Embed Size (px)
Citation preview

111
Department of Biology, 371 Serra Mall, Stanford University, Stanford, CA94305-5020, [email protected]
OONN TTHHEE UUSSEE OOFF TTAAXXOONN SSUUBBSSTTIITTUUTTEESS IINN RREEWWIILLDDIINNGGPPRROOJJEECCTTSS OONN IISSLLAANNDDSS
DENNIS M. HANSEN
Islands and Evolution (2010). Pérez-Mellado, V. and Ramon, C. (eds.) Institut Menorquí d’Estudis. Recerca,19: 111-146
AAbbssttrraacctt.. Recent proposals to rewildcontinental ecosystems with taxonsubstitutes –replacing extinct species withextant analogues– have met withcontroversy, chiefly because the involvedspecies are extinct Pleistocene mega-fauna and their still-extant counterparts,and because of the massive areasrequired. In contrast, I argue that islandrewilding offers ideal and much lesscontroversial scenarios to rapidlyadvance our empirical understanding oftaxon substitutions. The main reasonsare twofold: firstly, anthropogenicextinctions happened very recently onislands, often within the last fewcenturies; secondly, island species aremuch smaller and require less space toimplement rewilding projects. Islandrewilding offers hope for restoring evenhighly degraded island ecosystemswhere many species have been lost, andallows radically different restorationtrajectories to be pursued than thoseavailable with the surviving nativespecies pools. I here present an overviewof currently ongoing, planned, andproposed taxon substitution projects onislands, discuss several salient points,
and highlight promising avenues forfuture research and restoration mana-gement.
KKeeyy wwoorrddss:: naturalness, extinction,restoration, ecological analogues, plant-animal interactions, ecosystem functio-ning, assisted migration.
RReessuummeenn.. Las recientes propuestaspara repoblar ecosistemas continentalescon taxones sustitutivos (reemplazandolas especies extinguidas con otras vi-vientes análogas) han sido objeto decontroversia, especialmente porque lasespecies involucradas formaban partede la megafauna extinguida del Pleisto-ceno, al igual que sus todavía vivientescontrapartes y también porque se preci-san áreas de una gran extensión. Por elcontrario, propongo que la repoblaciónde las islas ofrece escenarios ideales ymucho menos controvertidos, paraavanzar rápidamente en nuestra com-prensión empírica de la sustitución de ta-xones. Las razones principales sondobles: en primer lugar, las extincionesantropogénicas han sucedido en las is-las muy recientemente, frecuentementeen los últimos siglos; en segundo lugar,
on the use of taxon:2 columnas 15/11/2010 13:53 Página 111

las especies insulares son mucho máspequeñas y requieren menos espaciopara implementar los proyectos de re-población. La repoblación insular ofreceuna esperanza para restaurar inclusoecosistemas insulares intensamente de-gradados, donde muchas especies sehan perdido y permite seguir trayectoriasde restauración radicalmente diferentesde aquéllas disponibles con el conjuntode especies nativas supervivientes. Pre-sento aquí el panorama de los proyec-tos de sustitución en islas que se hallanen curso de realización, en planifica-ción o propuestos, discuto numerosospuntos a resaltar y señalo las promete-doras líneas de futura investigación yde manejo de la restauración.
PPaallaabbrraass ccllaavvee.. Naturalidad, extin-ción, restauración, análogos ecológicos,interacciones planta-animal, funciona-miento de los ecosistemas, migraciónasistida.
RReessuumm.. Les recents propostes perrepoblar ecosistemes continentals ambtaxons substitutius (reemplaçant lesespècies extingides amb altres viventsanàlogues) han estat objecte decontrovèrsia, especialment perquè lesespècies involucrades formaven part dela megafauna extingida del Plistocè,igual que els seus encara vivents
contrapartes i també perquè es precisenàrees d'una gran extensió. Per contra,proposo que la repoblació de les illesofereix escenaris ideals i molt menyscontrovertits, per avançar ràpidamenten la nostra comprensió empírica de lasubstitució de taxons. Les raonsprincipals són dobles: en primer lloc, lesextincions antropogèniques han succeïta les illes molt recentment, freqüentmentals últims segles; en segon lloc, lesespècies insulars són molt més petites irequereixen menys espai per imple-mentar els projectes de repoblació. Larepoblació insular ofereix una espe-rança per restaurar fins i tot ecosistemesinsulars intensament degradats, onmoltes espècies s'han perdut i permetseguir trajectòries de restauració radi-calment diferents d'aquelles disponiblesamb el conjunt d'espècies nativessupervivents. Presento aquí el panoramadels projectes de substitució en illes quees troben en curs de realització, enplanificació o proposats, discuteixonombrosos punts a ressaltar i assenyaloles prometedores línies de futurainvestigació i de maneig de larestauració.
PPaarraauulleess ccllaauu.. Naturalitat, extinció,restauració, anàlegs ecològics, inter-accions planta-animal, funcionamentdels ecosistemes, migració assistida.
HANSEN
112
on the use of taxon:2 columnas 15/11/2010 13:53 Página 112

Many studies have documentedhow the majority of island ecosystemsare badly damaged after catastrophicanthropogenic impacts that mainlyoccurred within the last two millennia,depending on the date of humancolonisation. However, adding to thealready overwhelming litany ofecosystem destruction on islands is notthe topic of the present contribution. AsDonlan et al. (2005) succinctly put it,more than enough has sadly beenwritten about the “doom and gloom”facing Earth’s biodiversity.
Between the two opening quotes byJohn Lawton, one could say, lies theemerging field of rewilding with taxonsubstitutes, resurrecting the lostecosystem functions of recently extinctspecies by substituting them with extantspecies (unless stated otherwise,‘rewilding’ hereafter refers to restorationpractices involving taxon substitutes).Even though extinctions on islandshappened so recently, and are beingincreasingly well documented throughthe discovery of subfossil material, thereis still a widespread disregard of theecological and evolutionary roles played
by many of these ‘ghost species’–yetthere is no doubt that on many islandsthey fulfilled important functional rolesuntil very recently, for example asherbivores, frugivores and pollinators(e.g. Cox and Elmqvist, 2000;McConkey and Drake, 2002; Godfreyet al., 2008; Hansen and Galetti, 2009).As essential as modern-day conser-vation of endangered species andhabitats is in halting the sixth massextinction, some ecologists andpractitioners have come to realise thatwe ignore the ecological andevolutionary consequences of thesefunctional gaps at our own peril.
For continental ecosystems, theseviews have resulted in several recentsuggestions to rewild large tracts ofcontinental ecosystems, in NorthAmerica, South America and Eurasia(Galetti, 2004; Donlan et al., 2005;Zimov, 2005; Donlan et al., 2006), allof which aim to replace extinctPleistocene megafauna and restore lostecosystem dynamics. The notion that itmay be possible to get one up on thatmost ultimate of anthropogenicdisturbance regimes, extinction, may
113
ISLAND REWILDING
IINNTTRROODDUUCCTTIIOONN
“...stars, comets and extinct species share another convenient property thatappeals to people. Not only are they very interesting, but there is nothing we can
actually do about them either. By this I mean that we are powerless to intervene tochange what happens to them or what they do (or did). So discovering more aboutthem carries no responsibility; they are the ultimate, free, intellectual curiosity trip,
devoid of any need to 'do' anything when we discover more about them”
Lawton, 1995
“The ecology of the afterlife is alive and well”Lawton, 1995
on the use of taxon:2 columnas 15/11/2010 13:53 Página 113

seem heretic to some, and certainly hassparked controversy (Rubenstein et al.,2006; Caro, 2007; Caro and Sherman,2009; Donlan and Greene, 2010).
In contrast to this, I believe islandsoffer some of the best suited scenariosto rapidly advance our empiricalunderstanding of rewilding andexploring the use of taxon substitutionsin conservation and restoration (see alsoGriffiths and Harris, 2010; Griffiths etal., 2010; Hansen et al., 2010; Kaiser-Bunbury et al., 2010). This is chieflybecause the relevant temporal andspatial scales are much smaller onislands. Temporally, the majority ofisland species went extinct very recently,often within the last five centuries. Thiscontrasts greatly with the peak ofcontinental megafauna extinctionsthousands of years ago, in thePleistocene, and puts most island ghostssquarely in the youngest category ofLawton’s (1995) poetically termed‘noveau-dead’. Spatially, the involvedspecies are physically smaller thancontinental megafauna, making it morefeasible to manage them in restorationprojects. As a consequence, the size ofthe areas required to establishecologically meaningful rewildingprojects is also much smaller on islands,compared to the grand vistas requiredfor continental rewilding.
My point of view is rooted in thelong-standing tradition of evolutionarybiologists and ecologists to regardislands as natural laboratories: due totheir relative simple ecosystems, islandsoffer ideal setups to answer complexquestions that are difficult to address in
more complex mainland settings.However, taking this metaphor at facevalue, Holocene extinction events onislands worldwide have left thelaboratories more than half empty andsorely in need of equipment―from aplethora of pipet-tips and test tubes tolumbering gas-chromatographs. Pro-perly applied, rewilding is an explicit,hypothesis-driven and adaptive mana-gement option that adds a promisingtool to the growing arsenal available toisland restoration projects―re-stockingthe laboratories, so to speak.
Island rewilding is inherently part ofa whole-ecosystem view of restoration(Steadman and Martin, 2003; Lazell,2005; Hutton et al., 2007; Jones,2008), and is closely allied to theconcept of ‘inter-situ’ conservationchampioned by Burney and Burney(2007), which they define as: “Theestablishment of species by reintro-duction to locations outside the currentrange but within the recent past rangeof the species. In some cases, closestliving relatives or ecological surrogatesmay be substituted for globally extinctspecies that are regarded as essential tomaintain a process believed critical tothe function of the target ecosystem.Inter-situ conservation, in effect, bridgesthe gaps between in-situ and ex-situconservation.”. In both rewilding andinter-situ conservation, the emphasis ison maintaining or restoring ecosystemfunctioning rather than on speciesconservation, even though the two areoften intimately linked. In essence, forhighly degraded island ecosystemswhere many species have been lost,rewilding allows radically different
114
HANSEN
on the use of taxon:2 columnas 15/11/2010 13:53 Página 114

restoration trajectories to be pursuedthan those available with the survivingnative species pools. Depending on howdegraded the targeted ecosystem is,island rewilding spans the scale between‘ecological restoration’ and ‘ecologicalengineering’ on the managementcontinuum visualised by Atkinson(2001), and should be considered asconservation translocations (sensuSeddon et al., 2007; Seddon et al., inpress) within an expanded definition ofthe IUCN reintroduction guidelines(Hansen et al., 2010).
Opponents of rewilding with taxonsubstitutions claim that these schemesare doomed to create more problemsthan they can solve. It is true that islandshave suffered more havoc at the handsof introduced species than mostecosystems, and the utmost cautionshould be taken in any rewilding project.Examples of necessary cautionarymeasures are covered elsewhere(Donlan et al., 2006; Caro, 2007;Griffiths et al., 2010; Hansen et al., in2010), and I will not be elaborating onthis point in great detail here. Instead, Ihope the examples below, as well as thecase studies covered in greater detail inBoxes 1–3, will illustrate the potential fortaxon substitution to rapidly movebeyond the status of gimmick andbecome an integral part of restoration
In this contribution, I aim to firstlypresent an overview of selectedongoing, planned and suggestedrewilding projects on islands around theworld; secondly, to discuss how we canderive the information needed to assessthe appropriateness of a taxon substitutecandidate for a rewilding project; andthirdly to argue some salient points thatarise about how island rewilding canoffer fruitful research and managementavenues.
schemes for some of the most degradedhabitats on our planet. Encouragingly, avital first step in island rewilding―toaddress and manage or remove theknown or suspected causal agents thatled to extinctions in the first place―isincreasingly being achieved on islandsaround the world, e.g. eradications ofinvasive mammals (Veitch and Clout,2002; Nogales et al., 2004), setting thestage for the potential use of taxonsubstitutions in many dysfunctionalisland ecosystems around the world.
AAnniimmaall rreewwiillddiinngg
Probably reflecting the traditionalhigh visibility of research on islandavifauna, birds have until now been the
115
ISLAND REWILDING
IIssllaanndd rreewwiillddiinngg:: ccuurrrreenntt ssttaattee aanndd ffuuttuurree ooppttiioonnss
“Reintroduction of the 'missing' forest fauna [of Hong Kong] would bring ecological,conservation and educational benefits, but depending, as at present, on accidental
escapes and kind-hearted Buddhists to do this is a recipe for disaster”
Corlett, 2002
on the use of taxon:2 columnas 15/11/2010 13:53 Página 115

