Embed Size (px)
Citation preview

1
Ausarbeitung des Vortrags
EKG, EEG und EMG
Im Rahmen des Hauptseminars in Experimentalphysik
Universität Duisburg-Essen
Campus Duisburg
„Physikalische Grundlagen der medizinischen Diagnostik“
im SS 2006
Datum des Vortrags 22.06.2006
Von Nathalie Reckers
Betreuer: Dr. C. Meier

2
Inhaltsverzeichnis 1. Motivation
2. Begriffserklärung
3. Geschichte
3.1. Geschichte des EKG
3.2. Geschichte des EEG
3.3. Geschichte des EMG
4. Theorie
4.1. Membrankanäle für Ionen
4.2. Na-K-Pumpe
4.3. Ruhepotenzial
4.4. Aktionspotenzial
5. EKG
5.1. Theorie EKG
5.2. Anatomie des Herzen
5.3. Schema eines Herzmuskels
5.4. Normales EKG und seine Entstehung
5.5. Art der Ableitungen
5.6. Beispiele
5.7. Anwendungen
6. EEG
6.1. Theorie EEG
6.2. Elektrodenpositionen und ihre Verschaltung
6.3. Probleme bei der Messung
6.4. Verstärker
6.5. Wellenarten und ihre Bedeutungen
6.6. Beispiele
6.7. Anwendungen
7. EMG
7.1. Reizpunkte
7.2. Anwendungen
7.3. Beispiele
Quellenangaben

3
1. Motivation
Im Rahmen dieses Vortrags wird das Thema „EKG, EEG und EMG“ behandelt. Bei diesen
drei Methoden handeln es sich um diagnostische Verfahren, welche es schon sehr lange gibt
und die einen wichtigen Stellenwert in der Medizin haben.
2. Begriffserklärung
Um eine kurze Übersicht zu geben, welche Art der Diagnostik betrachtet wird, soll zunächst
eine Erklärung der einzelnen Begriffe gegeben werden.
EKG, bedeutet Elektrokardiographie. Dies beinhaltet das Messen von Aktionsströmen des
Herzens durch Anlegen von Elektroden an den Körper.
EEG, bedeutet Elektroenzephalographie. Hier werden Aktionsströme des Gehirns gemessen,
indem man Elektroden an der Kopfhaut befestigt.
EMG, bedeutet Elektromyographie. Diese Art der Diagnostik dient zur Feststellung des
elektrischen Aktionspotentials der Muskulatur.
3. Geschichte
3.1. Geschichte des EKG
Das erste EKG wurde 1882 von Waller (Physiologe) durchgeführt. Dabei war sein
Versuchsobjekt sein Hund, von diesem wurden alle vier Pfoten in eine leitfähige
Silberchloridlösung getaucht. Anschließend wurde ein EKG gemessen. Fünf Jahre später,
1887, gelang ihm diese Messung auch an einem Menschen. Jene Messung wurde über eine
Elektrode auf Brust und einer auf dem Rücken bewerkstelligt.
Im Jahre 1895 nahm Willem Einthoven die Versuche Waller wieder auf und wiederholte diese.
Diese Forschung dauerte bis 1901, in dieser Zeit verbesserte er die Versuche Wallers
beständig. 1924 bekam Einthoven für seine Forschung am EKG den Nobelpreis für Medizin
und Physiologie.
Anfangs gestaltete sich die Untersuchung sehr schwierig, da der Aufbau sehr aufwändig war
oder relativ groß. In den 20er Jahren kamen jedoch die ersten fahrbaren EKG-Geräte und in
ab den 30er Jahren bereits die ersten tragbaren Geräte.

4
3.2. Geschichte des EEG
1875 wurde das erste Mal durch Richard Caton beschrieben, dass elektrische Phänomene am
Gehirn festgestellt werden konnten. Bei ihm waren die Versuchsobjekte Affen und Kaninchen.
Es dauerte lange, bis die Untersuchungen am Gehirn weitergeführt wurden.
Abb.1: Erstes EEG von Hans Berger.
Daher musste Berger die Messmethode noch verbessern.
Ein Jahr später (1925) wurde das erste EEG am intakten Schädel aufgenommen.
1926-1929 nahm man die ersten gut ausgebildeten α-Wellen auf. Dies ist ein Wellentyp, dem
ein bestimmter Zustand des Patienten zugeordnet werden kann.
3.3. Geschichte des EMG
Als Grundstein für diese Art der Diagnostik und eigentlich auch der beiden anderen
diagnostischen Verfahren war die Entdeckung der Bioelektrizität durch Galvani Ende des
18.Jahrhunderts. Tiefer in die Materie ging die Entdeckung von durch willkürlichen
Kontraktionen erzeugten Strömen in Muskeln durch DuBois-Reymond im Jahre 1851. 15
Jahre später wurde der Terminus „Aktionspotenzial“ eingeführt, dessen Bedeutung im
weiteren Verlauf geklärt wird. Erst 1925 wurde das Konzept der „motorischen Einheit“ durch
Sherrington und Liddell eingeführt. Als motorische Einheit wird eine Muskeleinheit im
Körper angesehen, z.B. der Bizeps.
Erst 1924 wurde von Hans Berger das erste EEG
am Menschen durchgeführt. Dieses EEG wurde
jedoch an einem Menschen mit
Schädeldeckendefekten durchgeführt. Der Patient
hatte Löcher in der Schädeldecke und das EEG
wurde über Nadelelektroden, die in das Gehirn
gestochen wurden, gemessen.
Dieses erste EEG ist in Abbildung 1 dargestellt..
Man kann hier erkennen, dass die Signale stark
verrauscht sind.

5
Vier Jahre später (1929) konnte man mit Nadelelektroden einzelne motorische Einheiten
registrieren, dies gelang Adrian und Bonk.
4. Theorie
Damit verständlich werden kann, wie es dazu kommt, dass man diese Aktionsströme messen
kann, muss man zurückgehen bis zu den Grundlagen der Zellphysiologie. Speziell muss man
dabei auf den Stoffaustausch einer Zelle mit ihrer Umgebung eingehen. Hierbei ist es wichtig
zu sehen, dass eine Zelle kein statisches Gebilde ist. Die verschiedenen Räume einer Zelle
stehen untereinander im ständigen Stoffaustausch, genauso verhält es sich auch mit der Zelle
nach außen. Alle Strukturen der Zelle stehen in einem dynamischen Gleichgewicht
miteinander, jedoch erst durch Austausch mit ihrer Umgebung und untereinander entsteht ein
funktionierender Organismus. Im Folgenden sollen zwei wichtige Austauschmechanismen
betrachtet werden.
4.1. Membrankanäle für Ionen
Abb.2: Schema eines K+-Kanals (oben) und seines Energieprofils (unten).
Wie man hier erkennen kann, handelt es sich um
eine Membran. Links davon befindet sich das
Zelleninnere und rechts davon das Zellenäußere.
Durch diese Kanäle in der Membran ist es
möglich, dass Ionen transportiert werden. Auch
lassen sich bei der Permeation von Ionen Ströme
für die einzelnen Kanäle messen. Es hat sich
gezeigt, dass diese Kanäle spontan und
hochfrequent zwischen offenem und geschlossenen
Zustand hin- und herschalten. Wie hier in dem
Beispiel für einen K+-Kanal kann man Stromstöße
von etwa 2 pA (10-12A) Amplitude und einer
mittleren Dauer von einigen Millisekunden
messen. Während einer solchen
„Kanalöffnung“ fließen einige 10000 Ionen.

6
In Abbildung 2 sieht man oben den schematischen Aufbau eines K+-Kanals. Dieser enthält
vier negative Festladungen in der Kanalwand, welche hellblau gekennzeichnet sind. Zudem
sieht man ein K+-Ion am inneren Kanaleingang, dieses ist rot dargestellt. Unter der
schematischen Darstellung ist ein Energieprofil des Kanals dargestellt. Hierbei ist die
notwendige kinetische Energie des Ions über den Weg des Ions von der Innenseite zur
Außenseite der Membran aufgetragen. Die Energiemaxima entsprechen
Diffusionshindernissen innerhalb des Kanals. Man nimmt an, dass die Deformationen durch
das Kanalprotein an (1) und (2) spontan oszilliert und das Energieprofil abwechselnd die
ausgezogenen und die gestrichelten Profile annehmen kann. Dies vereinfacht die
Überwindung der Energiebarriere für vor der Barriere gebundene Ionen ungemein.
Dieser Prozess, der hier von innen nach außen dargestellt ist, funktioniert natürlich auch in die
andere Richtung und für andere Ionen.
Dadurch, dass die Ionen durch die Membrankanäle diffundieren können, sollte es zum
Ausgleich der Konzentrationsdifferenz zwischen innen und außen kommen. Es verhält sich
jedoch so, dass die Konzentrationen innen und außen bestehen bleiben. Daher muss es ein
Gleichgewicht zwischen Diffusions- und anderen Transportprozessen über die Membran
geben. Zudem bewegen sich die Ionen mit ihrem natürlichen Konzentrationsgradienten.
4.2. Na-K-Pumpe
Dieses Gleichgewicht wird durch den aktiven Ionentransport gewährleistet. Der wichtigste
Transportprozess ist hierbei die Na-K-Pumpe. Diese wird benötigt, da die intrazelluläre
Konzentration nicht ohne weiteres stabil bleibt. Dies liegt daran, dass das Membranpotential
etwas weniger negativer ist als das Kaliumpotential und erheblich negativer als das
Natriumpotential ist. Durch Diffusion würde sich letztlich die intrazelluläre Konzentration
zumindest für K+ und Na+ sich an die extrazelluläre Konzentration angleichen. Für die
Stabilität des natürlichen Ionengradienten ist die Na-K-Pumpe zuständig. Diese vollführt
einen aktiven Ionentransport. Dabei verhält es sich so, dass Membranproteine Ionen über die
Membran transportieren, dies geschieht entgegengesetzt des Konzentrations- und/ oder dem
elektrischen Gradienten. Hierbei wird Stoffwechselenergie verbraucht.

7
Abb.3: Schematischer Aufbau einer Na-K-Pumpe
In Abbildung 3 sieht man den schematischen Aufbau einer Na-K-Pumpe. Hier ist wieder eine
Membran abgebildet, dabei ist oben das Äußere der Zelle und unten das Innere der Zelle. Der
Na+-Gradient zeigt von außen nach innen, in die entgegengesetzte Richtung weist der K+-
Gradient. Wie man deutlich erkennen kann, wird Na+ aus der Zelle heraustransportiert und K+
hineintransportiert. Damit wird sichergestellt, dass in der Zelle eine niedrige Na+- und eine
hohe K+-Konzentration befindet. Der so erzielte Na-Konzentrationsgradient an der Membran
wird funktionell für die elektrische Informationsleitung genutzt.
Hierbei ist zu erwähnen, dass das Na-K-Transportprotein eine ATPase ist. Bei ATP handelt es
sich um Adenosintriphosphat. Dies ist eine Zusammensetzung aus Adenin, Ribose (Zucker)
und 3 Phosphatresten. Hierbei handelt es sich um einen Energiespeicher.
Das Transportprotein ist also eine ATPase, diese spaltet sich an der Innenseite der
Zellmembran in ADP (Adenosindiphosphat) und ein Phosphat. Mit Hilfe der von einem ATP
übernommenen Energie werden netto drei Na+ aus der Zelle und zwei K+ in die Zelle
transportiert. Dies hat zur Folge, dass bei jedem Pumpzyklus tatsächlich eine Ladung aus der
Zelle entfernt wird. Somit treibt die Na-K-Pumpe also einen elektrischen Strom über die
Membran aus der Zelle, dieser macht das Membranpotential um etwa 10 mV negativer. Das
Protein hat eine hohe Transportgeschwindigkeit, es werden 150 bis 600 Na+ pro Sekunde
umgesetzt.
Als mehrstufige chemische Reaktion ist die Na-K-Pumpe stark temperaturabhängig. Dies
zeigt das nachfolgende Experiment in Abbildung 4. Hier wurde der Na+-Ausstrom einer
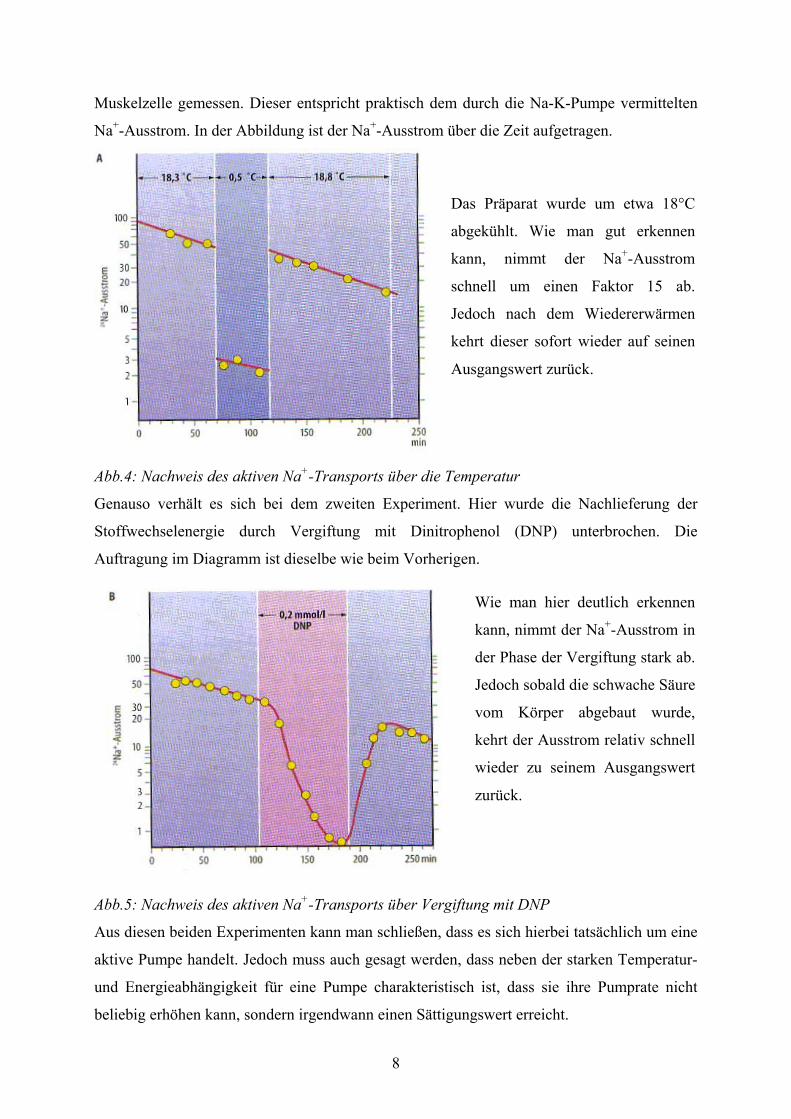
8
Muskelzelle gemessen. Dieser entspricht praktisch dem durch die Na-K-Pumpe vermittelten
Na+-Ausstrom. In der Abbildung ist der Na+-Ausstrom über die Zeit aufgetragen.
Abb.4: Nachweis des aktiven Na+-Transports über die Temperatur
Genauso verhält es sich bei dem zweiten Experiment. Hier wurde die Nachlieferung der
Stoffwechselenergie durch Vergiftung mit Dinitrophenol (DNP) unterbrochen. Die
Auftragung im Diagramm ist dieselbe wie beim Vorherigen.
Abb.5: Nachweis des aktiven Na+-Transports über Vergiftung mit DNP
Aus diesen beiden Experimenten kann man schließen, dass es sich hierbei tatsächlich um eine
aktive Pumpe handelt. Jedoch muss auch gesagt werden, dass neben der starken Temperatur-
und Energieabhängigkeit für eine Pumpe charakteristisch ist, dass sie ihre Pumprate nicht
beliebig erhöhen kann, sondern irgendwann einen Sättigungswert erreicht.
Das Präparat wurde um etwa 18°C
abgekühlt. Wie man gut erkennen
kann, nimmt der Na+-Ausstrom
schnell um einen Faktor 15 ab.
Jedoch nach dem Wiedererwärmen
kehrt dieser sofort wieder auf seinen
Ausgangswert zurück.
Wie man hier deutlich erkennen
kann, nimmt der Na+-Ausstrom in
der Phase der Vergiftung stark ab.
Jedoch sobald die schwache Säure
vom Körper abgebaut wurde,
kehrt der Ausstrom relativ schnell
wieder zu seinem Ausgangswert
zurück.

