Embed Size (px)
Citation preview

Der Einsatz mykorrhizierter Gehölze in biologischen Sanierungsverfahren
unter dem Aspekt TNT-belasteter Böden
Dissertation zur Erlangung des akademischen Grades Doktor der Naturwissenschaften
- Dr. rer. nat. -
vorgelegt dem Fachbereich 2 Biologie/Chemie
Universität Bremen
von
Ingo Dobner
Bremen 2003

Gutachter: Prof. Dr. Wolfgang Heyser Prof. Dr. Gunter-Otto Kirst

Inhalt
1. Einleitung...........................................................................................................1
1.1 Der Sprengstoff TNT.........................................................................................1 1.1.1 Herstellung und Vorkommen von TNT und Nebenprodukten ....................1 1.1.2 Toxikologische Eigenschaften von TNT.........................................................3 1.1.3 Abbau von TNT unter natürlichen Bedingungen ..........................................5 1.2 Bodenreinigung durch Pilze und Pflanzen.....................................................7 1.2.1 Bodenreinigung durch Weißfäulepilze ............................................................7 1.2.2 Phytoremediation von Schadstoffen – Bodenreinigung durch Pflanzen .................................................................................................10 1.2.3 Strukturen der Ektomykorrhiza; Bodenreinigung durch Mykorrhizapilze .....................................................................................11 1.3 Fragestellung und Zielsetzung des Arbeitsvorhabens ...............................13 2. Material und Methode...................................................................................16
2.1 Organismen in den Experimenten.................................................................16 2.1.1 Pilze ...................................................................................................................16 2.1.1.1 Kultivierung der Mykorrhizapilze auf Agarmedium.....................................16 2.1.1.2 Kultivierung der Weißfäulepilze auf Agarmedium ......................................16 2.1.1.3 Kultivierung der Weißfäulepilze auf Stroh und Weichholzschredder ......17 2.1.2 Pflanzen.............................................................................................................18 2.1.2.1 Anzucht der Kiefern.........................................................................................18 2.1.2.2 Herkunft der Pappeln ......................................................................................18 2.1.2.3 Mykorrhizierung der Kiefern in Rhizotronkulturen ......................................19 2.1.2.4 Mykorrhizierungen an Pappeln......................................................................19 2.2 Experimente......................................................................................................20 2.2.1 Kultivierung von Kiefern auf TNT-belasteten Boden (Kiefernexperiment).........................................................................................20 2.2.2 Kultivierung von Pappeln auf TNT-belasteten Boden (Pappelexperiment) .........................................................................................22 2.2.3 Aufnahme des Radiotracers 14C-TNT in Kiefernsämlinge ........................25 2.2.4 Phytotoxizitätstest mit Kiefern........................................................................26 2.2.4.1 Vorbereitung der Pflanzen..............................................................................27 2.2.4.2 Vorbereitung des Bodensubstrates ..............................................................27 2.2.4.3 Homogenitätskontrolle ....................................................................................27 2.2.4.4 Versuchsdesign und Probennahme..............................................................27 2.3 Methoden ..........................................................................................................29 2.3.1 Qualitativer Nachweis ligninolytischer Enzyme auf Agar-Festmedien ....29 2.3.1.1 Nachweis der Mangan-Peroxidase-Aktivität................................................30 2.3.1.2 Nachweis der Lignin-Peroxidase-Aktivität ...................................................30 2.3.2 Extraktionsverfahren .......................................................................................30 2.3.2.1 Nitroaromatenextraktion aus den Bodenmischproben ..............................30 2.3.2.2 Nitroaromatenextraktion aus Pflanzengeweben.........................................31 2.3.3 Nitroaromaten-Analytik ...................................................................................32 2.3.3.1 HPLC-Analytik ..................................................................................................32 2.3.3.2 GC/MS-Analytik................................................................................................33 2.3.4 Quantifizierung der Radioaktivität durch Szintillationszählung .................35

2.3.4.1 Vorbereitung der Pflanzenproben.................................................................35 2.3.4.2 Aufnahme der Nährlösung .............................................................................36 2.3.4.3 Auswaschung des Kohlepapiers ...................................................................36 2.3.4.4 Ermittlung der Radioaktivität ..........................................................................36 2.3.5 Angewandte Methoden für die Mikroautoradiographie..............................36 2.3.5.1 Gefriertrocknung der präparierten Wurzeln.................................................36 2.3.5.2 Kunststoffinfiltration.........................................................................................38 2.3.5.3 Schneiden der Proben am Ultramikrotom....................................................39 2.3.5.4 Mikroautoradiographie ....................................................................................39 2.3.6 Rasterelektronenmikroskopie (REM)............................................................39 2.3.7 Bestimmung des pH-Wertes von Bodenproben .........................................40 2.3.8 Fotografische Dokumentation........................................................................40 2.3.9 Ergebnisauswertung und Statistik.................................................................41 3. Ergebnisse.......................................................................................................42
3.1 Qualitativer Nachweis ligninolytischer Enzyme ..........................................42 3.2 TNT-Abbau-Experimente mit Kiefern ...........................................................43 3.2.1 Entwicklung der TNT-Belastung im Boden..................................................43 3.2.2 Entwicklung der ADNT-Belastung im Boden...............................................46 3.2.3 Biomasseentwicklung und Mykorrhizierungsgrad der Kiefern..................48 3.2.4 Zusammenfassung ..........................................................................................48 3.3 TNT-Abbauexperimente mit Pappeln als Phytoremediatoren ..................49 3.3.1 Bodenanalytik ...................................................................................................49 3.3.1.1 Entwicklung der TNT-Belastung im Boden..................................................50 3.3.1.2 Entwicklung der ADNT-Belastung im Boden...............................................53 3.3.1.3 Dinitrotoluole .....................................................................................................55 3.3.2 Biomasseentwicklung und Mykorrhizierungsgrad der Pappeln................56 3.3.3 Zusammenfassung ..........................................................................................56 3.3.4 Nachweis des Nitroaromatentransfers aus dem Boden in die Versuchspflanzen ............................................................................................57 3.3.4.1 Quantitativer Nachweis von Nitroaromaten mittels GC/MS ......................59 3.3.4.2 Wiederfindungsraten für TNT in Pflanzenproben.......................................61 3.3.4.3 Zusammenfassung ..........................................................................................62 3.4 Aufnahme und Verteilung von 14C-TNT/14C-TNT-Metaboliten in Pinus sylvestris Sämlingen ........................................................................62 3.4.1 Mikroautoradiographische Untersuchungen von nicht-mykorrhizierten und mykorrhizierten Kiefernwurzeln .............................................................63 3.4.2 Bestimmung der Radioaktivität in den einzelnen Pflanzenorganen durch Szintillationszählung.............................................................................69 3.4.3 Bilanzierung ......................................................................................................72 3.4.3.1 Wiederfindung nach Dotierung einer hohen 14C-TNT-Konzentration......72 3.4.3.2 Wiederfindung nach Dotierung einer niedrigen 14C-TNT-Konzentration ...................................................................................73 3.4.3.3 Verbleibende Radioaktivität in Kohlepapier und Düngelösung ................73 3.4.4 Zusammenfassung ..........................................................................................73 3.5 Toxizitätstest mit mykorrhizierten Kiefern in TNT-haltigem Bodensubstrat ..................................................................................................74 3.5.1 Wiederfindung der TNT-Kontamination........................................................74 3.5.2 Ermittlung der Nitroaromatengehalte im Boden und in der Bodenlösung ..................................................................................75

3.5.3 Ermittlung des Pflanzenwachstums in den Dotierungskategorien...........79 3.5.3.1 Gesamtüberblick ..............................................................................................80 3.5.3.2 Pflanzenwachstum in Bezug zur Nitroaromatenkonzentration in der Bodenlösung..........................................................................................82 3.5.4 Zusammenfassung ..........................................................................................87 4. Diskussion.......................................................................................................88
4.1 Wechselwirkungen zwischen Nitroaromaten und Bodenmatrix...............88 4.1.1 Adsorption an Tonmineralien.........................................................................88 4.1.2 Wechselwirkung mit Huminstoffen................................................................89 4.1.3 Reduktive TNT-Transformation in den Experimenten und Wechselwirkungen mit Bodenbestandteilen........................................90 4.1.4 Einfluss von Bodenfaktoren auf die Bindungskinetik .................................91 4.1.5 Hydroxylaminodinitrotoluole (HADNT) als Intermediate der TNT-Reduktion ..........................................................................................93 4.1.6 Entstehung von Azoxyverbindungen bei der TNT-Transformation..........94 4.2 Dekontamination sprengstoffbelasteter Böden im Kiefern- und Pappelexperiment; ein Variantenvergleich ..........................................95 4.2.1 Abreicherung der Nitroaromatenkonzentration im Boden der Kontrollvariante (Variante K)...................................................................98 4.2.2 Abreicherung der Nitroaromatenkonzentration im Boden der Weißfäulepilzvariante (Variante C) ........................................................99 4.2.3 Abreicherung der Nitroaromatenkonzentration im Boden der Versuchsvarianten mit mykorrhizierten Pflanzen (Varianten A und B) 100 4.2.3.1 Verwendete Organismen und ihre Fähigkeit zur Nitroaromatentransformation...................................................................... 101 4.2.3.2 Mykorrhizapilze und ihr Einfluss auf die mikrobielle Lebensgemeinschaft im Boden .................................................................. 102 4.2.4 Nitroaromatenabreicherung in Versuchsvariante D mit Weiß- fäulepilzen und mit nicht-mykorrhizierten Pflanzen................................. 104 4.3 Nitroaromaten in Pflanzengeweben........................................................... 106 4.3.1 Aufnahme von Nitroaromaten aus dem Boden........................................ 106 4.3.2 Kiefern als Versuchspflanzen bei der Applikation mit 14C-TNT............. 109 4.3.3 Nitroaromatenaufnahme in die Wurzeln.................................................... 110 4.3.4 Apoplastischer Transport............................................................................. 111 4.3.5 Symplastischer Transport ............................................................................ 113 4.3.6 Metabolismus der aufgenommenen Nitroaromaten................................ 114 4.3.6.1 Orte der Metabolisierung ............................................................................. 114 4.3.6.2 Einbau von 14C-TNT und Metaboliten in die Zellwände........................ 117 4.3.6.3 Einbau von TNT-Metaboliten in die Ligninfraktion................................... 118 4.3.6.4 Festlegung an Zellwandbestandteilen der Wurzeloberfläche ................ 119 4.3.7 Extraktion von Nitroaromaten aus Pflanzengeweben............................. 120 4.4 Toxizitätstest mit mykorrhizierten Kiefern in TNT-haltigem Bodensubstrat ............................................................................................... 122 5. Zusammenfassung..................................................................................... 128

6. Phytoremediation am Standort ‚Werk Tanne’; Durchführung und Beurteilung unter Berücksichtigung der Laborergebnisse.... 131
6.1 Kurze Beschreibung des Standortes ‚Werk Tanne’ ................................ 131 6.2 Verfahrenskonzept........................................................................................ 132 6.3 Einrichtung der Versuchsflächen................................................................ 132 6.4 Grenzen des Versuchsansatzes ................................................................ 133 6.5 Beurteilung des Phytoremediationsverfahrens in Hinblick seines Sanierungerfolges unter Berücksichtigung der Ergebnisse aus den Laborversuchen............................................................................. 134 6.6 Sanierung TNT-belasteter Flächen durch Phytoremediation; Anwendungsbereiche und Machbarkeit.................................................... 137 6.6.1 Zeitrahmen und Art des Verfahrens ........................................................... 138 6.6.2 Auswahl der Organismen............................................................................ 138 7. Literatur ......................................................................................................... 140 8. Anhang .......................................................................................................... 159

Abkürzungen ADNT Aminodinitrotoluol 2-ADNT 2-Aminodinitrotoluol 4-ADNT 4-Aminodinitrotoluol 4-A-2-NT 4-Amino-2-nitrotoluol 4-A-2-HA-6NT 4-Amino-2-hydroxylamino-6-nitrotoluol bar Bar BIA Berufsgenossenschaftliches Institut für Arbeitssicherheit bidest bidestillata (bidestilliert) BM Biomasse Bq Becquerel °C Grad/Celsius CAS Chemical Abstract Service cm Zentimeter cpm counts per minute Da Dalton 2,4-DAHAT 2,4-Diaminohydroxylaminotoluol DANT Diaminonitrotoluol 2,4-DANT 2,4-Diaminonitrotoluol 2,6-DANT 2,6-Diaminonitrotoluol 2,4-DAT 2,4-Diaminotoluol DC Dünnschichtchromatographie DCM Dichlormethan DNT Dinitrotoluol 2,4-DNT 2,4-Dinitrotoluol 2,6-DNT 2,6-Dinitrotoluol dpm disintegrations per minute (Zerfälle pro Minute) Fa Firma FG Frischgewicht g Gramm GCP Gelpermeationschromatographie h Stunde HADNT Hydroxylaminodinitrotoluol HPLC High Performance Liquid Chromatography (Hochleistungsflüssigkeitschromatographie) k Kilo KAKeff effektive Kationenaustauschkapazität Kow Octanol-Wasser-Verteilungskoeffizient l Liter LUFA Landwirtschaftliche Untersuchungs- und Forschungsanstalt Speyer m Meter mg Milligramm min Minute mm Millimeter mV Millivolt

MS Massenspektrometer PAK Polyzyklische aromatische Kohlenwasserstoffe rpm rounds per minute (Umdrehungen pro Minute) s Sekunde t Zeit TAT 2,4,6-Triaminotoluol TB Trockenboden TM Trockenmasse TNT 2,4,6-Trinitrotoluol W Watt µ Mikro

Abstract The use of mycorrhizal plants in the bioremediation of TNT -contaminated soils The production and improper handling of the explosive 2,4,6-trinitrotoluene (TNT) led
to the contamination of soil and groundwater at many former ammunition and military
sites in germany. This persistent organopollutant is very dangerous for the
environment, because of its toxicity to a large variety of organisms including animals
and humans.
Conventional ex situ remediation of TNT sites by chemico-physical treatment is costly
and an energy-intensive process. Alternatively, phytoremediation is an
environmentally acceptable in situ method and holds promise for low-cost
remediation. Phytoremediation is the use of plants to clean up soils contaminated
with a number of harzadous substances. This technique is also practical to remediate
TNT-contaminated soils.
The present work concentrates on the potential application especially of mycorrhizal
plants in remediation of TNT-contaminated soils. Provided that plants clean up soils
by a) release of exudates (sugars, organic acids etc.) and enzymes, that stimulate
microbial activity and biochemical dregadation in the rhizosphere ; b) furnishing the
host of the symbiosis with carbon sources, while mycorrhizal fungi are capable of
TNT biotransformation by extracellular enzymes; c) direct uptake of TNT and
degradation products and subsequent accumulation of non-phytotoxic products into
plant tissue, the following experiments were carried out.
The degradation of TNT in rhizosphere soil was tested in experiments with
ectomycorrhizal associations of Pinus sylvestris/Pisolithus tinctorius and Populus
tremula/Hebeloma spec., cultivated in rhizotron systems. The rhizotrons were filled
with soil from the ammunition site ‘Tanne’ at Clausthal Zellerfeld/Lower Saxony. In
addition, the investigations involved the ligninolytic white rot basidiomycete Pleurotus
ostreatus which is able to degrade TNT via excretion of lignin and manganese
peroxidase. The results indicated, that mycorrhizal plants can play an important role
for biotransformation of TNT and other nitroaromatic compounds. Contamination
levels were lowest in rhizotrons with mycorrhizal plants. In this context, it was
negligible whether rhizotrons were inoculated with white rot fungi or not.

Furthermore, uptake and translocation activity of TNT in mycorrhizal and non-
mycorrhizal plants (Pinus sylvestris/Pisolithus tinctorius) were examined using
ringlabelled 14C-TNT. Most of the radioactivity was detected in the roots with only
small levels detected in stems and needles. A mycorrhiza effect was not observed,
mycorrhizal and non-mycorrhizal plants showed similar results.
Ectomycorrhiza can be used for effective remediation of soil provided plants and
fungi are tolerant or resistant to toxic contaminants. Because less is known of
phytotoxicity thresholds of TNT and its metabolites in soils. In this context the effects
of TNT and transformation products on growth and development of Pinus
sylvestris/Pisolithus tinctorius mycorrhiza were investigated. The results showed that
levels less than 50 mg TNT/kg1 soil had no effects on mycorrhizal plants. Higher TNT
or ADNT concentrations up to 50 mg /kg1 soil substantially delayed production of
shoot biomass. Although plant growth was inhibited, mycorrhizal plants might be able
to tolerate higher levels and kept a relevant potential of regeneration.
The studies allow the conclusion, that mycorrhizal plants have the capacity to
remediate TNT-contaminated soils. Most efficiency may be expected at sites with
shallow contamination, where TNT and metabolites were treated in the rhizosphere
by microbial activity and by root uptake. Uptake, accumulation and binding of TNT
and its transformation products into plants seem to be dependent on plant species
and plant organs. In this case, we need more information for a better understanding.
Therefore, further investigations are required.

Einleitung
1
1.Einleitung 1.1 Der Sprengstoff TNT 1.1.1 Herstellung und Vorkommen von TNT und Nebenprodukten 2,4,6-Trinitrotoluol (TNT) wurde erstmalig im Jahre 1863 von J. Wilbrand in unreiner Form
synthetisiert. Als reine Verbindung gelang P. Hepp 1880 die Herstellung. Nach ersten
Sprengversuchen 1884 in Hanau wurde schließlich um 1900 die großtechnische Produktion
aufgenommen. Vor allem während des ersten und zweiten Weltkrieges wurden große Mengen TNT
hergestellt. Allein im Deutschen Reich lag die Produktion im zweiten Weltkrieg nach Schätzungen
über 800000 Tonnen (Preuß et al. 1988). Die Synthese von TNT erfolgt durch schrittweise
Nitrierung des Toluols. Außer TNT fielen bei der Produktion größere Mengen verschiedener
Nebenprodukte, wie Mononitrotoluole, Dinitrotoluole und auch TNT-Isomere an. Diese
unerwünschten Nebenprodukte ließen sich durch mehrmaliges Waschen mit Natriumsulfit-Lösung
aus dem Rohprodukt entfernen. Nach Neutralisierung mit Kalk wurden die Abwässer in der Regel
vor Ort in Vorfluter und Schlammteiche geleitet oder in Schluckbrunnen verpresst. Unfälle bei der
Produktion und unsachgemäße Handhabung haben zudem zur Kontamination der Böden zahlreicher
ehemaliger Produktionsstätten beigetragen. So lassen sich TNT und Folgeprodukte aufgrund ihrer
hohen Persistenz auch gegenwärtig noch an vielen Stellen in kristalliner Form im Boden nachweisen.
Die größten Sprengstofffabriken mit ihren damaligen monatlichen Kapazitäten waren nach Wolff
(1989) Stadtallendorf (5000 Tonnen), Hessisch Lichtenau (3500 Tonnen), Clausthal-Zellerfeld
(2800 Tonnen) Krümmel (2800 Tonnen) und Elsnig (2600 Tonnen). Aus zahlreichen Unter-
suchungen geht hervor, dass neben dem TNT selbst auch verschiedene Neben- und
Zwischenprodukte aus der TNT-Herstellung sowie einige Transformationsprodukte, die aus der
TNT-Degradation hervorgehen, ein stark gesundheitsgefährdendes Potenzial aufweisen (BIA-Report
1998, Berufsgenossenschaftliches Institut für Arbeitssicherheit). Zahlreiche Verbindungen befinden
sich diesbezüglich derzeit noch in der Überprüfung. Häufig auf Rüstungsaltlasten nachgewiesene
Verbindungen sind u. a. 2,4-DNT, 2,6-DNT, 2-ADNT, 4-ADNT, 2,4-DANT, 2,6-DANT, 2-NT,
4-NT und 3-NT. Nachstehend sind einige von ihnen
mit Formel und allgemeinen Angaben aufgeführt (Abbildung 1).
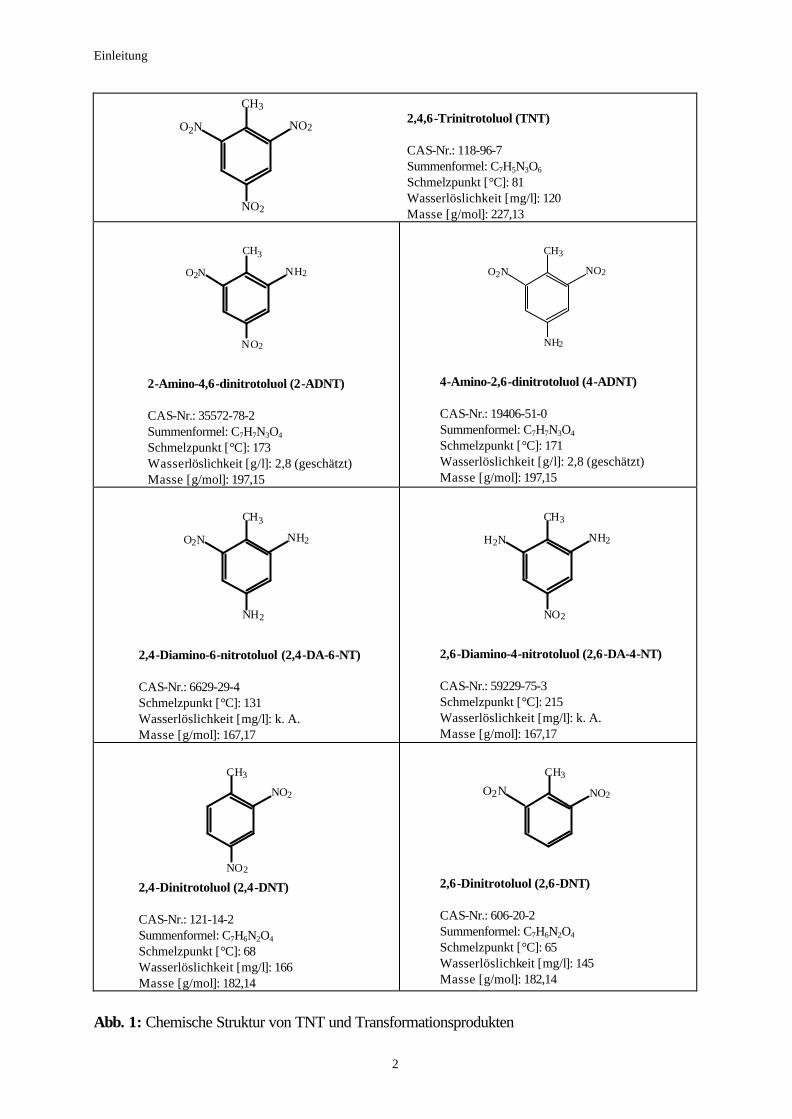
Einleitung
2
CH3
NO2
NO2
O2N
2,4,6-Trinitrotoluol (TNT) CAS-Nr.: 118-96-7 Summenformel: C7H5N3O6 Schmelzpunkt [°C]: 81 Wasserlöslichkeit [mg/l]: 120 Masse [g/mol]: 227,13
CH3
NH2
NO2
O2N
2-Amino-4,6-dinitrotoluol (2-ADNT) CAS-Nr.: 35572-78-2 Summenformel: C7H7N3O4 Schmelzpunkt [°C]: 173 Wasserlöslichkeit [g/l]: 2,8 (geschätzt) Masse [g/mol]: 197,15
CH3
NO2
NH2
O2N
4-Amino-2,6-dinitrotoluol (4-ADNT) CAS-Nr.: 19406-51-0 Summenformel: C7H7N3O4 Schmelzpunkt [°C]: 171 Wasserlöslichkeit [g/l]: 2,8 (geschätzt) Masse [g/mol]: 197,15
CH3
NH2
NH2
O2N
2,4-Diamino-6-nitrotoluol (2,4-DA-6-NT) CAS-Nr.: 6629-29-4 Schmelzpunkt [°C]: 131 Wasserlöslichkeit [mg/l]: k. A. Masse [g/mol]: 167,17
CH3
NH2
NO2
H2N
2,6-Diamino-4-nitrotoluol (2,6-DA-4-NT) CAS-Nr.: 59229-75-3 Schmelzpunkt [°C]: 215 Wasserlöslichkeit [mg/l]: k. A. Masse [g/mol]: 167,17
CH3
NO2
NO2 2,4-Dinitrotoluol (2,4-DNT) CAS-Nr.: 121-14-2 Summenformel: C7H6N2O4 Schmelzpunkt [°C]: 68 Wasserlöslichkeit [mg/l]: 166 Masse [g/mol]: 182,14
O2N
CH3
NO2
2,6-Dinitrotoluol (2,6-DNT) CAS-Nr.: 606-20-2 Summenformel: C7H6N2O4 Schmelzpunkt [°C]: 65 Wasserlöslichkeit [mg/l]: 145 Masse [g/mol]: 182,14
Abb. 1: Chemische Struktur von TNT und Transformationsprodukten
Einleitung
3
In den meisten Fällen sind aromatische Nitroverbindungen anthropogenen Ursprungs und haben
somit Fremdstoffcharakter. Als natürliche Verbindungen kommen nitrierte Aromaten dagegen sehr
selten und lediglich mit einer Nitrogruppe substituiert vor. Zu nennen sind hier die Antibiotika
Chloramphenicol und Pyrrolnitrin, die von Streptomyces- und Pseudomonas-Stämmen gebildet
werden, sowie die dem Fraßschutz dienende und von den Pflanzen über den Sekundärmetabolismus
gebildete Aristolochiasäure (Pailer 1960). Der Pilz Lepista diemii Singer synthetisiert das 4-
Nitroanisol als Metabolit (Thaller & Turner 1972). Stabilität und seltenes natürliches Vorkommen
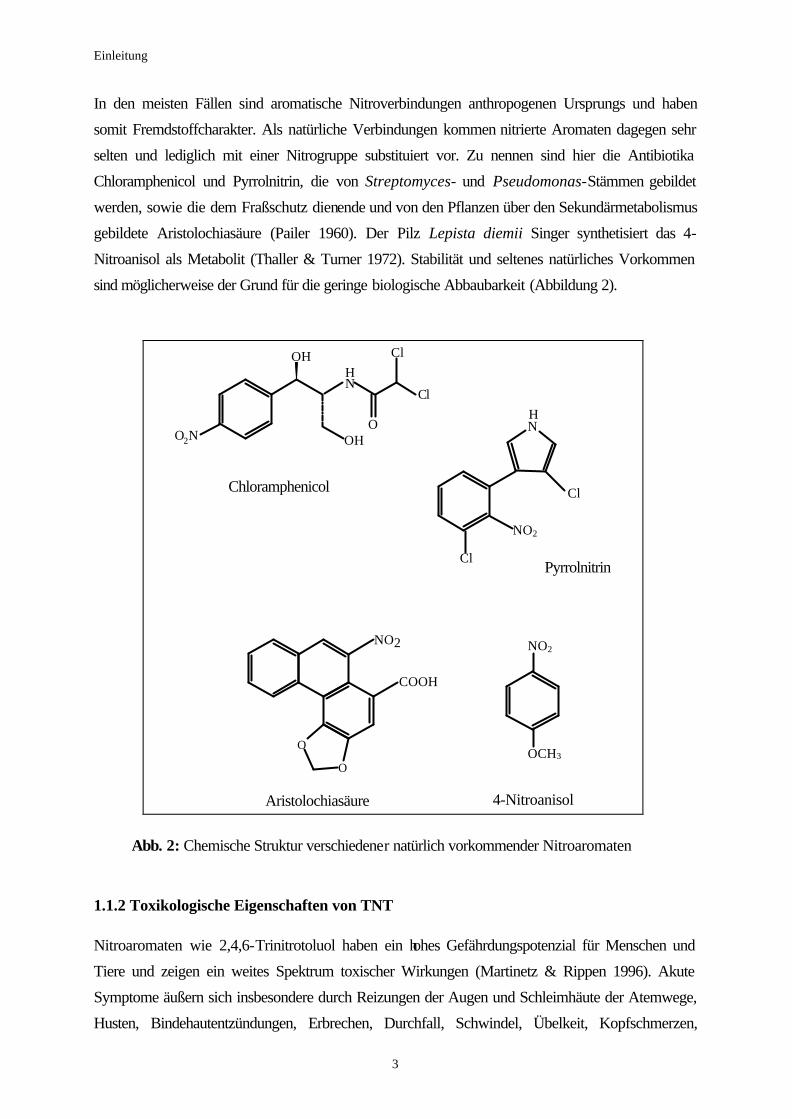
sind möglicherweise der Grund für die geringe biologische Abbaubarkeit (Abbildung 2).
O
O
NO2
COOH
N
O2N
OHH
Cl
Cl
OOH
NO2
OCH3
N
Cl
NO2
Cl
H
Chloramphenicol
Pyrrolnitrin
Aristolochiasäure 4-Nitroanisol
Abb. 2: Chemische Struktur verschiedener natürlich vorkommender Nitroaromaten 1.1.2 Toxikologische Eigenschaften von TNT Nitroaromaten wie 2,4,6-Trinitrotoluol haben ein hohes Gefährdungspotenzial für Menschen und
Tiere und zeigen ein weites Spektrum toxischer Wirkungen (Martinetz & Rippen 1996). Akute
Symptome äußern sich insbesondere durch Reizungen der Augen und Schleimhäute der Atemwege,
Husten, Bindehautentzündungen, Erbrechen, Durchfall, Schwindel, Übelkeit, Kopfschmerzen,

Einleitung
4
Atemstörungen bis hin zum Blutdruckabfall, Kollaps und Koma. Als Folgekrankheiten treten u. a.
Hepatitis, Gelbsucht, Anämie und Nierenschäden auf. Als Symptome nach chronischer
Inkorporation werden neben zahlreichen anderen Erkrankungen Methämoglobinanämie,
Leberzirrhose, Nierenschäden und Trübung der Augenlinsen (TNT-Star) beschrieben (Ludewig &
Lohs 1991, Henschler 1988, Moeschlin 1986). TNT wird gut resorbiert. Über die Haut des
Menschen können innerhalb von acht Stunden zwischen 2,6 und 3,8 µg TNT/ cm2 aufgenommen
werden (Henschler 1988). Weitere Aufnahmepfade sind Lunge, Schleimhäute und Verdauungstrakt.
Aufgrund einer zügigen Metabolisierung besteht trotz lipophiler Eigenschaften jedoch weniger die
Neigung zur Anreicherung im Fettgewebe (Koss et al. 1989). Die Halbwertzeit der Ausscheidung
bei Ratten nach gleichmäßiger Verteilung im Organismus beträgt 6,3 Stunden. Die Abgabe erfolgt
überwiegend mit dem Urin. Hauptmetaboliten im Harn sind 4-ADNT, 2-ADNT, 2,4-DANT, 4-
HADNT sowie das 2,4,6-TNT selbst (Henschler 1988). Von der Senatskommission zur Prüfung
gesund-heitsschädlicher Arbeitsstoffe der Deutschen Forschungsgemeinschaft wurde TNT bezüglich
seiner Kanzerogenität in Kategorie 3 eingestuft. Hiernach besteht die Möglichkeit einer
krebserregenden Wirkung beim Menschen. Jedoch liegen noch nicht genügend Informationen für
eine sichere Beurteilung vor (BIA-Report 1998). Allerdings lassen epidemiologische Studien
vermuten, dass Nitroaromaten starke Kanzerogene und Mutagene darstellen (Anderson et al. 1997).
Im Ames-Test ließ sich ein mutagenes Potenzial bei Konzentrationen zwischen 10 und 5000 µg TNT
bei Salmonella typhimurium TA 100 und anderen Stämmen feststellen (Spanggord et al. 1982).
Selbst Harnproben TNT-exponierter Arbeiter hatten bei Salmonella typhimurium TA 98 mutagene
Wirkung (Ahlborg et al. 1985). Als Luftgrenzwert wird eine Maximale Arbeitsplatzkonzentration von
0,1 mg TNT-Staub/m3 Luft (MAK-Wert) angegeben, wobei aufgrund hautresorptiver Eigenschaften
des TNT die Einhaltung des Luftgrenzwertes für den Schutz der Gesundheit nicht ausreicht und beim
Umgang zusätzliche arbeitshygienische Maßnahmen getroffen werden müssen (BIA-Report 1998).
Die teilweise reduzierten Metabolite des TNT, die ADNTs und die DANTs sind bisher nicht
ausreichend untersucht, um sichere Aussagen über ihre toxikologische Einstufung machen zu können.
Die wenigen vorliegenden Informationen deuten auf eine Abnahme der Toxizität durch Reduktion der
Nitrogruppen zu Aminogruppen am TNT-Molekül hin (Martinetz & Rippen 1996). Im
Biolumineszenztest mit dem Bakterium Vibrio fischeri nahm das Toxizitätspotenzial der untersuchten
Nitroaromaten in der Reihenfolge TNT > 4-ADNT > 2-ADNT ab (Frische 2002).
Für Dinitrotoluole ist ein breites Spektrum toxischer Wirkungen beschrieben (Martinetz & Rippen
1996). Aus verschiedenen Kanzerogenitätsstudien lässt sich eine kanzerogene Potenz für 2,4-DNT
und auch für 2,6-DNT ableiten, wobei 2,6-DNT eine stärkere krebsauslösende Wirkung ausübt. In
der Grenzwertliste 1998 des BIA wurde 2,6-DNT in der Rubrik Kanzerogene Stoffe in die
Kategorie 2 eingestuft. Kategorie 2 gilt für Stoffe, die als krebserregend für den Menschen

Einleitung
5
angesehen werden. Die Annahmen beruhen insbesondere auf Langzeit-Tierversuchen. Der MAK-
Wert beträgt 0,05 mg/m3 Luft.
1.1.3 Abbau von TNT unter natürlichen Bedingungen Chemische Stabilität, Persistenz und Toxizität sind die wesentlichen Eigenschaften von TNT und
zahlreichen anderen Nitroaromaten. Trotzdem konnte inzwischen bei Bakterien und Pilzen
verschiedener Gattungen die Fähigkeit, TNT zu transformieren oder biologisch abzubauen,
nachgewiesen werden. Die mikrobielle Spaltung des Aromaten mit nachfolgender Mineralisierung
wurde bisher allerdings nur selten beschrieben. Angaben hierzu konzentrieren sich auf ligninolytische
Pilze, welche mit Hilfe ihres extrazellulären Enzymsystems Nitroaromaten oxidativ angreifen und
mineralisieren können (Scheibner et al. 1997). Der Abbau erfolgt jedoch erst nach initialer Reduktion
der Nitrogruppen des TNTs zu Aminogruppen (Van Aken et al. 2000, Fritsche et al. 2000),
wodurch das Elektronendefizit am aromatischen Ring herabgesetzt wird. Über die weiteren
Abbauwege der reduzierten Transformationsprodukte ist bisher noch wenig bekannt. Das TNT-
Molekül wird sowohl unter aeroben als auch unter anaeroben Bedingungen durch schrittweise
Reduktion der Nitrogruppen transformiert. Als Zwischenprodukte entstehen Aminodinitrotoluole und
Diaminonitrotoluole (Abbildung 3).
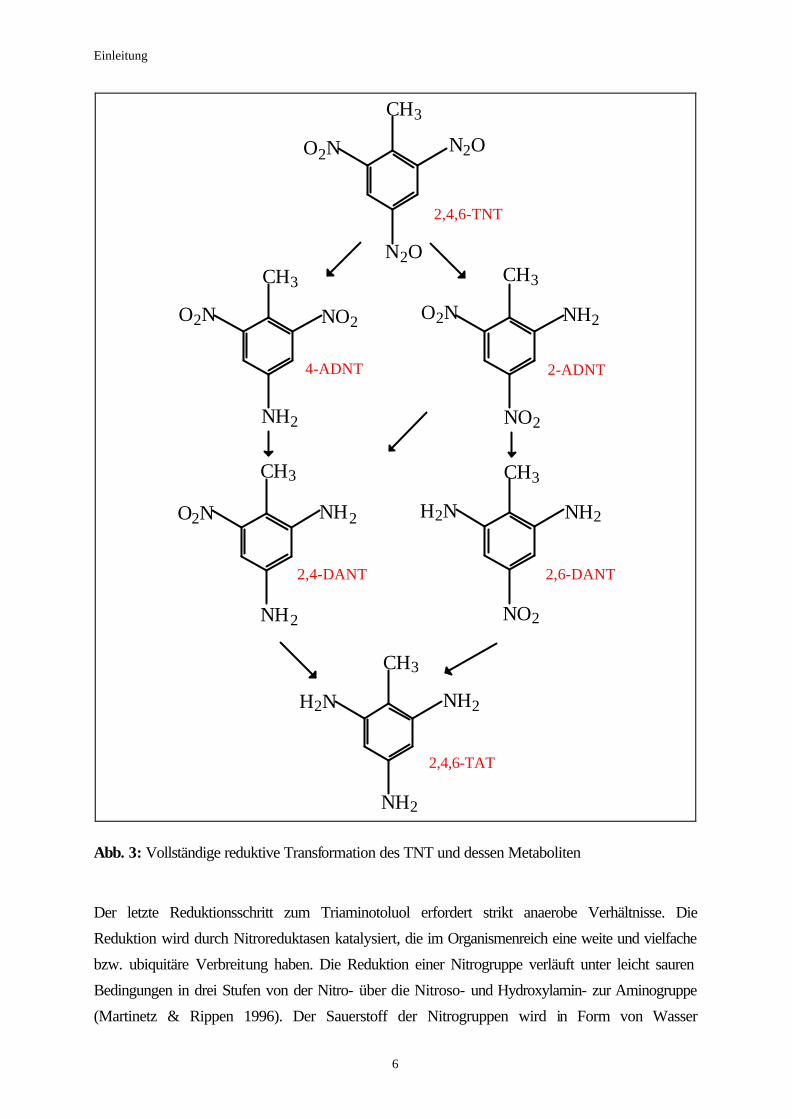
Einleitung
6
CH3
O2N N2O
N2OCH3 CH3
NO2O2N
NH2
NH2O2N
NO2
CH3 CH3
NH2 NH2O2N H2N
NH2 NO2
CH3
NH2H2N
NH2
2,4,6-TNT
2-ADNT4-ADNT
2,4-DANT 2,6-DANT
2,4,6-TAT
Abb. 3: Vollständige reduktive Transformation des TNT und dessen Metaboliten Der letzte Reduktionsschritt zum Triaminotoluol erfordert strikt anaerobe Verhältnisse. Die
Reduktion wird durch Nitroreduktasen katalysiert, die im Organismenreich eine weite und vielfache
bzw. ubiquitäre Verbreitung haben. Die Reduktion einer Nitrogruppe verläuft unter leicht sauren
Bedingungen in drei Stufen von der Nitro- über die Nitroso- und Hydroxylamin- zur Aminogruppe
(Martinetz & Rippen 1996). Der Sauerstoff der Nitrogruppen wird in Form von Wasser

Einleitung
7
abgespalten. Die nötigen Reduktionsäquivalente werden aus der Verwertung zusätzlicher C-Quellen
gewonnen. Nitroso- als auch Hydroxylaminoverbindungen reagieren schnell weiter, so dass die
Aminodinitrotoluole als erste relativ stabile Verbindungen anzusehen sind.
Für Mono- und Dinitroaromaten sind auch oxidative Initialangriffe in mikrobiellen Abbauprozessen
bekannt. Eine durch Bakterien erwirkte Dioxygenase-katalysierte Initialreaktion, die über den Weg
der Doppelhydroxylierung letztendlich zur Ringöffnung führt, wurde beim Abbau von 2,6
Dinitrotoluol festgestellt (Nishino et al. 2000b). Auch bei anderen Nitroaromaten, wie 1,3-
Dinitrobenzol, 2,4-Dinitrotoluol oder 3-Nitrobenzoat ließen sich Dioxygenase-katalysierte
Initialreaktionen nachweisen. Für 4-Nitrotoluol konnte über eine reduktive Initialreduktion der
Nitrogruppe zum 4-Hydroxylaminotoluol und nach-folgender Oxidation durch eine Dioxygenase
ebenfalls eine Ringöffnung mittels Bakterien beobachtet werden (verschiedene Referenzen in Lenke
2001).
Im Anschluss gilt die Aufmerksamkeit den Pilzen und Pflanzen, deren Remediationspotenzial beim
TNT-Abbau in der vorliegenden Arbeit untersucht wurde.
1.2 Bodenreinigung durch Pilze und Pflanzen 1.2.1 Bodenreinigung durch Weißfäulepilze Die saprophytischen Weißfäulepilze, wie beispielsweise die Arten Phanerochaete chrysosporium,
Trametes versicolor oder auch Pleurotus ostreatus leben im Holz und sind aufgrund ihrer
Enzymaustattung in der Lage, Lignin, Zellulose und Hemizellulose als Substrat zu nutzen und
abzubauen. Das Lignin als quantitativ wichtigste Substanz in der Natur kommt praktisch in allen
Pflanzenteilen vor und kann bis zu 30% der Zellwände ausmachen (Hüttermann et al. 1988, Reid
1995). Aufgrund seiner chemischen Struktur, es besteht überwiegend aus vielfach kondensierten
Benzolringen, gilt es als schwer angreifbar. Ligninpolymere setzen sich aus einer Mischung der drei
Monolignole p-Cumaryl-, Sinapyl- und Coniferylalkohol zusammen (Abbildung 4). Das Verhältnis
der drei Grundbausteine ist in verschiedenen Pflanzen sehr unterschiedlich. Während das Lignin der
Coniferen einen hohen Coniferylanteil hat, überwiegt in Getreidehalmen der Cumarylanteil. Die
Biosynthese erfolgt durch enzymatische Oxidation eines Monolignols unter Verwendung von H2O2 zu
einem Phenoxyradikal. Die freien Phenoxyradikale als Zwischenprodukte verknüpfen sich schließlich
untereinander auf vielfältige Weise zu einem hochmolekularen Netzwerk. Es entsteht ein sowohl über
Ether- als auch Kohlenstoff-Kohlenstoff-Bindungen zusammengesetztes Biopolymer.

Einleitung
8
OH
OH
OH
OH
OH
OH
OCH3 H3CO OCH3
p-Cumarylalkohol Coniferylalkohol Sinapylalkohol Abb. 4: Die Monolignole p-Cumaryl-, Sinapyl- und Coniferylalkohol dienen als Grundbau-steine der Ligninmakromoleküle Die wasserunlöslichen Lignin-Polymere können bei Abbauprozessen wegen ihrer Größe nicht von
den Zellen der Pilzhyphen aufgenommen werden. Zudem liegen sie oft fest eingebettet in Zellulose-
und Hemizellulosebestandteile der Zellwand vor (Reid 1995). Lignin ist kovalent gebunden mit
Zellulose verknüpft. Verschiedene Autoren vergleichen lignifizierte Zellwände mit Stahlbeton, wobei
die Zellulosefibrillen als Stahlgerüst und das Lignin als Beton angesehen werden. Deshalb sind seitens
der Weißfäulepilze besondere Strategien notwendig, um diesen weit verbreiteten Naturbaustein für
sich als Substrat erschließen zu können. Aus Studien mit den Weißfäuleerregern Trametes
versicolor und Phanerochaete chrysosporium geht hervor, dass bei der Biodegradation von
Weizenstroh zuerst Anteile weniger lignifizierter Gewebe angegriffen werden, während die in der
Regel stark lignifizierten Xylemelemente und Sklerenchymfasern einer verzögerten Attacke
unterliegen (Barrasa et al. 1995). Die extrazellulären Enzyme reagieren relativ substratunspezifisch
und können alle Verknüpfungen des Ligninmoleküls angreifen. In von Pilzhyphen infiziertem und sich
bereits im Holzfäuleprozess befindlichem Holzgewebe ließen sich lignocellulytische Lignin- und
Mangan-Peroxidasen am Plasmalemma, in der Zellwand und in den extrazellulären Schleimschichten
der Pilzhyphenzellen nachweisen. In den Sekundärwänden der Pflanzenzellen binden diese Enzyme
an bereits depolymerisierten Ligninfragmenten. Ähnlich verhält es sich mit den Laccasen. Zellulose
wird durch Cellulasen und ß-Glukosidasen angegriffen, welche, lokalisiert in intrazellulären Vesikeln,
in die äußere Schleimschicht des Pilzes diffundieren und an den Mikrofibrillen der pflanzlichen
Zellwand binden. Die lignocellulytischen Enzyme sind aufgrund ihrer Molekülgröße nicht fähig, durch
intakte pflanzliche Zellwände zu diffundieren. Eingeleitet wird der Abbauprozess durch
niedermolekulare, bewegliche Substanzen, wie Veratryalkohol-Kationradikale und Peroxid-radikale,
welche die Poren in den Zellwänden erweitern und somit das Eindringen der Enzyme ermöglichen

Einleitung
9
(Heinzkill 1995, nach Evans et al. 1994 u. 1991). Zu den wichtigsten von den Weißfäulepilzen ins
Substrat ausgeschiedenen Enzymen zählen Lignin-Peroxidase, Mangan-Peroxidase und Laccasen.
Lignin-Peroxidase (E.C. 1.11.1.14; Ligninase): das Enzym ist relativ substratunspezifisch und wird in
inaktivem Zustand durch H2O2 oxidiert, in aktiver Form katalysiert die Lignin-Peroxidase über Ein-
Elektronen-Oxidationen die Bildung von Phenoxyradikalen. Auch nicht-phenolische Verbindungen
können oxidiert werden. Die monomeren N-und O-glykosylierten Proteine besitzen ein
Eisenprotoporphyrin IX als prosthetische Gruppe. Das Molekulargewicht liegt zwischen 38 und 43
kDa (De Jong 1993).
Mangan-Peroxidase (E.C. 1.11.1.13): hierbei handelt es sich ebenfalls um N- und O-glykosylierte
Proteine mit einem Eisenprotoporphyrin IX. Die Molekulargewichte betragen zwischen 43 und 49
kDa (De Jong 1993). Das Enzym oxidiert zunächst ein zweiwertiges Mangan-Atom unter Verbrauch
von H2O2. Das dreiwertige Mangan oxidiert wiederum phenolische Substrate (Reid 1995). Die
enzymatische Reaktion erfolgt demnach im Gegensatz zur Lignin-Peroxidase indirekt über das
dreiwertige Manganion.
Laccase (E.C. 1.10.3.2): bei diesen Enzymen handelt es sich um kupferhaltige Glykoproteine.
Allerdings benötigen sie kein H2O2 als Cosubstrat (Reid 1995). Laccasen sind relativ
substratunspezifisch und katalysieren u. a. die Oxidation phenolischer Substrate unter Bildung freier
Phenoxyradikale.
Mit ihrer Enzymausstattung sind Weißfäulepilze auch zur Degradation aromatischer Xenobiotika
fähig. Eingesetzt werden sie beim Abbau von polyzyklischen aromatischen Kohlenwasserstoffen
(PAK), Dioxinen, Chlorphenolen und TNT (Majcherczyk et al. 1993, Stahl und Aust 1993a,
Scheibner et al. 1997). Die Metabolisierung des TNT ist anhand des Weißfäulepilzes
Phanerochaete chrysosporium mittlerweile genauer untersucht. Das TNT wird durch den Pilz
zunächst zu 4-ADNT bzw. 2-ADNT reduziert. Sich anschließende Acylierungsreaktionen führen zu
unterschiedlichen formylierten und acetylierten Zwischenprodukten (Hawari et al. 1999, Michels und
Gottschalk 1995). Eines dieser Zwischenprodukte, das formylierte 2-Amino-4-formamido-6-
nitrotoluol konnte als Substrat der Lignin-Peroxidase identifiziert werden (Michels und Gottschalk
1995). Auch die anderen Produkte werden unter ligninolytischen Bedingungen metabolisiert.
Scheibner et al. (1997a) gelang der Nachweis einer direkten Mineralisierung von 4-ADNT mit dem
Weißfäulepilz Nematoloma frowardii durch eine Mangan-Peroxidase.

Einleitung
10
1.2.2 Phytoremediation von Schadstoffen – Bodenreinigung durch Pflanzen Die Phytoremediation als Teilbereich der Bioremediation ist definiert als Entgiftung
schadstoffbelasteter Böden durch Pflanzen. Außer Bakterien und Pilzen verfügen auch Pflanzen über
Mechanismen, Schadstoffe abzubauen. Bisher kommen biologische in-situ Verfahren wenig zur
Anwendung (Trapp & Karlson 2000) und konzentrieren sich in Deutschland hauptsächlich auf
schwermetallbelastete Flächen. Fortgeschrittener ist die Entwicklung in den USA, vor allem bei der
Sanierung von Standorten, die mit organischen Schadstoffen belastet sind (Rock 1999, Newmann et
al. 1998). Einen Überblick über den Anwendungsbereich der Phytoremediation geben die Artikel
von Schnoor et al. (1995) sowie Trapp et al. (2000), wonach insbesondere die Kontaminanten
Nitrobenzol, Trichlorethen, TNT, PAK, Pentachlorphenol u. a. im Mittelpunkt stehen. In jüngerer
Zeit fanden auch Mineralöle immer mehr Beachtung (Coats 1999). Bei der Phytoremediation greifen
verschiedene Mechanismen seitens der Pflanzen. Zunächst besteht die Möglichkeit der Aufnahme
von Stoffen in die Pflanzen mit nachfolgender Metabolisierung im Gewebe. Reinigungstechniken, die
auf solchen Prozessen basieren, werden allgemein unter dem Begriff Phytodegradation
zusammengefasst. Auch TNT und TNT-Metaboliten werden von Pflanzen aufgenommen und
abgebaut. Dies wurde sowohl für zahlreiche krautige Pflanzen (Cataldo et al. 1989, Görge 1994,
Sens 1998, Scheidemann 1998) als auch für einige Gehölze nachgewiesen (Thompson et al. 1998,
Schönmuth und Pestemer 2000). Gewichtiger sind jedoch die Einflüsse der Pflanzen, die indirekt
zum Abbau der Schadstoffe beitragen. So kann über eine Veränderung des Bodenmilieus zugunsten
von Bakterien und Pilzen deren Aktivität und dadurch auch der Abbau schädlicher Substanzen
verbessert werden. Effiziente Phytoremediation beruht also im Wesentlichen auf eine Assoziation
oder Gemeinschaft von Pflanzen, Bakterien und Pilzen im Wurzeleinzugsbereich. Solche Verfahren,
die sich auf den Abbau von Kontaminanten im Wurzelraum konzentrieren, sind durch den Begriff
Rhizodegradation definiert. Ein Phytoremediationsverfahren, das auf Rhizodegradation beruht, wurde
mittlerweile auf dem TNT-belasteten Standort ‚Werk Tanne’ in Clausthal-Zellerfeld erprobt
(Warrelmann et al. 2000). Durch Beeinflussung der Wasserbilanz und des Gashaushaltes in der
Rhizosphäre, durch Abgabe von Wurzelexsudaten und durch Beeinflussung des Boden-pH-Wert
schaffen Pflanzen für Mikroorganismen günstige Milieubedingungen. Bis zu 40% des
photosynthetisch gebundenen Kohlenstoff kann von den Wurzeln in Form von organischen Säuren,
Zucker und Alkoholen wieder an die Umgebung abgegeben werden (Yoshitomi & Shann 2001) und
steht somit dem mikrobiellen Metabolismus zur Verfügung. Die besondere Gemeinschaft der
Rhizosphärenorganismen kann aufgrund vielfältiger enzymatischer Reaktionen vorhandene
Xenobiotika entgiften. Mit den Wurzeln assoziierten Pilzen ist es zudem möglich, den Boden
weiträumig und feinmaschig mit ihren dünnen Pilzhyphen für sich und für die
Rhizosphärengemeinschaft zu erschließen. Im Wurzeleinzugsraum mit einem Abstand < 1mm zur
Wurzel beträgt die Anzahl der geschätzten Organismen ca. 1,2 x 1011 Zellen je cm3, in 2 cm

Einleitung
11
Entfernung lediglich 1,3 x 1010 (Paul & Clark 1989). Etwa 5-10 % der Wurzeloberfläche sind von
Bakterien besiedelt. Außerdem sind die Pilzhyphen der Mykorrhiza mit einem bakteriellen Biofilm
überzogen (Romantschuk et al. 2000). Weiterhin können durch schnellwüchsige und starkwurzelnde
Pflanzen die aktiven Mikroorganismen verstärkt im Boden verteilt werden. Pflanzen tragen also auf
vielfältige Weise zur Reinigung kontaminierter Böden bei. Entsprechendes trifft auch auf die mit den
Wurzeln der Pflanzen in Symbiose lebenden Mykorrhizapilze zu, deren Rolle in der Bodensanierung
im folgenden Abschnitt dargestellt wird.
1.2.3 Strukturen der Ektomykorrhiza; Bodenreinigung durch Mykorrhizapilze Im Jahr 1885 prägte A. B. Frank den Begriff Mykorrhiza, mit dem er eine Symbiose zwischen Pilzen
und den Wurzeln verschiedener Landpflanzen beschrieb. Diese für beide Partner förderliche
Lebensgemeinschaft ist bei etwa 90% aller an Land lebenden Gefäßpflanzen verbreitet (Steffens et
al. 1994). Unterschieden wird heute aufgrund anatomischer Merkmale zwischen ekto-, ektendo- und
endotrophen Mykorrhizen. Die ektotrophe Form der Mykorrhiza bilden nur ca. 3% aller Arten der
Samenpflanzen aus. Die symbiontischen Pilze der Ektomykorrhiza gehören überwiegend zu den
Basidiomyceten und zu den Ascomyceten. Bei den Pflanzen mit ektotropher Mykorrhiza dominieren
die Bäume und Sträucher. Auch die im Rahmen der vorliegenden Arbeit in die Experimente
einbezogenen Pflanzen der Arten Pinus sylvestris und Populus tremula gehören zu den Vertretern
mit dieser Mykorrhiza-Form, wobei Populus tremula auch zu den endotrophen Mykorrhizabildnern
(VA-) gezählt wird (Harley & Harley 1987).
Auffällig ist das sehr einheitliche Bauprinzip von Ektomykorrhizastrukturen. Ein äußerer Mantel von
Pilzhyphen umschließt als dichtes Geflecht bevorzugt die Feinwurzeln der Wirtspflanzen. Folglich
müssen Stoffe aus dem Boden, wenn sie letztendlich in die Pflanzen gelangen sollen, den Weg über
die Pilzhyphen nehmen. Die durch die Ummantelung der Pilzhyphen allgemein in ihrem
Längenwachstum reduzierten und keulig verdickten Kurzwurzeln sind durch ein Geflecht von
Pilzhyphen zwischen ihren Rhizodermis- und Rindenzellen, dem sogenannten Hartigschen Netz,
gekennzeichnet. Das Hartigsche Netz entwickelt sich bei den verschiedenen Gattungen der
Wirtspflanzen unterschiedlich tief im Interzellularraum zwischen den Zellen, dringt aber nie in den
Zentralzylinder ein. Die Pilzhyphen mit ihren fingerförmigen Strukturen können die Zellen des
Wirtsgewebes nahezu vollständig umschließen und bilden mit den Wirtszellen durch engen Kontakt
ein ‘Interface’, eine gemeinsame Austauschzone für unterschiedlichste Nährstoffe (Kottke u.
Oberwinkler 1987, Massicotte et al. 1987).
Obwohl ektotroph mykorrhizierten Wurzeln aufgrund des sie umgebenden Hyphenmantels
Wurzelhaare fehlen, erfahren die Pflanzen durch den Pilzsymbionten eine verbesserte Nährstoff- und
Wasseraufnahme. Dem Mykorrhizapilz kommt hierbei eine Schlüsselrolle zu. Gegenüber nicht-

Einleitung
12
mykorrhizierten Feinwurzeln mit ihren Wurzelhaaren durchwachsen Pilzhyphen ein weitaus größeres
Bodenareal, aus dem sie Wasser und Nährstoffe entnehmen können. Die dünnen Hyphen haben
überdies den Vorteil, kleinste Interkapillarräume für ihren Wirtspartner erschließen zu können (Hilber
1992). Neben einer verbesserten Wasseraufnahme erfahren die mykorrhizierten Pflanzen vor allem
eine effizientere Versorgung mit Phosphor, Stickstoff und anderen Nährelementen (Werner 1987,
Hilber 1992). Besonders Phosphat steht den Pflanzen unter natürlichen Bedingungen im Boden oft
nur in sehr geringen Mengen zur Verfügung. Mykorrhizapilze besitzen die Fähigkeit, unter Abgabe
organischer Säuren fest im mineralischen Gestein gebundene Phosphate verstärkt in Lösung zu
bringen (Steffens et al. 1994). Über den Pilz gelangen die Phosphate schließlich in die Wurzeln.
Erschöpft sich die Phosphataufnahme durch die Mykorrhizen, können die Pflanzen von den seitens
der symbiontischen Pilze angelegten Phosphatspeichern partizipieren (Werner 1987). Im Gegenzug
profitiert der Pilz in erster Linie von Kohlenhydraten, aber auch von verschiedenen Aminosäuren und
Vitaminen, die er von der Pflanze erhält (Werner 1987, Steffens et al. 1994).
Die Bedeutung der Mykorrhiza in biologischen Verfahren zur Bodenreinigung wird zunehmend
erkannt. Das Spektrum an Xenobiotika, das durch den Einsatz dieser Pilz-Pflanze-Symbiose
wirksam reduziert werden kann, findet gegenwärtig eine ständige Erweiterung. Die
Nitroaromatenproblematik blieb hiervon allerdings weitgehend ausgeschlossen und das Potenzial
mykorrhizierter Pflanzen beim TNT-Abbau bisher unerforscht. Experimentelle Untersuchungen hierzu
wurden entweder mit Mykorrhizapilzen selbst oder mit Pflanzen, jedoch nicht unter dem Aspekt der
gemeinsamen Symbiose durchgeführt. Die Fähigkeit zur Nitroaromatendegradation ist u. a. für die
Mykorrhizapilzspezies Paxillus involutus, Pisolithus tinctorius und Suillus variegatus (Scheibner
et al. 1997, Meharg et al. 1997a, Meharg & Cairney 2000) nachgewiesen. Experimente mit
Paxillus involutus ergaben eine 100 %ige Metabolisierung von 250 µmol TNT innerhalb von 6
Tagen. Allerdings wurde nur etwa 1,6 % des dotierten 14C-TNT vollständig mineralisiert (Scheibner
et al. 1997). Auch andere persistente organische Schadstoffe lassen sich durch Mykorrhizapilze
abbauen. Zum Schadstoffspektrum zählen verschiedene PAK (Gramss et al. 1999), Polychlorierte
Biphenyle (Donnelly & Fletcher 1995), 2,4-Dichlorphenol (Meharg 1997b) und Chlorpropham
(Rouillon et al. 1989). Durchaus vergleichbar mit den Weißfäulepilzen beruht die Fähigkeit der
Biodegradation bei Mykorrhizapilzen auf dem Besitz unspezifischer, extrazellulärer Enzymsysteme,
die oxidative Angriffe am Kohlenstoffring katalysieren können. Unter den Phenol oxidierenden
Enzymen konnten bei Mykorrhizapilzen bisher Tyrosinase, Catechol-Oxidase, Ascorbat-Oxidase
und Laccasen nachgewiesen werden. Außerdem fanden sich Lignin-Peroxidase und Mangan-
Peroxidase (Colpaert & van Laere, Gramss 1997, Timonen & Sen 1998, Griffiths & Caldwell
1992, Cairney & Burke 1998).
In taxonomischer Hinsicht wurden bisher nur relativ wenige Spezies auf ihre Fähigkeit zur
Biodegradation organischer Kontaminanten gescreent. Die große Artenvielfalt sowie die Variationen

Einleitung
13
innerhalb einer Spezies in physiologischer Hinsicht rechtfertigen die Annahme, dass noch zahlreiche
Mykorrhizapilze, insbesondere Arten, die auch als Streuzersetzer in Frage kommen, bei
bodenbiologischen Sanierungsvorhaben sinnvoll eingesetzt werden können. Eine maßgebliche
Förderung im Remediationsprozess könnte zusätzlich durch die oft vorherrschende hohe Diversität
mykorrhizierender Pilze im Boden erfolgen. So ließen sich in einem Hektar Waldboden über hundert
Morphotypen nachweisen (Jonsson et al. 1999).
1.3 Fragestellung und Zielsetzung des Arbeitsvorhabens Vor dem Hintergrund des kurz dargestellten Wissensstand über die bestehenden Möglichkeiten,
Weißfäule- und Mykorrhizapilze sowie Pflanzen beim Abbau TNT-relevanter Verbindungen in
biologischen Bodensanierungsverfahren einzusetzen, blieb erstaunlicherweise ein gewichtiger Punkt
bisher unerforscht und unbeantwortet. Welches Sanierungspotenzial bietet eine Kombination dieser
Organismen in einem Phyto-remediationsverfahren zur Reinigung sprengstoffbelasteter Böden? Im
Mittelpunkt der hier durchgeführten Laboruntersuchungen standen daher Überlegungen für ein
modellhaftes Phytoremediationskonzept, das auf mikrobielle Degradation der
Sprengstoffverbindungen im Einzugsbereich ektotroph mykorrhizierter Wurzeln unter Beteiligung von
Bakterien, Pilzen und Pflanzen als Organismengemeinschaft beruht (Rhizosphärendegradation). In
mehreren Reaktionsschritten kommt es zunächst zu einer enzymatischen Reduktion des TNTs.
Abgesehen von den Rhizosphärenbakterien und Pilzen können an diesem Prozess auch die Pflanzen
beteiligt sein. Nach initialer Reduktion sollen enzymatische Oxidationen am Aromaten seitens der
Weißfäule- und Mykorrhizapilze letztendlich zu einer Ringspaltung in der Verbindung führen. Die
entstehenden stickstoffhaltigen Ringspaltungsprodukte können dann in weiteren Schritten von den
Bakterien verwertet (im Blickpunkt steht hier insbesondere der bakterielle Biofilm auf den Hyphen)
oder von den mit den Pflanzen assoziierten und im Boden weitverzweigten Mykorrhizapilzhyphen
aufgenommen und metabolisiert bzw. an die Wirtspflanze des Pilzes abgegeben werden. Ergänzend
hierzu erfährt die bakterielle Aktivität der Rhizosphäre durch Wurzelexsudation seitens der Pflanzen
und einer sich daraus ergebenden Verbesserung des Substratangebots für die Mikroorganismen eine
erhebliche Steigerung, so dass von einem zusätzlichen cometabolischen Umsatz der Kontaminanten
auszugehen ist. Reduzierte TNT-Metabolite können darüber hinaus eine Festlegung durch
Adsorption an Bodenbestandteilen erfahren. Von Adsorptionsprozessen sind neben den
Diaminonitrotoluolen hauptsächlich Triaminotoluole betroffen. Der Reduk-tionsschritt zum
Triaminotoluol erfolgt nur unter streng anaeroben Bedingungen. Infolge der mikroskopischen
Heterogenität des Rhizosphärenraumes kann allerdings schon über eine Distanz von wenigen
Millimetern ein Übergang von vollständiger Aerobie zu vollständiger Anaerobie erfolgen. Gerade eine

Einleitung
14
starke Mikroorganismentätigkeit an der Wurzeloberfläche kann mitunter zu semianaeroben
Bedingungen führen, in deren Folge anaerobe Prozesse ermöglicht werden (Gisi et al. 1990).
Im Blickpunkt der Versuche und bei der Wahl des experimentellen Versuchsdesigns steht die
Symbiose der Mykorrhiza. Wie bereits erwähnt wurde, können verschiedene Spezies der
Mykorrhizapilze TNT und TNT-Metabolite enzymatisch angreifen. Sie sind somit nicht nur
Verwerter der Ringspaltungsprodukte. Im Hinblick auf diese Tatsache erscheint die Frage berechtigt,
ob möglicherweise die ursprünglich in den Holzanteilen von Pflanzen und der Streuschicht
verbreiteten und wenig an das Bodenleben angepassten saprophytischen Weißfäulepilze von den
bodenbewohnenden Mykorrhizapilzen ergänzt oder ersetzt werden können. Die enge Assoziation
der Mykorrhizapilze mit den Pflanzen sowie deren Fähigkeit zur Ausbildung weiträumiger und
feinmaschiger Hyphengeflechte im Boden ermöglichen eine äußerst effiziente Besiedlung weiter
Bodenbereiche über den Wurzeleinzugsbereich hinaus. Außerdem ist eine Substratzugabe in den
Boden, wie es für Weißfäulepilze erforderlich ist, nicht notwendig. Dazu wird die Vitalität beider
Symbiosepartner durch die Mykorrhiza in der Regel gestärkt mit wiederum positiven Folgen für die
assoziierten Bakterien und deren metabolische Aktivität. Eine zentrale Bedeutung im angestrebten
Phytoremediationskonzept kommt zudem der Frage zu, ob und in welchem Ausmaß TNT und
Metaboliten von der Symbiose Pilz-Pflanze prinzipiell aufgenommen, akkumuliert oder metabolisiert
werden können. Die Voraussetzung für ein erfolgreiches Phytoremediationsverfahren ist entweder
durch eine umfassende Metabolisierung der Schadstoffe in der Rhizosphäre oder, falls es anteilig zur
Aufnahme kommt, durch eine grundlegende Detoxifizierung der Stoffe innerhalb der Symbionten
erfüllt.
Zusammenfassend sind die wesentlichen experimentellen Fragestellungen nachstehend aufgeführt:
Remediationsleistung: Können ektotroph mykorrhizierte Gehölze zur Reinigung sprengstoff-belasteter
Böden beitragen? Welche Bedeutung kommt dabei den jeweiligen Symbiose-partnern zu? Welche
Rolle spielen zubeimpfte Weißfäulepilze bei der Dekontamination TNT-belasteter Böden? Kann die
Aufgabe der Weißfäulepilze möglicherweise durch Mykor-rhizapilze übernommen werden?
Aufnahme und Akkumulationsverhalten: In welcher Größenordnung kommt es zur Aufnahme von
Nitroaromaten aus dem Boden in das System Mykorrhiza? Unterscheiden sich mykorrhizierte und
nicht-mykorrhizierte Pflanzen im Aufnahme- und Akkumulations-verhalten voneinander? Gibt es
einen Transfer aufgenommener Nitroaromaten in oberirdische Pflanzenteile? Liegt ein organ- oder
gewebespezifisches Akkumulationsverhalten vor, das Rückschlüsse auf Metabolisierungsvorgänge
innerhalb der Mykorrhiza zulässt?

Einleitung
15
Remediationspotenzial und Toxizität: Birgt der Kontakt der Wurzeln mit Nitroaromaten die Gefahr
toxischer Wirkungen auf die mykorrhizierten Pflanzen? Sind mit der Aufnahme der Schadstoffe in die
Pflanzen toxische Wirkungen auf die Pflanzen verbunden? Welche Schadstoffgehalte im Boden
können mykorrhizierte Gehölze tolerieren? Sind sie bezüglich ihres Toleranzverhaltens gegenüber
Nitroaromaten zur Remediation sprengstoffbelasteter Flächen geeignet?
Lassen sich die aus den Laborversuchen gewonnenen Erkenntnisse prinzipiell auf groß angelegte in-
situ Phytoremediationsverfahren zur Sanierung Sprenstoff-kontaminierter Böden übertragen? Diese
Frage wird in einem sich der Diskussion anschließenden Kapitel am Ende der Arbeit diskutiert. In
diesem Zusammenhang können die aus einem kürzlich maßstabsgerecht erprobten
Phytoremediationsverfahren gewonnenen Erkenntnisse (Warrelmann 2000, Koehler 2001a und b) in
die Diskussion einfließen, da Verfahrens-konzepte von Laborversuchen und in-situ Verfahren
weitgehende Übereinstimmungen zeigen. Das hier erwähnte in-situ Freilandexperiment wurde vom
UFT Bremen (Zentrum für Umweltforschung und Umwelttechnologie) in enger Kooperation mit der
Firma Umweltschutz Nord GmbH & Co Ganderkesee auf dem Gelände des ehemaligen
Sprengstoffwerks ‘Tanne’ bei Clausthal-Zellerfeld erprobt. Die Vergabe des Projektes erfolgte als
Unterauftrag von Umweltschutz-Nord im Rahmen eines Verbundvorhabens. Die finanzielle
Förderung wurde durch das BMBF (Projektträger Abfallwirtschaft und Altlastensanierung, Berlin)
getragen. Umfassende Arbeiten zum wissenschaftlichen Begleitmonitoring konnten darüber hinaus mit
finanzieller Unterstützung durch den Senator für Bildung und Wissenschaft der Freien und Hansestadt
Bremen durchgeführt werden.

