Embed Size (px)
Citation preview

U. Albrecht BC1
Glykogenstoffwechsel und Gluconeogenese
1. Glykogenabbau
2. Glykogensynthese
3. Kontrolle des Glykogenstoffwechsels
4. Gluconeogenese
5. Andere Biosynthesewege für Kohlenhydrate

U. Albrecht BC1
In Tieren: Gehrin und Erythrocyten -> Energieversorgung mit Glucose (Fettverbrennung in diesen Geweben nicht möglich). Leber speichert Glucose in Form von Glykogen -> Glucoseversorgung 1/2 Tag
Fasten -> Gluconeogenese (aus Aminosäuren).
Glykogenabbau defekt -> Muskelkrämpfe nach Anstrengung = McArdle-Krankheit
Glykogen Stärke
Speicherung von Glucose
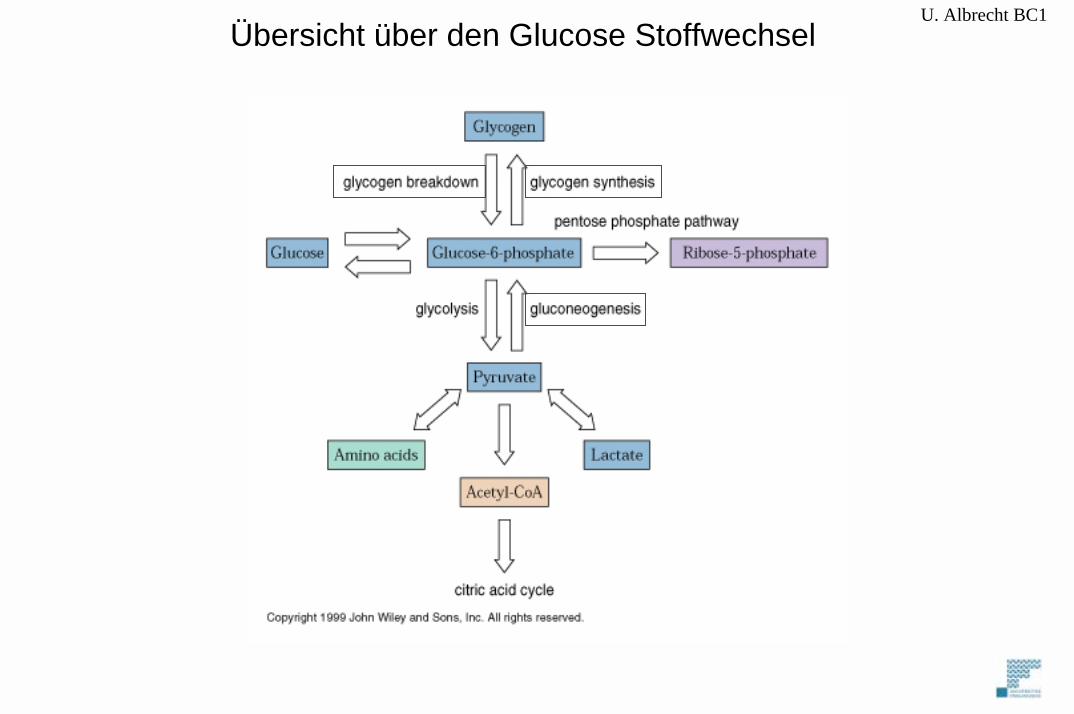
U. Albrecht BC1Übersicht über den Glucose Stoffwechsel
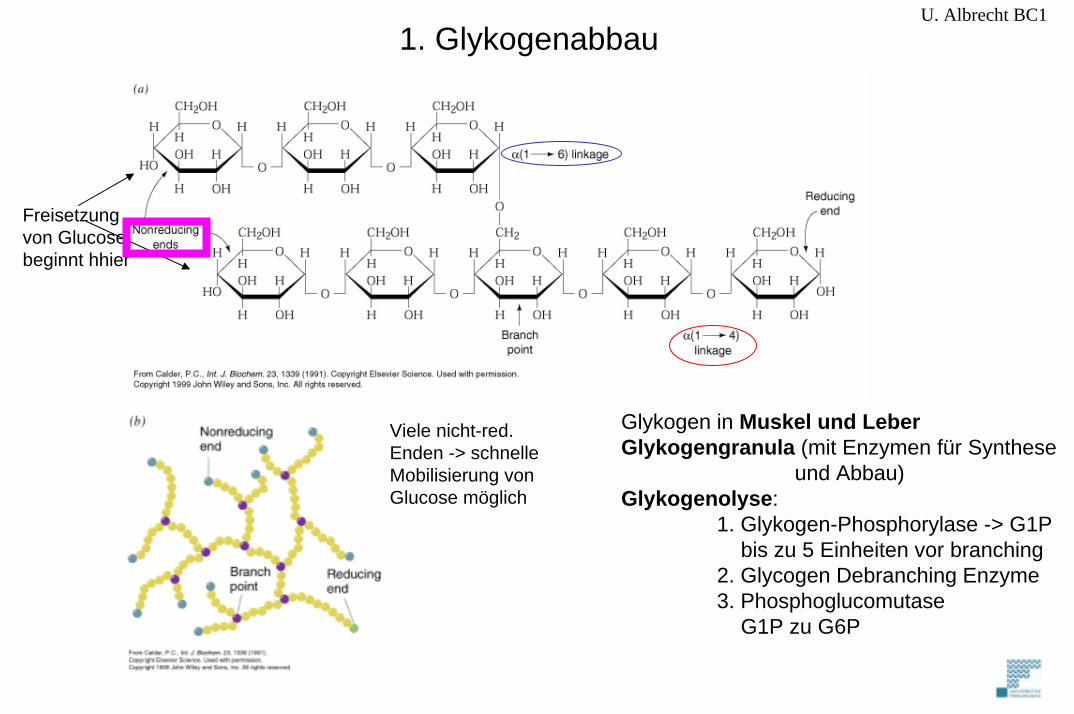
U. Albrecht BC1
Freisetzungvon Glucosebeginnt hhier
Viele nicht-red.Enden -> schnelleMobilisierung von Glucose möglich
Glykogen in Muskel und LeberGlykogengranula (mit Enzymen für Synthese
und Abbau)Glykogenolyse:
1. Glykogen-Phosphorylase -> G1Pbis zu 5 Einheiten vor branching
2. Glycogen Debranching Enzyme3. Phosphoglucomutase
G1P zu G6P
1. Glykogenabbau
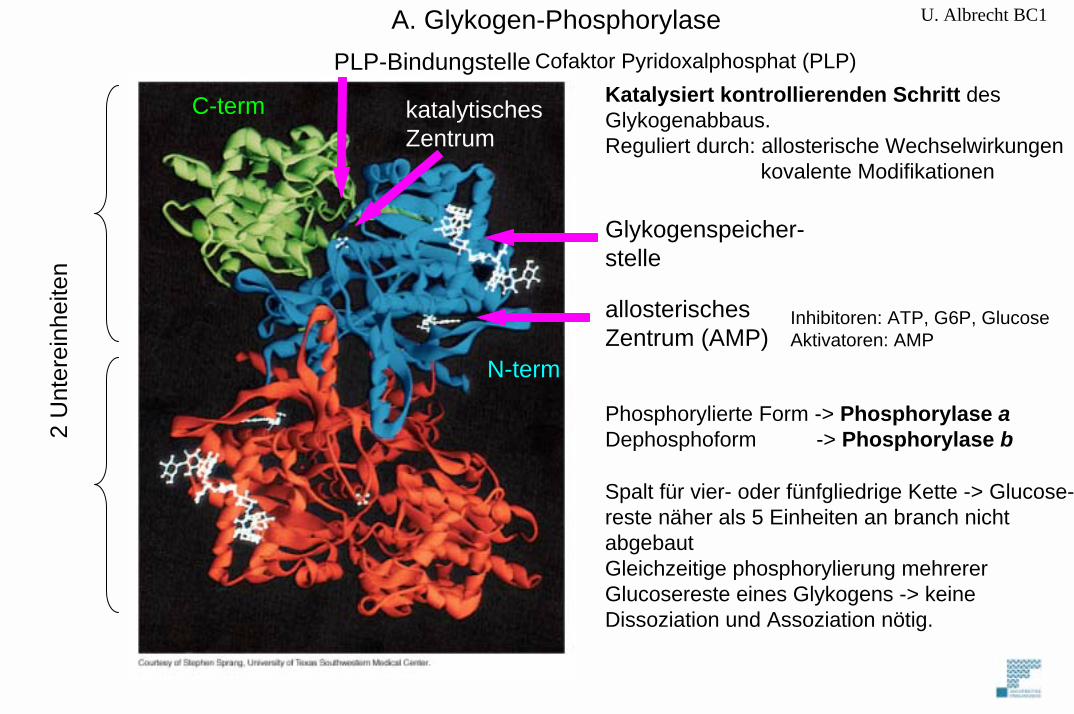
U. Albrecht BC1
2 U
nter
einh
eite
n
C-term
N-term
katalytisches Zentrum
PLP-BindungstelleKatalysiert kontrollierenden Schritt des Glykogenabbaus. Reguliert durch: allosterische Wechselwirkungen
kovalente Modifikationen
Phosphorylierte Form -> Phosphorylase aDephosphoform -> Phosphorylase b
Inhibitoren: ATP, G6P, GlucoseAktivatoren: AMP
Spalt für vier- oder fünfgliedrige Kette -> Glucose-reste näher als 5 Einheiten an branch nicht abgebaut Gleichzeitige phosphorylierung mehrerer Glucosereste eines Glykogens -> keine Dissoziation und Assoziation nötig.
Cofaktor Pyridoxalphosphat (PLP)
A. Glykogen-Phosphorylase
Glykogenspeicher-stelle
allosterischesZentrum (AMP)
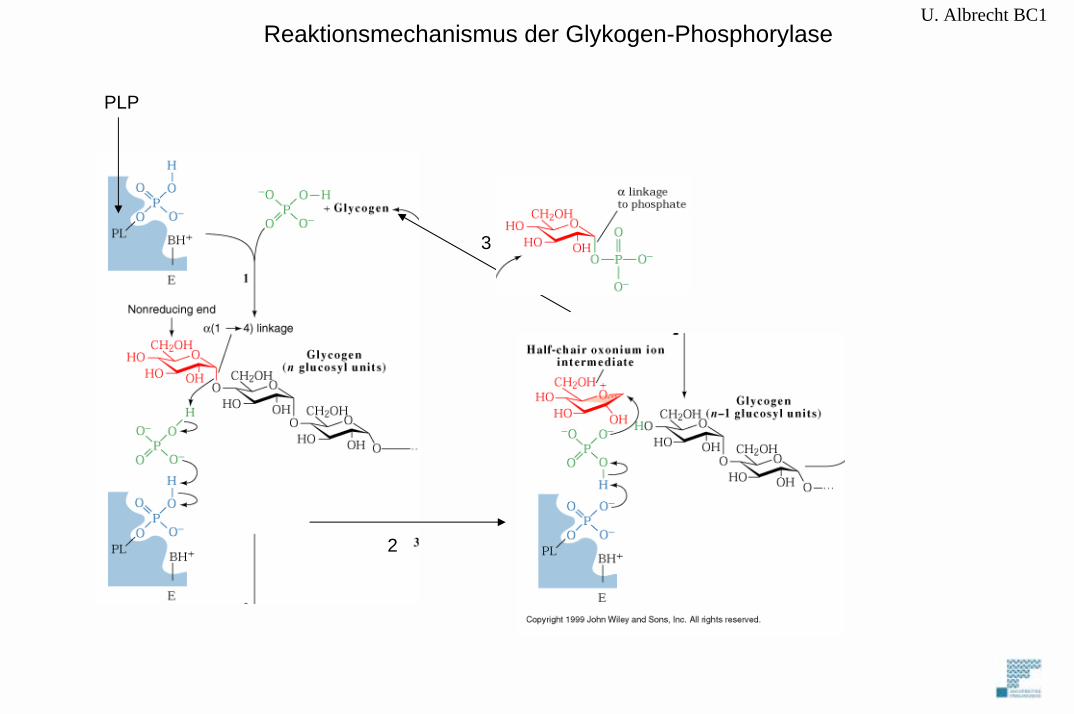
U. Albrecht BC1Reaktionsmechanismus der Glykogen-Phosphorylase
PLP
2
3
U. Albrecht BC1
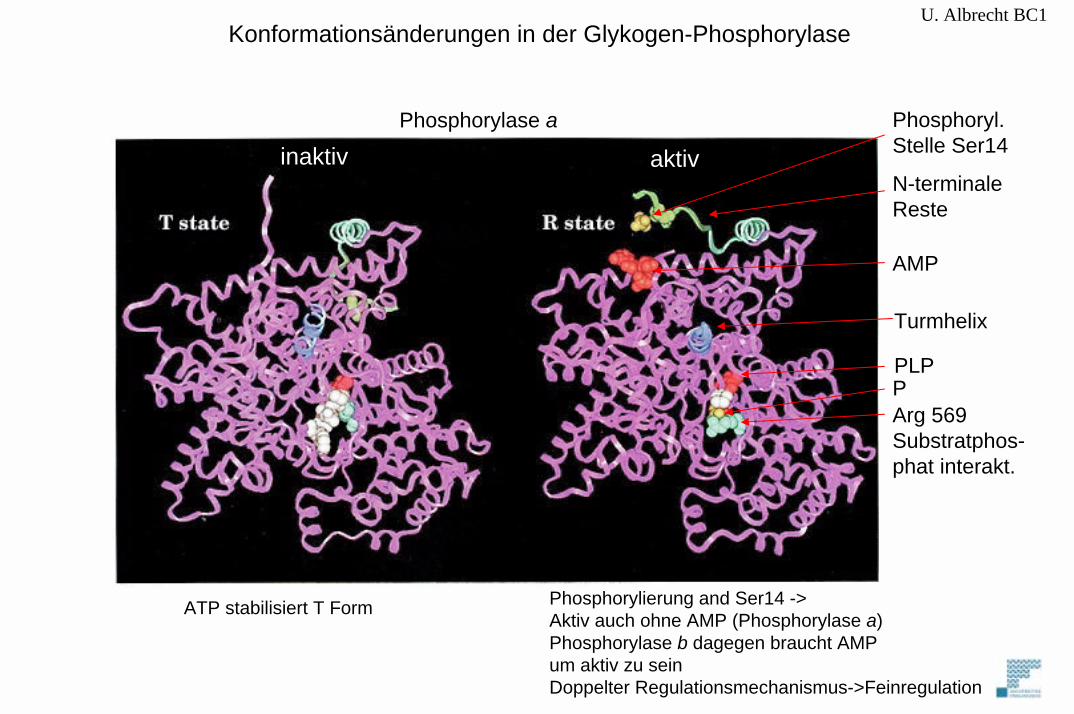
aktivinaktiv
Turmhelix
N-terminaleReste
AMP
Phosphoryl.Stelle Ser14
PLP
Arg 569Substratphos-phat interakt.
