Embed Size (px)
Citation preview

Einführung in die Biophysik!Schwerpunktstudium "Biophysik !
Prof. Dr. Joachim Rädler!!Lipide!!!!!!!!
LMU!
www.softmatter.physik.lmu.de/teaching!
• Plasmamembran!• Biochemie der Lipide!• Lipid Selbst-Assemblierung!• Permeabilität!• Diffusion in Membranen!
11. Juli 2001!
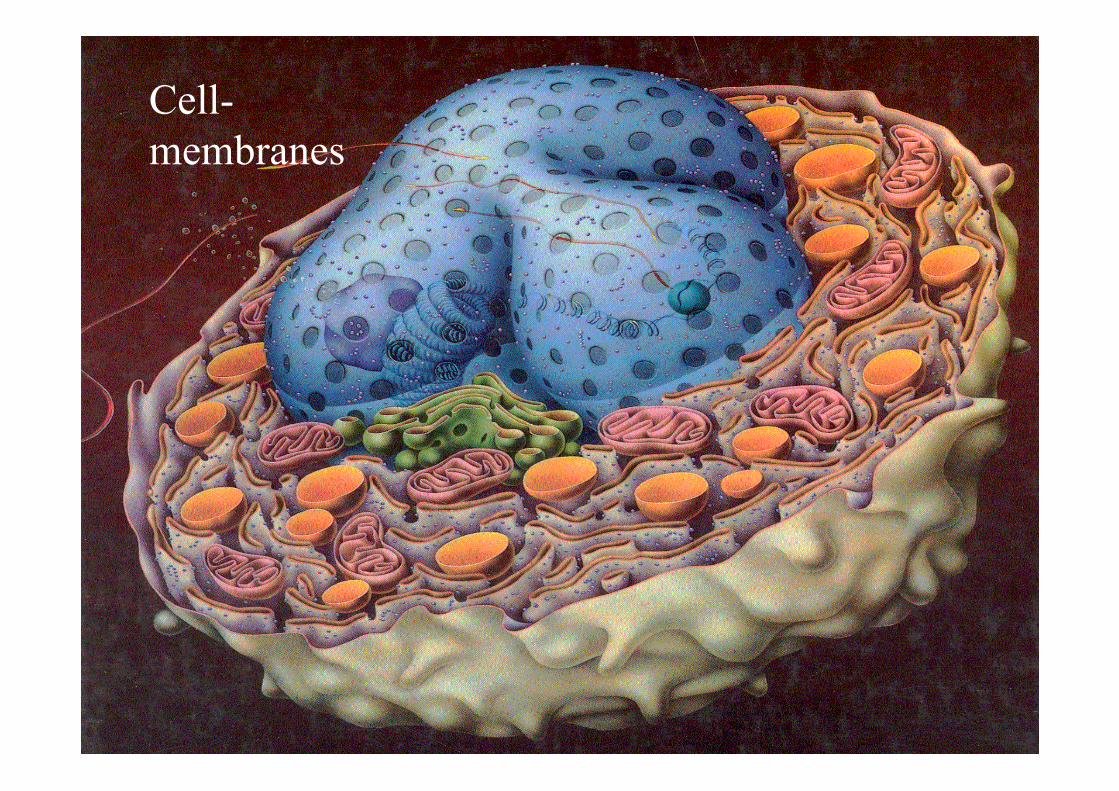
Cell- membranes
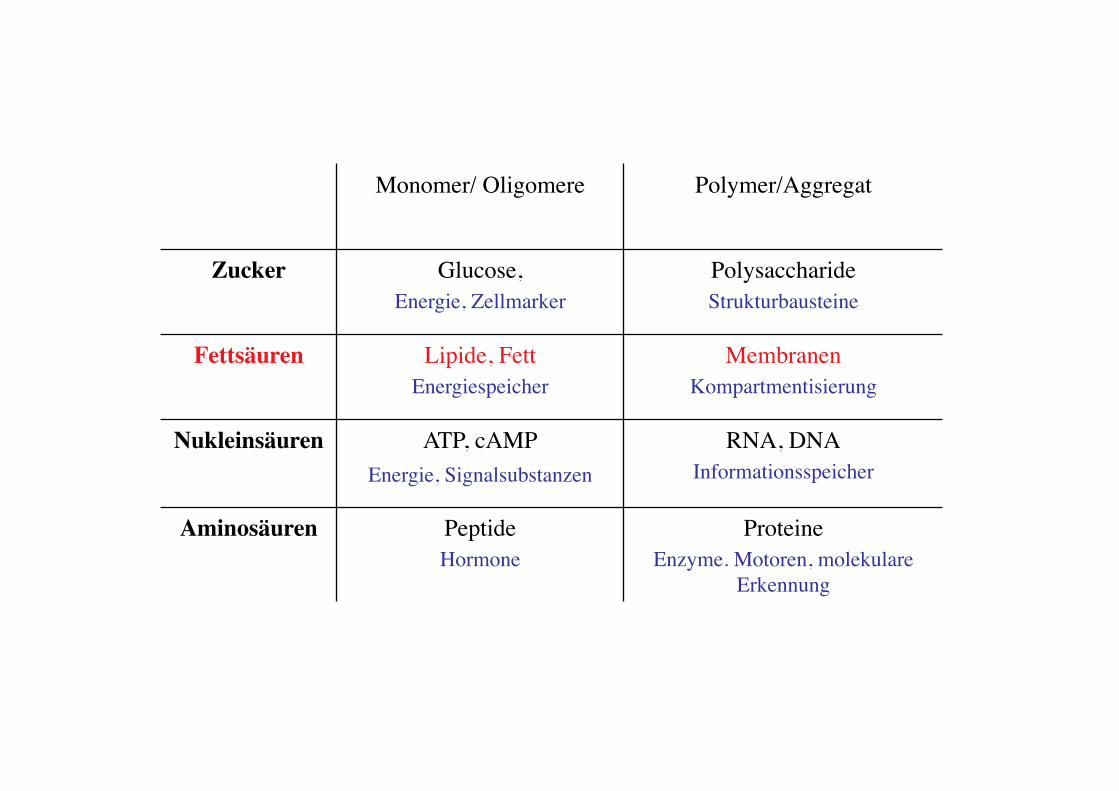
Monomer/ Oligomere! Polymer/Aggregat!
Zucker! Glucose, !Energie, Zellmarker!
Polysaccharide!Strukturbausteine!
Fettsäuren! Lipide, Fett!Energiespeicher!
Membranen!Kompartmentisierung!
Nukleinsäuren! ATP, cAMP!Energie, Signalsubstanzen !
RNA, DNA!Informationsspeicher!
Aminosäuren! Peptide!Hormone!
Proteine !Enzyme. Motoren, molekulare
Erkennung !
Gaub LMU! BPE§3.5! 4!
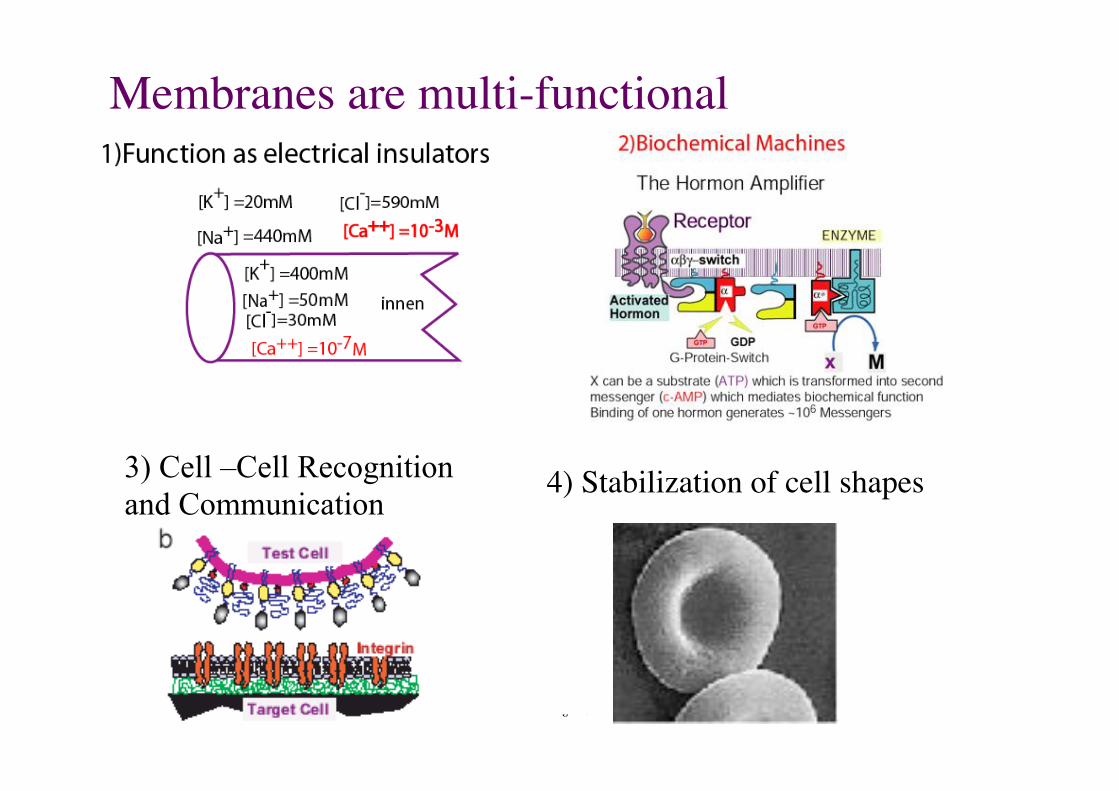
Membranes are multi-functional!
3) Cell –Cell Recognition and Communication
4) Stabilization of cell shapes!
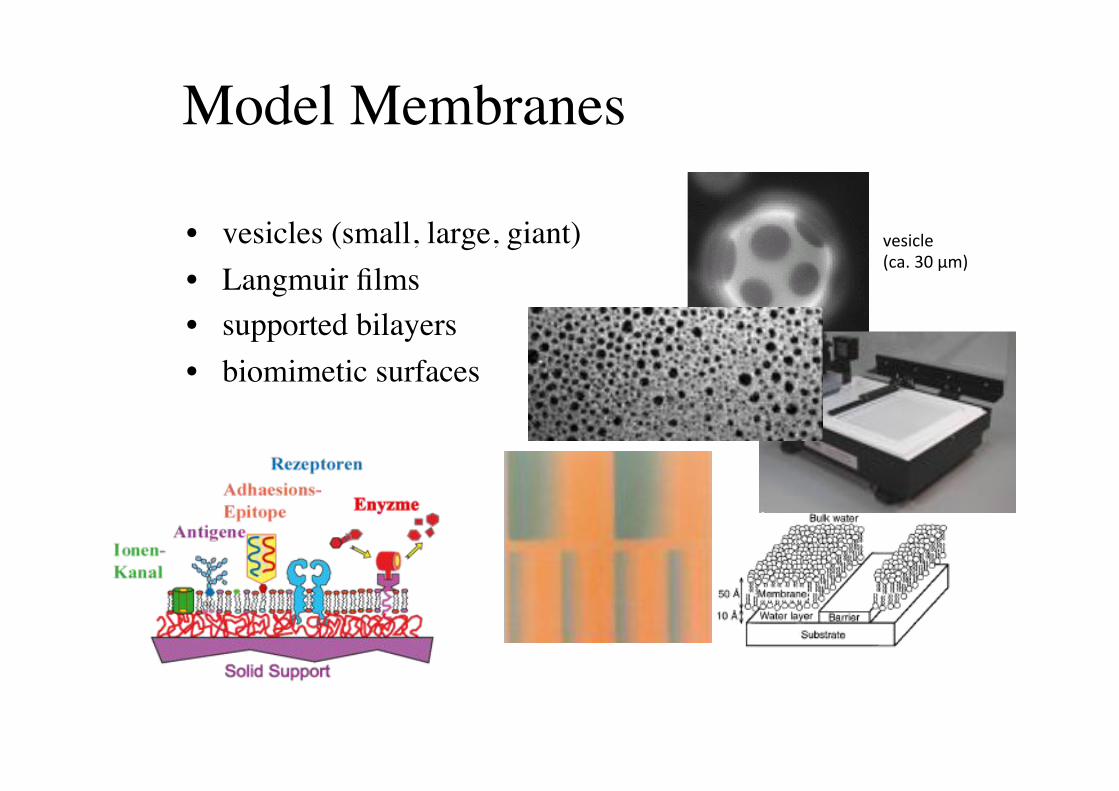
Model Membranes!
• vesicles (small, large, giant)!• Langmuir films!• supported bilayers!• biomimetic surfaces!
!"#$%&"''(%)*'+,'-./'
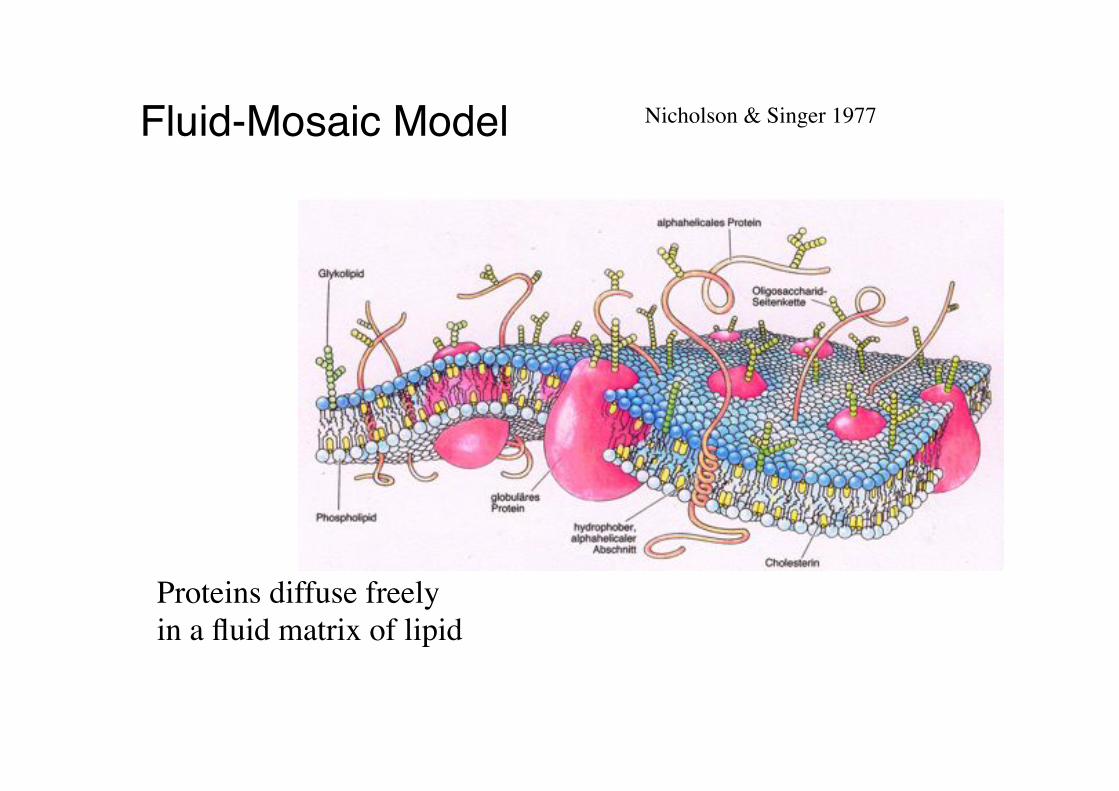
Fluid-Mosaic Model" Nicholson & Singer 1977!
Proteins diffuse freely!in a fluid matrix of lipid!
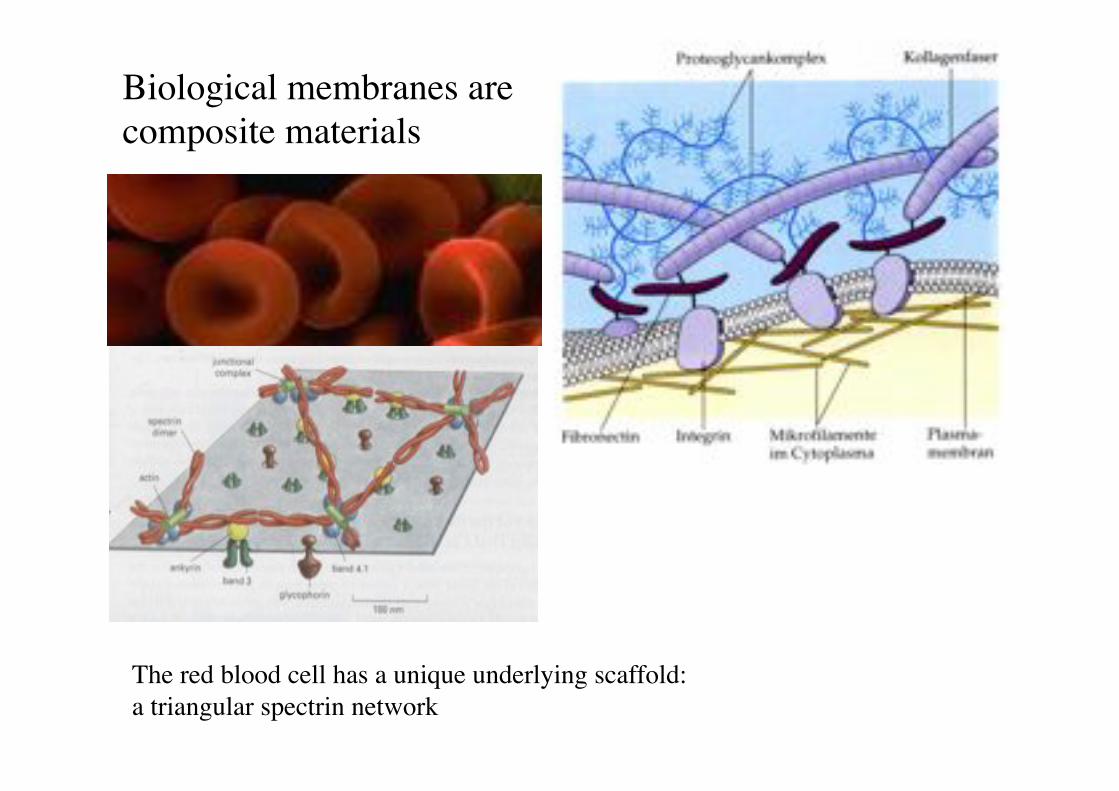
Biological membranes are composite materials!
The red blood cell has a unique underlying scaffold:"a triangular spectrin network!
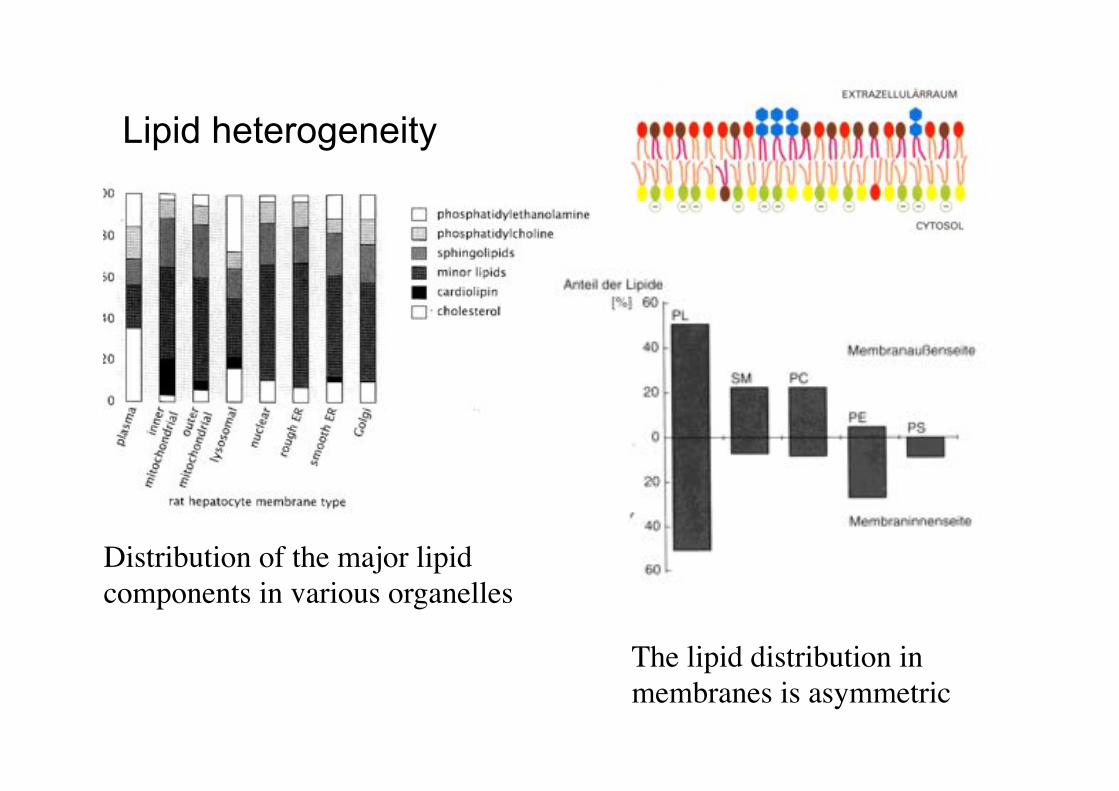
Lipid heterogeneity
Distribution of the major lipid components in various organelles!
The lipid distribution in membranes is asymmetric!
9!
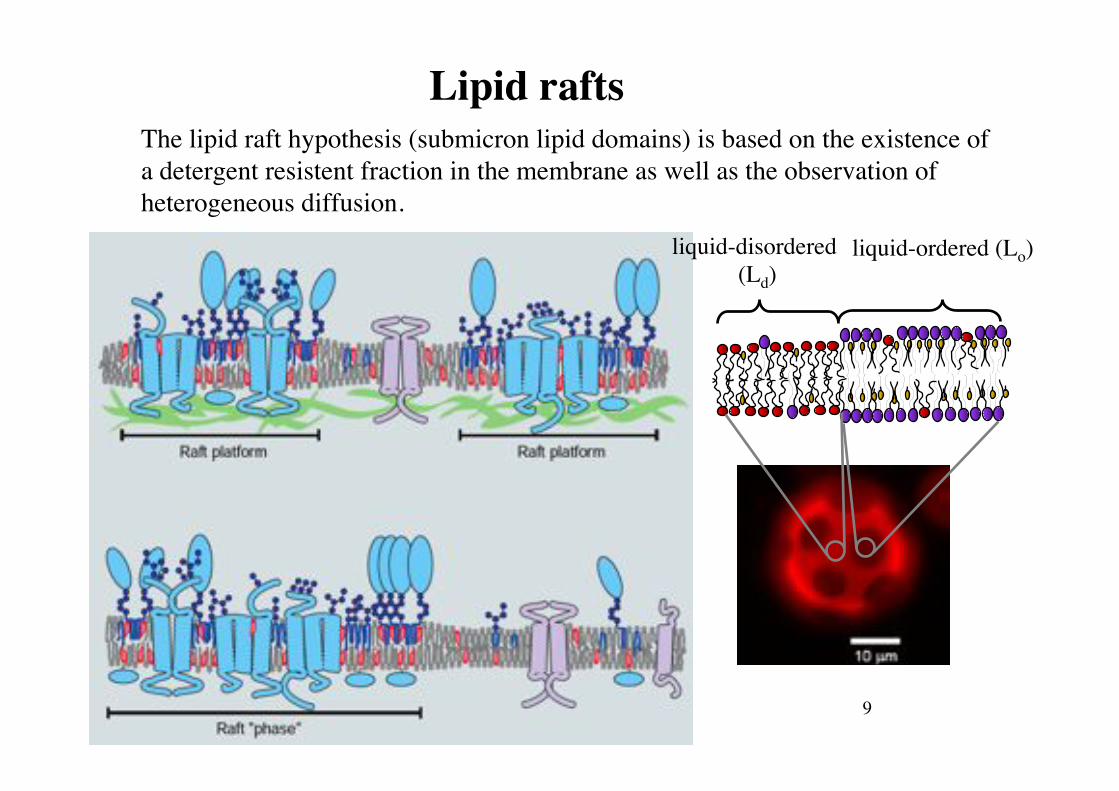
Lipid rafts!The lipid raft hypothesis (submicron lipid domains) is based on the existence of a detergent resistent fraction in the membrane as well as the observation of heterogeneous diffusion.!
liquid-disordered (Ld)!
liquid-ordered (Lo)!

