Embed Size (px)
Citation preview

MerkePrim�rstruktur: genetisch determinierteAminos�uresequenzSekund�rstruktur: a-Helix oder b-FaltblattTerti�rstruktur: dreidimensionale Strukturder ProteinketteQuart�rstruktur: r�umliche Anordnung meh-rerer Proteinuntereinheiten
Die Prim�rstruktur legt letztlich alle anderenStrukturen fest.
2.1.5 Die Nukleins�urenNukleins�uren sind der Speicher der genetischenInformation. Man unterscheidet Desoxyribonu-kleins�uren (DNA) von Ribonukleins�uren (RNA).
2.1.5.1 Die Grundbausteine: NukleotideNukleins�uren sind Polymere aus Nukleosid-monophosphaten, die aus Nukleosidtriphosphatenunter Pyorphosphatabspaltung synthetisiert wer-den.Nukleosidmonophosphate bestehen aus einer orga-nischen Base (Purin oder Pyrimidinbase), einemZucker (Ribose oder 2l-Desoxyribose) und einemPhosphatrest. Diese Komponenten sind charakte-ristisch miteinander verkn�pft. Dabei nennt man
die Verbindung aus Zucker und Base allein Nukleo-
sid. Kommen eine oder mehrere Phosphatgruppenhinzu, so spricht man von Nukleotiden.Die organischen Basen der DNA sind die Purinbasen
Adenin (A) und Guanin (G) sowie die Pyrimidin-
basen Cytosin (C) und Thymin (T). Die Zuckerkom-ponente in der DNA ist die Pentose 2’-Desoxyribose(s. S. 7). In der RNA kommen die gleichen Basen wiein der DNA vor, allerdings findet man an Stelle vonThymin die Base Uracil (U). Der Zucker der RNA istdie Pentose Ribose (s. S. 7).Die drei Bausteine Zucker, Base und Phosphatrestsind folgendermaßen verkn�pft: Am C-1 des Zu-ckers h�ngt die organische Purin- oder Pyrimidin-base, das C-5-Atom des Zuckers ist mit Phosphatverestert (Abb. 2.11a).
2.1.5.2 Der Aufbau und die Strukturder Nukleins�urenDie Verkn�pfung der NukleotideSchreibt man zwei Nukleotide �bereinander, sostellt man fest, dass �ber die Phosphatgruppe amC-5-Atom des einen Molek�ls die Ausbildungeiner Esterbindung mit der OH-Gruppe am C-3-Atom des anderen Molek�ls m�glich ist. In derDNA und RNA sind viele Nukleotide �ber diese C-3-
12 Die biologisch wichtigen Makromolek�le 2 Allgemeine Zellbiologie
�� ��
� ����
�
�
� ��
�
���
�
�
� ��
���
� � ��
���
�
��
��
�
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
�
��
�
��
�
�
�
�
�
��
�
�
�
�
�
�
�
�
�
�
�
�
��
�
�
���
�
��
�
�
� ���
��
� �
�
�
�
��
���
��
�
�
�
�
�
�Abb. 2.10 (a) Antiparalleles b-Faltblatt; (b)paralleles b-Faltblatt; farbig gestrichelt sinddie Wasserstoffbr�cken zwischen den ver-schiedenen Peptidstr�ngen.
Poeggel, Kurzlehrbuch Biologie (ISBN 3131409819), c Georg Thieme Verlag KG
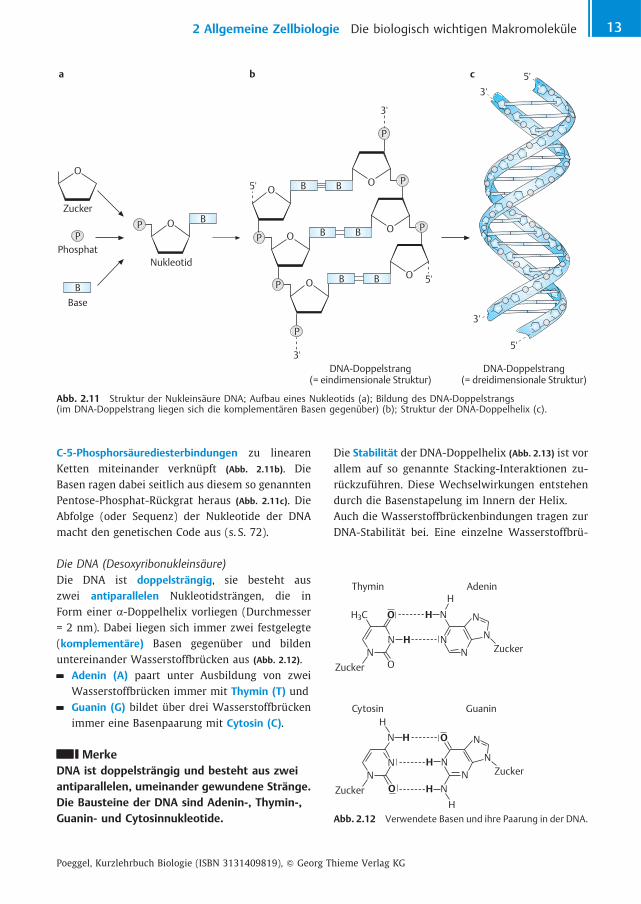
C-5-Phosphors�urediesterbindungen zu linearenKetten miteinander verkn�pft (Abb. 2.11b). DieBasen ragen dabei seitlich aus diesem so genanntenPentose-Phosphat-R�ckgrat heraus (Abb. 2.11c). DieAbfolge (oder Sequenz) der Nukleotide der DNAmacht den genetischen Code aus (s. S. 72).
