Embed Size (px)
Citation preview

Untersuchung der
Kälteanpassungsmechanismen
bakterieller Zellmembranen
Inaugural-Dissertation
zur
Erlangung des Grades
Doctor throphologiae
(Dr. troph.)
der Landwirtschaftlichen Fakultät
der
Rheinischen Friedrich-Wilhelms-Universität Bonn
vorgelegt am 14. August 2014
von
Julia Derichs geb. Krüger
aus Dormagen

Referent: Professor Dr. André Lipski
Korreferent: Professor Dr. Andreas Schieber
Tag der mündlichen Prüfung: 9. Februar 2015
Erscheinungsjahr: 2015

Inhaltsverzeichnis III
Inhaltsverzeichnis
Abkürzungsverzeichnis ................................................................................................ VI
Kurzfassung ............................................................................................................... VIII
Abstract ....................................................................................................................... IX
Veröffentlichungen ........................................................................................................ X
1. Einführung ................................................................................................................ 1
1.1 Definition Psychrophilie/Psychrotoleranz ............................................................. 2
1.1.1 Psychrophile und psychrotolerante Mikroorganismen in der
Lebensmittelindustrie ............................................................................................. 3
1.2 Aufbau der Plasmamembran / Das Flüssig-Mosaik-Modell / Funktionen ............. 4
1.3 Fettsäuren in der Zellmembran ............................................................................ 8
1.3.1 Nomenklatur der Fettsäuren ......................................................................... 8
1.3.2 Fettsäurezusammensetzung in der Kälteanpassung ..................................... 9
1.4 sonstige Lipide in der Zellmembran ................................................................... 11
1.4.1 Chinone ...................................................................................................... 12
1.4.2 Carotinoide ................................................................................................. 15
1.4.3 Weitere Mechanismen in der Kälteanpassung von Bakterien ...................... 18
1.5 Ziel der Arbeit .................................................................................................... 19
2. Material und Methoden ........................................................................................... 20
2.1 Herkunft der Bakterienisolate ............................................................................ 20
2.2 Verwendete Medien und Kultivierung der Bakterien .......................................... 23
2.3 Makroskopische und mikroskopische Morphologiebeschreibung ....................... 23
2.4 Temperaturtoleranztest ..................................................................................... 23
2.5 Salzkonzentrationstest ...................................................................................... 23
2.6 Molekularbiologische Methoden ........................................................................ 24
2.6.1 Molekularbiologische Identifizierungen anhand von 16S rRNA-Gen-
Sequenzanalysen ................................................................................................ 24

IV Inhaltsverzeichnis
2.6.2 Molekulare Phylogenie-Analyse .................................................................. 26
2.6.3 DNA-DNA Hybridisierung ............................................................................ 26
2.7 Chemotaxonomische Methoden ........................................................................ 27
2.7.1 Fettsäureanalyen ........................................................................................ 27
2.7.2 Chinonanalysen .......................................................................................... 28
2.7.3 Kombinierte polare Lipid- und Chinonanalyse ............................................. 30
2.7.4 Carotinoidextraktion .................................................................................... 31
2.8 Messungen des Zellertrages ............................................................................. 34
2.8.1 Messung des Wachstumsverhaltens mittels optischer Dichte ..................... 34
2.8.2 Messung des Proteingehaltes ..................................................................... 34
2.9 Kälteresistenztest .............................................................................................. 36
2.10 Überlebenstest - Zellertragsmessung bei 10°C und 30°C in einem
Modelllebensmittel ................................................................................................... 36
2.11 Physiologisches Reaktionsprofil ...................................................................... 37
3. Ergebnisse .............................................................................................................. 38
3.1 Makroskopische und mikroskopische Isolatbeschreibung .................................. 38
3.2 Identifizierung mittels 16S rRNA-Gen Sequenzierung ....................................... 45
3.2.1 Phylogenetische Beziehung der kälteangepassten Isolate .......................... 47
3.3 Neubeschreibung Isolat J22 .............................................................................. 50
3.3.1 Makroskopische und mikroskopische Zellmorphologie ................................ 50
3.3.2 Temperatur- und Salztoleranztest ............................................................... 50
3.3.3 Chemotaxonomische Beschreibung ............................................................ 51
3.3.4 physiologisches Reaktionsprofil .................................................................. 53
3.3.5 Molekulare Phylogenie-Analyse und DNA-DNA-Hybridisierung .................. 54
3.3.6 Namensgebung ........................................................................................... 54
3.4 Fettsäuremuster bei 10°C- und 30°C-Inkubation ............................................... 56
3.5 Chinonmuster und -gehalt bei 10°C- und 30°C-Inkubation ................................ 66
3.5.1 Qualitative Chinonanalyse ........................................................................... 66
3.5.2 Quantitative Chinonanalyse ........................................................................ 69

Inhaltsverzeichnis V
3.6 Carotinoidgehalt bei 10°C- und 30°C-Inkubation ............................................... 74
3.7 Wachstumsverhalten ......................................................................................... 79
3.7.1 Wachstumsverlauf einzelner Isolate bei 10°C- und 30°C-
Inkubationstemperatur ......................................................................................... 79
3.7.2 Wachstumsverhalten einzelner Isolate bei 10°C und 30°C im Vergleich zu
Typ- bzw. Referenzstämmen ............................................................................... 82
3.7.3. Exponentielle Wachstumsraten und maximale OD-Werte .......................... 85
3.8 Zelldichte ........................................................................................................... 86
3.9 Kälteresistenztest Listeria monocytogenes ........................................................ 88
3.10 Überlebenstest bei 10°C und 30°C in einem Modelllebensmittel ..................... 89
4 Diskussion ............................................................................................................... 91
4.1 Isolierung kälteangepasster Bakterien ............................................................... 91
4.2 Neubeschreibung Pedobacter nutrimenti J22T ................................................... 91
4.3 Fettsäurebewertung .......................................................................................... 93
4.3.1 Taxonomische Bedeutung .......................................................................... 93
4.3.2 Bewertung der Fettsäuremuster in der Kälteanpassung .............................. 94
4.4 Chinone ............................................................................................................. 96
4.4.1 Taxonomische Bedeutung der Chinone ...................................................... 96
4.4.2 Chinone in der Kälteanpassung .................................................................. 99
4.5 Carotinoide ...................................................................................................... 102
4.5.1 Carotinoide in Bakterien ............................................................................ 102
4.5.2 Carotinoide in der Kälteanpassung ........................................................... 103
4.6 Zelldichte und Wachstumsverhalten ................................................................ 106
4.7 Kälteresistenztest und Überlebenstest Listeria monocytogenes ...................... 108
5. Zusammenfassung ............................................................................................... 111
6. Literaturverzeichnis ............................................................................................... 113
8. Danksagung ......................................................................................................... 143

VI Abkürzungsverzeichnis
Abkürzungsverzeichnis
ATCC American Type Culture Collection
bp Basenpaare
CCUG Culture Collection, University of Göteborg, Sweden
CSP cold shock proteins
c/t cis/trans
DNA Desoxyribonukleinsäure
dNTP Desoxyribonukleosidtriphosphat
DSMZ Deutschen Sammlung von Mikroorganismen und Zellkulturen
ECL Equivalent Chain Length
EDTA Ethylendiamintetraacetat
g Einheit der Erdbeschleunigung (~ 9,81 m/s2)
GC Gaschromatographie
FS Fettsäure
h Stunden
HP Hewlett-Packard
HTA Hefeextrakt Trypton Soja Agar
HTB Hefeextrakt Trypton Soja Bouillon
HPLC High Pressure Liquid Chromatography
IAM IAM Culture Collection, University of Tokyo
IFO Institute for Fermentation Osaka
M molar
mM milli-Molar
mAU*s milli-Absorbance-Units pro Sekunde
MK Menachinon
MTK Methionachinon
MS Massenspektrometrie
N Normalität
OD Optische Dichte
PCR Polymerase Chain Reaction
pmol Pikomol
PQ Plastochinon
Q Ubichinon
rpm Rounds per minute

Abkürzungsverzeichnis VII
rRNA Ribosomale Ribonukleinsäure
spp. Species pluralis
TSA Trypton Soja Agar
TSB Trypton Soja Bouillon
U Unit (Enzymeinheit)
UV Ultraviolett

VIII
Kurzfassung
Die Lebensmittelindustrie versucht stetig, die Haltbarkeit ihrer Produkte zu verlängern,
um ihre Produktivität zu optimieren. Dafür verwendet sie auch Kühlungs- und
Gefriertechniken. Durch den Einsatz dieser Techniken sollen die für den Menschen
pathogenen Mikroorganismen im Lebensmittel absterben bzw. die Anzahl vermindert
werden, da ein Wachstum bei niedrigen Temperaturen nicht mehr möglich oder stark
verlangsamt ist. Es gibt jedoch auch psychrophile bzw. psychrotolerante Bakterien, die
sich in sehr kalten Regionen der Erde an die dort herrschenden Umwelt- und
Klimabedingungen evolutiv angepasst haben. Einige dieser kälteadaptierten Bakterien
finden optimale Wachstumsvoraussetzungen bei kühlschrankähnlichen Bedingungen
vor. Gelangen diese Bakterien nun mit Lebensmittelrohstoffen in ein Lebensmittel,
bietet sich ihnen ein nährstoffreiches Biotop, in dem sie sich gut vermehren können. Zu
diesen psychrophilen bzw. psychrotoleranten Bakterien zählen die Gattungen Listeria,
Clostridium, Yersinia und einige für den Menschen pathogene Bacillus-Stämme.
In dieser Dissertation werden die Kälteanpassungsmechanismen psychrophiler bzw.
psychrotoleranter Bakterien näher untersucht. Als Standardmechanismus der
Kälteanpassung wird in der herrschenden Literatur die Änderung im Fettsäuremuster
innerhalb der bakteriellen Membran definiert, welche sich qualitativ und quantitativ
verändern kann. Es gibt aber auch Bakteriengruppen, bei denen dieser
Anpassungsmechanismus bei Untersuchungen nicht nachgewiesen werden konnte.
Diese Feststellung führt zu der Annahme, dass es weitere Mechanismen der
Kälteanpassung bei Bakterien geben muss. Diese sollen im Rahmen der vorliegenden
Arbeit gefunden und erläutert werden. Sie konzentriert sich dabei auf die lipophilen
Substanzen Chinone und Carotinoide, die Bestandteile der bakteriellen Zellmembran
sind. Als Keimquellen dienten lebensmittelassoziierte Habitate.
Diese Arbeit konnte nachweisen, dass bei bestimmten Bakterienarten der
Chinongehalt und der Carotinoidgehalt durch Kälteeinfluss verändert wird. Zum Teil
kommt es zu einer Erhöhung des Gehaltes, durch den die Kältetoleranz dieser
Bakterien erklärt werden kann. Zusätzlich konnte durch diese Arbeit eine erhöhte
Zellausbeute bei einer Kühlinkubation gezeigt werden. Dieser Effekt trat nur bei
Wildisolaten auf, jedoch nicht bei Referenzstämmen. Die vorliegenden
Forschungsergebnisse können in der Lebensmittelindustrie bei der Haltbarmachung
von Produkten und in der Medizin angewendet werden. Sie erweitern den
Forschungsstand, um besser mit kälteangepassten, potenziell pathogenen
Mikroorganismen umzugehen und daraus entstehende Gefahren (für den Menschen)
besser beherrschbar zu machen.

IX
Abstract
To increase its productivity, food industry continuously tries to enhance the shelf life of
its products. To achieve this objective they employ refrigeration and cooling
techniques. These techniques make sure that human-pathogenic microorganisms in
food die off or their number decreases because they don’t grow at these low
temperatures or they grow very slowly. However, there are psychrophilic or
psychrotolerant bacteria that have adapted to cold temperatures because of
environmental and climate conditions on earth. Some of these cold-adapted bacteria
find their growth optimum at refrigerated conditions. Getting in food via e.g. food
ingredients these bacteria have a nutrient-rich habitat which allows them to proliferate.
These psychrophilic or psychrotolerant bacteria include the genera Listeria,
Clostridium, Yersinia and some human-pathogenic Bacillus-strains.
This graduate thesis examines the cold adaptation mechanisms of psychrophilic and
psychrotolerant bacteria more closely. In literature the qualitatively or quantitatively
change in fatty acid pattern inside the bacterial membrane is described as standard
mechanism. However, some groups of bacteria obviously don´t use this mechanism.
This leads to the assumption that other mechanisms exist. The present study tries to
find out and explain these mechanisms. The main focus is on the quinones and
carotenoids as lipophilic substances that can be components of the bacterial cell
membrane. As major source for bacterial isolates food-associated habitats were used.
The results show that quinone content and carotenoid content are altered by cold
influence in several types of bacteria. Partly, the content of quinones and carotenoids
increased. That affects the cold tolerance of bacteria. Additionally, the graduate thesis
resulted in an increased cell yield at cool incubation. This observation occurred only in
wild isolates but not in reference strains.
The present results can be applied in food industry and as well in medicine. They
deepen knowledge to optimize handling of cold-adapted, potentially human-pathogenic,
microorganisms and improve the management of emerging risks.

X
Veröffentlichungen
Beiträge bei Fachtagungen
Derichs, J. & Lipski, A. (2012): Fatty-acid independent adaptation of bacterial
membranes to cold temperatures. Posterpräsentation auf der VAAM Jahrestagung,
Tübingen, 18.-21.03.2012
Derichs, J., Falkenberg, F., Fritsch, C., Verbeek, J. & Lipski, A. (2013): Low growth
temperatures elevate cell yield for mesophilic isolates from food. Posterpräsentation
auf der VAAM Jahrestagung, Bremen, 10.-13.03.2013
Publikation
Derichs, J., Kämpfer, P. & Lipski, A. (2014): Pedobacter nutrimenti sp. nov., isolated
from chilled food. International Journal of Systematic and Evolutionary Microbiology 64,
1310-1316.

Einführung 1
1. Einführung
Organismen sind in der Lage, in vielen ökologischen Nischen zu überleben und sich zu
vermehren. Dafür notwendig ist die Bildung von Enzymen, die dem Organismus die
Synthese und den Abbau der Substrate ermöglichen, die für das Überleben in der
jeweiligen Umgebung notwendig sind. (Georlette et al. 2004)
In weiten Teilen der Erdbiosphäre, wie Ozeanböden, den Polarregionen oder
Hochgebirgsregionen herrschen das ganze Jahr über Temperaturen von unter 5°C.
Diese permanent kalten Lebensräume bilden eine Wachstumsnische für psychrophile
und psychrotolerante Mikroorganismen, die sich auf unterschiedliche Art und Weise
den dortigen Umweltbedingungen angepasst haben. Für eine lange Zeit wurden
Psychrophile als exotische Organismen bezeichnet, aber in den letzten Jahren wurden
immer mehr psychrophile Isolate untersucht (Helmke und Weyland 2004). So stieg die
Anzahl der Neubeschreibungen psychrophiler Mikroorganismen pro Jahr im
International Journal of Systematic and Evolutionary Microbiology von 25 im Jahr 2000
auf 73 im Jahr 2013 an.
Aufgrund des regen Interesses, z.B. aus der Biotechnologie, wurde die Erforschung
der Mechanismen der Kälteanpassung in den letzten Jahren vorangetrieben (Margesin
et al. 2008). Die Weiterentwicklung der industriellen Produktion von Lebensmitteln und
der immer häufigere Einsatz von Kühlung bzw. Tiefgefrieren zur Konservierung sorgen
dafür, dass psychrophile und pychrotolerante Mikroorganismen auch in Lebensmitteln
entsprechende Habitate vorfinden und an Bedeutung gewinnen (Hébraud und Potier
1999). Dies ist besonders im Zusammenhang mit den heutigen modernen
Anforderungen an die Lebensmittelproduktion wichtig, da Konservierungsmittel vom
Konsumenten nur teilweise toleriert werden, da natürlicher Geschmack und
Bekömmlichkeit an oberster Stelle stehen (Russell 2002).
Es gibt psychrophile und psychrotrophe Bakterienspezies, die als
Lebensmittelverderbniserreger oder sogar Lebensmittelpathogene bezeichnet werden.
Als Verderbniserreger gelten u.a. Brochothrix thermosphacta, Pseudomonas spp.,
Micrococcus spp. oder Lactobacillus-Arten. Zu den lebensmittelassoziierten
Pathogenen zählen beispielsweise Listeria monocytogenes, Clostridium botulinum oder
Yersinia enterocolitica (Laksanalasmai et al. 2011; Russell 2002). Diese nehmen
großen Einfluss auf die Lebensmittelsicherheit. Die Bakterien gelangen durch
Kontamination von ihren natürlichen Standorten in gekühlte Lebensmittelrohstoffe bzw.
–produkte und finden dort ein nährstoffreiches Habitat vor. Dadurch können sie sich

2 Einführung
vermehren und somit auch ihre Stoffwechselprodukte oder gegebenenfalls Toxine
ansammeln, welche letztlich die Eigenschaften des Produktes verändern oder die
Gesundheit des Lebensmittelkonsumenten schädigen können. (Kraft 1992)
1.1 Definition Psychrophilie/Psychrotoleranz
Neben der Toleranz von hohen Temperaturen, extremen pH-Werten und begrenzter
Verfügbarkeit von Sauerstoff oder Wasser, stellt die Kälteanpassung eine notwendige
Adaption für die Bakterien dar (Morita 1975). Eine niedrige Temperatur ist ein
wesentlicher Umweltfaktor und hat großen Einfluss auf das Wachstum und die
Entwicklung von lebenden Organismen (Suutari und Laakso 1994). Diese Organismen
werden als psychrophil und psychrotolerant bezeichnet. Durch die Fähigkeit
psychrophiler Organismen auch bei den niedrigsten Temperaturen überleben zu
können, sind sie in der Lage große Teile der Erdoberfläche zu besiedeln. Diese
extremophilen Organismen, entweder prokaryotisch oder eukaryotisch, stellen damit
eine wichtige Klasse der belebten Welt dar, wenn wir die Weitläufigkeit dieser
dauerhaft kalten Regionen der Welt, wie beispielweise polare und alpine Regionen,
betrachten. Die erfolgreiche Ansiedlung von Psychrophilen in solch extremen
ökologischen Nischen, wurde in den letzten Jahren immer intensiver erforscht. (Feller
und Gerday 1997)
Diese genannte Fähigkeit von Bakterien in kalter Umgebung zu überleben und sogar
sich vermehren zu können, zählt somit zu den bedeutsamsten Anpassungen an
unterschiedliche und teils extreme Umwelteinflüsse. Tiefe Temperaturen führen zu
einer Verlangsamung des bakteriellen Stoffwechsels bis hin zum Wachstumsstillstand
(Busta 1978). Im Normalfall beeinflusst die Temperatur die Wachstumsrate welche
durch die Enzymaktivität bestimmt wird. Im drastischen Fall würde es zu
Konformationsänderung zellulärer Makromoleküle und Denaturierung kommen, was
einen Zelltod zur Folge hätte. (Ratkowsky et al. 1982) Dieser Zusammenhang
zwischen Temperatur und Enzymaktivität wird durch die Arrhenius-Gleichung
beschrieben (Russell et al. 1990).
Mit Hilfe der unterschiedlichsten Mechanismen können Mikroorganismen Lebensräume
mit Temperaturen unter 5°C besiedeln. Es gibt zwei Arten von Kältetoleranz.
Psychrophile Mikroorganismen besitzen bezogen auf die Wachstumsgeschwindigkeit
ein Temperaturoptimum von unter 15°C, und können auch bei -5°C noch wachsen.
Psychrotolerante Bakterien haben ebenfalls ein Temperaturminimum bei -5°C, das
Wachstumsoptimum liegt dagegen zwischen 20 und 40°C. (Morita 1975; Gounot und
Russell 1999) Somit sind diese auch bei Kühlschranktemperaturen vermehrungsfähig.

Einführung 3
Mit sinkender Temperatur können jedoch teilweise die Generationszeiten im
Wachstumsverlauf ansteigen und die lag-Phase sich verlängern. (Krämer 2007).
Kältetolerante Vertreter der Gram-negativen Bakterien, der Gram-positiven Bakterien,
der Archaen, der Hefen, der Pilze und der Mikroalgen spiegeln sehr auffallend die
Merkmale der Kälteanpassung wieder. (Deming 2002; Gounot und Russell 1999;
Morita 1975; Russell et al. 1990). Beispielsweise können psychrophile Bakterien in
einem Vollmedium bei 4ºC Verdopplungszeiten aufweisen wie der mesophile
Escherichia coli bei 37°C; nur können psychrophile Bakterien bei dieser hohen
Temperatur nicht wachsen (Feller und Gerday 1997). Die meisten zellulären
Anpassungsmechanismen an niedrige Temperaturen sind noch nicht vollständig
verstanden und werden noch untersucht. Dazu gehören Studien über die Regulierung
der Membranfluidität, die Aufrechterhaltung der Proteinsynthese und die Produktion
von Kälte-Proteine und die speziellen Mechanismen der Kältetoleranz. (Saunders et al.
2003)
1.1.1 Psychrophile und psychrotolerante Mikroorganismen in der
Lebensmittelindustrie
Mehrere Studien zeigen, dass psychrophile und psychrotolerante Bakterien oft in der
Lebensmittelmatrix auftreten können. Diese Veröffentlichungen beschäftigen sich
entweder mit Lebensmittelverderbniserregern oder mit Pathogenen, die unter kalten
Bedingungen wachsen können. (Scherer und Neuhaus 2006)
Beispielsweise wurden psychrotolerante Milchsäurebakterien als Hauptursache für den
Verderb von Schweinefleisch-Wurstwaren verantwortlich gemacht (Hamasaki et al.
2003). Außerdem wurden als Hauptbesiedler von kühlgelagerten Schweinefleisch die
Gattungen Pseudomonas, Aeromonas und Acinetobacter identifiziert. (Olsson et al.
2003) Aber auch andere für die Lebensmittelindustrie wichtige Mikroorganismen sind in
der Lage bei niedrigen Temperaturen zu überleben und sich sogar zu vermehren. Die
Fähigkeit von Campylobacter jejuni Kühl- und Gefriertemperaturen zu überleben, hat
eine hohe Relevanz für die Lebensmittelsicherheit und die öffentliche Gesundheit
(Chan et al. 2001). Andere Beispiele für lebensmittelassoziierte Spezies, die sich bei
kalten Temperaturen vermehren können sind Bacillus cereus, Clostridium perfringens,
Staphylococcus aureus und Yersinia enterocolitica. (Lechner et al. 1998; Guenlcrl und
Linton 2003; Harrison et al. 2000; Kalinowski et al. 2003; Steele und Wright 2001; Zhao
et al. 2003).

4 Einführung
Die Reduzierung psychrophiler und psychrotoleranter Bakterien spielt daher eine sehr
wichtige Rolle in der Lebensmittelindustrie. Jedoch ist die Strategie, wie sich Bakterien
genau in einer Lebensmittelmatrix bei kalter Umgebungstemperatur vermehren können
nur wenig aufgeklärt. (Scherer und Neuhaus 2006)
1.2 Aufbau der Plasmamembran / Das Flüssig-Mosaik-Modell / Funktionen
In der Bakterienzelle ist neben der Proteinsynthese, Enzymaktivität und
Nährstoffaufnahme auch die Zusammensetzung, Integrität und Struktur der
Zellmembran temperaturabhängig (Suutari und Laakso 1994). Der Erhalt der
Membranfluidität ist von wesentlicher Bedeutung für die Funktion der
Transportvorgänge und die Signaltransduktion und daher überlebenswichtig (Berg et
al. 2003). Bei Senkung der Temperatur reagieren Bakterien in den meisten Fällen mit
einer Beibehaltung der Membranfluidität durch Veränderung im Fettsäureprofil. Der
Begriff „homeoviscous adaptation“ wurde im Jahr 1974 für diese Anpassung geprägt
(Sinensky 1974). Dadurch wird die Funktionsfähigkeit der bakteriellen Zellen in
bestimmten Stresssituationen wie Kälteeinfluss aufrechterhalten. (Heller et al. 1993)
Darüber hinaus gibt es spezielle Verbindungen zur Anpassung an einen Kälteschock,
die in der Zelle produziert werden. So ist beispielsweise die Anpassung nach
Kälteeinfluss bei Bakterien auch durch sogenannte „cold induced proteins“ (CSP)
reguliert. Bei diesen CSP´s handelt es sich um circa 7 KDa große Proteine, die den
generellen Metabolismus, die Translation und die Proteinfaltung beeinflussen können.
(Graumann et al. 1996). Neben den CSP´s können auch weitere Stoffgruppen zur
Akklimatisation der Zellen bei Kälte beitragen. Es ist bekannt, dass kompatible Solute
(z.B. Betain, Treholose und Ectoin) die Zellen bei Stress schützen können. Es wird
davon ausgegangen, dass kompatible Solute als chemische Chaperone gegen die
Denaturierung und Aggregation von Proteinen bei niedrigen Temperaturen und gegen
oxidativen Stress als Radikalfänger wirken. Weiterhin wird vermutet, dass sie die
Membran stabilisieren und schützen sowie als mögliches Frostschutzmittel fungieren
können. (Kandror et al. 2002) Kompatible Solute können entweder synthetisiert
(Kandror et al. 2002) oder mit Hilfe von spezifischen Transportern aus der Umgebung
aufgenommen werden. (Brigulla et al. 2003; Mendum und Smith 2002) Diese Stoffe
werden jedoch im Cytoplasma eingelagert und nicht in der Plasmamembran (Cleland
et al. 2004).
Für die optimale Funktionstüchtigkeit der Plasmamembran ist es notwendig, dass sich
die Membranlipide in einem sogenannten flüssig-kristallinen Zustand befinden. Unter
normalen physiologischen Bedingungen ist dies auch der Fall. Wenn allerdings die

Einführung 5
Umgebungstemperatur der Bakterien absinkt, wird durch Veränderung der
Membrankomponenten die normalerweise fluide Membran viskös; nun befindet sich sie
sich in der Gelphase. Diese Fluidität der Zellmembranen spielt aber eine sehr wichtige
Rolle bei den biologischen Prozessen, die zwischen dem Extrazellulär- und dem
Intrazellulärraum stattfinden. Dadurch wird unter anderem die normale Funktion der
Enzyme beeinträchtigt. Somit müssen einige Anpassungen zur Erhaltung dieser
Fluidität vorgenommen werden. (Beales 2004; Sinensky 1974).
Daher ist das Verständnis des Phasenüberganges von Gelphase (lipid-geordnet) zu
flüssig-kristallinen Phase (flüssig-ungeordnet) sehr wichtig (Abb.1). Dieser Zustand, in
dem sich die Kohlenwasserstoffketten der Lipide befinden und der die Fluidität der
Membran bestimmt, ist der wesentliche Parameter für die Funktion biologischer
Membranen. Bei der für eine Lipidsorte spezifischen Phasenübergangstemperatur TC
findet ein reversibler Phasenübergang statt. TC hängt dabei von der
Fettsäurezusammensetzung und der Kopfgruppenstruktur der Lipide sowie von
Umgebungsvariablen, wie dem pH-Wert und der Ionenkonzentration im umgebenden
Medium, ab. (Heller et al. 1993)
Abbildung 1: Simulation der Membranstruktur in der Gelphase (oben) und der flüssigkristallinen
Phase (unten) in Bezug auf die Temperatur (Heller et al. 1993).
Mit der Abnahme der Umgebungstemperatur, sind eine Reihe von Veränderungen im
Fettsäuremuster der Bakterienzellmembranen bekannt. Dadurch wird die laterale
Diffusion innerhalb der Membran aufrechterhalten. (Chattopadhyay 2006)
Viele Studien haben adaptive Veränderungen in der Fettsäurezusammensetzung mit
dem Hauptaugenmerk auf ungesättigte und verzweigtkettige Fettsäuren gezeigt. Die

6 Einführung
wichtigsten Anpassungsmechanismen werden durch Gounot & Russell (1999),
Chintalapati et al. (2004) und Scherer & Neuhaus (2006) beschrieben. Dazu zählen der
Ersatz von gesättigten durch ungesättigte Fettsäuren, das Abnehmen der Fettsäure-
Kettenlänge, der Ersatz von geradkettigen durch methylverzweigte Fettsäuren, der
Ersatz von iso-verzweigten durch anteiso-verzweigte Fettsäuren, die Isomerisierung
von trans- in cis-Doppelbindungen und der Ersatz von geradzahligen durch
ungeradzahlige Fettsäuren. Diese Anpassungen können mit den verschiedenen
Schmelzpunkten der jeweiligen Fettsäuren erklärt werden und somit kommt es zu einer
gewünschten Schmelzpunkterniedrigung. In Tabelle 1 sind Schmelzpunkte
verschiedener Fettsäuren aufgeführt.
Tabelle 1: Schmelzpunkte bestimmter Fettsäuren nach Guendouzi und Mekelleche (2012), Kaneda
(1991), Muranushi et al. (1981) und United States Environmental Protection Agency, Washington,
DC, USA (2012)
Fettsäure Schmelzpunkt [°C]
C 13:0 iso 89,3
C 14:1 trans 2 99,5
C 14:0 53,9
C 15:0 52,5
C 16:0 63,1
C 17:0 61,3
C 18:0 69,6
C 16:1 cis 9 0,5
C 18:1 trans 9 51,0
C 18:1 cis 9 8-10
C 18:1 trans 11 44,0
C 18:2 cis,cis 9,12 -5,0
15:0 iso 51,7
17:0 iso 60,2
15:0 anteiso 23,0
17:0 anteiso 36,8
Trotz der allgemein anerkannten dominierenden Funktion der Fettsäureprofile in der
Membrananpassung gibt es einige aktuelle Literaturquellen über mikrobielle
Gemeinschaften in arktischen und antarktischen Lebensräumen, die - teilweise
zusätzlich - andere Strategien verfolgen. Beispiele dafür sind das Verändern der
polaren Kopfgruppe, die Konzentrationsänderung von integralen Membranproteinen
und das Ändern der Carotinoidzusammensetzung und –gehaltes aus polaren und
unpolaren Carotinoiden (Chintalapati et al. 2004; Margesin et al. 2007).

Einführung 7
Im den folgenden Abschnitten wird nun detaillierter auf die Fettsäuren und andere
lipophile Komponenten der Zellmembran, die eine Rolle bei der Kälteanpassung auf
zellulärer Ebene spielen könnten, eingegangen.
8 Einführung
1.3 Fettsäuren in der Zellmembran
1.3.1 Nomenklatur der Fettsäuren
Fettsäuren können nach der Länge der Acylkette, Anzahl, Position und Konfiguration
der Doppelbindungen, sowie nach dem Vorkommen zusätzlicher funktioneller Gruppen
klassifiziert werden.
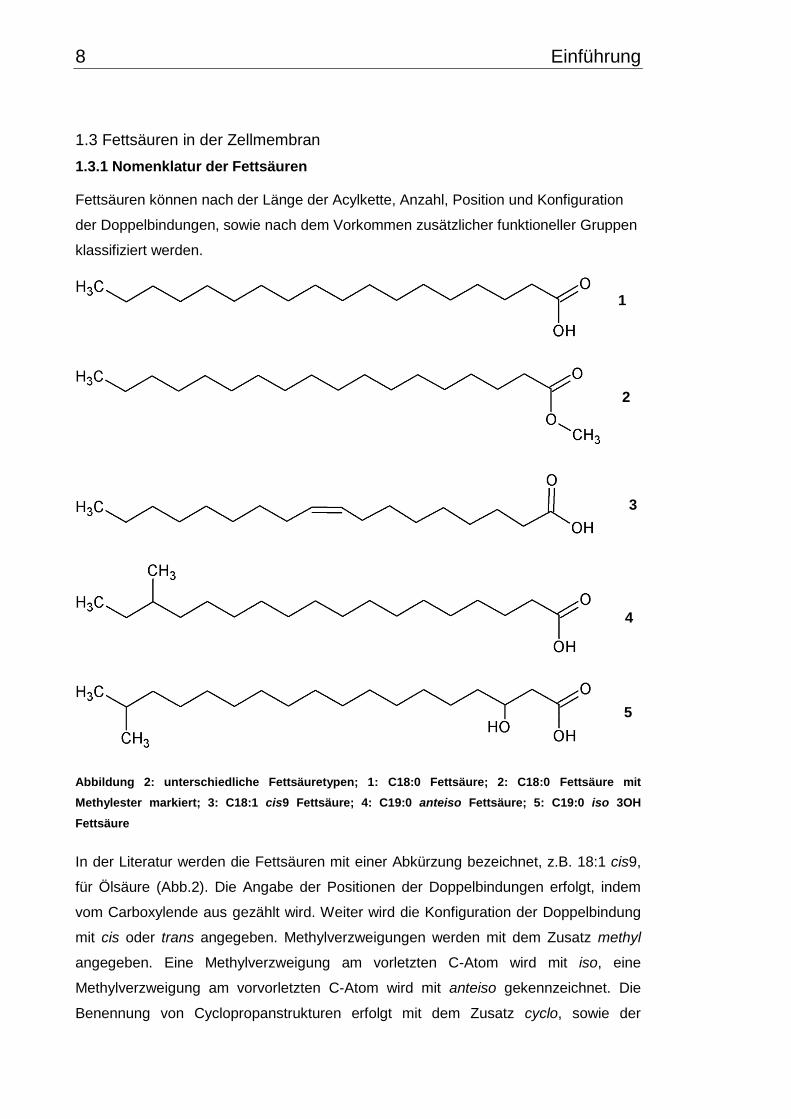
Abbildung 2: unterschiedliche Fettsäuretypen; 1: C18:0 Fettsäure; 2: C18:0 Fettsäure mit
Methylester markiert; 3: C18:1 cis9 Fettsäure; 4: C19:0 anteiso Fettsäure; 5: C19:0 iso 3OH
Fettsäure
In der Literatur werden die Fettsäuren mit einer Abkürzung bezeichnet, z.B. 18:1 cis9,
für Ölsäure (Abb.2). Die Angabe der Positionen der Doppelbindungen erfolgt, indem
vom Carboxylende aus gezählt wird. Weiter wird die Konfiguration der Doppelbindung
mit cis oder trans angegeben. Methylverzweigungen werden mit dem Zusatz methyl
angegeben. Eine Methylverzweigung am vorletzten C-Atom wird mit iso, eine
Methylverzweigung am vorvorletzten C-Atom wird mit anteiso gekennzeichnet. Die
Benennung von Cyclopropanstrukturen erfolgt mit dem Zusatz cyclo, sowie der
1
2
3
2
4
2
5
2

Einführung 9
Positionsangabe des C-Atoms. Hydroxylgruppen werden mit OH, sowie der
Positionsangabe gekennzeichnet. (Belitz et al. 2008; IUPAC-IUB-Commission on
Biochemical Nomenclature 1978)
1.3.2 Fettsäurezusammensetzung in der Kälteanpassung
Durch Schwankungen der Umgebungstemperatur der Mikroorganismen, sind die
Fettsäuren der Phospholipiddoppelschicht einer Anzahl von möglichen Änderungen
ausgesetzt (Chintalapati et al. 2004). Die Anpassungen wurden bereits im Abschnitt
1.2 aufgelistet. Fettsäuren unterschiedlicher Kettlängen und Sättigungsgrad befinden
sich in der Lipiddoppelschicht. Gesättigte Fettsäuren fördern die Aneinanderlagerung
benachbarter Acylketten, was wiederum stabilere Strukturen verursacht und die
Membran visköser macht. Diese Membranen haben wegen der optimalen, engen side
by side Packung und der daraus resultierenden höheren van-der-Waals-Kräfte einen
höheren Schmelzpunkt. (Freedman 1981; Hazel und Eugene Williams 1990)
Im Gegensatz dazu sind kürzere Fettsäuren, besonders die mit weniger als zwölf
Kohlenstoff-Atomen, nicht in der Lage in so hohem Maße hydrophobe
Wechselwirkungen mit anderen Lipiden oder Proteinen einzugehen, da sie zu kurz
sind, um durch die Lipiddoppelschicht der Membran zu reichen. Somit wird der fluide
Zustand der Membran gefördert. (Quinn 1981) Dieser Mechanismus der
Acylkettenlängenmodifizierung bei abfallender Temperatur ist allerdings an das
Wachstum der Zellen gekoppelt, das heißt die kürzeren Fettsäuren müssen de novo
synthetisiert werden. (Denich et al. 2003; Russell 1984).
Außerdem besitzen einige Bakterien membrangebundene Desaturasen. Diese sind in
der Lage in einem aeroben Prozess Doppelbindungen in Fettsäuren einzubauen, die
bereits in der Membran existieren. Ihre Funktion wird durch mehrere verknüpfte
Untereinheiten vermittelt, welche die Wasserstoffatome vom Substrat zu molekularem
Sauerstoff kanalisieren, um eine Doppelbindung und Wasser zu bilden. Die Fluidität
wird erhöht, indem die Doppelbindung in cis-Konfiguration in der Fettsäure mehr Platz
einnimmt und so für mehr Fluktuation in der Membran sorgt. Trans-Fettsäuren
dagegen reduzieren ähnlich wie gesättigte Fettsäuren die Fluidität der Membran.
(Aguilar und Mendoza 2006; Heipieper et al. 1992; Russell 1984) Aerobe Bakterien
erzeugen diese trans-Fettsäuren in ihrer Membran durch Umwandlung von bereits
bestehenden cis-Fettsäuren. Dies geschieht mit Hilfe der cis-trans-Isomerase. Sie ist in
der Lage eine einfach ungesättigte Doppelbindung in der cis-Konfiguration in einem
energie- und wachstumsunabhängigen Prozess in die trans-Konfiguration zu

10 Einführung
überführen. Die trans-Isomere der Fettsäuren haben daher Einfluss auf die
Membranfluidität. Sie weisen allerdings aufgrund ihrer langgestreckten, sterischen
Struktur, im Gegensatz zu den cis-Isomeren, eher die Eigenschaften von gesättigten
Fettsäuren auf und haben somit einen eher reduzierenden Effekt auf die
Membranfluidität. Die aufgelisteten Schmelzpunkte aus der Tabelle 1 können diesen
Effekt erklären. Zusätzlich sind Bakterien mit anaerobem Stoffwechsel in der Lage
trans-Fettsäuren de novo zu synthetisieren und in ihre Membran einzubauen.
(Heipieper et al. 2003; Keweloh und Heipieper 1996)
Als Beispiel für diesen Effekt konnten in einigen psychrophilen Bakterien der Gattung
Vibrio und Pseudomonas mit steigender Wachstumstemperatur ein Anstieg des trans-
Fettsäuregehaltes verzeichnet werden. Diese vermehrte Bildung der trans-Fettsäuren
kann den Effekt des Temperaturanstieges ausgleichen und somit die Membranfluidität
reduzieren. (Kiran et al. 2004; Okuyama et al. 1990)
Zusätzlich spielen in der Kälteanpassung die strukturellen Isomere der geradkettigen
Fettsäuren eine wichtige Rolle. Die strukturellen Isomere iso und anteiso sind die
häufigsten Formen der methylverzweigten Fettsäuren (Chintalapati et al. 2004). Diese
endständig verzweigten Fettsäuren werden de novo synthetisiert, das heißt die
Position der Methylverzweigung ist durch die jeweilige Aminosäure, aus der die
Fettsäure in der Fettsäuresynthese hervorgeht, determiniert und kann nicht mehr
verändert werden. Die zusätzliche Methylgruppe der verzweigten Fettsäuren benötigt
mehr Raum in der Plasmamembran und kann somit die Ordnung einer
Phospholipiddoppelschicht mehr beeinträchtigen als die iso-verzweigten Fettsäuren.
(Kaneda 1991; Russell 1984; Suutari und Laakso 1994).
Zusammengefasst kann gesagt werden, dass bei der Kälteadaptation von
Mikroorganismen das temperaturabhängige Fettsäuremuster der Membranen eine
wesentliche Rolle spielt. Durch Verzweigungen, die Variation der Kettenlänge, der
Anteils an Doppelbindungen und Cyclisierungen, können Fettsäuren verändert werden
(Suutari und Laakso 1994). Die Verschiebung zu verzweigten Fettsäuren (Annous
1997), die Verringerung des Sättigungsgrades und die Kürzung der Kettenlänge
erhöhen die Membranfluidität bei sinkenden Temperaturen (Quinn 1981).

Einführung 11
1.4 sonstige Lipide in der Zellmembran
Neben den membranbildenen Lipiden (Phospholipide, Sphingolipide) sind Isoprenoide
lipophile Strukturbestandteile der pro- und eukaryotischen Zellmembranen. Zu den
Isoprenoiden zählen u.a. die große Gruppe der Steroide (Zoo- und Phytosterine),
Carotinoide und Chinone.
Bezogen auf die Kälteanpassung übernimmt in eukaryotischen Zellen vor allem das
Cholesterin (Steroid) die Funktion zur Regulierung der Membranstabilität bzw.
Membranfluidität. In Eukaryoten ist Cholesterin daher ein wichtiger Bestandteil von
Membranen. Cholesterin ist bekannt für die Wirkung auf die physikalischen
Eigenschaften von Membranen. Das starre Steroidgerüst unterbricht die
Wechselwirkungen zwischen den Fettsäureketten. Durch die zusätzliche Bildung
spezifischer Komplexe mit den Phospholipiden wird die Membran weniger fluide und
die Wahrscheinlichkeit für Phasenübergänge wird gleichzeitig gesenkt (Berg et al.
2003). Aus diesem Grund kann es die Auswirkungen der Temperatur auf die
Membranstruktur und -funktion minimieren, d.h. die Phase des Übergangszustandes
wird verbreitert. Eine Änderung der Umgebungstemperatur hat unmittelbare Folgen für
die Membranen. Einige Eigenschaften der Membran sind aber besonders wichtig, um
die Funktionalität aufrechtzuerhalten. Dabei spielt, wie zuvor in Abschnitt 1.2
beschrieben, die Ordnung der Membran, die entsprechende Phase der
Lipiddoppelschicht sowie die Durchlässigkeit der Membran eine wichtige Rolle. Durch
Temperaturschwankungen ist eine Umstrukturierung bzw. Unordnung der Membran
erforderlich, um diese aufrecht zu erhalten. Wie bereits erwähnt, ist das Cholesterin
eine ideale Wahl für diese Umgestaltung der Membran, da die planare Struktur die
Ordnung der Akylketten im höheren Wachstumsbereich stabilisiert und die Ordnung
der Akylketten im niedrigen Bereich stört. (Crockett 1998)
Jedoch wird Cholesterin von Prokaryonten nicht gebildet bzw. exogen aufgenommen.
Eine Ausnahme sind dabei die Mykoplasmen. Mykoplasmen können das Steroid
exogen aufnehmen und dieses kann somit in die Membran diffundieren, um so u.a. die
Bildung einer Gelphase der Membran bei niedrigen Temperaturen zu verhindern.
(Razin et al. 1980; Rottem et al. 1973; Rottem 1981).
Diese Studien unterstützen die Arbeitshypothese, dass das Prinzip der Unordnung der
Acylketten der Phospholipide durch hydrophobe membranlösliche Moleküle in
kälteangepassten Bakterien als alternativer Mechanismus zur Fettsäuremodifikation
realisiert werden kann. Daher können auch andere Lipide an der Regulation der

12 Einführung
Membranfluidität beteiligt sein. In dieser Arbeit wurde versucht, Indizien dafür zu
finden, ob isoprenoide Strukturen wie Chinone und Carotinoide an der Regulation der
Fluidität beteiligt sind.
1.4.1 Chinone
Chinone sind in der Natur weit verbreitet und kommen größtenteils als fettlösliche
Komponenten membrangebunden vor. (Søballe und Poole 1999)
Der Begriff Chinon beschreibt eine chemische Verbindungsklasse, die häufig in
Naturstoffen vorkommt und vielfältige Eigenschaften aufweist. Er leitet sich von dem
Namen der Chinasäure ab, einer Zwischenstufe im Shikimatweg, welche wichtig für die
Biogenese von Aromaten ist. Im Allgemeinen werden Chinone nach dem jeweiligen
Aromaten benannt, von dem sie sich durch Oxidation ableiten. So erhält man aus
chemischer Sicht para-Benzochinon und ortho-Benzochinon aus Benzol, para-
Napthochinon und ortho-Naphthochinon aus Naphthalin sowie Anthrachinon aus
Anthracen. (Effenberger-Neidnicht 2011) Bakterielle Chinone können in zwei Gruppen
aufgeteilt werden. Die erste Gruppe umfasst die Benzochinone, die auch als
Ubichinone oder Coenzym Q bezeichnet werden. Beide Namen werden von der
IUPAC-Kommission für die Nomenklatur anerkannt (Nič et al. 2009). Der Begriff
Coenzym Q wird vermehrt im biochemischen und klinischen Bereich verwendet. Die
Abkürzung für Ubichinon, welche auch in dieser Arbeit verwendet wird, ist entweder Q
oder Q-n, wobei n für die Anzahl der Isopreneinheiten in der Seitenkette steht. Die
zweite Gruppe sind die Naphthochinone. Diese lassen sich in Phyllochinone (Vitamin
K1) und Menachinone (MK-n, Vitamin K2) unterscheiden. Phyllochinone tragen am 3.
Kohlenstoffatom des Naphtochinonrings eine Phytylgruppe und werden in grünen
Pflanzen gebildet. Dagegen sind Menachinone, die am C-3 eine ungesättigte
Polyprenylgruppe tragen, in Bakterien vorhanden. (Søballe und Poole 1999; Stein
2003)
Die verschiedenen Chinontypen unterscheiden sich in der Länge der
Isoprenseitenketten sowie dem Sättigungs- und Hydrierungsgrad der Isopreneinheiten.
Beispielsweise haben Menschen Chinone mit 10 Isoprenoidseitenketten, wohingegen
Nagetiere, das Bakterium Pseudomonas und Hefen überwiegend 6, 8 oder 9
Isoprenoideinheiten in der Seitenkette vorweisen. Die chemische Struktur von
Chinonen ist am Beispiel von Q-8 und MK-8 in Abbildung 3 gezeigt. (Collins und Jones
1981; Søballe und Poole 1999)

Einführung 13
Abbildung 3: chemische Struktur von Ubichinon (links) und Menachinon (rechts) am Beispiel von
Q-8 und MK-8 (Søballe und Poole 1999)
Die Klasse der Ubichinone ist in der Natur weit verbreitet und kommen in Zellen von
Tieren, Pflanzen und Mikroorganismen (v.a. Alpha-, Beta und Gammaproteobacteria)
vor. Gram-positive Bakterien und einige Gram-negative Bakterien wie Sulfatreduzierer
oder die Gattung Bacteroides und Cyanobakterien besitzen dagegen (zum Teil
zusätzlich) Menachinone. (Munk 2008)
Chinone sind Elektronenüberträger in der Atmungskette und spielen daher eine
wichtige Rolle bei der oxidativen Phosphorylierung. Zu den verbreitetsten
Atmungskettenchinonen zählen die Ubichinone, die Menachinone und die
Dimethylmenachinone. Der bekannteste Elektronencarrier ist das Coenzym Q, ein
Ubichinon. Das Chinon kann zwei Elektronen nacheinander aufnehmen, dabei entsteht
im ersten Schritt ein Semichinon-Radikal und bei Aufnahme des zweiten Elektrons ein
Hydrochinon. Chinone fungieren als Schalter zwischen Ein-und Zwei-Elektronen-
Übergängen. Bei der Reduktion des Chinons mit zwei Elektronen werden auf der
Membraninnenseite zwei Protonen aufgenommen, die bei der Oxidation auf der
Membranaußenseite wieder abgegeben werden. Wird das Hydrochinon durch einen
Elektronencarrier oxidiert, der nur eines der beiden Elektronen aufnimmt, wird das
zweite Elektron in den Chinonpool zurückgeführt, wo es ein festgebundenes Chinon
zum Semichinon-Radikal reduziert. Bei der Oxidation eines weiteren Hydrochinons
wird erneut ein Elektron auf den nachfolgenden Elektronencarrier übertragen, das
andere reduziert das gebundene Semichinon-Radikal zum Hydrochinon. Insgesamt
werden so in einem kompletten Chinonzyklus zwei Hydrochinone zum Chinon oxidiert,
ein gebundenes Chinon zu Hydrochinon reduziert und dabei zwei Elektronen und vier
Protonen transportiert. An den Elektronentransportketten der verschiedenen
Organismen sind wie oben beschrieben unterschiedliche Chinone beteiligt. (Madigan et
al. 2008; Madigan et al. 2013; Munk 2008)

14 Einführung
Bemerkenswert ist, dass das prokaryotische Atmungssystem ein flexibles Design
aufweist, welches der Zelle ermöglicht, Veränderungen der Wachstumsbedingungen
auszugleichen und sich an verschiedene verfügbare Elektronenakzeptoren wie z.B.
Sauerstoff oder Nitrat anzupassen. Dies wird dadurch bestätigt, dass Chinone eine
Rolle im Management von oxidativem Stress spielen. (Søballe und Poole 1999)
1.4.1.1 Chinone in der Taxonomie
Da sich die Art der Chinone und ihre Seitenkettenlänge je nach Bakteriengattung
unterscheiden, können diese zur chemotaxonomischen Einordnung von
Mikroorganismen herangezogen werden. Die Arbeiten von Jeffries et al. (1967), Collins
et al. (1979) und Collins & Jones (1981) wurden als neue Grundlage zur Nutzung von
Chinonstrukturen als taxonomische Kriterium genommen. Daraus geht hervor, dass
Ubichinone nur in Eukaryoten und bestimmten gramnegativen Bakterien vorkommen,
wogegen Menachinone in gramnegativen und grampositiven Bakterien sowie Archaea
enthalten sind.
1.4.1.2 Chinone in der Kälteanpassung
In der Literatur ist ein Einfluss von Chinonen auf die Kälteanpassung von
Zellmembranen bisher nicht beschrieben. Einer der Grundgedanken dieser Arbeit war
es daher, dass die membranständigen Menachinone und Ubichinone einen Einfluss
auf die Membranfluidität und damit gegebenenfalls auf die Kälteadaptation von
psychrophilen und psychrotoleranten Bakterien haben können. Dies ist in erster Linie
auf ihre molekulare Struktur, den dreidimensionalen, raumfordernden Aufbau und die
enthaltenen Doppelbindungen ungesättigter Isoprenoidseitenketten zurückzuführen.
Durch die Lokalisation in bakteriellen Membranen kommt es zur der Hypothese, dass
bei einer kälteinduzierten Restrukturierung der Membran ebenso das Chinonprofil
betroffen ist, welches unter Umständen sowohl qualitativ als auch quantitativ aktiv
angepasst werden kann, um die Adaption an die neuen Umweltbedingungen zu
gewährleisten.

Einführung 15
1.4.2 Carotinoide
Abbildung 4: Strukturformel unterschiedlicher Carotinoide; 1 -Carotin; 2: Bacterioruberin
Alle Carotinoide werden aus der gleichen Grundstruktur von 40 Kohlenstoffatomen
gebildet. Die aus acht Isopreneinheiten aufgebaute Polyen-Kohlenwasserstoffkette
bildet das Zentrum des Moleküls. Durch die Cyclisierung der Kettenenden,
Hydrierungen der Doppelbindungen sowie durch Addition von sauerstoffhaltigen
funktionellen Gruppen können Modifikationen entstehen (Abb. 4). Obwohl jede
Doppelbindung als cis oder trans Konfiguration auftreten kann, sind in der Natur
hauptsächlich die thermodynamisch stabileren all-trans-Carotinoide vertreten.
Carotinoide werden in die zwei Hauptgruppen Carotine und Xanthophylle unterteilt. Die
Carotine sind reine Polyene-Kohlenwasserstoffe, während Xanthophylle zusätzlich
mindestens eine sauerstoffhaltige funktionelle Gruppe (z.B. Hydroxylgruppe) enthalten.
(Belitz et al. 2008; Rodriguez-Amaya 2001).
Für die chemischen und physikalischen Eigenschaften der Carotinoide sind
hauptsächlich zwei Aspekte in der molekularen Struktur verantwortlich. Der strukturelle
Aufbau der Carotinoide (Größe, Gestalt, funktionellen Gruppen) ermöglicht die
funktionelle Lokalisation und Orientierung innerhalb der zellulären Strukturen. Das
System aus konjugierten Doppelbindungen ist entscheidend für die photochemischen
Eigenschaften und chemische Reaktivität. Aufgrund der konjugierten Doppelbindungen
können Carotinoide im sichtbaren Wellenlängenbereich von 400 bis 500 nm
Lichtenergie absorbieren. (Britton 1995)
Bei Mikroorganismen wird die Wirkung von Carotinoiden oft zwischen phototroph und
nicht-phototroph unterschieden. Phototrophe Bakterien benötigen Carotinoide für die
2
2
1
2

16 Einführung
Aufnahme von Lichtenergie zur Photosynthese und zum Schutz der
Chlorophyllmoleküle vor photooxidativen Schäden (Cogdell und Frank 1987). Bei den
nicht-phototrophen Bakterien wirken Carotinoide unter anderem schützend gegenüber
UV-Strahlung (Dieser et al. 2010).
1.4.2.1 Carotinoide in der Kälteanpassung
Carotinoide zeigen als lipophile Moleküle die Fähigkeit sich in biologische Membranen
einzulagern und somit einen Einfluss auf die Membranfluidität und Membranrigidität zu
nehmen. Bei Bakterien können daher die Carotinoide eine ähnliche regulierende
Funktion wie das Cholesterin bei Eukaryonten übernehmen. Damit kann die Regulation
der Membranfluidität in der Kälteadaptation bei Bakterien neben der unterschiedlichen
Fettsäurezusammensetzung in der Lipiddoppelschicht (Annous 1997; Quinn 1981;
Suutari und Laakso 1994) zusätzlich durch die Gruppe der Carotinoide übernommen
werden (Chattopadhyay et al. 1997; Jagannadham et al. 2000; Pintea et al. 2005;
Subczynski et al. 1991)
Die lipophilen Verbindungen sind meist in den Membranen lokalisiert, wobei die
Orientierung innerhalb der Lipiddoppelschicht von der Struktur und den funktionellen
Gruppen abhängt. Carotinoide mit polaren Substituenten (z.B. Zeaxanthin) können
Interaktionen mit den polaren Kopfgruppen der Phospholipide eingehen und somit die
Membran durchspannen. Hydrocarbone (β-Carotin) liegen dagegen innerhalb des
lipophilen Kerns der Membran vor. (Britton 2008)
Vor allem polare Carotinoide können einen Einfluss auf die Membranstruktur haben.
Oberhalb der Phasenübergangstemperatur erzielen sie über die Rigidisierung eine
Senkung der Membranfluidität (Subczynski et al. 1991). Indem die polaren Carotinoide
die Lipiddoppelschicht vollständig durchspannen, wird die Beweglichkeit der Lipide
innerhalb der Membran eingeschränkt und die Membranstabilität im fluiden Stadium
erhöht. Die apolaren Carotinoide wie β-Carotin ermöglichen dagegen eine verstärkte
Beweglichkeit der Membranlipide und erhöhen dadurch die Membranfluidität.
Carotinoide erfüllen somit ähnliche Funktionen wie das Cholesterin (Britton 2008).
In vitro Studien synthetischer Membranen mit Phosphatidylcholin haben gezeigt, dass
die Carotinoide Zeaxanthin, Cryptoxanthin und -Carotin in der Membran gebunden
wurden und damit die Fluidität der Membran verringert wurde (Jagannadham et al.
2000). In einer in vivo Studie konnte an dem psychrotoleranten Bakterienstamm
Micrococcus roseus 45R eine temperaturabhängige Synthese von Carotinoiden

Einführung 17
gezeigt werden. Bei der Wachstumstemperatur von 5°C stiegen der
Gesamtcarotinoidgehalt sowie der Anteil an den polaren Carotinoiden
Bacterioruberindiglycosid und Bacterioruberinmonoglycosid im Vergleich zu einer
Wachstumstemperatur von 25°C an (Chattopadhyay et al. 1997). Auch das
psychrotolerante Bakterium Sphingobacterium antarcticus steigerte bei 5°C die
Synthese an polaren Carotinoiden und ungesättigten Fettsäuren. Die ungesättigten
Fettsäuren erhöhen die Fluidität der Membran, während dagegen die polaren
Carotinoide die Stabilität erhalten können (Jagannadham et al. 2000).
In diesem beschriebenen Zusammenhang ist es sehr interessant, dass grampositive
Bakterien aus der Antarktis kollektiv eine Dominanz an pigmentierten Isolaten
vorweisen. Bei Arthrobacter agilis, ein Bakterium, welches aus dem antarktischen
Eismeer isoliert wurde, wird die starke Pigmentierung bei niedriger Temperatur durch
ein C-50 langes Carotinoidmolekül hervorgerufen, welches als Derivat von
Bacterioruberin identifiziert wurde. Diese Aussagen, dass solche Carotinoide die
Membran stabilisieren, werden auch von anderen extremophilen Bakterien und
Archaeen berichtet, die starken Salzkonzentrationen oder UV-Strahlenbelastung
ausgesetzt sind. (Fong et al. 2001)
Diese Aussagen passen auch zu der Beobachtung von Varkonyi et al. (2002), dass
einige polare Carotinoide nur bei einer niedrigen Temperatur in den
Thylakoidmembranen des Cyanobakteriums Cylondrospermopsis raciborskii induziert
wurden.
Jedoch scheint es auch so, dass die oben beschriebene Carotinoid-vermittelte
Stabilisierung von Membranen und die dadurch auftretende Abnahme der
Membranfluidität, durch Veränderungen im Fettsäuremuster wieder ausgeglichen
werden müssen. Dadurch würde die Membranfluidität wieder erhöht.
Allgemein kann gesagt werden, dass in Abhängigkeit von der Struktur und Polarität
Carotinoide unterschiedliche Effekte auf die Membran ausüben können und die
Membranfluidität oder die Rigidität erhöhen können. In Kombination mit den Fettsäuren
könnten Carotinoide eine regulierende Rolle bei der Anpassung an niedrige
Temperaturen spielen.
Jedoch sind Studien, die den Einfluss der Wachstumstemperatur auf die
Carotinoidsynthese bei Mikroorganismen und die damit verbundene Funktion der
Carotinoide bei der Kälteadaption liefern, insbesondere außerhalb der Psychrophilen
sehr selten. Es ist wichtig, das Zusammenspiel von Carotinoiden und Fettsäuren bei

18 Einführung
niedriger Temperatur verstehen zu können und damit aufzuklären. (Scherer und
Neuhaus 2006)
Daher war ein weiterer Grundgedanke dieser Arbeit, dass Carotinoide nicht nur einen
Einfluss auf die Membranstabilität haben, sondern auch auf die Membranfluidität, um
damit gegebenenfalls die Kälteadaptation von psychrophilen und psychrotoleranten
Bakterien erklären zu können. Carotinoide könnten wie Chinone oder ungesättigte
Fettsäuren durch ihre molekulare Struktur und die enthaltenen Doppelbindungen
raumfordernd auf die Membran wirken. Durch die Lokalisation in bakteriellen
Membranen kommt es zur erweiterten Arbeitshypothese, dass bei Kälteeinfluss die
Membran durch den vermehrten Einbau von Carotinoiden aktiv angepasst werden
kann, um die Funktionsfähigkeit der Membran zu gewährleisten.
1.4.3 Weitere Mechanismen in der Kälteanpassung von Bakterien
Neben den oben genannten Veränderungen können noch weitere membranständige
Substanzen einen Einfluss auf die Fluidität der Membranen nehmen und somit eine
Kälteadaption erklären, wovon beispielhaft zwei Gruppen kurz beschrieben werden.
Membranproteine tragen durch ihre Fähigkeit mit Lipiden zu interagieren zur
Gesamtstabilität der Phospholipiddoppelschicht bei (Epand 1998). Takeuchi et al.
zeigten bereits 1978 bei Escherichia coli, dass dessen Membranphospholipide mit den
Membranproteinen interagieren. In einer späteren Studie von Am Kropinski et al.
(1987) konnten auch temperaturabhängige Schwankungen im Proteingehalt der
Plasmamembran von Pseudomonas aeruginosa gezeigt werden. Diese Ergebnisse
untermauerten die Bedeutung der Membranproteine in der Kälteadaptation, auch wenn
sie im Vergleich zu den anderen Mechanismen eine untergeordnete Rolle spielen
(Shivaji et al. 2007).
Hopanoide stellen einem dem Cholesterin strukturell ähnlichen Stoff dar, der von
Bakterien gebildet werden kann. Somit können Hopanoide in Membranen von
Bakterien eine ähnliche Funktion besitzen, wie Cholesterin in Membranen von
Eukaryonten. Die Bildung von Hopanoiden scheint, wie auch die Bildung von
Carotinoiden und langkettigen, gesättigten Fettsäuren, die Membran als hydrophobe
Barriere zu stabilisieren und einer möglichen Erhöhung der Permeabilität
entgegenzuwirken. (Bringer et al. 1985; Kannenberg und Poralla 1999)

Einführung 19
1.5 Ziel der Arbeit
Am Ende dieser Arbeit soll die Erkenntnis stehen, ob und wie andere lipophile
Komponenten der Zellmembran von Bakterien einen Einfluss auf die Kälteanpassung
haben.
Mit dem Hintergrund der Bedeutsamkeit kälteangepasster Bakterien in der
Mikrobiologie, aber auch für die Lebensmittelindustrie im Bereich Biotechnologie und
Lebensmittelsicherheit, wurden für diese Arbeit lebensmittelassoziierte Kühlhabitate
und gekühlte Lebensmittel untersucht, um psychrophile bzw. psychrotolerante
Bakterien zu isolieren. Die erhaltenen Isolate werden morphologisch und
chemotaxonomisch charakterisiert und per 16S rRNA-Gensequenzierung identifiziert.
Bezogen auf die Kälteanpassung werden alle Isolate hinsichtlich ihres
Fettsäuremusters bei 10°C und 30°C analysiert und in Gruppen eingeordnet. Bei
ausgewählten Isolaten werden zusätzlich bei 10°C und 30°C zum einen die
Chinonanalytik und zum anderen die Carotinoidanalytik angewendet, um die unter
Abschnitt 1.4 beschriebenen weiteren Kälteanpassungsstrategien auf Membranebene
auszuarbeiten.

