Embed Size (px)
Citation preview

Aus dem Tierarztlichen Institut der Universitat Gottingen Direktor: Prof. Dr. E . Mitscherlich
Untersuchungen iiber die Ubertragbarkeit eines in leukotischem Tumormaterial vom Rind auftretenden
Agens auf weiBe Mause
Von
E. MITSCHERLICH
(Eingegangen am 3. Janrar 1969)
Die Erforschung der ktiologie und Epiderniologie der Rinderleukose triff t aus verschiedenen Griinden auf groi3e Schwierigkeiten. Es steht fur derar- tige Untersuchungen kein kleines Versuchstier zur Verfugung. Es ist bislang kein Kulturverfahren bekannt geworden, bei dem das Agens auf andere Weise als durch den langwierigen und kostspieligen Ruckiibertragungsversuch auf das Rind hatte nachgewiesen werden konnen (MONTEMAGNO, 1967). Es ist bis- her nicht gelungen, im Blute erkrankter Rinder erregerspezifische Abwehrstoff e nachzuweisen (LEHNERT, 1964; SCHMIDT, 1968). Die Leukose kann somit zur Zeit nur hamatologisch aufgrund der persistierenden Lymphozytose und morphologisch aufgrund der charakteristischen Organveranderungen diagnosti- ziert werden. Beide Verfahren erfassen wahrscheinlich nicht die Anfangssta- dien der Erkrankung. Experimentelle Untersuchungen uber die Ktiologie und Epidemiologie der Seuche lassen sich demzufolge nur am Rinde selbst durch- fiihren. Finanzielle Grunde beschranken diese Versuche allseitig auf ein unzu- reichendes Mass. Aus dieser Situation heraus schien der Versuch gerechtfertigt, nach anderen Wegen zum Nachweis des Erregers der Rinderleukose zu suchen.
In ausgedehnten Vorversuchen gelang es nicht, im Serum von Rindern, die an der enzootsichen Form der Rinderleukose erkrankt waren, mit Hilfe der Komplementbindungsreaktion oder dem Agargeldiffusionstest nach OUCHTERLONY erregerspezifische Antikorper nachzuweisen, obwohl der Modus der Reaktionen in der verschiedensten Weise variiert wurde. Als Antigene wurden verschiedenartige Extrakte sowie Kern- und Mitochondrienfraktionen aus leukotischem Tumorgew,ebe vom Rind oder auch Extrakte aus den Orga- nen von Mausen verwandt, auf die zuvor leukotisches Tumorgewebe vom Rind iibertragen worden war. Bei diesen Versuchen ergab sich, dai3 in den Lebern derart vorbehandelter Mause in einem wenige Tage umfassenden Zeit- raum, etwa 3 Tage p. i. beginnend, eine hamolysierende Substanz auftrat, die in den folgenden Ausfuhrungen als Hamolysin bezeichnet wird. Die vor- liegende Arbeit befaflt sich mit diesem Phanomen. Sie gliedert sich in die Ab- schnitte
l) Die Arbeit, die sich iiber 6 Jahre erstreckte, wurde in dankenswerter Weise durch eine finanzielle Unterstutzung der Milchwirtschaftlichen Arbeitsgemeinschaft Hannover- Braunschweig e. V. und des Herrn Niedersachsischen Mininsters fur Ernahrung, Landwirt- schaft und Forsten ermoglicht.

518 E. MITSCHERLICH
1. Nachweis des Hamolysins. 2. Art des Hamolysins. 3. Auftreten des Hamolysins in den Lebern von Mausen nach der intra-
abdominalen Injektion von Rinderleukosematerial. 3.1 Grundversuch 3.2 Auswertung gehaufter Hamolysinbefunde. 3.3 Wiederholung des Grundversuches.
4. Untersuchungen iiber die Bestandigkeit des mit Rinderleukosematerial mutmafllich auf Mause iibertragenen Agens, das in deren Lebern zur Ausbildung des Hamolysins fiihrt.
5. Versuche zur Neutralisation des betreffenden Agens mit dem Blut von einem leukosefreien und zwei leukoseinfizierten Rindern.
6. Riickiibertragung auf Rinder von hamolysinhaltigen Mauselebern aus Leukoseiibertragungsversuchen.
7. Diskussion 8. Zusammenfassung
1. Nachweis des Hamolysins Tiermaterial. Als Versuchstiere dienten 20-25 g schwere mannliche
NMRI-Mause. Ober diesen Stamm liegen im hiesigen Institut besonders hin- sichtlich des spontanen Auftretens von Adenokarzinomen und Leukamien um-
TOLLE, SCHMIDT u. SCHIMMELPFENNIG, 1967). Die Mause wurden von der Gesellschaft f i i v Versuchstierkunde in Hannover-Linden bezogen und spates- tens einen Tag nach ihrem Eintrefien im Institut in den Versuch genommen. Sie wurden mit einem Standardfutter der Fa. Altromin in Lage (Lippe) ge- futtert und fur die Dauer des jeweiligen Versuches zu 5 Tieren in Makrolon- kafigen gehalten.
Untersuchung dev Lebern auf Hamolysin. Die Mause wurden durch Genickschlag getotet, dekapitiert und vollig entblutet. Das Blut wurde zur Gewinnung der Erythrozyten in einer Natriumcitrat-Losung2) aufgefangen. Bei jeder Maus wurde die Leber bei der Sektion herausgenommen und zu einem Teil fur spatere Versuche bei - 70° C eingefroren. Der andere Teil wurde gewogen und nach Zugabe der vierfachen Menge (GewJVol.) Phosphat- puff erlosungs) mit einem elektrisch angetriebenen Glashomogenisator zu einer feinen Suspension verarbeitet. Die Suspensionen wurden anschlieflend 30 Mi- nuten in einem Ultraschallgerat bei 200 mA beschallt und waren damit zur Untersuchung, die stets am Totungstage erfolgte, fertig.
Zum Ansetzen des Testes wurden mit der Natriumcitrat-Losung 5 Rohr- chen beschickt, von denen das erste 0,75 ccm und die folgenden je 0,5 ccm er- hielten. In das erste Rohrchen wurde 0,25 ccm der zu untersuchenden Leber- suspension hinzugefiigt und nunmehr jeweils nach sorgfaltigem Durchmischen 0,5 ccm von Rohrchen zu Rohrchen iiberpipettiert. Aus dem letzten Rohrchen wurde 0,5 ccm entfernt. Damit enthielt jedes Rohrchen 0,5 ccm Lebersuspen- sion, deren Konzentration im ersten Rohrchen 1 : 20 und in den folgenden 1 : 40, 1 : 80, 1 : 160 und 1 : 320 betrug. Hierauf wurden in jedes Rohrchen 0,5 ccm einer 2 O/oigen Aufschwemmung von dreimal gewaschenen Schaf- oder Mauseerythrozyten in Natriumcitrat-Losung hinzugegeben.
z, Zusammensetzung der Natriumcitrat-Losung: tri-Natriumcitrat-2-hydrat 200,O; Natriumchlorid 50,O; Natriumcyanid 1,6; Aqua dest ad 10000,O. Die Losung wurde mit 10 O/oiger HCl auf p H 7,2 eingestellt.
s, Zusammensetzung der Phosphatpufferlosung: Natriumchlorid 17,6; KH,PO, 0,25; Na,HPO, * 2 H,O 3,25; Aqua dest. ad 2000,O.
fangreiche Erfahrungen VOn (SCHREIER U. SCHIMMELPFENNIG, 1965 ; SCHREIER,

Untersuchungen uber die Obertragbarkeit eines Agens auf weide Mause 519
Die Reaktionszeit betrug alternativ 5 Stunden bei 37' C im Wasserbad oder 18 Stunden bei Zimmertemperatur. Vielfach wurden die Teste parallel sowohl rnit der einen als auch mit der anderen Reaktionszeit angesetzt. Wah- rend der Reaktionszeit wurden die Rohrchen mehrmals aufgeschuttelt. Zuin Ablesen der Reaktionen wurden sie nach kurzem Aufschutteh abzentrifugiert.
Die Reaktion galt als positiv, wenn eine wolle Lyse der Erythrozyten eingetreten war: bei einer Bebrutungszeit von 5 Stunden bei 37' C wenigstens in dem 1. Rohr- chen (Verdunnung 1 : 20) und bei einer Reaktionszeit von 18 Stunden bei Zimmertemperatur wenigstens in den ersten beiden Rohrchen (Verdunnung 1 : 20 und 1 : 40).
Die bei der Untersuchung der Lebersuspensionen erreichte maximale Titerhohe betrug 1 : 640. Prozonenphanomene wurden nicht beobachtet.
Es ist wichtig, dafl das Ansetzen des Testes am Totungstage erfolgt. In Lebern, die in der Gefriertruhe aufbewahrt werden, laufen die enzymatischen Prozesse weiter. Dabei kommt es haufig zu einer Steigerung, mitunter aber auch zu einem Abfall des Hamolysingehaltes. Die Ergebnisse, die rnit eingefro- renen Lebern oder auch Lebersuspensionen erhalten wurden, stimmten deshalb oft nicht mit denen uberein, die mit frischem Material erzielt wurden. Im folgenden wird nur uber Ergebnisse mit frischem Material berichtet.
Die zum Ansetzen des Testes verwandte Natriumcitrat-Losung wurde unter mehreren vergleichend angewandten Losungen gewahlt, weil der bei der Hamolyse freiwerdende rote Blutfarbstoff in ihr wahrend der Reaktionszeit farblich nicht verandert wird, und die Losung eine bakteriostatische Wirkung besitzt.
2. Art des Hamolysins Das Hamolysin 1ai3t sich aus positiv reagierenden Mauselebern nach einem
Verfahren extrahieren, das von MARINETTI und Mitarbeiter (1959) fur die Extraktion von Lysolecithin angegeben wurde. Es nutzt die Tatsache aus, dai3 Lysolecithin in Methylalkohol und Chloroform loslich, in Aether und Aceton hingegen unloslich ist (DEUEL, 195 1). Das untersuchte Hamolysin verhalt sich gleichartig. Zur Extraktion wird 1 ccm der mit Phosphatpufferlosung bereite- ten 20 O/oigen Lebersuspension lyophilisiert und anschliei3end mit 12 ccm er- warmtem Methanol (60 O C) und 4 ccm Chloroform 2 Minuten lang bei 60 O C extrahiert. Die Suspension wird abzentrifugiert, der Oberstand aufgehoben und der Bodensatz noch zweimal mit je 10 ccm einer Mischung von Chloro- form und Methylalkohol ZZ jeweils 2 Minuten lang bei 60° C extrahiert. Nach jeder Extraktion werden die Rohrchen abzentrifugiert. Die 3 auf diese Weise erhaltenen Extrakte werden vereint, durch Zentrifugieren geklart, mit 3 ccm Benzol versetzt und bei 40 O C getrocknet. Der trockene Ruckstand wird in 2 ccm Phosphatpuff erlosung aufgenommen. Es entsteht hierbei eine homo- gene, leicht milchige Emulsion. Diese Emulsion wird im Hamolysetest genau so ausgewertet wie Lebersuspensionen.
