Embed Size (px)
Citation preview

M. Siniatchkin1 · K. Gröning2 · F. Moeller2 · N. Japaridze2 · U. Stephani2
1 Klinik für Psychiatrie, Psychosomatik und Psychotherapie des Kindes- und Jugendalters,
Johann-Wolfgang Goethe-Universität zu Frankfurt am Main, Frankfurt2 Klinik für Neuropädiatrie, Christian-Albrechts-Universität, Kiel
Bildgebung bei epileptischen Enzephalopathien
Trotz einzelner Studien ist das Wis-sen über epileptische Enzephalopa-thien (EE) lückenhaft, und es besteht nach wie vor dringender Bedarf, die Mechanismen kognitiver Defizite bei EE zu untersuchen. Besonders we-nig ist über neuronale Netzwerke be-kannt, die den EE zugrunde liegen, und über Mechanismen, wie die epi-leptische Aktivität mit kognitiven Funktionen interferiert und auf diese Art und Weise kognitive Defizite ver-ursacht. Die Bildgebung kognitiver Defizite kann hier einen wichtigen Beitrag leisten.
Hintergrund
Epileptische Enzephalopathien (EE) ge-hören zu der Gruppe von Epilepsien, die mit kognitiven Defiziten und Verhal-tensstörungen infolge epileptischer Akti-vität einhergehen [3, 26]. Die EE stellen ein großes gesundheitliches Problem dar, da sie häufig schwer auf eine antiepilep-tische Behandlung ansprechen und mit einer ungünstigen Prognose für Entwick-lung des Kindes assoziiert sind [14], Trotz großer Bedeutung sind die Vorstellun-gen über pathogenetische Mechanismen von EE unzureichend untersucht. Beson-ders wenig ist über neuronale Netzwer-ke bekannt, die den EE zugrunde liegen, und über Mechanismen, wie die epilepti-sche Aktivität mit kognitiven Funktionen interferiert und auf diese Art und Weise kognitive Defizite verursacht [26, 43].
Der vorliegende Überblick fasst Studien zur funktionellen Bildgebung zusammen, die zum Ziel hatten, spezifische neurona-le Netzwerke bei EE zu beschreiben.
Bildgebung kognitiver Defizite
Ein möglicher Mechanismus, wie die epi-leptische Aktivität kognitive Defizite ver-ursachen könnte, bezieht sich auf die Interaktion zwischen epileptischen und kognitiven neuronalen Netzwerken. Es ist beispielsweise möglich, dass die epi-leptische Aktivität die Aktivität im Ruhe-netzwerk unterbricht („Default-mode“-Netzwerk, DMN, [19]). Dieser Mechanis-mus wurde am häufigsten diskutiert. Das DMN besteht aus eng interagierenden Hirnarealen wie Praecuneus, lateraler pa-rietaler und anteriorer frontaler sowie re-trosplenialer Kortex [7]. Man nimmt an, dass das DMN eine günstige neurometa-bolische Umgebung für jegliche kogniti-ve Prozesse bildet und die physiologische Grundfunktion darstellt, die für die kog-nitive Leistung sowie die Integration ko-gnitiver und emotionaler Erfahrungen wichtig ist [36]. Veränderte Aktivität im DMN oder gestörte Konnektivität zwi-schen Strukturen des DMN könnte die Leistungsbereitschaft beeinträchtigen, die Performanz reduzieren, zur Störung der Aufmerksamkeit und des Arbeitsge-dächtnisses führen sowie zur Entstehung neuropsychiatrischer Erkrankungen wie Aufmerksamkeitsdefizit- und Hyperak-tivitätssyndrom oder Autismus beitragen
