Embed Size (px)
Citation preview

Auswirkungen des Klimawandels auf den Wald 33.6 Mortalität von Waldbäumen: Ursachen und
TrendsSophia Etzold1, Jan Wunder1, Sabine Braun2, Brigitte Rohner1, Christof Bigler3, Meinrad Abegg1 und Andreas Rigling1
1 Eidg. Forschungsanstalt für Wald, Schnee und Landschaft WSL, Birmensdorf2 Institut für angewandte Pflanzenbiologie IAP, Schönenbuch3 Eidg. Technische Hochschule ETH Zürich, Zürich Korrespondenz: [email protected]
Mortalität oder das Absterben von Bäumen ist ein natürliches Phänomen im Entwick-lungszyklus eines Waldes. Die Gründe für Mortalität sind in der Regel nicht monokausal, sondern durch komplexe Interaktionen von verschiedenen Faktoren und Ereignissen ge-steuert. Auf Schweizer Waldstandorten sterben im langjährigen Mittel pro Jahr 0,1 bis 1,9 Prozent der Bäume. Während der letzten Jahrzehnte lässt sich bei der Mortalität von fünf Hauptbaumarten der Schweiz (Fichte, Föhre, Tanne, Buche, Eiche) kein genereller Trend ableiten. Einzig in den tief gelegenen Föhrenstandorten in den Walliser und Bünd-ner Trockentälern und den Edelkastanienwäldern des Tessins sind einige «Hotspots» mit erhöhter Mortalität als Folge von Trockenheit zu beobachten. Sowohl die zeitliche und räumliche Variabilität von Mortalität auf Bestandesebene als auch der Absterbeprozess von Einzelbäumen lässt sich mit einer Kombination aus endogenen Faktoren wie Topo-grafie, Bestandesstruktur, Baumgrösse, jährlichem Zuwachs oder Schattentoleranz und exogenen Faktoren wie Klimabedingungen erklären. Die Relevanz der einzelnen Faktoren ist dabei je nach Baumart und Standort unterschiedlich. Einige Baumarten (Föhre, Fichte, Tanne, Buche) zeigen erhöhte Mortalität unter trockenen Bedingungen. Der Einfluss von Trockenheit auf die Baummortalität wird dabei durch Interaktionen mit biotischen Fakto-ren, wie beispielsweise Borkenkäferbefall oder Neobiota, zusätzlich verstärkt. Obwohl in der Schweiz und weltweit eine erhöhte Mortalität in gewissen Waldtypen, vor allem auf Trockenstandorten, auftritt, ist es sehr schwierig abzuschätzen, wie sich mit zuneh-mender Klimaveränderung die Mortalitätsraten entwickeln werden. Mit zunehmender Frequenz von Ereignissen wie dem Jahrhundertsommer 2003 ist jedoch mit erhöhter Mortalität zu rechnen.
< Abgestorbener Laubbaum in Hochfelden, ZH. Foto: P. Brang. 177

Mortalität
men. 3) Optimalphase: Der Bestand dünnt weiter aus, und der Deckungsgrad der Oberschicht nimmt ab. Durch den stärkeren Lichteinfall können sich schattentolerante Arten verjüngen. 4) Altbestand/Zerfallsphase: Mit erhöhtem Alter steigt die Mor-talität durch erhöhte Störungsanfälligkeit gegen-über externen Faktoren wie Trockenheit, Pilz- oder Insektenbefall wieder an (goFF und WeSt 1975; lorimer et al. 2001; coomeS und allen 2007; lineS et al. 2010; hurSt et al. 2011). Gleichzeitig profi-tiert der Jungwuchs von den besseren Lichtver-hältnissen und wächst in die Kronenschicht auf. Mit Beginn der Zerfallsphase des Altbestandes wird also die Verjüngungsphase der Folgegene-ration eingeleitet.
Generell ist Mortalität in einem Jungwald kon-kurrenzbedingt sehr hoch, nimmt mit zunehmen-dem Bestandesalter durch Selbstausdünnung ab und im Alter wieder zu (Abb. 3.6.1). In Wäldern, die seit vielen Jahrhunderten nicht bewirtschaftet wur-den, überlappen sich Generationen mit Bäumen al-ler Höhen- und Durchmesserklassen (korPel 1995). Da die Dauer des Entwicklungszyklus baumarten-spezifisch ist, überlagern sich im Mischwald auch die Generationen der einzelnen Baumarten. Das natürliche Alter der Bäume beziehungsweise die Dauer ihres Entwicklungszyklus kann dabei je nach Baumart und Standort mehrere Hundert Jahre um-fassen. Die Produktionszyklen im Wirtschaftswald dauern dagegen meistens nicht länger als 160 bis 200 Jahre und erreichen somit gerade mal die Selbstausdünnungs- oder Optimalphase.
Aus menschlicher Perspektive wird Baummor-talität zum Problem, wenn die Ökosystemfunktio-nen und -leistungen des Waldes wie zum Beispiel die Holzproduktion, Schutzfunktion oder der Was-ser- und Kohlenstoffhaushalt nicht mehr gewähr-leistet sind (anderegg et al. 2013). Mortalität kann in den Folgegenerationen einen Wechsel der do-minanten Arten und der mit ihnen assoziierten Pflanzengesellschaften einleiten, was zu einer Ver-änderung der Biodiversität führen kann. Nicht zu-letzt kann durch Mortalität und die damit einher-gehenden Veränderungen der Bestandesstruktur
Baummortalität – wenn ein natürlicher Prozess zum Problem wird
Baummortalität oder Absterben ist ein natürli-cher Prozess der Walddynamik und findet in allen Phasen der Bestandesentwicklung statt. Sowohl die physiologischen Prozesse als auch die Ursa-chen, die zu erhöhter Mortalität führen, sind bis heute noch nicht alle verstanden. Für eine effizi-ente Waldbewirtschaftung und einen erfolgreichen Waldschutz ist es aber essenziell, die treibenden Faktoren zu verstehen, die zu Baummortalität füh-ren (Franklin et al. 1987).
Je nach Alter und Entwicklungstand eines Waldes kann Mortalität verschieden stark ausge-prägt sein. Ausgehend von einem gleichaltrigen Baumbestand gibt es gemäss oliver und larSon (1996) vier Phasen der Bestandesentwicklung (Abb. 3.6.1): 1) Verjüngungsphase: Etablierung der Verjüngung in einer Lücke, die durch eine Stö-rung (Holzschlag, Windwurf usw.) oder das Ab-sterben der Oberschicht entstanden ist. 2) Jung-wald/Selbstausdünnung: Der Jungwuchs wächst hoch und durch das sich allmählich schließende Kronendach wird der Konkurrenzdruck um Licht, Kronen- und Wurzelraum, Wasser und Nährstoffe grösser. In der Folge sterben schwache Individuen ab, während starke Individuen an Biomasse zuneh-
Abbildung 3.6.1. Bestandeszyklus eines gleichförmigen Hoch-waldes mit den Entwicklungsphasen nach oliver und larSon (1996). Die Phasendauern sind in der Realität nicht gleich lang. Die rote Linie stellt den theoretischen Verlauf der Mortalität dar. Verändert aus KiMMinS (2004).
Verjü
ngun
gs-
phas
e
Verjü
ngun
gs-
phas
e
Jung
wald/
Selbs
taus
dünn
ung
Optim
alpha
se
Altbes
tand
/
Zerfa
llsph
ase
Mor
taliä
tsra
te
178

Auswirkungen des Klimawandels auf den Wald 3oder Vegetation das ästhetische Empfinden des Menschen gestört werden, der den Wald als Erho-lungsraum und kulturelle Stätte nutzt.
Mortalität ist meistens die Folge von mehre-ren interagierenden Ursachen, was die Ursachen-forschung sowie die Folgeabschätzung als auch Handlungsempfehlungen sehr schwierig macht. Die heute mehrheitlich akzeptierte «Decline Di-sease» Theorie (Sinclair 1967; manion 1981; houSton 1984) unterscheidet drei Faktorengruppen, die in komplexem Beziehungsgeflecht zum Absterben der Bäume führen (Abb. 3.6.2): Die schwächen-den Faktoren («predisposing factors») vermindern die Baumvitalität über lange Zeit. Dazu gehören beispielsweise genetische Veranlagung, das Alter, Klima oder Bodenfaktoren. Die auslösenden Fak-toren («inciting factors») sind kurzfristige Ereig-nisse wie Frost, Stürme, Entlaubung durch Insek-ten, Trockenheit oder mechanische Verletzungen. Die beitragenden Faktoren («contributing factors») können langfristiger Natur sein, wie zum Beispiel Pilzbefall oder Viruserkrankungen. Sie können aber auch zu einem sehr schnellen Absterben des ge-schwächten Baumes führen, wenn ein bestimm-ter Schwellenwert überschritten wird (millar und StePhenSon 2015), wie zum Beispiel bei Borkenkä-
ferbefall. Während in den 1980er-Jahren vor allem Schwefeldioxid- und Stickstoffeinträge («saurer Regen») für das regionale Waldsterben in Mitteleu-ropa verantwortlich gemacht wurden (ulrich 1986; Schulze 1989), sind diese Prozesse nach der in den neunziger Jahren erreichten starken Reduktion der Schwefelemissionen in den Hintergrund getreten. Heute auftretende Mortalitätsereignisse werden vor allem mit den sich verändernden Klimabedin-gungen und ihren möglichen Auswirkungen auf die Waldgesundheit in Zusammenhang gebracht (allen et al. 2010; millar und StePhenSon 2015). In diesem Kapitel liegt der Hauptfokus auf der kli-mainduzierten Mortalität, die vor allem nach Ereig-nissen wie Trockenperioden sichtbar wird.
Mortalität in Schweizer Wäldern – wie viel Sterben ist normal?
Jährliche Mortalitätsraten für einheimische Baumarten in der Schweiz
Mortalitätsraten sind schwierig zu ermitteln, denn dazu sind lange Beobachtungszeiträume und ein-zelbaumbasierte Messungen nötig. Dementspre-
Vita
lität
Zeit
auslösender Faktorbeitragende Faktoren
schwächende Faktoren
Abbildung 3.6.2. Schematische Darstellung der «Decline Disease»-Theorie am Beispiel von zwei Bäumen. Die obere Linie kennzeich-net den überlebenden Baum, die untere Linie den absterbenden Baum. In diesem Beispiel haben die schwächenden Faktoren («pre-disposing factors») grösseren Einfluss auf die Vitalität des absterbenden Baumes. Der auslösende Faktor («inciting factor») führt bei beiden Bäumen zu einem Einbruch der Vitalität. Es überlebt nur derjenige Baum, welcher weniger stark durch die schwächenden Faktoren beeinträchtigt wurde, sich von dem auslösenden Faktor wieder erholen kann und durch die beitragenden Faktoren («contri-buting factors») kaum beeinflusst wird. Der absterbende Baum aber wird durch die beitragenden Faktoren stark beeinflusst und stirbt schliesslich ab. Gradienten von schwarz zu grau bedeuten abnehmende Effekte der Mortalitätsfaktoren. Verändert aus Bigler (2003).
179