Wird der zum Test verwandten 2 O/oigen Schaferythrozytensuspension Rinderserumalbumin zugefugt, so erfolgt eine Hemmung der Hamolyse. Diese Hemmung kann bei Sstundiger Bebrutung im Wasserbad bei 37' C gerade noch vollstandig sein, wenn 0,5 ccm einer Emulsion, die Hamolysin in ein- facher Titerdosis enthalt, mit 0,5 ccm einer 2 O/oigen Schaferythrozytensuspen- sion zusammengebracht wird, der zuvor O,I O / o Rinderserumalbumin zuge- fugt wurde. KLIBANSKY und De VRIES (1963) haben darauf hingewiesen, dai3 das gleiche fur Lysolecithin zutrifft. Sie zeigten, dafl an die Membran von
Zbl. Vet. Med., Reihe B, Bd. 16, Heft 6 14

520 E. MITSCHERLICH
Erythrozyten gebundenes Lysolecithin wieder abgelost werden kann, wenn die Erythrozyten in einer physiologischen Kochsalzlosung gewaschen werden, die 4,s O / o menschliches Serumalbumin enthalt. Menschliches Fibrinogen, gamma- Globulin und Hamoglobin dagegen besitzen diese Hemmwirkung nicht.
Unter der Einwirkung des Hamolysins erfolgt eine Formveranderung der Erythrozyten. Sie blahen sich auf und nehmen dabei zunachst die Gestalt eines eingebeulten Balles, dann die einer vollen Kugel an, um schliei3lich einen schei- benformigen, langsam vergehenden Schatten zu hinterlassen. Dieselben Form- veranderungen erleiden Erythrozyten unter der Einwirkung von Lysolecithin. Nach KLIBANSKY und DE VRIes (1 963) besteht eine direkte Abhangigkeit zwischen der in dem Medium vorhandenen Lysolecithinkonzentration, der an die Erythrozytenmembran gebundenen Lysolecithinmenge und der Form bzw. Lysis der Erythrozyten.
Das gleichartige Verhalten von Hamolysin und Lysolecithin gegeniiber Methylalkohol, Chloroform, Aether, Aceton und Rinderserumalbumin sowie die gleichartigen Formveranderungen der Erythrozyten unter der Einwirkung von Hamolysin und Lysolecithin sprechen dafiir, dai3 beide Substanzen nahe miteinander verwandt oder identisch sind.
Von GROSS (1947, 1948 a, 1948 b) wurde in 10-20 O/oigen Tumorsuspen- sionen von Adenokarzinomen der Mause eine hamolysierende Substanz nach- gewiesen. Fur sie war kennzeichnend, dai3 sie nach dem Abzentrifugieren der Gewebssuspensionen auch in den iiberstehenden zellfreien Extrakten vorhan- den war und dai3 sie in ganz spezifischer Weise mit Mauseerythrozyten rea- gierte jedoch nicht rnit Erythrozyten von Mensch, Kaninchen oder Meer- schweinchen. Die in den vorliegend beschriebenen Versuchen wirksame Substanz lief3 sich in 10 Minuten bei 4000U/min mit den zelli en Bestandteilen der Lebersuspensionen abzentrifugieren. In dem zellfreien 8berstand war sie da- nach nicht mehr enthalten. Sie reagierte zudem nicht nur mit Mauseerythrozy- ten sondern auch rnit Meerschweinchen-, Kaninchen- und Schaferythrozyten. Sie wird deshalb als nicht identisch mit der von GROSS gefunden Substanz an- gesehen.
3. Auftreten des Himolysins in den Lebern von Mausen nach der intraabdominalen In jektion von Rinderleukosematerial
Den Untersuchungen liegt die Frage zugrunde, ob zwischen der intraabdo- minalen Injektion von Rinderleukosematerial in Mause und dem Auftreten des Hamolysins in deren Lebern eine Beziehung besteht. Hierzu sol1 zunachst eiii Grundversuch beschrieben werden, dessen Ergebnis diese Frage berechtigt er- scheinen lie&. Es folgt die Auswertung gehaufter Hamolysinbefunde in den Lebern der Versuchstiere in Hinblick auf die Fragestellung. Im Anschlui3 daran werden Wiederholungen des Grundversuches beschrieben.
3.1 Grundversuch Leukotisches Tumormaterial vom Rind war wenige Stunden nach der
Schlachtung bei - 30' C eingefroren worden. 52 Tage spater wurde aus ihm mit physiologischer Kochsalzlosung eine 20 O/oige Suspension bereitet. Sie wurde am gleichen Tage zum Ansetzen der 1. Mausepassage verwandt.
1. Passage. Von 100 Mausen wurden 10 als Kontrollen getotet, den ubri- gen 90 wurden je Tier 0,3 ccm der Suspension, das sind etwa 0,06 g leuko- tisches Rindertumorgewebe, intraabdominal injiziert. Die Tiere zeigten im Anschlui3 an die Injektion keinerlei Krankheitserscheinungen. Vom 3. Tage p. inoc. ab wurden von den Versuchstieren taglich 10 Tiere getotet. Ihre Lebern sowie die Lebern der Kontrolltiere wurden an dem jeweiligen Totungstage auf
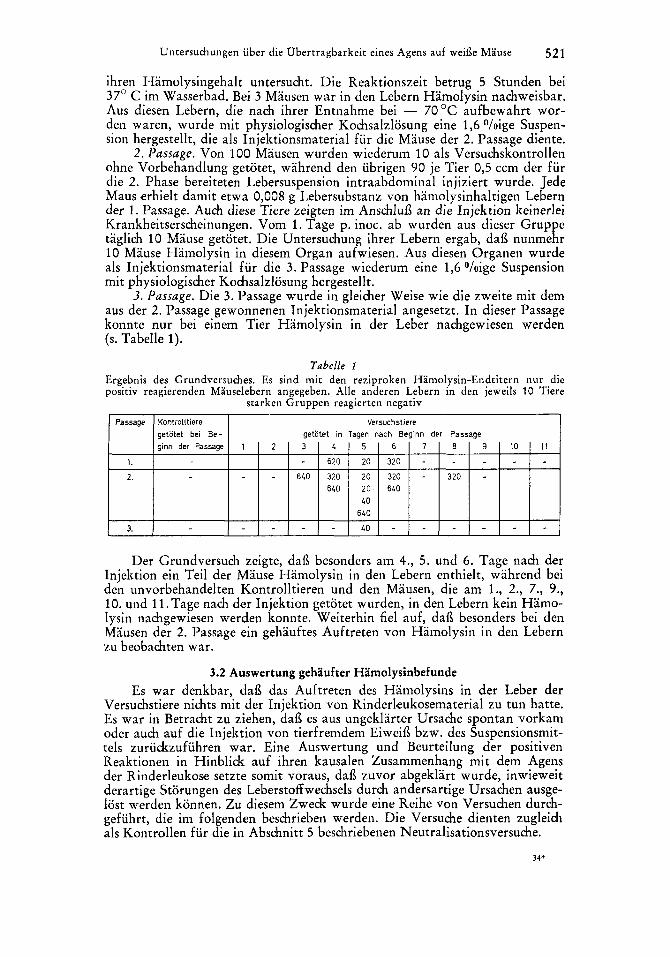
Untersuchungen iiber die Obertragbarkeit eines Agens auf weide Mause 521
Passage Kontrolltiere Versuchstiere
1.
2.
3.
Der Grundversuch zeigte, dai3 beson Iniektion ein Teil der Mause Hamolysin
ginn der Passage 1 2 3 4
- 620
- 640 320 640
5 6
20 320
20 320 20 640 LO 640
LO -
lers am 4., 5 . und 6 . Tage nach der n den Lebern enthielt. wahrend bei
7 8 9 10 I 1 - - 320 -
deh unvorbehandelten Kontrolltiere; und den Mausen, die am' l., 2., 7., 9., 10. und 11. Tage nach d.er Injektion getotet wurden, in den Lebern kein Hamo- lysin nachgewiesen werden konnte. Weiterhin fie1 auf, dai3 besonders bei den Mausen der 2 . Passage eiii gehauftes Auftreten von Hamolysin in den Leberii zu beobachten war.
3.2 Auswertung gehiufter Himolysinbefunde Es war denkbar, dai3 das Auftreten des Hamolysins in der Leber der
Versuchstiere nichts mit der Injektion von Kinderkukosematerial zu tun hatte. Es war in Betracht zu ziehen, dai3 es aus ungeklarter Ursache spontan vorkam oder auch auf die Injektion von tierfremdem Eiweifl bzw. des Suspensionsmit- tels zuruckzufiihren war. Eine Auswertung und Beurteilung der positiven Reaktionen in Hinblick auf ihren kausalen Zusammenhang mit dem Agens der Rinderleukose setzte somit voraus, dai3 zuvor abgeklart wurde, inwieweit derartige Storungen des Leberstofiwechsels durch andersartige Ursachen ausge- lost werden konnen. Zu diesem Zweck wurde eine Reihe von Versuchen durch- gefuhrt, die im folgenden beschrieben werden. Die Versuche dienten zugleich als Kontrollen fur die in Abschnitt 5 beschriebenen Neutralisationsversuche.
34*
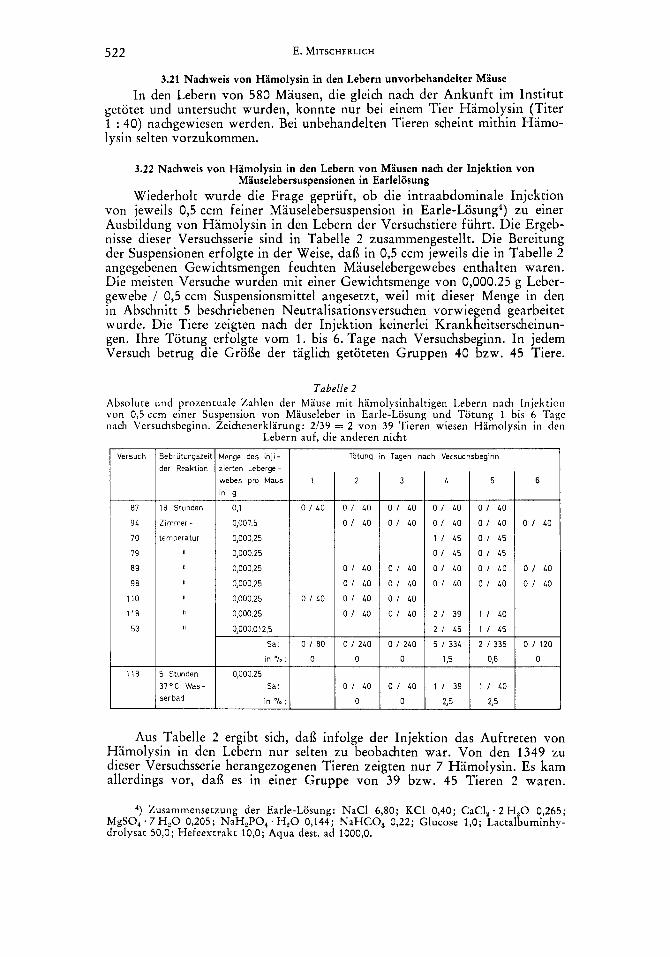
522 E. MITSCHERLICH
3.21 Nachweis von Himolysin in den Lebern unvorbehandelter Mluse In den Lebern von 580 Mausen, die gleich nach der Ankunft im Institut
getotet und untersucht wurden, konnte nur bei einem Tier Hamolysin (Titer 1 : 40) nachgewiesen werden. Bei unbehandelten Tieren scheint mithin Hamo- lysin selten vorzukommen.