20 Material & Methoden
2. Material und Methoden
Die verwendeten Chemikalien und Lösungsmittel stammen, sofern nicht zusätzlich
angegeben, von den Firmen Sigma-Aldrich (Taufkirchen, Deutschland) und Merck
KGaA (Darmstadt, Deutschland).
2.1 Herkunft der Bakterienisolate
Als Isolationsquellen dienten zum einen Oberflächen von verschiedenen
Kühlschrankeninnenseiten, Kühlräumwänden sowie ein gekühlter Milchtank und zum
anderen verschiedene, gekühlte Lebensmittelproben.
Die Isolate wurden entweder mit dem Rodac-Plattenverfahren oder dem
Tupferverfahren von der jeweiligen Oberfläche isoliert oder mithilfe des
Oberflächenverfahrens aus dem jeweiligen Lebensmittel isoliert (Bast 1999). Die
Isolate wurden als kälteadaptiert bezeichnet, sobald ein sichtbares Wachstum bei 10°C
nach 7 Tagen stattfand.
In Tabelle 2 werden die jeweiligen kälteadaptierten Bakterienisolate mit Herkunft
aufgelistet.
Tabelle 2: untersuchte Bakterienisolate und deren Herkunft
Isolat Isolationsquelle
J1
Oberfläche der Innenwand eines Kühlschrankes
J2
J3
J5
J6
J7
J8
J9
J10
J11
J12
J16
J17

Material & Methoden 21
J20 frische Tortellini mit Schweinefleischfüllung (gekühlt) J21
J22
J23
J29 Vanillespeiseeis aus Eisdiele J32
J34
J39
Oberfläche der Innenseite eines Kühlschrankes
J40
J44
J45
J48
J50
J52 Schlagsahne aus Sahneaufschlagmaschine einer Eisdiele J53
J54 Bananenspeiseeis von einer Eisdiele J55
J57
J58
Schokoladenspeiseeis von einer Eisdiele J59
J60
J61
J62
Rohmilch aus Milchtank (Forschungsgut Frankenforst der Universität
Bonn)
J63
J64
J65
J66
J67
J68
J69
J70
J71
J72
Tupfer-Abstrich Milchtankablass (Forschungsgut Frankenforst der
Universität Bonn)
J73
J74
J75
J76
J77
J78
J79
J80 Rohmilch Milchtank (Forschungsgut Frankenforst der Universität Bonn) J81
J82 Emmentaler Biomarkt J83
J84 Eichenblattsalat Biomarkt
22 Material & Methoden
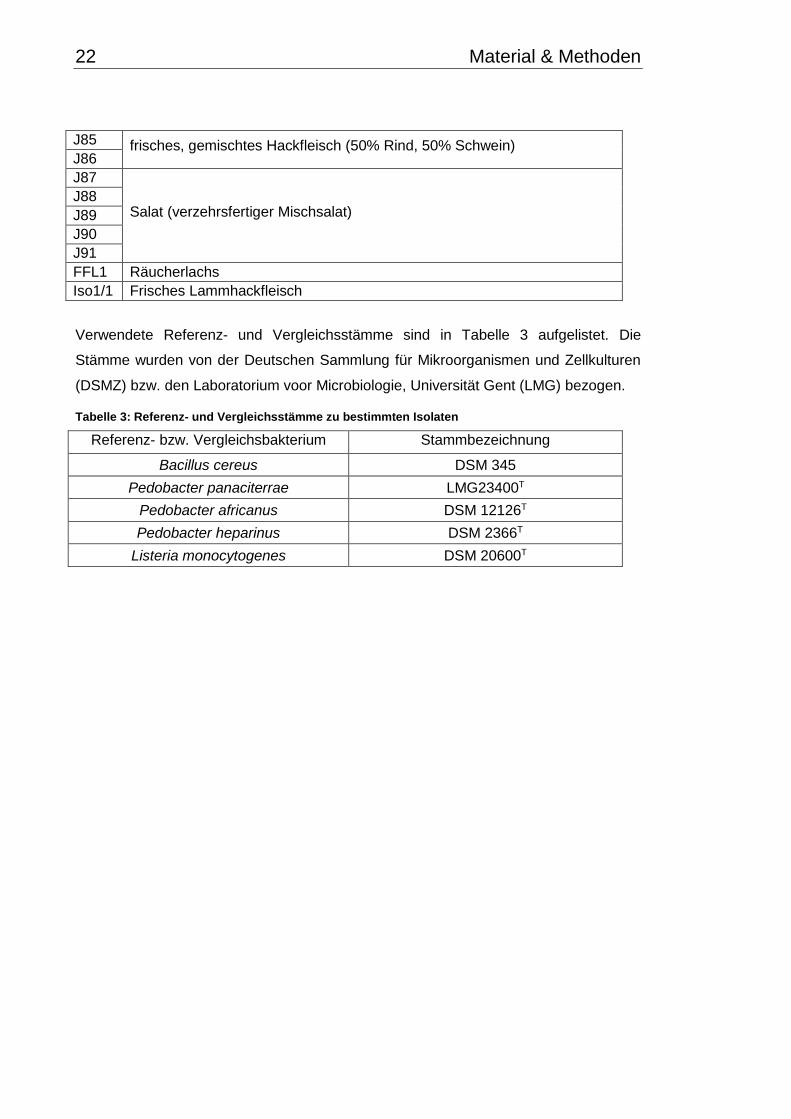
J85 frisches, gemischtes Hackfleisch (50% Rind, 50% Schwein) J86
J87
Salat (verzehrsfertiger Mischsalat) J88
J89
J90
J91
FFL1 Räucherlachs
Iso1/1
11
Frisches Lammhackfleisch
Verwendete Referenz- und Vergleichsstämme sind in Tabelle 3 aufgelistet. Die
Stämme wurden von der Deutschen Sammlung für Mikroorganismen und Zellkulturen
(DSMZ) bzw. den Laboratorium voor Microbiologie, Universität Gent (LMG) bezogen.
Tabelle 3: Referenz- und Vergleichsstämme zu bestimmten Isolaten
Referenz- bzw. Vergleichsbakterium Stammbezeichnung
Bacillus cereus DSM 345
Pedobacter panaciterrae LMG23400T
Pedobacter africanus DSM 12126T
Pedobacter heparinus DSM 2366T
Listeria monocytogenes DSM 20600T

Material & Methoden 23
2.2 Verwendete Medien und Kultivierung der Bakterien
Die Isolate wurden für die Untersuchungen und für die Stammhaltung in Trypton-Soja-
Bouillon (TSB) (Merck KGaA, Darmstadt, Deutschland; Zusammensetzung: Pepton
aus Casein 17,0 g/L, Pepton aus Sojamehl 3,0 g/L, D(+)Glucose-Monohydrat 2,5 g/L,
Natriumchlorid 5,0 g/L di-Kaliumhydrogenphosphat 2,5g/L) oder auf Trypton-Soja-Agar
(TSA) (Merck KGaA, Darmstadt, Deutschland; Zusammensetzung: Pepton aus Casein
15,0g/L, Pepton aus Sojamehl 5,0 g/L, Natriumchlorid 5,0 g/L Agar-Agar 15,0 g/L)
angezogen. Bei der Gattung Listeria wurde Hefeextrakt-Trypton-Soja-Bouillon (HTB)
(Zusammensetzung TSB und 6g/L Hefeextrakt (Oxoid, Hampshire, UK) oder
Hefeextrakt-Trypton-Soja-Agar (HTA) (Zusammensetzung: TSA und 6g/L Hefeextrakt)
zur Kultivierung verwendet.
2.3 Makroskopische und mikroskopische Morphologiebeschreibung
Zur Differenzierung der gewonnenen Reinkulturen wurde neben der
Koloniemorphologie auch die Zellmorphologie mit einem Lichtmikroskop (Modell BH-2,
Olympus, Hamburg, Deutschland untersucht. Die Zellgröße wurde mit einem
Phasenkontrastmikroskop (Axio, Zeiss, Deutschland) bestimmt. Eine weitere
Differenzierung der Isolate erfolgte mittels Gram-Färbung mit anschließender
mikroskopischer Betrachtung (Gram & Friedländer 1884). Die Ergebnisse des
Gram-Färbeverhaltens wurden mit dem KOH-Test (Buck 1982) und dem L-Alanin-
Aminopeptidase-Nachweis mit Bactident® Aminopeptidase-Teststreifen (Merck
KGaA, Darmstadt, Deutschland) überprüft. Außerdem wurde die Katalasereaktion
nach Gerhardt et al. (1981) getestet. Die Oxidasereaktion wurde mittels
Oxidaseteststreifen (Merck KGaA, Darmstadt, Deutschland) bestimmt.
2.4 Temperaturtoleranztest
Für einen Temperaturtoleranztest wurde bei 4°C, 8°C, 10°C, 17°C, 28°C, 30°C, 37°C
und 42°C inkubiert und auf Wachstum untersucht. Als Nährmedium diente Trypton-
Soja-Agar.
2.5 Salzkonzentrationstest
Um die Salztoleranz festzustellen, wurde bei verschiedenen Salzkonzentrationen bei
30°C inkubiert. Die Salzkonzentrationen lagen bei 0,5%, 1%, 1,5%, 2% 3% und 4%.
Als Nährmedium diente R2A-Bouillon (LAB MLimited, Lancashire, UK).

24 Material & Methoden
2.6 Molekularbiologische Methoden
2.6.1 Molekularbiologische Identifizierungen anhand von 16S rRNA-Gen-
Sequenzanalysen
Um die Isolate molekularbiologisch zu identifizieren, wurde das 16S rRNA-Gen
sequenziert. Dafür erfolgte die DNA-Extraktion mit dem Qiagen DNeasy® Blood and
Tissue Kit (Qiagen, Hilden, Deutschland) nach Herstellerangaben.
Die Bakterienbiomasse wurde als Zellpellet aus 1 mL Bakteriensuspension durch
Zentrifugation bei 7.500 x rpm extrahiert, anschließend in 180 µL Lysepuffer (20 mg/mL
Lysozym, 20 mM Tris, pH 8,0, 2 mM EDTA, 1,2% Triton X-100) resuspendiert und 30
Minuten bei 37°C inkubiert.
Durch Zugabe von 200 µL des Puffers AL sowie 25 µl Proteinase K und
anschließender Inkubation für 30 Minuten bei 70°C setzte die Lyse der Zellen ein.
Nach Zusatz von 200 µL Ethanol und gründlichen Vortexen wurde die Probe auf die
Säule pipettiert. Durch einen 1-minütigen Zentifugationsschritt bei 8.000 rpm wurde die
Probe durch den Filter in ein Sammelgefäß gedrückt. Die Nukleinsäuren werden dabei
durch Anwesenheit der chaotropen Salze an die Glasfasern des Filters gebunden. Um
PCR-störende Komponenten, Salze, Proteine sowie zelluläre Verunreinigungen von
der Säule zu waschen, wurde die Säule in ein neues Sammelgefäß überführt, mit 500
µL AW1-Puffer versetzt und erneut bei 8.000 rpm eine Minute zentrifugiert. Ein zweiter
Waschschritt erfolgte auf gleicher Weise mit Waschpuffer AW2. Zum Schluss folgte
noch ein letzter Zentrifugationsschritt bei voller Drehzahl (13.200 rpm), um die Säule zu
trocknen. Die DNA wurde schließlich mit Hilfe von 200 µL AE Puffer und Zentrifugation
von der Säule eluiert und anschließend in einem 1,5 mL-Reaktionsgefäß bei -20°C
gelagert. Die Amplifikation der 16S rRNA-Gen-DNA wurde mit der Polymerase
Kettenreaktion (PCR) durchgeführt. Für den Mastermix wurden 0,5 µl (1U/Ansatz) Taq-
PolymeraseBioTherm, Ares Bioscience GmbH, Köln, Deutschland), 1 µl 10mM dNTP´s
(Bioline GmbH, Luckenwalde, Deutschland), 1,5 µL 50 mM MgCl (BioTherm, Ares
Bioscience GmbH, Köln), 1 µL (8pmol/µL) foward-Primer GM3 (VBC, Biotech, Wien,
Österreich), 1 µL (8pmol/µL) reverse-Primer GM4 (VBC, Biotech, Wien, Österreich),
0,75 µL Bovine Serum Albumin (Promega Corporation, Madison, USA) gemischt und
auf 50 µl Reaktionsvolumen mit hochreinem und autoklaviertem HPLC-Wasser
aufgefüllt. Die PCR wurde mit dem Thermocycler (Mastercycler gradient, Eppendorf,
Hamburg, Deutschland) durchgeführt. Das Temperaturprogramm umfasste 10 Zyklen
mit jeweils der Denaturierungsphase bei 95°C (1 Minute), der Annealingphase des
Primers bei 49°C (1 Minute) und der Polymerisation bei 72°C (1,5 Minuten).

Material & Methoden 25
Anschließend folgten 20 Zyklen mit jeweils einer Minute bei 95°C, einer Minute bei
44°C sowie 1,5 Minuten bei 72 °C.
Zur Überprüfung der erfolgreichen DNA-Isolierung und der PCR-Amplifikation erfolgte
eine Gelelektrophorese mit einem 1%igen Agarosegel (VWR International GmbH,
Darmstadt, Deutschland) bei 70 V für 40 Minuten. 2 µL des DNA-Extrakts wurden dafür
mit 1 µL DNA Loading Buffer (Bioline GmbH, Luckenwalde, Deutschland) vermischt
und in die Geltaschen gegeben. Als Längenstandard wurden 3 µL einer DNA-Leiter
(HyperLadder I, Bioline GmbH, Luckenwalde, Deutschland) mitgeführt. Das Gel wurde
anschließend mit dem 3-fach konzentrierten DNA-Farbstoff GelRed (Biotium, Hayward,
USA ) 20 Minuten gefärbt und konnte unter UV-Licht betrachtet werden.
Bei erfolgreicher Amplifikation wurde das PCR-Produkt mit dem QIAquick PCR
Purification Kit (Qiagen, Hilden, Deutschland) nach Herstellerangaben aufgereinigt.
Nach Zugabe von 250 µL PB Puffer wurde das Gemisch auf eine Säule pipettiert.
Durch Zentrifugation bei 8.000 rpm wurde die DNA an die Glasfasern der Säule
gebunden. Die Säule wurde anschließend in ein neues Sammelgefäß überführt und mit
PE-Puffer gewaschen, um störende Komponenten zu entfernen. Nach Zugabe von
30 µL EB Puffer und einminütiger Inkubationszeit bei Raumtemperatur wurde durch
Zentrifugation das aufgereinigte PCR-Produkt von der Säule gelöst. Dieses wurde
dann in einem 1,5 mL-Reaktionsgefäß bei -20°C gelagert.
5 µL des aufgereinigten PCR-Produktes wurde mit jeweils 2 µL der Primer GM8Rneu
(5‘-GGACTACCAGGGTATCTAAT-3‘) und GM1F (Muyzer et al. 1993) (jeweils 20
pmol/µL) versetzt und zur Sequenzierung der Isolate an die Firma SeqLab (Sequence
Laboratories GmbH, Göttingen, Deutschland) geschickt. Die erhaltenen
Basensequenzen wurden zunächst mit dem Programm Chromas Lite (Technelysium
Pty Ltd, Brisbane, Australien) geprüft und falls nötig manuell editiert. Die
Zusammensetzung der Basensequenzen erfolgte mit dem Programm MEGA 5.0
(Tamura et al. 2007). Zum Schluss wurden die erhaltenen Basensequenzen mit
bekannten Gensequenzen über die Datenbank EzTaxon-e server 2.1 (Kim et al. 2012)
abgeglichen.

26 Material & Methoden
2.6.2 Molekulare Phylogenie-Analyse
Die Konstruktion eines phylogenetischen Stammbaumes wurde ebenfalls mit dem
Programm MEGA 5.0 (Tamura et al. 2007) mittels „Neighbor-Joining“-Analyse (Saitou
und Nei 1987) durchgeführt. Im „Bootstrap“-Verfahren konnten durch wiederholte
Berechnung (1000x) der Baumtopologie mit dem „Neighbor-Joining“-Verfahren Werte
für die statistische Signifikanz ermittelt werden. Um dieses Ergebnis zu bestätigen,
wurde zusätzlich die „Maximum Liklihood“ Analyse und die „Maximum Parsimony“
Analyse durchgeführt.
2.6.3 DNA-DNA Hybridisierung
Durch die DNA-DNA-Hybridisierung kann man den Grad der Ähnlichkeit zwischen
genetischen Pools von DNA-Sequenzen messen. Die Hybridisierung wurde mit
genomischer DNA des zu testenden Isolates und mit DNA des nächstähnlichen
Typstammes durchgeführt. Es wurde die von Ziemke et al. (1998) beschriebene
Methode verwendet, mit der Ausnahme, dass für die Nick-Translation 2 mg DNA
während einer 3-stündigen Inkubation bei 15°C markiert wurde.
Diese Analyseexperimente wurden von Prof. Peter Kämpfer im Institut für angewandte
Mikrobiologie, Abteilung Mikrobiologie der Recyclingprozesse der Universität Giessen,
durchgeführt.

Material & Methoden 27
2.7 Chemotaxonomische Methoden
2.7.1 Fettsäureanalyen
Die Fettsäureprofile der kälteangepassten Isolate wurden gaschromatographisch
bestimmt. Dabei wurde nach der Methode der sauren Methanolyse von Sasser (1990)
und Miller (1982) vorgegangen. Die eingesetzten Lösemittel besaßen HPLC-
Reinheitsgrad. Zunächst wurden die Isolate auf TSA bzw. HT-Agar ausgestrichen und
bei 30°C für 48 Stunden oder bei 10°C für 168 Stunden inkubiert. Anschließend
wurden 40 mg Zellmasse in ein Pyrexröhrchen eingewogen und eine
Fettsäuremethylester-Präparation durchgeführt. Dafür wurde als erstes die
Verseifungsreagenz (15% NaOH in Methanol/bidest. Wasser 1:1 v/v) hinzugegeben
und anschließend bei 100°C gekocht, um die Fettsäuren abzuspalten. Die
anschließende Methylierung erfolgte durch das Methylierungsreagenz (6,0 N
HCl/Methanol 13:11 v/v) und einen weiteren Erhitzungsprozess bei 80°C. Durch
Zugabe von einer Extraktionsreagenz (1:1 Hexan:Methyl-tert-Butylether) wurde eine
Phasentrennung hervorgerufen, durch die die Fettsäuremethylester gewonnen werden
konnten. Nach einer Waschung mit 1,2% NaOH-bidest. H2O-Lösung wurden die
Fettsäuren in ein Glas-Vial überführt und mittels
Gaschromatographie/Massenspektrometrie bestimmt (GC 7890A, GC
Massenspektrometer 5975C, Agilent Technologies, Böblingen). Zur besseren
Detektion hydroxilierter Fettsäuren wurde der Gaschromatograph mit einen FID-
Detektor (GC 6890A, Agilent Technologies, Böblingen, Deutschland) verwendet.
Das Injektionsvolumen betrug 1 µL, welches mit dem Trägergas Helium 5.0 auf die 5%-
ige Phenylmethylsilicon-Kapillarsäule (0,25 mm x 30 m) überführt wurde. Der
Injektorblock besaß eine Temperatur von 250°C und der Säulenofen wurde mit einer
Heizrate von 5°C/min von 120°C auf 240°C erhitzt (Lipski & Altendorf, 1997). Für die
Auswertung wurden die Software ChemStation GCMS Data-Analysis (Agilent
Technologies, Böblingen) verwendet. Die Retentionszeiten der Fettsäuremethylester
wurden mit denen eines Standards (Microbial ID, Newark, USA) abgeglichen und mit
Hilfe des Equivalent-Chain-Length-Wertes (ECL-Wert) über eine hausinterne
Datenbank identifiziert. Zur Überprüfung der Ergebnisse wurde das Massenspektrum
der Peaks ausgewertet. Der prozentuale Gehalt einer Fettsäure wurde durch die
ChemStation Software bestimmt. Die Lagerung der Proben erfolgte bei -20°C.

28 Material & Methoden
2.7.2 Chinonanalysen
Die Analyse des Chinongehaltes in Bakterienzellen wurde nach einer modifizierten
Methode von Hiraishi et al. (1996) und Hu et al. (1999) durchgeführt. Die Gewinnung
der Bakterienbiomasse erfolgte aus 100 mL Zellsuspension durch Zentrifugation für 20
Minuten bei 10.000 g. Es folgte eine zweifache Waschung mit 0,9%-iger NaCl-Lösung.
Zum Teil wurden die Zellpellets in einer Gefriertrocknungsanlage (Christ, Alpha 1-2 LD,
Osterode) plus Vakuumpumpe (vacuubrand, Wertheim) lyophyllisiert, um den
potentiellen Fehlerfaktor durch das Gewicht des Wassers zu eliminieren. Alle
eingesetzten Lösemittel besaßen HPLC-Reinheitsgrad.
Die bakterielle Biomasse wurde mit 5 mL Phosphatpuffer (50 mM, pH 7,4), 10 mL
Methanol und 5 mL Chloroform versetzt. Während des Mischens im Überkopf-Schüttler
für 1,5 Stunden wurden die Chinone aus den Zellen extrahiert. Aufgrund der
Lichtempfindlichkeit der Chinone sowie der Flüchtigkeit der eingesetzten Lösungsmittel
erfolgte dieser Schritt in abgedeckten 50 mL-Pyrexröhrchen mit Teflon-Dichtung.
Anschließend wurden 5 mL bidestilliertes Wasser und 5 mL Chloroform hinzugegeben
und 5 Minuten bei 3.000 g zentrifugiert. Zum Wasserentzug und zur Trocknung des
Lösungsmittels wurde die untere Phase (Chloroform-Methanol-Phase) über
Filterflocken mit wasserfreiem Natriumsulfat filtriert und das Filtrat im
Rotationsverdampfer (CVC 3000 vakuubrand, IKA RV 10 digital, IKA, HB 10 digital,
VWR, Darmstadt) bei 45°C und 100 mbar bis zur Trockne eingeengt.
Zur Extraktion der Chinone wurde anschließend 5 mL bidestilliertes Wasser und 10 mL
Hexan zugesetzt und 5 Minuten geschüttelt. Die Hexanphase wurde in einen neuen
Spitzkolben überführt und die Extraktion im ersten Kolben mit 10 mL Hexan wiederholt.
Die Hexanphasen wurden vereinigt und wieder bis zur Trockne eingeengt. Danach
wurde das Extrakt in 1 mL Hexan wieder aufgenommen und auf eine vorkonditionierte
Silicagelsäule (1 g Sorbent, 6 mL Reservoir, Biotage, Uppsala, Schweden)
aufgetragen. Die Elution der Menachinone erfolgte mit 20 mL eines Hexan-
Diethylether-Gemisches (98:2, v/v). Ubichinone wurden mit 20 mL Hexan-Diethylether
(90:10, v/v) von der Säule gelöst. Die zusammengeführten Extrakte wurden bis zur
Trockne im Rotationsverdampfer eingeengt, in 300 µL Methanol aufgenommen und für
die Analyse in Glas-Vials abgefüllt.
Die Analyse der Chinongehalte erfolgte mittels HPLC (HP Series 1050, Böblingen,
Deutschland). 15 µL der Probe wurden auf die ODS Hypersil RP18-Säule (HP,
Böblingen/Thermo Fisher Scientific, Massachusetts, USA) injiziiert, die durch den Ofen
(Mistral, Spark Holland, Emmen, Niederlande) auf einer Temperatur von 30°C gehalten
wurde. Als isokratisches Laufmittel wurde ein Gemisch aus Methanol und

Material & Methoden 29
Diisopropylether (9:2, v/v) mit einer Flussrate von 1 mL/min eingesetzt. Die Detektion
erfolgte über einen Diodenarray-Detektor (HP Series 1050, Böblingen, Deutschland)
bei 275 nm. Bis zu ihrer Verwendung wurden die Proben bei -20°C abgedunkelt
gelagert.
Zur Auswertung wurde die Software ChemStation (HP, Böblingen, Deutschland)
verwendet. Die auftretenden Peaks konnten über die Retentionszeiten und ihre
charakteristischen UV-Spektren identifiziert werden. Nach der Identifizierung der Ubi-
und Menachinone über das charakteristische UV-Spektrum konnte über das Verhältnis
der Retentionszeiten von Probe zu mitgeführtem Standard und mit Hilfe einer internen
Auswertungstabelle die genaue Chinonart mit der Anzahl an Isopreneinheiten ermittelt
werden. Als Standard wurde Ubichinon Q10 sowie das Phyllochinon Vitamin K1 mit
jeweils einer Konzentration von 100 nmol/mL in Hexan Methanol (1:2, v/v) eingesetzt.
Für die quantitative Bestimmung wurden die Peakflächen von Probe und mitgeführtem
Standard zur Berechnung herangezogen. Die Fläche unter dem Peak gibt die Menge
an absorbiertem Licht in milli-Absorbance-Units mal Sekunde (mAU*s) an. Über die
bekannte Konzentration des Standards und die eingesetzten Biomassenmenge konnte
der Chinongehalt pro Gramm Biomasse bzw. Lyophylisat mit Hilfe einer Formel
berechnet werden.
𝐶ℎ𝑖𝑛𝑜𝑛𝑔𝑒ℎ𝑎𝑙𝑡 𝑖𝑛 𝑑𝑒𝑟 𝐵𝑖𝑜𝑚𝑎𝑠𝑠𝑒 [𝑛𝑚𝑜𝑙
𝑔]
= 𝑃𝑒𝑎𝑘𝑓𝑙ä𝑐ℎ𝑒 𝑑𝑒𝑟 𝑃𝑟𝑜𝑏𝑒 [𝑚𝐴𝑈 ∗ 𝑠] × 100
𝑛𝑚𝑜𝑙𝑚𝑙
× 0,3 𝑚𝑙
𝑃𝑒𝑎𝑘𝑓𝑙ä𝑐ℎ𝑒 𝑆𝑡𝑎𝑛𝑑𝑎𝑟𝑑 [𝑚𝐴𝑈 ∗ 𝑠] × 𝐵𝑖𝑜𝑚𝑎𝑠𝑠𝑒 [𝑔]
Biomasse in g: Nassgewicht bzw. Trockengewicht
0,3 mL: aufgenommen in 300µl Methanol

30 Material & Methoden
2.7.3 Kombinierte polare Lipid- und Chinonanalyse
Dieses Verfahren ist eine kombinierte Methode und nach Minnikin et al. (1984)
modifiziert. Alle Lösungsmittel hatten HPLC-Reinheitsgrad.
In einem ersten Schritt wurden aus lyophylisierter Biomasse mit einen wässrigem
Methanol/Petroleumether (60-80°C)-Gemisch (1:1, v/v) die Chinone extrahiert. Der
Rückstand wurde in Aceton aufgenommen und mittels Dünnschichtchromatographie
auf einer Kieselgelplatte 60F254 getrennt. Als Standard für Menachinone wurde Vitamin
K1, als Standard für Ubichinone wurde Coenzym Q10 mit aufgetragen. Als Laufmittel
diente Petroleumether/Diethylether (85:15, v/v). Das Ergebnis wurde unter einer UV-
Lampe betrachtet. Die getrennten Mena- und Ubichinone wurden dann aus der
Kieselgelplatte eluiert und in der HPLC nach Hu et al. (1999) analysiert. Als
Elutionsmittel für Menachinone diente Diethylether und für Ubichinone Aceton.
In einem zweiten Schritt wurde das Lyophylisat nach der Chinonextraktion weiter mit
Chloroform/Methanol/0,3% NaCl-Lösung (9:10:3, v/v) behandelt, um die polaren Lipide
aus der Zellmembran zu extrahieren. Das gewonnene Extrakt wurde auf eine
Silicagelplatte aufgetragen und zweidimensional entwickelt. Als erstes Laufmittel wurde
Chloroform/Methanol/Wasser (65:25:4, v/v) und als zweites Laufmittel
Chloroform/Essigsäure/Methanol/Wasser (80:15:12:4, v/v) verwendet. Die getrocknete
Kieselgelplatte wurde zuerst mit Ninhydrinsprühreagenz besprüht und 5 Minuten bei
100°C inkubiert, um die Lipide mit freier Aminogruppe sichtbar zu machen. Nach einer
kurzen Abkühlung wurden die Platten mit Molybdänblau-Reagenz besprüht und
ebenfalls für 5 Minuten bei 100°C inkubiert. Entstandene blaue Flecken
kennzeichneten Phospholipide.
Es wurde jeweils eine Dokumentationsplatte unter den gleichen Bedingungen
angefertigt. Diese Platte wurde jedoch nicht mit den verschiedenen Färbereagenzien
besprüht, sondern mit 30% Schwefelsäure behandelt und anschließend erhitzt. Die
sich hier bildenden braunen Flecken kennzeichnen alle auf der Platte aufgetrennten
polaren Lipide.

Material & Methoden 31
2.7.4 Carotinoidextraktion
Die Carotinoidextraktionsmethode modifiziert nach Kaiser et al. (2007) und Rezwan et
al. (2007) stellte sich in der Abschlussarbeit von Jacobsen (2013) als effektivste
Extraktionsmethode heraus und würde daher als Carotinoidanalysemethode
angewendet.
Der Zellaufschluss wurde mit Lysozym und Ultraschall durchgeführt, um anschließend
die Carotinoide mit den Lösungsmitteln Methanol und Chloroform zu extrahieren. Alle
verwendeten Lösemittel besaßen HPLC-Reinheitsgrad.
Die Herstellung der Lysozym- (Sigma-Aldrich, Taufkirchen, Deutschland, Artikelnr:
L6876) und Lipase- (Sigma-Aldrich, Taufkirchen, Deutschland, Artikelnr: L1754)-
Lösung mit einer Konzentration von 2,4 mg/mL erfolgte mit einem Phosphatpuffer mit
pH 7,0. Die zur Extraktion benötigten Lösungsmittel Chloroform und das Chloroform-
Methanol-Gemisch wurden mit der Zugabe von 0,1% Butylhydroxytoluol (Sigma-
Aldrich, Taufkirchen, Deutschland) stabilisiert. Es wurden 20-50 mg Zelllyophilisat in
50 mL-Pyrexröhrchen eingewogen und in 5 mL Lysozym- und Lipase-Lösung
resuspendiert. Für den Zellaufschluss wurde die Suspension für zwei Stunden bei 37°C
unter Schütteln inkubiert und anschließend für 60 Minuten im Ultraschallbad (Sonorex,
Bandelin electronic, Berlin) behandelt. Nach dem Zellaufschluss wurde die
Zellsuspension in Zentrifugenröhrchen überführt und das Zellpellet durch Zentrifugation
für 10 Minuten bei 12.000 g und 4°C für die Carotinoidextraktion gewonnen.
Für die Extraktion der Carotinoide wurden 4 mL Methanol/Chloroform-Gemisch (7:3,
v/v) zum Zellpellet gegeben und gevortext. Nach einer Zentrifugation für 10 Minuten bei
12.000 g und 4°C wurde der Überstand, bestehend aus dem Extrakt mit den
Carotinoiden, über Filterflocken in ein 50 mL-Pyrexröhrchen überführt. Die Extraktion
wurde je nach Isolat drei- bis viermal wiederholt, bis der Extrakt farblos erschien. Zur
Gewinnung der Chloroformphase wurde den vereinigten Extrakten 10 mL 10%ige
NaCl-Lösung und 3 mL Chloroform hinzugefügt und die Mischung gevortext. Über
einen Zentrifugationsschritt für 5 Minuten bei 4000 rpm und 4°C konnte eine
Phasentrennung erreicht werden. Die untere Chloroform-Phase wurde in einen 25 mL
Spitzkolben überführt. Die Extraktion der Carotinoide aus der wässrigen Phase wurde
wiederholt, bis beide Phasen farblos waren.
Aus den vereinigten Extrakten wurde mit dem Rotationsverdampfer bei 100 mbar und
20-35°C das Lösungsmittel entfernt und der Rückstand wurde für die photometrische
und chromatographische Carotinoidbestimmung in Hexan aufgenommen.

32 Material & Methoden
2.7.4.1 Quantitative Carotinoidbestimmung
Die quantitative Carotinoidbestimmung erfolgte über eine photometrische und eine
chromatographische Methode.
Für die photometrische Bestimmung der Carotinoidkonzentration in den Proben
wurden die Lösungsmittel nach der Extraktion mit dem Rotationsverdampfer bei
100 mbar bei 30°C entfernt und die Rückstände in 1-5 mL Hexan aufgenommen. Das
Hexanvolumen war von der Rückstandsmenge abhängig, um eine Absorption von <1,0
im Spektralphotometer zu erreichen. Nach Rodriguez-Amaya (2001) wurden die
Lösungen bei einer Wellenlänge von 450 nm mit dem Spektralphotometer (Genesys
10uv Scanning, Thermo Fisher Scientific, Massachusetts, USA) gemessen. Die
Berechnung der Carotinoidmenge in der Probe in µg und der Carotinoidkonzentration
als Carotinoidmenge pro Gramm Lyophylisat wurden nach Rodriguez-Amaya (2001)
berechnet. Für ein Gemisch aus verschiedenen unbekannten Carotinoiden wird nach
Britton (1995) die Verwendung eines spezifischen Absorptionskoeffizienten von
2500 x 100𝑚𝐿
𝑔∙𝑐𝑚 vorgeschlagen.
𝑥[µ𝑔] =𝐴 ∙ 𝑦[𝑚𝐿] ∙ 106
𝐴1𝑐𝑚1% ∙ 100
𝑥[µ𝑔
𝑔] =
𝑥[µ𝑔]
𝐸𝑖𝑛𝑤𝑎𝑎𝑔𝑒 [𝑔]
x = Carotinoidmenge in µg bzw. -konzentration in µg/g Trockengewicht
y = Volumen an Lösungsmittel in mL
A = Absorption bei 450 nm
𝐴1𝑐𝑚1% = spezifischer Absorptionskoeffizient (2500
100𝑚𝐿
𝑔∙𝑐𝑚)
Für die chromatographische Bestimmung des Carotinoidgehaltes und zur Auftrennung
des Carotinoidgemisches wurden die Extrakte aus der photometrischen Bestimmung
erneut mit dem Rotationsverdampfer eingedampft. Nach der Entfernung des
Lösungsmittels wurde der Rückstand in 200 µl Hexan aufgenommen, in Glas-Vials
überführt und bis zur Messung bei -20°C gelagert.
Die chromatographische Analyse der Carotinoide wurde nach der modifizierten
Methode von Rählert (2007) durchgeführt. Die Detektion erfolgte wieder über das
HPLC-System mit einen Diodenarray-Detektor (HP Series 1050, Böblingen). 20 µL der
Probe wurden in das HPLC-System injiziert. Die Auftrennung der Analyten erfolgte
über eine auf 20°C temperierte ODS Hypersil RP18-Säule (Thermo Fisher Scientific,

Material & Methoden 33
Massachusetts, USA). Als mobile Phase wurde ein Gemisch aus Acetonitril, Methanol
und Isopropanol im Verhältnis 85:10:5 mit einer Flussrate von 0,8mL/min eingesetzt.
Der Diodenarray-Detektor detektierte die aufgetrennten Analyten bei einer Wellenlänge
von 440 nm. Zur Auswertung wurde die Software ChemStation 2.0 (HP, Böblingen)
verwendet. Die erhaltenen Peaks wurden über ihre charakteristischen
Absorptionspektren als Carotinoide identifiziert und der Gesamtcarotinoidgehalt mithilfe
des mitgeführten β-Carotin-Standards berechnet.
Über den Vergleich der gemessen Peakfläche des Standards mit bekannter
Konzentration und der Gesamtpeakfläche der Carotinoide in der Probe wurde die
Carotinoidkonzentration des Extraktes in mg/L nach der nachstehenden Formel
berechnet (Matissek et al. 2010).
𝑋[𝑚𝑔
𝐿] =
𝐹𝑙ä𝑐ℎ𝑒𝑃𝑟𝑜𝑏𝑒 ∙ 𝑚𝑆𝑡𝑑
𝐹𝑙ä𝑐ℎ𝑒𝑆𝑡𝑑 ∙ 𝑉𝐷
X = Carotinoidkonzentration in mg/L
Fläche (Probe) = Gesamtpeakfläche der Probe
Fläche (Std) = Peakfläche Standard
VD = Verdünnung des Standards für die chromatographische Analyse
mStd = Konzentration des Standards in mg/L
Über die Carotinoidkonzentration in mg/L in dem Probenextrakt konnte der
Carotinoidgehalt in der Probe in µg und der Carotinoidgehalt bezogen auf die
Einwaage (Lyophilisat) berechnet werden.
𝑥[µ𝑔] =𝑋[
𝑚𝑔𝐿 ]
106 ∙ 𝑦[µ𝑙]∙ 103
𝑥[µ𝑔
𝑔] =
𝑥[µ𝑔]
𝐸𝑖𝑛𝑤𝑎𝑎𝑔𝑒 [𝑔]
X = Carotinoidkonzentration der Probe in mg/L
x = Carotinoidmenge in der Probe in µg bzw. -konzentration in µg/g
y = Volumen an Lösungsmittel in µl

34 Material & Methoden
2.8 Messungen des Zellertrages
2.8.1 Messung des Wachstumsverhaltens mittels optischer Dichte
Zur Bestimmung des bakteriellen Zellertrages wurde von ausgewählten Isolaten eine
Wachstumskurve erstellt. Somit konnte man den bakteriellen Wachstumsverlauf
verfolgen. Hierfür wurde die indirekte Methode der Trübungsmessung gewählt. Dabei
wird die optische Dichte (OD) einer Bakteriensuspension, die innerhalb bestimmter
Grenzwerte proportional zur Zellzahl ist, photometrisch gemessen (Madigan et al.
2008). Als Nährmedium wurde TSB bzw. HT-Bouillon verwendet. Die optische Dichte
wurde bei einer Wellenlänge von 625 nm im Spektralphotometer (Genesys 10uv
Scanning, Thermo Fisher Scientific, Massachusetts, USA) gemessen.
Im Verlauf dieser Messung wurde die optische Dichtung in regelmäßigen Abständen
bestimmt. Damit konnten die unterschiedlichen Wachstumsphasen möglichst genau
bestimmt werden. Besonders relevant waren die Zeitpunkte der spätexponentiellen
Wachstumsphase und der stationären Wachstumsphase, weil in diesen Phasen die
optische Dichte am höchsten war. Danach wurde die Messung beendet, da die
Bakterien die stationäre Phase erreicht hatten und kein Anstieg mehr erfolgte.
Die Wachstumsgeschwindigkeit während der exponentiellen Vermehrung wurde über
die Wachstumsrate µ berechnet (Schlegel et al. 2007)
µ = ln(𝑥𝑡) − ln(𝑥0)
𝑡 − 𝑡0
µ: exponentielle Wachstumsrate einer statischen Kultur [h-1]
xt: Zelldichte zum Zeitpunkt t [hier: OD]
x0: Zelldichte zum Zeitpunkt t0 [hier: OD]
t: Zeit am Ende der exponentiellen Wachstumsphase [h]
t0: Zeit zu Beginn der exponentiellen Wachstumsphase [h]
2.8.2 Messung des Proteingehaltes
Zur Messung des bakteriellen Zellertrages wurde zusätzlich neben der Ermittlung der
optischen Dichte eine Proteinbestimmung nach Biuret (Bast 1999) durchgeführt.
Die ausgewählten Isolate wurden in Flüssigkultur (TSB bzw. HT-Bouillon) bis zu dem
Zeitpunkt des Erreichens der spätexponentiellen Wachstumsphase bei der jeweiligen
Temperatur inkubiert.

Material & Methoden 35
5 mL dieser Flüssigkultur wurden abgenommen, abzentrifugiert, mit 0,9%-iger NaCl-
Lösung zweimal gewaschen und anschließend die Gesamtproteinmenge des
entstandenen Zellpellets bestimmt. Dafür wurde das jeweilige Zellpellet mit 3 molarer
Natronlauge versetzt und anschließend 5 Minuten bei 100°C gekocht. Danach wurde
2,5% Kupfersulfat-Lösung hinzupipettiert und der Ansatz bei Raumtemperatur für 30
Minuten stehengelassen. Der Ansatz wurde zentrifugiert und der Überstand bei einer
Wellenlänge von 555 nm im Spektralphotometer gemessen.
Zusätzlich zu den Proben wurde eine Eichgerade erstellt. Die gemessenen Extinktion
der entsprechend eingestellten Proteinmengen (in mg pro 5 mL Probenansatz) wurden
als Eichkurve graphisch dargestellt. Somit konnte die Proteinmenge pro Probenansatz
ermittelt werden. Das eingesetzte Volumen wurde dividiert, um den Proteingehalt pro
mL Ausgangssuspension zu erhalten.

36 Material & Methoden
2.9 Kälteresistenztest
Bei dem Kälteresistenztest wurde die Resistenz gegenüber Einfrier- und Auftaustress
bestimmter Listerienstämme untersucht.
Dafür würde eine Bakterienkultur des jeweiligen Listerienstammes in HT-Bouillon bis
zur spätexponentiellen Wachstumsphase bei 6 °C wachsen gelassen und die Zellzahl
kulturell im Oberflächenverfahren bestimmt. Danach würde im Dreifachansatz jeweils
1 mL dieser kälteadaptierten Bakteriensuspension bei -20°C für 20 Stunden
eingefroren, anschließend wieder aufgetaut und die Zellzahl erneut kulturell im
Oberflächenverfahren bestimmt. Ein zweiter Dreifachansatz wurde zusätzlich bei -20°C
für 20 Stunden eingefroren, aufgetaut, erneut wieder für 20 Stunden bei -20°C
eingefroren und aufgetaut und dann die Zellzahl kulturell im Oberflächenverfahren
bestimmt. Das Verhältnis der Lebendzellzahl pro mL vor und nach der Behandlung
wurde als Maß für die Stressresistenz gegenüber kalten Bedingungen verwendet.
2.10 Überlebenstest - Zellertragsmessung bei 10°C und 30°C in einem
Modelllebensmittel
Als Modelllebensmittel diente 3,5% ultrahocherhitzte Milch. Isolat Iso 1/11 wurde als
Versuchskeim verwendet. Als Vergleichskeim wurde der Typstamm Listeria
monocytogenes DSM 20600T ausgewählt.
Für den Versuch wurden für jeden Keim jeweils 1 Liter Milch mit 108 KbE/mL inokuliert
(Endkonzentration 105 KbE/mL Milch) und parallel bei 10°C und 30°C inkubiert. Als
Negativkontrolle diente bei beiden Temperaturen 1L Milch, die nicht inokuliert wurde.
Die Ausgangskeimzahl wurde kulturell im Oberflächenverfahren bestimmt. Danach
wurden in bestimmten Abständen die Keimzahlen bestimmt, bis die Absterbephase der
Wachstumskurve einsetzte.