Mortalität
Box 3.6.1. Erfassung und Analyse der Mortalitätsraten von fünf Hauptbaumarten in der Schweiz
Bewirtschaftungsintensität oder Höhenlage. Den-noch geben die Zahlen einen guten Überblick über Mortalitätsraten auf Schweizer Waldstandorten. Besser vergleichbar sind aber die zeitliche Ent-wicklung der Mortalität und ihre Abhängigkeit von Einflussfaktoren.
Für Buche (Fagus sylvatica), Eiche (Quercus robur, Q. petraea), Fichte (Picea abies), Föhre (Pi-nus sylvestris, P. mugo) und Tanne (Abies alba) mit einem Brusthöhendurchmesser (BHD) > 7 cm in nicht oder nur geringfügig bewirtschafteten Wäl-dern (Abb. 3.6.3) ergab sich über die letzten 100
chend wenige Daten liegen über Mortalität und ihre zeitliche Entwicklung in Wäldern der Schweiz und Zentraleuropas vor. Im Rahmen der in den Boxen 3.6.1 und 3.6.2 vorgestellten drei Projekte wurden Mortalitätsraten für verschiedene Wald-standorte in der Schweiz berechnet. Die absolu-ten Zahlen sind dabei nicht direkt miteinander zu vergleichen, da in den Projekten unterschied-liche Methoden, zum Beispiel bei der Wahl der Kluppschwelle, zur Anwendung kamen. Zudem unterscheiden sich die untersuchten Waldstand-orte erheblich in Bezug auf Entwicklungsstadium,
Projekt 1: Mortalitätsraten von fünf Hauptbaumarten Buche (Fagus sylvatica), Eiche (Quercus robur, Q. petraea), Fichte (Picea abies), Föhre (Pinus sylvestris, P. mugo) und Tanne (Abies alba) von 86 Ertragskundeflächen (EKF), 221 Kernflächen von 30 Naturwaldreservaten (NWR) und 13 Plots der Langfristigen Waldökosystemforschung (LWF) wurden über einen Zeitraum von mehr als 100 Jahren mit Inventurperioden von 5 bis 10 Jah-ren untersucht (Abb. 3.6.3 a–e). Nach Brang et al. (2011) befan-den sich 33 Standorte in der Verjüngungsphase, 52 Standorte in der Jungwaldphase, 227 Standorte in der Optimalphase und 8 Standorte in der Zerfallsphase (Abb. 3.6.1). Insgesamt wur-den Messungen von 247 529 Bäumen mit einem BHD > 7 cm verwendet. Die Untersuchungsflächen wurden pro Baumart in jeweils zwei sich standörtlich stark unterscheidende Grup-pen aufgeteilt. Das waren bei der Föhre und Fichte die Tief- und Hochlagenstandorte (Föhre: 900 m ü. M., Fichte: 1300 m ü. M.), bei Buche, Eiche und Tanne, die überwiegend in tieferen Lagen zu finden sind, feuchte respektive trockene Standorte (minimale Standortwasserbilanz («Site Water Balance») bei Tanne und Buche:, SWBmin = 100 mm/Jahr, Eiche SWBmin = 50 mm/Jahr).
Projekt 2 : Im Rahmen der Interkantonalen Walddauerbeobach-tung (Braun und flücKiger, 2013) wurden über etwa 30 Jahre jährliche Mortalitätsraten von Buchen und Fichten mit einem Brusthöhendurchmesser (BHD) > 16 cm in ausgewachsenen (= Optimalphase), bewirtschafteten Waldbeständen erhoben (Abb. 3.6.3 f ). Der Datensatz umfasst 131 118 Einzelbeobach-tungen bei den Buchen und 84 693 Einzelbeobachtungen bei den Fichten. Es wurden nur Bäume als «abgestorben» aufge-nommen, wenn sie bei der jährlichen Aufnahme tot angetroffen wurden oder wenn eine dokumentierte Zwangsnutzung wegen Borkenkäferbefalls vorlag.
Mortalitätsmodelle: Anhand der Einzelbaumdaten wurden jähr-liche Mortalitätsraten nach dem Modell von Sheil und May (1996) berechnet:
m = (1-(Nt /N0)1/t) × 100 (1)
Mit m = Mortalitätsrate in %, N0 = Anzahl Bäume zu Beginn der Inventurperiode, Nt = Anzahl der überlebenden Bäume am Ende der Inventurperiode, und t = Dauer der Inventurperiode in Jahren.
Die Mortalitätsraten wurden mit gemischten logistischen Regressionsmodellen, mit Standort als gruppierender Variable, der Kombination von Anzahl abgestorbener und überlebender Bäume als abhängiger Variable und diversen erklärenden Vari-ablen (siehe unten) ausgewertet.
In diesem Kapitel werden die Mortalitätsraten mit zwei Trockenheitsindikatoren in Verbindung gebracht, die auf stand-ortspezifischen Berechnungen mit dem hydrologischen Modell WaSiM-ETH (Schulla 2013) basieren und die im Vergleich ver-schiedener Indikatoren am besten abgeschnitten haben (Kap. 3.4, rohner et al. 2016; Braun 2016): 1) ETa/ETp: Verhältnis zwischen aktueller (ETa) und potenziel-
ler (ETp) Evapotranspiration. Je näher dieses Verhältnis bei eins liegt, umso geringer ist der Wasserstress.
2) SWBmin: minimale Standortwasserbilanz nach SPellMann et al. (2007); Aufsummierung der täglichen Differenzen zwi-schen Niederschlag und potenzieller Verdunstung, mit nutz-barer Feldkapazität als Startwert. Je tiefer die SWBmin ist, desto grösser ist der Trockenstress. In den Auswertungen wurde der während der Vegetationsperiode erreichte mini-male Wert verwendet.
180

Auswirkungen des Klimawandels auf den Wald 3
In Projekt 1 wurden als erklärende Variablen Bestandespara-meter (Bestandesdichte, mittlerer BHD als Schätzer für das Bestandesalter, drei BHD-Klassen [klein, mittel, gross]), Topo-grafie (Neigung, Exposition), Klimavariablen (Abweichung der SWBmin, sowie Temperatur vom langjährigen Standortmittel während der Vegetationsperiode) geprüft. BHD-Klassen wur-den für jeden Standort und jedes Zeitfenster so definiert, dass in jeder BHD-Klasse gleich viele Bäume enthalten waren. Me-teorologische Daten wurden von Meteotest für alle Standorte modelliert (reMund et al. 2011).
In Projekt 2 wurden als erklärende Variablen verschiedene Trockenheitsindikatoren sowie Meereshöhe, Bestandesalter, Bestandesstruktur, und Blattkonzentrationen von Kalium, Ma-gnesium und Phosphor sowie Stickstoffdeposition geprüft. Die Wahl der Parameter erfolgte in einer schrittweisen Regression rückwärts durch Vergleich der AIC-Werte («Akaike Information Criterion», hurvich und tSay 1989).
Resultate des Projektes 1 sind in den Abbildungen 3.6.4, 3.6.5, 3.6.7 und 3.6.8 dargestellt, Resultate des Projektes 2 in den Abbildungen 3.6.6, 3.6.12 und Tabellen 3.6.1, 3.6.2.
Box 3.6.1. Erfassung und Analyse der Mortalitätsraten von fünf Hauptbaumarten in der Schweiz
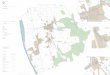
Abbildung 3.6.3. Beobachtungsflächen der EKF, NWR und LWF mit mindestens 20 % Basalflächenanteil der jeweiligen Baumart (a–e) und Beobachtungsflächen des Interkantonalen Walddauerbeobachtungsprogramms (f). In a–e liegen die Punkte der Stand-orte zum Teil über- oder sehr dicht nebeneinander. Rote Kreise zeigen Tieflagenflächen für Föhre und Fichte beziehungsweise trockene Standorte für Tanne, Buche und Eiche. Blaue Dreiecke stellen Hochlagenflächen beziehungsweise feuchte Standorte dar. Die Zahlen geben die jeweilige Anzahl an Beobachtungsflächen pro Baumart an.
BucheFichteBuche u. Fichte
Jahre im Mittel eine jährliche Mortalitätsrate von 1,9 %, das heisst, es starb ungefähr jeder 50. Baum pro Jahr ab (Abb. 3.6.4). Die Variabilität zwischen Baumarten und Untersuchungsflächen ist aber hoch. Die relativ hohe mittlere Mortalitätsrate er-gibt sich durch den Einbezug von teilweise sehr jungen Beständen (Box 3.6.1) und die niedrig an-gesetzte Kluppschwelle. Sie entspricht aber Morta-litätsraten von Studien auf ebenfalls nicht oder nur wenig bewirtschafteten Flächen (doBBertin et al. 2009; rohner et al. 2012; PretzSch et al. 2014). Die Mortalität nahm für Fichte, Föhre, Tanne und Eiche
mit zunehmender Höhe ab (Abb. 3.6.5). Dies bestä-tigen Daten aus den «Stillen Reservaten» der Stich-proben des Landesforstinventars (LFI, Brändli 2010) im hochmontanen bis subalpinen Bereich (Box 3.6.2; Wunder und aBegg in Vorb.). Hier lagen die jährlichen Mortalitätsraten für 35 einheimische Baumarten generell viel tiefer: Zwischen LFI2 und LFI3 starben pro Jahr nur etwa 0,7 % der Bäume mit BHD > 12 cm ab. Die jährlichen Mortalitätsraten von Buche und Fichte während der letzten etwa 30 Jahre auf den bewirtschafteten, ausgewachsenen Flächen des Interkantonalen Walddauerbeobach-
c) Tanne 11 41
b) Fichte 35 44
e) Eiche 37 41
f)
a) Föhre 22 58
d) Buche 55 45
50 km
181

