Embed Size (px)
Citation preview

23 Tiergeographie
Werner Peters
23.1 Allgemeines
Die Tiergeographie beschreibt die heutige und frühere Verteilung der Tiergruppen auf der Erde. Sieversucht, die Evolution in Raum und Zeit sowiedie Verbreitung der Tierarten und -gruppen zuerfassen und die komplexen Ursachen dieser Verbreitung zu deuten . Um diese Aufgaben lösen zukönnen, benötigt sie Erkenntnisse aus vielen benachbarten Wissenschaftszweigen: Taxonomie,Botanik , Genetik, insbes. Populati onsgenetik ,Ökologie, Bodenkunde, Geographie, Geologie, Paläont ologie und Klimakunde.Die vielseitigen Wechselbeziehungen dieser Wissenschaften haben immer wieder dazu geführt, die Tiergeographie entweder als Teilgebiet der Systematik, der Ökologieoder der Geographie aufzufassen. All diese Versuchesind aber stets im Sande verlaufen. Sie zeigen, dass dieTiergeographie offensichtlich ein eigenständiger Wissenschaftszweig ist. Tier- und Pflanzengeographie werdenals Biogeographie zusammengefasst. Sie haben vielerleiÜbereinstimmungen und ergänzen sich in vielfaltigerWeise.
Bisher wurde vielfach zwischen beschreibender oderdeskriptiver und kausaler Tiergeographie unterschieden.Dies hat wohl in erster Linie historische Gründe; beideRichtungen ergänzen einander und sind heute kaumstreng voneinander trennbar.
In neuerer Zeit ist, ausgehend von zwei ganz verschiedenen Entdeckungen, in wesentlichen Bereichen der Tiergeographie ein beachtlicher Wandeleingetreten:• Unter dem Einfluss der Ideen Hennig s zur phy
logenetischen Systematik entstand eine sta rkeTendenz zur Entwicklung einer phylogenetischor ientierten Tiergeographie, die im taxonomischen Bereich ausschließlich von monophyletischen Einheiten ausgeht, um deren Aufgliederung und Ausbreitung zu untersuchen . Von besonderer Bedeutung sind hierbei Gruppen, vondenen umfangrei ches fossiles Material aus verschiedenen Erdperioden und aus unterschiedlichen Gebieten bekannt ist.
• Einen enormen Auftrieb erfuhr die histori scheTiergeographie (s. 23.5) außerdem durch dieheute als weitgehend gesichert geltende Theorieder Kontinentalverschiebung und Plattentektonik. Dennoch gibt es namhafte Autoren auf demGebiet der Tiergeographie, die diese Fakten
noch nicht kannten oder sie nicht anerkennenoder sie skeptisch beurteilen . Ebenso gibt esAutoren, denen die Hennigsche Cladistik nichtals geeignetes Verfahren zur Aufklärung derEvolution und Verbreitung einer Tiergruppe erscheint. Bei der Lektüre von Arbeiten sollte mandaher die Grundauffassungen der betreffendenAutoren berücksich tigen.
Die Biogeographie der Insekten befindet sich imGrunde noch immer in den Anfängen. Die Ergebnisse älterer Arbeiten müssen vielfach unterBerücksichtigung der Methoden der phylogenetischen Systematik und neuerer Ergebnisse der Kontinenalverschiebung überpr üft werden.
23.2 Chorologie (Arealkunde)
Das Verbreitungsgebiet einer Tierart wird alsAreal bezeichnet. Diese einfach erscheinende Definition erfordert in vielen Fällen Anmerkungen.Als Verbreitungsgebiet kann nicht einfach das Gebiet aufgefasst werden, in dem jemals die betreffende Art gefunden wurde. Verdriftungen ,Wanderungen , Veränderungen bei den Nahrungsquellen usw. können Ursachen für Schwierigkeitenbei der Feststellung des Verbreitungsgebietes sein.
Grundlage der Chorologie ist die Faunenaufnahme kleiner Areale. Diese Arbe iten haben inneuerer Zeit dadurch erheblichen Auftrieb erhal ten, dass fauni stische und ökologische Untersuchungen in vordem ungekanntem Ausmaß kleinwie großr äumig erfolgen. Durch die Verwendungautomatischer Sammelverfahren wird bei derart igen Arbe iten ein riesiges Material aus zahlreichenTiergruppen zusammengetragen. Die sog. BarberFallen dienen dem Fang laufaktiver Insekten , Berlese-Trichter dem Sammeln versteckt lebender Bodenbewohner, Malaise-Fallen und Lichtfallen demFang fliegender Insekten sowie Emergenz-Fallendem Einsammeln frisch geschlüpfter Insekten ausAbschnitten von Fließgewässern usw. In allen Erdteilen kam auf diese Weise ein riesiges Materialzusammen, dessen Sichtung verblüffende Ergebnisse brachte. Viele neue Arten wurden gefundenund scheinbar seltene Arten erwiesen sich häufigals weiter verbreitet als ursprünglich angenommen

722 23 Tiergeographie
wurde. Die zunächst sporadischen Untersuchungsergebnisse wurden in den vergangenen Jahren in zunehmendem Maße mithilfe immer leistungsfähiger werdender Computer und der fürdiese Aufgaben besonders entwickelten Softwareausgewertet . Die Ergebnisse wurden in nationalenwie internationalen Datenbanken gesammelt. 1969wurde in Europa ein internationales Programmunter der Bezeichnung European Invertebrate Survey (EIS) gestartet, mit dessen Hilfe die Verbreitungsangaben gesammelt und kartiert werden. Diezunächst in nationalen Zentren gesammelten Daten werden seither an diese Zentrale weitergeleitet.Auf Wunsch werden von der Zentrale die erstelltenKarten ausgedruckt und verschickt, damit sie alsGrundlage für weitere Untersuchungen und Eintragungen dienen können .
Die Gittergröße der erstellten Rasterkarten variiert zwischen 10 km der regionalen Karten(Abb. 23-1), denen die Topografische Karte (Messtischblatt 1:25000) zugrunde liegt, und einem50 km-Gitter (UTM) für Europa. Bei den Angaben der Fundorte wird vielfach die Glaubwürdigkeit älterer Angaben berücksichtigt, bei denennoch andere taxonomische Auffassungen vorherrschten oder ein Bezug zu heute gültigen Art namen nicht möglich ist. In diesen Fällen könnenAngaben vor 1950 oder 1960 und danach getrenntaufgeführt sein.