focus of the majority of island rewildingprojects (see also Box 1). Lately, tortoiseshave joined the fray (Griffiths et al.,2010; Hansen, et al., 2010), and other‘typical’ island taxa like fruitbats andlizards offer similarly well-suited sce-
narios. Conservation and restoration ofinvertebrates on islands is sadly laggingfar behind that of vertebrates (New,2008) and the following overview isrestricted to examples of vertebraterewilding.
116
HANSEN
BBooxx 11:: NNeeww ZZeeaallaanndd―ttaakkiinngg tthhee lleeaadd iinn aavviiaann rreewwiillddiinngg??
In New Zealand, island restoration plans are strongly influenced by progress in thescientific understanding of pre-anthropogenic ecosystems. Most recently, a special issue ofthe New Zealand Journal of Ecology, “Feathers to Fur” (Sullivan et al., 2010), has brilliantlyhighlighted how the dramatic shift from avian- to mammal-dominated ecological guilds hashad large impacts on ecosystem dynamics, often to the detriment of native species (e.g. Forsythet al., 2010; Lee et al., 2010; see also Greenwood and Atkinson, 1977; Caughley, 1989;Atkinson and Cameron, 1993; Cooper et al., 1993; Bond et al., 2004; Wood et al., 2008).
In response, following the inspiring address by Atkinson (1988), a flurry of New Zealandisland restoration plans have advocated translocations of extant species within the NewZealand archipelago to replace extinct species or subspecies, focusing on birds. In relation totaxon substitutions New Zealand is therefore―as is the case with many aspects of islandconservation and restoration―ahead of much of the rest of the world. Most of these restorationplans stress the ecosystem approach behind the proposed taxon substitutions, aiming to restorelost trophic interactions as well as mutualistic interactions like pollination and seed dispersal.
An early example, perhaps far-flung to some, Meurk (1990, cited in Norton et al., 2005)suggested using extant ratites from outside New Zealand to replace extinct moas on QuailIsland. More realistically, Miskelly (1999) proposed to use the extant Chatham Island snipeCoenocorypha pusilla as a substitute for the extinct North Island snipe C. barrierensis. However,this could be complicated by politics if the Chatham Island community is resistant to endemictaxa being moved elsewhere. In this case, their approval could perhaps be secured after‘preciprocal’ translocations from elsewhere in New Zealand to replace extinct Chatham Islandtaxa (C. Miskelly, pers. comm.). For example, the Snares Island fernbird Bowdleria punctatacaudata, could be introduced to replace the extinct Chatham Island fernbird B. rufescens(Miskelly, 1999). A few years later, in 2001, the management plan for threatened birds onChatham Islands included a list with nine extinct birds that could be replaced by extant speciesand subspecies (Table 3 in Aikman et al., 2001), which was expanded to 11 species byAtkinson (Table 3 in Atkinson, 2001). Roberts and Miskelly (2003) developed further restorationand rewilding plans for snipe. Together with recent advances in New Zealand snipe taxonomy(Miskelly and Baker, 2009; Baker et al.,(2010), all these restoration plans provide an idealtemplate for scientifically rigorous implementation of snipe rewilding projects in New Zealand.
In 2005, the first thoroughly planned taxon substitution in New Zealand took place when30 Snares Island snipe C. huegeli were translocated to Putauhinu Island to replace the extinctStewart Island snipe C. iredalei (Charteris and Miskelly, 2005). The second New Zealand taxonsubstitution project, translocating North Island kokako Callaeas wilsoni to Secretary Island asa substitute for the extinct South Island kokako C. cinerea, took place in 2008 and 2009,
on the use of taxon:2 columnas 15/11/2010 13:53 Página 116

A fitting first stop on our round the(rewilded) world tour is the tiny island ofGuana in the British Virgin Islands. In theeminent edited volume on the naturalhistory of Guana, Lazell (2005) high-lights how conservation translocationsare a powerful tool in the restoration ofislands. The island has been the focus ofseveral reintroductions of locally extinctspecies, and Lazell (2005) argues thatthe logical next step is to use analogousspecies from elsewhere to replaceglobally extinct taxa. The list includesHawaiian monk seal Monachusschauinslandi, Guam rail Rallus owstoni,and Puerto Rican iguaca Amazoniavittata vittata as stand-ins for the extinctCaribbean monk seal M. tropicalis,DeBooy’s rail Nesotrochis debooyi, andVirgin Islands parrot Amazonia vittatassp. indet., respectively. Anothergroundbreaking example comes fromNonsuch Island near Bermuda, whererewilding spans from the introduction ofa scale-insect resistant genotype of theendemic Bermuda cedar Juniperusbermudiana to the replacement of theextinct night heron Nyctanassa carcino-catactes with the widespread N. violacea(Wingate, 2001; Olson and Wingate,2006).
On to the often heavily populatedarchipelagos of the Indomalayanregion, where, even on urban metropoleislands like Hong Kong, rewilding offersa way to drive restoration forward. Assuccinctly summarised in the openingquote of this section, Corlett (2002)highlights how a scientifically rigorousrewilding approach, rather than relyingon present-day haphazard non-con-servation introductions, is likely to resultin significant conservation benefits.
The islands in the Indian Oceanoffer yet another eminently suited labo-ratory for rewilding experiments.Recently, Cheke (2008) gave aninspiring overview of the vast potentialfor avifaunal reintroductions and taxonsubstitutions on the oceanic islands ofthe Western Indian Ocean (see also Box2). On the largest island in the WesternIndian Ocean, Madagascar, rewildingmay be the only way to resurrect someof the lost interactions of the extinctelephant birds, giant lemurs, and gianttortoises (Burney, 2003; Hansen et al.,2010).
117
ISLAND REWILDING
where a total of 17 birds were translocated. This substitution is part of a well-laid restorationplan for Secretary Island, aiming to restore lost herbivory, pollination and seed dispersalinteractions (Seddon et al., in press). Lastly, earlier translocations of South island takahePorphyrio mantelli to sites on North Island and some of its islets, which used to be home tothe extinct takahe P. hochstetteri (Lee and Jamieson, 2001), were in fact taxon substitutions eventhough they weren’t embedded in a rewilding framework but rather part of the species’recovery programme.
on the use of taxon:2 columnas 15/11/2010 13:53 Página 117

118
HANSEN
The Mascarene Islands―Mauritius, Rodrigues and Réunion, located between 600–1100km east of Madagascar―perfectly illustrate how, from an ecosystem function point-of-view,taxon substitutions can bring about a more holistic ecosystem restoration practice thanconservation of extant native species and control of alien invasive species on their own. Therehave been several proposals for the use of carefully selected taxon substitutes in the MascareneIslands (Jones, 1993; Jones, 1996; Jones, 2002; Cheke and Hume, 2008; Hansen et al.,2008; Jones, 2008 and refs therein). Potential rewilding options have recently been discussedin greater detail for two taxon groups: tortoises (Griffiths et al., 2010; Hansen et al., 2010)and birds (Cheke, 2008). Instead of repeating all of the proposed taxa, I here want to give asense of the kind of Mascarene restoration schemes, where taxon substitutes would likely bemost useful.
MMaauurriittiiuuss
On Mauritius, walking through the last remnants of native forests, one is surrounded byghosts. Since human colonisation in the 17th century, Mauritius has suffered the extinction ofmost large-bodied native vertebrates. The litany includes the famous flightless Dodo, but alsogiant tortoises, parrots, pigeons, fruitbats and giant lizards (Cheke and Hume, 2008). Manyof the extinct vertebrates were generalist herbivores and frugivores. The loss has consequentlyled native Mauritian habitats and species to suffer from disrupted herbivory and seed dispersalinteractions that once were pivotal for creating and maintaining ecosystem heterogeneity anddynamics on the island (Baider and Florens, 2006; Hansen et al., 2008; Virah-Sawmy et al.,2009; Griffiths et al., 2010). The real ghosts in the Mauritian forests are therefore not asmuch the extinct animals themselves, but more importantly the more or less extinct networksof interactions.
What about the few remaining extant endemic vertebrate species? Intense conservationefforts since the early 1980s have pulled back several endemic species from the literal brinkof extinction. The ‘Big Three’ in this regard are the Mauritius kestrel, Falco punctatus, the pinkpigeon, Nesoenas mayeri, and the echo parakeet, Psittacula echo; all three were down to orrapidly approaching single-digit numbers of surviving individuals. The rescue projects of thesethree species are-rightly so!-widely hailed as success stories that should inspire us to realisethat until the last breeding pair dies, there is hope to save a species from extinction.
However, a key question in the restoration of Mauritian habitats is whether theregeneration dynamics of native plant species in Mauritian forests ―themselves strugglingunder massive incursions by invasive alien plants and animals― are being restored as well?An example of a controversial but relevant question could be whether the Mauritian forests are‘better or worse off’ with the successful species conservation projects bringing, for example,the pigeons and parakeets back up in numbers? From the point of view of at least one critical
BBooxx 22:: TThhee MMaassccaarreennee IIssllaannddss:: aa ttaaxxoonn ssuubbssttiittuuttiioonn llaabboorraattoorryy iinn tthhee mmaakkiinngg
“Indeed the Mascarene Islands, in a nutshell, encapsulate both the fascination and frustration ofworking on the ecology of the afterlife; ghosts are not easy to pin down.”
Lawton, 1995
on the use of taxon:2 columnas 15/11/2010 13:53 Página 118

119
ISLAND REWILDING
functional component, seed dispersal, the answer may, perhaps surprisingly, be ‘worse’. Bothpink pigeons and echo parakeets feed on a variety of plant material, including substantialamounts of fruits and seeds. However, both birds are chiefly seed predators, rather than seeddispersers; the powerful gizzard of the pink pigeon crushes all but the hardest of seeds, whilethe echo parakeet mostly feeds destructively on fruits and seeds before they ripen. Anotherexample is the need to reinstate a suitable Mauritian herbivory regime that can promote habitatheterogeneity and native biodiversity. Currently, the lack of appropriate large herbivores inareas under restoration necessitates continuous weeding of invasive plants by humans; thisstate of affairs is certainly not restoring for the future, but rather only gardening for the present.
Therefore, in parallel to the conservation projects focusing on endangered species, theuse of suitable taxon substitutes to specifically resurrect efficient seed dispersal and herbivoryinteractions should be an equally important focus. This would likely not only directly benefitMauritian plant species, but also indirectly benefit long-term conservation of e.g. pink pigeonsand echo parakeets by ensuring the regeneration of biodiverse and productive native habitats.
Consequently, the situation in Mauritius is (over-)ripe for the use of taxon substitutes, but,as Lawton rightly pointed out, ‘ghosts are not easy to pin down’, much less to resurrect! Withso many ghosts to choose between, where do you start? Substitution of which species andresurrection of which functions should be attempted first? (Figure 1). The iconic dodo Raphuscucullatus would of course be an eye-catching start, but the world’s islands are sadly short ongiant, flightless pigeons. Also, despite early claims that dodos might be essential for forestregeneration (Temple, 1977), their role was likely more that of a seed predator than a seeddisperser (Witmer and Cheke, 1991; Hansen et al., 2008). Instead, the choice fell on theextinct Cylindraspis tortoises. Studies of extant species have painted an increasingly clearpicture of tortoises as keystone ecosystem components, in particular with respect to herbivoryand seed dispersal. Accordingly, Mauritius is emerging as a leader in rewilding of islandmegafauna, with successful tortoise taxon substitution projects already implemented on twooffshore islands, Round Island and Ile aux Aigrettes (Jones, 2002; Jones, 2008; Griffiths et al.,2010), and more are likely to follow.
Figure 1. How to select which functional island ghosts to resurrect? On Mauritius the dodo Raphus cucullatus(left) may be the most famous ghost, but empirical evidence strongly suggests that substituting the extinctCylindraspis tortoises (right) with extant counterparts will bring more benefits to habitat restoration (animal
illustrations by Carl Buell; Cylindraspis drawing modified from Hansen and Galetti 2009)
on the use of taxon:2 columnas 15/11/2010 13:53 Página 119

120
HANSEN
RRooddrriigguueess
Rodrigues has been hit harder than most islands; the island today is largely covered withgrassland and a few stands of exotic trees (Gade, 1985; Cheke and Hume, 2008). However,this also means that there is great scope for intelligently tinkering together novel ecosystemsthat can provide much-needed habitat for the remaining native Rodrigues biota. In fact,survival and recent population growth of all three surviving endemic vertebrates, a fruitbat andtwo passerines, has been largely attributed to the existence of (non-conservation) plantings ofexotic trees in the 1970s (Impey et al., 2002; Showler et al., 2002; Fox, 2003). Restorationprojects involving replanting forests with native species are on their way on Grande Montagneand in the Francois Leguat Giant Tortoise Reserve, as well as on some smaller islets (Jones,2008; Weaver and Griffiths, 2008). These will soon provide further habitat for the remainingnative species, as well as an ideal scenario for the introduction of taxon substitutes to re-startessential ecosystem functions like herbivory, pollination, and seed dispersal (Hansen et al.,2010; Kaiser-Bunbury et al., 2010).
RRééuunniioonn
On Réunion, the largest and youngest of the three Mascarene Islands, there are still fairlylarge tracts of native forests left in the mountainous interior of the island (Strasberg et al.,2005). Much of this region was included in a national park created in 2007. However, likeMauritius and Rodrigues, Réunion has lost the majority of its vertebrates (Cheke and Hume,2008). Also here there is evidence that plant species with fleshy fruits may be suffering fromseed dispersal limitation (Strasberg, 1996), especially on newly formed lava flows from the still-active volcano, Piton de la Fournaise (Thebaud and Strasberg, 1997).
Temple (1981) suggested that the large native forests on Réunion may be good sites tointroduce analogues from the neighbouring islands, particularly the Mauritian pink pigeonNesoenas mayeri and the echo parakeet P. echo because of empty ‘frugivorous pigeon’ and‘frugivorous parrot’ niches. There are ongoing discussions and nascent plans to replace theextinct Réunion parakeets, Psittacula eques and P. bensoni, with P. echo from Mauritius (A.Reuleaux, pers. comm). While these schemes make perfect sense from a species conservationpoint of view―literally putting valuable eggs in more than one basket―the case may not beas clear from the point of view of ecosystem function. The same argumentation as forMauritius, above, applies to the situation in Réunion. To facilitate forest regeneration―and forthis active, volcanic island in particular also to enable native plants to colonise recent lavaflows―it would make more sense to introduce generalist, efficient seed dispersers. Amongprime candidates could be blue pigeons (genus Alectroenas) from other islands in the WesternIndian Ocean, where species of this genus are still extant (Comoros, Madagascar, or theSeychelles). Tortoises are another excellent candidate group for taxon substitutions in Réunion.Radiated tortoises Astrochelys radiata from Madagascar have been introduced to Réunion aspets many years ago and are widely bred in captivity; as many as 40,000 A. radiata areestimated to be kept on the island (Boullay, 1995). While the species isn’t naturalised on theisland, animals could easily be sourced from the captive population and used as substitutesfor the extinct endemic species, Cylindraspis indica.
on the use of taxon:2 columnas 15/11/2010 13:53 Página 120