Mortalität
<1960 >1960 <1960 >1960 <1960 >1960 <1960 >1960 <1960 >1960Föhre Fichte Tanne Buche Eiche
0
2
4
6
8
10
Jähr
liche
Mor
talit
ätsr
ate
[%]
BHD−Klasse klein mittel gross
Abbildung 3.6.4. Jährliche Mortalitätsraten (± Standardfehler) pro Baumart und BHD-Klasse (klein, mittel, gross; siehe Box 3.6.1) für die Zeitperioden 1900 – 1959 (< 1960) und 1960 – 2013 (> 1960).
Box 3.6.2. Analyse von Mortalitätsprozessen auf Einzelbaumebene in unbewirtschafteten Waldbeständen
Projekt 3: Langfristige Datenreihen aus unbewirtschafteten Be-ständen des Schweizerischen Landesforstinventars (LFI) wur-den analysiert, um die Absterbeprozesse von 35 einheimischen Baumarten zu erfassen und diese Prozesse mit statistischen Mortalitätsmodellen zu analysieren.
«Stille Reservate»: Für die Mortalitätsanalysen wurden aus dem LFI-Netzwerk jene Plots ausgewählt, die in naturnahen, unbe-wirtschafteten Beständen liegen. Folgende Kriterien mussten erfüllt sein (Wunder und aBegg in Vorb.): 1) Klassifikation als «naturnahe Bestockung»; 2) Entstehung aus natürlicher Verjün-gung; 3) keine intensive Beweidung (früher und heute); und 4) keine Bewirtschaftung nach 1955. Diese Wälder entsprechen in vielen Kriterien (offiziellen) Waldreservaten (Brang et al. 2011). 530 Plots (etwa 8 % aller LFI-Plots) entsprechen den oben auf-geführten Kriterien. Diese Plots sind nicht gleichmässig über die Schweiz verteilt, vielmehr dominieren die hochmontanen und subalpinen Wälder der Alpen und der Südschweiz – das Mittelland ist kaum vertreten. Dennoch umspannen die «Stil-len Reservate» einen grossen klimatischen Gradienten von 345 bis 2840 Tagesgraden (berechnet nach BugMann 1994) bezie-hungsweise – 1,1 bis + 12 °C Jahresmitteltemperatur im unter-suchten Zeitraum.
Mortalitätsraten: In jedem LFI-Plot liegt die Kluppschwelle im grossen Probekreis (500 m2) bei 36 cm Brusthöhendurchmes-
ser (BHD), im kleinen Probekreis (200 m2) bei 12 cm BHD. Für die Mortalitätsraten wurden zunächst die Dichten der toten und lebenden Bäume pro Plot berechnet, wobei die unterschied-lichen Kluppschwellen der Probekreise entsprechend ihrer Fläche gewichtet wurden. Die jährliche Mortalitätsrate zwi-schen LFI2 (1993–1995) und LFI3 (2004–2006) ergibt sich aus dem Quotienten der toten und lebenden Bäume aller LFI-Plots nach Gleichung (1) in Box 3.6.1, mit mittlerer Dichte anstelle von N.
Mortalitätsmodelle: Mit logistischen Regressionsanalysen wurde untersucht, ob lebende Bäume (zum Zeitpunkt des LFI1 und LFI2) den Zeitraum zwischen LFI2 und LFI3 überleben und welche Faktoren den Absterbeprozess am besten erklären. Eine Reihe von Kandidaten-Modellen von unterschiedlicher Komplexität wurden getestet und das beste Modell mithilfe des korrigierten AICc («corrected Akaike Information Crite-rion»; hurvich und tSay 1989) ermittelt. Dieses finale Modell umfasste die erklärenden Variablen Baumwachstum, Baum-grösse, Schattentoleranz und Tagesgrade (nach BugMann 1994) des Standortes. Die Modellleistung wurde mit dem AUC («area under the receiver operating characteristic curve») er-mittelt. Für das finale Modell wurde ein AUC von 0,73 erreicht (entspricht guter Modelleistung, «good performance», nach hoSMer und leMeShoW 2000).
Resultate des Projektes 3 sind in Abbildung 3.6.9 dargestellt.
182

Auswirkungen des Klimawandels auf den Wald 3
0
2
4
6
8
10
500 1000 1500 2000
Jähr
liche
Mor
talit
ätsr
ate
[%]
Höhenlage [m ü. M.]
BucheEicheFichteFöhreTanne
�
Abbildung 3.6.5. Mittlere jährliche Mortalitätsraten von fünf Hauptbaumarten pro EKF, LWF und NWR Plot in Abhängigkeit zur Standorthöhe. Die Linien kennzeichnen den jeweiligen Erwartungswert pro Baumart (P < 0,05).
tungsprogramms lagen für die 30 bis 60 gemes-senen Bäume pro Plot mit einem BHD > 16 cm bei 0,1 bzw. 0,5 % (Abb. 3.6.6). Da Zwangsnutzungen nicht in der Mortalität eingeschlossen sind (aus-ser bei Borkenkäferbefall), ist die aufgenommene Mortalität eine konservative Schätzung. Ebenfalls niedrigere jährliche Mortalitätsraten von im Mittel 1 % (maximal 1,7 %) wurden auf bewirtschafteten österreichischen LFI-Flächen gemessen (monSerud und SterBa 1999). Im Waldzustandsbericht von Hessen (Deutschland) wird eine mittlere Mortali-tätsrate von 0,3 % über die Beobachtungszeit von 1984 bis 2013 erwähnt (Paar et al. 2013).
Zeitliche Trends der Mortalität
Betrachtet man die Mortalität der wichtigsten Baum arten in der Schweiz über die letzten Jahr-zehnte (Abb. 3.6.6) beziehungsweise die letzten hundert Jahre (Abb. 3.6.7), ist kein einheitlicher Trend zu erkennen. Generell kann man aber sa-gen, dass die Mortalität in Schweizer Wäldern
während der letzten Jahrzehnte nicht signifikant angestiegen ist, mit Ausnahme der tief gelege-nen Waldföhrenbestände, wo die Mortalität seit den 1980er-Jahren kontinuierlich zugenommen hat. Vergleicht man die zwei Zeitperioden vor und nach 1960, ergibt sich für die großen BHD-Klassen der Buchen, Fichten und Tannen eine Zunahme an Mortalität, während Mortalität in den kleinen bis mittleren BHD-Klassen meist abnahm (Abb. 3.6.4), wahrscheinlich auch bedingt durch Bestandesalte-rung und Bewirtschaftungsstrategien. Die zuneh-mende Mortalität alter Individuen ebnet demnach den Weg für einen Anstieg der Überlebenschancen der jüngeren.
Gründe für Mortalität – ein komplexes Wirkungs-gefüge
Mortalität ist in der Regel nicht monokausal, son-dern durch komplexe Interaktionen von verschie-denen Faktoren gesteuert (siehe Abschnitt zur «Decline Disease»-Theorie). Sowohl auf Bestan-
183

Mortalität
tragskundeflächen (EKF)1, Flächen der Langfristi-gen Waldökosystemforschung (LWF, SchauB et al. 2011) und der Naturwaldreservate (NWR, Brang et al. 2011) ist die Bestandesdichte denn auch für die meisten Baumarten und Standorte einer der wichtigsten erklärenden Faktoren. Erwartungsge-mäss nimmt die Mortalität konkurrenzbedingt mit zunehmender Bestandesdichte zu (Abb. 3.6.8). Der mittlere BHD eines Bestandes, als Mass für das Bestandesalter, hat je nach Entwicklungsstadium (Abb. 3.6.1) der jeweiligen Bestände einen unter-schiedlichen Effekt auf die Mortalität (Abb. 3.6.8). Generell weisen die kleinen BHD-Klassen eine hö-here Mortalität auf als die grösseren BHD-Klas-sen (Abb. 3.6.4), da kleine Bäume weniger kon-kurrenzstark sind als grosse Bäume (oliver und larSon 1996; reynoldS und Ford 2005; coomeS und allen 2007; Wunder et al. 2008; rohner et al. 2012). Im einzelbaumbasierten Mortalitätsmodell der LFI-Standorte (Box 3.6.2) geht der BHD als quadratischer Term ein, das heisst, mit zunehmen-der Grösse beziehungsweise Alter eines Baumes nimmt die Mortalität zunächst kontinuierlich ab, um dann ab etwa 70 cm BHD wieder anzusteigen (Abb. 3.6.9, vgl. auch Abb. 3.6.1).
1 www.wsl.ch/fe/waldressourcen/projekte/ertragskunde
desebene (Tab. 3.6.1, 3.6.2; Abb. 3.6.8) als auch auf Einzelbaumebene (Abb. 3.6.9) konnte die Va-riabilität der Mortalität in logistischen Regressi-onsmodellen mit Kombinationen aus endogenen Faktoren, wie Topografie und Bestandesstruktur, und exogenen Faktoren, wie Klimabedingungen, erklärt werden. Die Relevanz der einzelnen Fak-toren ist dabei je nach Baumart und Standort un-terschiedlich. Die statistischen Mortalitätsmodelle ermöglichen eine quantitative Erfassung der wich-tigsten Faktoren, welche zur Baummortalität bei-tragen. Man muss allerdings beachten, dass die Aussagekraft der Modelle sowohl von der Qualität der Daten als auch von der gewählten Modellstruk-tur abhängt. Prinzipiell können die Mortalitätsmo-delle jedoch zur Verbesserung von Waldsukzessi-onsmodellen beitragen und ermöglichen damit verlässlichere Projektionen der Waldentwicklung für die kommenden Dekaden (Kap. 3.5, Bircher et al. 2016).
Mortalität und Bestandesstrukturen
Mortalitätsraten eines Bestandes ergeben sich natürlicherweise aus dessen Struktur und Alter (Abb. 3.6.1). In den Mortalitätsmodellen für Er-
1990 1995 2000 2005 2010Jahr
0,4
0,8
1,2
1,6
2,0
0
5000
10 000
15 000
Anza
hl
Bork
enkä
ferb
efal
lshe
rde
Beobachtete MortalitätAnzahl Borkenkäferherde
1990 1995 2000 2005 2010Jahr
0,05
0,10
0,00
0,15
0,20
0,25
0,30
Mor
talit
ät p
ro J
ahr [
%]
Buche Fichtea) b)
0,0
Abbildung 3.6.6. Beobachteter Verlauf der Buchenmortalität (a) und Fichtenmortalität (b) auf den Interkantonalen Walddauerbeob-achtungsflächen. Dargestellt ist der Mittelwert ± Standardfehler. Gestrichelte Linie: Mittelwert über die Beobachtungszeit. Buchen und Fichten haben unterschiedlichen Skalen auf der Y-Achse. In der Fichtengrafik ist die Anzahl der Borkenkäferbefallsherde als gesamtschweizerischer Wert integriert (Quelle: Meier et al. [2014] und kantonale Borkenkäferdaten).
184