Die DNA (Desoxyribonukleins�ure)Die DNA ist doppelstr�ngig, sie besteht auszwei antiparallelen Nukleotidstr�ngen, die inForm einer a-Doppelhelix vorliegen (Durchmesser= 2 nm). Dabei liegen sich immer zwei festgelegte(komplement�re) Basen gegen�ber und bildenuntereinander Wasserstoffbr�cken aus (Abb. 2.12).
Adenin (A) paart unter Ausbildung von zweiWasserstoffbr�cken immer mit Thymin (T) undGuanin (G) bildet �ber drei Wasserstoffbr�ckenimmer eine Basenpaarung mit Cytosin (C).
MerkeDNA ist doppelstr�ngig und besteht aus zweiantiparallelen, umeinander gewundene Str�nge.Die Bausteine der DNA sind Adenin-, Thymin-,Guanin- und Cytosinnukleotide.
Die Stabilit�t der DNA-Doppelhelix (Abb. 2.13) ist vorallem auf so genannte Stacking-Interaktionen zu-r�ckzuf�hren. Diese Wechselwirkungen entstehendurch die Basenstapelung im Innern der Helix.Auch die Wasserstoffbr�ckenbindungen tragen zurDNA-Stabilit�t bei. Eine einzelne Wasserstoffbr�-
132 Allgemeine Zellbiologie Die biologisch wichtigen Makromolek�le
��
��
��
��
������ ������������ ���� ������� ����������
���
�
�� �
!�"� �
�#���#��
��
���� ����
��
�
�
�
� �
�
��
�
��
�
�
��
� ��
������ ���������� ����� ������� ����������
Abb. 2.11 Struktur der Nukleins�ure DNA; Aufbau eines Nukleotids (a); Bildung des DNA-Doppelstrangs(im DNA-Doppelstrang liegen sich die komplement�ren Basen gegen�ber) (b); Struktur der DNA-Doppelhelix (c).
$��
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
!�"� �
!�"� �
!�"� �
!�"� �
%#&��� � ���
'�����$&�����
Abb. 2.12 Verwendete Basen und ihre Paarung in der DNA.
Poeggel, Kurzlehrbuch Biologie (ISBN 3131409819), c Georg Thieme Verlag KG

ckenbindung hat nur eine sehr geringe Bindungs-energie, ihre hohe Anzahl jedoch tr�gt zum Zusam-menhalt der beiden DNA-Str�nge bei. Durch W�r-mezufuhr kann man diese Bindungen sprengen,die DNA liegt dann einzelstr�ngig vor.
Die RNA (Ribonukleins�ure)RNA ist einstr�ngig und bildet nur abschnittsweiseintramolekulare helikale Strukturen aus. Innerhalbder RNA findet man statt der Pyrimidinbase Thymindie Base Uracil, die sich durch das Fehlen einerCH3-Gruppe von Thymin unterscheidet. Kommt esunter RNA-Beteiligung zur Basenpaarung (z. B. w�h-rend der Transkription, s. S. 77), so paart ein Uracil-nukleotid U mit einem Adeninnukleotid A.
MerkeRNA ist einzelstr�ngig; durch intramolekulareBasenpaarungen k�nnen jedoch doppelhelikaleBereiche entstehen. Die Bausteine der RNA sindAdenin-, Uracil-, Guanin- und Cytosinnukleotide.
