Embed Size (px)
Citation preview

Arch. exper. Path. u. Pharmakol., Bd. 233, S. 531--540 (1958)
Aus dem Physiologischen Institut der Universit~t Heidelberg
~ber die Acetyleholin-Empfindlichkeit verschiedener Herzabschnitte ~
Von ROBERT F° SCHMIDT
Mi~ 4 Textabbildungen
(Eingegangen am 15..Februar 1958)
Seit LOEWI die humorale l~bertragung der Vaguswirkung (1921) und als t)bertr£gerstoff das Acetyleholin (Ach) entdeekte (LoEwI u. NAVRA~L 1926), ist oft versueht worden, die Wirkungen des Aeh auf die verschiede- nen Herzabsehnitte im einzelnen zu kl~ren. Am Froschherzen zeigte LXNCZOS (1930), daft die negativ inotrope Wirkung des Ach am Vorhof starker ist als an der Kammerbasis und an dieser starker als an der Kammerspitze. Dagegen fanden LISSi[K u. MARTIN (1940) keine unter- schiedliehe Empfindlichkeit innerha.lb der Frosehkammer und keine nennenswerte zwischen Kammer und Vorhof.
Am Vorhof des Warmblfiters werden als Regelfall negativ inotrope (BuRoE~ u. TERROUX 1953), unter gewissen Bedingungen aber aueh positiv inotrope Wirkungen des Aeh auf die Muskulatur beschrieben (McEwEN 1956). Keine Klarheit herrscht fiber die Effekte des Ach an der Kammermuskulatur des Warmbliiters. Zuletzt haben MIDDLETOlg et al. (1956) am Papillarmuskel der Katze in Abh~ngigkeit v o n d e r Aeh- Konzentration hemmende und fSrdernde Wirkungen gesehen.
Die Ver/~nderungen des Ruhe- und Aktionspotentials dureh Aeh sind in neuerer Zeit mit intracellul/~rer Potentialabteilung (LINe u. GERARD 1949) untersucht worden. M_it dieser Methode fanden DEL CASTILLO U. KATZ (1955a), HCTTE~ U. TRAUTW~,IN (1955) am Froschherzen, BURGE~ U. TERROCX (1953) am Vorhof der Katze eine ErhShung des Ruhepoten- rials, sowie eine sehnellere Repolarisation des Aktionspotentials (s. a. SCH~TZ 1936). _~hnliehe Ergebnisse besehrieben WEB]~ u. HOLLANDER (1956) am Rattenvorhof. An der Kammermuskulatur ver£ndert Aeh das Membranpotential nicht (HoFF~AN U. SUCKLING 1953).
Diese Befunde veranla~ten uns, die untersehiedliche Aeh-Empfmd- lichkeit der verschiedenen Herzabsehnitte vergleichend zu untersuehen, wobei wir am Frosehherzen nur die Amplitude der Kontraktion, an den Warmblfiterpr~paraten gleichzeitig auch Ruhe- und Aktionspotential registrierten.
* Ausgefiihrt mit Unterstiitzung der Deutschen Forschungsgemeinschaft.

532 R . F . SCI~IIDT:
Methodik _Frosch. Neben Herzen an der Straubkaniile wurden haupts~chlich isolierte
Vorh6fe und Streifenpr~parate der Kammer untersucht (Rana esculanta, Prepara- tion siehe ROT~SCHV~ 1948). Diese hingen in einer MeBkammer aus Plexiglas, die yon O2-gesattigter L6sung mit einer Geschwindigkeit yon 0,2--0,8 cm/sec durch- str6mt wurde. Es wurde mit Rechteekimpulsen von 0,4--2 msee gereizt. Die Kon- traktionen wurden mit einem Hebel auf ein Ru•kymographion iibertragen. Die Zusammensetzung der Ringer-L6sung war in mMol: Na 112,8; K2,7; Ca 1,8; C1 117,3; HCO 3 1,8. Die Raumtemperatur betrug 21°--24 ° C.
Ach (Ach-Chlorid der Firma Hoffmann-La Roche) wurde in Mengen yon 0,001 7--1,0mg vor der Me•kammer in den die Ringer-L6sung zufiihrenden Schlauch gespritzt. Dabei verdiinnt es sich in etwa 1 ml, der innerhalb yon 1--5 sec am Pri~parat vorbeiflie~t.