Karten, aus denen die Fundorte und damit dieVerbreitung einer Art hervorgehen, können alsRasterkarte (Abb. 23-1), Umrisskarte (Abb.23-2)oder Punktkarte (Inset in Abb. 23-3) ausgeführtsein. In eine Punktkarte werden die einzelnenFundorte eingetragen ; ein Punkt entspricht dahermindestens einem Fund. Bei einer Rasterkarte istdas Gitternetz der verwendeten Karte hervorgehoben; sie wird im Allgemeinen bei regionalenUntersuchungen verwendet . Eine Umrisskarteverallgemeinert diese Befunde und zeigt durchUmrandung oder zusätzliche Musterung der Fläche das Verbreitungsgebiet eines Taxons.
Die Größe der Areale kann sehr verschiedensein. Kosmopoliten oder Ubiquisten können in fastallen Erdteilen riesige Areale bewohnen . Zu denKosmopoliten gehören auch manche Kulturfolgersowie Arten, die durch den Menschen verbreitetwurden (s. 23.2).
Innerhalb eines Verbreitungsgebietes können die Lebensbedingungen mehr oder weniger günstig sein. In diesenZusammenhang gehört auch die auffallende Tatsache,dass bei zahlreichen Arten im Süden des europäischenVerbreitungsgebietes geschlechtliche Fortpfl anzung , imNorden dagegen parthenogenetische Fortpflanzung vorherrscht.
Andererseits können endemische Arten in sehrkleinen Arealen , beispielsweise Gebirgstälern,Bergrücken, Inseln, Höhlen, Seen, Bächen usw.
vorkommen. Entscheidend ist, wo diese Arten ausreichende Lebensbedingungen finden . Thermophile Arten können beispielsweise auf Binnendünen beschränkt sein. Grundsätzlich ist Ortstreuedurch ausreichende Nahrung, Brutmöglichkeitenusw. bedingt . Reliktareale (relictum, lat. = Übriggebliebenes) sind Reste eines früher größeren Areals. Die Einengung kann durch viele möglicheökologische Faktoren zustande gekommen sein.Als Refugium bezeichnet man ein Zufluchts- oderRückzugsgebiet. Dieses muss nicht für die betreffende Art optimale Bedingungen bieten . Zusammenhängende Areale nennt man kontinuierliche Areale. Ein diskontinuierliches Areal ist gekennzeichnet durch Lücken im Verbreitungsgebiet, die unter Umständen zeitweise überbrücktwerden, unter günstigen Umständen sogar für längere Zeit. Eine disjunkte Verbreitung liegt vor,wenn das Areal durch geographische Schrankenoder andere Hindernisse über längere Zeiträumein mehrere Teile gegliedert ist (Abb. 23-2).
Sind die Areale zweier nah e verwandter Artenvollständig voneinander getrennt, so spricht manvon allopatrischer Verbreitung, besiedeln sie dasgleiche Areal oder sind sie überwiegend nicht voneinander getrennt, so nennt man dies sympatrischeVerbreitung.
Ortstreue kann dadurch bedingt sein, dass ausreichend Nahrung, Brutmöglichkeiten usw. vorhanden sind. Wanderungen haben verschiedeneUrsachen und unterschiedlichen Umfang. Bei regelmäßig wandernden Arten unterscheidet manzwischen Wohn- und Wanderraum. Man kannauch Wohn- oder Fortpflanzungsraum sowieWander- oder Verkehrsraum angeben. Die Begrenzung des letzteren ist schwieriger festzulegenals die des Wohnraums. Die Grenzen zwischenWohn- und Wanderraum können ebenso schwanken wie die Ausdehnung des Wanderraums. ImWanderraum kann unter günstigen Bedingungeneine Fortpflanzung über 1-2 Generationen erfolgen.
Die Ausbreitung ist ein fundamentaler undüberaus vielschichtiger Aspekt der Tiergeographie. Innere Faktoren, die eine Ausbreitungstendenz zur Folge haben, können auf klimatischeVeränderungen, Übervölkerung und damit einhergehend auf eine Einengung des Lebensraumes,ständige Störungen, Nahrungsmangel u.a. ansprechen . Die Geschwindigkeit der Ausbreitungeiner Art hängt nicht allein von der Umwelt ab,sondern hat offensichtlich noch weitere Gründe.Auffallend ist, dass die einzelnen Arten eine unterschiedliche Ausbreitungsgeschwindigkeit zeigen,sodass bei konkurrierenden Arten diejenigen mitrascher Ausbreitungstendenz im Vorteil sind. DieAusbreitung von Aren kann behindert werden,wenn die klimatischen Bedingungen nicht zuträg-

23.2 Chorologie (Arealkunde) 723
A
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
343536373839404142434445464748495051525354555657
B
\ JC" - 1"J ~I"(
""'\ '\ U ·"J' r-- } J ,J'
1/ 1 I• ./ ~ r-;
"" '-- '\,-, j -i-- --, r. "'\. s:
i/" ...... N l o ;t
"- h .... ~ :. •_"l - " ./..... :-- -" • ,...'-:: • .. j
0 \- h ", .:...... 1' ....... \ Il."'l' 1
1" ~ (J • ..~
11 ~ (J. • (J(J (J .r- ~ c,/ ~ 1- • •• (J •• I. c ~ ....- IJ<' I<, •• • •I§ ... (J
~ •• . 1• •0 cl ..... • .~ r"" ~ 8 ~ (J
0 \ I..... (J<J o'<l .}l1) • ~
Cl • • (J l"- r-
0 ••.. r-, fif-• (J ri .-
r:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
Abb. 23-1: Rasterkarten zur Darstellung der regionalen Verbreitung von Köcherfliegen (Trichoptera)in Nordrhein-Westfalen. Als Rastereinheit dienen die Quadranten der Messtischblätter (Topographische Karte1:25000), die einer Kantenlänge von10km entsprechen. Das Bergland ist indieser Dasteilung dunkel hinterlegt, umes vom Tiefland unterscheiden zu können.O leerer Kreis Nachweis vor 1950,o halb ausgefüllter Kreis Nachweisnach 1950 glaubhaft, _ ausgefüllterKreis Nachweis nach 1950 geprüft.A Drusus annulatus (Limnephilidae) bewohnt als Larve Bergbäche. B Limnephilus marmoratus (Limnephilidae) ist inder Norddeutschen Tiefebene weit verbreitet. (Nach Robert und Wichard1994)
3435363738394041424344
45464748495051525354555657
\ Jc ,.J 1"( 1- .
~ .. ""\. ~ i:\ -./ '/~ -- ..... J J ,J'~
1/ l() 1<1 I"", ~ ~ -,.. KJ 1'-- 1'11"'l j • :--r-, .... "\ s:-...... ...,- I. • I...- •• ;l"0 h ... • v •
,~ • r-, ./I-""' - ,..... ~ ......." •n • 1- 1'1-- ""- 1\ '""• 0 ,.. I"- --,
~ e I\'-'~ (
1""0:,. , ( II '"- IJ,<' I.le
I't. Ie'Ir! I- e ~ r"" e- I""~1\ • • ;.J I
e .1) rc.. 'r ". ....... .....
i'\ ~...J r-, f.... ,....", ri
r
lieh sind, schwer überwindbare Hindernisse auftreten, geeignete Nahrung und Brutmöglichkeitenfehlen usw.
