Embed Size (px)
Citation preview

42PARACONULARIA BACHMANNI N. SP. FROM THE GERMANIC UPPER TRIASSIC,
THE FIRST FRESH-WATER OCCURRENCE OF A CONULARIID
GREGOR BARTH1, HEINZ W. KOZUR2, MATTHIAS FRANZ1 AND ROBERT E. WEEMS3
1 Technische Universität Bergakademie Freiberg - Institut für Geologie, Bernhard-von-Cotta-Str. 2, 09599 Freiberg, Germany,e-mail: [email protected] and [email protected];2 Rézsü u. 83, H-1029 Budapest, Hungary, e-mail: [email protected];
3 926A National Center, U.S. Geological Survey, Reston VA 20192, USA, e-mail: [email protected]
Abstract—For the first time a conulariid fossil, Paraconularia bachmanni n. sp., has been found in the GermanicUpper Triassic. All previously known conulariids were fully marine, but Paraconularia bachmanni n. sp. wasfound in a fresh water lake deposit. The presence of a specimen of this normally marine fossil group in a fresh waterlake deposit suggests that this species was a euryhaline marine conulariid that normally lived in a nearby largebrackish sea but occasionally penetrated into fresh water areas far from the marine realm.
Dedication—This paper is dedicated to Prof. Dr. Gerhard H. Bachmann, Halle, on the occasion of his 70thbirthday on July 3, 2013.
INTRODUCTION
The Conulariida Miller and Gurley, 1896 typically have beenregarded as a group of scyphozoan cnidarians or a group closely relatedto them (starting with Kiderlen, 1937 and thereafter). Some more recentworkers have concluded that conulariids represent their own phylum(e.g. Feldmann and Babcock, 1986; see discussion in de Moraes Leme etal., 2008; Van Iten et al., 2006; Lucas, 2012). Conulariids have beenfound in marine strata ranging in age from the latest Precambrian to thelatest Triassic (de Moraes Leme et al., 2008; Lucas, 2012). They werenever very diverse, but in some Paleozoic beds they can be common.Throughout the Triassic they were very rare and had very low diversity,but they managed to persist into and perhaps through the Rhaetian. VanIten (in Lucas, 2012) has stated that only one Triassic genus was present(Paraconularia Sinclair, 1940), but Flectoconularia Waterhouse, 1979from the Carnian of New Zealand represents a second genus. Despite thefact that only a few Triassic conulariid species are known, they do havea wide geographic distribution. They are known from Lower Triassicstrata of Japan (Sugiyama, 1942), Middle Triassic strata of Kashmir(Diener, 1913), Carnian and Rhaetian strata of New Zealand (Trechman,1918; Waterhouse, 1979), Norian and Rhaetian strata of the Alps (Bittner,1890; Osswald, 1918), and Rhaetian strata of New Caledonia (Hagdornand Campbell, 1990).
Two reports of Liassic and Late Cretaceous Conularia cannot beconfirmed. Argéliez (1856 a,b) mentioned a specimen he called Conulariacancellata Argéliez from the Liassic of Milhau, Aveyron, but this speci-men was never documented. If it still exists, it would be a junior hom-onym of the Silurian species Conularia cancellata Sandberger, 1847.This was pointed out by Osswald (1918), who regarded the presence ofany conulariids from the Liassic of Milhau as unproven. Since then, nocompelling evidence has been forthcoming for the existence of Liassicconulariids anywhere. Willard (1966, 1967) described a putativeconulariid fossil from the Late Cretaceous of Peru, but Waterhouse (1979)and Babcock (1985) later suggested that this fossil belongs to the bivalvePinna Linné. Lucas (2012) made a careful re-investigation of the materialfrom Peru and agreed with Waterhouse and Babcock that the Late Creta-ceous “conulariid” in fact is a Pinna. Thus, no verifiable post-Triassicconulariids are known and the youngest confirmed occurrences are fromthe Rhaetian of the Alps, New Zealand and New Caledonia.
All previously described conulariids have been found in marinedeposits. In the case of the conulariid here reported from the GermanicBasin, however, the occurrence is in a fresh-water deposit. Thus the
location and the depositional setting of this specimen both are remark-able.