Auswirkungen des Klimawandels auf den Wald 3
−6
−4
−2
0
0
2
4
6
8 Föhre
�
� � ��
� �
��
�� �
�
��
�
�� HochlageTieflage
SWBTrockenjahreStürme
−6
−4
−2
0
0
2
4
6
8
2 6 8 13 14 11 14 13 12 14 10 13 10 11 20 20 24 23 22 31
Fichte
�
� � ��
��
� ��
�� �
�
� � �
� HochlageTieflage
N:
−6
−4
−2
0
0
2
4
6
8
2 7 10 14 17 16 16 19 15 14 10 8 14 7 25 12 17 24 19 29
Tanne
��
� �� � � �
� ��
��
��
��
� �
�
Jähr
liche
Mor
talit
ätsr
ate
[%]
Nor
mal
isie
rte
SWB m
in
� feuchttrocken
N:
−6
−4
−2
0
0
2
4
6
8
4 8 10 10 9 13 13 13 8 14 8 9 4 16 22 40 37 37 45 27 48
Buche
� ��
��
��
��
�
�� �
�
��
�� � � �
� feuchttrocken
N:
−6
−4
−2
0
0
2
4
6
8
2 4 7 5 8 8 8 14 8 22 18 16 43 34 35 35 28 41
Eiche
�
�
�
�
�
��
�
�
� �
�
�
� �
� � �
� feuchttrocken
N:
1900 1920 1940 1960 1980 2000Jahr
2 3 4 2 10 10 3 8 4 19 18 27 15 31 21 23 23N:
Abbildung 3.6.7. Jährliche Mortalitätsraten pro Baumart und mittlere, normalisierte SWBmin an den entsprechenden Standorten. Bei Föhre und Fichte wurden Hoch- und Tieflagenstandorte unterschieden, für Tanne, Buche und Eiche feuchte und trockene Standorte (Box 3.6.1). Jeder Punkt stellt die mittlere Mortaliätsrate (± Standardfehler) für alle zu diesem Zeitpunkt gemessenen Plots dar. Da die Inventurperioden pro Plot über unterschiedlich lange Zeiträume integrieren (5–12 Jahre), ist die zeitliche Auf-lösung etwas verwischt. Ein Punkt stellt die Mortalität seit der letzten Inventur dar. Die Anzahl aller verfügbaren Plots (N) ist je-weils auf der x-Achse gegeben.
185

Mortalität
0
10
20FöhreTieflage
Bestandesdichtelocker dicht
mittlerer BHDdünn dick
∆SWBtrocken feucht
−0,94 (1,00)
∆Temperaturkalt warm
0,361 (0,90)
0
10
20FöhreHochlage
0,103 (1,00) 0,527 (1,00)
0
10
20FichteTieflage
0,184 (1,00) −0,022 (1,00)
0
10
20FichteHochlage
0,051 (1,00)
0
10
20Tannetrocken
0,443 (1,00) 0,067 (1,00) −0,078 (0,10) −0,368 (1,00)
0
10
20Tannefeucht
0,077 (1,00)
0
10
20Buchetrocken
0,053 (0,89) 0,388 (0,89)
0
10
20Buchefeucht
0,232 (1,00) −0,07 (1,00) 0,203 (0,83)
0
10
20Eichetrocken
0,665 (1,00) 0,064 (1,00) 2,007 (1,00) −1,288 (1,00)
0
10
20Eichefeucht
0 10 20 30 40
Bestandesdichte [m2/ha]
0,138 (1,00)
10 20 30 40 50
mittlerer BHD [cm]
−0,029 (1,00)
−2 −1 0
∆SWB
0,582 (1,00)
−1 0 1
∆Temperatur
0,594 (1,00)
Jähr
liche
Mor
talit
ätsr
ate
[%]
Abbildung 3.6.8. Responsekurven der jährlichen Mortalitätsraten in Abhängigkeit von signifikanten (P < 0,05) Parameterkoeffizienten der logistischen Regressionsmodelle für die fünf untersuchten Baumarten und Standortgruppen der EKF-, NWR- und LWF-Flächen (Box 3.6.1). Aus allen möglichen Modellkombinationen mit maximal fünf erklärenden Variablen wurden die besten Modelle basie-rend auf dem korrigierten AIC zu einem mittleren Modell gemittelt (Barton 2015). Dargestellt sind die Responsekurven der Morta-lität in Abhängigkeit von Bestandesdichte, dem mittleren BHD, Abweichung der SWBmin und Temperatur vom langjährigen Mittel. Die Responsekurven wurden über die BHD-Klassen (klein, mittel, gross; Box 3.6.1) und Exposition (Nord, Ost, Süd, West) gemittelt. Leere Felder bedeuten, dass der Parameter im Modell nicht signifikant ist. Für jede Kurve wurden die anderen Modellparameter konstant auf dem Mittelwert gehalten. Angegeben sind der Wert des Modellparameters und in Klammer der Wichtigkeitsfaktor des Koeffizienten (maximal 1) nach Barton (2015) innerhalb des gemittelten Modells. Der rote Balken verdeutlicht vor allem bei sehr flachen Kurvenverläufen den Zusammenhang zwischen Mortalität und Parameter (rot = hohe Mortalität, weiss = niedrige Mortalität).
186

Auswirkungen des Klimawandels auf den Wald 3
BHD [cm]
Jähr
l. M
orta
lität
sw'k
eit [
%]
<15 20 30 40 50 60 70 80 90 100 110 1200
1
2
3
4
5
Relativer Basalflächenzuwachssehr gering gering mittel gross
0
1
2
3
4
5
Jähr
l. M
orta
lität
sw'k
eit [
%]
300 600 900 1200 1500 1800 2100 2400 27000
1
2
3
4
5
Tagesgrade [°C] Schattentoleranz
0
1
2
3
4
5
tolerant intermediär intolerant
Abbildung 3.6.9. Jährliche Mortalitätswahrscheinlichkeiten in Abhängigkeit zu BHD, relativem Basalflächenzuwachs, Tagesgra-den (berechnet nach BugMann [1994]) und Schattentoleranz für 35 einheimische Baumarten in den «Stillen Reservaten» des LFI. Verändert aus Wunder und aBegg (in Vorb.).
Mortalität und sich verändernde Klimabedingungen
FöhreDie logistischen Regressionsmodelle zeigen bei den Waldföhren in tiefen Lagen den stärksten Tro-ckenheitseffekt: Hier steigt Mortalität unter trocke-nen, warmen Bedingungen stark an (Abb. 3.6.8). Der Klimaeffekt ist die wichtigste erklärende Vari-able für Mortalität und überlagert sogar den Ein-fluss der Bestandesdichte. Dies bestätigt die seit Jahren beobachteten, trockenheitsinduziert stei-genden Mortalitätsraten in den Trockentälern des Wallis (Rhonetal, Abb. 3.6.10; rigling et al. 2006, 2012, 2013) und des Churer Rheintals (Wohlgemuth et al. 2014). Während Waldföhren in der Lage sind, sich nach einzelnen Trockenjahren wieder zu erholen (eilmann et al. 2010; eilmann und rigling 2012), führen mehrere aufeinander folgende Trockenjahre zu einer stufenweisen Reduktion ih-rer Vitalität, und die Bäume werden anfälliger für Stressfaktoren wie Schädlinge, Pathogene oder
die Föhrenmistel (Viscum album ssp. austriacum, Abb. 3.6.11; Bigler et al. 2006; rigling et al. 2010). Auf den niederschlagsreicheren Hochlagenflächen ist bei der Föhre dagegen eine umgekehrte Bezie-hung von Mortalität und Trockenheit zu sehen: Die Mortalität nimmt unter relativ trockenen Be-dingungen ab (Abb. 3.6.8). Auch auf das Baum-wachstum wurde in Hochlagen kein oder sogar ein positiver Effekt des Jahrhundertsommers 2003 be-ziehungsweise von erhöhten Temperaturen festge-stellt (Jolly et al. 2005; etzold et al. 2011; manuSch et al. 2014; Wohlgemuth et al. 2014). In subalpinen Lagen können vor allem bereits kleine Änderungen in der Dauer der Schneebedeckung grosse, noch nicht abschätzbare Auswirkungen auf die Mortali-tät der Föhre haben (BarBeito et al. 2012).
FichteBei der Fichte auf den EKF, LWF und NWR hat das Klima in Hoch- sowie Tieflagen keinen direkten si-gnifikanten Einfluss auf die Mortalitätsraten (Abb.
187

