Embed Size (px)
Citation preview

BIOCHIMICA ET BIOPKYSICA ACTA 249
BBA 25 072
A C E T A T A K T I V I E R U N G IN G R I ) N A L G E N
II. DIE BILDUNG VON ACETYLPHOSPHAT IN STICHOCOCCUS BACILLARIS
UND ANDEREN ARTEN DER CHLOROCOCCALES
E R I C H OHMANN
Insti tut f~ir A llgemeine Botanik der Universitdt und A kademieinstitut f~r Biochemie der Pflanzen, Halle, Saale (Deutschland) (Eingegangen am 4. Dezember, I963)
SUMMARY
Acetate activation in green algae
II. The formation of acetyl phosphate in Stichococcus baciIlaris and other species of Chloroeoccales
I. Resting cells of Stichoeoccus baeillaris rapidly metabolise acetate. In contrast to former experiments with Euglena gracilis there was no lag phase in O2-uptake. Immediately after addition of acetate to respiring cells O,-uptake rose to maximum level. Only about 50 % of the added acetate is oxidised, the remainder is assimilated.
2. Although in crude ceU-free preparations acethydroxamic acid formation occurred also after CoA had been removed, there was a small stimulative effect upon adding CoA to the mixture. In reaction mixtures, however, which had been heated for 20 min to 55 °, CoA had no influence on the formation of hydroxamic acid.
3. In short time experiments authentic acetyl phosphate will be metabolised by crude extracts only if ADP as a phosphate acceptor is present. Exact stoichiometric relations exist between the disappearance of acetyl phosphate and ADP and the synthesis of ATP.
3. All experiments performed provide strong evidence, that in Stichococcus bacillaris and some other species of Chlorococcales acetate activation is initiated by acetyl phosphate formation. The enzyme responsible for this reaction, acetate-kinase (ATP:acetate phosphotransferase, EC 2.7.2.I), has been purified. The requirements for the reaction were studied, and the Michaelis constants determined.
EINLEITUNG
Heterotrophie ist bei den Algen eine verbreitete Erscheinung 1. In ganz besonderem Masse gilt das ftir einige Flagellaten. Viele ihrer Formen sind stAndig auf die Zuftihrung organischer Substrate angewiesen. Andere dagegen verm6gen sowohl rein autotroph im Licht wie heterotroph im Dunkeln auf geeigneten C-Quellen zu wachsen, z.B. Euglena gracilis auf Acetat. Die Gruppe der Chlorococcales hat bei den Anzuchten im Labor ihre BefAhigung zum reinen autotrophen Wachstum seit Jahren immer wieder
Biochim. Biophys. Acta, 90 (1964) 249-259

250 E. OHMANN
bewiesen. Zu ihr geh6ren auch die Gattungen Chlorella, Scenedesmus und Sticho- coccus. Sie gelten als autotrophe Organismen par excellence. Jecloch unter den Be- dingungen ihres natiirlichen Standortes ist auch bei diesen Arten an die M6glichkeit einer gelegentlichen oder dauernden Nutzung vorgebildeter organischer Substanzen fiir die Synthese yon Zellbestandteilen zu denken. Jedenfalls ist ihre prinzipielle Bef~ihigung dazu in vielen Versuchen erwiesen. Acetat wird von Chlorella pyrenoidosa nicht nur veratmet 2, sondern der Kohlenstoff dieser S~iure geht rasch in viele Zell- bestandteile ein ~,4. Ein Mechanismus, dessen sich die Zellen bei der Assimilation des Acetats bedienen, ist der Glyoxylat-Cyclus 5. Das gilt sowohl fiir die heterotroph auf Acetat als C-Quelle wachsende Euglena gracilis 6-8 wie ftir einige Arten der Chloro- coccalesS, 9. Voraussetzung ftir die Wirksamkeit dieses Prozesses ist eine vorherige Aktivierung der freien Essigs~iure. Erst in der Form des Acetyl-CoA ist sie bef~higt, an den durch Citrat-Synthase (EC 4.1.3.7) und Malat-Synthase (EC 4.1.3.2) kataly- sierten Aldolkondensationen dieses cyclischen Geschehens teilzuhaben. In den Einzel- heiten des enzymatischen Ablaufes der Bildung von Acetyl-CoA bestehen innerhalb tier Algen Unterschiede. In Euglena gracilis erfolgt die Aktivierung wie in tierischen Zellen 1°-15, in h6heren Pflanzen 16,17, in HefelS, .1 und Rhodospirillum rubrum ~, 23 an der Acetyl-CoA-Synthetase (EC 6.2.I.I)~4,~5:
Acetat + ATP + CoA--~ Acetyl-CoA + AMP + PPI (I)
In den in der vorliegenden Arbeit niiher untersuchten Arten der Chlorococcales wird Acetylphosphat gebildet, eine Reaktion, die bisher als charakteristisch ftir Bakterien angesehen Wllrde 26-29:
Acetat + ATP ~ Acetylphosphat + ADP (2)
MATERIAL UND METHODEN
Die zu den Untersuchungen verwendeten Algen Stichococcus bacillaris, Chlorella pyrenoidosa sowie zwei nicht n~ther identifizierte Scenedesmus-St~mme H I und H 2 wurden in einer MineralsalzlSsung nach KANDLER 3° unter frtiher 25 beschriebenen Bedingungen kultiviert.
Zur Herstellung zellfreier Extrakte wurden m6glichst dichte Aufschwemmungen mehrfach gewaschener Zellen in eisgektihltem 0.05 M Tris- oder Kaliumphosphat- Puffer, denen noch 0.005 M Cystein zugeffigt wtirde, in der Vibrogen Zellmtihle der Firma Btihler, Tiibingen, mechanisch mit Glaskugeln geschfittelt. Der ~bers tand nach einer Zentrifugation von 30 rain bei 20000 × g wird als Rohextrakt bezeichnet und nicht sp/iter als 12o rain nach dem Homogenisieren zur Fraktionierung oder direkt zu den Versuchen verwendet. Die Entfernung yon zelleigenem CoA erfolgte durch Behandlung mit Dowex-I nach CHANTRENNE UND LIPMANN 31.
