Embed Size (px)
Citation preview

Planta (Ber].) 76, 359--366 (1967)
Eine neue Beweisf'fihrung fiir die Hypothese einer differentiellen Genaktivierung
durch Phytochrom 730
HEI~BERT LANGE, ILSE BIENGEIr u n d HANS Mol-m
Botanisches Ins t i tu t der Universi t~t Freiburg i. Br.
Eingegangen am 15. Jul i 1967
New Evidence in Favour o/the Hypothesis o/Di//erential Gene Activation by Phytochrome 730
Summary. 1)hytochrome-mediated anthocyanin synthesis of the mustard seed- ling (Sinapis alba L.) can be regarded as a prototype of a "posi t ive" photoresponse. "Posi t ive" photeresponscs are those which are characterized by an ini t iat ion or an increase of biosynthetic or growth processes. The initial lag-phase of this type of photorcsponse is relatively long. In the case of anthocyanin synthesis of the mustard seedling it takes about 3 hours after the initial formation of t),30, the active phyto- chrome, unti l anthocyanin synthesis can be measured (Fig.). This period of t ime has been called initial (or primary) lag-phase.
In earlier papers (e.g. LANGE and MORR, 1965; MOR~, 1966a, b; SC~OeFE~, 1967) we have advanced and supported the hypothesis t ha t the positive photo- responses (in the above sense) of the mustard seedling can be explained by a dif- ferential gene act ivat ion through 1)730. Those genes of which the act ivi ty can be s tar ted or increased by 1)730 have been called potentially active genes (1)7s0).
I n order to verify this hypothesis fur ther we have been trying to characterize the processes which occur during the initial lag-phase after the onset of continuous far-red light. There is good evidence (HA~TMAN~, 1966; CLA~KSO~ and HrLLMAN, 1967) t h a t under continuous far-red a low bu t virtually s ta t ionary concentration of the active 1)~30 can be maintained in the seedling's tissues over a period of many hours. Consequently we are dealing with s teady state conditions of 1)730 when we investigate the process of anthocyanin accumulation under continuous far-red.
In the present paper we present data which indicate t ha t P 30 exerts two func- tions during the initial lag-phase. First, i t eliminates a barrier before the potentially active genes insofar as i t makes these genes accessible for the act ivat ing action of P:a0. Secondly, P~a0 starts the act ivi ty of potentially active genes. To mainta in gene act ivi ty the continuous presence of 1)~a 0 is required. On the other hand, ii a gene has once been "opened" to the action of 1)7a 0 i t remains easily accessible for the acti- vat ing action of 1)73 o even in the case when 1)730 disappeared and gene act ivi ty ceased for an extended period of time. After the re-appearance of 1)v30 the gene can be act ivated almost immediately. These conclusions have been derived from the following facts.
1. The initial lag-phase after the onset of far-red is always in the order of 3 hours (Fig.). If, however, a seedling which was preirradiated with 12 hours of far-red is kept in darkness for an extended period and is then re-irradiated with far-red no lag-phase for the action of the second irradiation can be found even when antho- cyanin synthesis had already ceased during the preceding dark period (Fig.).
25*

360 H. LANGE, I. BIENGER und H. Mom~:
2. Since the action of the second irradiation as measured by anthocyanin syn- thesis can be completely inhibited by relatively low doses of Puromycin and Cyclo- heximide (Table) we conclude that the re-appearance of P780 leads to de novo syn- thesis of enzyme protein which is required for a resumption of anthocyanin synthe- sis. In a preceding paper (MOOR and S]~F, 1966) we have shown that Puromycin has no direct influence on anthocyanin production.
3. Application of Actinomycin D (10 ~g/ml) only partially inhibits the action of the second irradiation as measured by anthocyanin synthesis (Table). This find- ing was to be expected. In a preceding paper (Mom~ and BIENGER, 1967) we have shown that far-red mediated anthocyanin synthesis of the mustard seedling can be blocked by moderate concentrations of Actinomycin I) (10 ~g/ml) if the substance is applied before the onset of far-red. If, however, the seedlings are irradiated for several hours (well within the lag-phase!), transferred to the dark for one hour, in- cubated with Actinomycin I) and then reirradiated with continuous far-red, antho- cyanin synthesis can only partially be inhibited. I t has been concluded that genes which have once been activated by P~a0 remain less sensitive towards Actinomycin D even when PTao has disappeared. I t seems that during the initial lag-phase P~3o leads to stable structural changes in the neighbourhood of potentially active genes.