9
Das Membranpotenzial hängt mit der Elektrolytkonzentration zusammen. Dies wird mit der
Nernst-Gleichung beschrieben. Diese drückt das Gleichgewichtspotenzial EIon aus, bei dem
der Nettostrom für dieses Ion verschwindet. Die Formel lautet:
R: Gaskonstante [Ion]a: Konzentration des Ions extrazellulär
T: absolute Temperatur [Ion]i: Konzentration des Ions intrazellulär
z: Wertigkeit des Ions
F: Faraday-Konstante
Zudem wurde es einmal bei Körpertemperatur (T=320K) und für das Kalium-Ion
ausgerechnet:
4.3. Ruhepotenzial
Jede Zelle weist ein Ruhepotenzial auf. Dies ist gleichbedeutetend mit dem
Membranpotenzial , welches die Zelle ohne Erregung zeigt. Für eine Membran liegen die
Werte für das Ruhepotenzial zwischen -55 mV und -100 mV. Nur bei glatten Muskelzellen ist
es größer und liegt bei bis zu -30 mV. Das Ruhepotenzial ist hauptsächlich durch das
Gleichgewichtspotenzial des Kalium-Ions bestimmt. Dies liegt daran, dass die
Membrankanäle vorwiegend für das Kalium-Ion offen sind. Das Ruhepotenzial kann
gemessen werden. Dafür gibt es zwei verschiedenen Messmethoden.
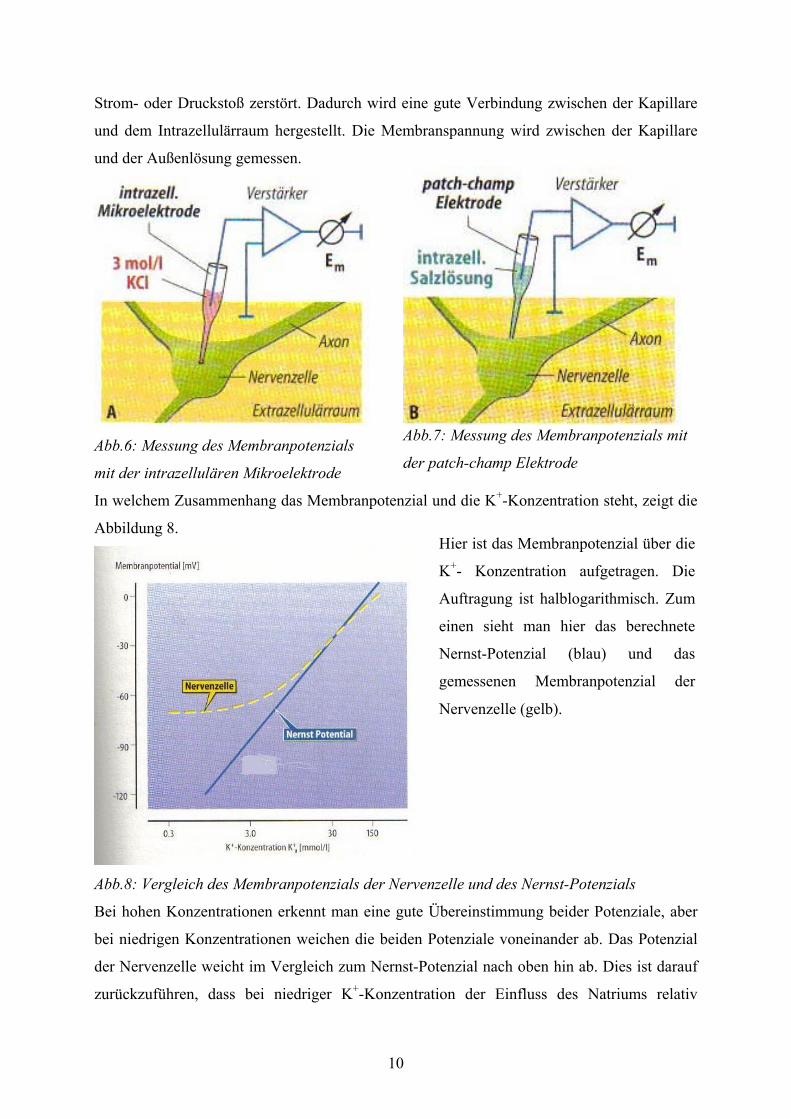
Eine Messmethode verwendet eine intrazellulären Mikroelektrode (Abb.6). Hierbei wird die
Mikroelektrode durch die Membran in die Zelle gestochen. Der potenzialfreie Kontakt ist nur
möglich, da die Elektrode mit 3 mol/l KCl gefüllt ist. Das Membranpotenzial wird nun
zwischen Elektrode und dem Extrazellulärraum gemessen.
Bei der anderen Messmethode wird eine sogenannte patch-champ Elektrode (Abb.7)
verwendet. Die Spitze der Elektrode hat einen Durchmesser von 1 µm. Die Spitze wird auf die
Zellmembran aufgesetzt. Dann bildet sich eine Dichtung aus zwischen Glasrand und der
Membran. Anschließend wird die innerhalb der Kapillare liegende Membran durch einen
[ ][ ]i
aIon Ion
IonFzTRE ln⋅⋅
=
[ ][ ]a
iK K
KE +
+
⋅−= logmV61
10
Strom- oder Druckstoß zerstört. Dadurch wird eine gute Verbindung zwischen der Kapillare
und dem Intrazellulärraum hergestellt. Die Membranspannung wird zwischen der Kapillare
und der Außenlösung gemessen.
Abb.6: Messung des Membranpotenzials
mit der intrazellulären Mikroelektrode
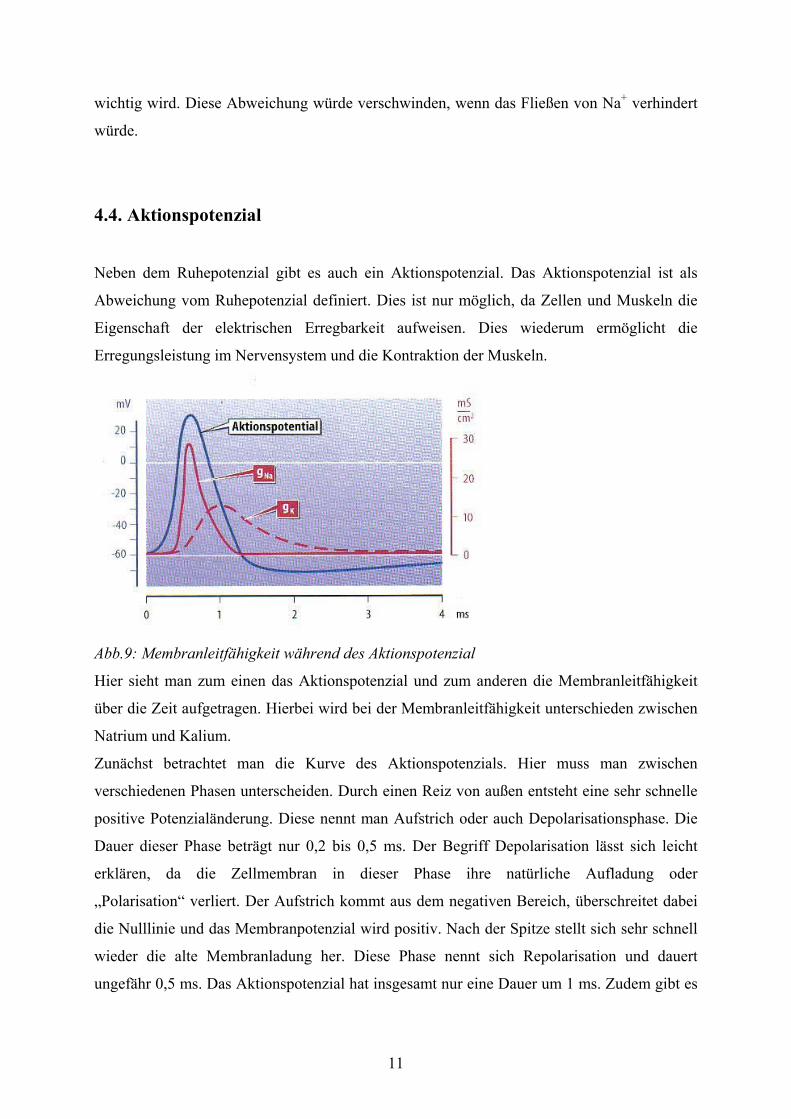
In welchem Zusammenhang das Membranpotenzial und die K+-Konzentration steht, zeigt die
Abbildung 8.
Abb.8: Vergleich des Membranpotenzials der Nervenzelle und des Nernst-Potenzials
Bei hohen Konzentrationen erkennt man eine gute Übereinstimmung beider Potenziale, aber
bei niedrigen Konzentrationen weichen die beiden Potenziale voneinander ab. Das Potenzial
der Nervenzelle weicht im Vergleich zum Nernst-Potenzial nach oben hin ab. Dies ist darauf
zurückzuführen, dass bei niedriger K+-Konzentration der Einfluss des Natriums relativ
Abb.7: Messung des Membranpotenzials mit
der patch-champ Elektrode
Hier ist das Membranpotenzial über die
K+- Konzentration aufgetragen. Die
Auftragung ist halblogarithmisch. Zum
einen sieht man hier das berechnete
Nernst-Potenzial (blau) und das
gemessenen Membranpotenzial der
Nervenzelle (gelb).
11
wichtig wird. Diese Abweichung würde verschwinden, wenn das Fließen von Na+ verhindert
würde.
4.4. Aktionspotenzial
Neben dem Ruhepotenzial gibt es auch ein Aktionspotenzial. Das Aktionspotenzial ist als
Abweichung vom Ruhepotenzial definiert. Dies ist nur möglich, da Zellen und Muskeln die
Eigenschaft der elektrischen Erregbarkeit aufweisen. Dies wiederum ermöglicht die
Erregungsleistung im Nervensystem und die Kontraktion der Muskeln.
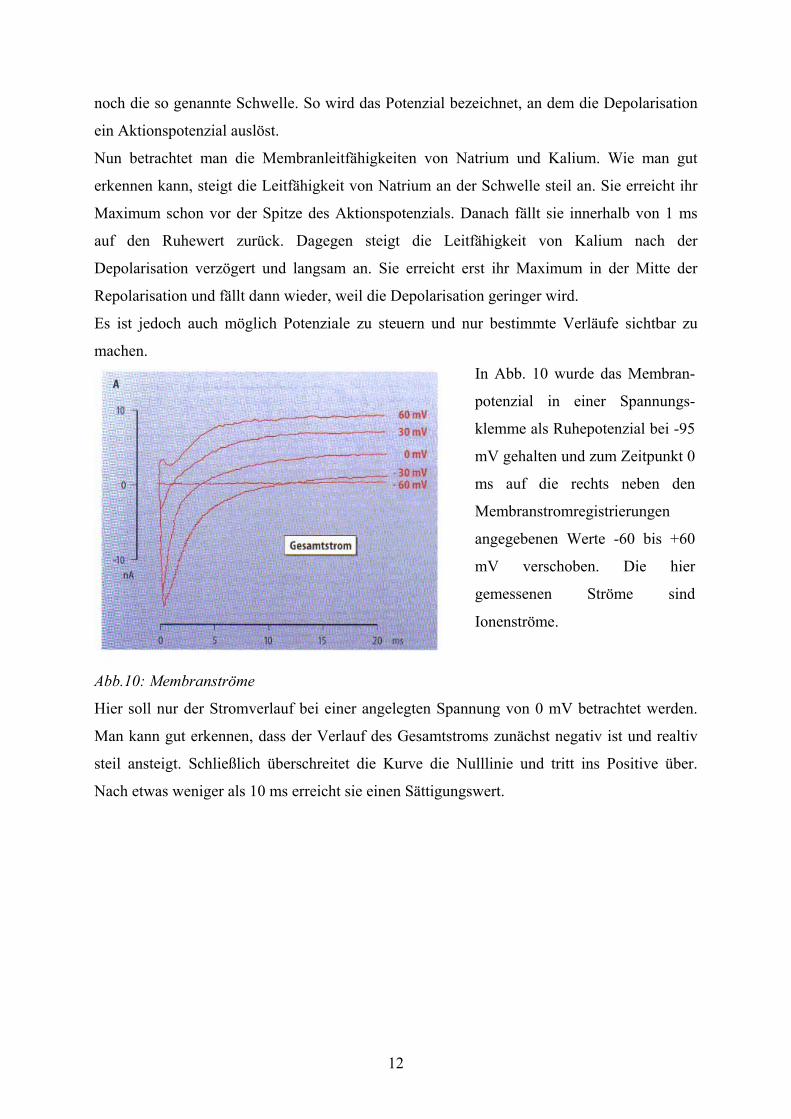
Abb.9: Membranleitfähigkeit während des Aktionspotenzial
Hier sieht man zum einen das Aktionspotenzial und zum anderen die Membranleitfähigkeit
über die Zeit aufgetragen. Hierbei wird bei der Membranleitfähigkeit unterschieden zwischen
Natrium und Kalium.
Zunächst betrachtet man die Kurve des Aktionspotenzials. Hier muss man zwischen
verschiedenen Phasen unterscheiden. Durch einen Reiz von außen entsteht eine sehr schnelle
positive Potenzialänderung. Diese nennt man Aufstrich oder auch Depolarisationsphase. Die
Dauer dieser Phase beträgt nur 0,2 bis 0,5 ms. Der Begriff Depolarisation lässt sich leicht
erklären, da die Zellmembran in dieser Phase ihre natürliche Aufladung oder
„Polarisation“ verliert. Der Aufstrich kommt aus dem negativen Bereich, überschreitet dabei
die Nulllinie und das Membranpotenzial wird positiv. Nach der Spitze stellt sich sehr schnell
wieder die alte Membranladung her. Diese Phase nennt sich Repolarisation und dauert
ungefähr 0,5 ms. Das Aktionspotenzial hat insgesamt nur eine Dauer um 1 ms. Zudem gibt es
12
noch die so genannte Schwelle. So wird das Potenzial bezeichnet, an dem die Depolarisation
ein Aktionspotenzial auslöst.
Nun betrachtet man die Membranleitfähigkeiten von Natrium und Kalium. Wie man gut
erkennen kann, steigt die Leitfähigkeit von Natrium an der Schwelle steil an. Sie erreicht ihr
Maximum schon vor der Spitze des Aktionspotenzials. Danach fällt sie innerhalb von 1 ms
auf den Ruhewert zurück. Dagegen steigt die Leitfähigkeit von Kalium nach der
Depolarisation verzögert und langsam an. Sie erreicht erst ihr Maximum in der Mitte der
Repolarisation und fällt dann wieder, weil die Depolarisation geringer wird.
Es ist jedoch auch möglich Potenziale zu steuern und nur bestimmte Verläufe sichtbar zu
machen.
Abb.10: Membranströme
Hier soll nur der Stromverlauf bei einer angelegten Spannung von 0 mV betrachtet werden.
Man kann gut erkennen, dass der Verlauf des Gesamtstroms zunächst negativ ist und realtiv
steil ansteigt. Schließlich überschreitet die Kurve die Nulllinie und tritt ins Positive über.
Nach etwas weniger als 10 ms erreicht sie einen Sättigungswert.
In Abb. 10 wurde das Membran-
potenzial in einer Spannungs-
klemme als Ruhepotenzial bei -95
mV gehalten und zum Zeitpunkt 0
ms auf die rechts neben den
Membranstromregistrierungen
angegebenen Werte -60 bis +60
mV verschoben. Die hier
gemessenen Ströme sind
Ionenströme.
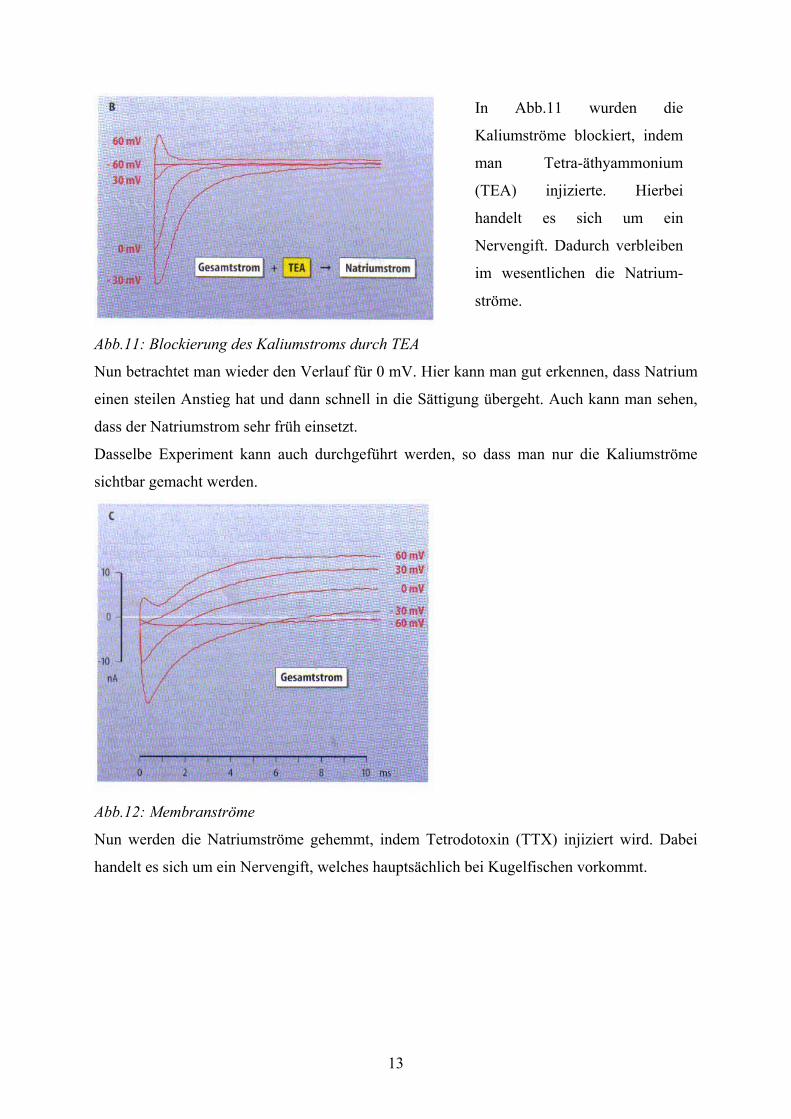
13
Abb.11: Blockierung des Kaliumstroms durch TEA
Nun betrachtet man wieder den Verlauf für 0 mV. Hier kann man gut erkennen, dass Natrium
einen steilen Anstieg hat und dann schnell in die Sättigung übergeht. Auch kann man sehen,
dass der Natriumstrom sehr früh einsetzt.
Dasselbe Experiment kann auch durchgeführt werden, so dass man nur die Kaliumströme
sichtbar gemacht werden.
Abb.12: Membranströme
Nun werden die Natriumströme gehemmt, indem Tetrodotoxin (TTX) injiziert wird. Dabei
handelt es sich um ein Nervengift, welches hauptsächlich bei Kugelfischen vorkommt.
In Abb.11 wurden die
Kaliumströme blockiert, indem
man Tetra-äthyammonium
(TEA) injizierte. Hierbei
handelt es sich um ein
Nervengift. Dadurch verbleiben
im wesentlichen die Natrium-
ströme.
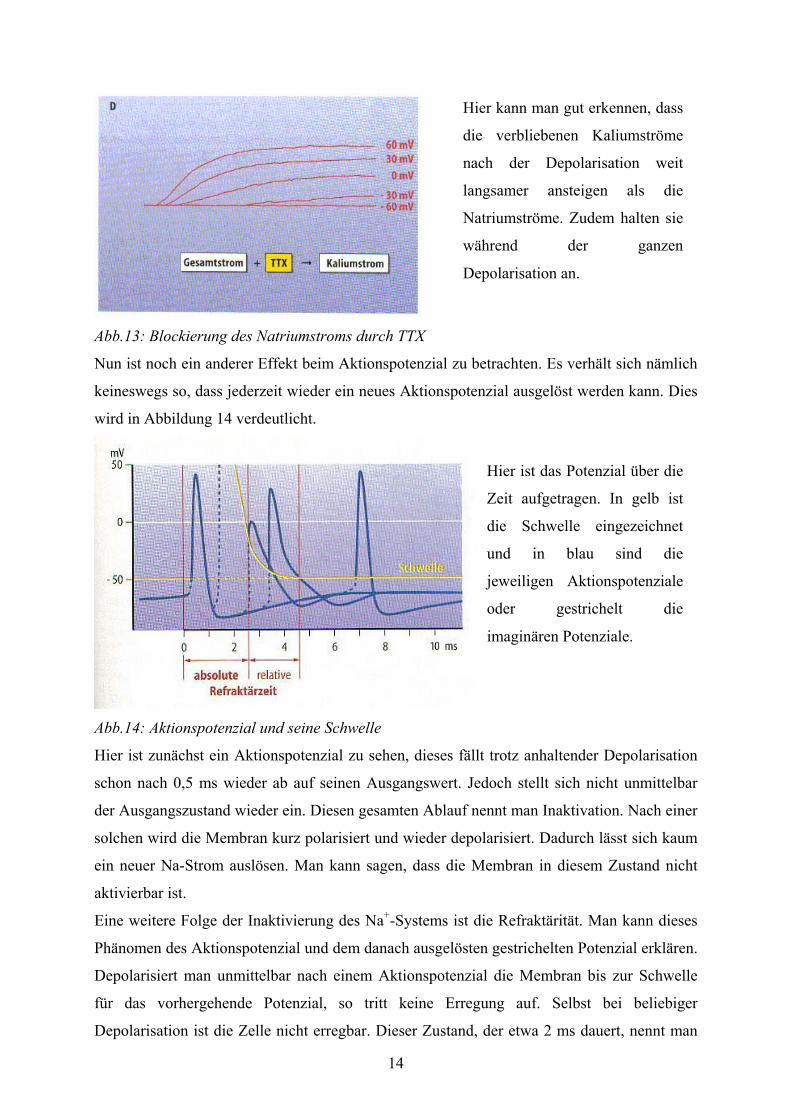
14
Abb.13: Blockierung des Natriumstroms durch TTX
Nun ist noch ein anderer Effekt beim Aktionspotenzial zu betrachten. Es verhält sich nämlich
keineswegs so, dass jederzeit wieder ein neues Aktionspotenzial ausgelöst werden kann. Dies
wird in Abbildung 14 verdeutlicht.
Abb.14: Aktionspotenzial und seine Schwelle
Hier ist zunächst ein Aktionspotenzial zu sehen, dieses fällt trotz anhaltender Depolarisation
schon nach 0,5 ms wieder ab auf seinen Ausgangswert. Jedoch stellt sich nicht unmittelbar
der Ausgangszustand wieder ein. Diesen gesamten Ablauf nennt man Inaktivation. Nach einer
solchen wird die Membran kurz polarisiert und wieder depolarisiert. Dadurch lässt sich kaum
ein neuer Na-Strom auslösen. Man kann sagen, dass die Membran in diesem Zustand nicht
aktivierbar ist.
Eine weitere Folge der Inaktivierung des Na+-Systems ist die Refraktärität. Man kann dieses
Phänomen des Aktionspotenzial und dem danach ausgelösten gestrichelten Potenzial erklären.
Depolarisiert man unmittelbar nach einem Aktionspotenzial die Membran bis zur Schwelle
für das vorhergehende Potenzial, so tritt keine Erregung auf. Selbst bei beliebiger
Depolarisation ist die Zelle nicht erregbar. Dieser Zustand, der etwa 2 ms dauert, nennt man
Hier kann man gut erkennen, dass
die verbliebenen Kaliumströme
nach der Depolarisation weit
langsamer ansteigen als die
Natriumströme. Zudem halten sie
während der ganzen
Depolarisation an.
Hier ist das Potenzial über die
Zeit aufgetragen. In gelb ist
die Schwelle eingezeichnet
und in blau sind die
jeweiligen Aktionspotenziale
oder gestrichelt die
imaginären Potenziale.

