Embed Size (px)
Citation preview

Aus dem Institut für Kreislaufforschung und Sportmedizin
Abteilung für Molekulare und Zelluläre Sportmedizin
Deutsche Sporthochschule Köln
Geschäftsführender Leiter: Univ.-Prof. Dr. med. Wilhelm Bloch
Der Einfluss einer sporttherapeutischen Trainingsintervention
auf die Dichte der Monocarboxylattransporter in Skelettmuskulatur und
Erythrozyten bei nicht-insulinpflichtigen
Typ 2 Diabetikern
von der Deutschen Sporthochschule Köln
zur Erlangung des akademischen Grades
Doktor der Sportwissenschaft
genehmigte Dissertation
vorgelegt von
Thorsten Kreutz
aus
Hagen
Köln 2010

Erster Referent: Univ.-Prof. Dr. med. Wilhelm Bloch
Zweiter Referent: Prof. Dr. rer. nat. Klara Brixius
Vorsitzende des Promotionsausschusses: Univ.-Prof. Dr. I. Hartmann-Tews
Tag der Disputation: 29.11.2010
Erklärung „Hierdurch versichere ich: Ich habe diese Arbeit selbständig und nur unter Benutzung der
angegebenen Quellen und technischen Hilfen angefertigt; sie hat noch keiner anderen Stelle
zur Prüfung vorgelegen. Wörtlich übernommene Textstellen, auch Einzelsätze oder Teile
davon, sind als Zitate kenntlich gemacht worden.
Hierdurch erkläre ich, dass ich die Leitlinien guter wissenschaftlicher Praxis’ der Deutschen
Sporthochschule Köln in der aktuellen Fassung eingehalten habe.“
Köln, den 28. Juli 2010 ___________________ Thorsten Kreutz

Für meine Frau und meinen Sohn Berke
und in Gedenken an meinen Vater

Inhaltsverzeichnis
Inhaltsverzeichnis
1 Einleitung 1
1.1 Laktat: Energiereiches Stoffwechselprodukt und Signalmolekül 1
1.2 Laktattransport 4
1.2.1 Monocarboxylattransporter 5
1.3 Physiologie des Laktattransports in Muskulatur und Erythrozyten 6
1.4 Veränderungen des Laktatstoffwechsels bei Typ 2 Diabetikern 11
1.5 Diabetes mellitus und körperliche Aktivität 12
1.6 Fragestellungen 15
2 Material und Methoden 17
2.1 Studiendesign 17
2.1.1 Probandenbeschreibung 17
2.1.2 Untersuchungsablauf 18
2.2 Untersuchungsverfahren 19
2.2.1 Erhebung der anthropometrischen Parameter 19
2.2.2 Laborparameter 20
2.2.3 Muskelbiopsie 25
2.3 Immunhistochemie 25
2.3.1 Immunhistochemischer Nachweis des MCT1 und MCT4 im Muskelgewebe 26
2.3.2 Immunhistochemischer Nachweis des MCT1 im Erythrozyten 33
2.3.3 Durchflusszytometrische Analyse des CD147 im Erythrozyten 38
2.3.4 Immunfluoreszenz-Nachweis des MCT1 und CD147 im Erythrozyten 39
2.4 Muskelfasertypisierung 41
2.5 Leistungsphysiologische Parameter 43
2.5.1 Fahrradergometrische Belastungsuntersuchung 43
2.5.2 Kraftmessungen 44

Inhaltsverzeichnis
2.6 Bewegungsinterventionen 45
2.6.1 Sporttherapeutisches Krafttraining 45
2.6.2 Sporttherapeutisches Ausdauertraining 48
2.7 Statistische Auswertung 49
3 Ergebnisse 50
3.1 Basischarakterisierung des Probandenkollektivs 50
3.2 Leistungsphysiologische Parameter 51
3.3 Stoffwechselparameter 52
3.4 Muskelfasertypisierung 54
3.5 Immunhistochemischer Nachweis des MCT1 und MCT4 im Muskelgewebe 56
3.6 Immunhistochemischer Nachweis des MCT1 im Erythrozyten 59
3.7 Durchflusszytometrische Analyse des CD 147 im Erythrozyten 62
3.8 Fluoreszenzmikroskopischer Nachweis des MCT1 und CD147 im Erythrozyten 65
4 Diskussion 67
4.1 Langfristig trainingsbedingte Veränderungen der MCT1-Dichte im Muskelgewebe 67
4.2 Langfristig trainingsbedingte Veränderungen der MCT4-Dichte im Muskelgewebe 71
4.3 Langfristig trainingsbedingte Veränderungen der MCT1- und CD147- Dichte im Erythrozyten 75
4.4 Kurzfristig trainingsbedingte Veränderungen der MCT1- und CD147- Dichte im Erythrozyten nach einer Maximalbelastung 79
4.5 Der Einfluss des sporttherapeutischen Ausdauer- und Krafttrainings auf den Stoffwechsel und die Leistungsfähigkeit bei nicht-insulin- pflichtigen Typ 2 Diabetikern 83
5 Zusammenfassung 89

Inhaltsverzeichnis
6 Literaturverzeichnis 92
7 Anhang 112
7.1 Abbildungsverzeichnis 112
7.2 Tabellenverzeichnis 115
7.3 Abstract 116
7.4 Lebenslauf 119

Abkürzungsverzeichnis
Abkürzungsverzeichnis
Abb. Abbildung
AG Ausdauergruppe
ATP Adenosintriphosphat
BEKG Belastungselektrokardiogramm
BMI Body Mass Index
BSA Bovine Serum Albumin
DAB Diaminobenzidin
dest. destilliert
DU Density Unit
EKG Elektrokardiogramm
et al. und andere
FACS Fluorescence Activated Cell Scanning
FSC Vorwärtsstreulicht
G Zentifugalbeschleunigung
g Gramm
g/l Gramm pro Liter
GLUT 4 Glukosetransporter Typ 4
H+ Hydrogen-Ion
HCL Chlorwasserstoffsäure (Salzsäure)
HDL High Density Lipoprotein
H2O Wasser
H2O2 Wasserstoffperoxid
HRP Horseradish Peroxidase
Hz Hertz
IHC Immunhistochemische Kontrollfärbung
J Jahre
KG Kraftgruppe
Kg Kilogramm
Km Michaelis-Menten-Konstante
l Liter
La Laktat

Abkürzungsverzeichnis
LDH Laktatdehydrogenase
LDL Low Density Lipoprotein
M Molarität
m Meter
m2 Meter zum Quadrat
max. Maximum
MCT Monocarboxylattransporter
mg Milligramm
mg/dl Milligramm pro Deziliter
ml/kg/min Milliliter pro Kilogramm pro Minute
ml/min Milliliter pro Minute
mm Millimeter
mmHg Millimeter Quecksilbersäule
mM Milli-Molarität
mmol/l Millimol pro Liter
n.s. nicht signifikant
min. Minimum
N Newton
NAD Nicotinamid-Adenin-Dinukleotid
n Anzahl
μ Micro (10-6)
μl Mikroliter (10-6)
μU/ml Mikrounit pro Milliliter
p Signifikanzniveau
PBS Phosphate buffered saline solution
PFA Paraformaldehyd
pH-Wert Negativer dekadischer Logarithmus der H+ -Ionenkonzentration
pK-Wert Negativer Logarithmus der Dissoziationskonstante eines Elektrolyten
pmol/l Pikomol pro Liter
RM Repitition Maximum
RPE Received Perception of Exertion
SD Standard Deviation
Sek. Sekunden
Std. Stunde

Abkürzungsverzeichnis
SSC Seitwärtsstreulicht
t Zeit
t0 Zeitpunkt vor der Belastungsuntersuchung
t1 Zeitpunkt nach der Belastungsuntersuchung
Tab. Tabelle
TBS Tris-Buffered Saline
TM Transmembrane domain
U1 Eingangsuntersuchung
U2 Ausgangsuntersuchung
U/min. Umdrehungen pro Minute
VO2max maximale Sauerstoffaufnahme
WHO World Health Organisation
°C Grad Celsius

Einleitung
1
1 Einleitung
1.1 Laktat: Energiereiches Stoffwechselprodukt und Signalmolekül
Über viele Jahre hinweg galt das Molekül Laktat als ein Abfallprodukt des
glykolytischen Stoffwechsels und wurde darüber hinaus für die zelluläre Azidose
verantwortlich gemacht. Diese älteren Ansichten wurden in den letzten Jahren
revidiert und die Bedeutung des energiereichen Moleküls Laktat hat für den
Stoffwechsel und die Diagnostik eine Wandlung erfahren.
Das Molekül Laktat ist ein Salz der Milchsäure, das in zwei verschiedenen
stöchiometrischen Konfigurationen je nach Ausrichtung als L-Laktat und D-Laktat
vorkommt (vgl. Abb. 1).
Abb. 1: Strukturformel L-Laktat
Laktat entsteht im Zytoplasma bei der anaeroben Glykolyse durch den Abbau von
einem Glukosemolekül mit einer Nettoenergieausbeute von zwei Molekülen ATP pro
Glukosemolekül. Durch die Laktatbildung unter anaeroben Bedingungen wird ein
Mangel des Elektronencarriers NAD+ im Zytoplasma kompensiert, der sonst die
Glykolyse zum Erliegen bringen würde (Horn et al. 2005).
Bei der Verstoffwechslung von einem Glukosemolekül entstehen nach zehn
katalysierten Reaktionsschritten zwei Pyruvatmoleküle, die in einer nachfolgenden
Laktatdehydrogenase-Reaktion zu zwei Laktatmolekülen reduziert werden. Aufgrund
des niedrigen pK-Wertes der Milchsäure (pK=3.86) kommt es zu einer sofortigen
Dissoziation und zur Anlagerung eines Natriumions, sodass im physiologischen
System überwiegend das Natriumlaktat vorliegt (vgl. Abb. 2).

Einleitung
2
𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃 + 𝑁𝑁𝑁𝑁𝑁𝑁𝑁𝑁 + 𝑁𝑁+ 𝐿𝐿𝑁𝑁𝑁𝑁�� 𝑀𝑀𝑀𝑀𝑀𝑀𝑀𝑀ℎ𝑠𝑠ä𝑃𝑃𝑃𝑃𝑢𝑢 + 𝑁𝑁𝑁𝑁𝑁𝑁+
𝐿𝐿𝑁𝑁𝑁𝑁�⎯� [𝐿𝐿𝑃𝑃𝐿𝐿𝑃𝑃𝑃𝑃𝑃𝑃−𝑁𝑁𝑃𝑃+]
+ 𝑁𝑁𝑁𝑁𝑁𝑁+ + 𝑁𝑁+
Abb. 2: Reaktionsgleichung für die Bildung von Natriumlaktat
Bei der Milchsäurebildung lagert sich sowohl ein H+-Ion vom NADH als auch ein
H+-Ion aus dem Zytoplasma an das Pyruvatmolekül an, wodurch die Gesamt-
protonenkonzentration im Zytoplasma reduziert wird (vgl. Abb. 3). Für die verstärkte
Protonenfreisetzung unter körperlicher Belastung wird hauptsächlich die verstärkte
ATP-Hydrolyse sowie eine erhöhte glykolytische Flussrate verantwortlich gemacht
und nicht die Laktatdehydrogenase-Reaktion (Robergs et al. 2003).
Abb. 3: Laktatdehydrogenase-Reaktion (Robergs et al. 2003)
Das energiereiche Stoffwechselzwischenprodukt Laktat wird sowohl im Ruhe-
zustand, als auch unter Belastung ständig produziert und abgebaut. Der Normbereich
des Ruhelaktatwertes liegt bei < 1.8 mmol/l (Biedler et al. 1995). Zu Beginn der
körperlichen Belastung kommt es zu einem akut erhöhten ATP-Bedarf innerhalb der
Muskelzelle, der nur kurzfristig durch Steigerung der glykolytischen Energie-
bereitstellung und einer damit verbundenen erhöhten Laktatproduktion bereitgestellt
werden kann. Ebenso zeigt sich auch bei längeren Belastungen mit höherer Intensität
eine verstärkte Anhäufung von Laktat in den Skelettmuskelzellen. Dies führt
langfristig zu einer Überforderung der oxidativen Kapazität, weshalb das benötigte
ATP nach Ausschöpfung des oxidativen Stoffwechsels über die glykolytische
Energiegewinnung zur Verfügung gestellt wird (Juel, 2004c).

Einleitung
3
Einerseits dient das entstandene Laktat als Energiequelle im Rahmen der aeroben
Energiegewinnung für Herz, Gehirn und Skelettmuskulatur (Chatham et al.1999;
Galdden, 2000; Halestrap et al. 2004; Waniewski et al. 2004), anderseits kann es in
der Leber, Nieren und Muskulatur über die Glukoneogenese als Glykogen
gespeichert werden (Donovan et al. 1990; McDermott et al. 1992; Bonen et al. 1994).
Das Monocarboxylat Laktat dient nicht nur als Energiequelle, sondern nimmt bei
zellulären Gewebeanpassungen die Rolle eines Signalmoleküls ein (Gladden et al.
2001; Philp et al. 2005; Brooks et al. 2008).
Nachgewiesen werden konnte dies u.a. bei der Wundheilung (Trabold et al. 2003),
bei der Kollagenbildung (Green et al. 1964) und bei Regulationsprozessen von
VEGF (Constant et al. 2000). Ebenso konnte erstmalig an Ratten gezeigt werden,
dass Laktat potentielle Bindungsstellen für ROS-reaktive Transkriptionsfaktoren
besitzt und somit vermutlich bei körperlicher Belastung an der mitochondrialen Bio-
genese im Muskel und der MCT1 Genexpression auf Transkriptionsebene beteiligt
ist (Hashimoto et al. 2007). Zudem fördert der belastungsbedingte Laktatanstieg die
Vasodilatation der Gefäße und die Katecholaminfreisetzung (Fattor et al. 2005).
Innerhalb der Schmerzregulation konnte der Einfluss des Laktats auf die säure-
empfindlichen Rezeptoren (ASIC) der Neuronen nachgewiesen werden (Immke et al.
2001). Bei der Umwandlung von Pyruvat zu Laktat ließ sich zellulär eine nachhaltige
Beeinflussung des Redoxstatus der Zelle beobachten (Brooks, 2002) und bei Ratten
bewirkten erhöhte Ruhelaktatwerte eine Verschlechterung des Glukosetransportes
innerhalb der Muskelzellen (Lombardi et al. 1999).

Einleitung
4
1.2 Laktattransport
Der Transport von Laktat durch die Membranen der unterschiedlichen Zellen und
Geweben ist essentiell für den Energiestoffwechsel und der intrazellulären
pH-Wertregulation aller Organismen (Juel et al. 1999). Der Transport und die
schnellere Verteilung von Laktat in die Körperzellen werden durch die Familie der
Monocarboxylattransporter (MCT) katalysiert.
Bei körperlicher Belastung kommt es im kontrahierenden Muskel zu einer
Akkumulation von Laktat und H+-Ionen. Das entstandene Laktat wird zusammen mit
einem H+-Ion durch die MCT aus der Muskelzelle und durch die Endothelschicht ins
Blutplasma transportiert. In Abhängigkeit von den metabolischen Bedingungen kann
das Laktat einerseits mittels der MCT-Proteine in die Erythrozyten transportiert
werden oder im Blutplasma verbleiben. Anderseits kann das Laktat im Rahmen der
Glukoneogenese als Glykogen gespeichert werden bzw. als Energiequelle für den
oxidativen Energiestoffwechsel der Zielzellen dienen (vgl. Abb. 4).
Erythrozyt
Laktat/H+Laktat/H+Laktat/H+
Laktat/H+
Laktat/H+
Laktat/H+
Oxidative Energiegewinnung
Laktatproduktion
Laktatverwertung
Glukoneogenese
Laktat/H+
MCT1
MCT1/4
Kontrahierender Skelettmuskel
Blutgefäß
MCT1/4
Laktat/H+ Laktat/H+
Laktat/H+
MCT1Intrazellularraum
MCT1
Abb. 4: Vereinfachtes Schema zum Laktattransport bei körperlicher Belastung

Einleitung
5
1.2.1 Monocarboxylattransporter
Über viele Jahre hinweg wurde die Auffassung vertreten, dass der Laktattransport
über die biologischen Membranen zwischen den Geweben und Zellen durch
vereinfachte Diffusion erfolgte. Erst im Jahre 1994 konnte das erste Laktat-
transportprotein aus Zellen eines chinesischen Hamster (Garcia et al. 1994a) und
nachfolgend von Menschen (Garcia et al. 1994b) kloniert werden. Da diese
Transporterproteine überwiegend Monocarboxylate wie Laktat, Pyruvat und Keton-
körper transportieren, wurden sie von ihren Entdecker als Monocarboxylattransporter
(MCT) bezeichnet.
Die MCT gehören zu der SCL16 Genfamilie (Hediger et al. 2004). Bisher konnten
insgesamt 14 Isoformen der MCT mit einem charakteristischen Gewebeverteilungs-
muster und unterschiedlicher Substratspezifität detektiert werden.
Die Monocarboxylattransporter bestehen aus zwölf transmembranären α -helikalen
Domänen (TM) und weisen einen charakteristischen Aufbau aus einer großen
Schleife zwischen TM 6 und TM 7 und einem intrazellulären N- und C-Terminus auf
(vgl. Abb. 5). Es wird angenommen, dass der N-Terminus für die energetischen
Kopplungsvorgänge (Na+- oder H+-Kotransport), den Membraneinbau und für die
Strukturerhaltung verantwortlich ist. Der C-Terminus hingegen für die spezifische
Substratbindung (Saier, 1994). Die 14 Isoformen unterscheiden sich strukturell vor
allem in der Aminosäurensequenz des intrazellulären N- bzw. C-Terminus und der
Schleifengröße voneinander (Halestrap et al. 1999).
Abb. 5: Aufbau des MCT1-Proteins (modifiziert nach Halestrap et al. 2004)

Einleitung
6
Die ersten vier Isomere (MCT1-MCT4) wurden bisher funktionell am intensivsten
untersucht. Hier konnte ein protonengekoppelter Transport durch die Membranen der
Gewebe von Laktat, Pyruvat und Ketonkörper nachgewiesen werden (Halestrap et al.
2004). Der zugrunde liegende Transportmechanismus ist ein gekoppelter Protonen-
/Monocarboxylat-Symport im Sinne der erleichterten Diffusion (Enerson et al. 2003).
Der Transport erfolgt im Verhältnis 1:1, elektroneutral (Roth et al. 1990; Juel et al.
2004c) und verläuft ohne Energieverbrauch stets in Richtung des negativen
Konzentrationsgradienten (Merezhinskaya et al. 2009). Der Transport erfolgt nach
dem Konzentrationsgefälle und unterliegt den Gesetzmäßigkeiten des folgenden
Gleichgewichts: [lactate]in/[lactate]out=[H+]out/[H+]in. Die kinetischen Parameter
sind allerdings gemäß dem Gleichgewicht nach Haldane (Vmax/Km)influx=
(Vmax/Km)efflux thermodynamisch beschränkt (Halestrap et al. 1999). Somit besitzt
der MCT die Fähigkeit auf die vorliegenden metabolischen Zustände in der Zelle zu
reagieren und in beide Richtungen, d.h. entweder in oder aus der Zelle heraus zu
transportieren.
1.3 Physiologie des Laktattransports in Muskulatur und Erythrozyten
Hauptverantwortlich für den stereoselektiven Transport von L-Laktat (Poole et al.
1993; Juel et al. 1997; Broer et al. 1998; Halestrap et al. 2004) in der Muskulatur
sind die beiden Transporterproteine MCT1 und MCT4, die coexprimiert werden und
eine muskelfasertypische Verteilung aufweisen (Pilegaard et al. 1999a).
Es zeigt sich eine erhöhte Expressionsrate des MCT1-Proteins in den langsamen,
oxidativen Typ 1 Muskelfasern (McCullagh et al. 1996; Bonen et al. 2001), wohin-
gegen sich in den schnellen, glykolytischen Typ 2 Muskelfasern eine geringere
MCT1-Dichte und erhöhte Expression des MCT4-Proteins beobachten lässt
(Mc Cullagh et al. 1997; Bonen et al. 2000a). Da sich diese Verteilung des MCT1
und MCT4 auch in anderen Zellen, wie u.a. in den oxidativen Herzmuskelzellen
(Halestrap et al. 1997) oder in glykolytischen weißen Blutzellen (Wilson et al. 1998)
feststellen lässt, wird angenommen, dass der MCT1 vermehrt für die Laktataufnahme
in die Zelle und der MCT4 überwiegend für den Laktatabfluss aus Zellen mit hoher
glykolytischer Kapazität verantwortlich ist (McCullagh et al. 1996; Bonen et al.
2000b; Manning et al. 2000). Hinzu kommt, dass der MCT1 eine größere Affinität zu

Einleitung
7
L-Laktat (Km=5 mM) besitzt als der MCT4 (Km=20-30 mM), so dass der MCT1
schon früher als der MCT4 bei niedrigeren Laktatkonzentrationen zu transportieren
beginnt (Juel et al. 1999; Dimmer et al. 2000).
In Ruhe und bei geringen körperlichen Belastungen sind für Aufrechterhaltung des
konstanten pH-Wertes hauptsächlich der Na/H+-Austauscher und der
Na/Bicarbonat-Austauscher verantwortlich (Juel, 1998a). Unter körperlicher
Belastung führt die erhöhte Laktatproduktion zu intrazellulären Ionenverschiebungen
mit einer Zunahme der H+-Ionenkonzentration und einer daraus resultierenden
Abnahme des pH-Wertes. Für den schnellen Transport von Laktat und H+-Ionen
unter Belastung sind der MCT1 und MCT4 zuständig. Die beiden MCT-Proteine
übernehmen somit bei intensiven körperlichen Belastungen eine wichtige Funktion
bei der intrazellulären pH-Wertregulation der Muskelzelle (Juel et al. 2004a). Es
konnte gezeigt werden, dass eine Korrelation zwischen der Zeitspanne bei der
pH-Wertnormalisierung und des Laktatwertes in der Regenerationszeit vorlag
(Juel, 1998b). Die Transportrichtung ist dabei von dem Laktatgradienten und somit
vom intrazellulären pH-Wert der Zelle abhängig (Roth et al. 1990; Halestrape et al.
1999).
In der Muskulatur ist die Expressionsrate des MCT-Proteins abhängig von der
Muskelaktivität, also von den metabolischen und energetischen Anforderungen der
Muskulatur. Zahlreiche Studien an Menschen und Tieren belegen, dass sowohl
Ausdauer- (Dubouchaud et al. 2000; Bonen et al. 2000c, Yoshida et al. 2004) und
Krafttraining (Juel et al. 2004b) als auch die elektrische Stimulation (McCullagh
et al. 1997; Bonen et al. 2000b) der Muskulatur langfristig den MCT1- und MCT4-
Gehalt erhöhen.
Die Laktatbeseitigung aus der Zelle nach körperlicher Belastung korreliert positiv
mit dem MCT-Gehalt (Donovan et al. 1983; McCullagh et al. 1996; Bonen et al.
1998; Dubouchaud et al. 2000) und dem Trainingszustand (Pilegaard et al. 1994).
Hierbei ist aber anzumerken, dass nicht nur alleine die Zunahme des MCT-
Gehaltes, sondern auch andere Ursachen wie u.a. ein verbesserter Blutfluss und eine
Vermehrung der Kapillarendichte für die erhöhte Laktattransportkapazität mit-
verantwortlich sind (Juel et al. 2004a).
Es ist davon auszugehen, dass sich in der Muskulatur die Proteinexpression des
MCT1 und MCT4 unterschiedlich an körperliche Belastung adaptiert. Verschiedene
Trainingsstudien zeigten, dass ein körperliches Training zu unterschiedlichen

Einleitung
8
Expressionsraten des MCT1 und MCT4 in der Muskulatur führte (Pilegaard et al.
1999a; Bonen et al. 2000b; Tonouchi et al. 2002; Juel et al. 2004b; Mohr et al. 2007).
Dabei kam es schon bei moderaten Trainingsbelastungen zu einer erhöhten MCT1-
Proteinexpression, wohingegen der MCT4 erst bei höheren Belastungen vermehrt
zusammen mit dem MCT1 exprimiert wurde (Juel et al. 2006). Körperliche
Inaktivität bewirkt dagegen eine Reduzierung der MCT-Dichte sowohl im humanen
(Pilegaard et al. 1998) als auch tierischen Skelettmuskel (McCullagh et al. 1995;
Dubouchaud et al. 1996; Wilson et al. 1998). Es wird kontrovers diskutiert, ob die
trainingsinduzierte Genexpression der MCT-Proteine in der Muskulatur nach
transkriptionellen, posttranskriptionellen oder translationellen Mechanismen erfolgt
(Jackson et al. 1997; Sachs et al. 1997; Bonen et al. 2000a; Coles et al. 2004;
Halestrape et al. 2004).
Eine erhöhte, aktivitätsbedingte „Up-Regulation“ der MCT-Proteine zeigt sich dabei
nicht nur nach langfristig angelegten Trainingsprogrammen, sondern auch kurzfristig
unmittelbar nach einer einzelnen Trainingseinheit (Zhou et al. 2000; Green et al.
2002; Coles et al. 2004). Dies könnte auf eine aktivitätsbedingte Translokation der
MCT, ähnlich wie bei den Glut4-Transportern in der Muskelzelle, zurückzuführen
sein (Hirshman et al. 1988; Phillips et al. 1996; Richter et al. 1998).
Die mögliche Translokation der MCT1- und MCT4-Proteine in die Zellmembran ist
dabei von dem Glykoprotein CD147 (Emmprin, Basigin) abhängig (Philp et al.
2003b; Wilson et al. 2005; Jiang et al. 2007). Eine Kolokalisation des CD147- und
MCT1-Proteins konnte bereits für eine Vielzahl von Geweben, wie u.a. Herz- und
Skelettmuskel, Leber, Niere, Darm und Hoden nachgewiesen werden (Nackai et al.
2006).
Das CD147-Protein ist ein ubiquitär exprimiertes Membranglykoprotein, das an
vielen physiologischen und pathologischen Prozessen (Zhou et al. 2005; Gallagher
et al. 2007) beteiligt ist und der Immunglobin-Superfamilie angehört. Das
Glykoprotein (vgl. Abb. 6) gliedert sich in zwei extrazelluläre Immun-
globulindomainen, einer Transmembranregion und einem kurzen zytoplasmatischen
Domän (Fossum et al. 1991). Es wird angenommen, dass der zytoplasmatische und/ -
oder transmembranäre Anteil des CD147 für die Verbindung mit zwei MCT-
Proteinen verantwortlich ist (Wilson et al. 2002).

Einleitung
9
SS
SS
C
CN
N
MCT1
CD147
Abb. 6: MCT1-Protein und sein Chaperon CD147 (modifiziert nach Halestrap et al. 2004)
Das CD147-Protein ist ein essentielles Chaperon für den Transport des MCT1 und
MCT4 in die Zellmembran und ist für die korrekte Lokalisation (Kirk et al. 2000),
Expression und Funktionalität der beiden MCT-Proteine in der Muskulatur
verantwortlich (Wilson et al. 2002; Halestrap et al. 2004).
Für den Laktattransport in den Erythrozyten sind das Bande 3-Protein, die nicht-
ionische Diffusion und der MCT1 verantwortlich (Deuticke et al. 1982a; De Bruijne
et al. 1983; Poole et al. 1993). Im humanen Erythrozyten konnte bisher nur die
Isoform MCT1 nachgewiesen werden (Deuticke et al. 1982b; Poole et al. 1996;
Halestrap et al. 1999; Juel et al. 2003). Der erythroide MCT1 transportiert
stereoselektiv L-Laktat zusammen mit einem Hydrogen-Ion im Verhältnis 1:1, wobei
die Transportrichtung von dem Konzentrationsgradienten der Plasmalaktat-
konzentration abhängig ist.
Die reifen Erythrozyten besitzen keine Zellorganellen und können deshalb ihren
Energiebedarf nur über die anaerobe Energiebereitstellung decken. Die Glukose ist
der Hauptenergieträger des Erythrozyten, welches mit einer Nettoenergieausbeute
von zwei Molekülen ATP zu Laktat abgebaut wird. Im Ruhezustand erfolgt über-
wiegend ein Ausstrom von Laktat aus dem Erythrozyten in das Blutplasma, während
es unter körperlicher Belastung, aufgrund des Konzentrationsgefälles, zu einem

Einleitung
10
Einstrom von Laktat in den Erythrozyten kommt (Väihkönen et al. 2001). Bei
aktivitätsbedingten hohen Laktatkonzentrationen erfolgt der Laktattransport vom
Blutplasma in die humanen Erythrozyten überwiegend durch den MCT1 (90%),
während die Bande 3-Proteine (6%) und die nicht ionische Diffusion (4%) eher eine
untergeordnete Rolle einnehmen (Deuticke et al. 1989; Skelton et al. 1998). Im
Vergleich verschiedener Tierspezies ließen sich Unterschiede in der Laktat-
transportkapazität und Michaelis-Menten-Konstanten der MCT1-Proteine feststellen
(Väihkönen et al. 2001). Dabei besitzen Menschen, Pferde und Hunde artspezifisch
eine höhere Lakattransportkapazität als Rentiere, Ziegen oder Schafe. Bei der
geringeren Laktattransportkapazität erfolgt der Laktattransport überwiegend durch
die nicht-ionische Diffusion und das Band 3-Protein (Skelton et al. 1995). Bei
Pferden ließen sich auch interindividuelle Unterschiede hinsichtlich der Laktat-
transportaktivität in den Erythrozyten feststellen, wofür genetische Faktoren
verantwortlich gemacht werden (Väihkönen et al. 2002).
Der Trainingszustand scheint dabei positiv mit der erythroiden Laktataufnahme zu
korrelieren, da sich bei trainierten Pferden nach einem Rennen höhere Laktatwerte in
den Erythrozyten zeigten als bei untrainierten Pferden (Räsänen et al. 1995;
Väihkönen et al. 1999). Ebenso konnte bereits in Trainingsstudien gezeigt werden,
dass die Laktattransportkapazität vom Plasma in die Erythrozyten bei trainierten
Tieren (Skelton et al. 1995; Koho et al. 2002) sowie Menschen (Connes et al. 2004b)
erhöht und bei körperlicher Inaktivität reduziert ist (Skelton et al. 1998). Jedoch ist
diese Laktataufnahme durch die Ionenverteilung im intra- und extraerythrozytärem
Raum eingeschränkt, da ein konstantes Laktat-Erythrozyten-Plasma-Verhältnis
im Erythrozyten von 0,6 bis 0,4 vorliegt (Harris et al. 1989; Smith et al.1997;
Hildebrand et al. 2000).
Unter Hypoxie-Bedingungen konnte eine erhebliche Steigerung der Expressionsrate
des MCT1 und einer Verbesserung der Laktattransportkapazität im humanen
Erythrozyten beobachtet werden (Juel et al. 2003). Eine Verbesserung der Kapazität
des Laktatransportes und ein geringeres Ansteigen der Laktatwerte bei körperlicher
Belastung ließen sich auch durch die Injektion des Medikamentes Erythropoetin bei
humanen Erythrozyten erreichen. (Connes et al. 2004a). Langfristig ließ sich nach
einem dreiwöchiges Schwimmtraining eine Steigerung der MCT1-Proteinexpression
in den Erythrozyten bei Ratten beobachten (Aoi et al. 2004), wobei körperliche
Inaktivität (Skelton et al. 1998) und Mutationen in der DNA des MCT1

Einleitung
11
(Merezhinskaya et al. 2000) zu einer Verschlechterung des Laktattransportes in den
Erythrozyten führt. Das CD147-Protein scheint wie in der Muskulatur eine wichtige
Funktion für den erythroiden Lakatttransport einzunehmen. Die Untersuchungs-
ergebnisse weniger Studien deuten darauf hin, dass das CD147-Protein als Chaperon
des MCT1 in den Erythrozyten fungiert. Langfristig konnte bei Pferden eine positive
Korrelation zwischen der Lakattransportaktivität und der erythroiden CD147-Dichte
sowie dem Trainingszustand nachgewiesen werden (Koho et al. 2002; Koho et al.
2006). Die zugrundeliegenden aktivitätsbedingten Regulationsmechanismen wurden
bisher noch nicht an den Erythrozyten untersucht.
1.4 Veränderungen des Laktatstoffwechsels bei Typ 2 Diabetikern
Bei Typ 2 Diabetikern ist die Aufnahme der Energieträger Laktat und Glukose in die
Zielzellen gestört. Der Glukosetransport durch die Zellmembran in die verschiedenen
Körperzellen wird durch die Familie der Glukosetransporter (GLUT) reguliert. Die
Glukoseaufnahme in die Muskelzelle kann einerseits durch die insulinabhängige und
andererseits durch die insulinunabhängige Translokation der GLUT4-Transporter aus
intrazellulären Speichervesikel an die Zellmembran erfolgen (Marette et al. 1992;
Hayashi et al. 1997; Zierath et al. 2002; Jessen et al. 2005). Bei Typ 2 Diabetikern
konnte festgestellt werden, dass eine Insulinresistenz u.a. durch Defekte innerhalb
der einzelnen Signalwege der Insulin stimulierten GLUT4-Translokation verursacht
werden (Eriksson et al. 1992; Czech et al. 1995; Kennedy et al. 1999; Krook et al.
2000).
Zudem konnte beobachtet werden, dass Typ 2 Diabetiker nicht nur erhöhte Glukose-,
sondern auch erhöhte Ruhelaktatwerte aufweisen (Reaven et al. 1988; Lovejoy et al.
1992). Dabei wiesen Patienten mit Typ 2 Diabetes in Ruhe höhere Laktatwerte als
fettleibige Personen auf (Reaven et al. 1988) und es ließ sich eine Korrelation
zwischen erhöhten HBA1c-Werten und erhöhten Ruhelaktatwerten feststellen (Chen
et al. 1999). Die Ursachen für die erhöhten Ruhelaktatwerte werden kontrovers
diskutiert. Es wird vermutet, dass das Fettgewebe für die verstärkte Laktatproduktion
mitverantwortlich ist (Hagström et al. 1990; DiGirolamo et al. 1992). Sowohl durch
die vermehrte Anzahl als auch den vergrößerten Durchmesser der Adipozyten wird
die Laktatproduktion in Ruhe entscheidend erhöht (Crandall et al. 1983). Ebenso
konnte ein signifikanter Einfluss der Skelettmuskulatur auf die Gesamtlaktat-

Einleitung
12
produktion aufgrund des verschlechterten oxidativen Glukosestoffwechsels bei
Typ 2 Diabetikern nachgewiesen werden (Consoli et al. 1990; Avogaro et al. 1996).
Möglicherweise führen die erhöhten Fettwerte zu einer Verminderung der
Pyruvatdehydrogenase-Aktivität (Randle et al. 1963), welches eine Verschiebung des
Pyruvat-/Laktat-Gleichgewichtes in Richtung des Laktats bedingt (Mondon et al.
1992).
Erhöhte Ruhelaktatwerte gelten als unabhängiger Risikofaktor für die Entstehung
von Typ 2 Diabetes (Ohlson et al. 1988), da gezeigt werden konnte, dass erhöhte
Laktatkonzentrationen den Insulin stimulierten GLUT-4 Transport am tierischen
Skelett- und Herzmuskel verschlechtern (Vettor et al. 1997; Lombardi et al. 1999;
Choi et al. 2002).
1.5 Diabetes mellitus und körperliche Aktivität
Die Erkrankung Diabetes mellitus wird infolge körperlicher Inaktivität,
Fehlernährung und Überalterung der Bevölkerung zu einem der führenden
Gesundheitsprobleme des 21. Jahrhunderts werden. In Europa leiden derzeit über 50
Millionen Betroffene an der Erkrankung, weltweit insgesamt schätzungsweise 285
Millionen Menschen (International Diabetes Federation, 2010). Es wird davon
ausgegangen, dass sich die Zahl der Erkrankungen bis ins Jahr 2030 weltweit auf 366
Millionen erhöht (Wild et al. 2004). Weltweit sterben jährlich etwa 3,2 Millionen
Menschen an den diabetischen Folgeerkrankungen. In Deutschland sind 7,1
Millionen Menschen von der Erkrankung betroffen (Hauner et al. 2007), wobei
zusätzlich noch von einer Dunkelziffer mit einem nicht-diagnostizierten Diabetes
von zwei bis drei Millionen Personen innerhalb der Bevölkerung ausgegangen wird
(Rathmann et al. 2003).
Der Typ 2 Diabetes ist mit etwa 90 % die häufigste Diabetesform und stellt eine
chronisch, progrediente Erkrankung dar, die sich aus einer vererbten und erworbenen
Insulinresistenz mit relativen Insulinmangel entwickelt und im Krankheitsverlauf zu
einem sekretorischen Defekt mit Insulinresistenz führt (Kerner et al. 2004; Matthaei
et al. 2008). Der Typ 2 Diabetes wird zusammen mit den Erkrankungen von
abdominaler Adipositas, Dyslipidämie und arterieller Hypertonie dem Symptomen-
komplex des Metabolischen Syndroms zugeordnet (Grundy et al. 2004). Treten
mehrere Komponenten des metabolischen Syndroms gemeinsam auf liegt für den

Einleitung
13
Patienten ein erhöhtes Risiko für kardiovaskuläre Erkrankungen vor, welches durch
große Bevölkerungsstudien wie u.a. der Framingham-Studie und PROCAM-Studie
(Hense et al. 2003; D'Agostino et al. 2002) belegt wurde.
Kardiovaskuläre Erkrankungen stellen mit 75% der Gesamtmortalität die häufigste
Todesursache bei Typ 2 Diabetikern dar, da die Erkrankten ein zwei bis vierfach
erhöhtes kardiovaskuläres Risiko gegenüber Nichtdiabetikern aufweisen (Wilson et
al. 1992; Geiss et al. 1995). Die chronische Hyperglykämie gilt als ein eigenständiger
Risikofaktor für die Entstehung von pathologischen Gefäßveränderungen (Hanefeld
et al. 1997). Langzeitstudien (ADVANCE, UKPDS) bei Typ 2 Diabetikern belegen,
dass ein langfristig gut eingestellter Blutzuckerspiegel das kardiovaskuläre Risiko bei
den Betroffenen nachhaltig reduzieren kann (Advance et al. 2008; Holman et al.
2008).
Die körperliche Aktivität hat innerhalb des Therapiekonzeptes des Typ 2 Diabetes
einen hohen therapeutischen Nutzen, weil durch die zusätzliche Muskelarbeit eine
Verbesserung der peripheren Insulinresistenz und eine Reduzierung der im Rahmen
des Metabolischen Syndroms auftretenden kardiovaskulären Risikofaktoren erreicht
werden kann.
Im Rahmen der Primärprävention untermauern große randomisierte Inter-
ventionsstudien, dass sich in der Krankheitsphase der beginnenden Insulinresistenz
der Blutzuckerspiegel durch eine Lebensstilveränderung, im Sinne einer
Ernährungsumstellung sowie regelmäßiger körperlicher Bewegung, normalisieren
lässt und somit eine Manifestation des Diabetes Typ 2 verzögert bzw. verhindert
werden kann (Eriksson et al. 1991; Manson et al. 1991; Tuomilehto et al. 2001; Hu et
al. 2002; Knowler et al. 2002; Krook et al. 2003; Petrella et al. 2005; Lindstrom et al.
2006; Ramachandran et al. 2006).
In der Sekundärprävention sieht die Deutsche Diabetes Gesellschaft die körperliche
Aktivität neben einer Ernährungsmodifikation als die erste Therapiemaßnahme bei
der Behandlung eines neu manifesten Typ-2-Diabetes an (Halle et al. 2008). In
zahlreichen randomisierten Studien wurde der sekundärtherapeutische Nutzen von
körperlicher Aktivität innerhalb der Therapie bei manifesten Typ-2-Diabetes
nachgewiesen (Castaneda et al. 2002; Knowler et al. 2002; Boule et al. 2003; Cauza
et al. 2005; Snowling et al. 2006). Auf der Basis kontrollierter Studien hat die
amerikanische Diabetes Gesellschaft sowohl dem Ausdauer- als auch dem

Einleitung
14
Krafttraining den „Evidenzgrad A“ für die Verbesserung der glykämischen Stoff-
wechsellage verliehen (Sigal et al. 2004; König et al.2006).
In der Grundlagenforschung gibt es auf molekularer und zellulärer Ebene
gegenwärtig nur vereinzelte Studien zum diabetischen Laktatstoffwechsel, welche
die langfristigen und kurzfristigen Dichteveränderungen und Regulations-
mechanismen der Monocarboxylattransporter im Muskel und Erythrozyten bei Typ 2
Diabetikern untersucht haben.
Im Tiermodell konnte ein verringerter MCT1-Gehalt und ein verschlechterter
Lakattransport in die Adipozyten von diabetischen (Hajduch et al. 2000) sowie von
übergewichtigen Ratten (Py et al. 2001) im Skelettmuskel festgestellt werden. In
einer weiteren Studie zeigte sich auch eine Verschlechterung des Laktattransportes in
der Skelettmuskulatur bei diabetischen Ratten, jedoch blieb der MCT1-Gehalt
unverändert (Py et al. 2002).
In einer Trainingsstudie mit diabetischen Ratten konnte nachgewiesen werden, dass
durch ein Ausdauertraining eine Erhöhung des krankheitsbedingten reduzierten
MCT1- und MCT4-Gehalts im Herz- und Skelettmuskel sowie eine Verbesserung
des Laktattransports erreicht werden kann (Enoki et al. 2003). Diese Beobachtungen
wurden durch die Ergebnisse einer weiteren Trainingsstudie bekräftigt, bei der die
Laktattransportkapazität und MCT1-Expression nach einem Ausdauertraining erhöht
werden konnte (Metz et al. 2005).
In der bisher einzigen mit Typ 2 Diabetikern durchgeführten Trainingsstudie konnte
eine um 35% reduzierte MCT1-Dichte im Vergleich zu der gesunden Kontroll-
gruppe festgestellt werden. Mittels eines sechswöchigen Krafttrainings konnte der
MCT1-Gehalt wieder normalisiert werden (Juel et al. 2004b). Die Auswirkungen
eines Ausdauertrainings auf die MCT-Dichte sowohl im Skelettmuskel und als auch
in den Erythrozyten sowie die zugrundeliegenden aktivitätsbedingten Regulations-
mechanismen wurden bisher bei Typ 2 Diabetikern noch nicht untersucht.