Man unterscheidet funktionell drei wichtige Typenvon RNA:
Die Messenger-RNA (mRNA): Sie fungiert als Bo-ten-RNA bei der Synthese von Proteinen. Die ge-netische Information der DNA wird w�hrend derTranskription (s. S. 77) in mRNA umgeschriebenund ins Zytoplasma der Zelle transportiert. Daes viele verschieden große Proteine gibt, gibtes auch viele mRNA-Molek�le unterschiedlicherL�nge. Die mRNA ist die vielf�ltigste RNA.Die ribosomale RNA (rRNA): Sie ist eine Struk-
tur-RNA und baut gemeinsam mit Proteinendie Ribosomen auf (s. S. 33). In prokaryontischenZellen gibt es drei, in eukaryontischen Zellengibt es vier verschiedene rRNA-Molek�le.Die Transfer-RNA (tRNA): Sie bindet im Zyto-plasma die Aminos�uren und transportiert siezur Proteinsynthese (Translation, s. S. 82) zuden Ribosomen. Da es 21 proteinogene Amino-s�uren gibt, muss es auch mindestens 21 ver-schiedene tRNA-Molek�le geben. Tats�chlichist die Zahl jedoch h�her. Aufgrund des so ge-nannten degenerierten genetischen Codes gibtes f�r viele Aminos�uren mehrere tRNAs. Injeder Zelle finden sich daher mindestens 60 ver-schiedene tRNA-Molek�le.
Bringt man ein tRNA-Molek�l zweidimensionalin eine Ebene, sieht es aus wie ein Kleeblatt(Abb. 2.14). Durch posttranskriptionale Modifikationwerden nach der Synthese der tRNA viele Basennachtr�glich ver�ndert. Es entstehen so genannteseltene Basen, die zu ungew�hnlichen Wechselwir-kungen f�hren. Im Bereich der Stege dieses Klee-blattes kommt es durch intramolekulare Basen-paarungen zu doppelhelikalen Abschnitten.Die Bindung der Aminos�uren erfolgt am letztenNukleotid an der 3’-OH-Gruppe des Zuckers (Adeno-sin). Dieses Ende ist bei allen tRNA-Molek�len iden-tisch (CCA-Ende). Die richtige Position auf dermRNAwird �ber das Anticodon nach dem Prinzip derBasenpaarung gefunden (s. „Translation“ S. 82).Die beiden anderen Schleifen dienen der Wechsel-
14 Die biologisch wichtigen Makromolek�le 2 Allgemeine Zellbiologie
��
��
��(��
�#���#��
��
���) �*��"#
��
��
+,-��
�� �� �*��"#
�,.��
Abb. 2.13 Ausschnitt aus einer DNA Doppelhelix. Manbeachte, dass die beiden Str�nge antiparallel vorliegen.
Poeggel, Kurzlehrbuch Biologie (ISBN 3131409819), c Georg Thieme Verlag KG

wirkung mit dem Ribosom und der Aminoacyl-tRNA-Synthetase (das Enzym, welches die passendetRNA mit der passenden Aminos�ure verkn�pft).
2.1.6 Klinische Bez�ge2.1.6.1 PrionenEinige gef�hrliche und in ihrem Verlauf t�dlicheKrankheiten werden wahrscheinlich durch eine In-fektion mit reinen Proteinen ausgel�st. Diese infek-ti�sen Proteine nennt man Prionen (von: protein-artige infekti�se Partikel). Sie enthalten keine
Nukleins�uren. Es sind also keine lebenden Erreger
f�r diese Erkrankungen verantwortlich wie bei an-deren Infektionskrankheiten. Dennoch sind Prio-nenkrankheiten �bertragbar, wobei die �bertra-gungswege noch nicht ganz aufgekl�rt sind. M�gli-cherweise erfolgt nach Aufnahme des infekti�senProteins �ber die Nahrung eine aufsteigende Infek-tion �ber Nervenendigungen des autonomen Ner-vensystems zum Zentralnervensystem.Prionen sind Ausl�ser von Rinderwahnsinn (BSE),
Katzen- und Nerzwahnsinn, Scrapie (bei Schafen)und der Creutzfeldt-Jakob-Krankheit (CJD) beimMenschen.
Die Erkrankung beruht auf einer Akkumulation vonfehlgefalteten Proteinen im Gehirn, durch die dasNervensystem zerst�rt wird:Offensichtlich kommen die Gene von Prion-Prote-inen in den S�ugetieren selbst vor. Sie kodierenf�r Proteine, die im Gehirn bestimmte Funktionenerf�llen. Wenn sich ein solches Protein fehlfaltetund eine unphysiologische Raumstruktur ein-nimmt, wird es zum Prion und induziert (�hnlicheiner Kettenreaktion) die Fehlfaltung weiterer nor-maler Proteine, die so ihre urspr�ngliche Funktionnicht mehr erf�llen k�nnen. Es wirkt also wie einKristallisationskeim und wird zu einem infekti�senAgens. Diese fehlgefalteten Proteine sind außerdemextrem stabil gegen�ber Proteasen, k�nnen also vonden erkrankten Organismen nicht abgebaut werden.�ber Scrapie weiß man, dass es zwei stabile Kon-formationen des betroffenen Proteins gibt:
PrPc ist normal gefaltet und kommt nat�rlicher-weise im Gehirn von Schafen vor.PrPsc ist fehlgefaltet und somit krankheitsaus-l�send.
