Embed Size (px)
Citation preview

Zeitschrift ffir
Lebensmittel-Untersudmng und-Forschung 102. BAND DEZEMBER 1955 H E F T 6
Uber die Proteolyse der Fleischproteine H I . Mitteilung
Der Einflull yon Bindegewebs-Kollagen auf die Proteolyse des Fleisches*
Von
REINER HAMM
MitteiIung aus dem Chemisch-Physikalischen Institut der Bundes/orschungsanstalt ]i~r Fleischwirtscha/t, Kulmbach
Mit 8 Textabbildungen
(Eingegangen am 5. Juli 1955)
Es ist bekannt, dab Fleisch durch proteolytische Enzyme um so schwerer abgebaut wird, je mehr Bindegewebe es enth~l~, da die Proteine des Bindegewebes, Kollagen und Elastin, gegen Proteinasen resistenter sind Ms die des Muskelgewebes. Auf Grund dieser Tatsache war yon uns eine Methode zur Bindegewebsbestimmung in Fleisch und Fleischwaren entwickelt worden 1.
I m AnschluB an diese Arbeiten interessierte uns die Frage, ob die Verdaulichkeit 4er Muskelproteine dureh Pepsin-Salzs~ure yon Sehnenkollagen in irgendeiner Weise beeinfluBt wird. In Anbetracht dessen, dab die Enzyme des Verdauungstraktes st~ndig vor die Aufgabe gestellt werden, Fleisch in Gegenwart yon Bindegewebe zu verdauen, n immt as Wunder, dab in der Literatur eine experimentelle Bearbeitung dieses Problems nicht zu linden ist.
Die L5sung dieser Frage wird durch die sehr komplexe Natur des Systems Kol lagen- -F le i sch- -Peps in - -S~lzs~ure erschwert: Die Sehnen quellen w~hrend des Fleischabbaues, wobei ein Tell der Enzyml5sung, aber auch ein Tell der Abbau- produkte yon dem Bindegewebe unter Quellung aufgenommen werden. Gleiehzeitig wird durch Sehnenzusatz der p~-Wert der Pepsinl5sung nach h5heren Werten hin verschoben. Neben dem allgemeinen EinfluB der Quellung und der p.-Verschiebung eine spezifische Wirkung der Sehnen auf die Proteolyse des Muskelgewebes erkennen zu k5nnen, erscheint um so sehwieriger, Ms auch das Kollagen einen gewissen Abbau erleidet, werm dieser aueh welt langsamer als derjenige des Fleisches vonstat ten geht.
Zun~chst kam es darauf an, die einzelnen Effekte, n~mlich die Proteolyse der Einzelsubstanzen (Fleiseh und Sehnen), die Quellung und die p.-Versehiebung getrennt voneinander zu untersuchen. Ers~ dann konnte man an die Untersuchung yon Gemischen aus Fleisch und Sehnen herangehen, wobei zuni~ehst durch Wahl
niedriger Substr~tkonzentration die Wirkung der Quellung weitgehend auszusch~lten war, um schlieBlich such zu h5heren Konzentr~tionen fiberzugehen. I m letzteren Falle liegt ein System vor, bei dem durch den hohen Gehal~ an Sehnen-Koll~gen der
* Fr~u Dr. A ~ L I ~ S BSm~ danke ich ffir ihre wertvolle Mitarbeit. G~Av, R., u. R. I-IA~xta: Diese Z. 93, 201 (1951); 98, 425 (1954).
Z. Lebensmittel-Unters. u. -Forsch., Band 102 27

418 R. H A ~ :
grSl]te Teil des Wassers durch Quel lung , , immobfl is ier t" ist. Ob Pro te inasen un te r solchen U m s t ~ n d e n ebenso wirken, wie in ,,frei bewegl ichem" Wasser , da r i ibe r scheint wenig b e k a n n t zu sein. Phys io logiseh is t Pine solche F rage yon a l lgemeinerem Interesse , da z. B. be i den enzymat i s chen Prozessen im Zel lp lasma Pin groBer Teil der Fl i i ss igkei t durch Quellung der Kol lo ide gebunden ist.
F i i r unsere Un te r suchungen w~hl ten wir EnzymlSsungen yon re l a t iv n iedr iger K o n z e n t r a t i o n (0,1~o Pepsin) , u m eine e twaige A d s o r p t i o n oder t t e m m u n g des E n z y m s u m so deut l icher in Ersche inung t r e t en zu lassen. Die Peps in -Hydro ly se geh t bekann t l i eh in zwei Phasen vor sich 1. W~hre nd der e rs ten Phase werden die Pro te inmolek i i le zu grol~en, durch Trichloressigs~ure noch f~l lbaren Ol igopept iden abgebau t . Eine t t y d r o l y s e zu freien Aminos~uren wird d u t c h Peps in-Salzs~ure n icht bewirk t . Die ) [nderung des Res t -S t i cks to f f s s te l l t ein wesent l ich gi inst igeres Mal~ ffir den Grad des Peps inabbaues dar , als die re la t iv geringe )~nderung des Amino-S t i ck - stoffs. Offenbar wird bere i ts dureh die Aufspa l tung weniger P e p t i d b i n d u n g e n die F~ l lba rke i t du reh Trichloressigs~ure aufgehoben.
Methodik Untersuchungsmaterial
F1eisch: Von anh~ngenden Sehnen und Fet t befreiter, 5 Tage abgeh~ngter l~indermuskel, zweimal durch den Wolf gedreht.
Sehnen: Von anh~ngendem Fet t und Muskelgewebe befreite Achillessehne des Rindes, Pin- real durch den Wolf gedreht.
In je 1 g des MateriMs wurde nach Veraschung mit Selen-Schwefels~ure der Stiekstoffgehalt nach K J E L ] ) ~ ermittelt,
Pepsin- Proteol yse ]:)us Substrat wurde auf 1 mg genau in 100 mLErlenmeyerkolben eingewogen und mit Enzym-
lSsung versetzt. Diese enthielt, wenn nicht im folgenden ausdrficldich anders vermerkt, 0,1% Pepsin (Merck) in 0,05 n-HCI (p~ 1,05). Naeh Inkubation im Brutschrank bei 37 ° C wurde abfiltriert und das Filtratvolumen gemessen, ttierzu wurde das in Vorversuehen ermittelte, yore Papierfilter aufgenommene Fliissigkeitsvolumen addiert. Der p~-Wert yon Filtrat und Rfiekstand wurde elektrometrisch (Glaselektrode) gemessen.