Chemistry and self-assembly!
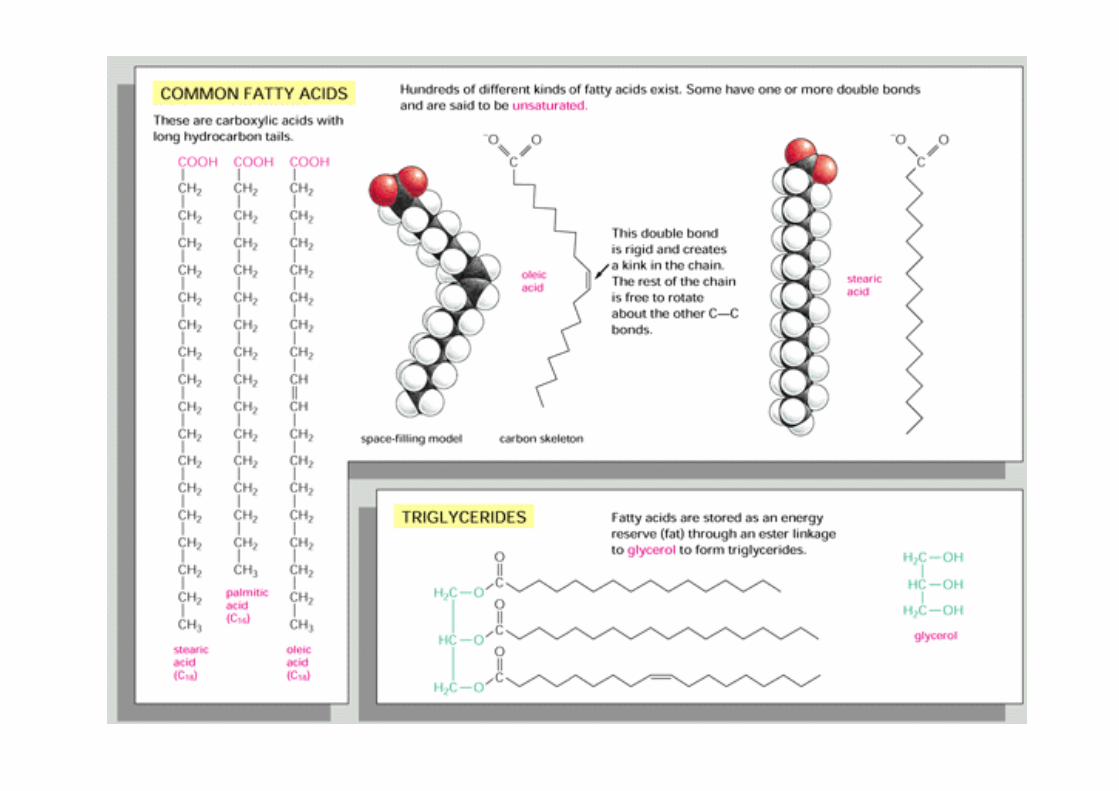
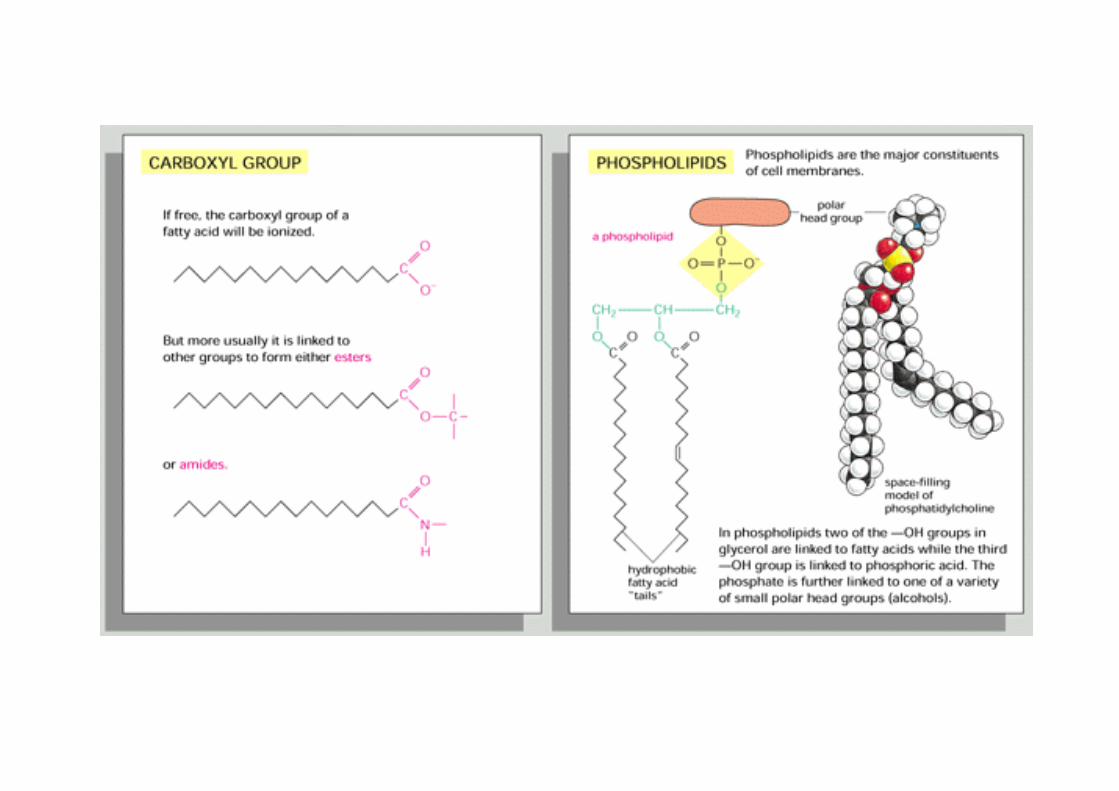
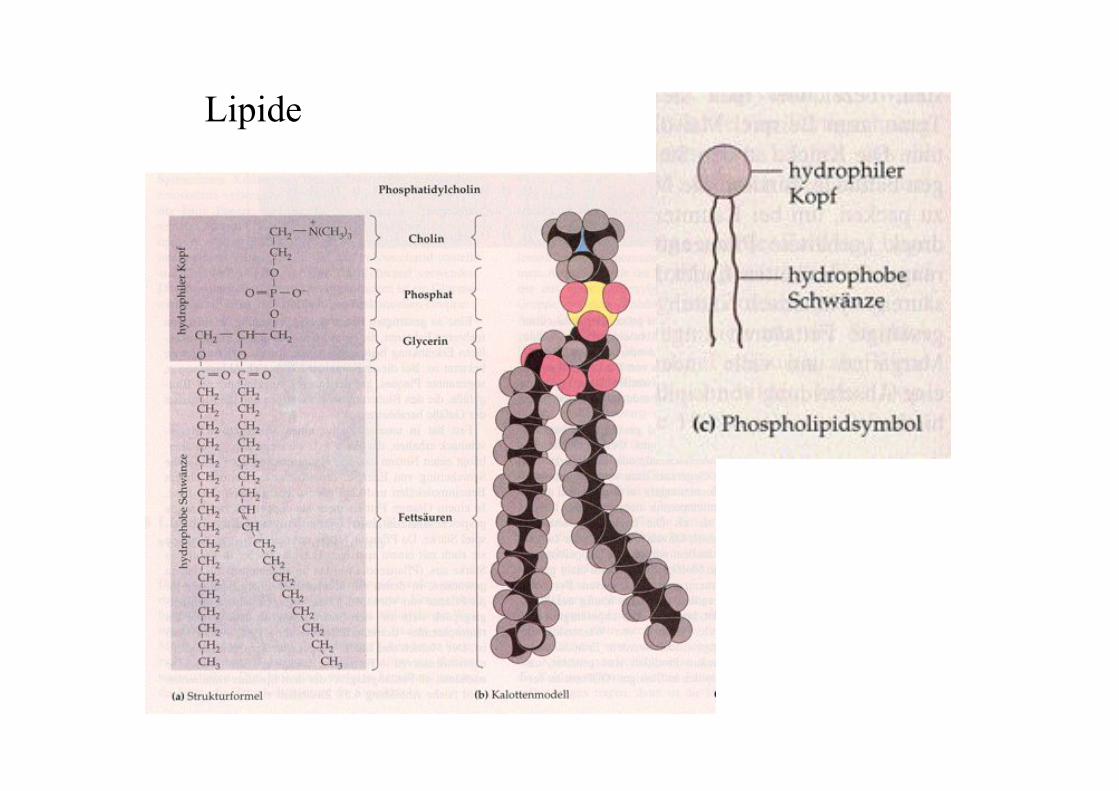
Lipide
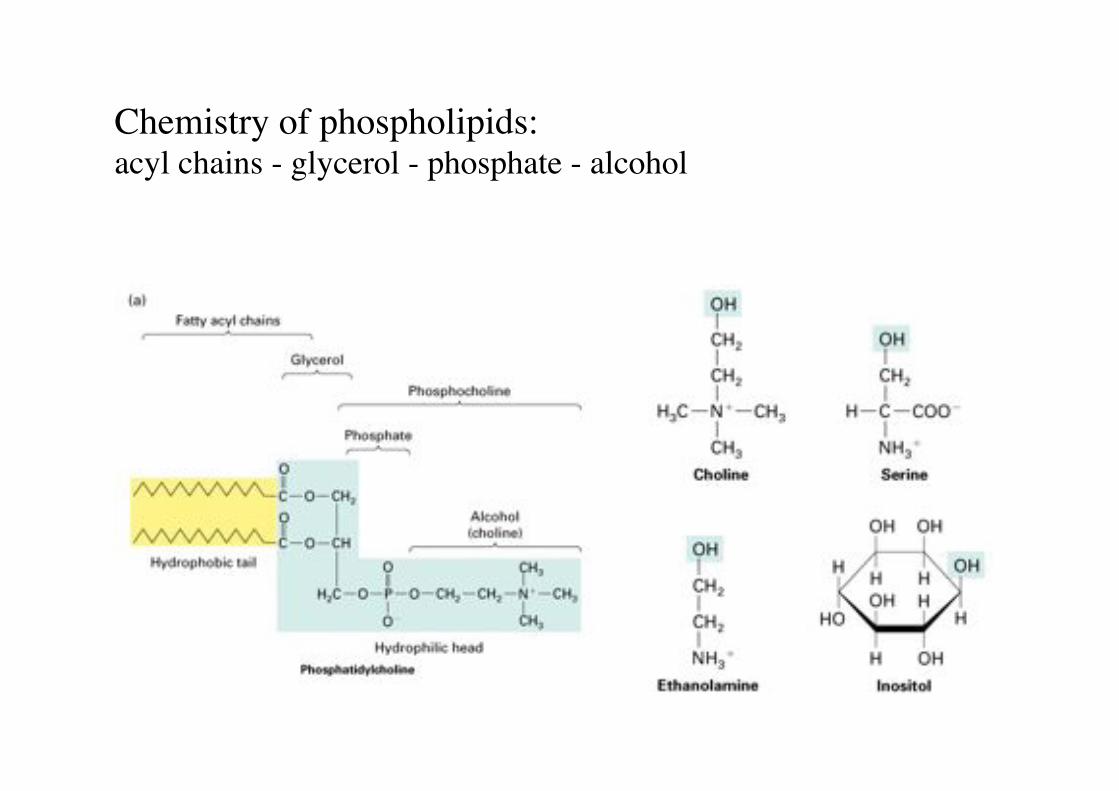
Chemistry of phospholipids:"acyl chains - glycerol - phosphate - alcohol!
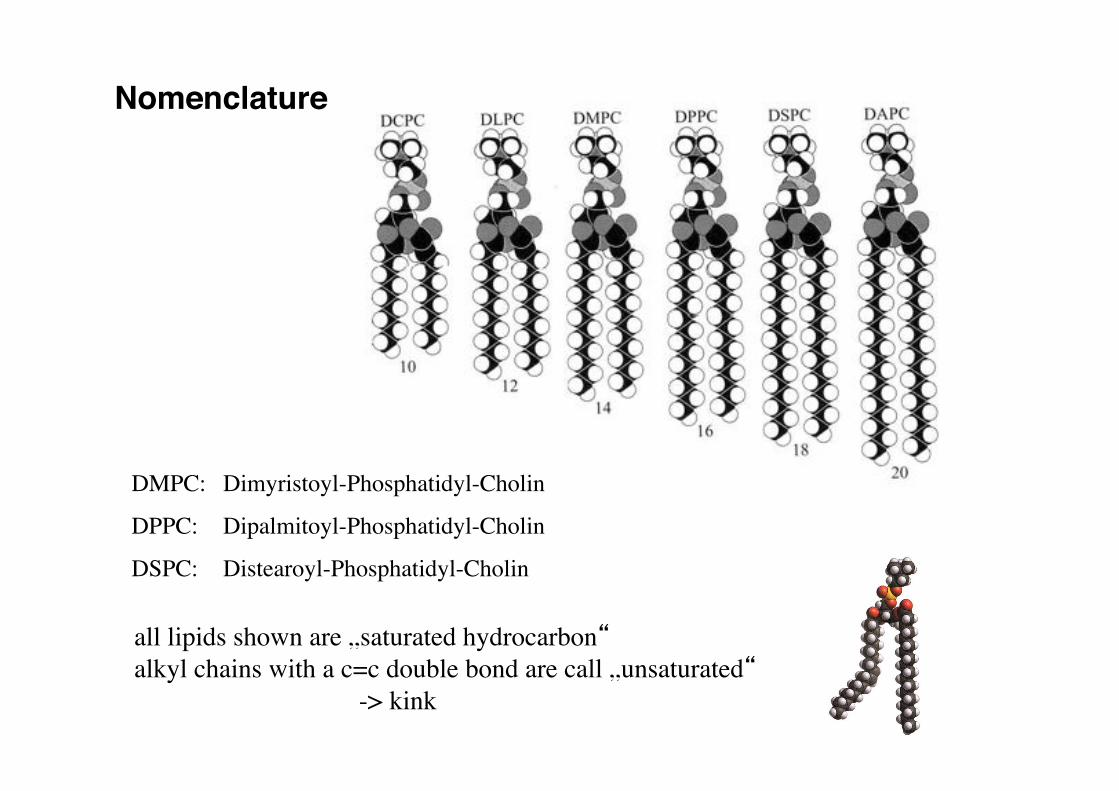
Nomenclature!
DMPC: !Dimyristoyl-Phosphatidyl-Cholin!
DPPC: !Dipalmitoyl-Phosphatidyl-Cholin!
DSPC: !Distearoyl-Phosphatidyl-Cholin!
all lipids shown are „saturated hydrocarbon“!alkyl chains with a c=c double bond are call „unsaturated“ !
-> kink!
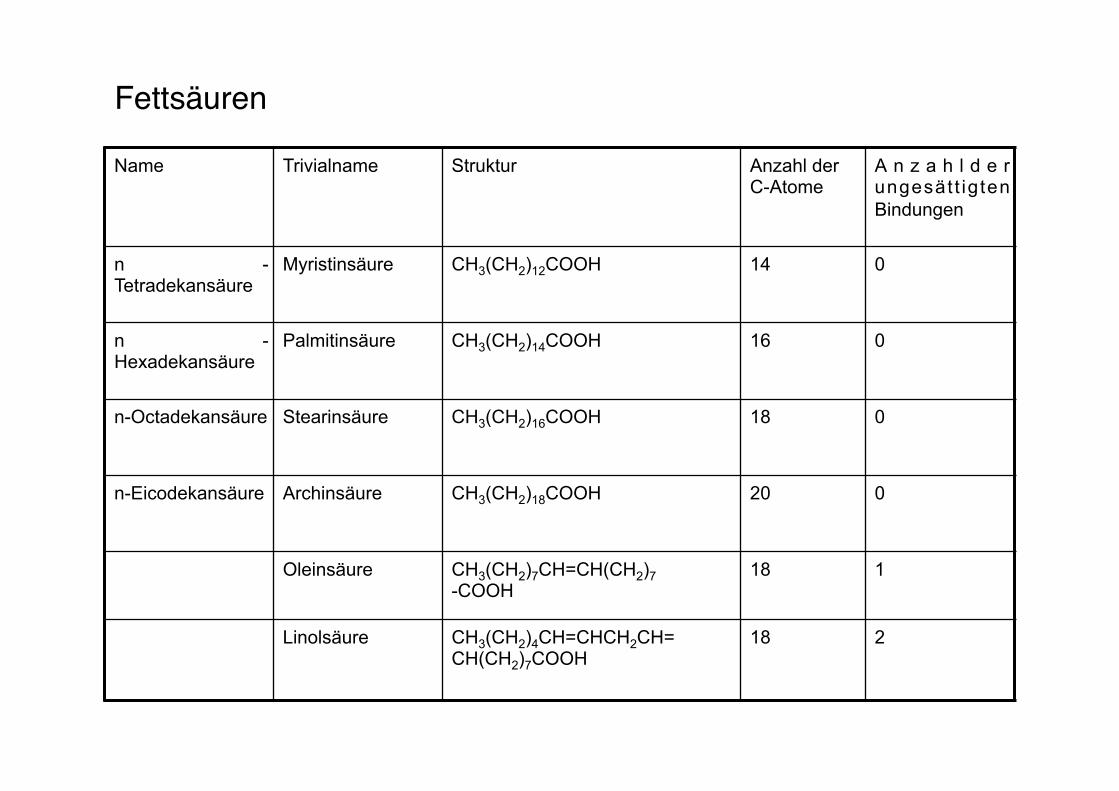
Fettsäuren"
Name Trivialname Struktur Anzahl der C-Atome
A n z a h l d e r ungesät t ig ten Bindungen
n -Tetradekansäure
Myristinsäure CH3(CH2)12COOH 14 0
n -Hexadekansäure
Palmitinsäure CH3(CH2)14COOH 16 0
n-Octadekansäure Stearinsäure CH3(CH2)16COOH 18 0
n-Eicodekansäure Archinsäure CH3(CH2)18COOH 20 0
Oleinsäure CH3(CH2)7CH=CH(CH2)7 -COOH
18 1
Linolsäure CH3(CH2)4CH=CHCH2CH= CH(CH2)7COOH
18 2
17!
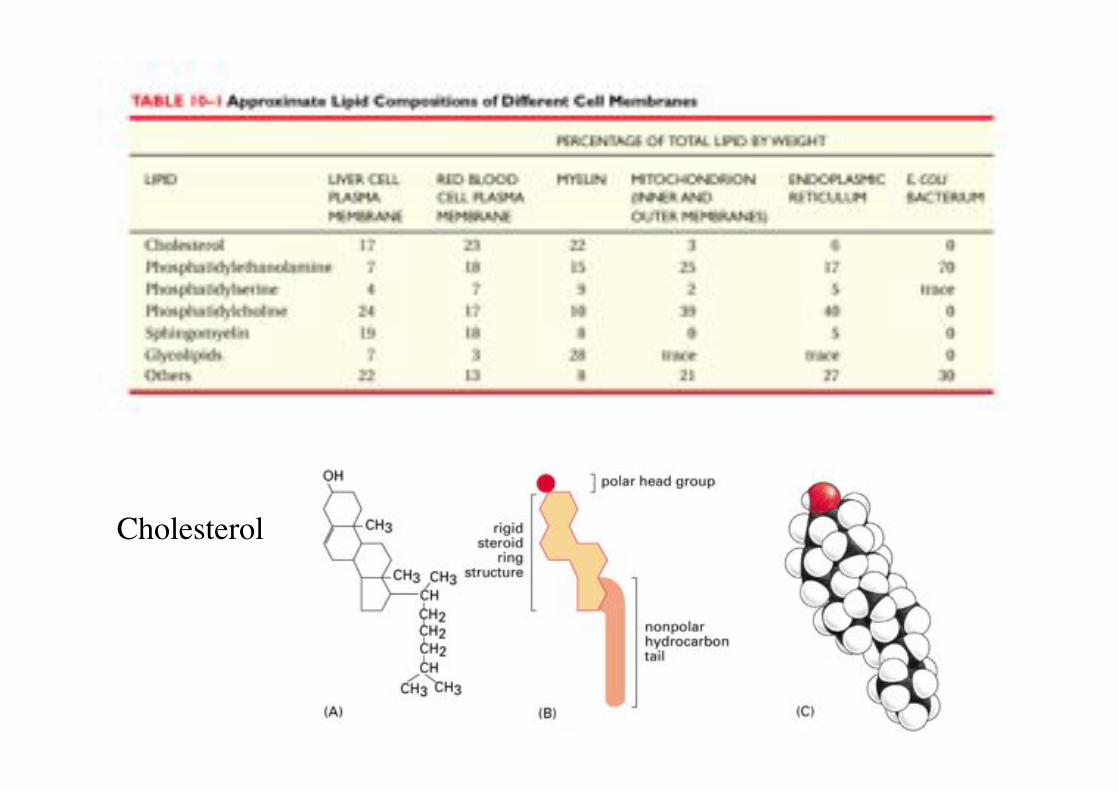
Cholesterol!
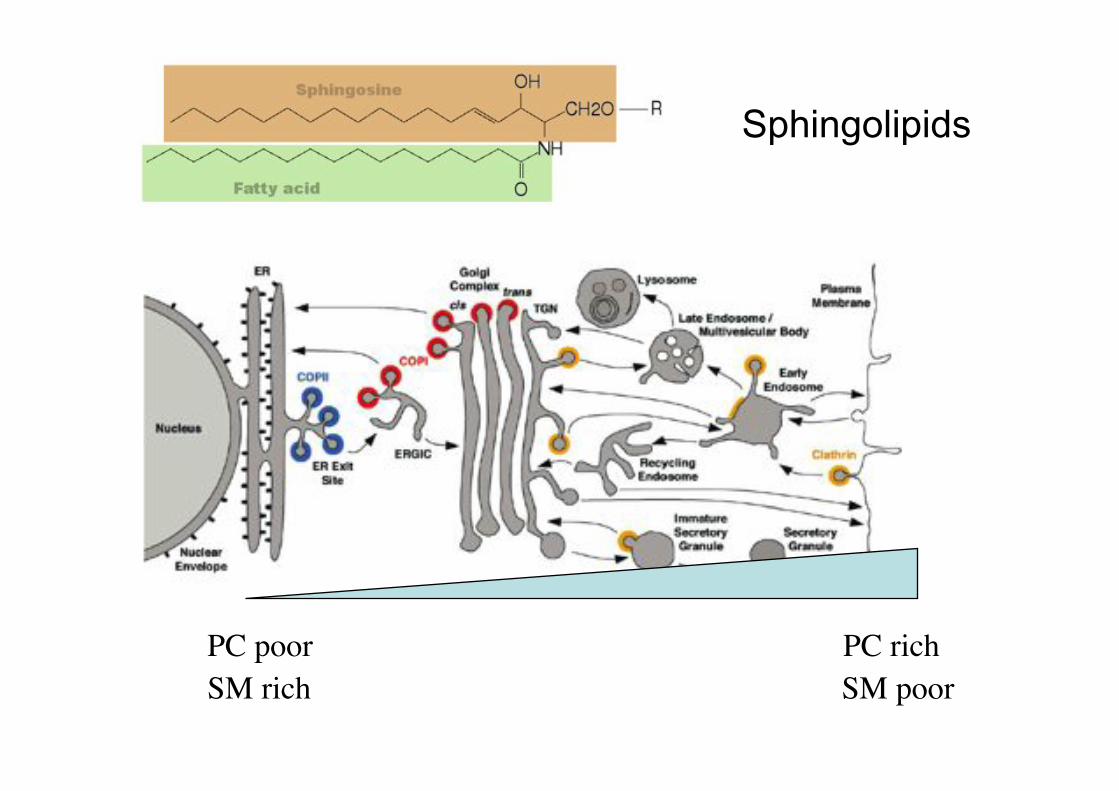
PC rich!PC poor!SM rich! SM poor!
Sphingolipids
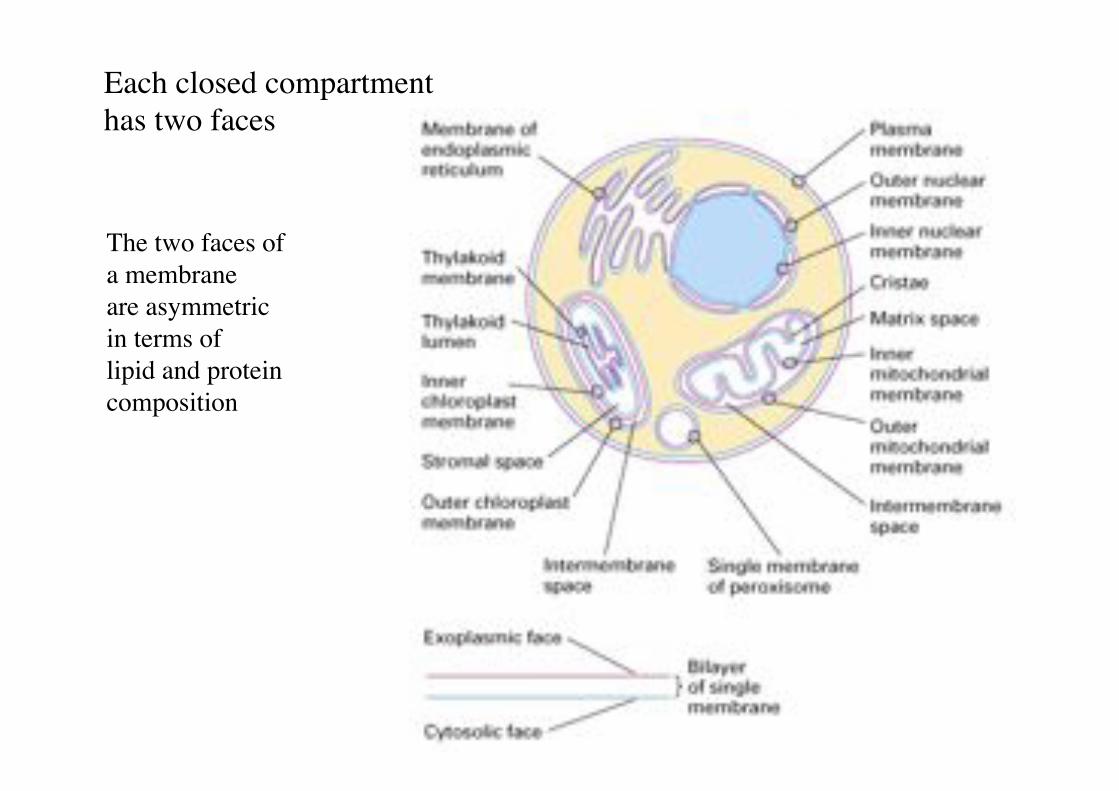
Each closed compartment "has two faces!
The two faces of "a membrane !are asymmetric "in terms of !lipid and protein composition!
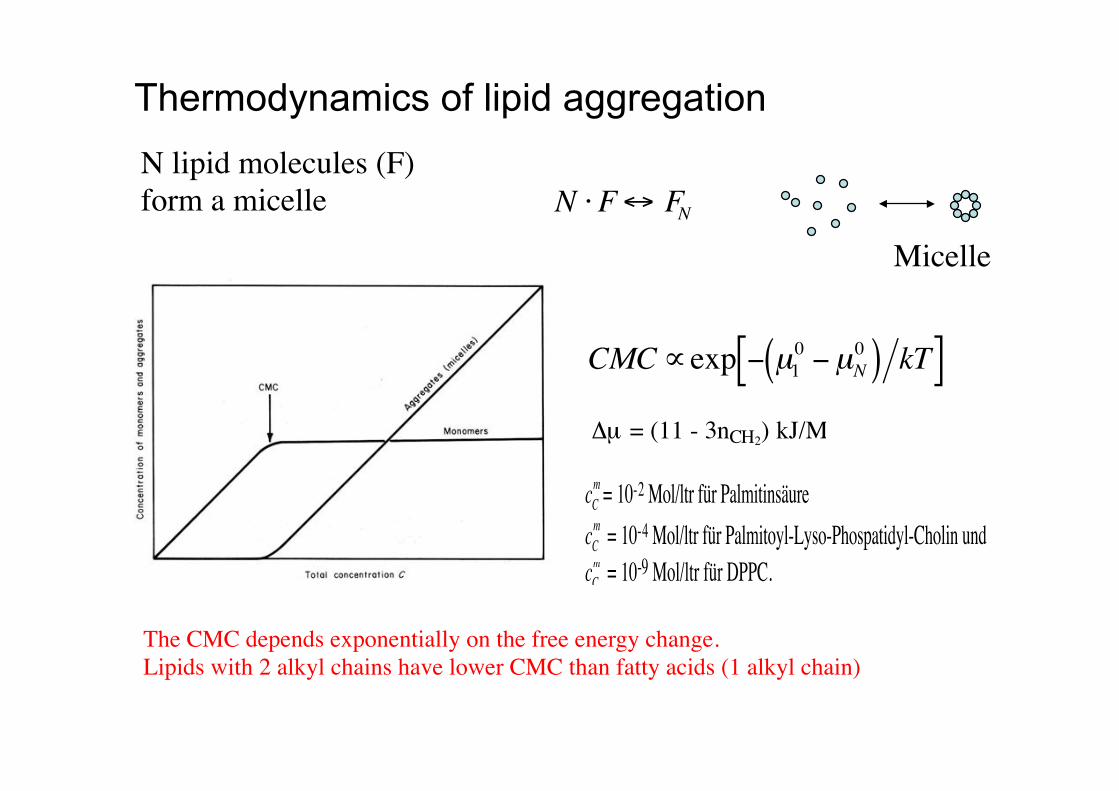
N !F" FN
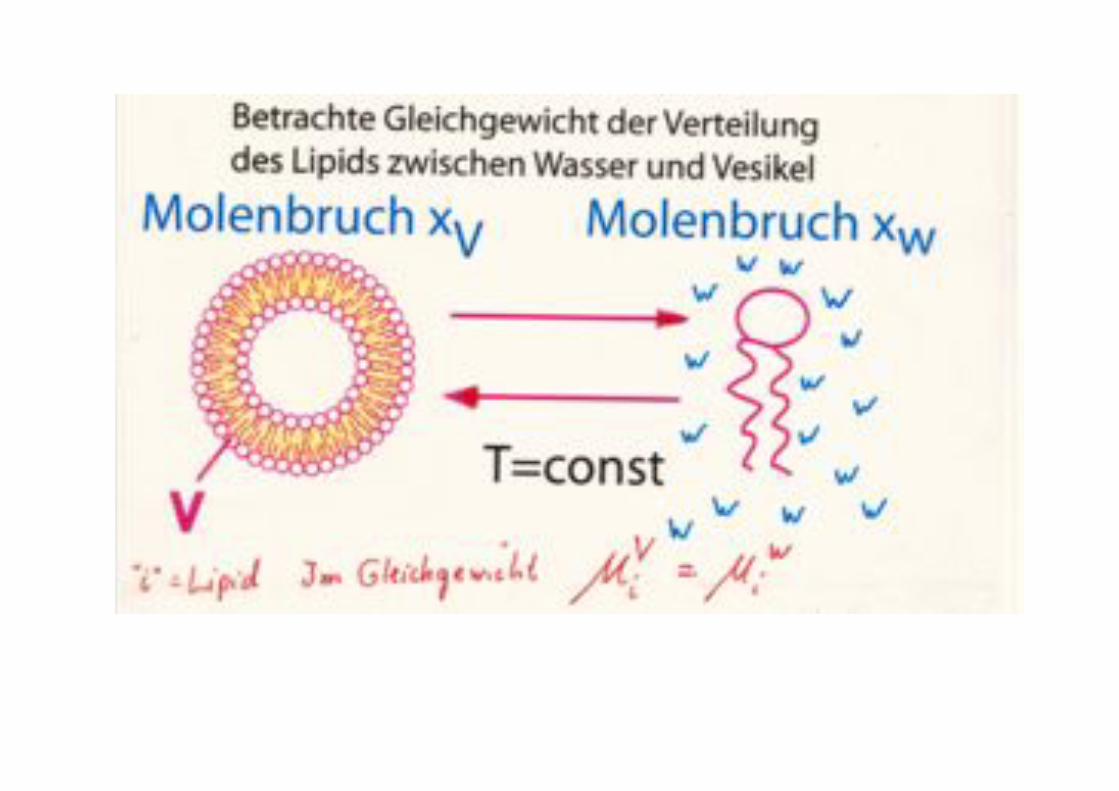
Thermodynamics of lipid aggregation
cCm = 10-2 Mol/ltr für PalmitinsäurecCm = 10-4 Mol/ltr für Palmitoyl-Lyso-Phospatidyl-Cholin undcCm = 10-9 Mol/ltr für DPPC.
!
CMC"exp # µ10 #µN
0( ) kT[ ]!µ = (11 - 3nCH2) kJ/M
Micelle!
N lipid molecules (F)!form a micelle!
The CMC depends exponentially on the free energy change.!Lipids with 2 alkyl chains have lower CMC than fatty acids (1 alkyl chain) !

Pulling a single lipid out of a membrane!
The unbinding force of PC-C14:0 f*# 12 pN. !This corresponds to a binding energy per molecule of !g*~5 kBT !(which is about factor 5 smaller than expected from the hydrophopic effect)!Note: thermal energy = kTRT = 4.1 nm pN!

0$12#2."#3'4 5.)&&'67$&)."&&)8'9"#$%&"'(569/':';-.'4 0)8<"'67$&)."&&)8'9"#$%&"'(069/':';,-.'4 =$)7>'67$&)."&&)8'9"#$%&"'(=69/':';,,-.'
Lipids are hygroscopic: They swell in water forming multilamellar (myelinlike) or unilamellar !liposomes (vesicles) !

