Embed Size (px)
Citation preview

Mathematische Modelle fur
Chemotaxis gemischter
Zellsysteme
Bachelorarbeitzur Erlangung des akademischen Grades
Bachelor of Science
Westfalische Wilhelms-Universitat Munster
Fachbereich Mathematik und Informatik
Institut fur Numerische und Angewandte Mathematik
Betreuung:
Prof. Dr. Martin Burger
Dr. Frank Wubbeling
Eingereicht von:
Lena Frerking
Munster, August 2010

i
Zusammenfassung
In dieser Arbeit entwickeln wir Kontinuumsmodelle fur Chemotaxis am Beispiel des
zellularen Schleimpilzes Dictyostelium discoideum. Wir leiten partielle Differentialglei-
chungen her, die die gerichtete Bewegung von Zellen aufgrund eines chemischen Gradi-
enten beschreiben. Dabei berucksichtigen wir die Existenz einer maximalen Zelldichte,
welcher eine erhebliche Bedeutung fur die Existenz von globalen Losungen solcher Mo-
delle zukommt.
Der Großteil der Modelle fur Chemotaxis beschrankt sich ausschließlich auf den Fall,
dass in einem Gebiet nur ein Typ von Zellen vorkommt. Wir betrachten auch Gebiete
mit verschiedenen Populationen, die sich gegenseitig beeinflussen. Hierfur zeigen wir,
dass die Entropie der Zellen mit der Zeit abnimmt. Die unterschiedlichen Populationen
sortieren sich durch die Wirkung der Chemotaxis auf eine bestimmte Art und Weise
und bilden so ein spezifisches Muster.

ii
Eidesstattliche Erklarung
Hiermit versichere ich, Lena Frerking, dass ich die vorliegende Arbeit selbststandig
verfasst und keine anderen als die angegebenen Quellen und Hilfsmittel verwendet
habe. Gedanklich, inhaltlich oder wortlich Ubernommenes habe ich durch Angabe von
Herkunft und Text oder Anmerkung belegt bzw. kenntlich gemacht. Dies gilt in gleicher
Weise fur Bilder, Tabellen, Zeichnungen und Skizzen, die nicht von mir selbst erstellt
wurden.
Das auf der CD beigefugte Programm ist von mir selbst programmiert worden.
Munster, 23. August 2010
Lena Frerking

iii
Inhaltsverzeichnis
1. Einleitung 1
2. Ein-Zell-Populationen 5
2.1. Minimalmodell . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
2.1.1. Herleitung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
2.1.2. Existenz von globalen Losungen . . . . . . . . . . . . . . . . . . 9
2.2. Modellvariationen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
2.3. Volume-Filling Modell . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
2.3.1. Herleitung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
2.3.2. Existenz von globalen Losungen . . . . . . . . . . . . . . . . . . 13
3. Zwei-Zell-Populationen 14
3.1. Herleitung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
3.2. Eigenschaften des Modells . . . . . . . . . . . . . . . . . . . . . . . . . 18
3.3. Numerische Ergebnisse . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
4. Fazit und Ausblick 29
A. Quellcode 31
Literaturverzeichnis 33

1
1. Einleitung
In dieser Arbeit widmen wir uns der mathematischen Modellierung der Chemotaxis von
Zellen. Zunachst werden wir in diesem Kapitel den Prozess der Chemotaxis biologisch
erlautern und anschließend einen kurzen Uberblick uber den Aufbau der Arbeit geben.
Taxis bezeichnet allgemein die Reaktion von Zellen oder Organismen auf ein externes
Signal. Dabei wird aufgrund einer Veranderung der Umwelt eine Bewegung ausgefuhrt.
Dies kann zum Beispiel eine Anderung der Temperatur sein, oder aber auch veranderte
Licht- oder Windverhaltnisse.
Chemotaxis ist eine von chemischen Reizen ausgeloste gerichtete Bewegung. Die Veran-
derung der Umwelt ist hier somit die Veranderung eines Konzentrationsgradienten ei-
nes chemischen Stoffes. Beim Schleimpilz Dictyostelium discoideum, auf den wir spater
noch naher eingehen werden, ist dieses chemische Signal cyclisches Adenosinmonophos-
phat (cAMP).
Chemotaktische Bewegungen werden unterschieden in positiv und negativ. Eine positi-
ve chemotaktische Bewegung ist hierbei die Bewegung hin zu einer hoheren chemischen
Konzentration. Eine negative chemotaktische Bewegung ist eine Bewegung in die Ge-
genrichtung der hoheren Konzentration.
Anhand einiger Beispiele werden wir nun die enorme Bedeutung der Chemotaxis in der
Biologie und in der Medizin erlautern.
Zunachst wird die Bewegung von Tieren direkt durch Chemotaxis beeinflusst. Ist bei-
spielsweise ein Tier auf Nahrungssuche, wird es sich, sofern es eine Futterquelle wittert,
darauf zubewegen. Hier findet eine positive chemotaktische Bewegung statt. Anders-
herum wird sich ein Tier, wenn es eine Gefahr wittert, zum Beispiel in Form eines
anderen Tieres, im Normalfall davon wegbewegen. Es bewegt sich negativ chemotak-
tisch.
Auch in der Medizin ist Chemotaxis von großer Bedeutung. So konnen sich auch im
menschlichen Korper allein durch Chemotaxis die Leukozyten, also die Immunzellen,
direkt zu bakteriell entzundeten Gebieten hinbewegen, um so moglichst schnell eine
Heilung herbeizufuhren.

1 Einleitung 2
(a) Fruhe Aggregationsphase [1] (b) Pseudoplasmodium [2] (c) Fruchtkorper [3]
Abbildung 1.1.: Entwicklungsstadien des Schleimpilzes Dictyostelium discoideum
Allerdings spielt Chemotaxis auch in Bezug auf Krankheiten eine Rolle. Tumorzellen
sind durch diese Form von Bewegung in der Lage, umgebende Gebiete zu befallen und
sich somit weiter auszubreiten.
Auf ein weiteres Beispiel werden wir jetzt etwas genauer eingehen. Der nur aus einer
einzigen Zelle bestehende Schleimpilz Dictyostelium discoideum schafft es nur mit Hilfe
von Chemotaxis, den Fortbestand seiner Art auch bei schlechten außeren Bedingun-
gen zu sichern. Solange noch genugend Nahrung vorhanden ist, pflanzen sich die Zellen
durch Zellteilung fort; die Population wachst. Im Fall einer Nahrungsknappheit beginnt
eine Zelle, die sogenannte Grunderzelle, cyclisches Adenosinmonophosphat (cAMP)
abzusondern. Es kann hierbei jede beliebige Zelle zur Grunderzelle werden. Von der
erhohten Konzentration von cAMP in der Umgebung der Grunderzelle werden nun die
anderen Zellen angezogen. Das cAMP wirkt also als Lockstoff. Ist eine Zelle in der
Nahe der Grunderzelle angekommen, so beginnt sie ebenfalls cAMP abzusondern um
so noch mehr Zellen anzulocken. Auf diese Art und Weise versammeln sich nahezu
alle Zellen an einer Stelle. Nun aggregieren immer 50.000 bis 100.000 Zellen zu ein-
zelnen Haufen, was in Abbildung 1.1 (a) zu sehen ist. Sind diese Haufen entstanden,
beginnen sich die Zellen eines jeden Haufens, wie in Abbildung 1.1 (b), zu einem soge-
nannten Pseudoplasmodium zu vereinigen. Dieses Pseudoplasmodium bewegt sich in
Richtung Licht. Hat es eine helle Stelle erreicht, sortieren sich die beiden Untergruppen
Prasporenzellen, zu denen 80 Prozent der gesamten Zellen gehoren, und Prastielzellen,
die die restlichen 20 Prozent bilden, nach ihrem Typ. Danach beginnen sie mit der
Ausbildung eines Fruchtkorpers. Dieser besteht, wie in Abbildung 1.1 (c) zu sehen ist,
aus einer Spore und einem Stiel, welche sich aus den Prasporenzellen beziehungsweise
aus den Prastielzellen zusammensetzen. Ist der Fruchtkorper fertig ausgebildet, setzt
er die Spore frei. Die Stielzellen sterben dann ab. Wenn die Bedingungen irgendwann

1 Einleitung 3
Abbildung 1.2.: Entwicklungszyklus des Schleimpilzes Dictyostelium discoideum. Diese Grafik ent-stammt [4].
wieder gunstiger sind, keimt die Spore und setzt wieder neue Zellen frei. Der Kreislauf
kann nun wieder von Neuem beginnen.
Das Besondere an dem Schleimpilz Dictyostelium discoideum ist, dass die einzelnen
Zellen wahrend des gesamten Prozesses ihre Plasmamembran und damit ihre eigene
Identitat behalten. Jede Zelle ist immer noch selbststandig lebensfahig. Der große Orga-
nismus Pseudoplasmodium setzt sich somit aus vielen kleinen Organismen zusammen.
Er ist ein multizellularer Organismus. Außerdem ist der Schleimpilz ein besonders in-
teressantes Forschungsobjekt fur Biologen, da er nur aus zwei verschiedenen Zelltypen,
namlich den Sporenzellen und den Stielzellen besteht. Musterbildung und Morpho-
genese, also die Entwicklung von Organen, finden gleichzeitig wie bei vielen hoheren
Organismen statt.
Abbildung 1.2 zeigt im Uhrzeigersinn eine Zusammenfassung des gesamten Lebenszy-
klus von Dictyostelium discoideum. Chemotaxis tritt hier gleich in mehreren Stadien
auf. Zu Beginn ist Chemotaxis dafur verantwortlich, dass die Zellen zu Haufen einer
bestimmten Große aggregieren. Sie sorgt sowohl dafur, dass die Zellen sich uberhaupt
an einem Ort versammeln als auch dafur, dass die Haufen, die gebildet werden, eine
bestimmte Große einhalten. Letzeres geschieht dadurch, dass die Zellen aufgrund der
unterschiedlichen Konzentration des chemischen Signals in ihrer Umgebung abschatzen,
wie viele andere Zellen sich bereits auf einem Haufen befinden. Außerdem ist Chemo-

1 Einleitung 4
taxis der Grund fur die Sortierung der Zellen im Stadium des Pseudoplasmodiums
kurz vor der Ausbildung des Fruchkorpers. Hier fuhren unterschiedliche Reaktionen
der verschiedenen Zellpopulationen auf das chemische Signal zu einer Musterbildung.
Unser Ziel ist es, ein mathematisches Modell zu entwickeln, das Chemotaxis in einem
Gebiet mit unterschiedlichen Zelltypen darstellen kann. Wir beginnen damit, in Kapi-
tel 2 das Keller-Segel Modell als ein Modell fur Chemotaxis herzuleiten, welches nur
Gebiete mit einer einzigen Population berucksichtigt. Insbesondere gehen wir dabei
auf die Teilmodelle Minimalmodell und Volume-Filling Modell ein, zu denen wir auch
die Existenz von globalen Losungen betrachten werden. In Kapitel 3 erweitern wir das
Volume-Filling Modell zu einem Modell fur zwei Populationen und charakterisieren
sowohl analytisch als auch numerisch dessen Eigenschaften in Bezug auf die Sortierung
von Zellen. Die Arbeit endet mit einer Diskussion und einem Ausblick auf zukunftige
Forschungsaspekte.