Messung der Acetataktivierung
Die Acetataktivierung wurde durch Spaltung des in der Reaktion entstandenen Acetylphosphats mit Hydroxylamin und anschliessender Reaktion der Acethydroxam- s~ure mit dem Trichloressigs/iure-FeCl~-Reagens nach LIPMANN UND TUTTLE 32 erfasst. Nach deln Abzentrifugieren des Proteins wurde der rotgef/irbte Hydroxam- s / iure-Fe-Komplex im Eppendorf-Photometer bei 54 6 mr,, d = I em, gemessen. Als Standard diente Succinhydroxamat, das nach STADTMAN 33 aus Bernsteins~urean-
Biochim. Biophys. dcta, 9o (1964) 249-259

ACETATAKTIVIERUNG IN GRUNALGEN 251
hydrid hergestellt wurde. Die Acethydroxams~ure wurde nach STAI)TMAN UND BARKER m extrahiert und chromatographiert.
Die rtickl~ufige Reaktion, die t3bertragung des Phosphatrestes von Acetyl- phosphat auf ADP wurde an Hand der Abnahme des Acetylphosphates erfasst. Diese Reaktion wurde durch Zugabe yon 2 ml I M Hydroxylaminl6sung zu den AnsAtzen yon I ml abgestoppt. Dadurch wurde das noch nicht umgesetzte Acetylphosphat sofort durch Hydroxams~turebildung aus der enzymatischen Reaktion genommen. Nach IO min bei 30 ° erfolgte die Zugabe von Trichloressigs~ure-FeC13-Reagens.
Als Enzymeinheit* wird die Enzymmenge definiert, die in einer Minute einen Substratumsatz von I/~Mol katalysiert 34.
Weitere Methoden
ATP wurde mit Phosphoglycerat-Kinase (EC 2.7.2.3) und Glycerinaldehyd- phosphat-Dehydrogenase (EC 1.2.I.I2), ADP mit Pyruvat-Kinase (EC 2.7.1.4o) und Lactat-Dehydrogenase (EC 1.1.1.27) in der jeweiligen Testzusammensetzung nach ADAM s5 bestimmt. Protein wurde in den zellfreien Extrakten nach LOWRY et al. ~ bestimmt. Die Messung der RadioaktivitAt des aus [~2P!Acetylphosphat und ADP an der Acetat-Kinase (ATP:Acetat-Phosphotransferase, EC 2.7.2.1 ) gebildeten [32PIATP geschah nach der yon LYNEN et a l F modifizierten Methode von BERG zz.
Die Messung der respiratorischen O~-Aufnahme erfolgte mit konventioneller I-Gef~tssmethode in 15 ml Kegelgef~ssen mit 20 % KOH im Mitteleinsatz 3.. Vor der Manometrie ~ wurden die Zellen 20 h i m Luftstrom an zelleigenen Speicherstoffen verarmt.
P r i~'p ar ate
ATP, ADP, CoA, Phosphoenolpyruvat, 3-Phosphoglycerat, NADH 2, Phospho- glycerat-Kinase, Pyruvat-Kinase, Glycerinaldehydphosphat-Dehydrogenase und Lactat-Dehydrogenase waren Handelsprodukte der Firma Boehringer, Mannheim. Acetylphosphat wurde als Di-Lithiumsalz dutch Umsetzung von Essigs~ureanhydrid mit K2HPO 4 in Pyridin bei o ° nach AVlSON 39 synthetisiert. Nach Umf~llung mit Athanol in der K~lte betrug die Reinheit 95 %. In einem Mikroansatz wurde nach dem gleichen Prinzip [32p~Acetylphosphat hergestellt 4°.
EXPERIMENTE UND ERGEBNISSE
Oxydative Assimilation des Acetats durch intakte Zd len yon Stichococcus bacillaris
Wird zu einer Suspension verarmter Zellen von Stichococcus Acetat gegeben, ist sofort ein Anstieg der O~-Aufnahme zu verzeichnen, der so lange anh~tlt, bis das Acetat verbraucht ist (Fig. I). Im Unterschied zu den VerhAltnissen bei autotroph gezogenen Zellen von Euglena gracilis, wo in Form eines induktiven Vorganges die maximalen Werte der 02-Aufnahme erst nach einigen Stunden erreicht werden .5, setzt hier der Umsatz sofort mit voller Geschwindigkeit ein. Dabei zeigt sich jedoch, dass die respiratorische 02-Aufnahme schon dann wieder auf die Werte der sub- stratfreien Kontrolle zurtickgefallen ist, wenn erst ein Bruchteil der be'i vollstitndiger Oxydation des gegebenen Acetats theoretisch zu erwartenden O ~-Menge aufgenommen
* Abkiirzung: E, Enzymeinhe i t hier definiert.