3.22 Nachweis von Hamolysin in den Lebern von Mausen nach der Injektion von Mauselebersuspensionen in Earlelosung
Wiederholt wurde die Frage gepriift, ob die intraabdominale Injektion von jeweils 0,5 ccm feiner Mauselebersuspension in Earle-Losung4) zu einer Ausbildung von Hamolysin in den Lebern der Versuchstiere fuhrt. Die Ergeb- nisse dieser Versuchsserie sind in Tabelle 2 zusammengestellt. Die Bereitung der Suspensionen erfolgte in der Weise, dai3 in 0,5 ccm jeweils die in Tabelle 2 angegebenen Gewichtsmengen feuchten Mauselebergewebes enthalten waren. Die meisten Versuche wurden mit einer Gewichtsmenge von 0,000.25 g Leber- gewebe / 0,5 ccm Suspensionsmittel angesetzt, weil mit dieser Menge in den in Abschnitt 5 beschriebenen Neutralisationsversuchen vorwiegend gearbeitet wurde. Die Tiere zeigten nach der Injektion keinerlei Krankheitserscheinun- gen. Ihre Totung erfolgte vom 1. bis 6. Tage nach Versuchsbeginn. In jedem Versuch betrug die Groi3e der taglich getoteten Gruppen 40 bzw. 45 Tiere.
Tabelle 2 Absolute und prozentuale Zahlen der Mause mit hamolysinhaltigen Lebern nach Injektion von 0,s ccm einer Suspension von Mauseleber in Earle-Losung und Totung 1 bis 6 Tagc nach Versuchsbeginn. Zeichenerklarung: 2/39 = 2 von 39 Tieren wiesen Hamolysin in den
Lebern auf, die anderen nicht
Versuch
87
9 L
70
79
89
98
110
118
53
118
-
3ebrutungszeit 3er Reaktion
I 8 Stunden
Zimrner - emperatur
11
I1
II
I/
I/
I1
5 Stunden 37OC Was- ser bad
lenge des in j i - ierten Leberge - vebes pro Maus
7 9
0,l
0,007.5
0,000.25
0,000.25
0,000.25
0,000.25
0.000.25
0,000.25
0,000.012.5
Sa:
in '1. :
0,000.2s Sa:
in 'lo :
1
0 I LO
0 I LO
0 I 8 0
0
Totung in Tagen nach Versuchsbeginn
2
0 1 40
0 1 LO
0 1 LO
0 1 LO
0 1 40
0 1 LO
0 I 2 L O
0
0 1 LO
0
0 1 LO
0 1 LO
0 1 LO
0 1 LO
0 1 LO
0 1 LO
0 I 260 T O I L 0 1 / 3 9 1 / 4 0
6
0 1 LO
0 1 LO
0 1 LO
0 I 120
0
Aus Tabelle 2 ergibt sich, dai3 infolge der Injektion das Auftreten von Hiimolysin in den Lebern nur selten zu beobachten war. Von den 1349 ZLI dieser Versuchsserie herangezogenen Tieren zeigten nur 7 Hamolysin. Es kam allerdings vor, dai3 es in einer Gruppe von 39 bzw. 45 Tieren 2 waren.
4, Zusarnmensetzung der Earle-Losung: NaCl 6,80; KCI 0,40; CaCI, . 2 H,O 0,265; MgSO, .7 H,O 0,205; NaH,PO, . H,O 0,144; NaHCO, 0,22; Glucose 1,O; Lactalbuminhy- drolysat 50,O; Hefeextrakt 10,O; Aqua dest. ad 1000,O.
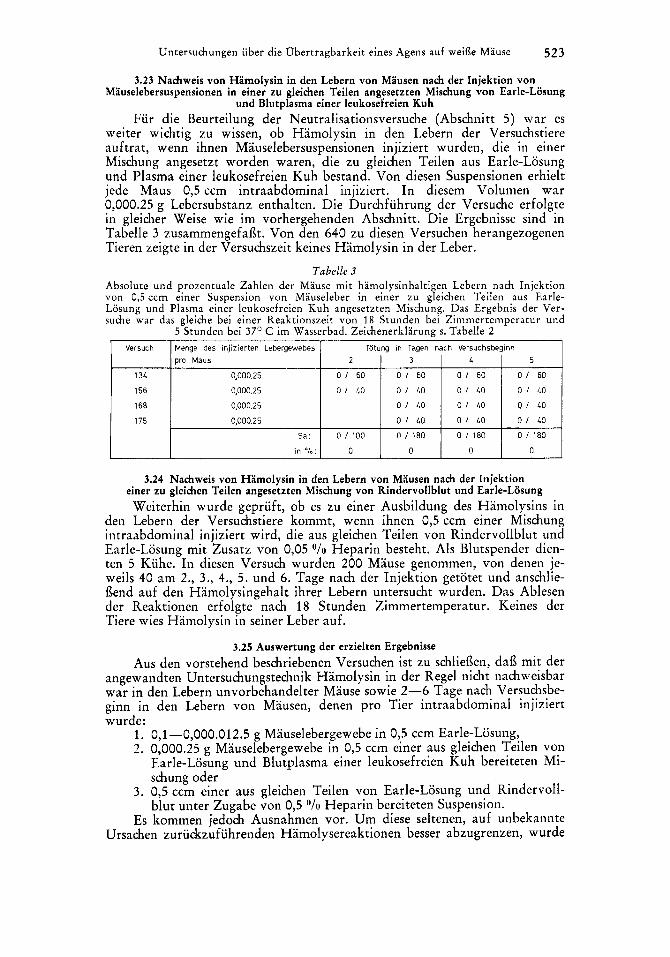
Untersuchungen uber die Obertragbarkeit eines Agens auf weilk Mause 523
Versuch Menge des injizierten Lebergewebes Totung in Tagen nach Versuchsbeginn pro Maus 2 1 3 1 4 1 5
I34
IS6 168
175
0,000.25 0 1 60 0 1 60 0 1 60 0 1 60
0,000.25 0 1 40 0 I L O 0 1 40 0 1 40
0,000.25 0 1 40 0 1 40 0 1 40
0,000.25 0 1 40 0 1 40 0 1 LO
Sa: 0 1 1 0 0 0 I 180 0 I 180 0 I 1 8 0
in % : 0 0 0 0

E. MITSCHERLICH 524
in den folgenden Versuchen so vorgegangen, dai3 stets mit Gruppen von 30 bzw. 40 Tieren gearbeitet wurde. Fur die Beurteilung der Versuche wurde das Gesamtergebnis aller Tiere einer Gruppe herangezogen. Es wurde dabei beurteilt ein nachweisbarer Hamloysingehalt der Leber bei
0- 3,4 O / o aller Tiere als: - 3,5- 7,4 O / o aller Tiere als: +/- 7,5-14,9 O/o aller Tiere ah: +
15,O-29,9 O/o aller Tiere als: + + uber 30,O O/o aller Tiere als: + + +
3.3 Wiederholung des Grundversuches In 8 weiteren Versuchen wurde nunmehr versucht, die Ergebnisse des
Grundversuches zu reproduzieren und zu erweitern. Die Resultate dieser Versuche sind in Tabelle 4 zusammengefai3t.
Das zu diesen Versuchen verwandte leukotische Tumormaterial stammte von 19 Kuhen. Es wurde entweder wenige Stunden nach der Schlachtung so- fort verarbeitet oder bei - 30' C eingefroren.
Im einzelnen betrug die Aufbewahrungszeit des Tumormateriales in der Gefriertruhe vor seiner Verarbeitung im 1. Versuch 4 Tage-5 Monate, im 2 . Versuch 1 Tag-3 Jahre, im 3. Versuch 14 Tage--13 Monate, im 4. Versuch 70 Tage-4 Monate, im 5. Versuch 14 Tage, im 6. Versuch 10-58 Tage, im 7. Versuch 29 Tage und im 8. Versuch wurde es frisch verarbeitet.
Aus dem Tumormaterial wurde mit Hilfe eines elektrisch betriebenen Glashomogenisators mit Earle-Losung eine 10-20 O/oige Grundsuspension hergestellt. Jede Versuchsmaus erhielt von dieser Suspension 0,5 ccm intraabb- dominal unjiziert.
Die Mause wurden vom 2. bis 6. Tage nach der Injektion taglich in Grup- pen von mehreren Tieren getotet. Sie zeigten bis dahin keinerlei Krankheits- erscheinungen. Ihre Lebern wurden am Totungstage auf Hamolysin untersucht. Sofort nach der Totung wurde von jeder Maus ein groi3eres Leberstuck ent- nommen und fur weitere Passagen in verschlossenen Glasrohrchen bei - 70' C aufbewahrt. Passagiert wurde Lebermaterial, das positive Hamolysereaktio- lien ausgelost hatte. Ausgehend von einer 20 O/oigen Grundsuspension wurde dieses Material so stark weiterverdunnt, dai3 in dem fur jede Maus der folgen- den Passage bestimmten Injektionsvolumen von 0,5 ccm die in der Tabelle 4 angegebene Gewichtsmenge hamolysinhaltiger Lebersubstanz enthalten war. Die Ergebnisse der Passagierungsversuche sind Tabelle 4 zu entnehmen.
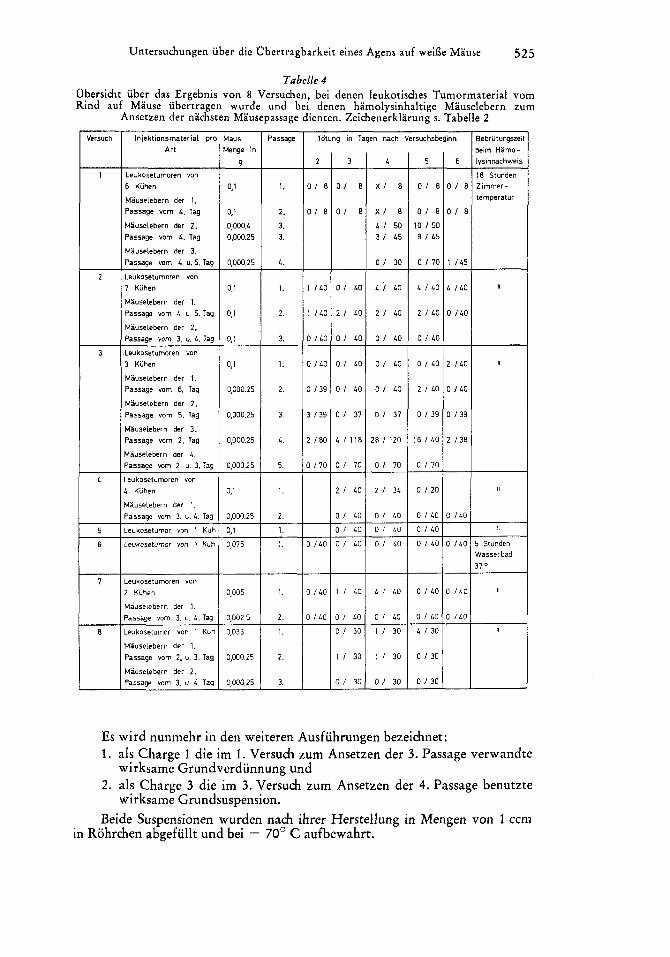
Aus Tab.elle 4 geht hervor, dai3 im I., 2., 3., 4., 7. und 8. Versuch Lebern mit Hamolysingehalt festgestellt wurden. Im 5. und 6. Versuch war das nicht der Fall. Die Zahl der hamolysinhaltigen Lebern war uber das auf Grund ,,unspezifischer" Ursachen mogliche Mai3 uber mehrere Passagen nur im 1. und 3. Versuch erhoht. Im 1. Versuch hauften sich die Falle in der 3. Passage, im 3. Versuch in der 4. Passage an. In beiden Versuchen waren die folgenden Passagen wieder frei von hamolysinhaltigen Lebern.