Einleitung
15
1.6 Fragestellungen
Der Schwerpunkt der zahlreichen Trainingsstudien zum Laktatstoffwechsel befasste
sich überwiegend mit den kinetischen Parametern zum Laktattransport. Bisher gibt es
nur wenige Trainingsstudien, die den langfristigen Einfluss von körperlicher
Aktivität auf die MCT-Transporter in der Muskulatur und Erythrozyten bei Typ 2
Diabetikern und die zugrundeliegenden kurzfristigen zellulären Regulations-
mechanismen untersucht haben. Analog zu der aktivitätsbedingten GLUT4-
Translokation in der Muskelzelle kann aus den gegenwärtigen Studienergebnissen
vermutet werden, dass möglicherweise körperliche Aktivität zu einer kurzfristigen
Translokation der MCT-Proteine und des Chaperons CD147 in die Muskelmembran
führt. Untersuchungen zu den kurzfristigen aktivitätsbedingten Regulations-
mechanismen der MCT1- und CD147-Proteine im Erythrozyten liegen nach gegen-
wärtigem Wissensstand nicht vor.
Vor diesem Hintergrund ergaben sich für die DiabetesAktiv-Studie folgende
Fragestellungen:
1. Kommt es durch ein sporttherapeutisches Ausdauer- und Krafttraining bei nicht-
insulinpflichtigen Typ 2 Diabetikern zu langfristigen Veränderungen der MCT1- und
MCT4-Dichte in der Muskulatur?
Für die Bestimmung dieser trainingsinduzierten Veränderungen der MCT1- und
MCT4-Dichten wurden immunhistochemische Färbungen an den Muskelschnitten
aus der Eingangs- und Ausgangsuntersuchung durchgeführt.
2. Zeigen sich im Erythrozyten neben den langfristigen auch kurzfristige
Veränderungen der MCT1- und CD147-Dichte unmittelbar nach dem Belastungs-
test?
Die Bestimmung der trainingsinduzierten Veränderungen der MCT1-Dichte erfolgte
anhand von immunhistochemischen Färbungen der Erythrozyten aus den venösen
Blutproben vor und unmittelbar nach dem Belastungstest der Eingangs- und
Ausgangsuntersuchung. Die CD147-Dichteveränderungen an den Erythrozyten
wurden durchflusszytometrisch aus den venösen Blutproben vor und unmittelbar
nach dem Belastungstest aus der Eingangs- und Ausgangsuntersuchung analysiert.

Einleitung
16
3. Zeigt sich unter körperlicher Belastung kurzfristig eine trainingsinduzierte
verstärkte Translokation der MCT1- und CD147-Proteine analog zur Glut4-
Translokation in der Erythrozytenmembran nach einem dreimonatigen Ausdauer-
und Krafttraining?
Der Nachweis einer möglichen aktivitätsbedingten Translokation der MCT1- und
CD147-Proteine im Erythrozyten erfolgte anhand von Immunfluoreszenz-Färbungen
aus den venösen Blutproben vor und unmittelbar nach dem Belastungstest aus der
Eingangs- und Ausgangsuntersuchung
4. Welchen Einfluss haben die beiden Trainingsformen auf den diabetischen
Stoffwechsel und die körperliche Leistungsfähigkeit der Patienten?
Die Auswirkungen des Ausdauer- und Krafttrainings wurden durch eine
Gegenüberstellung von ausgewählten metabolischen und leistungsphysiologischen
Parametern aus der Eingangs- und der Ausgangsuntersuchung aufgezeigt.
Material und Methoden
17
2 Material und Methoden
2.1 Studiendesign 2.1.1 Probandenbeschreibung
Eingeschlossen in die Studie wurden Männer über 30 Jahre, die einen nicht-
insulinpflichtigen Typ 2 Diabetes aufwiesen und zuvor nicht regelmäßig körperlich
aktiv waren. Ausgeschlossen waren insulinpflichtige Typ 1- und Typ 2 Diabetiker
und ebenso Patienten mit einem akuten Koronarsyndrom, limitierende orthopädische
Begleiterkrankungen, schweren mikro- und makrovaskuläre Folgeerkrankungen des
Diabetes mellitus, schwerer COPD, Alkohol- und Drogenmissbrauch oder weiteren
bösartigen Grunderkrankungen.
Insgesamt wurden 32 männliche Patienten mit einem nicht-insulinpflichtigen Typ 2
Diabetes in die Studie eingeschlossen. Davon haben insgesamt 27 Patienten an der
Eingangs- und Ausgangsuntersuchung teilgenommen und mindestens 90% der
gesamten Trainingseinheiten absolviert. Vier Studienteilnehmer sind aus gesund-
heitlichen Gründen und ein Patient wegen unregelmäßiger Teilnahme aus der Studie
ausgeschlossen worden. Dadurch reduzierte sich die Anzahl der Teilnehmer bei der
Ausdauergruppe auf dreizehn Probanden und bei der Kraftgruppe auf vierzehn
Probanden. Die Manifestation der Erkrankung lag bei den Patienten im Durchschnitt
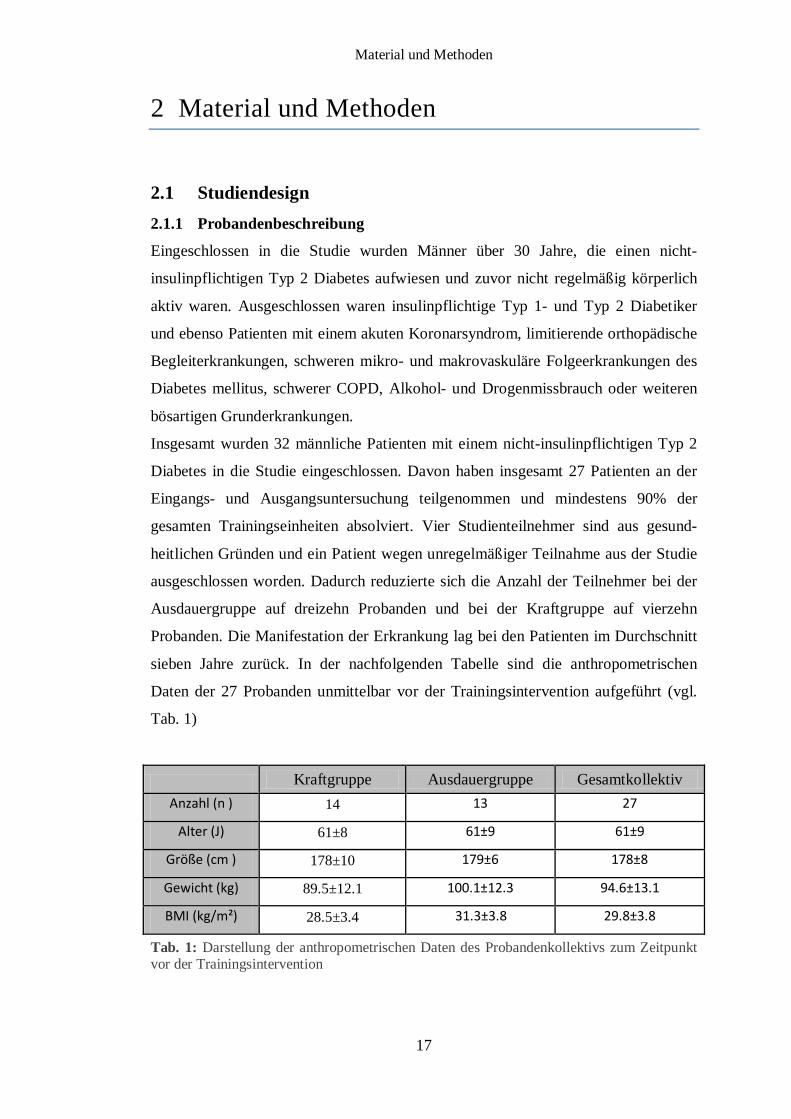
sieben Jahre zurück. In der nachfolgenden Tabelle sind die anthropometrischen
Daten der 27 Probanden unmittelbar vor der Trainingsintervention aufgeführt (vgl.
Tab. 1)
Tab. 1: Darstellung der anthropometrischen Daten des Probandenkollektivs zum Zeitpunkt vor der Trainingsintervention
Kraftgruppe Ausdauergruppe Gesamtkollektiv Anzahl (n ) 14 13 27
Alter (J) 61±8 61±9 61±9
Größe (cm ) 178±10 179±6 178±8
Gewicht (kg) 89.5±12.1 100.1±12.3 94.6±13.1
BMI (kg/m²) 28.5±3.4 31.3±3.8 29.8±3.8

Material und Methoden
18
2.1.2 Untersuchungsablauf
Die Rekrutierung erfolgte mittels zweier Zeitungsanzeigen und eines Informations-
briefes an die ortsansässigen Diabetologen. Im Rahmen eines Informationsabends
wurden die interessierten Diabetiker ausführlich über die Studienablauf und dessen
Inhalte sowie die möglichen Risiken bei den Untersuchungen aufgeklärt.
Die Eingangs- als auch Ausgangsuntersuchung wurde im Institut für Kreislauf-
forschung und Sportmedizin der Sporthochschule Köln durchgeführt. Zu den Unter-
suchungszeitpunkten fanden jeweils eine ausführliche Anamnese und eine Nüchtern-
Blutabnahme sowie eine klinische Untersuchung der Patienten statt. Nach der
Eingangsuntersuchung (U1) wurden die Teilnehmer in die Studie ein- bzw.
ausgeschlossen und anschließend in eine Kraftgruppe und Ausdauergruppe
randomisiert unterteilt. Vor Interventionsbeginn wurde bei achtzehn Patienten im
Institut für Motorik und Bewegungstechnik eine Muskelbiopsie durchgeführt.
Danach begann die dreimonatige sporttherapeutische Intervention, in dem beide
Trainingsgruppen jeweils zweimal pro Woche ihr Trainingsprogramm an der
Sporthochschule Köln und noch zusätzlich eigenverantwortlich einmal pro Woche
ein Heimtrainingsprogramm absolvierten. Zudem wurden bei beiden Trainings-
gruppen in der ersten und letzten Trainingswoche isometrische Kraftmessungen
durchgeführt. Nach der letzten Trainingseinheit erfolgten zeitnah die zweite Muskel-
biopsie und anschließend die Abschlussuntersuchung (U2) der Studienteilnehmer
(vgl. Abb. 7).
Der Ethikantrag wurde vor Studienbeginn von der Ethikkommission der Deutschen
Sporthochschule genehmigt und entsprach den Richtlinien der Deklaration von
Helsinki. Die Studienteilnehmer wurden ausführlich über die Studieninhalte und
mögliche Risiken bei den Untersuchungen aufgeklärt und haben vor der Eingangs-
untersuchung ihre schriftliche Einverständniserklärung zu den Untersuchungen und
Studienbedingungen abgegeben.

Material und Methoden
19
ScreeningVorauswahl der Patienten (n=32)
Studienablauf der DiabetesAktiv-Studie
Randomisierung / Gruppeneinteilung
Nov./Dez.2006
U1 / EingangsuntersuchungRuhe EKG, Labor, Ärztl. Untersuchung,
Echokardiographie, Bel. EKG, Muskelbiopsien, Isometrische Kraftmessungen
Ausdauergruppe 3 Monate
+ Heimtrainingsprogramm
Kraftgruppe 3 Monate
+ Heimtrainingsprogramm
19. März-13. Juni
2007
U2 / Abschlussuntersuchung Ruhe EKG, Labor, Ärztl. Untersuchung,
Echokardiographie, Bel. EKG, Muskelbiopsien, Isometrische Kraftmessungen
14-17.Juni2007
ZEITLAUF
8-11. März 2007
Abb. 7: Studienablaufschema der DiabetesAktiv-Studie
2.2 Untersuchungsverfahren
2.2.1 Erhebung der anthropometrischen Daten
Alter und Gewicht
Das Alter der Probanden wurde aus dem Geburtsdatum und dem Zeitpunkt der
Eingangsuntersuchung berechnet. Die Messung des Körpergewichtes erfolgte gemäß
den standardisierten Messvorgaben der IDIS (Laaser et al. 1989) auf einer Stand-
waage Typ Seca761 (Vogel & Halke, Hamburg, D) ohne Schuhe und in leichter
Kleidung.
Die Körpergröße wurde mittels eines geeichten Messbandes Typ Seca 225 (Vogel &
Halke, Hamburg, D) an einer senkrechten Wand, ohne Schuhe in aufrechter Position
erfasst.
Body-Mass-Index
Die Bestimmung des Body-Mass-Index (BMI) wurde aus den ermittelten Daten
anhand folgender Formel errechnet:
BMI (kg/m²) = Körpergewicht (kg) / Körpergröße zum Quadrat (m²)

Material und Methoden
20
Die Klassifikation des BMI-Wertes erfolgte entsprechend den Richtlinien der
Deutschen Adipositas-Gesellschaft (Hauner et al. 2007). Übergewicht ist definiert als
BMI ≥ 25 kg/m2, Adipositas als BMI ≥ 30 kg/m2.
Blutdruckmessung
Die arterielle Ruheblutdruckmessung erfolgte mit der indirekten Methode nach Riva
Rocci mittels eines vollautomatischen Messverfahrens (ELAG BE 227,
Schwarzhaupt, D) am Oberarm der Probanden gemäß den Richtlinien der Deutschen
Hochdruckliga (Deutsche Hochdruckliga et al. 1997).
Ruhe-EKG und Echokardiographie
Vor der Belastungsuntersuchung wurde das Ruhe-EKG der Brustwand nach
WILSON sowie der Extremitäten nach EINTHOVEN und GOLDBERGER
abgeleitet und mit dem MAC 1200 (Medical Systems, Freiburg, D) durchgeführt. Bei
der Echokardiographie wurden die Probanden in linker Halbseitenlage mit dem
VIVD TM 3 (Medical Systems, Solingen, D) untersucht.
2.2.2 Laborparameter
Cholesterin
Zur quantitativen Bestimmung des Cholesterins wurde das ABX Pentra Cholesterol
CP Reagenzsystem (ABX Diagnostics, Montpellier, F) verwendet. Der Nachweis des
Cholesterins im Serum erfolgte mittels eines enzymatischen, photometrischen Tests.
Bei dieser CHOD-PAP-Methode dient der rote Farbstoff Chinonimin als
kolorimetrischer Indikator. Die photometrischen Messungen wurden am Zentrifugal-
analyser Corbas Mira Plus System (Hoffmann-La Rouche, Basel, Schweiz)
durchgeführt.
HDL-Cholesterin
Die quantitative Bestimmung der Cholesterin-Lipoproteine von hoher Dichte im
Serum erfolgte mit Hilfe des ABX Pentra HDL Direct CP Reagenzsystems (ABX
Diagnostics, Montpellier, F) mittels Kolorimetrie. Bei dieser „ Accelerator Selective
Detergent-Methode“ wird das HDL über zwei Reagenzien einer Enzymreaktion
ausgesetzt. Mit Hilfe dieser homogenen Methode kann die HDL-Cholesterin-

Material und Methoden
21
konzentration direkt über eine Farbentwicklung mit Hilfe des Corbas Mira Plus
System bestimmt werden.
LDL-Cholesterin
Die Bestimmung der LDL-Cholesterinkonzentration wurde mittels der
FRIEDEWALD-Formel berechnet:
LDL-Cholesterin = Gesamtcholesterin - Triglyzeride/5 – HDL Cholesterin
Triglyzeride
Für die quantitative Messung der Triglyzeridkonzentration wurde das ABX Pentra
Triglycerides CP Reagenzsystem (ABX Diagnostics, Montpellier, F) eingesetzt. Bei
dieser enzymatischen, kolorimetrischen Methode wurde die Triglyzeridkonzentration
mittels des roten Farbstoffs Chinonimin durch das Zentrifugalanalysegerät Corbas
Mira Plus System bestimmt.
Insulin
Zur quantitativen Bestimmung der Insulinkonzentration im Serum wurde das
Elektrochemilumineszenz Immunoassay Elecsys 1010/2010 und Modular Analytics
E170 (Roche, Mannheim, Deutschland) verwendet. Das Grundprinzip dieser Mess-
methode beruht auf einem Sandwichprinzip, bei dem zwei monoklonale Insulin-
spezifische Antikörper nach der ersten Inkubation mit dem Insulin ein Sandwich-
Komplex bilden. Nach der zweiten Inkubation wird nach Zugabe von Streptavidin
beschichteten Mikropartikeln der Komplex über Biotin-Streptavidin-
Wechselwirkung an die Festphase gebunden. Anschließend wird das Reaktions-
gemisch in eine Messzelle gegeben, bei denen die Mikropartikel mittels der
magnetischen Wirkung auf der Oberfläche der Elektrode fixiert werden.
Ungebundene Substanzen wurden danach mit ProCell entfernt. Die durch Spannung
induzierte Chemilumineszenzemission wurde mit einem Photomultiplier gemessen.
Abschließend wurden anhand von Kalibrationskurven die Ergebnisse ermittelt.
Proinsulin
Die Messung der Proinsulinkonzentration im Humanserum erfolgte mit dem
Radioimmunoassay HPI-15K (Linco Research, Charles, USA). Diese radio-
immunologische Hormonbestimmung wurde erstmalig 1959 von Berson und Yalow

Material und Methoden
22
zur Insulinbestimmung durchgeführt. Nach einer viertägigen Inkubationszeit bei
Raumtemperatur wurde die Radioaktivität der gebildeten Antigen-Antikörper-
komplexe bestimmt, woraus sich die gesuchte Proinsulinkonzentration berechnen
ließ. Als Tracer wurde ein 125 J markiertes Humanproinsulin verwendet.
Glukose
Die quantitative Bestimmung von Glukose im Serum erfolgte mit Hilfe des ABX
Pentra Glukose HK CP Reagenzsystem (ABX Diagnostics, Montpellier, F) mittels
Kolorimetrie. Bei dieser enzymatischen Methode wurde die Glukosekonzentration
durch den Zentrifugalanalyser Corbas Mira Plus System bestimmt.
Homa-Index
Die Insulinresistenz der Probanden wurde über den Homa-Index berechnet. Dieser
Index ist ein mathematisches Modell, der den Grad der Insulinresistenz aus dem
Nüchterninsulin- und blutzuckerwert nach folgender Orginalformel (Matthews et al.
1985) berechnet:
Homa-Index = Insulin (nüchtern, μU/ml) x Glukose (nüchtern, mg/dl) : 405
Normwertbereich : ≤ 1 normal
> 2 Hinweis auf eine Insulinresistenz
> 2.5 Insulinresistenz sehr wahrscheinlich
> 5 Werte bei Typ 2 Diabetikern
Laktat
Bei den fahrradergometrischen Belastungsuntersuchungen wurden den Probanden
nach dem Einstich mit einer sterilen Einmallanzette (ASID Bonz, Herrenberg, D)
vor, in den letzten 10 Sekunden jeder Belastungsstufe, bei Testabbruch sowie fünf
und zehn Minuten in der Erholungsphase 20 μl arterialisiertes Kapillarblut aus dem
hyperämisierten Ohrläppchen mit Hilfe einer End-to-End-Kapillare (EKF-
Diagnostik, Barleben, D) entnommen. Nach der Entnahme wurde die Kapillare in ein
Eppendorfcup (Eppendorf, Hamburg, D) mit 1ml Systemlösung gegeben und zehn
Sekunden geschüttelt und vermischt und anschließend bis zur Analyse bei 4° C im
Kühlschrank gelagert.
Die Bestimmung der Laktatkonzentration erfolgte mit dem Laktatmeßgerät Ebio-plus
(Eppendorf, Hamburg, D). Vor den einzelnen Analysen erfolgte eine Nullpunkt-

Material und Methoden
23
bestimmung und Gerätekalibrierung mit anschließenden vier weiteren Kontroll-
messungen. Danach wurde der Eppendorfcup in den Probenteller gesteckt und 200 μl
Probenlösung wurden mittels automatisch gesteuerter Hohlnadel in die Messzelle
gesaugt und nach dem enzymatischen-amperometrischen Messprinzip analysiert.
Blutentnahme und Blutverarbeitung
Bei den zwei Untersuchungsterminen wurden den Studienteilnehmern zwei venöse
Blutproben aus einer Armvene mittels einer Butterfly-Nadel in eine EDTA-Vakuette
abgenommen (Becton Dichinson Vacutainer Systems, Heidelberg, D). Die erste
venöse Blutentnahme (to) wurde im nüchternen Zustand vor dem BEKG durch-
geführt, da aus dieser Blutprobe u.a. die Laborparameter mitbestimmt wurden. Die
zweite Blutabnahme erfolgte unmittelbar nach dem BEKG (t1).
Anschließend wurde das gewonnene Blut mit einer Zentrifuge des Typs Rotixa 1200
(Hettich Zentrifugen, Tuttlingen, D) bei einer Temperatur von 20 ° C bei
3400 U/Min. zehn Minuten lang zentrifugiert, um das benötigte Serum für die Unter-
suchungen der Laborparameter zu gewinnen.
Konservierung der Blutproben für die MCT1-Bestimmung der Erythrozyten
Zur Fixierung der Erythrozyten wurden aus jeder Probe 200 μl EDTA-Vollblut in ein
Eppendorfcup (Eppendorf, Hamburg, D) abpipettiert und dazu 200 μl vierprozentige
Paraformaldehydlösung mit einem pH-Wert von 7,4 gegeben. Nach einer
Inkubationszeit von zwanzig Minuten erfolgte eine dreiminütige Zentrifugation
(Hettich Zentrifugen, Tuttlingen, D) bei 800 U/Min. Nach der Abpipettierung des
Überstandes wurden 200 μl Phosphatpuffer (0.1 M) hinzugegeben, um die im
Sediment befindlichen Erythrozyten zu waschen. Nach einer dreiminütigen
Inkubationszeit erfolgte eine erneute dreiminütige Zentrifugation bei 800 U/Min. und
abschließend wurden die Proben nach dem Entfernen des Überstandes mit 800 μl
PBS-Puffer (0.1 M) gemischt. Die Proben wurden anschließend im Kühlschrank bei
4° C bis zu den immunhistochemischen Untersuchungen aufbewahrt.

Material und Methoden
24
Verwendete Lösungen und Puffer
Phosphatpuffer (PBS) 0.2 M [pH 7.4]:
28.8 g/l Dinatrium-Hydrogenphosphat-Dihydrat (Merck-Schuchardt,
Hohenbrunn, D) in 250 ml Aqua dest. lösen
5.2 g/l Natrium-Dihydrogenphosphat.Monohydrat (Merck, Darmstadt, D)
dazugeben
17.532 g/l Natriumchlorid (Merck, Darmstadt, D) hinzufügen
mit dem pH-mV-Meter (Mettler Toledo, Schwerzenbach, CH) mit HCL
(Merck, Darmstadt, D) auf pH 7.4 einstellen
mit Aqua dest. auf 1000 ml auffüllen
4% Paraformaldehydlösung:
• 4 g Paraformaldehyd (Merck, Darmstadt, D) in 40 ml Aqua dest. (60° C)
lösen
• mit 1n NaOH (Merck, Darmstadt, D) klären und filtrieren
mit 50 ml 0.2 M mischen und mit dem pH-mV-Meter (Mettler Toledo,
Schwerzenbach, CH) mit HCL (Merck, Darmstadt, D) auf pH 7.4 einstellen
mit Aqua dest. auf 100 ml auffüllen
Phosphatpuffer (PB) 0.1 M:
14.4 g/l Dinatrium-Hydrogenphosphat-Dihydrat in 250 ml Aqua dest. lösen
(Merck-Schuchardt, Hohenbrunn, D)
2.6 g/l Natrium-Dihydrogenphosphat.Monohydrat (Merck, Darmstadt, D)
dazugeben
mit dem pH-mV-Meter (Mettler Toledo, Schwerzenbach, CH) mit HCL auf
pH 7.4 einstellen
mit Aqua dest. auf 1000 ml auffüllen

Material und Methoden
25
2.2.3 Muskelbiopsie
Den Studienteilnehmern wurden vor und unmittelbar nach der Trainingsintervention
mit einer modifizierten Biopsienadel nach Evans et al. (1982) 50-100 mg an Muskel-
gewebe aus dem M. vastus lateralis entnommen.
Abb. 8 : Modifizierte Biopsienadel (Evans et al. 1982)
Über die Risiken und möglichen Komplikationen dieses freiwilligen Eingriffes
wurden die Probanden mündlich und schriftlich aufgeklärt. Die Technik der
Nadelbiopsie nach Bergstöm ist mit einer geringen Komplikationsrate behaftet
(Bergström, 1975), da der Muskel selbst keine Schmerzrezeptoren und große Gefäße
besitzt. Nach der Desinfektion und Lokalanästhesie durch eine 2 ml Lösung aus 2%
Xylocain 1:200000 mit Adrenalin wurde die Haut und Muskelfascie mit Hilfe eines
Skalpells etwa um 1 cm geöffnet. Anschließend wurde die Hohlnadel in den Muskel-
bauch eingeführt. Das durch den Unterdruck in die Biopsienadel gesaugte Muskel-
gewebe wurde mittels eines Stanzzylinders abgetrennt. Im Anschluss wurde die
Wunde mit einem Klammerpflaster versorgt sowie ein Kompressionsverband für 24
Stunden angelegt. Die entnommenen Muskelproben wurden unmittelbar danach in
Tissue Tek (Sakura, Zoeterwoude, NL) eingebettet. Anschließend wurden die Proben
in flüssigem Stickstoff tiefgefroren und bis zur Auswertung im Gefrierschrank bei
-80° C im Labor aufbewahrt.
2.3 Immunhistochemie In der Immunhistologie können zelluläre Antigene mit Hilfe von hochspezifischen
Antigen-Antikörper-Reaktionen nachgewiesen werden. Es wird zwischen zwei
Detektionssystemen unterschieden. Bei der direkten Methode wird der Primär-
antikörper direkt oder mit einem Enzym Fluorchrom markiert und mittels einer
Farbreaktion nachgewiesen. Dagegen bei der indirekten Methode bindet zuerst ein

Material und Methoden
26
unkonjugierter Primärantikörper an das zu untersuchende Antigen, welches mittels
von einem gegen den Primärantikörper gerichteten Sekundärantikörper indirekt über
eine Farbreaktion nachgewiesen werden kann (Luttman et al. 2004).
In der vorliegenden Arbeit wurde für die immunhistochemischen Untersuchungen
die indirekte StreptABC-Methode verwendet. Bei dieser Methode wurde die hohe
Bindungsaffinität von Avidin und Streptavidin zu Biotin ausgenutzt. Avidin ist ein
aus dem Hühnereiweiß stammendes basisches Glykoprotein, das an der Oberfläche
vier spezifische Biotin-Bindungsstellen aufweist. Streptavidin stammt aus dem
Bakterium Streptomyces und besitzt vier Bindungsstellen für das Biotin. Das wasser-
lösliche Vitamin Biotin besitzt gegenüber dem Avidin eine hohe Affinitätskonstante
und geht deshalb sehr schnell eine nicht kovalente Bindung mit dem Avidin ein. Zu
Beginn wurde ein unkonjugierter Primärantikörper auf die Proben gegeben, der an
die vorhandenen Zielantigene bindet. Im zweiten Schritt erfolgte die Inkubation mit
einem gegen die Tierspezies des Primärantikörpers gerichteten biotinylierten
Sekundärantikörper. Im Anschluss gehen die Moleküle des zugegebenen Enzym-
Konjugates aus Streptavidin und Meerretich-Peroxidase (HRP) mit dem Biotin des
Sekundärantikörpers eine Bindung ein. Abschließend erfolgt durch die Zugabe des
Substrates DAB (3,3 Diamino-Benzidin) eine Enzym-Substrat-Reaktion, bei der
mittels einer Farbreaktion das Zielantigen visualisiert und nachgewiesen werden
konnte (Hsu et al. 1981).
2.3.1 Immunhistochemischer Nachweis des MCT1 und MCT4 im
Muskelgewebe
Herstellung der Kryostatschnitte
Die Kryostatschnitte wurden mit dem Kryostat-Schneidegerät Leica CM 1900
(Reichert-Jung, Nussloch, D) angefertigt. Dazu wurden die mit Tissue-Tek (Sakura,
Zoeterwoude, NL) eingebetteten Muskelbiopsieproben auf dem Schneideteller fest-
gefroren und anschließend in das Mikrotom eingespannt. Die Gefrierkammer wurde
vor dem Schneiden auf -25° C herunter gekühlt. Die Dünnschnitte hatten eine Dicke
von 7μm und wurden anschließend auf einen Objektträger (Menzel, Braunschweig,
D) aufgezogen. Nach der lichtmikroskopischen Kontrolle (Optech, München, D)
wurden die Schnitte bis zu den immunhistochemischen Untersuchungen bei -80° C
im Gefrierschrank (Heraeus Instruments, Osterode, D) wieder eingefroren.

Material und Methoden
27
Verwendete Puffer und Lösungen
Trispuffer (TBS) 0.05 M:
6.057 g Tris(hydroxymethyl)-aminomethan (Merck, Darmstadt, D) in 250 ml
Aqua dest. lösen
8.766 g NaCl (=150 mmol) dazugeben (Merck, Darmstadt, D)
mit dem pH-mV-Meter (Mettler Toledo, Schwerzenbach, CH) mit HCL
(Merck, Darmstadt, D) auf pH 7.6 einstellen
mit Aqua dest. auf 1000 ml auffüllen
Phosphatpuffer (PB) 0.1 M [pH 7.4]:
14.4 g/l Dinatrium-Hydrogenphosphat-Dihydrat (Merck, Darmstadt, D) in
250 ml Aqua dest. lösen
2.6 g/l Natrium-Dihydrogenphosphat.Monohydrat (Merck, Darmstadt, D)
dazugeben
mit dem pH-mV-Meter (Mettler Toledo, Schwerzenbach, CH) mit HCL
(Merck, Darmstadt, D) auf pH 7.4 einstellen
mit Aqua dest. auf 1000 ml auffüllen
Wasserstoffperoxidlösung:
20 ml Methanol (Merck, Darmstadt, D)
4.5 ml Aqua dest.
0.5 ml 30% H2O2 (Merck, Darmstadt, D)
5% Bovine Serum Albumin (BSA)- Lösung:
5g /100 ml BSA (PAA, Cölbe, D) in 0.05 M TBS
Ammoniumchlorid-/ TritonX-Lösung:
0.59 g 0,5 M Ammoniumchlorid (Merck, Darmstadt, D)
50 µl 0.25% Triton X-100 (Serva, Heidelberg, D)
in 20 ml 0.05 M TBS lösen
Diaminobenzidin (DAB)-Lösung:
15 ml 0.1M PB [pH 7,4]:
150 µl Ammoniumchlorid = 6.0 mg (Sigma-Aldrich, Steinheim, D)
300 µl Nickel-II-Sulfat = 0.05 mol (Merck, Darmstadt, D)
300 µl 10% β-D-Glucose (Sigma-Aldrich, Steinheim, D)

Material und Methoden
28
50 µl Glucoseoxidase (Sigma-Aldrich, Steinheim, D)
150 µl Diaminobenzidin (DAB) = 7.5 mg (Merck, Darmstadt, D)
Filtrieren!
Entwässerung in einer aufsteigenden Alkoholreihe:
2 Min. in 70% Ethanol (Hoffmann, Düsseldorf, D)
2 Min. in 96% Ethanol
2 Min. in 100% Ethanol
2 Min. in Xylol (Quadflieg, Gelsenkirchen, D)
Immunhistochemisches Färbeprotokoll Die immunhistochemischen Untersuchungen am Muskelgewebe wurden gemäß eines
modifizierten Färbeprotokolls aus der Abteilung für molekulare und zelluläre
Sportmedizin des Institutes für Kreislaufforschung und Sportmedizin durchgeführt.
Zu Beginn wurden zwei Areale mit Muskelgewebe auf den Objektträgern (Menzel,
Braunschweig, D) mit einem hydrophoben PAP-Pen Markerstift (Dako, Hamburg,
D) umkreist. Auf dem oberen Areal erfolgte der Nachweis des MCT1 und auf dem
unteren Areal der des MCT4. Zudem dienten zwei Objektträger als immun-
histochemische Kontrolle (IHC), die keinen Kontakt mit dem Primärantikörper
erfuhren.
Danach wurden die Gewebeschnitte dreimal jeweils für fünf Minuten mit 0.05 M
TBS gewaschen. Auch die weiteren zwischen den einzelnen Arbeitsschritten
durchgeführten Waschvorgänge hatten jeweils eine Dauer von fünf Minuten. Im
nachfolgenden Arbeitsschritt wurde die endogene Peroxidaseaktivität durch eine
zwanzigminütige Inkubation der Gewebeschnitte mit einer frisch aufgesetzten
Wasserstoffperoxidlösung in TBS blockiert. Anschließend erfolgten zwei Wasch-
vorgänge bevor die Membranen für zehn Minuten mittels einer Ammoniumchlorid-
/TritonX-Lösung permeabilisiert wurden. Das Ammoniumchlorid bewirkt eine
Auflösung der Aldehydvernetzungen und Demaskierung des Gewebes, wohingegen
das Detergenz Triton X die Membran auflöst, so dass der Antikörper leichter zu dem
Antigen gelangen kann. Nach zwei weiteren Waschvorgängen erfolgte eine
Blockierung der unspezifischen Antikörperbindungsstellen durch 5%-ige BSA-
Lösung für eine Dauer von 60 Minuten. Direkt im Anschluss ohne weiteren Spül-

Material und Methoden
29
schritt wurde ein in 0.8%-iger BSA-Lösung verdünnter Primärantikörper des MCT1
(Rabbit Anti- Monocarboxylate Transporter 1, Chemicon, California, U.S.A.,
Verdünnung 1:1000) und MCT4 (Rabbit Anti- Monocarboxylate Transporter 4,
Santa Cruz Biotechnology, U.S.A., Verdünnung 1:500) auf die Gewebeschnitte
gegeben und über Nacht bei 4° C im Kühlschrank inkubiert. Hierbei erfolgt die
spezifische Bindung des Primärantikörpers an das Antigen. Bei der immunhisto-
chemischen Kontrollfärbung (IHC) wurde anstatt des Primärantikörpers lediglich die
0.8%-ige BSA-Lösung auf die Proben aufgetragen. Nach der Inkubation mit dem
Primärantikörper wurde die Gewebeschnitte dreimal mit TBS gespült bevor im
nachfolgenden Arbeitsschritt der in TBS verdünnte Biotin konjugierte Sekundär-
antikörper (Goat-Anti Rabbit, Dako, Hamburg, D, Verdünnung 1:400) auf die beiden
Areale des MCT1 und MCT4 appliziert wurde. Dieser Sekundärantikörper war gegen
die Tierspezies des Primärantikörpers gerichtet und wurde auf den Gewebeschnitten
für 60 Minuten inkubiert.
Nach drei weiteren Waschvorgängen wurde der Sekundärantikörper durch ein im
Verhältnis von 1:150 in TBS angesetzter Streptavidin-Hoseradish-Peroxidase-
Komplex (Healthcare, Chalfont St. Giles, Bucks Großbritannien) für die nach-
folgende Farbreaktion markiert. Die Inkubationszeit betrug 60 Minuten. Für die
anschließende Farbreaktion wurden die Schnitte mit einer frisch angesetzten 3.3´
Diaminobenzidin (DAB)-Lösung in Phosphatpuffer versetzt. Die Färbung wurde bei
wiederholter Sichtkontrolle bis zum Einsetzen einer bräunlichen Färbung unter dem
Mikroskop kontrolliert. Danach wurde die Reaktion direkt durch Abklopfen der
Substratlösung und drei weiteren Waschvorgängen abgestoppt. Im Anschluss
erfolgte eine Dehydrierung der Gewebeschnitte durch eine aufsteigende Alkohol-
reihe aus 70-, 96- und 100-prozentigem Ethanol und Xylol. Abschließend wurden die
Gewebeschnitte mit Entellan (Merck, Darmstadt, D) eingedeckt und mit einem
Deckgläschen (VWR, Langenfeld, D) versiegelt.

Material und Methoden
30
Abb. 9: MCT4-Färbung der Muskelzellen bei 200-facher Vergrößerung
Abb. 10: IHC-Kontrolle der Muskelzellen bei 200-facher Vergrößerung
Auswertung
Bildliche Darstellung der Gewebeschnitte
Für die bildliche Dokumentation der Gewebeschnitte wurde das Mikroskop KS 300
Axiophot (Carl Zeiss Vision GmbH, Hallbergmoos, D) verwendet und bei 200-
facher Vergrößerung ausgewertet. Die digitalen Bilder wurden mit der digitalen CCD
Kamera (Axio Cam, Jena, Deutschland) aufgenommen und mittels des Software-
programms „ KS 300 3.0“ auf den PC übertragen und als JPG- Datei abgespeichert.
Für den MCT1 und MCT4 wurden jeweils acht Bilder mit einer einheitlichen
Hintergrundgraustufenwert von 215±3 Density Unit (DU) mittels des Bildanalyse-
programmes ImageJ 137v (National Institute of Health, USA) angefertigt. Die
einzelnen Bilder wurden unter standardisierten Bedingungen bei identischer
Vergrößerung, Belichtung und Bildeinstellungen aufgenommen.