15
absolute Refraktärphase. Nach dieser Phase können in der relativen Refraktärphase durch
große Polarisationen Aktionspotenziale ausgelöst werden. Jedoch haben diese im Vergleich
zu dem ersten Aktionspotenzial eine erheblich niedrigere Amplitude. Erst nach mehreren ms
kann mit normaler Schwellendepolarisation ein Aktionspotenzial mit normaler Amplitude
ausgelöst werden. Damit ist die relative Refraktärphase abgeschlossen.
5. EKG
5.1. Theorie EKG Zunächst wird auch hier eine möglichst kleine Dimension betrachtet. In diesem Fall wird eine
Myokardfaser (Herzmuskelfaser) im Querschnitt einer näheren Betrachtung unterzogen.
Abb.15: Ionenpumpe
Abb.16: Faser vor Erregung und ihr Membranpotenzial
Hier sieht man die Faser vor der Erregung. Daher hat die Zelle ein Ruhepotenzial von ca. -80
mV. Dies ist im rechten Teil der Abbildung 16 gezeigt.. Hier ist auch angegeben wo das
Nullpotenzial und das Schwellenpotenzial liegt. Die Ladungsangaben innen und außen an der
Zelle beziehen sich nicht auf die tatsächliche Ladung, sondern auf die Potenziale der
Ionenkonzentrationen. Da innen in der Zelle eine höhere Kaliumkonzentration ist und dieses
ein negatives Potenzial besitzt, wirkt das Zelleninnere negativ nach außen. Dagegen ist die
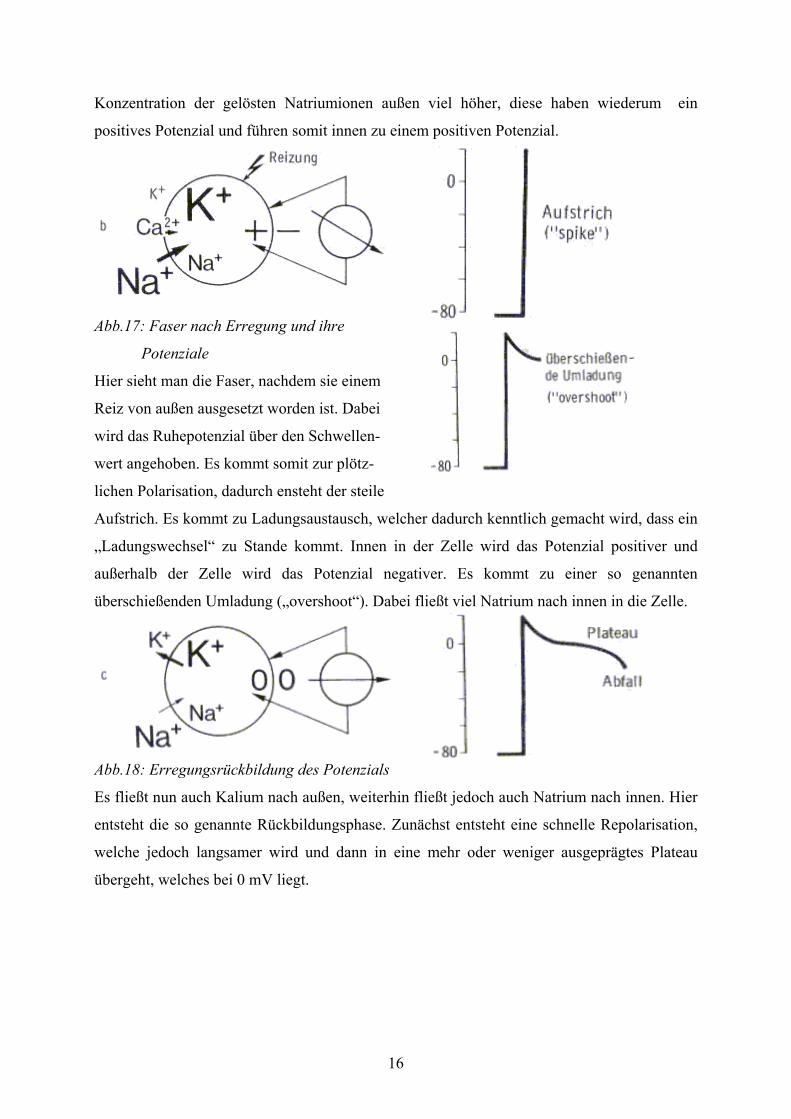
16
Konzentration der gelösten Natriumionen außen viel höher, diese haben wiederum ein
positives Potenzial und führen somit innen zu einem positiven Potenzial.
Abb.17: Faser nach Erregung und ihre
Potenziale
Hier sieht man die Faser, nachdem sie einem
Reiz von außen ausgesetzt worden ist. Dabei
wird das Ruhepotenzial über den Schwellen-
wert angehoben. Es kommt somit zur plötz-
lichen Polarisation, dadurch ensteht der steile
Aufstrich. Es kommt zu Ladungsaustausch, welcher dadurch kenntlich gemacht wird, dass ein
„Ladungswechsel“ zu Stande kommt. Innen in der Zelle wird das Potenzial positiver und
außerhalb der Zelle wird das Potenzial negativer. Es kommt zu einer so genannten
überschießenden Umladung („overshoot“). Dabei fließt viel Natrium nach innen in die Zelle.
Abb.18: Erregungsrückbildung des Potenzials
Es fließt nun auch Kalium nach außen, weiterhin fließt jedoch auch Natrium nach innen. Hier
entsteht die so genannte Rückbildungsphase. Zunächst entsteht eine schnelle Repolarisation,
welche jedoch langsamer wird und dann in eine mehr oder weniger ausgeprägtes Plateau
übergeht, welches bei 0 mV liegt.
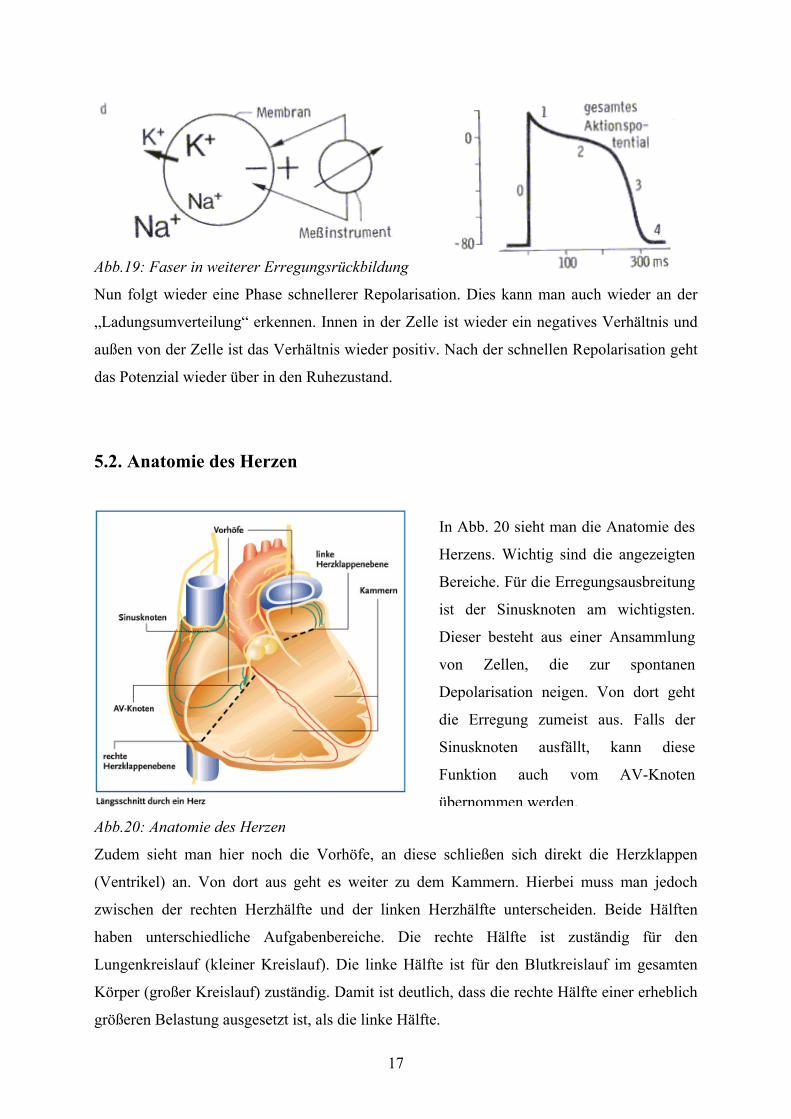
17
Abb.19: Faser in weiterer Erregungsrückbildung
Nun folgt wieder eine Phase schnellerer Repolarisation. Dies kann man auch wieder an der
„Ladungsumverteilung“ erkennen. Innen in der Zelle ist wieder ein negatives Verhältnis und
außen von der Zelle ist das Verhältnis wieder positiv. Nach der schnellen Repolarisation geht
das Potenzial wieder über in den Ruhezustand.
5.2. Anatomie des Herzen
Abb.20: Anatomie des Herzen
Zudem sieht man hier noch die Vorhöfe, an diese schließen sich direkt die Herzklappen
(Ventrikel) an. Von dort aus geht es weiter zu dem Kammern. Hierbei muss man jedoch
zwischen der rechten Herzhälfte und der linken Herzhälfte unterscheiden. Beide Hälften
haben unterschiedliche Aufgabenbereiche. Die rechte Hälfte ist zuständig für den
Lungenkreislauf (kleiner Kreislauf). Die linke Hälfte ist für den Blutkreislauf im gesamten
Körper (großer Kreislauf) zuständig. Damit ist deutlich, dass die rechte Hälfte einer erheblich
größeren Belastung ausgesetzt ist, als die linke Hälfte.
In Abb. 20 sieht man die Anatomie des
Herzens. Wichtig sind die angezeigten
Bereiche. Für die Erregungsausbreitung
ist der Sinusknoten am wichtigsten.
Dieser besteht aus einer Ansammlung
von Zellen, die zur spontanen
Depolarisation neigen. Von dort geht
die Erregung zumeist aus. Falls der
Sinusknoten ausfällt, kann diese
Funktion auch vom AV-Knoten
übernommen werden.