Wird unterstellt, dai3 die Ursache des Hamolysingehaltes der Mauselebern ein in dem Ausgangsmaterial vorhandenes Agens war, so ist zu folgern:
1. Der Nachweis dieses Agens gelang befriedigend nur in 2 von 8 Ver- suchen, bei denen das Tumormaterial der Rinder nach der Schlachtung ent- nommen und bis zu seiner Verarbeitung 4 Tage-5 Monate bzw. 14 Tage bis 13 Monate bei - 70' C eingefroren aufbewahrt worden war.
2 . Es gelang nicht, dieses Agens uber die 4. Passage hinaus in MY usen fortzufuhren.
Untersuchungen iiber die Obertragbarkeit eines Agens auf weide Mause 525
0 1 8
0 1 8
1 1 L O
1 / L O
0 1 4 0
O I L 0
0 1 3 9
3 I 3 9
2 I 8 0
0 I 7 0
0 / L O
0 110
0 1 4 0
Tabelle 4 Obersicht iiber das Ergebnis von 8 Versuchen, bei denen leukotisches Tumormaterial vom Rind auf Mause iibertragen wurde und bei denen hamolysinhaltige Miiuselebern zum
Ansetzen der nachsten Mausepassage dienten. Zeichenerklarung s. Tabelle 2
2 3
0 1 8
0 1 8
0 1 LO
2 I LO
0 I LO
0 1 LO
0 I LO
0 I 37
L I l l 8
0 I 7 0
2 1 LO
0 1 LO
0 1 LO
0 1 LO
1 I LO
0 I LO
0 1 30
I I 30
0 1 30
Versuch
- 1
laus lenge in
9 - 0.1
O,! 0,OOO.L 0,000.25
0,000.25
0, 1
0,1
0, 1
0.1
-
-
0,000.25
0,000.25
0,000.25
0,000.25
Passage
-
1.
2.
3. 3.
L.
I beim Harno- lnjektionsmaterial pri
Art L -
X I 8
X I 8
L I 50 3 1 L5
0 1 30
5 - 0 1 E
0 1 E
10 1 % 9 I LE
0 I 7c
Leukosetumoren van 6 Kuhen
Mauselebern der I , Passage vorn L. l a g
Mauselebern der 2. Passage vom L. l a g
Mauselebern der 3. Passage vorn L. u. 5. lag
Leukosetumoren von 7 Kiihen
Mauselebern der I. Passage vom L, u. 5. Tag
Mauselebern der 2. Passage vom 3. u. 4. lag
Leukosetumoren von 3 Kuhen
Mauselebern der 1. Passage vom 6. Tag
Mauselebern der 2. Passage vorn 5. lag
Mauselebern der 3. Passage vom 2. lag
Mauselebern der L. Passage vom 2. u. 3. lag
ternperatur
O'
1 l L 5
2 L I LO
2 1 LO
0 1 LO
L I LC
2 I LC
0 I LC
3 I .
2.
3.
4.
5. - 1.
2.
I . -
0 1 LO
0 1 LO
0 1 37
28 I i20
0 1 7 0 -
2 1 3L
0 1 LO
0 1 LO -
0 I 40
2 I LO
0 1 3 9
16 I LO
0 1 7 0 - 0 1 2 0
0 I LO
L
- 5
0,l
0,000.25 - 0.1 - 0,075
-
0,005
0,002.5
0,033 -
O,W0.25
0,000.25
Leukosetumoren von L Kuhen
Mauselebern der 1. Passage vom 3. u. L Tag
Leukosetumor von I Kut 0 I 4 0
0 I LO -
6
- 7
- 8
Leukoseturnor von 1 K u t 1.
- I .
2.
0 1 40 Wasserbad
L l LO
0 1 LO
0 I LO
0 1 LO k Leukosetumoren van 2 Kuhen
Mauselebern der 1, Passage vom 3. u. L. Tag
Leukosetumor von 1 Kut
Mauselebern der 1. Passage vorn 2. u. 3 . l a g
Mauselebern der 2. Passage vom 3. u. 4. Tag
1.
2.
3. -
1 I 30
1 I 30
0 1 30 -
4 I 30
0 I 3 0
0 1 3 0 -
Es wird nunmehr in den weiteren Ausfuhrungen bezeichnet : 1. aIs Charge 1 die im 1. Versuch zum Ansetzen der 3. Passage verwandte
2. als Charge 3 die im 3. Versuch zum Ansetzen der 4. Passage benutzte
Beide SusDensionen wurden nach ihrer Herstellung in Mengen von 1 ccm
wirksame Grundverdunnung und
wirksame Grundsuspension.
" " in Rohrchen a6gefullt und bei - 70' C aufbewahrt.

526 E. MITSCHERLICH
Versuchsgruppe
Suspendierung der Charge 3 i n :
Untersuchunq am Tage p. I. :
4. Untersuchungen uber die Bestandigkeit des mit Rinderleukosematerial mutmai3lich auf Mause ubertragenen Agens, das in deren Lebern
zur Ausbildung von Hamolysin fuhrt 4.1 Resistenz bei - 7OoC
Die als Charge 1 und 3 (s. Abschnitt 3.3) bezeichneten Suspensionen hamolysinhaltiger Mauselebern wurden in der Gefriertruhe bei - 70 O C auf- bewahrt und in gewissen Zeitabstanden durch intraabdominale Obertragung auf Mause wie in den bisher dargestellten Versuchen auf ihre Wirksamkeit iiberpruft. Zur Obertragung wurden sie derart verdunnt, dai3 jede Maus in 0,5 ccm Injektionsvolumen 0,000.25 g hamolysinhaltiges Lebermaterial erhielt.
Charge 1 war noch nach einem Aufenthalt von 224 Tagen in der Gefrier- truhe wirksam und danach verbraucht. Charge 3 war nach 143 Tagen Aufbe- wahrung in der Gefriertruhe voll wirksam. Nach einem Aufenthalt von 184 Tagen hatte sie jedoch ihre Aktivitat vollig eingebui3t.
1 2
Earle - Lasung Earle - Losung und BLut - plasma P I gernischt
2 I 3. I 4. 3. I 4
4.2 Resistenz bei Zimmertemperatur Fur die nachfolgend beschriebenen Neutralisationsversuche (Abschnitt 5)
war es wichtig zu wissen, welche Resistenz das Agens in verschiedenen Suspen- sionsmitteln bei Zimmertemperatur hat.
Aus der Charge 3 wurden zwei Verdunnungen hergestellt, davon die erste mit Earle-Losung und die zweite mit einer Mischung, die zu gleichen Teilen aus Earle-Losung und dem Blutplasma einer leukosefreien Kuh bestand. Beide Verdunnungen enthielten in 0,5 ccm 0,000.25 g hamolysinhaltiges Leber- material. Sie wurden 1 Stunde unter leichter Bewegung bei Zimmertemperatur gehalten und danach Miusen in der Menge von 0,s ccm pro Tier intraabdomi- nal injiziert. Die Mause wurden in Gruppen am 3., 4. und 5. Tage nach der Injektion getotet. Die Untersuchung der Lebern auf Hamolysin erfolgte am jeweiligen Totungstag in zwei parallel angesetzten Testen. In dem ersten Test wurden die Reaktionen nach 5stundiger Bebrutung bei 37 O C im Wasserbad und im zweiten nach 1 8stundigem Aufenthalt bei Zimmertemperatur abgele- sen. Die Ergebnisse dieses Versuches sind Tabelle 5 zu entnehmen.
Reaktionszeit Beurteilung
18 Stunden
Beurteilunq
Tabelle 5 zeigt, dai3 das Agens innerhalb einer Stunde bei Zimmertempe- ratur seine Wirksamkeit einbiiflt, wenn es in Earle-Losung suspendiert ist, dai3 es hingegen seine Wirksamkeit behalt, wenn als Suspensionsmittel eine Mischung verwandt wird, die zu gleichen Teilen aus Earle-Losung und Plasma einer leukosefreien Kuh besteht. Diese Beobachtungen weisen darauf hin, dai3 die Bestandigkeit des Agens in der Umwelt gering ist.
Untersuchungen iiber die Ubertragbarkeit eines Agens auf weiBe Mause 527
5. Versuche zur Neutralisation des mit Rinderleukosematerial mutmaBlich auf Mause iibertragenen Agens rnit dem Blut von einem leukosefreien
und zwei leukoseinfizierten Rindern Die Befunde, dai3 in den Lebern von Versuchsmausen, auf die Rinderleu-
kosematerial ubertragen wurde, ein Agens auftreten kann, das bei den Ver- suchstieren zu einer anormalen Steigerung des Hamolysingehaltes der Lebern fuhrt, lassen nicht den Schlui3 zu, dai3 dieses Agens etwas mit dem Erreger der Rinderleukose zu tun hat. Eine derartige Annahme wurde jedoch an Wahr- scheinlichkeit gewinnen, wenn es gelingt, die Wirksamkeit dieses Agens mit dem Blut von Rindern zu neutralisieren, die mit Leukose infiziert sind.
5.1 Material und Methoden Fur die Durchfuhrung dieser Neutralisationsversuche wurden verwandt: I . Die Charge 3 als Material, das das wirksame Agens enthalt. 2. Blut von Rindern, die an Leukose erkrankt waren. Zu diesem Zweck
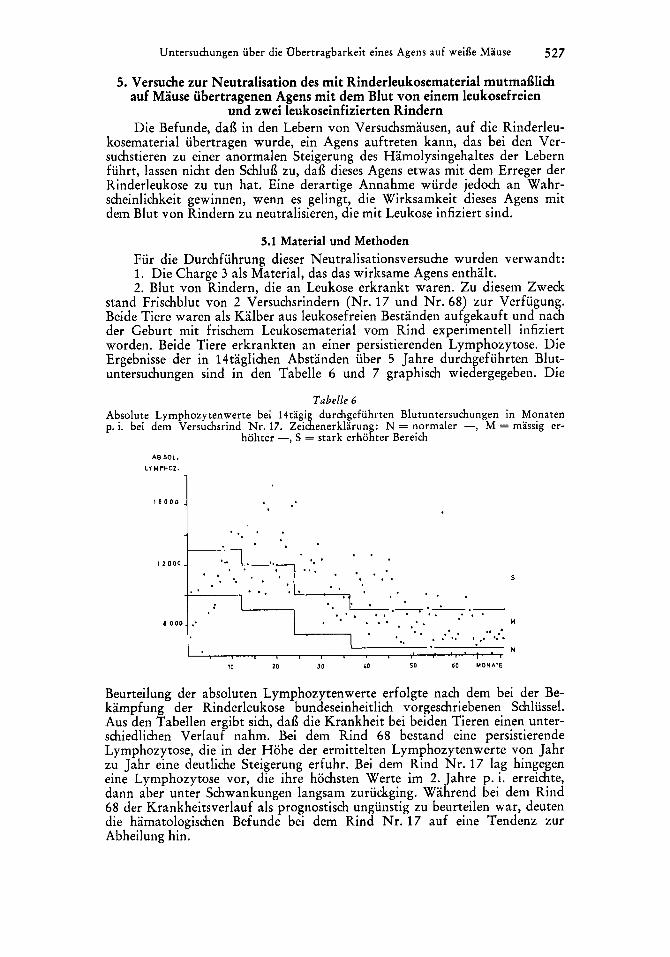
stand Frischblut von 2 Versuchsrindern (Nr. 17 und Nr. 68) zur Verfugung. Beide Tiere waren als Kalber aus leukosefreien Bestanden aufgekauft und nach der Geburt mit frischem Leukosematerial vom Rind experimentell infiziert worden. Beide Tiere erkrankten an einer persistierenden Lymphozytose. Die Ergebnisse der in 14taglichen Abstanden uber 5 Jahre durchgefuhrten Blut- untersuchungen sind in den Tabelle 6 und 7 graphisch wiedergegeben. Die
Tabelle 6 Absolute Lymphozytenwerte bei 14tagig durchgefiihrten Blutuntersuchungen in Monaten p. i. bei dem Versuchsrind Nr. 17. Zeichenerklarung: N = normaler -, M = massig er-
hohter -, S = stark erhohter Bereich
AB SOL.
