Embed Size (px)
Citation preview

Aus dem
INSTITUT FUR VIROLOGIE
der TIERARZTLICHEN HOCHSCHULE HANNOVER
Untersuchung der Bedeutung derSialinsaure-bindenden Aktivitat fur die
Enteropathogenitat des Virus der ubertragbarenGastroenteritis der Schweine
INAUGURAL-DISSERTATION
Zur Erlangung des Grades einer Doktorin der Veterinarmedizin
(Dr. med. vet.)
durch die Tierarztliche Hochschule Hannover
Vorgelegt von
Christel Elisabeth Schwegmann-Weßels, geb. Schwegmann
aus Lingen (Ems)
Hannover 2002

Wissenschaftliche Betreuung: Prof. Dr. Georg Herrler
1. Gutachter: Prof. Dr. Herrler
2. Gutachter: PD Dr. Fischer
Tag der mundlichen Prufung: 30.05.2002

Meinen Eltern und
meinem Mann Peter
So eine Arbeit wird eigentlich nie fertig,
man muss sie fur fertig erklaren,
wenn man nach Zeit und Umstanden
das moglichste getan hat.
(J. W. Goethe, Italienische Reise, 16. Marz 1787)


Inhaltsverzeichnis
1 Einleitung 17
1.1 Coronaviren . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
1.1.1 Taxonomie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
1.1.2 Strukturproteine . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
1.1.2.1 N-Protein . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
1.1.2.2 M-Protein . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
1.1.2.3 E-Protein . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
1.1.2.4 S-Protein . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
1.1.2.5 HE-Protein . . . . . . . . . . . . . . . . . . . . . . . . . . 25
1.2 Das Virus der ubertragbaren Gastroenteritis (TGEV) . . . . . . . . . . . . 26
1.2.1 Pathogenitat und Epidemiologie . . . . . . . . . . . . . . . . . . . . 26
1.2.2 Eigenschaften des S-Proteins von TGEV . . . . . . . . . . . . . . . 29
1.2.2.1 Epitope . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29
1.2.2.2 Bindungsaktivitaten . . . . . . . . . . . . . . . . . . . . . 31
1.2.2.3 Tropismus . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
1.3 Escherichia coli F5/K99 (E. coli F5/K99) . . . . . . . . . . . . . . . . . . 33
1.4 Virusrezeptoren . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34

2 Zielsetzung 39
3 Material 41
3.1 Zelllinien . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
3.2 Zellkulturmedien . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
3.3 Viren . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
3.4 Bakterien . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
3.5 Erythrozyten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
3.6 Darmmaterial . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43
3.7 Enzyme . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43
3.8 Antikorper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43
3.9 Kits . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
3.10 Substrate . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
3.11 Chemikalien . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
3.12 Puffer und Losungen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
4 Methoden 53
4.1 Zellkulturen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53
4.1.1 Passagierung von ST-Zellen . . . . . . . . . . . . . . . . . . . . . . 53
4.1.2 Passagierung von LLC-PK1-Zellen . . . . . . . . . . . . . . . . . . 53
4.2 Bakterien . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 54
4.2.1 Kultivierung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 54
4.2.2 Fimbrientest . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 54
4.3 Virusanzucht . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 54
4.4 Plaquetest . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55

4.5 Virusreinigung mittels Dichtegradientenzentrifugation . . . . . . . . . . . . 56
4.6 Isolierung des S-Proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . 56
4.7 Erythrozyten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57
4.7.1 Isolierung aus Vollblut . . . . . . . . . . . . . . . . . . . . . . . . . 57
4.7.2 Fixierung von Erythrozyten . . . . . . . . . . . . . . . . . . . . . . 57
4.7.3 Hamagglutinationstest . . . . . . . . . . . . . . . . . . . . . . . . . 58
4.7.4 Hamagglutinationsinhibition . . . . . . . . . . . . . . . . . . . . . . 58
4.8 Desialylierung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 59
4.8.1 Desialylierung von Viren . . . . . . . . . . . . . . . . . . . . . . . . 59
4.8.2 Desialylierung von Zellen . . . . . . . . . . . . . . . . . . . . . . . . 59
4.8.3 Desialylierung von Muzinen, MP und BSMV . . . . . . . . . . . . . 59
4.8.4 Desialylierung von immobilisierten Muzinen . . . . . . . . . . . . . 60
4.8.5 Desialylierung von Zelllysaten . . . . . . . . . . . . . . . . . . . . . 60
4.8.6 Desialylierung von Erythrozyten . . . . . . . . . . . . . . . . . . . . 60
4.9 Membranproteinisolierung . . . . . . . . . . . . . . . . . . . . . . . . . . . 61
4.10 Fallung von Oberflachenproteinen und Glykoproteinen . . . . . . . . . . . 61
4.10.1 Biotinylierung und Streptavidinfallung von Oberflachenproteinen . . 61
4.10.2 Fallung von Glykoproteinen mit immobilisiertem WGA . . . . . . . 62
4.11 Muzine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
4.11.1 Isolierung von Darmmuzinen . . . . . . . . . . . . . . . . . . . . . . 62
4.11.2 Reinigung der extrazellularen Muzine . . . . . . . . . . . . . . . . . 63
4.11.3 Kohlenhydratbestimmung mittels PAS-Farbung . . . . . . . . . . . 63
4.12 Burstensaummembranvesikel (BSMV) . . . . . . . . . . . . . . . . . . . . . 64

4.12.1 Herstellung von BSMV . . . . . . . . . . . . . . . . . . . . . . . . . 64
4.12.1.1 Darm von Absetzferkeln . . . . . . . . . . . . . . . . . . . 64
4.12.1.2 Darm von Saugferkeln . . . . . . . . . . . . . . . . . . . . 65
4.12.2 Enzymbestimmungen von BSMV . . . . . . . . . . . . . . . . . . . 65
4.12.2.1 Bestimmung der Aktivitat der AP . . . . . . . . . . . . . 65
4.12.2.2 Bestimmung der Aktivitat der Na/K-ATPase . . . . . . . 66
4.13 Proteinbestimmung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 66
4.13.1 Proteinbestimmung nach Bradford . . . . . . . . . . . . . . . . . . 66
4.13.2 BCA-Test . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 67
4.13.3 Messung bei 280 nm . . . . . . . . . . . . . . . . . . . . . . . . . . 68
4.14 Analyse von Sialinsauren . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68
4.15 SDS-Polyacrylamidgel-Elektrophorese . . . . . . . . . . . . . . . . . . . . . 68
4.16 Detektion von Proteinen in Polyacrylamidgelen . . . . . . . . . . . . . . . 69
4.16.1 Coomassie-Farbung . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
4.16.2 Silberfarbung zur Detektion von Glykoproteinen . . . . . . . . . . . 69
4.17 Western Blot . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
4.18 Immunchemische Detektion von Proteinen . . . . . . . . . . . . . . . . . . 70
4.19 Untersuchung der Virusbindung an Sialoglykokonjugate . . . . . . . . . . . 70
4.19.1 ELISA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70
4.19.2 Virusbindung in Zellkultur . . . . . . . . . . . . . . . . . . . . . . . 71
4.19.3 Virusbindung auf Nitrozellulose . . . . . . . . . . . . . . . . . . . . 72
4.19.4 Virusbindungstest mittels Immunfluoreszenz . . . . . . . . . . . . . 72
4.20 Analyse von Glykoproteinen . . . . . . . . . . . . . . . . . . . . . . . . . . 73

4.20.1 Triton X-114-Phasenseparierung . . . . . . . . . . . . . . . . . . . . 73
4.20.2 Glykoproteindetektion . . . . . . . . . . . . . . . . . . . . . . . . . 74
4.20.2.1 Kohlenhydraterkennung . . . . . . . . . . . . . . . . . . . 74
4.20.2.2 Selektive Sialinsaureerkennung . . . . . . . . . . . . . . . 74
4.20.3 Analyse von Glykoproteinen mit Lektinen . . . . . . . . . . . . . . 75
4.20.3.1 Lektintest auf dem Blot . . . . . . . . . . . . . . . . . . . 75
4.20.3.2 Fallung von BSMV mit Lektinen . . . . . . . . . . . . . . 75
5 Ergebnisse 77
5.1 Vergleich der Sialinsaurebindungsaktivitat von TGEV und E. coli F5 . . . 77
5.1.1 Interaktion mit Erythrozyten verschiedener Tierarten . . . . . . . . 77
5.1.2 Neuraminidase-Sensitivitat der Erythrozytenrezeptoren . . . . . . . 79
5.1.3 Untersuchung der Hamagglutinationshemmung . . . . . . . . . . . . 80
5.1.3.1 Praparation intestinaler Muzine . . . . . . . . . . . . . . . 80
5.1.3.2 Analyse des Sialinsauregehalts der Inhibitoren . . . . . . . 81
5.1.3.3 Hemmung der Hamagglutination . . . . . . . . . . . . . . 82
5.2 Bindung von TGEV an Zellkulturen . . . . . . . . . . . . . . . . . . . . . . 84
5.3 Untersuchung von Zelloberflachenproteinen . . . . . . . . . . . . . . . . . . 86
5.3.1 Bindung an Membranproteine . . . . . . . . . . . . . . . . . . . . . 86
5.3.2 Bindung an Zelloberflachenproteine . . . . . . . . . . . . . . . . . . 88
5.3.3 Darstellung der Zelloberflachenproteine . . . . . . . . . . . . . . . . 91
5.3.4 Sialinsaureabhangigkeit der Bindung von TGEV . . . . . . . . . . . 92
5.4 Charakterisierung des Sialoglykoproteins . . . . . . . . . . . . . . . . . . . 93
5.4.1 Triton X-114-Phasenseparierung . . . . . . . . . . . . . . . . . . . . 94

5.4.2 Prazipitation mit WGA . . . . . . . . . . . . . . . . . . . . . . . . 95
5.5 Interaktion von TGEV mit intestinalen Muzinen . . . . . . . . . . . . . . . 96
5.5.1 Bindung von TGEV an immobilisierte Darmmuzine . . . . . . . . . 97
5.5.2 Bedeutung der Sialinsauren fur die Bindung . . . . . . . . . . . . . 99
5.5.3 Vergleich der Muzinbindung von TGEV mit Mutanten . . . . . . . 99
5.5.4 Interaktion von TGEV mit einem hochmolekularen Protein . . . . . 101
5.6 Untersuchung von Burstensaummembranen . . . . . . . . . . . . . . . . . . 102
5.6.1 Praparation von Burstensaummembranvesikeln . . . . . . . . . . . 102
5.6.2 Analyse des Sialinsauregehalts der BSMV . . . . . . . . . . . . . . 103
5.6.3 Bindung an Proteine aus BSMV . . . . . . . . . . . . . . . . . . . . 103
5.7 Charakterisierung des BSMV-Proteins . . . . . . . . . . . . . . . . . . . . 107
5.7.1 Triton X-114-Phasenseparierung . . . . . . . . . . . . . . . . . . . . 107
5.7.2 Charakterisierung des Kohlenhydratanteils . . . . . . . . . . . . . . 108
5.7.2.1 Nachweis der Kohlenhydrate und Sialinsauren . . . . . . . 108
5.7.2.2 Charakterisierung der Kohlenhydratreste mit Lektinen . . 109
5.8 Interaktion von TGEV mit Darmepithel . . . . . . . . . . . . . . . . . . . 112
6 Diskussion 117
6.1 Bedeutung der Desialylierung von TGEV fur die Interaktion mit Sialogly-
kokonjugaten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 117
6.2 Bewertung des Vergleichs von TGEV mit E. coli F5 . . . . . . . . . . . . . 120
6.3 Interaktion von TGEV mit Zellkulturzellen . . . . . . . . . . . . . . . . . . 122
6.4 Interaktion von TGEV mit intestinalen Muzinen und mit Darmepithel . . 124
6.5 Bedeutung der Sialinsaure-bindenden Aktivitat fur die Enteropathogenitat
von TGEV . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 131

7 Zusammenfassung 135
8 Summary 137
Literaturverzeichnis 139
Abbildungsverzeichnis 161
Tabellenverzeichnis 163

Abkurzungsverzeichnis
ABTS 2,2’-Azino-di-[3-ethylbenzthiazolinsulfonat(6)]
a.E. absorbierende Einheiten
AP Alkalische Phosphatase
APN Aminopeptidase N
APS Ammoniumpersulfat
AS Aminosaure
ATCC”American Tissue Culture Collection“
ATP Adenosintriphosphat
BCA Bicinchonininsaure
BCoV Bovines Coronavirus
BSA Bovines Serumalbumin
BSMV Burstensaummembranvesikel
C Cystein
CCoV Coronavirus der Hunde
CCR Chemokinrezeptor
CD”cluster of differentiation“
CEA carcinoembryonales Antigen
cpE zytopathischer Effekt
D Asparaginsaure
DFP Diisopropylfluorophosphat
DIG Digoxigenin
DMB 4,5-Methylendioxy-1,2-phenylendiamin × 2 H2O
DMEM Dulbecco’s modified Eagle medium
DNA Desoxyribonukleinsaure

DTT Dithiothreitol
E Extinktion
ECL Enzym-Chemolumineszenz
E. coli Escherichia coli
EDTA Ethylendiamin-tetraessigsaure
EGTA Ethylenglykol-bis(β-Aminoethylether)N,N,N’,N’-Tetraessigsaure
ELISA”enzyme-linked immuno sorbent assay“
E-Protein Envelope-Protein
ETEC enterotoxinogene E. coli
F Phenylalanin
F5 Fimbrientyp 5
FCoV Felines Coronavirus
FIPV Virus der felinen infektiosen Peritonitis
FITC Fluoresceinisothiocyanat
FKS Fetales Kalberserum
G Glycin
gp/g Glykoprotein
HA Hamagglutination
HAD hamagglutinationsdefizient
hAPN humane Aminopeptidase N
HAU hamagglutinierende Einheiten
HCoV Humanes Coronavirus
HE Hamagglutinin-Esterase
HEF Hamagglutinin-Esterase-Fusions-Protein
HEPES N-(2-Hydroxyethyl)piperazin-N’-(2-Ethansulfonsaure)
HEV Hamagglutinierendes Enzephalomyelitis Virus
HIU hamagglutinationsinhibierende Einheiten
HIV Humanes Immundefizienzvirus
H2O Reinstwasser
HPLC Hochdruck-Flussigkeitschromatographie
HRP Peroxidase aus Meerrettich
HSV Herpes-simplex-Virus

IBV Virus der aviaren infektiosen Bronchitis
ICAM”intercellular adhesion molecule“
ICTV internationales Komitee fur Virus-Taxonomie
Ig Immunglobulin
IP isoelektrischer Punkt
kDa Kilo-Dalton
L Leucin
MAA Maackia amurensis Agglutinin
MAD Madrid
mAk monoklonaler Antikorper
MES 2(N-Morpholino)ethansulfonsaure
MHV Maus-Hepatitis-Virus
m.o.i.”multiplicity of infection“
MP Zellmembranen/Membranproteine
M-Protein Membran-Protein
mRNA”messenger RNA“
-NA neuraminidasebehandelt
Neu5Ac N-Acetylneuraminsaure
Neu5,9Ac2 N-Acetyl-9-O-Acetylneuraminsaure
Neu5Gc N-Glykolylneuraminsaure
Neu5Gc-GM3 N-Glykolylneuraminyl-neolaktotetraosyl-ceramid
NHS N-Hydroxysuccinimid
N-Protein Nukleokapsid-Protein
OG n-Oktyl-β-D-Glukopyranosid/Oktylglukosid
ORF offener Leserahmen
P Prolin
PAGE Polyacrylamidgel-Elektrophorese
pAPN porzine Aminopeptidase N
PAR Paris
PBS Phosphat-gepufferte Kochsalzlosung
PBSM PBS ohne Calcium und Magnesium
PEDV Virus der epidemischen Diarrhoe

PFU Plaque-bildende Einheiten
PK”porcine kidney“
PNA Peanut Agglutinin
PRCoV Porzines respiratorisches Coronavirus
PSM Muzine aus der Glandula submandibularis des Schweines
PTV”Purdue-type virus“ (fruher: NEB72)
PUR Purdue
PVDF Polyvinylidendifluorid
PVR Poliovirus-Rezeptor
R Arginin
RNA Ribonukleinsaure
RT Raumtemperatur
RtCoV”rabbit coronavirus“
S Serin
SDS Natriumdodecylsulfat
SNA Sambucus nigra Agglutinin
S-Protein Spike-/Surface-Protein
ST”swine testicular“
TBS TRIS-gepufferte Kochsalzlosung
TCoV Coronavirus des Truthahns
TEMED N,N,N’,N’-Tetramethylethylendiamin
TGE ubertragbare/transmissible Gastroenteritis
TGEV Virus der ubertragbaren Gastroenteritis
TiHo Tierarztliche Hochschule
TRIS Tris-Hydroxymethylaminomethan
VCNA Neuraminidase aus Vibrio cholerae
WGA Weizenkeimagglutinin (Wheat Germ Agglutinin)
w/o”without“


1. Einleitung
1.1 Coronaviren
1.1.1 Taxonomie
Das Virus der ubertragbaren Gastroenteritis der Schweine (TGEV) gehort zur Familie
der Coronaviridae, Genus Coronavirus. Diese Virusfamilie wurde 1975 durch das interna-
tionale Komitee fur Virus-Taxonomie (ICTV) eingefuhrt (Tyrrell et al., 1975). Bereits
1968 wurden Coronaviren aufgrund ihres charakteristischen Erscheinungsbildes im Elek-
tronenmikroskop als eigenstandige Virusgruppe definiert (Tyrrell et al., 1968).
Der Familie der Coronaviridae werden zwei Genera, Coronavirus und Torovirus, zuge-
ordnet (Pringle, 1992). Diese Zuordnung erfolgte aufgrund der Ahnlichkeiten in der
Organisation und Expression der Genome und in der Struktur der Genprodukte dieser
Viren. Im August 1996 wurde die Familie der Coronaviridae mit der Familie der Arte-
riviridae durch das ICTV zu der zweiten viralen Ordnung, den Nidovirales zusammen-
gefasst (Cavanagh et al., 1997; Enjuanes et al., 2000). Die Familien dieser Ordnung
enthalten umhullte Viren mit einem unsegmentierten einzelstrangigen RNA-Genom posi-
tiver Polaritat. Der Name”Nido“ (lat., deutsch: Nest) bezieht sich auf den einzigartigen
Transkriptions- und Replikationsmechanismus dieser Virusfamilien. Wahrend der Infekti-
on entsteht in der Zelle ein 3’ coterminales”nested set“ von bis zu 8 mRNAs, von denen
nur das Gen am 5’-Ende der jeweiligen mRNA translatiert wird (Ubersicht s. van der
Most und Spaan, 1995; de Vries et al., 1997).
Bei den Coronaviren handelt es sich um umhullte, pleomorphe, haufig spharische Partikel
17

18 1. Einleitung
mit einem Großendurchmesser von 60 bis 220 nm. Sie tragen charakteristische ca. 20 nm
lange keulenformige Oberflachenprojektionen, die die Viruspartikel im Elektronenmikros-
kop wie von einem Strahlenkranz (lat. Corona) umgeben erscheinen lassen (Siddell et al.,
1983). Diese Oberflachenprojektionen werden von dem Spike- bzw. Surface-Protein (S-
Protein) gebildet. Das positiv-strangige, nichtsegmentierte RNA-Genom besteht aus ca.
30000 Nukleotiden und beinhaltet ein ungewohnlich großes RNA-Polymerase-Gen (20000
Nukleotide), ein großes Oberflachenglykoprotein(S-Protein)-Gen, ein integrales Membran-
Protein(M-Protein)-Gen und ein Nukleokapsid-Protein(N-Protein)-Gen. Diese Gene sind
in der oben beschriebenen Reihenfolge im Genom angeordnet (5’ → 3’ Richtung), konnen
aber von zusatzlichen Genen, die fur weitere Struktur- oder Nichtstrukturproteine kodie-
ren, unterbrochen sein (Siddell, 1995a). Die RNA tragt ein Cap, ist polyadenyliert und
bildet zusammen mit dem N-Protein das Nukleokapsid (Lai und Cavanagh, 1997). Die
Morphologie des Nukleokapsids wurde als helikal beschrieben (Macnaughton et al.,
1978), es gibt jedoch auch Arbeitsgruppen, die eine ikosaedrische Struktur diskutieren
(Risco et al., 1996). In die das Nukleokapsid umgebende Virusmembran sind die viralen
Glykoproteine, das S-Protein, das M-Protein und das nur bei Viren der Serogruppe II vor-
kommende Hamagglutinin-Esterase(HE)-Protein eingelagert. Ein weiteres Strukturprote-
in ist das unglykosylierte Envelope-Protein (E-Protein), welches in der Virusmembran in
geringer Kopienzahl vorkommt (s. Abb. 1.1). Die Virusreifung findet an den intrazellularen
Membranen des intermediaren Kompartiments zwischen Endoplasmatischem Retikulum
und Golgi-Apparat statt.
Aufgrund serologischer und phylogenetischer Untersuchungen des Verwandschaftsgrades
werden die Coronaviren in drei Gruppen unterteilt (Tabelle 1.1). Bei dem porzinen respi-
ratorischen Coronavirus (PRCoV) und dem felinen Coronavirus (FCoV) handelt es sich
allerdings eher um Varianten des transmissiblen Gastroenteritisvirus (TGEV) bzw. des
Virus der felinen infektiosen Peritonitis (FIPV) als um eigene Coronavirusspezies. Die
meisten Coronaviren infizieren die Epithelien des Respirations- und/oder des Gastroin-
testinaltraktes von Saugetieren, Vogeln oder dem Menschen. Eine Ubersicht uber die von
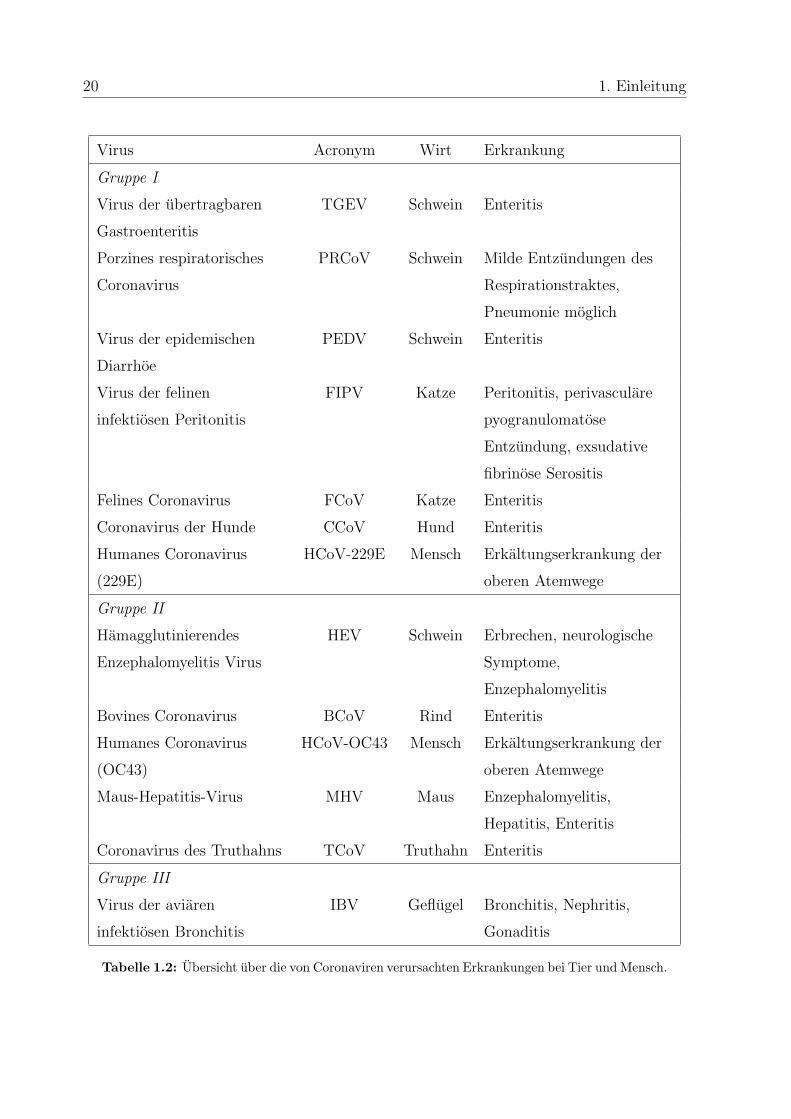
Coronaviren verursachten Erkrankungen bei den jeweiligen naturlichen Wirten gibt Ta-
belle 1.2.

1.1. Coronaviren 19
N
S
M
RNA
Membran
CAP
AA
A
E
HE
Gruppe IGruppe III
Gruppe II
Abbildung 1.1: Schematische Darstellung eines Coronaviruspartikels. S=Spike-Protein,
M= Membran-Protein, E= Envelope-Protein, HE=Hamagglutinin-Esterase, N =Nukleokapsid-
Protein. Die rechte Seite des Schemas zeigt die Viren der Serogruppe II, die als zusatzliches Struk-
turprotein das HE-Protein besitzen.
Gruppe I Gruppe II Gruppe III
HCoV-229E HCoV-OC43 IBV
TGEV MHV
PRCoV BCoV
PEDV HEV
CCoV TCoV
FIPV RtCoV
FCoV
Tabelle 1.1: Serologische und phylogenetische Verwandschaft der Coronaviren. RtCoV steht fur
”rabbit coronavirus“. Die Bedeutung der ubrigen Abkurzungen ist in Tabelle 1.2 aufgefuhrt.
20 1. Einleitung
Virus Acronym Wirt Erkrankung
Gruppe I
Virus der ubertragbaren
Gastroenteritis
TGEV Schwein Enteritis
Porzines respiratorisches
Coronavirus
PRCoV Schwein Milde Entzundungen des
Respirationstraktes,
Pneumonie moglich
Virus der epidemischen
Diarrhoe
PEDV Schwein Enteritis
Virus der felinen
infektiosen Peritonitis
FIPV Katze Peritonitis, perivasculare
pyogranulomatose
Entzundung, exsudative
fibrinose Serositis
Felines Coronavirus FCoV Katze Enteritis
Coronavirus der Hunde CCoV Hund Enteritis
Humanes Coronavirus
(229E)
HCoV-229E Mensch Erkaltungserkrankung der
oberen Atemwege
Gruppe II
Hamagglutinierendes
Enzephalomyelitis Virus
HEV Schwein Erbrechen, neurologische
Symptome,
Enzephalomyelitis
Bovines Coronavirus BCoV Rind Enteritis
Humanes Coronavirus
(OC43)
HCoV-OC43 Mensch Erkaltungserkrankung der
oberen Atemwege
Maus-Hepatitis-Virus MHV Maus Enzephalomyelitis,
Hepatitis, Enteritis
Coronavirus des Truthahns TCoV Truthahn Enteritis
Gruppe III
Virus der aviaren
infektiosen Bronchitis
IBV Geflugel Bronchitis, Nephritis,
Gonaditis
Tabelle 1.2: Ubersicht uber die von Coronaviren verursachten Erkrankungen bei Tier und Mensch.

1.1. Coronaviren 21
1.1.2 Strukturproteine
1.1.2.1 N-Protein
Die N-Proteine der Coronaviren sind 377 bis 455 Aminosauren lang mit einem Molekular-
gewicht von 45 bis 63 kDa. Mit einem Isoelektrischen Punkt (IP) von 10,3 bis 10,7 sind
N-Proteine stark basisch aufgrund eines hohen Gehaltes an Lysin- und Argininresten. Im
Gegensatz dazu steht ein saurer Abschnitt von 45 Aminosauren am Carboxylende mit
einem IP von 4,3 bis 5,5. N-Proteine besitzen viele Serinreste (7–11%), die posttransla-
tional phosphoryliert werden (Laude und Masters, 1995). Trotz dieser Ahnlichkeiten
sind die N-Proteine der Coronavirusspezies untereinander nur begrenzt homolog (Lapps
et al., 1987). Basierend auf dem Vergleich der Aminosauresequenzen konnten 3 struktu-
relle Domanen des N-Proteins identifiziert werden (Parker und Masters, 1990). Von
diesen Domanen stellt Domane II die potenzielle RNA-Bindungsstelle dar (Masters,
1992; Nelson und Stohlman, 1993). Das N-Protein kann sowohl RNA von Coronaviren
als auch andere RNA binden (Stohlman et al., 1988; Masters, 1992), scheint jedoch
virale RNA effizienter zu binden als nicht-virale RNA (Cologna et al., 2000). Die mogli-
che Funktion der Domanen I und III ist nicht bekannt, jedoch wurde in Domane III des
N-Proteins aller drei Coronavirusgruppen vor kurzem ein Nucleolus-Lokalisationssignal
(NuLS) identifiziert (Hiscox et al., 2001; Wurm et al., 2001).
Dem N-Protein der Coronaviren werden mehrere Funktionen wahrend des viralen Lebens-
zyklus zugeschrieben (Laude und Masters, 1995). Zum einen formt es mit der genomi-
schen RNA den Ribonukleoproteinkomplex (Davies et al., 1981), zum anderen bildet es
gemeinsam mit dem Membranprotein (M-Protein) das innere Core (Risco et al., 1996;
Escors et al., 2001b). Es wurde gezeigt, dass das N-Protein mit der leader RNA, die im
5’-Bereich der genomischen RNA lokalisiert ist, assoziiert ist (Baric et al., 1988; Nelson
et al., 2000). Ebenso wurde gezeigt, dass das N-Protein an RNA-Sequenzen im 3’-Bereich
des Genoms bindet (Zhou et al., 1996). Da die erwahnten RNA-Regionen an der Synthese
coronaviraler RNA beteiligt sind, wird postuliert, dass das N-Protein eine Rolle in der
Replikation genomischer RNA (Compton et al., 1987; Chang und Brian, 1996), in der
Transkription subgenomischer RNAs (Baric et al., 1988; Stohlman et al., 1988) und
in der Tranlation subgenomischer RNAs (Tahara et al., 1994) spielt. Bei den Arterivi-

22 1. Einleitung
ren wurde jedoch gezeigt, dass Replikation und Transkription auch in Abwesenheit des
N-Proteins stattfinden konnen (Molenkamp et al., 2000).
1.1.2.2 M-Protein
Bei den M-Proteinen handelt es sich um Glykoproteine mit einem Molekulargewicht von
20 bis 38 kDa (Ubersicht s. Rottier, 1995). Das M-Protein eines Coronavirus tragt ent-
weder N- oder O-glykosidisch gebundene Oligosaccharide. So kommen z.B. bei TGEV,
FIPV, CCoV, IBV und HCoV-229E nur N-glykosidisch gebundene Zuckerseitenketten
vor, bei BCoV, MHV und HCoV-OC43 ist das M-Protein hingegen O-glykosyliert. Das
M-Protein enthalt 44–51% hydrophobe Aminosauren. Allen M-Proteinen der Coronavi-
ren ist gemeinsam, dass sie drei hydrophobe Domanen alternierend zu kurzen hydrophilen
Regionen besitzen. Mit diesen hydrophoben Regionen durchspannt das Protein die Virus-
membran (Laude et al., 1990). Am N-Terminus befindet sich ein hydrophiler Bereich, der
die glykosylierte Ektodomane des M-Proteins bildet. Die carboxyterminale Halfte aller M-
Proteine ist amphiphil mit einer hydrophilen Domane am carboxyterminalen Ende. Mit
dem amphiphilen Bereich ist das Protein moglicherweise mit der Innenseite der Virusmem-
bran assoziiert (Rottier et al., 1986; Laude et al., 1987, 1990). Neuere Untersuchungen
mit monoklonalen Antikorpern, die spezifisch gegen den C-Terminus des M-Proteins von
TGEV gerichtet sind, zeigen, dass es zusatzlich zur Nexo-Cendo-Topologie (N-Terminus
nach außen, C-Terminus ins Virusinnere gerichtet) auch eine Nexo-Cexo-Topologie (N-
und C-Terminus nach außen gerichtet) des M-Proteins in der Virusmembran gibt (Risco
et al., 1995; Escors et al., 2001a). In der Nexo-Cendo-Topologie scheint das M-Protein
eine wichtige Rolle im Assembly des Nukleokapsids zu spielen (Narayanan et al., 2000;
Escors et al., 2001b). Die Funktion des M-Proteins in der Nexo-Cexo-Topologie ist noch
nicht bekannt.
Das M-Protein ist das einzige virale Genprodukt, das bei seinem Transport an intrazel-
lularen Membranen zuruckgehalten wird. In infizierten Zellen akkumuliert das Protein
an den Membranen des Endoplasmatischen Retikulums und des Intermediaren Kompar-
timents und wird nicht bis zur Plasmamembran transportiert. Auch einzeln exprimier-
tes M-Protein gelangt nur bis zum Trans-Golgi-Network (Klumperman et al., 1994;
Krijnse-Locker et al., 1994, 1995). Die Lokalisation des M-Proteins in virusinfizierten

1.1. Coronaviren 23
Zellen weist darauf hin, dass das M-Protein eine wichtige Funktion fur die Virusreifung
besitzt. Coexpressionsstudien mit M- und E-Protein zeigten, dass diese zwei Proteine
ausreichend sind fur die Bildung von”virus-like particles“ (Bos et al., 1996; Vennema
et al., 1996; Kim et al., 1997). In virusinfizierten Zellen interagiert das M-Protein mit
dem S-Protein und dem HE-Protein (Opstelten et al., 1995; Nguyen und Hogue,
1997). Diese Oberflachenproteine assoziieren mit dem M-Protein in großen heteromeren
Komplexen, wodurch sie im Kompartiment der Virussreifung zuruckgehalten werden.
1.1.2.3 E-Protein
Das Gen des kleinen Envelope-Proteins ist im Virusgenom aller Coronavirusspezies strom-
aufwarts des M-Protein-Gens zu finden. Das Gen kodiert Polypeptide mit einem vorher-
sagbaren Molekulargewicht von 9,1 bis 12,4 kDa (Siddell, 1995b). Im Viruspartikel
konnte das E-Protein zu Beginn der 90er Jahre bei IBV (Liu und Inglis, 1991), bei
TGEV (Godet et al., 1992) und bei MHV (Yu et al., 1994) als integrales Membran-
protein nachgewiesen werden. Fur TGEV konnte gezeigt werden, dass das E-Protein in
Cexo-Nendo-Konfiguration vorliegt. Monoklonale Antikorper, die spezifisch Epitope des
C-Terminus erkennen, ergaben in fixierten, TGEV-infizierten ST-Zellen ein positives Si-
gnal bei der Oberflachenimmunfluoreszenz-Analyse (Godet et al., 1992). Fur das E-
Protein von MHV wurde gezeigt, dass es posttranslational acyliert wird (Yu et al., 1994).
Das E-Protein kommt im Viruspartikel generell nur in geringer Kopienzahl vor (z.B. bei
TGEV 20 Molekule pro Partikel (Godet et al., 1992)).
Das E-Protein spielt eine essentielle Rolle bei der Virusreifung. Die Coexpression von M-
und E-Protein ist fur die Bildung von”virus-like particles“ notwendig und ausreichend
(Vennema et al., 1996). Es wird vermutet, dass das E-Protein die spharische Morphologie
und die Abschnurung der Partikel bestimmt. Die Schlusselrolle des E-Proteins fur die
Virusreifung konnte ebenfalls durch die Erstellung und Untersuchung von Virusmutanten
durch gezielte Rekombination gezeigt werden (Fischer et al., 1998).

24 1. Einleitung
1.1.2.4 S-Protein
Das Surface- oder Spike-Protein (S-Protein) bildet die charakteristischen 20 nm langen,
keulenformigen Oberflachenprojektionen, denen die Coronaviren ihren Namen verdanken.
Das S-Protein besitzt bei der Neusynthese eine N-terminale Signalsequenz. Nahe dem hy-
drophilen C-Terminus befindet sich eine hydrophobe Region, mit der das Protein in der
Lipidmembran der Viruspartikel verankert ist. Die Ektodomane stellt den Hauptteil des
Proteins dar. Die Große des Polypeptids variiert je nach Coronavirusspezies zwischen 1160
Aminosauren (IBV) und 1452 Aminosauren (FIPV). Das Molekulargewicht von ungespal-
tenem S-Protein liegt zwischen 170 und 220 kDa. Die S-Proteine der Coronaviren besitzen
eine große Zahl potentieller N-Glykosylierungsstellen (bis zu 35), von denen die meisten
auch genutzt zu werden scheinen. Abhangig von Virustyp und -stamm und von der infizier-
ten Zielzelle werden die S-Proteine der Coronaviren zu unterschiedlichen Graden in eine
S1- und eine S2-Untereinheit gespalten. Gespaltenes S wurde nie bei TGEV und FIPV be-
obachtet, hingegen kamen die S-Proteine bei IBV-Stammen von Huhnerembryonen oder
Huhnernierenzellen uberwiegend in gespaltener Form vor. Bei MHV variiert der Grad
der Spaltung je nach Wirtszelltyp von 0–100%. Die S2-Untereinheit stellt den Teil des
S-Proteins dar, der in der Virusmembran verankert ist, und besitzt innerhalb der Virusfa-
milie die hochste Konservierung bezuglich der Aminosauresequenzen (fur eine Ubersicht
s. Cavanagh, 1995). Analysen der Aminosauresequenzen von S-Proteinen verschiede-
ner Coronavirusspezies haben die Anwesenheit von zwei Regionen mit”heptad repeats“
in der S2-Untereinheit ergeben, die auf eine α-helikale”coiled-coil“ Struktur hinweisen
(de Groot et al., 1987; Rasschaert und Laude, 1987). Daher wird das S-Protein als
Multimer, moglicherweise Homotrimer, angesehen, wobei die S2-Untereinheit einen Stiel
mit einer dem Influenza-Hamagglutinin-Trimer ahnlichen Struktur bildet (Laude et al.,
1990; Cavanagh, 1995). Es wurde gezeigt, dass die S-Protein-Monomere erst nach Erhalt
ihrer Konformation zu stabilen Trimeren zusammengelagert werden. Die Trimerbildung
findet jedoch vor den letzten Glykosylierungsschritten im Golgi-Apparat statt (Delmas
und Laude, 1990). Im Gegensatz zur S2-Untereinheit zeigen die S1-Untereinheiten der
Coronaviren wenig Aminosaureidentitat. Selbst innerhalb einer Spezies wie z.B. IBV sind
die S1-Untereinheiten der verschiedenen Stamme sehr heterogen. Die S1-Untereinheit be-
sitzt ein hydrophobes Profil und bildet den globularen Kopf des Virusspikes.

1.1. Coronaviren 25
Dem S-Protein kommt in der Anfangsphase der Infektion eine entscheidende Bedeutung
zu, denn es vermittelt die Bindung des Virions an den zellularen Rezeptor und die an-
schließende Fusion der viralen Lipidhulle mit der Zellmembran. Fur die Fusion scheint
das S-Protein alleine ausreichend zu sein (Vennema et al., 1990). Viele Coronaviren fu-
sionieren mit der Plasmamembran der Wirtszelle bei einem pH-Wert von 7 oder hoher.
Fur TGEV wurde aber gezeigt, dass die Infektion nach rezeptorvermittelter Endozytose
und nachfolgender saureabhangiger Fusion mit einem intrazellularen Kompartiment statt-
findet (Hansen et al., 1998). Auch fur bestimmte IBV-Stamme wurde der Eintritt des
Virions in die Zelle durch Endozytose beschrieben.
Das S-Protein induziert uberwiegend die Bildung neutralisierender Antikorper im Ver-
lauf einer Infektion. Die antigenen Epitope der Coronaviren befinden sich vor allem im
aminoterminalen Bereich (S1-Untereinheit) des S-Proteins (Delmas et al., 1990).
1.1.2.5 HE-Protein
Die Coronaviren der Gruppe II besitzen ein zusatzliches Oberflachenprotein, die Ham-
agglutinin-Esterase (HE), die als rezeptorzerstorendes Enzym wirkt (fur eine Ubersicht
s. Brian et al., 1995). Im Elektronenmikroskop ist bei den Coronaviren der Gruppe II
ein zweiter Ring kurzerer Spikes zu sehen, der vom HE-Protein gebildet wird. Das Gen
des HE-Proteins befindet sich auf der 5’ Seite des S-Protein-Gens. Das HE-Protein ist ein
N-glykosyliertes Typ I-Membranprotein, welches als uber Disulfidbrucken verbundenes
Homodimer vorkommt. Ein Monomer hat ein Molekulargewicht von 60–65 kDa, das Dimer
von 130–140 kDa.
Eine Hauptfunktion des HE-Proteins ist seine Enzymaktivitat in Form einer 9-O-Acetyl-
esterase wie sie auch bei Influenza-C-Viren vorkommt (Vlasak et al., 1988a,b). Das
HE-Protein zeigt betrachtliche Homologie zum HEF-Protein des Influenza-C-Virus (30%
Homologie auf Aminosaureebene) und weist in seiner Aminosauresequenz ein F-G-D-S-
Motiv auf, welches identisch mit dem katalytischen Zentrum der Acetylesterase des HEF-
Proteins bei Influenza-C ist. Bei dieser Esterase handelt es sich um eine klassische Serines-
terase, die durch DFP (Diisopropylfluorophosphat) gehemmt wird. Eine solche Hemmung
der Enzymaktivitat wirkt auch inhibierend auf die Virusvermehrung, was vermuten ließ,

26 1. Einleitung
dass eine aktive virale Esterase essentiell fur den Viruseintritt in die Zelle ist (Vlasak
et al., 1988a).
Dem HE-Protein wurde zusatzlich eine Hamagglutinationsaktivitat uber die Bindung an
N-Acetyl-9-O-Acetylneuraminsaure (Neu5,9Ac2) zugeschrieben. Das HE-Protein wurde
aus diesem Grund als Bindungsprotein zur Initiation einer Infektion diskutiert (Vlasak
et al., 1988b). Es wurde jedoch gezeigt, dass das S-Protein von BCoV und HCoV-OC43
eine hohere Bindungsaktivitat fur Neu5,9Ac2 besitzt und damit das effizientere Hamag-
glutinin ist (Schultze et al., 1991a,b; Schultze und Herrler, 1992; Kunkel und
Herrler, 1993). Die Funktion des HE-Proteins scheint somit vor allem darin zu be-
stehen, die Virusoberflache als rezeptorzerstorendes Enzym frei von Glykokonjugaten zu
halten. Die Funktion der Bindung (S-Protein) und der rezeptorzerstorenden Aktivitat
(HE-Protein) scheinen sich somit bei den Coronaviren der Gruppe II wie bei Influenza-
A-Viren (Hamagglutinin, Neuraminidase) auf zwei Proteine zu verteilen.
1.2 Das Virus der ubertragbaren Gastroenteritis
(TGEV)
1.2.1 Pathogenitat und Epidemiologie
Falle der ubertragbaren Gastroenteritis der Schweine wurden vermutlich bereits in den
30er Jahren beobachtet (Smith, 1956). Die virale Atiologie der transmissiblen Gastro-
enteritis (TGE) wurde 1946 durch einen Bericht von Doyle und Hutchings (1946) in
den Vereinigten Staaten bestatigt. Sie konnten zeigen, dass ein filtrierbares Agens auf
Schweine ubertragen werden konnte und daraufhin eine haufig fatale Gastroenteritis her-
vorrief. In den folgenden Jahrzehnten wurde die TGE in allen Kontinenten beschrieben.
Von Bedeutung wurde die ubertragbare Gastroenteritis fur die Schweineindustrie mit ih-
rer Intensivierung (Woode, 1969). Den Kriterien der Familie der Coronaviridae folgend
(Tyrrell et al., 1968) wurde TGEV von Tajima (1970) in die Gruppe der Coronaviren
eingeordnet.
Bis in die 70er Jahre traten TGEV-Infektionen in den Vereinigten Staaten und in Europa

1.2. Das Virus der ubertragbaren Gastroenteritis (TGEV) 27
vor allem im Winter als akute Epizootien in Aufzuchtbetrieben auf. Nach einem Aus-
bruch der TGE hielt die Immunitat der Herde ca. 2 bis 3 Jahre an, bis es dann durch
Einschleppung des Virus zu einem neuen Ausbruch kam. Durch die Intensivierung der
Schweinezucht wandelte sich das Bild der TGEV-Infektion zu einer Enzootie vor allem
in großen Zuchtbetrieben, in denen das Virus nach einem Erstausbruch in Absetzferkeln
persistierte. In Europa und in den Vereinigten Staaten ist das Auftreten der ubertragba-
ren Gastroenteritis rucklaufig (Enjuanes und van der Zeijst, 1995). Es gibt jedoch
auch Berichte von schweren TGE-Ausbruchen in den 90er Jahren. So starben z.B. 1996
bei einem schweren Ausbruch in England 750 Ferkel und 5 Sauen (Jones und Paton,
1996; McGoldrick et al., 1999).
TGEV ist ein typisch enteropathogener Vertreter der Coronaviren, kann aber auch im Res-
pirationstrakt und in der Milchdruse laktierender Sauen replizieren. Das Virus wird mit
infektiosem Material z.B. den Faces oder der Sauenmilch auf oralem Wege aufgenommen.
Es erreicht uber die Magen-Darm-Passage den Dunndarm und infiziert die Epithelzellen
im apikalen Bereich der Zotten von Jejunum und Ileum. Das Virus breitet sich uber den
Villus auf die weiter abwarts liegenden Epithelzellen aus, wurde aber nie in den Zellen an
der Zottenbasis und in den Zellen der Krypten gefunden (Garwes, 1995). Die infizierten
Epithelzellen losen sich vom Villus ab und werden von neuen Zellen aus den Krypten
ersetzt. Dieser Vorgang beendet die Infektion nach ca. 5–7 Tagen. Die neuen Zellen wer-
den nicht infiziert. Durch die Ablosung der Epithelzellen kommt es zur Villusatrophie
und dadurch bedingt zu einer wassrigen Diarrhoe (Hooper und Haelterman, 1969;
Pensaert et al., 1970). Epizootische Ausbruche der ubertragbaren Gastroenteritis be-
treffen Schweine aller Altersgruppen und breiten sich in 2–3 Tagen auf die gesamte Herde
aus. Der Schweregrad der Erkrankung ist jedoch je nach Alter der Tiere unterschiedlich.
Bei Saugferkeln bis zum Alter von zwei Wochen kommt es haufig zu Erbrechen und wass-
riger Diarrhoe, die zu Dehydratation, starkem Gewichtsverlust und Tod in 2 bis 5 Tagen
fuhren. Die Mortalitatsrate in dieser Altersgruppe liegt bei bis zu 100%. Bei Absetzferkeln
(3–8 Wochen alt) betragt die Mortalitatsrate weniger als 10–20%. Es kommt jedoch zu
Wachstumsverzogerungen und somit zu wirtschaftlichen Verlusten. Erwachsene Tiere ent-
wickeln ebenfalls einen wassrigen Durchfall, der 2–4 Tage anhalt, aber die Mortalitatsrate
in dieser Altersgruppe liegt unter 5%.