P
Phosphorylierung and Ser14 ->Aktiv auch ohne AMP (Phosphorylase a)Phosphorylase b dagegen braucht AMPum aktiv zu seinDoppelter Regulationsmechanismus->Feinregulation
Konformationsänderungen in der Glykogen-Phosphorylase
Phosphorylase a
ATP stabilisiert T Form
U. Albrecht BC1
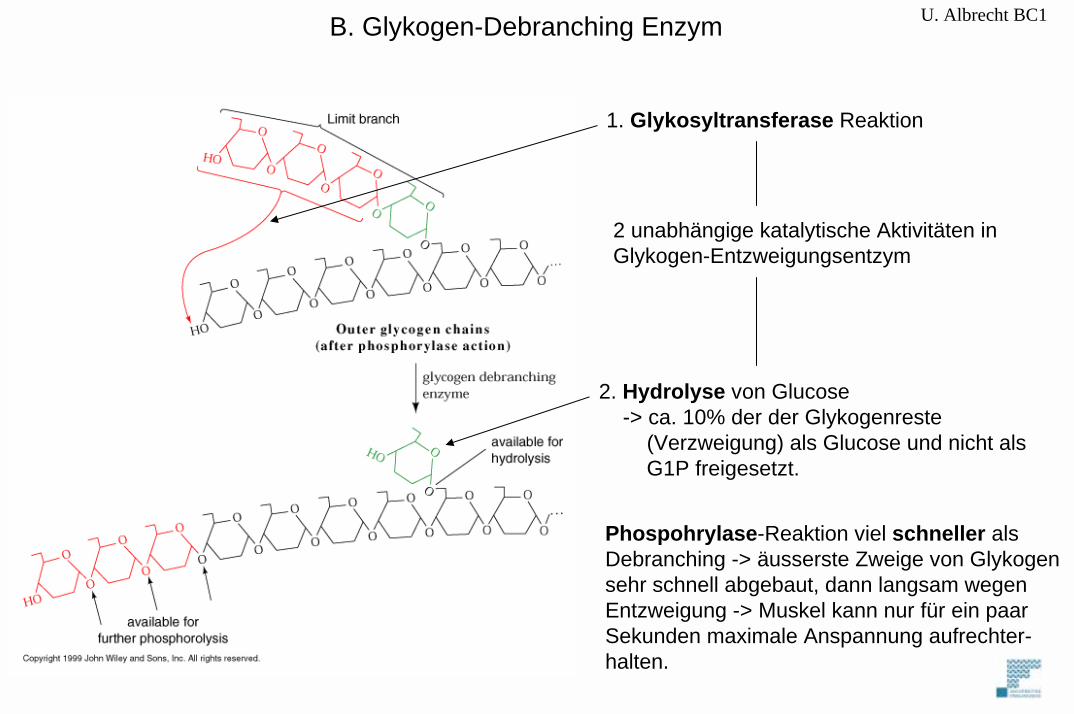
1. Glykosyltransferase Reaktion
2. Hydrolyse von Glucose-> ca. 10% der der Glykogenreste
(Verzweigung) als Glucose und nicht als G1P freigesetzt.
2 unabhängige katalytische Aktivitäten inGlykogen-Entzweigungsentzym
Phospohrylase-Reaktion viel schneller als Debranching -> äusserste Zweige von Glykogensehr schnell abgebaut, dann langsam wegenEntzweigung -> Muskel kann nur für ein paarSekunden maximale Anspannung aufrechter-halten.
B. Glykogen-Debranching Enzym
U. Albrecht BC1
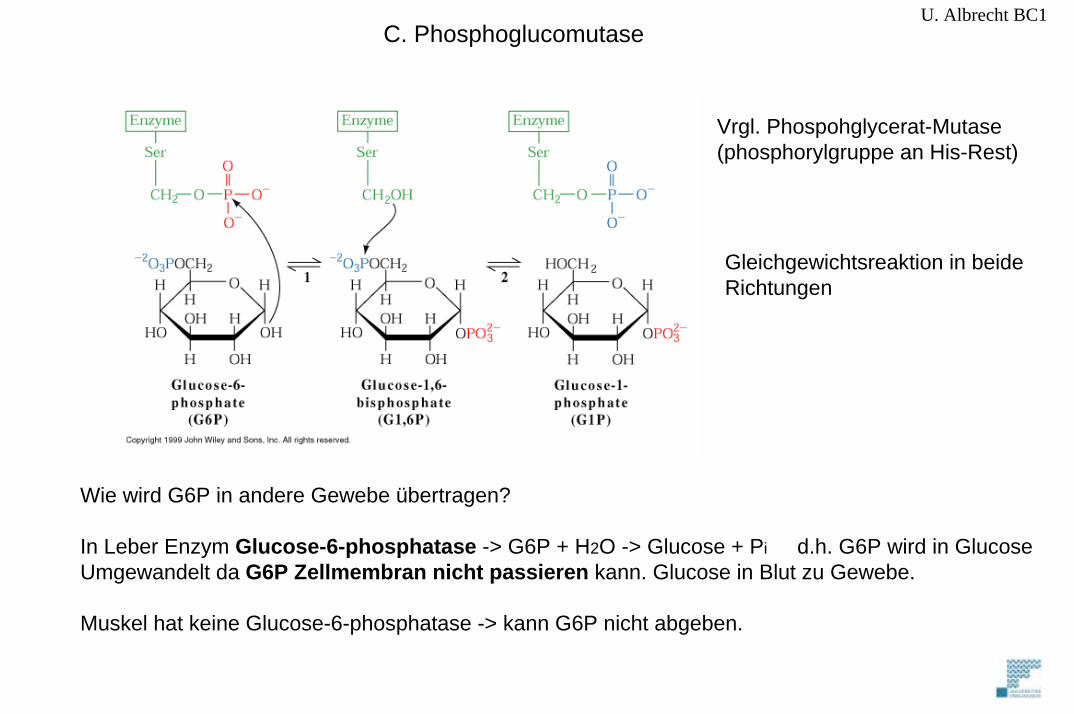
Vrgl. Phospohglycerat-Mutase(phosphorylgruppe an His-Rest)
Gleichgewichtsreaktion in beideRichtungen
Wie wird G6P in andere Gewebe übertragen?
In Leber Enzym Glucose-6-phosphatase -> G6P + H2O -> Glucose + Pi d.h. G6P wird in GlucoseUmgewandelt da G6P Zellmembran nicht passieren kann. Glucose in Blut zu Gewebe.
Muskel hat keine Glucose-6-phosphatase -> kann G6P nicht abgeben.
C. Phosphoglucomutase
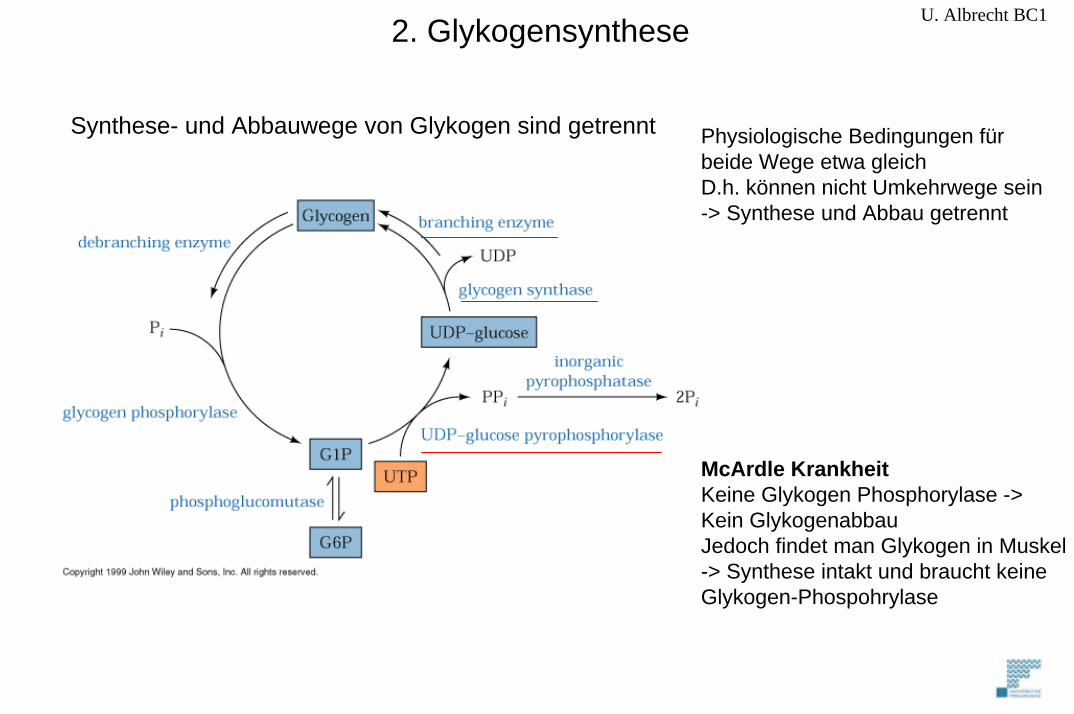
Physiologische Bedingungen fürbeide Wege etwa gleichD.h. können nicht Umkehrwege sein-> Synthese und Abbau getrennt
McArdle KrankheitKeine Glykogen Phosphorylase ->Kein GlykogenabbauJedoch findet man Glykogen in Muskel-> Synthese intakt und braucht keineGlykogen-Phospohrylase
U. Albrecht BC12. Glykogensynthese
Synthese- und Abbauwege von Glykogen sind getrennt
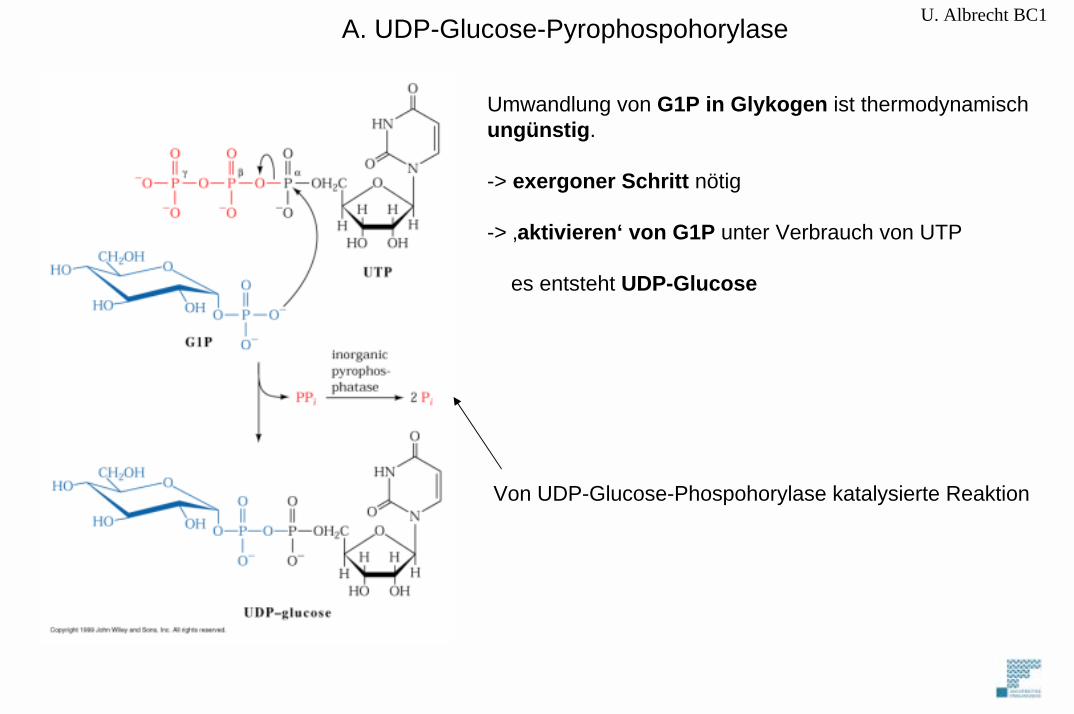
U. Albrecht BC1
Umwandlung von G1P in Glykogen ist thermodynamischungünstig.