[7, 16]. Zudem resultiert die Beeinträchti-gung der Aktivität im DMN in Vigilanz-schwankungen und Bewusstseinsstörun-gen [6]. Es kann angenommen werden, dass die Störung der Aktivität im DMN, z. B. dessen Deaktivierung infolge epilep-tischer Aktivität, kognitive Defizite ver-ursachen kann [19]. Eine derartige De-aktivierung wurde sowohl bei primär als auch sekundär generalisierten Paro-xysmen und bei Absencen beschrieben (. Abb. 1; [1, 19, 20, 30, 31]). Sie könn-te die Beeinträchtigung des Bewusstseins oder der Vigilanz bei Absencen und ge-neralisierten Paroxysmen widerspiegeln [19]. Die gestörte Konnektivität im DMN wurde bei verschiedenen epileptischen Syndromen beschrieben [46, 47], u. a. auch bei typischen EE wie dem „Continu-ous spike-and-wave syndrome“ (CSWS) und dem Lennox-Gastaut-Syndrom (LGS; [29, 35, 40]). Die Interaktion zwi-schen dem DMN und der epileptischen Aktivität ist jedoch nicht der einzige Me-chanismus der EE. Epileptische Aktivität kann darüber hinaus auch die Konnekti-vität zwischen kognitiven Netzwerken so-wie innerhalb kognitiver Netzwerke be-einträchtigen und dadurch kognitive De-fizite verursachen [4]. Diese Hypothese muss jedoch in zukünftigen Studien de-taillierter untersucht werden. Besonders mangelt es an den Studien, die direkt die Interaktion zwischen kognitiven Leistun-gen, neuronalen Netzwerken und epilep-sietypischen Entladungen untersuchen [43]. Andererseits kann auch versucht
Leitthema
Z Epileptol 2014 DOI 10.1007/s10309-013-0354-y© Springer-Verlag Berlin Heidelberg 2014
1Zeitschrift für Epileptologie 2014 |

werden, indirekt Schlüsse über Mecha-nismen kognitiver Beeinträchtigung bei EE zu ziehen, indem neuronale Netzwer-ke untersucht werden, die mit EE assozi-iert sind.
West-Syndrom
Das West-Syndrom stellt einen Prototyp der schweren EE des Säuglingsalters dar, die mit Blitz-Nick-Salaam-Anfällen, psy-chomotorischer Entwicklungsverzöge-rung und einem charakteristischen EEG-Muster von Hypsarrhythmie einhergeht [14, 15, 21]. Frühere Positronen-Emis-sions-Tomographie(PET)-Studien ha-ben ein spezifisches Netzwerk, bestehend aus einem fokalen Hypometabolismus in verschiedenen kortikalen Arealen und einem Hypermetabolismus in Putamen und Hirnstamm, beschrieben, das mit der Hypsarrhythmie assoziiert ist [9, 10, 27]. Basierend auf den vorgenannten Studien war jedoch nicht klar, ob dieses Netzwerk auf epilepsietypische Potenzia-
le im Rahmen der Hypsarrhythmie oder auf die hochamplitudige langsame Hirn-aktivität zurückgeführt werden kann. In einer kombinierten Studie aus EEG und „functional magnetic resonance imaging“ (fMRI, [39]) untersuchten Siniatchkin et al. Säuglinge mit dem West-Syndrom und zeigten, dass die epilepsietypischen Entla-dungen die Aktivierung in den kortikalen Arealen (besonders im okzipitalen Kor-tex) und die hochamplitudige δ-Aktivi-tät die Veränderungen des „Blood-oxy-genation-level-dependent“(BOLD)-Sig-nals in subkortikalen Strukturen wie Pu-tamen, Hirnstamm und Thalamus verur-sachen (. Abb. 1). Es wurden keine De-aktivierungen, insbesondere im DMN, assoziiert mit den pathologischen Eigen-schaften der Hypsarrhythmie festgestellt. Dieses beschriebene Muster der Aktivie-rung scheint spezifisch für das West-Syn-drom zu sein, da bei keinem der Säuglin-ge mit fokalen Epilepsien aus der Kont-rollgruppe ähnliche Aktivierungsmuster beschrieben werden konnten. Interessan-