Einleitung Die durch P730, das physiologisch aktive Phytochrom, ausgel6ste
Anthocyansyn these des Senfkeimlings k a n n als P ro to typ einer ,,posi- r iven Photomorphose" ( = ,,positive photoresponse") angesehen werden (MoH~, 1966a). Posit ive Photomorphosen sind solche phytochrom- abhi~ngigen Ver~nderungen einer Keimpflanze, die durch eine AuslSsung oder Steigerung von Biosynthesen oder Wachs tumsvorg~ngen ausge- zeichnet sind. Charakterist isch ist die relat iv lange lag-Phase. Bei der Anthocyansyn these des Senfkeimlings dauer t es etwa 3 Std, bis auf die erstmalige Bildung yon PTa0 hin An thocyan meBbar wird. I)iesen Zei t raum nennen wit ,,primi~re lag-Phase".
Wir haben auf Grund vieler experimenteller Befunde die Auftassung vertreten (MOHn, 1966a, b; SCHOPFSR, 1967), dab die positiven Photo- morphosen auf eine differentielle Genaktivierung durch P~30 zurfickzu- ffihren seien.
Bei der weiteren Verifizierung dieser Hypothese kommt es wesent- lieh darauf an, die Vorgi~nge, die sich w~hrend der mehrere Stunden langen prim~ren lag-Phase abspielen, kennenzulernen. -- In der vorlie- genden Arbeit werden wit Daten behandeln, die darauf hinweisen, dab das Pva0 w~hrend der prim~ren lag-Phase zwei versehiedene Funktionen ausfibt. Ers~ens beseitigt das Pwo eine Barriere an den potentiell aktiven Genen; es macht diese Gene fiir die aktivierende Wirkung yon PTso zu- g~nglieh. Zweitens 15st P~ao die Aktivit~t dieser Gene aus. W~hrend fiir die Aufreehterhaltung der Genaktivit~t die best~ndige Anwesenheit yon PTao erforderlieh ist, bleiben die yon P73o zug~ng]ich gemachten Gene auch naeh der Entfernung yon Pvs0 ,,often", d.h. sic b]eiben fiir die akti- vierende Wirkung des P~ao ]eicht zugi~nglich. Offenbar bewirkt das P~ao

Differentielle Genaktivierung durch Phytochrom 361
w/~hrend dcr prim/~ren lag-Phase eine strukturel le Ver/~nderung im Be- reich der potentiel l ak t iven Gene. Hinweise darauf haben wit bereits in einer vorangegangenen Arbei t publizier~ (MoH~ und B I ~ G E ~ , 1967). Wi t haben dort den Sehlul~ gezogen, ,,dal~ die dureh PTao in F u n k t i o n gesetzten Gene aueh naeh dem Erl6sehen der P730-Wirkung l~icht wieder in den Zus tand zuriiekgehen, den sie vor der ersten Bel iehtung ha t t en . "
Material und Methoden Als experimentelles System benutzen wir wiederum die fiber P~o ausgelSste
Anthocyansynthese des Senfkeimlings (Sinapis alba L.). Eine relativ niedrige, aber langfristig station~re Konzentration des wirksamen P~a0 wird mit Dauer-Dunke]rot eingestellt (HA~T~A~N, 1966; MOKa, 1966a, b). Das Samenmaterial yon Sinapis alba L. (Ernte 1965), die Standard-Bestrahlungsanlage fiir Dauer-Dunkelrot (= DR), der allgemeine Versuchsansatz (25 ~ C; erster Lichtbeginn 24 Std nach Aussaat) und die Auswertung (Extraktion und Messung des Anthocyans) sind bei MOOR (1966a) und bei WAGN]~R und lYlom~ (1966a, b) beschrieben. Die Zeiten, zu denen die Zweitbelichtung einsetzte und die Zeitpunkte des Lichtbeginns bei der Bestim- mung der prim~ren lag-Phase verschieden alter Keimlinge, gehen aus der Abb. hervor. Die Kurvenpunkte sind Mittelwerte aus 4--8, die Tabellenwerte sind Mittel- werte aus acht unabh~ngigen Einzelbestimmungen.