Eine Fehlfaltung dieser Proteine kann jedoch auchohne Infektion genetisch bedingt stattfinden.Durch eine Mutation im normalen Gen entstehtein defektes Protein, welches sich z. B. bei der klas-sischen Creutzfeldt-Jakob-Krankheit fehlfaltet, mitder Zeit akkumuliert und in fortgeschrittenemAlter zu einer Zerst�rung der Hirnsubstanz f�hrt.Bei der neuen Variante der CJD (nvCJD), die bereitsin fr�hen Lebensjahren auftreten kann, geht manjedoch davon aus, dass sie durch die Aufnahmeinfekti�ser Partikel �ber die Nahrung (BSE-konta-miniertes Rindfleisch) ausgel�st wird.
Check-up4 Rekapitulieren Sie den Aufbau einer
Aminos�ure.4 Wiederholen Sie die chemischen Reaktio-
nen zwischen zwei OH-Gruppen (Zucker undNukleins�uren), zwischen COOH- und NH2-Gruppen (Peptidbindung) sowie zwischenCOOH- und OH-Gruppen (Triglyceride).
4 Vergegenw�rtigen Sie sich den Aufbau vonNukleotiden und ihre Polymerisation zuNukleins�uren.
4 Machen Sie sich die Unterschiede zwischenRNA und DNA klar.
152 Allgemeine Zellbiologie Die biologisch wichtigen Makromolek�le
'
'��
'' '
''/$
$$'
0'
/$
���"����
���� �����12���3�� �
���� �����4���3�� �
�����5�� ��������� ��
$$$'$//
'
'$''///
$ $
$ /
'
$�%
$ / ' / '$/''
$ / $ '
' ' $
�
Abb. 2.14 Kleeblattstruktur eines tRNA-Molek�ls.
Poeggel, Kurzlehrbuch Biologie (ISBN 3131409819), c Georg Thieme Verlag KG

2.2 Die Zytoplasmamembran
LerncoachDie chemischen Eigenschaften der Zyto-plasmamembran und die von ihr gebilde-ten Strukturen sind f�r viele zellul�reFunktionen wichtig. Ihr Hauptbestandteilsind die Phospholipide. Wiederholen Siedeshalb ggf. deren Aufbau.
2.2.1 �berblick und FunktionDas Zytoplasma der Zellen ist von der Zytoplasma-membran (Plasmalemma, Durchmesser 5–10 nm)umgeben. Das Plasmalemma grenzt die Zellennach außen ab und verhindert einen freien un-kontrollierten Stoffaustausch mit der Umgebung.Dadurch ist jede Zelle „relativ“ isoliert.Der prinzipiell gleiche Typ von Membran umgibtauch viele Zellorganellen, kompartimentiert alsodie Zelle und schafft so relativ unabh�ngige Reak-tionsr�ume. Diese Abgrenzungsfunktion wird er-g�nzt durch eine Kontrollfunktion, da der Stoffaus-tausch durch die Membran �ber eine Vielzahl vonspezifischen Transportmechanismen reguliert wird.Außerdem ist die Zytoplasmamembran bei der Aus-bildung von Zell-Zell-Kontakten beteiligt. Diesesind wichtig f�r die Stabilit�t von Zellen und Gewe-
ben, sie k�nnen der Abdichtung von Zellzwischen-
r�umen dienen und erm�glichen Stoffaustausch
zwischen benachbarten Zellen.
2.2.2 Der Aufbau der Zytoplasmamembran2.2.2.1 Das „Unit-Membrane“-ModellDavson und Danielli erkannten Mitte der 30er Jah-re, dass es sich bei der Zytoplasmamembran umein einheitliches Gebilde („unit membrane“) han-delt, das aus einer regelm�ßigen Anordnung vonPhospholipiden besteht (s. S. 8). Phospholipidehaben aufgrund ihres amphiphilen Charakters dieTendenz in w�ssriger L�sung Doppelschichten zubilden. Die hydrophilen K�pfe sind dem w�ssrigenMedium zugewandt, die hydrophoben Schw�nzewenden sich im Inneren der Doppelschicht einan-der zu (Abb. 2.15). Weitere Strukturen, die gebildetwerden k�nnen (Mizellen, Monolayer und Liposo-men), werden ebenfalls in Abb. 2.15 gezeigt.
2.2.2.2 Das „Fluid-Mosaik“-ModellDie physikochemischen Eigenschaften von Zyto-plasmamembranen ließen sich jedoch allein durchdas „Unit-Membrane“-Modell nicht erkl�ren. 1972stellten Singer und Nicholson daher ihr „Fluid-Mosaik“-Modell vor. Die Grundstruktur der Zyto-plasmamembran entspricht auch in diesem Modell
16 Die Zytoplasmamembran 2 Allgemeine Zellbiologie
�����#�� #&����#��
���#��#�� 6������� �7�
85����� �9�� �����
6�:�;<�
=��� �
6�������� ��"#�"#�
>�? ��
6������
>�����& �
Abb. 2.15 Strukturierung von Lipidmolek�len an Grenzfl�chen.