The countless islands of the PacificOcean are also ideally suited candidateecosystems for rewilding. For theGalapagos Islands, we have recentlydiscussed options for tortoises elsewhere(Hansen et al., 2010). More outlandishPacific rewilding proposals that involvetortoises include some of the recentlyextinct meiolaniid tortoises in theAustral-Pacific region and the giantflightless anatids of Hawaii (Box 3). Forthe Australian Lord Howe Island, Huttonet al. (2007) provide a candidate listwith eight extinct animals and theirpotential substitutes in their Table 2. Asa good example of socio-culturalbarriers to rewilding, they caution thatone driver of extinction of the parakeetCyanoramphus novaezelandiae subfla-vescens was people killing the parakeetto protect their fruit crop―underscoringthe need for acceptance by locals incases where they may otherwise causea newly rewilded species to go extinct.
In their evocatively titled paperabout paleoecology and “the futureresurrection of birds on Pacific islands”,Steadman and Martin (2003) advocatedthe use of Didunculus strigirostris asreplacement for an extinct Didunculus onthe island of Tofua, Tonga, and theGuam rail Gallirallus owstoni as
replacement for an extinct undescribedGallirallus on Aguiguan. These twoexamples were not suggested with aspecific emphasis on ecosystem dyna-mics, but rather on conservation of theextant species. However, in the thirdexample, replacing the extinct Duculaaurorae of Mangaia, Cook Islands, theyreferred to the candidate D. pacifica asan “ecological analog” for the extinctspecies, hinting at the possibleresurrection of lost interactions here,too–a point elaborated on later bySteadman (2006).
Hawaii is another potential Pacificrewilding laboratory. Being another ofthe world’s island extinction hotspots,there are sadly also more than enoughcandidate ghosts that could beresurrected here. Progress is beingmade on the island of Kaua`i, which ishome to one of the most promisinghabitat recreation projects anywhere(Burney and Burney, 2007) that couldbenefit greatly from using taxonsubstitutes (see Box 3). Some of thesmall, northwestern islands in thearchipelago also represent goodscenarios for rewilding projects thatcould be rapidly implemented. Forexample, on the once-barren butrecuperating Laysan Island, Sincock and
121
ISLAND REWILDING
Another candidate taxon would be Pteropus fruitbats. Réunion once harboured twospecies, P. niger and P. subniger; while the former is still extant and common on Mauritius, thelatter is extinct. Interestingly, Réunion is currently the scene of a natural re-colonisation event:a few individuals of P. niger, have recently found their way to Réunion from Mauritius (MauritianWildlife Foundation, 2009). Should they fail to establish here, conservationists have a verystrong case to translocate P. niger, or potentially use P. rodricensis from Rodrigues as a taxonsubstitute for P. subniger. Inter-island movements of fruit bats have been observed in severalarchipelagos (Wiles and Glass, 1990; McConkey and Drake, 2007), and within-archipelagotaxon substitution with fruitbats would thus closely mimic natural colonisation events.
on the use of taxon:2 columnas 15/11/2010 13:53 Página 121

Kridler (1977 cited in Morin andConant, 1998) suggested introducingNihoa Millerbirds Acrocephalusfamiliaris kingi to replace an extinctsubspecies of A. familiaris. Indeed, thefirst translocation is scheduled to goahead in 2011 (H. Freifeld and S.Plentovich, pers. comm.). Additionally,Morin and Conant (1998) proposedtranslocating the main island honey-creeper Himatione sanguinea as a
substitute for the extinct subspecies H. s.freethii, and replacing the extinctflightless Laysan Island rail Porzanapalmeri with the widely distributed,flighted and closely related rail Porzanapusilla. The difference in flight capa-bilities of the taxa in the final examplehints at a promising avenue of research:can rewilding projects “re-start” evo-lutionary trajectories or states that wereoccupied by extinct species? (see below).
122
HANSEN
BBooxx 33.. OOuuttllaannddiisshh ttoorrttooiissee ttaaxxoonn ssuubbssttiittuuttiioonnss??
In two recent papers, we have highlighted how replacing recently extinct large and gianttortoises with extant tortoises represent a low-risk, high-impact restoration scenario for manyecosystems where tortoises once occurred, especially on islands (Griffiths et al., 2010; Hansenet al., 2010). In the following two case studies, I illustrate how the use of extant tortoises astaxon substitutes could also be extended beyond the taxonomic realm of the true land tortoises,Testudinidae.
TThhee ssttrraannggee ccaassee ooff tthhee mmeeiioollaanniiiiddss
Meiolaniid tortoises―an ancient reptilian lineage that went extinct within the last fewthousand years―were bizarre–looking creatures with horned heads and spiked, club–liketails. Some species grew to gigantic proportions with likely total lengths of more than threemeters. Their diet was probably that of a highly generalised herbivore. While not closely relatedto testudinid tortoises, meiolaniids were also Eucryptodires (Gaffney, 1996). They most recentlyinhabited continental and insular regions in the Australian-Pacific region (Gaffney, 1996;Worthy et al., 1999; White et al, 2010). In New Caledonia there is evidence that humans co-occurred with meiloaniids, and may have caused their demise (Gaffney, 1984). In Vanuatu,a recently discovered early Lapita settler site dating to between 3000–2800 BP (Bedford et al.,2006) includes extensive midden deposits containing many subfossil remains of a meiolaniidtortoise; remains that have not been found at any later sites throughout the archipelago todate, suggesting a rapid anthropogenic ‘blitzkrieg’ extinction of this species (White et al. 2010;S. Bedford, pers. comm.), as so often documented for birds on isolated islands (Steadman,2006). However, in at least one case, Lord Howe Island, M. platyceps apparently went extinctbefore human arrival for unknown reasons (Gaffney, 1996).
Because virtually nothing is known about the biology of the meiolaniids, suggestions toreplace them with extant taxa could be deemed highly speculative with no practical application.For islands where meiolaniids went extinct very recently, the case may be different. Here, giantextant testudinid tortoises may well be able to act as ecological replacements for extinctmeiolaniids, restoring recently lost megafaunal plant-animal interactions. Intriguingly, in his
on the use of taxon:2 columnas 15/11/2010 13:53 Página 122

123
ISLAND REWILDING
description of the extinct giant flightless pigeon of Fiji, Worthy (2001) mentions that, “Fiji hasan abundance of trees with large fruits, whose dropped fruit probably composed the bulk ofthe diet [of the giant pigeon]”. It is not inconceivable that the extinct Fijian meiolaniid tortoisesonce had a sizeable share of this bounty, too―and that extant testudinid tortoises thereforewould be excellent substitute frugivores.
HHaawwaaiiii:: ffrroomm ttoorrttooiissee--bbeeaakkeedd ggiiaanntt dduucckkss ttoo ttoorrttooiisseess??
In Hawaii, restoration ecologists have suggested to examine the use of giant tortoises asrestoration tools under controlled circumstances in enclosures (D.A. Burney, pers. comm.),even though tortoises never ocurred on Hawaiian islands. What may initially sound like aneccentric proposal has in fact got solid arguments behind it. Giant tortoises are likely to beclosely aligned with the type of pre-human grazer/browser regime once present on Hawaii.This was dominated by now-extinct large flightless anatids, often referred to as moa-nalo,many of which had beaks with tortoise-like features–so much so that one of the moa-nalogenera was named Chelychelynechen or ‘tortoise-jawed goose’ (Olson and James, 1991).Furthermore, some endemic Hawaiian plant lineages have traits such as spines andheterophylly, which have been suggested to have evolved as anti-browser adaptations againstmoa-nalos (Givnish et al., 1984); similar traits have been associated with anti-browsingadaptations against giant tortoises in the Mascarene Islands (Eskildsen et al., 2004). A studyof coprolites from the moa-nalo Thambetochen chauliodous showed that it was a browser,feeding on coarse material (James and Burney, 1997). It is thus not inconceivable that largeor giant tortoises could play key roles in Hawaiian restoration projects, e.g. the MakauwahiCave project on Kaua`i, that aims to reconstruct a native ecosystem more or less from scratchin former agricultural lands (Burney and Burney, 2007). Moreover, using giant tortoises ratherthan large extant exotic geese in restoration projects in Hawaii has the important addedadvantage of completely removing any risk of avifaunal disease transfer to endangeredendemic birds, such as the ne-ne goose (D.A. Burney, pers. comm).
Table 1. Recently extinct meioloaniid tortoises
on the use of taxon:2 columnas 15/11/2010 13:53 Página 123

PPllaanntt rreewwiillddiinngg
Native island plants are beset byhabitat loss, invasive plants, introducedseed- and seedling predators, andintroduced herbivores (Strahm, 1994;Maunder et al., 1998; Sakai et al.,2002; Meyer and Butaud, 2009). Eventhough there have been comparativelyfew documented extinctions of plants onislands (Sax and Gaines, 2008), there isno doubt that many native island florastoday are composed of the living dead(sensu Janzen, 1986)―species, whichare reduced to such pitiful numbers that
natural regeneration is impossible, andextinction without active intervention isextremely likely. While species conser-vation and habitat restoration clearlyhas the most important role to play inconserving island plants, the role ofplant rewilding should not be under-estimated, particularly when recreatingnative ecosystems more or less fromscratch in severely degraded islands(Burney and Burney, 2007).
Like in animals, it is perhaps easierfor large plants to get the attention ofresearchers and practitioners. A goodexample is the extinct giant palm
124
HANSEN
In summary, because of their great versatility and ease of management, I envisage thatlarge and giant tortoises could be used as substitutes for extinct non-chelonians. In fact,tortoises could be used as a generalised restoration tool in many island habitats, where othernon-native browsers or grazers, such as cattle, sheep or goats, would be too disruptive or toodifficult to manage. Rewilding with large and giant tortoises thus has the potential to be amultifunctional ‘Swiss army knife’ of ecosystem restoration in numerous degraded insular (andcontinental) ecosystems, where generalist herbivores and frugivores are deemed necessaryto achieve specific, desired restoration trajectories.
Figure 2. Outlandish tortoise taxon substitutions. Even across high-order taxonomical borders, ecologicallysound arguments can be made for the use of taxon substitutes. Two examples would be substituting the extinctHawaiian flightless anatid moa-nalo Chelychelynechen quassus or meiloaniid tortoises Meioloania spp. fromislands in the Austral-Pacific region (right) with extant giant tortoises, e.g. Aldabrachelys gigantea (left)
(illustrations of extinct animals by Carl Buell)
on the use of taxon:2 columnas 15/11/2010 13:53 Página 124

Paschalococos disperta (Arecaceae)from the now largely denuded EasterIsland (Dransfield et al., 1984), wheretheories have abounded on itsdistribution, usage by humans, and whyit went extinct (Hunt, 2007; Hunt andLipo, 2009 and references therein). Evenif recent studies have questioned to whatextent Easter Island was forested beforehuman arrival (Rull et al., 2010), therecan be no doubt that this huge, endemicpalm was a keystone component of theecosystem, and thus a prime candidatefor rewilding. A good substitute could bethe equally giant and likely closelyrelated Chilean palm Jubaea chilensis.Some people believe that the EasterIsland palm is indeed identical with J.chilensis (even though no clear suppor-ting evidence exists), and have plantedseveral small populations of J. chilensison Easter Island already (Moreno andFernández, 2004). Such replantingcould easily be expanded within aproper rewilding framework.
Another example comes from thenorthwesternmost Hawaiian Islands,where the restoration plan for LaysanIsland (Morin and Conant, 1998)proposes that several extinct species orsubspecies from this island could bereplaced with extant taxa from nearbyislands―in concert with the suggestionsto rewild the Laysan avifauna (seeabove). The restoration plan stresses theneed for careful studies to determine thesuitability of the substitutions, andincludes detailed information on stepsthat should be taken to assess if andhow the extinct Achyranthes atollensis
(Amaranthaceae) could be replaced byA. splendens var. rotundata, andCenchrus agrimonioides var. laysanensis(Poaceae) by C. agrimonioides var.agrimonioides, respectively. It alsomentions the possibility to replace anextinct Pritchardia palm, once animportant component of the Laysanecosystem (Athens et al., 2007), with anextant Pritchardia species, potentially P.remota from Nihoa (Rauzon, 2001). Afinal example comes from Lord HoweIsland, where Hutton et al. (2007)speculated that the extinct Solanumbauerianum (Solanaceae) could bereplaced with S. viride.
However, apart from obvious caseslike the Pritchardia palm on Laysan, orthe giant wine palm on EasterIsland―i.e. large plants in an otherwisedenuded landscape―it is very difficult toconfirm when/if a plant has gone extinct(Maunder, 1992), and thus couldbecome a ‘candidate ghost’ to berewilded. Indeed, some island plantsthat were thought to be extinct havebeen rediscovered (Strahm, 1983;IUCN, 2001; Simbaña and Tye, 2009),underscoring the need for extremecaution when determining the appro-priateness of plant rewilding projects.However, once a project is deemed to bedesirable, its implementation can benefita lot from the lessons learnt in plantreintroduction and translocation projects(Maunder, 1992; Allen, 1994). On theplus side of things, rewilded plant indi-viduals or populations are obviouslyeasier to monitor than rewilded animals(Maunder, 1992).
125
ISLAND REWILDING
on the use of taxon:2 columnas 15/11/2010 13:53 Página 125