Material & Methoden 37
2.11 Physiologisches Reaktionsprofil
Um das biochemische Reaktionsprofil zu bestimmen, wurden verschiedene
biochemische Tests durchgeführt.
Als erster Test wurde der API 20 E®-Test durchgeführt. Es ist ein biochemischer Test,
der zur Identifizierung von Enterobacteriaceae und gram-negativen Stäbchenbakterien
(BioMèrieux, Marcy l'Etoile, Frankreich) verwendet wird. Der Test wurde nach Angaben
des Herstellers durchgeführt.
Zur Analyse der Kohlenhydratverwertung wurde der API50CH Test (BioMèrieux, Marcy
l'Etoile, Frankreich) durchgeführt. Der Test wurde nach Angaben des Herstellers
durchgeführt. Jedoch wurde das CHB-Medium, welches laut Hersteller für die Gattung
Bacillus verwendet werden soll auch für gramnegative Stäbchenbakterien benutzt. Ein
positives Ergebnis wird durch einen Farbumschlag nach Gelb angezeigt; dann wurde in
Folge der Verwertung des Kohlenhydrats Säure gebildet.
Der GN2 Microplate Test (Biolog, Hayward, USA) diente zur Identifikation und
Charakterisierung von aeroben gramnegativen Bakterien. Er beruht auf der Reduktion
von Tetrazolium, welches bei einem aktiven Metabolismus reduziert wird. Es wurden in
einer 96-well Mikrotiterplatte 95 verschiedene Kohlenstoffquellen angeboten. Bei
Verstoffwechselung einer Kohlenstoffquelle kommt es zu einem Farbumschlag, der im
Mikrotiterplattenspektralphotometer (Epoch BioTekR, Winooski, USA) bei 550 nm
gemessen wird.
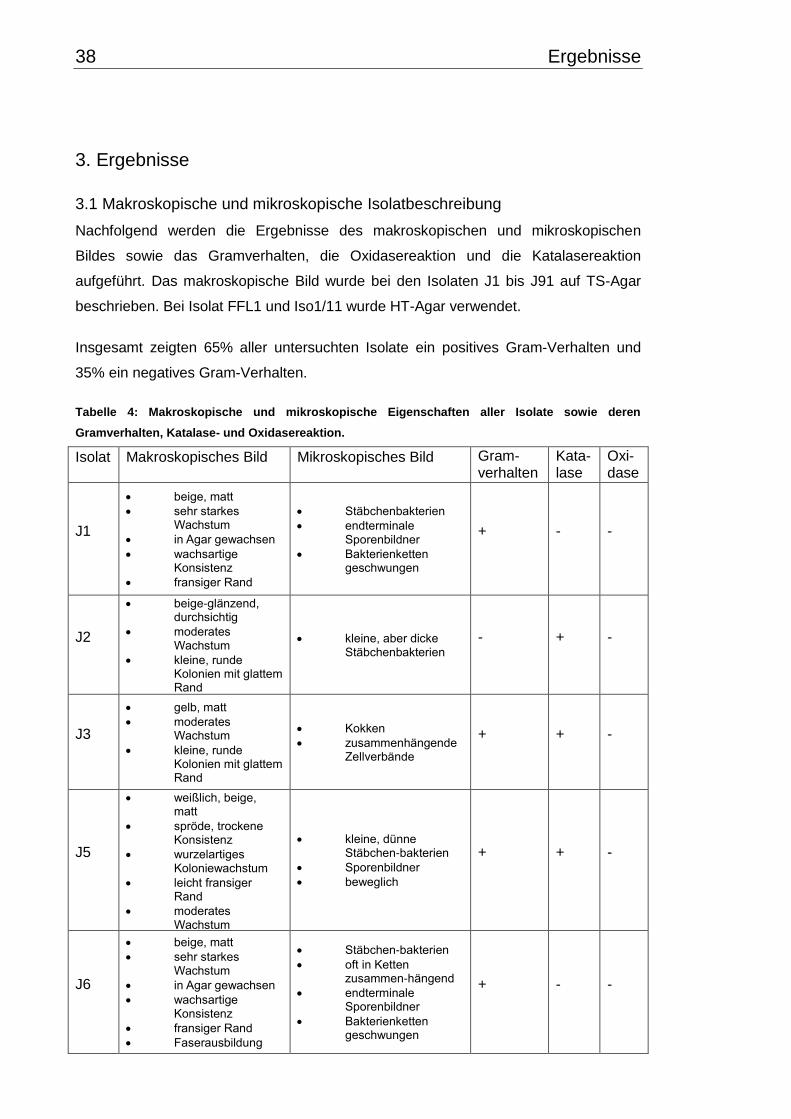
38 Ergebnisse
3. Ergebnisse
3.1 Makroskopische und mikroskopische Isolatbeschreibung
Nachfolgend werden die Ergebnisse des makroskopischen und mikroskopischen
Bildes sowie das Gramverhalten, die Oxidasereaktion und die Katalasereaktion
aufgeführt. Das makroskopische Bild wurde bei den Isolaten J1 bis J91 auf TS-Agar
beschrieben. Bei Isolat FFL1 und Iso1/11 wurde HT-Agar verwendet.
Insgesamt zeigten 65% aller untersuchten Isolate ein positives Gram-Verhalten und
35% ein negatives Gram-Verhalten.
Tabelle 4: Makroskopische und mikroskopische Eigenschaften aller Isolate sowie deren
Gramverhalten, Katalase- und Oxidasereaktion.
Isolat Makroskopisches Bild Mikroskopisches Bild Gram-verhalten
Kata-lase
Oxi-dase
J1
beige, matt
sehr starkes Wachstum
in Agar gewachsen
wachsartige Konsistenz
fransiger Rand
Stäbchenbakterien
endterminale Sporenbildner
Bakterienketten geschwungen
+ - -
J2
beige-glänzend, durchsichtig
moderates Wachstum
kleine, runde Kolonien mit glattem Rand
kleine, aber dicke Stäbchenbakterien
- + -
J3
gelb, matt
moderates Wachstum
kleine, runde Kolonien mit glattem Rand
Kokken
zusammenhängende Zellverbände
+ + -
J5
weißlich, beige, matt
spröde, trockene Konsistenz
wurzelartiges Koloniewachstum
leicht fransiger Rand
moderates Wachstum
kleine, dünne Stäbchen-bakterien
Sporenbildner
beweglich
+ + -
J6
beige, matt
sehr starkes Wachstum
in Agar gewachsen
wachsartige Konsistenz
fransiger Rand
Faserausbildung
Stäbchen-bakterien
oft in Ketten zusammen-hängend
endterminale Sporenbildner
Bakterienketten geschwungen
+ - -
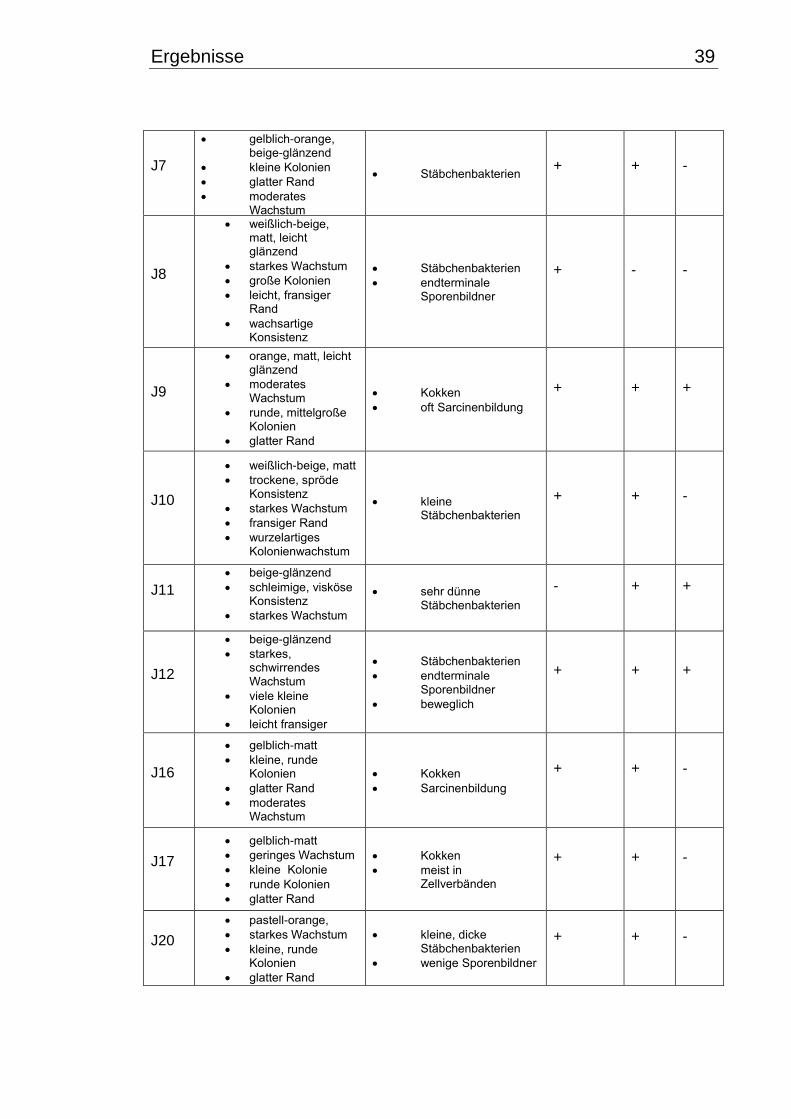
Ergebnisse 39
J7
gelblich-orange, beige-glänzend
kleine Kolonien
glatter Rand
moderates Wachstum
Stäbchenbakterien + + -
J8
weißlich-beige, matt, leicht glänzend
starkes Wachstum
große Kolonien
leicht, fransiger Rand
wachsartige Konsistenz
Stäbchenbakterien
endterminale Sporenbildner
+ - -
J9
orange, matt, leicht glänzend
moderates Wachstum
runde, mittelgroße Kolonien
glatter Rand
Kokken
oft Sarcinenbildung
+ + +
J10
weißlich-beige, matt
trockene, spröde Konsistenz
starkes Wachstum
fransiger Rand
wurzelartiges Kolonienwachstum
kleine Stäbchenbakterien
+ + -
J11
beige-glänzend
schleimige, visköse Konsistenz
starkes Wachstum
sehr dünne Stäbchenbakterien
- + +
J12
beige-glänzend
starkes, schwirrendes Wachstum
viele kleine Kolonien
leicht fransiger Rand
Stäbchenbakterien
endterminale Sporenbildner
beweglich
+ + +
J16
gelblich-matt
kleine, runde Kolonien
glatter Rand
moderates Wachstum
Kokken
Sarcinenbildung
+ + -
J17
gelblich-matt
geringes Wachstum
kleine Kolonie
runde Kolonien
glatter Rand
Kokken
meist in Zellverbänden
+ + -
J20
pastell-orange,
starkes Wachstum
kleine, runde Kolonien
glatter Rand
kleine, dicke Stäbchenbakterien
wenige Sporenbildner
+ + -
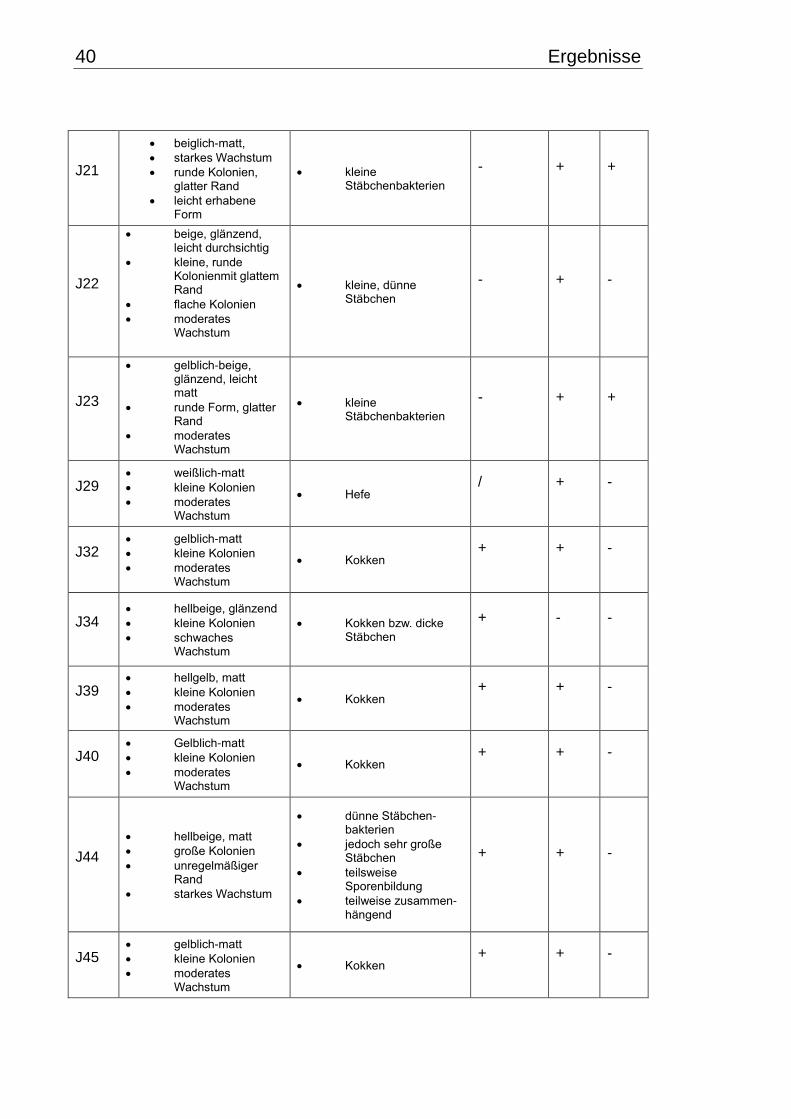
40 Ergebnisse
J21
beiglich-matt,
starkes Wachstum
runde Kolonien, glatter Rand
leicht erhabene Form
kleine Stäbchenbakterien
- + +
J22
beige, glänzend, leicht durchsichtig
kleine, runde Kolonienmit glattem Rand
flache Kolonien
moderates Wachstum
kleine, dünne Stäbchen
- + -
J23
gelblich-beige, glänzend, leicht matt
runde Form, glatter Rand
moderates Wachstum
kleine Stäbchenbakterien
- + +
J29 weißlich-matt
kleine Kolonien
moderates Wachstum
Hefe / + -
J32 gelblich-matt
kleine Kolonien
moderates Wachstum
Kokken + + -
J34 hellbeige, glänzend
kleine Kolonien
schwaches Wachstum
Kokken bzw. dicke Stäbchen
+ - -
J39 hellgelb, matt
kleine Kolonien
moderates Wachstum
Kokken + + -
J40 Gelblich-matt
kleine Kolonien
moderates Wachstum
Kokken + + -
J44
hellbeige, matt
große Kolonien
unregelmäßiger Rand
starkes Wachstum
dünne Stäbchen-bakterien
jedoch sehr große Stäbchen
teilsweise Sporenbildung
teilweise zusammen-hängend
+ + -
J45 gelblich-matt
kleine Kolonien
moderates Wachstum
Kokken + + -
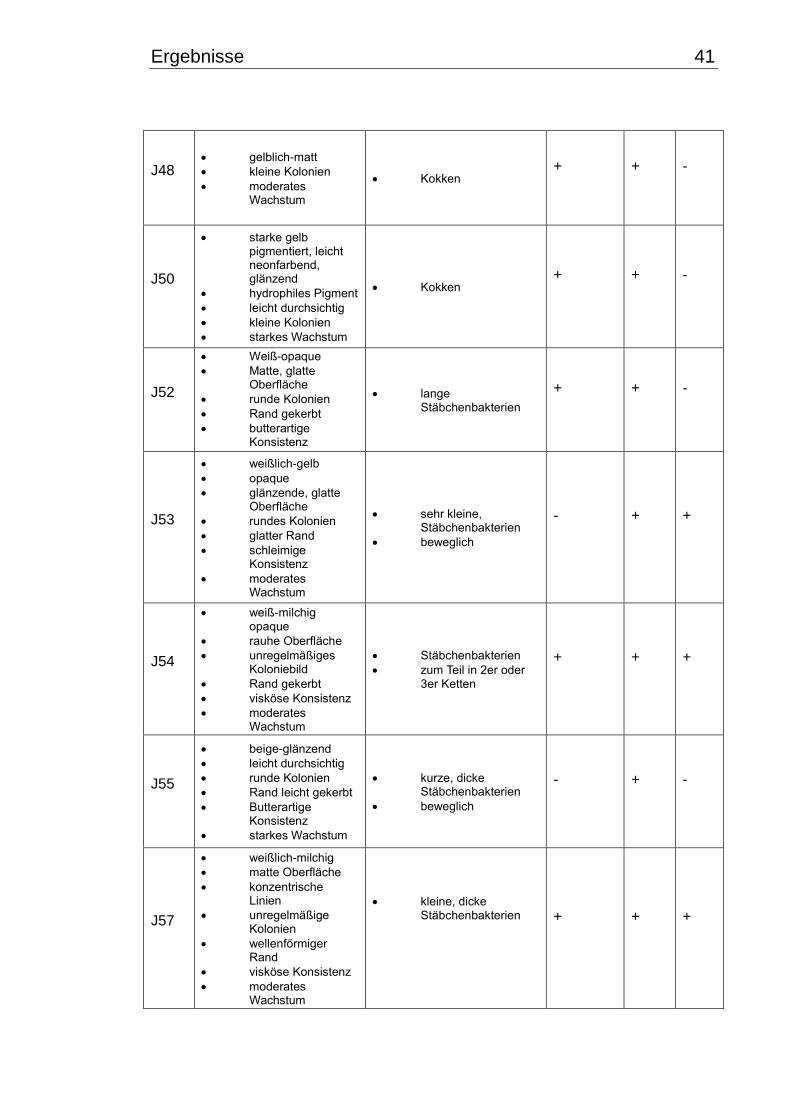
Ergebnisse 41
J48 gelblich-matt
kleine Kolonien
moderates Wachstum
Kokken + + -
J50
starke gelb pigmentiert, leicht neonfarbend, glänzend
hydrophiles Pigment
leicht durchsichtig
kleine Kolonien
starkes Wachstum
Kokken + + -
J52
Weiß-opaque
Matte, glatte Oberfläche
runde Kolonien
Rand gekerbt
butterartige Konsistenz
moderates Wachstum
lange Stäbchenbakterien
+ + -
J53
weißlich-gelb
opaque
glänzende, glatte Oberfläche
rundes Kolonien
glatter Rand
schleimige Konsistenz
moderates Wachstum
sehr kleine, Stäbchenbakterien
beweglich
- + +
J54
weiß-milchig opaque
rauhe Oberfläche
unregelmäßiges Koloniebild
Rand gekerbt
visköse Konsistenz
moderates Wachstum
Stäbchenbakterien
zum Teil in 2er oder 3er Ketten
+ + +
J55
beige-glänzend
leicht durchsichtig
runde Kolonien
Rand leicht gekerbt
Butterartige Konsistenz
starkes Wachstum
kurze, dicke Stäbchenbakterien
beweglich
- + -
J57
weißlich-milchig
matte Oberfläche
konzentrische Linien
unregelmäßige Kolonien
wellenförmiger Rand
visköse Konsistenz
moderates Wachstum
kleine, dicke Stäbchenbakterien
+ + +
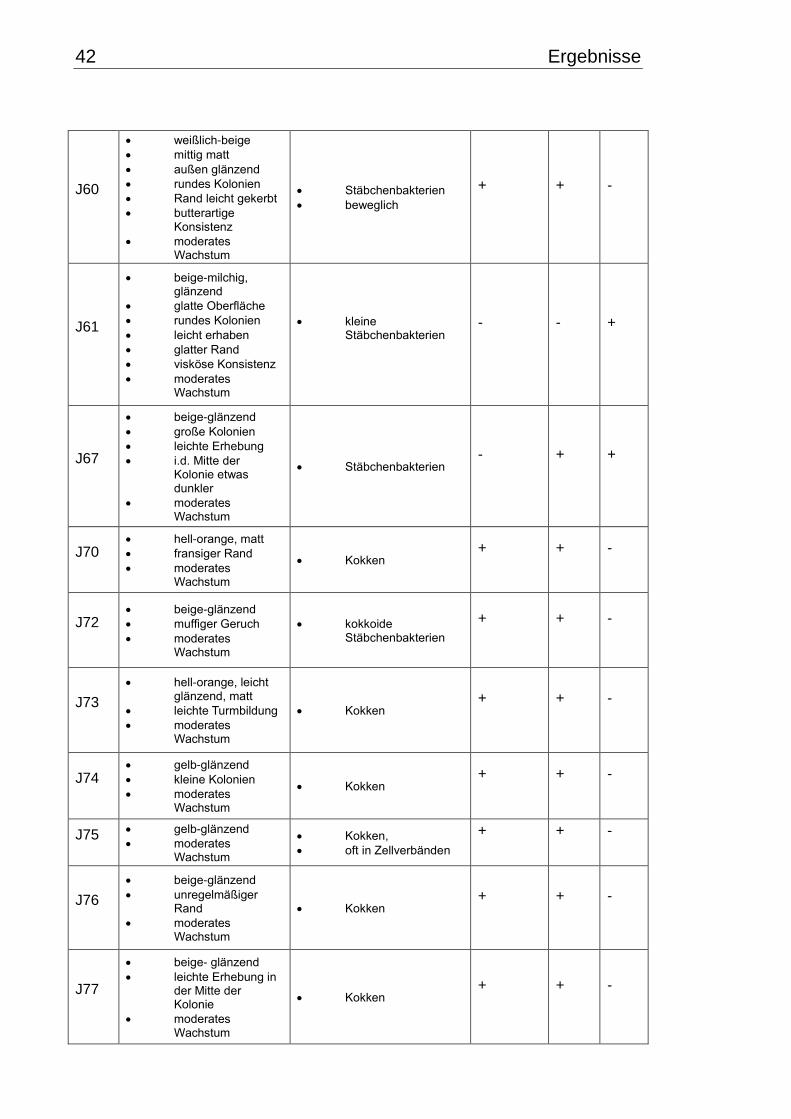
42 Ergebnisse
J60
weißlich-beige
mittig matt
außen glänzend
rundes Kolonien
Rand leicht gekerbt
butterartige Konsistenz
moderates Wachstum
Stäbchenbakterien
beweglich
+ + -
J61
beige-milchig, glänzend
glatte Oberfläche
rundes Kolonien
leicht erhaben
glatter Rand
visköse Konsistenz
moderates Wachstum
kleine Stäbchenbakterien
- - +
J67
beige-glänzend
große Kolonien
leichte Erhebung
i.d. Mitte der Kolonie etwas dunkler
moderates Wachstum
Stäbchenbakterien - + +
J70 hell-orange, matt
fransiger Rand
moderates Wachstum
Kokken + + -
J72 beige-glänzend
muffiger Geruch
moderates Wachstum
kokkoide Stäbchenbakterien
+ + -
J73
hell-orange, leicht glänzend, matt
leichte Turmbildung
moderates Wachstum
Kokken + + -
J74 gelb-glänzend
kleine Kolonien
moderates Wachstum
Kokken + + -
J75 gelb-glänzend
moderates Wachstum
Kokken,
oft in Zellverbänden
+ + -
J76
beige-glänzend
unregelmäßiger Rand
moderates Wachstum
Kokken + + -
J77
beige- glänzend
leichte Erhebung in der Mitte der Kolonie
moderates Wachstum
Kokken + + -
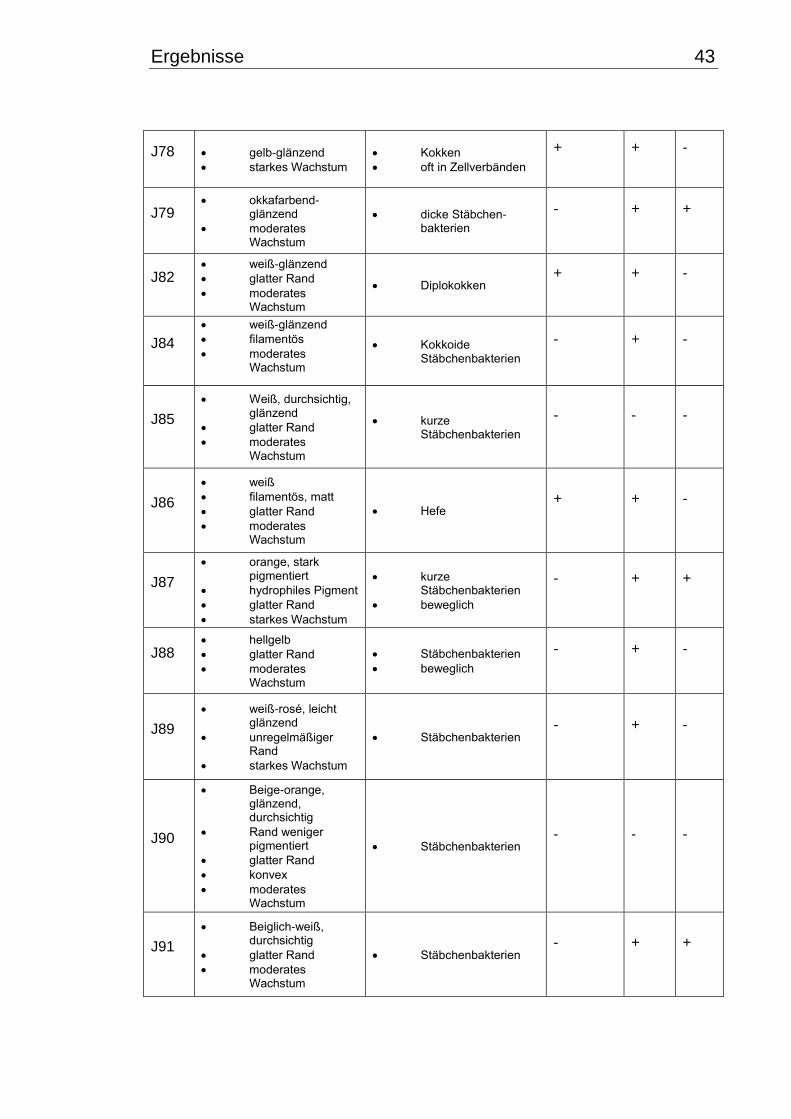
Ergebnisse 43
J78 gelb-glänzend
starkes Wachstum
Kokken
oft in Zellverbänden
+ + -
J79 okkafarbend-
glänzend
moderates Wachstum
dicke Stäbchen-bakterien
- + +
J82 weiß-glänzend
glatter Rand
moderates Wachstum
Diplokokken + + -
J84
weiß-glänzend
filamentös
moderates Wachstum
Kokkoide Stäbchenbakterien
- + -
J85
Weiß, durchsichtig, glänzend
glatter Rand
moderates Wachstum
kurze Stäbchenbakterien
- - -
J86
weiß
filamentös, matt
glatter Rand
moderates Wachstum
Hefe + + -
J87
orange, stark pigmentiert
hydrophiles Pigment
glatter Rand
starkes Wachstum
kurze Stäbchenbakterien
beweglich
- + +
J88 hellgelb
glatter Rand
moderates Wachstum
Stäbchenbakterien
beweglich
- + -
J89
weiß-rosé, leicht glänzend
unregelmäßiger Rand
starkes Wachstum
Stäbchenbakterien - + -
J90
Beige-orange, glänzend, durchsichtig
Rand weniger pigmentiert
glatter Rand
konvex
moderates Wachstum
Stäbchenbakterien - - -
J91
Beiglich-weiß, durchsichtig
glatter Rand
moderates Wachstum
Stäbchenbakterien - + +
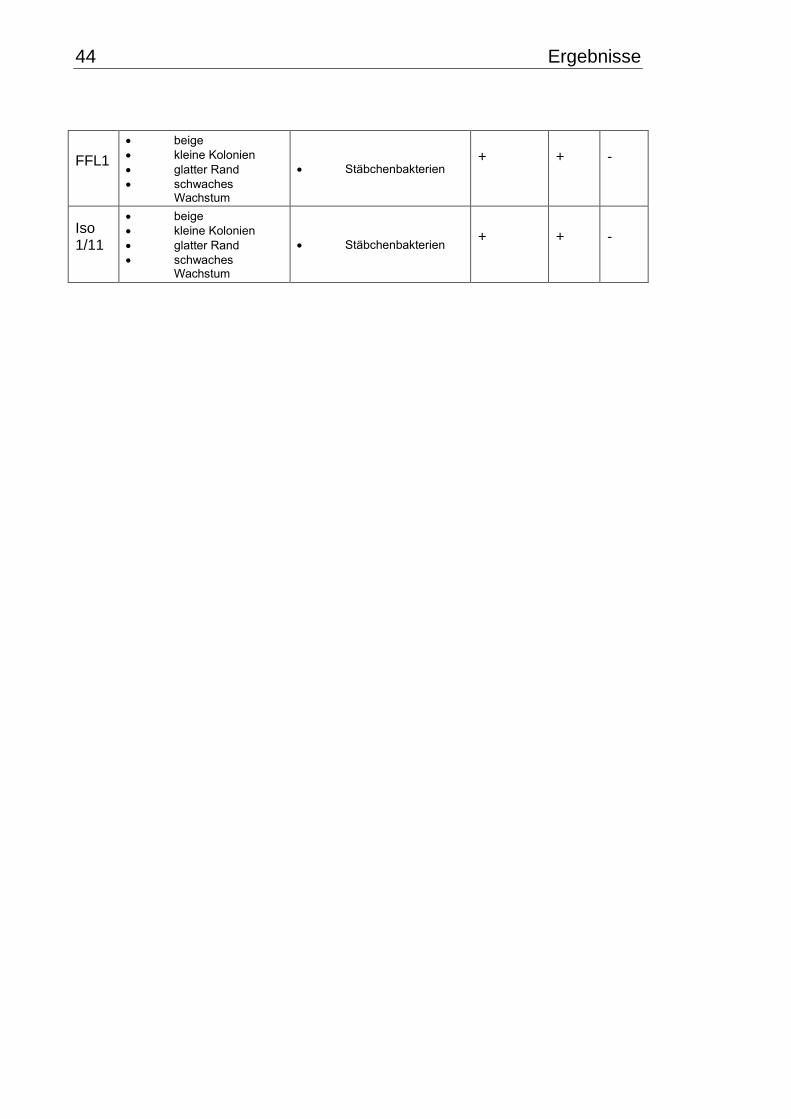
44 Ergebnisse
FFL1
beige
kleine Kolonien
glatter Rand
schwaches Wachstum
Stäbchenbakterien + + -
Iso 1/11
beige
kleine Kolonien
glatter Rand
schwaches Wachstum
Stäbchenbakterien + + -
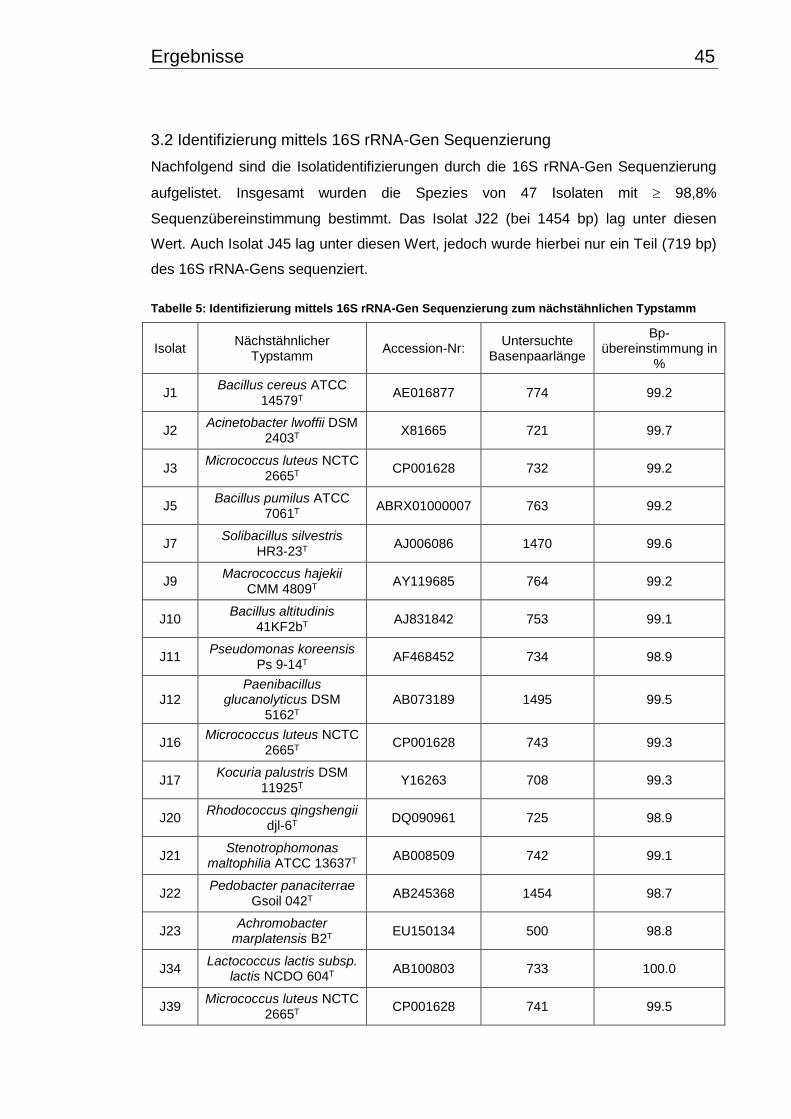
Ergebnisse 45
3.2 Identifizierung mittels 16S rRNA-Gen Sequenzierung
Nachfolgend sind die Isolatidentifizierungen durch die 16S rRNA-Gen Sequenzierung
aufgelistet. Insgesamt wurden die Spezies von 47 Isolaten mit 98,8%
Sequenzübereinstimmung bestimmt. Das Isolat J22 (bei 1454 bp) lag unter diesen
Wert. Auch Isolat J45 lag unter diesen Wert, jedoch wurde hierbei nur ein Teil (719 bp)
des 16S rRNA-Gens sequenziert.
Tabelle 5: Identifizierung mittels 16S rRNA-Gen Sequenzierung zum nächstähnlichen Typstamm
Isolat Nächstähnlicher
Typstamm Accession-Nr:
Untersuchte Basenpaarlänge
Bp-übereinstimmung in
%
J1 Bacillus cereus ATCC
14579T AE016877 774 99.2
J2 Acinetobacter lwoffii DSM
2403T X81665 721 99.7
J3 Micrococcus luteus NCTC
2665T CP001628 732 99.2
J5 Bacillus pumilus ATCC
7061T ABRX01000007 763 99.2
J7 Solibacillus silvestris
HR3-23T AJ006086 1470 99.6
J9 Macrococcus hajekii
CMM 4809T AY119685 764 99.2
J10 Bacillus altitudinis
41KF2bT AJ831842 753 99.1
J11 Pseudomonas koreensis
Ps 9-14T AF468452 734 98.9
J12 Paenibacillus
glucanolyticus DSM 5162T
AB073189 1495 99.5
J16 Micrococcus luteus NCTC
2665T CP001628 743 99.3
J17 Kocuria palustris DSM
11925T Y16263 708 99.3
J20 Rhodococcus qingshengii
djl-6T DQ090961 725 98.9
J21 Stenotrophomonas
maltophilia ATCC 13637T AB008509 742 99.1
J22 Pedobacter panaciterrae
Gsoil 042T AB245368 1454 98.7
J23 Achromobacter
marplatensis B2T EU150134 500 98.8
J34 Lactococcus lactis subsp.
lactis NCDO 604T AB100803 733 100.0
J39 Micrococcus luteus NCTC
2665T CP001628 741 99.5
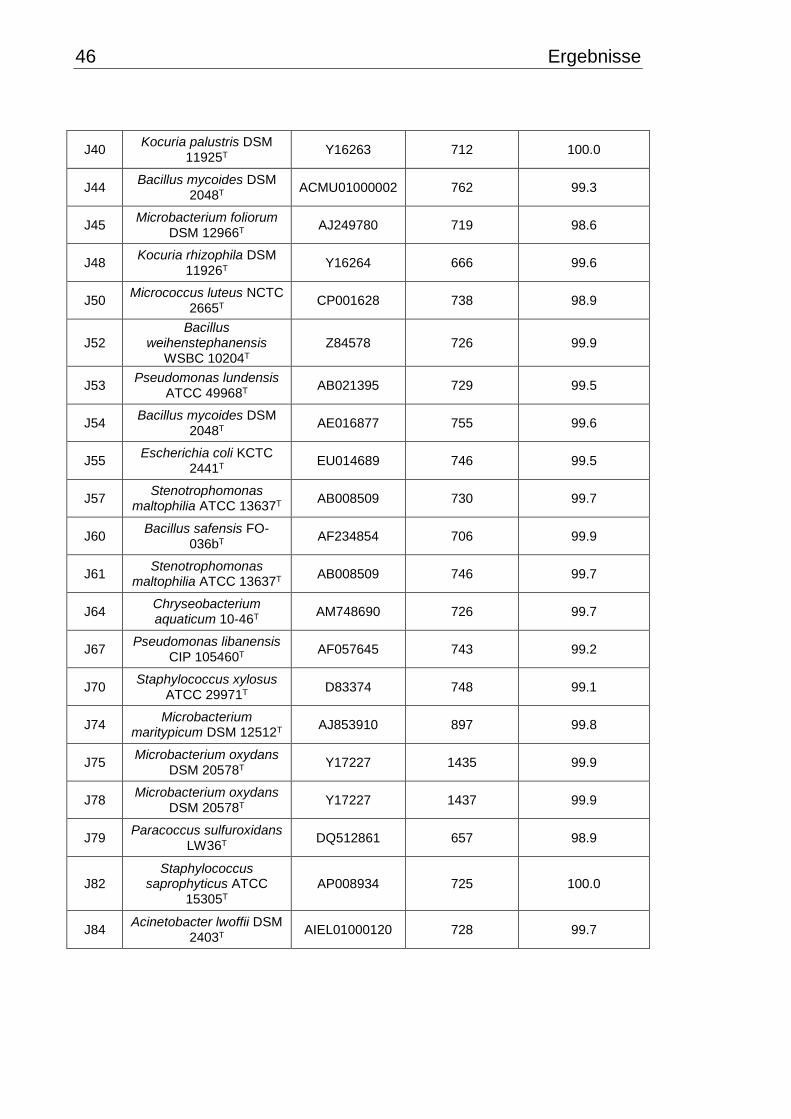
46 Ergebnisse
J40 Kocuria palustris DSM
11925T Y16263 712 100.0
J44 Bacillus mycoides DSM
2048T ACMU01000002 762 99.3
J45 Microbacterium foliorum
DSM 12966T AJ249780 719 98.6
J48 Kocuria rhizophila DSM
11926T Y16264 666 99.6
J50 Micrococcus luteus NCTC
2665T CP001628 738 98.9
J52 Bacillus
weihenstephanensis WSBC 10204T
Z84578 726 99.9
J53 Pseudomonas lundensis
ATCC 49968T AB021395 729 99.5
J54 Bacillus mycoides DSM
2048T AE016877 755 99.6
J55 Escherichia coli KCTC
2441T EU014689 746 99.5
J57 Stenotrophomonas
maltophilia ATCC 13637T AB008509 730 99.7
J60 Bacillus safensis FO-
036bT AF234854 706 99.9
J61 Stenotrophomonas
maltophilia ATCC 13637T AB008509 746 99.7
J64 Chryseobacterium aquaticum 10-46T
AM748690 726 99.7
J67 Pseudomonas libanensis
CIP 105460T AF057645 743 99.2
J70 Staphylococcus xylosus
ATCC 29971T D83374 748 99.1
J74 Microbacterium
maritypicum DSM 12512T AJ853910 897 99.8
J75 Microbacterium oxydans
DSM 20578T Y17227 1435 99.9
J78 Microbacterium oxydans
DSM 20578T Y17227 1437 99.9
J79 Paracoccus sulfuroxidans
LW36T DQ512861 657 98.9
J82 Staphylococcus
saprophyticus ATCC 15305T
AP008934 725 100.0
J84 Acinetobacter lwoffii DSM
2403T AIEL01000120 728 99.7
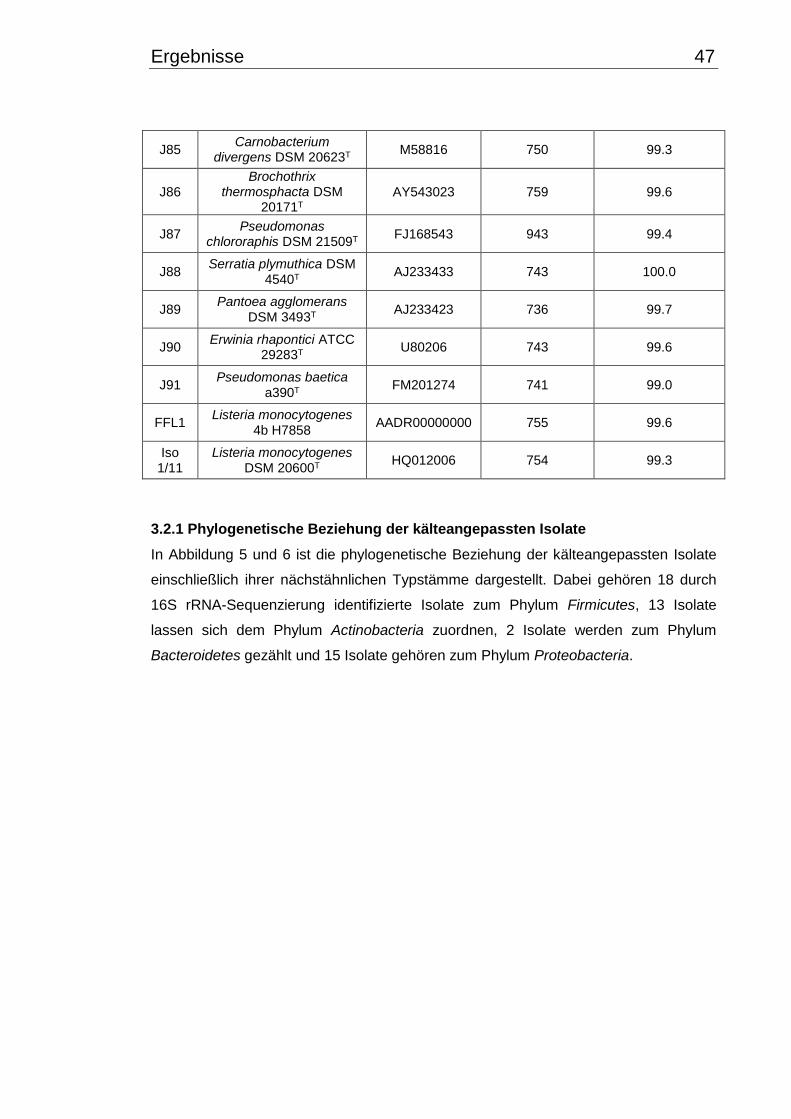
Ergebnisse 47
J85 Carnobacterium
divergens DSM 20623T M58816 750 99.3
J86 Brochothrix
thermosphacta DSM 20171T
AY543023 759 99.6
J87 Pseudomonas
chlororaphis DSM 21509T FJ168543 943 99.4
J88 Serratia plymuthica DSM
4540T AJ233433 743 100.0
J89 Pantoea agglomerans
DSM 3493T AJ233423 736 99.7
J90 Erwinia rhapontici ATCC
29283T U80206 743 99.6
J91 Pseudomonas baetica
a390T FM201274 741 99.0
FFL1 Listeria monocytogenes
4b H7858 AADR00000000 755 99.6
Iso 1/11
Listeria monocytogenes DSM 20600T
HQ012006 754 99.3
3.2.1 Phylogenetische Beziehung der kälteangepassten Isolate
In Abbildung 5 und 6 ist die phylogenetische Beziehung der kälteangepassten Isolate
einschließlich ihrer nächstähnlichen Typstämme dargestellt. Dabei gehören 18 durch
16S rRNA-Sequenzierung identifizierte Isolate zum Phylum Firmicutes, 13 Isolate
lassen sich dem Phylum Actinobacteria zuordnen, 2 Isolate werden zum Phylum
Bacteroidetes gezählt und 15 Isolate gehören zum Phylum Proteobacteria.
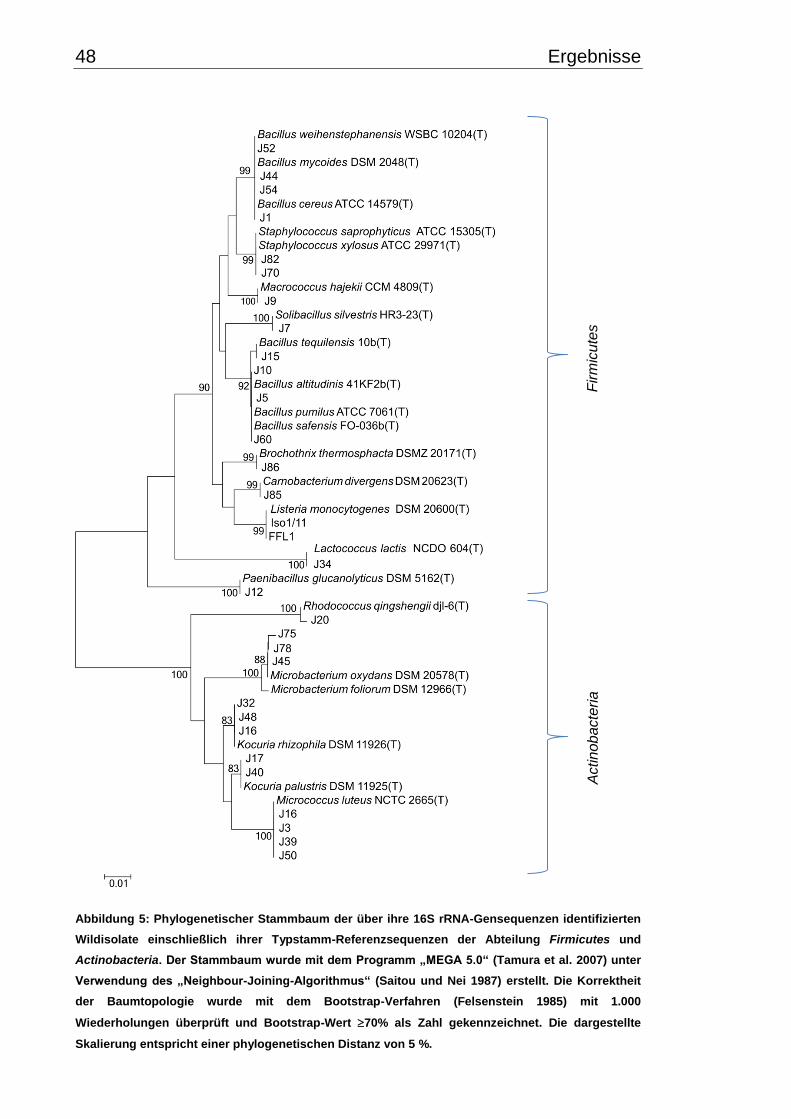
48 Ergebnisse
Abbildung 5: Phylogenetischer Stammbaum der über ihre 16S rRNA-Gensequenzen identifizierten
Wildisolate einschließlich ihrer Typstamm-Referenzsequenzen der Abteilung Firmicutes und
Actinobacteria. Der Stammbaum wurde mit dem Programm „MEGA 5.0“ (Tamura et al. 2007) unter
Verwendung des „Neighbour-Joining-Algorithmus“ (Saitou und Nei 1987) erstellt. Die Korrektheit
der Baumtopologie wurde mit dem Bootstrap-Verfahren (Felsenstein 1985) mit 1.000
Wiederholungen überprüft und Bootstrap-Wert 70% als Zahl gekennzeichnet. Die dargestellte
Skalierung entspricht einer phylogenetischen Distanz von 5 %.
Firm
icu
tes
Actin
ob
acte
ria

Ergebnisse 49
Abbildung 6: Phylogenetischer Stammbaum der über ihre 16S rRNA-Gensequenzen identifizierten
Wildisolate einschließlich ihrer Typstamm-Referenzsequenzen der Abteilung Bacteriodetes und
Proteobacteria. Der Stammbaum wurde mit dem Programm „MEGA 5.0“ (Tamura et al. 2007) unter
Verwendung des „Neighbour-Joining-Algorithmus“ (Saitou und Nei 1987) erstellt. Die Korrektheit
der Baumtopologie wurde mit dem Bootstrap-Verfahren (Felsenstein 1985) mit 1.000
Wiederholungen überprüft und ein Bootstrap-Wert in 70% als Zahl gekennzeichnet. Die
dargestellte Skalierung entspricht einer phylogenetischen Distanz von 5 %.
Pro
teo
bacte
ria
Ba
cte
rio
de
tes

50 Ergebnisse
3.3 Neubeschreibung Isolat J22
Im Verlauf dieser Arbeit wurde das Isolat J22 aus frischen Fleischtortellini isoliert,
welches bei der 16S rRNA-Gen Sequenzierung bei 1454 Basenpaaren nur eine 98,7%
Übereinstimmung mit dem nächstähnlichen Typstamm Pedobacter panaciterrae
LMG23400T aufwies. Durch diese niedrige Sequenzübereinstimmung war eine
Differenzierung auf Speziesebene nicht möglich und muss daher überprüft werden. Um
diesen Verdacht zu bestätigen, wurde eine DNA-DNA Hybridisierung mit dem
nächstähnlichen Typstamm auf 16S rRNA-Gensequenzebene, nämlich Pedobacter
panaciterrae LMG23400T durchgeführt. Hierbei ergab sich ein Wert von weniger als
36%. Der Schwellenwert für eine neu zu beschreibende Spezies wird mit einem
Grenzwert von weniger als 70% angegeben (Wayne et al. 1987), d.h. Isolat J22 gehört
nicht zur Spezies Pedobacter panaciterrae und kann als Vertreter einer neuen Spezies
neu beschrieben werden.
3.3.1 Makroskopische und mikroskopische Zellmorphologie
Isolat J22 wächst unter aeroben Bedingungen und bildet keine Sporen. Das Gram-
negative Stäbchenbakterium hat eine Länge von 2 µm und einem Durchmesser von
0,6 µm. Die Kolonien hatten auf TS-Agar einen Durchmesser von 2 mm nach zwei
Tagen Inkubation bei 30°C. Außerdem war es Oxidase- und Katalase-positiv.
3.3.2 Temperatur- und Salztoleranztest
Die Ergebnisse zum Wachstumstemperaturprofil und der Salztoleranz von Isolat J22
und drei Referenzstämmen sind in Tabelle 6 dargestellt.
Tabelle 6: Wachstumstemperaturprofil und Salztoleranz von Isolat J22 (1) sowie den
Referenzstämmen Pedobacter panaciterrae LMG 23400T (2) , Pedobacter africanus DSM12126T (3)
und Pedobacter heparinus DSM2366T (4)
1 2 3 4
Wachstumsbereich in °C 4-37 4-37 4-37 8-37
NaCl-Toleranz in R2A-Medium (in
%)
0-2 0-3 0-2 0-2
Isolat J22 und alle drei Referenzstämme zeigen Wachstum bis zur Maximaltemperatur
von 37°C. Die Minimaltemperatur für Wachstum liegt, mit der Ausnahme von
Pedobacter heparinus DSM2366T, bei 4°C. Alle vier untersuchten Stämme tolerieren
NaCl-Konzentrationen bis 2%, der Stamm Pedobacter panaciterrae LMG23400T sogar
bis 3%.

Ergebnisse 51
3.3.3 Chemotaxonomische Beschreibung
Als Hauptfettsäuren wurden bei Isolat J22 C15:0 iso und C16:1 cis 9 detektiert. Eine
genaue Auflistung des kompletten Fettsäuremusters von Isolat J22 sowie der drei
weiteren Referenzstämmen zum Vergleich befinden sich in Tabelle 7.
Als einziges respiratorisches Chinon wurde bei Isolat J22 und den Referenzstämmen
Pedobacter panaciterrae LMG23400T, Pedobacter africanus DSM12126T und
Pedobacter heparinus DSM2366T das Menachinon MK-7 detektiert.
Tabelle 7: Fettsäuremuster mittels GC-FID Analyse des Isolates J22 sowie der Referenzstämme
Pedobacter panaciterrae LMG23400T, Pedobacter africanus DSM12126T und Pedobacter heparinus
DSM2366T bei der Wachstumstemperatur von 30°C; Angaben in %.
J22 P. panaciterrae P. africanus P. heparinus
ECLa 12,880 2,4 0,8 0,7 1,0
ECLa 13,530 4,6 4,8 5,6 6,3
ECLa 13,600 - - - 1,3
C 14:0 4,8 3,0 1,5 1,0
C 15:0 iso 34,8 37,5 33,8 31,6
C 15:0 anteiso - 0,7 0,8 1,2
C 15:1 cis 9 0,7 0,8 - 2,5
C 15:0 2,6 1,3 0,5 2,2
C 14:0 3-OH 1,8 - - -
C 16:0 iso 0,7 - - -
C 16:1 cis 9 24,9 35,5 28,6 25,6
C 16:1 cis 11 4,1 3,3 1,9 1,7
C 16:0 7,3 1,6 2,3 1,2
C 15:0 iso 3-OH 2,3 1,4 2,3 3,3
C 17:1 iso cis 9 0,6 3,5 6,5 3,3
C 17:1 iso cis 11 - 1,0 1,5 2,1
C 17:0 cyclo 0,5 - 0,9 0,6
ECLa 17,115 0,7 - - 0,8
C 16:0 3-OH 3,0 0,9 1,5 1,3
ECLa17,875 - - 1,1 0,8
C 17:0 iso 3-OH 4,4 4,1 9,8 8,4
ECLa18,225 - - - 1,5
Verhältnis S/Ub 1,66 1,00 0,99 1,05 a ECL: Equivalent Chain Length b S: gesättigte Fettsäuren C 14:0, C 15:0 iso, C 15:0 anteiso, C 15:0, C 16:0 und C16:0 iso b U: ungesättigte Fettsäuren C 15:1 cis 9, C 16:1 cis 9, C 16:1 cis 11, C 17:1 iso cis 9 und C 17:1 iso cis 11
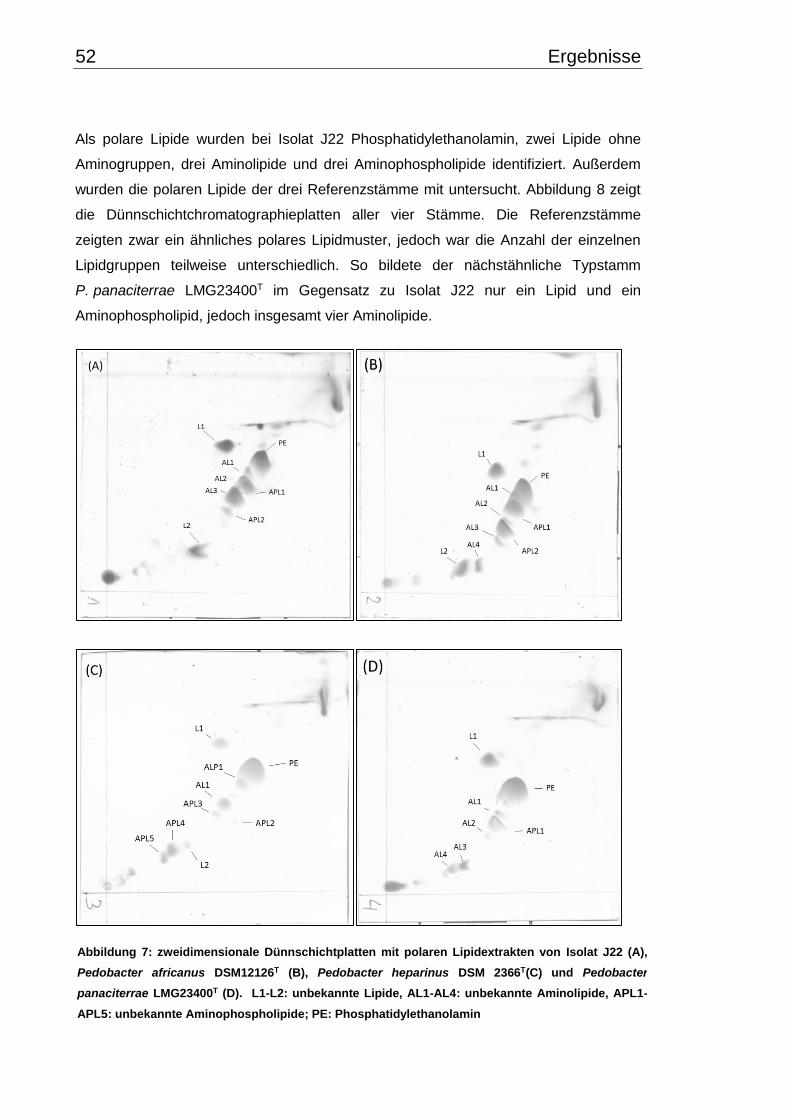
52 Ergebnisse
Als polare Lipide wurden bei Isolat J22 Phosphatidylethanolamin, zwei Lipide ohne
Aminogruppen, drei Aminolipide und drei Aminophospholipide identifiziert. Außerdem
wurden die polaren Lipide der drei Referenzstämme mit untersucht. Abbildung 8 zeigt
die Dünnschichtchromatographieplatten aller vier Stämme. Die Referenzstämme
zeigten zwar ein ähnliches polares Lipidmuster, jedoch war die Anzahl der einzelnen
Lipidgruppen teilweise unterschiedlich. So bildete der nächstähnliche Typstamm
P. panaciterrae LMG23400T im Gegensatz zu Isolat J22 nur ein Lipid und ein
Aminophospholipid, jedoch insgesamt vier Aminolipide.
Abbildung 7: zweidimensionale Dünnschichtplatten mit polaren Lipidextrakten von Isolat J22 (A),
Pedobacter africanus DSM12126T (B), Pedobacter heparinus DSM 2366T(C) und Pedobacter
panaciterrae LMG23400T (D). L1-L2: unbekannte Lipide, AL1-AL4: unbekannte Aminolipide, APL1-
APL5: unbekannte Aminophospholipide; PE: Phosphatidylethanolamin
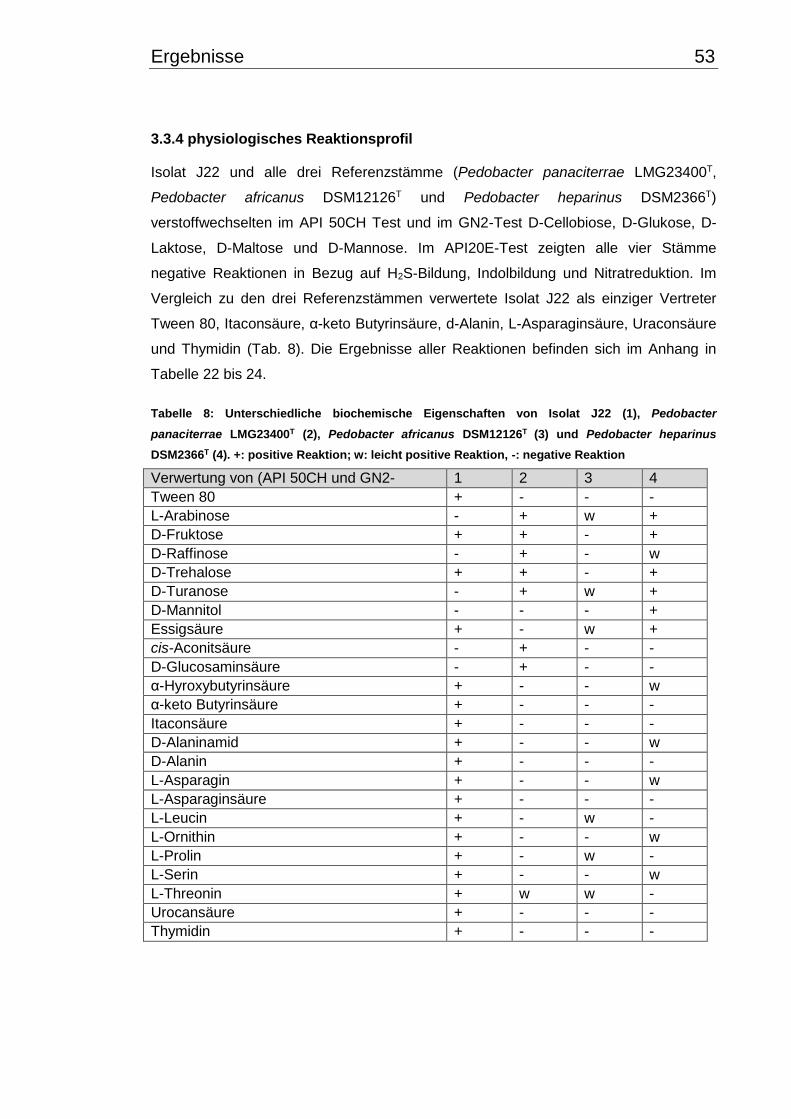
Ergebnisse 53
3.3.4 physiologisches Reaktionsprofil
Isolat J22 und alle drei Referenzstämme (Pedobacter panaciterrae LMG23400T,
Pedobacter africanus DSM12126T und Pedobacter heparinus DSM2366T)
verstoffwechselten im API 50CH Test und im GN2-Test D-Cellobiose, D-Glukose, D-
Laktose, D-Maltose und D-Mannose. Im API20E-Test zeigten alle vier Stämme
negative Reaktionen in Bezug auf H2S-Bildung, Indolbildung und Nitratreduktion. Im
Vergleich zu den drei Referenzstämmen verwertete Isolat J22 als einziger Vertreter
Tween 80, Itaconsäure, α-keto Butyrinsäure, d-Alanin, L-Asparaginsäure, Uraconsäure
und Thymidin (Tab. 8). Die Ergebnisse aller Reaktionen befinden sich im Anhang in
Tabelle 22 bis 24.
Tabelle 8: Unterschiedliche biochemische Eigenschaften von Isolat J22 (1), Pedobacter
panaciterrae LMG23400T (2), Pedobacter africanus DSM12126T (3) und Pedobacter heparinus
DSM2366T (4). +: positive Reaktion; w: leicht positive Reaktion, -: negative Reaktion
Verwertung von (API 50CH und GN2-
Microplate)
1 2 3 4
Tween 80 + - - -
L-Arabinose - + w +
D-Fruktose + + - +
D-Raffinose - + - w
D-Trehalose + + - +
D-Turanose - + w +
D-Mannitol - - - +
Essigsäure + - w +
cis-Aconitsäure - + - -
D-Glucosaminsäure - + - -
α-Hyroxybutyrinsäure + - - w
α-keto Butyrinsäure + - - -
Itaconsäure + - - -
D-Alaninamid + - - w
D-Alanin + - - -
L-Asparagin + - - w
L-Asparaginsäure + - - -
L-Leucin + - w -
L-Ornithin + - - w
L-Prolin + - w -
L-Serin + - - w
L-Threonin + w w -
Urocansäure + - - -
Thymidin + - - -