Poeggel, Kurzlehrbuch Biologie (ISBN 3131409819), c Georg Thieme Verlag KG

dem Modell der „unit membrane“. Der bimoleku-lare Phospholipidfilm wird jedoch als ein visk�ses
L�sungsmittel betrachtet, in das verschiedene peri-phere und integrale Proteine eingelagert sind, dieinnerhalb der Membran lateral beweglich sind.
MerkeDie Lipide bilden ein zweidimensionales visk�sesL�sungsmittel, in das sowohl integrale als auchperiphere Proteine eingebettet sind. Die Kom-ponenten der Zytoplasmamembran sind lateralbeweglich.
Die Lipiddoppelschicht ist asymmetrisch. Die Phos-pholipide (s. S. 8) sind in dieser Doppelschicht un-gleichm�ßig verteilt. Die innere (intrazellul�re)
Schicht wird von einem h�heren Anteil Phosphati-dylserin, Phosphatidylinositol und Phosphatidyl-ethanolamin gebildet, w�hrend die �ußere (extra-
zellul�re) Schicht mehr Sphingomyelin und Phos-phatidylcholin enth�lt.Die Fluidit�t der Membran h�ngt von ihrem Gehaltan unges�ttigten Fetts�uren ab, je mehr unges�t-tigte Fetts�uren vorhanden sind, desto fluider istdie Membran. Cholesterin (s. S. 9), das auf beidenMembranseiten gleichm�ßig verteilt ist, erh�ht inMembranen mit �berwiegend ges�ttigten Fetts�u-ren die Fluidit�t. In Membranen, die viele unges�t-tigte Fetts�uren enthalten f�llt es die L�cken, diedurch das Abknicken unges�ttigter Fetts�ure-schw�nze entstehen, und senkt damit ihre Fluidit�t.
Eingelagerte Proteine k�nnen die Membran einmaloder mehrfach in Form von a–Helices durchziehenoder kovalent an Lipide der �ußeren oder der inne-ren Schicht gebunden sein. Die in der Membran lie-genden Teile der a–Helices bestehen aus hydropho-ben Aminos�uren, deren Seitenketten nach außengerichtet sind (in das hydrophobe Innere der Mem-bran). Die Proteine und Lipide der Membran sindaußerdem h�ufig mit Kohlenhydraten (Oligosac-chariden) verkn�pft (Glykoproteine, Glykolipide).Diese Kohlenhydratanteile nennt man in ihrer Ge-samtheit Glykokalix. Sie zeigen immer nach außen(Abb. 2.16).Zwischen den Membranen einzelner Zellen bestehtnormalerweise ein Abstand von 10–20 nm (Inter-
zellularspalt).
MerkeDie Kohlenhydratanteile der Zytoplasmamem-bran liegen immer auf der Extrazellularseite!Sie werden in ihrer Gesamtheit als Glykokalixbezeichnet.
2.2.3 Die Funktionen derZytoplasmamembran2.2.3.1 Die Zytoplasmamembran alsPermeationsschrankeDie physikochemischen Eigenschaften der Zyto-plasmamembran haben zur Folge, dass nur kleine,nicht polare Stoffe (z. B. Gase) und sehr kleine, po-lare Stoffe (wie z. B. Wasser) hindurch diffundieren
172 Allgemeine Zellbiologie Die Zytoplasmamembran
��������� �������
�������""#����
'�&������������
������
$#�� �� ���
� ���# � �> �(������� ��
��� ���� �> �(������� ��
���������������������
'�&������ ��
Abb. 2.16 Aufbau der Zytoplasma-membran.
Poeggel, Kurzlehrbuch Biologie (ISBN 3131409819), c Georg Thieme Verlag KG

k�nnen. Die Membran wirkt somit als Barriere f�rgr�ßere polare Substanzen und Ionen.
2.2.3.2 Die ErkennungsfunktionDie Glykoproteine und Glykolipide der Zytoplasma-membran dienen den Zellen der gegenseitigenIdentifizierung, so sind sie z. B. ein chemischer Aus-
weis gegen�ber dem k�rpereigenen Immunsystem.Die Glykokalix erm�glicht also die Erkennung vonZellen und nichtzellul�re Strukturen. Diese Erken-nung wird auch f�r gezielte Wanderbewegungengenutzt (z. B. w�hrend der Embryonalentwicklung).Selektine sind z. B. Proteine, die bei Entz�ndungenauf der Oberfl�che von Endothelzellen exprimiertund von Lymphozyten erkannt werden. Auf dieseArt werden die Zellen des Immunsystems zueinem Entz�ndungsherd „gelockt“.