Mortalität
3.6.8). Bei der Fichte spielen vermehrt die sekun-dären Faktoren eine Rolle – wie die Borkenkäfer, die zwar an Störungen und trockene Jahre gekop-pelt sind, jedoch zeitlich nicht so eng wie das bei-spielsweise bei der Föhre der Fall ist (siehe Ab-schnitt zu «Mortalität und biotische Faktoren»). Im Vergleich verschiedener Trockenheitsindizes war auf den Flächen des Interkantonalen Walddauer-beobachtungsprogramms das Verhältnis von aktu-eller zu potenzieller Evapotranspiration (ETa/ETp), gemittelt über die ersten 80 Tage der letzten drei Vegetationsperioden, der beste Prädiktor für die Mortalität der Fichte. Fichten mit K- und Mg-Man-gel gemäss göttlein et al. (2011) haben eine er-höhte Mortalität (Tab. 3.6.1).
TanneDie Tanne zieht gut wasserversorgte Standorte vor. Hier ist die Bestandesdichte die wichtigste erklä-rende Variable für Mortalität, vor allem auf den feuchteren Standorten, wo das Klima keinen we-sentlichen Einfluss auf die Mortalität zeigt (Abb. 3.6.8). Dieses Muster bestätigen auch Untersu-
chungen aus dem Churer Rheintal, wo der nega-tive Einfluss von Trockenheit auf das Wachstum der Tannen durch tiefgründige Böden und Nordex-position stark abgeschwächt wird (Wohlgemuth et al. 2014). Auf den eher trockenen Standorten dagegen ergibt sich die Mortalität aus einem Fak-torengefüge von Bestandesdynamik, Neigung und Klimabedingungen. Hier ist die Mortalität nach tro-ckenen Jahren leicht erhöht (Abb. 3.6.8). Zudem ist die Tanne sehr frostempfindlich (leBourgeoiS et al. 2010), was sich in einer erhöhten Mortalität nach kalten Jahren (Abb. 3.6.8), wie zum Beispiel infolge des extremen Frostereignisses im Winter 1956 (leBourgeoiS 2007), widerspiegelt.
BucheDie Buche zeigt auf trockenen ebenso wie auf fri-schen Standorten eine erhöhte Mortalität unter feuchteren Bedingungen (Abb. 3.6.8). Dieser eher unerwartete Zusammenhang wurde auch von rohner et al. (2012) auf Schweizer und von nothdurFt (2013) auf deutschen Buchenstandorten gefunden. Mögliche Erklärungen sind eine Ver-
Tabelle 3.6.1. Bestes Regressionsergebnis für Mortalität von Fichten unter Einschluss eines über drei Jahre berechneten Mittel-wertes des Verhältnisses zwischen aktueller und potenzieller Evapotranspiration (ETa/ETp, über die ersten 80 Tage der Vegetati-onsperiode gemittelt). T.S., Trockensubstanz; SE, Standardfehler.
Parameter Koeffizient SE PWert
(Achsenabschnitt) 45,135 10,830 0,0000
K in Nadeln, 2 Gruppen: ≤ 2,8, > 2,8 mg/g T.S. –37,524 10,537 0,0004
Mg in Nadeln, 2 Gruppen: ≤ 0,8, > 0,8 mg/g T.S. –0,813 0,172 0,0000
ETa/ETp Gleitmittel über 3 Jahre –62,958 14,168 0,0000
K in Nadeln, 2 Gruppen × ETa/ETp Gleitmittel 45,978 13,771 0,0008
Tabelle 3.6.2. Bestes Regressionsergebnis für Mortalität von Buchen unter Einbezug eines über zwei Jahre berechneten Mittel-wertes der minimalen Standortwasserbilanz (SWBmin über die ersten 80 Tage der Vegetationsperiode gemittelt). SE, Standardfehler.
Parameter Koeffizient SE PWert
(Achsenabschnitt) –1,864 1,031 0,0705
Meereshöhe [km] –2,924 1,077 0,0066
SWBmin (2-Jahresmittel) –4,676 1,074 0,0000
188

Auswirkungen des Klimawandels auf den Wald 3zunimmt. Ähnliche Muster fanden auch rohner et al. (2012). Die Mortalität der Eiche kann zudem durch Faktoren wie den Befall von Wurzelpathoge-nen bei Staunässe oder Frost (gaertig et al. 2005) ausgelöst werden, was bei den trockenen Eichen-standorten am negativen Temperatureffekt zu se-hen ist (Abb. 3.6.8).
Mortalität und biotische Faktoren
Der Einfluss von Trockenheit auf die Baummortali-tät wird durch Interaktionen mit biotischen Fakto-ren verstärkt. So erhöht Mistelbefall (Abb. 3.6.11) die Anfälligkeit von Waldföhren gegenüber Tro-ckenheit durch zusätzlichen Wasser- und Kohlenhy-dratentzug (doBBertin und rigling 2006; zWeiFel et al. 2012). Die mit einer Trockenperiode meist einhergehenden hohen Temperaturen können die Entwicklung von Insekten beschleunigen (Wermelinger und SeiFert 1999). Trockenheit kann die Bäume für Wurzel- und Rindenkrankheiten an-fälliger machen (rouault et al. 2006). Bei vielen Pilzkrankheiten gilt Ähnliches (guillaumin et al. 1985; Blodgett et al. 1997; deSPrezlouStau et al. 2006; Jung 2009). Bei ausgewachsenen Waldbäu-men sind diese sekundären Trockenheitseffekte wahrscheinlich bedeutender als ein primäres aku-tes «Verdursten». Bekanntestes Beispiel hierfür ist der Zusammenhang zwischen Borkenkäferbe-fall und den Stürmen Vivian und Lothar und der Trockenheit 2003 bei der Fichte (meier et al. 2014; Kap. 3.9, JakoBy et al. 2016). So war die jährliche Mortalitätsrate der Fichte auch auf den Flächen des Interkantonalen Walddauerbeobachtungspro-gramms sehr eng an die gesamtschweizerische Buchdruckersituation gekoppelt (Abb. 3.6.6 b), und zwischen 2001 und 2006 als Folge der Borkenkä-ferinvasion nach dem Sturm Lothar 1999 deut-lich erhöht. Auch die erhöhten Mortalitätsraten in den 1950er-Jahren auf den tief gelegenen Fich-tenstandorten sind auf Borkenkäferbefall infolge der Trockenjahre 1947 und 1949 zurückzuführen (Abb. 3.6.7). Da Borkenkäferbefall oft durch vor-
grösserung der Blattfläche in feuchten Jahren und damit ein zunehmender Konkurrenzdruck auf un-terdrückte, kleine Bäume (rohner et al. 2012), was zu einer erhöhten Mortalität vor allem der kleinen BHD-Klasse führt. Die gute Durchwurzelungsrate der Buche beziehungsweise ihre effiziente Sto-matakontrolle (nothdurFt 2013) sowie ihre hohe evolutive Anpassungsfähigkeit wie die reversible Veränderung von Gestaltungsmerkmalen (roloFF und grundmann 2008) sind weitere Erklärungsan-sätze für die Trockenheitstoleranz der Buche. Nach den extremen Trockenjahren von 1976 und 2003 war die Mortalität der Buche dennoch erhöht, vor allem auf den trockenen Standorten (Abb. 3.6.7). Auch in den jährlichen Daten des Interkantonalen Walddauerbeobachtungsprogramms (Abb. 3.6.6 a) konnte eine zunehmende Buchenmortalität un-ter trockenen Bedingungen nachgewiesen wer-den (Tab. 3.6.2). Das Regressionsmodell schätzt die trockenheitsbedingte Erhöhung der Mortalität im Folgejahr des Hitzesommers 2003 auf 110 %. Absterbende Buchen zeigten mehrere Jahre vor dem Tod im Mittel eine erhöhte Kronenverlichtung (Abb. 3.6.12 a). Das heisst, dass sich der Absterbe-prozess über mehrere Jahre erstreckt und die Tro-ckenheit als auslösender Faktor vor allem bereits geschwächte Bäume trifft. Dementsprechend war die Mortalität nicht nur mit der Trockenheit des Vor-jahres, sondern auch des vorausgehenden Jahres signifikant korreliert (Tab. 3.6.2).
EicheAuch die Eiche zeigt sowohl auf trockenen wie auf frischen Standorten eine signifikant erhöhte Mor-talität unter feuchten Bedingungen (Abb. 3.6.8). Die Eiche ist eine sehr trockenheitstolerante, aber lichtbedürftige Art. Sie kann sich vor allem auf tro-ckenen Standorten etablieren, da sie unter frischen Bedingungen von konkurrenzstärkeren Arten wie der Buche verdrängt wird. In feuchten Perioden kann die Buche auf mittleren Standorten (hier die feuchten Eichenstandorte) im Wachstum zulegen, sodass der Konkurrenzdruck auf die Eiche steigt und deren Mortalität in feuchten, warmen Jahren
189

Mortalität
krebs (Cryphonectria parasitica) verursacht und durch klimatische Extremereignisse wie den Hit-zesommer 2003 verstärkt wurden (conedera et al. 2010). Dies zeigt sich auch in erhöhten Mortalitäts-raten (1,5 % für Edelkastanien > 12 cm BHD in den «Stillen Reservaten» des LFI). Derart geschwächte Edelkastanienwälder werden anfälliger gegenüber dem Eindringen weiterer Neobiota, wie der Edel-kastanien-Gallwespe (Dryocosmus kuriphilus; ForSter et al. 2009) oder dem aus China stam-menden Götterbaum (Ailanthus altissima, Abb. 3.6.13; Wunder et al. 2014; knüSel et al. 2015). Der Klimawandel fördert die Zunahme der Neobiota insofern, als dass mildere Winter ihr Überleben oft erleichtern und Sommertrockenheit die Wider-standskraft ihrer Wirtspflanzen und potenziellen Konkurrenten herabsetzt (Wermelinger 2014).
angehende, plötzlich auftretende Störungen wie Stürme oder Schneebruch ausgelöst wird, ist die Kronenverlichtung der Fichte in den Jahren vor dem Absterben, anders als bei der Buche, nicht erhöht (Abb. 3.6.12 b).
Zunehmend sind invasive Schadorganis-men (Neobiota) wie die Krankheiten Ulmenwelke (Ophiostoma ulmi, bzw. O. novo-ulmi) und Eschen-triebsterben (Hymenoscyphus pseudoalbidus) oder Schadinsekten wie der Asiatische Laubholz-bockkäfer (Anoplophora glabripennis; rigling und SchaFFer 2015) Ursache für höhere Mortalitäts-raten. Die Edelkastanie (Castanaea sativa) weist seit der Aufgabe vieler Kastanienselven in den 1950er-Jahren zunehmend Stresssymptome auf, die durch den 1948 in die Schweiz eingeschlepp-ten und aus China stammenden Kastanienrinden-
Abbildung 3.6.10. Schutzwald zwischen Stalden und Visp (VS) im Jahre 1999. Die extremen Trockenjahre 1996 und 1998 lös-ten ein grossflächiges Föhrensterben aus. (Foto: B. Werme-linger, WSL).
Abbildung 3.6.11. Von Misteln befallene Föhre bei St. German (VS; Foto: A. Rigling, WSL).
190