54 Ergebnisse
3.3.5 Molekulare Phylogenie-Analyse und DNA-DNA-Hybridisierung
Der mit der Neighbour-Joining-Analyse erstellte phylogenetische Stammbaum der 16S
rRNA-Gen Sequenz (Abb. 9) zeigt, dass Isolat J22 zu der Gattung Pedobacter
zugeordnet werden kann. Das gleiche Ergebnis wurde auch mit der Maximun-
Likelihood-Methode und der Maximum-Parsimony-Methode erzielt. Diese
phylogenetischen Stammbäume sind im Anhang in Abbildung 42 und 43 dargestellt.
Vergleichende 16S rRNA-Gen-Sequenzanalysen von Isolat J22 (1553 bp) zeigten
Sequenzübereinstimmung mit der Bakterienstämmen Pedobacter panaciterrae
LMG23400T (98,7%), gefolgt von Pedobacter africanus DSM 12126T (98,5%) und
Pedobacter heparinus DSM2366T (98,3%).
Zusätzlich wurde eine DNA-DNA Hybridisierung mit dem nächstähnlichen Typstamm
Pedobacter panaciterrae LMG23400T durchgeführt. Hierbei ergab sich ein Wert von
35,2% und einem reziprokalen Wert von 7,4%.
3.3.6 Namensgebung
Isolat J22 wurde aus frischen Fleischtortellini isoliert, welche bei 6°C gekühlt gelagert
wurden. Dieses Lebensmittelprodukt wurde in Deutschland hergestellt und vertrieben.
Aus diesem Grund wurde für das Isolat J22 der Speziesname Pedobacter nutrimenti
(nu.tri´men.ti L. gen. n. nutrimenti von nutriment (engl.); Nahrungsmittel) vorgeschlagen
(Derichs et al. 2014).
Der Typstamm J22T ist in der Stammsammlung der DSMZ unter der Nummer DSM
27373T und in der Stammsammlung der CCUG mit der Nummer CCUG 64422T
hinterlegt.
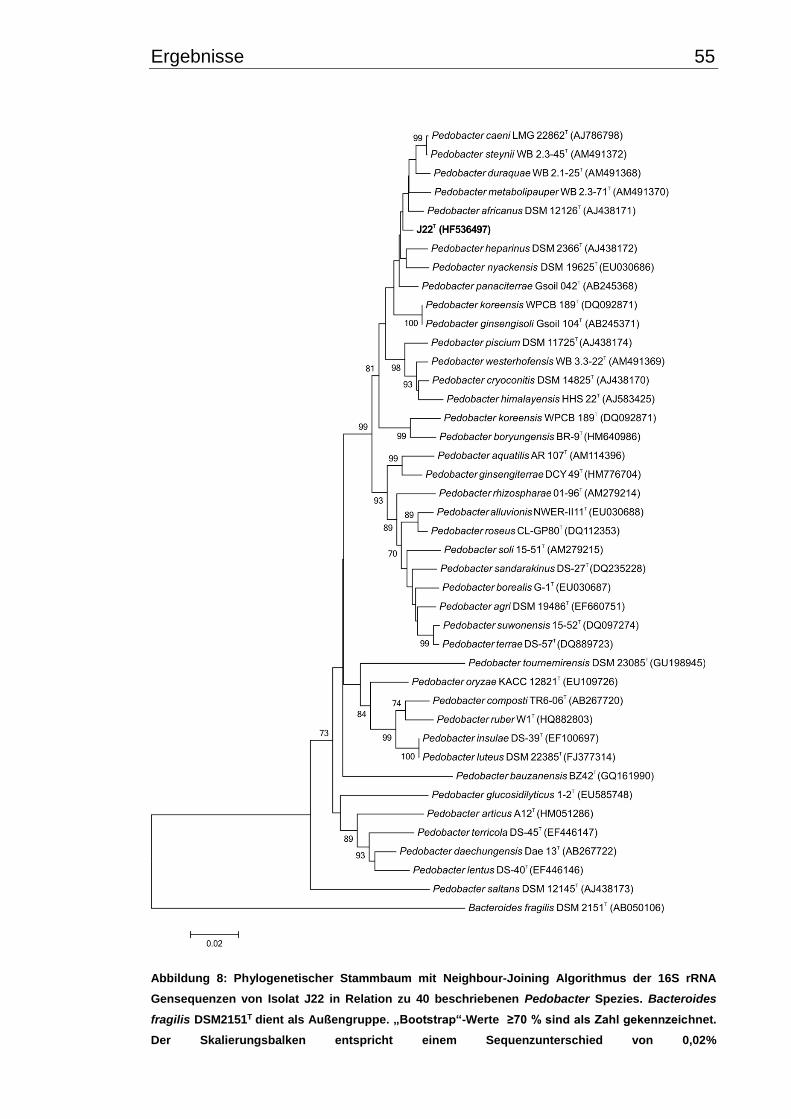
Ergebnisse 55
Abbildung 8: Phylogenetischer Stammbaum mit Neighbour-Joining Algorithmus der 16S rRNA
Gensequenzen von Isolat J22 in Relation zu 40 beschriebenen Pedobacter Spezies. Bacteroides
fragilis DSM2151T dient als Außengruppe. „Bootstrap“-Werte ≥70 % sind als Zahl gekennzeichnet.
Der Skalierungsbalken entspricht einem Sequenzunterschied von 0,02%
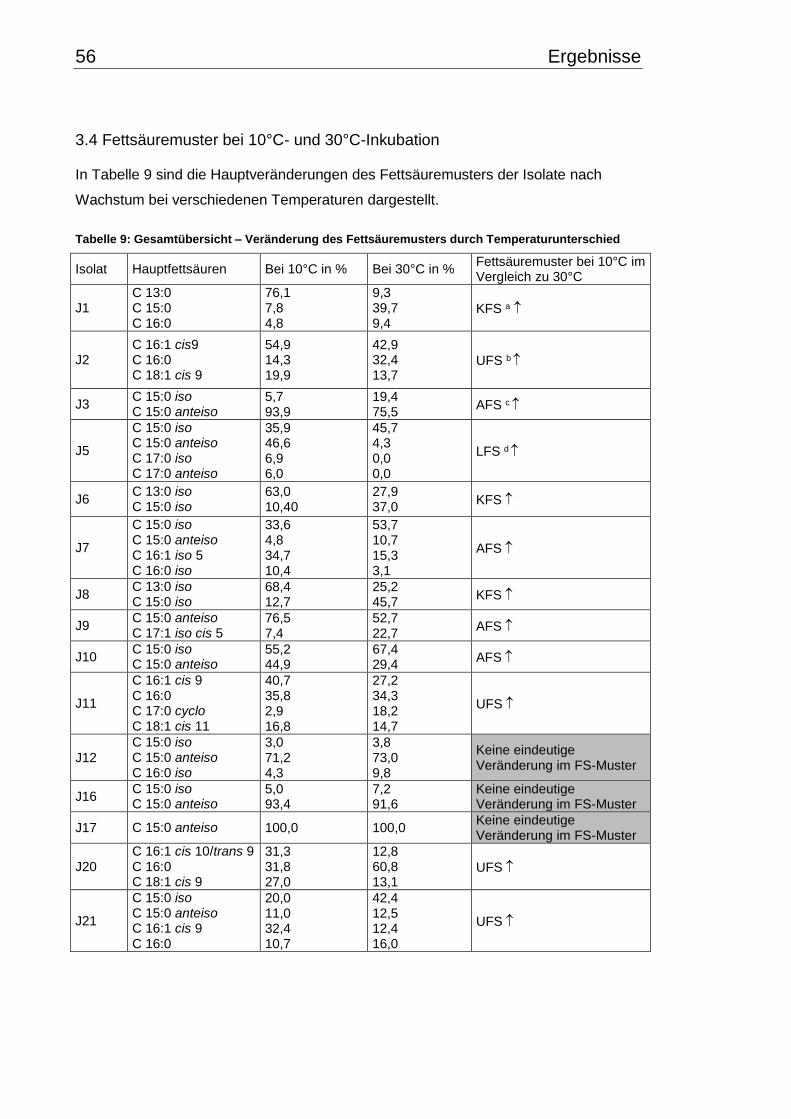
56 Ergebnisse
3.4 Fettsäuremuster bei 10°C- und 30°C-Inkubation
In Tabelle 9 sind die Hauptveränderungen des Fettsäuremusters der Isolate nach
Wachstum bei verschiedenen Temperaturen dargestellt.
Tabelle 9: Gesamtübersicht – Veränderung des Fettsäuremusters durch Temperaturunterschied
Isolat Hauptfettsäuren Bei 10°C in % Bei 30°C in % Fettsäuremuster bei 10°C im Vergleich zu 30°C
J1 C 13:0 C 15:0 C 16:0
76,1 7,8 4,8
9,3 39,7 9,4
KFS a
J2 C 16:1 cis9 C 16:0 C 18:1 cis 9
54,9 14,3 19,9
42,9 32,4 13,7
UFS b
J3 C 15:0 iso C 15:0 anteiso
5,7 93,9
19,4 75,5
AFS c
J5
C 15:0 iso C 15:0 anteiso C 17:0 iso C 17:0 anteiso
35,9 46,6 6,9 6,0
45,7 4,3 0,0 0,0
LFS d
J6 C 13:0 iso C 15:0 iso
63,0 10,40
27,9 37,0
KFS
J7
C 15:0 iso C 15:0 anteiso C 16:1 iso 5 C 16:0 iso
33,6 4,8 34,7 10,4
53,7 10,7 15,3 3,1
AFS
J8 C 13:0 iso C 15:0 iso
68,4 12,7
25,2 45,7
KFS
J9 C 15:0 anteiso C 17:1 iso cis 5
76,5 7,4
52,7 22,7
AFS
J10 C 15:0 iso C 15:0 anteiso
55,2 44,9
67,4 29,4
AFS
J11
C 16:1 cis 9 C 16:0 C 17:0 cyclo C 18:1 cis 11
40,7 35,8 2,9 16,8
27,2 34,3 18,2 14,7
UFS
J12 C 15:0 iso C 15:0 anteiso C 16:0 iso
3,0 71,2 4,3
3,8 73,0 9,8
Keine eindeutige Veränderung im FS-Muster
J16 C 15:0 iso C 15:0 anteiso
5,0 93,4
7,2 91,6
Keine eindeutige Veränderung im FS-Muster
J17 C 15:0 anteiso 100,0 100,0 Keine eindeutige Veränderung im FS-Muster
J20 C 16:1 cis 10/trans 9 C 16:0 C 18:1 cis 9
31,3 31,8 27,0
12,8 60,8 13,1
UFS
J21
C 15:0 iso C 15:0 anteiso C 16:1 cis 9 C 16:0
20,0 11,0 32,4 10,7
42,4 12,5 12,4 16,0
UFS
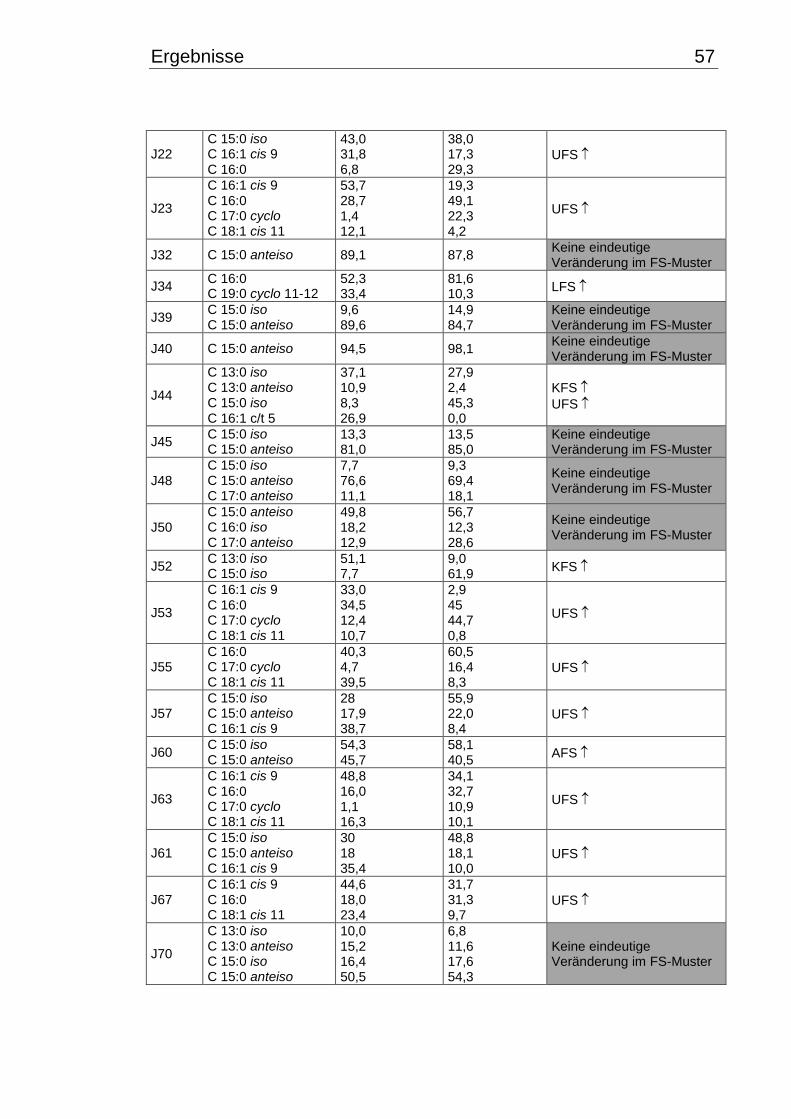
Ergebnisse 57
J22 C 15:0 iso C 16:1 cis 9 C 16:0
43,0 31,8 6,8
38,0 17,3 29,3
UFS
J23
C 16:1 cis 9 C 16:0 C 17:0 cyclo C 18:1 cis 11
53,7 28,7 1,4 12,1
19,3 49,1 22,3 4,2
UFS
J32 C 15:0 anteiso 89,1 87,8 Keine eindeutige Veränderung im FS-Muster
J34 C 16:0 C 19:0 cyclo 11-12
52,3 33,4
81,6 10,3
LFS
J39 C 15:0 iso C 15:0 anteiso
9,6 89,6
14,9 84,7
Keine eindeutige Veränderung im FS-Muster
J40 C 15:0 anteiso 94,5 98,1 Keine eindeutige Veränderung im FS-Muster
J44
C 13:0 iso C 13:0 anteiso C 15:0 iso C 16:1 c/t 5
37,1 10,9 8,3 26,9
27,9 2,4 45,3 0,0
KFS
UFS
J45 C 15:0 iso C 15:0 anteiso
13,3 81,0
13,5 85,0
Keine eindeutige Veränderung im FS-Muster
J48 C 15:0 iso C 15:0 anteiso C 17:0 anteiso
7,7 76,6 11,1
9,3 69,4 18,1
Keine eindeutige Veränderung im FS-Muster
J50 C 15:0 anteiso C 16:0 iso C 17:0 anteiso
49,8 18,2 12,9
56,7 12,3 28,6
Keine eindeutige Veränderung im FS-Muster
J52 C 13:0 iso C 15:0 iso
51,1 7,7
9,0 61,9
KFS
J53
C 16:1 cis 9 C 16:0 C 17:0 cyclo C 18:1 cis 11
33,0 34,5 12,4 10,7
2,9 45 44,7 0,8
UFS
J55 C 16:0 C 17:0 cyclo C 18:1 cis 11
40,3 4,7 39,5
60,5 16,4 8,3
UFS
J57 C 15:0 iso C 15:0 anteiso C 16:1 cis 9
28 17,9 38,7
55,9 22,0 8,4
UFS
J60 C 15:0 iso C 15:0 anteiso
54,3 45,7
58,1 40,5
AFS
J63
C 16:1 cis 9 C 16:0 C 17:0 cyclo C 18:1 cis 11
48,8 16,0 1,1 16,3
34,1 32,7 10,9 10,1
UFS
J61 C 15:0 iso C 15:0 anteiso C 16:1 cis 9
30 18 35,4
48,8 18,1 10,0
UFS
J67 C 16:1 cis 9 C 16:0 C 18:1 cis 11
44,6 18,0 23,4
31,7 31,3 9,7
UFS
J70
C 13:0 iso C 13:0 anteiso C 15:0 iso C 15:0 anteiso
10,0 15,2 16,4 50,5
6,8 11,6 17,6 54,3
Keine eindeutige Veränderung im FS-Muster
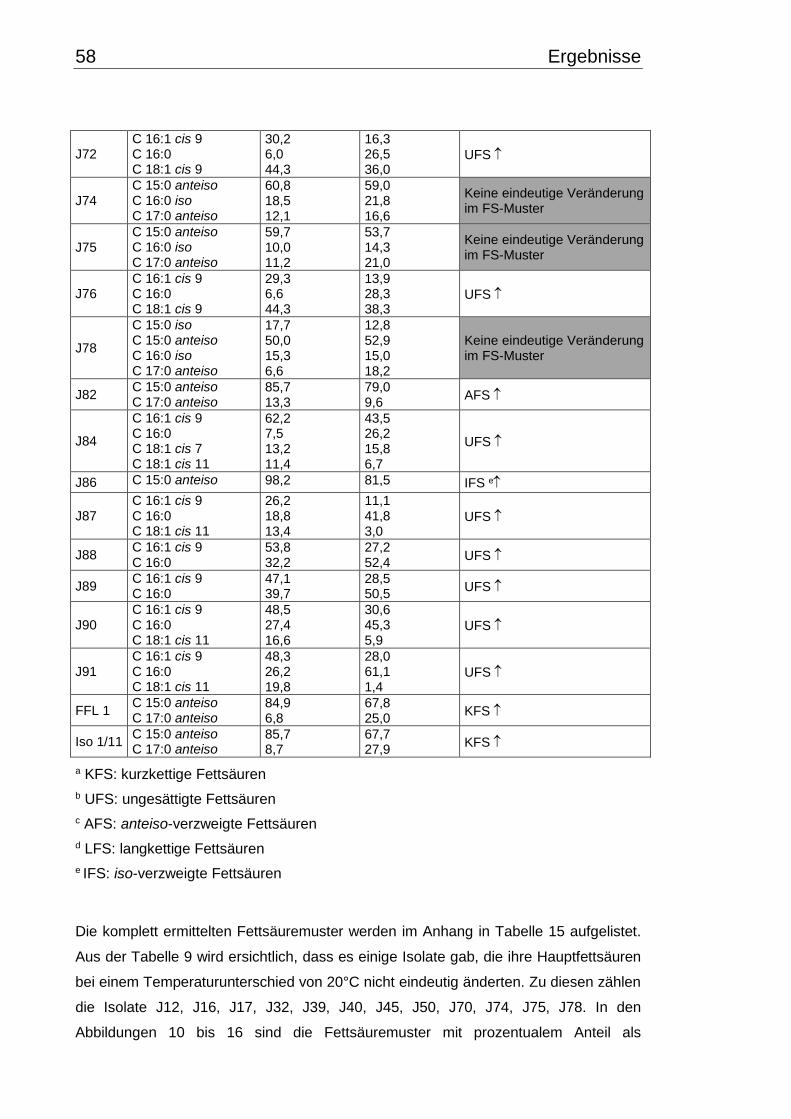
58 Ergebnisse
a KFS: kurzkettige Fettsäuren
b UFS: ungesättigte Fettsäuren
c AFS: anteiso-verzweigte Fettsäuren
d LFS: langkettige Fettsäuren
e IFS: iso-verzweigte Fettsäuren
Die komplett ermittelten Fettsäuremuster werden im Anhang in Tabelle 15 aufgelistet.
Aus der Tabelle 9 wird ersichtlich, dass es einige Isolate gab, die ihre Hauptfettsäuren
bei einem Temperaturunterschied von 20°C nicht eindeutig änderten. Zu diesen zählen
die Isolate J12, J16, J17, J32, J39, J40, J45, J50, J70, J74, J75, J78. In den
Abbildungen 10 bis 16 sind die Fettsäuremuster mit prozentualem Anteil als
J72 C 16:1 cis 9 C 16:0 C 18:1 cis 9
30,2 6,0 44,3
16,3 26,5 36,0
UFS
J74 C 15:0 anteiso C 16:0 iso C 17:0 anteiso
60,8 18,5 12,1
59,0 21,8 16,6
Keine eindeutige Veränderung im FS-Muster
J75 C 15:0 anteiso C 16:0 iso C 17:0 anteiso
59,7 10,0 11,2
53,7 14,3 21,0
Keine eindeutige Veränderung im FS-Muster
J76 C 16:1 cis 9 C 16:0 C 18:1 cis 9
29,3 6,6 44,3
13,9 28,3 38,3
UFS
J78
C 15:0 iso C 15:0 anteiso C 16:0 iso C 17:0 anteiso
17,7 50,0 15,3 6,6
12,8 52,9 15,0 18,2
Keine eindeutige Veränderung im FS-Muster
J82 C 15:0 anteiso C 17:0 anteiso
85,7 13,3
79,0 9,6
AFS
J84
C 16:1 cis 9 C 16:0 C 18:1 cis 7 C 18:1 cis 11
62,2 7,5 13,2 11,4
43,5 26,2 15,8 6,7
UFS
J86 C 15:0 anteiso 98,2 81,5 IFS e
J87 C 16:1 cis 9 C 16:0 C 18:1 cis 11
26,2 18,8 13,4
11,1 41,8 3,0
UFS
J88 C 16:1 cis 9 C 16:0
53,8 32,2
27,2 52,4
UFS
J89 C 16:1 cis 9 C 16:0
47,1 39,7
28,5 50,5
UFS
J90 C 16:1 cis 9 C 16:0 C 18:1 cis 11
48,5 27,4 16,6
30,6 45,3 5,9
UFS
J91 C 16:1 cis 9 C 16:0 C 18:1 cis 11
48,3 26,2 19,8
28,0 61,1 1,4
UFS
FFL 1 C 15:0 anteiso C 17:0 anteiso
84,9 6,8
67,8 25,0
KFS
Iso 1/11 C 15:0 anteiso C 17:0 anteiso
85,7 8,7
67,7 27,9
KFS
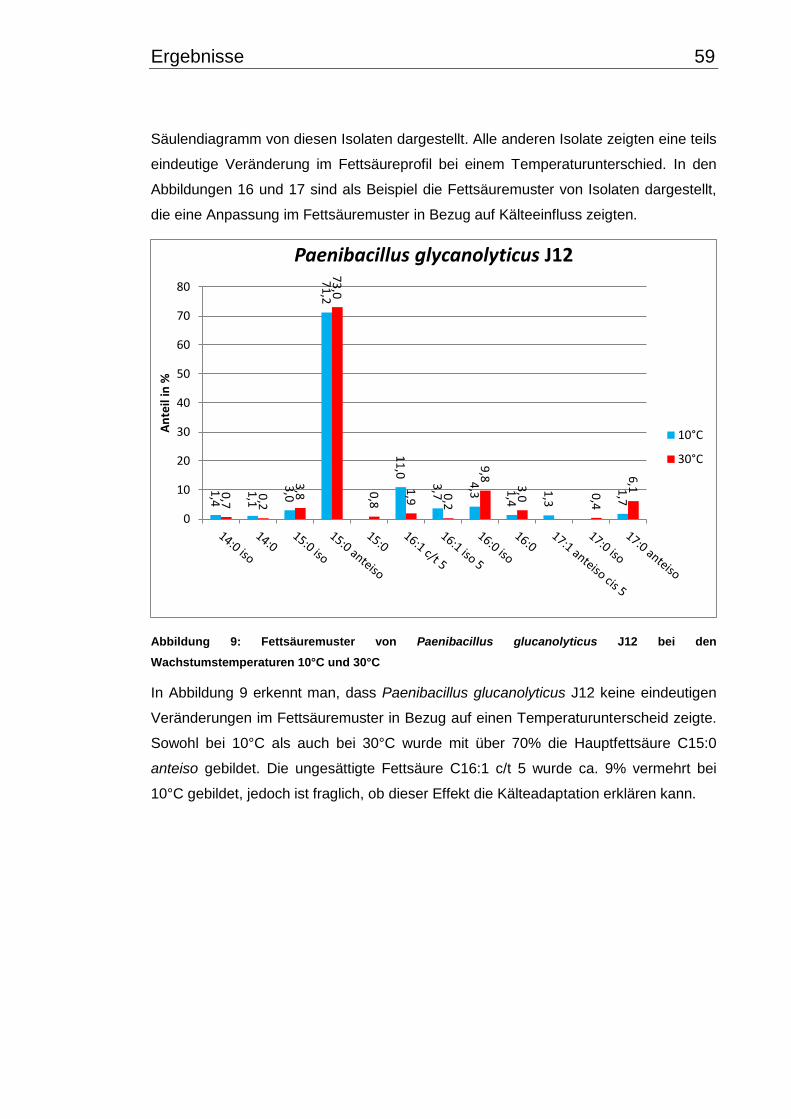
Ergebnisse 59
Säulendiagramm von diesen Isolaten dargestellt. Alle anderen Isolate zeigten eine teils
eindeutige Veränderung im Fettsäureprofil bei einem Temperaturunterschied. In den
Abbildungen 16 und 17 sind als Beispiel die Fettsäuremuster von Isolaten dargestellt,
die eine Anpassung im Fettsäuremuster in Bezug auf Kälteeinfluss zeigten.
Abbildung 9: Fettsäuremuster von Paenibacillus glucanolyticus J12 bei den
Wachstumstemperaturen 10°C und 30°C
In Abbildung 9 erkennt man, dass Paenibacillus glucanolyticus J12 keine eindeutigen
Veränderungen im Fettsäuremuster in Bezug auf einen Temperaturunterscheid zeigte.
Sowohl bei 10°C als auch bei 30°C wurde mit über 70% die Hauptfettsäure C15:0
anteiso gebildet. Die ungesättigte Fettsäure C16:1 c/t 5 wurde ca. 9% vermehrt bei
10°C gebildet, jedoch ist fraglich, ob dieser Effekt die Kälteadaptation erklären kann.
1,4
1,1
3,0
71
,2
11
,0
3,7
4,3 1
,4
1,3
1,7
0,7
0,2
3,8
73
,0
0,8
1,9 0,2
9,8
3,0 0
,4
6,1
0
10
20
30
40
50
60
70
80
An
teil
in %
Paenibacillus glycanolyticus J12
10°C
30°C
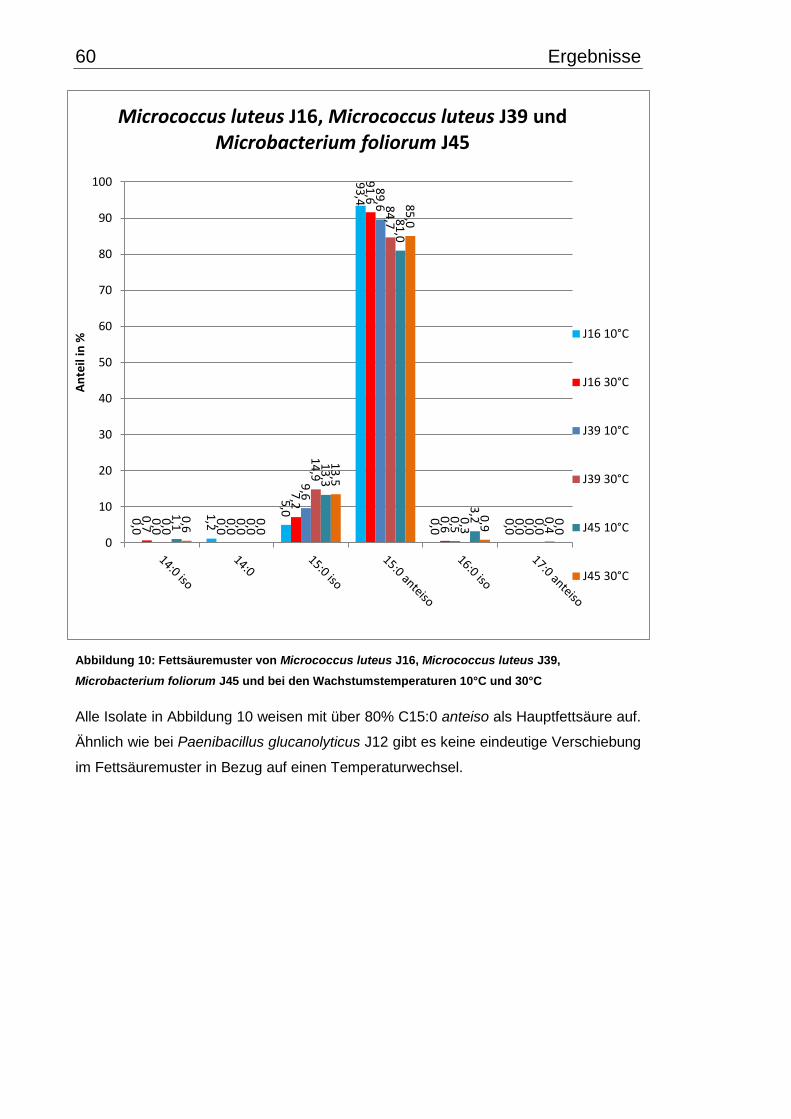
60 Ergebnisse
0,0
1,2
5,0
93
,4
0,0
0,0
0,7
0,0
7,2
91
,6
0,6
0,0
0,0
0,0
9,6
89
,6
0,5
0,0
0,0
0,0
14
,9
84
,7
0,3
0,0
1,1 0,0
13
,3
81
,0
3,2 0
,4
0,6
0,0
13
,5
85
,0
0,9
0,0
0
10
20
30
40
50
60
70
80
90
100
An
teil
in %
Micrococcus luteus J16, Micrococcus luteus J39 und Microbacterium foliorum J45
J16 10°C
J16 30°C
J39 10°C
J39 30°C
J45 10°C
J45 30°C
Abbildung 10: Fettsäuremuster von Micrococcus luteus J16, Micrococcus luteus J39,
Microbacterium foliorum J45 und bei den Wachstumstemperaturen 10°C und 30°C
Alle Isolate in Abbildung 10 weisen mit über 80% C15:0 anteiso als Hauptfettsäure auf.
Ähnlich wie bei Paenibacillus glucanolyticus J12 gibt es keine eindeutige Verschiebung
im Fettsäuremuster in Bezug auf einen Temperaturwechsel.
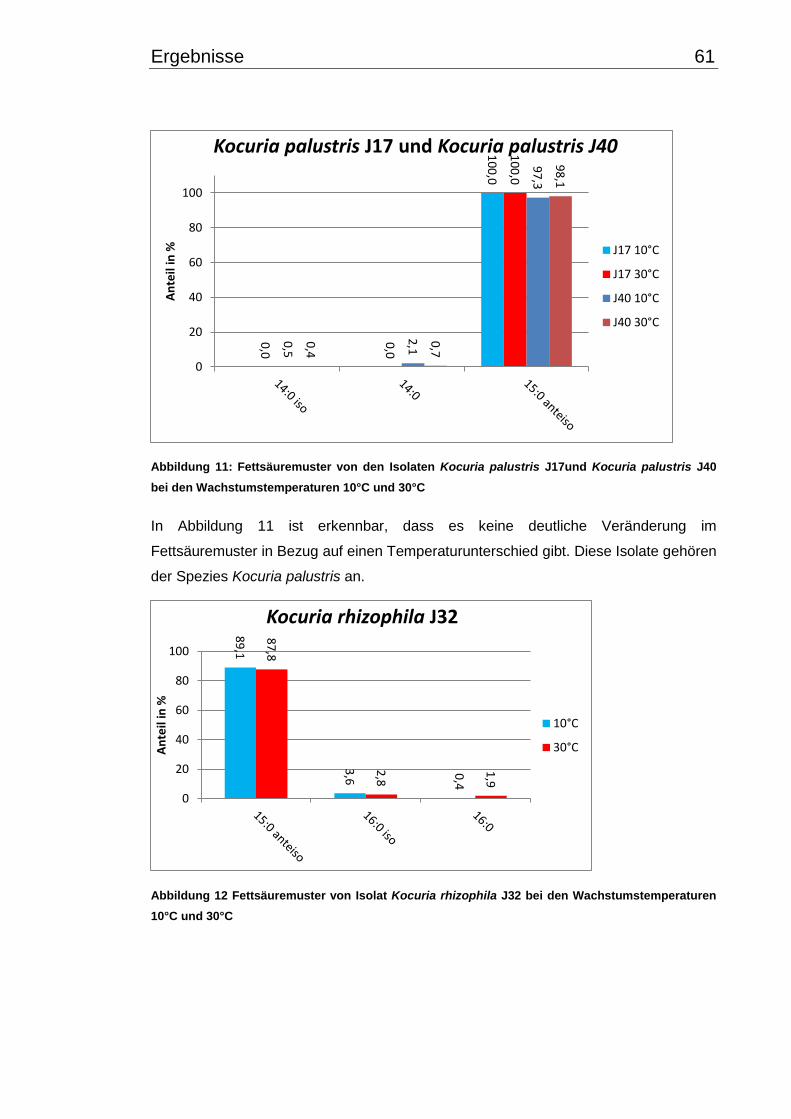
Ergebnisse 61
Abbildung 11: Fettsäuremuster von den Isolaten Kocuria palustris J17und Kocuria palustris J40
bei den Wachstumstemperaturen 10°C und 30°C
In Abbildung 11 ist erkennbar, dass es keine deutliche Veränderung im
Fettsäuremuster in Bezug auf einen Temperaturunterschied gibt. Diese Isolate gehören
der Spezies Kocuria palustris an.
Abbildung 12 Fettsäuremuster von Isolat Kocuria rhizophila J32 bei den Wachstumstemperaturen
10°C und 30°C
10
0,0
0,0
0,0
10
0,0
0,5
2,1
97
,3
0,4
0,7
98
,1
0
20
40
60
80
100
An
teil
in %
Kocuria palustris J17 und Kocuria palustris J40
J17 10°C
J17 30°C
J40 10°C
J40 30°C
89
,1
3,6 0,4
87
,8
2,8
1,9
0
20
40
60
80
100
An
teil
in %
Kocuria rhizophila J32
10°C
30°C
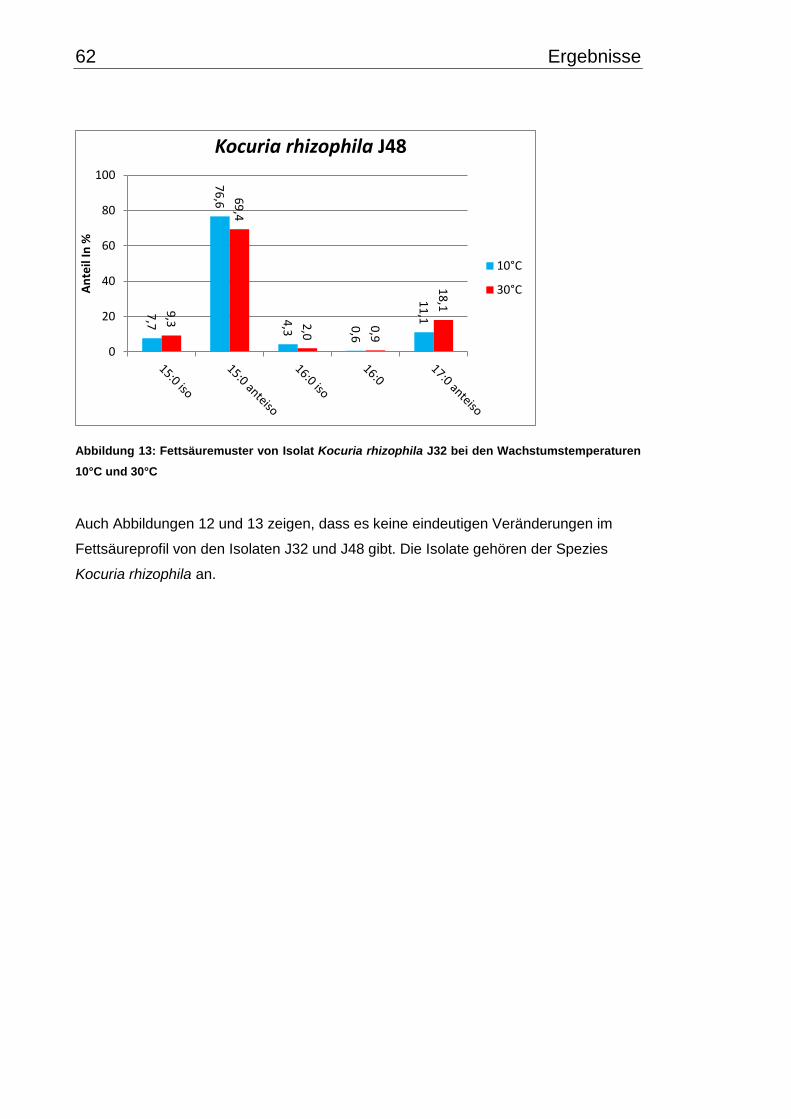
62 Ergebnisse
Abbildung 13: Fettsäuremuster von Isolat Kocuria rhizophila J32 bei den Wachstumstemperaturen
10°C und 30°C
Auch Abbildungen 12 und 13 zeigen, dass es keine eindeutigen Veränderungen im
Fettsäureprofil von den Isolaten J32 und J48 gibt. Die Isolate gehören der Spezies
Kocuria rhizophila an.
7,7
76
,6
4,3 0
,6
11
,1
9,3
69
,4
2,0
0,9
18
,10
20
40
60
80
100
An
teil
In %
Kocuria rhizophila J48
10°C
30°C
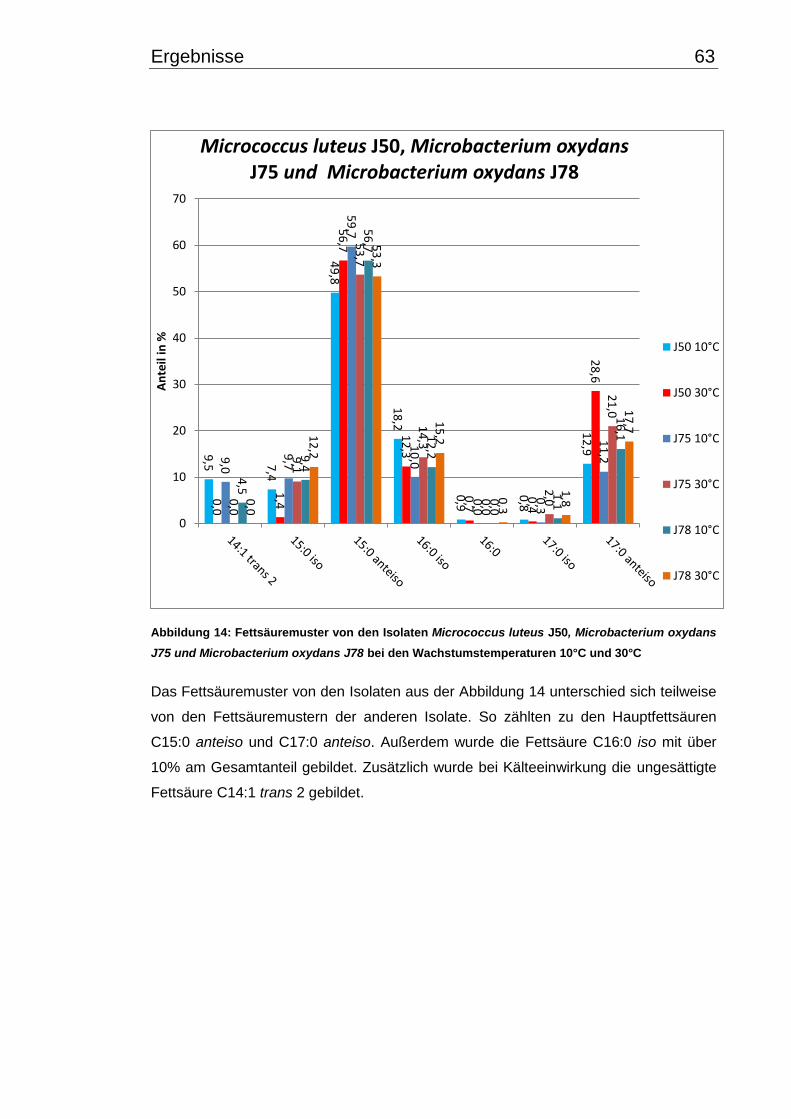
Ergebnisse 63
Abbildung 14: Fettsäuremuster von den Isolaten Micrococcus luteus J50, Microbacterium oxydans
J75 und Microbacterium oxydans J78 bei den Wachstumstemperaturen 10°C und 30°C
Das Fettsäuremuster von den Isolaten aus der Abbildung 14 unterschied sich teilweise
von den Fettsäuremustern der anderen Isolate. So zählten zu den Hauptfettsäuren
C15:0 anteiso und C17:0 anteiso. Außerdem wurde die Fettsäure C16:0 iso mit über
10% am Gesamtanteil gebildet. Zusätzlich wurde bei Kälteeinwirkung die ungesättigte
Fettsäure C14:1 trans 2 gebildet.
9,5 7
,4
49
,8
18
,2
0,9
0,8
12
,9
0,0
1,4
56
,7
12
,3
0,7
0,4
28
,6
9,0
9,7
59
,7
10
,0
0,0
0,3
11
,2
0,0
9,1
53
,7
14
,3
0,0
2,0
21
,0
4,5
9,4
56
,7
12
,2
0,0
1,1
16
,1
0,0
12
,2
53
,3
15
,2
0,3
1,8
17
,7
0
10
20
30
40
50
60
70
An
teil
in %
Micrococcus luteus J50, Microbacterium oxydans
J75 und Microbacterium oxydans J78
J50 10°C
J50 30°C
J75 10°C
J75 30°C
J78 10°C
J78 30°C
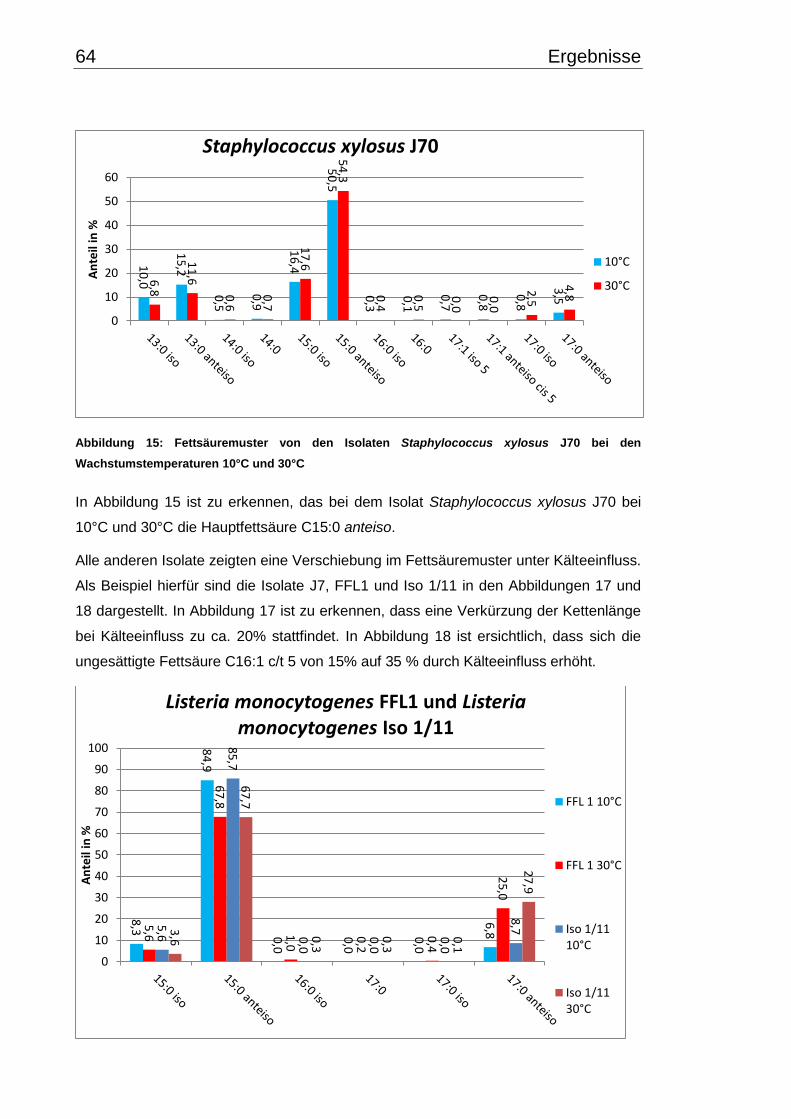
64 Ergebnisse
Abbildung 15: Fettsäuremuster von den Isolaten Staphylococcus xylosus J70 bei den
Wachstumstemperaturen 10°C und 30°C
In Abbildung 15 ist zu erkennen, das bei dem Isolat Staphylococcus xylosus J70 bei
10°C und 30°C die Hauptfettsäure C15:0 anteiso.
Alle anderen Isolate zeigten eine Verschiebung im Fettsäuremuster unter Kälteeinfluss.
Als Beispiel hierfür sind die Isolate J7, FFL1 und Iso 1/11 in den Abbildungen 17 und
18 dargestellt. In Abbildung 17 ist zu erkennen, dass eine Verkürzung der Kettenlänge
bei Kälteeinfluss zu ca. 20% stattfindet. In Abbildung 18 ist ersichtlich, dass sich die
ungesättigte Fettsäure C16:1 c/t 5 von 15% auf 35 % durch Kälteeinfluss erhöht.
10
,0
15
,2
0,5
0,9
16
,4
50
,5
0,3
0,1
0,7
0,8
0,8
3,5
6,8
11
,6
0,6
0,7
17
,6
54
,3
0,4
0,5
0,0
0,0
2,5
4,8
0
10
20
30
40
50
60
An
teil
in %
Staphylococcus xylosus J70
10°C
30°C
8,3
84
,9
0,0
0,0
0,0
6,8
5,6
67
,8
1,0
0,2
0,4
25
,0
5,6
85
,7
0,0
0,0
0,0
8,73
,6
67
,7
0,3
0,3
0,1
27
,9
0
10
20
30
40
50
60
70
80
90
100
An
teil
in %
Listeria monocytogenes FFL1 und Listeria monocytogenes Iso 1/11
FFL 1 10°C
FFL 1 30°C
Iso 1/1110°C
Iso 1/1130°C
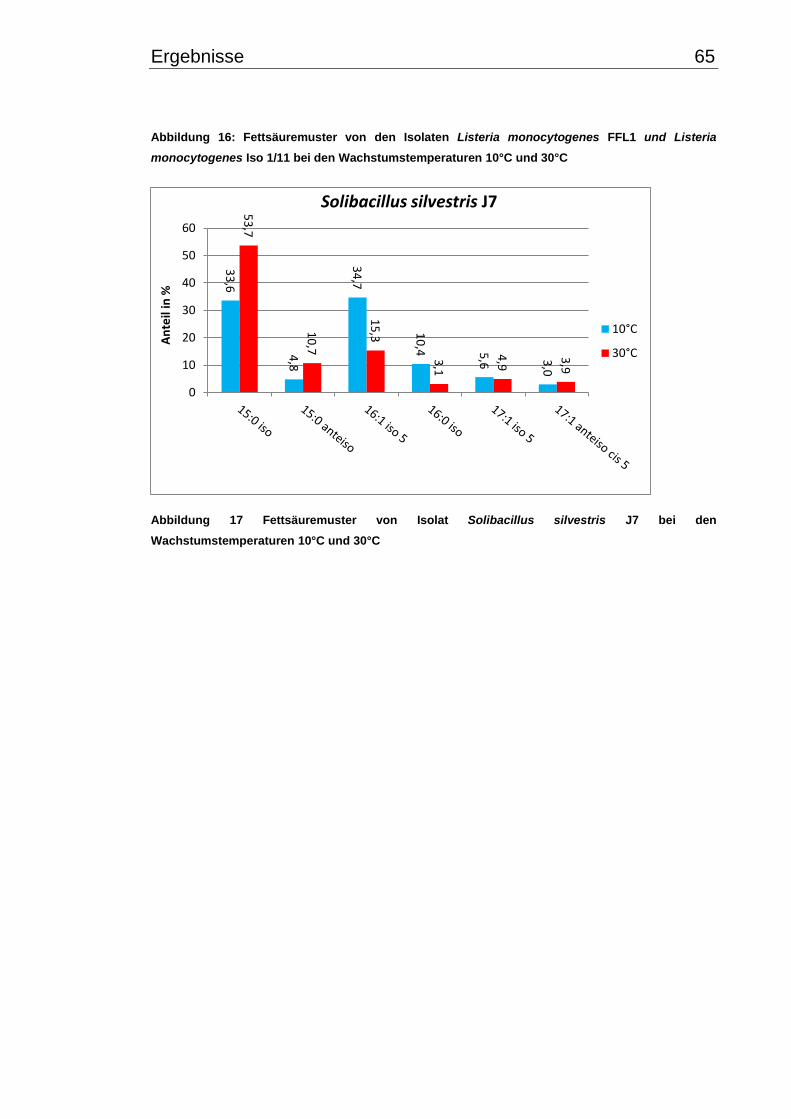
Ergebnisse 65
Abbildung 16: Fettsäuremuster von den Isolaten Listeria monocytogenes FFL1 und Listeria
monocytogenes Iso 1/11 bei den Wachstumstemperaturen 10°C und 30°C
Abbildung 17 Fettsäuremuster von Isolat Solibacillus silvestris J7 bei den
Wachstumstemperaturen 10°C und 30°C
33
,6
4,8
34
,7
10
,4 5,6 3
,0
53
,7
10
,7
15
,3
3,1
4,9
3,9
0
10
20
30
40
50
60
An
teil
in %
Solibacillus silvestris J7
10°C
30°C
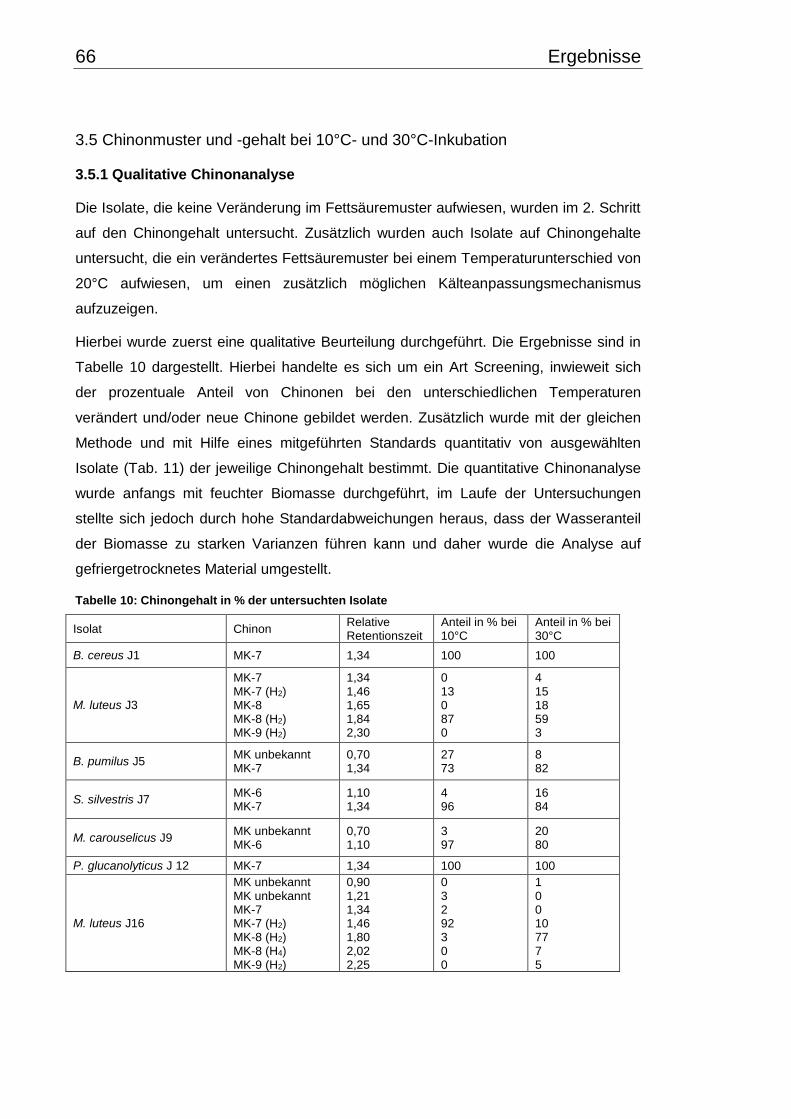
66 Ergebnisse
3.5 Chinonmuster und -gehalt bei 10°C- und 30°C-Inkubation
3.5.1 Qualitative Chinonanalyse
Die Isolate, die keine Veränderung im Fettsäuremuster aufwiesen, wurden im 2. Schritt
auf den Chinongehalt untersucht. Zusätzlich wurden auch Isolate auf Chinongehalte
untersucht, die ein verändertes Fettsäuremuster bei einem Temperaturunterschied von
20°C aufwiesen, um einen zusätzlich möglichen Kälteanpassungsmechanismus
aufzuzeigen.
Hierbei wurde zuerst eine qualitative Beurteilung durchgeführt. Die Ergebnisse sind in
Tabelle 10 dargestellt. Hierbei handelte es sich um ein Art Screening, inwieweit sich
der prozentuale Anteil von Chinonen bei den unterschiedlichen Temperaturen
verändert und/oder neue Chinone gebildet werden. Zusätzlich wurde mit der gleichen
Methode und mit Hilfe eines mitgeführten Standards quantitativ von ausgewählten
Isolate (Tab. 11) der jeweilige Chinongehalt bestimmt. Die quantitative Chinonanalyse
wurde anfangs mit feuchter Biomasse durchgeführt, im Laufe der Untersuchungen
stellte sich jedoch durch hohe Standardabweichungen heraus, dass der Wasseranteil
der Biomasse zu starken Varianzen führen kann und daher wurde die Analyse auf
gefriergetrocknetes Material umgestellt.
Tabelle 10: Chinongehalt in % der untersuchten Isolate
Isolat Chinon Relative Retentionszeit
Anteil in % bei 10°C
Anteil in % bei 30°C
B. cereus J1 MK-7 1,34 100 100
M. luteus J3
MK-7 MK-7 (H2) MK-8 MK-8 (H2) MK-9 (H2)
1,34 1,46 1,65 1,84 2,30
0 13 0 87 0
4 15 18 59 3
B. pumilus J5 MK unbekannt MK-7
0,70 1,34
27 73
8 82
S. silvestris J7 MK-6 MK-7
1,10 1,34
4 96
16 84
M. carouselicus J9 MK unbekannt MK-6
0,70 1,10
3 97
20 80
P. glucanolyticus J 12 MK-7 1,34 100 100
M. luteus J16
MK unbekannt MK unbekannt MK-7 MK-7 (H2) MK-8 (H2) MK-8 (H4) MK-9 (H2)
0,90 1,21 1,34 1,46 1,80 2,02 2,25
0 3 2 92 3 0 0
1 0 0 10 77 7 5
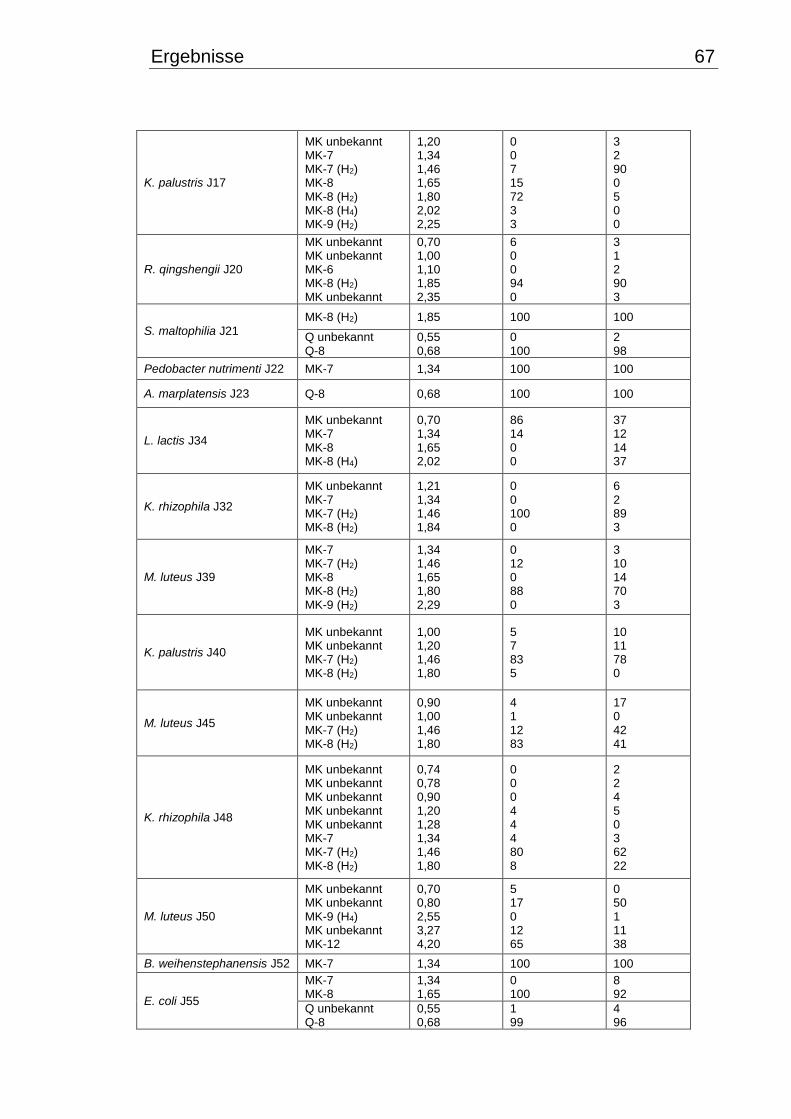
Ergebnisse 67
K. palustris J17
MK unbekannt MK-7 MK-7 (H2) MK-8 MK-8 (H2) MK-8 (H4) MK-9 (H2)
1,20 1,34 1,46 1,65 1,80 2,02 2,25
0 0 7 15 72 3 3
3 2 90 0 5 0 0
R. qingshengii J20
MK unbekannt MK unbekannt MK-6 MK-8 (H2) MK unbekannt
0,70 1,00 1,10 1,85 2,35
6 0 0 94 0
3 1 2 90 3
S. maltophilia J21 MK-8 (H2) 1,85 100 100
Q unbekannt Q-8
0,55 0,68
0 100
2 98
Pedobacter nutrimenti J22 MK-7 1,34 100 100
A. marplatensis J23 Q-8 0,68 100 100
L. lactis J34
MK unbekannt MK-7 MK-8 MK-8 (H4)
0,70 1,34 1,65 2,02
86 14 0 0
37 12 14 37
K. rhizophila J32
MK unbekannt MK-7 MK-7 (H2) MK-8 (H2)
1,21 1,34 1,46 1,84
0 0 100 0
6 2 89 3
M. luteus J39
MK-7 MK-7 (H2) MK-8 MK-8 (H2) MK-9 (H2)
1,34 1,46 1,65 1,80 2,29
0 12 0 88 0
3 10 14 70 3
K. palustris J40
MK unbekannt MK unbekannt MK-7 (H2) MK-8 (H2)
1,00 1,20 1,46 1,80
5 7 83 5
10 11 78 0
M. luteus J45
MK unbekannt MK unbekannt MK-7 (H2) MK-8 (H2)
0,90 1,00 1,46 1,80
4 1 12 83
17 0 42 41
K. rhizophila J48
MK unbekannt MK unbekannt MK unbekannt MK unbekannt MK unbekannt MK-7 MK-7 (H2) MK-8 (H2)
0,74 0,78 0,90 1,20 1,28 1,34 1,46 1,80
0 0 0 4 4 4 80 8
2 2 4 5 0 3 62 22
M. luteus J50
MK unbekannt MK unbekannt MK-9 (H4) MK unbekannt MK-12
0,70 0,80 2,55 3,27 4,20
5 17 0 12 65
0 50 1 11 38
B. weihenstephanensis J52 MK-7 1,34 100 100
E. coli J55
MK-7 MK-8
1,34 1,65
0 100
8 92
Q unbekannt Q-8
0,55 0,68
1 99
4 96
68 Ergebnisse
S. maltophilia J57 Q unbekannt Q-8
0,55 0,68
0 100
2 98
B. safensis J60 MK-6 MK-7
1,10 1,34
0 100
1 99
M. oxydans J75
MK unbekannt MK unbekannt MK-11 MK-12
0,70 0,80 3,27 4,20
10 11 9 70
7 25 23 45
M. oxydans J78
MK unbekannt MK unbekannt MK-11 MK-12
0,70 0,80 3,27 4,20
4 15 11 70
0 9 30 61
A. lwoffii J 84
Q unbekannt Q-8 Q-9 Q-10
0,56 0,65 0,80 1,00
1 31 65 1
1 30 68 1
B. thermosphacta J86 MK-7 1,34 100 100
P. agglomerans J89
MK-7 MK-8
1,34 1,65
4 96
6 94
Q unbekannt Q unbekannt Q-8
0,48 0,56 0,65
0 2 98
1 6 93
P. baetica J91 Q-8 Q-9 Q-10
0,65 0,80 1,00
3 97 0
5 94 1
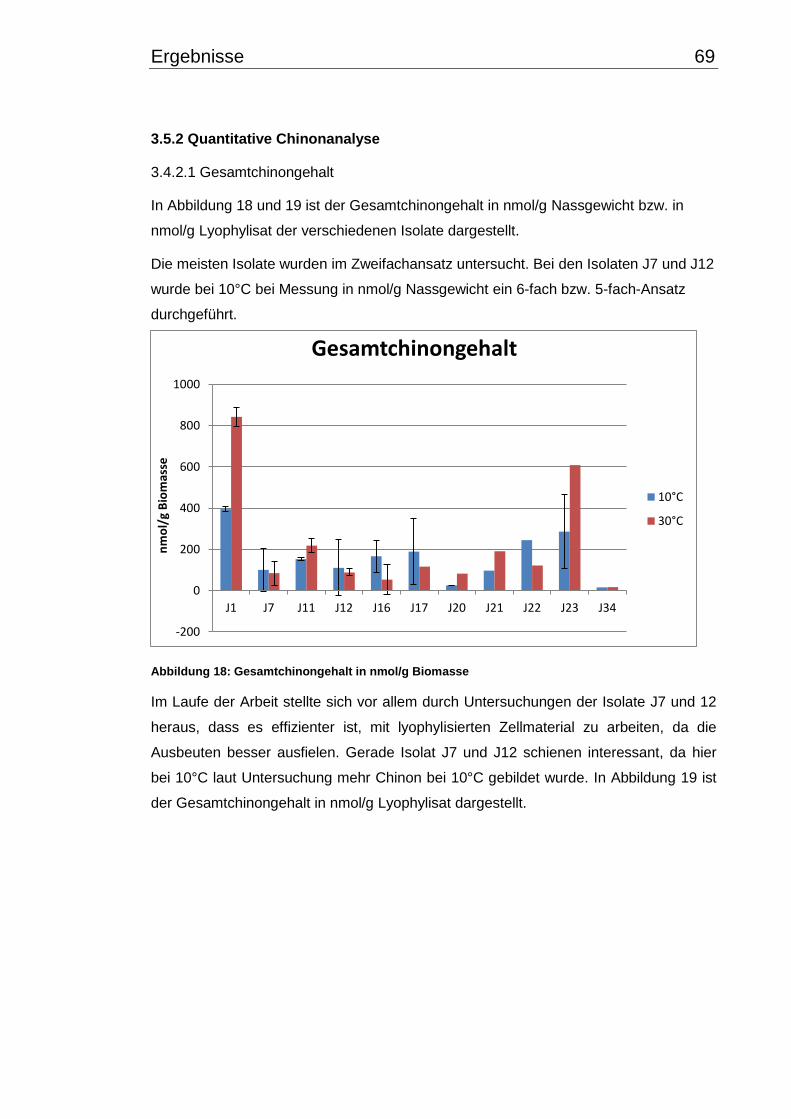
Ergebnisse 69
3.5.2 Quantitative Chinonanalyse
3.4.2.1 Gesamtchinongehalt
In Abbildung 18 und 19 ist der Gesamtchinongehalt in nmol/g Nassgewicht bzw. in
nmol/g Lyophylisat der verschiedenen Isolate dargestellt.
Die meisten Isolate wurden im Zweifachansatz untersucht. Bei den Isolaten J7 und J12
wurde bei 10°C bei Messung in nmol/g Nassgewicht ein 6-fach bzw. 5-fach-Ansatz
durchgeführt.
Abbildung 18: Gesamtchinongehalt in nmol/g Biomasse
Im Laufe der Arbeit stellte sich vor allem durch Untersuchungen der Isolate J7 und 12
heraus, dass es effizienter ist, mit lyophylisierten Zellmaterial zu arbeiten, da die
Ausbeuten besser ausfielen. Gerade Isolat J7 und J12 schienen interessant, da hier
bei 10°C laut Untersuchung mehr Chinon bei 10°C gebildet wurde. In Abbildung 19 ist
der Gesamtchinongehalt in nmol/g Lyophylisat dargestellt.
-200
0
200
400
600
800
1000
J1 J7 J11 J12 J16 J17 J20 J21 J22 J23 J34
nm
ol/
g B
iom
asse
Gesamtchinongehalt
10°C
30°C
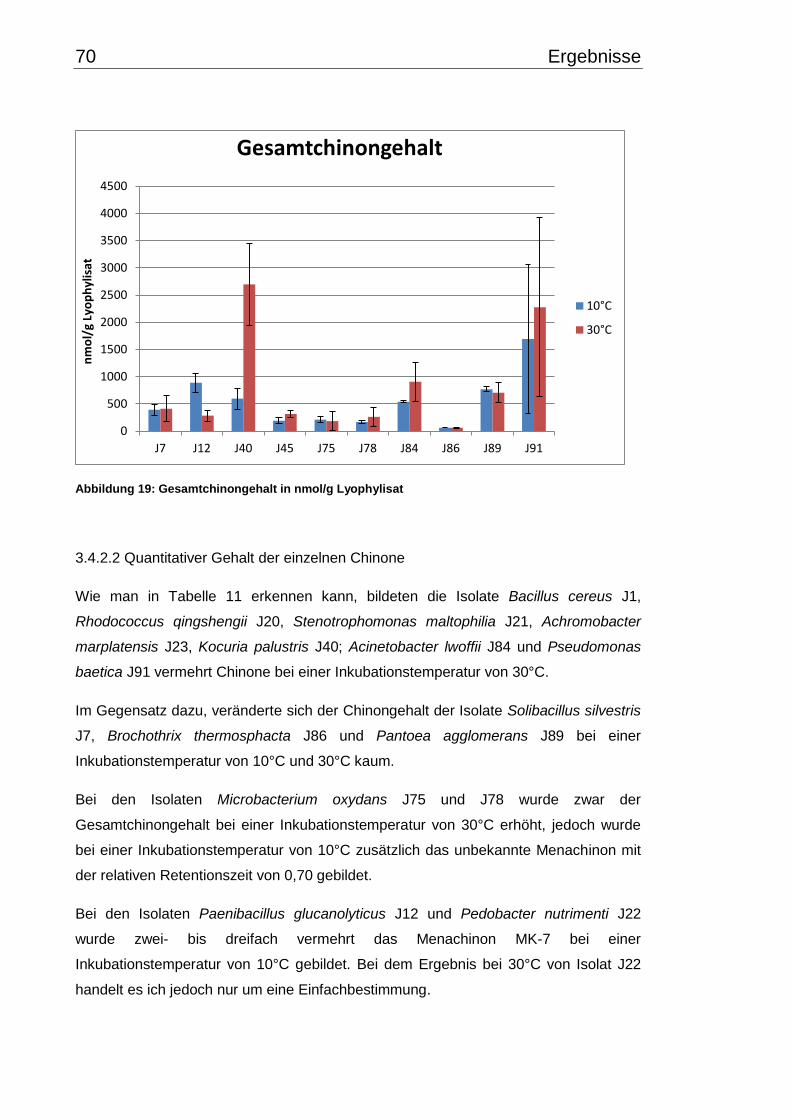
70 Ergebnisse
Abbildung 19: Gesamtchinongehalt in nmol/g Lyophylisat
3.4.2.2 Quantitativer Gehalt der einzelnen Chinone
Wie man in Tabelle 11 erkennen kann, bildeten die Isolate Bacillus cereus J1,
Rhodococcus qingshengii J20, Stenotrophomonas maltophilia J21, Achromobacter
marplatensis J23, Kocuria palustris J40; Acinetobacter lwoffii J84 und Pseudomonas
baetica J91 vermehrt Chinone bei einer Inkubationstemperatur von 30°C.
Im Gegensatz dazu, veränderte sich der Chinongehalt der Isolate Solibacillus silvestris
J7, Brochothrix thermosphacta J86 und Pantoea agglomerans J89 bei einer
Inkubationstemperatur von 10°C und 30°C kaum.
Bei den Isolaten Microbacterium oxydans J75 und J78 wurde zwar der
Gesamtchinongehalt bei einer Inkubationstemperatur von 30°C erhöht, jedoch wurde
bei einer Inkubationstemperatur von 10°C zusätzlich das unbekannte Menachinon mit
der relativen Retentionszeit von 0,70 gebildet.
Bei den Isolaten Paenibacillus glucanolyticus J12 und Pedobacter nutrimenti J22
wurde zwei- bis dreifach vermehrt das Menachinon MK-7 bei einer
Inkubationstemperatur von 10°C gebildet. Bei dem Ergebnis bei 30°C von Isolat J22
handelt es ich jedoch nur um eine Einfachbestimmung.
0
500
1000
1500
2000
2500
3000
3500
4000
4500
J7 J12 J40 J45 J75 J78 J84 J86 J89 J91
nm
ol/
g Ly
op
hyl
isat
Gesamtchinongehalt
10°C
30°C
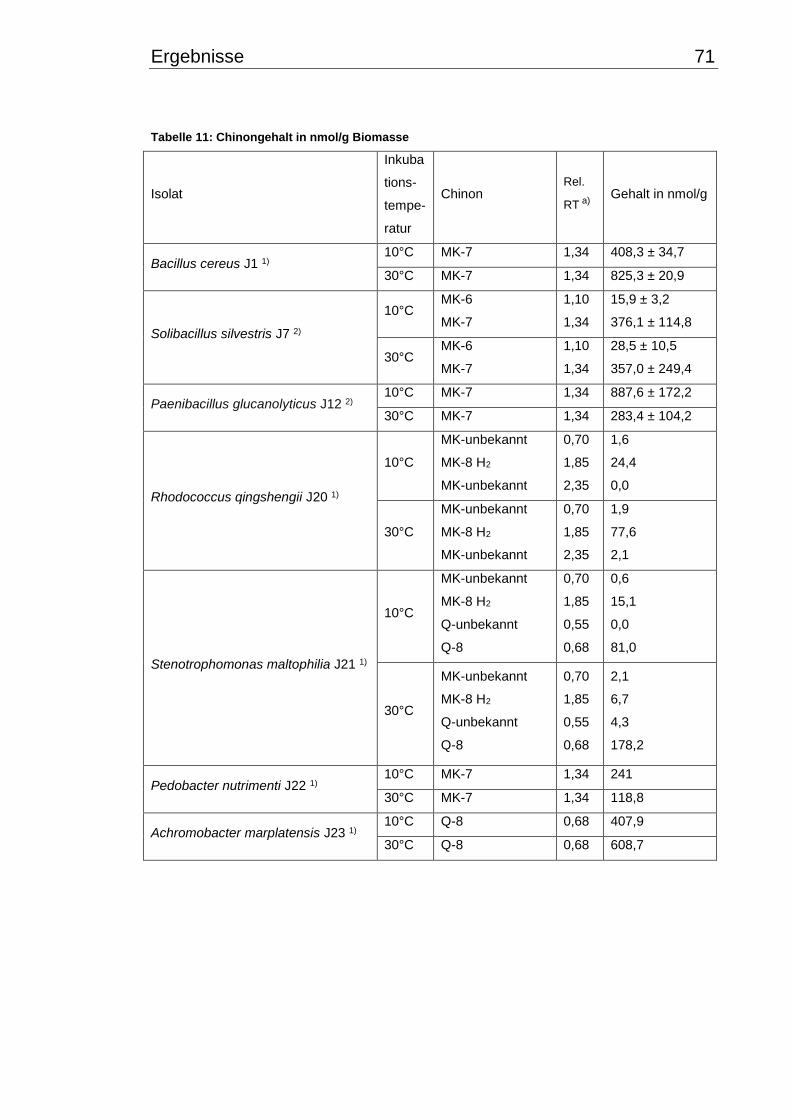
Ergebnisse 71
Tabelle 11: Chinongehalt in nmol/g Biomasse
Isolat
Inkuba
tions-
tempe-
ratur
Chinon Rel.
RT a) Gehalt in nmol/g
Bacillus cereus J1 1) 10°C MK-7 1,34 408,3 ± 34,7
30°C MK-7 1,34 825,3 ± 20,9
Solibacillus silvestris J7 2)
10°C MK-6
MK-7
1,10
1,34
15,9 ± 3,2
376,1 ± 114,8
30°C MK-6
MK-7
1,10
1,34
28,5 ± 10,5
357,0 ± 249,4
Paenibacillus glucanolyticus J12 2) 10°C MK-7 1,34 887,6 ± 172,2
30°C MK-7 1,34 283,4 ± 104,2
Rhodococcus qingshengii J20 1)
10°C
MK-unbekannt
MK-8 H2
MK-unbekannt
0,70
1,85
2,35
1,6
24,4
0,0
30°C
MK-unbekannt
MK-8 H2
MK-unbekannt
0,70
1,85
2,35
1,9
77,6
2,1
Stenotrophomonas maltophilia J21 1)
10°C
MK-unbekannt
MK-8 H2
Q-unbekannt
Q-8
0,70
1,85
0,55
0,68
0,6
15,1
0,0
81,0
30°C
MK-unbekannt
MK-8 H2
Q-unbekannt
Q-8
0,70
1,85
0,55
0,68
2,1
6,7
4,3
178,2
Pedobacter nutrimenti J22 1) 10°C MK-7 1,34 241
30°C MK-7 1,34 118,8
Achromobacter marplatensis J23 1) 10°C Q-8 0,68 407,9
30°C Q-8 0,68 608,7
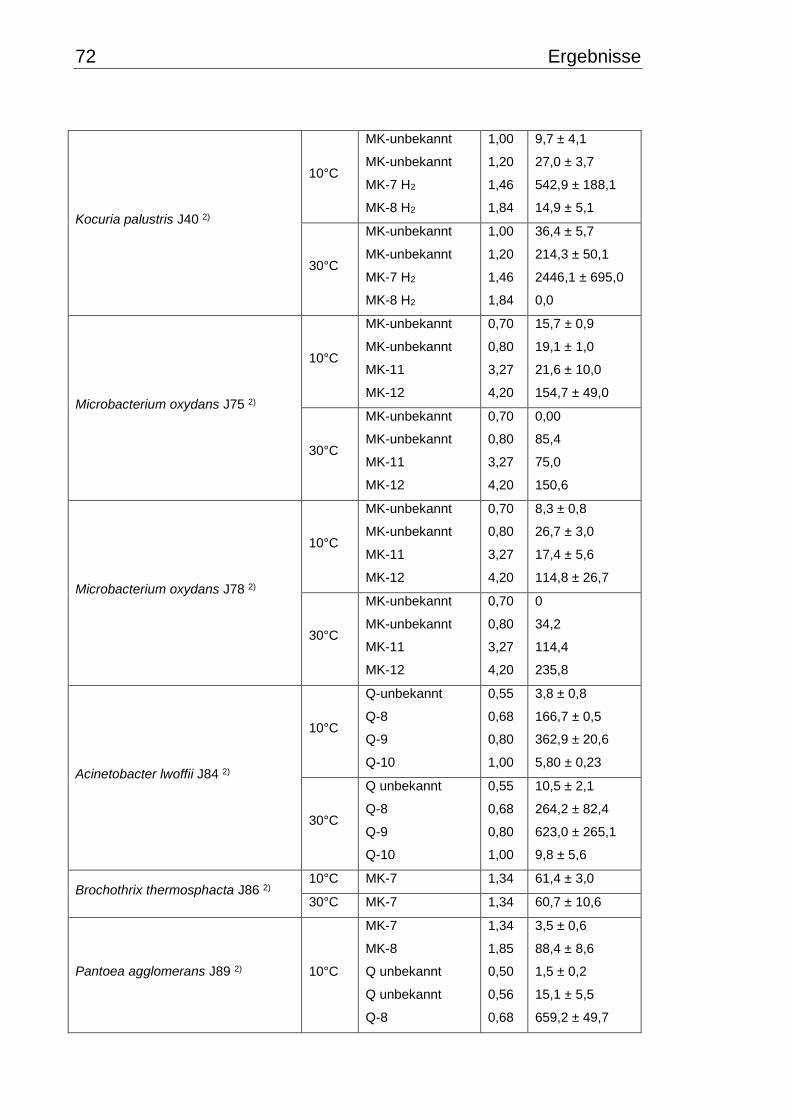
72 Ergebnisse
Kocuria palustris J40 2)
10°C
MK-unbekannt
MK-unbekannt
MK-7 H2
MK-8 H2
1,00
1,20
1,46
1,84
9,7 ± 4,1
27,0 ± 3,7
542,9 ± 188,1
14,9 ± 5,1
30°C
MK-unbekannt
MK-unbekannt
MK-7 H2
MK-8 H2
1,00
1,20
1,46
1,84
36,4 ± 5,7
214,3 ± 50,1
2446,1 ± 695,0
0,0
Microbacterium oxydans J75 2)
10°C
MK-unbekannt
MK-unbekannt
MK-11
MK-12
0,70
0,80
3,27
4,20
15,7 ± 0,9
19,1 ± 1,0
21,6 ± 10,0
154,7 ± 49,0
30°C
MK-unbekannt
MK-unbekannt
MK-11
MK-12
0,70
0,80
3,27
4,20
0,00
85,4
75,0
150,6
Microbacterium oxydans J78 2)
10°C
MK-unbekannt
MK-unbekannt
MK-11
MK-12
0,70
0,80
3,27
4,20
8,3 ± 0,8
26,7 ± 3,0
17,4 ± 5,6
114,8 ± 26,7
30°C
MK-unbekannt
MK-unbekannt
MK-11
MK-12
0,70
0,80
3,27
4,20
0
34,2
114,4
235,8
Acinetobacter lwoffii J84 2)
10°C
Q-unbekannt
Q-8
Q-9
Q-10
0,55
0,68
0,80
1,00
3,8 ± 0,8
166,7 ± 0,5
362,9 ± 20,6
5,80 ± 0,23
30°C
Q unbekannt
Q-8
Q-9
Q-10
0,55
0,68
0,80
1,00
10,5 ± 2,1
264,2 ± 82,4
623,0 ± 265,1
9,8 ± 5,6
Brochothrix thermosphacta J86 2) 10°C MK-7 1,34 61,4 ± 3,0
30°C MK-7 1,34 60,7 ± 10,6
Pantoea agglomerans J89 2) 10°C
MK-7
MK-8
Q unbekannt
Q unbekannt
Q-8
1,34
1,85
0,50
0,56
0,68
3,5 ± 0,6
88,4 ± 8,6
1,5 ± 0,2
15,1 ± 5,5
659,2 ± 49,7
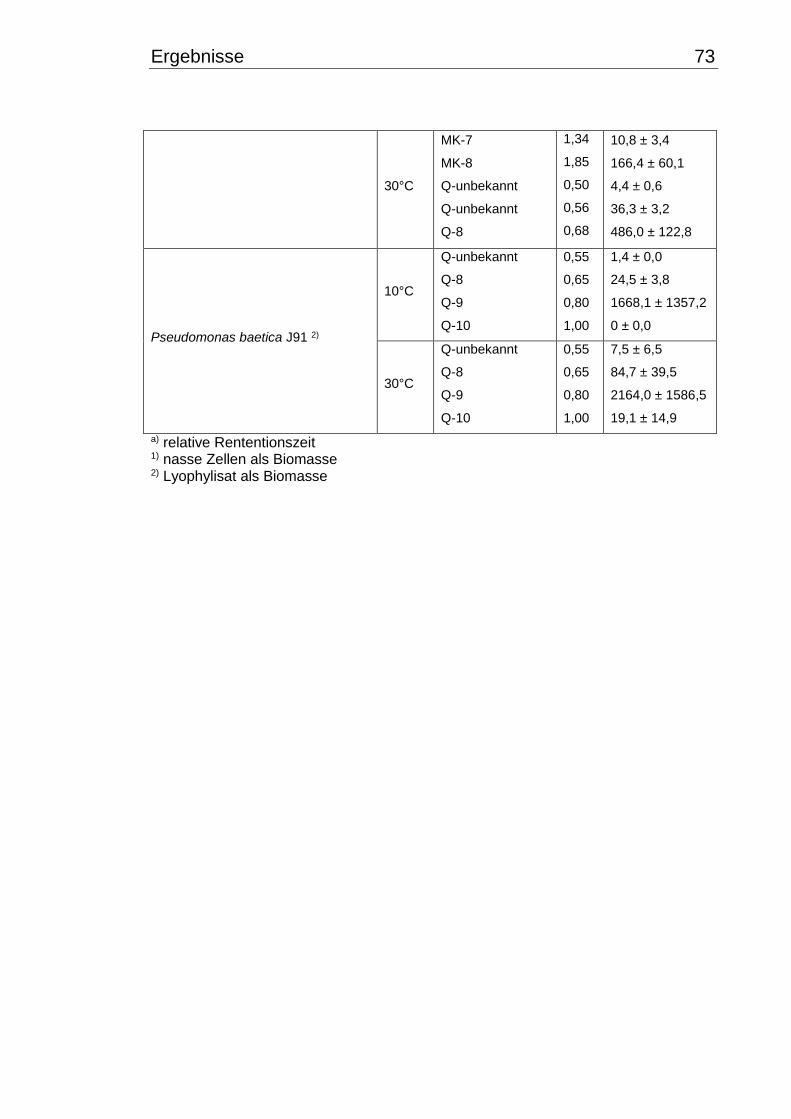
Ergebnisse 73
30°C
MK-7
MK-8
Q-unbekannt
Q-unbekannt
Q-8
1,34
1,85
0,50
0,56
0,68
10,8 ± 3,4
166,4 ± 60,1
4,4 ± 0,6
36,3 ± 3,2
486,0 ± 122,8
Pseudomonas baetica J91 2)
10°C
Q-unbekannt
Q-8
Q-9
Q-10
0,55
0,65
0,80
1,00
1,4 ± 0,0
24,5 ± 3,8
1668,1 ± 1357,2
0 ± 0,0
30°C
Q-unbekannt
Q-8
Q-9
Q-10
0,55
0,65
0,80
1,00
7,5 ± 6,5
84,7 ± 39,5
2164,0 ± 1586,5
19,1 ± 14,9
a) relative Rententionszeit 1) nasse Zellen als Biomasse 2) Lyophylisat als Biomasse