Flugfähige Insekten können sich aktiv ausbreiten, doch hat sich gezeigt, dass sie dabei insbesondere Luftströmungen oder in geringerem MaßeWasserströmungen zusätzlich nutzen . Als Luft-
strömungen sind nicht nur die horizontalen, sondern auch die vertikalen, durch Thermik bedingten Luftströmungen von Bedeutung . In einer Luftsäule mit einem Querschnitt von einer Quadratmeile wurden im Pazifik Millionen von Insektennachgewiesen. Aber nicht nur in tropischen odersubtropischen Gebieten erfolgen derartige Verdrif-

724 23 TIergeographie
Abb. 23-2: Disjunkte Verbreitung als Folge der Eiszeit am Beispiel von Zegris eupheme (Lepidoptera, Pieridae). ImWesten lebt die Unterart meridionalis, in Mitteleuropa ssp. eupheme (nicht eingezeichnet) und imOstarealdie ssp. sulphurea. (Nachde Lattin 1967)
tungen durch Th ermik. Auch in den Alpen kannman zahllose Insekten auf dem Gletscherschneeantreffen , die von den Almen dorthin verdriftetwurden; dies sind in ers ter Linie die gut fliegendenSyrphidae. Neuere Untersuchungen haben gezeigt,dass Insekten in ganz anderem Maße als Wirbeltiere zu größeren Wanderungen über See befähigtsind. Fliegende Insekten müssen nicht die Gesamtstrecke in einem Zuge bewält igen , sondern könnenim Meer eine Zwischenstation einlegen, indem sieauf der Wasseroberfläch e landen, eine Weile au sruhen und dann wieder weiterfliegen. "Island hopping" im Bereich der Molukken sowie im Pazifischen Ozean dürfte mehr als früher angenommenbei der Verbreitung von Insekten eine Rolle gespielt haben. Dabei geht es nicht allein um flugfähige Insekten, die mit günstigen Luftströmungenüber teilweise erstaunliche Entfernungen verdriftetwerden können, sondern teilweise auch um wenigoder gar nicht flugfähige Insekten, die von derThermik pas siv bis in große H öhen getragen undan schließend als "Luftplankton" verdriftet werden .
Als Beispiel für die erstgenannte Gruppe sei eine Bremsenart erwähnt, Tabanus ceylonicus, die von Ceylon biszu den Philippinen, den Salomon-Inseln und im nördlichen Queensland in Australien verbreitet ist. Beispielefür die zweite Gruppe liefern die Collembola, ungeflügelte Blattläuse und manche Mückenarten aus derFamilie Chironomidae. Bis zu einem gewissen Gradekann eine Ausbreitung auch auf driftenden Baumstämmen, anderen natürlichen "Flößen", Eisbergen und dergleichen unter Ausnutzung von Meeresströmungen oderdurch Nutzung der Schwimmfähigkeit aufgrund der
nicht benetzbaren Cuticula erfolgen. Es hat sich abergezeigt, dass Luftströmungen von wesentlich größererBedeutung für die Ausbreitung von Insekten sind alsMeeresströmungen.
In den vergangenen Jahrzehnten wurden die Möglichkeiten der Ausbreitung von Insekten durchaktiven Flug wie durch Verdriftung in vertika lenund horizontalen Luftströmungen weltweit sehrgründlich untersucht, sodass man heute nichtmehr nur Vermutungen an stellen kann, sondernfundi ert e Belege zur Verfügung hat.
Der Ausbreitung von Arten kann deren Ansiedlung folgen. Ein gelegentliches Einschleppen oderdas Erreichen eines vom ursprünglichen Siedlung sgebiet entfernten Gebietes reicht dafür nochnicht au s. Entscheidend ist, da ss neben au sreichender Ernähru ng auch Möglichkeiten zur Fortpflanzung gegeben sind, sodass ein kontinuierlicher Aufenthalt möglich wird. Das Vorhandenseinmöglich st zahlreicher ökologischer Nischen för dert die zunehmende Evolution neuer Arten ausden ursprünglich eingewanderten Arten. Das Gegenteil, die Auslöschung , kann durch etliche Fak toren wie ökologische Bedingungen, überm ächtigeKonkurrenz oder übertriebene Spezialisierung bedingt sein.
Manche Autoren halten die Möglichkeit einer Ausbreitung von einer Landrnasse zur anderen oder zwischenInseln über größere Meeresgebiete hinweg nur dann fürmöglich, wenn in früheren Zeiten Landbrücken existierten, wie beispielsweise die Landbrücken in der BeringSee sowie im Bereich der Molukken (s. aber 23.5 undAbb. 23-8).

23.2 Chorologie (Arealkunde) 725
600_-'-__-lI km
Abb. 23·3: Verbreitung der thermophilen Schmetterlingsart Ammoconia senex (Noctuidae). Das Verbreitungsgebietdieser impontisch-mediterranen Bereich weitverbreiteten Art ist durch Graurasterung angedeutet. ImGebiet des Mittelrheins sowiederunteren Mosel und Nahe istein durch einen Pfeil gekennzeichnetes Reliktvorkommen in warmen Gebieten vorhanden . ImInsetsind die Fundorte in Form einer Punktkarte dargestellt. Postglazial haben sich diese Populationen zu einer Unterart (ssp.mediorhenana) entwickelt. (Nach de Lattin 1967)
Das WanderverhaIten der Insekten scheint eineökologische Anpassungserscheinung zu sein, dennals Ursachen werden immer wieder knappe oderschlechter werdende Ernährungsbedingungen sowie Übervölkerung gesehen. Im Einzelnen sind diegenetischen Hintergründe wie auch die auslösenden Faktoren noch wenig bekannt und nur beiwenigen Arten nachgewiesen und näher analysiertworden.