LVMPHOZ.
10 60 MONATE 30 10 so
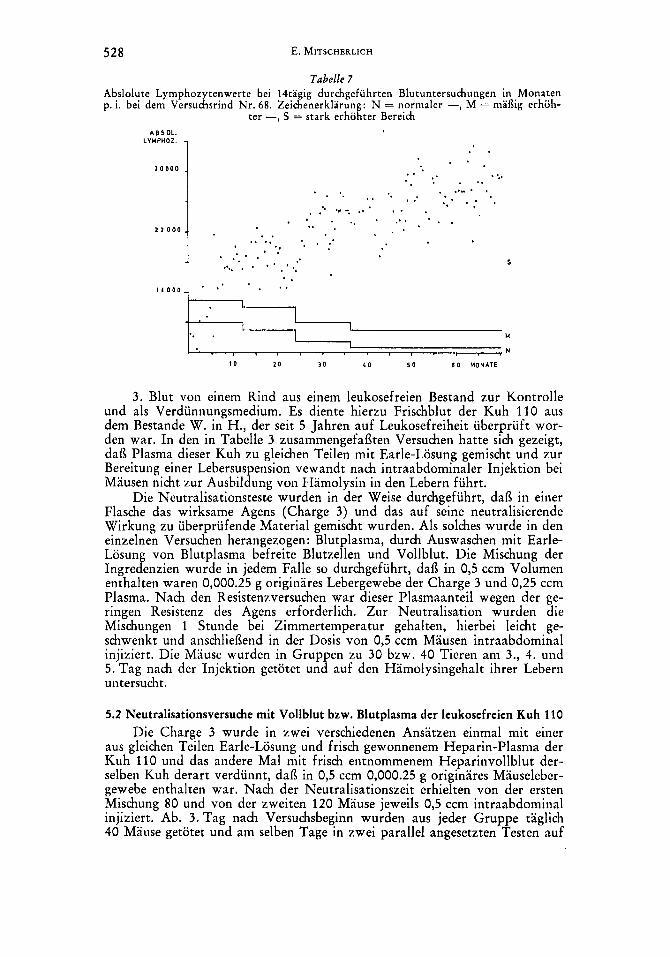
Beurteilung der absoluten Lymphozytenwerte erfolgte nach dem bei der Be- kampfung der Rinderleukose bundeseinheitlich vorgeschriebenen Schlussel. Aus den Tabellen ergibt sich, dai3 die Krankheit bei beiden Tieren einen unter- schiedlichen Verlauf nahm. Bei dem Rind 68 bestand eine persistierende Lymphozytose, die in der Hohe der ermittelten Lymphozytenwerte von Jahr zu Jahr eine deutliche Steigerung erfuhr. Bei dem Rind Nr . 17 lag hingegen eine Lymphozytose vor, die ihre hochsten Werte im 2. Jahre p. i. erreichte, dann aber unter Schwankungen langsam zuruckging. Wahrend bei dem Rind 68 der Krankheitsverlauf als prognostisch ungunstig zu beurteilen war, deuten die hamatologischen Befunde bei dem Rind Nr. 17 auf eine Tendenz zur Abheilung hin.
528 E. MITSCHERLICH
3 0 0 0 0 .
1 2 0 0 0 .
i ( o o a -
Tabelle 7 Abslolute Lymphozytenwerte bei 14tagig durchgefuhrten Blutuntersuchungen in Monaten p. i. bei dem Versuchsrind Nr. 68. Zeichenerklarung: N = normaler -, M = rnaflig erhoh-
ter -, S = stark erhohter Bereich
. . . * - :. .. - . . . .. . .... - . . . . . . : . . . 6 . * . . . * . . .. .Z -. .. . . . . . . . . ... ' . . '. .
. . . . .. .. - . . ." . ...* . . - * ' . s
* . * . . a .
* . ' *
lL*" . . I
N
A B S O L .
LYHPHoZ. 1 . .
3. Blut von einem Rind aus einem leukosefreien Bestand zur Kontrolle und als Verdunnungsmedium. Es diente hierzu Frischblut der Kuh 110 aus dem Bestande W. in H., der seit 5 Jahren auf Leukosefreiheit uberpruft wor- den war. In den in Tabelle 3 zusammengefaflten Versuchen hatte sich gezeigt, daf3 Plasma dieser Kuh zu gleichen Teilen mit Earle-Losung gemischt und zur Bereitung einer Lebersuspension vewandt nach intraabdominaler Injektion bei Mausen nicht zur Ausbildung von Hamolysin in den Lebern fuhrt.
Die Neutralisationsteste wurden in der Weise durchgefuhrt, dai3 in einer Flasche das wirksame Agens (Charge 3) und das auf seine neutralisierende Wirkung zu uberprufende Material gemischt wurden. Als solches wurde in den einzelnen Versuchen herangezogen: Blutplasma, durch Auswaschen mit Earle- Losung von Blutplasma befreite Blutzellen und Vollblut. Die Mischung der Ingredenzien wurde in jedem Falle so durchgefuhrt, dafl in 0,5 ccm Volumen enthalten waren 0,000.25 g originares Lebergewebe der Charge 3 und 0,25 ccm Plasma. Nach den Resistenzversuchen war dieser Plasmaanteil wegen der ge- ringen Resistenz des Agens erforderlich. Zur Neutralisation wurden die Mischungen 1 Stunde bei Zimmertemperatur gehalten, hierbei leicht ge- schwenkt und anschlieflend in der Dosis von 0,5 ccm Mausen intraabdominal injiziert. Die Mause wurden in Gruppen zu 30 bzw. 40 Tieren am 3., 4. und 5. Tag nach der Injektion getotet und auf den Hamolysingehalt ihrer Lebern untersucht.
5.2 Neutralisationsversuche mit Vollblut bzw. Blutplasma der leukosefreien Kuh 110 Die Charge 3 wurde in zwei verschiedenen Ansatzen einmal mit einer
aus gleichen Teilen Earle-Losung und frisch gewonnenem Heparin-Plasma der Kuh 110 und das andere Ma1 mit frisch entnommenem Heparinvollblut der- selben Kuh derart verdunnt, dai3 in 0,5 ccrn 0,000.25 g originares Mauseleber- gewebe enthalten war. Nach der Neutralisationszeit erhielten von der ersten Mischung 80 und von der zweiten 120 Mause jeweils 0,5 ccm intraabdominal injiziert. Ab. 3. Tag nach Versuchsbeginn wurden aus jeder Gruppe taglich 40 Mause getotet und am selben Tage in zwei parallel angesetzten Testen auf
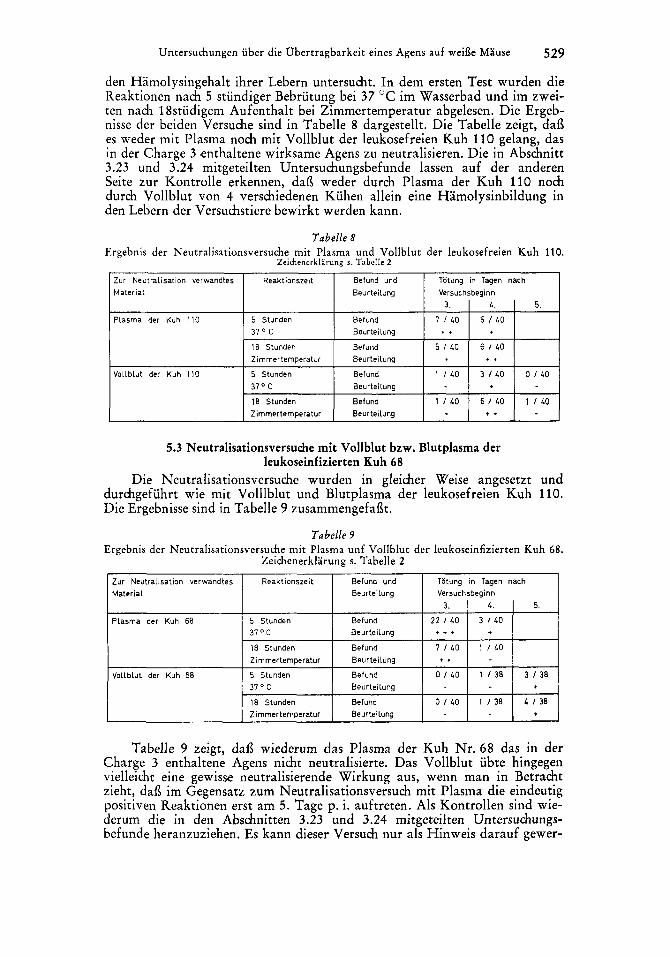
Untersuchungen iiber die Ubertragbarkeit eines Agens auf weile Mause 529
Zur Neutralisation verwandtes Reaktionszeit Befund und Material Beurteilung
Plasma der Kuh 110 5 Stunden Befund 31' C Beurteilung
18 Stunden Befund Zimrnertemperatur Beurteilung
Vollblut der Kuh I10 5 Stunden Befund 31" C Beurteilung
den Hamolysingehalt ihrer Lebern untersucht. In dem ersten Test wurden die Reaktionen nach 5 stundiger Bebrutung bei 37 "C im Wasserbad und im zwei- ten nach 18studigem Aufenthalt bei Zimmertemperatur abgelesen. Die Ergeb- nisse der beiden Versuche sind in Tabelle 8 dargestellt. Die Tabelle zeigt, dai3 es weder mit Plasma noch mit Vollblut der leukosefreien Kuh 110 gelang, das in der Charge 3 enthaltene wirksame Agens zu neutralisieren. Die in Abschnitt 3.23 und 3.24 mitgeteilten Untersuchungsbefunde lassen auf der anderen Seite zur Kontrolle erkennen, dai3 weder durch Plasma der Kuh 110 noch durch Vollblut von 4 verschiedenen Kuhen allein eine Hamolysinbildung in den Lebern der Versuchstiere bewirkt werden kann.
Totung in Tagen nach Versuchsbeginn
3. L. 5.
7 1 4 0 5 / 4 0 t f
5 1 L O 6 / L O . *
1 \ 4 0 3 / L O O I L 0
I8 Stunden Zimmerternperatur
Befund 1 / 4 0 6 / 4 0 I I L O Beurteilung + *
Zur Neutralisation verwandtes Reaktionszeit Befund und Material Beurteilung
Plasma der Kuh 68 5 Stunden Befund 3 I 0 C Beurteilung
18 Stunden Befund Z immertemperatur Beurteilung
Totung in Tagen nach Versuchsbeginn
3. L. 5.
3 / LO 2 2 I LO + + +
I I L O 1 1 L O * *
Tabelle 9 zeigt, dai3 wiederum das Plasma der Kuh Nr. 68 das in der Charge 3 enthaltene Agens nicht neutralisierte. Das Vollblut ubte hingegen vielleicht eine gewisse neutralisierende Wirkung aus, wenn man in Betracht zieht, dai3 im Gegensatz zum Neutralisationsversuch mit Plasma die eindeutig positiven Reaktionen erst am 5. Tage p. i. auftreten. Als Kontrollen sind wie- derum die in den Abschnitten 3.23 und 3.24 mitgeteilten Untersuchungs- befunde heranzuziehen. Es kann dieser Versuch nur als Hinweis darauf gewer-
Vollblut der Kuh 68 I 5 Stunden 31 C
18 Stunden Zimrnerternperatur
Befund O I L 0 1 / 3 8 3 / 3 8 Beurteilung
Befund O I L 0 1 / 3 8 L / 3 8 Beurteilung

530 E. MITSCHERLICH
Zur Neutralisation verwandtes Reaktionszeit Befund und Material Beurteilung
tet werden, dai3, wenn uberhaupt eine Neutralisation der Charge 3 moglich ist, die Blutzellen hierbei nicht auaer acht gelassen werden durfen.