74 Ergebnisse
3.6 Carotinoidgehalt bei 10°C- und 30°C-Inkubation
Im Laufe der Arbeit stellte sich heraus, dass einige Bakterienisolate eine stärkere
Pigmentbildung bei 10°C Inkubationstemperatur zeigten als bei 30°C
Inkubationstemperatur. Aus den Abbildungen 20 bis 24 ist beispielhaft ersichtlich, dass
sich die Intensität der Pigmentbildung der Isolate J7, J48, J50 und J70 bei 10°C im
Vergleich zu 30°C unterscheidet.
Abbildung 20: Isolat Solibacillus silvestris J7 bei 10°C (links) und 30°C (rechts)
Inkubationstemperatur
Abbildung 21: Isolat Kocuria rhizophila J48 bei 10°C (links) und 30°C (rechts)
Inkubationstemperatur
Ergebnisse 75
Abbildung 22: Isolat Micrococcus luteus J50 bei 10°C (links) und 30°C (rechts)
Inkubationstemperatur
Abbildung 23: Isolat Staphylococcus xylosus J70 bei 10°C (links) und 30°C (rechts)
Inkubationstemperatur
Abbildung 24: Isolat Microbacterium maritypicum J74 bei 10°C (links) und 30°C (rechts)
Inkubationstemperatur
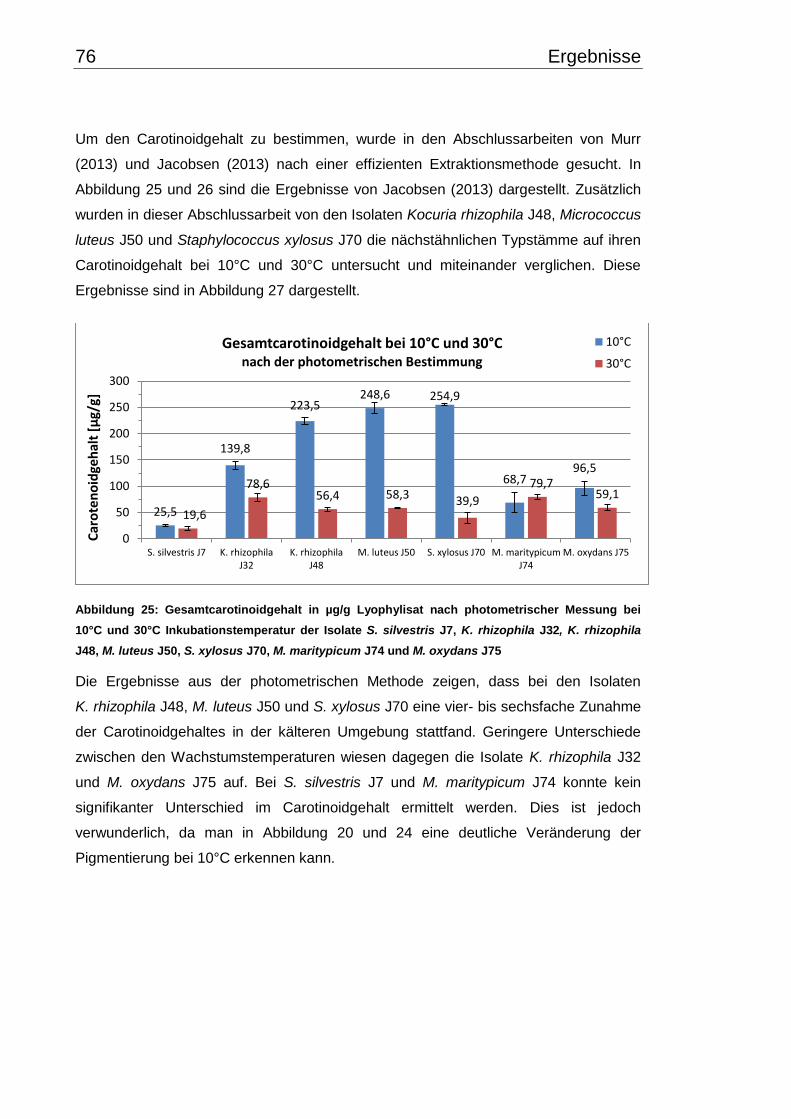
76 Ergebnisse
Um den Carotinoidgehalt zu bestimmen, wurde in den Abschlussarbeiten von Murr
(2013) und Jacobsen (2013) nach einer effizienten Extraktionsmethode gesucht. In
Abbildung 25 und 26 sind die Ergebnisse von Jacobsen (2013) dargestellt. Zusätzlich
wurden in dieser Abschlussarbeit von den Isolaten Kocuria rhizophila J48, Micrococcus
luteus J50 und Staphylococcus xylosus J70 die nächstähnlichen Typstämme auf ihren
Carotinoidgehalt bei 10°C und 30°C untersucht und miteinander verglichen. Diese
Ergebnisse sind in Abbildung 27 dargestellt.
Abbildung 25: Gesamtcarotinoidgehalt in µg/g Lyophylisat nach photometrischer Messung bei
10°C und 30°C Inkubationstemperatur der Isolate S. silvestris J7, K. rhizophila J32, K. rhizophila
J48, M. luteus J50, S. xylosus J70, M. maritypicum J74 und M. oxydans J75
Die Ergebnisse aus der photometrischen Methode zeigen, dass bei den Isolaten
K. rhizophila J48, M. luteus J50 und S. xylosus J70 eine vier- bis sechsfache Zunahme
der Carotinoidgehaltes in der kälteren Umgebung stattfand. Geringere Unterschiede
zwischen den Wachstumstemperaturen wiesen dagegen die Isolate K. rhizophila J32
und M. oxydans J75 auf. Bei S. silvestris J7 und M. maritypicum J74 konnte kein
signifikanter Unterschied im Carotinoidgehalt ermittelt werden. Dies ist jedoch
verwunderlich, da man in Abbildung 20 und 24 eine deutliche Veränderung der
Pigmentierung bei 10°C erkennen kann.
25,5
139,8
223,5248,6 254,9
68,796,5
19,6
78,656,4 58,3 39,9
79,759,1
0
50
100
150
200
250
300
S. silvestris J7 K. rhizophilaJ32
K. rhizophilaJ48
M. luteus J50 S. xylosus J70 M. maritypicumJ74
M. oxydans J75
Car
ote
no
idge
hal
t [µ
g/g]
Gesamtcarotinoidgehalt bei 10°C und 30°Cnach der photometrischen Bestimmung
10°C
30°C
C
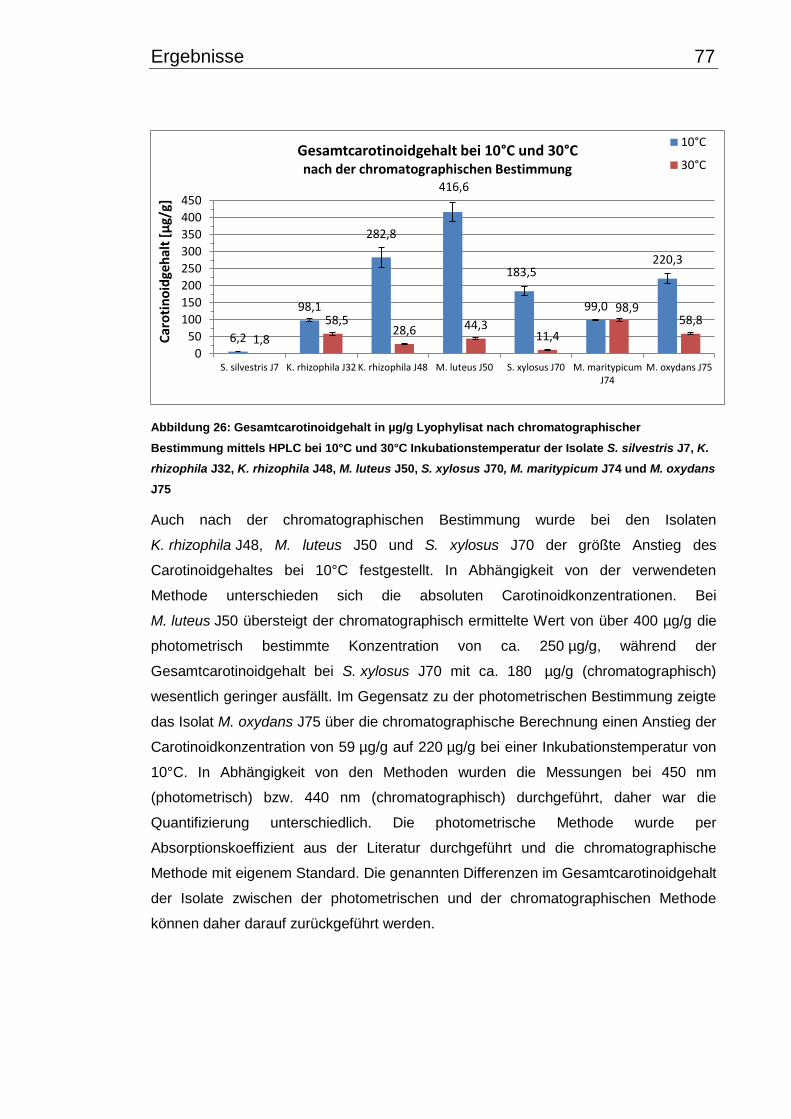
Ergebnisse 77
Abbildung 26: Gesamtcarotinoidgehalt in µg/g Lyophylisat nach chromatographischer
Bestimmung mittels HPLC bei 10°C und 30°C Inkubationstemperatur der Isolate S. silvestris J7, K.
rhizophila J32, K. rhizophila J48, M. luteus J50, S. xylosus J70, M. maritypicum J74 und M. oxydans
J75
Auch nach der chromatographischen Bestimmung wurde bei den Isolaten
K. rhizophila J48, M. luteus J50 und S. xylosus J70 der größte Anstieg des
Carotinoidgehaltes bei 10°C festgestellt. In Abhängigkeit von der verwendeten
Methode unterschieden sich die absoluten Carotinoidkonzentrationen. Bei
M. luteus J50 übersteigt der chromatographisch ermittelte Wert von über 400 µg/g die
photometrisch bestimmte Konzentration von ca. 250 µg/g, während der
Gesamtcarotinoidgehalt bei S. xylosus J70 mit ca. 180 µg/g (chromatographisch)
wesentlich geringer ausfällt. Im Gegensatz zu der photometrischen Bestimmung zeigte
das Isolat M. oxydans J75 über die chromatographische Berechnung einen Anstieg der
Carotinoidkonzentration von 59 µg/g auf 220 µg/g bei einer Inkubationstemperatur von
10°C. In Abhängigkeit von den Methoden wurden die Messungen bei 450 nm
(photometrisch) bzw. 440 nm (chromatographisch) durchgeführt, daher war die
Quantifizierung unterschiedlich. Die photometrische Methode wurde per
Absorptionskoeffizient aus der Literatur durchgeführt und die chromatographische
Methode mit eigenem Standard. Die genannten Differenzen im Gesamtcarotinoidgehalt
der Isolate zwischen der photometrischen und der chromatographischen Methode
können daher darauf zurückgeführt werden.
6,2
98,1
282,8
416,6
183,5
99,0
220,3
1,8
58,528,6 44,3
11,4
98,958,8
0
50
100
150
200
250
300
350
400
450
S. silvestris J7 K. rhizophila J32 K. rhizophila J48 M. luteus J50 S. xylosus J70 M. maritypicumJ74
M. oxydans J75
Car
oti
no
idge
hal
t [µ
g/g]
Gesamtcarotinoidgehalt bei 10°C und 30°Cnach der chromatographischen Bestimmung
10°C
30°C
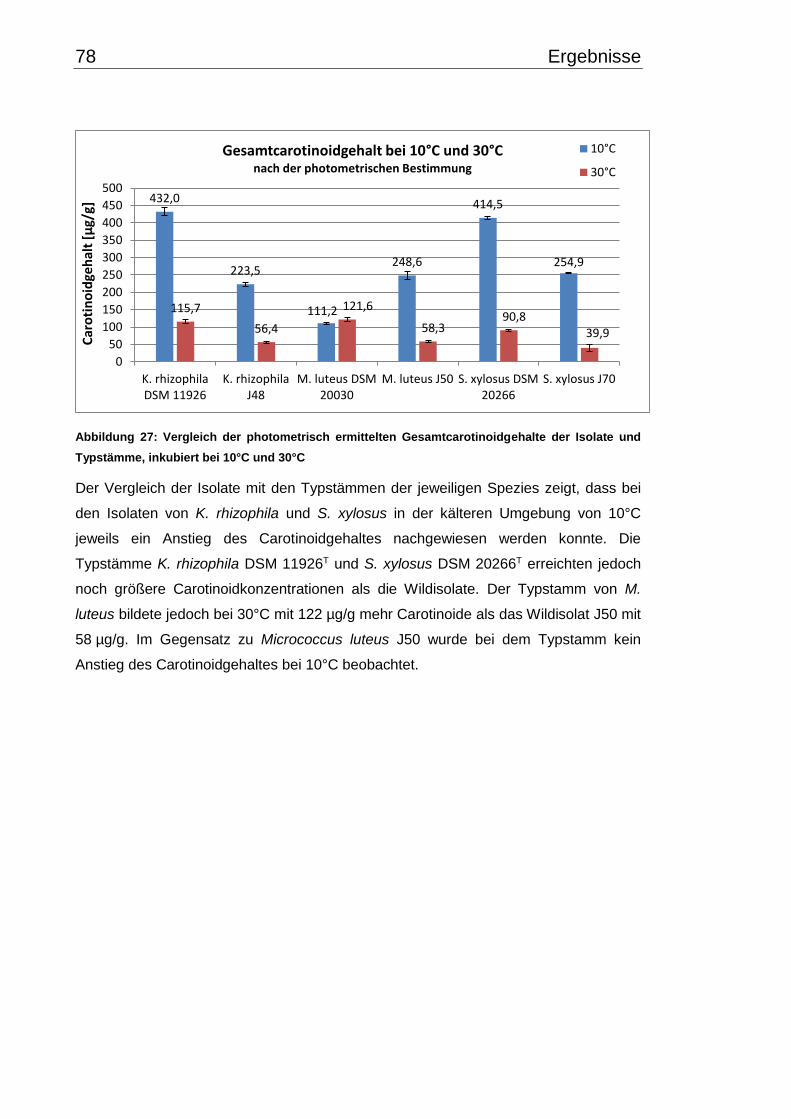
78 Ergebnisse
Abbildung 27: Vergleich der photometrisch ermittelten Gesamtcarotinoidgehalte der Isolate und
Typstämme, inkubiert bei 10°C und 30°C
Der Vergleich der Isolate mit den Typstämmen der jeweiligen Spezies zeigt, dass bei
den Isolaten von K. rhizophila und S. xylosus in der kälteren Umgebung von 10°C
jeweils ein Anstieg des Carotinoidgehaltes nachgewiesen werden konnte. Die
Typstämme K. rhizophila DSM 11926T und S. xylosus DSM 20266T erreichten jedoch
noch größere Carotinoidkonzentrationen als die Wildisolate. Der Typstamm von M.
luteus bildete jedoch bei 30°C mit 122 µg/g mehr Carotinoide als das Wildisolat J50 mit
58 µg/g. Im Gegensatz zu Micrococcus luteus J50 wurde bei dem Typstamm kein
Anstieg des Carotinoidgehaltes bei 10°C beobachtet.
432,0
223,5
111,2
248,6
414,5
254,9
115,7
56,4
121,6
58,390,8
39,9
0
50
100
150
200
250
300
350
400
450
500
K. rhizophilaDSM 11926
K. rhizophilaJ48
M. luteus DSM20030
M. luteus J50 S. xylosus DSM20266
S. xylosus J70
Car
oti
no
idge
hal
t [µ
g/g]
Gesamtcarotinoidgehalt bei 10°C und 30°C nach der photometrischen Bestimmung
10°C
30°C

Ergebnisse 79
3.7 Wachstumsverhalten
Bei einigen Isolaten zeigte sich, dass der Zellertrag in der stationären Phase bei einer
Inkubationstemperatur von 10°C deutlich höher lag als bei der Inkubationstemperatur
von 30°C. Um diesen Effekt näher untersuchen zu können, wurden Wachstumskurven
erstellt und der maximale Zellertrag mittels Trübungsmessung und Proteingehalt
bestimmt. Im folgenden Abschnitt sind einige Isolate als Beispiel für einen erhöhten
Zellertrag dargestellt. Zum Teil werden die Wachstumsverläufe bzw. Zellgehalte der
Isolate mit Wachstumsverläufen bzw. Zellgehalten von Typ- bzw. Referenzstämmen
verglichen.
3.7.1 Wachstumsverlauf einzelner Isolate bei 10°C- und 30°C-
Inkubationstemperatur
Die Abbildungen 28 bis 34 zeigen die Wachstumsverläufe der untersuchten Isolate. Auf
der X-Achse ist jeweils die Dauer der Messung in Stunden dargestellt und auf der
Ordinate die Werte der Optischen Dichte (OD) bei einer Wellenlänge von 625 nm.
Die Zellkulturen aller untersuchten Isolate hatte bei 10°C Inkubationstemperatur eine
deutlich längere lag-Phase als bei 30°C. Auch die stationäre Phase wurde wesentlich
später erreicht als bei 30°C Inkubationstemperatur, nämlich nach mindestens 100 h,
bei den Isolaten FFL1 und Iso1/11 nach 80 h. Die stationäre Phase bei 30°C wurde
bereits nach ca. 24 h erreicht.
Der maximale Zellertrag fällt bei allen Isolaten gemessen anhand der ODmax bei 10°C
deutlich höher aus als bei 30°C Inkubationstemperatur. Bei Isolat J70 wird sogar eine
ODmax von 25 bei 10°C gemessen. Bei 30°C Inkubationstemperatur liegt der
Maximalwert 5-fach niedriger. Alle anderen Isolate zeigten eine 0,5 bis 3-fache
Erniedrigung des Zellertrages bei 30°C.
80 Ergebnisse
Abbildung 28: Wachstumskurven von Isolat Bacillus cereus J1 bei 10°C und 30°C Inkubation
Abbildung 29: Wachstumskurven von Isolat Paenibacillus glucanolyticus J12 bei 10°C und 30°C
Inkubation
Abbildung 30: Wachstumskurven von Isolat Pedobacter nutrimenti J22 bei 10°C und 30°C
Inkubation
0123456789
0 50 100 150
OD
be
i 62
5 n
m
Zeit in h
Bacillus cereus J110°C
10°C
10°C
30°C
30°C
30°C
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 50 100 150
OD
be
i 62
5 n
m
Zeit in h
Paenibacillus glucanolyticus J12
10°C
10°C
10°C
30°c
30°C
30°C
0
1
2
3
4
5
6
7
8
9
0 50 100 150 200
OD
be
i 62
5 n
m
Zeit in h
Pedobacter nutrimenti J22
10°C
10°C
10°C
30°C
30°C
30°C
Ergebnisse 81
Abbildung 31: Wachstumskurven von Isolat Escherichia coli J55 bei 10°C und 30°C Inkubation
Abbildung 32: Wachstumskurven von Isolat Staphylococcus xylosus J70 bei 10°C und 30°C
Inkubation
Abbildung 33: Wachstumskurven von Isolat Listeria monocytogenes FFL1 bei 10°C und 30°C
Inkubation
0
2
4
6
8
10
12
0 50 100 150 200
OD
be
i 62
5 n
m
Zeit in h
Escherichia coli J55
10°C
10°C
10°C
30°C
30°C
30°C
0
5
10
15
20
25
30
0 50 100 150
OD
be
i 62
5 n
m
Zeit in h
Staphylococcus xylosus J70
10°C
10°C
10°C
30°C
30°C
30°C
0
0,5
1
1,5
2
2,5
3
3,5
4
0 20 40 60 80 100
Od
be
i 62
5 n
m
Zeit in h
Listeria monocytogenes FFL1
10°C
10°C
10°C
30°C
30°C
30°C
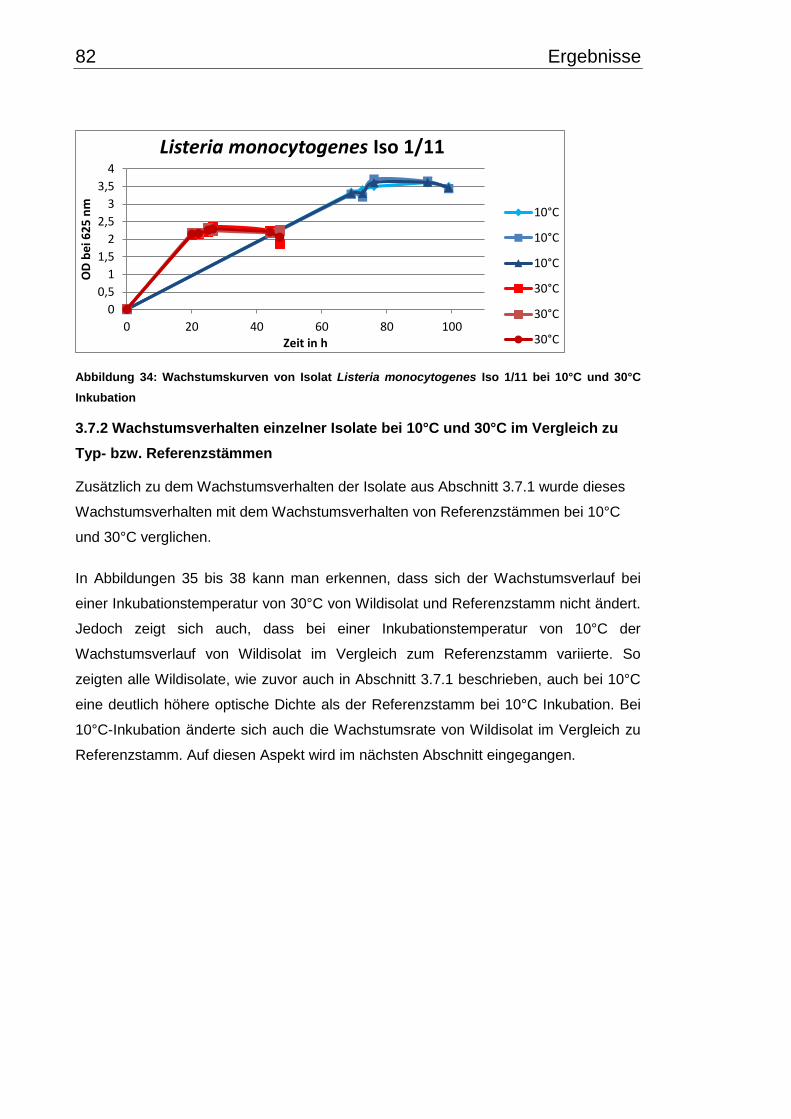
82 Ergebnisse
Abbildung 34: Wachstumskurven von Isolat Listeria monocytogenes Iso 1/11 bei 10°C und 30°C
Inkubation
3.7.2 Wachstumsverhalten einzelner Isolate bei 10°C und 30°C im Vergleich zu
Typ- bzw. Referenzstämmen
Zusätzlich zu dem Wachstumsverhalten der Isolate aus Abschnitt 3.7.1 wurde dieses
Wachstumsverhalten mit dem Wachstumsverhalten von Referenzstämmen bei 10°C
und 30°C verglichen.
In Abbildungen 35 bis 38 kann man erkennen, dass sich der Wachstumsverlauf bei
einer Inkubationstemperatur von 30°C von Wildisolat und Referenzstamm nicht ändert.
Jedoch zeigt sich auch, dass bei einer Inkubationstemperatur von 10°C der
Wachstumsverlauf von Wildisolat im Vergleich zum Referenzstamm variierte. So
zeigten alle Wildisolate, wie zuvor auch in Abschnitt 3.7.1 beschrieben, auch bei 10°C
eine deutlich höhere optische Dichte als der Referenzstamm bei 10°C Inkubation. Bei
10°C-Inkubation änderte sich auch die Wachstumsrate von Wildisolat im Vergleich zu
Referenzstamm. Auf diesen Aspekt wird im nächsten Abschnitt eingegangen.
0
0,5
1
1,5
2
2,5
3
3,5
4
0 20 40 60 80 100
OD
be
i 62
5 n
m
Zeit in h
Listeria monocytogenes Iso 1/11
10°C
10°C
10°C
30°C
30°C
30°C
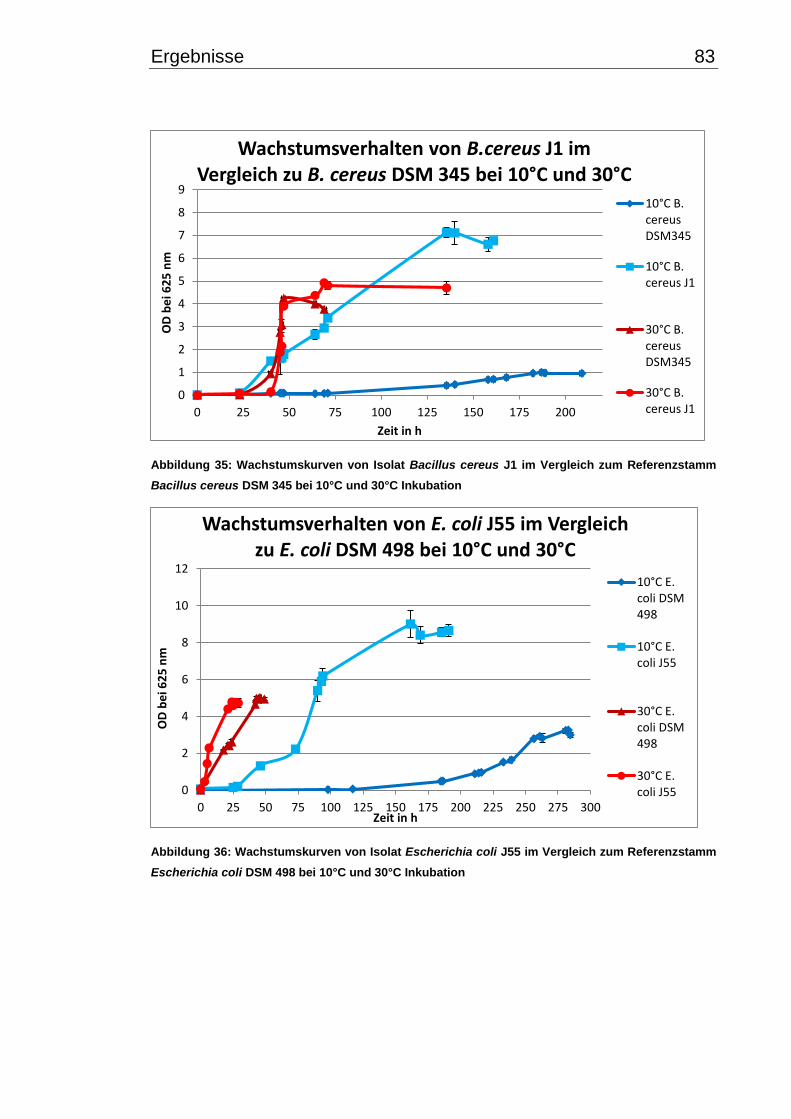
Ergebnisse 83
Abbildung 35: Wachstumskurven von Isolat Bacillus cereus J1 im Vergleich zum Referenzstamm
Bacillus cereus DSM 345 bei 10°C und 30°C Inkubation
Abbildung 36: Wachstumskurven von Isolat Escherichia coli J55 im Vergleich zum Referenzstamm
Escherichia coli DSM 498 bei 10°C und 30°C Inkubation
0
1
2
3
4
5
6
7
8
9
0 25 50 75 100 125 150 175 200
OD
be
i 62
5 n
m
Zeit in h
Wachstumsverhalten von B.cereus J1 im Vergleich zu B. cereus DSM 345 bei 10°C und 30°C
10°C B.cereusDSM345
10°C B.cereus J1
30°C B.cereusDSM345
30°C B.cereus J1
0
2
4
6
8
10
12
0 25 50 75 100 125 150 175 200 225 250 275 300
OD
be
i 62
5 n
m
Zeit in h
Wachstumsverhalten von E. coli J55 im Vergleich zu E. coli DSM 498 bei 10°C und 30°C
10°C E.coli DSM498
10°C E.coli J55
30°C E.coli DSM498
30°C E.coli J55
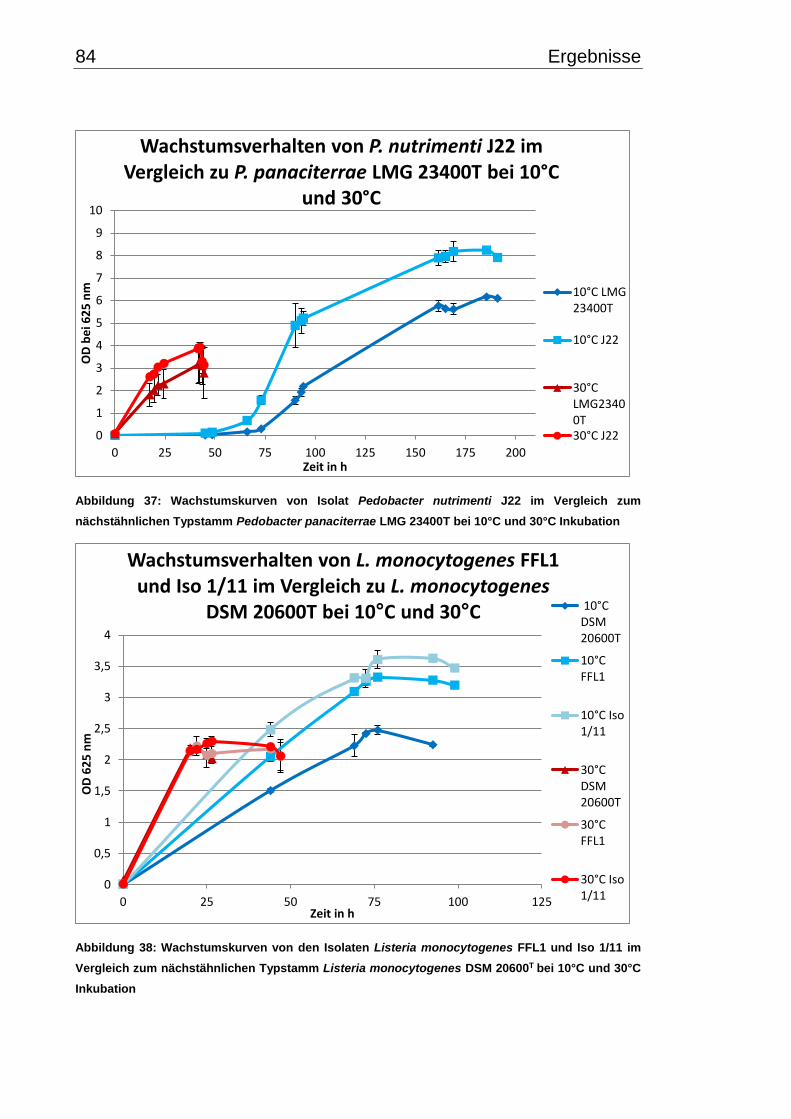
84 Ergebnisse
Abbildung 37: Wachstumskurven von Isolat Pedobacter nutrimenti J22 im Vergleich zum
nächstähnlichen Typstamm Pedobacter panaciterrae LMG 23400T bei 10°C und 30°C Inkubation
Abbildung 38: Wachstumskurven von den Isolaten Listeria monocytogenes FFL1 und Iso 1/11 im
Vergleich zum nächstähnlichen Typstamm Listeria monocytogenes DSM 20600T bei 10°C und 30°C
Inkubation
0
1
2
3
4
5
6
7
8
9
10
0 25 50 75 100 125 150 175 200
OD
be
i 62
5 n
m
Zeit in h
Wachstumsverhalten von P. nutrimenti J22 im Vergleich zu P. panaciterrae LMG 23400T bei 10°C
und 30°C
10°C LMG23400T
10°C J22
30°CLMG23400T30°C J22
0
0,5
1
1,5
2
2,5
3
3,5
4
0 25 50 75 100 125
OD
62
5 n
m
Zeit in h
Wachstumsverhalten von L. monocytogenes FFL1 und Iso 1/11 im Vergleich zu L. monocytogenes
DSM 20600T bei 10°C und 30°C 10°CDSM20600T
10°CFFL1
10°C Iso1/11
30°CDSM20600T
30°CFFL1
30°C Iso1/11
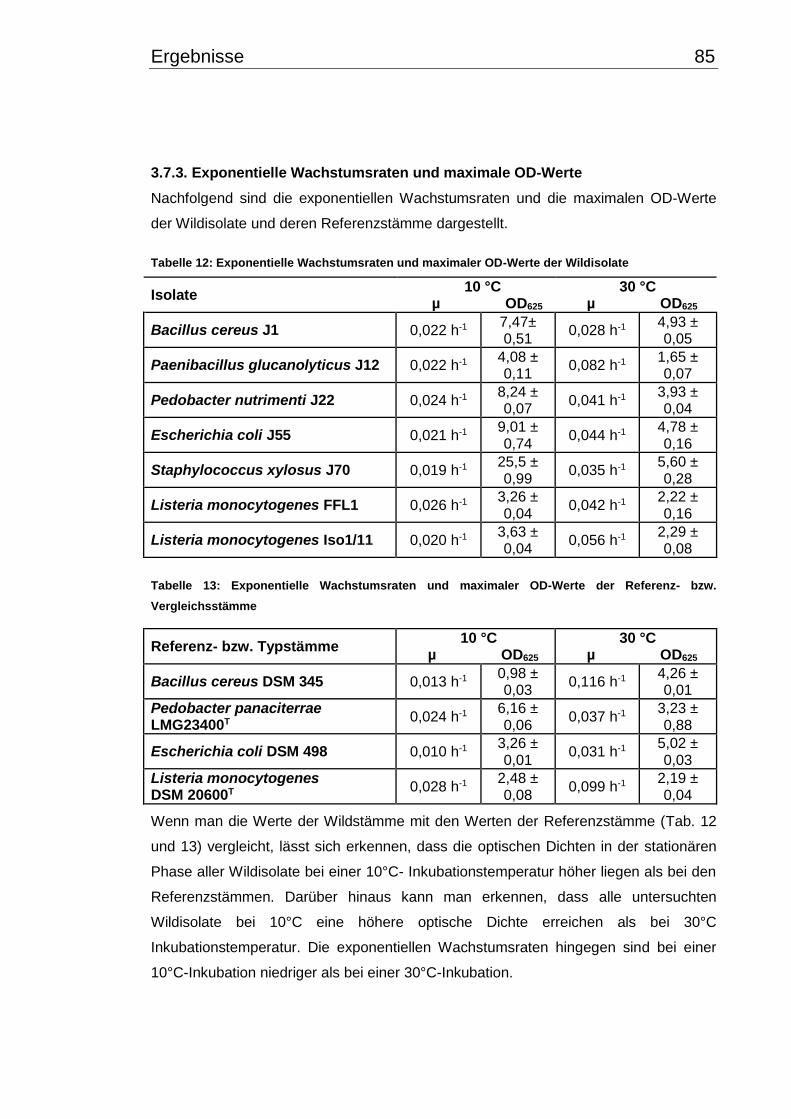
Ergebnisse 85
3.7.3. Exponentielle Wachstumsraten und maximale OD-Werte
Nachfolgend sind die exponentiellen Wachstumsraten und die maximalen OD-Werte
der Wildisolate und deren Referenzstämme dargestellt.
Tabelle 12: Exponentielle Wachstumsraten und maximaler OD-Werte der Wildisolate
Isolate 10 °C µ OD625
30 °C µ OD625
Bacillus cereus J1 0,022 h-1 7,47± 0,51
0,028 h-1 4,93 ± 0,05
Paenibacillus glucanolyticus J12 0,022 h-1 4,08 ± 0,11
0,082 h-1 1,65 ± 0,07
Pedobacter nutrimenti J22 0,024 h-1 8,24 ± 0,07
0,041 h-1 3,93 ± 0,04
Escherichia coli J55 0,021 h-1 9,01 ± 0,74
0,044 h-1 4,78 ± 0,16
Staphylococcus xylosus J70 0,019 h-1 25,5 ± 0,99
0,035 h-1 5,60 ± 0,28
Listeria monocytogenes FFL1 0,026 h-1 3,26 ± 0,04
0,042 h-1 2,22 ± 0,16
Listeria monocytogenes Iso1/11 0,020 h-1 3,63 ± 0,04
0,056 h-1 2,29 ± 0,08
Tabelle 13: Exponentielle Wachstumsraten und maximaler OD-Werte der Referenz- bzw.
Vergleichsstämme
Referenz- bzw. Typstämme 10 °C µ OD625
30 °C µ OD625
Bacillus cereus DSM 345 0,013 h-1 0,98 ± 0,03
0,116 h-1 4,26 ± 0,01
Pedobacter panaciterrae LMG23400T
0,024 h-1 6,16 ± 0,06
0,037 h-1 3,23 ± 0,88
Escherichia coli DSM 498 0,010 h-1 3,26 ± 0,01
0,031 h-1 5,02 ± 0,03
Listeria monocytogenes DSM 20600T
0,028 h-1 2,48 ± 0,08
0,099 h-1 2,19 ± 0,04
Wenn man die Werte der Wildstämme mit den Werten der Referenzstämme (Tab. 12
und 13) vergleicht, lässt sich erkennen, dass die optischen Dichten in der stationären
Phase aller Wildisolate bei einer 10°C- Inkubationstemperatur höher liegen als bei den
Referenzstämmen. Darüber hinaus kann man erkennen, dass alle untersuchten
Wildisolate bei 10°C eine höhere optische Dichte erreichen als bei 30°C
Inkubationstemperatur. Die exponentiellen Wachstumsraten hingegen sind bei einer
10°C-Inkubation niedriger als bei einer 30°C-Inkubation.
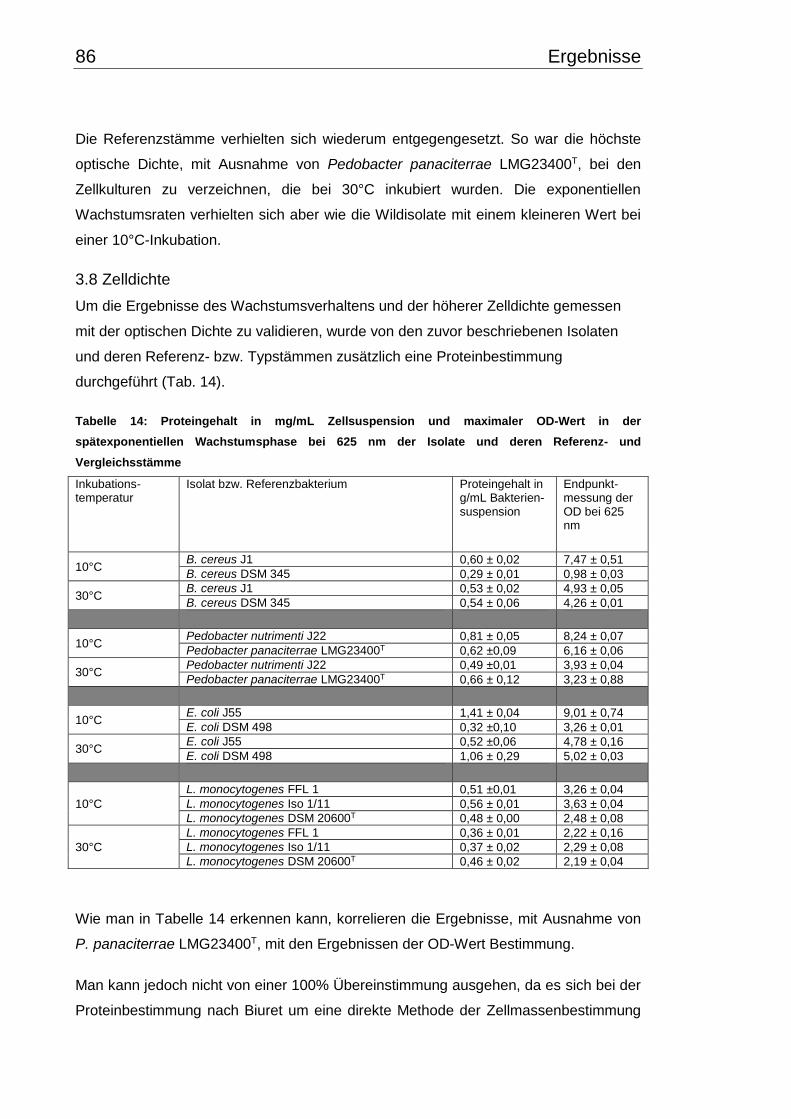
86 Ergebnisse
Die Referenzstämme verhielten sich wiederum entgegengesetzt. So war die höchste
optische Dichte, mit Ausnahme von Pedobacter panaciterrae LMG23400T, bei den
Zellkulturen zu verzeichnen, die bei 30°C inkubiert wurden. Die exponentiellen
Wachstumsraten verhielten sich aber wie die Wildisolate mit einem kleineren Wert bei
einer 10°C-Inkubation.
3.8 Zelldichte
Um die Ergebnisse des Wachstumsverhaltens und der höherer Zelldichte gemessen
mit der optischen Dichte zu validieren, wurde von den zuvor beschriebenen Isolaten
und deren Referenz- bzw. Typstämmen zusätzlich eine Proteinbestimmung
durchgeführt (Tab. 14).
Tabelle 14: Proteingehalt in mg/mL Zellsuspension und maximaler OD-Wert in der
spätexponentiellen Wachstumsphase bei 625 nm der Isolate und deren Referenz- und
Vergleichsstämme
Inkubations- temperatur
Isolat bzw. Referenzbakterium Proteingehalt in g/mL Bakterien-suspension
Endpunkt-messung der OD bei 625 nm
10°C B. cereus J1 0,60 ± 0,02 7,47 ± 0,51
B. cereus DSM 345 0,29 ± 0,01 0,98 ± 0,03
30°C B. cereus J1 0,53 ± 0,02 4,93 ± 0,05
B. cereus DSM 345 0,54 ± 0,06 4,26 ± 0,01
10°C Pedobacter nutrimenti J22 0,81 ± 0,05 8,24 ± 0,07
Pedobacter panaciterrae LMG23400T 0,62 ±0,09 6,16 ± 0,06
30°C Pedobacter nutrimenti J22 0,49 ±0,01 3,93 ± 0,04
Pedobacter panaciterrae LMG23400T 0,66 ± 0,12 3,23 ± 0,88
10°C E. coli J55 1,41 ± 0,04 9,01 ± 0,74
E. coli DSM 498 0,32 ±0,10 3,26 ± 0,01
30°C E. coli J55 0,52 ±0,06 4,78 ± 0,16
E. coli DSM 498 1,06 ± 0,29 5,02 ± 0,03
10°C
L. monocytogenes FFL 1 0,51 ±0,01 3,26 ± 0,04
L. monocytogenes Iso 1/11 0,56 ± 0,01 3,63 ± 0,04
L. monocytogenes DSM 20600T 0,48 ± 0,00 2,48 ± 0,08
30°C
L. monocytogenes FFL 1 0,36 ± 0,01 2,22 ± 0,16
L. monocytogenes Iso 1/11 0,37 ± 0,02 2,29 ± 0,08
L. monocytogenes DSM 20600T 0,46 ± 0,02 2,19 ± 0,04
Wie man in Tabelle 14 erkennen kann, korrelieren die Ergebnisse, mit Ausnahme von
P. panaciterrae LMG23400T, mit den Ergebnissen der OD-Wert Bestimmung.
Man kann jedoch nicht von einer 100% Übereinstimmung ausgehen, da es sich bei der
Proteinbestimmung nach Biuret um eine direkte Methode der Zellmassenbestimmung