Material und Methoden
2. Material und Methoden 2.1 Organismen in den Experimenten 2.1.1 Pilze In den Experimenten kamen sowohl saprophytische Weißfäulepilze als auch mit Pflanzen in Symbiose lebende Mykorrhizapilze zum Einsatz. Alle hier verwendeten Arten werden den Basidiomyceten zugeordnet. 2.1.1.1 Kultivierung der Mykorrhizapilze auf Agarmedium Als Mykobionten für die Testpflanzen wurden folgende Pilzkulturen verwendet: Pisolithus tinctorius (Pers.) Coker & Couch; Kultur von I. Kottke, Universität Tübingen Paxillus involutus (Batsch ex Fr.); Kultur von R. D. Finlay, Universität Lund Paxillus involutus (Batsch ex Fr.) Fr. Paxi 2; Kultur von T. Günther, Universität Jena Suillus bovinus (L.ex Fr.) Kuntze; Kultur von R. D. Finlay, Universität Lund Die Kultivierung der Mykorrhizapilze erfolgte auf einem modifizierten Melin-Norkans-Medium (MMN) nach Marx (1969) unter Zugabe von 2% Agar Agar (high gel strengh) in Kunststoffpetrischalen (Durchmesser 9 cm) im Dunkeln und bei Raumtemperatur. Das Rezept ist im Anhang aufgeführt (Tabelle 25). Zur Herstellung von Pilzinokulaten wurde mit einem Korkbohrer aus dem Randbereich der Schalen Pilzmyzel ausgestochen (Durchmesser 1 cm) und auf frisches MMN-Agar-Medium übertragen. Nach 5-10 Tagen entwickelten sich aus den Pellets wattebauschartige Myzelien. Diese konnten schließlich als Inokulate für die Beimpfung der Pflanzenwurzeln verwendet werden. 2.1.1.2 Kultivierung der Weißfäulepilze auf Agarmedium Im Vorfeld der Arbeit und während der Versuche wurde mit insgesamt 4 verschiedenen Weißfäulepilzarten experimentiert. Sämtliche Kulturen stammen von der DSM (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig). Pleurotus ostreatus (Jacquin ex Fries), Stamm Nr.1833 Trametes versicolor (Linnè ex Fries), Stamm Nr.1977 Phanerochaete chrysosporium (Burdsall & Eslyn), Stamm Nr.1556 Heterobasidion annosum (Fries ex Fries), Stamm Nr.2728
16

Material und Methoden
Die Kultivierung der verschiedenen Pilzstämme erfolgte auf einem besonderen von der DSM empfohlenen Malz-Pepton-Agar (Tabelle 26 im Anhang). Nach dem Beimpfen der Agar-Platten mit zuvor ausgestochenen Pellets, wurden die Pilzkulturen im Dunkeln und bei Zimmertemperatur aufbewahrt. Die vollständige Besiedlung der Oberflächen der Kulturplatten durch die Myzelien erfolgte je nach Pilzart in unterschiedlichen Zeiträumen. P. ostreatus und T. versicolor benötigten etwa 2-3, P. chrysosporium und H. annosum 3-4 Wochen für einen flächendeckenden Bewuchs in den Schalen. Die Pilzkulturen wurden alle 4-6 Wochen erneuert, dabei die Inokulate (Durchmesser 1cm) mit dem Korkbohrer aus dem Randbereich des Myzels ausgestochen und auf frisches Agar-Medium überimpft. 2.1.1.3 Kultivierung der Weißfäulepilze auf Stroh und Weichholzschredder Weißfäulepilze sind bezüglich ihrer Lebensraumansprüche keine typischen Bodenbewohner. Sie leben hauptsächlich in Holzanteilen von Bäumen und Sträuchern und in der Streuschicht. Dementsprechend galt es zunächst einmal für die Weißfäulepilze geeignete Bedingungen zu schaffen, damit sie im Boden überleben können, um dort Nitroaromaten abzubauen. Gesucht wurde ein lignin- und zellulosehaltiges Material, das sich von den Pilzen schnell besiedeln ließ und nach Einarbeitung in den Boden in seiner Eigenschaft längerfristig als Ernährungs-grundlage dienen würde. Als Substrat für die Pilze getestet wurden verschiedene geschredderte Weichlaubhölzer und Weizenstroh. Letztendlich fiel die Wahl unter den Pilzen auf Pleurotus ostreatus. Diese Pilzspezies zeigte die günstigsten Kultivierungs- und Vermehrungseigenschaften. Von den Substraten erwies sich Weizenstroh am geeignetsten. Die zerkleinerten und 60 min autoklavierten Strohhalme wurden etwa 6-8 Wochen nach Animpfen von den Hyphen des Pilzes dicht überwachsen. Das Stroh/Pilzsubstrat konnte nun mit dem kontaminierten Boden vermischt werden. Die Beimpfung des Strohs erfolgte mit Pilzinokulaten, die mittels eines Korkbohrers aus den Myzelien der Kulturplatten ausgestochen wurden. Als Gefäße für die Kultivierung fanden Weckgläser Verwendung.
17
Material und Methoden
Tabelle 1: Wachstum der Myzelien verschiedener Weißfäulepilze auf unterschiedlichen Kultursubstraten; der Anteil des bewachsenen Substrats ist in % angegeben
Pilzkultur auf Weichholz
Wachstum nach 8 Wochen
Wachstum nach 16 Wochen
P. ostreatus 10 20 T. versicolor 10 10
P. chrysosporium >5 >5 H. annosum >5 >5
Pilzkultur auf Weizenstroh
P. ostreatus 80 100 T. versicolor 30 50
P. chrysosporium 10 10 H. annosum 10 10
2.1.2 Pflanzen In den Experimenten wurden Pflanzen von Pinus sylvestris L. (Gemeine Kiefer oder Föhre) und Populus tremula L. (Zitter-Pappel) verwendet. 2.1.2.1 Anzucht der Kiefern Samen von Pinus sylvestris L. (Fa. Rahte, Wietze) wurden einen Tag in Leitungswasser vorgequollen, davon die ersten 60 Minuten unter fließendem Wasser, und danach 30 min mit H2O2 (30%) oberflächensterilisiert. Anschließend wurden die Samen nochmals kurz mit sterilem Leitungswasser nachgespült. Die Aussaat der keimungsfähigen Samen erfolgte in einem feuchten und autoklavierten Sand/Perlite Gemisch (Perlite ist ein künstliches Tonmineral). Unter kontrollierten, nicht-sterilen Bedingungen wuchsen die Pflanzen bis zur weiteren Verwendung in besonderen Phytotron-Klimakammern heran (Tages-/Nachtrhythmus: 14 h hell mit Lichtstärke 45 W m-2/ 10 h dunkel, 20°C/16°C, rel. Luftfeuchtigkeit 70%). Gegossen wurde nach Bedarf mit Leitungswasser. Die Düngung erfolgte etwa alle 4 Wochen mit einer besonderen Nährlösung nach Ingestad (1960), deren Bestandteile im Anhang genannt sind. 2.1.2.2 Herkunft der Pappeln Reife Pappelsamen bleiben nur wenige Tage keimfähig. Eine Erfolg versprechende Aussaat ist kompliziert und mit aufwendigen Arbeitsschritten verbunden. Aus diesen Gründen wurden die für die Experimente benötigten Pappeln nicht selbst angezogen, sondern als einjährige Pflänzchen von der Pflanzenhandlung Bunk (Elmshorn/ Schleswig-Holstein) bezogen. Die
18

Material und Methoden
etwa handbreit hohen aus Stecklingen gezogenen Pappeln wurden dankenswerter Weise unentgeldlich zur Verfügung gestellt. Die Kultivierung der kleinen Pappeln erfolgte bis zur weiteren Verwendung in den Experimenten in einem Sand/Torf Gemisch in Petrischalen (Durchmesser 9 cm). Die Pflanzen standen ohne künstliche Beleuchtung in einer geschlossenen Gewächshauskabine im Tageslicht (natürlicher Tag/Nacht Rhythmus), wobei die Tag/Nacht Temperatur 20°C/16°C betrug. Die rel. Luftfeuchtigkeit lag bei 70%. Gegossen wurde mit Leitungswasser nach Bedarf. Das Substrat wurde feucht, jedoch nicht nass gehalten. Gedüngt wurde mit flüssigem Volldünger (Tabelle 30 im Anhang) gelöst in Gieß-wasser während des Blattaustriebes einmal in 14 Tagen, sonst während der Vegetations-periode einmal in 4 Wochen. 2.1.2.3 Mykorrhizierung der Kiefern in Rhizotronkulturen Ab einem Alter von 8 Wochen erfolgte die weitere Kultivierung der jungen Kiefern in den Gewächshäusern des Biologischen Gartens der Universität wie unter Punkt 2.1.2.2 beschrieben. Nach einer Eingewöhnungszeit an die neuen Klimaverhältnisse von 4-6 Wochen konnten die Pflanzen schließlich mit den entsprechenden Pilzsymbionten beimpft werden. Für die Mykorrhizierung wurden die inzwischen 3-4 Monate alten Kiefern nach gründlichem Spülen ihrer Wurzeln unter fließendem Leitungswasser in spezielle Rhizotrone überführt (modifizierte Petrischalen, Durchmesser 9cm). Eine Lage steriles Aktivkohlepapier (Fa. Schleicher und Schüll) trennte die Wurzeln in diesem Verfahren (abgewandelt nach Kottke et al. 1987) von dem autoklavierten Perlite-Substrat. In den Petrischalen bildeten die kleinen Kiefern nach 2-3 Wochen genügend junge Seiten- oder Feinwurzeln mit wachsender Wurzelspitze aus. In der Regel sind im Wachstum befindliche Wurzeln leichter zu mykorrhizieren. Die Pilzinokulate wurden vorsichtig an die auswachsenden Wurzeln angelegt. Mykorrhizen etablierten sich nach 10-14 Tagen. 2.1.2.4 Mykorrhizierungen an Pappeln Auf eine selbst durchgeführte Beimpfung der Pappeln mit Mykorrhizapilzen wurde verzichtet, da die Feinwurzeln bei einem Teil der Setzlinge bereits unmittelbar nach Erhalt aus der Baumschule etablierte Mykorrhizen aufwiesen. Die taxonomische Zugehörigkeit des an der Mykorrhiza beteiligten Pilzes war nicht bekannt und sollte deshalb festgestellt werden. Grundsätzlich bestand die Möglichkeit einer Mehrfachinfektion der Pappeln mit verschiedenen Pilzen. Eine Identifizierung des oder der pilzlichen Symbionten lediglich anhand charakteristischer äußerer Merkmale des Hyphenmantels durchzuführen ist schwierig und unsicher. Die Bestimmung erfolgte daher mit genetischen Bestimmungsmethoden im Labor für Bioanalytik der Universität Bremen, AG Prof. Hildebrandt. Für dieses Vorhaben
19

Material und Methoden
wurden mykorrhizierte Wurzelproben von etwa 1 cm Länge von den Pappeln geerntet, lyophilisiert, homogenisiert und anschließend unter sterilen Bedingungen aus den Proben die DNA extrahiert. Nach durchgeführter PCR (Polymerase Kettenreaktion) wurden die PCR-Amplifikationen (Summe der in der PCR erzeugten DNA-Stränge) durch nachfolgende Gelelektrophorese in ein charakteristisches DNA-Bandenmuster aufgetrennt. Durch einen Abgleich mit den Bandenmustern bekannter Mykorrhizapilze bestand nun die Möglichkeit, die unbekannte Spezies mit hoher Zuverlässigkeit taxonomisch einzuordnen (Methoden-beschreibung nach persönlicher Mitteilung von Christoph Kulmann und Dr. Carsten Harms, AG Prof. Hildebrandt, Universität Bremen). Aus dem DNA-Bandenvergleich ging hervor, dass die untersuchten Pappelwurzeln mit zwei Spezies der Gattung Hebeloma eine Symbiose eingegangen waren. Bei dem einen Pilz handelte es sich entweder um H. helodes oder um H. cavipes. Der andere Pilz war entweder mesophaeum oder H. collariatum zugehörig. 2.2 Experimente 2.2.1 Kultivierung von Kiefern auf TNT-belastetem Boden (Kiefernexperiment) Mit der Durchführung dieses Experimentes sollte überprüft werden, ob und inwieweit durch den Einsatz von mykorrhizierten Kiefern (Pinus sylvestris / Pisolithus tinctorius) unter Einbeziehung des Weißfäulepilzes Pleurotus ostreatus die Transformation von Nitroaromaten im Boden gefördert werden kann. Etwa 3-4 Monate alte Kiefernsetzlinge wurden in vier Versuchsvarianten (mit jeweils 32 Parallelansätzen) auf einem TNT-belasteten Boden vom Standort des ehemaligen Sprengstoffwerkes „Tanne“ (Clausthal-Zellerfeld/ Harz) kultiviert und die Entwicklung der Nitroaromatengehalte im Boden untersucht. Vor dem Befüllen der Rhizotrone mit jeweils 70 g Bodensubstrat wurde der durch einen hohen Schluffanteil charakterisierte und skelettreiche Standortboden in luftgetrocknetem Zustand mehrmals gesiebt (Maschenweite 2 mm) und anschließend mit einem Drittel seines Gewichtes mit getrocknetem Sand vermischt. Verschiedene bodenkundliche Angaben zum Substrat können den Tabellen 31 und 32 im Anhang entnommen werden. In den entsprechenden Varianten wurde der Boden in den Rhizotronen dann mit jeweils 3 Gramm Weißfäulepilz-Stroh-Substrat beimpft. Die Kultivierung der Pflanzen in den Rhizotronen erfolgte in einer Gewächshaus-kabine wie unter Punkt (2.1.2.2) beschrieben.
20

Material und Methoden
Das experimentelle Design beinhaltete folgendes Variantenspektrum: Variante A: Kultivierung von mykorrhizierten Kiefern auf belastetem Boden unter Zugabe von Weißfäulepilz-Stroh-Substrat Variante B: Kultivierung von mykorrhizierten Kiefern auf belastetem Boden, auf die Zugabe von Weißfäulepilz-Stroh-Substrat wurde verzichtet Variante C: ohne Pflanzen, für diesen Ansatz wurde der kontaminierte Boden lediglich mit Weißfäulepilz-Stroh-Substrat vermischt Variante K: in diesem Kontrollansatz wurde das Bodensubstrat lediglich mechanisch bearbeitet (Homogenisieren durch Sieben), mykorrhizierte Kiefern und Weißfäulepilze fehlten in dieser Variante
Abb. 5: Rhizotron der Versuchsvariante A mit mykorrhizierter Kiefer und entwickeltem Weiß-fäulepilzmyzel.
Kiefernsämling (Pinus sylvestris)
Petrischale seitlich mit Öffnung
Weißfäulepilz (Pleurotus ostreatus) auf Weizenstroh
TNT-belasteter Boden
Der Versuchszeitraum erstreckte sich insgesamt über 24 Wochen. Nach 8, 12, 20 und 24 Wochen wurden 8 Parallelansätze jeder Versuchsvariante beprobt. Die Beprobung beinhaltete in diesem Experiment die Entnahme einer Bodenmischprobe (nach Lufttrocknung 5 g) für die Nitroaromatenextraktion aus jedem Rhizotron sowie die Entnahme einer 10 g Bodenprobe aus
21
Material und Methoden
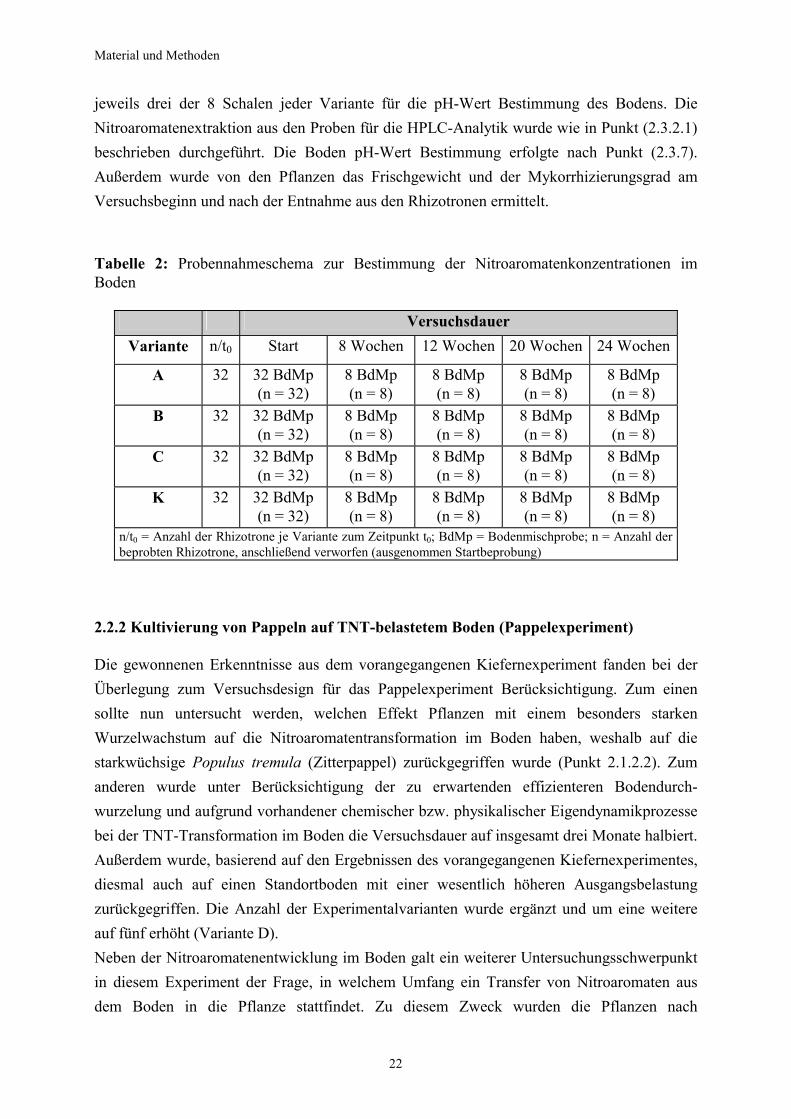
jeweils drei der 8 Schalen jeder Variante für die pH-Wert Bestimmung des Bodens. Die Nitroaromatenextraktion aus den Proben für die HPLC-Analytik wurde wie in Punkt (2.3.2.1) beschrieben durchgeführt. Die Boden pH-Wert Bestimmung erfolgte nach Punkt (2.3.7). Außerdem wurde von den Pflanzen das Frischgewicht und der Mykorrhizierungsgrad am Versuchsbeginn und nach der Entnahme aus den Rhizotronen ermittelt. Tabelle 2: Probennahmeschema zur Bestimmung der Nitroaromatenkonzentrationen im Boden
Versuchsdauer Variante n/t0 Start 8 Wochen 12 Wochen 20 Wochen 24 Wochen
A 32 32 BdMp (n = 32)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
B 32 32 BdMp (n = 32)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
C 32 32 BdMp (n = 32)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
K 32 32 BdMp (n = 32)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
n/t0 = Anzahl der Rhizotrone je Variante zum Zeitpunkt t0; BdMp = Bodenmischprobe; n = Anzahl der beprobten Rhizotrone, anschließend verworfen (ausgenommen Startbeprobung)
2.2.2 Kultivierung von Pappeln auf TNT-belastetem Boden (Pappelexperiment) Die gewonnenen Erkenntnisse aus dem vorangegangenen Kiefernexperiment fanden bei der Überlegung zum Versuchsdesign für das Pappelexperiment Berücksichtigung. Zum einen sollte nun untersucht werden, welchen Effekt Pflanzen mit einem besonders starken Wurzelwachstum auf die Nitroaromatentransformation im Boden haben, weshalb auf die starkwüchsige Populus tremula (Zitterpappel) zurückgegriffen wurde (Punkt 2.1.2.2). Zum anderen wurde unter Berücksichtigung der zu erwartenden effizienteren Bodendurch-wurzelung und aufgrund vorhandener chemischer bzw. physikalischer Eigendynamikprozesse bei der TNT-Transformation im Boden die Versuchsdauer auf insgesamt drei Monate halbiert. Außerdem wurde, basierend auf den Ergebnissen des vorangegangenen Kiefernexperimentes, diesmal auch auf einen Standortboden mit einer wesentlich höheren Ausgangsbelastung zurückgegriffen. Die Anzahl der Experimentalvarianten wurde ergänzt und um eine weitere auf fünf erhöht (Variante D). Neben der Nitroaromatenentwicklung im Boden galt ein weiterer Untersuchungsschwerpunkt in diesem Experiment der Frage, in welchem Umfang ein Transfer von Nitroaromaten aus dem Boden in die Pflanze stattfindet. Zu diesem Zweck wurden die Pflanzen nach
22

Material und Methoden
Versuchsende geerntet und die Nitroaromaten aus den Geweben extrahiert (Methode abgewandelt nach Görge et al. 1994). Die Herstellung und Bestückung der Rhizotrone erfolgte wie bereits in Punkt (2.2.1) beschrieben. Die für das Pappelexperiment eingesetzten Versuchsvarianten sind nachstehend aufgeführt. Variante A: Kultivierung von mykorrhizierten Pappeln auf belastetem Boden unter Zugabe von Weißfäulepilz-Stroh-Substrat Variante B: Kultivierung von mykorrhizierten Pappeln auf belastetem Boden, auf die Zugabe von Weißfäulepilz-Stroh-Substrat wurde verzichtet Variante C: ohne Pflanzen, für diesen Ansatz wurde der kontaminierte Boden lediglich mit Weißfäulepilz-Stroh-Substrat vermischt Variante D: wie Variante A, allerdings waren die Rhizotrone mit nicht-mykorrhizierten Pappeln bestückt Variante K: in diesem Kontrollansatz wurde das Bodensubstrat wie in den übrigen Varianten lediglich mechanisch bearbeitet (Homogenisieren durch Sieben), mykorrhizierte Pappeln und Weißfäulepilze fehlten in dieser Variante
23

Material und Methoden
TNT-belastete
Abb. 6: Rhizotron der Variante A mit mykorrhizierter Pappel und entwickeltem Weißfäulepilzmycel.
Zitterpappel (Populus tremula)
Petrischale seitlich mit Öffnung
Weißfäulepilz (Pleurotus ostreatus) auf Weizenstroh
r Boden
Der Versuch wurde nach 12 Wochen beendet. Beprobung und Ernte erfolgten nach 2, 4, 8 und 12 Wochen. Die Entnahme der Bodenproben für die Nitroaromatenbestimmung und die pH-Wert-Messung ist in den Punkten 2.3.2.1 und 2.3.7 beschrieben. Zur Erfassung der Nitroaromatenmengen in den Pflanzengeweben erfolgte eine Auftrennung in Wurzel- und Sprossanteile (Stamm/Blatt). Für die sich anschließende Extraktion wurden die Proben aus jeweils vier Rhizotronen zu einer Mischprobe zusammengefasst und wie in Punkt 2.3.2.2 beschrieben extrahiert.
24
Material und Methoden
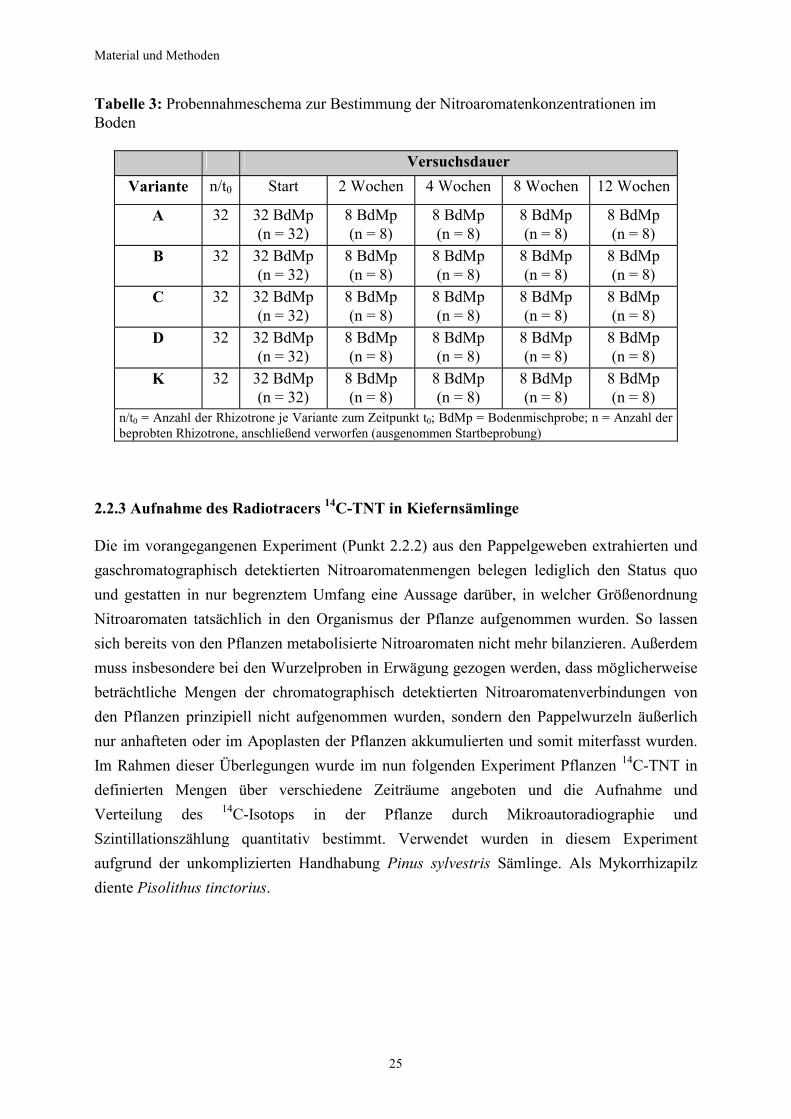
Tabelle 3: Probennahmeschema zur Bestimmung der Nitroaromatenkonzentrationen im Boden
Versuchsdauer Variante n/t0 Start 2 Wochen 4 Wochen 8 Wochen 12 Wochen
A 32 32 BdMp (n = 32)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
B 32 32 BdMp (n = 32)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
C 32 32 BdMp (n = 32)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
D 32 32 BdMp (n = 32)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
K 32 32 BdMp (n = 32)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
8 BdMp (n = 8)
n/t0 = Anzahl der Rhizotrone je Variante zum Zeitpunkt t0; BdMp = Bodenmischprobe; n = Anzahl der beprobten Rhizotrone, anschließend verworfen (ausgenommen Startbeprobung)
2.2.3 Aufnahme des Radiotracers 14C-TNT in Kiefernsämlinge Die im vorangegangenen Experiment (Punkt 2.2.2) aus den Pappelgeweben extrahierten und gaschromatographisch detektierten Nitroaromatenmengen belegen lediglich den Status quo und gestatten in nur begrenztem Umfang eine Aussage darüber, in welcher Größenordnung Nitroaromaten tatsächlich in den Organismus der Pflanze aufgenommen wurden. So lassen sich bereits von den Pflanzen metabolisierte Nitroaromaten nicht mehr bilanzieren. Außerdem muss insbesondere bei den Wurzelproben in Erwägung gezogen werden, dass möglicherweise beträchtliche Mengen der chromatographisch detektierten Nitroaromatenverbindungen von den Pflanzen prinzipiell nicht aufgenommen wurden, sondern den Pappelwurzeln äußerlich nur anhafteten oder im Apoplasten der Pflanzen akkumulierten und somit miterfasst wurden. Im Rahmen dieser Überlegungen wurde im nun folgenden Experiment Pflanzen 14C-TNT in definierten Mengen über verschiedene Zeiträume angeboten und die Aufnahme und Verteilung des 14C-Isotops in der Pflanze durch Mikroautoradiographie und Szintillationszählung quantitativ bestimmt. Verwendet wurden in diesem Experiment aufgrund der unkomplizierten Handhabung Pinus sylvestris Sämlinge. Als Mykorrhizapilz diente Pisolithus tinctorius.
25

Material und Methoden
Den Pflanzen wurde das14C-TNT mit einer radiochemischen Reinheit > 98% in einer Ingestad Nährlösung in zwei verschiedenen Konzentrationen 3, 7 und 14 Tage angeboten: 0,85 x 104 Bq in 2 ml Düngelösung je Schale, (0,00075 mg TNT ml-1 oder 0,75 mg TNT l-1) 17 x 104 Bq in 2 ml Düngelösung je Schale, (0,015 mg TNT ml-1 oder 15mg TNT l-1). Tabelle 4 gibt eine Übersicht über die verschiedenen Applikationszeiten des Radiotracers: Tabelle 4: Übersicht über die Anzahl der untersuchten Kiefern nach entsprechender Applikation; myk (-) (nicht- mykorrhiziert), myk (+) (mykorrhiziert)
Applikation niedriges 14C Angebot hohes 14C Angebot
myk (-) myk (+) myk (-) myk (+)
3 Tage 3 Pflanzen 3 Pflanzen 3 Pflanzen 3 Pflanzen
7 Tage 3 Pflanzen 3 Pflanzen 3 Pflanzen 3 Pflanzen
14 Tage 3 Pflanzen 3 Pflanzen 3 Pflanzen 3 Pflanzen
Als Rhizotrone dienten für die Pflanzen wiederum Petrischalen ( 9 cm Durchmesser), allerdings wurde diesmal auf eine Füllung mit Perlite verzichtet. Die Kohlepapierscheiben lagen den Schalenböden plan auf und wurden im weiteren Versuchsverlauf mit Leitungswasser stets feucht gehalten. Die Auflage von Glasobjektträgern gewährleistete bei Bedarf den direkten Kontakt der Wurzeln mit dem Kohlepapier. Nach der Ernte wurden die Pflanzenproben entsprechend ihrer weiteren Bestimmung entweder mit schmelzenden Stickstoff kryofixiert und anschließend in flüssigem Stickstoff aufbewahrt (für die Mikroautoradiographie, siehe Punkt 2.3.5) oder im Ofen bei 70° C getrocknet (für die Szintillationszählung, siehe Punkt 2.3.4). Durch die Kryofixierung konnten die Transportvorgänge in den Pflanzen schlagartig unterbrochen werden. Somit blieb das Verteilungsmuster des aufgenommenen 14C-TNT in den Pflanzen zum Erntezeitpunkt für die Mikroautoradiographie erhalten.
26

Material und Methoden
2.2.4 Phytotoxizitätstest mit Kiefern Die toxische Wirkung von TNT auf höhere Pflanzen ist bisher noch wenig untersucht. Toxizitätsdaten liegen größtenteils für krautige Pflanzen und Algen vor, wobei sich diese Daten nicht ohne weiteres auf Gehölzpflanzen übertragen lassen. Voraussetzung für den Erfolg eines Phytoremediationsverfahrens ist die Toleranz der eingesetzten Pflanzen gegenüber bodenrelevanten Schadstoffgehalten. Entsprechendes gilt auch für Mykorrhiza-pilze, wenn die Pflanzen mykorrhiziert sind. Mit einem Toxizitätstest sollte der Einfluss des TNTs auf mykorrhizierte Gehölzpflanzen untersucht werden. Die Kultivierung der kleinen mykorrhizierten Kiefernsämlinge erfolgte bei verschiedenen TNT-Konzentrationen im Bodensubstrat. 2.2.4.1 Vorbereitung der Pflanzen Als Testpflanzen dienten 4-5 Monate alte Kiefern (Pinus sylvestris). Die Anzucht erfolgte unter semisterilen Bedingungen wie in Punkt 2.1.2.1 beschrieben. Mykorrhiziert wurden die Pflanzen, wie in Punkt 2.1.2.3 erläutert. 2.2.4.2 Vorbereitung des Bodensubstrates Luftgetrockneter LUFA-Boden vom Typ 2.2 wurde durch sorgfältiges Sieben (Maschenweite 2 mm) homogenisiert und anschließend gezielt mit TNT angereichert. Dazu wurde kristallines TNT (Reinheitsgrad > 99%) zuvor in Aceton gelöst und zusammen mit dem Lösungsmittel mit Quarzsand vermengt. Das Aceton verdampfte bei ständigem Umrühren unter dem Abzug rasch, wobei sich das zurückgebliebene und getrocknete TNT-Quarzsand-Gemisch anschließend bequem mit dem vorbereiteten LUFA-Boden vermischen ließ. Es wurden Substrate mit Ausgangskonzentrationen von 3, 300 und 3000 mg TNT/kg Boden hergestellt. 2.2.4.3 Homogenitätskontrolle Im Rahmen einer Homogenitätskontrolle wurde überprüft, ob das zudotierte TNT in den Testböden in gleichmäßiger Verteilung vorlag. Von jeder Bodenvariante wurden fünf Mischproben (5 g) auf ihren TNT-Gehalt untersucht. Die Nitroaromatenextraktion erfolgte wie in Punkt 2.3.2.1 beschrieben. Zur Nitroaromatenanalytik siehe Punkt 2.3.3.1.
27

Material und Methoden
2.2.4.4 Versuchsdesign und Probennahme Für die Durchführung des Phytotoxizitätstest kamen als Rhizotrone modifizierte Petrischalen (Durchmesser 9 cm) zur Anwendung. Diese Pflanzenkammern erhielten im Rand eine kleine Öffnung für die Sprossachse und wurden mit 70 g kontaminiertem Boden gefüllt. Die kleinen mykorrhizierten Kiefern wurden zur Bestimmung des Frischgewichtes mit einer Analysenwaage (Sartorius) gewogen. Die Ermittlung des Mykorrhizierungsgrades der Feinwurzeln erfolgte mit der Stereolupe. Anschließend wurden die Setzlinge in den angefeuchteten Boden gepflanzt. Um Sickerverluste von TNT durch austretendes Wasser zu vermeiden, wurden die Schalen zusätzlich mit einem Streifen Parafilm abgedichtet. Die Exposition der Pflanzen erfolgte in Böden mit TNT-Konzentrationen von 3, 300 und 3000 mg/kg Boden (lufttrocken) sowie in einem unbelasteten Kontrollboden für maximal 12 Wochen. Nach einer Dauer von 4, 8 und 12 Wochen wurden jeweils fünf Parallelansätze von jeder Bodenvariante geerntet, siehe dazu auch Tabelle 5. Nach Ermittlung von Frischgewicht und Mykorrhizierungsgrad der Pflanzen sind die Nadeln auf Nekrosen und Verfärbungen untersucht worden, außerdem wurde der allgemeine Entwicklungszustand und das Erscheinungsbild der Wurzeln kontrolliert. Da aufgrund chemischer Einflüsse und mikrobieller Abbautätigkeit mit einem stetigen Rückgang der TNT-Belastung im Boden während der Versuchszeit zu rechnen war, musste parallel zum Pflanzenmonitoring die Entwicklung der Schadstoffkonzentration im Bodensubstrat im zeitlichen Verlauf bestimmt werden. Deshalb wurde zur Ermittlung des TNT-Gehaltes aus jeder Petrischale nach 2, 4, 8 und 12 Wochen eine Bodenmischprobe von 5 g (lufttrocken) entnommen und wie in Punkt 2.3.2.1 beschrieben extrahiert und analysiert. Als zusätzliche Begleituntersuchung wurde außerdem die TNT-Konzentration in der Bodenlösung bestimmt und die hierfür erforderliche Probenentnahme mit der Probenentnahme zur Bodenuntersuchung verbunden (Tabelle 5). Die Bodenlösung wurde aus 2,5 Gramm feuchtem bis nassem Boden 10 Minuten bei 14000 rpm abzentrifugiert (Centrifuge 5415 C, Fa. Eppendorf) und anschließend filtriert ( Micropure Filter, 22µm, Fa. Amicon). Aufgrund der geringen Probenvolumina von etwa 200-300 µl gewonnener Bodenlösung aus jeder Bodenprobe wurden die filtrierten Proben vor der HPLC Analytik in 175 µl Mikroeinsätze aus Glas für 2 ml Headspacevials überführt.
28
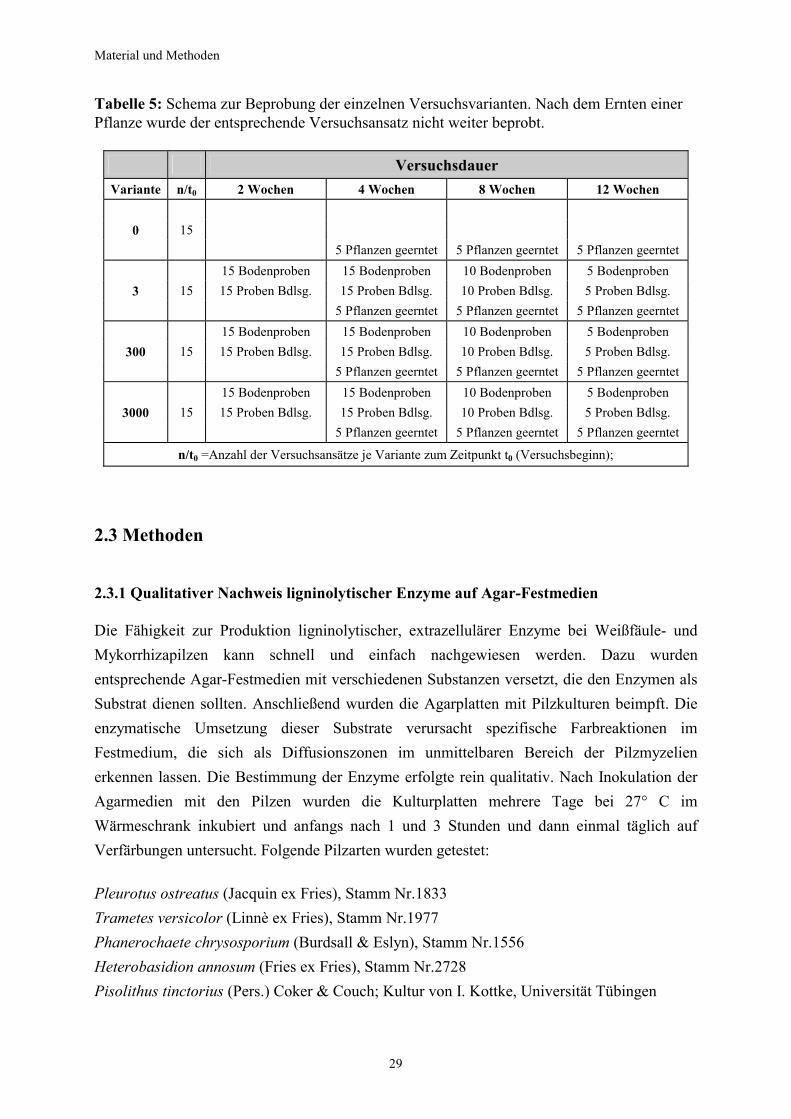
Material und Methoden
Tabelle 5: Schema zur Beprobung der einzelnen Versuchsvarianten. Nach dem Ernten einer Pflanze wurde der entsprechende Versuchsansatz nicht weiter beprobt.
Versuchsdauer Variante n/t0 2 Wochen 4 Wochen 8 Wochen 12 Wochen
0 15 5 Pflanzen geerntet 5 Pflanzen geerntet 5 Pflanzen geerntet 15 Bodenproben 15 Bodenproben 10 Bodenproben 5 Bodenproben
3 15 15 Proben Bdlsg. 15 Proben Bdlsg. 10 Proben Bdlsg. 5 Proben Bdlsg. 5 Pflanzen geerntet 5 Pflanzen geerntet 5 Pflanzen geerntet 15 Bodenproben 15 Bodenproben 10 Bodenproben 5 Bodenproben
300 15 15 Proben Bdlsg. 15 Proben Bdlsg. 10 Proben Bdlsg. 5 Proben Bdlsg. 5 Pflanzen geerntet 5 Pflanzen geerntet 5 Pflanzen geerntet 15 Bodenproben 15 Bodenproben 10 Bodenproben 5 Bodenproben
3000 15 15 Proben Bdlsg. 15 Proben Bdlsg. 10 Proben Bdlsg. 5 Proben Bdlsg. 5 Pflanzen geerntet 5 Pflanzen geerntet 5 Pflanzen geerntet
n/t0 =Anzahl der Versuchsansätze je Variante zum Zeitpunkt t0 (Versuchsbeginn);
2.3 Methoden 2.3.1 Qualitativer Nachweis ligninolytischer Enzyme auf Agar-Festmedien Die Fähigkeit zur Produktion ligninolytischer, extrazellulärer Enzyme bei Weißfäule- und Mykorrhizapilzen kann schnell und einfach nachgewiesen werden. Dazu wurden entsprechende Agar-Festmedien mit verschiedenen Substanzen versetzt, die den Enzymen als Substrat dienen sollten. Anschließend wurden die Agarplatten mit Pilzkulturen beimpft. Die enzymatische Umsetzung dieser Substrate verursacht spezifische Farbreaktionen im Festmedium, die sich als Diffusionszonen im unmittelbaren Bereich der Pilzmyzelien erkennen lassen. Die Bestimmung der Enzyme erfolgte rein qualitativ. Nach Inokulation der Agarmedien mit den Pilzen wurden die Kulturplatten mehrere Tage bei 27° C im Wärmeschrank inkubiert und anfangs nach 1 und 3 Stunden und dann einmal täglich auf Verfärbungen untersucht. Folgende Pilzarten wurden getestet: Pleurotus ostreatus (Jacquin ex Fries), Stamm Nr.1833 Trametes versicolor (Linnè ex Fries), Stamm Nr.1977 Phanerochaete chrysosporium (Burdsall & Eslyn), Stamm Nr.1556 Heterobasidion annosum (Fries ex Fries), Stamm Nr.2728 Pisolithus tinctorius (Pers.) Coker & Couch; Kultur von I. Kottke, Universität Tübingen
29

Material und Methoden
2.3.1.1 Nachweis der Mangan-Peroxidase-Aktivität MnP-Agar Assay (nach Heinzkill 1995): Für den qualitativen Nachweis der Mangan-Peroxidase (MnP) wurden mehrere Pilzinokulate (Agarplättchen, Durchmesser 4 mm) auf das Festmedium A (Tabellen 27 und 28 Anhang) mit 0,1 mM MnSO4 gelegt und bei 27°C inkubiert. Der Nachweis von Mangan-Peroxidase-Aktivität erfolgte durch das Vorkommen orangebrauner Verfärbungen (Oxidation von Mn2+ zu Mn3+) um die Myzelstücke. 2.3.1.2 Nachweis der Lignin-Peroxidase-Aktivität 3-NA-Agar-Assay (nach Heinzkill 1995): Für den qualitativen Nachweis der Lignin-Peroxidase (LiP) wurden mehrere Pilzinokulate (Agarplättchen, Durchmesser 4 mm) auf das Festmedium A mit einem Anteil von 0,01 % w/v 3-Nitroanilin (3-Na) gelegt und bei 27° C inkubiert. Die Zugabe des gelben Substrates 3-Na (gelöst in 50 %igem Ethanol) erfolgte nach dem Autoklavieren. Der Nachweis von LiP-Aktivität ergab sich durch das Vorkommen brauner Diffusionszonen (Oxidation von 3-NA) um die Myzelstücke. Das Screening nach Laccasen bei den entsprechenden Pilzstämmen erfolgte durch Herrn Dr. Jens Dittmann im Rahmen seiner Promotionsarbeit (Dittmann 1999). 2.3.2 Extraktionsverfahren 2.3.2.1 Nitroaromatenextraktion aus den Bodenmischproben Für die Extraktion der Nitroaromaten aus den Bodenproben wurde das Bodensubstrat in den geöffneten Rhizotronen etwa 1 Woche im Dunkeln bei 27° C an der Luft getrocknet, nachdem zuvor sorgfältig die Pflanzen und das Stroh aus dem Boden entfernt wurden. Anschließend erfolgte die Entnahme einer 5 g Bodenmischprobe (lufttrocken) aus den Petrischalen. In verschlossenen 20 ml Rollrandflaschen aus Glas wurden die Bodenproben in 10 ml Methanol für 5 Stunden im Ultraschallbad (Sonorex Super RK 106, 35 Khz, Fa. Bandelin) bei 20° C extrahiert. Um eine zu starke Erwärmung der Proben im Ultraschall-Dauerbetrieb zu vermeiden, wurde das Wasser über einen geschlossenen Kreislauf in einem Kühlaggregat (Fa. Haake) auf Raumtemperatur gehalten. Der Überstand aus den Probenfläschchen wurde nach Filtration durch Membranfilter (Polypropylen, 0,22 µm Porendurchmesser, Fa. Macherey & Nagel) in 2 ml Braunglas Autosamplergefäße aufgenommen und bis zur HPLC-Analytik bei -18° C im Kühlschrank aufbewahrt.
30

Material und Methoden
2.3.2.2 Nitroaromatenextraktion aus Pflanzengeweben Eine effiziente Methode zur Nitroaromatenextraktion aus unterschiedlichen Pflanzengeweben stellt das in einigen Schritten modifizierte Extraktionsverfahren nach Görge et al. (1994b) dar. Nach sehr sorgfältiger Reinigung der Wurzeln von anhaftenden Bodenpartikeln mit Wasser wurden die Pflanzen grob zerkleinert und in speziellen Gazé-Netzen gefriergetrocknet (modifizierte Gefriertrocknungsanlage Gamma 1a, Fa. Christ). Die Mahlung des entwässerten Materials zu Feinpulver erfolgte mit einer Kugelmühle (Pulverisette 6 und Mahlbecher aus Sinterkorund von Fa. Fritsch, Idar Oberstein) für 2 x 3 min. bei 600 U/min. Die sich anschließenden Extraktionsschritte sind zur besseren Übersicht in einzelnen Punkten dargestellt: DCM-Aufschluss Jeweils 100 mg gemahlenes Pflanzenmaterial wurden in verschraubbare Zentrifugengläser aus Glas eingewogen (Schraubverschlüsse mit Teflondichtung) und nach Zugabe von 4 ml DCM bei 20°C für 60 min im Ultraschallbad extrahiert. Anschließend wurden die Proben 10 min bei 2000 g zentrifugiert (Minifuge RF, Fa. Heraeus) und die DCM-Fraktion dann vorsichtig mit einer Spritze abgesaugt. Dieser Extrakt soll fortan als DCM-Aufschluss bezeichnet werden. Hydrolyse mit Schwefelsäure Das in den Zentrifugengläsern verbleibende Pflanzenmaterial wurde mit 2 ml 25 %iger Schwefelsäure versetzt und für 90 min bei 80°C im Wasserbad durch Säurehydrolyse weiter aufgeschlossen. Dann erfolgte eine erneute Extraktion für 30 min. im Ultraschallbad bei 20°C nach Zugabe von 2 ml DCM. Dieser Extraktionsschritt mit DCM wurde zweimal durchgeführt, wobei die jeweiligen DCM-Extrakte nach Zentrifugation aus den Gläsern abgesaugt und vereint wurden. Alkalisierung mit NaOH Unter vorsichtiger Zugabe (tropfenweise) von 8 M NaOH wurde das Pflanzenmaterial mit der Schwefelsäure bis zu einem pH-Wert von 11-12 alkalisiert (Kontrolle mit Indikatorpapier, Fa. Merck). Nochmals erfolgte eine 30minütige Extraktion im Ultraschallbad in 2 ml DCM. Die Lösungsmittelfraktion konnte nach dem Zentrifugieren vorsichtig abgesaugt und mit dem Extrakt aus der Säurehydrolyse vereint werden. Dieser vereinte Extrakt wird nachfolgend als Hydrolyse-Aufschluss bezeichnet.
31

Material und Methoden
2.3.3 Nitroaromaten-Analytik 2.3.3.1 HPLC-Analytik Die Auftrennung der Extrakte aus den Bodenproben für den Nitroaromatennachweis erfolgte mit der High Performance Liquid Chromatographie (HPLC). Die Integration der Peaks wurde mit der Software (GAT-Chrom, Fa. GAT) durchgeführt. Mittels Regressionsgeraden der Eichreihen ließen sich über die Peakflächen die Nitroaromatenkonzentrationen in den Proben errechnen. Die Bodenproben wurden auf insgesamt 15 Nitroaromaten getestet. Die Konzentrationen der Standardlösungen lagen zwischen 1 und 100 mg/l. Pumpe: Merck-Hitachi L-6200, 3-Kanal Gradientenpumpe Autosampler: Perkin-Elmer ISS-100, variable volume injection system Detektor: LDC Analytical SM 4000 programmable wavelength detector, Perkin-Elmer LC 480 Dioden Array Detektor Integrator: GATchrom PC-based LC/GC Chromatography Data System Säule: Macherey & Nagel ET 250/4 Nucleosil 120-5, C18, 5 µm, 250x4 mm Vorsäule: Merck LiChroCart 4-4 mm, RP-18, Teilchengröße 5µm Mobile Phase: 47% Methanol/ 53% Wasser, Flussrate 1,0 ml/min isokratisch Detektion: 230 nm Wellenlänge, 0,1AUFS Injektion: 20 µl Probenschleife Thermostat: 35° C Säulentemperatur Verwendete Standardsubstanzen: 1. 2,6-Diamino-4-nitrotoluol 9. 4-Amino-2,6-dinitrotoluol 2. 2,4-Diamino-6-nitrotoluol 10. 2-Amino-4,6-dinitrotoluol 3. 6-Amino-2-nitrotoluol 11. 2,6-Dinitrotoluol 4. 4-Amino-2-nitrotoluol 12. 2,4-Dinitrotoluol 5. 2-Amino-4-nitrotoluol 13. 2-Nitrotoluol 6. 1,2-Dinitrobenzol 14. 4-Nitrotoluol 7. 1,3-Dinitrobenzol 15. 3-Nitrotoluol 8. 2,4,6-Trinitrotoluol (TNT)
32
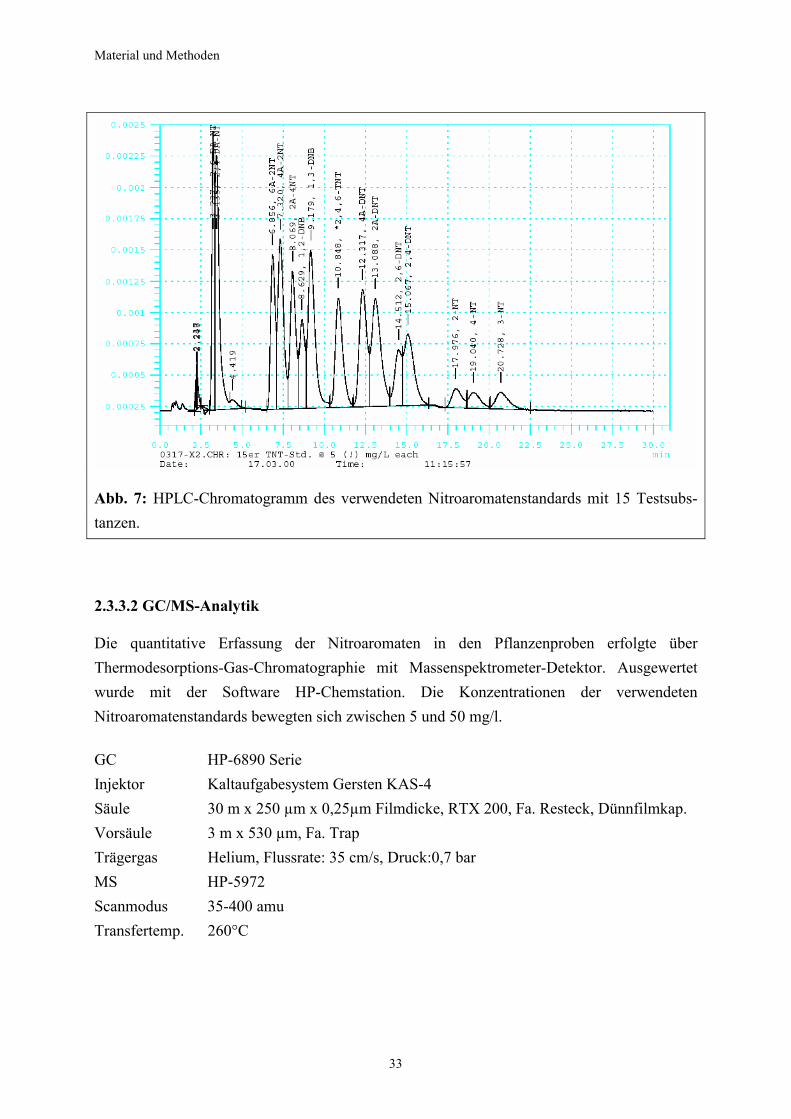
Material und Methoden
Abb. 7: HPLC-Chromatogramm des verwendeten Nitroaromatenstandards mit 15 Testsubs-tanzen.
2.3.3.2 GC/MS-Analytik Die quantitative Erfassung der Nitroaromaten in den Pflanzenproben erfolgte über Thermodesorptions-Gas-Chromatographie mit Massenspektrometer-Detektor. Ausgewertet wurde mit der Software HP-Chemstation. Die Konzentrationen der verwendeten Nitroaromatenstandards bewegten sich zwischen 5 und 50 mg/l. GC HP-6890 Serie Injektor Kaltaufgabesystem Gersten KAS-4 Säule 30 m x 250 µm x 0,25µm Filmdicke, RTX 200, Fa. Resteck, Dünnfilmkap. Vorsäule 3 m x 530 µm, Fa. Trap Trägergas Helium, Flussrate: 35 cm/s, Druck:0,7 bar MS HP-5972 Scanmodus 35-400 amu Transfertemp. 260°C
33

Material und Methoden
Thermodesorption Die Proben wurden im Glasröhrchen (Länge 15 cm, Durchmesser innen 4 mm) direkt vor dem Injektor im kontinuierlichem Trägergasstrom erhitzt (5°C - 230°C, für 8 - 10 min.). Hierdurch ließen sich die flüchtigen Nitroaromaten aus den Proben heraustreiben und mit dem Gasstrom in den Injektor transportieren. Im stark heruntergekühlten Injektor (-100°C) kondensierten sie, so dass darin schließlich alle Nitroaromaten vollständig konzentriert werden konnten. Durch schnelles und starkes Erhitzen des Injektors (12°C/s bis auf 260°C) wurden die Nitroaromaten im gasförmigen Zustand auf die Säule transportiert. Verwendete Standardsubstanzen: 1. 2,6-Diamino-4-Nitrotoluol 9. 4-Amino-2,6-Dinitrotoluol 2. 2,4-Diamino-6-Nitrotoluol 10. 2-Amino-4,6-Dinitrotoluol 3. 6-Amino-2-Nitrotoluol 11. 2,6-Dinitrotoluol 4. 4-Amino-2-Nitrotoluol 12. 2,4-Dinitrotoluol 5. 2-Amino-4-Nitrotoluol 13. 2-Nitrotoluol 6. 1,2-Dinitrobenzol 14. 4-Nitrotoluol 7. 1,3-Dinitrobenzol 15. 3-Nitrotoluol 8. 2,4,6-Trinitrotoluol (TNT)
34
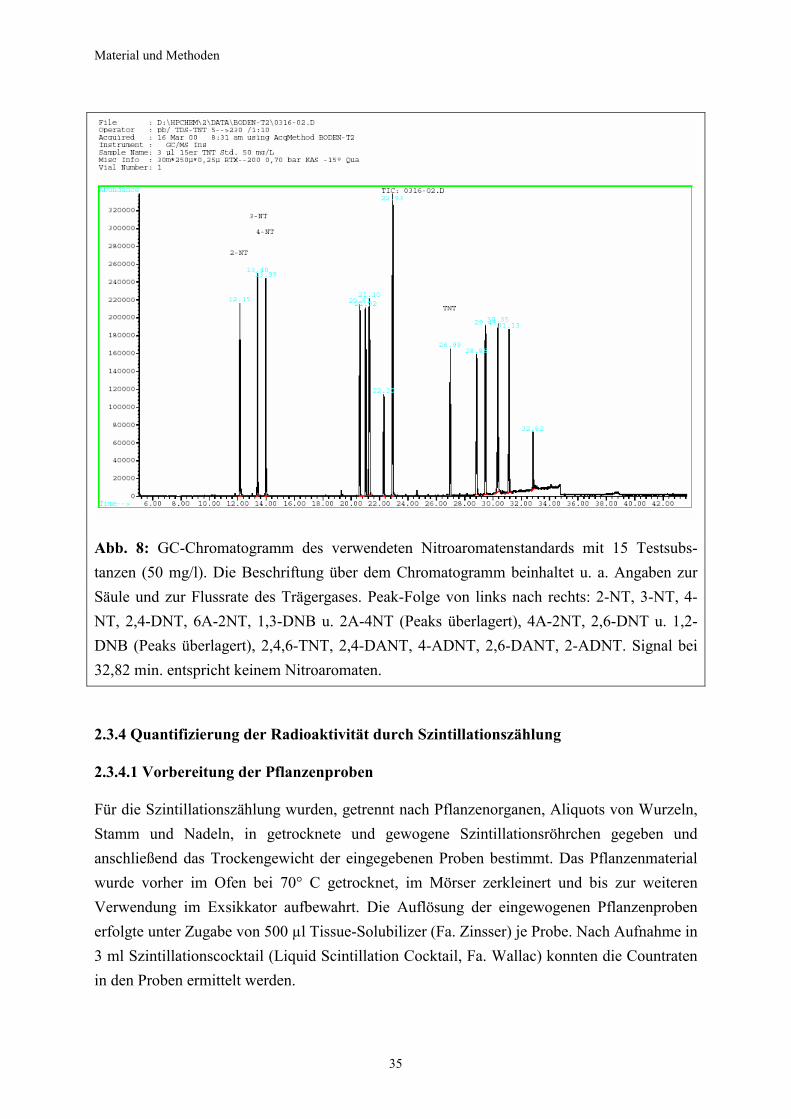
Material und Methoden
Abb. 8: GC-Chromatogramm des verwendeten Nitroaromatenstandards mit 15 Testsubs-tanzen (50 mg/l). Die Beschriftung über dem Chromatogramm beinhaltet u. a. Angaben zur Säule und zur Flussrate des Trägergases. Peak-Folge von links nach rechts: 2-NT, 3-NT, 4-NT, 2,4-DNT, 6A-2NT, 1,3-DNB u. 2A-4NT (Peaks überlagert), 4A-2NT, 2,6-DNT u. 1,2-DNB (Peaks überlagert), 2,4,6-TNT, 2,4-DANT, 4-ADNT, 2,6-DANT, 2-ADNT. Signal bei 32,82 min. entspricht keinem Nitroaromaten.
2.3.4 Quantifizierung der Radioaktivität durch Szintillationszählung 2.3.4.1 Vorbereitung der Pflanzenproben Für die Szintillationszählung wurden, getrennt nach Pflanzenorganen, Aliquots von Wurzeln, Stamm und Nadeln, in getrocknete und gewogene Szintillationsröhrchen gegeben und anschließend das Trockengewicht der eingegebenen Proben bestimmt. Das Pflanzenmaterial wurde vorher im Ofen bei 70° C getrocknet, im Mörser zerkleinert und bis zur weiteren Verwendung im Exsikkator aufbewahrt. Die Auflösung der eingewogenen Pflanzenproben erfolgte unter Zugabe von 500 µl Tissue-Solubilizer (Fa. Zinsser) je Probe. Nach Aufnahme in 3 ml Szintillationscocktail (Liquid Scintillation Cocktail, Fa. Wallac) konnten die Countraten in den Proben ermittelt werden.
35

Material und Methoden
2.3.4.2 Aufnahme der Nährlösung Die in den Rhizotronen verbleibende Düngelösung wurde nach erfolgter Ernte der Kiefern vorsichtig mit einer Eppendorfpipette abgenommen und in Szintillationsröhrchen überführt. In diese wurden nach Einengung der Proben durch vollständiges Trocknen im Ofen bei 70° C jeweils 3 ml Szintillationscocktail dazugegeben und anschließend die Countraten ermittelt. 2.3.4.3 Auswaschung des Kohlepapiers Die nach der Ernte verbliebenen Kohlepapierscheiben wurden im Ofen getrocknet. Danach wurden von den Scheiben je fünf Papierstücke (5mg pro Stück) von unterschiedlichen Stellen mit einer Schere entfernt und zusammen mit 3ml Szintillationscocktail in vorgewogene und trockene Szintillationsröhrchen gegeben. Über einen Zeitraum von 14 Tagen wurden die 14C-Anteile nun unter zwischenzeitigem kräftigem Schütteln (Vortex) herausgespült und anschließend gemessen. 2.3.4.4 Ermittlung der Radioaktivität Nach Messung der Proben mit einem Szintillationszähler (1450 Microbeta, Fa. Wallac) erfolgte die Ermittlung der Countraten (cpm). Hierbei wurde der Mittelwert aus fünf Einzelmessungen genommen. Um mögliche Quencheffekte auf die Countrate in der Probe auszugleichen, wurden je nach Stärke der Radioaktivität in den Proben verschiedene interne Standards verwendet. Die Berechnung der tatsächlichen Kernzerfälle je Zeiteinheit (dpm) erfolgte schließlich unter Berücksichtigung der Countraten (cpm-Werte) und der Wirksamkeit der Zählung in Anlehnung an Dyer (1974) und Fox (1976). 2.3.5 Angewandte Methoden für die Mikroautoradiographie 2.3.5.1 Gefriertrocknung der präparierten Wurzeln Die Infiltration der Wurzeln mit dem Epoxidharz Spurr (nach Spurr 1969) erfordert eine völlige Entwässerung der Proben aufgrund der Nichtmischbarkeit dieses Kunststoffes mit Wasser. Bei konventionellen Entwässerungsverfahren wird das Wasser in den Proben vor der Kunststoffeinbettung schrittweise durch Lösungsmittel wie Ethanol und Aceton ersetzt. Aufgrund ihrer guten Löslichkeit in solchen Lösungsmitteln würden in mykorrhizierten Wurzeln befindliche Nitroaromaten während der Entwässerung durch Auswaschung vermutlich verloren gehen. Auf eine herkömmliche Einbettungsmethode musste deshalb verzichtet werden.
36

Material und Methoden
Eine schonende Entwässerung durch Kryofixierung und nachfolgende Gefriertrocknung der Proben verhindert zum einen eine Auswaschung und eine nachträgliche Verlagerung der Nitroaromaten in den Geweben und ermöglicht zum anderen einen weitestgehenden Erhalt der Gewebe- und Zellstrukturen. Anfangstemperaturen von etwa -100 C° verhindern bei der Gefriertrocknung eine Rekristallisation des Wassers. Verwendet wurde ein Gerät der Firma Leica (Leica EM CFD) mit einer speziellen Zeolithkammer. Das darin befindliche und zuvor ausgeheizte Granulat erhält und verbessert ein zuvor erzeugtes Vakuum, indem die aus den Proben freiwerdenden Gase absorbiert werden. Cryofixierung und Gefriertrocknung wurden wie folgend durchgeführt: �� Schockgefrierung der frisch abpräparierten Wurzelproben durch Überführung in schmelzenden Stickstoff. �� Bestückung der Gefriertrockung mit den tiefgefrorenen Proben. Der Probenteller wurde zuvor mit flüssigem Stickstoff auf -180°C heruntergekühlt. Erzeugung eines Vorvakuums von 16 mbar in der Probenkammer mit einer Rotationspumpe. Durch die Bindeeigenschaften des Zeolith-Granulates erhöhte sich das Vakuum anschließend auf 5 x 10-4 mbar. �� Gefriertrocknung der Wurzelproben in mehreren aufeinander folgenden Teilschritten. Reihenfolge und Dauer der einzelnen Teilschritte sind mit den dazugehörigen Temperaturen in Tabelle 6 dargestellt. �� Nach Beendigung der Gefriertrocknung wurde über ein Ventil in der Probenkammer Normaldruck erzeugt. Die Aufbewahrung der Wurzelproben erfolgte bis zur weiteren Verarbeitung im Exsikkator über CaCl2.
37
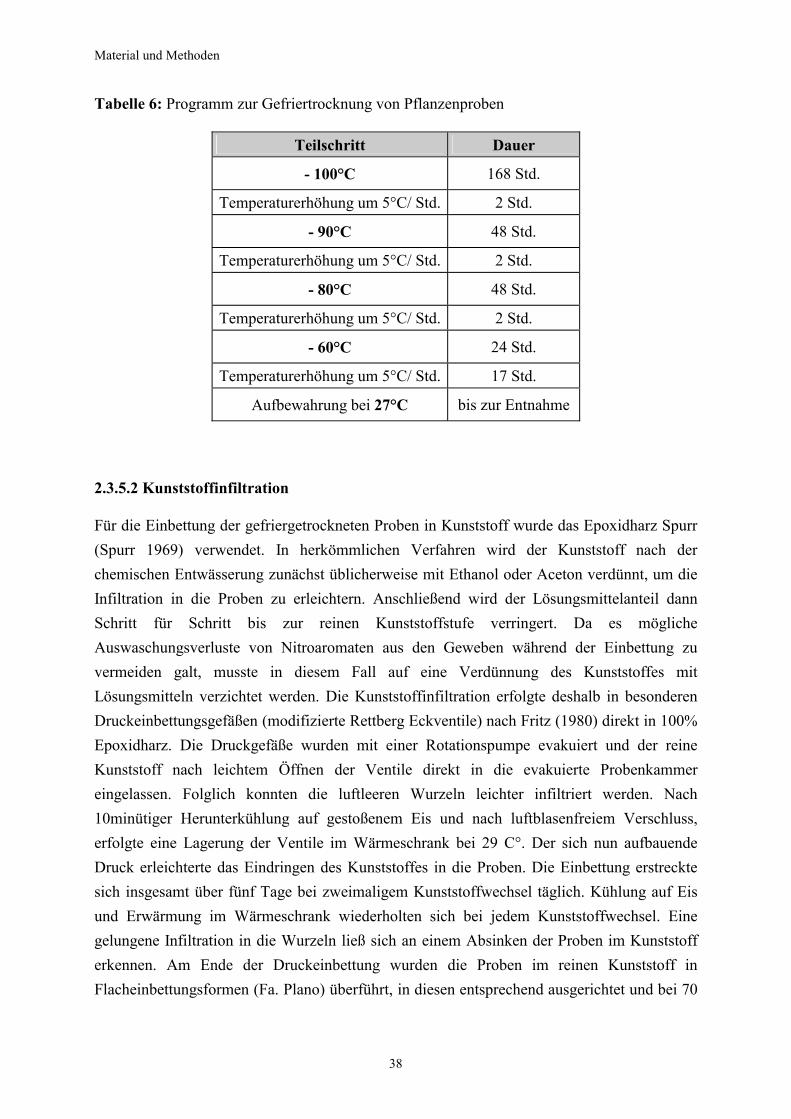
Material und Methoden
Tabelle 6: Programm zur Gefriertrocknung von Pflanzenproben
Teilschritt Dauer
- 100°C 168 Std.
Temperaturerhöhung um 5°C/ Std. 2 Std.
- 90°C 48 Std.
Temperaturerhöhung um 5°C/ Std. 2 Std.
- 80°C 48 Std.
Temperaturerhöhung um 5°C/ Std. 2 Std.
- 60°C 24 Std.
Temperaturerhöhung um 5°C/ Std. 17 Std.
Aufbewahrung bei 27°C bis zur Entnahme
2.3.5.2 Kunststoffinfiltration Für die Einbettung der gefriergetrockneten Proben in Kunststoff wurde das Epoxidharz Spurr (Spurr 1969) verwendet. In herkömmlichen Verfahren wird der Kunststoff nach der chemischen Entwässerung zunächst üblicherweise mit Ethanol oder Aceton verdünnt, um die Infiltration in die Proben zu erleichtern. Anschließend wird der Lösungsmittelanteil dann Schritt für Schritt bis zur reinen Kunststoffstufe verringert. Da es mögliche Auswaschungsverluste von Nitroaromaten aus den Geweben während der Einbettung zu vermeiden galt, musste in diesem Fall auf eine Verdünnung des Kunststoffes mit Lösungsmitteln verzichtet werden. Die Kunststoffinfiltration erfolgte deshalb in besonderen Druckeinbettungsgefäßen (modifizierte Rettberg Eckventile) nach Fritz (1980) direkt in 100% Epoxidharz. Die Druckgefäße wurden mit einer Rotationspumpe evakuiert und der reine Kunststoff nach leichtem Öffnen der Ventile direkt in die evakuierte Probenkammer eingelassen. Folglich konnten die luftleeren Wurzeln leichter infiltriert werden. Nach 10minütiger Herunterkühlung auf gestoßenem Eis und nach luftblasenfreiem Verschluss, erfolgte eine Lagerung der Ventile im Wärmeschrank bei 29 C°. Der sich nun aufbauende Druck erleichterte das Eindringen des Kunststoffes in die Proben. Die Einbettung erstreckte sich insgesamt über fünf Tage bei zweimaligem Kunststoffwechsel täglich. Kühlung auf Eis und Erwärmung im Wärmeschrank wiederholten sich bei jedem Kunststoffwechsel. Eine gelungene Infiltration in die Wurzeln ließ sich an einem Absinken der Proben im Kunststoff erkennen. Am Ende der Druckeinbettung wurden die Proben im reinen Kunststoff in Flacheinbettungsformen (Fa. Plano) überführt, in diesen entsprechend ausgerichtet und bei 70
38

Material und Methoden
C° im Ofen über Nacht ausgehärtet. Bis zur weiteren Bearbeitung am Ultramikrotom konnten die Proben nun problemlos aufbewahrt werden. 2.3.5.3 Schneiden der Proben am Ultramikrotom Die im Kunststoff eingebetteten Wurzelproben wurden mit Teflon beschichteten Single-edge-Klingen (Fa. Plano) entsprechend zurechtgetrimmt und mit selbst angefertigten Glasmessern (Messerbrecher von Fa. LKB Bromma, Schweden) trocken am Ultramikrotom (Ultracut E, Fa. Reichert und Jung) geschnitten. Die Glasmesser erhielten zuvor einen dünnen Teflonüberzug (Teflonspray, Fa. Reichelt), um Stauchungen der Wurzellängs- und -querschnitte auf der Messeroberfläche während des Schneidevorgangs zu vermeiden. Anschließend konnten die 1-1,5 µm dicken Schnitte mit sehr feinen Nadeln vorsichtig von den Messern abgehoben und auf mit Gelatine beschichteten Glasobjektträgern überführt werden. 2.3.5.4 Mikroautoradiographie Mit jeweils 100 µl Röntgenfilmemulsion ( Ilford, L4, in Gelform, im Verhältnis 1:2 mit bidest verdünnt, w/v, 15 min bei 40° C im Wasserbad gerührt) erhielten die Objektträger im Dunkelkammerlicht (Ilford, Filter 902) einen gleichmäßig dünnen Überzug. Anschließend wurden die beschichteten Objektträger 24 Stunden in einer lichtdicht verschließbaren Holzbox neben einer kleinen Schale mit CaCl2 getrocknet und hinterher bis zur Entwicklung in verschließbaren Kunststoffboxen im Dunkeln bei 4° C im Kühlschrank aufbewahrt. Die Entwicklung des Röntgenfilms erfolgte nach verschiedenen Expositionszeiten zwischen 2 Tagen und 20 Wochen mit dem Feinkornentwickler D 19 A/S (Sanderson 1981). Nach Fixierung des Röntgenfilms (Express-Fixiersalz, Fa. Tetanal) und sorgfältiger Spülung der Objektträger mit entmineralisiertem Wasser konnten die ungefärbten Wurzelschnitte (Mikroautoradiographien) nun an einem Universalmikroskop für Durchlicht (Fa. Zeiss) ausgewertet werden. Die fotografische Dokumentation der Mikroautoradiographien wurde an einem Interferenzmikroskop mit differentiellem Interferenzkontrast (Fa. Olympus) durchgeführt. 2.3.6 Rasterelektronenmikroskopie (REM) Die Präparation der Proben für die Rasterelektronenmikroskopie wurde folgenderweise durchgeführt: frische und feuchte Bodenaggregate mit den anhaftenden Weißfäulepilzhyphen wurden mit einer Federstahlpinzette direkt aus den Rhizotronen entnommen und 12-15 Stunden über Nacht bei 4°C in 2% Glutaraldehyd in 0,05 M Na-Cacodylatpuffer pH-Wert 7,0 vorfixiert. Nach etwa 90minütigem Auswaschen der Proben in 0,05M Cacodylatpuffer (Puffer
39

Material und Methoden
alle zehn Minuten ausgetauscht) konnten die Proben in einer aufsteigenden Ethanolreihe entwässert werden. 15% Ethanol 15 min. 30% Ethanol 15 min. 50% Ethanol 15 min. 70% Ethanol 15 min. 90% Ethanol 30 min. 100% Ethanol 2x15 min. (gewechselt) Um die Pilzhyphen in ihrer Struktur möglichst gut zu erhalten, wurde nach dem Entwässern der Proben eine Kritisch-Punkt-Trocknung angewendet (Gerät CPD 010, Balzers Union, Liechtenstein). Hierbei wurde das Entwässerungsmedium Ethanol gegen Kohlendioxid als Übergangsmedium oder Kritisch-Punkt-Medium ausgetauscht. Nach der Trocknung wurden die Proben mit beidseitig selbstklebenden Leit-Tabs (Plano, Marburg) auf Aluminium-Probenteller befestigt, mit Gold bedampft (Gerät SCD 040, Balzers Union, Liechtenstein) und im Rasterelektronenmikroskop ISI 100B (Leitz, Wetzlar) betrachtet. 2.3.7 Bestimmung des pH-Wertes von Bodenproben Die Bestimmung der Boden pH-Werte erfolgte nach Scheffer und Schachtschabel (1984). Je Probe wurden 10 g luftgetrockneter Boden mit 25 ml 0,01 M CaCl2 vermengt und 24 Stunden geschüttelt. Anschließend wurde der pH-Wert im Überstand mit einer pH-Elektrode gemessen (WTW pH 525). 2.3.8 Fotografische Dokumentation Beobachten und Fotografieren erfolgte am Universalmikroskop für Durchlicht und am Stereomikroskop DV4 (Stereolupe), Aufsetzkameras: MC 63 und 45 60 70 - 3, Belichtungs-steuerung: Gerät M und.45 93 40. Alle Geräte von Zeiss, Oberkochen. Für fotografische Aufnahmen wurden Kleinbildfilme von Ilford Pan F50 18DIN/50ASA, schwarz/weiß und Kodak Ektachrome 160T 23DIN/160ASA, Kunstlicht Farbdiafilme verwendet. Herstellung von schwarz/weiß Vergrößerungen auf Fotopapier von Ilford, Multigrade IV und von Farbvergrößerungen im Digital-Printverfahren. Für die weitere Bearbeitung mit der Computer-Software Adobe Photoshop 5.0 wurden die Aufnahmen mit einem Agfa Snap Scan 600 Flachbettscanner eingescannt.
40

Material und Methoden
2.3.9 Ergebnisauswertung und Statistik Die Berechnung der Mittelwerte, Mediane, Standardabweichungen, Maxima und Minima erfolgte mit Microsoft Excel für Windows 97 NT Version 7.0 oder Excel für Windows 2000 Professional, Version 2002. Insbesondere die ermittelten Messdaten zur Erfassung der Nitroaromatenbelastung im Boden wiesen keine Normalverteilung auf. Aus diesem Grund wurde als Signifikanztest der nichtparametrische U-Test nach Mann & Whitney mit dem Statistikprogramm Unistat Version 5.1 durchgeführt. Der H-Test nach Kruskal und Wallis fand als Ausweitung des U-Testes Anwendung. Als Signifikanzgrenze wurde die Irrtumswahrscheinlichkeit p (α) ≤ 0,05 unter zweiseitiger Testanwendung gewählt. Die Graphischen Darstellungen wurden mit Excel für Windows 97 NT oder Windows 2000 Professional (Versionen 7.0 und 2002) erstellt.
41
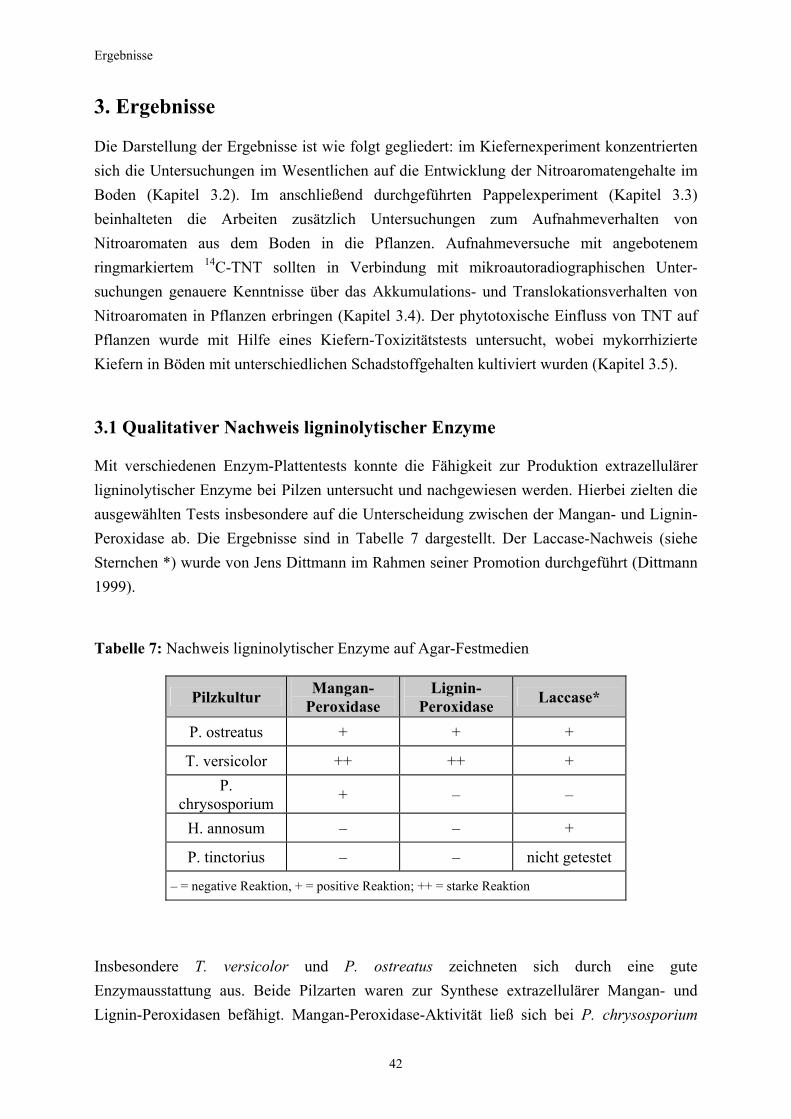
Ergebnisse
42
3. Ergebnisse Die Darstellung der Ergebnisse ist wie folgt gegliedert: im Kiefernexperiment konzentrierten sich die Untersuchungen im Wesentlichen auf die Entwicklung der Nitroaromatengehalte im Boden (Kapitel 3.2). Im anschließend durchgeführten Pappelexperiment (Kapitel 3.3) beinhalteten die Arbeiten zusätzlich Untersuchungen zum Aufnahmeverhalten von Nitroaromaten aus dem Boden in die Pflanzen. Aufnahmeversuche mit angebotenem ringmarkiertem 14C-TNT sollten in Verbindung mit mikroautoradiographischen Unter-suchungen genauere Kenntnisse über das Akkumulations- und Translokationsverhalten von Nitroaromaten in Pflanzen erbringen (Kapitel 3.4). Der phytotoxische Einfluss von TNT auf Pflanzen wurde mit Hilfe eines Kiefern-Toxizitätstests untersucht, wobei mykorrhizierte Kiefern in Böden mit unterschiedlichen Schadstoffgehalten kultiviert wurden (Kapitel 3.5). 3.1 Qualitativer Nachweis ligninolytischer Enzyme Mit verschiedenen Enzym-Plattentests konnte die Fähigkeit zur Produktion extrazellulärer ligninolytischer Enzyme bei Pilzen untersucht und nachgewiesen werden. Hierbei zielten die ausgewählten Tests insbesondere auf die Unterscheidung zwischen der Mangan- und Lignin-Peroxidase ab. Die Ergebnisse sind in Tabelle 7 dargestellt. Der Laccase-Nachweis (siehe Sternchen *) wurde von Jens Dittmann im Rahmen seiner Promotion durchgeführt (Dittmann 1999). Tabelle 7: Nachweis ligninolytischer Enzyme auf Agar-Festmedien
Pilzkultur Mangan- Peroxidase
Lignin- Peroxidase Laccase*
P. ostreatus + + +
T. versicolor ++ ++ + P.
chrysosporium + – –
H. annosum – – +
P. tinctorius – – nicht getestet
– = negative Reaktion, + = positive Reaktion; ++ = starke Reaktion Insbesondere T. versicolor und P. ostreatus zeichneten sich durch eine gute Enzymausstattung aus. Beide Pilzarten waren zur Synthese extrazellulärer Mangan- und Lignin-Peroxidasen befähigt. Mangan-Peroxidase-Aktivität ließ sich bei P. chrysosporium