Hund. Trabekel aus Vorhof und Kammerwand, Papillarmuskeln der Kammer und Purkinje-F~den des Reizleitungssystems wurden aus noch schlagenden Herzen excidiert und in einer MeBkammer aus Plexiglas an einem Ende durch ein Zangchen fixiert, am anderen durch efll. Haar mit der Anode der zur isometrisehen Registrie- rung der Kontraktionsamplitude verwendeten transducer-RShre (RCA 5734) ver- bunden. Alle Pr~parate wurden mit Rechteckimpulsen yon 0,1 msec angetrieben. Die Mel3kammer wurde yon Tyrode-LSsung durchstr6mt (37 ° C), die mit einem Gas- gemisch ges~ttigt war (95% 02, 5~o CO2). Die Zusammensetzung der Tyrode-L6sung war in mMol: Na 153; K2,7; Ca 1,8; Mg 1,05; Cl145; HCOa 13,5; H2POa2,4; Traubenzucker 5,2. Das Aeh wurde wie oben geschildert appliziert.
Die Ableitung des Ruhe- un4 Aktionspotentials erfolgte mit Glascapillaren, die mit 3 Mol KC1-L6sung gefiillt waren. Ihr Spitzendurchmesser war nieht grSBer als 0,5/~, der Widerstand betrug 30--60 M~2. Das abgeleitete Potential wurde fiber einen Kathodenfolger dureh den einen Strahl eines Cossor-Oscillographen registriert (weitere Einzelheiten siehe DUDEL U. TRAUTWEIN 1954). Der zweite Strahl wurde zur Aufzeichnung der Kontraktionsamplitude verwendet.
Ergebnisse Versuche a m Froschherzen
Am Herzen an der Straubkanfi le ] i~ t sich die unterschiedliche Emp-
findlichkeit yon Vorhof und K a m m e r nur schlecht untersuchen, da die U m g e b u n g des Sinusknotens und der AV-Kno ten schon durch sehr nied- rige Ach-Konzen t ra t ionen (10 -1° bis 10 -12 g/ml) geblockt werden. Das
Herz bleibt stehen, obwohl der Schr i t tmacher noch t~t ig ist, s ichtbar an den schwankenden Lichtref lexen in der Sinusgegend. Der Vorhof wird aufgebl~ht und atonisch. In diesem Zustand bleiben Vorhof und K a m m e r
er regbar : h£ufig 15sen K a m m e r e i g e n r h y t h m e n oder ein part iel ler Block den St i l ls tand ab (Abb. 1). Elektr ische Reizung ffihrt immer zu Kon- t r ak t ionen ; erst bei Ach-Konzen t ra t ionen yon mehr als 10 -4 g/ml kann an der K a m m e r kein Reizerfolg mehr registr ier t werden. Chronotrope Effekte t r e t en nie vor dem Block auf, so dal~ die Frequenzminderung nur
am Sinus mi t der Lupe festgestel l t werden kann. Wegen dieser untiber- sichtl ichen Verhiiltnisse ~m ganzen Herzen sind zum Studium der ino- t ropen Effekte des Ach isolierte Pr~para te von Vorhof und K a m m e r besser geeignet.

Acetyleholin-Empfindlichkeit verschiedener Herzabschnitte 533
Abb. 2A zeigt die A c h - W i r k u n g an e inem spon tan schlagenden iso- l ie r ten S inus -Vorhof -Pr~para t ; 0,1 y b r ing t e inen S t i l l s t and fiir e twa eine Minute , obwohl d a s A c h nur wenige Sekunden am P r ~ p a r a t vorbeifloS. E in Sinus-Vorhofblock h a t die Tg t igke i t des P r~pa ra t e s un te rbunden . W i r d es dagegen e lekt r i sch gereizt , so k o m m t es weder bei der gleichen (0,1 y), noch bei der 10fach grSl3eren Dosis zu einem S t i l l s t and (Abb. 2, C und D). Kle ine Mengen Ach, die noch n ieh t zu e inem Block ffihren, w i rken a m spon tan schlagenden und a m elekt r i sch gere iz ten P r g p a r a t gleich s t a rk nega t iv inot rop .
Abb. 1. Spontan schlagendes Herz an der Straubkanfile, Kont rak t ion registr ier t mi t Hebel auf RuB- kymograph. A: zwischen den Pfeilen Ersatz der Ringer-LSsung dutch Ach 10 -23 g/ml in Ringer, minimal wirksame Konzentrat ion. 10 - ~ g/nil Ach wirkt nicht. B : 5 rain spi~ter nach 3nlal igem Wechsel der Ringer-LSsung bei Pfeil Ach 10 - ~ g/ml in Ringer. Zei tmarken alle 15 sec. Versuch yore 23. 1. 1957
Der isol ierte gereizte K a m m e r s t r e i f e n is t gegen Ach 10 - -100fach unempf indl icher als das Vorhofpr~para t (Abb. 2, E--G). U n e r r e g b a r k e i t des P r£pa ra t e s war auch nach 1 mg Ach n ich t zu erzielen. W u r d e n die Versuche in n ich t z i rku l ie render BadelSsung durchgeff ihr t , so pr~gte sich der Unte r sch ied zwischen Vorhof und K a m m e r noch s t a rke r aus. Die min ima l wi rksame Konzen t r a t i on , die eine nega t ive Ino t rop ie hervor- rief, lag be im Vorhofp r~para t regelm~lt ig bei 10 -12, be im K a m m e r s t r e i f e n bei 10 -9 bis 10 -s g/ml.