18
5.3. Schema eines Herzmuskels
Zunächst betrachten wir den Erregungszyklus einer einzelnen Herzmuskelfaser. Dafür wird
im folgenden der Muskel nur schematisch dargestellt. Dabei ist zu beachten, dass die
Spannung auf zweierlei Weise abgegriffen wird.
Zum einen transmembranär, dies geschieht zwischen Punkt a und b mittels einer
intrazellulären Mikroelektrode.
Zum anderen extrazellulär zwischen den Punkten a und c, dies geschieht im Außenmedium
längs der Faser.
Abb.21: Herzmuskelfaser vor der Erregung
An der unerregten Faser zeigt sich im transmembrären Abgriff das Ruhepotenzial.
Extrazellulär dagegen wird keine Spannung gemessen. Die Kurve verläuft auf der Nulllinie.
Abb.22: Herzmuskelfaser nach der Erregung
In diesem Beispiel erfolgt die Erregung der Herzmuskelfaser von rechts her. Die
transmembräre Messung ergibt den Aufstrich des Aktionspotenzials. Extrazellulär dagegen
ergibt sich eine Potenzialdifferenz zwischen dem erregten und dem noch unerregten Abschnitt.
Punkt a ist negativ gegenüber Punkt c. Von außen betrachtet wirkt die Faser in stark
vereinfachter Darstellung wie ein elektrischer Dipol.

