
60
80
100
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 28: Rohfett-Verschwindensrate (%) bei KT und PL-Tieren ohne und unter Einfluss von Enzymzulagen der Dosierung D1
A
B
B
B BC BC
C
B
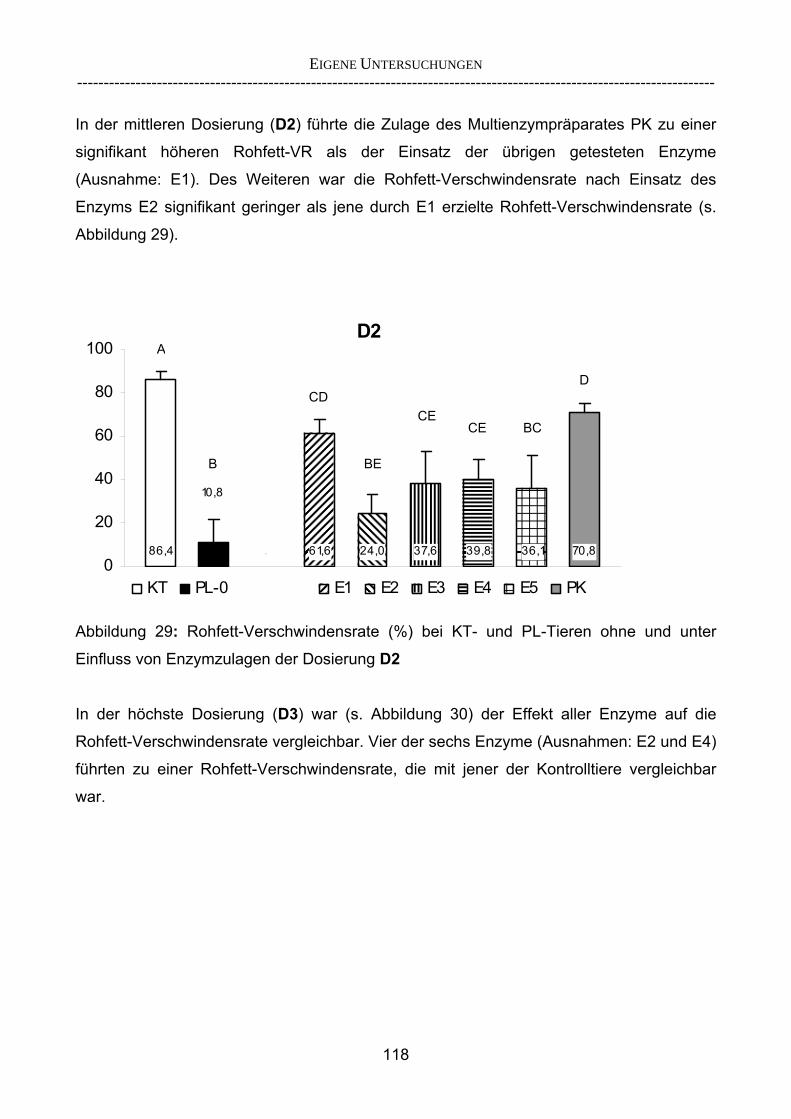
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
118
In der mittleren Dosierung (D2) führte die Zulage des Multienzympräparates PK zu einer
signifikant höheren Rohfett-VR als der Einsatz der übrigen getesteten Enzyme
(Ausnahme: E1). Des Weiteren war die Rohfett-Verschwindensrate nach Einsatz des
Enzyms E2 signifikant geringer als jene durch E1 erzielte Rohfett-Verschwindensrate (s. Abbildung 29).
D2
86,4 0 61,6 24,0 37,6 39,8 36,1 70,8
10,8
0
20
40
60
80
100
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 29: Rohfett-Verschwindensrate (%) bei KT- und PL-Tieren ohne und unter
Einfluss von Enzymzulagen der Dosierung D2 In der höchste Dosierung (D3) war (s. Abbildung 30) der Effekt aller Enzyme auf die
Rohfett-Verschwindensrate vergleichbar. Vier der sechs Enzyme (Ausnahmen: E2 und E4)
führten zu einer Rohfett-Verschwindensrate, die mit jener der Kontrolltiere vergleichbar
war.
A
B BE
CDCE
CE
D
BC
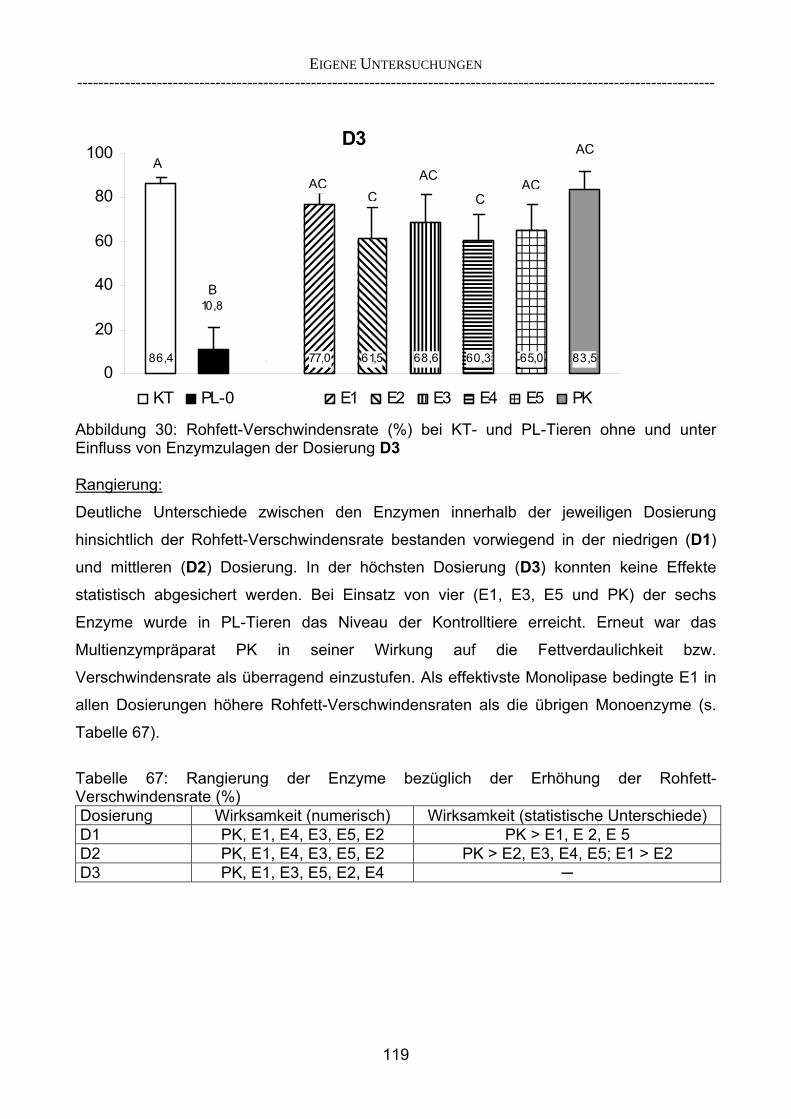
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
119
D3
86,4 0 77,0 61,5 68,6 60,3 65,0 83,5
10,8
0
20
40
60
80
100
KT PL-0 Reihe3 E1 E2 E3 E4 E5 PK
Abbildung 30: Rohfett-Verschwindensrate (%) bei KT- und PL-Tieren ohne und unter Einfluss von Enzymzulagen der Dosierung D3 Rangierung:
Deutliche Unterschiede zwischen den Enzymen innerhalb der jeweiligen Dosierung
hinsichtlich der Rohfett-Verschwindensrate bestanden vorwiegend in der niedrigen (D1)
und mittleren (D2) Dosierung. In der höchsten Dosierung (D3) konnten keine Effekte
statistisch abgesichert werden. Bei Einsatz von vier (E1, E3, E5 und PK) der sechs
Enzyme wurde in PL-Tieren das Niveau der Kontrolltiere erreicht. Erneut war das
Multienzympräparat PK in seiner Wirkung auf die Fettverdaulichkeit bzw.
Verschwindensrate als überragend einzustufen. Als effektivste Monolipase bedingte E1 in
allen Dosierungen höhere Rohfett-Verschwindensraten als die übrigen Monoenzyme (s.
Tabelle 67).
Tabelle 67: Rangierung der Enzyme bezüglich der Erhöhung der Rohfett-Verschwindensrate (%) Dosierung Wirksamkeit (numerisch) Wirksamkeit (statistische Unterschiede) D1 PK, E1, E4, E3, E5, E2 PK > E1, E 2, E 5 D2 PK, E1, E4, E3, E5, E2 PK > E2, E3, E4, E5; E1 > E2 D3 PK, E1, E3, E5, E2, E4 ─
B
CAC AC
C
AC
AC A

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
120
3.3.3.9 Relative Anflutung von Trockensubstanz
Wird die ileal bei PL-0-Tieren innerhalb von 8 h n. F. angeflutete Chymusmenge (TS-
Menge) mit 100% gleichgesetzt, fluteten im Chymus des terminalen Ileums der
Kontrolltiere im gleichen Zeitraum lediglich 35,2 % der Chymusmenge der PL-0-Tiere an
(s. Tabelle 68).
Tabelle 68: Absolute und relative Trockensubstanz-Anflutung (%) am terminalen Ileum bei KT- und PL-0 –Tieren (Zeitraum: über 8 h n. F.; Werte der PL-0 = 100) Absolut (g) Relativ (PL-0 = 100) KT 24,2 35,2 PL-0 68,8 100 A
Vergleich der verschiedenen Enzyme innerhalb einer Dosierung:
Bereits in der niedrigsten Dosierung (D1) hatte der Einsatz des Multienzympräparates PK
eine signifikante Verringerung der relativen TS-Anflutung um fast 50 % im Vergleich zu
den PL-0-Tieren zur Folge; alle eingesetzten Monolipasen bewirkten hingegen lediglich
eine Verringerung der innerhalb von 8 Stunden n. F. anflutenden Chymusmenge (TS) auf
67,8 bis 86,6 % der Werte der PL-0-Tiere (s. Tabelle 69).
In der mittleren Dosierung (D2) wurde die relative Trockensubstanz-Anflutung durch den
Einsatz von PK und E1 signifikant reduziert.
In der höchsten Dosierung (D3) der Enzyme wurde die Trockensubstanz-Anflutung durch
alle eingesetzten Enzyme im Vergleich zu den PL-0-Tieren signifikant verringert, wobei
sich die verschiedenen Enzyme in ihrer Wirkung jedoch nicht voneinander abgrenzen
ließen (s. Tabelle 69).
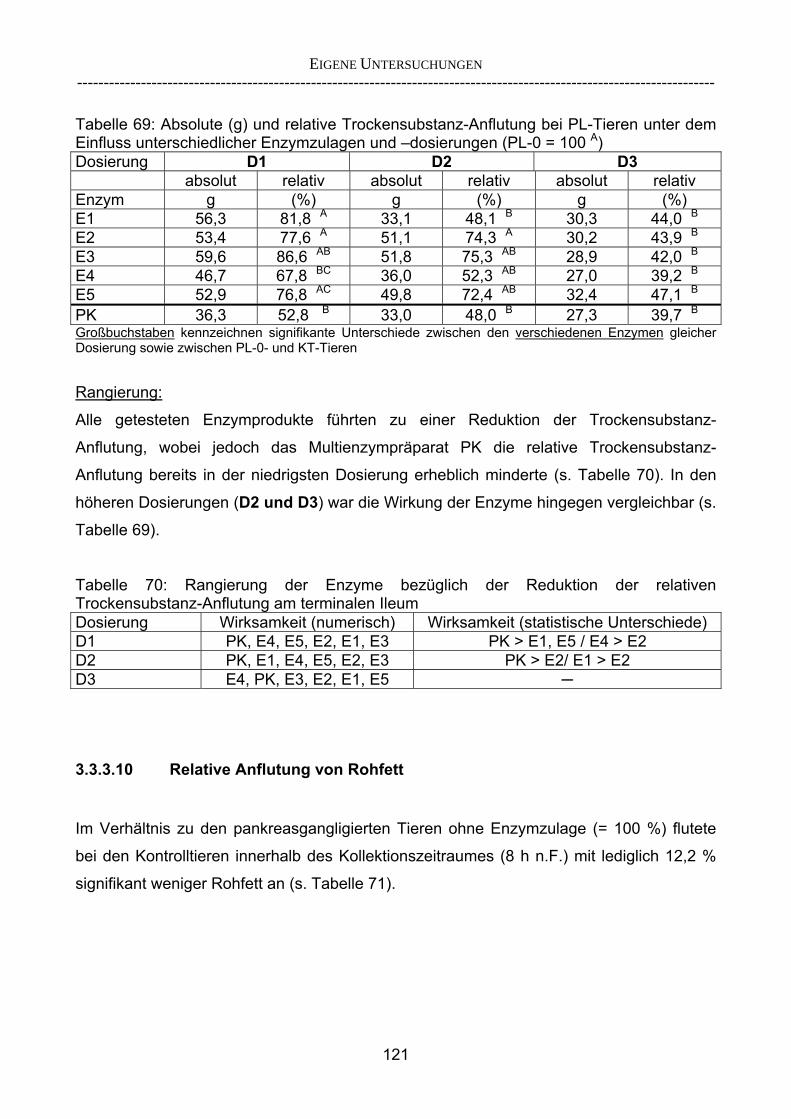
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
121
Tabelle 69: Absolute (g) und relative Trockensubstanz-Anflutung bei PL-Tieren unter dem Einfluss unterschiedlicher Enzymzulagen und –dosierungen (PL-0 = 100 A) Dosierung D1 D2 D3 absolut relativ absolut relativ absolut relativ Enzym g (%) g (%) g (%) E1 56,3 81,8 A 33,1 48,1 B 30,3 44,0 B E2 53,4 77,6 A 51,1 74,3 A 30,2 43,9 B E3 59,6 86,6 AB 51,8 75,3 AB 28,9 42,0 B E4 46,7 67,8 BC 36,0 52,3 AB 27,0 39,2 B E5 52,9 76,8 AC 49,8 72,4 AB 32,4 47,1 B PK 36,3 52,8 B 33,0 48,0 B 27,3 39,7 B Großbuchstaben kennzeichnen signifikante Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung sowie zwischen PL-0- und KT-Tieren
Rangierung:
Alle getesteten Enzymprodukte führten zu einer Reduktion der Trockensubstanz-
Anflutung, wobei jedoch das Multienzympräparat PK die relative Trockensubstanz-
Anflutung bereits in der niedrigsten Dosierung erheblich minderte (s. Tabelle 70). In den
höheren Dosierungen (D2 und D3) war die Wirkung der Enzyme hingegen vergleichbar (s.
Tabelle 69).
Tabelle 70: Rangierung der Enzyme bezüglich der Reduktion der relativen Trockensubstanz-Anflutung am terminalen Ileum Dosierung Wirksamkeit (numerisch) Wirksamkeit (statistische Unterschiede) D1 PK, E4, E5, E2, E1, E3 PK > E1, E5 / E4 > E2 D2 PK, E1, E4, E5, E2, E3 PK > E2/ E1 > E2 D3 E4, PK, E3, E2, E1, E5 ─
3.3.3.10 Relative Anflutung von Rohfett
Im Verhältnis zu den pankreasgangligierten Tieren ohne Enzymzulage (= 100 %) flutete
bei den Kontrolltieren innerhalb des Kollektionszeitraumes (8 h n.F.) mit lediglich 12,2 %
signifikant weniger Rohfett an (s. Tabelle 71).

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
122
Tabelle 71: Absolute und relative Rohfett-Anflutung am terminalen Ileum bei KT- und PL-0 –Tieren (Zeitraum: über 8 h n. F.; Werte der PL-0-Tiere = 100) Absolut (g) Relativ (PL-0 = 100) KT 4,05 12,2 acA PL-0 33,3 100 bB
Vergleich der verschiedenen Enzyme innerhalb einer Dosierung:
Eine dosisabhängige Verringerung der relativen Rohfett-Anflutung konnte bei allen
eingesetzten Testenzymen beobachtet werden (s. Tabelle 72).
Die relative Rohfett-Anflutung wurde durch Substitution des Enzyms PK bereits in der
niedrigsten Dosierung (D1) auf weniger als 50 % der bei PL-0-Tieren beobachteten
Rohfett-Anflutung reduziert, während die Testenzyme E1 bis E5 in der niedrigsten
Dosierung (D1) nur eine moderate Reduktion – auf Werte zwischen 69,7 und 86,8 %-
verglichen mit den PL-0-Tieren-bewirkten. Gegenüber den PL-0-Tieren signifikant war
dieser Effekt lediglich für PK und E4. Die für E1, E2 und E3 ermittelten Werte ließen sich –
aufgrund hoher Standardabweichung - nicht von den Kontrolltieren abgrenzen.
In der mittleren Dosierung (D2) sank auch nach Einsatz der Enzyme E1 und E4 die
Rohfett-Anflutung signifikant auf weniger als 50 % der Referenzwerte (Anflutung bei PL-0-
Tieren), während PK in der mittleren Dosierung (D2) sogar eine signifikante Reduktion
gegenüber den PL-0-Tieren auf 27,5 % bewirkte. Die Enzyme E2, E3 und E5 vermochten
bei vergleichbarer Dosierung die Rohfett-Anflutung hingegen lediglich auf ca. 60 % der
Werte der PL-0-Tiere (nicht signifikant) zu senken.
In der höchsten Dosierung (D3) wurde bei Verwendung aller Enzyme eine signifikante
Verringerung der relativen Rohfett-Anflutung im Vergleich zu den PL-0-Tieren ermittelt. Bei
Einsatz von E2 bis E5 wurden Ergebnisse zwischen 26,6 und 29,2 % erreicht. Dies
bedeutete, dass die Rohfett-Anflutung der PL-0-Tiere um ca. 2 / 3 reduziert werden
konnte, das Niveau der Kontrolltiere jedoch von E1, E2 und E4 nicht erreicht wurde. E1
und PK bedingten sogar eine Rohfett-Anflutung von ca. 20 % bezogen auf die PL-0-Tiere.
Die Steigerung der Enzymdosis von D1 auf D2 des Testenzyms E1 bedingte eine
signifikante Reduktion der Rohfett-Anflutung. Der Einsatz der Enzyme in der höchsten
Dosierung führte bei allen Enzymen zu einer signifikanten Verringerung der Rohfett-
Anflutung gegenüber dem Einsatz der geringsten Dosierung (D1). Zwischen den
Dosierungen D3 und D2 bestand lediglich für das Enzym E5 ein signifikanter Effekt der
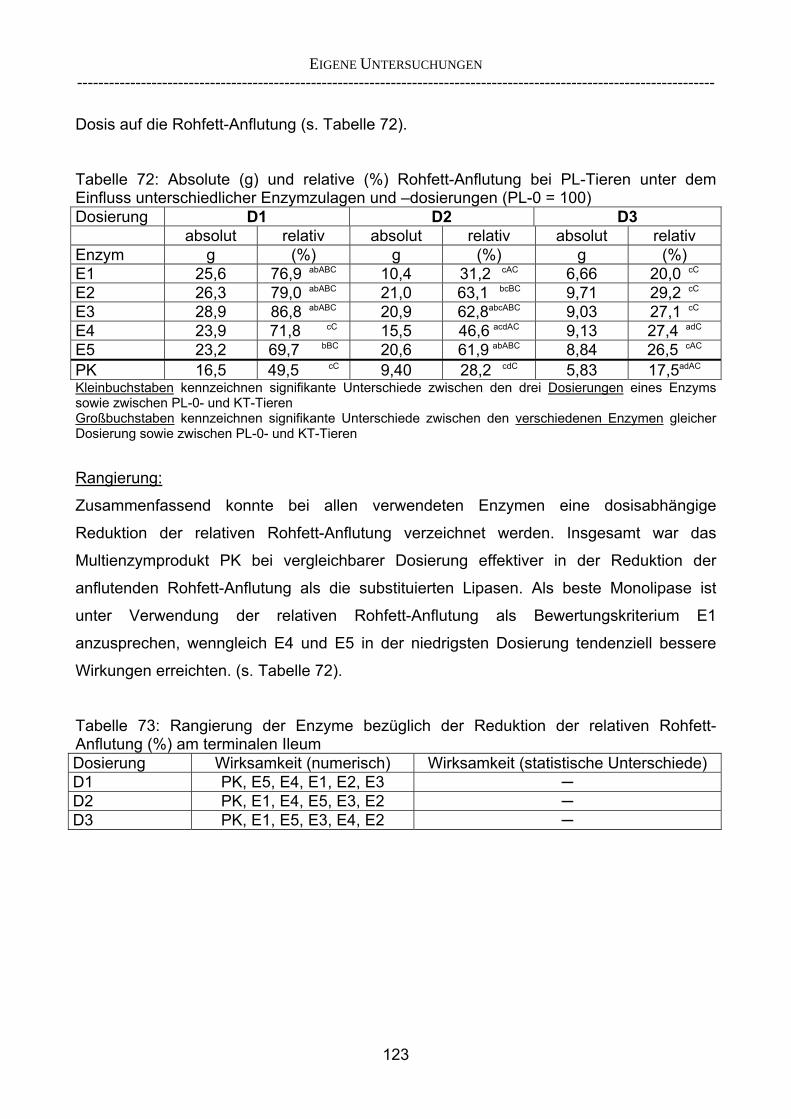
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
123
Dosis auf die Rohfett-Anflutung (s. Tabelle 72).
Tabelle 72: Absolute (g) und relative (%) Rohfett-Anflutung bei PL-Tieren unter dem Einfluss unterschiedlicher Enzymzulagen und –dosierungen (PL-0 = 100) Dosierung D1 D2 D3 absolut relativ absolut relativ absolut relativ Enzym g (%) g (%) g (%) E1 25,6 76,9 abABC 10,4 31,2 cAC 6,66 20,0 cC E2 26,3 79,0 abABC 21,0 63,1 bcBC 9,71 29,2 cC E3 28,9 86,8 abABC 20,9 62,8abcABC 9,03 27,1 cC E4 23,9 71,8 cC 15,5 46,6 acdAC 9,13 27,4 adC E5 23,2 69,7 bBC 20,6 61,9 abABC 8,84 26,5 cAC PK 16,5 49,5 cC 9,40 28,2 cdC 5,83 17,5adAC Kleinbuchstaben kennzeichnen signifikante Unterschiede zwischen den drei Dosierungen eines Enzyms sowie zwischen PL-0- und KT-Tieren Großbuchstaben kennzeichnen signifikante Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung sowie zwischen PL-0- und KT-Tieren
Rangierung:
Zusammenfassend konnte bei allen verwendeten Enzymen eine dosisabhängige
Reduktion der relativen Rohfett-Anflutung verzeichnet werden. Insgesamt war das
Multienzymprodukt PK bei vergleichbarer Dosierung effektiver in der Reduktion der
anflutenden Rohfett-Anflutung als die substituierten Lipasen. Als beste Monolipase ist
unter Verwendung der relativen Rohfett-Anflutung als Bewertungskriterium E1
anzusprechen, wenngleich E4 und E5 in der niedrigsten Dosierung tendenziell bessere
Wirkungen erreichten. (s. Tabelle 72).
Tabelle 73: Rangierung der Enzyme bezüglich der Reduktion der relativen Rohfett-Anflutung (%) am terminalen Ileum Dosierung Wirksamkeit (numerisch) Wirksamkeit (statistische Unterschiede) D1 PK, E5, E4, E1, E2, E3 ─ D2 PK, E1, E4, E5, E3, E2 ─ D3 PK, E1, E5, E3, E4, E2 ─

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
124
3.3.3.11 Relative Trockensubstanz-Verschwindensrate
Wird die Trockensubstanz-Verschwindensrate der PL-0-Tiere (35,0 %) mit 100
gleichgesetzt, so beträgt die Trockensubstanz-Verschwindensrate der Kontrolltiere 205 %
(s. Tabelle 74). Die Pankreasgangligatur bedingt demnach eine Reduktion der
Trockensubstanz-Verschwindensrate um ca. 50 %.
Tabelle 74: Absolute und relative Trockensubstanz-Verschwindensrate am terminalen Ileum bei KT und PL-0 –Tieren (Zeitraum: über 8 h n. F.) Absolut (%) Relativ (PL-0 = 100) KT 71,6 205 PL-0 35,0 100 A
Vergleich der verschiedenen Enzyme innerhalb einer Dosierung:
Alle eingesetzten Enzyme bewirkten eine dosisabhängige Steigerung der relativen
Trockensubstanz-Verschwindensrate, für deren Kalkulation die Werte der PL-Tiere ohne
Enzymzulage gleich 100 gesetzt wurden. Ausnahmen hierzu bildeten lediglich die Enzyme
E5 in der niedrigsten sowie E2 in der niedrigsten (D1) und mittleren (D2) eingesetzten
Dosierung, bei denen eine numerische Abnahme der Trockensubstanz-Verschwindensrate
gegenüber den Tieren ohne Enzymzulage zu verzeichnen war.
Bereits in der niedrigsten Dosierung (D1) erreichte PK mit der 1,37fache
Verschwindensrate eine signifikante Steigerung der relativen Trockensubstanz-
Verschwindensrate verglichen mit pankreasgangligierten Tieren ohne Enzymzulage. Die
effektivste Lipase (E4) bedingte eine Steigerung um den Faktor 1,29 und war damit
statistisch von E3 abzugrenzen.
Auch in der mittleren Dosierung (D2) ergaben sich durch den Einsatz des Produktes PK
die höchsten Werte (182 %), die signifikant gegenüber den Werten der PL-0-Tiere, aber
auch von E1 und E2 abgrenzbar waren. Die Werte der besten Monolipase E1 konnten von
denen der PL-0-Tiere abgegrenzt werden. Nach Einsatz der übrigen Testenzyme wurden
Werte ermittelt, die zwischen 99,1 und 163 % lagen (verglichen mit den PL-0-Tiere; s. Tabelle 72).
Infolge der Zulage von PK wurden in der höchsten Dosierung (D3) signifikant höhere
relative Trockensubstanz-Verschwindensraten ermittelt als für alle Monolipasen
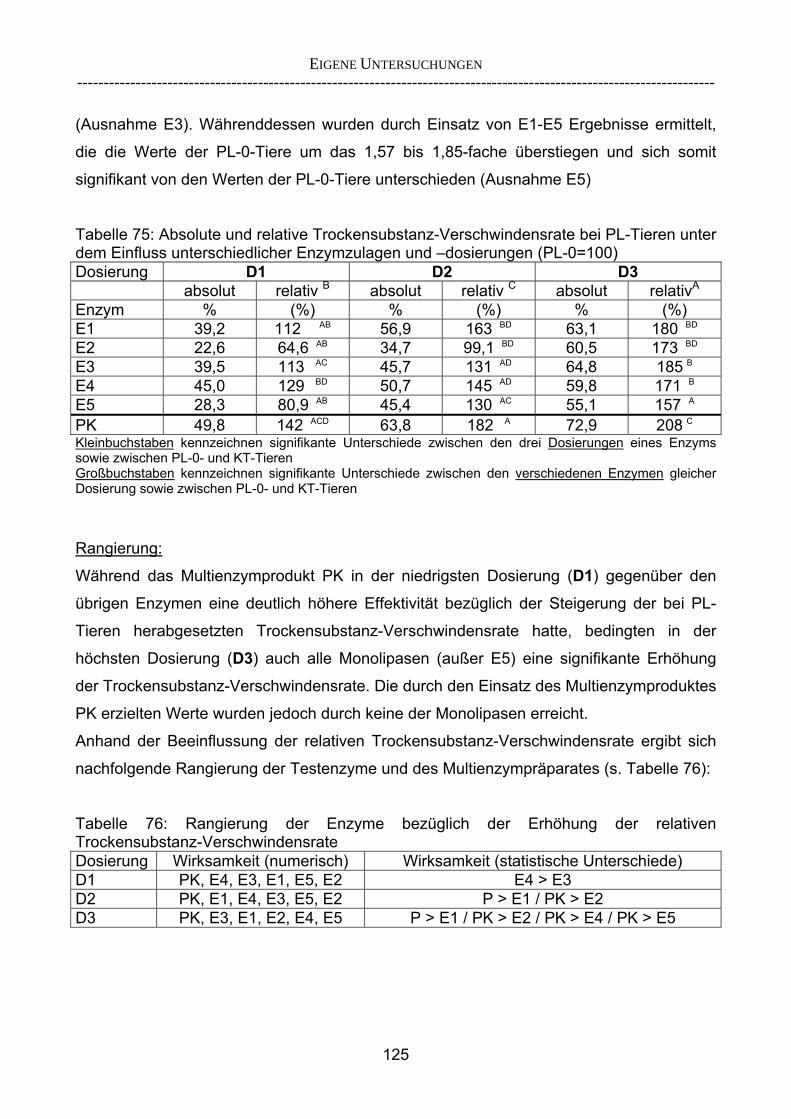
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
125
(Ausnahme E3). Währenddessen wurden durch Einsatz von E1-E5 Ergebnisse ermittelt,
die die Werte der PL-0-Tiere um das 1,57 bis 1,85-fache überstiegen und sich somit
signifikant von den Werten der PL-0-Tiere unterschieden (Ausnahme E5)
Tabelle 75: Absolute und relative Trockensubstanz-Verschwindensrate bei PL-Tieren unter dem Einfluss unterschiedlicher Enzymzulagen und –dosierungen (PL-0=100) Dosierung D1 D2 D3 absolut relativ B absolut relativ C absolut relativA
Enzym % (%) % (%) % (%) E1 39,2 112 AB 56,9 163 BD 63,1 180 BD E2 22,6 64,6 AB 34,7 99,1 BD 60,5 173 BD E3 39,5 113 AC 45,7 131 AD 64,8 185 B E4 45,0 129 BD 50,7 145 AD 59,8 171 B E5 28,3 80,9 AB 45,4 130 AC 55,1 157 A PK 49,8 142 ACD 63,8 182 A 72,9 208 C Kleinbuchstaben kennzeichnen signifikante Unterschiede zwischen den drei Dosierungen eines Enzyms sowie zwischen PL-0- und KT-Tieren Großbuchstaben kennzeichnen signifikante Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung sowie zwischen PL-0- und KT-Tieren
Rangierung:
Während das Multienzymprodukt PK in der niedrigsten Dosierung (D1) gegenüber den
übrigen Enzymen eine deutlich höhere Effektivität bezüglich der Steigerung der bei PL-
Tieren herabgesetzten Trockensubstanz-Verschwindensrate hatte, bedingten in der
höchsten Dosierung (D3) auch alle Monolipasen (außer E5) eine signifikante Erhöhung
der Trockensubstanz-Verschwindensrate. Die durch den Einsatz des Multienzymproduktes
PK erzielten Werte wurden jedoch durch keine der Monolipasen erreicht.
Anhand der Beeinflussung der relativen Trockensubstanz-Verschwindensrate ergibt sich
nachfolgende Rangierung der Testenzyme und des Multienzympräparates (s. Tabelle 76):
Tabelle 76: Rangierung der Enzyme bezüglich der Erhöhung der relativen Trockensubstanz-Verschwindensrate Dosierung Wirksamkeit (numerisch) Wirksamkeit (statistische Unterschiede) D1 PK, E4, E3, E1, E5, E2 E4 > E3 D2 PK, E1, E4, E3, E5, E2 P > E1 / PK > E2 D3 PK, E3, E1, E2, E4, E5 P > E1 / PK > E2 / PK > E4 / PK > E5
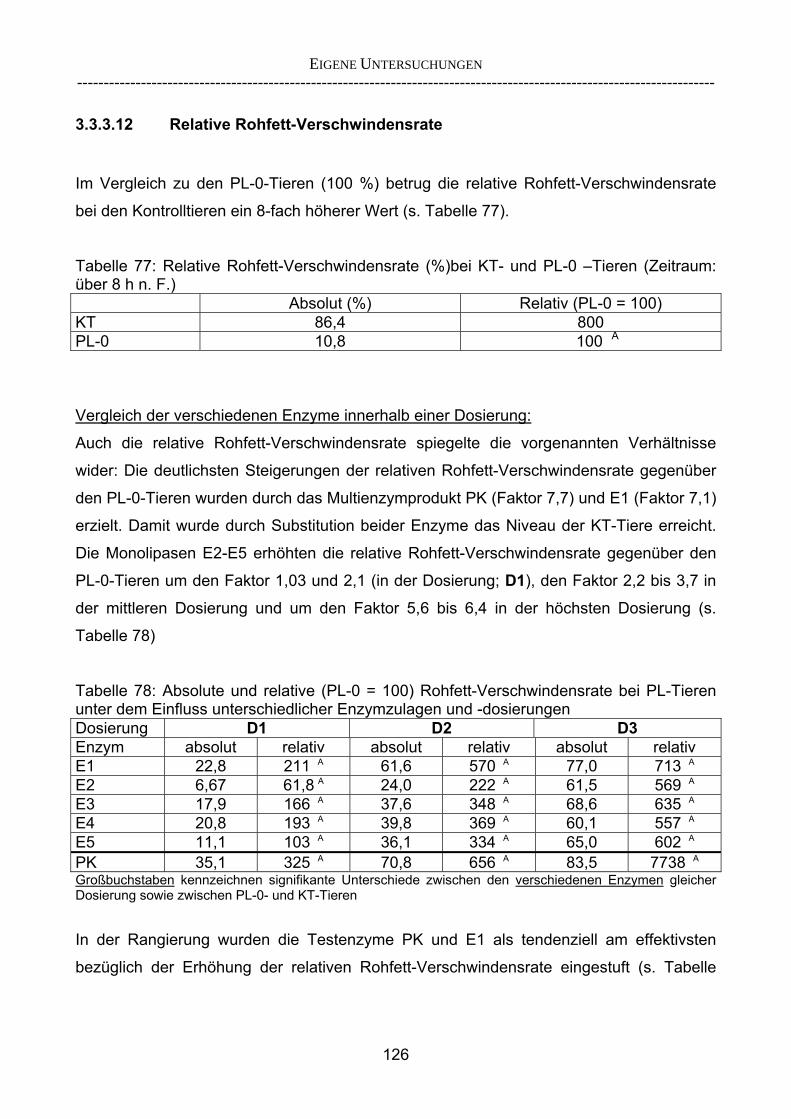
EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
126
3.3.3.12 Relative Rohfett-Verschwindensrate
Im Vergleich zu den PL-0-Tieren (100 %) betrug die relative Rohfett-Verschwindensrate
bei den Kontrolltieren ein 8-fach höherer Wert (s. Tabelle 77).
Tabelle 77: Relative Rohfett-Verschwindensrate (%)bei KT- und PL-0 –Tieren (Zeitraum: über 8 h n. F.)
Absolut (%) Relativ (PL-0 = 100) KT 86,4 800 PL-0 10,8 100 A
Vergleich der verschiedenen Enzyme innerhalb einer Dosierung:
Auch die relative Rohfett-Verschwindensrate spiegelte die vorgenannten Verhältnisse
wider: Die deutlichsten Steigerungen der relativen Rohfett-Verschwindensrate gegenüber
den PL-0-Tieren wurden durch das Multienzymprodukt PK (Faktor 7,7) und E1 (Faktor 7,1)
erzielt. Damit wurde durch Substitution beider Enzyme das Niveau der KT-Tiere erreicht.
Die Monolipasen E2-E5 erhöhten die relative Rohfett-Verschwindensrate gegenüber den
PL-0-Tieren um den Faktor 1,03 und 2,1 (in der Dosierung; D1), den Faktor 2,2 bis 3,7 in
der mittleren Dosierung und um den Faktor 5,6 bis 6,4 in der höchsten Dosierung (s. Tabelle 78)
Tabelle 78: Absolute und relative (PL-0 = 100) Rohfett-Verschwindensrate bei PL-Tieren unter dem Einfluss unterschiedlicher Enzymzulagen und -dosierungen Dosierung D1 D2 D3 Enzym absolut relativ absolut relativ absolut relativ E1 22,8 211 A 61,6 570 A 77,0 713 A E2 6,67 61,8 A 24,0 222 A 61,5 569 A E3 17,9 166 A 37,6 348 A 68,6 635 A E4 20,8 193 A 39,8 369 A 60,1 557 A E5 11,1 103 A 36,1 334 A 65,0 602 A PK 35,1 325 A 70,8 656 A 83,5 7738 A Großbuchstaben kennzeichnen signifikante Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung sowie zwischen PL-0- und KT-Tieren
In der Rangierung wurden die Testenzyme PK und E1 als tendenziell am effektivsten
bezüglich der Erhöhung der relativen Rohfett-Verschwindensrate eingestuft (s. Tabelle