Beim Vergleieh der wirksamen Konzentrationen an Vorhof und Kammer (z. B. Abb. 2, C u. D mit Abb. 2, E u. F) ist zu bedenken, dab die Kammerwand etwas dicker als die des Vorhofs ist. Der durch Diffusion und Spaltung verursachte Diffusionsgradient kSnnte im Inneren der diekeren Pr~parate zu re]ativ geringerer Aeh-Konzentration fiihren, ein solches Pr~parat seheint dann weniger empfindlich. Die Diffusions- und Spaltungsverh~ltnisse sind nicht zu fibersehen. In der i~ul]ersten Sehicht herrsch~ jedoch unabh~ngig vonder Dieke fast die Aul~enkonzentration. Bei der minimal wirksamen Konzentration wird nur die Kontraktion der ~uBersteu Schichten vermindert werden, diese Konzentrationen lassen sich also vergleiehen. Es ist aber durchaus mSglich, dal3 in Abb. 2, E bei einem diinneren Pr~parat die Hemmung etwas ausgepri~gter wi~re.
Die beschr iebenen Befunde decken sich nur tei lweise m i t dem yon LISS~K u. MARTIN (1940), die zwischen isolie1~em Vorhof und

534 R.F. SCHMIDT:
,,Kammersegment" nur einen Unterschied yon ungef/~hr einer Zehner- potenz beschrieben haben. Ebenso wie sie, im Gegensatz zu L£~czos (1930), fanden wir keine verschiedene Empfindlichkeit des Streifens der Kammerbasis und der Kammerspitze. Allerdings wurden die Befunde von L~.Nczos am Straubherzen mit Hilfe yon Ligaturen erhoben. Es ist mSglich, dal~ bei dieser Methode die Ausgangsbedingungen so stark ver~ndert wurden, da~ der beschriebene Effekt entstand.
Abb. 2. Die Wi rkung yon Ach a u f den spontan schlagenden (A u. B) und den elektrisch gereizten Vorhof (C u. D) , sowie den gereizten Kammers t r e i f en (E--G) des gleichen Herzens. Pr~iparatc in du rchs t r6mte r ~Iel ]kammer, S t rSmungsgeschwindigkei t der Ringer -LSsung 0,4 cm/sec. Bei A, C, E jeweils In j ek t ion von 0,1 ~' Ach, bei B, D, F 1,0 ~ Ach und bei G 10 y Ach. Ze i tmarken alle 15 sec.
Versuch v o m 12. 2. 1957
Versuche am Myokard des Hundes
Die Versuche am Froschherzen lieBen erwarten, daI~ auch die ver- schiedenen Abschnitte des Warmblfiterherzens gegenfiber Ach verschie- dene Empfindlichkeit besitzen. Um neben der inotropen Wirkung des Ach auch seinen Einflul~ auf die die Erregung auslSsenden Vorgiinge zu beobachten, registrierten wir bei den folgenden Versuchen neben der isometrischen Kontraktion auch das Ruhe- und Aktionspotential.