Ergebnisse
43
nachweisen. H. annosum besaß weder Mangan- noch Lignin-Peroxidase-Aktivität. Negativ war die Reaktion für diese beiden Enzyme auch bei dem Mykorrhizapilz P. tinctorius. 3.2 TNT-Abbau-Experimente mit Kiefern Dieses Experiment zielte in erster Linie darauf ab, Kenntnisse darüber zu erlangen, in welcher Größenordnung die Biodegradation von TNT und verschiedenen TNT-Metaboliten im Boden durch Bepflanzung mit mykorrhizierten Kiefern gefördert wird. Die Versuchsanleitung und Versuchsdurchführung ist aus Punkt 2.2.1 zu entnehmen. Der vom ehemaligen Sprengstoffwerk „Tanne“ entnommene Standortboden enthielt zahlreiche TNT-Kristalle und Aggregate unterschiedlicher Größe. Diese kleinen Splitter verursachten zum Teil eine sehr inhomogene Verteilung des TNT. Durch sorgfältiges Vermischen und Sieben des Bodens konnte die Inhomogenität zwar gemindert, jedoch nicht vollständig behoben werden. Von der Inhomogenität im Boden war allerdings nur das TNT selbst betroffen. Die ADNTs als unmittelbare Reduktionsprodukte des TNT zeigten einheitlichere Belastungswerte. Demzufolge war das vereinzelte Auftreten von stark abweichenden Extremwerten bezüglich der ermittelten Schadstoffkonzentrationen im Boden, hervorgerufen durch das Erfassen solcher Splitter bei der Beprobung, auf das TNT begrenzt. Entsprechende Werte mit extremer Abweichung konnten bei der weiteren statistischen Auswertung der Messergebnisse nicht berücksichtigt werden, da sonst der tatsächliche Abreicherungsvorgang im Boden nicht korrekt wiedergegeben worden wäre. Im Anhang sind die nicht in die Auswertung eingegangenen Werte in den Tabellen 33 und 34 fettgedruckt aufgeführt. Als Kontaminanten wurden TNT und dessen unmittelbaren Reduktionsprodukte 2-ADNT und 4-ADNT gefunden. Weitere Nitroaromatenverbindungen konnten mit Hilfe der eingesetzten Extraktionsmethoden nicht detektiert werden. Die Ausstattung der einzelnen Versuchsvarianten mit den entsprechenden Organismen ist in Punkt 2.2.1 beschrieben. Die beobachtete Transformationsdynamik bezüglich des TNT und der beiden ADNTs ist in verschiedenen Abschnitten dargestellt. In einem weiteren Kapitel soll kurz auf die Biomasseentwicklung der Pflanzen und auf den Mykorrhizierungsgrad ihrer Wurzeln eingegangen werden. 3.2.1 Entwicklung der TNT-Belastung im Boden In Abbildung 9 sind die Ergebnisse der Bodenanalysen dargestellt. Mit Ausnahme von Variante K war das TNT bis zur ersten Probennahme bereits größtenteils eliminiert. Auch in der Kontrollvariante K fiel der Rückgang deutlich aus, war allerdings zeitlich verzögert. Bis zur letzten Probennahme pendelte sich das Belastungsniveau der analysierten Bodenproben in allen Varianten auf einem niedrigen Level ein. Der ermittelte zwischenzeitliche Anstieg in den Varianten A und C ist vermutlich eine Folge der inhomogenen Verteilung des TNT
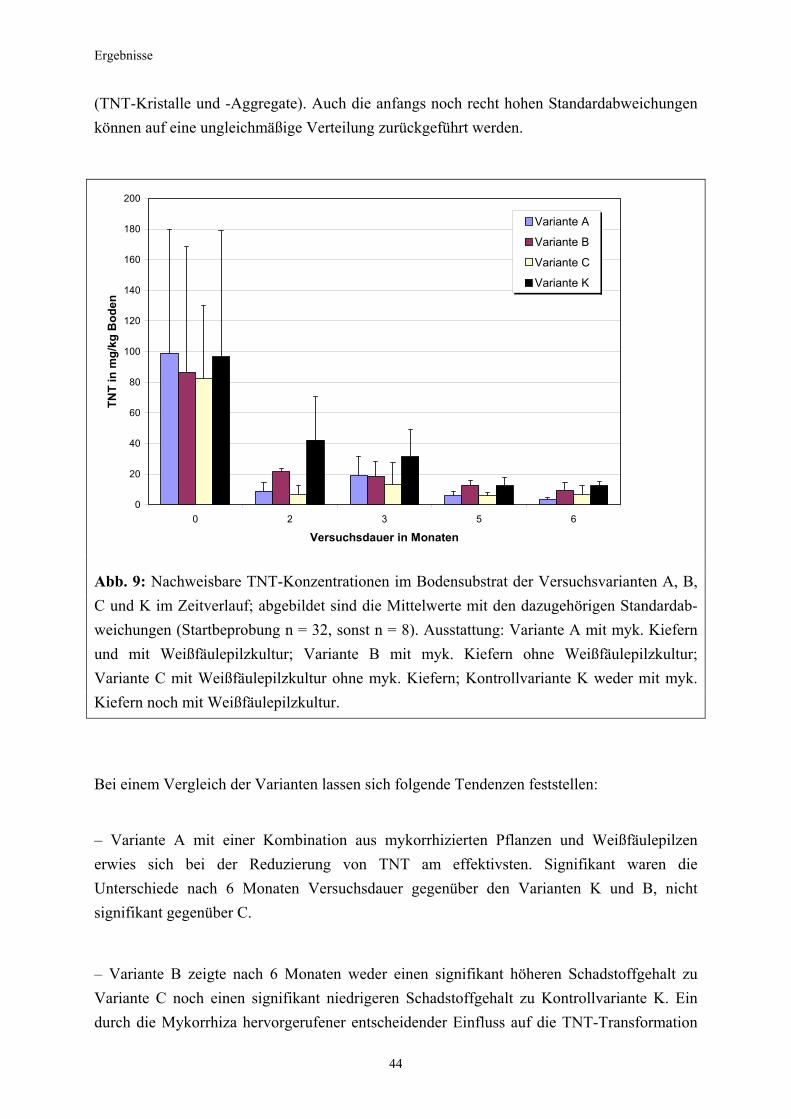
Ergebnisse
44
(TNT-Kristalle und -Aggregate). Auch die anfangs noch recht hohen Standardabweichungen können auf eine ungleichmäßige Verteilung zurückgeführt werden.
0
20
40
60
80
100
120
140
160
180
200
0 2 3 5 6
Versuchsdauer in Monaten
TNT
in m
g/kg
Bod
en
Variante A
Variante B
Variante C
Variante K
Abb. 9: Nachweisbare TNT-Konzentrationen im Bodensubstrat der Versuchsvarianten A, B, C und K im Zeitverlauf; abgebildet sind die Mittelwerte mit den dazugehörigen Standardab-weichungen (Startbeprobung n = 32, sonst n = 8). Ausstattung: Variante A mit myk. Kiefern und mit Weißfäulepilzkultur; Variante B mit myk. Kiefern ohne Weißfäulepilzkultur; Variante C mit Weißfäulepilzkultur ohne myk. Kiefern; Kontrollvariante K weder mit myk. Kiefern noch mit Weißfäulepilzkultur. Bei einem Vergleich der Varianten lassen sich folgende Tendenzen feststellen: – Variante A mit einer Kombination aus mykorrhizierten Pflanzen und Weißfäulepilzen erwies sich bei der Reduzierung von TNT am effektivsten. Signifikant waren die Unterschiede nach 6 Monaten Versuchsdauer gegenüber den Varianten K und B, nicht signifikant gegenüber C. – Variante B zeigte nach 6 Monaten weder einen signifikant höheren Schadstoffgehalt zu Variante C noch einen signifikant niedrigeren Schadstoffgehalt zu Kontrollvariante K. Ein durch die Mykorrhiza hervorgerufener entscheidender Einfluss auf die TNT-Transformation
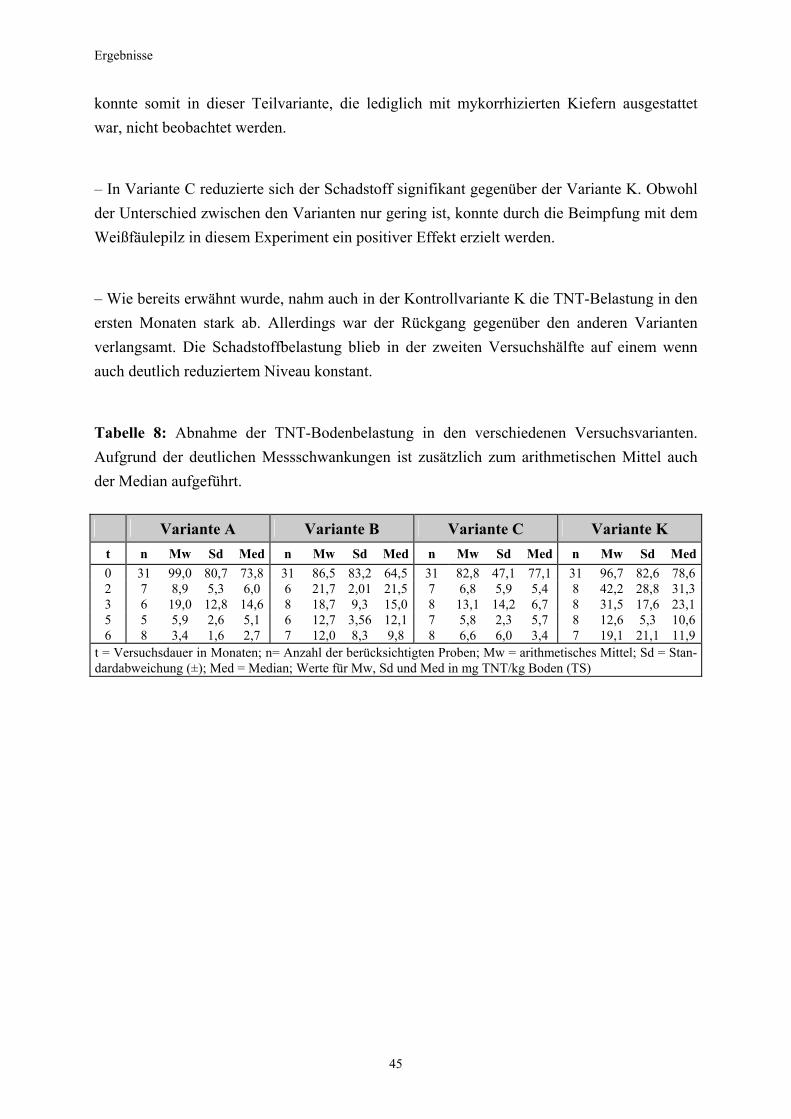
Ergebnisse
45
konnte somit in dieser Teilvariante, die lediglich mit mykorrhizierten Kiefern ausgestattet war, nicht beobachtet werden. – In Variante C reduzierte sich der Schadstoff signifikant gegenüber der Variante K. Obwohl der Unterschied zwischen den Varianten nur gering ist, konnte durch die Beimpfung mit dem Weißfäulepilz in diesem Experiment ein positiver Effekt erzielt werden. – Wie bereits erwähnt wurde, nahm auch in der Kontrollvariante K die TNT-Belastung in den ersten Monaten stark ab. Allerdings war der Rückgang gegenüber den anderen Varianten verlangsamt. Die Schadstoffbelastung blieb in der zweiten Versuchshälfte auf einem wenn auch deutlich reduziertem Niveau konstant. Tabelle 8: Abnahme der TNT-Bodenbelastung in den verschiedenen Versuchsvarianten. Aufgrund der deutlichen Messschwankungen ist zusätzlich zum arithmetischen Mittel auch der Median aufgeführt.
Variante A Variante B Variante C Variante K t n Mw Sd Med n Mw Sd Med n Mw Sd Med n Mw Sd Med0 31 99,0 80,7 73,8 31 86,5 83,2 64,5 31 82,8 47,1 77,1 31 96,7 82,6 78,62 7 8,9 5,3 6,0 6 21,7 2,01 21,5 7 6,8 5,9 5,4 8 42,2 28,8 31,33 6 19,0 12,8 14,6 8 18,7 9,3 15,0 8 13,1 14,2 6,7 8 31,5 17,6 23,15 5 5,9 2,6 5,1 6 12,7 3,56 12,1 7 5,8 2,3 5,7 8 12,6 5,3 10,66 8 3,4 1,6 2,7 7 12,0 8,3 9,8 8 6,6 6,0 3,4 7 19,1 21,1 11,9
t = Versuchsdauer in Monaten; n= Anzahl der berücksichtigten Proben; Mw = arithmetisches Mittel; Sd = Stan-dardabweichung (±); Med = Median; Werte für Mw, Sd und Med in mg TNT/kg Boden (TS)
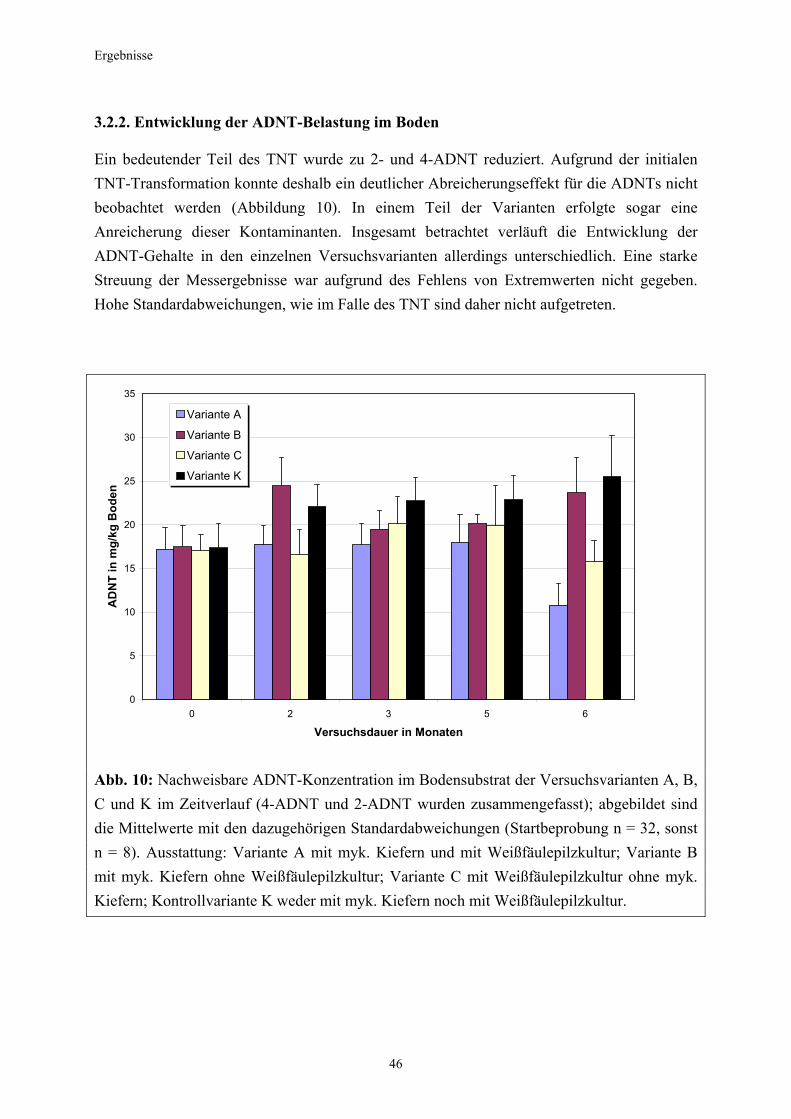
Ergebnisse
46
3.2.2. Entwicklung der ADNT-Belastung im Boden Ein bedeutender Teil des TNT wurde zu 2- und 4-ADNT reduziert. Aufgrund der initialen TNT-Transformation konnte deshalb ein deutlicher Abreicherungseffekt für die ADNTs nicht beobachtet werden (Abbildung 10). In einem Teil der Varianten erfolgte sogar eine Anreicherung dieser Kontaminanten. Insgesamt betrachtet verläuft die Entwicklung der ADNT-Gehalte in den einzelnen Versuchsvarianten allerdings unterschiedlich. Eine starke Streuung der Messergebnisse war aufgrund des Fehlens von Extremwerten nicht gegeben. Hohe Standardabweichungen, wie im Falle des TNT sind daher nicht aufgetreten.
0
5
10
15
20
25
30
35
0 2 3 5 6
Versuchsdauer in Monaten
AD
NT
in m
g/kg
Bod
en
Variante A
Variante B
Variante C
Variante K
Abb. 10: Nachweisbare ADNT-Konzentration im Bodensubstrat der Versuchsvarianten A, B, C und K im Zeitverlauf (4-ADNT und 2-ADNT wurden zusammengefasst); abgebildet sind die Mittelwerte mit den dazugehörigen Standardabweichungen (Startbeprobung n = 32, sonst n = 8). Ausstattung: Variante A mit myk. Kiefern und mit Weißfäulepilzkultur; Variante B mit myk. Kiefern ohne Weißfäulepilzkultur; Variante C mit Weißfäulepilzkultur ohne myk. Kiefern; Kontrollvariante K weder mit myk. Kiefern noch mit Weißfäulepilzkultur.
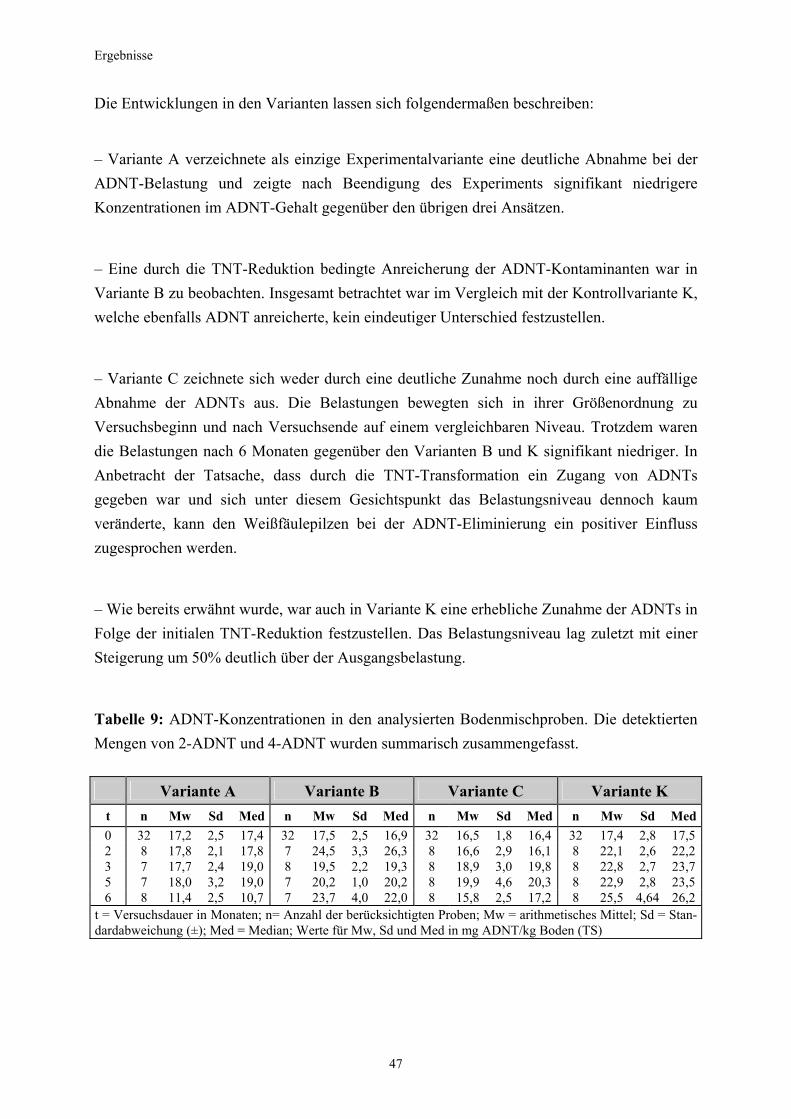
Ergebnisse
47
Die Entwicklungen in den Varianten lassen sich folgendermaßen beschreiben: – Variante A verzeichnete als einzige Experimentalvariante eine deutliche Abnahme bei der ADNT-Belastung und zeigte nach Beendigung des Experiments signifikant niedrigere Konzentrationen im ADNT-Gehalt gegenüber den übrigen drei Ansätzen. – Eine durch die TNT-Reduktion bedingte Anreicherung der ADNT-Kontaminanten war in Variante B zu beobachten. Insgesamt betrachtet war im Vergleich mit der Kontrollvariante K, welche ebenfalls ADNT anreicherte, kein eindeutiger Unterschied festzustellen. – Variante C zeichnete sich weder durch eine deutliche Zunahme noch durch eine auffällige Abnahme der ADNTs aus. Die Belastungen bewegten sich in ihrer Größenordnung zu Versuchsbeginn und nach Versuchsende auf einem vergleichbaren Niveau. Trotzdem waren die Belastungen nach 6 Monaten gegenüber den Varianten B und K signifikant niedriger. In Anbetracht der Tatsache, dass durch die TNT-Transformation ein Zugang von ADNTs gegeben war und sich unter diesem Gesichtspunkt das Belastungsniveau dennoch kaum veränderte, kann den Weißfäulepilzen bei der ADNT-Eliminierung ein positiver Einfluss zugesprochen werden. – Wie bereits erwähnt wurde, war auch in Variante K eine erhebliche Zunahme der ADNTs in Folge der initialen TNT-Reduktion festzustellen. Das Belastungsniveau lag zuletzt mit einer Steigerung um 50% deutlich über der Ausgangsbelastung. Tabelle 9: ADNT-Konzentrationen in den analysierten Bodenmischproben. Die detektierten Mengen von 2-ADNT und 4-ADNT wurden summarisch zusammengefasst.
Variante A Variante B Variante C Variante K t n Mw Sd Med n Mw Sd Med n Mw Sd Med n Mw Sd Med0 32 17,2 2,5 17,4 32 17,5 2,5 16,9 32 16,5 1,8 16,4 32 17,4 2,8 17,52 8 17,8 2,1 17,8 7 24,5 3,3 26,3 8 16,6 2,9 16,1 8 22,1 2,6 22,23 7 17,7 2,4 19,0 8 19,5 2,2 19,3 8 18,9 3,0 19,8 8 22,8 2,7 23,75 7 18,0 3,2 19,0 7 20,2 1,0 20,2 8 19,9 4,6 20,3 8 22,9 2,8 23,56 8 11,4 2,5 10,7 7 23,7 4,0 22,0 8 15,8 2,5 17,2 8 25,5 4,64 26,2
t = Versuchsdauer in Monaten; n= Anzahl der berücksichtigten Proben; Mw = arithmetisches Mittel; Sd = Stan-dardabweichung (±); Med = Median; Werte für Mw, Sd und Med in mg ADNT/kg Boden (TS)
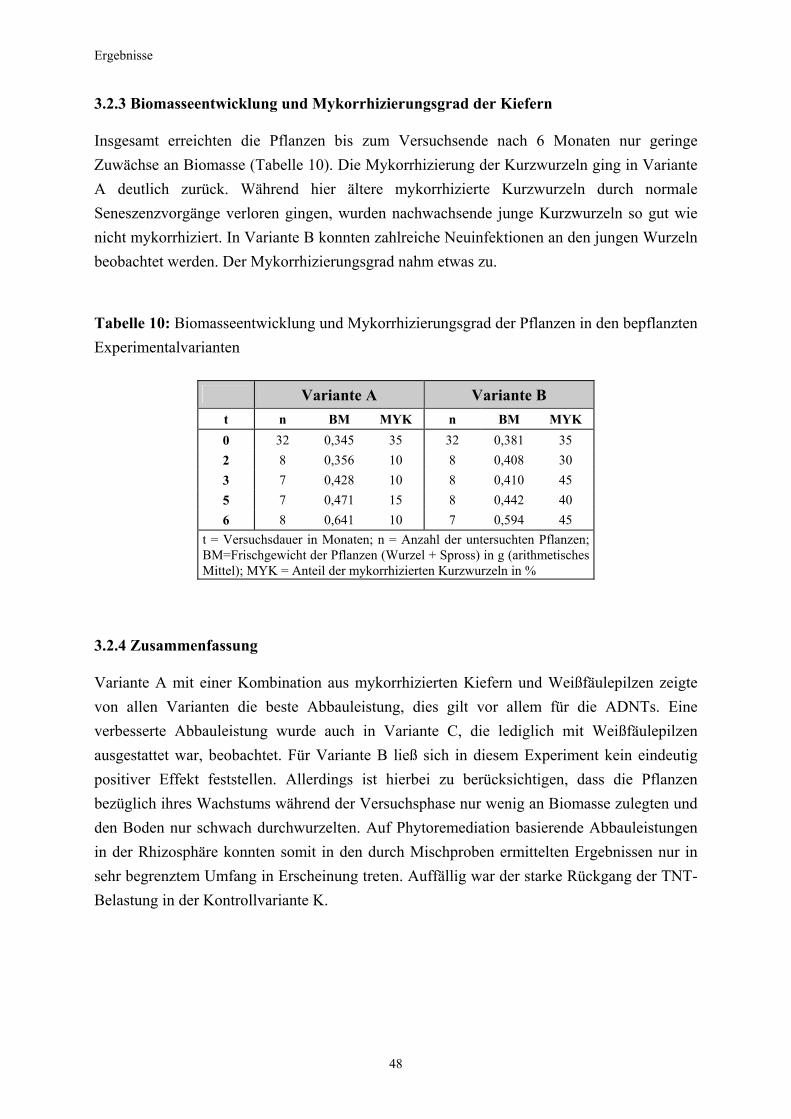
Ergebnisse
48
3.2.3 Biomasseentwicklung und Mykorrhizierungsgrad der Kiefern Insgesamt erreichten die Pflanzen bis zum Versuchsende nach 6 Monaten nur geringe Zuwächse an Biomasse (Tabelle 10). Die Mykorrhizierung der Kurzwurzeln ging in Variante A deutlich zurück. Während hier ältere mykorrhizierte Kurzwurzeln durch normale Seneszenzvorgänge verloren gingen, wurden nachwachsende junge Kurzwurzeln so gut wie nicht mykorrhiziert. In Variante B konnten zahlreiche Neuinfektionen an den jungen Wurzeln beobachtet werden. Der Mykorrhizierungsgrad nahm etwas zu. Tabelle 10: Biomasseentwicklung und Mykorrhizierungsgrad der Pflanzen in den bepflanzten Experimentalvarianten
Variante A Variante B t n BM MYK n BM MYK 0 32 0,345 35 32 0,381 35 2 8 0,356 10 8 0,408 30 3 7 0,428 10 8 0,410 45 5 7 0,471 15 8 0,442 40 6 8 0,641 10 7 0,594 45
t = Versuchsdauer in Monaten; n = Anzahl der untersuchten Pflanzen; BM=Frischgewicht der Pflanzen (Wurzel + Spross) in g (arithmetisches Mittel); MYK = Anteil der mykorrhizierten Kurzwurzeln in %
3.2.4 Zusammenfassung Variante A mit einer Kombination aus mykorrhizierten Kiefern und Weißfäulepilzen zeigte von allen Varianten die beste Abbauleistung, dies gilt vor allem für die ADNTs. Eine verbesserte Abbauleistung wurde auch in Variante C, die lediglich mit Weißfäulepilzen ausgestattet war, beobachtet. Für Variante B ließ sich in diesem Experiment kein eindeutig positiver Effekt feststellen. Allerdings ist hierbei zu berücksichtigen, dass die Pflanzen bezüglich ihres Wachstums während der Versuchsphase nur wenig an Biomasse zulegten und den Boden nur schwach durchwurzelten. Auf Phytoremediation basierende Abbauleistungen in der Rhizosphäre konnten somit in den durch Mischproben ermittelten Ergebnissen nur in sehr begrenztem Umfang in Erscheinung treten. Auffällig war der starke Rückgang der TNT-Belastung in der Kontrollvariante K.

Ergebnisse
49
3.3 TNT-Abbauexperimente mit Pappeln als Phytoremediatoren Aus dem vorangegangenen Kiefernexperiment wurden wichtige Erkenntnisse gewonnen, die bei den Planungen für die Durchführung des sich nun anschließenden Pappelexperimentes berücksichtigt wurden. Zum einen wurde aufgrund zu erwartender Abbauraten in der vorgesehenen Kontrollvariante (Punkt 3.2.1) auf einen deutlich stärker belasteten Stand-ortboden zurückgegriffen, um das Experiment mit einer höheren Ausgangsbelastung starten zu können. Zum anderen wurden die Versuchszeit und die Zeitintervalle zwischen den jeweiligen Beprobungen verkürzt, um der voraussichtlich zügigen Abbaukinetik der Kontaminanten gerecht zu werden. Unter Berücksichtigung der im Kiefernexperiment beobachteten Langsamwüchsigkeit der Pflanzen und einer damit verbundenen geringen Durchwurzelung des Substrates in den Rhizotronen wurde nun auf die schnellwüchsige Zitterpappel (Populus tremula) zurückgegriffen. Besonders Pappeln zeichnen sich durch ein starkes Wurzelwachstum aus. Verbunden mit einer intensiven Durchwurzelung des kontaminierten Bodens sollte ein möglicherweise auftretender und auf die Pflanzen zurückzuführender Phytoremediationseffekt (v. a. Rhizosphärendegradation) mit diesem Experiment besser fassbar werden. Ergänzend wurde die Anzahl der Versuchsvarianten um eine weitere auf fünf erhöht. Die unterschiedliche Bestückung der Rhizotrone ist in Punkt 2.2.2 beschrieben. Zusätzlich zur Rückstandsanalytik des Bodensubstrates, wurde untersucht, ob und in welcher Größenordnung eine Aufnahme und Akkumulation von TNT und Metaboliten in die Versuchspflanzen stattgefunden hat. Versuchsanleitung und Versuchsdurchführung sind im Methodenteil Punkt 2.2.2 zu ent-nehmen. 3.3.1 Bodenanalytik In den Bodenproben wurden regelmäßig TNT und dazu die unmittelbaren Reduktionsprodukte 2-ADNT und 4-ADNT detektiert. In sehr geringen Konzentrationen ließen sich in einem Teil der untersuchten Proben außerdem 2,4- und 2,6-DNT über die Rückstandsanalytik nach-weisen. Weitere Nitroaromaten konnten nicht gefunden werden.
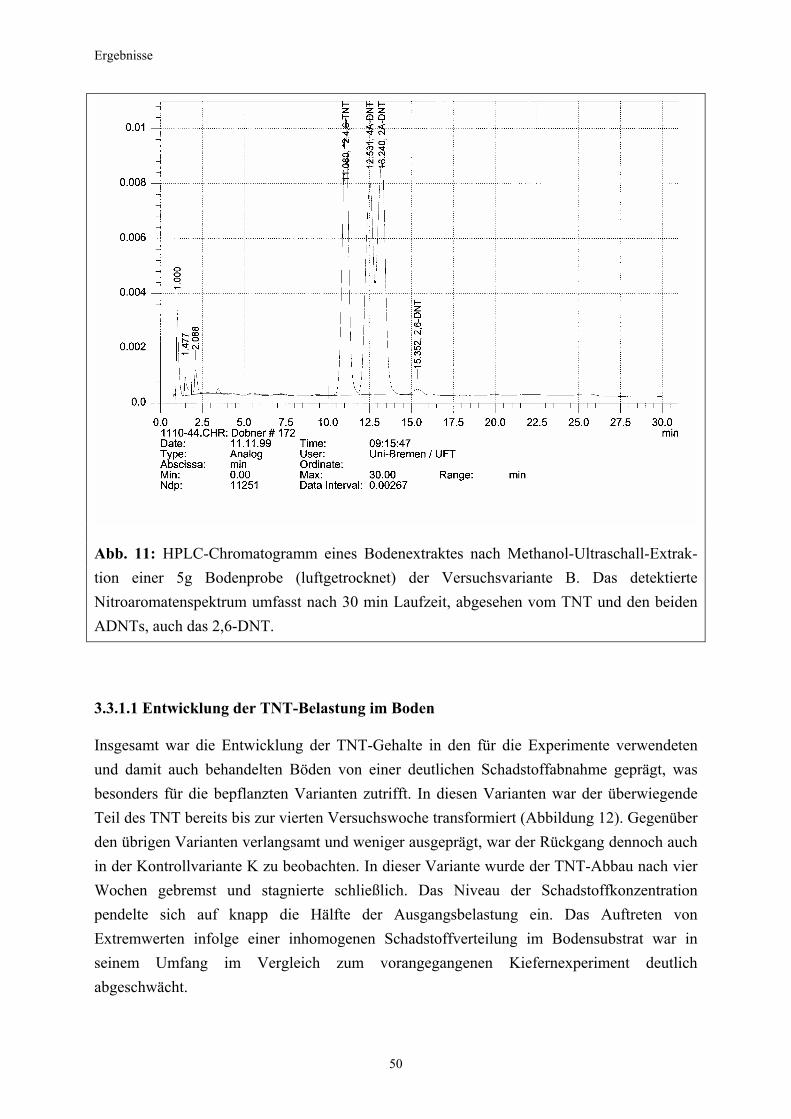
Ergebnisse
50
Abb. 11: HPLC-Chromatogramm eines Bodenextraktes nach Methanol-Ultraschall-Extrak-tion einer 5g Bodenprobe (luftgetrocknet) der Versuchsvariante B. Das detektierte Nitroaromatenspektrum umfasst nach 30 min Laufzeit, abgesehen vom TNT und den beiden ADNTs, auch das 2,6-DNT. 3.3.1.1 Entwicklung der TNT-Belastung im Boden Insgesamt war die Entwicklung der TNT-Gehalte in den für die Experimente verwendeten und damit auch behandelten Böden von einer deutlichen Schadstoffabnahme geprägt, was besonders für die bepflanzten Varianten zutrifft. In diesen Varianten war der überwiegende Teil des TNT bereits bis zur vierten Versuchswoche transformiert (Abbildung 12). Gegenüber den übrigen Varianten verlangsamt und weniger ausgeprägt, war der Rückgang dennoch auch in der Kontrollvariante K zu beobachten. In dieser Variante wurde der TNT-Abbau nach vier Wochen gebremst und stagnierte schließlich. Das Niveau der Schadstoffkonzentration pendelte sich auf knapp die Hälfte der Ausgangsbelastung ein. Das Auftreten von Extremwerten infolge einer inhomogenen Schadstoffverteilung im Bodensubstrat war in seinem Umfang im Vergleich zum vorangegangenen Kiefernexperiment deutlich abgeschwächt.
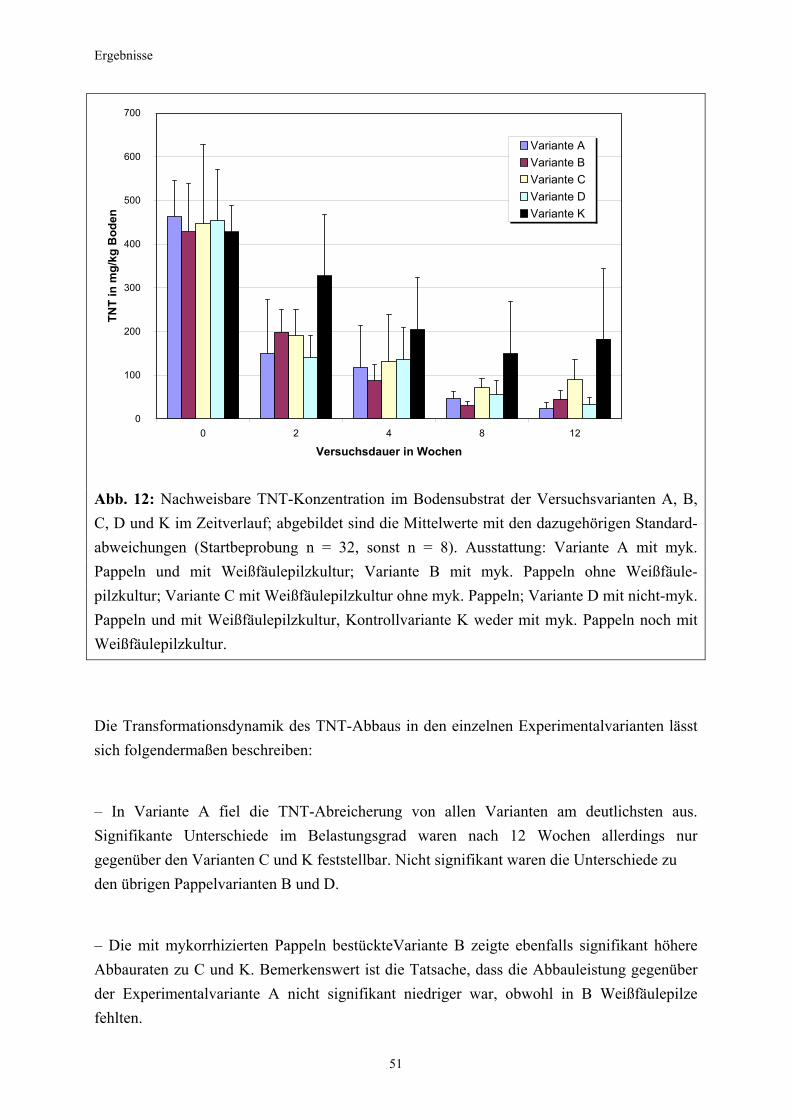
Ergebnisse
51
0
100
200
300
400
500
600
700
0 2 4 8 12
Versuchsdauer in Wochen
TNT
in m
g/kg
Bod
en
Variante AVariante BVariante CVariante DVariante K
Abb. 12: Nachweisbare TNT-Konzentration im Bodensubstrat der Versuchsvarianten A, B, C, D und K im Zeitverlauf; abgebildet sind die Mittelwerte mit den dazugehörigen Standard-abweichungen (Startbeprobung n = 32, sonst n = 8). Ausstattung: Variante A mit myk. Pappeln und mit Weißfäulepilzkultur; Variante B mit myk. Pappeln ohne Weißfäule-pilzkultur; Variante C mit Weißfäulepilzkultur ohne myk. Pappeln; Variante D mit nicht-myk. Pappeln und mit Weißfäulepilzkultur, Kontrollvariante K weder mit myk. Pappeln noch mit Weißfäulepilzkultur. Die Transformationsdynamik des TNT-Abbaus in den einzelnen Experimentalvarianten lässt sich folgendermaßen beschreiben: – In Variante A fiel die TNT-Abreicherung von allen Varianten am deutlichsten aus. Signifikante Unterschiede im Belastungsgrad waren nach 12 Wochen allerdings nur gegenüber den Varianten C und K feststellbar. Nicht signifikant waren die Unterschiede zu den übrigen Pappelvarianten B und D. – Die mit mykorrhizierten Pappeln bestückteVariante B zeigte ebenfalls signifikant höhere Abbauraten zu C und K. Bemerkenswert ist die Tatsache, dass die Abbauleistung gegenüber der Experimentalvariante A nicht signifikant niedriger war, obwohl in B Weißfäulepilze fehlten.
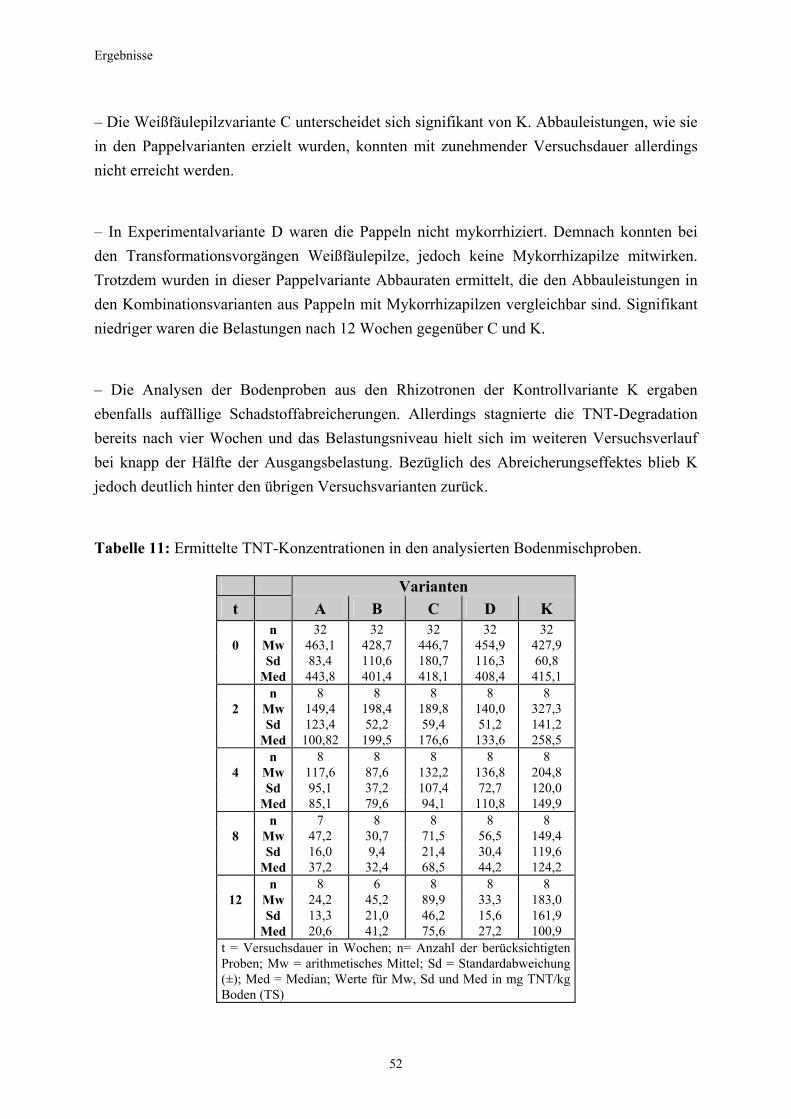
Ergebnisse
52
– Die Weißfäulepilzvariante C unterscheidet sich signifikant von K. Abbauleistungen, wie sie in den Pappelvarianten erzielt wurden, konnten mit zunehmender Versuchsdauer allerdings nicht erreicht werden. – In Experimentalvariante D waren die Pappeln nicht mykorrhiziert. Demnach konnten bei den Transformationsvorgängen Weißfäulepilze, jedoch keine Mykorrhizapilze mitwirken. Trotzdem wurden in dieser Pappelvariante Abbauraten ermittelt, die den Abbauleistungen in den Kombinationsvarianten aus Pappeln mit Mykorrhizapilzen vergleichbar sind. Signifikant niedriger waren die Belastungen nach 12 Wochen gegenüber C und K. – Die Analysen der Bodenproben aus den Rhizotronen der Kontrollvariante K ergaben ebenfalls auffällige Schadstoffabreicherungen. Allerdings stagnierte die TNT-Degradation bereits nach vier Wochen und das Belastungsniveau hielt sich im weiteren Versuchsverlauf bei knapp der Hälfte der Ausgangsbelastung. Bezüglich des Abreicherungseffektes blieb K jedoch deutlich hinter den übrigen Versuchsvarianten zurück. Tabelle 11: Ermittelte TNT-Konzentrationen in den analysierten Bodenmischproben.
Varianten t A B C D K n 32 32 32 32 32
0 Mw 463,1 428,7 446,7 454,9 427,9 Sd 83,4 110,6 180,7 116,3 60,8 Med 443,8 401,4 418,1 408,4 415,1 n 8 8 8 8 8
2 Mw 149,4 198,4 189,8 140,0 327,3 Sd 123,4 52,2 59,4 51,2 141,2 Med 100,82 199,5 176,6 133,6 258,5 n 8 8 8 8 8
4 Mw 117,6 87,6 132,2 136,8 204,8 Sd 95,1 37,2 107,4 72,7 120,0 Med 85,1 79,6 94,1 110,8 149,9 n 7 8 8 8 8
8 Mw 47,2 30,7 71,5 56,5 149,4 Sd 16,0 9,4 21,4 30,4 119,6 Med 37,2 32,4 68,5 44,2 124,2 n 8 6 8 8 8
12 Mw 24,2 45,2 89,9 33,3 183,0 Sd 13,3 21,0 46,2 15,6 161,9 Med 20,6 41,2 75,6 27,2 100,9
t = Versuchsdauer in Wochen; n= Anzahl der berücksichtigten Proben; Mw = arithmetisches Mittel; Sd = Standardabweichung (±); Med = Median; Werte für Mw, Sd und Med in mg TNT/kg Boden (TS)
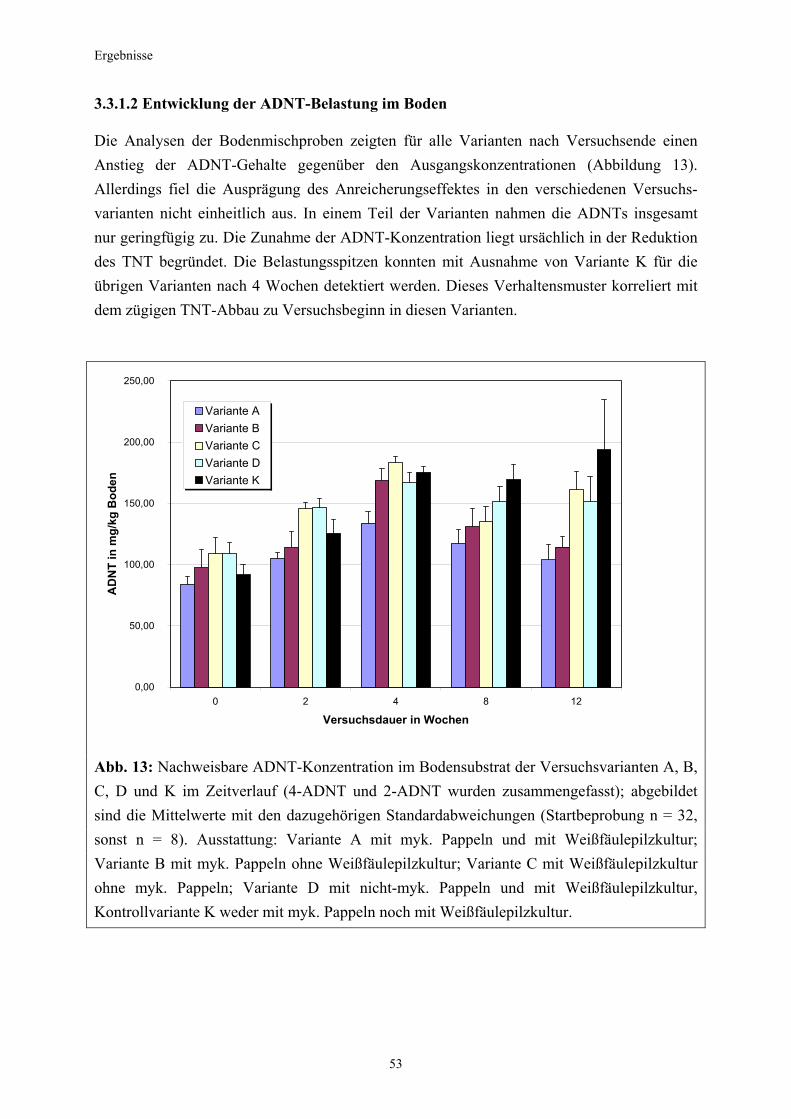
Ergebnisse
53
3.3.1.2 Entwicklung der ADNT-Belastung im Boden Die Analysen der Bodenmischproben zeigten für alle Varianten nach Versuchsende einen Anstieg der ADNT-Gehalte gegenüber den Ausgangskonzentrationen (Abbildung 13). Allerdings fiel die Ausprägung des Anreicherungseffektes in den verschiedenen Versuchs-varianten nicht einheitlich aus. In einem Teil der Varianten nahmen die ADNTs insgesamt nur geringfügig zu. Die Zunahme der ADNT-Konzentration liegt ursächlich in der Reduktion des TNT begründet. Die Belastungsspitzen konnten mit Ausnahme von Variante K für die übrigen Varianten nach 4 Wochen detektiert werden. Dieses Verhaltensmuster korreliert mit dem zügigen TNT-Abbau zu Versuchsbeginn in diesen Varianten.
0,00
50,00
100,00
150,00
200,00
250,00
0 2 4 8 12
Versuchsdauer in Wochen
AD
NT
in m
g/kg
Bod
en
Variante AVariante BVariante CVariante DVariante K
Abb. 13: Nachweisbare ADNT-Konzentration im Bodensubstrat der Versuchsvarianten A, B, C, D und K im Zeitverlauf (4-ADNT und 2-ADNT wurden zusammengefasst); abgebildet sind die Mittelwerte mit den dazugehörigen Standardabweichungen (Startbeprobung n = 32, sonst n = 8). Ausstattung: Variante A mit myk. Pappeln und mit Weißfäulepilzkultur; Variante B mit myk. Pappeln ohne Weißfäulepilzkultur; Variante C mit Weißfäulepilzkultur ohne myk. Pappeln; Variante D mit nicht-myk. Pappeln und mit Weißfäulepilzkultur, Kontrollvariante K weder mit myk. Pappeln noch mit Weißfäulepilzkultur.

Ergebnisse
54
Für die einzelnen Varianten konnten folgende Beobachtungen gemacht werden: – In Variante A hat die ADNT-Konzentration nach dem Erreichen der Belastungsspitze bis zur Beendigung des Versuchs stetig abgenommen und erreichte nach 12 Wochen von allen Varianten das niedrigste Belastungsniveau. Signifikant niedriger war der ADNT-Gehalt zu diesem Zeitpunkt gegenüber den Varianten C, D und K. Nicht signifikant war der Unterschied gegenüber Variante B. – In Variante B zeigte die Belastung in ihrer Entwicklung ein ähnliches Verhaltensmuster wie in Variante A. Nach Erreichen der Belastungsspitze war ein deutlicher Rückgang der ADNT-Konzentration zu verzeichnen. Das Belastungsniveau war gegenüber A in der zweiten Versuchshälfte nicht signifikant höher. Signifikant reduziert war in der Variante B nach Ablauf des Versuches die ADNT-Belastung gegenüber den Varianten C, D und K. – Die Weißfäulepilzvariante C grenzte sich insbesondere in der zweiten Versuchshälfte aufgrund eines signifikant niedrigeren Schadstoffgehaltes von der Kontrollvariante K ab. Abbauleistungen, wie sie in den Pappelvarianten A und B erzielt wurden, konnten bei zunehmender Versuchsdauer in C allerdings nicht erreicht werden. – In der Pappelvariante D waren die Pflanzen nicht mykorrhiziert. Bei einem Vergleich mit den beiden übrigen Pappelvarianten A und B, in denen Mykorrhizapilze zugegen waren, zeigte sich in D ein deutlich geringerer Effekt bei der Eliminierung der ADNTs, so dass der Schadstoffgehalt nach 12 Wochen signifikant höher war. Ein signifikant niedrigerer ADNT-Gehalt konnte letztendlich nur gegenüber der Kontrollvariante K festgestellt werden. – In der Kontrollvariante K wurden die maximalen ADNT-Konzentrationen am Ende des Experimentes nach zwölf Wochen detektiert. Das Auftreten einer Belastungsspitze nach 4 Wochen mit einem sich anschließendem Schadstoffrückgang konnte nicht beobachtet werden, so dass der ADNT-Gehalt in dieser Variante zum Schluss die höchsten Werte erreichte.
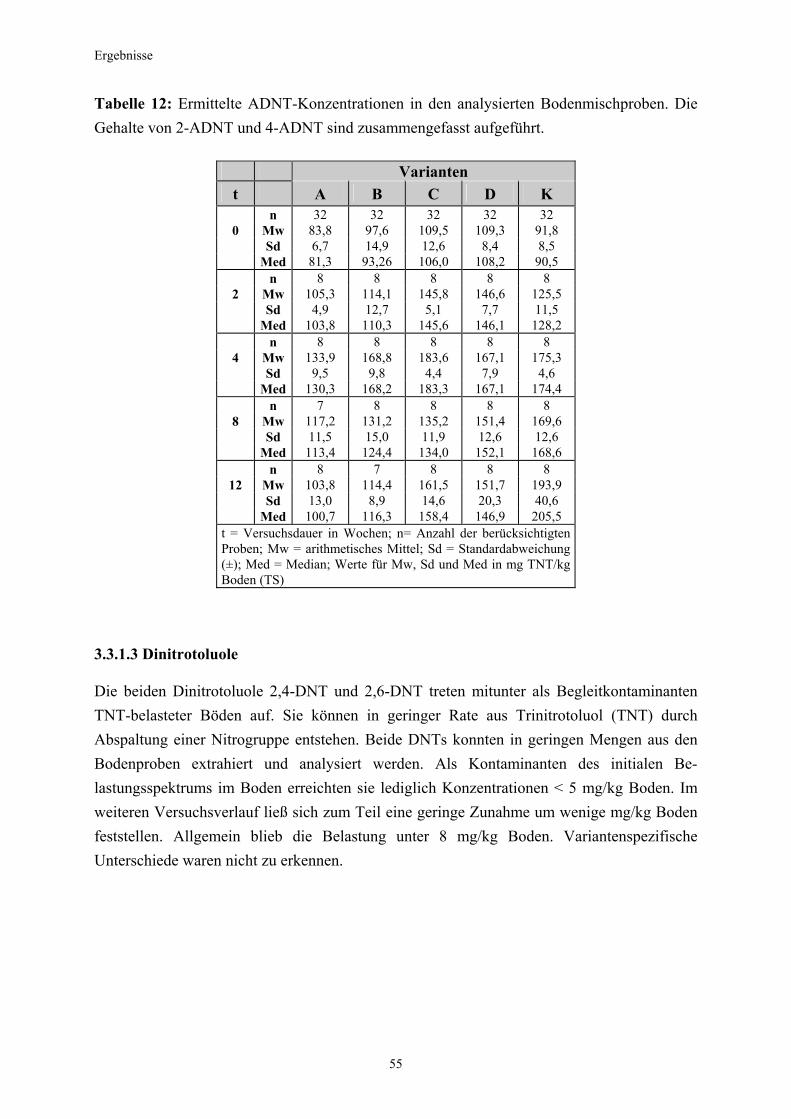
Ergebnisse
55
Tabelle 12: Ermittelte ADNT-Konzentrationen in den analysierten Bodenmischproben. Die Gehalte von 2-ADNT und 4-ADNT sind zusammengefasst aufgeführt.
Varianten t A B C D K n 32 32 32 32 32
0 Mw 83,8 97,6 109,5 109,3 91,8 Sd 6,7 14,9 12,6 8,4 8,5 Med 81,3 93,26 106,0 108,2 90,5 n 8 8 8 8 8
2 Mw 105,3 114,1 145,8 146,6 125,5 Sd 4,9 12,7 5,1 7,7 11,5 Med 103,8 110,3 145,6 146,1 128,2 n 8 8 8 8 8
4 Mw 133,9 168,8 183,6 167,1 175,3 Sd 9,5 9,8 4,4 7,9 4,6 Med 130,3 168,2 183,3 167,1 174,4 n 7 8 8 8 8
8 Mw 117,2 131,2 135,2 151,4 169,6 Sd 11,5 15,0 11,9 12,6 12,6 Med 113,4 124,4 134,0 152,1 168,6 n 8 7 8 8 8
12 Mw 103,8 114,4 161,5 151,7 193,9 Sd 13,0 8,9 14,6 20,3 40,6 Med 100,7 116,3 158,4 146,9 205,5
t = Versuchsdauer in Wochen; n= Anzahl der berücksichtigten Proben; Mw = arithmetisches Mittel; Sd = Standardabweichung (±); Med = Median; Werte für Mw, Sd und Med in mg TNT/kg Boden (TS)
3.3.1.3 Dinitrotoluole Die beiden Dinitrotoluole 2,4-DNT und 2,6-DNT treten mitunter als Begleitkontaminanten TNT-belasteter Böden auf. Sie können in geringer Rate aus Trinitrotoluol (TNT) durch Abspaltung einer Nitrogruppe entstehen. Beide DNTs konnten in geringen Mengen aus den Bodenproben extrahiert und analysiert werden. Als Kontaminanten des initialen Be-lastungsspektrums im Boden erreichten sie lediglich Konzentrationen < 5 mg/kg Boden. Im weiteren Versuchsverlauf ließ sich zum Teil eine geringe Zunahme um wenige mg/kg Boden feststellen. Allgemein blieb die Belastung unter 8 mg/kg Boden. Variantenspezifische Unterschiede waren nicht zu erkennen.
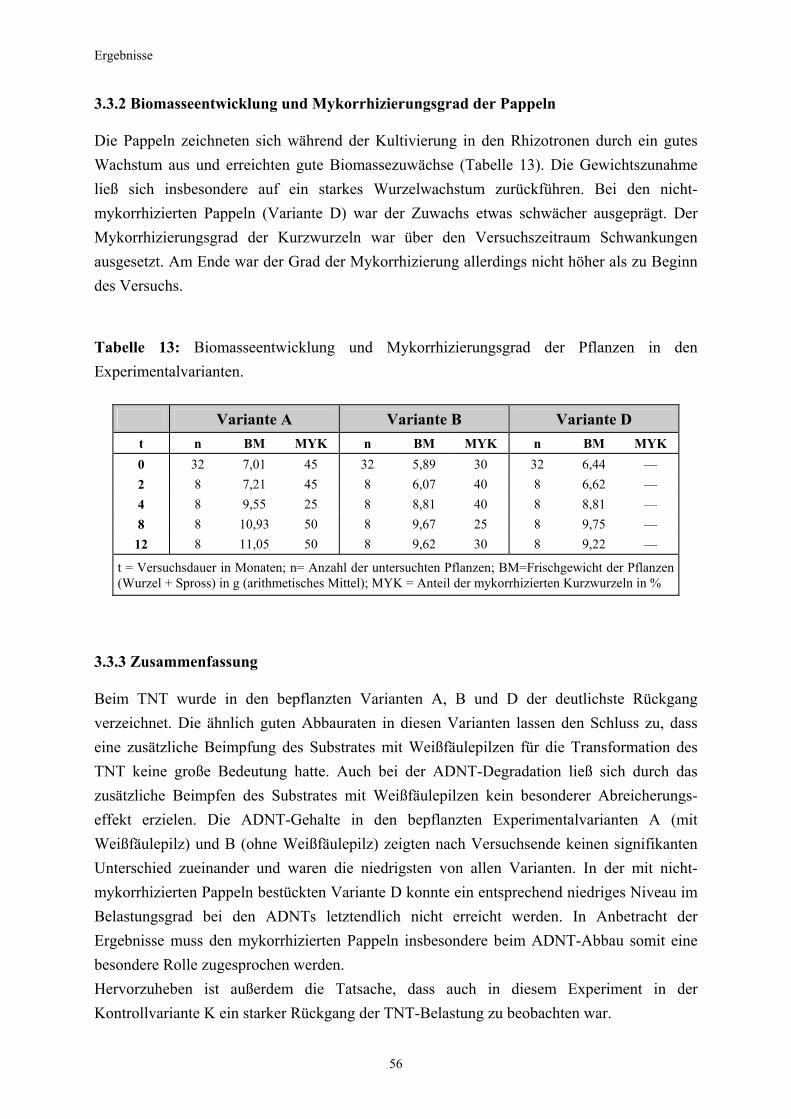
Ergebnisse
56
3.3.2 Biomasseentwicklung und Mykorrhizierungsgrad der Pappeln Die Pappeln zeichneten sich während der Kultivierung in den Rhizotronen durch ein gutes Wachstum aus und erreichten gute Biomassezuwächse (Tabelle 13). Die Gewichtszunahme ließ sich insbesondere auf ein starkes Wurzelwachstum zurückführen. Bei den nicht-mykorrhizierten Pappeln (Variante D) war der Zuwachs etwas schwächer ausgeprägt. Der Mykorrhizierungsgrad der Kurzwurzeln war über den Versuchszeitraum Schwankungen ausgesetzt. Am Ende war der Grad der Mykorrhizierung allerdings nicht höher als zu Beginn des Versuchs. Tabelle 13: Biomasseentwicklung und Mykorrhizierungsgrad der Pflanzen in den Experimentalvarianten.
Variante A Variante B Variante D t n BM MYK n BM MYK n BM MYK 0 32 7,01 45 32 5,89 30 32 6,44 — 2 8 7,21 45 8 6,07 40 8 6,62 — 4 8 9,55 25 8 8,81 40 8 8,81 — 8 8 10,93 50 8 9,67 25 8 9,75 —
12 8 11,05 50 8 9,62 30 8 9,22 —
t = Versuchsdauer in Monaten; n= Anzahl der untersuchten Pflanzen; BM=Frischgewicht der Pflanzen (Wurzel + Spross) in g (arithmetisches Mittel); MYK = Anteil der mykorrhizierten Kurzwurzeln in %
3.3.3 Zusammenfassung Beim TNT wurde in den bepflanzten Varianten A, B und D der deutlichste Rückgang verzeichnet. Die ähnlich guten Abbauraten in diesen Varianten lassen den Schluss zu, dass eine zusätzliche Beimpfung des Substrates mit Weißfäulepilzen für die Transformation des TNT keine große Bedeutung hatte. Auch bei der ADNT-Degradation ließ sich durch das zusätzliche Beimpfen des Substrates mit Weißfäulepilzen kein besonderer Abreicherungs-effekt erzielen. Die ADNT-Gehalte in den bepflanzten Experimentalvarianten A (mit Weißfäulepilz) und B (ohne Weißfäulepilz) zeigten nach Versuchsende keinen signifikanten Unterschied zueinander und waren die niedrigsten von allen Varianten. In der mit nicht-mykorrhizierten Pappeln bestückten Variante D konnte ein entsprechend niedriges Niveau im Belastungsgrad bei den ADNTs letztendlich nicht erreicht werden. In Anbetracht der Ergebnisse muss den mykorrhizierten Pappeln insbesondere beim ADNT-Abbau somit eine besondere Rolle zugesprochen werden. Hervorzuheben ist außerdem die Tatsache, dass auch in diesem Experiment in der Kontrollvariante K ein starker Rückgang der TNT-Belastung zu beobachten war.

Ergebnisse
57
Bezüglich ihrer Biomasseentwicklung zeigten die Pappeln ein deutliches Wachstum. Dieser Umstand spiegelte sich vor allem in einer guten Durchwurzelung des Substrates wieder. 3.3.4 Nachweis des Nitroaromatentransfers aus dem Boden in die Versuchspflanzen Durch die Schadstoffanalysen der Bodenproben konnte in den bepflanzten Rhizotronen eine besonders effiziente Nitroaromatenabreicherung nachgewiesen werden. Deshalb sollte nun untersucht werden, ob dieser Effekt auf einen verstärkten Abbau im Wurzelraum (Rhizosphäreneffekte) oder möglicherweise auch auf eine Aufnahme der Schadstoffe aus dem Boden in die Pflanzen zurückzuführen ist. Wie bereits aus Abschnitt 2.3.2.2 hervorgeht, wurden die Pflanzenproben mit Dichlormethan (DCM) extrahiert. Außerdem beinhaltete die Extraktion eine Säurehydrolyse mit anschließender Alkalisierung, damit auch die fest in die pflanzliche Matrix eingebundenen Metabolite herausgelöst werden konnten. Abbildung 14 zeigt den Chromatogrammausschnitt eines Wurzelextraktes nach Säurehydrolyse. Die analytische Erfassung der Nitroaromaten erfolgte über eine thermische Desorption gekoppelt mit nachfolgender GC/MS. Im Anhang sind die Nitroaromaten-Peaks einer Probe herausgefiltert dargestellt (Abbildung 44). Außerdem ist dort ein Massenspektrum für TNT aufgeführt (Abbildung 45).
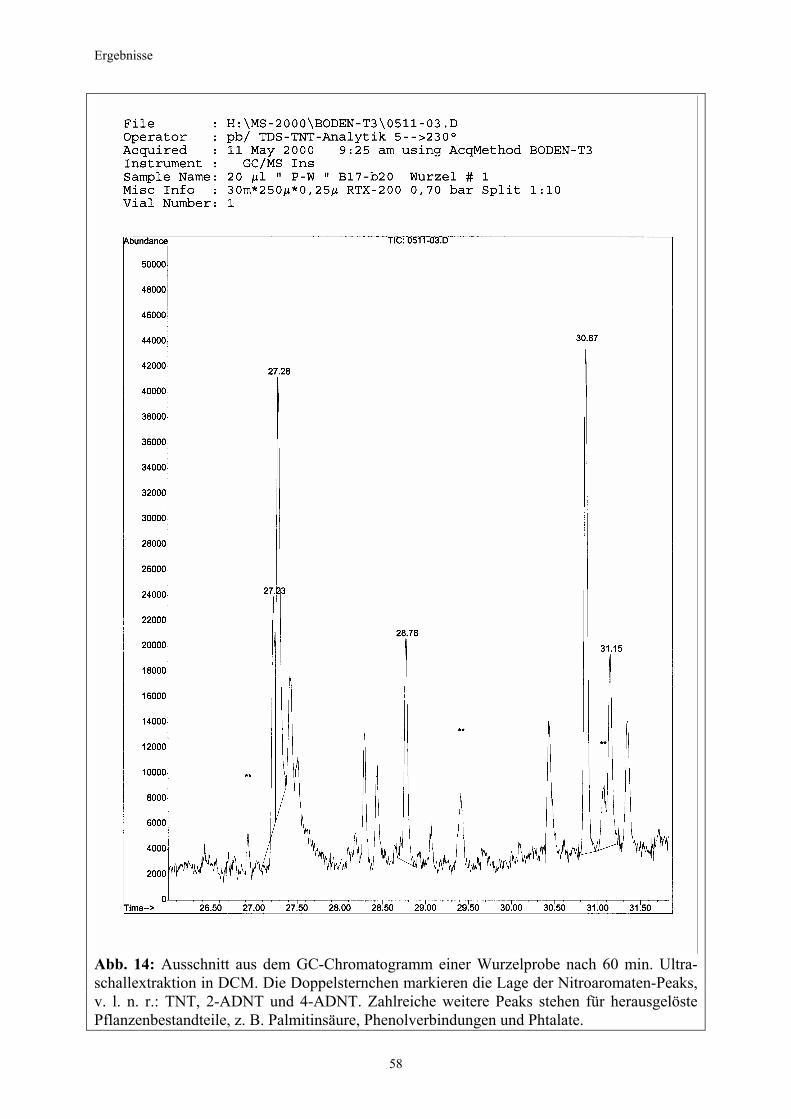
Ergebnisse
58
Abb. 14: Ausschnitt aus dem GC-Chromatogramm einer Wurzelprobe nach 60 min. Ultra-schallextraktion in DCM. Die Doppelsternchen markieren die Lage der Nitroaromaten-Peaks, v. l. n. r.: TNT, 2-ADNT und 4-ADNT. Zahlreiche weitere Peaks stehen für herausgelöste Pflanzenbestandteile, z. B. Palmitinsäure, Phenolverbindungen und Phtalate.
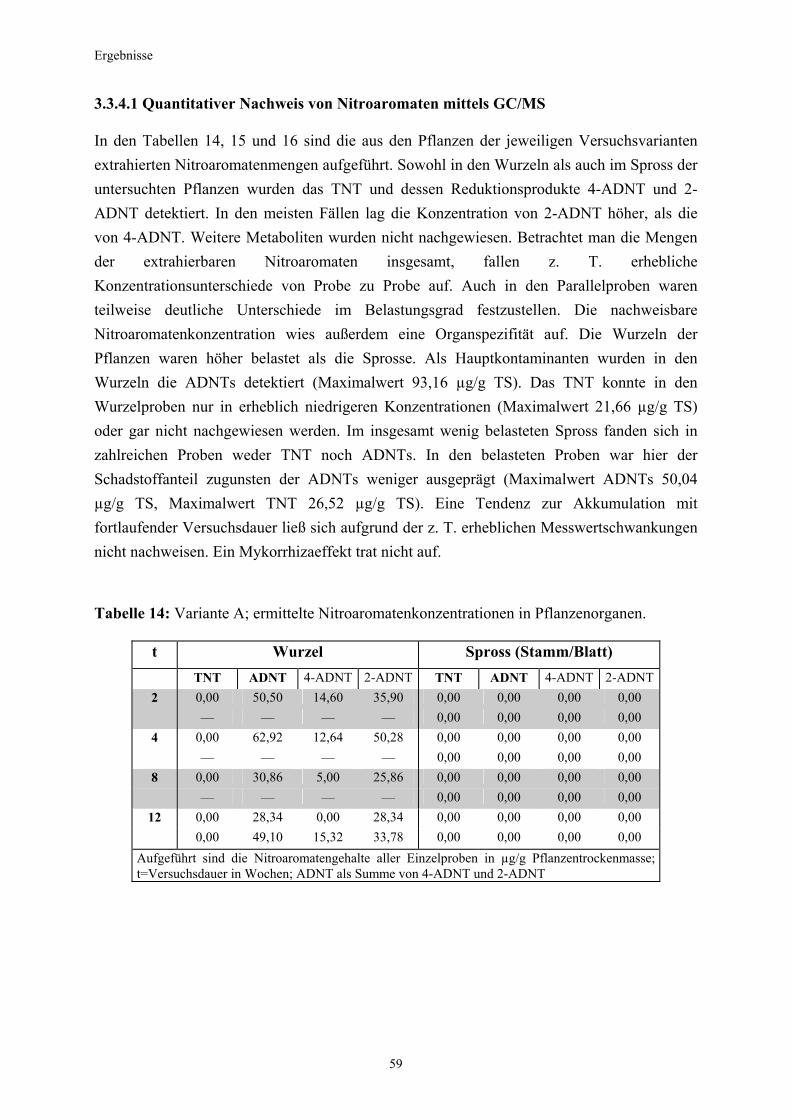
Ergebnisse
59
3.3.4.1 Quantitativer Nachweis von Nitroaromaten mittels GC/MS In den Tabellen 14, 15 und 16 sind die aus den Pflanzen der jeweiligen Versuchsvarianten extrahierten Nitroaromatenmengen aufgeführt. Sowohl in den Wurzeln als auch im Spross der untersuchten Pflanzen wurden das TNT und dessen Reduktionsprodukte 4-ADNT und 2-ADNT detektiert. In den meisten Fällen lag die Konzentration von 2-ADNT höher, als die von 4-ADNT. Weitere Metaboliten wurden nicht nachgewiesen. Betrachtet man die Mengen der extrahierbaren Nitroaromaten insgesamt, fallen z. T. erhebliche Konzentrationsunterschiede von Probe zu Probe auf. Auch in den Parallelproben waren teilweise deutliche Unterschiede im Belastungsgrad festzustellen. Die nachweisbare Nitroaromatenkonzentration wies außerdem eine Organspezifität auf. Die Wurzeln der Pflanzen waren höher belastet als die Sprosse. Als Hauptkontaminanten wurden in den Wurzeln die ADNTs detektiert (Maximalwert 93,16 µg/g TS). Das TNT konnte in den Wurzelproben nur in erheblich niedrigeren Konzentrationen (Maximalwert 21,66 µg/g TS) oder gar nicht nachgewiesen werden. Im insgesamt wenig belasteten Spross fanden sich in zahlreichen Proben weder TNT noch ADNTs. In den belasteten Proben war hier der Schadstoffanteil zugunsten der ADNTs weniger ausgeprägt (Maximalwert ADNTs 50,04 µg/g TS, Maximalwert TNT 26,52 µg/g TS). Eine Tendenz zur Akkumulation mit fortlaufender Versuchsdauer ließ sich aufgrund der z. T. erheblichen Messwertschwankungen nicht nachweisen. Ein Mykorrhizaeffekt trat nicht auf. Tabelle 14: Variante A; ermittelte Nitroaromatenkonzentrationen in Pflanzenorganen.
t Wurzel Spross (Stamm/Blatt) TNT ADNT 4-ADNT 2-ADNT TNT ADNT 4-ADNT 2-ADNT
2 0,00 50,50 14,60 35,90 0,00 0,00 0,00 0,00 — — — — 0,00 0,00 0,00 0,00
4 0,00 62,92 12,64 50,28 0,00 0,00 0,00 0,00 — — — — 0,00 0,00 0,00 0,00
8 0,00 30,86 5,00 25,86 0,00 0,00 0,00 0,00 — — — — 0,00 0,00 0,00 0,00
12 0,00 28,34 0,00 28,34 0,00 0,00 0,00 0,00 0,00 49,10 15,32 33,78 0,00 0,00 0,00 0,00
Aufgeführt sind die Nitroaromatengehalte aller Einzelproben in µg/g Pflanzentrockenmasse; t=Versuchsdauer in Wochen; ADNT als Summe von 4-ADNT und 2-ADNT
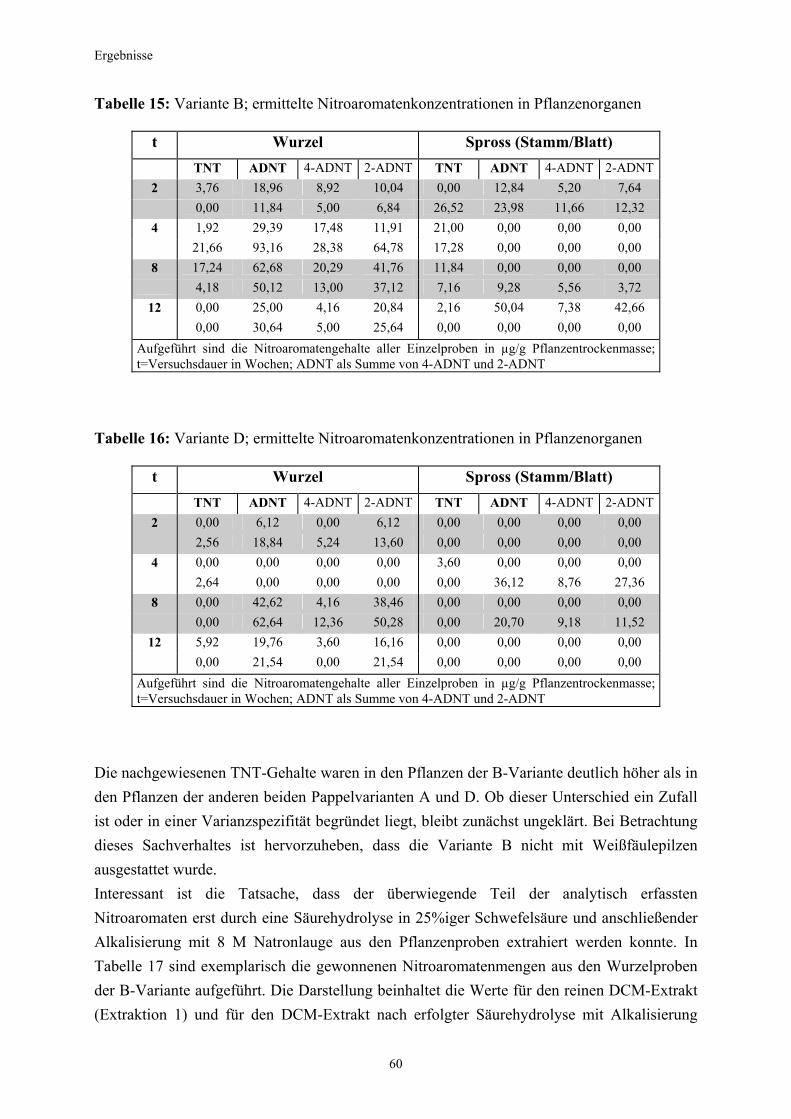
Ergebnisse
60
Tabelle 15: Variante B; ermittelte Nitroaromatenkonzentrationen in Pflanzenorganen
t Wurzel Spross (Stamm/Blatt) TNT ADNT 4-ADNT 2-ADNT TNT ADNT 4-ADNT 2-ADNT
2 3,76 18,96 8,92 10,04 0,00 12,84 5,20 7,64 0,00 11,84 5,00 6,84 26,52 23,98 11,66 12,32
4 1,92 29,39 17,48 11,91 21,00 0,00 0,00 0,00 21,66 93,16 28,38 64,78 17,28 0,00 0,00 0,00
8 17,24 62,68 20,29 41,76 11,84 0,00 0,00 0,00 4,18 50,12 13,00 37,12 7,16 9,28 5,56 3,72
12 0,00 25,00 4,16 20,84 2,16 50,04 7,38 42,66 0,00 30,64 5,00 25,64 0,00 0,00 0,00 0,00
Aufgeführt sind die Nitroaromatengehalte aller Einzelproben in µg/g Pflanzentrockenmasse; t=Versuchsdauer in Wochen; ADNT als Summe von 4-ADNT und 2-ADNT
Tabelle 16: Variante D; ermittelte Nitroaromatenkonzentrationen in Pflanzenorganen
t Wurzel Spross (Stamm/Blatt) TNT ADNT 4-ADNT 2-ADNT TNT ADNT 4-ADNT 2-ADNT
2 0,00 6,12 0,00 6,12 0,00 0,00 0,00 0,00 2,56 18,84 5,24 13,60 0,00 0,00 0,00 0,00
4 0,00 0,00 0,00 0,00 3,60 0,00 0,00 0,00 2,64 0,00 0,00 0,00 0,00 36,12 8,76 27,36
8 0,00 42,62 4,16 38,46 0,00 0,00 0,00 0,00 0,00 62,64 12,36 50,28 0,00 20,70 9,18 11,52
12 5,92 19,76 3,60 16,16 0,00 0,00 0,00 0,00 0,00 21,54 0,00 21,54 0,00 0,00 0,00 0,00
Aufgeführt sind die Nitroaromatengehalte aller Einzelproben in µg/g Pflanzentrockenmasse; t=Versuchsdauer in Wochen; ADNT als Summe von 4-ADNT und 2-ADNT
Die nachgewiesenen TNT-Gehalte waren in den Pflanzen der B-Variante deutlich höher als in den Pflanzen der anderen beiden Pappelvarianten A und D. Ob dieser Unterschied ein Zufall ist oder in einer Varianzspezifität begründet liegt, bleibt zunächst ungeklärt. Bei Betrachtung dieses Sachverhaltes ist hervorzuheben, dass die Variante B nicht mit Weißfäulepilzen ausgestattet wurde. Interessant ist die Tatsache, dass der überwiegende Teil der analytisch erfassten Nitroaromaten erst durch eine Säurehydrolyse in 25%iger Schwefelsäure und anschließender Alkalisierung mit 8 M Natronlauge aus den Pflanzenproben extrahiert werden konnte. In Tabelle 17 sind exemplarisch die gewonnenen Nitroaromatenmengen aus den Wurzelproben der B-Variante aufgeführt. Die Darstellung beinhaltet die Werte für den reinen DCM-Extrakt (Extraktion 1) und für den DCM-Extrakt nach erfolgter Säurehydrolyse mit Alkalisierung
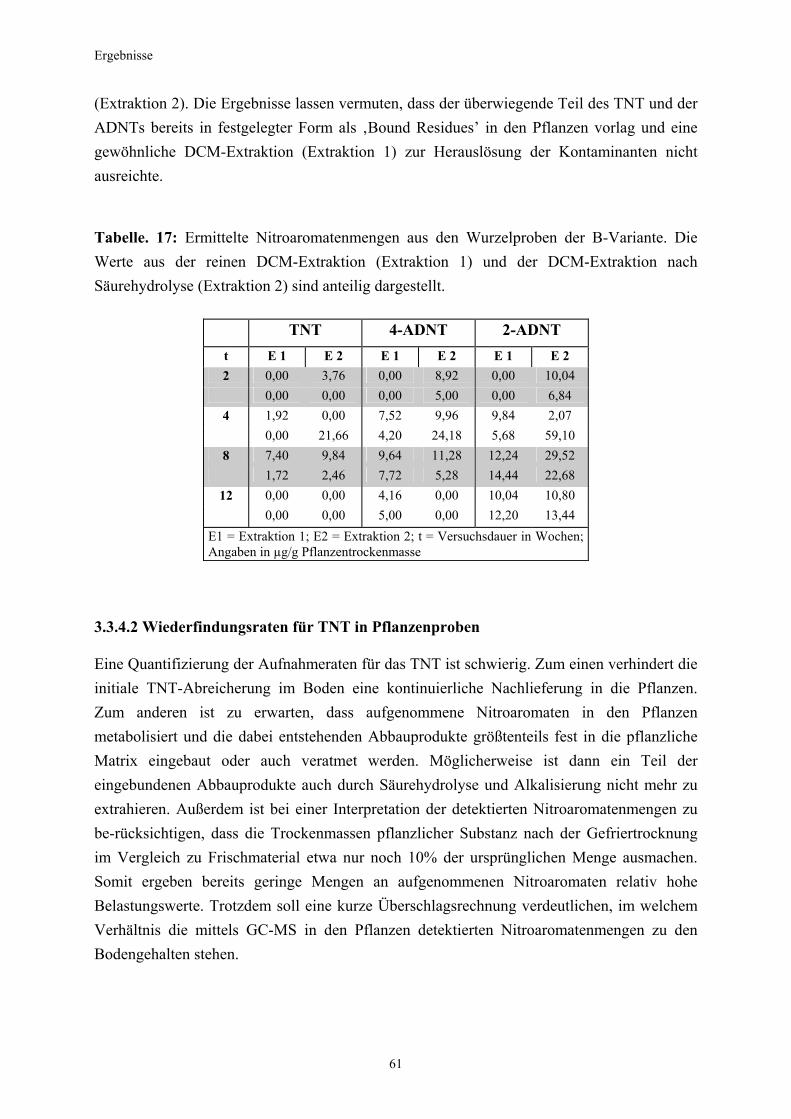
Ergebnisse
61
(Extraktion 2). Die Ergebnisse lassen vermuten, dass der überwiegende Teil des TNT und der ADNTs bereits in festgelegter Form als ‚Bound Residues’ in den Pflanzen vorlag und eine gewöhnliche DCM-Extraktion (Extraktion 1) zur Herauslösung der Kontaminanten nicht ausreichte. Tabelle. 17: Ermittelte Nitroaromatenmengen aus den Wurzelproben der B-Variante. Die Werte aus der reinen DCM-Extraktion (Extraktion 1) und der DCM-Extraktion nach Säurehydrolyse (Extraktion 2) sind anteilig dargestellt.
TNT 4-ADNT 2-ADNT t E 1 E 2 E 1 E 2 E 1 E 2 2 0,00 3,76 0,00 8,92 0,00 10,04 0,00 0,00 0,00 5,00 0,00 6,84
4 1,92 0,00 7,52 9,96 9,84 2,07 0,00 21,66 4,20 24,18 5,68 59,10
8 7,40 9,84 9,64 11,28 12,24 29,52 1,72 2,46 7,72 5,28 14,44 22,68
12 0,00 0,00 4,16 0,00 10,04 10,80 0,00 0,00 5,00 0,00 12,20 13,44
E1 = Extraktion 1; E2 = Extraktion 2; t = Versuchsdauer in Wochen; Angaben in µg/g Pflanzentrockenmasse
3.3.4.2 Wiederfindungsraten für TNT in Pflanzenproben Eine Quantifizierung der Aufnahmeraten für das TNT ist schwierig. Zum einen verhindert die initiale TNT-Abreicherung im Boden eine kontinuierliche Nachlieferung in die Pflanzen. Zum anderen ist zu erwarten, dass aufgenommene Nitroaromaten in den Pflanzen metabolisiert und die dabei entstehenden Abbauprodukte größtenteils fest in die pflanzliche Matrix eingebaut oder auch veratmet werden. Möglicherweise ist dann ein Teil der eingebundenen Abbauprodukte auch durch Säurehydrolyse und Alkalisierung nicht mehr zu extrahieren. Außerdem ist bei einer Interpretation der detektierten Nitroaromatenmengen zu be-rücksichtigen, dass die Trockenmassen pflanzlicher Substanz nach der Gefriertrocknung im Vergleich zu Frischmaterial etwa nur noch 10% der ursprünglichen Menge ausmachen. Somit ergeben bereits geringe Mengen an aufgenommenen Nitroaromaten relativ hohe Belastungswerte. Trotzdem soll eine kurze Überschlagsrechnung verdeutlichen, im welchem Verhältnis die mittels GC-MS in den Pflanzen detektierten Nitroaromatenmengen zu den Bodengehalten stehen.

Ergebnisse
62
angenommener Nitroaromatengehalt (TNT und ADNTs) je 1000 g Trockenboden, bezogen auf die Ausgangskon-zentration angenommener Nitroaromatengehalt (TNT und ADNTs) je 70 g Trockenboden (entspricht dem Bodenvolumen ei-nes Rhizotrons) angenommenes Frischgewicht von 10 g je Pflanze, dass entspricht etwa 1 g Trockenmasse angenommener Nitroaromatengehalt je 1 g Pflanzen-trockenmasse (ungefährer detektierter Wert) Anteil detektierter Nitroaromaten je Pflanze bezogen auf den initialen Schadstoffgehalt in 70 g Trockenboden
550 mg 38,5 mg 50 µg 0,1 - 0,2 %
3.3.4.3 Zusammenfassung In den Wurzel- und Sprossproben der Pappeln konnten die Nitroaromaten TNT, 4-ADNT und 2-ADNT detektiert werden. Als Hauptkontaminanten erwiesen sich die beiden ADNTs. Die nachweisbare Nitroaromatenkonzentration war organspezifisch. Die Wurzeln der Pflanzen waren höher belastet als der Spross. 3.4 Aufnahme und Verteilung von 14C-TNT/14C-TNT-Metaboliten in Pinus sylvestris Sämlingen In Experiment 2.2.2 konnten verschiedene Nitroaromaten in den Wurzeln und im Spross der Pflanzen gaschromatographisch nachgewiesen werden. Allerdings gaben die erzielten Ergebnisse keine genaue Auskunft darüber, in welchem Umfang die detektierten Schadstoffmengen von den Pflanzen tatsächlich aufgenommen wurden oder möglicherweise den Wurzeln nur äußerlich anhafteten. Um die Aufnahme und Verteilung von TNT und dessen Metaboliten in die Pflanzen quantitativ erfassen zu können, wurde nicht-mykorrhizierten und mykorrhizierten Pinus sylvestris Sämlingen 14C-TNT in zwei Konzentrationen angeboten. Die sich anschließende Untersuchung zur Aufnahme der Radiotracer erfolgte mikroautoradiographisch und durch Szintillationszählung. Die experimentellen Schritte können im einzelnen den Angaben in Kapitel 2.2.3 entnommen werden.

Ergebnisse
63
3.4.1 Mikroautoradiographische Untersuchungen von nicht-mykorrhizierten und mykorrhizierten Kiefernwurzeln Für die mikroautoradiographischen Untersuchungen wurden sowohl nicht-mykorrhizierte als auch mit Pisolithus tinctorius mykorrhizierte Wurzeln von Pinus sylvestris Sämlingen verwendet. Nach einem Angebot von 17 x 104 Bq in 2 ml Düngelösung (10,2 x 106 dpm) je Pflanze, erfolgte die Ernte der Wurzeln nach 3, 7 und 14 Tagen Applikation. Die Radioaktivität wurde durch Auflage einer speziellen Röntgenfilmemulsion auf die Semidünnschnitte nachgewiesen (Punkt 2.3.5.4). Bezüglich ihrer Körnung wurden Proben mit Expositionszeiten zwischen 2 Tagen und 20 Wochen nach Fixierung des Röntgenfilms lichtmikroskopisch untersucht. Aufgrund der schwachen Ausbildung von Kornstrukturen sind im folgenden Abbildungsteil ausschließlich beschichtete Präparate nach 20wöchiger Exposition dargestellt. Grundsätzlich ließ sich nach relativ hoher 14C-TNT-Applikation in den Wurzeln insgesamt nur wenig 14C nachweisen. Größtenteils wurde eine Schwärzung des Films durch den Wurzeln äußerlich anhaftendes 14C hervorgerufen . Eine Tendenz zur Akkumulation von Radioaktivität mit fortdauernder Applikation konnte in den Proben nicht beobachtet werden. Auffällige Unterschiede zwischen mykorrhizierten und nicht-mykorrhizierten Wurzeln bezüglich der Aufnahmemenge und der Verteilung der aufgenommenen Radioaktivität in den Pflanzen-zellen wurden nicht festgestellt. In Abbildung 15 ist eine nicht-mykorrhizierte Seitenwurzel nach 14tägiger Applikation im Längsschnitt (die dazugehörige Trägerwurzel im Querschnitt) als Übersicht dargestellt. Erkennbare Schwärzungen des Films befanden sich lediglich im Randbereich der Wurzeln (siehe Pfeile). Die großen Silberkornflecken im Bereich der Trägerwurzel wurden wahrscheinlich von kontaminierten Kohlepapierresten verursacht, die den Wurzeln anhafteten. In zahlreichen Schnitten konnten entsprechend großflächige Schwärzungen beobachtet werden. Innerhalb der Wurzeln waren die Markierungen durch eine geringe Silberkorndichte gekennzeichnet. Abbildung 16 zeigt einen Detailausschnitt mit Rhizodermis- und Rindenzellen der Seitenwurzel. Während sich die Silberkörner im Bereich der Rhizodermisaußenseite konzentrierten, blieb die Anzahl der Körner über der Rhizodermis und den Zellen der primären Rinde wesentlich niedriger. Die höhere Silberkorndichte an der Wurzeloberfläche könnte ein Indiz für einen erschwerten (gehemmten) Eintransport in die Wurzel sein oder auf eine verstärkte Adsorption hinweisen. Die aufgenommene Radioaktivität erfuhr eine relativ gleichmäßige Verteilung in Rhizodermis und primärer Rinde und gelangte schließlich bis in die Zellen des Zentralzylinders. So konnten zahlreiche Silberkörner in den Tracheiden des Xylems nachgewiesen werden (Abbildung 17). Allgemein war die Silberkorndichte in den tracheidalen Elementen etwas höher als in den anliegenden Zellen, was insbesondere für die ring- und schraubenartig verdickten Wandbereiche der Leitelemente zutraf. Radioaktivität im Xylem ließ sich bereits nach 3 Tagen Applikation

Ergebnisse
64
mikro-autoradiographisch nachweisen (nicht abgebildet). Eine auffällige Akkumulation mit fortschreitender Angebotsdauer war in den Tracheiden aufgrund der niedrigen Aufnahmerate insgesamt innerhalb der 14tägigen Applikation allerdings nicht erkennbar, wobei berücksichtigt werden muss, dass anteilig Radioaktivität mit dem Xylemstrom bereits abtransportiert wurde. Eine nennenswerte Verlagerung des aufgenommenen 14C in die apikalen Regionen der Wurzeln fand nicht statt. Über den Zellen der Wurzelspitze wurden kaum Silberkörner gefunden (Abbildung 18). Radioaktivität konnte auch im Trägerwurzelsystem der Kiefern nachgewiesen werden (Abbildung 19). Silberkörner fanden sich hier vor allem über den tracheidalen Leitelementen des Zentralzylinders. Der Aufnahmepfad des in den Trägerwurzeln vorgefundenen 14C verlief vermutlich überwiegend über die Leitelemente der Seitenwurzeln, die eine direkte Verbindung mit den Tracheiden der Trägerwurzeln herstellen. Über den Zellen der primären Rinde ließen sich kaum Silberkörner nachweisen. Wie bereits erwähnt wurde, konnten zwischen nicht-mykorrhizierten und mykorrhizierten Wurzeln kaum Unterschiede bezüglich der Aufnahmemenge und der Verteilung der aufgenommenen Radioaktivität in den Wurzeln festgestellt werden. Abbildung 20 zeigt eine mykorrhizierte Seitenwurzel im medianen Längsschnitt mit dazugehöriger Trägerwurzel. Letztere ist quer geschnitten. Der Hyphenmantel umgibt die Seitenwurzel vollständig. Im apikalen Bereich der Wurzel noch locker und verhältnismäßig dünn, nimmt der Mantel in proximaler Richtung an Stärke und Kompaktheit zu. Das Hartigsche Netz ist nur sporadisch ausgebildet und bleibt auf wenige Bereiche der Rinde begrenzt. Abgesehen von durch anhaftende Kohlepapierfasern verursachte Schwärzungen, die an zahlreichen mykorrhizierten Wurzeln beobachtet wurden, waren Markierungen im Hyphenmantel allgemein wenig zu finden. Silberkörner traten nur ganz vereinzelt auf (Abbildung 21). Die Markierung über den Zellen der primären Rinde, des Zentralzylinders und des Apikalmeristems ist in ihrer Intensität der Markierung vergleichbar, wie sie bereits für entsprechende Regionen nicht-mykorrhizierter Wurzeln nachgewiesen wurde. In den Rindenzellen war die Verteilung der Aktivität relativ gleichmäßig (Abbildung22). Im Zentralzylinder traten Silberkörner insbesondere über den Tracheiden und in diesen vor allem im Bereich der Wände in Erscheinung (Abbildungen 23 und 24). Auch in mykorrhizierten Kiefern ließ sich Radioaktivität in den tracheidalen Leitbahnen der Trägerwurzeln nachweisen (Abbildung 24). Der apikale Wurzelabschnitt blieb von einem 14C-Eintransport weitgehend ausgenommen. Radioaktivität war hier kaum zu finden (Abbildung 25).