EIGENE UNTERSUCHUNGEN ------------------------------------------------------------------------------------------------------------------------
127
79). Durch Zulage der Enzyme E2 - E5 konnten vergleichbare Werte verzeichnet werden,
so dass eine weitergehende Rangierung dieser Enzyme nicht möglich war.
Tabelle 79: Rangierung der Enzyme bezüglich der Erhöhung der relativen Rohfett-Verschwindensrate (%) Dosierung Wirksamkeit (numerisch) Wirksamkeit (statistische Unterschiede) D1 PK, E1, E4, E3, E5, E2 ─ D2 PK, E1, E4, E3, E5, E2 ─ D3 PK, E1, E3, E5, E2, E4 ─

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
128
4 Diskussion
Ziel der vorliegenden Arbeit war es, unter Verwendung pankreasgangligierter
Miniaturschweine einen In-vivo-Screening-Test für die Überprüfung der Effektivität
substituierter Lipasen zu entwickeln. Nachdem von BECKER (2005) ein solcher Test für
Amylasen und Proteasen entwickelt wurde, sollte in der hier vorliegenden Studie überprüft
werden, ob dieser Ansatz auch für ein Lipase-Screening-Verfahren umzusetzen sei.
Nach FUENTE-DEGE (2003) ist die Bestimmung der Rohfett-Verdaulichkeit über den
gesamten Verdauungstrakt geeignet, die Wirksamkeit lipolytischer Enzyme zu
quantifizieren, da die im postilealen Bereich erfolgenden mikrobiellen Umsetzungen
bezüglich des Rohfettes – im Unterschied zu Kohlenhydraten und Protein - zu
vernachlässigen sind. Als große Nachteile dieser etablierten Methode sind nach
GREGORY (persönliche Mitteilung, 2006) jedoch ein hoher zeitlicher Aufwand sowie der
Verbrauch relativ großer Mengen des zu prüfenden Enzyms zu nennen. Trotz Verzichts
auf eine mehrtägige Anfütterungsphase wird von einem Screening-Test die Möglichkeit
einer vergleichenden Quantifizierung der Effizienz lipolytischer Monopräparate verlangt.
Die im Screening-Test ermittelten Ergebnisse sollten also eine erste Einschätzung
ermöglichen, so dass nur „erfolgversprechende“ Neuentwicklungen in größerem Umfang
für weitere Untersuchungen produziert werden müssten. Ein Schnelltest ist jedoch nur
sinnvoll, wenn die Ergebnisse mit denen aus klassischen Verdaulichkeitsstudien
korrelieren. Eine Fehleinschätzung hätte gravierende Folgen wie beispielsweise
Investitionen in eine Weiterentwicklung von Lipasen unzureichender Wirksamkeit oder
einen Abbruch der Entwicklung effektiver Enzyme.
4.1 Kritik der Methoden
4.1.1 Anzahl der Tiere
Die Versuche im Rahmen der Entwicklung eines Screening-Tests sowie die Überprüfung
der Wirksamkeit verschiedener lipolytischer Enzyme wurden lediglich an einer geringen

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
129
Tierzahl durchgeführt. Die geringe Tierzahl (n=3 bzw. 4) in der Phase der Entwicklung des
Screening-Tests begründete sich zunächst einmal mit den räumlichen und personellen
Gegebenheiten, die parallele Chymussammlungen lediglich bei 3, ausnahmsweise bei 4,
Tieren je Tag erlaubten. In der Phase der Entwicklung einer optimierten Versuchsmahlzeit
wurden insgesamt 7 PL-Tiere eingesetzt, die in zwei Gruppen abwechselnd zum Einsatz
kamen. Dieses Vorgehen war trotz eingeschränkter Vergleichbarkeit notwendig, um den
Tieren eine „Regenerationsphase“ zwischen den Versuchen im Stoffwechselkäfig zu
gewähren. Abschließend wurde die optimierte Testmahlzeit unter identischen
Versuchsbedingungen an allen 7 Tieren überprüft, so dass durch eine größere Anzahl
vergleichbarer Ergebnisse die Reproduzierbarkeit abgesichert werden konnte.
In der Testphase bislang ungeprüfter Lipasen kam nur eine Tiergruppe, bestehend aus
drei identischen Tieren pro Enzymdosierung, zum Einsatz, so dass - trotz des z. T.
unterschiedlichen Niveaus der Werte der Tiere - die Vergleichbarkeit der Ergebnisse
gegeben war. Dennoch verstärkte sich das Gewicht von „Ausreißern“ dadurch, dass
anstelle einer dreitägigen Chymussammlung mit 3 - 6 Tieren pro Dosierung (entsprechend
9 bzw. 18 Ergebnissen pro Dosierung) in etablierten Verdaulichkeitsuntersuchungen von
TABELING 1998; FASSMANN 2001; HELDT 2001; MANDISCHER 2002; FUENTE-DEGE
2003; KAMMLOTT 2003; KARTHOFF 2004) im Screening-Test lediglich bei drei Tieren
eine eintägige Kollektion pro Enzymdosierung stattfand (3 Ergebnisse pro Dosierung).
Dementsprechend wäre auch im Screening-Verfahren eine Erhöhung der Tierzahl
wünschenswert. Aufgrund der räumlichen, technischen und personellen Ausstattung (s. o.)
wäre die Testung einer weiteren Tiergruppe jedoch nicht zeitgleich, sondern nur
anschließend durchführbar. Außerdem würde die Erhöhung der Tierzahl auch einen
erhöhten Enzymverbrauch bedeuten. Der sich demnach etwa verdoppelnde Bedarf an Zeit
und Material schien jedoch nicht gerechtfertigt: Erstens war als entscheidende
Anforderung an ein Lipase-Screening ein möglichst geringer Zeit- und Enzymaufwand
definiert worden. Auch wenn die Ergebnisse der einzelnen Tiere durchaus in der Höhe der
Werte variierten, so war zweitens festzuhalten, dass die dosisabhängigen Effekte bei allen
Tieren vergleichbar waren. Drittens wurden zur Minimierung tagesabhängiger Einflüsse
die drei zu testenden Dosierungen jedes Testenzyms randomisiert im 3 x 3 Lateinischen
Quadrat an die drei Tiere verabreicht. Darüber hinaus besteht für ein Lipase-Screening die
Anforderung, lediglich eine erste Rangierung und keine Absolutwerte zu erlangen.

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
130
Der Verzicht auf eine Ausweitung des Versuchsumfanges war hier somit tolerierbar.
Idealer Weise sollte aber bei entsprechender Verfügbarkeit der Enzyme der Einsatz einer
größeren Tierzahl erfolgen.
4.1.2 Validierung des Schnelltests
In einer Vergleichsstudie zur Validierung des Lipase-Screenings wurde auf bereits
vorhandene Daten klassischer Verdaulichkeitsversuche zurückgegriffen, in denen die
beiden Testlipasen E3 und E4 zum Einsatz kamen.
Jedoch konnte im Screening lediglich in der höchsten Dosierung ein deutlicher, jedoch
nicht abzusichernder Unterschied verzeichnet werden. Allerdings bestätigte sich dadurch
die in den Ergebnissen der Gesamtverdaulichkeitsstudie nachgewiesene Überlegenheit
der Effekte der Lipase E3 gegenüber dem Produkt E4.
Die Rangierung der Testlipasen im Screening-Test ergab für beide Enzyme eine
Bewertung im Mittelfeld. Es wäre jedoch wünschenswert gewesen, wenn zur Validierung
des Testverfahrens zwei Lipasen hätten gegenübergestellt werden können, die deutlichere
Differenzen aufwiesen, z.B. das als bestes und das als schlechtestes eingeschätzte
Enzym der Rangierung. Dieses war infolge mangelnder Verfügbarkeit entsprechender
Enzymmengen jedoch nicht realisierbar.
4.1.3 Eignung der erstmalig angewandten Methode zur Fettbestimmung (AnkomHCl Hydrolysis System®)
Durch die Veränderungen in Versuchsablauf und Anzahl der Versuche pro Woche infolge
der Entwicklung eines Lipase-Screenings stieg die Zahl der Proben erheblich.
Die Einführung des AnkomHCl Hydrolysis System® bot gegenüber der bisher in diesem
Projekt verwendeten Methode folgende Vorteile:
• wesentliche Steigerung des täglichen Probenumsatzes
• entscheidende Verringerung der benötigten Probenmasse
• Senkung der Arbeitsaufwandes infolge weitgehender Automatisierung

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
131
Durch Nutzung dieser Methode wurde es möglich, das erhöhte Probenaufkommen zu
bewältigen. Außerdem konnte auf diese Art und Weise - trotz zum Teil geringer
Probenmengen - der Rohfett-Gehalt der Proben bestimmt werden, so dass die Bildung
von Sammelproben mehrerer Kollektionsintervalle lediglich in Ausnahmefällen notwendig
war. Bei Verwendung der Ankom Geräte ergaben sich methodisch bedingte, gerichtete,
Abweichungen gegenüber der herkömmlichen VDLUFA-Methode (Soxhlet) von ca. 10 %.
Diese Differenz könnte eine Folge von methodischen Unterschieden sein: mit dem
Soxhlet-Verfahren wird der Rohfettgehalt durch Quantifizierung des Rückstandes nach
Etherextraktion und Trocknung bestimmt, während bei Verwendung der Ankom-Methode
die Bestimmung des Fettgehaltes über Differenzbildung (Masse vor Extraktion − Masse
nach Extraktion) erfolgt. Somit werden im Gegensatz zum Soxhlet-Verfahren bei
Bestimmung durch das Ankom-Verfahren alle flüchtigen lipophilen Substanzen in den
Rohfett-Gehalt mit einbezogen. Innerhalb des Screening-Tests fand ein Vergleich der
Ergebnisse lediglich anhand der mit der neuen Methode ermittelten Werte statt, so dass
die Vergleichbarkeit der Werte gegeben war. Da bei dem Vergleich der Screening-Test-
Ergebnisse mit denen aus Gesamtverdaulichkeiten aufgrund mehrerer Unterschiede keine
direkte Vergleichbarkeit vorlag, sollte lediglich die Rangierung begutachtet werden.
Demzufolge fand auch hier kein direkter Vergleich der beiden Methoden statt, so dass
innerhalb der jeweiligen Versuche alle eingesetzten Enzyme unter denselben
Bedingungen geprüft wurden.
Unter Verwendung der neuen Methode (AnkomHCl Hydrolysis System® (Filterbag-
Säurehydrolyse-Technik®)) wurde eine sehr gute Reproduzierbarkeit der Ergebnisse
erzielt. Die neu etablierte Ankom-Methode konnte demnach als zuverlässig und geeignet
zur Bestimmung der Rohfett-Konzentration im Chymus eingestuft werden.

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
132
4.2 Erörterung der eigenen Ergebnisse
4.2.1 Wie muss eine Testmahlzeit beschaffen sein, um In-vivo die Wirksamkeit von Lipasen in einem Schnelltest überprüfen zu können?
Die in dieser Studie zu prüfenden Lipasen waren bisher lediglich mittels In-vitro-Versuchen
auf ihre hydrolytische Aktivität und ihre pH-Stabilität untersucht worden (Ausnahme: E3
und E4, s. o.). Zur Überprüfung ihrer tatsächlichen Effektivität war eine In-vivo-
Untersuchung unerlässlich. Da in dieser Phase der Entwicklung jedoch nur sehr begrenzte
Enzymmengen zur Verfügung standen, war die Durchführung herkömmlicher
Verdaulichkeitsuntersuchungen unmöglich. Demzufolge bestand die Notwendigkeit zur
Entwicklung eines zeit- und materialsparenden In-vivo-Verfahrens, das dennoch eine erste
Einschätzung der In-vivo-Wirksamkeit der Lipasen ermöglichte.
Wie in Kapitel 3.1.6 – 3.1.8 (Material- und Methoden) beschrieben, stand bei der
Entwicklung eines Lipase-Screenings die Optimierung einer geeigneten Testmahlzeit im
Vordergrund: Trotz zu befürchteter Passagestörungen wurde ein hoher Fettanteil
angestrebt, insbesondere unter Verwendung langkettiger Fettsäuren, um auch im unteren
Dosierungsbereich eine ausreichende Differenzierung der Wirksamkeit zwischen den
verschiedenen Lipase-Produkten zu forcieren. Anderenfalls wären Wirkungen der
substituierten Enzyme nicht von denen endogener, z. B. gastrischer Lipasen
differenzierbar.
Die entscheidende Anforderung an die Beschaffenheit einer geeigneten Testmahlzeit
war jedoch die Erzielung eines parallelen Flusses von Trockensubstanz, Rohfett, Enzym
und Marker. Dieses Kriterium entstand aus folgender Problematik:
⇒ Ist es Ausdruck einer hohen Rohfett-Verdaulichkeit oder einer geringen
Chymusflussrate, wenn lediglich geringe Mengen Rohfett am terminalen Ileum
anfluten?
⇒ Ist es vorstellbar, dass lediglich Trockensubstanz und Marker an der ilealen Fistel
aufgefangen werden, Rohfett jedoch selektiv zurückgehalten wird?

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
133
Um sicher zu stellen, dass eine verringerte Rohfett-Anflutung Folge der Verdauung
(Resorption der Hydrolysespaltprodukte infolge der Enzymsubstitution) und kein Effekt von
Passagestörungen war, wurde eine fraktionierte Sammlung und Analyse des Chymus
durchgeführt, anhand derer relative parallele Flussraten kalkuliert wurden. Des Weiteren
wurde zur Überprüfung der Rohfett-Verschwindensrate eine Vielzahl von Parametern
eingesetzt.
Die Diskussion der Optimierungsvorgänge der Testmahlzeit zur Verbesserung paralleler
Flussraten erfolgt anhand folgender Gesichtspunkte:
• Modifikation des Trockensubstanz- und Rohfasergehaltes (4.2.1.1)
• Verringerung der Chromoxidmenge pro Mahlzeit (s. 4.2.1.2)
• Einsatz einer flüssigen Mahlzeit am Vorabend der Verabreichung der Testmahlzeit
(4.2.1.3)
• Stabilisierung der Testmahlzeit (Homogenisierung; 4.2.1.4)
4.2.1.1 Modifikation des Trockensubstanz- und Rohfasergehaltes
Bei der Optimierung der Testmahlzeit wurde von der Idee einer flüssigen Mahlzeit
ausgegangen. Die auf Basis eines Milchshakes nach verschiedenen Modifikationen
entwickelte endgültige Variante wies einen Trockensubstanz-Gehalt von ca. 40 % auf (147
g TS in 372 g uS). Die Testmahlzeit war eine Öl-in-Wasser-Emulsion, in der die feinst
verteilten unlöslichen Bestandteile für einen bestimmten Zeitraum eine stabile Mischung
bildeten. Im Gegensatz dazu sedimentierten die in der herkömmlichen Standard-Fettdiät
vorhandenen festen Bestandteile umgehend, so dass sich eine wässrige Schicht über den
festen Bestandteilen bildete. Hingegen blieb die flüssige „optimierte“ Testmahlzeit über
Stunden stabil, so dass von der berechtigte Annahme ausgegangen werden konnte, dass
auch im GIT des Tieres keine Entmischungen stattfanden. Trotz des höheren
Trockensubstanz-Anteils der Screening-Test-Diät gegenüber der Standard-Fettdiät konnte
deshalb von einem Flüssigfutter gesprochen werden, von dem zu erwarten war, dass sich
die passageregulierenden Aspekte einer flüssigen Mahlzeit insbesondere auf die
Magenentleerung auswirken könnten (BRAUDE et al.1970; HUNT u. STUBBS 1975;
READ 1984; DECUYPERE et al. 1985). Im 5. Versuch zur Optimierung einer Testmahlzeit

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
134
wurde erstmals zusätzlich Rohfaser (9,88 g Methocel®) eingesetzt, um Entmischungen
des Markers von der Testmahlzeit entgegenzuwirken. In den folgenden Versuchen traten
deutlichere dosisabhängige Effekte auf. Durch diese Maßnahmen konnten die erwarteten
Probleme infolge des hohen Fettgehaltes in der Diät reduziert und der parallele Fluss der
Mahlzeitkomponenten verbessert werden.
4.2.1.2 Verwendung von Chromoxid als Marker
Der eingesetzte Marker Chromoxid diente neben der optischen Abgrenzung des aus der
Testmahlzeit stammenden Chymus zur Berechnung der „Verschwindensrate“, der die
Formel der Indikatormethode zu Grunde lag. Eine geringe Anflutung von Chymus (bzw.
Rohfett) infolge hoch wirksamer lipolytischer Enzyme konnte damit von einer geringen
Anflutung infolge einer geringen Passagerate abgegrenzt werden.
Für die Verwendung dieser Formel ist ein paralleler Fluss von Trockensubstanz, Rohfett
und Marker von zentraler Bedeutung. In den ersten Ansätzen zur Optimierung einer
geeigneten Testmahlzeit konnten jedoch keine parallelen relativen Flussraten ermittelt
werden.
Dennoch wurde am Einsatz von Cr2O3 aufgrund folgender Gesichtspunkte festgehalten:
• Die Bestimmung des Markers war etabliert und es lagen entsprechende
Erfahrungen aus vorangegangen Studien des Projektes vor.
• Der Marker sollte eine optische Abgrenzung ermöglichen, um mit dem ersten
Erscheinen des gefärbten Chymus an der Fistel mit der Kollektion beginnen zu
können.
• Die Vergleichbarkeit mit vorangegangenen Studien des Projektes sollte erhalten
bleiben.
• Nach Zubereitung der Mahlzeit als Emulsion fand über Stunden keine
Entmischung von wässriger und fettiger Phase statt.
Infolge der durchgeführten Modifikationen gelang bei Verwendung der endgültigen
Testmahlzeit ein weitestgehend paralleler Fluss von Nährstoffen und Chromoxid im
Chymus (s. Abbildung 31):

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
135
• Anpassung der Chromoxidmenge an die verringerte Trockensubstanz-Menge (2,5 g
/ kg TS entsprechen 0,368 g Cr2O3)
• Verbesserte Homogenisierung der Emulsion durch mehrminütige Vermischung
• der Komponenten mittels Ultra-turrax 900® anstelle eines Handmixers
4.2.1.3 Einsatz einer flüssigen Mahlzeit am Vorabend des Screening-Tests
Um Interferenzen der dem Versuch vorangegangenen Abendfütterung mit der
Testmahlzeit im GIT infolge von „Verschleppungen“ (TS- und Rfe-Übertrag des
Haltungsfutters in den Kollektionszeitraum) zu vermeiden, wurde eine fettfreie
Flüssigmahlzeit (ProvideXtra®) 12 Stunden vor Fütterung der Testmahlzeit verabreicht.
Der Hintergrund dieses Vorgehens war das Bestreben, evtl. Reste vorheriger Mahlzeiten
in Magen oder Dünndarm weiter zu transportieren und eine zügige Passage der
Abendmahlzeit selbst sicherzustellen. Die im Lipase-Screening verwendete Testmahlzeit
war besonders empfindlich gegenüber einem Nährstoffeintrag aus vorherigen Mahlzeiten,
da sie lediglich 150 g TS (im Gegensatz zum üblicherweise verabreichten Mischfutter-
Mahlzeiten: ca. 225 g TS) beinhaltete, so dass selbst relativ geringe Mengen des sonst
verwendeten Mischfutters verhältnismäßig starken Einfluss auf Trockensubstanz-
Anflutung und -Verschwindensrate ausüben würden. Des Weiteren bestand zwischen
herkömmlichem Mischfutter und Testmahlzeit eine große Differenz hinsichtlich des
Rohfett-Gehaltes (Mischfutter ca. 2 % vs. Testmahlzeit ca. 35 % der TS), wodurch bei
Vermischung mit der Testmahlzeit eine erhöhte Rohfett-Verschwindensrate im
aufgefangenen Chymus vorgetäuscht würde.
Bei Einsatz von ProvideXtra® war jedoch davon auszugehen, dass es zu keinen
Interferenzen mit der Testmahlzeit im Magen und Dünndarm kam und an der ilealen Fistel
nach Farbwechsel lediglich aus der Testmahlzeit stammender (und damit enzymhaltiger)
Chymus gesammelt wurde, so dass eine Verbesserung in den parallelen Flussraten
verzeichnet werden konnte.

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
136
4.2.1.4 Stabilisierung der Testmahlzeit (Homogenisierung)
Zur Erzielung einer stabilen Testmahlzeit wurde die neu entwickelte Testmahlzeit
zusätzlich mit dem Homogenisator Ultra-turrax 900® für insgesamt sechs Minuten
homogenisiert.
Durch diese intensive Vermischung gewann die Emulsion an Stabilität, so dass ab dem
ersten Einsatz des Ultra-turrax 900® im sechsten Optimierungsversuch ein paralleler Fluss
von Trockensubstanz, Rohfett und Cr2O3 (s. Abbildung 31) erreicht wurde. Außerdem
wurden nicht nur dosisabhängige Effekte der Trockensubstanz- und Rohfett-
Verschwindensraten sondern auch der Trockensubstanz- und Rohfett-Anflutung
festgestellt.
Fazit: Trotz des hohen Fettgehaltes in der Testmahlzeit, der flüssigen Darreichungsform
der Testmahlzeit und der Reduktion der Chromoxidmenge pro Mahlzeit gelang es, durch
Zusatz einer Rohfaserquelle sowie Verwendung eines Homogenisators eine stabile
Emulsion zu schaffen. Zusätzlich wurde ein Übertrag von Chymus vorheriger Mahlzeiten
durch den Einsatz einer flüssigen Trinknahrung am Vorabend der Versuche bedeutend
vermindert. Mit Durchführung dieser Modifikationen stand ein Verfahren zur Verfügung, in
dem zuverlässig parallele relative Flussraten von Trockensubstanz, Rohfett und
Chromoxid ermittelt werden konnten, wie in nachstehender Abbildung (s. Abbildung 31)
exemplarisch dargestellt wird. Zur Ermittlung und Überprüfung der parallelen Flussraten
war der Einsatz des Markers Chromoxid entscheidend. Aufgrund dieser nachgewiesenen
Parallelität war es möglich, die Formel der Indikatormethode zur Kalkulation der
Verschwindensrate einzusetzen.
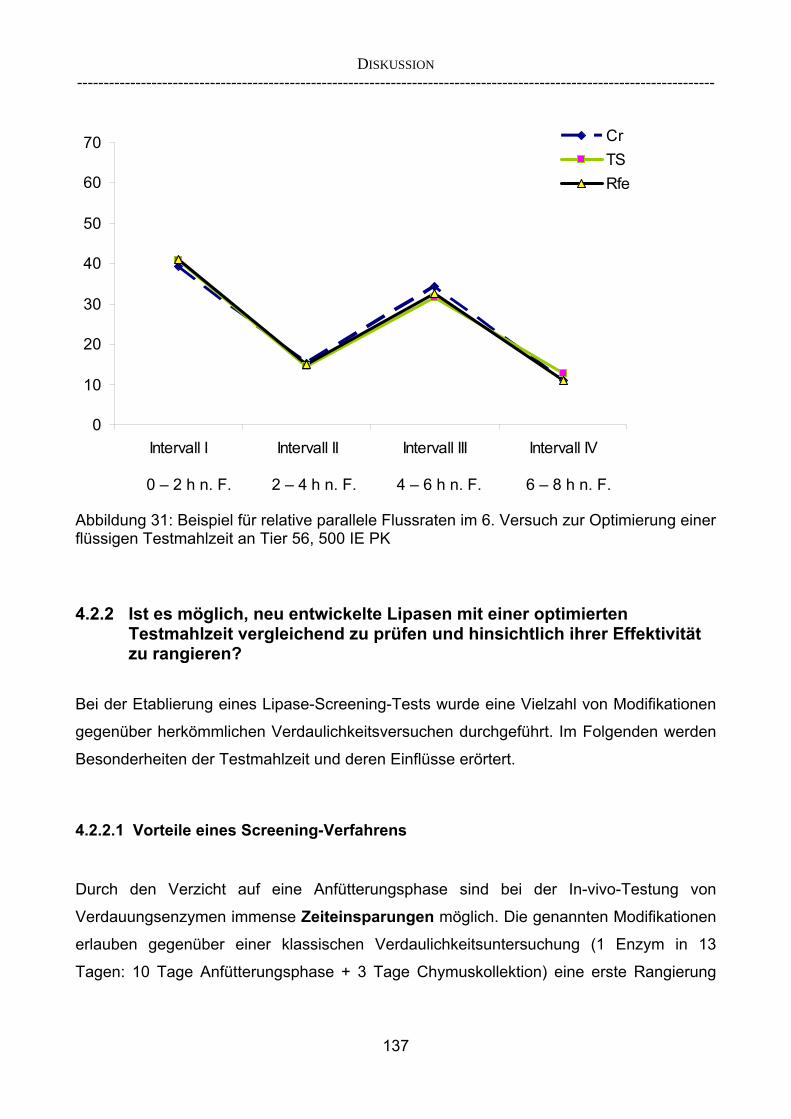
DISKUSSION ------------------------------------------------------------------------------------------------------------------------
137
0
10
20
30
40
50
60
70
Intervall I Intervall II Intervall III Intervall IV
CrTSRfe
0 – 2 h n. F. 2 – 4 h n. F. 4 – 6 h n. F. 6 – 8 h n. F.
Abbildung 31: Beispiel für relative parallele Flussraten im 6. Versuch zur Optimierung einer flüssigen Testmahlzeit an Tier 56, 500 IE PK
4.2.2 Ist es möglich, neu entwickelte Lipasen mit einer optimierten Testmahlzeit vergleichend zu prüfen und hinsichtlich ihrer Effektivität zu rangieren?
Bei der Etablierung eines Lipase-Screening-Tests wurde eine Vielzahl von Modifikationen
gegenüber herkömmlichen Verdaulichkeitsversuchen durchgeführt. Im Folgenden werden
Besonderheiten der Testmahlzeit und deren Einflüsse erörtert.
4.2.2.1 Vorteile eines Screening-Verfahrens
Durch den Verzicht auf eine Anfütterungsphase sind bei der In-vivo-Testung von
Verdauungsenzymen immense Zeiteinsparungen möglich. Die genannten Modifikationen
erlauben gegenüber einer klassischen Verdaulichkeitsuntersuchung (1 Enzym in 13
Tagen: 10 Tage Anfütterungsphase + 3 Tage Chymuskollektion) eine erste Rangierung

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
138
von 6 Enzymen bzw. 6 verschiedenen Enzymdosierungen im gleichen Zeitraum.
Außerdem werden bedeutend geringere Enzymmengen benötigt: 1 statt 26
Enzymapplikationen pro Enzym, so dass erste In-vivo-Studien zu einem deutlich früheren
Zeitpunkt im pharmazeutischen Entwicklungsprozess möglich werden. Dies ist von
besonderer Bedeutung, da die ersten In-vivo-Tests oftmals durch die in dieser
Entwicklungsphase nur in geringeren Mengen verfügbaren neuen Substanzen limitiert
werden.
4.2.2.2 Bedeutung der relativen Flussraten für die Berechnung der „Verschwindensrate“
Zur Berechnung der Verschwindensrate wird die Formel der Indikatormethode (s. 3.1.5.8)
herangezogen. Voraussetzung zur Bestimmung der Nährstoffverdaulichkeit mit der
Indikatormethode ist der parallele Fluss von Trockensubstanz, Rohfett und Chromoxid.
Durch diesen ist die Verwendung der Formel zu rechtfertigen, die in der Regel nur im
Zustand des „steady state“ nach mehrtägiger Anfütterungsphase (klassische
Verdaulichkeitsuntersuchung) gilt. Im Rahmen eines Screening-Verfahrens ohne
Anfütterungsphase kann die Indikatormethode also nur dann angewandt werden, wenn ein
paralleler Fluss der Parameter nachweisbar ist (BECKER 2005). Um den relativen Fluss
von Trockensubstanz, Rohfett und Chromoxid zu ermitteln, wurde die absolute Anflutung
dieser Parameter in 2-Stunden-Intervallen fraktioniert gesammelt und analysiert.
Anschließend wurden die angefluteten Mengen an Trockensubstanz, Rohfett und
Chromoxid auf den gesamten Kollektionszeitraum bezogen.
Erst mit Einsatz der optimierten Testmahlzeit konnte der parallele Fluss der Parameter
über den Kollektionszeitraum von 8 Stunden sichergestellt werden. Während der
Durchführung der Wirksamkeitsprüfung der Testenzyme wurde der parallele relative Fluss
weiterhin bestätigt (s. Abbildung 32).
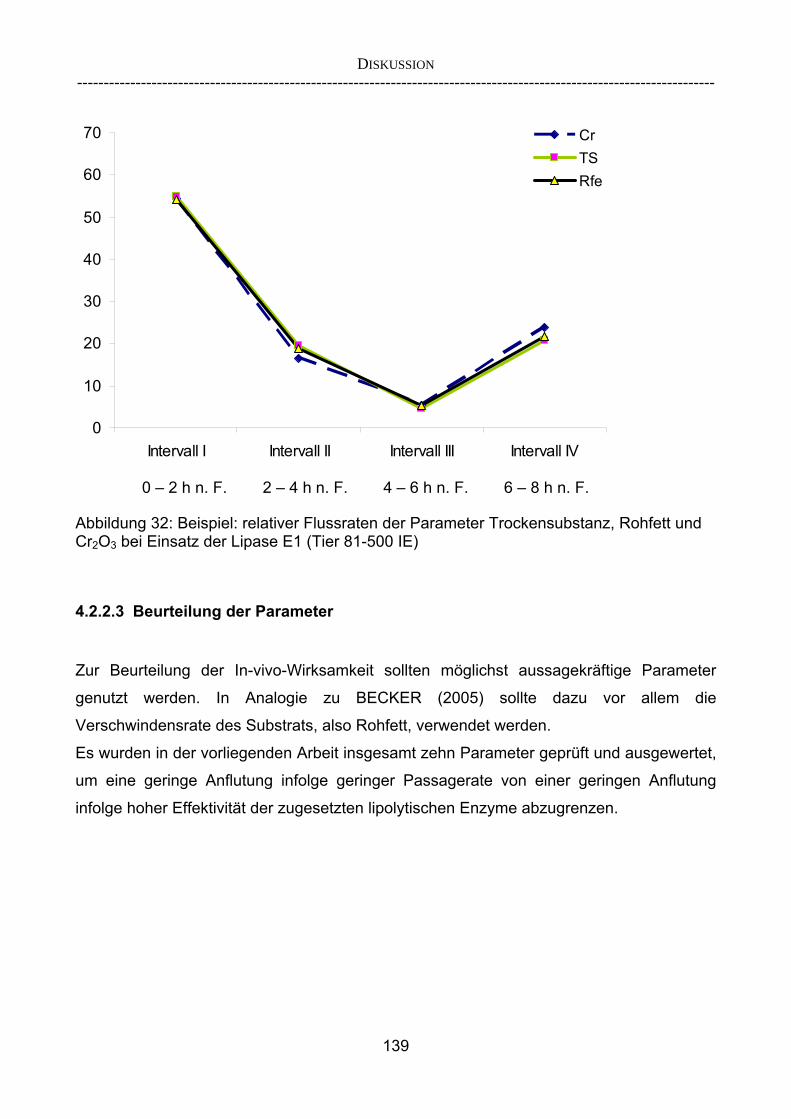
DISKUSSION ------------------------------------------------------------------------------------------------------------------------
139
0
10
20
30
40
50
60
70
Intervall I Intervall II Intervall III Intervall IV
CrTSRfe
0 – 2 h n. F. 2 – 4 h n. F. 4 – 6 h n. F. 6 – 8 h n. F.
Abbildung 32: Beispiel: relativer Flussraten der Parameter Trockensubstanz, Rohfett und Cr2O3 bei Einsatz der Lipase E1 (Tier 81-500 IE)
4.2.2.3 Beurteilung der Parameter
Zur Beurteilung der In-vivo-Wirksamkeit sollten möglichst aussagekräftige Parameter
genutzt werden. In Analogie zu BECKER (2005) sollte dazu vor allem die
Verschwindensrate des Substrats, also Rohfett, verwendet werden.
Es wurden in der vorliegenden Arbeit insgesamt zehn Parameter geprüft und ausgewertet,
um eine geringe Anflutung infolge geringer Passagerate von einer geringen Anflutung
infolge hoher Effektivität der zugesetzten lipolytischen Enzyme abzugrenzen.

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
140
Folgende Parameter wurden erhoben:
⇒ Trockensubstanz- und Rohfett-Anflutung (g):
(Keine Differenzierung zwischen hoher Passagerate und schlechter Wirksamkeit
des Enzyms möglich)
⇒ Rohfett- und Chromoxid-Konzentration (g / kg TS):
(Maß für die Wirksamkeit der substituierten Enzyme. Bei zunehmender Wirksamkeit
der Enzyme: Rfe-Konz. ↓ und Cr2O3 ↑)
⇒ Chromoxid-Wiederfindung (%):
(Berechnung der Verschwindensrate mittels Indikatormethode, enzym- und
dosisunabhängige Wiederfindung)
⇒ Verhältnis der Konzentrationen von Rohfett zu Chromoxid, Trockensubstanz- und
Rohfett-Verschwindensrate (%), Relative Anflutung und Verschwindensrate von
Trockensubstanz und Rohfett:
(Differenzierung zwischen der Wirksamkeit der Lipasen und Einflüssen der
Passageraten durch Einbeziehung der Cr2O3-Wiederfindung)
Für alle Parameter waren dosisabhängige Effekte erkennbar. Einzige Ausnahme bildete
die Chromoxid-Wiederfindung: bezüglich dieser Werte ließen sich weder dosis- noch
enzymabhängige Zusammenhänge nachweisen, womit Cr2O3 in diesen Versuchen seiner
Aufgabe als unverdaulicher, farblicher Marker gerecht wurde.