GEOLOGIC SETTING
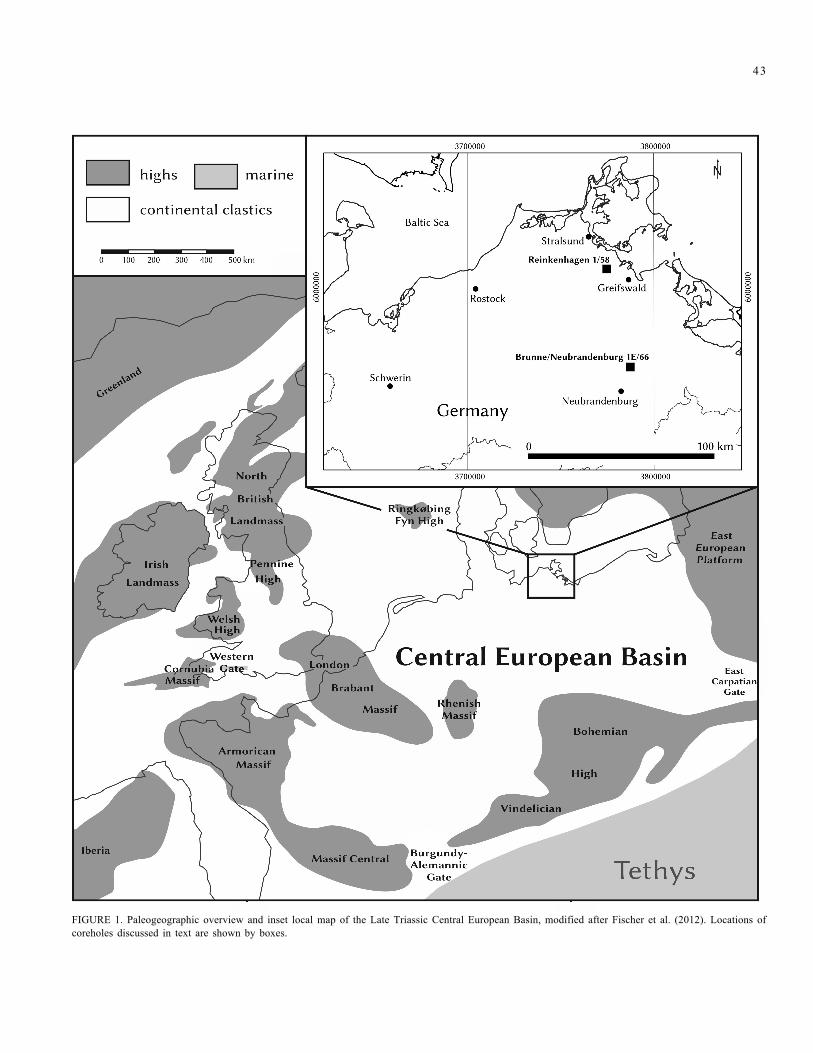
Paraconularia bachmanni n. sp. was found within the Norian-Rhaetian succession of the Brunne/Neubrandenburg 1E/66 (BruN 1E/66)core, located approximately 15 km northeast of the city of Neubranden-burg, Germany. During the Triassic this locality was part of the CentralEuropean Basin (Germanic Basin), an epicontinental basin linked to thenorthwestern Tethyan realm (Fig. 1). The Norian-Rhaetian basin fill ischaracterized by an upward transition from predominantly terrestrialplaya-like environments in the Norian to a generally shallow, brackish tobrachyhaline marine epicontinental sea that covered much of the basin inthe Rhaetian (Rhaetian Sea of Fischer et al., 2012). According to theoriginal descriptions of this borehole, the Norian-Rhaetian successioncan been subdivided into five informal units: the NorianSteinmergelkeuper, Saurichthys Beds, and Postera Beds, the middleRhaetian Contorta Beds, and the upper Rhaetian Triletes Beds (Wehland,1970; Schichtenverzeichnis Geologischer Dienst, LUNG M-V, Kb BruN1E/66, Landesamt für Umwelt, Naturschutz und Geologie Mecklenburg-Vorpommern, 2 Figs., distributed by Dr. K. Obst, Güstrow). A modernlithostratigraphic subdivision of the Triassic of the BruN 1E/66 core byBarth and Franz is in preparation and therefore is beyond the scope ofthe present paper.