Als "Wanderheuschrecken" bezeichnet man Feldheuschreckenarten , die unter bestimmten Bedingungen dieTendenz haben, Schwärme zu bilden, die hüpfend oderfliegend teilweise sehr ausgedehnte Wanderzüge unternehmen. Innerhalb einer Gattung kann es wanderndeund nicht wandernde Arten geben. Die Bezeichnung
"Wanderheuschrecke" ist daher kein systematischer, sondern ein biologischer Begriff. Wanderheuschrecken haben Brutgebiete (Fortpflanzungsräume) (Abb. 23-4), indenen ein für die Entwicklung aller Stadien günstigesMikroklima herrscht und ausreichend Nahrung vorhanden ist. Nach der Phasentheorie von Uvarov, die ursprünglich an Locusta migratoria entwickelt wurde, aberoffensichtlich auch für die übrigen Wanderheuschreckengilt, kann man eine solitäre und eine Schwarmphaseunterscheiden, die jeweils ineinander übergegehen können. Wenn in den Brutgebieten in günstigen Jahren eineMassenvermehrung stattgefunden hat, anschließend abereine zunehmende Trockenheit und Einengung des Lebensraumes zur Übervölkerung im immer kleiner werdenden Brutgebiet kommt , führt dies zur Induktion derSchwarmphase. Die Tiere sind in diesem Zustand durcheine starke Melanisierung der Cuticula, eine gegenüber

726 23 Tiergeographie
o Brutgebiete
~ Wanderungen
Abb. 23-4: Brutgebiete und größere Wanderungen von Schwärmen der Wüstenheuschrecke Schistocerca gregariaim Jahre 1968. Während der Wanderungen werden Pausen zur Fortpflanzung in geeigneten Gebieten eingelegt. Somit sind dieIndividuen eines Schwarmes während der gesamten, unter Umständen sich über Jahre ausdehnenden Wanderungen nicht mehrdiejenigen aus der Anfangszeit.
der solitären Form um 5-8 °C höhere Körpertemperatur,größere Flügel, größere Bewegungsaktivität und ein auffallendes gegenseitiges Nachahmungsve rhalten gekennzeichnet. Erreicht die Bevölkerungsdichte der Schwarmtiere ein bestimmtes Maß, so bilden sich Wanderschwärme, die nach und nach riesige Gebiete durchwandern können und in den Befallsgebieten ungeheureNahrun gsmengen vertilgen. Die Wanderzüge könnenviele Jahre andauern (Abb. 23-4). Sie werden zeitweiseunterbrochen , wenn in günstigen Gebieten eine Fortpflanzung möglich ist. Gerat en die Schwärme in Wüstenoder fliegen sie aufs offene Meer hinaus, so gehen sie imallgemeinen zugrunde. Heuschreckenschwärme werdenin schriftlichen Überlieferungen seit dem Altertum alsfurchtbare Plagen beschrieben, denen man einst hilflosausgeliefert war (Abb, 23-5). Die riesigen, kilometerlangen, bis zu mehrere Milliarden Tiere enthaltendenSchwärme verdunkelten die Sonne. Wanderheuschreckensind als Plage vor allem in Afrika (u.a. Locusta migratoria, Dociostaurusmaroccanus, Schistocerca gregaria;Locustana parda/ina in Südafrika) und Vorderasien (u. a.S. gregaria sowie L. migratoria) bekannt, kommen aberauch in Nordamerika (u.a. Melanoplus mexicanus) undSüdamerika (u.a. Schistocerca paranensis), Australien(u.a. Chortoicetes terminifera, Gastrimargus musicus, L.migratoria) und Ostasien (L. migratoria) vor. In Mitteleuropa fielen im 14.-18 .Jh. immer wieder Schwärme vonWanderheuschrecken ein, deren Brutgebiete am Unterlauf der Donau oder in Südrussland lagen (Abb. 23-5).
In Europa und Nord amerika werden alle Jahre wieder im Frühjahr eine Reihe von Schmetterlingsarten mit
den warmen, nordwärts gerichteten Winden verdriftet ,pflanzen sich mit 1-2 Generationen während des Sommers fort. Sie sind aber, von einzelnen Ausnahmen abgesehen, nicht in der Lage, in den nördlichen Gebieten,d. h. in Europa nördlich der Alpen, zu überwintern . Einbekanntes Beispiel ist der Admiral Vanessa ata/anta, derim Frühjahr von Italien her Mittel- und Nordeuropasowie England besiedelt. Ein Teil der 2. Sommergeneration fliegt wieder nach Italien zurück, ein weiterer Teilgeht im Winter zugrunde. Das Wanderverhalten vonSchmetterlingen der Gattung Danaus in Nordamerikawird in Abschnitt 9.5.2.3 geschildert (Abb. 9-35).
Die Cicadellide Circulifer tenellus kommt im Mittelmeeraum und in den semiariden westlichen Gebieten derUSA an zahlreichen Kulturpflanzen und wilden Chenopodiaceen vor. Aus den Brutgebieten im Südwesten derUSA wandern im Frühling und vielfach auch im Sommer, wenn die Nährpflan zen vertrocknen und Übervölkerung einsetzt, Teile dieser Populationen mehrerehundert Kilometer nord- bis ostwärt s. Im Oktober bisDezember erfolgt in den USA die Rückwanderung der 3.und 4. Generation zur Überwinterung in den Ausgangsgebieten.
Eine andere wirtschaftlich wichtige Zikadenart, Empoascafabae, wandert regelmäßig im Frühling aus ihrenÜberwinterungsgebieten in den Staaten am Golf vonMexico nach Norden bis nach New York. Die Tierefliegen in großen Schwärmen unter Ausnutzung der vorherrschenden Windrichtung bis zu 150 km pro Tag undkönnen dabei bis in Höhen von 5000 m getragen werden.

23.3 Faunistik 727
Eine in Ostasien vorkommende Zikade, Nilaparvatalugens, die als Reisschädling gefürchtet ist, gelangt imFrühjahr mithilfe der stetigen Südwestwinde aus denBrut- und Überwinterungsgebieten im Süden Chinasweit nach Nordchina und kehrt im Herbst mithilfe derdann vorherrschenden Nordost-Winde wieder in dieAusgangsgebietezurück.
Die anholozyklische an Mais lebende Blattlaus Rhopalosiphum maidis lebt in tropischen und subtropischenGebieten und dringt im Sommer weit nach Norden vor,in Amerika bis nach Kanada, in Europa bis nach Skandinavien. Eine Rückwanderung findet in diesem Fallenicht statt und eine Überwinterung ist nicht möglich. Diein den Norden gelangten Populationen sterben imHerbst aus.
Inseln im Meer, Höhlen, aus einer Ebene aufragende Gebirgsstöcke oder Oasen in der Wüste sindIsolationsräume und haben gemeinsame Merkmale. In manchen dieser Räume wird die Evolution neuer Arten erheblich gefördert. Ein bekanntes Beispiel ist Neu-Guinea, dessen Artenfülle bedingt ist durch die zahllosen Möglichkeiten zurEinnischung in Bergregionen von mehr als 5000 mHöhe, deren Hänge wegen des feucht-tropischenKlimas bis in die höchsten Regionen besiedeltwerden können.