In general, for rewilding projects onislands it is possible to suggest at leasttwo prioritised lists for selecting extantproxy taxa. First, in relation to sourceareas a reasonable initial list of options,in order of most preferred to leastpreferred, could be: 1) translocationwithin an island (including smallersurrounding islets), 2) translocationbetween islands within an archipelago,3) translocations between archipelagoswithin a region, and 4) translocationsfrom islands in other regions or fromcontinental regions. Second, in relationto the relevance of genetic relatednesswhen selecting analogue species, Huttonet al. (2007) argue that a similarapproach should be taken. That is,preferably 1) reintroducing the samespecies, before 2) substituting with adifferent subspecies, or 3) a congeneric.In their opinion, substitutions that involvetaxa above subspecies level or that arebased on ecological function should beavoided. However, both lists may be toorestrictive and indeed fundamentallyflawed when considering islands, whichare home to many of the world’s mostspectacular examples of adaptiveradiation―speciation events, whereclose relatives on neighbouring islandsare often the ones most morpho-logically, and thus also potentiallyecologically, dissimilar. Classical exam-ples of this include Galapagos finches
and Hawaiian honeycreepers. On theother hand, there are also severalgroups of closely related animals withinand between archipelagos that see-mingly fulfill the same function, forexample congeneric species of frugi-vorous pigeons and fruitbats acting asseed dispersers. Moreover, it is con-ceivable that extinct taxa could bereplaced with functionally similar extanttaxa even across genera or higher-ordertaxonomical borders (see Box 3). Whatis clear is that the more we know aboutthe extinct species, and the pre-anthropogenic islands they inhabited,the better we will be able to judge therelative merits of specific taxon substitutecandidates.
LLeeaarrnniinngg aabboouutt tthhee gghhoossttss
Lawton (1995) said that endeavoursto learn about ghosts were interesting,but ultimately of little practical use.However, both paleoecologists andconservation biologists now realise thatconservation and restoration projectsstand to gain tremendously from preciseand detailed information about extinctspecies and their pre-anthropogenicecosystems (e.g. Lyman, 2006; Froydand Willis, 2008; Frazier, 2010; Hadleyand Barnosky, 2009; Stewart, 2009)―and it is clear that rewilding projectswould, too.
126
HANSEN
HHooww ttoo sseelleecctt ttaaxxoonn ssuubbssttiittuutteess:: hheellpp ffrroomm bbeeyyoonndd tthhee ggrraavvee
“Most of the research about species extinction has been conductedon islands because islands are controlled environments
and scientists can get drinks with little umbrellas in them there”
O’Rourke, 1994
on the use of taxon:2 columnas 15/11/2010 13:53 Página 126

An obvious first step is to discoverand identify as many of the recentlyextinct species on an island as possible.While there are historical records oreven scientific studies available for someof the most recent island ghosts, mostoftenly paleobiologists have to dig a bitdeeper. Tremendous progress in thisfield has been made on many islandsaround the world in recent decades. Thepioneering work of island paleoeco-logists working in Hawaii and otherPacific islands, the Mascarenes, NewZealand, Madagascar, and islands inthe Caribbean, have unearthed inva-luable subfossil material and increasedthe known tally of recently extinct speciesmanifold (e.g. Olson and James, 1991;Burney, 1997; Worthy et al., 1999;Burney et al., 2001; Worthy and Holda-way, 2002; Burney et al., 2004;Stead-man, 2006; Steadman et al.,2007; Cheke and Hume, 2008).
After building a reasonablycomplete understanding of an island’spre-anthropogenic species communitiescomes the need to reconstruct theevolution and ecology of the species. Inmany cases, DNA can be extracted frommuseum specimens or subfossilremains, and used for molecularphylogenetic analyses to shed light one.g. relationships, origins, and rates ofspeciation (e.g. Cooper et al., 1996;Sorenson et al., 1999; Austin andArnold, 2001; Shapiro et al., 2002;James, 2004; Karanth et al., 2005;Austin and Arnold, 2006). Again, whilesuch studies often provide importantinformation for conservation of still-extant species (Leonard, 2008), it is clearthat they offer similarly central insights
for identifying suited taxon substitutecandidates.
Subfossil remains can also yieldtreasure troves of data on a centralquestion in re-assembling a partially lostisland ecosystem: what was the diet ofthe extinct species? Such insights can begained from studying dentition andpatterns of dental wear (Godfrey et al.,2004), stable isotopes (Burleigh andArnold, 1986; Koch et al., 2009;Crowley et al., 2010), and coprolitecontent (James and Burney, 1997;Wood et al., 2008). For community-levelinformation, Burney et al. (2003) usedan innovative approach to establishbody mass indices of extinct herbivoresin Madagascar by analysing abun-dances of fungal spores associated withdung from large herbivores. Pre-anthropogenic plant communities canalso be studied via analyses of seedsand pollen in sediments from e.g.wetlands and lakes (Colinvaux andSchofield, 1976; Burney et al., 2004;Athens et al., 2007; Rijsdijk et al.,2009).
A final, more indirect line of inquiryis examining putative links between traitsin extant island species and the causalevolutionary pressures once exerted byextinct island species; for example, howherbivores may have shaped the plantcommunities (Greenwood and Atkinson,1977; Caughley, 1989; Givnish et al.,1994; Bond et al., 2004; Eskildsen etal., 2004; Hansen et al., 2004; Burnsand Dawson, 2006; Bond and Silander,2007; Godfrey et al., 2008; Fadzly etal., 2009; Griffiths et al., 2010), howfrugivores may have interacted with fruit-
127
ISLAND REWILDING
on the use of taxon:2 columnas 15/11/2010 13:53 Página 127

bearing plants (Lord et al., 2002;McConkey and Drake, 2002; Godfreyet al., 2008), or assessing which flowersand pollinators may once haveinteracted (Lammers and Freeman,1986; Cox and Elmqvist, 2000).
Equipped with all of these detailedtools, paleoecologists can envision
In this section I will highlight somequestions raised, and opportunitiesoffered, by island rewilding. Some havedirect bearing on larger-scale con-tinental rewilding, while others arerelevant only for islands.
AAlliieenn eexxoottiicc oorr ddee ffaaccttoo ssuubbssttiittuuttee??
While negative impacts of intro-duced species are often readilyapparent, it can be more complex todisentangle their overall function,including potential positive roles theymay play in their new home. It is thusnot always a clear-cut case thateradication or control is the cure-all forintroduced species, and effects ofremoval may be difficult to predict(Zavaleta et al., 2001). This becomesespecially important to understand incases where native species may havecome to rely on the introduced species
entire prehistoric island ecosystems witha great deal of detail (Burney, 2003;Cheke and Hume, 2008; Godfrey et al.,2008), and rewilding practitioners arearmed with a solid scientific basis onwhich to progress when selecting specifictaxon substitute candidates.
for survival or reproduction (Westman,1990; Ewel and Putz, 2004). Warren(2007) therefore suggests to rather focuson problematic versus non-problematicspecies. Such deliberations raise animportant point in relation to rewilding:in ecosystems where introduced specieshave far more positive than negativeeffects and where they fulfill roles ofextinct species, should they be toleratedand indeed be considered de factotaxon substitutes?
Continental restoration projects andnature reserve managers have longmade use of livestock to providecontrollable, desired levels of herbivory(Posada et al., 2000; Papanastasis,2009). Even if not employed within anexplicit rewilding framework, such largemammals are, in fact, often de factotaxon substitutes, replacing recentlyextinct, local mammalian herbivoremegafauna. While it may be difficult to
128
HANSEN
FFuuttuurree rreesseeaarrcchh aanndd mmaannaaggeemmeenntt ddiirreeccttiioonnss
“The pragmatic reality is probably that restoration is moreabout creating a new future than recreating the past”
Macdonald, 2009
“I have found a strong inverse correlation between peoples’ecologicalpurity and the spatial extent of restoration they have personally achieved”
Cabin, 2007
on the use of taxon:2 columnas 15/11/2010 13:53 Página 128

assess if, and to what extent, e.g. anintroduced herbivore benefits thefunctioning of an island ecosystem (Leeet al., 2010), it is easier to understandthe advantage of an introduced speciesresurrecting extinct mutualisticinteractions, such as pollination andseed dispersal.
In relation to seed dispersal, somestudies have illustrated how introducedbirds can efficiently disperse nativeseeds on islands (Cole et al., 1995;Ferguson and Drake, 1999; Foster andRobinson, 2007; Kawakami et al.,2009), while other studies found theopposite pattern, with introduced birdslargely dispersing seeds of introducedand invasive plants (Lewin and Lewin,1984; Linnebjerg et al., 2010; Chimeraand Drake, 2010; Staddon et al.,2010). In the Balearic Islands, Riera etal. (2002) highlighted how introducedmammals did consume native fruits, butthat they were not providing as good aseed dispersal service to the endangeredCneorum tricoccon (Cneoraceae) as theendemic Podarcis lizards. In NewZealand, introduced rodents and pos-sums may feed on fruits and seeds ofsome native plant species, but destroythe majority of seeds and should thus beconsidered inefficient substitutes(Williams et al., 2000; Kelly et al.,2006). Likewise for pollination inter-actions, where introduced species canbe functional substitutes for extinctpollinators (Cox, 1983; Lord, 1991), butwhere they more oftenly disrupt nativeinteractions, to the detriment of nativeplants and/or animal mutualists (Tra-veset and Richardson, 2006).
These examples stress how the rolesof introduced species in native islandecosystems are idiosyncratic, and howdetailed case-by-case studies arerequired to determine whether already-present exotic species should beregarded as unwanted pests, or, inspecific cases, as welcome additions todegraded habitats (Rodriguez, 2006).
CCoonnsseerrvvaattiioonn ppoolliittiiccss,, hhuummaann ccuullttuurree,,aanndd ttaaxxoonn ssuubbssttiittuuttiioonnss oonn iissllaannddss
Many islands form, or are part of,small independent states, especially inthe Caribbean and in the Pacific Ocean.Tourism often forms a major part of theeconomy of these states, and pictures ofthe ‘unspoiled paradise island’ areamong the most frequently usedadvertisement gimmicks. With an in-creasing emphasis on the environment,it is in the interest of such islands tomaintain or enhance their ‘green’ eco-tourism image, and restoration of nativehabitats is a good way to achieve this.Therefore, parallel to the ecological andevolutionary arguments about islandsbeing relatively simple ecosystems,stakeholders in such island states,including parliamentary politicians, maybe more likely to support ecosystemrewilding with taxon substitutes thanpoliticians from more economicallycomplex continental countries. Therepublic of Mauritius in the MascareneIslands offers a good example of this(Box 2). In general, conservation mana-gement on islands is often very hands-on, and adaptive management isfrequently used. Both governmental and
129
ISLAND REWILDING
on the use of taxon:2 columnas 15/11/2010 13:53 Página 129

non-governmental conservation agen-cies are thus well-poised to capitalise onthe development of rewilding with taxonsubstitutes, and indeed to contributesignificantly to the maturation of rewil-ding as a management option and to itsmainstream acceptance elsewhere.
Within a small island nation thereis a great need to ensure local supportfor taxon substitution projects. This couldbe achieved via economical or culturalpathways. While economical benefitsare easy to envision, cultural ones maybe more difficult to imagine. A goodexample of the latter comes fromMangaia, Cook Islands, where peopleexpressed their “desire to establish andprotect populations of pigeons anddoves, thereby restoring some of theauditory and visual enchantment thatthese forests once offered” (Steadmanand Martin, 2003). An even morecharismatic example is linked to a snipetaxon substitution project in NewZealand (see Box 1). After an inves-tigation worthy of Agatha Christie,Miskelly (1987) identified the call of alegendary nocturnal bird, the hakawai,as the sounds created by the night-timeaerial displays of endemic snipe(Miskelly, 1987). Two of the male SnaresIsland snipe Coenocorypha huegeliintroduced to Putauhinu Island in 2005as substitutes for the extinct C. iredaleihad a pattern of tail feather wear sug-gestive of hakawai displays, causingelation among the people involved thatthey had resurrected not only a real butalso a legendary bird! (Charteris andMiskelly, 2005). To hear the hakawaiwas earlier considered a bad omen
(Miskelly, 1987), but in an interestingtwist to the story, local muttonbirdersenthusiastic about the snipe’s return maynow consider hearing the hakawai aboon―demonstrating another way inwhich rewilding can garner localsupport.
Of course, many island specieswent extinct very soon after humancolonisation; these taxa do not oftenpersist in the memory of local folklore.For such species, though, a creative useof nomenclature by the scientists thatdiscover the remnants may have animportant role to play in paving the wayfor cultural acceptance of taxon subs-titutes to replace them. A good exampleof this is could be the evocative namecoined by Olson and James (1991) forthe extinct giant, flightless anatids ofHawaii, the moa-nalo, “from Hawaiianmoa, fowl, and nalo, meaning lost,vanished, forgotten”―after all, whowouldn’t want to be the one to find long-lost fowls? (see Box 3).
MMaannaaggiinngg eeccoossyysstteemmss ffoorr tthhee ffuuttuurree
Despite early calls for an integrationof ecosystem dynamics in the then-emerging field of restoration ecology(Pickett and Parker, 1994), the goal ofmany restoration projects is still toachieve a ‘natural’ state where therestored ecosystem is as close aspossible to the way it was beforeanthropogenic impacts (Hilderbrand etal., 2005). There has been a longdiscussion of what is natural, andseveral schemes and metrics have been
130
HANSEN
on the use of taxon:2 columnas 15/11/2010 13:53 Página 130