28 1. Einleitung
Die Empfanglichkeit fur eine TGEV-Infektion setzt sich aus unterschiedlichen Faktoren
zusammen: Alter der Tiere, Umweltbedingungen, Virusdosis und Virulenz des jeweiligen
Virusstammes. Auch die Moglichkeit zur Kolostrumaufnahme scheint bei neugeborenen
Ferkeln die Empfanglichkeit gegenuber einer TGEV-Infektion zu beeinflussen. So konn-
te gezeigt werden, dass Neugeborene, die kein Kolostrum erhielten, nach einer Infektion
mit TGEV (PUR46-MAD) starben (100% Mortalitat). Ihre Wurfgeschwister, die fur 7 h
Zugang zum Kolostrum hatten, zeigten hingegen keinerlei Krankheitszeichen. In welcher
Weise die Kolostrumaufnahme die Empfanglichkeit fur eine TGEV-Infektion beeinflusst,
ob z.B. dadurch die Epithelzelldifferenzierung verandert ist oder ob Inhaltsstoffe des Ko-
lostrums mit den Virionen interagieren, ist noch nicht bekannt (Enjuanes und van der
Zeijst, 1995). Als Ursache fur den Schweregrad des Krankheitsverlaufs epizootischer
TGE-Ausbruche bei Saugferkeln wurden ebenfalls unterschiedliche Faktoren diskutiert:
eine geringe Enterozytenerneuerungsrate bei neugeborenen Ferkeln (Moon, 1973); das
Vorkommen von Enterozyten fetalen Ursprungs auf dem Villus, welche durch ihr tubulo-
vakuolares System die Virusreplikation erleichtern (Wagner et al., 1973); die fehlende
Aktivitat von naturlichen Killerzellen in den intraepithelialen Lymphozyten neugebore-
ner Ferkel (Cepica und Derbyshire, 1984) sowie das Vorkommen eines zusatzlichen
Rezeptors auf den Zotten Neugeborener (Weingartl und Derbyshire, 1993).
Wahrend der serologischen Routineuberwachung von Schweinebestanden in Großbritan-
nien, Belgien, den Niederlanden und Frankreich vermerkte man Mitte der 80er Jahre eine
Erhohung der Anzahl an Schweinebestanden, die Antikorper gegen TGEV besaßen. Dieser
Anstieg korrelierte nicht mit einer Erhohung der Erkrankungsrate an TGE. 1986 konnte
schließlich eine TGEV-Variante, das porzine respiratorische Coronavirus (PRCoV), aus
dem Respirationstrakt infizierter Schweine in Belgien isoliert werden (Pensaert et al.,
1986). Diese Variante replizierte mit hoher Effizienz im Respirationstrakt, jedoch nicht
oder nur in geringem Ausmaß im Gastrointestinaltrakt von Schweinen (Cox et al., 1990).
Die meisten Beobachter registrierten in Verbindung mit einer PRCoV-Infektion nur we-
nige oder keine Anzeichen einer respiratorischen Erkrankung (Enjuanes und van der
Zeijst, 1995). Serologische Untersuchungen ergaben, dass TGEV und PRCoV nahe ver-
wandt sind (Callebaut et al., 1988; Sanchez et al., 1992). Aus diesem Grund wird
PRCoV auch als TGEV-Variante und nicht als eigene Coronavirusspezies betrachtet. Da
das porzine respiratorische Coronavirus aerogen ubertragen wird, besitzt es eine hohe

1.2. Das Virus der ubertragbaren Gastroenteritis (TGEV) 29
Ausbreitungsrate. So waren bis Ende der 80er Jahre fast alle Schweinebestande in Belgien
infiziert (Pensaert und Cox, 1989). Diese Ausbreitung von PRCoV wird als Grund
fur den Ruckgang der TGEV-Infektionen angesehen. So wirkt eine PRCoV-Infektion
aufgrund der serologischen Verwandtschaft haufig als naturlicher Impfschutz gegen eine
TGEV-Infektion. Die Analyse der Sequenzen beider Viren ergab eine Homologie von 96%.
Die Hauptunterschiede in der Sequenz bestehen in drei Deletionen des PRCoV-Genoms
(Rasschaert et al., 1990; Wesley et al., 1991; Sanchez et al., 1992). Durch zwei der
Deletionen wird der ORF3a, der fur ein Nicht-Strukturprotein unbekannter Funktion ko-
diert, in ein Pseudogen verwandelt. Die dritte Deletion betrifft das Gen des S-Proteins.
Durch eine Deletion von 672 Nukleotiden (224 Aminosauren) bei europaischen Isolaten
bzw. 681 Nukleotiden (227 Aminosauren) bei nordamerikanischen Isolaten besitzt PRCoV
ein in seinem aminoterminalen Bereich stark verkurztes S-Protein.
1.2.2 Eigenschaften des S-Proteins von TGEV
Das Gen des S-Proteins von TGEV kodiert ein Polypeptid mit einer Lange von 1447 Ami-
nosauren. Nach Abspaltung des Signalpeptids (16 Aminsauren) im Endoplasmatischen Re-
tikulum werden an die meisten 32 potentiellen N-Glykosylierungsstellen Mannose-reiche
Kohlenhydratketten angehangt. Nach einem zusatzlichen Glykosylierungsschritt im Golgi-
Apparat wird das reife S-Protein (220 kDa) in die Viruspartikel eingebaut (Laude et al.,
1990). Die Spikes auf der Oberflache des Virions werden durch Homotrimere des S-Proteins
gebildet. Diese Trimere bilden sich, bevor die komplexen Glykosylierungsschritte ablaufen
(Delmas und Laude, 1990). Eine proteolytische Spaltung des S-Proteins in eine S1- und
eine S2-Untereinheit findet nicht statt.
1.2.2.1 Epitope
Die Antigen-Struktur des S-Proteins wurde von den Arbeitsgruppen Laude in Frankreich
und Enjuanes in Spanien mit Hilfe von monoklonalen Antikorpern eingehend untersucht.
Beschrieben wurden vier Epitope, die alle im aminoterminalen Bereich des S-Proteins
liegen (Delmas et al., 1986; Jimenez et al., 1986; Correa et al., 1988). Durch die
Isolierung von Escape-Mutanten wurden die an der Bildung der Epitope beteiligten Ami-

30 1. Einleitung
D 224
COOH
COOH
C B
D
D
C
A
A-B
NH2
NH2
PRCoV
TGEV-PUR46 Madrid
TGEV-PUR115 Paris
5051
97144
163165
385
383384
538 (Aa)543 (Ac)586 (Aa/b)591 (Ab)
549586
4 229
Abbildung 1.2: Schematische Darstellung der Epitope des S-Proteins von TGEV (Madrid) nach
Gebauer et al. (1991) und TGEV (Paris) nach Delmas et al. (1990). Die Zahlen unter den
einzelnen Epitopen bezeichnen die Positionen der an der Epitopbildung beteiligten Aminosaur-
en. Vergleichend ist das S-Protein von PRCoV mit seiner Deletion im aminoterminalen Bereich
dargestellt.
nosauren identifiziert (Correa et al., 1990; Delmas et al., 1990; Gebauer et al., 1991).
Bezeichnung und Lage der Epitope sind in Abbildung 1.2 dargestellt.
Das von der Arbeitsgruppe Enjuanes zusatzlich gefundene Epitop C ist nicht auf der
Oberflache nativer Virionen exponiert. Innerhalb der TGEV-Stamme sind die Epitope A
und B (nach Enjuanes) bzw. A/B und D (nach Laude) stark konserviert. Das Epitop A
(nach Enjuanes) bzw. A/B (nach Laude) ist der Hauptverursacher der Bildung neutra-
lisierender Antikorper. Monoklonale Antikorper gegen dieses Epitop reagieren ebenfalls
mit FIPV. Ein Vergleich der Aminosauresequenzen beider Viren im Bereich dieses Epi-
tops ergab eine Homologie von 92%. Im Bereich des Epitops D (nach Laude) bzw. B
(nach Enjuanes) lag hingegen nur eine Homologie von 25% zwischen TGEV und FIPV
vor (Delmas et al., 1990). Die korrekte Faltung der Epitope A und B (nach Enjuanes)
bzw. A/B und D (nach Laude) ist abhangig von der Glykosylierung. Nach vollstandigem
Zusammenbau des Virions ist der Kohlenhydratanteil nicht bedeutsam fur die Antigenitat
der Epitope (Gebauer et al., 1991).

1.2. Das Virus der ubertragbaren Gastroenteritis (TGEV) 31
1.2.2.2 Bindungsaktivitaten
Das S-Protein von TGEV besitzt zwei Bindungsaktivitaten. Mit einer Bindungsstelle
im Bereich zwischen Aminosaure 522 und 744 wird der zellulare Rezeptor, die porzine
Aminopeptidase N (pAPN) erkannt (Delmas et al., 1992; Godet et al., 1994). Eine
zweite Bindungsaktivitat erlaubt es TGEV Erythrozyten zu agglutinieren (Noda et al.,
1987, 1988). Die Hamagglutinationsaktivitat des Virus wird durch die Bindung an Sia-
linsauren verursacht (Schultze et al., 1993). Genauere Untersuchungen zeigten, dass
N-Glykolylneuraminsaure (Neu5Gc) der haufig vorkommenden N-Acetylneuraminsaure
(Neu5Ac) vorgezogen wird (Schultze et al., 1995, 1996). Die Sialinsaurebindungsstel-
le befindet sich im Bereich des Epitops B (nach Enjuanes) bzw. D (nach Laude) des
S-Proteins von TGEV (Schultze et al., 1996).
PRCoV besitzt wie TGEV die Eigenschaft an pAPN zu binden (Delmas et al., 1993).
Eine Hamagglutinationsaktivitat und damit Sialinsaurebindungsaktivitat besitzt das por-
zine respiratorische Coronavirus jedoch nicht, da die Bindungsstelle fur Sialinsauren in
dem bei PRCoV deletierten Bereich liegt (Schultze et al., 1996). Die S-Proteine von
TGEV und PRCoV sind zum Vergleich schematisch in Abbildung 1.2 dargestellt.
Die Sialinsaurebindungsaktivitat des transmissiblen Gastroenteritisvirus bewirkt, dass in
Zellkultur wachsendes Virus mit Hamagglutinationsinhibitoren zellularen Ursprungs um-
geben wird. Dieser Maskierung der Sialinsaurebindungsaktivitat kann durch Neuraminida-
sebehandlung entweder der Zellen vor der Infektion oder der Virionen nach der Virusernte
entgegengewirkt werden (Krempl et al., 1997). Eine nahere Analyse der Sialinsaure-
bindungseigenschaft von TGEV mit Hilfe von hamagglutinationsdefizienten Mutanten
(HAD-Mutanten) zeigte, dass das Virus durch die Bindung an zellulare Sialoglykokon-
jugate eine erhohte Resistenz gegenuber dem Detergenz Oktylglukosid besitzt (Krempl
et al., 2000). Da Epitop-D-Mutanten, die keine Hamagglutinationsaktivitat zeigten, auch
eine stark reduzierte Enteropathogenitat aufwiesen, wird vermutet, dass die Sialinsaure-
bindungsaktivitat von TGEV einen Pathogenitatsfaktor darstellt (Bernard und Laude,
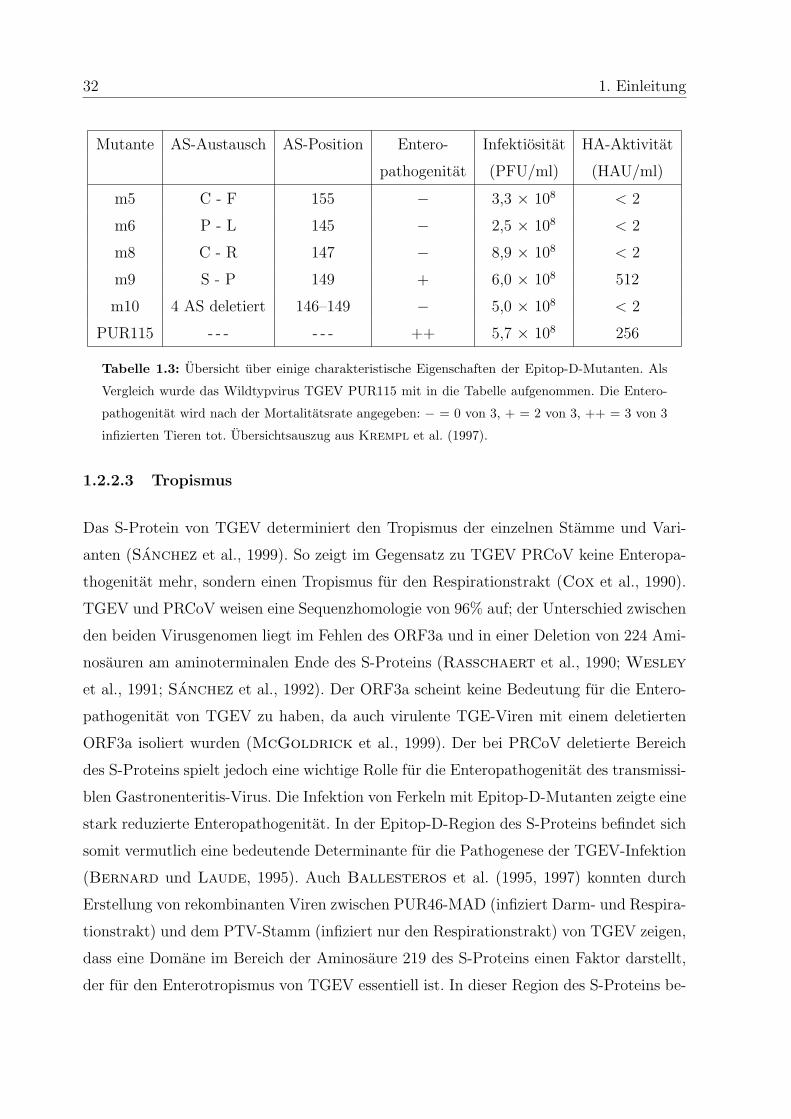
1995; Krempl et al., 1997). Eine Ubersicht uber einige der Epitop-D-Mutanten und ihre
Eigenschaften gibt Tabelle 1.3.
32 1. Einleitung
Mutante AS-Austausch AS-Position Entero- Infektiositat HA-Aktivitat
pathogenitat (PFU/ml) (HAU/ml)
m5 C - F 155 − 3,3 × 108 < 2
m6 P - L 145 − 2,5 × 108 < 2
m8 C - R 147 − 8,9 × 108 < 2
m9 S - P 149 + 6,0 × 108 512
m10 4 AS deletiert 146–149 − 5,0 × 108 < 2
PUR115 - - - - - - ++ 5,7 × 108 256
Tabelle 1.3: Ubersicht uber einige charakteristische Eigenschaften der Epitop-D-Mutanten. Als
Vergleich wurde das Wildtypvirus TGEV PUR115 mit in die Tabelle aufgenommen. Die Entero-
pathogenitat wird nach der Mortalitatsrate angegeben: − = 0 von 3, + = 2 von 3, ++ = 3 von 3
infizierten Tieren tot. Ubersichtsauszug aus Krempl et al. (1997).
1.2.2.3 Tropismus
Das S-Protein von TGEV determiniert den Tropismus der einzelnen Stamme und Vari-
anten (Sanchez et al., 1999). So zeigt im Gegensatz zu TGEV PRCoV keine Enteropa-
thogenitat mehr, sondern einen Tropismus fur den Respirationstrakt (Cox et al., 1990).
TGEV und PRCoV weisen eine Sequenzhomologie von 96% auf; der Unterschied zwischen
den beiden Virusgenomen liegt im Fehlen des ORF3a und in einer Deletion von 224 Ami-
nosauren am aminoterminalen Ende des S-Proteins (Rasschaert et al., 1990; Wesley
et al., 1991; Sanchez et al., 1992). Der ORF3a scheint keine Bedeutung fur die Entero-
pathogenitat von TGEV zu haben, da auch virulente TGE-Viren mit einem deletierten
ORF3a isoliert wurden (McGoldrick et al., 1999). Der bei PRCoV deletierte Bereich
des S-Proteins spielt jedoch eine wichtige Rolle fur die Enteropathogenitat des transmissi-
blen Gastronenteritis-Virus. Die Infektion von Ferkeln mit Epitop-D-Mutanten zeigte eine
stark reduzierte Enteropathogenitat. In der Epitop-D-Region des S-Proteins befindet sich
somit vermutlich eine bedeutende Determinante fur die Pathogenese der TGEV-Infektion
(Bernard und Laude, 1995). Auch Ballesteros et al. (1995, 1997) konnten durch
Erstellung von rekombinanten Viren zwischen PUR46-MAD (infiziert Darm- und Respira-
tionstrakt) und dem PTV-Stamm (infiziert nur den Respirationstrakt) von TGEV zeigen,
dass eine Domane im Bereich der Aminosaure 219 des S-Proteins einen Faktor darstellt,
der fur den Enterotropismus von TGEV essentiell ist. In dieser Region des S-Proteins be-

1.3. Escherichia coli F5/K99 (E. coli F5/K99) 33
findet sich die Sialinsaurebindungsaktivitat von TGEV, so dass eine Korrelation zwischen
der Enteropathogenitat und der Sialinsaurebindungsaktivitat postuliert wurde (Krempl
et al., 1997).
1.3 Escherichia coli F5/K99 (E. coli F5/K99)
Die durch enterotoxinogene E. coli (ETEC) verursachten Diarrhoen zahlen zu den wich-
tigsten Infektionskrankheiten bei Absetz- und Saugferkeln. Die Erkrankungen werden
durch E. coli -Stamme verschiedener Serogruppen hervorgerufen, die den Darm des Tieres
besiedeln und durch Toxinfreisetzung den Wirtsorganismus schadigen. Fur die Besiedelung
des Darms sind spezielle Adhasionsmechanismen verantwortlich. Die Bakterien besitzen
auf ihrer Oberflache Fimbrien, die das Haften an der Darmschleimhaut ermoglichen und
somit den Abtransport mit dem Darminhalt verhindern. Bei porzinen ETEC-Stammen
sind verschiedene Fimbrien-Typen beschrieben, die aus Protein bestehen und sich mor-
phologisch und antigenetisch voneinander unterscheiden (fur eine Ubersicht s. Stamm
und Sorg (1993)). Die Fimbrien dienen den Bakterienzellen als Kolonisationsfaktor, da
sie mit ihnen an spezifische Rezeptoren auf der Darmschleimhaut binden und somit den
Darm kolonisieren konnen.
Epidemiologische Untersuchungen in Spanien ergaben, dass das Vorhandensein von F5-
Fimbrien statistisch mit Durchfallerkrankungen von Ferkeln, die junger als 15 Tage waren,
assoziiert war (Garabal et al., 1997). Bei Kalbern und Lammern ist E. coli F5 ein
haufiger Vertreter der enterotoxinogenen E. coli -Stamme (Guinee et al., 1976). Kalber
und Lammer sind nur bis zum zweiten Lebenstag empfanglich fur eine Infektion mit dem
Bakterium; bei Ferkeln konnten nach dem Absetzen keine enterotoxinogenen E. coli F5
mehr bei Durchfallerkrankungen nachgewiesen werden (Gaastra und de Graaf, 1982).
Fimbrien besitzen die Eigenschaft Erythrozyten zu agglutinieren. Bei E. coli F5 geschieht
dies uber die Bindung an Sialinsauren von Zelloberflachenkomponenten. Durch diese Sia-
linsaurebindungseigenschaft konnen die Bakterien auch an Sialoglykokonjugate im Darm
binden und so diesen kolonisieren (Gaastra und de Graaf, 1982). E. coli F5 bevorzugt
Neu5Gc als Rezeptordeterminante auf der Erythrozytenoberflache (Lindahl et al., 1987;
Ono et al., 1989). Rezeptoren fur F5 wurden einerseits als Glykolipide, andererseits als

34 1. Einleitung
Glykoproteine beschrieben. So isolierten Teneberg et al. (1990) selektiv Glykolipide aus
dem Dunndarm neugeborener und erwachsener Schweine und identifizierten als rezep-
toraktives Glykolipid Neu5Gc-GM3 (N-Glykolylneuraminyl-neolaktotetraosyl-ceramid),
welches nur bei der Gruppe der Neugeborenen vorkam. Andere Arbeitsgruppen beschrie-
ben die Bindung der F5-Fimbrien an Muzine auf der Darmschleimhaut des Ferkeldunn-
darms oder die Reduzierung der Adhasion von E. coli F5 an das Darmepithel durch
Kohlenhydratanteile von Glykoproteinen aus Rinderseren (Lindahl und Carlstedt,
1990; Mouricout et al., 1990). Zusatzlich konnte nachgewiesen werden, dass das Auf-
treten von adhasiven und nicht-adhasiven Phanotypen bei der Infektion mit E. coli F5
mit dem Gehalt bestimmter Sialoglykolipide korreliert (Seignole et al., 1991).
1.4 Virusrezeptoren
Viren besitzen keinen eigenen Stoffwechselapparat und sind somit außerhalb lebender
Zellen nicht vermehrungsfahig. Um sich vermehren zu konnen, nutzen Viren die Synthe-
semaschinerie von Zellen und funktionieren sie fur ihre Zwecke um. Die Infektion einer
Zelle beginnt mit der Anlagerung der Viruspartikel an die Plasmamembran. In diesem
ersten Schritt binden die Virionen uber virale Oberflachenproteine an Molekule auf der
Zellmembran, die in diesem Zusammenhang als Virusrezeptoren bezeichnet werden. Die
Bindung an die Zelle ist notwendig, um in einem zweiten Schritt die Plasmamembran
penetrieren und damit die virale Erbinformation in die Zelle einschleusen zu konnen. Bei
behullten Viren ist hierzu die Fusion der Virusmembran mit der Plasmamembran bzw.
nach Endozytose mit der Endosomenmembran notwendig.
Das Vorkommen eines viralen Rezeptors kann den Tropismus und die Pathogenitat von
Viren bestimmen. Als Rezeptoren konnen sowohl Proteine und Lipide als auch Kohlen-
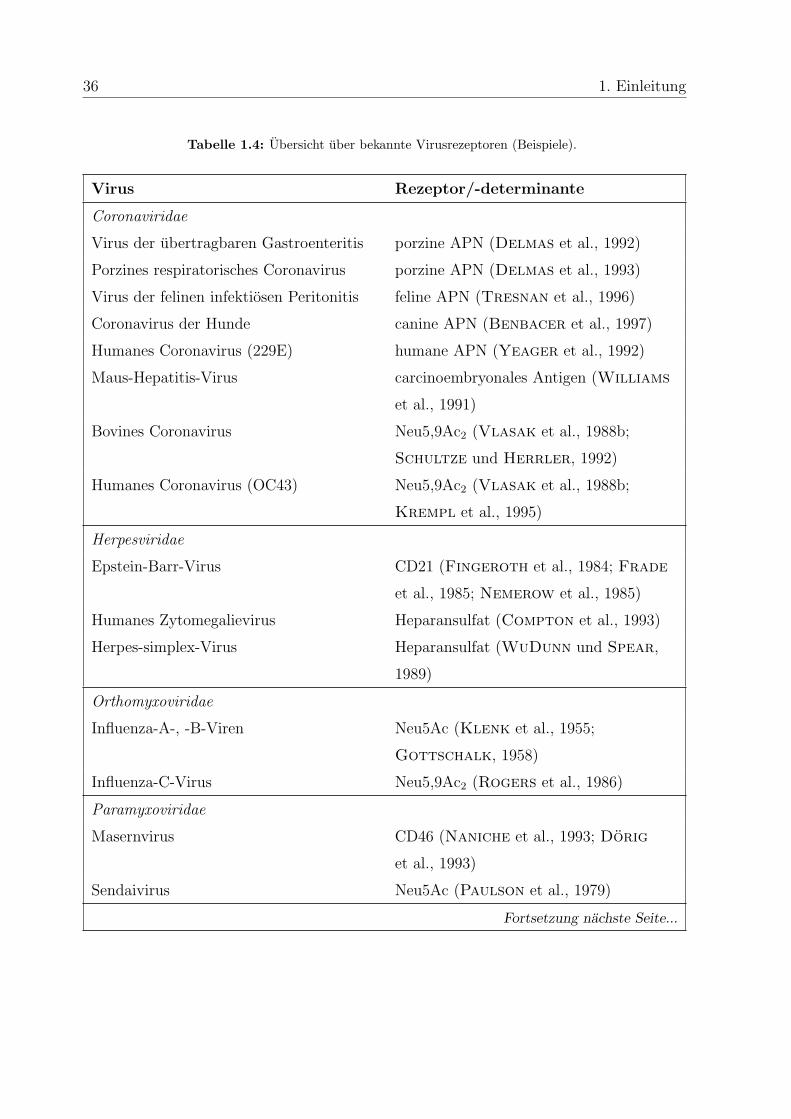
hydrate dienen. Eine beispielhafte Ubersicht uber bekannte Virusrezeptoren gibt Tabelle
1.4. Viele Viren erkennen mehr als einen Rezeptor oder benotigen zusatzlich zu ihrem
zellularen Rezeptor weitere Faktoren zur Etablierung einer Infektion in vivo. So sind z.B.
bei der Infektion von HIV-1 neben CD4 auch Proteine aus der Chemokinrezeptorfamilie
beteiligt (Lapham et al., 1996; Trkola et al., 1996; Wu et al., 1996). Die Bindung
an CD4 bewirkt eine Konformationsanderung des HIV-Spikeproteins (gp120), wodurch

1.4. Virusrezeptoren 35
die Bindung an den Corezeptor aus der Chemokinfamilie (CCR5) ermoglicht wird. Diese
zweite Interaktion bewirkt bei dem Transmembranprotein gp41 die Umwandlung in eine
fusionsaktive Konformation. Erst dann lauft die Fusion zwischen Virus- und Zellmembran
ab.
Der erste Virusrezeptor, der identifiziert wurde, war N-Acetylneuraminsaure (Neu5Ac),
die von Influenza-A- und -B-Viren sowie von einigen Paramyxoviren erkannt wird (Klenk
et al., 1955; Gottschalk, 1958). Dieses Beispiel fur einen Kohlenhydratrezeptor ist auf-
grund der jahrzehntelangen Forschungen gut charakterisiert. Neu5Ac bindet uber Wasser-
stoffbrucken und van-der-Waals-Krafte in einer Rezeptortasche, die sich im globularen Teil
des Influenza-Hamagglutinins befindet (Weis et al., 1988). Verschiedene Influenzastamme
haben eine Praferenz fur die α2,3- bzw. α2,6-gebundene Form der Sialinsaure (Rogers
und Paulson, 1983). Eine andere Sialinsaure, die N-Acetyl-9-O-Acetylneuraminsaure
(Neu5,9Ac2), wird z.B. von Influenza-C-Viren als Rezeptordeterminante erkannt (Rogers
et al., 1986). Inzwischen kennt man sowohl einige umhullte als auch nicht-umhullte Vi-
ren, die Sialinsauren als Rezeptordeterminante fur die Einleitung einer Infektion nutzen
(Tab. 1.4). Unter diesen Viren besitzen die umhullten Viren i.d.R. zusatzlich ein rezep-
torzerstorendes Enzym. Bei Influenza-A- und -B-Viren sowie Paramyxoviren handelt es
sich hierbei um eine Neuraminidase, die die Sialinsaure von der Galaktose abspaltet. Die
Viren, die Neu5,9Ac2 als Rezeptordeterminante nutzen, besitzen eine 9-O-Acetylesterase,
die die Acetylgruppe am C9 hydrolysiert.
Bei den Coronaviren sind bisher fur einige Vertreter Proteinrezeptoren entdeckt worden.
Andere Coronaviren nutzen Sialinsauren als Rezeptordeterminante. Fur die meisten Ver-
treter der Serogruppe I wurde Aminopeptidase N (APN), eine Metalloprotease, die auf der
Oberflache von Enterozyten, Lungenfibroblasten und Nierenepithelzellen vorkommt, als
Rezeptor identifiziert (Delmas et al., 1992; Yeager et al., 1992; Tresnan et al., 1996;
Benbacer et al., 1997). Die Bindung an Aminopeptidase N scheint die Speziesspezifitat
der Viren zu determinieren (Delmas et al., 1994). Bei TGEV (Delmas et al., 1992) und
HCoV-229E (Yeager et al., 1992), fur die als erstes die Bindung an APN gezeigt wurde,
liegt die Bindungsstelle in unterschiedlichen Domanen des Enzyms. Fur die Bindung von
TGEV ist die Region zwischen Aminosaure 717 und 813 von APN essentiell (Delmas
et al., 1994). Hingegen ist bei der HCoV-229E-Infektion ein Bereich zwischen Aminosaure
36 1. Einleitung
Tabelle 1.4: Ubersicht uber bekannte Virusrezeptoren (Beispiele).
Virus Rezeptor/-determinante
Coronaviridae
Virus der ubertragbaren Gastroenteritis porzine APN (Delmas et al., 1992)
Porzines respiratorisches Coronavirus porzine APN (Delmas et al., 1993)
Virus der felinen infektiosen Peritonitis feline APN (Tresnan et al., 1996)
Coronavirus der Hunde canine APN (Benbacer et al., 1997)
Humanes Coronavirus (229E) humane APN (Yeager et al., 1992)
Maus-Hepatitis-Virus carcinoembryonales Antigen (Williams
et al., 1991)
Bovines Coronavirus Neu5,9Ac2 (Vlasak et al., 1988b;
Schultze und Herrler, 1992)
Humanes Coronavirus (OC43) Neu5,9Ac2 (Vlasak et al., 1988b;
Krempl et al., 1995)
Herpesviridae
Epstein-Barr-Virus CD21 (Fingeroth et al., 1984; Frade
et al., 1985; Nemerow et al., 1985)
Humanes Zytomegalievirus Heparansulfat (Compton et al., 1993)
Herpes-simplex-Virus Heparansulfat (WuDunn und Spear,
1989)
Orthomyxoviridae
Influenza-A-, -B-Viren Neu5Ac (Klenk et al., 1955;
Gottschalk, 1958)
Influenza-C-Virus Neu5,9Ac2 (Rogers et al., 1986)
Paramyxoviridae
Masernvirus CD46 (Naniche et al., 1993; Dorig
et al., 1993)
Sendaivirus Neu5Ac (Paulson et al., 1979)
Fortsetzung nachste Seite...

1.4. Virusrezeptoren 37
... Fortsetzung von letzter Seite
Virus Rezeptor/-determinante
Picornaviridae
Poliovirus PVR (Mendelsohn et al., 1989)
CD44 (Shepley und Racaniello,
1994)
Rhinovirus ICAM-1 (Greve et al., 1989; Staunton
et al., 1989; Tomassini et al., 1989)
Enzephalomyokarditis-Virus Neu5Ac (Burness und Pardoe, 1981)
Papovaviridae
Polyomavirus Neu5Ac (Fried et al., 1981)
Retroviridae
HIV-1 CD4 (Dalgleish et al., 1984;
Klatzmann et al., 1984)
CXCR4 (T-Zell-trope Stamme) (Berson
et al., 1996; Feng et al., 1996)
CCR5 (Makrophagen und TH-Zell-trope
Stamme) (Alkhatib et al., 1996; Choe
et al., 1996; Deng et al., 1996; Doranz
et al., 1996; Dragic et al., 1996)
Mause-Leukamie-Virus basischer Aminosauretransporter
(Albritton et al., 1989)
Phosphattransporter (O’Hara et al.,
1990)
260 und 353 in der Nahe des katalytischen Zentrums von hAPN von Bedeutung (Yeager
et al., 1992; Kolb et al., 1996). FIPV und CCoV erkennen Domanen, die dem von TGEV
genutzten Bereich auf dem Enzym entsprechen (Tresnan et al., 1996; Benbacer et al.,
1997). Die feline Aminopeptidase N wird nicht nur von FIPV und FCoV, sondern auch
von CCoV, TGEV und HCoV-229E erkannt. Die felinen und caninen Coronaviren erken-
nen jedoch die porzine und humane Form der Aminopeptidase nicht (Tresnan et al.,
1996; Benbacer et al., 1997). So konnen Katzen z.B. mit TGEV infiziert werden; der

38 1. Einleitung
Verlauf ist jedoch asymptomatisch (Reynolds und Garwes, 1979).
Die Maus-Hepatitis-Viren (MHV) als Vertreter der Serogruppe II der Coronaviren nut-
zen Glykoproteine aus der Familie der carcinoembryonalen Antigene (CEA), die der
Immunglobulin-Superfamilie zugeordnet werden, als Rezeptoren (Dveksler et al., 1991;
Williams et al., 1991; Dveksler et al., 1993b). Dieses von MHV genutzte CEA-Gly-
koprotein befindet sich auf Zelloberflachen von Leber, Dunn- und Dickdarm. Diese Or-
gane stellen Ziele der MHV-Replikation dar. Das Virus bindet an die aminoterminale
Immunglobulin-artige Domane seines Rezeptors (Dveksler et al., 1993a). Zusatzlich
erkennen einige MHV, die ein HE-Protein besitzen, Neu5,9Ac2. Diese zusatzliche Re-
zeptordeterminante ist aber fur die Virulenz zumindest in Zellkultur nicht notwendig
(Gagneten et al., 1995).
Die Coronaviren BCoV, HEV und HCoV-OC43 nutzen Neu5,9Ac2 als Rezeptordetermi-
nante (Vlasak et al., 1988b; Schultze et al., 1990, 1991a; Schultze und Herrler,
1992). Entsprechend besitzen sie mit ihrem HE-Protein ein rezeptorzerstorendes Enzym,
die 9-O-Acetylesterase (Vlasak et al., 1988a; Schultze et al., 1991b).

2. Zielsetzung
Das ubertragbare Gastroenteritis-Virus der Schweine (TGEV) ist ein typisch enteropatho-
genes Coronavirus. Das S-Protein von TGEV besitzt zwei Bindungsaktivitaten. Die eine
vermittelt die Bindung an den zellularen Rezeptor, porzine Aminopeptidase N (pAPN),
der fur die Infektion in Zellkultur unerlasslich ist. Bei der zweiten Bindungsaktivitat
handelt es sich um eine Sialinsaure-bindende Aktivitat. Virusvarianten wie das porzine
respiratorische Coronavirus (PRCoV) und bestimmte Mutanten besitzen die Bindungs-
aktivitat an pAPN, binden jedoch nicht an Sialinsauren. Diese Varianten und Mutanten
konnen Zellkulturen effizient infizieren, haben aber in vivo ihre Enteropathogenitat verlo-
ren. Aus diesem Grund wird vermutet, dass die Sialinsaurebindungsaktivitat eine wichtige
Rolle fur die Enteropathogenitat von TGEV spielt. Die Sialinsaurebindungsstelle auf dem
S-Protein von TGEV wurde bereits naher charakterisiert. Uber die Bindungspartner auf
zellularer Ebene — in Zellkultur und im Darmtrakt der Schweine — ist jedoch bisher
noch nichts bekannt.
Ziel der vorliegenden Arbeit war es, durch die Untersuchung der Interaktion von TGEV
mit intestinalen Sialoglykokonjugaten wie Muzinen und Oberflachenkomponenten von
Darmepithelzellen, die Bedeutung der Sialinsaurebindungsaktivitat fur die Enteropatho-
genitat von TGEV besser zu verstehen. TGEV eignet sich gut als Modell fur die Unter-
suchung molekularer Mechanismen, die zur virusbedingten Gastroenteritis fuhren, da es
Mutanten und Varianten gibt, bei denen die Enteropathogenitat verloren gegangen ist.
Durch vergleichende Untersuchungen ist es so moglich, die Faktoren, die fur den Entero-
tropismus des Virus von Bedeutung sind, zu erkennen und zu analysieren.
Nahere Informationen uber die Bindungspartner, die TGEV uber seine Sialinsaurebin-
dungsaktivitat erkennen kann, sollten uber drei Ansatze gewonnen werden:
39

40 2. Zielsetzung
1. Der Vergleich von TGEV und E. coli F5 sollte Anhaltspunkte liefern, ob die beiden
Krankheitserreger gleiche oder unterschiedliche Bindungspartner im Darm erken-
nen. E. coli F5 wurde fur den Vergleich ausgewahlt, da dieses Bakterium Sialo-
glykokonjugate des Darmtrakts zur Kolonisation nutzt, also wie TGEV uber eine
Sialinsaurebindungsaktivitat verfugt.
2. Die Untersuchung der Interaktion von TGEV mit Zellkulturzellen sollte klaren, ob
es neben pAPN noch andere Strukturen gibt, die uber die Sialinsaurebindungsak-
tivitat von TGEV erkannt und fur die Bindung an Zellen genutzt werden konnen.
Bei diesem Ansatz sollten die Mutanten HAD3 und m10, die nicht an Sialinsauren
binden konnen, vergleichend eingesetzt werden. Die bei der Untersuchung der Zell-
kulturzellen erarbeiteten Methoden sollten auf die weiterfuhrenden Untersuchungen
mit Komponenten des Dunndarms ubertragen werden.
3. Intestinale Muzine und Oberflachenstrukturen des Darmepithels sollten auf poten-
tielle Bindungspartner fur TGEV untersucht werden. Durch den Vergleich mit den
Mutanten HAD3 und m10 sollten die Strukturen, die TGEV aufgrund seiner Sia-
linsaurebindungsaktivitat erkennt, gefunden werden.
Mit diesen drei Ansatzen wurde im Rahmen dieser Arbeit die Bedeutung der Sialinsaure-
bindenden Aktivitat fur die Enteropathogenitat des Virus der ubertragbaren Gastroente-
ritis der Schweine naher untersucht.

3. Material
3.1 Zelllinien
ST-Zellen (swine testicular): isoliert aus dem trypsinierten Hoden eines Schweinefoten
(80–90 d alt) (McClurkin und Norman, 1966).
LLC-PK1-Zellen: Schweinenierenzellen (Ref.: ATCC CRL-1392, 1994).
3.2 Zellkulturmedien
Dulbecco’s modified Eagle medium (DMEM) mit
4500 mg/l Glucose, Pyridoxin und Glutamax-I
GIBCO BRL Life Technologies
PBS Dulbecco’s w/o Calcium und Magnesium GIBCO BRL Life Technologies
Fetales Kalberserum (FKS), auf Mykoplasmen
und Viren untersucht
GIBCO BRL Life Technologies
Trypsin-EDTA-Losung GIBCO BRL Life Technologies
Penicillin-Streptomycin Antibiotikalosung GIBCO BRL Life Technologies
3.3 Viren
TGEV PURDUE 46-MAD: Das Ausgangsisolat wurde 1946 an der Purdue Universitat
in Indiana gewonnen (Doyle und Hutchings, 1946; Bohl, 1972). Die verwendeten
Virusstocks wurden seit Isolierung 115-mal in Dr. Bohls, 9-mal in Dr. Enjuanes (Sanchez
et al., 1990, 1992), 2-mal in Dr. Herrlers Labor in Marburg (Krempl, 1998) und 2-mal
in unserem Labor in ST-Zellen passagiert. Die letzte Plaque-Reinigung liegt 7 bzw. 8
41

42 3. Material
Passagen zuruck (Bezeichnung: C1P7 bzw. C1P8).
HAD-Mutanten: Diese Mutanten wurden als hamagglutinationsdefiziente Mutanten von
Dr. Krempl in Dr. Herrlers Labor aus PUR 46-MAD isoliert (Krempl et al., 1998,
2000). In dieser Arbeit wurde die HAD3-Mutante verwendet (L → P, Position 209 des
S-Proteins).
Epitop-D-Mutanten: Die Mutanten wurden als Escape-Mutanten mit dem monoklonalen
Antikorper 40.1 (Epitop D) aus PUR 115-PAR isoliert (Delmas et al., 1986; Bernard
und Laude, 1995). In dieser Arbeit wurde die von Dr. Laude zur Verfugung gestellte
Mutante PUR 115-PAR m10 verwendet (4 AS deletiert, Position 146–149 des S-Proteins).
Die verwendeten Mutanten HAD3 und m10 wurden aufgrund ihrer hohen Infektiositat in
Zellkultur bei nicht vorliegender Hamagglutinationsaktivitat ausgewahlt. Sie wurden in
unserem Labor 2-mal in ST-Zellen passagiert.
3.4 Bakterien
E. coli F5/K99 2 Feldstamme aus Kalbern: A 408/86 (F5), A 131/95 (O9:K35, F5) zur
Verfugung gestellt von Dr. Amtsberg, Institut fur Mikrobiologie der TiHo Hannover.
3.5 Erythrozyten
Huhnererythrozyten Geflugelklinik, TiHo Hannover
Rindererythrozyten Klinik fur Reproduktionsmedizin,
TiHo Hannover
Pferdeerythrozyten Klinik fur Reproduktionsmedizin,
TiHo Hannover
Schweineerythrozyten Klinik fur Reproduktionsmedizin,
TiHo Hannover

3.6. Darmmaterial 43
3.6 Darmmaterial
Darmmuzinisolierung: Jejunum aus 2 Saugferkeln, 12 und 14 Tage alt, und 2 Absetzfer-
keln, 10 und 15 Wochen alt.
Burstensaummembranvesikel-Praparation: Jejunum aus 2 Saugferkeln, 1 Tag alt, 2 Saug-
ferkeln, 3 Tage alt, 2 Absetzferkeln, 12 Wochen alt, 2 Absetzferkeln, 6–8 Wochen alt.
Darmschnitte: Jejunum eines Saugferkels, 3 Tage alt.
Schweinerasse (wo bekannt): Muttersau F1 Deutsche Landrasse × Deutsches Edelschwein,
Eber F1 Hampshire × Pietrain.
3.7 Enzyme
Acylneuraminyl-Hydrolase (Neuraminidase) aus
Vibrio cholerae
DADE Behring
Streptavidin-Peroxidase(HRP)-Komplex Amersham/Pharmacia
3.8 Antikorper
Monoklonaler Antikorper gegen TGEV (S-Protein, Epitop A.c) 6A.C3 von L. Enjuanes,
Madrid. Monoklonaler Antikorper gegen porzine Aminopeptidase N G43 (Delmas et al.,
1992) von H. Laude, Jouys-en-Josas. Die verwendeten monoklonalen Antikorper stammten
aus Mausen und lagen als Zellkulturuberstand vor.
Anti-Maus-Immunglobuline vom Schaf,
biotinyliert
Amersham/Pharmacia
Anti-Maus-Immunglobuline vom Schaf,
FITC-gekoppelt
Amersham/Pharmacia
Anti-Maus-Immunglobuline vom Kaninchen,
HRP-gekoppelt
DAKO
Anti-Maus-Immunglobuline aus der Ziege,
HRP-gekoppelt
Institut fur Virologie, TiHo

44 3. Material
Monovalentes diagnostisches Kalberkoliruhrserum
K99 (F5)
BgVV
3.9 Kits
Silver Stain Kit BIO-RAD
DIG Glykan Differentiation Kit Roche
ECL glycoprotein detection module Amersham/Pharmacia
BCA-Protein-Assay Pierce
BIO-RAD Protein-Assay BIO-RAD
3.10 Substrate
ABTS Losung Roche
BM Chemoluminescence Blotting Substrate
(POD)
Roche
Super Signalr West Dura Extended Duration
Substrate
Pierce
3.11 Chemikalien
Acetonitril, isocratic grade fur
Flussigkeitschromatographie
Merck
Acrylamidlosung 30%”rotiphoreser Gel 30“ Roth
Aminocapronsaure Sigma
Ammoniumpersulfat (APS) BIO-RAD
Blocking-Reagenz Roche
Bovines Serumalbumin Sigma
Casaminoacids Difco
Complete, Proteaseinhibitorcocktail Roche
Coomassie-Brilliantblau Merck
1,4-Dithiotreitol (DTT) Roth

3.11. Chemikalien 45
Essigsaure Roth
Ethanol Merck
Glucose Roth
Glutaraldehyd Sigma
Glycerin Roth
Glycin Roth
Leupeptin Roche
2-Mercaptoethanol Fluka
Methanol Roth
4,5-Methylendioxy-1,2-phenylendiamin × 2 H2O
(DMB)
Fluka
Muzin aus der Glandula submandibularis des
Schweines
R. Schauer, Kiel
N-Acetylneuraminsaure Sigma
Natriumacetat Merck
Natriumdesoxycholat Roth
Natriumdodecylsulfat (SDS) Roth
Neutralrot Merck
N-Glykolylneuraminsaure Sigma
2(N-Morpholino)ethansulfonsaure (MES) Sigma
N,N,N’,N’-Tetramethylethylendiamin (TEMED) Roth
n-Oktyl-β-D-Glukopyranosid (OG) Sigma
Nonidetr P40 (4-Nonylphenolpolyethylenglykol) Roche
Paraformaldehyd Fluka
PefablockrSC
(4-(2-Aminoethyl)benzolsulfonylfluorid,
Hydrochlorid)
Roche
Pepstatin Roche
PolyViteX bioMerieux
Proteinmarker (Rainbow) Amersham/Pharmacia
Saccharose Roth
Salzsaure Roth

46 3. Material
Sea Plaquer Agarose FMC Bio Products
Streptavidin-Agarose Pierce
Sulfo-NHS-Biotin Pierce
Tris-Hydroxymethylaminomethan (TRIS) Roth
Triton X-114 (Octylphenolpolyethylenglycolether) Serva
Tween 20 Roth
WGA-Agarose (Wheat Germ Agglutinin) Sigma
Alle nicht weiter aufgefuhrten Chemikalien waren ebenfalls vom Reinheitsgrad p.A. oder
gleichwertiger Qualitat und stammten von den bereits erwahnten Firmen.
3.12 Puffer und Losungen
10fach Laufpuffer SDS 10 g
fur Polyacrylamidgele: TRIS 30 g
Glycin 144 g
mit H2O auf 1 l auffullen
Sammelgellosung H2O 3,4 ml
fur Polyacrylamidgele TRIS-HCl 1 M pH 6,8 0,63 ml
(5 ml Ansatz): Acrylamidlosung 30% 0,83 ml
SDS 10% (in H2O) 50 µl
APS 10% (in H2O) 50 µl
TEMED 10 µl
Trenngellosung (8%) H2O 4,6 ml
fur Polyacrylamidgele TRIS-HCl 1,5 M pH 8,8 2,5 ml
(10 ml Ansatz): Acrylamidlosung 30% 2,7 ml
SDS 10% (in H2O) 100 µl
APS 10% (in H2O) 100 µl
TEMED 24 µl

3.12. Puffer und Losungen 47
Trenngellosung (10%) H2O 4,0 ml
fur Polyacrylamidgele TRIS-HCl 1,5 M pH 8,8 2,5 ml
(10 ml Ansatz): Acrylamidlosung 30% 3,3 ml
SDS 10% (in H2O) 100 µl
APS 10% (in H2O) 100 µl
TEMED 16 µl
Probenpuffer 2fach: TRIS-HCl pH 6,8 100 mM
SDS 4%
Glycerin 20%
Bromphenolblau 0,02%
(DTT 200 mM)
Coomassie-Farbelosung: Coomassie-Brilliantblau 0,5 g
Ethanol 200 ml
Essigsaure (konz.) 50 ml
H2O 250 ml
Entfarbelosung/ Ethanol 400 ml
Fixierlosung Essigsaure (konz.) 100 ml
fur Proteingele: H2O 500 ml
Anodenpuffer I (pH 9,0): TRIS 1 M 300 ml
Ethanol 200 ml
H2O 500 ml
mit HCl auf pH 9,0 einstellen
Anodenpuffer II (pH 7,4): TRIS 1 M 25 ml
Ethanol 200 ml
H2O 770 ml
mit HCl auf pH 7,4 einstellen

48 3. Material
Kathodenpuffer (pH 9,0): Aminocapronsaure 5,25 mg
TRIS 1M 25 ml
Ethanol 200 ml
H2O 770 ml
mit HCl auf pH 9,0 einstellen
NP40-Lysispuffer Natriumdesoxycholat 0,5%
(pH 7,5): TBS pH 7,5 50 ml
Nonidetr P40 1%
bei Bedarf Complete 1 Tabl.
Phosphat-gepufferte NaCl 8 g
Kochsalzlosung KCl 0,2 g
pH 7,5 (PBS): Na2HPO4 1,15 g
MgCl2 × 6H2O 0,1 g
KH2PO4 0,2 g
CaCl2 × 2H2O 0,13 g
mit H2O auf 1 l auffullen
PBS ohne Calcium NaCl 8 g
und Magnesium KCl 0,2 g
pH 7,5 (PBSM): Na2HPO4 1,15 g
KH2PO4 0,2 g
mit H2O auf 1 l auffullen
PBSM/0,1% Tween: PBSM 2 l
Tween-20 2 ml
PBSM/0,05% Tween: PBSM 2 l
Tween-20 1 ml
3.12. Puffer und Losungen 49
TRIS-gepufferte TRIS 0,05 M
Kochsalzlosung NaCl 0,15 M
pH 7,5 (TBS): mit HCl auf pH 7,5 einstellen
TRIS-Puffer: TRIS-HCl pH 7,4 10 mM
Pepstatin 1 µM
Leupeptin 1 µM
PefablockrSC 1 mM
DIG Glycan TBS pH 7,5
Differentiation Kit MgCl2 1 mM
Puffer 1 (pH 7,5): MnCl2 1 mM
CaCl2 1 mM
DIG Glycan TRIS 0,1 M
Differentiation Kit MgCl2 0,05 M
Puffer 2 (pH 9,5): NaCl 0,1 M
mit HCl auf pH 9,5 einstellen
MES-Puffer (pH 6,5): MES 0,05 M
NaCl 0,1 M
CaCl2 0,02 M
mit 1 M NaOH auf pH 6,5 einstellen
Natriumacetat-Puffer, Natriumacetat 0,2 M
einfach (pH 5,5): mit Essigsaure auf pH 5,5 einstellen
Natriumacetat-Puffer Natriumacetat 0,05 M
(pH 5,5): NaCl 0,154 M
CaCl2 0,009 M
mit Essigsaure auf pH 5,5 einstellen
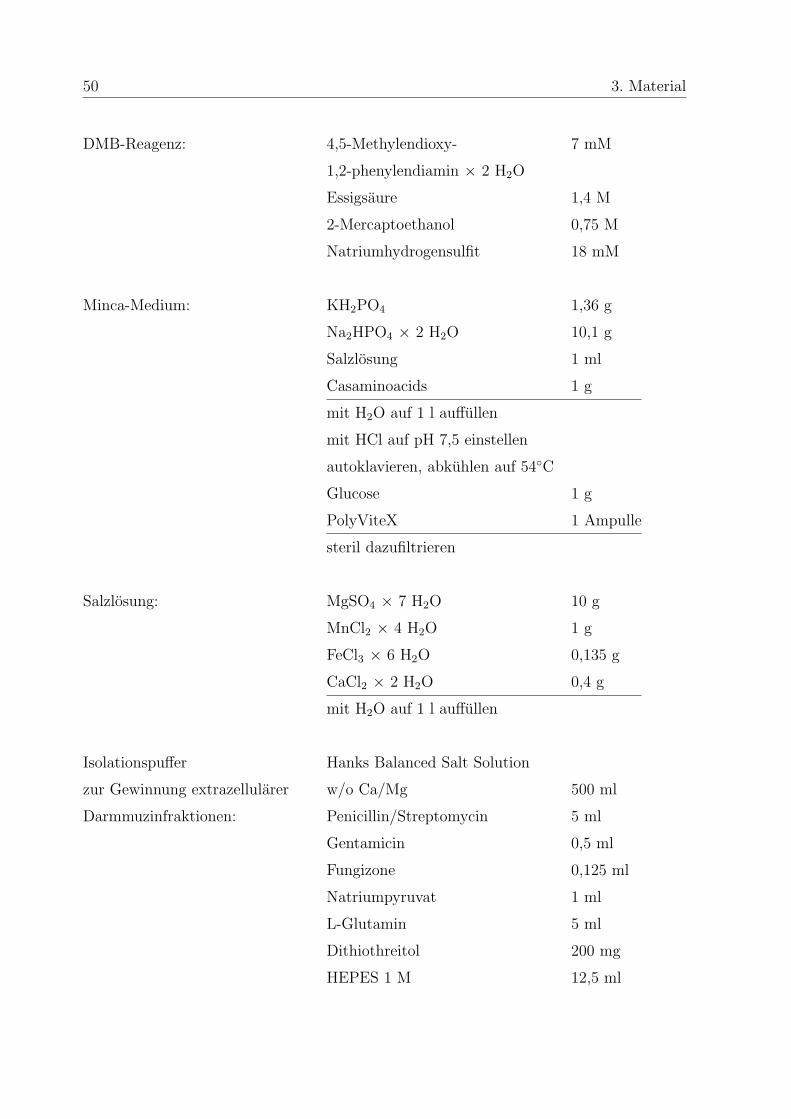
50 3. Material
DMB-Reagenz: 4,5-Methylendioxy- 7 mM
1,2-phenylendiamin × 2 H2O
Essigsaure 1,4 M
2-Mercaptoethanol 0,75 M
Natriumhydrogensulfit 18 mM
Minca-Medium: KH2PO4 1,36 g
Na2HPO4 × 2 H2O 10,1 g
Salzlosung 1 ml
Casaminoacids 1 g
mit H2O auf 1 l auffullen
mit HCl auf pH 7,5 einstellen
autoklavieren, abkuhlen auf 54◦C
Glucose 1 g
PolyViteX 1 Ampulle
steril dazufiltrieren
Salzlosung: MgSO4 × 7 H2O 10 g
MnCl2 × 4 H2O 1 g
FeCl3 × 6 H2O 0,135 g
CaCl2 × 2 H2O 0,4 g
mit H2O auf 1 l auffullen
Isolationspuffer Hanks Balanced Salt Solution
zur Gewinnung extrazellularer w/o Ca/Mg 500 ml
Darmmuzinfraktionen: Penicillin/Streptomycin 5 ml
Gentamicin 0,5 ml
Fungizone 0,125 ml
Natriumpyruvat 1 ml
L-Glutamin 5 ml
Dithiothreitol 200 mg
HEPES 1 M 12,5 ml