Acetylcholin-Empfindlichkeit verschiedener Herzabschnitte 535
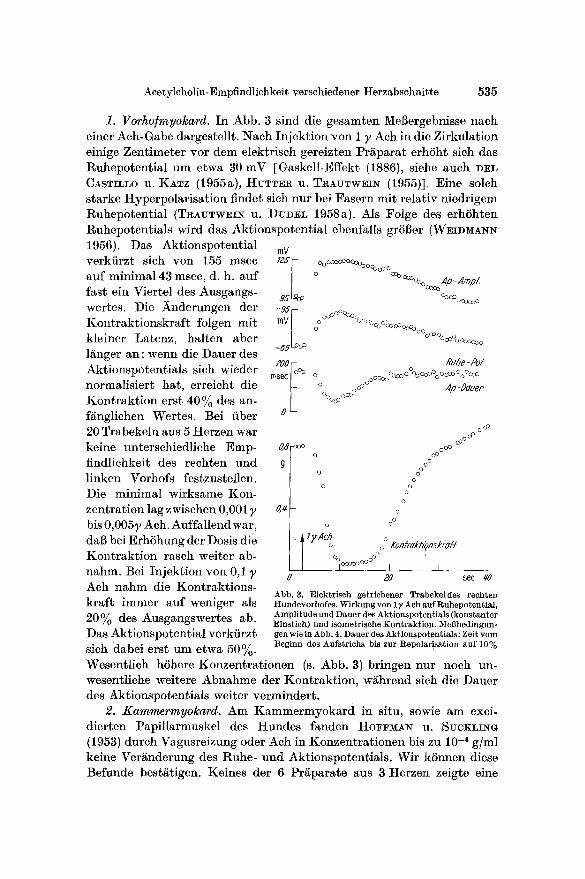
1. Vorho/myolcard. In Abb. 3 sind die gesamten MeSergebnisse hack einer Ach-Gabe dargestellt. Nach Injektion yon I }, Ach in die Zirkulation einige Zentimeter vor dem elektrisch gereizten Pr£para t erhSht sich das Ruhepotential um etwa 30 mV [Gaskell-Effekt (1886), siehe auch ])EL CASTILLO u. KATZ (1955a), HUTTER U. TRAUTWEIN (1955)]. Eine solch starke Hyperpolarisation finder sich nur bei Fasern mit relativ niedrigem l~uhep0tential (TlCAUTWEIN U. DUD]EL 1958a). Als Folge des erhShten Ruhepotentials wird das Aktionspotential ebenfalls gr58er (WEIDMA~I~ t956). Das Aktionspotential verkiirzt sich von 155 msec auf minimal 43 msec, d. h. auf fast ein Viertel des Ausgangs- wertes. Die ~nderungen der Kontrakt ionskraf t folgen mit kleiner Latenz, halten aber l&nger an: wenn die Dauer des Aktionspotentials sich wieder normalisiert hat, erreicht die Kontrakt ion erst 40% des an- f/inglichen Wertes. Bei fiber 20 Trabekeln aus 5 Herzen war keine unterschiedliche Emp- findlichkeit des rechten und linken Vorhofs festzustellen. Die minimal wirksame Kon- zentration lag zwischen 0,0017 bis 0,0057 Ach. Auffallend war, dab bei ErhShung der Dosis die Kontrakt ion rasch weiter ab- nahm. Bei Injektion yon 0,1 7 Ach nahm die Kontraktions- kraf t immer auf weniger als 20% des Ausgangswertes ab. Das Aktionspotential verkfirzt sich dabei erst um etwa 50%.
lnV 7Z5 ~ oOOO~OOoooooooeo~oo~o o~ooooo Ap_Amp Z
95 ~vo %00000000
-95 1 o °°°°°%° I/1V o %COocoOcooo
-65 c° °°°°°°°°°°°°°%°°°°°°
200 Ruhe - P# msec I °° o OoooO%oOOO%ooco%°coo
o o °° °°°°°° ,#p -DG'/./'ep °Oooo oCrJ
O L
~0
g
I I--- I I g 20 sec
o oo coo o°
o o o°°° o o
o o o
Q o ° o
o
o
o o o
7yAch o o o Konlraktl&nskra~
% o o ° ,
I c°° ' j~°° I I J
Abb. 3. Elektrisch getriebener Trabekeldes rechten Hundevorhofes. Wirkung von 17 Ach auf Ruhepotential, Amplitude und Dauer des Aktionspot entials (konstanter Einstich) und isometrisehe Kontraktion. MeBbedingun- gen wie in Abb. 4. Dauer des Ak$ionspotentials: Zeit vom Beginn des Aufstrichs bis zur Repolar!sation auf 10%
Wesentlich h6here Konzentrat ionen (s. Abb. 3) bringen nur noch un- wesentliche weitere Abnahme der Kontraktion, w/ihrend sich die Dauer des Aktionspotentials weiter vermindert.
2. Kammermyokard. Am Kammermyokard in situ, sowie am exci- dierten Papillarmuskel des Hundes fanden HOrFMAI¢ u. SUCKLINO (1953) durch Vagusreizung oder Ach in Konzentrat ionen bis zu 10 -4 g/ml keine Ver/inderung des Ruhe- und Aktionspotentials. Wir kSnnen diese Befunde best/~tigen. Keines der 6 Pr/iparate aus 3 Herzen zeigte eine
536 R.F. SCHMIDT:
meBbare VerKnderung des Ruhe- und Aktionspotentials bei Injektion yon 0,01 7 bis 1 mg Ach.