23.3 Faunistik
Die Faunistik stellt die einzelnen Tierformen alsFaunenelemente bestimmter Gebiete zusammen,untersucht sie taxonomisch und versucht, ihr Verhältnis zu den anderen Tierformen innerhalb derBiozönosen zu ermitteln. Sie charakterisiert KernAreale, in denen diese Arten leben. Beispielsweisebreiten sich wärmeliebende Arten, deren KernAreal im nördlichen Mittelmeerraum liegt , überFlußtäler bis weit in nördliche Gegenden aus. Aufdiese Weise gelangen mediterrane Faunenelementebeispielsweise über das Rheintal bis in das Moselund Ahrtal oder über Gebirgstäler aus dem Mittelmeerraum bis tief in die Schweiz . PannonischeFaunenelemente erreichen über das Flusssystemder Donau bestimmte Gebiete in Deutschlandoder über die Po-Ebene Gebirgstäler in derSchweiz .
Die vielfältigen ökologischen Aspekte der Faunistik sollen hier nicht erörtert werden.

728 23 Tiergeographie
Tab. 23·1: Die tiergeographischen Regionen der landfauna. (Nach de Lattin, 1967)
Region
1. Holarktische Region
2. Orientalische (= Indische) Region3. Äthiopische Region
4. Neotropische Region5. Australische Region
23.4 TiergeographischeRegionen
Im Jahre 1858 erschienen in den .Proceedings ofthe Linnean Society of London" nicht nur dieberühmten Abhandlungen von Darwin und Wallace über die Evolution der Lebewesen durch na türliche Selektion, sondern auch die erste Definition tiergeographischer Regionen du rch Slater.Wallace erweiterte diese Ergebnisse. Slater (1858)und Wallace (1876) erkannten, dass die Landfauna der Erde in großen Gebieten Übereinstimmungen aufwe ist. Daher konzipierten sie ein Sy-
Subregion
Paläarktische SubregionNearktische Subregion
Afrikanische SubregionMadegassische Subregion
Kontinental-australische SubregionNeuseeländische SubregionPolynesische Subregion
stem von 5 Regionen, das im Laufe der Zeit inmancherlei Hinsicht Veränderungen erforderte,aber im Prinzip erh alten blieb (Abb. 23-6). DieCharakteris ierung tiergeographischer Regionenerfolgt anhand der nur in diesen Gebieten vorkommenden endemischen Arten . Zunächst dientenzur Charakterisierung dieser Regionen und ihrerAusdehnung vorwiegend das Vorhandensein oderFehlen bestimmter Wirbeltierarten und -gruppen,in erster Linie Säugetiere und Vögel. Je mehrTiergruppen im Laufe der Zeit für diese Charakterisierung herangezogen wurden, desto mehrMeinungsverschiedenheiten gab es im einzelnen.Dennoch blieb die regionale Gliederung der Land-
Abb. 23·6: Die tiergeographischen Regionen der Erde. Nur bei der Holarktis sind die Subregionen (Paläarktis und Nearktis),genannt. Die Ausdehnung der übrigen (s. Text) istlediglich durch gestrichelte Linien angedeutet. Misch- oder Übergangsgebiete sinddurch Punktierung hervorgehoben. Sie können beträchtliche Größe erreichen. Das im Bereich der Sundainseln und Molukkenliegende, als Wallacea bezeichnete Übergangsgebiet zwischen Orientalis und Australis ist um der Übersicht willen nicht punktiert.Die Antarktis ist nicht eingezeichnet. (Nach Ziswiler 1981 , verändert)

fauna in allen wesentlichen Aspekten erhalten.Bemerkenswert ist die Übereinstimmung von Faunen- und Florenregionen.
Problematisch bleibt die Grenzziehung zwischen zwei Regionen, wenn in umfangreichen Gebieten eine Durchmischung der für die benachbarten Kerngebiete charakteristischen Fauna erfolgt. Beispiele hierfür sind die sehr weiträumigeMischzone zwischen holarktischer und orientalischer Region in Ostasien, die fast ganz Chinabetrifft (Abb. 23-6) sowie die sehr ausgedehntenordafrikanische Mischzone zwischen Paläarktisund Äthiopis und die mittelamerikanische Mischzone zwischen Nearktis und Neotropis.
Die Antarktis wird wegen der geringen Zahl der Artennicht als 6. Region gezählt. Sie ist neuerdings von besonderem Interesse im Zu sammenhang mit der Besiedelung der Südkontinente während des Mesozoikums (s.23.5).
Problematisch sind auch die Faunen küstenferner,kleiner Inseln, deren Endemiten dort auf unterschiedliche Weise eintreffen und einander auchdezimieren können. Nachträglich durch denSchiffsverkehr eingeschleppte Tiere, vor allemRatten und Katzen, können zumindest Teile derursprünglich angesiedelten Fauna vernichten. Daher werden Faunen küstenferner Inseln weder beider Abgrenzung der großen Faunenregionen nochals gesonderte Bereiche berücksichtigt.
Wegen der beachtlichen Fähigkeit der Insektenzu Wanderungen über See (s. 23.2) wurde neuerdings vorgeschlagen, Papua, Melanesien, Micronesien und Polynesien der Orientalischen Region zuzuordnen und anstelle der umstrittenenSubregion Wallacea eine Mischzone zwischen Orientalischer und Australischer Region im Bereichdes südlichen Neuguinea und des nordöstlichenAustralien anzunehmen.
23.5 Historische Tiergeographie
Dieses Teilgebiet der Tiergeographie der Insektenerhielt starken Auftrieb durch die Entdeckung undSicherung der Vorstellungen von der Kontinentalverschiebung und der Plattentektonik (Abb. 23-7)sowie durch die Entdeckung fossilen Materials,das zeigte, dass viele Insektengruppen älter alserwartet sind. Von ganz besonderer Bedeutungsind in dieser Hinsicht die Neuentdeckungen vonFossilien im Bereich der Südkontinente, einschließlich Antarctica. Die Insekten dürften mindestens im Devon , wahrscheinlich aber schon früher im damals tropischen Laurentia (s.u.) entstanden sein. Die ersten geflügelten Insekten fand
23.5 HistorischeTiergeographie 729
man im Devon von Angara (Sibirien) (s. u.) sowieim mittleren Karbon von Laurentia, Nordamerikaund Grönland. Im Allgemeinen reichen die Fossilfunde aber noch keineswegs aus, um gesicherteVorstellungen über den Ort und Zeitraum derEntstehung einzelner Gruppen zu gewinnen . InTab. 23-2 sind die Erdperioden und ihre jeweiligeDauer aufgeführt.