3.12. Puffer und Losungen 51
Bradford-Reagenz, konz.: Coomassie-Brilliantblau 100 mg
Ethanol, 97% 50 ml
Phosphorsaure, 85% 100 ml
Bradford-Gebrauchslosung: Konzentrat 3 ml
H2O 17 ml
Schiff-Reagenz: Fuchsin (basisch)in H2O 1%
HCl 1 M 20%
Aktivkohle 0,5%
Aktivkohle 2-mal abfiltrieren
Schiff-Reagenz, entfarbt: Natriummetabisulfit 100 mg
Schiff-Reagenz 6 ml
Perjodessigsaure: Perjodsaure 10 mg
Essigsaure, 7% 10 ml
Burstensaummembranvesikel Mannit 300 mM
(BSMV) Praparationspuffer 1: TRIS 12 mM
EGTA 5 mM
mit HCl auf pH 7,4/4◦C einstellen
BSMV Mannit 60 mM
Praparationspuffer 2: TRIS 12 mM
EGTA 5 mM
mit HCl auf pH 7,4/4◦C einstellen
BSMV Mannit 100 mM
Praparationspuffer A: HEPES 10 mM
mit TRIS auf pH 7,4/4◦C einstellen

52 3. Material
BSMV Mannit 100 mM
Praparationspuffer B: HEPES 10 mM
MgSO4 × 7 H2O 0,1 mM
mit TRIS auf pH 7,4/4◦C einstellen
BSMV Mannit 100 mM
Vesikelpuffer: KCl 100 mM
HEPES 10 mM
mit TRIS auf pH 7,4/4◦C einstellen
Alkalische Phosphatase-Puffer: Diethanolamin 1,02 M
MgCl2 × 6 H2O 0,51 M
mit HCl auf pH 9,8 einstellen
Na-Nitrophenylphosphat 10 mM
Assaymix (2fach konz.): MgCl2 × 6 H2O 10 mM
NaCl 200 mM
KCl 20 mM
TRIS 200 mM
mit HCl auf pH 7,4 einstellen
Assaymix-Puffer (f. 40 Proben): Assaymix 10 ml
H2O 8 ml
ATP 20 mM pH 7,4 (TRIS) 2 ml
EDTA 100 mM PH 7,4 (NaOH) 0,6 ml
(Ouabain 0,5%)
Farbreagenz: Schwefelsaure 1,15 M 100 ml
Ammoniumheptamolybdat 1,1 g
FeSO4 × 7 H2O 4 g
Trichloressigsaure, 5% 50 ml

4. Methoden
4.1 Zellkulturen
4.1.1 Passagierung von ST-Zellen
ST-Zellen wurden in DMEM, welches mit 10% FKS und Antibiotikalosung versetzt war,
kultiviert. Die Passagierung erfolgte im Drei- bzw. Viertagesrhythmus in 75 cm2-Zell-
kulturflaschen (Costar). Der adharente Zellrasen wurde nach zweimaligem Waschen mit
PBS Dulbecco’s (ohne Calcium und Magnesium) mit 1 ml Trypsin/EDTA-Losung bei
RT inkubiert. Nach ca. 1 min wurde die uberschussige Trypsinlosung abgeschuttet und
die Zellen bis zur Ablosung vom Flaschenboden bei 37◦C im Brutschrank inkubiert (ca.
10 min). Die Zellsuspension wurde in 10 ml DMEM/10% FKS resuspendiert und 1 ml
bzw. 0,75 ml davon wurden mit 20 ml neuem Kultivierungsmedium weitergefuhrt (1:10
bzw. 1:15 verdunnt).
4.1.2 Passagierung von LLC-PK1-Zellen
Die Passagierung dieser adharent wachsenden Nierenzelllinie erfolgte analog zu den ST-
Zellen. Die LLC-PK1-Zellen wurden in einer 1:20 Verdunnung weitergefuhrt.
Beide Zelllinien wurden in einem 37◦C Brutschrank mit 5% CO2 Begasung kultiviert.
53

54 4. Methoden
4.2 Bakterien
4.2.1 Kultivierung
E. coli F5 (K99) wurde auf Minca-Agar kultiviert, um die Fimbrienausbildung zu erhalten
(Mouricout et al., 1990). In regelmaßigen Abstanden (3–4 Wochen) wurden Subkulturen
erstellt (ca. 18 h, 37◦C), die bei 4◦C gelagert wurden. Zur Herstellung einer Flussigkul-
tur wurden 3–5 ml Minca-Medium mit 1–2 Kolonien angeimpft und bei 37◦C fur 18 h
geschuttelt. Die Bakterien wurden bei 3000 × g, 15 min, 4◦C pelletiert und zweimal mit
PBS/1% Mannose gewaschen. Zur weiteren Verwendung, z.B. fur Hamagglutinationstests,
wurden die Bakterien in PBS/1% Mannose bei 4◦C aufbewahrt.
4.2.2 Fimbrientest
Auf einen Objekttrager wurde ein Tropfen Kalberkoliruhrserum F5 (K99) gegeben, und
mit einer Ose eine Bakterienkolonie nach und nach mit dem Tropfen verstrichen. Als Kon-
trolle diente ein Tropfen physiologische Kochsalzlosung, die ebenfalls mit einer Kolonie
vermischt wurde. Die Vermischung mit dem Kalberkoliruhrserum bewirkte eine Aggluti-
nation mit den Bakterienfimbrien und zeigte somit durch weiße Klumpchenbildung ein
positives Testergebnis an.
4.3 Virusanzucht
Zur Herstellung von Virusstocks wurden zu einem dichten Monolayer (75 cm2 Zellkultur-
flasche) gewachsene ST-Zellen zweimal mit PBS gewaschen und mit Stockvirus in einer
m.o.i. von 1 PFU/Zelle in 3 ml DMEM fur 1 h bei 37◦C inkubiert. Anschließend wurden
12 ml DMEM hinzugefugt, und die Zellen im CO2-begasten Brutschrank bei 37◦C inku-
biert. Die Virusernte erfolgte nach 20–24 h, sobald ein deutlicher cpE zu erkennen war.
Zur Virusernte wurde der virushaltige Zellkulturuberstand abgenommen, mit 1% FKS
versetzt, und die Zelltrummer durch eine Zentrifugation bei 1900 × g fur 10 min bei 4◦C
entfernt. Der Uberstand wurde in sterile Polyethylenrohrchen (1,8 ml, Greiner) a 1 ml

4.4. Plaquetest 55
aliquotiert, in flussigem Stickstoff schockgefroren und bei −80◦C gelagert. Der Virustiter
des Stocks wurde von einem Aliquot mittels Plaquetest bestimmt (ca. 108 PFU/ml). Die
Virusanzucht erfolgte fur alle TGEV-Isolate nach dem oben beschriebenen Vorgang.
4.4 Plaquetest
Der Plaquetest dient zur Quantifizierung der infektiosen Viruspartikel einer Suspension.
Durch das halbfeste Agarmediumgemisch wird die Ausbreitung der Viren durch Diffusion
verhindert, und die infektiosen Partikel konnen sich nur von Zelle zu Zelle ausbreiten.
Dadurch entsteht ein sogenannter”Plaque“ im Zellrasen, gebildet von virusinfizierten
Zellen. Diese Lasion wird durch Farbung sichtbar gemacht.
Zur Bestimmung des Virustiters mittels Plaque-Test wurde von der Virussuspension ei-
ne Verdunnung in 10er Schritten in PBS angelegt. Mit den verschiedenen Verdunnungen
wurden zu einem dichten Monolayer gewachsene ST-Zellen nach zweimaligem Waschen in-
fiziert. Lagen die Zellen in 60 mm Zellkulturschalen vor, wurden 500 µl Inokulum verwen-
det; bei 35 mm Schalen wurde ein Volumen von 300 µl eingesetzt. Nach einer Inkubation
von 1 h bei 37◦C wurde das Inokulum entfernt, die Zellen zweimal mit PBS gewaschen
und mit 0,8%iger Sea-Plaquer-Agarose (44◦C) in DMEM/2% FKS uberschichtet (5 bzw.
3 ml). Nach Erhartung des Agarosemediumgemisches bei Raumtemperatur (RT), wurde
der Ansatz bei 37◦C unter CO2 Begasung inkubiert. Nach zwei Tagen wurde zur Dar-
stellung der”Plaques“ der Agar mit 0,8%iger Sea-Plaquer-Agarose/0,0125% Neutralrot
uberschichtet (2 bzw. 1 ml). Nach Erkalten des Gemisches bei RT wurden die Schalen
uber Nacht bei 37◦C inkubiert, so dass das Neutralrot durch die Agarschicht diffundie-
ren und die intakten Zellen rot anfarben konnte. Die virusinfizierten Zellen bildeten die
ungefarbten”Plaques“ im Zellrasen, die fur die Titerbestimmung gezahlt wurden. Der
Virustiter wurde in Plaque-Forming-Units (Plaque-bildende Einheiten) pro ml (PFU/ml)
angegeben.

56 4. Methoden
4.5 Virusreinigung mittels Dichtegradientenzentrifu-
gation
ST-Zellmonolayer in 145 mm Zellkulturschalen (Greiner) wurden mit einer m.o.i. von 1
bzw. 0,1 PFU/Zelle infiziert. Nach ein bzw. zwei Tagen wurde der Virus-haltige Zellkul-
turuberstand geerntet und bei 1900 × g fur 10 min bei 4◦C zur Entfernung der Zell-
trummer zentrifugiert. Die Viruspartikel wurden aus dem Uberstand durch Ultrazentrifu-
gation bei 120000 × g uber 1 h bei 4◦C sedimentiert. Der Uberstand wurde verworfen, und
das Sediment in PBSM resuspendiert. Nach Bedarf wurden die resuspendierten Viruspar-
tikel bzw. ein Aliquot mit Neuraminidase aus Vibrio cholerae, wie unter 4.8 beschrieben,
behandelt. Zur Reinigung wurde die Virussuspension uber einen kontinuierlichen Saccha-
rosegradienten (20–50% w/w) geschichtet. Dem Gradienten war ein 0,5 ml Kissen aus
60%iger Saccharose unterschichtet. Nach einer Zentrifugation von 3 h bei 150000 × g und
4◦C wurde die sichtbare Virusbande mit einer 1 ml Spritze (Kanule 1,2 × 40) abgenom-
men. Nach Verdunnung mit PBSM wurden die Viruspartikel bei 150000 × g (1 h, 4◦C)
pelletiert, in ein geringes Volumen PBSM aufgenommen, aliquotiert und bei −20◦C zur
Aufbewahrung eingefroren. Zur Analyse der gereinigten Viren wurden eine SDS-PAGE
mit Coomassie-Farbung des Gels, ein Hamagglutinationstest und die Proteinkonzentrati-
onsbestimmung durch Messung im Photometer bei 280 nm durchgefuhrt.
4.6 Isolierung des S-Proteins
Gereinigtes und neuraminidasebehandeltes TGEV wurde in 400 µl PBSM mit 1% OG
resuspendiert und fur 10 min bei RT inkubiert. M-Protein und RNP-Komplex wurden
durch eine 30-minutige Zentrifugation (16000 × g, 4◦C) pelletiert und der Uberstand auf
einen Saccharosegradienten (10–30% w/w, 0,5 ml Kissen aus 60% Saccharose) in PBS/1%
OG geschichtet. Nach Zentrifugation fur 18 h bei 214000 × g, 4◦C wurden durch Anste-
chen des Zentrifugenrohrchenbodens 0,5 ml Aliquots gesammelt und in der SDS-PAGE
mit anschließender Silberfarbung auf ihren Gehalt an S-Protein untersucht. Als weitere
Kontrolle wurde ein Hamagglutinationstest durchgefuhrt, da neuramindidasebehandeltes
S-Protein eine hohe Hamagglutinationsaktivitat aufweist.

4.7. Erythrozyten 57
Fur die weitere Verwendung, z.B. in Bindungstests, wurden die S-Protein-haltigen Fraktio-
nen vereinigt und gegen H2O:PBS (4:1) dialysiert. Die dialysierte Probe wurde in Aliquots
bei −20◦C aufbewahrt.
4.7 Erythrozyten
4.7.1 Isolierung aus Vollblut
Zur Isolierung der Erythrozyten wurde mit EDTA versetztes Vollblut der entsprechen-
den Tierart bei 2000 × g fur 10 min bei 4◦C zentrifugiert. Das Plasma und die Leu-
kozytenschicht wurden abgesaugt, und die sedimentierten Erythrozyten in 0,9% NaCl-
Losung resuspendiert. Nach erneutem Zentrifugieren und zweimaliger Wiederholung die-
ses Waschvorgangs wurden die Erythrozyten als 10%ige Suspension in physiologischer
Kochsalzlosung bei 4◦C aufbewahrt.
Huhnererythrozyten wurden i.d.R. als frische Suspension verwendet. Die Erythrozyten
der anderen Tierarten wurden nach der Isolierung einem Fixationsvorgang zur Haltbar-
machung unterzogen.
4.7.2 Fixierung von Erythrozyten
Einer 10%igen Erythrozytensuspension wurde zur Haltbarmachung 25%iges Glutaralde-
hyd tropfenweise unter Schwenken bis zu einer Endkonzentration von 1% hinzugefugt.
Diese Suspension wurde uber Nacht bei 4◦C kontinuierlich geruhrt. Am nachsten Tag
wurde zur Inaktivierung des uberschussigen Glutaraldehyds 0,1 M Glycin eingewogen,
und die Suspension erneut uber Nacht bei 4◦C geruhrt. Anschließend wurde sie viermal
wie unter 4.7.1 beschrieben gewaschen und als 10%ige Erythrozytensuspension in 0,9 %
NaCl-Losung bei 4◦C aufbewahrt.
Als Kontrolle fur die Vollstandigkeit der Fixierung wurde eine Inkubation mit H2O (Reinst-
wasser) durchgefuhrt, wobei die Erythrozyten nicht lysiert werden durften.

58 4. Methoden
4.7.3 Hamagglutinationstest
Viren und Bakterien, die eine Sialinsaurebindungseigenschaft besitzen, konnen aufgrund
dieser Fahigkeit auch an Erythrozyten binden. Durch die polyvalente Adsorptionsmoglich-
keit der Viruspartikel (Bakterienzellen)kommt es bei entsprechend hoher Partikelkonzen-
tration zum Quervernetzen der Erythrozyten, so dass diese sich nicht mehr am Boden
eines Napfes als Sediment absetzen konnen. Der Hamagglutinationstest dient somit zur
qualitativen Bestimmung der Sialinsaurebindungsaktivitat eines Virus/ eines Bakteriums
und kann daruberhinaus bei Viren mit sialinsaurebindenden Eigenschaften zur Quantifi-
zierung der relativen Virusmenge einer Suspension dienen.
Hamagglutinationstests wurden in Mikrotiterplatten mit U-formigen Vertiefungen (Grei-
ner) durchgefuhrt. Von der zu testenden Virussuspension (Bakteriensuspension) wurde
eine Verdunnungsreihe in Zweierpotenzen in PBSM angelegt (50 µl/Vertiefung). Zu jeder
Verdunnungstufe wurde das gleiche Volumen einer 0,5%igen Erythrozytensuspension ge-
geben. Nach Inkubation von ca. 1 h bei 4◦C (bei fixierten Erythrozyten ca. 3 h)war eine
Agglutination durch gleichmassige Verteilung der Erythrozyten zu sehen. Nicht aggluti-
nierte Erythrozyten sedimentierten als scharf umrissener Knopf am Boden der Vertiefung.
Der in Hamagglutinationseinheiten/ml (HAU/ml, haemagglutinating units/ml) angege-
bene Titer stellt den reziproken Wert der hochsten Verdunnungsstufe dar, die noch eine
Agglutination erkennen lasst.
4.7.4 Hamagglutinationsinhibition
Die Agglutination von Erythrozyten durch Viruspartikel (Bakterienzellen) kann durch
bestimmte Substanzen gehemmt werden.
Von den Inhibitorsubstanzen wurde eine Verdunnungsreihe in Zweierpotenzen in PBSM
hergestellt (25 µl/Vertiefung, Mikrotiterplatten wie in 4.7.3). Zu jeder Verdunnungsstufe
wurde das gleiche Volumen einer Virussuspension (Bakteriensuspension) mit 8 HAU/ml
gegeben. Anschließend wurden zu jeder Vertiefung 50 µl einer 0,5%igen Erythrozytensus-
pension hinzugefugt. Nach einer Inkubation von ca. 2 h bei 4◦C ließ sich die Hemmung
der Agglutination als am Boden der Vertiefung sedimentierte Erythrozyten in Form ei-

4.8. Desialylierung 59
nes Knopfes ablesen. Der reziproke Wert der hochsten Hemmstoffverdunnung, die eine
Hamagglutination verhindert, wird als Hemmtiter der entsprechenden Substanz bezeich-
net und in hamagglutinationshemmenden Einheiten/ml (HIU/ml, haemagglutination in-
hibiting units/ml) angegeben.
4.8 Desialylierung
4.8.1 Desialylierung von Viren
Sedimentiertes und in PBSM resuspendiertes Virus wurde mit Neuraminidase aus Vibrio
cholerae (VCNA, 1 U/ml) in einer Endkonzentration von 50 mU/ml versetzt, fur 30 min
bei 37◦C inkubiert und anschließend auf Eis gekuhlt. Die so behandelten konzentrierten
Viruspartikel wurden, wie in 4.5 beschrieben, uber einen Saccharosegradienten gereinigt.
4.8.2 Desialylierung von Zellen
Zu einem konfluenten Monolayer gewachsene ST- bzw. LLC-PK1-Zellen wurden dreimal
mit PBS gewaschen und mit VCNA (1 U/ml) fur 1 h bei 37◦C inkubiert. Es wurde eine
Enzymaktivitat von 250 mU/ml MES-Puffer verwendet. Die entsprechenden Kontrollzel-
len wurden in gleicher Weise jedoch mit MES-Puffer allein behandelt. Nach der Inkubation
wurden die Zellen zwei- bzw. dreimal mit PBS gewaschen und fur die jeweiligen Experi-
mente benutzt.
4.8.3 Desialylierung von Muzinen, Zellmembranen (MP) und
Burstensaummembranvesikeln (BSMV)
Muzine wurden bei einer Konzentration von 1 mg/ml (Muzinkonzentration), MP und
BSMV bei einer Proteinkonzentration von 4 mg/ml 1:2 mit VCNA (1 U/ml) versetzt.
Die Kontrollproben wurden statt mit VCNA mit Natriumacetat-Puffer, in dem auch die
Neuraminidase gelost vorliegt, behandelt. Nach einer Inkubation von 1 h bei 37◦C unter
Schutteln wurde SDS-Probenpuffer (2fach) zu den Proben gegeben. Diese wurden fur

60 4. Methoden
5 min bei 96◦C erhitzt und fur die SDS-Polyacrylamidgelelektrophorese (SDS-PAGE)
verwendet.
4.8.4 Desialylierung von immobilisierten Muzinen
Auf Mikrotiterplatten (Maxisorpr, Nunc) immobilisierte Muzine wurden zwecks Desia-
lylierung mit VCNA (1 U/ml) in einer Konzentration von 50 mU/Napf fur 1 h bei 37◦C
inkubiert. Als Kontrolle diente die Inkubation mit Natriumacetat-Puffer. Anschließend
wurden die Platten fur einen indirekten ELISA zur Detektion der Virusbindung an Mu-
zine verwendet (4.19.1).
4.8.5 Desialylierung von Zelllysaten
Zellen wurden, wie unter 4.10.1 beschrieben, lysiert und mit Streptavidin- oder WGA-
Agarose prazipitiert. Zu den Agarosepellets (100 µl) wurden nach dreimaligem Waschen
50 µl VCNA (1 U/ml) in Natriumacetatpuffer mit Proteaseinhibitoren (1 µM Pepsta-
tin, 1 µM Leupeptin, 1 mM PefablockrSC) hinzugegeben und die Pellets fur 1 h bei
37◦C geschuttelt. Die Kontrolllysate wurden ohne VCNA, nur in Natriumacetatpuffer
mit Proteaseinhibitoren inkubiert. Anschließend wurden die Proteine durch die zusatzli-
che Zugabe von je 50 µl Probenpuffer (2fach) fur 10 min bei 96◦C eluiert.
4.8.6 Desialylierung von Erythrozyten
Pro Ansatz wurden 0,5 ml einer 10%igen Rindererythrozytensuspension (fixiert, siehe
4.7.2) verwendet. Die bei 1400 × g, 3 min, RT sedimentierten Erythrozyten (ca. 50 µl)
wurden im 10fachen Volumen MES-Puffer (450 µl), dem die gewunschte Aktivitat an
VCNA (1 U/ml) bereits zugesetzt worden war, resuspendiert und fur 30 min bei 37◦C
geschwenkt. Um die Neuraminidase wieder zu entfernen, wurden die Erythrozyten bei
1400 × g abzentrifugiert und zweimal mit PBSM/1% BSA gewaschen. Von den desialy-
lierten Erythrozyten wurde eine 1%ige Suspension in PBSM hergestellt, die fur Hamag-
glutinationstests (4.7.3) verwendet wurde.

4.9. Membranproteinisolierung 61
4.9 Membranproteinisolierung
Aus der Plasmamembran von ST- und LLC-PK1-Zellen sollten Membranproteine isoliert
werden, um sie im Virusbindungstest einzusetzen.
In 145 mm Zellkulturschalen zu einem dichten Monolayer gewachsene Zellen wurden zwei-
mal mit PBSM gewaschen und in je 10 ml PBSM mit einem Zellschaber abgelost. Die
Zellen wurden bei 1400 × g pelletiert und das Pellet in 1 ml PBSM resuspendiert. Das
20fache Volumen an TRIS-Puffer wurde vorgelegt und die Zellsuspension tropfenweise un-
ter Schwenken hinzugefugt. Nach einer Inkubation von 20 min wurde die Zellsuspension
zwanzigmal im Dounce-Homogenisator geschert. Die Kerne wurden bei 500 × g, 5 min
pelletiert und der Uberstand vorsichtig abpipettiert. Das Kernsediment wurde in TRIS-
Puffer resuspendiert und ein weiteres Mal bei 500 × g zentrifugiert. Der Uberstand wurde
mit dem ersten Uberstand vereinigt und bei 100000 × g, 1 h sedimentiert. Das Pellet,
welches die Membranfraktion darstellt, wurde in 200 µl Phosphatpuffer (50 mM, pH 7,0)
aufgenommen und bei −20◦C aufbewahrt. Alle Schritte wahrend der Praparation wurden
auf Eis bzw. bei 4◦C durchgefuhrt. Von einem Aliquot wurde die Proteinkonzentration
mit dem BCA-Protein-Assay bestimmt.
4.10 Fallung von Oberflachenproteinen und Glyko-
proteinen
4.10.1 Biotinylierung und Streptavidinfallung von Oberflachen-
proteinen
Konfluente, auf 94 mm Zellkulturschalen gewachsene Zellrasen wurden dreimal mit kaltem
PBS gewaschen. Die Markierung der Oberflachenproteine erfolgte durch Inkubation der
Zellen mit 3 ml Sulfo-NHS-Biotin (0,5 mg/ml PBS) fur 30 min bei 4◦C. Zur Inaktivierung
uberschussigen Biotins wurden die Zellen anschließend mit 10 ml PBS/0,1 M Glycin
gewaschen und fur 15 min bei 4◦C mit diesem Puffer inkubiert.
Zur Fallung der biotinmarkierten Oberflachenproteine wurden die Zellen erneut mit kal-

62 4. Methoden
tem PBS gewaschen und in 10 ml PBS mit einem Zellschaber abgeschabt. Nach Zen-
trifugation (693 × g, 5 min, 4◦C) wurde das Zellpellet in 1 ml NP40-Lysispuffer mit
dem Proteaseinhibitorcocktail Complete resuspendiert und fur 15 min auf Eis inkubiert.
Unlosliche Zellbestandteile wurden durch eine Zentrifugation bei 16000 × g (30 min, 4◦C)
entfernt. Das Zelllysat wurde mit 100 µl 50%iger Streptavidin-Agarose (dreimal mit NP40-
Lysispuffer vorgewaschen) uber Nacht bei 4◦C unter Schutteln inkubiert. Am nachsten
Tag wurde die Streptavidin-Agarose pelletiert (16000 × g, 3 min, 4◦C) und dreimal mit
NP40-Lysispuffer gewaschen. Es folgte die Desialylierung eines Teils der Pellets wie unter
4.8.5 beschrieben. Die Proteine wurden durch Erhitzen der Pellets in SDS-Probenpuffer
(zweifach) bei 96◦C fur 10 min eluiert. Die Agarose wurde sedimentiert (16000 × g, 5 min,
RT) und der Uberstand mit den eluierten Oberflachenproteinen bis zur weiteren Verwen-
dung bei −20◦C aufbewahrt. Fur reduzierende Bedingungen wurde den Eluaten 100 mM
DTT zugesetzt und diese erneut bei 96◦C fur 5 min erhitzt.
4.10.2 Fallung von Glykoproteinen mit immobilisiertem Wei-
zenkeimagglutinin (WGA)
Dicht gewachsene Zellmonolayer wurden dreimal mit kaltem PBS gewaschen, wie unter
4.10.1 beschrieben, lysiert und mit 100 µl 50%iger WGA-Agarose gefallt. Die folgenden
Schritte erfolgten analog zu 4.10.1.
4.11 Muzine
4.11.1 Isolierung von Darmmuzinen
Darmstucke aus dem mittleren Jejunum 12–14 Tage alter Saug- bzw. 10–15 Wochen alter
Absetzferkel (frischtot entnommen, mit physiologischer Kochsalzlosung gespult und in
flussigem Stickstoff eingefroren; Lagerung bei −80◦C) wurden aufgetaut, gewogen (15–
20 g) und in kleine Stucke geschnitten. Diese Stucke wurden in ca. 50 ml Isolationspuffer
gegeben und 20 min bei 37◦C im Wasserbad inkubiert. Alle 5 min wurde unter kraftigem
Schutteln die extrazellulare Muzinschicht abgelost. Anschließend wurde die Suspension

4.11. Muzine 63
durch ein Sieb gegeben und der schleimhaltige Uberstand in einem Becherglas aufgefangen.
Dieser Schritt wurde mit dem im Sieb verbliebenen Darmgewebe mit 50 ml Isolationspuffer
noch einmal wiederholt. Nach Abspulen der aufgefangen Gewebestucke mit 50 ml PBSM
wurde der Uberstand zur Entfernung grober Stucke bei 2000× g, 15 min, 4◦C zentrifugiert.
4.11.2 Reinigung der extrazellularen Muzine
Zur Reinigung der Muzine wurden grobe Partikel durch Zentrifugation bei 14000 × g,
15 min, 4◦C entfernt. Die gewonnenen Proben wurden nun uber eine Membran mit ei-
ner Molekulausschlussgroße von 30 kD auf ein Volumen von ca. 4–8 ml eingeengt (bei
Gewebestucken von 15–20 g). Die so konzentrierten Proben wurden auf ein Sepharo-
se 4B-Chromatographie-Gel gegeben (Saule: 61 × 2,7 cm) und die Proteine ihrer Mo-
lekulgroße entsprechend aufgetrennt. Von den einzelnen Fraktionen (je 7,5 ml) wurde der
Kohlenhydrat- und Proteingehalt bestimmt und anhand dessen ermittelt, welche Fraktio-
nen als Muzinfraktionen definiert werden konnten. Diese wurden vereinigt, konzentriert,
gegen H2O dialysiert und lyophilisiert.
4.11.3 Kohlenhydratbestimmung mittels PAS-Farbung
Die Kohlenhydratbestimmung der Fraktionen erfolgte in Mikrotiterplatten (Greiner) mit
H2O als Leerwert. Es wurden pro Vertiefung 150 µl Probe bzw. H2O vorgelegt, und
jeweils 50 µl Perjodessigsaure hinzugefugt. Die Platte wurde in den Brutschrank (37◦C)
gestellt, bis das mit Natriummetabisulfit versetzte Schiff-Reagenz entfarbt war. Von dem
entfarbten Schiff-Reagenz wurden je 50 µl zu jeder Vertiefung gegeben und die Platte
30 min bei RT abgedeckt stehen gelassen. Die Extinktion wurde bei einer Wellenlange
von 550 nm gemessen. Die Multiplikation mit vorher berechneten Umrechnungsfaktoren
ergab die Kohlenhydratkonzentration der einzelnen Fraktionen.

64 4. Methoden
4.12 Burstensaummembranvesikel (BSMV)
4.12.1 Herstellung von BSMV
4.12.1.1 Darm von Absetzferkeln
Die Darmproben wurden von Dr. Schroder, Institut fur Physiologie, zur Verfugung ge-
stellt. Alle Schritte der Praparation von BSMV erfolgten auf Eis bzw. bei 4◦C. Der ge-
frorene Darm (Absetzferkel, mittleres Jejunum) wurde gewogen, mit der gleichen Menge
an Praparationspuffer 1 (isoton) ubergossen und auf Eis aufgetaut. Durch 10-minutiges
Schutteln im”Sinusquirl“ wurden die Enterozyten von der Darmwand gelost. Die Suspen-
sion mit gequirltem Darm wurde durch ein Sieb geseiht und mit der 4fachen Menge an
H2O (bezogen auf das Volumen des Praparationspuffers 1) nachgespult. Die Zellverbande
wurden in einem Standmixer (zweimal fur 1,5 min, hochste Stufe) weiter aufgeschlossen.
Anschließend wurde die Enterozytensuspension 2 min stehen gelassen, um den entstan-
denen Schaum absaugen zu konnen. Zur Enzym- und Proteinbestimmung wurden Kon-
trollproben P1 (4 × 250 µl, −20◦C) entnommen. Um die apikalen von den basolateralen
Enterozytenmembranen zu trennen, wurden letztere mit 10 mM MgCl2 gefallt. Hierzu
wurde 1 M MgCl2 dem Homogenat bis zu einer Endkonzentration von 10 mM tropfenwei-
se unter Ruhren zugefugt und fur 15 min auf Eis inkubiert. Durch eine Zentrifugation bei
5019 × g fur 18 min wurden die basolateralen Membranen sedimentiert. Aus dem Uber-
stand wurden nachfolgend die apikalen Membranen pelletiert (14740 × g, 40 min). Nach
Aufschwemmen der Pellets in 35 ml Praparationspuffer 2 und 10maligem Homogenisieren
im Potter erfolgte eine zweite Fallung mit 10 mM MgCl2. Es folgten die zwei Zentrifu-
gationsschritte, und die Pellets wurden in insgesamt 35 ml Vesikelpuffer aufgeschwemmt.
Durch erneutes Scheren im Potter wurden die Vesikel weiter homogenisiert. Die apikalen
Membranvesikel sedimentierten bei einer Zentrifugation von 19950 × g fur 45 min, und
das Pellet wurde in 1–3 ml Vesikelpuffer resuspendiert. Zum weiteren Homogenisieren
wurden die Membranen mit der Insulinspritze (Kanule: 0,45 × 23) mehrmals aufgezogen
(10–20-mal). Abschließend wurden Kontrollproben P2 (3 × 100 µl) fur die Enzym- und
Proteinbestimmung entnommen. Die BSMV wurden in flussigem Stickstoff eingefroren
und bei −80◦C gelagert.

4.12. Burstensaummembranvesikel (BSMV) 65
4.12.1.2 Darm von Saugferkeln
Die Herstellung von BSMV aus dem Jejunum von Saugferkeln erfolgte prinzipiell wie unter
4.12.1.1 beschrieben. Die bereits von der Serosa abgeschabte Mukosa wurde gewogen, mit
150 ml Praparationspuffer A versetzt und zweimal fur 1 min im Standmixer homogenisiert.
Die weiteren Schritte sind bis zur Zentrifugation mit 4.12.1.1 identisch. Die erste Zentrifu-
gation erfolgte bei 3687 × g, die apikalen Membranen wurden bei 16000 × g pelletiert und
anschließend in Praparationspuffer B aufgeschwemmt. Nach der zweiten Fallung wurde
die Suspension fur 10 min bei 3293 × g zentrifugiert. Die apikalen Membranen wurden aus
dem Uberstand bei 26430 × g, 30 min sedimentiert. Der letzte Zentrifugationsschritt nach
der Resuspendierung im Vesikelpuffer und dem Pottern wurde bei 31300× g durchgefuhrt.
4.12.2 Enzymbestimmungen von BSMV
Die Alkalische Phosphatase ist ein spezifisches Enzym der Burstensaummembran (Leit-
enzym), wahrend die Na/K-ATPase ein Leitenzym der basolateralen Membran ist. Bei
der BSMV-Praparation wurde eine Kontrollprobe (P1) vor der Fallung und Differential-
zentrifugation entnommen und eine Kontrollprobe der Vesikel (P2). Um den Gehalt der
Vesikel an Burstensaummembranen bzw. deren Anreicherung zu bestimmen, wurden die
Konzentrationen der beiden Leitenzyme in den entnommenen Kontrollproben bestimmt
(bezogen auf die Proteinkonzentration). Die Anreicherung ergab sich aus dem Verhaltnis
Enzymaktivitat/Proteinmenge von P2 zu P1.
Enzymaktivitat [U/l]
Proteingehalt [g/l]= U/g Protein
Anreicherung =U/g Protein P2
U/g Protein P1
4.12.2.1 Bestimmung der Aktivitat der Alkalischen Phosphatase (AP)
Die Alkalische Phosphatase spaltet p-Nitrophenylphosphat hydrolytisch in p-Nitrophenol
und Phosphat. Die entstehende Menge an p-Nitrophenol ist proportional zur Phosphata-
seaktivitat und kann aufgrund ihrer gelben Farbe photometrisch bestimmt werden.

66 4. Methoden
Je 50 µl Probe wurden mit 3 ml AP-Puffer/Na-Nitrophenylphosphat gemischt und sofort
im Photometer bei 405 nm gemessen. Die Extinktion wurde nach 1, 2 und 3 min abgelesen.
Die Messung erfolgte gegen Luft als Leerwert. Die Berechnung erfolgte nach folgender
Formel:
AP-Aktivitat [U/l] = 3300×∆E/min× Verdunnung
4.12.2.2 Bestimmung der Aktivitat der Na/K-ATPase
Die Aktivitat der Na/K-ATPase wird photometrisch als Differenz der ATPase-Aktivitat
mit und ohne Ouabainzusatz gemessen. Ouabain hemmt spezifisch die Na/K-ATPase.
Durch die ATPase-Aktivitat wird anorganisches Phosphat frei, welches in saurer Losung
mit Molybdat Phosphomolybdansaure bildet, die zu Molybdanblau reduziert wird. Die
Extinktion dieses blauen Farbkomplexes kann bei 690 nm gemessen werden.
0,5 ml Assaymix-Puffer wurden mit und ohne Ouabainzusatz fur 5 min bei 37◦C im Was-
serbad inkubiert. Es wurden jeweils 20 µl Probe bzw. H2O als Leerwert hinzugegeben
und das Gemisch 30 min bei 37◦C inkubiert. Die Reaktion wurde mit je 1,5 ml Farbrea-
genz gestoppt. Nach einer weiteren Inkubationszeit von 30 min, RT wurde die Extinktion
bei 690 nm gegen den Leerwert gemessen. Die Berechnung der Na/K-ATPase-Aktivitat
erfolgte nach der Formel:
∆E = Mittelwert E ohne Ouabainzusatz−Mittelwert E mit Ouabainzusatz
Na/K-ATPase-Aktivitat [U/l] = ∆E× 862
4.13 Proteinbestimmung
4.13.1 Proteinbestimmung nach Bradford
Durch die Bindung von Coomassie-Brilliantblau an Proteine verschiebt sich das Absorp-
tionsmaximum des Farbstoffs (465 nm ohne Protein, 595 nm mit Protein). Die Zunahme
der Absorption bei 595 nm ist ein Maß fur die Proteinkonzentration der Losung.

4.13. Proteinbestimmung 67
Die Proteinbestimmung der gelfiltrierten Fraktionen der Muzinreinigung erfolgte in Mi-
krotiterplatten (Greiner). Als Leerwert diente H2O. Es wurden je 50 µl Probe bzw. H2O
pro Vertiefung vorgelegt und mit 200 µl Bradford-Gebrauchslosung (s. Material 3.12) ver-
setzt. Nach 10-minutiger Inkubation bei RT (abgedeckt) wurde die Extinktion bei 580 nm
gemessen. Die Proteinkonzentration wurde mit Hilfe von Umrechnungsfaktoren bestimmt.
Zur Proteinbestimmung der BSMV wurde ebenfalls der Bradford-Test verwendet, hier
jedoch in Form des kauflichen BIO-RAD Protein-Assays. Um alle Proteinbindungsstellen
der BSMV zuganglich zu machen, wurde oberflachenaktives Saponin zugegeben. 50 µl
Probe bzw. BIO-RAD Proteinstandard (Bovines Plasma-Gamma-Globulin) wurden mit
50 µl Saponin (1%) versetzt und fur 20 min bei RT inkubiert. Zu dem Gemisch wurden je
2,5 ml verdunntes BIO-RAD-Farbstoffkonzentrat hinzugefugt. Es folgte eine Inkubation
von 10 min bei RT. Zur anschließenden Messung im Photometer wurden die Proben in
Kuvetten umgefullt. Die Proteinkonzentration der Proben wurde mit Hilfe der Standards
berechnet.
4.13.2 BCA-Test
Zur Bestimmung des Proteingehalts von praparierten Zellmembranen wurde der BCA-
Assay von Pierce nach den Angaben der Herstellers verwendet.
Die Proteine bilden bei diesem Test in alkalischer Losung einen Komplex mit Cu2+-Ionen
(Biuret-Reaktion). Die Cu2+-Ionen werden zu Cu+-Ionen reduziert, die mit Bicinchonin-
insaure (BCA) einen violetten Farbkomplex bilden.
Der Test wurde in Mikrotiterplatten (Greiner) durchgefuhrt. Je 10 µl der Proben, der
Standards (BSA) und des Leerwerts (H2O) wurden in die Vertiefungen gegeben und mit
200 µl des Substrats gemischt. Nach einer Inkubation von 30 min bei 37◦C wurde die
Extinktion bei 570 nm im Photometer gemessen. Die Proteinkonzentration wurde mit
Hilfe der Standards berechnet.

68 4. Methoden
4.13.3 Messung bei 280 nm
Die Proteinkonzentration der gereinigten Viren wurde durch Messung der Extinktion bei
einer Wellenlange von 280 nm (UV-Absorption) bestimmt. Zur Berechnung diente eine
Eichgerade, die mit Transferrin erstellt wurde.
4.14 Analyse von Sialinsauren
Losungen der Glykokonjugate in H2O (i.d.R. 1 mg/ml) wurden 1:2 mit 0,2 M HCl ver-
setzt. Durch eine Inkubation uber 50 min bei 80◦C wurden die Sialinsauren hydroly-
siert (Schauer, 1987). Die freigesetzten Sialinsauren wurden mit einer fluoreszieren-
den Verbindung derivatisiert, indem 50 µl des Hydrolysats mit dem gleichen Volumen
DMB-Reagenz fur 150 min bei 65◦C im Dunkeln inkubiert wurden (Hara et al., 1989).
Als Referenzsubstanzen dienten N-Acetyl- und N-Glykolylneuraminsaure der Firma Sig-
ma. Diese Sialinsauren wurden ebenfalls mit DMB-Reagenz (1 µl/50 µl) derivatisiert.
Die Auftrennung der Sialinsaurederivate erfolgte auf einer C-18”Reversed Phase“-Saule
(4 × 250 mm, Kornung 4 µm, Porengrosse 100 A, Merck) mit Acetonitril/Methanol/H2O
(9:7:84, v/v/v) als mobile Phase. Es wurden 20 µl des Derivats aufgetragen und bei einer
Flussrate von 1 ml/min isokratisch innerhalb von 15 min eluiert (L-6200A HPLC-Pumpe,
Merck). Die Detektion erfolgte uber einen Fluoreszenzdetektor (F-1050, Merck) bei ei-
ner Anregungswellenlange von 375 nm und einer Emissionswellenlange von 445 nm. Die
Daten wurden mit Hilfe eines elektronischen Datenverarbeitungssystems (D-6000 HPLC
Manager, Merck) gespeichert und ausgewertet.
4.15 SDS-Polyacrylamidgel-Elektrophorese
Proteine wurden mit Hilfe der diskontinuierlichen Natriumdodecylsulfat-Polyacrylamid-
gel-Elektrophorese (SDS-PAGE) aufgetrennt (Laemmli, 1970). Die Trenngele besaßen
die Maße 50 × 80 × 0,75 mm und eine Acrylamidkonzentration von 8 oder 10%. Die Auf-
trennung der Proteine erfolgte bei konstant 150 V in Miniproteingel-Kammern der Firma
Keutz. Die Elektrophorese wurde, wenn nicht anders angegeben, unter nicht-reduzierenden

4.16. Detektion von Proteinen in Polyacrylamidgelen 69
Bedingungen durchgefuhrt. Als Marker wurden vorgefarbte Proteine mit Molekularge-
wichten zwischen 14,3 und 220 kDa verwendet. Die aufgetrennten Proteine wurden ent-
weder im Gel gefarbt oder auf Nitrozellulose transferiert.
4.16 Detektion von Proteinen in Polyacrylamidgelen
4.16.1 Coomassie-Farbung
Der Farbstoff Coomassie-Brilliantblau bindet an Proteine, die im Gel auf diese Weise
sichtbar gemacht werden konnen. Die Gele wurden zu diesem Zweck 30 min in Coomassie-
Farbelosung geschwenkt und solange in Entfarbelosung inkubiert (ca. 1 h) bis der Hin-
tergrund entfarbt und die Proteinbanden gut sichtbar waren. Nach Wasserung der Gele
konnten diese uber Nacht zwischen zwei Zellophanfolien gespannt und getrocknet werden.
4.16.2 Silberfarbung zur Detektion von Glykoproteinen
Die Coomassie-Farbung von Gelen ist fur Proteinmengen unter 1 µg/Bande sowie fur
reich sialylierte Proteine ungeeignet. Daher wurde fur geringere Proteinmengen und vor
allem fur den Nachweis von Sialoglykoproteinen im Gel eine Silberfarbung durchgefuhrt
(Dzandu et al., 1984; Merril et al., 1979, 1981). Zu diesem Zweck wurde das Silver
Stain Kit der Firma BIO-RAD, das speziell zur selektiven Farbung von Glykoproteinen
im Handel ist, verwendet. Die Anwendung erfolgte entsprechend den Anweisungen des
Herstellers.
4.17 Western Blot
Die Elution der durch SDS-PAGE aufgetrennten Proteine mit anschließender Immobili-
sierung auf Nitrozellulose erfolgte im”Semi-Dry-Blot“ nach Kyhse-Anderson (1984)
mit modifizierten pH-Werten der Anodenpuffer (von pH 10,4 auf pH 9,0 bzw. auf pH 7,4).
Der Transfer erfolgte bei konstanter Stromstarke von 0,8 mA/cm2 uber einen Zeitraum

70 4. Methoden
von 75 min. Anschließend wurden noch freie Bindungsstellen auf der Membran durch
Inkubation mit”Blocking Reagenz“ (0,5% in PBSM) uber Nacht bei 4◦C abgesattigt.
4.18 Immunchemische Detektion von Proteinen
Auf Nitrozellulosemembranen immobilisierte Proteine wurden in einem indirekten immu-
nologischen Nachweis mit spezifischen monoklonalen Antikorpern detektiert. Alle Inku-
bationsschritte wurden bei 4◦C uber 1 h durchgefuhrt. Nach dem Blockierschritt (siehe
4.17) und nach jedem Inkubationsschritt wurde die Membran dreimal fur je 10 min mit
PBSM/0,1% Tween gewaschen. Es wurden 500 µl einer Verdunnung des entsprechenden
monoklonalen Antikorpers (in PBSM/0,05% Tween) pro Membran (6 cm × 8 cm) ein-
gesetzt. Dabei wurde die Antikorperlosung auf die Membran getraufelt und als dunner
Flussigkeitsfilm auf der Nitrozellulose verteilt, indem diese mit einem Stuck Parafilm
bedeckt wurde. Anschließend folgte eine Inkubation mit 10 ml Peroxidase-gekoppelten
Anti-Maus-Immunglobulinen (1:5000 bzw. 1:2000 in PBSM/0,05% Tween). Die gebun-
dene Peroxidase konnte mit BM Chemoluminescence Blotting Substrate (Roche) bzw.
Super Signalr (Pierce), beides auf Chemolumineszenz basierende Nachweissysteme, auf
Rontgenfilmen dargestellt werden.
4.19 Untersuchung der Virusbindung an Sialoglyko-
konjugate
4.19.1 ELISA
Die Bindung von TGEV bzw. TGEV-Mutanten an isolierte Darmmuzine (4.11) und an
Muzine aus der Glandula submandibularis des Schweines wurde untersucht, indem die
Viruspartikel mit den immobilisierten Testsubstanzen inkubiert und anschließend immu-
nologisch nachgewiesen wurden.
Die Muzine wurden auf Mikrotiterplatten (Maxisorpr, Nunc) uber Nacht bei 4◦C im-
mobilisiert (je 50 µl der entsprechenden Verdunnung in PBSM). Durch Ausschlagen der

4.19. Untersuchung der Virusbindung an Sialoglykokonjugate 71
Platten wurden die Losungen entfernt und noch freie Bindungsstellen mit”Blocking Rea-
genz“ abgesattigt (200 µl/Napf, 2 h, 4◦C). Nach der Absattigung und zwischen jedem
Inkubationsschritt wurden die Platten dreimal mit PBSM/0,05% Tween gewaschen. Die
Muzine jedes Napfes wurden mit je 50 µl einer Virussuspension (1 µg Protein/Napf) fur
1 h bei 4◦C inkubiert. Es wurden gereinigtes TGEV und die gereinigten TGEV-Mutanten
HAD3 und m10 verwendet. Der Nachweis gebundener Viren erfolgte indirekt mit einem
gegen das S-Protein gerichteten monoklonalen Antikorper (mAk 6A.C3, 1:50 in PBSM,
50 µl/Napf, 1 h, 4◦C) und HRP-gekoppelten Anti-Maus-Immunglobulinen vom Kaninchen
(DAKO, 1:1000 in PBSM, 50 µl/Napf, 1 h, 4◦C). Die Enzymreaktion wurde mit dem Zu-
satz von je 50 µl ABTS-Losung (Roche) gestartet und nach einigen Minuten (ca. 5 min)
mit dem gleichen Volumen einer 1%igen SDS-Losung gestoppt. Die Extinktion des Tests
wurde in einem ELISA-Messgerat bei einer Wellenlange von 405 nm bestimmt.
Zur Bestimmung des Grades der sialinsaurebedingten Bindung der Viruspartikel an die
Muzine wurden diese vor dem Virusinkubationsschritt mit Neuraminidase aus Vibrio cho-
lerae behandelt (siehe 4.8.4).
4.19.2 Virusbindung in Zellkultur
In Mikrotiterplatten (Greiner) gewachsene ST- bzw. LLC-PK1-Monolayer wurden, wie
unter 4.8.2 beschrieben, desialyliert bzw. mit MES-Puffer inkubiert. Nach dreimaligem
Waschen mit PBS wurden die Zellen mit gereinigtem TGEV oder TGEV-Mutanten fur
1 h bei 4◦C inkubiert (11 µg/Napf in PBS). Nach einem weiteren Waschschritt mit PBS
wurden die Zellen durch eine Behandlung mit 3% Paraformaldehyd fur 20 min bei Raum-
temperatur fixiert. Uberschussiges Paraformaldehyd wurde durch Waschen und 5-minuti-
ge Inkubation mit 0,1 M Glycin in PBSM inaktiviert. Zum Nachweis der gebundenen
Virusmenge folgten eine Inkubation mit dem gegen das S-Protein gerichteten mAk 6A.C3
(1:50 in PBSM, 1 h, 4◦C) und dann eine Inkubation mit Peroxidase-gekoppelten Anti-
Maus-Immunglobulinen vom Kaninchen (DAKO, 1:1000 in PBSM, 1 h, 4◦C). Zwischen
allen Inkubationsschritten wurden die Platten dreimal mit PBSM gewaschen. Der Nach-
weis des gebundenen Enzyms und damit indirekt des gebundenen Virus erfolgte, wie in
4.19.1 beschrieben. Die Reaktion mit der ABTS-Losung wurde nach 5 min mit 1% SDS
gestoppt und die Extinktion bei 405 nm im Photometer gemessen.