Die Applikation der Antibiotica (Actinomycin D = Act. D, ein Geschenk yon Merck, Sharp & Dohme, Rahway N.J., U.S.A; Puromycin-Dihydrochlorid = Pu, yon Nutritional Bioehemicals Corporation, Cleveland, Ohio U.S.A. ; Aetidion purum = Cycloheximid = Cyc, yon Fluka AG, Buchs, Schweiz) erfolgte eine Stunde vor Beginn der Zweitbelichtung. Die Vorbehandlung bis zu diesem Zeitpunkt war fol- gende (vgl. Abb.): Eachdem die Keimlinge 24 Std im Dunkeln gekeimt waren, warden sie 12 Std mit DR bestrahlt und anschlieBend flit 17 Std verdunkelt. Dann wurden je 25 Keimlinge einer Keimdose im sehr schwachen griinen Sicherheitslicht in kleine Petrisehalen fibefffihrt und mit 10 ml Antibiotica-LSsung - - (Konzen- trationen, vgl. Tabelle) - - bzw. mit 10 ml Aqua dest. iibergossen. Each einstfindiger submerser Inkubation im Dunkeln wurden die Keimlinge im DR-Standardfeld wieder in die urspriinglichen Keimdosen fibertragen. Die 10 ml InkubationslSsung wurde in die Keimdosen gegeben, deren Keimpapier in der Zwischenzeit getrock- net worden war. Die ttydraturbedingungen in der Dose sind denen beim Standard- ansatz recht ~hnlich (gequollenes Keimpapier plus 3,5 ml Aqua dest.; vgl. MOHR, 1966a). Die Auswertung erfolgte nach 18 Std Zweitbelichtung mit Standard-DR. Ffir den zum Vergleieh dienenden Ausgangswert wurden die Keimlinge nach der einstfindigen Inkubation im Dunkeln, also zum Zeitpunkt des Einsetzens der Zweit- belichtung extrahiert. - - Die mit den Antibiotica behandelten Keimlinge zeigten auch im langffistigen Experiment keinerlei unspezifische Ver~nderungen.
Experimentelle Daten 1. Einige l~ekapitulationen. Es gibt gute Griinde fiir die Auffassung (WAo~CER
und Morn% 1966a), dab die physiologische Wirkung der geringen station~Lren Kon- zentration an PTs0, die yon dem Standard-DR in der Pflanze aufrecht erhalten wird, bei Abschalten des DR rasch aufhSrt. Die Tatsache, dal3 die Anthocyansynthese nach dem Abschalten des DR noch etwa 6 Std mit gleichbleibender Intensit~Lt weitergeht (vgl. Abb.), ist darauf zuriickzuffihren, da$ das kurzlebigste unter den an der Anthocyansynthese beteiligten Enzymen eine Lebensdauer in dieser GrS~en- ordnung hat (Mom~ und S~NF, 1966). Die postulierte PTao-abhiingige raRES, welche
25b Plan~a (:BerI.), Bd. 76
362 It. LANCE, I. BIENGEI~ und It. MGI~:
die limitierenden Enzyme der Anthoeyansynthese eodiert, dfirf~e sehr kurzlebig sein (Mom~ und SENF, 1966; DlmS~ und Mom~, 1966).