terweise konnten in einer nachfolgenden Quellenanalyse in der Frequenzdomä-ne („dynamic imaging of coherent sour-ces“) ähnliche Quellen für die hocham-plitudige pathologische δ-Aktivität wie in der vorgenannten EEG-fMRT-Studie beschrieben werden (okzipitaler Kor-tex, Putamen, Hirnstamm). Zusätzlich ist es mithilfe der Quellenanalyse mög-lich, die Hierarchie zwischen den Quel-len zu untersuchen. So ergab die spezi-fische Analyse des Informationsflusses zwischen den Quellen, dass der Hirn-stamm in dem beschriebenen Netzwerk eine dominierende Rolle spielt: Der In-formationsfluss scheint vom Hirnstamm zu den anderen kortikalen und subkor-tikalen Strukturen zu laufen [22]. Da-mit scheint der Hirnstamm eine zentra-le Rolle in den Mechanismen der Hyps-arrhythmie zu spielen. Es kann angenom-men werden, dass die pathologische Akti-vierung im Hirnstamm dafür verantwort-lich ist, dass es während der BNS-Anfälle bei Patienten mit West-Syndrom zu einer
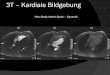
Abb. 1 8 Aktivierungsmuster bei diskutierten Syndromen, die mit kognitiven Defiziten einhergehen: Absence-Epilepsie, West-Syndrom, Lennox-Gastaut-Syndrom (nach [30, 39, 41], mit freundlichen Genehmigung von Wiley & Sons) und „Continu-ous spike-and-wave syndrome“ (CSWS; nach [40], mit freundlicher Genehmigung von Oxford University Press)
2 | Zeitschrift für Epileptologie 2014
Leitthema

Enthemmung spinaler Reflexe kommt, die zu myoklonieähnlichen reflexartigen bilateralen Anfällen führen kann. Die Hypsarrhythmie und die hiermit ver-bundene multifokale Aktivität resultie-ren wahrscheinlich auch aus der Hyper-aktivierung aufsteigender Verbindungen vom Hirnstamm, hauptsächlich aus den diffusen Einflüssen der retikulären For-mation auf den zerebralen Kortex [21]. Deswegen scheint die Hypsar rhythmie beim West-Syndrom infolge der Inter-aktion zwischen kortikalen und subkor-tikalen Strukturen zu entstehen; vermut-lich resultiert sie nicht aus einer kortika-len Struktur mit Schrittmacherfunktion [3, 8].
Lennox-Gastaut-Syndrom
Der Hirnstamm scheint eine entschei-dende Komponente im epileptischen neuronalen Netzwerk bei Patienten mit LGS zu repräsentieren. Das LGS entwi-ckelt sich oft aus dem West-Syndrom. Es ist eine EE, die einerseits mit einem spezi-fische Typus von Anfällen einhergeht (to-nische, tonisch-klonische und atonische Anfälle sowie atypische Absencen), an-dererseits auch mit den typischen EEG-Veränderungen (1- bis 2,5/s-“slow-spike-wave“-Komplexe und „polyspikes“) so-wie mentaler Retardierung verbunden ist [2]. Eine EEG-fMRT-Studie an Kindern mit kryptogenem und symptomatischem LGS beschrieb eine Aktivierung in Hirn-stamm und Thalamus in einem Grup-penvergleich (. Abb. 1; [41]). Die ande-re EEG-fMRT-Studie, die an Kindern und Erwachsenen mit LGS durchgeführt wor-den ist, zeigte, dass die paroxysmale Po-lyspike-Aktivität („paroxysmal fast activi-ty“) mit dem Anstieg hämodynamischer Veränderungen in Arealen des assozia-tiven Kortex, aber auch in Hirnstamm, Thalamus und Basalganglien assoziiert ist [34]. Der Hirnstamm spielt eine wesent-liche Rolle bei der Generierung tonischer Anfälle (diese sind ein wichtiger Bestand-teil der Symptomatologie bei LGS):F Tonische Anfälle wurden auch bei
Patienten mit einer Hydranenzepha-lie beobachtet [44].
F Diese Anfälle können durch eine Kallosotomie nicht beeinflusst wer-den [37].
F Die tonischen Anfälle verschwinden nicht nach der Resektion der mögli-chen kortikalen epileptogenen Zone [5].