Material und Methoden
31
Bestimmung der zellulären MCT1- und MCT4-Dichte
Die quantitative Auswertung der digitalen Bilder erfolgte mit dem Bildanalyse-
programm ImageJ 137v. Bei dieser Auswertemethode lässt sich die Expression der
gesuchten Proteine über die Intensität der Färbung im Zytoplasma der einzelnen
Muskelzellen berechnen. Für die Bestimmung der trainingsinduzierten Veränder-
ungen der MCT1- und MCT4-Dichte im Zytoplasma wurden jeweils für jeden
Probanden sowohl vor und nach der Intervention acht Bilder mit je fünf Muskel-
zellen ausgewertet. Dazu wurde für jedes Bild ein mittlerer Hintergrundwert und aus
den fünf zytoplasmatischen Messpunkten ein Mittelwert für die Färbungsintensität
berechnet. Anschließend wurde für jedes Bild der Wert für die mittlere
Färbungsintensität von dem mittleren Hintergrundwert abgezogen, um so die
bildspezifischen geringen Abweichungen der Hintergrundwerte bei der Auswertung
zu berücksichtigen. Abschließend wurde die zelluläre MCT1- und MCT4-Dichte
mittels einer Mittelwertbildung aus diesen berechneten Werten der acht Bilder
ermittelt.
Bestimmung der absoluten und der relativen membranösen MCT1- und MCT4-
Dichte
Zu Beginn der Auswertung wurden zunächst die Gefäße und Artefakte aus dem
Orginalbild (vgl. Abb. 11, Bild A) unter Verwendung des Bildbearbeitungs-
programms (Adobe Photoshop CS3 Extended, Version10.0) weiß überblendet, so
dass ausschließlich nur die Membran und das Muskelgewebe ausgewertet wurde
(vgl. Abb. 11, Bild B).
Abb. 11: Orginalbild (Bild A) Bearbeitetes Bild (Bild B)

Material und Methoden
32
Anschließend wurde das zu analysierende digitale Farbbild mit dem ImageJ Bild-
analyseprogram in ein Graubild 8 Bit konvertiert. Für die weitere Auswertung wurde
eine Schwelle mit einer Farbintensität zwischen 0 und 175 DU festgelegt, um die
nachzuweisenden MCT-Proteine ausschließlich in der Muskelmembran zu erfassen
(vgl. Abb. 12). Höhere Schwellenwerte würden zu falschen Ergebnissen hinsichtlich
der Proteindichte in der Muskelmembran führen, da sonst auch die Muskelzellen-
innenfläche mit in die Auswertung einginge. Abschließend wurde der Gesamt-
mittelwert der Färbungsintensität der acht ausgewerteten Bilder vom Gesamt-
hintergrundwert abgezogen, so dass die bildspezifischen geringen Unterschiede der
Hintergrundwerte bei der Bestimmung der membranösen MCT1-und MCT4-Dichte
mitberücksichtigt wurden.
Abb. 12: Markierung des MCT1-Proteins in der Zellmembran der Muskelzellen bei einer
festgelegten Schwelle von 0-175 DU
Im anschließenden Schritt muss die absolute MCT-Dichte und die Messfläche in
Relation zu der Gesamtmuskelfläche gesetzt werden, um vergleichbare Aussagen zu
den MCT-Proteinveränderungen treffen zu können. Dazu wurden für die
Bestimmung der mittleren Gesamtmuskelfläche die acht Bilder erneut mittels des
Adobe Photoshop Programmes bearbeitet (vgl. Abb. 13). Alles außer der Muskel-
fläche wurde hell überblendet und nachfolgend mit einem festgelegten Schwellen-
wert von 250 DU mit dem Bildbearbeitungsprogramm Image J ausgewertet
(vgl. Abb. 14).

Material und Methoden
33
Abb. 13: Hintergrundflächenmarkierung mit dem Adobe Photoshop Programmes
Abb. 14: Gesamtmuskelflächenberechnung bei einer festgelegten Schwelle von 0-250 DU
Für die Berechnung der relativen MCT-Dichte wurde die Messfläche der MCT-
Proteine durch die mittlere Gesamtmuskelfläche geteilt und anschließend das
Ergebnis mit der zuvor berechneten absoluten MCT-Dichte multipliziert. Danach
wurden die Messdaten in eine Microsoft Excel-Tabelle übertragen und die absoluten
und die relativen membranösen MCT1- und MCT4-Dichten statistisch ausgewertet
und visualisiert.
2.3.2 Immunhistochemischer Nachweis des MCT1 im Erythrozyten
Verwendete Puffer und Lösungen
Trispuffer (TBS) 0.05 M:
6.057 g Tris(hydroxymethyl)-aminomethan (Merck, Darmstadt, D) in 250 ml
Aqua dest. lösen
8.766 g NaCl (=150 mmol) dazugeben (Merck, Darmstadt, D)
mit dem pH-mV-Meter (Mettler Toledo, Schwerzenbach, CH) mit HCL
(Merck, Darmstadt, D) auf pH 7.6 einstellen
mit Aqua dest. auf 1000 ml auffüllen
Phosphatpuffer (PB) 0.1 M:
14.4 g/l Dinatrium-Hydrogenphosphat-Dihydrat in 250 ml Aqua dest. lösen
(Merck-Schuchardt, Hohenbrunn, D)
2.6 g/l Natrium-Dihydrogenphosphat.Monohydrat (Merck, Darmstadt, D)
dazugeben

Material und Methoden
34
mit dem pH-mV-Meter (Mettler Toledo, Schwerzenbach, CH) mit HCL
(Merck, Darmstadt, D) auf pH 7.4 einstellen
mit Aqua dest. auf 1000 ml auffüllen
Wasserstoffperoxidlösung:
20 ml Methanol (Merck, Darmstadt, D)
4.5 ml Aqua dest.
0.5 ml 30% H2O2 (Merck, Darmstadt, D)
0.03 % Tween 20-Lösung
0.03g Tween 20 (Sigma, Taufkirchen, D)/100ml in TBS lösen
Diaminobenzidin (DAB)-Lösung:
15 ml 0.1M PB [pH 7.4]:
150 µl Ammoniumchlorid = 6.0 mg (Sigma-Aldrich, Steinheim, D)
300 µl Nickel-II-Sulfat = 0.05 mol (Merck, Darmstadt, D)
300 µl 10% β-D-Glucose (Sigma-Aldrich, Steinheim, D)
50 µl Glucoseoxidase (Sigma-Aldrich, Steinheim, D)
150 µl Diaminobenzidin (DAB) = 7.5 mg (Merck, Darmstadt, D)
Filtrieren!
Entwässerung in einer aufsteigenden Alkoholreihe:
2 Min. in 70% Ethanol (Hoffmann, Düsseldorf, D)
2 Min. in 96% Ethanol
2 Min. in 100% Ethanol
2 Min. in Xylol (Quadflieg, Gelsenkirchen, D)
Immunhistochemisches Färbeprotokoll
Die immunhistochemischen Untersuchungen am Erythrozyten wurden gemäß eines
modifizierten Färbeprotokolls aus der Abteilung für molekulare und zelluläre
Sportmedizin des Institutes für Kreislaufforschung und Sportmedizin durchgeführt.
Zu Beginn wurden 20 μl Blut aus den in PFA fixierten Erythrozyten der Probanden
auf einen beschrifteten Objektträger (Menzel, Braunschweig, D) gleichmäßig
ausgestrichen und über dem Bunsenbrenner durch mehrmaliges Schwenken hitze-
fixiert. Anschließend wurde die Zelldichte und die Unversehrtheit der Erythrozyten
unter dem Mikroskop (Optech, München, D) kontrolliert. Mittels eines Fettstiftes

Material und Methoden
35
(Dako, Hamburg, D) wurden zwei Areale auf dem Objektträger umrandet. Dabei
stellt das erste Areal die immunhistochemisch zu behandelnde Fläche dar und das
zweite Areal die immunhistochemische Kontrollfärbung (IHC), die keinen Kontakt
mit dem primären Antikörper erfährt. Zeigt die IHC-Kontrolle bei der Auswertung
keine charakteristische Farbreaktion können somit unspezifische Reaktionen
während der immunhistochemischen Färbung ausgeschlossen werden.
Die zu behandelnden Areale auf den Objektträgern werden zu Beginn mit einer
0.05 M Tris-Pufferlösung (TBS) für fünf Minuten gewaschen, um mögliche Verun-
reinigungen zu beseitigen. Die Dauer für die folgenden Waschvorgänge zwischen
den einzelnen Arbeitsschritten wurde auf fünf Minuten festgelegt. Im Anschluss
erfolgt eine bis 45-minütige Inkubation bei 37° C mit dem Enzym Trypsin (Sigma-
Aldrich, Steinheim, D), um eine Permeabilisierung der Zellmembranen und somit ein
leichteres Eindringen der Antikörper in den Erythrozyten zu erreichen. Anschließend
wurde die Reaktion durch einen Waschvorgang mit Leitungswasser abgestoppt. Im
nachfolgenden Arbeitsschritt wurde die endogene Peroxidaseaktivität durch
Wasserstoffperoxid blockiert. Dazu wurde zweimal eine neu aufgesetzte
Wasserstoffperoxidlösung für jeweils 20 Minuten auf die Areale der Objektträger
pipettiert. Nach einem weiteren Waschvorgang erfolgte eine Inkubation mit einer
dreiprozentigen in TBS gelösten Milchpulverlösung für 60 Minuten, um un-
spezifische Antikörperbindungsstellen zu blockieren. Anschließend, ohne weiteren
Spülgang, wurde der primäre Antikörper (Rabbit Anti- Monocarboxylate
Transporter1, Chemicon, California, U.S.A., Verdünnung 1:1000) in einer Lösung
bestehend aus 0.3%-igen Milchpulver und 0.03%-igen Tween 20 gelöst und für 60
Minuten auf das erste Areal des Objektträgers gegeben. Das für die IHC-Kontrolle
vorgesehene zweite Areal wurde nur mit der 0,3%-igen Milchpulverlösung ohne den
primären Antikörper inkubiert. Anschließend erfolgte ein Waschvorgang bevor für
30 Minuten ein 3%-iges Normal-Goat-Serum (Dako, Hamburg, D) für die Block-
ierung der unspezifischen Antikörperbindungsstellen auf die Proben aufgetragen
wurde. Unmittelbar nach dem Absaugen kam es ohne weiteren Waschvorgang zur
Inkubation mit dem sekundären Antikörper (Goat- Anti Rabbit, Dako, Hamburg, D,
Verdünnung 1:400). Dieser biotinylierte Antikörper war gegen die Tierspezies des
primären Antikörpers gerichtet. Nach 30 Minuten wurde die Inkubation durch einen
weiteren Waschvorgang abgestoppt.

Material und Methoden
36
Für die Vorbereitung für die enzymatische Färbereaktion wurde ein im Verhältnis
von 1:150 in TBS angesetzter Streptavidin-Hoseradish- Peroxidase- Komplex
(Healthcare, Chalfont St. Giles, Bucks Großbritannien) auf die Proben aufgetragen,
um den sekundären Antikörper für den nachfolgenden Arbeitsschritt zu markieren.
Nach einem weiteren Spülvorgang erfolgte anschließend durch die Zugabe einer
frisch angesetzten 3,3´ Diaminobenzidin (DAB)-Lösung in Phosphatpuffer die
enzymatische Färbereaktion.
Die Dauer der Färbung ist abhängig vom Färbeergebnis, welches ständig unter dem
Mikroskop beobachtet wurde. Die Färbereaktion wurde unmittelbar beim Einsetzen
einer bräunlichen Anfärbung der Proben durch ein Abklopfen der Substratlösung und
dreimaliges Waschen mit TBS abgestoppt. Anschließend erfolgte ein Equilibrieren
der Proben in einer aufsteigenden Alkoholreihe aus 70-, 96- und 100-prozentigem
Ethanol und Xylol. Abschließend wurden die Proben mit Entellan eingedeckt und
mit einem Deckgläschen versiegelt.
Abb. 15: Immunhistochemische MCT1-Färbung bei 400-facher Vergrößerung
Abb. 16: Immunhistochemische IHC-Kontrolle bei 400-facher Vergrößerung

Material und Methoden
37
Auswertung
Bildliche Darstellung der Erythrozyten
Die bildliche Darstellung der Präparate erfolgte am Mikroskop KS 300 Axiophot
(Carl Zeiss Vision GmbH, Hallbergmoos, D) bei 400-facher Vergrößerung. Die
Bilder von den Erythrozyten wurden mittels der digitalen CCD Kamera (Axio Cam,
Jena, Deutschland) abfotografiert und unter Verwendung des Softwareprogramms
„KS 300 3.0“ standardisiert als JPG- Datei abgespeichert. Dabei wiesen die Hinter-
gründe der aufgenommenen Bilder eine einheitliche Farbintensität von 215 ± 5 DU
auf, um eine Vergleichbarkeit der Proben zu erreichen. Die Bestimmung der Farb-
intensität jedes Bildes erfolgte mit dem Bildanalyseprogramm ImageJ 137v. Die
Aufnahmen der einzelnen Bilder wurden unter standardisierten Bedingungen bei
identischer Vergrößerung, Belichtung und Bildeinstellungen durchgeführt.
Quantitative Auswertung der immunhistochemischen Färbung am Erythrozyten
Für die quantitative Auswertung der digitalen Bilder wurde die Intensität der
Färbung von den Erythrozyten mittels des Bildanalyseprogramms densitometrisch
gemessen. Bei dieser Auswertemethode korreliert die Färbungsintensität mit der
untersuchten MCT1-Dichte der Erythrozyten. Desto dunkler die Erythrozyten
angefärbt waren, umso höher war die Proteinexpression des MCT1 im Erythrozyten.
Dazu wurden von jedem Ausstrich 40 Erythrozyten der Färbung sowie zehn Zellen
der IHC-Kontrolle zum Zeitpunkt U1 und U2 vor und unmittelbar nach der
Belastungsuntersuchung mittels des Bildanalyseprogramms bestimmt. Anschließend
wurden die Messdaten in eine Microsoft Excel-Tabelle übertragen und statistisch
ausgewertet und graphisch dargestellt.
Die Gesamtfärbung der 40 ausgewerteten Erythrozyten ergab sich aus der Differenz
des arithmetischen Mittelwertes der gemessenen Färbung der einzelnen Erythrozyten
und dessen Hintergrundwertes:
Gesamtfärbung des Erythrozyten = Erythrozytenfärbung - Hintergrundwert
Von diesem errechneten Wert der Gesamtfärbung der Erythrozyten wurde
anschließend der arithmetrische Mittelwert der Gesamtfärbungen der IHC-Kontrolle
abgezogen, so dass man als Ergebnis den Wert für die spezielle Antigen-Antikörper-

Material und Methoden
38
färbung erhält. Dieser ermittelte Wert steht für den gesuchten Antigengehalt der
immunhistochemischen Färbung.
Antigengehalt = Gesamtfärbung des Erythrozyten – Gesamtfärbung der IHC-Kontrolle
2.3.3 Durchflusszytometrische Analyse des CD147 im Erythrozyten
Bei der Durchflusszytometrie (FACS: Fluorescence-activated-cell-sorter) handelt es
sich um ein optisch-elektronisches Mess- und Analyseverfahren, das eine Einzelzell-
typisierung nach Fluoreszenz- und Streulichteigenschaften von den zuvor mit
fluoreszenzgekoppelten Antikörpern markierten Zellpopulationen ermöglicht. Bei
dieser quantitativen Messmethode werden die zu untersuchenden Zellen in
Suspension an einen Laserstrahl vorbeigeleitet und die aus spezifischen Zell-
eigenschaften resultierende individuelle Streuung des Laserlichtes von Photo-
detektoren und Photomultipliern detektiert und digitalisiert. Die Auswertung des
Vorwärtsstreulichtes (FSC, forward scatter) gilt als Maß für die Größe und die des
Seitwärtsstreulichtes (SSC, side scatter) als Indikator für die Granularität der
Einzelzelle. Bei der Fluoreszenzwertung werden die durch das Laserlicht hervor-
gerufenen verschiedenen Wellenlängen des emittierenden Lichtes der Zellen mit den
fluoreszenzmarkierten Antikörper analysiert (Sack et al. 2007).
Probenvorbereitung für die FACS-Analyse des CD147
Insgesamt wurden drei beschriftete Röhrchen-Tubes (Sarstedt, Stadt, D) für die Blut-
aufbereitung vorbereitet. Im ersten Schritt wurden 200 µl von den zuvor in PFA
fixierten und bei 4° C gelagerten Erythrozyten in ein Röhrchen gegeben und danach
zweimal zum Lösen der Erythrozyten mit 1.5 ml Cellwash (Becton Dickinson,
Heidelberg, Deutschland) gewaschen. Dazwischen erfolgte ein kurzes Mischen mit
dem Vortex-Mischer (Scientific Industries, Bohemia, USA). Anschließend wurde die
Probe für zwei Minuten bei 2500 G in der Rotixa-Zentrifuge (Hettich Zentrifugen,
Tuttlingen, Deutschland) zentrifugiert und der Überstand abgekippt. Im zweiten
Schritt wurde das Pellet mit 1,5 ml Cellwash versetzt. Nach kurzem Durchmischen
wurde die Erythrozytenkonzentration mit Hilfe des Blutbildanalyser (Sysmex,
Norderstedt, Deutschland) bestimmt. Bei der nachfolgenden Verdünnungsreihe
wurden die einzelnen Proben in ein neues zweites beschriftetes Röhrchen auf

Material und Methoden
39
insgesamt 10.000.000 Erythrozyten/ml mit Cellwash auf 1.0 ml verdünnt und wieder
mit dem Vortex-Mixer vermischt. Anschließend wurden aus der Probe 100 µl in das
dritte vorbereitete Röhrchen abpipettiert und 20 µl von dem monoklonalen
Antikörper CD147 (Phycoerythrin (PE) anti-human CD147 (EMMPRIN, Basigin),
eBioscience, Inc., San Diego, USA) dazugegeben und für 60 Minuten bei Raum-
temperatur im Dunkeln inkubiert. Nach zwei weiteren Wasch- und Mischvorgängen
mit 1,5 ml Cellwash erfolgte eine zweiminütige Zentrifugation bei 1000 G. Die übrig
gebliebenen Zellen wurden daraufhin mit 220 µl Cellwash angelöst, gemischt und
anschließend für drei Sekunden bei 15 G kurz an zentrifugiert. Im letzten Arbeits-
schritt wurden die Proben auf eine 96-Wellplate übertragen.
Die Messungen erfolgten mit dem Durchflusszytometer (ArrayPlate, 96 Well, U-
Bottom, Becton Dickinson, Heidelberg) und die statistische Auswertung und
Digitalisierung wurde mit Hilfe dem Tabellenkalkulationsprogramms Excel vor-
genommen.
2.3.4 Immunfluoreszenz-Nachweis des MCT1 und CD147 im Erythrozyten
Das Verfahren der Immunfluoreszenz ermöglicht die Lokalisation und den Nachweis
von Gewebestrukturen mit Hilfe von fluoreszenzmarkierten Antikörpern. Bei der
hier angewendeten indirekten Methode wird zunächst ein unkonjugierter Primär-
antikörper auf die Proben gegeben und anschließend der mit einem Fluoreszfarbstoff
gekoppelte Sekundärantikörper. Unter dem Fluoreszenzmikroskop kommt es zur
Anregung der Fluorezenzfarbstoffe und somit zu einer fluorometrischen Detektion
des gesuchten Antigens (Luttmann et al. 2004).
Die Vorbereitung der Objektträger und die Zusammensetzung der verwendeten
Puffer und Lösungen sind analog zum immunhistochemischen Färbeprotokoll des
MCT1 im Erythrozyten. Zu Beginn wurden die zu behandelnden Areale auf den
Objektträgern für eine Dauer von fünf Minuten mit 0.05 M TBS gewaschen.
Anschließend erfolgte eine bis zu 45-minütige Permeabilisierung der Zellmembranen
bei 37° C durch Zugabe des Enzyms Trypsin. Anschließend erfolgte ein Wasch-
vorgang mit Leitungswasser, bevor die unspezifischen Antikörperbindungssstellen
durch eine dreiprozentige in TBS gelöste Milchpulverlösung für 45 Minuten
blockiert wurden. Direkt im Anschluss ohne erneutes Waschen wurde ein in 0.3%-
iger Milchpulverlösung mit 0.03% Tween 20 verdünnter Primärantikörper des MCT1

Material und Methoden
40
(Rabbit Anti- Monocarboxylate Transporter1, Chemicon, California, U.S.A.,
Verdünnung 1:1000) für 60 Minuten auf die Areale des Objektträgers aufgetragen.
Nach dem Entfernen der ungebundenen Antikörper durch einen weiteren Wasch-
vorgang wurde für 30 Minuten ein 3%-iges Normal-Goat-Serum (Dako, Hamburg, D)
für die Neutralisation der unspezifischen Antikörperbindungsstellen auf die Proben
appliziert. Ohne weiteren Waschvorgang erfolgte anschließend die Inkubation mit
einem fluorochrommarkierten Sekundärantikörper Cy3 (FluoroLinkTM CyTM 3
labelled streptavidin, Amersham Biosciences, UK, Verhältnis 1:1000). Die
Inkubation erfolgte im Dunkeln und dauerte 60 Minuten.
Anschließend wurde die Reaktion mit drei fünfminütigen Waschvorgängen
abgestoppt. Im nächsten Arbeitsschritt erfolgte wiederum eine 45-minütige Block-
ierung der unspezifischen Antikörperbindungssstellen durch eine dreiprozentige in
TBS gelöste Milchpulverlösung. Danach erfolgte unmittelbar die Inkubation mit dem
Primärantikörper des CD147 (Purified anti-human CD147, Biozol, Eching, D, 1:400),
der in einer 0,3%igen Milchpulverlösung mit Tween 20 verdünnt wurde. Nach
60-miütiger Inkubationszeit wurden die Proben gewaschen und anschließend die
unspezifischen Antikörperbindungsstellen für 30 Minuten mit einem 3%-igen
Normal-Goat-Serum blockiert. Anschließend wurde für 60 Minuten der Sekundär-
antikörper des CD147 (CyTM2-conjugated*AffiniPure Goat Anti-Rabbit lgG, Jackson
Immuno Research, USA, Verhältnis 1:500) auf die Proben im Dunkeln appliziert.
Das Abstoppen der Reaktion erfolgte durch drei weitere Spülvorgänge. Im Anschluss
erfolgten das Equilibrieren der Proben in einer aufsteigenden Alkoholreihe und das
Eindecken mit Entellan.
Auswertung
Die Visualisierung der immunfluoreszenzischen Färbung erfolgte mit Hilfe des
Mikroskop LSM 510 Meta (Carl Zeiss Vision GmbH, Hallbergmoos, D). Die Bilder
wurden mit einer an das Fluoreszenzmikroskop angeschlossene Digitalkamera
aufgenommen.

Material und Methoden
41
2.4 Muskelfasertypisierung Basierend auf der Heterogenität der verschiedenen Myosin-Schwerketten und den
damit verbundenen biochemischen Eigenschaften kann histochemisch über die unter-
schiedliche pH-Wertstabilität der ATPasen, nach der Methode von Brooke und
Kaiser, eine Muskelfasertypisierung vorgenommen werden (Brooke et al. 1970).
Herstellung der Kryostatschnitte
Die Muskelbiopsieproben wurden mit dem Kryostat-Schneidegerät Leica CM 1900
(Reichert-Jung Nussloch, Deutschland) bei einer Temperatur -25° C geschnitten.
Dazu wurden die Proben mit einem Fixierkleber Tissue Tek (Sakura, Zoeterwoude,
NL) auf dem Schneideteller festgefroren und anschließend in die Haltevorrichtung
des Mikrotoms eingespannt. Die Schnitte wurden mit einer Dicke von 7 μm an-
gefertigt und anschließend auf einen Objektträger (Menzel, Braunschweig, D)
aufgezogen. Anschließend wurden die Gewebeschnitte unter dem Lichtmikroskop
(Optech, München, D) kontrolliert und bis zur Myosin-ATPase-Färbung bei -80° C
im Gefrierschrank (Heraeus Instruments, Osterode, D) wieder eingefroren.
Verwendete Lösungen
Inkubationslösung I:
6 mM Natriumacetat (1.3g) (Merck, Darmstadt, D)
mit dem pH-mV-Meter (Mettler Toledo, Schwerzenbach, CH) mit HCL
(Merck, Darmstadt, D) auf pH-Wert von 4.55 kalibrieren
das Volumen auf 160 ml mit Aqua dest. auffüllen
Inkubationslösung II:
60 mM Glycin, (0.8g) (Merck, Darmstadt, D)
25 mM Calciumchlorid-2-hydrat krist., (0.66g) (Merck, Darmstadt, D)
55 mM Natriumchlorid, (0.58g) (Merck, Darmstadt, D)
ATP, (0.306g) (Sigma-Aldrich, Steinheim, D)
mit dem pH-mV-Meter (Mettler Toledo, Schwerzenbach, CH) den pH-Wert
mit HCL (Merck, Darmstadt, D) auf 9.4 kalibrieren
das Volumen auf 180 ml mit Aqua dest. auffüllen

Material und Methoden
42
2% Cobaltchloridlösung:
4g Cobaltchloridlösung- Hexahydrat (Merck, Darmstadt, D) in 200 ml Aqua dest. lösen
1% Ammoniumsulfidlösung:
2g Ammoniumsulfid (Merck, Darmstadt, D) in 200 ml Aqua dest. Lösen
Myosin-ATPase-Färbeprotokoll
Zu Beginn der Myosin-ATPase-Färbung wurden die beschrifteten Objektträger
(Menzel, Braunschweig, D) mit den konsequtiv geschnitten Querschnittpräparaten in
ein Schiffchen (VWR, Langenfeld, D) einsortiert. Die einzelnen Inkubationslösungen
wurden jeweils frisch angesetzt. Zunächst erfolgt die Inkubation in Lösung I bei
einem pH-Wert von 4,55 über eine Dauer von zehn Minuten. Bei diesem Schritt
erfolgt die Perforation und Vorbereitung für die nachfolgende ATP-Lösung. Im
Anschluss wurde die Inkubation durch einen dreißig Sekunden dauernden
Waschvorgang in destilliertem Wasser gestoppt. Im nachfolgenden Arbeitsschritt
werden die Proben fünfundzwanzig Minuten einer basischen ATP-Lösung ausgesetzt.
Nach einem weiteren Waschvorgang von dreißig Sekunden mit destilliertem Wasser
erfolgt die Inkubation in Kobaltchlorid über ein Zeitraum von drei Minuten.
Anschließend wird auch diese durch sechs aufeinanderfolgenden, jeweils dreißig
Sekunden dauernde, Waschschritte abgestoppt. Die nachfolgende Inkubation in
Ammoniumsulfid erfolgt bis zu einem makroskopisch erkennbaren Farbumschlag,
welcher durch die Umsetzung des ATP mit dem hieran gebunden Kobaltchlorid
entsteht. Im Anschluss wurde die Reaktion mit einem fünfminütigen Waschschritt
mit Leitungswasser abgestoppt. Abschließend wurden die Proben mit Glycerin-
Gelatin (Merck, Darmstadt, D) eingedeckt und mit einem Deckgläschen versiegelt.
Auswertung
Für die bildliche Darstellung der Muskelschnitte wurde ein Mikroskop KS 300
Axiophot (Carl Zeiss Vision GmbH, Hallbergmoos, D) bei 200-facher Vergrößerung
mit einem zehnfach vergrößerten Okular benutzt. Mit Hilfe der digitalen CCD
Kamera (Axio Cam, Jena, Deutschland) wurden die Bilder aufgenommen und für die
Übertragung auf den PC das Softwareprogramm „KS 300 3.0“ verwendet. Im
Anschluss wurden die Bilder als JPG-Datei auf den PC abgespeichert. Bei der
mikroskopischen Auswertung ließen sich durch den Farbumschlag drei voneinander

Material und Methoden
43
unterscheidbare Farbnuancen erkennen. Die Typ I-Fasern zeigten eine dunkle
Färbung, die Typ IIX-Fasern eine intermediäre und die Typ IIA-Fasern eine helle
Farbe. Für jedes Präparat wurden unter dem Mikroskop 100 Zellen optisch aus-
gezählt und die Werte in eine Excel-Tabelle übertragen. Anschließend wurde der
mittlere prozentuale Anteil des jeweiligen Muskelfasertyps berechnet und visualisiert.
2.5 Leistungsphysiologische Parameter 2.5.1 Fahrradergometrische Belastungsuntersuchung
Die Bestimmung der Ausdauerleistungsfähigkeit der Probanden erfolgte anhand
eines Stufentests auf dem Ergometer nach dem WHO-Schema. Auswertungsrelevant
war die letzte Wattstufe, die der Proband vollständig durchfahren konnte. Die
Anfangsbelastung von 25 Watt wurde alle zwei Minuten stufenweise um 25 Watt bis
zur subjektiven Ausbelastung unter Berücksichtigung der allgemeinen Abbruch-
kriterien (Rost et al. 1995; Steinacker et al. 2002) gesteigert. Die Durchführung
orientierte sich an den Leitlinien der Deutschen Gesellschaft für Kardiologie (Trappe
et al. 2000). Die Untersuchung fand auf einem drehzahlunabhängigen Ergometer
ERGO-METRICS 900 (Ergoline, Bitz, D) statt. Der Blutdruck der Probanden wurde
über ein angeschlossenes Blutdruckmessgerät, basierend auf der Riva Rocci
Methode, gemessen. Die Aufzeichnung der Herzströme erfolgte mit dem
ERGOSCRIPT EK3012 (Ergoline, Bitz, D). Dabei wurden die Brustwand-
ableitungen nach WILSON und die Extremitätenableitungen nach EINTHOVEN und
GOLDBERGER angelegt. Am Ende jeder Belastungsstufe wurden Blutdruck,
Herzfrequenz und das EKG mittels Computer aufgezeichnet. Vor der Belastung, in
den letzten zehn Sekunden jeder Belastungsstufe, bei Testabbruch sowie nach fünf
und zehn Minuten in der Erholungsphase wurde den Probanden 20 μl arterialisiertes
Kapillarblut aus dem Ohrläppchen entnommen. Das subjektive Belastungsempfinden
der Probanden wurde nach jeder Belastungsstufe über die Borg-Skala erfragt und
dokumentiert (Borg, 1998; Fletcher et al. 2001; Löllgen, 2004).

Material und Methoden
44
2.5.2 Kraftmessungen
Isometrische Maximalkraftmessungen
Vor der isometrischen Maximalkraftmessung erfolgte eine Voraktivierung der
Muskulatur auf dem Vibrationskrafttrainingsgerät „Power-Plate Classic“ (Power-
Plate, Frankfurt/Main, D) bei einer Frequenz von 30 Hz und einer Amplitude von
2 mm. Dies erfolgte 30 Sekunden im schulterbreiten Stand bei einem Kniegelenks-
winkel von 130°.
Anschließend wurde die isometrischen Maximalkraft an der desmodromischen
Beinpresse „Desmotronic“ (Schnell, Peutenhausen, D) gemessen. Bei der Eingangs-
und Ausgangsuntersuchung wurde die Testung in normierter Position durchgeführt,
um eine Reliabilität der Testergebnisse zu ermöglichen:
- aufrechte Sitzposition, Gesäß und Rücken liegen am Polster auf
- Füße und Knie in schulterbreiter Position
- Zehenspitzen zeigen nach vorne-oben
- 130° Kniegelenkswinkel
- Position der Fußplattform bei Loch 4
- Schiene des Rückenpolsters steht bei 8 cm
Die Studienteilnehmer beider Trainingsgruppen mussten auf ein akustisches Signal
hin aus einer vorgespannter Position mit der höchst möglichen Kraft aus beiden
Beinen zwei Sekunden gegen die Kraftmessplatte drücken. Es wurden drei Durch-
gänge durchgeführt, wobei jeweils nur der höchst gemessene Wert in die Aus-
wertung einging. Die Pausenzeit zwischen den Versuchen betrug 30 Sekunden.
Bestimmung des Einer-Wiederholungs-Maximums
Bei den Teilnehmern der Krafttrainingsgruppe wurde zur Ermittlung der maximalen
Kraftveränderungen unmittelbar vor und nach der Trainingsintervention an den
beiden Krafttrainingsgeräten (Gym 80, Gelsenkirchen, D) Brustpresse und dem
Beinstrecker eine konzentrische Maximalkraftmessung (engl.1RM) gemäß dem
Protokoll von Kraemer (Kraemer et al. 1991) durchgeführt. Vor der Krafttestung
erfolgte ein Eingewöhnungssatz mit einer Gewichtsbelastung von 15% des Körper-
gewichts an beiden Testgeräten, um die neuromuskuläre Leistungsbereitschaft zu
erhöhen und das Verletzungsrisiko zu mindern (Schlumberger et al. 2004). Zudem
wurden die individuellen Geräteeinstellungen protokolliert und auf die richtige
Bewegungsausführung der Teilnehmer geachtet. In maximal fünf Versuchen erfolgte

Material und Methoden
45
eine ständige Gewichtsannäherung zur maximalen Last, welche die Teilnehmer
gerade einmal über die volle Bewegungsamplitude bei einer gleichmäßigen
Geschwindikeit von vier Sekunden bewegen konnten. Die Pausenzeit zwischen den
einzelnen Sätzen betrug dabei zwei Minuten.
2.6 Bewegungsinterventionen
Die dreimonatige Bewegungsintervention beinhaltete ein zweimal wöchentlich von
Diplom-Sportlehrern betreutes Kraft- und Ausdauertraining an der Sporthochschule
Köln. Zudem wurde zusätzlich einmal pro Woche selbständig und eigen-
verantwortlich ein Heimtrainingsprogramm durchgeführt, um eine langfristige
Verhaltensänderung zu einem aktiveren Lebensstil bei den Teilnehmern zu bewirken.
Die Teilnehmer erhielten während der Studie keine begleitenden Ernährungs-
empfehlungen und wurden angewiesen nicht noch zusätzlich mit anderen sportlichen
Aktivitäten im Interventionszeitraum zu beginnen, damit eine kausale Zuordnung der
Trainingseffekte zu der Trainingsform und den erhobenen Parametern erfolgen
konnte.
2.6.1 Sporttherapeutisches Krafttraining
Das dreimonatige dynamische Muskelaufbautraining fand zweimal wöchentlich mit
einem Trainingsumfang von 60 Minuten im Krafttraum der Leichtathletikhalle der
Sporthochschule Köln statt.
Gestaltung und Durchführung des Trainings orientierten sich an den Empfehlungen
der Deutschen Gesellschaft für Prävention und Rehabilitation von Herz-
Kreislauferkrankungen sowie nach den Richtlinien des gesundheitsorientierten
Krafttrainings (Boeckh-Behrens et al. 2002; Bjarnason-Wehrens et al. 2004).
Bei dieser Studie wurde auf ein allgemeines Aufwärmen, Cool down und Dehnungs-
übungen verzichtet, um weitere Einflüsse auf die Untersuchungsergebnisse aus-
zuschließen. Zur Verletzungsprophylaxe führten die Teilnehmer vor jedem Gerät
einen muskelspezifischen Aufwärmsatz mit fünfzehn Wiederholungen und 30% des
Trainingsgewichtes durch, um die Muskulatur auf die nachfolgende Trainings-
belastung vorzubereiten.
Das Trainingsgewicht wurde zu Beginn der Intervention über das 1RM und im
weiteren Verlauf über das subjektive Belastungsempfinden der Teilnehmer

Material und Methoden
46
festgelegt. Die subjektive Belastungseinschätzung wurde über eine modifizierte
zehnstufige Borg-Skala für das Krafttraining operationalisiert (vgl. Abb. 17).
1
2 sehr leicht
3
4 Leicht
5
6 mittelschwer
7
8 schwer
9
10 sehr schwer
Abb. 17: Modifizierte Borg-Skala für die Beurteilung des subjektiven Belastungsempfindens während des Krafttrainings
Zur Vermeidung von zu hohen Blutdruckanstiegen trainierten die Teilnehmer nie bis
zur maximalen Ausbelastung, sondern nur im mittleren bis maximal schweren
Bereich ihres subjektiven Belastungsempfindens. Die Pausenzeiten zwischen den
einzelnen Trainingssätzen dauerten zwischen ein und zwei Minuten. Bei der Durch-
führung des Krafttrainings wurde das Trainingsprinzip der progressiven Belastungs-
steigerung angewendet, damit sich die Trainingsbelastung kontinuierlich dem neuen
Kraftniveau des Teilnehmers anpasst.
Der methodische Aufbau des Trainings unterteilt sich in drei Phasen:
1. Vortraining:
In der ersten bis zweiten Woche absolvierten die Teilnehmer ein Gewöhnungs- und
Anpassungstraining, um die richtige Bewegungsausführung an den Trainingsgeräten
kennen zu lernen. Die Ziele dieser Phase lagen in der Verbesserung der inter-
muskuläre Koordination und Körperwahrnehmung sowie Vorbereitung des
Bewegungsapparates auf das nachfolgende Muskelaufbautraining. Die Teilnehmer
wurden angehalten auf die richtige Atemtechnik und auf ein gleichmäßiges
Bewegungstempo von vier Sekunden pro Wiederholung zu achten.
Das Training wurde nach dem Aufwärmsatz bei einer geringen Belastungsintensität
(<30% 1RM) mit 2 Sätzen a` 20 Wiederholungen durchgeführt. Das subjektive
Belastungsempfinden der Teilnehmer lag in dieser Phase noch im leichten Bereich.