5
2. Ein-Zell-Populationen
In diesem Kapitel werden wir die Chemotaxis in Gebieten betrachten, in denen nur
ein einziger Typ von Zellen vorkommt. Hierzu werden wir das Keller-Segel Modell
einfuhren und ausfuhrlich auf die Herleitung des Minimalmodells als Teilmodell des
Keller-Segel Modells eingehen. Außerdem werden wir auch kurz die Existenz von globa-
len Losungen in diesem Modell betrachten. Anschließend werden wir eine Verfeinerung
des Minimalmodells, das Volume-Filling Modell, herleiten und analysieren.
Es gibt zwei Perspektiven, um den Prozess der Chemotaxis mathematisch zu betrach-
ten: die mikroskopische und die makroskopische Perspektive. Bei der mikroskopischen
Perspektive werden die einzelnen Zellen betrachtet. Dafur wird davon ausgegangen,
dass die Zellen sich in einem Random-Walk bewegen. Das heißt, eine Zelle kann sich in
einer Dimension nur nach rechts oder nach links bewegen. Außerdem wird in der Regel
angenommen, dass die Zellen nur Sprunge einer konstanten Lange h machen konnen.
Gesucht ist dann die Wahrscheinlichkeit dafur, dass sich eine Zelle zu einer bestimmten
Zeit an einem bestimmten Ort befindet. Befindet sich eine Zelle zum Zeitpunkt t + 1
an der Stelle x, so muss sie sich also zum Zeitpunkt t entweder an der Stelle x−h oder
bei x+ h befunden haben.
Bei der makroskopischen Perspektive wird ein Kontinuumsmodell betrachtet. Man lasst
die Anzahl der Zellen gegen unendlich laufen. Es werden also nicht mehr die einzelnen
Zellen betrachtet, sondern die Populationsdichte.
Das popularste Modell fur Chemotaxis ist das Keller-Segel Modell. Dies beschreibt
die Zelldichte und die Konzentration des chemischen Signals. Es ist also ein makro-
skopisches Modell. Das Besondere an diesem Modell ist, dass es die Modellierung von
raumlicher Musterbildung zum Beispiel bei Dictyostelium discoideum ermoglicht.
Es werden jedoch einige Voraussetzungen fur das Keller-Segel Modell benotigt. Zum
einen muss das chemische Signal, das betrachtet wird, als auto-attractant wirken, das
heißt als ein Lockstoff, von dem ein Organismus angezogen wird und den derselbe
Organismus auch produziert. cAMP ist fur Dictyostelium discoideum ein solcher auto-
attractant. Die Produktion dieses Stoffes ist damit abhangig von den Zellen. Eine

2 Ein-Zell-Populationen 6
weitere Vorraussetzung ist, dass das chemische Signal linear abgebaut wird. Außerdem
sollen Zellwachstum und Zelltod unabhangig von dem chemischen Signal sein [13].
In den verschiedenen Teilmodellen des Keller-Segel Modells werden die Parameter Dif-
fusionsvermogen und chemotaktische Sensitivitat der Zellen, Zellwachstum, Zelltod und
Produktion des chemischen Signals berucksichtigt und unterschiedlich festgelegt.
2.1. Minimalmodell
Das Keller-Segel Modell besteht aus mehreren Teilmodellen, die es ermoglichen, ver-
schiedene Aspekte zu berucksichtigen. Das am meisten erforschte Teilmodell ist das
Minimalmodell. Hier wird angenommen, dass alle Parameter, die im Keller-Segel Mo-
dell zu wahlen sind, konstant sind. Außerdem sollen die Zellen nicht direkt miteinander
interagieren. Jegliche Beeinflussung der Zellen untereinander soll indirekt durch das
chemische Signal stattfinden [13]. Es werden homogene Neumann-Randbedingungen
vorausgesetzt.
2.1.1. Herleitung
Der folgende Abschnitt orientiert sich an [13]. Zur Herleitung des Minimalmodells be-
trachten wir zunachst die mikroskopische Perspektive. Die Zellbewegung wird also als
Random-Walk mit konstanter Sprunglange h auf einem beschrankten, diskreten 1D-
Gitter angenommen. Dies fuhrt zu der sogenannten Mastergleichung
∂u(x, τ)
∂τ= T+
x−hu(x− h, τ) + T−x+hu(x+ h, τ)− (T+x + T−x )u(x, τ), (1)
wobei u(x, τ) die Zelldichte in x zur Zeit τ ist, T+x die Wahrscheinlichkeit von x um
h nach rechts und T−x die Wahrscheinlichkeit von x um h nach links zu springen. Der
erste Term gibt also die Wahrscheinlichkeit an, dass zum Zeitpunkt τ eine Zelle von
x− h nach x springt, der zweite Term steht fur die Wahrscheinlichkeit, dass eine Zelle
zur Zeit τ von x+h nach x springt und der letzte Term spiegelt die Wahrscheinlichkeit
wider, dass eine Zelle zur Zeit τ von x nach x+ h oder x− h springt.
Nun ist an der Chemotaxis gerade das Besondere, dass die Sprunge der Zellen durch den
raumlichen Gradienten eines chemischen Signals v beeinflusst werden. Daher besitzt T

2 Ein-Zell-Populationen 7
die folgende Form:
T±x = a+ b(v(x± h, τ)− v(x, τ)
)(2)
Das Einsetzen von T in die Mastergleichung ergibt zunachst
∂u
∂τ=(a+ b
(v(x, τ)− v(x− h, τ)
))u(x− h, τ)
+(a+ b
(v(x, τ)− v(x+ h, τ)
))u(x+ h, τ)
−((
a+ b(v(x+ h, τ)− v(x, τ)
))+(a+ b
(v(x− h, τ)− v(x, τ)
)))u(x, τ)
(3)
und eine anschließende Erweiterung der rechten Seite mit h2 und die Skalierung von τ
zu λt fuhrt zu
ut = λh2
h2
(a(u(x− h, λt) + u(x+ h, λt)− 2u(x, λt)
)− b((− v(x, λt) + v(x− h, λt)
)u(x− h, λt)
−(v(x, λt)− v(x+ h, λt)
)u(x+ h, λt)
+(v(x+ h, λt)− v(x, λt)
)u(x, λt)
+(v(x− h, λt)− v(x, λt)
)u(x, λt)
))= λ
h2
h
(a(u(x+ h, λt)− u(x, λt)
h− u(x, λt)− u(x− h, λ)
h
)− b(− v(x, λt)− v(x− h, λt)
hu(x− h, λt)
+v(x+ h, λt)− v(x, λt)
hu(x+ h, λt)
+v(x+ h, λt)− v(x, λt)
hu(x, λt)
− v(x, λt)− v(x− h, λt)h
u(x, λt)))
.
(4)

2 Ein-Zell-Populationen 8
Wir fuhren eine Taylor-Entwicklung bis zur zweiten Ordnung durch:
ut = λh2
(a(u(x+ h, λt)− u(x, λt)
h2− u(x, λt)− u(x− h, λt)
h2
)− b(v(x+ h, λt)− v(x, λt)
h2u(x+ h, λt)− v(x, λt)− v(x− h, λt)
h2u(x, λt)
+v(x+ h, λt)− v(x, λt)
h2u(x, λt)− v(x, λt)− v(x− h, λt)
h2u(x− h, λt)
))= λh2
(auxx − b
(2(vxu)x
)+O(h2)
)= λh2
(auxx − 2b(vxu)x
)+O(h4)
= λh2(aux − 2bvxu)x +O(h4)
(5)
Nach einer Umskalierung der Zeit mit Hilfe der Grenzwerte
limh→0,λ→∞
aλh2 = Du, limh→0, λ→∞
2bλh2 = χu (6)
folgt dann
ut = (Duux − χuuvx)x. (7)
Interessant ist das Modell nicht nur in einer, sondern auch in den hoheren Dimensionen.
Die Herleitung erfolgt aquivalent zu der fur eine Dimension:
ut = ∇ · (Du∇u− χuu∇v)
vt = ∆v + µu− δv(8)
Zum Schluss wird zur Vereinfachung des Modells noch eine Entdimensionalisierung
durchgefuhrt. Dazu werden die Parameter wie folgt gewahlt:
x∗ =
√δ
Dv
x, t∗ = δt, u∗ =u
u′, v∗ =
δ
µu′, D =
Du
Dv
, χ =χuµu
′
δDv
(9)
mit u′ =∫
Ωu(x, 0) dx/|Ω|. Daraus ergibt sich durch Einsetzen in (8) und nach Weglas-
sen der Sternchen das Minimalmodell :
ut = ∇ · (D∇u− χu∇v)
vt = ∆v + u− v(10)