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
141
Anhand der übrigen zehn Parameter waren folgende Ergebnisse hinsichtlich der Testung
und Rangierung der In-vivo-Wirksamkeit der zu prüfenden Enzyme feststellbar:
• Für das Multienzymprodukt PK sowie die Monolipase E1 wurden in allen drei
geprüften Dosierungen die höchste Effizienz ermittelt.
• In der höchsten Dosierung wurde von drei der sechs Enzyme das Niveau der
Kontrolltiere bei allen Parametern erreicht.
• In der niedrigsten und mittleren Dosierung war infolge höherer
Standardabweichungen oftmals keine statistische Abgrenzung gegenüber den
Kontrolltieren zu verzeichnen.
• Beim Vergleich der Enzyme miteinander in einer einzigen Dosierung konnten
nur wenige signifikante Differenzen in der Wirksamkeit abgegrenzt werden: Das
Multienzym PK und E1 als beste Monolipase waren jedoch in mehreren Fällen
von den weniger wirksamen Lipasen (z.B. E2) signifikant abzugrenzen.
Als Ursache für die häufig nicht abzusichernden Effekte könnte die Auswahl der
Dosierungen in Betracht kommen: In vorangegangenen Arbeiten dieses Projektes wurden
112 000 bis 1 120 000 IE lipolytischer Aktivität des Multienzymproduktes PK unter
Verwendung der breiförmigen Standard-Fettdiät zur Bestimmung der scheinbaren Rohfett-
Verdaulichkeit eingesetzt. Die ermittelten Werte der KT-Tiere lagen in den
Untersuchungen von TABELING 1998; FASSMANN 2001; HELDT 2001; MANDISCHER
2002; FUENTE-DEGE 2003) bei 88,2 – 97,6 %, während von den PL-Tieren in der
höchsten Dosierung (336 000 IE) des Multienzymproduktes PK eine scheinbare
Rohfettverdaulichkeit (Rfe-sV) von 71,4 – 83,3 % erreicht wurde. Im Verlauf der
Optimierung der Testmahlzeit zur Entwicklung eines Lipase-Screenings wurden die
Dosierungen wesentlich reduziert. Mit Einsatz einer flüssigen Mahlzeit wurden bereits in
geringsten Dosierungen vergleichbare Effekte erzielt. Schon bei Einsatz von nur 75 000 IE
wurden die Werte der maximalen Wirksamkeit von PK ermittelt (87,8 % bzw. 85,0 % Rfe-
VR im 6. bzw. 7. Versuch zur Optimierung einer Testmahlzeit). Ebenfalls konnte bereits
bei 5 000 IE ein vergleichbares Niveau erreicht werden: 77,1 % bzw. 81,9 % Rfe-VR im 3
bzw. 4. Versuch zur Optimierung einer Testmahlzeit. Ein Erklärungsansatz könnte in den
Unterschieden der eingesetzten Rationen liegen: In den vorherigen Studien wurde die
Standard-Fettdiät des Projektes eingesetzt. Diese wurde im Gegensatz zur Testmahlzeit

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
142
(stabile Emulsion s. o.) als Suspension (Brei) verabreicht, die sich eher entmischte (s. o.:
verbesserte Bedingungen für die Enzyme in flüssiger, homogener Umgebung).
Schließlich wurden bei der optimierten Testmahlzeit deutliche dosisabhängige Effekte in
den Dosierungen D1 = 500, D2 = 5 000 und D3 = 75 000 IE lipolytischer Aktivität erreicht.
Es ist zum derzeitigen Zeitpunkt nicht abschließend zu bewerten, ob die hohe Effektivität
der substituierten Enzyme (inkl. des etablierten Multienzymproduktes PK) eine Folge der
speziellen Zusammensetzung der Testmahlzeit oder aber der Zubereitung (flüssige,
homogenisierte Diät) ist. Es ist zu berücksichtigen, dass unterschiedliche Fettquellen in
den verschiedenen Versuchsmahlzeiten verwendet wurden: während in den klassischen
Gesamtverdaulichkeitsstudien Sojaöl zum Einsatz kamen, wurde der Screening-Test-Diät
Olivenöl und Milchfett zugesetzt. Eine besonders hohe Effektivität der zugesetzten
Monolipasen kann ausgeschlossen werden, da sich das bekannt sehr wirksame
Multienzymprodukt PK ebenfalls im Screening-Test durch eine vergleichsweise höhere
Effektivität als in früheren Studien auszeichnete (71,4 – 83,3 % Rfe-sV vs. 85,0 – 87,8 %
Rfe-VR).
Fazit: Insgesamt kann der Schluss gezogen werden, dass alle erhobenen Parameter zur
Rangierung der Lipasen geeignet sind. Die im Screening-Verfahren mittels Rohfett-
Verschwindensrate aufgestellte Rangierung war demnach anhand aller verglichenen
Parameter (exklusive Cr2O3-Wdf.) nachzuvollziehen.
4.2.3 Korrelieren die Ergebnisse aus dem Schnelltest für Lipasen mit denen aus etablierten Verdaulichkeitsuntersuchungen?
Bei Vergleich der Ergebnisse des Lipase-Screenings mit Ergebnissen aus herkömmlichen
Gesamtverdaulichkeitsversuchen ausgewählter Enzyme (KARTHOFF (2004) und
BECKER (2005)) wurde lediglich der Parameter Rfe-Verschwindensrate ausgewertet.
Anschließend wurde die im Screening aufgestellte Rangierung der Testenzyme E3 und E4
anhand weiterer Parameter überprüft.

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
143
4.2.3.1 Vergleich der Ergebnisse des Screening-Tests mit denen herkömmlicher Gesamtverdaulichkeitsstudien
In einem ersten Schritt (s. 3.1.6 bis 3.1.8) wurde ein Lipase-Screening-Test unter
Verwendung des bekannten Multienzymproduktes PK entwickelt.
In einem zweiten Schritt wurde das neue Testverfahren an fünf Lipasen fungaler Herkunft
überprüft.
Da lediglich geringe Mengen der Testenzyme vorlagen, war es jedoch unmöglich,
klassische Verdaulichkeitsversuche, die eine mehrtägige Anfütterungsphase erfordern,
durchzuführen. Deshalb kamen für den Vergleich der Methoden nur die Monolipasen E3
und E4 in Betracht, die bereits in früheren herkömmlichen Verdaulichkeitsstudien
untersucht worden waren (KARTHOFF 2004; BECKER 2005). Bei dem Vergleich mit
Ergebnissen klassischer Verdaulichkeitsversuche sind mehrere Unterschiede im
Versuchsablauf zu beachten: In den vorangegangenen Studien des Projektes
„pankreasgangligiertes Minischwein“ wurden die Enzyme E3 und E4 in ähnlicher, jedoch
nicht identischer Dosierung eingesetzt: Die Lipase E3 wurde in 100 000 IE und die Lipase
E4 in 110 000 IE lipolytischer Einheiten dosiert. Des Weiteren befand sich zu dem
damaligen Zeitpunkt noch keine Flüssigmahlzeit als Testdiät im Einsatz. Stattdessen
wurde die Standard-Fettdiät des Projektes mit einem Fettanteil von 30 % (2x täglich, 250g
uS, ca. 70 g Rfe / Mahlzeit) verabreicht. Die angegebenen Werte sind Mittelwerte von
Einzelwerten, die an je sechs pankreasgangligierten Tieren im Rahmen einer fünftägigen
Gesamtverdaulichkeitsuntersuchung ermittelt wurden. Da aber sowohl Versuchsdesign als
auch Versuchsdiät in den beiden Gesamtverdaulichkeitsstudien absolut vergleichbar
waren und nahezu identische Tiergruppen verwendet wurden, ist ein direkter Vergleich der
Ergebnisse und eine Rangierung der beiden Enzyme anhand ihres Einflusses auf die
Fettverdaulichkeit trotz zeitlicher Differenz zwischen den Versuchen zulässig.
Dieser Versuch einer Validierung kann und soll daher keinen direkten Vergleich zwischen
Screening-Verfahren und etablierter Gesamtverdaulichkeitsstudie darstellen, sondern
lediglich der Überprüfung dienen, inwiefern die anhand der Ergebnisse im Screening-Test
vorgenommene Rangierung mit der aus klassischen Verdaulichkeitsuntersuchungen
übereinstimmt.
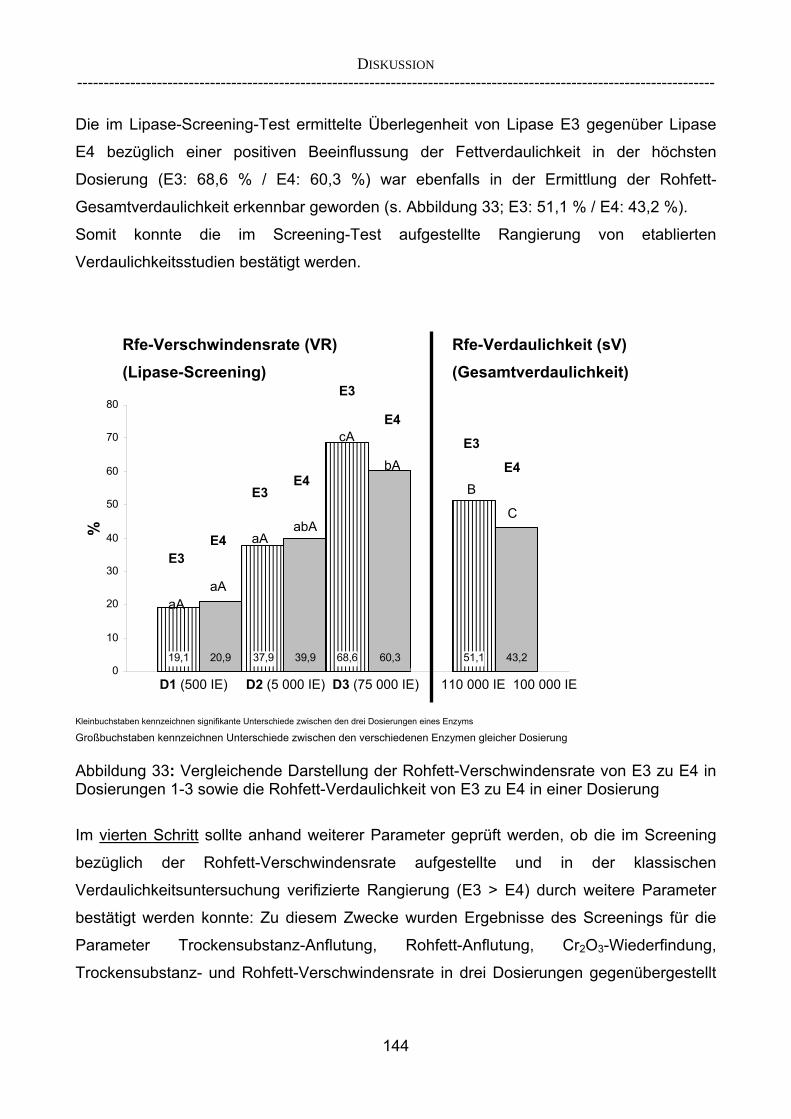
DISKUSSION ------------------------------------------------------------------------------------------------------------------------
144
Die im Lipase-Screening-Test ermittelte Überlegenheit von Lipase E3 gegenüber Lipase
E4 bezüglich einer positiven Beeinflussung der Fettverdaulichkeit in der höchsten
Dosierung (E3: 68,6 % / E4: 60,3 %) war ebenfalls in der Ermittlung der Rohfett-
Gesamtverdaulichkeit erkennbar geworden (s. Abbildung 33; E3: 51,1 % / E4: 43,2 %).
Somit konnte die im Screening-Test aufgestellte Rangierung von etablierten
Verdaulichkeitsstudien bestätigt werden.
Rfe-Verschwindensrate (VR) Rfe-Verdaulichkeit (sV) (Lipase-Screening) (Gesamtverdaulichkeit)
19,1 20,9 37,9 39,9 68,6 60,3 1 51,1 43,20
10
20
30
40
50
60
70
80
%
Kleinbuchstaben kennzeichnen signifikante Unterschiede zwischen den drei Dosierungen eines Enzyms
Großbuchstaben kennzeichnen Unterschiede zwischen den verschiedenen Enzymen gleicher Dosierung
Abbildung 33: Vergleichende Darstellung der Rohfett-Verschwindensrate von E3 zu E4 in Dosierungen 1-3 sowie die Rohfett-Verdaulichkeit von E3 zu E4 in einer Dosierung
Im vierten Schritt sollte anhand weiterer Parameter geprüft werden, ob die im Screening
bezüglich der Rohfett-Verschwindensrate aufgestellte und in der klassischen
Verdaulichkeitsuntersuchung verifizierte Rangierung (E3 > E4) durch weitere Parameter
bestätigt werden konnte: Zu diesem Zwecke wurden Ergebnisse des Screenings für die
Parameter Trockensubstanz-Anflutung, Rohfett-Anflutung, Cr2O3-Wiederfindung,
Trockensubstanz- und Rohfett-Verschwindensrate in drei Dosierungen gegenübergestellt
E3
aA
E4
aA
E3
aA
E4
abA
E3
cA E4
bA E3
B E4 C
D1 (500 IE) D2 (5 000 IE) D3 (75 000 IE) 110 000 IE 100 000 IE
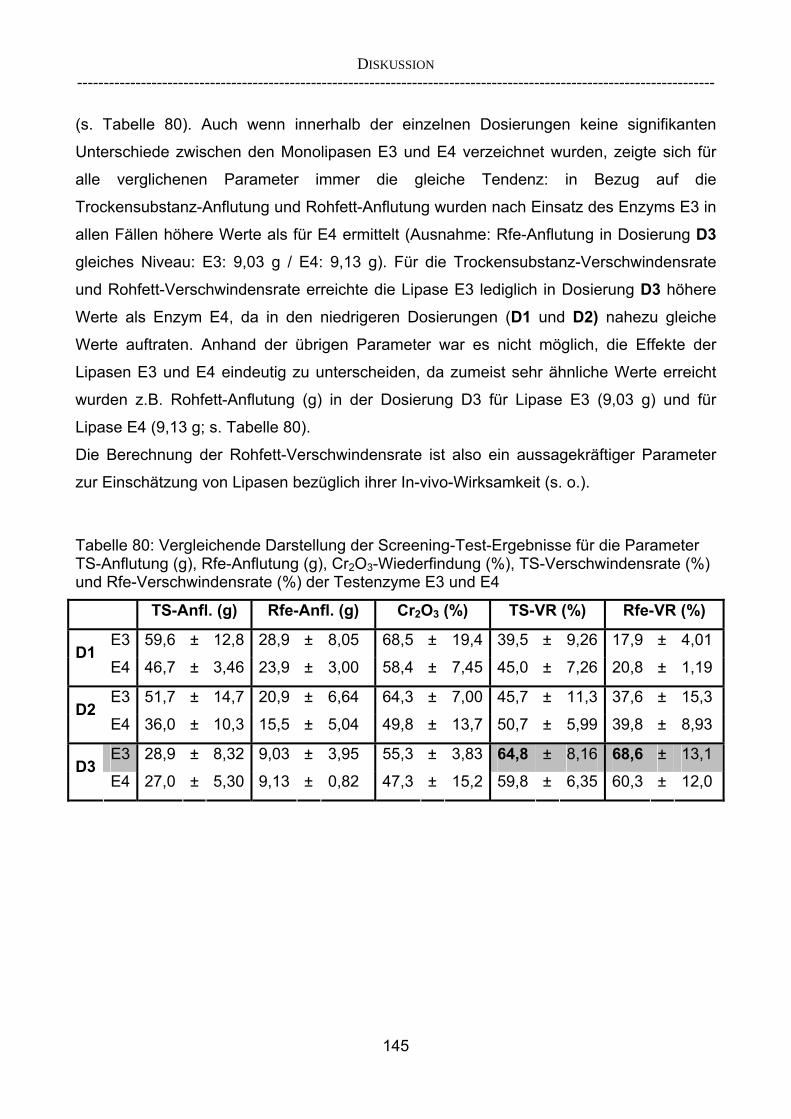
DISKUSSION ------------------------------------------------------------------------------------------------------------------------
145
(s. Tabelle 80). Auch wenn innerhalb der einzelnen Dosierungen keine signifikanten
Unterschiede zwischen den Monolipasen E3 und E4 verzeichnet wurden, zeigte sich für
alle verglichenen Parameter immer die gleiche Tendenz: in Bezug auf die
Trockensubstanz-Anflutung und Rohfett-Anflutung wurden nach Einsatz des Enzyms E3 in
allen Fällen höhere Werte als für E4 ermittelt (Ausnahme: Rfe-Anflutung in Dosierung D3
gleiches Niveau: E3: 9,03 g / E4: 9,13 g). Für die Trockensubstanz-Verschwindensrate
und Rohfett-Verschwindensrate erreichte die Lipase E3 lediglich in Dosierung D3 höhere
Werte als Enzym E4, da in den niedrigeren Dosierungen (D1 und D2) nahezu gleiche
Werte auftraten. Anhand der übrigen Parameter war es nicht möglich, die Effekte der
Lipasen E3 und E4 eindeutig zu unterscheiden, da zumeist sehr ähnliche Werte erreicht
wurden z.B. Rohfett-Anflutung (g) in der Dosierung D3 für Lipase E3 (9,03 g) und für
Lipase E4 (9,13 g; s. Tabelle 80).
Die Berechnung der Rohfett-Verschwindensrate ist also ein aussagekräftiger Parameter
zur Einschätzung von Lipasen bezüglich ihrer In-vivo-Wirksamkeit (s. o.).
Tabelle 80: Vergleichende Darstellung der Screening-Test-Ergebnisse für die Parameter TS-Anflutung (g), Rfe-Anflutung (g), Cr2O3-Wiederfindung (%), TS-Verschwindensrate (%) und Rfe-Verschwindensrate (%) der Testenzyme E3 und E4
TS-Anfl. (g) Rfe-Anfl. (g) Cr2O3 (%) TS-VR (%) Rfe-VR (%)
E3 59,6 ± 12,8 28,9 ± 8,05 68,5 ± 19,4 39,5 ± 9,26 17,9 ± 4,01 D1
E4 46,7 ± 3,46 23,9 ± 3,00 58,4 ± 7,45 45,0 ± 7,26 20,8 ± 1,19
E3 51,7 ± 14,7 20,9 ± 6,64 64,3 ± 7,00 45,7 ± 11,3 37,6 ± 15,3 D2
E4 36,0 ± 10,3 15,5 ± 5,04 49,8 ± 13,7 50,7 ± 5,99 39,8 ± 8,93
E3 28,9 ± 8,32 9,03 ± 3,95 55,3 ± 3,83 64,8 ± 8,16 68,6 ± 13,1 D3
E4 27,0 ± 5,30 9,13 ± 0,82 47,3 ± 15,2 59,8 ± 6,35 60,3 ± 12,0

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
146
Die im Screening-Test aufgestellte Rangierung bezüglich der Rohfett-Verschwindensrate
wurde auch durch die Ergebnisse der Gesamtverdaulichkeitsuntersuchungen bestätigt.
Für die Enzyme PK, E3 und E4 war beim Vergleich der Ergebnisse der
Gesamtverdaulichkeitsuntersuchungen mit denen der Screening-Tests folgendes
beobachtet worden (s. Abbildung 34):
• Die Ergebnisse beider Verfahren ergaben eine übereinstimmende Rangierung
der Enzyme beobachtet: PK ≥ E3 ≥ E4.
• Generell überstieg die Rohfett-Verschwindensrate die Werte der Rohfett-
Verdaulichkeit (Effekt der flüssigen Darreichungsform gegenüber der
herkömmlichen breiförmigen Standard-Fettdiät, s. o.). Es ist jedoch zu
berücksichtigen, dass es sich um ein gänzlich anderes Versuchsdesign handelte
und somit identische Werte auch nicht zu erwarten waren.
• Die Gesamtheit der Kurven des einen Verfahrens war der des anderen
Verfahrens im Verlauf geometrisch ähnlich, d. h. dass sich das Verhältnis der
Gesamtheit der Kurven beider Verfahren in der Anordnung der Kurven
zueinander ähnelte.
• Wie bereits beschrieben, unterschieden sich die Effekte der Lipasen E3 und E4
in der niedrigen und mittleren Dosierung des Screening-Tests nur unwesentlich.
Obwohl die Testlipase E4 in der Gesamtverdaulichkeitsstudie aufgrund von
Umrechnungsfaktoren geringgradig höher dosiert war als das Multienzymprodukt PK
und die Testlipase E3, wurden für E4 niedrigere Rohfett-Verdaulichkeiten ermittelt.
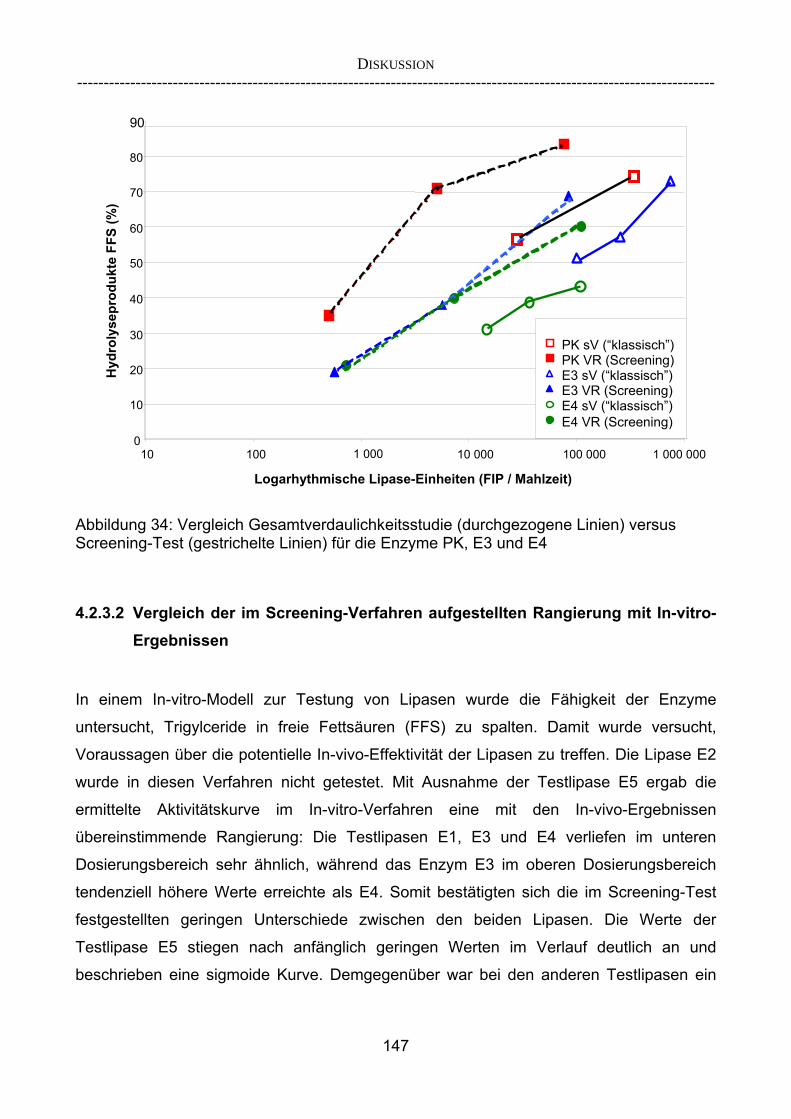
DISKUSSION ------------------------------------------------------------------------------------------------------------------------
147
Abbildung 34: Vergleich Gesamtverdaulichkeitsstudie (durchgezogene Linien) versus Screening-Test (gestrichelte Linien) für die Enzyme PK, E3 und E4
4.2.3.2 Vergleich der im Screening-Verfahren aufgestellten Rangierung mit In-vitro-Ergebnissen
In einem In-vitro-Modell zur Testung von Lipasen wurde die Fähigkeit der Enzyme
untersucht, Trigylceride in freie Fettsäuren (FFS) zu spalten. Damit wurde versucht,
Voraussagen über die potentielle In-vivo-Effektivität der Lipasen zu treffen. Die Lipase E2
wurde in diesen Verfahren nicht getestet. Mit Ausnahme der Testlipase E5 ergab die
ermittelte Aktivitätskurve im In-vitro-Verfahren eine mit den In-vivo-Ergebnissen
übereinstimmende Rangierung: Die Testlipasen E1, E3 und E4 verliefen im unteren
Dosierungsbereich sehr ähnlich, während das Enzym E3 im oberen Dosierungsbereich
tendenziell höhere Werte erreichte als E4. Somit bestätigten sich die im Screening-Test
festgestellten geringen Unterschiede zwischen den beiden Lipasen. Die Werte der
Testlipase E5 stiegen nach anfänglich geringen Werten im Verlauf deutlich an und
beschrieben eine sigmoide Kurve. Demgegenüber war bei den anderen Testlipasen ein
0
10
20
30
40
50
60
70
80
90
10 100 1 000 10 000 100 000 1 000 000
Logarhythmische Lipase-Einheiten (FIP / Mahlzeit)
Hyd
roly
sepr
oduk
te F
FS (%
)
PK sV (“klassisch”) PK VR (Screening)E3 sV (“klassisch”) E3 VR (Screening)E4 sV (“klassisch”)E4 VR (Screening)
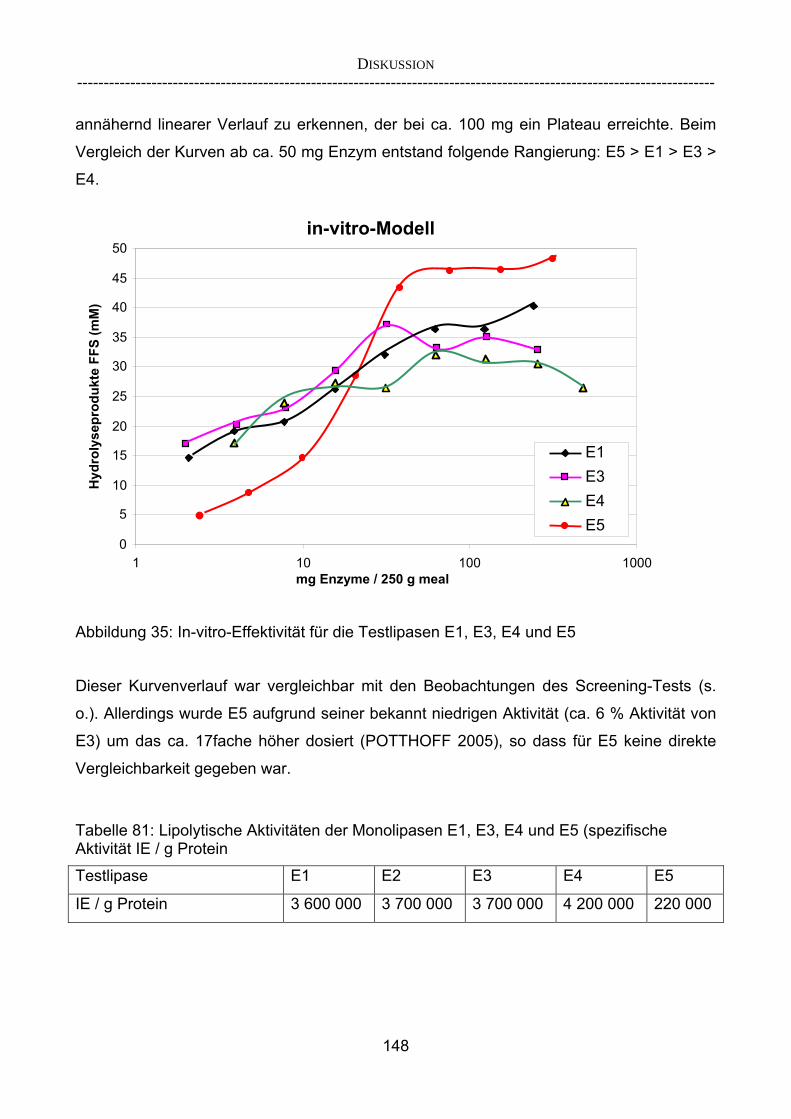
DISKUSSION ------------------------------------------------------------------------------------------------------------------------
148
annähernd linearer Verlauf zu erkennen, der bei ca. 100 mg ein Plateau erreichte. Beim
Vergleich der Kurven ab ca. 50 mg Enzym entstand folgende Rangierung: E5 > E1 > E3 >
E4.
Abbildung 35: In-vitro-Effektivität für die Testlipasen E1, E3, E4 und E5
Dieser Kurvenverlauf war vergleichbar mit den Beobachtungen des Screening-Tests (s.
o.). Allerdings wurde E5 aufgrund seiner bekannt niedrigen Aktivität (ca. 6 % Aktivität von
E3) um das ca. 17fache höher dosiert (POTTHOFF 2005), so dass für E5 keine direkte
Vergleichbarkeit gegeben war.
Tabelle 81: Lipolytische Aktivitäten der Monolipasen E1, E3, E4 und E5 (spezifische Aktivität IE / g Protein
Testlipase E1 E2 E3 E4 E5
IE / g Protein 3 600 000 3 700 000 3 700 000 4 200 000 220 000
in-vitro-Modell
0
5
10
15
20
25
30
35
40
45
50
1 10 100 1000mg Enzyme / 250 g meal
Hyd
roly
sepr
oduk
te F
FS (m
M)
E1 E3 E4 E5

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
149
Die Monolipase E5 wurde trotz geringer Hydrolyseaktivität im In-vivo-Screening in einer
mit den anderen Testenzymen vergleichbaren Dosierung eingesetzt, da die Lipase sich in
den In-vitro-Tests als besonders gallensalz- und pepsinstabil gezeigt hatte (POTTHOFF
2005). Allerdings konnte auch In-vivo die hohe Stabilität nicht die niedrige Aktivität der
Lipase ausgleichen. Ob der Einsatz des Enzyms E5 in wesentlich höheren Dosierungen
sinnvoll ist, muss allerdings hinterfragt werden, da einer der besonderen Vorteile des
Einsatzes mikrobieller Produkte die drastische Reduktion der Enzymmengen (SUZUKI et
al. 1997, 1999) sein sollte. Es also zu bedenken, inwiefern E5 unter diesen Zielvorgaben
gegenüber den anderen Testlipasen konkurrenzfähig ist (GRIFFIN et al. 1989;
MORRISON et al. 1992; SANTINI et al. 2000). Andererseits ist die außerordentliche
Gallensalz- und Pepsinstabilität von E5 im Rahmen einer Weiterentwicklung zukünftiger
Optimierungen ein interessanter Ansatz.
4.2.3.3 Kombinationsversuch
Eine im Screening-Test erfolgreich getestete Lipase sollte auch auf ihre Stabilität
gegenüber Proteasen geprüft werden: Bei isolierter Prüfung der Wirksamkeit von Lipasen
besteht die Problematik, dass eine eventuelle Empfindlichkeit gegenüber proteolytisch
wirksamen Enzymen nicht beurteilt werden kann. Die bekannte Sensitivität von Lipasen
gegenüber Proteasen, verstärkt durch die bei EPI häufig vorliegenden tiefen pH-Werte,
wurde bereits von THIRUVENGADAM u. DIMAGNO (1988) u. LAYER et al. (1990)
beschrieben.
Da das Ziel des Gesamtprojektes die Entwicklung eines Multienzymproduktes ist, sollte
die Lipase in Kombination mit Proteasen und Amylasen geprüft werden. Zu diesem
Zwecke wurden eine Protease und eine Amylase der Wirksamkeitsstudien von BECKER
(2005) mit der Testlipase E3 des hier vorgestellten Lipase-Screenings gemeinsam
eingesetzt. Das Auswahlkriterium der Lipase war vor allem die Verfügbarkeit
ausreichender Mengen, so dass hier leider nicht die effektivste Monolipase verwendet
werden konnte. Auch in diesem Versuch konnten dosisabhängige Effekte hinsichtlich der
Anflutungen und Verschwindensraten von Trockensubstanz und Rohfett ermittelt werden.
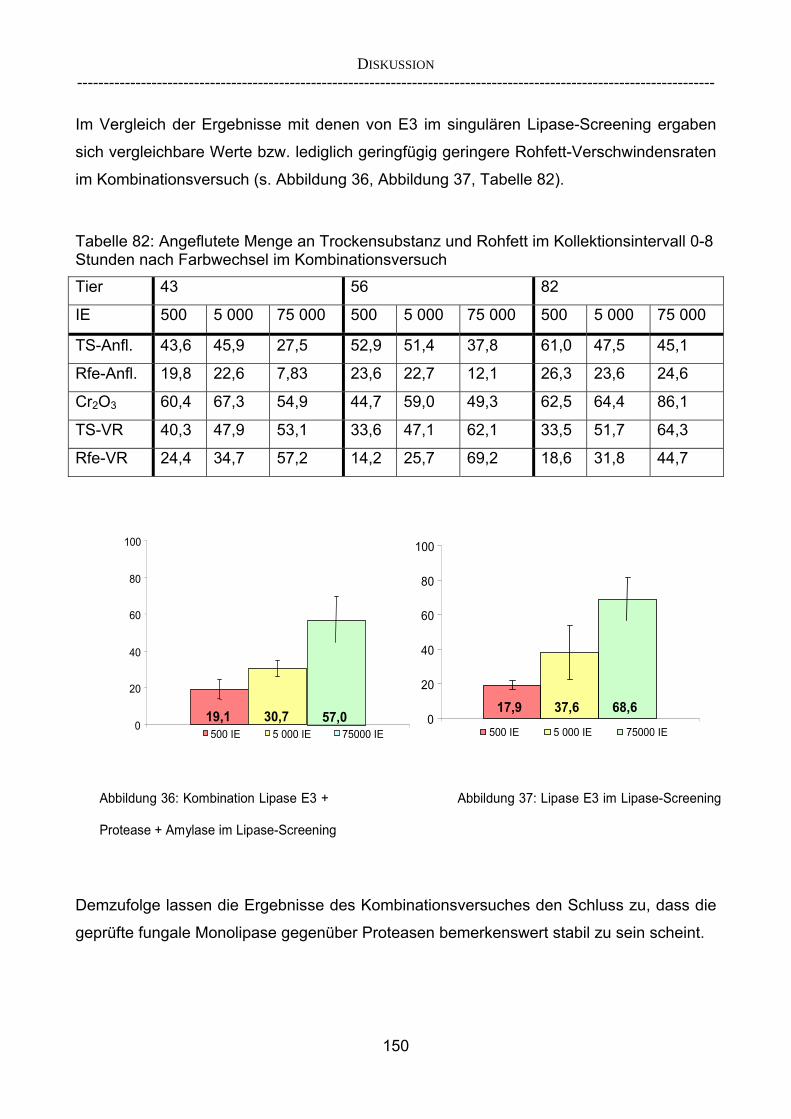
DISKUSSION ------------------------------------------------------------------------------------------------------------------------
150
Im Vergleich der Ergebnisse mit denen von E3 im singulären Lipase-Screening ergaben
sich vergleichbare Werte bzw. lediglich geringfügig geringere Rohfett-Verschwindensraten
im Kombinationsversuch (s. Abbildung 36, Abbildung 37, Tabelle 82).
Tabelle 82: Angeflutete Menge an Trockensubstanz und Rohfett im Kollektionsintervall 0-8 Stunden nach Farbwechsel im Kombinationsversuch
Tier 43 56 82
IE 500 5 000 75 000 500 5 000 75 000 500 5 000 75 000
TS-Anfl. 43,6 45,9 27,5 52,9 51,4 37,8 61,0 47,5 45,1
Rfe-Anfl. 19,8 22,6 7,83 23,6 22,7 12,1 26,3 23,6 24,6
Cr2O3 60,4 67,3 54,9 44,7 59,0 49,3 62,5 64,4 86,1
TS-VR 40,3 47,9 53,1 33,6 47,1 62,1 33,5 51,7 64,3
Rfe-VR 24,4 34,7 57,2 14,2 25,7 69,2 18,6 31,8 44,7
Abbildung 36: Kombination Lipase E3 + Abbildung 37: Lipase E3 im Lipase-Screening Protease + Amylase im Lipase-Screening
19,1 30,7 57,00
20
40
60
80
100
500 IE 5 000 IE 75000 IE
17,9 37,6 68,6 0
20
40
60
80
100
500 IE 5 000 IE 75000 IE
Demzufolge lassen die Ergebnisse des Kombinationsversuches den Schluss zu, dass die
geprüfte fungale Monolipase gegenüber Proteasen bemerkenswert stabil zu sein scheint.