Ermittlung des Eiweifl-Abbaues Im Ffltrat wurde der Gesamt-N und - - nach F~llung mit einem gleichen Volumen 15%iger
Trichloressigs~ure - - der Rest-N in tier fib]ichen Weise bestimmt. Amino-N: 2--5 ml des Filtrates win, den in ein Zentrifugenglas abpipettiert und mit einer
Kupferphosphat-Suspension 2 auf 10 ml aufgefiillt. Naeh 5 rain wurde zentrifugiert, 2 ml des Zentrffugates in pine LSsung yon 2 g K J in 20 ml 10%ige Essigs~ure pipettiert und sofort mit 0,005 n-Natrinmthiosu]fatlSsung titriert miter Verwendung Pines besonderen St~rkeindicators 3.
Untersuchung der QueUungs/li~ssig]~eit Aussalzen: ]:)us abfiltrierte, gequollene Material wurde in einem Zentrifugenglas mit festem
NaC1 durchgeknetet, nach 1/: Std. die ausgeschiedene Flfissigkeit abzentrifugiert, der Zentrffugen- rfickstand mit 5 n-NaC1-LSsung durchgeknetet, nochmals zentrifugiert und der Rfiekstand lest ausgeprefit. Zentrifugate und Prel~saft wurden vereinigt, auf 50 ml aufgefiillt, filtriert und der N-Gehalt des Filtrates nach Veraschen mit Selen-Schwefe]si~ure naeh KZELI)~I~L ermittelt.
Extraktion mit heiBem H30: Das abfiltrierte, gequollene Material wurde mit etwa 200 ml tt~O 2 Std. unter Rfickfinl~ gekoeht, der Rfickstand abffltriert, in ein Zentrifugenglas fibergeffihrt, zentrifugiert, mit H20 gewaschen, nochmals zentrifugiert und im Zentrffugierriiekstand der Stick- stoffgehalt ermittelt.
Berechnungsweise Die proteolytisch nicht ~bgebauten Fleischproteine (a) blieben naeh dieser Extraktion
ungelSst, au~erdem allerdings noch ein sehr kleiner, aus dem Bindegewebe stammender Anteil, der
1 DESNUELLE, P., M. ROVER:EY U. :B. BONJOUR: Biochim. et Biophysica Acta 5, 116 (1950). SC~OEDEE, W. A., L. M. KAY u. R. S. MILLS: Analyt. Chemistry 22, 760 (1950).
3 POPE, C. G., u. M. F. STEVENS: Bioehemic. J. 38, 1070 (1939).
Uber die Proteolyse der Fleischproteine. I I I 419
sich durch entsprechende Behandlung des Riickstandes einer Sehnenproteolyse (ohne Fleiseh) ermitteln liel~ (b). Auf Grund dieser Daten und des in Pepsin-Salzsaure nieht, in heil]em Wasser aber fast quantitativ 15sliehen Bindegewebs-Stiekstoffes des Fleisehes (ermittelt dutch Pepsin- abbau yon Fleisch allein) (c) l~l~t sich die Menge des in einem Fleisch-Sehnen-Gemisch nicht abgebauten FleiseheiweiBes (d) berechnen:
c -7 (a - - b) = d (a, b, c, d = Stiekstoffwerte).
Dureh Subtraktion dieses d-Wertes vom Gesamt-Stickstoffgehalt des eingesetzten Fleisehes erhalt man den in LSsung gegangenen Fleisch-ST (e). Hierzu kommt nun noeh der aus den Sehnen proteolytiseh gelSste Stickstoff (/), der sich dureh Proteolyse der Sehnen allein ermitteln lgl~t. Der in der Quellungsflfissigkeit vorhandene Gesamt-N (g) erreelmet sich unter I-Iinzuziehung des experimentell gefundenen Ffltrat-Stiekstoffs (Filtrat der Pepsin-Hydrolyse) (h):
(e ~-/) - - h = g (e, f, g, h = Stickstoffwerte).
Bezeichnungen ,,LSslichkeit" ~ Filtrat-N in % des eingesetzten Substrat-N, ,,Peptidabbau" = Rest-N in % des Filtrat-N, ,,Amino-N" ~ Amino-N in % des Ffltrat-N, ,Substratkonzentration" in % = g Substrat/100 ml Enzyml6sung.
Ergebnisse 1. Die Proteolyse des Muskelgewebes durch Pepsin-Salzsiiure
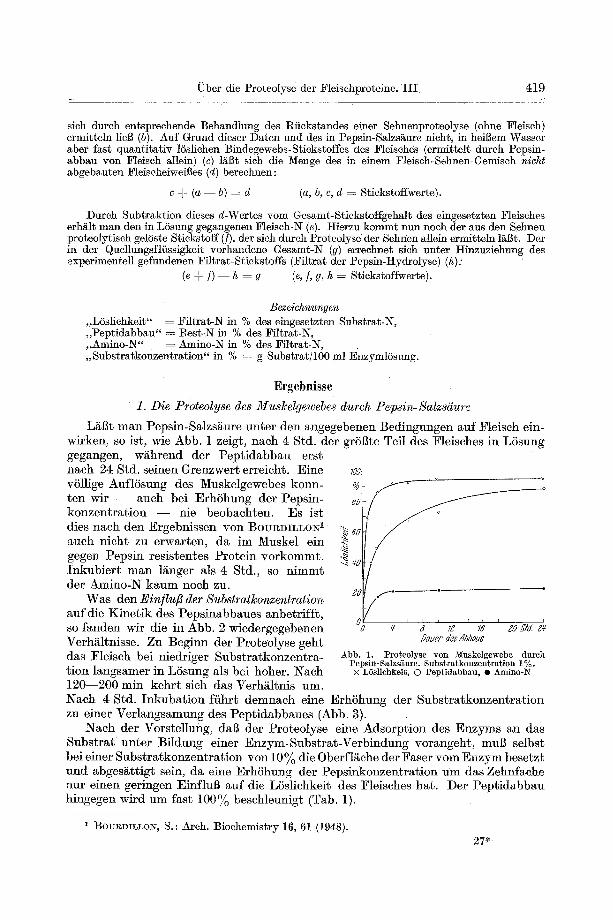
L£1~t m a n Peps in-Salzs~ure un te r den angogebenen Bedingungen au f F le isch ein- wirken, so ist, wio Abb. 1 zeigt, naeh 4 Std. der grSl~te Toil des Fle isches in LSsung gegangon, wghrend dor P e p t i d a b b a u erst nach 24 Std. soinen Grenzwort erreieht . E ine vSllige Auf l6sung des Muskelgewebes konn- t en wi t - - auch boi E rh6hung dot Popsin- konzen t r a t i on - - nie beobachton. Es is t dies nach den Ergebnissen yon BOURDILLON 1 auch n icht zu erwarton, da im Muskel oin gegen Peps in res is ten tes P ro t e in v o r k o m m t . I n k u b i e r t m a n 1/~nger als 4 Std. , so n i m m t der Amino -N k a u m noch zu.