Da der Hirnstamm über die retikulä-re Formation einen diffusen Einfluss auf die kortikale Aktivität ausübt, kann ange-nommen werden, dass diese diffuse Ak-tivierung die Multifokalität bei Patienten mit LGS miterklären kann [21]. In der Tat fand eine vor Kurzem durchgeführte Stu-die eine erhöhte Konnektivität zwischen
Arealen des DMN bei Kindern mit mul-tifokalen Epilepsien; mehrere von ihnen litten unter dem LGS [28, 29]. Diese er-höhte funktionelle Konnektivität könnte drauf hinweisen, dass das Gehirn von Pa-tienten mit LGS zu der multifokalen epi-leptischen Aktivität prädisponiert. Neben dem Hirnstamm scheint auch der Thala-mus eine besondere Rolle bei den Me-chanismen epileptischer Entladungen in Patienten mit LGS zu spielen. Die Studie von Siniatchkin et al. [41] demonstrierte, dass der zentromediane und der anterio-
Zusammenfassung · Abstract
Z Epileptol 2014 · [jvn]:[afp]–[alp] DOI 10.1007/s10309-013-0354-y© Springer-Verlag Berlin Heidelberg 2014
M. Siniatchkin · K. Gröning · F. Moeller · N. Japaridze · U. StephaniBildgebung bei epileptischen Enzephalopathien
ZusammenfassungEpileptische Enzephalopathien (EE) gehören zu der Gruppe von Epilepsien, die mit ausge-prägten kognitiven Defiziten und Verhaltens-störungen infolge epileptischer Aktivität ein-hergehen. Trotz großer Bedeutung von EE für die Entwicklung des betroffenen Kindes und dessen psychosozialer Anpassung sind bis-lang Mechanismen kognitiver Defizite bei EE unzureichend untersucht worden. Der vor-liegende Überblick fasst Studien zusammen, die zum Ziel hatten, spezifische neuronale Netzwerke bei EE zu beschreiben. Diese zei-gen, dass, obwohl epileptische Aktivität bei EE in unterschiedlichen Gehirnregionen ge-neriert werden kann, spezifische Propaga-tionswege und Netzwerke existieren, die sehr charakteristisch für die jeweilige Art der En-zephalopathie sind. Bei einigen EE scheint
die epileptische Aktivität die Integrität des Ruhe(„Default-mode“)-Netzwerks zu beein-trächtigen und möglicherweise über diesen Mechanismus mit kognitiven Funktionen zu interferieren. Darüber hinaus liegen den EE weitere Mechanismen zugrunde, z. B. eine gestörte Konnektivität zwischen Gehirnarea-len, die an der Steuerung kognitiver Funktio-nen beteiligt sind, oder Beeinträchtigungen der Verbindungen bzw. Aktivität in thalamo-kortikalen Regelkreisen.
SchlüsselwörterWest-Syndrom · Lennox-Gastaut-Syndrom · Dravet- Syndrom · „Continuous spike-and-wave syndrome“ · Funktionelle Magnetresonanztomographie
Imaging of epileptic encephalopathies
AbstractEpileptic encephalopathies (EE) belong to the group of epilepsies which are associated with expressed cognitive and behavioral distur-bances subsequent to epileptic activity. De-spite the great importance of EE in the devel-opment and psychosocial adaptation of af-fected children, the mechanisms of cognitive deficits in EE have so far been insufficiently investigated. This review article summarizes the various neuroimaging studies which have tried to describe specific neuronal networks in EE. The results show that although epilep-tic activity in EE can be generated in different brain regions, specific propagation pathways and networks exist which are very character-istic for each different form of encephalopa-thy. In some forms of EE the epileptic activi-
ty seems to impair the integrity of the default mode network and possibly to interfere with cognitive function through this mechanism. Furthermore, there are further mechanisms underlying EE, e.g. abnormal connectivity patterns between brain regions which partic-ipate in the control of cognitive functions and impairment of the connections and activities in the thalamocortical network.