Material und Methoden
47
2. Kraftausdauertraining
Ab der dritten bis sechsten Woche erfolgte das Kraftausdauertraining mit dem Ziel
der Verbesserung der lokalen aeroben Ausdauer der Muskulatur. Nach dem Auf-
wärmsatz wurden in der dritten und vierten Woche 3 Sätze mit 20 Wiederholungen
durchgeführt und ab der fünften bis sechsten Woche die Wiederholungszahl auf
15 Wiederholungen reduziert. Das Gewicht wurde auf 30-60% des 1RM gesteigert.
Die Teilnehmer trainierten im mittelschweren Bereich ihres subjektiven Belastungs-
empfindens.
3. Muskelaufbautraining
Von der siebten bis zur zwölften Woche wurde das Muskelaufbautraining mit dem
Ziel der Muskelquerschnittsvergrößerung durchgeführt. Dabei wurde die Wieder-
holungszahl nach einem Aufwärmsatz in der siebten bis neunten Woche auf 3 Sätze
mit 12 Wiederholungen reduziert und in der zehnten bis zwölften Woche nochmals
auf 10 Wiederholungen pro Satz. Die Teilnehmer trainierten in dieser Phase bei einer
Belastungsintensität von 60-80% des 1RM und sollten ihr Training nach ihrem
subjektiven Belastungsempfinden im schweren Bereich absolvieren.
Zu Beginn der Studie erfolgte für jeden Teilnehmer eine ausführliche Einweisung an
den Krafttrainingsgeräten (Gym 80, Gelsenkirchen, D). Die individuellen Ein-
stellungen und das Trainingsgewicht wurden in einem Trainingsplan nach jeder
Trainingseinheit protokolliert. Folgende Übungen wurden von den Probanden durch-
geführt:
Brustpresse (M. pectoralis major); Latissimuszug (M. latissimus dorsi, M. bizeps
brachii); Beinpresse (M. quadrizeps femoris, M. glutaeus maximus); Beinstrecker
(M. quadrizeps femoris); Beinbeuger (M. bizeps femoris) ; Hyperextensionsbank (M.
erector spinae); Crunches (M. rectus abdominis).
Die Übungen für die Bauchmuskulatur und Rückenmuskulatur wurden mit dem
eigenen Körpergewicht bis zur muskulären Erschöpfung durchgeführt.
Nach sechs Wochen wurden die folgenden beiden Trainingsgeräte noch zusätzlich in
den Trainingsplan der Probanden aufgenommen, um das Training noch vielseitiger
und abwechslungsreicher für die Teilnehmer zu gestalten:
Ruderzug (M. latissimus dorsi, M. bizeps brachii, Mm. rhomboidei); Schulterpresse
(M. deltoideus, M.trapezius).
Des Weiteren führten die Teilnehmer zusätzlich einmal pro Woche ein aus acht

Material und Methoden
48
Übungen bestehenden Heimtrainingsplan durch. Jeder Teilnehmer bekam eine aus-
führliche Einführung für die einzelnen Übungen und einen Dokumentationsplan zur
Protokollierung der einzelnen Trainingseinheiten ausgehändigt. In Abhängigkeit von
den individuellen Voraussetzungen der Teilnehmer wurden 3 Sätze mit
10-15 Wiederholungen pro Übung absolviert.
Das Heimtraining setzte sich aus den folgenden acht Kräftigungsübungen zusammen:
Theraband: oberer Rücken; Theraband: unterer Rücken; Bauchmuskulatur;
Rückenstrecker; Schultergürtelmuskulatur; Liegestütze auf den Knien/Wand; Knie-
beugen; Wadenmuskulatur.
2.6.2 Sporttherapeutisches Ausdauertraining
Das sporttherapeutische Ausdauertraining fand an zwei Tagen in der Woche im
Ergometerraum der Sporthochschule Köln statt und wurde von Diplom-Sportlehrern
überwacht und betreut. Vor und kurz nach Ende der Trainingseinheit erfolgten bei
jedem Teilnehmer eine Blutdruckmessung und die Einschätzung des subjektiven
Belastungsempfindens. Das Ziel des Ausdauertrainings lag in der Verbesserung der
Grundlagenausdauer und deshalb wurde die Trainingsbelastung basierend auf dem
Untersuchungsprotokoll des BEKG`s bei einer Intensität von 2 mmol/l Laktat fest-
gelegt. Bei dem monitorkontrollierten Ergometertraining erfolgte eine ständige
Herzfrequenz- und EKG-Überwachung der Teilnehmer.
Die einzelne Trainingseinheit lässt sich in vier Phasen unterteilen:
1. Aufwärmphase I
In dieser Aufwärmphase I trainierten die Teilnehmer bei <50% der empfohlenen
Trainingswattbelastung und einer Belastungsdauer von zwei Minuten.
2. Aufwärmphase II
In der anschließenden Phase II erfolgte eine allmähliche Belastungssteigerung bis zur
vorgegebenen Trainingswattbelastung. Dabei wurde die Belastung innerhalb von
fünf Minuten kontinuierlich gesteigert.
3. Trainingsphase
In dieser Trainingsphase erreichten die Teilnehmer ihre individuelle
Trainingswattbelastung- und -herzfrequenz. Die Dauer dieser Phase wurde bei den

Material und Methoden
49
Teilnehmern ausgehend von fünfzehn Minuten progressiv auf 40 Minuten bis zum
Interventionsende gesteigert. Das subjektive Belastungsempfinden sollte zu Beginn
als leicht und später bis maximal anstrengend (Orginal-Borg-Skala, RPE Werte
11-15) empfunden werden.
4.Erholungsphase
In der Erholungsphase wurde die Belastung allmählich auf null Watt
heruntergefahren und die Trainingsdaten im Trainingsprotokoll festgehalten.
Zudem absolvierten die Teilnehmer ebenfalls ein Heimtrainingsprogramm, bei dem
sie die Art der körperlichen Belastung frei auswählen und nach ihrem individuellen
Trainingspuls trainierten sollten. Die Dauer der zusätzlichen Trainingseinheit betrug
45 Minuten und die Trainingsdaten (Dauer, Herzfrequenz, Borg-Wert) wurden in
einem Trainingsprotokoll eigenständig aufgeschrieben. Im Laufe der Studie wurden
die Grundtechniken der Ausdauersportarten Walken und Nordic Walking vermittelt,
damit sie ihren Heimtrainingsplan abwechslungsreicher gestalten konnten.
2.7 Statistische Auswertung
Die deskriptive Darstellung erfolgte anhand der Berechnung von Mittelwerten und
Standardabweichungen mittels des Tabellenkalkulationsprogramms Excel 2007
(Microsoft Corporation, Redmond, USA). Der statistische Mittelwertvergleich für
die einzelnen Parameter innerhalb der Trainingsgruppen wurde mit dem Statistik-
programm SPSS 18.0 (SPSS Inc., Chicago, USA) durchgeführt. Die graphische
Darstellung der Ergebnisse erfolgte wiederum mit Excel 2007.
Voraussetzung für einen Mittelwertvergleich ist die Normalverteilung der
Untersuchungsergebnisse. Die Überprüfung erfolgte durch den Kolmogorov-
Smirnov-Test mit Lilliefors-Korrektur. Die Signifikanzüberprüfung der einzelnen
Untersuchungsparameter innerhalb der einzelnen Trainingsgruppen wurde per t-Test
mit einem festgelegten Signifikanzniveau von p<0.05 durchgeführt (Bühl, 2006;
Sachs, 2009).

Ergebnisse
50
3 Ergebnisse
3.1 Basischarakterisierung des Probandenkollektivs
Zur detaillierten Analyse des Probandenkollektivs erfolgte eine Gegenüberstellung
ausgewählter Daten aus der Eingangs- und Ausgangsuntersuchung. Dazu wurden die
anthropometrischen Daten, Blutdruck- und Ruhelaktatwerte, leistungsphysiologische
Parameter sowie Stoffwechselparameter von den Studienteilnehmern erhoben.
Anthropometrische Daten
Entsprechend den Richtlinien der Deutschen Adipositas Gesellschaft ließ sich die
Ausdauergruppe als leicht adipös und die Kraftgruppe als stark übergewichtig vor
der Trainingsintervention klassifizieren. Nach der sporttherapeutischen Intervention
zeigte sich in beiden Trainingsgruppen eine geringe Reduktion des Körpergewichtes
und des BMI-Wertes, wobei die Abnahme bei der Ausdauergruppe signifikant war
(vgl. Tab. 1).
Blutdruck
Bei Diabetikern gelten erhöhte Ruheblutdruckwerte als Risikofaktor für das
Auftreten von diabetischen Folgeerkrankungen. Nach dem Trainingsprogramm kam
es sowohl in der Ausdauergruppe als auch Kraftgruppe zu einer Reduktion des
systolischen und diastolischen Blutdruckes in den optimalen Blutdruckbereich von
unter 140/85 mmHg (Bretzel et al. 2008). Der systolische Blutdruck wurde in beiden
Gruppen signifikant gesenkt, wohingegen der diastolische Blutdruck nur in der
Ausdauergruppe signifikant verringert werden konnte (vgl. Tab. 1).
Ruhelaktatwerte
Es gibt Hinweise, dass bei Diabetikern die Ruhelaktatkonzentration im Plasma
erhöht ist (Lovejoy et al. 1992). Der Normbereich des Ruhelaktatwertes liegt
bei < 1.8 mmol/l (Biedler et al. 1995). Beide Gruppen befanden sich vor der
Intervention im Normbereich des Ruhelaktatwertes. Nach dem Trainingsprogramm
konnten in beiden Gruppen keine trainingsinduzierten Veränderungen der
Ruhelaktatkonzentration im Plasma festgestellt werden (vgl. Tab.1)

Ergebnisse
51
3.2 Leistungsphysiologische Parameter
BEKG
Die Bestimmung der Ausdauerleistungsfähigkeit erfolgte anhand eines standard-
isierten Stufentests nach dem WHO-Schema auf dem drehzahlunabhängigen
Ergometer. Beide Trainingsgruppen konnten ihre maximale Wattleistung durch die
Trainingsintervention steigern. Diese Verbesserungen der Ausdauerleistungs-
fähigkeit zeigten sowohl in der Ausdauergruppe als auch in der Kraftgruppe ein
signifikantes Ergebnis (vgl. Tab. 1).
Isometrische Maximalkrafttestung
Die Kraftleistungsfähigkeit der Probanden wurde anhand einer standardisierten
isometrischen Maximalkraftmessung an der desmodromischen Beinpresse bestimmt.
Durch das Ausdauertraining kam es zu einem Zuwachs der beidbeinigen und
rechtsbeinigen isometrischen Maximalkraft. Die linksbeinige isometrische Maximal-
kraft blieb nach dem Ausdauerprogramm unverändert. Diese Ergebnisse der
Ausdauergruppe verfehlten jedoch die statistische Signifikanz. Bei der Kraftgruppe
ließ sich eine beidbeinige, rechtsbeinige und linksbeinige Zunahme der iso-
metrischen Maximalkraft feststellen. Ein signifikantes Ergebnis konnte dabei nur für
die beidbeinige und rechtsbeinige Zunahme der Maximalkraft ermittelt werden
(vgl. Tab. 1).
1 RM
Für die Darstellung der maximalen Kraftveränderungen der Probanden wurde eine
konzentrische Maximalkraftmessung vor und unmittelbar nach dem Krafttrainings-
programm an den beiden Krafttrainingsgeräten Beinstrecker und Brustpresse
durchgeführt. Das Krafttrainingsprogramm bewirkte sowohl am Beinstrecker (U1:
65±22 kg; U2: 84±25 kg) als auch an der Brustpresse (U1: 46±11 kg; U2: 59±12 kg)
eine Verbesserung des 1 RM. Für diese Kraftzuwächse an beiden Krafttrainings-
geräten konnte ein signifikanter Einfluss durch das Training nachgewiesen werden.
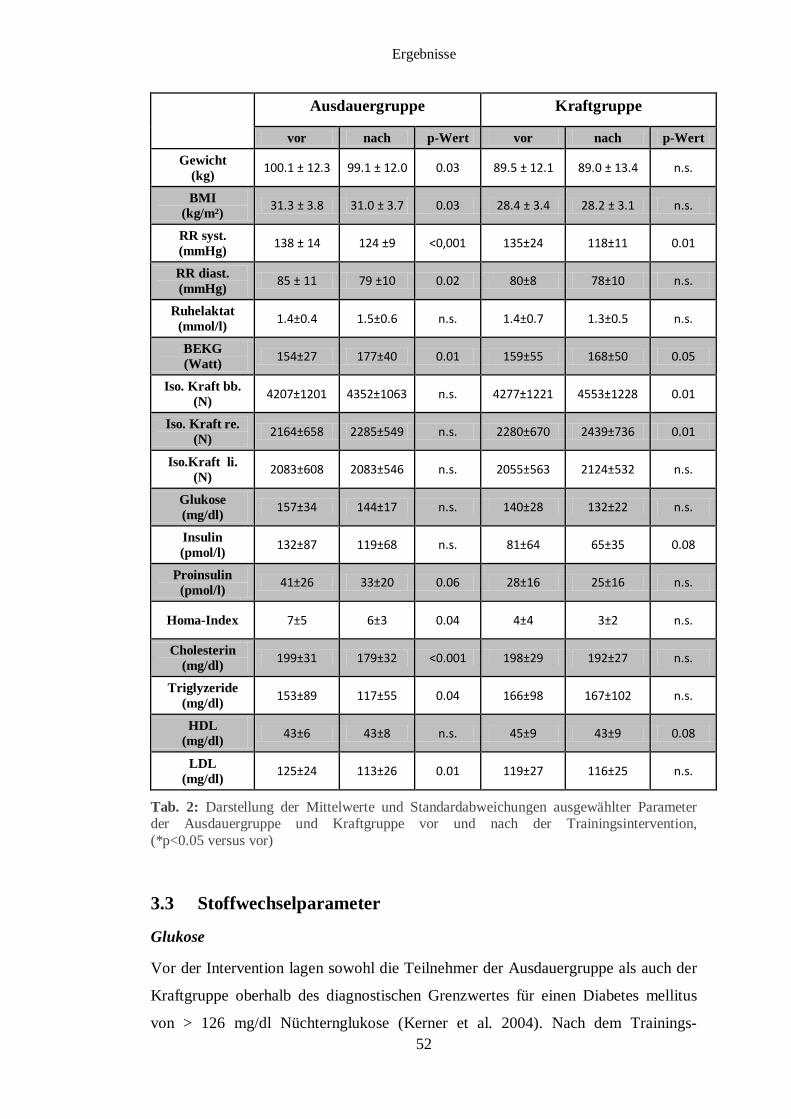
Ergebnisse
52
Ausdauergruppe Kraftgruppe
vor nach p-Wert vor nach p-Wert
Gewicht (kg) 100.1 ± 12.3 99.1 ± 12.0 0.03 89.5 ± 12.1 89.0 ± 13.4 n.s.
BMI (kg/m²) 31.3 ± 3.8 31.0 ± 3.7 0.03 28.4 ± 3.4 28.2 ± 3.1 n.s.
RR syst. (mmHg) 138 ± 14 124 ±9 <0,001 135±24 118±11 0.01
RR diast. (mmHg) 85 ± 11 79 ±10 0.02 80±8 78±10 n.s.
Ruhelaktat (mmol/l) 1.4±0.4 1.5±0.6 n.s. 1.4±0.7 1.3±0.5 n.s.
BEKG (Watt) 154±27 177±40 0.01 159±55 168±50 0.05
Iso. Kraft bb. (N) 4207±1201 4352±1063 n.s. 4277±1221 4553±1228 0.01
Iso. Kraft re. (N) 2164±658 2285±549 n.s. 2280±670 2439±736 0.01
Iso.Kraft li. (N) 2083±608 2083±546 n.s. 2055±563 2124±532 n.s.
Glukose (mg/dl) 157±34 144±17 n.s. 140±28 132±22 n.s.
Insulin (pmol/l) 132±87 119±68 n.s. 81±64 65±35 0.08
Proinsulin (pmol/l) 41±26 33±20 0.06 28±16 25±16 n.s.
Homa-Index 7±5 6±3 0.04 4±4 3±2 n.s.
Cholesterin (mg/dl) 199±31 179±32 <0.001 198±29 192±27 n.s.
Triglyzeride (mg/dl) 153±89 117±55 0.04 166±98 167±102 n.s.
HDL (mg/dl) 43±6 43±8 n.s. 45±9 43±9 0.08
LDL (mg/dl) 125±24 113±26 0.01 119±27 116±25 n.s.
Tab. 2: Darstellung der Mittelwerte und Standardabweichungen ausgewählter Parameter der Ausdauergruppe und Kraftgruppe vor und nach der Trainingsintervention, (*p<0.05 versus vor)
3.3 Stoffwechselparameter
Glukose
Vor der Intervention lagen sowohl die Teilnehmer der Ausdauergruppe als auch der
Kraftgruppe oberhalb des diagnostischen Grenzwertes für einen Diabetes mellitus
von > 126 mg/dl Nüchternglukose (Kerner et al. 2004). Nach dem Trainings-

Ergebnisse
53
programm ließ sich in beiden Gruppen eine Reduktion des mittleren Blutzucker-
wertes feststellen, jedoch verfehlten diese Abnahmen die statistische Signifikanz
(vgl. Tab. 1).
Insulin
Die erhobenen Insulinwerte liegen vor dem Interventionsbeginn bei beiden
Trainingsgruppen im Zielwertbereich zwischen 17,8 und 174 pmol/l. Im Rahmen der
Trainingsintervention kam es sowohl in der Ausdauergruppe als auch in der
Kraftgruppe zu einer Verringerung der Insulinplasmakonzentration. Bei der
Kraftgruppe konnte dabei eine tendenzielle Abnahme beobachtet werden
(vgl. Tab. 1).
Proinsulin
Der Proinsulinspiegel beider Trainingsgruppen lag vor Trainingsbeginn außerhalb
des Normwertbereiches von 6,4-9,4 pmol/l. Durch das Trainingsprogramm kam es in
beiden Trainingsgruppen zu einer Abnahme des mittleren Proinsulinwertes, wobei
lediglich in der Ausdauergruppe eine tendenzielle Verringerung festgestellt werden
konnte (vgl. Tab. 1).
Homa-Index
Der Grad der Insulinresistenz wurde anhand des Homa-Indexes berechnet. Nach den
Beurteilungskriterien des Homa-Indexes wies die Ausdauergruppe vor Trainings-
beginn eine Insulinresistenz auf, wohingegen für die Kraftgruppe eine Insulin-
resistenz als sehr wahrscheinlich anzunehmen war. Beide Gruppen konnten ihren
mittleren Homa-Index durch das Trainingsprogramm reduzieren. Bei der Ausdauer-
gruppe war dabei ein signifikanter Einfluss des Trainings auf die Insulinresistenz
nachgewiesen worden (vgl. Tab. 1).
Cholesterin
Die diabetische Dyslipoproteinämie geht mit einem höheren kardiovaskulären Risiko
einher und daher gelten für Typ 2-Diabetiker insgesamt engere Normwerte
(American Diabetes Association 2009). Die Gesamt-Cholesterinkonzentration lag bei
beiden Interventionsgruppen am oberen Grenzwert des Normwertbereiches von
200 mg/dl. Bei der Ausdauergruppe ließ sich ein signifikantes Sinken der

Ergebnisse
54
Cholesterinwerte beobachten, wohingegen für die Abnahme in der Kraftgruppe kein
signifikantes Ergebnis ermittelt werden konnte (vgl. Tab. 1).
HDL
Die mittlere HDL-Konzentration beider Trainingsgruppen befand sich vor
Interventionsbeginn innerhalb des Normwertbereiches von > 40 mg/dl für männliche
Typ 2-Diabetiker (American Diabetes Association 2009). Die HDL-Werte der
Ausdauergruppe änderten sich durch das Trainingsprogramm nicht, wohingegen sich
bei der Kraftgruppe eine tendenzielle Abnahme des HDL-Wertes beobachten ließ
(vgl. Tab. 1).
LDL
Beide Interventionsgruppen zeigten vor Trainingsbeginn eine erhöhte atherogene
LDL-Konzentration außerhalb des Zielwertbereiches von < 100 mg/dl (American
Diabetes Association 2009). Nach dem Trainingsprogramm konnten beide Trainings-
gruppen die LDL-Werte reduzieren, wobei nur für die Ausdauergruppe ein
signifikantes Ergebnis festgestellt werden konnte (vgl. Tab. 1).
Triglyzeride
Der Zielwert für die Triglyzeride liegt für Typ 2 Diabetiker bei <150 mg/dl
(American Diabetes Association 2009). Vor Trainingsbeginn lagen sowohl die
Ausdauergruppe als auch die Kraftgruppe außerhalb des Normwertbereiches. Das
Ausdauertraining erbrachte eine signifikante Verringerung der Triglyzerid-
konzentration in den Zielwertbereich. Bei der Kraftgruppe blieben die Werte eher
unverändert (vgl. Tab. 1).
3.4 Muskelfasertypisierung
Zur Charakterisierung der trainingsinduzierten Veränderungen des Muskelfaser-
typenprofils wurde eine Myosin-ATPase-Färbung mit den Muskelbiopsieproben der
Eingangs- und Ausgangsuntersuchung durchgeführt. Die Auszählung der einzelnen
Muskelzellen zeigte, dass das Ausdauertraining keinen signifikanten Einfluss auf die
prozentuale Muskelfasertypenzusammensetzung hatte (vgl. Abb. 18).
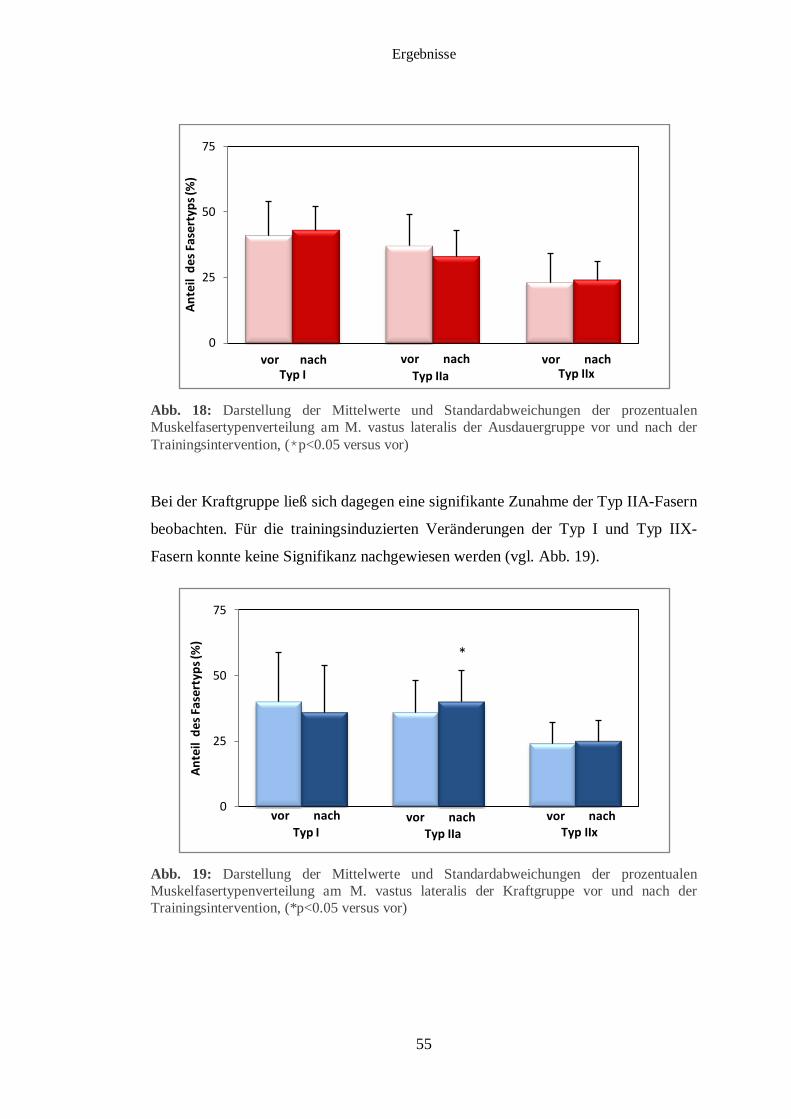
Ergebnisse
55
Abb. 18: Darstellung der Mittelwerte und Standardabweichungen der prozentualen Muskelfasertypenverteilung am M. vastus lateralis der Ausdauergruppe vor und nach der Trainingsintervention, (*p<0.05 versus vor)
Bei der Kraftgruppe ließ sich dagegen eine signifikante Zunahme der Typ IIA-Fasern
beobachten. Für die trainingsinduzierten Veränderungen der Typ I und Typ IIX-
Fasern konnte keine Signifikanz nachgewiesen werden (vgl. Abb. 19).
Abb. 19: Darstellung der Mittelwerte und Standardabweichungen der prozentualen Muskelfasertypenverteilung am M. vastus lateralis der Kraftgruppe vor und nach der Trainingsintervention, (*p<0.05 versus vor)
0
25
50
75
Ante
il d
es F
aser
typs
(%)
Typ IIaTyp I Typ IIxvor nach vor nach vor nach
0
25
50
75
Ante
il d
es F
aser
typs
(%)
vor nach Typ IIa
vor nachTyp I
vor nach Typ IIx
*
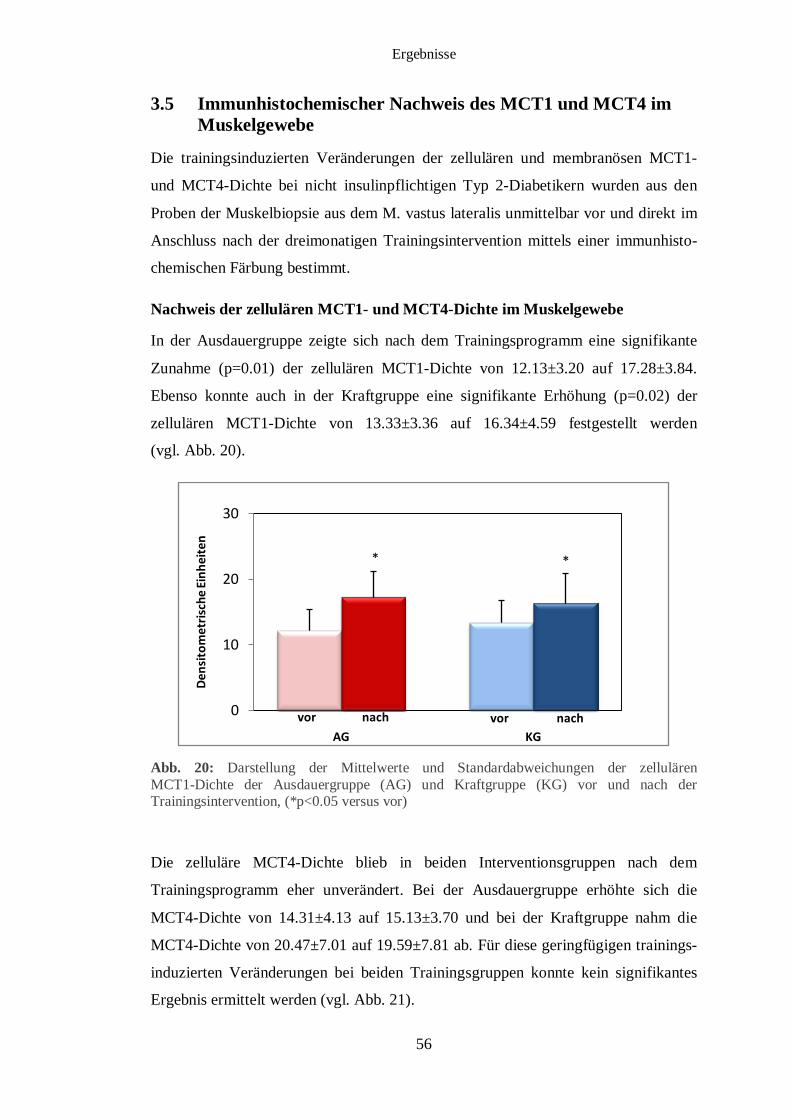
Ergebnisse
56
3.5 Immunhistochemischer Nachweis des MCT1 und MCT4 im Muskelgewebe
Die trainingsinduzierten Veränderungen der zellulären und membranösen MCT1-
und MCT4-Dichte bei nicht insulinpflichtigen Typ 2-Diabetikern wurden aus den
Proben der Muskelbiopsie aus dem M. vastus lateralis unmittelbar vor und direkt im
Anschluss nach der dreimonatigen Trainingsintervention mittels einer immunhisto-
chemischen Färbung bestimmt.
Nachweis der zellulären MCT1- und MCT4-Dichte im Muskelgewebe
In der Ausdauergruppe zeigte sich nach dem Trainingsprogramm eine signifikante
Zunahme (p=0.01) der zellulären MCT1-Dichte von 12.13±3.20 auf 17.28±3.84.
Ebenso konnte auch in der Kraftgruppe eine signifikante Erhöhung (p=0.02) der
zellulären MCT1-Dichte von 13.33±3.36 auf 16.34±4.59 festgestellt werden
(vgl. Abb. 20).
Abb. 20: Darstellung der Mittelwerte und Standardabweichungen der zellulären MCT1-Dichte der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention, (*p<0.05 versus vor)
Die zelluläre MCT4-Dichte blieb in beiden Interventionsgruppen nach dem
Trainingsprogramm eher unverändert. Bei der Ausdauergruppe erhöhte sich die
MCT4-Dichte von 14.31±4.13 auf 15.13±3.70 und bei der Kraftgruppe nahm die
MCT4-Dichte von 20.47±7.01 auf 19.59±7.81 ab. Für diese geringfügigen trainings-
induzierten Veränderungen bei beiden Trainingsgruppen konnte kein signifikantes
Ergebnis ermittelt werden (vgl. Abb. 21).
0
10
20
30
Dens
itom
etris
che
Einh
eite
n
vor nachvor nach AG KG
** *
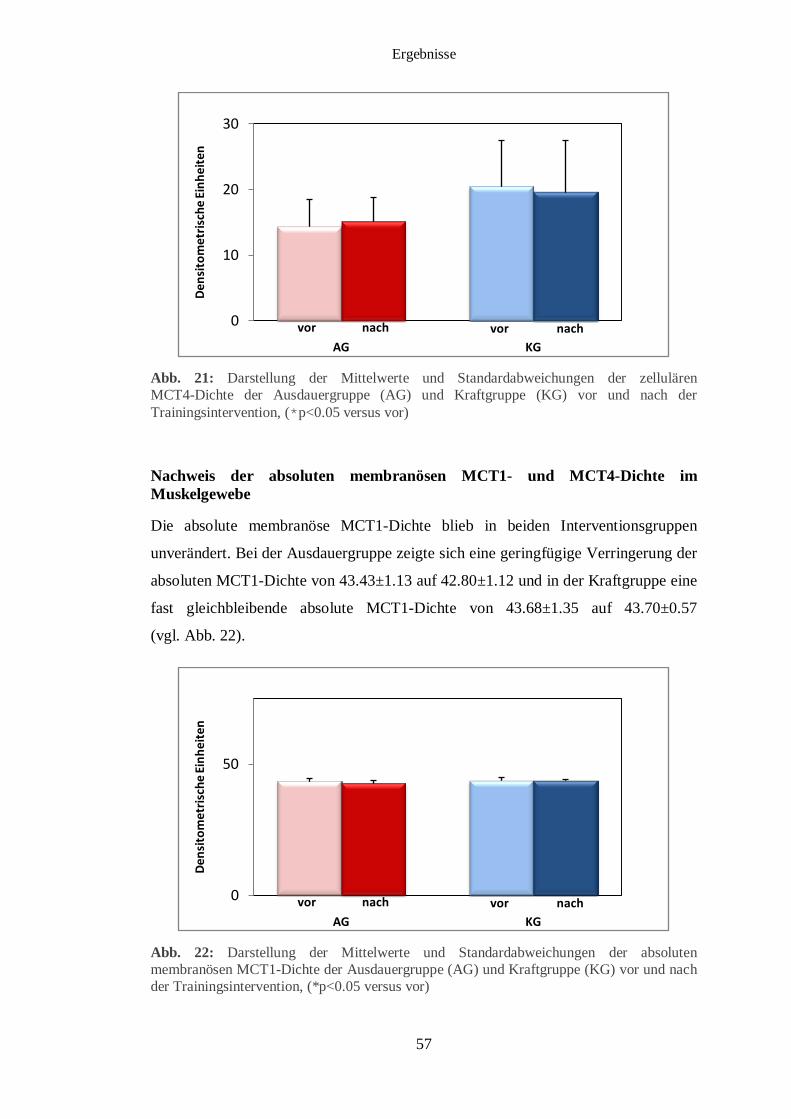
Ergebnisse
57
Abb. 21: Darstellung der Mittelwerte und Standardabweichungen der zellulären MCT4-Dichte der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention, (*p<0.05 versus vor)
Nachweis der absoluten membranösen MCT1- und MCT4-Dichte im Muskelgewebe
Die absolute membranöse MCT1-Dichte blieb in beiden Interventionsgruppen
unverändert. Bei der Ausdauergruppe zeigte sich eine geringfügige Verringerung der
absoluten MCT1-Dichte von 43.43±1.13 auf 42.80±1.12 und in der Kraftgruppe eine
fast gleichbleibende absolute MCT1-Dichte von 43.68±1.35 auf 43.70±0.57
(vgl. Abb. 22).
Abb. 22: Darstellung der Mittelwerte und Standardabweichungen der absoluten membranösen MCT1-Dichte der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention, (*p<0.05 versus vor)
0
10
20
30
Dens
itom
etris
che
Einh
eite
n
vor nachvor nach AG KG
0
50
Dens
itom
etris
che
Einh
eite
n
vor nachvor nach AG KG
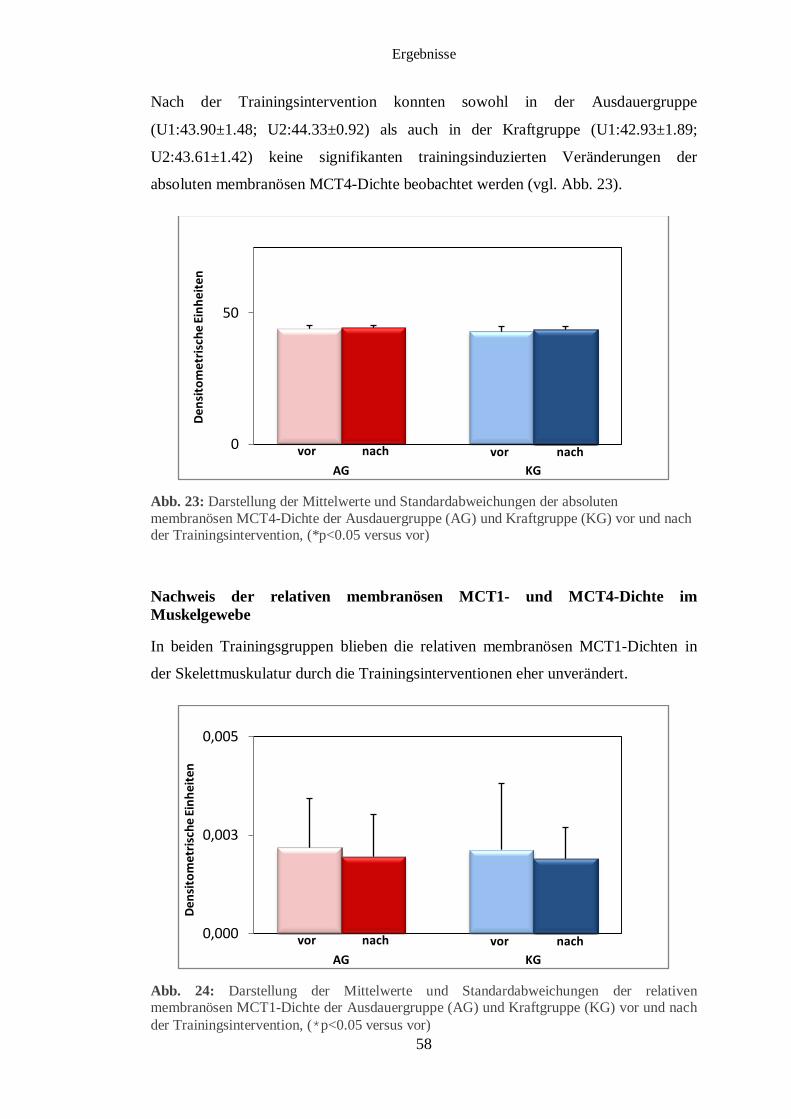
Ergebnisse
58
Nach der Trainingsintervention konnten sowohl in der Ausdauergruppe
(U1:43.90±1.48; U2:44.33±0.92) als auch in der Kraftgruppe (U1:42.93±1.89;
U2:43.61±1.42) keine signifikanten trainingsinduzierten Veränderungen der
absoluten membranösen MCT4-Dichte beobachtet werden (vgl. Abb. 23).
Abb. 23: Darstellung der Mittelwerte und Standardabweichungen der absoluten membranösen MCT4-Dichte der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention, (*p<0.05 versus vor)
Nachweis der relativen membranösen MCT1- und MCT4-Dichte im Muskelgewebe
In beiden Trainingsgruppen blieben die relativen membranösen MCT1-Dichten in
der Skelettmuskulatur durch die Trainingsinterventionen eher unverändert.
Abb. 24: Darstellung der Mittelwerte und Standardabweichungen der relativen membranösen MCT1-Dichte der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention, (*p<0.05 versus vor)
0
50
Dens
itom
etris
che
Einh
eite
n
vor nachvor nach AG KG
0,000
0,003
0,005
Dens
itom
etris
che
Einh
eite
n
vor nachvor nach AG KG
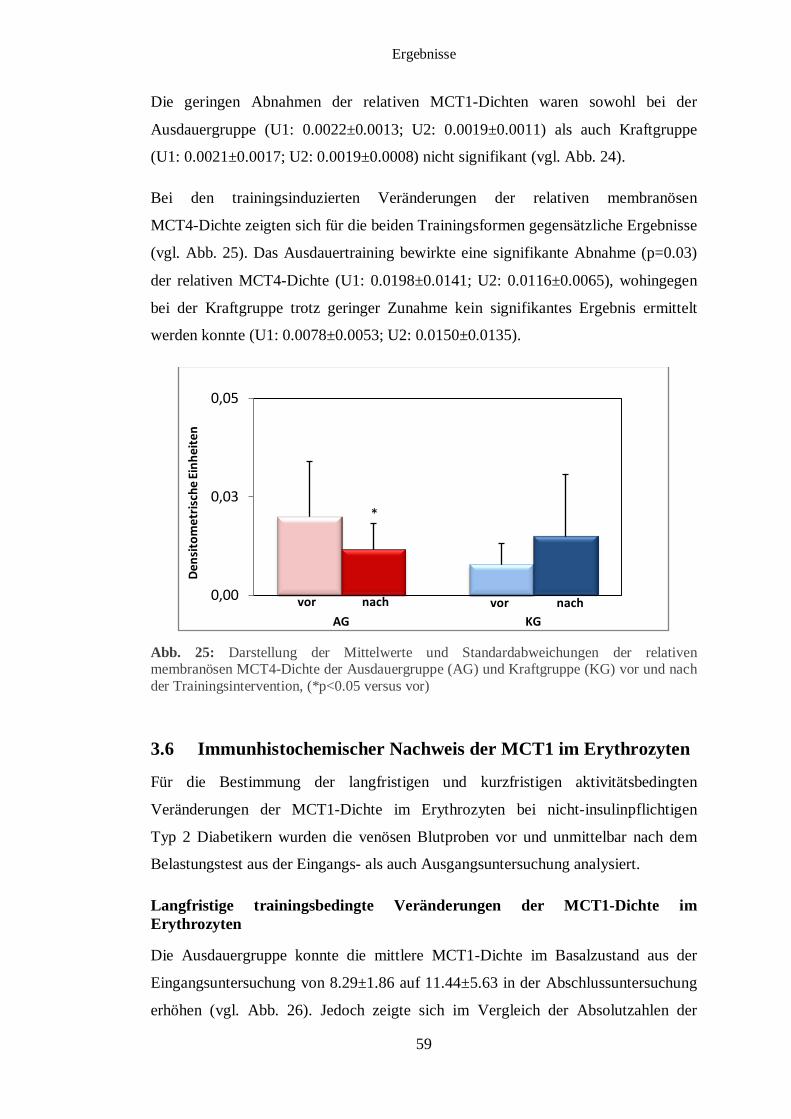
Ergebnisse
59
Die geringen Abnahmen der relativen MCT1-Dichten waren sowohl bei der
Ausdauergruppe (U1: 0.0022±0.0013; U2: 0.0019±0.0011) als auch Kraftgruppe
(U1: 0.0021±0.0017; U2: 0.0019±0.0008) nicht signifikant (vgl. Abb. 24).
Bei den trainingsinduzierten Veränderungen der relativen membranösen
MCT4-Dichte zeigten sich für die beiden Trainingsformen gegensätzliche Ergebnisse
(vgl. Abb. 25). Das Ausdauertraining bewirkte eine signifikante Abnahme (p=0.03)
der relativen MCT4-Dichte (U1: 0.0198±0.0141; U2: 0.0116±0.0065), wohingegen
bei der Kraftgruppe trotz geringer Zunahme kein signifikantes Ergebnis ermittelt
werden konnte (U1: 0.0078±0.0053; U2: 0.0150±0.0135).
Abb. 25: Darstellung der Mittelwerte und Standardabweichungen der relativen membranösen MCT4-Dichte der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention, (*p<0.05 versus vor)
3.6 Immunhistochemischer Nachweis der MCT1 im Erythrozyten
Für die Bestimmung der langfristigen und kurzfristigen aktivitätsbedingten
Veränderungen der MCT1-Dichte im Erythrozyten bei nicht-insulinpflichtigen
Typ 2 Diabetikern wurden die venösen Blutproben vor und unmittelbar nach dem
Belastungstest aus der Eingangs- als auch Ausgangsuntersuchung analysiert.
Langfristige trainingsbedingte Veränderungen der MCT1-Dichte im Erythrozyten
Die Ausdauergruppe konnte die mittlere MCT1-Dichte im Basalzustand aus der
Eingangsuntersuchung von 8.29±1.86 auf 11.44±5.63 in der Abschlussuntersuchung
erhöhen (vgl. Abb. 26). Jedoch zeigte sich im Vergleich der Absolutzahlen der
0,00
0,03
0,05
Dens
itom
etris
che
Einh
eite
n
vor nachvor nach AG KG
*
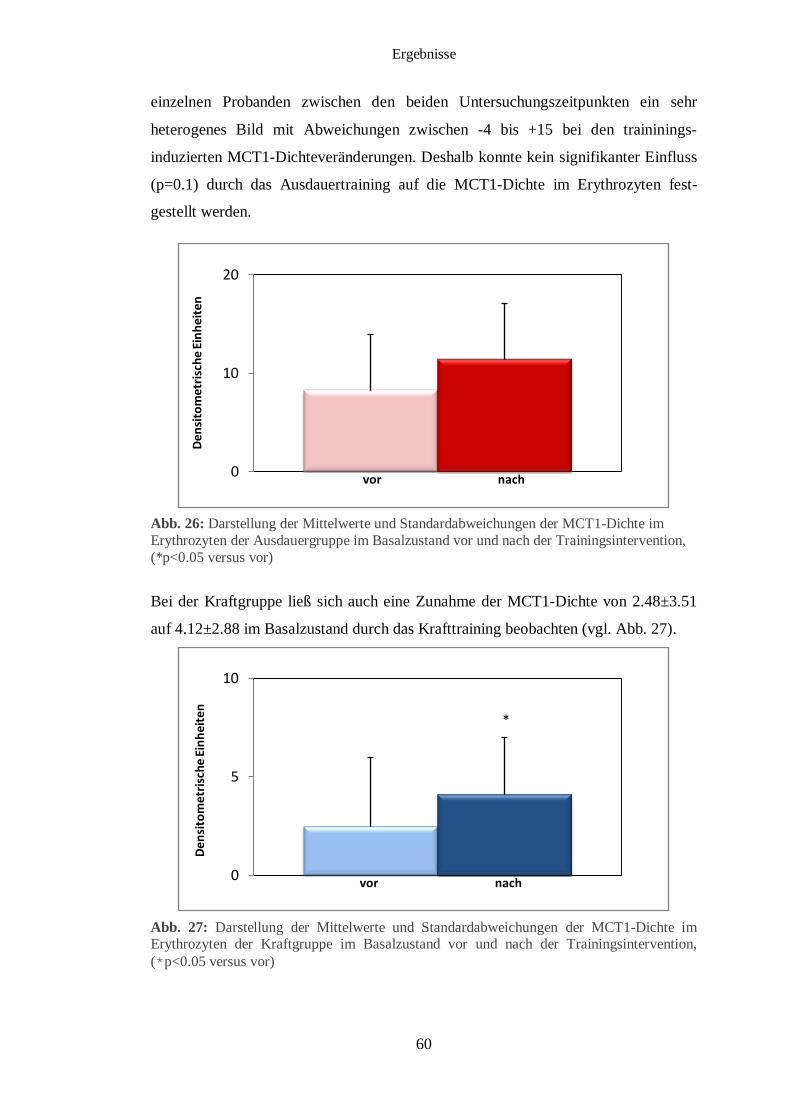
Ergebnisse
60
einzelnen Probanden zwischen den beiden Untersuchungszeitpunkten ein sehr
heterogenes Bild mit Abweichungen zwischen -4 bis +15 bei den traininings-
induzierten MCT1-Dichteveränderungen. Deshalb konnte kein signifikanter Einfluss
(p=0.1) durch das Ausdauertraining auf die MCT1-Dichte im Erythrozyten fest-
gestellt werden.
Abb. 26: Darstellung der Mittelwerte und Standardabweichungen der MCT1-Dichte im Erythrozyten der Ausdauergruppe im Basalzustand vor und nach der Trainingsintervention, (*p<0.05 versus vor)
Bei der Kraftgruppe ließ sich auch eine Zunahme der MCT1-Dichte von 2.48±3.51
auf 4.12±2.88 im Basalzustand durch das Krafttraining beobachten (vgl. Abb. 27).
Abb. 27: Darstellung der Mittelwerte und Standardabweichungen der MCT1-Dichte im Erythrozyten der Kraftgruppe im Basalzustand vor und nach der Trainingsintervention, (*p<0.05 versus vor)
0
10
20
Dens
itom
etris
che
Einh
eite
n
vor nach
0
5
10
Dens
itom
etris
che
Einh
eite
n
vor nach
*