Biochim. Biophys. Acta, 90 (1964) 249-259
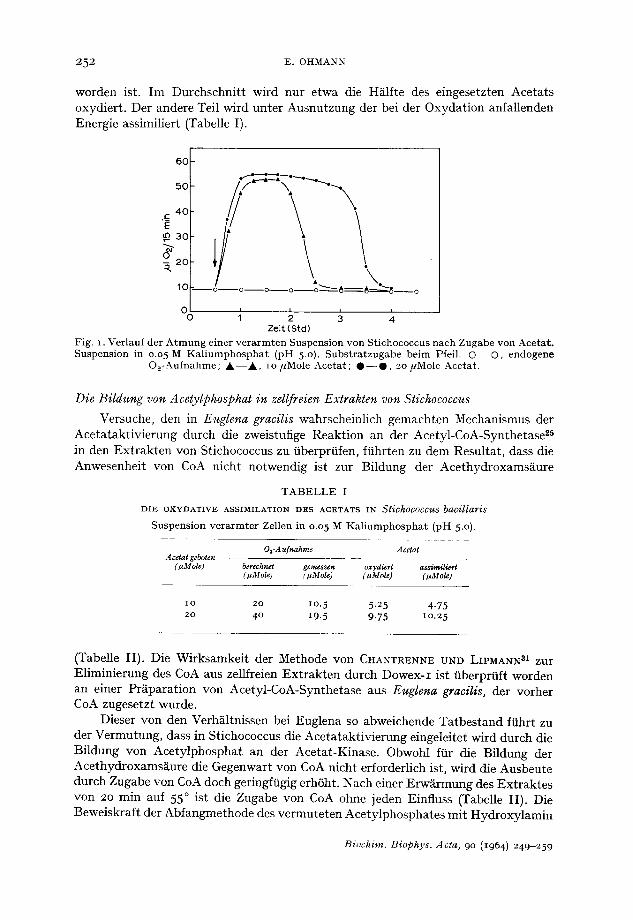
252 E. OKMANN
worden ist. Im Durchschnitt wird nur etwa die H~ilfte des eingesetzten Acetats oxydiert. Der andere Tell wird unter Ausnutzung der bei der Oxydation anfallenden Energie assimiliert (Tabelle I).
60
--I 5 0
.G 40
E 3O
d~ ~. 2o
1 0 o o o o o 6 ~=~.--" 8 o
I I I I
0 1 2 3 4 Zeit (Std)
Fig. I. Ver lauf der A t m u n g einer v e r a r m t e n Suspens ion von St ichococcus nach Zugabe von Ace ta t . Suspens ion in 0.05 M K a l i u m p h o s p h a t (pH 5.0). S u b s t r a t z u g a b e be im Pfeil. © - - O , endogene
O2-Aufnahme ; ~ k - - A , io /zMole Ace ta t ; 0 - - 0 , 20/*Mole Aceta t .
Die Bildung von Acetylphosphat in zellfreien Extrakten yon Stichococcus
Versuche, den in Euglena gracilis wahrscheinlich gemachten Mechanismus der Acetataktivierung durch die zweistufige Reaktion an der Acetyl-CoA-Synthetase ~5 in den Extrakten -con Stichococcus zu fiberpriifen, fiihrten zu dem Resultat, dass die Anwesenheit yon CoA nicht notwendig ist zur Bildung der Acethydroxams~.ure
T A B E L L E I
D I E O X Y D A T I V E A S S I M I L A T I O N D E S A C E T A T S IN Stichococcus bacillaris
Suspens ion v e r a r m t e r Zellen in o.o 5 M K a l i u m p h o s p h a t (pH 5.0).
O,- A ufnahme A ceta~ Acetat geboten
(#Mole) berechnet gemessen oxydiert assimiliert ( l~ M ol 0 ( it Mole) (#Mole) (l,Mole)
I0 20 10"5 5"25 4"75 20 4 ° 19.5 9"75 10"25
(Tabelle II) . Die Wirksamkeit der Methode von CHANTRENNE UND LIPMANN 81 zur Eliminierung des CoA aus zellfreien Ext rakten durch Dowex-I ist fiberprfift worden an einer Pr~iparation von Acetyl-CoA-Synthetase aus Euglena gracilis, der vorher CoA zugesetzt wurde.
Dieser yon den VerhAltnissen bei Euglena so abweichende Tatbestand fiihrt zu der Vermutung, dass in Stichococcus die Acetataktivierung eingeleitet wird durch die Bildung von Acetylphosphat an der Acetat-Kinase. Obwohl ftir die Bildung der Acethydroxamsliure die Gegenwart von CoA nicht erforderlich ist, wird die Ausbeute durch Zugabe yon CoA doch geringftigig erh6ht. Nach einer Erw~irmung des Extraktes yon 20 rain auf 55 ° ist die Zugabe yon CoA ohne jeden Einfluss (Tabelle II). Die Beweiskraft der Abfangmethode des vermuteten Acetylphosphates mit Hydroxylamin
B i o c h i m . B i o p h y s . A c t a , 90 (1964) 249-259

ACETATAKTIVIERUNG IN GR/JNALGEN 253
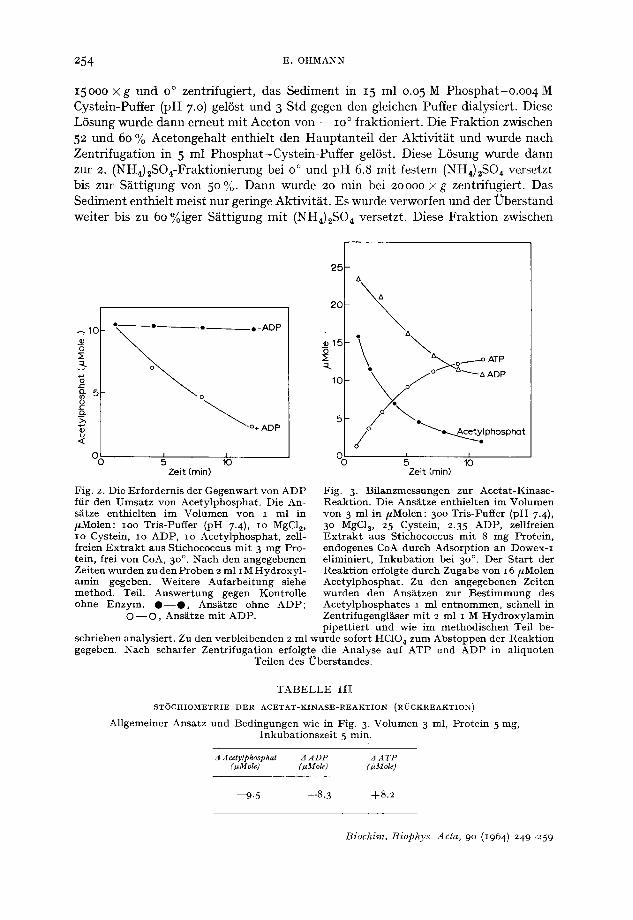
in der H in reak t ion ist allein n icht gross genug ftir die E r m i t t l u n g des Mechanismus der A c e t a t a k t i v i e r u n g in Stichococcus. Daher wurden Versuche mi t au then t i schem A c e t y l p h o s p h a t durchgeft ihrt . Sie f t ihrten zu dem Ergebnis , dass im zellfreien Ex- t r ak t , der von endogenem CoA befrei t wurde, A c e t y l p h o s p h a t nur dann in nennens- wer tem Masse umgese tz t wird, wenn A D P als Phospha t accep to r gegeben wird (Fig. 2).