Creating Vesicles
'?8")@7<'1A2#1A2&$1$B#'27')'C)>"8D2$&'$7>"8E)%"3'4 F$##2&!"'&$1$B#'$7'.$7"8)&'2$&*'4 G28.')7'".H&#$27'C$>A'C)>"8'4 I&)%"'>A"'".H&#$27'27'>21'2E'C)>"8'(E""B$7<'#2&H@27/'
'''''''')7B'%"7>8$EH<"'>A"'B821&">#'B2C7'9"#$%&"#'
J2$8")HK')7B'0$L%A)L"8'
! '#C"&&$7<'M'#27$%)@27'(569/'! '>A"8.)&'#C"&&$7<'(569N'=69/'! '"&"%>82'#C"&&$7<'(=69/'! '#2&!"7>'"K%A)7<"'.">A2B#'! '8"!"8#"'1A)#"'"K>8)%@27#'
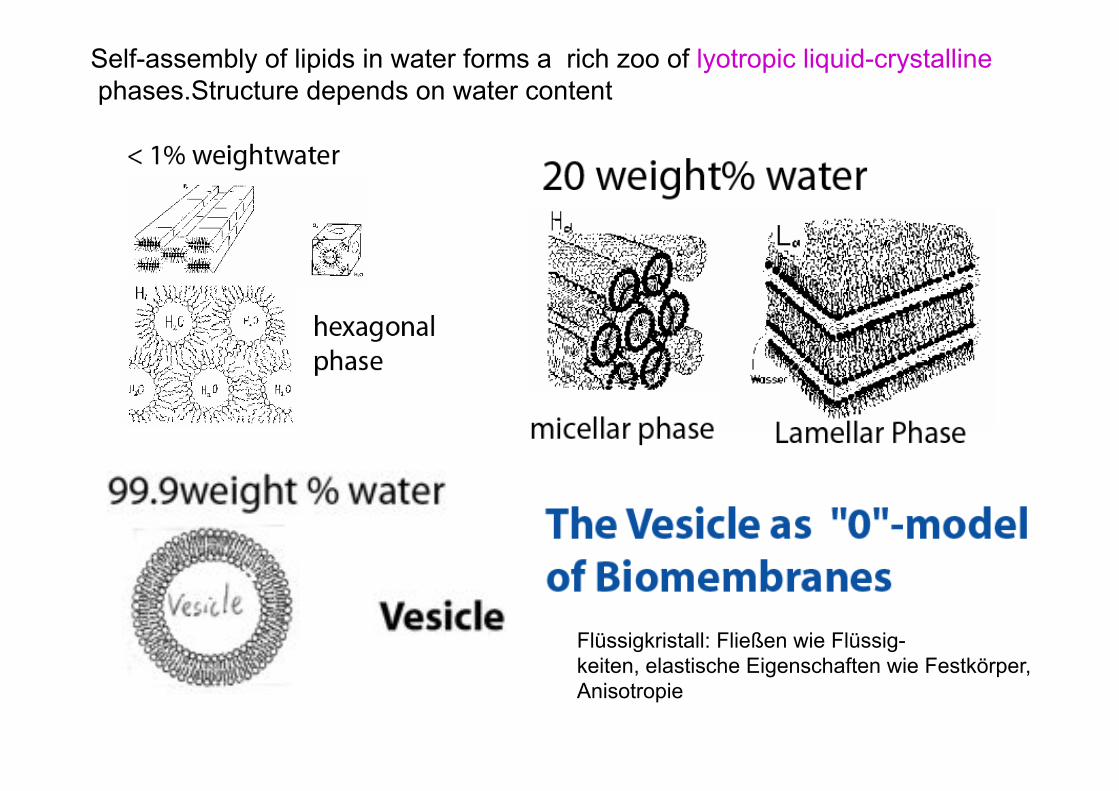
Self-assembly of lipids in water forms a rich zoo of lyotropic liquid-crystalline phases.Structure depends on water content
Flüssigkristall: Fließen wie Flüssig- keiten, elastische Eigenschaften wie Festkörper, Anisotropie
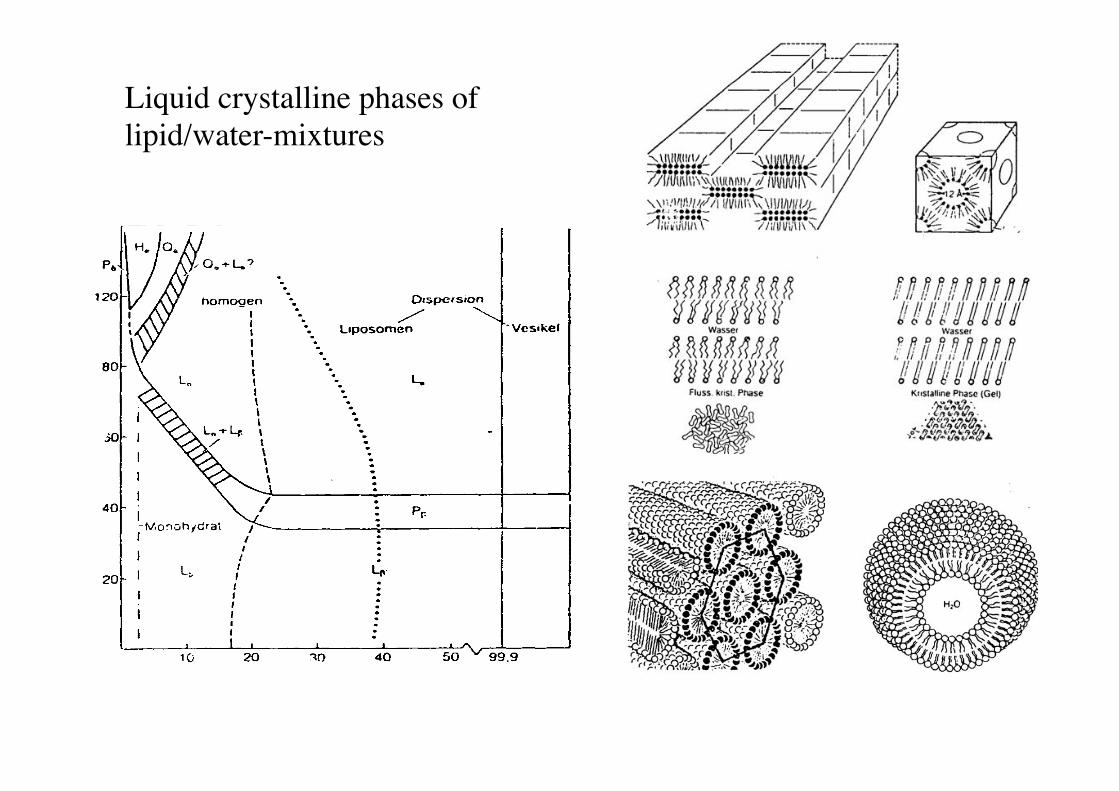
Liquid crystalline phases of lipid/water-mixtures!
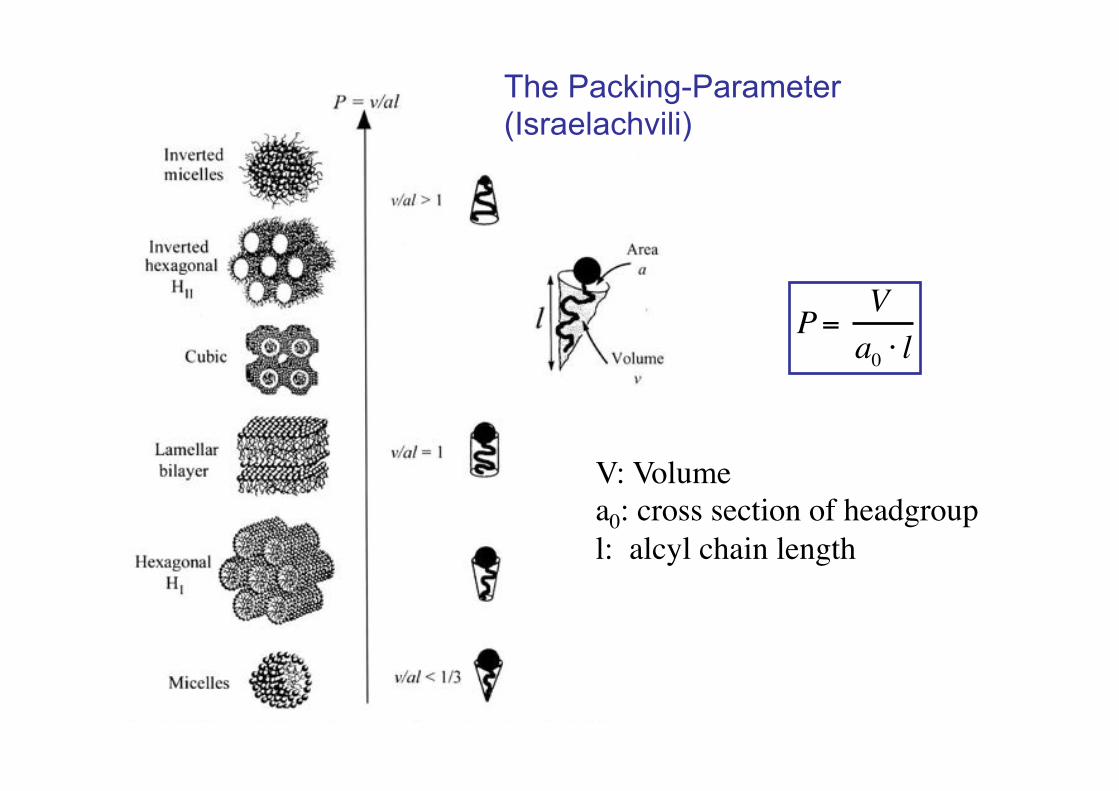
P= Va0 ! l
V: Volume!a0: cross section of headgroup!l: alcyl chain length!
The Packing-Parameter (Israelachvili)
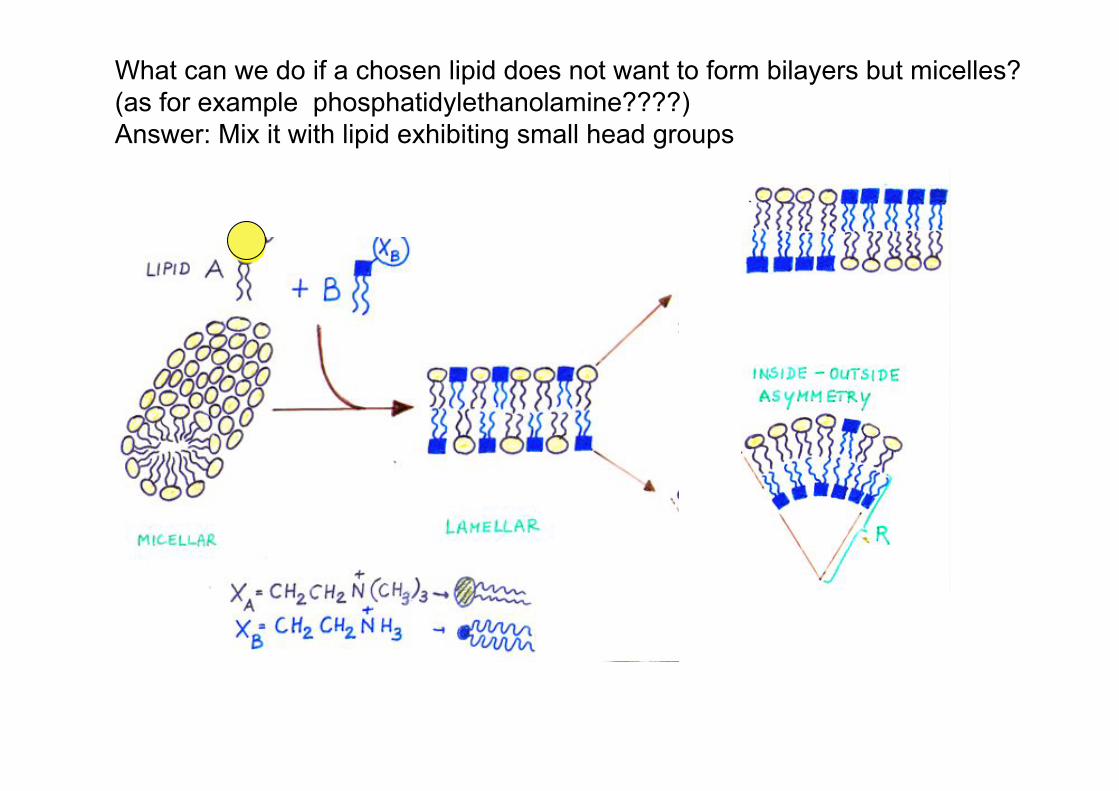
What can we do if a chosen lipid does not want to form bilayers but micelles? (as for example phosphatidylethanolamine????) Answer: Mix it with lipid exhibiting small head groups
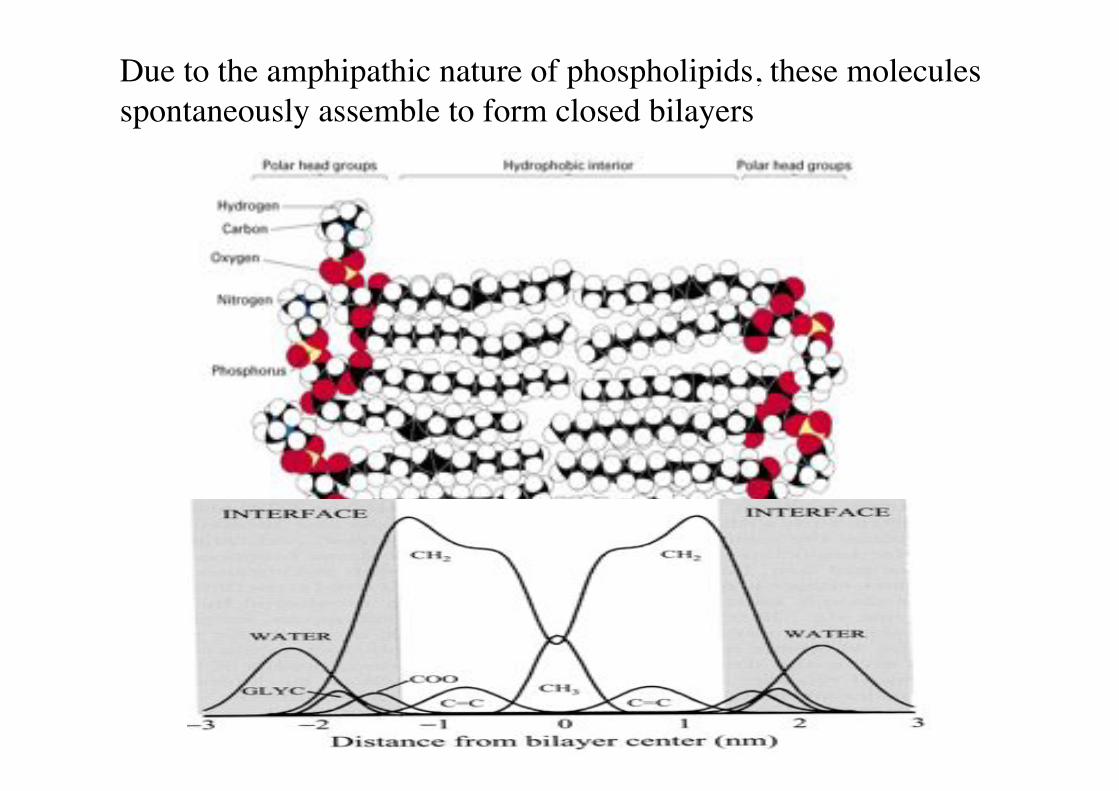
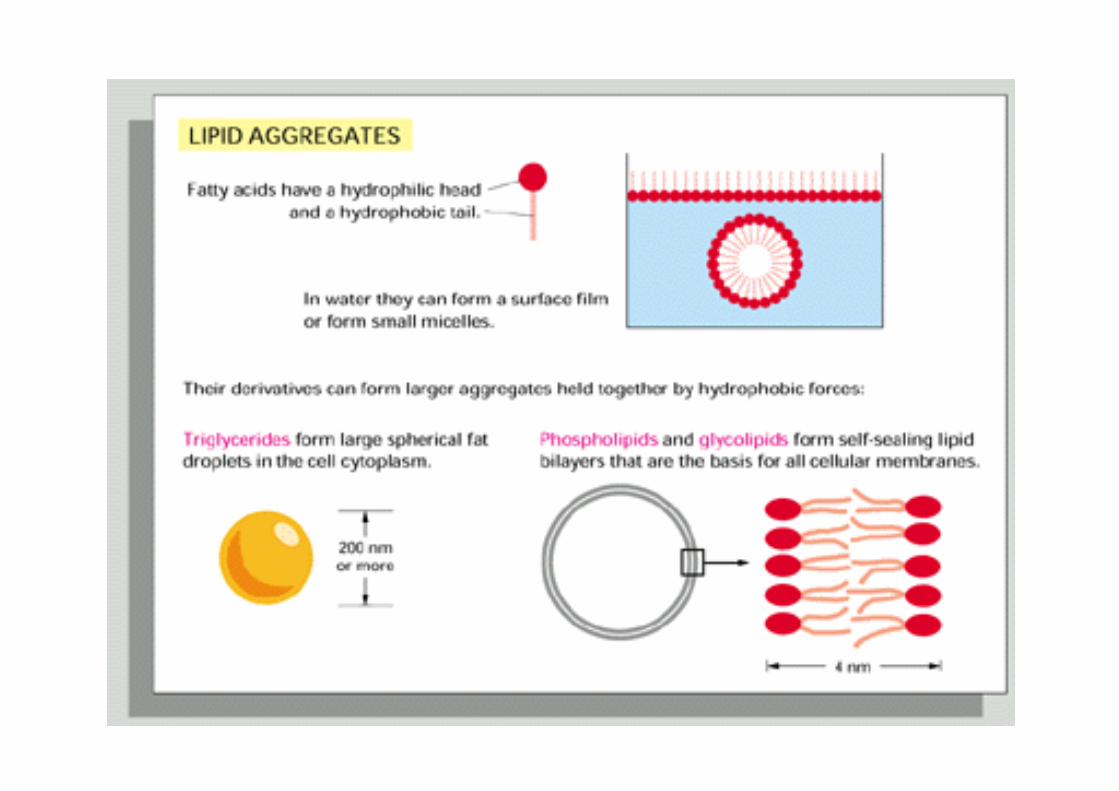
Due to the amphipathic nature of phospholipids, these molecules spontaneously assemble to form closed bilayers!
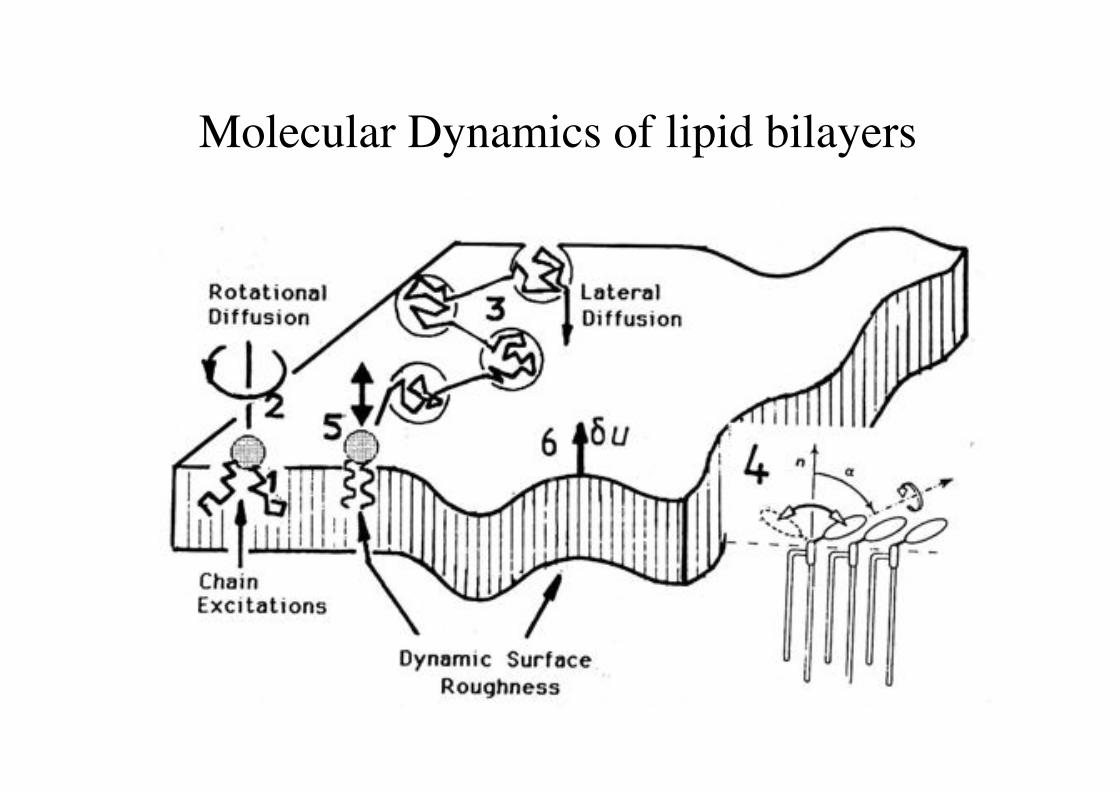
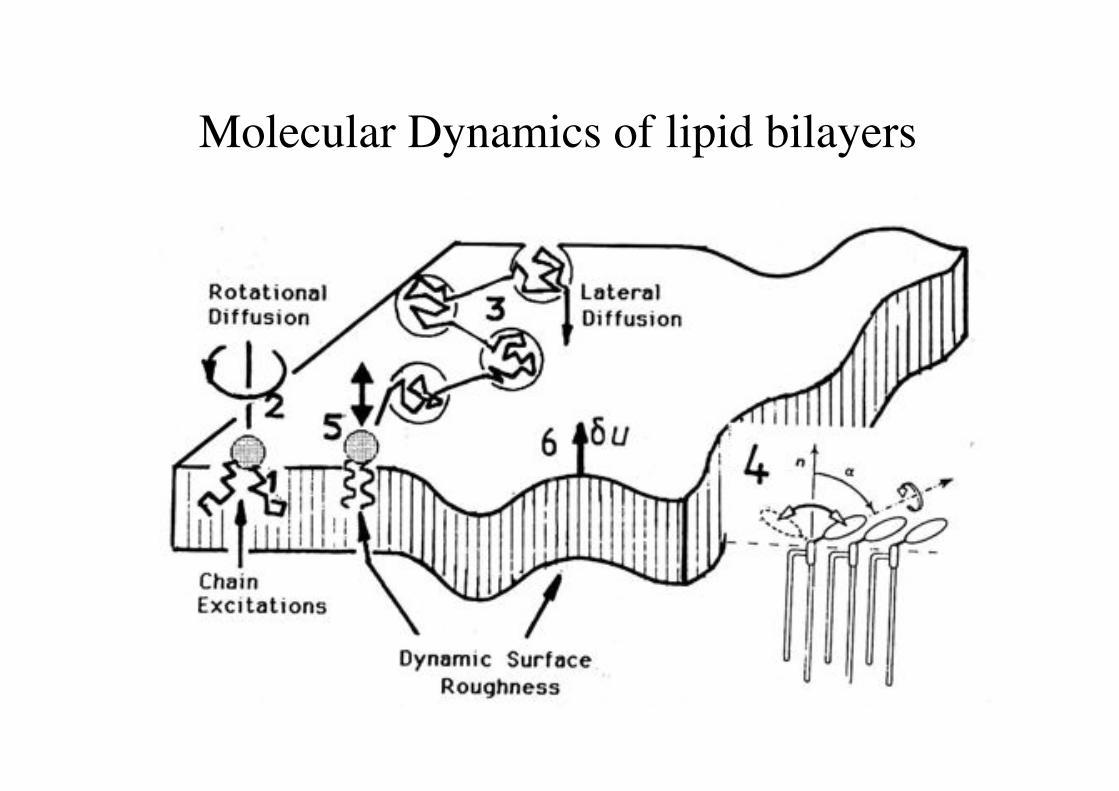
Molecular Dynamics of lipid bilayers!
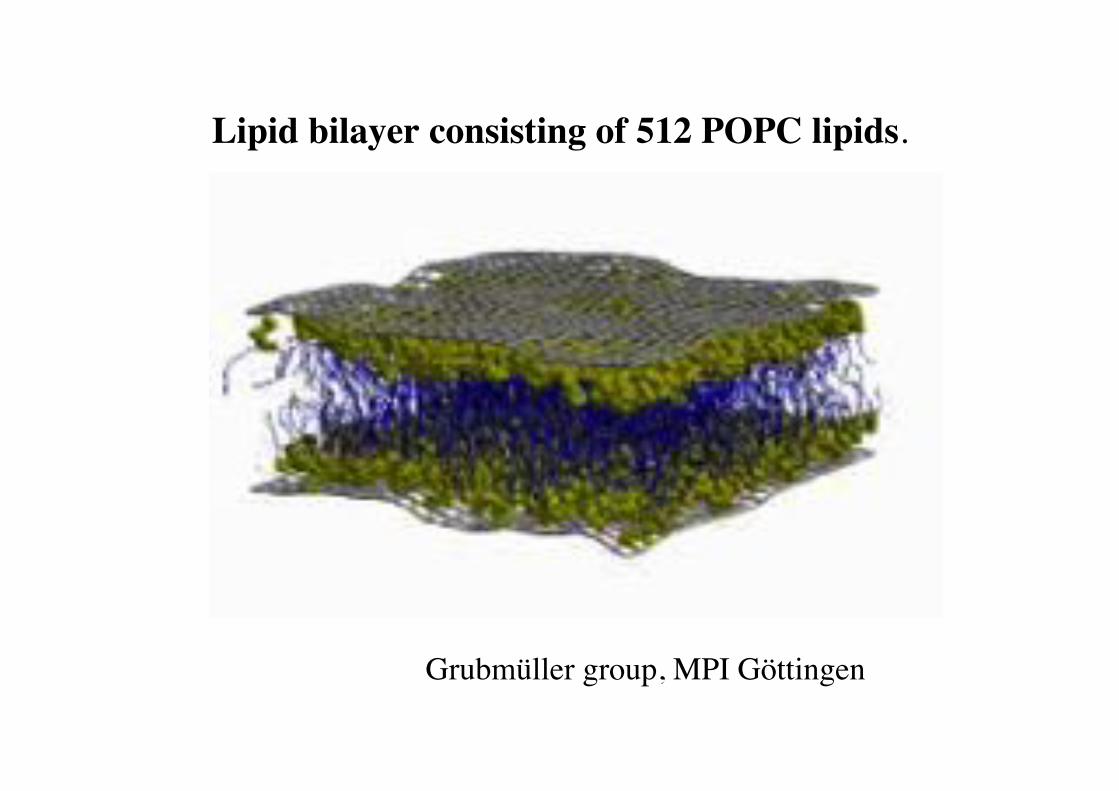
Lipid bilayer consisting of 512 POPC lipids.!
Grubmüller group, MPI Göttingen!

H. Grubmüller, Göttingen!

Snap shots!
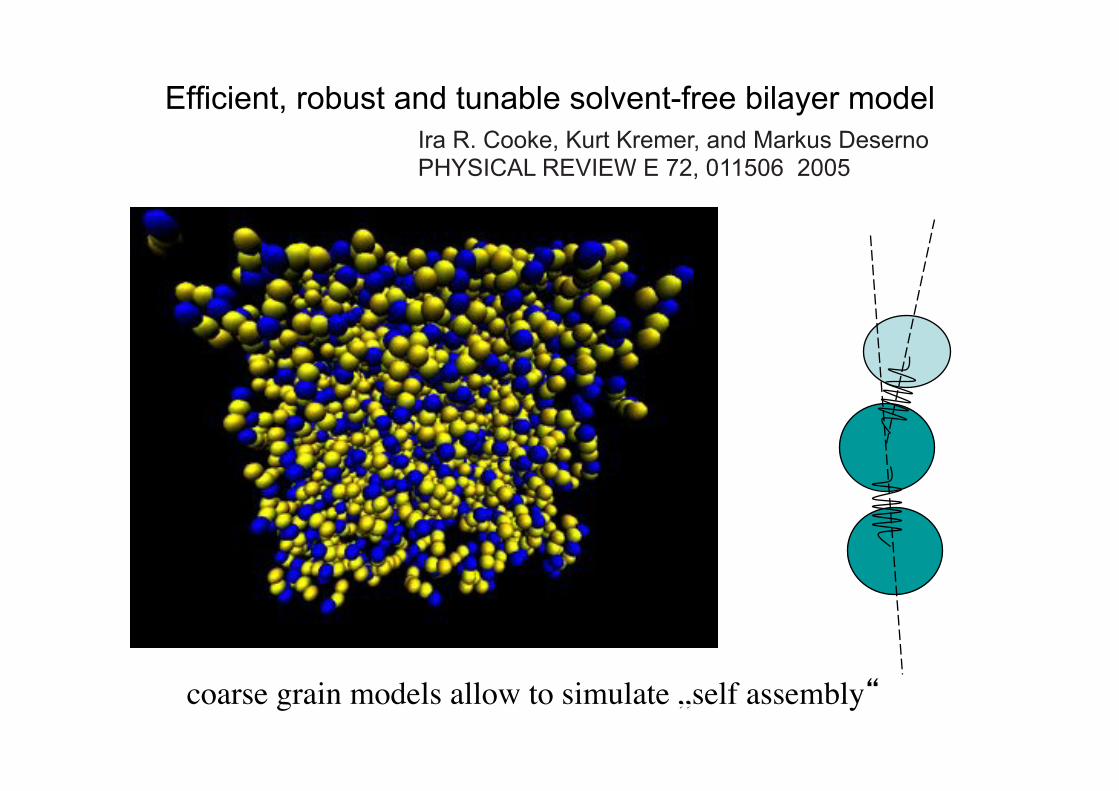
Efficient, robust and tunable solvent-free bilayer model Ira R. Cooke, Kurt Kremer, and Markus Deserno PHYSICAL REVIEW E 72, 011506 2005
coarse grain models allow to simulate „self assembly“!