2 Ein-Zell-Populationen 9
2.1.2. Existenz von globalen Losungen
Das Minimalmodell hat interessante Eigenschaften in Bezug auf die Existenz von globa-
len Losungen. Es ist namlich so, dass in einer Dimension fur alle Anfangswerte globale
Losungen existieren. Das heißt, egal welche Werte eingesetzt werden, es gibt zu jeder
beliebigen Zeit t feste Werte fur die Zelldichte und die Population des chemischen Si-
gnals. Dies resultiert aus
Lemma 1 (nach Hillen und Painter [13]). Losungen fur die 1D-Version von (10) sind
global in der Zeit.
Allerdings kann fur t→∞ ein sogenannter blow-up auftreten. Das bedeutet, dass die
Zelldichte gegen unendlich geht. Biologisch gesehen ist es jedoch unmoglich, dass die
Zelldichte unendlich groß ist. Dies ist ein Zeichen dafur, dass das Modell zu sehr ver-
einfacht wurde. Es spiegelt nicht den exakt realistischen Verlauf der Populationsdichte
wider.
In zwei Dimensionen gibt es eine Art Grenze fur die Anfangswerte bezuglich der Exis-
tenz von globalen Losungen. Liegt ein Anfangswert darunter, so gibt es eine globale
Losung. Dieses Ergebnis spiegelt sich in folgendem Satz wider:
Satz 1 (nach Jager und Luckhaus [17]). Wir nehmen an, dass Ω ein beschranktes
Gebiet im R2 ist, δΩ ist ein C1-Rand, u0 ist C1 und erfullt die Randbedingung. Dann
existiert ein kritischer Wert c(Ω), so dass
1
|Ω|
∫Ω
u(x, 0) dxχ < c(Ω)
impliziert, dass eine eindeutige, glatte, positive Losung von (10) fur alle Zeiten existiert.
Liegt er jedoch daruber, so tritt nach [13] ein blow-up schon in endlicher Zeit auf. Das
bedeutet, dass ein t0 existiert mit limt→t0 u(x, t) = ∞. Fur ein großeres t ist u dann
nicht mehr definiert. Tritt so ein blow-up auf, so liegt er in jedem Fall auf dem Rand
des Gebiets. Biologisch sinnvoll fur das Auftreten von Aggregation sind allerdings nur
solche Anfangswerte, die uber der Grenze liegen, fur die also in diesem Modell ein
blow-up auftritt.
In drei oder mehr Dimensionen tritt fur jeden beliebigen Anfangswert ein blow-up schon
in endlicher Zeit auf. Es existieren also uberhaupt keine globalen Losungen mehr.

2 Ein-Zell-Populationen 10
Satz 2 (nach Perthame [23]). Fur d > 2 setzen wir m0 :=∫Rdu(x, 0) dx. Es gibt eine
Konstante C klein genug, so dass wenn
χ
∫Rd
|x|2
2u(x, 0) dx ≤ C(χm0)
dd−2 ,
dann gibt es keine globale glatte Losung mit ausreichend Abfall in x in unendlich fur
das Keller-Segel Modell (10).
2.2. Modellvariationen
Außer dem Minimalmodell gibt es noch viele weitere Modellvariationen. Diese beruck-
sichtigen jeweils einen zusatzlichen Parameter. Dieser wird realistischer gewahlt, also
nicht mehr wie im Minimalmodell linear. Im Normalfall regularisiert dieser Parameter
das Modell so weit, dass globale Losungen fur alle Anfangswerte existieren. Aus diesem
Grund heißt dieser jeweilige Parameter regularisation parameter [13]. In jedem dieser
Modelle wird auf diese Weise ein beliebiger Aspekt der Chemotaxis genauer untersucht.
Jede der Modellvariationen lasst sich durch bestimmte Grenzwerte ganz einfach wieder
auf das Minimalmodell zuruckfuhren. Das Minimalmodell ist also ein Spezialmodell fur
alle diese Modellvariationen.
Allgemein haben alle hier betrachteten Modellvariationen die Form
ut = ∇ ·(D(u)∇u− h(u, v)u∇v
)+ f(u)
vt = ∆v + ug(u)− v(11)
mit homogenen Neumann-Randbedingungen
n ·(D(u)∇u− h(u, v)u∇v
)= n · ∇v = 0 (12)
und festgelegten Anfangswerten
u(x, 0) = u0(x), v(x, 0) = v0(x), (13)
wobei u wieder die Zelldichte auf einem beschrankten Gebiet Ω ⊂ R2 und v die Konzen-

2 Ein-Zell-Populationen 11
tration des chemischen Signals ist. D(u) steht fur das Diffusionsvermogen der Zellen.
Weiter ist h(u, v) die chemotaktische Sensitivitat, also die Eigenschaft, wie schnell eine
Zelle auf die Veranderung der Konzentration eines chemischen Signals in ihrer Umge-
bung reagiert. f(u) steht fur das Zellwachstum und den Zelltod. g(u) beschreibt die
Produktion und den Abbau des chemischen Signals. Des Weiteren ist n der außere
Normalenvektor zu δΩ.
2.3. Volume-Filling Modell
Zellen befinden sich im Normalfall nicht alleine in einem Raum. In der Aggregations-
phase des Schleimpilzes Dictyostelium discoideum befinden sich zum Beispiel bis zu
100.000 Zellen auf engstem Raum. Dadurch werden die einzelnen Zellen in ihrer Bewe-
gungsfreiheit stark eingeschrankt. Es kann vorkommen, dass der Ort, zu dem sich eine
Zelle aufgrund des chemischen Gradienten hinbewegen wurde, bereits von einer ande-
ren Zelle belegt ist. Die Bewegung einer Zelle ist also abhangig von dem verfugbaren
Platz in ihrer Umgebung. Dieser Aspekt wird im Volume-Filling Modell berucksichtigt.
Fur ein bestimmtes Gebiet wird eine maximale Zelldichte γ vorausgesetzt. Diese hat
Einfluss auf die chemotaktische Sensitivitat der Zellen. Ist namlich in dem Raum zwi-
schen einer Zelle und der hochsten Konzentration des chemischen Signals die maxi-
male Zelldichte schon erreicht, so kann sich die Zelle nicht weiter in Richtung des
chemischen Gradienten bewegen. Die chemotaktische Sensitivitat h der Zellen wird im
Volume-Filling Modell nach [13] h(u, v) = χ(
1− uγ
)gesetzt. Zellwachstum und Zell-
tod werden in diesem Modell vernachlassigt. Es gilt damit f(u) = 0. Die Produktion
und der Abbau g des chemischen Signals werden des Weiteren gleich Eins gesetzt:
ut = ∇ ·(D∇u− χu
(1− u
γ
)∇v)
vt = ∆v + u− v(14)
Die chemotaktische Reaktion wird bei hoher Zelldichte praktisch ausgeschaltet. Dieses
Modell spiegelt damit die Tatsache wider, dass Zellen in der Lage sind, die Große
einer Aggregation zu bestimmen. Im weiteren Verlauf dieses Abschnitts folgen wir der
Darstellung von [22].

2 Ein-Zell-Populationen 12
2.3.1. Herleitung
Fur die Herleitung des Volume-Filling Modells wird die Wahrscheinlichkeit T , dass eine
Zelle von einem Ort x auf x± h springt, wie folgt gewahlt:
T±x = q(u(x± h, τ)
)(a+ b
(v(x± h, τ)− v(x, τ)
)), (15)
wobei q(u) die Wahrscheinlichkeit dafur widerspiegelt, dass die Zelle einen freien Platz
in ihrer Umgebung findet. q wird gewahlt als
q(u) = 1− u
γ> 0, 0 ≤ u ≤ γ, γ ≥ 1. (16)
Damit nimmt die Wahrscheinlichkeit, dass eine Zelle zu einem bestimmten Ort springt,
mit steigender Zelldichte linear ab. Weiter erfolgt die Herleitung analog zu der des
Minimalmodells. Das Einsetzen von T in die Mastergleichung fuhrt zunachst zu
ut = q(u(x, τ)
)(a+ b
(v(x, τ)− v(x− h, τ)
))u(x− h, τ)
+ q(u(x, τ)
)(a+ b
(v(x, τ)− v(x+ h, τ)
))u(x+ h, τ)
−(q(u(x+ h, τ)
)(a+ b
(v(x+ h, τ)− v(x, τ)
))+ q(u(x− h, τ)
)(a+ b
(v(x− h, τ)− v(x, τ)
)))u(x, τ)
= a
(q(u(x, τ)
)u(x− h, τ) + q
(u(x, τ)
)u(x+ h, τ)
−(q(u(x+ h, τ)
)+ q(u(x− h, τ)
))u(x, τ)
)+ b
(q(u(x, τ)
)(v(x, τ)− v(x− h, τ)
)u(x− h, τ)
+ q(u(x, τ)
)(v(x, τ)− v(x+ h, τ)
)u(x+ h, τ)
− q(u(x+ h, τ)
)(v(x+ h, τ)− v(x, τ)
)u(x, τ)
− q(u(x− h, τ)
)(v(x− h, τ)− v(x, τ)
)u(x, τ)
).
(17)
Aus dem Erweitern der rechten Seite und der Definition der Grenzwerte folgt
ut = ∇ ·(Du
(q(u)− q′(u)u
)∇u− χuq(u)∇v
), (18)

2 Ein-Zell-Populationen 13
wobei q′(u) die Ableitung von q nach u ist. Fur nichtansteigendes q(u) ist hierbei
der Diffusionskoeffizient Du
(q(u)− q′(u)
)immer positiv und damit (18) wohldefiniert.
Die Einbeziehung der Zellbewegung und der Signaldynamik sowie das Einsetzen von
q(u) = 1− uγ
fuhrt zum Volume-Filling Modell. Dabei ist zu beachten, dass in diesem
Fall gilt: q(u)− q′(u)u = 1
2.3.2. Existenz von globalen Losungen
Das Volume-Filling Modell schließt durch die Einfuhrung einer maximalen Zelldichte
das Auftreten von blow-ups, also Orten mit unendlicher Zelldichte, fur sinnvolle An-
fangswerte aus. Damit existieren globale Losungen ohne jegliche Einschrankung durch
Dimensionen.
Satz 3 (nach Hillen und Painter [12]). Seien p, σ und α gegeben durch
1 < σ < 2, 1 < α < 2, 2 < σ + α < 3, max
n
σ − 1,
n
σ − 2
< p.
Fur alle Anfangswerte u0 ∈ W σ,p, v0 ∈ W σ+α,p mit(u0(x), v0(x)
)∈ Ω fur alle
x ∈ [0, 1] mit periodischen Randbedingungen existiert eine globale Losung (u, v) ∈C([0,∞),W σ,p ×W σ+α,p
)von (14).
Oft wird die maximale Zelldichte im Volume-Filling Modell zu Eins skaliert. Dann gilt
also 0 ≤ u ≤ 1.