proposed on how to classify andquantify ‘naturalness’ e.g. (Anderson,1991; Machado, 2004). It should bynow be clear, though, that there is not asingle non-anthropogenic ecosystem leftanywhere on this planet. Moreover, ourknowledge of ecosystems haveprogressed to an understanding thatecosystems are not static, but rather arefluid and dynamic (Wu and Loucks,1995; Bengtsson et al., 2003; Manninget al., 2009), an especially importantpoint in relation to conservation andrestoration (Gillson and Willis, 2004;Manning et al., 2009; Bellingham et al.,2010; Hall, 2010). In face of anuncertain future, a strong case cantherefore be made for also restoringtowards future resilience rather than onlyaccording to past history (Swetnam etal., 1999; Choi, 2007; Hayward, 2009;Jackson and Hobbs, 2009). Ideally,rewilding combines key aspects of boththese approaches by aiming to restorelost ecosystem functions and dynamics,based on recent ecological history,rather than aiming to achieve ormaintain a specific ecosystem state orspecies roster. Rewilders and restorationecologists should work together asmediators and creators of desiredecosystem trajectories and optimallandscape fluidity (sensuManning et al.,2009).
An interesting parallel has recentlybeen drawn between how we viewhuman health and ecosystem health,and specifically how approaches toecosystem restoration ideally couldmirror human medicine in how todiagnose ‘unhealthy’ ecosystems, andhow to ‘heal’ them (Temperton, 2007;Keulartz, 2010; Lowenthal, 2010). Justlike human health is easier to maintainvia prevention than to restore via cure,rewilding with taxon substitutes thereforehas potential as a ‘future’ managementoption in another sense. Cole et al.(2008) argued that, “Although contro-versial, [practitioners could] considerfunctional substitutes for species thatcannot survive under current con-ditions”. This could become importantfor many oceanic island ecosystems,where even the most focusedconservation efforts may not be enoughto save a species, or where climatechange will transform a restrictedhabitat type in a way that renderssurvival impossible (Harris et al. 2006;see also Box 4). In such cases, inves-tigating the possible use of taxonsubstitutions ‘ahead of time’ wouldmean that if a species goes extinct, taxonsubstitution as a restoration measurecould be undertaken fast enough toprevent undesirable trajectories ofecosystem dynamics.
131
ISLAND REWILDING
on the use of taxon:2 columnas 15/11/2010 13:53 Página 131

AA mmeettaappooppuullaattiioonn aapppprrooaacchh ttoo iissllaannddrreewwiillddiinngg
Even more so than in manycontinental ecosystems, current conser-vation or restoration areas on islandsare often very small and disjunct. Theuse of the same taxon substitute speciesin a network of small intra- or inter-
island conservation management areas(CMAs) could take place within a‘managed metapopulation’ strategy(Davies-Mostert et al., 2009), whichwould have important implications forlong-term management. For example,human-mediated dispersal of indi-viduals between CMAs could beemployed to mimic natural levels of
132
HANSEN
BBooxx 44.. KKiinnddrreedd ssppiirriittss:: rreewwiillddiinngg aanndd aassssiisstteedd mmiiggrraattiioonn
Models and projections increasingly demonstrate how the distributions of many specieswill have to change to respond to climate change. However, some species or groups of specieswill likely not be able to respond and migrate fast enough, or even to disperse acrossanthropogenically altered and disconnected landscapes to new, suitable habitats (Walther etal., 2002; Colwell et al., 2008; Devictor et al., 2008).
In response, some scientists and practitioners have proposed to proactively translocatepopulations of plants and animals―confusingly using four different names for the sameprocess: assisted migration (Keel, 2007; McLachlan et al., 2007), assisted colonisation (Hunter,2007; Hoegh-Guldberg et al., 2008), facilitated range shift (Manning et al., 2009), ormanaged relocation (Schwartz et al., 2009). All these terms describe an approach aiming atidentifying species that are particularly at risk due to climate change and moving them toother, more suited, parts of the ‘same broad biogeographic region’ (Hoegh-Guldberg et al.,2008). The concerns raised against this practise largely mirror the ones against taxonsubstitutions, mainly the inherent risk of creating new invasive species or pests (Ricciardi andSimberloff, 2009). However, this risk is very likely small. Mueller and Hellmann (2008) foundthat introduced species that had been moved within a continent were unlikely to becomeinvasive, compared to species that had been moved to another continent. Moving specieswithin one island, or between islands in an archipelago, would in all likelihood be even lessrisky, given the much smaller geographical ranges involved.
In parallel to the arguments in favour of rewilding islands with taxon substitutes, islandsthus also appear eminently suitable as both donors and recipients of species targeted forassisted migration. Within their relatively small areas, many islands harbour very differentclimatic regimes. Even within a dramatic future scenario for climate change, it is thusconceivable that slowly migrating native species, e.g. plants and invertebrates, can betranslocated within an island. Moreover, within or between archipelagos, or even betweenregions, there may be opportunities to combine taxon substitutions and assisted migration, byusing ‘climatic refugees’ (Hunter, 2007) from one island as substitutes for extinct, similarspecies on another. Carter et al. (2001) take a conservative view and argue that adaptationoptions are limited to broadscale measures such as slowing invasion rates, optimisingconservation policies, and increasing public awareness. For species at risk of extinction, suchmeasures will appear more than a little ‘fluffy’, with little, if any, practical actions before it’stoo late.
on the use of taxon:2 columnas 15/11/2010 13:53 Página 132

gene flow or simply to maintain geneticdiversity, or to prevent populations fromfluctuating towards zero. The meta-population management approach maybe more prudent on small oceanicislands than on larger continentalislands, where larger areas can berewilded and ideally harbour long-termself-sustaining populations of thesubstitute species. Obviously, though,the metapopulation metaphor shouldnot be taken too far; for example, itwould be undesirable to let populationsin one CMA go extinct before supple-menting them with individuals fromanother.
IIssllaanndd rreewwiillddiinngg aass ((rree--))eevvoolluuttiioonnaarryyeexxppeerriimmeennttss
With proper planning and moni-toring, introducing taxon substitutes ontoone or several islands may function aslong term, replicable evolutionary‘re–speciation’ or ‘re–evolution’experiments―as Atkinson (1988)
Public perception and under-standing of concepts in ecology andconservation is usually lagging behindscientific research (Ladle and Gillson,
asked: “Can carefully planned speciestranslocations reinstate the evolutionaryprocesses formerly acting on related butextinct species?”. Extant and extinctisland rails (Rallidae), most but not all ofwhom are/were flightless, and theirislands offer an ideal candidate setup.Until recently these birds werewidespread, especially in the Pacificregion (Steadman, 2006), andSteadman and Martin (2003) envisionedthe need for an “innovative, proactiveconservation strategy... to restartevolution of some of the lineages thathave suffered the most loss, such asflightless rails” (see also Lazell, 2005).We know that traits of island species canrespond evolutionarily to changes intheir environment within a very shorttime (Grant, 1998). Initial stages of ‘re-evolution’ of flightlessness in flightedsubstitutes of island birds, such asbehavioural modifications and extendedground feeding, could perhaps bedetected within relatively fewgenerations.
2009). Rewilding with taxon substitutionsrepresents a case where public andpolitical perception―and hence priori-tisation and funding―could well be
133
ISLAND REWILDING
CCoonncclluuddiinngg rreemmaarrkkss
“those who repair the earth should ultimately marvel atnature’s ways even more than lament nature’s losses”
Hall, 2010
“Restoration is fencing, planting, fertilizing, tilling, and weeding the wildland garden:succession, bioremediation, reforestation, aforestation, fire control, proscribed burning,
crowd control, biological control, reintroduction, mitigation, and much more” Janzen, 1998
on the use of taxon:2 columnas 15/11/2010 13:53 Página 133

ahead of ecologists in the decadesahead. Even among scientists, thefascination with the ‘ecology of theafterlife’, and debating the relativemerits of tinkering with species andecosystems for restoration purposes,causes creative coining of terms like‘resurrection ecology’ (Pyle, 2000), or‘recombinant ecology’ (Soulé, 1990).
Interestingly, many of the positiveresponses to the suggestions to rewildNorth America with substitutemegafauna have come from a perhapsunexpected side: landowners andranchers, who are willing to put theirlands at the disposal of rewilders(Donlan and Greene, 2010). Thus,advances in applied rewilding may alsobe driven by private conservationentrepreneurs, one example of which isthe recent rewilding of the Bolsontortoise in New Mexico on lands ownedby the Turner Endangered Species Fund(Truett and Phillips, 2009). Similarly,many small islands or parts of largerislands are privately owned, and incases where the owners are sympatheticto the rewilding concept these could beused for taxon substitutions.
Opponents against taxon substi-tutions most frequently invoke the fearof creating new invasive species pro-blems, often proceeding to listing themany well-known examples from intro-duced biocontrol agents gone out ofcontrol. While this concern is valid andimportant, in some cases it makes littlesense, especially in relation to rewildingon islands. For example, Ricciardi andSimberloff (2009) recount how “human-assisted introductions of predators
have...caused species loss in insularhabitats worldwide”―but noone in theirright mind would advocate replacing anextinct island species with a mainlandpredator! More drastically, in a recentcomment, Caro and Sherman (2009) goas far as to say there should be “aninternational moratorium” on the use ofnon-native megafauna in rewildingprojects. Quite apart from lackingsuggestions as to how such a mora-torium could be achieved, the questionremains who should enforce it? Anunderlying thread in the argumentationof rewilding opponents is to say that “weshouldn’t allow any rewilding projectsbecause there is too little empiricalevidence from rewilding projects”.However, as rightly pointed out byGriffiths and Harris (2010), this is quitean unconstructive catch-22.
Instead of blanket condemnation oftaxon substitution as a restoration toolon islands, we need case-by-casejudgments based on the particularnatural, economical, political, andcultural idiosyncrasies of each case.Moreover, to avoid creating eco-touristgimmicks only, rewilding projects shouldcomprise explicit and quantifiable short-and long-term objectives. Properlyapplied, rewilding with taxon substitutesrepresents a very clear case of evidence-based conservation (sensu Sutherland etal., 2004).
I conclude this contribution with acall to nations and organisations thatare custodians of appropriate species orpopulations to consider sharing thisessentially global natural heritage withnations or organisations that are pla-
134
HANSEN
on the use of taxon:2 columnas 15/11/2010 13:53 Página 134

nning restoration projects, where thebest available evidence demonstrateshow these species could be successfullyused as taxon substitutes.
AACCKKNNOOWWLLEEDDGGEEMMEENNTTSS
My greatest gratitude goes toValentín Pérez-Mellado, M.M. Ramon,and the organising committee of theIsland Evolution Symposium in Menorcafor the opportunity to join the Sym-posium and to present my thoughts andviews here. I am also grateful to DavidBurney, Trevor Worthy, Stuart Bedford,Colin Miskelly, and Anthony Cheke forsharing information and unpublisheddata, to Christine Griffiths, Josh Donlan,Karl Campbell, Rachel Atkinson, andChristopher Kaiser-Bunbury for manyinspiring discussions, and to the manyother colleagues and friends in theMascarene Islands and elsewhere, withwhom I have discussed island rewildingduring the last few years.
RREEFFEERREENNCCEESS
Aikman, H., Davis, A., Miskelly, C.,O'Connor, S. and Taylor, G. (2001).Chatham Islands threatened birds.Recovery and management plans. NewZealand Department of Conservation,Wellington, New Zealand.
Allen, W. H. (1994). Reintroduction ofendangered plants. Bioscience 44: 65–68.
Anderson, J. E. (1991). A conceptual frame-work for evaluating and quantifyingnaturalness. Cons. Biol. 5: 347–352.
Athens, J. S., Ward, J. V. and Blinn, D. W.(2007). Vegetation history of LaysanIsland, Northwestern Hawaiian Islands.Pac. Sci. 61: 17–37.
Atkinson, I. A. E. (1988). Opportunities forecological restoration. NZ J. Ecol.11: 1–12.
Atkinson, I. A. E. and Cameron, E. K. (1993).Human influence on the terrestrial biotaand biotic communities of NewZealand. Trends Ecol. Evol. 8: 447–451.
Atkinson, I. A. E. (2001). Introducedmammals and models for restoration.Biol. Cons. 99: 81–96.
Austin, J. J. and Arnold, E. N. (2001). Ancientmitochondrial DNA and morphologyelucidate an extinct island radiation ofIndian Ocean giant tortoises(Cylindraspis). Proc. R. Soc. Lond. B268: 2515–2523.
Austin, J. J. and Arnold, E. N. (2006). Usingancient and recent DNA to explorerelationships of extinct and endangeredLeiolopisma skinks (Reptilia : Scincidae)in the Mascarene islands. Mol. Phyl.Evol. 39: 503–511.
Baider, C. and Florens, F.B.V. (2006). Currentdecline of the "Dodo Tree": a case ofbroken-down interactions with extinctspecies or the result of new interactionswith alien invaders? In: W. F. Lauranceand C. A. Peres (eds), Emerging threatsto tropical forests, 199–214. ChicagoUniversity Press, Chicago.
Baker, A. J., Miskelly, C.M. and Haddrath, O.(2010). Species limits and populationdifferentiation in New Zealand snipes(Scolopacidae: Coenocorypha). Cons.Genet., 11: 1363–1374.
135
ISLAND REWILDING
on the use of taxon:2 columnas 15/11/2010 13:53 Página 135