Ergebnisse
65
Abb.15: Mikroautoradiographie einer längsgeschnittenen nicht-mykorrhizier-ten Seitenwurzel (Sw) mit quergeschnit-tener Trägerwurzel (Tw) von Pinussylvestris; angeboten wurden 17 x 104
Bq 14C-TNT für einen Zeitraum von 14 Tagen; mit Silberkornflecken im Rand-bereich außerhalb der Wurzel; erhöhte Silberkorndichte im äußeren Randbe-reich der Rhizodermis (siehe Pfeile); Detailausschnitte sind markiert; Apikal-meristem (Am), Rinde (Ri), Zentral-zylinder (Zz), Maßstab 100 µm.
Abb.16: Detail aus Abbildung 15; Ver-teilung der Radioaktivität (Silberkörner) in Rhizodermis (Rh), primärer Rinde (Ri) und im Randbereich der Seitenwurzel (Pfeile), Maßstab 25 µm.Rh
Ri
Tw
Rh
Am
Sw
Ri
Zz
Abb. 19
Abb.:17
Abb.:16
Abb.:18

Ergebnisse
66
Abb.17: Detail aus Abbildung 15; Verteilung der Radioaktivität (Sil-berkörner) im Zentralzylinder (Zz) der Seitenwurzel, Tracheide (Tr); Maßstab 15 µm.
Abb.18: Detail aus Abbildung 15; Verteilung der Radioaktivität (Sil-berkörner) im apikalen Wurzel-abschnitt; Apikalmeristem (Am), Rhizodermis (Rh), Rinde (Ri); Maßstab 25 µm.
Abb.19: Detail aus Abbildung 15; Aus-schnitt aus dem Zen-tralzylinder der quer angeschnittenen Trä-gerwurzel; Verteilung der Radioaktivität (Sil-berkörner); Trachei-den (Tr) mit sich ausdifferenzierenden Zellwänden (Pfeile); Maßstab 15 µm.
Zz
Tr
Am
Rh
Ri
Tr

Ergebnisse
67
Abb. 20: Mikroautoradiographie einer mykorrhizierten Seitenwurzel (Sw) mit Trägerwurzel (Tw) als Übersicht (Pinus sylvestris/Piso-lithus tinctorius Mykorrhiza); ange-boten wurden 17 x 104 Bq 14C-TNT für einen Zeitraum von 14 Tagen; Detailausschnitte sind markiert; Hyphenmantel (Hm), HartigschesNetz (Hn), Apikalmeristem (Am), Rinde (Ri), Zentralzylinder (Zz); Maßstab 100 µm.
Abb. 21: Ausschnitt aus einer quer angeschnittenen mykorrhizierten Kie-fernwurzel (Pinus sylvestris/Pisolithustinctorius); angeboten wurden 17 x 104
Bq 14C-TNT für einen Zeitraum von 14 Tagen; Silberkörner (Pfeile) nur ganz vereinzelt im Bereich des Hyphenman-tels (Hm), Hartigsches Netz (Hn), Rinde (Ri); Maßstab 20 µm.
Hm
Hn
Ri
Am
Zz
Ri
Hn
Sw
Tw
Abb.: 23
Abb.: 22
Abb.: 25
Abb.: 24
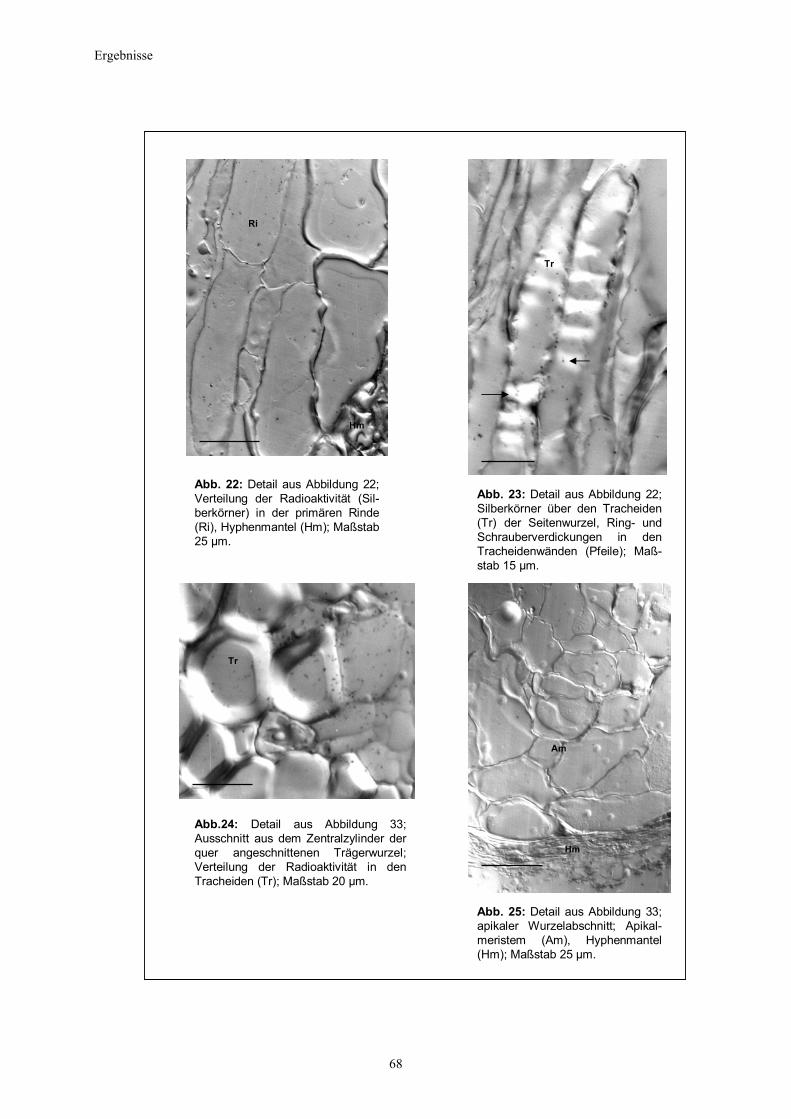
Ergebnisse
68
Abb. 22: Detail aus Abbildung 22; Verteilung der Radioaktivität (Sil-berkörner) in der primären Rinde (Ri), Hyphenmantel (Hm); Maßstab 25 µm.
Abb. 23: Detail aus Abbildung 22; Silberkörner über den Tracheiden(Tr) der Seitenwurzel, Ring- und Schrauberverdickungen in den Tracheidenwänden (Pfeile); Maß-stab 15 µm.
Abb.24: Detail aus Abbildung 33; Ausschnitt aus dem Zentralzylinder der quer angeschnittenen Trägerwurzel; Verteilung der Radioaktivität in den Tracheiden (Tr); Maßstab 20 µm.
Abb. 25: Detail aus Abbildung 33; apikaler Wurzelabschnitt; Apikal-meristem (Am), Hyphenmantel(Hm); Maßstab 25 µm.
Ri
Hm
Tr
Tr
Am
Hm

Ergebnisse
69
3.4.2 Bestimmung der Radioaktivität in den einzelnen Pflanzenorganen durch Szintillationszählung Die Quantifizierung der aufgenommenen Aktivität mittels Szintillationszählung erfolgte für Wurzeln, Spross und Nadeln getrennt. Untersucht wurden sowohl nicht-mykorrhizierte als auch mit Pisolithus tinctorius mykorrhizierte Pinus sylvestris Sämlinge. Angeboten wurde das 14C-TNT in zwei Konzentrationen: 0,85 x 104 Bq in 2 ml Düngelösung (0,501 x 106 dpm) 17 x 104 Bq in 2 ml Düngelösung (10,2 x 106 dpm) Die ermittelten Aktivitäten zeigten für die verschiedenen Organe der Pflanzen, besonders für die Wurzelproben, zum Teil hohe Schwankungsbreiten in ihren Werten. Die Ursache für das Auftreten solcher Streuungen liegt vermutlich darin begründet, dass für die Auswertung nur Aliquots von den jeweiligen Pflanzenteilen entnommen wurden, wobei die Organe der Pflanzen allgemein durch Bereiche unterschiedlicher Aufnahme- und Transportaktivitäten charakterisiert sind. Bei den Wurzeln dürften zu den zum Teil erheblichen Schwankungen außerdem sehr feine anhaftende kontaminierte Kohlepapierfasern beigetragen haben. Aufgrund der inhomogenen Verteilung der Radioaktivität sind daher in den Diagrammen die Mediane mit den dazugehörigen Maximal- und Minimalwerten aufgeführt. Ein Mykor-rhizaeffekt trat beim Anreicherungsverhalten nicht auf. Grundsätzlich ließen sich zwischen nicht-mykorrhizierten und mykorrhizierten Pflanzen keine auffälligen Unterschiede bezüglich der aufgenommenen radioaktiv markierten Substanzen nachweisen.
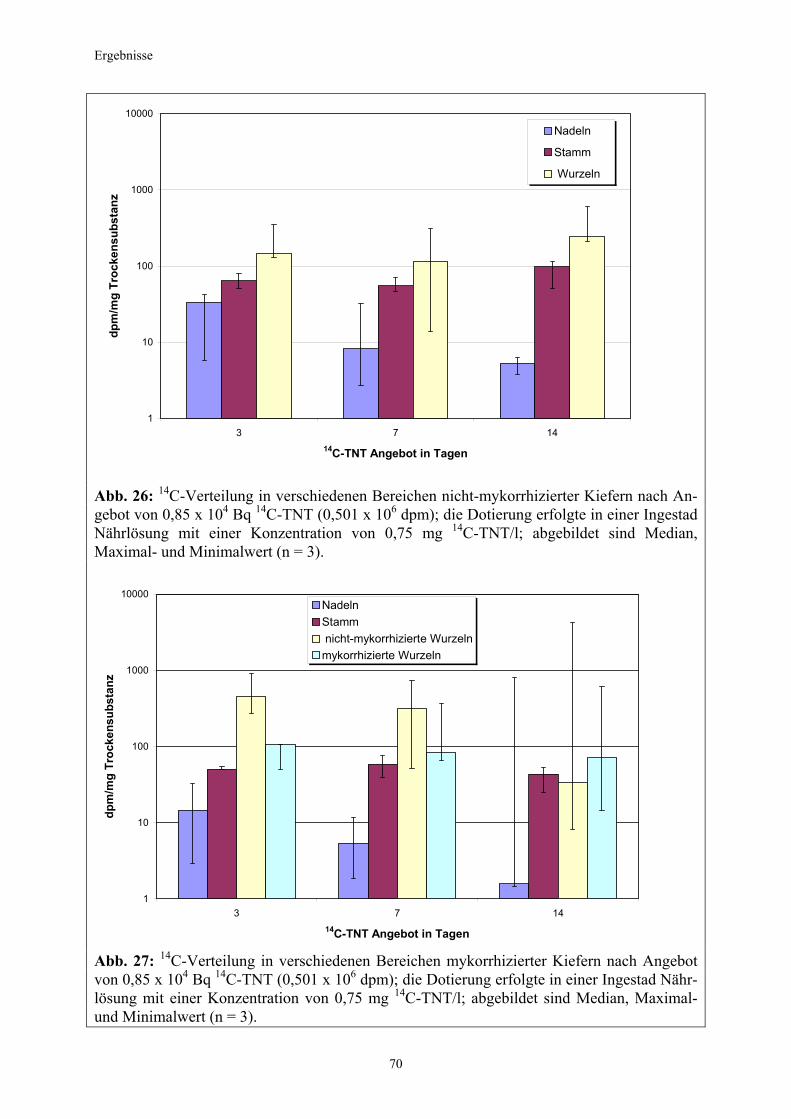
Ergebnisse
70
1
10
100
1000
10000
3 7 1414C-TNT Angebot in Tagen
dpm
/mg
Troc
kens
ubst
anz
Nadeln
Stamm
Wurzeln
Abb. 26: 14C-Verteilung in verschiedenen Bereichen nicht-mykorrhizierter Kiefern nach An-gebot von 0,85 x 104 Bq 14C-TNT (0,501 x 106 dpm); die Dotierung erfolgte in einer Ingestad Nährlösung mit einer Konzentration von 0,75 mg 14C-TNT/l; abgebildet sind Median, Maximal- und Minimalwert (n = 3).
1
10
100
1000
10000
3 7 1414C-TNT Angebot in Tagen
dpm
/mg
Troc
kens
ubst
anz
NadelnStamm nicht-mykorrhizierte Wurzelnmykorrhizierte Wurzeln
Abb. 27: 14C-Verteilung in verschiedenen Bereichen mykorrhizierter Kiefern nach Angebot von 0,85 x 104 Bq 14C-TNT (0,501 x 106 dpm); die Dotierung erfolgte in einer Ingestad Nähr-lösung mit einer Konzentration von 0,75 mg 14C-TNT/l; abgebildet sind Median, Maximal- und Minimalwert (n = 3).
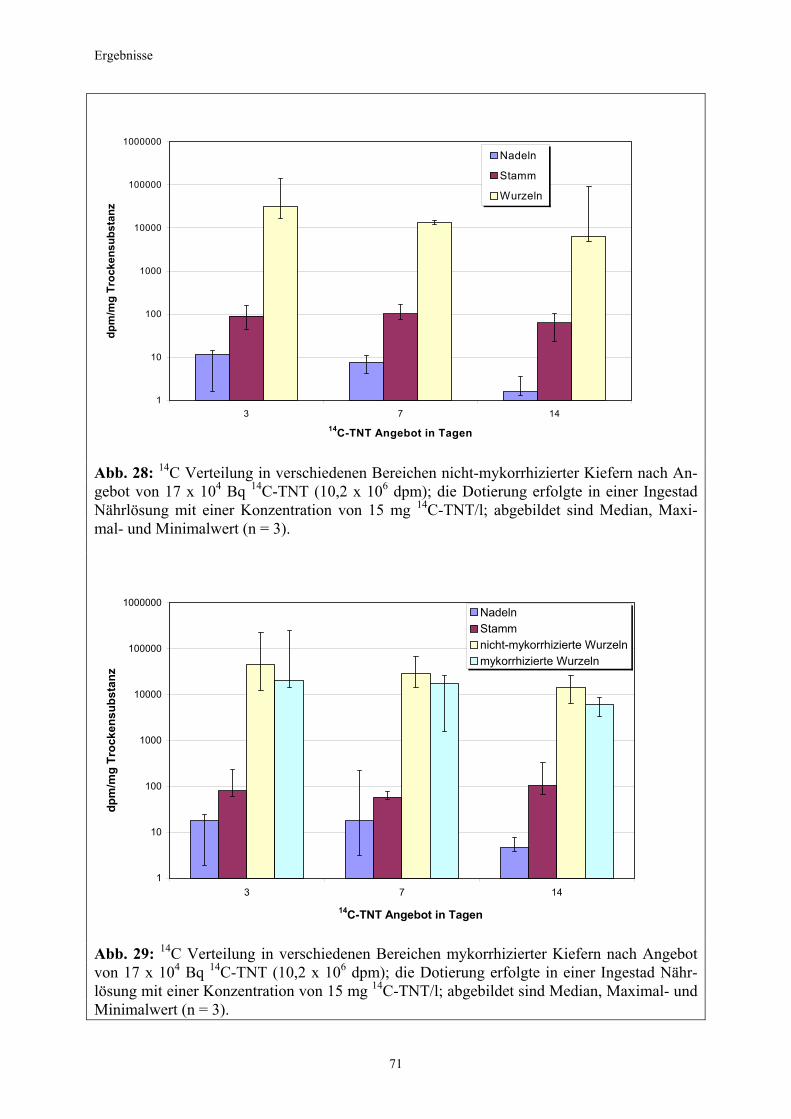
Ergebnisse
71
1
10
100
1000
10000
100000
1000000
3 7 1414C-TNT Angebot in Tagen
dpm
/mg
Troc
kens
ubst
anz
Nadeln
Stamm
Wurzeln
Abb. 28: 14C Verteilung in verschiedenen Bereichen nicht-mykorrhizierter Kiefern nach An-gebot von 17 x 104 Bq 14C-TNT (10,2 x 106 dpm); die Dotierung erfolgte in einer Ingestad Nährlösung mit einer Konzentration von 15 mg 14C-TNT/l; abgebildet sind Median, Maxi-mal- und Minimalwert (n = 3).
1
10
100
1000
10000
100000
1000000
3 7 14
14C-TNT Angebot in Tagen
dpm
/mg
Troc
kens
ubst
anz
Nadeln Stammnicht-mykorrhizierte Wurzelnmykorrhizierte Wurzeln
Abb. 29: 14C Verteilung in verschiedenen Bereichen mykorrhizierter Kiefern nach Angebot von 17 x 104 Bq 14C-TNT (10,2 x 106 dpm); die Dotierung erfolgte in einer Ingestad Nähr-lösung mit einer Konzentration von 15 mg 14C-TNT/l; abgebildet sind Median, Maximal- und Minimalwert (n = 3).

Ergebnisse
72
In den Aliquots der Wurzelproben ließen sich stets die größeren Mengen an 14C nachweisen. Unter Berücksichtigung der mikroautoradiographischen Untersuchungsergebnisse muss allerdings angenommen werden, dass der wesentlich größere Teil der ermittelten Radioaktivität auf Kontaminationen zurückzuführen ist, die den Wurzeln zum Zeitpunkt der Probennahme tatsächlich von außen anhafteten. Nur der geringere Teil dürfte demnach von den Pflanzen aufgenommen worden sein. Eine Tendenz zur Akkumulation der Aktivität in den Wurzeln mit fortschreitender Versuchsdauer bzw. zunehmender Angebotsdauer ließ sich nicht feststellen. Allerdings wäre bei insgesamt niedrigen 14C-Aufnahmeraten in die Wurzeln eine Überlagerung zeitlich bedingter Anreicherungen durch anteilig größere Anhaftungen an den Wurzeloberflächen möglich. Diesbezüglich konnte eine Zunahme der Silberkörner in den Mikroautoradiographien mit fortschreitender Angebotsdauer jedoch nicht beobachtet werden. In den Stämmchen blieb die nachweisbare Radioaktivität im gesamten Versuchsverlauf auf einem sehr niedrigen Niveau konstant. Weder die Angebotsdauer noch eine Erhöhung des extern applizierten 14C bewirkten hier eine Zunahme der Aktivität. Eine Verlagerung des von den Wurzeln aufgenommenen 14C in den Stamm der Pflanze war also kaum von Bedeutung. Dies ist insoweit bemerkenswert, da selbst die von den Xylemleitbahnen der Wurzeln aufgenommenen 14C-Anteile kaum in die oberirdischen Teile der Pflanzen transportiert wurden. Die Nadelfraktion war im Vergleich zu den Wurzel- und Sprossanteilen am geringsten belastet. Ein nennenswerter Eintransport von markierten Substanzen in die Nadeln konnte im Allgemeinen in nur ganz niedrigen Konzentrationen nachgewiesen werden. Mit fortlaufender Versuchsdurchführung nimmt die Radioaktivität hier ab. Dieser Befund könnte auf eine Veratmung radioaktiver Verbindungen hindeuten. 3.4.3 Bilanzierung 3.4.3.1 Wiederfindung nach Dotierung einer hohen 14C-TNT-Konzentration Abgesehen von wenigen Ausnahmen lag die ermittelte Countrate für die Wurzelfraktion (nicht-mykorrhizierte und mykorrhizierte Wurzeln) zwischen 10 x103 und 100 x 103 dpm je mg Trockensubstanz. Die nachweisbare Aktivität für die Stamm- und Nadelfraktion ist aufgrund der niedrigen Werte bezüglich der Berechnung der aufgenommenen Gesamtaktivität in der Pflanze kaum von Bedeutung. Wird für eine Bilanzierung eine Aktivität von 30 x 103 dpm je mg Wurzelfraktion (TS) angenommen, beträgt die Aktivität für das Gesamtwurzelwerk einer Pflanze bei einer Wurzeltrockenmasse von 15 mg (ermittelter Durchschnittswert) insgesamt 450 x 103 dpm bzw. 7,5 x 103 Bq. Bei einer Dotierung von 10,2 x 106 dpm bzw. 17 x 104 Bq wurden demzufolge 4,4 % des angebotenen 14C in der Pflanze mittels Szintillationszählung wiedergefunden. Unter Berücksichtigung der mikroautoradio-graphischen Untersuchungen muss allerdings bedacht werden, dass erhebliche Mengen 14C

Ergebnisse
73
den geernteten Wurzeln äußerlich anhafteten und nicht von der Pflanze aufgenommen wurden. Demzufolge dürfte die Wiederfindungsrate für tatsächlich aufgenommenes 14C erheblich niedriger sein und unter 1 % liegen. 3.4.3.2 Wiederfindung nach Dotierung einer niedrigen 14C-TNT-Konzentration Nennenswerte Mengen Radioaktivität konnten lediglich in der Wurzelfraktion (nicht-mykorrhizierte und mykorrhizierte Wurzeln) detektiert werden. Die Countrate je mg Wurzeltrockenmasse lag zwischen 0,1 x 103 und 1 x 103 dpm. Für das Gesamtwurzelwerk einer Pflanze mit einer durchschnittlichen Wurzeltrockenmasse von 15 mg (ermittelter Wert) und einer angenommenen Aktivität von 0,6 x 103 dpm je mg Trockensubstanz lässt sich somit eine Gesamtaktivität von 9x103 dpm bzw. 0,15 x 103 Bq errechnen. Dotiert wurden 0,501 x 106 dpm bzw. 0,85 x 104 Bq. Demnach konnten 1,8 % des angebotenen 14C in den Wurzelproben wieder gefunden werden. Nach Abzug der Radioaktivität, die den Wurzeln von außen anhaftete, dürfte die tatsächliche Wiederfindungsrate für aufgenommenes 14C weit unter 1 % liegen. 3.4.3.3 Verbleibende Radioaktivität in Kohlepapier und Düngelösung Nach erfolgter Ernte der Pflanzen wurden die Kohlepapierscheiben und die nicht aufgenommenen Reste der Düngelösung aus den Rhizotronen entfernt und auf ihre Radioaktivität untersucht. Hierbei ließen sich zusammengenommen im Mittel etwa 75% der ursprünglich eingesetzten Radioaktivität wieder finden. Unter Berücksichtigung der von den Pflanzen aufgenommenen 14C-Anteile (inklusive Anhaftungen an den Wurzeloberflächen) lässt sich eine Bilanzlücke von etwa 20% feststellen, die vermutlich größtenteils durch Anhaftungen an Wänden und Böden der Petrischalen erklärt werden kann. 3.4.4 Zusammenfassung Allgemein ließ sich nach 14tägiger Applikation nur wenig Radioaktivität in den Pflanzen nachweisen. Die lokale 14C-Verteilung war organspezifisch und konzentrierte sich auf die Wurzeln. Insbesondere die Tracheiden waren hier deutlich markiert. Oberirdische Organe waren nur gering belastet. Eine Akkumulation in den Geweben mit zunehmender Angebotsdauer wurde nicht beobachtet. In den Kiefernnadeln als Endpunkt des Transportes wurden möglicherweise radioaktive Substanzen anteilig veratmet, da dort die ermittelte 14C-Menge mit der Zeit abnahm. Ein Mykorrhizaeffekt trat nicht auf.

Ergebnisse
74
3.5 Toxizitätstest mit mykorrhizierten Kiefern in TNT-haltigem Boden-substrat Die erfolgreiche Durchführung eines Phytoremediationsverfahren setzt Kenntnisse über den Einfluss der toxischen Bodenkontaminanten voraus, denen die Pflanzen ausgesetzt werden. Schädliche Wirkungen sind in der Regel mit einer Verminderung der Vitalität verbunden und setzen die Effizienz der Pflanzen bei der Sanierung herab. Oft sind Toleranzspielräume gegenüber Schadstoffgehalten im Boden gegeben. Allerdings müssen Resistenzverhalten und Regenerationsfähigkeit ermittelt werden um herauszufinden, bis zu welchen Kontaminations-graden Phytoremediation möglich ist. Unter diesem Aspekt wurden in einem Toxizitätstest mykorrhizierte Kiefernsämlinge auf einem TNT-belasteten Standardboden kultiviert und die Pflanzen auf ihre Vitalität und Entwicklung geprüft. Die genaue Versuchsbeschreibung ist im Methodenteil in Kapitel 2.2.4 zu finden. Als Maß für die Toxizität wurde in erster Linie der Effekt von Nitroaromaten auf die Biomasseentwicklung berücksichtigt. Weitere Größen zur Bewertung der Phytotoxizität waren die Entwicklung des Mykorrhizierungsgrades der Pflanzen sowie das Auftreten morphologischer Veränderungen an Nadeln und Wurzeln. Die inhomogene Verteilung von TNT in Standortböden würde Rückschlüsse auf den Zusammenhang zwischen Belastungsgrad des Bodens und Vitalität der Pflanzen kaum zulassen. Aus diesem Grund wurde ein Standardboden gleichmäßig in drei unterschiedlichen Belastungskategorien mit TNT angereichert. niedrig belastet 3 mg TNT/ kg Trockenboden
mäßig belastet 300 mg TNT/ kg Trockenboden
hoch belastet 3000 mg TNT/ kg Trockenboden
3.5.1 Wiederfindung der TNT-Kontamination Etwa 24 Stunden nach der Dotierung des Bodens mit dem TNT wurden vor der Bepflanzung der Rhizotrone Bodenproben zur Überprüfung der Kontamination entnommen. Abgesehen von der Erfassung der Wiederfindungsraten, sollte zusätzlich kontrolliert werden, ob eine gleichmäßige Verteilung der TNT-Kontamination im Boden vorlag. In Tabelle 18 sind die ermittelten Messwerte für die jeweiligen Dotierungsklassen dargestellt. Auffällig sind die deutlichen Verluste an TNT in den Dotierungsklassen 300 mg /kg und 3000 mg/kg Boden. Durch Extraktion konnten hier lediglich 73% der Ausgangsmengen wieder gefunden werden. ADNTs ließen sich nicht nachweisen und gehörten zum Zeitpunkt der Bepflanzung der Rhizotrone somit noch nicht zum Schadstoffspektrum des Bodens. Die geringen Standard-
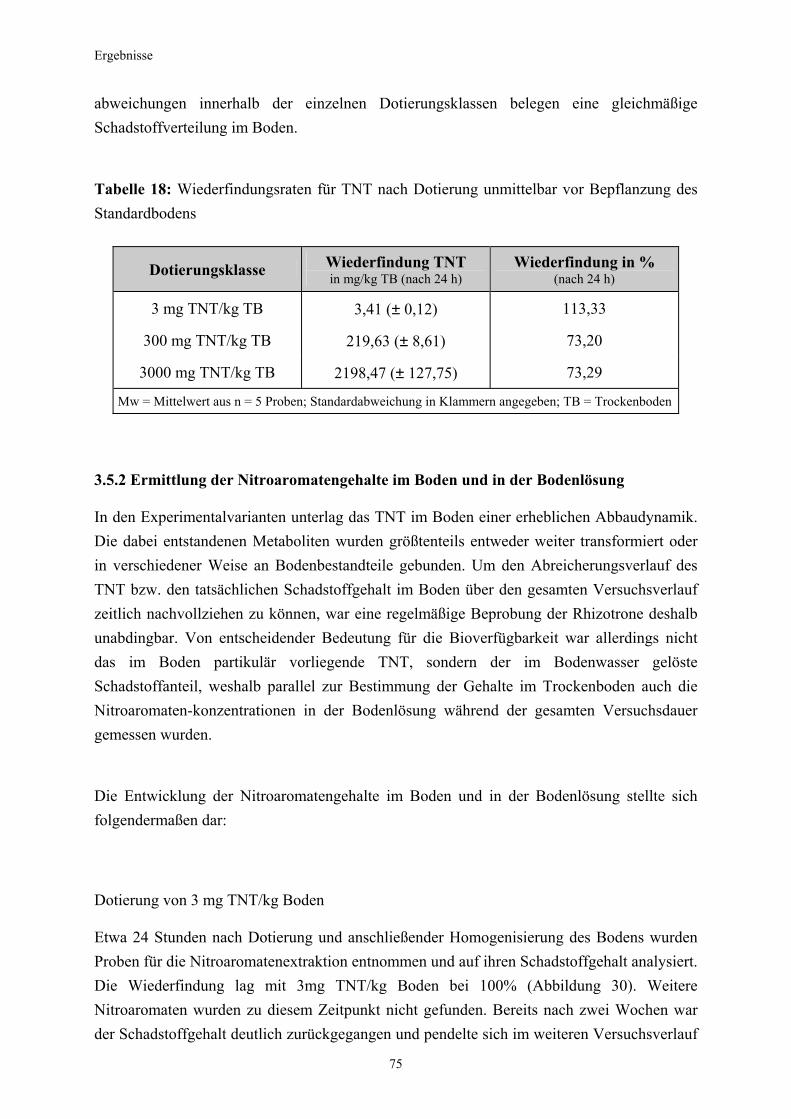
Ergebnisse
75
abweichungen innerhalb der einzelnen Dotierungsklassen belegen eine gleichmäßige Schadstoffverteilung im Boden. Tabelle 18: Wiederfindungsraten für TNT nach Dotierung unmittelbar vor Bepflanzung des Standardbodens
Dotierungsklasse Wiederfindung TNT in mg/kg TB (nach 24 h)
Wiederfindung in % (nach 24 h)
3 mg TNT/kg TB 3,41 (± 0,12) 113,33
300 mg TNT/kg TB 219,63 (± 8,61) 73,20
3000 mg TNT/kg TB 2198,47 (± 127,75) 73,29
Mw = Mittelwert aus n = 5 Proben; Standardabweichung in Klammern angegeben; TB = Trockenboden
3.5.2 Ermittlung der Nitroaromatengehalte im Boden und in der Bodenlösung In den Experimentalvarianten unterlag das TNT im Boden einer erheblichen Abbaudynamik. Die dabei entstandenen Metaboliten wurden größtenteils entweder weiter transformiert oder in verschiedener Weise an Bodenbestandteile gebunden. Um den Abreicherungsverlauf des TNT bzw. den tatsächlichen Schadstoffgehalt im Boden über den gesamten Versuchsverlauf zeitlich nachvollziehen zu können, war eine regelmäßige Beprobung der Rhizotrone deshalb unabdingbar. Von entscheidender Bedeutung für die Bioverfügbarkeit war allerdings nicht das im Boden partikulär vorliegende TNT, sondern der im Bodenwasser gelöste Schadstoffanteil, weshalb parallel zur Bestimmung der Gehalte im Trockenboden auch die Nitroaromaten-konzentrationen in der Bodenlösung während der gesamten Versuchsdauer gemessen wurden. Die Entwicklung der Nitroaromatengehalte im Boden und in der Bodenlösung stellte sich folgendermaßen dar: Dotierung von 3 mg TNT/kg Boden Etwa 24 Stunden nach Dotierung und anschließender Homogenisierung des Bodens wurden Proben für die Nitroaromatenextraktion entnommen und auf ihren Schadstoffgehalt analysiert. Die Wiederfindung lag mit 3mg TNT/kg Boden bei 100% (Abbildung 30). Weitere Nitroaromaten wurden zu diesem Zeitpunkt nicht gefunden. Bereits nach zwei Wochen war der Schadstoffgehalt deutlich zurückgegangen und pendelte sich im weiteren Versuchsverlauf
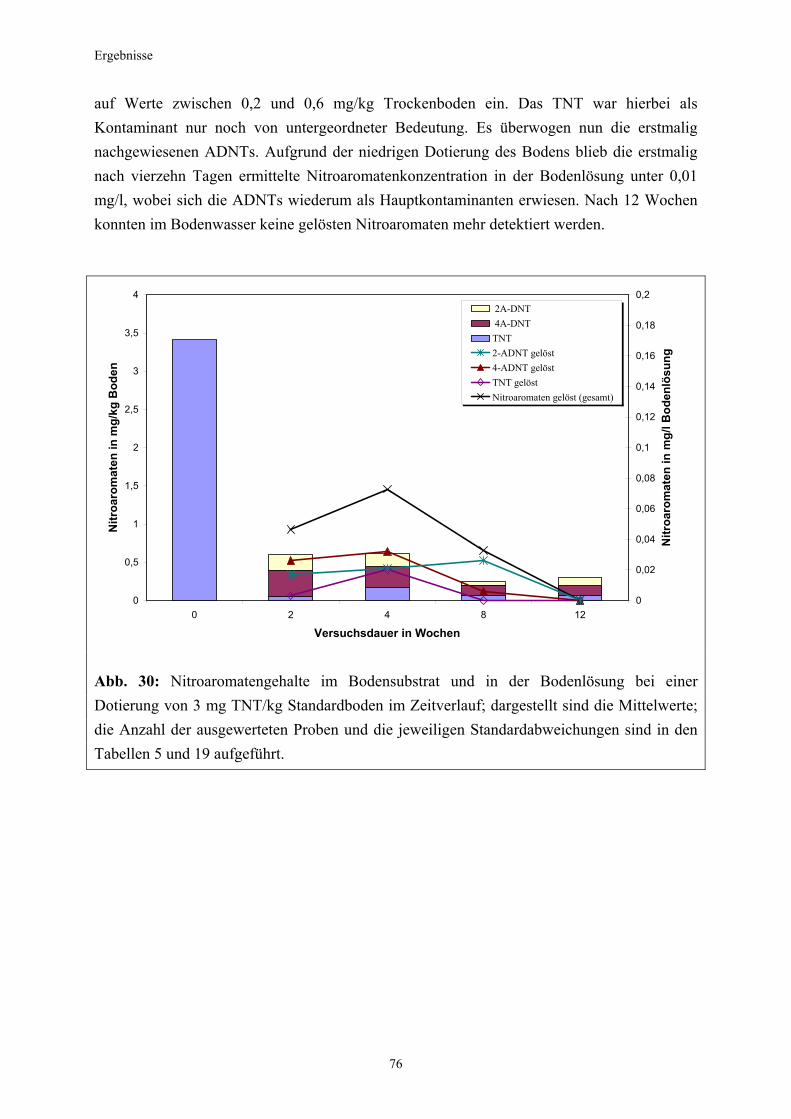
Ergebnisse
76
auf Werte zwischen 0,2 und 0,6 mg/kg Trockenboden ein. Das TNT war hierbei als Kontaminant nur noch von untergeordneter Bedeutung. Es überwogen nun die erstmalig nachgewiesenen ADNTs. Aufgrund der niedrigen Dotierung des Bodens blieb die erstmalig nach vierzehn Tagen ermittelte Nitroaromatenkonzentration in der Bodenlösung unter 0,01 mg/l, wobei sich die ADNTs wiederum als Hauptkontaminanten erwiesen. Nach 12 Wochen konnten im Bodenwasser keine gelösten Nitroaromaten mehr detektiert werden.
0
0,5
1
1,5
2
2,5
3
3,5
4
0 2 4 8 12
Versuchsdauer in Wochen
Nitr
oaro
mat
en in
mg/
kg B
oden
0
0,02
0,04
0,06
0,08
0,1
0,12
0,14
0,16
0,18
0,2
Nitr
oaro
mat
en in
mg/
l Bod
enlö
sung
2A-DNT 4A-DNTTNT2-ADNT gelöst4-ADNT gelöstTNT gelöstNitroaromaten gelöst (gesamt)
Abb. 30: Nitroaromatengehalte im Bodensubstrat und in der Bodenlösung bei einer Dotierung von 3 mg TNT/kg Standardboden im Zeitverlauf; dargestellt sind die Mittelwerte; die Anzahl der ausgewerteten Proben und die jeweiligen Standardabweichungen sind in den Tabellen 5 und 19 aufgeführt.
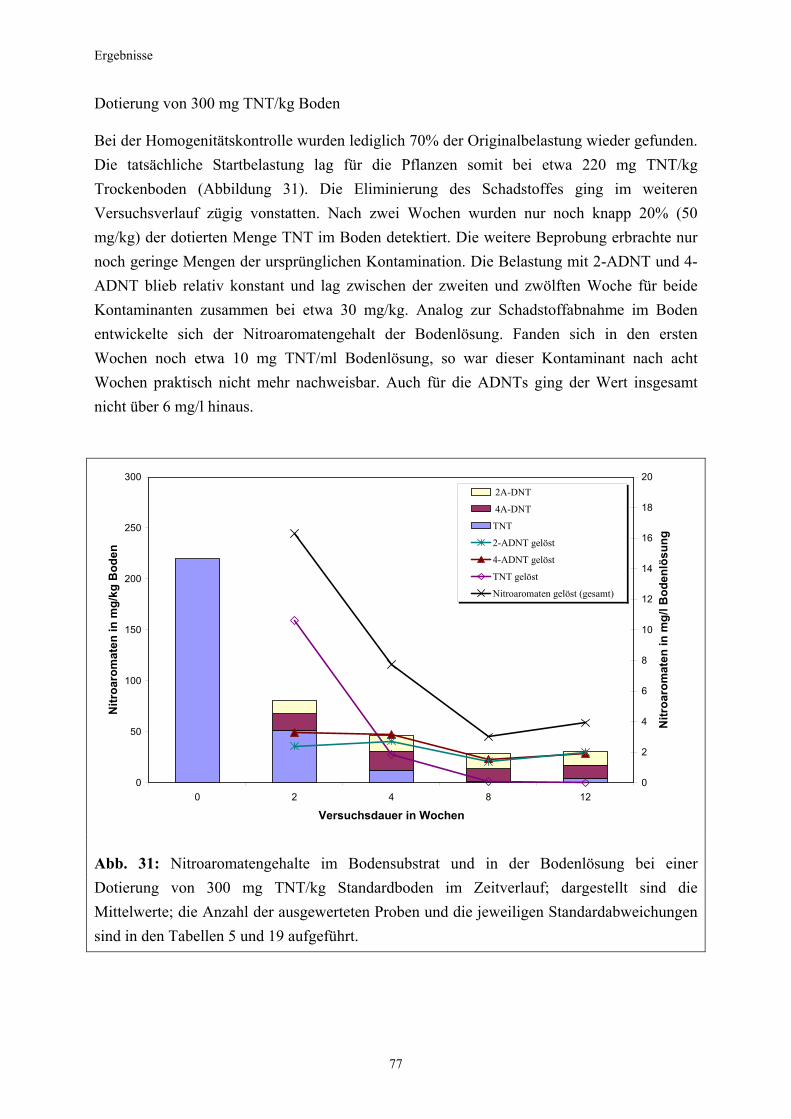
Ergebnisse
77
Dotierung von 300 mg TNT/kg Boden Bei der Homogenitätskontrolle wurden lediglich 70% der Originalbelastung wieder gefunden. Die tatsächliche Startbelastung lag für die Pflanzen somit bei etwa 220 mg TNT/kg Trockenboden (Abbildung 31). Die Eliminierung des Schadstoffes ging im weiteren Versuchsverlauf zügig vonstatten. Nach zwei Wochen wurden nur noch knapp 20% (50 mg/kg) der dotierten Menge TNT im Boden detektiert. Die weitere Beprobung erbrachte nur noch geringe Mengen der ursprünglichen Kontamination. Die Belastung mit 2-ADNT und 4-ADNT blieb relativ konstant und lag zwischen der zweiten und zwölften Woche für beide Kontaminanten zusammen bei etwa 30 mg/kg. Analog zur Schadstoffabnahme im Boden entwickelte sich der Nitroaromatengehalt der Bodenlösung. Fanden sich in den ersten Wochen noch etwa 10 mg TNT/ml Bodenlösung, so war dieser Kontaminant nach acht Wochen praktisch nicht mehr nachweisbar. Auch für die ADNTs ging der Wert insgesamt nicht über 6 mg/l hinaus.
0
50
100
150
200
250
300
0 2 4 8 12
Versuchsdauer in Wochen
Nitr
oaro
mat
en in
mg/
kg B
oden
0
2
4
6
8
10
12
14
16
18
20
Nitr
oaro
mat
en in
mg/
l Bod
enlö
sung
2A-DNT
4A-DNT
TNT
2-ADNT gelöst
4-ADNT gelöst
TNT gelöst
Nitroaromaten gelöst (gesamt)
Abb. 31: Nitroaromatengehalte im Bodensubstrat und in der Bodenlösung bei einer Dotierung von 300 mg TNT/kg Standardboden im Zeitverlauf; dargestellt sind die Mittelwerte; die Anzahl der ausgewerteten Proben und die jeweiligen Standardabweichungen sind in den Tabellen 5 und 19 aufgeführt.
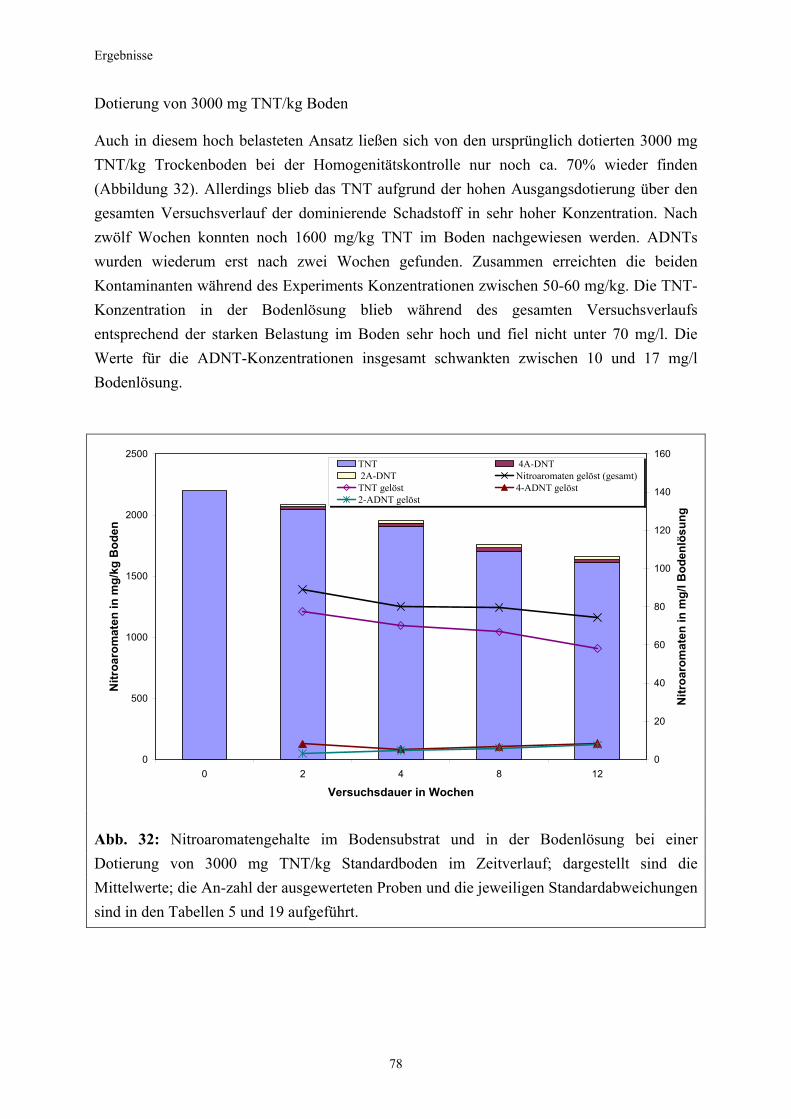
Ergebnisse
78
Dotierung von 3000 mg TNT/kg Boden Auch in diesem hoch belasteten Ansatz ließen sich von den ursprünglich dotierten 3000 mg TNT/kg Trockenboden bei der Homogenitätskontrolle nur noch ca. 70% wieder finden (Abbildung 32). Allerdings blieb das TNT aufgrund der hohen Ausgangsdotierung über den gesamten Versuchsverlauf der dominierende Schadstoff in sehr hoher Konzentration. Nach zwölf Wochen konnten noch 1600 mg/kg TNT im Boden nachgewiesen werden. ADNTs wurden wiederum erst nach zwei Wochen gefunden. Zusammen erreichten die beiden Kontaminanten während des Experiments Konzentrationen zwischen 50-60 mg/kg. Die TNT-Konzentration in der Bodenlösung blieb während des gesamten Versuchsverlaufs entsprechend der starken Belastung im Boden sehr hoch und fiel nicht unter 70 mg/l. Die Werte für die ADNT-Konzentrationen insgesamt schwankten zwischen 10 und 17 mg/l Bodenlösung.
0
500
1000
1500
2000
2500
0 2 4 8 12
Versuchsdauer in Wochen
Nitr
oaro
mat
en in
mg/
kg B
oden
0
20
40
60
80
100
120
140
160
Nitr
oaro
mat
en in
mg/
l Bod
enlö
sung
TNT 4A-DNT 2A-DNT Nitroaromaten gelöst (gesamt)TNT gelöst 4-ADNT gelöst 2-ADNT gelöst
Abb. 32: Nitroaromatengehalte im Bodensubstrat und in der Bodenlösung bei einer Dotierung von 3000 mg TNT/kg Standardboden im Zeitverlauf; dargestellt sind die Mittelwerte; die An-zahl der ausgewerteten Proben und die jeweiligen Standardabweichungen sind in den Tabellen 5 und 19 aufgeführt.
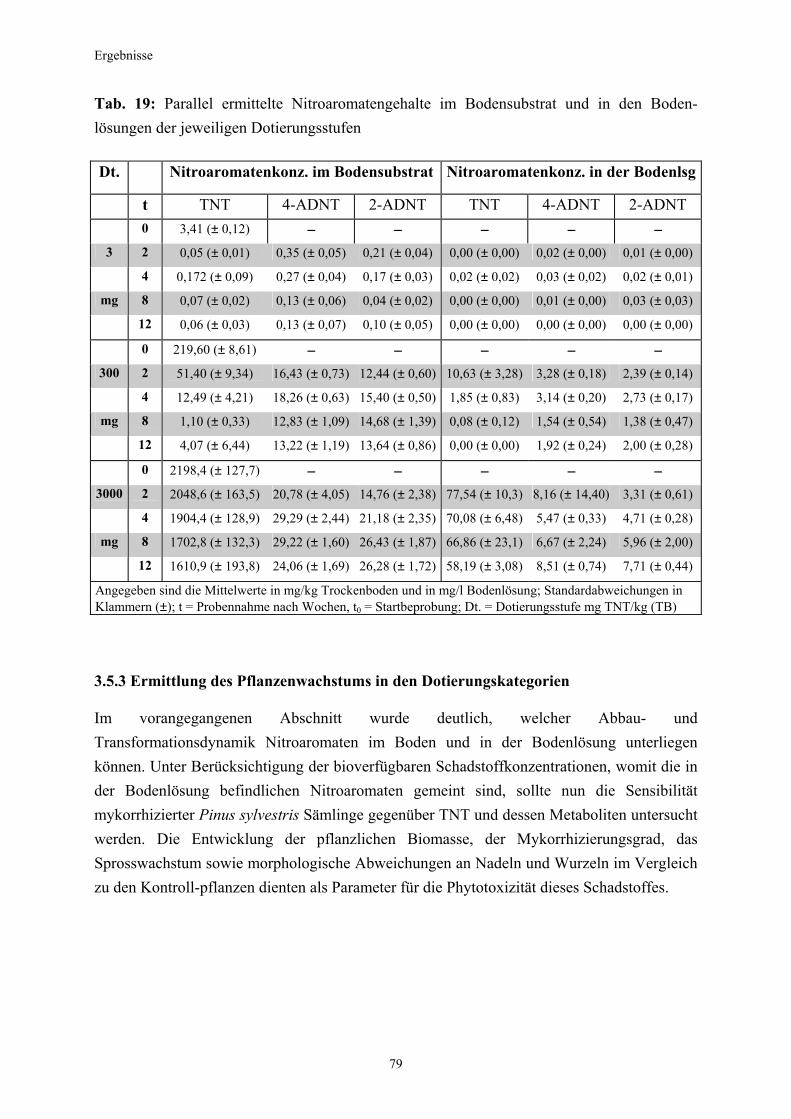
Ergebnisse
79
Tab. 19: Parallel ermittelte Nitroaromatengehalte im Bodensubstrat und in den Boden-lösungen der jeweiligen Dotierungsstufen Dt. Nitroaromatenkonz. im Bodensubstrat Nitroaromatenkonz. in der Bodenlsg
t TNT 4-ADNT 2-ADNT TNT 4-ADNT 2-ADNT 0 3,41 (± 0,12) − − − − −
3 2 0,05 (± 0,01) 0,35 (± 0,05) 0,21 (± 0,04) 0,00 (± 0,00) 0,02 (± 0,00) 0,01 (± 0,00)
4 0,172 (± 0,09) 0,27 (± 0,04) 0,17 (± 0,03) 0,02 (± 0,02) 0,03 (± 0,02) 0,02 (± 0,01)
mg 8 0,07 (± 0,02) 0,13 (± 0,06) 0,04 (± 0,02) 0,00 (± 0,00) 0,01 (± 0,00) 0,03 (± 0,03)
12 0,06 (± 0,03) 0,13 (± 0,07) 0,10 (± 0,05) 0,00 (± 0,00) 0,00 (± 0,00) 0,00 (± 0,00)
0 219,60 (± 8,61) − − − − − 300 2 51,40 (± 9,34) 16,43 (± 0,73) 12,44 (± 0,60) 10,63 (± 3,28) 3,28 (± 0,18) 2,39 (± 0,14)
4 12,49 (± 4,21) 18,26 (± 0,63) 15,40 (± 0,50) 1,85 (± 0,83) 3,14 (± 0,20) 2,73 (± 0,17)
mg 8 1,10 (± 0,33) 12,83 (± 1,09) 14,68 (± 1,39) 0,08 (± 0,12) 1,54 (± 0,54) 1,38 (± 0,47)
12 4,07 (± 6,44) 13,22 (± 1,19) 13,64 (± 0,86) 0,00 (± 0,00) 1,92 (± 0,24) 2,00 (± 0,28)
0 2198,4 (± 127,7) − − − − − 3000 2 2048,6 (± 163,5) 20,78 (± 4,05) 14,76 (± 2,38) 77,54 (± 10,3) 8,16 (± 14,40) 3,31 (± 0,61)
4 1904,4 (± 128,9) 29,29 (± 2,44) 21,18 (± 2,35) 70,08 (± 6,48) 5,47 (± 0,33) 4,71 (± 0,28)
mg 8 1702,8 (± 132,3) 29,22 (± 1,60) 26,43 (± 1,87) 66,86 (± 23,1) 6,67 (± 2,24) 5,96 (± 2,00)
12 1610,9 (± 193,8) 24,06 (± 1,69) 26,28 (± 1,72) 58,19 (± 3,08) 8,51 (± 0,74) 7,71 (± 0,44)
Angegeben sind die Mittelwerte in mg/kg Trockenboden und in mg/l Bodenlösung; Standardabweichungen in Klammern (±); t = Probennahme nach Wochen, t0 = Startbeprobung; Dt. = Dotierungsstufe mg TNT/kg (TB) 3.5.3 Ermittlung des Pflanzenwachstums in den Dotierungskategorien Im vorangegangenen Abschnitt wurde deutlich, welcher Abbau- und Transformationsdynamik Nitroaromaten im Boden und in der Bodenlösung unterliegen können. Unter Berücksichtigung der bioverfügbaren Schadstoffkonzentrationen, womit die in der Bodenlösung befindlichen Nitroaromaten gemeint sind, sollte nun die Sensibilität mykorrhizierter Pinus sylvestris Sämlinge gegenüber TNT und dessen Metaboliten untersucht werden. Die Entwicklung der pflanzlichen Biomasse, der Mykorrhizierungsgrad, das Sprosswachstum sowie morphologische Abweichungen an Nadeln und Wurzeln im Vergleich zu den Kontroll-pflanzen dienten als Parameter für die Phytotoxizität dieses Schadstoffes.
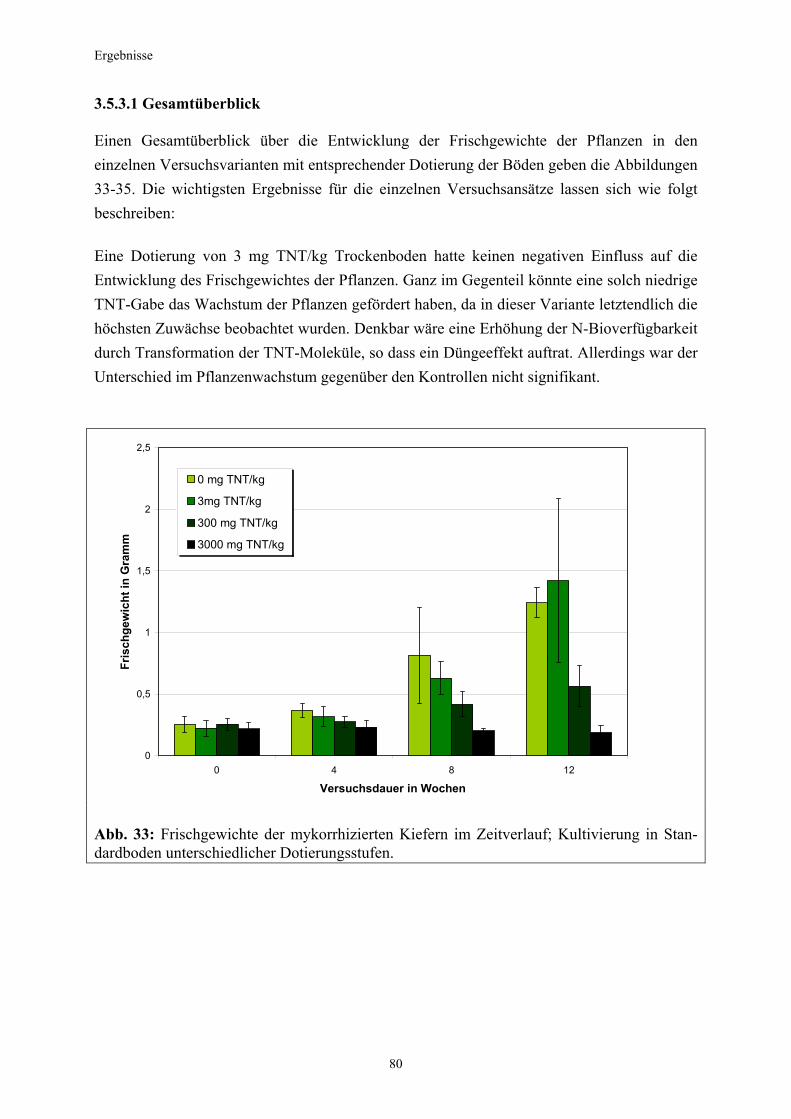
Ergebnisse
80
3.5.3.1 Gesamtüberblick Einen Gesamtüberblick über die Entwicklung der Frischgewichte der Pflanzen in den einzelnen Versuchsvarianten mit entsprechender Dotierung der Böden geben die Abbildungen 33-35. Die wichtigsten Ergebnisse für die einzelnen Versuchsansätze lassen sich wie folgt beschreiben: Eine Dotierung von 3 mg TNT/kg Trockenboden hatte keinen negativen Einfluss auf die Entwicklung des Frischgewichtes der Pflanzen. Ganz im Gegenteil könnte eine solch niedrige TNT-Gabe das Wachstum der Pflanzen gefördert haben, da in dieser Variante letztendlich die höchsten Zuwächse beobachtet wurden. Denkbar wäre eine Erhöhung der N-Bioverfügbarkeit durch Transformation der TNT-Moleküle, so dass ein Düngeeffekt auftrat. Allerdings war der Unterschied im Pflanzenwachstum gegenüber den Kontrollen nicht signifikant.
0
0,5
1
1,5
2
2,5
0 4 8 12
Versuchsdauer in Wochen
Fris
chge
wic
ht in
Gra
mm
0 mg TNT/kg
3mg TNT/kg
300 mg TNT/kg
3000 mg TNT/kg
Abb. 33: Frischgewichte der mykorrhizierten Kiefern im Zeitverlauf; Kultivierung in Stan-dardboden unterschiedlicher Dotierungsstufen.

Ergebnisse
81
Eine Dotierung von 300 mg TNT hemmte das Pflanzenwachstum. Reduziert waren sowohl Spross- als auch Wurzelwachstum. Mit zunehmender Versuchsdauer erholten sich die Pflanzen und legten an Biomasse zu. Die verlangsamte Gewichtszunahme machte sich allerdings erst nach acht Wochen in einem signifikanten Unterschied gegenüber der Kontrollvariante und der Variante mit 3 mg TNT/kg Boden bemerkbar. Eine hohe TNT-Belastung (Dotierung 3000 mg/kg Trockenboden) hemmte das Wachstum der Pflanzen vollständig. Ein Biomassezuwachs konnte somit nicht beobachtet werden. Signifikant waren die Unterschiede gegenüber der Kontrolle nach 4 Wochen und gegenüber den übrigen Varianten nach 8 Wochen.
Abb. 34: Spross- und Wurzelwachstum nach 12 Wochen Versuchsdauer; Dotierungsstufen von l. n. r, in mg TNT/kg Standardboden: 3000, 300, 3 und unbelastete Kontrollvariante.
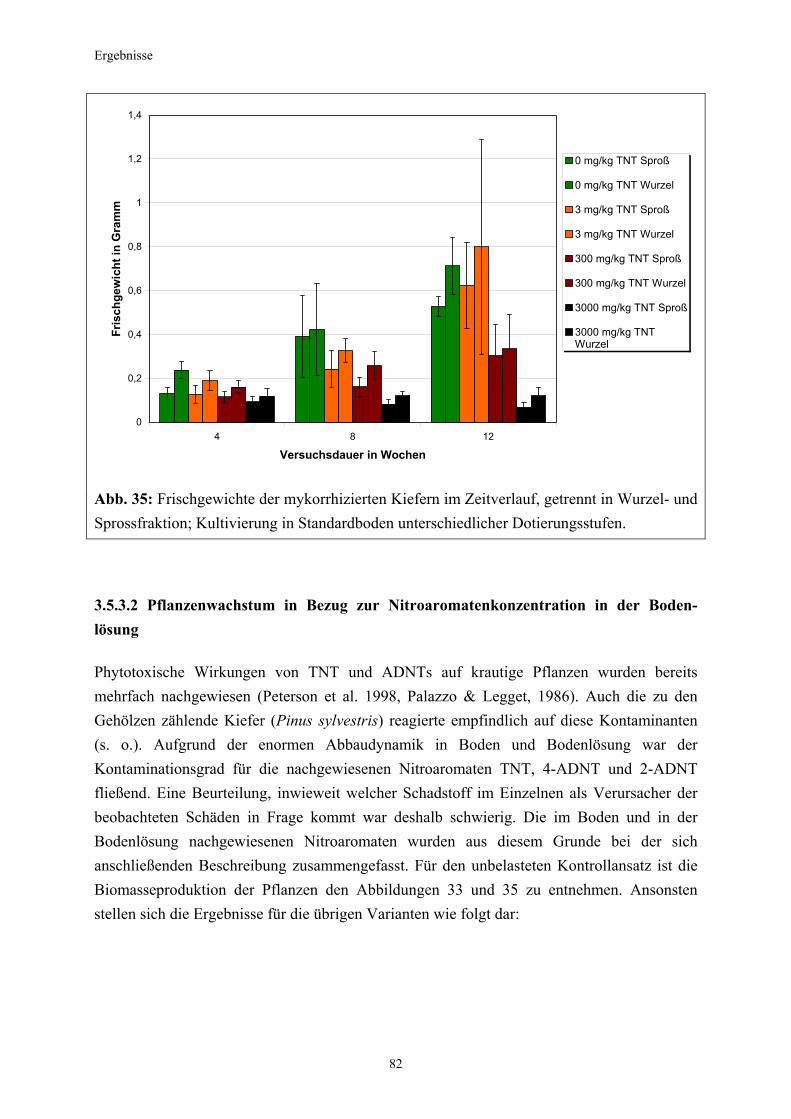
Ergebnisse
82
0
0,2
0,4
0,6
0,8
1
1,2
1,4
4 8 12
Versuchsdauer in Wochen
Fris
chge
wic
ht in
Gra
mm
0 mg/kg TNT Sproß
0 mg/kg TNT Wurzel
3 mg/kg TNT Sproß
3 mg/kg TNT Wurzel
300 mg/kg TNT Sproß
300 mg/kg TNT Wurzel
3000 mg/kg TNT Sproß
3000 mg/kg TNTWurzel
Abb. 35: Frischgewichte der mykorrhizierten Kiefern im Zeitverlauf, getrennt in Wurzel- und Sprossfraktion; Kultivierung in Standardboden unterschiedlicher Dotierungsstufen. 3.5.3.2 Pflanzenwachstum in Bezug zur Nitroaromatenkonzentration in der Boden-lösung Phytotoxische Wirkungen von TNT und ADNTs auf krautige Pflanzen wurden bereits mehrfach nachgewiesen (Peterson et al. 1998, Palazzo & Legget, 1986). Auch die zu den Gehölzen zählende Kiefer (Pinus sylvestris) reagierte empfindlich auf diese Kontaminanten (s. o.). Aufgrund der enormen Abbaudynamik in Boden und Bodenlösung war der Kontaminationsgrad für die nachgewiesenen Nitroaromaten TNT, 4-ADNT und 2-ADNT fließend. Eine Beurteilung, inwieweit welcher Schadstoff im Einzelnen als Verursacher der beobachteten Schäden in Frage kommt war deshalb schwierig. Die im Boden und in der Bodenlösung nachgewiesenen Nitroaromaten wurden aus diesem Grunde bei der sich anschließenden Beschreibung zusammengefasst. Für den unbelasteten Kontrollansatz ist die Biomasseproduktion der Pflanzen den Abbildungen 33 und 35 zu entnehmen. Ansonsten stellen sich die Ergebnisse für die übrigen Varianten wie folgt dar:
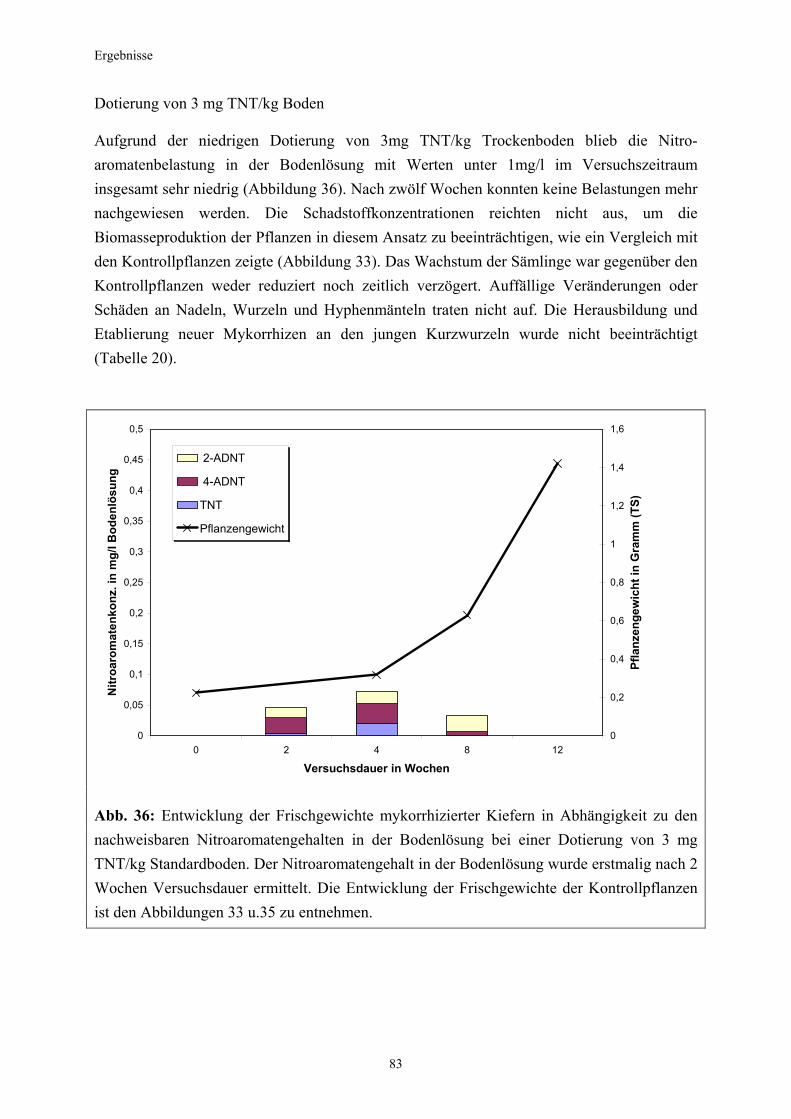
Ergebnisse
83
Dotierung von 3 mg TNT/kg Boden Aufgrund der niedrigen Dotierung von 3mg TNT/kg Trockenboden blieb die Nitro-aromatenbelastung in der Bodenlösung mit Werten unter 1mg/l im Versuchszeitraum insgesamt sehr niedrig (Abbildung 36). Nach zwölf Wochen konnten keine Belastungen mehr nachgewiesen werden. Die Schadstoffkonzentrationen reichten nicht aus, um die Biomasseproduktion der Pflanzen in diesem Ansatz zu beeinträchtigen, wie ein Vergleich mit den Kontrollpflanzen zeigte (Abbildung 33). Das Wachstum der Sämlinge war gegenüber den Kontrollpflanzen weder reduziert noch zeitlich verzögert. Auffällige Veränderungen oder Schäden an Nadeln, Wurzeln und Hyphenmänteln traten nicht auf. Die Herausbildung und Etablierung neuer Mykorrhizen an den jungen Kurzwurzeln wurde nicht beeinträchtigt (Tabelle 20).
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,45
0,5
0 2 4 8 12
Versuchsdauer in Wochen
Nitr
oaro
mat
enko
nz. i
n m
g/l B
oden
lösu
ng
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
Pfla
nzen
gew
icht
in G
ram
m (T
S)
2-ADNT
4-ADNT
TNT
Pflanzengewicht
Abb. 36: Entwicklung der Frischgewichte mykorrhizierter Kiefern in Abhängigkeit zu den nachweisbaren Nitroaromatengehalten in der Bodenlösung bei einer Dotierung von 3 mg TNT/kg Standardboden. Der Nitroaromatengehalt in der Bodenlösung wurde erstmalig nach 2 Wochen Versuchsdauer ermittelt. Die Entwicklung der Frischgewichte der Kontrollpflanzen ist den Abbildungen 33 u.35 zu entnehmen.

Ergebnisse
84
Dotierung von 300 mg TNT/kg Boden Eine Nitroaromatenkonzentration um 16 mg/l Bodenlösung wirkte sich noch hemmend auf die Biomasseentwicklung der Pflanzen aus, wobei die zeitgleiche Belastung der entsprechenden Kontaminanten im Boden bei etwa 80 mg/kg lag. Der Beginn des Wachstums war zeitlich verzögert und die Gewichtszunahme verlangsamt (Abbildung 37). Morphologische Schädigungen traten allerdings wenig in Erscheinung. Nur vereinzelt kam es zu Braunfärbungen an den Nadelspitzen. Wurzeln und Hyphenmäntel des Mykobionten zeigten keine auffälligen Veränderungen, die auf Schädigungen zurückzuführen waren. Neue Kurzwurzeln differenzierten sich anfangs jedoch nicht aus, weshalb aufgrund fehlender Neuetablierungen zu Beginn keine Steigerung der Mykorrhizierungrate zu beobachten war. Erst bei einem Rückgang der Nitroaromatenkonzentration unter 10 mg/l Bodenlösung (4. Woche), das entsprach einer Bodensubstratkonzentration von etwa 50 mg/kg Nitroaromaten, erholten sich die Kiefern sichtbar. Die Pflanzen reagierten nun mit einer verstärkten Biomasseproduktion. Es differenzierten sich zahlreiche neue Nadeln und Wurzeln aus. Einhergehend mit der Ausbildung neuer Kurzwurzeln war parallel eine zunehmende Mykorrhizierung der jungen Wurzeln durch den Mykobionten zu beobachten. Dieser Trend verstärkte sich im weiteren Versuchsverlauf. Die Ergebnisse zeigten, dass mäßige Kontaminationsgrade mit Nitroaromaten phytotoxisch wirken. Belastungswerte zwischen 10 und 15 mg/l Bodenlösung (50 bis 100 mg/kg Bodensubstrat) verminderten das Wachstum der Pflanzen und verursachten leichte Schäden an den Nadeln, hatten jedoch keine letale Wirkung. Möglicherweise können auch höhere Schadstoffgehalte toleriert werden. Die Pflanzen behielten ihr Regenerationsvermögen und zeigten sich mit abnehmender Nitroaromatenbelastung schnell erholt. Schwierig ist die Bewertung eines möglichen phytotoxischen Effektes auf den Pilzsymbionten. So könnte die herabgesetzte Mykorrhizierungsrate in einer verminderten Seitenwurzelbildung und nicht in einer Schwächung des Mykorrhizapilzes begründet liegen.
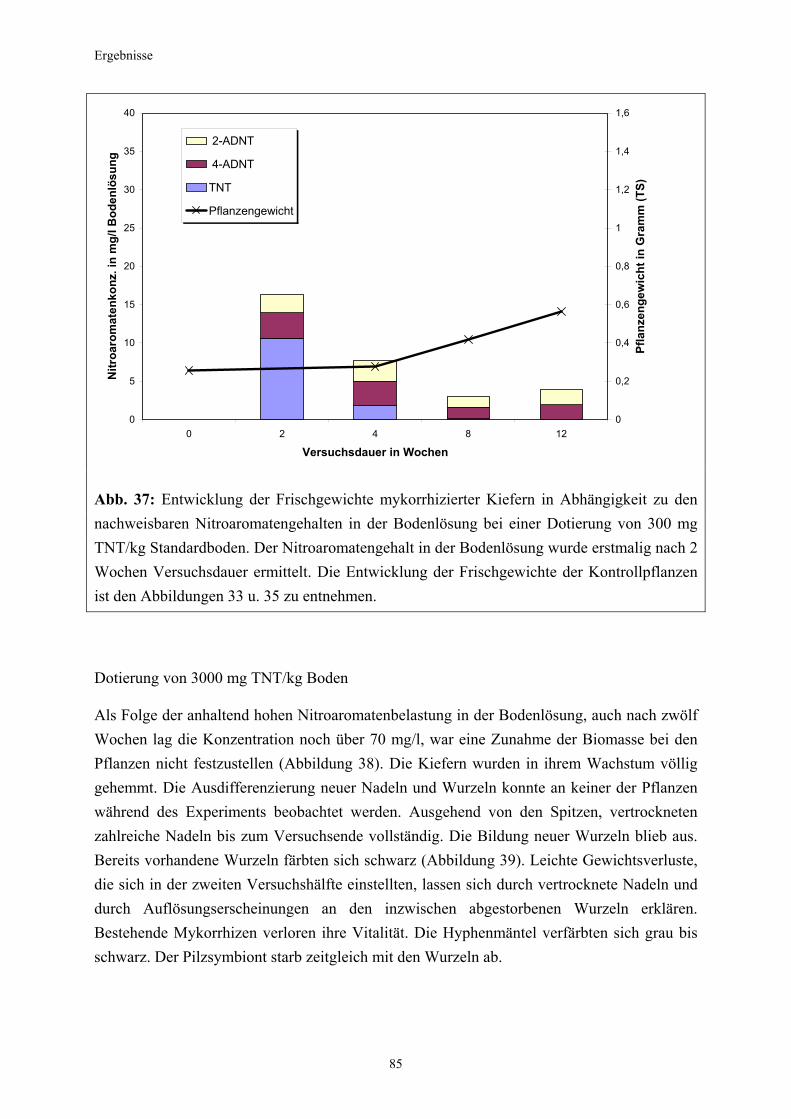
Ergebnisse
85
0
5
10
15
20
25
30
35
40
0 2 4 8 12
Versuchsdauer in Wochen
Nitr
oaro
mat
enko
nz. i
n m
g/l B
oden
lösu
ng
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
Pfla
nzen
gew
icht
in G
ram
m (T
S)
2-ADNT
4-ADNT
TNT
Pflanzengewicht
Abb. 37: Entwicklung der Frischgewichte mykorrhizierter Kiefern in Abhängigkeit zu den nachweisbaren Nitroaromatengehalten in der Bodenlösung bei einer Dotierung von 300 mg TNT/kg Standardboden. Der Nitroaromatengehalt in der Bodenlösung wurde erstmalig nach 2 Wochen Versuchsdauer ermittelt. Die Entwicklung der Frischgewichte der Kontrollpflanzen ist den Abbildungen 33 u. 35 zu entnehmen. Dotierung von 3000 mg TNT/kg Boden Als Folge der anhaltend hohen Nitroaromatenbelastung in der Bodenlösung, auch nach zwölf Wochen lag die Konzentration noch über 70 mg/l, war eine Zunahme der Biomasse bei den Pflanzen nicht festzustellen (Abbildung 38). Die Kiefern wurden in ihrem Wachstum völlig gehemmt. Die Ausdifferenzierung neuer Nadeln und Wurzeln konnte an keiner der Pflanzen während des Experiments beobachtet werden. Ausgehend von den Spitzen, vertrockneten zahlreiche Nadeln bis zum Versuchsende vollständig. Die Bildung neuer Wurzeln blieb aus. Bereits vorhandene Wurzeln färbten sich schwarz (Abbildung 39). Leichte Gewichtsverluste, die sich in der zweiten Versuchshälfte einstellten, lassen sich durch vertrocknete Nadeln und durch Auflösungserscheinungen an den inzwischen abgestorbenen Wurzeln erklären. Bestehende Mykorrhizen verloren ihre Vitalität. Die Hyphenmäntel verfärbten sich grau bis schwarz. Der Pilzsymbiont starb zeitgleich mit den Wurzeln ab.
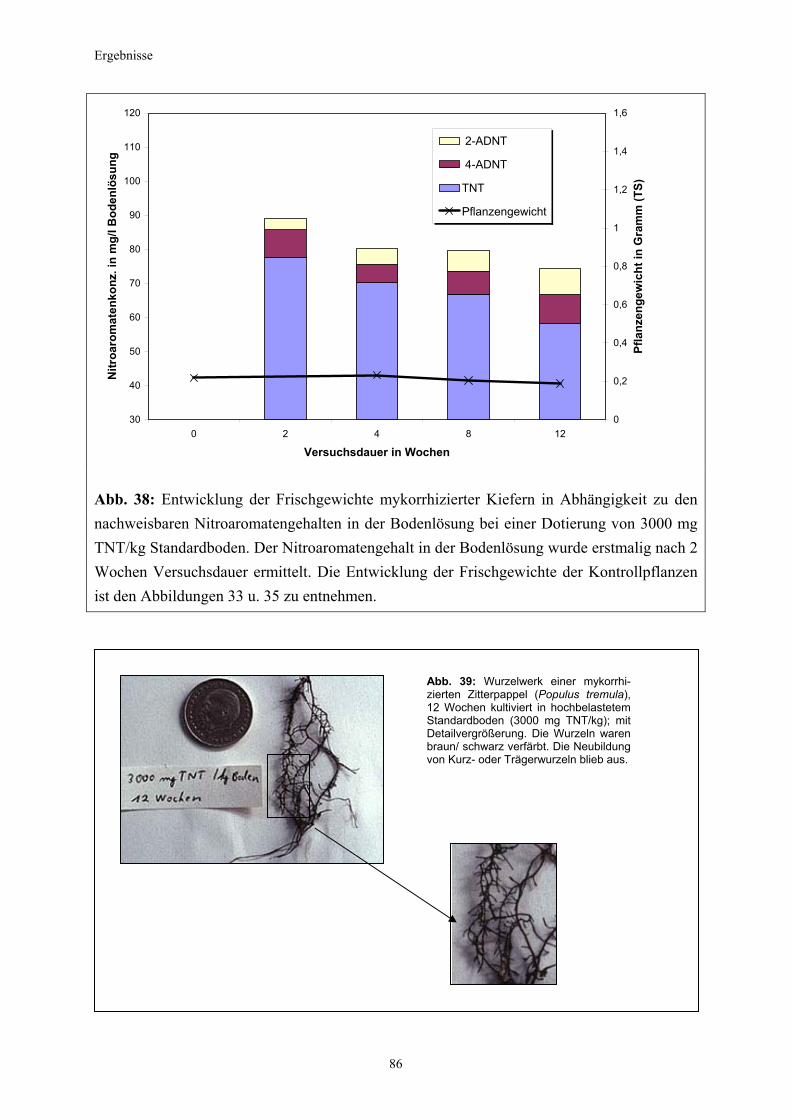
Ergebnisse
86
30
40
50
60
70
80
90
100
110
120
0 2 4 8 12
Versuchsdauer in Wochen
Nitr
oaro
mat
enko
nz. i
n m
g/l B
oden
lösu
ng
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
Pfla
nzen
gew
icht
in G
ram
m (T
S)
2-ADNT
4-ADNT
TNT
Pflanzengewicht
Abb. 38: Entwicklung der Frischgewichte mykorrhizierter Kiefern in Abhängigkeit zu den nachweisbaren Nitroaromatengehalten in der Bodenlösung bei einer Dotierung von 3000 mg TNT/kg Standardboden. Der Nitroaromatengehalt in der Bodenlösung wurde erstmalig nach 2 Wochen Versuchsdauer ermittelt. Die Entwicklung der Frischgewichte der Kontrollpflanzen ist den Abbildungen 33 u. 35 zu entnehmen.
Abb. 39: Wurzelwerk einer mykorrhi-zierten Zitterpappel (Populus tremula), 12 Wochen kultiviert in hochbelastetem Standardboden (3000 mg TNT/kg); mit Detailvergrößerung. Die Wurzeln waren braun/ schwarz verfärbt. Die Neubildung von Kurz- oder Trägerwurzeln blieb aus.
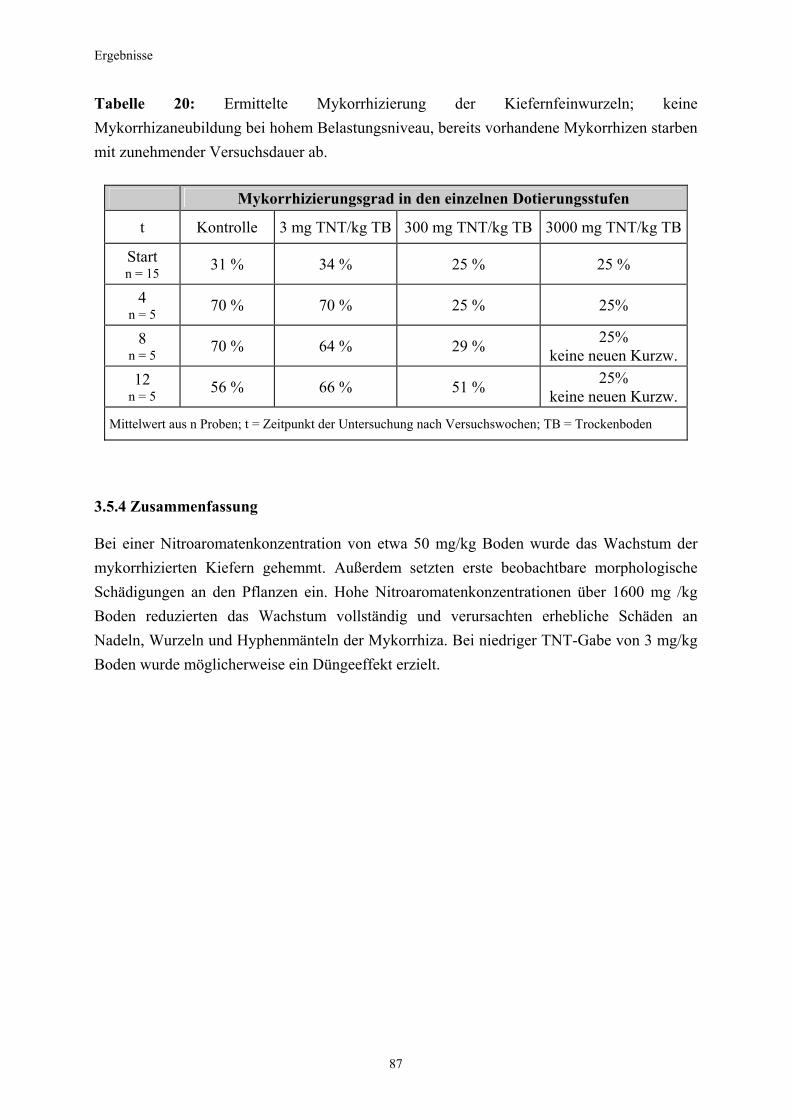
Ergebnisse
87
Tabelle 20: Ermittelte Mykorrhizierung der Kiefernfeinwurzeln; keine Mykorrhizaneubildung bei hohem Belastungsniveau, bereits vorhandene Mykorrhizen starben mit zunehmender Versuchsdauer ab.
Mykorrhizierungsgrad in den einzelnen Dotierungsstufen
t Kontrolle 3 mg TNT/kg TB 300 mg TNT/kg TB 3000 mg TNT/kg TB
Start n = 15 31 % 34 % 25 % 25 %
4 n = 5 70 % 70 % 25 % 25%
8 n = 5 70 % 64 % 29 % 25%
keine neuen Kurzw. 12
n = 5 56 % 66 % 51 % 25% keine neuen Kurzw.
Mittelwert aus n Proben; t = Zeitpunkt der Untersuchung nach Versuchswochen; TB = Trockenboden
3.5.4 Zusammenfassung Bei einer Nitroaromatenkonzentration von etwa 50 mg/kg Boden wurde das Wachstum der mykorrhizierten Kiefern gehemmt. Außerdem setzten erste beobachtbare morphologische Schädigungen an den Pflanzen ein. Hohe Nitroaromatenkonzentrationen über 1600 mg /kg Boden reduzierten das Wachstum vollständig und verursachten erhebliche Schäden an Nadeln, Wurzeln und Hyphenmänteln der Mykorrhiza. Bei niedriger TNT-Gabe von 3 mg/kg Boden wurde möglicherweise ein Düngeeffekt erzielt.