2.2.3.3 Die RezeptorfunktionViele Membranproteine fungieren als Rezeptoren.Sie erkennen chemische Signale anderer Zellen(z. B. Hormone, Neurotransmitter) und leiten dieseInformation �ber verschiedene Mechanismen (s. S.64) in die Zelle hinein. Rezeptoren k�nnen per-
manent vorhanden sein (wie z. B. der Insulin-rezeptor auf der Oberfl�che von Hepatozyten)oder tempor�r ausgebildet werden.
2.2.3.4 Pumpstation, Reizperzeption undReizleitungF�r den geregelten Ablauf zellul�rer Vorg�nge isth�ufig eine Ionenungleichverteilung zwischen Zell-innerem und Zell�ußerem n�tig. Dieses Ungleich-
gewicht realisieren in der Membran liegende trans-membran�se Proteine (Ionenpumpen, z. B. dieNa+-K+-ATPase).Das durch Ionenpumpen erzeugte Ionenungleich-gewicht von Na+-, K+-, Ca2+- und Cl–-Ionen bildetbeispielsweise die Grundlage f�r Erkennung undWeiterleitung elektrischer Signale �ber die Mem-branoberfl�che innerhalb des Nervensystems.
2.2.3.5 Die elektrische IsolationWenn Information durch den Fluss von elektri-schen Str�men �bertragen wird, m�ssen nat�rlichauch verschiedene Informationsleiter voneinanderelektrisch isoliert werden. Diese elektrische Iso-lation wird im Nervensystem von Membranenrealisiert, die in vielfachen Lagen �bereinandergewickelt sind (Myelinscheiden).
2.2.3.6 Die Zell-Zell-KontaktbildungF�r den Zusammenhalt und die Kommunikationuntereinander bilden Zellen spezifische Zell-Zell-Kontakte zwischen ihren Zytoplasmamembranenaus.
Bei der Ausbildung von Zell-Zell-Kontaktenspielen verschiedene Komponenten desZytoskeletts eine wichtige Rolle. Ausf�hrlicheInformationen zum Zytoskelett finden Sieab S. 26.
Die Zonula occludensDie Zonula occludens (tight junction) dient demVerschluss von Zellzwischenr�umen. Sie ist eine
18 Die Zytoplasmamembran 2 Allgemeine Zellbiologie
!�������""��� ������#��@��"�����
!��������#� � ��
� A��������@��"�����
� ������
$��# ���
>����:���� ��
$��� A����( �� # ������2�$��� A��/�� � ��# �� ��
$��# ��� ?&�����������"# ����B� %���:���� ��
����:�� �� ?&�����������"# ���B� 9�� ���� :���� ��C������
����������� ��� ������
%���:���� ��
Abb. 2.17 �bersicht�ber Zell-Zell-Kontakte.
Poeggel, Kurzlehrbuch Biologie (ISBN 3131409819), c Georg Thieme Verlag KG

g�rtelf�rmige Struktur, die einen Interzellularspaltvon weniger als 1 nm zwischen benachbarten Zel-len bel�sst. Praktisch ist also kein Interzellularspaltmehr vorhanden. Proteine der benachbarten Zyto-plasmamembranen r�cken in so enge Nachbar-schaft, dass es zur Ausbildung von so genanntenVerschlussn�hten kommt. Diese Form des Zell-Zell-Kontaktes soll verhindern, dass Stoffe unkontrol-liert zwischen den Zellen hindurch diffundierenk�nnen. Tight junctions findet man daher �beralldort, wo K�rperinneres gegen K�rper�ußeres abge-dichtet werden muss (z. B. im Darmepithel) oderwo besonders empfindliche Organe gesch�tzt wer-den m�ssen (Endothelien der Hirnkapillaren). Eshandelt sich also um Diffusionsbarrieren. Eine wei-tere Funktion dieser Kontakte ist die Fixierung von
Membranproteinen auf bestimmte Bereiche der Zy-toplasmamembran, da durch die „Verschweißung“benachbarter Zellen die laterale Diffusion von Pro-teinen in der Membran behindert wird. Die Prote-ine k�nnen diese „N�hte“ nicht �berwinden undwerden auf bestimmte Dom�nen der Epithelzellenfixiert (apikal oder basolateral, vgl. (Abb. 2.17).
Die Zonula adhaerensDie Zonula adhaerens (Haftzone) ist ebenfalls eineStruktur, die g�rtelf�rmig um Epithelzellen heruml�uft. Sie dient der mechanischen Stabilisierung
dieser Zellen in einem Zellverband. An diesen Stel-len erscheint die Zytoplasmamembran optisch ver-dickt. Dieser Eindruck entsteht durch dicke Aktin-
faserb�ndel (Mikrofilamente, Bestandteile des Zy-toskeletts, s. S. 27), die auf der zytoplasmatischenSeite aufgelagert und �ber Anheftungsproteine(z. B. a-Aktinin) mit transmembran�sen Proteinen(Cadherinen) verbunden sind. Die Cadherine �ber-lappen im Interzellularspalt und verhindern somitein Auseinandergleiten der miteinander verbunde-nen Zellen (Abb. 2.17).