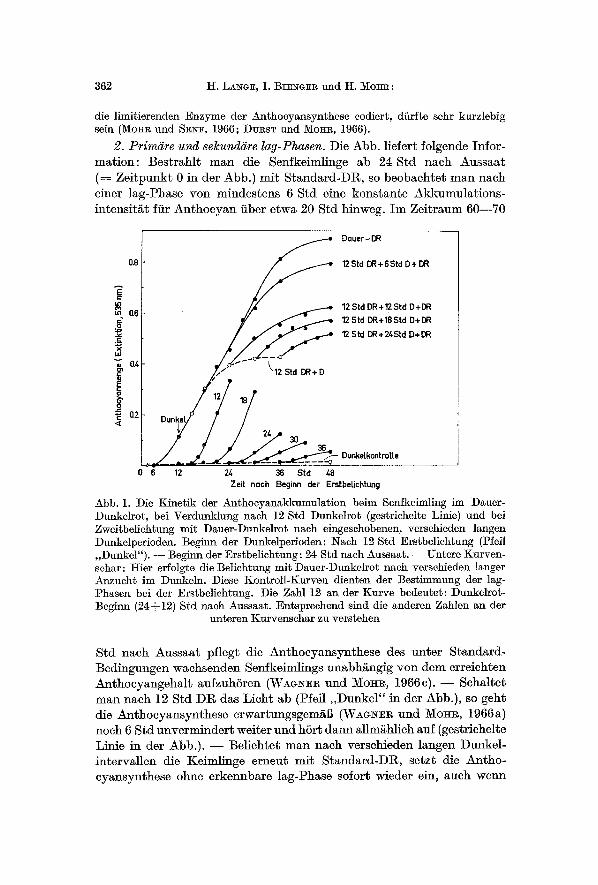
2. Primi~re und selcundgre lag-Phasen. Die Abb. l iefert folgende Infor- ma t ion : Bes t r ah l t m a n die Senfkeimlinge ab 24 S t d nach Aussaa t ( = Z e i t p u n k t 0 in der Abb.) mi t S t a n d a r d - D R , so beobach te t m a n nach einer lag-Phase yon mindes tens 6 S td eine kons t an t e Akkumula t i ons - in tens i t / i t ftir A n t h o c y a n fiber e~wa 20 S td hinweg. I m Ze i t r aum 60 - -70
Dauer- DR
0.8 12 Std DR + 6 Std D + DR
7 12 Std DR§ D+DR
m (16 E 12 Std DR+18 Std D+ DR
~= 12 Std DR+2/.Std D+DR
~ a2
Ou~etkontrotte
0 6 12 24 36 Std 48 Zeit noch Beginn der ErstbeUchtung
Abb. 1. Die Kinetik der Anthocyanakkumulation beim Senfkeimling im Dauer- Dunkelro~, bei Verdunklung nach 12 Std Dunkelrot (gestriehelte Linie) und bei Zweitbelichtung mit Dauer-Dunkelrot nach eingeschobenen, versehieden ]angen Dunkelperioden. Beginn der Dunkelperioden: 7Nach 12 Std Erstbeliehtung (Pfeil ,,Dunkel"). - - Begilm der Erstbelichtung: 24 Std nach Aussaat. - - Untere Kurven- schar: t i ier effolgte die Belichtung mit Dauer-Dunkelrot nach versehieden langer Anzuchb im Dunkeln. Diese Kontroll-Kurven dienten der Bestimmung der lag- Phasen bei der Erstbelichtung. Die Zahl 12 an der Kurve bedeutet: Dunkelrot- Begilm (243 12) Std nach Aussaat. Entspreehend sind die anderen Zahlen an der
unteren Kurvenschar zu verstehen
S t d nach Aussaa t pf legt die A n t h o e y a n s y n t h e s e des u n t e r S t anda rd - Bed ingungen wachsenden Senfkeimlings unabh~ngig yon dem er re ichten A n t h o e y a n g e h a l t aufzuhSren (WAG~]~R u n d M o i l , 1966 c). - - Seha l t e t m a n nach 12 S td D R das L ich t ab (Pfeil , ,Dunkel" in der Abb.) , so geh t die An~hocyansynthese e rwar tungsgem~6 (WAa~wR und MOl~, 1966a) noch 6 S td u n v e r m i n d e r t wei te r und h6r t d a n n al lm~hlich anf (gestr ichelte Linie in der Abb.) . - - Be l ich te t m a n nach versckieden langen Dunkel - in te rva l l en die Keiml inge e rneut mi~ S t a n d a r d - D R , se tz t die An tho- cy~nsynthese ohne e rkennba re lag-Phase sofort wieder ein, aueh wenn

Differentielle Genaktivierung dutch Phytochrom 363
die Anthocyanbildung im Dunkeln bereits vSllig zum Erliegen gekom- men war. Die reduzierte Intensitat der Anthoeyansynthese unter dem Einflul~ der Zweitbeliehtung ditrfte mat dem Fortschreiten der ,,primaren Differenzierung" ( W A G ~ und MOHR, 1966e) wahrend der Dunkel- intervalle zusammenhangen. Darauf soll jetzt nieht weiter eingegangen werden. Das ffir unsere Fragestellung nnmittelbar wiehtige Resultat ist, dal~ bei der Zweitbeliehtung keine lag-Phase erfal]t werden kann, wah- rend bei der Erstbelichtung (unterer Tefl der Abb.) die lag-Phase stets deutlich ansgepragt ist nnd - - zumindest ab 30 Std nach Aussaat - - in der Gr6Benordnung yon 3 Std liegt. Kurz zusammengefal3t: Die ,,pri- mare" lag-Phase, d.h. die lag-Phase nach Erstbelichtung, liegt in der Gr6Benordnung yon 3 Std; die ,,seknndare" lag-Phase, d.h. die lag- Phase naeh Einsetzen der Zweitbeliehtung, ist hingegen nieht mel~bar. Die Extrapolation der Kinetik-Kurven naeh Zweitbelichtung weist direkt auf den Zeitpunkt des Lichtbeginns.