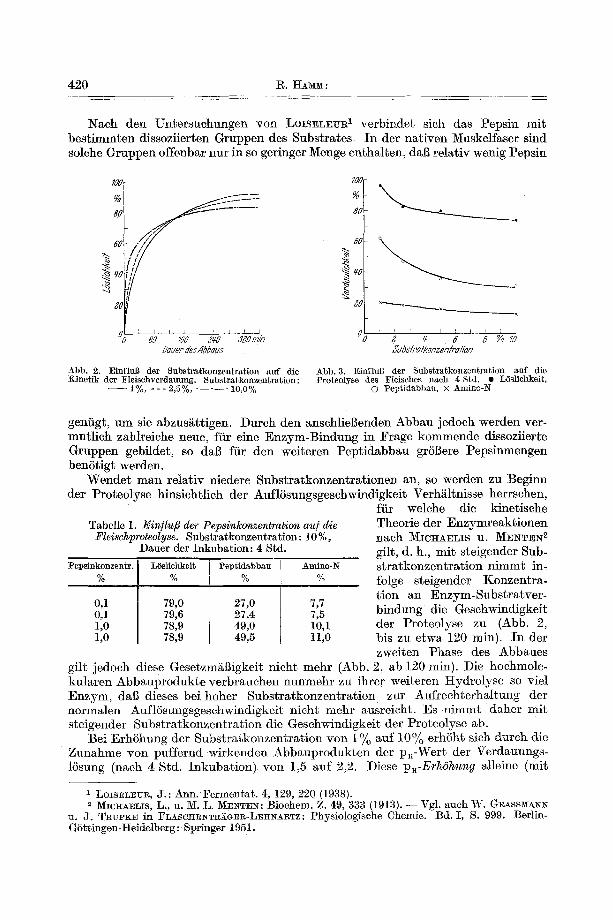
W a s don Einflufi der Substratkonzentration a u f d i e K i n e t i k dos Peps inabbaues anbet r i f f t , so fanden wir die in Abb. 2 wiodergegebenen Verhifltnisse. Zu Beginn der Pro teo lyse geh t das F le iseh bei n iedr iger Subs t r a tkonzen t r a - Lion l angsamer in LSsung als bei hoher. Nach 120--200 rain keh r t sieh das Vorh/~ltnis um.
7O5
o o t/ 3 72 7# g0 Sld ZL/ Dauer des /hbau~
Abb. 1. Proteotyse yon .~uskelgewebe dutch Pepsin-Salzsiiure. Substratkonzen&ration 1%, × LSslichkeit, O Peptidabbau, • Amino-~
Nach 4 Std. I n k u b a t i o n fiihrL demnaeh eine ErhShung der Subs t r~ tkonzen t r a t ion zu einer Ver l~ngsamung des P e p t i d a b b a u e s (Abb. 3).
Naeh der Vorstel lung, dab der Pro teo lyse eine Adsorp t ion des E n z y m s an das S u b s t r a t un te r Bi ldung einer E n z y m - S u b s t r a t - V e r b i n d u n g vorangeht , muB selbst bei e iner S u b s t r a t k o n z e n t r a t i o n von 10% die Oberf lgehe der Fase r vom E n z y m bese tz t und abgesi~ttigt sein, da eine ErhShung der Peps inkonzen t r a t ion Um das Zehnfache nur einen ger ingen EinfluB auf die L6sl ichkei t des Fleisches hat . Der P e p t i d a b b a u hingegen wird um fast 100~o beschleunigt (Tab. 1).
1 BO~RDILLO~, S. : Arch. Biochemistry 16, 61 (1948). 27*
420 R. I-I.A~M:
N a e h den Un te r suehungen von LOISELEUg x ve rb inde t sieh das Peps in mi t b e s t i m m t e n dissozi ier ten Gruppen des Subs t ra tes . I n der na t iven Muskelfaser s ind solehe G r u p p e n offenbar nur in so ger inger Menge en tha l t en , dab re la t iv wenig Peps in
700[ 70~
% . . . . . . . :---= %
60- /, ~. 85
~ . ~,u " I . ~ ~'~
'~ so I ' ~ zo
/71 ; I ~ I I I I I. Oi -0 80 7#0 ZgO 3ZO FI7t/7
Dauer des/thbaus
~ B
,.~ub."t/'a/konzen/m//o,7
Abb. 2. Einflufl der Substratkonzentration auf die Kinetik der Fleischverdauung. Substratkonzentration:
- - 1%, - - - 2,5% - . . . . 3_0,0%
Abb. g. EinfluB der Sttbstratkonzentration auf die Proteolyse des Fleisches nach 4 Std. • LSslichkelt,
© Peptidabbau, x Amino-N
gentigt , u m sic abzus~t t igen. Durch den anschl ieBenden A b b a u jedoeh werden ver- mut l i eh zahlre iehe neue, fi ir e ine E n z y m - B i n d u n g in F r a g e k o m m e n d e dissozi ier te Gr uppen gebildet , so dab fiir den wei te ren P e p t i d a b b a u grSgere Peps inmengen benSt ig t werden.
W e n d e t m a n r e l a t i v n iedere S u b s t r a t k o n z e n t r a t i o n e n an, so werden zu Beg inn der P ro teo lyse hinsieht l ieh der Auf lSsungsgeschwindigkei t Verh/i l tnisse herrsehen,
ffir welehe die k inet isehe Tabelle 1. Ein/luf i der Pepsinkonzentration au/ die
Eleischproteolyse. Substratkonzentration: 10%, Dauer der Inkubation: 4 Std.