Bedford, S., Spriggs, M. and Regenvanu, R.(2006). The Teouma Lapita site and theearly human settlement of the PacificIslands. Antiquity 80: 812–828.
Bellingham, P.J., Towns, D.R., Cameron, E.K.,Davis, J. J., Wardle, D. A., Wilmshurst,J. M., Mulder, C. P. H. (2010). NewZealand island restoration: seabirds,predators, and the importance ofhistory. NZ J. Ecol. 34: 115–136.
Bengtsson, J., Angelstam, P., Elmqvist, T.,Emanuelsson, U., Folke, C., Ihse, M.,Moberg, F., Nyström, M. (2003).Reserves, Resilience and DynamicLandscapes. Ambio 32: 389–396.
Bond, W. J., Lee, W. G. and Craine, J. M.(2004). Plant structural defencesagainst browsing birds: a legacy ofNew Zealand's extinct moas. Oikos104: 500–508.
Bond, W.J. and Silander, J.A. (2007). Springsand wire plants: anachronistic defencesagainst Madagascar’s extinct elephantbirds. Proc. R. Soc. Lond. B 274: 1985–1992.
Boullay, S. (1995). Repatriation of radiatedtortoises, Geochelone radiata, fromRéunion Island to Madagascar. Chelon.Cons. Biol. 1: 319–320.
Burleigh, R. and Arnold, E. N. (1986). Ageand dietary differences of recentlyextinct Indian Ocean tortoises(Geochelone s. lat.) revealed by carbonisotype analysis. Proc. R. Soc. Lond. B227: 137–144.
Burney, D. A. (1997). Tropical islands aspaleoecological laboratories: Gaugingthe consequences of human arrival.Hum. Ecol. 25: 437–457.
Burney, D. A., James, H. F., Burney, L. P.,Olson, S .L., Kikuchi, W., Wagner, W. L.,Burney, M., McCloskey, D., Kikuchi, D.,Grady, F. V., Gage, R., Nishek, R.(2001). Fossil evidence for a diversebiota from Kaua'i and its transformationsince human arrival. Ecol. Monogr. 71:615–641.
Burney, D. A. (2003). Madagascar’sprehistoric ecosystems. In: S. M.Goodman and J. P. Benstead (eds), 47–51. Natural history of Madagascar,University of Chicago Press, Chicago.
Burney, D. A., Robinson, G. S. and Burney,L. P. (2003). Sporormiella and the lateHolocene extinctions in Madagascar.PNAS 100: 10800–10805.
Burney, D. A., Burney, L. P., Godfrey, L. R.,Jungers, W. L., Goodman, S.M., Wright,H. T., Jull, A. J. T. (2004). A chronologyfor late prehistoric Madagascar. J. Hum.Evol. 47: 25–63.
Burney, D. A. and Burney, L. P. (2007).Paleoecology and "inter-situ" restorationon Kaua'i, Hawai'i. Front. Ecol. Environ.5: 483–490.
Burns, K. C. and Dawson, J. W. (2006). Amorphological comparison of leafheteroblasty between New Caledoniaand New Zealand. NZ J. Bot. 44: 387–396.
Cabin, R. J. (2007). Science-driven resto-ration: a square grid on a round earth?Rest. Ecol. 15: 1–7.
Caro, T. (2007). The Pleistocene re-wildinggambit. Trends Ecol. Evol. 22: 281–283.
Caro, T. and Sherman, P. (2009). Rewildingcan cause rather than solve ecologicalproblems. Nature 462: 985.
Carter, L. M., Shea, E., Hamnett, M.,Anderson, C., Dolcemascolo, C.,
136
HANSEN
on the use of taxon:2 columnas 15/11/2010 13:53 Página 136

Guard, C., Taylor, M., Barnston, T., He,Y., Larsen, M., Loope, L., L., M., Meehl,G. (2001). Potential consequences ofclimate variability and change for theUS-affiliated islands of the Pacific andCaribbean. –In: Climate ChangeImpacts on the United States: ThePotential Consequences of ClimateVariability and Change. CambridgeUniversity Press: Cambridge, UK., pp.315–349.
Caughley, G. (1989). New Zealand plant-herbivore systems: past and present.NZ J. Ecol. 12: 3–10.
Charteris, M. and Miskelly, C. (2005). SnaresIsland snipe (tutukiwi) translocation toPutauhinu Island, April 2005. Depart-ment of Conservation, Wellington, NewZealand.
Cheke, A. S. (2008). Bird reintroduction andanalogue options in the oceanic islandsof the western Indian Ocean. In: J. G.Ewen, D. P. Armstrong, K. A. Parker, P.J. Seddon (eds), Avian reintroductionbiology: current issues for science andmanagement, 27–50. Avian BiologyResearch 1, London, United Kingdom.
Cheke, A. S. and Hume, J. P. (2008). Lostland of the Dodo. Christopher Helm,London, UK.
Chimera, C. G. and Drake, D. R. (2010).Patterns of seed dispersal and dispersalfailure in a Hawaiian dry forest havingonly introduced birds. Biotropica 42:493–502.
Choi, Y. D. (2007). Restoration ecology to thefuture: a call for new paradigm. Rest.Ecol. 15: 351–353.
Cole, D. N., Yung, L., Zavaleta, E. S., Aplet,G. H., III, F. S. C., Graber, D. M., Higgs,E. S., Hobbs, R. J., Landres, P. B., Millar,C. I., Parsons, D. J., John M. Randall,Stephenson, N. L., Tonnessen, K. A.,
White, P. S., Woodley, S. (2008).Naturalness and beyond: protectedarea stewardship in an era of globalenvironmental change. The GeorgeWright Forum 25: 36–56.
Cole, F. R., Loope, L. L., Medeiros, A. C.,Raikes, J. A. and Wood, C. S. (1995).Conservation implications of introducedgame birds in high-elevation Hawaiianshrubland. Cons. Biol. 9: 306–313.
Colinvaux, P. A. and Schofield, E. K. (1976).Historical ecology in the GalapagosIslands: I. A Holocene pollen recordfrom El Junco Lake, Isla San Cristobal.J. Ecol. 64: 989–1012.
Colwell, R. K., Brehm, G., Cardelus, C. L.,Gilman, A.C. and Longino, J.T. (2008).Global warming, elevational rangeshifts, and lowland biotic attrition in thewet tropics. Science 322: 258–261.
Cooper, A., Atkinson, I. A. E., Lee, W. G. andWorthy, T.H. (1993). Evolution of theMoa and their effect on the NewZealand flora. Trends Ecol. Evol. 8:433–437.
Cooper, A., Rhymer, J., James, H. F., Olson,S. L., McIntosh, C.E., Sorenson, M. D.,Fleischer, R. C. (1996). Ancient DNAand island endemics. Nature 381:484–484.
Corlett, R. T. (2002). Reintroduction of"missing" vertebrates to Hong Kong:Benefits, problems and prospects. In: J.Hodgkiss (ed) Challenges of natureconservation in the face of developmentpressure, 175–180. 2001 IUCN WorldCommission on Protected Areas, EastAsia Conference, AFCD, Hong Kong.
Cox, P. A. (1983). Extinction of the Hawaiianavifauna resulted in a change ofpollinators for the ieie, Freycynetiaarborea. Oikos 41: 195–199.
137
ISLAND REWILDING
on the use of taxon:2 columnas 15/11/2010 13:53 Página 137

Cox, P. A. and Elmqvist, T. (2000). Pollinatorextinction in the Pacific Islands. Cons.Biol. 14: 1237–1239.
Crowley, B. E., Godfrey, L. R. and Irwin, M. T.(2010). A glance to the past: subfossils,stable isotopes, seed dispersal, andlemur species loss in SouthernMadagascar. Am. J. Primatol. 71:1–13
Davies-Mostert, H. T., Mills, M. G. L. andMacdonald, D. W. (2009). A criticalassessment of South Africa's managedmetapopulation recovery strategy forAfrican wild dogs. In: M. W. Haywardand M. J. Somers (eds), Reintroductionof top-order predators, 10–42. Wiley-Blackwell, Oxford.
Devictor, V., Julliard, R., Couvet, D. andJiguet, F. (2008). Birds are trackingclimate warming, but not fast enough.Proc. R. Soc. Lond. B 275: 2743–2748.
Donlan, J., Greene, H. W., Berger, J., Bock,C. E., Bock, J. H., Burney, D. A., Estes,J. A., Foreman, D., Martin, P.S., Roemer,G. W., Smith, F. A., Soulé, M. E. (2005).Re-wilding North America. Nature 436:913–914.
Donlan, C. J., Berger, J., Bock, C. E., Bock, J.H., Burney, D. A., Estes, J. A., Foreman,D., Martin, P. S., Roemer, G. W., Smith,F. A., Soule, M. E., Greene, H. W.(2006). Pleistocene rewilding: Anoptimistic agenda for twenty-firstcentury conservation. Am. Nat. 168:660–681.
Donlan, C. J.and Greene, H. W. (2010).NLIMBY: No lions in my backyard. In:M. Hall (ed) Restoration and history: thesearch for a usable environmental past,293–305. Routledge, New York.
Dransfield, J., Flenley, J. R., King, S. M.,Harkness, D. D. and Rapu, S. (1984). Arecently extinct palm from Easter Island.Nature 312: 750–752.
Eskildsen, L. I., Olesen, J. M. and Jones, C.G. (2004). Feeding response of theAldabra giant tortoise (Geochelonegigantea) to island plants showingheterophylly. J. Biogeogr. 31: 1785–1790.
Ewel, J. J. and Putz, F. E. (2004). A place foralien species in ecosystem restoration.Frontiers Ecol. Environ. 2: 354–360.
Ewen, J. G., Armstrong, D., Parker, K. andSeddon, P. (2008). Avian reintroductionbiology: current issues for science andmanagement. Avian Biology Research1: 27–50
Fadzly, N., Jack, C., Schaefer, H. M. andBurns, K. C. (2009). Ontogenetic colourchanges in an insular tree species:signalling to extinct browsing birds?New Phytol. 184: 495–501.
Ferguson, R. N. and Drake, D. R. (1999).Influence of vegetation structure onspatial patterns of seed deposition bybirds. NZ J. Bot. 37: 671–677.
Forsyth, D. M., Wilmshurst, J. M., Allen, R. B.and Coomes, D. A. (2010). Impacts ofintroduced deer and extinct moa onNew Zealand ecosystems. NZ J. Ecol34: 48–65.
Foster, J. T. and Robinson, S. K. (2007).Introduced birds and the fate ofHawaiian rainforests. Cons. Biol. 21:1248–1257.
Fox, D. (2003). Using exotics as temporaryhabitat: an accidental experiment onRodrigues island. Conservation inPractice 4.
Frazier, J. (2010). The call of the wild. In: R.M. Dean (ed) The archaeology ofanthropogenic environments, 341–369.Center for Archaeological Investi-gations, Occasional Paper No. 37,Southern Illinois University, Car-bondale.
138
HANSEN
on the use of taxon:2 columnas 15/11/2010 13:53 Página 138