Within the Norian-Rhaetian succession, a distinct lithologic changefrom grey and greenish dolomitic marls to grey claystones can be ob-served. The dolomitic marls show various features reflecting intensepedogenesis, thus comprising mature petrocalcic horizons (nodules andcrusts) that have been termed “Steinmergel” in Thuringia and elsewhere.At the top of this sequence the pedogenic overprint culminates in a thickdolomite crust that is the correlative equivalent of the “Mühlburgbank”in Thuringia (Franz, 2008). Above the grey to dark grey claystones andgreyish siltstones, pedogenic features are rare to absent (Fig. 2). Theselatter deposits, which yielded the herein described specimen ofParaconularia bachmanni n. sp., are very rich in conchostracans andplant detritus. Almost all beds within this interval have yielded theostracod Darwinula and conchostracans, which are a typical fresh waterassociation. In slightly brackish, low-salinity oligohaline deposits of thisage, small Rhombocythere are common. Rhombocythere is absent at theParaconularia bachmanni horizon and also absent immediately belowand above it. About 55 km toward the north-northwest, Rhombocytheregracilis Kozur, 1968 (= Notocythere media prima Will, 1969), a small
Tanner, L.H., Spielmann, J.A. and Lucas, S.G., eds., 2013, The Triassic System. New Mexico Museum of Natural History and Science, Bulletin 61.
43
FIGURE 1. Paleogeographic overview and inset local map of the Late Triassic Central European Basin, modified after Fischer et al. (2012). Locations ofcoreholes discussed in text are shown by boxes.

44
species of Rhombocythere, occurs in the Reinkenhagen 1/58 core (Fig. 1)in very low salinity oligohaline brackish beds (Kozur, 1968) of latestmiddle Norian (?) to early late Norian age (formerly regarded as lowerRhaetian). Thus, the common occurrence of Darwinula andconchostracans, in the absence of the very low salinity brackish waterostracod Rhombocythere gracilis and any other indicators of brackishwater in all samples around the occurrence of Paraconularia bachmanni,clearly indicate it was found in a fresh water deposit.
The conchostracan age for the Paraconularia horizon is veryprecisely known. In the Paraconularia bed (and even on theParaconularia bedding surface at 1082.3 m), are the youngest occur-rences in borehole BruN 1E/66 of Anyuanestheria lucasi Kozur andWeems. Thus, Paraconularia was found at the very top of the A. lucasiZone. According to Kozur and Weems (2011), the A. lucasi Zone in thesouthwestern United States belongs to the uppermost middle Norian.The oldest certain A. lucasi in BruN 1E/66 was found at 1084.4 m, and at1085.1 m a transitional form is found that is morphologically intermedi-ate between Anyuanestheria n. sp. (forerunner of A. lucasi) and A. lucasi.The oldest questionable A. lucasi was found slightly lower at 1087.60 m.Below this level is the Shipingia hebaozaiensis Zone and below that theShipingia mcdonaldi Zone is recognized. The Shipingia hebaozaiensisZone occurs from China through Middle Europe to the Newark Super-group basins and the southwestern United States within the upper middleNorian. The Shipingia mcdonaldi Zone occurs from Middle Europe tothe Newark Supergroup and the southwestern United States in the up-per part of the middle Norian below the S. hebaozhaiensis Zone. Thelatter index species is a long-ranging form that persists into the twooverlying conchostracan zones of the upper middle Norian. This succes-sion also is found in the BruN 1E/66 core. The S. mcdonaldi Zone and theS. hebaozhaiensis Zone occur as well in south and central Germany instratigraphic order within the upper part of the middle Norian (Kozur
FIGURE 2. Position of Paraconularia bachmanni n. sp. and associated fossils within the borehole BruN 1E/66.
and Weems, 2010). The A. lucasi Zone previously was known only fromthe southwestern United States (Kozur and Weems, 2011) but now hasbeen identified in Germany. This conchostracan succession therefore isapplicable over a very wide area and shows that Paraconularia bachmanniwas found within the middle/upper Norian boundary interval.