KeywordsWest syndrome · Lennox-Gastaut syndrome · Dravet syndrome · Continuous spike-and-wave syndrome · Functional magnetic resonance imaging
3Zeitschrift für Epileptologie 2014 |

re thalamische Kern in das mit dem LGS verbundene Netzwerk involviert sind. Die Beteiligung des Thalamus an der Ge-nerierung epileptischer Aktivität bei Pa-tienten mit LGS wurde auch durch die Studie von Pillay et al. [34] bestätigt. Auf die Bedeutung thalamokortikaler Regel-kreise in der Pathogenese von LGS wur-de schon mehrmals hingewiesen:F Die tiefe Hirnstimulation des zentro-
medianen thalamischen Kerns führte zu einer deutlichen Verbesserung des klinischen Verlaufs bei Patienten mit medikamentenrefraktärem LGS [45].
F Der anteriorer Thalamus wurde häu-fig und erfolgreich zur Reduktion der Anfallsbereitschaft durch die tie-fe Hirnstimulation bei multifokalen Epilepsien genutzt [38].
Diese therapeutischen Optionen unter-mauern die Bedeutung des Thalamus in der Pathogenese des LGS.
„Continuous spike-and-wave syndrome“
Das CSWS ist mit dem Erscheinen in-teriktaler epileptischer Aktivität in mehr als 85% des Slow-wave-Schlafs verbun-den. Dieses EEG-Merkmal kann sich aus unterschiedlichen Epilepsieformen ent-wickeln und geht häufig mit kognitiven Defiziten einher. Unabhängig von der Ätiologie sind Patienten mit CSWS durch eine Aktivierung des gleichen neurona-len Netzwerks charakterisiert [40], be-stehend aus perisylvischer Region, Insel und Gyrus cinguli (. Abb. 1). Zudem geht mit dem CSWS auch die Deaktivie-rung von Regionen des DMN einher [12, 40]. In einer Quellenanalyse konnte ge-zeigt werden, dass diese Aktivierungs-muster syndromspezifische Propaga-tionswege widerspiegeln. In verschiede-nen Studien wurde darauf hingewiesen, dass die epileptische Aktivität bei Patien-ten mit CSWS entweder in der perisylvi-schen Region generiert oder in diese Re-gion propagiert [32, 33, 42]. Dies steht in Übereinstimmung mit PET- und „Sing-le-photon-emission-computed-tomogra-phy“(SPECT)-Studien, die Hirnareale des Hypermetabolismus und der Hypoperfu-sion in der perisylvischen Region und im temporoparietalen Kortex bei Patienten
mit CSWS beschrieben [11, 13, 18, 24, 25]. Wenn auch der perisylvische Kortex eine bedeutende Rolle in der Generierung epileptischer Aktivität bei Patienten mit Landau-Kleffner-Syndrom (LKS) zu spie-len scheint, kann die epileptsche Aktivi-tät bei Patienten mit CSWS ohne LKS in anderen Gehirnregionen entstehen und erst dann in die perisylvische Region pro-pagieren [11, 18, 24, 25, 32]. Das bedeu-tet, dass zwar der Generator bei CSWS individuell in verschiedenen kortikalen Regionen lokalisiert sein kann, die Pro-pagationswege jedoch bei unterschiedli-chen Patienten die gleichen sind [18, 25]. Hierzu muss erwähnt werden, dass die-ses Muster der Propagation spezifischer für CSWS zu sein scheint als die zugrun-de liegende Ätiologie.
Die beschriebene Deaktivierung im DMN kann möglicherweise erklären, wa-rum Patienten mit CSWS kognitive De-fizite aufweisen. Es kann vermutet wer-den, dass die epileptische Aktivität bei diesen Patienten die normale Funktion und Konnektivität im DMN beeinträch-tigt und dadurch auf kognitive Funktio-nen negativen Einfluss nimmt [19]. Ar-gumente für die Behauptung können in den Studien von De Tiege et al. [12, 13] gefunden werden: Mithilfe longitudinaler PET-Untersuchungen haben die Autoren gezeigt, dass eine erfolgreiche Behand-lung des CSWS zu einer signifikanten Reduktion des ursprünglich vorliegen-den Hypometabolismus in Arealen des DMN führen kann. Je besser die Norma-lisierung des Metabolismus im DMN war, desto größer war der klinische Effekt.