Ergebnisse 87
handelt und die Trübungsmessung mittels optischer Dichte eine indirekte
Bestimmungsmethode der Biomasse ist. Chemische Zellbestandteile wie zum Beispiel
Gesamtstickstoff, Protein oder DNA lassen sich zur direkten Bestimmung der
Biomasse heranziehen, da der Bestandteil einen genügend großen Anteil der
Zellmasse ausmacht und zum größten Teil in einem gleichbleibenden Verhältnis zur
Gesamtbiomasse steht. (Bast 1999)
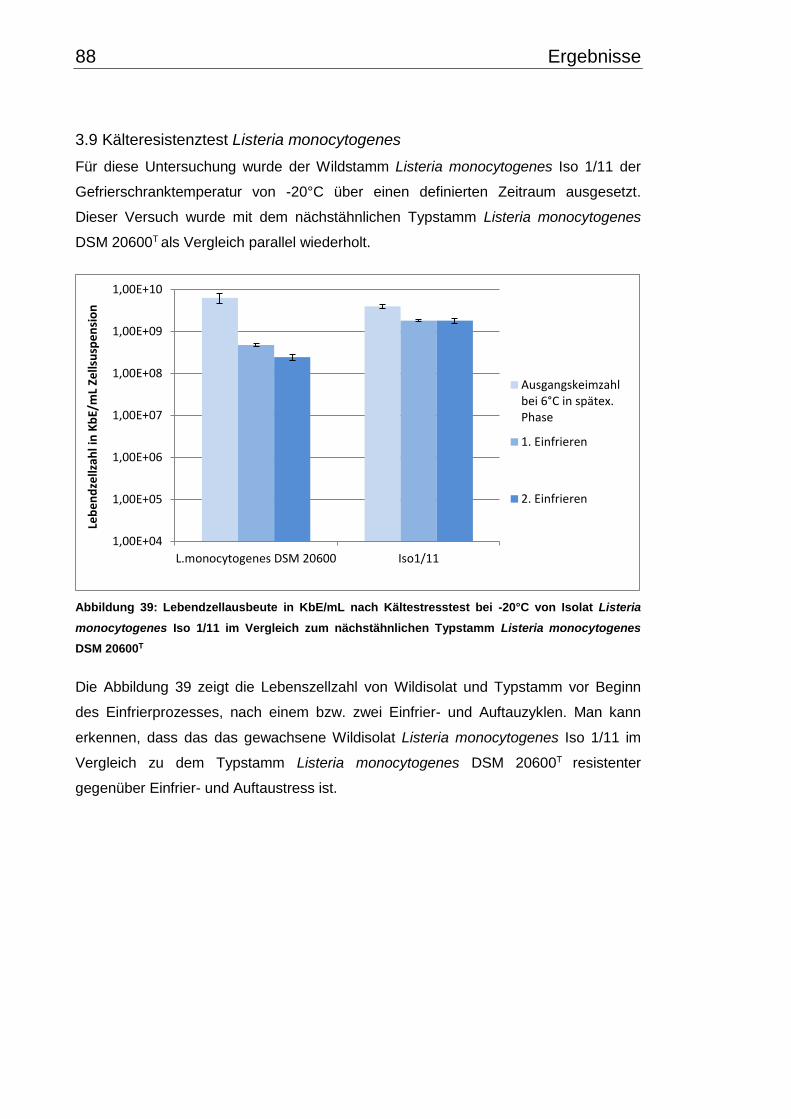
88 Ergebnisse
3.9 Kälteresistenztest Listeria monocytogenes
Für diese Untersuchung wurde der Wildstamm Listeria monocytogenes Iso 1/11 der
Gefrierschranktemperatur von -20°C über einen definierten Zeitraum ausgesetzt.
Dieser Versuch wurde mit dem nächstähnlichen Typstamm Listeria monocytogenes
DSM 20600T als Vergleich parallel wiederholt.
Abbildung 39: Lebendzellausbeute in KbE/mL nach Kältestresstest bei -20°C von Isolat Listeria
monocytogenes Iso 1/11 im Vergleich zum nächstähnlichen Typstamm Listeria monocytogenes
DSM 20600T
Die Abbildung 39 zeigt die Lebenszellzahl von Wildisolat und Typstamm vor Beginn
des Einfrierprozesses, nach einem bzw. zwei Einfrier- und Auftauzyklen. Man kann
erkennen, dass das das gewachsene Wildisolat Listeria monocytogenes Iso 1/11 im
Vergleich zu dem Typstamm Listeria monocytogenes DSM 20600T resistenter
gegenüber Einfrier- und Auftaustress ist.
1,00E+04
1,00E+05
1,00E+06
1,00E+07
1,00E+08
1,00E+09
1,00E+10
L.monocytogenes DSM 20600 Iso1/11
Leb
en
dze
llzah
l in
Kb
E/m
L Ze
llsu
spe
nsi
on
Ausgangskeimzahlbei 6°C in spätex.Phase
1. Einfrieren
2. Einfrieren
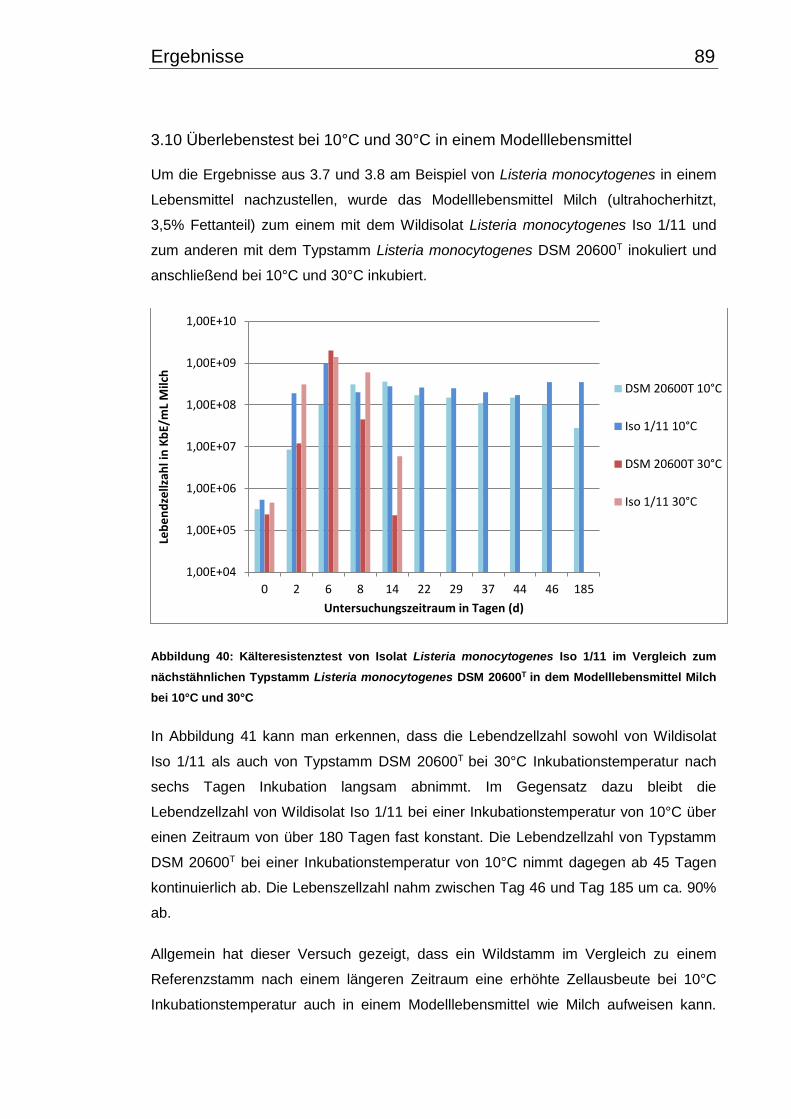
Ergebnisse 89
3.10 Überlebenstest bei 10°C und 30°C in einem Modelllebensmittel
Um die Ergebnisse aus 3.7 und 3.8 am Beispiel von Listeria monocytogenes in einem
Lebensmittel nachzustellen, wurde das Modelllebensmittel Milch (ultrahocherhitzt,
3,5% Fettanteil) zum einem mit dem Wildisolat Listeria monocytogenes Iso 1/11 und
zum anderen mit dem Typstamm Listeria monocytogenes DSM 20600T inokuliert und
anschließend bei 10°C und 30°C inkubiert.
Abbildung 40: Kälteresistenztest von Isolat Listeria monocytogenes Iso 1/11 im Vergleich zum
nächstähnlichen Typstamm Listeria monocytogenes DSM 20600T in dem Modelllebensmittel Milch
bei 10°C und 30°C
In Abbildung 41 kann man erkennen, dass die Lebendzellzahl sowohl von Wildisolat
Iso 1/11 als auch von Typstamm DSM 20600T bei 30°C Inkubationstemperatur nach
sechs Tagen Inkubation langsam abnimmt. Im Gegensatz dazu bleibt die
Lebendzellzahl von Wildisolat Iso 1/11 bei einer Inkubationstemperatur von 10°C über
einen Zeitraum von über 180 Tagen fast konstant. Die Lebendzellzahl von Typstamm
DSM 20600T bei einer Inkubationstemperatur von 10°C nimmt dagegen ab 45 Tagen
kontinuierlich ab. Die Lebenszellzahl nahm zwischen Tag 46 und Tag 185 um ca. 90%
ab.
Allgemein hat dieser Versuch gezeigt, dass ein Wildstamm im Vergleich zu einem
Referenzstamm nach einem längeren Zeitraum eine erhöhte Zellausbeute bei 10°C
Inkubationstemperatur auch in einem Modelllebensmittel wie Milch aufweisen kann.
1,00E+04
1,00E+05
1,00E+06
1,00E+07
1,00E+08
1,00E+09
1,00E+10
0 2 6 8 14 22 29 37 44 46 185
Leb
en
dze
llzah
l in
Kb
E/m
L M
ilch
Untersuchungszeitraum in Tagen (d)
DSM 20600T 10°C
Iso 1/11 10°C
DSM 20600T 30°C
Iso 1/11 30°C

90 Ergebnisse
Außerdem konnte eine unterschiedliche Überlebensdauer und einen unterschiedlichen
Wachstumsverlauf bei niedrigen Temperaturen detektiert werden. kann.

Diskussion 91
4 Diskussion
4.1 Isolierung kälteangepasster Bakterien
Insgesamt wurden 54 Bakterienstämme aus gekühlten Lebensmitteln und
lebensmittelassoziierten Kalthabitaten isoliert. Bei 48 Isolaten wurden das 16S rRNA-
Gen sequenziert, um sie zu identifizieren. Diese Methode hat sich zur Beschreibung
neuer Prokaryoten-Spezies sowie zur Identifikation von Bakterienisolaten bewährt
(Janda und Abbott 2007; Stackebrandt und Goebel 1994).
Die Phyla Proteobacteria, Actinobacteria und Firmicutes waren unter den für die
Identifizierung ausgewählten Isolaten zu etwa gleichen Anteilen vertreten, während
das Phylum Bacteroidetes den geringsten Anteil an identifizierten Isolaten
ausmachte. Dies stellt ein phylogenetisches Muster für kälteliebende Bakterien dar,
wie Simon et al. (2009) bei Proben von Gletschereis feststellten.
4.2 Neubeschreibung Pedobacter nutrimenti J22T
Isolat J22 konnte der durch 16S rRNA-Gensequenz, Fettsäuremuster, Chinonmuster
und polaren Lipidmuster der Gattung Pedobacter zugeordnet werden, die zuerst von
Steyn et al. (1998) beschrieben wurde. Die Gattung umfasste zum Zeitpunkt der
Beschreibung Pedobacter heparinus (vorher: Flavobacterium heparinum (Takeuchi und
Yokota 1992); Sphingobacterium heparinum (Payza und Korn 1956) und die zwei
neuen Spezies Pedobacter africanus und Pedobacter saltans. Seitdem wurden
mehrere Pedobacter-Spezies neu beschrieben und zurzeit umfasst die Gattung
Pedobacter 40 Spezies. Hauptsächlich wurde die Gattung aus Bodenproben isoliert
((Yoon et al. 2007), (Zhou et al. 2012), (Luo et al. 2010), (Oh et al. 2013)) aber auch
aus Wasserproben (Baik et al. 2007), Gletscher Kyroconit (Margesin 2003), Kompost
(Lee et al. 2009) oder der Pflanzenrhizosphäre (Kwon et al. 2007).
Obwohl vermehrt die verschiedenen Spezies der Gattung Pedobacter aus Sediment-
und Umweltproben isoliert wurden, gibt es Beschreibungen, in denen vereinzelt
Pedobacter-Stämme aus Lebensmitteln isoliert wurden. So isolierte Herranen et al.
(2010) einen Pedobacter-Stamm aus Cornflakes und Tsukamoto et al. (2002) aus dem
Speisepilz Clitocybe sp. Die 16S rRNA-Gen Analysen in diesen Arbeiten könnten
aufgrund von Sequenzübereinstimmungen unter 98% auf eine neue Spezies
hinweisen, jedoch wurde dieser Verdacht nicht weiter untersucht. Daher ist es sehr
wichtig die potentielle Rolle der Gattung Pedobacter im Lebensmittelbereich weiterhin
zu untersuchen.

92 Diskussion
Die Gattung Pedobacter besitzt eine gewisse Affinität zu kalten Habitaten. Es gibt
mehrere Pedobacter-Spezies, die auch wie Isolat J22 aus kalten Habitaten isoliert
wurden, wie beispielsweise Pedobacter articus aus dem Erdboden der Kältesteppe
(Zhou et al. 2012) und Pedobacter himalayensis aus einem Gletscher des Himalaya-
Gebirges (Shivaji 2005). Dadurch, dass auch Isolat J22 aus einem kalten Habitat
isoliert wurde, spielt dessen Temperaturwachstumsbereich eine wichtige Rolle. Es
wurde ein Temperaturbereich von 4°C bis 37°C ermittelt, mit einem Optimum bei 30°C.
Beim Test der biochemischen Reaktionsprofile mit den Testsystemen API 20E und API
50CH zeigte sich, dass im Vergleich zu den drei Referenzstämmen nur das
Pedobacter-Isolat J22 die Substrate Tween 80, Itaconsäure, α-Keto-Butylsäure, D-
Alanin, L-Asparaginsäure, Urocansäure und Thymindin verstoffwechselte. Damit
konnte dieses Isolat von den anderen nächstähnlichen Typstämmen abgegrenzt
werden.
Bei der phylogenetischen Analyse mit den drei verschiedenen Algorithmen Neighbour-
Joining, Maximum-Likelihood und Maximum-Parsimony zeigte sich, dass Isolat J22 in
die Gattung Pedobacter eingeordnet werden konnte. Bei der DNA-DNA Hybridisierung
wurde ein Wert von 35,2% (reziprok 7,4%) zwischen dem Isolat J22 und dem
nächstähnlichen Typstamm Pedobacter panaciterrae LMG23400T gezeigt. Wayne et al.
(1987) definierten den Wert von weniger als 70% als Kriterium zur Abgrenzung und
Beschreibung einer neuen Spezies in der bakteriellen Taxonomie.
Die Fettsäureanalyse ermittelte die Hauptfettsäuren C14:0, C15:0 iso, C16:1 cis 9,
C16:0 und C17:0 iso 3-OH, welche im Vergleich zu anderen Pedobacter-Spezies
typisch für diese Gattung sind. (Kim et al. (2013); Luo et al. (2010); Steyn et al. (1998);
Yoon et al. (2007); Zhou et al. (2012)). Jedoch gab es teilweise Unterschiede zu den
Referenzstämmen. So konnte Isolat J22 durch den höheren Gehalt an der Fettsäure
C16:0 und dem niedrigeren Gehalt an C17:1 iso cis 9 von den anderen drei Referenz-
Pedobacter-Stämmen abgegrenzt werden. Außerdem wurde die für die Gattung
Pedobacter typische Fettsäure C17:1 iso cis 11 nicht detektiert. Bei den polaren
Lipiden ergab sich das typische Pedobacter-Muster aus Phosphatidylethanolamin, zwei
Lipiden, zwei Aminophospholipiden und drei Aminolipide. Dieses Muster ist im
Vergleich zu den drei Referenzstämmen teilweise übereinstimmend. Menachinon MK-7
wurde als einziges Chinon sowohl bei Isolat J22 als auch bei den drei
Referenzstämmen detektiert. Dieses Ergebnis ist aber übereinstimmend mit allen
Vertretern der Familie Sphingobacteriaceae (Margesin und Shivaji 2011).

Diskussion 93
Die oben aufgeführten chemotaxischen Daten und die phylogenetische Inferenz lässt
die Schlussfolgerung zu, dass Isolat J22 zu der Gattung Pedobacter gehört. Es kann
aber durch die unterschiedlich phänotypische Charakteristik und durch die DNA-DNA
Hybridisierung mit dem nächstähnlichen Typstamm Pedobacter panaciterrae
LMG23400T mit einem Wert von weniger als 40% abgegrenzt werden.
Bezogen auf die hier beschriebenen Daten, konnte Isolat J22 als Repräsentant einer
neuen Spezies in der Gattung Pedobacter eingeordnet werden, für welche der Name
Pedobacter nutrimenti sp. nov. vorgeschlagen wurde (Derichs et al. 2014).
4.3 Fettsäurebewertung
4.3.1 Taxonomische Bedeutung
Die Fettsäureanalytik spielt in der Chemotaxonomie und damit auch in der
Neubeschreibung von Bakterienarten eine wichtige Rolle. Viele Bakteriengattungen
weisen ein unterschiedliches Fettsäuremuster auf und können damit von anderen
Bakteriengattungen abgegrenzt werden. Die hier aus kalten Habitaten stammenden
beschriebenen Isolate weisen alle ihr gattungstypisches Fettsäuremuster auf. Die am
häufigsten vertretenden Gattungen sind Bacillus, Micrococcus und Pseudomonas. So
sind die Fettsäuren C13:0 iso, C15:0 iso, C16:0, C17:0 iso, C15:0 anteiso, C16:0 iso
typisch für die Gattung Bacillus (Guinebretiere et al. 2013). Die Gattung Micrococcus
hat mit 15:0 iso und 15:0 anteiso zwei Hauptfettsäuren (Wieser et al. 2002) und die
Gattung Pseudomonas besitzt neben den für gramnegative Bakterien typische
hydroxylierten Fettsäuren noch die Fettsäuren C16:0 und C18:1 cis 11 (Yu et al. 2013).
Durch die Inkubation bei 10°C veränderten sich das Fettsäuremuster und die
Fettsäurezusammensetzung von vielen Isolaten; zum Teil kam es sogar zu starken
Veränderungen. Somit erkennt man, dass die Veränderung im Fettsäuremuster eine
wichtige Rolle in der bakteriellen Kälteanpassung spielt. Dies ist eine bereits bekannte
und in der Literatur oft beschriebene Strategie zur Änderung der Membranfluidität und -
beweglichkeit in der Zellmembran (Russell 1984).

94 Diskussion
4.3.2 Bewertung der Fettsäuremuster in der Kälteanpassung
Die Grundstruktur jeder Zellmembran ist die Lipiddoppelschicht. Diese
Membrandoppelschicht muss die richtige Fluidität aufweisen, um die Permeabilität und
somit die Beweglichkeit der Membran zu gewährleisten. Der funktionelle Zustand ist
immer die flüssig-kristalline Phase, jedoch kann es beim Absenken der Temperatur zu
einem Übergang in die Gelphase kommen, wodurch essentielle
Membraneigenschaften verloren gehen können. Die Temperatur, bei der dieser
Phasenübergang stattfindet, ist abhängig von der Lipidzusammensetzung der
Membran, aber zum größten Teil von den Fettsäureketten, die in der
Lipiddoppelschicht verankert sind. Daher ist eine der wichtigsten Aufgabe von
psychrophilen und psychrotoleranten Bakterien, diese Fettsäurezusammensetzung an
die Umgebungstemperatur anzupassen. (Feller und Gerday 2003)
Anhand der Fettsäureanalyse nach Sasser (1990) ist dieser aus der Literatur bekannte
Kälteanpassungsmechanismus bei verschiedenen Isolaten nachgewiesen geworden.
Zu den häufigsten Modifizierungen im Fettsäuremuster zählen die Erhöhung des
Anteils von ungesättigten Fettsäuren, der Ersatz von langkettigen durch kurzkettige
Fettsäuren, die Substitution von geradkettigen durch methyl-verzweigte Fettsäuren, die
Substitution von iso-verzweigten durch anteiso-verzweigte Fettsäuren und die
Isomerisierung von trans in cis-Doppelbindungen (Scherer und Neuhaus 2006). Alle
diese Modifizierungen bzw. Veränderungen, mit der Ausnahme von der Isomerisierung
von trans in cis-Doppelbindungen und die Substitution von geradkettigen durch methyl-
verzweigte Fettsäuren, wurden in dieser Arbeit nachgewiesen.
Viele Isolate erhöhten bei Kälteeinfluss ihren Anteil an ungesättigten Fettsäuren.
Dieser Effekt wurde vermehrt von gramnegativen Bakterienisolaten angewendet, wie
Isolat J2, J11, J21, J22, J23, J52, J55, J57, J61, J63, J67, J84, J88, J86, J90 und J91.
Diese Isolate zählten alle zu der Familie der Pseudomonadaceae, Moraxellaceae oder
Enterobacteriaceae. Dies kann man jedoch auch darauf zurückzuführen, dass
ungesättigte Fettsäuren die Hauptfettsäuren dieser Bakterienfamilien darstellen (Zelles
1999).
Die Substitution von iso-verzweigten Fettsäuren durch anteiso-verzweigte Fettsäuren
wurde vermehrt durch Bakterienisolate der Familien Bacillaceae, Listeriaceae und
Staphylococcaceae aus der Ordnung Bacillales gezeigt. Eine Erhöhung des Gehaltes
an der verzweigten, kurzkettigen Fettsäuren C13:0 iso wurde bei Isolaten der Spezies
Bacillus cereus nachgewiesen. Der Anteil an langkettigeren Fettsäuren nahm dabei ab.

Diskussion 95
Die Veränderungen im Fettsäuremuster führen zu einer Herabsetzung des
Schmelzpunktes (Tab.1), wodurch die Membranfluidität auch bei niedrigen
Temperaturen aufrechterhalten werden kann (Russell 1984). Laut Tabelle 1 müsste die
Membran jedoch durch den Ersatz mit ungesättigten Fettsäuren wie beispielsweise
C16:1 cis 9 oder C18:1 cis 9 stärker fluid gehalten werden als durch den Ersatz von
anteiso-verzweigten Fettsäuren. Damit kann man auch erklären werden, dass das
Ausmaß der Veränderung je nachdem welche Fettsäure eingebaut wird, stärker oder
schwächer ausfällt.
Auch die Übersicht aus Tabelle 9 verdeutlicht, dass die Erhöhung der ungesättigten
Fettsäuren zu Lasten der gesättigten Fettsäuren eine bevorzugte Strategie zur
Kälteadaptation darstellt. Somit ist ableitbar, dass dieser Mechanismus gegenüber
anderen eine effektivere Methode darstellen könnte, um sich an kältere
Umgebungstemperaturen anzupassen. Die Einführung von Doppelbindungen wird
daher bevorzugt angewendet.
Doppelbindungen können mit zwei verschiedenen Mechanismen in die Fettsäuren
eingebaut werden (Russell 1984). Der Einbau und die dadurch entstehende
Veränderungen im Fettsäuremuster werden gattungsspezifisch entweder durch die
Sauerstoff-unabhängige Synthese ungesättigter Fettsäuren erreicht oder durch die
sauerstoffabhängige Desaturierung von gesättigten Fettsäuren. Der
sauerstoffunabhängige Mechanismus wurde nicht nur in anaeroben Bakterien
gefunden (Clostridium, Lactobacillus), sondern auch im aeroben Bakterien, wie
Escherichia oder Pseudomonas. Der sauerstoffabhängige Mechanismus wurde bei
Spezies von Bacillus, Mycobacterium und Micrococcus entdeckt. (Erwin und Bloch
1964; Härtig et al. 1999; Magnuson et al. 1993)
Der Einbau über den anaeroben Weg geschieht während des
Kettenverlängerungsprozess während der de-novo Fettsäurebiosynthese. Hierbei wird
die Doppelbindung in der Regel bei Position 7 eingeführt (Magnuson et al.1993). Der
Einbau der Doppelbindung über den sogenannten aeroben Weg der
Fettsäurebiosynthese kann mittels Desaturase eingefügt werden. Dieser Mechanismus
ist dadurch charakterisiert, dass die Doppelbindung bei Position C9 der
entsprechenden Fettsäure spezifisch eingeführt wird (Fulco 1974). Da C18:1 cis 9 die
Hauptfettsäure bei Acinetobacter darstellt, ist dies beispielsweise ein Indikator für die
Gegenwart dieses Desaturase-Systems und daher auch für den aeroben
Stoffwechselweg der Fettsäurebiosynthese in der Gattung Acinetobacter. (Härtig et al.

96 Diskussion
1999) Bei der Gattung Pseudomonas (Wada et al. 1989) und Vibrio (Morita et al. 1993)
scheinen beide Biosynthesewege zu existieren.
Allgemein lässt sich aus den Ergebnissen erkennen, dass drei auf Fettsäureebene
bekannte Adaptationsvorgänge an niedrige Umgebungstemperaturen vorliegen. Es gibt
jedoch auch Isolate, die keinen eindeutigen Unterschied bezüglich dieser
Anpassungsstrategien zeigen. Dies gibt Hinweise darauf, dass diese kälteangepassten
Stämme andere Anpassungsmechanismen besitzen müssen. Bei diesen Stämmen ist
es sehr interessant, die zuvor aufgestellten Hypothesen der fettsäureunabhängigen
Kälteanpassung auf andere lipophile Komponenten der Zellmembran zu untersuchen.
Somit könnten andere und unbekannte Strategien aufgeklärt werden. Diese
Hypothesen werden in den folgenden Abschnitten untersucht und diskutiert.
4.4 Chinone
4.4.1 Taxonomische Bedeutung der Chinone
Die chemisch-analytische Erfassung der Membranzusammensetzung
(Fettsäuremuster, polare Lipide, Mycolsäuren, Chinone) dient unter anderem der
Gattungs- und Speziesbestimmung von Bakterien (Kämpfer 1996). Chinone sind in
allen Arten von aeroben und photosynthetischen Mikroorganismen vorhanden und
zeigen deutliche Strukturvariationen in Abhängigkeit von der mikrobiellen Taxonomie.
Daher werden Chinone nicht nur als Marker in der chemotaxonomischen mikrobiellen
Systematik, sondern auch als Maßstab von mikrobiellen Populationen in Bezug auf
Quantität, Qualität und Aktivität verwendet. Insbesondere die Analyse von
Chinonprofilen ist nützlich für die Überwachung von mikrobiellen
Populationsverschiebungen z.B. in einem Ökosystem, soweit die Verschiebungen nicht
durch konventionelle Kulturverfahren und molekulare Techniken detektiert werden
können. (Hiraishi 1999)
Chinone stellen Coenzyme dar und werden für den Elektronentransfer in der
mikrobiellen Zelle verwendet. Ein bakterielles Phylum hat in der Regel eine dominante
Art von respiratorischen Chinonen. So stellen Ubichinone gute Marker für Alpha-, Beta-
und Gammaproteobacteria dar. (Kunihiro et al. 2014)
Das fakultativ anaerobe, gram-positive Isolat Bacillus cereus J1 bildete die beiden
Menachinone MK-6 und MK-7, wobei MK-7 den Hauptanteil der Chinonkonzentration in
den Zellen ausmachte. Dies entspricht dem Chinonmuster des nächstähnlichen
Typstamms IAM 1074T (Watanuki und Aida 1972).

Diskussion 97
Die Isolate Acinetobacter lowffii J2 und J84 bildeten als Hauptubichinon Q-9 und zu
geringen Anteilen Q-8 und Q-10. Laut Malhotra et al. (2012) bildet die Gattung
Acinetobacter das Hauptubichinon Q-9.
Bei Isolat Solibacillus silvestris J7 wurden die Menachinone MK-6 und MK-7
nachgewiesen. In der Umbenennung von Krishnamurthi et al. (2009) des Bakteriums
Bacillus silvestris (Rheims et al. 1999) in Solibacillus silvestris wurden ebenfalls MK-7
als Hauptchinon nachgewiesen.
In der Zellmembran von Isolat Paenibacillus glucanolyticus J12 wurde als einziges
Menachinon MK-7 detektiert. Dieses Ergebnis deckt sich mit der Erstbeschreibung von
Shida et al. (1997).
Mehrere Menachinonarten wurden bei Isolat Micrococcus luteus J16 detektiert. Als
Hauptmenachinon wurde bei 30°C das hydrierte Menachinon MK-8 H2 analysiert.
Zusätzlich wurden auch die Menachinone MK-7, MK-8 und die hydrierten Isoprene MK-
7 H2 und MK-9 H2 nachgewiesen. Laut Wieser et al. (2002) werden vorwiegend die
Menachinone MK-8 H2 und MK-8 von der Spezies Micrococcus luteus gebildet. Die
phylogenetisch-verwandte Spezies Micrococcus yunnanensis und Micrococcus flavus
bilden MK-7 H2 und MK-8 H2 (Rieser et al. 2013). Zusätzlich wurden aber auch noch
andere hydrierte Menachinone in der Zellmembran detektiert. Auf diesen Aspekt wird
jedoch in der angegebenen Literatur nicht eingegangen. .
Bei Kocuria palustris J17 wurden wie bei Isolat Micrococcus luteus J16 wieder mehrere
Menachinone, teils auch hydrierte Menachinone analysiert. Dieses Ergebnis stimmt mit
dem Ergebnis von Kovacs et al. (1999) überein.
Im Isolat Rhodococcus qingshengii J20 wurde genauso wie bei der Erstbeschreibung
von Xu et al. (2007) als dominierendes Chinon das hydrierte Menachinon MK-8 H2
nachgewiesen.
Die Menachinone MK-8 und MK-9 wurden im aerotoleranten Isolat Lactococcus lactis
J34 analysiert. Andere Isolate dieser Spezies wiesen zusätzlich zu kleinen Anteilen die
Chinone MK-7 oder MK-10 auf (Morishita et al. 1999). Auch die taxonomisch
benachbarte Gattung Streptococcus weist als Hauptchinon MK-9, gefolgt von MK-8
und zu kleinen Anteilen MK-7 und MK 10 auf. (Collins et al. 1983)
Wie auch in der Literatur für die Gattung Stenotrophomonas beschrieben, bildet Isolat
Stenotrophomonas maltophilia J21, als aerobes und gramnegatives Bakterium als
Hauptchinon das Ubichinon Q-8 (Palleroni und Bradbury 1993).

98 Diskussion
Bei Isolat Pedobacter nutrimenti J22 wird als einziges Menachinon MK-7 bestimmt.
Diese Aussage deckt sich mit der Speziesbeschreibung des nächstähnlichen
Typstammes Pedobacter panaciterrae (Yoon et al. 2007) und kann auf die komplette
Gattung Pedobacter bezogen werden (Steyn et al. 1998).
In der Zellmembran von den Isolaten Microbacterium oxydans J75 und J78 wurden
neben zwei unbekannten Chinonen vorwiegend die Menachinone MK-11 und MK-12
identifiziert. Die Bakteriengattung Microbacterium bildet als Hauptchinone MK-11 und
MK-12. (Takeuchi und Hatano 1998). Auch Yu et al. (2013) detektierte unbekannte
Chinone zu geringen Anteilen. Auf die Identifikation wurde jedoch in dieser Arbeit nicht
eingegangen. Interessant ist jedoch, dass es sich bei dem von Yu et al (2013)
untersuchten Bakterium um ein psychrotolerantes Microbacterium handelte.
Wie bei den meisten hier beschriebenen gram-negativen und aeroben Bakterien bilden
Achromobacter marplatensis J23 (Yabuuchi und Yano 1981) und Pantoea
agglomerans J89 (Kageyama et al. 1992) als Hauptchinon Ubichinon Q-8.
In Isolat Brochothrix thermosphacta J86 wird das Menachinon MK-7 als
Hauptmenachinon detektiert (Collins et al. 1979).
Das für die Gattung Pseudomonas typische Ubichinon Q-9 (Lin et al. 2013) wurde auch
als Hauptchinon in Isolat Pseudomonas baetica J91 nachgewiesen. Zusätzlich wurden
zu kleinen Anteilen Q-8 und Q-10 detektiert. Das Ubichinon Q-8 wurde auch zu 4% von
(Ramos et al. 2013) in der Neuschreibung von der phylogenetisch-benachbarten
Spezies Pseudomonas punonensis detektiert.

Diskussion 99
4.4.2 Chinone in der Kälteanpassung
Arbeiten über das Thema Temperaturabhänigkeit bei Chinonprofilen und
Chinongehalten in Bakterien sind nicht vorhanden. 1975 wurde von Ashcroft und
Haddock jedoch eine veränderte Chinonsynthese in Abhängigkeit von der
Sauerstoffverfügbarkeit ermittelt. So wurde bei der Spezies Escherichia coli mehr
Ubichinon Q-8 unter aeroben Wachstum und mehr Menachinon MK-8 unter
anaerobem Wachstum detektiert. Dieser Effekt kann jedoch auf die unterschiedlichen
Redoxpotentiale zurückgeführt werden.
Durch Absenken der Umgebungstemperatur wurden in der vorliegenden Arbeit bei
bestimmten Bakterienisolaten Veränderungen im Chinongehalt und Chinonmuster der
Bakterienzellen festgestellt. Bei einigen Isolaten stieg die Chinonkonzentration an oder
es wurden zusätzliche Chinone gebildet. Dadurch wird die Hypothese bestärkt, dass
Chinone eine Rolle bei der Kälteanpassung spielen können.
Auch Hiraishi et al. (1999) detektierten bei einer Temperaturänderung eine
Veränderung im Chinongehalt von Bakterien. Jedoch bezog sich dieses Ergebnis auf
Bakterien, die aus heißen Quellen isoliert wurden und daher starker Hitze ausgesetzt
waren. Die interessante Beobachtung war, dass die Chinonprofile der untersuchten
Bakteriengemeinschaften unterschiedlich in Abhängigkeit von der Temperatur waren
und dass diese auftretenden Unterschiede unabhängig von der untersuchten
geographischen Lage waren. Es wurden vorwiegend folgende Strukturtypen von
Chinonen in Abhängigkeit der jeweiligen Umgebungstemperatur erkannt. Bei 68°C war
das Hauptchinon Methionachinon MTK-7 vertreten, bei 61 bis 65°C wurde
hauptsächlich Menachinon MK-10 detektiert, bei 45 bis 56°C Quellentemperatur das
Plastochinon PQ-9 und das Vitamin K1 und bei unter 40°C Umgebungstemperatur das
Ubichinon Q-8.
Bei vielen hier untersuchten Bakterienisolaten führte das Wachstum in kalter
Umgebung zu einem reduzierten Gehalt an bakterieneigenen Chinonen. Zu erklären ist
dies mit einer geringeren Biosyntheserate bei niedrigen Temperaturen. In Bakterien
sind an der Biosynthese von Ubi- und Menachinonen verschiedene Enzyme beteiligt
(Bentley und Meganathan 1982; Meganathan 2001). Da normalerweise die Aktivität
von Enzymen temperaturabhängig ist und innerhalb bestimmter Grenzen mit der
Temperatur ansteigt (Feller et al. 1994), liegt bei tiefen Temperaturen folglich eine
niedrigere Enzymaktivität vor. Der verringerte Chinongehalt kann daher auf einen
Verminderung der Biosyntheserate zurückgeführt werden. Zusätzlich werden dadurch

100 Diskussion
vermutlich auch weniger Chinone in der Atmungskette benötigt, da bei weniger
Substratumsatz auch weniger Elektronen eingespeist werden müssen.
Im Gegensatz zu dieser allgemeinen Aussage wurde in Bezug auf den
Gesamtchinongehalt im Zellmaterial der Isolate Paenibacillus glucanolyticus J12,
Pedobacter nutrimenti J22 und Pantoea agglomerans J89 bei 10°C ein erhöhter
Chinongehalt gemessen. Zusätzlich dazu wurde in den Isolaten Kocuria palustris J40,
Microbacterium oxydans J75 und J78 ein neues Chinon unter Kälteeinfluss gebildet. Es
kam demzufolge zu einer Veränderung im Chinonprofil und Chinongehalt. Dadurch
kann die aufgestellte Hypothese unterstützt werden, dass diese Bakterienisolate u.a.
durch diese erhöhte bzw. geänderte Chinonkonzentrationen in der Lage waren, in
dieser kalten Umgebung sich zu vermehren. Diese Hypothese wird durch die Arbeit
von Hiraishi et al. (1999) unterstützt, bei der es auch zu einer Änderung im Chinonprofil
bei einem Temperaturunterschied kommt.
Da die Sauerstofflöslichkeit bei niedrigen Temperaturen besser ist, könnten aerobe
Bakterien aber auch eine verlängerte bzw. verstärkt ablaufende Atmungskette und
anschließende oxidative Phosphorylierung betreiben. Durch einen größeren
Chinonvorrat in der Zellmembran wären die Bakterien in der Lage, schneller Energie in
Form von ATP zu synthetisieren und für biosynthetische Reaktionen zu nutzen
(Madigan et al. 2013).
In der Einleitung wurden die grundsätzlichen Mechanismen der Erhaltung der
mikrobiellen Membranfluidität, z.B. durch Modifikation des Fettsäuremusters bzw.
Erhöhung des Cholesteringehaltes bei Eukaryonten als Reaktion auf niedrige
Umgebungstemperaturen geschildert. Da auch Chinone in das Membransystem
integriert sind, wird die Vermutung aufgestellt, dass für Chinone ähnliche Annahmen
bezüglich der Anpassung an Kälte gelten und somit die Unordnung der Membran
beibehalten wird. Damit könnte die Aufrechterhaltung der Membranfluidität durch
vermehrten Einbau von Chinonen in die Zellmembran erklärt werden. Die Ergebnisse
von Asai und Watanabe (1999), in denen ein Anstieg von Chinonen eine fluidere
Modellmembran ergibt, würden diese Theorie bekräftigen. Dabei wurden Chinone in
eine Modellmembran eingebaut, dadurch die Temperatur der Phasenumwandlung
gesenkt und folglich die Membranfluidität erhöht. Damit konnte ein fluidisierender Effekt
von Chinonen an der Modellmembran beobachtet werden.
Um diese Hypothese zusätzlich zu überprüfen, wurde in der Abschlussarbeit von
Fritsch (2012) Bakterienzellen Vitamin K1 beim Wachstum zugesetzt, diese dann
detektiert und in die Bakterienzelle aufgenommene Vitamin K1-Menge berechnet. Die

Diskussion 101
Hypothese dieser Arbeit lag in der Annahme, dass Chinone als eine Art Schutzstoff in
die Membran aufgenommen werden und somit die Stresstoleranz von Bakterien bei
niedrigen Temperaturen erhöhen. Ein Zusammenhang zwischen der Synthese bzw.
Aufnahme von kompatiblen Soluten und der Kälteadaption konnte in verschiedenen
Studien bereits nachgewiesen werden. In Untersuchungen von Ko et al. (1994) mit
Listeria monocytogenes wurde beispielsweise ein erhöhter Gehalt an Glycinbetain bei
Wachstum in kalter Umgebung (4°C) entdeckt. Anhand weiterer Versuche konnte
bestätigt werden, dass die Anreicherung des osmotisch aktiven Soluts dem Bakterium
auch die Kältetoleranz verleiht. Ebenfalls wurde durch kompatible Solute eine
induzierte Kältetoleranz in überwinternden Insekten bewiesen (Storey und Storey
1986).
Der genaue Mechanismus, wie Glycinbetain als Gefrierschutzmittel wirkt, ist
unbekannt. Allerdings ist zu berücksichtigen, dass Betain ein Stabilisierungsmittel
darstellt und daher eine nicht gewünschte Ordnung der Lipiddoppelschicht verhindern
kann. Außerdem kann Betain die Löslichkeit von Zellproteinen oder die physikalischen
Eigenschaften der Zellmembran positiv verändern (Rudolph et al. 1986). Osmotischer
Stress scheint das Phasenverhalten der Zellmembran zu verändern (Sanderson et al.
1991) und Glycinbetain kann sowohl osmotischen Stress als auch Kältestress durch
einen ähnlichen Mechanismus entgegenwirken.
Fast alle untersuchten Bakterienisolate nahmen Vitamin K1 aus einer mit Vitamin K1
versetzten Flüssigbouillon während des Wachstums auf und bauten es zu
unterschiedlichen Anteilen vermutlich in ihre Plasmamembran ein, da es im
analysierten Zellpellet nachgewiesen wurde. Dabei wurden jedoch größere Mengen in
nmol/g Biomasse detektiert, wenn die Bakterienisolate bei 30°C inkubiert wurden.
Allerdings zeigte sich eine Ausnahme: Bei Isolat Lactococcus lactis J34 war der
Vitamin K1-Gehalt in den Zellen nach 10°C-Anzucht um etwa 60% höher als nach
Anzucht bei 30°C. Die Ergebnisse von Fritsch (2012) deuten auf eine erhöhte
Chinonproduktion und eine erhöhte Chinonaufnahme bei steigender Temperatur hin,
obwohl ein untersuchtes Isolat ein umgekehrtes Verhalten zeigt.
Dieses Isolat J34 aus der Masterarbeit von Fritsch (2012) und die untersuchten Isolate
J12, J22, J75 und J78 aus dieser Arbeit, die eine erhöhte Chinonkonzentration bzw.
eine Veränderung im Chinonmuster bei Kälteeinfluss zeigen, geben Anlass dazu, einen
weiteren Kälteanpassungsmechanismus mit lipophilen Substanzen der Zellmembran
zu untersuchen. Wie es scheint, können Chinone bei Kälteeinfluss zwar nicht allgemein
aber teilweise eine Rolle spielen. Um dieses Ergebnis abzusichern, wäre es sinnvoll,

102 Diskussion
die temperaturabhängigen Chinonversuche auf eine komplette Gattung zu beziehen
und mit Referenzstämmen zu vergleichen.
4.5 Carotinoide
4.5.1 Carotinoide in Bakterien
22 der in Tabelle 4 aufgelisteten Isolate zeigten eine tyisch durch Carotinoide
hervorgerufene Pigmentierung der Kolonie.
Carotinoide spielen innerhalb der Bakterienzellmembran eine wichtige Rolle. Sie
dienen zum Beispiel als Schutz gegen durch Singulett-Sauerstoff verursachte
Schäden. Bakterien und Pilze bedienen sich des Schutzes von Carotinoiden, wenn sie
unter Licht und Sauerstoffanwesenheit wachsen. So können Bakterien diese
membrangebundenen Carotinoide als Antioxidans gegen oxidativen Stress nutzen.
Schon 1957 konnte Swart-Füchtbauer beobachten, dass der Prozentsatz der
pigmentierten Bakterien in der Luft etwa dreimal so hoch war wie in der Erde. Daraus
wurde geschlossen, dass pigmentierte Bakterien gegenüber Sonnenstrahlen
widerstandsfähiger sind als farblose Bakterien. Carotinoide sind außerdem beteiligt am
Lichtsammelkomplex, eine Ansammlung von Membranproteinen in den
photosynthetischen Membranen von Mikroorganismen, die Photosynthese betreiben,
wie beispielsweise Purpurbakterien (Guedes et al. 2011). Damit besitzen mikrobielle
Carotinoide biochemisch wie auch pflanzliche Carotinoide als sekundärer Pflanzenstoff
zwei charakteristische Eigenschaften: 1. Radikalfänger (Antioxidans) und 2. bei der
UV-VIS-Absorption (Lichtsammelpigmente).
Eine vergleichsweise wenig geprüfte Hypothese zur Funktionalität bakterieller
Carotinoide ist die Funktion eines Membranstabilisierers, welche in Eukaryoten von
Cholesterin übernommen wird (Mitchell et al. 1986; Ourisson und Nakatani 1994).
Durch ihre Lipophilie und Molekülstruktur können sich Carotinoide in biologische
Membranen einlagern und in der Folge die Fluidität verändern (Pintea et al. 2005). Bei
thermophilen Bakterien wird beispielsweise eine funktionelle Membranverstärkung
durch Carotinoide als Überlebensstrategie bei extrem hohen Temperaturen postuliert
(Havaux 1998; Subczynski et al. 1992).
Diese Aussage der Hitzeresistenz könnte jedoch auch auf andere Umwelteinflüsse wie
osmotischer Stress oder Kälteeinfluss übertragen werden. So bildet beispielsweise das
pathogene Bakterium Staphylococcus aureus ein goldschimmerndes Pigment aus,
welches laut Liu (2005) dazu benutzt wird, sich gegen äußere Umwelteinflüsse zu

Diskussion 103
schützen. Bei dem Pigment könnte es sich das Staphylokokken-typische Carotinoid
Staphyloxanthin (Marshall und Wilmoth 1981) handeln. Auf diesen Aspekt wird jedoch
bei Liu (2005) nicht eingegangen.
4.5.2 Carotinoide in der Kälteanpassung
Jacobsen (2013) zeigte bei einer 10°C-Inkubation vier- bis sechsfache Zunahmen der
Carotinoidkonzentration bei den Isolaten Kocuria rhizophila J48, Micrococcus luteus
J50 und Staphylococcus xylosus J70. Die stärkste Zunahme wurde bei S. xylosus J70
ermittelt, die Carotinoidkonzentration stieg von 40 µg/g bei 30°C auf 255 µg/g in der
kälteren Umgebung an. Die ermittelten Carotinoidmengen bezogen sich auf das
Lyophilisat und können nicht mit Literaturangaben verglichen werden, da
entsprechende Daten für die analysierten Bakterienstämme fehlen. Jagannadham et
al. (2000) und Chattopadhyay et al. (1997) bezogen sich lediglich auf die prozentualen
Carotinoidanteile in der Biomasse in Abhängigkeit von der Wachstumstemperatur und
nicht auf absolute Carotinoidgehalte. So stieg beispielsweise der Gehalt in der
Biomasse von Zeaxanthin bei Sphingobacterium antarcticus von ca. 50 % bei 25°C auf
ca. 80 % bei 5°C an.
Bei dem psychrotrophen Bakterium Arthrobacter agilis konnten
Carotinoidkonzentrationen zwischen 250 µg/g bei 30°C Inkubation bis zu über
1000 µg/g bei 5°C Inkubation bezogen auf das Trockengewicht detektiert werden
(Fong et al. 2001). Die Mikroalge Chlorella protothecoides zeigte unter
Stressbedingungen, wie hoher Belichtung, Salzstress oder Nährstoffmangel höhere
Carotinoidgehalte pro Gramm Lyophilisat (Campenni’ et al. 2013). Ebenfalls zeigte der
Schimmelpilz Mucor strictus bei einer Inkubationstemperatur von 0°C eine erhöhte ß-
Carotin Produktion im seiner Biomasse im Gegensatz zu 25°C Inkubationstemperatur
(Dexter und Cooke 1984).
Die temperaturabhängigen Veränderungen der Carotinoidmengen deuten auf einen
Zusammenhang zwischen der Carotinoidsynthese und der Kälteanpassung hin. Die
Erhöhung der Carotinoidkonzentration unter kälteren Wachstumsbedingungen bei den
Isolaten K. rhizophila J48, M. luteus J50 und S. xylosus J70 werden durch die
Ergebnisse von Chattopadhyay et al. (1997) bei Micrococcus roseus und
Jagannadham et al. (2000) bei Sphingobacterium anarticus bestätigt.
Die mangelhafte Auftrennung der Peaks über die HPLC-Messung nach der Methode
von Rählert (2007) und die fehlende Bestimmung der Absorptionsmaxima verhinderte

104 Diskussion
leider die zusätzliche Ermittlung der prozentualen Carotinoidprofile. Die Angabe von
Chattopadhyay et al. (1997) und Jagannadham et al. (2000), dass bei niedrigeren
Wachstumstemperaturen ein höherer Anteil an polaren Carotinoiden gebildet wird,
kann ohne die prozentualen und die Identifizierung der Carotinoide nicht bestätigt
werden. Bei der chromatographischen Analyse des Isolates M. maritypicum J74
wurden die isolierten Carotinoide nach 10°C-Inkubation bei kürzeren Retentionszeiten
detektiert als nach 30°C-Inkubation. Da für die Auftrennung eine C18-Säule und ein
polares Lösungsmittel verwendet wurden, kann dies ein Indiz auf eine vermehrte
Synthese von polaren Carotinoiden sein. Doch auch die Molekülgröße hat einen
Einfluss auf die Retentionszeit, z.B. durchlaufen kleinere Verbindungen die stationäre
Phase schneller.
Neben den beschriebenen Wildisolaten wurden zusätzlich in der Abschlussarbeit von
Jacobsen (2013) die Referenzstämme K. rhizophila DSM 11926T, M. luteus DSM
20030T und S. xylosus DSM 20266T auf den Carotinoidgehalt analysiert. Bereits in der
Literatur wird auf die Pigmentierung der Typstämme hingewiesen. Nach Kovacs et al.
(1999) wächst Kocuria rhizophila DSM 11926T bei 28°C in gelb gefärbten Kolonien und
auch Micrococcus luteus DSM 20030T bildet Kolonien mit cremig gelben Pigmenten bei
37°C (Wieser et al. 2002). Einige Stämme von Staphylococcus xylosus zeigen nur bei
Wachstumstemperaturen unter 20°C eine orange-gelbe Pigmentierung und sind bei
30-37°C farblos (Schleifer und Kloos 1975). Dieser Aspekt unterstützt die zweite
Hypothese dieser Arbeit, dass Carotinoide einen Einfluss auf die Kälteanpassung
haben. Nach der Anzucht bei 10°C und 30°C konnten bei diesen drei Typstämmen
Carotinoide isoliert werden. Die Typstämme von K. rhizophila DSM 11926T und
S. xylosus DSM 20266T wiesen entsprechend der Wildisolate bei 10°C einen höheren
Carotinoidgehalt auf. Der Typstamm von M. luteus zeigte jedoch keine Veränderung im
Gesamtcarotinoidgehalt (Abb. 28).
Dieser zuvor beschriebene Effekt der Erhöhung der Carotinoidkonzentration bei
Kälteeinfluss wurde auch bereits in einer angewandten Studie der
Lebensmittelindustrie postuliert. Dort wurde beschrieben, dass es Bakterien gibt, die
Pigmente ausbilden und zu einem Verderb bei Lebensmitteln führen können. So
konnte Kröckel (2007) beobachten, dass es in einer vorgepackten, kühl gelagerten
Weißwurst zu einer Gelbfärbung kam. Es stellte sich heraus, dass es sich um das
gelbpigmentierte Milchsäurebakterium Leuconostoc gelidum handelte, welches diese
Gelbfärbung verursachte. Kröckel (2007) stellte in weiteren Versuchen fest, dass diese
Gelbfärbung jedoch nur bei kalten Temperaturen vorkam, wie beispielsweise der

Diskussion 105
gekühlten Lagertemperatur. Die Identifizierung des Carotinoids erfolgte mittels UV-VIS
Spektroskopie und Größenausschlusschromatographie. Während ein gelbes Pigment
von Leuconostoc citreum spektrophotometrisch eindeutig als 4,4’-Diaponeurosporin
identifiziert werden konnte, welches temperaturunabhängig produziert wird, handelte
es sich bei dem nur in der Kälte produzierten gelben Pigment von Leuconostoc
gelidum um eine Vorstufe dieses Triterpenoids. Leuconostoc gelidum ist zwar für den
Menschen nicht pathogen, jedoch wird es als Lebensmittelverderber angesehen, da
durch die Verfärbung des Lebensmittelproduktes es zu einer Ablehnung des
Konsumenten gegenüber dem Produkt kommt.
Wie die Ergebnisse der Carotinoidbestimmung zeigen, liegt die Vermutung nahe, dass
es durch Kälteeinfluss zu einer erhöhten Carotinoidproduktion in der Zellmembran
kommen kann. Dieser Effekt wird jedoch, wie bei Kröckel (2007) bevorzugt bei
Wildisolaten beobachtet. Somit kann davon ausgegangen werden, dass dieser erhöhte
Gehalt an Carotinoiden einen Schutzmechanismus gegenüber Kältestress darstellen
kann.
Der verstärkte Einbau von Carotinoiden in die Zellmembran bei Kälteeinfluss kann im
Gegensatz zu Fettsäuren nicht mit der physikalischen Eigenschaft des
Schmelzpunktes erklärt werden. Die Schmelzpunkte verschiedener Carotinoide liegen
bei über 150°C (Rothblat et al. 1964; Zscheile und White 1940), somit kann der
Mechanismus nicht mit einer Verschiebung des Schmelzpunktes begründet werden.
Vielmehr kann hier die molekulare Struktur der Carotinoide eine Rolle spielen. Die
genaue Funktion von unterschiedlichen Carotinoiden in Membranen ist immer noch
unbekannt. Es gibt jedoch Hinweise, dass das bakterielle Carotinoid Bacterioruberin
sich an Membranvesikel bindet und somit die Rigidität der Membran ansteigt (Strand et
al. 1997). Die Daten von Dieser et al. (2010) tragen zum Verständnis der
Funktionsweise von Carotinoiden bei. Carotinoid-pigmentierte heterotrophe Bakterien
tolerieren niedrige Temperaturen und episodisches Einfrieren erfolgreicher als nicht-
pigmentierte Bakterien. Diese Ergebnisse deuten stark darauf hin, dass Carotinoide bei
der Modulation der Membranfluidität mitwirken müssen und zusätzlich auf die
Membranstabilität einen Einfluss haben. Daher ist dies eine notwendige Anpassung an
physikalischen Stress, der unter niedrigen oder Gefriertemperaturen entsteht. Laut
Donato et al. (1997) kann die Anwesenheit von Carotinoiden, eine höhere
Packungsdichte der Phospholipide in der Membran verursachen, womit diese
stabilisiert wird.