EXPLANATION FOR THE OCCURRENCE OF ACONULARIID WITHIN FRESH WATER DEPOSITS
There are two possible explanations for the occurrence of aconulariid in fresh water sediments: (1) A conulariid evolved in the LateTriassic that habitually lived in a fresh water environment, or (2) aeuryhaline marine conulariid, which normally lived in a large brackishsea, penetrated into a fresh water area far from its normal marine habitat.We think it is improbable (but not impossible) that close to the extinctionof conulariids, when both the taxonomic diversity and number of indi-vidual specimens had become very low, that a totally new ecologicalbehavior developed. Therefore we prefer the second explanation, espe-cially as modern marine taxa are known to migrate into fresh waterhabitats. For example, one author (H.W. Kozur) has observed marinefaunal elements in the fluvial environments of the river Don close to thecity of Rostov/Don, about 70 km upstream from the Asov Sea, which isonly narrowly connected to the Black Sea. The surface waters of theBlack Sea are brackish, having about 1.7 % salt content. In the southernAsov Sea the salt content is lower, around 1.1 %, and in the northern partclose to the Don River the salt content is only 0.4-0.5 %, indicatingoligohaline brackish conditions. In the Don River itself, close to Rostov/Don and about 70 km upstream from its mouth, brackish-marine mol-lusks including Cardium are common in this fluvial freshwater environ-ment. These brackish-marine mollusks obviously can and do survive in afreshwater environment, but what remains unclear is how they manage

45to migrate upstream 70 km against a river current. Larval freshwatermollusks (glochidia) attach themselves to the gills, fins, or skin of fishand use them as transportation upstream (Burch, 1975). Fish, someother mobile aquatic animal, or perhaps the feet of birds could providethe mechanism for upstream transport of the normally brackish watermollusks found in fresh water near Rostov/Don, and some sort of eco-logically similar animal likely carried Paraconularia bachmanni into fresh-water environments as well.
In the lower Rhombocythere gracilis Zone of the upper middleNorian(?) and lower upper Norian (Fig. 2), R. gracilis is common in thelower Postera Beds of northwestern Germany (Will, 1969). At that timea large, slightly brackish (oligohaline) inland sea covered northwesternGermany that may have been a precursor ingression of the later wide-spread Rhaetian ingression. The easternmost occurrence of theseoligohaline brackish beds containing R. gracilis was found by Kozur(1968) in the Reinkenhagen borehole, about 55 km north-northwest ofborehole BruN 1E/66. Paraconularia bachmanni probably migrated fromthis oligohaline brackish environment into the fresh water lake environ-ment found in the BruN 1E/66 core.
SYSTEMATIC DESCRIPTION
Phylum COELENTERATA Frey and Lenckart, 1847Class SCYPHOZOA Götte, 1887
Subclass CONULATA Moore and Harrington, 1956Order CONULARIIDA Miller and Gurley, 1896
Family CONULARIIDAE Walcott, 1886Subfamily PARACONULARIINAE Sinclair, 1952
Genus Paraconularia Sinclair, 1940
Type species: Conularia inaequicostata de Koninck, 1883.
Paraconularia bachmanni n. sp.Figs. 3-4
Derivatio nominis: In honor of Prof. Dr. Gerhard H. Bachmann,Halle, for his contributions to the stratigraphy, sequence stratigraphy,cyclic stratigraphy, and facies of the Germanic Triassic.
Holotype: The specimens shown in Figs. 3 and 4 are deposited inthe Technische Universität Bergakademie Freiberg, Institut für Geologie,Freiberg, Germany.
Locus typicus: Borehole Brunne/Neubrandenburg 1E/66 (BruN1E/(66), northeast of Neubrandenburg, northeastern Germany (CentralEuropean Basin).
Stratum typicum: Sample 11-03a at 1082.30 m (core box no.150), grey siltstone containing abundant plant detritus, conchostracans,specimens of Darwinula sp., and fish scales and teeth that collectivelyconstitute a typical freshwater lake assemblage.
Material: Part and counterpart of one specimen.Diagnosis: Specimen broken close to the apical end, about 35
FIGURE 3. Paraconularia bachmanni n. sp., holotype, borehole BruN 1E/66, 1082.3 m, in a grey siltstone fresh water deposit with plant debris,Darwinula sp. (ostracod), and Anyuanestheria lucasi Kozur and Weems,part and counterpart of the specimen.
FIGURE 4. Higher magnification of the left part of the specimen in Fig. 3.

46mm long as preserved, its widest point (13.3 mm) close to the aperture,width of the apical part about 5 mm. Apical half is distinctly curved,apertural half straight. Two of the four faces are visible. One face ispreserved over its full width or nearly its full width. The other face isvisible across nearly half of its width close to the aperture, but in itsapical region it is not visible. The flattened corner between these twofaces has in its apical region (about 40 % of the shell length) a distinctcorner groove; in the apertural region (about 60 % of the shell length) nocorner groove is present. The transverse ribs are wide and generallysmooth, but locally a very faint striation parallel to the margin of thetransverse ribs is visible. Within some transverse ribs short narrow ver-tical bars are visible, and the shell is thickest beneath the transverse ribs.The shell is preserved only as a secondary replacement composed of ironoxide (probably oxidized pyrite). The integument is either not preservedor preserved only as thin remnants.