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
151
Der Vergleich verschiedener Verfahren mit dem Lipase-Screening-Test erlaubt
folgendes Fazit:
• Zwischen den im In-vitro- und im Screening-Verfahren erhobenen Werten konnte
eine enge Korrelation bezüglich der Rangierung von Enzymen festgestellt werden.
o Die im Screening-Verfahren aufgestellte Rangierung der Monolipasen
bestätigte demzufolge die Ergebnisse der In-vitro-Untersuchungen.
• Es konnte eine gute Übereinstimmung der Screening-Test-Ergebnisse mit
herkömmlichen Gesamtverdaulichkeitsuntersuchungen ermittelt werden.
• Die Kombinationsversuche bestätigen zudem, dass nur unwesentliche Abstriche
bezüglich der Lipase-Wirksamkeit der eingesetzten Lipasen Amylase und Protease
zu erwarten sind.
4.3 Ausblick
Nach der Entwicklung einer geeigneten Testmahlzeit wurde ein Lipase-Screening-
Verfahren etabliert, mit welchem fünf mikrobielle Monolipasen vergleichend auf ihre In-
vivo-Wirksamkeit hin untersucht und entsprechend rangiert wurden. Werden die
Ergebnisse der neu entwickelten Methode mit Ergebnissen aus In-vitro-Studien ergänzt,
ist es möglich, erfolgversprechende Lipasen frühzeitig und gezielt auszuwählen. Somit
können die zeitaufwändigen herkömmlichen Verdaulichkeitsstudien auf ein Minimum
beschränkt werden, da eine weitergehende Testung lediglich für die effektivsten und
stabilsten Enzyme durchgeführt werden muss. Außerdem können die ersten In-vivo-
Untersuchungen der Enzyme zu einem viel früheren Entwicklungszeitpunkt eines neuen
Produktes stattfinden, da für den Screening-Test vergleichsweise geringere
Enzymmengen benötigt werden. Da mit der Entwicklung des Lipase-Screenings nun
Schnelltest-Verfahren für alle drei Gruppen der Verdauungsenzyme vorliegen (Amylase-
und Protease-Screening siehe BECKER 2005), besteht die Möglichkeit, die Effektivität von
Monopräparaten beispielsweise für die Auswahl eines mikrobielles Multienzymproduktes
innerhalb eines wesentlich verkürzten Zeitraumes zu beurteilen. Durch das beschriebene
Verfahren des Lipase-Screening-Tests wird eine Rangierung der untersuchten Enzyme im
direkten Vergleich ermöglicht. Die Ergebnisse sind dabei nicht als absolute Werte zu

DISKUSSION ------------------------------------------------------------------------------------------------------------------------
152
verstehen, sondern stellen lediglich eine vergleichende Bewertung der Testenzyme
untereinander dar.
Als Ziel bleibt die Entwicklung eines Schnell-Test-Verfahrens zur gleichzeitigen Prüfung
von Amylasen, Proteasen und Lipasen bestehen (BECKER 2005). Der unter Punkt 4.2.3.3
beschrieben Kombinationsversuch, in welchem die Testmahlzeit des Lipase-Sreenings
genutzt wurde, könnte hierzu als Ansatz dienen. Die Versuchsmahlzeit müsste dann mit
einem höheren Stärke- und Proteingehalt modifiziert werden, um die Effektivität von
Proteasen und Amylasen hinreichend quantifizieren zu können.
4.4 Schlussfolgerungen
Im Rahmen der vorliegenden Arbeit wurde ein Screening-Test zur zeit-, kosten- und
arbeitssparenden Untersuchung von Lipasen entwickelt. Bei Anwendung dieser Methode
werden bedeutend geringere Enzymmengen (lediglich ca. 2 %) im Vergleich zu etablierten
Verdaulichkeitsstudien benötigt. Deshalb ist es mit dieser Methode möglich, erste In-vivo-
Untersuchungen zu einem früheren Zeitpunkt in der Entwicklungsphase neuer
Verdauungsenzyme durchzuführen. Aufgrund des beschleunigten Verfahrens kann
darüber hinaus der Einsatz von Versuchstieren reduziert werden.
Durch die Entwicklung der Lipase-Screening-Methode besteht nun die Möglichkeit,
verschiedene Lipasen mit bisher unbekannter In-vivo-Wirkung durch Applikation einer
einzigen Versuchsmahlzeit auf ihre Effektivität hinsichtlich ihres Einflusses auf die
Rohfettverdaulichkeit vergleichend zu untersuchen.
Die Testergebnisse des Screenings stimmen mit den Ergebnissen aus In-vitro- und
klassischen (aufwändigeren) Verdaulichkeitsversuchen hinsichtlich einer Rangierung der
getesteten Enzyme überein. Nach einer Vorselektion durch das In-vivo-Screening-
Verfahren sollten „erfolgversprechende“ Lipasen mit weiterführenden klassischen
Verdaulichkeitsstudien näher quantifiziert werden.
Infolge des hier etablierten Screening-Verfahrens besteht in Zukunft die Möglichkeit, die
vorklinische Entwicklungsphase pharmazeutischer Neuentwicklungen deutlich zu
verkürzen und aufgrund der frühzeitigen Vorselektion die Ausgaben für abgebrochene
Projekte (s. 2.4.1: entsprechen 70 % der Gesamtkosten) zu reduzieren.

ZUSAMMENFASSUNG ------------------------------------------------------------------------------------------------------------------------
153
5 Zusammenfassung
Sonka Zantz
Untersuchungen zur Entwicklung eines Screening-Tests zur Beurteilung substituierter Lipasen im Modell pankreasgangligiertes Miniaturschwein
Die Substitution von Verdauungsenzymen stellt die entscheidende Möglichkeit für die
Behandlung einer exokrinen Pankreasinsuffizienz (EPI) dar. Vor diesem Hintergrund
verständlich zielten die eigenen Untersuchungen auf die Entwicklung eines In-vivo-
Schnelltests zur Wirksamkeitsprüfung neuer Enzyme bei Verfügbarkeit nur geringer
Enzymmengen. Basierend auf dem von BECKER (2005) erfolgreich etablierten Screening-
Test für Amylasen und Proteasen sollte in der vorliegenden Arbeit ein Schnelltest für Lipasen entwickelt werden. Hierbei ging es zunächst um die Entwicklung einer geeigneten
Testmahlzeit, darauf folgten vergleichende Untersuchungen zur Wirksamkeit neuer
Lipasen, in denen ein bekannt wirksames Multienzymprodukt als Positivkontrolle zum
Einsatz kam. Ziel des Schnelltests war eine vergleichende Bewertung verschiedener
Lipasen, um Weiterentwicklung und Produktion größerer Enzymmengen nur auf die
erfolgreichsten Neuentwicklungen beschränken zu können.
Für die In-vivo-Untersuchungen standen insgesamt 10 adulte weibliche Göttinger
Miniaturschweine mit ileocaecaler Umleitungsfistel zur Verfügung. Zur Auslösung einer
EPI war bei 7 Tieren der Ausführungsgang des Pankreas ligiert worden (PL-Tiere),
wohingegen die übrigen 3 Tiere als Kontrolltiere (KT) dienten. Die Analyse des
Rohnährstoffgehaltes der Futtermittel erfolgte nach den Vorschriften zur Weender-Analyse
von NAUMANN u. BASSLER (2004). Die Chromoxidbestimmung wurde nach PETRY u.
RAPP (1970) durchgeführt. Die Analyse des Fettgehaltes der Futter- und Chymusproben
fand unter Verwendung der Geräte der Firma Ankom Technology Corporation® (AnkomHCl
Hydrolysis System und AnkomXT15 Extraktor) statt, wobei zuvor eine Validierung der
Methoden durchgeführt wurde.
Basis der Testmahlzeit war ein flüssiges Nahrungsergänzungsmittel, das im Rahmen der
Optimierung einer einmalig zu applizierenden markerhaltigen Testmahlzeit modifiziert

ZUSAMMENFASSUNG ------------------------------------------------------------------------------------------------------------------------
154
wurde. Die Versuchsmahlzeit wurde insbesondere hinsichtlich des Fettgehaltes verändert,
so dass sie schließlich aus (g / kg Trockensubstanz (TS)): 351 Rohfett (Rfe); 76,9
Rohprotein (Rp); 46,3 Rohfaser (Rfa); 22,8 Rohasche (Ra); 2,5 Cr2O3 bestand. Diese
flüssige Testmahlzeit (ca. 150 g TS) wurde lediglich am Morgen des Versuchstages
verabreicht, während sonst das übliche Mischfutter (g / kg TS: 49,8 Rfe / 189 Rp / 477
Stärke / 30,3 Rfa / 53 Ra) ohne Enzymzulage zum Einsatz kam. Am Vorabend des
Versuchstages wurden anstelle des o. g. Mischfutters 400 ml einer fettfreien Trinkmahlzeit
(g /100 ml: 525 KJ / 3,75 Rp / 27,5 KH / 5,5 Zucker / 0 Rfe / 1 Rfa / 80 ml H2O) angeboten.
Die Chymuskollektion begann mit der Anflutung der durch den Marker farblich
gekennzeichneten Testmahlzeit an der ilealen Fistel und endete 8 Stunden später. In
diesem neu entwickelten Lipase-Screening wurden fünf verschiedene Monolipasen
mikrobiellen Ursprungs (E1 - E5) in drei verschiedenen Dosierungen (500, 5 000 und
75 000 IE) geprüft. Hierzu wurde das von BECKER (2005) entwickelte Versuchsdesign
(Fütterungs-, Kollektions- und Versuchsintervalle) angewandt.
Die Beurteilung der Wirksamkeit der zugelegten Lipasen nach einmaliger Verabreichung
der Testmahlzeit erfolgte anhand nachstehender Parameter: absolute TS- und Rfe-
Anflutung, Rfe-Konzentration im Chymus, Rfe / Cr2O3-Verhältnis und kalkulierte
Verschwindensraten (VR) für TS und Rfe. Eine Korrelation zwischen Ergebnissen im
Schnelltest und denen klassischer Verdaulichkeitsstudien wurde anhand von Daten bereits
vorgenommener Untersuchungen geprüft.
1. Die optimierte Testmahlzeit führte zu einer homogenen Chymusanflutung am
terminalen Ileum. Anhand paralleler relativer Flussraten von Trockensubstanz,
Rohfett und Marker konnte eine Entmischung der Komponenten weitestgehend
ausgeschlossen werden. Der „grüne“ Chymus der Testmahlzeit flutete regelmäßig
4 - 6 Stunden ppr. am Ileumende an, so dass von einer zügigen Magen-
Dünndarmpassage auszugehen war. Infolge des hohen Fettgehaltes (34,6 % der
Testmahlzeit) war es trotz der flüssigen Darreichungsform möglich, PL-0-Tiere von
gesunden Kontrolltieren zu unterscheiden (Rfe-VR: PL-0-Tiere 10,8 %; KT 86,4 %).
2. Der Lipase-Screening-Test wurde eingesetzt, um fünf Monolipasen vergleichend
einer ersten In-vivo-Wirksamkeitsprüfung zu unterziehen. Dabei konnten für alle
Enzyme bezüglich der Rfe-VR (und weiterer erhobener Parameter) dosisabhängige
Effekte ermittelt werden. In der höchsten Dosierung erreichten mehrere Enzyme

ZUSAMMENFASSUNG ------------------------------------------------------------------------------------------------------------------------
155
das Niveau des bekannt hochwirksamen Multienzymproduktes PK, wobei die
Rangierung PK, E1, E3, E5, E2, E4 entstand. In den verschiedenen Dosierungen
waren produktabhängige Unterschiede zu erkennen, wobei in allen
Untersuchungen PK und E1 tendenziell den anderen Testenzymen überlegen
waren (Rfe-VR (%): PK 83,5; E1 77,0; E3 68,6; E5 65,0; E2 61,5; E4 60,1). Für die
meisten Parameter (s. o.) konnte eine übereinstimmende Rangierung der fünf
Testlipasen und des Multienzymproduktes PK erstellt werden.
3. Um die im Lipase-Screening-Test ermittelte Rangierung abzusichern, wurde auf
Ergebnisse aus früheren Studien zur Gesamtverdaulichkeit zurückgegriffen, in
denen zwei (E3 und E4) der fünf Testlipasen eingesetzt wurden. Zusätzlich wurden
Ergebnisse aus In-vitro-Verfahren mit denen des Screening-Tests verglichen. Die
verschiedenen Verfahren ergaben dabei eine übereinstimmende Rangierung; stets
war das Testenzym E3 in seiner Wirksamkeit dem Produkt E4 überlegen.
Schlussfolgerung: Der im Rahmen dieser Studie neu entwickelte In-vivo-Screening-Test
zur vergleichenden Beurteilung von Lipasen erlaubte eine Differenzierung der Enzyme
anhand ihrer Wirksamkeit.
Diese ergänzende Methode stellt dabei keinen vollständigen Ersatz für herkömmliche
Verdaulichkeitsstudien dar, sondern ein Verfahren, mit Hilfe dessen innerhalb kürzester
Zeit (nach einmaliger Fütterung der Testmahlzeit mit Enzymzulage) und unter minimalem
Enzymverbrauch eine erste Einschätzung der Wirksamkeit neu entwickelter Lipasen
möglich ist. Die in dem Screening-Test ermittelte erste In-vivo-Bewertung erlaubt damit
eine gezielte Vorauswahl effektiver Enzyme, deren weitere Entwicklung und Produktion
forciert werden kann.

SUMMARY ------------------------------------------------------------------------------------------------------------------------
156
6 Summary
Sonka Zantz
Development of a novel In-vivo screening test to assess the efficacy of substitute lipases in pancreatic duct ligated minipigs.
The substitution of digestive enzymes is an essential treatment of pancreatic exocrine
insufficiency (PEI). The aim of the present investigation was to develop an In-vivo lipase screening test, which requires only small amounts of enzymes to estimate enzyme
efficacy. The test which was developed was based on BECKER’s (2005) successfully
established protease and amylase screening tests. At first it was necessary to select a
suitable test diet. The assay was then established using different doses of a well known
and highly potent standard multienzyme product (PK). In a third step the efficacy of 5
microbial lipases were compared to each other and to that of the standard product.
The studies were performed in 10 female Göttingen Minipigs all fitted with an ileo-caecal-
reentrant fistula. Additional pancreatic exocrine insufficiency was induced by ligation of the
pancreatic duct (PL-animals) in 7 of them. The remaining 3 pigs served as controls (KT-
animals). The test diet and the chyme samples were analysed for crude nutrients
according to standard VDLUFA methods (NAUMANN u. BASSLER 2004). Chromic oxide
Cr2O3 was analysed after PETRY u. RAPP (1970). For the analysis of crude fat Ankom
Technology Corporation® was used, a method which was validated in this study.
The test diet was based on a liquid nutritional supplement called “Calshake®”, which was
optimised for single lipase dose application mainly with respect to crude fat content and
homogeneity. The final test diet contained (g / kg) 351 crude fat (cfa); 76,9 crude protein
(cp); 46,3 crude fibre (cfi); 22,8 crude ash (ca); 2,5 Cr2O3. This liquid test meal (ca. 150 g
DM), into which the test enzymes were mixed, was given only once, on the morning of
each trial day, while in the evenings of the trials and on all other days the pigs were fed a
regular diet (g / kg DM: 49,8 cfa / 189 cp / 477 st / 30,3 cfi / 53 ca) without enzymes. An
exception was the evening meal given 12 hours before starting each trial: instead of the
regular diet a fat-free liquid meal was offered (ProvideXtra® g /100 ml: 525 KJ / 3,75 cp /
27,5 carbohydrates / 5,5 sugar / 0 cfa / 1 cfi / 80 ml H2O). The chyme collection was

SUMMARY ------------------------------------------------------------------------------------------------------------------------
157
started individually for each pig with the arrival of the Cr2O3 (green) labelled chyme of the
test diet at the ileal fistula and was finished after an 8-hour-chyme collection. The dose-
dependency of a standard multienzyme product (PK) on fat digestibility was investigated
and three doses (500, 5000, 75000 IU) selected. Five unknown microbial lipases (E1 – E5)
were then assayed in this newly developed lipase screening test using the same three
doses (500, 5000 and 75000 IU). Therefore the same trial design used by BECKER (2005)
(feeding, collection and time schedule for testing) was used in this screening test.
After single dose application of the test diet the efficacy of the lipases was estimated
according to the following parameters: absolute DM- and cfa- appearance and
disappearance rate and the relationships between them. The results of the screening test
with two of the microbial and the standard lipase supplement were compared to results in
previously performed standard total digestibility trials.
1. By calculating parallel flow rates it could be excluded that there was separation of
the diet’s components (DM, cfa and marker). The “green” chyme of the test diet
flowed regularly 4 to 6 hour postprandial at the ileal fistula. Although the cfa
digestibility was improved by using a liquid test diet, marked differences were
observed between PL- and KT-animals (Rfe-VR: PL-0-Tiere 10,8 %; KT 86,4 %),
due to the high fat content of the diet (34,6 %). The conspicuous green colour of the
marker, Cr2O3, was used as a signal to determine the time of arrival of the test meal
at the ileal fistula and to separate this from earlier meals (brown chyme). Thus the
marker indicated the ileal arrival of the test diet and the beginning of the 8 hour
chyme collection.
2. The standard multienzyme product PK caused a dose-dependent improvement in
cfa digestibility. Subsequently, the screening test was used to compare the in-vivo-
efficacy of five microbial lipases. For all five enzymes dose-dependent responses
were measured. Several lipases approached the level of efficacy of PK at the
highest dosage used, although differences were observed at the low and medium
dosages. Nevertheless in all trials PK and E1 tended to show higher potency than
the other enzymes (Rfe-VR: PK 83,5 %; E1 77,0 %; E3 68,6 %; E5 65,0 %; E2 61,5
%; E4 60,1 %;). The five test lipases and PK showed a corresponding ranking of
most parameters. The following tendentious ranking demonstrates the precaecal
cfa disappearance rate (highest dosage): PK, E1, E3, E5, E2, E4

SUMMARY ------------------------------------------------------------------------------------------------------------------------
158
3. The ranking of selected lipases in the screening test compared well to results
observed in standard efficacy studies by KARTHOFF (2004) and BECKER (2006,
personal communication). Thus for PK and the test lipases E3 and E4 the same
ranking was ascertained in total digestibility studies. Furthermore, an in-vitro-
digestibility model yielded a similar ranking of lipases (exception: E5). Because of
different trial conditions E5’s in-vivo-results were not directly comparable.
Conclusion: A screening test has been established that allows differentiation and
selection of lipases according to their efficacy in-vivo. The results of the screening test
compare well to findings in standard in-vivo-digestibility trials and additional in-vitro-
assays. This lipase screening test is not a replacement for standard digestibility trials, but
is to be used as a way to estimate the in-vivo-efficacy of substituted enzymes with a
minimum of time and enzyme consumption. This allows preliminary selection of effective
enzymes, whose efficacy has still to be properly quantified in standard digestibility trials.

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
159
7 Literaturverzeichnis
ABELLO J., X. PASCAUD, C. SIMONES-NUNES, J. C. CUBER, J. L. JUNIEN u. C. ROZE (1989): Total pancreatic insufficiency in pigs: A model to study intestinal enzymes and plasma levels of digestive hormones after pancreatic supplementation by a whole pancreas preparation. Pancreas 4, Nr. 5, 556-564 ABRAMS, J., M. HAMOSH, S. K. DUTTA, V. S. HUBBARD u. P. HAMOSH (1987): Role of nonpancreatic lipolytic activity in exocrine pancreatic insufficiency. Gastroenterol. 92, 125-129 ALLIANCE PHARMACEUTICAL CORPERATION (2006): Phases of Product development 2006 AMMAN, R. W., H. BUEHLER, R. MUENCH, A. W. FREIBURGHAUS u. W. SIEGENTHALER (1987): Differences in the natural history of idiopathic (nonalcoholic) and alcoholic chronic pancreatitis. Pancreas 2, 368-377 ANDERSON, D. M. u. R. W. ASH (1971): The effect of ligating the pancreatic duct on digestion in the pig. Proc. Nutr. Soc. 30 (1), 34A-35A ARGENZIO, R. A., J. E. LOWE, D. W. PICKARD u. C. E. STEVENS (1974): Digesta passage and water exchange in the equine large intestine. Am. J. Physiol. 226 (5), 1035-1042 BECKER, C. (2005): Untersuchungen zur Entwicklung eines Screening-Tests zur Beurteilung der Wirksamkeit substituierter Enzyme (Proteasen und Amylasen) am Modelltier pankreasgangligiertes Minischwein Hannover, Tierärztl. Hochschule, Diss. BEITZ, D. C. u. R. S. ALLEN (1993): Lipid Metabolism. In: M. J. SWENSON u. W. O. REECE (Hrsg.) Duke’s Physiology of domestic animals. 11th ed. Cornell University Press, Ithaca, New York, 453-460

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
160
BERKHOFF, M., D. GROßNER, R. KLAPDOR, G. KLÖPPEL u. H. v. KROGE (1988): Die intravenös stimulierte Insulinreserve des Pancreas nach experimenteller Pancreasgangligatur beim Schwein – Untersuchungen 2, 4 und etwa 60 Tage nach Ligatur. Langenbecks Arch. Chir. 373, 151-158 BOIVIN, M., S. J. LANSPA, A. R. ZINSMEISTER, V. L. GO u. E. P. DIMAGNO (1990): Are diets associated with different rates of human interdigestive and postprandial pancreatic enzyme secretion? Gastroenteol. 99, 1763-71 BOOK, S. A. u. L. K. BUSTAD (1974): The fetal and neonatal pig in biomedical research. J. Anim. Sci. 38 (5), 997-1002. BRAUDE, R., M. H. NEWPORT u. J. W. G. PORTER (1970): Artificial rearing of pigs. Br. J. Nutr. 24 (3), 827-842 BROWNING, T. (1998): Exocrine pancreatic insufficiency in a cat. Austr. Vet. J. 76 (2), 104-6. BRUNO, M. J., E. B. HAVERKORT, G. N. TYTGAT u. D. J. VAN LEEUWEN (1995): Maldigestion associated with exocrine pancreatic insufficiency: implications of gastrointestinal physiology and properties of enzyme preparations for a cause-related and patient-tailored treatment. Am. J. Gastroenterol. 90 (9), 1383-93. CARRIERE, F., J. A. BARROWMAN, R. VERGER u. R. LAUGIER (1993): Secretion and contribution to lipolysis of gastric and pancreatic lipases during a test meal in humans. Gastroenterol. 105, 876-888 CARRIERE, F., P. GRANDVAL, P. C. GREGORY, C. RENOU, F. HENNIGES, S. SANDER-STRUCKMEIER u. R. LAUGIER (2005): Does the pancreas really produce much more lipase than required for fat digestion? JOP. 6 (3), 206-15 CRANWELL, P. D. u. P. J. MOUGHAN (1989): Biological limitations imposed by the digestive system to the growth performance of weaner pigs. in: J. L. BARNETT u D. P. HENNESSY (Hrsg.).: Manipulating Pig Production II. Australasian Pig Science Association, Werribee, Australia, S. 140-159

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
161
DECUYPERE, J. A., R. M. DENDOOVEN u. H. K. HENDERICKX (1985): Stomach emptying of milk diets in pigs – A mathematical model allowing description and comparison of the emptying patterns. Prod. 3rd Int. Sem. Dig. Physiol. in the Pig, Copenhagen, 16. – 18.5.1985, S. 51-57 DEGEN, L., D. MATZINGER, J. DREWE u. C. BEGLINGER (2001): The effect of cholecystokinin in controlling appetite and food intake in humans. Peptides 22 (8), 1265-9. DESPORT, J. C., C. JUSTE, B. BEAUFRÈRE u. T. CORRING (1997): The pig as a model in clinical investigation in the exocrine pancreatic insufficiency and of cholesterol crystallisation from bile. in: J. P. LAPLACE, C. FÉVRIER u. A BARBEAU (Hrsg.): 7th Int. Sympos. Dig. Physiol. In Pigs, Saint Malo, 26.-28.5.1997, Proc., S.17-27 DIMAGNO, E. P., V. L. W. GO u. W. H. J. SUMMERSKILL (1973): Relations between pancreatic enzyme outputs and malabsorption in severe pancreatic insufficiency. New Engl. J. Med. 288, 813-815 DIMAGNO, E. P., J. R. MALAGELADA, V. L. W. GO u. C. G. MOERTEL (1977): Fare of orally ingested enzymes in pancreatic insufficiency: comparison of two dosage schedules. New Eng. J. Med. 9, 1318-1322 DIMAGNO, E. P., H. A. REBER u. M. A. TEMPERO (1999): AGA technical review on the epidemiology, diagnosis, and treatment of pancreatic ductal adenocarcinoma. American Gastroenterological Association. Gastroenterology 117, (6), 1464-84 DIMAGNO, E. P. (2001): Gastric acid suppression and treatment of severe exocrine pancreatic insufficiency. Best Pract Res Clin Gastroenterol 15, 477-86 DIMAGNO, E. P. (2003): Autoimmune chronic panceatitis: a plea for simplification and consistency. Clin. Gastroenterol. Hepatol. 1 (6), 421-2. DODGE, J. A., J. M. CUSTANCE, M. C. GOODCHILD, S. C. LAING u. M. VAUGHAN (1990): Paradoxical effects of essential fatty acid supplementation on lipid profiles and sweat electrolytes in cystic fibrosis. Br. J. Nutr. 63 (2), 259-271

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
162
DOBRILLA, G. (1989): Management of chronic pancreatitis. Focus on enzyme replacement therapy. Int. J. Pancreatol. 5 (Suppl.), 17-29. DREYER, J. (1990): In-vitro-Untersuchungen mit der Colon-Simulations-Technik (COSITEC) zum mikrobiellen Stoffwechsel im Dickdarm von Schweinen. Göttingen, Univ., Fachber. Agrarwiss., Diss. DROCHNER, W. (1984): Einfluss wechselnder Rohfaser- und Pektingehalte im Futter auf einige praecaecale und postileale Verdauungsvorgange beim wachsenden Schwein. Fortschr. Tierphysiol. Tierernähr., Band 14 DROCHNER, W. (1987): Untersuchungen zur Verdauung im Dickdarm beim Schwein. Fortschr. Tierphysiol. Tierernähr., Beih. 17 DROCHNER, W. u. H. MEIER (1991): Verdauung organischer Substanzen im Dickdarm verschiedener Haustierarten. Fortschr. Tierphysiol. Tierernähr., Beih. 22 DUTTA, S. K., M. P. BUSTIN, R. M. RUSSELL u. B. S. COSTA (1982): Deficiency of fat-soluble vitamins in treated patients with pancreatic insufficiency. Ann. Intern. Med. 97 (4), 549-52. EHRLEIN, H. J. (2000a): Motorik des Dünndarms. in: W. v. ENGELHARDT u. G. BREVES (Hrsg.): Physiologie der Haustiere. Enke im Hippokrates Verlag GmbH, Stuttgart, S. 323-327 EHRLEIN, H. J. (2000b): Motorik des Dickdarms. in: W. v. ENGELHARDT u. G. BREVES (Hrsg.): Physiologie der Haustiere. Enke im Hippokrates Verlag GmbH, Stuttgart, S. 327-333 FASSMANN, C. (2001): Untersuchungen zum Umfang und Lokalisation der Nährstoffverdaulichkeit von fettreichen Rationen bei pankreasgangligierten Schweinen. Hannover, Tierärztl. Hochschule., Diss. FLEMMING, S. E. u. M. M. WASILEWSKI (1984): Using the pig as a tool for studying fermentation in the gut. Nutr. Rep. Int. 30, 825-834

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
163
FLORES, C. A., P. M. BRANNON, S. A. BUSTAMANTE, J. BEZERRA, K. T. BUTLER, T. GODA u. O. KOLDOVSKY (1988): Effect of diet on intestinal and pancreatic enzyme activities in the pig. J. Pediatr. Gastroenterol. Nutr. 7, 914-921 FREUDIGER, U. (1971): Die Erkrankung des exokrinen Pankreas des Hundes. Kleintier-Prax. 16, 210-228 FREUDIGER, U. (1991): Physiologie, Pathologie, Labor und Therapie der exokrinen Erkrankung der Bauchspeicheldrüse beim Hund. Kleintier-Prax. 36, 5-16 FRIESS, H., J. DING, J. KLEEFF, Q. LIAO, P. O. BERBERAT, J. HAMMER u. M. W. BUCHLER (2001): Identification of disease-specific genes in chronic pancreatitis using DNA array technology. Ann. Surg. 234 (6), 769-78 FUENTE-DEGE, A. (2003): Untersuchungen an pankreasgangligierten Schweinen zu Beziehungen zwischen der Verdaulichkeit von Nährstoffen im praecaecalen Bereich und der Verdaulichkeit über den gesamten Verdauungstrakt. Hannover, Tierärztl. Hochsch., Diss. GOODCHILD, M. C. u. J. A. DODGE (1986): Biochemical basis of cystic fibrosis. Nature 323 (6087), 400 GRABOWSKI, H. (2004): Are the economics of pharmaceutical research and development changing?: productivity, patents and political pressures. Pharmacoeconomics 22 (Suppl 2), 15-24. GRAHAM, D. Y. (1977): Enzyme replacement therapy of exocrine pancreatic insufficiency in man Relation between In-vitro Enzyme activities and In-vivo Potency in Comercial Pancreatic Extracts. New Engl. J. Med. 296, Nr. 23, 1314-1317 GREGORY, P. C. (2006): Persönliche Mitteilung. Solvay Pharmaceuticals Deutschland, Hannover

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
164
GRIFFIN, S. M., D. ALDERSON u. J. R. FARNDON (1989): Acid resistant lipase as replacement therapy in chronic pancreatic exocrine insufficiency: a study in dogs. Gut 30, 1012-1015 HASHOLT, J. (1972): Atrophy of the pancreas in budgerigars. Nord. Vet. Med. 24 (9), 458-61. HAYAKAWA, T., T. KONDO, T. SHIBATA, Y. SUGIMOTO u. M. KIAGAWA (1989): Chronic alcoholism and evolution of pain and prognosis in chronic pancreatitis. Dig. Dis. Sci. 34, 33-8 HELDT, D. (2001): Untersuchungen am pancreasgangligierten Schwein zu Effekten zweier unterschiedlicher Enzympräparationen auf die Verdaulichkeit (praecaecal/ in toto) einer fettreichen Diät. Hannover, Tierärztl. Hochsch., Diss. HERTEL, J., H. J. ALTMANN u. K. DREPPER (1970): Nutritional physiology studies of the horse. II. Raw nutrient studies of the gastrointestinal tract of slaughtered horses. Tierphysiol. Tierernahr. Futtermittelkd. 26 (3), 169-74. HEYMANS,H. S. (1989): Gastrointestinal dysfunction and its effects on nutrition in CF. Acta Paediatr. Scand. Suppl. 363, 74-8 HINZE, S., T. REISS, I. DOMINGUEZ-LACASA u. S. WÖRNER (2001): Einfluss der Biotechnologie auf das Innovationssystem der pharmazeutischen Industrie Bericht an das Bundesministerium für Bildung und Forschung, Referat Z25, Frauenhofer Institut für Systemtechnik und Innovationsforschung, Karlsruhe HUNT, J. N. u. D. F. STUBBS (1975): The volume and energy content of meals as determinants of gastric emptying. J. Physiol. 245, 209-25 IFFLÄNDER, F. (2006): Persönliche Mitteilung. Solvay Pharmaceuticals, Hannover JENSEN, M. S., S. K. JENSEN u. K. JAKOBSEN (1997): Development of digestive enzymes in pigs with emphasis on lipolytic activity in the stomach and pancreas. J. Anim. Sci. 75, 437-445

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
165
JONES, T. (1999): The changes of market success in pharmaceutical research and development. In: Sussex, J.; Marchant, N (eds.): Risk and return in the pharmaceutical industry. Springer Verlag, Berlin KAMMLOTT, E. (2003): Untersuchungen am pankreasgangligierten Schweinen zur Nährstoffverdaulichkeit (praecaecal/ in toto) unter dem Einfluss einer Enzympräperation sowie zur Wirksamkeit ergänzender Maßnahmen (Einsatz von Omeprazol oder Lecithin bzw. fraktionierte Futter- und/oder Enzymgabe). Hannover, Tierärztl. Hochsch., Diss. KAMPHUES, J. (1987): Untersuchungen zu Verdauungsvorgängen bei Absatzferkeln in Abhängigkeit von Futtermengen und –zubereitungen sowie Futterzusätzen Hannover, Tierärztl. Hochsch., Habil.-Schr. KAMPHUES, J., M. COENEN, E. KIENZLE, J. PALLAUF, O. SIMON u. J. ZENTEK (Hrsg.; 2004): Supplemente zu Vorlesungen und Übungen in der Tierernährung. 10. überarbeitete Aufl., Verlag M.&H. Schaper, Alfeld, Hannover KARTHOFF, J. (2004): Beurteilung der Wirksamkeit einer Enzymsubstitution am Modell des pankreasgangligierten Minischweins mittels unterschiedlicher Methoden bzw. Parameter (Nährstoffverdaulichkeit, Konzentration absorbierter Nährstoffe und unterschiedlicher Testsubstanzen im Blut). Hannover, Tierärztl. Hochsch., Diss. KAWCHAK, D. A., H. ZHAO, T. F. SCANLIN, J. L. TOMEZSKO, A. CNAAN u. V. A. STALLINGS (1996): Longitudinal, prospective analysis of dietary intake in children with cystic fibrosis. J. Pediatr. 129 (1), 119-29. KEYS, J. E. u. J. V. DE BARTHE (1974): Site and extend of carbohydrate, dry matter, energy and protein digestion and the rate of passage of grain diets in swine. J. Anim. Sci. 39, 57-62 KIDDER, D. E., u. M. J. MANNERS (1978): Digestion in the pig. Scientechnica, Bristol, UK KIES, A. K., P. J. MOUGHAN u. W. C. SMITH (1986): The apparent and truth ileal digestibility of nitrogen and amino acids in lactic casein for the growing pig. Anim. FEED Sci. Technol. 16, 169-178