19
Abb.23: Vollständig erregte Herzmuskelfaser
Die Faser ist vollständig erregt. Dabei kommt es wie schon zuvor erklärt zu dem
„overshoot“ und der anschließenden Repolarisation, welches bei der intrazellulären Messung
auch zu sehen ist. Jedoch extrazellulär dagegen ist keine Potenzialdifferenz mehr nachweisbar
und der Ausschlag kehrt auf die Nulllinie zurück.
Abb.24: Erregungsrückbildung der Hezmuskelfaser
Hier beginnt die Erregungsrückbildung. Nun tritt auch extrazellulär wieder eine
Potenzialdifferenz auf. Diese ist jedoch nun entgegengesetzt gepolt, dies verursacht auch den
Abfall nach unten. Transmembrär fällt die Repolarisation noch steiler ab.
Abb.25: Muskel wieder in Ruhe
Sowohl transmembrär als auch extrazellulär geht das Potenzial wieder in den
„Ruhezustand“ über.
Hier sollte noch erwähnt werden, dass für das EKG nur die extrazelluläre Messung relevant
ist.
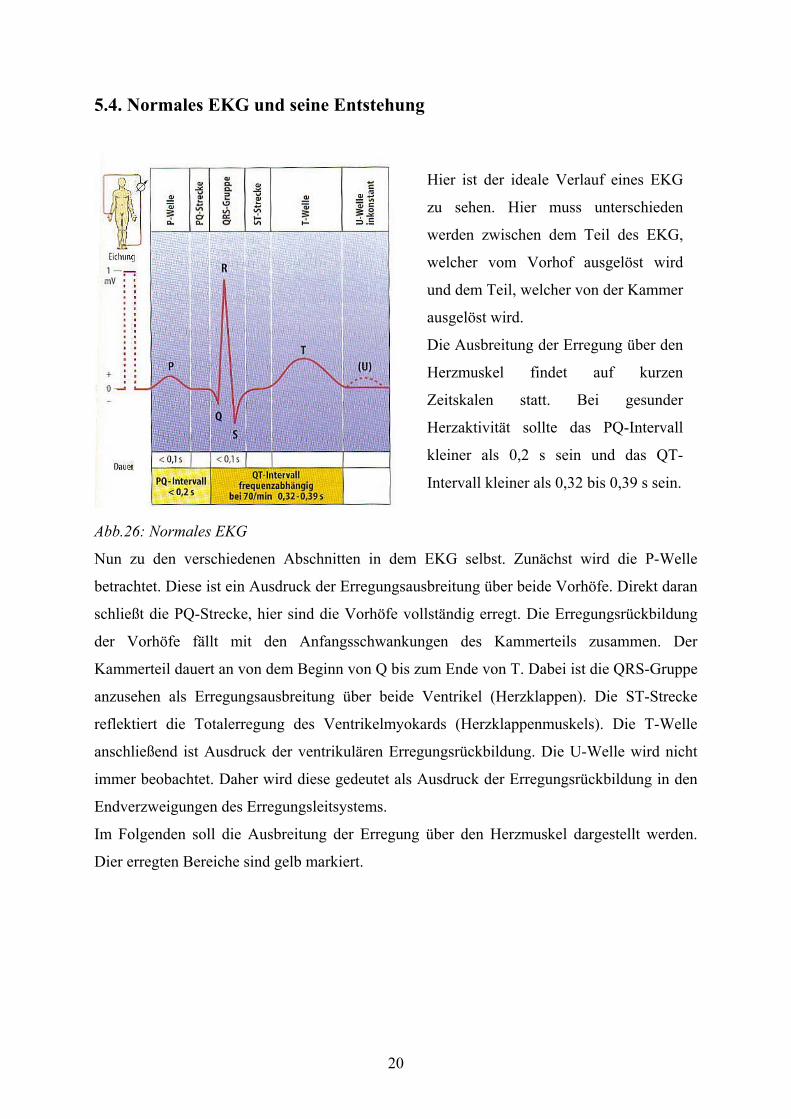
20
5.4. Normales EKG und seine Entstehung
Abb.26: Normales EKG
Nun zu den verschiedenen Abschnitten in dem EKG selbst. Zunächst wird die P-Welle
betrachtet. Diese ist ein Ausdruck der Erregungsausbreitung über beide Vorhöfe. Direkt daran
schließt die PQ-Strecke, hier sind die Vorhöfe vollständig erregt. Die Erregungsrückbildung
der Vorhöfe fällt mit den Anfangsschwankungen des Kammerteils zusammen. Der
Kammerteil dauert an von dem Beginn von Q bis zum Ende von T. Dabei ist die QRS-Gruppe
anzusehen als Erregungsausbreitung über beide Ventrikel (Herzklappen). Die ST-Strecke
reflektiert die Totalerregung des Ventrikelmyokards (Herzklappenmuskels). Die T-Welle
anschließend ist Ausdruck der ventrikulären Erregungsrückbildung. Die U-Welle wird nicht
immer beobachtet. Daher wird diese gedeutet als Ausdruck der Erregungsrückbildung in den
Endverzweigungen des Erregungsleitsystems.
Im Folgenden soll die Ausbreitung der Erregung über den Herzmuskel dargestellt werden.
Dier erregten Bereiche sind gelb markiert.
Hier ist der ideale Verlauf eines EKG
zu sehen. Hier muss unterschieden
werden zwischen dem Teil des EKG,
welcher vom Vorhof ausgelöst wird
und dem Teil, welcher von der Kammer
ausgelöst wird.
Die Ausbreitung der Erregung über den
Herzmuskel findet auf kurzen
Zeitskalen statt. Bei gesunder
Herzaktivität sollte das PQ-Intervall
kleiner als 0,2 s sein und das QT-
Intervall kleiner als 0,32 bis 0,39 s sein.
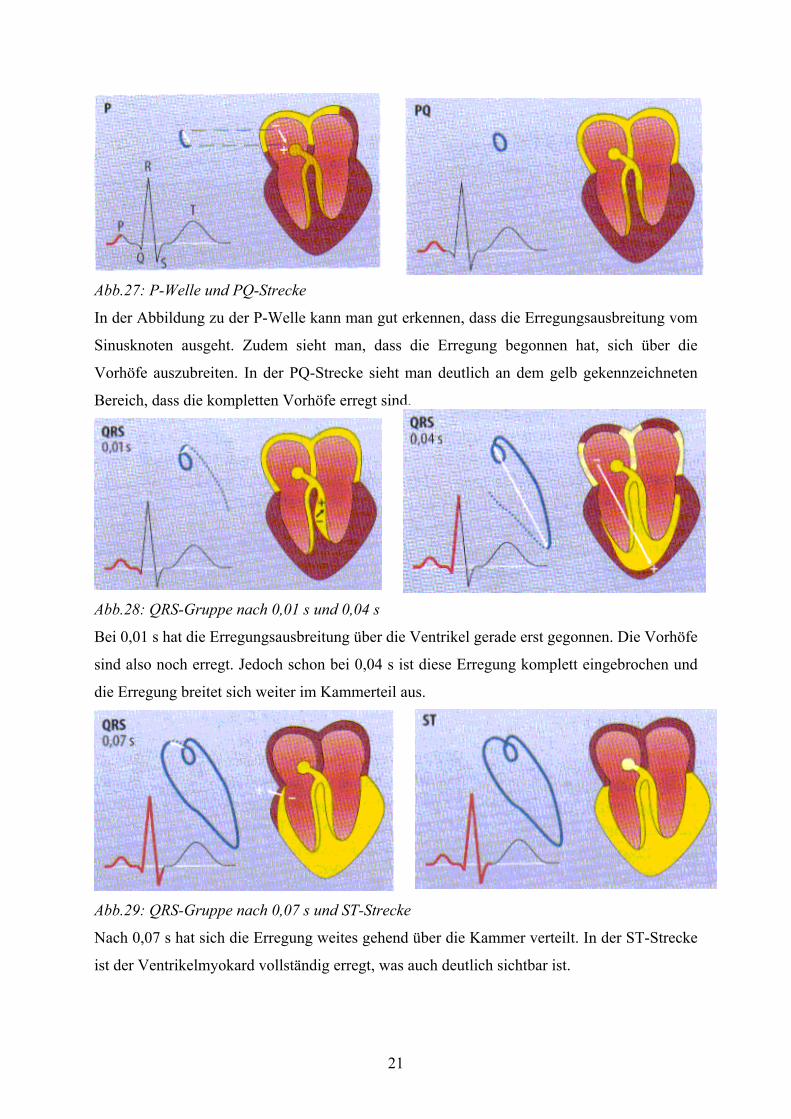
21
Abb.27: P-Welle und PQ-Strecke
In der Abbildung zu der P-Welle kann man gut erkennen, dass die Erregungsausbreitung vom
Sinusknoten ausgeht. Zudem sieht man, dass die Erregung begonnen hat, sich über die
Vorhöfe auszubreiten. In der PQ-Strecke sieht man deutlich an dem gelb gekennzeichneten
Bereich, dass die kompletten Vorhöfe erregt sind.
Abb.28: QRS-Gruppe nach 0,01 s und 0,04 s
Bei 0,01 s hat die Erregungsausbreitung über die Ventrikel gerade erst gegonnen. Die Vorhöfe
sind also noch erregt. Jedoch schon bei 0,04 s ist diese Erregung komplett eingebrochen und
die Erregung breitet sich weiter im Kammerteil aus.
Abb.29: QRS-Gruppe nach 0,07 s und ST-Strecke
Nach 0,07 s hat sich die Erregung weites gehend über die Kammer verteilt. In der ST-Strecke
ist der Ventrikelmyokard vollständig erregt, was auch deutlich sichtbar ist.
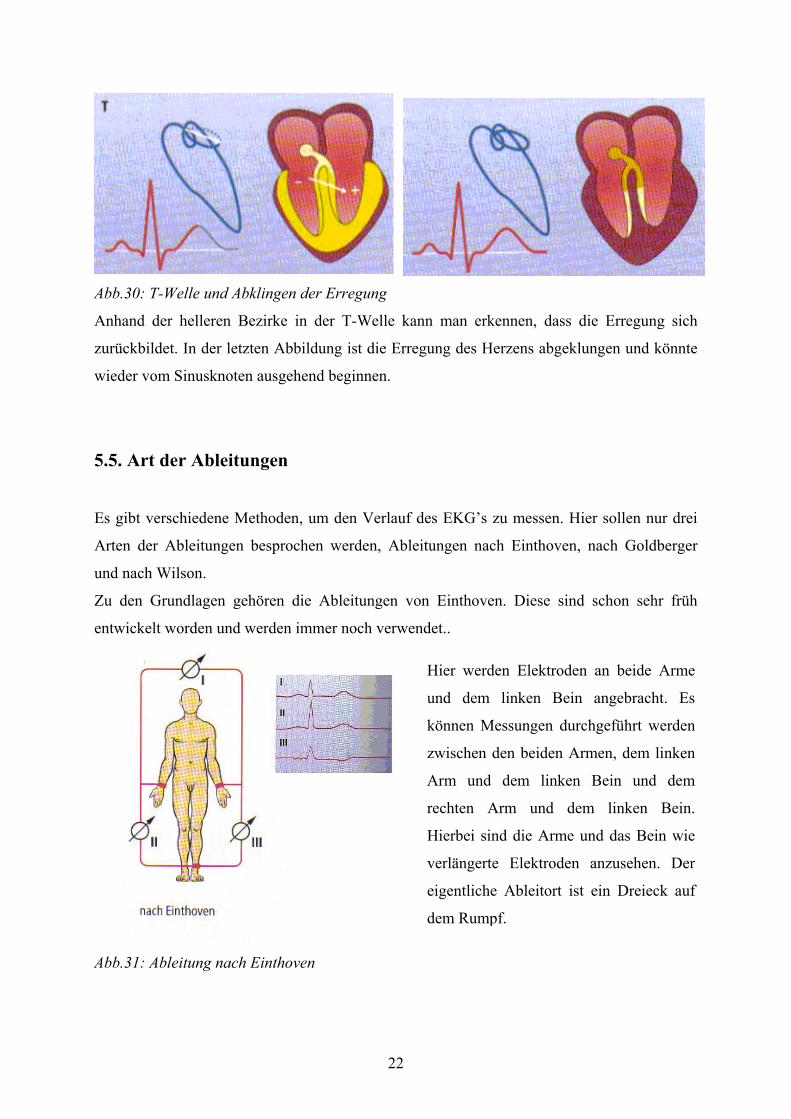
22
Abb.30: T-Welle und Abklingen der Erregung
Anhand der helleren Bezirke in der T-Welle kann man erkennen, dass die Erregung sich
zurückbildet. In der letzten Abbildung ist die Erregung des Herzens abgeklungen und könnte
wieder vom Sinusknoten ausgehend beginnen.
5.5. Art der Ableitungen
Es gibt verschiedene Methoden, um den Verlauf des EKG’s zu messen. Hier sollen nur drei
Arten der Ableitungen besprochen werden, Ableitungen nach Einthoven, nach Goldberger
und nach Wilson.
Zu den Grundlagen gehören die Ableitungen von Einthoven. Diese sind schon sehr früh
entwickelt worden und werden immer noch verwendet..
Abb.31: Ableitung nach Einthoven
Hier werden Elektroden an beide Arme
und dem linken Bein angebracht. Es
können Messungen durchgeführt werden
zwischen den beiden Armen, dem linken
Arm und dem linken Bein und dem
rechten Arm und dem linken Bein.
Hierbei sind die Arme und das Bein wie
verlängerte Elektroden anzusehen. Der
eigentliche Ableitort ist ein Dreieck auf
dem Rumpf.
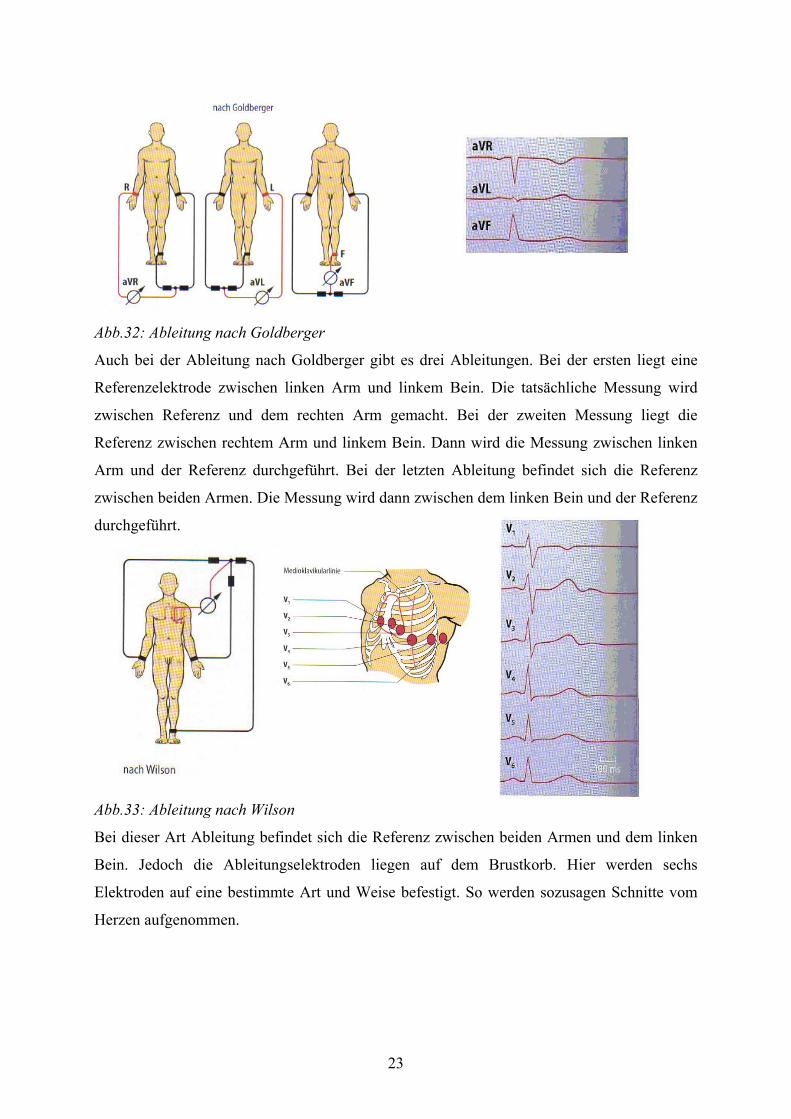
23
Abb.32: Ableitung nach Goldberger
Auch bei der Ableitung nach Goldberger gibt es drei Ableitungen. Bei der ersten liegt eine
Referenzelektrode zwischen linken Arm und linkem Bein. Die tatsächliche Messung wird
zwischen Referenz und dem rechten Arm gemacht. Bei der zweiten Messung liegt die
Referenz zwischen rechtem Arm und linkem Bein. Dann wird die Messung zwischen linken
Arm und der Referenz durchgeführt. Bei der letzten Ableitung befindet sich die Referenz
zwischen beiden Armen. Die Messung wird dann zwischen dem linken Bein und der Referenz
durchgeführt.
Abb.33: Ableitung nach Wilson
Bei dieser Art Ableitung befindet sich die Referenz zwischen beiden Armen und dem linken
Bein. Jedoch die Ableitungselektroden liegen auf dem Brustkorb. Hier werden sechs
Elektroden auf eine bestimmte Art und Weise befestigt. So werden sozusagen Schnitte vom
Herzen aufgenommen.
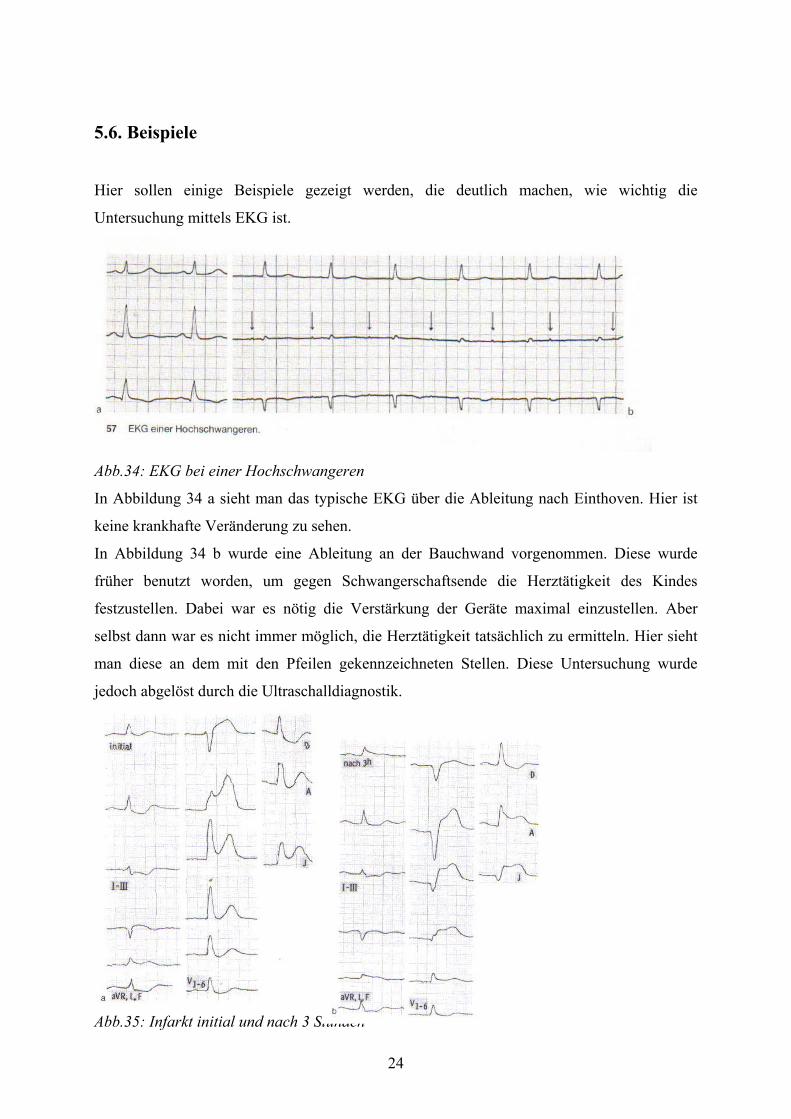
24
5.6. Beispiele
Hier sollen einige Beispiele gezeigt werden, die deutlich machen, wie wichtig die
Untersuchung mittels EKG ist.
Abb.34: EKG bei einer Hochschwangeren
In Abbildung 34 a sieht man das typische EKG über die Ableitung nach Einthoven. Hier ist
keine krankhafte Veränderung zu sehen.
In Abbildung 34 b wurde eine Ableitung an der Bauchwand vorgenommen. Diese wurde
früher benutzt worden, um gegen Schwangerschaftsende die Herztätigkeit des Kindes
festzustellen. Dabei war es nötig die Verstärkung der Geräte maximal einzustellen. Aber
selbst dann war es nicht immer möglich, die Herztätigkeit tatsächlich zu ermitteln. Hier sieht
man diese an dem mit den Pfeilen gekennzeichneten Stellen. Diese Untersuchung wurde
jedoch abgelöst durch die Ultraschalldiagnostik.
Abb.35: Infarkt initial und nach 3 Stunden