-> exergoner Schritt nötig
-> ‚aktivieren‘ von G1P unter Verbrauch von UTP
es entsteht UDP-Glucose
Von UDP-Glucose-Phospohorylase katalysierte Reaktion
A. UDP-Glucose-Pyrophospohorylase
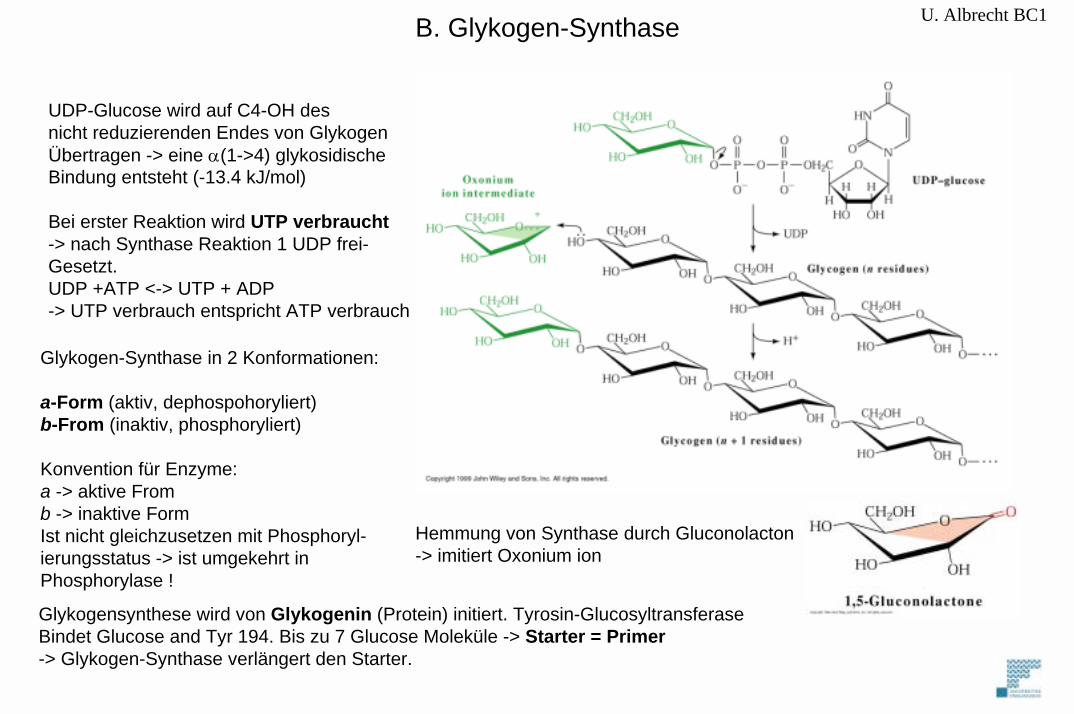
U. Albrecht BC1B. Glykogen-Synthase
UDP-Glucose wird auf C4-OH desnicht reduzierenden Endes von GlykogenÜbertragen -> eine α(1->4) glykosidischeBindung entsteht (-13.4 kJ/mol)
Bei erster Reaktion wird UTP verbraucht-> nach Synthase Reaktion 1 UDP frei-Gesetzt.UDP +ATP <-> UTP + ADP -> UTP verbrauch entspricht ATP verbrauch
Glykogen-Synthase in 2 Konformationen:
a-Form (aktiv, dephospohoryliert)b-From (inaktiv, phosphoryliert)
Konvention für Enzyme: a -> aktive Fromb -> inaktive FormIst nicht gleichzusetzen mit Phosphoryl-ierungsstatus -> ist umgekehrt in Phosphorylase !
Hemmung von Synthase durch Gluconolacton-> imitiert Oxonium ion
Glykogensynthese wird von Glykogenin (Protein) initiert. Tyrosin-GlucosyltransferaseBindet Glucose and Tyr 194. Bis zu 7 Glucose Moleküle -> Starter = Primer-> Glykogen-Synthase verlängert den Starter.
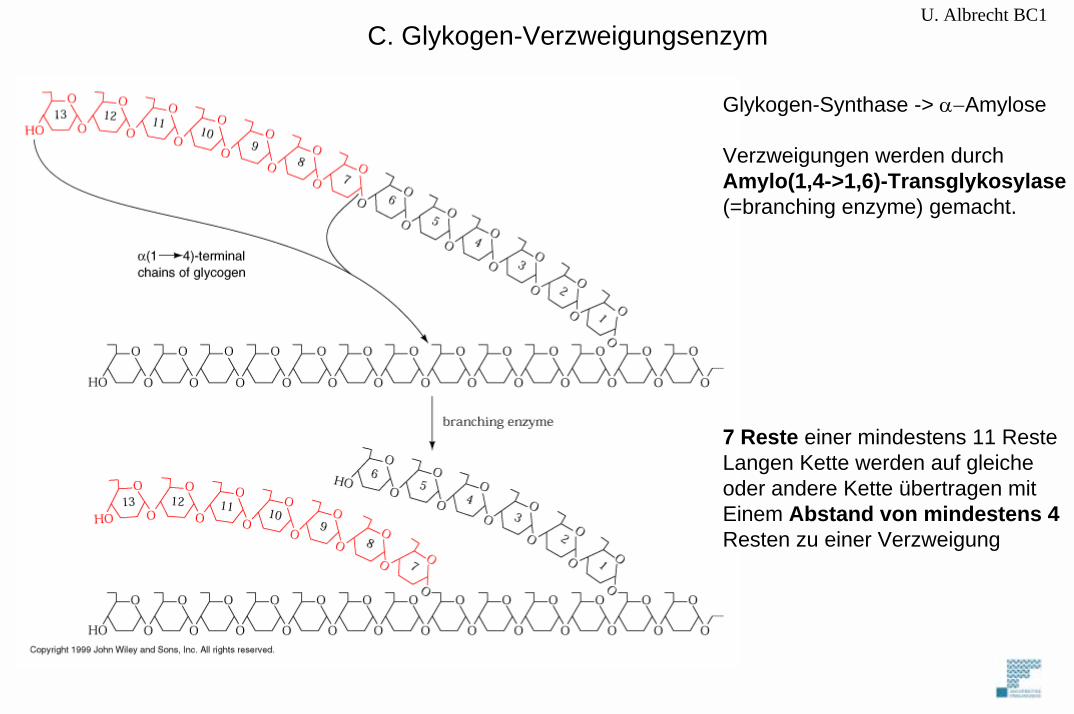
U. Albrecht BC1C. Glykogen-Verzweigungsenzym
Glykogen-Synthase -> α−Amylose
Verzweigungen werden durch Amylo(1,4->1,6)-Transglykosylase(=branching enzyme) gemacht.
7 Reste einer mindestens 11 ResteLangen Kette werden auf gleicheoder andere Kette übertragen mit Einem Abstand von mindestens 4Resten zu einer Verzweigung

U. Albrecht BC1
Glykogensynthese und Abbau nicht gleichzeitig, da sonst verschwendung von UTP-hydrolyse.
Regulation des Glykogenstoffwechsels auf 2 Ebenen: 1. Allosterische Kontrolle (AMP,ATP, G6P)2. Hormonelle Kontrolle (Glucagon, Adrenalin)
führt zu kovalenter Modifikation
A. Direkte allostersiche Kontrolle von Glykogen-Phosphorylase und -Synthase
Nettofluss = V for - V rück Gleichgewichtsreaktion -> lim 0 -> nicht kontrollierbar
-> 2 Reaktionen -> V for und V rück verändern sich unabhängig voneinander-> Kontrolle der Reaktionsrate und der Reaktionsrichtung.
Phosphorylase aktiviert durch AMPgehemmt durch ATP
G6P aktiviert Synthase
-> hoher Bedarf and ATP -> Aktivierung der Phosphorylase und Hemmung der Synthase
Zusätzlich überlagertes Kontrollsystem das Ansprechempfindlichkeit reguliert (Phosphorylierung)
3. Kontrolle des Glykogenstoffwechsels
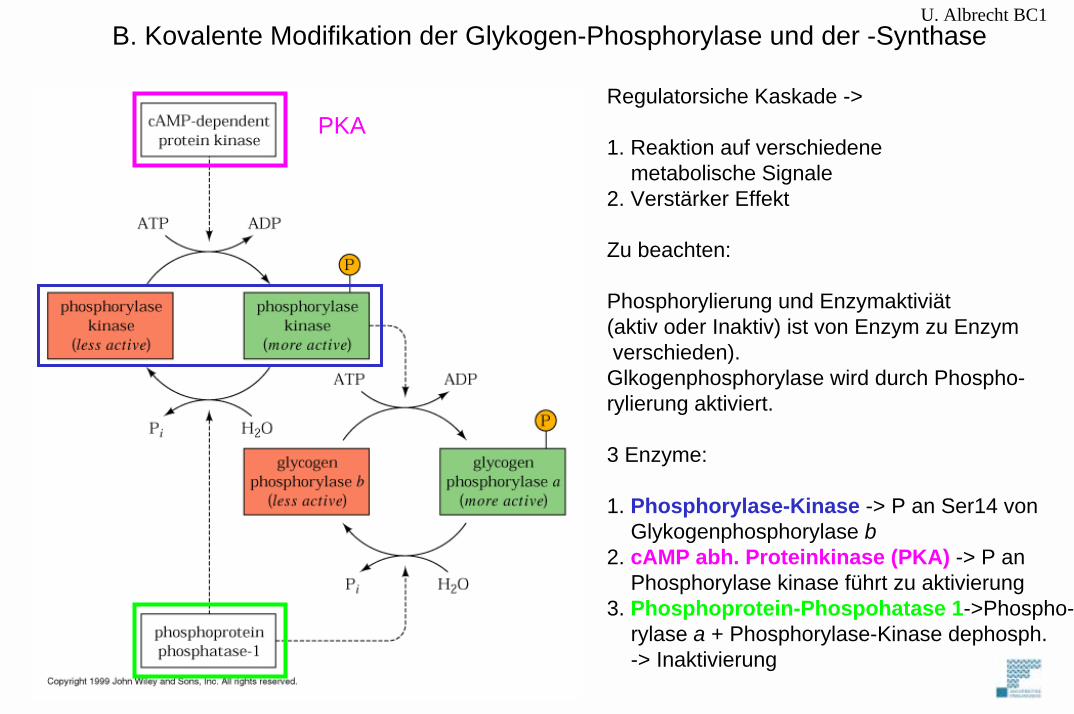
Regulatorsiche Kaskade ->
1. Reaktion auf verschiedene metabolische Signale
2. Verstärker Effekt
Zu beachten:
Phosphorylierung und Enzymaktiviät(aktiv oder Inaktiv) ist von Enzym zu Enzymverschieden).Glkogenphosphorylase wird durch Phospho-rylierung aktiviert.
3 Enzyme:
1. Phosphorylase-Kinase -> P an Ser14 vonGlykogenphosphorylase b
2. cAMP abh. Proteinkinase (PKA) -> P an Phosphorylase kinase führt zu aktivierung
3. Phosphoprotein-Phospohatase 1->Phospho-rylase a + Phosphorylase-Kinase dephosph. -> Inaktivierung
B. Kovalente Modifikation der Glykogen-Phosphorylase und der -SynthaseU. Albrecht BC1
PKA
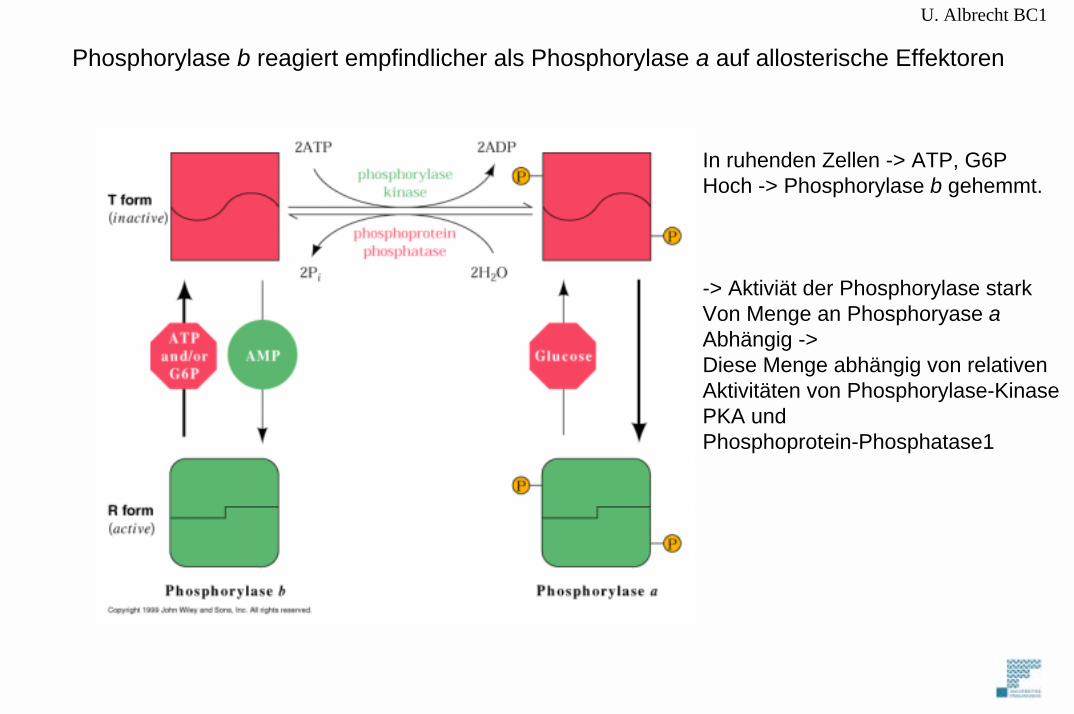
U. Albrecht BC1
Phosphorylase b reagiert empfindlicher als Phosphorylase a auf allosterische Effektoren
In ruhenden Zellen -> ATP, G6PHoch -> Phosphorylase b gehemmt.