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
166
KIRCHGESSNER, M. (2004): Tierernährung. 11. Aufl., DLG-Verlag, Frankfurt (Main) KNOX, M. T. u. C. N. MALLINSON (1971): Gastric emptying of fats in patients with pancreatitis. Rendiconti 3, 115 KOLBEL, C., P. LAYER, J. HOTZ u. H. GOEBELL (1986): Effect of an acid protected, micro-encapsulated pancreatin preparation on pancreatogenic steatorrhea. Med. Klin. (Munich) 81 (3), 85-6. KUHEL, D. G., S. ZHENG, P. TSO u. D. Y. HUI (2000): Adenovirus-mediated human pancreatic lipase gene transfer to rat bile: gene therapy of fat Malabsorption. Am. J. Physiol. Gastrointes. Liver Physiol. 279, G1031-G1036 KUMAR, G., S. FARR-JONES u. M. SCHUPPENHAUER (2001): Value chain management. In: BioCentury 9, No. 6, A1-A5 LADAS, S. D., K. GIORGIOTIS u. S. A. RAPITS (1993): Complex carbohydrate malabsorption in exocrine pancreatic insufficiency. Gut 34, 984-987 LANKISCH, P. G. (1993): Function tests in the diagnosis of chronic pancreatitis. Critical evaluation. Int. J. Pancreatol. 14 (1), 9-20. LANKISCH, P. G. (2001): Natural course of chronic pancreatitis. Pancreatology 1 (1), 3-14. LAYER, P., A. R. ZINSMEISTER u. E. P. DIMAGNO (1986): Effects of decreasing intraluminal activitiy on starch digestion and postprandial gastrointestinal function in humans. Gastroenteol. 91, 41-48 LAYER, P., J. B. JANSEN, L. CHERAN, C. B. LAMERS u. H. GOEBELL (1990): Feedback regulation of human pancreatic secretion. Gastroenterol. 98, 1311-1319 LAYER, P. u. G. HOLTMANN (1994): Pancreatic enzymes in chronic pancreatitis (state-of-the-art). Int. J. Pancreatol. 15, 1-11

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
167
LAYER, P., H. YAMAMOTO, L. KALTHOFF et al. (1994): The different courses of early- and late-onset idiopathic and alcoholic chronic pancreatitis. Gastroenterol 107, 1481-7 LAYER, P., M. RÜNZI u. U. ROSIEN (1997): Klinik und Verlauf der chronischen Pankreatitis. Z. Gastroenterol. (Suppl. 1) LAYER, P. u. E. P. DIMAGNO (1999): Early and late onset in idiopathic and alcoholic chronic pancreatitis. Different clinical courses. Surg. Clin. North. Am. 79 (4), 847-60. LAYER, P. u. J. KELLER (1999): Pancreatic Enzymes: Secretion and Luminal Nutrient Digestion in Health and Disease Clinical Reviews: Pancreatic and Biliary Diseases. J. Clin. Gastroenterol. 28, 1-15 LAYER, P., J. KELLER u. P. G. LANKISCH (2001): Pancreatic enzyme replacement therapy. Curr. Gastroenterol. Rep. 3 (2), 101-8. LAYER, P. u. J. KELLER (2003): Lipase supplementation therapy: standards, alternatives, and perspectives. Pancreas 26, 1-7. LECHOWSKI, R., J. PISARSKI, J. GOSHAWSKI u. M. LENARCIK (1991): Exocrine pancreatic insufficiency-like syndrome in giraffe. J. Wildl. Dis. 27, 728-730 LEBENTHAL, E., D. D. ROLSTON u. D. S. HOLSCLAW jr. (1994): Enzyme therapy for pancreatic insufficiency: present status and future needs. Pancreas 9 (1), 1-12 LEIBNIZ GEMEINSCHAFT (2005): Zwischenruf: Umweltforschung für politische Praxis Heft 2 / 2005 LEIBOLZ, J., R. J. LOVE, Y. MOLLAH u. R. R. CARTER (1986): The absorption of dietary L-Lysine and extruded L-Lysine in pigs. Anim. FEED Sci. Technol. 15, 141-148 LEIDINGER, E. (1997): Exokrine Pankreasinsuffizienz bei Hund und Katze. Wien. tierärztl. Monatsschr. 84, 355-358

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
168
LIDE, D. R., u. H. P. R. FREDERIKSE (1995): CRC Handbook of Chemistry and Physics. CRC Press, Boca Raton, Florida LÖSCHER, W. (2006): Generelle Aspekte bei der Entwicklung neuer Arzneimittel. Vortrag i. Rahmen des pharmakolog.-toxikolog. Seminars, Stiftung Tierärztliche Hochschule Hannover, 03.05.06 LÖSER, CH. u. U. R. FÖLSCH (1991): Klinische und pharmakologische Aspekte der Pankreasenzymsubstitution. Leber Magen Darm 2, 56-65 LÖSER, CH. u. U. R. FÖLSCH (1995): Differentialtherapie der exokrinen Pankreasinsuffizienz – Aktuelle Aspekte und zukünftige Perspektiven der Substitutionstherapie mit Pankreasenzymen. Z. Gastroenterol. 33, 715-722 LOW, A. G. (1982): Digestibility and availability of amino acids from feedstuffs for pigs: a review. Livest. Prod. Sci. 9, 511-520 LÜTHEN, R. u. C. NIEDERAU (1994): Die Magenlipase übernimmt einen wesentlichen Teil der Fettverdauung. Z. Gastroenterol. 32, 519-520 MAEDA, H., C. DANEL u. R. G. CRYSTAL (1994): Adenovirus-mediated transfer of human lipase complementary DNA to the gallbladder. Gastroenterology 106, 1638-44 MALLINSON, C. N. (1968): Effect of pancreatic insufficiency and intestinal lactase deficiency on the gastric emptying of starch and lactose. Gut 9 (6), 737 MANDISCHER, C. (2002): Untersuchungen am pankreasgangligierten Schwein zur Verdaulichkeit (praecaecal / in toto) eines stärkereichen Mischfutters unter dem Einfluss zwei verschiedener oral verabreichter Enzympräparationen. Hannover, Tierärztl. Hochsch., Diss. MAYERLE, J. u. M. M. LERCH (2001): Course of illness in acute pancreatitis. Kongressbd. Dtsch. Ges. Chir. Kongr. 118, 296-300

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
169
MOREAU, H., Y. GARGOURI, D. LECAT, J. L. JUNIEN u. R. VERGER (1988): Screening of preduodenal lipases in several mammals. Biochim. biophys. Acta 959, 247-252 MORRISON, G., J. M. MORRISON, A. O. REDMOND, C. A. BYERS, K. J. MCCRACKEN, J. A. DODGE, S. A. GUILFORD u. M. W. BOWDEN (1992): Comparison between a standard pancreatic supplement and a high enzyme preparation in cystic fibrosis. Alimentary pharmacology & therapeutics 6 (5), 549-55 MÖSSELER, A. K., P. C. GREGORY u. J. KAMPHUES (2005): Influence of ligation of the pancreatic duct on praecaecal / total digestibility of starch in minipigs In: Proceedings of the 9th ESVCN-conference, Grufliasco (Torino), 22. – 24.09.2005 MÖSSELER, A. K., P. C. GREGORY u. J. KAMPHUES (2006 a): Compensatory digestion of fat, protein and starch (rates and amounts) in the large intestine of pigs in cases of reduced prececal digestion 10th International Symposium on Digestive Physiology in Pigs, Danish Institute of Agricultural Science, Vejle (Denmark), 25. – 27.06.06, 55 MÖSSELER, A. K., P. C. GREGORY u. J. KAMPHUES (2006 b): Bedeutung des exokrinen Pankreas für die Verdauung beim Schwein. Übersichten zur Tierernährung 1, 57-103 MÖSSNER, J. (1994): Sense and nonsense in the treatment of exocrine pancreatic insufficiency. Schweiz Rundsch 9, 873-6 MOUGHAN, P. J., P. D. CRANWELL, A. J. DARRAGH u. A. M. ROWAN (1994): The domestic pig as a model: animal for studying in humans. in: W. B. SOUFFRANT und HAGEMEISTER (eds.): Proc. 6th Int. Sympos. Dig. Physiol. In Pigs, Bad Doberan, 4-6.10.1994, S. 389-396 NAKAMURA, T., H. KIKUCHI, K TABEKE, M. ISHII, K. IMAMURA, N. YAMADA, K. KUDOH u. A. TERADA (1994): Correlation between bile acid malabsorption and pancreatic exocrine dysfunction in patients with chronic pancreatitis. Pancreas 9, 580-584 NAKAMURA, T., K. TAKEBE, M. ISHII et al. (1996): Study of gastric emptying in patients with pancreatic diabetes (chronic pancreatitis) using acetaminophen and isotope. Acta Gastroenterol. Belg. 59, 177

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
170
NAUMANN, C., u. R. BASSLER (2004): Die chemische Untersuchung von Futtermitteln. 3. Aufl., 5. Ergänzungslieferung 2004 VDLUFA-Verlag, Darmstadt OECD, ANBERD DATABASE (2000): Industrial structure statistics, Vol. 1, 1999 edition. Paris OELFKE, S. (1978): Untersuchungen zur Verdaulichkeit verschiedener Stärkearten im Magen-Dünndarmbereich des Schweins. Hannover, Tierärztl. Hochsch. (Inst. Tierernähr.), Staatsexamensarbeit PAP, A., M. KOROM, E. MAROSI u. V. VARRO (1990): Requirements for successful pancreatic enzyme replacement therapy ( comparative study of Kreon and Panpur). Orv. Hetil. 131 (5), 241-4. PARRA, R. (1978): Comparison of foregut and hindgut fermentation in herbivores. in: G. G. MONTGOMERY (Hrsg.): The ecology of arboreal folivores. Smithsonian Inst., Washington D. C., S. 205-229 PAYNE, W. L., G. F. COMBS, R. R. KIFER u. D. G. SNYDER (1968): Investigation of protein quality – ileal recovery of amino acids. Fed. Proc. 27, 1190-1203 PEKAS, J. C., V. W. HAYS u. A. M. THOMPSON (1964): Exclusion of the exocrine pancreatic secretion: effect on digestibility of soybean and milk protein by baby pigs at various ages. J. Nutr. 82, 277-286 PEREZ, M. M., A. D. NEWCOMER, C. G. MOERTEL, V. L. GO u. E. P. DIMAGNO (1983): Assessment of weight loss, food intake, fat metabolism, malabsorption, and treatment of pancreatic insufficiency in pancreatic cancer. Cancer 15, 346-52 PITKÄRANTA, P., L. KIVISAARI, S. NORDLING, A. SAARI u. T. SCHRÖDER (1989): Experimental chronic pancreatitis in the pig. Scand. J. Gastroenterol. 24, 987-992 POINTER, H. u. FLEGEL (1975): Die Behandlung der exokrinen Pankreasinsuffizienz mit einer säurestabilen Pilzlipase. Arzneim. Forsch. 25, Nr. 11, 1833-1835

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
171
POND, W. G. u. K A. HOUPT (1978): The biology of the pig. Cornell University Press, Ithaca, New York POTTHOFF, A. (2005): Persönliche Mitteilung. Solvay Pharmaceuticals Deutschland, Hannover PROESMANS, M. u. K. DE BOECK (2003): Omeprazole, a proton pump inhibitor, improves residual steatorrhea in cystic fibrosis patients treated with high dose pancreatic enzymes. Eur. J. Pediatr. 162 (11), 760-3. RADUN, D. u. P. MALFERTHEINER (1996): Chronic pancreatitis: conservative therapy. Ther. Umsch. 53 (5), 359-64. RADUN, D., J. E. DOMINGUEZ-MUNOZ u. P. MALFERTHEINER (1997): Chronische Pankreatitis: Konservative Therapie. Z. Gastroenterol. Suppl. 1, 130-134 RAHKO, T., T. V. KALIMA u. H. SALONIEMI (1987): Pancreatic Duct Obstruction in the Pig: Light Microscopy of chronic Pancreatitis. Acta vet. scand. 28, 285-289 RAIMONDO, M. u. E. P. DIMAGNO (1994): Lipolytic activity of bacterial lipase survives better than that of porcine lipase in human gastric and duodenal content. Gastroenteology 107, 231-5 READ, N. W. (1984): The control of gastrointestinal transit by luminal nutrients. in: E. SKADHAUGE u. K. HEINTZE (Hrsg.): Intestinal absorption and secretion. MTP Press, 153-160 READ, N. W., A. MCFARLANE, R. I. KINSMAN et al. (1984): Effect of infusion of nutrient solutions into the ileum on gastrointestinal transit and plasma levels of neurotensin and enteroglucagon. Gastroenterology 86, 274-80 REGAN, P. T., J. R. MALAGELADA, E. P. DIMAGNO u. V. L. GO (1977): Reduced intraluminal bile acid concentration and fat maldigestion in pancreatic insufficiency: correction by treatment. Gastroenterology 77, 285-9

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
172
RERAT, A. (1983): Function and dysfunction of the small intestine. in: R. M. BATT u. T. L. J. LAWRENCE (eds.): Proc. 2nd George Durant Memorial Symposium, 22 RIMAILA-PARNANEN, E. u. E. WESTERMARCK (1982): Pancreatic degenerative atrophy and chronic pancreatitis in dogs. A comparative study of 60 cases. Acta vet. scand. 23 (3), 400-6. RINDERKNECHT, H. (1993): Pancreatic secretory enzymes. in: V. L. W. GO, E. P. DIMAGNO, J. D. GARDNER, E. LEBENTHAL, H. A. REBER, G. A. SCHEELE (Hrsg.): The pancreas: Biology, Pathobiology and Disease. Raven press, New York, S. 219-251 RITCHEY, J. W., L. A. DEGERNES u. T. T. BROWN (1997): Exocrine pancreatic insufficiency in a yellow-naped Amazon with pancreatic adenocarcinoma. Vet. Pathol. 34, 55-57 ROTHENBACHER, D., M. LOW, P. D. HARDT, H. U. KLOR, H. ZIEGLER u. H. BRENNER (2005): Prevalence and determinants of exocrine pancreatic insufficiency among older adults: results of a population-based study. Scand J Gastroenterol 40, 697-704 RUTZ, G. M., J. M. STEINER u. J. HIRSCHBERGER (2000): Exokrine Pankreasinsuffizienz des Hundes. Tierärztl. Prax. 28, 138-144 SANTINI, B., M. ANTONELLI, A. BATTISTINI, S. BERTASI, M. COLLURA, I. ESPOSITO, L. DI FEBBRARO, R. FERRARI, L. FERRERO, L. IAPICHINO, V. LUCIDI, A. MANCA, C. L. PISCONTI, G. PISI, V. RAIA, L. ROMANO, P. ROSATI, I. GRAZIOLI u. G. MELZI (2000): Comparison of two enteric coated microsphere preparations in the treatment of pancreatic exocrine insufficiency caused by cystic fibrosis. Dig. Liver. Dis. 32 (5), 406-11 SCHARRER, E. u. S. WOLFFRAM (2000): Funktionen des Dünndarms und seiner Anhangdrüsen. in: W. v. ENGELHARDT u. G. BREVES (Hrsg.): Physiologie der Haustiere. Enke Verlag, Stuttgart, S. 369-394

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
173
SCHNEIDER, M. U., S. KNOLL-RUZICKA, S. DOMSCHKE, G. HEPTNER u. W. DOMSCHKE (1985): Pancreatic enzyme replacement therapy: Comparative effects of conventional and enteric-coated microspheric pancreatin and acid-stable fungal enzyme preparations on steatorrhea in chronic pancreatitis. Hepato-Gastroenterol. 32, 97-102 SCHÜRCH, A. (1969): Die Verdaulichkeit der Nahrung bzw. Nahrungskomponenten. In: LENKEIT, W., K. BREIREM u. E. CRASEMANN: Handbuch der Tierernährung, Bd. 1 Paul Parey Verlag, Berlin und Hamburg, S. 272-294 SHAW, D. W. u. G. O. BARBEZAT (1982): Comparative In-vitro enzyme activities of five commercially available pancreatic supplements. N. Z. Med. J. 95 (710), 424-5 SHIAU, Y. - F. (1987): Lipid digestion and absorption. in: L. R. JOHNSON (Hrsg.): Physiology of the Gastrointestinal Tract. Vol. II., 2.ed. Raven press, New York, U.S.A. SINGER, M. V. u. M. K. MÜLLER (1995): Epidemiologie, Ätiologie und Pathogenese der chronischen Pankreatitis. in: J. MÖSSNER, G. ADLER, U. R. FÖLSCH u. M. V. SINGER (Hrsg.): Erkrankungen des exkretorischen Pancreas. 2. Auflage, Gustav Fischer Verlag, Jena, Stuttgart, S. 313-324 SLAFF, J., D. JACOBSON, C. R. TILLMANN, C. CURINGTON u. P. TOSKES (1984): Protease-specific suppression of pancreatic exocrine secretion. Gastroenterol. 87 (1), 44-52 STEINBERG, W. M., S. T. CHARI, C. E. FORSMARK, S. SHERMAN, H. A. REBER, E. L. BRADLEY 3rd u. E. DIMAGNO (2003): Controversies in clinical pancreatology: management of acute idiopathic recurrent pancreatitis. Pancreas 27 (2), 103-17 STERNBY, B., G. HOLTMANN, D. G. KELLY u. E. P. DIMAGNO (1992): Effect of gastric or duodenal nutrient infusion on gastric and pancreatic lipase secretion. Gastroenterol. 102, A292

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
174
STEVENS, C. E. (1977): Comparative physiology of the digestive system. in: M. J. SWENSON (Hrsg.): Duke's Physiology of Domestic Animals. 9th ed. Cornell University Press, Ithaca, New York SUZUKI A., A. MIZUMOTO, M. G. SARR u. E. P. DIMAGNO (1997): Bacterial lipase and high-fat diets in canine exocrine pancreatic insufficiency: a new therapy of steatorrhea? Gastroenterol. 112, 2048-55 SUZUKI, A., A. MIZUMOTO, R. RERKNIMITR, M. G. SARR u. E. P. DIMAGNO (1999): Effect of bacterial or porcine lipase with low- or high-fat diets on nutrient absorption in pancreatic-insufficient dogs. Gastroenterol. 116, 431-437 TABELING, R. (1998): Untersuchungen am pankreasgangligierten Schwein zu Effekten einer Enzympräparation auf die Nährstoffverdaulichkeit (praecaecal / in toto). Hannover, Tierärztl. Hochsch. Diss. THIRUVENGADAM, R. u. E. P. DIMAGNO (1988): Inactivation of human lipase by proteases. Am. J. Physiol. 255, G476-81 TUFTS CENTER FOR THE STUDY OF DRUG DEVELOPMENT (1999): European and U.S. approval times for new drugs are virtually identical. Impact Report Vol. 1, November 1999, Boston VFA STATISTICS - VERBAND FORSCHENDER ARZNEIMITTELHERSTELLER e.V. (2005): Die Arzneimittelindustrie in Deutschland www.vfa.de, Berlin VILLE, E., F. CARRIERE, C. RENOU u. R. LAUGIER (2002) Physiological study of pH stability and sensitivity to pepsin of human gastric lipase. Digestion 65, 73-81 WALKOWIAK, J., H. WITMANOWSKI, K. STRZYKALA, B. BYCHOWIEC, T. SONGIN, K. BORSKI u. K. H. HERZIG (2003): Hemmung der endogenen Sekretion von Pankreasenzymen durch orale Therapie mit Pankreasenzymen. Eur. J. Clin.Invest. 33, 65-69 WATSON, A. D. J., D. B. CHURCH, D. J. MIDDLETON u. T. L. W. ROTHWELL (1981): Weight loss in cats which eat well. J. Small. Anim. Pract. 22,473-482