25
Abb.36: Infarkt nach 24 Stunden und 48 Stunden
Hier wurden alle drei Ableitungen aufgenommen, die vorgestellt worden sind. Wie man
deutlich erkennen kann, ist bei jeder Ableitung das EKG stark verändert. Dies ist auf den
Infarkt zurückzuführen. Diese EKG’s wurden an einem 57-jährigen Mann durchgeführt.
5.7. Anwendungen
Es gibt verschiedene Arten der Anwendung für das EKG. Die häufigste Methode ist das
Ruhe-EKG. Dies wird meist im Liegen angefertigt. Der große Vorteil bei dieser Art der
Diagnose ist, dass sie auch im Notfall durchführbar ist. Dies ist möglich, da die Untersuchung
nur eine Dauer von einigen Sekunden hat. Das Ruhe-EKG ist somit die schnellste
Diagnosemethode.
Eine weitere Methode ist das Langzeit-EKG. Diese Art der Diagnose wird eingesetzt bei
Beschwerden, die nur ab und zu oder zu bestimmten Anlässen auftreten. Daher ist die Dauer
auch relativ lang, das EKG wird über 24 Stunden aufgenommen und anschließend
ausgewertet.
Eine weitere bekannte Diagnosemethode ist das Belastungs-EKG. Dabei fährt der Patient
Fahrrad, währenddessen wird sein EKG aufgenommen.
Zuletzt soll noch die Methode des intrakardialen EKG vorgestellt werden. Diese Art der
Untersuchung kann während einer Herzkatheteruntersuchung im Krankenhaus durchgeführt

26
werden. Hierbei wird eine Sonde zur Registrierung des EKG’s durch den Katheter bis zum
Herzen geschoben und es wird im Inneren des Herzen ein EKG aufgenommen.
6. EEG
6.1. Theorie EEG
Nun soll auf das Thema Elektroenzephalographie eingegangen werden. Dabei ist es wichtig,
dass man im Gehirn keine Muskeln betrachtet, sondern Synapsen. Synapsen sind
Kontaktstellen zwischen Nervenzellen bzw. Nervenzellen und anderen Zellen (wie Sinnes-,
Muskel- oder Drüsenzellen). An ihnen findet die Erregungsübertragung von einer Zelle auf
die andere statt.
Dabei unterscheidet man hauptsächlich zwischen axosomatischen und axodendritischen
Synapsen. Diese Unterscheidung richtet sich danach, ob der synaptische Kontakt am
Zellkörper (axosomatisch) oder an den Dendriten hergestellt wird (axodentritisch). Zudem
können Synapsen erregend oder hemmend wirken.
Abb.37: Schematische Darstellung der verschiedenen Strukturen eines Neurons
Hier wird zunächst ein Neuron dargestellt. Eine Nervenzelle oder ein Neuron ist eine auf
Erregungsleitung spezialisierte Zelle. Das Neuron steht über Synapsen in Verbindung mit
anderen Nervenzellen. Durch ihre elektrische Erregbarkeit und Leitfähigkeit sind
Nervenzellen in der Lage Nervenimpulse selektiv weiterzuleiten und im Verbund sind sie
27
befähigt, Informationen zu verarbeiten und gegebenenfalls zu speichern. Das menschliche
Gehirn enthält zwischen 30 und 100 Milliarden von Neuronen.
6.2. Elektrodenpositionen und ihre Verschaltung
Nun soll geklärt werden, in welcher Art und Weise diese Potenziale gemessen werden. Dazu
werden die Elektroden in einer bestimmten Weise auf dem Kopf befestigt.
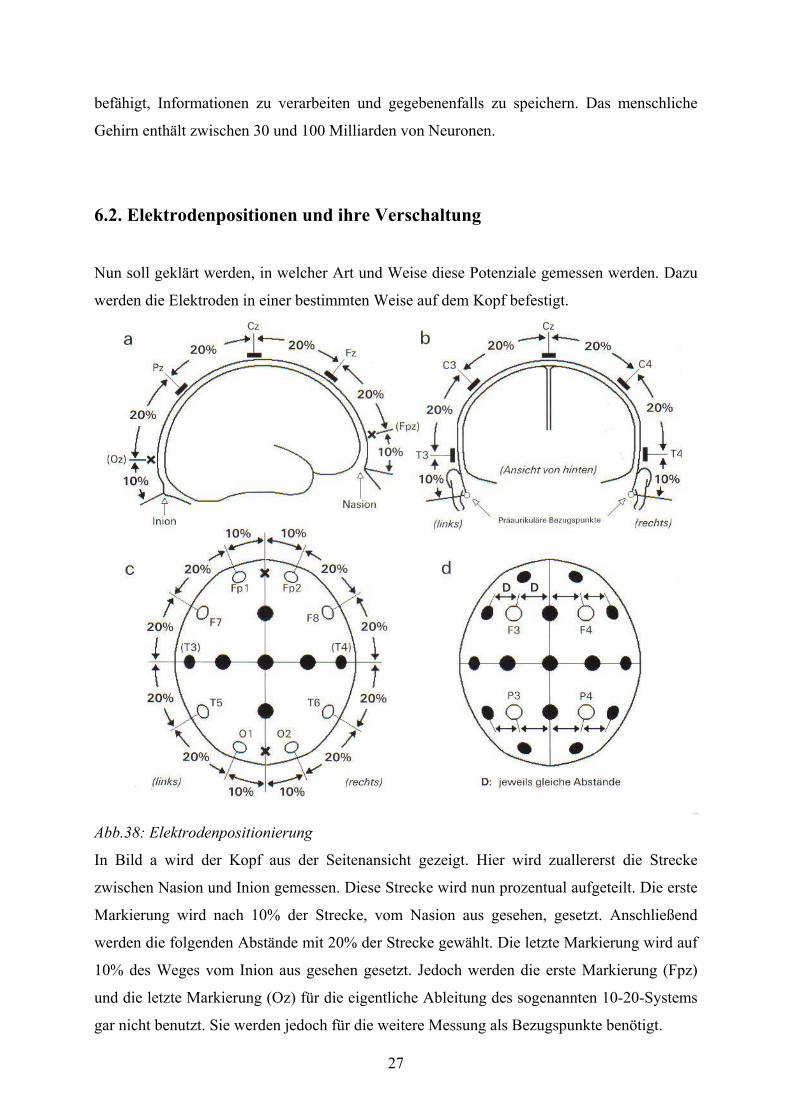
Abb.38: Elektrodenpositionierung
In Bild a wird der Kopf aus der Seitenansicht gezeigt. Hier wird zuallererst die Strecke
zwischen Nasion und Inion gemessen. Diese Strecke wird nun prozentual aufgeteilt. Die erste
Markierung wird nach 10% der Strecke, vom Nasion aus gesehen, gesetzt. Anschließend
werden die folgenden Abstände mit 20% der Strecke gewählt. Die letzte Markierung wird auf
10% des Weges vom Inion aus gesehen gesetzt. Jedoch werden die erste Markierung (Fpz)
und die letzte Markierung (Oz) für die eigentliche Ableitung des sogenannten 10-20-Systems
gar nicht benutzt. Sie werden jedoch für die weitere Messung als Bezugspunkte benötigt.

28
In Bild b wird der Kopf von hinten abgebildet. Auch hier wird die Strecke gemessen, als
Bezugspunkte dienen Punkte an den Ohren. Die Strecke wird ebenfalls in feste prozentuale
Schritte unterteilt. Der erste und letzte Punkt werden nach 10% der Strecke gesetzt und die
anderen werden wieder nach 20% der Strecke fest gemacht. Als Hilfe dient die schon in Bild
a gesetzte Markierung von Cz.
In Bild c sieht man den Kopf von oben. Hier sieht man schon die gesetzten Markierungen. Als
Bezugpunkte dienen hier die Markierungen von Fpz und Oz. Die neuen Markierungen werden
wieder nach einer bestimmten prozentualen Einteilung gesetzt.
In Bild d sieht man wieder den Kopf von oben und die schon gesetzten Markierungen. Die
letzten Markierungen werden gemacht, indem man Die Abstände zwischen zwei
Markierungen gleich groß wählt. Anschließend können die Elektroden gesetzt werden.
Nun zu den verschiedenen Verschaltungsarten der Elektroden. Hier sollen nur drei
verschiedene Arten aufgezeigt werden.
Abb.39: Verschaltungen der Elektroden
In Bild a sieht man bipolare Längsreihen mit alternierender Links-Rechts-Zuordnung der
Schreiberkanäle.
In Bild b sieht man bipolare Längsreihen mit blockförmiger Zusammenfassung der linken und
rechten Seite.
In Bild c sieht man bipolare Querreihen.
6.3. Probleme bei der Messung Es ist mit Schwierigkeiten verbunden, diese meist sehr kleinen Potenzialveränderungen zu
messen. Durch Hornhaut, Epidermis, Fettgewebe sowie andere Zwischenschichten werden die
Signale abgeschwächt. Es ergeben sich aus den verschiedenen Schichten unterschiedliche
Widerstände und Kapazitäten. Aus allen zusammen ergibt sich ein Übergangswiderstand, der
sich aus zwei Arten von Widerständen zusammensetzt. Zum einen dem
Gleichstromwiderstand (ohmscher Widerstand), dieser behindert jedoch lediglich die freie