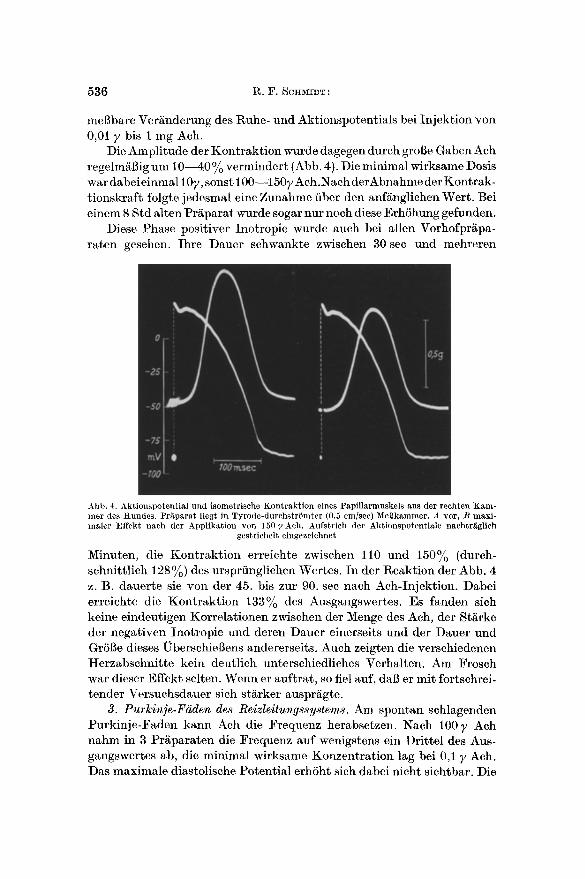
Die Amplitude der Kontrakt ion wurde dagegen durch groBe Gaben Ach regelmi~Big um 1 0 4 0 % vermindert (Abb. 4). Die minimal wirksame Dosis war dabei einmal 10y, sonst 100--1507 Ach.Naeh derAbnahme der Kontrak- tionskraft folgte jedesmal eine Zunahme fiber den anfKnglichen Wert. Bei einem 8 Std alten PrKparat wurde sogar nur noch diese ErhShung gefunden.
Diese Phase positiver Inotropie wurde auch bei allen VorhofprKpa- raten gesehen. Ihre Dauer schwankte zwischen 30 see und mehreren
Abb. 4. Akt iouspoteut ia l und isometr ische Kon t rak t ion eines Papi l larmuskels aus der rechten K a m - m e r des t tundes . Pr~iparat l iegt in Tyrode-durchs t r6mte r (0,5 cm/see) MeBkammer. A vor, /~ max i - maler Effekt nach der Appl ikat ion yon 150 y Ach. Aufs t r ich der Aktionspotent ia le nachtr~glich
gestr ichel t eingezeichnet
Minuten, die Kontrakt ion erreichte zwischen l l 0 und 150% (durch- schnittlich 128 %) des ursprfingliehen Wertes. In der Reaktion der Abb. 4 z. B. dauerte sie yon der 45. bis zur 90. see nach Aeh-Injektion. Dabei erreichte die Kontrakt ion 133~o des Ausgangswertes. Es fanden sich keine eindeutigen Korrelationen zwischen der Menge des Ach, der St~rke der negativen Inotropie und deren Dauer einerseits und der Dauer und GrSl~e dieses l~berschieI~ens andererseits. Auch zeigten die verschiedenen Herzabschnitte kein deutlich unterschiedliches Verhalten. Am Frosch war dieser Effekt selten. Wenn er auftrat, so fiel auf, dab er mit fortschrei- tender Versuchsdauer sich st£rker auspr£gte.
3. Purkin]e-FSden des Reizleitungssystems. Am spontan schlagenden Purkinje-Faden kann Ach die Frequenz herabsetzen. Nach 100 7 Ach nahm in 3 Pr£paraten die Frequenz auf wenigstens ein Drittel des Aus- gangswertes ab, die minimal wirksame Konzentration lag bei 0,1 7 Ach. Das maximale diastolische Potential erhSht sich dabei nicht sichtbar. Die

Acetylcholin-Empfindtichkeit verschiedener Herzabsctmitte 537
Dauer des Aktionspotentials - - gemessen bei durch Reiz kons tan t ge- haltener Frequenz - - v e r k f i r z t sich nach 100 ? Ach um etwa 10~o. Diese Aeh-Wirkungen am Purkinje-Faden sind nieht konstant : an 2 weiteren Pr£paraten £nderte sich auf Aeh selbst die Spontanfrequenz nicht. Entgegen der Darstellung yon TRAUTWEn¢ (1957) wirkt Aeh also zu- mindest bei einem Teil der Purkinje-F/iden. Bei der schwaehen Reaktion der Purkinje-Fiiden wird deutlieh, dab von den Erregnngsvorg/ingen die spontane diastolische Depolarisation auf Ach am empfindlichsten reagiert (s. auch T~AUTWmN U. DUDrL 1958b).