TABELLE II
DIE BILDUNG VON ACETHYDROXAMSAURE IM EXTRAKT VON Stichococcus bacillaris
Die Ans~.tze enthielten im Volumen yon i ml in #Molen: IOO Tris-Puffer (pH 7.4), 20 Kalium- acetat, io MgCI~, io Cystein, io ATP, 0.2 CoA, 200 Hydroxylamin, zellfreien Extrakt mit 3 mg Protein, frei yon endogenem CoA. Die AnsAtze 5 und 6 enthielten zellfreien Extrakt, der 20 rain im Wasserbad auf 55 ° erhitzt und durch nachfolgende Zentrifugation von ausgefallenen Proteinen befreit wurde. Nach 3 ° min Inkubation bei 3 o° wurde mit Trichloressigs~.ure-FeClz-Reagens
abgestoppt.
Hydroxams~ure Nr. A nsatz gebildet
(#Mole)
I komplett i. 5 2 --CoA t .25 3 --ATP o 4 --Acetat 0.02 5 komplett, Extrakt 2o rain auf 55 ° erhitzt I. I 6 --CoA, Extrakt 2o rain auf 55 ° erhitzt i . i
In Bi lanzversuchen zur E r m i t t l u n g der St6chiometr ie , in denen die Reakt ions- pa r t ne r Ace ty lphospha t , A D P und ATP nebene inander b e s t i m m t wurden, ha t sich gezeigt, dass zwischen der A b n a h m e von A D P und A c e t y l p h o s p h a t auf der einen Seite und der Zunahme von A T P auf der anderen Seite eine d i rekte Beziehung bes teh t (Fig. 3). Pro Mol ATP, das gebi ldet wird, werden je ein Mol A D P und Ace ty l - p h o s p h a t ve rb rauch t (Tabelle I I I ) .
A n r e i c h e r u n g der A c e t a t - K i n a s e
Ft i r wei terf t ihrende Versuche war eine Anre icherung des E n z y m s notwendig. Eine E r p r o b u n g anderer , ebenfal ls zur sys temat i schen Gruppe der Chlorococcales geh6render Algen ergab, dass in Chlorella p y r e n o i d o s a und zwei n ich t n~.her charak- te r i s ier ten St~tmmen von Scenedesmus die Ace t a t ak t iv i e rung ebenfalls wie bei St icho- coccus an der Ace t a t -K inase erfolgt. Als Ausgangsmate r i a l ftir die Anre icherung d ienten 260 ml eines zellfreien E x t r a k t e s aus Scenedesmus H I. Der Aufschluss der Zellen erfolgte in o.o 5 M Ka l iumphospha t -Pu f f e r (pH 7.o), dem 0.004 M Cystein beigesetz t war. Die Frakt ionieI -ang verl~uft hier besser als bei Stichococcus. Zu dem E x t r a k t wurde l angsam unte r Umrt ihren kal tes Ace ton von - - 1 5 ° gegeben, bis ein Gehal t von 4 ° % erreicht war. Die T e m p e r a t u r der Mischung be t rug danach - - i o °. Das grobflockig ausgefallene Pro te in wurde darauf bei dieser T e m p e r a t u r 15 min bei 25o0 × g zentr i fugier t , der ~ b e r s t a n d dann auf einen Ace tongeha l t von 55 % gebrach t , 15 min spXter e rneut 15 min bei 5000 • g zentr i fugier t . Das Sediment wurde sorgf~tltig in 25 ml (NH4)2SO4-L6sung von 55 %iger S~tttigung + 0 . 0 0 4 M Cystein bei o ° gel6st. Es enthie l t e twa die H~lfte der Gesamtak t iv i t / i t . Du tch Zugabe von kris t . (NH4)2SO 4 wurde die L6sung auf 65 %ige Si i t t igung gebracht , dann 20 min bei
Biochim. Biophys. Acta, 90 (1964) 249-259
254 E. OHMANN
15ooo × g und o ° zentrifugiert, das Sediment in 15 ml o.o5 M Phosphat-o.oo4 M Cystein-Puffer (pH 7.0) gel6st und 3 Std gegen den gleichen Puffer dialysiert. Diese L6sung wurde dann erneut mit Aceton yon --IO ° fraktioniert. Die Fraktion zwischen 52 und 60 % Acetongehalt enthielt den Hauptanteil der Aktivit/it und wurde nach Zentrifugation in 5 ml Phosphat-Cystein-Puffer gel6st. Diese L6sung wurde dann zur 2. (NH4)2SO4-Fraktionierung bei o ° und pH 6.8 mit festem (NH4)2S0 4 versetzt bis zur S/ittigung von 50 %. Dann wurde 20 rain bei 20000 × g zentrifugiert. Das Sediment enthielt meist nur geringe Aktivit/it. Es wurde verworfen und der 13berstand weiter bis zu 60 %iger S/ittigung mit (NH4)2S0 4 versetzt. Diese Fraktion zwischen
~10 o
0
%
• ~ i • -ADP
~ - o + ADP
Zeit (rain) Fig. 2. Die Erfordernis der Gegenwar t yon ADP fiir den Umsa tz von Acetylphosphat . Die An- siitze enthiel ten im Volumen von I ml in #Molen: ioo Tris-Puffer (pH 7.4), io MgC12, io Cystein, IO ADP, IO Acetylphosphat , zell- freien E x t r a k t aus Stichococcus mi t 3 mg Pro- tein, frei yon CoA, 3 o°. Nach den angegebenen Zeiten wurden zu den Proben 2 ml i M Hydroxyl - amin gegeben. Weitere Aufarbei tung siehe method. Teil. Auswer tung gegen Kontrolle ohne Enzym. @ - - @ , Ans~tze ohne ADP;
O - - O , Ansiitze mi t ADP.