Pepsinkonzentr. LSslichkeit Peptidabbau Amino-N % % % %
0,1 0,1 1,0 1,0
79,0 79,6 78,9 78,9
27,0 27,4 49,0 49,5
7,7 7,5 10,1 11,0
Theorie de r E n z y m r e a k t i o n e n naeh M~ICI-IAELIS n. ~¢[ENTEN 2 gilt , d. h., m i t s te igender Sub~ s t r a t k o n z e n t r a t i o n n i m m t in , folge s te igender K onz e n t r a - t ion an E n z y m - S u b s t r a t v e r - b i n d u n g die Gesehwindigkei t der P ro teo lyse zu (Abb. 2, bis zu e twa 120 min). I n der zwei ten Phase des A bba ue s
gri t jedoch diese GesetzmaBigkei t n ich t mehr (Abb. 2, ab 120 rain). Die hochmole- ku l a r en A b b a u p r o d u k t e ve rb rauchen nunmehr zu ihrer wei te ren H y d r o l y s e so viel E n z y m , dab dieses be i hoher S u b s t r a t k o n z e n t r a t i o n zur Auf rech te rha l tung der no rma len Auf lSsungsgeschwindigkei t n ich t mehr ausreicht . Es n i m m t dahe r mi t s te igender S u b s t r a t k o n z e n t r a t i o n die Geschwindigkei t der P ro teo lyse ab.
Bei E rhShung der S u b s t r a t k o n z e n t r a t i o n y o n 1 °/o ~uf 10% erhSht sieh durch die Zunahme yon puf fe rnd w i r k e n d e n A b b a u p r o d u k t e n der p n - W e r t der Verdauungs- 15sung (nach 4 Std. I n k u b a t i o n ) yon 1,5 au f 2,2. Diese p H - E r h S h u n g alleine (mit
1 LOISELEUR, J. : _Ann. Fermentat. 4, 129, 220 (1938). 2 MIC~ELIS, L., u. M. L. MENTE~: Biochem. Z. 49, 333 (1913). - - Vgl. auch W. Gt~ASS~AN~ ~
U. J. TI~UPKE in FLASCHENTRXGER-LEHNAt~TZ: Physiologische Chemie. Bd. I, S. 999. Berlin- GOt~ingen-tteidelberg: Springer 1951.
Ober die Protoolyse der Fleischproteine. III 421
NaOIt eingestellt) hat, wie Tab. 2 zeigt, auf die Proteo- lyse nur einen geringen Ein- flug.
2. Die Proteolyse des Bindegewebes ( Sehnen) durch
Pepsin- Salzsiiure
Bei niedriger Substratkon- zentration (1%), bei welcher
Tabelle 2. Ein/lufi des pK-Wertes au/ den Pepsin-Abbau des Muskels. Substratkonzentr~tion: 2,5 %,
Bauer der Inkubation: 4 Std. Filtrat-p, L6slichkeit Peptidabbau
naeh Inkubation % %
1,50 1,83 2,20
91,8 91,2 87,9
44,6 42,0 39,6
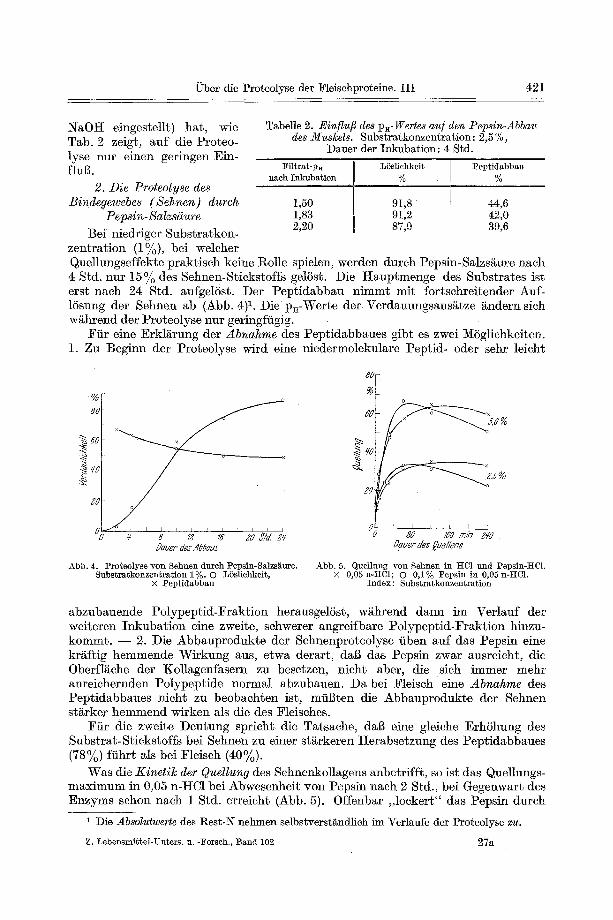
Quelhmgseffekte praktiseh keine l~olle spielen, werden durch Pepsin-Salzs/~ure naeh 4 Std. nur 15% des Sehnen-Stiekstoffs gelSst. Die I-Iauptmenge des Substrates ist erst nach 24 Std. aufgelSst. Der Peptidabbau nimmt mit fortschreitender Auf- 15sung der Sehnen ab (Abb. 4)L Die p~-Werte der Verdauungsans~tze ~ndern sich w/thrend der Proteo]yse nur geringfiigig.
Fiir eine Erklarung der Abnahme des Peptidabbaues gibt es zwei M6glichkeiten. 1. Zu Beginn der Proteolyse wird eine niedermolekulare Peptid- oder sehr leicht
% 80
ZU
#auer des Abbaus
Abb. 4. :Proteolyse yon Sehnen dureh Pepsin-Salzs~ttre. Substratkonzentration 1%. O LSslichkeit,
x Peptidabbau
~o o
<,4,o %
5% 20
i r [ I
80 1#0 rain Jr# Dauer des ~ue//ens
Abb. 5. Quellung yon Sehnen in HCl und Pepsin-]tCL × 0,05 n-tiC1; O 0,1% Pepsin in 0,05 n-tiC1.
Index: Substratkonzentration
abzubauende Polypeptid-Fraktion herausgelSst, w~hrend darm im Verlauf der weiteren Inkubation eine zweite, schwerer angreifbare Polypeptid-Fraktion hinzu- kommt. - - 2. Die Abbauprodukte der Sehnenproteolyse iiben auf das Pepsin eine kr~ftig hemmende Wirkung aus, etwa derart, dag das Pepsin zwar ausreicht, die Oberfi~che der Kollagenfasern zu besetzen, nicht aber, die sich immer mehr anreichernden Polypeptide normal abzubauen. Da bei F]eisch eine Abnahme des Peptidabbaues nicht zu beobachten ist, miil3ten die Abbauprodukte der Sehnen starker hemmend wirken als die des Fleisches.
F/Jr die zweite Deutung spricht die Tatsache, dab eine gleiche Erh6hung des Substrat-Stickstoffs bei Sehnen zu einer st~rkeren tIerabsetzung des Peptidabbaues (78%) fiihrt als bei Fleisch (40%).