5.4 Neutralisationsversuche rnit Blutplasma, Blutzellen und Vollblut der leukoseinfizierten Kuh Nr. 17
Die Neutralisationsversuche wurden zunachst mit Plasma und Vollblut der Kuh Nr. 17 in gleicher Weise angesetzt und durchgefuhrt, wie es rnit Plasma und Vollblut der Kuh Nr . 110 (Tab. 8) geschehen war. Die Ergebnisse dieses Versuches sind Tabelle 10 zu entnehmen.
Tdtung in Tagen nach Versuchsbeginn
3. I L. I 5.
5 Stunden
37 o c
18 Stunden Z immertemperatur
5 Stunden
3 1 O C
I8 Stunden Zimmertemperatur
[P lasma der Kuh 17 Befund 5 / 3 0 1 / 3 0 Beurteilung + *
Beurteilung * + Befund 6 / 3 0 0 / 3 0
Befund 0 / 3 9 0 / 4 0 0 / 3 8 Beurteilung
Befund 0 / 3 9 0 1 4 0 0 1 3 8 Beurteilung
I Vollblut der Kuh 17
I
Die Ergebnisse dieses Versuches stehen in Obereinstimmung mit denen, die rnit Vollblut und Blutplasma der Kuh 68 erzielt wurden. Mit Blutplasma war eine Neutralisation nicht zu erzielen, wohl aber mit Vollblut.
Damit ergab sich die Frage, ob der Neutralisationseffekt allein auf die Blutzellen zuriickzufuhren war oder auf ein Zusammenwirken von Blutzellen und bestimmten Plasmafraktionen. Um diese Frage zu klaren wurde Frisch- blut der Kuh 17 mit gleichen Teilen Earle-Losung unter Zugabe von 0,05 " / o Heparin gemischt und bei + 10' C abzentrifugiert. Die Blutkorperchen wurden dreimal mit Earle-Losung ausgewaschen.
Ein Teil der Blutkorperchen wurde hierauf derart in unverdunntem Plasma der leukosefreien Kuh 110 aufgenommen, dai3 das zuvor naturlicher- weise im Tier gegebene Zell-Plasma-Verhaltnis wieder hergestellt war. Dem Plasma der Kuh 110 war dabei soviel von der Charge 3 zugefugt worden, dai3 in 0,5 ccm Blutzellen-Plasma-Mischung 0,000.25 g hamolysinhaltiges Le- bergewebe enthalten war (Ansatz A).
Ein weiterer Teil der Blutkorperchen wurde ebenfalls mit Blutplasma in der Weise wieder aufgenommen, dai3 das ursprungliche, im Tier gegebene Zell- Plasma-Verhaltnis wieder hergestellt war. In dem Gesamtvolumen der Blut- zellen-Plasma-Mischung war jedoch abweichend von dem zuvor beschriebenen Ansatz 2 O / o unverdunntes Plasma der leukoseinfizierten Kuh 17 und im ubri- gen unverdunntes Plasma der leukosefreien Kuh 110 enthalten. Daruber hinaus enthielt die Mischung in 0,5 ccm 0,000.25 g hamolysinhaltiges Leberge- webe der Charge 3 (Ansatz B).
Nach einer einstundigen Neutralisationszeit wurde 120 Mausen je 0,5 ccm des Ansatzes ,,A" und weiteren 120 Mausen je 0,5 ccm des Ansatzes ,,B" intra- abdominal injiziert. Ab 3. Tag p. i. wurden taglich aus jeder Gruppe 40 Mause getotet und auf den Hamolysingehalt ihrer Lebern untersucht.
Die Ergebnisse der beiden Versuche sind in Tabelle 11 zusammengestellt. Es ergibt sich aus der Tabelle, dai3 eine Neutralisation des in der Charge 3

Zur Neutralisation verwandtes Material
BLutzellen der Kuh 17 in Plasma der Kuh 110
Blutzellen der Kuh 17 in 2 "10 Plasma der Kuh 17 ad 100% Plasma der Kuh 110
enthaltenen Agens mit Blutzellen der leukoseinfizierten Kuh 17 nicht gelang, wenn diese Zellen in Plasma der leukosefreien Kuh 110 resuspendiert waren. Enthielt jedoch die Mischung aui3erdem 2 O/o unverdunntes Plasma der leukose- infizierten Kuh 17, so war eine Neutralisation moglich.
Die Ergebnisse der Neutralisationsversuche lassen es insgesamt betrachtet moglich erscheinen, dai3 besonders im Blute von Rindern, bei denen die Leu- kose einen prognostisch giinstigen Verlauf nimmt, bestimmte Verbindungen auftreten, die allein fur sich unwirksam sind. Im Zusammenwirken mit den Blutzellen dieser Tiere sind sie jedoch in der Lage, das in der Charge 3 ent- haltene Agens zu neutralisieren, welches bei Mausen zu einer Hamolysinbil- dung in der Leber fiihrt. Damit gewinnt die Annahme an Wahrscheinlichkeit, dai3 dieses Agens in direktem Zusammenhang mit dem Erreger der Rinder- leukose steht oder mit ihm identisch ist.
Diese Schlui3folgerungen wurden weiterhin eine gewisse Absicherung er- fahren, wenn es sich in Ruckiibertragungsversuchen auf Rinder nachweisen liefie, dai3 in den hamolysinhaltigen aus Leukoseiibertragungsversuchen stam- ineiiden Mauselebern das Agens der Rinderleukose enthalten ist. Zur Klarung dieser Frage wurde eine Reihe von Ruckubertragungsversuchen angesetzt.
6. Riickiibertragung auf Rinder von hamolysinhaltigen Miuselebern aus Leukoseiibertragungsversuchen
In den vorher beschriebenen Versuchen war frisches Rinderleukosematerial auf Mause des NMRI-Stammes ubertragen worden. Aus den Lebern der Mause dieser I . Passage waren, sofern sie Hamolysin nachweisbar enthielten, Suspen- sionen hergestellt worden, um Mause der nachsten Passa e zu infizieren und SO fort. Es ergab sich dabei, dai3 es in einigen von diesen 6bertragungsversuchen nach wenigen Passagen zum Auftreten von Hamolysin in einer groi3eren Zahl von Mauselebern kam. Um nachzuweisen, ob diese Lebern den Erreger der Rinderleukose enthielten, wurde in 2 Ruckiibertragungsversuchen derartiges Lebermaterial Versuchkalbern injiziert. Die zu diesen Versuchen verwandten Kalber wurden gleich nach der Geburt aus Betrieben aufgekauft, die seit mehreren Jahren durch Blutuntersuchung aller uber 2 Jahre alten Rinder auf Leukosefreiheit uberpriift worden waren. Die Kalber wurden in einem separa- ten Versuchsstall gehalten.
6.1 Riidciibertragungsversuch Zu diesem Versuch dienten 6 Kalber. Ihnen wurden dreimal Suspensionen
aus hamolysinhaltgigen Mauselebern injiziert: zum ersten Ma1 am Tage der
Reaktionszeit Befund und Totung in Tagen nach Beurteilung Versuchsbeginn
3 4 5
5 Stunden Befund 5 / 3 0 0 1 3 0 1 / 3 0 37oc Beurteilung + +
Z immertemperatur Beurteilung * + * 18 Stunden Befund 9 / 3 0 0 1 3 0 1 / 3 0
5 Stunden Befund 0 / 3 0 0 / 3 0 1 / 3 0 37OC Beurteilung
18 Stunden Befund 0 / 3 0 1 / 3 0 Zimmertemperatur Beurteilung
E. MITSCHERLICH 532
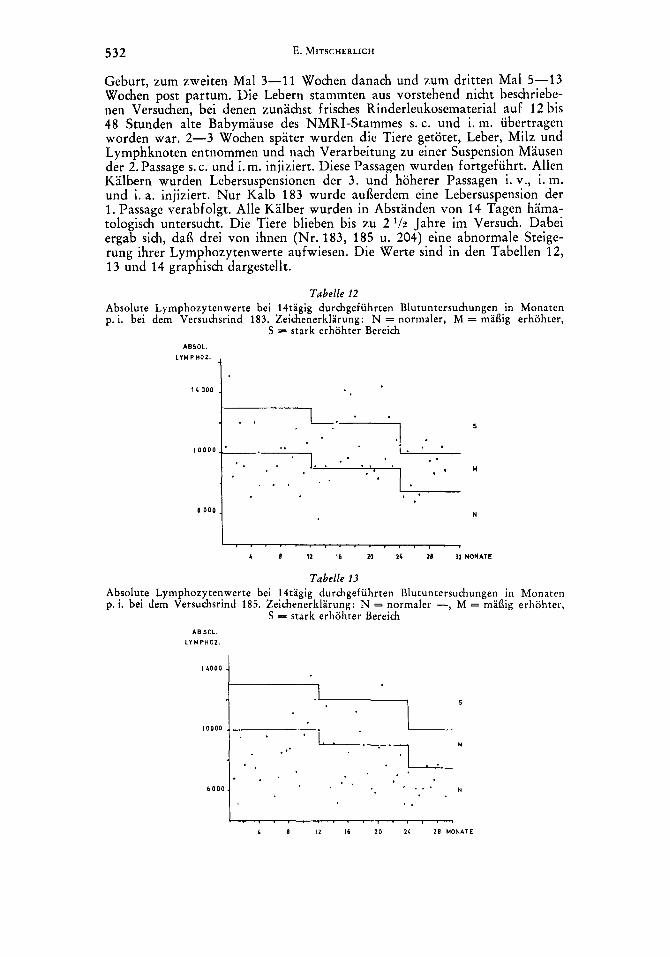
Geburt, zum zweiten Ma1 3-11 Wochen danach und zum dritten Ma1 5-13 Wochen post partum. Die Lebern stammten aus vorstehend nicht beschriebe- nen Versuchen, bei denen zunachst frisches Rinderleukosematerial auf 12 bis 48 Stunden alte Babymause des NMRI-Stammes s. c. und i. m. ubertragen worden war. 2-3 Wochen spater wurden die Tiere getotet, Leber, Milz und Lymphknoten entnommen und nach Verarbeitung zu einer Suspension Mausen der 2.Passage s. c. und i. m. injiziert. Diese Passagen wurden fortgefuhrt. Allen Kalbern wurden Lebersuspensionen der 3. und hoherer Passagen i. v., i. m. und i. a. injiziert. Nur Kalb 183 wurde aui3erdem eine Lebersuspension der 1. Passage verabfolgt. Alle Kalber wurden in Abstanden von 14 Tagen hama- tologisch untersucht. Die Tiere blieben bis zu 2 l/n Jahre im Versuch. Dabei ergab sich, dai3 drei von ihnen (Nr. 183, 185 u. 204) eine abnormale Steige- rung ihrer Lymphozytenwerte aufwiesen. Die Werte sind in den Tabellen 12, 13 und 14 graphisch dargestellt.
Tabelle 12 Absolute Lymphozytenwerte bei l4tagig durchgefuhrten Blutuntersuchungen in Monaten p. i. bei dem Versuchsrind 183. Zeichenerklarung: N = normaler, M = maflig erhohter,
S = stark erhohter Bereich ASSOL.