Innerhalb der einzelnen Gruppen der Insekten kann manin einigen Untergruppen Phasen starker Aufgliederung(Radiation) zu verschiedenen Zeiten feststellen. Unterden Diptera entwickelten sich beispiel sweise die Nematocera und die primitiven Brachycera bereits während desmittleren Mesozoikums, während die CyclorrhaphaSchizophora erst im Terti är eine starke Aufgliederungzeigten. So kommt es, dass eng verwandte Arten derNematocera sowohl in Süd amerika wie in Australienvorkommen. Bei manchen dieser Arten konnte wahrscheinlich gemacht werden , dass sie auf dem Wege überAntarctica entweder in der einen oder in de r anderenRichtung diese Verbreitung erreichten. Für Cyclorrhapha ist dergleichen nichts nachgewiesen.
Nach einer übertriebenen Vorstellung hätten die Insekten nach ihrer Entstehung im Devon wegen ihrerBeweglichkeit hinreichend Zeit gehabt, sich weltweit aus zubreiten, sodass ein Einfluss der erdgeschichtlichen Veränderungen und der Kontinentalverschiebung bedeutungslos sei. Alle bisher gewonnenen, einigermaßen gesichert erscheinenden Ergebnisse spre chen gegen einederart radikale und zugleich na ive Auffa ssung.
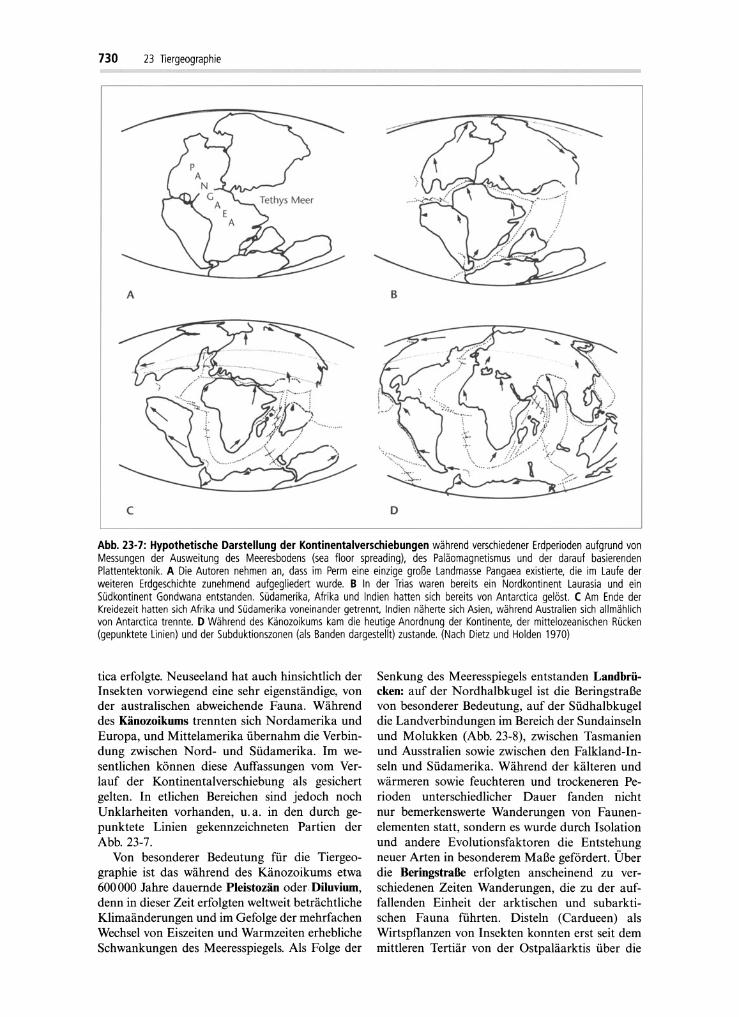
Abb. 23-7 zeigt eine Darstellung der Veränderungen auf der Erde in 4 Erdperioden, die für diePhylogen ie der Lebewesen von besonderer Bedeutung sind. In diesen Zeiten erfolgte eine zunehmende Aufgliederung der ursprünglichen, bis zumEnde des Perm vorhandenen riesigen, Pangaeagenannten Landrnasse durch sog. "seafloor spreading" und einer dadurch zustande kommendenKontinentalverschiebung. In der Trias war auf derNordhalbkugel eine riesige Landrnasse, Laurasia,bereits abgegliedert. Auf der Südhalbkugel wurdeeine ähnlich große Landrnasse, Gondwana-Land,abgetrennt und zunehmend zergliedert. Dabei lösten sich von Antarctica sowohl Südamerika alsauch Afrika und das damit noch verbundene Madagascar. Außerdem trennten sich von Antarcticanacheinander auch Indien und Australien. Allediese Teile drifteten nordwärts. Während des Mesozoikums begann die Aufgliederung von Laurasiain Laurentia und Angara (Abb. 23-7). Letztereswar seit dem Kambrium nicht von Wasser bedeckt. Auf der Südhalbkugel trennten sich Südamerika und Afrika sowie Afrika und Madagaskar. Indien driftete während des Tertiär gegen dieasiatische .Landmasse, wodurch es zur Auffaltungdes Himalaja und der benachbarten Gebirge kam.Vermutlich verlor Neuseeland erst in der UnterenKreide die Verbindung zu Antarctica, und zwarspäter als die Trennung Australiens von Antarc-
730 23 Tiergeographie
A B
C D
Abb. 23-7: Hypothetische Darstellung der Kontinentalverschiebungen während verschiedener Erdperioden aufgrund vonMessungen der Ausweitung des Meeresbodens (sea floar spreading), des Paläomagnetismus und der darauf basierendenPlattentektonik. A Die Autoren nehmen an, dass im Perm eine einzige große Landmasse Pangaea existierte, die im Laufe derweiteren Erdgeschichte zunehmend aufgegliedert wurde. B In der Trias waren bereits ein Nordkontinent Laurasia und einSüdkontinent Gondwana entstanden. Südamerika, Afrika und Indien hatten sich bereits von Antarctica gelöst. C Am Ende derKreidezeit hatten sich Afrika und Südamerika voneinander getrennt, Indien näherte sich Asien, während Australien sich allmählichvon Antarctica trennte. D Während des Känozoikums kam die heutige Anordnung der Kontinente, der mittelozeanischen Rücken(gepunktete Lin ien) und der Subduktionszonen (als Banden dargestellt) zustande. (Nach Dietz und Holden 1970)
tica erfolgte. Neuseeland hat auch hinsichtlich derInsekten vorwiegend eine sehr eigenständige, vonder australischen abweichende Fauna . Währenddes Känozoikums trennten sich Nordamerika undEuropa, und Mittelamerika übernahm die Verbindung zwischen Nord- und Südamerika. Im wesentlichen können diese Auffassungen vom Verlauf der Kontinentalverschiebung als gesichertgelten. In etlichen Bereichen sind jedoch nochUnklarheiten vorhanden, u.a. in den durch gepunktete Linien gekennzeichneten Partien derAbb. 23-7.