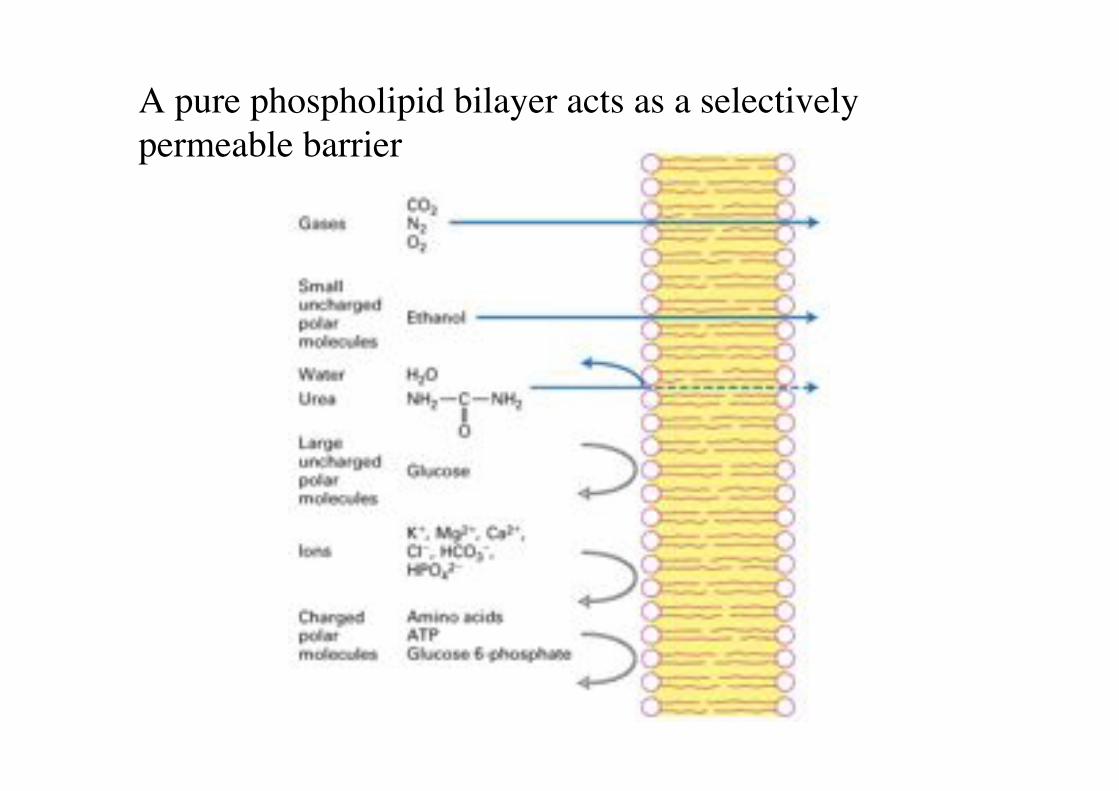
A pure phospholipid bilayer acts as a selectively permeable barrier!
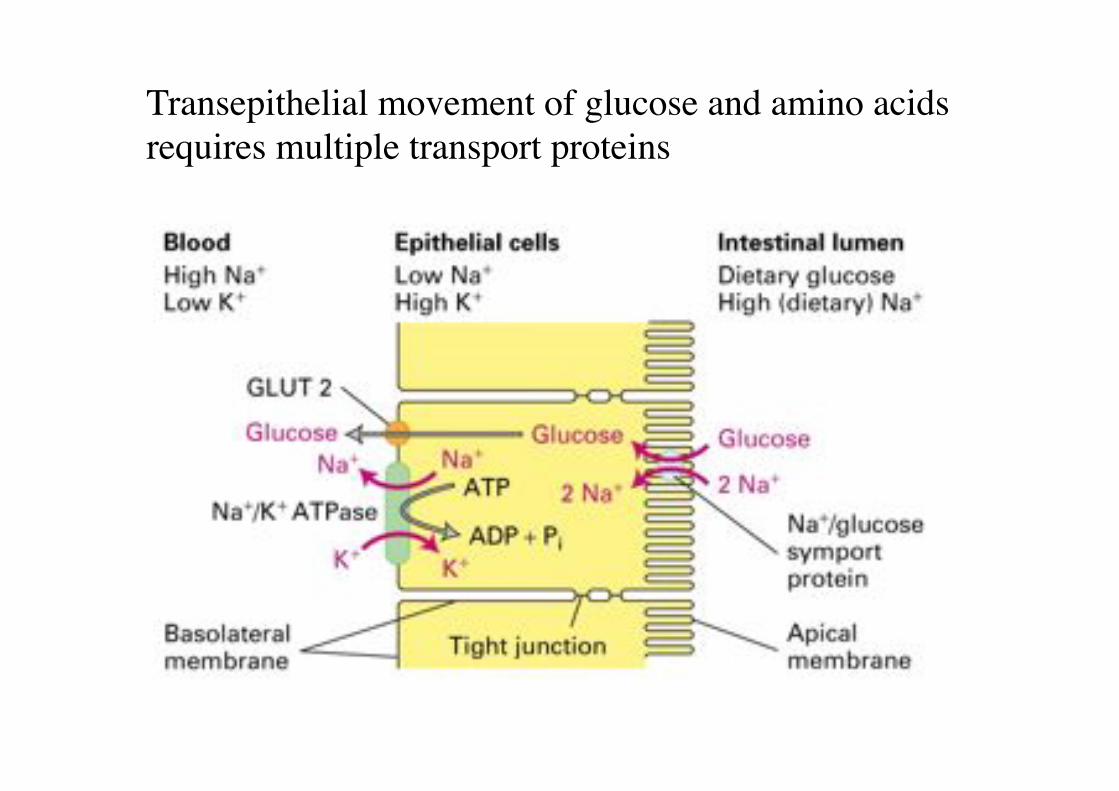
Transepithelial movement of glucose and amino acids requires multiple transport proteins!
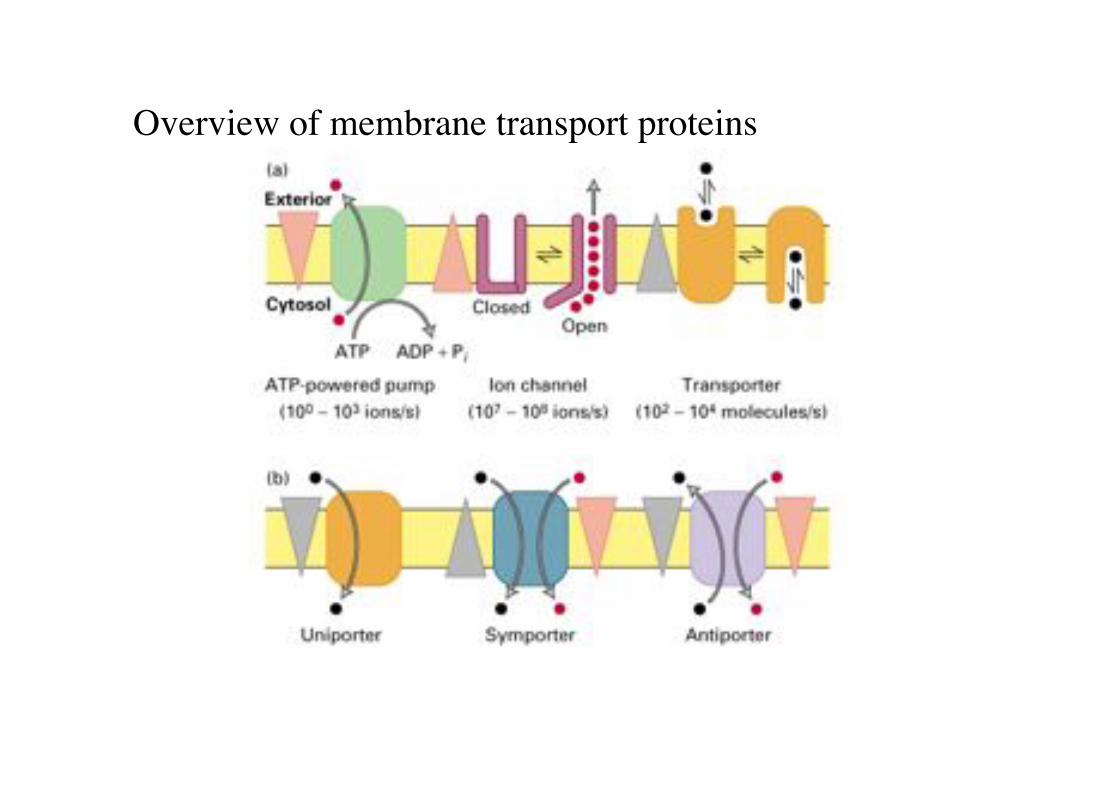
Overview of membrane transport proteins!
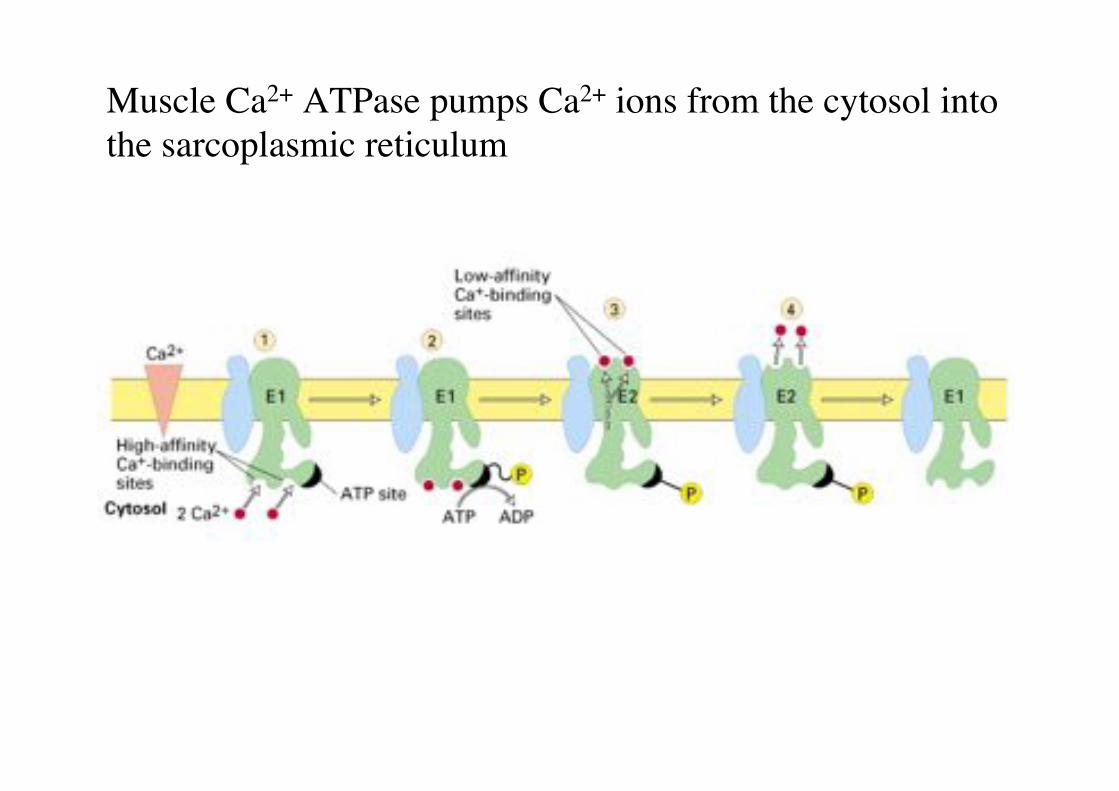
Muscle Ca2+ ATPase pumps Ca2+ ions from the cytosol into the sarcoplasmic reticulum"
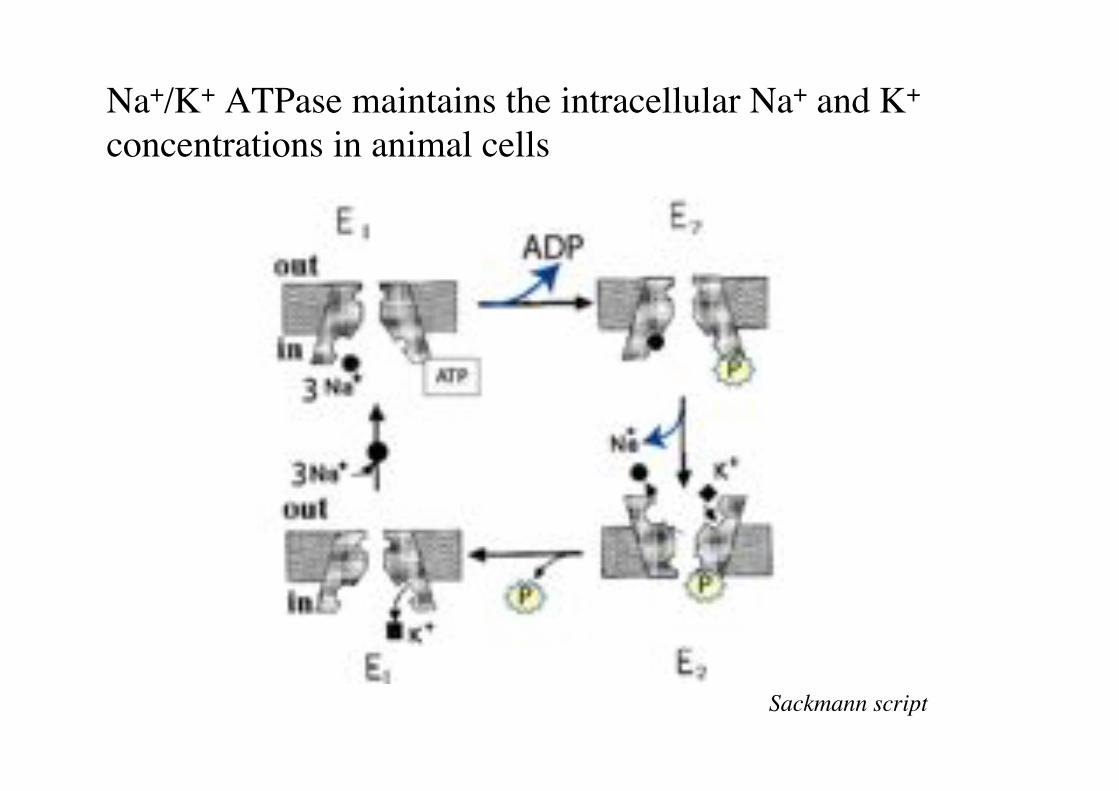
Na+/K+ ATPase maintains the intracellular Na+ and K+ concentrations in animal cells"
Sackmann script!
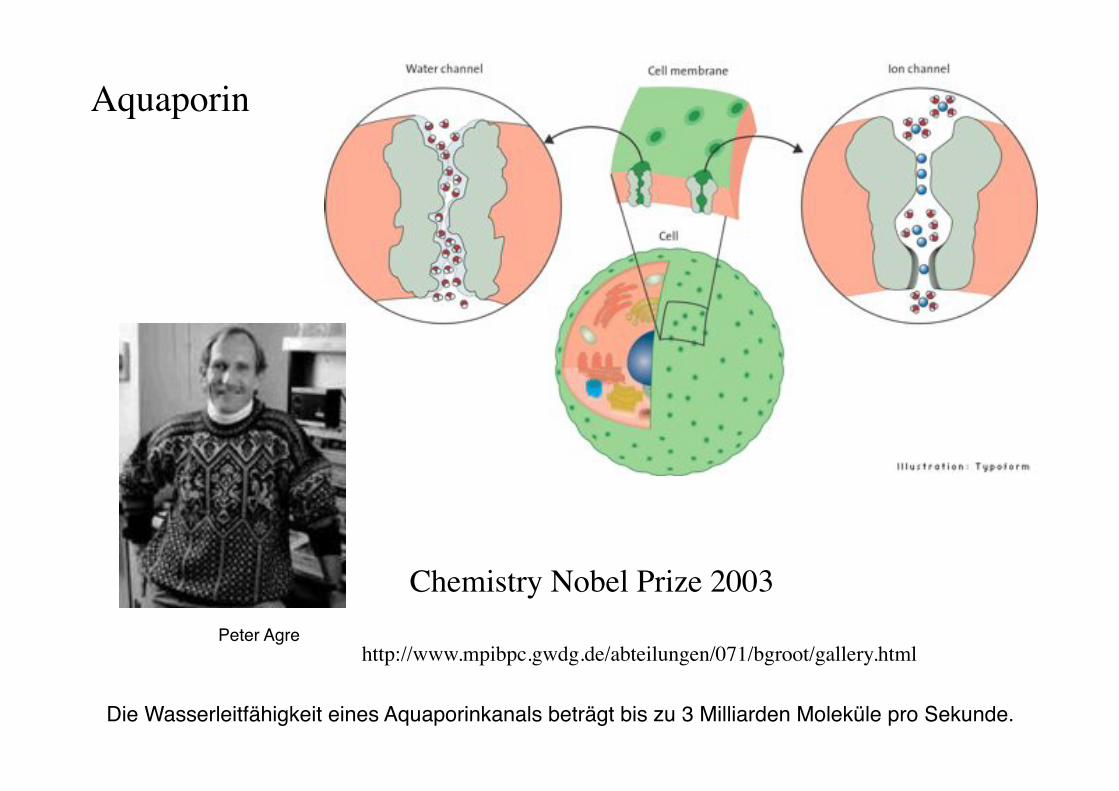
Aquaporin!
http://www.mpibpc.gwdg.de/abteilungen/071/bgroot/gallery.html!
Chemistry Nobel Prize 2003!
Die Wasserleitfähigkeit eines Aquaporinkanals beträgt bis zu 3 Milliarden Moleküle pro Sekunde. "
Peter Agre"


Aquaporin-1 in action. Simulation fragment of 200 ps of aquaporin-1. !Generated using Molscript/Bobscript and raster3d.!

„Real time“ molecular dynamics simulation!
Aquaglyceroporin tetramer (blue, cyan, orange, magenta), embedded within a POPE lipid bilayer(yellow head groups and green tails) surrounded by water (red, white).The total system consists of about 101,000 atoms.!!Overlaid snapshots from a trajectory of a water molecule passing through AQP1 (left and middle panels, surface and ribbon representation of the same protein structure) and hydrogen bond energies per water molecule (right). The permeation event shown on the left lasted 3.3 ns.!

Thermotropic and lyotropic "lipid phase behavior!
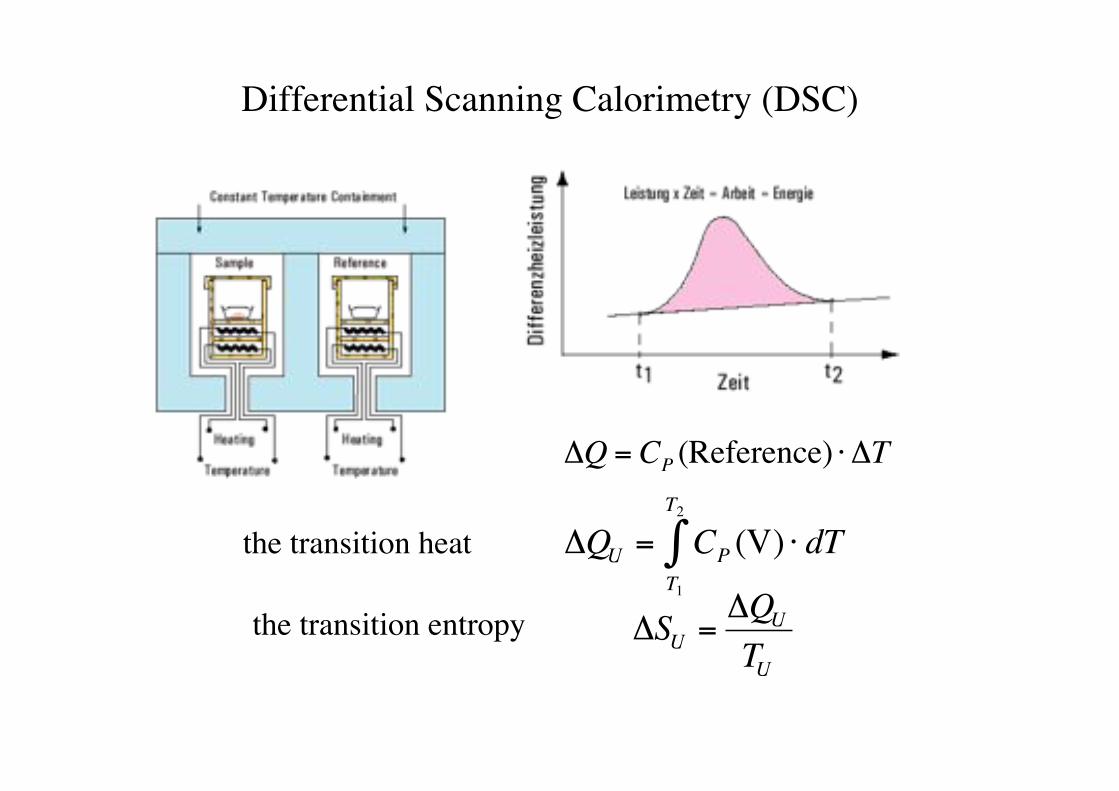
Differential Scanning Calorimetry (DSC)!
!
"Q = CP (Reference) # "T
!
"QU = CPT1
T2
# (V) $ dTthe transition heat!
the transition entropy!
!
"SU ="QU
TU
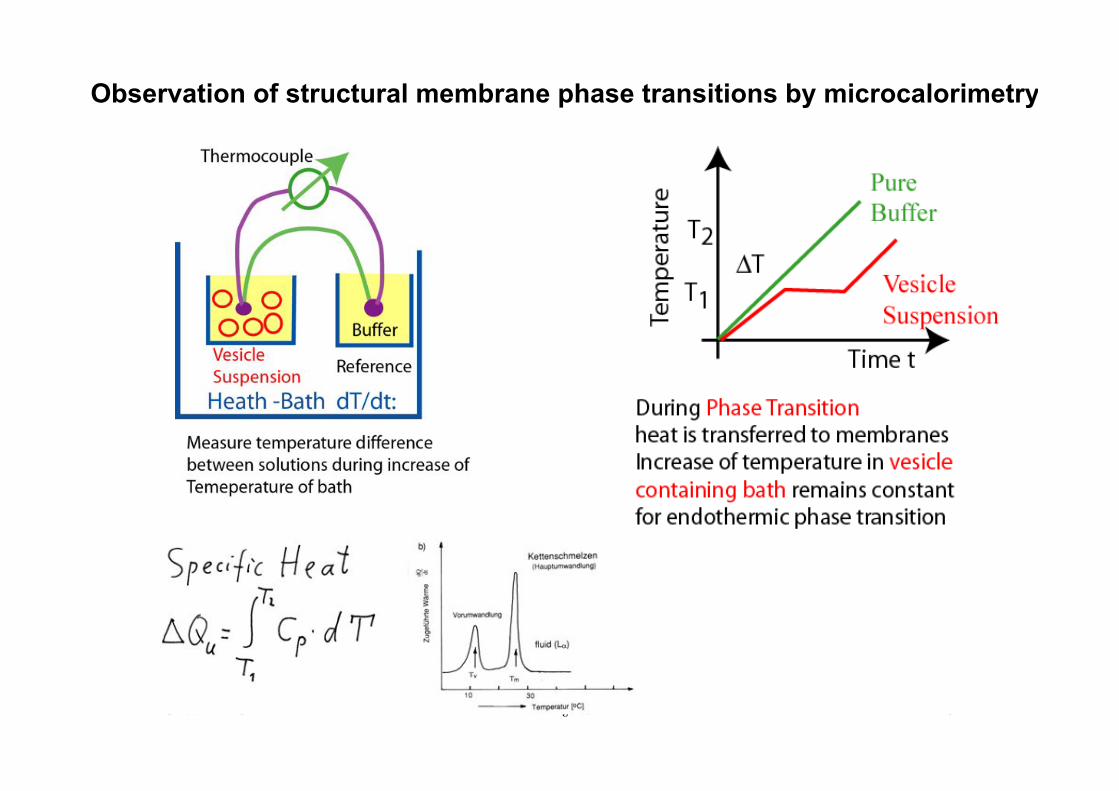
Gaub LMU! BPE§3.5! 49!
Observation of structural membrane phase transitions by microcalorimetry
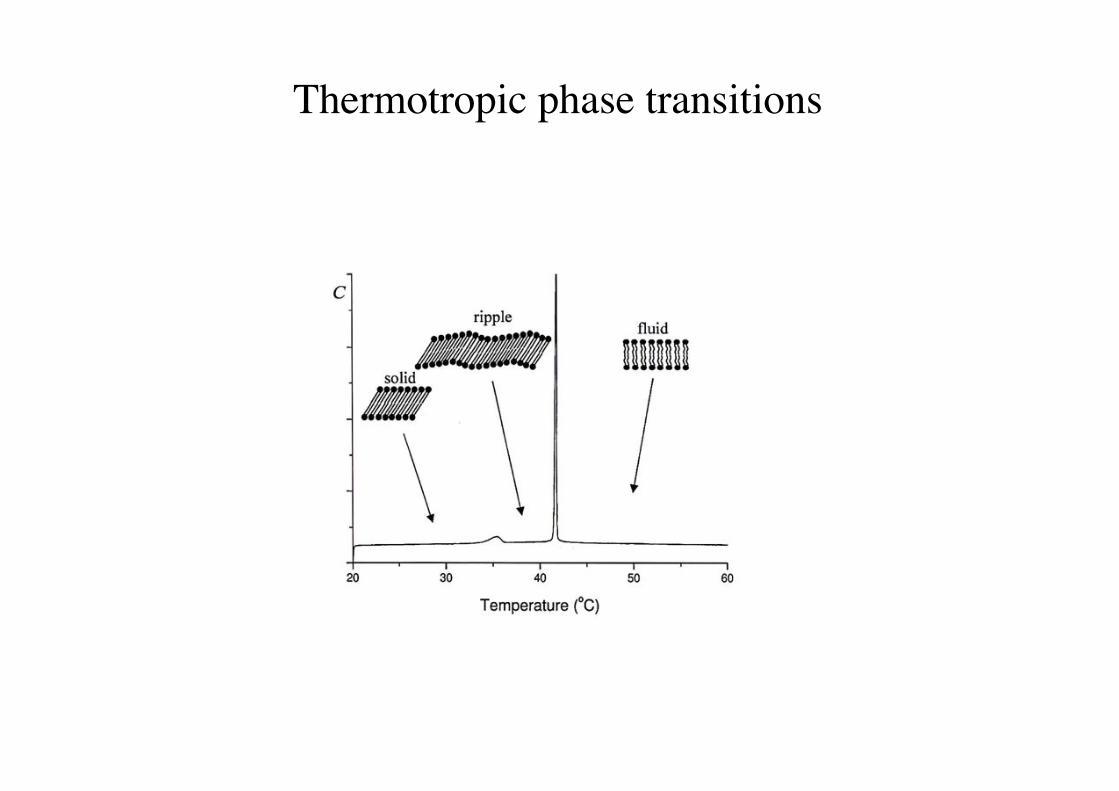
Thermotropic phase transitions!
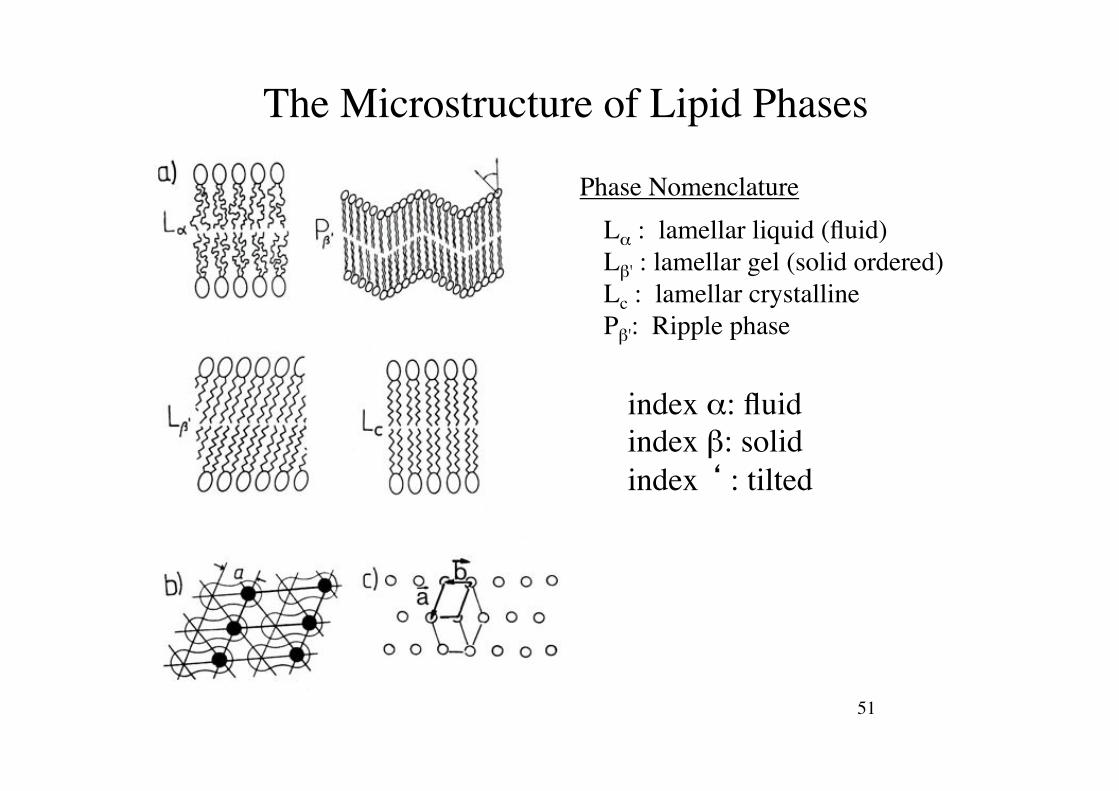
51!
The Microstructure of Lipid Phases!
index ": fluid!index #: solid!index ‘ : tilted!
L" : lamellar liquid (fluid)!L#' : lamellar gel (solid ordered)!Lc : lamellar crystalline!P#': Ripple phase!
Phase Nomenclature!
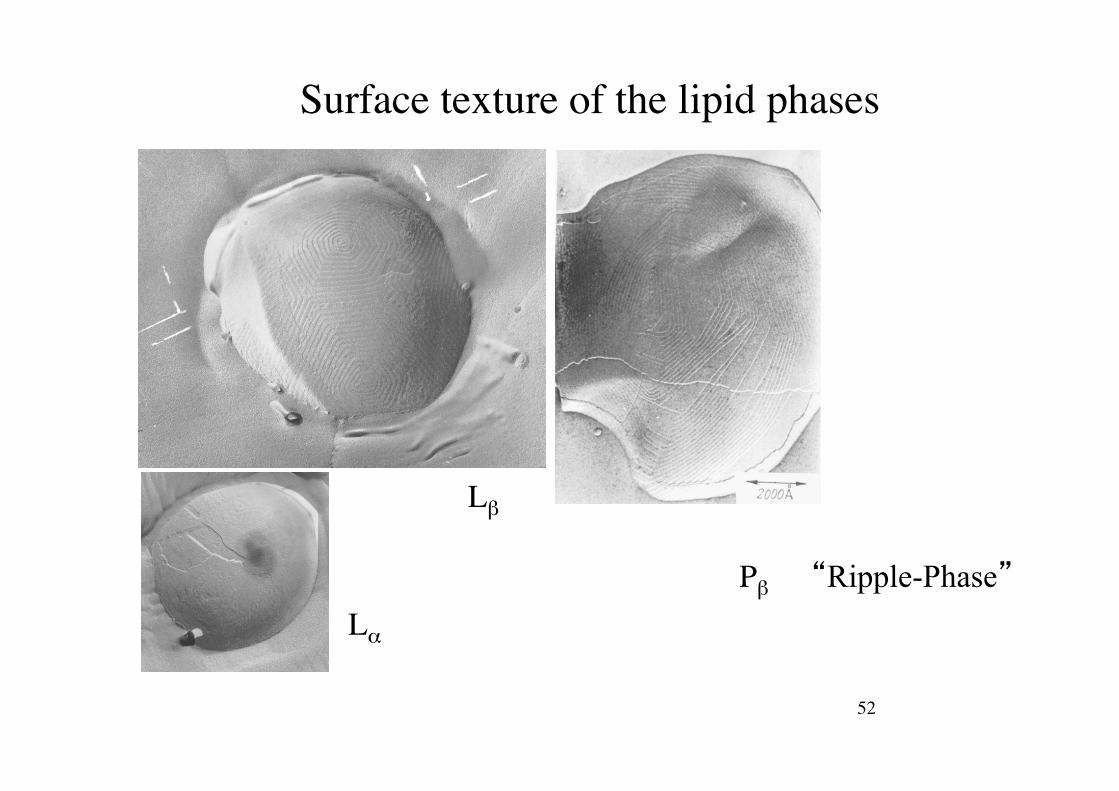
52!
Surface texture of the lipid phases!
L"$
L#$
P#$ “Ripple-Phase”
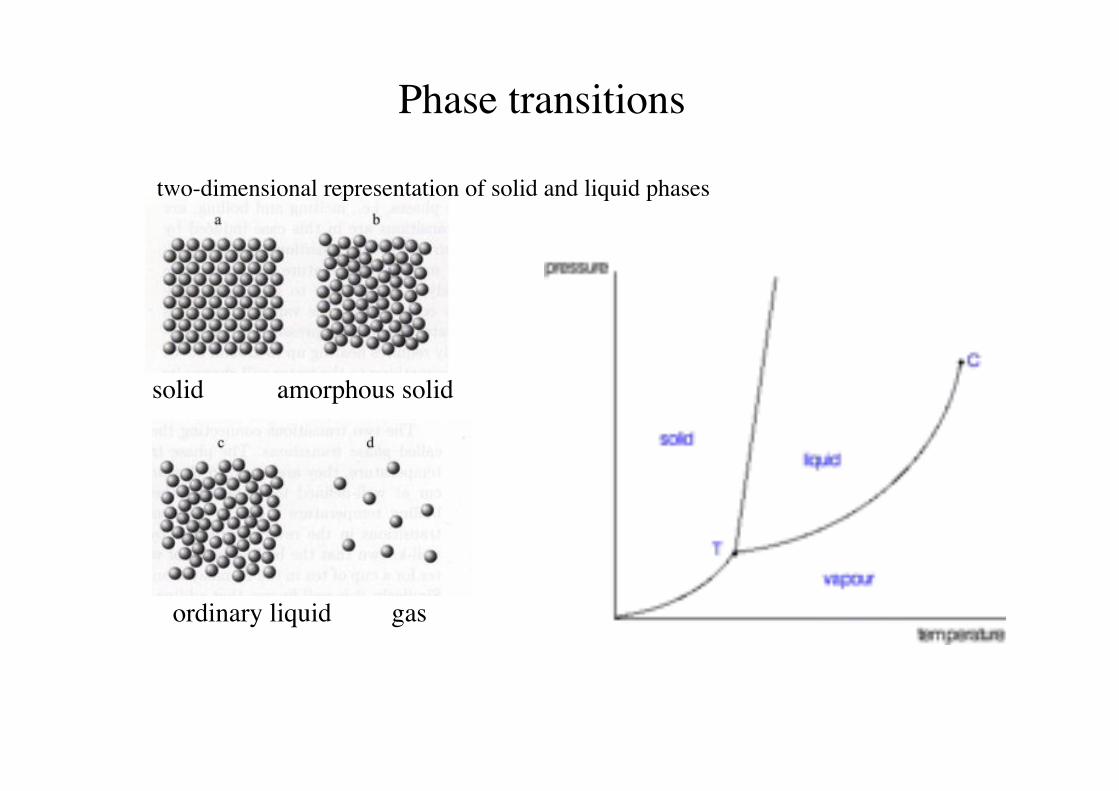
Phase transitions!
two-dimensional representation of solid and liquid phases!
solid amorphous solid!
ordinary liquid gas!
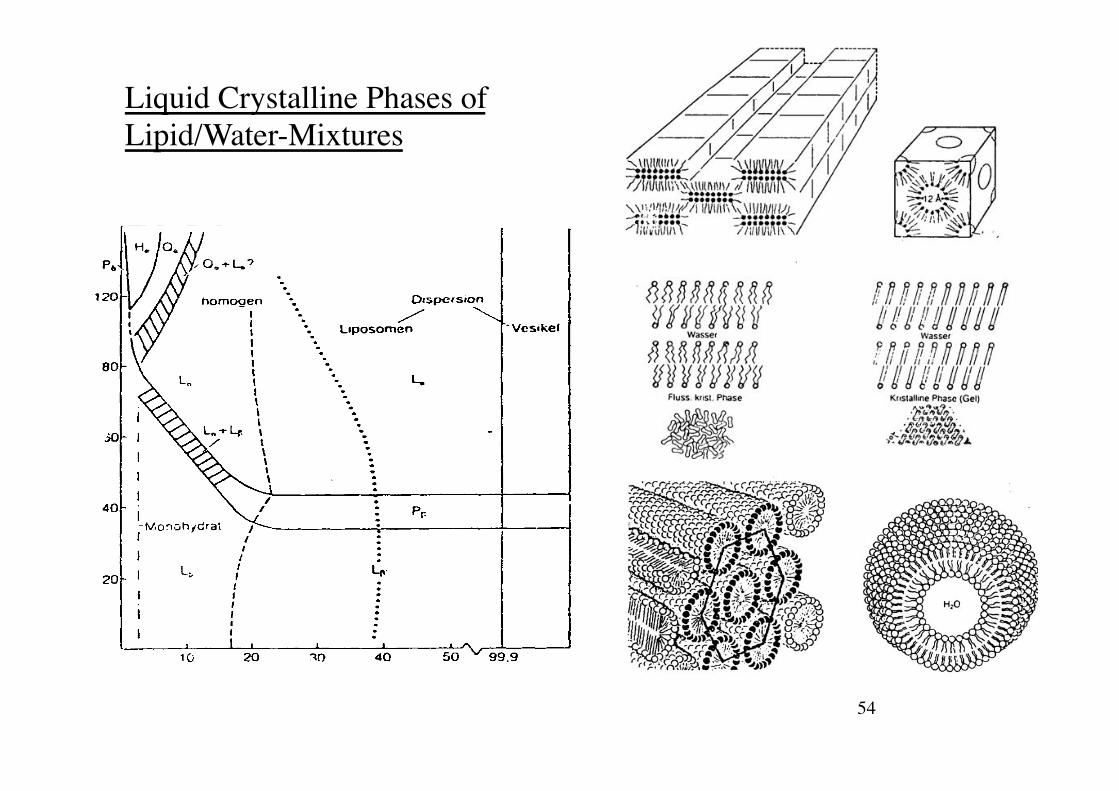
54!
Liquid Crystalline Phases of Lipid/Water-Mixtures!