Diskussion
88
4. Diskussion Die nun folgende Diskussion ist in vier wesentliche Abschnitte untergliedert. Neben den Wechselwirkungen und Bindungen von Nitroaromaten mit Bestandteilen der Bodenmatrix sollen in einem zweiten Abschnitt unter Berücksichtigung der eingesetzten Experimentvarianten mikrobielle Aspekte und die Funktion mykorrhizierter Pflanzen beim Nitroaromatenabbau näher beleuchtet werden. In einem weiteren Kapitel wird über das Aufnahme-, Akkumulations- und Abbauverhalten von Nitroaromaten in Pflanzen diskutiert. Im vierten Abschnitt wird das Toleranzverhalten mykorrhizierter Kiefern gegenüber Nitroaromaten bewertet. Im Anschluss an die Diskussion sollen die im Rahmen dieser Arbeit erzielten Laborergebnisse mit den Ergebnissen eines maßstabsgerecht erprobten in situ-Phytoremediationsverfahren zur Sanierung TNT-kontaminierter Böden am Standort ‘Werk Tanne’ verglichen und das Phytoremediationspotenzial mykorrhizierter Gehölze beim Abbau von TNT und dessen Metaboliten diskutiert werden. 4.1 Wechselwirkungen zwischen Nitroaromaten und Bodenmatrix Aromatische Verbindungen wie das TNT und dessen Transformationsprodukte können auf unterschiedliche Weise mit den Bestandteilen des Bodens in Wechselwirkung treten. Dabei kann es zu reversiblen oder irreversiblen Bindungen mit der Bodenmatrix kommen. Als Hauptadsorbentien des Bodens gelten die Tonmineralien, die organische Bodenfraktion sowie die Ton- Humuskomplexe. Bei den Bindungsmechanismen unterscheidet man zwischen Physisorption, (Van der Waals-Kräfte, Wasserstoffbrückenbindungen und Ionenbindungen) und Chemiesorption (kovalente Bindungen). Allgemein wird die Art der Bindungsweise von der chemischen Struktur der Verbindungen, der Beschaffenheit der Bodenstrukturen, von den Umweltbedingungen und gegebenenfalls von den Inkubationsbedingungen bestimmt (Lenke 2001). 4.1.1 Adsorption an Tonmineralien Tonmineralien bestehen aus Aggregaten kleinster anorganischer Partikel, den mit einer negativen Oberflächenladung versehenen Tonplättchen. Die Tonplättchen werden durch einen Belag von Kationen, z. B. Ca2+, zusammengehalten und bilden dadurch Feinporen. Aufgrund ihrer negativen Oberflächenladung fungieren die Tonplättchen als Kationenaustauscher. Adsorbierte Kationen können durch gleichsinnig geladene Ionen aus der Bodenlösung ausgetauscht werden. Generell verlaufen die Austauschvorgänge schnell und sind in der Regel reversibel (Gisi et al. 1990). Organische Kationen mit aromatischen Ringsystemen werden besonders stark gebunden (Martinetz & Rippen 1996). Die Adsorption verschiedener

Diskussion
89
Nitroaromaten erfolgt größtenteils an der Oberfläche der Tonmineralien. Dabei nimmt die Affinität und das Adsorptionsvermögen verschiedener Tonmineralien gegenüber Nitroaromaten in der Reihenfolge Kaolinit < Illit < Montmorillinit zu (Haderlein et al. 1996). Der Sorptionsprozess verläuft sehr schnell innerhalb weniger Minuten. Allerdings sind die Substanzen einem weiteren biologischen Abbau nicht dauerhaft entzogen, da die Bindung überwiegend von kurzer Dauer und reversibel bleibt. Bei einer länger andauernden Bindung können die Kationen allerdings in die Tonoberfläche hinein diffundieren, was eine sehr starke und nahezu irreversible Adsorption zur Folge hat (Martinetz & Rippen 1996). Vor allem für das TNT und dessen unmittelbaren Transformationsprodukte 4-ADNT und 2-ADNT konnte eine vollständig reversible Bindung an Montmorillonit nachgewiesen werden, während die Sorption von 2,4-DANT, welches aufgrund seiner basischen Aminogruppen und seiner höheren Polarität stärker an die negative Oberfläche der Tonmineralien adsorbiert, nur zum Teil reversibel blieb (Daun et al. 1998). Von weitaus geringerer Bedeutung ist vermutlich die Einlagerung von Nitroaromaten in die Feinporen der Tonmineralien, da die Oberflächen primär schneller als Kontaktzone erreichbar sind und dort eine Adsorption dann in der Regel schnell vonstatten geht (Haderlein et al. 1996). 4.1.2 Wechselwirkung mit Huminstoffen Die organischen Huminstoffe spielen bei den Wechselwirkungen zwischen Nitroaromaten und Bodenmatrix vermutlich eine weitaus bedeutendere Rolle als die anorganischen Tonmineralien. Chemisch sind Huminstoffe schwierig zu charakterisieren. Sie sind ein Gemisch hochmolekularer Substanzen mit vielfältigen Strukturen und sind überwiegend aus aromatischen Bestandteilen (als Grundgerüst von Lignin) zusammengesetzt. Ein Abbau ist schwierig und erfolgt sehr langsam. Unterteilt werden die Huminstoffe in Humin, Fulvinsäuren und Huminsäuren. Als Bindungsstellen für aromatische Amine werden u. a. Chinone und Carbonylstrukturen diskutiert (Thorn et al. 1996). Allgemein sind die Wechselwirkungen zwischen Niroaromaten und den organischen Bodenbestandteilen bisher wenig untersucht. Einerseits wäre theoretisch eine reversible Sorption von TNT und seinen Metaboliten an die Huminstoffmatrix denkbar. Andererseits findet ein mikrobieller Einbau der Kontaminanten in die Huminstoffe statt (Martinetz & Rippen 1996). Die letztere Möglichkeit sollte als dominierend angesehen werden, wobei die Schadstoffe mit zunehmender Reduktion der Nitrogruppen durch Chemisorption irreversibel binden. Elovitz et al. (1999) stellten fest, dass unter aeroben Verhältnissen 2,4-DANT irreversibel in Sedimenten gebunden wurde, während dieses für TNT und 4-ADNT nicht zutraf.

Diskussion
90
4.1.3 Reduktive TNT-Transformation in den Experimenten und Wechselwirkungen mit Bodenbestandteilen In allen Experimenten war allgemein ein deutlicher Rückgang der originalen TNT-Belastung festzustellen. Die Menge des durch methanolische Extraktion aus dem Bodensubstrat desorbierbaren TNT variierte dabei je nach Versuchsvariante (siehe Abschnitte 3.2 und 3.3). Über die Transformationswege des TNT im Bodensubstrat und das Bindungsverhalten des TNT und möglicher Metabolite mit Bodenbestandteilen können nur Vermutungen angestellt werden. Falls es anteilig zu Wechselwirkungen zwischen TNT und der Bodensubstanz kam, kann in diesem Fall in der Regel von einer reversiblen Sorption des Schadstoffes an die Bodenmatrix ausgegangen werden. Prinzipiell bleibt auch die reversibel gebundene Fraktion einer Umsetzung nicht dauerhaft entzogen. Der weitaus bedeutendere Teil des TNT dürfte einem schnell einsetzenden und umfassenden Reduktionsprozess zum ADNT ausgesetzt gewesen sein. Kurzzeitversuche mit Boden als Bakterieninokulum zeigten, dass unter aeroben und anaeroben Bedingungen die Transformation von TNT zu 2- ADNT und 4-ADNT innerhalb von 24 Stunden abläuft (Pennington & Pattrick 1990). In den hier durchgeführten Experimenten akkumulierten die ADNT-Gehalte in einigen Versuchsvarianten, und hier insbesondere in den Kontrollvarianten, zum Teil erheblich. Im Pappelexperiment hat sich die extrahierbare ADNT-Menge in der Kontrollvariante K letztendlich verdoppelt. Bezogen auf die umgesetzte TNT-Menge ließen sich nach Versuchsende immerhin noch 42 % als ADNT wieder finden. Die relativ hohe ADNT-Wiederfindungsrate in der Kontrollvariante zeigte, dass abgesehen vom TNT auch das ADNT entweder weniger zur Sorption neigte oder, falls doch, in größeren Mengen nicht fest an die Bodenmatrix gebunden wurde. Ähnliche Beobachtungen machten bereits Elovitz et al. (1999). Waren die Versuchsvarianten mit Bäumchen bestückt, ließ sich trotz einer hohen Abnahme der TNT-Konzentration lediglich ein verhältnismäßig geringer Anstieg der ADNT-Konzentration ermitteln. So wurden in Variante A nach Versuchsende nur noch 5 % des umgesetzten TNT als ADNT nachgewiesen. In Anbetracht der relativ geringen bzw. reversiblen Sorptionsneigung von ADNT zur Bodenmatrix ist demzufolge von einem umfassenden Fortgang der Transformation auszugehen. Sei es anteilig durch Reduktion einer weiteren Nitrogruppe zu DANT oder über andere mikrobiell vermittelte Reaktionswege, z. B. durch Oxidationsprozesse zu verschiedensten anderen nicht identifizierten Verbindungen. Obwohl sich die Reduktionssequenz bis zum DANT nicht nachweisen ließ, DANT wurde in keiner Versuchsvariante gefunden, kann davon ausgegangen werden, dass in den Bodenproben sämtlicher Varianten relevante Mengen DANT gebildet wurden, anschließend schnell und irreversibel an Bodenbestandteile adsorbierten und sich deshalb nicht extrahieren ließen. Die Reduktion bis zum DANT unter aeroben Bedingungen ist bereits belegt (Schackmann und Müller 1991). Das in mäßig belasteten Böden ungebundenes DANT während der Abbaukinetik unter aeroben Verhältnissen nicht akkumuliert und unter der Nachweisgrenze

Diskussion
91
bleibt, ist nicht ungewöhnlich. Selbst in hochkontaminierten TNT-Böden bleibt die DANT-Konzentration allgemein niedrig und kurzzeitig nachweisbar. So fanden Achtnich et al. (2000), wenn auch in einem alternierend anaeroben/ aeroben Prozess, bei einer Ausgangskonzentration von nahezu 5000 mg TNT kg-1 Boden (TS) maximal zwischen 50 und 60 mg DANT kg-1 Boden, wobei diese Konzentrationen lediglich wenige Tage nach Inkubationsbeginn gemessen werden konnten und danach sehr schnell abnahmen. Für eine weitere Reduktion vom DANT zum TAT sind anaerobe Verhältnisse und ein Redoxpotenzial (EH) von < -200 mV erforderlich (Lenke et al. 1994 in Martinetz & Rippen 1996). Im aeroben Milieu der jeweiligen Versuchsvarianten dürfte die reduktive Trans-formation deshalb auf der Stufe des DANT ihr Ende gefunden haben. TAT wurde in keinem Fall nachgewiesen. Neben TNT und ADNT wurden aus den Bodenproben des Pappelexperimentes zusätzlich geringe Mengen DNT extrahiert und analytisch erfasst. Aufgrund fehlender funktioneller Gruppen ist generell von einer geringen Bindungsneigung dieser Verbindungen mit Bodenbestandteilen auszugehen. So beobachteten Pörschmann und Stottmeister (1993) keine Bindung von 2,6-DNT an in Grundwasser gelöste Huminstoffe. Die aerobe Mineralisierung des 2,4-DNT durch Bakterien ist inzwischen gut untersucht (Martinetz & Rippen 1996). Die Ringspaltung wird durch Oxigenasereaktionen unter Bildung verschiedener aromatischer Zwischenprodukte katalysiert. Auch für 2,6-DNT ließ sich mittlerweile ein oxidativer Abbau über Dioxygenase-vermittelte Initialreaktionen nachweisen (Nishino et al. 2000b). 4.1.4 Einfluss von Bodenfaktoren auf die Bindungskinetik Kenntnisse über den Einfluss verschiedener Milieufaktoren im Boden auf die Bindungskinetik der Nitroaromaten mit Bodenbestandteilen sind bisher stark limitiert. Neben dem Redox-potenzial sind insbesondere der pH-Wert und der Gehalt organischer Bodensubstanzen von entscheidender Bedeutung für Bindungsreaktionen. Der optimale pH-Wert-Bereich für die biologische Aktivität zahlreicher Mikroorganismen liegt zwischen 4 und 7. Entsprechende Bereiche gelten z. B. für die bei vielen Pilzen vorkommenden und beim mikro-organismenvermittelten Einbau von Xenobiotika wichtigen Enzyme wie Peroxidasen und Laccasen. In einem für optimale Aktivität geeigneten Bereich befanden sich auch die in den TNT-Abbau-Experimenten verwendeten Bodensubstrate (siehe Abbildungen 41 und 42 im Anhang). Abgesehen von geringen Schwankungen lag der pH-Wert des Bodens im Pappelexperiment bei 4,2 und im Kiefernexperiment bei 5,5. Auch für die physiko-chemischen und in der Regel reversiblen Sorptionsprozesse zwischen Nitroaromaten und Bodenbestandteilen spielt der pH-Wert eine maßgebliche Rolle. Mit sinkendem pH-Wert in der Bodenlösung erhöht sich der Anteil protonierter Amine. Folglich kommt es zu einer zunehmenden Bindung reduzierter Verbindungen an die Bodenmatrix. In Tabelle 21 sind die pKa-Werte für verschiedene Nitroaromaten dargestellt. Der pKa-Wert gibt

Diskussion
92
als Maß für die Ionisation infolge Protonierung des Amins denjenigen pH-Wert an, bei dem sich 50 % der in einer Lösung vorhandenen Moleküle in einem ionisierten Zustand befinden. Tabelle 21: pKa-Werte verschiedener TNT-Metaboliten (aus Martinetz & Rippen 1996; berechnet nach Harris und Hayes 1990)
4-ADNT 1,21 2,4-DANT 3,47 TAT 5,79 4-A-2-NT 3,67 2,4-DAT 5,52 4-A-2-HA-6-NT 3,28 2,4-DAHAT 5,47
Werden beim TNT die Nitrogruppen zunehmend durch Aminogruppen ersetzt, steigt auch das Maß der Ionisierung infolge Protonierung der Aminogruppen unter umweltrelevanten Bedingungen. So hat 2,4-DANT einen pKa-Wert von 3,47, für 2,6-DANT ist er mit 3,37 nur geringfügig niedriger. Von den in den Experimentvarianten anfallenden TNT-Metaboliten dürften sich die DANTs daher durch deutlich höhere Bindungseigenschaften ausgezeichnet haben als vergleichsweise die ADNTs, was sich auch in ihrer Nachweisbarkeit ausdrückte. Für 2,6-DANT liegen Untersuchungen über das Bindungsverhalten bei verschiedenen pH-Werten vor. In Abhängigkeit von der Huminsäurekonzentration in einer Testlösung wurde bei einem pH-Wert von 4,6 eine um bis zu 25 % höhere Bindung an Huminsäure beobachtet, als bei pH 6,8 (Li et al. 1997). Unter dem Aspekt, dass zum einen der mikrobiell vermittelte Einbau reduzierter Nitroaromaten bevorzugt bei niedrigen pH-Werten abläuft (niedriger pH-Wert erhöht ja auch Anteil protonierter Amine) und zum anderen die Sorption reduzierter Verbindungen aufgrund zunehmender Protonierung der Aminogruppen bei abnehmenden pH-Wert zunimmt, ist zu berücksichtigen, dass im unmittelbaren Rhizosphärenbereich von mykorrhizierten Pflanzenwurzeln der pH-Wert um mehrere Einheiten niedriger sein kann als im umgebenden Boden. Dass entsprechende Beobachtungen mit deutlichen pH-Wert-Differenzen zwischen bepflanzten und unbepflanzten Varianten in den Experimenten allerdings nicht gemacht werden konnten, dürfte darin begründet liegen, dass die Bodenprobenahme für die pH-Wert-Bestimmung in den bepflanzten Ansätzen als Bodenmischprobe aus dem gesamten Bereich der Rhizotrone erfolgte, und nicht nur aus dem Rhizosphärenbereich wenige mm um die Wurzeln herum. Eventuelle pH-Wert-Unterschiede zwischen Rhizosphärenboden und umgebenem Boden könnten somit möglicherweise verwischt worden sein. Über den Einfluss

Diskussion
93
mykorrhizierter Pflanzen bei der Transformation von Nitroaromaten wird an späterer Stelle ausführlich diskutiert. Eine wichtige Rolle bei der TNT-Transformation im Boden spielt auch der C-Gehalt. Zur Reduktion der Nitrogruppen werden aus dem Abbau von Kohlenstoffquellen stammende Reduktionsäquivalente benötigt (kometabolische Reaktion). Da Pflanzen über ihre Wurzeln erhebliche Mengen verschiedener niedermolekularer Substanzen durch Exsudation an ihre Umgebung abgeben, dürfte insbesondere in den bepflanzten Experimentalvarianten von einer Erhöhung des bioverfügbaren C-Pools auszugehen sein, was schließlich eine verstärkte mikrobielle metabolische Aktivität zur Folge hatte. Außerdem wurden mit den Pflanzen allgemein vermehrt komplexere Kohlenstoffverbindungen mit den abgestorbenen Wurzeln in den Boden eingebracht (vor allem durch die starkwüchsigen Pappeln), die für anfallende Transformationsprodukte als geeignete Reaktionspartner zur Festlegung fungieren können. Möglicherweise wurde der C-Gehalt im Bodensubstrat auch durch das Einbringen des Weizenstrohs beeinflusst und erhöht. Allerdings ist eine Beurteilung darüber, inwieweit Strohanteile zwischenzeitlich biochemisch zersetzt und somit für Bindungsprozesse verfügbar wurden, nicht möglich. Ein durch Strohzugabe hervorgerufener Abreicherungseffekt bei der Nitroaromatenkonzentration ließ sich nicht beobachten. Letztendlich kann über die Entwicklung der C-Konzentrationen in den Böden der jeweiligen Versuchsvarianten nur spekuliert werden, da sich die Untersuchungen zur Gehaltsbestimmung auf die Ausgangssubstrate beschränken. Die Messergebnisse können aus Tabelle 31 im Anhang entnommen werden. 4.1.5 Hydroxylaminodinitrotoluole (HADNT) als Intermediate der TNT-Reduktion Ein Nachweis der sehr hydrophilen HADNT-Isomere in den Bodenproben konnte mit der im Rahmen dieser Arbeit zur Anwendung gekommenen HPLC-Methode nicht geführt werden. Auf den Einsatz zusätzlicher Nachweismethoden zur Quantifizierung der HADNTs wurde verzichtet, weil diese Transformationsprodukte in Gegenwart von Bodenbestandteilen gewöhnlich einer zügigen Umwandlung unterliegen. Dafür spricht zum einen die deutliche Zunahme der ADNTs. In einigen Experimentalvarianten konnte ein Anstieg um nahezu 100% nachgewiesen werden. Zum anderen besteht die Möglichkeit zur schnellen Kondensation mit Nitroso-Derivaten zu Azoxyverbindungen (s. u.). In den Experimenten ist deshalb lediglich von sehr geringen Mengen längerfristig akkumulierender HADNTs im Boden auszugehen. Entsprechende Beobachtungen machten Achtnich et al. (2000) in einem Kompostierungs-versuch im Labormaßstab bei schneller Transformation des in großer Menge dotierten TNT (4500 mg kg -1 TS). Eine im Verhältnis zur dotierten TNT-Menge sehr geringe Konzentration von 0,33 mg kg-1 (TS) isomerer HADNTs ließ sich lediglich bis zum zweiten Tag nach Versuchsbeginn nachweisen.

Diskussion
94
Für eine Bewertung des ökotoxikologischen Potentials der Hydroxylaminoverbindungen liegen bisher nur wenige Daten vor. Intermediate wie 2-Hydroxylamino-4,6-Dinitrotoluol (2HA46DNT) inhibierten das Wachstum der Grünalge Selenastrum capricornutum bei einer Konzentration von 1,4 mg l-1 um etwa 50 %, wobei die Wachstumshemmung deutlich niedriger ausfiel als bei vergleichbarer TNT-Konzentration (Tadros et al. 2000). Für eine abschließende Beurteilung des Toxizitätsprofils von HADNTs sind noch weitere Studien notwendig. Da allerdings von einer geringen Neigung zur Akkumulation im Boden auszu-gehen ist, dürfte das toxische Wirkungspotenzial auf Pflanzen und andere Organismen eher eine untergeordnete Rolle spielen. 4.1.6 Entstehung von Azoxyverbindungen bei TNT-Abbauprozessen Unter aeroben Bedingungen, kaum jedoch unter anaeroben Bedingungen, kann es im Verlauf der Reduktion der Nitrogruppe zu einer abiotischen Kondensationsreaktion von Nitroso- und Hydroxylaminoaromaten zu Azoxyverbindungen kommen (Abbildung 40). Mikroorganismen spielen dabei lediglich eine Rolle als Lieferanten der Kopplungssubstrate, entweder beim bakteriellen Abbau durch Kompostierung oder bei der Transformation des TNT durch Pilze (Garg et al. 1991, Isbister et al. 1984, Michels u. Gottschalk 1994; in Martinetz & Rippen 1996).
CH3
O2N N=O
NO2
CH3
N OH
NO2
O2NH
O2N CH3
O2N
N=N
H3C NO2
NO2
+
4,4',6,6'-Tetranitro-2,2'-azoxytoluol
+ H2O
O
Abb. 40: Entstehung von 2,2’-Azoxy aus TNT-Metaboliten (aus Martinetz & Rippen 1996, nach Layton et al. 1987)

Diskussion
95
Azoxyverbindungen gelten als hochresistent. Ihre toxikologische Relevanz ist bisher ungeklärt. In welcher Größenordnung diese Verbindungen entstehen und dann möglicherweise Wechselwirkungen mit der Huminstofffraktion im Boden eingehen und dabei fest gebunden werden oder im Boden akkumulieren, ist bisher ebenfalls wenig untersucht. Sollte es tatsächlich zu einer Akkumulation kommen, könnte dies bei Sanierungsvorhaben so ein Problem darstellen. In welchem Umfang Azoxyverbindungen durch Kondensation in den hier durchgeführten Experimenten im Bodensubstrat anfielen, wurde im Rahmen dieser Arbeit nicht näher untersucht. Eine Akkumulation kann grundsätzlich nicht ausgeschlossen werden. Achtnich et al. (2000) fanden in einem generell aeroben Prozess bei einer Ausgangsdotierung von 4500 mg TNT kg-1 Boden (TS) 1-2 Tage nach Inkubationsbeginn lediglich Maximalwerte von 7-13 mg kg-1 (TS). Im Laufe der weiteren Behandlung lag die Konzentration nach 9 Behandlungstagen schließlich unter der Nachweisgrenze, wobei nicht ausgeschlossen wurde, dass durch mikrobielle Sauerstoffzehrung anaerobe Mikrobereiche entstanden sein könnten und die Azoxy-Bildung somit unterbunden wurde. In anderen Versuchen wurden bei ähnlich hoch TNT-belasteten Böden dagegen wesentlich höhere Azoxy-Konzentrationen nach-gewiesen (Achtnich et al. 1999). 4.2 Dekontamination sprengstoffbelasteter Böden im Kiefern- und Pappel-experiment; ein Variantenvergleich Die Eignung verschiedener Kraut- und Gehölzpflanzen beim Abbau von Nitroaromaten im Boden wurde bereits mehrmals erfolgreich getestet und in der Literatur beschrieben (Cataldo et al. 1989, Görge et al. 1994a und 1995, Scheidemann 1998, Schönmuth und Pestemer 2000). Der Aspekt der Mykorrhiza fand in den hierbei erwähnten Experimenten allerdings wenig Beachtung. Böhler et al. (2000) erprobten im kleintechnischen Maßstab in Gefäßversuchen eine Kombination von verschiedenen krautigen Pflanzen (Medicago sativa, Carex buchananii, Miscanthus sinensis) mit dem sowohl saprophytisch als auch Mykorrhiza-bildend lebenden Basidiomyzeten Stropharia rugosoannulata. Die Inokulation des Bodens mit dem Pilz erfolgte durch Einbringung von Stroh, welches als Wuchssubstrat diente. Der Mykor-rhizaeffekt beim TNT-Abbau wurde nicht näher beleuchtet. Ein weiteres Freilandexperiment speziell mit mykorrhizierten Gehölzen zur Phytoremediation TNT-belasteter Flächen wurde in den vergangenen Jahren auf dem Gelände einer ehemaligen Sprengstoffproduktionsstätte im Harz (‘Werk Tanne’/Clausthal-Zellerfeld) durchgeführt (Warrelmann et al. 2000, Koehler et al. 2001 a und 2001 b). Die während der Erprobung dieses in situ-Phytoremediations-verfahrens gewonnenen Erkenntnisse werden noch an späterer Stelle mit den Ergebnissen aus den Laborexperimenten der vorliegenden Arbeit verglichen und diskutiert. Weitere Experimente, die bei der Sanierung TNT-belasteter Böden in ihrem Schwerpunkt insbesondere auf den Einsatz mykorrhizierter Pflanzen basieren, sind bisher nicht bekannt.

Diskussion
96
Weißfäulepilze in ihrer Funktion als ligninolytische Organismen sind in der Lage, TNT enzymatisch zu reduzieren und mittels Peroxidasen eine Ringspaltung des Aromaten herbeizuführen (Stahl und Aust 1993a, Spiker et al. 1992). Verbunden mit einem enzymatischen Angriff auf TNT im Boden durch Weißfäulepilze ist allerdings die Einbringung organischer Zuschlagstoffe in den Boden, welche als Kultursubstrate für den Pilz dienen. Dabei haben über das Substrat eingebrachte Weißfäulepilze in der Regel Schwierigkeiten, sich im Boden zu etablieren (Böhler et al. 2000; eigene Beobachtungen), da sie nicht zu den eigentlichen Bodenorganismen zählen, sondern überwiegend in der Holzfraktion von Bäumen und in der Streuschicht verbreitet sind. Im Gegensatz zu Weißfäulepilzen ist bei einer Beimpfung des Bodens mit Mykorrhizapilzen eine Substratzugabe in Form von Zuschlagstoffen nicht erforderlich, da eine kontinuierliche Versorgung mit Kohlenstoffverbindungen über Photoassimilate durch die Pflanzen erfolgt. Des Weiteren fungieren zahlreiche Mykorrhizapilze als Saprophyten (z. B. Paxillus und Stropharia) und können sich Kohlenstoffquellen aus dem Material der Streuschicht erschließen. Der Besitz reduktiver Enzymsyteme und verschiedener ringspaltender Peroxidasen konnte inzwischen für zahlreiche Mykorrhizapilze festgestellt werden (Scheibner et al. 1997, Gramss et al. 1998, Meharg et al. 1997a). Als Bodenorganismen ist es Mykorrhizapilzen zudem möglich, sich im Boden gut zu etablieren und diesen mit ihrem feinen und weit verzweigten Hyphensystem weiträumig zu durchwachsen und als Nahrungsquelle zu erschließen. Basierend auf diesen Tatsachen sollten mit Hilfe der Variantenauswahl in den beiden Experimenten mit mykorrhizierten Kiefern und Pappeln zur Untersuchung der TNT-Abreicherung im Boden zwei wesentliche Fragen näher beleuchtet werden. • Führt eine Kombination aus Beimpfung mit Weißfäulepilzen und Bepflanzung mit mykorrhizierten Gehölzen generell zu einer Dekontamination TNT-belasteter Böden (Variante A)? • Sind in den Boden eingebrachte Mykorrhizapilze aufgrund ihrer Eigenschaften dazu in der Lage, Weißfäulepilze in ihrer Funktion beim TNT-Abbau zu ergänzen oder sogar zu ersetzen (Varianten A und B)? Darüber hinaus wurde die Anzahl der Versuchsvarianten im Pappelexperiment um eine weitere ergänzt und das Spektrum der Varianten um eine Kombination aus Weißfäulepilzen und nicht-mykorrhizierten Pflanzen erweitert (Variante D). Dabei sollte die Einflussgröße der Pflanzen auf die TNT-Abreicherung im Boden ermittelt werden.

Diskussion
97
• Inwieweit lässt sich ein Einfluss der nicht-mykorrhizierten Pflanzen auf die TNT-Abreicherung im Boden erkennen (Variante D) bei einem Vergleich mit mykorrhizierten Pflanzen (Variante A)? In der vorliegenden Arbeit zeigte sich in den Varianten des Kiefernexperimentes ein anderes Abbauverhalten der Bodenkontaminationen als in den gleichartig angesetzten Varianten des Pappelexperimentes. Ein geringer Biomassezuwachs der Kiefern, einhergehend mit einer schwachen Durchwurzelung des Substrates in den Rhizotronen insgesamt, resultierte in einer Ausprägung der Rhizosphäreneffekte auf niedrigem Niveau. Ein lediglich kleiner Einflussbereich der Wurzeln mit einer gerade dort zu erwartenden erhöhten mikrobiellen Biomasse mit hoher metabolischer Aktivität aufgrund einer großen Bioverfügbarkeit dort freigesetzter organischer Nährstoffe durch die Wurzeln ließ pflanzeninduzierte Transformationsvorgänge kaum in Erscheinung treten. Auffällige Abreicherungseffekte, insbesondere bei den ADNTs, ließen sich überwiegend in den mit Weißfäulepilzen beimpften Varianten beobachten und sind demnach vermutlich hauptsächlich auf Abbauleistungen der Weißfäulepilze zurückzuführen. Allerdings müssen auch die Mykorrhizapilze eine positive Wirkung auf die Abreichungsprozesse gehabt haben, da in der Variante A die Belastungen am stärksten zurückgingen. Im Pappelexperiment stellte sich die Situation grundlegend anders dar. Hier hatte die Bestückung der Rhizotrone mit den starkwüchsigen mykorrhizierten Gehölzen erhebliche Effekte auf die Entwicklung der Nitroaromatenbelastung im Substrat. Die mykorrhizierten Pappeln erwiesen sich bei der Dekontamination nicht nur als sinnvolle Ergänzung zu den Weißfäulepilzen, viel mehr scheinen die Untersuchungen zu belegen, dass die Funktion der Weißfäulepilze durch eine Assoziation Mykorrhizapilz/Pflanze übernommen werden konnte. Eine zusätzliche Beimpfung der bepflanzten Varianten mit Weißfäulepilzen erbrachte keine wesentlich verbesserte Abbauleistung (vergleiche Varianten A und B). Als erstaunlich effizient erwiesen sich außerdem nicht-mykorrhizierte Pappeln beim TNT-Abbau. Das Fehlen der Mykorrhizapilze wirkte sich nicht nachteilig auf die TNT-Degradation aus. Eine entsprechend positive Tendenz konnte beim Abbau der ADNTs allerdings nicht beobachtet werden (Variante D). Verglichen mit den Kiefern hatten die starkwüchsigen Pappeln nicht nur wesentlich mehr Biomasse zu Versuchsbeginn aufzuweisen, sondern legten auch während des Experimentes erheblich mehr an Biomasse zu (vergleiche Tabellen 10 und 13). Ein besserer Durchwurzelungsgrad im Substrat der Rhizotrone ließ dementsprechend ein deutlich stärkeres Auftreten von Rhizosphäreneffekten erwarten. Mehr Lebendbiomasse bzw. Wurzelbiomasse der Pflanzen hatte vermutlich eine höhere Exsudatfreisetzung in die Rhizosphäre zur Folge und ein größeres Angebot freiverfügbarer organischer Nährstoffe für die Mikroflora ermöglichte dann eine Steigerung der Rhizosphärendegradation aufgrund einer erhöhten metabolischen Aktivität. In Verbindung mit dem guten Durchwurzelungsgrad der Pappeln kann außerdem von einer besseren Verteilung der Mykorrhizapilzhyphen im Substrat

Diskussion
98
ausgegangen werden, die mit den Wurzeln der Wirtspflanze assoziiert waren. Ein effizienteres Durchwachsen der Bodenbereiche wird den Abreicherungsprozess somit ebenfalls positiv beeinflusst haben. Im Hinblick auf die unterschiedliche Ausstattung der einzelnen Versuchsvarianten sollen die erzielten Ergebnisse im Folgenden genauer betrachtet werden. 4.2.1 Abreicherung der Nitroaromatenkonzentration im Boden der Kontrollvariante (Variante K) Sowohl im Kiefern- als auch im Pappelexperiment zeigten die Bodenanalysen der Kontrollansätze eine erhebliche Transformation des TNT, obwohl dem Boden zubeimpfte Mykorrhiza- und Weißfäulepilze sowie Pflanzen fehlten. Eine möglicherweise durch mechanische Bearbeitung des Bodens herbeigerufene Initiierung des TNT-Abbaus wurde schon mehrmals beobachtet (Koehler et al. 2001a, Kraß 1998, Thomas et al. 2001). Im Allgemeinen zeigt TNT eine jahrzehntelange Persistenz in ungestörten Standortböden. Diese Eigenschaft kann vermutlich auf eine Selbsteinkapselung der unterschiedlich großen TNT-Partikel und Kristalle zurückgeführt werden (Seifert und Schecker 1993), so dass die Bioverfügbarkeit dieser Verbindung herabgesetzt wird. Bodenstörungen in der Startphase der Experimente könnten somit als Anschub für Abbauvorgänge in Frage kommen. Eine mechanische Bearbeitung des Bodens (Substrat wurde gesiebt) zerkleinerte das in partikulärer Form vorliegende TNT, wodurch zwischenzeitlich die Einkapselung aufgehoben wurde und mehr TNT in Lösung gehen konnte. Einhergehend mit der Bearbeitung des Bodens könnte eine damit verbundene Belüftung eine pulsartige Aktivierung der Bodenmikroflora bewirkt und die TNT-Transformation initiiert haben (Koehler et al. 2001b). Mit der Zeit kann allerdings der Wiedereinkapselungsprozess an den Kristallen einsetzen (Seifert und Schecker 1993). In diesem Zusammenhang lässt sich die nach einigen Wochen beobachtete Stagnation des TNT-Abbaus, welche insbesondere im Kontrollansatz des Pappelexperimentes beobachtet wurde, erklären. Die lange anhaltende Persistenz von TNT und Metaboliten in Standortböden ist außerdem darauf zurückzuführen, dass die Schadstoffe mit der Zeit in für Bakterien nicht erreichbare Mikroporen des Bodengefüges diffundieren. In gealterter Erde sind sie damit für die Bakterien kaum bioverfügbar (Martinetz und Rippen 1996). Auch in diesem Fall wäre eine Initiierung der Biotransformation durch Freisetzung der Kontaminanten nach Störung des Bodengefüges während der mechanischen Einwirkung vorstellbar. Als abiotische Mechanismen bei der Transformation von TNT können möglicherweise auch Photolyseprozesse mitgewirkt haben, wobei die Bedeutung dieses Abbauweges in den abgedunkelten Rhizotronen allerdings eher gering gewesen sein dürfte. Durch Lichtenergie erfolgt eine Umwandlung des TNTs in verschiedene aromatische Verbindungen. Diese photoinstabilen Zwischenprodukte sind zum Teil in der Lage, Dimere zu bilden (Layton et al.

Diskussion
99
1987, in Martinetz und Rippen 1996). Des Weiteren können sie einer mikrobiellen Transformation unterliegen. Nach Zepp et al. (1985) können Photolyseprozesse durch die Anwesenheit von Huminstoffen erheblich beschleunigt werden, da sie als Photosensibilatoren und Bindungspartner für Metabolite wirken. 4.2.2 Abreicherung der Nitroaromatenkonzentration im Boden der Weißfäulepilz-variante (Variante C) Weißfäulepilze sind im Boden normalerweise nicht konkurrenzfähig, weil die dort vorherrschenden Bedingungen für sie ungünstig sind (Hüttermann et al. 1988). Als Besiedler der Holzanteile von Pflanzen und der Streuschicht sind sie im Boden dem Wettkampf mit den dort lebenden Mikroorganismen nicht gewachsen. Ihre Möglichkeiten, den Bodenbereich in den Rhizotronen mit ihren Hyphen von den Zuschlagstoffen ausgehend intensiv zu besiedeln, blieben deshalb begrenzt. Ein dichtes Pilzmyzel des zugeimpften Weißfäulepilzes ließ sich in der Regel nur in Bodenbereichen mit direktem Kontakt zu den überwachsenen Strohanteilen beobachten. Lediglich Rhizomorphen durchzogen vereinzelt das Substrat. Die Möglichkeit zur Ausbildung eines feinen, engmaschigen und weit verzweigten Hyphengeflechtes, um damit möglichst effizient und ganzräumig die Bodenbereiche durchwachsen zu können, blieb daher den Mykorrhizapilzen in den anderen Varianten vorbehalten. Zwischen der Besiedlungsdichte transformierender Pilze im Boden einerseits und der Transformationsrate andererseits besteht jedoch ein enger Zusammenhang. So konnte für den Weißfäulepilz Phanerochaete chrysosporium eine direkte Korrelation der TNT-Reduktion mit der Myzelmasse nachgewiesen werden (Stahl und Aust 1993a und b). Demzufolge kann davon ausgegangen werden, dass eine eingeschränkte Besiedlung des Bodensubstrates mit dem zubeimpften Weißfäulepilz und das Fehlen von Mykorrhizapilzen in Versuchsvariante C aufgrund einer reduzierten Pilzbiomasse und Pilzaktivität im Boden insgesamt zu einer Verlangsamung des Degradationsprozesses im Variantenvergleich geführt hat. Diese Entwicklung im Abreicherungsprozess zeichnete sich insbesondere im Pappelexperiment ab. In der lediglich mit dem Weißfäulepilz beimpften Variante C blieben die Abbauleistungen beim TNT und bei den ADNTs signifikant niedriger als in den Varianten A und B, welche zusätzlich mit starkwüchsigen, mykorrhizierten Pappeln bestückt waren und in Verbindung mit einer intensiven Durchwurzelung deutliche Abreichungseffekte (auch durch eine damit geförderte Mikroflora) sichtbar werden ließen. Obwohl der verwendete Weißfäulepilz Pleurotus ostreatus in Folge seiner eingeschränkten Etablierungsmöglichkeiten im Boden in seiner Leistungsfähigkeit dem Abbaupotenzial mykorrhizierter Pappeln nachstand, konnte immerhin noch ein verbesserter Abreicherungsverlauf gegenüber den weder mit Pilzen beimpften noch mit Pflanzen bestückten Kontrollvarianten (Variante K) beobachtet werden. Beim Abbau nitroaromatischer Verbindungen durch Weißfäulepilze sind sehr unspezifische, auf viele Aromaten reduktiv und

Diskussion
100
oxidativ einwirkende Enzyme wirksam. Bereits Anfang der 80er Jahre wurden hauptsächlich mit dem Weißfäulepilz Phanerochaete chrysosporium für zahlreiche Xenobiotika gute Abbauergebnisse in Flüssigkultur erzielt (Bumpus et al. 1985). Majcherczyk et al. (1993) gelang der Abbau verschiedener Xenobiotika, u. a. TNT, durch Weißfäulepilze später auch unter Bodenbedingungen. Neben niederkondensierten Aromaten können verschiedene Weißfäulepilze außerdem hochkondensierte Verbindungen wie das Benz[a]pyren angreifen (Hüttermann et al 1988). Allerdings erfordert eine erfolgreiche Biodegradation solcher schwerstabbaubarer Bodenkontaminanten die unmittelbare Nähe des belasteten Bodens zum bewachsenen Substrat damit die Schadstoffe für den Metabolismus des Weißfäulepilzes erschlossen werden können. Durch das Einbringen eines aktiven Pilz-/Stroh-Gemisches in den Boden konnte diese Voraussetzung nur unzureichend erfüllt werden, wie die eigenen Experimente zeigten, da der Pilz mit seinen Hyphen nur wenig in den Boden vordrang. Gerade in Hinblick auf die Verwendung von Pilzen in in-situ-Sanierungsverfahren ist deshalb die Fähigkeit zur Ausbildung sehr feiner und weiträumiger Hyphengeflechte im Boden von vorrangiger Bedeutung, um eine umfassende Biodegradation, sei es durch den Pilz selbst oder durch eine Förderung der autochthonen Bakterienflora wie auch der Pflanzen zu erreichen. Diesbezüglich bieten sich bestimmte Mykorrhizapilz-Pflanzen-Assoziationen zur Dekonta-mination TNT-belasteter Böden an, da sie in der Lage sind den Bodenbereich äußerst effizient zu durchwachsen, so dass eine Nitroaromatentransformation nicht lokal begrenzt bleibt. 4.2.3 Abreicherung der Nitroaromatenkonzentration im Boden der Versuchsvarianten mit mykorrhizierten Pflanzen (Varianten A und B) Im Mittelpunkt der experimentellen Fragestellungen stand die Überlegung, ob Weißfäulepilze in ihrer Funktion als TNT-Abbauer durch den Einsatz mykorrhizierter Gehölze sinnvoll ergänzt oder möglicherweise sogar ersetzt werden können. Es zeigte sich, dass unter Verwendung mykorrhizierter Gehölze (Variante B) vergleichbar gute Abbauleistungen erzielt wurden wie unter Verwendung einer Kombination aus mykorrhizierten Gehölzen und Weißfäulepilzen (Variante A). Die eingebrachten Weißfäulepilzkulturen bewirkten keine signifikante Verbesserung der Abbauleistung, wenn die Varianten mit mykorrhizierten Gehölzen bestückt waren. Die Voraussetzung für einen durchgreifenden Einfluss der Mykorrhiza auf den Abreicherungseffekt war eine intensive Durchwurzelung des Substrates, da eine höhere Wurzeldichte ein höheres externes Myzelium in der wurzelfreien Zone zur Folge hat. Ausgehend von den ummantelten Feinwurzeln konnten die Hyphen mit ihrem Wuchs das Substrat feinmaschig und weiträumig erschließen und somit die Mikroflora des Bodens nachhaltig stimulieren bzw. den mikrobiellen Abbau fördern. Gleichzeitig beinhaltete eine dichte Besiedlung des Substrates mit dem Mykorrhizapilz eine bessere Verteilung transformierender Enzyme, die von den Pilzhyphen selbst ausgeschieden werden. Auch die Pflanzen dürften einen erheblichen Einfluss auf die Abnahme der Nitroaromaten gehabt

Diskussion
101
haben. Zum einen sind Pflanzen in der Lage die metabolische Aktivität der Rhizosphärenorganismen durch Exsudation organischer Substanzen anzukurbeln, was eine gesteigerte Schadstoffdegradation nach sich zieht. Zum anderen können auch von den Wurzeln Enzyme abgegeben werden, so dass die Transformation beschleunigt wird. Über die Bedeutung der Aufnahme und Translokation von TNT und Abbauprodukten in die Pflanzen beim Abreicherungsprozess im Boden wird noch an späterer Stelle diskutiert. 4.2.3.1 Verwendete Organismen und ihre Fähigkeit zur Nitroaromatentransformation Es wurde bereits erwähnt, dass neben den holzzersetzenden Weißfäulepilzen auch zahlreiche Arten streuabbauender Mykorrhizapilze aufgrund ihrer enzymatischen Ausstattung die Fähigkeit zur Biotransformation von Nitroaromaten haben (Scheibner et al. 1997, Meharg et al. 1997a, Gramss et al. 1998). In der vorliegenden Arbeit wurde im Kiefernexperiment der Mykorrhizapilz Pisolithus tinctorius als Symbiont für die Kiefernsämlinge verwendet. Pisolithus tinctorius ist als Mykobiont für Kiefern hervorragend geeignet und bildet gemeinsam mit der Wirtspflanze in kurzer Zeit etablierte Mykorrhizen aus. Ein qualitativer Nachweis oxidativer Enzyme mittels Platten-Enzymtests gelang für diese Pilzspezies allerdings nicht. Die durchgeführten Nachweisreaktionen blieben negativ (siehe Tabelle 7). Die im Pappelexperiment verwendeten Pflanzen zeichneten sich an ihren Wurzeln durch Mehrfachinfektionen mit mindestens zwei unterschiedlichen Hebeloma-Arten aus (siehe Abschnitt 2.1.2.4). Die Isolierung steriler Kulturen einzelner dieser Pilze auf Agar-Nährmedien von den mykorrhizierten Pappelwurzelfragmenten gelang trotz mehrerer Versuche nicht. Sämtliche produzierten Pilzmischisolate auf den Agarplatten waren außerdem mit Kolonien verschiedener Bakterien kontaminiert. Isolierungsversuche aus Pilzfruchtkörpern konnten nicht vorgenommen werden, da solche nicht vorlagen. Aufgrund nicht bestehender Zuordnungsmöglichkeiten eventuell auftretender positiver Nachweis-reaktionen für den einzelnen Organismus wurde auf Plattenenzymtests mit den Mykobionten der Pappeln verzichtet. Gramss et al. (1998) gelang der Nachweis für den Besitz verschiedener oxidativer Enzyme für Hebeloma crustuliniforme (Bull.) Quél. und weiterer Spezies dieser Gattung in Reinkultur. Auch für eine Reihe anderer Ektomykorrhizapilzgattungen gelang dieser Nachweis. Zahlreiche Pilze, u. a. Stropharia und Paxillus ließen Aktivität von Phenoloxidasen, Laccasen, Tyrosinasen und Peroxidasen beobachten (Scheibner et al. 1997, Gramss et al. 1998). Die Fähigkeit aromatische Verbindungen mit Hilfe oxidativer Enzyme anzugreifen, scheint unter den Mykorrhizapilzen also eine weit verbreitete Eigenschaft zu sein. Diesbezüglich ist die Annahme berechtigt, dass auch die hier zum Einsatz gekommenen Pilzstämme der Gattung Hebeloma durchaus dazu in der Lage waren, aromatische Verbindungen wie das TNT und dessen Metaboliten oxidativ anzugreifen. Oxidative Enzyme gelangen nicht nur durch Weißfäule- und Mykorrhizapilze in

Diskussion
102
den Boden. Auch Pflanzen produzieren zahlreiche Peroxidasen, welche über die Wurzeloberfläche in den Rhizos-phärenboden freigesetzt werden (Adler et al. 1994). Bevor über oxidative Angriffe durch extrazelluläre Enzyme letztendlich eine Ringspaltung erfolgen kann und damit der weitere Mineralisierungsprozess eröffnet wird, muss das TNT zuvor in einer Initialreaktion enzymatisch reduziert werden. Reduktaseaktivitäten sind ubiquitär bei Organismen (Bakterien, Pilze, Pflanzen) vorhanden. Viele Untersuchungen belegen inzwischen die enzymkatalysierte Reduktion von Nitroaromaten durch zahlreiche Prokaryonten und Eukaryonten. So konnte inzwischen auch für verschiedene Weißfäule- und Ektomykorrhizapilze, u. a. für Pisolithus tinctorius, eine entsprechende Fähigkeit nachgewiesen werden (Stahl und Aust 1993b, Meharg et al. 1997a und 2000). Von pflanzlichen Reduktasen, welche in Verbindung mit Laccasen zum Aufbrechen des Aromatenringes führen sollen, berichteten Schnoor et al. (1995). Die partielle Nitrogruppenreduktion des TNT mittels Reduktasen läuft unspezifisch ab und stellt nicht die eigentliche Funktion dieser Enzyme dar, da natürliche aromatische Nitroverbindungen nur selten vorkommen. Zu erwähnen sind hier vor allem die Antibiotika Chloramphenicol und Pyrrolnytrin, sowie die in Pflanzen synthetisierte und dem Fraßschutz dienende Aristolochiasäure (Pailer 1960, Rastrick 1949). In Anbetracht der Tatsache, dass die bei der TNT-Transformation wirksamen Enzyme nicht durch TNT oder dessen Metaboliten induziert werden und ein weites Substratspektrum aufweisen, ist von einer allgemein weit verbreiteten Fähigkeit zur reduktiven Nitroaromatentransformation unter den Organismen auszugehen. Daher ist zu vermuten, dass auch die hier zur Anwendung gekommenen Pilz- und Pflanzenspezies entsprechende reduktive Fähigkeiten besaßen. 4.2.3.2 Mykorrhizapilze und ihr Einfluss auf die mikrobielle Lebensgemeinschaft im Boden Einerseits können Mykorrhizapilze einen direkten Einfluss auf die autochthone Lebens-gemeinschaft der Mikroorganismen ausüben und so ihre metabolische Aktivität im Boden verändern. Andererseits ist eine Modifizierung der mikrobiellen Abbauvorgänge auch indirekt über die Einflussnahme auf Pflanzen möglich, so dass letztendlich die Pflanzen als Reaktion auf den Mykobionten die Bedingungen in der Rhizosphäre ändern und auf diesem Wege die mikrobiologische Aktivität beeinflussen. Beispielsweise wird durch die Mykorrhizierung der Anteil an Photoassimilaten und Kohlenhydraten in den Wurzeln und damit schließlich auch im Rhizosphärenbereich deutlich erhöht. Dies belegen verschiedene Studien. So fanden sich am selben Individuum von Eucalyptus pilularis/Pisolithus tinctorius Mykorrhizen im Vergleich zu nicht-mykorrhizierten Wurzeln etwa 18 Mal mehr 14C-markierte Photoassimilate (Cairney et al. 1989 in Smith und Read 1997). Da sich der TNT-Abbau im Boden prinzipiell als schwierig darstellt, gilt es als unwahrscheinlich, dass dieser lediglich durch einige wenige Bakterienspezies erfolgt. Vielmehr kann von der Beteiligung ganzer Bakterienkonsortien an

Diskussion
103
der kometabolischen Nitroaromatendegradation ausgegangen werden (Breitung et al. 1995). Gerade solche Bakterienkonsortien können durch die Besiedlung des Bodens mit Mykorrhizapilzhyphen sowohl qualitativ als auch quantitativ entscheidend verändert werden. In letzter Konsequenz erfährt aus diesem Grunde auch die mikrobielle Aktivität eine Veränderung. Mit einem Größendurchmesser zwischen 2-15 µm ist es Hyphen von Mykorrhizapilzen möglich auch in sehr kleine Bodenporen einzuwachsen, welche für die im Durchmesser größeren Wurzeln und Wurzelhaare unerreichbar bleiben (O’Keefe und Sylvia 1992). Prinzipiell ist daher auch in diesen Kleinsträumen eine Einflussnahme auf die mikrobielle Aktivität durch die vermehrte Freisetzung von Stoffen aus den Hyphen in den Boden möglich, die dann dort von den Bakterien als Wachstumssubstrat genutzt werden können (Joner et al. 2000 und 2001). So geben lebende Pilzhyphen verwertbare Proteine wie Glomalin und extrazelluläre Enzyme in die Umgebung ab. Des Weiteren können Pilzhyphen größere Mengen an P und N in ihren in der Regel kurzlebigen Zellen akkumulieren (Johansen et al. 1993, Zhao et al. 1997), welche nach dem Absterben verfügbar werden. Dies gilt auch für die Biomasse der abgestorbenen Pilzmyzelien selbst. Üben Mykorrhizapilze auf die mikrobielle Stoffwechselaktivität im rhizosphärenfernen Bereich einen direkten Einfluss aus, erfolgt die Einflussnahme auf die Aktivität der Mikroflora im Rhizosphärenbereich auch indirekt über die Wirtspflanzen. Hierbei äußert sich der Mykorrhizaeffekt in einer qualitativ als auch quantitativ veränderten Exsudation der mykorrhizierten Wurzeln. So ließ sich bei Wurzeln mykorrhizierter krautiger Pflanzen eine deutlich höhere Exsudation von Zuckern, Aminosäuren und noch anderen organischen Säuren nachweisen als bei Wurzeln nicht-mykorrhizierter Pflanzen (Schwab et al. 1983). Ähnliches wäre auch für mykorrhizierte Gehölze vorstellbar. Eine erheblich modifizierte Wurzelexsudation kann entscheidend zu einer Veränderung der mikrobiellen Gemeinschaft in ihrer Zusammensetzung und somit auch in ihrer Aktivität im Rhizosphärenbereich führen. In diesem Zusammenhang verweisen Joner et al. (2001) auf positive Effekte während der mikrobiellen PAK-Degradation. Es ist daher zu vermuten, dass auch bei der Nitroaromatendegradation in den Experimentalvarianten möglicherweise positive Einflüsse zum Tragen kamen, die auf einer modifizierten Wurzelexsudation basieren. Vermutlich verursachen Infektionen mit Mykorrhizapilzen nicht nur eine Stimulierung der mikrobiellen Nitroaromatentransformation, sei es direkt oder indirekt über die Pflanzen. Vielmehr können Pflanzen nach erfolgreicher Mykorrhizierung auch selbst ihre Abbauleistung effizient verbessern, indem sie nach der Infektion mit dem Pilzpartner zu einer vermehrten Expression extrazellulärer reduktiv und oxidativ wirksamer Enzyme angeregt werden. Criquet et al. (2000) beobachteten eine Erhöhung der Peroxidaseaktivität bei mykorrhizierten Luzernen, McArthur und Knowles (1992) fanden eine erhöhte extrazelluläre Oxidoreduktaseaktivität bei mykorrhizierten Kartoffelwurzeln. Die Infektion mit einem Mykorrhizapilz induziert in der Wirtspflanze allgemein verschiedene Abwehrreaktionen (Gianinazzi-Pearson et al. 1996). In diesem Zusammenhang könnte auch das verstärkte

Diskussion
104
Einsetzen der Peroxidaseaktivität in Pflanzen nach erfolgreicher Mykorrhizierung eine Abwehrreaktion auf die Pilzinfektion darstellen (Salzer et al. 1999), was letztendlich als positiven Nebeneffekt eine verbesserte enzymatische Nitroaromatendegradation im Rhizosphärenbereich zur Folge hat. 4.2.4 Nitroaromatenabreicherung in Versuchsvariante D mit Weißfäulepilzen und mit nicht-mykorrhizierten Pflanzen Das Versuchsdesign für das Kiefernexperiment beinhaltete vier Versuchsvarianten, im anschließend durchgeführten Pappelexperiment wurde die Anzahl der Experimentalvarianten um Variante D ergänzt und somit auf insgesamt fünf erhöht. Bei Experimentalvariante D handelte es sich um eine Kombination von nicht-mykorrhizierten Pflanzen mit dem zubeimpften Weißfäulepilz Pleurotus ostreatus. Vordergründig sollte unter Hinzunahme dieser Variante näher untersucht werden, welchen Beitrag Pflanzen selbst am Abreicherungsvorgang zu leisten im Stande sind, ohne Einfluss eines Pilzsymbionten. Es zeigte sich, dass bei der Abreicherung der TNT-Konzentration mit nicht-mykorrhizierten Pappeln vergleichbar gute Ergebnisse erzielt wurden wie unter Verwendung mykorrhizierter Pappeln (siehe Abbildung 12). So war nach Ablauf des Versuchs kein signifikanter Unterschied des TNT-Gehaltes in den Bodenproben der Varianten A, B und D festzustellen. Signifikant höher blieb die TNT-Konzentration in Variante C, welche unbepflanzt war und lediglich mit dem Weißfäulepilz Pleurotus ostreatus beimpft wurde. Grundsätzlich anders war die Situation bei den ADNTs. Die 2-ADNT- und 4-ADNT-Konzentrationen blieben in Variante D auf einem signifikant höheren Niveau als in den zwei mit mykorrhizierten Pappeln bestückten Varianten A und B (Abbildung 13). Im Vergleich zu Variante C (nur Weißfäulepilz) ließ die ADNT-Konzentration in Variante D keinen signifikanten Abreicherungseffekt erkennen. Die Ergebnisse in den einzelnen Varianten zeigten, dass nicht-mykorrhizierte Pflanzen bei der initialen TNT-Reduktion zum ADNT im Boden einen effektiven Beitrag leisten konnten. Im fortschreitenden Transformationsverlauf konnte dieser Trend bei der Eliminierung der ADNTs dagegen nicht mehr beobachtet werden. Hier ließen die Mykorrhizapilze ein größeres Einflusspotenzial auf die Abreicherungsprozesse erkennen. Es stellt sich die Frage, in welcher Weise Pflanzen die Abreicherungsvorgänge im Rhizosphärenboden zu beeinflussen vermögen. Als wesentlicher Faktor ist hier die Abgabe von Exsudaten aus den Wurzeln in ihre unmittelbare Umgebung zu nennen. In Folge der Wurzelexsudation und auch der Wurzeldeposition wird der organische C-Gehalt des Bodens in Wurzelnähe erheblich erhöht (Shimp et al. 1993). So kann über 40% des photosynthetisch fixierten C durch Exsudation von Zucker, Aminosäuren u. a. wieder in die Rhizosphäre abgegeben werden. Für Zea mays wurde ein Anteil täglich produzierter wasserlöslicher Exsudate von 0,4-0,6 mg C je Gramm Wurzeltrockenmasse ermittelt (Yoshitomi und Shann 2001). Cheng et al. (1996) ermittelten einen täglichen Kohlenstoffeintrag von über 1500 µg je

Diskussion
105
Gramm Boden (TS) in die Rhizosphäre. Entsprechend dem verbesserten Angebot freiverfügbarer Nährstoffe zieht eine hohe Mikrobiomasse mit dazugehöriger erhöhter metabolischer Aktivität eine deutlich verstärkte Biotransformation organischer Schadstoffe nach sich, wovon somit auch TNT und Metaboliten betroffen wären. Da TNT-Metabolite die Neigung haben, an die organische Matrix des Bodens als “bound residues” zu binden, werden durch den verstärkten Eintrag organischer Substanz aus den Wurzeln außerdem Adsorptions- und Bindemöglichkeiten verbessert, was wiederum Einfluss auf die Konzentration freier Nitroaromaten im Boden hat. Aus den Pflanzenwurzeln können zudem zahlreiche reduktive und oxidative Enzyme in den Boden gelangen (Adler et al. 1994). Hierbei haben sie eine gewichtige Funktion beim Streuabbau und bei der Humusbildung. Solche in die Rhizosphäre freigesetzten und in der Regel substratunspezifischen Enzyme könnten somit eine weitere Ursache für die TNT-Abreicherung in den Rhizotronen der D-Variante sein. Dass diese Möglichkeit mit hoher Wahrscheinlichkeit bestand hat, belegen inzwischen vereinzelte Hinweise aus der Literatur. So erwähnten Schnoor et al. (1995) eine nicht genauer beschriebene pflanzliche Nitroreduktase, die im Zusammenspiel mit einer Laccase letztendlich sogar zum Aufbrechen des Aromatenringes führen kann. Sens (1998) gelang der Nachweis von ADNTs in Pflanzenwurzeln einer sterilen mit 14C-TNT dotierten Hydrokultur. Ein weiterer Beleg dafür, dass Pflanzen die Fähigkeit besitzen, TNT zu reduzieren. Eine weitere mögliche Ursache für die Abreicherung des extrahierbaren TNT im Boden könnte auch die Adsorption von Nitroaromaten an die Wurzeloberfläche bzw. ihre Aufnahme in die Pappelwurzeln sein. Diesbezüglich bleibt allerdings festzustellen, dass solche Adsorptions- und Aufnahmeprozesse im 14C-Fütterungsversuch mit Kiefern insgesamt nur eine untergeordnete Rolle bei der Abreicherung spielten, wie im Rahmen dieser Arbeit durchgeführte Untersuchungen zeigten (siehe Abschnitt 3.4). Von einem Transfer in die Pflanze oder einer Bindung an die Wurzeloberfläche waren hier nur geringe Mengen Nitroaromaten betroffen. Jedoch vermögen allgemein schnellwüchsige Pappeln dem Boden möglicherweise in wesentlich deutlicherem Ausmaß bioverfügbares TNT bzw. dessen Metaboliten zu entziehen und in ihren Geweben aufzunehmen. Über die Aufnahme und Festlegung von Nitroaromaten in Pflanzen wird ausführlich im folgenden Kapitel diskutiert.

Diskussion
106
4.3 Nitroaromaten in Pflanzengeweben 4.3.1 Aufnahme von Nitroaromaten aus dem Boden Grundsätzlich können nur in der Bodenlösung gelöste Nitroaromaten von den Pflanzen aufgenommen werden. Liegt das TNT ursprünglich in kristalliner Form im Boden vor, wird es aufgrund seiner persistenten Eigenschaften nur langsam im Bodenwasser gelöst. Die Wasserlöslichkeit von TNT beträgt bei 25° C 140 mg/l (Martinetz & Rippen 1996). Für den Lösungsvorgang mitentscheidend ist dabei auch die Größe der TNT-Partikel. Von kleineren Partikeln geht im Verhältnis mehr TNT in Lösung als von großen Partikeln. Je kleiner der Durchmesser der Teilchen, desto größer ist im Verhältnis zur Partikelgröße die Oberfläche und somit auch die zur Bodenlösung exponierten Fläche (Voudrias & Assaf 1996). Dissoziieren mehr TNT-Moleküle, erhöht sich dementsprechend auch der Anteil dissoziierter Transformationsprodukte. Solche Metaboliten, genannt seien hier insbesondere 2-ADNT und 4-ADNT, können für die Pflanzen dann ebenfalls besser verfügbar sein. Bei den im Rahmen dieser Arbeit durchgeführten Untersuchungen über das Transferverhalten von Nitroaromaten in Pflanzen wurden abgesehen vom TNT außerdem 2-ADNT und 4-ADNT im Schadstoffspektrum der Pflanzenproben nachgewiesen. Weitere Verbindungen wurden nicht gefunden. Auch in der Literatur wurden bisher ähnliche Befunde beschrieben. Unabhängig vom Kultivierungssystem wurden neben dem TNT die beiden ADNTs als dominierende Kontaminanten in Pflanzen identifiziert (Görge et al. 1994 und 1995, Thompson et al. 1998, Scheidemann et al. 1998, Schönmuth und Pestemer 2000, Schneider et al. 1996, Palazzo & Leggett 1986, Hughes et al. 1997, Harvey et al. 1990). DANTs als unmittelbare Reduktionsprodukte der ADNTs ließen sich in Pflanzengeweben bisher nur in zwei Fällen in sehr geringen Mengen nachweisen (Thompson et al. 1998, Sens 1998). In einigen Fällen konnten 2,6-DNT und 2,4-DNT gefunden werden (Görge 1995, Scheidemann 1998, Thompson et al. 1998). Allen Untersuchungen gemeinsam ist die deutliche Dominanz der ADNTs gegenüber dem TNT im Nitroaromatenspektrum der Pflanzen. Eine weitere Gemeinsamkeit liegt in der Tatsache begründet, dass die nachweisbare Nitroaromaten-konzentration stets eine hohe Organspezifität aufwies. Die Wurzeln der untersuchten Pflanzen waren mit deutlichem Abstand anteilig am höchsten belastet. Die Sprosse waren auffällig niedriger, z. T. gar nicht belastet. In den Kiefernnadeln wurde für den Gehalt der 14C-Radioaktivität mit zunehmender Versuchsdauer eine rückläufige Tendenz beobachtet. Möglicherweise könnte dieses Ergebnis als ein Indiz dafür angesehen werden, dass 14C-TNT-Metaboliten in den Nadeln zum Teil veratmet und als 14CO2 in die Atmosphäre freigesetzt wurde. Zu berücksichtigen ist hierbei allerdings das insgesamt sehr niedrige Belastungsniveau in den Blattorganen der Kiefern über den gesamten Versuchsverlauf. In den Nadeln wurde bei hoher Dotierung anteilig unter 0,1% und in den Stämmchen maximal 0,5% der

Diskussion
107
Gesamtaufnahmemenge nachgewiesen. Thompson et al. (1998) konnten nach 14C-TNT-Applikation bei Hybridpappeln keine respiratorische 14CO2-Freisetzung detektieren. In Abschnitt 3.4 sind Verteilung und Wiederfindung der eingesetzten Radioaktivität nach 14-tägiger Fütterung in den jeweiligen Fraktionen aus dem 14C-Dotierungsversuch aufgeführt. Nur der geringste Teil des ursprünglich applizierten 14C konnte mittels Szintillationszählung später in den mykorrhizierten und nicht-mykorrhizierten Wurzelfraktionen der Kiefern detektiert werden. Der weitaus größte Teil des 14C befand sich im getränkten Kohlepapier, im nicht von den Pflanzen aufgenommenen Gießwasser oder als Anhaftungen an den Wänden der Petrischalen. Mikroautoradiographisch konnte zudem gezeigt werden, dass große Anteile der mittels Szintillationszählung in den Wurzeln detektierten 14C-TNT und 14C-Metaboliten nicht tatsächlich von den Zellen aufgenommen waren, sondern den Wurzeln äußerlich nur anhafteten. Demzufolge lag der Anteil von den Wurzeln inkorporierter 14C-Mengen noch wesentlich niedriger, als mittels Szintillationszählung ermittelt wurde. Für die untersuchten Kiefern dürfte die tatsächliche Aufnahmerate nach 14-tägiger Applikation unter 1% gelegen haben, wobei es keine Rolle spielte, ob die Wurzeln mykorrhiziert oder nicht-mykorrhiziert waren. Aufnahmeexperimente mit 14C-TNT liegen auch von anderen Pflanzenspezies vor. Schönmuth und Pestemer (2000) fanden nach 60tägiger Applikation in Topfkulturen mit Altlastenboden in Fichten (Picea abies) immerhin 39% der eingesetzten Radioaktivität wieder. In Sandboden betrug die Wiederfindungsrate für Fichten sogar 77%. Auch Weiden (Salix-Hybride EW-20) erreichten in Sandkultur mit 72% ähnlich hohe Aufnahmeraten. Thompson et al. (1998) erreichten mit Pappelhybriden in Flüssigkultur nach 20tägiger Inkubation 14C-TNT Aufnahmeraten von über 80%, in Bodenkultur war die Aufnahmerate mit 25% dagegen wesentlich niedriger. In Anbetracht dieser Ergebnisse sind mehrere Rückschlüsse erlaubt. Zum einen haben verschiedene Pflanzenspezies vermutlich ein unterschiedliches Aufnahmevermögen für TNT und Metaboliten. Schnellwüchsige Pappel- und Weiden-Hybride mit einem besonders starken Wurzelwachstum vermögen dem Boden wesentlich mehr bioverfügbares TNT zu entziehen und aufzunehmen als weniger schnellwüchsige Arten, wie z. B. Kiefern mit einem mäßigen Wurzelwachstum. Zum anderen ist die Aufnahme in Pflanzen stark von der Bioverfügbarkeit im Boden abhängig. Im Vergleich zu künstlich hergestellten Quarzsandsubstraten oder Flüssigkulturen waren die Aufnahmeraten aus den ton- und humushaltigen Standortböden deutlich niedriger. In ihrer Ursache dürfte die herabgesetzte Pflanzenverfügbarkeit in den verschiedenen Wechselwirkungen zwischen TNT und seinen Metaboliten mit den Bestandteilen der Bodenmatrix begründet liegen. Kritisch angemerkt werden muss allerdings, dass weder bei Schönmuth und Pestemer (2000) noch bei Thompson et al. (1998) die Wurzeln genauer daraufhin untersucht wurden, in welchen Anteilen die gemessene Radioaktivität tatsächlich in den Wurzeln oder nur äußerlich anhaftend lokalisiert war. Demzufolge könnten die tatsächlichen Aufnahmeraten wesentlich niedriger liegen als angenommen wurde. Im Hinblick auf Phytoremediationsverfahren gilt es deshalb in Zukunft noch genauer zu

Diskussion
108
untersuchen, in welcher Größenordnung Pflanzen zur Aufnahme von TNT und dessen Metaboliten aus dem Boden befähigt sind, wobei Wachstumsaktivität und Transpirationsfähigkeit noch stärker berücksichtigt werden müssen. Wichtig ist außerdem die Frage, ob aufgenommene Nitroaromaten in Pflanzengeweben akkumulieren oder zügig metabolisiert bzw. in die Pflanzenmatrix eingebaut werden. Nach Schönmuth und Pestemer (2000) ließ sich etwa 80% der aufgenommenen TNT-Radioaktivität selbst mit Essigsäure nicht extrahieren. Entsprechende Beobachtungen machten Thompson et al. (1998) mit Hybrid-Pappeln. In eigenen Versuchen ließen sich große Teile der analytisch erfassten Nitroaromaten erst nach Säurehydrolyse mit anschließender Alkalisierung aus den Pflanzengeweben der Pappeln extrahieren (siehe Tabelle 17), wobei auch hier von nicht extrahierten Resten auszugehen ist. Vergleichbare Erfahrungen wurden auch von Görge et al. (1994 und 1995), Sens (1998) und Scheidemann (1998) beschrieben. Insgesamt sprechen diese Ergebnisse für umfassende Metabolisierungs- und Festlegungsvorgänge in Pflanzen. Untermauert wird dies durch die Feststellung, dass bei 14C-TNT-Versuchen große Teile des in den Pflanzen gefundenen 14C in Form nicht-identifizierter polarer Bestandteile, bzw. als gebundene Rückstände, den so genannten “bound residues”, vorzufinden waren (Sens 1998, Thompson et al. 1998). Des Weiteren sind die Transformationsvorgänge in den Pflanzen artspezifisch. So waren bei Phaseolus vulgaris und bei Triticum aestivum die Anzahl der polaren Metaboliten und ihr Anteil an der ermittelten Gesamtradioaktivität verschieden (Sens 1998). Artspezifisch sind möglicherweise auch Akkumulation und Festlegung in den unterschiedlichen Organen. Während in den Kiefernnadeln kaum erhöhte Radioaktivität nachgewiesen wurde (siehe Abbildungen 26-29), fanden Thompson et al., (1998) bereits nach wenigen Tagen in den Blättern von Hybrid-Pappeln teilweise über 10% der applizierten Radioaktivität wieder und konnten somit einen Transfer von 14C-TNT/ 14C-TNT-Metaboliten in die Blätter nachweisen. In Fichtennadeln wurden nach 60 Tagen Applikation zwischen 2 und 5% der Radioaktivität wieder gefunden (Schönmuth und Pestemer 2000). In den hier gaschromatographisch untersuchten Pappelproben (Populus tremula) ließen sich teilweise TNT und die beiden ADNTs im oberirdischen Sprossteil finden. Da Blatt- und Stammanteile nicht getrennt sondern als Mischproben analysiert wurden, konnte eine weiterführende organspezifische Aufgliederung nicht durchgeführt und ein gesicherter Nachweis von Nitroaromaten in den Pappelblättern nicht mit letzter Sicherheit beschrieben werden. Bei der Erprobung von Pflanzen bezüglich ihrer Rolle in der Nitroaromatentransformation wurde der Aspekt der Mykorrhiza bisher kaum berücksichtigt. Angaben über das Aufnahme- und Akkumulationsverhalten von Nitroaromaten in den Pilzpartner der Mykorrhiza sind in der Literatur nicht zu finden. Für andere aromatische Xenobiotika ist das Aufnahmeverhalten in Mykorrhizapilze inzwischen besser untersucht. So gelang Dittmann (1999) röntgenmikro-analytisch 3-Chlorbenzoesäure intrazellulär im Hyphenmantel von Suillus bovinus/Pinus sylvestris Mykorrhizen nachzuweisen. Hier kam es insbesondere zu einer Akkumulation in den Vakuolen der Hyphen. Die im Rahmen dieser Arbeit durchgeführten mikroauto-

Diskussion
109
radiographischen Untersuchungen an mykorrhizierten Kiefernwurzeln lassen Aussagen über Aufnahme und Verteilung von 14C-TNT und 14C-TNT-Metaboliten auf zellulärer Ebene im Hyphenmantel nicht zu. Allerdings konnte eine erhöhte Radioaktivität aufgrund akkumulierender Silberkörner im Bereich der Hyphenmäntel insgesamt nicht beobachtet werden. Grundsätzlich hatte die Mykorrhizierung keinen deutlichen Effekt auf den Nitroaromatengehalt in den Wurzeln. So ließen sich weder mittels Szintillationszählung in den mykorrhizierten Kiefernwurzeln noch durch gaschromatographische Untersuchungen in den mykorrhizierten Pappelwurzeln erhöhte Werte gegenüber nicht-mykorrhizierten Wurzeln detektieren, wobei im letzteren Fall bereits festgelegte Bestandteile von der GC-MS-Analytik nicht erfasst wurden und somit von der Bilanzierung ausgenommen waren. Insgesamt betrachtet gilt es, den anteiligen Mykorrhizaeffekt bezüglich der Nitroaromatenaufnahme in die Pflanze noch genauer zu untersuchen, insbesondere unter dem Aspekt der artspezifischen Wurzelwüchsigkeit. Möglicherweise müssen auch seitens des Pilzsymbionten hervorgerufene artspezifische Mykorrhizaeffekte mehr Berücksichtigung finden. 4.3.2 Kiefern als Versuchspflanzen bei der Applikation mit 14C-TNT Die mittels Gaschromatographie detektierten Nitroaromatenkonzentrationen in den Pappelproben (siehe Pappelexperiment 3.3.4) erlauben letztendlich keine genaue Quantifizierung des tatsächlichen Schadstofftransfers in die Pflanzen, da analytisch nur ein Teil der vielen möglichen Transformationsprodukte erfasst bzw. identifiziert werden konnte. Außerdem sind von der GC-MS-Analyse bereits in den Pflanzenkörper fest inkorporierte Rückstände in Teilen ausgenommen. Deshalb wurde im Anschluss an das Pappelexperiment in einem gesonderten Versuch mit Kiefern eine Applikation mit einer definierten Menge 14C-TNT durchgeführt, um Aufnahmeraten und den 14C-Gehalt in den Pflanzenproben abschließend genauer bilanzieren zu können. Da die Ergebnisse bisher durchgeführter Versuche zur TNT-Aufnahme in die Pflanze Hinweise auf ein artspezifisches Transfer-, Akkumulations- und Transformationsverhalten geben, wäre es im Hinblick auf die Verwendung einer einheitlichen Pflanzenlinie in den jeweiligen Versuchen zu wünschen gewesen, auch im 14C-Experiment für eine bessere Vergleichbarkeit auf Pappeln zurückzugreifen. Aus mehreren praktischen Gründen konnte dieses Vorhaben nicht verwirklicht werden. Im 14C-Experiment wurde auf die Inokulation und Infektion der Pflanzen mit nur einem definierten Mykorrhizapilz abgezielt. Deshalb war es unabdingbar, die Versuchspflanzen aus zuvor oberflächensterilisiertem Saatgut anzuziehen, damit Fremdinfektionen mit anderen Pilzen weitgehend ausgeschlossen werden konnten. Da Pappelsamen nur für kurze Zeit nach der Reife keimungsfähig bleiben ist eine Anzucht von Pflanzen aus Samen allgemein sehr schwierig. Auf die Anzucht von Pappeln aus Saatgut wurde daher verzichtet. Zudem ließen sich in eigens durchgeführten Vorversuchen junge Pappelpflänzchen nach gezielter Beimpfung mit unterschiedlichen Mykorrhizapilzen bei

Diskussion
110
niedriger Erfolgsquote kaum erfolgreich infizieren, was möglicherweise allgemein für Arten mit einem starken Wurzelwachstum zutrifft. Letztendlich wurde deshalb bei der Durchführung der Radiotracer-Untersuchung auf die in der Arbeitsgruppe etablierte Pinus sylvestris/ Pisolithus tinctorius Mykorrhiza zurückgegriffen. Beide Organismen dieser Symbiose ließen sich problemlos in den bewährten Rhizotronsystemen halten und garantierten hohe Mykorrhizierungsraten. 4.3.3 Nitroaromatenaufnahme in die Wurzeln Prinzipiell kann die Aufnahme organischer Stoffe aus der Bodenlösung in die Pflanzen passiv durch Diffusion oder Massenfluss erfolgen (Marschner 1995, Trapp 1995). Entsprechendes dürfte auch für Pilze gelten. Allerdings wird die Aufnahme stark von den lipophilen Eigenschaften des Moleküls bestimmt. Als Maß für die Lipophilie eines Stoffes kann der Octanol-Wasser-Verteilungskoeffizient (KOW) herangezogen werden. Dieser Verteilungs-koeffizient definiert das Verteilungsverhältnis einer Substanz zwischen der Octanol- bzw. der Wasserphase, wobei das Octanol als Modell-Lösungsmittel zur Simulierung des Lipidanteils biologischer Membranen dient. Aufgenommen werden lipophile Substanzen mit einem log KOW zwischen 0,5 und 3,0 (Briggs et al. 1982 in Schnoor et al. 1995). In der Regel adsorbiert das Schadstoff-Molekül zuerst an der Wurzeloberfläche, um dann über die Biomembran in die Wurzelzelle passiv einzudringen (Shone & Wood 1972). Sehr lipophile Substanzen mit einem log KOW > 3,0 sind dagegen kaum bioverfügbar, da sie zu stark sorbiert sind. Zum einen erschwert die starke Bindung an die Wurzeloberfläche eine Aufnahme durch die Zellmembran erheblich, zum anderen adsorbieren sie stark an die organische Bodenmatrix. Letzteres ist nicht verwunderlich, da die organische Bodensubstanz (Humus) im Wesentlichen aus pflanzlichen Materialien entsteht und die Sorptionseigenschaften von Boden und Wurzeln für sehr lipophile organische Stoffe deshalb als ähnlich anzusehen sind. Sehr polare, hydrophile Stoffe mit einem log KOW < 0,5 sorbieren nicht genügend an Wurzeln. Außerdem wird die Aufnahme durch den Widerstand der Zellmembran gehemmt (Briggs et al. 1982 in Schnoor et al. 1995, Trapp 2000). Der Octanol-Wasser-Verteilungskoeffizient beträgt für TNT log KOW 1,65 (Palazzo & Legget 1983), für 2-ADNT log KOW 1,59 und für 4-ADNT log KOW 1,31 (Görge 1994 in Martinetz & Rippen 1996). Aufgrund ihrer lipophilen Eigenschaften und mit den entsprechenden Koeffizienten ist es diesen Nitroaromaten also möglich, Biomembranen zu durchdringen. In Anbetracht des nachgewiesenen Schadstoffspektrums der Wurzeln mit einer auffälligen Dominanz von 2-ADNT und 4-ADNT gegenüber dem TNT wäre einerseits eine Aufnahme des transformierten TNT nach mikrobieller Reduktion im wurzelnahen Raum als ADNT durchaus vorstellbar, zumal sich auch in anderen Untersuchungen die ADNTs stets als dominante Schadstoffgruppe in den Wurzeln nachweisen ließen (u. a. Görge et al. 1995, mit verschiedenen krautigen Pflanzen; Schönmuth und Pestemer 2000, mit Salix spec. und Picea abies; Thompson et al. 1998, mit