Was die Kinetil~ der Quellung des Sehnenkollagens anbetrffft, so ist das Que]lungs- maximum in 0,05 n-tiC1 bei Abwesenheit yon Pepsin nach 2 Std., bei Gegenwart des Enzyms schon nach 1 Std. erreicht (Abb. 5). Offenbar ,,10ekert" das Pepsin durch
1 Die Absolutwerte des Rest-N nehmen selbstverst~ndlich im Verlaufe der Proteolyse zu.
Z. Lebensmi~tel-Unters. u. -Forsch., Band 102 2 7 a
422 1~. H x ~ :
beginnende Proteolyse die Struktur des Sehnenkollagens auf und bewirkt so eine r~sehere Quel]nng. Das Ausmal~ der Quellung jedoeh wird dureh Pepsin nieht ver- ~ndert, Die bei l~ngerer Inkubation erfolgende Quellungsabnahme (d. h. die Bildung eines Maximums) ist in Gegenwart yon Pepsin welt deutlieher als ohne ~ und auf die enzym~tisehe ,Verflfissigung" zuriickzuffihren.
Die p~-Werte des gequollenen Kollagens und der Flfissigkeit (HC1 bzw. Pepsin- HC1) gleiehen sich im Verlaufe der Que]lung raseh aus und sind n~eh 4 Std. einander gleieh.
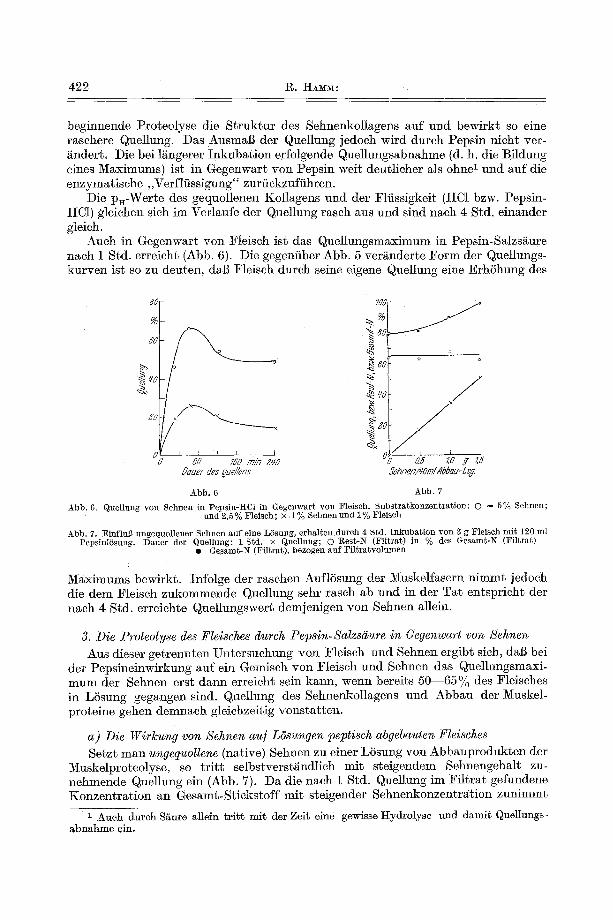
Auch in Gegenwart yon Fleiseh ist das Quellungsm~ximum in Pepsin-Salzs~ure nach 1 Std. erreieht (Abb. 6). Die gegen/iber Abb. 5 ver~nderte Form der Quellungs- kurven ist so zu deuten, d~l~ Fleisch dureh seine eigene Quellung eine ErhShung des
8O
~y
706
~ 65
7~'0 m/k/ #YO D~uer d~ ~udl~ns
Abb. 6
.____...¢.J/'J"
o o
7// I P i
O.5 gO g 7,5 Se~en/¢Om/Abbau-l~y
Abb. 7
Abb. 6. Quellung yon Sehnen in Peps in-HCl in Gegenwar t yon Fleisch. Subs t ra tkonzen t ra t ion : O = 5% Sehnen; trod 2 ,5% Fleisch; x . 1% Sehnen und 1 % Fleisch
Abb. 7. ]~influl3 ungequol]ener Sehnen h u f eine LSsung, erhal~en d t t rch 4 Std. I n k u b a t i o n yon 3 g Fleisch m i t 120 ml :PepsinlSsung. Dauer der Quel lung: 1 Std. × Quellung; O l~est-N (Fi l t ra t ) in % des Gesamt -N (Filtra~)
I Gesamt -N (Fi l t ra t ) , bezogea a u f :Fi l t ra tvolumen
M~ximums bewirkt. Infolge der rasehen AuflSsung der Muskelfasern nimmt jedoeh die dem Fleiseh zukommende Quellung sehr raseh ab und in der Tat entsprieht der n~eh 4 Std. erreiehte Quellungswert demjenigen yon Sehnen allein.
3. Die Proteolyse des Fleisches durch Pepsin-Salzsiiure in Gegenwart yon Sehnen Aus dieser getrennten Untersuehung yon Fleiseh und Sehnen ergibt sieh, dal3 bei
der Pepsineinwirkung auf ein Gemiseh yon Fleiseh und Sehnen das Quellungsmaxi- mum der Sehnen erst dann erreieht sein kann, wenn bereits 50--65% des Fleisehes in L5sung gegangen sind. Quellung des Sehnenko]lagens und Abbau der Muske]- proteine gehen demnaeh gleiehzeitig vonstatten.
a) Die Wirlcung yon Sehnen au/ LSsungen peptiseh abgebauten Fleisehes Setzt man ungequollene (native) Sehnen zu einer L6sung von Abbauprodukten tier
Muskelproteolyse, so tr i t t selbstverst/~ndlieh mit steigendem Sehnengehalt zu- nehmende Quellung ein (Abb. 7). Da die naeh 1 Std. Quellung im Filtrat gefundene Konzentration an Gesamt-Stiekstoff mit steigender Sehnenkonzentration zunimmt
1 Auch dutch S~ure ~llein trier mi~ der Zeit eine gewisse ttydrolyse und dami~ Quellungs- abnahme ein.

iJber die Proteolyse der Fleischpr0teine. III 423
(Abb. 7), muB die yon den Sehnen aufgenommene Fliissigkeit ein geringeres LSsungs- vermSgen ftir Abbauprodukte besitzen als die ,,freie" Flfissigkeit. Es handelt sich hier um die bekannte Erseheinung des ,,nieht 16senden Raumes", d. h. das yon dem Kolloid hydratisch gebundene Wasser hat ein vermindertes LSsungsvermSgen. Aueh bei l~ngerer Quellungsdauer (4 Std.) ist dieser Effekt zu beobaehten. In diesem Falle wurde das Pepsin der Fleisehproteolyse-LSsung vor dem Sehnenzusatz dutch kurzes Erhitzen auf 85 ° C inaktiviert.