LITERATURVERZEICHNIS ------------------------------------------------------------------------------------------------------------------------
175
WESTERMARCK, E. (1980): The hereditary nature of canine pancreatic degenerative atrophy in the German Shepherd dog. Acta vet. scand. 21, 389-394 WESTERMARCK, E., V. MYLLYS u. M. AHO (1993): Effect of treatment on the jejunal and colonic bacterial flora of dogs with exocrine pancreatic insufficiency. Pancreas 8, 559-562 WIBERG, D. A. (2004): Pancreatic acinar atrophy in german Shepherd dogs and rough-coated collies. A review. Veterinary Quarterley 26 (2), 61-75 WILLIAMS, D. A. u. F. MINNICH (1990): Canine exocrine pancreatic insufficiency – a survey of 640 cases diagnosed by assay of serum trypsin like immunoreactivity. J. Vet. Int. Med. 4, 123 YIN, Y.-L., J. D. G. MCEVOY, H. SCHULZE u. K. J. MCCRACKEN (2000): Studies on cannulation method and alternative indigestible markers and the effects of food enzyme supplementation in barley-based diets on ileal and overall apparent digestibility in growing pigs. Animal Science 70, 63-72 ZABIELSKI, R., T. ONAGA, H. MINRO u. S. KATO (1993): Periodic fluctuations in pancreatic secretion and duodenal motility investigated in neonatal calves. Exp. Physiol. 78 (5), 675-84 ZABIELSKI, R., P. KIELA, V. LESNIEWSKA, R. KREMINSKI, M. MIKOLAICZYK u. W. BAREJ (1997): Kinetics of pancreatic juice secretion in relation to duodenal migrating myoelectric complex in preruminant and ruminant calves fed twice daily. Br. J. Nutr. 78 (3), 427-42 ZEBROWSKA, T., O. SIMON, R. MÜNCHMEYER u. H. BERGER (1980): Untersuchungen zur Sezernierung endogener Aminosäuren in den Verdauungstrakt und zur Aminosäureresorption beim Schwein. Arch. Tierernähr. 26, 69-82 ZENTLER-MUNRO, P. L., B. A. ASSOUFI, K. BALASUBRAMANIAN, S. CORNELL, D. BENOLIEL, T. C. NORTHFIELD u. M. E. HODSON (1992): Therapeutic potential and clinical efficacy of acid-resistant fungal lipase in the treatment of pancreatic steatorrhea due to Cystic Fibrosis. Pancreas 7, Nr. 3, 311-319
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
176
8 Tabellenanhang
Anhang 1.1: Rohfett- Konzentration des Chymus (g/ kg TS) sowie Relation Rohfettkonzentration / Chromoxidkonzentration im Kollektionsintervall 0-8 h n. F. (1.-3. Versuch zur Optimierung einer flüssigen Testmahlzeit) Nr. Enzym Dosierung (IE)] Tier Fett Konz. Fett / Cr Konz.
63 344 59,7 82 361 51,1 0 83 462 50,7 82 173 23,6 63 210 17,7 10 000 83 110 7,41 82 90,7 7,30 63 234 13,5
1. PK
75 000 83 132 6,40 63 450 104 83 363 65,2 0 82 529 89,8 63 420 74,4 82 480 57,6 5 000 83 271 24,7 63 306 28,6 83 88,5 7,00
2. PK
55 000 82 145 7,52 43 477 41,9 0 93 479 37,8 43 410 54,3 83 543 60,7 500 93 531 87,9 82 481 41,0 43 296 30,6 93 360 27,8 5 000
83 138 10,2 82 323 20,2
3. PK
55 000 83 162 6,57
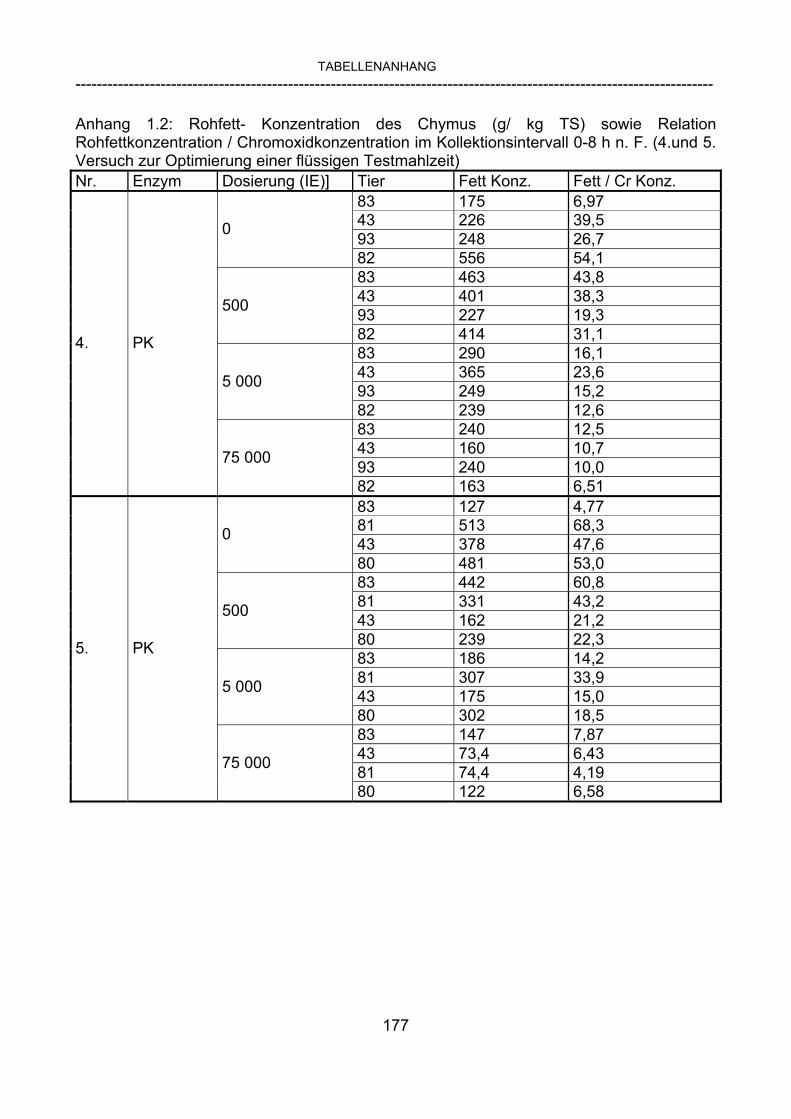
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
177
Anhang 1.2: Rohfett- Konzentration des Chymus (g/ kg TS) sowie Relation Rohfettkonzentration / Chromoxidkonzentration im Kollektionsintervall 0-8 h n. F. (4.und 5. Versuch zur Optimierung einer flüssigen Testmahlzeit) Nr. Enzym Dosierung (IE)] Tier Fett Konz. Fett / Cr Konz.
83 175 6,97 43 226 39,5 93 248 26,7 0
82 556 54,1 83 463 43,8 43 401 38,3 93 227 19,3 500
82 414 31,1 83 290 16,1 43 365 23,6 93 249 15,2 5 000
82 239 12,6 83 240 12,5 43 160 10,7 93 240 10,0
4. PK
75 000
82 163 6,51 83 127 4,77 81 513 68,3 43 378 47,6 0
80 481 53,0 83 442 60,8 81 331 43,2 43 162 21,2 500
80 239 22,3 83 186 14,2 81 307 33,9 43 175 15,0 5 000
80 302 18,5 83 147 7,87 43 73,4 6,43 81 74,4 4,19
5. PK
75 000
80 122 6,58
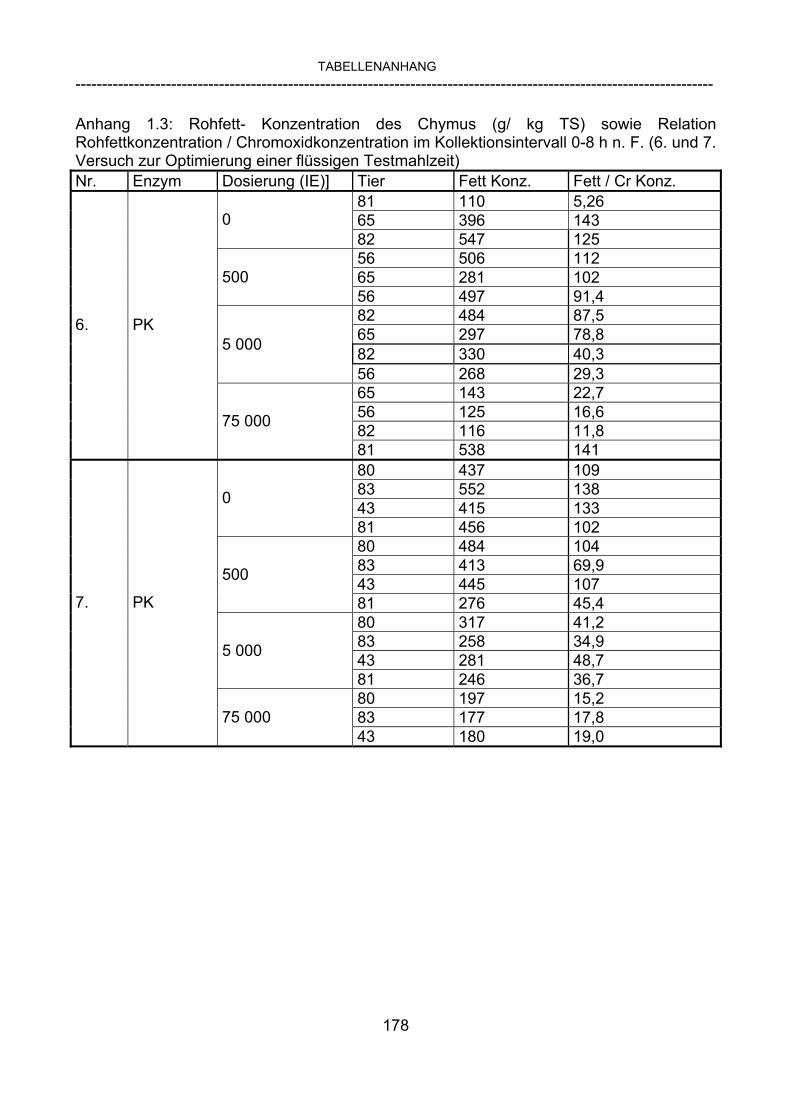
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
178
Anhang 1.3: Rohfett- Konzentration des Chymus (g/ kg TS) sowie Relation Rohfettkonzentration / Chromoxidkonzentration im Kollektionsintervall 0-8 h n. F. (6. und 7. Versuch zur Optimierung einer flüssigen Testmahlzeit) Nr. Enzym Dosierung (IE)] Tier Fett Konz. Fett / Cr Konz.
81 110 5,26 65 396 143 0 82 547 125 56 506 112 65 281 102 500 56 497 91,4 82 484 87,5 65 297 78,8 82 330 40,3 5 000
56 268 29,3 65 143 22,7 56 125 16,6 82 116 11,8
6. PK
75 000
81 538 141 80 437 109 83 552 138 43 415 133 0
81 456 102 80 484 104 83 413 69,9 43 445 107 500
81 276 45,4 80 317 41,2 83 258 34,9 43 281 48,7 5 000
81 246 36,7 80 197 15,2 83 177 17,8
7. PK
75 000 43 180 19,0
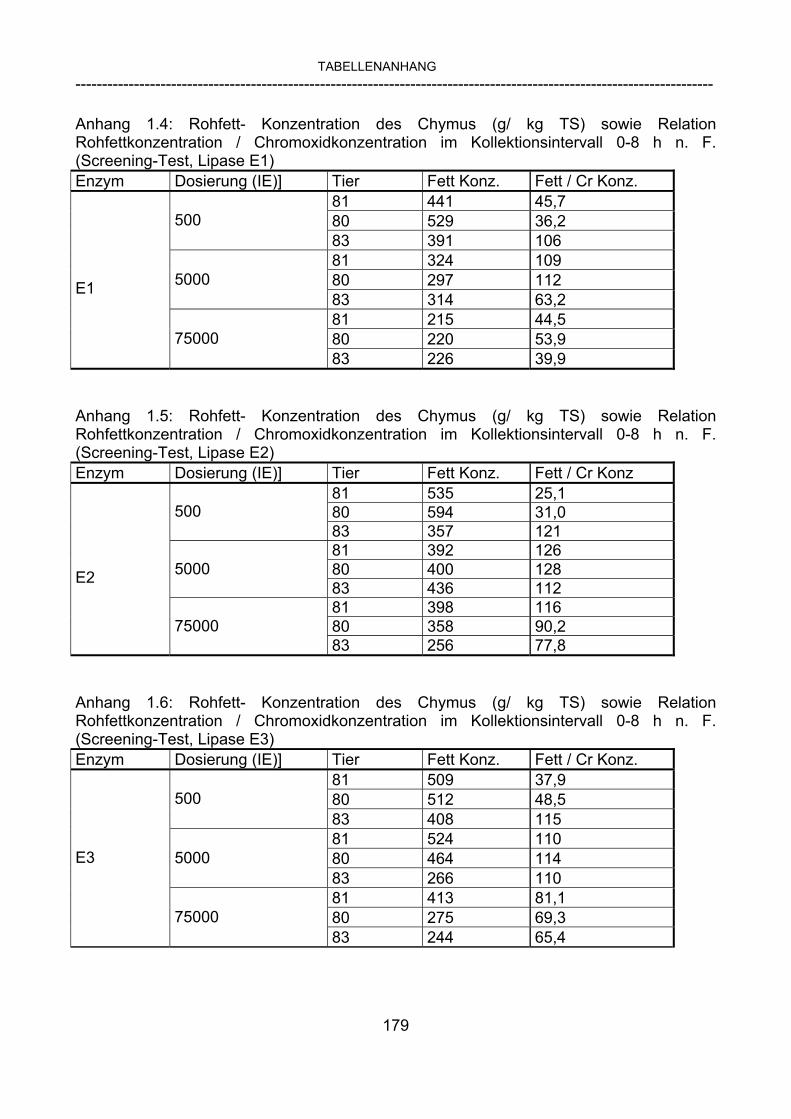
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
179
Anhang 1.4: Rohfett- Konzentration des Chymus (g/ kg TS) sowie Relation Rohfettkonzentration / Chromoxidkonzentration im Kollektionsintervall 0-8 h n. F. (Screening-Test, Lipase E1) Enzym Dosierung (IE)] Tier Fett Konz. Fett / Cr Konz.
81 441 45,7 80 529 36,2 500 83 391 106 81 324 109 80 297 112 5000 83 314 63,2 81 215 44,5 80 220 53,9
E1
75000 83 226 39,9
Anhang 1.5: Rohfett- Konzentration des Chymus (g/ kg TS) sowie Relation Rohfettkonzentration / Chromoxidkonzentration im Kollektionsintervall 0-8 h n. F. (Screening-Test, Lipase E2) Enzym Dosierung (IE)] Tier Fett Konz. Fett / Cr Konz
81 535 25,1 80 594 31,0 500 83 357 121 81 392 126 80 400 128 5000 83 436 112 81 398 116 80 358 90,2
E2
75000 83 256 77,8
Anhang 1.6: Rohfett- Konzentration des Chymus (g/ kg TS) sowie Relation Rohfettkonzentration / Chromoxidkonzentration im Kollektionsintervall 0-8 h n. F. (Screening-Test, Lipase E3) Enzym Dosierung (IE)] Tier Fett Konz. Fett / Cr Konz.
81 509 37,9 80 512 48,5 500 83 408 115 81 524 110 80 464 114 5000 83 266 110 81 413 81,1 80 275 69,3
E3
75000 83 244 65,4
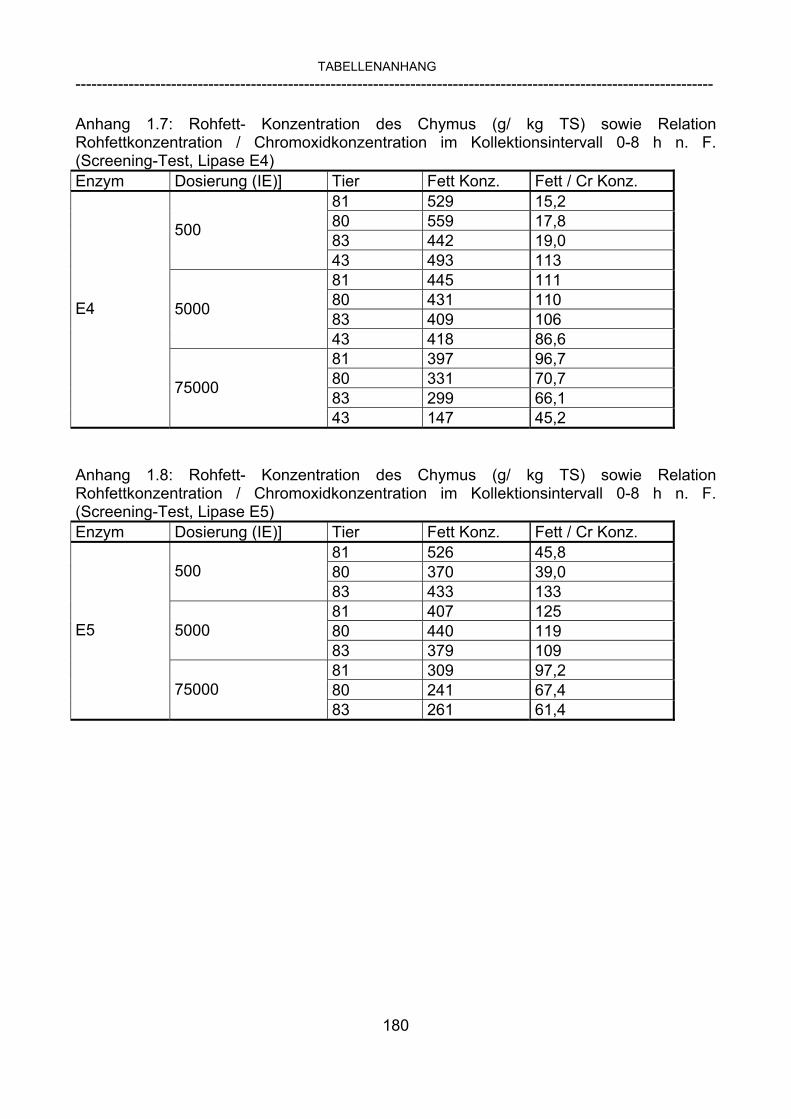
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
180
Anhang 1.7: Rohfett- Konzentration des Chymus (g/ kg TS) sowie Relation Rohfettkonzentration / Chromoxidkonzentration im Kollektionsintervall 0-8 h n. F. (Screening-Test, Lipase E4) Enzym Dosierung (IE)] Tier Fett Konz. Fett / Cr Konz.
81 529 15,2 80 559 17,8 83 442 19,0
500
43 493 113 81 445 111 80 431 110 83 409 106 5000
43 418 86,6 81 397 96,7 80 331 70,7 83 299 66,1
E4
75000
43 147 45,2 Anhang 1.8: Rohfett- Konzentration des Chymus (g/ kg TS) sowie Relation Rohfettkonzentration / Chromoxidkonzentration im Kollektionsintervall 0-8 h n. F. (Screening-Test, Lipase E5) Enzym Dosierung (IE)] Tier Fett Konz. Fett / Cr Konz.
81 526 45,8 80 370 39,0 500 83 433 133 81 407 125 80 440 119 5000 83 379 109 81 309 97,2 80 241 67,4
E5
75000 83 261 61,4
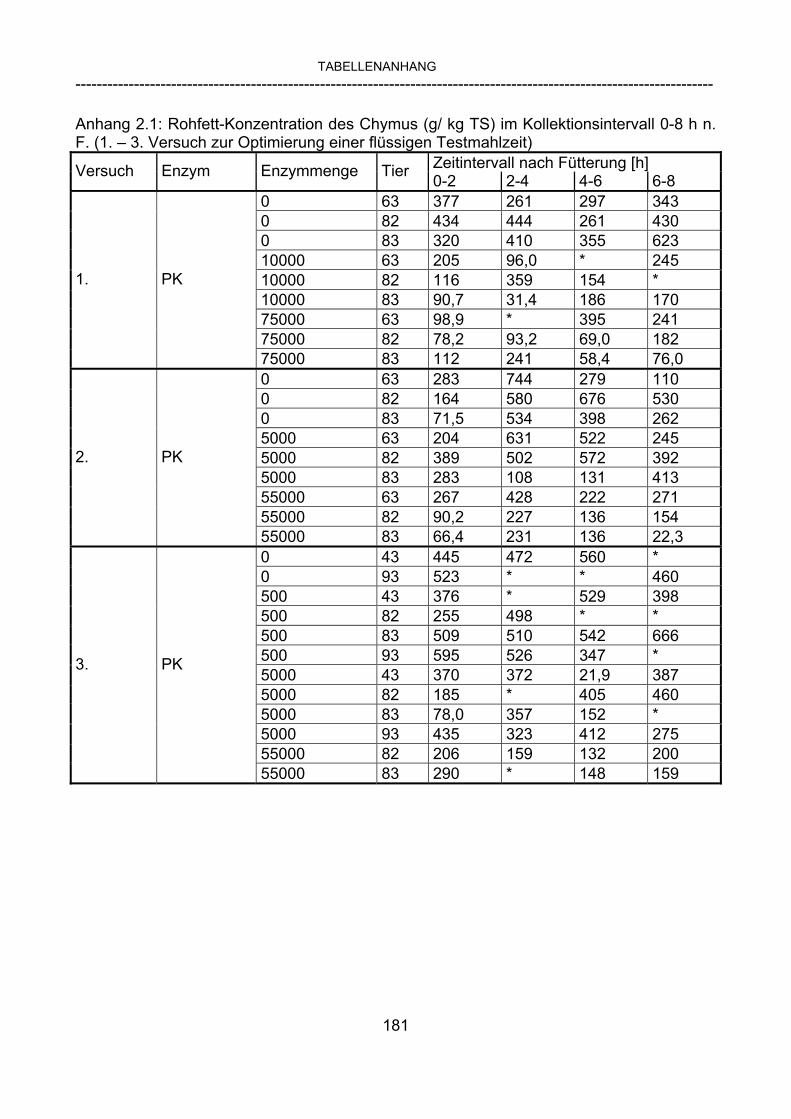
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
181
Anhang 2.1: Rohfett-Konzentration des Chymus (g/ kg TS) im Kollektionsintervall 0-8 h n. F. (1. – 3. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Zeitintervall nach Fütterung [h] Versuch Enzym Enzymmenge Tier 0-2 2-4 4-6 6-8 0 63 377 261 297 343 0 82 434 444 261 430 0 83 320 410 355 623 10000 63 205 96,0 * 245 10000 82 116 359 154 * 10000 83 90,7 31,4 186 170 75000 63 98,9 * 395 241 75000 82 78,2 93,2 69,0 182
1. PK
75000 83 112 241 58,4 76,0 0 63 283 744 279 110 0 82 164 580 676 530 0 83 71,5 534 398 262 5000 63 204 631 522 245 5000 82 389 502 572 392 5000 83 283 108 131 413 55000 63 267 428 222 271 55000 82 90,2 227 136 154
2. PK
55000 83 66,4 231 136 22,3 0 43 445 472 560 * 0 93 523 * * 460 500 43 376 * 529 398 500 82 255 498 * * 500 83 509 510 542 666 500 93 595 526 347 * 5000 43 370 372 21,9 387 5000 82 185 * 405 460 5000 83 78,0 357 152 * 5000 93 435 323 412 275 55000 82 206 159 132 200
3. PK
55000 83 290 * 148 159
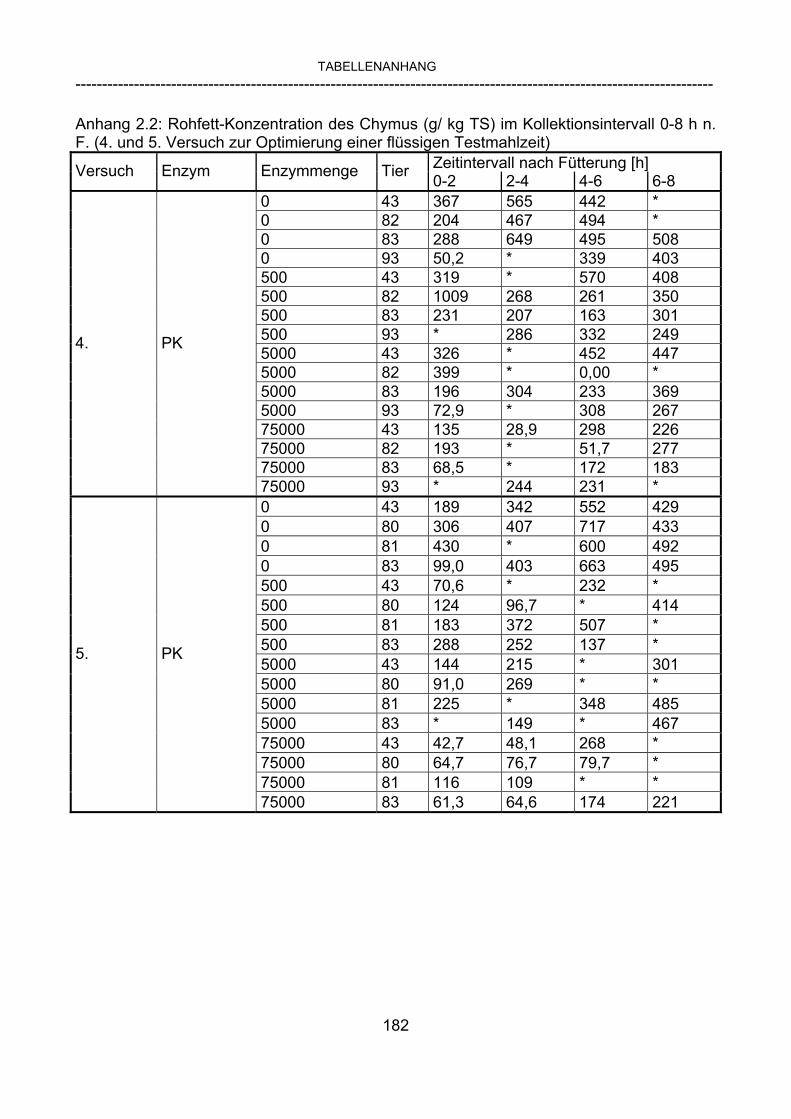
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
182
Anhang 2.2: Rohfett-Konzentration des Chymus (g/ kg TS) im Kollektionsintervall 0-8 h n. F. (4. und 5. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Zeitintervall nach Fütterung [h] Versuch Enzym Enzymmenge Tier 0-2 2-4 4-6 6-8 0 43 367 565 442 * 0 82 204 467 494 * 0 83 288 649 495 508 0 93 50,2 * 339 403 500 43 319 * 570 408 500 82 1009 268 261 350 500 83 231 207 163 301 500 93 * 286 332 249 5000 43 326 * 452 447 5000 82 399 * 0,00 * 5000 83 196 304 233 369 5000 93 72,9 * 308 267 75000 43 135 28,9 298 226 75000 82 193 * 51,7 277 75000 83 68,5 * 172 183
4. PK
75000 93 * 244 231 * 0 43 189 342 552 429 0 80 306 407 717 433 0 81 430 * 600 492 0 83 99,0 403 663 495 500 43 70,6 * 232 * 500 80 124 96,7 * 414 500 81 183 372 507 * 500 83 288 252 137 * 5000 43 144 215 * 301 5000 80 91,0 269 * * 5000 81 225 * 348 485 5000 83 * 149 * 467 75000 43 42,7 48,1 268 * 75000 80 64,7 76,7 79,7 * 75000 81 116 109 * *
5. PK
75000 83 61,3 64,6 174 221
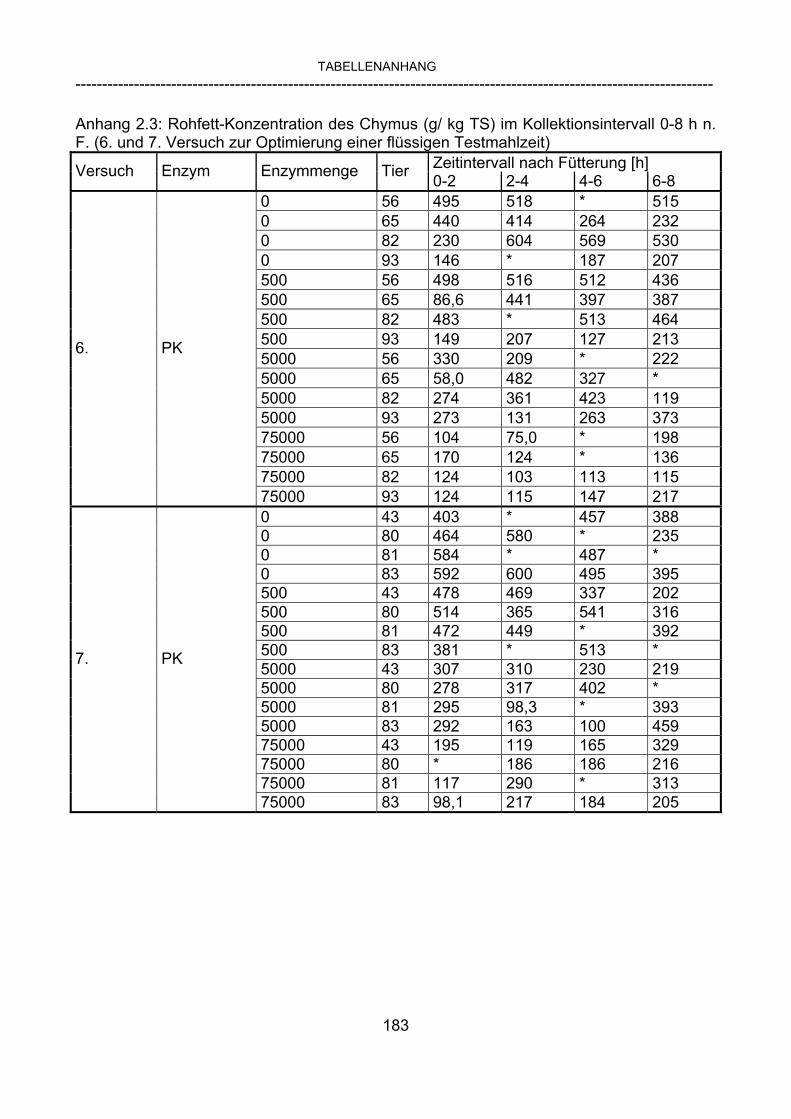
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
183
Anhang 2.3: Rohfett-Konzentration des Chymus (g/ kg TS) im Kollektionsintervall 0-8 h n. F. (6. und 7. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Zeitintervall nach Fütterung [h] Versuch Enzym Enzymmenge Tier 0-2 2-4 4-6 6-8 0 56 495 518 * 515 0 65 440 414 264 232 0 82 230 604 569 530 0 93 146 * 187 207 500 56 498 516 512 436 500 65 86,6 441 397 387 500 82 483 * 513 464 500 93 149 207 127 213 5000 56 330 209 * 222 5000 65 58,0 482 327 * 5000 82 274 361 423 119 5000 93 273 131 263 373 75000 56 104 75,0 * 198 75000 65 170 124 * 136 75000 82 124 103 113 115
6. PK
75000 93 124 115 147 217 0 43 403 * 457 388 0 80 464 580 * 235 0 81 584 * 487 * 0 83 592 600 495 395 500 43 478 469 337 202 500 80 514 365 541 316 500 81 472 449 * 392 500 83 381 * 513 * 5000 43 307 310 230 219 5000 80 278 317 402 * 5000 81 295 98,3 * 393 5000 83 292 163 100 459 75000 43 195 119 165 329 75000 80 * 186 186 216 75000 81 117 290 * 313
7. PK
75000 83 98,1 217 184 205
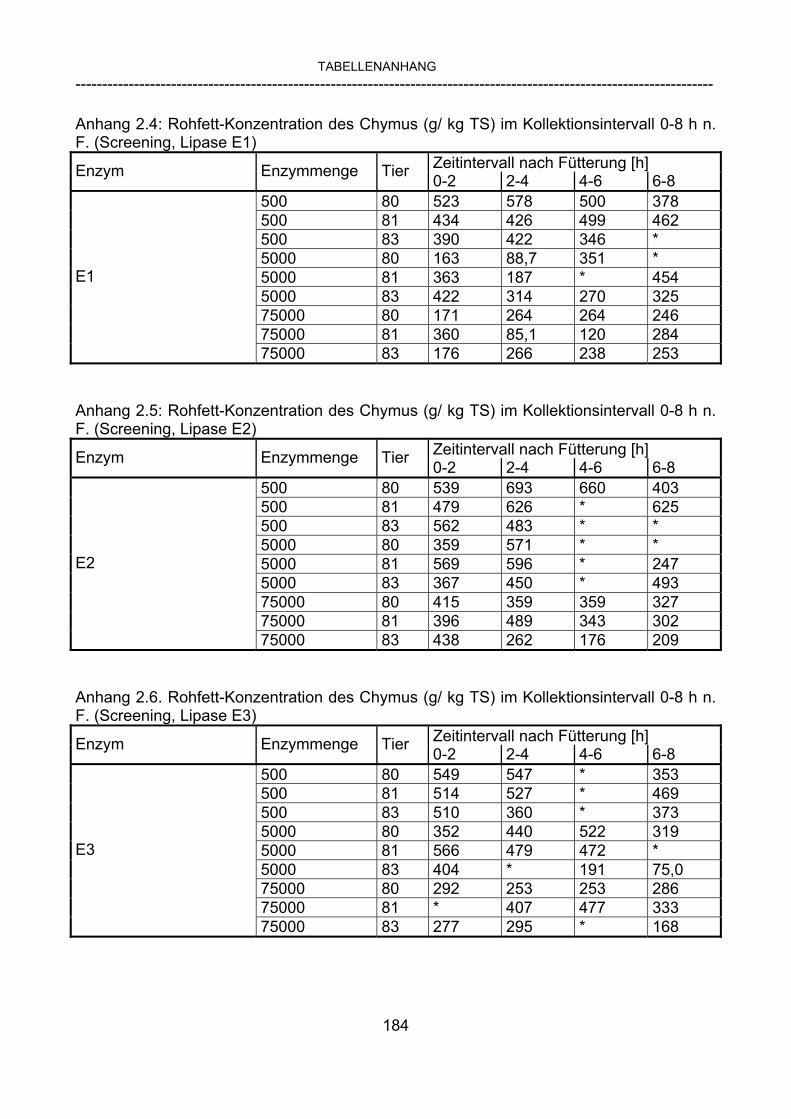
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
184
Anhang 2.4: Rohfett-Konzentration des Chymus (g/ kg TS) im Kollektionsintervall 0-8 h n. F. (Screening, Lipase E1)
Zeitintervall nach Fütterung [h] Enzym Enzymmenge Tier 0-2 2-4 4-6 6-8 500 80 523 578 500 378 500 81 434 426 499 462 500 83 390 422 346 * 5000 80 163 88,7 351 * 5000 81 363 187 * 454 5000 83 422 314 270 325 75000 80 171 264 264 246 75000 81 360 85,1 120 284
E1
75000 83 176 266 238 253 Anhang 2.5: Rohfett-Konzentration des Chymus (g/ kg TS) im Kollektionsintervall 0-8 h n. F. (Screening, Lipase E2)
Zeitintervall nach Fütterung [h] Enzym Enzymmenge Tier 0-2 2-4 4-6 6-8 500 80 539 693 660 403 500 81 479 626 * 625 500 83 562 483 * * 5000 80 359 571 * * 5000 81 569 596 * 247 5000 83 367 450 * 493 75000 80 415 359 359 327 75000 81 396 489 343 302
E2
75000 83 438 262 176 209 Anhang 2.6. Rohfett-Konzentration des Chymus (g/ kg TS) im Kollektionsintervall 0-8 h n. F. (Screening, Lipase E3)
Zeitintervall nach Fütterung [h] Enzym Enzymmenge Tier 0-2 2-4 4-6 6-8 500 80 549 547 * 353 500 81 514 527 * 469 500 83 510 360 * 373 5000 80 352 440 522 319 5000 81 566 479 472 * 5000 83 404 * 191 75,0 75000 80 292 253 253 286 75000 81 * 407 477 333
E3
75000 83 277 295 * 168
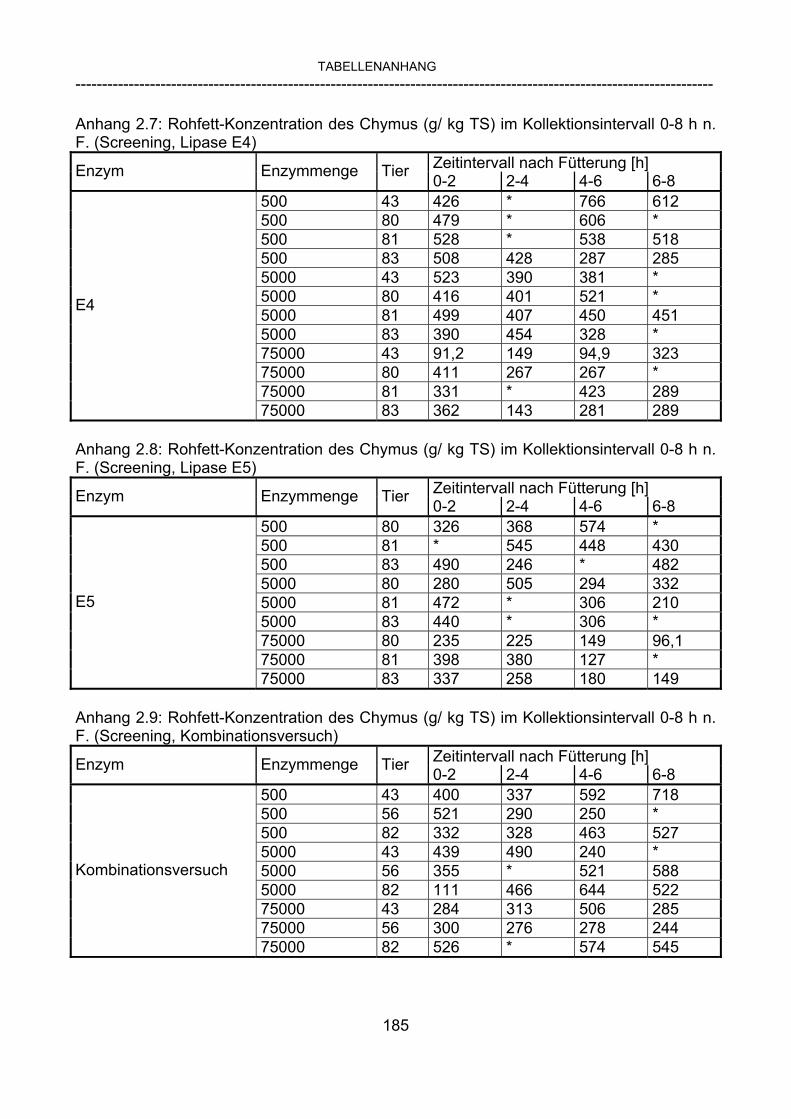
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
185
Anhang 2.7: Rohfett-Konzentration des Chymus (g/ kg TS) im Kollektionsintervall 0-8 h n. F. (Screening, Lipase E4)
Zeitintervall nach Fütterung [h] Enzym Enzymmenge Tier 0-2 2-4 4-6 6-8 500 43 426 * 766 612 500 80 479 * 606 * 500 81 528 * 538 518 500 83 508 428 287 285 5000 43 523 390 381 * 5000 80 416 401 521 * 5000 81 499 407 450 451 5000 83 390 454 328 * 75000 43 91,2 149 94,9 323 75000 80 411 267 267 * 75000 81 331 * 423 289
E4
75000 83 362 143 281 289 Anhang 2.8: Rohfett-Konzentration des Chymus (g/ kg TS) im Kollektionsintervall 0-8 h n. F. (Screening, Lipase E5)
Zeitintervall nach Fütterung [h] Enzym Enzymmenge Tier 0-2 2-4 4-6 6-8 500 80 326 368 574 * 500 81 * 545 448 430 500 83 490 246 * 482 5000 80 280 505 294 332 5000 81 472 * 306 210 5000 83 440 * 306 * 75000 80 235 225 149 96,1 75000 81 398 380 127 *
E5
75000 83 337 258 180 149 Anhang 2.9: Rohfett-Konzentration des Chymus (g/ kg TS) im Kollektionsintervall 0-8 h n. F. (Screening, Kombinationsversuch)
Zeitintervall nach Fütterung [h] Enzym Enzymmenge Tier 0-2 2-4 4-6 6-8 500 43 400 337 592 718 500 56 521 290 250 * 500 82 332 328 463 527 5000 43 439 490 240 * 5000 56 355 * 521 588 5000 82 111 466 644 522 75000 43 284 313 506 285 75000 56 300 276 278 244
Kombinationsversuch
75000 82 526 * 574 545
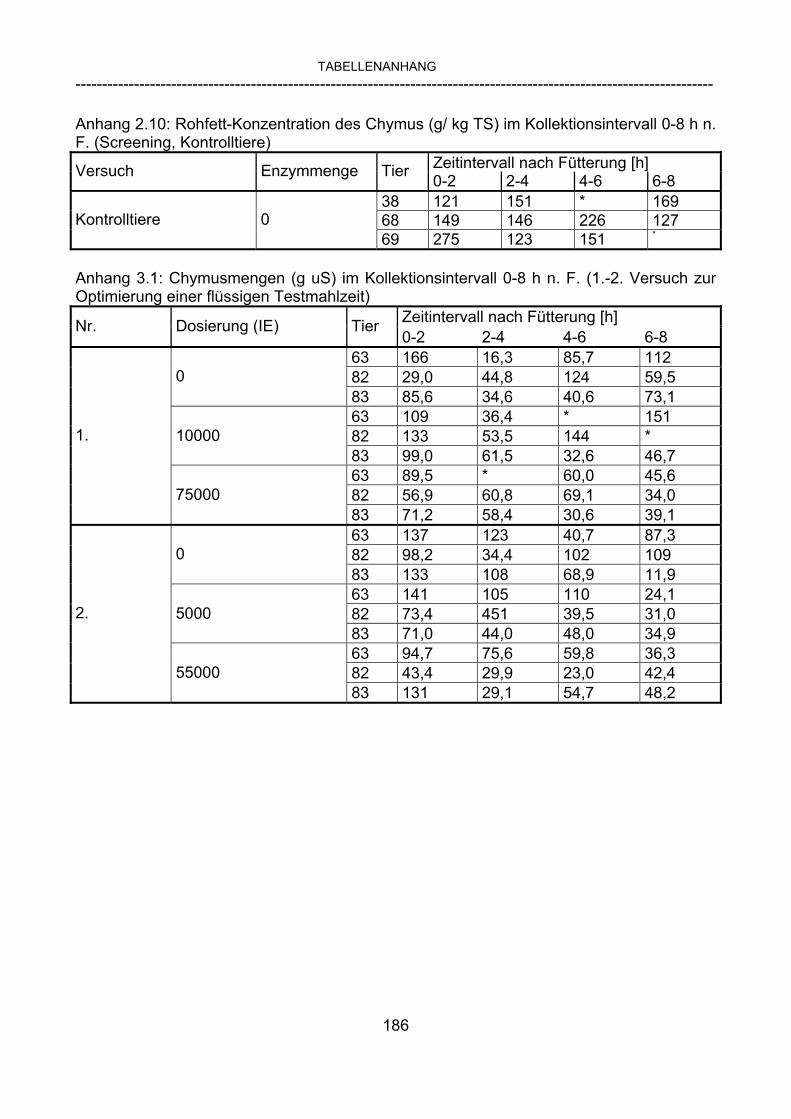
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
186
Anhang 2.10: Rohfett-Konzentration des Chymus (g/ kg TS) im Kollektionsintervall 0-8 h n. F. (Screening, Kontrolltiere)
Zeitintervall nach Fütterung [h] Versuch Enzymmenge Tier 0-2 2-4 4-6 6-8 38 121 151 * 169 68 149 146 226 127 Kontrolltiere 0 69 275 123 151 *
Anhang 3.1: Chymusmengen (g uS) im Kollektionsintervall 0-8 h n. F. (1.-2. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Zeitintervall nach Fütterung [h] Nr. Dosierung (IE) Tier 0-2 2-4 4-6 6-8
63 166 16,3 85,7 112 82 29,0 44,8 124 59,5 0 83 85,6 34,6 40,6 73,1 63 109 36,4 * 151 82 133 53,5 144 * 10000 83 99,0 61,5 32,6 46,7 63 89,5 * 60,0 45,6 82 56,9 60,8 69,1 34,0
1.
75000 83 71,2 58,4 30,6 39,1 63 137 123 40,7 87,3 82 98,2 34,4 102 109 0 83 133 108 68,9 11,9 63 141 105 110 24,1 82 73,4 451 39,5 31,0 5000 83 71,0 44,0 48,0 34,9 63 94,7 75,6 59,8 36,3 82 43,4 29,9 23,0 42,4
2.
55000 83 131 29,1 54,7 48,2
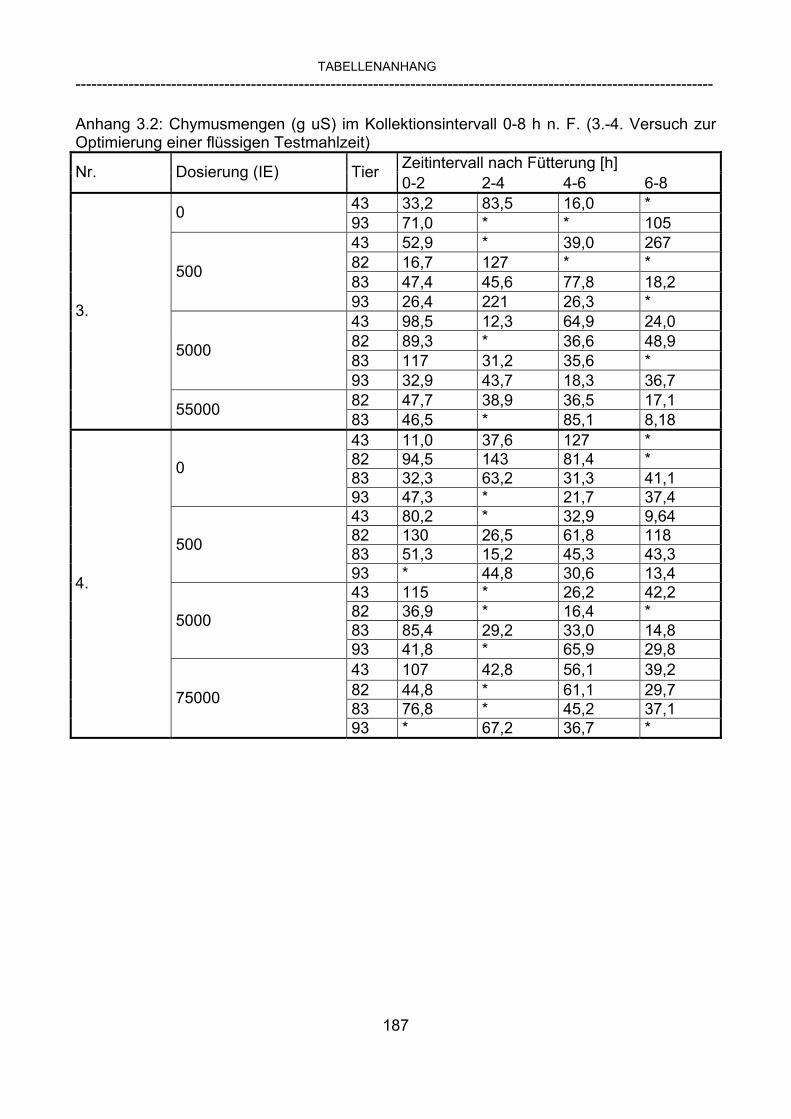
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
187
Anhang 3.2: Chymusmengen (g uS) im Kollektionsintervall 0-8 h n. F. (3.-4. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Zeitintervall nach Fütterung [h] Nr. Dosierung (IE) Tier 0-2 2-4 4-6 6-8
43 33,2 83,5 16,0 * 0 93 71,0 * * 105 43 52,9 * 39,0 267 82 16,7 127 * * 83 47,4 45,6 77,8 18,2 500
93 26,4 221 26,3 * 43 98,5 12,3 64,9 24,0 82 89,3 * 36,6 48,9 83 117 31,2 35,6 * 5000
93 32,9 43,7 18,3 36,7 82 47,7 38,9 36,5 17,1
3.
55000 83 46,5 * 85,1 8,18 43 11,0 37,6 127 * 82 94,5 143 81,4 * 83 32,3 63,2 31,3 41,1 0
93 47,3 * 21,7 37,4 43 80,2 * 32,9 9,64 82 130 26,5 61,8 118 83 51,3 15,2 45,3 43,3 500
93 * 44,8 30,6 13,4 43 115 * 26,2 42,2 82 36,9 * 16,4 * 83 85,4 29,2 33,0 14,8 5000
93 41,8 * 65,9 29,8 43 107 42,8 56,1 39,2 82 44,8 * 61,1 29,7 83 76,8 * 45,2 37,1
4.
75000
93 * 67,2 36,7 *
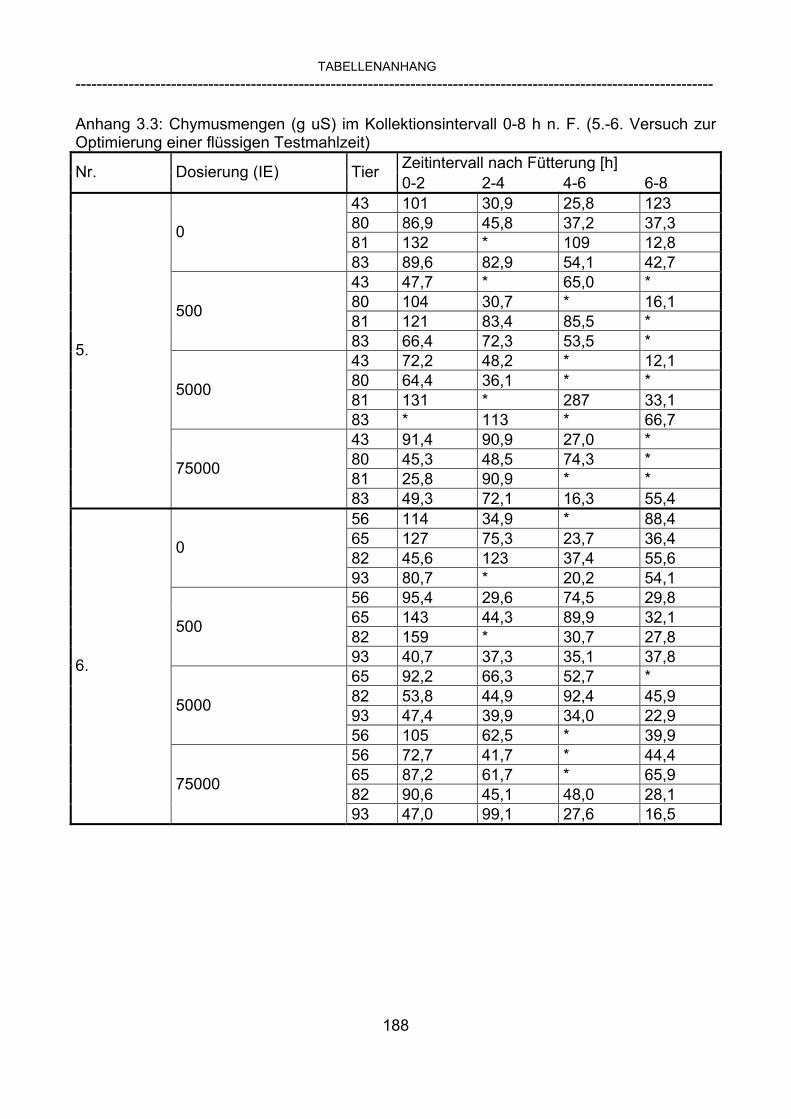
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
188
Anhang 3.3: Chymusmengen (g uS) im Kollektionsintervall 0-8 h n. F. (5.-6. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Zeitintervall nach Fütterung [h] Nr. Dosierung (IE) Tier 0-2 2-4 4-6 6-8
43 101 30,9 25,8 123 80 86,9 45,8 37,2 37,3 81 132 * 109 12,8
0
83 89,6 82,9 54,1 42,7 43 47,7 * 65,0 * 80 104 30,7 * 16,1 81 121 83,4 85,5 * 500
83 66,4 72,3 53,5 * 43 72,2 48,2 * 12,1 80 64,4 36,1 * * 81 131 * 287 33,1 5000
83 * 113 * 66,7 43 91,4 90,9 27,0 * 80 45,3 48,5 74,3 * 81 25,8 90,9 * *
5.
75000
83 49,3 72,1 16,3 55,4 56 114 34,9 * 88,4 65 127 75,3 23,7 36,4 82 45,6 123 37,4 55,6
0
93 80,7 * 20,2 54,1 56 95,4 29,6 74,5 29,8 65 143 44,3 89,9 32,1 82 159 * 30,7 27,8 500
93 40,7 37,3 35,1 37,8 65 92,2 66,3 52,7 * 82 53,8 44,9 92,4 45,9 93 47,4 39,9 34,0 22,9 5000
56 105 62,5 * 39,9 56 72,7 41,7 * 44,4 65 87,2 61,7 * 65,9 82 90,6 45,1 48,0 28,1
6.
75000
93 47,0 99,1 27,6 16,5
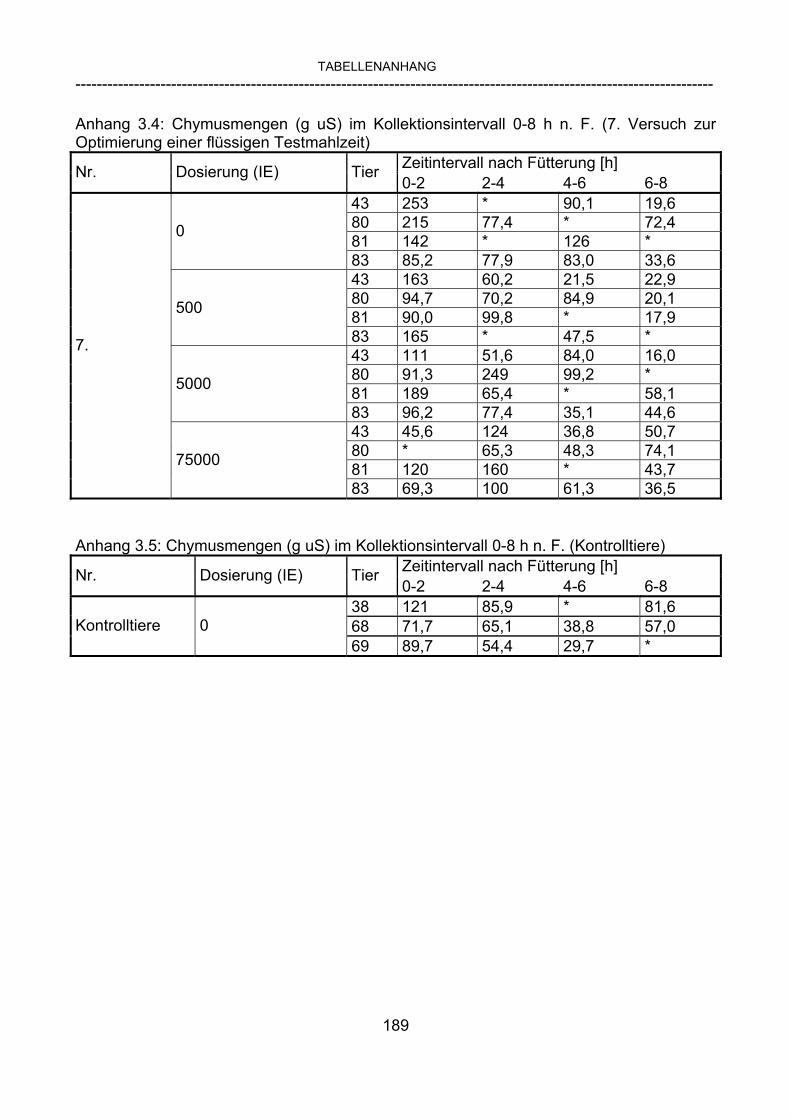
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
189
Anhang 3.4: Chymusmengen (g uS) im Kollektionsintervall 0-8 h n. F. (7. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Zeitintervall nach Fütterung [h] Nr. Dosierung (IE) Tier 0-2 2-4 4-6 6-8
43 253 * 90,1 19,6 80 215 77,4 * 72,4 81 142 * 126 * 0
83 85,2 77,9 83,0 33,6 43 163 60,2 21,5 22,9 80 94,7 70,2 84,9 20,1 81 90,0 99,8 * 17,9 500
83 165 * 47,5 * 43 111 51,6 84,0 16,0 80 91,3 249 99,2 * 81 189 65,4 * 58,1 5000
83 96,2 77,4 35,1 44,6 43 45,6 124 36,8 50,7 80 * 65,3 48,3 74,1 81 120 160 * 43,7
7.
75000
83 69,3 100 61,3 36,5 Anhang 3.5: Chymusmengen (g uS) im Kollektionsintervall 0-8 h n. F. (Kontrolltiere)
Zeitintervall nach Fütterung [h] Nr. Dosierung (IE) Tier 0-2 2-4 4-6 6-8
38 121 85,9 * 81,6 68 71,7 65,1 38,8 57,0 Kontrolltiere 0 69 89,7 54,4 29,7 *
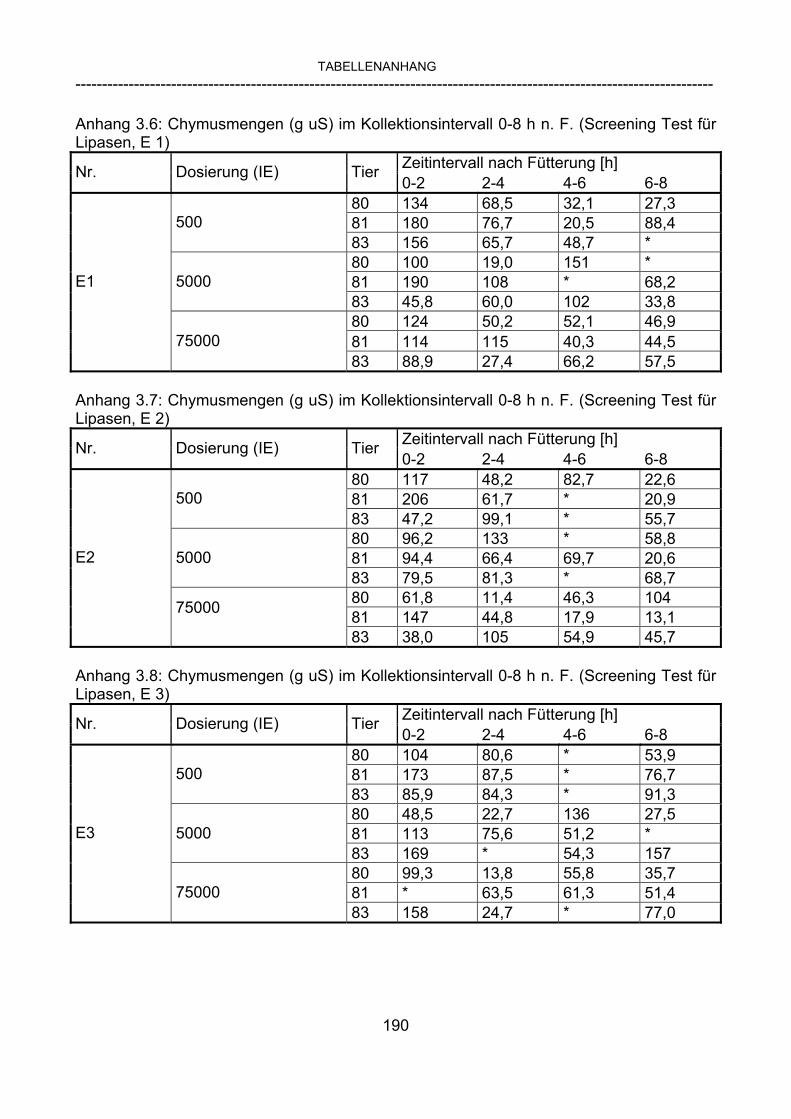
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