25 A
2°1
5 / ~~'phosphoi
Zeit (rain) Fig. 3. Bilanzmessungen zur Aeetat-Kinase- Reaktion. Die AnsAtze enthielten im Volumen yon 3 ml in #Molen: 3oo Tris-Puffer (pH 7-4), 3 ° MgC12, 25 Cystein, 2.35 ADP, zellfreien E x t r a k t aus Stichococcus mi t 8 mg Protein, endogenes Co& dutch Adsorpt ion an Dowex-I eliminiert, I nkuba t ion bei 3 o°. Der Star t der :Reaktion erfolgte durch Zugabe von 16 #Molen Acetylphosphat . Zu den angegebenen Zeiten wurden den Ansiitzen zur Bes t immung des Acetylphosphates i ml en tnommen, schnell in Zentrifugengl~ser mi t 2 ml I M Hydroxy lamin pipet t ier t und wie im methodischen Teil be-
schrieben analysiert . Zu den verbleibenden 2 ml wurde sofort HC104 zum Abs toppen der Reakt ion gegeben. Nach scharfer Zentr i fugat ion erfolgte die Analyse auf ATP und ADP in al iquoten
Teilen des ?3berstandes.
T A B E L L E I I I
STOCHIOMETRIE DER ACETAT-KINASE-REAKTION (R/,)CKREAKTION)
Allgemeiner Ansatz und Bedingungen wie in Fig. 3. Volumen 3 ml, Protein 5 rag, Inkubat ionsze i t 5 rain.
A Acetylphosphat A A D P A A T P (taMole) (l~Mote) (l, Mole)
--9.5 --8.3 +8 .2
Biochim. Biophys. Acta, 9 ° (1964) 249-259
ACETATAKTIVIERUNG IN GRUNALGEN 255
5o und 6o %iger S~tttigung enthielt den Hauptanteil der Aktivit~tt und wurde nach Samm|ung durch 20 min Zentrifugation bei 20000 × g in 2 ml Phosphat-Cystein- Puffer gelSst. Sie enthielt 5 mg Protein und hatte eine spez. AktivitAt yon lO5 E/rag Protein. Die Gegenwart yon Cystein w~hrend des Aufarbeitungsganges ist notwendig. Zur Vermeidung grSsserer Aktivit~itsverluste sind die Produkte der Acetonfraktio- nierung mSglichst schnell weiter zu verarbeiten.
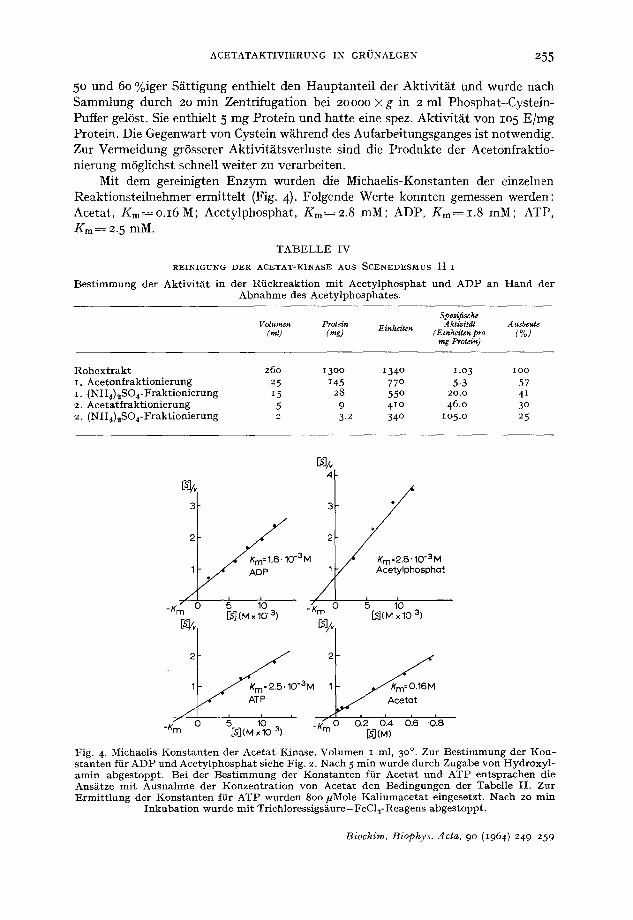
Mit dem gereinigten Enzym wurden die Michaelis-Konstanten der einzelnen Reaktionsteilnehmer ermittelt (Fig. 4). Folgende Werte konnten gemessen werden: Acetat, Km~O.I6M; Acetylphosphat, Kin----2.8 raM; ADP, Km=I .8 mM; ATP, Km--~ 2.5 raM.
T A B E L L E IV
R ] ~ I N I G U N G D E R A C I ~ T A T - K I N A S E A U S S C E N E D ] ~ S M U S H I
B e s t i m m u n g der Aktivi t~t in der Riickreaktion mi t Ace ty lphospha t und ADP an Hand der A bnahm e des Acetylphosphates .