LYWPHOL. 1
6 0 0 0 1 N
I 1 B 12 I 6 10 11 18 32 MONATE
Tabelle 13 Absolute Lymphozytenwerte bei 14tagig durchgefuhrten Blutuntersuchungen in Monaten p. i. bei dem Versuchsrind 185. Zeichenerklarung: N = normaler -, M = maflig erhohter,
S = stark erhohter Bereich A050L.
L V M P H O Z
I4000
10000
6 0 0 0
. .
L 8 12 16 2 0 2 4 1 8 HOUATE
Untersuchungen iiber die Obertragbarkeit eines Agens auf we& Mause 533
Tabelle 14 Absolute Lymphozytenwerte bei 14tagig durchgefiihrten Blutuntersuhungen in Monaten p. i. bei dem Versuchsrind 204. Zeichenerklarung: N = normaler -, M - mBi3ig erhohter -,
S = stark erhohter Bereich 48ML.
LYM P noz.
-I 1 LOO0
.I- S
I I
6oooL----- b 8 12 16 20 14 28 HONATE
Die Lymphozytenzahlen des Versuchstieres 83 bewegten sich vom 16. Monat p. i. an fast ausschliei3lich im mai3ig bis stark erhohten Bereich. Die Lymphozytenzahlen des Rindes 204 waren vom 13. Monat p. i. an in iiber der Halfte der Falle mai3ig erhoht und die des Versuchstieres 185 zeigten gleichfalls sporadisch abnormale Stei erungen in den mai3ig bis stark erhohten Bereich hinein. Bei der Schlachtung d er Tiere wurden keine tumorosen Veran- derungen festgestellt.
6.2 Riickiibertragungsverswh Zum 2. Ruckiibertragungsversuch wurden 7 Kalber verwandt. Als In-
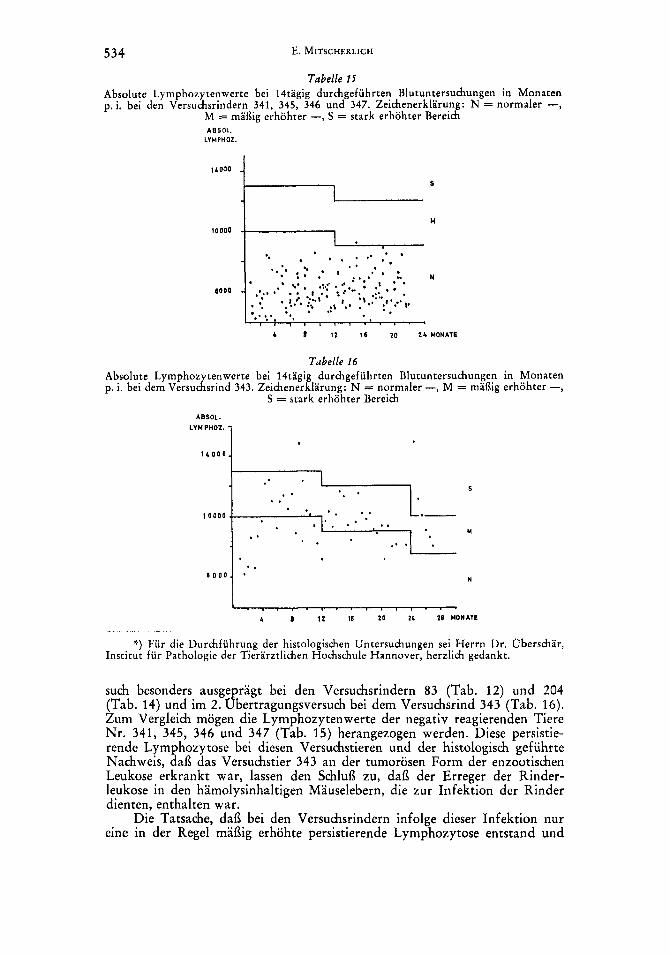
fektionsmaterial dienten bei allen Tieren Suspensionen hamolysinhaltiger Lebern der 3. Passage des I . Leukoseubertragungsversuches (s. Tab. 4). Jedes Tier erhielt 30-40 ccm einer 20 O/oigen Suspension zum groi3eren Teil intraabdominal und zum kleineren intramuskular injiziert. Die Tiere wurden alle 14 Tage hamatologsich untersucht. Das Versuchsrind 343 blieb 2 Jahre, 4 Monate im Versuch, die ubrigen 1 Jahr, 10 Monate.
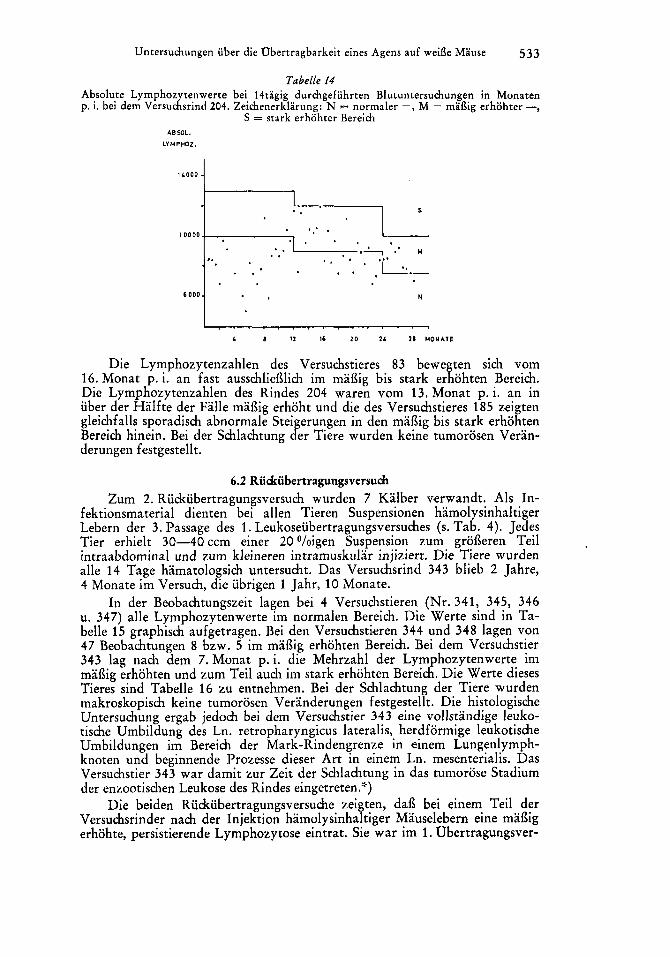
In der Beobachtungszeit lagen bei 4 Versuchstieren (Nr. 341, 345, 346 u. 347) alle Lymphozytenwerte im normalen Bereich. Die Werte sind in Ta- belle 15 graphisch aufgetragen. Bei den Versuchstieren 344 und 348 lagen von 47 Beobachtungen 8 bzw. 5 im mai3ig erhohten Bereich. Bei dem Versuchstier 343 lag nach dem 7.Monat p. i. die Mehrzahl der Lymphozytenwerte im mai3ig erhohten und zum Teil auch im stark erhohten Bereich. Die Werte dieses Tieres sind Tabelle 16 zu entnehmen. Bei der Schlachtung der Tiere wurden makroskopisch keine tumorosen Veranderungen festgestellt. Die histologische Untersuchung ergab jedoch bei dem Versuchstier 343 eine vollstandige leuko- tische Umbildung des Ln. retropharyngicus lateralis, herdformige leukotische Umbildungen im Bereich der Mark-Rindengrenze in ,einem Lungenlymph- knoten und beginnende Prozesse dieser Art in einem Ln. mesenterialis. Das Versuchstier 343 war damit zur Zeit der Schlachtung in das tumorose Stadium der enzootischen Leukose des Rindes eingetreten.'>)
Die beiden Riidrubertragungsversuche zeigten, dai3 bei einem Teil der Versuchsrinder nach der Injektion hamolysinhaltiger Mauselebern eine mai3ig erhohte, persistierende Lymphozytose eintrat. Sie war im 1. Obertragungsver-
E. MITSCHERLICH 534
Tabelle 15 Absolute Lymphozytenwerte bei 14tagig durchgefuhrten Blutuntersuchungen in Monaten p. i. bei den Versuchsrindern 341, 345, 346 und 347. Zeichenerklarung: N = normaler -,
M = maflig erhohter -, S = stark erhohter Bereich ABSOL.
LVHPHOZ.
I4000
l0000
l o o 0
Absolute Lymphozytenwerte p. i. bei dem Versuchsrind 343
-I
I I *
ABSOL.
lVM PHoZ. 1
Tabelle 16 bei 14tagig durchgefuhrten Blutuntersuchungen in Monaten
. Zeichenerklarung: N = normaler -, M = maflig erhohter -, S = stark erhohter Bereich
1 0 0 0 0 .- . .. L .- ' . . H .. . . . *
SN 4 8 12 16 20 21 28 HONATE
+) Fur die Durchfuhrung der histologischen Untersuchungen sei Herrn Dr. Oberschar, Institut fur Pathologie der Tierarztlichen Hochschule Hannover, herzlich gedankt.
such besonders ausge ragt bei den Versuchsrindern 83 (Tab. 12) und 204 (Tab. 14) und im 2. gbertragungsversuch bei dem Versuchsrind 343 (Tab. 16). Zum Vergleich mogen die Lymphozytenwerte der negativ reagierenden Tiere Nr. 341, 345, 346 und 347 (Tab. 15) herangezogen werden. Diese persistie- rende Lymphozytose bei diesen Versuchstieren und der histologisch gefuhrte Nachweis, dai3 das Versuchstier 343 an der tumorosen Form der enzootischen Leukose erkrankt war, lassen den Schlui3 zu, dai3 der Erreger der Rinder- leukose in den hamolysinhaltigen Mauselebern, die zur Infektion der Rinder dienten, enthalten war.
Die Tatsache, dai3 bei den Versuchsrindern infolge dieser Infektion nur eine in der Regel mai3ig erhohte persistierende Lymphozytose entstand und

Untersuchungen iiber die Ubertragbarkeit eines Agens auf weide Mause 535
sehr stark erhohte Werte wie bei den Versuchsrindern Nr. 17 (Tab. 6) und Nr. 68 (Tab. 7) ausblieben, konnte darauf zuruckzufuhren sein, dai3 der Erreger der Rinderleukose in den Mausen bezuglich seiner Virulenz fur das Rind eine Abschwachung erfuhr.
Da auch das in leukotischem Tumormaterial vom Rind enhaltene Agens, das bei seiner Obertragung auf die weii3e Maus zu einer Hamolysinbildung in der Leber fuhrte, sich nur wenige Passagen in den Mausen fortfuhren liei3, besteht hier eine Parallelitat der Beobachtungen.
Insgesamt gesehen erfahrt die Annahme, dai3 dieses Agens mit dem Er- reger der Rinderleukose in direktem Zusammenhang steht oder mit ihm identisch ist, durch den Ausgang der Ruckubertragungsversuche auf das Rind eine weitere Stutze.
7. Diskussion Die vorliegenden Untersuchungen hatten zum Ziel, die Moglichkeiten
fur den Nachweis des Erregers der Rinderleukose zu erweitern. Er wird bis- lang nur durch die Feststellung pathologischer Symptome beim Rinde selbst gefuhrr.