Von besonderer Bedeutung für die Tiergeographie ist das während des Känozoikums etwa600000 Jahre dauernde Pleistozän oder Diluvium,denn in dieser Zeit erfolgten weltweitbeträchtlicheKlimaänderungen und im Gefolge der mehrfachenWechselvon Eiszeiten und Warmzeiten erheblicheSchwankungen des Meeresspiegels. Als Folge der
Senkung des Meeresspiegels entstanden Landbrücken: auf der Nordhalbkugel ist die Beringstraßevon besonderer Bedeutung, auf der Südhalbkugeldie Landverbindungen im Bereich der Sundainselnund Molukken (Abb. 23-8), zwischen Tasmanienund Ausstralien sowie zwischen den Falkland-Insein und Südamerika. Während der kälteren undwärmeren sowie feuchteren und trockeneren Perioden unterschiedlicher Dauer fanden nichtnur bemerkenswerte Wanderungen von Faunenelementen statt, sondern es wurde durch Isolationund andere Evolutionsfaktoren die Entstehungneuer Arten in besonderem Maße gefördert . Überdie Beringstraße erfolgten anscheinend zu verschiedenen Zeiten Wanderungen , die zu der auffallenden Einheit der arktischen und subarktischen Fauna führten . Disteln (Cardueen) alsWirtspflanzen von Insekten konnten erst seit demmittleren Tertiär von der Ostpaläarktis über die

23.5 HistorischeTiergeographie 731
I
.:-
•~ .~...~:.
... ...\
\ ...\ ...
'.
"" .
~.,f>. ~~" :•'c::::::::r
,
I
If
f,,/ Lyd;kker-Linie
. " "',1 "
'I' 0 /f II I
I I'-O._~_
-f • (~I~~''''\ ~.....\ ...
Abb. 23-8: Die Übergangszone zwischen Orientalis und Australis. DerVersuch einer Abgrenzung diesesüberauskomplexen,vielfach und gründlich untersuchten Übergangsbereichs durch dieWallace-LinieimWesten, dieWeber-Linie inder MittesowiederimOsten verlaufenden Lydekker-Linie und die Benennung dieses Gebietes als Wallacea erwies sich im Laufe der Zeit als nichtbefriedigend. (Nach de Lattin 1967)
Bering-Landbrücke nach Nordamerika einwandern. So bildete beispielsweise die Gattung Cirsium dort 50 endemische Arten, die inzwischenvorwiegend im Westen Nordamerikas verbreitetsind . Von den Phytophagen dieser Disteln erreichten Bohrfliegen des Tribus Terellini die Ostpaläarktis erst , als die Beringstraße nicht mehr pas sierbar war. Sie konnten aber mit ihren Wirtspflanzen nach Nordamerika einwandern, als imPliozän eine weitere Hebung erneut eine BeringLandbrücke ergab. Im Pleistozän dürfte über dieseLandbrücke auch die Bohrfliegen-Gattung Chaestome/la mit Distel-Arten der Gattung Cirsiumnach Nordamerika gelangt sein.
Ein intensiver Austau sch der Faunen und Floren dürfte auch über die breite, bis zum Ende derKre idezeit existierende Verbindung zwischenNordeuropa und dem heutigen Kan ad a erfolgtsein. Die Beringstraße war außerdem auch für dieBesiedelung Mittel- und Süd amerikas von Bedeutung, denn die Insektenfauna der Nearktis und derNeotropis weisen viele Übereinstimmungen auf.Andererseits lässt die Fauna des südlichen Süd amerika deutlich erkennen, da ss sie in erheblichemMaße durch einen Austausch von Faunenanteilenauf dem Wege über Antarctica entstanden seindürfte.
Während der Eiszeiten in Europa drangen, ausgehend von den skandinavischen Hochgebirgeneinerseits und den Alpen andererseits, gewaltigeEismassen vor. In Norddeutschland unterscheidetman 3 Eiszeiten (Elster-, Saale-, Weichseleiszeit) ,im Gebiet der Alpen 4 Eiszeiten (Günz-, Mindel-,Riss- und Würmeiszeit). Diese Eismassen zogensich in Norddeutschland in 4, in den Alpen in 5Stufen wieder zurück. Zahlreiche Faunenelemente
Tab. 23-2: Die Erdzeitalter. (Nach Hohl, 1981)
Periodein
ambnum 70OrdovlZIum 70Silur 35Devon 55Karbon 65Perm 60
esozoi um Trias 30Jura 55Kreide 75
Kano-, eozoikum Iertiär 65Quartär 1,5
Plelstozan (Diluvium)Holoz än (Alluvium)

732 23 TIergeographie
I,,
.. ..
i/
II
/./ : -.
.1'. . :: " e.. .
"-J-'-'- : J) .- ~.. .: ."'--" ' ' . .. ........ "'-.-._)
»>:
•••• Elster-Vereisung------ Weichsel -Vereisung
Saale-Vereisungnicht vereiste Gebiete
Abb. 23·9: Die Ausdehnung der nordeuropäischen und alpinen Eismassen im Pleistozän, dieAbwanderung von Arten inwärmere Gebiete Südwest- und Südosteuropas und die Rückwanderung nach dem Ende der Eiszeiten. DieWanderungsrich tungensind durch die Pfeil spitzen angedeutet. (Nach versch . Autoren, verändert)
wichen vor dem vordringenden Eis und dem damiteinsetzenden kälteren Klima einerseits nach Südwesten und andererseits nach Südosten aus(Abb. 23-9). Anscheinend fanden viele Arten keineAusweichmöglichkeit in wärm ere Gebiete und gingen zugrunde.
In der Nacheiszeit, dem Holozän oder Alluvium, das etwa 12000 Jahre andauerte, zogen dieausgewanderten Faunenanteile wieder in die zunehmend wärmer werdenden Gebiete ein. Je nachWanderungsgeschwindigkeit und anderen Faktoren sind die heutigen Bcsiedelungsgrenzen der einzelnen Arten in unte rschiedlichen Bereichen zufinden . Vielfach haben sich bereits Subspezies ge-
bildet (Abb. 23-2). Ein Refugium während der Eiszeit wurde zum Ausbreitungsareal nach der Eiszeit.