72 4. Methoden
4.19.3 Virusbindung auf Nitrozellulose
Proteine von Membranpraparationen, Zelllysaten, Muzinen und BSMV wurden nach der
Auftrennung durch eine SDS-PAGE auf Nitrozellulose geblottet. Nach Absattigung un-
spezifischer Bindungsstellen durch die Inkubation mit”Blocking-Reagenz“ (uber Nacht,
4◦C) wurde die Membran mit gereinigtem TGEV, HAD3 oder m10 (ca. 8 µg Protein/Blot,
Parafilm) fur 1 h bei 4◦C inkubiert. Die Detektion der gebundenen Viruspartikel erfolg-
te immunologisch uber den oben erwahnten mAk 6A.C3 (1:300 in PBSM/0,05% Tween,
Parafilm, 1 h, 4◦C) und im Fall der Membranproteine und Zelllysate uber HRP-gekoppelte
Anti-Maus-Immunglobuline aus der Ziege bzw. aus dem Kaninchen (1:5000 bzw. 1:2000
in 10 ml PBSM/0,05% Tween, 1 h, 4◦C). Handelte es sich bei den Ausgangssubstanzen
um Muzine bzw. BSMV wurde die Membran nach der Inkubation mit dem monoklo-
nalen Antikorper gegen das virale S-Protein mit biotinylierten polyklonalen Anti-Maus-
Immunglobulinen vom Schaf (1:1000 in 10 ml PBSM/0,05% Tween/Membran, 1 h, 4◦C)
und mit Streptavidin-Peroxidase (1:5000 in 10 ml PBSM/0,05% Tween/Membran, 1 h,
4◦C) inkubiert. Zwischen allen Schritten wurde die Membran dreimal fur 10 min mit
PBSM/0,1% Tween gewaschen. Die Detektion gebundener Peroxidase erfolgte wie unter
4.18 beschrieben uber Chemolumineszenz.
4.19.4 Virusbindungstest mittels Immunfluoreszenz
Von frisch entnommenen und in Isopenthan/flussigem Stickstoff eingefrorenen Jejunum-
proben wurden Kryoschnitte (ca. 7 µm dick) angefertigt. Mit 3% Paraformaldehyd (20 min,
RT) wurde das Gewebe fixiert. Uberschussiges Paraformaldehyd wurde durch Inkubati-
on mit 0,1 M Glycin/PBSM inaktiviert. Zur Klarung, ob und wo TGEV bzw. TGEV-
Mutanten am Darmepithel binden, wurden die Schnitte mit den gereinigten Viren in
konzentrierter Form (ca. 6 µg/µl, ca. 15 µl/Schnitt unter Parafilm) fur 1 h bei 4◦C inku-
biert. Die Detektion der gebundenen Viren erfolgte durch nachfolgende Inkubation mit
dem mAk gegen das S-Protein (6A.C3, 1:50 in PBSM, 50 µl unter Parafilm) und FITC-
gekoppelten Anti-Maus-Immunglobulinen aus dem Schaf (Amersham, 1:200 in PBSM,
50 µl unter Parafilm). Zwischen allen Inkubationsschritten wurden die Gewebeschnitte
jeweils dreimal mit PBSM gewaschen. Vor dem Eindeckeln der Proben mit Mowiol wur-

4.20. Analyse von Glykoproteinen 73
den diese mit H2O gewaschen, um Salzreste zu entfernen.
Die Auswertung der Virusbindung erfolgte durch Betrachtung der Gewebeschnitte mit Hil-
fe eines Fluoreszenzmikroskops (Anregungslicht 450–490 nm, Emissionslicht 520–560 nm)
bei verschiedenen Vergroßerungen. Zum Vergleich wurden die Darmschnitte im Durchlicht
betrachtet.
4.20 Analyse von Glykoproteinen
4.20.1 Triton X-114-Phasenseparierung
Die Oberflachenproteine konfluenter ST- bzw. LLC-PK1-Zellen wurden mit Biotin mar-
kiert (siehe 4.10.1) und die Zellen in 1 ml TBSE (TBS/1 mM EDTA), 1% Triton X-114
lysiert. Nach einer Inkubation von 1 h auf Eis wurde unlosliches Zellmaterial durch Zen-
trifugation entfernt (14000 × g, 10 min, 4◦C). Der Uberstand wurde fur 10 min bei 37◦C
inkubiert. Bei dieser Temperatur wird die kritische Micellenkonzentration uberschritten,
so dass sich eine wassrige Phase und eine Detergenzphase bilden, die durch Zentrifuga-
tion (1000 × g, 5 min, RT) voneinander getrennt werden konnen (Lisanti et al., 1988).
Die Detergenzphase (ca. 100 µl) wurde mit 900 µl TBSE, 0,06% Triton X-114, die obere
wassrige Phase (ca. 900 µl) mit 100 µl 10% Triton X-114 extrahiert. Die Proben wur-
den fur 5 min auf Eis gestellt, zentrifugiert (s.o.) und erneut einer temperaturinduzierten
Phasentrennung unterworfen. Die beiden extrahierten Phasen wurden anschließend mit
TBS auf 1 ml verdunnt und mit WGA- oder Streptavidin-Agarose gefallt (siehe 4.10.2
bzw. 4.10.1). Die gefallten Proteine wurde durch eine SDS-PAGE aufgetrennt und zur Un-
tersuchung hinsichtlich Virusbindung auf Nitrozellulose geblottet. Alternativ wurde das
Polyacrylamidgel einer Silberfarbung unterzogen.

74 4. Methoden
4.20.2 Glykoproteindetektion
4.20.2.1 Kohlenhydraterkennung
BSMV-Proben (mit und ohne vorherige Desialylierung, siehe 4.8.3) wurden elektropho-
retisch aufgetrennt und auf eine PVDF-Membran geblottet. Nach einem 10-minutigen
Waschschritt mit PBSM wurden die Kohlenhydratgruppen der Proteine mit 10 mM Na-
triummetaperjodat (in 100 mM Natriumacetat-Puffer pH 5,5, 20 min, RT, im Dunkeln)
oxidiert. Durch eine anschließende Inkubation mit 0,125 mM Biotinhydrazid (in 100 mM
Natriumacetat-Puffer, 30 min, RT) wurden die Glykoproteine im Bereich ihrer oxidier-
ten Kohlenhydratreste markiert. An dieser Stelle folgte die Inkubation mit”Blocking-
Reagenz“ (4◦C, uber Nacht), um uberschussige Bindungsstellen der Membran abzusatti-
gen. Durch Schwenken in Streptavidin-Peroxidase (1:6000 in PBSM, 30 min) konnten
uber die Enzymreaktion mittels Chemolumineszenz die Kohlenhydrate im Rontgenfilm
sichtbar gemacht werden (4.17). Zwischen allen Inkubationsschritten wurde die Membran
dreimal fur 10 min in PBSM gewaschen. Es wurde das ECL glycoprotein detection module
(Amersham) verwendet.
4.20.2.2 Selektive Sialinsaureerkennung
Zur exklusiven Markierung der Sialinsaurereste wurde ebenfalls das Kit von Amersham
verwendet. Die 10-minutige Inkubation der Membran in PBSM erfolgte hier auf Eis. Der
Oxidationsschritt mit 1 mM Natriummetaperjodat (in 100 mM Natriumacetat-Puffer,
20 min) wurde ebenso wie der anschließende Waschschritt (dreimal PBSM fur 10 min)
auf Eis durchgefuhrt. Die oxidierten Sialinsaurereste wurden durch eine Inkubation mit
5 mM Biotinhydrazid (in 100 mM Natriumacetat-Puffer, 30 min, RT) markiert. Nach
dem Blocken wurde die Membran mit Streptavidin-Peroxidase (1:1000 in PBSM, 30 min)
inkubiert. Zwischen allen Inkubationen wurde mit PBSM gewaschen (dreimal 10 min).
Die Detektion erfolgte ebenfalls mittels Chemolumineszenz (4.17, 4.20.2.1).

4.20. Analyse von Glykoproteinen 75
4.20.3 Analyse von Glykoproteinen mit Lektinen
4.20.3.1 Lektintest auf dem Blot
Zur naheren Charakterisierung der Kohlenhydratgruppen wurden in der SDS-PAGE auf-
getrennte und auf Nitrozellulose transferierte BSMV mit Lektinen inkubiert. Hierzu wurde
das DIG Glycan Differentiation Kit der Firma Roche verwendet. Nach Absattigung der
Membranbindungsstellen in Blockierungslosung wurden die Membranen zweimal mit TBS
und einmal mit Puffer 1 gewaschen (je 10 min). Es folgte eine Inkubation mit Digoxigenin-
markierten Lektinen nach den Anweisungen des Herstellers in Puffer 1. Fur die Untersu-
chung wurden die Lektine MAA (Maackia amurensis Agglutinin), SNA (Sambucus nigra
Agglutinin) und PNA (Peanut Agglutinin) verwendet. Nach dreimaligem Waschen (TBS,
je 10 min) wurden die Membranen mit AP-gekoppelten Anti-Digoxigenin-Fab-Fragmenten
inkubiert (1:1000 in TBS, 1 h, RT). Die Detektion erfolgte nach einem weiteren Wasch-
schritt uber die Zugabe einer Losung aus Nitroblautetrazoliumchlorid/X-phosphat. Die
Reaktion wurde nach einigen Minuten mit H2O gestoppt. Das Vorhandensein der jewei-
ligen Zucker war durch Graufarbung der Membranen an den entsprechenden Stellen zu
erkennen.
4.20.3.2 Fallung von BSMV mit Lektinen
Zellen wurden wie unter 4.10.2 beschrieben zur Glykoproteinanalyse mit WGA-Agarose
gefallt.
Die Fallung von BSMV mit Lektinen erfolgte auf ahnliche Weise. Hierzu wurden die
BSMV nach Inkubation mit VCNA bzw. Natriumacetat-Puffer (siehe 4.8.3) in NP40-
Lysispuffer unter Zusatz von 1 mM MgCl2, 1 mM CaCl2 und 1 mM Mn2Cl aufgenom-
men. Nach einer Lyse von 15 min auf Eis wurden die Proben fur 30 min bei 16000 × g
und 4◦C zentrifugiert. Die so von Zelltrummern befreiten Uberstande wurden auf gewa-
schene WGA-, PNA- oder Jacalin-Agarose gegeben und uber Nacht bei 4◦C geschuttelt.
Die uber ihre jeweiligen Zucker gebundenen Glykoproteine wurden mit SDS-Probenpuffer
(zweifach) eluiert (10 min, 96◦C). Die Proben wurden fur Virusbindungstests auf Nitro-
zellulose verwendet.


5. Ergebnisse
5.1 Vergleich der Sialinsaurebindungsaktivitat von
TGEV und E. coli F5
Die Fimbrien von E. coli F5 besitzen wie TGEV eine Sialinsaurebindungseigenschaft.
Aufgrund dieser Eigenschaft konnen sie an intestinale Sialoglykokonjugate binden, und
E. coli kann auf diese Weise den Darm kolonisieren (Gaastra und de Graaf, 1982).
TGEV agglutiniert bevorzugt Erythrozyten, die Neu5Gc auf ihrer Oberflache prasentieren
(Schultze et al., 1996). Auch E. coli F5 bevorzugt Neu5Gc als Rezeptordeterminante auf
der Erythrozytenoberflache (Lindahl et al., 1987; Ono et al., 1989). Im Schweinedarm
scheint E. coli F5 ebenfalls bevorzugt an Neu5Gc als Sialinsaure bestimmter Glykolipide
zu binden (Teneberg et al., 1990, 1994).
Zur Bindung von E. coli F5 an Sialoglykokonjugate des Darmes sind bereits einige Un-
tersuchungen von verschiedenen Arbeitsgruppen durchgefuhrt worden (s. Einleitung 1.3).
Der Vergleich der Sialinsaurebindungsaktivitat von TGEV und E. coli F5 sollte klaren,
ob die beiden Krankheitserreger die gleichen Sialoglykokonjugate erkennen und somit die
gleichen Rezeptoren fur eine Darminfektion benutzen.
5.1.1 Interaktion mit Erythrozyten verschiedener Tierarten
Eine einfache Moglichkeit, die Sialinsaurebindungsaktivitat verschiedener Erreger zu ver-
gleichen, ist ein Hamagglutinationstest. Um Unterschiede und Gemeinsamkeiten zwischen
TGEV und E. coli F5 in der Bevorzugung bestimmter Sialinsauretypen zu erkennen, wur-
77

78 5. Ergebnisse
den Erythrozyten von verschiedenen Tierarten vergleichend eingesetzt.
ST-Zellen wurden mit TGEV, Stamm PURDUE 46 (PUR46), infiziert. Die im Zellkul-
turuberstand enthaltenen Virionen wurden durch Sedimentation konzentriert. Um eine
optimale Hamagglutinationsaktivitat zu erhalten, wurden die Viruspartikel mit Neurami-
nidase aus Vibrio cholerae desialyliert (TGEV-NA) und uber einen Saccharosegradienten
gereinigt. E. coli F5 wurde uber Nacht in Minca-Medium kultiviert, wodurch die Fimbri-
enausbildung gewahrleistet wurde. Anschließend wurden die Bakterien mit PBS/1% Man-
nose gewaschen und im gleichen Puffer resuspendiert. Um die Hamagglutinationsaktivitat
vergleichen zu konnen, wurden die beiden Erreger in einer Konzentration eingesetzt, die
bei Pferdeerythrozyten einen ubereinstimmenden Hamagglutinationstiter ergab.
Erythrozyten Hamagglutinationstiter
(Tierart) (HAU/ml)
TGEV-NA E. coli F5
Huhn 128 ≤2
Rind 64 32
Pferd 128 128
Schwein 256 64
Tabelle 5.1: Vergleich der Hamagglutinationsaktivitat von TGEV und E. coli F5. Es wurden
Erythrozyten verschiedener Tierarten eingesetzt.
Wie in Tabelle 5.1 zu sehen ist, konnte E. coli F5 Pferde-, Schweine- und Rindererythro-
zyten agglutinieren. Huhnererythrozyten wurden dagegen nur geringgradig, meist aber
nicht agglutiniert. Neu5Gc ist der Sialinsauretyp, der mehr als 90% der Sialinsauren auf
der Oberflache von Pferde-, Schweine- und Rindererythrozyten ausmacht (Reuter et al.,
1988). Auf Huhnererythrozyten kommt hingegen keine Neu5Gc, sondern uberwiegend
Neu5Ac vor. Das Hamagglutinationsverhalten von E. coli F5 stimmte somit mit fruhe-
ren Berichten uberein, dass dieser Erreger spezifisch Neu5Gc erkennt (Ono et al., 1989;
Teneberg et al., 1990). TGEV-NA konnte, im Gegensatz zu E. coli F5, Erythrozyten
aller verwendeten Spezies agglutinieren. Die Bindung an bestimmte Sialinsauretypen wie
Neu5Gc ist somit bei desialyliertem TGEV weniger spezifisch als bei E. coli F5.
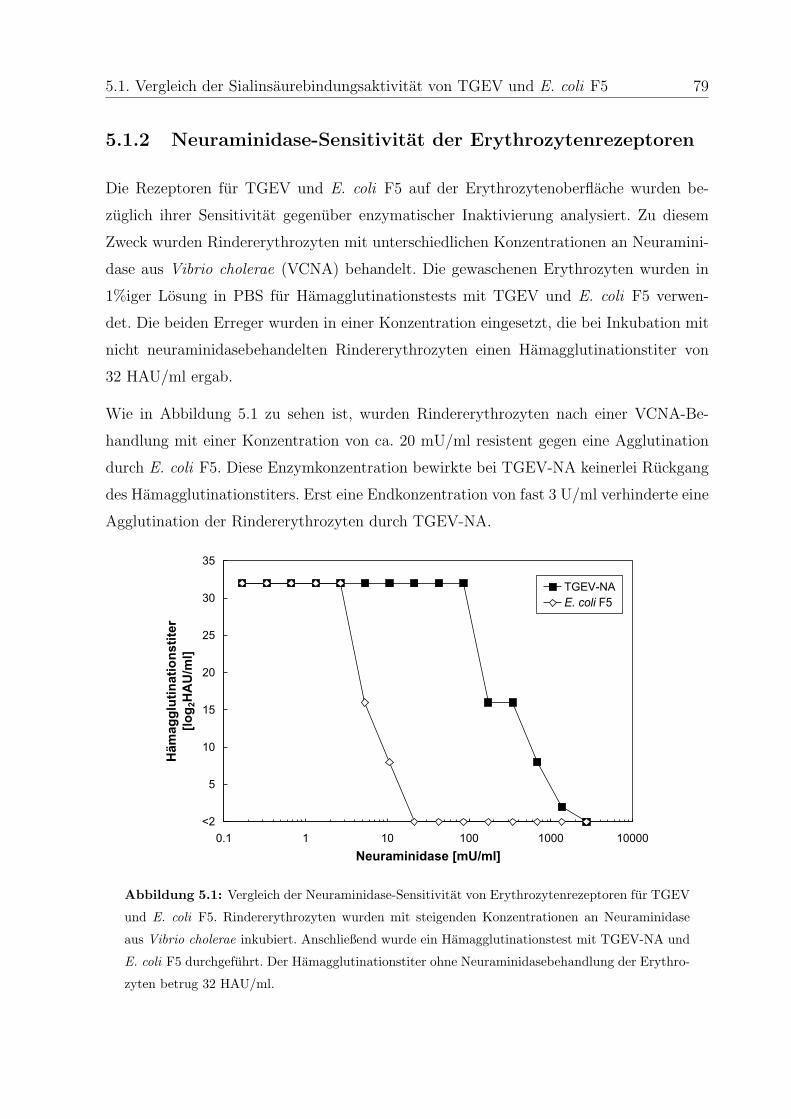
5.1. Vergleich der Sialinsaurebindungsaktivitat von TGEV und E. coli F5 79
5.1.2 Neuraminidase-Sensitivitat der Erythrozytenrezeptoren
Die Rezeptoren fur TGEV und E. coli F5 auf der Erythrozytenoberflache wurden be-
zuglich ihrer Sensitivitat gegenuber enzymatischer Inaktivierung analysiert. Zu diesem
Zweck wurden Rindererythrozyten mit unterschiedlichen Konzentrationen an Neuramini-
dase aus Vibrio cholerae (VCNA) behandelt. Die gewaschenen Erythrozyten wurden in
1%iger Losung in PBS fur Hamagglutinationstests mit TGEV und E. coli F5 verwen-
det. Die beiden Erreger wurden in einer Konzentration eingesetzt, die bei Inkubation mit
nicht neuraminidasebehandelten Rindererythrozyten einen Hamagglutinationstiter von
32 HAU/ml ergab.
Wie in Abbildung 5.1 zu sehen ist, wurden Rindererythrozyten nach einer VCNA-Be-
handlung mit einer Konzentration von ca. 20 mU/ml resistent gegen eine Agglutination
durch E. coli F5. Diese Enzymkonzentration bewirkte bei TGEV-NA keinerlei Ruckgang
des Hamagglutinationstiters. Erst eine Endkonzentration von fast 3 U/ml verhinderte eine
Agglutination der Rindererythrozyten durch TGEV-NA.
<2
5
10
15
20
25
30
35
0.1 1 10 100 1000 10000
Neuraminidase [mU/ml]
Häm
ag
glu
tin
ati
on
sti
ter
[lo
g2H
AU
/ml]
TGEV-NA
E. coli F5
Abbildung 5.1: Vergleich der Neuraminidase-Sensitivitat von Erythrozytenrezeptoren fur TGEV
und E. coli F5. Rindererythrozyten wurden mit steigenden Konzentrationen an Neuraminidase
aus Vibrio cholerae inkubiert. Anschließend wurde ein Hamagglutinationstest mit TGEV-NA und
E. coli F5 durchgefuhrt. Der Hamagglutinationstiter ohne Neuraminidasebehandlung der Erythro-
zyten betrug 32 HAU/ml.

80 5. Ergebnisse
Dieser große Unterschied in der Neuraminidase-Sensitivitat der Erythrozytenrezeptoren
beider Erreger deutet darauf hin, dass TGEV und E. coli F5 verschiedene Rezeptoren
zur Bindung an Erythrozyten nutzen. Es konnte aber auch sein, dass E. coli F5 eine
wesentlich hohere Anzahl von Sialinsaureresten auf der Erythrozytenoberflache benotigt,
um uberhaupt eine Hamagglutinationsreaktion hervorrufen zu konnen.
5.1.3 Untersuchung der Hamagglutinationshemmung durch
Muzine und Glykolipide
Um nahere Informationen uber die Sialinsaurebindungsaktivitaten von TGEV und E. co-
li F5 zu erhalten, wurde ihre Sensitivitat gegenuber Hamagglutinationsinhibitoren ge-
testet. Die Agglutination von Erythrozyten kann durch bestimmte Substanzen gehemmt
werden. Die Starke der Hemmung gibt einen Hinweis, wie effizient der betreffende Bin-
dungspartner mit dem viralen bzw. bakteriellen Erreger interagieren kann.
Intestinale Muzine sind hoch sialylierte Glykokonjugate und somit potentielle Liganden
fur Mikroorganismen wie TGEV und E. coli F5, die uber die Magen-Darm-Passage in den
Organismus eindringen. Da die Infektionen mit beiden Erregern v.a. Ferkel, die junger
als 15 Tage sind, betreffen, wurden Muzine aus zwei unterschiedlichen Altersgruppen
gewonnen (ca. 14 Tage und ca. 10 bis 15 Wochen alte Tiere). So konnten durch den
Vergleich der Bindung an diese Muzine mogliche altersbedingte Unterschiede erkannt
werden.
E. coli F5 besitzt eine Affinitat fur Ganglioside, die auf den Enterozyten junger Schwei-
ne exprimiert werden (Teneberg et al., 1990). Aus diesem Grund wurden neben den
Muzinen zwei verschiedene Gangliosidtypen, Rinderhirnganglioside und Hamatosid aus
Pferdeerythrozyten, auf ihre Hemmaktivitat untersucht.
5.1.3.1 Praparation intestinaler Muzine
Muzine wurden aus dem Jejunum 12–14 Tage alter Saug- und 10–15 Wochen alter Ab-
setzferkel isoliert und anschließend gelfiltriert (s. Methodenteil 4.11). Nach Bestimmung
des Protein- und Kohlenhydratgehalts konnten aus den gesammelten Fraktionen diejeni-

5.1. Vergleich der Sialinsaurebindungsaktivitat von TGEV und E. coli F5 81
gen bestimmt werden, die einen hohen Kohlenhydratanteil besaßen und somit die Muzine
beinhalteten. Die Muzinfraktionen wurden vereinigt und lyophilisiert.
5.1.3.2 Analyse des Sialinsauregehalts der Inhibitoren
Zur qualitativen und quantitativen Bestimmung des Sialinsauregehalts der Inhibitoren
wurden je 0,5 mg der Muzine und je 0,1 mg der Glykolipide in 1 ml H2O gelost und, wie
unter 4.14 beschrieben, behandelt. Die Analyse erfolgte mittels HPLC. Um den Gehalt
an Neu5Gc und Neu5Ac quantifizieren zu konnen, wurden mit unterschiedlichen Kon-
zentrationen der Referenzsubstanzen (von 0,5 µg/ml bis 500 µg/ml) Eichgeraden erstellt.
Anhand der Steigung der Geraden konnte die Konstante zur Berechnung der Sialinsaure-
menge bestimmt werden. Der Gehalt Gx an N-Glykolyl- bzw. N-Acetylneuraminsaure
berechnete sich nach folgenden Formeln:
GNeu5Gc [µg/20µl] =F
56558884bzw. GNeu5Ac [µg/20µl] =
F
52348368,
wobei F die Flache unter dem zu der jeweiligen Sialinsaure gehorenden HPLC-Peak ist.
Den Gehalt in µg/mg erhalt man unter Berucksichtigung der Anfangskonzentrationen der
Proben und der durch die Hydrolyse und Derivatisierung bedingten Verdunnungsschritte
(s. 4.14).
Der Gehalt der intestinalen Muzine an Neu5Gc und Neu5Ac ist in Abbildung 5.2 dar-
gestellt. Alle vier Proben enthielten beide Typen von Sialinsauren, jedoch in einem un-
terschiedlichen Verhaltnis und in unterschiedlichen Mengen. Der Gehalt an Neu5Gc und
Neu5Ac der Muzine aus dem Saugferkel A stimmte annahernd mit dem Gehalt der Muzi-
ne aus Absetzferkel D uberein. Die intestinalen Muzine des Saugferkels B besaßen zu fast
gleichen Teilen Neu5Gc und Neu5Ac. Der Gesamtsialinsauregehalt betrug jedoch mehr
als das Vierfache der Muzine von Tier A und D. Die Muzine des Absetzferkels C hingegen
unterschieden sich von den anderen Muzinen v.a. durch einen 10fach hoheren Gehalt an
Neu5Ac im Vergleich zu Neu5Gc. Diese Ergebnisse weisen darauf hin, dass es eine Variati-
on in der Verteilung und Menge an Neu5Gc und Neu5Ac in Darmmuzinen gibt und somit
keine klaren Unterschiede zwischen den Proben von Saug- und Absetzferkeln bestehen.
Da die Inhibitoren bei der Hamagglutinationshemmung auf einen ubereinstimmenden Sia-
linsauregehalt eingestellt werden sollten, wurde auch von den Rinderhirngangliosiden und
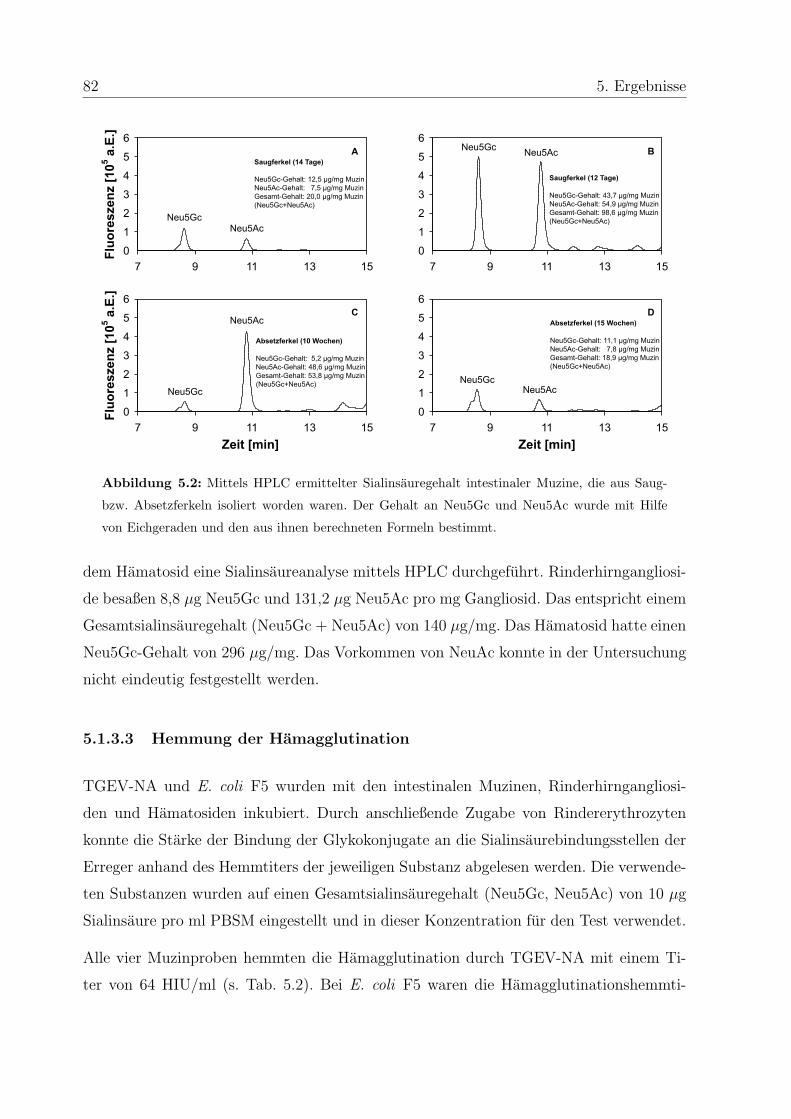
82 5. Ergebnisse
0
1
2
3
4
5
6
7 9 11 13 15
Flu
ore
szen
z[1
05
a.E
.]
Saugferkel (14 Tage)
Neu5Gc-Gehalt: 12,5 µg/mg Muzin
Neu5Ac-Gehalt: 7,5 µg/mg Muzin
Gesamt-Gehalt: 20,0 µg/mg Muzin
(Neu5Gc+Neu5Ac)
Neu5GcNeu5Ac
A
0
1
2
3
4
5
6
7 9 11 13 15
Flu
ore
szen
z[1
05
a.E
.]
Saugferkel (12 Tage)
Neu5Gc-Gehalt: 43,7 µg/mg Muzin
Neu5Ac-Gehalt: 54,9 µg/mg Muzin
Gesamt-Gehalt: 98,6 µg/mg Muzin
(Neu5Gc+Neu5Ac)
Neu5GcNeu5Ac B
0
1
2
3
4
5
6
7 9 11 13 15
Zeit [min]
Flu
ore
szen
z[1
05
a.E
.]
Absetzferkel (10 Wochen)
Neu5Gc-Gehalt: 5,2 µg/mg Muzin
Neu5Ac-Gehalt: 48,6 µg/mg Muzin
Gesamt-Gehalt: 53,8 µg/mg Muzin
(Neu5Gc+Neu5Ac)Neu5Gc
Neu5AcC
0
1
2
3
4
5
6
7 9 11 13 15
Zeit [min]
Flu
ore
szen
z[1
05
a.E
.]
Absetzferkel (15 Wochen)
Neu5Gc-Gehalt: 11,1 µg/mg Muzin
Neu5Ac-Gehalt: 7,8 µg/mg Muzin
Gesamt-Gehalt: 18,9 µg/mg Muzin
(Neu5Gc+Neu5Ac)
Neu5GcNeu5Ac
D
Abbildung 5.2: Mittels HPLC ermittelter Sialinsauregehalt intestinaler Muzine, die aus Saug-
bzw. Absetzferkeln isoliert worden waren. Der Gehalt an Neu5Gc und Neu5Ac wurde mit Hilfe
von Eichgeraden und den aus ihnen berechneten Formeln bestimmt.
dem Hamatosid eine Sialinsaureanalyse mittels HPLC durchgefuhrt. Rinderhirngangliosi-
de besaßen 8,8 µg Neu5Gc und 131,2 µg Neu5Ac pro mg Gangliosid. Das entspricht einem
Gesamtsialinsauregehalt (Neu5Gc + Neu5Ac) von 140 µg/mg. Das Hamatosid hatte einen
Neu5Gc-Gehalt von 296 µg/mg. Das Vorkommen von NeuAc konnte in der Untersuchung
nicht eindeutig festgestellt werden.
5.1.3.3 Hemmung der Hamagglutination
TGEV-NA und E. coli F5 wurden mit den intestinalen Muzinen, Rinderhirngangliosi-
den und Hamatosiden inkubiert. Durch anschließende Zugabe von Rindererythrozyten
konnte die Starke der Bindung der Glykokonjugate an die Sialinsaurebindungsstellen der
Erreger anhand des Hemmtiters der jeweiligen Substanz abgelesen werden. Die verwende-
ten Substanzen wurden auf einen Gesamtsialinsauregehalt (Neu5Gc, Neu5Ac) von 10 µg
Sialinsaure pro ml PBSM eingestellt und in dieser Konzentration fur den Test verwendet.
Alle vier Muzinproben hemmten die Hamagglutination durch TGEV-NA mit einem Ti-
ter von 64 HIU/ml (s. Tab. 5.2). Bei E. coli F5 waren die Hamagglutinationshemmti-

5.1. Vergleich der Sialinsaurebindungsaktivitat von TGEV und E. coli F5 83
Hamagglutinationshemmtiter
(HIU/ml)
TGEV-NA E. coli F5
intestinale Muzine
Saugferkel A 64 16
Saugferkel B 64 8
Absetzferkel C 64 8
Absetzferkel D 64 32
Glykolipide
Rinderhirnganglioside <2 16
Hamatosid <2 8
Tabelle 5.2: Vergleich von porzinen intestinalen Muzinen und Glykolipiden bezuglich ihrer Fahig-
keit, die Agglutination von Rindererythrozyten durch TGEV und E. coli F5 zu hemmen.
ter 2 bis 8fach niedriger. Anders verhielten sich die Ganglioside in ihrer Hemmung. Die
Hamagglutination durch E. coli F5 wurde bis zu einem Titer von 16 HIU/ml gehemmt.
Bei TGEV-NA war es den Glykolipiden dagegen nicht moglich, die Hamagglutination zu
hemmen.
Dieses Ergebnis zeigt, dass TGEV-NA Muzinen gegenuber sensitiver ist als E. coli F5. Bei
Gangliosiden dagegen ist E. coli F5 sensitiver als das Virus. Vergleicht man die Aktivitat
der intestinalen Muzine mit der Aktivitat der Glykolipide bezuglich der Hemmung der
TGEV-NA-bedingten Hamagglutination, scheinen Muzine im Gegensatz zu Glykolipiden
potentere Inhibitoren und damit moglicherweise auch Bindungspartner bei Infektionen zu
sein. In der Hemmung der Hamagglutination ist bei TGEV-NA kein Unterschied zwischen
den Muzinen aus den zwei Altersgruppen zu erkennen. Da der mittels HPLC-Analyse
bestimmte Gesamtsialinsauregehalt aller eingesetzten Hemmsubstanzen ubereinstimmte,
hatte nur durch das unterschiedliche Verhaltnis an Neu5Gc zu Neu5Ac oder durch zusatz-
liche sich unterscheidende Oberflachenstrukturen ein Unterschied in der Hemmaktivitat
der Muzine auftreten konnen. Die im Hamagglutinationshemmtest gemessene Interaktion
mit Muzinen erklart nicht, warum die TGEV-Infektion bei Saugferkeln soviel schwerer
verlauft als bei alteren Tieren.
Die vergleichenden Untersuchungen von TGEV und E. coli F5 zeigen, dass sich diese bei-

84 5. Ergebnisse
den Krankheitserreger in der Spezifitat ihrer Sialinsaurebindungsaktivitat unterscheiden.
Somit ist es wahrscheinlich, dass die beiden Mikroorganismen auch im Tier verschiede-
ne Sialoglykokonjugate als Liganden nutzen, um eine intestinale Infektion zu etablieren.
Falls Sialoglykokonjugate auf der Oberflache der Enterozyten an der Replikation bzw.
Pathogenitat von TGEV beteiligt sein sollten, spricht dieses Ergebnis dafur, dass eher
Glykoproteine als Glykolipide Bindungspartner fur das Virus darstellen.
5.2 Bindung von TGEV an Zellkulturen
Porzine Aminopeptidase N (pAPN) ist als Rezeptor fur die Infektion von Zellkulturen
durch TGEV unerlasslich (Delmas et al., 1992). Die Sialinsaurebindungsaktivitat wird
hingegen fur das Wachstum in Zellkultur nicht benotigt. Mutanten und Varianten, die
keine Sialinsaurebindungsaktivitat besitzen, konnen zu gleichen Titern in Zellkultur re-
plizieren wie das Wildtypvirus (Krempl et al., 1997, 2000). Da die Sialinsaurebindungs-
aktivitat somit scheinbar keinen Einfluss auf die Infektiositat der Virionen in Zellkultur
hat, wurde genauer untersucht, ob auch die Bindung an die Zelloberflache nur durch
die Interaktion mit pAPN erfolgt oder ob die Sialinsaurebindungsaktivitat von TGEV
ebenfalls zur Bindung an Zellkulturzellen beitragt.
Zu diesem Zweck wurden in Mikrotiterplatten ausgesate ST- und LLC-PK1-Zellen mit ge-
reinigtem Virus bei 4◦C inkubiert. Nachdem nicht gebundene Viruspartikel durch Waschen
mit PBS von der Zelloberflache entfernt worden waren, wurden die Zellen mit Paraformal-
dehyd fixiert. Gebundene Virionen wurden mit Hilfe eines gegen das S-Protein gerichteten
monoklonalen Antikorpers und eines enzymgebundenen Zweitantikorpers nachgewiesen.
Um die Sialinsaurebindungsstellen auf den S-Proteinen der Virionen von Faktoren zel-
lularen Ursprungs zu demaskieren, wurden die Viruspartikel vor ihrer Saccharosereinigung
desialyliert (TGEV-NA). Vergleichend wurden nicht desialylierte Virionen eingesetzt. Ne-
ben TGEV-PUR46 wurde die Bindung von zwei Mutanten getestet. Eingesetzt wurden die
Mutante HAD3, die mit Hinblick auf ihre Hamagglutinationsdefizienz von Krempl et al.
(2000) selektiert worden war, und die Epitop D-Mutante m10, die als Escape-Mutante
mit dem monoklonalen Antikorper 40.1 (Epitop D) aus PUR 115-PAR isoliert worden
war (Delmas et al., 1986). Beide Mutanten binden nicht an Sialinsauren, was sich in

5.2. Bindung von TGEV an Zellkulturen 85
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
TGEV
- NA
TGEV HAD3
- NA
HAD3 m10
- NA
m10
DE
40
5
ST
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
TGEV
- NA
TGEV HAD3
- NA
HAD3 m10
- NA
m10
DE
40
5
LLC-PK1
Abbildung 5.3: Interaktion von TGEV sowie der Mutanten HAD3 und m10 mit ST- und LLC-
PK1-Zellen. Die Viren wurden in desialylierter (-NA) und in unbehandelter Form eingesetzt. Die
weißen Saulen stehen fur die nicht desialylierten Zellen. Die grauen Saulen reprasentieren die
mit 25 mU Neuraminidase fur 1 h bei 37◦C behandelten Zellen. Die Starke der Bindung wurde
immunologisch mit Hilfe eines enzymgebundenen Zweitantikorpers gemessen und ist in Extinkti-
onseinheiten angegeben.
der fehlenden Hamagglutinationsaktivitat dieser Viren ausdruckt (Krempl et al., 1997,
2000). Um zusatzlich zum Vergleich des Wildtypvirus mit sialinsaurebindungsdefizienten
Mutanten einen weiteren Beweis der sialinsaurebedingten Bindung an die Oberflache der
Zellkulturzellen zu bekommen, wurden die Zellen vor der Inkubation mit Virus fur 1 h
bei 37◦C mit Neuraminidase aus Vibrio cholerae desialyliert. Kontrollzellen wurden an-
stelle des Enzyms mit MES-Puffer inkubiert. Unterschiede in der Virusbindung zwischen
diesen so vorbehandelten Zellen beruhen somit auf einer Bindung an Sialinsauren auf der
Zelloberflache.
Wie in Abbildung 5.3 zu sehen ist, gab es eine messbare Bindung von TGEV an ST- und
LLC-PK1-Zellen (weiße Saulen). Wurde der Test mit desialylierten Virionen durchgefuhrt
(TGEV-NA), stieg die Menge gebundener Viruspartikel um etwa das Sechsfache an.
Die Behandlung der Virionen mit Neuraminidase entfernt Sialoglykokonjugate von der Vi-
rusoberflache, die sich im Laufe der Infektion dort ansammeln. Diese Sialoglykokonjugate
stammen von der Zelloberflache der infizierten Zellen und bewirken, dass zu einem spaten
Zeitpunkt geerntetes TGEV keine Hamagglutinationsaktivitat mehr besitzt. Nur in sehr
hohen Viruskonzentrationen ist noch eine Hamagglutinationsaktivitat nachweisbar. Der
quantitative Unterschied in der Bindung von TGEV und TGEV-NA an die Zelloberflachen
ist dadurch bedingt, dass bei unbehandeltem Virus (TGEV) die Bindungsstellen fur Sia-
linsaure auf dem S-Protein durch Inhibitoren besetzt sind und somit nicht mit den Zellen

86 5. Ergebnisse
des Testansatzes interagieren konnen. Die niedrigen Bindungswerte des unbehandelten
TGEV stimmen mit den Bindungswerten der sialinsaurebindungsdefizienten Mutanten
uberein. Bei den Mutanten unterschieden sich die niedrigen Werte an gebundenen Virio-
nen (HAD3 und m10) kaum von denen, die mit desialylierten Virionen (HAD3-NA und
m10-NA) erzielt wurden. Die Bindung der Mutanten an die Zelloberflache war ebenso wie
die TGEV-Bindung mit Werten < 0,1 deutlich geringer als die mit TGEV-NA erzielte
Bindung (> 0,4). Eine weitere Bestatigung fur die Bedeutung der Sialinsaurebindungsak-
tivitat von TGEV fur die Bindung an Zellen lieferte die Reduktion der Bindung durch die
vorherige Desialylierung der Zellen. Die Menge an gebundenem TGEV-NA nahm nach
enzymatischer Entfernung der Sialinsauren von der Zelloberflache bei ST-Zellen um mehr
als das Vierfache und bei LLCPK-1-Zellen fast um das Sechsfache ab (graue Saulen). Bei
TGEV, welches nicht neuraminidasebehandelt war, kam es nur zu einer geringen Abnahme
der Bindung. Die Mutanten ließen keinerlei Abnahme ihrer Bindung an Zellen erkennen.
Diese Ergebnisse zeigen, dass bei hamagglutinationsaktivem TGEV die Sialinsaurebin-
dungsaktivitat effizient zur Bindung an Zellen in Zellkultur beitragt.
5.3 Untersuchung von Zelloberflachenproteinen
5.3.1 Bindung an Membranproteine
Die Bindung von TGEV an Sialoglykokonjugate von ST- und LLC-PK1-Zellen wurde
naher untersucht. Da der Vergleich der Sialinsaurebindungsaktivitaten von TGEV und
E. coli F5 darauf hinwies, dass Glykoproteine im Gegensatz zu Glykolipiden die potenteren
Bindungspartner fur TGEV darstellen, wurden Membranproteine von ST- und LLC-PK1-
Zellen untersucht.
Membranproteine wurden, wie unter 4.9 beschrieben, aus den Zellen isoliert. Um spezifi-
sche Proteine als Bindungspartner identifizieren zu konnen, wurden die Membranproteine
(20 µg/Spur) in einem SDS-Polyacrylamidgel elektrophoretisch aufgetrennt und anschlie-
ßend auf Nitrozellulose geblottet. Nachdem die freien Bindungsstellen auf der Nitrozel-
lulose uber Nacht mit”Blocking Reagenz“ abgesattigt worden waren, erfolgte eine Inku-
bation mit desialyliertem gereinigten Virus (TGEV-NA bzw. m10-NA) fur 1 h bei 4◦C.