Diskussion
Naeh den erhobenen Befunden reagiert das ganze Froschherz auf Ach wesentlich empfindlicher als das isolierte gereizte Kammermyokard . Eine negative Inotropie der Kammermyokardfaser kann daher, entgegen der bisherigen Interpretat ion, nicht die haupts£chliehste Ursaehe der ver- minderten Kontrakt ion des Iterzens an der Straub-Kaniile bei niedrigen Aeh-Konzentrationen sein. Eine negative Inotropie der Vorhoffasern, die bei diesen Konzentrat ionen mSglieh ist, reicht ebenfalls zur Erkl£rung der Kontrakt ionsabnahme nieht aus, da die Vorh5fe an der Kontrakt ion einen sehr geringen Anteil haben. Es miissen vielmehr die die Kontrakt ion auslSsenden Vorg£nge dureh das Aeh so beeinfluBt werden, dab es zur Kon- t rakt ionsabnahme kommt. So ffihrt z. B. eine teilweise Unterdrfickung der Reizleitung zu einer sehw/~eheren Kontraktion, da die Zahl der er- regten Fasern abnimmt. Wahrscheinlieh wird auf diese Weise die Kon- t rakt ion am Straubherzen bei niedrigenAeh-Konzentrationenvermindert , da wir fanden, dab die Kontrakt ion meist sprunghaft abnimmt und der Stillstand des Herzens bei h5heren Konzentrationen durch vollkommene Unterdrfiekung der l~eizleitung zustande kommt. Dutch die gleiehzeitige Messung des Ruhe- und Aktionspotentials 1/~Bt sieh dieser Einflul3 der Reizleitung auf die Kontrakt ion noeh besser verfolgen: Vagusreiz ffihrt am Vorhof zu abortiven Aktionspotentialen in der Sinusgegend, die nicht oder nur schlecht geleitet werden (HuTTER u. TRAUTWEI~ 1956). Bei dieser Verkniipfung der kontraktionsausl5senden Vorg/inge mit der Kon- t rakt ion des ganzen Herzens stellt sieh die Frage, ob es aueh an der Einzel- laser unter Ach eine Kopplung zwischen der Form des Aktionspotentials und der Kontrakt ion gibt.
BUR(~EN U. TEI~I~OUX (1953) nahmen einen solchen Zusammenhang an: sie waren der Ansicht, dab die verminderte Kontrakt ion unter Aeh durch das verkiirzte Aktionspotential verursaeht sei. Unser Befund, Abnahme der Kontrakt ion des Papillarmuskels bei relativ grol~en Ach- Dosen ohne Ver/inderung des Ruhe- und Aktionspotentials, I/~St eine solehe Annahme zweifelhaft erseheinen, Der gleiche Befund widerlegt aueh die Ansicht yon HOFFMAN U. SVCKL~O (1953), dab die fehlende

538 R . F . SC][~IIDT ."
Ach-Empf ind l i chke i t der K a m m e r m y o k a r d m e m b r a n durch eine Imper - meabilit /~t der Membran fiir Ach bedingt sei. Vielmehr mug es sich um ein Feh len jener spezifischen Ach-Recep to ren (DI~L CASTILLO n. KATZ 1955 b) der M e m b r a n handeln, die z. B. am Vorhof die Ka l ium-Pe rmeab i - l i t i i t erhShen. Eine enge K o p p l u n g der Ak t ionspo ten t i a ldaue r und der K o n t r a k t i o n wird auch dadurch unwahrscheinl ich, dab der Phase posi t i - ver Ino t ropie , die bei unseren Versuchen auf die negat ive folgt, keine Ver/i, nderung des Ruhe- und Akt ionspo ten t i a l s an Vorhof und K a m m e r entspr icht .
Bisher sind diese Phasen positiver Inotropie durch Ach nur ohne vorhergehende negative oder unter dem Einflul~ yon Pharmaka, insbesondere Atropin, beschrieben worden (MIDDLETON et al. 1956). Ohne Einflul3 yon Pharmaka erreichte McEwu~¢ (1956) am Rattenvorhof durch kurzfristige Vagusreizung ehle zweiphasisch ver- laufende Inotropie, verbunden mit Frequenzsteigerungen in der positiven Phase. Er vermutet mit JOURDAC¢ U. NOWAK (1936), KABAT (1939) undMIDDLETON et al. (1949) Acceleransfasern in den Nn. vagi. Da wir aber auch nach Ach positive Inotropie finden, liegt es nahe anzunehmen, dab der Effekt dutch Freisetzung yon Adrenalin aus cholinergen Ganglien verursacht wird. Allerdings haben M~DDLETO:N eta | . (1956) in Katzenpapillarmuskeln, die positive Inotropie zeigten, bei anschliel3ender histo- logischer Untersuchung kein chromaffines Gewebe gefunden. Eine andere Erkl~- rungsmSglichkeit bietet die ErhShung des Ruhepotentials nach Ach durch die erhShte Kalium-Leitf/~higkeit (TRAUTWEIN U. DUDEL 1958a): Fasern, die dutch niedriges Ruhepotential unerregbar geworden sind, kSnnen durch die Hyperpolarisation unter Ach wieder erregbar werden und anschlieSend die Inotropie verst/~rken. Solche ,,erregende" Wirkungen des Ach sind auch yon BuRet u. VANE (1949), MARSHALL U. VAUGttAlvlWILLIAMS(1956) und TRAUTWEIN U. DUDEL(1958b) (friihere Literatur siehe ROTHSCHU]~ 1954) beschrieben worden.