-> Aktiviät der Phosphorylase starkVon Menge an Phosphoryase aAbhängig -> Diese Menge abhängig von relativen Aktivitäten von Phosphorylase-KinasePKA und Phosphoprotein-Phosphatase1
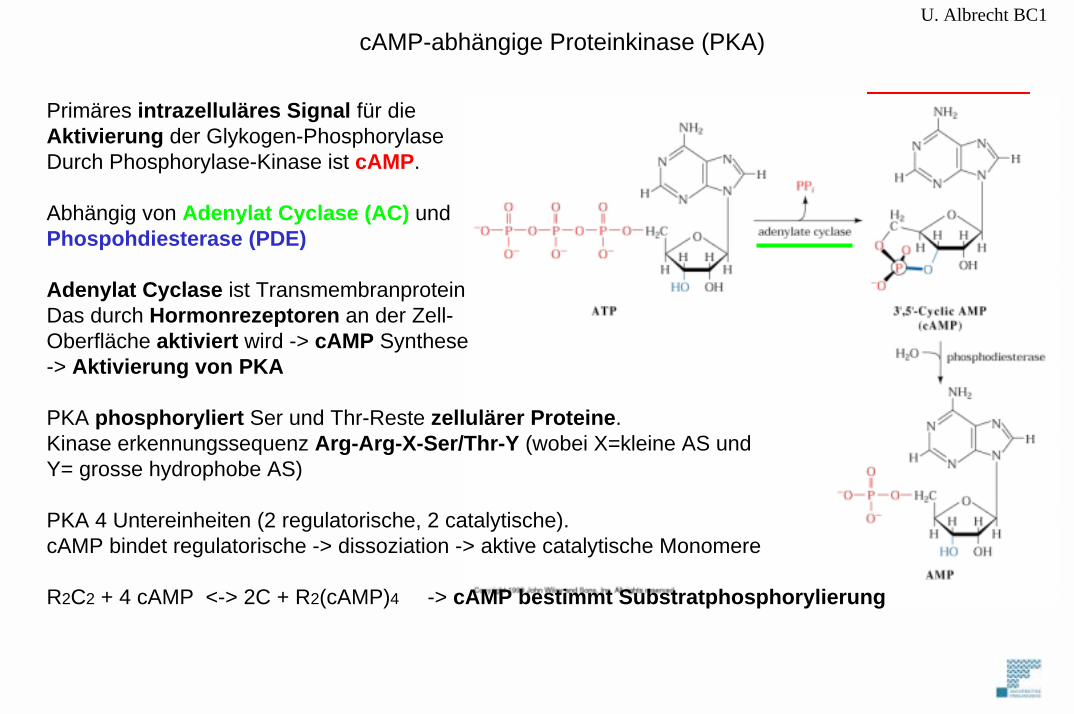
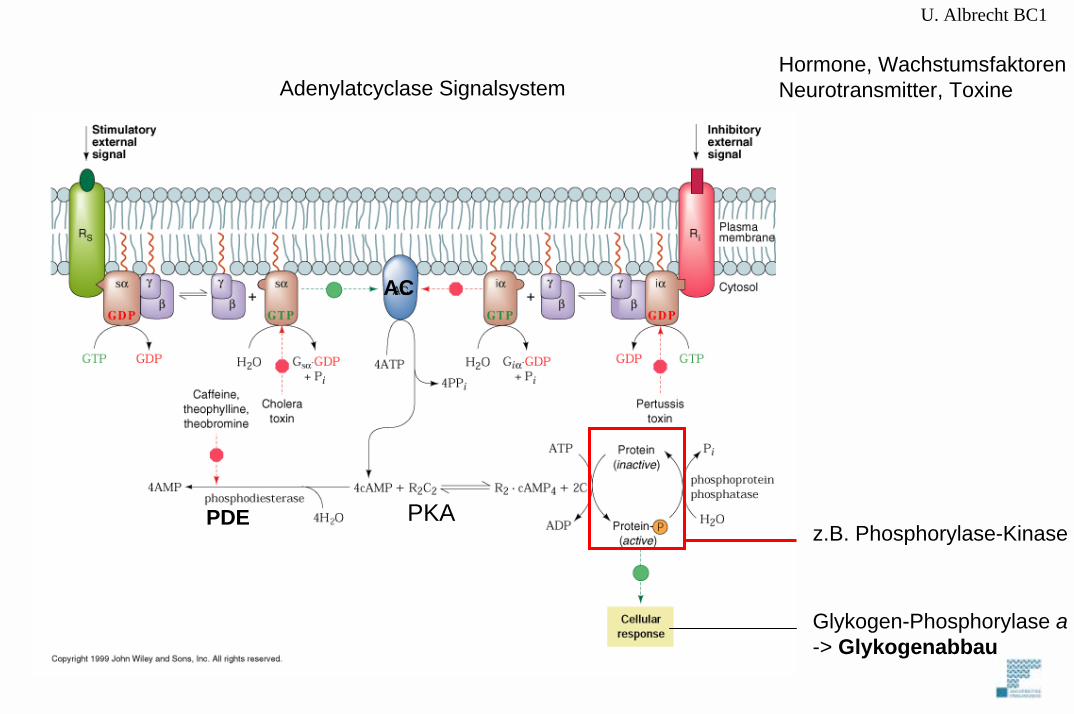
U. Albrecht BC1cAMP-abhängige Proteinkinase (PKA)
Primäres intrazelluläres Signal für dieAktivierung der Glykogen-PhosphorylaseDurch Phosphorylase-Kinase ist cAMP.
Abhängig von Adenylat Cyclase (AC) und Phospohdiesterase (PDE)
Adenylat Cyclase ist TransmembranproteinDas durch Hormonrezeptoren an der Zell-Oberfläche aktiviert wird -> cAMP Synthese-> Aktivierung von PKA
PKA phosphoryliert Ser und Thr-Reste zellulärer Proteine.Kinase erkennungssequenz Arg-Arg-X-Ser/Thr-Y (wobei X=kleine AS und Y= grosse hydrophobe AS)
PKA 4 Untereinheiten (2 regulatorische, 2 catalytische). cAMP bindet regulatorische -> dissoziation -> aktive catalytische Monomere
R2C2 + 4 cAMP <-> 2C + R2(cAMP)4 -> cAMP bestimmt Substratphosphorylierung
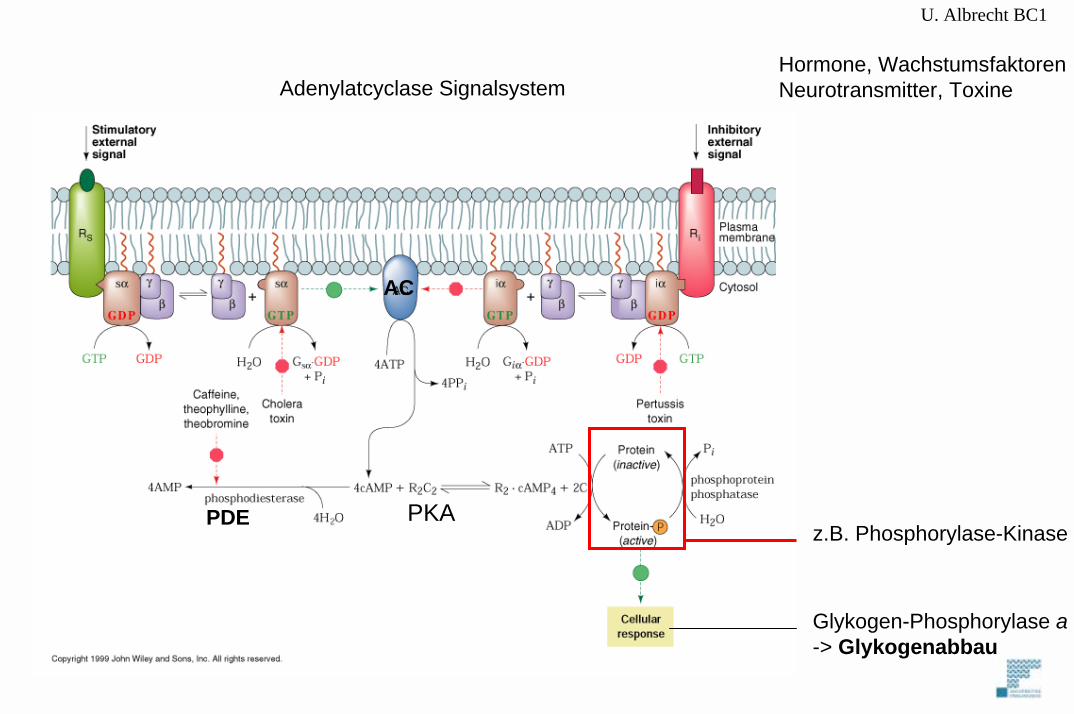
PKA
Glykogen-Phosphorylase a-> Glykogenabbau
PDE
AC
U. Albrecht BC1
Hormone, WachstumsfaktorenNeurotransmitter, ToxineAdenylatcyclase Signalsystem
z.B. Phosphorylase-Kinase
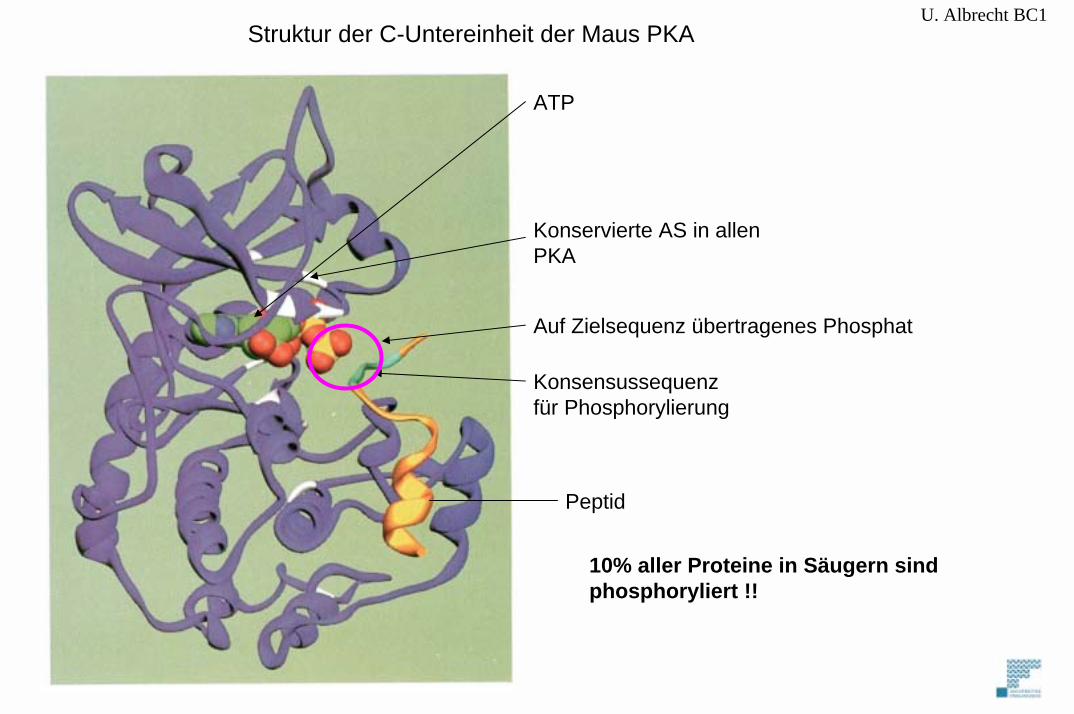
U. Albrecht BC1
Auf Zielsequenz übertragenes Phosphat
10% aller Proteine in Säugern sind phosphoryliert !!