Dravet-Syndrom
Das Dravet-Syndrom gehört zu den schweren myoklonischen Epilepsien des Säuglingsalters („severe myoclonic epi-lepsy of infancy“, SMEI). Es handelt sich um eine resistente EE, die in etwa 80% der Fälle mit einer Mutation im SCN1A-Gen einhergeht. In einer vor Kurzem pu-blizierten Studie [29] wurden 10 Patien-ten mit Dravet-Syndrom und nachgewie-sener Mutationen im SCN1A-Gen mithil-fe der EEG-fMRT untersucht. Trotz der ähnlichen Ätiologie konnte in dieser Stu-die kein syndromspezifisches neuronales Netzwerk beschrieben werden. Das ein-
zige gemeinsame Merkmal der Hirnak-tivität während epileptischer Entladun-gen bei Patienten mit Dravet-Syndrom war die Einbeziehung der Areale des DMN in das epileptische Netzwerk bei 7 der 10 Patienten. Die Gruppenanalyse er-brachte jedoch kein schlüssiges Ergebnis. Möglicherweise kann die Tatsache, dass kein Dravet-Syndrom-spezifisches Netz-werk gefunden wurde, so gedeutet wer-den, dass das Dravet Syndrom, wie in an-deren Studien nachgewiesen, pathogene-tisch heterogen ist [17, 23].
Fazit
F Verschiedene epileptische Syndro-me und EE sind durch ein spezifisches „activation pattern“ charakterisiert. Das Wissen darüber, wie die kogniti-ven Defizite bei EE entstehen, ist je-doch immer noch unzureichend.
F Die Unterbrechung der Aktivität im DMN durch epileptische Aktivität lie-fert eine mögliche Erklärung für kog-nitive Einbußen bei Patienten mit EE. Da sich allerdings bei vielen EE keine Deaktivierung im DMN findet (LGS, West-Syndrom), kann diese Erklärung nicht pauschal für jede EE gelten.
F Nach wie vor existieren zu wenige Studien, die direkt das Verhältnis zwi-schen kognitiver Funktion, epilepti-scher Aktivität und strukturellen bzw. funktionellen Veränderungen im Ge-hirn untersucht haben. Der momen-tane Wissensstand ermöglicht, Hypo-thesen über pathogenetische Me-chanismen von EE zu bilden, die eine Grundlage für weitere Studien zu Ef-fekten epileptischer Aktivität auf die Kognition bilden sollten.
Korrespondenzadresse
Prof. Dr. M. SiniatchkinKlinik für Psychiatrie, Psychosomatik und Psychotherapie des Kindes- und Jugendalters, Johann-Wolfgang Goethe-Universität zu Frankfurt am Main,Deutschordenstr. 50, 60528 [email protected]
4 | Zeitschrift für Epileptologie 2014
Leitthema

Einhaltung ethischer Richtlinien
Interessenkonflikt. M. Siniatchkin, K. Gröning, F. Moeller, N. Japaridze und U. Stephani geben an, dass kein Interessenkonflikt besteht.
Der Beitrag enthält keine Studien an Menschen oder Tieren.