Naeh unseren Versuchen kann die yon don Sehnen unter Quellung aufgenommene S~ure nicht mehr als 60--80 mg N/100 ml aufnehmen.
Von Interesse ist der Befund, dab das Verh~Itnis l~est-N/gel6stem N im Filtrat nach der Quellung das gleiche ist wie vorher (Abb. 7). Es werden demnaeh die hoch- molekularen Abbauprodukte ebenso ]eicht yon der Quellungsfliissigkeit gelSst wie die niedermolekularen.
Da das Kollagen Protonen binder, verschiebt sich der p . -Wert des Filtrates (und der Sehnen) bei steigendem Sehnenzusatz naeh der alkalisehen Seite (z. B. pa 1,50 bei 1% Sehnenzusatz, pa 1,89 bei 2% Sehnenzusatz).
Setzt man einer Fleisehproteolyse-LSsung start nativem Gewebe vorher 1 Std. (37 ° C) in 0,05 n-HC1 gequollene Sehnen zu, so wirct natfirlieh keine Fliissigkeit mehr unter Quelhmg aufgenommen. Die in der AbbaulSsung vorhandenen Stickstoff- substanzen verteilen sich iimerhalb yon 4 Std. fast vSllig gleiehm~13ig zwisehen Quellungsfliissigkeit und ,,freier" LSsung (Tab. 3).
Tabelle 3. Einwirkung gequollener Sehnen (1 Std. in 0,05 n-HC], 37 ° G) au] 40 ml einer f~6sung, erhalten durch Inkubation yon 4 g tZleisch mit 160 ml Enzyml6sung. Ges~mt-N ist bezogen auf Filtratvolumen. - - L6sungsvermSgen der Que]lungsfliissigkeit in % derjenigen des Filtrates
In den Sehnen I enthaRene I Dauer der
Sehnenzusatz Quellungs- I Einwirkung f l t i s~ke i t
g S~d.
o - 0
2,0 18,6 2,0 17,6 1,0 69,2 44
Gesamt-N im ~i l t ra t
mg
32,39 28,02 23,12 26,49
Gesam~-N im Fil trat
%
83 70 58 6 6
Rest-N LSsungs- bezogen auf verm6gen Filtrat-lg in
% % I
] 54,0 55,3
94 51,1 98 55,1
:Filtrat-pK
1,60 1,60 1,60 1,60
DaB hier nieht, wie im FMle der oben erSrterten Versuehe (Abb. 7) eine ,,Konzen- trierung" des Filtrat-N eintritt, liegt daran, dab unter diesenVersuehsbedingungen die Grenze des LSsungsvermSgens der Quellungsflfissigkeit (etwa 60 mg N/100 ml) noeh nicht erreieht ist. Aueh hier werden hoeh- und niedermolekulare Peptide yon der Quellungsflfissigkeit gleieh gut gelSst (Rest-N in °/o des Gesamt-N bleibt fast unver~ndert). Da gequollene Sehnen bereits mit Protonen abgesEttigt sind, t r i t t keine p~-Versehiebung ein (Tab. 3).
b) Proteolyse von Fleisch bei Zusatz gequollener Sehnen
Der peptisehe Abbau yon Fleisch in Gegenwart yon Sehnen, die ihren maximalen Quellungszustand erreicht haben, ist tibersichtlicher, als der in Gegenwart unge- quollener Sehnen, da der Abbau nicht yon gleichzeitiger starker Quellung begleitet wir4. Auch hier verteilt sieh nach 4 Std. Pepsinverdauung der gelSste Stiekstoff gleichm~gig zwisehen Quellungsfliissigkeit und Filtrat, wie es nach dem in Tab. 3 angegebenen Versuch nicht anders zu erwarten war. Der Peptidabbau ist geringer
424 R. Hx~M:
Ms es auf Grund der Proteolyse der Einzelkomponenten (Fleiseh und Sehnen) zu erwarten war, und zwar wurde er durch den Sehnenzusatz um 22% herabgesetzt (Tab. 4).
Tabelle 4. Proteolyse yon Fleisch durch Pepsin-Salzs~iure in Gegenwart gequollener Sehnen. Substr~tkonzentration yon Sehnen und Fleisch je 2,5 %. Dauer der Inkubation: 4 Std
LSslichkeit %
• I Experimentell ermittelt . . . . . . . . 43,0 berechnet aus der Proteolyse der
Komponenten . . . . . . . . . . . 46,9
Peptidabbau %
44,4
56,7
Amino-~ %
13,0
14,7
Da sich durch den Sehnenzusatz die Konzentrat ion an gelSsten Peptiden von 34 mg N (Fleiseh ohne Sehnen) auf nut 4g mg N/40 ml LSsung erhSht hat, wird naeh den oben angeffihrten Versuehen der Hemmungseffekt weniger auf der Zunahme an Abbauprodukten, als vielmehr auf der Wirkung des ungelSsten Sehnenmaterials beruhen. Offenbar wird yon dem Sehnenkollagen Pepsin adsorbiert. Dal~ dies in der Tat zutrifft, ]i~Bt sich durch die Wirkung einer Vorbehandlung der PepsinlSsung mit Sehnen auf die proteolytisehe Aktivit~t nachweisen.
Tabelle 5. Wirkung einer Vorbehandlung der Pepsinl6sung mit Sehnen au/ die proteolytische Aktivit~it des Enzyms. 26 rnl der nach Quellung yon den Sehnen abfiltrierten EnzymlSsung wurden mit
1 g Fleisch 4 Std. inkubiert
Vorbehandlung der EnzymlSsung
k e i n e . . . . . . . . . . . . . . 1 Std. Quellen mit 2 % S e h n e n . . . keine . . . . . . . . . . . . . . 1 Std. Quellen mit 2 % S e h n e n . . .