Gegen diesen Mechanismus sprieht , dab wit die Phase pos i t iver Ino t rop ie an allen P r~pa ra t en finden, auch den friseh excid ier ten und besonders am Papi l la rmuske l , dessen Ruhepo ten t i a l durch Ach n icht hyperpo la r i s i e r t wird. Es bl iebe also zu prfifen, ob n icht Aeh an der Herzmuske l faser nach der nega t iven eine , ,eehte" posi t ive Ino t rop ie bewirk t , die aber nur nach kurzfr is t iger Aeh-Appl ika t ion au f t r i t t . Mit den vor l iegenden Versuehen ]£Bt sieh diese F rage nicht endgii l t ig ent- scheiden.
Zusammeniassung An Pr/~paraten verschiedener t t e r zabschn i t t e des Frosch- und Hunde-
herzens wurde die Ach -Wi rkung auf die K o n t r a k t i o n und das Membran- po ten t i a l untersucht . Dabe i f anden wir:
1. Die min imal wi rksame K o n z e n t r a t i o n lag beim ganzen Frosch- herzen bei 10 -14 bis 10 -12, bei den isol ier ten Vorh6fen bei 10 -12 und beim K a m m e r s t r e i f e n bei 10 -9 bis 10 -s g/m] Ach.
2. Bei 10fach h5herer als der min imal wi rksamen K onz e n t r a t i on ble iben das isolierte Froschherz und das isol ierte S inus-Vorhof -Pr~para t regelm/£i3ig du tch l~eizlei tungsblock stehen.

Acetylcholin-Empfindliehkeit verschiedener Herzabschnitte 539
3. Am Vorhoftr~bekel des Hundes t re ten gleichzeitig mi t der nega t iven Inotropie Hyperpolar i sa t ion des Ruhe- und Verkiirzung des Aktions- potent ia ls auf. Am Papi l la rmuskel wird die Kont r~kbion erst dureh 104 bis 105lath h5here Konzen t r a t i on verminder t , ohne dab sieh Ruhe- u n d Akt ionspoten t ia l /£ndern .
4. Bei allen Warmbli i terpr /~paraten ist die negat ive Inot ropie yon einer Phase posit iver Inot ropie gefolgt. Diese dauer t 30 sec bis mehrere Minuten, die K o n t r a k t i o n erreicht dabei 110- -150% des Ausgangswertes.
Herrn Dozent Dr. TRA~TW~.I~¢ danke ich herzlich f'tir die Themenstellung, seine Unterstiitzung und die ~berlassung des Versuchsstandes.
Literatur BURGEN, A. S. V., and K. G. TERROUX: On the negativ inotropic effect in the cat's
auricle. J. Physiol. (Lond.) 120, 449--464 (1953). BURN, J. H., and J. R. VANE: The relation between the motor and the inhibitor
action of acetyleholine. J. Physiol. (Lond.) 108, 104--115 (1949). DEL CASTILLO, J., and B. KATz: The membrane potential changes in the frog's heart
produced by inhibitory nerve impulses. Nature (Lond.) 175, 1035 (1955a). " DEL CASTILLO, J., and B. KATZ: On the localization of acetyleholine receptors. J.
Physiol. (Lond.) 128, 157--181 (1955b). :DUDEL, J., u. ~V. TRAUTWEIN: Das Aktionspotential und Mechanogramm des Herz-
muskels unter dem EinfiuB der ])ehnung. Cardiologia (Basel) 25, 344--362, (1954).
GASKELL, ~V. H. : The electrical changes in the quiescent cardiac muscle which accompany stimulation of the vagus nerve. J. Physiol. (Lond.) 7, 451--452 (1886).
HOFFI~N, B.F., and E. E. SUCKLING: Cardiac cellular potentials: effect of vagal stimulation and acetyleholine. Amer. J. Physiol. 173, 312--320 (1953).