Froyd, C. A. and Willis, K. J. (2008).Emerging issues in biodiversity &conservation management: The needfor a palaeoecological perspective.Quatern. Sci. Rev. 27: 1723–1732.
Gade, D. W. (1985). Man and nature onRodrigues: tragedy of an islandcommon. Environ. Cons. 12: 207–215.
Gaffney, E. S. (1984). New occurrences ofextinct meiolaniid turtles in NewCaledonia. Am. Mus. Nov. 2800: 1–6.
Gaffney, E. S. (1996). The postcranialmorphology of Meiloania platycepsand a review of the Meiloaniidae. Bull.Am. Mus. Nat. His. 229: 1–166.
Galetti, M. (2004). Parks of the Pleistocene:recreating the Cerrado and thePantanal with megafauna. Natureza &Conservação 2: 93–100.
Gillson, L. and Willis, K. J. (2004). ‘As earthtestimonies tell’: wilderness conser-vation in a changing world. Ecol. Lett.7: 990–998.
Givnish, T. J., Sytsma, K. J., Smith, J. F. andHahn, W. J. (1994). Thorn-like pricklesand heterophylly in Cyanea: adapta-tions to extinct avian browsers onHawaii? PNAS 91: 2810–2814.
Godfrey, L. R., Semprebon, G. M., Jungers,W. L., Sutherland, M. R., Simons, E. L.,Solounias, N. (2004). Dental use wearin extinct lemurs: evidence of diet andniche differentiation. J. Hum. Evol. 47:145–169.
Godfrey, L. R., Jungers, W. L., Schwartz, G. T.and Irwin, M. T. (2008). Ghosts andorphans. - In: Elwyn Simons: a searchfor origins. pp. 361–395.
Grant, P. R. (1998). Evolution on islands.Oxford University Press, Oxford, UK.
Greenwood, R. M. and Atkinson, I. A. E.(1977). Evolution of divaricating plantsin New Zealand in relation to moabrowsing. Proc. NZ Ecol. Soc. 24: 21–33.
Griffiths, C. J., Jones, C. G., Hansen, D. M.,Puttoo, M., Tatayah, R.V., Müller, C. B.,Harris, S. (2010). The use of extantnon-indigenous tortoises to replaceextinct ecosystem engineers: arestoration tool. Rest. Ecol. 18: 1–7.
Griffiths, C. J. and Harris, S. (2010) Preven-tion of secondary extinctions throughtaxon substitution. Cons. Biol. 24, 645–646.
Hadley, E. A. and Barnosky, A. D. (2009).Vertebrate fossils and the future ofconservation biology. In: G. P. Dietl andK. W. Flessa (eds.), Conservationpaleobiology: using the past to managefor the future, 39–59. The Paleon-tological Society, Lubbock, Texas.
Hall, M. (2010). Restoration and history: thesearch for a usable environmental past.Routledge, New York.
Hansen, D. M., Kaiser, C. N. and Müller, C.B. (2008). Seed dispersal and esta-blishment of endangered plants onoceanic Islands: the Janzen-Connellmodel, and the use of ecologicalanalogues. PLoS ONE 3(5): e2111.doi:10.1371/journal.pone.0002111
Hansen, D. M. and Galetti, M. (2009). Theforgotten megafauna. Science 324:42–43.
Hansen, D. M., Donlan.C. J, Griffiths, C. J.and Campbell, K. (2010). Ecologicalhistory and latent conservation poten-tial: Large and giant tortoises as amodel for taxon substitutions. Eco-graphy 33: 272–284.
139
ISLAND REWILDING
on the use of taxon:2 columnas 15/11/2010 13:53 Página 139

Hansen, I., Brimer, L. and Molgaard, P.(2004). Herbivore-deterring secondarycompounds in heterophyllous woodyspecies of the Mascarene Islands.Perspect. Plant Ecol. Evol. Syst. 6: 187–203.
Harris, J. A., Hobbs, R. J., Higgs, E. andAronson, J. (2006). Ecologicalrestoration and global climate change.Rest. Ecol. 14: 170–176.
Hayward, M. W. (2009). Conservationmanagement for the past, present andfuture. Biodiv. Cons. 18: 765–775.
Hilderbrand, R. H., Watts, A. C. and Randle,A. M. (2005). The myths of restorationecology. Ecol. Soc. 10: 19 [online] URL:http://www.ecologyandsociety.org/vol10/iss1/art19/
Hoegh-Guldberg, O., Hughes, L., McIntyre,S., Lindenmayer, D. B., Parmesan, C.,Possingham, H. P., Thomas, C. D.(2008). Assisted colonization and rapidclimate change. Science 321: 345–346.
Hunt, T. L. (2007). Rethinking Easter Island'secological catastrophe. J. Archaeol. Sci.34: 485–502.
Hunt, T. L. and Lipo, C. P. (2009). RevisitingRapa Nui (Easter Island) ‘‘ecocide’’.Pac. Sci. 63: 601–616.
Hunter, M. L., Jr. (2007). Climate changeand moving species: Furthering thedebate on assisted colonization. Cons.Biol. 21: 1356–1358.
Hutton, I., Parkes, J. P. and Sinclair, A. R. E.(2007). Reassembling island eco-systems: the case of Lord Howe Island.Anim. Cons. 10: 22–29.
Impey, A. J., Côte, I. M. and Jones, C. G.(2002). Population recovery of thethreatened endemic Rodrigues fody(Foudia flavicans) (Aves, Ploceidae)
following reforestation. Biol. Rest. 107:299–305.
IUCN (2001). Rediscovery of "extinct" specieson Mauritius spurs joint action to secureits future. IUCN News, June 21.
Jackson, S. T., Hobbs, R. J. (2009). Ecologicalrestoration in the light of ecologicalhistory. Science 325: 567–569.
James, H. F. and Burney, D. A. (1997). Thediet and ecology of Hawaii’s extinctflightless waterfowl: evidence fromcoprolites. Biol. J. Linn. Soc. 62: 279–297.
James, H. F. (2004). The osteology andphylogeny of the Hawaiian finchradiation (Fringillidae : Drepanidini),including extinct taxa. Zool. J. Linn. Soc.141: 207–255.
Janzen, D. H. (1986). The future of tropicalecology. Ann. Rev. Ecol. Syst. 17: 305–324.
Janzen, D. (1998). Gardenification ofwildland nature and the humanfootprint. Science 279: 1312–1313.
Jones, C. G. (1993). The ecology andconservation of Mauritian skinks. Proc.Roy. Soc. Arts Sci. Maur. 5: 71–95.
Jones, C. G. (1996). Bird introductions toMauritius; status and relationships withnative birds. In: J. S. Holmes and J. R.Simons (eds), The introduction andnaturalisation of birds, 113–123. JointNature Conservation Committee.London.
Jones, C. G. (2002). Reptiles and amphi-bians. In: M. R. Perrow and A. J. Davy(eds), Handbook of ecologicalrestoration, 355–375. CambridgeUniversity Press, Cambridge.
Jones, C. G. (2008). Practical conservationon Mauritius and Rodrigues: steps
140
HANSEN
on the use of taxon:2 columnas 15/11/2010 13:53 Página 140

towards the restoration of devastatedecosystems. In: A. S. Cheke and J. P.Hume (eds), Lost land of the Dodo,226–259. Christopher Helm, London.
Kaiser-Bunbury, C. N., Traveset, A. andHansen, D. M. (2010). Conservationand restoration of plant-animalmutualisms on oceanic islands.Perspect. Plant Ecol. Evol. Syst. 12:131–143.
Karanth, K. P ., Delefosse, T., Rakotosami-manana, B., Parsons, T. J. and Yoder, A.D. (2005). Ancient DNA from giantextinct lemurs confirms single origin ofMalagasy primates. PNAS 102: 5090–5095.
Kawakami, K., Mizusawa, L. and Higuchi, H.(2009). Re-established mutualism in aseed-dispersal system consisting ofnative and introduced birds and plantson the Bonin Islands, Japan. Ecol. Res.24: 741–748.
Keel, B. G. (2007). Assisted migration as aconservation strategy for rapid climatechange: investigating extended photo-period and mycobiont distributions forHabenaria repens Nuttall (Orchida-ceae) as a case study. PhD-Thesis,Antioch University, New England, USA.
Kelly, D., Robertson, A. W., Ladley, J. J.,Anderson, S. H. and McKenzie, R. J.(2006). Relative (un)importance ofintroduced animals as pollinators andseed dispersers of native plants. In: R. B.Allen and W. G. Lee (eds), Biologicalinvasions in New Zealand, 227–245.Springer, New York.
Keulartz, J. (2010). Toward a multiple visionof ecological restoration. In: M. Hall(ed) Restoration and history: the searchfor a usable environmental past.Routledge, New York, pp. 236–252.
Koch, P. L., Fox-Dobbs, K. and Newsome, S.D. (2009). The isotopic ecology of fossilvertebrates and conservation paleo-biology. In: G. P. Dietl and K. W. Flessa
(eds.), Conservation paleobiology: usingthe past to manage for the future, 95–112. The Paleontological Society,Lubbock, Texas.
Ladle, R. J. and Gillson, L. (2009). The(im)balance of nature: a publicperception time-lag? Public Underst.Sci. 18: 229–242
Lammers, T. G. and Freeman, C. E. (1986).Ornithophily among the HawaiianLobelioideae (Campanulaceae): Evi-dence from floral nectar sugarcompositions. Am. J. Bot. 73: 1613–1619.
Lazell, J. (2005). Island: fact and theory innature. University of California Press,Berkeley, California.
Lee, W. G. and Jamieson, I. G. (2001). Thetakahe: Fifity years of conservationmanagement and research. Universityof Otago Press, Dunedin, NewZealand.
Lee, W. G., Wood, J. R. and Rogers, G. M.(2010). Legacy of avian-dominatedplant–herbivore systems in NewZealand. NZ J. Ecol. 28–47.
Leonard, J. A. (2008). Ancient DNAapplications for wildlife conservation.Mol. Ecol. 17: 4186–4196.
Lewin, V. and Lewin, G. (1984). The Kalijpheasant, a newly established gamebird on the island of Hawaii. WilsonBull. 96: 634–646.
Linnebjerg, J. F., Hansen, D. M., Bunbury, N.and Olesen, J. M. (2010). Dietcomposition of the invasive red-whiskered bulbul Pycnonotus jocosus inMauritius. J. Trop. Ecol. 26: 347–350.
141
ISLAND REWILDING
on the use of taxon:2 columnas 15/11/2010 13:53 Página 141

Lord, J. M. (1991). Pollination and seeddispersal in Freycinetia baueriana, adioecious liane that has lost its batpollinator. NZ J. Bot. 29: 83–86.
Lord, J. M., Markey, A. and Marshall, J.(2002). Have frugivores influenced theevolution of fruit traits in New Zealand?In: D. Levey, A. Markey, J. Marshall(eds), Frugivory and seed dispersal:ecological, evolutionary and conser-vation perspectives, 55–68. CABInternational, Wallingford.
Lowenthal, D. (2010). Reflections onHumpty-Dumpty ecology. In: M. Hall(ed) Restoration and history: The searchfor a usable environmental past,13–34.Routledge, New York.
Lyman, R. L. (2006). Paleozoology in theservice of conservation biology. Evol.Anthropol. 15: 11–19.
Macdonald, D. W. (2009). Lessons learntand plans laid: seven awkwardquestions for the future of rein-troductions. In: M. W. Hayward and M.J. Somers (eds), Reintroduction of top-order predators, 411–448. Wiley-Blackwell, New York.
Machado, A. (2004). An index ofnaturalness. J. Nat. Cons. 12: 95–110.
Manning, A. D., Fischer, J., Felton, A.,Newell, B., Steffen, W., Lindenmayer,D.B. (2009). Landscape fluidity – aunifying perspective for understandingand adapting to global change. J.Biogeogr. 36: 193–199.
Maunder, M. (1992). Plant reintroduction: anoverview. Biodiv. Cons. 1: 51–61.
Maunder, M., Culham, A. and Hankamer, C.(1998). Picking up the pieces: botanicalconservation on degraded oceanicislands. In: P. L. Fiedler and P. M.Kareiva (eds), Conservation biology: for
the coming decade, 2nd ed, 317–344.Chapman and Hall, New York.
McConkey, K. R. and Drake, D. R. (2002).Extinct pigeons and declining batpopulations: are large seeds still beingdispersed in the tropical Pacific? In:D.Levey, A. Markey, J. Marshall (eds),Frugivory and seed dispersal: evo-lutionary and conservation perspectives,381–395.CAB International, Walling-ford, UK.
McConkey, K. R. and Drake, D. R. (2007).Indirect evidence that flying foxes trackfood resources among islands in aPacific archipelago. Biotropica 39:436–440.
McLachlan, J. S., Hellmann, J. J. andSchwartz, M. W. (2007). A frameworkfor debate of assisted migration in anera of climate change. Cons. Biol. 21:297–302.
Meyer, J.-Y. and Butaud, J.-F. (2009). Theimpacts of rats on the endangerednative flora of French Polynesia (PacificIslands): drivers of plant extinction orcoup de grâce species? Biol. Invas. 11:1569–1585.
Miskelly, C. (1987). The identity of thehakawai. Notornis 34: 95–116.
Miskelly, C. (1999). Mana Island ecologicalrestoration plan. New Zealand Depart-ment of Conservation, Wellington, pp.1–136.
Miskelly, C. M. and Baker, A. J. (2009).Description of a new subspecies ofCoenocorypha snipe from subantarcticCampbell Island, New Zealand.Notornis 56: 113–123.
Moreno, M., and Fernández, M. P. (2004).Managing the world’s longest-livingpalms: Chile. In: Examples of thesuccessful conservation and sustainableuse of dryland biodiversity, 35–40.
142
HANSEN
on the use of taxon:2 columnas 15/11/2010 13:53 Página 142