Auswirkungen des Klimawandels auf den Wald 3und veränderte Bewirtschaftungsstrategien. Einzig in den tief gelegenen Waldföhrenwäldern in den Walliser oder Bündner Trockentälern und in den Edelkastanienwäldern des Tessins gibt es einige «Hotspots», wo erhöhte Mortalität bedingt durch Trockenheit, gefolgt von dem Einwandern einhei-mischer oder invasiver Arten, tatsächlich zu beob-achten ist (rigling et al. 2013; Wunder et al. 2014; Wunder und aBegg in Vorb.). Ein wichtiger Grund für das gegenwärtige Eindringen neuer Arten in diese Ökosysteme ist aber auch die Aufgabe der traditionellen Bewirtschaftung dieser Wälder nach 1950 (conedera et al. 2000; gimmi et al. 2010). So waren Kastanie und Föhre durch menschlichen Ein-fluss über ihre ökologische Nische hinaus verbreitet und werden nun zunehmend zurückgedrängt. Die anderen untersuchten Baumarten scheinen sich an den meisten Standorten noch relativ gut an herr-schende Klimabedingungen anpassen zu können. Bei einigen Baumarten (Buche, Föhre, Fichte, Tanne) besteht allerdings ein signifikanter direkter oder in-direkter Zusammenhang zwischen Mortalität und Trockenheitsereignissen und/oder Temperatur.
Höhere Mortalität in einer wärmeren Welt?
Klimainduzierte Waldmortalität ist ein globales Phänomen geworden. Grossflächige Baummorta-lität durch Trockenheit und heisse Klimabedingun-gen wurde auf allen Kontinenten und in den meis-ten Bioregionen in den letzten zwei Jahrzehnten beobachtet (anderegg et al. 2013). Obwohl auch ohne Klimawandel episodische Mortalitätsereig-nisse dokumentiert sind, deuten zahlreiche Stu-dien darauf hin, dass zumindest gewisse Wälder (z. B. Trockenstandorte, Arten an ihrer Verbreitungs-grenze) schon heute auf Klimaänderungen reagie-ren und dass das Risiko von grossflächigem Baum-sterben und entsprechenden Waldveränderungen in Zukunft zunehmen dürfte (allen et al. 2010).
Die Mortalitätsraten der meisten Schweizer Wälder sind allerdings während der letzten Jahr-zehnte nicht signifikant gestiegen. Jedoch hat sich im Vergleich zu früheren Dekaden Mortalität von den dünnen zu den dickeren Bäumen verlagert, vermutlich auch bedingt durch Bestandesalterung
–5 –4 –3 –2 –1 0Jahre vor Absterben
–5 –4 –3 –2 –1 0Jahre vor Absterben
20
40
60
80
100
20
40
60
80
100
Kron
enve
rlich
tung
[%]
a) Buche Fichteb)
Abbildung 3.6.12. Kronenverlichtung von Buche (a) und Fichte (b) in den dem Absterben vorausgehenden Jahren. Balken: 95 %-Ver-trauensbereich.
191

Mortalität
steigen, da sich der biologische Umsatz der Wäl-der erhöhen wird? In Deutschland wurden abneh-mende Mortalitätsraten der Buche unter höheren Temperaturen beobachtet und mit der höheren Produktivität der Wälder und daher dem früheren Erreichen des Selbstausdünnungsstadiums (Abb. 3.6.1), das abnehmende Mortalität impliziert, er-klärt (PretzSch et al. 2014). Allerdings wird sich unter höheren Temperaturen nicht nur der Umsatz der Bäume steigern, sondern auch derjenige von Schadinsekten (Kap. 3.9, JakoBy et al. 2016). Damit kann also auch mit einer erhöhten Baummortalität gerechnet werden.
Auf gut wasserversorgten Standorten werden sich die Vegetationsgürtel unter wärmeren Tempe-raturen wahrscheinlich in die Höhe verschieben (Walther et al. 2002; gehringFaSel et al. 2007). Da die einzelnen Waldelemente, wie beispielsweise die langlebigen Bäume und die kurzlebigen Insek-ten, individuelle Klimaabhängigkeiten haben, wird die Höhenverschiebung vermutlich nicht simultan erfolgen. Die in den tiefen Lagen erhöhten Morta-litätsraten nach vergangenen extremen Hitzepe-rioden (Abb. 3.6.6, 3.6.7) und die teils signifikan-ten Beziehungen von Mortalität und Trockenheit (Tab. 3.6.1, 3.6.2; Abb. 3.6.8, 3.6.9) lassen vermu-ten, dass mit steigender Frequenz von extremen Trockenperioden auch, zumindest kurzfristig, mit steigenden Mortalitätsraten in Schweizer Wäldern zu rechnen ist. Wie hoch die Resilienz der jewei-ligen Baumarten nach solchen Trockenepisoden ist, bedarf weiterer Untersuchungen (millar und StePhenSon 2015). Verschiedene Studien deuten aber darauf hin, dass es auf trockenen Standorten schon bei moderaten Klimaszenarien zu Baumar-tenverschiebungen hin zu trockenheitstolerante-ren Baumarten kommen wird, da die heute stand-ortgerechten Baumarten an ihre physiologischen Grenzen stossen werden (rigling et al. 2012; léveSque et al. 2013, 2014; siehe auch Kap. 3.7, zimmermann et al. 2016). Neben der Trockentoleranz gilt es, auch die artspezifischen Anfälligkeiten ge-genüber Schadinsekten und Krankheiten zu be-rücksichtigen, die von entscheidender Bedeutung
Bei voraussichtlich weiter steigenden Tempe-raturen bestehen nach wie vor Unsicherheiten in Bezug auf die regionale und jahreszeitliche Nie-derschlagsverteilung (CH2014-Impacts 2014) und dementsprechend auch für die Vegetationsentwick-lung. Die Mortalität weist auf den untersuchten Schweizer Waldstandorten einen Höhen- bezie-hungsweise Temperaturgradienten auf (Abb. 3.6.5; Tab. 3.6.2). Auch die Mortalitätswahrscheinlichkeit eines einzelnen Baumes in den «Stillen Reserva-ten» des LFI hängt stark von den Tagesgraden sei-nes Standortes ab (Abb. 3.6.9). Erwartungsgemäss zeigen hierbei langsam wachsende, kleine, schat-ten-intolerante Bäume an warmen Standorten höchste Mortalität (Wunder und aBegg in Vorb.). Wird Mortalität also unter wärmeren Bedingungen
Abbildung 3.6.13. Vitale Götterbäume und gestresste Edelkas-tanien im Hitzesommer 2003 (San Vittore GR; Foto: M. Cone-dera, WSL).
192