Speaifiscke Volumen Protein E inkeiten A ktivitiit A usbeute
(ml) (rag) (Einheiten pro ( % ) mg Protein)
Rohex t r ak t 26o i . Acetonfrakt ionierung 25 I. (NH~) ~SO4-Fraktionierung 15 2. Aceta t f rakt ionierung 5 2. (NHa) 2SO4-Fraktionierung 2
13oo 134 ° 1.o 3 IOO 145 77 ° 5-3 57
28 55 ° 2o.o 41 9 4 lo 46.o 3 ° 3.2 34 ° lO5.O 25
3
2
1 ~ 1 o -3 M f ADP
[S]•l 3
2
1
/ / Acetylphl::;~Mt
_ ~ " / 0 ~S] (M x ll0 10 3) /
_Km 0 [S](M x 10 3)
1 O-3M 1 / q [ / ~ ATP ~ Acetat
/ ( " I I I I f l I I I I
.Kin O 5 10 _K~ n O 0.2 0.4 0.6 0.8 [s](M xlO 3) Es](M)
Fig. 4. Michael is-Konstanten der Acetat-Kinase. Volumen i ml, 3 o°. Zur Best in lmung der K o n - s t an ten fiir ADP und Acety lphospha t siehe Fig. 2. Nach 5 rain wurde du tch Zugabe von Hydroxyl - amin abgestoppt . Bei der Bes t immung der Kons tan ten {iir Acctat und ATP entsprachen die Ans~itze mi t Ausnahme der Konzent ra t ion von Acetat den Bedingungen der Tabelle I I . Zur E rmi t t l ung der Kons t an t en fiir ATP wurden 8o0 #Mole Kal iumace ta t eingesetzt. Nach 2o min
Inkuba t ion wurde mi t Trichloressigsiiure-FeC13-Reagens abgestoppt .
B i o c h i m . B i o p h y s . Ac ta , 90 (1964) 249-259

256 E. OKMANN
Diese Ergebnisse lassen eine befriedigende i3bereinstimmung mit den an Bak- terien gewonnenen Werten erkennen ~9. Der sehr hohe Wert ftir Acetat ist auffiillig. Er fibertrifft den ffir die Acetyl-CoA-Synthetase aus Euglena ermittelten um zwei Zehnerpotenzen 25. In vergleichenden Experimenten hatte sich ergeben, dass diese schon bei Acetatkonzentrationen nahezu v611ig gehemrnt ist, die ffir die Acetat-Kinase den Bereich der Halbs/ittigung darstellen.
Der Einsatz von [32p]Acetylphosphat in die Reaktion der Acetat-Kinase
Der t,)bergang der Phosphatgruppe des Acetylphosphates auf das ADP ist unter Verwendung yon [32p]Acetylphosphat leicht darstellbar. Die Tabelle V enth~ilt die Darstellung eines Versuches dieser Art. Er bringt noch einma! eine Best~itigung der auf andere Weise gewonnenen Ergebnisse. Radioaktivit~tt tr i t t nur dann im ATP auf, wenn das Enzym aktiv ist und wenn ADP in den Ansatz gegeben wird.
T A B E L L E V
I)IE BILOUNG VON [32p]ATP Aos A D P OND ~3gp~ACETYLPHOSPI-IAT
Dureh f i i h rung in Zentrifugengli~sern, die AnsO.tze en th i e l t en im Volumen von i ml in /zMolen: ioo Tr is -Puffer (pH 7.4), IO MgC12, i o Cystein, 3 ADP, 3 [32PlAcety lphosphat (etwa IOOOOO Im- pulse /min) , 20 E i n h e i t e n Ace ta t -Kinase . N a c h den angegebenen Ze i ten wurde die R e a k t i o n durch Zugabe von HC10 4 abges topp t . Nach E n t f e r n u n g des P ro t e i n Adsorp t ion der Nucleo t ide an
Akt ivkoh le .
Nr. A nsatz lnkubationszeit Impulse/rain in A TP
I k o m p l e t t o i oo 2 k o m p l e t t 5 4 °60 3 k o m p l e t t IO 8220 4 gekoch tes E n z y m i o i oo 5 ohne A D P IO 16 5
DISKUSSION
Voraussetzung fiir die ]3eteiligung der Essigs/iure an biochemischen Reaktionen ist ihre vorherige Aktivierung. Erst in der Form des Acetyl-CoA nimmt sie die bekannte dominierende Position als Schltisselsubstanz fiir viele Bezirke des Stoffwechsels ein. Wenn Mikroorganismen autotroph wachsen oder ihren Energiebedarf und ihre C-Ske- lette ftir Neusynthesen im wesentlichen aus dem Abbau von Kohlenhydraten oder Fetten beziehen, ist die im Zuge dieser Vorg/tnge entstehende Essigs/iure bereits immer an CoA gebunden. Die Notwendigkeit eines besonderen Mechanismus ftir die Akti- vierung der freien S/iure besteht daher hier nicht in dem Masse wie unter Bedingungen, wo Acetat den Zellen von aussen zur Verftigung gestellt wird.