United Nations Development Pro-gramme.
Morin, M. and Conant, S. (1998). LaysanIsland ecosystem restoration plan.University of Hawaii at Manoa.
Mueller, J. M. and Hellmann, J. J. (2008). Anassessment of invasion risk fromassisted migration. Cons. Biol. 22: 562–567.
New, T. R. (2008). A special issue on insectconservation and islands. J. InsectCons. 12: 195–195.
Nogales, M., Martin, A., Tershy, B. R.,Donlan, C. J., Witch, D., Puerta, N.,Wood, B., Alonso, J. (2004). A reviewof feral cat eradication on islands.Conserv. Biol. 18: 310–319.
Norton, D. A., Leighton, A. and Phipps, H.(2005). Otamahua/Quail Islandrestoration plan. University of Canter-bury, Christchurch, New Zealand.
Olson, S. L. and James, H. F. (1991).Descriptions of thirty-two new species ofbirds from the Hawaiian Islands. Part I.Non-Passeriformes. Ornithol. Monogr.45: 1–91.
Olson, S. L. and Wingate, D. B. (2006). Anew species of night-heron (Ardeidae:Nyctanassa) from Quaternary depositson Bermuda. Proc. Biol. Soc. Wash.119: 326–337.
O'Rourke, P. (1994). All the trouble in theworld: the lighter side of overpo-pulation, famine, plague, ecologicaldisaster, ethnic hatred, and poverty.Picador.
Papanastasis, V. P. (2009). Restoration ofdegraded grazing lands throughgrazing management: Can it work?Rest. Ecol. 17: 441–445.
Pickett, S. T. A. and Parker, V. T. (1994).Avoiding the old pitfalls: opportunitiesin a new discipline. Rest. Ecol. 2: 75–79.
Posada, J. M., Aide, T. M. and Cavelier, J.(2000). Cattle and weedy shrubs asrestoration tools of tropical montanerainforest. Rest. Ecol. 8: 370–379.
Pyle, R. M. (2000). Resurrection ecology:bring back the Xerces blue. Wild Earth10: 30–34.
Rauzon, M. J. (2001). Isles of refuge.University of Hawai'i Press, Honolulu.
Ricciardi, A. and Simberloff, D. (2009).Assisted colonization is not a viableconservation strategy. Trends Ecol. Evol.24: 248–253.
Riera, N., Traveset, A. and García, O.(2002). Breakage of mutualisms byexotic species: the case of Cneorumtricoccon L. in the Balearic Islands(Western Mediterranean Sea). J.Biogeogr. 29: 713–719.
Rijsdijk, K. F., Hume, J. P., Bunnik, F., Florens,F. B. V., Baider, C., Shapiro, B., van derPlicht, J., Janoo, A., Griffiths, O.,Ostende, L., Cremer, H., Vernimmen,T., De Louw, P. G. B., Bholah, A.,Saumtally, S., Porch, N., Haile, J.,Buckley, M., Collins, M., Gittenberger,E. (2009). Mid-Holocene vertebratebone Concentration-Lagerstatte onoceanic island Mauritius provides awindow into the ecosystem of the dodo(Raphus cucullatus). Quatern. Sci. Rev.28: 14–24.
Roberts, A. and Miskelly, C. (2003). Recoveryplan for the snipe species of NewZealand and the Chatham Islands(Coenocorypha spp.) Tutukiwi. 2003–2013. Department of Conservation,New Zealand.
143
ISLAND REWILDING
on the use of taxon:2 columnas 15/11/2010 13:53 Página 143

Rodriguez, L. (2006). Can invasive speciesfacilitate native species? Evidence ofhow, when, and why these impactsoccur. Biol. Invas. 8: 927–939.
Rubenstein, D. R., Rubenstein, D. I.,Sherman, P. W. and Gavin, T. A. (2006).Pleistocene park: does re-wilding NorthAmerica represent sound conservationfor the 21st century? Biol. Cons. 132:232–238.
Rull, V., Cañellas-Boltà, N., Sáez, A., Giralt,S., Pla, S., Margalef, O. (2010).Paleoecology of Easter Island: Evidenceand uncertainties. Earth-Sci. Rev. 99:50–60.
Sakai, A. K., Wagner, W. L. and Mehrhoff, L.A. (2002). Patterns of endangerment inthe Hawaiian flora. Syst. Biol. 51: 276–302.
Sax, D. F. and Gaines, S. D. (2008). Speciesinvasions and extinction: the future ofnative biodiversity on islands. PNAS105: 11490–11497.
Schwartz, M. W., Hellmann, J. J. andMcLachlan, J. S. (2009). Theprecautionary principle in managedrelocation is misguided advice. TrendsEcol. Evol. 24: 474–474.
Seddon, P. J., Armstrong, D. P. and Maloney,R.F. (2007). Developing the science ofreintroduction biology. Cons. Biol. 21:303–312.
Seddon, P. J., Strauss and W. M., Innes, J. (inpress). Animal translocations: what arethey and why do we do them? In: J. G.Ewen, D. P. Armstrong, K.A. Parker, P. J.Seddon (eds), The science andmanagement of translocations forconservation. Blackwell Publishing.
Shapiro, B., Sibthorpe, D., Rambaut, A.,Austin, J., Wragg, G. M., Bininda-Emonds, O. R. P., Lee, P. L. M., Cooper,A. (2002). Flight of the dodo. Science295: 1683–1683.
Showler, D. A., Côté, M., I. and Jones, C. G.(2002). Population census and habitatuse of Rodrigues warbler Acrocephalusrodericanus. Bird Cons. Int. 12: 211–230.
Simbaña, W. and Tye, A. (2009). Repro-ductive biology and responses to threatsand protection measures of the totalpopulation of a critically endangeredGalapagos plant, Linum cratericola(Linaceae). Bot. J. Linn. Soc. 161: 89–102.
Sorenson, M. D., Cooper, A., Paxinos, E. E.,Quinn, T. W., James, H. F., Olson, S. L.,Fleischer, R. C. (1999). Relationships ofthe extinct moa-nalos, flightlessHawaiian waterfowl, based on ancientDNA. Proc. R. Soc. Lond. B 266: 2187–2193.
Soulé, M. E. (1990). The onslaught of alienspecies, and other challenges in thecoming decades. Cons. Biol. 4: 233–239.
Staddon, S. C., Compton, S. G. and Portch,A. (2010). Dispersal of fig seeds in theCook Islands: introduced frugivores areno substitutes for natives. Biodivers.Cons. 19: 1905–1916.
Steadman, D. W. and Martin, P. S. (2003).The late Quaternary extinction andfuture resurrection of birds on Pacificislands. Earth-Sci. Rev. 61: 133–147.
Steadman, D. W. (2006). Extinction andbiogeography of tropical Pacific birds.University of Chicago Press, Chicago.
144
HANSEN
on the use of taxon:2 columnas 15/11/2010 13:53 Página 144

Steadman, D. W., Franz, R., Morgan, G. S.,Albury, N. A., Kakuk, B., Broad, K.,Franz, S. E., Tinker, K., Pateman, M. P.,Lott, T. A., Jarzen, D. M., Dilcher, D. L.(2007). Exceptionally well preservedlate Quaternary plant and vertebratefossils from a blue hole on Abaco, TheBahamas. PNAS 104: 19897–19902.
Stewart, J. R. (2009). The Quaternary fossilrecord as a source of data for evidence-based conservation: is the past the keyto the future? In: S. T. Turvey (ed)Holocene extinctions, 249–261. OxfordUniversity Press, Oxford.
Strahm, W. (1983). Rodrigues: can its florabe saved? Oryx 17: 122–125.
Strahm, W. (1994). The conservation andrestoration of the flora of Mauritius andRodrigues. PhD Thesis, University ofReading, UK.
Strasberg, D. (1996). Diversity, sizecomposition and spatial aggregationamong trees on a 1-ha rain forest plotat La Réunion. Biodivers. Cons. 5: 825–840.
Strasberg, D., Rouget, M., Richardson, D. M.,Baret, S., Dupont, J., Cowling, R. M.(2005). An assessment of habitatdiversity and transformation on LaReunion Island (Mascarene Islands,Indian Ocean) as a basis for identifyingbroad-scale conservation priorities.Biodivers. Cons. 14: 3015–3032.
Sullivan, J. J., Kelly, D. and Ladley, J. J.(2010). Feathers to fur: the status ofNew Zealand ecological research in2009. NZ J. Ecol. 34: 1–5.
Sutherland, W. J., Pullin, A.S., Dolman, P. M.and Knight, T.M. (2004). The need forevidence-based conservation. TrendsEcol. Evol. 19: 305–308.
Swetnam, T. W., Allen, C. D. and Betancourt,J. L. (1999). Applied historical ecology:using the past to manage for the future.Ecol. Appl. 9: 1189–1206.
Temperton, V. M. (2007). The recent doubleparadigm shift in restoration ecology.Rest. Ecol. 15: 344–347.
Temple, S. A. (1977). Plant-animal mutua-lism: coevolution with Dodo leads tonear extinction of plant. Science 97:885–886.
Temple, S. A. (1981). Applied island biogeo-graphy and the conservation ofendangered island birds in the Indianocean. Biol. Cons. 20: 147–161.
Thebaud, C. and Strasberg, D. (1997). Plantdispersal in fragmented landscapes: afield study of woody colonization inrainforest remnants of the MascareneArchipelago. - In: W. F. Laurance and R.O. Bierregaard, Jr. (eds), Tropical forestremnants. University of Chicago Press,Chicago, pp. 321–332.
Traveset, A. and Richardson, D. M. (2006).Biological invasions as disruptors ofplant reproductive mutualisms. TrendsEcol. Evol. 21: 208–216.
Truett, J. and Phillips, M. (2009). Beyondhistoric baselines: restoring bolsontortoises to Pleistocene range. Ecol.Rest. 27: 145–151.
Veitch, C. R. and Clout, M. N. (2002).Turning the tide: the eradication ofinvasive species. Occasional Paper ofthe IUCN Species Survival CommissionNo. 27, Auckland, New Zealand.
Virah-Sawmy, M., Mauremootoo, J., Marie,D., Motala, S. and Sevathian, J.-C.(2009). Rapid degradation of aMauritian rainforest following 60 yearsof plant invasion. Oryx 43: 599–607.
145
ISLAND REWILDING
on the use of taxon:2 columnas 15/11/2010 13:53 Página 145

Walther, G. R., Post, E., Convey, P., Menzel,A., Parmesan, C., Beebee, T. J. C.,Fromentin, J. M., Hoegh-Guldberg, O.,Bairlein, F. (2002). Ecological responsesto recent climate change. Nature 416:389–395.
Warren, C. R. (2007). Perspectives on the‘alien’ versus ‘native’ species debate: acritique of concepts, language andpractice. Prog. Hum. Geogr. 31: 427–446.
Weaver, E. A. and Griffiths, O. L. (2008). Aguide to La Vanille Réserve desMascareignes (Mauritius). BioculturePress, Rivière des Anguilles, Mauritius.
Westman, W. E. (1990). Park managementof exotic plant species: problems andissues. Cons. Biol. 4: 251–260.
White, A. W., Worthy, T. H., Hawkins, S.,Bedford, S. and Spriggs, M. (2010).Megafaunal meiolaniid horned turtlessurvived until early human settlement inVanuatu, Southwest Pacific. PNAS 107:15512–15516.
Wiles, G. J. and Glass, P. O. (1990).Interisland movements of fruit bats(Pteropus mariannus) in the MarianaIslands. Atoll Res. Bull. 323: 1–6.
Williams, P. A., Karl, B. J., Bannister, P. andLee, W. G. (2000). Small mammals aspotential seed dispersers in NewZealand. Austral Ecol. 25: 523–532.
Wingate, D. B. (2001). Strategies forsuccessful biodiversity conservation andrestoration on small oceanic islands:some examples from Bermuda. - In: M.Pienkowski (ed) Calpe 2000: Linking thefragments of paradise. UK OverseasTerritories Conservation Forum, pp. 16–24.
Witmer, M. C. and Cheke, A. S. (1991). Thedodo and the tambalacoque tree: anobligate mutualism reconsidered. Oikos61: 133–137.
Wood, J. R., Rawlence, N. J., Rogers, G. M.,Austin, J. J., Worthy, T. H., Cooper, A.(2008). Coprolite deposits reveal thediet and ecology of the extinct NewZealand megaherbivore moa (Aves,Dinornithiformes). Quatern. Sci. Rev.27: 2593–2602.
Worthy, T. H., Anderson, A. J. and Molnar, R.E. (1999). Megafaunal expression in aland without mammals - the first fossilfaunas from terrestrial deposits in Fiji.Senckenb. Biol. 79: 237–242.
Worthy, T. H. (2001). A giant flightless pigeongen. et sp. nov. and a new species ofDucula (Aves: Columbidae), fromQuaternary deposits in Fiji. J. Roy. SocNZ 31: 763–794.
Worthy, T. H. and Holdaway, R. N. (2002).The lost world of the moa: prehistoriclife of New Zealand. Indiana UniversityPress, Bloomington.
Wu, J. and Loucks, O. L. (1995). Frombalance of nature to hierarchical patchdynamics: a paradigm shift in ecology.Quart. Rev. Biol. 70: 439–466.
Zavaleta, E. S., Hobbs, R. J. and Mooney, H.A. (2001). Viewing invasive speciesremoval in a whole-ecosystem context.Trends Ecol. Evol. 16: 454–459.
Zimov, S. A. (2005). Pleistocene park: Returnof the mammoth's ecosystem. Science308: 796–798.
146
HANSEN
on the use of taxon:2 columnas 15/11/2010 13:53 Página 146



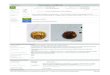



![-- I - . 't~scipp.ucsc.edu/~hartmut/Radiobiology/pCT_Lit/Schneider U 1996.pdf · 2 U Schneider et al, , proton stopping power and Hounsfie]d values, Tissue substitutes are usually](https://img.pdfslide.org/doc/110x75/5e823455102bdb23d80aaa90/i-tscippucsceduhartmutradiobiologypctlitschneider-u-1996pdf-2-u.jpg)





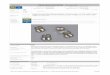



![Signatur Notizen Titel Autor Verlag Ort Jahr - uni-due.de · 1.2 SIT [1] SIT Stanford - Binet Intelligenz-Test Terman, Hogrefe Göttingen 1965 1.2 SIT [2] SIT Stanford Intelligenz-Test](https://img.pdfslide.org/doc/110x75/5b5442987f8b9add3a8cdbee/signatur-notizen-titel-autor-verlag-ort-jahr-uni-duede-12-sit-1-sit-stanford.jpg)