Diskussion
111
Populus spec.). Andererseits besteht auch die Möglichkeit einer raschen Transformation des aufgenommenen TNT zu ADNT innerhalb der Pflanzen, wonach die Dominanz der ADNTs letztendlich auf eine Metabolisierungsleistung der Pflanzen zurückzuführen wäre. Dass das TNT in die Wurzeln transferiert wird, zeigten u. a. auch die Schadstoffspektren dieser Arbeit und zahlreiche weitere in der Literatur beschriebene Untersuchungen, und dass TNT von Pflanzen reduziert werden kann, belegen Experimente mit Triticum aestivum unter sterilen Bedingungen (Sens et al. 1999). So ließen sich ADNTs in den Wurzeln aus Pflanzen nachweisen, denen unter sterilen Bedingungen in Hydrokultur 14C-TNT appliziert wurde. Bezogen auf die hier durchgeführten Versuche kann letztendlich nicht beantwortet werden, in welchem Umfang das TNT direkt oder nach Transformation im Boden als ADNT in die Pflanzen gelangt ist. Zu berücksichtigen bleibt die Tatsache, dass vermutlich beide Möglichkeiten bestehen. Berücksichtigt werden muss außerdem die Tatsache, dass neben TNT und den ADNTs vermutlich noch weitere im Boden gebildete unpolare und lipophile TNT-Metaboliten mit einem logKOW zwischen 0,5 und 3,0 aufgrund ihrer Membrangängigkeit ebenfalls diffusiv ihren Weg aus der Bodenlösung in die Wurzeln fanden. Für geladene Metaboliten mit einem zu niedrigen Octanol–Wasser–Verteilungskoeffizienten ist eine Diffusion durch Membranen dagegen kaum möglich. 4.3.4 Apoplastischer Transport Wie bereits in der Einleitung beschrieben wurde, bildet der Pilzsymbiont einer Mykorrhiza ein mehr oder weniger dichtes Hyphengeflecht, den so genannten Hyphenmantel, um die ektotroph mykorrhizierten Feinwurzeln der Wirtspflanze aus. Ein direkter Kontakt der Wurzelzellen mit dem umgebenden Bodenmilieu wird durch diesen Hyphenmantel unterbunden. Verursacht durch eine interzelluläre Matrix diskutieren verschiedene Autoren außerdem eine apoplastische Blockade und somit eine Impermeabilität des Hyphenmantels aufgrund einer interzellularen Matrix (Ashford et al. 1988 und 1989, Bücking 1995, Smith und Read 1997). Demzufolge wäre die mykorrhizierte Pflanze in ihrer Nährstoffaufnahme vollständig vom pilzlichen Symbiosepartner abhängig. Dass diese Annahme nicht uneingeschränkt gilt, zeigten Versuche mit verschiedenen Pinus sylvestris-Mykorrhizen, bei denen eine apoplastische Permeabilität des Hyphenmantels für mehrere Tracer nachgewiesen werden konnte (Behrmann 1995, Behrmann und Heyser 1992). Es ist deshalb davon auszugehen, dass die Wasseraufnahme und damit verbunden auch die Nährstoffaufnahme in die Wurzel nicht gänzlich vom Pilzsymbionten kontrolliert wird. Eine apoplastische Permeabilität des Hyphenmantels konnte auch an Picea abies/Hebeloma crustuliniforme Ektomykorrhizen nachgewiesen werden (Scheidegger und Brunner 1995). Insofern dürfte deshalb grundsätzlich die Möglichkeit eines apoplastischen Transportes von TNT/TNT-Metaboliten durch den Hyphenmantel für die in dieser Arbeit untersuchten Ektomykorrhizen nicht auszuschließen sein. Behrmann (1995) konnte eine unterschiedliche apoplastische

Diskussion
112
Permeabilität des Hyphenmantels entlang der Wurzelachse feststellen. Im jüngeren Wurzelbereich zeigte sich der Hyphenmantel durchlässiger als im basalen Abschnitt, so dass die Stoffe hier bevorzugt in das Hartigsche Netz gelangten, was vermutlich auf den kompakteren Aufbau des Hyphenmantels an der Wurzelbasis zurückzuführen war. Im Interface, das als Kontaktzone gemeinsam von den Zellwänden beider Symbionten gebildet wird, treten die Stoffe auch in die Rhizodermis- und Rindenzellwände der Wirtspflanze über. Apoplastisch können Stoffe wie TNT und seine Metabolite nun bis zur Endodermis gelangen. Bei nicht mykorrhizierten Wurzeln beginnt der apoplastische Weg der im Bodenwasser gelösten niedermolekularen Stoffe in den Zellwänden der Rhizodermis. Auch hier erfolgt das Eindringen in den leicht zugänglichen „Freien Raum“ (Apparent Free Space, AFS) passiv und nicht selektiv durch Diffusion. Sind Metabolite wie ADNTs und DANTs im sauren Milieu des Apoplasten positiv geladen, kann eine Bindung an Zellwandbestandteile erfolgen. Für solche Substanzen muss der AFS dann in den „Wasserfreiraum“ (Water Free Space, WSF) und den „Donnan-Freiraum“ (Donnan Free Space, DFS) unterteilt werden. Im DFS wäre insbesondere eine Bindung durch die Carboxylgruppen der Polygalacturonsäure des Protopektins möglich (Marschner 1995), während der WFS für Ladungsträger frei permeierbar bleibt. Die mikroautoradiographischen Untersuchungen zeigten, dass ein ausschließlich apoplastischer Transport von TNT und dessen Metaboliten bis zur Endodermis beim Aufnahmeprozess in die Wurzel vermutlich kaum eine Bedeutung hat. Nur sehr vereinzelt ließ sich Radioaktivität, verursacht durch mobile oder festgelegte TNT/TNT-Metaboliten mikroautoradiographisch im Bereich der Zellwände der Rindenzellen detektieren. Wahrscheinlicher ist eine Aufnahme durch die Plasmamembran ins Zellinnere nach kurzer apoplastischer Diffusion bereits im Bereich der Rhizodermis. Diese Annahme wird durch das gehäufte Auftreten von Radioaktivität im Inneren der Rhizodermiszellen erhärtet. Trotzdem kann die Möglichkeit für eine apoplastische Diffusion bis zur Endodermis prinzipiell nicht ausgeschlossen werden. Nach gängiger Lehrmeinung wird der apoplastische Transport von Substanzen über die primäre Rinde in den Zentralzylinder der Wurzel durch den Casparyschen Streifen in den Radialwänden der Endodermis effektiv unterbunden. Demzufolge müssten die Teilchen spätestens an der Endodermis das Plasmalemma überwinden, um letztendlich symplastisch durch die Plasmodesmen in den Zentralzylinder gelangen zu können. Allerdings ist eine grundsätzliche Sperrwirkung möglicherweise nicht gegeben. Untersuchungen von Behrmann (1995) zeigten eine gewisse Durchlässigkeit Casparyscher Streifen primär ausdifferenzierter Endodermen verschiedener Kiefernmykorrhizen für mehrere Fluoreszenzfarbstoffe und das Schwermetall Lanthan. Außerdem bleibt festzustellen, dass auch in jungen Bereichen der Wurzel nahe der Wurzelspitze, dort, wo in den Radialwänden der Endodermis der Casparysche Streifen noch fehlt oder auch über das Apikalmeristem ein apoplastischer Transport molekularer Teilchen in den Zentralzylinder grundsätzlich möglich ist (Steudtle et al. 1987, Peterson et al. 1986, Behrmann 1995). Jedoch ließ sich mikroautoradiographisch im apikalen und subapikalen

Diskussion
113
Bereich mykorrhizierter als auch nicht-mykorrhizierter Wurzeln kaum nennenswerte Radioaktivität weder in den Zellwänden noch innerhalb der Zellen nachweisen. Ein Aufnahmeweg für TNT/TNT-Metaboliten über die Wurzelspitze in den Zentralzylinder spielt deshalb vermutlich kaum eine Rolle. Dass TNT/TNT-Metaboliten in die Xylem-Leitungsbahnen von Pflanzen transportiert werden, belegen neben den eigenen Untersuchungen auch Experimente mit 14C-TNT von Sens (1998). Dass der Weg dorthin zumindest bei den hier untersuchten Kiefern hauptsächlich symplastisch über das Cytoplasma und durch die Plasmodesmen erfolgt, kann aufgrund der nachgewiesenen Radioaktivität im Zellinnern angenommen werden. 4.3.5 Symplastischer Transport TNT sowie 2-ADNT und 4-ADNT sind aufgrund ihrer lipophilen Eigenschaften gut membrangängig. Selektive Aufnahmeprozesse von TNT und dessen Metaboliten in die Pilz- bzw. Pflanzenzellen sind unwahrscheinlich. Die Aufnahme dieser Substanzen in die Zellen erfolgt passiv durch Diffusion aufgrund einer Gleichgewichtseinstellung zwischen der Bodenlösung und den mykorrhizierten bzw. nicht-mykorrhizierten Wurzeln. In den Pilzhyphen mykorrhizierter Wurzeln ist prinzipiell ein passiver Transport mit der cytoplasmatischen Strömung bis zur Interface denkbar. Die Löslichkeit von TNT in Wasser ist mit etwa 130 mg/l relativ gut. Möglichkeiten zur Wechselwirkung mit Wasser bietet das Molekül über die freien Elektronenpaare der Sauerstoffatome in den Nitrogruppen, die als Wasserstoff-Brücken-Akzeptoren fungieren. TNT besitzt keinen pKa-Wert, da die funktionellen Gruppen nicht protoniert oder deprotoniert werden. Eine Ionisierung durch Protonierung mit sinkendem pH-Wert wie im Fall aromatischer Amine tritt also nicht ein. Das TNT-Molekül behält sowohl beim Eintritt ins neutrale Cytoplasma (pH-Wert 6,8-7,4; nach Smith & Raven 1979), als auch im sauren Milieu des Apoplasten beim Übertritt in die gemeinsame Austauschzone, der pH-Wert des Apoplasten steriler und mykorrhizierter Wurzeln variiert zwischen 3 und 4 (Nenninger 1995 und 1999), seine membrangängigen Eigenschaften und kann ungehindert ins Zellinnere der Wurzelzellen diffundieren. Wiederum mit der cytoplasmatischen Strömung wäre in der Wurzel ein symplastischer Transport über die Plasmodesmen durch die Endodermis in den Zentralzylinder denkbar. Aufgrund seiner lipophilen Eigenschaften ist letztendlich ein Durchtritt des Moleküls durch die Plasmamembran der Xylemparenchymzellen und somit in den Apoplasten der Xylem-Tracheiden möglich. Auch die ADNTs büßen beim Eintritt ins Cytoplasma des Zellinneren nichts von ihrem lipophilen Charakter und ihrer Membrangängigkeit ein und können relativ gut bis zu den Wasserleitelementen des Xylems diffundieren. Bei 2-ADNT und 4-ADNT üben jeweils die beiden Nitrogruppen einen starken Elektronenzug auf das restliche Molekül aus. Das freie Elektronenpaar am Stickstoffatom der Aminogruppe steht deshalb für eine Protonierung kaum zur Verfügung (Kraß 1998). 4-ADNT besitzt einen entsprechend

Diskussion
114
niedrigen pKa-Wert von 1,21 (für 2-ADNT liegt kein pKa-Wert vor), d. h. erst bei einem sehr niedrigen pH-Wert von 1,21 sind 50% der 4-ADNT-Moleküle protoniert. Ist der pH-Wert um drei Einheiten auf 4,21 erhöht, beträgt der Anteil protonierter Moleküle lediglich noch 0,1%. Von wesentlicher Bedeutung für den symplastischen Transport von Zelle zu Zelle ist die Durchlässigkeit der Plasmodesmen für Substanzen. Durch Mikroinjektion fluores-zenzmarkierter Moleküle ließ sich die Durchgängigkeit für Stoffe bis etwa 1 kDa Molekulargewicht nachweisen (Robards und Lucas 1990, Lucas 1997). Hiernach ist nicht nur eine intrazelluläre Diffusion von Zuckern, verschiedenen Signalstoffen und Ionen möglich, sondern auch der Durchtritt von TNT und dessen Metaboliten. Bisher blieb die molekulare Architektur dieser symplastischen Verbindungen wenig untersucht, wobei inzwischen feststeht, dass Plasmodesmen mit unterschiedlicher und bisweilen komplizierter Struktur vorkommen (Blackman und Overall 2001). Mittlerweile belegen neuere Untersuchungen die Möglichkeit zum Durchtritt weitaus größerer Moleküle, u. a. für Proteine mit Größenordnungen über 50 kDa, wobei die Öffnungen hierzu erweitert werden können (Crawford und Zambryski 2000). 4.3.6 Metabolismus der aufgenommenen Nitroaromaten 4.3.6.1 Orte der Metabolisierung Außer TNT, 2- und 4-ADNT konnten in wenigen Fällen geringe Mengen 2,4- und 2,6-DNT bisher aus Pflanzen extrahiert und identifiziert werden (Görge et al. 1994 und 1995, Thompson et al. 1998, Scheidemann et al. 1998, Schönmuth und Pestemer 2000, Schneider et al. 1996 u.a.). Thompson et al. (1998) gelang es, 2,4-DANT in jungen Hybrid-Pappeln nachzuweisen. Des Weiteren ließen sich noch zahlreiche überwiegend polare nicht identifizierte aus dem TNT-Metabolismus hervorgegangene Verbindungen in Pflanzen finden. Aus Phaseolus vulgaris wurden insgesamt 10 unpolare, 17 polare und 3 sehr polare, aus Triticum aestivum 3 unpolare und 13 polare Metaboliten gewonnen und dünnschichtchromatographisch aufgetrennt (Sens 1998). Auch in Geweben von Hybridpappeln wurden mehrere nicht identifizierte polare TNT-Metaboliten gefunden (Thompson et al. 1998). Die Aufnahme von Xenobiotika in Zellen von Pflanzen und Pilzen setzt lipophile Eigenschaften dieser Stoffe voraus. Bei Pflanzen erfolgt die zelluläre Detoxifizierung aufgenommener Schadsubstanzen entweder durch chemische Transformation oder durch Kompartimentierung (Coleman et al. 1997). Für Pilze sind vergleichbare Prozesse anzunehmen. So wird die Vakuole von Pilzen beispielsweise als Kompartiment zur Ablagerung von Schwermetallen verwendet. Die Detoxifizierung wird durch eine Bindung der Schwermetalle über SH-Gruppen der Cystein-Bausteine an Phytochelatine oder Metallothionine erreicht (Richter 1998). Die aus der Transformation hervorgehenden

Diskussion
115
Folgemetaboliten haben in der Regel ein geringeres toxisches Potential für den Organismus, außerdem sind sie häufig polarer und damit hydrophiler als die Ausgangssubstanzen. In Phaseolus und Triticum sowie in Hybrid-Pappeln ließen sich zahlreiche polare Metaboliten nachweisen (Sens 1998 und Thompson et al. 1998). In Tomatenpflanzen konnte für 4-Nitrophenol die Bildung eines 4-Nitrophenyl-ß-D-Glucopyranosid und damit eine Erhöhung der Polarität der entstandenen Verbindung beobachtet werden (Wallnöfer et al. 1996). Mit zunehmender Polarität wird eine weitere Ausbreitung aufgrund einer herabgesetzten Durchlässigkeit für Membranen erschwert und somit eine gewisse “Festlegung“ durch Kompartimentierung erreicht. Diese Möglichkeit der Detoxifikation aufgenommener Nitroaromaten kann auch für die hier untersuchten mykorrhizierten und nicht-mykorrhizierten Kiefern angenommen werden. Insbesondere in den Rindenzellen konnte mikro-autoradiographisch eine Verteilung der Radioaktivität über das gesamte Zelllumen, folglich auch eine Verteilung in den Vakuolen nachgewiesen werden. Allerdings sei an dieser Stelle noch mal darauf hingewiesen, dass die in den Pflanzen detektierten Mengen an TNT/TNT-Metaboliten stets auf niedrigem Niveau blieben. Auch in den gaschromatographisch untersuchten Pappelproben wurden, abgesehen von den ADNTs, relevante Mengen weiterer polarer Verbindungen nicht gefunden. Allgemein wird die Biotransformation von aufgenommenen Xenobiotika in Phase-I- und Phase-II-Reaktionen unterteilt (Coleman et al. 1997, Zhang et al. 1996). Eine entsprechende Einteilung ist auch auf Metabolisierungsvorgänge von Nitroaromaten übertragbar. Die Phase-I-Reaktionen sind überwiegend durch enzymatisch katalysierte Reduktions- und Oxidationsvorgänge an den Nitroaromaten geprägt. Entstehende Metaboliten können dann in Phase-II-Reaktionen über Konjugationen an Gluthathion, Glukose und andere Zucker, Aminosäuren und weiteren Verbindungen verknüpft werden. Phase-I-Mechanismen Zahlreiche Enzyme katalysieren die Reduktion von Nitroaromaten in prokaryotischen als auch in eukaryotischen Zellen. In Frage kommen hierfür insbesondere Cytochrom c-Oxidoreduktasen und Cytochrom P-450-Oxidoreduktasen (Cerniglia und Somerville 1995). In Arabidopsis thaliana konnte die Reduktion von TNT durch eine NADPH-abhängige Thioredoxin-Reduktase nachgewiesen werden (Miskiniene et al. 1998). Insgesamt bleibt festzustellen, dass die eigentliche Funktion dieser Enzyme nicht die Reduktion von Nitroaromaten ist, da natürlich vorkommende Nitroverbindungen in sehr geringer Zahl auftreten. Als wesentlich dominante Kontaminanten treten in Pflanzen in der Regel 2-ADNT und 4-ADNT auf. DANT als reduktives Folgeprodukt des ADNT konnte bisher nur in sehr wenigen Ausnahmefällen in Hybrid-Pappeln bzw. in Phaseolus gefunden werden (Thompson et al. 1998, Sens et al. 1999). Die reduktive Transformation bis zum DANT dürfte demnach in Pflanzen eine untergeordnete Rolle spielen. Eine vollständige Reduktion des TNTs bis zur

Diskussion
116
Stufe des Triaminotoluols kann unterdessen ausgeschlossen werden, da eine vollständige Reduktion aller drei Nitrogruppen nur im strikt anaeroben Milieu erfolgt und entsprechende Bedingungen in Pflanzen nicht vorliegen. Bedeutsamer und verbreiteter im Transformationsverlauf von Nitroaromaten sind möglicherweise durch Mono- und Dioxygenasen katalysierte Metabolisierungsschritte. Oxidative Angriffe als Transformationsreaktionen an Aminonitrotoluolen sind durchaus vorstellbar. Für den weiteren oxidativen Angriff kommen zudem diverse extrazellulär transformierte und anschließend in die Zellen aufgenommene TNT-Metaboliten in Frage. Membrangebundene Monooxygenasen katalysieren den Einbau nur eines O-Atoms in das Kohlenstoffgerüst und kommen zahlreich in Pflanzenzellen vor. Als Beispiel sei hier die Cytochrom P450 Monooxygenase genannt. Mit Cytochrom P450 als O2-Bindungsstelle katalysiert dieses Enzym die Oxidation von Phenylpropanen, welche als Ligninvorstufen in die Ligninsynthese eingehen. Eine unter zahlreichen Bakterien nachgewiesene Monooxygenasereaktion ist die durch oxidativen Angriff hervorgerufene Nitritabspaltung beim 4-Nitrophenol (Kadiyala und Spain 1998), wobei das dabei entstehende Trihydroxyphenol letztendlich einer Ringspaltung unterliegt und weiter metabolisiert wird. Auch über Hydrochinon als Zwischenprodukt konnte eine mikrobielle Metabolisierung des 4-Nitrophenols (Gattung Arthrobacter) beobachtet werden (Hanne et al. 1993). Auch Dioxygenasen sind in Pflanzen zahlreich verbreitet. Sie katalysieren eine Doppelhydroxylierung des Aromaten. Die häufig vorkommende Lipooxygenase katalysiert die Dioxygenierung der mehrfach ungesättigten Fettsäuren Linol- und Linolensäure. Die Hydroperoxylinolensäure wird durch die Hydroperoxid-Ligase gespalten (Heldt 1996). Für verschiedene Nitroaromaten, wie 1,3-Dinitrobenzol, 2,4-Dinitrotoluol, 2,6-Dinitrotoluol u.a., konnte inzwischen die mikrobielle Transformation durch Dioxygenasen nachgewiesen werden (Nishino et al. 2000a und 2000b, Spanggord et al. 1991). Dinitrotoluole wurden in Phaseolus vulgaris (Görge et al. 1994, Scheidemann 1998 und Sens 1998) sowie in Medicago sativa und Phleum spec. (Scheidemann 1998) nachgewiesen. Phase-II-Mechanismen Durch Phase I-Reaktionen entstandene TNT-Metaboliten können vermutlich über Phase-II-Reaktionen weiter metabolisiert und über Konjugationen gebunden werden. Die Konjugate liegen entweder gebunden oder mobil in der Pflanze vor. Eine feste Bindung von TNT und dessen Metabolite ist unter anderem mit der Ligninfraktion sehr wahrscheinlich. Mobile Konjugate können vorerst kompartimentiert werden und erfahren somit eine gewisse zwischenzeitliche Festlegung. Aus dem Ergebnisteil dieser Arbeit geht hervor, dass ein großer Teil der ADNTs erst nach Säurehydrolyse mit anschließender Alkalisierung aus dem pflanzlichen Material extrahiert werden konnte, was auf eine Konjugation mit pflanzlichen Bestandteilen schließen lässt. Dass sowohl 2-ADNT als auch 4-ADNT durch

Diskussion
117
Konjugationsprozesse in Pflanzen detoxifiziert werden, zeigten Untersuchungen von Bhadra et al. (1999) mit sterilen Pflanzenwurzeln von Catharanthus roseus. NMR-spektroskopisch gelang der Nachweis von Konjugationsbindungen der beiden unmittelbaren TNT-Metaboliten an einen unbekannten C6-Körper im intrazellären Milieu. Da die sterilen Pflanzenkulturen ausschließlich mit TNT dotiert wurden, mussten die Pflanzen außerdem über reduzierende Systeme verfügen. Es ist bekannt, dass auch zahlreiche Pestizide, Herbizide, Fungizide und andere Xenobiotika über Konjugationen in der Pflanze detoxifiziert werden. Verschiedene Autoren beschreiben die vakuoläre und die vesikuläre Deposition von Konjugations-intermediaten als einen Zwischenschritt vor ihrer endgültigen Festlegung als so genannte “bound residues”. Verknüpft mit zukünftigen Zellwandpolymeren, wie Glukosiden, werden die vakuolären Konjugate letztendlich ins Cytosol und ins apoplastische Milieu transportiert und dort in der Zellwand festgelegt (Sandermann 1994, Schmidt et al. 1997, Laurent und Scalla 1999). Ein weiteres Beispiel für die Detoxifizierung aufgenommener Xenobiotika durch Konjugatbildung und Kompartimentierung stellt die Synthese von Gluthathion-konjugaten und der Transport dieser Verbindungen in die Vakuolen dar (Edwards et al. 2000, Coleman et al. 1997). Über diese Reaktion werden übrigens auch von der Pflanze selbst gebildete Verbindungen unschädlich gemacht. Die Konjugation wird durch Glutathion-S-Transferasen katalysiert. Dabei wird die sehr reaktive SH-Gruppe des Gluthations über Thioether an Carbonylgruppen und anderen funktionellen Gruppen der aufgenommenen Schadstoffe gebunden. Durch einen im Tonoplasten lokalisierten spezifischen Glutathion-Translokator werden die Glutathionkonjugate unter ATP-Verbrauch gegen einen Konzentrationsgradienten in die Vakuole transportiert und dort deponiert oder weiter metabolisiert. 4.3.6.2 Einbau von 14C-TNT und Metaboliten in die Zellwände Unabhängig von den Zelltypen der Pflanze ist vor allem die Sekundärwandbildung durch eine erhebliche Cellulosesynthese geprägt. In Geweben mit Stütz- und Festigungsfunktion ist ferner die Lignifizierung (Verholzung) der Sekundärwand von wesentlicher Bedeutung. Zusammen mit Suberin und Kutin, die zusätzlich in verschiedene Zellwände eingelagert werden, kommt dem Lignin außerdem eine gewisse Bedeutung bei der Imprägnierung von Zellwänden und bei dem Schutz vor Pflanzenpathogenen zu. Lignin entsteht durch radikalische Polymerisation der drei Monolignole Sinapyl-, Coniferyl- und p-Cumarylalkohol (Phenylpropane mit einem Aromaten als Grundkörper) zu Makromolekülen. Der Mechanismus ist in wesentlichen Teilen noch unklar. Die Synthese erfolgt außerhalb der Zellen (Heldt 1996). Bei der Verkettung der einzelnen Monolignole sind Peroxidasen beteiligt. Auch Suberin ist ein Polymerisat, das neben Fettsäuren, Fettalkoholen und anderen Bestandteilen aus Phenylpropanen zusammengesetzt wird. Bei der Verknüpfung der Phenylpropane beim Wandaufbau ist eine kovalente Einbindung verschiedener TNT-

Diskussion
118
Metabolite in das Gerüst der entstehenden Makromoleküle durchaus zu vermuten. Auch andere Formen der Festlegung oder Einlagerungen im Wandbereich sind denkbar. Auffällige Konzentrationen an 14C-TNT/Metaboliten konnten mikroautoradiographisch an den Rhizodermisaußenwänden nicht-mykorrhizierter Wurzeln sowie im Bereich des Xylems in den Leitbündeln nicht-mykorrhizierter als auch mykorrhizierter Wurzeln nachgewiesen werden. Zahlreiche Seitenwurzeln der Kiefern waren durch Spitzenwachstum gekennzeichnet und befanden sich noch im Stadium der Ausdifferenzierung. Bemerkenswert war die deutlich erhöhte Silberkorndichte über den ring- und schraubenförmigen Versteifungen in den tracheidalen Elementen. Möglicherweise lässt sich diese Häufung als Hinweis auf die Einbindung von TNT-Metaboliten in die Ligninfraktion deuten. Andererseits könnten die Silberkornansammlungen anteilig auch ein Präparationsartefakt des Entwässerungsvorganges darstellen, wobei sich 14C-TNT/14C-TNT-Metaboliten als Transportsubstanz des Xylems bevorzugt in den Aussteifungen der Zellwände sammelten. 4.3.6.3 Einbau von TNT-Metaboliten in die Ligninfraktion Hinweise für einen kovalenten Einbau von Xenobiotika in die Ligninfraktion ergaben Versuche mit 2,4-Dichlorphenoxyessigsäure (Scheel und Sandermann 1981) und mit 14C- Phenoxyessigsäure (Laurent und Scalla 1999) in Zellsuspensionskulturen von Glycine max (Sojabohne) und Triticum aestivum (Weizen). Letztere genannte Autoren konnten außerdem erhebliche Anteile der applizierten 14C-Phenoxyessigsäure fest gebunden in der Hemicellulosefraktion der Zellwände von Glycine max lokalisieren. Versuche mit ringmarkiertem 14C-TNT von Cataldo et al. (1989) zeigten, dass die tatsächlich von den Pflanzen aufgenommene Schadstoffmenge größer war als nach chemischer Extraktion wieder gefunden werden konnte. Bereits hier ergab sich der Hinweis auf eine feste Einbindung von TNT-Metaboliten als „bound residues“ in Pflanzen. Deutliche Hinweise für einen kovalenten Einbau von TNT/TNT-Metaboliten in das Lignin, vergleichbar dem angenommenen festen Einbau verschiedener Xenobiotka, ergaben umfangreiche Untersuchungen mit ringmarkiertem 14C-TNT an Phaseolus vulgaris und Triticum aestivum (Sens 1998). Gelpermeationschromatographische Untersuchungen (GPC) des Lignins zeigten, dass der wesentliche Anteil der 14C-markierten Ligninbestandteile ein Molekulargewicht >1000 g/mol hatten. Dagegen ließen sich nur geringe Mengen an Radioaktivität im Molekulargewichtsbereich von TNT und TNT-Transformationsprodukten finden. Der Autor schließt allerdings nicht aus, dass TNT-Metabolite in “käfigartige Strukturen” eingeschlossen wurden und sich daher in der GPC nicht herauslösen und somit nicht nachweisen ließen. Wie bereits erwähnt wurde, könnte die mikroautoradiographisch nachgewiesene Ansammlung von 14C-Radioaktivität in Xylem des Zentralzylinders ein weiteres Indiz für eine mögliche Einbindung von TNT-Metaboliten in das Lignin sein. Vor allem im Bereich der ring- und schraubenartig verholzten Wandverdickungen der Tracheiden konnten vermehrt radioaktive

Diskussion
119
Einlagerungen durch eine erhöhte Silberkorndichte aufgezeigt werden. Gewöhnlich wird für den Einbau des Lignins in die Zellwand die Beteiligung von Peroxidasen und Laccasen angenommen (Heldt 1996). Voraussetzung für eine Polymerisation ist der Besitz von Hydroxylgruppen am aromatischen Ring der Monolignole. Sens (1998) zieht in Erwägung, dass möglicherweise ebenso hydroxylgruppentragende TNT-Metaboliten für einen Einbau in die Ligninfraktion in Frage kämen. Die Einführung von Hydroxylgruppen in den aromatischen Ring der Nitroaromaten könnte durch Monooxygenasen mit Cytochrom P450 als O2-Bindungsstelle synthetisiert werden, ebenso, wie solche Monooxygenasen die Einführung von Hydroxylgruppen in den Phenylring von Ligninvorstufen synthetisieren. In der Regel sind alle P450-Monooxygenasen an die Membran des Endoplasmatischen Reticulums gebunden (Heldt 1996). Der Transport hydroxylierter TNT-Metabolite durch die Zellmembran in den Apoplasten könnte vergleichbar dem Export von Monolignolen aus dem Zellinnern erfolgen (Sens 1998). Nach Lewis und Yamamoto (1990) gibt es Befunde, dass die Monolignole als Glucoside die Zellmembran überwinden und im Apoplasten durch Glucosidasen wieder freigesetzt werden. Diese Annahme ist allerdings noch umstritten (Heldt 1996). Insgesamt bleibt festzustellen, dass über den Mechanismus des Exports bisher noch wenig bekannt ist. Auch der Mechanismus der Ligninbildung selbst ist bisher nur lückenhaft untersucht. An der Verknüpfung der Monolignole sind neben Peroxidasen noch Laccasen und andere Oxidasen beteiligt (Boudet 2000), wobei angenommen wird, dass die Polymerisierung unter Verbrauch von H2O2 erfolgt. Allerdings konnte im Xylem von Tabak eine Polyphenol-Oxidase vom Laccasetyp lokalisiert werden, welche Coniferylalkohol ohne H2O2 zu polymerisieren vermag (Richardson und McDougall 1997). 4.3.6.4 Festlegung an Zellwandbestandteilen der Wurzeloberfläche Mikroautoradiographisch ließen sich erhöhte Konzentrationen von 14C-TNT/TNT-Metaboliten vor allem an der Oberfläche nicht-mykorrhizierter Wurzeln detektieren. Möglicherweise handelt es sich bei der Sorption um kovalente Verbindungen mit verschiedenen Zellwandbestandteilen wie Suberinen und Tanninen. Die Vermutung, dass die Bindungen in fester Form vorliegen, wird durch die Tatsache erhärtet, dass sich selbst nach mehrmaligem Kunststoffwechsel während der Einbettung die radioaktiven Bestandteile nicht von der Wurzeloberfläche ablösten. Ähnliche Beobachtungen machte Sens (1998) mit radioaktiv markierten Nitroaromaten, die fest an die Wurzeln von Phaseolus vulgaris und Triticum aestivum sorbierten und sich selbst nach mehrmaligem Spülen mit Methanol nicht loslösen ließen. Vor allem die Tannine oder Gerbstoffe sind in Pflanzen weit verbreitet und oft sehr fest mit Proteinen verbunden. Über den genauen Syntheseweg ist nur anteiliges bekannt. Vermutlich sind radikalische Prozesse beteiligt (Heldt 1996). Die gebräuchliche Bezeichnung Polyphenole deutet auf den Ursprung dieser heterogenen Stoffgruppe hin. Neben phenolischen Hydroxylgruppen besitzen diese Monomere weitere Funktionelle

Diskussion
120
Gruppen (–OH, –NH, –NH2). Sie sind in der Lage zu hochmolekularen Verbindungen zu kondensieren. Solche Kondensationsreaktionen können durch Stress für die Pflanze stimuliert werden und sind als eine Abwehrreaktion gegen Mikroorganismen oder toxische Stoffe zu verstehen. Durch die Bindung an Tanninen können von Mikroorganismen freigesetzte, sezernierende Enzyme für die Pflanzen unschädlich gemacht werden. Ein vergleichbarer Schutzmechanismus wäre auch für toxisch wirkende Nitroaromaten vorstellbar, nachdem die Pflanze zuvor durch den Kontakt in eine Stresssituation gebracht wurde. Hinweise auf eine umfangreiche Tanninproduktion in der Rhizodermis und in den äußeren Rindenzellen liegen in einer dunkelbraunen Verfärbung der Kiefernwurzeln in den Experimenten begründet. Entsprechende Verfärbungen ließen sich insbesondere nach hohen TNT-Applikationen beobachten (14C-Versuch und Kiefern-Toxizitätstest). Anteilig könnte die Bildung einer Tanninschicht auf der Wurzeloberfläche auch auf tanninhaltige, kollabierte Kalyptrazellen zurückzuführen sein, die noch in Resten den Wurzeln anhafteten. 4.3.7 Extraktion von Nitroaromaten aus Pflanzengeweben Alle bisher in der Literatur beschriebenen Extraktionsmethoden zur Isolierung von Nitroaromaten aus Pflanzengeweben basieren letztendlich auf einem sauren Extraktionsschritt mit einer nachfolgenden Alkalisierung des sauren Hydrolysats. Durch die Verwendung unpolarer Lösungsmittel (Dichlormethan, Diethylether u. a.) ließen sich bisher die relativ "unpolaren" Kontaminanten TNT, 2-ADNT, 4-ADNT sowie vereinzelt in sehr geringen Mengen 2,4-DNT und 2,6-DNT extrahieren ( Palazzo & Leggett 1986, Cataldo et al. 1989, Harvey et al. 1990, Görge et al. 1994 , Schneider et al. 1996). Auch im Rahmen dieser Arbeit wurde auf ein entsprechendes Verfahren, modifiziert nach Görge et al. (1994a) zurückgegriffen. Über dünnschichtchromatographische Methoden konnten inzwischen zahlreiche weitere überwiegend polare nicht identifizierte TNT-Metaboliten in Pflanzengeweben nachgewiesen werden (Sens 1998). Etwa 70 % der detektierten Nitroaromaten ließen sich erst nach Säurehydrolyse und nachfolgender Alkalisierung mit Natronlauge aus den Pflanzen extrahieren, nachdem zuvor nach einfacher DCM-Extraktion lediglich 30 % der Nitroaromaten gewonnen werden konnten (siehe Tabelle 17). Vergleichbare Ergebnisse lieferten auch die Versuche von Cataldo et al. (1989), Görge et al. (1994 ), Scheidemann (1998) u. a. Autoren. Vermutlich liegt der überwiegende Teil der von den Pflanzen aufgenommenen Nitroaromaten demnach in einer Form vor, die eine Herauslösung allein durch eine DCM-Extraktion nicht zulässt. Schon Catoldo et al. (1989) belegten anhand von Experimenten mit radioaktiv markiertem 14C-TNT eine Festlegung großer Teile des 14C als gebundene Rückstände (bound residues) in den Pflanzen. Bestätigt wurden diese Beobachtungen durch Versuche von Sens et al. (1999). Gelpermeationschromatographisch ließen sich 14C-TNT-Metaboliten in der Ligninfraktion von Phaseolus und Triticum nachweisen. Aufgrund der hohen Molekulargewichte von über

Diskussion
121
1000 g/mol dieser Metaboliten gilt eine kovalente Bindung mit Ligninbestandteilen als wahrscheinlich. Diese Annahme wird durch eigene mikroautoradiographische Untersuchungen gestützt, denn vor allem im Bereich der ring- und schraubenartig lignifizierten Wandverdickungen der Tracheiden konnten vermehrt radioaktive Einlagerungen durch eine erhöhte Silberkorndichte festgestellt werden. Eine starke Adsorption von 14C-TNT/14C-TNT-Metaboliten an der Wurzeloberfläche der Kiefernwurzeln deutet außerdem auf eine Bindung an Tanninen, welche in ihrer Grundstruktur, ebenso wie die Lignine, aus phenolischen Verbindungen synthetisiert werden. Von den analytisch in den Pflanzen nachgewiesenen Nitroaromaten bildete das TNT gegenüber den ADNTs einen Anteil von weniger als 10 % der Gesamtmenge. Mit ca. 90 % Anteil waren 2-ADNT und 4-ADNT die Hauptkontaminanten. Vergleichbare Ergebnisse zeigten u. a. die Untersuchungen von Görge et al. (1994 und 1995) und Scheidemann (1998). Die Dominanz der ADNTs im identifizierten Schadstoffspektrum ist möglicherweise auf die Tatsache zurückzuführen, dass allgemein weniger TNT als vermehrt ADNT in die Wurzeln aufgenommen wird. Denkbar wäre andererseits eine zügige Metabolisierung des aufgenommenen TNT innerhalb der Wurzeln, so dass der Anteil des TNT am Nitro-aromatengehalt in der Wurzel insgesamt unbedeutend wird. Die deutliche Dominanz der ADNTs gegenüber dem TNT unter den extrahierbaren Nitroaromaten könnte eventuell auch auf eine unterschiedlich starke Bindung an Bestandteile der Pflanzen zurückzuführen sein. Schon mehrmals wurde die Bindung von TNT und Metaboliten mit Ligninverbindungen in den Zellwänden diskutiert (s. o.). Vorstellbar wären u. a. starke π-π-Donor-Akzeptor-Wechselwirkungen (Charge-Transfer-Wechselwirkungen) zwischen aromatischen π-Systemen. So könnte das in seiner Struktur planare TNT-Molekül (abgesehen von der Methylgruppe) aufgrund des hohen Elektronendefizits an seinem aromatischen π-System (verursacht durch die elektronenziehenden Nitrogruppen) als Elektronenakzeptor wirken, während Phenylstrukturen in der Ligninfraktion als Elektronendonor fungieren. Möglicherweise waren die TNT-Moleküle über Charge-Transfer-Wechselwirkungen so stark am Lignin gebunden, dass sie durch eine Säurehydrolyse mit anschließender Alkalisierung nur schwer herauszulösen waren. Im Falle der ADNTs führt die Reduktion der Elektronen ziehenden Nitrogruppen zu Elektronen gebenden Aminogruppen zu einer Erniedrigung des Elektronendefizits am aromatischen π-System mit der Folge deutlich schwächer ausgeprägter Charge-Transfer-Wechselwirkungen zwischen den Akzeptor-Donor-Komplexen, so dass eine Extraktion möglicher wurde.

Diskussion
122
4.4 Toxizitätstest mit mykorrhizierten Kiefern in TNT-haltigem Boden-substrat Über die toxikologische Wirkung von Nitroaromaten auf den Menschen ist in der Einleitung (Abschnitt 1.1.2) bereits ausführlich berichtet worden. Relativ begrenzt ist das Wissen über die phytotoxische Wirkung dieser Schadstoffe. Bisherige Untersuchungen beschränken sich hauptsächlich auf Algen (Bringmann und Kühn 1977) und verschiedene krautartige Pflanzen (Peterson et al. 1996 u. 1998; Gong et al. 1999 u. 2001 und Krishnan et al. 2000). Dabei zeigte sich eine unterschiedliche Anfälligkeit der Pflanzen gegenüber diesen Schadstoffen. Kenntnisse über die Empfindlichkeit von Gehölzpflanzen gegenüber TNT und dessen Metaboliten liegen bisher so gut wie nicht vor. Gerade im Hinblick auf eine erfolgreiche pflanzenbasierte Sanierung TNT-kontaminierter Böden, ist für den Sanierungserfolg jedoch von ganz wesentlicher Bedeutung, in welcher Größenordnung TNT und dessen Metaboliten von den Pflanzen toleriert werden können. Das Phytoremediationspotenzial ist also ganz wesentlich vom Resistenz- bzw. Toleranzvermögen der Pflanzen gegenüber diesen Kontaminanten im Boden abhängig. Verlieren Pflanzen aufgrund von toxischen Einflüssen der Bodenkontaminanten an Vitalität, würde somit sicherlich der Phytoremediationseffekt beeinträchtigt werden. Wie Gehölze mit Nitroaromatenkonzentrationen verschiedener Größenordnung umzugehen vermögen, sollte daher an mykorrhizierten Kiefern untersucht werden. Im Rahmen der vorliegenden Arbeit wurden mit Pisolithus tinctorius mykorrhizierte Kiefernsämlinge (Pinus sylvestris) über einen längeren Zeitraum unterschiedlichen TNT-Bodenkonzentrationen ausgesetzt, um festzustellen, inwieweit die Pflanzen auf toxikologische Einflüsse der Kontaminanten reagieren. Das toxische Potential von TNT und dessen Wirkung auf die Kiefern wurde in erster Linie über den Biomassezuwachs als Maß für die Vitalität ermittelt. Weitere Parameter waren eventuell auftretende morphologische Schäden an Nadeln und Wurzeln und die Entwicklung des Mykorrhizierungsgrades der Wurzeln. Biomasseentwicklung und morphologische Abnormitäten an Pflanzenorganen als wichtige Parameter zur Bestimmung der Phytotoxizität von Nitroaromaten wurden bereits mehrmals verwendet (Peterson et al. 1998, Gong et al. 1999, Krishnan et al. 2000). Da von der Annahme auszugehen war, dass das TNT im mechanisch bearbeiteten Bodensubstrat einer erheblichen Abbaudynamik unterliegt (siehe Schadstoffentwicklung im Kiefern- und Pappelexperiment) und der Kontaminationsgrad während der Experimentierphase somit nicht konstant bleiben, sondern stetig abnehmen würde, musste außerdem die Belastung in Boden und Bodenlösung über den gesamten Versuchszeitraum ermittelt werden. So war es möglich, die Schadstoffentwicklung im Boden in zeitlicher Abhängigkeit nachzuvollziehen und in diesem Zusammenhang einen Einfluss auf die Vitalität der Pflanzen herzustellen. Für die Bioverfügbarkeit ist allerdings in erster Linie der Nitroaromatengehalt in der wässrigen Phase

Diskussion
123
entscheidend, deshalb wurde parallel der Schadstoffgehalt in der Bodenlösung bestimmt. Als Kontaminanten konnten neben dem dotierten TNT auch 4-ADNT und 2-ADNT als unmittelbare Reduktionsprodukte des TNT im Bodensubstrat und in der Bodenlösung detektiert werden. Weitere Nitroaromaten ließen sich nicht finden. Die DANTs als reduktive Folgeprodukte der ADNTs binden unter aeroben Bedingungen irreversibel an Bodenbestandteile (Elovitz und Weber 1999). Vermutlich konnten die DANTs deshalb nicht nachgewiesen werden. Phytotoxische Eigenschaften für TNT, 4-ADNT und 2-ADNT sind bereits von Palazzo und Legget (1986) belegt worden. So reduzierten in Flüssigkultur alle drei Nitroaromaten das Wachstum von Cyperus esculentus L. signifikant. Allerdings wird die Toxizität der unmittelbaren TNT-Metabolite allgemein geringer eingestuft als für das TNT selbst (Martinetz und Rippen 1996). Dies konnte u. a. im Leuchtbakterien- Hemmtest festgestellt werden, wobei allgemein die Biolumineszenz als Maß für die Vitalität angenommen wird. In Tabelle 22 sind die jeweiligen Schadstoffkonzentrationen ausgewählter Nitroaromaten aufgeführt, welche die Biolumineszenz um 20% reduzieren (EC20). Tabelle 22: Wichtige Nitroaromaten und ihre Giftigkeit gegen Leuchtbakterien (Angaben aus Martinetz & Rippen 1996, nach Schäfer 1992)
Verbindung EC20 (in mg/l)
2,4,6-TNT 1
4-Amino-2,6-dinitro- toluol
115
2-Amino-4,6-dinitro- toluol
38
2,4-Dinitrotoluol 22
2,6-Dinitrotoluol 1,6
2,3-Dinitrotoluol 4,8
3,4-Dinitrotoluol 3,6
2-Amino-4-nitrotoluol 20
2-Nitrotoluol 2,1
3-Nitrotoluol 4,0
4-Nitrotoluol 12
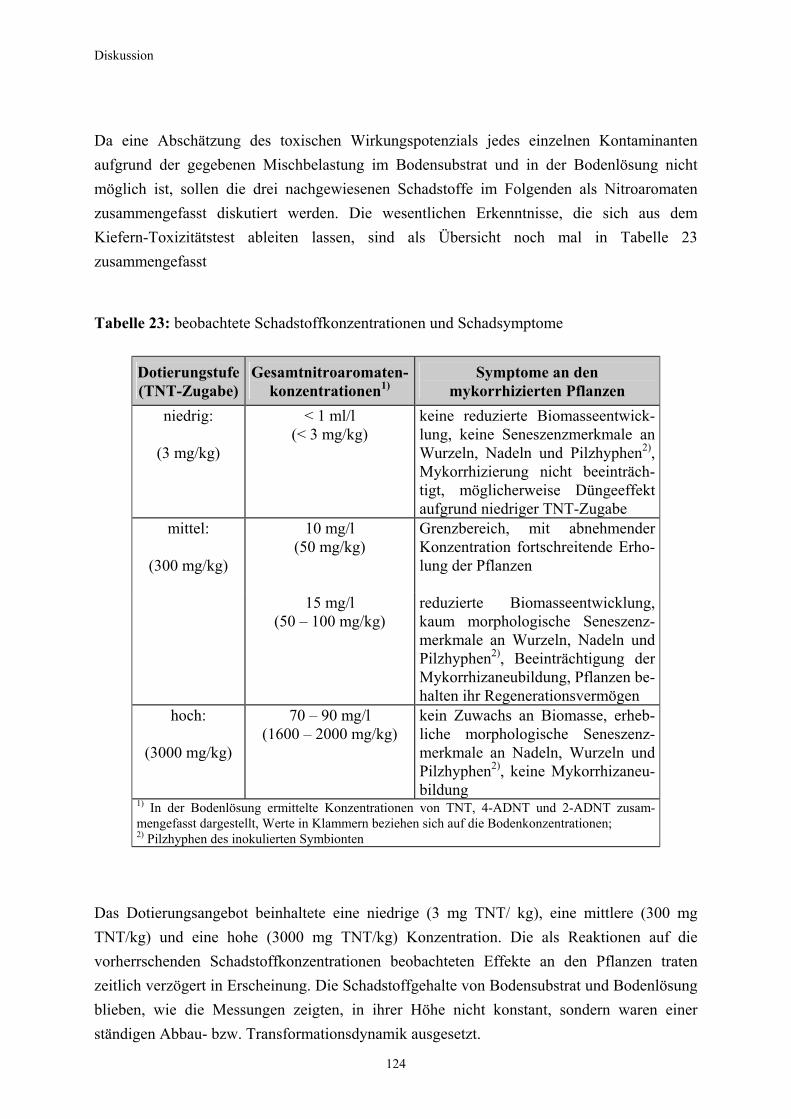
Diskussion
124
Da eine Abschätzung des toxischen Wirkungspotenzials jedes einzelnen Kontaminanten aufgrund der gegebenen Mischbelastung im Bodensubstrat und in der Bodenlösung nicht möglich ist, sollen die drei nachgewiesenen Schadstoffe im Folgenden als Nitroaromaten zusammengefasst diskutiert werden. Die wesentlichen Erkenntnisse, die sich aus dem Kiefern-Toxizitätstest ableiten lassen, sind als Übersicht noch mal in Tabelle 23 zusammengefasst Tabelle 23: beobachtete Schadstoffkonzentrationen und Schadsymptome
Dotierungstufe(TNT-Zugabe)
Gesamtnitroaromaten-konzentrationen1)
Symptome an den mykorrhizierten Pflanzen
niedrig:
(3 mg/kg)
< 1 ml/l (< 3 mg/kg)
keine reduzierte Biomasseentwick-lung, keine Seneszenzmerkmale an Wurzeln, Nadeln und Pilzhyphen2), Mykorrhizierung nicht beeinträch-tigt, möglicherweise Düngeeffekt aufgrund niedriger TNT-Zugabe
mittel:
(300 mg/kg)
10 mg/l (50 mg/kg)
Grenzbereich, mit abnehmender Konzentration fortschreitende Erho-lung der Pflanzen
15 mg/l (50 – 100 mg/kg)
reduzierte Biomasseentwicklung, kaum morphologische Seneszenz-merkmale an Wurzeln, Nadeln und Pilzhyphen2), Beeinträchtigung der Mykorrhizaneubildung, Pflanzen be-halten ihr Regenerationsvermögen
hoch:
(3000 mg/kg)
70 – 90 mg/l (1600 – 2000 mg/kg)
kein Zuwachs an Biomasse, erheb-liche morphologische Seneszenz-merkmale an Nadeln, Wurzeln und Pilzhyphen2), keine Mykorrhizaneu-bildung
1) In der Bodenlösung ermittelte Konzentrationen von TNT, 4-ADNT und 2-ADNT zusam-mengefasst dargestellt, Werte in Klammern beziehen sich auf die Bodenkonzentrationen; 2) Pilzhyphen des inokulierten Symbionten
Das Dotierungsangebot beinhaltete eine niedrige (3 mg TNT/ kg), eine mittlere (300 mg TNT/kg) und eine hohe (3000 mg TNT/kg) Konzentration. Die als Reaktionen auf die vorherrschenden Schadstoffkonzentrationen beobachteten Effekte an den Pflanzen traten zeitlich verzögert in Erscheinung. Die Schadstoffgehalte von Bodensubstrat und Bodenlösung blieben, wie die Messungen zeigten, in ihrer Höhe nicht konstant, sondern waren einer ständigen Abbau- bzw. Transformationsdynamik ausgesetzt.

Diskussion
125
Während Pflanzen bei hohen Nitroaromatenkonzentrationen allgemein mit einer reduzierten Biomasseproduktion reagieren oder ihr Wachstum ganz einstellen, können gering dotierte Mengen durchaus stimulierend auf das Wachstum wirken und letztendlich sogar zu einer gesteigerten Biomasseproduktion führen. Solche stimulierenden Effekte, verursacht durch geringe TNT-Zugaben, wurden bereits für die Dikotyledonen Lepidium sativum L. und Brassica rapa Metzg. sowie für die Monokotyledonen Avena sativa L. und Triticum aestivum L. beobachtet (Gong et al. 1999). Auch die in der vorliegenden Arbeit untersuchten Kiefern zeigten bei einer Dotierung von 3 mg TNT kg-1 eine entsprechende Tendenz. So lag die Biomasseproduktion der Pflanzen nach 12 Wochen um etwa 15 % höher als bei den Kiefern des unbelasteten Kontrollbodens (Abbildung 33). Zwar war der Unterschied nicht signifikant, trotzdem ist zu vermuten, dass mit der Dotierung des TNT und dem damit verbundenen zusätzlichen Stickstoffeintrag in den Boden ein gewisser Düngeeffekt bei den Pflanzen ausgelöst wurde und das TNT bzw. Metabolite als Stickstoffquelle dienten. Wie bereits erwähnt wurde, stellt die Ermittlung des EC-Wertes eine allgemein verbreitete Methode zur Bestimmung des toxischen Wirkungspotenzials von Schadstoffen gegenüber Organismen dar. Anwendung findet diese Methode vor allem im Leuchtbakterien- Hemmtest. Beträgt die Inkubationszeit, wie in solchen Tests allgemein üblich, nur Stunden oder maximal ein bis zwei Tage, kann der zwischenzeitliche Transformationsvorgang als beeinflussende Größe in der Regel vernachlässigt werden. Problematischer stellt sich die Lage für Wachstumshemmtests mit Pflanzen dar. Solche Untersuchungen erstrecken sich zeitlich über mehrere Wochen oder sogar Monate. Durch die zwischenzeitlich ablaufenden Transformationsvorgänge kann der Schadstoffabbau während der Versuchsdurchführung erhebliche Ausmaße annehmen. Dies belegen die eigenen Studien. Die mykorrhizierten Kiefern waren in den Rhizotronen keiner konstanten, sondern einer ständig in Abnahme begriffenen Belastung ausgesetzt. An den Pflanzen auftretende toxische Wirkungen konnten in diesem Fall dann nicht auf die dotierten Ausgangswerte bezogen werden, wie das sonst bei der EC-Wert-Bestimmung üblich ist. Ein auftretender Effekt konnte nur mit einem etwas weiter gefassten Kontaminationsbereich in Beziehung gebracht werden, zumal durch Schadstoffeinwirkung auftretende Effekte wie Wachstumshemmung und morphologische Schäden an den Pflanzen zeitlich verzögert in Erscheinung traten. Bezüglich des Toxizitätstest mit den mykorrhizierten Kiefern lassen sich deshalb folgende Aussagen machen: Belastungen zwischen 50 und 200 mg Nitroaromaten kg-1 Boden, das entspricht etwa einem Kontaminationsbereich zwischen 10 und 30 mg l-1, hemmten einerseits die Bio-masseproduktion der Pflanzen nahezu vollständig, bewirkten aber andererseits so gut wie keine Schadeffekte an Nadeln, Wurzeln und Hyphenmänteln bezüglich ihrer morphologischen Ausgestaltung. Schäden ließen sich nur sehr vereinzelt durch das Auftreten brauner Nadelspitzen feststellen. Die Kiefern als auch der Pilzsymbiont waren zum einen in der Lage,

Diskussion
126
Nitroaromatengehalte dieser Größenordnung mehrere Wochen mit Wachstumseinbußen relativ unbeschadet auszuhalten, zum anderen reagierten Pflanze und Pilz mit abnehmender Schadstoffkonzentration rasch mit einer deutlichen Erhöhung der Biomasseproduktion bzw. mit einer vermehrten Mykorrhizainfektion an den Kurzwurzeln. Das erstaunliche Regenerationspotenzial der mykorrhizierten Kiefern zeigte sich insbesondere bei Nitroaromatengesamtkonzentrationen unter 50 mg kg-1 Boden, das entspricht etwa 10 mg l-1 Bodenlösung. Die Fähigkeit, ungünstige Bedingungen zu überdauern, beobachteten auch Trapp und Karlson (2000) in ihren Studien mit Salix spec. in Laborversuchen in Verbindung mit mineralischen Kohlenwasserstoffen und Cyanid. Die Pflanzen konnten bis zu 5g Dieselöl, fast 1 g Superbenzin oder 0,5 g Gesamtzyanid je kg Boden mit Wachstumseinbußen aushalten. Verbesserte Überlebenschancen gegenüber hohen TNT-Belastungen hatten transgene Tabakpflanzen der Art Nicotiana tabacum CV Xanthi im Vergleich zum entsprechenden Wildtyp (Hannink et al. 2001). Durch den Besitz eines modifizierten Bakteriengens (Enterobacter cloacae NCIMB 10101) zur Kodierung von Nitroreduktase (nFsI) ließ sich das Toleranzvermögen erheblich steigern. Während das Schadstoffspektrum an aufgenommenen Nitroaromaten beim Wildtyp abgesehen von den ADNTs auch noch das TNT beinhaltete, wurden beim transgenen Typ lediglich ADNTs gefunden. Der Schluss liegt nahe, dass im letzteren Typ durch zügige reduktive Transformation des aufgenommenen TNT zum weniger toxischen ADNT die Schadwirkung insgesamt herabgesetzt und das Toleranzpotenzial somit letztendlich erhöht wird. Trotz solcher Erfolge müssen allerdings die Chancen und Risiken der Gentechnik beim Einsatz transgener Pflanzen in zukünftigen in-situ- Phytoremediationsverfahren gründlich überdacht und kritisch diskutiert werden. In hohem Maße toxisch für die mykorrhizierten Kiefern waren Bodenbelastungswerte zwischen 1600 und 2000 mg Nitroaromaten kg-1 Boden, das entsprach Konzentrationen zwischen 70 und 90 mg Nitroaromaten l-1 Bodenlösung, denen die Pflanzen in der hohen Belastungskategorie ausgesetzt wurden. Die hohen Schadstoffkonzentrationen verursachten nicht nur erhebliche morphologische Schäden an den Nadeln und Wurzeln, sie führten zum Teil auch mitsamt dem Mykobionten gänzlich zum Absterben der Pflanzen. Neue Nadeln und Wurzeln wurden nicht ausgebildet. Bereits ausdifferenzierte Mykorrhizen zeigten bald Auflösungserscheinungen. Die Pilzhyphen pigmentierten zunehmend und gaben dem Hyphenmantel bald eine dunkelbraune Färbung. Dunkel verfärbte Hyphen in alternden oder seneszenten Mykorrhizen sind bereits mehrfach von Autoren beschrieben worden (Downes et al. 1992, Behrmann 1995). Ein Biomassezuwachs konnte bei den Pflanzen nicht verzeichnet werden. Kontaminationsgrade dieser Größenordnung lagen demnach jenseits jeglicher Toleranzgrenzen für die Kiefern, wie das Auftreten der starken phytotoxischen Symptome zeigte. Die Fähigkeit, eine Schadstoffbelastung unter Beibehaltung eines bestimmten Regenerationspotenzials für einen gewissen Zeitraum zunächst mit einer Wachstumseinbuße auszuhalten - bei mittleren Belastungen unter 300 mg kg-1 Boden waren die Kiefern dazu in

Diskussion
127
der Lage - ging bei einem Nitroaromatengehalt über 1600 mg kg-1 Boden verloren. Auch die Literatur liefert Hinweise dafür, dass hohe Nitroaromatenkonzentrationen von Pflanzen nur selten vertragen werden. So ist der Stechapfel Datura innoxia Mill. dazu in der Lage, etwa 1000 mg TNT kg-1 Boden zu tolerieren (Müller et al. 1993 in Peterson et al. 1996). Der im Vergleich zu anderen Pflanzen als relativ TNT-tolerant geltende Saat-Hafer Avena sativa L. vermochte noch bei Konzentrationen zwischen 800 und 1600 mg TNT kg-1 Boden zu wachsen, wobei das Spross- und Wurzelwachstum bereits in erheblichem Maße verzögert und deutlich weniger ausgeprägt war (Gong et al., 1999). Die verkümmerten Wurzeln blieben kürzer und besaßen weniger Wurzelhaare. Die meisten bisher untersuchten Pflanzen, vor allem Gräser und dikotyle krautartige Pflanzen, reagieren schon bei wesentlich niedrigeren TNT-Konzentrationen sehr empfindlich (Peterson et al. 1996 und 1998, Krishnan et al. 2000, Gong et al 1999 u. 2001). Obwohl sich die Empfindlichkeit gegenüber Nitroaromaten für die verschiedenen Pflanzenarten unterschiedlich darstellt, dürften Toleranzwerte für Expositionsgrade, die ein gewisses Erholungspotential erlauben, somit allgemein deutlich unter 1000 mg TNT oder Nitroaromaten kg -1 Boden liegen. Bei hohen Belastungen werden jedoch nicht nur Pflanzen in ihrem Wachstum stark beeinträchtigt. Auch die mikrobielle Abbautätigkeit kann bei hohen TNT-Konzentrationen stark vermindert werden oder gänzlich zum erliegen kommen (Bradley und Chapelle 1995) mit wiederum weitreichenden Konsequenzen für die Pflanzen. Stagniert der Abbau oder ist er stark verlangsamt, bietet sich den Pflanzen nicht die Möglichkeit, ungünstige Bedingungen mit Wachstumseinbußen auszuhalten, wie dies im Ansatz der mittleren Belastungskategorie beobachtet werden konnte. Resümierend bleibt festzustellen, dass sich für Phytoremediationsverfahren daher Flächen mit niedrigen und mittleren Belastungsgraden anbieten, deren Schadstoffgehalte von Pflanzen toleriert werden können, während in Bereichen hoher Belastungskategorien dem Sanierungspotenzial durch Pflanzen aufgrund des zunehmend toxischen Einflusses der Nitroaromaten Grenzen gesetzt sind.

Zusammenfassung
128
5. Zusammenfassung
Ziel der vorliegenden Arbeit war es, das Phytoremediationspotenzial mykorrhizierter Gehölze für
TNT und Begleitkontaminanten bei der biologischen Bodensanierung zu untersuchen. Hierzu wurden
bei der Ausführung der Experimente drei wesentliche Schwerpunkte gebildet:
1) Untersuchungen zur Abreicherung der Nitroaromatenkonzentrationen im Boden.
Mit der Durchführung dieser Versuche wurde überprüft, ob und inwieweit durch den Einsatz
mykorrhizierter Kiefern und Pappeln (Pinus sylvestris/Pisolithus tinctorius, Populus
tremula/Hebeloma spec.) unter Einbeziehung des Weißfäulepilzes Pleurotus ostreatus in
besonderen Rhizotronsystemen der Abbau von TNT und Metaboliten gefördert werden kann. Als
kontaminiertes Substrat diente ein nachträglich mit Sand vermischter TNT-belasteter Standortboden
vom ehemaligen Sprengstoffwerk ‘Tanne’ bei Clausthal-Zellerfeld. Die Ausgangskonzentrationen für
TNT in den Bodensubstraten lagen bei 80-100 mg/kg (Experiment mit Kiefern) bzw. 400-450
mg/kg (Experiment mit Pappeln). Die Detektion der Nitroaromatenkonzentrationen in den
Bodenproben erfolgte mit der HPLC.
2) Untersuchungen zur Aufnahme und Verteilung von Nitroaromaten im Organismus Pflanze.
Mittels GC/MS-Analytik wurden verschiedene Nitroaromaten in den Geweben nicht-mykorrhizierter
und mykorrhizierter Pappeln erfasst. Allerdings gestattet eine gaschro-matographische Detektion in
nur begrenztem Umfang Aussagen darüber, in welcher Größenordnung Nitroaromaten tatsächlich
von den Pflanzen aufgenommen werden. Bereits in den Geweben metabolisierte oder festgelegte
TNT-/TNT-Metaboliten-Anteile lassen sich nicht mehr bilanzieren. Im Rahmen dieser Überlegungen
wurde in einem gesonderten Experiment nicht-mykorrhizierten und mykorrhizierten Kiefern 14C-TNT
in definierten Mengen angeboten und die Aufnahme und Verteilung des 14C-Radiotracers in den
Pflanzen durch mikroautoradiographische Verfahren und Szintillationszählung quantitativ bestimmt.
3) Untersuchungen zur Nitroaromatentoleranz bei Gehölzen
Voraussetzung für den Erfolg eines Phytoremediationsverfahrens ist die Toleranz der eingesetzten
Pflanzen gegenüber bodenrelevanten Schadstoffgehalten. Entsprechendes gilt auch für

Zusammenfassung
129
Mykorrhizapilze, wenn die Pflanzen mykorrhiziert sind. Daher wurde mit einem Toxizitätstest der
Einfluss des TNTs auf mykorrhizierte Kiefern untersucht (Pinus sylvestris/Pisolithus tinctorius).
Die Kultivierung der 4-5 Monate alten Pflanzen erfolgte bei verschiedenen TNT-
Ausgangskonzentrationen (3, 300 und 3000 mg TNT/kg Boden) in einem dotierten LUFA-
Standardboden (Typ 2.2). Das toxische Potenzial von TNT und dessen Wirkung auf die Kiefern
wurde in erster Linie über den Biomassezuwachs als Maß für die Vitalität ermittelt. Weitere
Parameter waren auftretende morphologische Schäden an Nadeln und Wurzeln und die Entwicklung
des Mykorrhizierungsgrades der Wurzeln.
Die wichtigsten Ergebnisse der kurz dargestellten Experimente lassen sich wie folgt beurteilen:
zu 1) Als erstaunlich effizient beim Nitroaromatenabbau (TNT und ADNTs) im Boden erwiesen sich
insbesondere mykorrhizierte als auch nicht-mykorrhizierte Pappeln, wobei sich die guten
Abbauleistungen nicht-mykorrhizierter Pappeln auf die TNT-Transformation konzentrieren. Die
Untersuchungen scheinen zu belegen, dass die Funktion der Weißfäule-pilze bei der
Nitroaromatendegradation durch eine Assoziation Mykorrhizapilz/Pflanze übernommen werden
kann. Voraussetzung hierfür ist allerdings ein guter Durchwur-zelungsgrad des Bodens, so dass eine
umfassende Rhizosphärendegradation ermöglicht wird. Die eingesetzten Pappeln erfüllten diese
Voraussetzung. Sie zeichneten sich durch ein starkes Wurzelwachstum aus. In den
Abreicherungsexperimenten mit Kiefern zeigte sich ein anderes Abbauverhalten der
Bodenkontaminationen. Ein geringer Biomassezuwachs der Kiefern, einhergehend mit einer
schwachen Durchwurzelung des Substrates insgesamt, resultierte in einer Ausprägung der
Rhizosphäreneffekte auf niedrigem Niveau und ließ pflanzeninduzierte Transformationsvorgänge
daher kaum in Erscheinung treten.
zu 2) Im Rahmen der durchgeführten Untersuchungen zum Transferverhalten von Nitroaromaten aus
dem Boden in die Pflanzen ließen sich TNT, 2-ADNT und 4-ADNT dem aufgenommenen
Schadstoffspektrum zuordnen. Die nachweisbare Nitroaromaten-konzentration in den Pflanzen zeigte
stets eine hohe Organspezifität. Die Wurzeln waren mit deutlichem Abstand anteilig am höchsten
belastet. Aufnahmeexperimente mit 14C-TNT ergaben bei Kiefern für tatsächlich inkorporierte 14C-
TNT/14C-TNT-Metaboliten lediglich eine Wiederfindung unter 1% nach 14 Tagen Applikation. Eine
Akkumulation in den Geweben mit zunehmender Angebotsdauer wurde nicht beobachtet. Durch
mikro-autoradiographische Verfahren ließ sich eine teilweise Bindung von 14C-TNT/14C-TNT-
Metaboliten an der Oberfläche der Seitenwurzeln feststellen. Aufgenommene 14C-Anteile fanden sich

Zusammenfassung
130
bevorzugt in den Tracheiden des Xylems der Seiten- und Trägerwurzeln und dort häufig im Bereich
der ring- und schraubenartig verstärkten Zellwandbereiche. Erhebliche Teile aufgenommener
Nitroaromaten konnten erst nach Säurehydrolyse mit nachfolgender Alkalisierung aus den Geweben
der Pflanzen (Populus tremula) extrahiert werden. Insgesamt sprechen die Ergebnisse für
umfassende Metabolisierungs- und Festlegungsvorgänge in den Pflanzen bezüglich der
aufgenommenen Schadstoffe. Ein Mykorrhizaeffekt hinsichtlich des Aufnahmeverhaltens trat nicht
auf.
zu 3) Die zu Beginn dotierten TNT-Mengen waren während der Experimentierphase einer
erheblichen Abbaudynamik unterlegen, so dass der Kontaminationsgrad im Substrat stetig abnahm.
Parallel zum Vitalitätszustand der Pflanzen wurde daher die Nitroaromaten-konzentration im Boden
und in der Bodenlösung über den gesamten Versuchszeitraum ermittelt. Das nachgewiesene
Schadstoffspektrum beinhaltete abgesehen vom TNT auch 2-ADNT und 4-ADNT. Beide
Metaboliten stellen Transformationsprodukte des TNT dar. Bei der Abschätzung des toxischen
Wirkungspotenzials wurden die nachgewiesenen Konta-minanten aufgrund der gegebenen
Mischbelastung als Nitroaromaten zusammengefasst diskutiert. Eine Dotierung von 3 mg TNT/kg
Boden wirkte sich nicht beeinträchtigend auf die Vitalität der Mykorrhiza aus. Die gesteigerte
Biomasseproduktion gegenüber den Pflanzen aus den unbelasteten Kontrollen legt einen gewissen
Düngeeffekt verbunden mit einem zusätzlichen Stickstoffeintrag in den Boden nahe. Als
Toleranzgrenze für Beeinträchtigungen der Vitalität sollten Nitroaromatenbelastungswerte um 50
mg/kg angesehen werden. Bei dieser Konzentration traten Wachstumsminderungen gegenüber den
Kontrollen ein, allerdings behielten die Pflanzen ihr Regenerationsvermögen und reagierten bei
abnehmender Schadstoffkonzentration mit steigender Biomasseproduktion. Unter Berücksichtigung
einer zügigen Abbaukinetik der Schadstoffe sollten die untersuchten Pflanzen aufgrund ihres
Erholungspotenzials daher in der Lage sein, Expositionsgrade im mäßigen Belastungsbereich mit
Werten deutlich über 50 mg TNT/kg Boden auszuhalten. Belastungswerte zwischen 1600-2000 mg
Nitroaromaten/kg Boden waren in hohem Maße toxisch für die mykorrhizierten Kiefern und
brachten zahlreiche Pflanzen mitsamt des Mykobionten zum absterben.

131
6. Phytoremediation am Standort ’Werk Tanne’; Durchführung und
Beurteilung unter Berücksichtigung der Laborergebnisse Zwischen 1998 und 2000 wurden auf dem ehemaligen Sprengstoffwerk ‘Tanne’ in Clausthal-
Zellerfeld verschiedene biologische Verfahren zur Sanierung TNT-belasteter Böden durchgeführt.
Die maßstabsgerechte Erprobung beruhte hierbei im Wesentlichen auf herkömmlichen ex-situ
Verfahren, die eine Auskofferung des kontaminierten Bodens und eine Aufschüttung als Mieten
voraussetzten (Walter et al. 1998, Winterberg et al. 1998 und Spreinat et al 1998). Darüber hinaus
führte das Zentrum für Umweltforschung und Umwelttechnologie (UFT) in Kooperation mit der
Firma ‘Umweltschutz Nord’ aus Ganderkesee parallel ein in-situ Phytoremediationsverfahren zur
Dekontamination großflächiger und oberflächennaher Bodenbelastungen an gleicher Stelle durch. Die
Idee des Verfahrens zielte auf einen mikrobiellen Abbau von Sprengstoffverbindungen in der
pflanzlichen Rhizosphäre unter Verwendung von Weißfäulepilzen und mykorrhizierten Gehölzen ab.
Im Hinblick auf weitreichende methodische Übereinstimmungen des in-situ Verfahrenskonzeptes am
Standort ‘Tanne’ mit den Modellansätzen mehrerer Labor-experimente der vorliegenden Arbeit
(Kiefern- und Pappelexperiment) sollen die gewonnenen Erkenntnisse sowohl aus dem
Freilandversuch als auch aus den Laborversuchen zueinander in Beziehung gesetzt werden (siehe
Tabelle 24). Im Mittelpunkt des Interesses steht insbesondere die Frage nach einer Übertragbarkeit
der positiven Laborergebnisse auf das hier dargestellteVerfahrensprinzip im Feld.
6.1 Kurze Beschreibung des Standortes ‘Werk Tanne’ Das ehemalige Sprengstoffwerk ‘Tanne’ in Clausthal-Zellerfeld zählte zu den bedeutendsten
Produktionsstätten für TNT im ehemaligen Deutschen Reich während des zweiten Weltkrieges.
Zwischen den Jahren 1939 und 1944 wurden dort 105.358 Tonnen dieses Sprengstoffes produziert
und verarbeitet (Braedt et al. 1999). Fabrikation und Verarbeitung auf dem etwa 120 Hektar großen
Werksgelände verursachten Bodenkontaminationen mit TNT sowie mit Zwischen- und
Nebenprodukten aus dem Herstellungsprozess. Verschiedene Abbauprodukte des TNT,
hauptsächlich Aminodinitrotoluole, zählen ebenso zu den verbreiteten Kontaminanten im Boden.
Nach Abholzung des Geländes Ende der vierziger Jahre erfolgte kurz darauf eine nahezu
flächendeckende Aufforstung mit Fichten (Warrelmann et al. 2000), so dass der Standort
gegenwärtig durch einen etwa 50jährigen Fichtenbestand (Picea abies) gekennzeichnet ist. Das
Versuchsareal schloss sich in seiner Lage dem ehemaligen Tonsilerdelager des Werkes an. Die
Tonsilerde wurde zur Rückgewinnung von TNT aus Fehlproduktionen und Produktionsverlusten
verwendet. Großflächige Verteilung der Belastung in ausreichender räumlicher Ausdehnung,
einheitliche Geländebeschaffenheit und gute Erreichbarkeit waren ausschlaggebende Faktoren für die

132
Wahl der Fläche. Innerhalb der Probefläche zeigte sich die Belastungssituation im Boden teilweise
sehr heterogen.
6.2 Verfahrenskonzept: Die weitreichenden methodischen Übereinstimmungen des in-situ Verfahrenskonzeptes am Standort
‘Tanne’ mit den Modellansätzen verschiedener Experimente im Labormaßstab wurden bereits
erwähnt (siehe auch Einleitung Abschnitt 1.3). Konzeptvorstellungen des in-situ
Phytoremediationsverfahrens sollen an dieser Stelle deshalb nur in den wesentlichen Grundzügen
erläutert werden. Den Kernpunkt des Feldversuches bildet der mikrobielle Abbau von
Sprengstoffverbindungen im Wurzelraum mykorrhizierter Gehölze (Rhizosphären-degradation).
Mykorrhizapilze mit ligninolytischen Fähigkeiten als Streuzersetzer in Symbiose mit Pflanzen,
Weißfäulepilze und dazu Bakterien, die in Assoziation mit den genannten Pilzen und Pflanzen leben,
erwirken als Organismengemeinschaft über einen Mehrstufenprozess letztendlich eine Ringspaltung
der Nitroaromaten. Nach reduktivem Initialangriff an den Nitrogruppen kommt insbesondere den
Mykorrhizapilzen in zweierlei Hinsicht eine Schlüsselrolle zu. Zum einen sollen sie durch den Besitz
verschiedener extrazellulärer und oxidativer Enzyme die Mineralisierung der Schadstoffe initiieren,
zum anderen partizipieren die Wirtspflanzen von stickstoffhaltigen Folgeprodukten, die durch die
metabolische Weiterverwertung der Ringspaltungsprodukte durch den Mykorrhizapilz seitens des
Mykobionten bereitgestellt werden. Bei der Durchführung des Verfahrens war außerdem von
Interesse, ob die Funktion der ligninolytischen Weißfäulepilze möglicherweise von den verwendeten
Mykorrhizapilzen übernommen werden kann, da nur letztere eigentliche Bodenbewohner sind und
zudem kontinuierlich von den Pflanzen mit Kohlen-stoffverbindungen versorgt werden. Im Gegensatz
zu den Weißfäulepilzen sind die Mykorrhizapilze somit nicht von Substratzugaben abhängig.
6.3 Einrichtung der Versuchsflächen: Auf dem Versuchsareal stehende Bäume wurden gefällt und die im Erdreich verbleibenden
Wurzelteller in einem Schutzwall auf dem Gelände integriert. Auflockerung und eine horizontale
Vermischung der oberen 30 cm Boden mit einer Bodenfräse brachten eine teilweise Aufhebung der
erheblichen Belastungsheterogenitäten mit sich. Trotz mechanischer Verteilung des TNTs ließen sich
Unterschiede im Kontaminationsgrad auf den Versuchs-varianten allerdings nicht vollständig
aufheben. Hierauf wird an späterer Stelle noch einzugehen sein.

133
Auf dem Areal (Nitroaromatenbelastung bis 1000 mg TNT/kg Boden TS) wurden nach
mechanischer Bearbeitung vier Versuchsparzellen mit einer Ausdehnung von jeweils 4,5 x 6,5 m
eingerichtet:
Variante U: unbelastet, bepflanzt mit ektotroph mykorrhizierten Fichten (Picea abies) und
mykorrhizierten Pappeln (Populus tremula) sowie mit Holunder (Sambucus nigra, möglicherweise
mit VA-Mykorrhiza) im 0,5-Meter-Raster (Mengenverhältnis 6:1:1), als ektotrophe Mykorrhizapilze
wurden Pisolithus tinctorius und Paxillus involutus Kulturen verwendet, Fläche bis etwa 20-30
cm Bodentiefe beimpft mit Weißfäulepilz-Pappelholz- Weizenstroh-Schreddersubstrat (Pleurotus
ostreatus und Trametes, versicolor), Abdeckung aus Kiefernrindenmulch.
Variante B: belastet, sonst wie Fläche U. Variante P: wie Fläche B, allerdings nicht mit Weißfäulepilz-Schredder-Substrat beimpft. Variante M: wie Fläche P, allerdings ohne Bepflanzung, lediglich mechanische Bearbeitung der
Bodenoberfläche.
In einem angrenzendem Fichtenforst (belastet, mit etwa 50jährigen Picea abies bestanden) wurden
parallel ebenfalls Bodenproben entnommen und auf ihren Nitroaromatengehalt überprüft; Variante K
ohne jeden Eingriff.
6.4 Grenzen des Versuchsansatzes Für die Durchführung des in-situ Phytoremediationsverfahrens wurde als Versuchsfläche auf dem
ehemaligen Werksgelände ein Areal ausgewählt, das in seiner Ausdehnung an das ehemalige
Tonsilerdelager anschloss (siehe oben). Gründe für die Wahl des Standortes waren neben einer
relativ einheitlichen morphologischen Beschaffenheit und günstigen Lage bzw. guten Erreichbarkeit
die deutliche Belastung der Fläche. Resultierend aus umfangreichen Voruntersuchungen erwies es
sich im Nachhinein als schwierig, replizierbare Flächen mit einer geeigneten Belastung in einem
Bereich von 100-1000 mg TNT/kg Boden (TS) mit ausreichender räumlicher Ausdehnung zu finden.
Wie es für Altlastenstandorte typisch ist, trat die Verteilung der Kontamination im Boden sehr
inhomogen in Erscheinung. Auch nach Bearbeitung der schließlich ausgewählten Fläche durch den
Einsatz der Bodenfräse ließen sich die vorhandenen Inhomogenitäten nur teilweise aufheben.