Literatur
1. Aghakhani Y, Bagshaw AP, Benar CG et al (2004) fMRI activation during spike and wave dischar-ges in idiopathic generalized epilepsy. Brain 127:1127–1144
2. Arzimanoglou A, French J, Blume WT et al (2009) Lennox-Gastaut syndrome: a consensus approach on diagnosis, assessment, management, and trial methodology. Lancet Neurol 8:82–93
3. Avanzini G, Manganotti P, Meletti S et al (2012) The system epilepsies: a pathophysiological hypo-thesis. Epilepsia 53:771–778
4. Bettus G, Guedj E, Joyeux F et al (2009) Decreased basal fMRI functional connectivity in epileptoge-nic networks and contralateral compensatory me-chanisms. Hum Brain Mapp 30:1580–1591
5. Bladin PF (1985) Adult Lennox-Gastaut syndrome: features and diagnostic problems. Clin Exp Neurol 21:93–104
6. Boly M, Phillips C, Tshibanda L et al (2008) Intrinsic brain activity in altered states of consciousness: how conscious is the default mode of brain functi-on. Ann N Y Acad Sci 1129:119–129
7. Broyd SJ, Demanuele C, Debener S et al (2009) Default-mode brain dysfunction in mental disor-ders: a systematic review. Neurosci Biobehav Rev 33:279–296
8. Capovilla G, Berg AT, Cross JH et al (2009) Concep-tual dichotomies in classifying epilepsies: partial versus generalized and idiopathic versus sympto-matic (April 18–20, 2008, Monreale, Italy). Epilep-sia 50:1645–1649
9. Chiron C, Dulac O, Bulteau C et al (1993) Study of regional cerebral blood flow in West syndrome. Epilepsia 34:707–715
10. Chugani HT, Shewmon DA, Sankar R (1992) Infan-tile spasms: II. Lenticular nuclei and brain stem ac-tivation on positron emission tomography. Ann Neurol 31:212–219
11. De Tiege X, Goldman S, Laureys S et al (2004) Re-gional cerebral glucose metabolism in epilepsies with continuous spikes and waves during sleep. Neurology 63:853–857
12. De Tiege X, Harrison S, Laufs H et al (2007) Im-pact of interictal epileptic activity on normal brain function in epileptic encephalopathy: an electro-encephalography-functional magnetic resonance imaging study. Epilepsy Behav 11:460–465
13. De Tiege X, Ligot N, Goldman S et al (2008) Meta-bolic evidence for remote inhibition in epilepsies with continuous spike-waves during sleep. Neu-roImage 40:802–810
14. Dulac O (2001) Epileptic encephalopathy. Epilep-sia 42:23–26
15. Dulac O (2001) What is West syndrome? Brain Dev 23:447–452
16. Eichele T, Debener S, Calhoun VD et al (2008) Pre-diction of human errors by maladaptive changes in event-related brain networks. Proc Natl Acad Sci U S A 105:6173–6178
17. Ferrie CD, Marsden PK, Maisey MN, Robinson RO (1997) Cortical and subcortical glucose metabo-lism in childhood epileptic encephalopathies. J Neurol Neurosurg Psych 63:181–187
18. Gaggero R, Caputo M, Fiorio P et al (1995) SPECT and epilepsy with continuous spike waves during slow-wave sleep. Childs Nerv Syst 11:154–160
19. Gotman J, Grova C, Bagshaw A et al (2005) Gene-ralized epileptic discharges show thalamocorti-cal activation and suspension of the default state of the brain. Proc Natl Acad Sci U S A 102:15236–15240
20. Hamandi K, Salek-Haddadi A, Laufs H et al (2006) EEG-fMRI of idiopathic and secondary generalized epilepsies. NeuroImage 31:1700–1710
21. Hrachovy RA, Frost JD (2003) Infantile epileptic encephalopathy with hypsarrhythmia. J Clin Neu-rophysiol 20:408–425
22. Japaridze N, Muthuraman M, Moeller F et al (2013) Neuronal networks in West syndrome as revealed by source analysis and renormalized par-tial directed coherence. Brain Topogr 26:157–170
23. Korinthenberg R, Bauer-Scheid C, Burkart P et al (2004) 18FDG-PET in epilepsies of infantile onset with pharmacoresistant generalised tonic-clonic seizures. Epilepsy Res 60:53–61
24. Luat AF, Asano E, Juhasz C et al (2005) Relation-ship between brain glucose metabolism positron emission tomography (PET) and electroencepha-lography (EEG) in children with continuous spike-and-wave activity during slow-wave sleep. J Child Neurol 20:682–690