14
3. Zwei-Zell-Populationen
In diesem Kapitel werden wir unsere Betrachtungen der Chemotaxis auf Gebiete aus-
weiten, in denen mehr als nur ein Typ von Zellen vorkommt. In der Praxis treten oft
viele verschiedene Zellen auf einmal auf. Der Einfachheit halber beschranken wir uns
auf den Fall, dass sich in einem Gebiet zwei verschiedene Zelltypen befinden. Hier-
zu werden wir zunachst ein Modell herleiten und anschließend dessen numerische und
analytische Eigenschaften erortern.
Die Bewegungen einer Population konnen die Verteilung einer anderen Population, ge-
rade in einem Gebiet mit sehr hoher Zelldichte, stark beeinflussen. Hierbei ist vor allem
entscheidend, wie sehr sich die Reaktionen der beiden Populationen auf chemische Gra-
dienten unterscheiden. Allgemein werden nach [21] zwei Falle unterschieden.
So kann zunachst nur ein einziges chemisches Signal auftreten, das von beiden Popu-
lationen produziert wird. Die beiden Populationen konnen dabei dennoch eine unter-
schiedliche Dichte aufweisen und vor allem auch verschieden auf dieses Signal reagieren.
Dies ist auch beim Schleimpilz Dictyostelium discoidem der Fall. So versammeln sich
vor der Ausbildung des Fruchtkorpers alle Prastielzellen an einem Ende des Pseudo-
plasmodiums und die Prasporenzellen am anderen Ende. Die Zellen sortieren sich also
nach ihrem Typ. Nur aufgrund dieser Musterbildung konnen dann anschließend die
Prasporenzellen die Spore und die Prastielzellen den Stiel ausbilden. Wie sehr sich die
Reaktionen auf ein chemisches Signal unterscheiden ist nicht festgelegt. Es kann so-
wohl sein, dass nur die Starke anders ist als auch, dass eine Population von dem Signal
angezogen und die andere abgestoßen wird.
Der zweite mogliche Fall ist, dass die verschiedenen Populationen unterschiedliche che-
mische Signale produzieren, auf die beide jeweils unterschiedlich reagieren konnen. Das
heißt, es ist auch nicht notwendig, dass die Population u auf das von v produzierte
Signal genauso reagiert wie die Population v auf das von u produzierte Signal. Wieder
konnen auch verschiedene Konzentrationen der Populationen angenommen werden.

3 Zwei-Zell-Populationen 15
3.1. Herleitung
Wir werden hier nun ein Chemotaxismodell fur zwei Populationen u und v in einer
Dimension herleiten. Dabei orientieren wir uns an [21] und betrachten nur den Fall,
dass beide Populationen dasselbe Signal produzieren.
Wir setzen zunachst ein diskretes Gitter mit einheitlichen Abstanden h und eine kon-
tinuierliche Zeit voraus, bis wir spater auch den Raum zu einem Kontinuum skalieren.
Außerdem nehmen wir wie im Volume-Filling Modell eine konstante maximale Zell-
dichte Wmax fur die Punkte im Gitter an. Dabei ist es nicht erforderlich, dass sich in
einem Punkt auf dem Gitter nur eine einzige Zelle befinden kann. Die Zelldichten der
beiden Populationen im Punkt x bezeichnen wir mit Ux bzw Vx. Wx = Ux+Vx gibt die
gesamte Zelldichte in x an. Die Anteile der durch die verschiedenen Zelltypen belegten
Platze in x definieren wir als ux = Ux
Wmax, vx = Vx
Wmaxund wx = Wx
Wmax.
Die Bewegung einer Zelle kann auf zwei unterschiedliche Arten durchgefuhrt werden.
Zunachst ist es moglich, dass eine Zelle, die sich in x befindet, vollig ungehindert an
einen freien benachbarten Platz springt, also zu x ± h. Dies geschieht mit der Rate
P 1±x ux(1−wx±h) fur eine Zelle vom Typ u und mit Rate Q1±
x vx(1−wx±h) fur eine Zelle
vom Typ v. Der Faktor (1− wx±h) spiegelt jeweils den freien Platz in x± h wider.
Eine zweite Moglichkeit fur die Bewegung von Zellen ist, dass eine Zelle, die sich in
x befindet, mit einer Zelle in x ± h den Platz tauscht. In diesem Fall ist es also nicht
erforderlich, dass sich in den benachbarten Punkten noch freie Platze befinden. Eine
solche Bewegung wird normalerweise nur durch eine der beiden Zellen aktiv ausgefuhrt.
Diese aktive Zelle druckt dann eine passive Zelle weg. Hierbei ist noch zu unterscheiden,
ob eine Zelle den Platz mit einer Zelle desselben oder des anderen Typs tauscht. Im
ersten Fall bewegt sich eine Zelle vom Typ u beziehungsweise v von x nach x± h und
gleichzeitig eine weitere Zelle vom Typ u beziehungsweise v von x ± h nach x. Dies
passiert mit der Rate P 2±x uxux±h fur Zellen vom Typ u beziehungsweise Q2±
x vxvx±h fur
Zellen vom Typ v. Im letzten Fall bewegt sich eine Zelle vom Typ u von x nach x± hund eine Zelle vom Typ v von x± h nach x oder umgekehrt. Hierfur werden die Raten
P 3±x uxvx±h und Q3±
x vxux±h definiert. Unter Berucksichtigung dieser Bewegungstypen
ergibt sich die folgende Mastergleichung:

3 Zwei-Zell-Populationen 16
duxdτ
= + P 1−x+hux+h(1− wx) + P 1+
x−hux−h(1− wx)
− P 1−x ux(1− wx−h)− P 1+
x ux(1− wx+h)
+ P 3−x+hux+hvx + P 3+
x−hux−hvx − P3−x uxvx−h − P 3+uxvx+h
−Q3−x+hvx+hux −Q3+
x−hvx−hux +Q3−vxux−h +Q3+x vxux+h
dvxdτ
= + P 1−x+hvx+h(1− wx) + P 1+
x−hvx−h(1− wx)
− P 1−x vx(1− wx−h)− P 1+
x vx(1− wx+h)
+ P 3−x+hvx+hux + P 3+
x−hvx−hux − P3−x vxux−h − P 3+vxux+h
−Q3−x+hux+hvx −Q3+
x−hux−hvx +Q3−uxvx−h +Q3+x uxvx+h
(19)
Hierbei spiegeln jeweils die ersten beiden Zeilen die Bewegungen der Zellen vom Typ
u beziehungsweise v an einen freien benachbarten Ort wider. Die dritte Zeile der Glei-
chung fur u und die vierte Zeile der Gleichung fur v stehen fur den Platztausch zwi-
schen einer aktiven Zelle vom Typ u mit einer passiven Zelle vom Typ v. Die vierte
Zeile der Gleichung fur u und die dritte Zeile der Gleichung fur v stehen entsprechend
fur das entgegengesetzte Ereignis. Bewegungen, bei denen eine Zelle den Platz mit ei-
ner Zelle desselben Typs tauscht, tauchen hier nicht auf, da sie keinen Einfluss auf die
Veranderung der Verteilung haben. Sie kurzen sich in den Gleichungen einfach weg.
Fur die Ratenfunktionen P 1±,3± und Q1±,3± wird zur Vereinfachung angenommen, dass
fur die gerichtete Bewegung der Zellen allein Chemotaxis eine Rolle spielt, also nicht
etwa Zell-Zell-Interaktionen oder Ahnliches. Die Zellen bewegen sich somit, abgesehen
von Diffusion, nur aufgrund eines lokalen chemotaktischen Gradienten. Außerdem sol-
len beide Zelltypen grundsatzlich dieselben Bewegungseigenschaften haben. Sie sollen
also gleich auf chemische Signale reagieren. Damit ergibt sich fur die Ratenfunktionen
P 1±,3±x ≡ Q1±,3±
x ≡ k1,3u,v
(1 + κu,v(ax±h − ax)
). (20)
a steht hier fur die Verteilung des chemischen Signals. k1 ist eine Konstante, die die
Kompetenz der Zellen zur Bewegung zu freien Orten widerspiegelt und k3 entsprechend
die Bewegung zu voll besetzten Orten. Durch die Konstante κ wird die ungerichtete
Bewegung aufgrund von Diffusion miteinbezogen. Sie stellt den Anteil der Chemotaxis
an der Bewegung dar. κ kann hierbei auch negativ sein. In diesem Fall wirkt das
chemische Signal abstoßend auf die Zellen. Sie bewegen sich also weg von der hochsten

3 Zwei-Zell-Populationen 17
Konzentration des Signals.
Die weitere Herleitung des Modells erfolgt nun wie bei den vorherigen Modellen. Die
Ratenfunktionen (20) werden zunachst in die Mastergleichungen (19) eingesetzt. An-
schließend werden die rechten Seiten der Gleichungen mit h erweitert und τ zu λt sowie
Wmax zu 1 skaliert. Zum Schluss werden folgende Grenzwerte definiert:
limλ→∞, h→0
k1u,vλh
2 = αu,v , limλ→∞, h→0
k3u,vλh
2 = βu,v , limλ→∞, h→0
2κu,vλh2 = φu,v (21)
Hierbei stehen αu und αv fur die zufallige Bewegung, also die Diffusion der Populationen
in freien Regionen und βu und βv fur die Diffusion in voll besetzten Regionen. αuΦu
und αvΦv geben die chemotaktische Effizienz der Populationen in leeren Gebieten an
und βuΦu und βvΦv entsprechend in vollen Gebieten.
Durch die beschriebenen Schritte entsteht das folgende System von Gleichungen:
∂u
∂t=
(1u)︷ ︸︸ ︷αu∇ ·
((1− w)∇u+ u∇w
)+
(2u)︷ ︸︸ ︷(βu + βv)∇ · (v∇u− u∇v)
− αuφu∇ ·((1− w)u∇a
)︸ ︷︷ ︸(3u)
− (βuφu − βvφv)∇ · (uv∇a)︸ ︷︷ ︸(4u)
∂v
∂t=
(1v)︷ ︸︸ ︷αv∇ ·
((1− w)∇v + v∇w
)+
(2v)︷ ︸︸ ︷(βu + βv)∇ · (u∇v − v∇u)
− αvφv∇ ·((1− w)v∇a
)︸ ︷︷ ︸(3v)
− (βvφv − βuφu)∇ · (uv∇a)︸ ︷︷ ︸(4v)
(22)
mit w = u+ v. Die Terme (1u) und (1v) beschreiben die zufallige Bewegung der Zellen
an einen freien Platz und (2u) und (2v) den zufalligen Platztausch mit einer Zelle des
jeweils anderen Typs. Genauso folgen (3u) und (3v) aus der chemotaktischen Bewegung
zu einem freien Punkt und (4u) und (4v) daraus, dass Zellen unterschiedlichen Typs
aufgrund von Chemotaxis den Platz tauschen.
Um die Verteilung aller Zellen zusammen zu betrachten ist es notwendig die Gleichun-
gen (22) zu addieren. Daraus resultiert folgende Gleichung fur beide Populationen:
∂w
∂t= ∇ ·
((1− w)(αu∇u+ αv∇v) + (αuu+ αvv)∇w
)−∇ ·
((1− w)(αuφuu+ αvφvv)∇a
) (23)
Die Terme (2u), (2v), (4u) und (4v) kurzen sich dabei gegenseitig weg. Dies ist auch