134
Diesbezüglich konnte nicht vermieden werden, dass die Versuchsvarianten durch erhebliche
Unterschiede in ihren Ausgangskontaminationen charakterisiert waren. Diesem Sachverhalt muss
beim Vergleich der Versuchsvarianten Rechnung getragen werden. Des Weiteren war aufgrund der
Anwendung umfangreicher Monitoring- und Testsysteme die wissenschaftliche Versuchsplanung nur
mit Abstrichen zu bewältigen. Hier wäre insbesondere eine Verkürzung der Zeitintervalle zwischen
den Beprobungen wünschenswert gewesen.
6.5 Beurteilung des Phytoremediationsverfahrens im Hinblick seines Sa-
nierungserfolges unter Berücksichtigung der Ergebnisse aus den Labor-
versuchen Das Begleitmonitoring zur Erfolgskontrolle des Verfahrens beinhaltete umfangreiche chemische
(Nitroaromatenrückstandsanalytik in Boden und Pflanzen), bodenkundliche und bodenbiologische
(Biotest-Batterie, Bodentiere, Streuabbau) Untersuchungen. Vordergründig von Interesse im
vorliegenden Abschnitt sind die Erhebungen zur Entwicklung der Nitroaromatengehalte in Böden und
Pflanzengeweben unter variantenspezifischem Aspekt (Tabelle 24). Weitere Angaben zur
Flächeneinrichtung und zu methodischen Ausführungen sowie eine detaillierte, umfassende
Ergebnisdokumentation bezüglich des Gesamtmonitorings sind den Publikationen Warrelmann et al.
(2000) bzw. Koehler et al. (2001a) zu entnehmen.
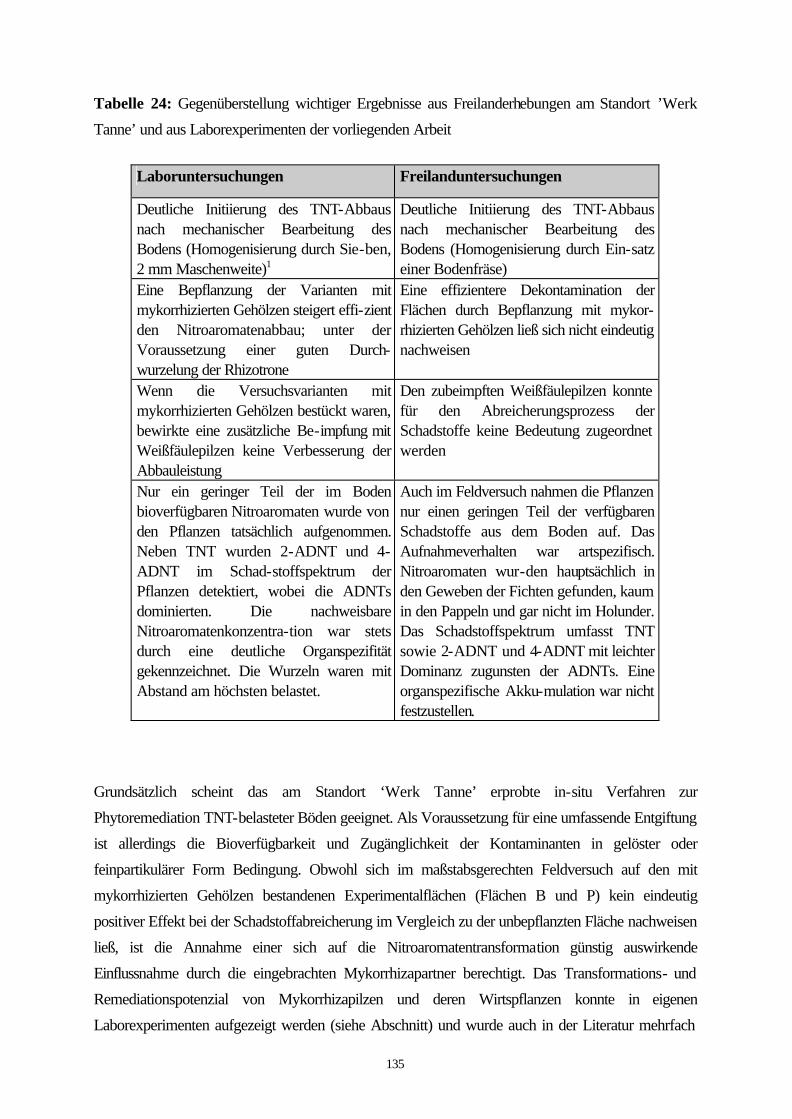
135
Tabelle 24: Gegenüberstellung wichtiger Ergebnisse aus Freilanderhebungen am Standort ’Werk
Tanne’ und aus Laborexperimenten der vorliegenden Arbeit
Laboruntersuchungen Freilanduntersuchungen
Deutliche Initiierung des TNT-Abbaus nach mechanischer Bearbeitung des Bodens (Homogenisierung durch Sie-ben, 2 mm Maschenweite)1
Deutliche Initiierung des TNT-Abbaus nach mechanischer Bearbeitung des Bodens (Homogenisierung durch Ein-satz einer Bodenfräse)
Eine Bepflanzung der Varianten mit mykorrhizierten Gehölzen steigert effi-zient den Nitroaromatenabbau; unter der Voraussetzung einer guten Durch-wurzelung der Rhizotrone
Eine effizientere Dekontamination der Flächen durch Bepflanzung mit mykor-rhizierten Gehölzen ließ sich nicht eindeutig nachweisen
Wenn die Versuchsvarianten mit mykorrhizierten Gehölzen bestückt waren, bewirkte eine zusätzliche Be-impfung mit Weißfäulepilzen keine Verbesserung der Abbauleistung
Den zubeimpften Weißfäulepilzen konnte für den Abreicherungsprozess der Schadstoffe keine Bedeutung zugeordnet werden
Nur ein geringer Teil der im Boden bioverfügbaren Nitroaromaten wurde von den Pflanzen tatsächlich aufgenommen. Neben TNT wurden 2-ADNT und 4-ADNT im Schad-stoffspektrum der Pflanzen detektiert, wobei die ADNTs dominierten. Die nachweisbare Nitroaromatenkonzentra-tion war stets durch eine deutliche Organspezifität gekennzeichnet. Die Wurzeln waren mit Abstand am höchsten belastet.
Auch im Feldversuch nahmen die Pflanzen nur einen geringen Teil der verfügbaren Schadstoffe aus dem Boden auf. Das Aufnahmeverhalten war artspezifisch. Nitroaromaten wur-den hauptsächlich in den Geweben der Fichten gefunden, kaum in den Pappeln und gar nicht im Holunder. Das Schadstoffspektrum umfasst TNT sowie 2-ADNT und 4-ADNT mit leichter Dominanz zugunsten der ADNTs. Eine organspezifische Akku-mulation war nicht festzustellen.
Grundsätzlich scheint das am Standort ‘Werk Tanne’ erprobte in-situ Verfahren zur
Phytoremediation TNT-belasteter Böden geeignet. Als Voraussetzung für eine umfassende Entgiftung
ist allerdings die Bioverfügbarkeit und Zugänglichkeit der Kontaminanten in gelöster oder
feinpartikulärer Form Bedingung. Obwohl sich im maßstabsgerechten Feldversuch auf den mit
mykorrhizierten Gehölzen bestandenen Experimentalflächen (Flächen B und P) kein eindeutig
positiver Effekt bei der Schadstoffabreicherung im Vergleich zu der unbepflanzten Fläche nachweisen
ließ, ist die Annahme einer sich auf die Nitroaromatentransformation günstig auswirkende
Einflussnahme durch die eingebrachten Mykorrhizapartner berechtigt. Das Transformations- und
Remediationspotenzial von Mykorrhizapilzen und deren Wirtspflanzen konnte in eigenen
Laborexperimenten aufgezeigt werden (siehe Abschnitt) und wurde auch in der Literatur mehrfach

136
beschrieben (Kreslavski et al. 1999, Scheidemann 1999). Relevante Gründe für einen fehlenden
Nachweis im Feld dürften in mehrfacher Weise vorliegen und sollen nun kurz erläutert werden.
Nachteilig für eine sichere Beurteilung des Mykorrhizaeffektes ist in jedem Fall die sehr heterogen
verteilte Ausgangskontamination im Boden nach der Flächeneinrichtung. So verhielt sich die
Belastung zum Zeitpunkt t0 auf den belasteten und bearbeiteten Flächen P>B>M im Verhältnis
10:4:2. Erreichte die Nitroaromatenbelastung auf Fläche P im Mittel anfangs etwa 110 mg/kg Boden
(TS), konnten für die lediglich mechanisch bearbeitete Kontrollfläche M zu Versuchsbeginn nur 25
mg/kg Boden (TS) detektiert werden. Damit war die verhältnismäßig gering belastete Fläche M für
einen Variantenvergleich letztendlich ungeeignet. Durch den mechanischen Eingriff initiierter
pulsartiger starker TNT-Abbau, wie er auch in den Laborexperimenten beobachtet werden konnte
(siehe Abschnitt Variante K Pappelversuch und Kiefernversuch) bewirkte bis zur ersten
Messkampagne nach 6 Monaten einen nahezu vollständigen Abbau der Belastung auf M. Auch die
Flächen B und P waren diesbezüglich bis zur ersten Messung von einer weitreichenden
Dekontamination betroffen. Vergleichende Aussagen zurAbbaukinetik während der schnellen
Abreicherung sind aufgrund fehlender weiterer Messungen innerhalb des ersten Halbjahres daher
kaum zulässig. Genauere Hinweise für eine allgemein schnell einsetzende Abbaukinetik nach
Bodenbearbeitung ergaben die Laborversuche mit drastischen Rückgängen der TNT-Belastung
innerhalb weniger Wochen nach Versuchsbeginn (siehe Abschnitte 3.2 und 3.3). Nachteilig für die
Beurteilung von Mykorrhizaeffekten wirkte sich auch der insgesamt kurze Versuchszeitraum über
lediglich zwei Vegetationsperioden aus. Für in ihrer Durchführung längerfristig ausgelegte
Phytoremediationsverfahren war dieser Zeitraum im Nachhinein zu knapp bemessen. Eine
flächendeckende Durchwurzelung des Bodens auf den bepflanzten Varianten B und P ließ sich nach
eingehender Kontrolle selbst nach Beendigung des Feldexperiments nicht bestätigen. Somit dürfte
sich die Ausbildung einer Rhizosphäre mit einer assoziierten metabolisch hoch aktiven Mikroflora
lokal begrenzt auf das unmittelbare Umfeld der gesetzten Bäumchen mit ihren Wurzeln konzentriert
haben, während Bereiche außerhalb des Wurzelraums weniger von den Einflüssen der Pflanzen
partizipierten. In diesem Zusammenhang würden sich insbesondere die mit den Pflanzen verbundenen
Mykorrhizapilze mit ihren sich weit verzweigenden Hyphensystemen mittelfristig für einen Abbau der
Restkontaminationen anbieten, wie sie vor allem für Fläche P detektiert wurden.
Keine große Bedeutung bei der Nitroaromatendekontamination konnte den zubeimpften
Weißfäulepilzen im Freilandversuch zugeordnet werden. Bereits die Ergebnisse der
Laborexperimente ließen erkennen, dass die Weißfäulepilze in ihrer Funktion mit Erfolg durch
Mykorrhizapilze und deren Wirtspflanzen ersetzt werden können (siehe Variante A und B). Es stellt
sich somit die Frage nach dem Verzicht von Weißfäulepilzen in entsprechenden zukünftigen
Phytoremediationsverfahren, zumal deren Verwendung mit aufwendiger und langwieriger Anzucht auf
Zuschlagstoffen verbunden ist, welche anschließend in den Boden eingebracht werden müssen.

137
Unter dem Aspekt eines eventuell bestehenden Risikopotenzials für die Umwelt durch Verlagerung
relevanter Mengen Nitroaromaten aus dem Boden über die Wurzeln in oberirdische Sprossteile
wurde über den gesamten Versuchszeitraum ein umfangreiches Schadstoffmonitoring an den Pflanzen
durchgeführt. Hierbei ließ sich insgesamt ein nur sehr niedriger Schadstofftransfer in die gepflanzten
Gehölze nachweisen, wobei zu berücksichtigen ist, dass vermutlich nur ein Teil der inzwischen weiter
metabolisierten Nitroaromaten detektiert werden konnte, da sich die Nachweisanalytik auf GC/MS-
Untersuchungen konzentrierte. Eigene Laborexperimente mit 14C-TNT und verschiedene Ergebnisse
aus der Literatur (Sens 1998, Schönmuth und Pestemer 2000) deuten auf eine zügige und
umfassende Metabolisierung aufgenommener Schadstoffe mit nachfolgendem Einbau der gebildeten
Verbindungen in die pflanzliche Matrix hin, so dass auch bei einer Aufnahme größerer Mengen
Nitroaromaten aufgrund einer rückhaltenden Festlegung vermutlich kaum von einer Gefährdung der
Umwelt durch einen Schadstoffaustrag über die Pflanzenbiomasse auszugehen ist. Untersuchte
Freiland- und Laborpflanzen zeigten ein übereinstimmendes Belastungsspektrum mit den
Nitroaromaten TNT, 2- ADNT und 4-ADNT. Weitere Nitroaromaten wurden nicht gefunden. Der
Feldversuch ließ artspezifische Unterschiede im Aufnahmeverhalten deutlich werden. Ein Indiz dafür,
dass niedrige Aufnahmeraten nicht generell auf alle Pflanzen übertragbar sind (Schönmuth und
Pestemer 2000). Kontaminationen fanden sich fast ausschließlich in den Fichten. Im Gewebe der
Holunderpflanzen konnten keine Schadstoffe detektiert werden. Zusammenfassend betrachtet muss
über fortführende Remobilisierungsexperimente das Reversibilitäts- und Festlegungsverhalten von
Nitroaromaten in Pflanzen weiter untersucht werden, um eine Gefährdung der Umwelt bei der
Anwendung von Phytoremediationsverfahren mit letzter Sicherheit ausschließen zu können.
6.6 Sanierung TNT-belasteter Flächen durch Phytoremediation;
Anwendungsbereiche und Machbarkeit Für eine sichere und abschließende Beurteilung über den Erfolg des durchgeführten
Phytoremediationsverfahrens war der knapp zweijährige Versuchszeitraum insgesamt zu kurz. Somit
besteht der Bedarf zusätzlicher Freilandexperimente auf entsprechenden Problemstandorten, zumal
weitere vergleichbare Testverfahren bisher nicht durchgeführt wurden. Unter Berücksichtigung der
eigenen und aus der Literatur bekannten positiven Laborergebnisse soll dennoch modellhaft skizziert
werden, bei welchen Bedingungen Phytoremediation auf TNT-Altlasten einen grundlegenden
Sanierungserfolg verspricht.

138
6.6.1 Zeitrahmen und Art des Verfahrens Grundsätzlich sollten zur Anwendung kommende Verfahren bis zum Erreichen des
Sanierungserfolges auf einen Zeitraum über mehrere Jahre ausgelegt sein, da die mykorrhizierten
Pflanzen mit ihren Wurzeln und Mykorrhizapilze mit ihren Hyphensystemen mehrere
Vegetationsperioden benötigen, um den Boden flächendeckend und feinmaschig zu erschließen. Ein
guter Durchwurzelungsgrad ist Voraussetzung für eine durchgreifende Schadstofftransformation. Als
dominierende Technik beim Nitroaromatenabbau ist die Rhizodegradation anzusehen, wobei
entstehende Transformationsprodukte in der Huminstoffmatrix dauerhaft festgelegt oder von
Rhizosphärenorganismen (einschließlich Mykorrhizapilze) weiter abgebaut werden. Auch der
Phytodegradation muss Beachtung geschenkt werden. Diesbezüglich müssen Aufnahmeverhalten,
Translokation und Metaboli-sierungsvorgänge mit anschließender Festlegung der Abbauprodukte im
Pflanzenkörper noch genauer untersucht werden. Im Hinblick auf die Toxizität von Nitroaromaten
und deren schädlichen Wirkung auf Organismen sind insbesondere Flächen mit einem schwachen bis
mittleren Kontaminationsgrad für Phytoremediationsverfahren geeignet, wobei die Höhe der
Kontamination 400-500 mg TNT/ kg Boden (TS) nicht überschreiten sollte. Im Blickpunkt stehen
hierbei die großflächigen heterogen belasteten und zum Teil bewaldeten Areale, die mit
herkömmlichen ex-situ Verfahren nicht gereinigt werden können.
6.6.2 Auswahl der Organismen Eine weitere Bedingung für den Sanierungserfolg eines Verfahrens ist die richtige Auswahl von
Pflanzen- und Mykorrhizapilzspezies. Abgesehen von einer guten Wüchsigkeit der eingesetzten
Pflanzen spielt insbesondere eine gute Anpassung an die jeweiligen Standortgegebenheiten der zum
Einsatz kommenden Arten eine wichtige Rolle. So sind möglicherweise natürlich am Standort
verbreitete Pflanzen bei der Flächeneinrichtung vorzuziehen. Als Mykobionten bieten sich Isolate von
belasteten Flächen an, wobei eventuell optimierte Detoxifizierungsmechanismen nicht nur die
Abbauleistung, sondern auch das Toleranzvermögen gegenüber der Toxizität der Nitroaromaten
erhöhen. Als Gehölzpflanzen sind außerdem Arten mit einem weiten Wirtsspektrum für verschiedene
Mykorrhizapilze vorzuziehen. Eine hohe Artendiversität an Ektomykorrhizapilzen im Boden dürfte für
eine rege Enzymaktivität im Boden nur förderlich sein. Bisher sind erst wenige Ektomykor-rhizapilze
auf ihre Fähigkeit zur Biodegradation untersucht. Unter Berücksichtigung der Artenvielfalt und auch
der Variationen innerhalb einer Spezies ergibt sich möglicherweise ein enormes
Degradationspotenzial (Meharg & Cairney 2000). Von 42 getesteten ECM-Arten besaßen 33 Arten
die Fähigkeit, eine oder mehrere Klassen von Schadstoffen zu degradieren.
Durch die Verwendung tief wurzelnder Gehölze könnte zudem die Schadstoffdegradation in tieferen
Bodenschichten gesteigert werden. Vor allem Bakterien könnten von den Bedingungen in der

139
Rhizosphäre profitieren, verrottende Wurzeln als Substrat nutzen und vermehrt Schadstoffe
cometabolisch umsetzen. Entstehende Wurzelgänge wiederum erleichtern den Gasaustausch mit der
Erdoberfläche. Positiv wirkt sich möglicherweise auch aus, dass Pflanzen über die Wasseraufnahme
ihrer Wurzeln eine Verdriftung der Nitroaromaten mit dem versickernden Bodenwasser in tiefere
grundwasserführende Bodenschichten verringern. Außerdem verbessern sie die
Wasserhaltekapazität des Bodens und vermindern auch auf diesem Wege die Verlagerung von
Nitroaromaten in das Grundwasser.
Umstritten ist bisher die Verwendung genetisch modifizierter Pflanzen in der Phytoremediation, wobei
sich Befürworter der Gentechnik hiervon einen verbesserten Abbau der Schadstoffe versprechen.
Der Nachweis vielversprechender Erfolge im Freiland blieb bisher allerdings aus. Im Hinblick auf
mögliche Risiken der Gentechnik sollte bedacht werden, ob sich genetische Modifikationen in der
Phytoremediation nicht als unnötig erweisen. Nur ein sehr kleiner Teil aller vorkommenden
Gefäßpflanzen (400000 Arten bekannt) wurde bisher hinsichtlich des Abbaupotenzials für organische
Schadstoffe untersucht. Erwartet wird, dass im Laufe der Zeit zahlreiche weitere für die
Phytoremediation geeignete Pflanzen gefunden werden (Trapp & Karlson 2000).

Literatur
140
Literatur Achtnich, C.; Fernandes, E.; Bolldag, J.-M.; Knackmuss, H.-J.; Lenke, H. 1999 Covalent binding of reduced metabolites of [15N3]-TNT to soil organic matter during a bioremediation process analyzed by 15N NMR spectroscopy Environ. Sci. Technol. 33, 4448-4456 Achtnich, C.; Peters, D.; Knackmuss, H.-J.; Lenke, H. 2000 Alternierender Anaerob-/Aerob-Prozess: Analyse der Bindungsstrukturen von metabolisiertem und humifiziertem TNT im Boden Verbundvorhaben: Biologische Verfahren zur Bodensanierung Teilverbund 5: Langzeit-Remobilisierungsverhalten von Schadstoffen bei der biologischen Bodensanierung Gefördert vom BMBF 2. Statusseminar, 22. Februar 2000 an der Hochschule Bremen Adler, P. R.; Arora, R. ; Ghaouth, A. E.; Glenn, D. M.; Solar, J. M. 1994 Bioremediation of phenolic compounds from water with plant root surface peroxidases J. Environ. Qual. 23, 1113-1117 Ahlborg, G.; Bergstroem, B.; Hogstedt, C.; Einisto, P.; Sorsa, M. 1985 Urinary screening for potentially genotoxic exposures in a chemical industry Brit. J. Ind. Med. 42, 691-699 Anderson, K. E.; Hammons, G. J.; Kadlubar, F. F.; Potter, J. D.; Kaderlik, K. R.; Ilett, K. F.; Minchin, R. R.; Teitel, C. H.; Chou, H.-C.; Martin, M. V.; Guengerich, F. P.; Barone, G. W.; Lang, N. P.; Peterson, L. A. 1997 Metabolic activation of aromatic amines by human pancreas Carcinogenesis. 18, 1085-1092 Ashford, A. E.; Peterson, C. A.; Carpenter, J. L.; Cairney, J. W. G.; Allaway, W. G. 1988 Structure and permeability of the fungal sheath in the Pisonia mycorrhiza Protoplasma 147, 149-161 Ashford, A. E.; Allaway, W. G.; Peterson, C. A.; Cairney, J. W. G.; 1989 Nutrient transfer and the fungus-root interface Aust. J. Plant Physiol. 16, 85-97 Barrasa, J M.; Camarero, S.; Martinez, A. T.; Ruel, K. 1995 Ultrastructural aspects of wheat straw degradation by Phanerochaete chrysosporium and Trametes versicolor Appl. Microbiol. Biotechnol. 43, 766-770 Behrmann, P. und Heyser, W. 1992 Apoplastic transport through the fungal sheath of Pinus sylvestris/Suillus bovinus ectomycorrhizae Bot. Acta 105, 427-434

Literatur
141
Behrmann, P. 1995 Entwicklung, Struktur und Funktion der Endodermis in mykorrhizierten und nicht-mykorrhizierten Baumwurzeln unter besonderer Berücksichtigung der Kiefer (Pinus sylvestris L. ) Dissertation, Fachbereich 2 Biologie/Chemie, Universität Bremen Bending, G. D.; Read, D. J. 1997 Lignin and soluble phenolic degradation by ectomycorrhizal and ericoid mycorrhizal fungi Mycol. Res. 101, 1348-1354 Bhadra, R.; Wayment, D. G.; Hughes, J. B.; Shanks, J. V. 1999 Confirmation of Conjugation Processes during TNT-Metabolism by Axenic Plant Roots Environ. Sci. Technol. 33, 446-452 BIA-Report 7/98, Grenzwertliste 1998 Sicherheit und Gesundheitsschutz bei der Arbeit Hauptverband der gewerblichen Berufsgenossenschaften (HVBG) Blackman, L. M.; Overall, R. L. 2001 Structure and function of plamodesmata Aust. J. Plant Physiol. 28, 709-727 Böhler, A.; Gerth, A.; Eulering, B.; Thomas, H. 2000 Biologische In-situ-Sanierung TNT-kontaminierter Böden TerraTech 4/2000, 61-63 Boudet, A.-M. 2000 Lignins and lignification: Select issues Plant Physiol. Biochem. 38 (1/2), 81-96 Bradley, P. M.; Chapelle, F. H. 1995 Factors Affecting Microbial 2,4,6-Trinitrotoluene Mineralization in Contaminated Soil Environ. Sci. Technol. 29, 802-806 Braedt, W.; Horseljau, H.; Jacobs, F.; Knolle, F. 1999 Die Sprengstofffabrik ‘Tanne’ in Clausthal-Zellerfeld. Geschichte und Perspektive einer Harzer Rüstungsaltlast Papierflieger, Clausthal-Zellerfeld, S. 166 Breitung, J.; Bruns-Nagel, D.; von Löw, E.; Steinbach, K.; Haas, R.; Gemsa, D. 1995 Mikrobielle Sanierung von 2,4,6-Trinitrotoluol (TNT) kontaminierten Böden UWSF-Z. Umweltchem. Ökotox. 7 (4), 195-200 Briggs, G.; Bromilow, R. H.; Evans, A. A. 1982 Relationship between lipophilicity and root uptake and translocation of non-ionised chemicals by barley Pestic. Sci. 13, 495-504

Literatur
142
Bringmann, G.; Kühn, R. 1977 Befunde der Schadwirkung wassergefährdender Stoffe gegen Bakterien (Pseudomonas putida) und Grünalgen (Scenedesmus quadricauda) im Zellvermehrungshemmtest Z. Wasser- Abwasserforschung 10, 87-98 Bumpus, J. A.; Tien, M.; Wright, D.; Aust, S. D. 1985 Oxidation of persistent environmental pollutants by a white-rot fungus Science 228, 1434-1436 Cairney, J.W.G.; Ashford, A. E.; Allaway, W. G. 1989 Distribution of photosynthetically fixed carbon within root systems of Eucalyptus pilularis plants ectomycorrhizal with Pisolithus tinctorius New Phytol. 112, 495-500 Cairney, J. W. G.; Burke, R. M. 1998 Do ecto- and ericoid mycorrhizal fungi produce peroxidase activity? Mycorrhiza 8, 61-65 Cataldo, D. A.; Harvey, S. D.;Fellows, R. J.; Bean, R. M.; McVeety, B. D. 1989 An evaluation of the environmental fate and behavior of munitions material (TNT, RDX) in soil and plant systems Pacific Northwest laboratories, Richland, Report AD-A223 546 Cerniglia, C. E.; Somerville, C. C. 1995 Reductive metabolism of nitroaromatic and nitropolycyclic aromatic hydrocarbons In: Biodegradation of nitroaromatic compounds J. Spain (eds), Plenum Press New York, 99-115 Cheng, W.; Zhang, Q.; Coleman, D. C.; Carroll, C. R.; Hoffman, C. A. 1996 Is available carbon limiting microbiol growth in the rhizosphere? Soil Biol. Biochem. 28, 1283-1288 Coats, J. R. 1999 Phytoremediation of soils polluted with organic contaminants In: Umweltbundesamt (Hrsg.): Pflanzenbelastung auf kontaminierten Standorten Erich Schmidt, Berlin, S. 198-199 Coleman, J. O. D.; Blake-Kalff, M. M. A.; Davies, T. G. E. 1997 Detoxification of xenobiotics by plants: chemical modification and vacuolar compartmentation Trends in Plant Science 2 (4), 144-151 Colpaert, J. V.; van Laere, A. 1996 A comparison of the extracellular enzyme activities of two ectomycorrhizal and leaf-saprotrophic basidiomycetes colonizing beech leaf litter New Phytol. 134, 133-141

Literatur
143
Crawford, K. M. und Zambryski, P. C. 2000 Subcellular localization determines the availability of non-targeted proteins to pasmodesmatal transport Current Biology 10, 1032-1040 Criquet, S.; Joner, E.; Leglize, P.; Leyval, C. 2000 Anthracene and mycorrhiza affect the activity of oxidoreductases in the roots and the rhizosphere of lucerne (Medicago sativa L.) Biotechnol. Letters 22, 1733-1737 Daun, G.; Lenke, H.; Reuss, M.; Knackmuss, H.-J. 1998 Biological treatment of TNT-contaminated soil. 1. Anaerobic cometabolic reduction and interaction of TNT and metabolites with soil components Environ. Sci. Technol. 32, 1956-1963 De Jong, E. 1993 Physiological roles and metabolism of fungal aryl alcohols Dissertation, Landbau-Universität Wageningen Dittmann, J. 1999 Akkumulation und Biodegradation von im Bioschlamm vorhandener 3-Chlorbenzoesäure durch Pilze, insbesondere Mykorrhizapilze und durch Wurzeln höherer Pflanzen Dissertation FB 2 Biologie/Chemie Universität Bremen Donnelly, P. K.; Fletcher, J. S. 1995 PCB metabolism by ectomycorrhizal fungi Bull. Environ. Contam. Toxicol. 54, 507-513 Donner, B.; Heyser, W. 1992 Buchenmykorrhizen: Möglichkeiten der Elementselektion unter besonderer Berücksichtigung einiger Schwermetalle Forstw. Cbl. 108, 150-163 Downes, G. M.; Alexander, I. J.; Cairney, J. W. G. 1992 A study of ageing of spruce (Picea sitchensis (Bong.) Carr.) ectomycorrhizas. 1. Morphological and cellular changes in mycorrhizas formed by Tylospora fibrillosa (Burt.) Donk and Paxillus involutus (Batsch. ex Fr.) Fr.. New Phytol. 122, 141-152 Durall, D. M.; Todd, A. W.; Trappe, J. M. 1994 Decomposition of 14C-labelled substrates by ectomycorrhizal fungi in association with Douglas fir New Phytol. 127, 725-729 Dyer, A. 1974 An introduction to liquid szintillation counting Heyden, London

Literatur
144
Edwards, R.; Dixon, D. P.; Walbot, V. 2000 Plant glutathione S-transferases: enzymes with multiple functions in sickness and in health Trends in Plant Science 5 (5), 193-198 Elovitz, M. S.; Weber, E. J. 1999 Sediment-mediated reduction of 2,4,6-trinitrotoluene and fate of the resulting (poly)amines Environ. Sci. Technol. 33, 2617-2625 Evans, C. S.; Dutton, M. V.; Guillén, F.; Veness, R. G. 1994 Enzymes and small molecular mass agents involved with lignocellulose degradation FEMS Microbiol. Rev. 13, 235-240 Evans, C. S.; Gallagher, I. M.; Atkey, P. T.; Wood, D. 1991 Localisation of degradative enzymes in white.rot decay of lignocellulose Biodegradation 2, 93-106 Fox, B. W. 1976 Techniques of sample preparation for liquid scintillation counting. In: Laboratory techniques in biochemistry and molecular biology Hrsg. Work, T. S. & Work, E. North-Holland Publishing Company, Amsterdam Frank, A. B. 1885 Über die auf Wurzelsymbiose beruhende Ernährung gewisser Bäume durch unterirdische Pilze Ber. Deutsch. Bot. Ges. 3, 128-145 Frische, T. 2002 Screening for Soil Toxicity and Mutagenicity Using Luminescent Bacteria-A Case Study of the Explosive 2,4,6-Trinitrotoluene (TNT) Ecotoxicol. Environ. Safety 51, 133-144 Fritsche, W.; Scheibner, K.; Herre, A.; Hofrichter M. 2000 Fungal degradation of explosives: TNT and related nitroaromatic compounds In: Spain, J. C.; Hughes, J. B.; Knackmuss, H.-J. (Hrsg.) Biodegradation of nitroaromatic compounds and explosives, 213-237 CRC Press: Boca Raton, Fl., USA Fritz, E. 1980 Microautoradiographic localisation of assimilates in phloem: problems and new methods Ber. Dtsch. Bot. Ges. 93, 109-121 Garg, R.; Grasso, D.; Hoag, G. 1991 Treatment of explosives contaminated lagoon sludge Hazardous Waste & Hazardous Materials 8, 319-340

Literatur
145
Gianinazzi-Pearson, V.; Dumas-Gaudot, E.; Gollotte, A.; Tahiri-Alaoui, A.; Gianinazzi, S. 1996 Cellular and molecular defence-related root responses to invasion by arbuscular mycorrhizal fungi New Phytol. 133, 45-57 Gisi, U.; Schenker, R.; Schulin, R.; Stadelmann, F. X.; Sticher, H 1990 Bodenökologie Thieme Verlag, Stuttgart Görge, E. 1994 Aufnahme von Trinitrotoluol durch Pflanzen CPM-Seminar „Bewertung von sprengstoffspezifischen Schadstoffen auf Rüstungs- und militärischen Standorten“ 25.-26. April, 1994, Bonn Bad Godesberg, Tagungsband, 13-1 - 13-10 Görge, E.; Brandt, S.; Werner, D. 1994 Uptake and Metabolism of 2,4,6-Trinitrotoluene in Higher Plants Environ. Sci. & Pollut. Res. 1 (4), 229-233 Görge, E.; Brandt, S.; Werner, D. 1995 Aufnahme von 2,4,6-Trinitrotoluol in Pflanzen - Freilandversuche auf dem Gelände einer ehemaligen Sprengstoffabrik in Stadtallendorf UWSF - Z. Umweltchem. Ökotox. 7 (3), 139-148 Gong, P.; Wilke, B.-M.; Fleischmann, S. 1999 Soil-Based Phytotoxicity of 2,4,6-Trinitrotoluene (TNT) to Terrestrial Higher Plants Arch. Environ. Contam. Toxicol. 36, 152-157 Gong, P.; Wilke, B.-M.; Strozzi, E.; Fleischmann, S. 2001 Evaluation and refinement of a continuous seed germination and early seedling growth test for the use in the ecotoxicological assessment of soils Chemosphere 44, 491-500 Gramss, G. 1997 Activity of oxidative enzymes in fungal mycelia from grassland and forest soils J. Bas. Microbiol. 37, 407-423 Gramss, G.; Günther, T.; Fritsche, W. 1998 Spot tests for oxidative enzymes in ectomycorrhizal, wood-, and litter decaying fungi Mycol. Res. 102 (1), 67-72 Gramss, G.; Kirsche, B; Voigt, K. D.; Günther, T.; Fritsche, W. 1999 Conversion rates of five polycyclic aromatic hydrocarbons in liquid cultures in fifty-eight fungi and the concomitant production of oxidativ enzymes Mycol. Res. 103, 1009-1018

Literatur
146
Griffiths, R. P.; Caldwell, B. A. 1992 Mycorrhizal mat communities in forest soil In: Read, D. J.; Lewis, D. H.; Fitter, A. H.; Alexander, I. J. (Eds.), Mycorrhizas in Ecosystems CAB International, Wallingford, 98-105 Haderlein, S. B.; Weissmahr, K. W.; Schwarzenbach, R. P. 1996 Specific Adsorption of Nitroaromatic Explosives and Pesticides to Clay Minerals Environ. Sci. Technol. 30, 612-622 Hankenson, K.; Schaefer, D. J. 1991 Microtox assay of trinitrotoluene, diaminonitrotoluene and dinitromethylaniline mixtures Bull. Environ. Contam. Toxicol 46, 550-553 Hanne, L. F.; Kirk, L. L.; Appel, S. M.; Narayan, A. D.; Bains, K. K. 1993 Degradation and induction specificity in actinomycetes that degrade p-nitrophenol Appl. Environ. Microbiol. 59, 3505-3508 Hannink, N.; Rosser, S. J.; French, C. E.; Basran, A.; Murray, J. A. H.; Nicklin, S., Bruce, N. C. 2001 Phytodetoxification of TNT by transgenic plants expressing a bacterial nitroreductase Nature Biotechnol. 19, 1168-1172 Harley, J. L.; Harley, E. L. 1987 A Check-List of Mycorrhiza in the British Flora New. Phytol. 105, 1-102 Harris, J. C.; Hayes, M. J. 1990 Acid dissociation constant. In: W. J. Lyman; W. F. Reehl; D. H. Rosenblatt (Hrsg.) Handbook of chemical Property Estimation Methods Amer. Chem. Soc., Wash. D.C., Chapt. 6 Hartig, T. 1851 Vollständige Naturgeschichte der forstlichen Culturpflanzen Deutschlands Berlin Harvey, S. D.; Fellows, R. J.; Cataldo, D. A.; Bean, R. M. 1990 Analysis of 2,4,6-trinitrotoluene and ist transformation products in soils and plant tissues by high-performance liquid chromatography J. Chrom. 518, 361-374 Hawari, J.; Halasz, A.; Beaudet, S.; Ampleman, G.; Thiboutot, S. 1999 Biotransformation of 2,4,6-trinitrotoluene (TNT) with Phanerochaete chrysosporium in agitated cultures at pH 4.5 Appl. Environ. Microbiol. 65, 2977-2986

Literatur
147
Heinzkill, M. 1995 Isolierung und Charakterisierung von Laccasen und Peroxidasen aus Basidiomyceten der Ordnung Agaricales Dissertation, Fachbereich Biologie, Universität Kaiserslautern Heldt, H.-W. 1996 Pflanzenbiochemie Spektrum Akademischer Verlag GmbH Heidelberg Henschler, D. 1988 Gesundheitsschädliche Arbeitsstoffe VCH, Weinheim Hilber, O. 1992 Angewandte Mykorrhizaforschung an der Versuchsanstalt für Pilzanbau AFZ 8, 366 Hüttermann, A.; Loske, D.; Majcherczyk A 1988 Der Einsatz von Weißfäulepilzen bei der Sanierung von besonders problematischen Altlasten In: Thomé-Kozmiensky, K. J. (ed.) Altlasten 2 EF-Verlag, Berlin Hughes, J. B.; Shanks, J.; Vanderford, M.; Lauritzen, J.; Bhadra, R. 1997 Transformation of TNT by aquatic plants and plant tissue cultures Environ. Sci. Technol. 31, 266-271 Ingestad, T. 1960 Studies on the nutrition of forest tree seedlings. III Mineral nutrition of pine Physiol. Plant. 13, 513-533 Isbister, J. D.; Anspach, G. L.; Kitchens, J. F.; Doyle, R. C. 1984 Composting for decontamination of soils containing explosives Microbiologica 7, 47-73 Johansen, A.; Jakobsen, I.; Jensen, E. S. 1993 External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifolium subterraneum L. New Phytol. 124, 61-68 Joner, E. J.; Ravnskov, S.; Jakobsen, I. 2000 Arbuscular mycorrhizal phosphate transport under monoxenic conditions using radio-labelled inorganic and organic phosphate Biotechnol. Letters 22, 1705-1708

Literatur
148
Joner, E. J.; Johansen, A.; Loibner, A. P.; Dela Cruz, M. A.; Szolar, O. H. J.; Portal, J. M.; Leyval, C. 2001 Rhizosphere Effects on Microbial Community Structure and Dissipation and Toxicity of Polycyclic Aromatic Hydrocarbons (PAHs) in Spiked Soil Environ. Sci. Technol. 35, 2773-2777 Jonsson, L.; Dahlberg, A.; Nilsson, M.-C.; Zackrisson, O.; Karen, O. 1999 Ectomycorrhizal fungal communities in late-successional Swedish boreal forests and composition following wildfire Mol. Ecol. 8, 205-215 Kadiyala, V.; Spain, J. C. 1998 A two-component monooxygenase catalyzes both the hydroxylation of p-nitrophenol and the oxidative release of nitrite from 4-nitrocatechol in Bacillus sphaericus JS905 Appl. Environ. Microbiol. 64, 2479-2484 Koehler, H.; Frische, T.; Dobner, I.; Behrend, P.; Schaefer, M.; Taubner, H.; Jastorff, B.; Warrelmann, J.; Walter, U. 2001a Erprobung und Erfolgskontrolle eines Phytoremediationsverfahrens zur Sanierung Sprengstoff-kontaminierter Böden Teil II: Ergebnisse eines Freilandexperimentes UWSF-Z. Umweltchem. Ökotox. 13 (5), 291-300 Koehler, H.; Warrelmann, J.; Behrend, P.; Dobner, I.; Frische, T.; Heyser, W.; Jastorff, B.; Walter, U. 2001b Erprobung und Erfolgskontrolle eines Phytoremediationsverfahrens zur Sanierung Sprengstoff-kontaminierter Böden Teil III: Beurteilung des Verfahrens UWSF-Z. Umweltchem. Ökotox. 13 (6), 359-368 Koss, G.; Lommel, A.; Ollroge, I.; Tesseraux, I.; Haas, R.; Kappos, A. D. 1989 Zur Toxikologie der Nitrotoluole und weiterer Nitroaromaten aus rüstungsbedingten Altlasten Bundesgesundheitsblatt 32, 527-536 Kottke, I.; Guttenberger, M.; Hampp, R.; Oberwinkler, S. 1987 An in vitro method for establishing mycorrhizae on coniferous tree seedlings Trees 1, 191-194 Kottke, I.; Oberwinkler, F. 1987a The Cellular structure and function of the Hartig net: coenocytic and transfer cell-like organization Nord. J. Bot. 2, 85-95 Kraß, J. D. 1998 2,4,6-Trinitrotoluol und relevante Metaboliten: Untersuchungenzu chemischen Eigenschaften und biologischen Wirkungen unter besonderer Berücksichtigung des Mediums Boden Wissenschaft & Technik Verlag Berlin

Literatur
149
Kreslavski, V. D.; Vasilyeva, G. K.; Comfort, S. D.; Drijber, R. A.; Shea, P. J. 1999 Accelerated Transformation and Binding of 2,4,6-trinitrotoluene in Rhizosphere Soil Bioremed. J. 2, 59-67 Krishnan, G.; Horst, G. L.; Darnell, S.; Powers, W. L. 2000 Growth and development of smooth bromegrass and tall fescue in TNT-contaminated soil Environmental Pollution 107, 109-116 Laurent, F. M. G.; Scalla, R. 1999 Metabolism and cell wall incorporation of phenoxyacetic acid in soybean cell suspension culture Pesticide Science 55, 3-10 Layton, D.; Mallon, B.; Mitchell, W.; Hall, L.; Fish, R.; Perry, L.; Snyder, G.; Bogen, K.; Malloch, W.; Ham, C.; Dowd, P. 1987 Conventional weapons demilitarization: A health and environmental effects data-base assessment. Explosives and their co-contaminants. Final Report, Phase II. U. S. Army Medical Research & Development Laboratory. Frederick, Maryland, USA, Dec.1987, UCRL-21109 Lenke, H.; Wagener, B.; Daun, G.; Knackmuss, H.-J. 1994 TNT-contaminated soil: A sequential anaerob/aerob process for bioremediation. In: Symposium on Biodegradation of Nitroaromatic Compounds 21.-23. May, San Diego, Abstract Booklet Lenke, H. 2001 Mikrobieller Abbau von aromatischen Nitroverbindungen und Explosivstoffen und dessen Relevanz für einen technischen Einsatz Habilitationsschrift, Fakultät Geo- und Biowissenschaften Universität Stuttgart, Fraunhofer IRB Verlag Lewis, N. G.; Yamamoto, E. 1990 Occurrence, biogenesis and biodegradation Annual Review of Plant Physiology and Plant Molecular Biology 41, 455-496 Li, A. Z.; Marx, K. A.; Walker, J.; Kaplan, D. L. 1997 Trinitrotoluene and metabolites binding to humic acid Environ. Sci. Technol. 31, 584-589 Lucas, W. J. 1997 Application of microinjection techniques to plant nutrition Plant and Soil 196, 175-189 Ludewig, R.; Lohs, K. 1991 Akute Vergiftungen Gustav Fischer, Jena, 8. Auflage, S. 328-333

Literatur
150
Majcherczyk, A.; Zeddel, A.; Kelschebach, M.; Loske, D.; Hüttermann, A. 1993 Abbau von Polyzyklischen Aromatischen Kohlenwasserstoffen durch Weißfäulepilze BioEngineering 9 (2), 27-31 Marschner, H. 1995 Mineral nutrition of higher plants 2th Ed., Academic Press, London Martinetz, D.; Rippen, G. 1996 Handbuch der Rüstungsaltlasten Ecomed Landsberg/Lech Marx, D. H. 1969 The influence of ectotrophic mycorrhizal fungi on the resistance of pine roots to pathogenic infections. 1. Antagonism of mycorrhizal fungi to root pathogenic fungi and soil bacteria Phytpathology 59, 159-163 Massicotte, H. B.; Ackerley, C. A.; Perterson, R. L. 1987 The root-fungus interface as an indicator of symbiont interaction in ectomycorrhizae Can. J. For. Res. 17, 846-854 McArthur, D. A. J.; Knowles, N. R. 1992 Resistance responses of potato to vesicular-arbuscular mycorrhizal fungi under varying abiotic phosphorus levels Plant Physiol. 100, 341-351 Meharg, A. A.; Dennis, G. R.; Cairney, J. W. G. 1997a Biotransformation of 2,4,6-trinitrotoluene (TNT) by ectomycorrhizal basidiomycetes Chemosphere 35 (3), 513-521 Meharg, A. A.; Cairney, J. W. G.; Maguire, N. 1997b Mineralization of 2,4-dichlorophenol by ectomycorrhizal fungi in axenic culture and in symbiosis with pine Chemosphere 34, 2495-2504 Meharg, A. A.; Cairney, W. G. 2000 Ectomycorrhizas - extending the capabilities of rhizosphere remediation? Soil Biol. Biochem. 32, 1475-1484 Michels, J.; Gottschalk, G. 1994 Inhibition of the lignin peroxidase of Phanerochaete chrysosporium by hydrxylamino-dinitrotoluene, an early intermediate in the degradation of 2,4,6-trinitrotoluene Appl. Environ. Microbiol. 60, 187-194

Literatur
151
Michels, J.; Gottschalk. G. 1995 Pathway of 2,4,6-trinitrotoluene (TNT) degradation by Phanerochaete chrysosporium In: Spain, J. C. (ed.) 1995, 135-149 Biodegradation of nitroaromatic compounds Environmental Science Research Series, Vol.49, Plenum Press Miskiniene, V.; Sarlauskas, J.; Jacquot, J. P.; Cenas, N. 1998 Nitroreductase reactions of Arabidopsis thaliana thioredoxin reductase Biochemica et Biophysica Acta 1366, 275-283 Moeschlin, S 1986 Klinik und Therapie der Vergiftungen Georg Thieme, Stuttgart, 7. Auflage, S. 408-409 Müller, W. F.; Bedell, G. W.; Baker, B; Shojaee, S. & Jackson, P. J. 1993 Bioremediation of TNT wastes by higher Plants Proc. WERC Technology Development Conference, pp 192-202 Nenninger, A. 1995 Methode zur Abschätzung des apoplastischen pH-Wertes am Interface von Pinus sylvestris / Suillus bovinus (Fr.) O. Kuntze Ektomykorrhizen über die Aufnahme von Fluoreszenzfarbstoffen Diplomarbeit, Universität Bremen Nenninger, A. 1999 Ermittlung apoplastischer pH-Werte in ektotroph mykorrhizierten Wurzeln der Kiefer mit Hilfe der konfokalen Laser Scan Mikroskopie (CLSM) Dissertation, Uni-Bremen, Fachbereich Biologie/Chemie Newman, L. A.; Doty, S. L.; Gery, K. L.; Heilman, P. E.; Muiznieks, I.; Shang, T. Q.; Siemieniec, S. T.; Strand, S. E.; Wang, X.; Wilson, A. M.; Gordon, M. P. 1998 Phytoremediation of organic contaminants: A review of phytoremediation research at the university of Washington J. Soil Contam. 7, 531-542 Nishino, S. F.; Spain, J.C.; He, Z. 2000a Strategies for aerobic degradation of nitroaromatic compounds by bacteria: Process discovery to field application. In Spain, J. C.; Hughes, J. B.; Knackmuss, H. J. (Hrsg.) Biodegradation of nitroaromatic compounds and explosives CRC Press: Boca Raton, USA, 7-61 Nishino, S. F.; Paoli, G.; Spain, J.C. 2000b Aerobic biodegradation of dinitrotoluenes and pathway for bacterial degradation of 2,6-dinitrotoluene Appl. Environ. Microbiol. 66, 2139-2147

Literatur
152
O’Keefe, D. M.; Sylvia, D. M. 1992 Chronology and mechanisms of P uptake by mycorrhizal sweet potato plants New Phytol. 122, 651-659 Pailer, M. 1960 Natürlich vorkommende Nitroverbindungen Fortschritte der Chemie organischer Naturstoffe 18, 55-82 Palazzo, A. J. & Legget, D. C. 1983 Toxicity, uptake, translocation and metabolism of TNT by plants Technical Report, US Army Cold Regions Research and Engineering Laboratory, Hannover Palazzo, A. J. & Legget, D. C. 1986 Effect and disposition of TNT in a terrestrial plant Journal of Environmental Quality 15, 49-52 Paul, E. A.; Clark, F. E. 1989 Soil microbiology and biochemistry Academic Press, San Diego Pennington, J. C.; Patrick, W. H. 1990 Adsorption and desorption of 2,4,6-trinitrotoluene by soils J. Environ. Qual. 19, 559-567 Peterson, M. M.; Horst, G. L.; Shea, P. J.; Comfort, S. D.; Peterson, R. K. D. 1996 TNT and 4-Amino-2,6-Dinitrotoluene influence on germination and early seedling development of Tall Fescue Environmental Pollution 93 (1), 57-62 Peterson, M. M.; Horst, G. L.; Shea, P. J.; Comfort, S. D., 1998 Germination and seedling development of switchgrass and smooth bromegrass exposed to 2,4,6-trinitrotoluene Environmental Pollution 99, 53-59 Peterson, T. A.; Swanson, E. S.; Hull, R. J. 1986 Use of lanthanum to trace apoplastic solute transport in intact plants J. Exp. Bot. 37, 807-822 Pörschmann, J.; Stottmeister, U. 1993 Methodical investigation of interactions between organic pollutants and humic organic material in coal wastewaters Chromatographia 36, 207-211 Preuß, J.; Haas, R.; Koss, G. 1988 Altstandorte, Altablagerungen, Altlasten Geogr. Rdsch. 6, 31-38

Literatur
153
Rastrick, H. 1949 Chloromycetin: Its structure and synthesis Nature 163, 553-554 Reid, I. D. 1995 Biodegradation of lignin Can. J. Bot. 73, 1011-1018 Richardson, A.; McDougall, G. J. 1997 A laccase-type polyphenol oxidase from lignifying xylem of tobacco Phytochemistry 44, 229-235 Richter,G. 1998 Stoffwechselphysiologie der Pflanzen 5. Auflage, Georg Thieme Verlag Stuttgart Rippen, G. 1994 Handbuch Umweltchemikalien - Stoffdaten, Prüfverfahren, Vorschriften (3. Aufl.) ecomed Verlag, Landsberg. Stand: 26. Erg.- Lfg., 10/94 Robards, A. W.; Lucas, W. J. 1990 Plasmodesmata Annual Review of Plant Physiology and Molecular Biology 41, 369-419 Rock, S. 1999 Possibilities and limitations of phytoremediation/ Möglichkeiten und Grenzen der Phytoremediation In: Umweltbundesamt (Hrsg.): Pflanzenbelastung auf kontaminierten Standorten Erich Schmidt, Berlin, S. 144-156 Romantschuk, M.; Sarand, I.; Petanen, T.; Peltola, R.; Jonsson-Vihanne, M.; Koivula, T.; Yrjala, K.; Haahtela, K. 2000 Means to improve the effect of in situ bioremediation of contaminated soil: an overview of novel approaches Environ. Pollut. 107, 179-185 Rouillon, R.; Poulain, C.; Bastide, J.; Coste, C. M. 1989 Degradation of the herbicide chlorpropham by some ectomycorrhizal fungi in pure culture Agricul. Ecosys. Environ. 28, 421-424 Salzer, P.; Corbiere, H.; Boller, T. 1999 Hydrogen peroxide accumulation in Medicago truncatula roots colonized by the arbuscular mycorrhiza-forming Glomus intraradices Planta 208, 319-325

Literatur
154
Sandermann, H. Jr. 1994 Higher plant metabolism of xenobiotics: the ´green liver` concept Pharmacogenetics 4, 225-241 Sanderson, J. 1981 Modified development to improve the performance of AR-10 stripping emulsions for use with the more energetic isotopes J. Microsc. 124, 177-182 Schackmann, A. V.; Müller, R. 1991 Reduction of nitroaromatic compounds by different Pseudomonas species under aerobic conditions Appl. Microbiol. Biotechnol. 34, 809-813 Schäfer, H 1992 Ableitung von Sanierungsrichtwerten für Böden mit spezifischen Belastungen aus der TNT-Produktion. In: Spyra, W.; Lohs, K.; Preussner, M.; Rüden, H.; Thomé-Kozmiensky, K. J. (Hrsg.): Verfahren zur Sanierung von Rüstungsaltlasten EF-Verlag, Berlin, S. 45-57 Scheel, D., Sandermann, H. Jr. 1981 Metabolism of 2,4-dichlorophenoxyacetic acid in cell suspension cultures of soybean (Glycine max L) and wheat (Triticum aestivum L.) II. Evidence for incorporation into lignin Planta 152, 248-252 Scheffer, F; Schachtschabel, P. 1984 Lehrbuch der Bodenkunde Ferdinand Enke Verlag Stuttgart, 11. Auflage Scheibner, K.; Hofrichter, M.; Herre, A.; Michels, J.; Fritsche, W. 1997 Screening for fungi intensively mineralizing 2,4,6-trinitrotoluene Appl. Microbiol. Biotechnol. 47, 452-457 Scheibner, K.; Hofrichter, M.; Fritsche, W. 1997a Mineralization of 2-amino-4,6-dinitrotoluene by manganese peroxidase of the white-rot fungus Nematoloma frowardii Biotechnol. Lett. 19, 835-839 Scheidegger, C.; Brunner, I. 1995 Electron microscopy of ectomycorrhiza: methods, applications and findings In: Varma, A.; Hock, B. (Hrsg.): Mycorrhiza: structure, function, molecular biology and biotechnology Springer Verlag Berlin, Heidelberg, S. 205-228 Scheidemann, P. 1998 Aufnahme und Metabolisierung von Trinitrotoluol (TNT) im System Pflanze/Boden Dissertation, Philipps-Universität Marburg

Literatur
155
Scheidemann, P.; Klunk, A.; Sens, C.; Werner, D. 1998 Species Dependent Uptake and Tolerance of Nitroaromatic Compounds by Higher Plants J. Plant Physiol. Vol. 152, 242-247 Schmidt, B.; Breuer, J; Thiede, B., Shuphan, I. 1997 Metabolism of the pesticide metabolite 4-[U-14C] nitrophenol in Datura stramonium (L.) cell suspension culture - A model for processing of xenobiotic carbohydrate conjugates and formation of nonextractable residues Pestic. Biochem. Physiol. 57, 109-118 Schneider, K.; Oltmanns, J.; Radenberg, T.; Schneider, T.; Pauly-Mundegar, D. 1996 Uptake of nitroaromatic compounds in plants - Implications for risk assesment of ammunition sites Environ. Sci. Pollut. Res. 3, 135-138 Schnoor, J. L.; Licht, L. A.; McCutcheon, S. C.; Wolfe, N. L.; Carreira, L. H. 1995 Phytoremediation of Organic and Nutrient Contaminants Pilot and full-scale studies are demonstrating the promise and limitations of using vegetation for remediating hazardous wastes in soils and sediments. Environmental Science & Technology 29 (7), 318A-323A Schnoor, J. L. 1997 Phytoremediation. Technology Evaluation Report to the Ground-Water Remediation Technologies Center Schönmuth, B.; Pestemer, W. 2000 Aufnahme und Verbleib von 14[C]-Trinitrotoluol in Weiden und Fichten 2. Symposium, Natural Attenuation 7.-8.12.2000, Dechema, Tagungsband Schwab, S. M.; Leonard, R. T.; Menge, J. A. 1983 Quantitative and qualitative comparison of root exudates of mycorrhizal and nonmycorrhizal plant species Can. J. Bot. 62, 1227-1231 Seifert, U.; Schecker, H.G. 1993 Selbsteinkapselung von organischen Schadstoffgemischen unterschiedlich flüchtiger Komponenten in Böden UWSF-Z. Umweltchem. Ökotox. 5 (2), 81-89 Shimp, J. F.; Tracy, J. C.; Davis, L. C.; Lee, E.; Huang, W.; Erickson, L. E. 1993 Beneficial effects of plants in the remediation of soil and groundwater contaminated with organic materials Crit. Rev. Environ. Sci. Technol. 23 (1), 41-77 Shone, M.G.T.; Wood, A.V. 1972 A comparison of the uptake and translocation of some organic herbicides and a systemic fungicide by barley. I. Absorbtion in relation to physico-chemical properties J. Exp. Bot. 25, 390-400

Literatur
156
Sens, C. 1998 Metabolisierung und Lokalisierung von 14C-Trinitrotoluol (TNT) in Phaseolus vulgaris (Buschbohne) und Triticum aestivum (Weizen) Dissertation, Fachbereich Biologie, Philipps-Universität Marburg Sens, C.; Scheidemann, P.; Werner, D. 1999 The distribution of 14C-TNT in different biochemical compartments of the monocotyledonous Triticum aestivum Environmental Pollution 104, 113-119 Smith, F. A.; Raven, J. A. 1979 Intracellular pH and ist regulation Ann. Rev. Plant Physiol. 30, 289-311 Smith, S. E.; Read, D. J. 1997 Mycorrhizal Symbiosis Second Edition, Academic Press, London Spanggord, R.J.; Mortelmans, K. E.; Griffin, A. F.; Simmon, V. F. 1982 Mutagenicity in Salmonella typhimurium and structure-activity relationships of wastewater components emanating from the manufacture of trinitrotoluene Environ. Mutag. 4, 163 Spanggord, R. J.; Spain, J. C.; Nishino, S. F.; Mortelmans, K. E. 1991 Biodegradation of 2,4-dinitrotoluene by a Pseudomonas sp. Appl. Environ. Microbiol. 57, 3200-3205 Spiker, J. K.; Crawford, D. L.; Crawford, R. L. 1992 Influence of 2,4,6-Trinitrotoluene (TNT) Concentration on the Degradation of TNT in Explosive-Contaminated Soils by the White Rot Fungus Phanerochaete chrysosporium Appl. Environ. Microbiol. 58 (9), 3199-3202 Spreinat, A.; Abken, H. J.; Tölle, K.; Gottschalk, G. 1998 Sanierung von Rüstungsaltlasten durch Pilze Terra Tech 3/98, Vereinigte Fachverlage, Mainz, S. 46-49 Spurr, A. R. 1969 A low-viscosity epoxy resin embedding medium for electron microscopy J. Ultrastr. Res. 26, 31-43 Stahl, J. D.; Aust, S. D. 1993a Metabolism and detoxification of TNT by Phanerochaete chrysosporium Biochem. Biophy. Res. Com. 192 (2), 477-482 Stahl, J. D.; Aust, S. D. 1993b Plasma membrane dependent reduction of 2,4,6-trinitrotoluene by Phanerochaete chrysosporium Biochem. Biophys. Res. Communicat. 192 (2), 471-476

Literatur
157
Steffens, F.; Arendholz, W. R.; Storrer, J. G. 1994 Die Ektomykorrhiza: Eine Symbiose unter der Lupe BIUZ, 24 Jahrg., Nr.4, 211-218 Steudle, E.; Oren, R.; Schulze, E. D. 1987 Water transport in maize roots Plant Physiol. 84, 1220-1232 Tadros, M. G.; Crawford, A.; Mateo-Sullivan, A.; Zhang, J.; Hughes, J. B. 2000 Toxic Effects of Hydroxylamino Intermediates from Microbial Transformation of Trinitrotoluene and Dinitrotoluenes on Algae Selenastrum capricornutum Bull. Environ. Contam. Toxicol. 64, 579-585 Thaller, V.; Turner, J. L. 1972 Natural acetylenes. XXXV. Polyacetylenic acid and benzenoid metabolites from cultures of the fungus Lepista diemii Singer J. Chem. Soc. Perkin 1 16, 2032-2034 Thomas, H.; Gerth, A.; Eulering, B.; Böhler, A. 2001 Neue Erkenntnisse zur biologischen In-situ-Sanierung TNT-kontaminierter Böden TerraTech 10, 52-54 Thompson, P. L.; Ramer, L. A.; Schnoor, J. L. 1998 Uptake and Transformation of TNT by Hybrid Poplar Trees Environ. Sci. Technol. 32, 975-980 Thorn, K. A.; Pettigrew, P. J.; Goldenberg, W. S.; Weber, E. J. 1996 Covalent binding to humic substances. 2. 15N NMR studies of nucleophilic addition reactions Environ. Sci. Technol. 30, 2764-2775 Timonen, S.; Sen, R 1998 Heterogeneity of fungal and plant enzyme expression in intact Scots pine-Suillus bovinus and -Paxillus involutus mycorrhizospheres developed in natural forest humus New Phytol. 138, 355-366 Trapp, S.; Matthies, M.; Kaune, A. 1994 Transfer von PCDD/F und anderen organischen Umweltchemikalien im System Boden-Pflanze-Luft: I. Modellierung des Transferverhaltens Z. Umweltchem. Ökotox. 6 (1), 31-40 Trapp, S. 1995 Model for uptake of xenobiotics into plants In: Trapp, S.; McFarlane, C (Hrsg.): Plant Contamination: Modeling and Simulation of Organic Chemical Processes Lewis Pub., Boca Raton, 107-151

Literatur
158
Trapp, S. 2000 Modelling uptake into roots and subsequent translocation of neutral and ionisable organic compounds Pest Management Science 56, 767-778 Trapp, S.; Karlson, U. 2000 Phytoremediation organischer Schadstoffe UWSF- Z. Umweltchem. Ökotox. 12 (5), 245-255 Van Aken, B.; Stahl, J. D.; Naveau, H.; Spiros, N. A.; Aust, S. 2000 Transformation of 2,4,6-Trinitrotoluene (TNT) Reduction Products by Lignin Peroxidase (H8) from the White Rot Basidiomycete Phanerochaete chrysosporium Biorem. J. 4 (2), 135-145 Voudrias, E. A.; Assaf, K. S. 1996 Theoretical evaluation of dissolution and biochemical reduction of TNT for phytoremediation of contaminated sediments Journal of Contaminant Hydrology 23, 245-261 Wallnöfer, P. R.; Preiß, U.; Ziegler, W.; Engelhardt, G. 1996 Konjugatbildung organischer Schadstoffe in Pflanzen UWSF-Z. Umweltchem. Ökotox. 8, 43-46 Walter U.; Fischer, D.; Warrelmann, J.; Heyser, W.; Henke, G.A. 1998 Anaerob/Aerob-Verfahren zur biologischen Behandlung sprengstoffkontaminierter Böden Terra Tech 3/98, Vereinigte Fachverlage, Mainz, S. 42-45 Warrelmann, J.; Koehler, H.; Frische, T.; Dobner, I.; Walter, U.; Heyser, W. 2000 Erprobung und Erfolgskontrolle eines Phytoremediationsverfahrens zur Sanierung Sprengstoff-kontaminierter Böden Teil I: Konzeption und Einrichtung eines Freilandexperimentes UWSF-Z. Umweltchem. Ökotox. 12 (6), 351-357 Werner, D. 1987 Pflanzliche und mikrobielle Symbiosen Thieme Verlag Stuttgart Winterberg, R.; von Löw, E.; Held, T. 1998 Dynamisches Mietenverfahren zur Sanierung von Rüstungsaltlasten Terra Tech 3/98, Vereinigte Fachverlage, Mainz, S. 39-41 Wolff, H.-J. 1989 Die Allendorfer Sprengstoffwerke DAG und WASAG 2. Aufl., Magistrat der Stadt Stadtallendorf, Stadtallendorf