Ergebnisse
61
Hier ließen sich im Vergleich der Absolutzahlen der einzelnen Probanden aus der U1
zu der U2-Untersuchung fast einheitliche Erhöhungen für die MCT1-Dichte mit
geringeren Abweichungen von -2 bis +3 feststellen, woraus eine signifikante
Erhöhung (p=0.02) durch das Krafttraining auf die MCT1-Dichte im Erythrozyten
resultiert.
In der nachfolgenden Abbildung ist die verstärkte Graufärbung der Erythrozyten
nach der Trainingsintervention exemplarisch bei einem Probanden aus der
Kraftgruppe dargestellt. Die intensivere Graustufenintensität deutet dabei auf eine
verstärkte MCT1-Dichte in den Erythrozyten nach der Trainingsintervention hin
(vgl. Abb. 28).
vor der Trainingsintervention nach der Trainingsintervention
Abb. 28: Immunhistochemischer Nachweis einer verstärkten MCT1-Färbung an den Erythrozyten eines Probanden der Kraftgruppe im Basalzustand vor und nach Trainingsintervention bei 400-facher Vergrößerung
Kurzfristige trainingsbedingte Veränderungen der MCT1-Dichte nach körperlicher Belastung im Erythrozyten
Im untrainierten Zustand blieb in beiden Trainingsgruppen unmittelbar nach dem
Belastungstest die MCT1-Dichte eher unverändert, welches sich optisch in einer
gleich bleibenden Graustufenfärbung der Erythrozyten zeigte (vgl. Abb. 29; Bild A).
Bei der Ausdauergruppe nahm die mittlere MCT1-Dichte geringfügig von 8.29±1.86
nach Belastungsende auf 8.59±2.55 zu. Bei der Kraftgruppe ließ sich eine minimale
Reduktion der MCT1-Dichte nach dem Belastungstest von 2.48±3.51 auf 2.28±3.10
beobachten.
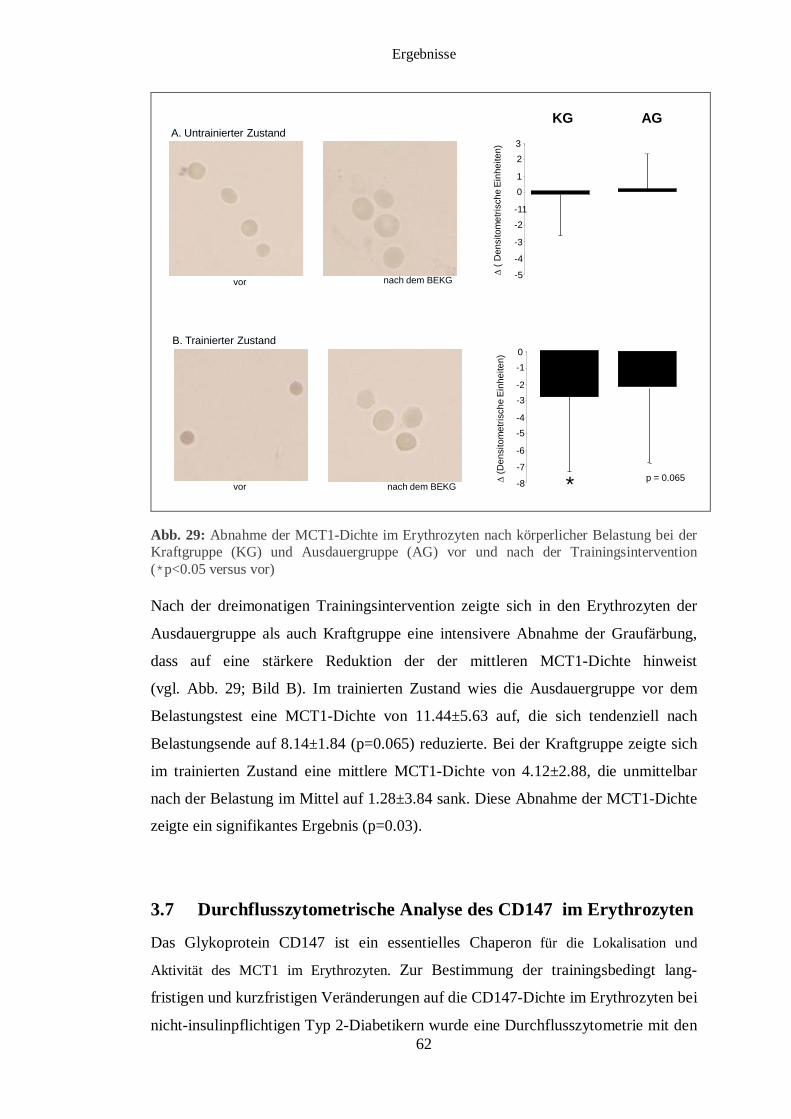
Ergebnisse
62
vor nach dem BEKG
vor
-5
-4
-3
-2
-11
01
23
∆( D
ensi
tom
etris
che
Einh
eite
n)-8
-7
-6
-5
-4
-3-2
-10
∆(D
ensi
tom
etris
che
Einh
eite
n)nach dem BEKG *
A. Untrainierter Zustand
B. Trainierter Zustand
KG AG
p = 0.065
Abb. 29: Abnahme der MCT1-Dichte im Erythrozyten nach körperlicher Belastung bei der Kraftgruppe (KG) und Ausdauergruppe (AG) vor und nach der Trainingsintervention (*p<0.05 versus vor) Nach der dreimonatigen Trainingsintervention zeigte sich in den Erythrozyten der
Ausdauergruppe als auch Kraftgruppe eine intensivere Abnahme der Graufärbung,
dass auf eine stärkere Reduktion der der mittleren MCT1-Dichte hinweist
(vgl. Abb. 29; Bild B). Im trainierten Zustand wies die Ausdauergruppe vor dem
Belastungstest eine MCT1-Dichte von 11.44±5.63 auf, die sich tendenziell nach
Belastungsende auf 8.14±1.84 (p=0.065) reduzierte. Bei der Kraftgruppe zeigte sich
im trainierten Zustand eine mittlere MCT1-Dichte von 4.12±2.88, die unmittelbar
nach der Belastung im Mittel auf 1.28±3.84 sank. Diese Abnahme der MCT1-Dichte
zeigte ein signifikantes Ergebnis (p=0.03).
3.7 Durchflusszytometrische Analyse des CD147 im Erythrozyten
Das Glykoprotein CD147 ist ein essentielles Chaperon für die Lokalisation und
Aktivität des MCT1 im Erythrozyten. Zur Bestimmung der trainingsbedingt lang-
fristigen und kurzfristigen Veränderungen auf die CD147-Dichte im Erythrozyten bei
nicht-insulinpflichtigen Typ 2-Diabetikern wurde eine Durchflusszytometrie mit den
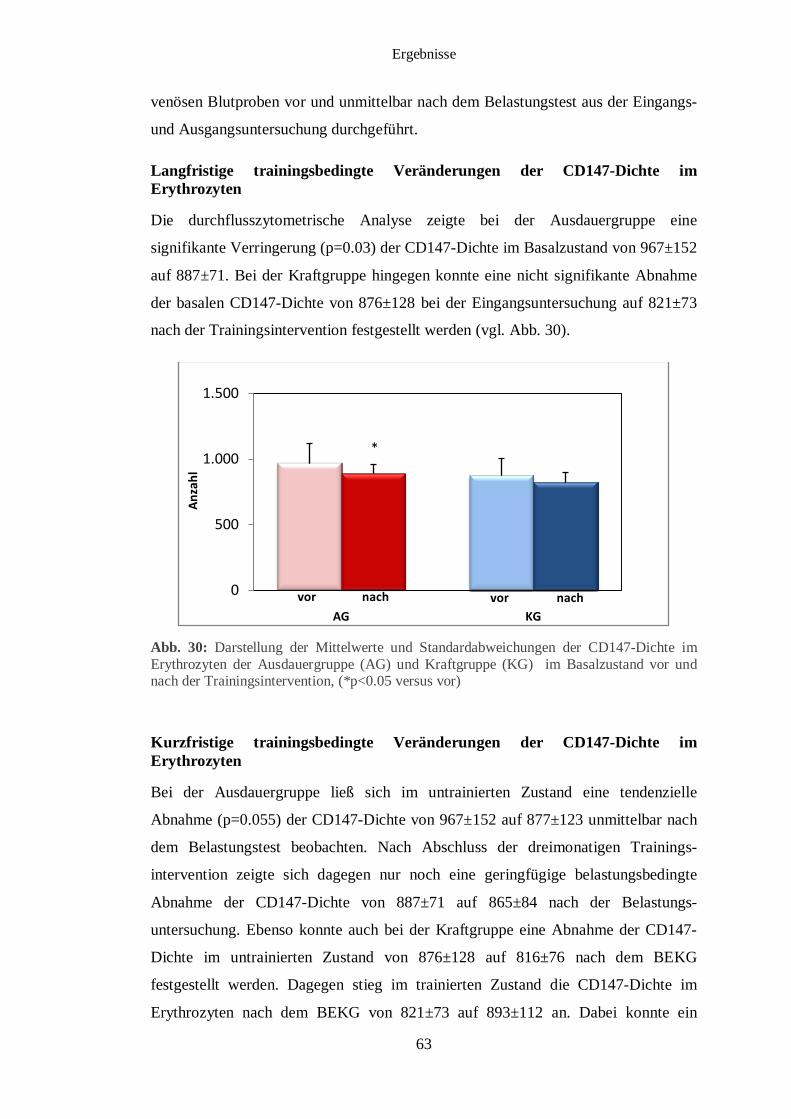
Ergebnisse
63
venösen Blutproben vor und unmittelbar nach dem Belastungstest aus der Eingangs-
und Ausgangsuntersuchung durchgeführt.
Langfristige trainingsbedingte Veränderungen der CD147-Dichte im Erythrozyten
Die durchflusszytometrische Analyse zeigte bei der Ausdauergruppe eine
signifikante Verringerung (p=0.03) der CD147-Dichte im Basalzustand von 967±152
auf 887±71. Bei der Kraftgruppe hingegen konnte eine nicht signifikante Abnahme
der basalen CD147-Dichte von 876±128 bei der Eingangsuntersuchung auf 821±73
nach der Trainingsintervention festgestellt werden (vgl. Abb. 30).
Abb. 30: Darstellung der Mittelwerte und Standardabweichungen der CD147-Dichte im Erythrozyten der Ausdauergruppe (AG) und Kraftgruppe (KG) im Basalzustand vor und nach der Trainingsintervention, (*p<0.05 versus vor)
Kurzfristige trainingsbedingte Veränderungen der CD147-Dichte im Erythrozyten
Bei der Ausdauergruppe ließ sich im untrainierten Zustand eine tendenzielle
Abnahme (p=0.055) der CD147-Dichte von 967±152 auf 877±123 unmittelbar nach
dem Belastungstest beobachten. Nach Abschluss der dreimonatigen Trainings-
intervention zeigte sich dagegen nur noch eine geringfügige belastungsbedingte
Abnahme der CD147-Dichte von 887±71 auf 865±84 nach der Belastungs-
untersuchung. Ebenso konnte auch bei der Kraftgruppe eine Abnahme der CD147-
Dichte im untrainierten Zustand von 876±128 auf 816±76 nach dem BEKG
festgestellt werden. Dagegen stieg im trainierten Zustand die CD147-Dichte im
Erythrozyten nach dem BEKG von 821±73 auf 893±112 an. Dabei konnte ein
0
500
1.000
1.500
Anza
hl
vor nachvor nach AG KG
*
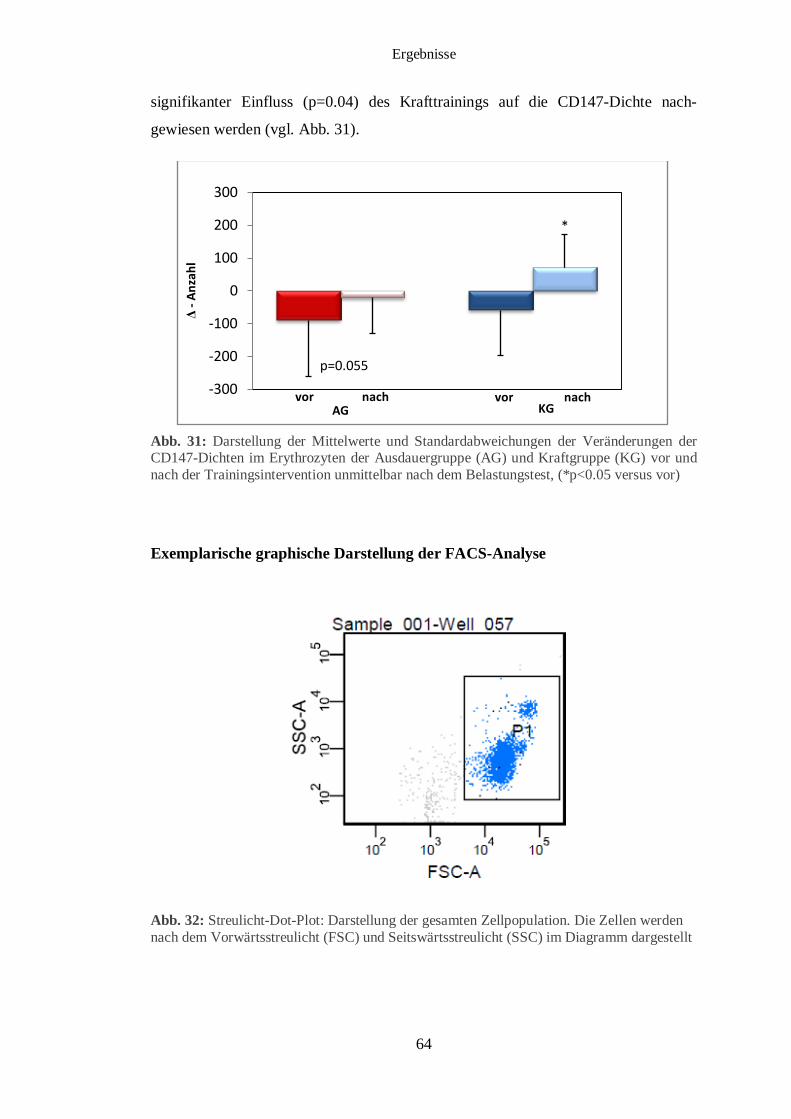
Ergebnisse
64
signifikanter Einfluss (p=0.04) des Krafttrainings auf die CD147-Dichte nach-
gewiesen werden (vgl. Abb. 31).
Abb. 31: Darstellung der Mittelwerte und Standardabweichungen der Veränderungen der CD147-Dichten im Erythrozyten der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention unmittelbar nach dem Belastungstest, (*p<0.05 versus vor)
Exemplarische graphische Darstellung der FACS-Analyse
Abb. 32: Streulicht-Dot-Plot: Darstellung der gesamten Zellpopulation. Die Zellen werden nach dem Vorwärtsstreulicht (FSC) und Seitswärtsstreulicht (SSC) im Diagramm dargestellt
-300
-200
-100
0
100
200
300Δ
-Anz
ahl
vor nach vor nach AG KG
*
p=0.055
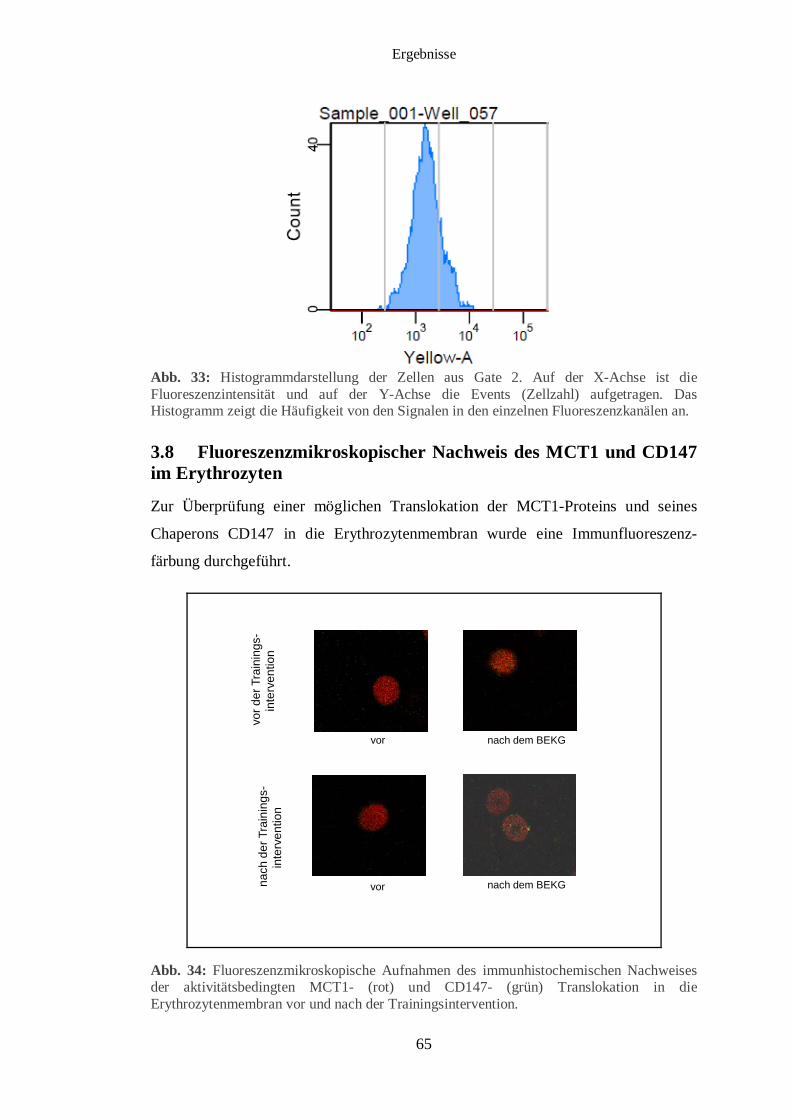
Ergebnisse
65
Abb. 33: Histogrammdarstellung der Zellen aus Gate 2. Auf der X-Achse ist die Fluoreszenzintensität und auf der Y-Achse die Events (Zellzahl) aufgetragen. Das Histogramm zeigt die Häufigkeit von den Signalen in den einzelnen Fluoreszenzkanälen an. 3.8 Fluoreszenzmikroskopischer Nachweis des MCT1 und CD147 im Erythrozyten Zur Überprüfung einer möglichen Translokation der MCT1-Proteins und seines
Chaperons CD147 in die Erythrozytenmembran wurde eine Immunfluoreszenz-
färbung durchgeführt.
vor nach dem BEKG
vor nach dem BEKG
nach
der
Tra
inin
gs-
inte
rven
tion
vor d
er T
rain
ings
-in
terv
entio
n
Abb. 34: Fluoreszenzmikroskopische Aufnahmen des immunhistochemischen Nachweises der aktivitätsbedingten MCT1- (rot) und CD147- (grün) Translokation in die Erythrozytenmembran vor und nach der Trainingsintervention.

Ergebnisse
66
Im untrainierten Zustand konnten keine Veränderungen der hier rot markierten
MCT1-Transporter und der grün markierten CD147-Proteine in die Erythrozyten-
membran beobachtet werden. Dagegen nach dem dreimonatigen Trainingsprogramm
ließ sich eine Verlagerung der MCT1- und CD147-Proteine unmittelbar nach dem
Belastungstest von innen nach außen in die Erythrozytenmembran nachweisen
(vgl. Abb. 34).

Diskussion
67
4 Diskussion
4.1 Langfristig trainingsbedingte Veränderungen der MCT1-Dichte im Muskelgewebe
Die immunhistochemischen Auswertungen am Muskel zeigten eine signifikante
Erhöhung der zellulären MCT1-Dichte in beiden Trainingsgruppen, wohingegen
keine trainingsinduzierten Veränderungen der absoluten und relativen membranösen
MCT1-Dichte in der Ausdauergruppe und Kraftgruppe beobachtet werden konnten.
Einfluss des Ausdauertrainings auf die MCT1-Dichte
In zahlreichen Studien konnte bisher nachgewiesen werden, dass ein langfristiges
Ausdauertraining sowohl die Proteinexpression des MCT1-Proteins im menschlichen
(Bonen et al. 1998; Dubouchaud et al. 2000) als auch im tierischen Skelettmuskel
(McDermott et al. 1993a; Baker et al. 1998; Yoshida et al. 2004) erhöht. Dabei
wurde eine enge Korrelation zwischen der Proteinmenge des MCT1 und der
oxidativen Kapazität (Mc Cullagh et al. 1996; Baker et al. 1998; Pilegaard
et al. 1999a; Bonen et al. 2001) sowie einer verbesserten Laktattransportkapazität
in der Muskulatur festgestellt (Pilegaard et al. 1993; Mc Cullagh et al. 1997;
Baker et al. 1998; Bonen et al. 2000c). Die Auswirkungen eines Ausdauertrainings
auf den MCT1-Gehalt bei Typ 2 Diabetikern wurden bisher noch nicht untersucht.
Lediglich in zwei Trainingsstudien mit Tieren konnte bisher gezeigt werden, dass ein
tägliches dreiwöchiges Laufbandtraining zu einer Erhöhung der Proteinexpression
des reduzierten MCT1-Gehaltes im Skelett- und Herzmuskel bei diabetischen Ratten
führte (Enoki et al. 2003). Dieser reduzierte MCT1-Gehalt geht mit einem
verschlechterten Laktattransport einher (McCullagh et al. 1997; Baker et al. 1998;
Hajduch et al. 2000). In der zweiten Trainingsstudie mit übergewichtigen Ratten
konnte auch eine Zunahme des MCT1-Gehaltes nach einem zehnwöchigen
Laufbandtraining mit fünf Trainingseinheiten in der Woche im Skelettmuskel
beobachtet werden (Metz et al. 2005).

Diskussion
68
In den bisher durchgeführten Studien wurde methodisch zur Detektion des MCT1-
Proteins ein aus Muskelzellen, Endothelzellen und Satellitenzellen bestehendes
Zellgemisch mittels eines Western Blot-Verfahren quantitativ analysiert. Nur in
wenigen Studien wurde gezielt versucht die Muskelmembran zu isolieren
(Dubouchaud et al. 2000; Juel et al. 2004b), um so eine detailliertere Aussage über
die lokale Proteinexpression des MCT-Proteins zu erlangen. Aus diesem Grund
wurde in der DiabetesAktiv-Studie für die Auswertung der Muskelschnitte das
immunhistochemische Analyseverfahren angewendet, welches eine qualitative
Unterscheidung zwischen zellulärer und membranöser MCT-Dichteveränderung
ermöglicht. Durch die unterschiedlichen Auswertungsverfahren ist deshalb ein
direkter Vergleich zwischen den Untersuchungsergebnissen der Trainingsstudien nur
bedingt möglich, da in der DiabetesAktiv-Studie eine Unterscheidung zwischen
zellulärer und membranöser MCT-Dichte erfolgte und nicht die Gesamtprotein-
menge des MCT-Proteins quantitativ aus einem Homogenat bestimmt wurde. Im
weiteren Methodenvergleich der Studien zeigten sich auch Differenzen zwischen den
Trainingsformen und der Belastungsteuerung der einzelnen Trainingsprogramme.
Aus der Analyse der Untersuchungsergebnisse der verschiedenen Studien kann
vermutet werden, dass die Belastungsintensität und die Trainingshäufigkeit sowie
Trainingsdauer der Belastung eine bedeutsame Rolle bei der Expression des MCT1-
Proteins spielt. In der DiabetesAktiv-Studie lag die Belastungsintensität des
Ausdauertrainings bei einer Trainingsherzfrequenz an der 2 mmol/l Lakatschwelle
und muss deshalb als moderat eingestuft werden.
Im Tiermodell zeigte sich nach einem moderaten Lauftraining mit gesunden Ratten
keine Veränderung der Proteingesamtmenge des MCT1 im Skelettmuskel (Baker et
al. 1998). Dies konnte auch in der DiabetesAktiv-Studie beobachtet werden, in der
die absolute und membranöse MCT1-Dichte nach dem moderaten Ergometertraining
unverändert blieb. Erst bei einer höheren Trainingsintensität ließ sich eine
signifikante Erhöhung der Proteinexpression des MCT1 ausschließlich in den
oxidativen Muskelfasern der Ratten feststellen (Baker et al. 1998). Dieser intensitäts-
bedingte Anstieg der MCT1- Proteinexpression zeigte sich auch im M. soleus nach
einem intensiven fünfwöchigen intervallartigen Laufradtraining bei Ratten (Thomas
et al. 2007). Hier muss jedoch kritisch hinterfragt werden, ob sich die Belastungs-
intensitäten bei menschlichen Probanden mit denen des Tiermodells problemlos
vergleichen lassen. Da ein Vergleich der Trainingsherzfrequenzen und der Laktat-

Diskussion
69
werte von Mensch und Tier nur bedingt möglich sind, bleiben Vergleiche und
Aussagen zur Belastungsintensität und den daraus resultierenden Veränderungen
schwierig.
Bei trainierten Skiläufern zeigte sich auch eine trainingsintensitätsabhängige
Zunahme (80 - 90 % der VO2max) der Expression des MCT1-Proteins nach einem
fünfmonatigen Laufbandtraining, wohingegen sich bei moderater Belastungs-
intensität (60 - 70 % der VO2max) eine Abnahme des MCT1-Gehaltes beobachten ließ
(Evertsen et al. 2001). Aus diesen Beobachtungen kann vermutet werden, dass der
Trainingszustand der Probanden auch einen Einfluss auf eine erhöhte Expressions-
rate des MCT1-Proteins hat. Bei untrainierten Probanden führen schon geringere
Belastungsintensitäten zu einer Zunahme der MCT1-Dichte (Bonen et al. 1998;
Dubouchaud et al. 2000), während sich bei hohen Trainingsintensitäten die MCT1-
Dichte bei trainierten Sportlern nicht weiter steigern ließ (Evertsen et al. 2001). In
zwei weiteren hochintensiven Trainingsstudien auf einem Ergometer in liegender
Position ließen sich bei gesunden Probanden bei einer Belastungsintensität von
150 % der VO2max eine signifikante Erhöhung der MCT1-Dichte um 15% (Juel et al.
2004a) und um 76% nach einem achtwöchigen Training mit fünf Trainingstagen in
der Woche beobachten (Pilegaard et al. 1999b). Die hochintensiven Intervalle lagen
bei 2 x 30 Sekunden und 3 x 1 Minute und wurden in diesen Untersuchungen bis zur
muskulären Erschöpfung durchgeführt. In der DiabetesAktiv-Studie zeigte sich bei
den Probanden ein signifikanter Anstieg der zellulären MCT1-Dichte, wohingegen
die absolute und relative membranöse MCT1-Dichte unverändert blieb. Als
möglicher Erklärungsansatz für diesen Anstieg der zellulären MCT1-Dichte könnte
ein trainingsinduzierter Anstieg der Mitochondriendichte sein. Es konnte bereits
nachgewiesen werden, dass MCT1-Proteine in der Mitochondrienmembran bei
Ratten lokalisiert sind, welche möglicherweise Laktat in das Mitochondrium
transportieren, wo es anschließend im Citratzyklus verstoffwechselt wird (Brooks et
al. 1999; Benton et al. 2004; Butz et al. 2004). In Anlehnung an diese Hypothese
würde mit einem trainingsinduzierten Anstieg der Mitochondriendichte auch die
zelluläre MCT1-Dichte ansteigen, wie es in der DiabetesAktiv-Studie nach dem
Ausdauertraining beobachtet wurde.
Nicht nur die Trainingsintensität, sondern auch die Trainingshäufigkeit und
Trainingsdauer scheinen für eine erhöhte Expression des MCT1-Proteins bei einem

Diskussion
70
Ausdauertraining verantwortlich zu sein. Bei untrainierten, männlichen Probanden
konnte eine trainingsbedingte Erhöhung der MCT1-Dichte (+18%) nach einem
täglichen Ergometertraining mit einer moderaten Belastungsintensität von 65% der
Vo2max in der Skelettmuskulatur festgestellt werden (Bonen et al. 1998). In einer
weiteren Trainingsstudie mit untrainierten Männern zeigte sich nach einem
sechstägigen einstündigem Ergometertraining über einen Zeitraum von neun Wochen
bei 75% der Vo2max eine Erhöhung des Proteingesamtgehaltes des MCT1 im
M. vastus lateralis (+90%), im Sarkolemm (+60%) und in der Mitochondrien-
membran (+78%). Zudem bewirkte das Ausdauertraining eine signifikante Erhöhung
der Typ I-Fasern und eine tendenzielle Abnahme der Typ IIa- und Typ IIx-Fasern
(Dubouchaud et al. 2000). Diese Korrelation zwischen den Veränderungen der
MCT1-Dichte und dem Muskelfasertyp konnte auch in anderen Untersuchungen
bisher nachgewiesen werden (Mc Cullagh et al. 1996; Baker et al. 1998; Bonen et al.
2000b; Kobayashi et al. 2004). Im Vergleich dazu kam es in der DiabetesAktiv-
Studie bei der Auswertung der Myosin-ATPase-Färbung zu keiner signifikant
trainingsinduzierten Verschiebung des prozentualen Typ I-Muskelfasertyps, welches
als weitere mögliche Ursache für die unveränderte absolute und relative MCT1-
Dichte angeführt werden kann. Ebenso konnte im Tiermodell nach einem täglichen
moderaten Laufradtraining nach sechs Wochen eine erhöhte Proteinexpression des
MCT1-Proteins bei Mäusen beobachtet werden (Yoshida et al. 2004). Diese
trainingsdauerbedingte erhöhte Expressionsrate zeigte sich auch nach elektrischer
Dauerstimulation bei geringer Intensität (Mc Cullagh et al. 1997; Bonen et al.
2000b). In der DiabetesAktiv-Studie muss die Trainingshäufigkeit mit zwei betreuten
einstündigen Trainingseinheiten und einer freiwilligen Ausdauereinheit in der Woche
als zu gering eingestuft werden, um eine erhöhte Expressionsrate der membranösen
und relativen MCT1-Dichte über die Trainingshäufigkeit zu erreichen. Zudem
scheint auch die Dauer der einzelnen Trainingseinheiten in der DiabetesAktiv-Studie
zu kurz gewesen zu sein, die ausgehend von 25 Minuten progressiv auf 50 Minuten
zum Interventionsende gesteigert wurde.
Einfluss des Krafttrainings auf die MCT1-Dichte
Gegenwärtig gibt es nur eine vergleichbare Krafttrainingsstudie mit Typ 2
Diabetikern, die den Einfluss eines Krafttrainings auf die MCT1-Proteinexpression
untersucht hat. Vor Trainingsbeginn wiesen die Typ 2 Diabetiker einen 35%igen

Diskussion
71
reduzierten membranösen MCT1-Gehalt gegenüber der gesunden Kontrollgruppe
auf. Durch ein dreimal in der Woche stattfindendes Muskelaufbautraining mit
progressiver Trainingsgewichtsanpassung konnte der reduzierte membranöse MCT1-
Gehalt wieder nach sechs Wochen normalisiert werden (Juel et al. 2004b). In der
DiabetesAktiv-Studie konnte dagegen eine Erhöhung der absoluten und relativen
membranösen MCT1-Dichte nicht erreicht werden. Hier lag die Trainingshäufigkeit
für das progressive Muskelaufbautraining bei zweimal pro Woche mit vergleichbarer
Belastungsintensität. Zum Interventionsende wurde das Training nach dem
subjektiven Belastungsempfinden im schweren Bereich mit drei Sätzen mit jeweils
zehn Wiederholungen an den Kraftgeräten durchgeführt. Offensichtlich muss die
Trainingshäufigkeit mit zwei Trainingseinheiten in der Woche im Vergleich zur
Studie von Juel et al. (2004b) als zu gering angesehen werden, um eine signifikante
Zunahme der absoluten und relativen membranösen MCT1-Dichte zu erreichen.
4.2 Langfristig trainingsbedingte Veränderungen der MCT4-Dichte im Muskelgewebe
In der Ausdauergruppe ließ sich eine Abnahme der relativen membranösen MCT4-
Dichte feststellen, wobei die relative membranöse MCT4-Dichte der Kraftgruppe
unverändert blieb. Für die zelluläre und absolute membranöse MCT4-Dichte zeigten
sich in beiden Interventionsgruppen keine Veränderungen nach dem dreimonatigen
Trainingsprogramm.
Einfluss des Ausdauertrainings auf die MCT4-Dichte
Das MCT4-Protein wurde bisher überwiegend in Muskelzellen mit einer hohen
glykolytischen Aktivität nachgewiesen (Mc Cullagh et al.1997; Wilson et al. 1998;
Pilegaard et al. 1999a). Dabei konnte eine hohe Korrelation zwischen dem MCT4-
Gehalt und den Typ II-Fasern beobachtet werden (Pilegaard et al. 1999a; Bonen et al.
2000a, Kobayashi et al. 2004). In verschiedenen Trainingsstudien konnte bisher
beobachtet werden, dass ein moderates Ausdauertraining zu unterschiedlichen
Expressionsraten des MCT4- und MCT1-Proteins in der Muskulatur führte. Beim
Menschen zeigte sich nach einem neunwöchigen Ergometertraining bei 75% der
Vo2max eine Zunahme des MCT1-Gehaltes in der Muskelmembran, wohingegen der
MCT4-Gehalt unverändert blieb (Dubouchaud et al. 2000). Ebenso ließ sich im Tier-

Diskussion
72
modell nach einer dreiwöchigen Dauerstimulation eine Erhöhung der MCT1-
Proteinexpression in den roten und weißen Muskelzellen des M. tibialis anterior bei
Ratten feststellen, wobei es zu keinen trainingsinduzierten Veränderungen des
MCT4-Gehaltes kam (Bonen et al. 2000b). Die gleichen intensitätsbedingten
Veränderungen des MCT1- und MCT4-Gehaltes konnten auch nach einem sieben-
tägigen moderaten Laufradtraining mit Mäusen (Yoshida et al. 2004) und nach
elektrischer Dauerstimulation bei Ratten beobachtet werden (Wilson et al. 1998). In
der DiabetesAktiv-Studie zeigte sich in der Ausdauergruppe in Übereinstimmung zu
den vorher beschriebenen Untersuchungsergebnissen eine erhöhte zelluläre MCT1-
Dichte, wohingegen die zelluläre MCT4-Dichte unverändert blieb. Aufgrund der
unterschiedlichen intensitätsbedingten Proteinexpression des MCT1 und MCT4 wird
angenommen, dass beide Proteine unterschiedlich reguliert werden. Es wird in der
gegenwärtigen Literatur kontrovers diskutiert, ob die aktivitätsbedingte
Genexpression der MCT-Proteine nach transkriptionellen, posttranskriptionellen oder
posttranslationellen Mechanismen erfolgt (Jackson et al. 1997; Sachs et al. 1997;
Bonen et al. 2000b; Wang et al. 2003; Coles et al. 2004; Halestrape et al. 2004).
Zudem konnten weitere Studien den Einfluss verschiedener Transkriptionsfaktoren
auf das MCT-Protein in unterschiedlichen Geweben nachweisen. Im Skelettmuskel
bei Ratten zeigte sich ein Einfluss des Transkriptionsfaktors HIF1α auf das MCT4-
Protein (Ullah et al. 2006) wie auch in den Endothelzellen und Astrozyten des
Gehirns auf das MCT1-Protein (Zhang et al. 2005). Im Darm konnte eine durch den
Transkriptionsfaktor AP2 und PKC induzierte MCT1-Genexpression beobachtet
werden (Saksena et al. 2008). Zudem scheint ein belastungsbedingter Laktatanstieg
den wichtigen Transkriptionsfaktor PGC1α für die mitochondriale Biogenese zu
aktivieren, der an der trainingsinduzierten erhöhten MCT1-Proteinexpression
beteiligt ist (Hashimoto et al. 2007; Benton et al. 2008).
Entscheidend für eine erhöhte Expressionsrate des MCT4 scheint die Belastungs-
intensität des Trainings zu sein, da der MCT4 erst bei höherer Trainingsintensität
zusammen mit dem MCT1 exprimiert wird (Juel, 2006). Dies ließ sich nach einem
sechstägigen einstündigem Ergometertraining mit untrainierten Männern über einen
Zeitraum von neun Wochen bei 75% der Vo2max beobachten, wonach sich der
Gesamtproteingehalt des MCT4 (+47%) und MCT1 (+90%) erhöhte (Dubouchaud
et al. 2000). Eine gleichzeitige Proteinexpression des MCT1- und MCT4-Proteins
zeigte sich auch nach einem progressiv gesteigerten Knie-Extensoren-Training auf

Diskussion
73
einem Ergometer in liegender Position bei 150 % der VO2max, bei dem sich der
MCT4-Gehalt um 34% und der MCT1-Gehalt um 76% im trainierten Bein der männ-
lichen Probanden erhöhte (Pilegaard et al. 1999b). Bei einer vergleichbaren Studie
mit ähnlicher Trainingsdurchführung bei gleicher Trainingsdauer ließ sich dagegen
keine signifikante Zunahme des MCT4-Gehaltes (11 %) nach acht Wochen im
trainierten Bein der männlichen Probanden feststellen (Juel et al. 2004a). Diese
beiden unterschiedlichen Ergebnisse können einerseits in der großen inter-
individuellen Variation des Muskelfasertyps innerhalb der Stichprobe (Pilegaard et
al. 1999a) und anderseits in der geringen Stichprobengröße (Juel n=6; Pilegaard n=7)
begründet sein. Eine gleichzeitige erhöhte Expressionsrate des MCT1- und MCT4-
Proteins konnte nicht nur nach langfristigen Trainingsprogrammen, sondern auch
kurzfristig nach einer einzelnen Trainingseinheit im Skelettmuskel bei Menschen
(Green et al. 2002) und Raten (Coles et al. 2004) beobachtet werden. Die
kurzfristigen Regulationsmechanismen der MCT-Proteine in der Skelettmuskulatur
wurden in der DiabetesAktiv-Studie nicht untersucht.
In der DiabetesAktiv-Studie zeigte sich eine signifikante Abnahme der relativen
membranösen MCT4-Dichte bei der Ausdauergruppe. In Übereinstimmung mit den
zuvor beschriebenen Untersuchungsergebnissen war offensichtlich die Belastungs-
intensität des moderaten Ausdauertrainings in der DiabetesAktiv-Studie zu gering,
um über die Trainingsintensität eine erhöhte relative membranöse Proteinexpression
des MCT4 zu erreichen. Ebenso kann auch die unveränderte absolute membranöse
und zelluläre MCT4-Dichte auf die geringe Intensität des Ausdauertrainings für die
intensitätsbedingte Proteinexpression des MCT4-Proteins zurückgeführt werden.
Zudem ließ sich in der Ausdauergruppe eine nicht signifikante geringe Abnahme der
Typ II-Muskelfasern beobachten, dass als weiterer Erklärungsansatz für die
Abnahme des überwiegend in den Typ II-Muskelfasern vorkommenden MCT4-
Proteins angeführt werden kann (Bonen et al. 2000a; Dimmer et al. 2000). Aber auch
im trainierten Zustand scheint eine Steigerung der Expressionsrate des MCT4 durch
hochintensive Trainingsprogramme limitiert zu sein. Sowohl nach einem hoch-
intensiv sechswöchigen Sprinttraining mit ausdauertrainierten Läufern (Bickham et
al. 2006) als auch nach einem intensiv fünfwöchigen Intervalltraining auf einem
Ergometer mit Sportstudentinnen zeigten sich keine weiteren signifikanten
Erhöhungen des MCT4-Gehaltes im trainierten Muskel (Bishop et al. 2008). Dies
konnte auch nach einem fünfmonatigen intensiven Laufbandtraining mit trainierten

Diskussion
74
Skiläufern beobachtet werden, wo es in der Vergleichsgruppe nach einer Reduktion
der Belastungsintensität sogar zu einer Abnahme des MCT4-Gehaltes bei den
trainierten Skiläufern kam (Evertsen et al. 2001).
Im untrainierten Zustand führt die fehlende Muskelaktivität zu einer Verringerung
des MCT4-Gehaltes in der Skelettmuskulatur (Wilson et al. 1998). Der Einfluss eines
Ausdauertrainings auf den MCT4-Gehalt wurde bisher noch nicht bei Typ 2
Diabetikern untersucht. Bisher konnte nur im Tiermodell eine reduzierte Laktat-
transportkapazität (Py et al. 2002) und geringerer MCT4-Gehalt nachgewiesen
werden, der sich nach einem täglichen Laufbandtraining nach drei Wochen bei
diabetischen Ratten wieder erhöhen ließ (Enoki et al. 2003). Beim Typ 2 Diabetiker
konnte eine weitere diabetesinduzierte Abnahme des MCT4-Gehaltes im Vergleich
zu der gesunden Kontrollgruppe nicht festgestellt werden (Juel et al. 2004b).
Einfluss des Krafttrainings auf die MCT4-Dichte
In der einzigen mit Typ 2 Diabetikern durchgeführten Krafttrainingsstudie zeigten
sich vor Studienbeginn keine Unterschiede hinsichtlich des MCT4-Gehaltes
zwischen den Typ 2 Diabetikern und der gesunden Kontrollgruppe im M. vastus
lateralis. Nach einem dreimal in der Woche stattfindenden progressiven Muskel-
aufbautraining konnte die Kontrollgruppe ihren MCT4-Gehalt signifikant erhöhen
(+32%), wohingegen bei den Typ 2 Diabetikern der MCT4-Gehalt nach
sechswöchigem Training eher unverändert (+6%) blieb (Juel et al. 2004b). Als
mögliche Ursache für die verschlechterte Proteinexpression des MCT4 bei den
Typ 2 Diabetikern kann deshalb nicht die Belastungsintensität angeführt werden, da
beide Trainingsgruppen ein vergleichbares Trainingsprogramm absolviert haben.
Vielmehr werden die Ursachen für die verschlechterte Proteinexpression des MCT4
in den diabetesbedingten Störungen des intrazellulären Laktatstoffwechsels gesehen
(Avogaro et al. 1996). Dieser pathologische Laktatstoffwechsel könnte auch für den
unverändert zellulären und membranösen MCT4-Gehalt in der DiabetesAktiv-Studie
verantwortlich gewesen sein. Für die Überprüfung dieser allgemeinen Annahme
bedarf es jedoch weiterer Trainingsstudien, die die Zusammenhänge zwischen einem
pathologischen Laktattransport bei Typ 2 Diabetikern und dessen Trainierbarkeit
detaillierter untersuchen.