3. Hiingt der El/elct der Zweitbelichtung mit einer Enzym-Neusynthese zusammen ? - - Diese Frage kann dnrch Inhibitor-Experimente beant- wortet werden. Die Keimlinge warden 12 Std im DR-Feld gehalten, dann 18 Std (bis zum ErlSschen der Anthocyansynthese) verdunkelt und an- schliel]end nochmals 18 Std mat DR belichtet. 1 Std vor der Zweitbe- lichtung erfolgte die Applikation yon Act. D, Pu bzw. Cyc (vgl. Metho- den). Act. D ist bekanntlieh ein hochgradig spezffischer Inhibitor der Transkription; Pu und Cyc hemmen die Proteinsynthese.
Wie die Tabelle zeigt, hemmen bereits relativ geringe Konzentrationen an Pu and Cyc die dureh die Zweitbelichtung ausgelSste Anthoeyan- synthese vSllig. Es sei hier daran erinnert, dal~ Pu keinen direkten Ein- flul~ auf die Anthocyansynthese hat (MoH~ und SE~F, 1966). SeineWir- knng auf die Anthocyansynthese kann nur fiber eine t temmung der relevanten Enzymsynthese verstanden werden. Fiir Cyc gelten ver- mutlich dieselben Argumente. - - Der Schlul~ erseheint berechtigt, dal] P~ao auch bei der Zweitbeliehtung dadurch wirkt, dal~ es die de novo Synthese yon Enzymen ermSglieht, die ffir die Anthoeyanbildung ge- braucht werden. - - Die Resnltate mat Act. D (nnr partielle Itemmung) best~tigen und erweitern die Befunde yon MOHR und BIE~Gv~ (1967): Eine Konzentration an Act. D (10 ~g/ml), die bei einer Applikation vor der Erstbelichtung die PTa0-abhangige Anthocyansynthese vsnig hemmt, fibt nur noch eine partielle t temmung aus, wenn sic vor einer Zweit- belichtung appliziert wird. Dieses Ergebnis kann wahrscheinlich nur so aufgefa~t werden (Argumente bei M o ~ und BIs.~Gv,~, 1967), dal~ die einmal yon P730 in Aktion gesetzten Gene eine reduzierte ,,Empfindlich- keit" fiir Act. D zeigen; offenbar deshalb, weft in der 57achbarschaft dieser Gene dureh das P73o bleibende strnkturelle Veranderungen ver- ursacht wurden. Diese Veranderungen bleiben auch erhalten, wenn P~so
364 H. L~G]:, I. BIE~G~ und H. tV[oHR:
nicht mehr vorhanden ist, z.B. 17 Std naeh Beendigung der Erstbe- lichtung.