5.3. Untersuchung von Zelloberflachenproteinen 87
VCNA+_ _
+
a a b b
TGEV-NA
__++
aa bb
m10-NA
__++
aa bb
antiAPN
220
97
[kDa]
pAPN
Abbildung 5.4: Bindung von desialyliertem TGEV (TGEV-NA) und der Mutante m10 (m10-NA)
an Membranproteine aus ST- (a) und LLC-PK1-Zellen (b). Die Membranproteine wurden vor der
elektrophoretischen Auftrennung mit Neuraminidase aus Vibrio cholerae (VCNA, +) desialyliert
bzw. mit Natriumacetat-Puffer behandelt (−, s. auch unter Methoden 4.8.3). Auf der rechten
Seite ist der immunchemische Nachweis der zellularen pAPN dargestellt. Die beiden Striche ganz
links kennzeichnen die Position zweier Markerproteine mit den Angaben der Molekulargewichte
in kDa. Der untere Pfeil weist auf die pAPN-Bande hin, der obere Pfeil markiert eine zusatzliche
hochmolekulare Proteinbande, die von TGEV-NA erkannt wurde.
Gebundene Viruspartikel wurden, wie unter 4.19.3 beschrieben, immunchemisch nachge-
wiesen. Zur Unterscheidung des zellularen Rezeptors porzine Aminopeptidase N (pAPN)
von moglichen anderen Bindungspartnern wurde parallel ein Western Blot mit nachfolgen-
der immunchemischer Detektion von pAPN durchgefuhrt (s. auch 4.18). Als monoklonaler
Antikorper (mAk) wurde bei diesem Nachweis der gegen pAPN gerichtete aus der Maus
stammende mAk G43 verwendet.
Abbildung 5.4 zeigt die Proteinbanden, an die die Viren bzw. der gegen pAPN gerich-
tete mAk G43 gebunden haben. Der gegen den zellularen Rezeptor pAPN gerichtete
mAk G43 erkannte eine Bande mit einem Molekulargewicht von ca. 150 kDa. An diese
Bande konnten sowohl desialylierte TGEV- als auch desialylierte m10-Virionen binden.
Von TGEV-NA wurde jedoch im Gegensatz zu m10-NA noch eine weitere Proteinban-
de erkannt. Sowohl ST- (a) als auch LLC-PK1-Zellen (b) besaßen dieses hochmolekulare
Protein, das von TGEV-NA bei beiden Zellarten annahernd gleich stark gebunden wurde.
Anders als das hochmolekulare Protein wurde der zellulare Rezeptor pAPN sowohl von
TGEV-NA als auch von m10-NA erkannt. Die Bindung der Viren an pAPN fuhrte bei
ST-Zellen zu einem starkeren Signal als bei LLC-PK1-Zellen.
Wurden die Membranproteine vor der elektrophoretischen Auftrennung mit Neuramini-

88 5. Ergebnisse
dase aus Vibrio cholerae, wie unter 4.8.3 beschrieben, behandelt (+ VCNA), kam es zu
einer starken Verringerung bis zu volligem Verlust der Virusbindung an das hochmoleku-
lare Protein. Es handelt sich somit bei der Bindung von TGEV-NA an die hochmolekulare
Proteinbande um eine sialinsaureabhangige Bindung. Wie die Inkubation mit dem mAk
G43 zeigte, stellt das hochmolekulare Protein keine Form der pAPN, sondern einen zweiten
Bindungspartner von TGEV dar. Auch die Tatsache, dass dieses zusatzliche Protein nicht
von m10-NA erkannt wird, belegt die Bedeutung der Sialinsaurebindungsaktivitat von
TGEV fur die Erkennung eines zweiten Proteins neben dem zellularen Rezeptor pAPN.
Die Bindung an pAPN wurde durch die Neuraminidasebehandlung der Membranproteine
nicht beseitigt. Die Effizienz der Neuraminidasebehandlung außerte sich in einem etwas
veranderten elektrophoretischen Laufverhalten der pAPN-Bande.
Der Sialinsauregehalt der Membranproteine wurde in einer HPLC-Analyse untersucht. Die
Proteine enthielten relativ viel Neu5Ac (13,5 µg/mg Protein), jedoch nur wenig Neu5Gc
(0,5 µg/mg Protein).
5.3.2 Bindung an Zelloberflachenproteine
Da die unter 4.9 beschriebene Praparation von Membranproteinen aus Zellkulturen keine
absolut reinen Plasmamembranen liefert und somit auch intrazellulare Strukturen enthal-
ten sein konnen, sollte ein zweiter Versuchsansatz die Lokalisation des Bindungspartners
von TGEV auf der Zelloberflache bestatigen.
ST- und LLC-PK1-Zellen wurden mit dem nicht membrangangigen Sulfo-NHS-Biotin
markiert und anschließend lysiert (s. Methodenteil 4.10.1). Die biotinylierten Oberflachen-
proteine wurden mit Streptavidin-Agarose aus den Zelllysaten gefallt, elektrophoretisch
aufgetrennt und auf Nitrozellulose geblottet. Die Bindungstests mit desialylierten Viren
erfolgten in der gleichen Art und Weise wie bei der Untersuchung der Bindung an Mem-
branproteine (5.3.1). Zusatzlich zu TGEV-NA und m10-NA wurde die hamagglutinati-
onsdefiziente Mutante HAD3 in desialylierter gereinigter Form (HAD3-NA) eingesetzt.
Abbildung 5.5 (obere Reihe) zeigt die Bindung der Viren an Oberflachenproteine von
ST- (a) und LLC-PK1-Zellen (b). Die Proteine wurden zusatzlich unter reduzierenden
Bedingungen (+DTT, untere Reihe) aufgetrennt, um daraus resultierende Unterschiede
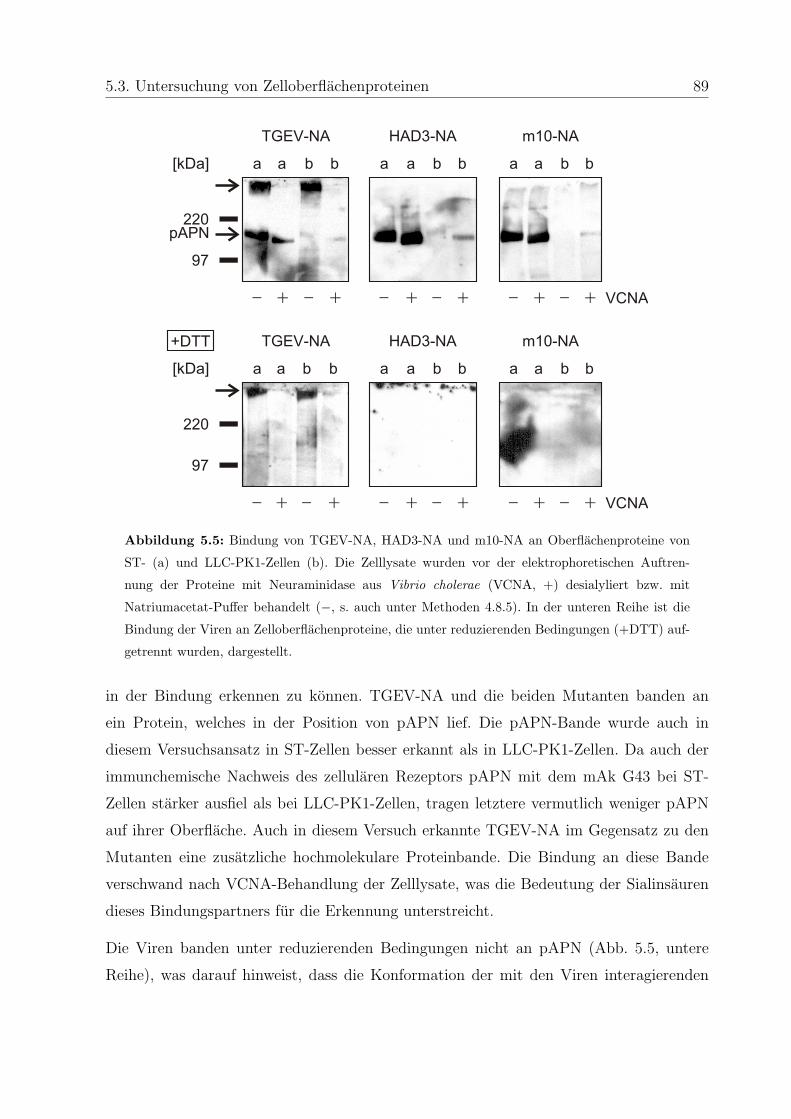
5.3. Untersuchung von Zelloberflachenproteinen 89
__++ VCNA
a a b b
TGEV-NA
aa bb
HAD3-NA
__++
aa bb
m10-NA
220
97
[kDa]
pAPN
+_ _
+
__++ VCNA
a a b b
TGEV-NA
aa bb
HAD3-NA
__++
aa bb
m10-NA
220
97
[kDa]
+_ _
+
+DTT
Abbildung 5.5: Bindung von TGEV-NA, HAD3-NA und m10-NA an Oberflachenproteine von
ST- (a) und LLC-PK1-Zellen (b). Die Zelllysate wurden vor der elektrophoretischen Auftren-
nung der Proteine mit Neuraminidase aus Vibrio cholerae (VCNA, +) desialyliert bzw. mit
Natriumacetat-Puffer behandelt (−, s. auch unter Methoden 4.8.5). In der unteren Reihe ist die
Bindung der Viren an Zelloberflachenproteine, die unter reduzierenden Bedingungen (+DTT) auf-
getrennt wurden, dargestellt.
in der Bindung erkennen zu konnen. TGEV-NA und die beiden Mutanten banden an
ein Protein, welches in der Position von pAPN lief. Die pAPN-Bande wurde auch in
diesem Versuchsansatz in ST-Zellen besser erkannt als in LLC-PK1-Zellen. Da auch der
immunchemische Nachweis des zellularen Rezeptors pAPN mit dem mAk G43 bei ST-
Zellen starker ausfiel als bei LLC-PK1-Zellen, tragen letztere vermutlich weniger pAPN
auf ihrer Oberflache. Auch in diesem Versuch erkannte TGEV-NA im Gegensatz zu den
Mutanten eine zusatzliche hochmolekulare Proteinbande. Die Bindung an diese Bande
verschwand nach VCNA-Behandlung der Zelllysate, was die Bedeutung der Sialinsauren
dieses Bindungspartners fur die Erkennung unterstreicht.
Die Viren banden unter reduzierenden Bedingungen nicht an pAPN (Abb. 5.5, untere
Reihe), was darauf hinweist, dass die Konformation der mit den Viren interagierenden

90 5. Ergebnisse
pAPN-Domane und nicht eine lineare Aminosaurekette fur die Interaktion wichtig ist.
Das hochmolekulare Protein konnte hingegen auch nach Auftrennung unter reduzieren-
den Bedingungen von TGEV-NA gebunden werden. Der zusatzliche Bindungspartner von
TGEV-NA ist somit ein Zelloberflachenprotein und wird uber Sialinsaurereste vom Virus
erkannt.
Um herauszufinden, ob isoliertes S-Protein aus gereinigtem TGEV-NA das gleiche Bin-
dungsverhalten wie die Virionen selbst aufweist, wurde das S-Protein mit Detergenzien,
wie unter 4.6 beschrieben, aus der Virusmembran gelost und uber einen Saccharosegra-
dienten gereinigt. In einem Bindungstest mit Zelloberflachenproteinen aus ST-Zellen ver-
hielt sich isoliertes S-Protein wie vollstandige Viruspartikel. Wie in Abbildung 5.6 zu
sehen ist, erkannte aus TGEV-NA isoliertes S-Protein eine hochmolekulare Bande und
eine Bande mit einem Molekulargewicht von ca. 150 kDa (linkes Bild). Die Identifizierung
dieser kleineren Bande als pAPN mittels mAk G43 ist nochmal in der Mitte der Abbil-
dung dargestellt. Auch die Interaktion des isolierten S-Proteins mit dem hochmolekularen
Zelloberflachenprotein beruht auf Sialinsaureerkennung, wie das Verschwinden der Bande
nach vorheriger Desialylierung der aufgetragenen Zelllysate beweist.
_+ VCNA
TGEV-NAS-Protein antiAPN
_+
Kontrolle
220
97
[kDa]
pAPN
+_
Abbildung 5.6: Bindung des aus TGEV-NA isolierten S-Proteins an Oberflachenproteine von
ST-Zellen. Das mittlere Bild zeigt den Nachweis von pAPN durch mAk G43. Auf der rechten Seite
ist zur Kontrolle eine Inkubation der Membran mit den im Bindungstest verwendeten Antikorpern
dargestellt.
Die Desialylierung der Virionen war bei der Bindung stets von entscheidender Bedeutung.
Wie in Abbildung 5.7 zu sehen ist, erkannte das nicht neuraminidasebehandelte TGEV,
wenn es in der gleichen Proteinkonzentration wie die anderen Virionen (8 µg/Blot) einge-
setzt wurde, weder die hochmolekulare Proteinbande noch die pAPN-Bande (linkes Bild).
Erst bei einer 10fach hoheren Konzentration von 80 µg/Blot wurde eine schwache pAPN-

5.3. Untersuchung von Zelloberflachenproteinen 91
TGEV
_+ VCNA
HAD3
_+
m10
220
97
[kDa]
pAPN
+_
TGEV10x
_+
Abbildung 5.7: Bindung nicht desialylierter Viren an Oberflachenproteine von ST-Zellen. Der
Pfeil gibt die Position der pAPN-Bande an. Uber den Bildern ist angegeben mit welchen Virionen
der Bindungstest durchgefuhrt wurde.
Bande von TGEV erkannt. Dies spricht dafur, dass die Viruspartikel ohne Neuraminida-
sebehandlung nicht ausreichend mit den auf der Nitrozellulose immobilisierten Proteinen
interagieren konnen. Eine mogliche Ursache konnte sein, dass die Viren uber gebundene
Sialoglykokonjugate zur Bildung großer Virusaggregate neigen.
Die Mutanten HAD3 und m10 verhielten sich ohne vorherige Neuraminidasebehandlung
genauso wie in desialyliertem Zustand. Der zellulare Rezeptor pAPN wurde bei 150 kDa
von beiden Mutanten gut erkannt. Dies bestatigt, dass sie keinerlei Sialinsaurebindungsak-
tivitat besitzen und somit auch nicht uber Sialoglykokonjugate aneinander binden konnen,
was wie bei TGEV ihre Bindung an pAPN behindern wurde.
5.3.3 Darstellung der Zelloberflachenproteine
Zusatzlich zu den Bindungstests wurden die Zelloberflachenproteine noch mit anderen
Nachweisverfahren dargestellt (Abb. 5.8). Die Oberflachenproteine von ST-Zellen ließen
sich mit den drei Methoden in unterschiedlicher Anzahl und Starke nachweisen. Das hoch-
molekulare Oberflachenprotein, welches von TGEV-NA uber Sialinsaurebindung erkannt
wird, wurde bei der Inkubation der auf der Nitrozellulosemembran immobilisierten bioti-
nylierten Oberflachenproteine mit Streptavidin-Peroxidase nicht gefunden (Abb. 5.8, A,
oberer Pfeil). Moglicherweise ist die Biotinylierung dieses Proteins nicht sehr effektiv, so
dass ein Nachweis durch Streptavidin-Peroxidase bei dieser Proteinkonzentration nicht
moglich war. Das biotinylierte Protein konnte jedoch offensichtlich in fur den Virusbin-

92 5. Ergebnisse
VCNA_
+
B
_+
CA
220
97
[kDa]
pAPN
46
66
Abbildung 5.8: Darstellung der Oberflachenproteine von ST-Zellen mit verschiedenen Nachweis-
verfahren. A = Nachweis biotinylierter Oberflachenproteine mit Streptavidin-Peroxidase; B = Sil-
berfarbung der Oberflachenproteine; C = Coomassiefarbung aller Zellproteine. Zum Vergleich sind
links die Positionen der Bindungspartner von TGEV mit Pfeilen markiert.
dungsnachweis ausreichender Menge von Streptavidin-Agarose gefallt werden (s. 5.3.2).
Auch im Coomassie-Gel (Abb. 5.8, C) ließ sich die hochmolekulare Proteinbande nicht
darstellen. Bei der Coomassie-Farbung mussen die Proteine in einer Mindestkonzentration
von 1 µg/Bande vorliegen. Stark glykosylierte Proteine sind auch in hoheren Konzentratio-
nen nicht oder nur schwer nachweisbar. Das hochmolekulare Protein kam somit entweder
in zu geringer Menge in der aufgetragenen Probe vor oder ließ sich aufgrund eines hohen
Kohlenhydratanteils nicht anfarben.
Wurde eine fur Glykoproteine spezifische Silberfarbung der mit Streptavidin-Agarose
gefallten Zelloberflachenproteine durchgefuhrt (Abb. 5.8, B), konnte man eine hochmole-
kulare Proteinbande erkennen, die nach VCNA-Behandlung des Zelllysats etwas schwacher
wurde. Bei dieser hochmolekularen Bande handelt es sich wahrscheinlich um den zusatz-
lichen Bindungspartner von TGEV.
5.3.4 Sialinsaureabhangigkeit der Bindung von TGEV
Wie die vorherigen Untersuchungen schon gezeigt haben, war fur die Erkennung des hoch-
molekularen Proteins durch TGEV-NA das Vorhandensein von Sialinsaureresten auf dem
Bindungspartner essentiell. Wurden die Zelllysate vor der SDS-PAGE, wie unter 4.8.5

5.4. Charakterisierung des Sialoglykoproteins 93
+_ _
+ VCNA
1 1 2 2TGEV-NA
220
97
[kDa]
pAPN
Abbildung 5.9: Sialinsaureabhangigkeit der Bindung von TGEV-NA an ST-Zellen. Die von
TGEV-NA gebundene hochmolekulare Proteinbande (oberer Pfeil) wurde nach Neuraminidase-
behandlung des Zellrasens (1, + VCNA) bzw. der Zelllysate (2, + VCNA) nicht mehr erkannt.
beschrieben, desialyliert, konnte TGEV-NA nicht mehr an das hochmolekulare Protein
binden (s. Abb. 5.5).
Vergleichend wurde getestet, ob eine Desialylierung der intakten Zellmonolayer den glei-
chen Effekt auf die Virusbindung hat. ST-Zellen wurden mit Neuraminidase aus Vibrio
cholerae in einer Konzentration von 250 mU/ml MES-Puffer fur 1 h bei 37◦C inkubiert.
Dies entsprach einer Enzymaktivitat von 11 mU/cm2 Zellrasen. Die Kontrollzellen wur-
den mit MES-Puffer ohne Enzymzusatz behandelt. Es folgte nach einem Waschschritt mit
PBS die Biotin-Markierung der Zelloberflachenproteine. Alle weiteren Schritte wurden wie
ublich durchgefuhrt (s. Methodenteil 4.10.1).
Wie in Abbildung 5.9 zu sehen ist, verschwand die hochmolekulare Proteinbande, die von
TGEV-NA bei nicht enzymatisch behandelten ST-Zellen erkannt wurde, nach vorheriger
Desialylierung des Zellrasens (1). Somit verhielt sich das Virus in seiner Erkennung des
hochmolekularen Zelloberflachenproteins uber Sialinsaurereste bei desialyliertem Zellra-
sen ebenso wie bei desialylierten Zelllysaten (2).
5.4 Charakterisierung des Sialoglykoproteins
TGEV erkennt aufgrund seiner Sialinsaurebindungsaktivitat ein hochmolekulares Protein
auf der Oberflache von ST- und LLC-PK1-Zellen. Dieses Protein, bei dem es sich —
wie die vorhandenen Sialinsaurereste zeigen — um ein Sialoglykoprotein handelt, wurde
durch weitere Untersuchungen naher charakterisiert. Zum einen wurde die Assoziation

94 5. Ergebnisse
des Proteins zur Zellmembran untersucht. Zum anderen sollte festgestellt werden, ob es
sich bei diesem Protein um das einzige Sialoglykoprotein des Zelllysats handelt, das von
TGEV erkannt wird.
5.4.1 Triton X-114-Phasenseparierung
Die Assoziation des hochmolekularen Proteins mit der Plasmamembran wurde mit Hil-
fe einer Triton X-114-Phasenseparierung untersucht (s. Methoden 4.20.1). Proteine von
ST- und LLC-PK1-Zellen wurden mit Triton X-114 extrahiert. Nach einer Phasensepa-
rierung wurden die biotinylierten Oberflachenproteine mit Streptavidin-Agarose aus der
Detergenz- und aus der wassrigen Phase prazipitiert und mit der fur Glykoproteine spezi-
fischen Silberfarbung im SDS-Gel dargestellt (s. Abb. 5.10, A). Parallel wurde ein Ansatz
mit unmarkierten Zellproteinen durchgefuhrt. In diesem Ansatz wurden die Glykopro-
teine der beiden Phasen mit immobilisiertem Weizenkeimagglutinin (WGA) gefallt und
ebenfalls nach elektrophoretischer Auftrennung im SDS-Gel gefarbt (s. Abb. 5.10, B).
Das hochmolekulare Protein wurde bei beiden Ansatzen uberwiegend in der wassrigen
Phase (w) detektiert. Die Bande war bei der WGA-Fallung starker ausgepragt, da dort
auch die intrazellularen Glykoproteine nachgewiesen wurden. Neben der Silberfarbung
w
a a b bA
aa bbB
aa bbC
220
97
[kDa]
wwwwwd d dddd
Abbildung 5.10: Charakterisierung der Assoziation des Sialoglykoproteins mit der Plasmamem-
bran mittels Triton X-114-Phasenseparierung. Detergenz- (d) und wassrige Phase (w) von ST-
(a) und LLC-PK1-Zellen (b) wurden nach Streptavidinfallung (A) bzw. WGA-Fallung (B) im Sil-
bergel dargestellt. Ein Bindungstest mit WGA-gefallten Proteinen und aus TGEV-NA isoliertem
S-Protein ist auf der rechten Seite(C) dargestellt.
wurde das Protein nach WGA-Fallung in einem Bindungstest mit aus TGEV-NA iso-
liertem S-Protein nachgewiesen. Wie in Abbildung 5.10 (C) zu sehen ist, befand sich

5.4. Charakterisierung des Sialoglykoproteins 95
das Glykoprotein eindeutig in der wassrigen Phase. Dieses Verhalten zeigt, dass es sich
bei dem hochmolekularen Sialoglykoprotein eher um ein membranassoziiertes als um ein
Transmembranprotein handelt.
5.4.2 Prazipitation mit WGA
Die bisherigen Ergebnisse zeigen, dass unter den biotinylierten Oberflachenproteinen von
ST- und LLC-PK1-Zellen die hochmolekulare Bande das Hauptsialoglykoprotein, das von
TGEV erkannt wird, darstellt. Um herauszufinden, ob das auch fur die Gesamtheit der
zellularen Sialoglykoproteine gilt, wurden die Zellen lysiert und mit immobilisiertem Wei-
zenkeimagglutinin (WGA) gefallt. WGA ist ein Lektin, welches an N-Acetylglukosamin
und an Sialinsauregruppen bindet (Gallagher et al., 1985).
Wie in Abbildung 5.11 zu erkennen ist, band TGEV sowohl bei ST- (a) als auch bei
LLC-PK1-Zellen (b) bevorzugt an die hochmolekulare Proteinbande. Nach Behandlung
der Zelllysate mit Neuraminidase (+ VCNA) verschwand nur diese Bande, was bestatigt,
dass es sich um ein sialyliertes Makromolekul handelt und dass die Erkennung durch
TGEV uber Sialinsaurereste vermittelt wird.
+_ _
+ VCNA
a a b b
TGEV-NA
220
97
[kDa]
+DTT
Abbildung 5.11: Spezifische Bindung von TGEV an das hochmolekulare Sialoglykoprotein. Aus
Lysaten von ST- (a) und LLC-PK1-Zellen (b) wurden die Glykoproteine mit WGA gefallt und
unter reduzierenden Bedingungen (+DTT) in der SDS-PAGE aufgetrennt. Anschließend wurde
ein Bindungstest mit desialylierten TGE-Virionen (TGEV-NA) durchgefuhrt.
Der zellulare Rezeptor pAPN ist in dieser Abbildung nicht zu erkennen, da die Elektro-
phorese unter reduzierenden Bedingungen durchgefuhrt wurde. Unter nicht reduzierenden
Bedingungen band TGEV auch an pAPN, wie in Abbildung 5.10 (C) gezeigt ist. Bei pAPN

96 5. Ergebnisse
handelt es sich somit auch um ein Glykoprotein und, wie das veranderte Laufverhalten der
pAPN-Bande nach VCNA-Behandlung der Zelllysate in den vorherigen Versuchen gezeigt
hat, ist auch dieses Protein sialyliert. Die Bindung von TGEV an pAPN erfolgt jedoch
nicht uber Sialinsaurereste, sondern uber Interaktion mit einer Aminosauredomane, da
eine VCNA-Behandlung keine Abnahme der Bindung von TGEV an die pAPN-Bande
erkennen ließ.
5.5 Interaktion von TGEV mit intestinalen Muzinen
Die Bindung an den zellularen Rezeptor pAPN ist die Voraussetzung fur eine TGEV-
Infektion. Zusatzlich existieren aber vermutlich Faktoren, die die Enteropathogenitat von
TGEV bedingen. Die Sialinsaurebindungsaktivitat von TGEV scheint solch einen Patho-
genitatsfaktor darzustellen, da Varianten wie PRCoV und Mutanten wie die Epitop-D-
Mutanten, die keine oder nur eine stark reduzierte Enteropathogenitat aufweisen, ihre
Sialinsaurebindungsaktivitat verloren haben (Krempl et al., 1997).
Nachdem durch die Bindungstests mit TGEV und den Mutanten HAD3 und m10 in
Zellkultur bereits ein hochmolekulares Sialoglykoprotein als zweiter Bindungspartner fur
TGEV gefunden wurde, sollte auch die Interaktion von TGEV mit Komponenten des
Dunndarms wie extrazellularen Muzinen und Darmepithel getestet werden. Durch die
vergleichende Untersuchung von TGEV und E. coli F5 wurde bereits festgestellt, dass
Muzine Bindungspartner fur TGEV sein konnen (s. Abschnitt 5.1.3). In Abschnitt 5.1.3.2
ist die Charakterisierung des Sialinsauregehalts von aus vier Ferkeln unterschiedlichen
Alters isolierten intestinalen Muzinen (A–D) beschrieben. Die Interaktion von TGEV mit
diesen Muzinen wurde naher untersucht. Vergleichend wurden Muzine aus der Glandula
submandibularis des Schweines (PSM) mit in die Untersuchung einbezogen. Diese Muzine
zeigten bereits in fruheren Untersuchungen eine gute Bindung an desialyliertes TGEV
(Krempl et al., 2000). Der Sialinsauregehalt von PSM wurde ebenfalls mittels HPLC,
wie in Abschnitt 5.1.3.2 beschrieben, analysiert. Abbildung 5.12 zeigt, dass etwa 20-mal
mehr Neu5Gc als Neu5Ac in der Probe enthalten waren.
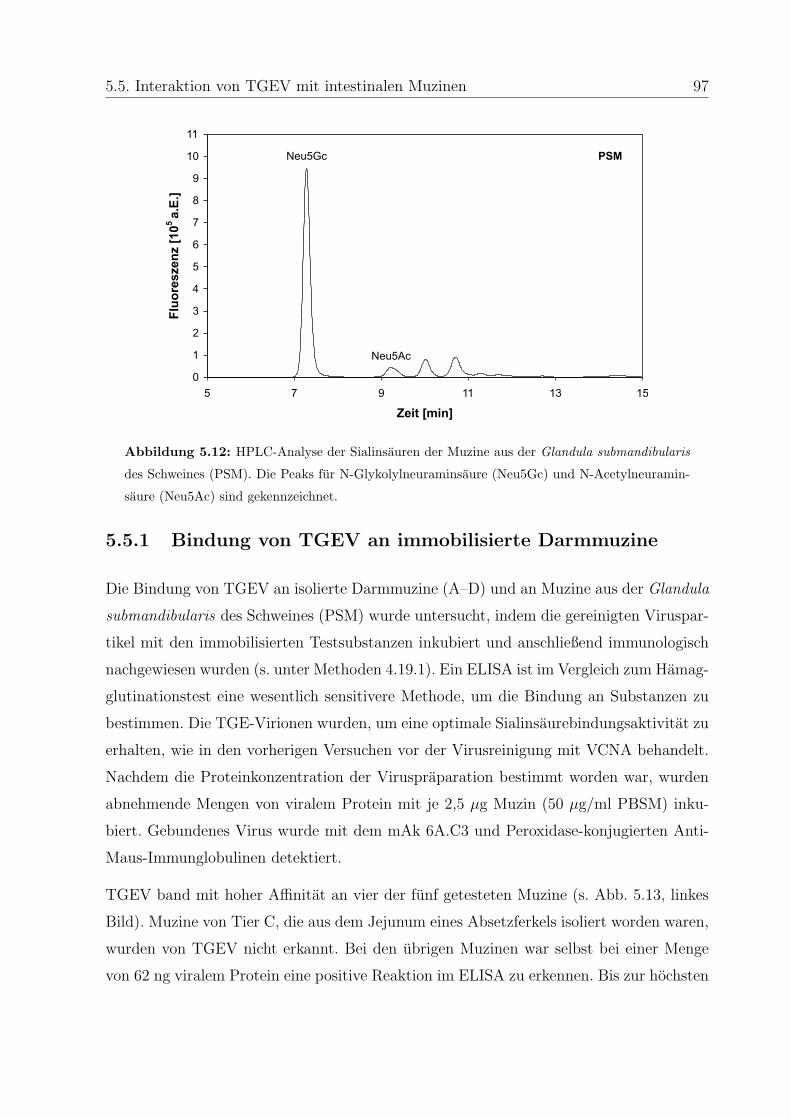
5.5. Interaktion von TGEV mit intestinalen Muzinen 97
0
1
2
3
4
5
6
7
8
9
10
11
5 7 9 11 13 15
Zeit [min]
Flu
ore
szen
z[1
05
a.E
.]
PSM
Neu5Ac
Neu5Gc
Abbildung 5.12: HPLC-Analyse der Sialinsauren der Muzine aus der Glandula submandibularis
des Schweines (PSM). Die Peaks fur N-Glykolylneuraminsaure (Neu5Gc) und N-Acetylneuramin-
saure (Neu5Ac) sind gekennzeichnet.
5.5.1 Bindung von TGEV an immobilisierte Darmmuzine
Die Bindung von TGEV an isolierte Darmmuzine (A–D) und an Muzine aus der Glandula
submandibularis des Schweines (PSM) wurde untersucht, indem die gereinigten Viruspar-
tikel mit den immobilisierten Testsubstanzen inkubiert und anschließend immunologisch
nachgewiesen wurden (s. unter Methoden 4.19.1). Ein ELISA ist im Vergleich zum Hamag-
glutinationstest eine wesentlich sensitivere Methode, um die Bindung an Substanzen zu
bestimmen. Die TGE-Virionen wurden, um eine optimale Sialinsaurebindungsaktivitat zu
erhalten, wie in den vorherigen Versuchen vor der Virusreinigung mit VCNA behandelt.
Nachdem die Proteinkonzentration der Viruspraparation bestimmt worden war, wurden
abnehmende Mengen von viralem Protein mit je 2,5 µg Muzin (50 µg/ml PBSM) inku-
biert. Gebundenes Virus wurde mit dem mAk 6A.C3 und Peroxidase-konjugierten Anti-
Maus-Immunglobulinen detektiert.
TGEV band mit hoher Affinitat an vier der funf getesteten Muzine (s. Abb. 5.13, linkes
Bild). Muzine von Tier C, die aus dem Jejunum eines Absetzferkels isoliert worden waren,
wurden von TGEV nicht erkannt. Bei den ubrigen Muzinen war selbst bei einer Menge
von 62 ng viralem Protein eine positive Reaktion im ELISA zu erkennen. Bis zur hochsten

98 5. Ergebnisse
0
0.1
0.2
0.3
0.4
0.5
0.6
1000 500 250 125 62 31 16
virales Protein [ng]
DE
40
5
Muzin A
Muzin B
Muzin C
Muzin D
PSM
0
0.1
0.2
0.3
0.4
0.5
0.6
1000 500 250 125 62 31 16 8
Muzinkonzentration [µg/ml]
DE
40
5
Abbildung 5.13: Interaktion von TGEV mit immobilisierten Muzinen. Auf der linken Seite ist
die Bindung abnehmender Mengen viralen Proteins an 2,5 µg Muzin pro Vertiefung dargestellt.
Das rechte Bild zeigt die Bindung von 1 µg viralen Proteins an abnehmende Muzinkonzentrationen.
Die gebundene Enzymmenge wurde anhand der Absorption des Substrats bei 405 nm gemessen.
eingesetzten Menge von 1 µg TGEV-Protein pro Vertiefung stieg die messbare Bindung
stetig an. Um in den folgenden Versuchen eindeutige Ergebnisse zu bekommen, wurden
somit bei allen ELISAs Viruskonzentrationen von 1 µg/Vertiefung eingesetzt.
Da Darmmuzine aus einem der Tiere nicht von TGEV gebunden wurden, wurde eine
großere Spanne an unterschiedlichen Muzinkonzentrationen auf Virusbindung getestet
(1000 µg/ml bis 8 µg/ml). Dadurch sollte ausgeschlossen werden, dass die nicht vorhan-
dene Bindung durch eine zu geringe Menge immobilisierten Muzins bedingt ist. Wie in
Abbildung 5.13 auf der rechten Seite zu sehen ist, wurden Muzine des Absetzferkels C in
keiner der eingesetzten Konzentrationen von TGEV erkannt. Die anderen Muzine hinge-
gen wurden sowohl bei einer Konzentration von 1000 µg/ml als auch bei einer Konzen-
tration von 8 µg/ml unverandert gut erkannt.
Ein Grund fur die fehlende Bindung von TGEV an Muzine des Tieres C konnte die Sia-
linsaurezusammensetzung der Muzine sein. Wie in Abbildung 5.2 (C) zu sehen ist, besitzen
diese Muzine zwar eine relativ große Gesamtmenge an Sialinsauren pro mg Muzin, diese
Sialinsauren kommen jedoch zu ca. 90% in Form der N-Acetylneuraminsaure (Neu5Ac)
vor. Da TGEV bevorzugt an Neu5Gc bindet (Schultze et al., 1995, 1996), konnte dieses
Bindungsverhalten die fehlende Interaktion mit diesen Muzinen erklaren. Die Muzine der
Saugferkel A und B wurden von TGEV gerringgradig besser erkannt als die des Absetzfer-
kels D und PSM. Welche Komponenten diese bessere Erkennung bedingten, war unklar,
da die Muzine der Ferkel A und D in ihrem Gehalt an Neu5Gc und Neu5Ac fast identisch
waren.

5.5. Interaktion von TGEV mit intestinalen Muzinen 99
0
0.1
0.2
0.3
0.4
0.5
0.6
DE
40
5
A B C D PSM
Abbildung 5.14: Sialinsaureabhangigkeit der Bindung von TGEV an intestinale Muzine. Die
weißen Saulen stellen die nicht enzymatisch behandelten Muzine dar. Die grauen Saulen reprasen-
tieren die mit 50 mU VCNA fur 1 h bei 37◦C behandelten Muzine. TGE-Virionen wurden wie
ublich in desialylierter gereinigter Form in einer Proteinmenge von 1 µg eingesetzt.
5.5.2 Bedeutung der Sialinsauren fur die Bindung
Es wurde untersucht, ob die nachgewiesene Interaktion von TGEV mit drei der vier
Darmmuzine und mit PSM tatsachlich durch die Sialinsaurebindungsaktivitat des Virus
verursacht wird. Daher wurden die Muzine vor der Virusinkubation mit 50 mU VCNA fur
1 h bei 37◦C desialyliert. Als Kontrolle diente die Inkubation mit Natriumacetatpuffer.
Wie in Abbildung 5.14 dargestellt ist, verschwand die Bindung von TGEV nach VCNA-
Behandlung der Muzine. Somit wurden die Muzine ausschließlich uber ihre Sialinsauren
von TGEV erkannt. Die Sialinsaurebindungsaktivitat von TGEV ist also essentiell fur die
Interaktion mit Darmmuzinen.
5.5.3 Vergleich der Muzinbindung von TGEV mit Mutanten
Als weiterer Beleg fur die Bedeutung der Sialinsaurebindungsaktivitat von TGEV bezug-
lich der Interaktion mit Muzinen aus dem Jejunum von Schweinen diente eine vergleichen-
de Untersuchung mit den Mutanten HAD3 und m10. Es wurde eine Muzinkonzentration
von je 50 µg/ml d.h. 2,5 µg/Vertiefung eingesetzt. Die Menge an viralem Protein betrug
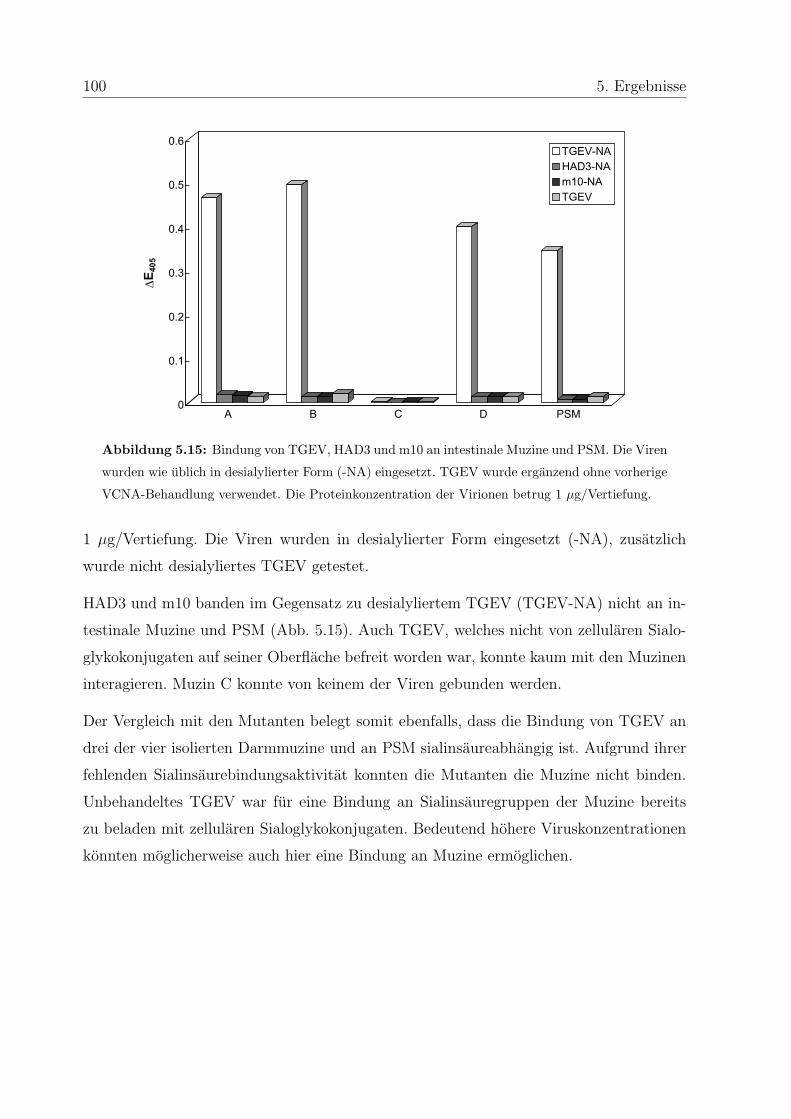
100 5. Ergebnisse
0
0.1
0.2
0.3
0.4
0.5
0.6D
E4
05
A B C D PSM
TGEV-NA
HAD3-NA
m10-NA
TGEV
Abbildung 5.15: Bindung von TGEV, HAD3 und m10 an intestinale Muzine und PSM. Die Viren
wurden wie ublich in desialylierter Form (-NA) eingesetzt. TGEV wurde erganzend ohne vorherige
VCNA-Behandlung verwendet. Die Proteinkonzentration der Virionen betrug 1 µg/Vertiefung.
1 µg/Vertiefung. Die Viren wurden in desialylierter Form eingesetzt (-NA), zusatzlich
wurde nicht desialyliertes TGEV getestet.
HAD3 und m10 banden im Gegensatz zu desialyliertem TGEV (TGEV-NA) nicht an in-
testinale Muzine und PSM (Abb. 5.15). Auch TGEV, welches nicht von zellularen Sialo-
glykokonjugaten auf seiner Oberflache befreit worden war, konnte kaum mit den Muzinen
interagieren. Muzin C konnte von keinem der Viren gebunden werden.
Der Vergleich mit den Mutanten belegt somit ebenfalls, dass die Bindung von TGEV an
drei der vier isolierten Darmmuzine und an PSM sialinsaureabhangig ist. Aufgrund ihrer
fehlenden Sialinsaurebindungsaktivitat konnten die Mutanten die Muzine nicht binden.
Unbehandeltes TGEV war fur eine Bindung an Sialinsauregruppen der Muzine bereits
zu beladen mit zellularen Sialoglykokonjugaten. Bedeutend hohere Viruskonzentrationen
konnten moglicherweise auch hier eine Bindung an Muzine ermoglichen.

5.5. Interaktion von TGEV mit intestinalen Muzinen 101
5.5.4 Interaktion von TGEV mit einem hochmolekularen
Protein
Die Bindung von TGEV an bestimmte Darmmuzine und PSM wurde naher untersucht.
Zu diesem Zweck wurden die Muzine in einer Konzentration von 1 mg/ml 1:2 mit VCNA
(1 U/ml) bzw. Natriumacetat-Puffer versetzt (s. auch unter Methoden 4.8.3). Nach einer
Inkubationszeit von 1 h bei 37◦C wurden die Proben 1:2 mit SDS-Probenpuffer verdunnt
und aufgekocht. Bei der folgenden SDS-PAGE wurden 5 µg Muzin pro Spur aufgetrennt.
Nach dem Blotten der Proteine auf Nitrozellulose wurde ein Bindungstest mit desialylier-
tem TGEV durchgefuhrt (s. Methoden 4.19.3).
Bei einer aufgetragenen Muzinmenge von 5 µg/Spur konnten nur bei den Muzinen des
Saugferkels B und PSM Proteinbanden im Virusbindungstest detektiert werden. Die
Interaktion von TGEV mit diesen Proteinbanden verschwand nach vorheriger VCNA-
Behandlung der Muzine (Abb. 5.16). Es wurden bei beiden Muzinarten zwei hochmole-
+_ _
+ VCNA
B PSM
TGEV-NA
220
97
[kDa]
Abbildung 5.16: Bindung von desialyliertem TGEV (TGEV-NA) an Muzine des Ferkels B und
PSM. Die Muzine wurden in einer SDS-PAGE mit und ohne vorherige Neuraminidasebehandlung
aufgetrennt (+/− VCNA). Die Pfeile in der linken Spalte kennzeichnen die Proteinbanden, die von
TGEV-NA gebunden wurden.
kulare Proteinbanden von TGEV erkannt. Diese sind in der Abbildung 5.16 bei Muzinen
des Saugferkels B besonders gut zu erkennen. Bei diesen beiden Proteinbanden konnte es
sich um das gleiche Protein in unterschiedlicher Glykosylierung handeln. In jedem Fall
handelte es sich in beiden Fallen um eine sialinsaureabhangige Bindung durch TGEV.
Das Protein, das TGEV in Muzinen uber seine Sialinsaurebindungsaktivitat erkennt, ist
somit ein Sialoglykoprotein.

102 5. Ergebnisse
5.6 Untersuchung von Burstensaummembranen
Wie die vorherigen Ergebnisse gezeigt haben, kann TGEV mit extrazellularen Darmmu-
zinen interagieren. Dies schließt die Anwesenheit eines zweiten Rezeptors neben pAPN
auf Zellen des Darmepithels aber nicht aus. Daher wurden Burstensaummembranen aus
Schweinedunndarm mit in die Untersuchungen einbezogen und auf ihre Interaktion mit
TGEV getestet.
5.6.1 Praparation von Burstensaummembranvesikeln
Burstensaummembranvesikel (BSMV) wurden aus dem Jejunum von vier Saug- und vier
Absetzferkeln prapariert (s. Methoden 4.12.1). Zur Kontrolle des Praparationserfolgs wur-
den die Aktivitaten der Alkalischen Phosphatase (AP), als Leitenzym fur die apikale Mem-
bran, und der Na/K-ATPase, als Leitenzym fur die basolaterale Membran, bestimmt und
auf die Proteinkonzentrationen der Praparationen bezogen (Berechnung s. 4.12.2). Die An-
reicherung beider Enzyme in den hergestellten BSMV zeigt Tabelle 5.3. Die Burstensaum-
membranen wurden in allen acht Praparationen angereichert, jedoch in unterschiedlichem
Maße. Wie in der Tabelle zu erkennen ist, gab es z.B. bei den BSMV von einem Saugferkel
AP (a) Na/K-ATPase (b) Verhaltnis a/b
Absetzferkel A1 4,2 1,4 3,0
Absetzferkel A2 6,1 0,5 12,2
Absetzferkel A3 11,9 2,2 5,4
Absetzferkel A4 8,6 2,1 4,1
Saugferkel S1 7,7 0,7 11,0
Saugferkel S2 5,6 0,7 8,0
Saugferkel S3 4,7 1,9 2,5
Saugferkel S4 7,1 1,3 5,5
Tabelle 5.3: Anreicherungen der beiden Leitenzyme AP und N/K-ATPase durch die BSMV-
Praparation. Es sind die Werte der Membranen aus dem Darmepithel von vier Absetzferkeln (A1–
A4) und vier Saugferkeln (S1–S4) angegeben (Berechnung der Werte s. 4.12.2). Die rechte Spalte
zeigt das Verhaltnis der beiden Anreicherungen zueinander.

5.6. Untersuchung von Burstensaummembranen 103
nur etwas mehr als doppelt so viele apikale Membranen wie basolaterale Membranen (S3).
Bei einem anderen Saugferkel (S1) hingegen reicherten sich die Burstensaummembranen
gut an und waren 11-mal starker vertreten als die basolateralen Membranen (gemessen an
den Enzymaktivitaten). Fur die folgenden Versuche wurden trotz der unterschiedlichen
Anreicherungen alle acht Praparationen verwendet.
5.6.2 Analyse des Sialinsauregehalts der BSMV
Der Sialinsauregehalt der BSMV wurde wie auch in den vorherigen Versuchen mittels
HPLC bestimmt. Die BSMV wurden in einer Proteinkonzentration von 1 mg/ml einge-
setzt. Der Gehalt an Neu5Gc und Neu5Ac ist in den Kurven in Abbildung 5.17 dargestellt.
Alle Proben besaßen einen hoheren Neu5Gc- als Neu5Ac-Gehalt. Die BSMV aus Saug-
ferkeln zeichneten sich durch einen insgesamt hoheren Sialinsauregehalt aus als die der
Absetzferkel (3–5-mal hoher). Der Gehalt an Neu5Gc war bei den Saugferkeln (S3, S4),
die einen Tag alt waren, hoher als bei den drei Tage alten Saugferkeln (S1, S2). Die Proben
der Absetzferkel A1 und A2 (12 Wochen) enthielten mehr Neu5Gc und Neu5Ac als die
der Absetzferkel A3 und A4 (6–8 Wochen).
5.6.3 Bindung an Proteine aus BSMV
Zur Untersuchung der Bindung von TGEV an Proteine aus den Burstensaummembranen
wurden die BSMV in einer Proteinkonzentration von 4 mg/ml 1:2 mit Neuraminidase
aus Vibrio cholerae (VCNA, 1 U/ml) versetzt und fur 1 h bei 37◦C desialyliert. Die
Kontrollproben wurden statt mit VCNA mit Natriumacetat-Puffer behandelt (Metho-
den 4.8.3). Anschließend wurden die Burstensaummembranen 1:2 mit SDS-Probenpuffer
versetzt und bei 96◦C fur 5 min denaturiert. Die Proteine wurden in einer SDS-PAGE
aufgetrennt (20 µg/Spur) und auf Nitrozellulose geblottet. Nach der Absattigung mit
”Blocking Reagenz“ wurde die Virusbindung, wie unter 4.19.3 beschrieben, untersucht.
Wie in Abbildung 5.18 (a) zu erkennen ist, wurde von TGEV-NA eine Bande auf der
Hohe von 150 kDa in allen Proben erkannt. Bei dieser Bande handelte es sich um pAPN,
was in Abbildung 5.18 (c) durch Interaktion mit dem mAk G43 gegen pAPN gezeigt ist.
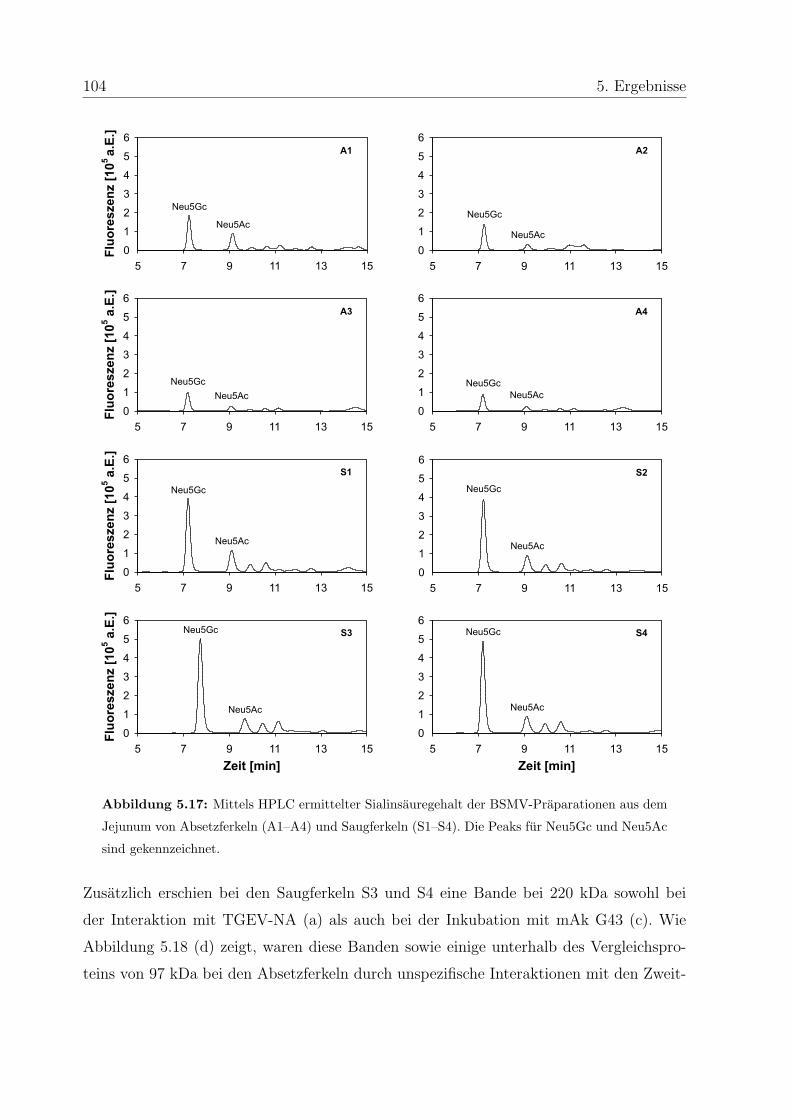
104 5. Ergebnisse
0
1
2
3
4
5
6
5 7 9 11 13 15
Flu
ore
szen
z[1
05a.E
.]
Neu5Gc
Neu5Ac
A1
0
1
2
3
4
5
6
5 7 9 11 13 15
Flu
ore
szen
z[1
05a.E
.]
Neu5Gc
Neu5Ac
A2
0
1
2
3
4
5
6
5 7 9 11 13 15
Flu
ore
szen
z[1
05
a.E
.]
Neu5Gc
Neu5Ac
A3
0
1
2
3
4
5
6
5 7 9 11 13 15
Flu
ore
szen
z[1
05
a.E
.]
Neu5Gc
Neu5Ac
A4
0
1
2
3
4
5
6
5 7 9 11 13 15
Flu
ore
szen
z[1
05
a.E
.]
Neu5Gc
Neu5Ac
S1
0
1
2
3
4
5
6
5 7 9 11 13 15
Flu
ore
szen
z[1
05
a.E
.]
Neu5Gc
Neu5Ac
S2
0
1
2
3
4
5
6
5 7 9 11 13 15
Zeit [min]
Flu
ore
szen
z[1
05
a.E
.]
Neu5Gc
Neu5Ac
S3
0
1
2
3
4
5
6
5 7 9 11 13 15
Zeit [min]
Flu
ore
szen
z[1
05
a.E
.]
Neu5Gc
Neu5Ac
S4
Abbildung 5.17: Mittels HPLC ermittelter Sialinsauregehalt der BSMV-Praparationen aus dem
Jejunum von Absetzferkeln (A1–A4) und Saugferkeln (S1–S4). Die Peaks fur Neu5Gc und Neu5Ac
sind gekennzeichnet.
Zusatzlich erschien bei den Saugferkeln S3 und S4 eine Bande bei 220 kDa sowohl bei
der Interaktion mit TGEV-NA (a) als auch bei der Inkubation mit mAk G43 (c). Wie
Abbildung 5.18 (d) zeigt, waren diese Banden sowie einige unterhalb des Vergleichspro-
teins von 97 kDa bei den Absetzferkeln durch unspezifische Interaktionen mit den Zweit-
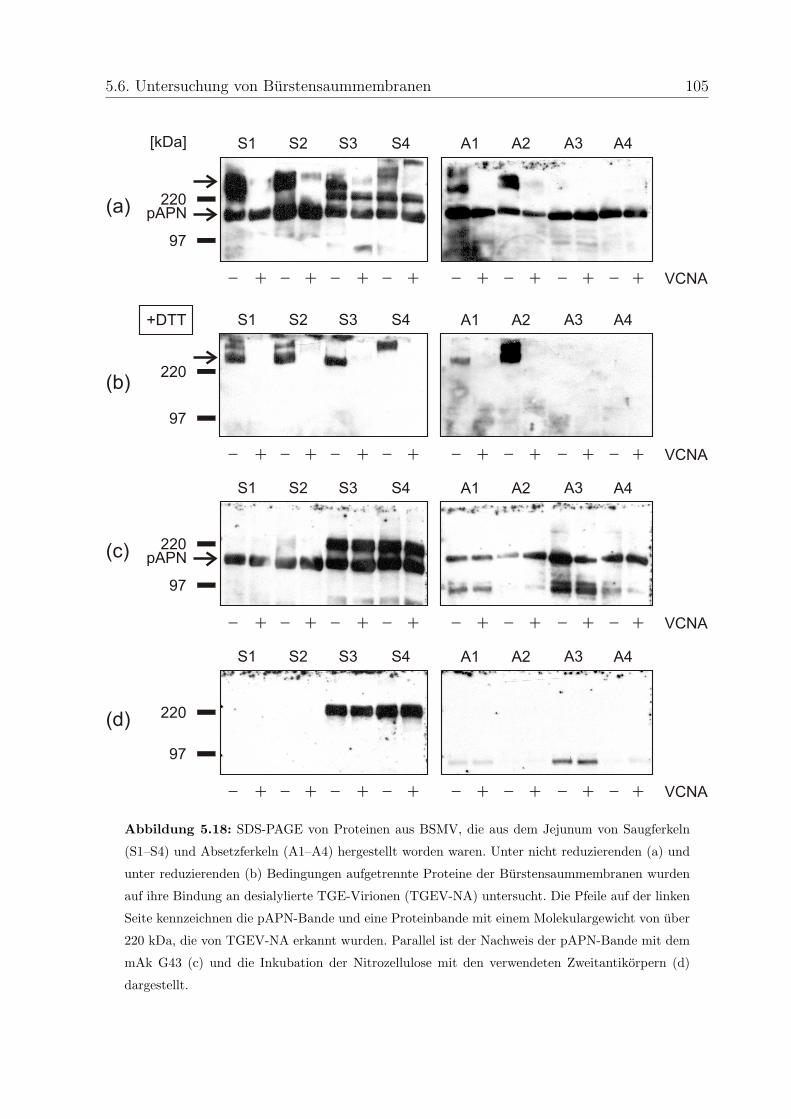
5.6. Untersuchung von Burstensaummembranen 105
VCNA+_ _
+
S1 S2
__++
S4S3
__++
220
97
[kDa]
pAPN
A1
__++
A4A3A2
(a)
VCNA+_ _
+
S1 S2
__++
S4S3
__++
220
97
A1
__++
A4A3A2
(b)
+DTT
VCNA+_ _
+
S1 S2
__++
S4S3
__++
220
97
pAPN
A1
__++
A4A3A2
(c)
VCNA+_ _
+
S1 S2
__++
S4S3
__++
220
97
A1
__++
A4A3A2
(d)
Abbildung 5.18: SDS-PAGE von Proteinen aus BSMV, die aus dem Jejunum von Saugferkeln
(S1–S4) und Absetzferkeln (A1–A4) hergestellt worden waren. Unter nicht reduzierenden (a) und
unter reduzierenden (b) Bedingungen aufgetrennte Proteine der Burstensaummembranen wurden
auf ihre Bindung an desialylierte TGE-Virionen (TGEV-NA) untersucht. Die Pfeile auf der linken
Seite kennzeichnen die pAPN-Bande und eine Proteinbande mit einem Molekulargewicht von uber
220 kDa, die von TGEV-NA erkannt wurden. Parallel ist der Nachweis der pAPN-Bande mit dem
mAk G43 (c) und die Inkubation der Nitrozellulose mit den verwendeten Zweitantikorpern (d)
dargestellt.