190
Anhang 3.6: Chymusmengen (g uS) im Kollektionsintervall 0-8 h n. F. (Screening Test für Lipasen, E 1)
Zeitintervall nach Fütterung [h] Nr. Dosierung (IE) Tier 0-2 2-4 4-6 6-8
80 134 68,5 32,1 27,3 81 180 76,7 20,5 88,4 500 83 156 65,7 48,7 * 80 100 19,0 151 * 81 190 108 * 68,2 5000 83 45,8 60,0 102 33,8 80 124 50,2 52,1 46,9 81 114 115 40,3 44,5
E1
75000 83 88,9 27,4 66,2 57,5
Anhang 3.7: Chymusmengen (g uS) im Kollektionsintervall 0-8 h n. F. (Screening Test für Lipasen, E 2)
Zeitintervall nach Fütterung [h] Nr. Dosierung (IE) Tier 0-2 2-4 4-6 6-8
80 117 48,2 82,7 22,6 81 206 61,7 * 20,9 500 83 47,2 99,1 * 55,7 80 96,2 133 * 58,8 81 94,4 66,4 69,7 20,6 5000 83 79,5 81,3 * 68,7 80 61,8 11,4 46,3 104 81 147 44,8 17,9 13,1
E2
75000
83 38,0 105 54,9 45,7 Anhang 3.8: Chymusmengen (g uS) im Kollektionsintervall 0-8 h n. F. (Screening Test für Lipasen, E 3)
Zeitintervall nach Fütterung [h] Nr. Dosierung (IE) Tier 0-2 2-4 4-6 6-8
80 104 80,6 * 53,9 81 173 87,5 * 76,7 500 83 85,9 84,3 * 91,3 80 48,5 22,7 136 27,5 81 113 75,6 51,2 * 5000 83 169 * 54,3 157 80 99,3 13,8 55,8 35,7 81 * 63,5 61,3 51,4
E3
75000 83 158 24,7 * 77,0
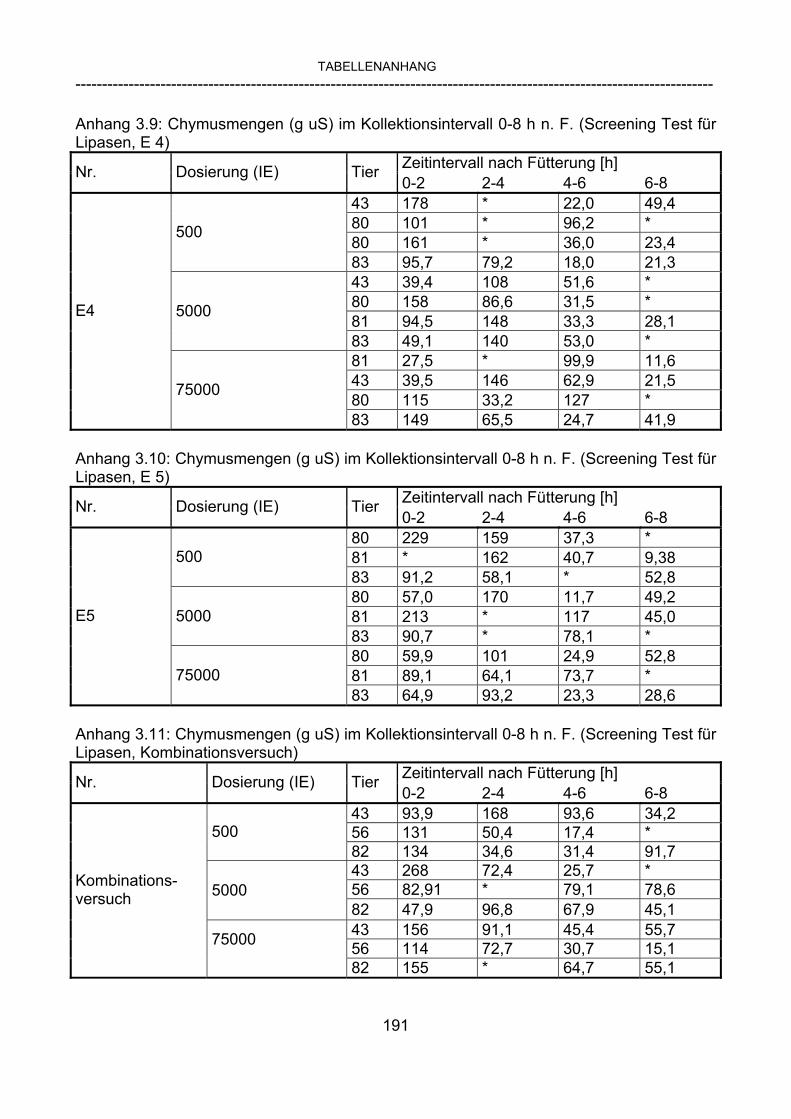
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
191
Anhang 3.9: Chymusmengen (g uS) im Kollektionsintervall 0-8 h n. F. (Screening Test für Lipasen, E 4)
Zeitintervall nach Fütterung [h] Nr. Dosierung (IE) Tier 0-2 2-4 4-6 6-8
43 178 * 22,0 49,4 80 101 * 96,2 * 80 161 * 36,0 23,4
500
83 95,7 79,2 18,0 21,3 43 39,4 108 51,6 * 80 158 86,6 31,5 * 81 94,5 148 33,3 28,1 5000
83 49,1 140 53,0 * 81 27,5 * 99,9 11,6 43 39,5 146 62,9 21,5 80 115 33,2 127 *
E4
75000
83 149 65,5 24,7 41,9 Anhang 3.10: Chymusmengen (g uS) im Kollektionsintervall 0-8 h n. F. (Screening Test für Lipasen, E 5)
Zeitintervall nach Fütterung [h] Nr. Dosierung (IE) Tier 0-2 2-4 4-6 6-8
80 229 159 37,3 * 81 * 162 40,7 9,38 500 83 91,2 58,1 * 52,8 80 57,0 170 11,7 49,2 81 213 * 117 45,0 5000 83 90,7 * 78,1 * 80 59,9 101 24,9 52,8 81 89,1 64,1 73,7 *
E5
75000 83 64,9 93,2 23,3 28,6
Anhang 3.11: Chymusmengen (g uS) im Kollektionsintervall 0-8 h n. F. (Screening Test für Lipasen, Kombinationsversuch)
Zeitintervall nach Fütterung [h] Nr. Dosierung (IE) Tier 0-2 2-4 4-6 6-8
43 93,9 168 93,6 34,2 56 131 50,4 17,4 * 500 82 134 34,6 31,4 91,7 43 268 72,4 25,7 * 56 82,91 * 79,1 78,6 5000 82 47,9 96,8 67,9 45,1 43 156 91,1 45,4 55,7 56 114 72,7 30,7 15,1
Kombinations-versuch
75000 82 155 * 64,7 55,1
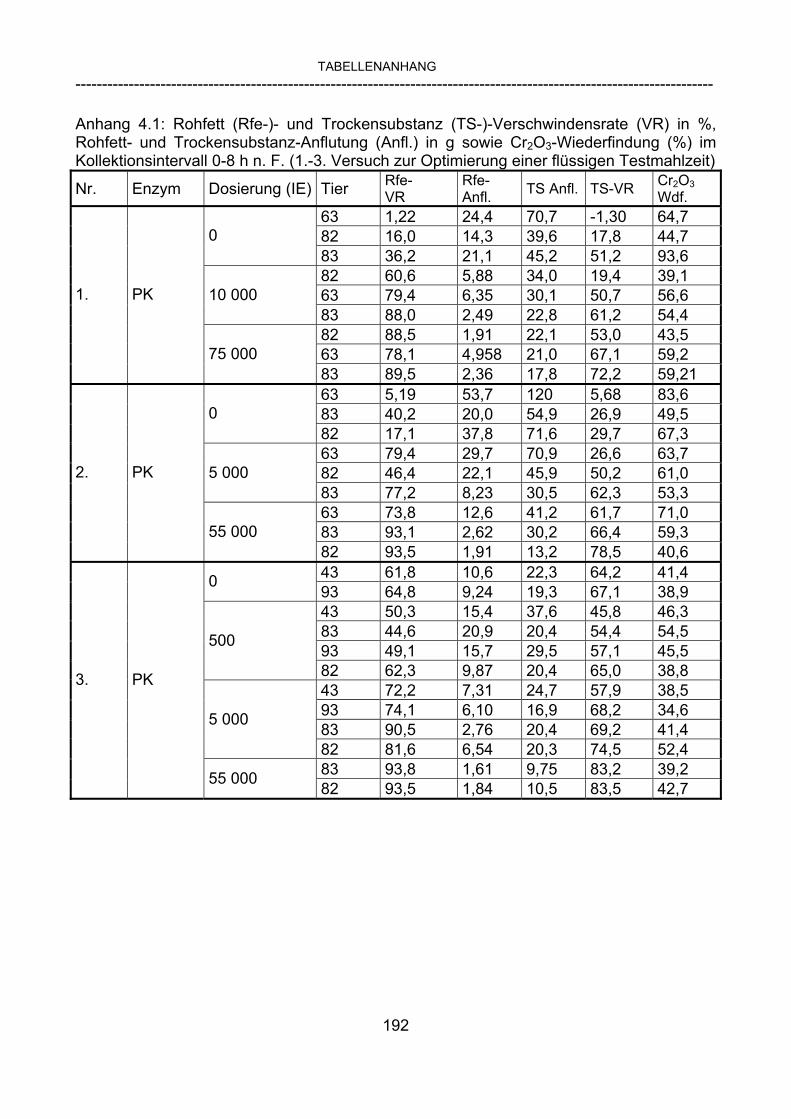
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
192
Anhang 4.1: Rohfett (Rfe-)- und Trockensubstanz (TS-)-Verschwindensrate (VR) in %, Rohfett- und Trockensubstanz-Anflutung (Anfl.) in g sowie Cr2O3-Wiederfindung (%) im Kollektionsintervall 0-8 h n. F. (1.-3. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Nr. Enzym Dosierung (IE) Tier Rfe- VR
Rfe-Anfl. TS Anfl. TS-VR Cr2O3
Wdf. 63 1,22 24,4 70,7 -1,30 64,7 82 16,0 14,3 39,6 17,8 44,7 0 83 36,2 21,1 45,2 51,2 93,6 82 60,6 5,88 34,0 19,4 39,1 63 79,4 6,35 30,1 50,7 56,6 10 000 83 88,0 2,49 22,8 61,2 54,4 82 88,5 1,91 22,1 53,0 43,5 63 78,1 4,958 21,0 67,1 59,2
1. PK
75 000 83 89,5 2,36 17,8 72,2 59,21 63 5,19 53,7 120 5,68 83,6 83 40,2 20,0 54,9 26,9 49,5 0 82 17,1 37,8 71,6 29,7 67,3 63 79,4 29,7 70,9 26,6 63,7 82 46,4 22,1 45,9 50,2 61,0 5 000 83 77,2 8,23 30,5 62,3 53,3 63 73,8 12,6 41,2 61,7 71,0 83 93,1 2,62 30,2 66,4 59,3
2. PK
55 000 82 93,5 1,91 13,2 78,5 40,6 43 61,8 10,6 22,3 64,2 41,4 0 93 64,8 9,24 19,3 67,1 38,9 43 50,3 15,4 37,6 45,8 46,3 83 44,6 20,9 20,4 54,4 54,5 93 49,1 15,7 29,5 57,1 45,5 500
82 62,3 9,87 20,4 65,0 38,8 43 72,2 7,31 24,7 57,9 38,5 93 74,1 6,10 16,9 68,2 34,6 83 90,5 2,76 20,4 69,2 41,4 5 000
82 81,6 6,54 20,3 74,5 52,4 83 93,8 1,61 9,75 83,2 39,2
3. PK
55 000 82 93,5 1,84 10,5 83,5 42,7
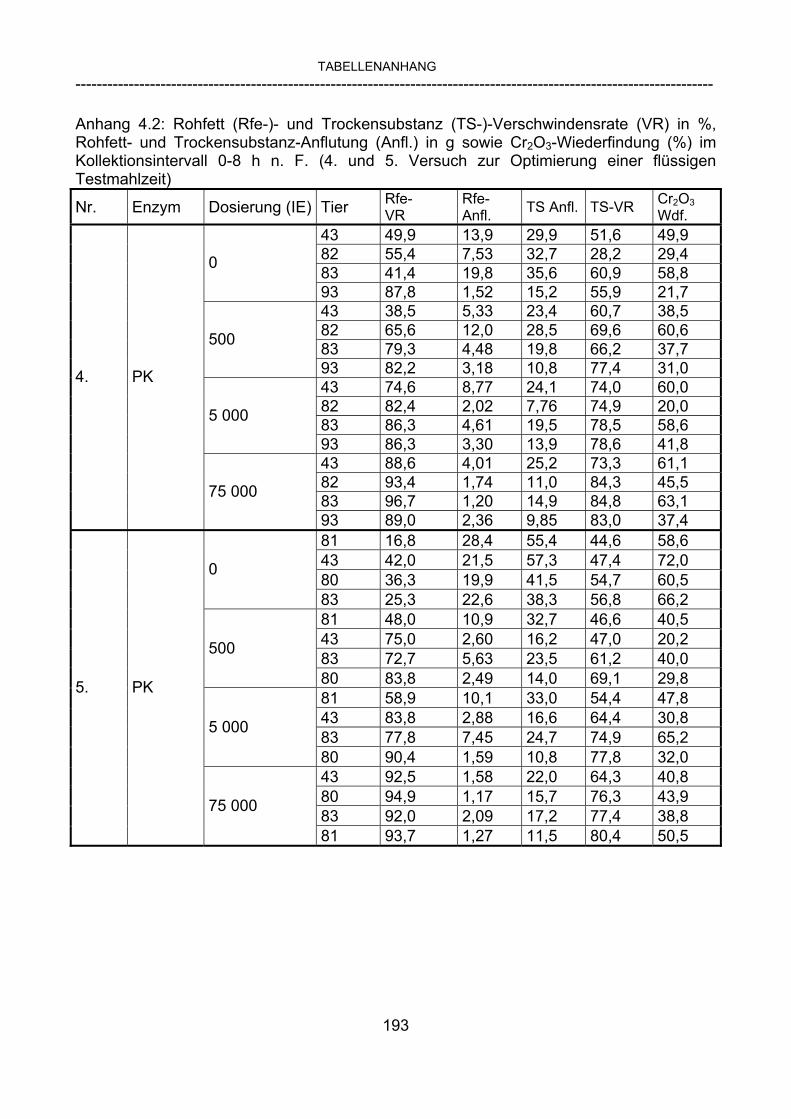
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
193
Anhang 4.2: Rohfett (Rfe-)- und Trockensubstanz (TS-)-Verschwindensrate (VR) in %, Rohfett- und Trockensubstanz-Anflutung (Anfl.) in g sowie Cr2O3-Wiederfindung (%) im Kollektionsintervall 0-8 h n. F. (4. und 5. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Nr. Enzym Dosierung (IE) Tier Rfe- VR
Rfe-Anfl. TS Anfl. TS-VR Cr2O3
Wdf. 43 49,9 13,9 29,9 51,6 49,9 82 55,4 7,53 32,7 28,2 29,4 83 41,4 19,8 35,6 60,9 58,8 0
93 87,8 1,52 15,2 55,9 21,7 43 38,5 5,33 23,4 60,7 38,5 82 65,6 12,0 28,5 69,6 60,6 83 79,3 4,48 19,8 66,2 37,7 500
93 82,2 3,18 10,8 77,4 31,0 43 74,6 8,77 24,1 74,0 60,0 82 82,4 2,02 7,76 74,9 20,0 83 86,3 4,61 19,5 78,5 58,6 5 000
93 86,3 3,30 13,9 78,6 41,8 43 88,6 4,01 25,2 73,3 61,1 82 93,4 1,74 11,0 84,3 45,5 83 96,7 1,20 14,9 84,8 63,1
4. PK
75 000
93 89,0 2,36 9,85 83,0 37,4 81 16,8 28,4 55,4 44,6 58,6 43 42,0 21,5 57,3 47,4 72,0 80 36,3 19,9 41,5 54,7 60,5
0
83 25,3 22,6 38,3 56,8 66,2 81 48,0 10,9 32,7 46,6 40,5 43 75,0 2,60 16,2 47,0 20,2 83 72,7 5,63 23,5 61,2 40,0 500
80 83,8 2,49 14,0 69,1 29,8 81 58,9 10,1 33,0 54,4 47,8 43 83,8 2,88 16,6 64,4 30,8 83 77,8 7,45 24,7 74,9 65,2 5 000
80 90,4 1,59 10,8 77,8 32,0 43 92,5 1,58 22,0 64,3 40,8 80 94,9 1,17 15,7 76,3 43,9 83 92,0 2,09 17,2 77,4 38,8
5. PK
75 000
81 93,7 1,27 11,5 80,4 50,5
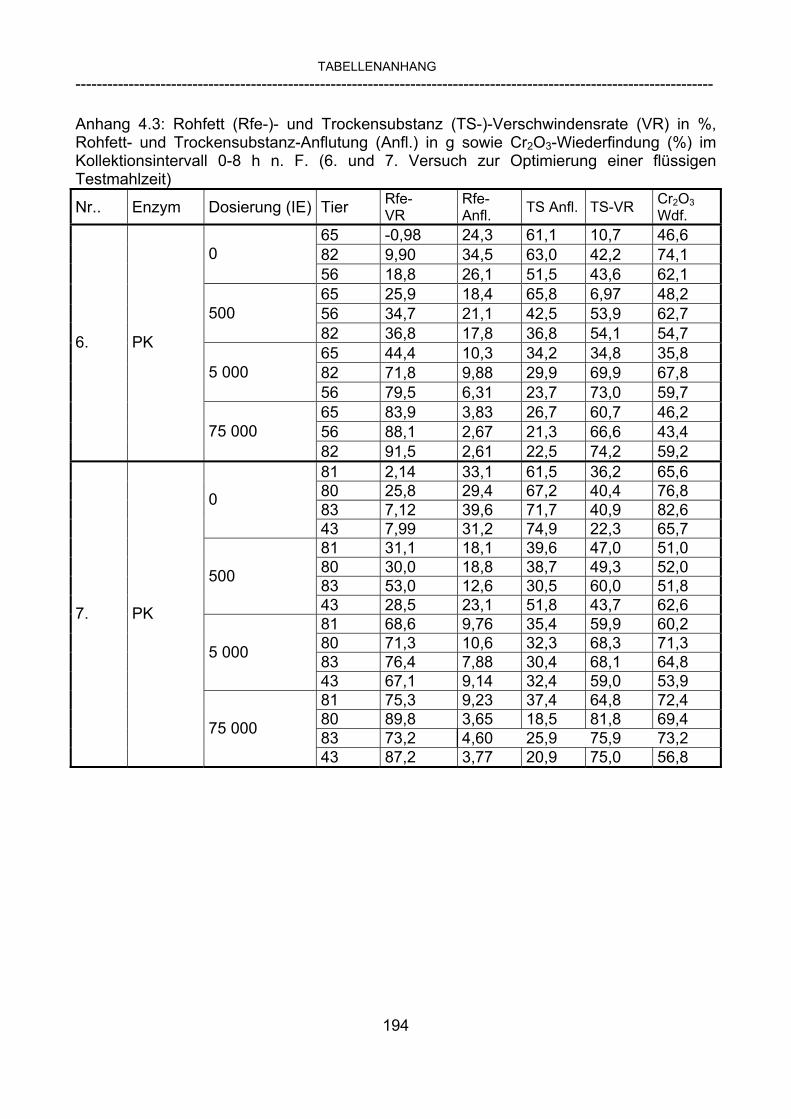
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
194
Anhang 4.3: Rohfett (Rfe-)- und Trockensubstanz (TS-)-Verschwindensrate (VR) in %, Rohfett- und Trockensubstanz-Anflutung (Anfl.) in g sowie Cr2O3-Wiederfindung (%) im Kollektionsintervall 0-8 h n. F. (6. und 7. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Nr.. Enzym Dosierung (IE) Tier Rfe- VR
Rfe-Anfl. TS Anfl. TS-VR Cr2O3
Wdf. 65 -0,98 24,3 61,1 10,7 46,6 82 9,90 34,5 63,0 42,2 74,1 0 56 18,8 26,1 51,5 43,6 62,1 65 25,9 18,4 65,8 6,97 48,2 56 34,7 21,1 42,5 53,9 62,7 500 82 36,8 17,8 36,8 54,1 54,7 65 44,4 10,3 34,2 34,8 35,8 82 71,8 9,88 29,9 69,9 67,8 5 000 56 79,5 6,31 23,7 73,0 59,7 65 83,9 3,83 26,7 60,7 46,2 56 88,1 2,67 21,3 66,6 43,4
6. PK
75 000 82 91,5 2,61 22,5 74,2 59,2 81 2,14 33,1 61,5 36,2 65,6 80 25,8 29,4 67,2 40,4 76,8 83 7,12 39,6 71,7 40,9 82,6 0
43 7,99 31,2 74,9 22,3 65,7 81 31,1 18,1 39,6 47,0 51,0 80 30,0 18,8 38,7 49,3 52,0 83 53,0 12,6 30,5 60,0 51,8 500
43 28,5 23,1 51,8 43,7 62,6 81 68,6 9,76 35,4 59,9 60,2 80 71,3 10,6 32,3 68,3 71,3 83 76,4 7,88 30,4 68,1 64,8 5 000
43 67,1 9,14 32,4 59,0 53,9 81 75,3 9,23 37,4 64,8 72,4 80 89,8 3,65 18,5 81,8 69,4 83 73,2 4,60 25,9 75,9 73,2
7. PK
75 000
43 87,2 3,77 20,9 75,0 56,8
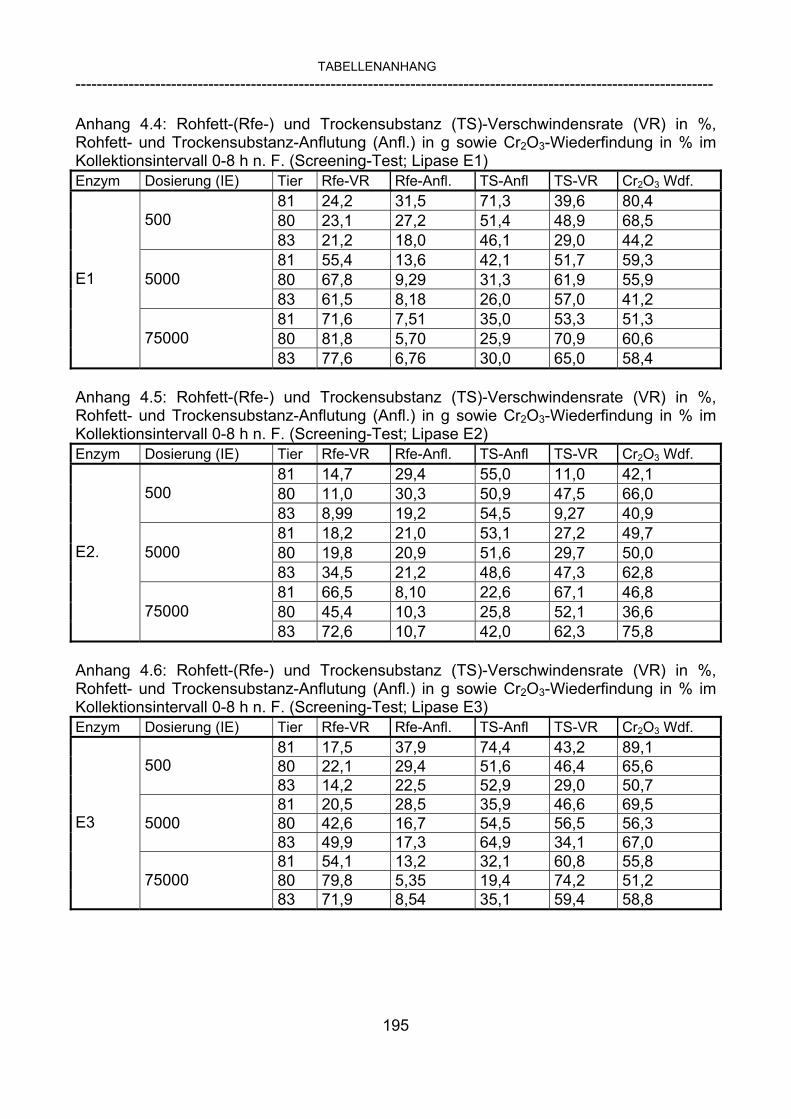
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
195
Anhang 4.4: Rohfett-(Rfe-) und Trockensubstanz (TS)-Verschwindensrate (VR) in %, Rohfett- und Trockensubstanz-Anflutung (Anfl.) in g sowie Cr2O3-Wiederfindung in % im Kollektionsintervall 0-8 h n. F. (Screening-Test; Lipase E1) Enzym Dosierung (IE) Tier Rfe-VR Rfe-Anfl. TS-Anfl TS-VR Cr2O3 Wdf.
81 24,2 31,5 71,3 39,6 80,4 80 23,1 27,2 51,4 48,9 68,5 500 83 21,2 18,0 46,1 29,0 44,2 81 55,4 13,6 42,1 51,7 59,3 80 67,8 9,29 31,3 61,9 55,9 5000 83 61,5 8,18 26,0 57,0 41,2 81 71,6 7,51 35,0 53,3 51,3 80 81,8 5,70 25,9 70,9 60,6
E1
75000 83 77,6 6,76 30,0 65,0 58,4
Anhang 4.5: Rohfett-(Rfe-) und Trockensubstanz (TS)-Verschwindensrate (VR) in %, Rohfett- und Trockensubstanz-Anflutung (Anfl.) in g sowie Cr2O3-Wiederfindung in % im Kollektionsintervall 0-8 h n. F. (Screening-Test; Lipase E2) Enzym Dosierung (IE) Tier Rfe-VR Rfe-Anfl. TS-Anfl TS-VR Cr2O3 Wdf.
81 14,7 29,4 55,0 11,0 42,1 80 11,0 30,3 50,9 47,5 66,0 500 83 8,99 19,2 54,5 9,27 40,9 81 18,2 21,0 53,1 27,2 49,7 80 19,8 20,9 51,6 29,7 50,0 5000 83 34,5 21,2 48,6 47,3 62,8 81 66,5 8,10 22,6 67,1 46,8 80 45,4 10,3 25,8 52,1 36,6
E2.
75000 83 72,6 10,7 42,0 62,3 75,8
Anhang 4.6: Rohfett-(Rfe-) und Trockensubstanz (TS)-Verschwindensrate (VR) in %, Rohfett- und Trockensubstanz-Anflutung (Anfl.) in g sowie Cr2O3-Wiederfindung in % im Kollektionsintervall 0-8 h n. F. (Screening-Test; Lipase E3) Enzym Dosierung (IE) Tier Rfe-VR Rfe-Anfl. TS-Anfl TS-VR Cr2O3 Wdf.
81 17,5 37,9 74,4 43,2 89,1 80 22,1 29,4 51,6 46,4 65,6 500 83 14,2 22,5 52,9 29,0 50,7 81 20,5 28,5 35,9 46,6 69,5 80 42,6 16,7 54,5 56,5 56,3 5000 83 49,9 17,3 64,9 34,1 67,0 81 54,1 13,2 32,1 60,8 55,8 80 79,8 5,35 19,4 74,2 51,2
E3
75000 83 71,9 8,54 35,1 59,4 58,8
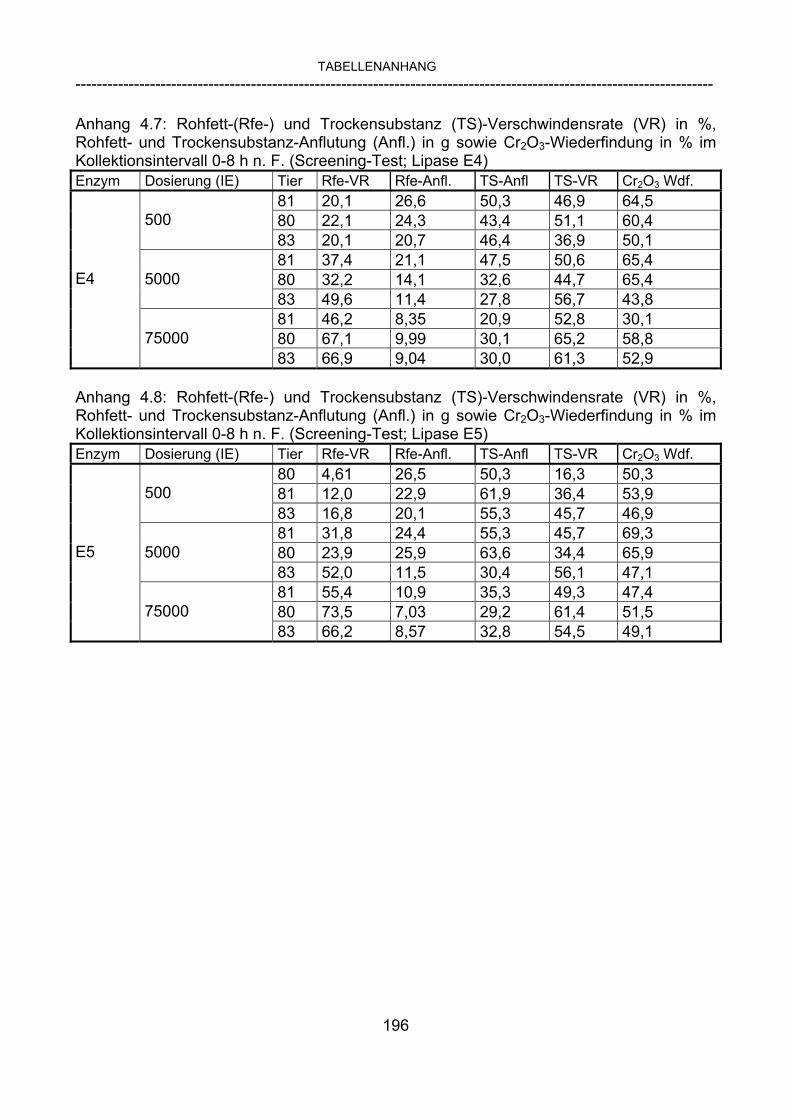
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
196
Anhang 4.7: Rohfett-(Rfe-) und Trockensubstanz (TS)-Verschwindensrate (VR) in %, Rohfett- und Trockensubstanz-Anflutung (Anfl.) in g sowie Cr2O3-Wiederfindung in % im Kollektionsintervall 0-8 h n. F. (Screening-Test; Lipase E4) Enzym Dosierung (IE) Tier Rfe-VR Rfe-Anfl. TS-Anfl TS-VR Cr2O3 Wdf.
81 20,1 26,6 50,3 46,9 64,5 80 22,1 24,3 43,4 51,1 60,4 500 83 20,1 20,7 46,4 36,9 50,1 81 37,4 21,1 47,5 50,6 65,4 80 32,2 14,1 32,6 44,7 65,4 5000 83 49,6 11,4 27,8 56,7 43,8 81 46,2 8,35 20,9 52,8 30,1 80 67,1 9,99 30,1 65,2 58,8
E4
75000 83 66,9 9,04 30,0 61,3 52,9
Anhang 4.8: Rohfett-(Rfe-) und Trockensubstanz (TS)-Verschwindensrate (VR) in %, Rohfett- und Trockensubstanz-Anflutung (Anfl.) in g sowie Cr2O3-Wiederfindung in % im Kollektionsintervall 0-8 h n. F. (Screening-Test; Lipase E5) Enzym Dosierung (IE) Tier Rfe-VR Rfe-Anfl. TS-Anfl TS-VR Cr2O3 Wdf.
80 4,61 26,5 50,3 16,3 50,3 81 12,0 22,9 61,9 36,4 53,9 500 83 16,8 20,1 55,3 45,7 46,9 81 31,8 24,4 55,3 45,7 69,3 80 23,9 25,9 63,6 34,4 65,9 5000 83 52,0 11,5 30,4 56,1 47,1 81 55,4 10,9 35,3 49,3 47,4 80 73,5 7,03 29,2 61,4 51,5
E5
75000 83 66,2 8,57 32,8 54,5 49,1
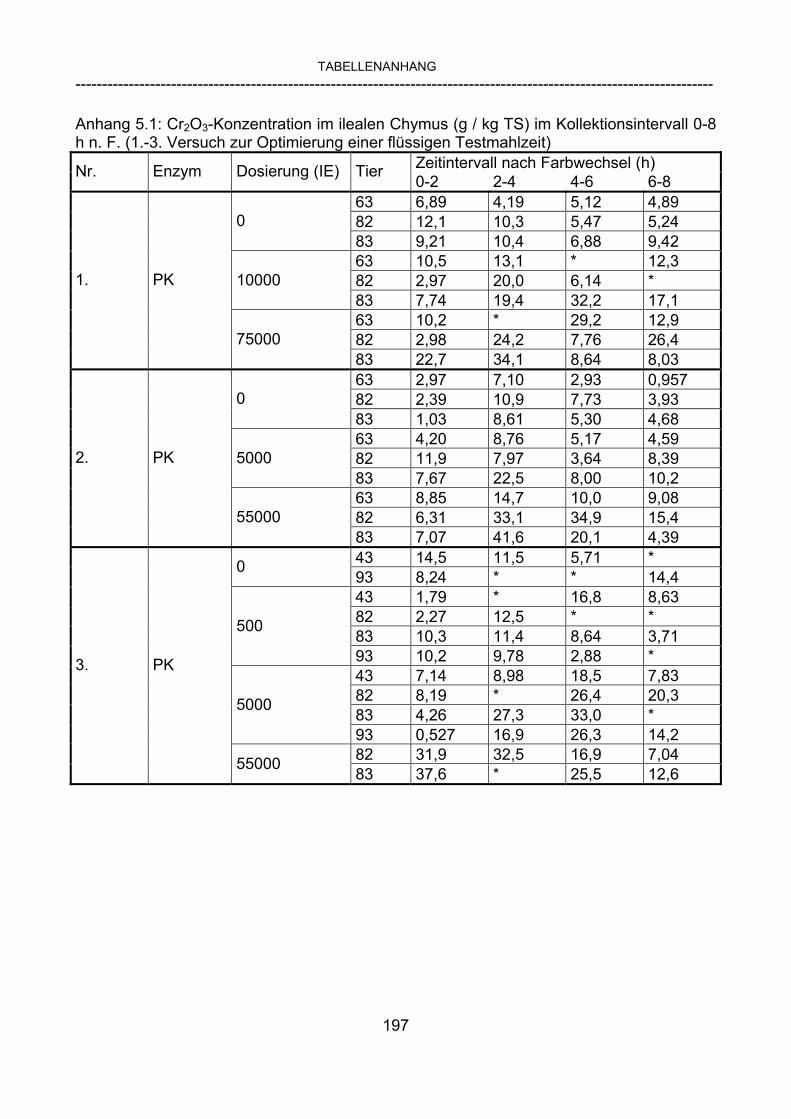
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
197
Anhang 5.1: Cr2O3-Konzentration im ilealen Chymus (g / kg TS) im Kollektionsintervall 0-8 h n. F. (1.-3. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Zeitintervall nach Farbwechsel (h) Nr. Enzym Dosierung (IE) Tier 0-2 2-4 4-6 6-8 63 6,89 4,19 5,12 4,89 82 12,1 10,3 5,47 5,24 0 83 9,21 10,4 6,88 9,42 63 10,5 13,1 * 12,3 82 2,97 20,0 6,14 * 10000 83 7,74 19,4 32,2 17,1 63 10,2 * 29,2 12,9 82 2,98 24,2 7,76 26,4
1. PK
75000 83 22,7 34,1 8,64 8,03 63 2,97 7,10 2,93 0,957 82 2,39 10,9 7,73 3,93 0 83 1,03 8,61 5,30 4,68 63 4,20 8,76 5,17 4,59 82 11,9 7,97 3,64 8,39 5000 83 7,67 22,5 8,00 10,2 63 8,85 14,7 10,0 9,08 82 6,31 33,1 34,9 15,4
2. PK
55000 83 7,07 41,6 20,1 4,39 43 14,5 11,5 5,71 * 0 93 8,24 * * 14,4 43 1,79 * 16,8 8,63 82 2,27 12,5 * * 83 10,3 11,4 8,64 3,71 500
93 10,2 9,78 2,88 * 43 7,14 8,98 18,5 7,83 82 8,19 * 26,4 20,3 83 4,26 27,3 33,0 * 5000
93 0,527 16,9 26,3 14,2 82 31,9 32,5 16,9 7,04
3. PK
55000 83 37,6 * 25,5 12,6
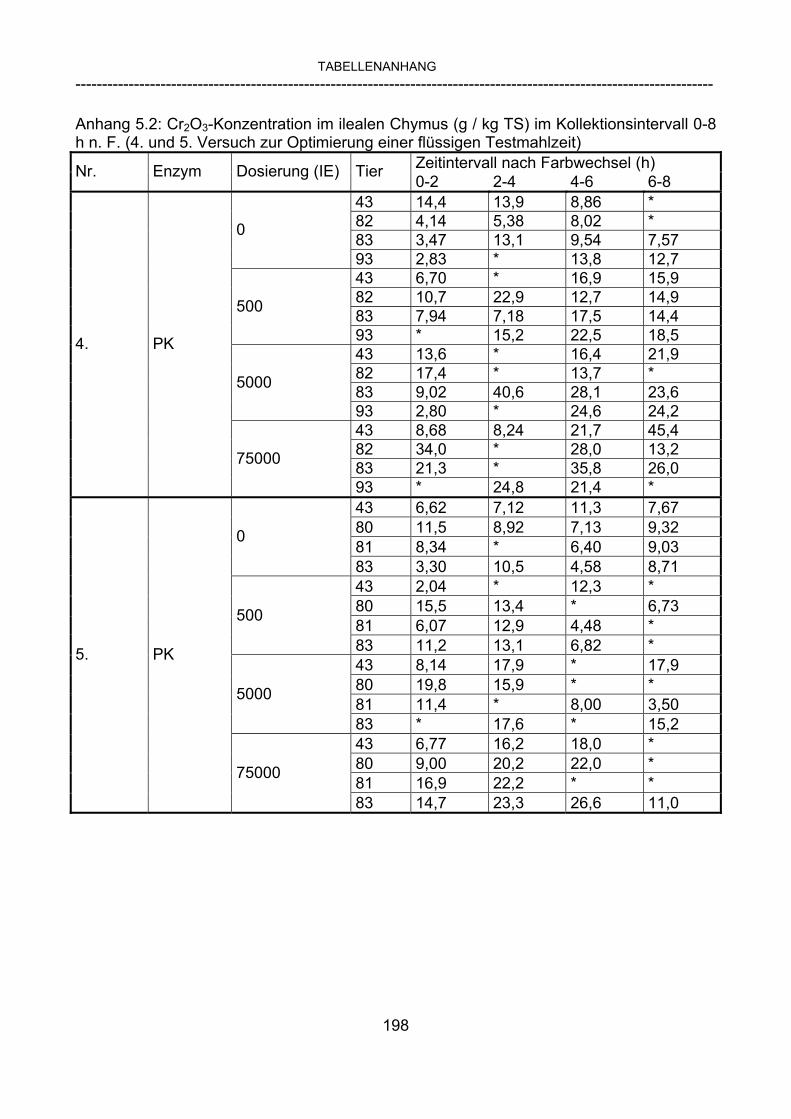
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
198
Anhang 5.2: Cr2O3-Konzentration im ilealen Chymus (g / kg TS) im Kollektionsintervall 0-8 h n. F. (4. und 5. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Zeitintervall nach Farbwechsel (h) Nr. Enzym Dosierung (IE) Tier 0-2 2-4 4-6 6-8 43 14,4 13,9 8,86 * 82 4,14 5,38 8,02 * 83 3,47 13,1 9,54 7,57 0
93 2,83 * 13,8 12,7 43 6,70 * 16,9 15,9 82 10,7 22,9 12,7 14,9 83 7,94 7,18 17,5 14,4 500
93 * 15,2 22,5 18,5 43 13,6 * 16,4 21,9 82 17,4 * 13,7 * 83 9,02 40,6 28,1 23,6 5000
93 2,80 * 24,6 24,2 43 8,68 8,24 21,7 45,4 82 34,0 * 28,0 13,2 83 21,3 * 35,8 26,0
4. PK
75000
93 * 24,8 21,4 * 43 6,62 7,12 11,3 7,67 80 11,5 8,92 7,13 9,32 81 8,34 * 6,40 9,03
0
83 3,30 10,5 4,58 8,71 43 2,04 * 12,3 * 80 15,5 13,4 * 6,73 81 6,07 12,9 4,48 * 500
83 11,2 13,1 6,82 * 43 8,14 17,9 * 17,9 80 19,8 15,9 * * 81 11,4 * 8,00 3,50 5000
83 * 17,6 * 15,2 43 6,77 16,2 18,0 * 80 9,00 20,2 22,0 * 81 16,9 22,2 * *
5. PK
75000
83 14,7 23,3 26,6 11,0
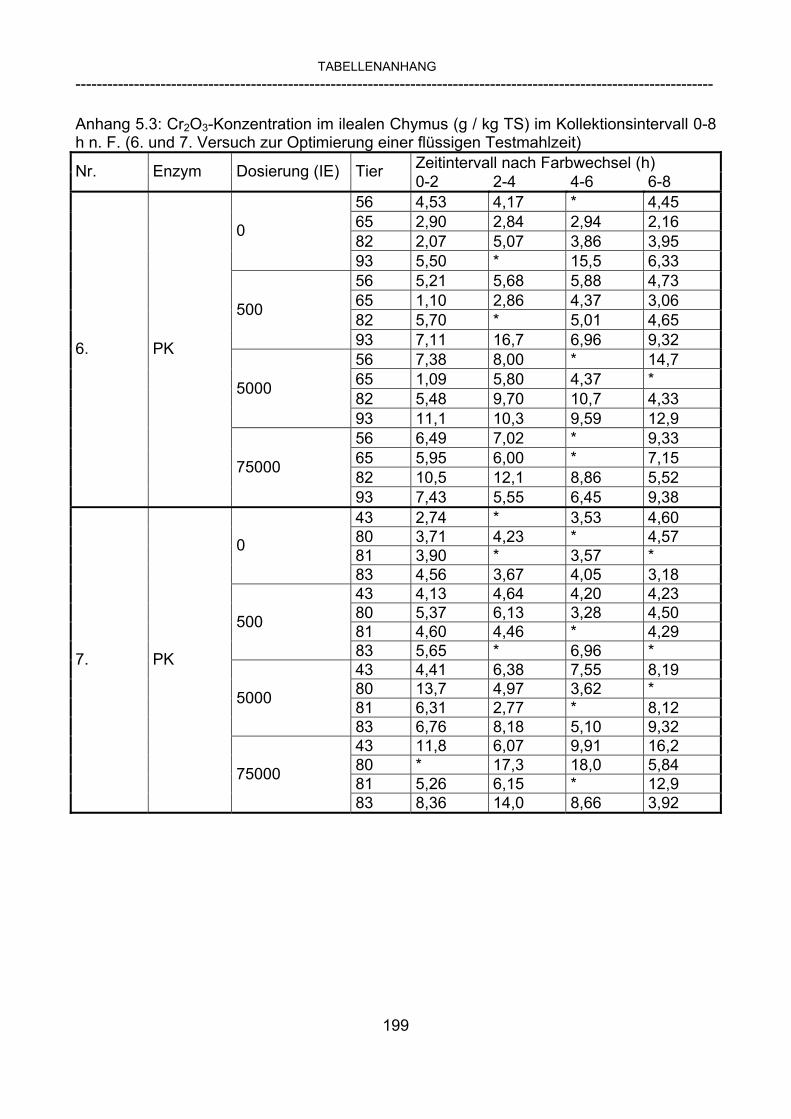
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
199
Anhang 5.3: Cr2O3-Konzentration im ilealen Chymus (g / kg TS) im Kollektionsintervall 0-8 h n. F. (6. und 7. Versuch zur Optimierung einer flüssigen Testmahlzeit)
Zeitintervall nach Farbwechsel (h) Nr. Enzym Dosierung (IE) Tier 0-2 2-4 4-6 6-8 56 4,53 4,17 * 4,45 65 2,90 2,84 2,94 2,16 82 2,07 5,07 3,86 3,95
0
93 5,50 * 15,5 6,33 56 5,21 5,68 5,88 4,73 65 1,10 2,86 4,37 3,06 82 5,70 * 5,01 4,65 500
93 7,11 16,7 6,96 9,32 56 7,38 8,00 * 14,7 65 1,09 5,80 4,37 * 82 5,48 9,70 10,7 4,33 5000
93 11,1 10,3 9,59 12,9 56 6,49 7,02 * 9,33 65 5,95 6,00 * 7,15 82 10,5 12,1 8,86 5,52
6. PK
75000
93 7,43 5,55 6,45 9,38 43 2,74 * 3,53 4,60 80 3,71 4,23 * 4,57 81 3,90 * 3,57 * 0
83 4,56 3,67 4,05 3,18 43 4,13 4,64 4,20 4,23 80 5,37 6,13 3,28 4,50 81 4,60 4,46 * 4,29 500
83 5,65 * 6,96 * 43 4,41 6,38 7,55 8,19 80 13,7 4,97 3,62 * 81 6,31 2,77 * 8,12 5000
83 6,76 8,18 5,10 9,32 43 11,8 6,07 9,91 16,2 80 * 17,3 18,0 5,84 81 5,26 6,15 * 12,9
7. PK
75000
83 8,36 14,0 8,66 3,92
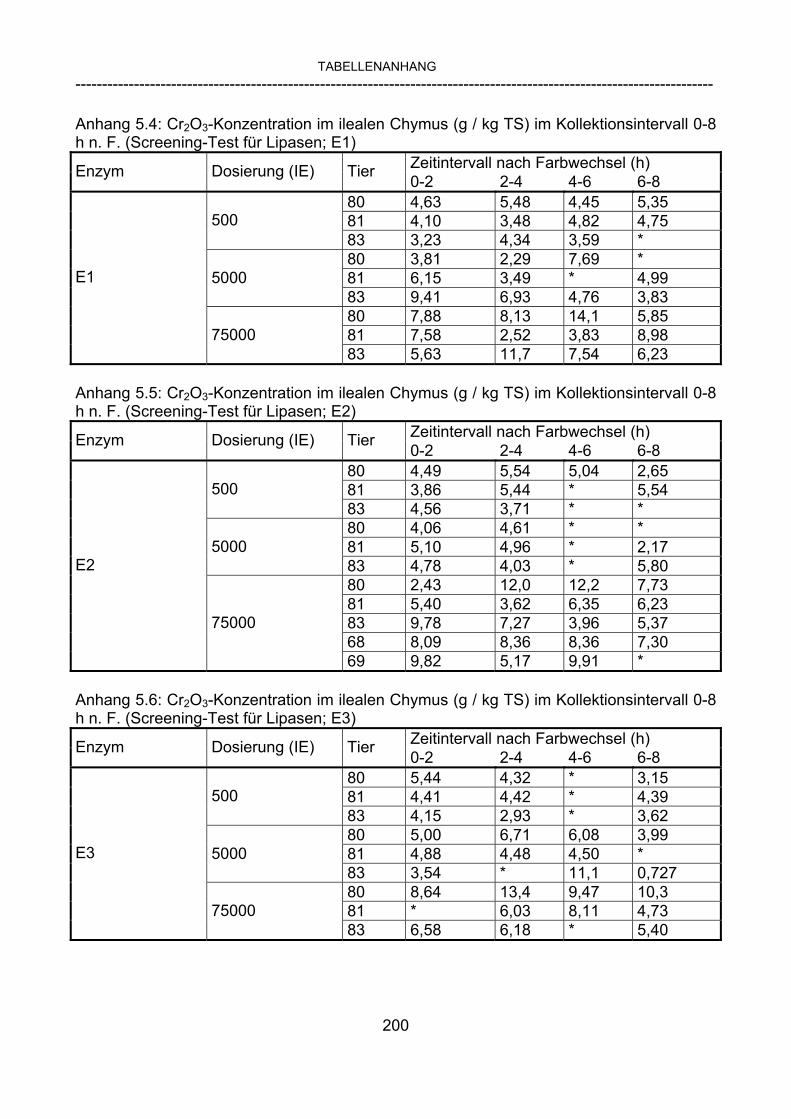
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
200
Anhang 5.4: Cr2O3-Konzentration im ilealen Chymus (g / kg TS) im Kollektionsintervall 0-8 h n. F. (Screening-Test für Lipasen; E1)
Zeitintervall nach Farbwechsel (h) Enzym Dosierung (IE) Tier 0-2 2-4 4-6 6-8 80 4,63 5,48 4,45 5,35 81 4,10 3,48 4,82 4,75 500 83 3,23 4,34 3,59 * 80 3,81 2,29 7,69 * 81 6,15 3,49 * 4,99 5000 83 9,41 6,93 4,76 3,83 80 7,88 8,13 14,1 5,85 81 7,58 2,52 3,83 8,98
E1
75000 83 5,63 11,7 7,54 6,23
Anhang 5.5: Cr2O3-Konzentration im ilealen Chymus (g / kg TS) im Kollektionsintervall 0-8 h n. F. (Screening-Test für Lipasen; E2)
Zeitintervall nach Farbwechsel (h) Enzym Dosierung (IE) Tier 0-2 2-4 4-6 6-8 80 4,49 5,54 5,04 2,65 81 3,86 5,44 * 5,54 500 83 4,56 3,71 * * 80 4,06 4,61 * * 81 5,10 4,96 * 2,17 5000 83 4,78 4,03 * 5,80 80 2,43 12,0 12,2 7,73 81 5,40 3,62 6,35 6,23 83 9,78 7,27 3,96 5,37 68 8,09 8,36 8,36 7,30
E2
75000
69 9,82 5,17 9,91 * Anhang 5.6: Cr2O3-Konzentration im ilealen Chymus (g / kg TS) im Kollektionsintervall 0-8 h n. F. (Screening-Test für Lipasen; E3)
Zeitintervall nach Farbwechsel (h) Enzym Dosierung (IE) Tier 0-2 2-4 4-6 6-8 80 5,44 4,32 * 3,15 81 4,41 4,42 * 4,39 500 83 4,15 2,93 * 3,62 80 5,00 6,71 6,08 3,99 81 4,88 4,48 4,50 * 5000 83 3,54 * 11,1 0,727 80 8,64 13,4 9,47 10,3 81 * 6,03 8,11 4,73
E3
75000 83 6,58 6,18 * 5,40
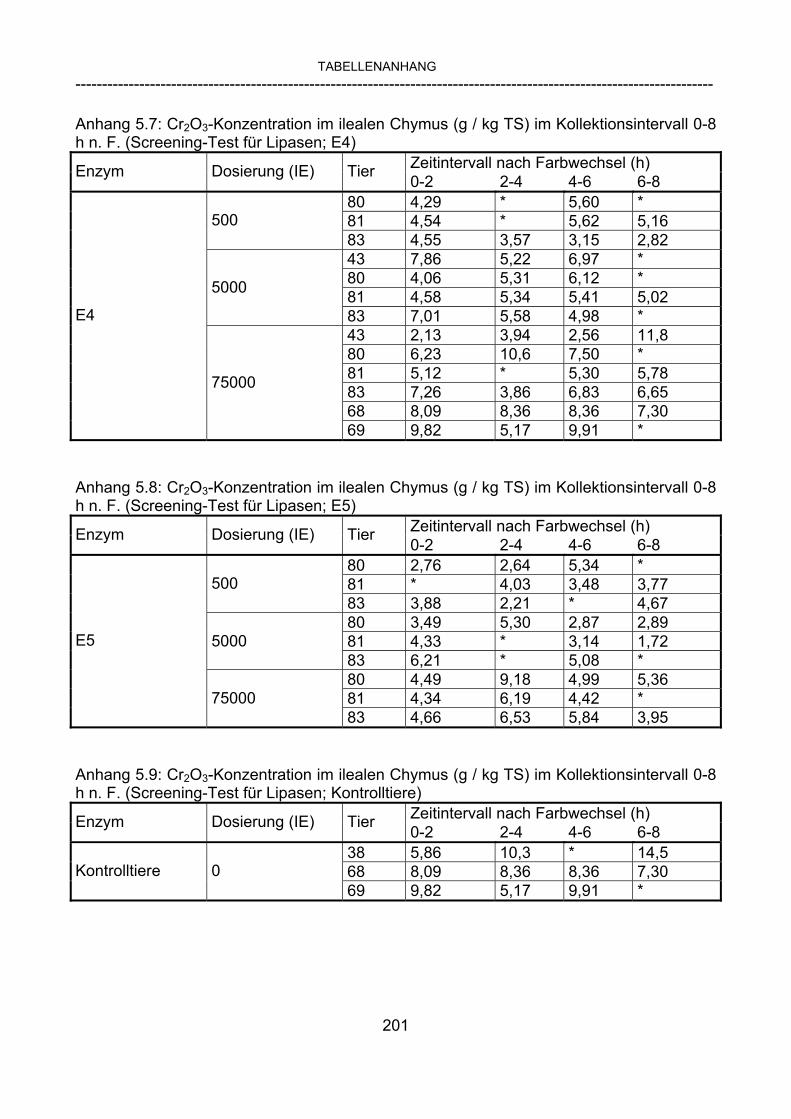
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
201
Anhang 5.7: Cr2O3-Konzentration im ilealen Chymus (g / kg TS) im Kollektionsintervall 0-8 h n. F. (Screening-Test für Lipasen; E4)
Zeitintervall nach Farbwechsel (h) Enzym Dosierung (IE) Tier 0-2 2-4 4-6 6-8 80 4,29 * 5,60 * 81 4,54 * 5,62 5,16 500 83 4,55 3,57 3,15 2,82 43 7,86 5,22 6,97 * 80 4,06 5,31 6,12 * 81 4,58 5,34 5,41 5,02 5000
83 7,01 5,58 4,98 * 43 2,13 3,94 2,56 11,8 80 6,23 10,6 7,50 * 81 5,12 * 5,30 5,78 83 7,26 3,86 6,83 6,65 68 8,09 8,36 8,36 7,30
E4
75000
69 9,82 5,17 9,91 * Anhang 5.8: Cr2O3-Konzentration im ilealen Chymus (g / kg TS) im Kollektionsintervall 0-8 h n. F. (Screening-Test für Lipasen; E5)
Zeitintervall nach Farbwechsel (h) Enzym Dosierung (IE) Tier 0-2 2-4 4-6 6-8 80 2,76 2,64 5,34 * 81 * 4,03 3,48 3,77 500 83 3,88 2,21 * 4,67 80 3,49 5,30 2,87 2,89 81 4,33 * 3,14 1,72 5000 83 6,21 * 5,08 * 80 4,49 9,18 4,99 5,36 81 4,34 6,19 4,42 *
E5
75000 83 4,66 6,53 5,84 3,95
Anhang 5.9: Cr2O3-Konzentration im ilealen Chymus (g / kg TS) im Kollektionsintervall 0-8 h n. F. (Screening-Test für Lipasen; Kontrolltiere)
Zeitintervall nach Farbwechsel (h) Enzym Dosierung (IE) Tier 0-2 2-4 4-6 6-8 38 5,86 10,3 * 14,5 68 8,09 8,36 8,36 7,30 Kontrolltiere 0 69 9,82 5,17 9,91 *
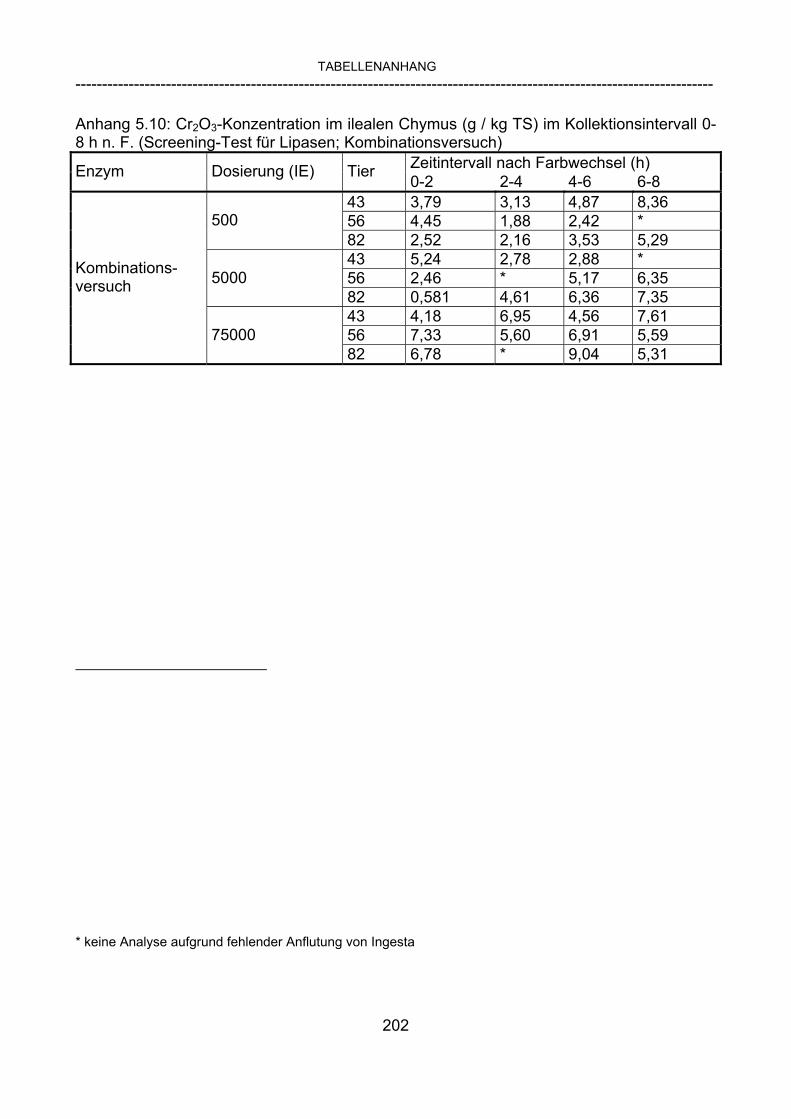
TABELLENANHANG ------------------------------------------------------------------------------------------------------------------------
202
Anhang 5.10: Cr2O3-Konzentration im ilealen Chymus (g / kg TS) im Kollektionsintervall 0-8 h n. F. (Screening-Test für Lipasen; Kombinationsversuch)
Zeitintervall nach Farbwechsel (h) Enzym Dosierung (IE) Tier 0-2 2-4 4-6 6-8 43 3,79 3,13 4,87 8,36 56 4,45 1,88 2,42 * 500 82 2,52 2,16 3,53 5,29 43 5,24 2,78 2,88 * 56 2,46 * 5,17 6,35 5000 82 0,581 4,61 6,36 7,35 43 4,18 6,95 4,56 7,61 56 7,33 5,60 6,91 5,59
Kombinations- versuch
75000 82 6,78 * 9,04 5,31
* keine Analyse aufgrund fehlender Anflutung von Ingesta