55!Rädler SS 09! Biophysik der Zelle!
Phase Determination using Small-Angle-X-ray scattering
1mm
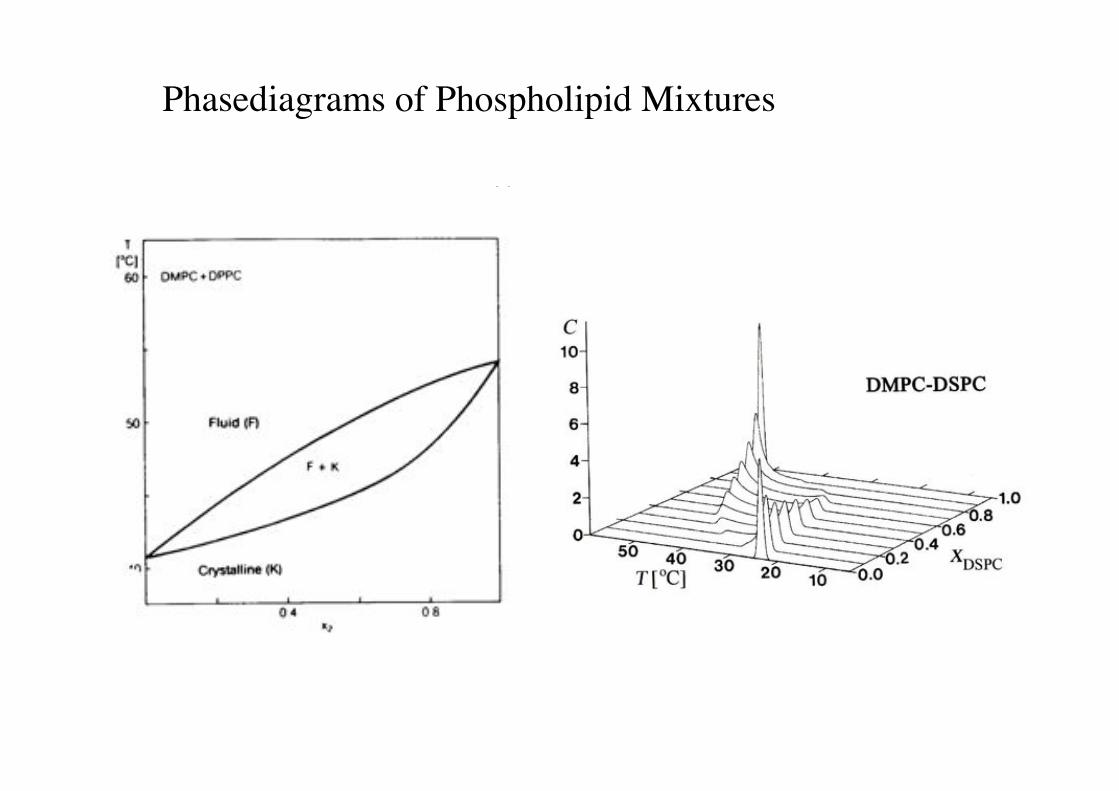
Phasediagrams of Phospholipid Mixtures!
a) b)
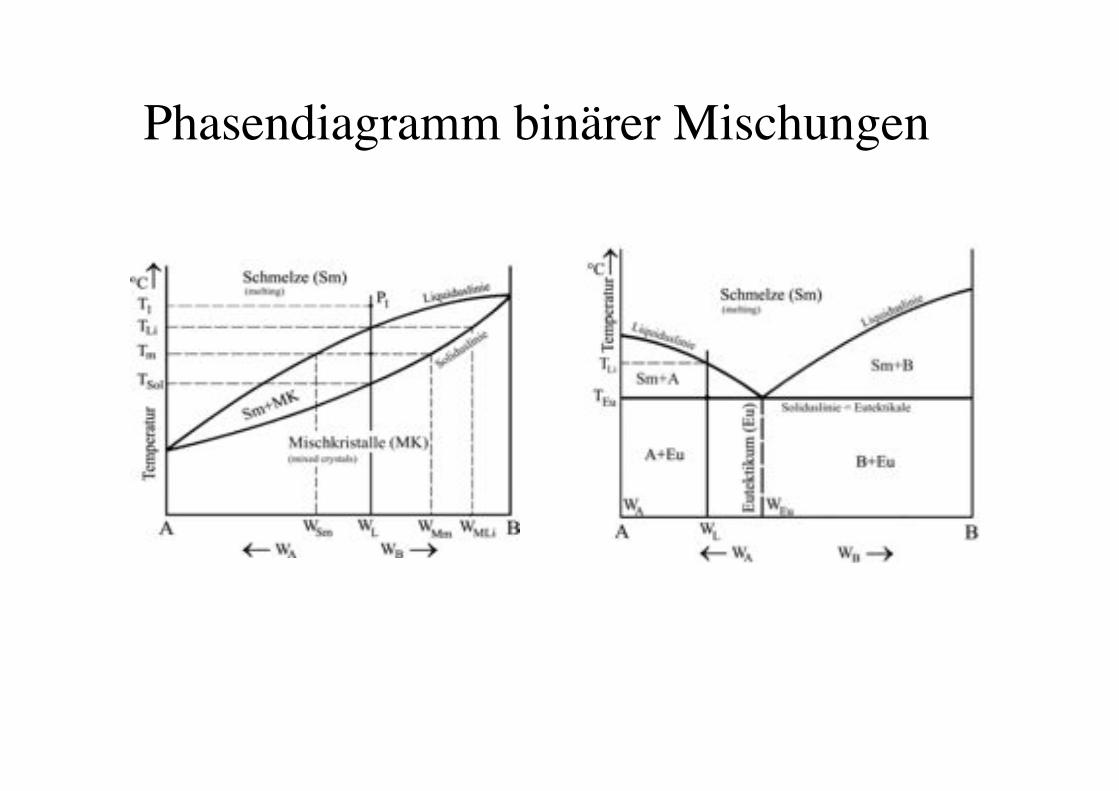
Phasendiagramm binärer Mischungen!
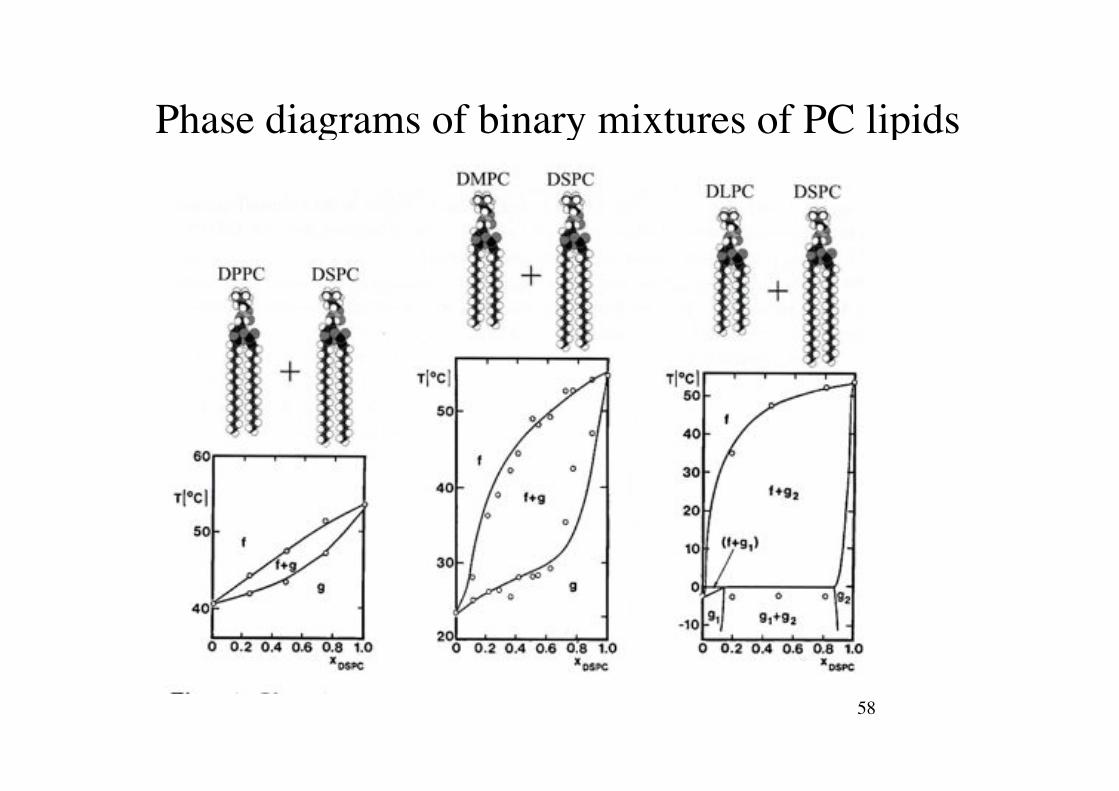
58!
Phase diagrams of binary mixtures of PC lipids!
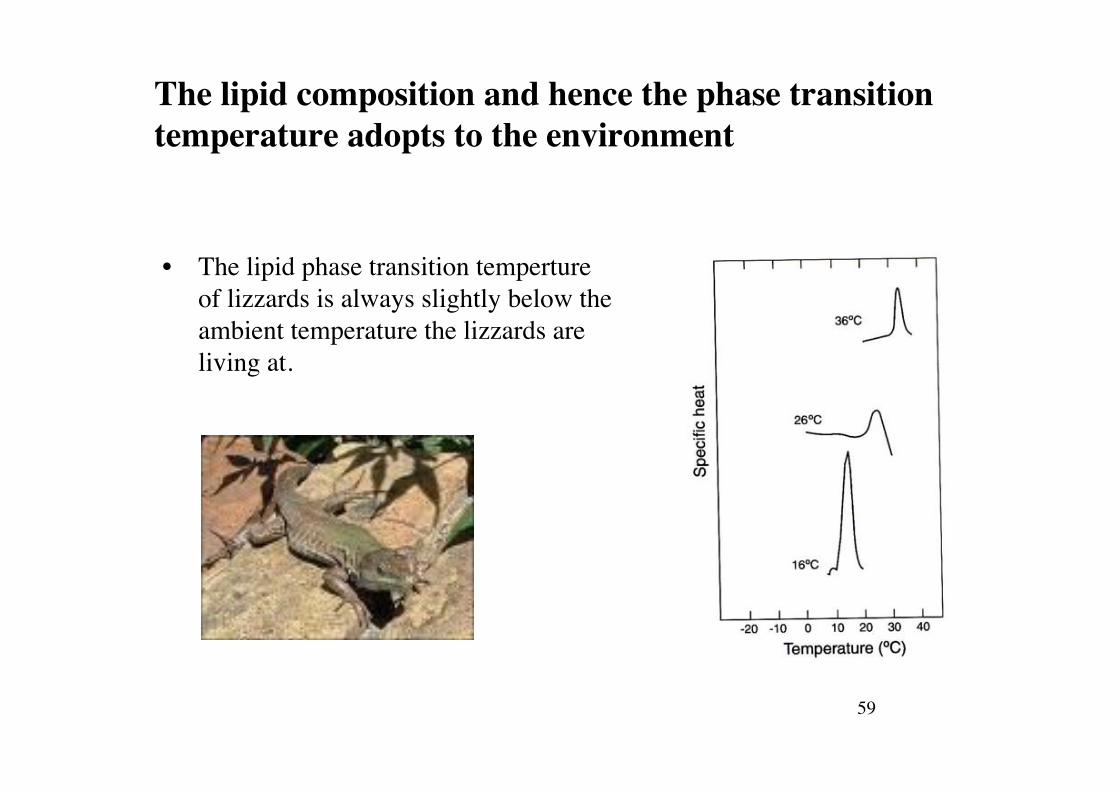
59!
The lipid composition and hence the phase transition temperature adopts to the environment!
• The lipid phase transition temperture of lizzards is always slightly below the ambient temperature the lizzards are living at.!

60!
Destabilization of the bilayer at the phase transition!
The permeability for ions is anomalously large at the phase separation region.!!Cholesterol suppresses the leakiness at the transition as well as suppresses anomalous binding at the transition.!
Molecular Dynamics of lipid bilayers!
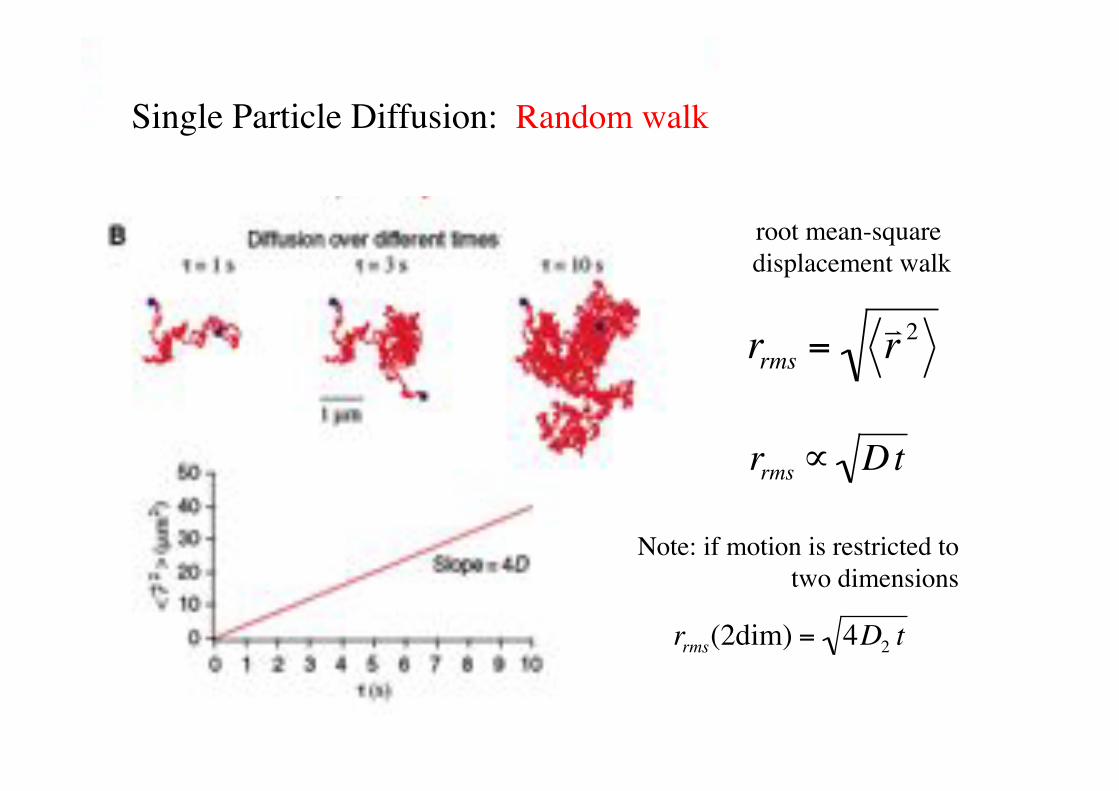
root mean-square displacement walk!
Single Particle Diffusion: Random walk !
!
rrms =! r 2
Note: if motion is restricted to two dimensions!
!
rrms(2dim) = 4D2 t!
rrms " Dt
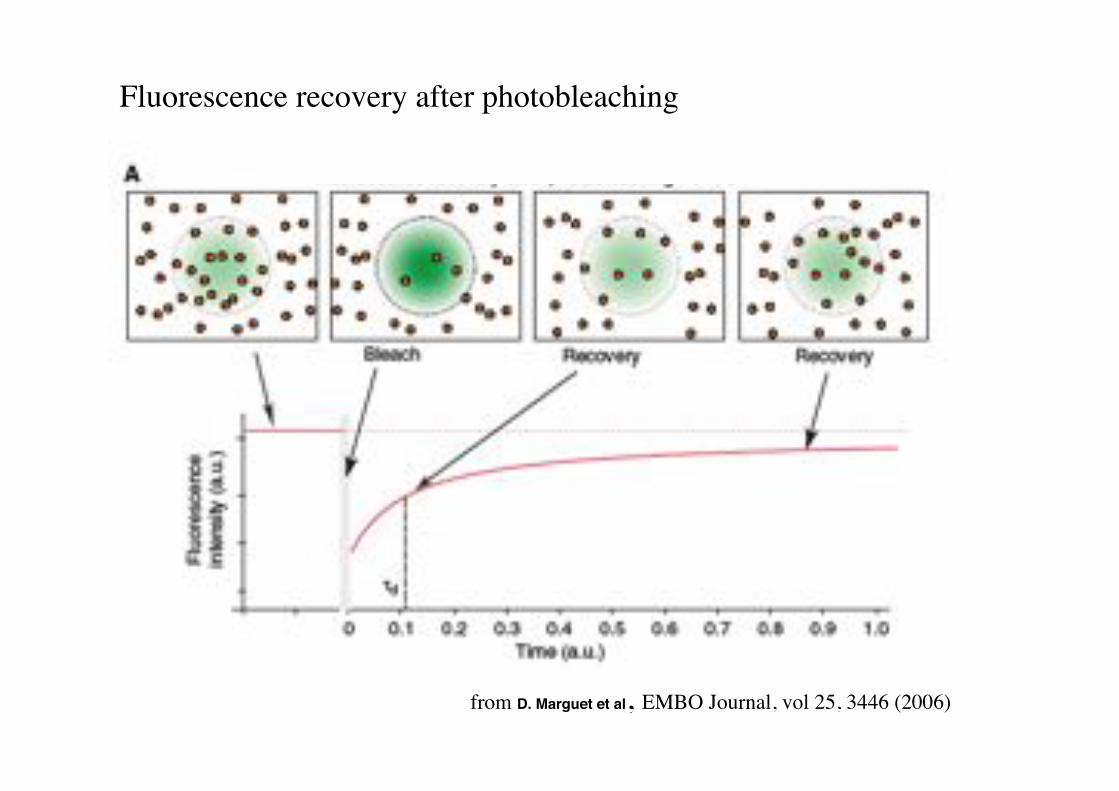
from D. Marguet et al, EMBO Journal, vol 25, 3446 (2006)!
Fluorescence recovery after photobleaching!!!
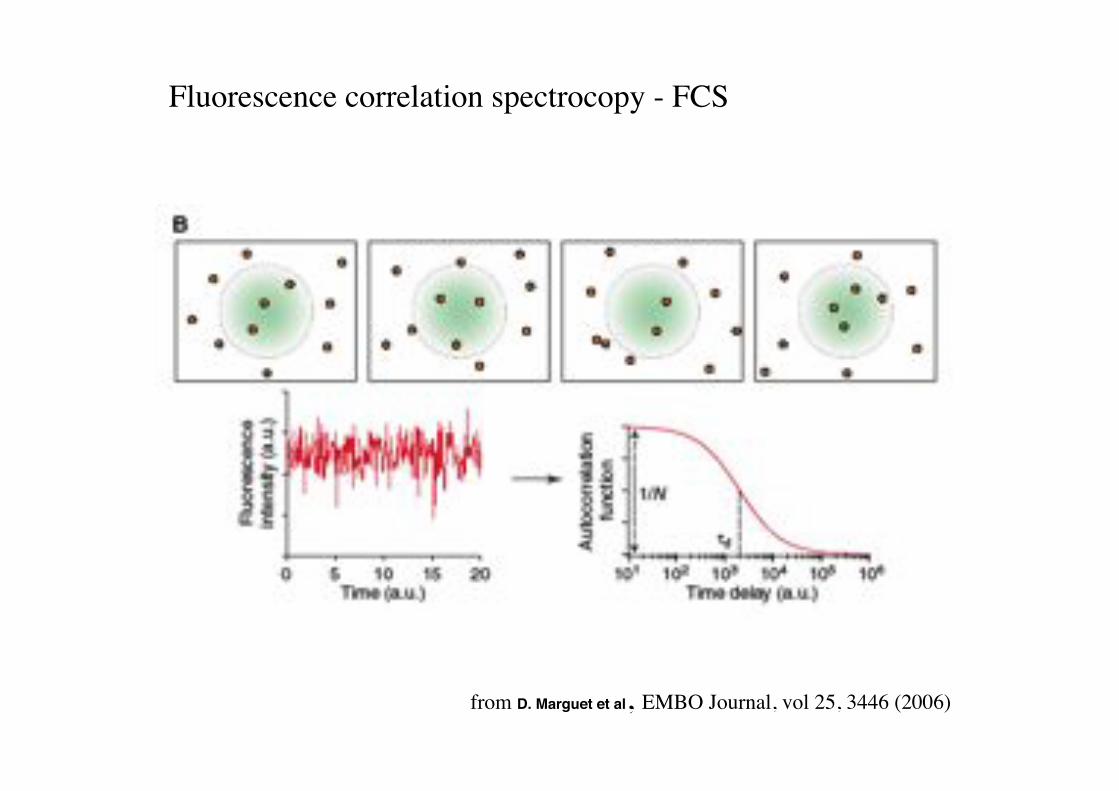
from D. Marguet et al, EMBO Journal, vol 25, 3446 (2006)!
!!
Fluorescence correlation spectrocopy - FCS!
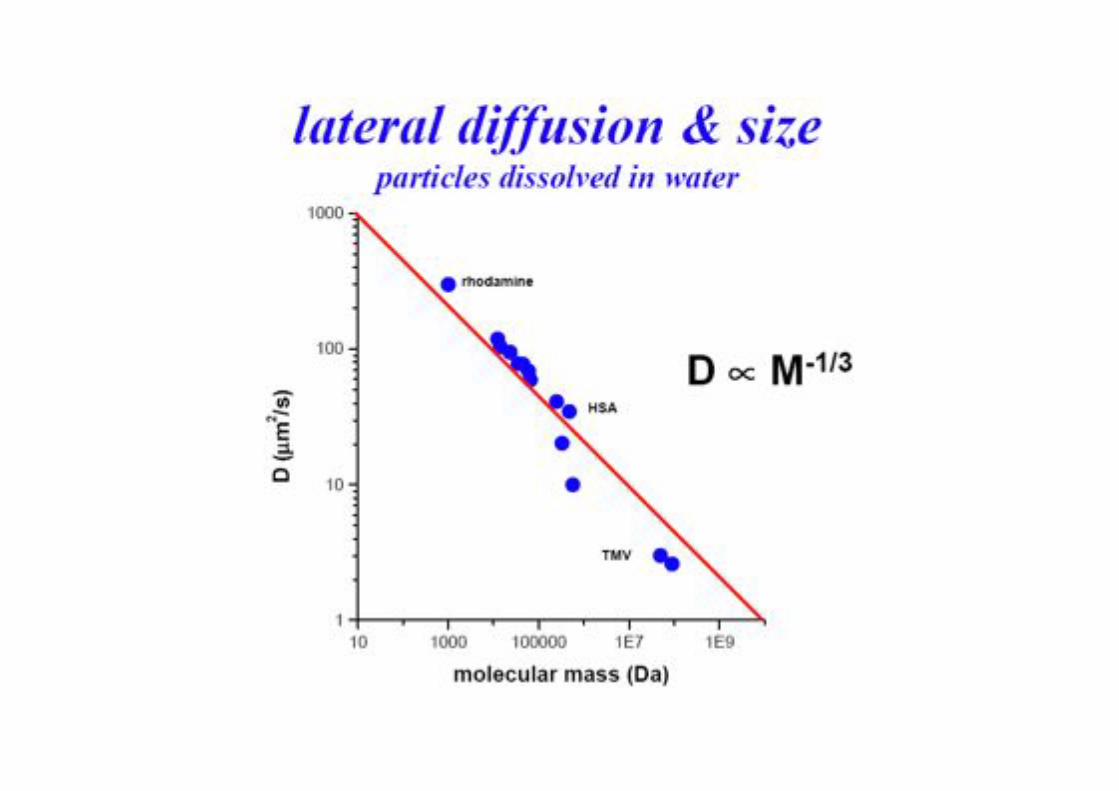