106 5. Ergebnisse
antikorpern (biotinylierte Anti-Maus-Immunglobuline, Streptavidin-Peroxidase) bedingt.
Spezifisch von TGEV-NA wurde jedoch eine Bande uber dem 220 kDa-Vergleichsprotein
bei allen vier Saugferkelproben und zwei der vier Absetzferkelproben (A1 und A2) erkannt.
Auch wenn die SDS-PAGE unter reduzierenden Bedingungen (+DTT) durchgefuhrt wur-
de (Abb. 5.18 b), wurden die hochmolekularen Banden von TGEV-NA gebunden. Wie
bei (b) gut zu erkennen ist, verhielten sich die von TGEV-NA gebundenen Proteinbanden
etwas heterogen. Bei S1, S2 und A2 schienen Doppelbanden in etwas unterschiedlicher
Große vorzuliegen; S3 und S4 wiesen jeweils nur einen Vertreter aus dieser Doppelban-
de auf. Moglicherweise handelt es sich bei diesen Proteinbanden um das gleiche Protein,
welches aber in unterschiedlich glykosylierter Form vorliegt.
Erfolgte die Auftrennung der BSMV-Proteine in desialylierter Form, wurden die hoch-
molekularen Proteinbanden nicht mehr bzw. kaum von TGEV-NA erkannt (VCNA, +).
Die Interaktion mit den hochmolekularen Proteinbanden begrundete sich somit auf die
Bindung von TGEV-NA an Sialinsauregruppen des Proteins. Die Sialinsaurebindungs-
aktivitat von TGEV bewirkt also auch in BSMV eine Interaktion mit hochmolekularen
Sialoglykoproteinen.
Die Virusbindungstests wurden ebenfalls mit den Mutanten HAD3 und m10 in desialylier-
ter und nicht desialylierter Form sowie mit unbehandelten TGE-Virionen durchgefuhrt.
In Abbildung 5.19 sind beispielhaft die Interaktionen von HAD3, m10 und TGEV mit
den Proteinen der Burstensaummembranen von Saugferkel S1 und Absetzferkel A1 dar-
gestellt. HAD3 und m10 banden an die pAPN-Bande. HAD3 erkannte keine zusatzlichen
Proteine. Die bei m10 sichtbare dunne 220 kDa-Bande trat nur bei dieser Mutante auf.
Worum es sich bei dieser Bande handelt, ist nicht bekannt. Die Mutanten verhielten sich in
desialylierter Form identisch. Nicht desialyliertes TGEV konnte in der eingesetzten Prote-
inkonzentration wie auch schon bei der Untersuchung der Bindung an Oberflachenproteine
von Zellkulturzellen (5.3.2) keine Proteinbanden binden. Da das spezifische Bindungsmus-
ter in den Proben aller acht Ferkel ubereinstimmte, wurde die Darstellung exemplarisch
auf S1 und A1 beschrankt. Die Mutanten konnten aufgrund ihrer fehlenden Sialinsaure-
bindungsaktivitat nicht an das hochmolekulare von TGEV-NA erkannte Protein binden.

5.7. Charakterisierung des BSMV-Proteins 107
VCNA+_ _
+
S1 A1
__++
A1S1
__++
220
97
[kDa]
pAPN
S1 A1
HAD3 m10 TGEV
Abbildung 5.19: Bindung von HAD3, m10 und TGEV (nicht neuraminidasebehandelt) an
BSMV-Proteine von Saugferkel S1 und Absetzferkel A1. Die Lokalisation der pAPN-Bande ist
mit einem Pfeil gekennzeichnet.
5.7 Charakterisierung des BSMV-Proteins
TGEV (desialyliert) erkennt aufgrund seiner Sialinsaurebindungsaktivitat ein hochmo-
lekulares Protein in BSMV-Praparationen von Saug- und Absetzferkeln. Dieses Protein
wurde durch weitere Untersuchungen naher charakterisiert. Die Assoziation des Proteins
zur Zellmembran und sein Kohlenhydratanteil wurden untersucht.
5.7.1 Triton X-114-Phasenseparierung
Die Assoziation des hochmolekularen Proteins zur Burstensaummembran wurde mit Hilfe
einer Triton X-114-Phasenseparierung untersucht (s. Methoden 4.20.1). Die Proteine der
Burstensaummembranen wurden mit Triton X-114 extrahiert. Nach einer Phasenseparie-
rung wurden die Proteine mit WGA aus der Detergenzphase und der wassrigen Phase
gefallt und nach einer SDS-PAGE fur einen Virusbindungstest mit TGEV-NA auf Ni-
trozellulose geblottet. Das hochmolekulare Protein wurde ausschließlich in der wassrigen
Phase (w) von TGEV erkannt (Abb. 5.20). Es handelt sich somit vermutlich bei dem
hochmolekularen BSMV-Protein um ein membranassoziiertes Protein und nicht um ein
Transmembranprotein.

108 5. Ergebnisse
w
S1 S2 A2A1
220
[kDa]
wwwd ddd
Abbildung 5.20: Charakterisierung der Assoziation des BSMV-Proteins mit der Burstensaum-
membran mittels Triton X-114-Phasenseparierung. Detergenz- (d) und wassrige Phase (w) der
BSMV-Proteine von den Ferkeln S1 und S2 bzw. A1 und A2 wurden nach WGA-Fallung fur einen
Bindungstest mit TGEV-NA durch SDS-PAGE aufgetrennt.
5.7.2 Charakterisierung des Kohlenhydratanteils
5.7.2.1 Nachweis der Kohlenhydrate und Sialinsauren
BSMV-Proteine wurden nach elektrophoretischer Auftrennung auf eine PVDF-Membran
geblottet und einer Perjodatoxidation unterzogen (s. Methoden 4.20.2.1). Nach Markie-
rung der oxidierten Kohlenhydratreste mit Biotinhydrazid, konnten diese uber Streptavi-
din-Peroxidase mittels Chemolumineszenz sichtbar gemacht werden. Abbildung 5.21 (a)
und (b) zeigen, dass es sich bei dem von TGEV erkannten Protein um ein Hauptglyko-
protein der Burstensaummembran handelt (Pfeil, teilweise als Doppelbande). Durch eine
milde Perjodatoxidation auf Eis konnten selektiv die Sialinsauregruppen der Proteine
dargestellt werden (Abb. 5.21 c). Das Hauptglykoprotein ist somit ein Sialoglykoprotein.
Die Absetzferkelproben A3 und A4 schienen auf der gleichen Hohe wie die anderen Proben
ebenfalls ein Hauptglykoprotein mit geringer Sialylierung zu besitzen, obwohl TGEV bei
diesen Proben kein hochmolekulares Protein gebunden hatte. Die vorherige Desialylierung
der Proben (VCNA, +) zeigte jedoch bei A3 und A4 keinen Unterschied im Laufverhalten
der Banden, wohingegen das Hauptsialoglykoprotein der anderen Proben mehr oder we-
niger stark nach oben verschoben wurde. Fur muzinartige sialinsaurereiche Glykoproteine
ist ein verlangsamtes elektrophoretisches Laufverhalten nach Desialylierung typisch. Wie
in Abbildung 5.21 (c) zu erkennen ist, befand sich die hochmolekulare Sialinsaurebande
bereits bei nicht desialylierten A3- und A4-Proben auf der Hohe der Banden der desialy-
lierten anderen Proben. Diese Banden stellen somit moglicherweise Sialinsauretypen dar,
die nicht durch VCNA abgespalten werden konnen und die ebenfalls nicht durch TGEV
gebunden werden.
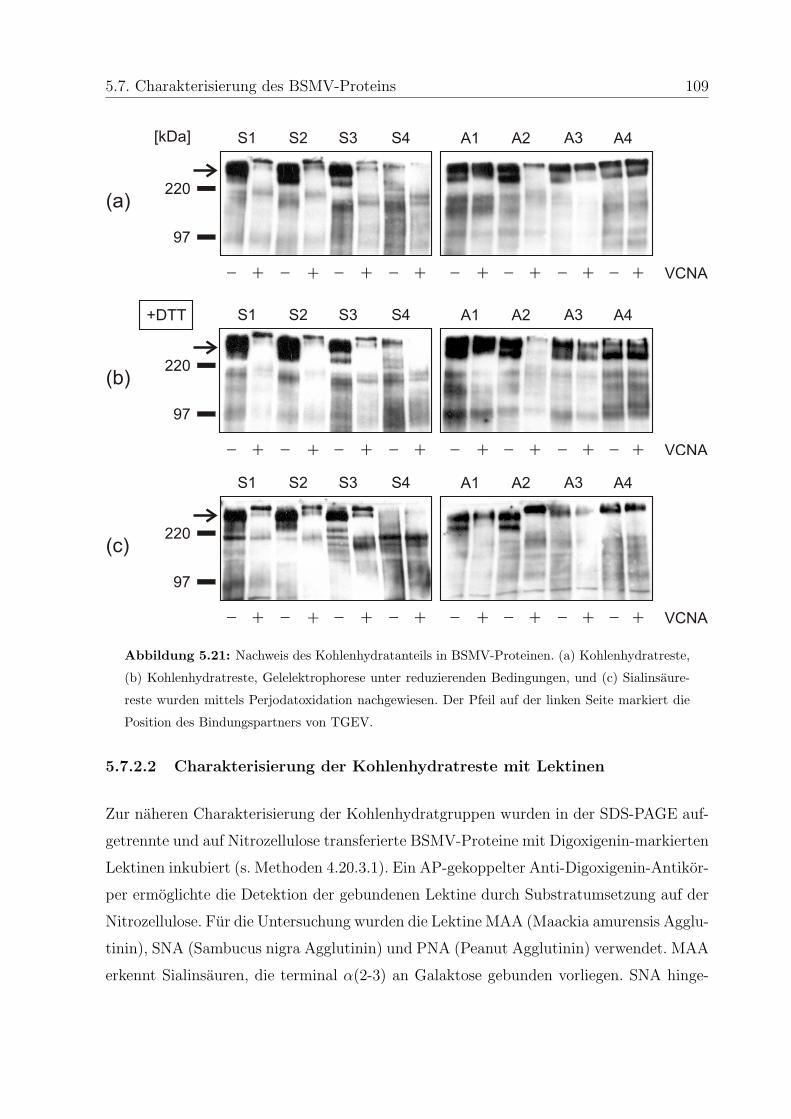
5.7. Charakterisierung des BSMV-Proteins 109
VCNA+_ _
+
S1 S2
__++
S4S3
__++
220
97
[kDa] A1
__++
A4A3A2
(a)
VCNA+_ _
+
S1 S2
__++
S4S3
__++
220
97
A1
__++
A4A3A2+DTT
(b)
VCNA+_ _
+
S1 S2
__++
S4S3
__++
97
A1
__++
A4A3A2
220(c)
Abbildung 5.21: Nachweis des Kohlenhydratanteils in BSMV-Proteinen. (a) Kohlenhydratreste,
(b) Kohlenhydratreste, Gelelektrophorese unter reduzierenden Bedingungen, und (c) Sialinsaure-
reste wurden mittels Perjodatoxidation nachgewiesen. Der Pfeil auf der linken Seite markiert die
Position des Bindungspartners von TGEV.
5.7.2.2 Charakterisierung der Kohlenhydratreste mit Lektinen
Zur naheren Charakterisierung der Kohlenhydratgruppen wurden in der SDS-PAGE auf-
getrennte und auf Nitrozellulose transferierte BSMV-Proteine mit Digoxigenin-markierten
Lektinen inkubiert (s. Methoden 4.20.3.1). Ein AP-gekoppelter Anti-Digoxigenin-Antikor-
per ermoglichte die Detektion der gebundenen Lektine durch Substratumsetzung auf der
Nitrozellulose. Fur die Untersuchung wurden die Lektine MAA (Maackia amurensis Agglu-
tinin), SNA (Sambucus nigra Agglutinin) und PNA (Peanut Agglutinin) verwendet. MAA
erkennt Sialinsauren, die terminal α(2-3) an Galaktose gebunden vorliegen. SNA hinge-

110 5. Ergebnisse
gen bindet an terminale Sialinsauren, die α(2-6) an Galaktose oder N-Acetylgalaktosamin
gebunden sind. Kommt es zu einer Bindung von PNA liegt das Disaccharid Galaktose-
β(1-3)-N-Acetylgalaktosamin vor, welches hauptsachlich die Core-Einheit von O-Glykanen
darstellt. Eine terminale Sialinsaure verhindert die PNA-Erkennung, in diesem Fall gelingt
der Nachweis mit PNA nur nach vorheriger Abspaltung der Sialinsauren mit Neuramini-
dase.
Die Lektine SNA und MAA eigneten sich nicht gut fur eine Kohlenhydratuntersuchung
auf dem Blot. Es gab nur sehr schwache Reaktionen, die ein Vorhandensein entsprechender
Sialinsauren erahnen ließen. Nur bei den Proben A1 und A2 gelang eine starkere Reaktion
mit MAA, die zum Nachweis einer Doppelbande auf Hohe des Hauptsialoglykoproteins
fuhrte (> 220 kDa, nicht gezeigt). Somit scheinen die Sialinsauren des Glykoproteins
zumindest zum Teil in α(2-3) an Galaktose gebundener Form vorzuliegen.
Abbildung 5.22 zeigt das Ergebnis der Inkubation mit PNA. Die Proben S2 und A2 wie-
sen beide nach vorheriger VCNA-Behandlung zwei Banden auf, die von PNA gebunden
wurden. Die obere dieser beiden Banden reprasentiert das Glykoprotein, das in nicht
+ +
97
VCNA_
S2 A2
220
[kDa]
_
Abbildung 5.22: Erkennung von BSMV-Proteinen durch PNA. Auf Hohe der oberen Bande von
S2 und A2 befindet sich das von TGEV gebundene Protein. Die Position ist durch einen Pfeil
markiert.
desialylierter Form von TGEV gebunden wird und nach Neuraminidasebehandlung ein
langsameres elektrophoretisches Laufverhalten aufweist. Die Proben S1, S3, S4 und A1
verhielten sich wie S2 und A2. Probe A3 zeigte die Reaktion mit PNA in Form der beiden
Banden bereits ohne Neuraminidasebehandlung. Bei Probe A4 ließ sich keine Bindung von
PNA nachweisen. Das von TGEV gebundene Sialoglykoprotein in sechs der acht Ferkelpro-
ben besitzt somit nachweislich das Disaccharid Galaktose-β(1-3)-N-Acetylgalaktosamin.
Daher handelt es sich bei dem Sialoglykoprotein um ein O-glykosyliertes Protein.
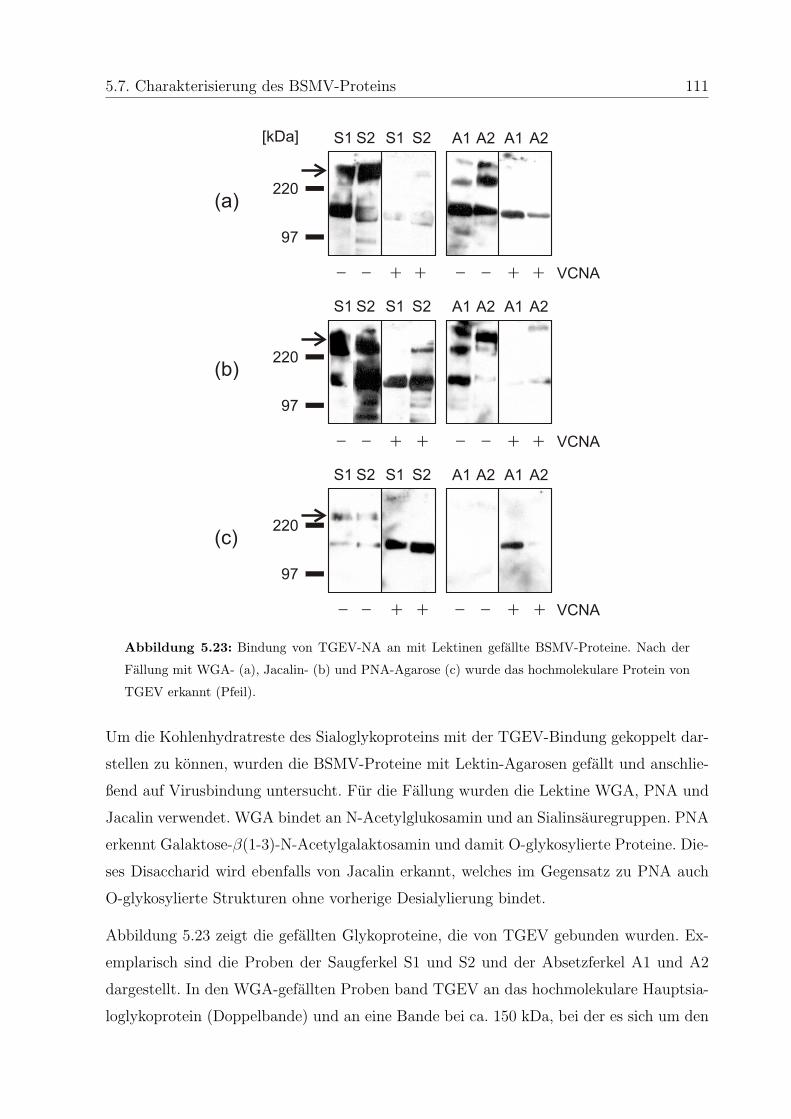
5.7. Charakterisierung des BSMV-Proteins 111
VCNA++
S1 S2
+ +
220
97
[kDa] A1 A2
(a)
S1 S2 A1 A2
____
VCNA++
S1 S2
+ +
220
97
A1 A2
(b)
S1 S2 A1 A2
____
VCNA
S1 S2
220
97
A1 A2
(c)
S1 S2 A1 A2
++____
++
Abbildung 5.23: Bindung von TGEV-NA an mit Lektinen gefallte BSMV-Proteine. Nach der
Fallung mit WGA- (a), Jacalin- (b) und PNA-Agarose (c) wurde das hochmolekulare Protein von
TGEV erkannt (Pfeil).
Um die Kohlenhydratreste des Sialoglykoproteins mit der TGEV-Bindung gekoppelt dar-
stellen zu konnen, wurden die BSMV-Proteine mit Lektin-Agarosen gefallt und anschlie-
ßend auf Virusbindung untersucht. Fur die Fallung wurden die Lektine WGA, PNA und
Jacalin verwendet. WGA bindet an N-Acetylglukosamin und an Sialinsauregruppen. PNA
erkennt Galaktose-β(1-3)-N-Acetylgalaktosamin und damit O-glykosylierte Proteine. Die-
ses Disaccharid wird ebenfalls von Jacalin erkannt, welches im Gegensatz zu PNA auch
O-glykosylierte Strukturen ohne vorherige Desialylierung bindet.
Abbildung 5.23 zeigt die gefallten Glykoproteine, die von TGEV gebunden wurden. Ex-
emplarisch sind die Proben der Saugferkel S1 und S2 und der Absetzferkel A1 und A2
dargestellt. In den WGA-gefallten Proben band TGEV an das hochmolekulare Hauptsia-
loglykoprotein (Doppelbande) und an eine Bande bei ca. 150 kDa, bei der es sich um den

112 5. Ergebnisse
zellularen Rezeptor pAPN handelte (Abb. 5.23 a). Das Hauptsialoglykoprotein und pAPN
konnten auch von Jacalin gefallt werden, wie der anschließende Nachweis der Virusbindung
zeigt (Abb. 5.23 b). Die Uberprufung des Erfolgs der PNA-Fallung durch Virusbindung
erwies sich als schwierig, da TGEV an Sialinsauregruppen des Proteins bindet, diese Sia-
linsauren aber fur eine erfolgreiche PNA-Fallung vorher abgespalten werden mussen. Wie
Abbildung 5.23 (c) zeigt, war es jedoch moglich, ohne vorherige VCNA-Behandlung der
S1/S2-Proben einen kleinen Teil des Sialoglykoproteins mit PNA zu fallen. Dieser Teil
konnte von TGEV erkannt werden (Pfeil). Auch von pAPN konnte ohne vorherige De-
sialylierung der S1/S2-Proben ein kleiner Teil mit PNA gefallt und von TGEV gebunden
werden (Abb. 5.23 c). Wurden die Proben mit VCNA behandelt, war die Fallung von
pAPN jedoch wesentlich effektiver. Die Starke der pAPN-Banden in Abbildung 5.23 (c)
bei den BSMV-Proben S1 und S2 zeigt, dass diese Banden gut von TGEV erkannt wur-
den. Die Virusbindung an Aminopeptidase N ist folglich nicht sialinsaureabhangig. Bei
den A1/A2-Proben konnten ohne vorherige VCNA-Behandlung keine fur die Virusbindung
ausreichenden Proteinmengen mit PNA gefallt werden (Abb. 5.23 c). Nach Neuraminida-
sebehandlung dieser Proben konnte bei A1 PNA-gefallte Aminopeptidase N von TGEV
gebunden werden. Bei der BSMV-Probe A2 ist die pAPN-Bande in Abbildung 5.23 (c)
trotz vorheriger Desialylierung der Probe nur schwach zu erkennen. Die Ursache hierfur
ist unbekannt. Die geschilderten Ergebnisse zeigen, dass das Hauptsialoglykoprotein, mit
dem TGEV durch seine Sialinsaurebindungsaktivitat interagiert, ein O-glykosyliertes Sia-
loglykoprotein ist.
5.8 Interaktion von TGEV mit Darmepithel
Nachdem in Burstensaummembranen ein Sialoglykoprotein gefunden worden war, das von
desialyliertem TGEV uber Sialinsauregruppen gebunden wurde, wurde die Bindung von
TGEV in Darmgefrierschnitten untersucht. Die Untersuchung der Bindung in Kryoschnit-
ten bietet den Vorteil, dass die Strukturen des Darmgewebes in ihrer naturlichen Zu-
sammensetzung weitestgehend erhalten bleiben, und so Aussagen uber die tatsachliche
Lokalisation des Bindungspartners gemacht werden konnen.
Gefrierschnitte des Jejunums eines 3 Tage alten Saugferkels wurden mit Paraformaldehyd
fixiert und anschließend mit konzentrierten Virussuspensionen von TGEV-NA, TGEV,

5.8. Interaktion von TGEV mit Darmepithel 113
HAD3-NA und m10-NA inkubiert (s. Methoden 4.19.4). Die Virusbindung wurde uber
den mAk 6A.C3 und einen FITC-gekoppelten Zweitantikorper nachgewiesen und im Fluo-
reszenzmikroskop beurteilt.
Die Fluoreszenz in Abbildung 5.24 (oben, unten) zeigt, dass TGEV-NA in umschriebenen
Bereichen an das Dunndarmepithel des Saugferkels band (weiße Pfeile). Es handelt sich
bei diesen Bereichen vermutlich um Becherzellen des Dunndarmepithels, was vor allem
in dem vergroßerten Bildausschnitt in Abbildung 5.24 (unten) bei der Gegenuberstellung
von Fluoreszenz und Durchlicht zu erkennen ist. Becherzellen sind modifizierte Saulen-
epithelzellen, die Schleim synthetisieren und sezernieren. Der apikale breitere Zellleib der
Becherzellen ist dicht mit Muzingranula gefullt, die durch Exozytose abgegeben werden
und mit Wasser den Darmschleim bilden.
Wurden die Kryoschnitte vor der Virusinkubation mit Neuraminidase aus Vibrio cholerae
behandelt, verschwand die Bindung von TGEV-NA in den Becherzellbereichen (Abb. 5.24
Mitte). Dies belegt die Sialinsaureabhangigkeit der Bindung von TGEV in diesen Regio-
nen. Die Bindung an den zellularen Rezeptor pAPN bewirkte nur ein schwaches Fluores-
zieren des gesamten Epithels (Nachweis mit mAk G43 nicht gezeigt.).
Zum Vergleich wurde die Interaktion von nicht desialyliertem TGEV und den Mutanten
HAD3 und m10 (in desialylierter Form) mit Gefrierschnitten des Jejunums des Saugferkels
untersucht. Die Ergebnisse dieser Bindungstests sind in Abbildung 5.25 dargestellt. Die
beiden Mutanten zeigten keine Bindung an die von TGEV-NA erkannten Becherzellberei-
che, was die Bedeutung der Sialinsaurebindungsaktivitat von TGEV fur diese Interaktion
unterstreicht. TGEV hingegen konnte auch ohne vorherige Neuraminidasebehandlung mit
Strukturen der Becherzellen interagieren, allerdings in abgeschwachter Form (Abb. 5.19
oben, weiße Pfeile).
TGEV bindet somit vermutlich an Sialinsauregruppen der Muzingranula in den Becher-
zellen und an den daraus entstehenden Darmschleim. Eine vorherige Desialylierung der
TGE-Virionen zur Optimierung der Sialinsaurebindungsaktivitat ist fur die Interaktion
mit Darmepithel im Kryoschnitt nicht unbedingt erforderlich. Entweder sind die Viruspar-
tikel aufgrund der eingesetzten hohen Konzentration auch ohne Desialylierung in der Lage
zu interagieren, oder die Affinitat zu den Strukturen im Becherzellbereich ist hoher als
die zu anderen Sialoglykokonjugaten.
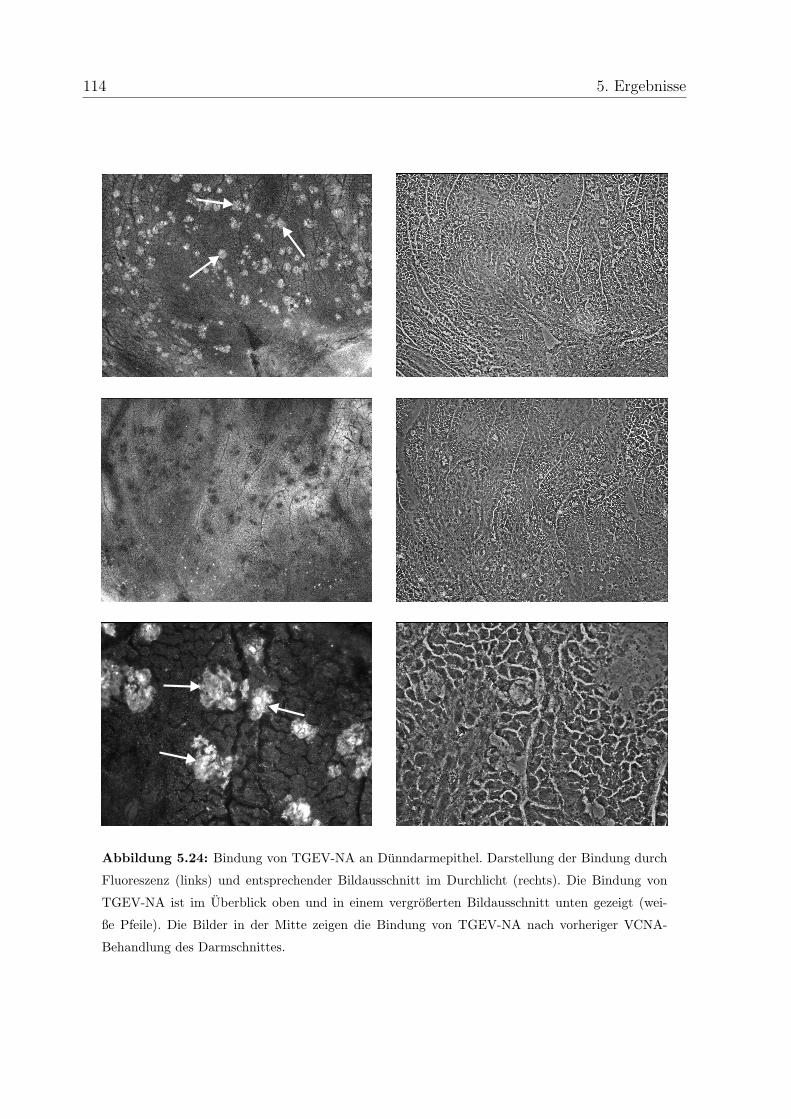
114 5. Ergebnisse
Abbildung 5.24: Bindung von TGEV-NA an Dunndarmepithel. Darstellung der Bindung durch
Fluoreszenz (links) und entsprechender Bildausschnitt im Durchlicht (rechts). Die Bindung von
TGEV-NA ist im Uberblick oben und in einem vergroßerten Bildausschnitt unten gezeigt (wei-
ße Pfeile). Die Bilder in der Mitte zeigen die Bindung von TGEV-NA nach vorheriger VCNA-
Behandlung des Darmschnittes.

5.8. Interaktion von TGEV mit Darmepithel 115
Abbildung 5.25: Bindung von nicht desialyliertem TGEV und den Mutanten HAD3 und m10 an
Dunndarmepithel. Inkubation der Kryoschnitte mit TGEV (oben), HAD3-NA (Mitte) und m10-
NA (unten).


6. Diskussion
6.1 Bedeutung der Desialylierung von TGEV fur die
Interaktion mit Sialoglykokonjugaten
Fur die Untersuchung der Sialinsaurebindungsaktivitat von TGEV wurden die Virionen
im Vorfeld stets mit Neuraminidase aus Vibrio cholerae desialyliert und anschließend uber
einen Saccharosegradienten gereinigt. Diese Vorbehandlung diente der Optimierung der
Bindung an Sialinsauren durch das S-Protein von TGEV und sollte klarere Ergebnisse
bezuglich potentieller Bindungspartner des Virus liefern. TGEV besitzt im Gegensatz zu
den Coronaviren der Gruppe II keine Rezeptor-inaktivierende Enzymaktivitat. Die Sia-
linsaurebindungsaktivitat von TGEV bewirkt, dass in Zellkultur wachsendes Virus mit
Sialoglykokonjugaten zellularen Ursprungs umgeben wird. Dieser Maskierung der Sia-
linsaurebindungsstellen des S-Proteins kann mangels viraler Enzymaktivitat nur durch
externe Neuraminidasebehandlung entweder der Virionen nach der Virusernte oder der
Zellen vor der Virusinfektion entgegengewirkt werden (Krempl et al., 1997).
Um zu sehen, wie sich die Maskierung der Sialinsaurebindungsstellen des S-Proteins durch
zellulare Sialoglykokonjugate auf die Bindungsaktivitaten von TGEV auswirkt, wurde
bei den Bindungsstudien in Zellkultur, mit intestinalen Muzinen, Burstensaummembra-
nen und Kryoschnitten von Darmepithelien parallel zu neuraminidasebehandeltem TGEV
auch nicht enzymbehandeltes Virus untersucht. Bei den Virusbindungstests auf Nitrozel-
lulose stellte sich heraus, dass TGEV-Partikel, die vor dem Reinigungsschritt uber einen
Saccharosegradienten nicht desialyliert worden waren, nicht nur in ihrer Sialinsaurebin-
dung sondern auch in ihrer Bindung an den zellularen Rezeptor Aminopeptidase N stark
117

118 6. Diskussion
beeintrachtigt waren (5.3.2, 5.6.3). Die Interaktion zellularer Sialoglykokonjugate mit den
S-Proteinen von TGEV-Partikeln konnte zu einer Vernetzung und somit zur Aggregatbil-
dung der Viruspartikel gefuhrt haben (Krempl und Herrler, 2001). Dies wurde auch
die Bindungsstellen fur pAPN zu großen Teilen verdecken und fur die Interaktion mit den
pAPN-Banden auf der Nitrozellulose nicht mehr genugend Kontaktstellen bieten. Selbst
der Einsatz unbehandelter TGE-Virionen in einer 10fach hoheren Proteinkonzentration
als die anderen Viruspartikel zeigte nur eine sehr schwache Bindung an die pAPN-Bande
bei 150 kDa (5.3.2). Fur einen Virusbindungstest auf Nitrozellulose ist somit der Ein-
satz nicht desialylierter TGEV-Partikel zur Untersuchung der Sialinsaurebindungsakti-
vitat und auch zur Untersuchung der Bindung an pAPN ungeeignet.
Bei der Untersuchung der Bindung von TGEV an Zellkulturen, die in Mikrotiterplatten
ausgesat worden waren, wurden ebenfalls VCNA-behandelte und unbehandelte Virionen
verwendet. Auch hier zeigte sich, dass die Sialinsaurebindungseigenschaft unbehandel-
ter Viruspartikel aufgrund bereits gebundener zellularer Sialoglykokonjugate stark einge-
schrankt war. Nicht desialyliertes TGEV kann aber sehr wohl noch Rindererythrozyten
agglutinieren, die zu 90% Neu5Gc auf ihrer Oberflache tragen. Dieser Sialinsauretyp wird
von TGEV bevorzugt erkannt und auf der Oberflache von Rindererythrozyten mogli-
cherweise auch in sterisch gunstiger Form prasentiert (Schultze et al., 1996). Auf ST-
und LLC-PK1-Zellen befindet sich aber uberwiegend Neu5Ac (5.3.1). Da die Affinitat
von TGEV zur Bindung an Neu5Ac wesentlich geringer ist als die Bindungsaffinitat zu
Neu5Gc (Schultze et al., 1996), findet eine effiziente Bindung von TGEV an die beiden
Zellkulturen erst nach Neuraminidasebehandlung der Viruspartikel statt.
Bei intestinalen Muzinen und Muzin aus der Glandula submandibularis des Schweines
(PSM) konnte man aufgrund des teilweise hohen Gehalts an Neu5Gc erwarten, dass die
Bindung von TGEV auch ohne vorherige Desialylierung der Viruspartikel nachweisbar
ist. Es zeigte sich jedoch, dass selbst bei den Muzinen des Saugferkels B und PSM, die
den hochsten Gehalt an Neu5Gc bei den eingesetzten Proben aufwiesen, nicht desialy-
liertes TGEV im Gegensatz zu desialyliertem TGEV nur wenig binden konnte (5.5.1).
Moglicherweise hatten hohere Viruskonzentrationen eine starkere Bindung bewirkt. Es
kann aber auch sein, dass durch die Immobilisierung der Muzine auf der ELISA-Platte
und die Vernetzung der Viruspartikel untereinander ahnlich ungunstige Voraussetzungen

6.1. Bedeutung der Desialylierung von TGEV fur die Interaktion... 119
geschaffen wurden wie bei dem Virusbindungstest auf Nitrozellulose.
Darmgewebe aus frischtoten Tieren, das direkt nach der Entnahme in Isopenthan/flussi-
gem Stickstoff eingefroren wurde, kann in relativ unverandertem Zustand gelagert werden
(−80◦C). Die Untersuchung der Virusbindung an Kryoschnitte, die aus solchen Jejunum-
proben angefertigt wurden, bietet somit die Moglichkeit, an relativ unveranderten Struk-
turen zu arbeiten. Da ein Schnitt nicht mehr das gesamte Gewebe in unveranderter Form
prasentiert, ist auch diese Untersuchungsmethode nur eine Annaherung an naturliche
Verhaltnisse. So sind durch die Anfertigung der Darmschnitte auch Strukturen ange-
schnitten, die im Tier vom Darmlumen aus nicht direkt zuganglich waren. Trotzdem ist
die Untersuchung der Bindung von TGEV an Kryoschnitte des Darmes, die Methode in
dieser Arbeit, die den naturlichen Bedingungen im Tier am nachsten kommt. Die Schnitte
wurden vor der Inkubation mit Virus mit Paraformaldehyd fixiert, da das Arbeiten mit
unfixierten Schnitten die zellularen Strukturen des Darmgewebes nach mehreren Inku-
bationsschritten schadigte bzw. schlechter erkennen ließ. Vorherige Tests mit unfixierten
Schnitten zeigten jedoch, dass sich das Bindungsverhalten von TGEV durch die Fixie-
rung nicht veranderte. Bei der Bindung von TGEV an Sialinsaurereste der Strukturen
des Darmepithels in diesen Kryoschnitten zeigte sich, dass die Vorbehandlung der Virio-
nen mit VCNA in dieser Untersuchungsmethode nicht notwendig war. Nicht desialylier-
te TGE-Virionen konnten uber ihre Sialinsaurebindungsaktivitat an Sialoglykokonjugate
des Darmepithels binden, wenn auch nicht so effizient wie TGEV-NA. Die Bindungspart-
ner, die TGEV uber seine Sialinsaure-bindende Aktivitat im Kryoschnitt des Jejunums
erkennt, sind somit offensichtlich fur eine Interaktion mit TGEV optimal sialyliert, so-
wohl was den Sialinsauretyp betrifft als auch hinsichtlich der raumlichen Prasentation
der Sialinsauren. Die Affinitat zu diesen Bindungspartnern scheint so stark zu sein, dass
die verbleibenden Sialinsaurebindungsstellen auf den nicht desialylierten S-Proteinen von
TGEV fur eine Bindung ausreichen. Denkbar ist auch, dass die Bindung der zellularen
Sialoglykokonjugate an TGEV aufgrund einer hoheren Affinitat zu den jejunalen Sialo-
glykokonjugaten gelost wird.
Die Notwendigkeit der vorherigen Desialylierung der Virionen ergibt sich also aus der Me-
thode, mit der man die TGEV-Bindung nachweisen mochte. Wahrend die enzymatische
Vorbehandlung bei der Bindung an immobilisierte Glykoproteine und Muzine, an Zellkul-

120 6. Diskussion
turen und bei der Agglutination von Huhnererythrozyten unbedingt erforderlich ist, kann
man bei der Agglutination von Rindererythrozyten und bei der Bindung an Kryoschnitte
des Darmes darauf verzichten.
Da fur die Interaktion mit Sialoglykokonjugaten des Darmepithels die Desialylierung von
TGEV nicht erforderlich ist, kann die Sialinsaurebindungsaktivitat fur die Enteropatho-
genitat von TGEV in vivo von großer Bedeutung sein. Ganz gleich, ob das Virus auf
seinem Weg in den Darm mit Sialoglykokonjugaten beladen ist oder nicht, es weist eine
hohe Bindungsaffinitat zu Sialoglykokonjugaten des Darmepithels auf und kann mit den
Enterozyten interagieren.
6.2 Bewertung des Vergleichs von TGEV mit
E. coli F5
E. coli F5 ist ein Bakterium, welches den Dunndarm uber die Bindung an Neu5Gc ko-
lonisiert. Die Fimbrien dieses Mikroorganismus besitzen somit wie die S-Proteine von
TGEV eine Sialinsaurebindungsaktivitat mit Bevorzugung von Neu5Gc. Die Infektion
von Schweinen mit E. coli F5 ist altersabhangig. Bei Saugferkeln kommt es zur Besiede-
lung des Darmes und zur Schadigung des Wirtsorganismus durch Toxinfreisetzung uber
die Bakterien (Stamm und Sorg, 1993). Mit zunehmendem Alter der Tiere haften immer
weniger Bakterien an den Burstensaummembranen. Bei 6 Wochen alten Tieren kommt es
nicht mehr zur Adhasion von F5-positiven E. coli, was als Entwicklung einer naturlichen
Resistenz der Enterozyten interpretiert wurde (Stamm und Sorg, 1993). Teneberg
et al. (1990) berichteten uber ein modifiziertes Rezeptormolekul, das keine Adhasion von
F5-Fimbrien ermoglicht. Auch die Infektion mit TGEV verlauft je nach Alter der Tiere un-
terschiedlich und betrifft Saugferkel besonders stark (Enjuanes und van der Zeijst,
1995). Im Gegensatz zu E. coli F5 war bei TGEV bisher aber weder bekannt, welche
Bedeutung die Sialinsaurebindungsaktivitat fur die Infektion besitzt, noch hatte man
ubereinstimmende Erklarungen fur den altersabhangigen Verlauf der Erkrankung. Der
Vergleich der Sialinsaurebindungsaktivitaten von TGEV und E. coli F5 sollte daher eine
Hilfe sein, aufgrund der Gemeinsamkeiten der beiden Mikroorganismen und des Wissens
uber E. coli F5, Anhaltspunkte fur die Bedeutung der Sialinsaurebindungseigenschaft von

6.2. Bewertung des Vergleichs von TGEV mit E. coli F5 121
TGEV zu bekommen.
Trotz einiger Gemeinsamkeiten bestehen zwischen einer Infektion mit E. coli F5 und einer
Infektion mit TGEV doch große Unterschiede. Bei E. coli F5 handelt es sich um ein nicht-
invasives Bakterium, welches die Mukosa des Dunndarms uber seine Adhasine kolonisieren
kann. Das bedeutet, dass die Bakterien einem Abtransport mit dem Darminhalt aufgrund
der Bindung ihrer Fimbrien (Adhasine) an Rezeptoren auf der Darmschleimhaut entge-
genwirken. Durch die Enterotoxine des Bakteriums werden die Epithelzellen des Darms zu
erhohter Flussigkeitssekretion stimuliert, die die eigentliche Diarrhoe bewirkt (Gaastra
und de Graaf, 1982). Die Epithelzellen selber bleiben intakt. Altere Tiere konnen wegen
einer Veranderung der Rezeptoren nicht infiziert werden. Bei einer TGEV-Infektion muss
das Virus, um sich vermehren zu konnen, in die Wirtszellen eindringen. Die Bindung an
das Epithel ist der erste Schritt, der den Replikationszyklus einleitet. Durch die Infektion
der Epithelzellen mit TGEV kommt es schließlich zu einer Zerstorung des Zottenepithels
und damit zu einer wassrigen Diarrhoe. Infiziert werden konnen Tiere jeden Alters, nur der
Verlauf der Erkrankung ist je nach Alter unterschiedlich. Somit gleichen sich diese beiden
Erreger nur in dem Punkt, dass sie durch die Bindung an Strukturen des Darmepithels
einen Abtransport durch die Darmperistaltik verhindern mussen, um den Fortbestand
ihrer Art sichern zu konnen. Diese erste Bindung erfolgt bei beiden uber Sialinsauren.
Bei den vergleichenden Untersuchungen der beiden Mikroorganismen wurde TGEV stets
in desialylierter Form eingesetzt, um moglichst viele freie Sialinsaurebindungsstellen zu
haben. Da bekannt ist, dass TGEV auch ohne vorherige Desialylierung Rindererythro-
zyten agglutinieren kann (Schultze et al., 1996), ware ein zusatzlicher Vergleich mit
nicht desialylierten Virionen durchaus interessant gewesen. Schultze et al. (1996) ha-
ben die Agglutination von Rinder-, Huhner- und Schweineerythrozyten durch TGEV und
TGEV-NA verglichen. Huhnererythrozyten werden wie bei E. coli F5 von nicht desialylier-
tem TGEV nicht agglutiniert. Interessanterweise werden Schweineerythrozyten, obwohl
sie wie Rindererythrozyten 90% der Sialinsauren in Form von Neu5Gc auf ihrer Oberflache
tragen, von nicht desialylierten TGE-Virionen ebenfalls nicht agglutiniert. Somit scheint
nicht nur der Gehalt an Neu5Gc, sondern auch die Art der raumlichen Prasentation der
Sialinsauren eine Rolle fur die Interaktion zu spielen.
Der Vergleich der Interaktion mit Hamagglutinationsinhibitoren sollte Anhaltspunkte lie-

122 6. Diskussion
fern, ob TGEV ahnliche Bindungspartner bevorzugt wie E. coli F5. Ein Hamagglutina-
tionstest ist nicht sehr sensitiv, bietet aber den Vorteil, dass man einfach und schnell
Anregungen fur die folgenden Untersuchungsrichtungen bekommt. Die Hamagglutination
durch E. coli F5 wurde sowohl von Muzinen als auch von Glykolipiden gehemmmt (5.1.3).
Dieses Ergebnis stimmt mit Berichten uberein, dass E. coli F5 Glykolipide wie Neu5Gc-
GM3 als Rezeptor zur Kolonisation des Darms nutzt (Teneberg et al., 1990; Seignole
et al., 1991), die Fimbrien aber ebenfalls an intestinale Muzine der Darmschleimschicht
binden (Lindahl und Carlstedt, 1990). Die Agglutination von Rindererythrozyten
durch TGEV-NA wurde durch Glykolipide nicht gehemmt. Intestinale Muzine hemmten
die Hamagglutination jedoch zu hoheren Titern als bei E. coli F5. Dieses Ergebnis lasst
somit den Schluss zu, dass TGEV Sialoglykoproteine gegenuber -lipiden als Bindungs-
partner bevorzugt.
6.3 Interaktion von TGEV mit Zellkulturzellen
Die Desialylierung von TGEV bewirkt eine bis zu 6fach hohere Bindung der Virionen
an die Oberflache von ST- und LLC-PK1-Zellen (5.2). Vergleicht man jedoch die Infek-
tiositat von TGEV-NA mit TGEV oder mit sialinsaurebindungsdefizienten Mutanten sind
die Virustiter annahernd gleich hoch (Schultze et al., 1996; Krempl et al., 1997, 2000).
Die Infektiositat in Zellkultur erhoht sich also nicht durch die Eigenschaft, zusatzlich an
Sialinsauren binden zu konnen. Die Bindung an den zellularen Rezeptor porzine Amino-
peptidase N (pAPN) hingegen ist unerlasslich fur die Einleitung einer Infektion. Durch
die Bildung von pAPN-TGEV-Komplexen kommt es zu einer erhohten Endozytoserate,
und auch fur die Fusion der Virus- mit der Endosomenmembran scheint die Interaktion
mit pAPN eine Rolle zu spielen (Hansen et al., 1998).
Die Bindung an mehr als einen Rezeptor ist bereits fur mehrere Viren demonstriert wor-
den und belegt die Komplexitat der Vorgange bei der Einleitung einer Virusinfektion. So
bewirkt z.B. bei einer HIV-Infektion die Bindung des externen Virus-Glykoproteins gp120
an zellulares CD4 eine Konformationsanderung des viralen Proteins, wodurch die Bindung
an den Corezeptor aus der Chemokinfamilie (CCR5) ermoglicht wird (Dalgleish et al.,
1984; Klatzmann et al., 1984; Deng et al., 1996). Diese zweite Interaktion bewirkt