In Untersuchungen an autotrophen Euglena-Zellen hatte sich ergeben 25, dass die Oxydation des Acetats anfangs gering ist und mit stetig zunehmender Geschwin- digkeit erst nach einigen Stunden ihr Maximum erreicht. Heterotroph auf Acetat als C-Quelle herangezogene Zellen sind dagegen sofort zur kr/iftigen Oxydation des Acetats bef/ihigt. Konform zum komplexen Vorgang des Gaswechsels der intakten Zelle ist im zellfreien Extrakt heterotropher Zellen yon Euglena auch die Aktivit/it der fiir die Aktivierung des Acetats verantwortlichen Acetyl-CoA-Synthetase um ein Mehrfaches erh6ht. Im Unterschied zu diesen Verh/iltnissen bei Euglena liegt bei
B i o c h i m . B iophys . Ac la , 9 ° (i964) 249-259

ACETATAKTIVIERUNG IN GRf3NALGEN 257
der Acetatoxydation in Stichococcus kein Induktionsprozess vor (Fig. I). ,~hnliches gilt auch f~r Chloretla pyrenoidosa und die beiden Scenedesmus-St~nlme. Die Experi- mente haben ergeben, dass der Mechanismus der Acetataktivierung in Stichococcus bacillaris und den anderen untersuchten St~nmen nicht wie bei Euglena gracilis ~5 durch Acetyl-CoA-Synthetase, sondern durch die bisher ausschliesslich in Bakterien nachgewiesene Acetat-Kinase 2~-~9 katalysiert wird. Als Folge dieser Reaktion wird Acetylphosphat gebildet. Da dieses aber nur in wenige biochemische Reaktionen einzutreten vermag und die universell verwendbare aktive Form des Acetats bei allen Organismen immer das Acetyl-CoA ist, wird in alien bisher bekannten Fiillen die Acetat-Kinase dutch eine von STADTMAN UND BARKER ~ zuerst in Clostridium kluyveri entdeckte Transacetylierungsreaktion komplettiert. Hierbei wird an der Phospho-Transacetylase (Acetyl-CoA:Orthophosphat-Acetyltransferase, EC 2.3.1.8) die Acetylgruppe des Acetylphosphates unter Bildung von Acetyl-CoA auf CoA fibertragen. P1 wird frei:
Acetylphosphat + CoA ~ Acetyl-CoA + PI (3)
Es bestehen Anhaltspunkte, dass diese Reaktion auch in Stichococcus abl~iuft. Da- durch w~ire auch die Tatsache, class die Gegenwart yon CoA eine geringftigige Stimu- lierung der Bildung yon Acethydroxams~ture in den Extrakten bewirkt, einer Er- kl~ung zug~nglich. In diesem Falle n~imlich k6nnte ein Teil des Acetylphosphates mit CoA zum Acetyl-CoA reagieren, noch bevor es restlos durch Hydroxylamin gebunden ist. Dann wiirden sowohl Acetylphosphat wie Acetyl-CoA durch Hydroxyl- amin gespalten. Dadurch k6nnte unter den gegebenen experimentellen Bedingungen die Bildung yon AcethydroxamsAure beschleunigt werden. Nach einer Erw~rmung des Ansatzes von 20 min auf 55 ° ist die M6glichkeit einer Beeinflussung der Bildung von Acethydroxams~iure durch CoA nicht mehr gegeben (Tabelle II). Es ist bekannt, dass bei dieser Temperatur die Phospho-Transacetylase vollst§mdig inaktiviert wird 41. Die Acetat-Kinase wird nut unwesentlich geschAdigt.
Obwohl die Evolution in Bezug auf die Bildung yon Acetyl-CoA aus freier Essigs~iure in Euglena und den bier untersuchten Arten der Chlorococcales zur Aus- bildung enzymatisch unterscbiedlicher Mechanismen innerhalb der Algen gefiihrt hat, sind doch die molekularen VorgAnge der durch die verscbiedenen Enzyme kataly- sierten Reaktionen sehr ~ihnlich. In beiden Fiillen besteht der primate Reaktions- schritt in einem nucleopbilen Angriff der ungepaarten Elektronen des Acetatcar- boxyls auf eine Pyrophosphatbindung im ATP 21. Im Falle der Acetat-Kinase erfolgt der Angriff auf die Bindung des terminalen P-Atoms, und es entsteht Acetylphosphat als frei dissoziables Zwischenprodukt, das dann in einer Folgereaktion an einem zweiten Enzym, der Phospho-Transacetylase, mit CoA zum Acetyl-CoA weiterrea- giert. Im Falle der Acetyl-CoA-Synthetase in Euglena erfolgt der Angriff in/ihnlicher Weise auf das AdenylsAure-P-Atom. Er ffihrt zur Bildung yon Adenylacetat und zur Abspaltung von Pyrophosphat. Das Adenylacetat bleibt am Enzym haften und kann mit CoA zum Acetyl-CoA weiterreagieren. Dieser zweite Teilprozess wird vom gleichen Enzym katalysiert.
Die weite Verbreitung der anorganischen Pyrophosphatase bedingt, dass die nach diesem Mechanismus ablaufende Acetataktivierung in vivo zu einem irrever- siblen, auf die Synthese von Acetyl-CoA ausgerichtetem Vorgang wird. Die Reaktion an der Acetat-Kinase ist auch in vivo reversibel. Bei einigen Bakterien ist die physio-
Biochim. Biophys. Acla, 90 (I964) 249-259

258 E. OHMANN
logische Bedeutung dieses Enzyms offenbar darin zu sehen, dass es im Dienste der Bildung yon ATP steht.
0 b sich aus der Ausbildung beider enzymatischer Mechanismen der Acetatakti- vierung innerhalb der Algen Gesichtspunkte ffir eine Chemotaxonomie gewinnen lassen, mfissen Untersuchungen ergeben, die im Gange sind.
DANK
Dank sagen m6chte ich an dieser Stelle Hern Professor Dr. K. MOTHES, Halle, ffir die Erm6glichung der Arbeit und ihre st~tndig f6rdernde Untersttitzung sowie Herrn Professor Dr. H. G. SCHLEGEL, G6ttingen, ffir beratende Diskussion und l~berlassung yon Chemikalien.
ZUSAMMENFASSUNG
I. Ruhende Zellen von Stichococcus bacillaris sind zu einem schnellen Umsatz von Acetat bef/ihigt. Im Gegensatz zu frfiheren Untersuchungen an Euglena gracilis liess sich bier keine lag-Phase der 02-Aufnahme beobachten. Die O~-Aufnahme er- reicht sofort nach Zugabe von Acetat zu atmenden Zellen ihr maximales Ausmass. Vom zugesetzten Acetat werden etwa 50 % oxydiert, der andere Teil wird assimiliert.
2. Obgleich in den zellfreien Extrakten die Bildung yon Acethydroxams~ure auch dann erfolgt, wenn diese frei sind yon CoA, ist nach Zugabe yon CoA doch eine geringe Stimulierung zu beobachten. In Reaktionsans/itzen, die 20 min auf 55 ° erhitzt wurden, hatte die Zugabe yon CoA keinen Einfluss auf die Bildung der Hydroxams/iure.