3 Zwei-Zell-Populationen 18
biologisch sinnvoll, da Bewegungen, die in Form von Platztauschen stattfinden, auf die
gesamte Verteilung aller Zellen keinen Einfluss haben. Gleichung (23) setzt sich also
nur aus Termen fur die Bewegung von Zellen an freie Orte zusammen.
Realistische Anfangswerte sowohl fur u und v als auch fur w liegen in jedem Fall
zwischen Null und Eins, denn die Zelldichten konnen weder negativ sein noch uber der
maximalen Zelldichte liegen, die wir zuvor auf Eins skaliert haben.
Die Gleichung fur die Verteilung des chemischen Signals im Volume-Filling Modell kann
ganz einfach zu einer Gleichung fur zwei Populationen erweitert werden:
∂a
∂t= ∆a+ u+ v − a (24)
3.2. Eigenschaften des Modells
Das Modell (22) besitzt eine Gradientenfluss-Struktur. Um dies zu zeigen, fuhren wir
zunachst eine Entropie-Funktion ein:
E(u, v) =
∫Ω
[u log u+ v log v + (1− w) log(1− w)− φuau− φvav] dx. (25)
Im Folgenden bezeichnen wir fur eine Funktion G(r, s) die partielle Ableitung von G
nach r mit G′(r) und die Ableitung von G nach s mit G′(s).
Zur Berechnung von E ′(u) und E ′(v) definieren wir die Funktionen
ϕ(u) = ut, ψ(x) = vt (26)
und das Funktional
f(u, v) = u log u+ v log v + (1− w) log(1− w)− φuau− φvav. (27)
Damit ist
E(u, v) =
∫Ω
f(u, v) dx. (28)
Fur die partielle Ableitung der Entropiefunktion nach u folgt, da ϕ aus dem Dualraum

3 Zwei-Zell-Populationen 19
von f ′(u) ist,
Euϕ = E ′(u(x)
)ϕ(x)
=
∫Ω
f ′(u(x)
)ϕ(x) dx
= 〈f ′(u), ϕ〉L2 .
(29)
Nach dem Riesz’schen Darstellungssatz gilt somit
Eu = f ′(u)
= log u− log(1− w)− φua,(30)
wobei zu beachten ist, dass w′(u) = 1.
Analoge Rechnungen ergeben fur die Ableitung von E nach v
Ev = log v − log(1− w)− φva. (31)
Hieraus folgt, dass die Gradienten von Eu und Ev die folgende Form besitzen:
∇Eu =∇uu
+∇w
1− w− φu∇a
∇Ev =∇vv
+∇w
1− w− φv∇a
(32)
Nun lasst sich ut wie folgt umformen:
ut = αu∇ ·((1− w)∇u+ u∇w
)+ (βu + βv)∇ · (v∇u− u∇v)
− αuφu∇ ·((1− w)u∇a
)− (βuφu − βvφv)∇ · (uv∇a)
= ∇ ·(αu((1− w)∇u+ u∇w − φu(1− w)u∇a
)+ (βu + βv)(v∇u− u∇v − φuuv∇a+ φvuv∇a)
)= ∇ ·
(αu(1− w)u
(∇uu
+∇w
1− w− φu∇a
)+ (βu + βv)uv
(∇uu
+∇w
1− w− φu∇a−
∇vv− ∇w
1− w+ φv∇v
))= ∇ ·
(αu(1− w)u∇Eu + (βu + βv)uv∇ · (Eu − Ev)
)= ∇ ·
((αu(1− w)u+ (βu + βv)uv
)∇Eu − (βu + βv)uv∇Ev
)
(33)

3 Zwei-Zell-Populationen 20
Durch aquivalente Umformungen erhalten wir fur vt
vt = ∇ ·((αv(1− w)v + (βu + βv)uv
)∇Ev − (βu + βv)uv∇Eu
)= ∇ ·
(− (βu + βv)uv∇Eu +
(αv(1− w)v + (βu + βv)uv
)∇Ev
).
(34)
Das Modell (22) lasst sich also schreiben in der Form
(u
v
)t
= ∇ ·
(M(u, v)∇ ·
(Eu
Ev
)), (35)
wobei
M =
(αu(1− w)u+ (βu + βv)uv −(βu + βv)uv
−(βu + βv)uv αv(1− w)v + (βu + βv)uv
). (36)
Hierbei gilt, wie beim Keller-Segel Modell, die Randbedingung
M(u, v)∇ ·
(Eu
Ev
)· n = 0. (37)
M ist eine positiv definite Matrix. Aufgrund der Symmetrie von M konnen wir diese
Eigenschaft uberprufen, indem wir zeigen, dass die beiden Hauptminoren großer als
Null sind.
Fur die erste Hauptminore gilt
αu(1− w)u+ (βu + βv)uv > 0, (38)
da die maximale Zelldichte auf Eins skaliert wurde und somit 1 − w ≥ 0 gilt. Alle
ubrigen Parameter, also αu, βu, βv, u und v, sind nach Voraussetzung großer als Null.
Die zweite Hauptminore sieht wie folgt aus:
(αu(1− w)u+ (βu + βv)uv
)(αv(1− w)v + (βu + βv)uv
)− (βu + βv)
2u2v2
= αuαv(1− w)2uv + (βu + βv)2u2v2 + (βu + βv)
(αu(1− w)u+ αv(1− w)v
)− (βu + βv)
2u2v2
= αuαv(1− w)2uv + (βu + βv)(αu(1− w)u+ αv(1− w)v
)> 0
(39)

3 Zwei-Zell-Populationen 21
Sie ist wie oben großer als Null, da alle Parameter großer als Null sind und w ≤ 1 ist.
Wir werden nun zeigen, dass die Entropiefunktion E(u, v) mit der Zeit abnimmt, also
dass Et(u, v) ≤ 0 gilt.
Zunachst einmal gilt fur die Ableitung von E nach t
Et = Euut + Evvt. (40)
Da ut wieder aus dem Dualraum von Eu ist, folgt aufgrund des Riesz’schen Darstel-
lungssatzes
Euut =
∫Ω
[Euut] dx
=
∫Ω
[(∇ · (M11∇Eu +M12∇Ev)
)Eu]dx
=
∫Ω
[M11∆Eu · Eu] dx+
∫Ω
[M12∆Ev · Eu] dx.
(41)
Durch partielle Integration ergibt sich
Euut = M11∇Eu · Eu −∫
Ω
[M11|∇Eu|2
]dx
+M12∇Ev · Eu −∫
Ω
[M12∇Eu∇Ev]dx.(42)
Die Randterme fallen wegen (37) weg. Daher gilt
Euut = −∫
Ω
[M11|∇Eu|2 +M12∇Eu∇Ev
]dx. (43)
Analog gilt
Evvt = −∫
Ω
[M22|∇Ev|2 +M12∇Eu∇Ev
]dx (44)
und daher
Et = −∫
Ω
[M11|∇Eu|2 +M22|∇Ev|2 + 2M12∇Eu∇Ev
]dx. (45)
Da M positiv definit ist, ist somit Et(u, v) ≤ 0.

3 Zwei-Zell-Populationen 22
Wir haben vorausgesetzt, dass die Zellen sich weder vermehren noch sterben. Daher
liegt eine Massenerhaltung vor, das heißt∫Ω
u(x) dx = c1 und
∫Ω
v(x) dx = c2, (46)
wobei c1 und c2 konstant sind.
Eine stationare Losung von (22) existiert dann, wenn E ein Extremum besitzt. Da wir
bereits gezeigt haben, dass Et ≤ 0 ist, wird in einem kritischen Punkt von E schon ein
Minimum angenommen. Um also E auf kritische Punkte zu uberprufen, konnen wir
wegen (46) die Nebenbedingungen
g1(x) =
∫Ω
u(x, t) dx− c1 = 0
g2(x) =
∫Ω
v(x, t) dx− c2 = 0
(47)
formulieren.
Wir wenden nun die Methode der Lagrange-Multiplikatoren an. Hierzu setzen wir
L(u, v) = E(u, v) + λ1
(∫Ω
u dx− c1
)+ λ2
(∫Ω
v dx− c2
), (48)
λ1 und λ2 sind positive Skalare. Ein kritischer Punkt von E liegt dann vor, wenn
L′(u, v) gleich Null ist, also wenn
Lu = Eu + λ1
= log u− log(1− w)− φua+ λ1
= 0
Lv = Ev + λ2
= log v − log(1− w)− φva+ λ2
= 0.
(49)
Fur den Fall, dass kein chemischer Gradient vorliegt, also dass φua und φva gleich
Null sind, lassen sich die Gleichungen eindeutig losen. Um dies zu zeigen, subtrahie-
ren wir die erste Gleichung mit λ1, die zweite mit λ2 und wenden anschließend die

3 Zwei-Zell-Populationen 23
Exponentialfunktion an:
u
1− w= e−λ1 ,
v
1− w= e−λ2 (50)
Wir addieren nun die beiden Gleichungen und losen das Ergebnis nach w auf:
w
1− w= e−λ1 + e−λ2
⇔ w = e−λ1 + e−λ2 − w(e−λ1 + e−λ2)
⇔ w(1 + e−λ1 + e−λ2) = e−λ1 + e−λ2
⇔ w =e−λ1 + e−λ2
1 + e−λ1 + e−λ2
(51)
Die Gleichungen (50) losen wir nach u und v auf und setzen w ein:
u = (1− w)e−λ1 =
(1− e−λ1 + e−λ2
1 + e−λ1 + e−λ2
)e−λ1
v = (1− w)e−λ2 =
(1− e−λ1 + e−λ2
1 + e−λ1 + e−λ2
)e−λ2
(52)
Es existiert somit eine eindeutige stationare Losung von (22). Diese ist außerdem kon-
stant, da λ1 und λ2 Konstanten sind, die sich mit (47) berechnen lassen.
Existiert jedoch ein chemischer Gradient, so muss, damit ein kritischer Punkt von E
vorliegt, zusatzlich gelten, dass
at = ∆a+ u(a) + v(a)− a = 0. (53)
Nur fur den Fall, dass diese Gleichung losbar ist, existiert ein Minimum von E und
somit eine stationare Losung von (22).
Die maximale Zelldichte haben wir in Modell (22) auf Eins skaliert. Es gilt also 0 ≤u, v ≤ 1. Wie im Volume-Filling Modell wird hierdurch auch das Auftreten von blow-
ups fur sinnvolle Anfangswerte verhindert.