The density of transverse ribs is 7 per 5 mm close to the aperturebut 8 per 5 mm close to the apex. For the most part they are nearlystraight or arched slightly towards the aperture. Close to the cornerbetween the faces, the transverse ribs are distinctly arched towards theapex. Close to the apex the transverse ribs are broader than the spacesbetween them. In the middle part of the test the ribs have either the samewidth as the distances between them or the ribs are somewhat narrowerthan the distances between them. Close to the aperture the transverseribs are as wide as the spaces between them or somewhat wider.
Occurrence: Siltstone, rich in plant detritus with some Darwinulaand conchostracans (Anyuanestheria lucasi Kozur and Weems). Fresh-water lake deposits of the uppermost A. lucasi Zone, within the Alaunian-Sevatian boundary interval.
Remarks: Paraconularia triadica (Bittner, 1890) from the lateNorian of the Alps is more slender, not curved, and has narrower trans-verse ribs that are distinctly narrower than the spaces between the ribs.
Paraconularia stromeri Osswald, 1918 from the Rhaetian of theAlps has a similar shape, but more strongly arched transverse ribs.
Paraconularia matauraensis Waterhouse, 1979 from the Carnianto Rhaetian of New Zealand is similar, but the transverse ribs are morestrongly arched. Additionally, the mid-line of the faces usually is raisedinto a ridge. The corner groove is distinct along the entire length of theshell.
CONCLUSIONS
For the first time, a conulariid specimen has been discovered in theGermanic Triassic. It represents a new species here named Paraconulariabachmanni. This specimen also is the first known freshwater occurrenceof a conulariid found anywhere. Because it was found far from anycoeval brackish or marine deposits, it apparently migrated or was carriedby a nektonic vector from adjacent very low salinity oligohaline brackishenvironments into the freshwater environment where it was found.
ACKNOWLEDGMENTS
We very much thank: Prof. Dr. J.B. Waterhouse, Oamaru (NewZealand); Dr. S.G. Lucas, Albuquerque; Dr. J. Ansorge, Greifswald; Prof.Dr. C. Brauckmann, Clausthal-Zellerfeld; Prof. Dr. O. Elicki, Freiberg forimportant help, discussions and literature on conulariids, Prof. Dr. G. H.Bachmann, Halle, Dr. K. Obst, Güstrow, for discussions of the stratigra-phy and Dr. M. Magnus, Freiberg, for the photograph of Fig. 4. We alsothank J.M. Self-Trail and B.R. Wardlaw for their very helpful reviews ofthis paper.
REFERENCES
Argéliez, M., 1856a, Letter to Èlie de Beaumont: Bulletin de la SocietéGéologique de France, v. 13, p. 186-188.
Argéliez, M., 1856b, Über den Lias von Milhau, Aveyron, und seineVersteinerungen: Neues Jahrbuch für Mineralogie, Geognosie, Geologieund Petrefaktenkunde, Jg. 1856, p. 732.
Babcock, L.E., 1985, A new Ordovician conulariid from Oklahoma?: Okla-homa Geology Notes, v.45(2), p. 66-70.
Bittner, A., 1890, Eine triadische Conularia: Verhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt Wien, Jg. 1890(9), p. 177-178.
Burch, J.B., 1975, Freshwater unionacean clams (Mollusca: Pelecypoda) ofNorth America: Biota of Freshwater Ecosystems Identification Manual11, p. 1-114.
De Moraes Leme, J., Simões, M.G., Rodrigues, S.C.Van Iten, H. and Marques,A.C., 2008, Major developments in conulariid research: Problems ofinterpretation and future perspectives: Ameghiniana (Rev. Asoc.Paleontol. Argent.), v. 45(2), p. 407-420.
Diener, C., 1913, Triassic faunae of Kashmir, Kannaur and Spiti:Palaeontologia Indica, v. 15, n.s. 5, Memoir, 1, p. 1-133.
Feldman, R.M. and Babcock, L.E., 1986, Exceptionally preserved conulariidsfrom Ohio – Reinterpretation of their anatomy: National GeographicResearch, v. 2(4), p. 464-472.