6.3. Interaktion von TGEV mit Zellkulturzellen 123
beim viralen Transmembranprotein gp41 die Umwandlung in eine fusionsaktive Konfor-
mation. Es existieren Virusvarianten, die statt CCR5 CXCR4 oder andere Rezeptoren
nutzen, um die Fusion zu bewirken (Berson et al., 1996; Feng et al., 1996). Auch Vi-
ren der Familie Herpesviridae wie das Herpes-simplex-Virus (HSV-1) benotigen mehrere
Zelloberflachenfaktoren zur Infektion einer Zelle. Mit dem Oberflachenglykoprotein gC
binden die Virionen an Heparansulfat (WuDunn und Spear, 1989). Der Viruseintritt
erfordert jedoch die Interaktion des viralen Glykoproteins gD mit Corezeptoren auf der
Zelloberflache (Campadelli-Fiume et al., 2000). Sowohl bei HIV als auch bei Herpes-
viren existieren Mutanten, die sich in Zellkultur effizient vermehren, obwohl sie nicht an
CD4 bzw. Heparansulfat binden konnen. Die Corezeptoren werden in diesem Fall auch
fur die Adsorption an die Zelle genutzt. Diese Mutanten konnen sich zwar in Zellkultur
vermehren, kommen aber nicht in naturlichen Isolaten vor. In vivo ist die Bindung an
CD4 bzw. Heparansulfat somit unerlasslich.
In ahnlicher Weise kann man die Bindung von TGEV an Sialinsauren betrachten. Unter
den kunstlichen Bedingungen in Zellkultur wird diese Bindungseigenschaft nicht benotigt,
sie ist fakultativ. Im Wirtsorganismus hingegen stellt die Sialinsaurebindungsaktivitat
einen Pathogenitatsfaktor dar, der es TGEV ermoglicht, effizient im Darmtrakt zu repli-
zieren. Im Gegensatz zu HIV und HSV-1 existiert bei TGEV auch in vivo eine naturliche
Variante — das porzine respiratorische Coronavirus (PRCoV), das keine Sialinsaurebin-
dungaktivitat besitzt (Schultze et al., 1996), sondern nur uber die Bindung an pAPN
eine Infektion einleitet (Delmas et al., 1993). PRCoV infiziert jedoch naturlicherweise nur
den Respirationstrakt und kann im Darmtrakt kaum replizieren (Cox et al., 1990). Somit
ist die Sialinsaurebindungsaktivitat von TGEV nicht generell fur eine naturliche Infektion
erforderlich, wohl aber fur eine erfolgreiche Replikation im Dunndarm des Schweines.
Die Sialinsaureanalyse der Membranproteine von ST- und LLC-PK1-Zellen ergab, dass
Neu5Ac den großten Anteil der Sialinsauren ausmacht (5.3.1). Dies ist wohl mit ein Grund,
warum TGEV erst nach Neuraminidasebehandlung an Sialoglykokonjugate auf der Zell-
kulturoberflache binden kann (s. auch 6.1). Bei dem Bindungspartner handelt es sich
um ein hochmolekulares Sialoglykoprotein. Einige Befunde weisen auf einen muzinartigen
Charakter dieses Sialoglykoproteins hin:

124 6. Diskussion
• Das Sialoglykoprotein besitzt ein Molekulargewicht von vermutlich mehreren hun-
dert kDa. Muzine sind Glykoproteine mit einem Molekulargewicht von mehreren
hundert bis mehreren tausend kDa (Roussel und Lamblin, 1996).
• Das Sialoglykoprotein ließ sich schlecht mit NHS-Biotin markieren (5.3.3). Da unter
anderem Hydroxyaminosauren mit dem N-Hydroxysuccinimidester des Biotins bei
einer Markierung reagieren, konnte die O-Glykosylierung von Serin- oder Threonin-
resten diese Interaktion verhindert haben. Dies hatte eine geringe Markierbarkeit
des Proteins mit Biotin zur Folge.
• Das Sialoglykoprotein ließ sich gut mit der fur Glykoproteine spezifischen Silberfar-
bung nachweisen (5.3.3). Es farbte sich in dieser Farbung gelb-braunlich, was fur
Proteine mit einem hohen Anteil an O-Glykanen typisch ist (Deh et al., 1985).
• Bei dem Sialoglykoprotein handelt es sich um ein membranassoziiertes Protein ohne
Transmembrananker (5.4.1). Viele Muzine werden von Epithelzellen sezerniert und
befinden sich danach in lockerer Assoziation zur Zellmembran.
• Das Sialoglykoprotein wurde von dem Lektin WGA gut erkannt. WGA bindet be-
vorzugt an N-Acetylglukosamin und hat eine eher schwache Affinitat fur Sialinsau-
ren (Gallagher et al., 1985). Die Kohlenhydratketten von Muzinen bilden haufig
sogenannte Cluster hoch glykosylierter Domanen (Roussel und Lamblin, 1996).
Diese Clusterbildung konnte bei dem hochmolekularen Sialoglykoprotein die gute
Erkennung durch WGA bewirkt haben.
6.4 Interaktion von TGEV mit intestinalen Muzinen
und mit Darmepithel
Die Agglutination von Rindererythrozyten durch TGEV wurde von den intestinalen Mu-
zinen aller Tiere zu gleichen Titern gehemmt (5.1.3). Im ELISA band TGEV gut an
intestinale Muzine von Saugferkel A und B und Absetzferkel D, konnte intestinale Muzi-
ne des Absetzferkels C jedoch nicht erkennen (5.5.1). Ein ELISA ist wesentlich sensitiver
als ein Hamagglutinationshemmtest, was erklart, dass die geringeren Bindungsunterschie-

6.4. Interaktion von TGEV mit intestinalen Muzinen und mit Darmepithel 125
de zwischen den Muzinen von Tier A, B und D im Hamagglutinationstest nicht erfassbar
waren. Die Muzine des Absetzferkels C verhielten sich in den beiden Testsystemen jedoch
vollig unterschiedlich. TGEV konnte diese Muzine nicht binden, was durch den spezifi-
schen Nachweis mittels Antikorper im ELISA gezeigt wurde. Trotzdem hemmten diese
Muzine die Hammagglutination durch TGEV zu gleichen Titern wie die Muzine, die auch
spezifisch mit TGEV interagieren. Eine mogliche Erklarung fur diese Hemmung sind un-
spezifische Interaktionen mit den Viruspartikeln, die nicht auf eine direkte Interaktion mit
der Sialinsaurebindungsstelle beruhen, aber die Interaktion von TGEV mit den Erythro-
zyten behindern. Eine spezifische Bindung von TGEV an Sialinsaurereste der Muzine des
Tieres C war vermutlich wegen des geringen Gehalts an Neu5Gc von nur 10% (bezogen
auf den Gesamtgehalt an Neu5Gc und Neu5Ac) nicht moglich. Desialyliertes TGEV kann
jedoch auch an Neu5Ac binden, was die hohen Hamagglutinationstiter beim Einsatz von
Huhnererythrozyten belegen (Schultze et al., 1996). Da die raumliche Prasentation der
Sialinsauren eine bedeutende Rolle fur die erfolgreiche Interaktion mit TGEV spielt, ist
die ungunstige Prasentation der Sialinsauren ein wahrscheinlicherer Grund fur die feh-
lende Bindung an Muzine des Absetzferkels C. In dem Bindungstest auf Nitrozellulose
wurde bei den Muzinen nur bei Tier B und PSM ein positives Signal detektiert. Es ist
wahrscheinlich, dass die aufgetragene Muzinmenge von 5 µg/Spur bei den anderen ELISA-
positiven Muzinen unterhalb der Nachweisgrenze lag. Die Sensitivitat des Bindungstests
auf einer Blotmembran erreicht nicht die eines ELISAs. Um auch die ubrigen Muzine im
Blot detektieren zu konnen, mussten vermutlich Muzinmengen von ≥ 20 µg/Spur gewahlt
werden.
Die Untersuchung der Bindung von TGEV an Burstensaummbranen ergab, dass bei al-
len vier Saugferkelproben (S1–S4) und zwei der vier Absetzferkelproben (A1, A2) neben
pAPN ein hochmolekulares Sialoglykoprotein erkannt wurde. Diese Interaktion erfolgte
uber Sialinsaurereste des Proteins mit der Sialinsaurebindungsstelle der S-Proteine von
TGEV, wie u.a. der Vergleich mit den sialinsaurebindungsdefizienten Mutanten HAD3
und m10 zeigte. Die HPLC-Analyse ergab, dass alle BSMV-Praparationen, auch die der
Absetzferkel A3 und A4, sowohl Neu5Gc als auch Neu5Ac enthielten (5.6.2). Obwohl der
Sialinsauregehalt der Burstensaummembranen von A3 und A4 geringer war als von den
anderen Tieren, erklart sich die fehlende Bindung an Sialinsaurereste des hochmolekularen
Proteins bei diesen beiden Tieren dadurch nicht. Beim Nachweis des Kohlenhydrat- und

126 6. Diskussion
Sialinsaureanteils der einzelnen BSMV-Proteine nach SDS-PAGE und Ubertragung auf
Nitrozellulose zeigte sich jedoch, dass sich die Proteine der Absetzferkel A3 und A4 in ih-
rer Kohlenhydratzusammensetzung von denen der anderen Tiere unterschieden (5.7.2.1).
Die beiden Proben enthielten zwar ebenfalls eine Hauptglykoproteinbande auf der Hohe
des Bindungspartners von TGEV, diese zeigte nach vorheriger Neuraminidasebehandlung
der Proben aber keine Veranderung in ihrem Laufverhalten und ihrer Intensitat. Bei den
ubrigen Proben hingegen kam es nach vorheriger Desialylierung zu einer Verschiebung der
Bande des Hauptglykoproteins nach oben und zu einer Abnahme der Bandenstarke. Die
Hauptglykoproteine der BSMV-Praparationen der Absetzferkel A3 und A4 unterscheiden
sich somit von denen der anderen Ferkel in ihrem Kohlenhydratanteil. Vor allem bei Pro-
be A3 scheinen kaum Sialinsauren an diesem Hauptglykoprotein vorzuliegen. Aus diesem
Grund konnte diese Glykoproteinbande auch ohne VCNA-Behandlung der Probe bereits
mit dem Lektin PNA reagieren. Das Hauptglykoprotein der Probe A4 hingegen schien
etwas starker sialyliert zu sein (5.7.2.1). Hier ließen sich die Sialinsauren jedoch nicht mit
VCNA abspalten, was auch die unterbliebene Bindung von PNA erklart. Diese Unterschie-
de in der Kohlenhydratzusammensetzung im Vergleich zu den anderen Proben erklaren
die fehlende Bindung von TGEV an das Hauptglykoprotein von A3 und A4. Erganzende
Untersuchungen konnten zeigen, ob sich die fehlende Bindung von TGEV an intestina-
le Muzine des Absetzferkels C durch ahnliche Unterschiede in der Kohlenhydratstruktur
erklart.
Die Unterschiede in der Kohlenhydratzusammensetzung der BSMV-Proben A3 und A4
und moglicherweise auch der Muzinprobe C zu den anderen Proben konnten durch die
Isolierungs- und Reinigungsschritte verursacht worden sein. Da aber die Muzinisolierung
und -reinigung sowie die BSMV-Praparation von Saugferkel- und Absetzferkeldarm inner-
halb der Gruppen immer gleich ablief, handelt es sich bei den Unterschieden in der Koh-
lenhydratzusammensetzung wohl eher um individuelle Unterschiede zwischen den Tieren.
Die Absetzferkel A3 und A4 waren 6–8 Wochen alt und kamen beide direkt vor der Totung
aus der gleichen Haltung in Mariensee. Absetzferkel A1 und A2 (12 Wochen alt) kamen
ursprunglich auch aus Mariensee, wurden jedoch bereits ca. 4 Wochen vor der Totung
im Institut fur Physiologie der TiHo Hannover eingestallt und unterlagen dort anderen
Haltungsbedingungen als in Mariensee (Futterumstellung, anderer Stall, Umweltfaktoren,
etc.). Diese Haltungsunterschiede und dabei vor allem eine andere Futterzusammenset-

6.4. Interaktion von TGEV mit intestinalen Muzinen und mit Darmepithel 127
zung konnten durchaus einen Einfluss auf die Kohlenhydratzusammensetzung der Gly-
koproteine des Darms haben. Die Saugferkelproben fur die BSMV-Praparationen kamen
aus dem Lehr- und Forschungsinstitut der TiHo Hannover in Ruthe. Auch die Tiere fur
die Muzinisolierungen kamen aus unterschiedlichen Haltungen. Der Darm der Absetzfer-
kel C und D und der Darm des Saugferkels A (14 Tage alt) wurden zu verschiedenen
Zeitpunkten vom Institut fur Pathologie zur Verfugung gestellt. Alle Tiere kamen aus
verschiedenen Betrieben mit verschiedenen Vorberichten. Der Darm des 12 Tage alten
Saugferkels wurde vom Institut fur Physiologie zur Verfugung gestellt, das Tier kam aus
Ruthe. Das Tiermaterial war somit sehr heterogen. Die unterschiedlichen Haltungs- und
Ernahrungsbedingungen der Tiere und auch genetisch bedingte individuelle Unterschie-
de konnten die Unterschiede im Bindungsverhalten von TGEV an die jeweiligen Proben
erklaren.
Die Probenauswahl aus jeweils zwei Altersgruppen (Saug- und Absetzferkel) sollte helfen,
die altersabhangigen Unterschiede im Verlauf einer TGEV-Infektion zu erklaren. Die Un-
tersuchungen ergaben, dass es Unterschiede in der Sialinsaurezusammensetzung und in
der Interaktion mit TGEV zwischen den Proben gab. Bei den intestinalen Muzinen wies
die Probe des Absetzferkels C mit einem hohen Neu5Ac- und einem geringen Neu5Gc-
Gehalt eine andere Sialinsaurezusammensetzung als die anderen Proben auf (5.1.3.2).
Diese Muzinprobe wurde von TGEV im ELISA nicht gebunden (5.5.1). Die Muzinprobe
des Absetzferkels D hatte eine fast identische Sialinsaurezusammensetzung (Neu5Gc +
Neu5Ac) wie die Muzinprobe des Saugferkels A (5.1.3.2), wurde im ELISA aber etwas
schlechter von TGEV erkannt als die intestinalen Muzine beider Saugferkel (5.5.1). Der
Sialinsauregehalt der BSMV-Praparationen aus Saugferkeldarm — speziell der Gehalt an
der fur die TGEV-Bindung wichtigen Sialinsaure Neu5Gc — war bezogen auf die Pro-
teinmenge ca. 4-mal hoher als der der Praparationen aus Absetzferkeldarm (5.6.2). Das
hochmolekulare Sialoglykoprotein wurde von TGEV in allen vier BSMV-Proben der Saug-
ferkel gut gebunden. Bei den Absetzferkeln wurden nur zwei von vier Proben von TGEV
erkannt (A1, A2), wobei die Bindung an die hochmolekulare Bande bei A1 schwacher war
als bei A2 (5.6.3). Die bessere Virusbindung an die Saugferkelproben im Vergleich zu den
Absetzferkelproben weist darauf hin, dass die hochgradige Gastroenteritis bei Saugferkeln
durch eine bessere Bindung des Virus an Sialoglykokonjugate im Darm bedingt sein kann.
Durch bessere Anheftungsmoglichkeiten der Virionen im Darm steigt die Wahrscheinlich-

128 6. Diskussion
keit fur eine erfolgreiche Virusreplikation in den Epithelzellen, so dass bei Saugferkeln
eine schwerere Erkrankung verursacht wird. Fur die altersabhangigen Unterschiede ei-
ner virusbedingten Gastroenteritis gibt es viele Erklarungen. Der schwerere Verlauf der
TGE konnte bedingt sein durch die geringe Enterozytenerneuerungsrate bei neugeborenen
Ferkeln (Moon, 1973). Die Malabsorption, die Maldigestion, der daraus folgende Flussig-
keitsverlust und somit der Durchfall dauern aufgrund dieser geringen Erneuerungsrate der
Epithelzellen bei Saugferkeln langer an als bei Absetzferkeln. Das Vorkommen von En-
terozyten fetalen Ursprungs auf dem Villus Neugeborener, die durch ihr tubulo-vakuolares
System die Virusreplikation erleichtern (Wagner et al., 1973), ist als ein weiterer Grund
fur den schweren Verlauf der Gastroenteritis bei Saugferkeln junger als 2 Wochen disku-
tiert worden. Auch haben Umweltfaktoren wie Kalte oder schwankende Temperaturen bei
Neugeborenen einen starkeren Einfluss auf den Schweregrad der Diarrhoe (Saif, 1990).
Das Immunsystem, vor allem die lokale Darmimmunitat, die bei der Bekampfung einer
TGEV-Infektion von großer Bedeutung ist, kann bei Neugeborenen ebenfalls noch nicht
ausreichend entwickelt sein. Diese Erklarungsversuche sind nicht durch experimentelle Da-
ten belegt. Die Ergebnisse der vorliegenden Arbeit sprechen dafur, dass die Bindung an
Sialoglykokonjugate die Infektion von Saugferkeln im Vergleich zu Absetzferkeln beguns-
tigen konnte.
Sowohl bei den Muzinen des Saugferkels B als auch bei den BSMV-Proben der Saugfer-
kel S1–S4 und der Absetzferkel A1 und A2 wurden hochmolekulare Proteinbanden uber
die Sialinsaurebindungsaktivitat von TGEV gebunden. Es ist moglich, dass dieses hoch-
molekulare Protein in den BSMV-Praparationen mit dem in den Muzinen identisch ist.
Bei dem Sialoglykoprotein der BSMV-Praparationen sprechen viele Anhaltspunkte dafur,
dass es sich wie bei den Muzinpraparationen ebenfalls um ein Muzin handelt:
• Bei dem von TGEV gebundenen Sialoglykoprotein handelt es sich um ein sehr hoch-
molekulares Protein. Ein Charakteristikum von Muzinen ist ihr Molekulargewicht
von mehreren hundert bis mehreren tausend kDa (Roussel und Lamblin, 1996).
• Bei dem Sialoglykoprotein handelt es sich um ein membranassoziiertes Protein ohne
Transmembrananker (5.7.1). Viele Muzine werden von speziellen Epithelzellen sezer-
niert und bilden zusammen mit Wasser und anderen Substanzen die Schleimschicht
auf der Oberflache von Epithelien wie z.B. im Darm, im Auge, im Respirationstrakt

6.4. Interaktion von TGEV mit intestinalen Muzinen und mit Darmepithel 129
etc. (Carlstedt et al., 1985).
• Der Nachweis der Kohlenhydratanteile in den BSMV-Proteinen weist durch die In-
tensitat der Kohlenhydratbande auf Hohe des hochmolekularen BSMV-Proteins da-
rauf hin, dass dieses Protein stark glykosyliert ist (5.7.2.1). Die Definition von Muzi-
nen basiert neben dem hohen Molekulargewicht auf einem hohen Kohlenhydratanteil
von 50–70% (Roussel und Lamblin, 1996).
• Die enzymatische Entfernung der Sialinsauren mit VCNA fuhrte zu keiner erhohten
Mobilitat des Proteins in der SDS-PAGE, sondern zu einem verlangsamten Lauf-
verhalten (5.7.2.1). Fur muzinartige sialinsaurereiche Glykoproteine ist solch ein
verlangsamtes elektrophoretisches Laufverhalten typisch.
• Das hochmolekulare Sialoglykoprotein wurde von den Lektinen Jacalin und PNA
erkannt (5.7.2.2), die an das fur O-glykosylierte Proteine charakteristische Disaccha-
rid Galaktose-β(1-3)-N-Acetylgalaktosamin binden (Sastry et al., 1986; Sueyoshi
et al., 1988). Die Kohlenhydratketten von Muzinen sind uber O-gykosidische Bin-
dungen zwischen N-Acetylgalaktosamin und den Hydroxygruppen der Aminosau-
ren Serin oder Threonin mit dem Apomuzin (Peptidanteil des Muzins) verankert
(Roussel und Lamblin, 1996).
• Die sialinsaurespezifischen Lektine MAA und SNA erkannten das sialylierte Glyko-
protein nur schwach bzw. gar nicht. Diese Lektine reagieren gut mit Sialinsauren
in N-Glykanen, konnen Sialinsauren in O-Glykanen jedoch nur ineffizient oder gar
nicht erkennen (Wang und Cummings, 1988; Haselbeck et al., 1990).
• Das Sialoglykoprotein wurde von dem Lektin WGA gut erkannt. WGA bindet bevor-
zugt an N-Acetylglukosamin und hat eine eher schwache Affinitat fur Sialinsauren
(Gallagher et al., 1985). Die Kohlenhydratketten von Muzinen bilden haufig so-
genannte Cluster hoch glykosylierter Domanen, und die Muzine erhalten dadurch
ein”flaschenburstenartiges“ Aussehen (Roussel und Lamblin, 1996). Diese Clus-
terbildung konnte bei dem hochmolekularen Sialoglykoprotein die gute Erkennung
durch WGA bewirkt haben.
TGEV erkannte bei einigen der Proben (Muzine, BSMV-Proben S1, S2, A2) zwei hochmo-
lekulare Banden in etwas unterschiedlicher Hohe, bei anderen (S3, S4, A1) nur jeweils eine

130 6. Diskussion
dieser Banden. Diese von TGEV erkannte Doppelbande konnte ein und dasselbe Prote-
in in unterschiedlicher Glykosylierung darstellen. Es kann aber auch nicht ausgeschlossen
werden, dass sich die beiden Banden in ihrer Aminosaurezusammensetzung unterscheiden.
Es ist bekannt, dass selbst in ein und derselben Zelle produzierte Muzine Unterschiede
aufweisen. Die Polydispersitat der Peptidvorlaufer von Muzinen zeigte sich in Großenun-
terschieden der Peptide von 100 bis uber 400 kDa (Roussel und Lamblin, 1996). Ob
sich die von TGEV erkannten zwei hochmolekularen Banden in ihrer Peptid- oder/und
in ihrer Kohlenhydratzusammensetzung unterscheiden, mussen weitere Untersuchungen
klaren. Die Isolierung und anschließende Sequenzierung der Proteine auf der Hohe dieser
Banden wird zeigen, ob es sich um ein unterschiedlich glykosyliertes Protein oder um zwei
verschiedene Proteine handelt.
Die sialinsaureabhangige Bindung von TGEV an Strukturen der Becherzellen des Darm-
epithels (5.8) spricht dafur, dass es sich hier um eine Bindung an Granula der in den
Becherzellen gebildeten Muzine handelt. Diese Muzine sind wahrscheinlich mit denen in
den BSMV-Praparationen und denen aus der Darmmuzinisolierung identisch. Da diese
Muzine in den Becherzellen gehauft vorkommen und die Becherzellen im Anschnitt fur
das Virus gut zuganglich waren, kam es in dem Bindungstest auf Kryoschnitten vor allem
dort zu einer starken Fluoreszenz. Diese Fluoreszenz uberstrahlte die Bindung von TGEV
an die sezernierten Muzine auf der Epitheloberflache, die nur schwach zu erkennen war. In
vivo sind vor allem die sezernierten Muzine fur eine Interaktion mit TGEV von Bedeutung,
die Muzingranula in den Becherzellen hingegen fur das Virus nicht zuganglich.
Die Bindung von TGEV an die Muzingranula war im Kryoschnitt sowohl im Bereich der
Zotten als auch im Bereich der Krypten vorhanden. Bei einer naturlichen TGEV-Infektion
werden die Epithelzellen im apikalen Bereich der Zotten von Jejunum und Ileum infiziert.
Das Virus breitet sich dann uber den Villus auf die weiter abwarts liegenden Epithelzellen
aus. Es werden somit die Zotten in ihrer gesamten Lange (bis auf die Basis), jedoch nicht
die Krypten infiziert (Pensaert et al., 1970; Saif, 1990). Der zellulare Rezeptor pAPN
wurde in Darmschnitten mit dem monoklonalen Antikorper G43 auf dem gesamten Vil-
lus nachgewiesen, in den Krypten hingegen nicht (Weingartl und Derbyshire, 1994).
Ohne pAPN ist eine erfolgreiche Virusreplikation nicht moglich, was erklart, wieso die
Krypten nicht infiziert werden konnen. In elektronenmikrographischen Scans des Darm-

6.5. Bedeutung der Sialinsaure-bindenden Aktivitat fur die Enteropathogenitat... 131
epithels ist die Dreidimensionalitat der Darmzotten gut zu erkennen. Das Darmepithel
ist von einer dicken Schleimschicht uberzogen, das Darmlumen mit dem im Abbau be-
findlichen Nahrungsbrei gefullt und der gesamte Inhalt ist durch die Darmperistaltik in
standiger Bewegung. Unter all diesen Gesichtspunkten ist es nicht verwunderlich, dass
die im Darmlumen befindlichen TGE-Virionen als erstes die Zottenspitzen infizieren und
sich erst dann auf die weiter abwarts liegenden Epithelzellen ausbreiten. Die Zottenspitze
ist am zuganglichsten und die Wahrscheinlichkeit der erfolgreichen Bindung an pAPN in
diesem Bereich am hochsten.
6.5 Bedeutung der Sialinsaure-bindenden Aktivitat
fur die Enteropathogenitat von TGEV
Die Sialinsaurebindungsaktivitat von TGEV stellt einen Pathogenitatsfaktor dar, der fur
die Infektion in Zellkultur nicht erforderlich, fur die Infektion des Gastrointestinaltrakts
hingegen unerlasslich ist. In den letzten Jahren wurden fur viele Viren Corezeptoren be-
schrieben, die fur eine erfolgreiche Infektion erforderlich sind. Das Coronavirus MHV z.B.
scheint einen zweiten Rezeptor zur Penetration der zellularen Membranen zu benotigen
(Yokomori et al., 1993). Corezeptoren, die den Zelltropismus bestimmen, wurden z.B.
fur HIV-1 gefunden. Nach der primaren Bindung an CD4 entscheidet die Bindungsaffinitat
fur CXCR4 bzw. fur CCR5, ob der jeweilige HIV-Stamm T-Zellen bzw. Makrophagen in-
fiziert (Berson et al., 1996; Deng et al., 1996; Dragic et al., 1996; Feng et al., 1996).
Im Gegensatz zu HIV bestimmt bei TGEV die Sialinsaurebindungsaktivitat im ersten
Anheftungsschritt den Enterotropismus. Im zweiten Schritt kommt es zur Bindung an
den zellularen Rezeptor pAPN. Die Bindung an pAPN ist ein gemeinsames Merkmal
der enteropathogenen und der apathogenen Isolate (Delmas et al., 1993). Auch Unter-
suchungen anderer Arbeitsgruppen ergaben, dass der N-terminale Bereich des S-Proteins
von TGEV eine bedeutende Rolle fur den Enterotropismus spielt (Bernard und Laude,
1995; Ballesteros et al., 1997; Sanchez et al., 1999). In diesem Bereich des S-Proteins
ist die Sialinsaurebindungsaktivitat lokalisiert (Schultze et al., 1996; Krempl et al.,
1997). Die Enteropathogenitat von TGEV kann naturlich zusatzlich von weiteren Fak-
toren beeinflusst werden. So sind Feldstamme von enteropathogenen TGEV wesentlich

132 6. Diskussion
virulenter als Zellkultur-adaptierte Virustamme wie TGEV-PUR46 und TGEV-PUR115,
obwohl letztere eine Sialinsaurebindungsaktivitat besitzen und den Darmtrakt infizieren
konnen (Bernard und Laude, 1995; Ballesteros et al., 1997). Die Sialinsaurebin-
dungsaktivitat stellt auch bei dem Bakterium E. coli F5 einen Pathogenitatsfaktor dar.
Durch die Interaktion mit Sialinsauren im Bereich der Glykokalix oder assoziierter Mu-
zinschichten kann das Bakterium den Darmtrakt kolonisieren und seinen Abtransport mit
dem Darminhalt verhindern.
Welche Funktion hat die Sialinsaure-bindende Aktivitat im Rahmen der Enteropathoge-
nitat von TGEV?
Eine mogliche Funktion ware der Schutz der Viruspartikel durch gebundene Sialoglykokon-
jugate bei der Magen-Darm-Passage. Coronaviren sind behullte Viren, die i.d.R. empfind-
licher gegenuber schadigenden Einflussen von außen reagieren als unbehullte Viren. Die
Enteropathogenitat der Viren setzt eine Resistenz gegenuber den niedrigen pH-Werten im
Magen, den Verdauungsenzymen und den detergenzartigen Gallensalzen voraus. Unter-
suchungen haben ergeben, dass sich TGEV in seiner Empfindlichkeit gegenuber Trypsin
und niedrigen pH-Werten nicht von sialinsaurebindungsdefizienten Mutanten unterschei-
det (Krempl et al., 2000). Auch die Stabilitat der Viruspartikel gegenuber dem Deter-
genz Oktylglukosid ist bei TGEV-PUR46 im Vergleich zu den Mutanten nur geringgradig
erhoht. Bei TGEV-PUR115 und den sialinsaurebindungsdefizienten Epitop-D-Mutanten
konnte auch hier kein Unterschied in der Stabilitat der Viruspartikel festgestellt werden
(Krempl et al., 2000). Auch wenn sich die Bedingungen im Magen-Darm-Trakt von den
Testbedingungen in diesen Versuchen unterscheiden, sprechen die Ergebnisse dafur, dass
die Sialinsaurebindungsaktivitat keine große Rolle fur die Stabilitat der Viruspartikel bei
der Magen-Darm-Passage spielt. Vielmehr scheinen andere Faktoren, die auch bei den
sialinsaurebindungsdefizienten Mutanten z.B. eine erstaunlich hohe Resistenz gegenuber
niedrigen pH-Werten bewirken, fur die Stabilitat der Virionen verantwortlich zu sein.
Die Sialinsaurebindungsaktivitat von TGEV scheint somit ihre Funktion als Pathoge-
nitatsfaktor in einer ersten Anlagerung der Viruspartikel an die Muzinschicht des Darms
zu haben. Sialinsaurebindungsdefiziente Mutanten wie die Mutante m10 konnen nicht
an diese Muzinschicht binden. Dies erklart die stark reduzierte Enteropathogenitat die-
ser Mutanten. Die Anlagerung an Muzine im Darmschleim erschwert ahnlich wie z.B.

6.5. Bedeutung der Sialinsaure-bindenden Aktivitat fur die Enteropathogenitat... 133
bei E. coli F5 den Abtransport der Virionen durch die Darmperistaltik. Der Mukus im
Darmtrakt hat die Aufgabe, die Epithelzellen vor Selbstverdau zu schutzen. Im Gegensatz
dazu muss das mukoziliare Transportsystem im Respirationstrakt die unteren Atemwege
von mit der Atemluft eingedrungenen Partikeln frei halten (Carlstedt et al., 1985). Die
Schleimhaut der unteren Atemwege ist steril. Im Darm hingegen gibt es eine physiologische
mikrobielle Flora, die u.a. die Mukosa vor pathogenen Mikroorganismen schutzt. Diese
unterschiedliche Funktion des Schleims zeigt schon, dass eine Bindung an Darmschleim fur
pathogene Mikroorganismen durchaus Vorteile fur eine erfolgreiche Infektion hat. Die Bin-
dung an die Schleimschicht des Atemtrakts hingegen ist eher von Nachteil, da es dadurch
zu einem schnelleren Abtransport der Mikroorganismen kommt. Auch der Darmschleim
kann einen Schutz vor potentiellen Krankheitserregern darstellen. Die Bindung an Muzi-
ne und andere kohlenhydratreiche Strukturen des Darmtrakts scheinen jedoch auch einige
Erreger fur sich zu nutzen.
Aus diesen Uberlegungen lasst sich fur eine Infektion des Darmtrakts mit TGEV folgendes
Modell ableiten:
1. Nach oraler Aufnahme und Passage durch den Magen binden die Viruspartikel uber
ihre Sialinsaurebindungsstellen an Muzine in der Schleimschicht, die die Darmmu-
kosa bedeckt.
2. Es kommt zu einem schrittweisen Durchdringen der Schleimschicht, wobei die Bin-
dung an Neu5Gc-tragende Muzine der dem Epithel nahen Schleimschicht eine große
Rolle spielt.
3. Die Viren gelangen zur Glykokalix, wo die Anheftung an Sialinsaure-tragende Mu-
zine ein weiteres Vordringen der Erreger bis zur Zellmembran ermoglicht.
4. Nach Erreichen der Epithelzelloberflache auf den Zottenspitzen kommt es zur Bin-
dung der Viruspartikel an pAPN. Dies bewirkt die Endozytose der Virionen (Hansen
et al., 1998).
5. Nach der Fusion der Endosomenmembran mit den viralen Membranen kommt es zur
Abgabe des Virusgenoms in die Zelle und zur Replikation (Hansen et al., 1998).
Die Virusreifung findet an den intrazellularen Membranen des intermediaren Kom-

134 6. Diskussion
partiments zwischen Endoplasmatischem Retikulum und Golgi-Apparat statt. Die
neuen Viruspartikel verlassen die Zelle.
6. Da TGEV spezifisch an bestimmte losliche Muzine bindet, kommt es trotz fehlen-
der Sialinsaure-spaltender Enzymaktivitat des Virus nicht zu einer Assoziation der
freigesetzten Virionen mit der Wirtszelle, sondern zur Abgabe der Partikel in den
Mukus des Darms.
7. Die Viruspartikel gelangen so zu Zotten, die noch nicht infiziert wurden, und konnen
ihren Replikationszyklus fortsetzen. Eine Infektion bereits erneuerter Epithelzellen
findet nicht statt, da die aus den Krypten nachwachsenden Zellen resistent gegenuber
einer TGEV-Infektion sind (Saif, 1990).
8. Die Virionen wandern, gebunden an losliche Muzine, mit dem Darmschleim Rich-
tung Dickdarm. Die Muzine werden im Rektum von verschiedenen Enzymen (Gly-
kosidasen, Sulfatasen, O-Acetylesterasen) abgebaut. Diese Enzyme werden von der
lokalen Bakterienflora sezerniert (Roussel und Lamblin, 1996). In den Faces ist
somit physiologischerweise kein Muzin mehr vorhanden. Die Viruspartikel werden
mit dem Faces abgegeben, konnen von anderen Tieren oral aufgenommen werden
und eine neue TGEV-Infektion verursachen.
Die Sialinsaurebindungsaktivitat stellt somit einen wichtigen Pathogenitatsfaktor des Vi-
rus der ubertragbaren Gastroenteritis der Schweine dar. Weitere Untersuchungen, wie
die Isolierung und Sequenzierung des hochmolekularen Sialoglykoproteins und genaue-
re Charakterisierung der Kohlenhydratanteile, konnen zur Identifizierung des Muzins im
porzinen Intestinaltrakt fuhren.

7. Zusammenfassung
Das ubertragbare Gastroenteritis-Virus (TGEV) ist ein typischer Vertreter der entero-
pathogenen Coronaviren. Es infiziert Schweine jeden Alters. Bei Saugferkeln kommt es
zu einer besonders schweren Gastroenteritis mit einer Mortalitatsrate von bis zu 100%.
Auf dem Oberflachenglykoprotein S von TGEV sind zwei Bindungsaktivitaten lokali-
siert. Die Bindung an den zellularen Rezeptor porzine Aminopeptidase N (pAPN) ist
fur die Infektion von Zellkulturen und auch im Wirtsorganismus unerlasslich. Uber die
zweite Bindungsaktivitat, deren aktives Zentrum im N-terminalen Bereich des S-Proteins
lokalisiert ist, interagiert TGEV mit den Sialinsaureresten von Glykokonjugaten, wobei
Neu5Gc bevorzugt gebunden wird. Diese Sialinsaurebindungsaktivitat ist zur Replikation
in Zellkultur nicht notwendig. Bei der Verursachung der ubertragbaren Gastroenteritis im
Schwein stellt sie jedoch einen wichtigen Pathogenitatsfaktor von TGEV dar, wie verglei-
chende Untersuchungen mit Mutanten zeigen.
Das Bakterium E. coli F5 bindet uber seine Fimbrien an Neu5Gc-tragende Strukturen im
Intestinaltrakt von Saugferkeln. Mit dieser Bindungsaktivitat kann es den Darm koloni-
sieren und eine schwere Diarrhoe verursachen. Als Rezeptor auf den Enterozyten fungiert
das Glykolipid Neu5Gc-GM3, welches nur bei neugeborenen Schweinen vorkommt. Der
Vergleich der Sialinsaurebindungsaktivitat von TGEV und E. coli F5 ergab, dass die Ag-
glutination von Rindererythrozyten durch diese beiden Krankheitserreger von intestinalen
Muzinen und Glykolipiden unterschiedlich gehemmt wird. Dieser Vergleich fuhrte zu dem
Schluss, dass TGEV und E. coli F5 auch im Darmtrakt des Schweines unterschiedliche
sialinsauretragende Strukturen zur Bindung nutzen. Da Glykolipide die Hamagglutinati-
on durch TGEV nicht messbar hemmen konnten, wurden die weiteren Untersuchungen
der Bindungspartner von TGEV auf Sialoglykoproteine konzentriert.
135

136 7. Zusammenfassung
TGEV wurde hinsichtlich seiner Interaktion mit Zellkulturzellen, intestinalen Muzinen
und Darmepithel untersucht. Die sialinsaurebindungsdefizienten Mutanten HAD3 und
m10 (stark reduzierte Enteropathogenitat) wurden bei diesen Untersuchungen verglei-
chend eingesetzt, um die Bedeutung der Sialinsaure-bindenden Aktivitat von TGEV bes-
ser verstehen zu konnen. TGEV erkannte zusatzlich zu pAPN ein hochmolekulares muzin-
artiges Sialoglykoprotein auf der Oberflache von ST- und LLC-PK1-Zellen. Die Bindung
der Virionen an die Zelloberflache war durch die Interaktion von TGEV mit diesem Pro-
tein um ca. das Sechsfache erhoht. Dies ließ der Vergleich mit den Mutanten HAD3 und
m10 erkennen, die mangels Sialinsaurebindungsaktivitat nur an pAPN binden konnten.
Um mit diesem muzinartigen Protein interagieren zu konnen, mussten die TGEV-Partikel
jedoch von auf der Oberflache haftenden Sialoglykokonjugaten zellularen Ursprungs be-
freit werden. Durch die Behandlung mit Neuraminidase aus Vibrio cholerae konnten die
Sialinsaurebindungsstellen auf den S-Proteinen von TGEV demaskiert und die Interaktion
mit dem hochmolekularen Sialoglykoprotein moglich gemacht werden.
TGEV konnte mit intestinalen Muzinen und Burstensaummembranen der Enterozyten des
Dunndarms interagieren. Die sialinsaurebindungsdefizienten Mutanten HAD3 und m10
banden nicht an die extrazellularen Muzine und konnten nur uber die Bindung an pAPN
mit den Burstensaummembranvesikeln (BSMV) interagieren. TGEV hingegen erkannte
uber seine Sialinsaurebindungsaktivitat zusatzlich ein hochmolekulares O-glykosyliertes
Sialoglykoprotein. Die Untersuchungen zu diesem Protein weisen darauf hin, dass es sich
um ein losliches Muzin handelt, welches in den Becherzellen des Jejunums produziert wird.
Dieses Muzin prasentiert die Sialinsaurereste in optimaler Form fur TGEV, so dass selbst
unbehandeltes, d.h. mit zellularen Sialoglykokonjugaten beladenes Virus an das Muzin
bindet. Die Unterschiede zwischen Saug- und Absetzferkeln in der Zusammensetzung die-
ser Muzine und in der Interaktion mit TGEV konnten eine Rolle fur den altersabhangigen
Verlauf der Gastroenteritis spielen.
Die Sialinsaurebindungsaktivitat von TGEV stellt einen wichtigen Pathogenitatsfaktor
dar, der fur die erfolgreiche Virusreplikation im Intestinaltrakt von Schweinen von ent-
scheidender Bedeutung ist. Die Funktion der Sialinsaure-bindenden Aktivitat scheint da-
rin zu bestehen, dass die Bindung von Virionen an Muzine im Darmschleim ermoglicht
und damit die Wahrscheinlichkeit, zum Rezeptor pAPN auf der Enterozytenoberflache
gelangen zu konnen, deutlich erhoht wird.

8. Summary
Christel Schwegmann-Weßels (2002):
Importance of the sialic acid binding activity for the enteropathogenicity of
transmissible gastroenteritis coronavirus (TGEV)
TGEV is a typical member of the enteropathogenic coronaviruses. It affects pigs of all ages
and causes gastroenteritis. Infection is most severe in suckling piglets with mortality rates
up to 100%. The surface glycoprotein S of TGEV has two binding activities. Binding to the
cellular receptor aminopeptidase N (pAPN) is required for the infection of both cultured
cells and pigs. The second binding activity, the active centre of which is localized in the
N-terminal region of the S protein, enables TGEV to interact with sialoglycoconjugates.
Neu5Gc is the type of sialic acid preferentially recognized by TGEV. The sialic acid
binding activity is dispensable for the replication in cultured cells. However, studies with
mutants indicated that binding to sialoglycoconjugates is an important pathogenicity
factor for the intestinal infection resulting in transmissible gastroenteritis of swine.
Escherichia coli F5 binds via its fimbriae to Neu5Gc-carrying structures in the intestinal
tract of suckling piglets. In this way, the bacterium can colonize the gut and cause severe
diarrhea. The receptor on enterocytes has been shown to be a glycolipid, Neu5Gc-GM3,
which is present only in newborn pigs. The comparison of the sialic acid binding activity of
TGEV and E. coli F5 showed that the agglutination of bovine erythrocytes by these two
pathogens is differentially inhibited by intestinal mucins and glycolipids. From the results
it was concluded that TGEV and E. coli F5 attach to different sialylated compounds in the
intestinal tract of pigs. As glycolipids could not inhibit the hemagglutination by TGEV,
the subsequent search for ligands recognized by TGEV was focussed on sialoglycoproteins.
TGEV was examined with respect to its interaction with cultured cells, intestinal mucins
137

138 8. Summary
and the intestinal epithelium. The mutants HAD3 and m10 that lack a sialic acid binding
activity have been used for comparison to better understand the importance of the viral
sialic acid binding activity. In addition to pAPN, TGEV recognized a high molecular
weight mucin-like sialoglycoprotein on the surface of ST and LLC-PK1 cells. The inter-
action with this protein enhanced the binding of the virions to the cell surface by a factor
of 6. This was evident from the comparison with the mutants HAD3 and m10, which
only bind to pAPN. For the efficient interaction with this mucin-like glycoprotein, TGEV
particles had to be separated from surface-bound sialoglycoconjugates of cellular origin.
The treatment of virions with neuraminidase of Vibrio cholerae unmasked the sialic acid
binding sites on the S proteins of TGEV and enabled strong interaction with the high
molecular weight sialoglycoprotein.
TGEV was found to interact with intestinal mucins and brush border membranes of
enterocytes derived from the small intestine. The mutants HAD3 and m10 did not bind
to extracellular mucins and could only interact with the brush border membrane vesicles
by binding to pAPN. TGEV on the other hand recognized a high molecular weight O-
glycosylated sialoglycoprotein. Analysis of this protein indicated that it is a soluble mucin,
which is produced in the goblet cells of the jejunum. The type, number, and steric presen-
tation of the sialic acid residues on this mucin appear to be optimal for interaction with
TGEV, because even virus not treated with neuraminidase, i.e. virions containing surface
bound cellular sialoglycoproteins, are able to bind to the mucin. The differences between
suckling and weaning piglets in the composition of these mucins and in their interaction
with TGEV may play a role for the age-dependent progression of the gastroenteritis.
The sialic acid binding activity of TGEV presents an important pathogenicity factor,
which is crucial for the successful viral replication in the intestinal tract of swine. Our
results indicate that the sialic acid binding activity allows the binding of virions to mucins
in the intestinal mucus. This interaction may enable TGEV to stay longer in the intestinal
tract, to get into contact with the epithelium, and to find the receptor pAPN on the
enterocyte surface.