HUTTER, 0. F., and W. T~UTWEIN: Effect of vagal stimulation on the sinus veno- sus of the frog's heart. Nature (Lond.) 176, 512 (1955).
HUTTER, O. F., and W. TRAUTWEIN: Vagal and sympathetic effects on the pace- maker fibers in the sinus venosus of the heart. J. gen. Physiol. 39, 715--733 (1956).
JOURnAl, F., and S. J. C. •OWAK: ]~tude experimental chez le chien des fibres eardio-ace61~ratriees du vague. Arch. int. Pharmaeodyn. 53, 121--135 (1936).
KARAT, M. : The ca~'diac accelerator fibers in the vagus nerve of the dog. Amer. J. Physiol. 128, 246--257 (1939).
L£~cczos, A. : ~ber die Wirkung des Vagusstoffes und Aeetylcholins ~uf die isolierte Kammer des Froschherzens. Pfltig. Arch. ges. Physiol. 225, 710---720 (1930).
LING, G., and R. W. GERARD: The normal membrane potential of frog sartorins fibers. J. cell. eomp. Physiol. 34, 383--396 (1949).
LISS~K,]~., u. J. ~RTIN: Beitr/ige zum Wirkungsmechanismus des Atropins. Naunyn-Schmiedeberg's Arch. exp. Path. Phaxmak. 196, 558--565 (1940).
Lo~wI, O.: Uber humorale Ubertragbarkeit der Herznervenwirkung. I. Mittlg. Pfliig. Arch. ges. Physiol. 189, 239--242 (1921).
Lo~wI, 0., u. E. lqAVRAT~L: Uber humorale t?bertragbarkeit der Herznerven- wirkung. X. Mittlg. Uber das Schicksal des Vagusstoffes. Pfliig. Arch. ges. Physiol. 214, 678--688 (1926).
McEw~¢, L. M. : The effect on the isolated rabbit heart of vagal stimulation and its modification by cocaine, hexamethonium and ouabaine. J. Physiol. (Lond.) 131, 678---689 {1956).

540 R . F . SC~IDT : Acetylcholin-Empfindlichkeit verschiedener Herzabschnitte
MARSHALL, J. M., and E. M. V.~VGI~AM WILLIAMS: Pacemaker potentials. The exci- tation of isolated rabbit auricles by Acetylcholinc at low temperatures. J. Physiol. (Lond.) 181, 186--199 (1956).
MIDDLETON, S., H. I-I. MIDDLETON and J. ToI~A: Adrenergic mechanism of vagal cardio-stimulation. Amer. J. Physiol. 158, 31--37 (1949).
MIDDLETON, S., C. OBERTI, R. PRAGER and H. H. MIDDLETON: Stimulation effect of acetylcholine on the papillary myocardium. Acta physiol, lat.-amer. 6, 82--89 (1956).
ROTHSCHVH, K. E. : Ober die Anwendbarkeit der Differenzanalyse und der Vektor- analyse auf das indirekt abgeleitete Elektrocardiogramm. Arch. Kreisl.-Forsch. 14, 155--204 (1948).
ROTHSCHVH, K. E.: Vorkommen und Funktionen des Acetylcholins im Herzen. ]Klin. Wschr. 32, 1--7 (1954).
SCRi2TZ, E. : Elektrophysiologie des Herzens bei cinphasischer Ableitung. Ergebn. Physiol. 88, 493--620 (1936).
T~AVTWEI~, W. : in K. St"ANt, RhythmusstSrungen des Herzens, S. 65. Stuttgart: Georg Thieme 1957.
TRAUTWEIN, W., u. J. DUDEL: Zunl Mechanismus der Membranwirkung des Acetyl- cholin an der Herzmuskelfaser. Pfliig. Arch. ges. Physiol. 266, 324--334 (1958 a).
TRAVTWEIN, W., u. J-. DUDEL: Hemmende und ,,erregende" ~rirkungen des Aeetyl- cholin am Warmbliiterherzcn. Zur l%age der spontanen Erregungsbildung.
Pfliig. Arch. ges. Physiol. 266, 653--664 (1958b). WE~]3, J. L., and P. B. HOLLANDER: The action of aeetylcholin and epinephrine on
the cellular membrane potentials and contractility of rat atrium. Circular. Res. 4, 332--336 (1956).
WEIDMANN, S. : Elektrophysiologie der Herzmuskclfaser. Bern, Stuttgart: Hans Huber 1956.
ROBERT F. SCm~IDT, Physiologisches Institut der Universit~t Heidelberg, Akademiestr. 3





