Fischer, J., Voigt, S., Franz, M., Schneider, J.W., Joachimski, M.M.,Tichomirowa, M., Götze, J. and Furrer, J., 2012, Palaeoenvironmentsof the late Triassic Rhaetian Sea: Implications from oxygen and stron-tium isotopes of hybodont shark teeth: Palaeogeography,Palaeoclimatology, Palaeoecology, v. 353-355, p. 60-72.
Franz, M., 2008, Litho- und Leitflächstratigraphie, Chronostratigraphie,Zyklo- und Sequenzstratigraphie des Keupers im östlichenZentraleuropäischen Becken (Deutschland, Polen) und Dänischen Becken(Dänemark, Schweden) [Dr. rer. nat. dissertation]: Martin-Luther-Universität Halle-Wittenberg, 196 p. and appendix.
Hagdorn, H. and Campbell, H.J., 1990, Paracomatula triadica sp. nov. – Anearly comatulid crinoid from the Otapirian (Late Triassic) of NewCaledonia: Alcheringa, v. 17, p. 1-17.
Kiderlen, H., 1937, Die Conularien. Über Leben und Bau der erstenScyphozoa: Neues Jahrbuch für Mineralogie, Beilage-Band, v. 77 Abt. B,p. 113-169.
Kozur, H., 1968, Einige seltene Ostrakoden-Arten aus der germanischenTrias: Monatsber, Deutsch Akad. Wiss. Berlin, v. 10(11), p. 848-872.
Kozur, H.W. and Weems, R.E., 2010, The biostratigraphic importance ofconchostracans in the continental Triassic of the northern hemisphere;in Lucas, S.G., ed., The Triassic timescale: Geological Society, London,Special Publication, v. 334, p. 315-417.
Kozur, H.W. and Weems, R.E., 2011, Additions to the uppermost Alaunianthrough Rhaetian (Triassic) conchostracan zonation of North America:New Mexico Museum of Natural History and Science, Bulletin 53, p.295-300.
Lucas, S.G., 2012, The extinction of the conulariids: Geosciences, v. 2012(2),p. 1-10; doi: 10.3390/geosciences2010001.
Miller, S.A. and Gurley, W.F.E., 1896, New species of Paleozoic inverte-brates from Illinois and other states: Illinois State Museum of NaturalHistory, Bulletin 11, 50 p.
Moore, R.C. and Harrington, H.J., 1956, Conulata; in Moore, R.C., ed.,Treatise on Invertebrate Paleontology, Part F: Coelenterata: GeologicalSociety of America and University of Kansas Press, F 54-F 66.
Osswald, K., 1918, Mesozoische Conulariiden: Centralblatt für Mineralogie,Geologie und Paläontologie, Jahrgang 1918, p. 337-344.
Sinclair, G.W., 1940, A discussion of the genus Metaconularia with descrip-tions of new species: Royal Soc. Canada, Trans., ser. 3, v. 34, sec. 4, p.101-121.
Sinclair, G.W., 1952, A classification of the Conularida; Chicago Nat. Hist.Mus, Fieldiana, Geology, v. 10(13), p. 135-145.

47Sugiyama, T., 1942, Studies on the Japanese Conularida: Geol. Soc. Japan, v.
49, p. 390-399.Trechman, C.T., 1918, The Trias of New Zealand: Quarterly Journal of the
Geological Society of London, v. 73, p. 165-246.Van Iten, H., de Moraes Leme, Simões, M.G., Marques, A.C. and Collins,
A.G., 2006, Reassessment of the phylogenetic position of conulariids(?Ediacaran-Triassic) within the subphylum Medusozoa (phylumCnidaria): Journal of Systematic Palaeontology, v. 4(2), p. 109-118.
Waterhouse, J.B., 1979, Permian and Triassic conulariid species from NewZealand: Journal of the Royal Society of New Zealand, v. 9(4), p. 475-489.
Wehland, H., 1970, Kurzbericht Nr. RGEE 122/70 über dieKartierungsbohrungen Brunn 1 und Brunn 1E. - 28. S., 18. Abb., 41 Anl.:ZGI Berlin. [LUNG Archiv-Signatur: UK0003].
Will, H.-J., 1969, Untersuchungen zur Stratigraphie und Genese desOberkeupers in Nordwestdeutschland: Geologisches Jahrbuch, Beihefte,v. 54, p. 1-240.
Willard, B., 1966, The Harvey Bassler collection of Peruvian fossils: LehighUniversity, Bethlehem, PA, USA, 255 p.
Willard, B., 1967, Cretaceous conularid from Peru: Pennsylvania Academyof Science, v. 40, p. 87-89.