Diskussion
75
Als weiteren möglichen Erklärungsansatz für den gleich bleibenden zellulären und
membranösen MCT4-Gehalt in der DiabetesAktiv Studie darf die geringe
Belastungsintensität trotz der obigen Studienergebnisse nicht ausgeschlossen werden,
wodurch möglicherweise die geringe Zunahme des Typ II-Muskelfaseranteil in der
DiabetesAktiv-Studie erklärt werden kann. Es ist anzunehmen, dass ein intensiveres
Krafttraining zu weiteren Veränderungen des Muskelfasertyps geführt hätte,
wodurch sich vermutlich auch der MCT4-Gehalt in Übereinstimmung zu vorherigen
Studienergebnissen erhöht hätte (Bonen et al. 2000a; Wilson et al. 1998). Zur
Analyse des Einflusses der Belastungsintensität beim Kraft- als auch Ausdauer-
training auf den MCT-Gehalt bedarf es jedoch weiterer Trainingsstudien sowohl mit
gesunden Kontrollgruppen als auch Typ 2 Diabetikern, um die Effizienz der
unterschiedlichen Belastungsintensitäten hinsichtlich der zellulären Anpassungen zu
überprüfen. Möglicherweise führen höhere Belastungsintensitäten mit längeren
Regenerationszeiten zu positiveren zellulären Anpassungserscheinungen als
geringere Belastungsintensitäten, woraus sich in Zukunft neue effizientere Trainings-
konzepte für das Kraft- und Ausdauertraining entwickeln ließen.
4.3 Langfristig trainingsbedingte Veränderungen der MCT1- und CD147-Dichte im Erythrozyten
Es konnte eine langfristig trainingsbedingte Zunahme der basalen MCT1-Dichte in
beiden Trainingsgruppen beobachtet werden, wobei nur für die Kraftgruppe ein
signifikantes Ergebnis ermittelt werden konnte. Parallel dazu nahm die basale
CD147-Dichte in der Erythrozytenmembran in beiden Trainingsgruppen ab, wobei
sich nur für die Ausdauergruppe ein trainingsbedingter signifikanter Einfluss zeigte.
Unter körperlicher Belastung ist hauptsächlich der MCT1 für den schnellen
Laktattransport vom Blutplasma in die Erythrozyten verantwortlich. Der prozentuale
Anteil des Laktattransports bei körperlicher Aktivität liegt für den MCT1 bei
ungefähr 90%, für die Bande-3-Proteine bei 6% und die nicht ionische Diffusion bei
4% (Skelton et al. 1998; Deuticke et al. 1989). Die zahlreichen Trainingsstudien
befassten sich bisher jedoch überwiegend mit den kinetischen Parametern zum
Laktattransport im Erythrozyten (Väihkönen et al. 1998; Connes et al. 2004b; Patillo
et al. 2005; Pösö, 2002; Sara et al. 2006). In der DiabetesAktiv-Studie wurden
dagegen die trainingsbedingt langfristigen und kurzfristigen MCT1- und CD147-

Diskussion
76
Dichteveränderungen und die zugrundeliegenden aktivitätsbedingten Regulations-
mechanismen im Erythrozyten untersucht. Zunahmen der MCT1-Proteinexpression
infolge von längerfristigen Trainingsinterventionen sind bereits in der Herz- und
Skelettmuskulatur bei Menschen und Tieren nachgewiesen worden. Dies zeigte sich
auch bei Typ 2 Diabetikern, bei denen der 35%ige geringere MCT1-Gehalt durch ein
Krafttraining wieder normalisiert werden konnte (Juel et al. 2004b). In den
Erythrozyten konnte bisher nur in einer einzigen vergleichbaren Trainingsstudie eine
Erhöhung der MCT1-Proteinexpression nach einem dreiwöchigen Schwimmtraining
mit fünf einstündigen Trainingseinheiten in der Woche bei männlichen Ratten
beobachtet werden. Zudem konnte dabei festgestellt werden, dass jüngere
Erythrozyten eine höhere MCT1-Dichte aufwiesen als ältere Erythrozyten. Basierend
auf den Beobachtungen wird angenommen, dass die MCT1-Proteinexpression
während der Erythropoese im Knochenmark abläuft (Aoi et al. 2004).
In Übereinstimmung mit den Ergebnissen von Aoi et al. (2004) ließ sich auch in der
DiabetesAktiv-Studie erstmalig beim Menschen eine trainingsbedingte Zunahme der
basalen MCT1-Dichte im Erythrozyten in beiden Trainingsgruppen beobachtet
werden. Dabei konnte nur für die Kraftgruppe ein signifikantes Ergebnis ermittelt
werden, weil sich in der Ausdauergruppe im Vergleich der Absolutzahlen der
einzelnen Probanden ein sehr heterogenes Bild mit großen Abweichungen zeigte.
Dies beeinflusst gerade bei kleinen Stichprobenumfängen die Aussagekraft des
ermittelten Ergebnisses. Als weiterer Erklärungsansatz für die nicht signifikanten
Veränderungen der MCT1-Dichte im Erythrozyten könnte die geringe Trainings-
intensität des Ausdauertrainings angeführt werden. Die Belastungsintensität des
Ausdauertrainings wurde bei der 2 mmol/l Laktatschwelle festgelegt. Es kann
vermutet werden, dass die Intensität des moderaten Ausdauertrainings nicht
ausreichend war, um eine stärkere Erythropoetinausschüttung und somit eine
Erhöhung der Erythrozytenanzahl zu bewirken. Nach der Annahme von Aoi et al.
(2004) würde der Anstieg der Erythrozytenanzahl mit einer Erhöhung der MCT1-
Dichte in den Erythrozyten einhergehen, welches in der DiabetesAktiv-Studie nicht
festgestellt werden konnte.
Die signifikante Zunahme der MCT1-Dichte in den Erythrozyten der Kraftgruppe
kann möglicherweise mit den höheren belastungsbedingten Blutlakatkonzentrationen
während des Krafttrainings gegenüber dem Ausdauertraining erklärt werden. Am

Diskussion
77
Ende des progressiv aufgebauten Krafttrainingsprogrammes trainierten die Pro-
banden in der Muskelaufbauphase sechs Wochen nach ihrem subjektiven
Belastungsempfinden im schweren Bereich. Aufgrund der höheren Intensität ist
davon auszugehen, dass die Probanden höhere Laktatwerte als 2 mmol/l wie beim
Ausdauertraining erreicht haben. Untersuchungen von vier verschiedenen Kraft-
trainingsmethoden bei männlichen Probanden haben nachgewiesen, dass in
Abhängigkeit von der jeweiligen Trainingsmethode ein mittlerer Laktatwert von
3.4 - 4.4 mmol/l während des Trainings erreicht werden kann (Gentil et al. 2006). Da
in der DiabetesAktiv-Studie die Laktatwerte während der einzelnen Trainings-
einheiten in beiden Interventionsgruppen nicht explizit untersucht worden sind,
können keine Aussagen über eine mögliche Korrelation zwischen trainingsbedingt
niedrigen und hohen Laktatwerten und der MCT1-Proteinexpression in den
Erythrozyten getroffen werden. In der Muskulatur konnte bereits der Einfluss des
Moleküls Laktat auf die MCT1-Proteinexpression in gezüchteten L6 Muskelzellen
von Ratten gezeigt werden, wo der belastungsbedingte Lakatanstieg intrazelluläre
Signalwege im Muskel aktiviert, welche für eine erhöhte MCT1-Expression und die
mitochondriale Biogenese verantwortlich sind (Hashimoto et al. 2007).
Aber nicht nur eine aktivitätsbedingte Hypoxie, sondern auch langfristig unter
Hypoxiebedingungen auf 4100 m Höhe konnte eine „Up-Regulation“ des MCT1-
Proteins (+330%) nach zwei Wochen in den Erythrozyten bei acht dänischen
Probanden beobachtet werden, wohingegen in der Muskulatur die MCT1- und
MCT4-Proteinexpression unverändert blieb (Juel et al. 2003). Connes et al. (2004b)
vermuten, dass für die trainingsinduzierte Regulation des MCT1-Proteins in den
Erythrozyten das Hormon Erythropoetin verantwortlich ist. Diese Vermutung konnte
in einer weiteren Studie bestätigt werden, wo sich nach Injektion von Erythropoetin
in humane Erythrozyten eine signifikante Verbesserung des Laktatflusses in die
Erythrozyten zeigte (Connes et al 2004a). Da die Erythrozyten keinen Zellkern und
keine Zellorganellen besitzen, wird momentan kontrovers diskutiert, wie die
trainingsbedingte Proteinexpression des MCT1 im Erythrozyten reguliert wird. Ob
die MCT1-Proteinexpression schon frühzeitig während der Erythropoese im
Knochenmark erfolgt (Aoi et al. 2004) oder möglicherweise aufgrund einer stabilen
mRNA nach posttranskriptionellen Mechanismen, muss in zukünftigen Studien
intensiver untersucht werden. Zudem bedarf es zur Überprüfung des Einflusses des
pathologischen Stoffwechsels der nicht-insulinpflichtigen Typ 2 Diabetiker auf die

Diskussion
78
trainingsbedingte Proteinexpression des MCT1-Proteins im Erythrozyten weiterer
Trainingsstudien mit gesunden Kontrollgruppen, um gezielt diabetes- und
belastungsbedingte Unterschiede aufzudecken.
Essentiell für die Aktivität und Lokalisation des MCT1-Proteins scheint das trans-
membranäre Glykoprotein CD147 zu sein (Halestrap et al. 1999; Kirk et al. 2000;
Wilson et al. 2005). Bei trainierten Pferden mit einer hohen Laktattransportaktivität
konnte eine hohe Dichte des CD147- wie auch des MCT1-Proteins in den
Erythrozyten beobachtet werden (Koho et al. 2006). Das Schlüsselprotein für die
Regulation eines verbesserten erythroiden Lakatttransports scheint dabei nicht das
MCT1-Protein, sondern vielmehr das CD147-Protein zu sein. Im interindividuellen
Vergleich von Pferden konnte gezeigt werden, dass die CD147-Dichte positiv mit
der erhöhten Laktattransportaktivität korreliert (Koho et al. 2002). In der
DiabetesAktiv-Studie ließ sich in der Ausdauergruppe eine signifikante Abnahme der
basalen CD147-Dichte nach dem dreimonatigem Trainingsprogramm beobachten,
wohingegen die basale CD147-Dichte in der Kraftgruppe nur geringfügig abnahm.
Für die trainingsbedingte Abnahme der basalen CD147-Dichte in beiden
Trainingsgruppen können möglicherweise Veränderungen der Membran-
Permeabilität durch die belastungsbedingte erhöhte Radikalenbildung mit einer damit
verursachten Peroxidation von ungesättigten Fettsäuren in der Erythrozytenmembran
verantwortlich sein (Davies et al. 1982). Als weiteren denkbaren Erklärungsansatz
könnten pathologische Störungen des Laktatstoffwechsels die aktivitätsbedingte
Proteinexpression des CD147-Proteins negativ beeinflusst haben. Bei dem
Probandenkollektiv handelte es sich um nicht-insulinpflichtige Typ 2 Diabetiker, die
wohl noch keine diabetischen Folgeerkrankungen aufwiesen, aber vermutlich schon
Störungen des intrazellulären Laktatstoffwechsels zeigten (Avogaro et al. 1996). Hier
besteht weiterer Forschungsbedarf, um die Auswirkungen des diabetischen Stoff-
wechsels und die der körperlichen Aktivität auf das CD147-Protein in Zukunft
detaillierter zu untersuchen.

Diskussion
79
4.4 Kurzfristig trainingsbedingte Veränderungen der MCT1- und CD147-Dichte im Erythrozyten nach einer Maximalbelastung
In der DiabetesAktiv-Studie wurden erstmalig die kurzfristig aktivitätsbedingten
Regulationsmechanismen der MCT1- und CD147-Proteine in humanen Erythrozyten
nach einer Maximalbelastung untersucht. Die Ergebnisse der Immunfluoreszenz
deuten darauf hin, dass es im trainierten Zustand kurzfristig zu einer möglichen
aktivitätsbedingten Translokation des MCT1-Proteins und seines Chaperons CD147
in die Erythrozytenmembran kommt, welches im untrainierten Zustand nicht
beobachtet werden konnte.
In der Skelettmuskulatur wurden die kurzfristig aktivitätsbedingten Regulations-
mechanismen für das GLUT4-Protein bereits nachgewiesen, wobei körperliche
Aktivität die Aufnahme von Glukose durch eine erhöhte Genexpression und insulin-
unabhängige Translokation der GLUT4-Transporter in die Skelettmuskelzellen
steigert (Kristiansen et al. 1996; Phillips et al. 1996; Chibalin et al. 2000; O'Gorman
et al. 2006). Zahlreiche Studien konnten zeigen, dass sowohl ein langfristiges
Ausdauertraining (Ren et al. 1994; Goodyear et al. 1998; Yu et al. 2001) und
Krafttraining (Tabata et al. 1999; Holten et al. 2004; Yaspelkis et al. 2006) sowie
eine einzelne Trainingseinheit (Lund et al. 1995; Oakes et al. 1997; Chibalin et al.
2000) den verschlechterten Glukosetransport in die Muskelzellen verbessern. Diese
aktivitätsbedingte erhöhte Proteinexpression des GLUT4-Proteins konnte auch nach
einer einzelnen Trainingseinheit für das MCT1-Protein in der Skelettmuskulatur
festgestellt werden. Beim Menschen ließ sich nach einer sechsstündigen Trainings-
einheit bei 60% der Vo2max nach zwei Tagen eine Zunahme des MCT1-Gehaltes
feststellen (Green et al. 2002). Diese erhöhte MCT1-Proteinexpression im
Skelettmuskel zeigte sich auch unmittelbar nach einem zweistündigen moderaten
Laufbandtraining bei Ratten (Coles et al. 2004). Keine Veränderungen des MCT1-
Gehaltes ließen sich nach einer einzelnen submaximalen Belastung nach einer
dreißigminütigen Laufbandbelastung in Typ I- und Typ II-Muskelfasern bei Ratten
beobachten (Eydoux et al. 2000). Bei sehr hohen Trainingsintensitäten zeigten sich
dagegen Abnahmen des MCT1- und MCT4-Gehaltes. Nach einer einzelnen hoch-
intensiven Trainingseinheit mit einer Dauer von 45 Sekunden bei 200% der Vo2max
konnte eine Abnahme des MCT1 (-24%) und MCT4-Gehaltes (-26%) in humanen
Skelettmuskelzellen beobachtet werden (Bishop et al. 2007). Dies konnte auch im
Tiermodell gezeigt werden, wo der MCT1 (-10%) und MCT4-Gehalt (-25%) nach

Diskussion
80
zehnminütiger hoch intensiver elektrischer Stimulation im Skelettmuskel bei Ratten
abnahm (Tonouchi et al. 2002). Diese gegensätzlichen Untersuchungsergebnisse sind
auf die teils unterschiedlichen methodischen Studiendesigns der Trainingsstudien
zurückzuführen. Insgesamt lässt sich aus den Beobachtungen eindeutig erkennen,
dass intensitätsabhängige Muskelkontraktionen kurzfristig eine „Up“ bzw. “Down“-
Regulation der MCT1- und MCT4-Proteine im Skelettmuskel nach einer einzelnen
Trainingseinheit bewirken.
In der DiabetesAktiv-Studie zeigte sich nach der dreimonatigen Trainings-
intervention in der Kraftgruppe eine signifikante und in der Ausdauergruppe eine
tendenzielle Abnahme der MCT1-Dichte unmittelbar nach dem Belastungstest,
wohingegen im untrainierten Zustand die MCT1-Dichte in beiden Trainingsgruppen
eher unverändert blieb. Die Ursache für die belastungsbedingte Abnahme der MCT1-
Dichten beider Trainingsgruppen nach Belastungsende könnte mit einer verstärkten
Translokation des MCT1-Proteins aus dem Zytoplasma in die Erythrozytenmembran
zusammenhängen. Dadurch ließe sich die optische Abnahme der Färbungsintensität
in den Erythrozyten erklären. Die gleichzeitige translokationsbedingte Dichte-
zunahme des MCT1-Proteins in der Membran kann bei einer immunhisto-
chemischen Auswertung nicht adäquat gemessen werden, sodass daraus insgesamt
die optisch festgestellte Dichteabnahme resultiert. Einen weiteren Nachweis für eine
mögliche aktivitätsbedingte Translokation des MCT1-Proteins liefern die
fluoreszenzmikroskopischen Aufnahmen der Erythrozyten vor und nach der
Trainingsintervention. Hier lässt sich gut erkennen, dass sich im trainierten Zustand
die MCT1- und CD147-Proteine unmittelbar nach dem Belastungstest von innen
nach außen in die Erythrozytenmembran verlagern. Diese optische Verlagerung der
CD147-Proteine und somit Zunahme der CD147-Dichte an der Erythrozyten-
membran werden durch die Ergebnisse der durchflusszytometrischen Analyse der
CD147-Dichte unterstützt. Bei der Kraftgruppe zeigte sich unmittelbar nach dem
Belastungstest ein signifikantes Ansteigen der CD147-Dichte, während die CD147-
Dichte im untrainierten Zustand noch abgenommen hatte. Bei der Ausdauergruppe
ließ sich vor dem Interventionsbeginn eine tendenzielle Verringerung der CD147-
Dichte nach dem Belastungstest feststellen, wohingegen sich nach der
Trainingsintervention nur noch eine geringe Abnahme der CD147-Dichte in der
Erythrozytenmembran zeigte. Im Vergleich der beiden Trainingsgruppen zeigen die
Veränderungen der CD147-Dichten die gleiche trainingsbedingte Tendenz, jedoch

Diskussion
81
lassen sich für die Kraftgruppe die eindeutigeren Reaktionen nachweisen. Es kann
vermutet werden, das das Chaperon CD147 auch im Erythrozyten eine Schlüssel-
funktion in der Proteinlokation des MCT1 einnimmt, da es mit dem aktivitäts-
bedingten Anstieg der CD147-Dichte in beiden Trainingsgruppen auch zu einer
verstärkten möglichen Translokation des MCT1-Proteins in die Erythrozyten-
membran im trainierten Zustand kam. Die essentielle Bedeutung des Zellober-
flächenproteins CD147 für die Lokalisation und eine mögliche Translokation des
MCT1 und MCT4 an die Zelloberfläche wurde bereits in-vitro-Studien herausgestellt
(Kirk et al. 2000; Wilson et al. 2005). Unter in-vivo-Bedingungen konnte an CD147-
Knockout-Mäusen gezeigt werden, dass das CD147-Protein für die Lokalisation des
MCT1 in Herz- und Skelettmuskelzellen verantwortlich ist (Nakai et al. 2006).
Ebenso konnte bei CD147-Knockout-Mäusen beobachtet werden, dass ein Fehlen
des CD147-Proteins zu einer reduzierten Proteinexpression des MCT1 und MCT4
am Pigmentepithelium der neuronalen Netzhaut führt (Philp et al. 2003a).
In der Skelettmuskelzelle konnte für das MCT4-Protein ein intrazellulärer Pool im
Zytoplasma entdeckt werden. Es wird vermutet, dass der MCT4 während
körperlicher Aktivität zur Muskelmembran transloziert und somit vermehrt Laktat
und H+-Ionen aus der Muskulatur heraus transportiert (Bonen et al. 2000a). Diese
intrazellulären Depots und die Translokation zur Muskelmembran wurden bereits für
das GLUT4-Protein nachgewiesen (Pessin et al. 1999). Sollte die Translokation der
MCT-Proteine analog zur GLUT4-Translokation ablaufen, könnte die Muskelzelle so
kurzfristig auf aktivitätsbedingte intrazelluläre pH-Wertveränderungen reagieren und
entscheidend in den Laktatmetabolismus eingreifen. Bei Typ 2 Diabetikern bewirkt
eine aktivitätsbedingte Translokation der GLUT4-Transporter langfristig eine
Reduktion der erhöhten Nüchternglukosewerte. Analog dazu könnte eine mögliche
aktivitätsbedingte Translokation der MCT1- und MCT4-Proteine in die Muskel-
membran die erhöhten Ruhelaktatwerte bei Typ 2 Diabetikern, durch einen
verbesserten Laktattransport sowie einer erhöhten Laktataufnahme in die
Körperzellen bei regelmäßiger körperlicher Aktivität, langfristig senken. Im
nachfolgenden hypothetischen Modell wird die mögliche aktivitätsbedingte
Translokation der MCT1- und CD147-Proteine bei körperlicher Belastung im
Erythrozyten dargestellt (vgl. Abb. 35). Die körperliche Belastung aktiviert über
bisher unbekannte Signalwege eine Translokation der MCT1- und CD147-Proteine
aus intrazellulären Speichervesikeln zur Erythrozytenmembran. Durch diese

Diskussion
82
mögliche aktivitätsbedingte Translokation der MCT1- und CD147-Proteine könnte
die ansteigende Plasmalaktatkonzentration während körperlicher Belastung durch die
vermehrte translokationsbedingte Laktataufnahme der MCT1-Transporter in die
Erythrozyten schneller reduziert werden. Dabei würden der Erythrozyten eine
Pufferfunktion einnehmen, weil durch die erythroide Laktataufnahme der intra-
zelluläre pH-Wert in der Muskulatur als auch das Laktatkonzentrationsgefälle
zwischen Muskel und Blutplasma bei körperlicher Belastung länger aufrechterhalten
werden kann.
MCT1- Vesikel
Körperliche Belastung
CD14
7CD
147
CD147
CD147
Erythrozytenmembran
Signalwege?
Abb. 35: Hypothetisches Modell einer möglichen aktivitätsbedingten Translokation im Erythrozyten des MCT1-und CD147-Proteins bei körperlicher Belastung
Zur Aufklärung der möglichen intrazellulären Signalwege und der zugrunde-
liegenden Regulationsmechanismen einer möglichen aktivitätsbedingten Trans-
lokation des MCT1- und CD147-Proteins im Erythrozyten bedarf es weiterer
Trainingsstudien im Bereich der Grundlagenforschung.

Diskussion
83
4.5 Der Einfluss des sporttherapeutischen Ausdauer- und Krafttrainings auf den Stoffwechsel und die Leistungsfähigkeit bei nicht-insulinpflichtigen Typ 2 Diabetikern Der körperlichen Bewegung wird innerhalb der diabetischen Therapie ein hoher
Stellenwert zugeschrieben (Boule et al. 2001; Sigal et al. 2006). Es konnte bisher in
zahlreichen Studien gezeigt werden, dass sowohl ein Ausdauer- (Boule et al. 2003;
Neil et al. 2006) als auch Krafttraining (Castaneda et al. 2002; Cauza et al. 2005) die
diabetische Stoffwechsellage verbessern und die Risikofaktoren bei der Entstehung
eines manifesten Typ 2 Diabetes reduzieren (Tuomilehto et al. 2001, Hu et al. 2002).
Dieses belegen auch die Ergebnisse der DiabetesAktiv-Studie, da beide Trainings-
formen die diabetische Stoffwechsellage und körperliche Leistungsfähigkeit der
Probanden verbessert haben. Aufgrund der eindeutigen Studienlage erfolgt im
Folgenden deshalb kein Vergleich zwischen den beiden Trainingsformen hinsichtlich
eines bereits bewiesenen gesundheitlichen Nutzen für den Typ 2 Diabetiker, sondern
eine erklärende Gegenüberstellung der Ergebnisse.
In der DiabetesAktiv-Studie bewirkte das Ausdauertraining eine geringe signifikante
Reduktion des Körpergewichtes und BMI-Wertes, wohingegen bei der Kraftgruppe
das Körpergewicht und der BMI-Wert unverändert blieb, welches durch die
krafttrainingsbedingte Zunahme an Muskelmasse erklärt werden kann. Ähnliche
Ergebnisse zeigten sich auch in anderen Trainingsstudien, wobei eine geringe
ausdauerbedingte Gewichtsabnahme (Boule et al. 2001; Dunstan et al. 1997; Cuff et
al. 2003) und ein unverändertes Körpergewicht nach einem Krafttraining (Ishii et al.
1998; Castaneda et al. 2002; Ibanez et al. 2005) beobachtet werden konnte. Es war zu
erwarten, dass in der dreimonatigen DiabetesAktiv-Studie ein zweimal in der Woche
stattfindendes Training zu gering war, um das Körpergewicht ohne eine ergänzende
Ernährungsumstellung effektiv zu reduzieren. Aus diesem Grund zählt innerhalb der
Diabetes-Therapie die Ernährungsumstellung neben der körperlichen Aktivität und
der medikamentösen Behandlung zu den drei Behandlungsschwerpunkten (Halle et
al. 2008).
Die Ruhelaktatwerte befanden sich vor Studienbeginn im Normbereich bei
1.4 mmol/l in beiden Trainingsgruppen und blieben nach der Bewegungsintervention
erwartungsgemäß eher unverändert. Inwieweit diabetesbedingt erhöhte Ruhe-

Diskussion
84
laktatwerte durch körperliche Aktivität reduziert werden können, muss in weiteren
Studien bei Typ 2 Diabetikern untersucht werden.
Eine signifikante Verbesserung des systolischen Ruheblutdrucks ließ sich in beiden
Trainingsgruppen beobachten, wohingegen für den diastolischen Ruheblutdruck nur
für die Ausdauergruppe ein signifikantes Ergebnis ermittelt werden konnte. Diese
ausdauerbedingten Ruheblutdruckveränderungen ließen sich auch in anderen
Trainingsstudien mit Typ 2 Diabetikern feststellen (Boule et al. 2001; Sigal et al.
2004). In einer Metaanalyse konnte eine signifikante Verbesserung des Ruhe-
blutdruckes bei Hypertonikern nach einem progressiv aufgebauten Krafttraining
nachgewiesen werden (Kelly et al. 2000). Bei Typ 2 Diabetikern ließen sich dagegen
nach kurzen Krafttrainingsprogrammen (< 8 Wochen) keine signifikanten
Verbesserungen des Ruheblutdrucks feststellen (Erikson et al. 1997; Dunstan et al.
1998). Bei länger angelegten Trainingsstudien mit Typ 2 Diabetikern zeigten sich in
Übereinstimmung zu den Ergebnissen mit der DiabetesAktiv-Studie signifikante
Verbesserungen sowohl des systolischen Blutdrucks (Castenada et al. 2002) als auch
des systolischen und diastolischen Ruheblutdrucks nach einem Krafttraining
(Dunstan et al. 2002; Cauza et al. 2005). Die signifikanten Veränderungen des Ruhe-
blutdrucks durch ein Krafttraining können vermutlich auf die längere Trainings-
dauer der Studien zurückgeführt werden.
Die amerikanische Diabetes Gesellschaft hat aufgrund der bisherigen Studien-
ergebnisse beiden Trainingsformen innerhalb der diabetischen Sporttherapie den
Evidenzgrad A für die Verbesserung der glykämischen Stoffwechsellage verliehen
(Sigal et al. 2004). Als Indikatoren für den Kohlenhydratstoffwechsel der Probanden
dienten in der DiabetesAktiv-Studie der Homa-Index, die Nüchternglukosewerte
sowie die Proinsulin- und Insulinwerte. In beiden Trainingsgruppen konnten der
Homa-Index und Nüchternblutzuckerwerte reduziert werden, wobei nur für die
ausdauertrainingsbedingte Abnahme der Insulinresistenz ein signifikantes Ergebnis
ermittelt werden konnte. Die Ursachen für die nicht signifikante Abnahme der Blut-
zuckerwerte können in der Trainingshäufigkeit und Trainingsintensität der
Interventionsprogramme der DiabetesAktiv-Studie liegen. Nach den Richtlinien der
amerikanischen Diabetes Gesellschaft sollte ein Ausdauertraining mindestens an drei
Tagen mit einem Trainingsumfang von 150 Minuten bei 40-60% der VO2max in der
Woche oder bei einer höheren Trainingsintensität >60% der VO2max mit einem

Diskussion
85
Wochenumfang von 90 Minuten erfolgen. Analog dazu werden für das Krafttraining
drei Sätze mit acht bis zehn Wiederholungen der großen Muskelgruppen auch
dreimal wöchentlich mit submaximaler Belastungsintensität empfohlen (Sigal et al.
2006). Durch die geringe Trainingshäufigkeit mit zwei betreuten Trainingseinheiten
pro Woche konnte möglicherweise der kontraktionsbedingte Effekt einer bis zu
48-stündigen erhöhten Insulinsensitivität an den Muskelzellen über die Woche in
beiden Trainingsgruppen nicht aufrechterhalten werden (König et al. 2006).
Eine signifikante Abnahme des Blutzuckerwertes konnte in der Metaanalyse von
Boule et al. (2001) beobachtet werden, wobei ein im Durchschnitt dreimal fünfzig-
minütiges Ausdauertraining bei 50-70% der VO2max. über achtzehn Wochen den
HBA1c unabhängig vom Körpergewicht um 0,66% reduzieren konnte. Diese
positiven Auswirkungen des Ausdauertrainings auf den HBA1c konnte in einer
weiteren Metaanalyse von Snowling et al. (2006) bestätigt werden. Darüber hinaus
führten höhere Belastungsintensitäten zu einer weiteren Verbesserung der Insulin-
sensitivität bei Typ 2 Diabetikern (O`Donovan et al. 2005; Coker et al. 2006;
Dipietro et al. 2006). Die Trainingsintensität für das Ausdauertraining wurde in der
DiabetesAktiv-Studie an der 2 mmol/l Laktatschwelle festgelegt. Diese moderate
Trainingsintensität war möglicherweise zu gering, um eine signifikante Abnahme der
Nüchternblutzuckerwerte zu bewirken. Diese Annahme wird von den Ergebnissen
zweier Metaanalysen bekräftigt, wo eine Korrelation zwischen Trainingsintensität
und der Abnahme des HBA1c-Wertes bei einem Ausdauertraining nachgewiesen
werden konnte (Boule et al. 2003; Zanuso et al. 2009).
In der Kraftgruppe zeigten sich keine signifikanten Veränderungen des Homa-
Indexes und der Nüchternblutzuckerwerte, welches auch in vereinzelten
Publikationen nach einem Krafttraining beobachtet werden konnte (Eriksson et al.
1997; Dunstan et al. 1998). In dem überwiegenden Anteil der mit Typ 2 Diabetikern
durchgeführten Krafttrainingsstudien ließen sich jedoch signifikante Verbesserungen
der Nüchternblutzuckerwerte und des HBA1c-Wertes feststellen (Castaneda et al.
2002; Baldi et al. 2003; Cauza et al. 2005; Dunstan et al. 2005). Für die richtige
Trainingsintensität beim Krafttraining muss eine Differenzierung hinsichtlich des
individuellen Trainingszieles des Patienten erfolgen. Einerseits führt ein Muskel-
aufbautraining mit einer geringen Wiederholungszahl bis zur muskulären
Erschöpfung zu einer Zunahme der Muskelmasse und andererseits bewirkt ein

Diskussion
86
Kraftausdauertraining durch die höhere Anzahl von Muskelkontraktionen für eine
höhere metabolische Umsatzrate von Glukose. Es konnte bisher gezeigt werden, das
sowohl ein intensives (Castaneda et al. 2002; Dunstan et al. 2002) als auch moderates
(Ishii et al. 1998; Maiorana et al. 2001; Baldi et al. 2003) Krafttraining für die
Verbesserung der glykämischen Stoffwechsellage in den diabetischen Sporttherapie
seine Berechtigung hat. Die nicht signifikanten Abnahmen des Homa-Indexes und
der Nüchternblutzuckerwerte in der DiabetesAktiv-Studie können deshalb aus
trainingswissenschaftlicher Sicht nur mit der kurzen Phasendauer des progressiv
aufgebauten Krafttrainingsprogrammes oder in der geringen Trainingshäufigkeit mit
zwei Krafttrainingseinheiten in der Woche begründet werden. Als weiteren
möglichen Erklärungsansatz für die nicht signifikanten Veränderungen der Nüchtern-
blutzuckerwerte in beiden Trainingsgruppen könnte das Ernährungsverhalten der
Teilnehmer angeführt werden. In der DiabetesAktiv-Studie erhielten die Probanden
keine weiteren Ernährungsempfehlungen, damit eine kausale Zuordnung der
Trainingseffekte zu der Trainingsform erfolgen kann. Trotzdem kann nicht
ausgeschlossen werden, dass eine einseitige kohlenhydratreiche Ernährung seitens
der Probanden die Ergebnisse der Nüchternblutzuckerwerte nachhaltig negativ
beeinflusst und somit die positiven Effekte der körperlichen Aktivität abschwächt.
Eine trainingsbedingte Verbesserung der Insulinresistenz an den Muskel- und
Fettzellen geht langfristig mit einer Abnahme des Insulin- und Proinsulinwertes
einher. In der DiabetesAktiv-Studie ließen sich in beiden Trainingsgruppen
Abnahmen des Insulinwertes feststellen, wohingegen nur für die Kraftgruppe ein
tendenzielles Ergebnis ermittelt werden konnte. Diese trainingsbedingte Abnahme
der Insulinwerte konnte zuvor bereits in anderen Ausdauer- (Wing et al. 1988;
Lehmann et al. 1995) und Krafttrainingsstudien (Dunstan et al. 1998; Baldi et al.
2003; Cauza et al. 2005) bei Typ 2 Diabetikern beobachtet werden. Zudem wurden in
der DiabetesAktiv-Studie erstmalig Proinsulinwerte erhoben, wodurch ein Vergleich
der Proinsulinwerte mit anderen Trainingsstudien nicht möglich ist. Es kann jedoch
angenommen werden, dass der Proinsulinwert durch ein körperliches Training
langfristig abnimmt, da sich mit einer langfristigen Senkung des HBA1c-Wertes
auch die Insulinproduktion in der Bauchspeicheldrüse anpasst. Diese Vermutung
wird durch die Ergebnisse der DiabetesAktiv-Studie teilweise unterstützt, da in der
Ausdauergruppe der Proinsulinwert tendenziell abnahm. In der Kraftgruppe nahm

Diskussion
87
dagegen der Proinsulinwert nur geringfügig ab, obwohl der Insulinwert eine
tendenzielle Abnahme zeigte.
Während es bei der Ausdauergruppe zu signifikanten Verbesserungen der
Cholesterin-, Triglyzerid-, und LDL-Konzentration kam, zeigte sich in der Kraft-
gruppe ein unverändertes Lipidprofil. In der Kraftgruppe konnte lediglich eine
tendenzielle Abnahme der HDL-Konzentration beobachtet werden. In der Meta-
analyse von Kelley et al. (2007) ließ sich durch ein Ausdauertraining eine
signifikante Abnahme des LDL-Cholesterins bei Typ 2 Diabetikern feststellen,
wobei sich bei den Verbesserungen des Gesamt- und HDL-Cholesterins und der
Triglyzeride keine signifikanten Veränderungen zeigten. Weitere Ausdauertrainings-
studien unterstützen die Ergebnisse der DiabetesAktiv-Studie, wo sich einheitliche
Verbesserungen des Lipidprofils zeigten, aber aufgrund kleiner Stichproben und
kurzer Interventionsdauer teilweise kein signifikantes Ergebnis ermittelt werden
konnte (Vanninen et al. 1992; Ligtenberg et al. 1997; Dunstan et al. 2002; Thomas et
al. 2006; Kadoglou et al. 2007). In Übereinstimmung mit den Ergebnissen der
DiabetesAktiv-Studie zeigten sich auch in anderen Studien keine Verbesserungen des
Lipidprofils durch ein Krafttraining (Erikson et al. 1997; Dunstan et al. 2002).
Obwohl einige Trainingsstudien krafttrainingsbedingte signifikante Verbesserungen
des Lipidprofils feststellten konnten (Honkola et al. 1997; Cauza et al. 2005; Misra et
al. 2008) scheint ein Ausdauertraining das Lipidprofil bei Typ 2 Diabetikers positiver
zu beeinflussen. Es konnte gezeigt werden, dass ein Ausdauertraining die
mitochondriale oxidative Enzymkapazität und die intrazellulären Fettsäure-
transportproteine erhöht und dadurch den Fettstoffwechsel und das Lipidprofil lang-
fristig verbessert (Bruce et al. 2004).
Die Effizienz der beiden Trainingsprogramme spiegelt sich in den trainings-
formspezifischen Verbesserungen der körperlichen Leistungsfähigkeit beider
Trainingsgruppen wieder. Sowohl bei der Ausdauergruppe als auch Kraftgruppe ließ
sich eine signifikante Verbesserung der Ausdauerleistungsfähigkeit feststellen,
welches sich auch in anderen Ausdauertrainingsstudien mit Typ 2 Diabetikern zeigte
(Boule et al. 2001; Cuff et al. 2003; Sigal et al. 2004; Cauza et al. 2005; Kadoglou et
al. 2007). Da in der DiabetesAktiv-Studie die Ausdauerleistungsfähigkeit über die
maximale Wattleistung bestimmt wurde, kann vermutet werden, dass für die
signifikante Verbesserung der Ausdauerleistungsfähigkeit der Kraftgruppe die

Diskussion
88
Zunahme der Muskelkraft im M. quadrizeps femoris mitverantwortlich ist. Cauza
et al. (2005) konnte ebenso nach einem progressiv aufgebauten Krafttraining eine
Verbesserung der Ausdauerleistungsfähigkeit in der Kraftgruppe feststellen. Zudem
konnten die Probanden der Kraftgruppe sowohl ihre konzentrische als auch
isometrische beidbeinige und rechtsbeinige Maximalkraft signifikant verbessern.
Diese Ergebnisse stehen im Einklang mit den Ergebnissen zahlreicher
Krafttrainingsstudien, bei denen sich signifikante Verbesserungen der Muskelkraft
bei Typ 2 Diabetikern zeigten (Smutok et al. 1994; Maiorana et al. 2002; Baldi et al.
2003; Fenicchia et al. 2004; Dunstan et al. 2005; Ibanez et al. 2005).