Vielfach ist der Mensch für die Ausbreitung von Artenverantwortlich. Bekannte Beispiele sind die StubenfliegeMusca domestica, die Stechmücke Cu/ex fat igans, dieKleiderlaus Pedicu/us humanus, die Filzlaus Pth irus pubis,zahlreiche Pflanzenschädlinge u. a. (s. Kap. 21). Europaund Nordamerika haben mindestens 5000 Tierart en gemeinsam. Die Hälfte davon dürfte durch den Menschenverschleppt worden sein, wobei etwa zehnmal so vielArten von Europa nach Nordameri ka gelangten wie inumgekehrter Richtung. 63 nach Nordamerika eingeschleppten Schmetterlingsarten, darunter 16 Pyralidae

Abb. 23-10: Die rasche Ausbreitungdes Kartoffelkäfers Leptinotarsadecemlineata in Europa. (Aus Sedlag1995)
Literatur 733
und 10 Tortricidae, stehen nur 3 nach Europa verfrachtete Arten gegenüber. Besonders beachtet und vielfachkartiert wurde die Ausbreitung von Schädlingen nachihrer erfolgreichen Einschleppung. Ein gutes Beispiel liefert ein gefürchteter Forstschädling, der Schwammspinner Lymantria dispar, der aus Europa in die USA importiert wurde. Besonders bekannt wurden die aus denUSA nach Europa eingeschleppten Schädlinge ReblausViteus vitifolii und der Kartoffelkäfer Leptinotarsa decemlineata (Abb. 23-10). Vielfach mussten nach der Einschleppung von Schädlingen geeignete Parasiten zur biologischen Bekämpfung importiert werden . Das gilt nichtnur für tierische Schädlinge, sondern auch für die Un krautbekämpfung (s. 21.1) . Nach der Einschleppung vonOpuntien wurde in Australien zur Bekämpfung derKlein schmetterling Cactoblastis cactorum eingeführt.Die Rinderzucht erforderte in Australien die Einfuhr vonMistkäfern, um den Rindermist auf den Weiden abzubauen (s. 21.2 .1.2). Einheimische Insekten waren nurzum Abbau von trockenem Rindermist fähig. Zahlreicheweitere Beispiele für die absichtliche und unabsichtlicheFaunenveränderung durch den Menschen könnten noc hangeführt werden .
Literatur
Avise, 1.c, Amold, 1., Ball, R. M., Bermingham, E.,Lamb, T., Neigel , 1.E., Reeb, C. A., Sanders, N.C.(1987): Intraspecific phylogeography: the mitochondrial DNA bridge between population genetics andsystematics. Ann . Rev. Entomo!. System. 18,489-522
Brundin, L. (1971): Phylogenetics and biogeography.Syst. Zoo!. 21: 69-79
Cooper, S.1., Ibrahim, K. M. , Hewitt, G. M. (1995): Post glacial expansion and genome subdivision in the European grasshopper Chorthippus parallelus. Mo!.Eco!. 4, 49-60
Dietz , R.S., Ho lden, 1.c. (1979): The breakup of Pan gaea . Sei. Amer. 223 (4): 3~1
Drake, V.A., Gatehouse, A.G. (1995): Insect migration.478 p. Cambridge University Press, Cambridge
Glemeroth, A. K . (1995): Paläoökologische Untersuchungen über die letzten 22000 Jahre in Europa.Akad. Wiss. Lit. Paläoklimaforschg. 18. Fischer Verlag, Stuttgart
Gressitt, 1.L. (1958): Zoogeography ofinsects. Ann , Rev.Entomo!. 3: 207-230
Gre ssitt , 1.L. (1974): Insect biogeography. Ann . Rev.Entomo!. 19: 293-321
Hallam, A. (1972): Continental drift and the fossil record. Sei. Amer. 227 (5): 56-66
Hennig, W. (1960): Die Dipteren-Fauna von Neu seelandals systematisches und tiergeografisches Problem.Beitr. Entomo!. 10: 221-329
Hennig, W. (1969): Die Stammesgeschichte der Insekten.436 S. Kramer, Frankfurt
Holdhaus, K. (1994): Die Spuren der Eiszeit in derTierwelt Europas. Abh . zoo!.-bot. Ges. Wien 18:1--493
Hurley, P. M. (1968): The confirmation of continentaldrift. Sei. Amer. 218 (4): 52-64
Munroe, E. (1965): Zoogeography of insects and alliedgroups. Ann . Rev. Entomo!. 10: 325-344
IlIies,1. (1983): Changing concepts in biogeography. Ann .Rev. Entomo!. 28: 391--406
Johnson, C. G. (1969): Migration and dispersal of insectsby t1ight. 763 p. Methuen, London
Keast, A. (1971): Continental drift and the evolution ofthe biota on southern continents. Quart. Rev. Bio!. 46(4): 335-378
De Lattin, G. (1967): Grundriß der Zoogeographie. Fischer, Stuttgart
Mackeras, r. M. (1970) : Composition and distribution ofthe fauna. In: Insects of Australia. r. M. Mackeras etal. (eds.): CSIRO, Melboume. 187-203
McKenzie, D. P. (1970): Plate tectonics and continentaldrift. Endeavour 29: 39--44
McKenzie, D.P. (1972): Plate tectonics and sea-t1oorspreading. Am. Sei. 60: 425--435
Müller, P. (1980): Biogeographie. Ulmer, StuttgartRoderick, G. K. (1996): Geographie structure of insect
populations: gene t1ow, phylogeography, and theiruses. Ann. Rev. Entomo!. 41, 325-352
Schlinger, E. r. (1974): Continental drift, Nothofagus, andsome ecologically associated insects. Ann. Rev. Entomo!. 19: 323-343

734 23 Tiergeographie
Taberlet , P., Fumagalli, L., Wust-Saucy, A.-G., Cosson,1.F. (1998): Comparative phylogeography and postglacial colonization routes in Europe. Mo!. Eco!. 7,453-464
Udvardy, M. D. F. (1969): Dynamic zoogeography, withspecial reference to land animals. 445 p. Van Nostrand & Reinhold, New York
Warnecke, G. (1961): Rezente Arealvergrößerung beiMakrolepidopteren in Mittel- und Nordeuropa.Bonn . Zoo!. Beitr. 12: 113-141
Williamson, M. (1981): Island populations. Oxford Uni versity Press, Oxford
Zeuner, F.E. (1959): The pleistocene period, its c1imate,chronology and faunal successions. 447 p. Hutehinson, London
Zwick, P.(1980): Plecoptera (Steinfliegen) . Handb. Zoo!.4 (2) 2/7: 1-115
Zwölfer, H. (1988): Evolutionary and ecological relationships of the insect fauna of thistles. Ann . Rev.Entomo!. 33: 103-122




![Lehrbuch Persönlichkeitspsychologie · : [Lehrbuch]__Schuetz__Persoenlichkeitspsychologie__[Druck-PDF]/04.05.2016/Seite 2 Lehrbuch Persönlichkeitspsychologie Astrid Schütz, Martin](https://img.pdfslide.org/doc/110x75/5e1ef9515ee7da6b5b6d41fc/lehrbuch-persnlic-lehrbuchschuetzpersoenlichkeitspsychologiedruck-pdf04052016seite.jpg)