Typische Werte für Diffusions -Koeffizienten!
Diffusion in Membranen
3 Gründe für Abweichungen: - 2D Flüssigkeit: Hydrodynamik - hydrodynamische Kopplung mit Wand (oder Bindung an das Zytoskelett) - mosaikartiger Membranaufbau aus fluiden und festen Bereichen
8)B$H#'(7./' O='(PF)/' 0$1$B'O)>8$K' F&)>'(-.Q#"%D;/'
1A2#1A2&$1$B' ,*R' ,*S' FOI?' T'
=&UP21A28$7' ,*R' R,' FOI?' T'
V?DW"X"1>28' ;NR' R,' 52U)D0"%*' +N+'
V?DW"#X"1>28' Y' Q,,' 52U)D0"%*' +N;'
>">8)."8'
WA2B21#$7' Q' R,' FOI?' +'
Z)7B"'[[[' +N+' \T,' ]8U>A82XU>' ,NQ'
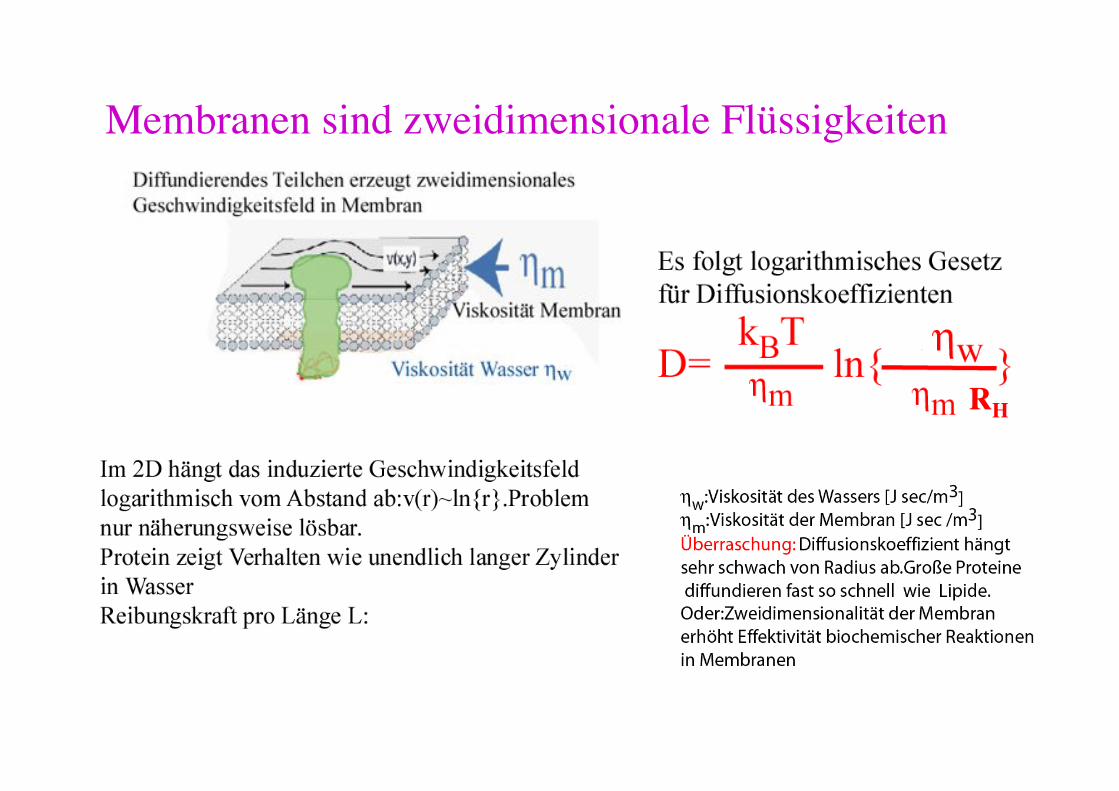
Membranen sind zweidimensionale Flüssigkeiten!
RH!
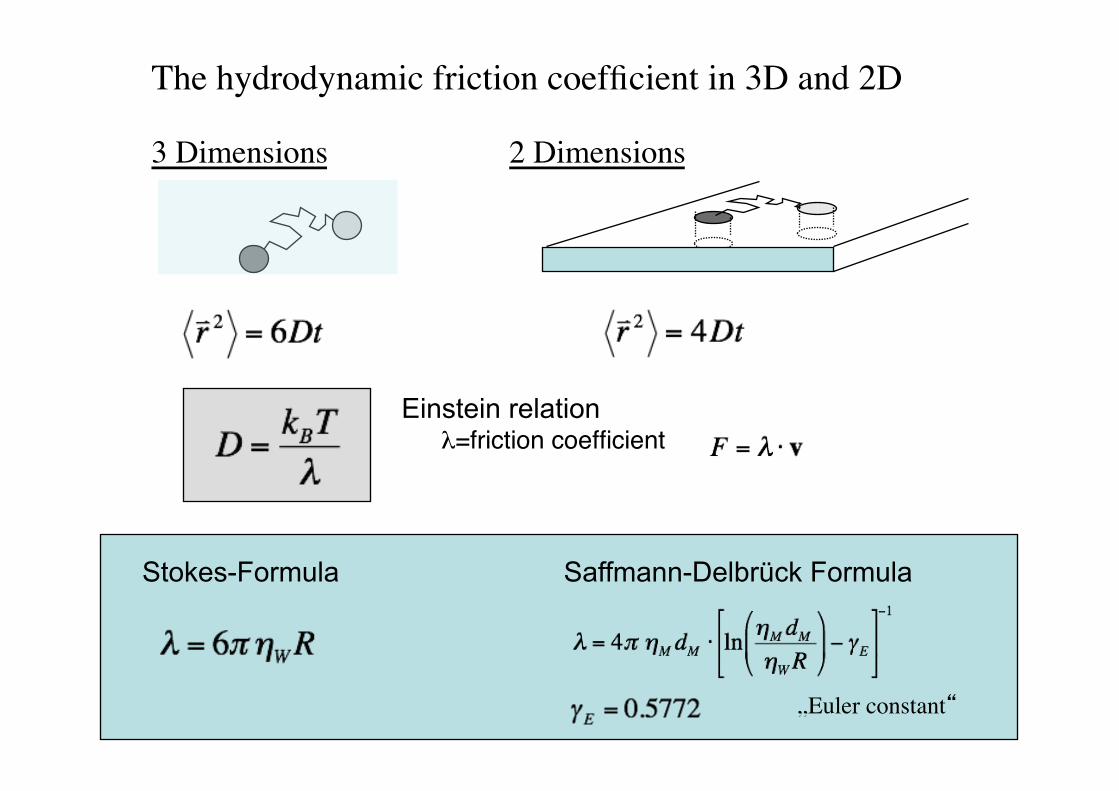
The hydrodynamic friction coefficient in 3D and 2D!
Saffmann-Delbrück Formula
„Euler constant“!
Stokes-Formula
Einstein relation
3 Dimensions! 2 Dimensions!
%=friction coefficient"
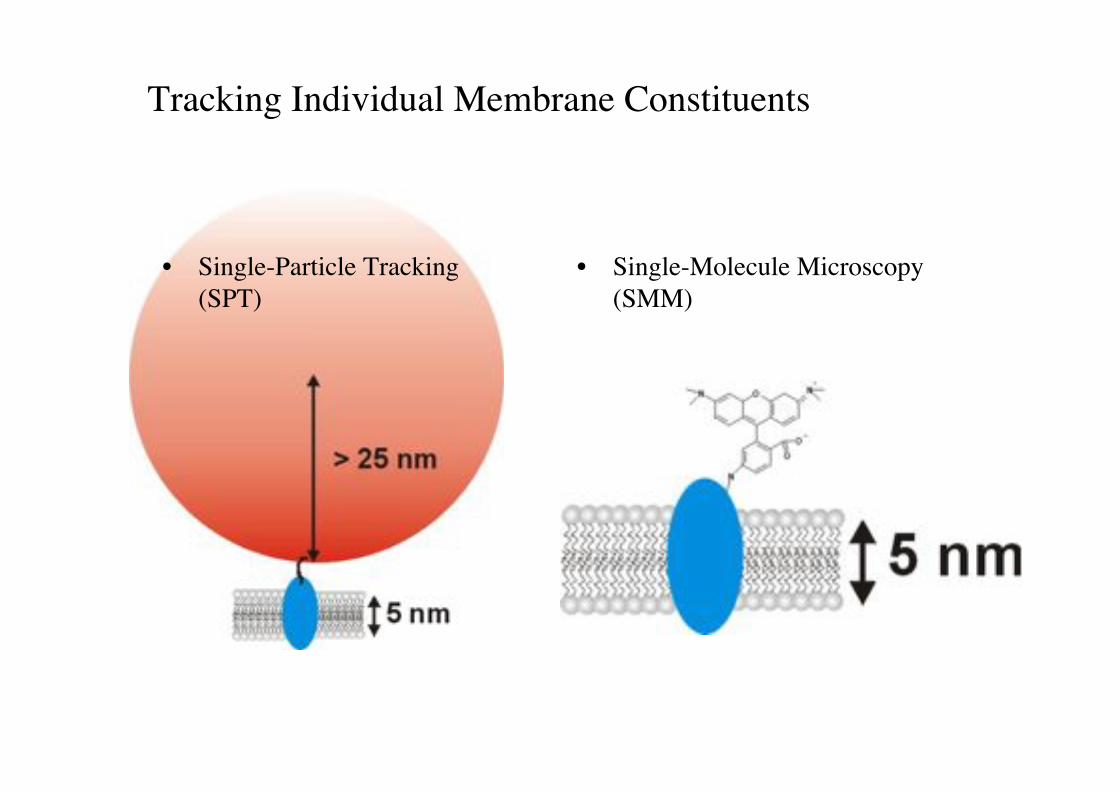
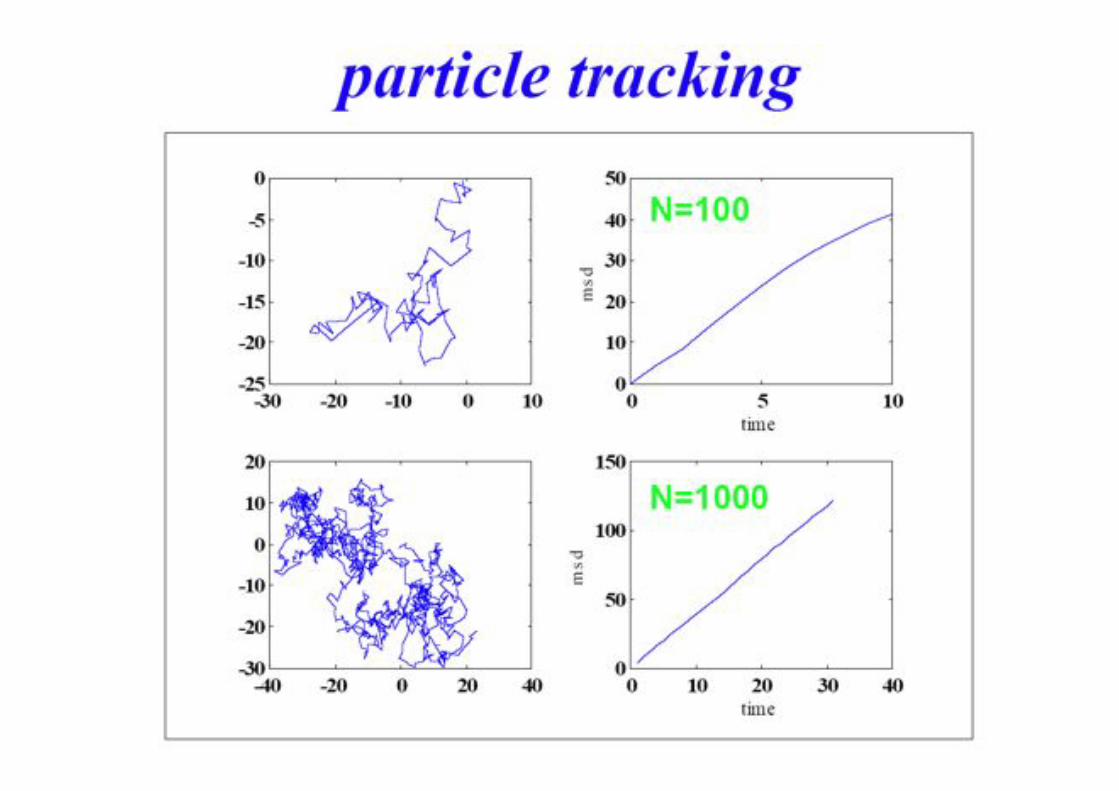
Tracking Individual Membrane Constituents!
• Single-Particle Tracking "(SPT)!
• Single-Molecule Microscopy (SMM)!
Brownian Motion!

Tinnefeld 2010!
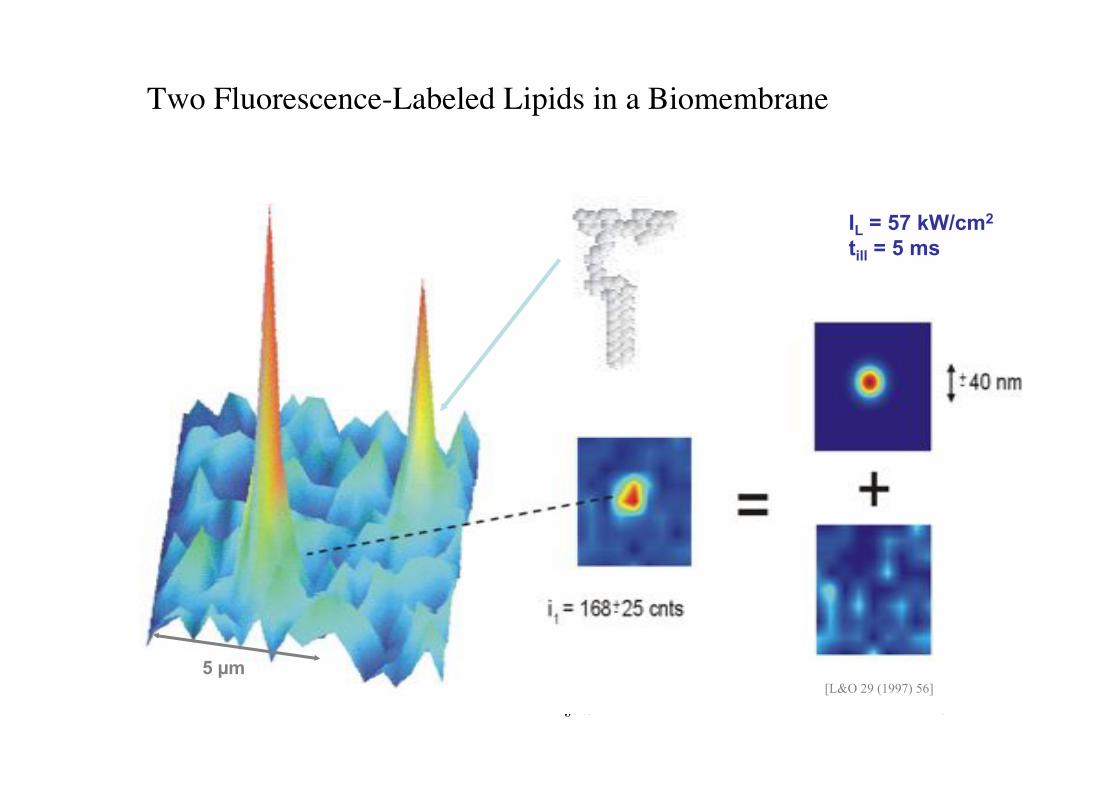
BPE §3.5! 72!
IL = 57 kW/cm2 till = 5 ms
5 !m [L&O 29 (1997) 56]
Two Fluorescence-Labeled Lipids in a Biomembrane!
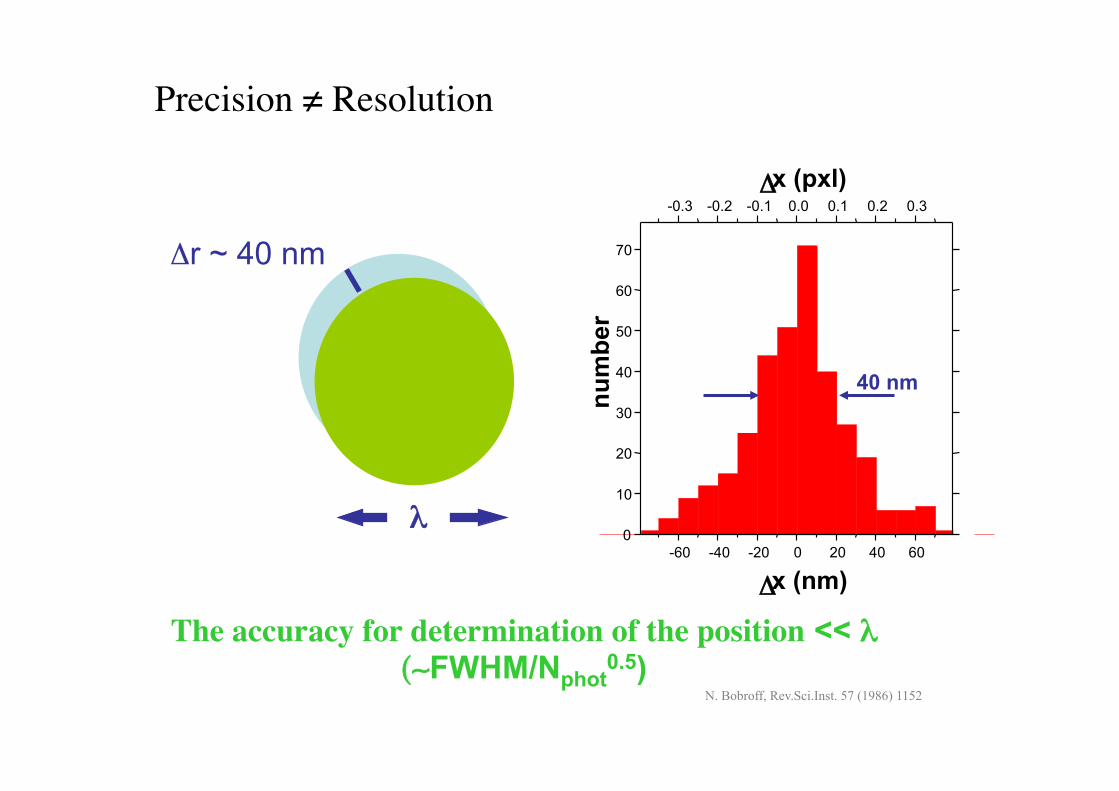
Precision $ Resolution!
The accuracy for determination of the position << % (&FWHM/Nphot
0.5)
%$
!r ~ 40 nm
N. Bobroff, Rev.Sci.Inst. 57 (1986) 1152
-60! -40! -20! 0! 20! 40! 60!0!10!20!30!40!50!60!70!
-0.3! -0.2! -0.1! 0.0! 0.1! 0.2! 0.3!
!
!!x (pxl)!
num
ber!
!!x (nm)!
40 nm
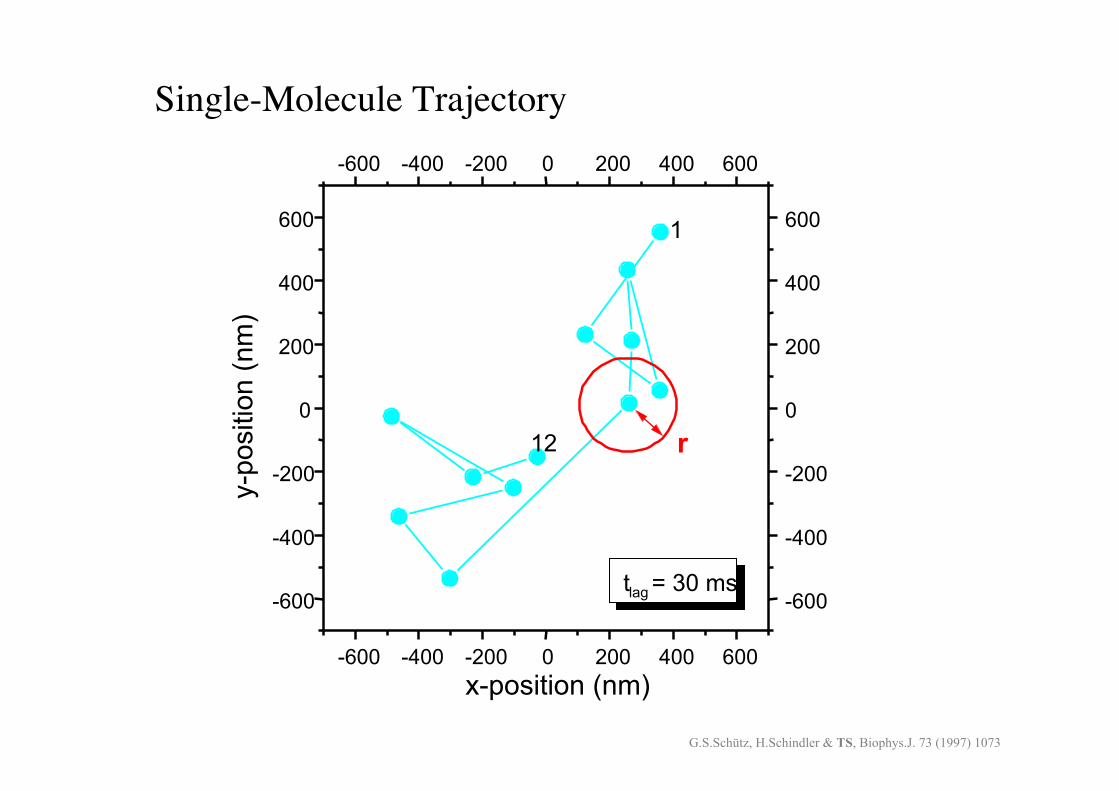
Single-Molecule Trajectory!
-600! -400! -200! 0! 200! 400! 600!
-600!
-400!
-200!
0!
200!
400!
600!
-600! -400! -200! 0! 200! 400! 600!
-600!
-400!
-200!
0!
200!
400!
600!
r!
t!lag! = 30 ms!
12!
1!y-
posi
tion
(nm
)!
x-position (nm)!G.S.Schütz, H.Schindler & TS, Biophys.J. 73 (1997) 1073
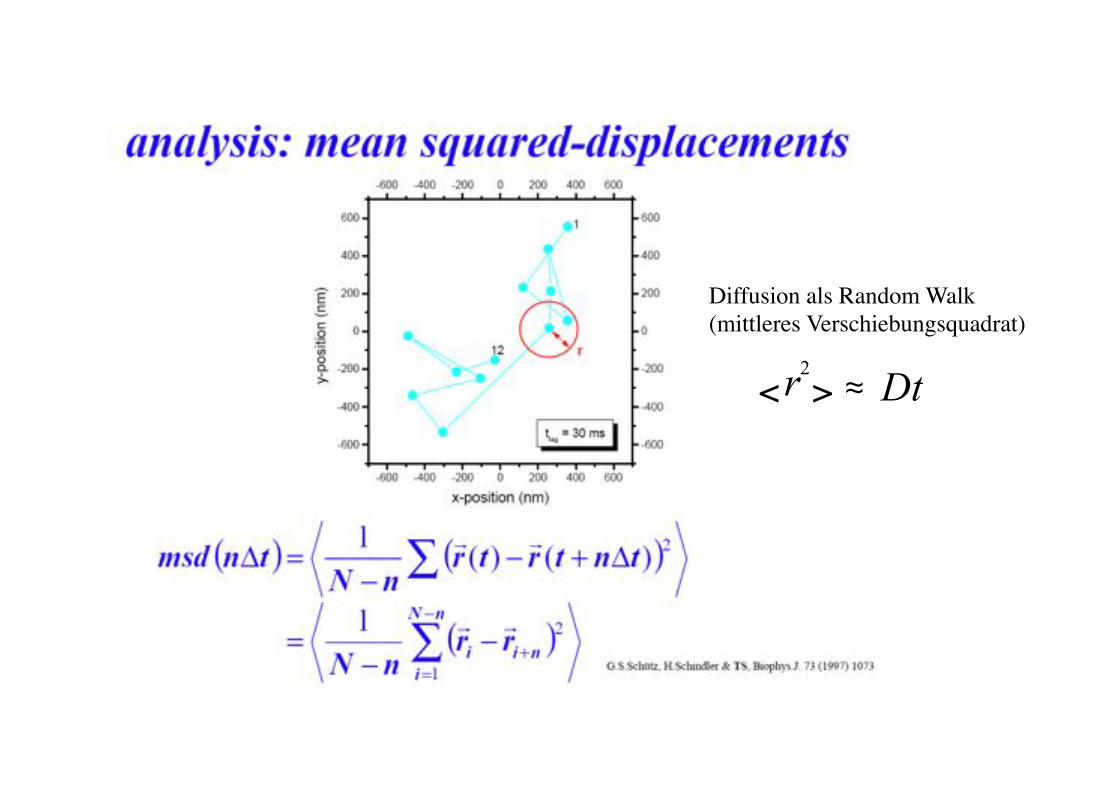
Diffusion als Random Walk!(mittleres Verschiebungsquadrat)!
2 r Dt < >$ #!
Tinnefeld, Gaub / SS2010!
BPE §3.5! 76!
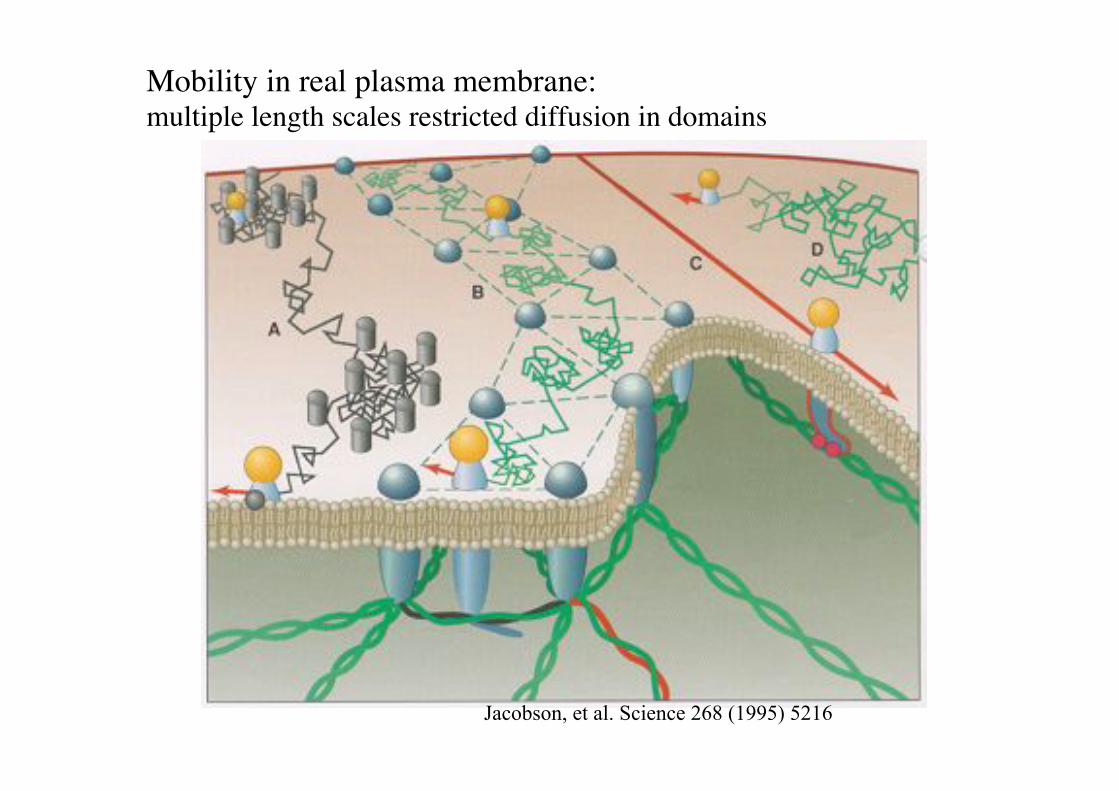
Mobility in real plasma membrane: "multiple length scales restricted diffusion in domains!
Jacobson, et al. Science 268 (1995) 5216
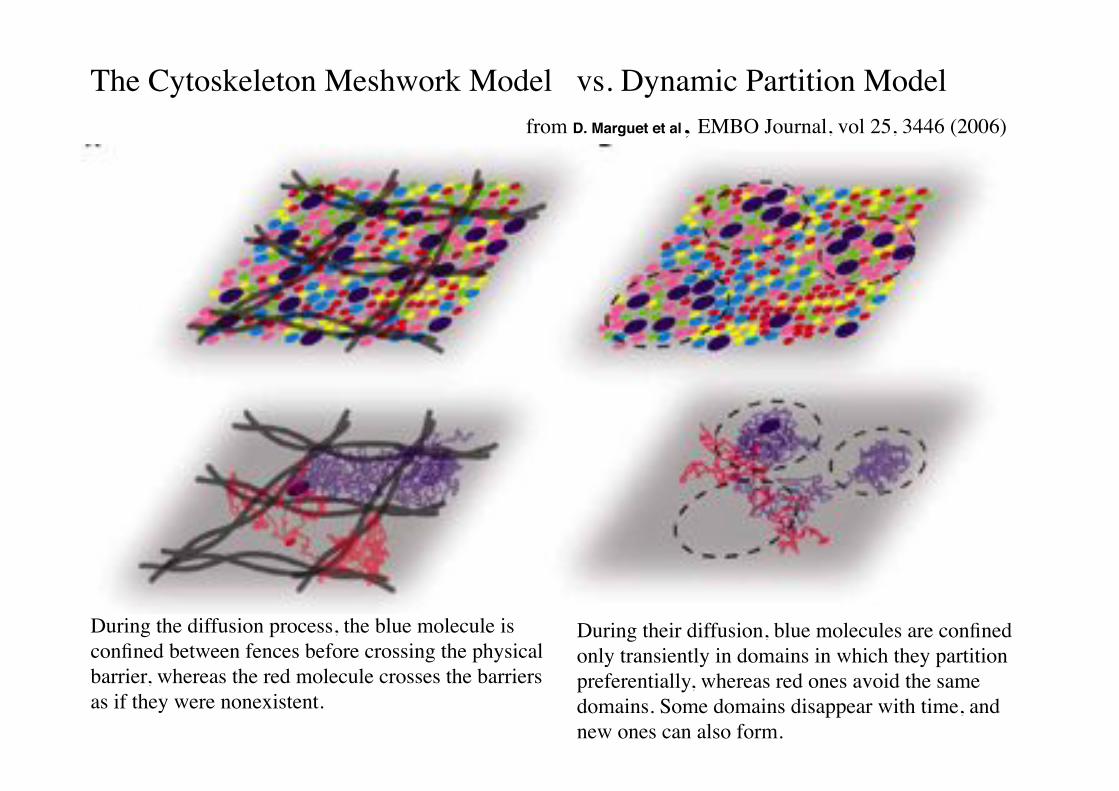
The Cytoskeleton Meshwork Model vs. Dynamic Partition Model!from D. Marguet et al, EMBO Journal, vol 25, 3446 (2006)!
During the diffusion process, the blue molecule is confined between fences before crossing the physical barrier, whereas the red molecule crosses the barriers as if they were nonexistent.!
During their diffusion, blue molecules are confined only transiently in domains in which they partition preferentially, whereas red ones avoid the same domains. Some domains disappear with time, and new ones can also form.!