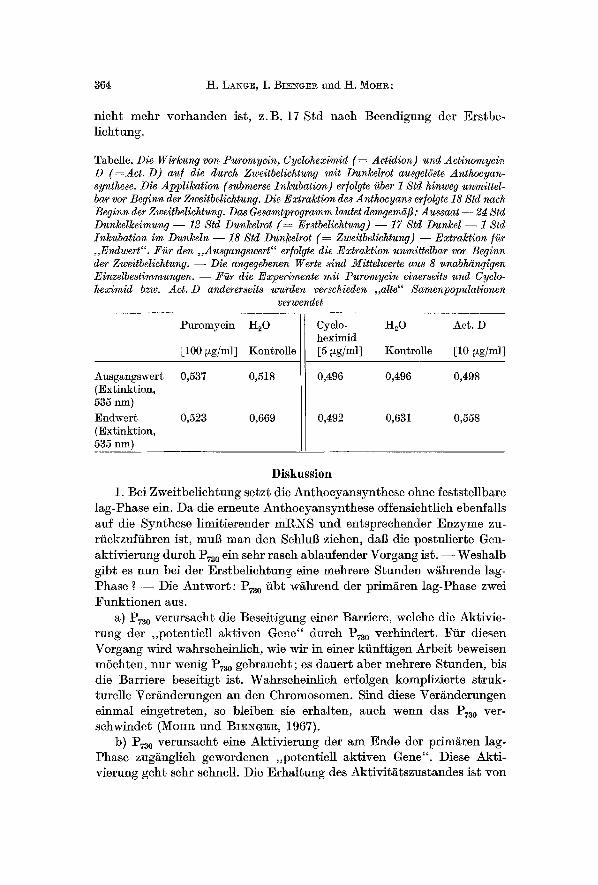
Tabelle. Die Wirkung yon Puromyein, Cycloheximid ( ~ Actidion) und Aetinomycin D ( : A c t . D) au/ die durch Zweitbeliehtung mit Dunkelrot ausgelgste Anthocyan- synthese. Die Applikation (submerse Inkubation) er/olgte i~ber 1 Std hinweg unmittel- bar vor Beginn der Zweitbelichtung. Die Extraktion des Anthocyans er/olgte 18 Std naeh Beginn der Zweitbelichtung. Das Gesamtprogramm lautet demgem~i[3 : Aussaat - - 24 Std Dunkel~eimung - - 12 Std Dun~elrot ( ~ Erstbelichtung) - - 17 Std Dunkel - - 1 Std Inl~ubation im Dun~eln - - 18 Std Dunkelrot ( ~ Zweitbelichtung) - - Extraktion liar ,,Endwert". Fi~r den ,,Ausgangswert ~ er/olgte die Extraktion unmittelbar vor Beginn der Zweitbetiehtung. - - Die angeffebenen Werte sind Mittelwerte aus 8 unabMingigen Einzelbestimmu~gen. - - ~ r die Experimente mit Puromycin einerselts und Cyclo- heximid bzw. Act. D andererseits wurden verschieden ,,alte" Samenpopulationen
verwendet
Ausgangswert (Extinktion, 535 nm) Endwert (Extinktion, 535 nm)
Puromycin H20
[100 ~g/ml] Kontrolle
0,537 0,518
0,523 0,669
Cyclo- H20 Act. D heximid [5 ~g/ml] Kontrolle [10 ~g/ml]
0,496 0,496 0,498
0,492 0,631 0,558
Diskussion
1. Bei Zweitbelichtung setz~ die Anthocyansynthese ohne feststellbare lag-Phase ein. Da die erneute Anthocyansynthese offensichtlieh ebenfalls auf die Synthese limitierender ml~NS und entsprechender Enzyme zu- rfickzuffihren ist, mul~ man den SchluB ziehen, dal~ die postulierte Gen- aktivierung durch PT~0 ein sehr raseh ablaufender Vorgang ist. - - WeshMb gibt es nun bei der Erstbelichtung eine mehrere Stunden w~hrende lag- Phase ? - - Die Antwort: P~30 fibt w~hrend der prim~ren lag-Phase zwei Funktionen aus.
a) P730 verursaeht die Beseitigung einer Barriere, welehe die Aktivie- rung der ,,po~entiell aktiven Gene" durch P~ao verhindert. Fiir diesen Vorgang wird wahrsehein]ich, wie ~ hi einer kfinftigen Arbeit beweisen InSchten, nur wenig P780 gebraucht; es dauert aber mehrere Stunden, bis die Barriere beseitigt ist. Wahrseheinlich erfolgen komplizierte struk- turelle Ver~nderungen an den Chromosomen. Sind diese Vers einmal eingetreten, so bleiben sie erhalten, aueh wenn das P~a0 ver- sehwindet (MOH~ und BIENG]~R, 1967).