MerkeDie Aktinfaserb�ndel reichen nicht von Zelle zuZelle. Die Verfestigung der Zellen untereinanderwird �ber Transmembranproteine (Cadherine)realisiert. Die Anbindung an das Zytoskeletterfolgt �ber Anheftungsproteine an Mikrofila-mente (Aktin).
Die Macula adhaerens (Desmosom, Haftplatte)ImUnterschied zur Zonula adhaerens handelt es sichbei den Desmosomen um punktf�rmige Zell-Zell-Kontakte. Sie dienen, vergleichbar mit Schweiß-punkten, ebenfalls der mechanischen Stabilisierungvon Zellen in einem Zellverband und wirken Scher-und Zugkr�ften entgegen. Auch bei Desmosomenerfolgt die Verfestigung zwischen den Zellen �berCadherine. Diese Cadherine sind jedoch intrazellu-l�r �ber Anheftungsproteine (zytoplasmatischePlaques) mit Tonofilamenten (Zytokeratin) verbun-den und nicht mit Mikrofilamenten. Die Tonofila-mente durchziehen die Zelle von Desmosom zuDesmosom und stabilisieren damit die gesamteZellstruktur.
Die HemidesmosomenHemidesmosomen bilden Kontakte von Zellen zunichtzellul�ren Strukturen. Sie dienen der Befesti-gung von Zellen auf einer Unterlage (Basallamina)und verhindern, dass Epithel- oder Endothelzellen�ber die Basallamina rutschen. Die Verbindungzwischen Membran und Basallamina wird �berbestimmte Proteine, die Integrine, hergestellt. DieAnbindung der Integrine an das Zytoskelett erfolgtwie bei Desmosomen �ber Anheftungsproteine(zytoplasmatische Plaques) an Zytokeratin.
Die fokalen KontakteFokale Kontakte sind ebenfalls punktf�rmig undvermitteln auf �hnliche Weise wie die Hemidesmo-somen einen Kontakt zwischen Zellen und extrazel-lul�rer Matrix. Im Unterschied zu Hemidesmoso-men sind die Integrine der Zytoplasmamembran je-doch �ber zytoplasmatische Plaques mit den Aktin-
fasern des Zytoskeletts verbunden. �ber kompli-zierte Mechanismen k�nnen durch extrazellul�reSignale Polymerisation und Depolymerisation die-ser Aktinfasern reguliert werden, sodass die Zelle�ber eine Unterlage „kriechen“ kann. Dabei werdenfokale Kontakte aufgel�st und wieder neu gekn�pftund dabei Filopodien oder Lamellopodien gebildet.Diese Form von Kontakten findet man weniger beiEpithelzellen, sondern �berall dort, wo Zellenaktiv wandern (Bewegung von Makrophagen, em-bryonale Zellbewegungen).
192 Allgemeine Zellbiologie Die Zytoplasmamembran
Poeggel, Kurzlehrbuch Biologie (ISBN 3131409819), c Georg Thieme Verlag KG

Die Kommunikationskontakte (Nexus, Gap junction)Kommunikationskontakte (Nexus, Gap junction)sind por�se Verbindungen des Zytoplasmas zweierbenachbarter Zellen. In die Zytoplasmamembranbeider Zellen sind Proteinrohre (Connexone) einge-lagert. Jedes Connexon besteht aus 6 transmem-bran�sen zylindrischen Proteinen (Connexinen),welche wiederum jeweils mit 4 a-Helices die Mem-bran durchqueren. Die Connexone zweier Zellen la-gern sich aneinander und bilden ein durchg�ngigesProteinrohr mit einem Durchmesser von ca. 1,5 nm.Dadurch ist ein Austausch kleiner Molek�le bis zueinem Molekulargewicht von 1000–1500 Daltonzwischen den Zellen m�glich (Disaccharide, Amino-s�uren, Vitamine, cAMP, Steroidhormonen, Wachs-tumsfaktoren). Der Interzellularspalt verringertsich an den Gap junctions auf 2–4 nm.Diese Kontakte dienen u. a. der elektrischen Kopp-lung von Zellen (z. B. im Herzmuskel) und w�hrendder Embryonalentwicklung der Synchronisation beider Gewebsdifferenzierung.
2.2.3.7 Die Regulation des Stoffaustausches
Machen Sie sich bei den nachfolgenddargestellten Transportvorg�ngen bewusst, wiediese angetrieben werden und in welchen F�llender Einsatz von ATP als Energielieferant erfor-derlich ist.