LOslichkei$ Pepsinabbau % %
I 93,4 J 50,0 94,5 [ 35,6 94,6 i 60,5 95,0 i 32,3
Amino-~ Filtra~-p K % nach Pro~eolyse
13,5 1,50 10,5 1,80
1,52 1,88
Durch eine solche Vorbehandlung wird die AuflSsungsgeschwindigkeit des Fleisches nicht beeintri~chtigt, der Pept idabbau hingegen um 26 bzw. 46 ~o (gegeniiber dam Abbau mit unbehandelter EnzymlSsung) herabgesetzt (Tab. 5). Diese Hemmung des Peptidabbaues ist nur durch eine Adsorption des Pepsins an das Sehnenkollagen zu erkli~ren. Das im Fil trat verbleibende Enzym reicht zwar - - wie oben erSrtert w u r d e - - noch aus, F]eisch mit unver~nderter Geschwindigkeit aufzulSsen, nicht abet, die Polypeptide normal abzubauen. Die durch den Sehnenzusatz bedingte p . - ErhShung des Verdauungsansatzes um 0,3 Einheiten (yon p . 1,50 auf p . 1,84) ist, wie Tab. 2 zeigt, ohne grS~eren Einflul~ auf die Proteolyse.
c) Proteolyse von Fleisch bei Zusatz nicht gequollener Sehnen Niedrige Substratkonzentration. Ffir die Untersuchung des proteolytischen Ab-
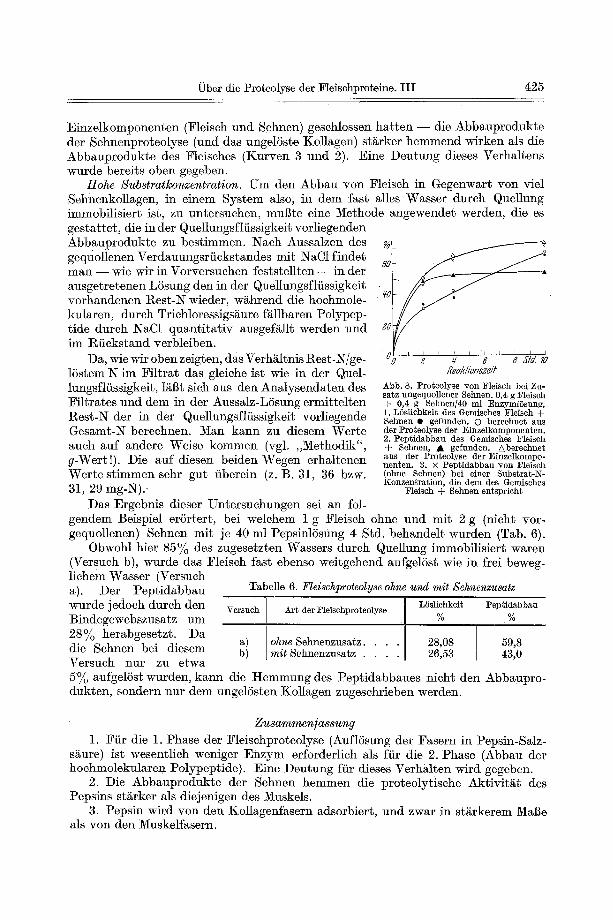
baues einer Mischung yon Fleisch und nativen, nicht vorgequollenen Sehnen wurden zun~ehst niedrige Substratkonzentrat ionen gew~hlt, um die durch Quellung beding- ten Effekte mSglichst auszusehalten. Aus Abb. 8 ist zu ersehen~ dab die hier beob- aehteten Effekte im gro•en und ganzen denen bei Zusatz vorgequollener Sehnen (Tab. 4) entspreehen. W~hrend die LSslichkeit unbeeinfluI~t bleibt (Kurve 1), wird der Pept idabbau durch Sehnenzusatz gehemmt. Er geht wesentlich langsamer vonsta t ten als es auf Grund der Proteolyse der Einzelkomponenten zu erwarten w~re (Kurve 2). Ferner zeigt sieh, dab - - w i e wir sehon oben aus dem A b b a u der
~lber die Proteolyse der Fleischproteine. I I I 425
Einzelkomponenten (Fleiseh und Sehnen) geschlossen bat ten - - die Abbauprodukte der Sehnenproteolyse (und das ungelSste Kollagen) starker hemmend wirken Ms die Abbauprodukte des Fleisches (Kurven 3 und 2). Eine Deutung dieses Verhaltens wurde bereits oben gegeben.
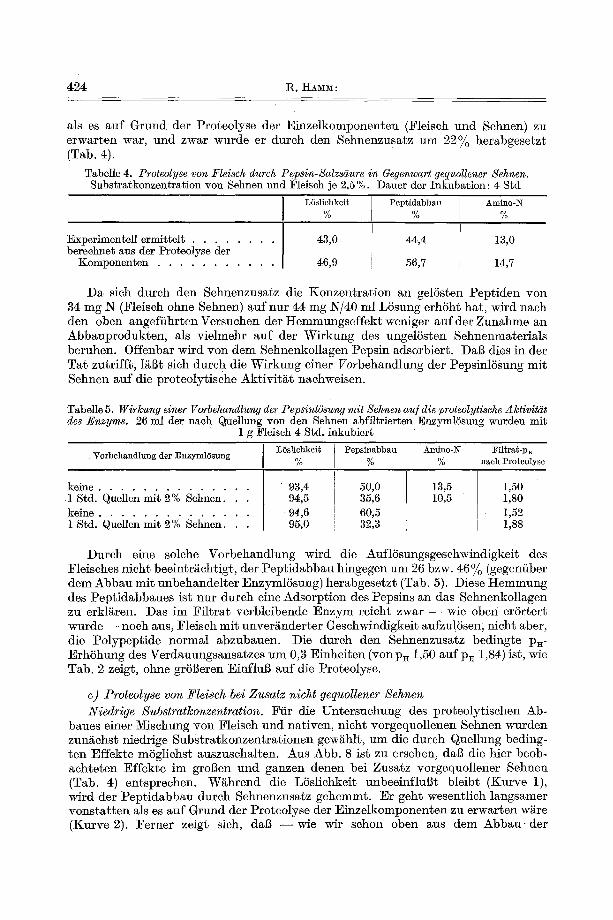
Hohe Substratlconzentration. Um den Abbau yon Fleisch in Gegenwart yon viel Sehnenkollagen, in einem System also, in dem fast ailes Wasser durch Quellung immobilisiert ist, zu untersuchen, muBte eine Methode angewendet werden, die es gestattet , die hi der Quellungsflfissigkeit vorliegenden Abbauprodukte zu bestimmen. Nach Aussalzen des gequollenen Verdauungsrfickstandes mit 2qaC1 finder man - - wie wir in Vorversuchen feststellten - - in der ausgetretenen LSsung den in der Quellungsflfissigkeit vorhandenen Rest-2q wieder, wghrend die hochmole- kularen, durch Triehloressigs~ure f~llbaren Polypep- tide (lurch NaC1 quanti tat iv ausgef~llt werden nnd im Rfickstand verbleiben.