3 Zwei-Zell-Populationen 24
3.3. Numerische Ergebnisse
Wir werden in diesem Abschnitt die Auswirkungen der Chemotaxis auf die Musterbil-
dung numerisch erforschen.
Damit mogliche andere Eigenschaften der Zellen keinen Einfluss auf die Sortierung ha-
ben konnen, nehmen wir hier an, dass die beiden Populationen, bis auf die Produktion
und die Reaktion auf die chemischen Signale, vollig identisch sind, also
αu = αv = βu = βv = α. (54)
Die Gleichungen (22) werden durch diese Annahme erheblich vereinfacht. Das Einsetzen
von (54) in (22) ermoglicht die folgenden Umformungen fur ut:
ut = α∇ ·((1− w)∇u+ u∇w
)+ 2α∇ · (v∇u− u∇v)
− αφu∇ ·((1− w)u∇a
)− (αφu − αφv)∇ · (uv∇a)
= α∇ ·((1− u− v)∇u+ u∇u+ u∇v + 2v∇u− 2u∇v
)− αφu∇ ·
((1− u− v)u∇a
)− αφu∇ · (uv∇a) + αφv∇ · (uv∇a)
= α∇ ·((1 + v)∇u− u∇v
)− αφu∇ ·
(u(1− u)∇a
)+ αφv∇ · (uv∇a)
= α(∇v∇u+ (1 + v)∆u− u∆v −∇u∇v
)− αφu∇ ·
(u(1− u)∇a
)+ αφv∇ · (uv∇a)
= α((1 + v)∆u− u∆v
)− αφu∇ ·
(u(1− u)∇a
)+ αφv∇ · (uv∇a)
(55)
Fur vt konnen die Umformungen analog durchgefuhrt werden, was zu
vt = α((1 + u)∆v − v∆u
)− αφv∇ ·
(v(1− v)∇a
)+ αφu∇ · (uv∇a)
(56)
fuhrt.
In den Abbildung 3.1, Abbildung 3.2, Abbildung 3.3 und Abbildung 3.4 sind die Zell-
dichten (55) und (56) der beiden Populationen sowie die gesamte Zelldichte w = u+ v
und die Dichte des chemischen Signals (24) zu verschiedenen Zeiten in einer Dimension
dargestellt. Als Gebiet ist hierbei das Einheitsintervall [0, 1] mit homogenen Neumann-
Randbedingungen vorgegeben.
3 Zwei-Zell-Populationen 25
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=0.5
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=1
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=2
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=20
t=0
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=300
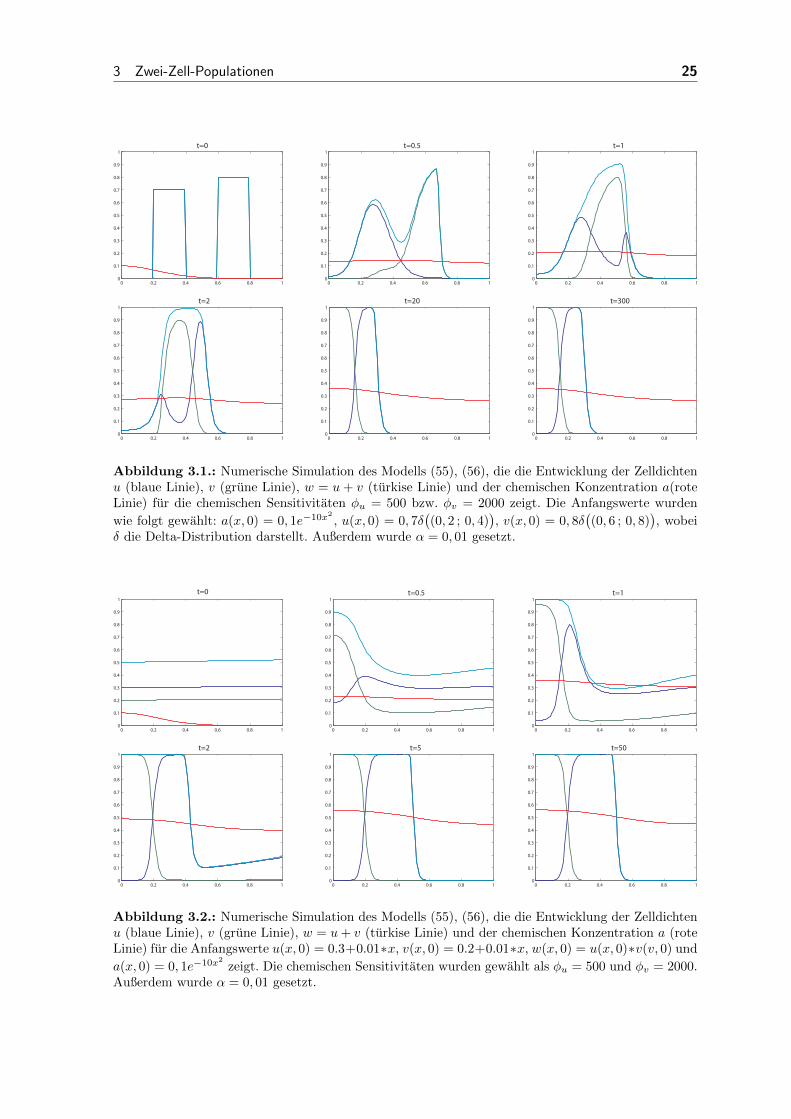
Abbildung 3.1.: Numerische Simulation des Modells (55), (56), die die Entwicklung der Zelldichtenu (blaue Linie), v (grune Linie), w = u+ v (turkise Linie) und der chemischen Konzentration a(roteLinie) fur die chemischen Sensitivitaten φu = 500 bzw. φv = 2000 zeigt. Die Anfangswerte wurden
wie folgt gewahlt: a(x, 0) = 0, 1e−10x2
, u(x, 0) = 0, 7δ((0, 2 ; 0, 4)
), v(x, 0) = 0, 8δ
((0, 6 ; 0, 8)
), wobei
δ die Delta-Distribution darstellt. Außerdem wurde α = 0, 01 gesetzt.
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=0
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=0.5
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=1
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=2
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=5
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=50
Abbildung 3.2.: Numerische Simulation des Modells (55), (56), die die Entwicklung der Zelldichtenu (blaue Linie), v (grune Linie), w = u+ v (turkise Linie) und der chemischen Konzentration a (roteLinie) fur die Anfangswerte u(x, 0) = 0.3+0.01∗x, v(x, 0) = 0.2+0.01∗x, w(x, 0) = u(x, 0)∗v(v, 0) und
a(x, 0) = 0, 1e−10x2
zeigt. Die chemischen Sensitivitaten wurden gewahlt als φu = 500 und φv = 2000.Außerdem wurde α = 0, 01 gesetzt.
3 Zwei-Zell-Populationen 26
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=0
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=0.5
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=1
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=2
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=5
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=50
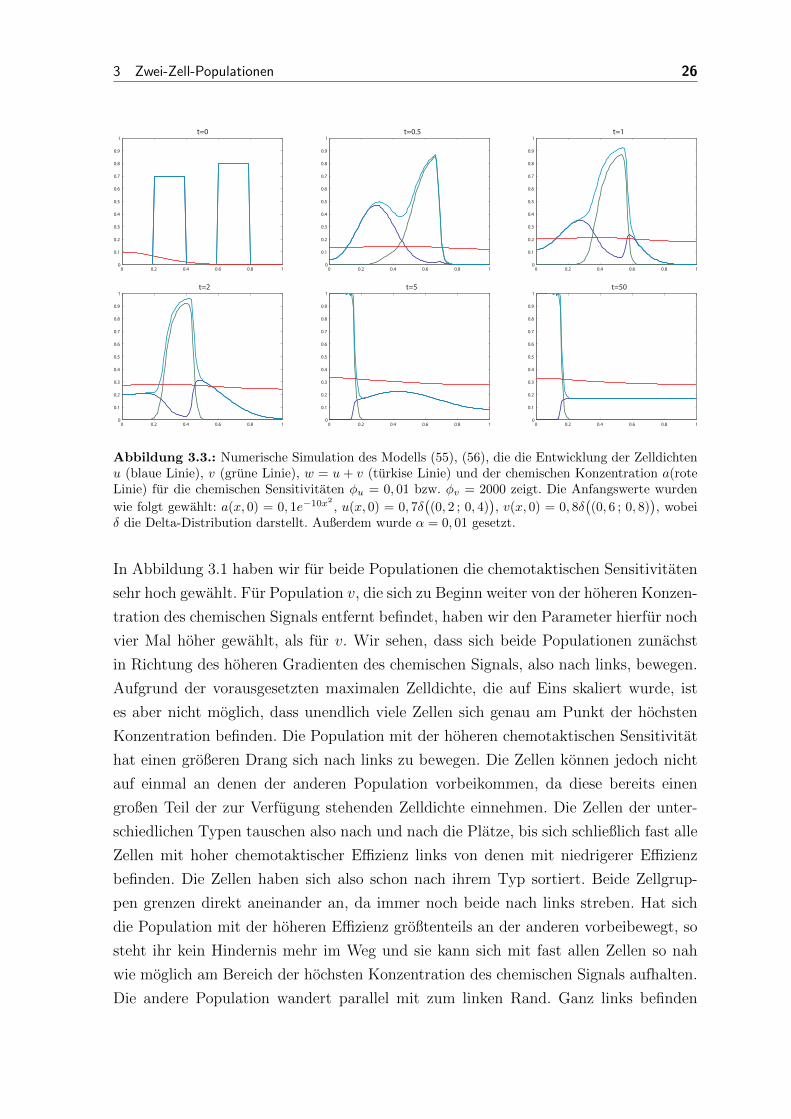
Abbildung 3.3.: Numerische Simulation des Modells (55), (56), die die Entwicklung der Zelldichtenu (blaue Linie), v (grune Linie), w = u+ v (turkise Linie) und der chemischen Konzentration a(roteLinie) fur die chemischen Sensitivitaten φu = 0, 01 bzw. φv = 2000 zeigt. Die Anfangswerte wurden
wie folgt gewahlt: a(x, 0) = 0, 1e−10x2
, u(x, 0) = 0, 7δ((0, 2 ; 0, 4)
), v(x, 0) = 0, 8δ
((0, 6 ; 0, 8)
), wobei
δ die Delta-Distribution darstellt. Außerdem wurde α = 0, 01 gesetzt.
In Abbildung 3.1 haben wir fur beide Populationen die chemotaktischen Sensitivitaten
sehr hoch gewahlt. Fur Population v, die sich zu Beginn weiter von der hoheren Konzen-
tration des chemischen Signals entfernt befindet, haben wir den Parameter hierfur noch
vier Mal hoher gewahlt, als fur v. Wir sehen, dass sich beide Populationen zunachst
in Richtung des hoheren Gradienten des chemischen Signals, also nach links, bewegen.
Aufgrund der vorausgesetzten maximalen Zelldichte, die auf Eins skaliert wurde, ist
es aber nicht moglich, dass unendlich viele Zellen sich genau am Punkt der hochsten
Konzentration befinden. Die Population mit der hoheren chemotaktischen Sensitivitat
hat einen großeren Drang sich nach links zu bewegen. Die Zellen konnen jedoch nicht
auf einmal an denen der anderen Population vorbeikommen, da diese bereits einen
großen Teil der zur Verfugung stehenden Zelldichte einnehmen. Die Zellen der unter-
schiedlichen Typen tauschen also nach und nach die Platze, bis sich schließlich fast alle
Zellen mit hoher chemotaktischer Effizienz links von denen mit niedrigerer Effizienz
befinden. Die Zellen haben sich also schon nach ihrem Typ sortiert. Beide Zellgrup-
pen grenzen direkt aneinander an, da immer noch beide nach links streben. Hat sich
die Population mit der hoheren Effizienz großtenteils an der anderen vorbeibewegt, so
steht ihr kein Hindernis mehr im Weg und sie kann sich mit fast allen Zellen so nah
wie moglich am Bereich der hochsten Konzentration des chemischen Signals aufhalten.
Die andere Population wandert parallel mit zum linken Rand. Ganz links befinden
3 Zwei-Zell-Populationen 27
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=0
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=0.5
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=1
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=2
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=5
0 0.2 0.4 0.6 0.8 10
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1t=50
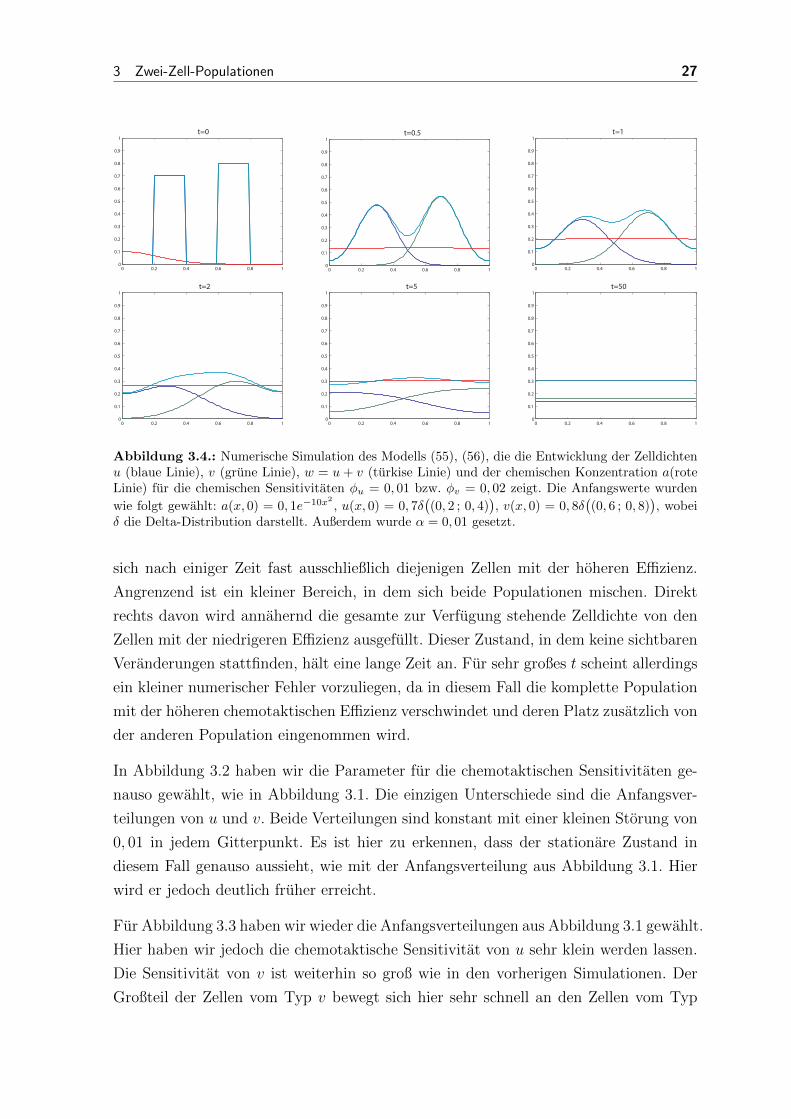
Abbildung 3.4.: Numerische Simulation des Modells (55), (56), die die Entwicklung der Zelldichtenu (blaue Linie), v (grune Linie), w = u+ v (turkise Linie) und der chemischen Konzentration a(roteLinie) fur die chemischen Sensitivitaten φu = 0, 01 bzw. φv = 0, 02 zeigt. Die Anfangswerte wurden
wie folgt gewahlt: a(x, 0) = 0, 1e−10x2
, u(x, 0) = 0, 7δ((0, 2 ; 0, 4)
), v(x, 0) = 0, 8δ
((0, 6 ; 0, 8)
), wobei
δ die Delta-Distribution darstellt. Außerdem wurde α = 0, 01 gesetzt.
sich nach einiger Zeit fast ausschließlich diejenigen Zellen mit der hoheren Effizienz.
Angrenzend ist ein kleiner Bereich, in dem sich beide Populationen mischen. Direkt
rechts davon wird annahernd die gesamte zur Verfugung stehende Zelldichte von den
Zellen mit der niedrigeren Effizienz ausgefullt. Dieser Zustand, in dem keine sichtbaren
Veranderungen stattfinden, halt eine lange Zeit an. Fur sehr großes t scheint allerdings
ein kleiner numerischer Fehler vorzuliegen, da in diesem Fall die komplette Population
mit der hoheren chemotaktischen Effizienz verschwindet und deren Platz zusatzlich von
der anderen Population eingenommen wird.
In Abbildung 3.2 haben wir die Parameter fur die chemotaktischen Sensitivitaten ge-
nauso gewahlt, wie in Abbildung 3.1. Die einzigen Unterschiede sind die Anfangsver-
teilungen von u und v. Beide Verteilungen sind konstant mit einer kleinen Storung von
0, 01 in jedem Gitterpunkt. Es ist hier zu erkennen, dass der stationare Zustand in
diesem Fall genauso aussieht, wie mit der Anfangsverteilung aus Abbildung 3.1. Hier
wird er jedoch deutlich fruher erreicht.
Fur Abbildung 3.3 haben wir wieder die Anfangsverteilungen aus Abbildung 3.1 gewahlt.
Hier haben wir jedoch die chemotaktische Sensitivitat von u sehr klein werden lassen.
Die Sensitivitat von v ist weiterhin so groß wie in den vorherigen Simulationen. Der
Großteil der Zellen vom Typ v bewegt sich hier sehr schnell an den Zellen vom Typ