Struktur der C-Untereinheit der Maus PKA
ATP
Konservierte AS in allenPKA
Konsensussequenzfür Phosphorylierung
Peptid
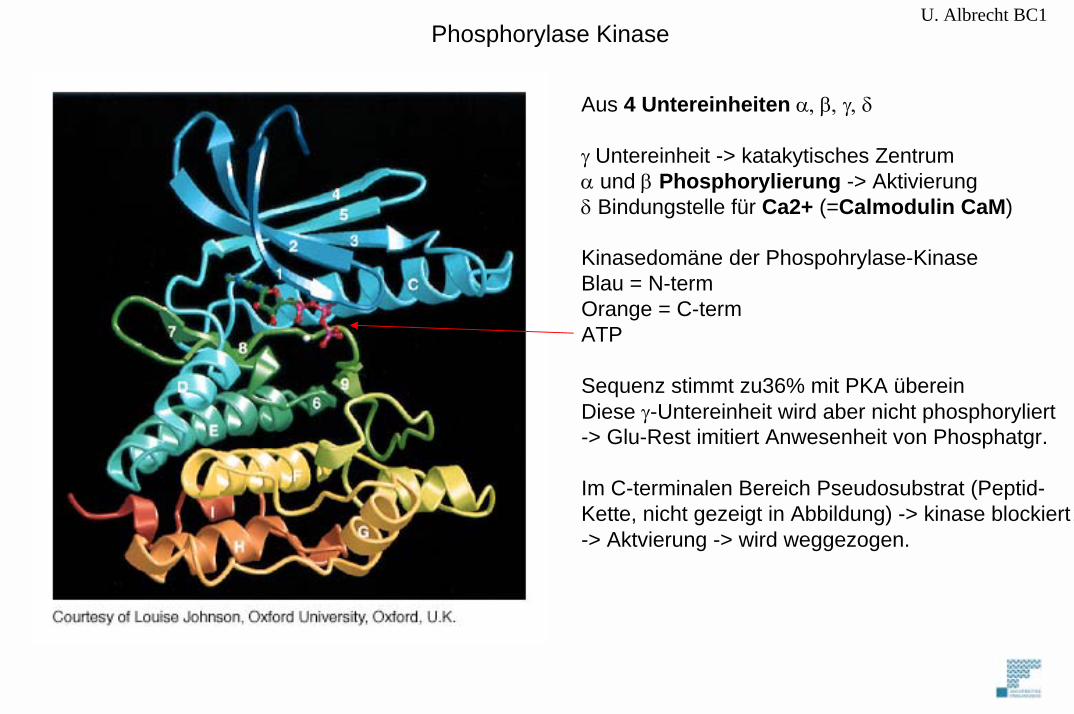
U. Albrecht BC1
Aus 4 Untereinheiten α, β, γ, δ
γ Untereinheit -> katakytisches Zentrumα und β Phosphorylierung -> Aktivierungδ Bindungstelle für Ca2+ (=Calmodulin CaM)
Kinasedomäne der Phospohrylase-KinaseBlau = N-termOrange = C-termATP
Sequenz stimmt zu36% mit PKA übereinDiese γ-Untereinheit wird aber nicht phosphoryliert-> Glu-Rest imitiert Anwesenheit von Phosphatgr.
Im C-terminalen Bereich Pseudosubstrat (Peptid-Kette, nicht gezeigt in Abbildung) -> kinase blockiert-> Aktvierung -> wird weggezogen.
Phosphorylase Kinase
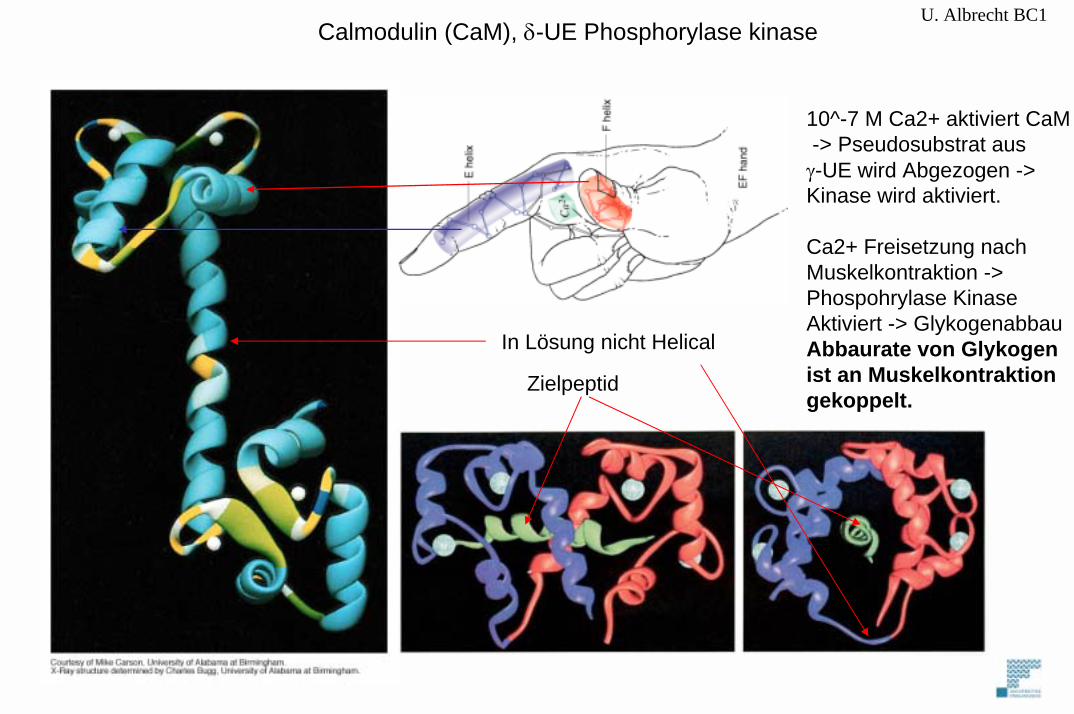
U. Albrecht BC1Calmodulin (CaM), δ-UE Phosphorylase kinase
10^-7 M Ca2+ aktiviert CaM-> Pseudosubstrat aus
γ-UE wird Abgezogen -> Kinase wird aktiviert.
Ca2+ Freisetzung nach Muskelkontraktion -> Phospohrylase KinaseAktiviert -> GlykogenabbauAbbaurate von Glykogenist an Muskelkontraktion gekoppelt.
In Lösung nicht Helical
Zielpeptid
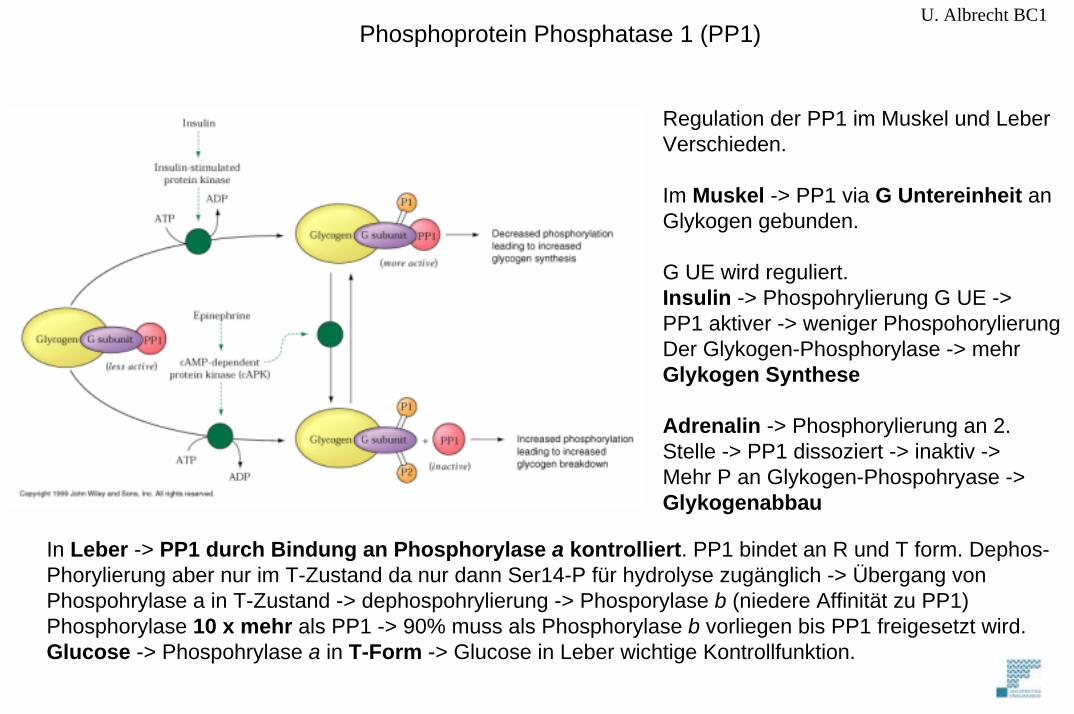
U. Albrecht BC1
Regulation der PP1 im Muskel und LeberVerschieden.
Im Muskel -> PP1 via G Untereinheit anGlykogen gebunden.
G UE wird reguliert. Insulin -> Phospohrylierung G UE ->PP1 aktiver -> weniger PhospohorylierungDer Glykogen-Phosphorylase -> mehrGlykogen Synthese
Adrenalin -> Phosphorylierung an 2.Stelle -> PP1 dissoziert -> inaktiv -> Mehr P an Glykogen-Phospohryase ->Glykogenabbau
In Leber -> PP1 durch Bindung an Phosphorylase a kontrolliert. PP1 bindet an R und T form. Dephos-Phorylierung aber nur im T-Zustand da nur dann Ser14-P für hydrolyse zugänglich -> Übergang vonPhospohrylase a in T-Zustand -> dephospohrylierung -> Phosporylase b (niedere Affinität zu PP1)Phosphorylase 10 x mehr als PP1 -> 90% muss als Phosphorylase b vorliegen bis PP1 freigesetzt wird.Glucose -> Phospohrylase a in T-Form -> Glucose in Leber wichtige Kontrollfunktion.
Phosphoprotein Phosphatase 1 (PP1)
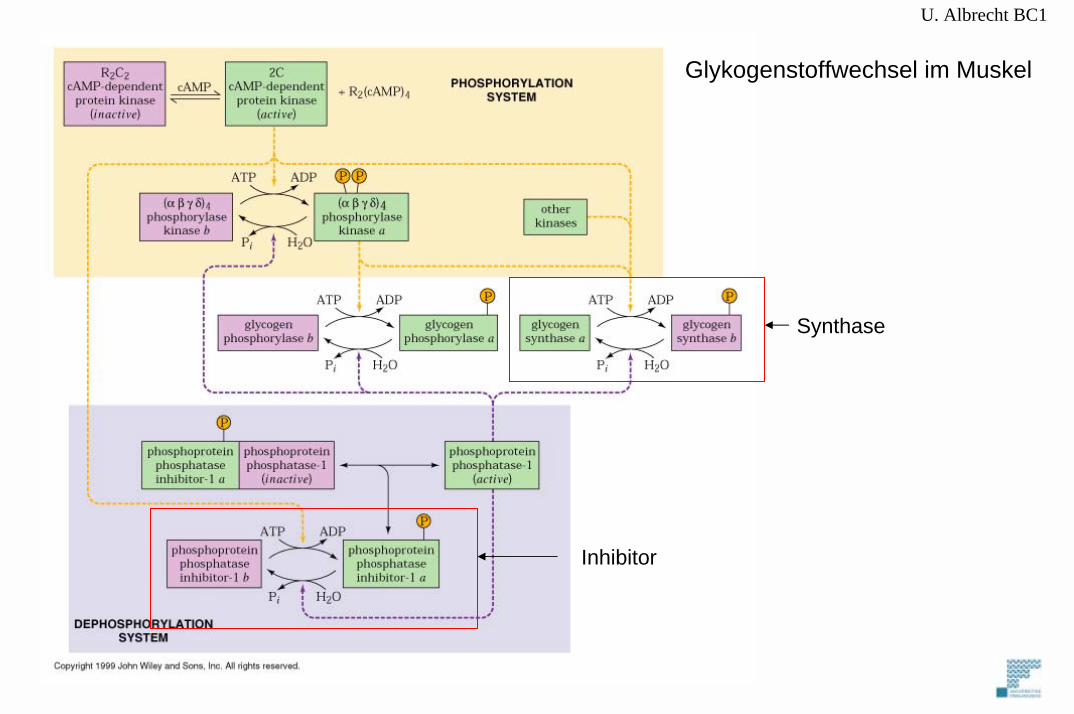
U. Albrecht BC1
Inhibitor
Glykogenstoffwechsel im Muskel
Synthase

U. Albrecht BC1
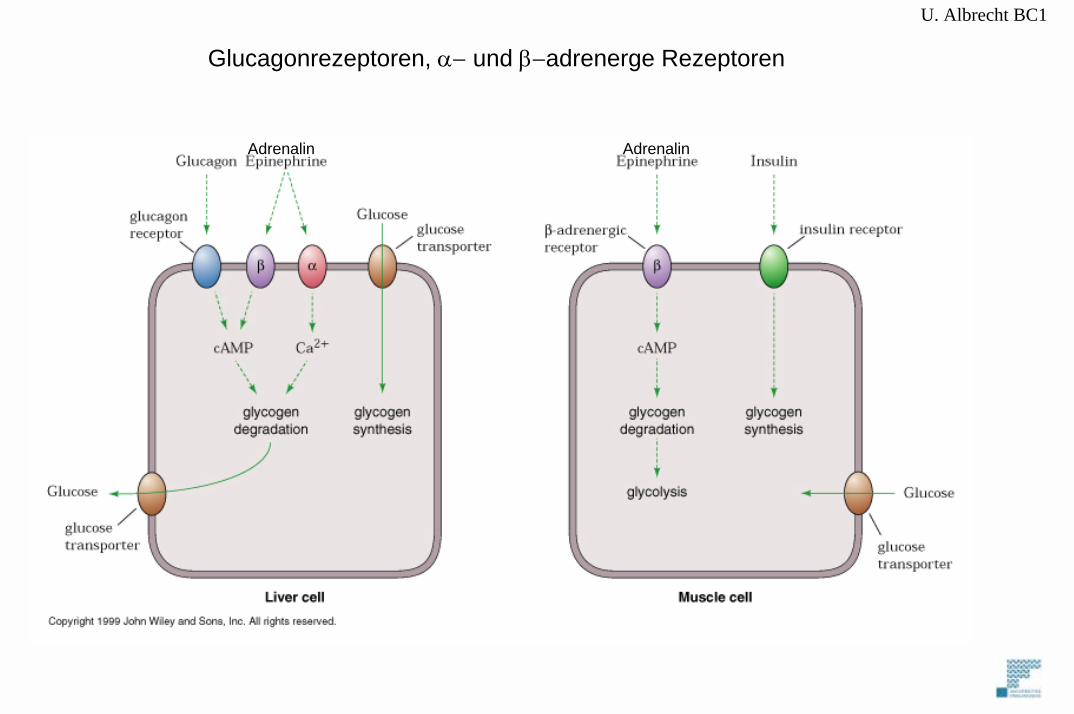
= Adrenalin,Noradrenalin
Wirken auf Membranrezeptoren und lösen Signalkaskaden überSekundäre Botenstoffe (second messengers) aus.