106 Diskussion
Somit kann festgestellt werden, dass Carotinoide eine wichtige Rolle bei Kältestress
von Bakterien auf Zellmembranebene spielen können. Sowohl für die Medizin als auch
für die Industrie ist es daher von großer Bedeutung genauer herauszufinden, warum
und zu welchem Zweck Carotinoide in Bakterien synthetisiert werden.
4.6 Zelldichte und Wachstumsverhalten
Um die Auswirkung von unterschiedlichen Temperaturen auf das jeweilige
Wachstumsverhalten von Bakterien zu untersuchen, wurde von ausgewählten Isolaten
der Wachstumsverlauf bei 10°C und 30°C bestimmt. An den beispielhaft in Abschnitt
3.7 beschriebenen Isolaten wurden temperaturabhängige Veränderungen bei den
maximalen Zellerträgen ermittelt. Es wurde jeweils eine höhere Zellausbeute bei 10°C
im Vergleich zu 30°C festgestellt. Dieses Verhalten wurde zuvor auch in den
Abschlussarbeiten von Fritsch (2012), Jacobsen (2013) und Murr (2013) beobachtet.
Bereits Margesin (2009) und Muranushi (2009) konnten diesen Effekt ebenfalls bei
ausgewählten Isolaten, die aus kalten Habitaten stammen, nachweisen.
Es fällt auf, dass dieser Effekt fast ausschließlich bei den Wildisolaten auftrat, nicht
aber bei verwandten Typ- oder Referenzstämmen. Dieses Verhalten konnte in der
Proteinbestimmung bestätigt werden.
Nach den Angaben aus der Literatur wachsen diese Isolate vorwiegend im mesophilen
Bereich. Das Wachstumsoptimum von Bacillus cereus liegt bei 37°C (Warth 1978) und
von Paenibacillus glucanolyticus liegt der Wachstumsbereich zwischen 17-37°C
(Alexander und Priest 1989). Pedobacter panaciterrae hat ein Wachstumsoptimum bei
25°C (Yoon et al. 2007) und Escherichia coli bei 37°C bis 42°C (Buchanan und La
Klawitter 1992). Staphylococcus xylosus wächst im Temperaturbereich zwischen 15°C
und 45°C (Schleifer und Kloos 1975). Listeria monocytogenes hat ein
Wachstumsoptimum zwischen 30°C und 37°C (Krämer 2007).
Bei mesophilen Bakterien liegt der maximale Wachstumsertrag meist in der Nähe des
Temperaturoptimums und bleibt zusammen mit der Wachstumsrate über einen großen
Temperaturbereich konstant. Manche psychrophile Bakterien (z.B. Psychrobacter)
versuchen dagegen sinkende Wachstumsraten bei Temperaturen unterhalb des
Wachstumsoptimums mit einer Erhöhung des Wachstumsertrages auszugleichen.
(Bakermans und Nealson 2004)
Jedoch zeigte sich bei diesen Isolaten, die laut Speziesbeschreibung ein
Wachstumsoptimum im mesophilen Bereich haben, eine deutlich höhere Zellausbeute

Diskussion 107
bei einer niedrigeren, im Kältebereich liegenden Temperatur. Da es sich bei allen
Isolaten laut Speziesbeschreibung eigentlich um mesophile Bakterien handelte, sind
diese Beobachtungen erstaunlich, da es eigentlich zu einer Reduzierung der
Zellausbeute hätte kommen müssen. Die erhöhte Zellausbeute wurde in dieser Arbeit
zusätzlich mit einer Proteinbestimmung bestätigt. Der potentielle Mechanismus dieses
Effektes sollte durch weiterführende Arbeiten ausführlicher untersucht werden, da
bislang wenige Literaturquellen zu diesem Thema existieren. Außerdem muss genauer
abgegrenzt werden, zu welchen Bakterientypen in Bezug auf das Temperaturverhalten
diese Wildisolate zählen. Außerdem sind sie nicht immer mit dem Verhalten der
Referenzstämme gleichzusetzen.
Die Ergebnisse der vorliegenden Arbeit zeigen, dass die Wachstumsausbeute ein sehr
wichtiger, jedoch auch oft übersehener, Faktor in der Temperaturanpassung von
Bakterien ist. Zu diesem Resultat kommen auch Knoblauch und Jørgensen (1999). Die
Korrelation zwischen einer hohe Zellausbeute und Wachstum bei niedrigen
Temperaturen kann ein Merkmal von kälteangepassten Bakterien sein und kann daher
den psychrophilen Bakterien einen Vorteil beim mikrobiellen Wettbewerb bieten.
Mikroorganismen, die über einen weiten Temperaturbereich leben, benötigen einen
Ausgleich für langsame Reaktionsgeschwindigkeiten bei niedrigen Temperaturen, um
wettbewerbsfähig zu bleiben zu können. So würde die langsamere Wachstumsrate und
die dadurch langsameren biochemischen Prozesse kompensiert werden. Einige
Psychrophile maximieren oder versuchen ihr Wachstum bei niedrigen Temperaturen zu
erhalten, um niedrige Wachstumsraten auszugleichen. Damit wären sie
konkurrenzfähig. Die Daten der vorliegenden Arbeit werden auch Harder & Veldkamp
(1968), Harder und Veldkamp (1971) Isaksen & Jorgensen (1996) und Knoblauch &
Jorgensen (1999) bestätigt, die u.a. Zellausbeute und spezifische Wachstumsrate von
unterschiedlichen psychrophilen Bakterienstämmen untersucht haben. So zeigte sich
beispielweise bei Knoblauch und Jorgensen (1999), dass der psychrophile Sulfat-
reduzierende Bakterienstamm LSv514 zwar die höchste spezifische Wachstumsrate
und die Sulfatreduktion im mesophilen Temperaturbereich aufwies, die höchste
Zellausbeute jedoch zwischen 0 und 4°C gemessen wurde. Außerdem kann damit die
längere Anpassungsphase erklärt werden. Der eigentlichen Vermehrung geht bei
kühlen Inkubationstemperaturen ebenso wie bei wärmeren Inkubationstemperaturen
zunächst immer diese Anpassungsphase voraus. Diese lag-Phase ist extrem
temperaturabhängig, wie Mossel et al. (1995) bereits festgestellt hatten. Die
Anpassungsphase von L. monocytogenes lag bei einem Temperaturbereich von 0 bis

108 Diskussion
1°C zwischen 3 und 33 Tagen, bereits eine geringfügige Erhöhung auf 3°C verkürzte
sie auf 2 bis 8 Stunden.
Insgesamt kann festgestellt werden, dass durch die Kälteanpassung das Wachstum
von Bakterien sich deutlich verändert. Es scheint, dass das Wachstum in der Kälte
einen interessanten jedoch auch bislang wenig untersuchten Effekt zur Folge hat. Es
zeigt sich ein teilweise stark erhöhter Zellertrag bei kälteangepassten Bakterien im
Vergleich mit mesophilen Bakterien bei ihrer optimalen Temperatur. Dies könnte darauf
hindeuten, dass kälteadaptierte Bakterien möglicherweise ihre Kohlenstoffquelle
effizienter nutzen, um somit die mehr verlangte Biomasse aufzubauen. Außerdem
muss der niedrige bzw. nicht auftretende Effekt des gesteigerten Zellertrages bei
Referenzstämmen, die meist aus Stammsammlungen bezogen werden, geklärt
werden.
4.7 Kälteresistenztest und Überlebenstest Listeria monocytogenes
Der Kälteresistenztest zeigt, dass ein Listeria monocytogenes-Wildstamm eine stärkere
Kälteresistenz aufweist als der vergleichende Referenzstamm. So konnte der
Wildstamm Listeria monocytogenes Iso 1/11 zum einen trotz Stress in Form von
Gefrieren seine Zelldichte aufrechterhalten und zum anderen über einen sehr langen
Zeitraum in dem Lebensmittel Milch bei niedriger Temperatur überleben und die
Zelldichte halten. Diese Effekte konnten bei dem Referenzstamm DSM 20600T nicht
gezeigt werden. Die Zelldichte des Referenzstammes nahm sowohl im
Kälteresistenztest als auch im Überlebenstest ab Tag 44 ab.
Diese Ergebnisse zeigen, dass man das Verhalten gegenüber einem Stressfaktor wie
in diesem Fall niedrige Temperaturen nicht immer auf eine komplette Spezies beziehen
kann, wie es bei der Spezies Listeria monocytogenes oft der der Fall ist (Tasara und
Stephan 2006). Unser Fall zeigt, dass nur das Wildisolat eine stärker ausgeprägte
Kälteresistenz, nicht jedoch der Referenzstamm DSM 20600T, aufweist.
In der Literatur ist der Bezug von der Spezies Listeria monocytogenes zu kalten
Umgebungstemperaturen ausführlich beschrieben. Bezogen auf die
Wachstumstemperatur ist der durchschnittliche minimale Wert bei 1,7°C anzusiedeln,
wie entsprechende Untersuchungen von Junttila et al. (1988) zeigen. Laut Guillier et al.
(2005) weist die Spezies Listeria monocytogenes eine innerhalb für Bakterien recht
weiten Spannbreite von 0°C bis 45°C auf.

Diskussion 109
Auch bezogen auf das Wachstumsverhalten von L. monocytogenes konnte Jiang et al.
(2004) ein anderes Wachstumsverhalten bei unterschiedlichen Temperaturen zeigen.
So waren in Erdboden eingebrachte L. monocytogenes-Stämme bei 21°C bis zu 3
Wochen lebensfähig und bei 5°C sogar bis zu 43 Tage. Aber auch bezogen auf den
Lebensmittelbereich werden in der Literatur besonders häufig die Spezies Listeria
monocytogenes in geräucherter und gekühlter Fischware beschrieben. Als Beispiel sei
hier Hudson und Mott (1993) genannt, wo sich in geräucherten, abgepackten Lachs
L. monocytogenes bei einer Lagertemperatur von 10°C innerhalb von 4 Tagen um ca.
3 Zehnerpotenzen vermehren konnte. Zusätzlich stellten Guyer und Jemmi (1991) bei
mit L. monocytogenes kontaminiertem Lachs fest, was die anschließende
Lagertemperatur bis zu 30 Tage bei 4°C und 10°C für eine wichtige Rolle spielen kann.
Die Anfangskontamination von 6,5 x 102 Zellen/g wurde bei 4°C nach 30 Tagen circa
verhundertfacht und zwar auf einen Wert von ca. 104 Zellen/g. Bei 10°C
Lagertemperatur lag dieser Wert nach 10 Tagen sogar schon bei knapp 107 Zellen/g.
Bradshaw et al. (1985) zeigte, dass bei einer Aufbewahrungstemperatur von Milch bei
8°C sich eine anfängliche Listerien-Population von ca. 10 Zellen innerhalb von 7 Tagen
auf ca. 104 KbE/mL und nach 10 Tagen auf 106 KbE/mL anstieg.
Wie man erkennen kann, spielen die extrinsischen Faktoren eines Lebensmittels
(Lagertemperatur, aber auch Gasatmosphäre) eine wichtige Rolle bezogen auf
Wachstumsverhalten und Vermehrungsfähigkeit von Listeria monocytogenes.
Wahrscheinlich mehr als die intrinsischen Faktoren (aw-Wert, pH-Wert,
Zusammensetzung) und die Struktur des Lebensmittels, da insbesondere der
extrinsische Faktor Kühlung in aller Regel nicht ausreicht, einmal ins Produkt gelangte
L. monocytogenes-Stämme im Wachstum einzuschränken oder gar zu reduzieren.
Man erkennt, dass L. monocytogenes somit in der Lage ist, in kühlpflichtigen
Lebensmitteln nicht nur zu überdauern, sondern auch zu wachsen. Die mögliche
Gefährdung für den Menschen steigt mit längerer Lagerungsdauer grundsätzlich an,
wenn das Lebensmittel erst einmal mit dem Erreger kontaminiert ist. Die Zellen
benötigen zwar nach der Besiedlung eine temperaturabhängige Anpassungszeit,
jedoch gehen die Zellen dann schnell in die exponentielle Vermehrung mit einer kurzen
Generationszeit über. (Bülte 2008b)
Für eine Risikobewertung sind die Temperatur und die Temperaturführung daher von
wesentlicher Bedeutung. Selbst bei geringer Belastung von Lebensmitteln mit Listeria
monocytogenes mit einigen wenigen Zellen, kann bei haushaltsüblichen

110 Diskussion
Kühlschranktemperaturen der Grenzwert von 100 KbE/mg bzw. mL schnell erreicht
und überschritten werden. (Bülte 2008a)
Wie in den zuvor erwähnten Bespielen beschrieben, wird in der Literatur von den
untersuchten Isolaten oder Bakterienstämmen auf die komplette Spezies geschlossen.
Gerade bei der Spezies Listeria monocytogenes ist dies der Fall, da sie ein gut
untersuchter Vertreter für kälteadaptierte Lebensmittelpathogene ist. Die vorliegenden
Versuche zeigen aber, dass es zu einem stammspezifisch starken Unterschied in der
Vermehrungsfähigkeit und im Wachstumsverhalten bei niedrigen Temperaturen
kommen kann. Insbesondere für die Qualitätssicherung der Lebensmittelindustrie ist
diese Erkenntnis von Bedeutung. So ist es interessant, wie sich in den Rückstellproben
bestimmte Bakterien während einer gewährleisteten kühlen Lagerung im Laufe des
Mindesthaltbarkeitsdatums verhalten und dabei ihren Zellgehalt verändern bzw. sich im
Gegensatz zu höheren Temperaturen erhöhen.
Zusätzlich ist es interessant und eigentlich unabdingbar zu testen, inwieweit sich das
Wachstum von Wildstämmen und den verwendeten Referenzstämmen in einem
Lebensmittel unterscheidet. Denn erst damit wird das Wachstumsverhalten von
Bakterienstämmen, die durch Umwelteinflüsse von der (erwarteten) Standardisierung
abweichen können, der Realität entsprechend abgebildet. Erfahrungen dieser
Testreihen könnten die Qualitätssicherung in der Lebensmittelindustrie voranbringen
und die Haltbarkeit der Produkte weiter optimieren.

Zusammenfassung 111
5. Zusammenfassung
In der vorliegenden Dissertation wurden verschiedene Kälteanpassungsmechanismen
bakterieller Zellmembranen untersucht. Die übergeordnete Leitfrage war, ob und wie
andere lipophile Komponenten der Zellmembran von Bakterien einen Einfluss auf die
Kälteanpassung haben.
Dafür wurden kälteadaptierte Bakterienisolate aus Lebensmitteln und
lebensmittelassozierten Habitaten isoliert und bei den Wachstumstemperaturen 10°C
und 30°C inkubiert. Untersucht wurden sie zuerst auf Unterschiede im Fettsäuremuster
und dann auf Unterschiede im Chinonmuster bzw. Chinongehalt sowie dem
Carotinoidgehalt. Zusätzlich wurde von ausgewählten Isolaten das jeweilige
Wachstumsverhalten bei verschiedenen Temperaturen untersucht und mit dem
Wachstumsverhalten bestimmter Referenzstämme verglichen. Darüber hinaus wurden
mit einem Wildstamm der Spezies Listeria monocytogenes ein Kälteresistenztest in
Bezug auf Gefriertemperaturen und ein Überlebenstest im Modelllebensmittel Milch
durchgeführt. Die Ergebnisse wurden mit Ergebnissen eines Referenzstammes aus der
Stammsammlung verglichen. Zudem konnte die Speziesneubeschreibung von
Pedobacter nutrimenti veröffentlicht werden.
Die Ergebnisse der Fettsäureanalyse entsprachen weitestgehend den Erwartungen: So
war bei Kälte bei vielen Bakterien eine deutliche Verschiebung des Fettsäuremusters
hin zu verzweigten, kurzkettigen, ungesättigten bzw. hydroxylierten Fettsäuren
erkennbar. Bei gram-positiven Vertretern zeigte sich diese Verschiebung nicht so
deutlich wie bei gram-negativen Vertretern. Der nächste Schritt dieser Arbeit bestand
deswegen darin, dieser Tendenz auf den Grund zu gehen und diese Isolate in Bezug
auf Chinone und Carotinoide weiter zu untersuchen.
Die Ergebnisse der Chinonanalyse zeigten dann oft eine Abnahme des Chinongehaltes
bei 10°C Inkubation, obwohl bei wenigen, ausgewählten Isolaten es entweder zu einem
Anstieg im Chinongehalt bzw. zu einer Veränderung des Chinonmusters bei
Kälteeinfluss kam.
Damit konnte gezeigt werden, dass unterschiedliche Chinongehalte bzw.
Chinonmuster einen potentiellen fettsäureunabhängigen
Kälteanpassungsmechanismus darstellen können.
Die Ergebnisse der Carotinoidanalyse ergaben, dass bestimmte Isolate bei 10°C eine
größere Carotinoidmenge aufwiesen als bei 30°C. Der Vergleich mit den
Fettsäuremustern unterstützte die Erkenntnis, dass es einen Zusammenhang zwischen

112 Zusammenfassung
Carotinoiden und einem fettsäureunabhängigen Kälteanpassungsmechanismus der
Bakterien gibt.
Eigentlich sollten in der vorliegenden Arbeit nur Fettsäuren, Chinone und Carotinoide
von Bakterien und ihr Einfluss auf die Kälteanpassung analysiert werden. Aber auch
das Wachstumsverhalten einiger der untersuchten Bakterien zeigte einen auffallenden
Effekt durch Kälteeinfluss. Somit brachte diese Arbeit eine weitere, nicht erwartete
Erkenntnis: Die Zellausbeute bei einer Inkubation von 10°C war bei einigen Isolaten
stark erhöht im Gegensatz zu einer Inkubation bei 30°C. Auffällig zudem: Dieser Effekt
trat nur bei Wildisolaten, nicht aber bei den Vergleichs- und Referenzstämmen auf.
Zusammenfassend kann gesagt werden, dass bei einigen Bakterienstämmen bei
sinkender Umgebungstemperatur eine Zunahme bzw. Änderung der Chinonsynthese
und Carotinoidsynthese stattfindet. Diese Ergebnisse unterstützen die in dieser Arbeit
voranstehende Hypothese, nämlich dass Chinone und Carotinoide als lipophile
Komponenten in der Zellmembran einen Einfluss auf die Membranstruktur bei
Kälteeinfluss haben können und die Kälteanpassung der Bakterienzelle auch
fettsäureunabhängig ablaufen kann. Somit wird die Fluidität der Zellmembran
beeinflusst, um auch bei niedrigeren Temperaturen in der flüssig-kristallinen und damit
funktionellen Phase zu bleiben.
In zukünftigen Arbeiten sollte der Einfluss dieser Substanzen auf die Membranfluidität
genauer untersucht und der Grad der Fluidität gemessen werden.
Außerdem sollte der Effekt der höheren Zellausbeute bei niedrigeren Temperaturen auf
ein Modelllebensmittel bezogen werden. Dieser Effekt und das Resultat, dass eine
erhöhte Zellausbeute bei einer niedrigeren Temperatur nachgewiesen wurde, obwohl
das Wachstumsoptimum dieser Isolate laut Literatur eher im mesophilen Bereich liegt,
kann auch für die Lebensmittelindustrie wichtig sein.
Zusätzlich sollte darauf geachtet werden, das Wachstumsverhalten eines Bakteriums
nicht immer auf eine komplette Spezies zu beziehen, sondern stammspezifisch zu
untersuchen. Gerade in der Lebensmittelindustrie wäre dies wichtig, da oft Versuche
zu Vermehrungsfähigkeit und Überdauerung in Lebensmitteln mit Referenzorganismen
aus Stammsammlungen durchgeführt werden, diese sich jedoch stark im
Wachstumsverhalten zu Wildisolaten unterscheiden können. Dieser Aspekt ist in der
Literatur kaum beschrieben und muss für die Qualitätssicherung stärker untersucht
werden.

Literatur 113
6. Literaturverzeichnis
Aguilar, P. S. & de Mendoza, D. (2006). Control of fatty acid desaturation: a
mechanism conserved from bacteria to humans. Molecular Microbiology 62, 1507–
1514.
Alexander, B. & Priest, F.G. (1989). Bacillus glucanolyticus, a new species that
degrades a variety of β-glucans. International Journal of Systematic Bacteriology 39,
112–115.
Annous, B. (1997). Critical role of anteiso-C15:0 fatty acid in the growth of Listeria
monocytogenes at low temperatures. Applied and Environmental Microbiology, 3887–
3894.
Asai, Y. & Watanabe, S. (1999). The interaction of ubiquinone-3 with phospholipid
membranes. FEBS Letters 446, 169–172.
Ashcroft, J.R. & Haddock, B.A. (1975). Synthesis of alternative membrane-bound
redox carriers during aerobic growth of Escherichia coli in the presence of potassium
cyanide. Biochemical Journal 148, 349–352.
Ashcroft, J.R. & Haddock, B.A. (1975). Synthesis of alternative membrane-bound
redox carriers during aerobic growth of Escherichia coli in the presence of potassium
cyanide. Biochemical Journal 148, 349–352.
Bakermans, C. & Nealson, K.H. (2004). Relationship of critical temperature to
macromolecular synthesis and growth yield in Psychrobacter cryopegella. Journal of
Bacteriology 186, 2340–2345.
Bast, E. (1999). Mikrobiologische Methoden. Eine Einführung in grundlegende
Arbeitstechniken. Heidelberg: Spektrum Akad. Verlag
Beales, N. (2004). Adaptation of microorganisms to cold temperatures, weak acid
preservatives, low pH, and osmotic stress: A review. Comprehensive Reviews in Food
Science and Food Safety 3, 1–20.
Belitz, H.D.; Grosch, W.; Schieberle, P. (2008). Lehrbuch der Lebensmittelchemie.
Mit 634 Tabellen. 6., vollst. überarb. Aufl., [Nachdr.]. Berlin: Springer
Bentley, R. & Meganathan, R. (1982). Biosynthesis of vitamin K (menaquinone) in
bacteria. Microbiological reviews 46, 241–280.

114 Literatur
Berg, J.M.; Tymoczko, J.L.; Stryer, L. (2003). Biochemie. 5. Aufl. Heidelberg:
Spektrum Akad. Verl (Spektrum-Lehrbuch).
Bradshaw, J.G.; Peeler, J.T.; Corwin, J. J.; Hunt, J. M.; Tierney, J. T.; Larkin, E. P.
& Twedt, R. M. (1985). Thermal resistance of Listeria monocytogenes in milk. Journal
of food protection 48.
Bringer, S.; Härtner, T.; Poralla, K. & Sahm, H. (1985). Influence of ethanol on the
hopanoid content and the fatty acid pattern in batch and continuous cultures of
Zymomonas mobilis. Archives of Microbiology 140, 312–316.
Britton, G. (1995). Structure and properties of carotenoids in relation to function. The
FASEB Journal, 1551–1558.
Britton, G. (2008). Functions of Intact Carotenoids. In: George Britton, Synnøve
Liaaen-Jensen und Hanspete Pfander (Hg.): Carotenoids, Bd. 4. Basel: Birkhäuser
Basel, 189–212.
Buchanan, R.L. & Klawitter L.A. (1992). The effect of incubation temperature, initial
pH, and sodium chloride on the growth kinetics of Escherichia coli O157: H7. Food
Microbiology 9, 185–196.
Buck, J.D. (1982). Nonstaining (KOH) method for determination of gram reactions of
marine bacteria. Applied and Environmental Microbiology 44, 992–993.
Bülte, Michael (2008a). Listeria monocytogenes Band 1. [Jubiläumsausg.]. Hamburg:
Behr (Pathogene Mikroorganismen, 1).
Bülte, Michael (2008b). Listeria monocytogenes Band 2. [Jubiläumsausg.]. Hamburg:
Behr (Pathogene Mikroorganismen, 2).
Busta, F. (1978). Introduction to injury and repair of microbial cells. In: Advances in
Applied Microbiology Volume 23, Bd. 23: Elsevier (Advances in Applied Microbiology),
195–201.
Campenni’, L.; Nobre, B. P.; Santos, C.A.; Oliveira, A.C.; Aires-Barros, M.R.;
Palavra, A.M.F. & Gouveia, L. (2013). Carotenoid and lipid production by the
autotrophic microalga Chlorella protothecoides under nutritional, salinity, and luminosity
stress conditions. Applied Microbiology and Biotechnology 97, 1383–1393.
Chan, K.F.; Le Tran, H.; Kanenaka, R.Y. & Kathariou, S. (2001). Survival of clinical
and poultry-derived isolates of Campylobacter jejuni at a low temperature (4 °C).
Applied and Environmental Microbiology 67, 4186–4191.

Literatur 115
Chattopadhyay, M.; Jagannadham, M.; Vairamani, M. & Shivaji, S. (1997).
Carotenoid pigments of an antarctic psychrotrophic bacterium Micrococcus roseus:
temperature dependent biosynthesis, structure, and interaction with synthetic
membranes. Biochemical and Biophysical Research Communications 239, 85–90.
Chattopadhyay, M.K. (2006). Mechanism of bacterial adaptation to low temperature.
Journal of Biosciences 31, 157–165.
Chintalapati, S.; Kiran, M.D. & Shivaji, S. (2004). Role of membrane lipid fatty acids
in cold adaptation. Cellular and Molecular Biology 50, 631–642.
Cleland, D.; Krader, P.; McCree, C.; Tang, J. & Emerson, D. (2004). Glycine betaine
as a cryoprotectant for prokaryotes. Journal of Microbiological Methods 58, 31–38.
Cogdell, R.J. & Frank, H.A. (1987). How carotenoids function in photosynthetic
bacteria. Biochimica et Biophysica Acta (BBA) - Reviews on Bioenergetics 895, 63–79.
Collins, M.D. & Jones, D. (1981). Distribution of isoprenoid quinone structural types in
bacteria and their taxonomic implications. Microbiological Reviews 45, 316–354.
Collins, M.D.; Jonse, D.; Goodfellow, M. & Minnikin, D.E. (1979). Isoprenoid
quinone composition as a guide to the classification of Listeria, Brochothrix,
Erysipelothrix and Caryophanon. Journal of General Microbiology 111, 453–457.
Collins, M.D.; Farrow, J.A.; Phillips, B.A. & Kandler, O. (1983). Streptococcus
garvieae sp. nov. and Streptococcus plantarum sp. nov. Journal of General
Microbiology 129, 3427–3431.
Crockett, E.L. (1998). Cholesterol function in plasma membranes from ectotherms:
membrane-specific roles in adaptation to temperature. American zoologist 38, 291–
304.
Deming, J.W. (2002). Psychrophiles and polar regions. Current opinion in microbiology
5, 301–309.
Denich, T.; Beaudette, L.; Lee, H. & Trevors, J. (2003). Effect of selected
environmental and physico-chemical factors on bacterial cytoplasmic membranes.
Journal of Microbiological Methods 52, 149–182.
Derichs, J.; Kämpfer, P. & Lipski, A. (2014). Pedobacter nutrimenti sp. nov., isolated
from chilled food. International Journal of Systematic and Evolutionary Microbiology 64,
1310-1316.

116 Literatur
Dexter, Y. & Cooke, R. (1984). Fatty acids, sterols and carotenoids of the psychrophile
Mucor strictus and some mesophilic Mucor species. Transactions of the British
Mycological Society 83, 455–461.
Dieser, M.; Greenwood, M. & Foreman, C.M. (2010). Carotenoid pigmentation in
Antarctic heterotrophic bacteria as a strategy to withstand environmental stresses.
Arctic, Antarctic and Alpine Research 42, 396–405.
Donato, M.M.; Antunes-Madeira, M.C.; Jurado, A.S. & Madeira, V.M. (1997).
Partition of DDT and DDE into membranes and extracted lipids of Bacillus
stearothermophilus. Bulletin of environmental contamination and toxicology 59, 696–
701.
Effenberger-Neidnicht, K. (2011). Biochemische Charakterisierung und
Wirkoptimierung potentieller Chemotherapeutika auf Basis der natürlichen Chinone
Doxorubicin und Thymochinon. Dissertation, Universität Bayreuth.
Epand, R.M. (1998). Lipid polymorphism and protein–lipid interactions. Biochimica et
Biophysica Acta (BBA)-Reviews on Biomembranes 1376, 353–368.
Feller, G. & Gerday, C. (1997). Review-Psychrophilic enzymes: molecular basis of
cold adaptation. Cellular and Molecular Life Sciences (CMLS) 53, 830–841.
Feller, G. & Gerday, C. (2003). Psychrophilic enzymes: hot topics in cold adaptation.
Nature reviews 1, 200–208.
Feller, G.; Narinx, E.; Arpigny, J. L.; Zekhnini, Z.; Swings, J. & Gerday, C. (1994).
Temperature dependence of growth, enzyme secretion and activity of psychrophilic
Antarctic bacteria. Applied Microbiology and Biotechnology 41, 477–479.
Felsenstein, J. (1985). Confidence limits on phylogenies: an approach using the
bootstrap. Evolution, 783–791.
Fong, N. J. C.; Burgess, M.L.; Barrow, K.D. & Glenn, D.R. (2001). Carotenoid
accumulation in the psychrotrophic bacterium Arthrobacter agilis in response to thermal
and salt stress. Applied Microbiology and Biotechnology 56, 750–756.
Freedman, R.B. (1981). Membrane-bound enzymes. New Comprehensive
Biochemistry 1, 161–214.
Fritsch, C. (2012). Beeinflussung des Wachstumsverhaltens von Bakterien bei
niedrigen Temperaturen durch Zugabe von Chinonen. Masterarbeit. Universität Bonn.

Literatur 117
Fulco, A. (1974). Metabolic alterations of fatty acids. Annual review of biochemistry 43,
215–241.
Georlette, D.; Blaise, V.; Collins, T.; D'Amico, S.; Gratia, E.; Hoyoux, A. et al.
(2004). Some like it cold: biocatalysis at low temperatures. FEMS Microbiology
Reviews 28, 25–42.
Gerhardt, P.; Murray, R.G.; Costilow, R.N.; Nester, E.W.; Wood, W.A.; Krieg, N.R.
& Phillips, G.B. (1981). Manual of methods for general bacteriology.
Gounot, A.M. & Russell, N.J. (1999). Physiology of cold-adapted microorganisms. In:
Rosa Margesin und Franz Schinner (Hg.): Cold-adapted organisms. Berlin, Heidelberg:
Springer Berlin Heidelberg, 33–55.
Gram, H.C.J.; Friedländer, C. (1884). Über die isolierte Färbung der Schizomyceten:
in Schnitt-und Trockenpräparaten: Theodor Fischer's medizinischer Buchhandlung.
Graumann, P.; Schröder, K.; Schmid, R. & Marahiel, M. A. (1996). Cold shock
stress-induced proteins in Bacillus subtilis. Journal of Bacteriology 178, 4611–4619.
Guedes, A.C.; Amaro, H.M. & Malcata, F.X. (2011). Microalgae as sources of
carotenoids. Marine Drugs 9, 625–644.
Guendouzi, A. & Mekelleche, S.M. (2012). Prediction of the melting points of fatty
acids from computed molecular descriptors: A quantitative structure–property
relationship study. Chemistry and Physics of Lipids 165, 1–6.
Guenlcrl, A.M. & Linton, R.H. (2003). Growth and survival of selected pathogens in
margarine-style table spreads.
Guillier, L.; Pardon, P. & Augustin, J.-C. (2005). Influence of stress on individual lag
time distributions of Listeria monocytogenes. Applied and Environmental Microbiology
71, 2940–2948.
Guinebretiere, M.-H.; Auger, S.; Galleron, N.; Contzen, M.; Sarrau, B. de; Buyser,
M.-L. de et al. (2013). Bacillus cytotoxicus sp. nov. is a novel thermotolerant species of
the Bacillus cereus group occasionally associated with food poisoning. International
Journal of Systematic and Evolutionary Microbiology 63, 31–40.
Guyer, S. & Jemmi, T. (1991). Behavior of Listeria monocytogenes during fabrication
and storage of experimentally contaminated smoked salmon. Applied and
Environmental Microbiology 57, 1523–1527.

118 Literatur
Hamasaki, Y.; Ayaki, M.; Fuchu, H.; Sugiyama, M. & Morita, H. (2003). Behavior of
psychrotrophic lactic acid bacteria isolated from spoiling cooked meat products.
Applied and Environmental Microbiology 69, 3668–3671.
Harder, W. & Veldkamp, H. (1968). Physiology of an obligatory psychrophilic marine
Pseudomonas species. Journal of Applied Microbiology 31, 12–23.
Harder, W. & Veldkamp, H. (1971). Competition of marine psychrophilic bacteria at
low temperatures. Antonie van Leeuwenhoek 37, 51–63.
Harrison, W.A.; Peters, A.C. & Fielding, L.M. (2000). Growth of Listeria
monocytogenes and Yersinia enterocolitica colonies under modified atmospheres at 4
and 8 °C using a model food system. Journal of Applied Microbiology 88, 38–43.
Härtig, C.; Loffhagen, N. & Babel, W. (1999). Glucose stimulates a decrease of the
fatty acid saturation degree in Acinetobacter calcoaceticus. Archives of Microbiology
171, 166–172.
Havaux, M. (1998). Carotenoids as membrane stabilizers in chloroplasts. Trends in
Plant Science 3, 147–151.
Hazel, J.R. & Eugene Williams, E. (1990). The role of alterations in membrane lipid
composition in enabling physiological adaptation of organisms to their physical
environment. Progress in lipid research 29, 167–227.
Hébraud, M. & Potier, P. (1999). Cold shock response and low temperature
adaptation in psychrotrophic bacteria. Journal of Molecular Microbiology and
Biotechnology 1, 211–219.
Heipieper, H.-J.; Diefenbach, R. & Keweloh, H. (1992). Conversion of cis
unsaturated fatty acids to trans, a possible mechanism for the protection of phenol-
degrading Pseudomonas putida P8 from substrate toxicity. Applied and Environmental
Microbiology 58, 1847–1852.
Heipieper, H.J.; Meinhardt, F. & Segura, A. (2003). The cis-trans isomerase of
unsaturated fatty acids in Pseudomonas and Vibrio. Biochemistry, molecular biology
and physiological function of a unique stress adaptive mechanism. FEMS Microbiology
Letters 229, 1–7.
Heller, H.; Schaefer, M. & Schulten, K. (1993). Molecular dynamics simulation of a
bilayer of 200 lipids in the gel and in the liquid crystal phase. The Journal of Physical
Chemistry 97, 8343–8360.

Literatur 119
Helmke, E. & Weyland, H. (2004). Psychrophilic versus psychrotolerant bacteria-
occurrence and significance in polar and temperate marine habitats. Cellular and
molecular Biology 50, 553–561.
Herranen, M.; Kariluoto, S.; Edelmann, M.; Piironen, V.; Ahvenniemi, K.; Iivonen,
V. et al. (2010). Isolation and characterization of folate-producing bacteria from oat
bran and rye flakes. International Journal of Food Microbiology 142, 277–285.
Hiraishi, A.; Ueda, Y.; Ishihara, J.B & Mori, T. (1996). Comparative lipoquinone
analysis of influent sewage and activated sludge by high-performance liquid
chromatography and photodiode array detection. The Journal of General and Applied
Microbiology 42, 457–469.
Hiraishi, A. (1999). Isoprenoid quinones as biomarkers of microbial populations in the
environment. Journal of Bioscience and Bioengineering 88, 449–460.
Hiraishi, A.; Umezawa, T.; Yamamoto, H.; Kato, K. & Maki, Y. (1999). Changes in
quinone profiles of hot spring microbial mats with a thermal gradient. Applied and
Environmental Microbiology 65, 198–205.
Hu, H.-Y.; Fujie, K. & Urano, K. (1999). Development of a novel solid phase extraction
method for the analysis of bacterial quinones in activated sludge with a higher
reliability. Journal of Bioscience and Bioengineering 87, 378–382.
Hudson, J. & Mott, S. (1993). Growth of Listeria monocytogenes, Aeromonas
hydrophila and Yersinia enterocolitica on cold-smoked salmon under refrigeration and
mild temperature abuse. Food Microbiology 10, 61–68.
Isaksen, M.F. & Jorgensen, B.B. (1996). Adaptation of psychrophilic and
psychrotrophic sulfate-reducing bacteria to permanently cold marine environments.
Applied and Environmental Microbiology 62, 408–414.
IUPAC-IUB, Commission on Biochemical Nomenclature (1978). The nomenclature
of lipids. Chemistry and Physics of Lipids 21, 159–173.
Jacobsen, M. (2013). Der Carotenoidgehalt bei Bakterien in Abhängigkeit von der
Wachstumstemperatur. Masterarbeit. Universität Bonn.
Jagannadham, M.V.; Chattopadhyay, M.K.; Subbalakshmi, C.; Vairamani, M.;
Narayanan, K.; Mohan Rao, C. & Shivaji, S. (2000). Carotenoids of an Antarctic
psychrotolerant bacterium, Sphingobacterium antarcticus, and a mesophilic bacterium,
Sphingobacterium multivorum. Archives of Microbiology 173, 418–424.

120 Literatur
Janda, J.M. & Abbott, S.L. (2007). 16S rRNA gene sequencing for bacterial
identification in the diagnostic laboratory: pluses, perils, and pitfalls. Journal of Clinical
Microbiology 45, 2761–2764.
Jeffries, L., Cawthrone, M.A., Harris, M., Diplock, A.T., Green, J. & Price, S.A.
(1967). Distribution of menaquinones in aerobic Micrococcaceae. Nature 215, 257–
259.
Jiang, X.; Islam, M.; Morgan, J. & Doyle, M. P. (2004). Fate of Listeria
monocytogenes in bovine manure-amended soil. Journal of Food Protection 67, 1676–
1681.
Junttila, J.R.; Niemelä, S.I. & Hirn, J. (1988). Minimum growth temperatures of
Listeria monocytogenes and non‐haemolytic listeria. Journal of Applied Microbiology
65, 321–327.
Kageyama, B.; Nakae, M.; Yagi, S. & Sonoyama, T. (1992). Pantoea punctata sp.
nov., Pantoea citrea sp. nov., and Pantoea terrea sp. nov. isolated from Fruit and Soil
Samples. International Journal of Systematic Bacteriology 42, 203–210.
Kaiser, P.; Surmann, P.; Vallentin, G. & Fuhrmann, H. (2007). A small-scale method
for quantitation of carotenoids in bacteria and yeasts. Journal of Microbiological
Methods 70, 142–149.
Kalinowski, R.M.; Tompkin, R.B.; Bodnaruk, P.W. & Pruett, W.P. (2003). Impact of
cooking, cooling, and subsequent refrigeration on the growth or survival of Clostridium
perfringens in cooked meat and poultry products. Journal of Food Protection 66, 1227–
1232.
Kämpfer, P. (1996). Klassische Methoden zur Charakterisierung von
Abwasserbakterien - Grenzen und Möglichkeiten. In: Ökologie der
Abwasserorganismen: Springer, 45–68.
Kandror, O.; DeLeon, A. & Goldberg, A.L. (2002). Trehalose synthesis is induced
upon exposure of Escherichia coli to cold and is essential for viability at low
temperatures. Proceedings of the National Academy of Sciences 99, 9727–9732.
Kaneda, T. (1991). Iso-fatty and anteiso-fatty acids in bacteria - Biosynthesis, function
and taxonomic significance. Microbiological Reviews 55, 288–302.
Kannenberg, E.L. & Poralla, K. (1999). Hopanoid biosynthesis and function in
bacteria. Naturwissenschaften 86, 168–176.

Literatur 121
Keweloh, H. & Heipieper, H.J. (1996). Trans unsaturated fatty acids in bacteria. Lipids
31, 129–137.
Kim, D.-U.; Kim, Y.-J.; Shin, D.-H.; Weon, H.-Y.; Kwon, S.-W.; Seong, C.-N. & Ka,
J.-O. (2013). Pedobacter namyangjuensis sp. nov. isolated from soil and
reclassification of Nubsella zeaxanthinifaciens Asker et al. 2008 as Pedobacter
zeaxanthinifaciens comb. nov. Journal of Microbiology 51, 25–30.
Kim, H.-B.; Srinivasan, S.; Sathiyaraj, G.; Quan, L.-H.; Kim, S.-H.; Bui, T. P. N. et
al. (2010). Stenotrophomonas ginsengisoli sp. nov., isolated from a ginseng field.
International Journal of Systematic and Evolutionary Microbiology 60, 1522–1526.
Kim, O.-S.; Cho, Y.-J.; Lee, K.; Yoon, S.-H.; Kim, M.; Na, H. et al. (2012).
Introducing EzTaxon-e: a prokaryotic 16S rRNA gene sequence database with
phylotypes that represent uncultured species. International Journal of Systematic and
Evolutionary Microbiology 62, 716–721.
Kiran, M. D.; Prakash, J.S.S.; Annapoorni, S.; Dube, S.; Kusano, T.; Okuyama, H.
et al. (2004). Psychrophilic Pseudomonas syringae requires trans-monounsaturated
fatty acid for growth at higher temperature. Extremophiles 8, 401–410.
Knoblauch, C. & Jorgensen, B.B. (1999). Effect of temperature on sulphate
reduction, growth rate and growth yield in five psychrophilic sulphate-reducing bacteria
from Arctic sediments. Environmental Microbiology 1, 457–467.
Ko, R.; Smith, L.T. & Smith, G.M. (1994). Glycine betaine confers enhanced
osmotolerance and cryotolerance on Listeria monocytogenes. Journal of Bacteriology
176, 426–431.
Kovacs, G.; Burghardt, J.; Pradella, S.; Schumann, P.; Stackebrandt, E. &
Marialigeti, K. (1999). Kocuria palustris sp. nov. and Kocuria rhizophila sp. nov.,
isolated from the rhizoplane of the narrow-leaved cattail (Typha angustifolia).
International Journal of Systematic Bacteriology 49, 167–173.
Kraft, A.A. (1992). Psychrotrophic Bacteria in Foods: Disease and Spoilage: Boca
Raton, Florida: CRC Press.
Krämer, J. (2007). Lebensmittel-Mikrobiologie. 6. Auflage, Stuttgart: Ulmer (UTB
Lebensmittelwissenschaften, Biologie).
Krishnamurthi, S.; Chakrabarti, T. & Stackebrandt, E. (2009). Re-examination of the
taxonomic position of Bacillus silvestris Rheims et al. 1999 and proposal to transfer it to

122 Literatur
Solibacillus gen. nov. as Solibacillus silvestris comb. nov. International Journal of
Systematic and Evolutionary Microbiology 59, 1054–1058.
Kropinski; A.M., Lewis, V. & Berry, D. (1987). Effect of growth temperature on the
lipids, outer membrane proteins, and lipopolysaccharides of Pseudomonas aeruginosa
PAO. Journal of Bacteriology 169, 1960–1966.
Kröckel, L. (2007). Ein bakterielles Carotinoid färbt vorverpackte, kühl gelagerte
Weißwurst gelb. Mitteilungsblatt der Fleischforschung Kulmbach 46, 223–230.
Kunihiro, T.; Veuger, B.; Vasquez-Cardenas, D.; Pozzato, L.; Le Guitton, M.;
Moriya, K. et al. (2014). Phospholipid-derived fatty acids and quinones as markers for
bacterial biomass and community structure in marine sediments. PloS one 9, e96219.
Kwon, S.-W.; Kim, B.-Y.; Lee, K.-H.; Jang, K.-Y.; Seok, S.-J.; Kwon, J.-S. et al.
(2007). Pedobacter suwonensis sp. nov., isolated from the rhizosphere of Chinese
cabbage (Brassica campestris). International Journal of Systematic and Evolutionary
Microbiology 57, 480–484.
Laksanalasmai, P.; Burall, L. & Datta, A.R. (2011). Adaptation Mechanisms of
Psychrotolerant Bacterial Pathogens. Extremophiles Handbook, 817–837.
Lechner, S.; Mayr, R.; Francis, K. P.; Prü, B. M.; Kaplan, T.; Wießner-Gunkel, E. et
al. (1998). Bacillus weihenstephanensis sp. nov. is a new psychrotolerant species of
the Bacillus cereus group. International Journal of Systematic Bacteriology 48, 1373–
1382.
Lee, H.-G.; Kim, S.-G.; Im, W.-T.; Oh, H.-M. & Lee, S.-T. (2009). Pedobacter
composti sp. nov., isolated from compost. International Journal of Systematic and
Evolutionary Microbiology 59, 345–349.
Lin, S.-Y.; Hameed, A.; Liu, Y.-C.; Hsu, Y.-H.; Lai, W.-A.; Chen, W.-M. et al. (2013).
Pseudomonas sagittaria sp. nov., a siderophore-producing bacterium isolated from oil-
contaminated soil. International Journal of Systematic and Evolutionary Microbiology
63, 2410–2417.
Liu, G. Y. (2005). Staphylococcus aureus golden pigment impairs neutrophil killing and
promotes virulence through its antioxidant activity. Journal of Experimental Medicine
202, 209–215.
Luo, X.; Wang, Z.; Dai, J.; Zhang, L.; Li, J.; Tang, Y. et al. (2010). Pedobacter
glucosidilyticus sp. nov., isolated from dry riverbed soil. International Journal of
Systematic and Evolutionary Microbiology 60, 229–233.

Literatur 123
Madigan, M. T.; Martinko, J. M.; Dunlap, P. V. & Clark, D. P. (2008). Brock Biology
of microorganisms 12th edn. International Microbiology 11, 65–73.
Madigan, M.T.; Martinko, J.M.; Stahl, D.A.; Clark, D.P.; Thomm, M.; Wirth, R.
(2013). Brock Mikrobiologie. 13., aktualisierte Aufl. München: Pearson Higher
Education (Bio Biologie).
Magnuson, K.; Jackowski, S.; Rock, C.O. & Cronan, J.E. (1993). Regulation of fatty
acid biosynthesis in Escherichia coli. Microbiological reviews 57, 522–542.
Malhotra, J.; Anand, S.; Jindal, S.; Rajagopal, R. & Lal, R. (2012). Acinetobacter
indicus sp. nov., isolated from a hexachlorocyclohexane dump site. International
Journal of Systematic and Evolutionary Microbiology 62, 2883–2890.
Marshall, J.H. & Wilmoth, G. J. (1981). Pigments of Staphylococcus aureus, a series
of triterpenoid carotenoids. Journal of Bacteriology 147, 900–913.
Margesin, R. (2003). Pedobacter cryoconitis sp. nov., a facultative psychrophile from
alpine glacier cryoconite. International Journal of Systematic and Evolutionary
Microbiology 53, 1291–1296.
Margesin, R. (2009). Effect of temperature on growth parameters of psychrophilic
bacteria and yeasts. Extremophiles 13, 257–262.
Margesin, R.; Neuner, G. & Storey, K.B. (2007). Cold-loving microbes, plants, and
animals - fundamental and applied aspects. Naturwissenschaften 94, 77–99.
Margesin, R. & Shivaji, S. (2011). Genus II. Pedobacter Steyn, Segers, Vancanneyt,
Sandra, Kersters and Joubert 1998, 171 VP. Bergey’s Manual of Systematic
Bacteriology 4, 339–347.
Margesin, R.; Gerday, C.; Marx, J.-C.; Schinner, F. (2008). Psychrophiles: from
biodiversity to biotechnology. Berlin, Heidelberg: Springer-Verlag Berlin Heidelberg.
Matissek, R.; Steiner, G.; Fischer, M. (2010). Lebensmittelanalytik: Springer
Germany.
Meganathan, R. (2001). Ubiquinone biosynthesis in microorganisms. FEMS
Microbiology Letters 203, 131–139.
Miller, L.T. (1982). Single derivatization method for routine analysis of bacterial whole-
cell fatty acid methyl esters, including hydroxy acids. Journal of Clinical Microbiology
16, 584–586.

124 Literatur
Mitchell, C.; Iyer, S.; Skomurski, J. F. & Vary, J. C. (1986). Red pigment in Bacillus
megaterium spores. Applied and Environmental Microbiology 52, 64–67.
Minnikin, D.; O'Donnell, A.; Goodfellow, M.; Alderson, G.; Athalye, M.; Schaal, A.
& Parlett, J. (1984). An integrated procedure for the extraction of bacterial isoprenoid
quinones and polar lipids. Journal of Microbiological Methods 2, 233–241.
Morishita, T.; Tamura, N.; Makino, T. & Kudo, S. (1999). Production of
menaquinones by lactic acid bacteria. Journal of Dairy Science 82, 1897–1903.
Morita (1975). Psychrophilic bacteria. Bacteriol. Rev, 144–167.
Morita, N.; Shibahara, A.; Yamamoto, K.; Shinkai, K.; Kajimoto, G. & Okuyama, H.
(1993). Evidence for cis-trans isomerization of a double bond in the fatty acids of the
psychrophilic bacterium Vibrio sp. strain ABE-1. Journal of Bacteriology 175, 916–918.
Mossel, D.A.A.; Corry, J.E.L.; Struijk, C.B.; Baird, R.M. (1995). Essentials of the
microbiology of foods: a textbook for advanced studies: John Wiley & Sons.
Munk, K. (2008). Biochemie. Zellbiologie. 1. Aufl. s.l: Georg Thieme Verlag KG.
Muranushi, N.; Takagi, N.; Muranishi, S. &Sezaki, H. (1981). Effect of fatty acids and
monoglycerides on permeability of lipid bilayer. Chemistry and Physics of Lipids 28,
269–279.
Murr, D. (2013). Carotinoidanalyse von Bakterien und deren Bedeutung in der
bakteriellen Kälteanpassung. Abschlussarbeit. Universität Bonn
Muyzer, G.; De Waal, E.C. & Uitterlinden, A.G. (1993). Profiling of complex microbial
populations by denaturing gradient gel electrophoresis analysis of polymerase chain
reaction-amplified genes coding for 16S rRNA. Applied and Environmental
Microbiology 59, 695–700.
Muyzer, G.; Teske, A.; Wirsen, C.O. & Jannasch, H.W. (1995). Phylogenetic
relationships of Thiomicrospira species and their identification in deep-sea
hydrothermal vent samples by denaturing gradient gel electrophoresis of 16S rDNA
fragments. Archives of Microbiology 164, 165–172.
Nič, M.; Jirát, J.; Košata, B.; Jenkins, A.; McNaught, A. (2009). IUPAC
Compendium of Chemical Terminology. Research Triagle Park, NC: IUPAC.
Oh, H.-W.; Kim, B.-C.; Park, D.-S.; Jeong, W.-J.; Kim, H.; Lee, K.H. & Kim, S.U.
(2013). Pedobacter luteus sp. nov., isolated from soil. International Journal of
Systematic and Evolutionary Microbiology 63, 1304–1310.

Literatur 125
Okuyama, H.; Sasaki, S.; Higashi, S. & Murata, N. (1990). A trans-unsaturated fatty-
acid in a psychrophilic bacterium, Vibrio sp. Strain ABE-1 Journal of Bacteriology, 172,
3515–3518.
Olsson, C.; Ahrné, S.; Pettersson, B. & Molin, G. (2003). The bacterial flora of fresh
and chill-stored pork: analysis by cloning and sequencing of 16S rRNA genes.
International Journal of Food Microbiology 83, 245–252.
Ourisson, G. & Nakatani, Y. (1994). The terpenoid theory of the origin of cellular life:
the evolution of terpenoids to cholesterol. Chemistry & biology 1, 11–23.
Palleroni, N. J.&Bradbury, J.F. (1993). Stenotrophomonas, a new bacterial genus for
Xanthomonas maltophilia (Hugh 1980) Swings et al. 1983. International Journal of
Systematic Bacteriology 43, 606–609.
Payza, A.N. & Korn, E.D. (1956). Bacterial degradation of heparin. Nature 177, 88–89.
Pintea, A.; Diehl, H. A.; Momeu, C.; Aberle, L.&Socaciu, C. (2005). Incorporation of
carotenoid esters into liposomes. Biophysical Chemistry 118, 7–14.
Quinn, P.J. (1981). The fluidity of cell membranes and its regulation. Progress in
biophysics and molecular biology 38, 1–104.
Rählert, N. (2007). Carotinoidbiosynthese in Flavobacterium spec P99-3.
Universitätsbibliothek Frankfurt
Ramos, E.; Ramírez-Bahena, M.-H.; Valverde, A.; Velázquez, E.; Zúñiga, D.;
Velezmoro, C. & Peix, A. (2013). Pseudomonas punonensis sp. nov., isolated from
straw. International Journal of Systematic and Evolutionary Microbiology 63, 1834–
1839.
Ratkowsky, D.A.; Olley, J.; McMeekin, TA. & Ball, A. (1982). Relationship between
temperature and growth rate of bacterial cultures. Journal of Bacteriology 149, 1–5.
Razin, S.; Kutner, S.; Efrati, H. & Rottem, S. (1980). Phospholipid and cholesterol
uptake by mycoplasma cells and membranes. Biochimica et Biophysica Acta (BBA)-
Biomembranes 598, 628–640.
Rezwan, M.; Lanéelle, M.-A.; Sander, P. & Daffé, M. (2007). Breaking down the wall:
fractionation of mycobacteria. Journal of Microbiological Methods 68, 32–39.
Rieser, G.; Scherer, S. & Wenning, M. (2013). Micrococcus cohnii sp. nov., isolated
from the air in a medical practice. International Journal of Systematic and Evolutionary
Microbiology 63, 80–85.

126 Literatur
Rheims, H.; Fruhling, A.; Schumann, P.; Rohde, M. & Stackebrandt, E. (1999).
Bacillus silvestris sp. nov., a new member of the genus Bacillus that contains lysine in
its cell wall. International Journal of Systematic Bacteriology 49, 795–802.
Rodriguez-Amaya, D.B. (2001). A guide to carotenoid analysis in foods: ILSI press
Washington, CDC.
Rothblat, G.H.; Ellis, D.S. & Kritchevsky, D. (1964). The carotenoid pigments of
Micrococcus lysodeikticus. Biochimica et Biophysica Acta (BBA)-Specialized Section
on Lipids and Related Subjects 84, 340–347.
Rottem, S. (1981). Cholesterol is required to prevent crystallization of Mycoplasma
arginini phospholipids at physiological temperature. FEBS Letters 133, 161–164.
Rottem,S.; Cirillo, V.P.; Kruyff, B. de; Shinitzky, M. & Razin, S. (1973). Cholesterol
in mycoplasm membranes. Correlation of enzymic and transport activities with physical
state of lipids in membranes of Mycoplasma mycoides var. capri adapted to grow with
low cholesterol concentrations. Biochimica et Biophysica Acta (BBA)-Biomembranes
323, 509–519.
Rucker, R.B.; Zempleni, J.; Suttie, J.W.; McCormick, D.B. (2010). Handbook of
vitamins: CRC Press.
Rudolph, A.S.; Crowe, J.H. & Crowe, L.M. (1986). Effects of three stabilizing
agents—proline, betaine, and trehalose—on membrane phospholipids. Archives of
biochemistry and biophysics 245, 134–143.
Russell, N.J. (1984). Mechanisms of thermal adaptation in bacteria: blueprints for
survival. Trends in Biochemical Sciences 9, 108–112.
Russell, N.J. (2002). Bacterial membranes: the effects of chill storage and food
processing. An overview. International Journal of Food Microbiology 79, 27–34.
Russell, N.J.; Harrisson, P.; Johnston, I.A.; Jaenicke, R.; Zuber, M.; Franks, F. &
Wynn-Williams, D. (1990). Cold adaptation of microorganisms Philosophical
Transactions of the Royal Society B: Biological Sciences 326, 595–611
Saitou, N. & Nei, M. (1987). The neighbor-joining method: a new method for
reconstructing phylogenetic trees. Molecular Biology and Evolution 4, 406–425.
Sanderson, P.W.; Lis, L. J.; Quinn, P.J. & Williams, W.P. (1991). The Hofmeister
effect in relation to membrane lipid phase stability. Biochimica et Biophysica Acta
(BBA)-Biomembranes 1067, 43–50.

Literatur 127
Sasser, M. (1990). Identification of bacteria by gas chromatography of cellular fatty
acids. Technical Note 101. Microbial Identification Applied Microbiology. 42, 457-469
Saunders, N.F.W.; Thomas, T.; Curmi, P. M. G.; Mattick, J. S.; Kuczek, E.; Slade, R.
et al. (2003). Mechanisms of thermal adaptation revealed from the genomes of the
Antarctic Archaea Methanogenium frigidum and Methanococcoides burtonii. Genome
Research 13, 1580–1588.
Scherer, S. & Neuhaus, K. (2006). Life at low temperatures. In: Martin Dworkin,
Stanley Falkow, Eugene Rosenberg, Karl-Heinz Schleifer und Erko Stackebrandt (Hg.):
The Prokaryotes. New York, NY: Springer New York, 210–262.
Schlegel, H.G.; Eitinger, T.; Fuchs, G. (2007). Allgemeine mikrobiologie: Georg
Thieme Verlag.
Schleifer, K.H. & Kloos, W.E. (1975). Isolation and characterization of Staphylococci
from human skin I. Amended descriptions of Staphylococcus epidermidis and
Staphylococcus saprophyticus and descriptions of three new species: Staphylococcus
cohnii, Staphylococcus haemolyticus, and Staphylococcus xylosus. International
Journal of Systematic Bacteriology 25, 50–61.
Shida, O.; Takagi, H.; Kadowaki, K.; Nakamura, L. K. & Komagata, K. (1997).
Transfer of Bacillus alginolyticus, Bacillus chondroitinus, Bacillus curdlanolyticus,
Bacillus glucanolyticus, Bacillus kobensis, and Bacillus thiaminolyticus to the genus
Paenibacillus and emended description of the genus Paenibacillus. International
Journal of Systematic Bacteriology 47, 289–298.
Shivaji, S. (2005). Pedobacter himalayensis sp. nov., from the Hamta glacier located in
the Himalayan mountain ranges of India. International Journal of Systematic and
Evolutionary Microbiology 55, 1083–1088.
Shivaji, S.; Kiran, M. D. & Chintalapati, S. (2007). Perception and transduction of low
temperature in bacteria. Physiology and biochemistry of extremophiles. ASM Press,
Washington, DC, 194–207.
Simon, C.; Wiezer, A.; Strittmatter, A.W. & Daniel, R. (2009). Phylogenetic diversity
and metabolic potential revealed in a glacier ice metagenome. Applied and
Environmental Microbiology 75, 7519–7526.
Sinensky, M. (1974). Homeoviscous Adaptation - a homeostatic process that regulates
the viscosity of membrane lipids in Escherichia coli. Proceedings of the National
Academy of Sciences 71, 522–525.