dp r t dt
D divgrad p(r,t)
p r t Dt
r Dt
( , )
( , ) exp
!"=$
=$ -$'$($)$
*$+$,$
2 2 1
4 4
R 2
P t p r t dr Dt
2 R
(R2, exp =$ =$ -$ -$'$($)$ *$+$,$-$2
0 1 4 ) ( , )
Particle Tracking in 2d! Ergodische Systeme:"(Ensemblemittel = Zeitmittel)!
W, innerhalb R2 für Zeit t zu bleiben!
W für Mean Square Displacement!r2 während der Zeit t!
W-dichte, Partikel zum Zeitpkt !t am Ort r zu finden!
Diffusion als Random Walk!(mittleres Verschiebungsquadrat)!
2 r Dt < >$ #! Zentraler Grenzwertsatz!
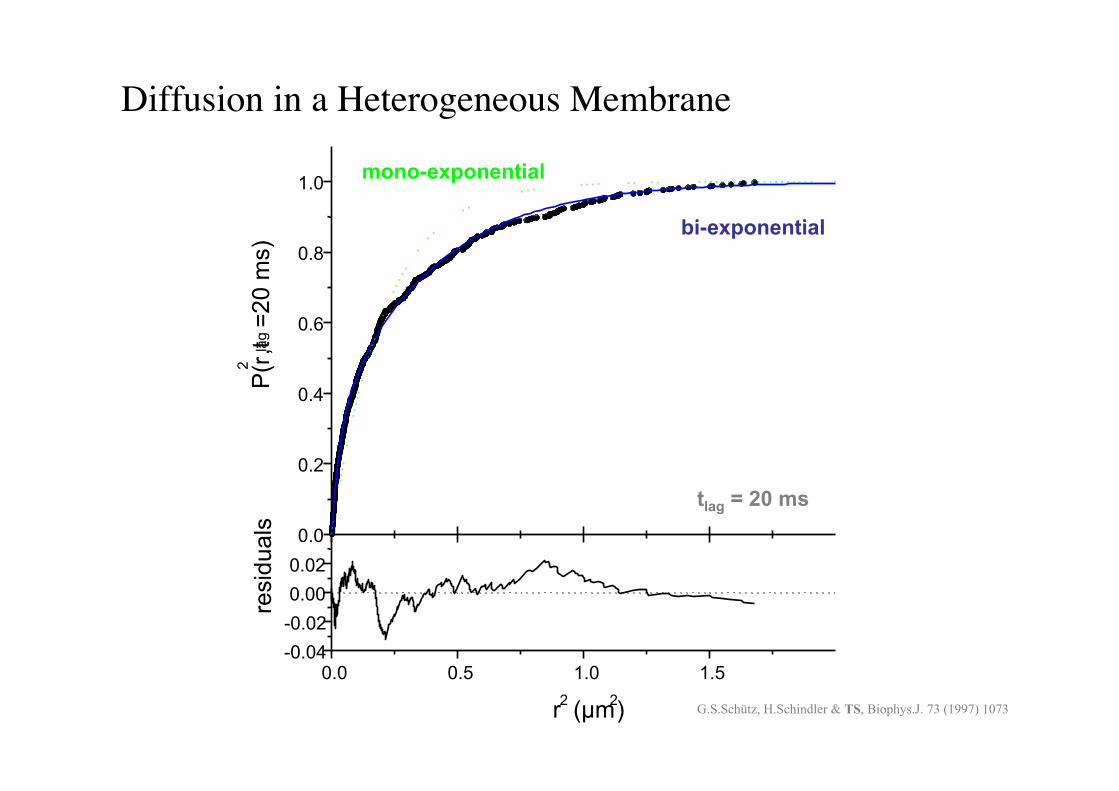
BPE §3.5! 80!
Diffusion in a Heterogeneous Membrane!
0.0!
0.2!
0.4!
0.6!
0.8!
1.0!
P(r!
2! ,t!lag! =2
0 m
s)!
0.0! 0.5! 1.0! 1.5!-0.04!-0.02!0.00!0.02!
resi
dual
s!
r!2! (!m!2!)!
bi-exponential
mono-exponential
tlag = 20 ms
G.S.Schütz, H.Schindler & TS, Biophys.J. 73 (1997) 1073
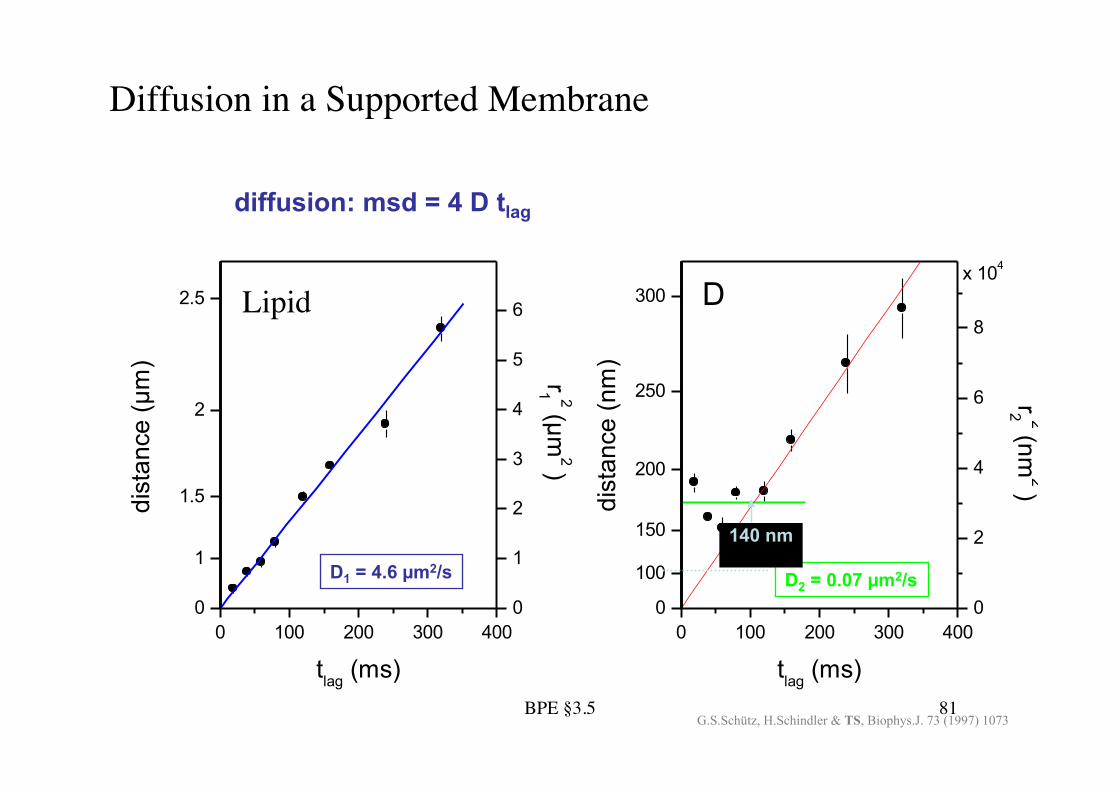
BPE §3.5! 81!
Diffusion in a Supported Membrane!
0 100 200 300 4000
2
4
6
8
x 104
r2 2 (nm2 )
dist
ance
(nm
)
tlag (ms)
0
100
150
200
250
300 D
0 100 200 300 4000
1
2
3
4
5
6
r1 2 (!m2 )
dist
ance
(!m
)
tlag (ms)
0
1
C
1.5
2
2.5
D1 = 4.6 !m2/s D2 = 0.07 !m2/s 140 nm
diffusion: msd = 4 D tlag
G.S.Schütz, H.Schindler & TS, Biophys.J. 73 (1997) 1073
Lipid!

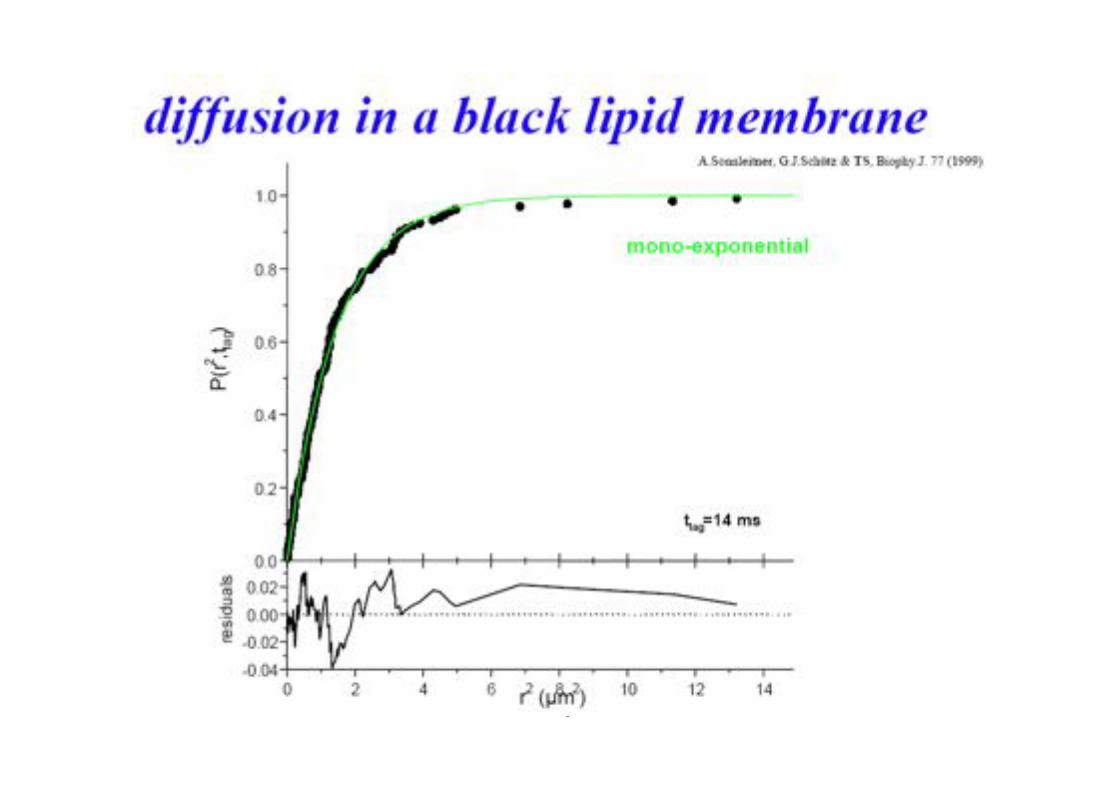
BPE §3.5! 83!
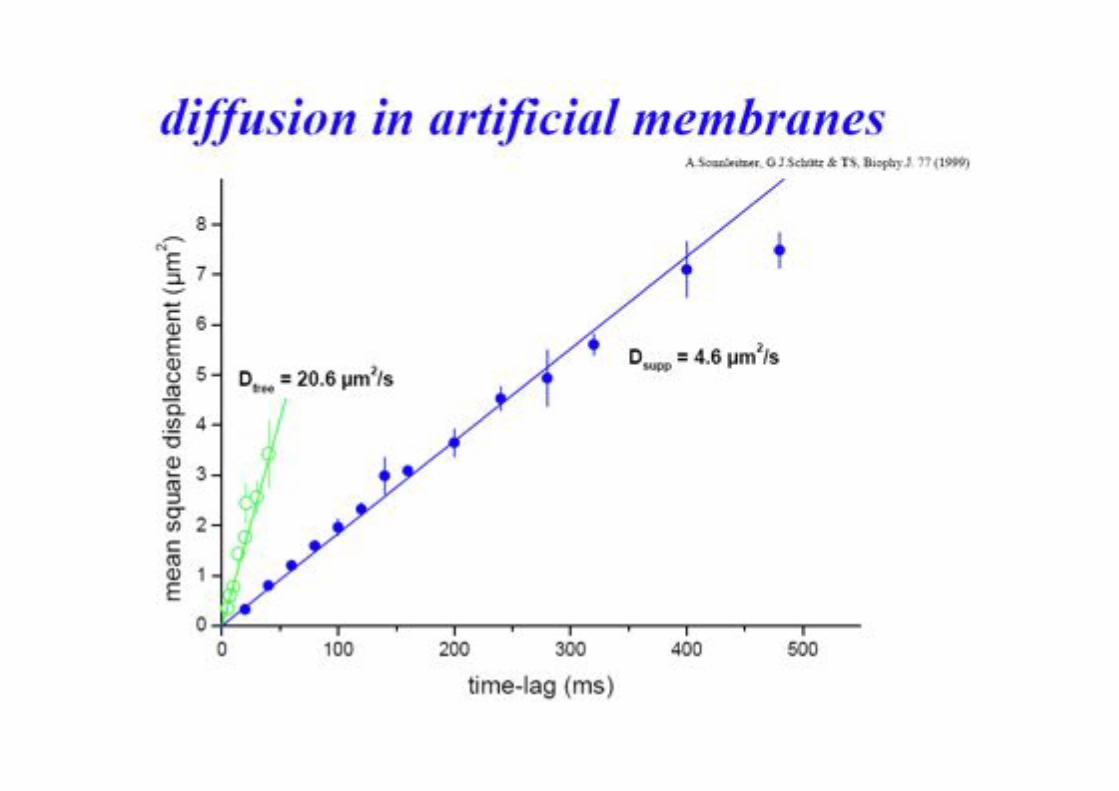
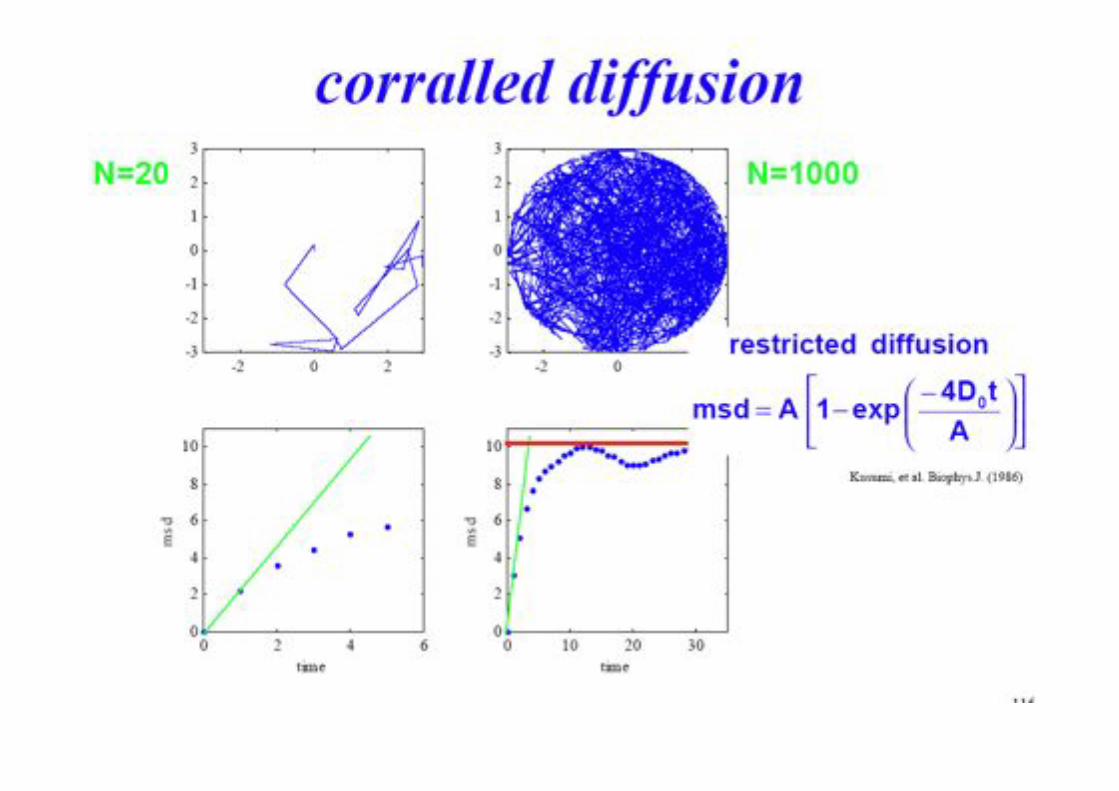
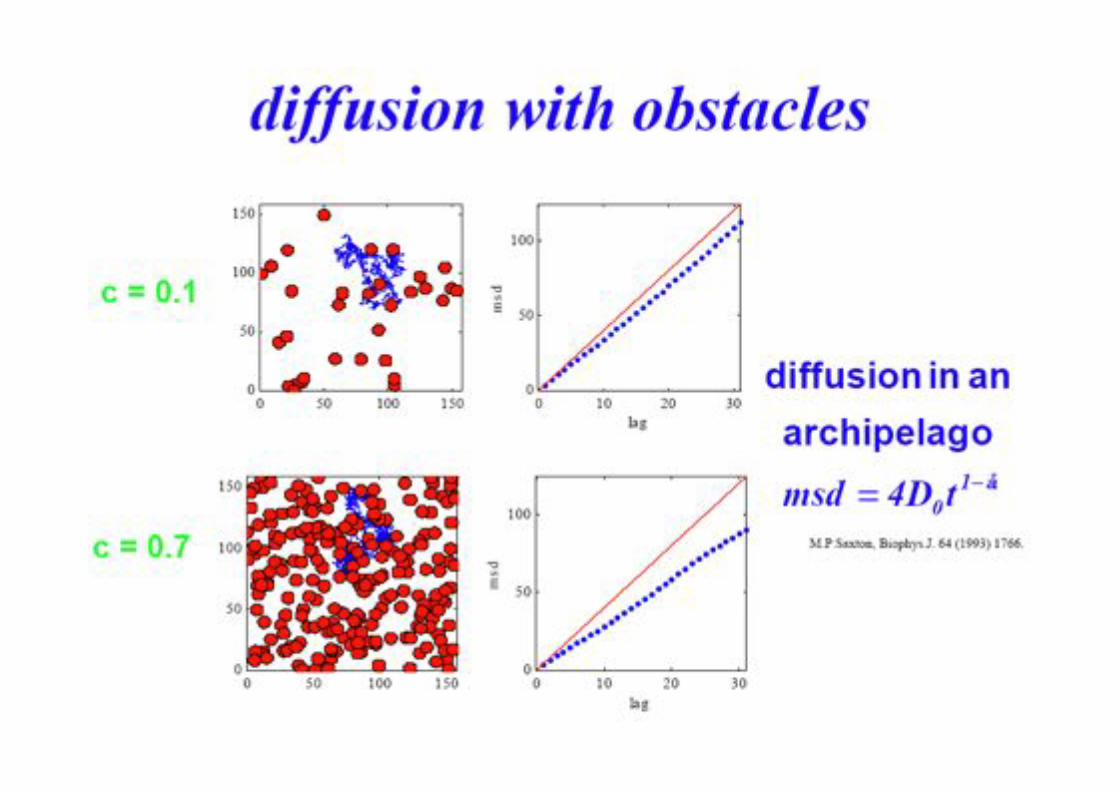
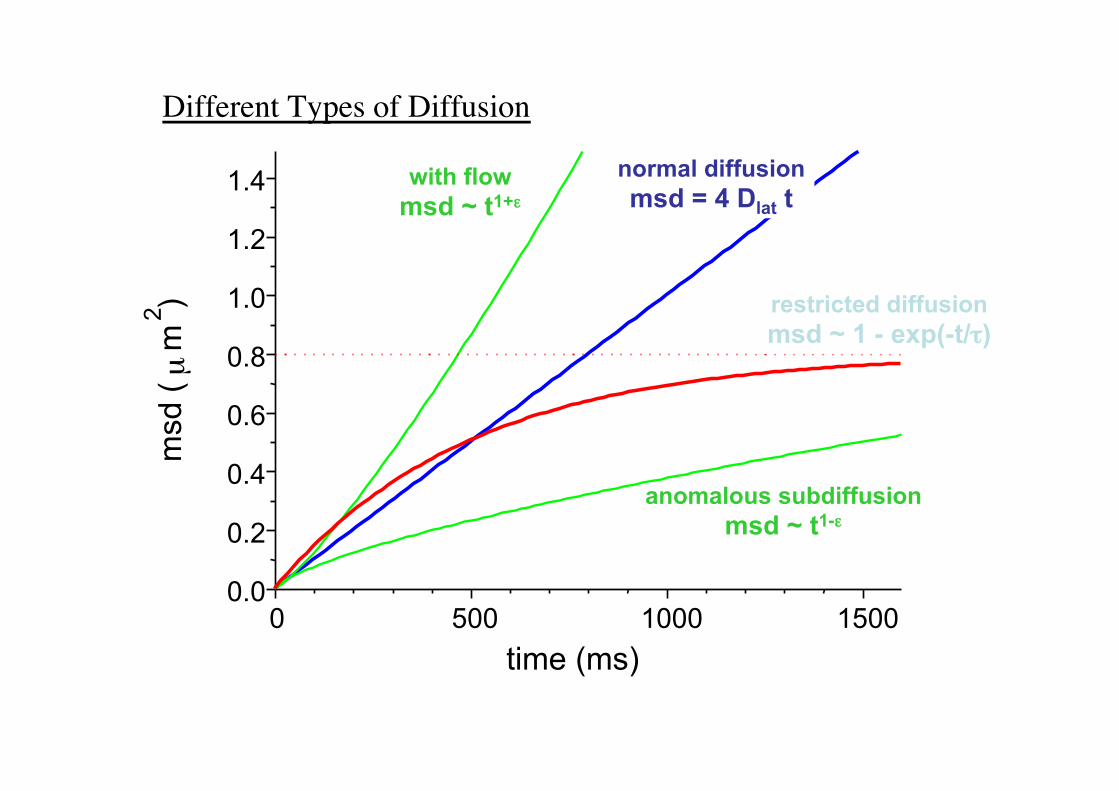
Different Types of Diffusion!
0! 500! 1000! 1500!0.0!
0.2!
0.4!
0.6!
0.8!
1.0!
1.2!
1.4!m
sd (!
µ!m!2! )!
time (ms)!
normal diffusion msd = 4 Dlat t
with flow msd ~ t1+.
anomalous subdiffusion msd ~ t1-.
restricted diffusion msd ~ 1 - exp(-t//)
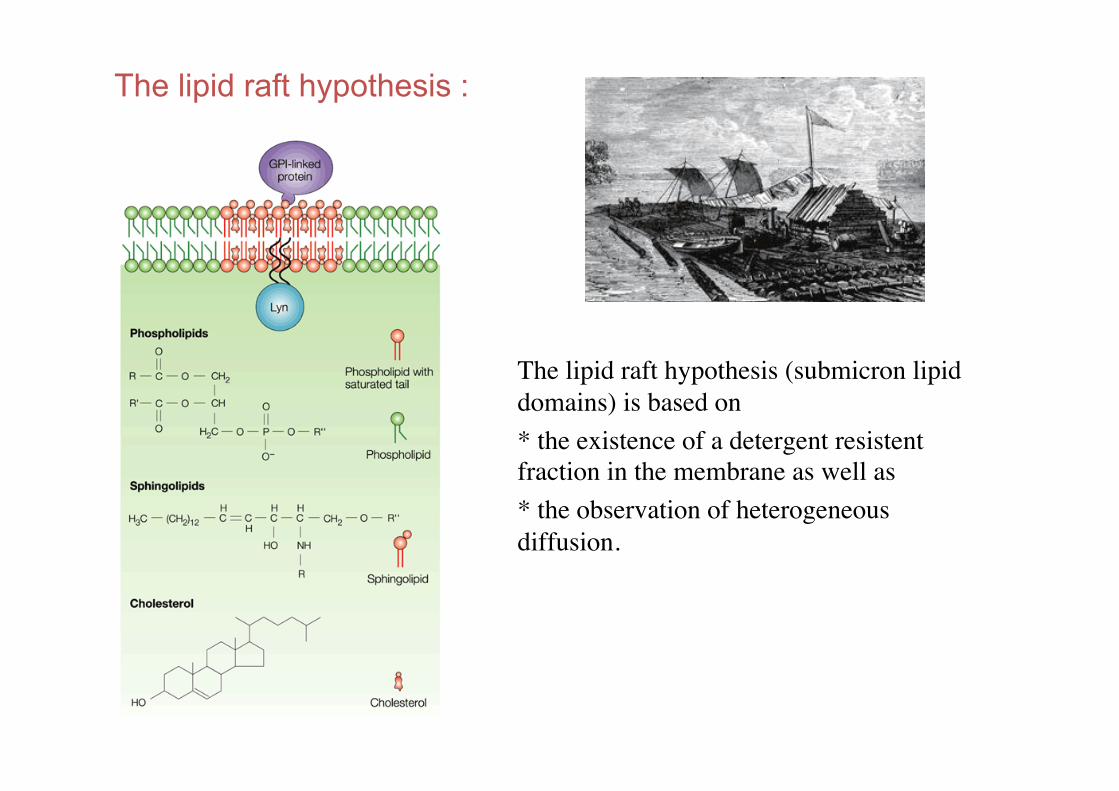
The lipid raft hypothesis :
The lipid raft hypothesis (submicron lipid domains) is based on !* the existence of a detergent resistent fraction in the membrane as well as !* the observation of heterogeneous diffusion.!
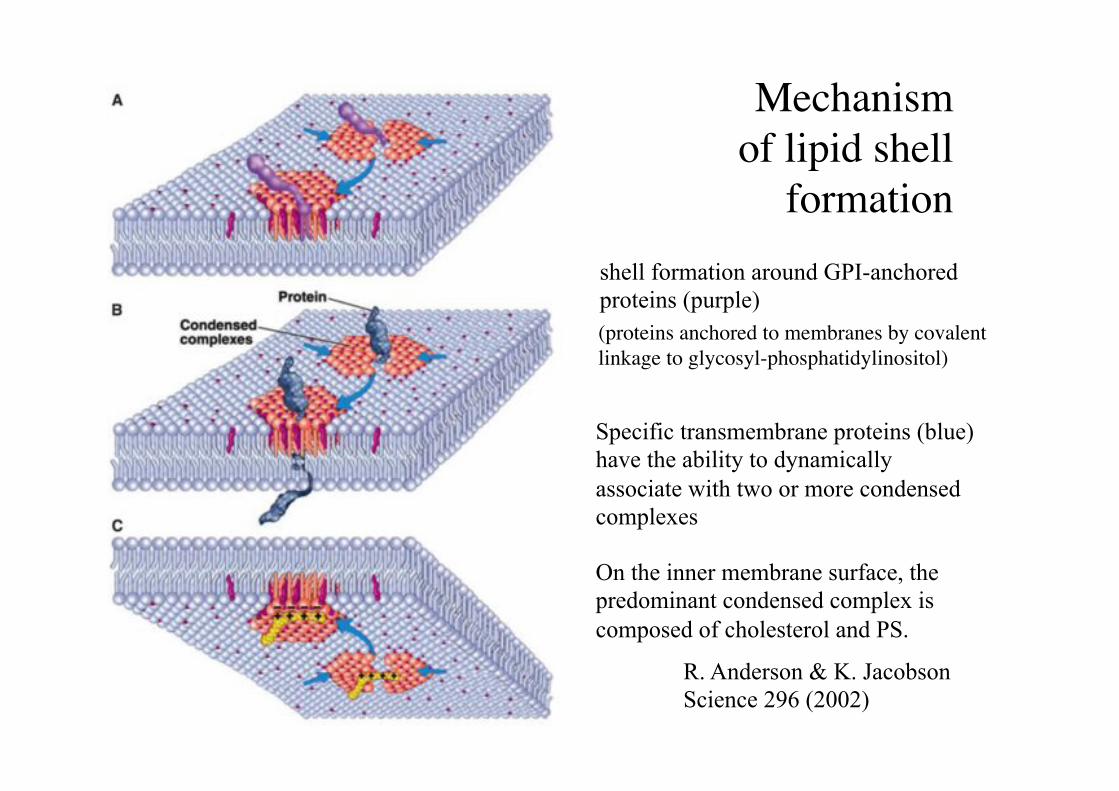
Mechanism "of lipid shell
formation !
On the inner membrane surface, the predominant condensed complex is composed of cholesterol and PS.
shell formation around GPI-anchored proteins (purple)
Specific transmembrane proteins (blue) have the ability to dynamically associate with two or more condensed complexes
R. Anderson & K. Jacobson Science 296 (2002)
(proteins anchored to membranes by covalent linkage to glycosyl-phosphatidylinositol)!
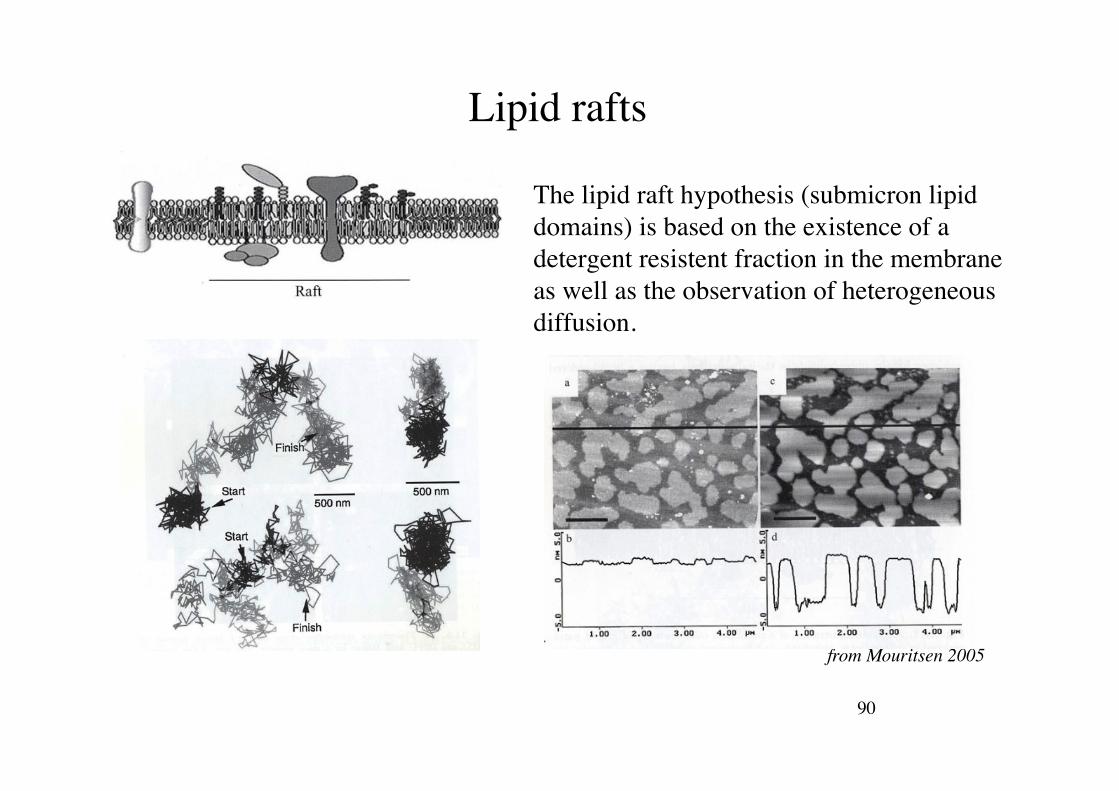
90!
Lipid rafts!
The lipid raft hypothesis (submicron lipid domains) is based on the existence of a detergent resistent fraction in the membrane as well as the observation of heterogeneous diffusion.!
from Mouritsen 2005!