Auswirkungen des Klimawandels auf den Wald 3Literaturallen, c.d.; Macalady, a.K.; chenchouni, h.; Bachelet, d.;
McdoWell, n.; vennetier, M.; KitZBerger, t.; rigling, a.; BreShearS, d.d.; hogg, e.h.; gonZaleZ, P.; fenShaM, r.; Zhang, Z.; caStro, J.; deMidova, n.; liM, J.-h.; allard, g.; running, S.W.; SeMerci, a.; coBB, n., 2010. A global over-view of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manage. 259, 660–684.
anderegg, W.r.l.; Kane, J.M.; anderegg, l.d.l., 2013. Conse-quences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 3, 30–36.
BarBeito, i.; daWeS, M.a.; rixen, c.; Senn, J.; Beri, P., 2012. Factors driving mortality and growth at treeline: a 30-year experiment of 92 000 conifers. Ecology 93, 389–401.
Barton, K., 2015. MuM. In: Model selection and model averaging based on information criteria (AICc and alike). https://cran.r-project.org/web/packages/MuMIn/index.html.
Bigler, c., 2003. Growth-dependent tree mortality: ecological processes and modeling approaches based on tree-ring data. Doctoral Thesis. Swiss Federal Institute of Technology Zurich. doi: 10.3929/ethz-a-004583451. 131 pp.
Bigler, c.; BraeKer, o.u.; BugMann, h.; doBBertin, M.; rigling, a., 2006. Drought as inciting mortality factor in Scots pine stands of the Valais, Switzerland. Ecosystems 9, 330–343.
Bircher, n.; cailleret, M.; Zingg, a.; BugMann, h., 2016. Poten-zielle Grundflächenveränderungen auf Bestandesebene im Klimawandel. In: PlueSS, a.r.; auguStin, S.; Brang, P. (Red.), Wald im Klimawandel. Grundlagen für Adaptations-strategien. Bundesamt für Umwelt BAFU, Bern; Eidg. For-schungsanstalt WSL, Birmensdorf; Haupt, Bern, Stuttgart, Wien. 157–175.
Blodgett, J.t.; Kruger, e.l.; StanoSZ, g.r., 1997. Effects of moderate water stress on disease development by Sphaeropsis sapinea on red pine. Phytopathology 87, 422–434.
Brändli, u.-B. (Red.), 2010. Schweizerisches Landesforst-inventar. Ergebnisse der dritten Erhebung 2004–2006. Bir-mens dorf, Eid. Forschungsanstalt für Wald, Schnee und Landschaft WSL. Bern, Bundesamt für Umwelt, BAFU. 312 S.
Brang, P.; heiri, c.; BugMann, h., 2011. Waldreservate. 50 Jahre natürliche Waldentwicklung in der Schweiz. Eidg. For-schungsanstalt WSL, Birmensdorf; ETH Zürich, Zürich. Haupt, Bern, Stuttgart, Wien. 272 S.
Braun, S., 2016. Die Eignung verschiedener Trockenheitsindizes zur Abschätzung des Trockenheitsrisikos für den Wald und ihre standortspezifische Entwicklung bis 2050/2100. Schlussbericht. Schlussbericht. Forschungsprogramm «Wald und Klimawandel» des Bundesamtes für Umwelt BAFU, Bern und der Eidg. Forschungsanstalt WSL, Birmensdorf. doi: 10.3929/ethz-a-010691682. 58 S.
sein werden für das erfolgreiche Bestehen der Baumarten in einem zukünftigen Klima.
In Deutschland und Österreich wird vor al-lem für tief gelegene Fichtenstandorte eine Zu-nahme des Mortalitätsrisikos prognostiziert, wogegen Buche und Eiche vermutlich vom Kli-mawandel profitieren werden (leXer et al. 2002; roloFF und grundmann 2008; nothdurFt 2013). Für eine zuverlässige Abschätzung von nationalen und weltweiten Trends sollte ein einheitliches Mo-nitoringsystem aufgebaut werden, damit aktuelle Mortalitätsraten direkt mit auslösenden Ereignis-sen korreliert werden können (allen et al. 2010; hartmann et al. 2015). Die Schweiz verfügt dabei schon heute über zahlreiche Monitoringssysteme (z. B. LFI, EKF, LWF, Interkantonales Walddauerbe-obachtungsprogramm, Sanasilva, TreeNet), die in das globale Netz integriert werden können.
DanksagungWir danken für die finanzielle Unterstützung der folgenden Pro-jekte (Projektleitung; Autorin, Mitautoren des Kap. 3.6) durch das Forschungsprogramm «Wald und Klimawandel» des Bun-desamtes für Umwelt BAFU und der Eidg. Forschungsanstalt WSL: «Relative Mortalität als Indikator für die Sensitivität von Waldbeständen» (M. Dobbertin / A. Rigling; S. Etzold); «Die Eig-nung verschiedener Trockenheitsindizes zur Abschätzung des Trockenheitsrisikos für den Wald und ihre standortspezifische Entwicklung bis 2050/2100» (S. Braun); «Retrospektive und pro-spektive Analyse der Sensitivität schweizerischer Waldbestan-destypen auf Klimaveränderungen» (H. Bugmann; J. Wunder, C. Bigler, M. Abegg). Die Erhebung der Daten des Interkantonalen Walddauerbeobachtungsprogramms wurde von den Forstäm-tern der Kantone AG, BE, BL, BS, FR, SO, TG, ZH und ZG und dem Bundesamt für Umwelt BAFU finanziert. Die langjährigen Aufnahmen auf den Monitoringstandorten der EKF, LFI, NWR und LWF erfordern ein hohes Mass an organisatorischem, so-wie technischem Aufwand. Wir danken den Verantwortlichen A. Zingg, U.-B. Brändli, P. Brang und A. Gessler / C. Hug für die Bereitstellung der Daten und den zahlreichen Mitarbeitenden für ihren Einsatz bei den Aufnahmen. M. Dobbertin, der im Jahr 2012 unerwartet verstorben ist, hat das Projekt «Relative Mor-talität als Indikator für die Sensitivität von Waldbeständen» ins Leben gerufen. J. Remund (Meteotest) danken wir für umfang-reiche meteorologische Daten sowie für die Modellierung von standortpezifischen Klimaszenarien.
193

Mortalität
giMMi, u.; WohlgeMuth, t.; rigling, a.; hoffMann, c.W.; Bürgi, M., 2010. Land-use and climate change effects in forest compositional trajectories in a dry Central-Alpine valley. Ann. For. Sci. 67, 701.
goff, f.g., WeSt, d., 1975. Canopy–understory interaction effects on forest population structure. For. Sci. 21, 98–108.
göttlein, a.; Baier, r.; Mellert, K.h., 2011. Neue Ernährungs-kenn werte für die forstlichen Hauptbaumarten in Mittel-europa – Eine statistische Herleitung aus van den Burg‘s Literaturzusammenstellung. Allg. Forst- Jagdztg. 182, 173–186.
guillauMin, J.J.; Bernard, c.; delatour, c.; Belgrand, M., 1985. Root pathology of Quercus robur dieback in the Troncais Forest (of Central France). Ann. Sci. For. 42, 1–22.
hartMann, h.; adaMS, h.d.; anderegg, W.r.l.; JanSen, S.; ZePPel, M.J.B., 2015. Research frontiers in drought-induced tree mortality: crossing scales and disciplines. New Phytol. 205, 965–969.
hoSMer, d.W.; leMeShoW, S., 2000. Applied logistic regression. Wiley, New York, NY, US, 383 pp.
houSton, d.r., 1984. Stress related to diseases. Arboric. J. 8, 137–149.
hurSt, J.M.; allen, r.B.; cooMeS, d.a.; duncan, r.P., 2011. Size-specific tree mortality varies with neighbourhood crowding and disturbance in a montane Nothofagus forest. PLoS ONE 6, e26670.
hurvich, c.M.; tSay, c.l., 1989. Regression and time series modeling in small samples. Biometrika 76, 297–307.
JaKoBy, o.; StadelMann, g.; liSchKe, h.; WerMelinger, B., 2016. Borkenkäfer und Befallsdisposition der Fichte im Klima-wandel. In: PlueSS, a.r.; auguStin, S.; Brang, P. (Red.), Wald im Klimawandel. Grundlagen für Adaptations stra tegien. Bundesamt für Umwelt BAFU, Bern; Eidg. For schungs anstalt WSL, Birmensdorf; Haupt, Bern, Stuttgart, Wien. 247–265.
Jolly, W.M.; doBBertin, M.; ZiMMerMann, n.e.; reichStein, M., 2005. Divergent vegetation growth responses to the 2003 heat wave in the Swiss Alps. Geophys. Res. Lett. 32, doi: 10.1029/2005gl023252. 1–4.
Jung, t., 2009. Beech decline in Central Europe driven by the interaction between Phytophthora infections and climatic extremes. For. Pathol. 39, 73–94.
KiMMinS, J.P., 2004. Forest ecology. Macmillan Publishing Com-pany, New York, NY, US. 611 pp.
KnüSel, S.; conedera, M.; rigling, a.; fonti, P.; Wunder, J., 2015. A tree-ring perspective on the invasion of Ailanthus altissima in protection forests. For. Ecol. Manage. 354, 334–343.
KorPel, S., 1995. Die Urwälder der Westkarpaten. Gustav Fischer Verlag, Stuttgart, Jena, New York. 310 S.
leBourgeoiS, f., 2007. Climatic signal in annual growth variation of silver fir (Abies alba Mill.) and spruce (Picea abies Karst.) from the French Permanent Plot Network (RENECOFOR). Ann. For. Sci. 64, 333–343.
Braun, S.; flücKiger, W., 2013. Wie geht es unserem Wald? Bericht 4. Institut für angewandte Pflanzenbiologie, Schö-nen buch. 127 S.
BugMann, h., 1994. On the ecology of mountainous forests in a changing climate: A simulation study. Doctoral Thesis. ETH Zürich. doi: 10.3929/ethz-a-000946508. 258 pp.
CH2014-Impacts, 2014. Toward quantitative scenarios of climate change impacts in Switzerland. OCCR, FOEN, MeteoSwiss, C2SM, Agroscope, ProClim, Bern. 136 pp.
conedera, M.; Stanga, P.; liScher, c.; StöcKli, v., 2000. Com-petition and dynamics in abandoned chestnut orchards in southern Switzerland. Ecol. Mediterr. 26, 101–112.
conedera, M.; Barthold, f., torriani, d., PeZZatti, g.B., 2010. Drought Sensitivity of Castanea sativa: Case Study of Summer 2003 in the Southern Alps. Acta Hortic. 866, 297–302.
cooMeS, d.a.; allen, r.B., 2007. Mortality and tree-size dis-tributions in natural mixed-age forests. J. Ecol. 95, 27–40.
deSPreZ-louStau, M.-l.; MarçaiS, B.; nageleiSen, l.-M.; Piou, d.; vannini, a., 2006. Interactive effects of drought and pathogens in forest trees. Ann. For. Sci. 63, 597–612.
doBBertin, M.; hug, c.; Waldner, P., 2009. Kronenverlichtung, Sterberaten und Waldwachstum in Langzeitstudien – Welche Indikatoren beschreiben den Waldzustand am besten? In: Langzeitforschung für eine nachhaltige Wald-nutzung. Forum für Wissen 2009, 7–20.
doBBertin, M.; rigling, a., 2006. Pine mistletoe (Viscum album ssp. austriacum) contributes to Scots pine (Pinus sylvestris) mortality in the Rhone valley of Switzerland. For. Pathol. 36, 309–322.
eilMann, B.; rigling, a., 2012. Using tree-growth analysis to estimate drought tolerance of different tree species. Tree Physiol. 32, 178–187.
eilMann, B.; BuchMann, n.; SiegWolf, r.; Saurer, M.; cheruBini, P.; rigling, a., 2010. Fast response to improved water availability of tree-ring width and δ13C in Scots pine. Plant Cell Environ. 33, 1351–1360.
etZold, S.; ruehr, n.K.; ZWeifel, r.; doBBertin, M.; Zingg, a.; PlueSS, P.; haeSler, r.; eugSter, W.; BuchMann, n., 2011. The carbon balance of two contrasting mountain forest ecosystems in Switzerland: Similar annual trends, but seasonal differences. Ecosystems 14, 1289–1309.
forSter, B.; engeSSer, r.; Meier, f., 2009. Die Kastaniengallwespe hat das Tessin erreicht. Waldschutz aktuell 2. Eidg. For-schungs anstalt WSL, Birmensdorf. 2 S.
franKlin, J.f.; Shugart, h.h.; harMon, M.e., 1987. Tree death as an ecological process. Bioscience 37, 550–556.
gaertig, t.; von WilPert, K.; SeeMann, d., 2005. Differentialdia–gnostische Untersuchungen zu Eichenschäden in Baden-Württemberg. Ber. Freibg. Forstl. Forsch. 61. 161 S.
gehring-faSel, J.; guiSan, a.; ZiMMerMann, n.e., 2007. Tree line shifts in the Swiss Alps: Climate change or land abandonment? J. Veg. Sci. 18, 571–582.
194