C. Hormonelle Einflüsse auf den Glykogenstoffwechsel
PKA
Glykogen-Phosphorylase a-> Glykogenabbau
PDE
AC
U. Albrecht BC1
Hormone, WachstumsfaktorenNeurotransmitter, ToxineAdenylatcyclase Signalsystem
z.B. Phosphorylase-Kinase
U. Albrecht BC1
Adrenalin Adrenalin
Glucagonrezeptoren, α− und β−adrenerge Rezeptoren
U. Albrecht BC1
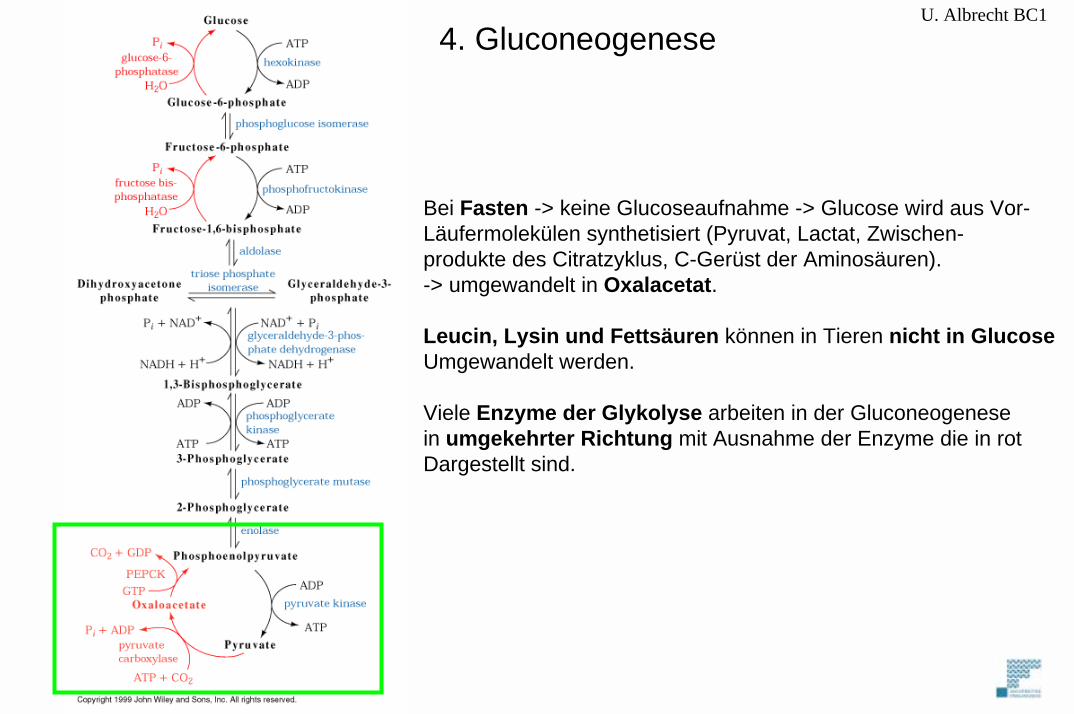
Bei Fasten -> keine Glucoseaufnahme -> Glucose wird aus Vor-Läufermolekülen synthetisiert (Pyruvat, Lactat, Zwischen-produkte des Citratzyklus, C-Gerüst der Aminosäuren). -> umgewandelt in Oxalacetat.
Leucin, Lysin und Fettsäuren können in Tieren nicht in GlucoseUmgewandelt werden.
Viele Enzyme der Glykolyse arbeiten in der Gluconeogenesein umgekehrter Richtung mit Ausnahme der Enzyme die in rotDargestellt sind.
4. Gluconeogenese
U. Albrecht BC1
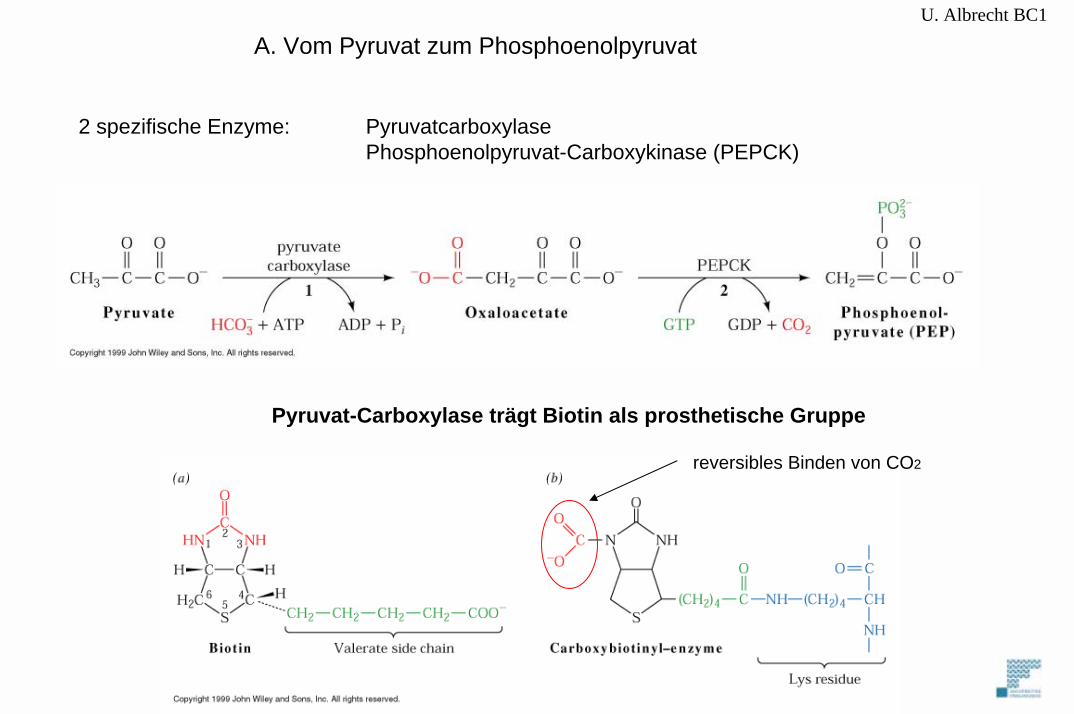
Pyruvat-Carboxylase trägt Biotin als prosthetische Gruppe
reversibles Binden von CO2
A. Vom Pyruvat zum Phosphoenolpyruvat
2 spezifische Enzyme: PyruvatcarboxylasePhosphoenolpyruvat-Carboxykinase (PEPCK)
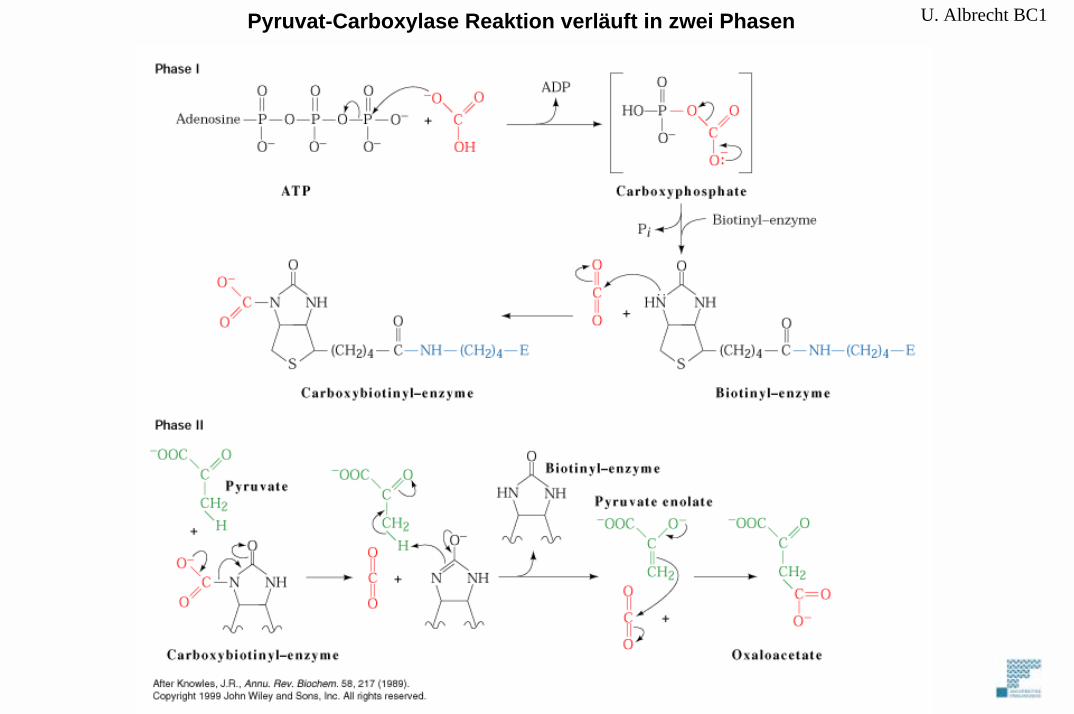
U. Albrecht BC1Pyruvat-Carboxylase Reaktion verläuft in zwei Phasen
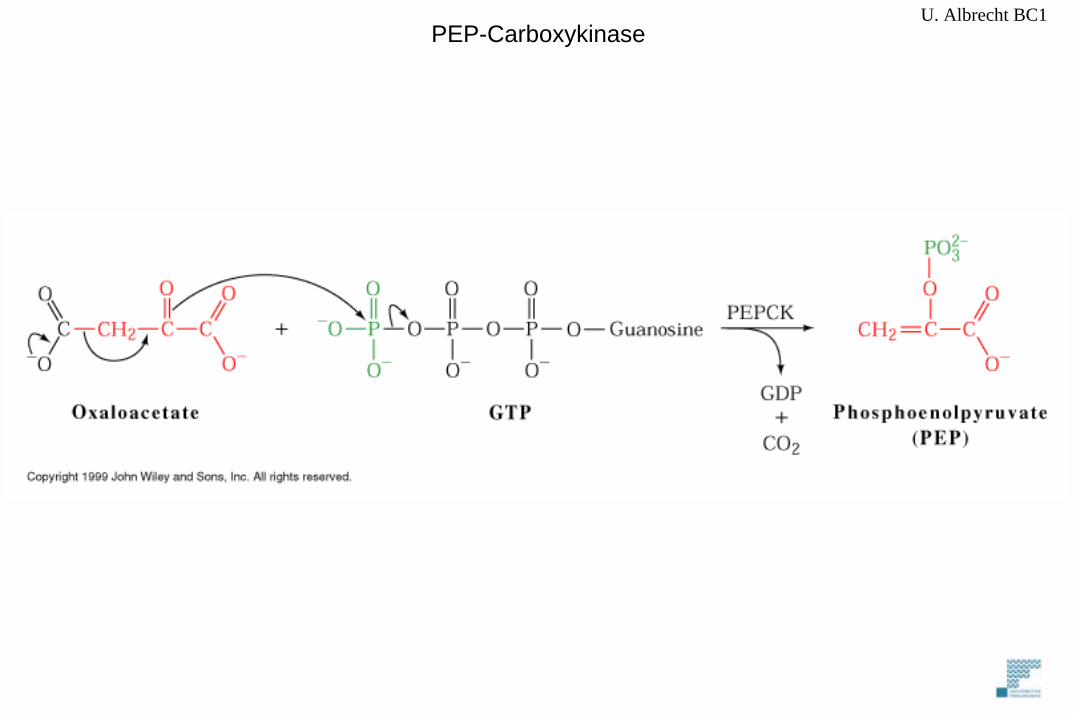
U. Albrecht BC1PEP-Carboxykinase
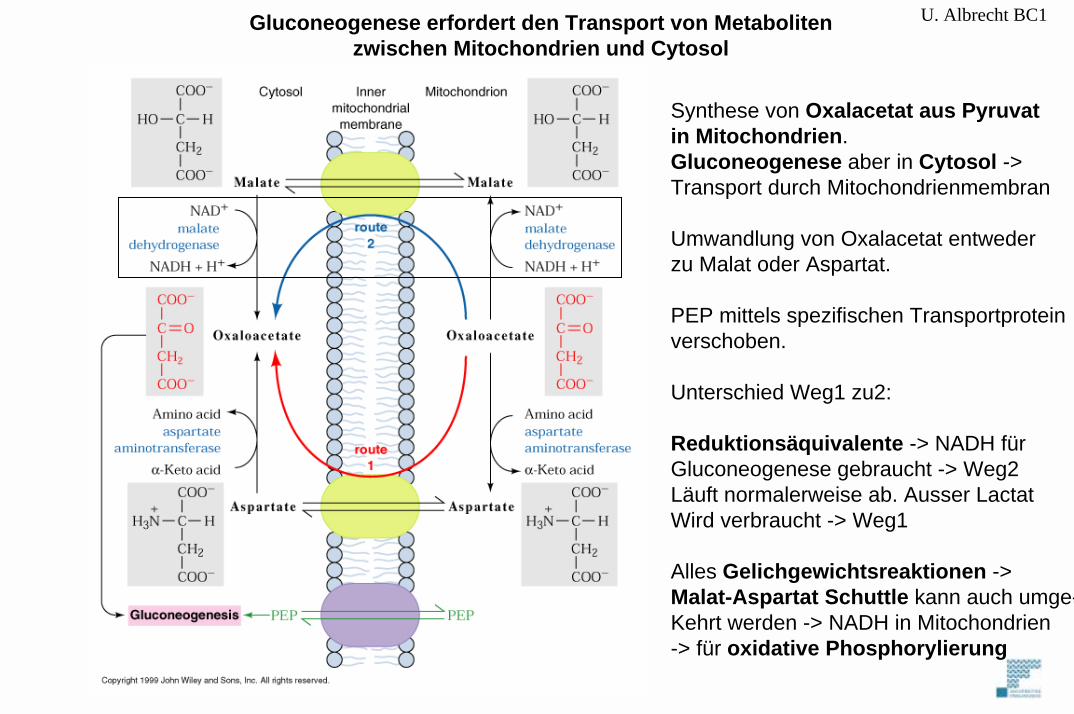
U. Albrecht BC1
Synthese von Oxalacetat aus Pyruvatin Mitochondrien.Gluconeogenese aber in Cytosol ->Transport durch Mitochondrienmembran
Umwandlung von Oxalacetat entwederzu Malat oder Aspartat.