3 Zwei-Zell-Populationen 28
u vorbei. Schon fur t = 5 sind fast alle dieser Zellen im Bereich zwischen 0 und 0, 2.
Die Population mit der geringen chemotaktischen Sensitivitat verteilt sich dagegen
gleichmaßig uber das gesamte Gebiet. Nach 50 Zeitschritten ist die Verteilung von u
konstant, mit Ausnahme des Bereichs mit der hochsten Konzentration des chemischen
Signals, der fast komplett von Population v eingenommen wird.
Abbildung 3.4 ist eine Simulation der Entwicklung der Zelldichten, in der fur bei-
de Populationen die chemotaktischen Sensitivitaten sehr niedrig gewahlt wurden. Der
chemische Gradient hat in diesem Fall keinen sichtbaren Einfluss auf das Verhalten der
Zellen. Die Populationen beginnen sofort sich gleichmaßig uber das gesamte Gebiet zu
verteilen, bis beide eine konstante Verteilung aufweisen.

29
4. Fazit und Ausblick
In dieser Arbeit haben wir ein Chemotaxismodell fur zwei Populationen entwickelt
und analysiert. Hierfur haben wir zunachst das Keller-Segel Modell fur eine Population
hergeleitet, indem wir anfangs nur das mikroskopische Verhalten der Zellen betrachtet
haben. Dadurch, dass wir die Ortsschrittweite gegen Null laufen lassen haben, haben
wir das makroskopische Modell erhalten.
Insbesondere fur das Minimalmodell haben sich interessante Eigenschaften in Bezug
auf die Existenz von globalen Losungen herausgestellt. Diese Existenz ist abhangig von
der Dimension des Modells. In einer Dimension existieren fur alle Anfangswerte globale
Losungen. In zwei Dimensionen existieren nur fur solche Anfangswerte Losungen, die
unter einer bestimmten Grenze liegen und in drei oder mehr Dimensionen gibt es gar
keine globalen Losungen mehr.
Durch die Einbeziehung einer maximalen Zelldichte fur jeden Punkt des Gebietes haben
wir das Volume-Filling Modell erhalten. Durch diese Verfeinerung des Minimalmodells
existiert hier in jeder Dimension eine globale Losung.
Das Volume-Filling Modell haben wir zuletzt zu einem Modell fur zwei Populationen
erweitert. Hierbei haben wir berucksichtigt, dass sich Zellen sowohl zu freien Orten
hinbewegen konnen als auch, dass sie den Platz mit einer Zelle desselben oder des
anderen Typs tauschen konnen. Der Fall, dass eine Zelle den Platz mit einer Zelle der
gleichen Population tauscht, spielt jedoch keine Rolle fur die Betrachtung der Zelldichte,
da sich hierdurch keine Veranderung ergibt. Beim Aufstellen einer Gleichung fur die
gesamte Zelldichte im betrachteten Gebiet fallt auch der Fall weg, dass eine Zelle mit
einer Zelle des anderen Typs den Platz tauscht, da dies auf die gesamte Zelldichte
keinen Einfluss hat.
Das hergeleitete Modell findet beispielsweise Anwendung bei der Betrachtung des
Schleimpilzes Dictyostelium disoideum. Wie sich herausgestellt hat, spiegelt es die
Eigenschaft des Pilzes wider, dass sich die unterschiedlichen Zellen nach ihrem Typ
sortieren. Diese Erkenntnis haben wir gewonnen, indem wir das Modell umgeschrieben
haben, sodass es abhangig von einer Entropiefunktion ist. Fur zwei Populationen, die