Die DiffusionEine Form des passiven Stoffaustausches mit derUmgebung ist die Diffusion durch die Zytoplasma-membran (Abb. 2.18). Sie wird durch die chemischenEigenschaften der Zellmembran beeinflusst und be-schr�nkt sich auf Gase, Wasser und unpolare lipo-phile Substanzen. Diffusion durch die Zytoplasma-membran kann prinzipiell in beide Richtungen er-folgen. Die Richtung wird jedoch durch das Konzen-trationsgef�lle des jeweiligen Stoffes festgelegt.Durch diesen Konzentrationsunterschied wird derVorgang der Diffusion �berhaupt erst angetrieben,da immer ein Konzentrationsausgleich auf beidenSeiten einer durchl�ssigen Membran angestrebtwird. Die Zelle muss daher bei der Diffusion keineEnergie aufwenden.Die Geschwindigkeit der Diffusion h�ngt u. a. vomAusmaß des Konzentrationsunterschiedes, von derMolek�lgr�ße, dem lipophilen Charakter und derGr�ße der Hydrath�lle einer Substanz ab.
Die facilitierte (erleichterte) DiffusionSie ist ebenfalls eine Form des passiven Transportsund findet, wie auch die Diffusion, nur in Richtungdes Konzentrationsgef�lles statt, verl�uft jedochwesentlich schneller. Die Barrierefunktion derZytoplasmamembran wird dabei durch spezifischetransmembran�se Proteine, so genannte Permea-
sen, herabgesetzt (Abb. 2.18). Diese Permeasen k�n-nen unterschiedlich funktionieren, z. B. durch Bil-
20 Die Zytoplasmamembran 2 Allgemeine Zellbiologie
6�������� ��"#�"#�
C��? �������������� ��
��:�"# ��::�����
��::���������"# �� ��C����
�� �"#� �� ���::��������"#�$���� �
�����D ��%�������������� ��C��? ���������� :5��
����D ��%������������"#�$���� �
���E��
%�
Abb. 2.18 Aktive und passive Transportsysteme.
Poeggel, Kurzlehrbuch Biologie (ISBN 3131409819), c Georg Thieme Verlag KG

dung eines Kanals (w�ssrige Pore) oder als Carrier
(Flip-flop-Modell oder Pendel). Einen solchen Car-rier kann man sich als einen nach einer Seite offe-nen Kanal vorstellen, der sich durch eine Konfor-mations�nderung (nach Bindung der zu transpor-tierenden Substanz) „umst�lpt“ und diese dannauf der anderen Seite der Membran wieder freigibt.Permeasen haben eine hohe Spezifit�t, transportie-ren also nur definierte Substanzen durch eineMembran. Der Transport kann – in Abh�ngigkeitvom Konzentrationsgradienten – auch hier prinzi-piell in beide Richtungen erfolgen. Die Konforma-tions�nderung der Permeasen verbraucht keineEnergie.Zucker und Aminos�uren sind Beispiele f�r Sub-stanzen, die h�ufig �ber facilitierte Diffusion trans-portiert werden.
MerkePermeasen funktionieren in beide Richtungen!Die effektive Transportrichtung h�ngt nur vonder Richtung des Konzentrationsgef�lles ab.
Der aktive TransportTransporte gegen ein Konzentrationsgef�lle durchdie Membranen ben�tigen immer entweder direkt
oder indirekt Energiezufuhr. Solche aktiven Trans-
porte werden ebenfalls �ber transmembran�se„Carrier“-Proteine realisiert (Abb. 2.18).Einer der bekanntesten Transporter ist derNa+-K+-Transporter: In den Axonen der Nervenzel-len ist das Verh�ltnis der Na+-Ionen innen zuaußen 1: 9, das der K+-Ionen 41:1. Dieses Ionen-ungleichgewicht muss f�r die elektrische Reizwei-terleitung immer aufrechterhalten werden. Dazupumpt die Na+-K+-ATPase jeweils gegen das Kon-zentrationsgef�lle gleichzeitig 3 Na+-Ionen nachaußen und 2 K+-Ionen nach innen. Der Energiebe-darf wird hierbei direkt durch Spaltung eines ATP-Molek�ls gedeckt. Daher wird dieser Transporterauch Na+-K+-ATPase genannt.
MerkeAktive Transporte ben�tigen Energie, meist inForm von ATP.
Der CotransportWerden zwei Substanzen gekoppelt durch die Zyto-plasmamembran transportiert, handelt es sich umCotransporte. Ist die Transportrichtung beider Sub-stanzen identisch, handelt es sich um einen Sym-
port; ist die Transportrichtung entgegengesetzt,
212 Allgemeine Zellbiologie Die Zytoplasmamembran
6�������� ��"#�"#�
/������ �&����� �������
$����������
Abb. 2.19 Cotransporte: Die Energie stammt aus dem Konzentrationsgef�lle der ersten Substanz (farbige Kreise), diezweite Substanz (Dreiecke) wird gegen ihr Konzentrationsgef�lle transportiert.
Poeggel, Kurzlehrbuch Biologie (ISBN 3131409819), c Georg Thieme Verlag KG