Zusammenfassung
89
5 Zusammenfassung
Der Forschungsschwerpunkt der gegenwärtigen Literatur befasste sich bisher
überwiegend mit den kinetischen Parametern zum Laktattransport im Erythrozyten
und in der Skelettmuskulatur. Die zugrundeliegenden aktivitätsbedingten
Regulationsmechanismen des MCT1- und CD147-Protein im Erythrozyten wurden
zuvor noch nicht an humanen Erythrozyten untersucht. Eine Zielsetzung dieser
Arbeit lag in der Aufdeckung dieser kurzfristigen aktivitätsbedingten Regulations-
mechanismen des MCT1- und CD147-Proteins in den Erythrozyten nach einer
Maximalbelastung. Neben diesen kurzfristigen Anpassungserscheinungen wurden
auch die langfristig trainingsbedingten Veränderungen der MCT1- und CD147-
Dichte im Erythrozyten und der MCT1- und MCT4-Dichte in der Skelettmuskulatur
bei nicht-insulinpflichtigen Typ 2 Diabetikern nach einem dreimonatigen sport-
therapeutischen Ausdauer- und Krafttraining immunhistochemisch untersucht.
Im Vergleich der beiden Trainingsformen zeigten sich hinsichtlich der langfristig als
auch kurzfristig trainingsbedingten MCT1- und CD147-Dichteveränderungen
ähnliche Ergebnisse, wobei das Krafttraining eindeutigere zelluläre Reaktionen im
Erythrozyten auslöste. Die Ergebnisse der fluoreszenzmikroskopischen Aufnahmen
an den Erythrozyten deuten darauf hin, dass es kurzfristig bei trainierten nicht-
insulinpflichtigen Typ 2 Diabetikern zu einer verstärkten aktivitätsbedingten
Translokation des MCT1-Proteins und des Chaperons CD147 in die Erythrozyten-
membran kommt. Langfristig konnte nur eine Erhöhung der MCT1-Dichte in den
Erythrozyten der Kraftgruppe beobachtet werden. Die Untersuchungen am
Skelettmuskel zeigten in beiden Trainingsgruppen eine signifikante Zunahme der
zellulären MCT1-Dichte, wohingegen die zelluläre MCT4-Dichte nach dem drei-
monatigem Trainingsprogramm eher unverändert blieb. Die absolute membranöse
MCT4-Dichte blieb in beiden Trainingsgruppen unverändert, wohingegen nur in der
Ausdauergruppe eine signifikante Abnahme der relativen membranösen MCT4-
Dichte beobachtet werden konnte.
Das nachfolgende hypothetische Regulationsmodell zum Laktattransport während
körperlicher Belastung veranschaulicht die funktionelle Bedeutung der MCT-
Proteine in Abhängigkeit vom Trainingszustand (vgl. Abb. 36). Im trainierten

Zusammenfassung
90
Zustand kommt es aufgrund einer intensitätsabhängigen Zunahme der MCT1- und
MCT4-Dichte und einer möglichen aktivitätsbedingten Translokation der MCT-
Proteine zu einer schnelleren Laktat/H+-Ionenbeseitigung aus dem kontrahierenden
Muskel in das Blutplasma. Durch eine trainingsbedingte Zunahme der MCT1-Dichte
und einer erhöhten translokationsbedingten Laktataufnahme der MCT1-Proteine in
den Erythrozyten kann die ansteigende Plasmalaktatkonzentration während der
körperlichen Belastung schneller reduziert werden. Hierbei fungieren die
Erythrozyten durch die schnellere Laktataufnahme als Puffer, da sie dadurch den
intrazellulären pH-Wert im Skelettmuskel und den konzentrationsabhängigen Laktat-
fluss vom Skelettmuskel ins Blutplasma aufrechterhalten. Diese erhöhte Laktat-
transportrate führt im Körper zu einer gleichmäßigeren Laktatverteilung und somit
kann das Laktat neben Glukose als Brennstoff für die aerobe Energiegewinnung in
den verschiedenen Zielzellen als Energieträger verwertet oder als Glykogen
gespeichert werden.
La/H+
La/H+
La/H+
La/H+
La/H+
La/H+
La/H+
La/H+
La/H+
Untrainierter Trainierter
La/H+La/H+La/H+
Muskel Herz Leber
Endothel
Muskel Herz Leber
Laktatproduktion
Laktatverwertung
Skelettmuskel
La/H+
La/H+La/H+
La/H+ La/H+
TranslokationLa/H+ Translokation
MCT1/4
MCT1Erythrozyt
La/H+La/H+
La/H+
La/H+
La/H+ La/H+ La/H+La/H+ La/H+ La/H+
La/H+
La/H+
La/H+La/H+ La/H+
MCT1/4
Abb. 36: Hypothetisches Regulationsmodell zum Laktattransport und der funktionellen
Bedeutung der MCT-Proteine im Muskel und Erythrozyten während körperlicher Belastung im trainierten und untrainierten Zustand

Zusammenfassung
91
In der diabetischen Sporttherapie nimmt die körperliche Aktivität einen hohen
Stellenwert ein. Epidemiologische Studien haben eindeutig gezeigt, dass körperliche
Aktivität in der Primär- und Sekundärprävention den diabetischen Stoffwechsel
positiv beeinflusst. Sowohl das Ausdauer- als auch das Krafttraining haben aufgrund
ihrer Trainingsauswirkungen ihre alleinige Berechtigung in der diabetischen
Sporttherapie. Dies konnte auch in der DiabetesAktiv-Studie bestätigt werden, da
beide Trainingsformen die diabetische Stoffwechsellage und die körperliche
Leistungsfähigkeit der Studienteilnehmer positiv beeinflusst haben. Dies wird durch
weitere Studienergebnisse bestätigt, da ein Kombinationstraining aus Ausdauer- und
Krafttrainingsanteilen für den Typ 2 Diabetiker am effizientesten zu sein scheint
(Maiorana et al. 2002; Balducci et al. 2004; Sigal et al. 2007).
Für die Zukunft besteht ein großer Forschungsbedarf hinsichtlich der zellulären
Auswirkungen von sporttherapeutischen Trainingsinterventionen auf den
diabetischen Stoffwechsel. Ein besseres Verständnis der aktivitätsbedingten
zellulären Anpassungserscheinungen stellt die Grundlage für die Arbeit des
qualifizierten Sportwissenschaftlers dar, um individuell dosiert die körperliche
Aktivität wie ein Medikament innerhalb der diabetischen Sporttherapie in Zukunft
einsetzen zu können und somit langfristig das Gesundheitssystem durch diese
kostengünstige und effiziente Therapieform nachhaltig zu entlasten.

Literaturverzeichnis
92
6 Literaturverzeichnis
ADVANCE Collaborative Group, et al.. Intensive Blood Glucose Control and Vascular Outcomes in Patients with Type 2 Diabetes. N Eng J Med 358: 2560-2572, 2008
American Diabetes Association. Standards of medical care in diabetes--2009. Diabetes Care Jan 32 Suppl 1: S13-61, 2009
Aoi W, Iwashita S, Fujie M, Suzuki M. Sustained swimming increases erythrocyte MCT1 during erythropoiesis and ability to regulate pH homeostasis in rat. Int J Sports Med. Jul 25(5): 339-44, 2004
Avogaro A, Toffolo G, Miola M, Valerio A, Tiengo A, Cobelli C, Del Prato S. Intracellular lactate- and pyruvate-interconversion rates are increased in muscle tissue of non-insulin-dependent diabetic individuals.J Clin Invest. Jul 1; 98(1): 108-15, 1996
Baker SK, McCullagh KJA, and Bonen A. Training intensity-dependent and tissue-specific increases in lactate uptake and MCT1 in heart and muscle. J Appl Physiol 84: 987–994, 1998
Baldi JC, Snowling N. Resistance training improves glycaemic control in obese type 2 diabetic man. Int J Sports Med 24: 419–423, 2003
Balducci S, Leonetti F, Di Mario U, Fallucca F. Is a long-term aerobic plus resistance training program feasible for and effective on metabolic profiles in type 2 diabetic patients? Diabetes Care 27: 841–842, 2004
Benton CR, Campbell SE, Tonouchi M, Hatta H, Bonen A. Monocarboxylate transporters in subsarcolemmal and intermyofibrillar mitochondria. Biochem Biophys Res Commun. Oct 8; 323(1): 249-53, 2004
Benton, C. R., Y. Yoshida, et al. "PGC-1alpha increases skeletal muscle lactate uptake by increasing the expression of MCT1 but not MCT2 or MCT4." Physiol Genomics 35(1): 45-54, 2008
Bergström J. Percutaneous needle biopsy of skeletal muscle in physiological and clinical research. In: Scand J Clin Invest 35: 609-616, 1975
Bickham DC, Bentley DJ, Le Rossignol PF, Cameron-Smith D. The effects of short-term sprint training on MCT expression in moderately endurance-trained runners. Eur J Appl Physiol 96: 636–643, 2006
Biedler A, Mertzlufft F. Die Messung der Laktatkonzentration mit Elektroden. Anaesthesiol. Intensivmed. Notfallmed. Schmerzther. 30 (Suppl.1): 24- 26, 1995

Literaturverzeichnis
93
Bishop D, Edge J, Thomas C, Mercier J. High-intensity exercise acutely decreases the membrane content of MCT1 and MCT4 and buffer capacity in human skeletal muscle. J Appl Physiol 102: 616–621, 2007
Bishop D, Edge J, Thomas C, Mercier J. Effects of high-intensity training on muscle lactate transporters and postexercise recovery of muscle lactate and hydrogen ions in women. Am J Physiol Regul Integr Comp Physiol. Dec; 295(6): R1991-8, 2008 Bjarnason-Wehrens B, Mayer-Berger W, Meister ER, Baum K, Hambrecht R, Gielen S. Einsatz von Kraftausdauertraining und Muskelaufbau in der kardiologischen Rehabilitation. Empfehlungen der Deutschen Gesellschaft für Prävention und Rehabilitation von Herz-Kreislauferkrankungen e.V. Z Kardiol; 93: 357–70, 2004 Boeckh-Behrens WU, Buskies W. Gesundheitsorientiertes Fitnesstraining. Lüneburg: Wehdemeier & Pusch, 2002
Bonen A, Homonko DA. Effects of exercise and glycogen depletion on glyconeogenesis in muscle. J Appl Physiol. Apr;76(4): 1753-8, 1994
Bonen A, McCullagh KJ, Putman CT, Hultman E, Jones NL, Heigenhauser GJ. Short-term training increases human muscle MCT1 and femoral venous lactate in relation to muscle lactate. Am J Physiol Endocrinol Metab 274: E102-E107, 1998
Bonen A, Miskovic D, Tonouchi M, Lemieux K, Wilson MC, Marette A, Halestrap AP. Abundance and subcellular distribution of MCT1 and MCT4 in heart and fast-twitch skeletal muscles. Am J Physiol Endocrinol Metab 278: E1067–E107, 2000a
Bonen A, Tonuchi M, Miskovic D, Heddle C, Heikkila JJ, Halestrap AP. Isoform-specific regulation of the lactate transporters MCT1 and MCT4 by contractile activity. Am J Physiol Endocrinol Metab 279: E1131-E1138, 2000b
Bonen A. Lactate transporters (MCT proteins) in heart and skeletal muscles. Med Sci Sports Exerc. Apr; 32(4): 778-89, 2000c
Bonen A. Expression of lactate transporters (MCT1, MCT4) in heart and muscle. Eur J Appl Physiol 86: 6–11, 2001
Borg G. Borg’s perceived exertion and pain scales. Human Kinetics, Champaign Il.: 1998
Boule NG, Haddad E, Kenny GP, Wells GA, Sigal RJ: Effects of exercise on glycemic control and body mass in type 2 diabetes mellitus: a meta-analysis of controlled clinical trials. J Am Med Ass 286: 1218-1227, 2001
Boule NG, Kenny GP, Haddad E, Wells GA, Sigal RJ: Meta-analysis of the effect of structured exercise training on cardiorespiratory fitness in Type 2 diabetes mellitus. Diabetol 46: 1071-1081, 2003

Literaturverzeichnis
94
Bretzel RG, et al.. Hypertonie beim Diabetes mellitus. In: Scherbaum WA (Hrsg.): Praxis-Leitlinien der Deutschen Diabetes-Gesellschaft (DDG). Diabetologie und Stoffwechsel, Band 3, Supplement 2: S 141-142, 2008
Broer S, Schneider HP, Broer A, Rahman B, Hamprecht B, Deitmer JW. Characterization of the monocarboxylate transporter 1 expressed in Xenopus laevis oocytes by changes in cytosolic pH. Biochem. J. Jul 1; 333 (Pt 1): 167–174, 1998
Brooke MH, Kaiser KK. Muscle fiber types: how many and what kind? Arch Neurol 23: 369-379, 1970
Brooks GA, Brown MA, Butz CE, Sicurello JP, Dubouchaud H. Cardiac and skeletal muscle mitochondria have a monocarboxylate transporter MCT1. J Appl Physiol. Nov; 87(5): 1713-8, 1999
Brooks GA. Lactate shuttles in nature. Biochem. Soc. Trans. 30: 258 -264, 2002
Brooks G et al.. Lactate as a metabolic signal of gene expression. Deutsche Zeitschrift für Sportmedizin, 59, 12: 280-286, 2008
Bruce CR, Hawley JA. Improvements in insulin resistance with aerobic exercise training: a lipocentric approach. Med Sci Sports Exerc 36: 1196-1201, 2004
Bühl A. SPSS 14 - Einführung in die moderne Datenanalyse. 10. überarbeitete Auflage. Pearson Education Deutschland GmbH: S.121-123, 2006
Butz CE, McClelland GB, Brooks GA. MCT1 confirmed in rat striated muscle mitochondria. J Appl Physiol. Sep; 97(3): 1059-66, 2004
Castaneda C, Layne JE, Munoz-Orians L, et al.. A randomized controlled trial of resistance exercise training to improve glycemic control in older adults with type 2 diabetes. Diabetes Care; 25: 2335-41, 2002
Cauza E, Hanusch-Enserer U, Strasser B, Kostner K, Dunky A, Haber P. Strength and endurance training lead to different post exercise glucose profiles in diabetic participants using a continuous subcutaneous glucose monitoring system. Eur J Clin Invest.Dec; 35(12): 745-51, 2005
Chatham JC, Gao Z-P, Bonen A, Forder JR. Preferential inhibition of lactate oxidation relative to glucose oxidation in the rat heart following diabetes. Cardiovasc Res 43: 96–106, 1999
Chen YD, Varasteh BB, Reaven GM. Plasma lactate concentration in obesity and type 2 diabetes. Diabetes & metabolism; Band 19; Heft 4: 348-54, 1999
Chibalin, AV, Yu M, Ryder JW, Song XM, Galuska D, Krook A, Wallberg-Henriksson H, Zierath JR. Exercise-induced changes in expression and activity of proteins involved in insulin signal transduction in skeletal muscle: differential effects on insulin receptor substrates 1 and 2. Proc Natl Acad Sci USA 97: 38-43, 2000

Literaturverzeichnis
95
Choi CS, Kim YB, Lee FN, Zabolotny JM, Kahn BB, Youn JH: Lactate induces insulin resistance in skeletal muscle by suppressing glycolysis and impairing insulin signaling. Am J Physiol Endocrinol Metab; 283(2): 233-40, 2002
Coker R, Hays NP, Williams RH, Brown AD, Freeling SA, Kortebein PM, Sullivan DH, Starling RD, Williams JE. Exercise-induced changes in insulin action and glycogen metabolism in elderly adults. Med Sci Sport Exerc 38: 433–438, 2006
Coles L, Litt J, Hatta H, Bonen A. Exercise rapidly increases expression of the monocarboxylate transporters MCT1 and MCT4 in rat muscle. J Physiol 561: 253–261, 2004
Consoli A, Nurjhan N, Reilly J, Bier D, Gerich J. Mechanism of increased gluconeogenesis in noninsulin-dependent diabetes mellitus: role of alteration in systemic, hepatic and muscle lactate and alanine metabolism. J. Clin. Invest. 86: 2038-204, 1990
Connes P, Caillaud C, Mercier J, Bouix D, Casties JF. Injections of recombinant human erythropoietin increases lactate influx into erythrocytes. J Appl Physiol 97: 326–332, 2004a
Connes P, Bouix D, Py G, Caillaud C, Kippelen P, Brun JF, Varray A, Prefaut C, Mercier J. Does exercise-induced hypoxemia modify lactate influx into erythrocytes and hemorheological parameters in athletes? J Appl Physiol. Sep; 97(3): 1053-8, 2004b
Constant, J. S., Feng, J. J., Zabel, D. D., Yuan, H., Suh, D. Y., Scheuenstuhl, H., Hunt, T. K. and Hussain, M. Z. Lactate elicits vascular endothelial growth factor from macrophages: a possible alternative to hypoxia. Wound Repair Regen. 8: 353-360, 2000
Crandall DL, Fried SK, Francendese AA, Nickel M, DiGirolamo M. Lactate release from isolated rat adipocytes: influence of cell size, glucose concentration, insulin and epinephrine. Horm Metab Res. Jul; 15(7): 326-9, 1983
Cuff DJ, Meneilly GS, Martin A, Ignaszewski A, Tildesley HD, Frohlich JJ. Effective exercise modality to reduce insulin resistance in women with type 2 diabetes. Diabetes Care; 26: 2977–2982, 2003
Czech MP. Molecular actions of insulin on glucose transport. Annu Rev Nutr.;15: 441-7, 1995
D'Agostino RB, Russell MW, Huse DM, Ellison RC, Silbershatz H, Wilson PW, Hartz SC. Primary and subsequent coronary risk appraisal: new results from the Framingham study. Am Heart J Jan; 143(1): 21, 2002
Davies KJ, et al.. Free radicals and tissue damage produced by exercise. Biochemical and biophysical research communications, 107: 1198-120, 1982

Literaturverzeichnis
96
De Bruijne AW, Vreeburg H, Van Steveninck J. Kinetic analysis of L-lactate transport in human erythrocytes via the monocarboxylate-specific carrier system. Biochim Biophys Acta. Aug 10; 732(3): 562-8, 1983
Deuticke B., Beyer E. and Forst B. Discrimination of three parallel pathways of lactate transport in the human erythrocyte membrane by inhibitors and kinetic properties. Biochem. Biophys. 684: 96–110, 1982a
Deuticke B. Monocarboxylate transport in erythrocytes. J Membr Biol 70: 89–103, 1982b
Deuticke B. Monocarboxylate transport in red blood cells: kinetics and chemical modification. Methods Enzymol.; 173: 300-29, 1989
Deutsche Liga zur Bekämpfung des hohen Blutdruckes e.V. Empfehlungen zur Blutdruckmessung. Heidelberg, 4. Aufl.: 1997b
DiGirolamo M, Newby FD, Lovejoy J. Lactate production in adipose tissue: a regulated function with extra-adipose implications. FASEB J. Apr; 6(7); Review: 2405-12, 1992
Dipietro L, Dziura L, Yeckel CW, Darrel Neufer P. Exercise and improved insulin sensitivity in older women: evidence of the enduring benefits of higher intensity training. J Appl Physiol 100: 142–149, 2006
Dimmer KS, Friedrich B, Lang F, Deitmer JW, Broer S. The low affinity monocarboxylate transporter MCT4 is adapted to the export of lactate in highly glycolytic cells. Biochem J 350: 219–227, 2000
Donovan C, Brooks GA. Endurance training affects lactate clearance, not lactate production. Am J Physiol 244: 83–92, 1983
Donovan CM, Pagliassotti MJ. Enhanced efficiency of lactate removal after endurance training. J Appl Physiol.Mar; 68(3): 1053-8, 1990
Dubouchaud, H., P. Granier, J. Mercier, C. Le Peuch, C. Préfaut. Lactate uptake by skeletal muscle sarcolemmal vesicles decreases after 4 wk of hindlimb unweighting in rats. J. Appl. Physiol. 80: 416-421, 1996
Dubouchaud H, Butterfield GE, Wolfel EE, Bergman BC, Brooks GA. Endurance training, expression, and physiology of LDH, MCT1, and MCT4 in human skeletal muscle. Am J Physiol Endocrinol Metab 278: 571–579, 2000
Dunstan DW, Mori TA, Puddey IB, Beilin LJ, Burke V, Morton AR, Stanton KG. The independent and combined effects of aerobic exercise and dietary fish intake on serum lipids and glycemic control in NIDDM: a randomized controlled study.Diabetes Care 20: 913–921, 1997
Dunstan DW, Puddey IB, Beilin LJ, Burke V, Morton AR, Stanton KG. Effects of a short-term circuit weight training program on glycaemic control in NIDDM. Diabetes Res Clin Pract; 40: 53-61, 1998

Literaturverzeichnis
97
Dunstan DW, Daly RM, Owen N, Jolley D, De Courten M, Shaw J, Zimmet P. High-intensity resistance training improves glycemic control in older patients with type 2 diabetes. Diabetes Care 25: 729-1736, 2002
Dunstan DW, Daly RM, Owen N, Jolley D, Vulikh E, Shaw J, Zimmet P. Home-based resistance training is not sufficient to maintain improved glycemic control following supervised training in older individuals with type 2 diabetes. Diabetes Care 28: 3–9, 2005 Enerson BE, Drewes LR. Molecular features, regulation, and function of monocarboxylate transporters: implications for drug delivery. J Pharm Sci. Aug; 92(8): 1531-44, 2003
Enoki T, Yoshida Y, Hatta H, and Bonen A. Exercise training alleviates MCT1 and MCT4 reductions in heart and skeletal muscles of STZ-induced diabetic rats. J Appl Physiol 94: 2433-2438, 2003
Eriksson K, Lindgrade F. Prevention of type 2 (non-insulin-dependent) diabetes mellitus by diet and physical exercises. The 6-year Malmö feasibility study. Diabetologia; 34: 891-898, 1991
Eriksson J, Koranyi L, Bourey R, Schalin-Jäntti C, Widén E, Mueckler M, Permutt AM, Groop LC. Insulin resistance in type 2 (non-insulin-dependent) diabetic patients and their relatives is not associated with a defect in the expression of the insulin-responsive glucose transporter (GLUT-4) gene in human skeletal muscle. Diabetologia. Feb; 35(2): 143-7. Erratum in: Diabetologia Jun;35(6): 594, 1992
Eriksson J, Taimela S, Eriksson K, Parviainen S, Peltonen J, Kujala U. Resistance training in treatment of non-insulindependent diabetes mellitus. Int J Sports Med 18: 242–246, 1997
Evans WJ, Phinney SD, Young VR. Suction applied to a muscle biopsy maximizes sample size. Med Sci Sports Exerc 14: 101-102, 1982
Eversten F, Medbo JI, Bonen A. Effect of training intensity on muscle lactate transporters and lactate threshold of cross-country skiers. Acta Physiol Scand 173: 195–205, 2001
Eydoux N, Dubouchaud H, Py G, Granier P, Prefaut C, Mercier J. Lactate transport in rat sarcolemmal vesicles after a single bout of submaximal exercise. Int J Sports Med 21: 393–399, 2000
Fattor JA, Miller BF, Jacobs KA, Brooks GA. Catecholamine response is attenuated during moderate-intensity exercise in response to the `lactate clamp'. Am J Physiol. 288: E143 -E147, 2005
Fenicchia LM, Kanaley JA, Azevedo JL, et al.. Influence of resistance exercise training on glucose control in women with type 2 diabetes, Metabolism 53 (3): 284–289, 2004

Literaturverzeichnis
98
Fletcher GF, Balady GJ, Amsterdam EA, et al.. Exercise standardsfor testing and training: a statement for healthcare professionals from the American Heart Association. Circulation; 104(14): 1694–740, 2001
Fossum S, Mallett S, Barclay AN. The MRC OX-47 antigen is a member of the immunoglobulin superfamily with an unusual transmembrane sequence. Eur J Immunol; 21(3): 671-679, 1991
Gallagher SM, Castorino JJ, Wang D, Philp NJ. Monocarboxylate transporter 4 regulates maturation and trafficking of CD147 to the plasma membrane in the metastatic breast cancer cell line MDA-MB-231. Cancer Res.; 67: 4182–4189, 2007
Garcia CK, Goldstein JL, Pathak RK, Anderson RG, Brown MS. Molecular characterization of a membrane transporter for lactate, pyruvate and other monocarboxylates: implications for the Cori cycle. Cell 76: 865–873, 1994a
Garcia CK, Li X, Luna J, Francke U. cDNA cloning of the human monocarboxylate transporter 1 andchromosomal localization of the SLC16A1 locus to 1p13.2-p12. Genomics 23: 500-503, 1994b
Geiss LS, Herman WH, Smith PJ. Mortality in non-insulin-dependent diabetes. In: National Diabetes Data Group, ed. Diabetes inAmerica. Bethesda, Md: US Department of Health and Human Services, Public HealthService, National Institutes of Health: 233–257, 1995
Gentil P, Oliveira E, Bottaro M. Time under tension and blood lactate response during four different resistance training methods. J Physiol Anthropol. Sep; 25(5): 339-44, 2006
Gladden LB. Muscle as a consumer of lactate. Med Sci Sports Exerc. Review Apr; 32(4): 764-71, 2000
Gladden B. Lactic acid: new roles in a new millennium. Proc Natl Acad Sci USA 98: 395–397, 2001
Goodyear, LJ, Kahn BB. Exercise, glucose transport, and insulin sensitivity. Annu Rev Med 49: 235-261, 1998
Green, H. Goldberg, B. Collagen and cell protein synthesis by established mammalian fibroblast line. Nature 204: 347-349, 1964
Green H, Halestrap A, Mockett C, O'Toole D, Grant S, and Ouyang J. Increases in muscle MCT are associated with reductions in muscle lactate after a single exercise session in humans. Am J Physiol Endocrinol Metab 282: E154–E160, 2002
Grundy SM, Cleeman JI, Merz CN, Brewer HB Jr, Clark LT, Hunninghake DB, Pasternak RC, Smith SC Jr, Stone NJ; National Heart, Lung, and Blood Institute; American College of Cardiology Foundation; American Heart Association. Implications of Recent Clinical Trials for theNational Cholesterol Education Program Adult Treatment Panel III Guidelines Circulation. Aug 10; 110(6): 763, 2004

Literaturverzeichnis
99
Hagström E., Arner P., Ungerstedt U., Bolinder J. Subcutaneous adipose tissue: a source of lactate production after glucose in humans. Am. J. Physiol. 258: E888-E89, 1990
Hajduch E, Heyes RR, Watt PW, and Hundal HS. Lactate transport in rat adipocytes: identification of monocarboxylate transporter 1 (MCT1) and its modulation during streptozotocin induced-diabetes. FEBS Lett 479: 89-92, 2000
Halestrap AP, Wang X, Poole RC, Jackson VN, Price NT. Lactate transport in heart in relation to myocardial ischemia. Am J Cardiol. Aug 4; 80(3A): 17A-25A, 1997
Halestrap AP, Price NT. The proton-linked monocarboxylate transporter (MCT) family: structure, function, and regulation, Biochemical Journal 343: 281–299, 1999
Halestrap AP, Meredith D. The SLC16 gene family-from monocarboxylate transporters (MCTs) to aromatic amino acid transporters and beyond, European Journal of Physiology 447: 619–628, 2004
Halle M, Kemmer F-W, Stumvoll M, Thurm U, Zimmer P. Körperliche Aktivität und Diabetes mellitus. Scherbaum WA, Haak T (Hrsg.): Evidenzbasierte Leitlinie der Deutschen Diabetes-Gesellschaft (DDG), 2008
Hanefeld M, Temelkova-Kurktschiev T. The postprandial state and the risk of atherosclerosis. Diabetic Med; 14: 6–11, 1997
Harris RT, Dudley GA. Exercise alters the distribution of ammonia and lactate in blood. J Appl. Physiol 66: 313-317, 1989
Hashimoto T, Hussien R, Oommen S, Gohil K, Brooks GA. Lactate sensitive transcription factor network in L6 cells: activation of MCT1 and mitochondrial biogenesis. FASEB J. Aug; 21(10): 2602-12, 2007
Hauner H, Köster I, Schubert I. Trends in der Prävalenz und ambulanten Versorgung von Menschen mit Diabetes mellitus. Eine Analyse der Versichertenstichprobe AOK Hessen/KV Hessen im Zeitraum von 1998 bis 2004. Deutsches Ärzteblatt 41: A2799-2805, 2007
Hauner et al.. Evidenzbasierte Leitlinen. Prävention und Therapie der Adipositas. Hrsg. Deutsche Adipositas-Gesellschaft, Deutsche Diabetes-Gesellschaft, DeutscheGesellschaft für Ernährung, Deutsche Gesellschaft für Ernährungsmedizin: S.6, 2007
Hayashi T, Wojtaszewski JF, Goodyear LJ. Exercise regulation of glucose transport in skeletal muscle. Am J Physiol Endocrinol Metab 273: E1039–E1051, 1997
Hediger MA, Romero MF, Peng JB, Rolfs A, Takanaga H, Bruford EA. The ABCs of solute carriers: physiological, pathological and therapeutic implications of human membrane transport proteinsIntroduction. Pflugers Arch. Review Feb; 447(5): 465-8, 2004

Literaturverzeichnis
100
Hense HW, Schulte H, Löwel H, Assmann G, Keil U. Framingham risk function overestimates risk of coronary heart disease in men and women from Germany--results from the MONICA Augsburg and the PROCAM cohorts. Eur Heart J.May; 24(10): 937-45, 2003
Hildebrand A, Lormes W, Emmert J, Liu Y, Lehmann M, Steinacker JM. Lactate concentration in plasma and red blood cells during incremental exercise. Int J Sports med 21: 463-468, 2000
Hildebrandt AL, Pilegaard H, Neufer PD. Differential transcriptional activation of select metabolic genes in response to variations in exercise intensity and duration. Am J Physiol Endocrinol Metab 285: E1021–E1027, 2003
Hirshman MF, Wallberg-Henriksson LJ, Wardzala FR, Horton ED, Horton ES. Acute exercise increases the number of plasma membrane glucose transporters in rat skeletal muscle. FEBS Lett. 238: 235–239, 1988
Holman RR, Paul SK, Bethel MA, Matthews DR, Neil HA.10-year follow-up of intensive glucose control in type 2 diabetes. N Engl J Med. Oct 9; 359(15): 1577-89, 2008
Holten MK, Zacho M, Gaster M, Juel C, Wojtaszewski JFP, Dela F. Strength training increases insulin-mediated glucose uptake, GLUT4 content and insulin signaling in skeletal muscle in patients with Type 2 diabetes. Diabetes 3: 394–305, 2004
Honkola A, Forsen T, Eriksson J. Resistance training improves the metabolic profile in individuals with type 2 diabetes. Acta Diabetol 34: 245–248, 1997
Horn F, et al.. Biochemie des Menschen. 3. grundlegend überarbeitete und erweiterte Auflage, Georg Thieme Verlag: S.85-100, 2005
Hsu SM, Raine L, Fanger H. Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques: a comparison between ABC and unlabeled antibody (PAP) procedures. J. Histochem. Cytochem. 29: 577-580, 1981 Hu FB, Stampfer MJ, Haffner SM, Solomon CG, Willett WC, Manson JE. Elevated risk of cardiovascular disease prior to clinical diagnosis of type 2diabetes. Diabetes Care 25: 1129-1134, 2002 Ibanez J, Izquierdo M, Arguelles I, Forga L, Larrion JL, Garcia- Unciti M, Idoate F, Gorostiaga EM. Twice weekly progressive resistance training decreases abdominal fat and improves insulin sensitivity in older man with type 2 diabetes. Diabetes Care 28: 662–667, 2005
Immke DC, McCleskey EW. Lactate enhances the acid-sensing Na+ channel on ischemia-sensing neurons. Nat. Neurosci. 4: 869-870, 2001
Ishii T, Yamakita T, Sato T, Tanaka S, Fujii S. Resistance training improves insulin sensitivity in NIDDM subjects without altering maximal oxygen uptake. Diabetes Care 21: 1353–1355, 1998

Literaturverzeichnis
101
Jackson VN, Price NT, Halestrap AP. Cloning of the monocarboxylate transporter isoform MCT2 from rat testis provides evidence that the expression is species specific and may involve posttranscriptional regulation. Biochem J 324: 447-453, 1997
Jessen N, Goodyear LJ. Contraction signaling to glucose transport in skeletal muscle. J Appl Physiol. Review Jul; 99(1): 330-7, 2005
Jiang JL, Tang J. CD147 and its interacting proteins in cellular functions. Sheng Li Xue Bao. Aug 25; 59(4): 517-23, 2007
Juel C. Lactate-proton cotransport in skeletal muscle. Physiol Rev 77: 321–358, 1997
Juel C. Skeletal muscle Na+/H+ exchange in rats: pH dependency and the effect of training. Acta Physiol Scand 164: 135–140, 1998a
Juel C. Muscle pH regulaion: role of training. Acta physiologica Scandinavica, 162: 359-366, 1998b
Juel C, Halestrap A. Lactate transport in skeletal muscle-role and regulation of the monocarboxylate transporter. J Physiol 517: 633–642, 1999
Juel C, Lundby C, Sander M, Calbet JA, and Hall G. Human skeletal muscle and erythrocyte proteins involved in acid-base homeostasis: adaptations to chronic hypoxia. J Physiol 548: 639–648, 2003
Juel C, Klarskov C, Nielsen JJ, Krustrup P, Mohr M, Bangsbo J. Effect of high-intensity intermittent training on lactate and H+ release from human skeletal muscle. Am J Physiol Endocrinol Metab 286: E245-51, 2004a
Juel C, Holten MK, Dela F. Effects of strength training on muscle lactate release and MCT1 and MCT4 content in healthy and type 2 diabetic humans. J Physiol 556: 297–304, 2004b
C. Juel. Laktattransport im Skelettmuskel: Trainingsinduzierte Anpassung und Bedeutung bei körperlicher Belastung, Deutsche Zeitschrift für Sportmedizin, Jahrgang 55, Nr. 6: S. 157-160, 2004c
Juel C. Training-induced changes in membrane transport proteins of human skeletal muscle. Eur J Appl Physiol. Apr; 96(6): 627-35, 2006
Kadoglou NP, Iliadis F, Angelopoulou N, Perrea D, Ampatzidis G, Liapis CD, Alevizos M. The anti-inflammatory effects of exercise training in patients with type 2 diabetes mellitus. Eur J Cardiovasc Prev Rehabil.; 14: 837– 843, 2007
Kelley GA, Kelley KS. Effects of aerobic exercise on lipids and lipoproteins in adults with type 2 diabetes a metaanalysis of randomized-controlled trials. Public Health 121(9): 643-655, 2007

Literaturverzeichnis
102
Kelly GA, Kelly KS. Progressive resistance exercise and resting blood pressure: a meta-analysis of randomized controlled trials. Hypertension 35: 838 –843, 2000
Kennedy JW, Hirshman MF, Gervino EV, Ocel JV, Forse RA, Hoenig SJ, Aronson D, Goodyear LJ, Horton ES. Acute exercise induces GLUT4 translocation in skeletal muscle of normal human subjects and subjects with Type 2 diabetes. Diabetes 48: 1192–1197, 1999
Kerner W, Brückel J, Böhm BO. Definition, Klassifikation und Diagnostik des Diabetes mellitus. Scherbaum WA, Kiess W (Hrsg.): Evidenzbasierte Leitlinie der Deutschen Diabetes-Gesellschaft (DDG), 2004
Kirk P, Wilson MC, Heddle C, Brown MH, Barclay AN, Halestrap AP. CD147 is tightly associated with lactate transporters MCT1 and MCT4 and facilitates their cell surface expression. EMBO J19: 3896–3904, 2000
Knowler W, Barrett-Connor E, Fowler SE, Hamman RF, Lachin JM. Reduction in the incidence of type 2 with lifestyle intervention or metformin. N Engl J Med; 346: 393-403, 2002
Kobayashi M. Fiber type-specific localization of monocarboxylate transporters MCT1 and MCT4 in rat skeletal muscle. Kurume Med J. 51(3-4): 253-61, 2004
Koho NM, Väihkönen LK, Pösö AR. Lactate transport in red blood cells by monocarboxylate transporters. Equine Vet J Suppl. Sep; (34): 555-9, 2002
Koho NM, Hyyppä S, Pösö AR. Monocarboxylate transporters (MCT) as lactate carriers in equine muscle and red blood cells. Equine Vet J Suppl. Aug; (36): 354-8, 2006
König D, Deibert P, Dickhuth H-H, Berg A. Bewegungstherapie bei Diabetes mellitus Typ II – metabolische Grundlagen und evidenzbasierte Empfehlungen. Deutsche Zeitschrift für Sportmedizin, Jahrgang 57, J 10: 242-247, 2006
Kraemer WJ, Fry AC. Strength testing: development and evaluation of methodology. In: Maud P.J. and Foster C., editors. Physiological Testing of Human Fitness, Champaign IL, Human Kinetics: 1991
Kristiansen S, Hargreaves M, Richter EA. Exercise induced increase in glucose transport, GLUT-4, and VAMP-2 in plasma membrane from human muscle. Am J Physiol.270 (Endocrinol. Metab. 33): E197–E201, 1996
Krook A, Björnholm M, Galuska D, Jiang XJ, Fahlman R, Myers MG, Jr, Wallberg-Henriksson H, Zierath JR. Characterization of signal transduction and glucose transport in skeletal muscle from type 2 diabetic patients. Diabetes 49: 284-292, 2000
Krook A, Holm I, Pettersson S, Wallberg-Henriksson H. Reduction of risk factors following lifestyle modification programme in subjects with type 2 (non-insulin dependent) diabetes mellitus. Clinical physiology and functional imaging; 23(1): 21-30, 2003