25. Maquet P, Hirsch E, Metz-Lutz MN et al (1995) Re-gional cerebral glucose metabolism in children with deterioration of one or more cognitive func-tions and continuous spike-and-wave discharges during sleep. Brain 118:1497–1520
26. McTague A, Cross JH (2013) Treatment of epileptic encephalopathies. CNS Drugs 27:175–184
27. Metsähonkala L, Gaily E, Rantala H et al (2002) Fo-cal and global cortical hypometabolism in pati-ents with newly diagnosed infantile spasms. Neu-rology 58:1646–1651
28. Moehring J, Spiczak S von, Moeller F et al (2013) EEG-fMRI study in Dravet syndrome patients with SCN1A mutations: one gene – individual results. Epilepsia (Epub ahead of print)
29. Moehring J, Kroeher B, Galka A et al (2013) Multi-focal epilepsy is associated with increased func-tional connectivity. Epilepsia (submitted)
30. Moeller F, Siebner H, Wolff S et al (2008) EEG-fMRI in children with untreated childhood absence epi-lepsy. Epilepsia 49:1510–1519
31. Moeller F, Siebner H, Wolff S et al (2008) Changes in activity of striato-thalamo-cortical network pre-cede generalized spike wave discharges. NeuroI-mage 39:1839–1849
32. Morrell F, Whisler WW, Smith MC et al (1995) Lan-dau-Kleffner syndrome: treatment with subpial intracortical transection. Brain 118:1529–1546
33. Paetau R (2009) Magnetoencephalography in Landau-Kleffner syndrome. Epilepsia 50(Suppl 7):51–54
34. Pillay N, Archer JS, Badawy RA et al (2013) Net-works underlying paroxysmal fast activity and slow spike and wave in Lennox-Gastaut syndro-me. Neurology 81:665–673
35. Pizoli CE, Shah MN, Snyder AZ et al (2011) Res-ting-state activity in development and mainte-nance of normal brain function. Proc Natl Acad Sci U S A 108:11638–11643
36. Raichle ME, Mintun MA (2006) Brain work and brain imaging. Annu Rev Neurosci 29:449–476
37. Rougier A, Claverie B, Pedespan JM et al (1997) Callosotomy for intractable epilepsy: overall out-come. J Neurosurg Sci 41:51–57
38. Samadani U, Baltuch GH (2007) Anterior thalamic nucleus stimulation for epilepsy. Acta Neurochir Suppl 97:343–346
39. Siniatchkin M, Baalen A van, Jacobs J et al (2007) Different neuronal networks are associated with spikes and slow activity in hypsarrhythmia. Epi-lepsia 48:2312–2321
40. Siniatchkin M, Groening K, Moehring J et al (2010) Neuronal networks in children with continu-ous spikes and waves during slow sleep. Brain 133:2798–2813
41. Siniatchkin M, Coropceanu D, Moeller F et al (2011) EEG-fMRI reveals activation of brainstem and thalamus in patients with Lennox-Gastaut syndrome. Epilepsia 52:766-774
42. Sobel DF, Aung M, Otsubo H, Smith MC (2000) Magnetoencephalography in children with Lan-dau-Kleffner syndrome and acquired epileptic aphasia. Am J Neuroradiol 21:301–307
43. Van Bogaert P, Urbain C, Galer S et al (2012) Im-pact of focal interictal epileptiform discharges on behavior and cognition in children. Neurophysiol Clin 42:53–58
44. Velasco M, Velasco F, Gardea G et al (1997) Poly-graphic characterization of the sleep epilepsy pat-terns in a hydranencephalic child with severe ge-neralized seizures of Lennox-Gastaut syndrome. Arch Med Res 28:297–302
45. Velasco AL, Velasco F, Jimenez F et al (2006) Neuromodulation of the centromedian thalamic nuclei in the treatment of generalized seizures and the improvement of the quality of life in pa-tients with Lennox-Gastaut syndrome. Epilepsia 47:1203–1212
46. Zhang Z, Lu G, Zhong Y et al (2009) Impaired at-tention network in temporal lobe epilepsy: a res-ting FMRI study. Neurosci Lett 458:97–101
47. Zhang Z, Lu G, Zhong Y et al (2010) Altered spon-taneous neuronal activity of the default-mode network in mesial temporal lobe epilepsy. Brain Res 1323:152–160
5Zeitschrift für Epileptologie 2014 |