4 Fazit und Ausblick 30
unterschiedlich auf ein chemisches Signal reagieren, wird diese Entropiefunktion mit
der Zeit immer kleiner. Das heißt also, die Zellen im betrachteten Gebiet sind immer
weniger vermischt. Sie sortieren sich nach ihrem Typ. Wie wir durch verschiedene nume-
rische Simulationen gezeigt haben, ist eine Voraussetzung fur die Sortierung der Zellen,
dass zumindest eine der Populationen eine relativ hohe chemotaktische Sensitivitat auf-
weist. Ist fur jeden Zelltyp die chemotaktische Sensitivitat zwar unterschiedlich, aber
sehr niedrig, so strebt jede Population fur sich eine gleichmaßige Verteilung uber das
gesamte Gebiet an.
In der Realitat sind die Bewegungen von Zellen nach [21] abhangig von einem soge-
nannten Adhasionskomplex. Diese Komplexe befinden sich an jeder Zelle. Zellen konnen
sich nur fortbewegen, indem sie sich mit ihren Adhasionskomplexen entweder an den
Rand des Gebietes oder an eine andere Zelle binden. Auch wenn es aussieht, als wurde
das von uns hergeleitete Modell diesen Aspekt nicht mit einbeziehen, so findet er doch
Berucksichtigung im Term k, der fur die Kompetenz der Zellen zur Bewegung steht.
Die Modellierung von Chemotaxis ist ein sehr aktuelles Forschungsgebiet, das noch
lange nicht ausgeschopft ist. Naheliegend ware es beispielsweise ein Modell fur den Fall
zu entwickeln, dass zwei Populationen verschiedene chemische Signale produzieren. Das
Modell (22) beinhaltet außerdem die Eingrenzung von u, v und w zwischen Null und
Eins. Denkbar sind aber auch Gebiete, in denen keine solche Beschrankung vorhan-
den ist. Diese und noch sehr viele weitere Aspekte konnen in zukunftigen Arbeiten
Berucksichtigung finden.

31
A. Quellcode
%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%% Modellierung von Chemotaxis fur zwei Populationen%% mit dem expliziten Eulerverfahren%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%close allclear all
T=300;n=100;m=T*25000;dt=T/m; %Schrittweite der Zeith=1/n; %Schrittweite des Ortest=0;x=0:h:1; %Intervall
%Anfangsverteilung von ufor i=1:n+1
if x(i)>=0.2 && x(i)<0.4u0(i)=0.7;
elseu0(i)=0;
endend
%Anfangsverteilung von vfor i=1:n+1
if x(i)>=0.6 && x(i)<0.8v0(i)=0.8;
elsev0(i)=0;
endend
a0=0.1*exp(-10*x.*x);
u0=u0’;v0=v0’;a0=a0’;
plot(x,u0,x,v0,x,a0,’LineWidth’,1.5)axis([0 1 0 1])

A Quellcode 32
%Parameteralpha=0.01;phi_u=500;phi_v=2000;
%Diskretisierung des Laplace Operatorse = ones(numel(u0), 1);Delta=1/(hˆ2) * spdiags([e -2*e e], -1:1, n+1, n+1);Delta(1, 1) = -1/h;Delta(1, 2) = 1/h;Delta(n+1, n) = -1/h;Delta(n+1, n+1) = 1/h;
%Diskretisierung des Gradientennabla=1/h * spdiags([-e e], 0:1, n+1, n+1);nabla(n+1, n)=-1/h;nabla(n+1, n+1) = 1/h;
%Losen der Gleichungfor i=1:m
u1=u0+dt*alpha*((1+v0).*(Delta*u0) - u0.*(Delta*v0));v1=v0+dt*alpha*((1+u0).*(Delta*v0) - v0.*(Delta*u0));u1=u1-dt*(alpha*phi_u*(nabla*((u0.*(1-u0)).*(nabla*a0))));v1=v1-dt*(alpha*phi_v*(nabla*((v0.*(1-v0)).*(nabla*a0))));u1=u1-dt*(-alpha*phi_v*(nabla*((u0.*v0).*(nabla*a0))));v1=v1-dt*(-alpha*phi_u*(nabla*((v0.*u0).*(nabla*a0))));a1=a0+dt*(Delta*a0+u0+v0-a0);
%homogene Randbedingungenu1(1)=u1(2);v1(1)=v1(2);a1(1)=a1(2);u1(n+1)=u1(n);v1(n+1)=v1(n);a1(n+1)=a1(n);
t=t+dt;
if(rem(i,100)==0)plot(x,u1,x,v1,x,a0,’LineWidth’,1.5)axis([0 1 0 1])title([’t=’ num2str(t)]);pause(0.0001)end;
u0=u1;v0=v1;a0=a1;
end

33
Literaturverzeichnis
[1] http://slimemold.uark.edu/fungi/images/NameImages/D.disc.agg2.smz1x5x.jpg.
gesichtet am 11.08.2010.
[2] http://www.uni-koeln.de/dictyostelium/images/interaptin.jpg. gesichtet am
12.08.2010.
[3] http://jcs.biologists.org/content/vol114/issue24/images/medium/joces11424cvf.
gif. gesichtet am 11.08.2010.
[4] http://www.nature.com/nature/journal/v408/n6815/images/408917aa.2.jpg. ge-
sichtet am 11.08.2010.
[5] C.J. Budd, R. Carretero-Gonzalez, and R.D. Russell. Precise computations of che-
motactic collapse using moving mesh methods. Journal of Computational Physics,
202:463–487, Jan. 2005.
[6] M. Burger, M. Di Francesco, J.-F. Pietschmann, and B. Schlake. Nonlinear cross-
diffusion with size exclusion. SIAM J. Math. Analysis, 2010.
[7] M. Burger, Y. Dolak-Struss, and C. Schmeiser. Asymptotic analysis of an
advection-dominated chemotaxis model in multiple spatial dimensions. Commun.
Math. Sci., 6(1):1–28, 2008.
[8] M. Di Francesco and J. Rosado. Fully parabolic Keller-Segel model for chemotaxis
with prevention of overcrowding. Nonlinearity, 21(11):2715–2730, 2008.
[9] Y. Dolak and C. Schmeiser. The Keller-Segel model with logistic sensitivity func-
tion and small diffusity. SIAM J. Appl. Math., 66:286–308, 2005.
[10] A. Early, T. Abe, and J. Williams. Evidence for positional differentiation of
prestalk cells and for a morphogenetic gradient in dictyostelium. Cell, 83:91–99,
1995.

Literaturverzeichnis 34
[11] F. Filbet. A finite volume scheme for the Patlak-Keller-Segel chemotaxis model.
Numer. Math., 104:457–488, 2006.
[12] T. Hillen and K.J. Painter. Global existence for a parabolic chemotaxis model
with prevention of overcrowding. Adv. Appl. Math., 26:280–301, 2001.
[13] T. Hillen and K.J. Painter. A user’s guide to PDE models for chemotaxis. J.
Math. Biol., 58:183–217, 2009.
[14] D. Horstmann. From 1970 until present: The Keller-Segel model in chemotaxis
and its consequences i. Jahresberichte DMV, 105(3):103–165, 2003.
[15] D. Horstmann. Generalizing Keller-Segel: Multi-species chemotaxis models with
attraction and repulsion between compatitive interaction species. Technical report,
Mathematisches Institut, Universitat zu Koln, Feb. 2009.
[16] D. Horstmann and M. Lucia. Nonlocal elliptic boundary value problems related to
chemotactic movement of mobile species. RIMS Kokyuroku Bessatsu, B15:39–72,
2009.
[17] W. Jager and S. Luckhaus. On explosions of solutions to a system of partial
differential equations modelling chemotaxis. Transactions of the american mathe-
matical society, 329(2):819–824, 1992.
[18] E.F. Keller and L.A. Segel. Model for chemotaxis. J. Theor. Biol., 30:225–234,
1971.
[19] A. Marrocco. 2d simulation of chemotactic bacteria aggregation. Research report,
INRIA, 2002. Project M3N.
[20] S. Matsukuma and A.J. Durston. Chemotactic cell sorting in Dictyostelium dis-
coideum. J. Embryol. exp. Morph., 50:243–251, 1979.
[21] K.J. Painter. Coninuous models for cell migration is tissues and applications to
cell sorting via differential chemotaxis. Bulletin of Mathematical Biology, 71:1117–
1147, 2009.
[22] K.J. Painter and T. Hillen. Volume-filling and quorum-sensing in models for
chemosensitive movement. Can. Appl. Math. Quart., 10(4):501–543, 2002.
[23] B. Perthame. Transport Equations in Biology. Birkhauser Basel, 2007.
[24] W.K. Purves, D. Sadava, G.H. Orians, and H.C. Heller. Biologie. Markl, J., 2007.

Literaturverzeichnis 35
[25] M. J. Simpson, K.A. Landman, and B.D. Hughes. Multi-species simple exclusion
processes. Physica A, 388:399–406, 2009.
[26] A. Stevens. The derivation of chemotaxis equations as limit dynamics of moderate-
ly interacting stochastic many-particle systems. SIAM J. Appl. Math., 61(1):183–
212, Jun.-Jul. 2000.
[27] R. Tyson, L.J. Stern, and R.J. LeVeque. Fractional step methods applied to a
chemotaxis model. J. Math. Biol., 41:455–475, 2000.
[28] T. Umeda and K. Inouye. Cell sorting by differential cell motility: A model for
pattern formation in Dictyostelium. J. Theor. Biol., 226:215–224, 2004.





