b) P~a0 verursaeht eine Aktivierung der am Ende der primgren lag- Phase zuggnglieh gewordenen ,,potentiell aktiven Gene". Diese Akti- vierung geht sehr schneU. Die Erhaltung des Aktivit~tszustandes ist yon

Differentielle Genaktivierung durch Phytochrom 365
der bestandigen Anwesenheit des PTs0 abh~ngig. Versehwindet P730, so hSrt die gen-abhiingige RNS- und Enzymsynthese sofort auf (DuRsT und MOHR, 1966). Mit einer VerzSgerung yon etwa 6 Std, die auf die betrs liche ,,Lebensdauer" des limitierenden Enzyms zurfickzuffihren ist, endet auch die Anthoeyansynthese. Wird Pva0 wieder gebildet, so kann die RNS- und Enzymsynthese sofort wieder einsetzen. Am Beispiel der Phenyl- alanindesaminase werden wir in einer folgenden Arbeit (RISSLAND und MOH~, in Vorbereitung) zeigen, dab die Untersuchung der Enzym- induktion durch Pv30 zu genau denselben Schlfissen ffihrt wie die Unter- suchung der P73o-abh~ngigen Anthocyansynthese.
2. Im Zusammenhang mit Arbeiten fiber phytochrom-abhi~ngige Blattbewegungen haben FO~DEWLLE, BORTItWICK und HENDRmS:S (1966) (vgl. Zitat) sowie ItrLLMAN und KOUXKARI (1967) darauf hinge- wiesen, dab photoresponses mit kurzer lag-Phase (ca. 5--10 min) nicht mit der Vorstelhing, Pv30 verursache bei positiven Photomorphosen eine Genaktivierung, in Einklang zu bringen seien. ("The rapidity of the response indicates that gene activation as currently suggested for phyto- chrome action by If. Moga and his coworkers is not involved"). Die Ergebnisse der vorliegenden Arbeit zeigen, dab dieser Einwand nicht berechtigt ist. Andererseits mSchten wir abermals betonen, dab wit durchaus der Feststellung zustimmen, die Funktion des Phytoehroms in den Pulvini yon Mimosa pudica L. habe vermutlich nichts mit differen- tietler Genaktivierung zu tun. Es erscheint durchaus mSglich, dab in diesem speziellen Fall das Phytoehrom seine Wirkung ausschlieBlich fiber Ver~nderungen der Membranpermeabilit~t entfaltet. Wir sehen aber keine MSglichkeit, die Funktion des Phytochroms im Zusammenhang mit Differenzierung und Morphogenese ohne das I(onzept einer differen- tiellen Genaktivierung zu diskutieren (MOHR, 1967).
Zusammenhssung Naeh Erstbelichtung des Senfkeimlings mit Dauer-Dunkelrot zeigt
die Anthocyansynthese, eine typische ,,positive Photomorphose", eine mehrere Stunden ws lag-Phase. Bei einer Zweitbeliehtung (Pro- gramm: Erstbelichtung - - liingere Dunkelperiode - - Zweitbeliehtung) fehlt die lag-Phase (Abb.). - - Auch die Zweitbelichtung dfirfte die Anthoeyansynthese fiber eine Genaktivierung, die eine Enzymsynthese zur Folge hat, auslSsen (Tabelle). - - Die wichtigsten Konsequenzen: P7a0, das aktive Phytochrom, kann die Enzymsynthese sehr rasch be- wirken, falls die Gene der Aktivierung dureh PTao zug~nglioh shad. Die lange lag-Phase nach Einsetzen der Erstbelichtung benStigt das 1)730 offenbar dazu, die potentiell aktiven Gene (Pvso) ffir das P7so zng~inglich zu maehen. Die diesbezfiglichen Xnderungen, vermutlich in der Umge- bung der Gene, bleiben aueh dann erhalten, wenn Pv30 verschwindet.

366 H. LANGE et al. : Differentielle Genaktivierung durch Phytochrom
P~80 scheint also zwei d i s t ink te F u n k t i o n e n zu haben, die m a n experi- mente l l t re rmen k a n n : Es m a c h t die Gene zug/~nglich und es ak t i v i e r t d ie be t ref fenden Gene.