91!
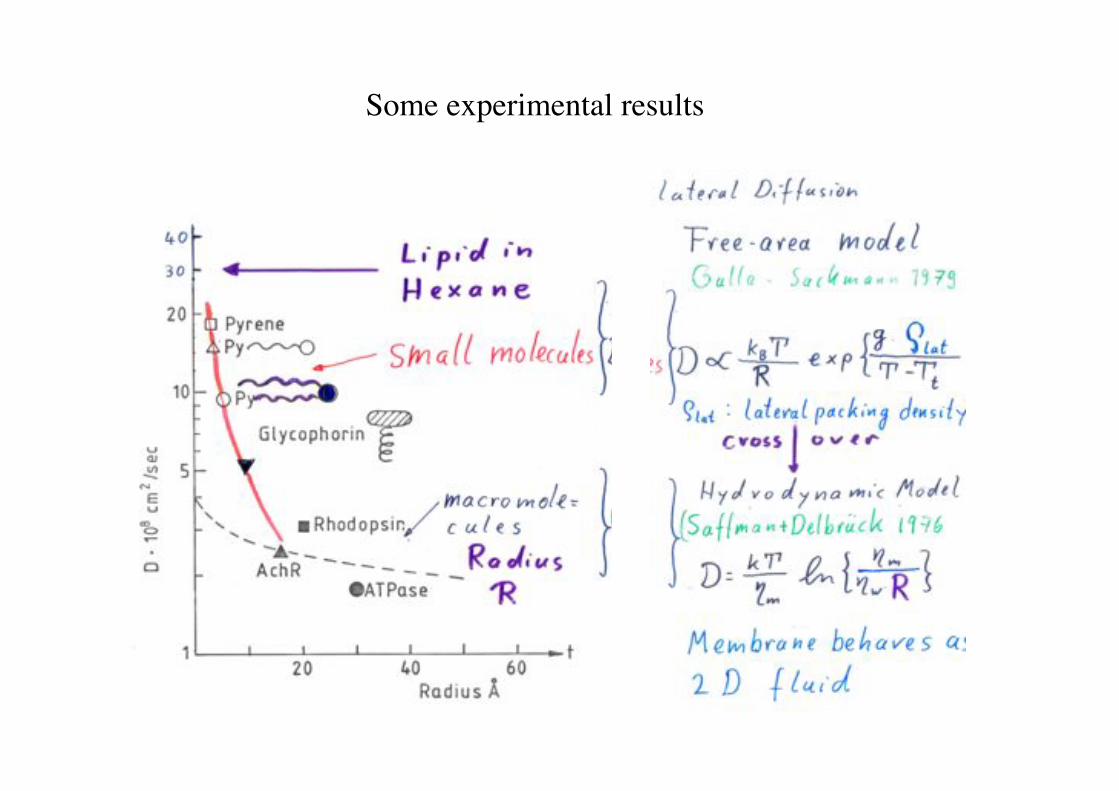
Das freie Volumen Modell!
- das hydrodynamische Modell (Saffman-Delbrück) beschreibt die Diffusion !
- auf kurzen Zeitskalen von Lipiden nicht gut.!
D =kBTfe!
"a*
a0# T!Tt( )
-Ein Molekül kann dann einen Diffusionssprung durch Fluktuation vollführen, wenn in der Nachbarschaft (durch thermische Fluktuationen) eine minimale freie Fläche a* entsteht (Fluktuation der Packungsdichte).!!
D0 : kbT/f a*: für Sprung benötigte freie Fläche a0: Fläche pro Molekül Lipid Tt : Umwandlungstemperatur
Gaub LMU! BPE§3.5! 92!
Some experimental results!

Lipid Monolayers!

Rädler SS 08! Biophysik der Zelle! 94!
- An der Oberfläche ist keine optimale Wechselwirkung mehr möglich!
- Das Erzeugen einer Oberfläche kostet Energie.!
- Die Energie pro Oberflächeneinheit heißt Oberflächenspannung!
- Moleküle wechselwirken miteinander in der Flüssigkeit. (Kohäsion)!
! ="E"A
[! ] = Nm
Oberflächenspannung und 2-D-Thermodynamik!
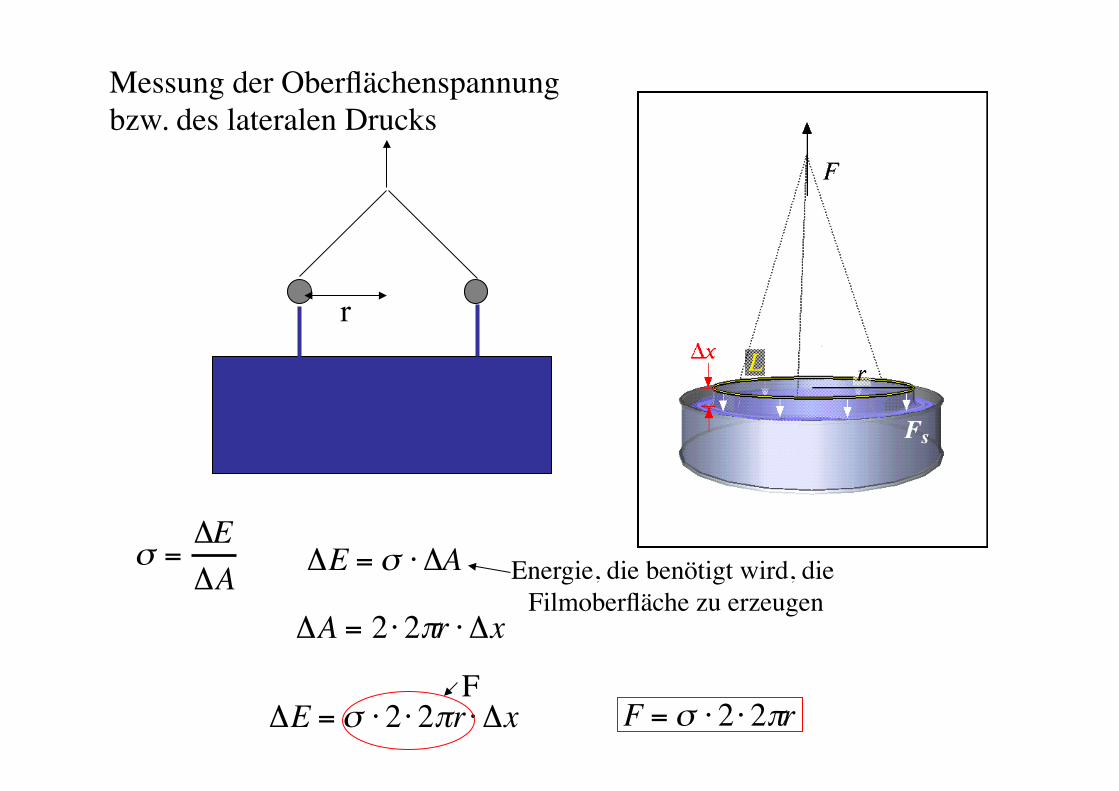
Rädler SS 08! Biophysik der Zelle! 95!
r!
! ="E"A
!A = 2 " 2#r " !x
!E = " # 2 # 2$r # !x F = ! " 2 " 2#rF!
!E = " # !A Energie, die benötigt wird, die Filmoberfläche zu erzeugen!
Messung der Oberflächenspannung "bzw. des lateralen Drucks!
Rädler SS 08! Biophysik der Zelle! 96!
F
Fl!ä!che A!B!a!r!r!i!e!r!e!
Brei
te"
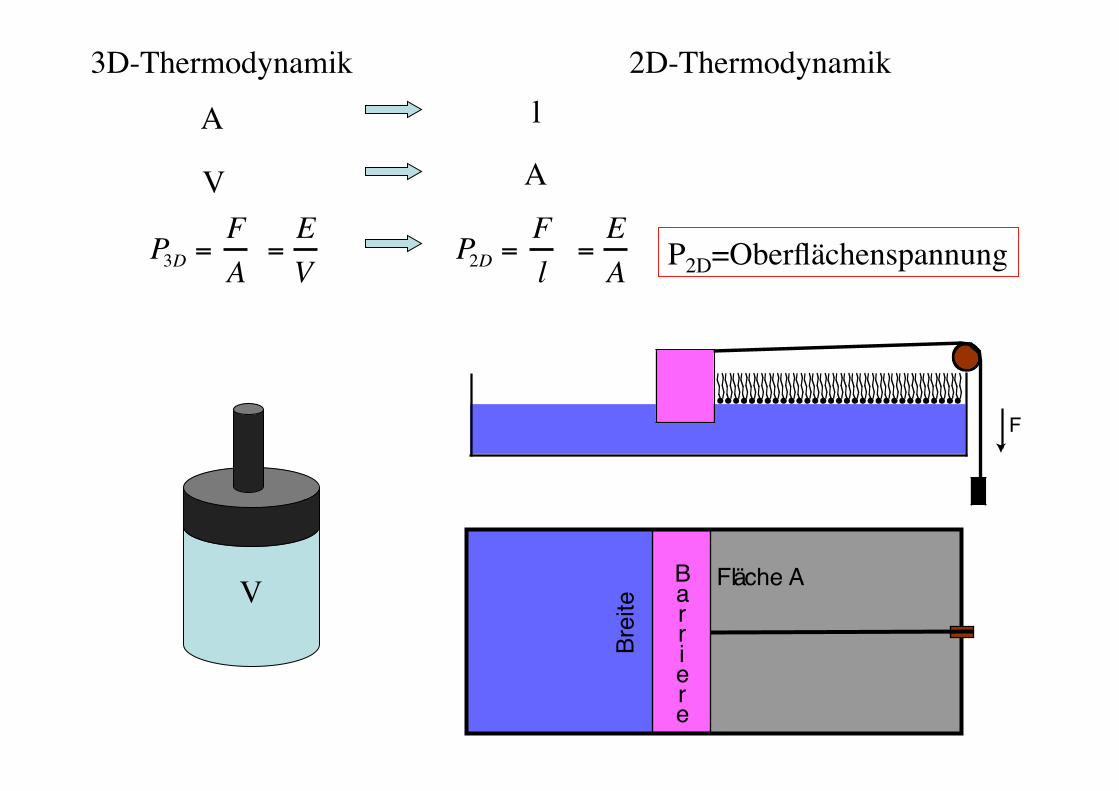
P3D =FA
=EV
P2D =Fl
=EA
2D-Thermodynamik!
V!
A!
V!
l!
A!
P2D=Oberflächenspannung!
3D-Thermodynamik!

98!
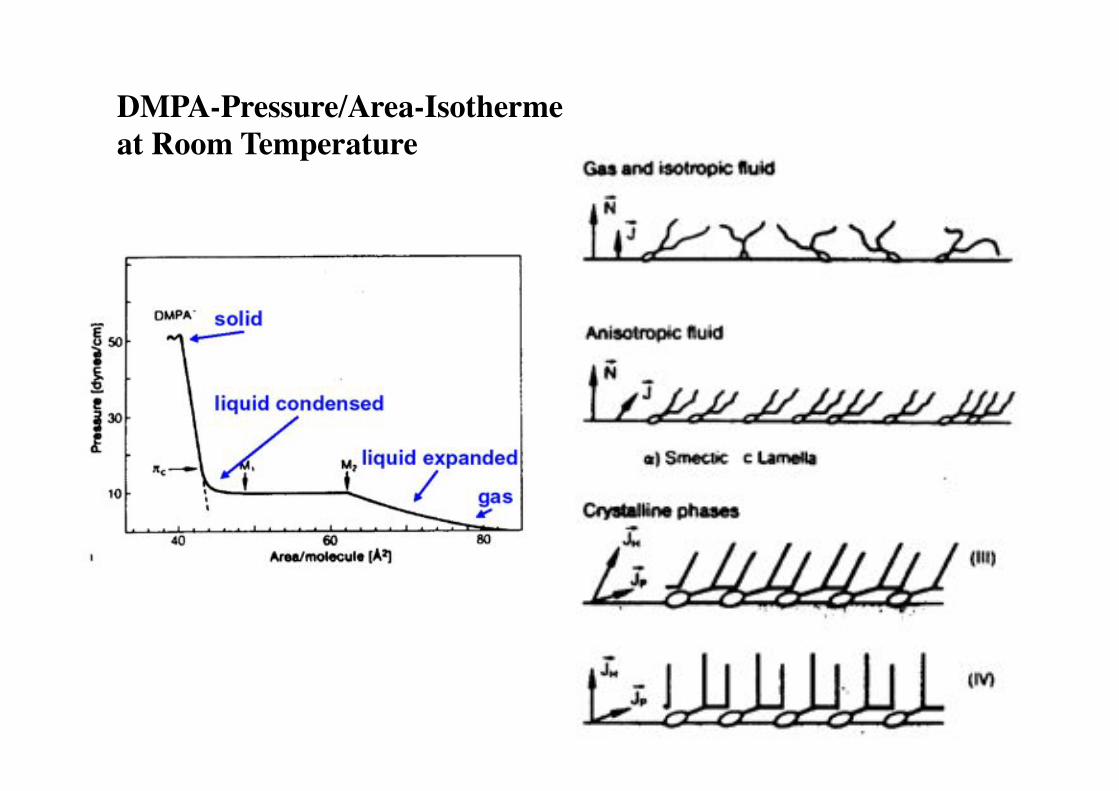
DMPA-Pressure/Area-Isotherme "at Room Temperature!

Dynamics of fluid phase coexistence!
DPPC domains!
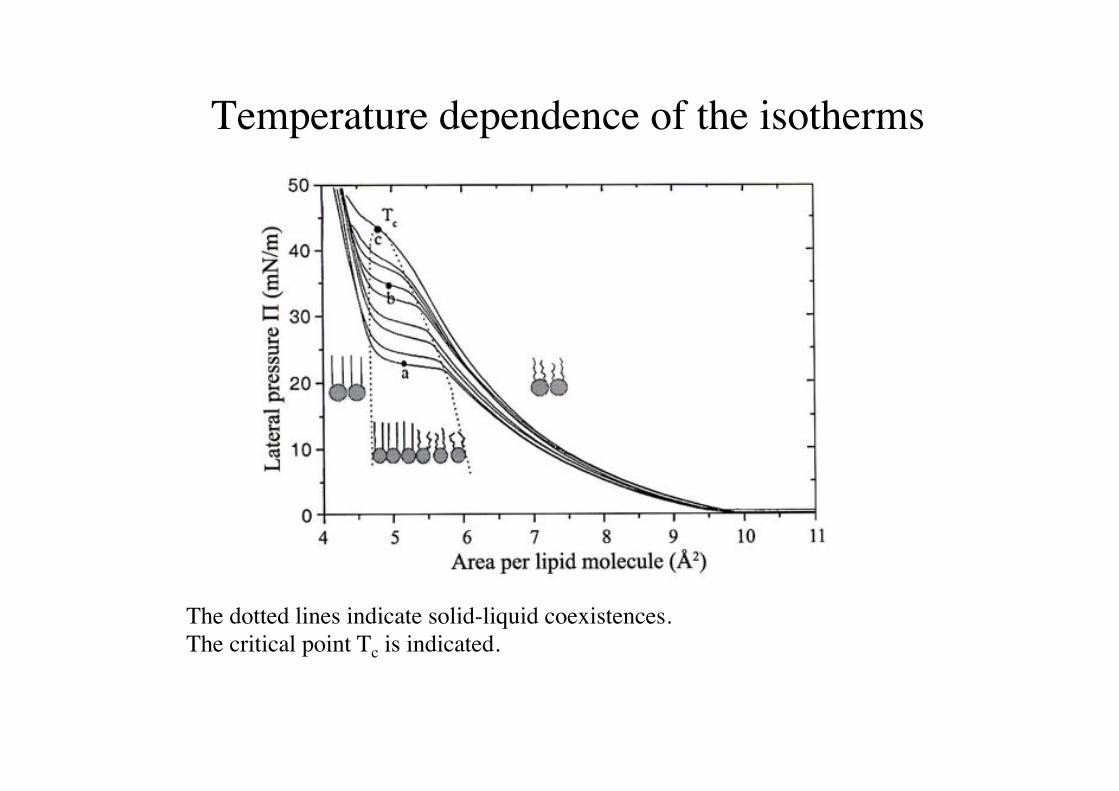
Temperature dependence of the isotherms!
The dotted lines indicate solid-liquid coexistences. !The critical point Tc is indicated.!
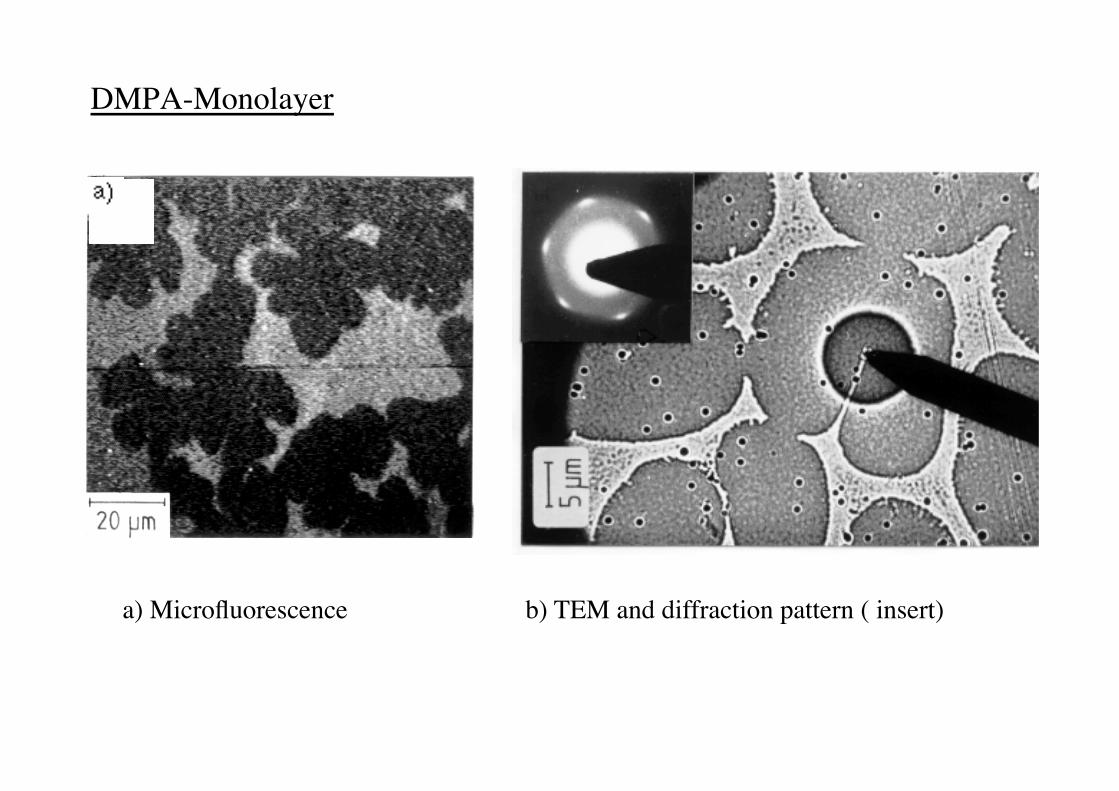
DMPA-Monolayer !
a) Microfluorescence ! ! b) TEM and diffraction pattern ( insert)!

Pattern!
Complex pattern arise as a consequence of the competition!between the line tension of the lipid domains and the long-range!electrostatic interaction between domains!
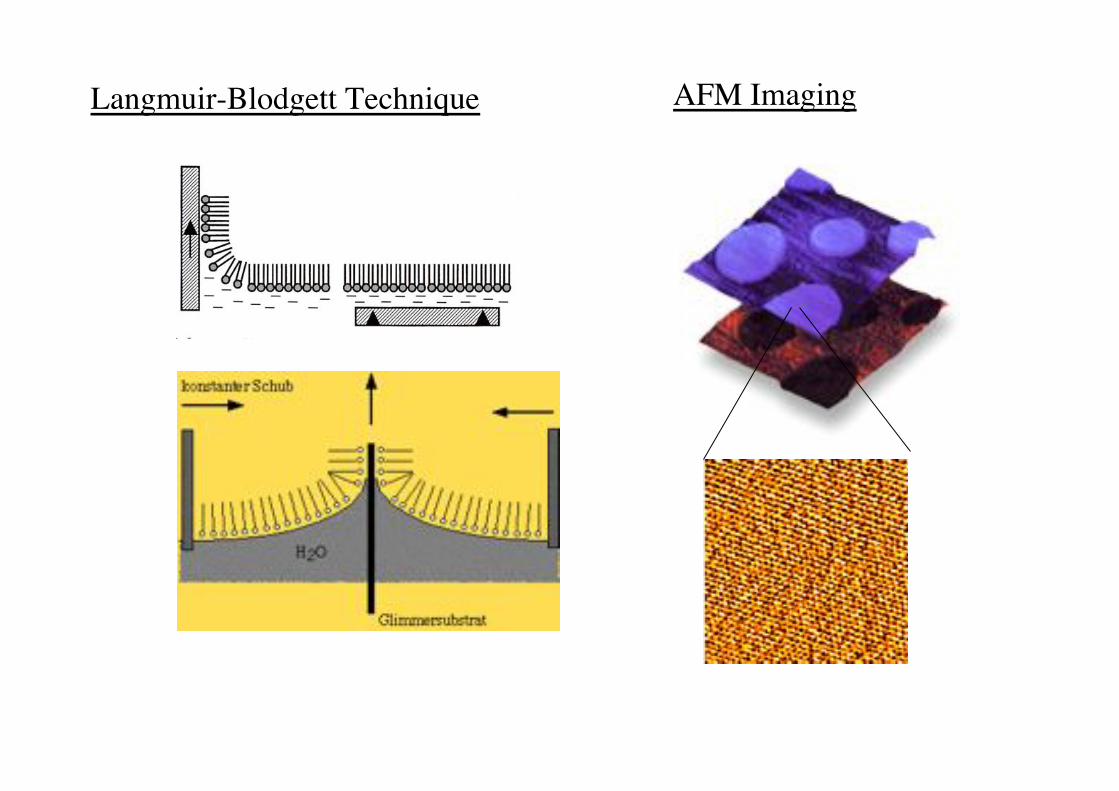
Langmuir-Blodgett Technique! AFM Imaging!
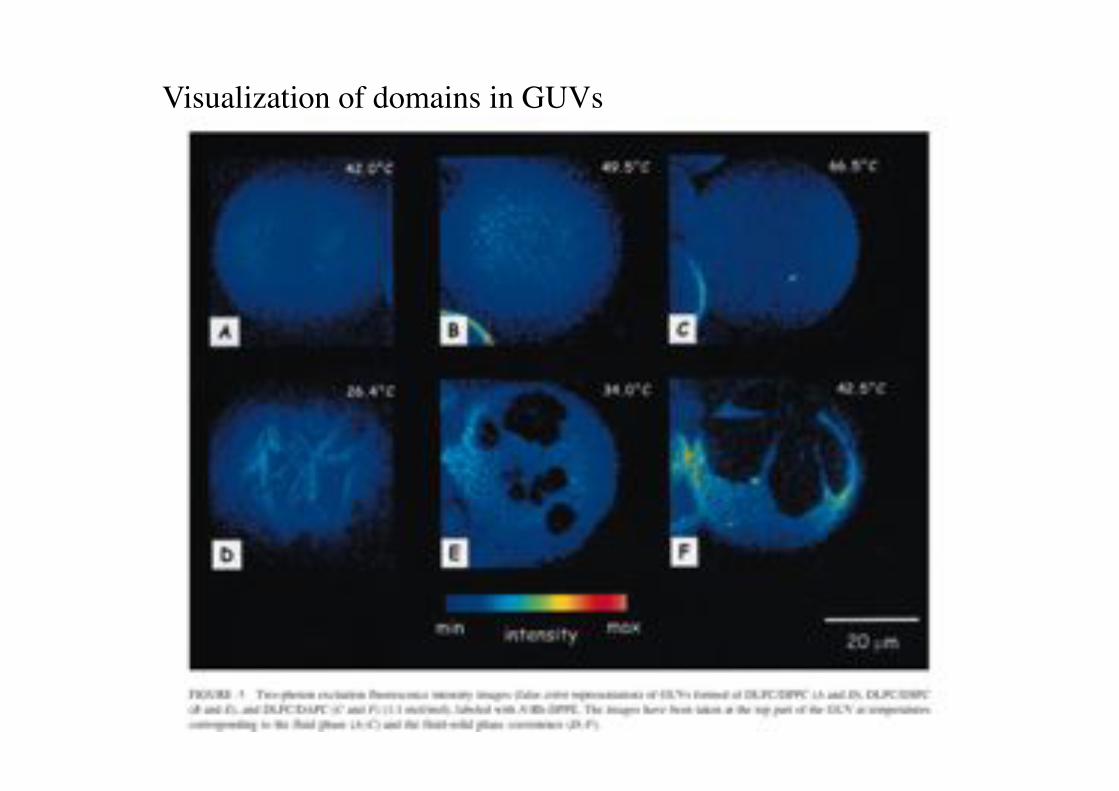
Visualization of domains in GUVs!
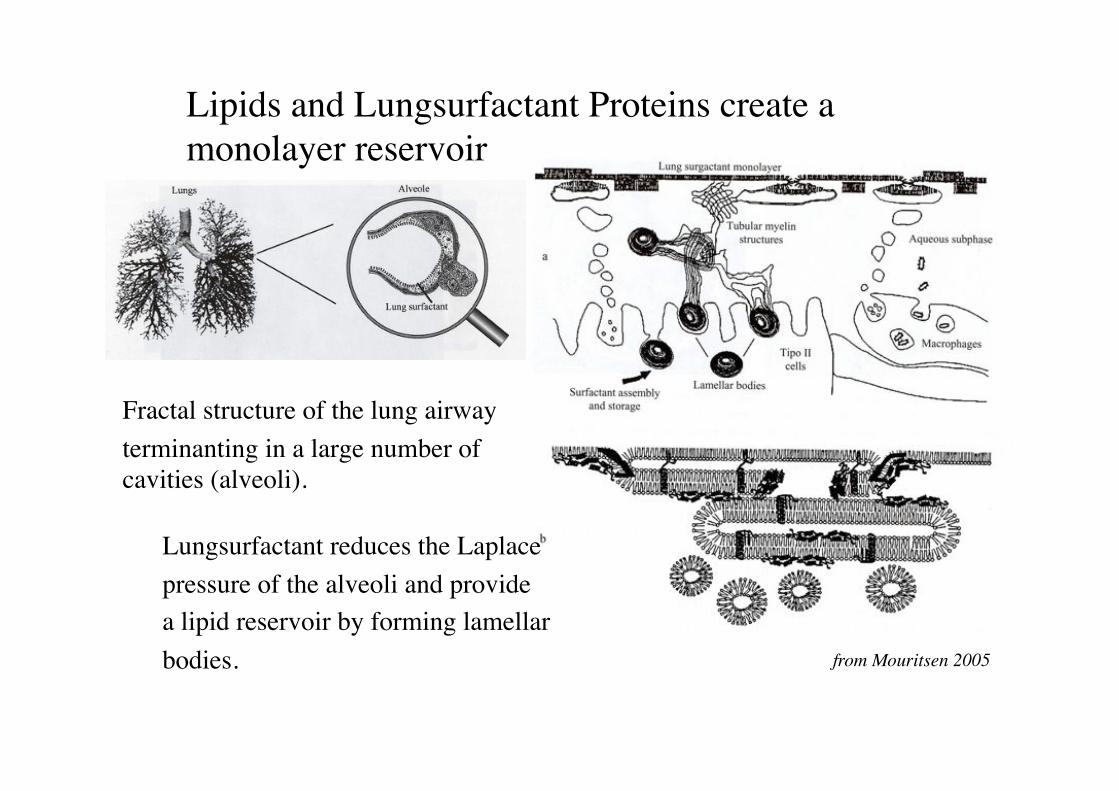
Lipids and Lungsurfactant Proteins create a monolayer reservoir!
Fractal structure of the lung airways.!terminanting in a large number of cavities (alveoli).!
Lungsurfactant reduces the Laplace!pressure of the alveoli and provide!a lipid reservoir by forming lamellar!bodies.! from Mouritsen 2005!





