PEP mittels spezifischen Transportproteinverschoben.
Unterschied Weg1 zu2:
Reduktionsäquivalente -> NADH fürGluconeogenese gebraucht -> Weg2Läuft normalerweise ab. Ausser LactatWird verbraucht -> Weg1
Alles Gelichgewichtsreaktionen -> Malat-Aspartat Schuttle kann auch umge-Kehrt werden -> NADH in Mitochondrien -> für oxidative Phosphorylierung
Gluconeogenese erfordert den Transport von Metabolitenzwischen Mitochondrien und Cytosol
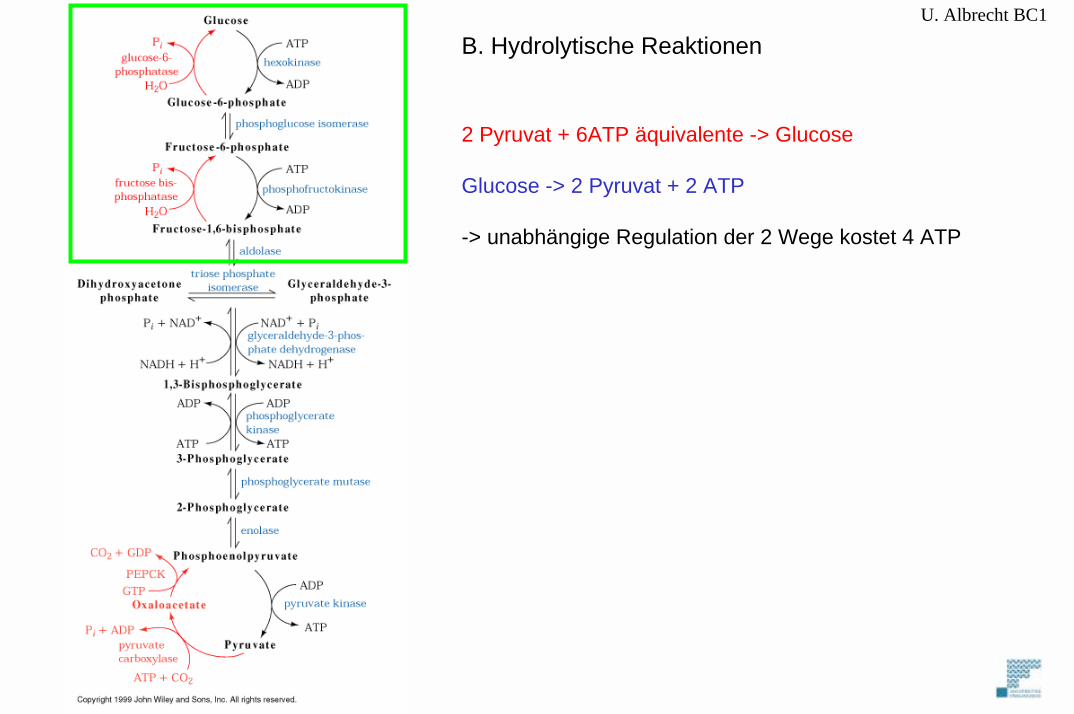
U. Albrecht BC1
2 Pyruvat + 6ATP äquivalente -> Glucose
Glucose -> 2 Pyruvat + 2 ATP
-> unabhängige Regulation der 2 Wege kostet 4 ATP
B. Hydrolytische Reaktionen
U. Albrecht BC1
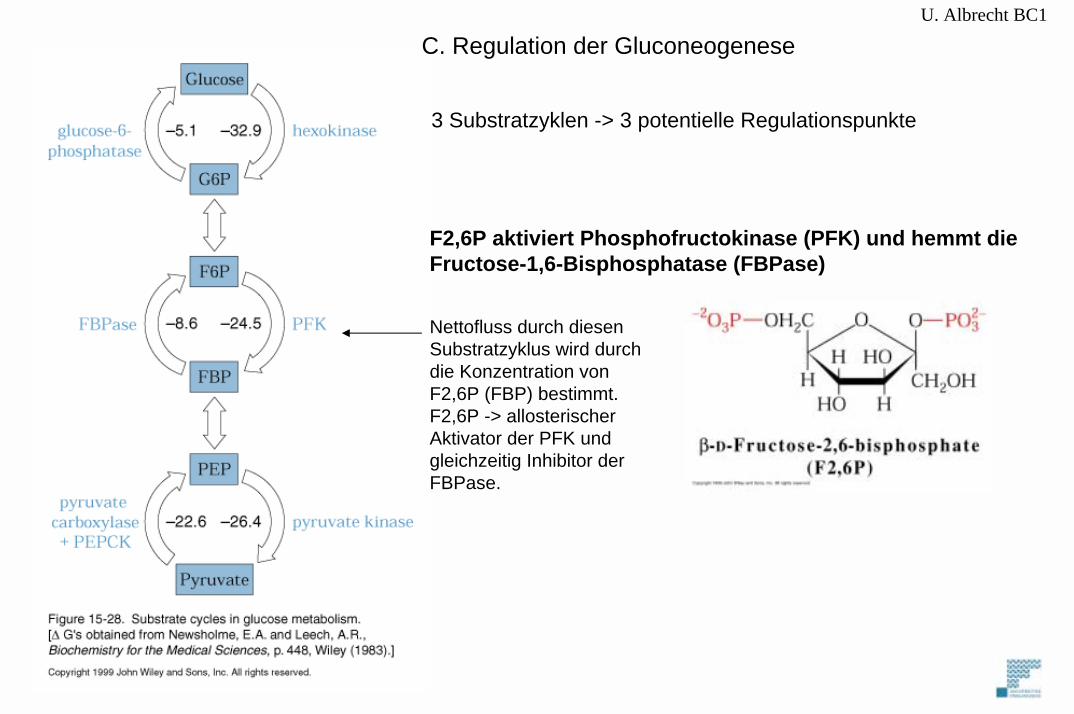
C. Regulation der Gluconeogenese
3 Substratzyklen -> 3 potentielle Regulationspunkte
F2,6P aktiviert Phosphofructokinase (PFK) und hemmt die Fructose-1,6-Bisphosphatase (FBPase)
Nettofluss durch diesenSubstratzyklus wird durchdie Konzentration von F2,6P (FBP) bestimmt.F2,6P -> allosterischerAktivator der PFK undgleichzeitig Inhibitor derFBPase.
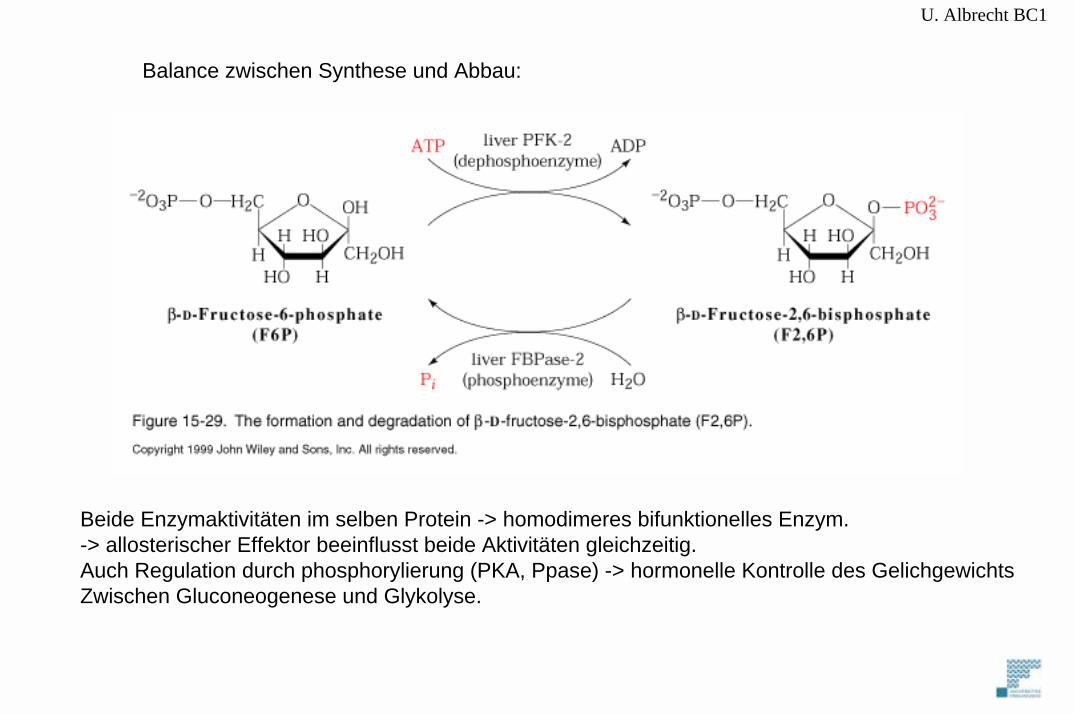
U. Albrecht BC1
Beide Enzymaktivitäten im selben Protein -> homodimeres bifunktionelles Enzym.-> allosterischer Effektor beeinflusst beide Aktivitäten gleichzeitig.Auch Regulation durch phosphorylierung (PKA, Ppase) -> hormonelle Kontrolle des GelichgewichtsZwischen Gluconeogenese und Glykolyse.
Balance zwischen Synthese und Abbau:

Achtung: In Muskel läuft keine Gluconeogenese ab, daanderes PFK-2 Isoenzym.Skelettmuskel: keine Phosphorylierungsstelle
-> keine cAMP abh. KontrolleHerzmuskel: andere Phosphorylierungsstelle
-> Aktivierung statt Hemmung derPFK-2 durch phosphorylierungals Antwort zu Hormonen dieGlykogenabbau im Herzmuskelfördern.