3. In kurzzeitigen Versuchen wird Acetylphosphat von den Rohextrakten nur dann umgesetzt, wenn ADP als Phosphatacceptor gegeben wird. Zwischen der Ab- nahme yon Acetylphosphat und ADP sowie der Bildung yon ATP bestehen st6chio- metrische Zusammenh/inge.
4- Alle Experimente sprechen sehr daffir, dass in Stichococcus bacillaris und einigen anderen Ar t ende r Chlorococcales die Acetataktivierung durch die Bildung yon Acetylphosphat eingeleitet wird. Die Acetat-Kinase (EC 2.7.2. I), das ftir diese Reaktion verantwortliche Enzym, ist gereinigt worden. Die Bedingungen der Reaktion wurden untersucht und die Michaelis-Konstanten bestimmt.
L I T E R A T U R
1 E. G. PRINGSHEIM, in W. RUHLAND, Handbuch der Pflanzenphysiologie, B a n d i i , Springer . Ber l in , 1959, S. 303 .
2 0 . KANDLER UND H. ERNST, Planta, 46 (1955) 46. 3 H. G. SCHLEGEL, Planta, 47 (1956) 51°. 4 H. G. SCHLEGEL, Z. Naturforsch., I 4b (1959) 246. 5 H. L. KORNBERG UND H. A. KREBS, Nature, 179 (1957) 988. 6 H. C. REEVERS, S. KADIS UND S. AJL, Biochim. Biophys. Acta, 57 (1962) 403 . 7 W. WIESSNER UND A. KUHL, Vortrage aus dem Gesamtgebiet der Botanik, Neue Folge, i (1962) lO2. $ E. OHMANN, in Vorbere i tung . 9 p. j . SYRETT, M. J. MERRETT UND S. M. BOCKS, f . Exptl. Bot., 14 (1963) 249.
10 F. LIPMANN, J. Biol. Chem., 16o (1945) 173. 11 T. C. CHou UND F. LIPMANN, f . Biol. Chem., 196 (1952) 89. 12 IX. BEINERT, D. E. GREEN, P. HELE, H. HIFT, lc{. W. MON KORFF UND C. V. RAMAKRISHNAN,
J. Biol. Chem., 203 (1953) 35. 13 W. P. JENCKS UND F. LIPMANN, J. Biol. Chem., 225 (1957) 207. 14 p. HELE, J. Biol. Chem., 2o6 (1954) 671. 15 F. CAMPAGNARI UND L. T. WEBSTER, J. Biol. Chem., 238 (1963) 1628.
Biochim. Biophys. Acta, 90 (1964) 249-259

ACETATAKTIVIERUNG IN GR/JNALGEN 259
is M. A. EISENBERG, f . Biol. Chem., 203 (1953) 815. 17 A. MILLERD UND J. BONNER, Arch. Biochem. Biophys., 49 (1954) 343. i8 F. LIPMANN, 31[. E. JONES, S. BLACK UND R. M. FLYNN, J. Am. Chem. Soc., 74 (1952) 2384. 19 M. E. JONES, Federation Proe., 12 (1953) 7 °8. z0 M. E. JONES, F. LIPMANN, H. HILZ UND F. LYNEN, J. Am. Chem. Soc., 75 (1953) 3285. 21 p. BERG, J. Biol. Chem., 222 (1956) 991. 22 M. A. EISENBERG, Biochim. Biophys. Aeta, 16 (1955) 58. 23 M. A. EISENBERG, Bioehim. Biophys. Ae/a, 23 (1957) 327 • 24 E. OHMANN, Naturwissenschaflen, 5 ° (1963) 578. 25 E. OHMANN, Biochim. Biophys. Acta, 82 (1964) 325. 26 F. LIPMANN, J. Biol. Chem., 155 (1944) 55. 27 N. O. KAPLAN UND F. LIPMANN, Federation Proc., 7 (1948) 163- 28 E. R. STADTMAN UND H. A. BARKER,.J. Biol. Chem., 184 (195o) 769. 29 j . A. RosE, M. GRUNBERG-MANAGO, S. R. KOREY UND S. OCI-IOA, J. Biol. Chem., 211 (1954) 737- 3o O. KANDLER, Z. Naturforsch., 9b (1954) 625. 21 H. CHANTRENNE UND F. LIPMANN, J. Biol. Chem., 187 (195 o) 757. 82 F. LIPMANN UND L. C. TUTTLE, J. Biol. Chem., 159 (1945) 21. 3 3 E. R. STADTMAN, in S. P. COLOWICK UND N. O. KAPLAN, Methods in Enzymology, Vol. 3,
Academic Press, New York, 1957, p. 231. 34 Report of/he Commission on Enzymes of/he International Union of Biochemistry, Pergamon Press,
Oxford, 1961. 35 H. ADAM, Biochem. Z., 335 (1961) 25. 26 O. H. LOWRY, N. J. ROSEBROUGH, A. L. FARR UND R. J. RANDALL, J. Biol. Chem., 193 (1951)
265 37 F. LYNEN, J. KNAPPE, E. LORCH, G. JOTTING, E. RINGELMANN UND J. P. LACHANCE, Biochem.
Z., 335 (1961) 123. 38 W. W. UMBREIT, R. H. BORRIS UND J. F. STAUFFER, Manometric Techniques, Burgess, Minnea-
polis, 1957. 39 A. W. D. AVlSON, J. Chem. Soc., (1955) 732. 4o A. KORNBERG, S. R. KORNBERG UND E. S. SIMMS, Biochim. Biophys. Acta, 20 (1956) 215. 41 J. R. STERN UND S. OCHOA, f . Biol. Chem., 191 (1951) I6x.
Biochim. Biophys. Acta, 90 (1964) 249-259