29
Bewegung der Ionen. Wichtiger hier ist der kapazitive Widerstand, dieser kann an
Gewebsstrukturen (-membranen), aber auch an elektrischen Doppelschichten unterhalb der
Elektrode auftreten. Dieser wirkt wie ein Kondensator, den die elektrischen Ladungsträger,
wie Ionen, nicht überwinden können.
Abb.40: Schematische Darstellung der Schichten und ihre Ersatzschaltbilder
Hierbei symbolisieren die Größe der ohmschen und der kapazitiven Widerstände ihren
Einfluss auf den Übergangswiderstand. Wie man gut erkennen kann,haben die elektrische
Doppelschicht und die Hornhaut den größten Einfluss.
Diese Einflüsse können auf zwei verschiedene Arten minimiert werden. Entweder wird der
Übergangswiderstand erniedrigt, indem die Hornhaut mit einem Elektrolyten durchfeuchtet.
Diese erhöht die Leitfähigkeit und minimiert so den Übergangswiderstand.
Die andere Möglichkeit ist, dass man die obere Hornhautschicht entfernt, indem man diese
abkratzt.
6.4. Verstärker
Bei der Messung müssen noch andere Aspekte berücksichtigt werden. Es werden bei allen
Diagnostikmethoden immer nur Potenziale über Strecken gemessen. Es wird nie ein Potenzial
nur über eine Elektrode gemessen. Dies lässt sich mit der folgenden Abbildung leicht erklären.
30
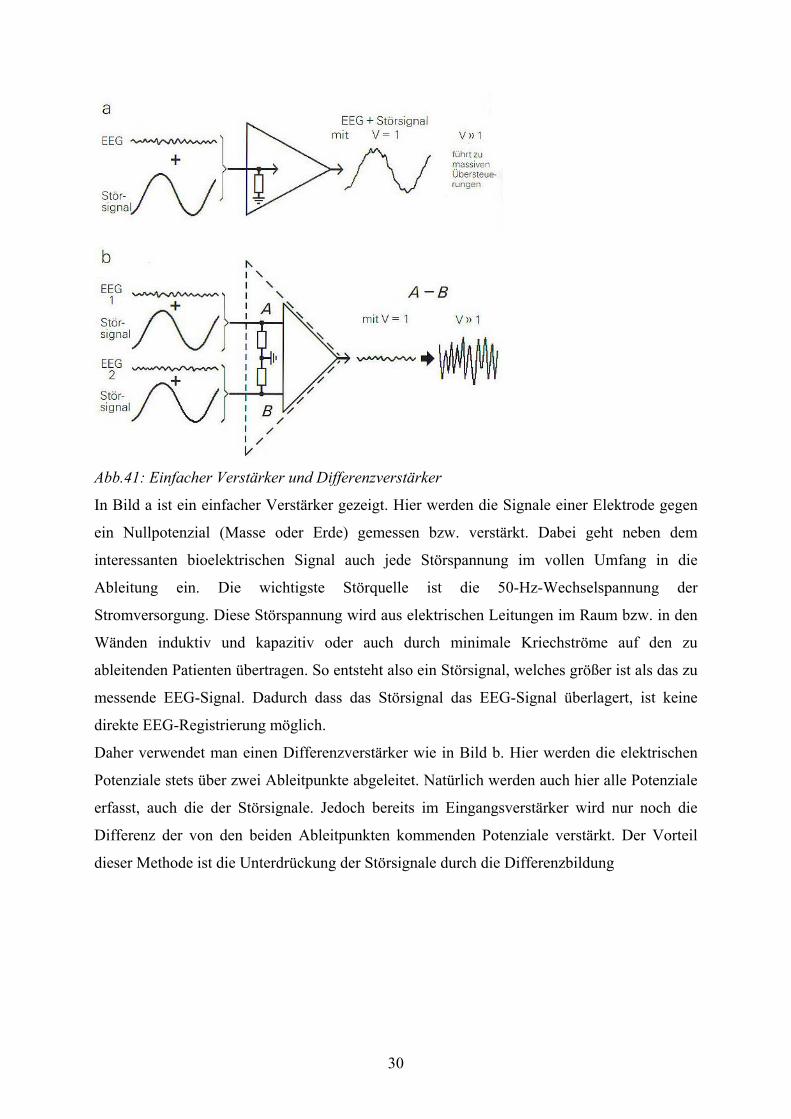
Abb.41: Einfacher Verstärker und Differenzverstärker
In Bild a ist ein einfacher Verstärker gezeigt. Hier werden die Signale einer Elektrode gegen
ein Nullpotenzial (Masse oder Erde) gemessen bzw. verstärkt. Dabei geht neben dem
interessanten bioelektrischen Signal auch jede Störspannung im vollen Umfang in die
Ableitung ein. Die wichtigste Störquelle ist die 50-Hz-Wechselspannung der
Stromversorgung. Diese Störspannung wird aus elektrischen Leitungen im Raum bzw. in den
Wänden induktiv und kapazitiv oder auch durch minimale Kriechströme auf den zu
ableitenden Patienten übertragen. So entsteht also ein Störsignal, welches größer ist als das zu
messende EEG-Signal. Dadurch dass das Störsignal das EEG-Signal überlagert, ist keine
direkte EEG-Registrierung möglich.
Daher verwendet man einen Differenzverstärker wie in Bild b. Hier werden die elektrischen
Potenziale stets über zwei Ableitpunkte abgeleitet. Natürlich werden auch hier alle Potenziale
erfasst, auch die der Störsignale. Jedoch bereits im Eingangsverstärker wird nur noch die
Differenz der von den beiden Ableitpunkten kommenden Potenziale verstärkt. Der Vorteil
dieser Methode ist die Unterdrückung der Störsignale durch die Differenzbildung
31
6.5. Wellenarten und ihre Bedeutungen
Im Folgenden sollen verschiedene EEG-Wellenarten diskutiert werden. Diese lassen
Rückschlüsse auf den Gemütszustand und auf bestimmte Krankheitsbilder zu.
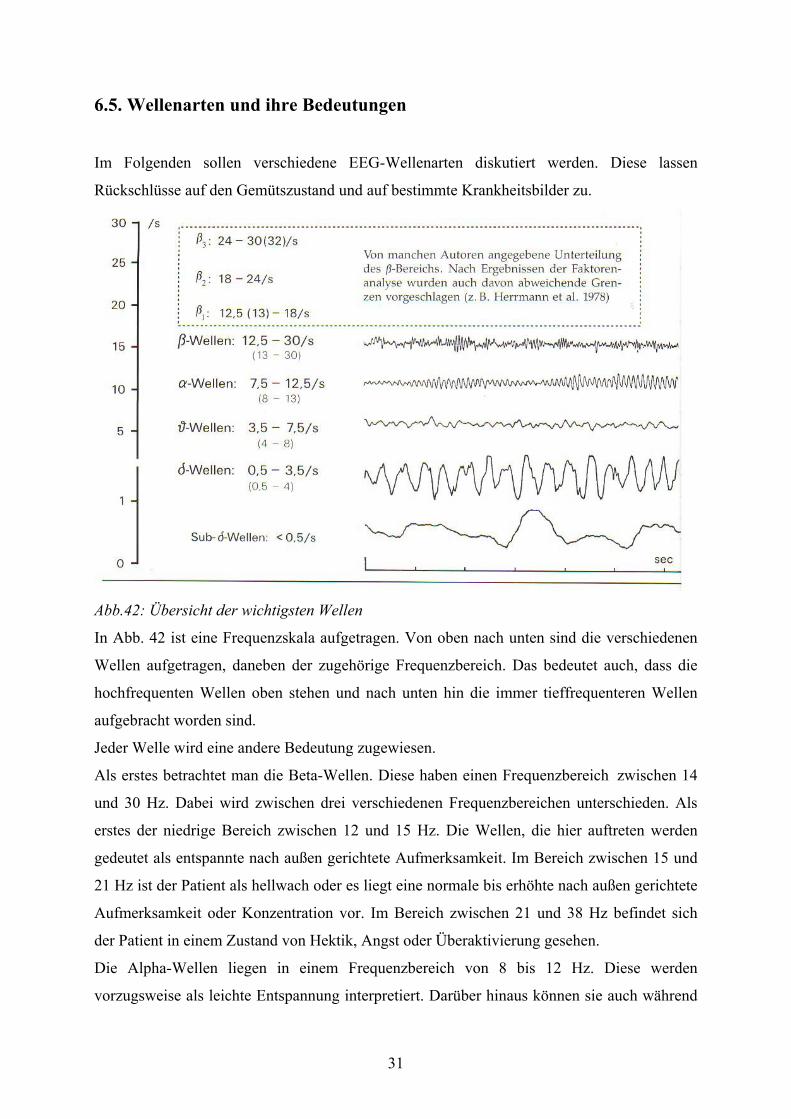
Abb.42: Übersicht der wichtigsten Wellen
In Abb. 42 ist eine Frequenzskala aufgetragen. Von oben nach unten sind die verschiedenen
Wellen aufgetragen, daneben der zugehörige Frequenzbereich. Das bedeutet auch, dass die
hochfrequenten Wellen oben stehen und nach unten hin die immer tieffrequenteren Wellen
aufgebracht worden sind.
Jeder Welle wird eine andere Bedeutung zugewiesen.
Als erstes betrachtet man die Beta-Wellen. Diese haben einen Frequenzbereich zwischen 14
und 30 Hz. Dabei wird zwischen drei verschiedenen Frequenzbereichen unterschieden. Als
erstes der niedrige Bereich zwischen 12 und 15 Hz. Die Wellen, die hier auftreten werden
gedeutet als entspannte nach außen gerichtete Aufmerksamkeit. Im Bereich zwischen 15 und
21 Hz ist der Patient als hellwach oder es liegt eine normale bis erhöhte nach außen gerichtete
Aufmerksamkeit oder Konzentration vor. Im Bereich zwischen 21 und 38 Hz befindet sich
der Patient in einem Zustand von Hektik, Angst oder Überaktivierung gesehen.
Die Alpha-Wellen liegen in einem Frequenzbereich von 8 bis 12 Hz. Diese werden
vorzugsweise als leichte Entspannung interpretiert. Darüber hinaus können sie auch während

32
des Super Learning (unterbewusstes Lernen) oder bei einer nach innen gerichtete
Aufmerksamkeit auftreten.
Die Theta-Wellen liegen in dem Frequenzbereich zwischen 3 und 8 Hz. Auch hier muss man
zwischen hoch- und niederfrequent unterscheiden. Der niedrige Bereich liegt zwischen 3 und
6,5 Hz wird als hypnagogisches Bewusstsein (Einschlafen), Hypnose oder Wachträumen
gedeutet. Der hohe Bereich liegt zwischen 6,5 und 8 Hz wird interpretiert als Entspannung,
Meditation, Hypnose oder Wachträumen.
Die Delta-Welle ist die niederfrequenteste Welle. Der Frequenzbereich liegt zwischen 0,5 und
4 Hz. Diese Art der Welle wird nur im Tiefschlaf oder in der Trance beobachtet.
Darüber hinaus gibt es noch zwei andere Wellen, die bedeutsam sind. Die Gamma-Welle liegt
in einem Frequenzbereich um 40 Hz. Hier erkennt man eine anspruchsvolle Tätigkeit mit
hohem Informationsfluss.
Die so genannten „Sharp-waves“ zeichnen sich durch besonders spitze, steile Verläufe,
welche eng beieinander liegen, aus. Sie sind typisch bei Epilepsie.
Abb.44: Beispiele für Sharp-waves
33
6.6. Beispiele
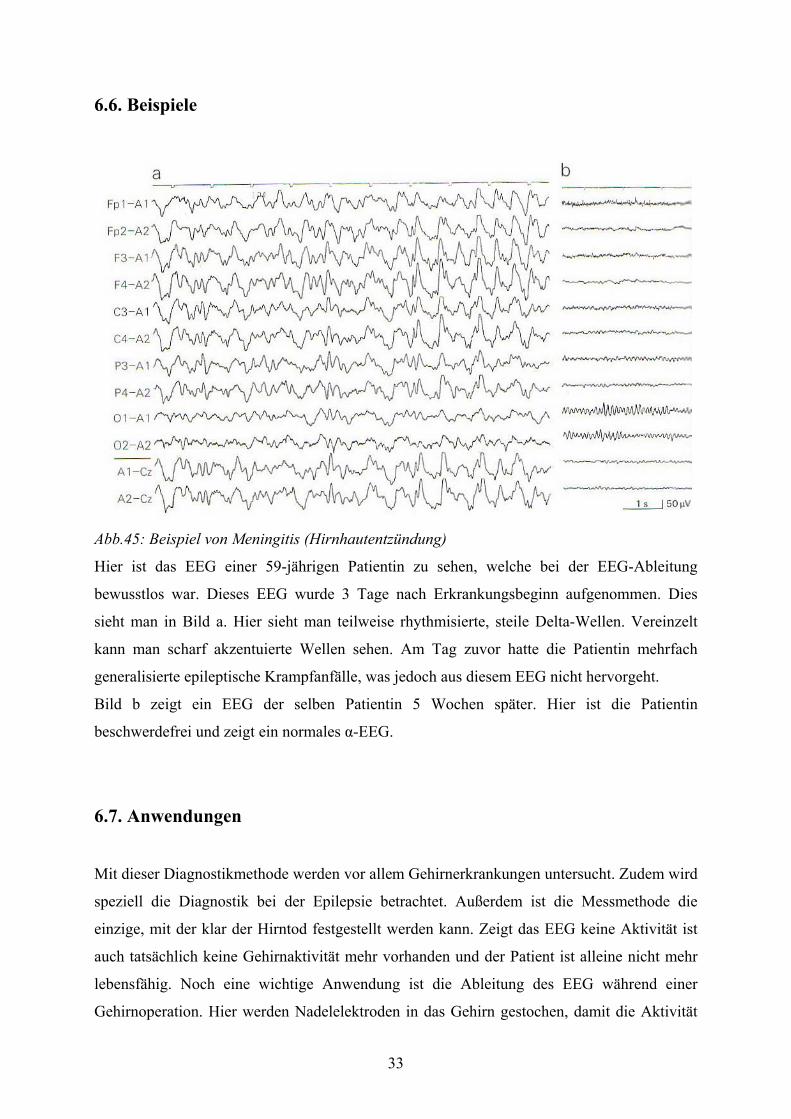
Abb.45: Beispiel von Meningitis (Hirnhautentzündung)
Hier ist das EEG einer 59-jährigen Patientin zu sehen, welche bei der EEG-Ableitung
bewusstlos war. Dieses EEG wurde 3 Tage nach Erkrankungsbeginn aufgenommen. Dies
sieht man in Bild a. Hier sieht man teilweise rhythmisierte, steile Delta-Wellen. Vereinzelt
kann man scharf akzentuierte Wellen sehen. Am Tag zuvor hatte die Patientin mehrfach
generalisierte epileptische Krampfanfälle, was jedoch aus diesem EEG nicht hervorgeht.
Bild b zeigt ein EEG der selben Patientin 5 Wochen später. Hier ist die Patientin
beschwerdefrei und zeigt ein normales α-EEG.
6.7. Anwendungen
Mit dieser Diagnostikmethode werden vor allem Gehirnerkrankungen untersucht. Zudem wird
speziell die Diagnostik bei der Epilepsie betrachtet. Außerdem ist die Messmethode die
einzige, mit der klar der Hirntod festgestellt werden kann. Zeigt das EEG keine Aktivität ist
auch tatsächlich keine Gehirnaktivität mehr vorhanden und der Patient ist alleine nicht mehr
lebensfähig. Noch eine wichtige Anwendung ist die Ableitung des EEG während einer
Gehirnoperation. Hier werden Nadelelektroden in das Gehirn gestochen, damit die Aktivität