128 Literatur
Søballe, B. & Poole, R. K. (1999). Microbial ubiquinones: multiple roles in respiration,
gene regulation and oxidative stress management. Microbiology 145, 1817–1830.
Sousa, E.T.; da Silva, M.M.; de Andrade, S.J.; Cardoso, M.P.; Silva, L.A. & de
Andrade, J.B. (2012). Evaluation of thermal stability of quinones by thermal analysis
techniques. Thermochimica Acta 529, 1–5.
Stackebrandt, E. & Goebel, B. M. (1994). Taxonomic Note: a place for DNA-DNA
reassociation and 16S rRNA sequence analysis in the present species definition in
bacteriology. International Journal of Systematic Bacteriology 44, 846–849.
Steele, F.M. & Wright, K.H. (2001). Cooling rate effect on outgrowth of Clostridium
perfringens in cooked, ready-to-eat turkey breast roasts. Poultry Science 80, 813–816.
Stein, Jürgen (2003). Praxishandbuch klinische Ernährung und Infusionstherapie,
Springer DE.
Steyn, P.L.; Segers, P.; Vancanneyt, M.; Sandra, P.; Kersters, K. & Joubert, J.J.
(1998). Classification of heparinolytic bacteria into a new genus, Pedobacter,
comprising four species: Pedobacter heparinus comb. nov., Pedobacter piscium comb.
nov., Pedobacter africanus sp. nov. and Pedobacter saltans sp. nov. proposal of the
family Sphingobacteriaceae fam. nov. International Journal of Systematic Bacteriology
48, 165–177.
Storey, K.B. & Storey, J.M. (1986). Freeze tolerance and intolerance as strategies of
winter survival in terrestrially-hibernating amphibians. Comparative Biochemistry and
Physiology Part A: Physiology 83, 613–617.
Strand, A.; Shivaji, S. & Liaaen-Jensen, S. (1997). Bacterial carotenoids 55. C50-
carotenoids 25.† revised structures of carotenoids associated with membranes in
psychrotrophic Micrococcus roseus Biochemical systematics and ecology 25, 547–552.
Subczynski, W.K.; Markowska, E. & Sielewiesiuk, J. (1991). Effect of polar
carotenoids on the oxygen diffusion-concentration product in lipid bilayers. An EPR
spin label study. Biochimica et Biophysica Acta (BBA) - Biomembranes 1068, 68–72.
Subczynski, W.K.; Markowska, E.; Gruszecki, W.I. & Sielewiesiuk, J. (1992).
Effects of polar carotenoids on dimyristoylphosphatidylcholine membranes: a spin-label
study. Biochimica et Biophysica Acta (BBA)-Biomembranes 1105, 97–108.
Suutari, M. & Laakso, S. (1994). Microbial fatty acids and thermal adaptation. Critical
Reviews in Microbiology 20, 285–328.

Literatur 129
Swart-Füchtbauer, H. (1957). Zur Strahlenresistenz pigmentierter Bakterien. Archiv
für Mikrobiologie 26, 132–150.
Takeuchi, A.; Inman, L. R.; O'Hanley, P.D.; Cantey, J.R. & Lushbaugh, W.B.
(1978). Scanning and transmission electron microscopic study of Escherichia coli O15
(RDEC-1) enteric infection in rabbits. Infection and immunity 19, 686–694.
Takeuchi, M. & Hatano, K. (1998). Proposal of six new species in the genus
Microbacterium and transfer of Flavobacterium marinotypicum ZoBell and Upham to
the genus Microbacterium as Microbacterium maritypicum comb. nov. International
Journal of Systematic Bacteriology 48, 973–982.
Takeuchi, M. & Yokota, A. (1992). Proposals of Sphingobacterium faecium sp. nov.,
Sphingobacterium piscium sp. nov., Sphingobacterium heparinum comb. nov.,
Sphingobacterium thalpophilum comb. nov. and two genospecies of the genus
Sphingobacterium, and synonymy of Flavobacterium yabuuchiae and
Sphingobacterium spiritivorum. The Journal of General and Applied Microbiology 38,
465–482.
Tamura, K.; Dudley, J.; Nei, M. & Kumar, S. (2007). MEGA4: Molecular evolutionary
genetics analysis (MEGA) Software Version 4.0. Molecular Biology and Evolution 24,
1596–1599.
Tani, Y. (1989). Microbial Production of Vitamin K2 (Menaquinone) and Vitamin K1
(Phylloquinone). In: Biotechnology of Vitamins, Pigments and Growth Factors:
Springer, 123–134.
Tasara, T. & Stephan, R. (2006). Cold stress tolerance of Listeria monocytogenes: a
review of molecular adaptive mechanisms and food safety implications. Journal of
Food Protection 69, 1473–1484.
Tsukamoto, T.; Murata, H. & Shirata, A. (2002). Identification of non-pseudomonad
bacteria from fruit bodies of wild agaricales fungi that detoxify tolaasin produced by
Pseudomonas tolaasii. Bioscience, Biotechnology, and Biochemistry 66, 2201–2208.
United States Environmental Protection Agency, Washington, DC, USA. (2012).
Estimation Programs Interface Suite™. Washington, DC, USA.: US EPA.
Varkonyi, Z.; Masamoto, K.; Debreczeny, M.; Zsiros, O.; Ughy, B.; Gombos, Z. et
al. (2002). Low-temperature-induced accumulation of xanthophylls and its structural
consequences in the photosynthetic membranes of the cyanobacterium

130 Literatur
Cylindrospermopsis raciborskii: An FTIR spectroscopic study. Proceedings of the
National Academy of Sciences 99, 2410–2415.
Wada, M.; Fukunaga, N. & Sasaki, S. (1989). Mechanism of biosynthesis of
unsaturated fatty acids in Pseudomonas sp. strain E-3, a psychrotrophic bacterium.
Journal of Bacteriology 171, 4267–4271.
Warth, A.D. (1978). Relationship between the heat resistance of spores and the
optimum and maximum growth temperatures of Bacillus species. Journal of
Bacteriology 134, 699–705.
Watanuki, M. & Aida, K.O. (1972). Significance of quinones in the classification of
bacteria. The Journal of General and Applied Microbiology 18, 469–472.
Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.; Kandler, O.; Krichevsky,
M.I. et al. (1987). Report of the ad hoc committee on reconciliation of approaches to
bacterial systematics. International Journal of Systematic Bacteriology 37, 463–464.
White, G.F.; Racher, K.I.; Lipski, A.; Hallett, F. & Wood, J. M. (2000). Physical
properties of liposomes and proteoliposomes prepared from Escherichia coli polar
lipids. Biochimica et Biophysica Acta (BBA) - Biomembranes 1468, 175–186.
Wieser, M.; Denner, E.B.M.; Kämpfer, P.; Schumann, P.; Tindall, B.; Steiner, U. et
al. (2002). Emended descriptions of the genus Micrococcus, Micrococcus luteus (Cohn
1872) and Micrococcus lylae (Kloos et al. 1974). International Journal of Systematic
and Evolutionary Microbiology 52, 629–637.
Xu, J.-L.; He, J.; Wang, Z.-C.; Wang, K.; Li, W.-J.; Tang, S.-K. & Li, S.-P. (2007).
Rhodococcus qingshengii sp. nov., a carbendazim-degrading bacterium. International
Journal of Systematic and Evolutionary Microbiology 57, 2754–2757.
Yabuuchi, E. & Yano, I. (1981). Achromobacter gen. nov. and Achromobacter
xylosoxidans (ex Yabuuchi and Ohyama 1971) nom. rev. International Journal of
Systematic Bacteriology 31, 477–478.
Yoon, M.-H.; Ten, L.N.; Im, W.-T. & Lee, S.-T. (2007). Pedobacter panaciterrae sp.
nov., isolated from soil in South Korea. International Journal of Systematic and
Evolutionary Microbiology 57, 381–386.
Yu, Z.; Chang, M.; Wu, M.; Yang, G.; Zhou, S. & Zhuang, L. (2013). Pseudomonas
oryzae sp. nov. isolated from a paddy soil in South China. Archives of Microbiology
195, 815–822.

Literatur 131
Yu, L.; Lai, Q.; Yi, Z.; Zhang, L.; Huang, Y.; Gu, L. & Tang, X. (2013).
Microbacterium sediminis sp. nov., a psychrotolerant, thermotolerant, halotolerant and
alkalitolerant actinomycete isolated from deep-sea sediment. International Journal of
Systematic and Evolutionary Microbiology 63, 25–30.
Zelles, L. (1999). Fatty acid patterns of phospholipids and lipopolysaccharides in the
characterisation of microbial communities in soil: a review. Biology and Fertility of Soils
29, 111–129.
Zhao, T.; Ezeike, G.O.I.; Doyle, M. P.; Hung, Y.-C. & Howell, R.S. (2003). Reduction
of Campylobacter jejuni on poultry by low-temperature treatment. Journal of Food
Protection 66, 652–655.
Zhou, Z.; Jiang, F.; Wang, S.; Peng, F.; Dai, J.; Li, W. & Fang, C. (2012).
Pedobacter arcticus sp. nov., a facultative psychrophile isolated from Arctic soil, and
emended descriptions of the genus Pedobacter, Pedobacter heparinus, Pedobacter
daechungensis, Pedobacter terricola, Pedobacter glucosidilyticus and Pedobacter
lentus. International Journal of Systematic and Evolutionary Microbiology 62, 1963–
1969.
Ziemke, F.; Höfle, M. G.; Lalucat, J. & Rossellö-Mora, R. (1998). Reclassification of
Shewanella putrefaciens Owen's genomic group II as Shewanella baltica sp. nov.
International Journal of Systematic Bacteriology 48, 179–186.
Zscheile, F.P. & White J.W. (1940). Microscope hot stage for determination of melting
points: application to carotenoid pigments. Industrial & Engineering Chemistry
Analytical Edition 12, 436–438.

7. Anhang
Tabelle 15: Fettsäuremuster aller Isolate bei 10°C und 30°C; Angaben in %
J1
J1
J2
J2
J3
J3
J5
J5
J7
J7
J8
J8
J9
J9
J10
J10
J11
J11
J12
J12
10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 10°C 30°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 10°C 30°c 30°C
10:0
0,00 0,05
11:0 iso
11:0 anteiso
11:0 3OH
12:0
1,90 5,30
12:0 cis 5
12:0 2OH
12:0 3OH
0,00 0,81
13:0
13:0 iso 76,14 9,32
68,38 25,22 0,00 1,26
13:0 anteiso 0,00 0,18
5,06 1,22
13:0 iso 3OH
14:0 iso 0,00 2,19
0,13 2,21
5,74 2,16 1,99 2,11
0,00 0,68 1,43 1,33 0,75 0,68
14:0 3OH
14:1 trans 2
14:0
0,00 0,12
0,00 1,15
1,18 0,0 1,23 0,97 0,28 0,18
15:0 iso 7,75 39,68
5,65 19,37 35,93 45,67 33,12 33,98 54,91 52,44 12,70 45,71 2,74 9,11 55,15 67,44 5,00 7,16 3,12 2,88 3,89 3,66
15:0 anteiso 0,00 1,91
93,91 75,48 46,64 54,33 4,58 5,00 10,85 10,6 0,00 5,20 76,53 52,70 44,85 29,44 93,41 91,60 70,46 71,96 71,75 74,33
15:0 2OH
15:0 iso 3OH
15:1 cis 10
15:0
0,00 0,00 0,99 0,66
16:1 cis 10 bzw. 16:1
trans 9
6,91 7,60
0,00 9,16
16:1 cis 9
54,89 42,90
0,41 0,00
16:1 cis 11
16:1 c/t 5 4,43 1,05
2,95 0,13 2,72 2,80
11,52 10,45 2,16 1,64
16:1 iso 5
35,76 33,64 15,03 15,63
3,59 3,78 0,33 0,16
16:0 iso 2OH
16:0 3OH
16:0 iso 0,00 5,55
0,25 2,49 1,71 0,00 9,8 10,97 2,88 3,3 0,00 0,50
0,00 0,57 4,28 4,22 9,91 9,63
16:0 4,77 9,37 14,34 32,37
1,70 0,00 4,16 4,61
1,56 1,2 3,11 2,86
17:1 iso cis 9
17:1 iso cis 6 bzw. cis
9
17:1 iso 5 0,00 9,53
5,9 5,31 4,73 5,13 0,00 1,07 7,35 22,70 0,00 1,48
1,15 1,39 0,00 0,00
17:1 (iso?) 10
0,00 4,92 0,00 1,53
17:1 anteiso cis 5 0,00 1,58
3,19 2,75 3,74 4,02 0,00 0,24 3,89 0,00
17:0 iso 3OH
17:0 cyclo
17:0 iso 0,00 11,45
6,93 0,00
0,00 1,82 0,00 1,50
0,39 0,32
17:0 anteiso 0,00 0,67
5,95 0,00
1,66 1,79 6,42 5,87
17:0
18:1 cis 7
4,29 3,07
18:1 cis 9
19,88 13,70
18:1 cis 11
8,99 4,28
18:1 trans 9 bzw. cis
11
18:0
0,00 0,44
130
An
ha
ng
J16 J17 J20 J21 J22 J23 J34 J39 J40
10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C
10:0
11:0 iso 1,59
4,13
11:0 anteiso
11:0 3OH
12:0
12:0 cis 5
12:0 2OH
12:0 3OH
13:0
13:0 iso
13:0 anteiso
13:0 iso 3OH 0,00 2,45
14:0 iso 0,00 0,68
0,52 0,36
14:0 3OH
0,99 1,81
14:1 trans 2
14:0 1,18 0,00 3,06 3,61 2,2
2,81
4,57 3,11 3,31 1,2 5,82 2,01 0,65
15:0 iso 5,00 7,16 20,03 42,43 42,9 37,98 9,62 14,87
15:0 anteiso 93,41 91,60 100 100 10,99 12,45 5,69 89,64 84,66 94,49 98,1
15:0 2OH
15:0 iso 3OH
1,58 2,6
15:1 cis 10
15:0 1,09 0,00
16:1 cis 10 bzw. 16:1 trans 9 31,32 12,81 6,29 0,00
16:1 cis 9 1,49 0,00 32,47 12,36 31,84 17,31 53,73 19,3
16:1 cis 11
6,99
16:1 c/t 5 3,99 0,00
16:1 iso 5
16:0 iso 2OH
16:0 3OH
2 3,94
16:0 iso
0,54 0,26
16:0 10,6 15,94
6,77 29,3 28,67 49,09 52,29 81,56
17:1 iso cis 9 1,49
2,51
17:1 iso cis 6 bzw. cis9
17:1 iso 5
17:1 (iso?) 10
17:1 anteiso cis 5
17:0 iso 3OH
2,23 4,31
17:0 cyclo 0,00 2,27 1,4 22,28
17:0 iso 0,69 2,64
17:0 anteiso
17:0
18:1 cis 7
18:1 cis 9 4,92 0,00
18:1 cis 11 4,42 0,00 12,1 4,22 13,08 2,36
18:1 trans 9 bzw. cis 11
18:0
19:0 cyclo 11-12
33,43 10,26
Anhang
13
1
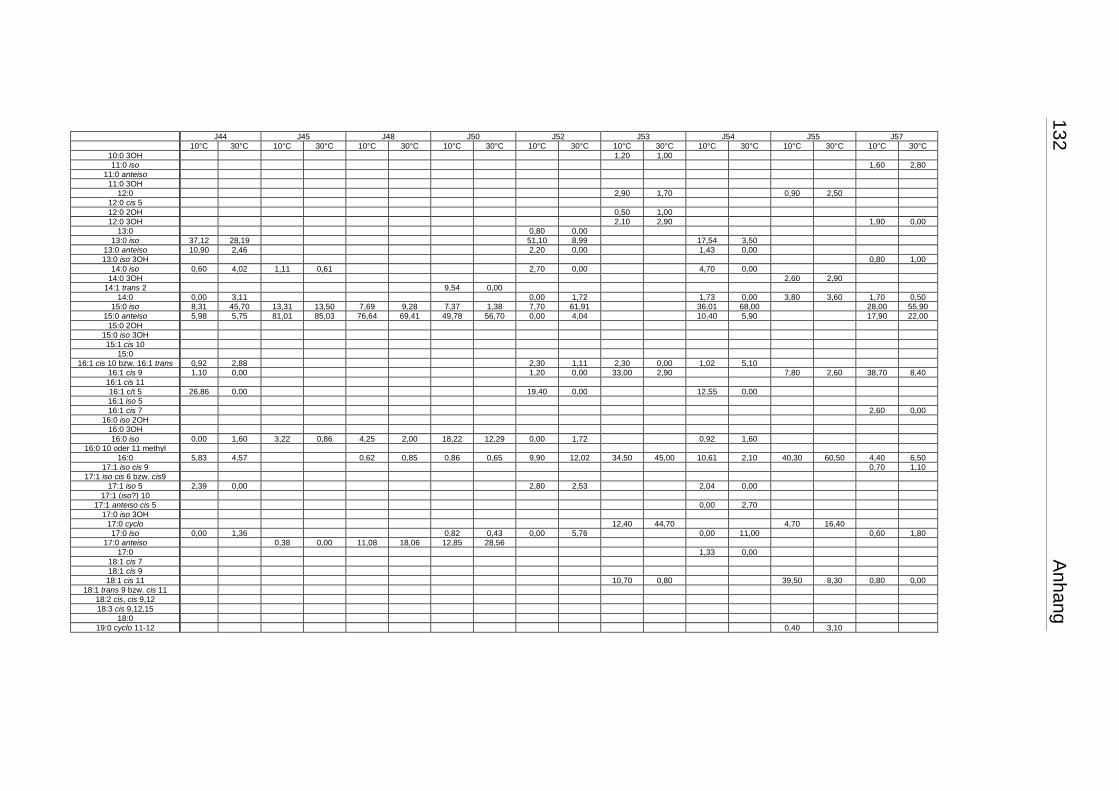
J44
J44
J45
J45
J48
J48
J50
J50
J52
J52
J53
J53
J54
J54
J55
J55
J57
J57
10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C
10:0 3OH
1,20 1,00
11:0 iso
1,60 2,80
11:0 anteiso
11:0 3OH
12:0
2,90 1,70
0,90 2,50
12:0 cis 5
12:0 2OH
0,50 1,00
12:0 3OH
2,10 2,90
1,90 0,00
13:0
0,80 0,00
13:0 iso 37,12 28,19
51,10 8,99
17,54 3,50
13:0 anteiso 10,90 2,46
2,20 0,00
1,43 0,00
13:0 iso 3OH
0,80 1,00
14:0 iso 0,60 4,02 1,11 0,61
2,70 0,00
4,70 0,00
14:0 3OH
2,60 2,90
14:1 trans 2
9,54 0,00
14:0 0,00 3,11
0,00 1,72
1,73 0,00 3,80 3,60 1,70 0,50
15:0 iso 8,31 45,70 13,31 13,50 7,69 9,28 7,37 1,38 7,70 61,91
36,01 68,00
28,00 55,90
15:0 anteiso 5,98 5,75 81,01 85,03 76,64 69,41 49,78 56,70 0,00 4,04
10,40 5,90
17,90 22,00
15:0 2OH
15:0 iso 3OH
15:1 cis 10
15:0
16:1 cis 10 bzw. 16:1 trans
9
0,92 2,88
2,30 1,11 2,30 0,00 1,02 5,10
16:1 cis 9 1,10 0,00
1,20 0,00 33,00 2,90
7,80 2,60 38,70 8,40
16:1 cis 11
16:1 c/t 5 26,86 0,00
19,40 0,00
12,55 0,00
16:1 iso 5
16:1 cis 7
2,60 0,00
16:0 iso 2OH
16:0 3OH
16:0 iso 0,00 1,60 3,22 0,86 4,25 2,00 18,22 12,29 0,00 1,72
0,92 1,60
16:0 10 oder 11 methyl
16:0 5,83 4,57
0,62 0,85 0,86 0,65 9,90 12,02 34,50 45,00 10,61 2,10 40,30 60,50 4,40 6,50
17:1 iso cis 9
0,70 1,10
17:1 iso cis 6 bzw. cis9
17:1 iso 5 2,39 0,00
2,80 2,53
2,04 0,00
17:1 (iso?) 10
17:1 anteiso cis 5
0,00 2,70
17:0 iso 3OH
17:0 cyclo
12,40 44,70
4,70 16,40
17:0 iso 0,00 1,36
0,82 0,43 0,00 5,76
0,00 11,00
0,60 1,80
17:0 anteiso
0,38 0,00 11,08 18,06 12,85 28,56
17:0
1,33 0,00
18:1 cis 7
18:1 cis 9
18:1 cis 11
10,70 0,80
39,50 8,30 0,80 0,00
18:1 trans 9 bzw. cis 11
18:2 cis, cis 9,12
18:3 cis 9,12,15
18:0
19:0 cyclo 11-12
0,40 3,10
132
An
ha
ng
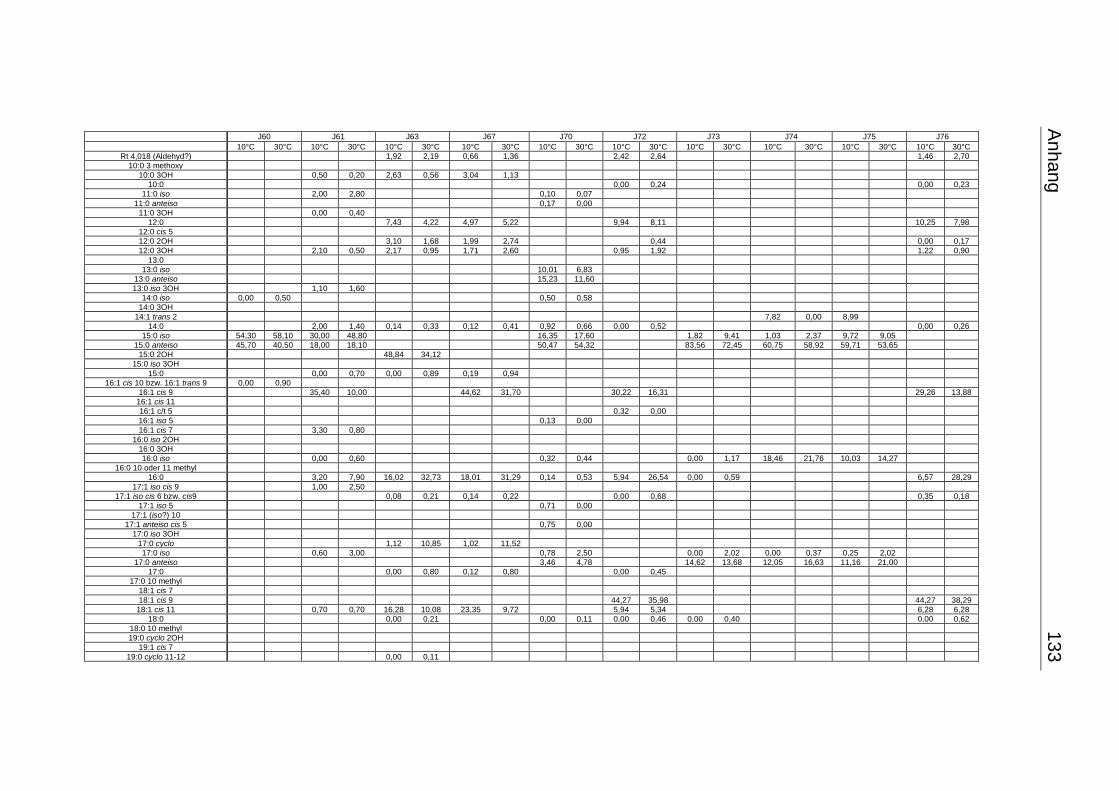
J60
J60
J61
J61
J63
J63
J67
J67
J70
J70
J72
J72
J73
J73
J74
J74
J75
J75
J76
J76
10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C 10°C 30°C
Rt 4,018 (Aldehyd?)
1,92 2,19 0,66 1,36
2,42 2,64
1,46 2,70
10:0 3 methoxy
10:0 3OH
0,50 0,20 2,63 0,56 3,04 1,13
10:0
0,00 0,24
0,00 0,23
11:0 iso
2,00 2,80
0,10 0,07
11:0 anteiso
0,17 0,00
11:0 3OH
0,00 0,40
12:0
7,43 4,22 4,97 5,22
9,94 8,11
10,25 7,98
12:0 cis 5
12:0 2OH
3,10 1,68 1,99 2,74
0,44
0,00 0,17
12:0 3OH
2,10 0,50 2,17 0,95 1,71 2,60
0,95 1,92
1,22 0,90
13:0
13:0 iso
10,01 6,83
13:0 anteiso
15,23 11,60
13:0 iso 3OH
1,10 1,60
14:0 iso 0,00 0,50
0,50 0,58
14:0 3OH
14:1 trans 2
7,82 0,00 8,99
14:0
2,00 1,40 0,14 0,33 0,12 0,41 0,92 0,66 0,00 0,52
0,00 0,26
15:0 iso 54,30 58,10 30,00 48,80
16,35 17,60
1,82 9,41 1,03 2,37 9,72 9,05
15:0 anteiso 45,70 40,50 18,00 18,10
50,47 54,32
83,56 72,45 60,75 58,92 59,71 53,65
15:0 2OH
48,84 34,12
15:0 iso 3OH
15:0
0,00 0,70 0,00 0,89 0,19 0,94
16:1 cis 10 bzw. 16:1 trans 9 0,00 0,90
16:1 cis 9
35,40 10,00
44,62 31,70
30,22 16,31
29,26 13,88
16:1 cis 11
16:1 c/t 5
0,32 0,00
16:1 iso 5
0,13 0,00
16:1 cis 7
3,30 0,80
16:0 iso 2OH
16:0 3OH
16:0 iso
0,00 0,60
0,32 0,44
0,00 1,17 18,46 21,76 10,03 14,27
16:0 10 oder 11 methyl
16:0
3,20 7,90 16,02 32,73 18,01 31,29 0,14 0,53 5,94 26,54 0,00 0,59
6,57 28,29
17:1 iso cis 9
1,00 2,50
17:1 iso cis 6 bzw. cis9
0,08 0,21 0,14 0,22
0,00 0,68
0,35 0,18
17:1 iso 5
0,71 0,00
17:1 (iso?) 10
17:1 anteiso cis 5
0,75 0,00
17:0 iso 3OH
17:0 cyclo
1,12 10,85 1,02 11,52
17:0 iso
0,60 3,00
0,78 2,50
0,00 2,02 0,00 0,37 0,25 2,02
17:0 anteiso
3,46 4,78
14,62 13,68 12,05 16,63 11,16 21,00
17:0
0,00 0,80 0,12 0,80
0,00 0,45
17:0 10 methyl
18:1 cis 7
18:1 cis 9
44,27 35,98
44,27 38,29
18:1 cis 11
0,70 0,70 16,28 10,08 23,35 9,72
5,94 5,34
6,28 6,28
18:0
0,00 0,21
0,00 0,11 0,00 0,46 0,00 0,40
0,00 0,62
18:0 10 methyl
19:0 cyclo 2OH
19:1 cis 7
19:0 cyclo 11-12 0,00 0,11
Anhang
133
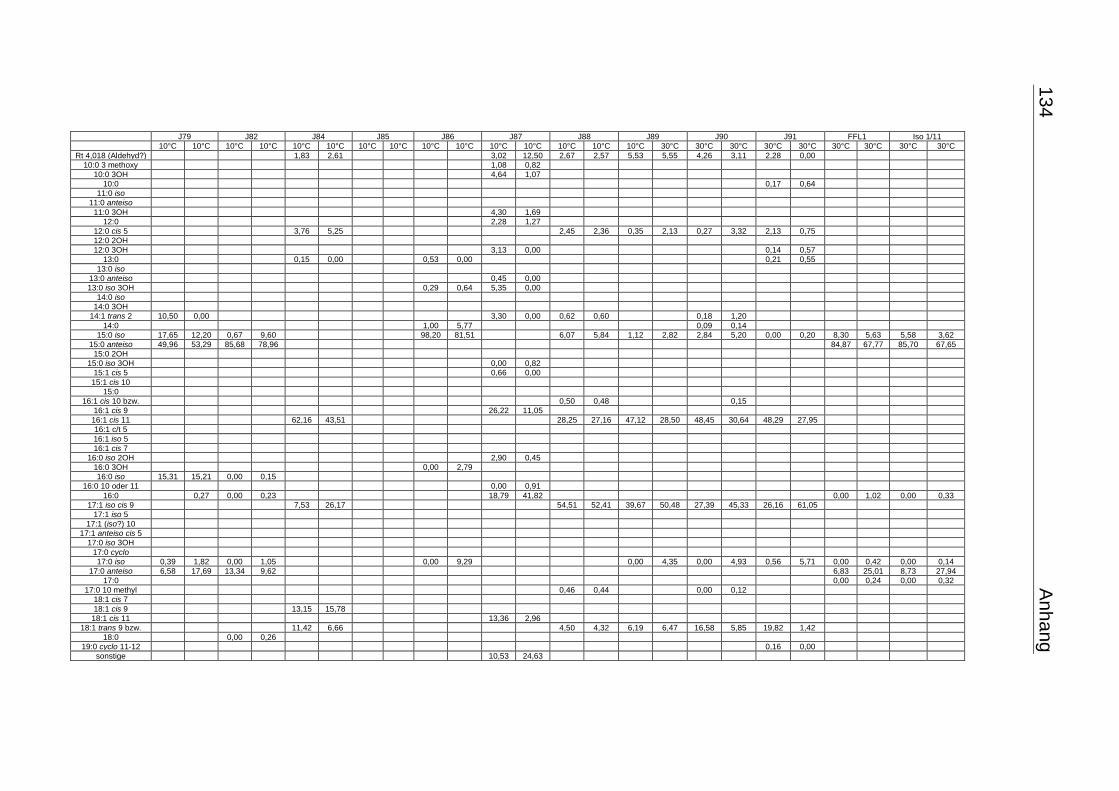
J79
J79
J82
J82
J82
J82
J84
J84
J84
J84
J85
J85
J85
J85
J86
J86
J86
J86
J87
J87
J87
J87
J88
J88
J88
J88
J89
J89
J89
J89
J90
J90
J90
J90
J91
J91
J91
J91
FFL1
FFL1
FFL1
FFL1
Iso 1/11
Iso 1/11
Iso 1/11
Iso 1/11
10°C 10°C 10°C 10°C 10°C 10°C 10°C 10°C 10°C 10°C 10°C 10°C 10°C 10°C 10°C 30°C 30°C 30°C 30°C 30°C 30°C 30°C 30°C 30°C
Rt 4,018 (Aldehyd?)
1,83 2,61
3,02 12,50 2,67 2,57 5,53 5,55 4,26 3,11 2,28 0,00
10:0 3 methoxy
1,08 0,82
10:0 3OH
4,64 1,07
10:0
0,17 0,64
11:0 iso
11:0 anteiso
11:0 3OH
4,30 1,69
12:0
2,28 1,27
12:0 cis 5
3,76 5,25
2,45 2,36 0,35 2,13 0,27 3,32 2,13 0,75
12:0 2OH
12:0 3OH
3,13 0,00
0,14 0,57
13:0
0,15 0,00
0,53 0,00
0,21 0,55
13:0 iso
13:0 anteiso
0,45 0,00
13:0 iso 3OH
0,29 0,64 5,35 0,00
14:0 iso
14:0 3OH
14:1 trans 2 10,50 0,00
3,30 0,00 0,62 0,60
0,18 1,20
14:0
1,00 5,77
0,09 0,14
15:0 iso 17,65 12,20 0,67 9,60
98,20 81,51
6,07 5,84 1,12 2,82 2,84 5,20 0,00 0,20 8,30 5,63 5,58 3,62
15:0 anteiso 49,96 53,29 85,68 78,96
84,87 67,77 85,70 67,65
15:0 2OH
15:0 iso 3OH
0,00 0,82
15:1 cis 5
0,66 0,00
15:1 cis 10
15:0
16:1 cis 10 bzw.
16:1 trans 9
0,50 0,48
0,15
16:1 cis 9
26,22 11,05
16:1 cis 11
62,16 43,51
28,25 27,16 47,12 28,50 48,45 30,64 48,29 27,95
16:1 c/t 5
16:1 iso 5
16:1 cis 7
16:0 iso 2OH
2,90 0,45
16:0 3OH
0,00 2,79
16:0 iso 15,31 15,21 0,00 0,15
16:0 10 oder 11
methyl
0,00 0,91
16:0
0,27 0,00 0,23
18,79 41,82
0,00 1,02 0,00 0,33
17:1 iso cis 9
7,53 26,17
54,51 52,41 39,67 50,48 27,39 45,33 26,16 61,05
17:1 iso 5
17:1 (iso?) 10
17:1 anteiso cis 5
17:0 iso 3OH
17:0 cyclo
17:0 iso 0,39 1,82 0,00 1,05
0,00 9,29
0,00 4,35 0,00 4,93 0,56 5,71 0,00 0,42 0,00 0,14
17:0 anteiso 6,58 17,69 13,34 9,62
6,83 25,01 8,73 27,94
17:0
0,00 0,24 0,00 0,32
17:0 10 methyl
0,46 0,44
0,00 0,12
18:1 cis 7
18:1 cis 9
13,15 15,78
18:1 cis 11
13,36 2,96
18:1 trans 9 bzw.
cis 11
11,42 6,66
4,50 4,32 6,19 6,47 16,58 5,85 19,82 1,42
18:0
0,00 0,26
19:0 cyclo 11-12
0,16 0,00
sonstige
10,53 24,63
134
A
nhang
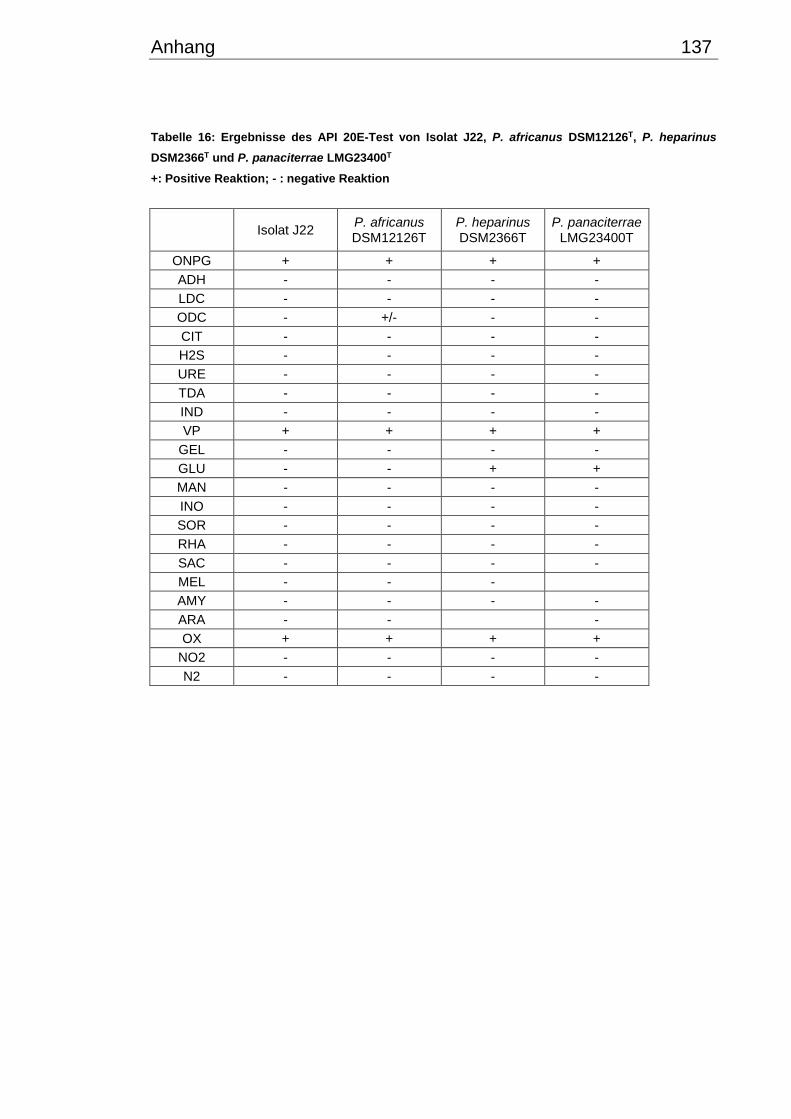
Anhang 137
Tabelle 16: Ergebnisse des API 20E-Test von Isolat J22, P. africanus DSM12126T, P. heparinus
DSM2366T und P. panaciterrae LMG23400T
+: Positive Reaktion; - : negative Reaktion
Isolat J22
P. africanus DSM12126T
P. heparinus DSM2366T
P. panaciterrae LMG23400T
ONPG + + + +
ADH - - - -
LDC - - - -
ODC - +/- - -
CIT - - - -
H2S - - - -
URE - - - -
TDA - - - -
IND - - - -
VP + + + +
GEL - - - -
GLU - - + +
MAN - - - -
INO - - - -
SOR - - - -
RHA - - - -
SAC - - - -
MEL - - -
AMY - - - -
ARA - -
-
OX + + + +
NO2 - - - -
N2 - - - -
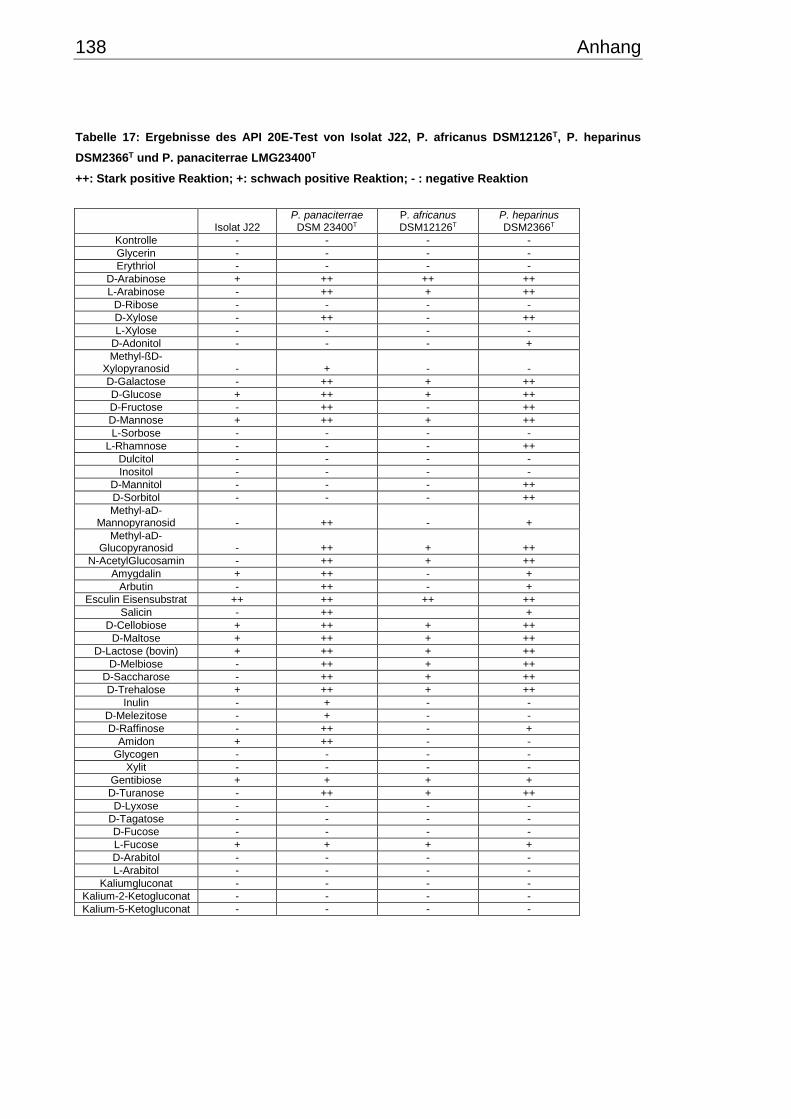
138 Anhang Anhang
Tabelle 17: Ergebnisse des API 20E-Test von Isolat J22, P. africanus DSM12126T, P. heparinus
DSM2366T und P. panaciterrae LMG23400T
++: Stark positive Reaktion; +: schwach positive Reaktion; - : negative Reaktion
Isolat J22 P. panaciterrae DSM 23400T
P. africanus DSM12126T
P. heparinus DSM2366T
Kontrolle - - - -
Glycerin - - - -
Erythriol - - - -
D-Arabinose + ++ ++ ++
L-Arabinose - ++ + ++
D-Ribose - - - -
D-Xylose - ++ - ++
L-Xylose - - - -
D-Adonitol - - - +
Methyl-ßD-Xylopyranosid - + - -
D-Galactose - ++ + ++
D-Glucose + ++ + ++
D-Fructose - ++ - ++
D-Mannose + ++ + ++
L-Sorbose - - - -
L-Rhamnose - - - ++
Dulcitol - - - -
Inositol - - - -
D-Mannitol - - - ++
D-Sorbitol - - - ++
Methyl-aD-Mannopyranosid - ++ - +
Methyl-aD-Glucopyranosid - ++ + ++
N-AcetylGlucosamin - ++ + ++
Amygdalin + ++ - +
Arbutin - ++ - +
Esculin Eisensubstrat ++ ++ ++ ++
Salicin - ++
+
D-Cellobiose + ++ + ++
D-Maltose + ++ + ++
D-Lactose (bovin) + ++ + ++
D-Melbiose - ++ + ++
D-Saccharose - ++ + ++
D-Trehalose + ++ + ++
Inulin - + - -
D-Melezitose - + - -
D-Raffinose - ++ - +
Amidon + ++ - -
Glycogen - - - -
Xylit - - - -
Gentibiose + + + +
D-Turanose - ++ + ++
D-Lyxose - - - -
D-Tagatose - - - -
D-Fucose - - - -
L-Fucose + + + +
D-Arabitol - - - -
L-Arabitol - - - -
Kaliumgluconat - - - -
Kalium-2-Ketogluconat - - - -
Kalium-5-Ketogluconat - - - -
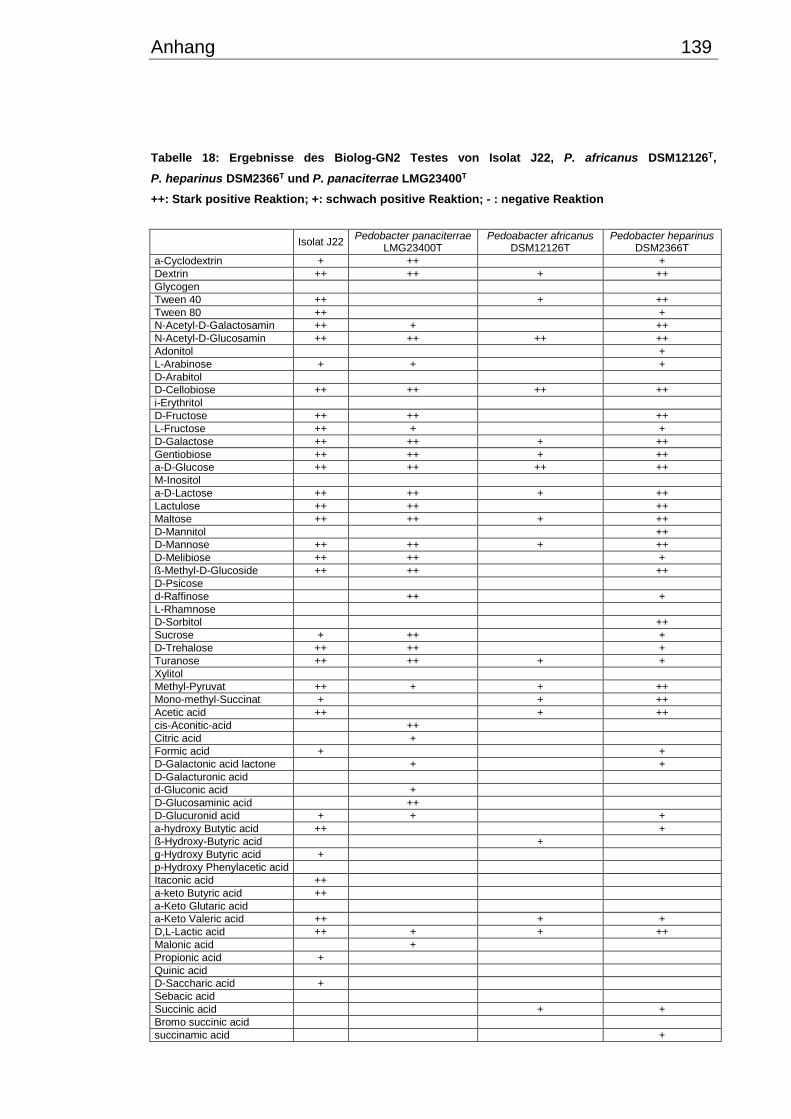
Anhang 139
Tabelle 18: Ergebnisse des Biolog-GN2 Testes von Isolat J22, P. africanus DSM12126T,
P. heparinus DSM2366T und P. panaciterrae LMG23400T
++: Stark positive Reaktion; +: schwach positive Reaktion; - : negative Reaktion
Isolat J22
Pedobacter panaciterrae LMG23400T
Pedoabacter africanus DSM12126T
Pedobacter heparinus DSM2366T
a-Cyclodextrin + ++ +
Dextrin ++ ++ + ++
Glycogen
Tween 40 ++ + ++
Tween 80 ++ +
N-Acetyl-D-Galactosamin ++ + ++
N-Acetyl-D-Glucosamin ++ ++ ++ ++
Adonitol +
L-Arabinose + + +
D-Arabitol
D-Cellobiose ++ ++ ++ ++
i-Erythritol
D-Fructose ++ ++ ++
L-Fructose ++ + +
D-Galactose ++ ++ + ++
Gentiobiose ++ ++ + ++
a-D-Glucose ++ ++ ++ ++
M-Inositol
a-D-Lactose ++ ++ + ++
Lactulose ++ ++ ++
Maltose ++ ++ + ++
D-Mannitol ++
D-Mannose ++ ++ + ++
D-Melibiose ++ ++ +
ß-Methyl-D-Glucoside ++ ++ ++
D-Psicose
d-Raffinose ++ +
L-Rhamnose
D-Sorbitol ++
Sucrose + ++ +
D-Trehalose ++ ++ +
Turanose ++ ++ + +
Xylitol
Methyl-Pyruvat ++ + + ++
Mono-methyl-Succinat + + ++
Acetic acid ++ + ++
cis-Aconitic-acid ++
Citric acid +
Formic acid + +
D-Galactonic acid lactone + +
D-Galacturonic acid
d-Gluconic acid +
D-Glucosaminic acid ++
D-Glucuronid acid + + +
a-hydroxy Butytic acid ++ +
ß-Hydroxy-Butyric acid +
g-Hydroxy Butyric acid +
p-Hydroxy Phenylacetic acid
Itaconic acid ++
a-keto Butyric acid ++
a-Keto Glutaric acid
a-Keto Valeric acid ++ + +
D,L-Lactic acid ++ + + ++
Malonic acid +
Propionic acid +
Quinic acid
D-Saccharic acid +
Sebacic acid
Succinic acid + +
Bromo succinic acid
succinamic acid +
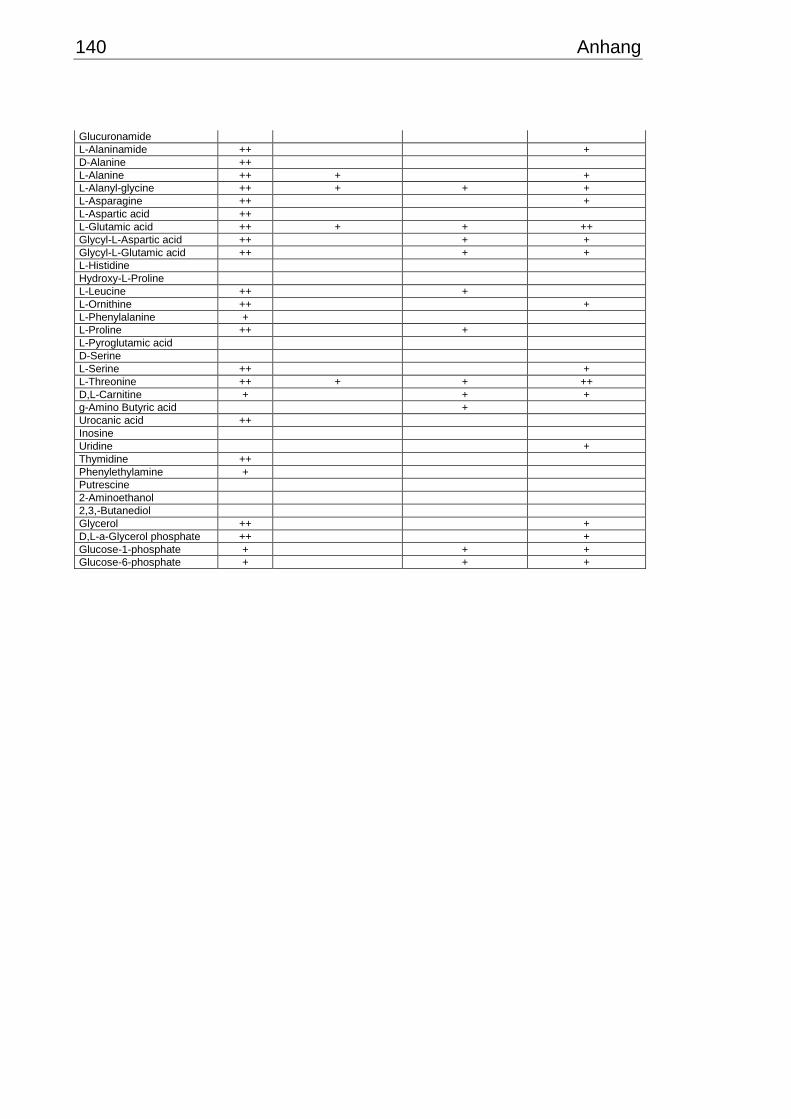
140 Anhang Anhang
Glucuronamide
L-Alaninamide ++ +
D-Alanine ++
L-Alanine ++ + +
L-Alanyl-glycine ++ + + +
L-Asparagine ++ +
L-Aspartic acid ++
L-Glutamic acid ++ + + ++
Glycyl-L-Aspartic acid ++ + +
Glycyl-L-Glutamic acid ++ + +
L-Histidine
Hydroxy-L-Proline
L-Leucine ++ +
L-Ornithine ++ +
L-Phenylalanine +
L-Proline ++ +
L-Pyroglutamic acid
D-Serine
L-Serine ++ +
L-Threonine ++ + + ++
D,L-Carnitine + + +
g-Amino Butyric acid +
Urocanic acid ++
Inosine
Uridine +
Thymidine ++
Phenylethylamine +
Putrescine
2-Aminoethanol
2,3,-Butanediol
Glycerol ++ +
D,L-a-Glycerol phosphate ++ +
Glucose-1-phosphate + + +
Glucose-6-phosphate + + +
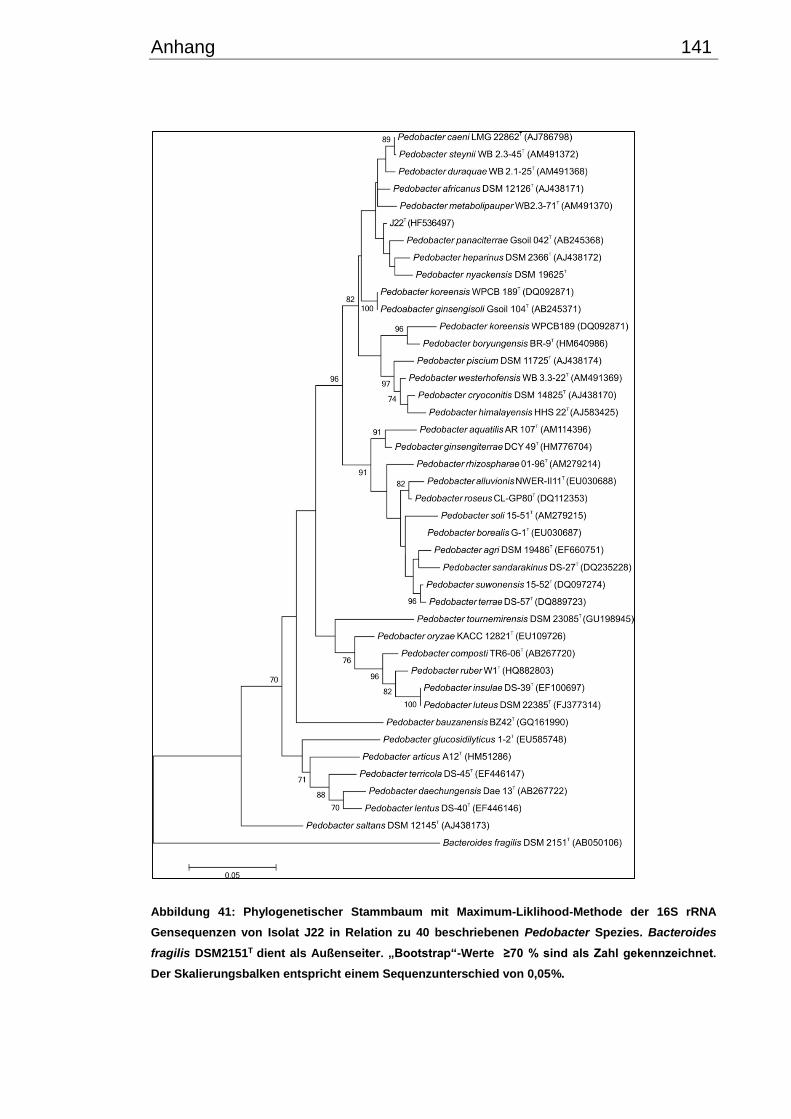
Anhang 141
Abbildung 41: Phylogenetischer Stammbaum mit Maximum-Liklihood-Methode der 16S rRNA
Gensequenzen von Isolat J22 in Relation zu 40 beschriebenen Pedobacter Spezies. Bacteroides
fragilis DSM2151T dient als Außenseiter. „Bootstrap“-Werte ≥70 % sind als Zahl gekennzeichnet.
Der Skalierungsbalken entspricht einem Sequenzunterschied von 0,05%.

142 Anhang Anhang
Abbildung 42: Phylogenetischer Stammbaum mit Maximum-Parsimony-Methode der 16S rRNA
Gensequenzen von Isolat J22 in Relation zu 40 beschriebenen Pedobacter Spezies. Bacteroides
fragilis DSM2151T dient als Außenseiter.

Danksagung 143
8. Danksagung
An dieser Stelle möchte ich mich bei allen bedanken, die mich während der letzten
Jahre mit Rat und Tat unterstützt haben:
Als erstes gilt mein großer Dank Prof. Dr. André Lipski für die Bereitstellung des
Themas, das entgegengebrachte Vertrauen und die wissenschaftliche Betreuung.
Außerdem möchte ich mich bei ihm für das Ermöglichen mehrerer Kongressteilnahmen
zur Veröffentlichung der Studienergebnisse und natürlich für die Unterstützung
bedanken.
Herr Prof. Dr. Andreas Schieber möchte ich herzlich für die Übernahme des
Korreferats danken.
Ein besonders herzlicher Dank gilt Eva Meyer. Bei Ihr möchte ich von Herzen für die
enorme Unterstützung und Hilfsbereitschaft, all die vielen schönen Pausen und die
unzähligen Gespräche während aller Höhen und Tiefen bedanken. Gleiches gilt für
Mareike Weber. Bei Ihr möchte ich mich für ihre (vor allem im Endspurt) enorme
Unterstützung in allen Bereichen der Arbeit, die unzähligen wissenschaftlichen und
nicht-wissenschaftlichen Gespräche ☺ und das Korrekturlesen der Arbeit danken.
Dieser Dank gilt natürlich auch Beate Silkens.
Frau Paula Nöthen danke ich für die schöne Zusammenarbeit und die vielen netten
und lustigen Gespräche.
Ich danke allen im Laufe der Zeit wechselnden Mitgliedern meiner Arbeitsgruppe für
die freundschaftliche und amüsante Zusammenarbeit. Ein besonders Dankeschön gilt
hierbei Nina Müller, Nicole Breuer, Rommy Schmidt, Sandra Szczepanski, Alina Meyer,
Jan Sieber, Daniela Reekers, Charlotte Weiß, Jonas Verbeek, Julian Weber, Caroline
Fritsch, Dorothee Murr, Marina Jacobsen und Waldemar Seel.
Herrn Dr. Benno Zimmermann danke ich für seine hilfreichen Ratschläge bei der
Carotinoidbestimmung.
Ich danke allen Mitarbeiterinnen und Mitarbeitern des Instituts für Ernährungs- und
Lebensmittelwissenschaften und dem Institut für Mikrobiologie und Biotechnologie.
Ganz zum Schluss möchte ich mich bei meiner Familie und vor allem bei meinem
Mann Marcus für die unendliche, liebevolle Unterstützung in allen Lebenslagen und
Euren festen Glaube an mich und meine Fähigkeiten bedanken.















![Modulhandbuch Wissenschaftszentrum Weihenstephan fur ... · Mikrobiologie / Organismische und Molekulare Mikrobiologie (Microbiology / Organismic and Molecular Microbiology) [WZ2503]](https://img.pdfslide.org/doc/110x75/5dd09306d6be591ccb61a6c0/modulhandbuch-wissenschaftszentrum-weihenstephan-fur-mikrobiologie-organismische.jpg)