Mit Unt~rst~tzung durch die Deutsche Forsehungsgemeinschaft. - - Herrn Dr. Cx. W. MuStrE~T bei der Firma Merck, Sharp und Dohme (Rahway, N.ff., U.S.A.) danken wir fiir die Uberlassung yon Actinomyein D.
L i te ra tu r
CLARXSO~, D. T., and W. S. H~L~A~: Stability of phytockrome concentration in dicotyledonous tissues under continuous far-red light. Planta (Berl.) 75, 286--290 (1967).
DV~ST, F., and H. Mon~: Half-life of phytochrome-induced pheny]alanine deami- nase in mustard seedlings (Sinapis alba L.). iYaturwissensehaften 53, 707 (1966).
FOI'qDEVILLE, J.C., H. A. BORTHWICK, and S. B. H:ENDI~[CKS: Leaflet movement of Mimosa 2udica L. indicative of phytochrome action. Planta (Berl.) 69, 359--364 (1966).
I-IA~T~AN~, K. M.: A general hypothesis to interpret "high energy phenomena" of photomorphogenesis on the basis of phytochrome. Photoehem. Photobio]. 5, 349--366 (1966).
Hmr~L~, W. S., and W. L. K O ~ K ~ I : Phytochrome effects in the nyctinastic leaf movements of Albizzia ]ulibrissin and some other legumes. Plant Physiol. (in press) (1967).
L.~G~, H., u. It. MO]tR: Die Hemmung der phytochrominduzierten Anthoeyan- synthese durch Actinomyein D und Puromycin. Planta (Berl.) 67, 107--121 (1965).
MOH~, H.: Untersuchungen zur phytochrominduzierten Photomorphogenese des Senfkeimlings (Sinapis alba L.). Z. Pflanzenphysiol. 54, 63--83 (1966a).
- - Differential gene activation as a mode of action of phytochrome 730. Photochem. Photobiol. 5, 469--483 (1966b).
- - Photomorphogenesis. In: The physiology of plant growth, development and responses (M. B. W~KI~s, ed.). Maidenhead: McGraw-Hill Book Co 1967.
--, u. I. BIE~v,R: Experimente zur Wirkung yon Actinomyein D auf die dureh Phytoehrom bewirkte Anthocyansynthese. Planta (Berl.) 75, 180--184 (1967).
- - , u. R. S E ~ : Die Hemmung der phytochrom-induzierten Anthoeyansynthese dureh Puromyein und 2-Thiouracil. Planta (Berl.) 71, 195--203 (1966).
Sc~oP]~]~R, P.: Die Hemmung der phytoehrominduzierten Photomorphogenese (,,positive" Photomorphosen) des Senfkeimlings (Sinapis alba L.) dutch Aetino- mycin D und Puromycin. Planta (BEE.) 72, 297--305 (1967).
WAO~ER, E., and If. Mong: Kinetic studies to interpret "high-energy phenomena" of photomorphogenesis on the basis of phytoehrome. Photochem. Photobiol. 5, 3 9 7 ~ 0 6 (1966a).
- - - - Kinetisehe Studien zur Interpretation der Wirkung yon Sukzedanbestrah- lungen mit Hellrot und Dunkelrot bei der Photomorphogenese (Anthocyan- synthese bei Sinapis alba L.). Planta (Berl.) 70, 34 41 (1966b).
- - - - ,,Prim~re" und ,,sekund~re" Differenzierung im Zusammenhang mit der Photomorphogenese yon Keimpflanzen (Sinapis alba L.). Planta (Berl.) 71, 20~--221 (1966e).
Prof. Dr. If. Mon~ Botanisehes Institut der Universit~t 78 Freiburg i.Br., Sch~nzlestraBe 9






![Ereignis-korrelierte fMRT-Studie zu differentiellen ... · perzeptuelles Lernen, sowie die klassische Konditionierung [127]. Das nicht-deklarative Gedächtnis ist entscheidend zur](https://img.pdfslide.org/doc/110x75/5b9f5d3d09d3f2d0208d0f85/ereignis-korrelierte-fmrt-studie-zu-differentiellen-perzeptuelles-lernen.jpg)





