Literaturverzeichnis
Albritton, L. M., L. Tseng, D. Scadden und J. M. Cunningham (1989): A puta-
tive murine ecotropic retrovirus receptor gene encodes a multiple membrane-spanning
protein and confers susceptibility to virus infection. Cell 57, 659–666
Alkhatib, G., C. Combadiere, C. C. Broder, Y. Feng, P. E. Kennedy, P. M.
Murphy und E. A. Berger (1996): CC CKR5: a RANTES, MIP-1alpha, MIP-1beta
receptor as a fusion cofactor for macrophage-tropic HIV-1. Science 272, 1955–1958
Ballesteros, M. L., C. M. Sanchez, J. Martin-Caballero und L. Enjuanes
(1995): Molecular bases of tropism in the PUR46 cluster of transmissible gastroenteritis
coronaviruses. Adv. Exp. Med. Biol. 380, 557–562
Ballesteros, M. L., C. M. Sanchez und L. Enjuanes (1997): Two amino acid
changes at the N-terminus of transmissible gastroenteritis coronavirus spike protein
result in the loss of enteric tropism. Virology 227, 378–388
Baric, R. S., G. W. Nelson, J. O. Fleming, R. J. Deans, J. G. Keck, N. Casteel
und S. A. Stohlman (1988): Interactions between coronavirus nucleocapsid protein
and viral RNAs: implications for viral transcription. J. Virol. 62, 4280–4287
Benbacer, L., E. Kut, L. Besnardeau, H. Laude und B. Delmas (1997): Interspe-
cies aminopeptidase-N chimeras reveal species-specific receptor recognition by canine
coronavirus, feline infectious peritonitis virus, and transmissible gastroenteritis virus.
J. Virol. 71, 734–737
Bernard, S. und H. Laude (1995): Site-specific alteration of transmissible gastroente-
ritis virus spike protein results in markedly reduced pathogenicity. J. Gen. Virol. 76,
2235–2241
139

140 Literaturverzeichnis
Berson, J. F., D. Long, B. J. Doranz, J. Rucker, F. R. Jirik und R. W. Doms
(1996): A seven-transmembrane domain receptor involved in fusion and entry of T-cell-
tropic human immunodeficiency virus type 1 strains. J. Virol. 70, 6288–6295
Bohl, E. H. (1972): Antibody in serum, colostrum, and milk of swine after infection or
vaccination with transmissible gastroenteritis virus. Infect. Immun. 6, 289–301
Bos, E. C., W. Luytjes, H. V. van der Meulen, H. K. Koerten und W. J. Spaan
(1996): The production of recombinant infectious DI-particles of a murine coronavirus
in the absence of helper virus. Virology 218, 52–60
Brian, D. A., B. G. Hogue und T. E. Kienzle (1995): The coronavirus hemagglutinin
esterase glycoprotein. In: S. G. Siddell (Hrsg.) The Coronaviridae, Plenum Press,
New York, S. 165–179
Burness, A. T. und I. U. Pardoe (1981): Effect of enzymes on the attachment of
influenza and encephalomyocarditis viruses to erythrocytes. J. Gen. Virol. 55, 275–288
Callebaut, P., I. Correa, M. Pensaert, G. Jimenez und L. Enjuanes (1988):
Antigenic differentiation between transmissible gastroenteritis virus of swine and a re-
lated porcine respiratory coronavirus. J. Gen. Virol. 69, 1725–1730
Campadelli-Fiume, G., F. Cocchi, L. Menotti und M. Lopez (2000): The novel re-
ceptors that mediate the entry of herpes simplex viruses and animal alphaherpesviruses
into cells. Rev. Med. Virol. 10, 305–319
Carlstedt, I., J. K. Sheehan, A. P. Corfield und J. T. Gallagher (1985):
Mucous glycoproteins: a gel of a problem. Essays Biochem. 20, 40–76
Cavanagh, D. (1995): The coronavirus surface glycoprotein. In: S. G. Siddell (Hrsg.)
The Coronaviridae, Plenum Press, New York, S. 73–113
Cavanagh, D., D. A. Brian, P. Britton, L. Enjuanes, M. C. Horzinek,
M. M. C. Lai, H. Laude, P. G. Plagemann, S. G. Siddell, W. J. Spaan
und P. J. Talbot (1997): Nidovirales: a new order comprising Coronaviridae and
Arteriviridae. Arch. Virol. 142, 629–633

Literaturverzeichnis 141
Cepica, A. und J. B. Derbyshire (1984): Antibody-dependent and spontaneus cell-
mediated cytotoxicity against transmissible gastroenteritis virus infected cells by lym-
phocytes from sows, fetuses and neonatal piglets. Can. J. Comp. Med. 48, 258–261
Chang, R. Y. und D. A. Brian (1996): Cis requirement for N-specific protein sequence
in bovine coronavirus defective interfering RNA replication. J. Virol. 70, 2201–2207
Choe, H., M. Farzan, Y. Sun, N. Sullivan, B. Rollins, P. D. Ponath,
L. Wu, C. R. Mackay, G. LaRosa, W. Newman, N. Gerard, C. Gerard
und J. Sodroski (1996): The beta-chemokine receptors CCR3 and CCR5 facilitate
infection by primary HIV-1 isolates. Cell 85, 1135–1148
Cologna, R., J. F. Spagnolo und B. G. Hogue (2000): Identification of nucleocapsid
binding sites within coronavirus-defective genomes. Virology 277, 235–249
Compton, S. R., D. B. Rogers, K. V. Holmes, D. Fertsch, J. Remenick und J. J.
McGowan (1987): In vitro replication of mouse hepatitis virus strain A59. J. Virol.
61, 1814–1820
Compton, T., D. M. Nowlin und N. R. Cooper (1993): Initiation of human cytome-
galovirus infection requires initial interaction with cell surface heparan sulfate. Virology
193, 834–841
Correa, I., G. Jimenez, C. Sune, M. J. Bullido und L. Enjuanes (1988): Antigenic
structure of the E2 glycoprotein from transmissible gastroenteritis coronavirus. Virus
Res. 10, 77–93
Correa, I., F. Gebauer, M. J. Bullido, C. Sune, M. F. Baay, K. A. Zwaagstra,
W. P. Posthumus, J. A. Lenstra und L. Enjuanes (1990): Localization of anti-
genic sites of the E2 glycoprotein of transmissible gastroenteritis coronavirus. J. Gen.
Virol. 71, 271–279
Cox, E., M. B. Pensaert, P. Callebaut und K. van Deun (1990): Intestinal rep-
lication of a porcine respiratory coronavirus closely related antigenically to the enteric
transmissible gastroeneteritis virus. Vet. Microbiol. 23, 237–243

142 Literaturverzeichnis
Dalgleish, A. G., P. C. Beverley, P. R. Clapham, D. H. Crawford, M. F.
Greaves und R. A. Weiss (1984): The CD4 (T4) antigen is an essential component
of the receptor for the AIDS retrovirus. Nature 312, 763–767
Davies, H. A., R. R. Dourmashkin und M. R. Macnaughton (1981): Ribonucleo-
protein of avian infectious bronchitis virus. J. Gen. Virol. 53, 67–74
de Groot, R. J., W. Luytjes, M. C. Horzinek, B. A. van der Zeijst, W. J.
Spaan und J. A. Lenstra (1987): Evidence for a coiled-coil structure in the spike
proteins of coronaviruses. J. Mol. Biol. 196, 963–966
de Vries, A. A., M. C. Horzinek, P. J. Rottier und R. J. de Groot (1997):
The genome organisation of the nidovirales: similarities and differences between arteri-,
toro-, and coronaviruses. Seminars Virol. 8, 33–47
Deh, M. E., J. K. Dzandu und G. E. Wise (1985): Sialoglycoproteins with a high
amount of O-glycosidically linked carbohydrate moieties stain yellow with silver in
sodium dodecyl sulfate - polyacrylamide gels. Anal. Biochem. 150, 166–173
Delmas, B. und H. Laude (1990): Assembly of coronavirus spike protein into trimers
and its role in epitope expression. J. Virol. 64, 5367–5375
Delmas, B., J. Gelfi und H. Laude (1986): Antigenic structure of transmissible gastro-
enteritis virus. II. Domains in the peplomer glycoprotein. J. Gen. Virol. 67, 1405–1418
Delmas, B., D. Rasschaert, M. Godet, J. Gelfi und H. Laude (1990): Four major
antigenic sites of the coronavirus transmissible gastroenteritis virus are located on the
amino-terminal half of spike glycoprotein S. J. Gen. Virol. 71, 1313–1323
Delmas, B., J. Gelfi, R. L’Haridon, L. K. Vogel, H. Sjostrom, O. Noren
und H. Laude (1992): Aminopeptidase N is a major receptor for the enteropathogenic
coronavirus TGEV. Nature 357, 417–420
Delmas, B., J. Gelfi, H. Sjostrom, O. Noren und H. Laude (1993): Further
characterization of aminopeptidase-N as a receptor for coronaviruses. Adv. Exp. Med.
Biol. 342, 293–298

Literaturverzeichnis 143
Delmas, B., J. Gelfi, E. Kut, H. Sjostrom, O. Noren und H. Laude (1994):
Determinants essential for the transmissible gastroenteritis virus-receptor interaction
reside within a domain of aminopeptidase-N that is distinct from the enzymatic site.
J. Virol. 68, 5216–5224
Deng, H., R. Liu, W. Ellmeier, S. Choe, D. Unutmaz, M. Burkhart, P. D.
Marzio, S. Marmon, R. E. Sutton, C. M. Hill, C. B. Davis, S. C. Peiper,
T. J. Schall, D. R. Littman und N. R. Landau (1996): Identification of a major
co-receptor for primary isolates of HIV-1. Nature 381, 661–666
Doranz, B. J., J. Rucker, Y. Yi, R. J. Smyth, M. Samson, S. C. Peiper,
M. Parmentier, R. G. Collman und R. W. Doms (1996): A dual-tropic primary
HIV-1 isolate that uses fusin and the beta-chemokine receptors CKR-5, CKR-3, and
CKR-2b as fusion cofactors. Cell 85, 1149–1158
Dorig, R. E., A. Marcil, A. Chopra und C. D. Richardson (1993): The human
CD46 molecule is a receptor for measles virus (Edmonston strain). Cell 75, 295–305
Doyle, L. P. und L. M. Hutchings (1946): A transmissible gastroenteritis in pigs.
Am. J. Vet. Med. Assoc. 108, 257–259
Dragic, T., V. Litwin, G. P. Allaway, S. R. Martin, Y. Huang, K. A.
Nagashima, C. Cayanan, P. J. Maddon, R. A. Koup, J. P. Moore und W. A.
Paxton (1996): HIV-1 entry into CD4+ cells is mediated by the chemokine receptor
CC-CKR-5. Nature 381, 667–673
Dveksler, G. S., M. N. Pensiero, C. B. Cardellichio, R. K. Williams, G. S.
Jiang, K. V. Holmes und C. W. Dieffenbach (1991): Cloning of the mouse hepa-
titis virus (MHV) receptor: expression in human and hamster cell lines confers suscep-
tibility to MHV. J. Virol. 65, 6881–6891
Dveksler, G. S., C. W. Dieffenbach, C. B. Cardellichio, K. McCuaig, M. N.
Pensiero, G. S. Jiang, N. Beauchemin und K. V. Holmes (1993a): Several mem-
bers of the mouse carcinoembryonic antigen-related glycoprotein family are functional
receptors for the coronavirus mouse hepatitis virus-A59. J. Virol. 67, 1–8

144 Literaturverzeichnis
Dveksler, G. S., M. N. Pensiero, C. W. Dieffenbach, C. B. Cardellichio,
A. A. Basile, P. E. Elia und K. V. Holmes (1993b): Mouse hepatitis virus strain
A59 and blocking antireceptor monoclonal antibody bind to the N-terminal domain of
cellular receptor. Proc. Nat. Acad. Sci. USA 90, 1716–1720
Dzandu, J. K., M. E. Deh, D. L. Barratt und G. E. Wise (1984): Detection of ery-
throcyte membrane proteins, sialoglycoproteins, and lipids in the same polyacrylamide
gel using a double-staining technique. Proc. Nat. Acad. Sci. USA 81, 1733–1737
Enjuanes, L. und B. A. van der Zeijst (1995): Molecular basis of transmissible ga-
stroenteritis virus epidemiology. In: S. G. Siddell (Hrsg.) The Coronaviridae, Plenum
Press, New York, S. 337–376
Enjuanes, L., D. A. Brian, D. Cavanagh, K. V. Holmes, M. M. C. Lai,
H. Laude, P. S. Masters, P. J. Rottier, S. G. Siddell, W. J. Spaan,
F. Taguchi und P. J. Talbot (2000): Coronaviridae. In: M. H. van Regenmortel,
C. M. Fauquet, D. H. Bishop, E. B. Carsten, M. K. Estes, S. M. Lemon,
D. J. McGeoch, J. Maniloff, M. A. Mayo, C. R. Pringle und R. B. Wickner
(Hrsg.) Virus taxonomy. Classification and nomenclature of viruses., Academic Press,
New York, S. 835–849
Escors, D., E. Camafeita, J. Ortego, H. Laude und L. Enjuanes (2001a): Orga-
nization of two transmissible gastroenteritis coronavirus membrane protein topologies
within the virion and core. J. Virol. 75, 12228–12240
Escors, D., J. Ortego, H. Laude und L. Enjuanes (2001b): The membrane M
protein carboxy terminus binds to transmissible gastroenteritis coronavirus core and
contributes to core stability. J. Virol. 75, 1312–1324
Feng, Y., C. C. Broder, P. E. Kennedy und E. A. Berger (1996): HIV-1 entry co-
factor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor.
Science 272, 872–877
Fingeroth, J. D., J. J. Weis, T. F. Tedder, J. L. Strominger, P. A. Biro und
D. T. Fearon (1984): Epstein-Barr virus receptor of human B lymphocytes is the
C3d receptor CR2. Proc. Nat. Acad. Sci. USA 81, 4510–4514

Literaturverzeichnis 145
Fischer, F., C. F. Stegen, P. S. Masters und W. A. Samsonoff (1998): Analysis
of constructed E gene mutants of mouse hepatitis virus confirms a pivotal role for E
protein in coronavirus assembly. J. Virol. 72, 7885–7894
Frade, R., M. Barel, B. Ehlin-Henriksson und G. Klein (1985): gp140, the C3d
receptor of human B lymphocytes, is also the Epstein-Barr virus receptor. Proc. Nat.
Acad. Sci. USA 82, 1490–1493
Fried, H., L. D. Cahan und J. C. Paulson (1981): Polyoma virus recognizes specific
sialyloligosaccharide receptors on host cells. Virology 109, 188–192
Gaastra, W. und F. K. de Graaf (1982): Host-specific fimbrial adhesins of noninva-
sive enterotoxigenic Escherichia coli strains. Microbiol. Rev. 46, 129–161
Gagneten, S., O. Gout, M. Dubois-Dalcq, P. J. Rottier, J. Rossen und K. V.
Holmes (1995): Interaction of mouse hepatitis virus (MHV) spike glycoprotein with
receptor glycoprotein MHVR is required for infection with an MHV strain that expresses
the hemagglutinin-esterase glycoprotein. J. Virol. 69, 889–895
Gallagher, J. T., A. Morris und T. M. Dexter (1985): Identification of two binding
sites for wheat-germ agglutinin on polylactosamine-type oligosaccharides. Biochem. J.
231, 115–122
Garabal, J. I., F. Vazquez, J. Blanco, M. Blanco und E. A. Gonzalez (1997):
Colonization antigens of enterotoxigenic Escherichia coli strains isolated from piglets
in Spain. Vet. Microbiol. 54, 321–328
Garwes, D. J. (1995): Pathogenesis of the porcine coronaviruses. In: S. G. Siddell
(Hrsg.) The Coronaviridae, Plenum Press, New York, S. 377–388
Gebauer, F., W. P. Posthumus, I. Correa, C. Sune, C. Smerdou, C. M.
Sanchez, J. A. Lenstra, R. H. Meloen und L. Enjuanes (1991): Residues in-
volved in the antigenic sites of transmissible gastroenteritis coronavirus S glycoprotein.
Virology 183, 225–238
Godet, M., R. L’Haridon, J. F. Vautherot und H. Laude (1992): TGEV corona
virus ORF4 encodes a membrane protein that is incorporated into virions. Virology
188, 666–675

146 Literaturverzeichnis
Godet, M., J. Grosclaude, B. Delmas und H. Laude (1994): Major receptor-
binding and neutralization determinants are located within the same domain of the
transmissible gastroenteritis virus (coronavirus) spike protein. J. Virol. 68, 8008–8016
Gottschalk, A. (1958): The influenza virus neuraminidase. Nature 181, 377–378
Greve, J. M., G. Davis, A. M. Meyer, C. P. Forte, S. C. Yost, C. W. Marlor,
M. E. Kamarck und A. McClelland (1989): The major human rhinovirus receptor
is ICAM-1. Cell 56, 839–847
Guinee, P. A., W. H. Jansen und C. M. Agterberg (1976): Detection of the K99 an-
tigen by means of agglutination and immunoelectrophoresis in Escherichia coli isolates
from calves and its correlation with entertoxigenicity. Infect. Immun. 13, 1369–1377
Hansen, G. H., B. Delmas, L. Besnardeau, L. K. Vogel, H. Laude,
H. Sjostrom und O. Noren (1998): The coronavirus transmissible gastroenteritis
virus causes infection after receptor-mediated endocytosis and acid-dependent fusion
with an intracellular compartment. J. Virol. 72, 527–534
Hara, S., M. Yamaguchi, K. Furuhata, H. Ogura und M. Nakamura (1989):
Determination of mono-O-acetylated N-acetylneuraminic acids in human and rat sera
by fluorometric High-Performance Liquid Chromatography. Anal. Biochem. 179, 162–
166
Haselbeck, A., E. Schickaneder, H. von der Eltz und W. Hosel (1990): Struc-
tural characterization of glycoprotein carbohydrate chains by using digoxigenin-labeled
lectins on blots. Anal. Biochem. 191, 25–30
Hiscox, J. A., T. Wurm, L. Wilson, P. Britton, D. Cavanagh und G. Brooks
(2001): The coronavirus infectious bronchitis virus nucleoprotein localizes to the nucleo-
lus. J. Virol. 75, 506–512
Hooper, B. E. und E. O. Haelterman (1969): Lesions of the gastrointestinal tract
of pigs infected with transmissible gastroenteritis. Can. J. Comp. Med. 33, 29–36
Jimenez, G., I. Correa, M. P. Melgosa, M. J. Bullido und L. Enjuanes (1986):
Critical epitopes in transmissible gastroenteritis virus neutralization. J. Virol. 60, 131–
139

Literaturverzeichnis 147
Jones, T. O. und D. J. Paton (1996): Classical transmissible gastroenteritis returns.
Vet. Rec. 138, 166–167
Kim, K. H., K. Narayanan und S. Makino (1997): Assembled coronavirus from com-
plementation of two defective interfering RNAs. J. Virol. 71, 3922–3931
Klatzmann, D., E. Champagne, S. Chamaret, J. Gruest, D. Guetard,
T. Hercend, J. C. Gluckman und L. Montagnier (1984): T-lymphocyte T4 mo-
lecule behaves as the receptor for human retrovirus LAV. Nature 312, 767–768
Klenk, E., H. Faillard und H. Lempfried (1955): Uber die enzymatische Wirkung
von Influenza-Virus. Z. Physiol. Chem. 301, 235–246
Klumperman, J., J. Krijnse-Locker, A. Meijer, M. C. Horzinek, H. J. Geuze
und P. J. Rottier (1994): Coronavirus M proteins accumulate in the Golgi complex
beyond the site of virion budding. J. Virol. 68, 6523–6534
Kolb, A. F., J. Maile, A. Heister und S. G. Siddell (1996): Characterization of
functional domains in the human coronavirus HCV 229E receptor. J. Gen. Virol. 77,
2515–2521
Krempl, C. (1998): Funktionelle Analyse des Oberflachenproteins S des Virus der
ubertragbaren Gastroenteritis . Dissertation, Fachbereich Humanmedizin der Philipps-
Universitat Marburg
Krempl, C. und G. Herrler (2001): Sialic acid binding activity of transmissible ga-
stroenteritis coronavirus affects sedimentation behavior of virions and solubilized gly-
coproteins. J. Virol. 75, 844–849
Krempl, C., B. Schultze und G. Herrler (1995): Analysis of cellular receptors for
human coronavirus OC43. Adv. Exp. Med. Biol. 380, 371–374
Krempl, C., B. Schultze, H. Laude und G. Herrler (1997): Point mutations in
the S protein connect the sialic acid binding activity with the enteropathogenicity of
transmissible gastroenteritis coronavirus. J. Virol. 71, 3285–3287

148 Literaturverzeichnis
Krempl, C., M. L. Ballesteros, L. Enjuanes und G. Herrler (1998): Isolation of
hemagglutination-defective mutants for the analysis of the sialic acid binding activity
of transmissible gastroenteritis virus. Adv. Exp. Med. Biol. 440, 563–568
Krempl, C., M. L. Ballesteros, G. Zimmer, L. Enjuanes, H. D. Klenk und
G. Herrler (2000): Characterization of the sialic acid binding activity of transmissible
gastroenteritis coronavirus by analysis of haemagglutination-deficient mutants. J. Gen.
Virol. 81, 489–496
Krijnse-Locker, J., M. Ericsson, P. J. Rottier und G. Griffiths (1994): Cha-
racterization of the budding compartment of mouse hepatitis virus: evidence that trans-
port from the RER to the Golgi complex requires only one vesicular transport step.
J. Cell Biol. 124, 55–70
Krijnse-Locker, J., D. J. Opstelten, M. Ericsson, M. C. Horzinek und P. J.
Rottier (1995): Oligomerization of a trans-Golgi/trans-Golgi network retained protein
occurs in the Golgi complex and may be part of its retention. J. Biol. Chem. 270, 8815–
8821
Kunkel, F. und G. Herrler (1993): Structural and functional analysis of the surface
protein of human coronavirus OC43. Virology 195, 195–202
Kyhse-Anderson, J. (1984): Electroblotting of mutiple gels: a simple apparatus with-
out buffer tank for rapid transfer of proteins from polyacrylamide to nitrocellulose.
J. Biochem. Biophys. Methods 10, 203–209
Laemmli, U. K. (1970): Cleavage of structural proteins during the assembly of the head
of bacteriophage T4. Nature 227, 680–685
Lai, M. M. C. und D. Cavanagh (1997): The molecular biology of coronaviruses. Adv.
Virus Res.. 48, 1–100
Lapham, C. K., J. Ouyang, B. Chandrasekhar, N. Y. Nguyen, D. S. Dimitrov
und H. Golding (1996): Evidence for cell-surface association between fusin and the
CD4-gp120 complex in human cell lines. Science 274, 602–605

Literaturverzeichnis 149
Lapps, W., B. G. Hogue und D. A. Brian (1987): Sequence analysis of the bovine
coronavirus nucleocapsid and matrix protein genes. Virology 157, 47–57
Laude, H. und P. S. Masters (1995): The coronavirus nucleocapsid protein. In: S. G.
Siddell (Hrsg.) The Coronaviridae, Plenum Press, New York, S. 141–163
Laude, H., D. Rasschaert und J. C. Huet (1987): Sequence and N-terminal proces-
sing of the transmembrane protein E1 of the coronavirus transmissible gastroenteritis
virus. J. Gen. Virol. 68, 1687–1693
Laude, H., D. Rasschaert, B. Delmas, M. Godet, J. Gelfi und B. Charley
(1990): Molecular biology of transmissible gastroenteritis virus. Vet. Microbiol. 23,
147–154
Lindahl, M. und I. Carlstedt (1990): Binding of K99 fimbriae of enterotoxigenic
Escherichia coli to pig small intestinal mucin glycopeptides. J. Gen. Microbiol. 136,
1609–1614
Lindahl, M., R. Brossmer und T. Waldstrom (1987): Carbohydrate receptor spe-
cificity of K99 fimbriae of enterotoxigenic Escherichia coli. Glycoconjugate J. 4, 51–58
Lisanti, M. P., M. Sargiacomo, L. Graeve, A. R. Saltiel und E. Rodriguez-
Boulan (1988): Polarized apical distribution of glycosyl-phosphatidylinositol-anchored
proteins in a renal epithelial cell line. Proc. Nat. Acad. Sci. USA 85, 9557–9561
Liu, D. X. und S. C. Inglis (1991): Association of the infectious bronchitis virus 3c
protein with the virion envelope. Virology 185, 911–917
Macnaughton, M. R., H. A. Davies und M. V. Nermut (1978): Ribonucleoprotein-
like structure from coronavirus particles. J. Gen. Virol. 39, 545–549
Masters, P. S. (1992): Localization of an RNA-binding domain in the nucleocapsid
protein of the coronavirus mouse hepatitis virus. Arch. Virol. 125, 141–160
McClurkin, A. W. und J. O. Norman (1966): Studies on transmissible gastroenteritis
of swine. II. Selected characteristics of a cytopathogenic virus common to five isolates
from transmissible gastroenteritis. Can. J. Comp. Med. 30, 190–198

150 Literaturverzeichnis
McGoldrick, A., J. P. Lowings und D. J. Paton (1999): Characterisation of a
recent virulent transmissible gastroenteritis virus from Britain with a deleted ORF 3a.
Arch. Virol. 144, 763–770
Mendelsohn, C. L., E. Wimmer und V. R. Racaniello (1989): Cellular receptor
for poliovirus: molecular cloning, nucleotide sequence, and expression of a new member
of the immunoglobulin superfamily. Cell 56, 855–865
Merril, C. R., R. C. Switzer und M. L. van Keuren (1979): Trace polypeptides
in cellular extracts and human body fluids detected by two-dimensional electrophoresis
and a highly sensitive silver stain. Proc. Nat. Acad. Sci. USA 76, 4335–4339
Merril, C. R., D. Goldman, S. A. Sedman und M. H. Ebert (1981): Ultrasensitive
stain for proteins in polyacrylamide gels shows regional variation in cerebrospinal fluid
proteins. Science 211, 1437–1438
Molenkamp, R., H. van Tol, B. C. Rozier, Y. van der Meer, W. J. Spaan und
E. J. Snijder (2000): The arterivirus replicase is the only viral protein required for
genome replication and subgenomic mRNA transcription. J. Gen. Virol. 81, 2491–2496
Moon, H. W. (1973): Epithelial cell migration in the alimentary mucosa of the suckling
pig. Proc. Soc. Exp. Biol. Med. 137, 151–154
Mouricout, M., J. M. Petit, J. R. Carias und R. Julien (1990): Glycoprotein
glycans that inhibit adhesion of Escherichia coli mediated by K99 fimbriae: treatment
of experimental colibacillosis. Infect. Immun. 58, 98–106
Naniche, D., G. Varior-Krishnan, F. Cervoni, T. F. Wild, B. Rossi,
C. Rabourdin-Combe und D. Gerlier (1993): Human membrane cofactor protein
(CD46) acts as a cellular receptor for measles virus. J. Virol. 67, 6025–6032
Narayanan, K., A. Maeda, J. Maeda und S. Makino (2000): Characterization of
the coronavirus M protein and nucleocapsid interaction in infected cells. J. Virol. 74,
8127–8134
Nelson, G. W. und S. A. Stohlman (1993): Localization of the RNA-binding domain
of mouse hepatitis virus nucleocapsid protein. J. Gen. Virol. 74, 1975–1979

Literaturverzeichnis 151
Nelson, G. W., S. A. Stohlman und S. M. Tahara (2000): High affinity interaction
between nucleocapsid protein and leader/intergenic sequence of mouse hepatitis virus
RNA. J. Gen. Virol. 81, 181–188
Nemerow, G. R., R. Wolfert, M. E. McNaughton und N. R. Cooper (1985):
Identification and characterization of the Epstein-Barr virus receptor on human B lym-
phocytes and its relationship to the C3d complement receptor (CR2). J. Virol. 55,
347–351
Nguyen, V. P. und B. G. Hogue (1997): Protein interactions during coronavirus
assembly. J. Virol. 71, 9278–9284
Noda, M., H. Yamashita, F. Koide, K. Kadoi, T. Omori, M. Asagi und Y. Inaba
(1987): Hemagglutination with transmissible gastroenteritis virus. Arch. Virol. 96, 109–
115
Noda, M., F. Koide, M. Asagi und Y. Inaba (1988): Physicochemical properties of
transmissible gastroenteritis virus hemagglutinin. Arch. Virol. 99, 163–172
O’Hara, B., S. V. Johann, H. P. Klinger, D. G. Blair, H. Rubinson, K. J.
Dunn, P. Sass, S. M. Vitek und T. Robins (1990): Characterization of a human
gene conferring sensitivity to infection by gibbon ape leukemia virus. Cell. Growth
Differ. 1, 119–127
Ono, E., K. Abe, M. Nakazawa und M. Naiki (1989): Ganglioside epitope recognized
by K99 fimbriae from enterotoxigenic Escherichia coli. Infect. Immun. 57, 907–911
Opstelten, D. J., M. J. Raamsman, K. Wolfs, M. C. Horzinek und P. J.
Rottier (1995): Envelope glycoprotein interactions in coronavirus assembly. J. Cell.
Biol. 131, 339–349
Parker, M. M. und P. S. Masters (1990): Sequence comparison of the N genes of
five strains of the coronavirus mouse hepatitis virus suggests a three domain structure
for the nucleocapsid protein. Virology 179, 463–468
Paulson, J. C., J. E. Sadler und R. L. Hill (1979): Restoration of specific myxovirus
receptors to asialoerythrocytes by incorporation of sialic acid with pure sialyltransfer-
ases. J. Biol. Chem. 254, 2120–2124

152 Literaturverzeichnis
Pensaert, M. B. und E. Cox (1989): Porcine respiratory coronavirus related to trans-
missible gastroenteritis virus. AGRI-PRACTICE 10, Nr. 2, 17–21
Pensaert, M. B., E. O. Haelterman und T. Burnstein (1970): Transmissible ga-
stroenteritis of swine: virus-intestinal cell interactions. I. Immunofluorescence, histopa-
thology and virus reproduction in the small intestine through the course of the infection.
Arch. gesamte Virusforsch. 31, 321–334
Pensaert, M. B., P. Callebaut und J. Vergote (1986): Isolation of a porcine
respiratory, non-enteric coronavirus related to transmissible gastroenteritis. Vet. Q. 8,
257–261
Pringle, D. A. (1992): Committee persues medly of virus taxonomic issues. ASM News
58, 475–476
Rasschaert, D. und H. Laude (1987): The predicted primary structure of the peplomer
protein E2 of the porcine coronavirus transmissible gastroenteritis virus. J. Gen. Virol.
68, 1883–1890
Rasschaert, D., M. Duarte und H. Laude (1990): Porcine respiratory coronavirus
differs from transmissible gastroenteritis virus by a few genomic deletions. J. Gen.
Virol. 71, 2599–2607
Reuter, G., S. Stoll, J. P. Kamerling, J. F. Vliegenthart und R. Schauer
(1988): Sialic acids on erythrocytes and in blood plasma of mammals. In: R. Schauer
und T. Yamakawa (Hrsg.) Sialic Acids 1988, Kieler Verlag Wissenschaft und Bildung,
Kiel, Germany, S. 88–89. Proceedings of the Japanese-German Symposium on sialic
acids
Reynolds, D. J. und D. J. Garwes (1979): Virus isolation and serum antibody respon-
ses after infection of cats with transmissible gastroenteritis virus. Brief report. Arch.
Virol. 60, 161–166
Risco, C., I. M. Anton, C. Sune, A. M. Pedregosa, J. M. Martin-Alonso,
F. Parra, J. L. Carrascosa und L. Enjuanes (1995): Membrane protein molecules
of transmissible gastroenteritis coronavirus also expose the carboxy-terminal region on
the external surface of the virion. J. Virol. 69, 5269–5277

Literaturverzeichnis 153
Risco, C., I. M. Anton, L. Enjuanes und J. L. Carrascosa (1996): The transmis-
sible gastroenteritis coronavirus contains a spherical core shell consisting of M and N
proteins. J. Virol. 70, 4773–4777
Rogers, G. N. und J. C. Paulson (1983): Receptor determinants of human and animal
influenza virus isolates: differences in receptor specificity of the H3 hemagglutinin based
on species of origin. Virology 127, 361–373
Rogers, G. N., G. Herrler, J. C. Paulson und H. D. Klenk (1986): Influenza C
virus uses 9-O-acetyl-N-acetylneuraminic acid as a high affinity receptor determinant
for attachment to cells. J. Biol. Chem. 261, 5947–5951
Rottier, P. J. (1995): The coronavirus membrane glycoprotein. In: S. G. Siddell
(Hrsg.) The Coronaviridae, Plenum Press, New York, S. 115–139
Rottier, P. J., G. W. Welling, S. Welling-Wester, H. G. Niesters, J. A.
Lenstra und B. A. van der Zeijst (1986): Predicted membrane topology of the
coronavirus protein E1. Biochemistry 25, 1335–1339
Roussel, P. und G. Lamblin (1996): Human mucosal mucins in diseases. In:
J. Montreuil, J. F. Vliegenthart und H. Schachter (Hrsg.) Glycoproteins
and disease. Elsevier Science, Amsterdam, S. 351–393
Saif, L. J. (1990): Comparative aspects of enteric viral infections. In: L. J. Saif und
K. W. Theil (Hrsg.) Viral diarrheas of man and animals. CRC Press, Boca Raton,
Florida, S. 10–31
Sanchez, C. M., G. Jimenez, M. D. Laviada, I. Correa, C. Sune, M. J. Bullido,
F. Gebauer, C. Smerdou, P. Callebaut, J. M. Escribano und L. Enjuanes
(1990): Antigenic homology among coronaviruses related to transmissible gastroenteritis
virus. Virology 174, 410–417
Sanchez, C. M., F. Gebauer, C. Sune, A. Mendez, J. Dopazo und L. Enjuanes
(1992): Genetic evolution and tropism of transmissible gastroenteritis coronaviruses.
Virology 190, 92–105

154 Literaturverzeichnis
Sanchez, C. M., A. Izeta, J. M. Sanchez-Morgado, S. Alonso, I. Sola,
M. Balasch, J. Plana-Duran und L. Enjuanes (1999): Targeted recombinati-
on demonstrates that the spike gene of transmissible gastroenteritis coronavirus is a
determinant of its enteric tropism and virulence. J. Virol. 73, 7607–7618
Sastry, M. V., P. Banarjee, S. R. Patanjali, M. J. Swamy, G. V. Swarnalatha
und A. Surolia (1986): Analysis of saccharide binding to Artocarpus integrifolia lectin
reveals specific recognition of T-antigen (beta-D-Gal(1–3)D-GalNAc). J. Biol. Chem.
261, 11726–11733
Schauer, R. (1987): Analysis of sialic acids. Methods Enzymol. 138, 132–161
Schultze, B. und G. Herrler (1992): Bovine coronavirus uses N-acetyl-9-O-acetyl-
neuraminic acid as a receptor determinant to initiate the infection of cultured cells.
J. Gen. Virol. 73, 901–906
Schultze, B., H. J. Gross, R. Brossmer, H. D. Klenk und G. Herrler (1990):
Hemagglutinating encephalomyelitis virus attaches to N-acetyl-9-O-acetylneuraminic
acid-containing receptors on erythrocytes: comparison with bovine coronavirus and in-
fluenza C virus. Virus Res. 16, 185–194
Schultze, B., H. J. Gross, R. Brossmer und G. Herrler (1991a): The S protein
of bovine coronavirus is a hemagglutinin recognizing 9-O-acetylated sialic acid as a
receptor determinant. J. Virol. 65, 6232–6237
Schultze, B., K. Wahn, H. D. Klenk und G. Herrler (1991b): Isolated HE-protein
from hemagglutinating encephalomyelitis virus and bovine coronavirus has receptor-
destroying and receptor-binding activity. Virology 180, 221–228
Schultze, B., L. Enjuanes, D. Cavanagh und G. Herrler (1993): N-acetyl-
neuraminic acid plays a critical role for the haemagglutinating activity of avian infec-
tious bronchitis virus and porcine transmissible gastroenteritis virus. Adv. Exp. Med.
Biol. 342, 305–310
Schultze, B., L. Enjuanes und G. Herrler (1995): Analysis of the sialic acid-binding
activity of the transmissible gastroenteritis virus. Adv. Exp. Med. Biol. 380, 367–370

Literaturverzeichnis 155
Schultze, B., C. Krempl, M. L. Ballesteros, L. Shaw, R. Schauer,
L. Enjuanes und G. Herrler (1996): Transmissible gastroenteritis coronavirus, but
not the related porcine respiratory coronavirus, has a sialic acid (N-glycolylneuraminic
acid) binding activity. J. Virol. 70, 5634–5637
Seignole, D., M. Mouricout, Y. Duval-Iflah, B. Quintard und R. Julien
(1991): Adhesion of K99 fimbriated Escherichia coli to pig intestinal epithelium: corre-
lation of adhesive and non-adhesive phenotypes with the sialoglycolipid content. J. Gen.
Microbiol. 137, 1591–1601
Shepley, M. P. und V. R. Racaniello (1994): A monoclonal antibody that blocks
poliovirus attachment recognizes the lymphocyte homing receptor CD44. J. Virol. 68,
1301–1308
Siddell, S. G. (1995a): The coronaviridae: An introduction. In: S. G. Siddell (Hrsg.)
The Coronaviridae, Plenum Press, New York, S. 1–10
Siddell, S. G. (1995b): The small-membrane protein. In: S. G. Siddell (Hrsg.) The
Coronaviridae, Plenum Press, New York, S. 181–189
Siddell, S. G., R. Anderson, D. Cavanagh, K. Fujiwara, H. D. Klenk, M. R.
Macnaughton, M. Pensaert, S. A. Stohlman, L. Sturman und B. A. van der
Zeijst (1983): Coronaviridae. Intervirology 20, 181–189
Smith, H. C. (1956): Advances made in swine practice. IX. Transmissible gastroenteritis.
Vet. Med. 51, 425–429
Stamm, M. und I. Sorg (1993): Intestinale Rezeptoren fur adhasive Fimbrien von Esche-
richia coli beim Schwein — eine Literaturubersicht. Schweiz. Arch. Tierheilkd. 135,
89–95
Staunton, D. E., V. J. Merluzzi, R. Rothlein, R. Barton, S. D. Marlin und
T. A. Springer (1989): A cell adhesion molecule, ICAM-1, is the major surface re-
ceptor for rhinoviruses. Cell 56, 849–853
Stohlman, S. A., R. S. Baric, G. N. Nelson, L. H. Soe, L. M. Welter und R. J.
Deans (1988): Specific interaction between coronavirus leader RNA and nucleocapsid
protein. J. Virol. 62, 4288–4295

156 Literaturverzeichnis
Sueyoshi, S., T. Tsuji und T. Osawa (1988): Carbohydrate-binding specificities of
five lectins that bind to O-glycosyl-linked carbohydrate chains. Quantitative analysis
by frontal-affinity chromatography. Carbohydr. Res. 178, 213–224
Tahara, S. M., T. A. Dietlin, C. C. Bergmann, G. W. Nelson, S. Kyuwa, R. P.
Anthony und S. A. Stohlman (1994): Coronavirus translational regulation: leader
affects mRNA efficiency. Virology 202, 621–630
Tajima, M. (1970): Morphology of transmissible gastroenteritis virus of pigs. Arch.
gesamte Virusforsch. 29, 105–108
Teneberg, S., P. Willemsen, F. K. de Graaf und K. A. Karlsson (1990):
Receptor-active glycolipids of epithelial cells of the small intestine of young and adult
pigs in relation to susceptibility to infection with Escherichia coli K99. FEBS Lett.
263, 10–14
Teneberg, S., P. T. Willemsen, F. K. de Graaf, G. Stenhagen, W. Pimlott,
P. A. Jovall, J. Angstrom und K. A. Karlsson (1994): Characterization of
gangliosides of epithelial cells of calf small intestine, with special reference to receptor-
active sequences for enteropathogenic Escherichia coli K99. J. Biochem. (Tokyo) 116,
560–574
Tomassini, J. E., D. Graham, C. M. DeWitt, D. W. Lineberger, J. A. Rodkey
und R. J. Colonno (1989): cDNA cloning reveals that the major group rhinovirus
receptor on HeLa cells is intercellular adhesion molecule 1. Proc. Nat. Acad. Sci. USA
86, 4907–4911
Tresnan, D. B., R. Levis und K. V. Holmes (1996): Feline aminopeptidase N serves
as a receptor for feline, canine, porcine, and human coronaviruses in serogroup I.
J. Virol. 70, 8669–8674
Trkola, A., T. Dragic, J. Arthos, J. M. Binley, W. C. Olson, G. P. Allaway,
C. Cheng-Mayer, J. Robinson, P. J. Maddon und J. P. Moore (1996): CD4-
dependent, antibody-sensitive interactions between HIV-1 and its co-receptor CCR-5.
Nature 384, 184–187

Literaturverzeichnis 157
Tyrrell, D. A., J. D. Almeida, D. M. Berry, C. H. Cunningham, D. Hamre,
M. S. Hofstad, L. Mallucci und K. McIntosh (1968): Coronaviruses. Nature
220, 650
Tyrrell, D. A., J. D. Almeida, C. H. Cunningham, W. R. Dowdle, M. S.
Hofstad, K. . McIntosh, M. Tajima, L. Y. Zakstelskaya, B. C. Easterday,
A. Kapikian und R. W. Bingham (1975): Coronaviridae. Intervirology 5, 76–82
van der Most, R. G. und W. J. Spaan (1995): Coronavirus replication, transcription,
and RNA recombination. In: S. G. Siddell (Hrsg.) The Coronaviridae, Plenum Press,
New York, S. 11–31
Vennema, H., L. Heijnen, A. Zijderveld, M. C. Horzinek und W. J. Spaan
(1990): Intracellular transport of recombinant coronavirus spike proteins: implications
for virus assembly. J. Virol. 64, 339–346
Vennema, H., G. J. Godeke, J. W. Rossen, W. F. Voorhout, M. C. Horzinek,
D. J. Opstelten und P. J. Rottier (1996): Nucleocapsid-independent assembly of
coronavirus-like particles by co-expression of viral envelope protein genes. EMBO J.
15, 2020–2028
Vlasak, R., W. Luytjes, J. Leider, W. J. Spaan und P. Palese (1988a): The
E3 protein of bovine coronavirus is a receptor-destroying enzyme with acetylesterase
activity. J. Virol. 62, 4686–4690
Vlasak, R., W. Luytjes, W. J. Spaan und P. Palese (1988b): Human and bovine
coronaviruses recognize sialic acid-containing receptors similar to those of influenza C
viruses. Proc. Nat. Acad. Sci. USA 85, 4526–4529
Wagner, J. E., P. D. Beamer und M. Ristic (1973): Electron microscopy of intestinal
epithelial cells of piglets infected with a transmissible gastroenteritis virus. Can. J.
Comp. Med. 37, 177–188
Wang, W. C. und R. D. Cummings (1988): The immobilized leukoagglutinin from the
seeds of Maackia amurensis binds with high affinity to complex-type Asn-linked oli-
gosaccharides containing terminal sialic acid-linked alpha-2,3 to penultimate galactose
residues. J. Biol. Chem. 263, 4576–4585

158 Literaturverzeichnis
Weingartl, H. M. und J. B. Derbyshire (1993): Binding of porcine transmissible
gastroenteritis virus by enterocytes from newborn and weaned piglets. Vet. Microbiol.
35, 23–32
Weingartl, H. M. und J. B. Derbyshire (1994): Evidence for a putative second
receptor for porcine transmissible gastroenteritis virus on the villous enterocytes of
newborn pigs. J. Virol. 68, 7253–7259
Weis, W., J. H. Brown, S. Cusack, J. C. Paulson, J. J. Skehel und D. C.
Wiley (1988): Structure of the influenza virus haemagglutinin complexed with its
receptor, sialic acid. Nature 333, 426–431
Wesley, R. D., R. D. Woods und A. K. Cheung (1991): Genetic analysis of porcine
respiratory coronavirus, an attenuated variant of transmissible gastroenteritis virus.
J. Virol. 65, 3369–3373
Williams, R. K., G. S. Jiang und K. V. Holmes (1991): Receptor for mouse hepatitis
virus is a member of the carcinoembryonic antigen family of glycoproteins. Proc. Nat.
Acad. Sci. USA 88, 5533–5536
Woode, G. N. (1969): Transmissible gastroenteritis of swine. Vet. Bull. 39, 239–248
Wu, L., N. P. Gerard, R. Wyatt, H. Choe, C. Parolin, N. Ruffing,
A. Borsetti, A. A. Cardoso, E. Desjardin, W. Newman, C. Gerard und
J. Sodroski (1996): CD4-induced interaction of primary HIV-1 gp120 glycoproteins
with the chemokine receptor CCR-5. Nature 384, 179–183
WuDunn, D. und P. G. Spear (1989): Initial interaction of herpes simplex virus with
cells is binding to heparan sulfate. J. Virol. 63, 52–58
Wurm, T., H. Chen, T. Hodgson, P. Britton, G. Brooks und J. A. Hiscox
(2001): Localization to the nucleolus is a common feature of coronavirus nucleoproteins,
and the protein may disrupt host cell division. J. Virol. 75, 9345–9356
Yeager, C. L., R. A. Ashmun, R. K. Williams, C. B. Cardellichio, L. H.
Shapiro, A. T. Look und K. V. Holmes (1992): Human aminopeptidase N is a
receptor for human coronavirus 229E. Nature 357, 420–422

Literaturverzeichnis 159
Yokomori, K., M. Asanaka, S. A. Stohlman und M. M. C. Lai (1993): A spike
protein-dependent cellular factor other than the viral receptor is required for mouse
hepatitis virus entry. Virology 196, 45–56
Yu, X., W. Bi, S. R. Weiss und J. L. Leibowitz (1994): Mouse hepatitis virus gene
5b protein is a new virion envelope protein. Virology 202, 1018–1023
Zhou, M., A. K. Williams, S. I. Chung, L. Wang und E. W. Collisson (1996): The
infectious bronchitis virus nucleocapsid protein binds RNA sequences in the 3’ terminus
of the genome. Virology 217, 191–199


Abbildungsverzeichnis
1.1 Schematische Darstellung eines Coronaviruspartikels . . . . . . . . . . . . . 19
1.2 Schematische Darstellung der Epitope des S-Proteins . . . . . . . . . . . . 30
5.1 Neuraminidase-Sensitivitat der Erythrozytenrezeptoren . . . . . . . . . . . 79
5.2 Qualitative und quantitative Analyse des Sialinsauregehalts intestinaler
Muzine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82
5.3 Interaktion von TGEV mit Zellkulturen . . . . . . . . . . . . . . . . . . . 85
5.4 Bindung an Membranproteine . . . . . . . . . . . . . . . . . . . . . . . . . 87
5.5 Bindung an Zelloberflachenproteine . . . . . . . . . . . . . . . . . . . . . . 89
5.6 Bindung von isoliertem S-Protein an Zelloberflachenproteine . . . . . . . . 90
5.7 Bindung nicht desialylierter Virionen an Zelloberflachenproteine . . . . . . 91
5.8 Oberflachenproteine von ST-Zellen . . . . . . . . . . . . . . . . . . . . . . 92
5.9 Sialinsaureabhangigkeit der Bindung von TGEV-NA an ST-Zellen . . . . . 93
5.10 Charakterisierung der Assoziation des Sialoglykoproteins mit der Plasma-
membran mittels Triton X-114-Phasenseparierung . . . . . . . . . . . . . 94
5.11 Spezifische Bindung von TGEV an das hochmolekulare Sialoglykoprotein . 95
5.12 Analyse der Sialinsauren der Muzine aus der Glandula submandibularis des
Schweines . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 97
5.13 Interaktion von TGEV mit immobilisierten Muzinen . . . . . . . . . . . . . 98
161

162 Abbildungsverzeichnis
5.14 Sialinsaureabhangigkeit der Bindung von TGEV an intestinale Muzine . . 99
5.15 Bindung an intestinale Muzine . . . . . . . . . . . . . . . . . . . . . . . . . 100
5.16 Bindung an Muzine des Ferkels B und PSM . . . . . . . . . . . . . . . . . 101
5.17 Qualitative Analyse des Sialinsauregehalts von BSMV . . . . . . . . . . . . 104
5.18 Bindung von TGEV-NA an Proteine aus BSMV . . . . . . . . . . . . . . . 105
5.19 Bindung von HAD3, m10 und TGEV an Proteine aus BSMV . . . . . . . . 107
5.20 Charakterisierung der Assoziation des Proteins mit der Burstensaummem-
bran mittels Triton X-114-Phasenseparierung . . . . . . . . . . . . . . . . 108
5.21 Nachweis des Kohlenhydratanteils in BSMV-Proteinen . . . . . . . . . . . 109
5.22 Erkennung von BSMV-Proteinen durch PNA . . . . . . . . . . . . . . . . . 110
5.23 Bindung von TGEV-NA an mit Lektinen gefallte BSMV-Proteine . . . . . 111
5.24 Bindung von TGEV-NA an Dunndarmepithel . . . . . . . . . . . . . . . . 114
5.25 Bindung von TGEV und Mutanten an Dunndarmepithel . . . . . . . . . . 115

Tabellenverzeichnis
1.1 Serologische und phylogenetische Verwandschaft der Coronaviren . . . . . . 19
1.2 Coronaviren . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
1.3 Ubersicht uber Eigenschaften der Epitop-D-Mutanten . . . . . . . . . . . . 32
1.4 Ubersicht uber bekannte Virusrezeptoren (Beispiele) . . . . . . . . . . . . . 36
5.1 Agglutination von Erythrozyten verschiedener Tierarten . . . . . . . . . . 78
5.2 Vergleich der Hemmaktivitaten von intestinalen Muzinen und Glykolipiden 83
5.3 Enzymanreicherungen durch die BSMV-Prapararation . . . . . . . . . . . . 102
163

Veroffentlichungen und Tagungsbeitrage
Teile der vorliegenden Arbeit wurden in folgenden Publikationsorganen veroffentlicht:
Schwegmann, C., G. Zimmer, T. Yoshino, M.-L. Enss und G. Herrler (2001):
Comparison of the sialic acid binding activity of transmissible gastroenteritis coronavirus
and E. coli K99. Virus Res. 75, 69–73.
Schwegmann, C., G. Zimmer und G. Herrler (2001):
Are intestinal mucins involved in the pathogenicity of transmissible gastroenteritis coro-
navirus? Adv. Exp. Med. Biol. 494, 219–223.
Schwegmann-Weßels, C., G. Zimmer, H. Laude, L. Enjuanes und G. Herrler (2002):
Binding of transmissible gastroenteritis coronavirus to cell surface sialoglycoproteins.
J. Virol. 76, 6037–6043.
Teile der vorliegenden Arbeit wurden auf folgenden Tagungen prasentiert:
Jahrestagung der Gesellschaft fur Virologie, Wien, 26.–29.04.2000.
Poster: Untersuchung der Sialinsaurebindungsaktivitat des ubertragbaren Gastroenteri-
tisvirus der Schweine (TGEV) — Vergleich zu E. coli F5 (K99).
Schwegmann, C., Zimmer, G. und Herrler, G.
VIIIth International Symposium on Nidoviruses (Coronaviruses and Arteriviruses),
Pennsylvania, USA, 20.–25.05.2000.
Kurzvortrag: Interaction of transmissible gastroenteritis coronavirus with intestinal sialo-
glycoconjugates.
Schwegmann, C. and Herrler, G.
6th International Symposium on Positive Strand RNA Viruses, Paris, 28.05.–02.06.2001.
Poster: Interaction of transmissible gastroenteritis coronavirus with glycoproteins of cul-
tured cells.
Schwegmann-Weßels, C., Zimmer, G. and Herrler, G.

Danksagung
Mein besonderer Dank gilt Herrn Prof. Dr. Georg Herrler fur die Uberlassung des Themas
und die hervorragende Betreuung und Unterstutzung wahrend der Durchfuhrung dieser
Arbeit.
Herrn Dr. Gert Zimmer danke ich fur die fachlichen Diskussionen und Anregungen und
fur seine Hilfe bei technischen Problemen im Labor.
Herrn Dr. B. Schroder und Herrn Prof. Dr. G. Breves aus dem Institut fur Physiolo-
gie mochte ich ganz herzlich fur die freundliche und hilfreiche Zusammenarbeit bei der
Besorgung von Darmproben sowie der Bereitstellung des Labors und der Technik zur Her-
stellung von BSMV danken. Frau M. Burmester und Frau G. Becker danke ich fur die
Unterstutzung bei der Arbeit im Labor des Instituts fur Physiologie.
Bei Frau Dr. M.-L. Enss aus dem zentralen Tierlabor der MHH bedanke ich mich fur
die gute Zusammenarbeit bei der Isolierung und Reinigung der intestinalen Muzine. Die
Bereitstellung des Labors und der Methodik waren sehr hilfreich. Frau R. Thrien danke ich
fur die tatkraftige Unterstutzung im Labor und die Durchfuhrung vieler Arbeitsschritte.
Durch ihre Hilfe blieb mir ein standiges Hin- und Herfahren zwischen TiHo und MHH
erspart.
Herrn PD Dr. A. Gruber aus dem Institut fur Pathologie mochte ich fur die Zusammen-
arbeit in histologischen Fragestellungen danken. Herrn K.-P. Kuhlmann danke ich fur die
Anfertigung der Kryoschnitte.
Ich mochte mich bei allen Mitarbeitern der AG Herrler und des gesamten Instituts fur
Virologie fur ihre Hilfsbereitschaft und das gute Arbeitsklima bedanken. Fur die gute
Stimmung im Labor danke ich vor allem meinen”Mit-Doktoranden“ Birthe, Brigitte,
Matthias und Wiebke. Des weiteren bedanke ich mich bei meinen”Ex-Kollegen“ Ina,
Linda, Holger und Oliver fur die schone Zeit im Labor.
Meinem Mann Peter gilt mein besonderer Dank fur seine moralische Unterstutzung
wahrend meiner Promotionszeit und seine technische Hilfe bei der Anfertigung der Dis-
sertation. Vor allem danke ich ihm dafur, dass er mich uberredet hat, diese Arbeit mit
LATEX zu schreiben.












![Kapitel2 Theorie - ULB Halle: Online-Publikationen · KAPITEL 2. THEORIE 4 in den Hepatozyten ausgeglichen [9]. Dort erfolgt die Bildung der prim¨aren Gallens ¨au-renChols¨aure](https://img.pdfslide.org/doc/110x75/5e0c193954287012ca4b301a/kapitel2-theorie-ulb-halle-online-publikationen-kapitel-2-theorie-4-in-den-hepatozyten.jpg)