DANKSAGUNG ------------------------------------------------------------------------------------------------------------------------
203
Danksagung
Mein besonderer Dank gilt Herrn Prof. Dr. J. Kamphues für die Überlassung des Themas
und die jederzeit gewährte freundliche und konsequente Unterstützung während der
Anfertigung dieser Arbeit.
Dr. Anne Katrin Mößeler möchte ich in freundschaftlicher Verbundenheit für ihre
unermüdliche und uneingeschränkte Hilfsbereitschaft und Unterstützung bei der Erstellung
dieser Arbeit während der letzten zwei Jahre herzlichst danken.
Bei der Firma Solvay Pharmaceutical GmbH bedanke ich mich nicht nur für die materielle
und finanzielle Unterstützung, sondern danke auch vielen Mitarbeitern für ihre freundliche
Hilfsbereitschaft. Besonders bedanke ich mich bei Dr. P. C. Gregory für die kollegiale
Zusammenarbeit. Bei dem „Minipig-Team“ (Petra, Bernd, Bianca, Ille und Aileen) danke
ich für die freundschaftliche Aufnahme, die umfassende und allgegenwärtige
Hilfsbereitschaft sowie das großartige Engagement.
Ein besonderer Dank gilt auch Katrin Kiesmann für die tatkräftige Unterstützung bei der
praktischen Durchführung der Versuche.
Ebenfalls möchte ich mich bei allen Mitarbeitern des Instituts für Tierernährung,
insbesondere bei Jutta und Katja, für die ausdauernde und herzliche Unterstützung
bedanken.
Des Weiteren gilt mein Dank dem Institut für Biometrie für die Beratung und Unterstützung
bei der statistischen Auswertung der Ergebnisse.
Meinen Eltern und meinem Bruder Tanno danke ich herzlichst für die umfassende
Unterstützung der letzten 28 Jahre, die mir letztendlich ermöglichte, diese Arbeit zu
erstellen.
Ganz besonders danke ich auch meinen Freunden, die mich in kleinen und großen Krisen
stets treu begleitet haben und immer für mich da waren.
Robert sei für die rettende Lösung meines Computerproblems gedankt.




