34
der jeweiligen Gehirnregion abgeleitet werden kann. Damit soll sicher gestellt werden, dass
keine wichtigen Bereiche zerstört werden.
7. EMG
Es gibt zwei verschiedene Arten der Elektromyographie. Entweder werden ganze motorische
Einheiten untersucht oder es lassen sich einzelne Muskelfasern erfassen.
Zunächst einmal ist jedoch wichtig, was man unter einer motorischen Einheit versteht. Dies
ist die kleinste funktionelle motorische Einheit. Die Kraftentwicklung der einzelnen Muskeln
wird über diese gesteuert. Hier wird noch zwischen kleinen und großen motorischen Einheiten
unterschieden. Kleine Einheiten versorgen etwa 5 bis 15 Muskelfasern (z.B. äußere
Augenmuskulatur) und erlauben eine feine Kraftabstufung. Eine große Einheit versorgt bis zu
1000 Muskelfasern (z.B. Haltemuskel des Rücken) und ist für die grobe Kraftabstufung
zuständig.
Die Potenzialschwankungen diese motorischen Einheiten lassen sich mit konzentrischen
Nadelelektroden erfassen. Diese haben eine Länge zwischen 2 und 6,5 cm und einen
Durchmesser zwischen 0,45 und 0,65 mm.
Abb.46: Konzentrische Nadelelektrode
Zur Registratur des Potenzials einer einzelnen Faser benötigt man spezielle Nadeln. Mit
diesen wird dann die Einzelfasermyographie durchgeführt.
35
7.1. Reizpunkte
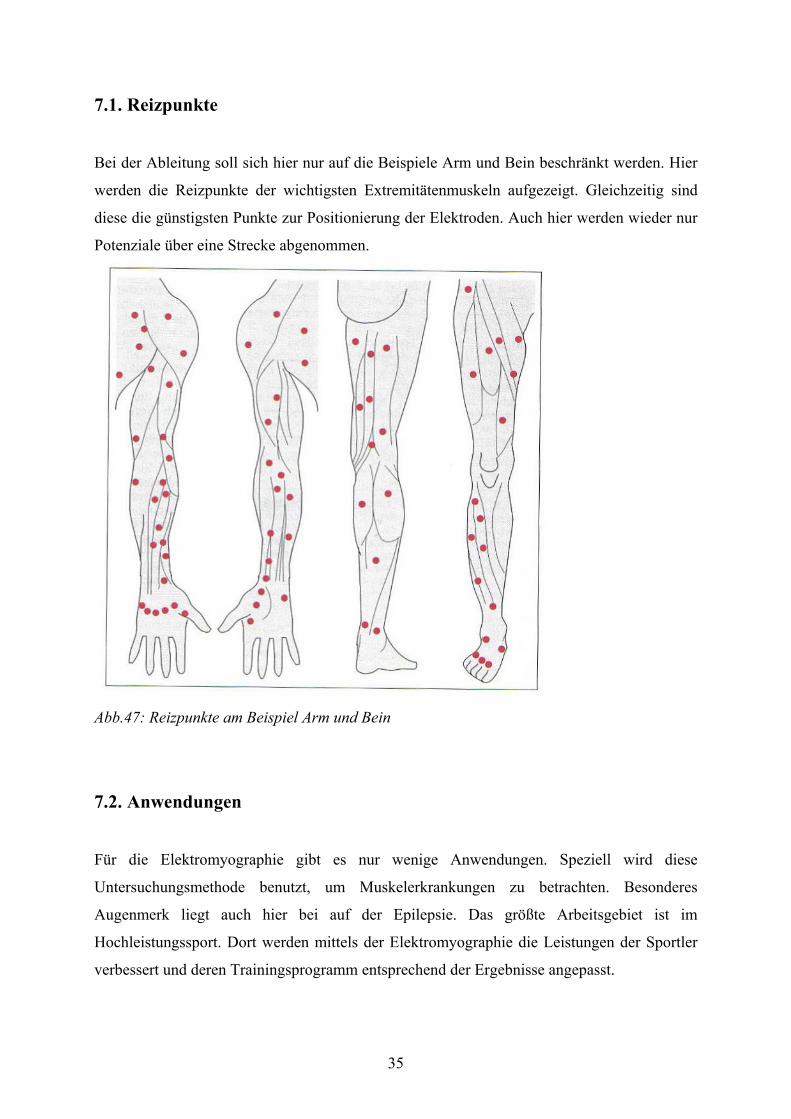
Bei der Ableitung soll sich hier nur auf die Beispiele Arm und Bein beschränkt werden. Hier
werden die Reizpunkte der wichtigsten Extremitätenmuskeln aufgezeigt. Gleichzeitig sind
diese die günstigsten Punkte zur Positionierung der Elektroden. Auch hier werden wieder nur
Potenziale über eine Strecke abgenommen.
Abb.47: Reizpunkte am Beispiel Arm und Bein
7.2. Anwendungen
Für die Elektromyographie gibt es nur wenige Anwendungen. Speziell wird diese
Untersuchungsmethode benutzt, um Muskelerkrankungen zu betrachten. Besonderes
Augenmerk liegt auch hier bei auf der Epilepsie. Das größte Arbeitsgebiet ist im
Hochleistungssport. Dort werden mittels der Elektromyographie die Leistungen der Sportler
verbessert und deren Trainingsprogramm entsprechend der Ergebnisse angepasst.
36
7.3. Beispiel
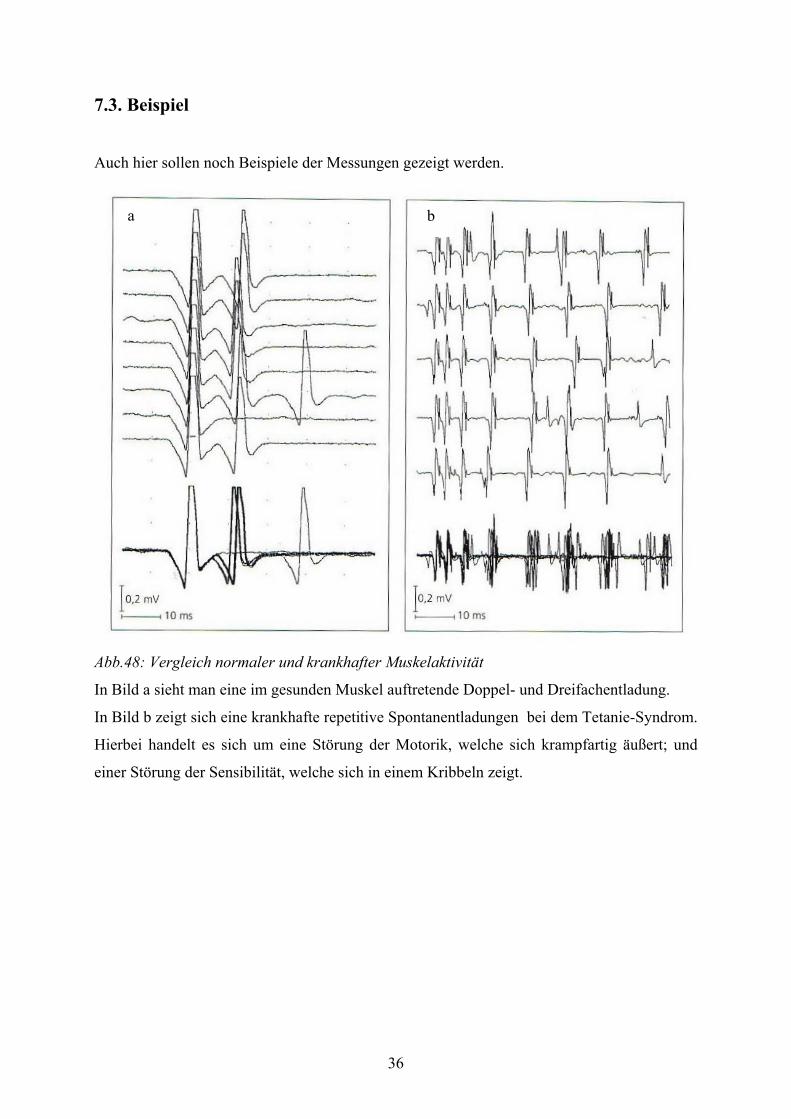
Auch hier sollen noch Beispiele der Messungen gezeigt werden.
Abb.48: Vergleich normaler und krankhafter Muskelaktivität
In Bild a sieht man eine im gesunden Muskel auftretende Doppel- und Dreifachentladung.
In Bild b zeigt sich eine krankhafte repetitive Spontanentladungen bei dem Tetanie-Syndrom.
Hierbei handelt es sich um eine Störung der Motorik, welche sich krampfartig äußert; und
einer Störung der Sensibilität, welche sich in einem Kribbeln zeigt.
a b
37
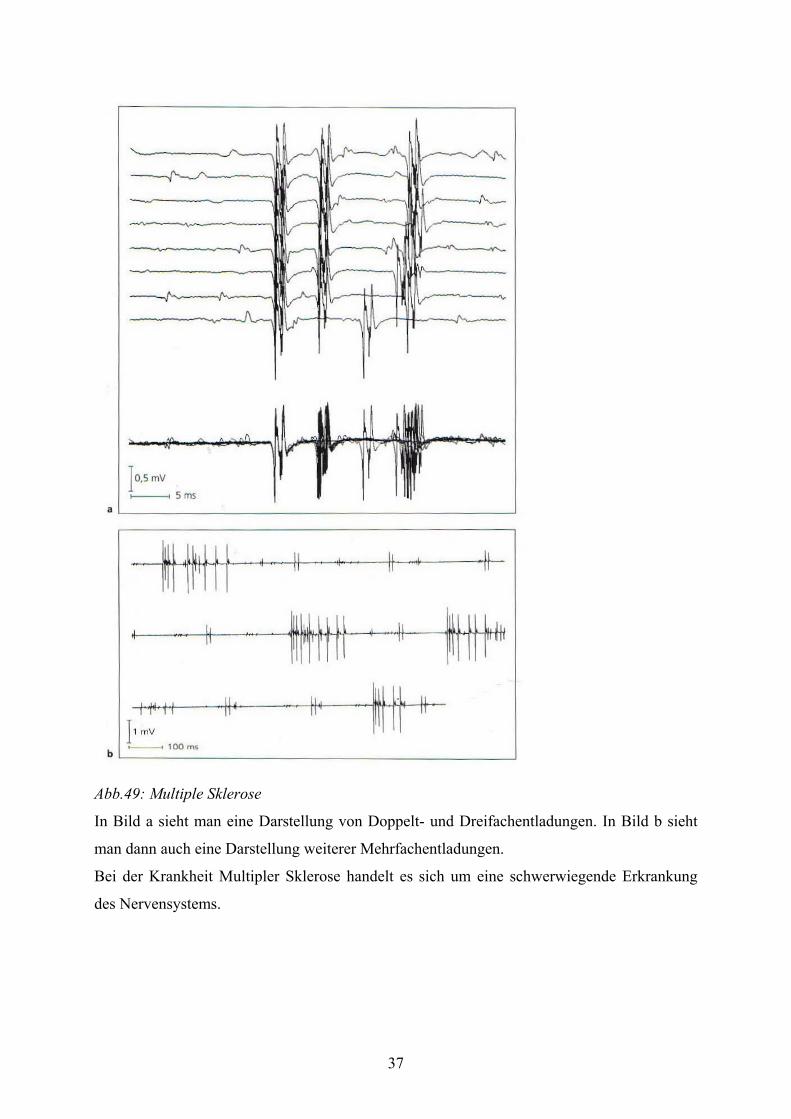
Abb.49: Multiple Sklerose
In Bild a sieht man eine Darstellung von Doppelt- und Dreifachentladungen. In Bild b sieht
man dann auch eine Darstellung weiterer Mehrfachentladungen.
Bei der Krankheit Multipler Sklerose handelt es sich um eine schwerwiegende Erkrankung
des Nervensystems.

38
Quellenangaben • www.ejk.de
• www.emgsrus.com
• www.owlnet.rice.edu
• www.neurocog.psy.tufts.edu
• www.neurobiologie.fu-berlin.de
• www.novafeel.de
• www.dr-schoenherr.at
• www.informatik.uni-bremen.de
• Sundoc.bibliothek.uni-halle.de
• Stephan Zschocke: Klinische Elektroenzephalographie
• Schmidt, Thews: Physiologie des Menschen
• Rolf Heinecker: EKG in Praxis und Klinik
• www.wikipedia.de
• Hopf, Dengler, Röder: Elektromyographie-Atlas
• www.ch-medizintechnik.de/ elektr.htm





