Auswirkungen des Klimawandels auf den Wald 3reMund, J.; frehner, M.; Walthert, l.; Kägi, M.; rihM, B., 2011.
Schätzung standortspezifischer Trockenstressrisiken in Schweizer Wäldern. Schlussbericht. Forschungsprogramm Wald und Klimawandel. Eidg. Forschungsanstalt WSL. doi: 10.3929/ethz-a-010693256. 56 S.
reynoldS, J.h.; ford, e.d., 2005. Improving competition re-presentation in theoretical models of self-thinning: a critical review. J. Ecol. 93, 362–372.
rigling, a.; Schaffer, h.P. (eds), 2015. Waldbericht 2015. Zustand und Nutzung des Schweizer Waldes. Bundesamt für Umwelt, Bern; Eidg. Forschungsanstalt WSL, Birmensdorf. 144 S.
rigling, a.; doBBertin, M.; Bürgi, M.; feldMeier-chriSte, e.; giMMi, u.; ginZler, c.; graf, u.; Mayer, P., 2006. Baumarten-wechsel in den Walliser Waldföhrenwäldern. Forum für Wissen 2006, 71 S.
rigling, a.; eilMann, B.; Köchli, r.; doBBertin, M., 2010. Mistletoe-induced crown degradation in Scots pine in a xeric environment. Tree Physiol. 30, 845–852.
rigling, a.; elKin, c.; doBBertin, M.; giuggiola, a.; WohlgeMuth, t.; BugMann, h., 2012. Waldentwicklung in zentral-alpinen Trockentälern unter fortschreitendem Klimawandel – die Fallstudie Region Visp. Schweiz. Z. Forstwes. 163, 481–492.
rigling, a.; Bigler, c.; eilMann, B.; Mayer, P.; ginZler, c.; vacchiano, g.; WeBer, P.; WohlgeMuth, t.; ZWeifel, r., 2013. Driving factors of a vegetation shift from Scots pine to pub escent oak in dry Alpine forests. Glob. Chang. Biol. 19, 229–240.
rohner, B.; Bigler, c.; Wunder, J.; Brang, P.; BugMann, h., 2012. Fifty years of natural succession in Swiss forest reserves: changes in stand structure and mortality rates of oak and beech. J. Veg. Sci. 23, 892–905.
rohner, B.; Braun, S.; WeBer, P.; thürig, e., 2016. Wachstum von Einzelbäumen: das Klima als Baustein im komplexen Wirkungsgefüge. In: PlueSS, a.r.; auguStin, S.; Brang, P. (Red.), Wald im Klimawandel. Grundlagen für Adaptations-strategien. Bundesamt für Umwelt BAFU, Bern; Eidg. For-schungsanstalt WSL, Birmensdorf; Haupt, Bern, Stuttgart, Wien. 137–155.
roloff, a.; grundMann, B., 2008. Klimawandel und Baumarten-Verwendung für Waldökosysteme. TU Dresden, Stiftung: Wald in Not, Tharandt, Deutschland, 1–46.
rouault, g.; candau, J.-n.; lieutier, f.; nageleiSen, l.-M.; Martin, J.-c.; WarZée, n., 2006. Effects of drought and heat on forest insect populations in relation to the 2003 drought in Western Europe. Ann. For. Sci. 63, 613–624.
SchauB, M.; doBBertin, M.; Kräuchi, n.; Kaennel doBBertin, M., 2011. Preface – long-term ecosystem research: under-standing the present to shape the future. Environ. Monit. Assess. 174, 1–2.
Schulla, J.; JaSPer, K., 2013. Model Description WaSiM-ETH (Zürich). http://www.wasim.ch/downloads/doku/wasim/wasim_2013_en.pdf.
leBourgeoiS, f.; rathgeBer, c.B.K.; ulrich, e., 2010. Sensitivity of French temperate coniferous forests to climate variability and extreme events (Abies alba, Picea abies and Pinus sylvestris). J. Veg. Sci. 21, 364–376.
léveSQue, M.; SiegWolf, r.; Saurer, M.; Brang, P.; eilMann, B.; BugMann, h.; rigling, a., 2013. Drought responses of five conifers at their xeric distribution limit in the Alps and under mesic conditions in Central Europe. Glob. Chang. Biol. 19, 3184–3199.
léveSQue, M.; rigling, a.; BugMann, h.; WeBer, P.; Brang, P., 2014. Growth response of five co-occurring conifers to drought across a wide climatic gradient in Central Europe. Agric. For. Meteorol. 197, 1–12.
lexer, M.J.; hoenninger, K.; Scheifinger, h.; Matulla, c.; groll, n.; KroMoP-KolB, h.; Schadauer, K.; Starlinger, f.; engliSch, M., 2002. The sensitivity of Austrian forests to scenarios and climatic change: a large-scale risk assessment based on a modified gap model and forest inventory data. For. Ecol. Manage. 162, 53–72.
lineS, e.r.; cooMeS, d.a.; PurveS, d.W., 2010. Influences of forest structure, climate and species composition on tree mortality across the eastern US. PLoS ONE 5, e13212.
loriMer, c.g.; dahier, S.e.; nordheiM, e.v., 2001. Tree mortality rates and longevity in mature and old-growth hemlock-hardwood forests. J. Ecol. 89, 960–971.
Manion, P.d., 1981. Tree disease concepts. Prentice-Hall Englewood Cliffs , New Jersey, US. 416 pp.
ManuSch, c.; BugMann, h.; Wolff, a., 2014. The impact of climate change and its uncertainty on carbon storage in Switzerland. Reg. Environ. Chang. 14, 1437–1450.
Meier, f.; forSter, B.; engeSSer, r., 2014. Borkenkäfer – Buchdrucker-Befall hat zugenommen. Waldschutz aktuell 1/2014.
Millar, c.l.; StePhenSon, n.l., 2015. Temperate forest health in an era of emerging megadisturbance. Science 349, 823–826.
MonSerud, r.a.; SterBa, h., 1999. Modeling individual tree mortality for Austrian forest species. For. Ecol. Manage. 113, 109–123.
nothdurft, a., 2013. Spatio-temporal prediction of tree mortality based on long-term sample plots, climate change scenarios and parameter frailty modeling. For. Ecol. Manage. 291, 43–54.
oliver, c.d.; larSon, B.c., 1996. Forest Stand Dynamics. John Wiley & Sons, Inc., New York; Chichester; Brisbane; Toronto; Singapore. 544 pp.
Paar, u.; daMMann, i.; WeyMar, J.; SPielMann, M.; eichhorn, J., 2013. Waldzustandsbericht 2013. Nordwestdeutsche Forstliche Versuchsanstalt. 40 S.
PretZSch, h.; BiBer, P.; SchütZe, g.; uhl, e.; rötZer, t., 2014. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 5, 4967.
195

Mortalität
tal. Schlussbericht Projekt Bündner Wald im Klimawandel. Eidg. Forschungsanstalt WSL, Birmensdorf; Amt für Wald und Naturgefahren, Kt. Graubünden. 81 S.
Wunder, J.; aBegg, M., in Vorb. Modelling individual tree mortality for Swiss forest species – predicting the un-predictable?
Wunder, J.; BrZeZiecKi, B.; ZyBura, h.; reineKing, B.; Bigler, c.; BugMann, h., 2008. Growth-mortality relationships as indicators of life-history strategies: a comparison of nine tree species in unmanaged European forests. Oikos 117, 815–828.
Wunder, J.; noBiS, M.; conedera, M., 2014. Der Götterbaum – eine Gefahr für den Schweizer Wald? Wald Holz 6, 40–43.
ZiMMerMann, n.e.; SchMatZ, d.r.; gallien, l.; Körner, c.; huBer, B.; frehner, M.; Küchler, M.; PSoMaS, a., 2016. Baum-artenverbreitung und Standorteignung. In: PlueSS, a.r.; auguStin, S.; Brang, P. (Red.), Wald im Klimawandel. Grund lagen für Adaptationsstrategien. Bundesamt für Umwelt BAFU, Bern; Eidg. Forschungsanstalt WSL, Bir-mensdorf; Haupt, Bern, Stuttgart, Wien. 199–221.
ZWeifel, r.; Bangerter, S.; rigling, a.; StercK, f.J., 2012. Pine and mistletoes: how to live with a leak in the water flow and storage system? J. Exp. Bot. 63, 2565–2578.
SchulZe, e.d., 1989. Air pollution and forest decline in a spruce (Picea abies) forest. Science 244, 776–783.
Sheil, d.; May, r.M., 1996. Mortality and recruitment rate evaluations in heterogeneous tropical forests. J. Ecol. 84, 91–100.
Sinclair, W.a., 1967. Decline of hardwoods: possible causes. In: International shade tree conference, 17–32.
SPellMann, h.; SutMöller, J.; MeeSenBurg, h., 2007. Risikovor-sorge im Zeichen des Klimawandels. AFZ/Wald 62, 1246–1249.
ulrich, B., 1986. Die Rolle der Bodenversauerung beim Wald-sterben: Langfristige Konsequenzen und forstliche Mög-lich keiten. Forstwiss. Cent.bl. 105, 421–435.
Walther, g.r.; PoSt, e.; convey, P.; MenZel, a.; ParMeSan, c.; BeeBee, t.J.c.; froMentin, J.M.; hoegh-guldBerg, o.; Bairlein, f., 2002. Ecological responses to recent climate change. Nature 416, 389–395.
WerMelinger, B., 2014. Invasive Gehölzinsekten: Bedrohung für den Schweizer Wald? Schweiz. Z. Forstwes. 165, 166–172.
WerMelinger, B.; Seifert, M., 1999. Temperature-dependent reproduction of the spruce bark beetle Ips typographus, and analysis of the potential population growth. Ecol. Entomol. 24, 103–110.
WohlgeMuth, t., rigling, a. (Red.), 2014. Kurz- und langfristige Auswirkungen des Klimas auf die Wälder im Churer Rhein-
Abgestorbene Nadelbäume in Conthey, VS. Foto: P. Brang. >196