U. Albrecht BC1
Niedriger Glucosespielgel in der Leber
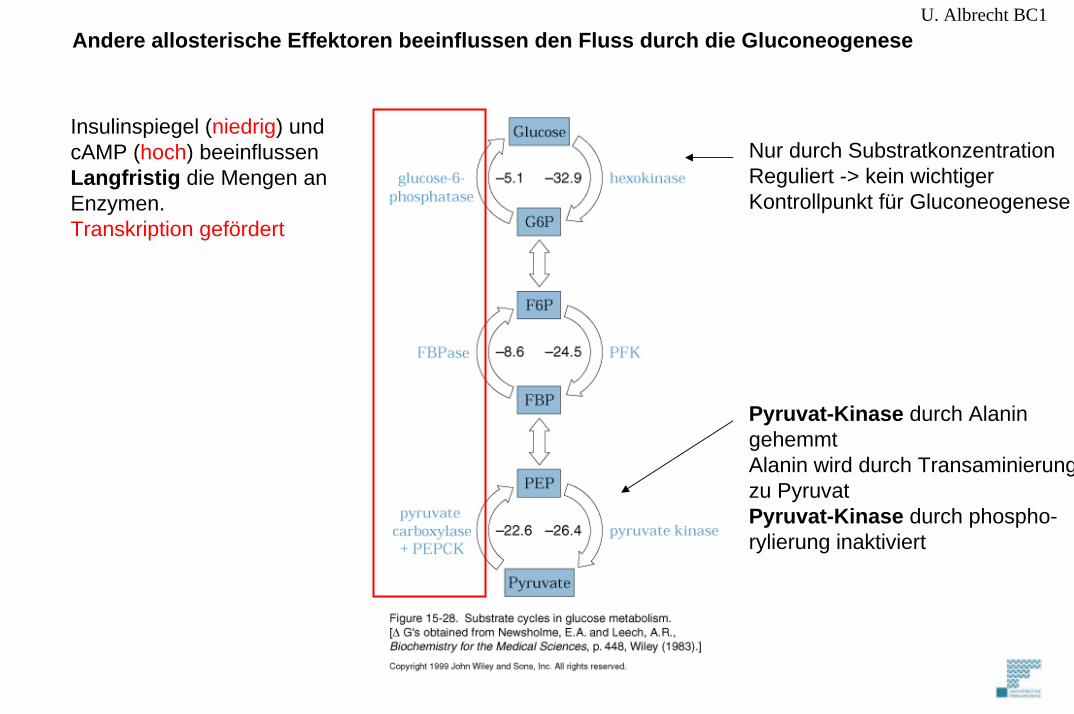
U. Albrecht BC1Andere allosterische Effektoren beeinflussen den Fluss durch die Gluconeogenese
Nur durch SubstratkonzentrationReguliert -> kein wichtigerKontrollpunkt für Gluconeogenese
Pyruvat-Kinase durch Alaningehemmt Alanin wird durch Transaminierungzu PyruvatPyruvat-Kinase durch phospho-rylierung inaktiviert
Insulinspiegel (niedrig) undcAMP (hoch) beeinflussenLangfristig die Mengen anEnzymen.Transkription gefördert
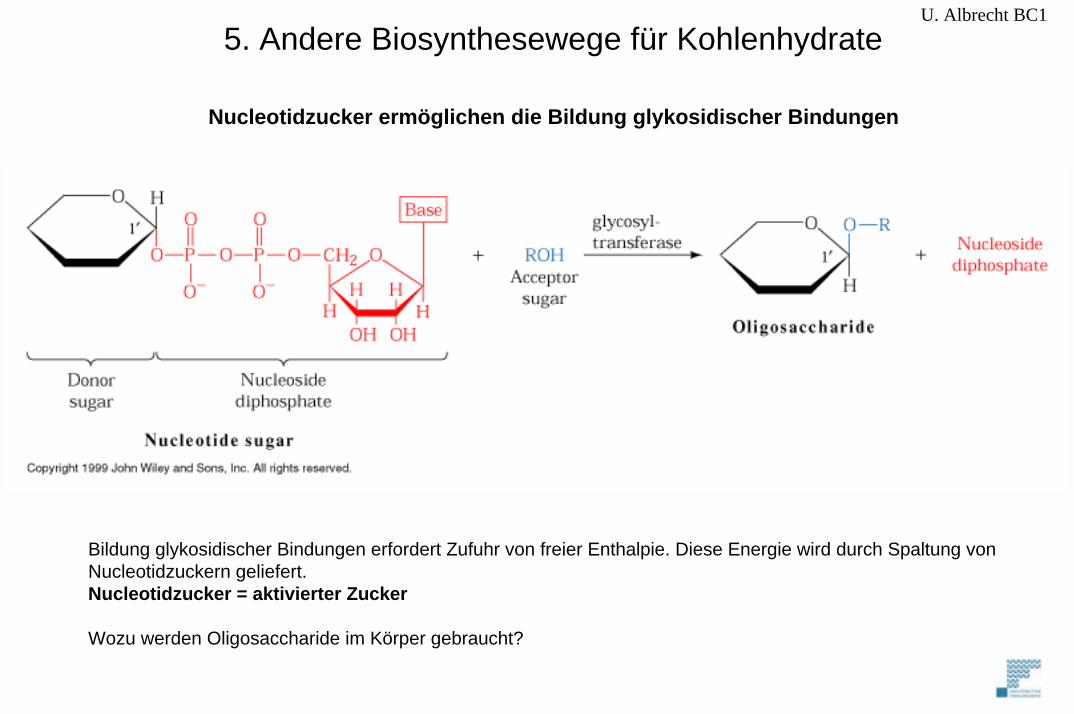
5. Andere Biosynthesewege für KohlenhydrateU. Albrecht BC1
Bildung glykosidischer Bindungen erfordert Zufuhr von freier Enthalpie. Diese Energie wird durch Spaltung vonNucleotidzuckern geliefert.Nucleotidzucker = aktivierter Zucker
Wozu werden Oligosaccharide im Körper gebraucht?
Nucleotidzucker ermöglichen die Bildung glykosidischer Bindungen
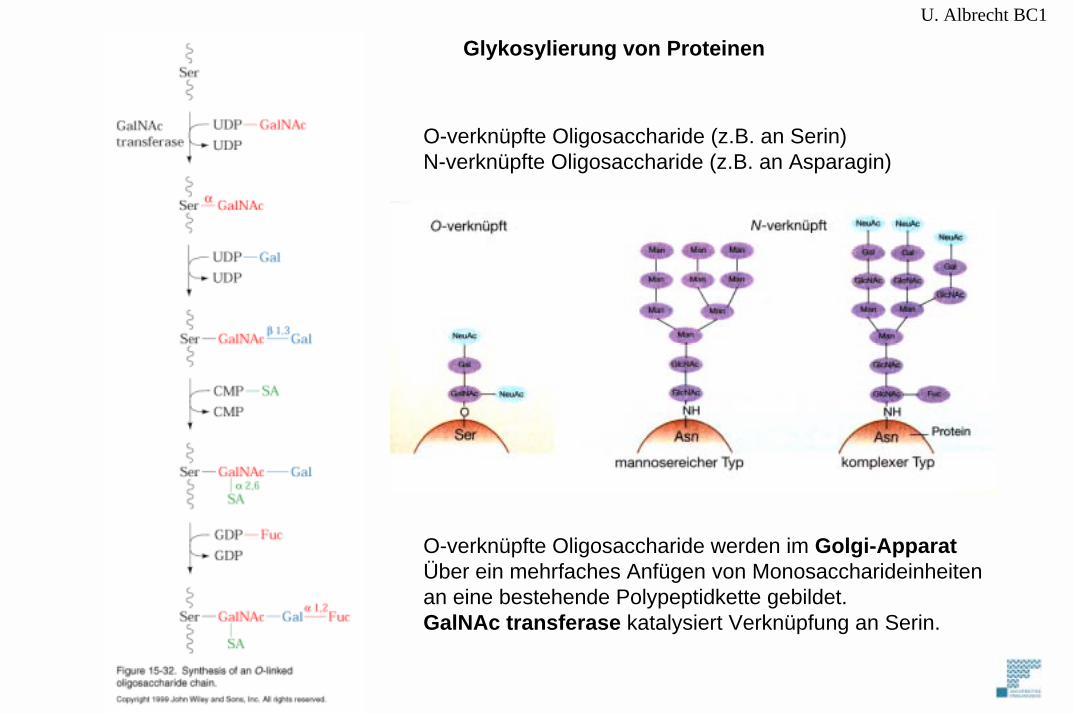
U. Albrecht BC1
O-verknüpfte Oligosaccharide (z.B. an Serin)N-verknüpfte Oligosaccharide (z.B. an Asparagin)
O-verknüpfte Oligosaccharide werden im Golgi-ApparatÜber ein mehrfaches Anfügen von Monosaccharideinheitenan eine bestehende Polypeptidkette gebildet.GalNAc transferase katalysiert Verknüpfung an Serin.
Glykosylierung von Proteinen
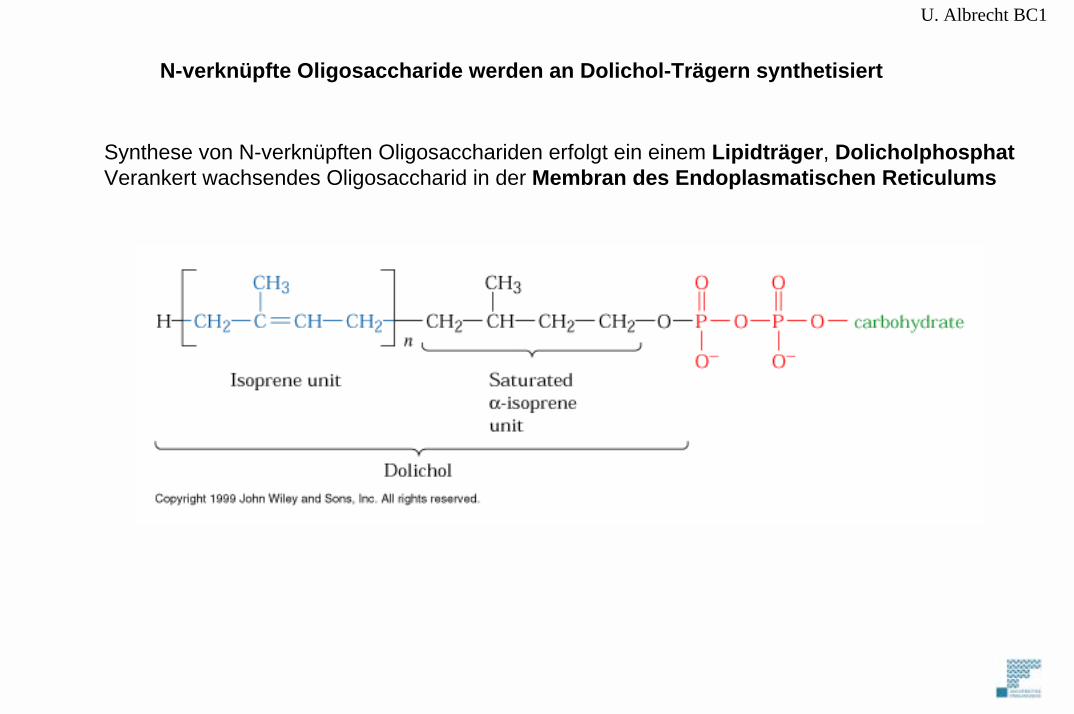
U. Albrecht BC1
Synthese von N-verknüpften Oligosacchariden erfolgt ein einem Lipidträger, DolicholphosphatVerankert wachsendes Oligosaccharid in der Membran des Endoplasmatischen Reticulums
N-verknüpfte Oligosaccharide werden an Dolichol-Trägern synthetisiert
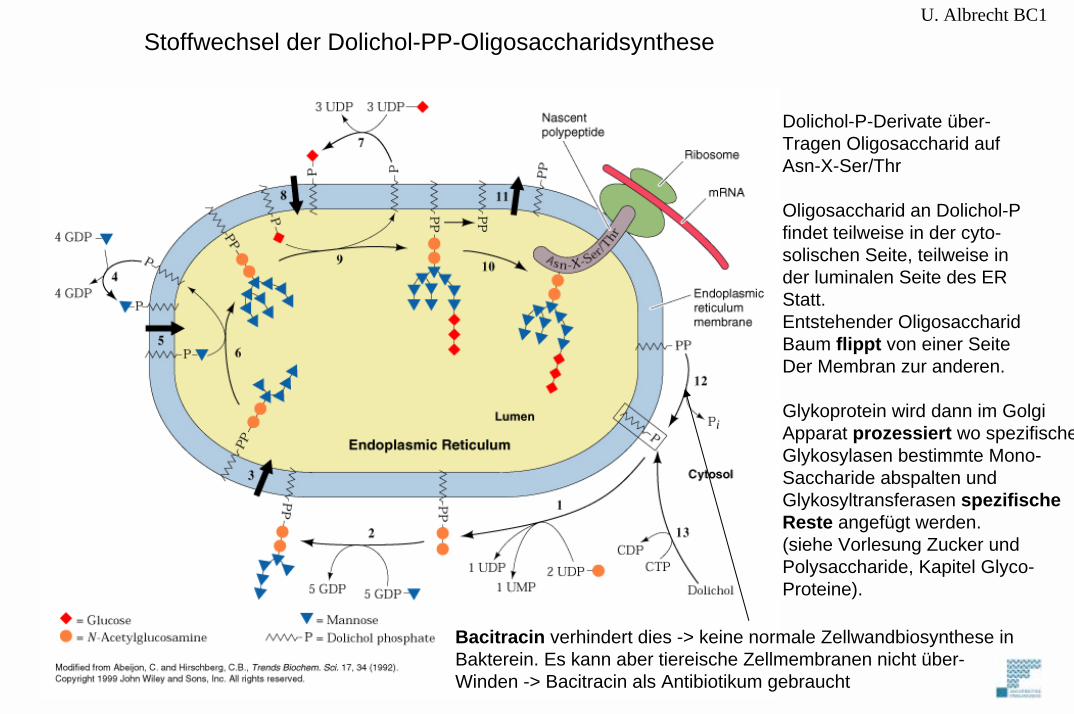
U. Albrecht BC1Stoffwechsel der Dolichol-PP-Oligosaccharidsynthese
Dolichol-P-Derivate über-Tragen Oligosaccharid aufAsn-X-Ser/Thr
Oligosaccharid an Dolichol-Pfindet teilweise in der cyto-solischen Seite, teilweise in der luminalen Seite des ERStatt.Entstehender OligosaccharidBaum flippt von einer SeiteDer Membran zur anderen.
Glykoprotein wird dann im GolgiApparat prozessiert wo spezifischeGlykosylasen bestimmte Mono-Saccharide abspalten und Glykosyltransferasen spezifischeReste angefügt werden.(siehe Vorlesung Zucker und Polysaccharide, Kapitel Glyco-Proteine).
Bacitracin verhindert dies -> keine normale Zellwandbiosynthese inBakterein. Es kann aber tiereische Zellmembranen nicht über-Winden -> Bacitracin als Antibiotikum gebraucht





