Literatur
159
Yoshitomi, K. J.; Shann, J. R. 2001 Corn (Zea mays L.) root exudates and their impact on 14C-pyrene mineralization Soil. Biol. Biochem. 33, 1769-1776 Zepp, R. G.; Schlotzhauer, P. F.; Sink, R. M. 1985 Photosensitized transformation involving electronic energy transfer in natural waters: role of humic substances Environ. Sci. Technol. 19, 74-81 Zhang, D.; Yang, Y; Leaky, E. A.; Cerniglia, C. E. 1996 Phase I and phase II enzymes produced by Cunninghamella elegans for the metabolism of xenobiotics FEMS Microbiology Letters 138, 221-226 Zhao, B.; Trouvelot, A.; Gianinazzi, S.; Gianinazzi-Pearson, V. 1997 Influence of two legume species on hyphal production and activity of two arbuscular mycorrhizal fungi Mycorrhia 7, 179-185
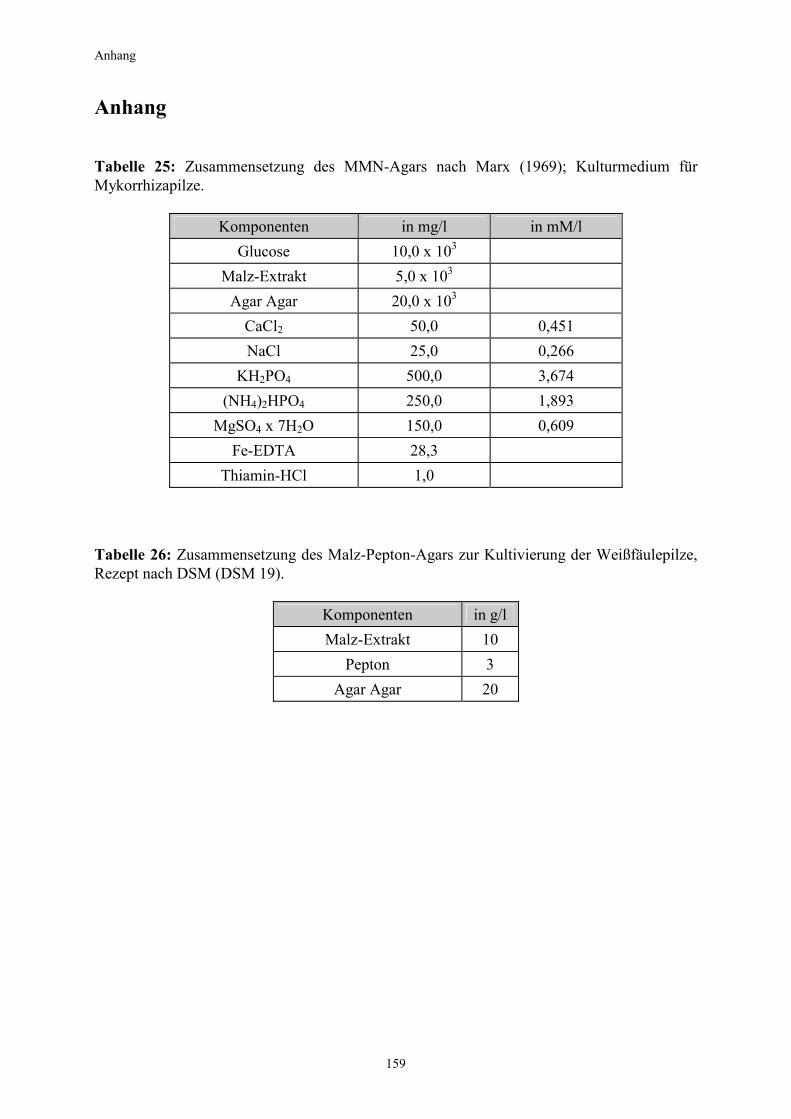
Anhang
Anhang Tabelle 25: Zusammensetzung des MMN-Agars nach Marx (1969); Kulturmedium für Mykorrhizapilze.
Komponenten in mg/l in mM/l Glucose 10,0 x 103
Malz-Extrakt 5,0 x 103 Agar Agar 20,0 x 103
CaCl2 50,0 0,451 NaCl 25,0 0,266
KH2PO4 500,0 3,674 (NH4)2HPO4 250,0 1,893
MgSO4 x 7H2O 150,0 0,609 Fe-EDTA 28,3
Thiamin-HCl 1,0 Tabelle 26: Zusammensetzung des Malz-Pepton-Agars zur Kultivierung der Weißfäulepilze, Rezept nach DSM (DSM 19).
Komponenten in g/l Malz-Extrakt 10
Pepton 3 Agar Agar 20
159
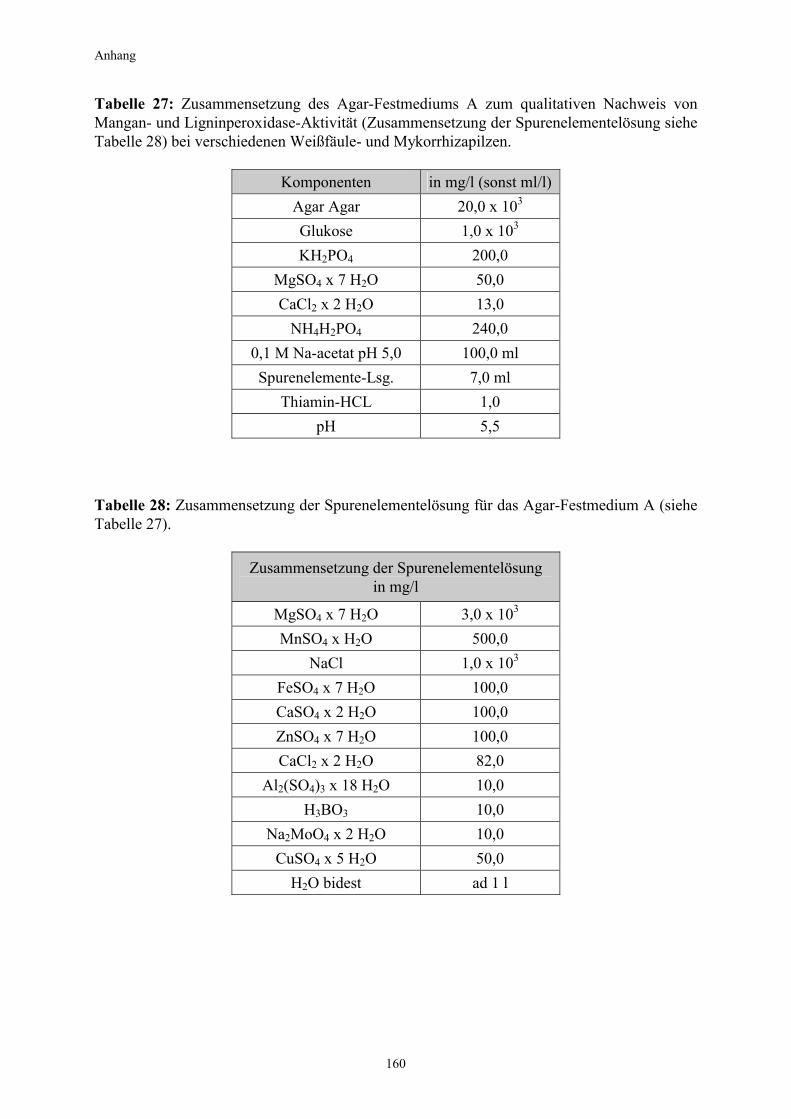
Anhang
Tabelle 27: Zusammensetzung des Agar-Festmediums A zum qualitativen Nachweis von Mangan- und Ligninperoxidase-Aktivität (Zusammensetzung der Spurenelementelösung siehe Tabelle 28) bei verschiedenen Weißfäule- und Mykorrhizapilzen.
Komponenten in mg/l (sonst ml/l) Agar Agar 20,0 x 103 Glukose 1,0 x 103 KH2PO4 200,0
MgSO4 x 7 H2O 50,0 CaCl2 x 2 H2O 13,0
NH4H2PO4 240,0 0,1 M Na-acetat pH 5,0 100,0 ml Spurenelemente-Lsg. 7,0 ml
Thiamin-HCL 1,0 pH 5,5
Tabelle 28: Zusammensetzung der Spurenelementelösung für das Agar-Festmedium A (siehe Tabelle 27).
Zusammensetzung der Spurenelementelösung in mg/l
MgSO4 x 7 H2O 3,0 x 103 MnSO4 x H2O 500,0
NaCl 1,0 x 103 FeSO4 x 7 H2O 100,0 CaSO4 x 2 H2O 100,0 ZnSO4 x 7 H2O 100,0 CaCl2 x 2 H2O 82,0
Al2(SO4)3 x 18 H2O 10,0 H3BO3 10,0
Na2MoO4 x 2 H2O 10,0 CuSO4 x 5 H2O 50,0
H2O bidest ad 1 l
160
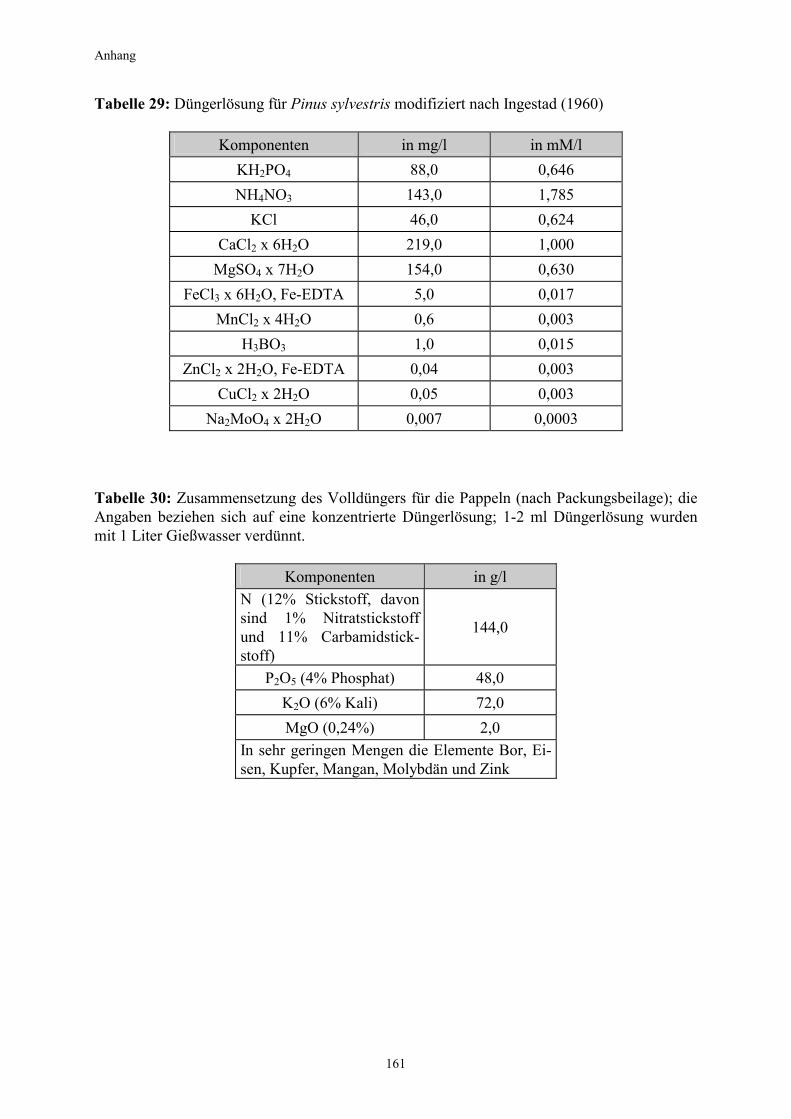
Anhang
Tabelle 29: Düngerlösung für Pinus sylvestris modifiziert nach Ingestad (1960)
Komponenten in mg/l in mM/l KH2PO4 88,0 0,646 NH4NO3 143,0 1,785
KCl 46,0 0,624 CaCl2 x 6H2O 219,0 1,000
MgSO4 x 7H2O 154,0 0,630 FeCl3 x 6H2O, Fe-EDTA 5,0 0,017
MnCl2 x 4H2O 0,6 0,003 H3BO3 1,0 0,015
ZnCl2 x 2H2O, Fe-EDTA 0,04 0,003 CuCl2 x 2H2O 0,05 0,003
Na2MoO4 x 2H2O 0,007 0,0003 Tabelle 30: Zusammensetzung des Volldüngers für die Pappeln (nach Packungsbeilage); die Angaben beziehen sich auf eine konzentrierte Düngerlösung; 1-2 ml Düngerlösung wurden mit 1 Liter Gießwasser verdünnt.
Komponenten in g/l N (12% Stickstoff, davon sind 1% Nitratstickstoff und 11% Carbamidstick-stoff)
144,0
P2O5 (4% Phosphat) 48,0 K2O (6% Kali) 72,0 MgO (0,24%) 2,0
In sehr geringen Mengen die Elemente Bor, Ei-sen, Kupfer, Mangan, Molybdän und Zink
161
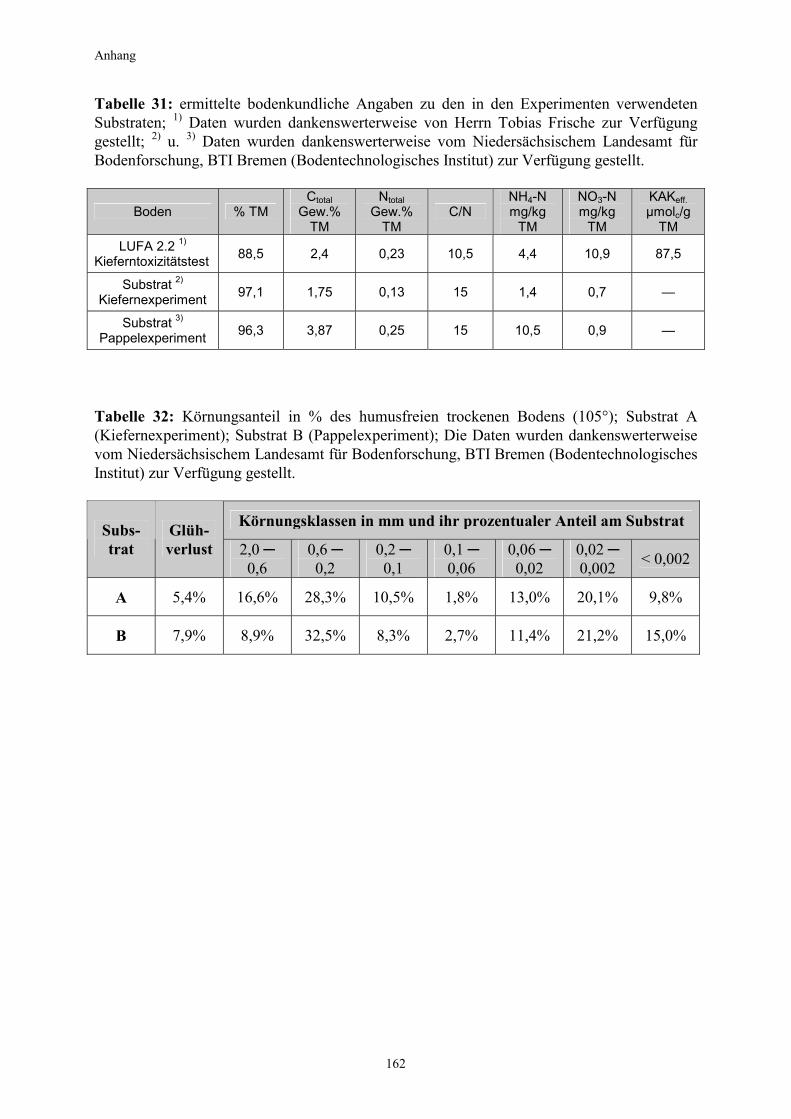
Anhang
Tabelle 31: ermittelte bodenkundliche Angaben zu den in den Experimenten verwendeten Substraten; 1) Daten wurden dankenswerterweise von Herrn Tobias Frische zur Verfügung gestellt; 2) u. 3) Daten wurden dankenswerterweise vom Niedersächsischem Landesamt für Bodenforschung, BTI Bremen (Bodentechnologisches Institut) zur Verfügung gestellt.
Boden % TM Ctotal
Gew.% TM
Ntotal Gew.%
TM C/N
NH4-N mg/kg
TM
NO3-N mg/kg
TM
KAKeff. µmolc/g
TM LUFA 2.2 1)
Kieferntoxizitätstest 88,5 2,4 0,23 10,5 4,4 10,9 87,5
Substrat 2) Kiefernexperiment 97,1 1,75 0,13 15 1,4 0,7 —
Substrat 3) Pappelexperiment 96,3 3,87 0,25 15 10,5 0,9 —
Tabelle 32: Körnungsanteil in % des humusfreien trockenen Bodens (105°); Substrat A (Kiefernexperiment); Substrat B (Pappelexperiment); Die Daten wurden dankenswerterweise vom Niedersächsischem Landesamt für Bodenforschung, BTI Bremen (Bodentechnologisches Institut) zur Verfügung gestellt.
Körnungsklassen in mm und ihr prozentualer Anteil am Substrat Subs- trat
Glüh- verlust 2,0 ─
0,6 0,6 ─ 0,2
0,2 ─ 0,1
0,1 ─ 0,06
0,06 ─ 0,02
0,02 ─ 0,002 < 0,002
A 5,4% 16,6% 28,3% 10,5% 1,8% 13,0% 20,1% 9,8%
B 7,9% 8,9% 32,5% 8,3% 2,7% 11,4% 21,2% 15,0%
162
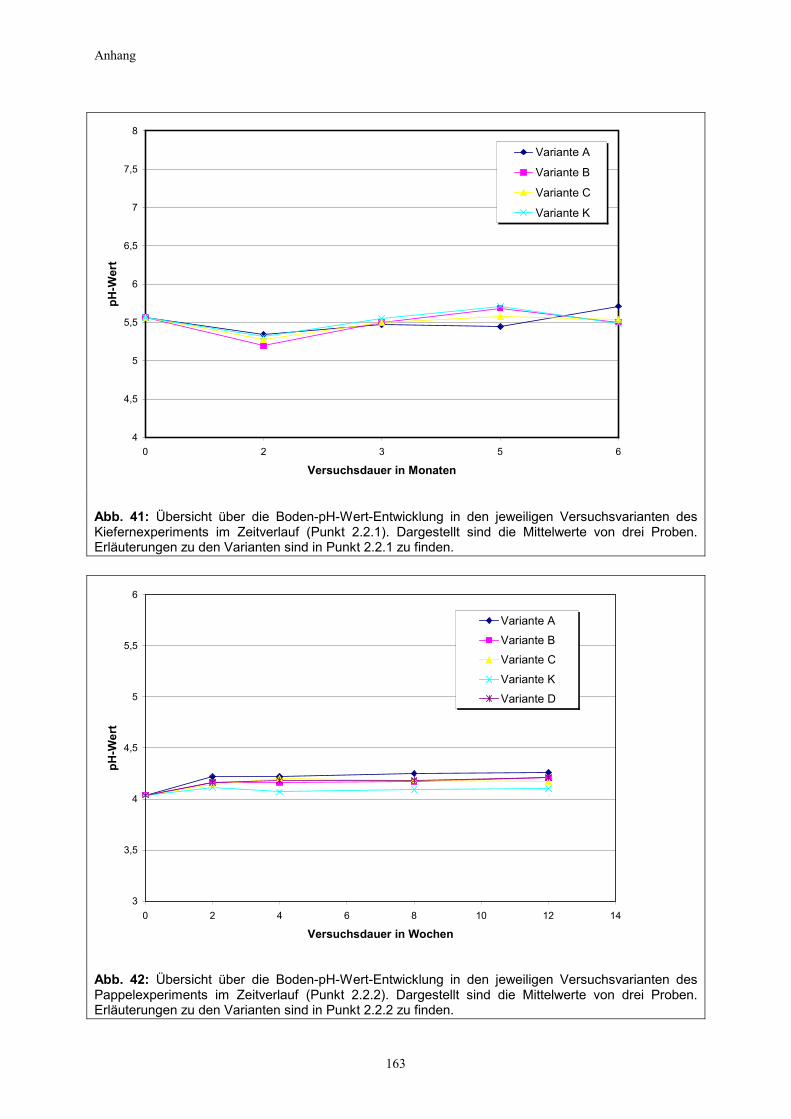
Anhang
4
4,5
5
5,5
6
6,5
7
7,5
8
0 2 3 5 6
Versuchsdauer in Monaten
pH-W
ert
Variante A
Variante B
Variante C
Variante K
Abb. 41: Übersicht über die Boden-pH-Wert-Entwicklung in den jeweiligen Versuchsvarianten des Kiefernexperiments im Zeitverlauf (Punkt 2.2.1). Dargestellt sind die Mittelwerte von drei Proben. Erläuterungen zu den Varianten sind in Punkt 2.2.1 zu finden.
3
3,5
4
4,5
5
5,5
6
0 2 4 6 8 10 12 14
Versuchsdauer in Wochen
pH-W
ert
Variante AVariante BVariante CVariante KVariante D
Abb. 42: Übersicht über die Boden-pH-Wert-Entwicklung in den jeweiligen Versuchsvarianten des Pappelexperiments im Zeitverlauf (Punkt 2.2.2). Dargestellt sind die Mittelwerte von drei Proben. Erläuterungen zu den Varianten sind in Punkt 2.2.2 zu finden.
163
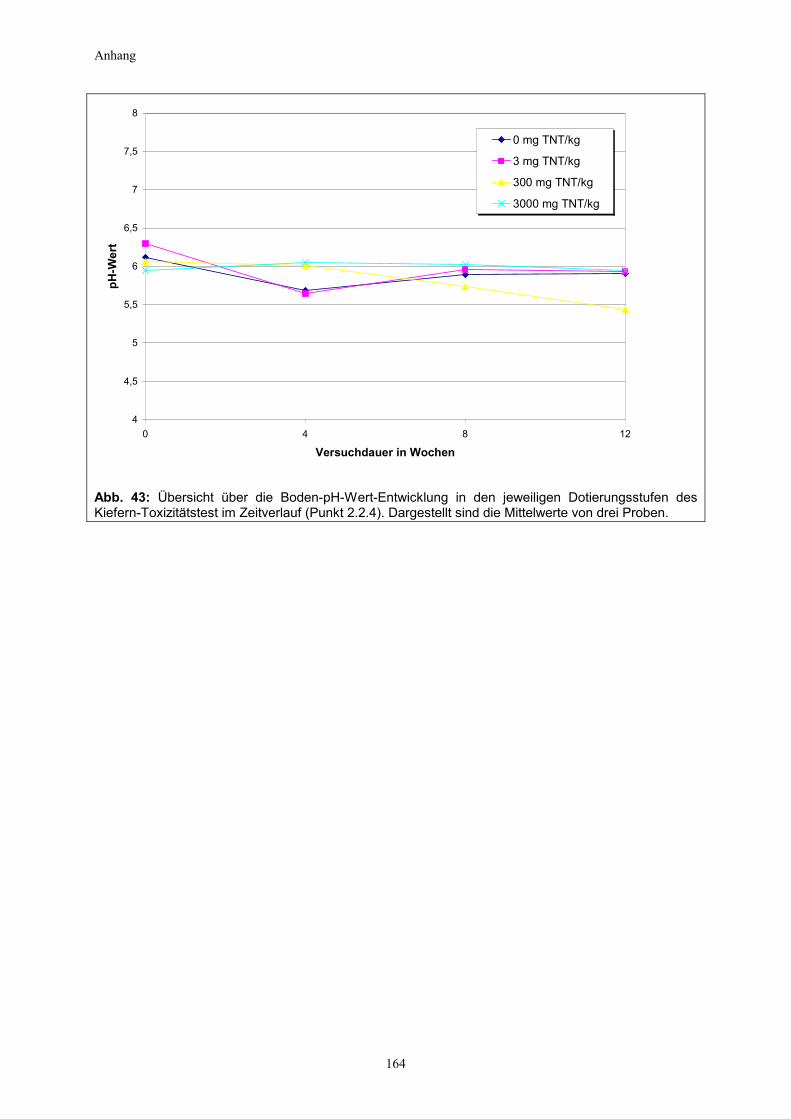
Anhang
4
4,5
5
5,5
6
6,5
7
7,5
8
0 4 8 1
Versuchdauer in Wochen
pH-W
ert
2
0 mg TNT/kg
3 mg TNT/kg
300 mg TNT/kg
3000 mg TNT/kg
Abb. 43: Übersicht über die Boden-pH-Wert-Entwicklung in den jeweiligen Dotierungsstufen des Kiefern-Toxizitätstest im Zeitverlauf (Punkt 2.2.4). Dargestellt sind die Mittelwerte von drei Proben.
164
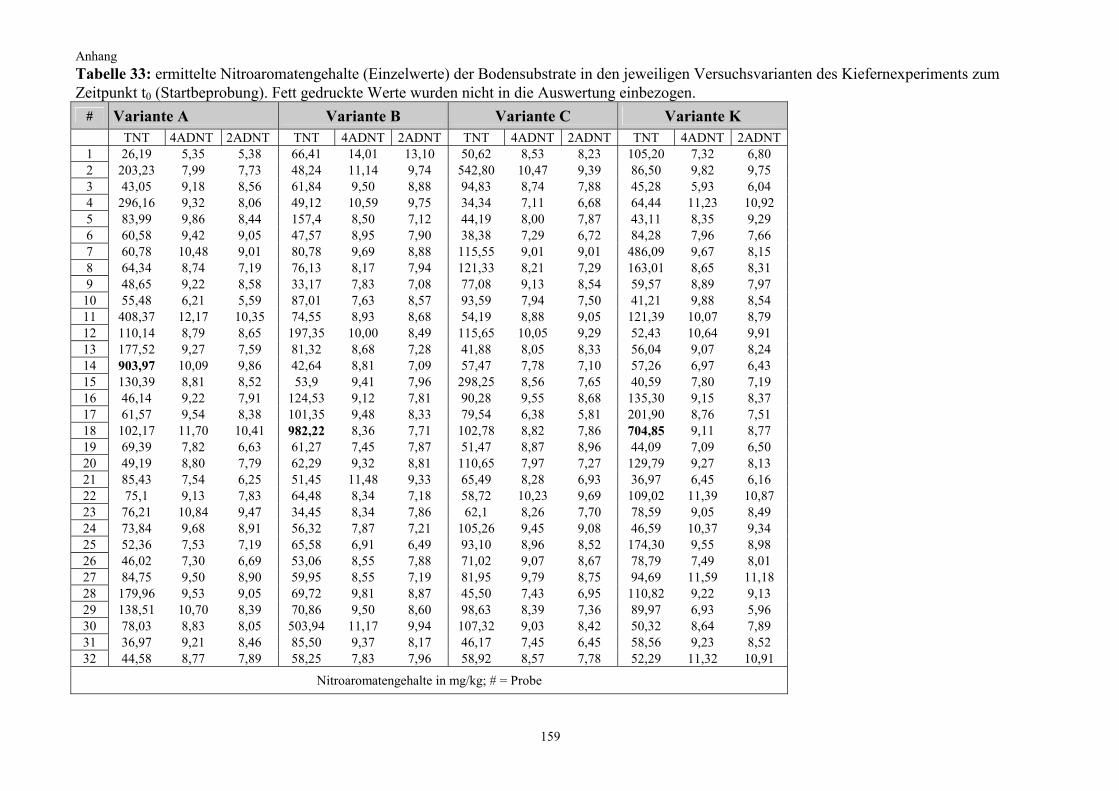
Anhang
159
Tabelle 33: ermittelte Nitroaromatengehalte (Einzelwerte) der Bodensubstrate in den jeweiligen Versuchsvarianten des Kiefernexperiments zum Zeitpunkt t0 (Startbeprobung). Fett gedruckte Werte wurden nicht in die Auswertung einbezogen.
# Variante A Variante B Variante C Variante K TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT
1 26,19 5,35 5,38 66,41 14,01 13,10 50,62 8,53 8,23 105,20 7,32 6,802 203,23 7,99 7,73 48,24 11,14 9,74 542,80 10,47 9,39 86,50 9,82 9,753 43,05 9,18 8,56 61,84 9,50 8,88 94,83 8,74 7,88 45,28 5,93 6,044 296,16 9,32 8,06 49,12 10,59 9,75 34,34 7,11 6,68 64,44 11,23 10,925 83,99 9,86 8,44 157,4 8,50 7,12 44,19 8,00 7,87 43,11 8,35 9,296 60,58 9,42 9,05 47,57 8,95 7,90 38,38 7,29 6,72 84,28 7,96 7,667 60,78 10,48 9,01 80,78 9,69 8,88 115,55 9,01 9,01 486,09 9,67 8,158 64,34 8,74 7,19 76,13 8,17 7,94 121,33 8,21 7,29 163,01 8,65 8,319 48,65 9,22 8,58 33,17 7,83 7,08 77,08 9,13 8,54 59,57 8,89 7,97
10 55,48 6,21 5,59 87,01 7,63 8,57 93,59 7,94 7,50 41,21 9,88 8,5411 408,37 12,17 10,35 74,55 8,93 8,68 54,19 8,88 9,05 121,39 10,07 8,7912 110,14 8,79 8,65 197,35 10,00 8,49 115,65 10,05 9,29 52,43 10,64 9,9113 177,52 9,27 7,59 81,32 8,68 7,28 41,88 8,05 8,33 56,04 9,07 8,2414 903,97 10,09 9,86 42,64 8,81 7,09 57,47 7,78 7,10 57,26 6,97 6,4315 130,39 8,81 8,52 53,9 9,41 7,96 298,25 8,56 7,65 40,59 7,80 7,1916 46,14 9,22 7,91 124,53 9,12 7,81 90,28 9,55 8,68 135,30 9,15 8,3717 61,57 9,54 8,38 101,35 9,48 8,33 79,54 6,38 5,81 201,90 8,76 7,5118 102,17 11,70 10,41 982,22 8,36 7,71 102,78 8,82 7,86 704,85 9,11 8,7719 69,39 7,82 6,63 61,27 7,45 7,87 51,47 8,87 8,96 44,09 7,09 6,5020 49,19 8,80 7,79 62,29 9,32 8,81 110,65 7,97 7,27 129,79 9,27 8,1321 85,43 7,54 6,25 51,45 11,48 9,33 65,49 8,28 6,93 36,97 6,45 6,1622 75,1 9,13 7,83 64,48 8,34 7,18 58,72 10,23 9,69 109,02 11,39 10,8723 76,21 10,84 9,47 34,45 8,34 7,86 62,1 8,26 7,70 78,59 9,05 8,4924 73,84 9,68 8,91 56,32 7,87 7,21 105,26 9,45 9,08 46,59 10,37 9,3425 52,36 7,53 7,19 65,58 6,91 6,49 93,10 8,96 8,52 174,30 9,55 8,9826 46,02 7,30 6,69 53,06 8,55 7,88 71,02 9,07 8,67 78,79 7,49 8,0127 84,75 9,50 8,90 59,95 8,55 7,19 81,95 9,79 8,75 94,69 11,59 11,1828 179,96 9,53 9,05 69,72 9,81 8,87 45,50 7,43 6,95 110,82 9,22 9,1329 138,51 10,70 8,39 70,86 9,50 8,60 98,63 8,39 7,36 89,97 6,93 5,9630 78,03 8,83 8,05 503,94 11,17 9,94 107,32 9,03 8,42 50,32 8,64 7,8931 36,97 9,21 8,46 85,50 9,37 8,17 46,17 7,45 6,45 58,56 9,23 8,5232 44,58 8,77 7,89 58,25 7,83 7,96 58,92 8,57 7,78 52,29 11,32 10,91
Nitroaromatengehalte in mg/kg; # = Probe
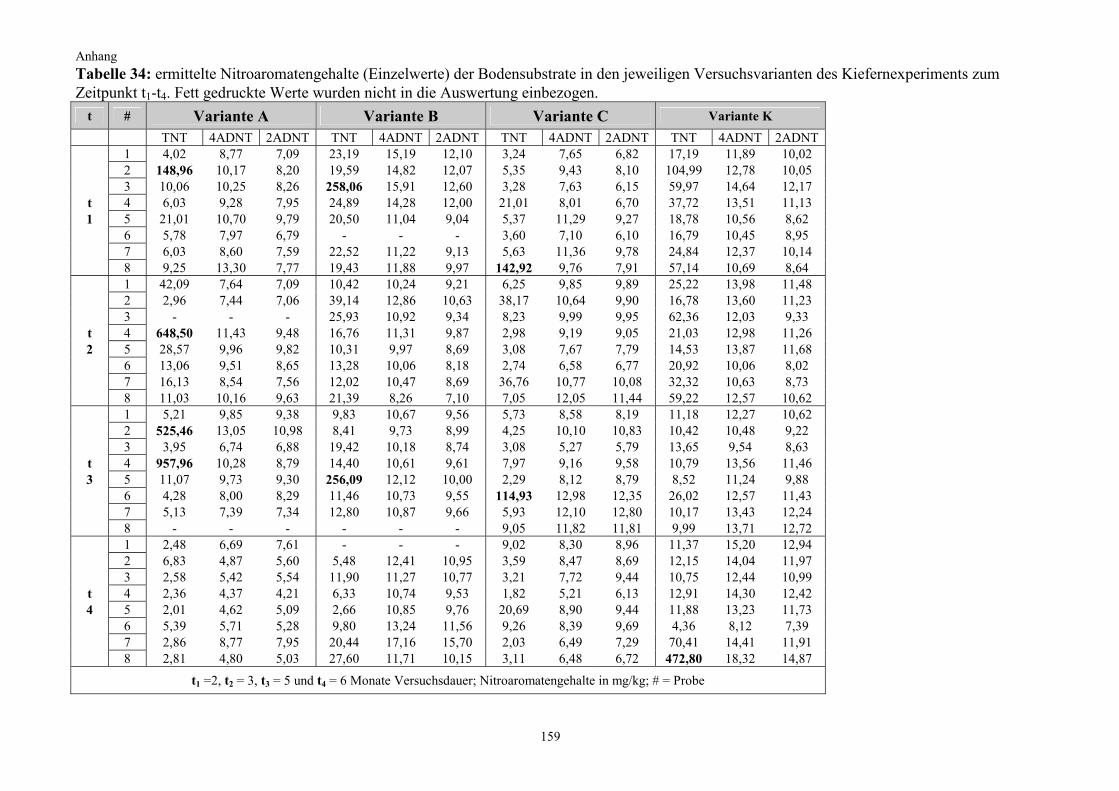
Anhang
159
Tabelle 34: ermittelte Nitroaromatengehalte (Einzelwerte) der Bodensubstrate in den jeweiligen Versuchsvarianten des Kiefernexperiments zum Zeitpunkt t1-t4. Fett gedruckte Werte wurden nicht in die Auswertung einbezogen.
t # Variante A Variante B Variante C Variante K TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT 1 4,02 8,77 7,09 23,19 15,19 12,10 3,24 7,65 6,82 17,19 11,89 10,02 2 148,96 10,17 8,20 19,59 14,82 12,07 5,35 9,43 8,10 104,99 12,78 10,05 3 10,06 10,25 8,26 258,06 15,91 12,60 3,28 7,63 6,15 59,97 14,64 12,17t 4 6,03 9,28 7,95 24,89 14,28 12,00 21,01 8,01 6,70 37,72 13,51 11,131 5 21,01 10,70 9,79 20,50 11,04 9,04 5,37 11,29 9,27 18,78 10,56 8,62 6 5,78 7,97 6,79 - - - 3,60 7,10 6,10 16,79 10,45 8,95 7 6,03 8,60 7,59 22,52 11,22 9,13 5,63 11,36 9,78 24,84 12,37 10,14 8 9,25 13,30 7,77 19,43 11,88 9,97 142,92 9,76 7,91 57,14 10,69 8,64 1 42,09 7,64 7,09 10,42 10,24 9,21 6,25 9,85 9,89 25,22 13,98 11,48 2 2,96 7,44 7,06 39,14 12,86 10,63 38,17 10,64 9,90 16,78 13,60 11,23 3 - - - 25,93 10,92 9,34 8,23 9,99 9,95 62,36 12,03 9,33t 4 648,50 11,43 9,48 16,76 11,31 9,87 2,98 9,19 9,05 21,03 12,98 11,262 5 28,57 9,96 9,82 10,31 9,97 8,69 3,08 7,67 7,79 14,53 13,87 11,68 6 13,06 9,51 8,65 13,28 10,06 8,18 2,74 6,58 6,77 20,92 10,06 8,02 7 16,13 8,54 7,56 12,02 10,47 8,69 36,76 10,77 10,08 32,32 10,63 8,73 8 11,03 10,16 9,63 21,39 8,26 7,10 7,05 12,05 11,44 59,22 12,57 10,62 1 5,21 9,85 9,38 9,83 10,67 9,56 5,73 8,58 8,19 11,18 12,27 10,62 2 525,46 13,05 10,98 8,41 9,73 8,99 4,25 10,10 10,83 10,42 10,48 9,22 3 3,95 6,74 6,88 19,42 10,18 8,74 3,08 5,27 5,79 13,65 9,54 8,63t 4 957,96 10,28 8,79 14,40 10,61 9,61 7,97 9,16 9,58 10,79 13,56 11,463 5 11,07 9,73 9,30 256,09 12,12 10,00 2,29 8,12 8,79 8,52 11,24 9,88 6 4,28 8,00 8,29 11,46 10,73 9,55 114,93 12,98 12,35 26,02 12,57 11,43 7 5,13 7,39 7,34 12,80 10,87 9,66 5,93 12,10 12,80 10,17 13,43 12,24 8 - - - - - - 9,05 11,82 11,81 9,99 13,71 12,72 1 2,48 6,69 7,61 - - - 9,02 8,30 8,96 11,37 15,20 12,94 2 6,83 4,87 5,60 5,48 12,41 10,95 3,59 8,47 8,69 12,15 14,04 11,97 3 2,58 5,42 5,54 11,90 11,27 10,77 3,21 7,72 9,44 10,75 12,44 10,99t 4 2,36 4,37 4,21 6,33 10,74 9,53 1,82 5,21 6,13 12,91 14,30 12,424 5 2,01 4,62 5,09 2,66 10,85 9,76 20,69 8,90 9,44 11,88 13,23 11,73 6 5,39 5,71 5,28 9,80 13,24 11,56 9,26 8,39 9,69 4,36 8,12 7,39 7 2,86 8,77 7,95 20,44 17,16 15,70 2,03 6,49 7,29 70,41 14,41 11,91 8 2,81 4,80 5,03 27,60 11,71 10,15 3,11 6,48 6,72 472,80 18,32 14,87
t1 =2, t2 = 3, t3 = 5 und t4 = 6 Monate Versuchsdauer; Nitroaromatengehalte in mg/kg; # = Probe
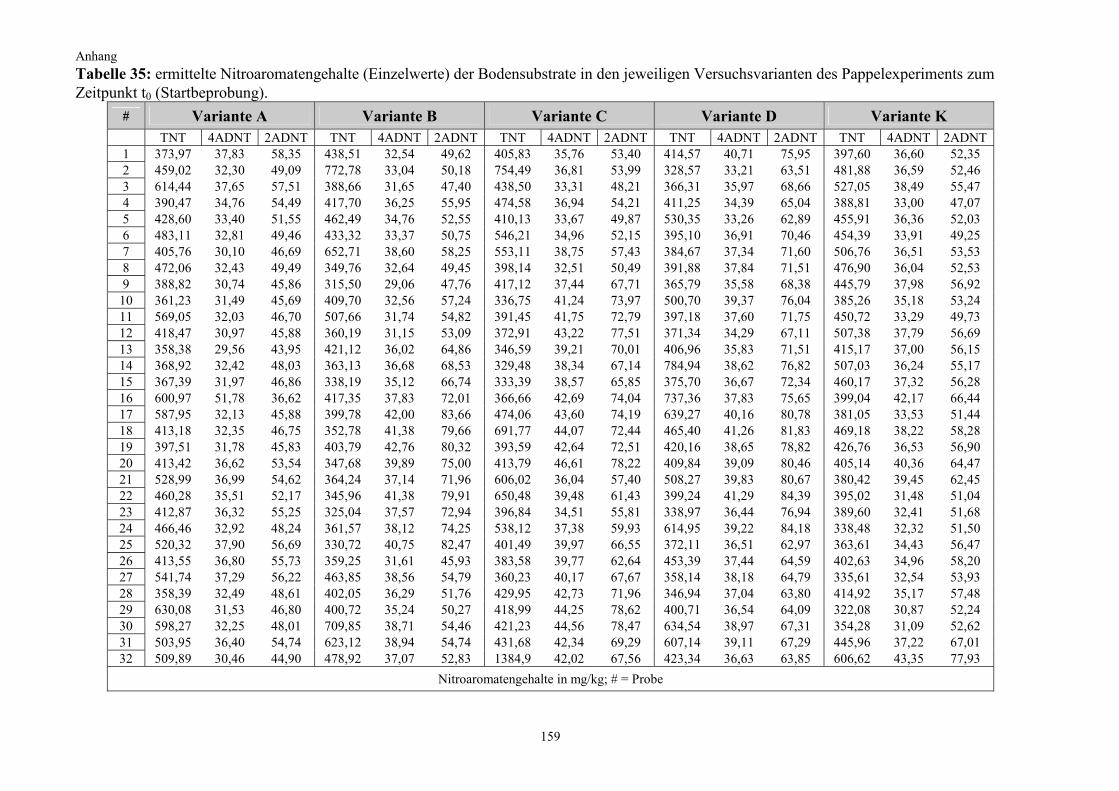
Anhang
159
Tabelle 35: ermittelte Nitroaromatengehalte (Einzelwerte) der Bodensubstrate in den jeweiligen Versuchsvarianten des Pappelexperiments zum Zeitpunkt t0 (Startbeprobung).
# Variante A Variante B Variante C Variante D Variante K TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT
1 373,97 37,83 58,35 438,51 32,54 49,62 405,83 35,76 53,40 414,57 40,71 75,95 397,60 36,60 52,352 459,02 32,30 49,09 772,78 33,04 50,18 754,49 36,81 53,99 328,57 33,21 63,51 481,88 36,59 52,463 614,44 37,65 57,51 388,66 31,65 47,40 438,50 33,31 48,21 366,31 35,97 68,66 527,05 38,49 55,474 390,47 34,76 54,49 417,70 36,25 55,95 474,58 36,94 54,21 411,25 34,39 65,04 388,81 33,00 47,075 428,60 33,40 51,55 462,49 34,76 52,55 410,13 33,67 49,87 530,35 33,26 62,89 455,91 36,36 52,036 483,11 32,81 49,46 433,32 33,37 50,75 546,21 34,96 52,15 395,10 36,91 70,46 454,39 33,91 49,257 405,76 30,10 46,69 652,71 38,60 58,25 553,11 38,75 57,43 384,67 37,34 71,60 506,76 36,51 53,538 472,06 32,43 49,49 349,76 32,64 49,45 398,14 32,51 50,49 391,88 37,84 71,51 476,90 36,04 52,539 388,82 30,74 45,86 315,50 29,06 47,76 417,12 37,44 67,71 365,79 35,58 68,38 445,79 37,98 56,92
10 361,23 31,49 45,69 409,70 32,56 57,24 336,75 41,24 73,97 500,70 39,37 76,04 385,26 35,18 53,2411 569,05 32,03 46,70 507,66 31,74 54,82 391,45 41,75 72,79 397,18 37,60 71,75 450,72 33,29 49,7312 418,47 30,97 45,88 360,19 31,15 53,09 372,91 43,22 77,51 371,34 34,29 67,11 507,38 37,79 56,6913 358,38 29,56 43,95 421,12 36,02 64,86 346,59 39,21 70,01 406,96 35,83 71,51 415,17 37,00 56,1514 368,92 32,42 48,03 363,13 36,68 68,53 329,48 38,34 67,14 784,94 38,62 76,82 507,03 36,24 55,1715 367,39 31,97 46,86 338,19 35,12 66,74 333,39 38,57 65,85 375,70 36,67 72,34 460,17 37,32 56,2816 600,97 51,78 36,62 417,35 37,83 72,01 366,66 42,69 74,04 737,36 37,83 75,65 399,04 42,17 66,4417 587,95 32,13 45,88 399,78 42,00 83,66 474,06 43,60 74,19 639,27 40,16 80,78 381,05 33,53 51,4418 413,18 32,35 46,75 352,78 41,38 79,66 691,77 44,07 72,44 465,40 41,26 81,83 469,18 38,22 58,2819 397,51 31,78 45,83 403,79 42,76 80,32 393,59 42,64 72,51 420,16 38,65 78,82 426,76 36,53 56,9020 413,42 36,62 53,54 347,68 39,89 75,00 413,79 46,61 78,22 409,84 39,09 80,46 405,14 40,36 64,4721 528,99 36,99 54,62 364,24 37,14 71,96 606,02 36,04 57,40 508,27 39,83 80,67 380,42 39,45 62,4522 460,28 35,51 52,17 345,96 41,38 79,91 650,48 39,48 61,43 399,24 41,29 84,39 395,02 31,48 51,0423 412,87 36,32 55,25 325,04 37,57 72,94 396,84 34,51 55,81 338,97 36,44 76,94 389,60 32,41 51,6824 466,46 32,92 48,24 361,57 38,12 74,25 538,12 37,38 59,93 614,95 39,22 84,18 338,48 32,32 51,5025 520,32 37,90 56,69 330,72 40,75 82,47 401,49 39,97 66,55 372,11 36,51 62,97 363,61 34,43 56,4726 413,55 36,80 55,73 359,25 31,61 45,93 383,58 39,77 62,64 453,39 37,44 64,59 402,63 34,96 58,2027 541,74 37,29 56,22 463,85 38,56 54,79 360,23 40,17 67,67 358,14 38,18 64,79 335,61 32,54 53,9328 358,39 32,49 48,61 402,05 36,29 51,76 429,95 42,73 71,96 346,94 37,04 63,80 414,92 35,17 57,4829 630,08 31,53 46,80 400,72 35,24 50,27 418,99 44,25 78,62 400,71 36,54 64,09 322,08 30,87 52,2430 598,27 32,25 48,01 709,85 38,71 54,46 421,23 44,56 78,47 634,54 38,97 67,31 354,28 31,09 52,6231 503,95 36,40 54,74 623,12 38,94 54,74 431,68 42,34 69,29 607,14 39,11 67,29 445,96 37,22 67,0132 509,89 30,46 44,90 478,92 37,07 52,83 1384,9 42,02 67,56 423,34 36,63 63,85 606,62 43,35 77,93
Nitroaromatengehalte in mg/kg; # = Probe
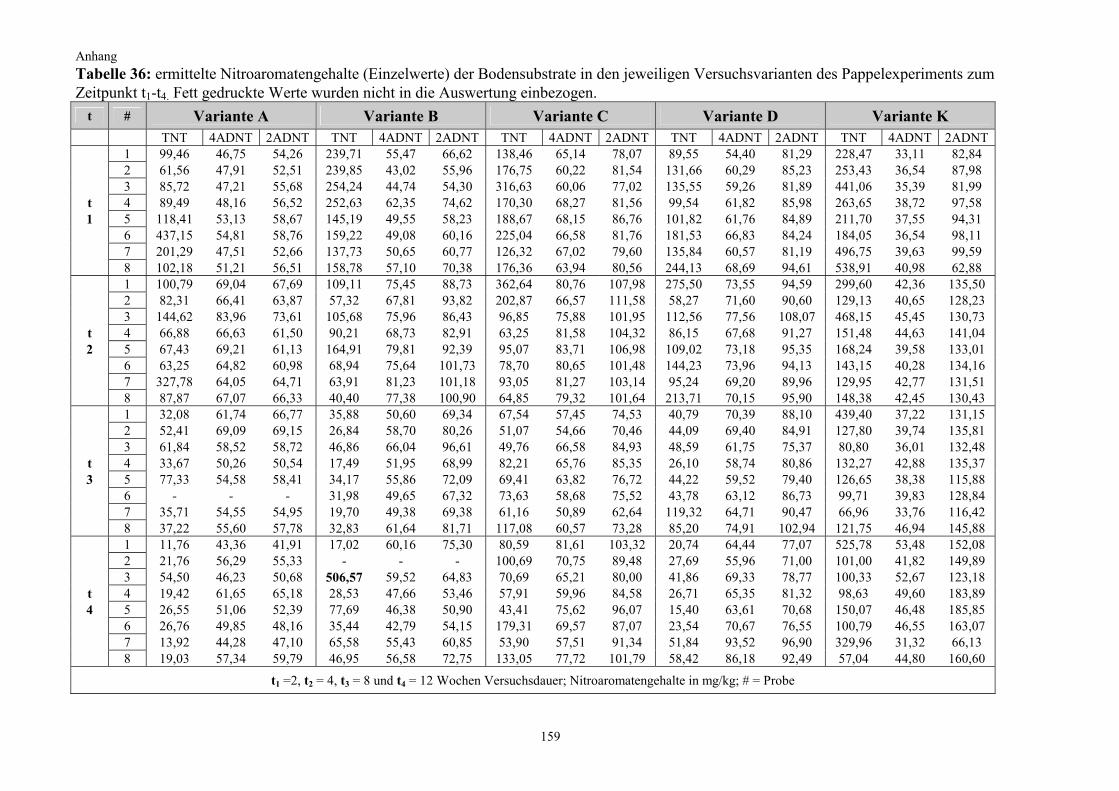
Anhang
159
Tabelle 36: ermittelte Nitroaromatengehalte (Einzelwerte) der Bodensubstrate in den jeweiligen Versuchsvarianten des Pappelexperiments zum Zeitpunkt t -t Fett gedruckte Werte wurden nicht in die Auswertung einbezogen. 1 4.
t # Variante A Variante B Variante C Variante D Variante K TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT TNT 4ADNT 2ADNT 1 99,46 46,75 54,26 239,71 55,47 66,62 138,46 65,14 78,07 89,55 54,40 81,29 228,47 33,11 82,84 2 61,56 47,91 52,51 239,85 43,02 55,96 176,75 60,22 81,54 131,66 60,29 85,23 253,43 36,54 87,98 3 85,72 47,21 55,68 254,24 44,74 54,30 316,63 60,06 77,02 135,55 59,26 81,89 441,06 35,39 81,99t 4 89,49 48,16 56,52 252,63 62,35 74,62 170,30 68,27 81,56 99,54 61,82 85,98 263,65 38,72 97,581 5 118,41 53,13 58,67 145,19 49,55 58,23 188,67 68,15 86,76 101,82 61,76 84,89 211,70 37,55 94,31 6 437,15 54,81 58,76 159,22 49,08 60,16 225,04 66,58 81,76 181,53 66,83 84,24 184,05 36,54 98,11 7 201,29 47,51 52,66 137,73 50,65 60,77 126,32 67,02 79,60 135,84 60,57 81,19 496,75 39,63 99,59 8 102,18 51,21 56,51 158,78 57,10 70,38 176,36 63,94 80,56 244,13 68,69 94,61 538,91 40,98 62,88 1 100,79 69,04 67,69 109,11 75,45 88,73 362,64 80,76 107,98 275,50 73,55 94,59 299,60 42,36 135,50 2 82,31 66,41 63,87 57,32 67,81 93,82 202,87 66,57 111,58 58,27 71,60 90,60 129,13 40,65 128,23 3 144,62 83,96 73,61 105,68 75,96 86,43 96,85 75,88 101,95 112,56 77,56 108,07 468,15 45,45 130,73t 4 66,88 66,63 61,50 90,21 68,73 82,91 63,25 81,58 104,32 86,15 67,68 91,27 151,48 44,63 141,042 5 67,43 69,21 61,13 164,91 79,81 92,39 95,07 83,71 106,98 109,02 73,18 95,35 168,24 39,58 133,01 6 63,25 64,82 60,98 68,94 75,64 101,73 78,70 80,65 101,48 144,23 73,96 94,13 143,15 40,28 134,16 7 327,78 64,05 64,71 63,91 81,23 101,18 93,05 81,27 103,14 95,24 69,20 89,96 129,95 42,77 131,51 8 87,87 67,07 66,33 40,40 77,38 100,90 64,85 79,32 101,64 213,71 70,15 95,90 148,38 42,45 130,43 1 32,08 61,74 66,77 35,88 50,60 69,34 67,54 57,45 74,53 40,79 70,39 88,10 439,40 37,22 131,15 2 52,41 69,09 69,15 26,84 58,70 80,26 51,07 54,66 70,46 44,09 69,40 84,91 127,80 39,74 135,81 3 61,84 58,52 58,72 46,86 66,04 96,61 49,76 66,58 84,93 48,59 61,75 75,37 80,80 36,01 132,48t 4 33,67 50,26 50,54 17,49 51,95 68,99 82,21 65,76 85,35 26,10 58,74 80,86 132,27 42,88 135,373 5 77,33 54,58 58,41 34,17 55,86 72,09 69,41 63,82 76,72 44,22 59,52 79,40 126,65 38,38 115,88 6 - - - 31,98 49,65 67,32 73,63 58,68 75,52 43,78 63,12 86,73 99,71 39,83 128,84 7 35,71 54,55 54,95 19,70 49,38 69,38 61,16 50,89 62,64 119,32 64,71 90,47 66,96 33,76 116,42 8 37,22 55,60 57,78 32,83 61,64 81,71 117,08 60,57 73,28 85,20 74,91 102,94 121,75 46,94 145,88 1 11,76 43,36 41,91 17,02 60,16 75,30 80,59 81,61 103,32 20,74 64,44 77,07 525,78 53,48 152,08 2 21,76 56,29 55,33 - - - 100,69 70,75 89,48 27,69 55,96 71,00 101,00 41,82 149,89 3 54,50 46,23 50,68 506,57 59,52 64,83 70,69 65,21 80,00 41,86 69,33 78,77 100,33 52,67 123,18t 4 28,53 19,42 61,65 65,18 47,66 53,46 57,91 59,96 84,58 26,71 65,35 81,32 98,63 49,60 183,894 5 26,55 51,06 52,39 77,69 46,38 50,90 43,41 75,62 96,07 15,40 63,61 70,68 150,07 46,48 185,85 6 26,76 49,85 48,16 35,44 42,79 54,15 179,31 69,57 87,07 23,54 70,67 76,55 100,79 46,55 163,07 7 13,92 44,28 47,10 65,58 55,43 60,85 53,90 57,51 91,34 51,84 93,52 96,90 329,96 31,32 66,13 8 19,03 57,34 59,79 46,95 56,58 72,75 133,05 77,72 101,79 58,42 86,18 92,49 57,04 44,80 160,60
t1 =2, t2 = 4, t3 = 8 und t4 = 12 Wochen Versuchsdauer; Nitroaromatengehalte in mg/kg; # = Probe
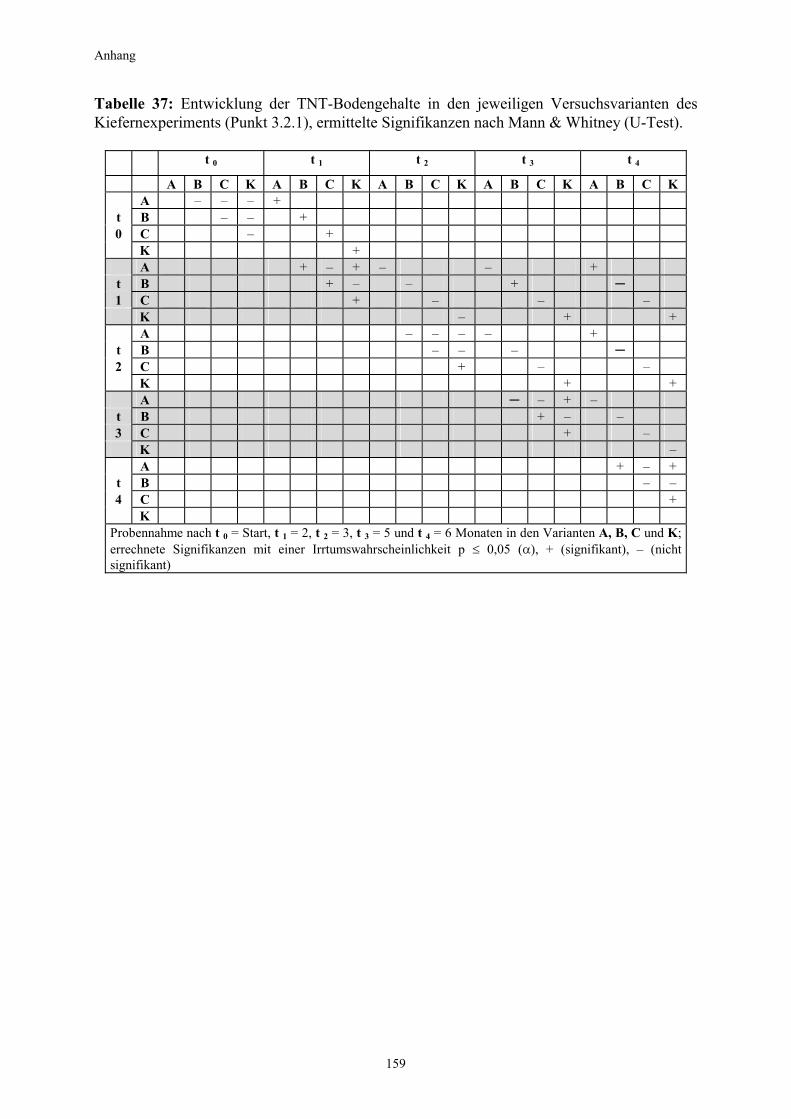
Anhang
Tabelle 37: Entwicklung der TNT-Bodengehalte in den jeweiligen Versuchsvarianten des Kiefernexperiments (Punkt 3.2.1), ermittelte Signifikanzen nach Mann & Whitney (U-Test).
t 0 t 1 t 2 t 3 t 4
A B C K A B C K A B C K A B C K A B C K A – – – + t B – – + 0 C – + K + A + – + – – + t B + – – + ─ 1 C + – – – K – + + A – – – – + t B – – – ─ 2 C + – – K + + A ─ – + – t B + – – 3 C + – K – A + – + t B – – 4 C + K
Probennahme nach t 0 = Start, t 1 = 2, t 2 = 3, t 3 = 5 und t 4 = 6 Monaten in den Varianten A, B, C und K; errechnete Signifikanzen mit einer Irrtumswahrscheinlichkeit p � 0,05 (�), + (signifikant), – (nicht signifikant)
159
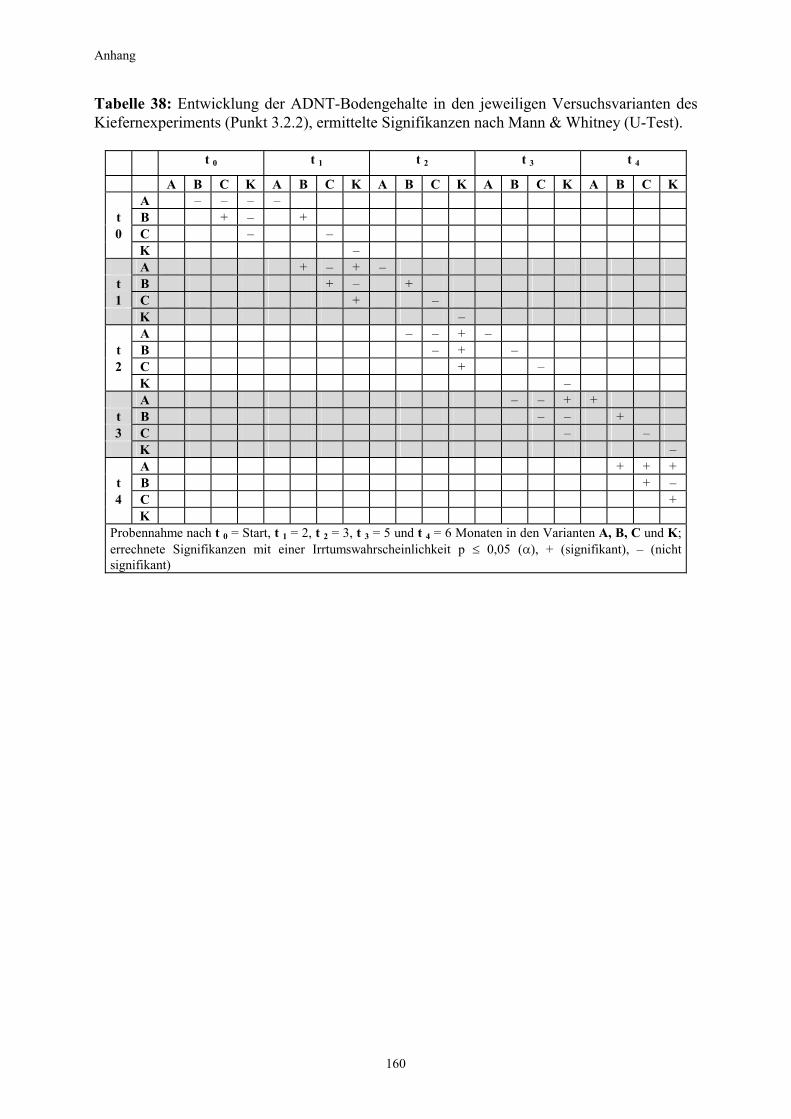
Anhang
Tabelle 38: Entwicklung der ADNT-Bodengehalte in den jeweiligen Versuchsvarianten des Kiefernexperiments (Punkt 3.2.2), ermittelte Signifikanzen nach Mann & Whitney (U-Test).
t 0 t 1 t 2 t 3 t 4
A B C K A B C K A B C K A B C K A B C K A – – – – t B + – + 0 C – – K – A + – + – t B + – + 1 C + – K – A – – + – t B – + – 2 C + – K – A – – + + t B – – + 3 C – – K – A + + + t B + – 4 C + K
Probennahme nach t 0 = Start, t 1 = 2, t 2 = 3, t 3 = 5 und t 4 = 6 Monaten in den Varianten A, B, C und K; errechnete Signifikanzen mit einer Irrtumswahrscheinlichkeit p � 0,05 (�), + (signifikant), – (nicht signifikant)
160
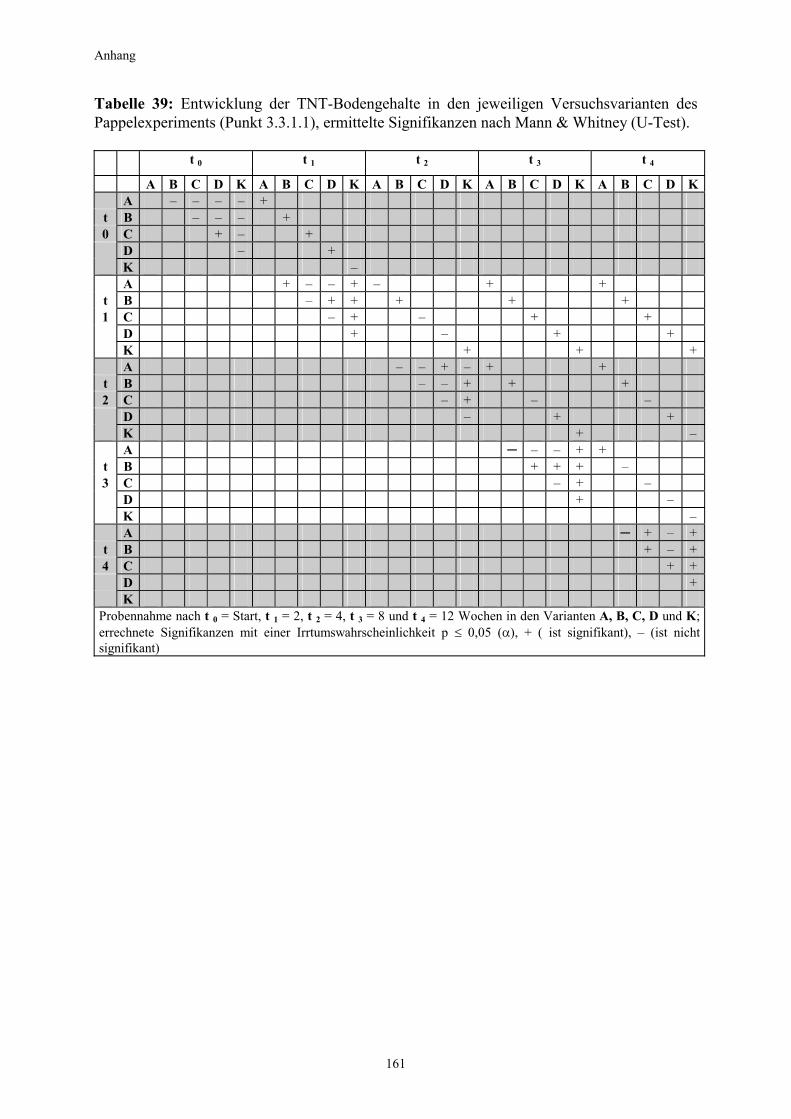
Anhang
Tabelle 39: Entwicklung der TNT-Bodengehalte in den jeweiligen Versuchsvarianten des Pappelexperiments (Punkt 3.3.1.1), ermittelte Signifikanzen nach Mann & Whitney (U-Test).
t 0 t 1 t 2 t 3 t 4
A B C D K A B C D K A B C D K A B C D K A B C D K A – – – – + t B – – – + 0 C + – + D – + K – A + – – + – + + t B – + + + + + 1 C – + – + + D + – + + K + + + A – – + – + + t B – – + + + 2 C – + – – D – + + K + – A ─ – – + + t B + + + – 3 C – + – D + – K – A ─ + – + t B + – + 4 C + + D + K
Probennahme nach t 0 = Start, t 1 = 2, t 2 = 4, t 3 = 8 und t 4 = 12 Wochen in den Varianten A, B, C, D und K; errechnete Signifikanzen mit einer Irrtumswahrscheinlichkeit p � 0,05 (�), + ( ist signifikant), – (ist nicht signifikant)
161
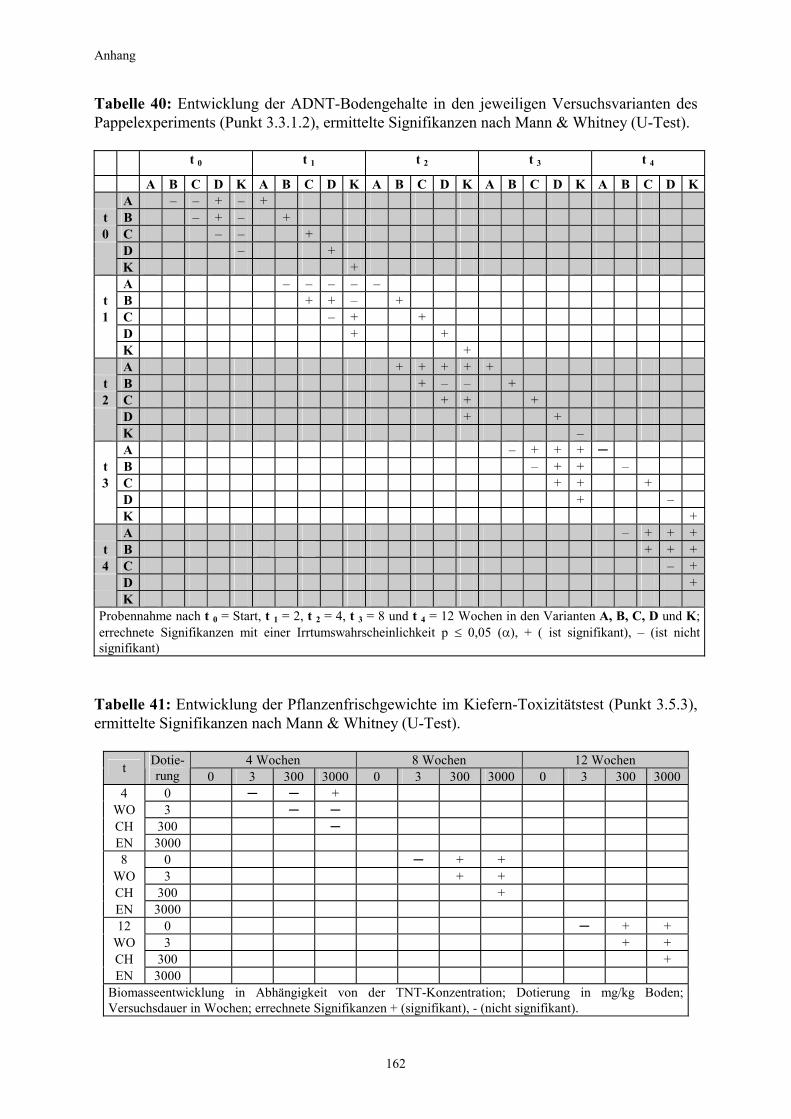
Anhang
Tabelle 40: Entwicklung der ADNT-Bodengehalte in den jeweiligen Versuchsvarianten des Pappelexperiments (Punkt 3.3.1.2), ermittelte Signifikanzen nach Mann & Whitney (U-Test).
t 0 t 1 t 2 t 3 t 4
A B C D K A B C D K A B C D K A B C D K A B C D K A – – + – + t B – + – + 0 C – – + D – + K + A – – – – – t B + + – + 1 C – + + D + + K + A + + + + + t B + – – + 2 C + + + D + + K – A – + + + ─ t B – + + – 3 C + + + D + – K + A – + + + t B + + + 4 C – + D + K
Probennahme nach t 0 = Start, t 1 = 2, t 2 = 4, t 3 = 8 und t 4 = 12 Wochen in den Varianten A, B, C, D und K; errechnete Signifikanzen mit einer Irrtumswahrscheinlichkeit p � 0,05 (�), + ( ist signifikant), – (ist nicht signifikant)
Tabelle 41: Entwicklung der Pflanzenfrischgewichte im Kiefern-Toxizitätstest (Punkt 3.5.3), ermittelte Signifikanzen nach Mann & Whitney (U-Test).
4 Wochen 8 Wochen 12 Wochen t Dotie- rung 0 3 300 3000 0 3 300 3000 0 3 300 3000
4 0 ─ ─ + WO 3 ─ ─ CH 300 ─ EN 3000 8 0 ─ + +
WO 3 + + CH 300 + EN 3000 12 0 ─ + +
WO 3 + + CH 300 + EN 3000
Biomasseentwicklung in Abhängigkeit von der TNT-Konzentration; Dotierung in mg/kg Boden; Versuchsdauer in Wochen; errechnete Signifikanzen + (signifikant), - (nicht signifikant).
162
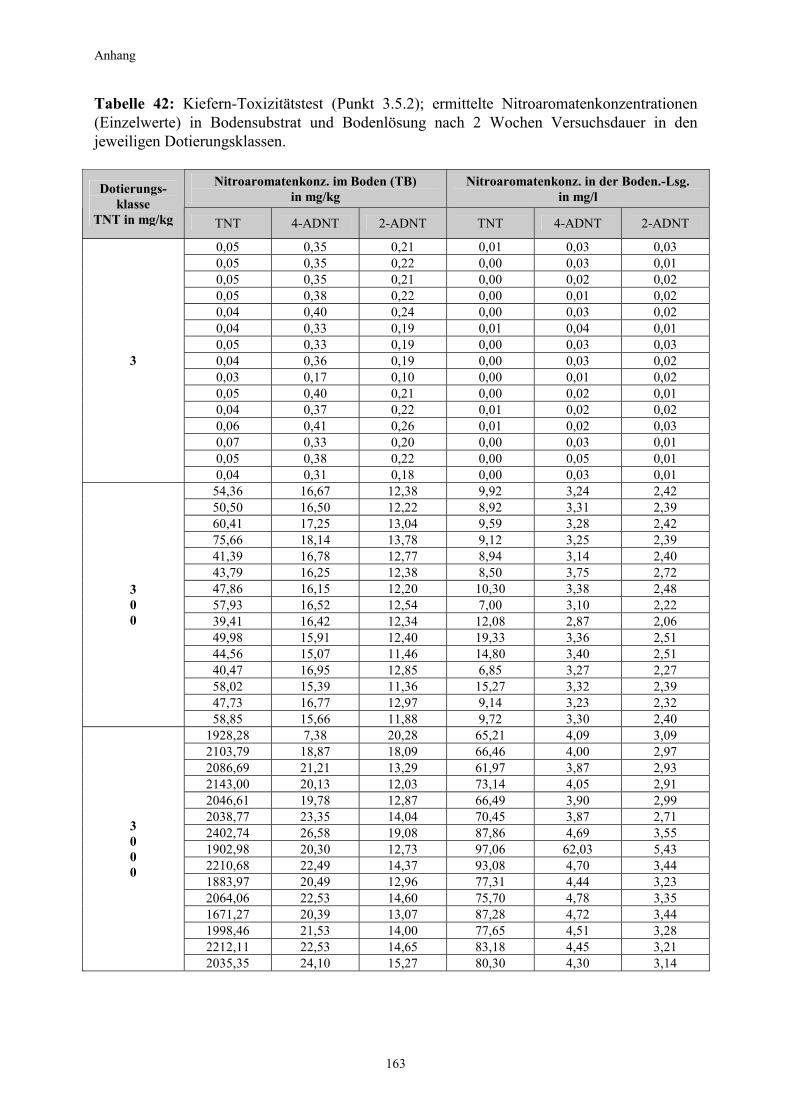
Anhang
Tabelle 42: Kiefern-Toxizitätstest (Punkt 3.5.2); ermittelte Nitroaromatenkonzentrationen (Einzelwerte) in Bodensubstrat und Bodenlösung nach 2 Wochen Versuchsdauer in den jeweiligen Dotierungsklassen.
Nitroaromatenkonz. im Boden (TB) in mg/kg
Nitroaromatenkonz. in der Boden.-Lsg. in mg/l Dotierungs-
klasse TNT in mg/kg TNT 4-ADNT 2-ADNT TNT 4-ADNT 2-ADNT
0,05 0,35 0,21 0,01 0,03 0,03 0,05 0,35 0,22 0,00 0,03 0,01 0,05 0,35 0,21 0,00 0,02 0,02 0,05 0,38 0,22 0,00 0,01 0,02 0,04 0,40 0,24 0,00 0,03 0,02 0,04 0,33 0,19 0,01 0,04 0,01 0,05 0,33 0,19 0,00 0,03 0,03 0,04 0,36 0,19 0,00 0,03 0,02 0,03 0,17 0,10 0,00 0,01 0,02 0,05 0,40 0,21 0,00 0,02 0,01 0,04 0,37 0,22 0,01 0,02 0,02 0,06 0,41 0,26 0,01 0,02 0,03 0,07 0,33 0,20 0,00 0,03 0,01 0,05 0,38 0,22 0,00 0,05 0,01
3
0,04 0,31 0,18 0,00 0,03 0,01 54,36 16,67 12,38 9,92 3,24 2,42 50,50 16,50 12,22 8,92 3,31 2,39 60,41 17,25 13,04 9,59 3,28 2,42 75,66 18,14 13,78 9,12 3,25 2,39 41,39 16,78 12,77 8,94 3,14 2,40 43,79 16,25 12,38 8,50 3,75 2,72 47,86 16,15 12,20 10,30 3,38 2,48 57,93 16,52 12,54 7,00 3,10 2,22 39,41 16,42 12,34 12,08 2,87 2,06 49,98 15,91 12,40 19,33 3,36 2,51 44,56 15,07 11,46 14,80 3,40 2,51 40,47 16,95 12,85 6,85 3,27 2,27 58,02 15,39 11,36 15,27 3,32 2,39 47,73 16,77 12,97 9,14 3,23 2,32
3 0 0
58,85 15,66 11,88 9,72 3,30 2,40 1928,28 7,38 20,28 65,21 4,09 3,09 2103,79 18,87 18,09 66,46 4,00 2,97 2086,69 21,21 13,29 61,97 3,87 2,93 2143,00 20,13 12,03 73,14 4,05 2,91 2046,61 19,78 12,87 66,49 3,90 2,99 2038,77 23,35 14,04 70,45 3,87 2,71 2402,74 26,58 19,08 87,86 4,69 3,55 1902,98 20,30 12,73 97,06 62,03 5,43 2210,68 22,49 14,37 93,08 4,70 3,44 1883,97 20,49 12,96 77,31 4,44 3,23 2064,06 22,53 14,60 75,70 4,78 3,35 1671,27 20,39 13,07 87,28 4,72 3,44 1998,46 21,53 14,00 77,65 4,51 3,28 2212,11 22,53 14,65 83,18 4,45 3,21
3 0 0 0
2035,35 24,10 15,27 80,30 4,30 3,14
163
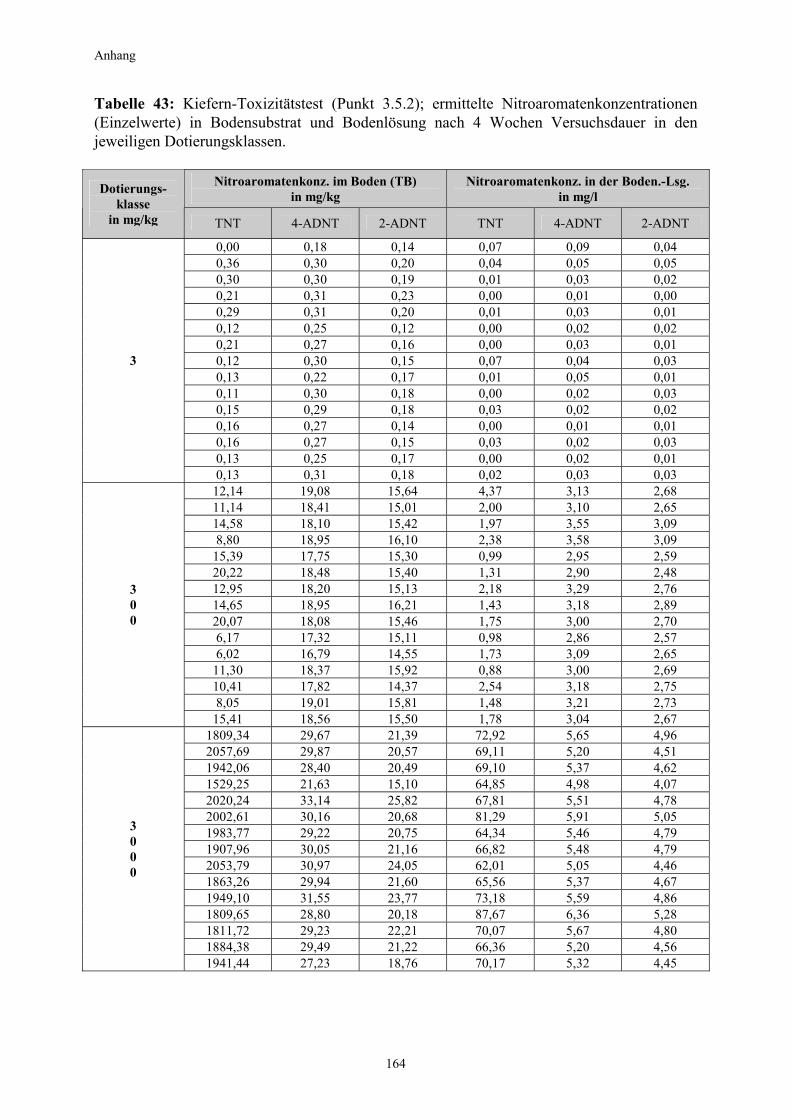
Anhang
Tabelle 43: Kiefern-Toxizitätstest (Punkt 3.5.2); ermittelte Nitroaromatenkonzentrationen (Einzelwerte) in Bodensubstrat und Bodenlösung nach 4 Wochen Versuchsdauer in den jeweiligen Dotierungsklassen.
Nitroaromatenkonz. im Boden (TB) in mg/kg
Nitroaromatenkonz. in der Boden.-Lsg. in mg/l Dotierungs-
klasse in mg/kg TNT 4-ADNT 2-ADNT TNT 4-ADNT 2-ADNT
0,00 0,18 0,14 0,07 0,09 0,04 0,36 0,30 0,20 0,04 0,05 0,05 0,30 0,30 0,19 0,01 0,03 0,02 0,21 0,31 0,23 0,00 0,01 0,00 0,29 0,31 0,20 0,01 0,03 0,01 0,12 0,25 0,12 0,00 0,02 0,02 0,21 0,27 0,16 0,00 0,03 0,01 0,12 0,30 0,15 0,07 0,04 0,03 0,13 0,22 0,17 0,01 0,05 0,01 0,11 0,30 0,18 0,00 0,02 0,03 0,15 0,29 0,18 0,03 0,02 0,02 0,16 0,27 0,14 0,00 0,01 0,01 0,16 0,27 0,15 0,03 0,02 0,03 0,13 0,25 0,17 0,00 0,02 0,01
3
0,13 0,31 0,18 0,02 0,03 0,03 12,14 19,08 15,64 4,37 3,13 2,68 11,14 18,41 15,01 2,00 3,10 2,65 14,58 18,10 15,42 1,97 3,55 3,09 8,80 18,95 16,10 2,38 3,58 3,09
15,39 17,75 15,30 0,99 2,95 2,59 20,22 18,48 15,40 1,31 2,90 2,48 12,95 18,20 15,13 2,18 3,29 2,76 14,65 18,95 16,21 1,43 3,18 2,89 20,07 18,08 15,46 1,75 3,00 2,70 6,17 17,32 15,11 0,98 2,86 2,57 6,02 16,79 14,55 1,73 3,09 2,65
0,88 3,00 2,69 10,41 17,82 14,37 2,54 3,18 2,75
19,01 15,81 1,48 3,21 2,73
3 0 0
15,41 18,56 15,50 1,78 3,04 2,67 1809,34 29,67 21,39 72,92 5,65 4,96 2057,69 29,87 20,57 69,11 5,20 4,51 1942,06 28,40 20,49 69,10 5,37 4,62 1529,25 21,63 15,10 64,85 4,98 4,07 2020,24 33,14 25,82 67,81 5,51 4,78
20,68 81,29 5,91 5,05 1983,77 29,22 20,75 64,34 5,46 4,79
21,16 66,82 5,48 4,79 2053,79 30,97 24,05 62,01 5,05 4,46 1863,26 29,94 21,60 65,56 5,37 4,67 1949,10 31,55 23,77 73,18 5,59 4,86 1809,65 28,80 20,18 87,67 6,36 5,28 1811,72 29,23 22,21 70,07 5,67 4,80 1884,38 29,49 21,22 66,36 5,20 4,56
3 0 0 0
1941,44 27,23 18,76 70,17 5,32 4,45
11,30 18,37 15,92
8,05
2002,61 30,16
1907,96 30,05
164
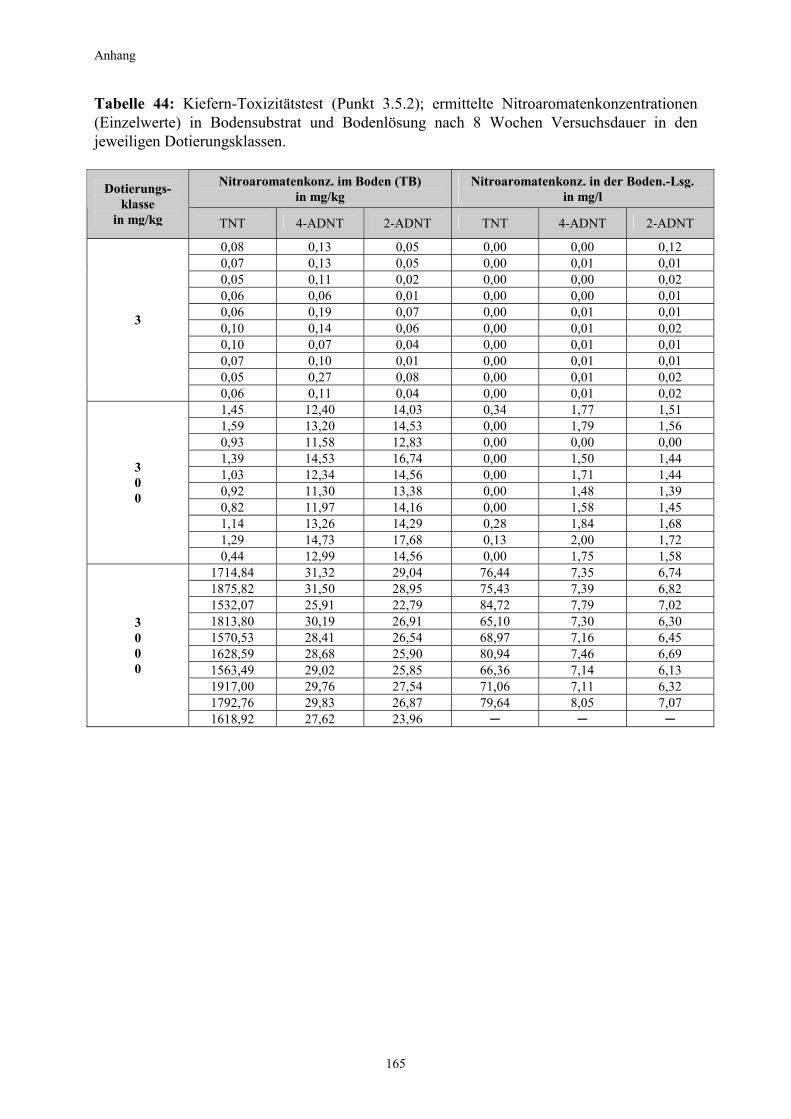
Anhang
Tabelle 44: Kiefern-Toxizitätstest (Punkt 3.5.2); ermittelte Nitroaromatenkonzentrationen (Einzelwerte) in Bodensubstrat und Bodenlösung nach 8 Wochen Versuchsdauer in den jeweiligen Dotierungsklassen.
Nitroaromatenkonz. im Boden (TB) in mg/kg
Nitroaromatenkonz. in der Boden.-Lsg. in mg/l Dotierungs-
klasse in mg/kg TNT 4-ADNT 2-ADNT TNT 4-ADNT 2-ADNT
0,08 0,13 0,05 0,00 0,00 0,12 0,07 0,13 0,05 0,00 0,01 0,01 0,05 0,11 0,02 0,00 0,00 0,02 0,06 0,06 0,01 0,00 0,00 0,01 0,06 0,19 0,07 0,00 0,01 0,01 0,10 0,14 0,06 0,00 0,01 0,02 0,10 0,07 0,04 0,00 0,01 0,01 0,07 0,10 0,01 0,00 0,01 0,01 0,05 0,27 0,08 0,00 0,01 0,02
3
0,06 0,11 0,04 0,00 0,01 0,02 12,40 14,03 0,34 1,77 1,51
1,59 13,20 14,53 0,00 1,79 1,56 0,93 11,58 12,83 0,00 0,00 0,00 1,39 14,53 16,74 0,00 1,50 1,44 1,03 12,34 14,56 0,00 1,71 1,44 0,92 11,30 13,38 0,00 1,48 1,39 0,82 11,97 14,16 0,00 1,58 1,45 1,14 13,26 14,29 0,28 1,84 1,68 1,29 14,73 17,68 0,13 2,00 1,72
3 0 0
14,56 0,00 0,44 12,99 1,75 1,58 1714,84 31,32 29,04 76,44 7,35 6,74 1875,82 31,50 28,95 75,43 7,39 6,82 1532,07 25,91 22,79 84,72 7,79 7,02 1813,80 30,19 26,91 65,10 7,30 6,30 1570,53 28,41 26,54 68,97 7,16 6,45 1628,59 28,68 25,90 80,94 7,46 6,69 1563,49 29,02 25,85 66,36 7,14 6,13 1917,00 29,76 27,54 71,06 7,11 6,32 1792,76 29,83 26,87 79,64 8,05 7,07
3 0 0 0
1618,92 27,62 23,96 ─ ─ ─
1,45
165
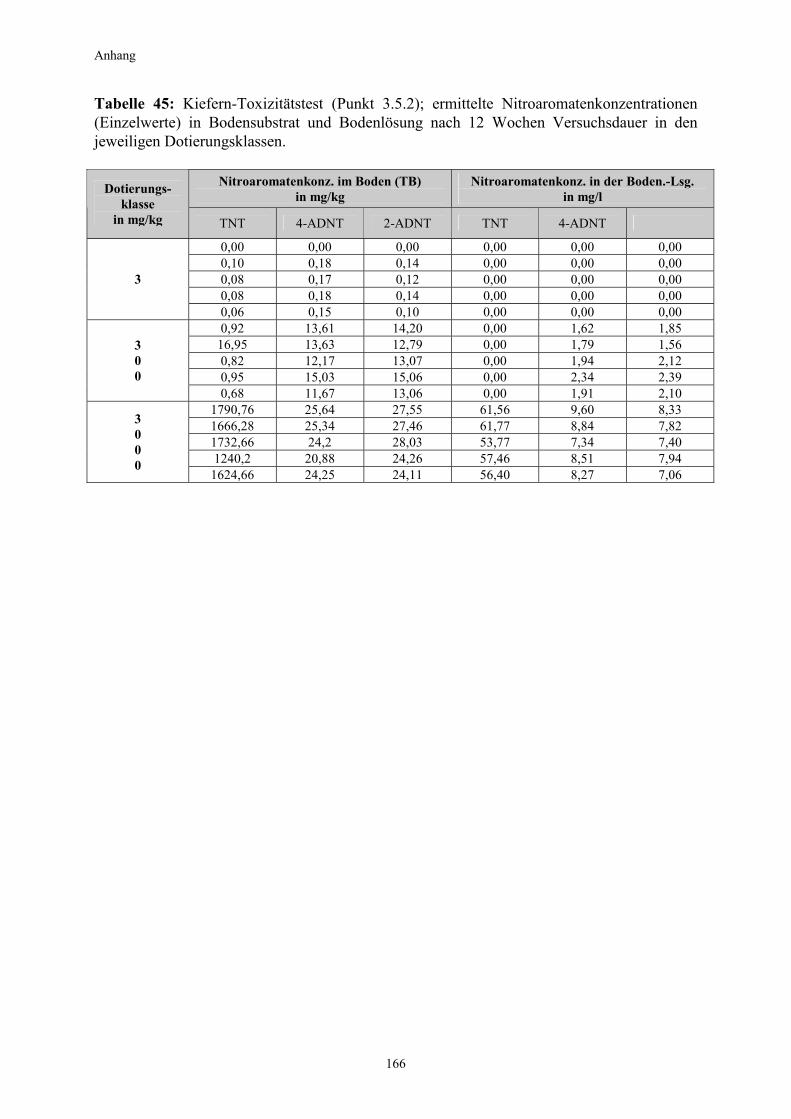
Anhang
Tabelle 45: Kiefern-Toxizitätstest (Punkt 3.5.2); ermittelte Nitroaromatenkonzentrationen (Einzelwerte) in Bodensubstrat und Bodenlösung nach 12 Wochen Versuchsdauer in den jeweiligen Dotierungsklassen.
Nitroaromatenkonz. im Boden (TB) in mg/kg
Nitroaromatenkonz. in der Boden.-Lsg. in mg/l Dotierungs-
klasse in mg/kg TNT 4-ADNT 2-ADNT 2-ADNT
0,00 0,00 0,00 0,00 0,00 0,00 0,10 0,18 0,14 0,00 0,00 0,00 0,08 0,17 0,12 0,00 0,00 0,00 0,08 0,18 0,14 0,00 0,00 0,00
3
0,06 0,15 0,10 0,00 0,00 0,00 13,61 14,20 0,00 1,62 1,85
16,95 13,63 12,79 0,00 1,79 1,56 12,17 13,07 0,00 1,94 2,12
0,95 15,03 15,06 0,00 2,34 2,39
3 0 0
0,68 11,67 13,06 0,00 1,91 2,10 1790,76 25,64 27,55 61,56 9,60 8,33 1666,28 25,34 27,46 61,77 8,84 7,82 1732,66 24,2 28,03 53,77 7,34 7,40 1240,2 20,88 24,26 57,46 8,51 7,94
3 0 0 0
1624,66 24,25 24,11 56,40 8,27 7,06
TNT 4-ADNT
0,92
0,82
166
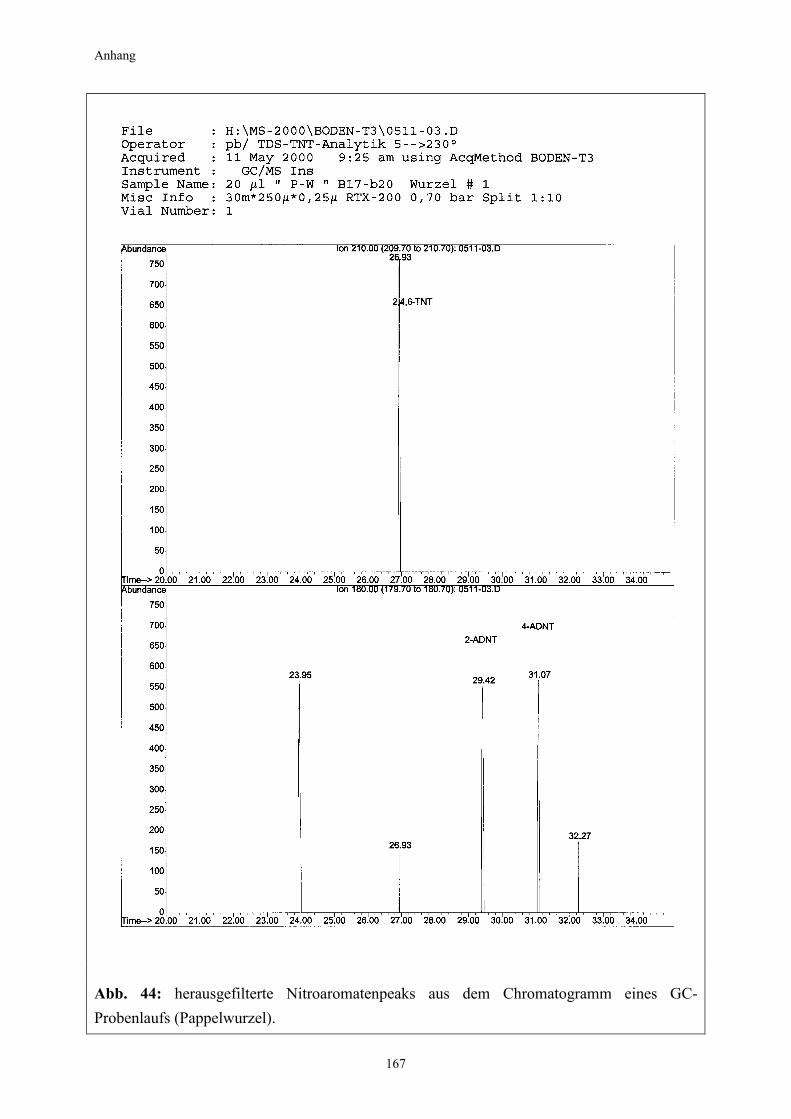
Anhang
Abb. 44: herausgefilterte Nitroaromatenpeaks aus dem Chromatogramm eines GC-Probenlaufs (Pappelwurzel).
167
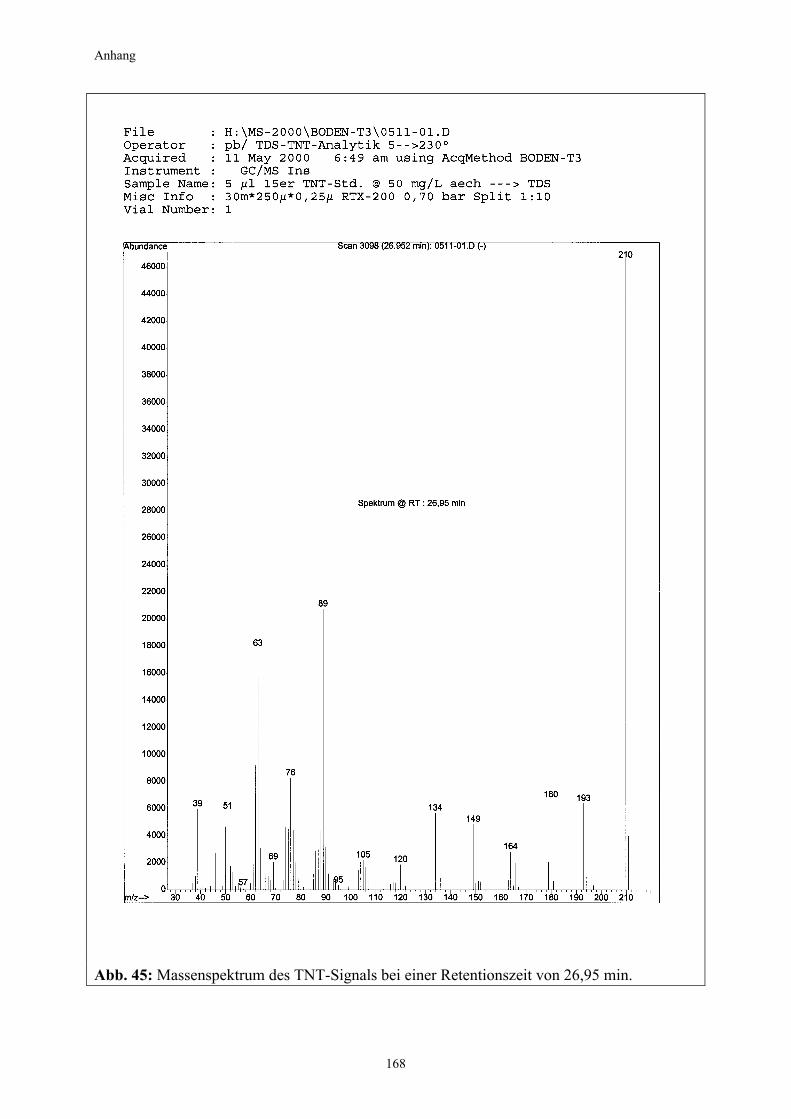
Anhang
Abb. 45: Massenspektrum des TNT-Signals bei einer Retentionszeit von 26,95 min.
168

Danksagung An dieser Stelle möchte ich allen Personen danken, die zum Gelingen der vorliegenden Arbeit beigetragen haben. In erster Linie gilt mein Dank Herrn Prof. Dr. Wolfgang Heyser für die interessante Aufgabenstellung, für die Bereitstellung einer Stelle als wissenschaftlicher Mitarbeiter, für den großen Freiraum bei der Bearbeitung des Themas und für die ständige Diskussions-bereitschaft. Herrn Prof. Dr. G. O. Kirst danke ich besonders für seine Bereitschaft sich als Gutachter dieser Arbeit zur Verfügung zu stellen. Auch bei Frau Prof. Dr. J. Filser, Herrn Dr. J. Warrelmann, Herrn. Dr. habil. Hartmut Koehler sowie Frau Diplom Biologin Antje Seebeck möchte ich mich für ihre Mitwirkung am Promotionsprüfungsausschuss bedanken. Ganz besonders danken möchte ich Herrn Peter Behrend für die Messung zahlreicher Boden- und Pflanzenproben mit der HPLC und GC/MS. Danke Peter! Mein ganz besonderer Dank gebührt auch Frau Anke Toltz und Frau Helga Wehrkamp für die große und freundliche Unterstützung im Laboralltag. Herrn Mario Kröger und Herrn Prof. Dr. G. Fels von der Universität Paderborn gilt mein besonderer Dank für die Bereitstellung des 14C-TNT-Radiotracers. Bei Frau Dr. habil. Heike Bücking möchte ich mich insbesondere für die Hilfestellung in den Experimenten mit Radiotracern bedanken. Bei Frau Antje Seebeck, Frau Myrna Landim, Frau Ulrike Jakob, Frau Dr. Maricel Alvarez, Herrn Dr. Seak Jin Kim, Herrn Rainer Hans und Herrn Dr. Jens Dittmann möchte ich mich für den Erfahrungsautausch und den besonders freundlichen Umgang miteinander bedanken. Frau Erika Beilfuß danke ich besonders für das Korrekturlesen dieser Arbeit. Ausdrücklich bedanken möchte ich mich bei allen Mitgliedern der Arbeitsgruppe für das freundliche, hilfsbereite und kooperative Arbeitsklima. Dank gilt dem ‘Tanne-Team’ für die besondere Zusammenarbeit und Durchführung des in-situ Projektes in Clausthal-Zellerfeld. Zum Schluss möchte ich mich bei meinen Eltern ganz herzlich bedanken, die mich während des Studiums und der Promotion immer nach Kräften unterstützt haben und meine Launen ertrugen.





