Literaturverzeichnis
103
Laaser U, Wolters P. Public health sciences graduate study at the Bielefeld University in the framework of comparative endeavours. Soz Praventivmed 34: 223-6, 1989
Lehmann R, Vokac A, Niedermann K, Agosti K, Spinas GA. Loss of abdominal fat and improvement of the cardiovascular risk profile by regular moderate exercise training in patients with NIDDM. Diabetologia 38: 1313-1319, 1995
Ligtenberg PC, Hoekstra JB, Bol E, Zonderland ML, Erkelens DW. Effects of physical training on metabolic control in elderly type 2 diabetes mellitus patients. Clin Sci 93: 127-135, 1997
Lindstrom J, Ilanne-Parikka P, Peltonen M, Aunola S, Eriksson JG, Hemio K, Hamalainen H, Harkonen P, Keinanen-Kiukaanniemi S, Laakso M, Louheranta A, Mannelin M, Paturi M, Sundvall J, Valle TT, Uusitupa M, Tuomilehto J. Sustainedreduction in the incidence of type 2 diabetes by lifestyle intervention: follow-up of theFinnish Diabetes Prevention Study. Lancet 368: 1673-1679, 2006
Löllgen H. Das Anstrengungsempfinden (RPE, Borg-Skala). Dtsch Z Sportmed 55: 299-300, 2004
Lombardi AM, Fabris R, Bassetto F, Serra R, Leturque A, Federspil G, Girad J, Vettor R. Hyperlactemia reduces muscle glucose uptake and GLUT-4 mRNA while increasing PDH gene expression in rat. Am J Physiol Endocrinol Metab 276: E922–E929, 1999
Lovejoy J, Newby FD, Gebhart SS, DiGirolamo M. Insulin resistance in obesity is associated with elevated basal lactate levels and diminished lactate appearance following intravenous glucose and insulin. Metabolism 41: 22–27, 1992
Lund, S, Holman GD, Schmitz O, Pedersen O. Contraction stimulates translocation of glucose transporter GLUT4 in skeletal muscle through a mechanism distinct from that of insulin. Proc Natl Acad Sci USA 92: 5817-5821, 1995
Luttmann W, Bratke K, Küpper M, Myrtek D. Der Experimentator. Immunologie. München: Spektrum akademischer Verlag, 1. Aufl.: 92-110, 2004
Maiorana A, O’Driscoll G, Cheetham C, Dembo L, Stanton K, Goodman C, Taylor R, Green D. The effect of combined aerobic and resistance exercise training on vascular function in type 2 diabetes. J Am Coll Cardiol 38: 860–866, 2001
Maiorana A, O’Driscoll G, Goodman C, Taylor R, Green D. Combined aerobic and resistance exercise improves glycemic control and fitness in type 2 diabetes. Diabetes Res Clin Pract 56: 115-123, 2002
Manning Fox JE, Meredith D, Halestrap AP. Characterisation of human monocarboxylate transporter 4 substantiates its role in lactic acid efflux from skeletal muscle. J Physiol 529: 285–293, 2000

Literaturverzeichnis
104
Manson JE, Colditz GA, Stampfer MJ, Willett WC, Krolewski AS, Rosner B, Arky RA, Speizer FE, Hennekens CH. A prospective study of maturity-onset diabetes mellitus and risk of coronary heart disease and stroke in women. Arch Intern Med
Matthews DR, Hosker JP, Rudenski AS, Naylor BA, Treacher DF, Turner RC.
. 151(1991): 1141-1147, 1991
Marette A, Richardson JM, Ramlal T, Balon TW, Vranic M, Pessin JE, and Klip A. Abundance, localization, and insulin induced translocation of glucose transporters in red and white muscle. Am J Physiol Cell Physiol 263: C443–C452, 1992
Matthaei, S. et al.. Behandlung des Diabetes mellitus Typ 2. Diabetologie; 3 Suppl 2: S157-S161, 2008
Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia. Jul; 28(7): 412-9, 1985
McCullagh KJA, Bonen A. Reduced lactate transport in denervated rat skeletal muscle. Am. J. Physiol. 269 (Regulatory Integrative Comp. Physiol. 38): R884-R888, 1995
McCullagh KJA, Poole RC, Halestrap AP, O'Brien M, Bonen A. Role of the lactate transporter (MCT1) in skeletal muscles. Am J Physiol Endocrinol Metab 271: E143-E150, 1996
McCullagh KJA, Poole RC, Halestrap AP, Tipton KF, O'Brien M, Bonen A. Chronic electrical stimulation increases MCT1 and lactate uptake in red and white skeletal muscle. Am J Physiol Endocrinol Metab 273: E239-E246, 1997
McDermott JC, Bonen A. Glyconeogenic and oxidative lactate utilization in skeletal muscle. Can J Physiol Pharmacol 70: 142–149, 1992
McDermott, JC, Bonen A. Endurance training increases skeletal muscle lactate transport. Acta Physiol. Scand. 147: 323-327, 1993a
McDermott JC, Bonen A. Lactate transport by skeletal muscle sarcolemmal vesicles. Mol Cell Biochem. May 26; 122(2): 113-21, 1993b
Merezhinskaya N, Fishbein WN, Davis JI, Foellmer JW. Mutations in MCT1 cDNA in patients with symptomatic deficiency in lactate transport. Muscle Nerve 23: 90–97, 2000
Merezhinskaya N, Fishbein WN. Monocarboxylate transporters: past, present, and future. Histol Histopathol. Feb; 24(2): 243-64, 2009
Metz L, Vermaelen M, Lambert K, Broca C, Sirvent P, Raynaud E, Mercier J. Endurance training increases lactate transport in male Zucker fa/fa rats. Biochem Biophys Res Commun. Jun 17; 331(4): 1338-45, 2005

Literaturverzeichnis
105
Misra A, Alappan NK, Vikram NK, Goel K, Gupta N, Mittal K, et al.. Effect of supervised progressive resistance-exercise training protocol on insulin sensitivity, glycemia, lipids, and body composition in Asian Indians with type 2 diabetes, Diabetes Care 31 (7): 1282–1287, 2008
Mohr M, Krustrup P, Nielsen JJ, Nybo L, Rasmussen MK, Juel C, Bangsbo J. Effect of two different intense training regimens on skeletal muscle ion transport proteins and fatigue development. Am J Physiol Regul Integr Comp Physiol 292: R1594–R1602, 2007
Mondon CE, Jones RI, Azhar S, Hollenbeck CB, Reaven GM. Lactate production and pyruvate dehydrogenase activity in fat skeletal muscle from diabetic rats. Diabetes 41: 1547-1554, 1992
Nakai M, et al.. Tissue Distribution of Basigin and Monocarboxylate Transporter 1 in the Adult Male Mouse: A Study Using the Wild- Type and Basigin Gene Knockout Mice. TheAnatomical Record Part A 288: 527-535, 2006
Oakes ND, Bell KS, Furler SM, Camilleri S, Saha AK, Ruderman NB, Chisholm DJ, Kraegen EW. Diet-induced muscle insulin resistance in rats is ameliorated by acute dietary lipid withdrawal or a single bout of exercise: parallel relationship between insulin stimulation of glucose uptake and suppression of long-chain fatty acyl-CoA. Diabetes 46: 2022-2028, 1997
O’Donovan G, Kearney EM, Nevill AM, Woolf-May K, Bird SR. The effects of 24 weeks of moderate- or high-intensity exercise on insulin resistance. Eur J Appl Physiol 95(5–6): 522–528, 2005
O'Gorman DJ, Karlsson HK, McQuaid S, Yousif O, Rahman Y, Gasparro D, Glund S, Chibalin AV, Zierath JR, Nolan JJ. Exercise training increases insulin-stimulated glucose disposal and GLUT4 (SLC2A4) protein content in patients with type 2 diabetes. Diabetologia. Dec; 49(12): 2983-92, 2006
Ohlson LO, Larsson B, Bjorntorp P, Eriksson H, Svardsudd K, Welin L, Tibblin G, Wilhelmsen L. Risk factors for type 2 (non-insulin-dependent) diabetes mellitus. Thirteen and one-half years of follow-up of the participants in a study of Swedish men born in 1913. Diabetologia 31: 798–805, 1988
Pattillo RE, Gladden LB. Red blood cell lactate transport in sickle disease and sickle cell trait. J Appl Physiol 99: 822–827, 2005
Pessin JE, Thurmond DC, Elmendorf JS, Coker KJ, Okada S. Molecular basis of insulin-stimulated GLUT4 vesicle trafficking. J Biol Chem 274: 2593-2596, 1999
Petrella,RJ, Lattanzio,CN, Demeray,A, Varallo,V, Blore,R. Can adoption of regular exercise later in life prevent metabolic risk for cardiovascular disease? Diabetes Care 28: 694-701, 2005
Philp NJ, et al.. Loss of MCT1, MCT3, and MCT4 Expression in the Retinal Pigment Epithelium and Neural Retina of the 5A11/ Basigin-Null Mouse. Investigative Ophthalmology and Visual Science. 44: 1305-1311, 2003a

Literaturverzeichnis
106
Philp NJ, Wang D, Yoon H, Hjelmeland LM. Polarized expression of monocarboxylate transporters in human retinal pigment epithelium and ARPE-19 cells. Invest Ophthalmol Vis Sci 44: 1716–1721, 2003b
Philp A, Macdonald AL, Watt PW. Lactate--a signal coordinating cell and systemic function. J Exp Biol. Review Dec; 208(Pt 24): 4561-75, 2005
Phillips S, Han X, Green HJ, Bonen A. Increments in skeletal muscle GLUT-1 and GLUT-4 after endurance training in humans. Am. J. Physiol. 270 (Endocrinol. Metab. 33): E456–E462, 1996
Pilegaard H, Juel C, Wibrand F. Lactate transport studied in sarcolemmal giant vesicles from rats: effect of training. Am. J. Physiol. 264 (Endocrinol. Metab. 27): E156-E160, 1993
Pilegaard H, Bangsbo J, Richter EA, Juel C. Lactate transport studied in sarcolemmal giant vesicles from human muscle biopsies: relation to training status. J Appl Physiol. Oct; 77(4): 1858-62, 1994
Pilegaard H, Juel C. Lactate transport studied in sarcolemmal giant vesicles from rat skeletal muscles: effect of denervation. Am. J. Physiol. 269 (Endocrinol. Metab. 32): E679-E682, 1995
Pilegaard H, Mohr T, Kjær M, Juel C. Lactate/H+ transport in skeletal muscle from spinal cord injured patients. Scand J Med Sci Sports 8: 98–101, 1998
Pilegaard H., Terzis G, Halestrap A, Juel C. Distribution of the lactate/H+ transporter isoforms MCT1 and MCT4 in human skeletal muscle. Am. J. Physiol. 276: 843–848, 1999a
Pilegaard H, Domino K, Noland T, Juel C, Hellsten Y, Halestrap AP, Bangsbo J. Effect of high-intensity exercise training on lactate/H+ transport capacity in human skeletal muscle. Am J Physiol Endocrinol Metab 276: E255–E261, 1999b
Pösö AR. Monocarboxylate transporters and lactate metabolism in equine athletes: a review. Acta Vet Scand. 43(2): 63-74, 2002
Poole RC, Halestrap AP. Transport of lactate and other monocarboxylates across mammalian plasma membranes. Am. J. Physiol. 264: C761-C782, 1993
Poole RC, Sansom CE, Halestrap AP. Studies of the membrane topology of the rat erythrocyte H+/lactate cotransporter (MCT1). Biochem J. Dec 15; 320 (Pt 3): 817-24, 1996
Py G, Lambert K, Perez-Martin A, Raynaud E, Prefaut C, Mercier J. Impaired sarcolemmal vesicle lactate uptake and skeletal muscle MCT1 and MCT4 expression in obese Zucker rats. Am J Physiol Endocrinol Metab 281: E1308-E1315, 2001

Literaturverzeichnis
107
Py G, Lambert K, Milhavet O, Eydoux N, Prefaut C, Mercier J. Effects of streptozotocin-induced diabetes on markers of skeletal muscle metabolism and monocarboxylate transporter 1 to monocarboxylate transporter 4 transporters. Metabolism 51: 807–813, 2002
Ramachandran A, Snehalatha C, Mary S, Mukesh B, Bhaskar AD, Vijay V, the Indian Diabetes Prevention Programme (IDPP). The Indian Diabetes Prevention Programme shows that lifestyle modification and metformin prevent type 2 diabetes in Asian Indian subjects with impaired glucose tolerance (IDPP-1). Diabetologia 49: 289–297, 2006
Räsänen LA, Lampinen KJ, Pösö AR. Responses of blood and plasma lactate and plasma purine concentrations to maximal exercise and their relation to performance in Standardbredtrotters. Am J Vet Res 56: 1651–1656, 1995
Randle PJ, Garland PB, Hales CN, Newsholme EA. The glucose fatty acid cycle: its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet 1: 785-789, 1963
Rathmann W, Haastert B, Icks A, Löwel H, Meisinger C, Holle R, Giani G for Diabetologia. High prevalence of undiagnosed diabetes mellitus in Southern Germany: Target populations for efficient screening. The KORA survey 2000 Diabetologia 46: 182-189, 2003
Reaven GM, Hollenbeck C, Jeng CY, Wu MS, Chen YD. Measurements of plasma glucose, free fatty acid, lactate and insulin for 24h in patients with NIDDM. Diabetes; 37: 1020-4, 1988
Ren, JM, Semenkovich CF, Gulve EA, Gao J, Holloszy JO. Exercise induces rapid increases in GLUT4 expression, glucose transport capacity, and insulin-stimulated glycogen storage in muscle. J Biol Chem 269: 14396-14401, 1994
Richter, EA, Jensen P, Kiens B, Kristiansen S. Sarcolemmal glucose transport and GLUT-4 translocation during exercise are diminished by endurance training. Am J Physiol Endocrinol Metab 274: E89-E95, 1998
Robergs R.A., Amann M. Belastungsbedingte metabolische Azidose: Woher kommen die Protonen? Öster J Sportmed 3: 11–25, 2003
Rost R. Diagnostische Methoden. In Rost R (Ed.), Sport- und Bewegungstherapie bei inneren Krankheiten, Deutscher Ärzte-Verlag GmbH, Köln: 276-277, 1995
Roth DA, Brooks GA. Lactate and pyruvate transport is dominated by a pH gradient-sensitive carrier in rat skeletal muscle sarcolemmal vesicles. Arch. Biochem. Biophys. 279: 386–394, 1990
Sachs AB, Sarnow P, Hentze MW. Starting at the beginning, middle and end: translation initiation in eukaryotes. Cell 89: 831-838, 1997
Sachs L, Hedderich J. Angewandte Statistik: Methodensammlung mit R. 13., aktualisierte und erw. Aufl. Berlin: Springer, 2009

Literaturverzeichnis
108
Sack U, Tarnok A, Rothe G. (Hrsg.). Zelluläre Diagnostik: Grundlagen, Methoden und klinische Anwendungen der Durchflusszytometrie. Basel, Karger: 27-70, 2007
Saksena SA, Dwivedi et al.. "PKC-dependent Stimulation of the Human MCT1 Promoter Involves Transcription Factor AP2." Am J Physiol Gastrointest Liver Physiol., 2008
Saier MH. Computer-aided analyses of transport protein sequences - gleaning evidence concerning function, structure, biogenesis, and evolution. Microbiological Reviews; 58: 71–93, 1994
Sara F, Connes P, Hue O, Montout-Hedreville M, Etienne-Julan M, Hardy-Dessources MD. Faster lactate transport across red blood cell membrane in sickle cell trait carriers. J Appl Physiol. Feb; 100(2): 427-32, 2006
Schlumberger A, Schmidtbleicher D. Grundlagen der Kraftdiagnostik. In: Banzer, W, Pfeifer K, Vogt L. Funktionsdiagnostik des Bewegungssystems in der Sportmedizin. Berlin: Springer: S. 87-106, 2004
Sigal RJ, Kenny GP, Wasserman DH, Castaneda-Sceppa C. Physical activity/exercise and type 2 diabetes. Diab Care 27: 2518-2539, 2004
Sigal RJ, Kenny GP, Wasserman DH, Castaneda-Sceppa C, White RD. Physical activity/exercise and type 2 diabetes: a consensus statement from the American Diabetes Association. Diabetes Care. Jun; 29(6): 1433-8, 2006
Sigal RJ, Kenny GP, Boulé NG, Wells GA, Prud'homme D, Fortier M, Reid RD, Tulloch H, Coyle D, Phillips P, Jennings A, Jaffey J. Effects of aerobic training, resistance training, or both on glycemic control in type 2 diabetes: a randomized trial. Ann Intern Med. Sep 18; 147(6): 357-69, 2007
Skelton MS, Kremer DE, Smith EW, Gladden LB. Lactate influx into red blood cells of athletic and nonathletic species. Am J Physiol Regul Integr Comp Physiol 268: R1121–R1128, 1995
Skelton MS, Kremer DE, Smith EW, Gladden LB. Lactate influx into red blood cells from trained and untrained human subjects. Med Sci Sports Exerc 30: 536–542, 1998
Smith EW, Skelton MS, Kremer DE, Pascoe DD, Gladden LB. Lactate distribution in the blood during progressive exercise. Med Sci Sports Exercise 29: 654-660, 1997
Smutok MA, Reece C, Kokkinos PF, Farmer CM, Dawson PK, DeVane J, Patterson J, Goldberg AP, Hurley BF. Effects of exercise training modality on glucose tolerance in men with abnormal glucose regulation. Int J Sports Med. Aug; 15(6): 283-9, 1994
Snowling NJ, Hopkins WG. Effects of different modes of exercise training on glucose control and risk factors for complications in type 2 diabetic patients: a meta-analysis. Diabetes Care. Nov; 29(11): 2518-27, 2006

Literaturverzeichnis
109
Steinacker JM, Liu Y, Reißnecker S. Abbruchkriterien bei der Ergometrie. Dtsch Z Sportmed 53: 228-229, 2002
Tabata I, Suzuki Y, Fukunaga T, Yokozeki T, Akima H, Funato K. Resistance training affects GLUT-4 content in skeletal muscle of humans after 19 days of head-down bed rest. J Appl Physiol 86 : 909-914, 1999
Thomas D, Elliott E, Naughton G. Exercise for type 2 diabetes mellitus. In: The cochrane library, Issue 3. Chichester, UK: Wiley: 2006
Thomas C, Bishop D, Moore-Morris T, Mercier J. Effects of high-intensity training on MCT1, MCT4, and NBC expressions in rat skeletal muscles: influence of chronic metabolic alkalosis. Am J Physiol Endocrinol Metab 293: E916–E922, 2007
Tonouchi M, Hatta H, Bonen A. Muscle contraction increases lactate transport while reducing sarcolemmal MCT4, but not MCT1. Am J Physiol Endocrinol Metab 282: E1062-E1069, 2002
Trabold O, Wagner S, Wicke C, Scheuenstuhl H, Hussain Z, Rosen N, Seremetiev A, Becker HD, Hunt TK. Lactate and oxygen constitute a fundamental regulatory mechanism in wound healing. Wound Rep. Reg. 11: 504-509, 2003
Trappe et al. 2000. Leitlinien zur Ergometrie. Zeitschrift für Kardiologie, Band 89. Z Kardiol 89: 821-837, 2000
Tuomilehto J, Lindström J, Eriksson JG, Valle TT, Hämäläinen H, Ilanne-Parikka P, Keinänen-Kiukaanniemi S, Laakso M, Louheranta A, Rastas M, Salminen V, Uusitupa M. Finnish Diabetes Prevention Study Group. Prevention of type 2 diabetes mellitus by changes in lifestyle among subjects with impaired glucose tolerance. N Engl J Med. May 3; 344(18): 1343-50, 2001
Ullah MS, Davies AJ, Halestrap AP. The plasma membrane lactate transporter MCT4, but not MCT1, is up-regulated by hypoxia through a HIF-1alpha-dependent mechanism. J Biol Chem 281: 9030-9037, 2006
Väihkönen LK, Pösö AR. Interindividual variation in total and carrier-mediated lactate influx into red blood cells. Am J Physiol Regul Integr Comp Physiol 274: R1025–R1030, 1998
Väihkönen LK, Hyyppä S, Pösö AR. Factors affecting accumulation of lactate in red blood cells. Equine Vet J Suppl 30: 443–447, 1999
Väihkönen LK, Heinonen OJ, Hyyppä S, Nieminen M, Pösö AR. Lactate-transport activity in RBCs of trained and untrained individuals from four racing species. Am J Physiol Regul Integr Comp Physiol 281: R19–R24, 2001
Väihkönen LK, Ojala M, Pösö AR. Age-related changes and inheritance of lactate transport activity in red blood cells. Equine Vet J Suppl. Sep;(34): 568-72, 2002

Literaturverzeichnis
110
Vanninen E, Uusitupa M, Siitonen O, Laitinen J, Lansimies E. Habitual physical activity, aerobic capacity and metabolic control in patients with newly-diagnosed type 2 (non-insulin-dependent) diabetes mellitus: effect of 1-year diet and exercise intervention. Diabetologia 35: 340-346, 1992
Vettor R, Lombardi AM, Fabris R, Pagano C, Cusin I, Rohner-Jeanrenaud F, Federspil G, Jeanrenaud B. Lactate infusion in anesthetized rats produces insulin resistance in heart and skeletal muscles. Metabolism; 46(6): 684-90, 1997
Wang Y, Tonouchi M, Miskovic D, Hatta H, Bonen A. T3 increases lactate transport and the expression of MCT4, but not MCT1, in rat skeletal muscle. Am J Physiol Endocrinol Metab 285: E622–E628, 2003
Waniewski RA, Martin DL. Astrocytes and synaptosomes transport and metabolize lactate and acetate differently. Neurochem. Res. 29, 209-217, 2004
Wild S, Roglic G, Green A, Sicree R, King H. Global Prevalence of Diabetes: Estimates for the year 2000 and projections for 2030. Diabetes Care; 27: 1047-1053, 2004
Wilson PWF, Kannel WB. Epidemiologyof hyperglycemia and atherosclerosis. In: Ruderman N, Williamson J, Brownlee M, eds.Hyperglycemia, diabetes, and vascular disease.New York: Oxford University Press: 21-9, 1992
Wilson MC, Jackson VN, Heddle C, Price NT, Pilegaard H, Juel C, Bonen A, Montgomery I, Hutter OF, Halestrap AP. Lactic acid efflux from white skeletal muscle is catalyzed by the monocarboxylate transporter isoform MCT3. J. Biol. Chem. 273: 15920-15926, 1998
Wilson MC, Meredith D, Halestrap AP. Fluorescence resonance energy transfer studies on the interaction between the lactate transporter MCT1 and CD147 provide information on the topology and stoichiometry of the complex in situ. J Biol Chem; 277: 3666–72, 2002
Wilson MC, Meredith D, Manning Fox JE, Manoharan C, Davies AJ, Halestrap AP. Basigin (CD147) is the target for organomercurial inhibition of monocarboxylate transporter isoforms 1 and 4: the ancillary protein for the insensitive MCT2 is embigin (gp70). J Biol Chem 280: 27213–27221, 2005
Wing RR, Epstein LH, Paternostro-Bayles M, Kriska A, Nowalk MP, Gooding W. Exercise in a behavioural weight control programme for obese patients with type 2 (non-insulin-dependent) diabetes. Diabetologia 31: 902-909, 1988
Yaspelkis BB. Resistance training improves insulin signaling and action in skeletal muscle. Exerc Sport Sci Rev. Jan; 34(1): 42-6, 2006
Yoshida Y, Hatta H, et al.."Relationship between skeletal muscle MCT1 and accumulated exercise during voluntary wheel running." J Appl Physiol 97(2): 527-34, 2004

Literaturverzeichnis
111
Yu M, Blomstrand E, Chibalin AV, Wallberg-Henriksson H, Zierath JR, Krook A. Exercise-associated differences in an array of proteins involved in signal transduction and glucose transport. J Appl Physiol 90: 29-34, 2001
Zanuso S, Jimenez A, Pugliese G, Corigliano G, Balducci S. Exercise for the management of type 2 diabetes: a review of the evidence. Acta Diabetol. Mar; 47(1): 15-22, 2010
Zhang F, Vannucci SJ, et al.. "Monocarboxylate transporter expression in the spontaneous hypertensive rat: effect of stroke." J Neurosci Res 79(1-2): 139-45, 2005
Zhou M, Lin B-Z, Coughlin S, Vallega G, Pilch PF. UCP-3 expression in skeletal muscle: effects of exercise, hypoxia and AMP-activated protein kinase. Am J Physiol Endocrinol Metab 279: E622–E629, 2000
Zhou S, Zhou H, Walian PJ, Jap BK. CD147 is a regulatory subunit of the gamma-secretase complex in Alzheimer’s disease amyloid beta-peptide production. Proc Natl Acad Sci U S A; 102: 7499-7504, 2005
Zierath JR et al.. Exercise training-induced changes in insulin signaling in skeletal muscle. J Appl Physiol. Aug; 93(2): 773-81, 2002
Internetquellen:
Halle M, Kemmer F-W, Stumvoll M, Thurm U, Zimmer P. Körperliche Aktivität und Diabetes mellitus. Oktober 2008. Scherbaum WA, Haak T (Hrsg.): Evidenzbasierte Leitlinie der Deutschen Diabetes-Gesellschaft (DDG). Zugriff am 12.05.2009 unter http://www.deutsche-diabetes gesellschaft.de/leitlinien/EBL Bewegung_2008. pdf
International Diabetes Federation. Zugriff am 21.03.2010 unter http://www.diabetesatlas.org/content/some-285-million-people-worldwide-will-live-diabetes-2010
Kerner W, Brückel J, Böhm BO. Definition, Klassifikation und Diagnostik des Diabetes mellitus. Oktober 2004. Scherbaum WA, Kiess W (Hrsg.): Evidenzbasierte Leitlinie der Deutschen Diabetes-Gesellschaft (DDG). Zugriff am 12.05.2009 unter http://www.deutsche-diabetes-gesellschaft.de/ leitlinien/EBL _Klassifikation_Update 2004. pdf

112
7 Anhang
7.1 Abbildungsverzeichnis Abb. 1: Strukturformel L- Laktat
1
Abb. 2: Reaktionsgleichung für die Bildung von Natriumlaktat
2
Abb. 3: Laktatdehydrogenase-Reaktion (Robergs et al. 2003)
3
Abb. 4: Vereinfachtes Schema zum Laktattransport bei körperlicher Belastung
4
Abb. 5: Aufbau des MCT1-Proteins (modifiziert nach Halestrap et al. 2004)
5
Abb. 6: MCT1-Protein und sein Chaperon CD147
9
Abb. 7: Studienablaufschema der DiabetesAktiv-Studie
19
Abb. 8: Modifizierte Biopsiennadel (Evans et al. 1982)
25
Abb. 9: MCT4-Färbung der Muskelzellen bei 200-facher Vergrößerung
30
Abb. 10: IHC-Kontrolle der Muskelzellen bei 200-facher Vergrößerung
30
Abb. 11: Bildbearbeitung bei der Bestimmung der absoluten und der relativen membranösen MCT1- und MCT4-Dichte
31
Abb. 12: Markierung des MCT1-Proteins in der Zellmembran der Muskelzellen bei einer festgelegten Schwelle von 0-175 DU
32
Abb. 13: Hintergrundflächenmarkierung mit dem Adobe Photoshop Programmes
33
Abb. 14: Gesamtmuskelflächenberechnung bei einer festgelegten Schwelle von 0- 250 DU
33
Abb. 15: Immunhistochemische MCT1-Färbung bei 400-facher Vergrößerung
36

113
Abb. 16: Immunhistochemische IHC-Kontrolle bei 400-facher
Vergrößerung
36
Abb. 17: Modifizierte Borg-Skala für die Beurteilung des subjektiven Belastungsempfindens während des Krafttrainings
46
Abb. 18: Darstellung der Mittelwerte und Standardabweichungen der prozentualen Muskelfasertypenverteilung am M. vastus lateralis der Ausdauergruppe vor und nach der Trainingsintervention, (*p<0.05 versus vor)
55
Abb. 19: Darstellung der Mittelwerte und Standardabweichungen der prozentualen Muskelfasertypenverteilung am M. vastus lateralis der Kraftgruppe vor und nach der Trainingsintervention, (*p<0.05 versus vor)
55
Abb. 21: Darstellung der Mittelwerte und Standardabweichungen der zellulären MCT4-Dichte der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention, (*p<0.05 versus vor)
57
Abb. 22: Darstellung der Mittelwerte und Standardabweichungen der membranösen MCT1-Dichte der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention, (*p<0.05 versus vor)
57
Abb. 23: Darstellung der Mittelwerte und Standardabweichungen der membranösen MCT4-Dichte der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention, (*p<0.05 versus vor)
58
Abb. 24: Darstellung der Mittelwerte und Standardabweichungen der relativen membranösen MCT1-Dichte der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention, (*p<0.05 versus vor)
58
Abb. 25: Darstellung der Mittelwerte und Standardabweichungen der relativen membranösen MCT4-Dichte der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention, (*p<0.05 versus vor)
59
Abb. 26: Darstellung der Mittelwerte und Standardabweichungen der MCT1-Dichte im Erythrozyten der Ausdauergruppe im Basalzustand vor und nach der Trainingsintervention, (*p<0.05 versus vor)
60
Abb. 27: Darstellung der Mittelwerte und Standardabweichungen der MCT1-Dichte im Erythrozyten der Kraftgruppe im Basalzustand vor und nach der Trainingsintervention, (*p<0.05 versus vor)
60

114
Abb. 28: Immunhistochemischer Nachweis einer verstärkten MCT1-Färbung an den Erythrozyten eines Probanden der Kraftgruppe im Basalzustand vor und nach Trainingsintervention bei 400-facher Vergrößerung
61
Abb. 29: Abnahme der MCT1-Dichte im Erythrozyten nach körperlicher Belastung bei der Kraftgruppe (KG) und Ausdauergruppe (AG) vor und nach der Trainingsintervention (*p<0.05 versus vor)
62
Abb. 30: Darstellung der Mittelwerte und Standardabweichungen der CD 147-Dichte im Erythrozyten der Ausdauergruppe (AG) und Kraftgruppe (KG) im Basalzustand vor und nach der Trainingsintervention, (*p<0.05 versus vor)
63
Abb. 31: Darstellung der Mittelwerte und Standardabweichungen der aktivitätsbedingten Veränderungen der CD147-Dichte im Erythrozyten der Ausdauergruppe (AG) und Kraftgruppe (KG) vor und nach der Trainingsintervention unmittelbar nach dem Belastungstest, (*p<0.05 versus vor)
64
Abb. 32: Streulicht-Dot-Plot: Darstellung der gesamten Zellpopulation. Die Zellen werden nach dem Vorwärtsstreulicht (FSC) und Seitswärtsstreulicht (SSC) im Diagramm dargestellt
64
Abb. 33: Histogrammdarstellung der Zellen aus Gate 2. Auf der X-Achse ist die Fluoreszenzintensität und auf der Y-Achse die Events (Zellzahl) aufgetragen. Das Histogramm zeigt die Häufigkeit von den Signalen in den einzelnen Fluoreszenzkanälen an
65
Abb. 34: Fluoreszenzmikroskopische Aufnahmen des immunhisto-chemischen Nachweises der aktivitätsbedingten MCT1- und CD147-Translokation in die Erythrozytenmembran vor und nach der Trainingsintervention
65
Abb. 35: Hypothetisches Modell einer möglichen aktivitätsbedingten Translokation im Erythrozyten des MCT1-und CD147-Proteins bei körperlicher Belastung
82
Abb. 36: Hypothetisches Regulationsmodell zum Laktattransport und der funktionellen Bedeutung der MCT-Proteine im Muskel und Erythrozyten während körperlicher Belastung im trainierten und untrainierten Zustand
90
Abb. 37: Hypothetical regulatory model of lactate transport and the function of the MCT proteins in the muscle and the erythrocytes during physical activity in trained and untrained condition
116

115
7.2 Tabellenverzeichnis Tab. 1: Darstellung der anthropometrischen Daten des Probanden-
kollektivs zum Zeitpunkt vor der Trainingsintervention
17
Tab. 2: Darstellung der Mittelwerte und Standardabweichungen ausgewählter Parameter der Ausdauergruppe und Kraftgruppe vor und nach der Trainingsintervention, (*p<0.05 versus vor)
52

116
7.3 Abstract
The focus of research in contemporary literature is mostly on the kinetic parameters
of lactate transport in erythrocytes and the skeletal muscles. The underlying activity-
induced regulatory mechanisms of the MCT1 and CD147 proteins in the erythrocytes
have never before been analyzed in human erythrocytes. One aim of this study was
the exposure of the short-term activity-induced regulatory mechanisms of the MCT1
and CD147 proteins in the erythrocytes after maximal physical exercise. Besides
these short-term adaption phenomena the long-term training-induced changes of the
MCT1 and CD147 density in the erythrocytes and the MCT1 and CD147 density in
the skeletal muscle of non insulin-dependent type 2 diabetics after a three month
sport-therapeutic endurance and strength training were analyzed by
immunohistochemistry.
In comparison, the two forms of training showed similar results regarding the long-
term and the short-term training-induced changes of the MCT1 and CD147 density,
whereas the strength training caused clearer cellular reactions in the erythrocytes.
The results of the fluorescence microscopy of the erythrocytes indicate that trained
non insulin-dependent type 2 diabetics show a short-term translocation of the MCT1
protein and the CD147 chaperone into the erythrocyte membrane. In long-term, only
an increase of the MCT1 density in the erythrocytes of the strength training group
was observed. Tests on the skeletal muscles in both training groups showed a
significant increase of the cellular MCT1 density, whereas the cellular MCT4 density
remained nearly unchanged. The absolute membranous MCT4 density remained
unchanged in both training groups, while a significant decrease of the relative
membranous MCT4 density was observed in the endurance group only.
The following hypothetical regulatory model of the lactate transport during physical
activity visualizes the function of the MCT proteins depending on the physical
condition (Figure 36). In trained individuals, a faster lactate/H+-ion removal from the
contracting muscle to the blood plasma occurs due to the intensity-dependent
increase of the MCT1 and MCT4 density and a possible activity-induced
translocation of the MCT proteins. Because of a training-induced increase of the
MCT1 density and an increased translocation-induced lactate reception of the MCT1
proteins in the erythrocytes, the rising plasma lactate concentration during physical
exercise can be reduced faster. Here, the erythrocytes with their faster lactate uptake

117
function as a buffer, keeping up the intra-cellular pH level in the skeletal muscle and
the concentration-dependent lactate flow from the skeletal muscle to the blood
plasma. This increased rate of lactate transport leads to a more steady dispersion of
lactate in the body and therefore the lactate, together with glucose, can be used as an
energy source for the aerobic energy generation in the different target cells or be
stored as glycogen.
La/H+
La/H+
La/H+
La/H+
La/H+
La/H+
La/H+
La/H+
La/H+
untrained trained
La/H+La/H+La/H+
muscle heart liver
endothelium
muscle heart liver
lactate production
lactate utilization
skeletal muscle
La/H+
La/H+La/H+
La/H+ La/H+
translocationLa/H+ translocation
MCT1/4
MCT1erythrocyte
La/H+La/H+
La/H+
La/H+
La/H+ La/H+ La/H+La/H+ La/H+ La/H+
La/H+
La/H+
La/H+La/H+ La/H+
MCT1/4
Abb. 37: Hypothetical regulatory model of lactate transport and the function of the MCT proteins in the muscle and the erythrocytes during physical activity in trained
and untrained condition
Physical activity plays an important role in diabetic sport therapy. Epidemiological
studies have clearly shown that physical activity in primary and secondary
prevention influence the diabetic metabolism positively. Endurance and strength
training are both qualified for diabetic sport therapy because of their effects. This
was also confirmed by the DiabetesAktiv study, since both forms of training have
positively influenced the diabetic metabolism and the physical working capacity of
the study participants. These findings are confirmed by other studies, where a
combination of endurance and strength training seem to be most efficient for type 2
diabetics (Maiorana et al. 2002; Balducci et al. 2004; Sigal et al. 2007).
For the future, a vast need for research exists regarding the cellular effects of sport-
therapeutic training on the diabetic metabolism. A better understanding of activity-

118
induced cellular adaption phenomena is a basic key for the professional work of a
qualified sport therapist in order to be able to implement individual doses of physical
activity just like a medication within diabetic sport therapy in the future and thereby
relieving the healthcare system in the long run with this low-cost and efficient form
of therapy in a sustainable way.

119
Lebenslauf Persönliche Daten:
Nachname: Kreutz
Vorname: Thorsten
Geburtsdatum: 28.03.1969
Geburtsort: Hagen
Familienstand: verheiratet
Schulbildung:
1975 – 1979 Grundschule Hestert in Hagen
1980 – 1985 Ernst-Eversbusch-Schule in Hagen
1985 – 1989 Ernst-Meister-Gymnasium in Hagen, Abitur
Hochschulausbildung
SS 1993 Immatrikulation an der Ruhr-Universität-
Bochum, Studiengang: Sportwissenschaften
und Pädagogik (Lehramt Sek. II)
SS 1995 Immatrikulation an der Deutschen
Sporthochschule Köln, Studiengang:
Sportwissenschaften (Diplom)
WS 1999/00 Abschluss: Diplom-Sportlehrer
seit SS 2004 Promotionsstudium an der Deutschen
Sporthochschule Köln
Weiterbildung
April 2000 – April 2001 Ausbildung zum Sporttherapeuten für
Innere Erkrankungen und Orthopädie (DVGS)
24.02.2006 1. Kolloquium „Laktatdiagnostik“ in Köln
27.- 29.09.2007 40. Deutscher Sportärztekongress in Köln
20.- 23.05.2009 44. Jahrestagung Deutschen Diabetes-
Gesellschaft in Leipzig (Posterpräsentation)
03.- 04.12.2009 High-Intensity-Training (HIT): Chancen und
Risiken in einem modernen Training in Köln

120
Berufserfahrungen
Oktober 1990 - Juli 2000 Trainertätigkeiten in verschiedenen
Gesundheitszentren
Dezember 2000 - Februar 2001 Praktikum im Rehabilitationszentrum
Orthomed von Borussia Dortmund
September 2001 - Februar 2002 Sporttherapeut in der Medizinischen
Trainingstherapie in einem Gesundheits-
zentrum in Goslar
März 2002 - April 2004 Freiberufliche Tätigkeiten in den Bereichen:
- Medizinische Trainingstherapie
- Präventions- und Koordinationsstrainer bei
Bayer 04 Leverkusen im Fußball Jugend-
bereich
- Personal Training
- Betrieblichen Gesundheitsförderung
- Halbjahrespraktikum als Sporttherapeut in der
Eifelhöhenklinik in Marmagen
- Mitarbeit und Entwicklung des M.O.B.I.L.I.S-
Bewegungsprogrammes der BEK des
Institutes für Kreislaufforschung und
Sportmedizin der Sporthochschule Köln
Mai 2004 - Februar 2010 Sporttherapeut in einem Therapiezentrum in
Solingen
- Referent in der ärztlichen Weiterbildung des
Sportärztebundes zu den Themenkomplexen:
Sport mit Diabetikern und über die Methodik
des Krafttraining
seit März 2010 Leitung eines Therapiezentrums in einem
medizinischen Versorgungszentrum in Hilden























![Immunhistochemische!Parameter!undderenprognostische ... · ! 2! Verfügbarkeit bildgebender! Verfahren! zurückzuführen! ist,! da hierdurch! die! Diagnose! einfacher!gestelltwerden!konnte![,2010].!Das](https://img.pdfslide.org/doc/110x75/5d59f61688c993c91f8bc1e9/immunhistochemischeparameterundderenprognostische-2-verfuegbarkeit.jpg)