Da, wie wir oben zeigten, das Verhgltnis Rest-BT/ge- 15stem B7 im Filtrat das gleiche ist wie in der Quel- lungsfliissigkeit, l~13t sich aus den Analysendaten des Filtrates und dem in der Aussa]z-LSsung ermittelten Rest-B7 der in tier Quellungsflfissigkeit vorliegende Gesamt-~7 berechnen. Man kann zu diesem Werte auch auf andere Weise kommen (vgl. , ,Methodik", g-Wertl). Die auf diesen beiden Wegen erhaltenen Werte st immen sehr gut iiberein (z. B. 3], 36 bzw. 31, 29 mg-/~).,
Das Ergebnis dieser Untersuchungen sei an fol-
80
feak//onsze# Abb. 8. Proteolyse yon F1eisch bei Zu- satz angequollener Sehnen. 0,4 g F1eisch ÷ 0,4 g Sehnen/40 m] :Enzyml~sung, 1. L6slichkeit des Oemisches Fleisch + Sehnen • gefunden. O berechnet aus der Prote01yse der Einzelkomponenten. 2. Pep~idabbau des Gemisches Fleisch + Sehnen, & gefunden. Aberechnet aus der Proteolyse der Einzelkompo- nen~en. 3. × Peptidabbau yon Fleisch (ohne Sehnen) bei einer Subs~l'at-N- Konzentration, die dem des Gemisches
Fleisch + Sehnen entspricht
lichem Wasser (Versuch a). Der Pept idabbau wurde jedoch dutch den Bindegewebszusatz um 28% herabgesetzt. Da
gendem Beispiel erSrtert, bei welchem 1 g Fleisch ohne und mit 2 g (nicht vor- gequo]lenen) Sehnen mit je 40 ml PepsinlSsung 4 Std. behandelt wurden (Tab. 6).
Obwohl hier 85% des zugesetzten Wassers dutch Quellung immobilisiert waren (Versuch b), wurde das Fleiseh fast ebenso weitgehend aufgelSst wie in frei beweg-
Tabelle 6. Fleischproteolyse ohne und mit Sehnenzusatz
L6slichkeit Peptidabbau Versuch AI'$ der Fleischproteolyse % %
a) ohne Sehnenzusatz . . . . 28,08 59,8 die Sehnen bei diesem b) mit Sehnenzusatz . . . . 26,53 43,0 Versuch nut zu etwa 5'~o aufgel6st wurden, kann die Hemmung des Peptidabbaues nicht den Abbaupro- dukten, sondern nur dem ungelSsten Ko]lagen zugeschrieben werden.
Zusammen/assung
1. Ffir die 1. Phase der Fleischproteolyse (AuflSsung der Fasern in Pepsin-S~lz- s~iure) ist wesentlich weniger Enzym erforderlich als ffir die 2. Phase (Abbau der hochmolekularen Polypeptide). Eine Deutung fiir dieses Verhalten wird gegeben.
2. Die Abbauprodukte der Sehnen hemmen die proteolytische Aktivit~t des Pepsins starker als diejenigen des Muskels.
3. Pepsin wird yon den Kollagenfasern adsorbiert, und zwar in st~rkerem Mal~e als yon den Muskelfasern.

426 H. T~u~TW~:
4. Bei der Proteolyse des Fleisches in Gegenwart nativer Sehnen gehen Quellung des Koll~gens und Abb~u des Muskeleiwei~es gleichzeitig vor sich.
5. Die yon den Sehnen unter Quellung aufgenommene Flfissigkeit hat ein geringeres LSsungsvermSgen fiir proteolytische Abbauprodukte als die ,,freie" Flfissigkeit. Ihr LSsungsvermSgen ffir hoehmolekulare Peptide ist d~s gleiehe wie ftir niedermolekulare.
6. In einem System, in welchem durch Kollagenquellung der grS~te Tell des Wassers immobilisiert ist, wird Fleisch fast ebenso rasch dutch 1)epsin-Salzs~ure auf- gel6st wie in einem sehnenfreien System, der Peptidabbau jedoch ist verlangsamt.
Bestimmung der Alterung des Fisches mittels jodometrischer Titration
I I1 . Mitteilung
Von HANS TRUTTWIN
Mitteilung aus der Avdelning ]6r ]6dviimneshygie~, Kungl. VeteriniirhSgskoIan, Stoclcholm*
1Kit 4 Textabbildungen
(Eingegangen am 16. Juni 1955)
Die frfiher an Dorsch, Schellfisch, Scholle und Witling durchgefiihrten Versuehe 1 zur Feststellung der Grenze der GenuBtaugliehkeit eines Fisches wurden nunmehr auf Fettfische ausgedehnt, zun~chst auf StrSmling, Hering und Lachs.
Arbeitsweise Die Methodik blieb grunds~tzlich die gieiche; es wurde zur Titration eine JodlSsung yon
0,800 g subl. Jod und 1,500 g Kaliumjodid in 1000 ml Wasser verwendet. Das feinverteil~e Fisch- fleisch, wiederum jeweils 1 g, wurde jedoch nicht wie bisher in 50 ml Wasser allein, sondern unter
Abb. 1. Sohfittelapparatur zm" Durchffihrung der jodometrischen Titration
Hinzuftigung yon Acegon (42 ml Wasser -~ 8 ml Aceton) duroh Schiig~eln suspendiert. Dadurch wurde ein glat~erer Reaktionsverlauf bei gleich guter Fests~ellung der UmscMagsfarbe (diesmM mit 1,5 ml 1%iger Sg~rkel6sung) erzielg.
* Abteilung fiir LebensmRte]hygiene der Kg]. Tier~rztlichen Hochschule Stockholm. 1 T~uTTWI~, It. : Diese Z. 97, 275 (1953) u. 99, 461 (1954).












![Beurteilung der Proteolyse von ausgewählten …othes.univie.ac.at/13606/1/2011-03-04_0503241.pdf · worden ist, hergestellt sind“ [Codexkapitel_B32], des weiteren aber auch Erzeugnisse](https://img.pdfslide.org/doc/110x75/5b9f6be609d3f2385c8bb4ad/beurteilung-der-proteolyse-von-ausgewaehlten-othes-worden-ist-hergestellt.jpg)