Der Gedanke, die weii3e Maus zu diesen Untersuchungen heranzuziehen, mui3te zunachst als Wagnis erscheinen. Die Verwendung der weii3en Maus als Versuchstier bei der Erforschung der Leukamien des Menschen (POLLARD, 1967) oder auch der Leukose des Rindes (STRAUB, 1966) erscheint untunlich, weil Mause selber Trager von Leukamieviren sind. Nach POLLARD sind prak- tisch in allen ,, k,eimfreien" Mausestammen diese oder andere Virusarten nachgewiesen worden. Wenn bei Mausen nach der Injektion von menschlichem Leukamie - oder auch Rinderleukosematerial eine Leukamie oder andere pathologische Symptome ausgelost werden, so lassen sich diese Befunde nicht ohne weiteres als Beweis dafur werten, dai3 es nunmehr gelungen sei, die Leukamie des Menschen oder die Leukose des Rindes auf die Maus zu ubertra- gen. Selbst wenn, wie in den Versuchen von VASILIJEV (1960, 1962 zit. n. WITTMANN, 1963) nach der Obertragung leukotischen Tumormaterials vom Rind auf Sauglingsmause bei diesen Tieren in der Folge erhohte Lymphozyten- zahlen und 4-7 Monate p. i. die Entstehung von Tumoren an verschiedenen Korperstellen beobachtet wurden, wenn es sich zeigte, dai3 sich diese Tumoren bei verkurzter Inkubationszeit in weiteren Passagen in Mausen fortzuchten liei3en und schliei3lich ein mit derartigen Mausetumoren infiziertes Rind leu- kamisch erkrankte, so kann daraus nur gefolgert werden, dai3 es gelang, den Erreger der Rinderleukose in Mausen fortzufuhren. Die bei den Mausen beob- achtete Lymphozytose und Tumorbildung konnen aber durchaus auf Virus- arten zuruckzufuhren sein, mit denen die Mause schon vor Beginn der Ver- suche latent infiziert waren und die durch die Passagen aktiviert wurden. Die Feststellung eines pathologischen Symptomes bei den Mausen nach der Ober- tragung von Rinderleukosematerial gewinnt erst dann fur den Erreger der Rinderleukose eine diagnostische Bedeutung, wenn sich nachweisen lafit, dai3 das in dem Leukosematerial enthaltene Agens dieses Symptom nicht mehr aus- losen kann, wenn es zuvor durch Abwehrkrafte immuner Rinder neutralisiert wurde.
Die vorstehend dargestellten Untersuchungen geben erste Hinweise dafur dai3:
1. In Rinderleukosematerial ein Agens vorhanden ist, das bei einer Ober- tragung auf die weii3e Maus zur Ausbildung von Hamolysin in den Lebern der Versuchstiere fuhrt.
2. Nach der Ubertragung derartiger hamolysinhaltiger Mauselebern auf Versuchsrinder bei einem Teil der Versuchstiere eine persistierende Lympho-
Zbl. Vet. Med., Reihe B, Bd. 16, Heft 6 35

536 E. MITSCHERLICH
zytose und die leukotische Umbildung von Lymphknoten auftreten, wie sie bei der Leukose des Rindes beobachtet werden.
3. Im Blute von Rindern, die das leukamische Stadium der Rinderleukose uberwinden spezifische Abwehrkrafte auftreten, die in Blutzellen und in be- stimmten Plasmafraktionen lokalisiert sind. Diese Abwehrkrafte sind jeder allein fur sich unwirksam, im engen Verbund miteinander jedoch in der Lage, das Agens t u neutralisieren, welches in der weii3en Maus zu einer Ausbildung von Hamolysin in der Leber Veranlassung gibt.
Die Untersuchungsergebnisse lassen es moglich erscheinen, dai3 das bei Mausen zu einer Ausbildung von Hamolysin in den Lebern fuhrende Agens mit dem Erreger der Rinderleukose in direktem Zusammenhang steht oder mit ihm identisch ist.
Aufgrund der in Abschnitt 2 wiedergegebenen Untersuchungsbefunde ist es weiterhin moglich, dai3 das in den Lebern der Versuchsmause auftretende Hamolysin Lysolecithin ist.
Lysolecithin kommt im Organismus in verschiedenen Organen in Spuren normalerweise vor. Es wurde in der Darmschleimhaut, Leber, Milz, Niere, Lunge und Herz der Ratte (MARINETTI u. Mitarb., 1957 a), in gereinigten Cytochrompraparaten des Schweineherzens (MARINETTI u. Mitarb., 1957 b), in den Nebennieren (TITUS u. Mitarb., 1956) und im Blutplasma des Menschen nachgewiesen (PHILIPS, 1958). Es ist nicht abgeklart welche Aufgabe es im Organismus erfullt. Es greift die Membranen der Zellen an und fuhrt auf- grund dieser Tatsache zu einer Hamolyse roter Blutkorperchen. BERGENHEIM und FAHRAEUS (1936) vertreten die Auffassung, dai3 Lysolecithin eine wichtige Rolle beim Abbau der roten Blutkorperchen in der Milz spielt. Nach TITUS (1956) besteht die Moglichkeit, dai3 es hilft, die Durchlassigkeit der Zellmem- branen fur Kaliumionen zu regulieren. Daruber hinaus wies der gleiche Autor nach, dai3 Lysolecithin eine digitalisahnliche Wirkung auf das isolierte Frosch- herz ausubt. Es ist nicht sicher, in welchen Organen Lysolecithin gebildet wird. Da es aber aus Lecithin entsteht und die Leber nach DEUEL (1955) die Quelle des Plasmalecithins ist, spricht vie1 dafur, dai3 die Leber eine Produktionsstatte des Lysolecithins ist (MARINETTI u. Mitarb., 1959). Der Lecithingehalt der Leber betragt beim Menschen 1,44O/0 und beim Rind 1,56O/o des Organfeucht- gewichtes (DEUEL, 1955). Lysolecithin entsteht aus Lecithin unter der Ein- wirkung der Lecithinase A. Dieses Enzym greift an der alpha-Stellung der Esterbindung des Lecithinmolekules an und bildet unter Abspaltung von Fett- saure Lysolecithin (HOLMAN, LUNDBERG u. MALKIN, 1957). Es ist verstandlich, dai3 diese fur die Zellen des Organismus gefahrliche Verbindung sich nicht in dem Organ anreichern darf. DAWSON (1956) konnte nachweisen, dai3 es in der Leber ein weiteres Enzym gibt, die Phospholipase B, die Lysolecithin in Glyceryl hosphorylcholin abbaut, das seinerseits durch eine Diesterase in
MARINETTI und Mitarbeiter (1958) zeigen, dai3 in der Leber noch ein weiteres Enzym vorkommt, das umgekehrt Lysolecithin in Lecithin zuriickiiberfiihrt und das durch Hitze und teilweise auch durch Cyanid inaktiviert werden kann. Die Menge des in der Leber vorkommenden Lysolecithins wird damit offenbar durch einen feinen Regelmechanismus gesteuert, an dem wahrschein- lich 3 Enzyme beteiligt sind, die den Aufbau bzw. Abbau dieser Substanz bewirken. Es erscheint denkbar, dai3 der die Lysolecithinbildung in der Leber steuernde Mechanismus durch eine Infektion mit dem Agens der Rinderleukose bei der Maus in der Weise gestort wird, dai3 es zu einer anormalen Erhohung des Lysolecithingehaltes in den Lebern der Versuchstiere kommt.
glycerop K osphorige Saure und Cholin gespalten wird. Daruber hinaus konnten

Untersuchungen iiber die Obertragbarkeit eines Agens auf weide Mause 537
Die Anwesenheit des Agens der Rinderleukose in der weii3en Maus, die auf die Infektion hin keinerlei sichtbare Krankheitserscheinungen erkennen lafit, wurde sich damit durch das patho-physiologische Symptom einer Ab- wandlung der normalen Enzymtatigkeit in ahnlicher Weise nachweisen lassen wie die des Kaninchenpapillomvirus im Menschen. Das Papillomvirus (SHOPE) induziert in den Epithelzellen der Kaninchen die Ausbildung eines neuen Enzymes, einer Arginase. Mit dem Virus infizierte Kaninchen weisen hierdurch bedingt eine geringere Argininkonzentration in ihrem Blute auf als gesunde Kontrolltiere (ROGERS, 1959; EVANS u. ROGERS, 1967). Personen, die mit dem Kaninchenpapillomvirus gearbeitet haben, tragen dieses Virus vielfach off en- bar als harmlosen Passanten in sich. Sie zeigen infolge der Infektion keinerlei Krankheitserscheinungen. Die Anwesenheit des Virus in ihrem Organismus gibt sich nach den Untersuchungen von ROGERS (1966) nur darin zu erkennen, dai3 sie wie infizierte Kaninchen in ihrem Blut eine geringere Argininkonzen- tration aufweisen als Kontrollpersonen.
Verschiedene Umstande haben dazu beigetragen, dai3 diese Untersuchun- gen nicht mehr als Hinweise der genannten Art zu geben vermogen.
Es zeigte sich, dai3 das zur Hamolysinbildung in den Lebern der Versuchs- mause fuhrende Agens durchaus nicht in jedem leuketischen Tumormaterial vom Rinde nachweisbar war. Den Grundversuch eingeschlossen war es in 9 Versuchen nur dreimal sicher nachweisbar.
Die Resistenz des Agens in der Umwelt ist offensichtlich gering. Epide- miologische Beobachtungen sprechen dafur, dai3 das in gleicher Weise fur den Erreger der Rinderleukose zutrifft. Da sich das Agens in der Gefriertruhe bei - 7OoC nur 5-7 Monate hielt, entstand sehr bald ein Materialmangel. Es war deshalb nicht moglich, zu den Neutralisationsversuchen mehr Blutprobeii sowohl von gesunden als auch von leukamisch erkrankten Rindern heranzu- ziehen und weitere Titrationen durchzufuhren.
Es mui3te bei den Mauseversuchen stets mit Gruppen von 30-40 Tieren gearbeitet werden, um zu verwertbaren Ergebnissen zu kommen. Der Grund hierfur lag darin, dai3 das Agens durchaus nicht bei allen infizierten Mausen zu einer Hamolysinbildung in den Lebern Anlai3 gab. Die Hoffnung diese Fahig- keit durch weitere Mausepassagen uber das in Tabelle 4 angegebene Mai3 zu erhohen, erfullte sich nicht. Beriicksichtigt man jedoch, dai3 bei der Ober- tragung von Rinderleukosematerial auf Versuchskalbern in der Regel auch nur etwa 3 O o / o der Kalber erkranken (BEDERKE u. TOLLE, 1964; TOLLE, 1966; BEDERKE, TOLLE u. LOPPNOW, 1967), so ergibt sich auch hier eine Parallelitat zu dem Erreger der Rinderleukose.
Zusammenfassung Es wird uber Versuche berichtet, bei denen leukotisches Tumorinaterial
vom Rind auf mannliche weii3e Mause des NMRI-Stammes ubertragen wurde. Bei 3 von derartigen Obertragungsversuchen kam es in der Leber der Mause in einem wenige Tage umfassenden, etwa 3 Tage p. i. beginnenden Zeitraum zur Ausbildung einer hamolysierenden Substanz, die mit Lysolecithin nahe ver- wandt oder identisch war. Das diese Stoffwechselstorung auslosende Agens konnte 3 bis 4 Passagen lang in Mausen fortgezuchtet werden. Suspensionen hamolysinhaltiger Mauselebern aus diesen Passagen wurden auf 13 Versuchs- kalber aus leukosefreien Betrieben ruckubertragen. Drei von ihnen erkrankten etwa 13 Monate p. i. an einer persistierenden Lymphozytose im mai3ig erhoh- ten Bereich. Bei einem konnte histologisch die tumorose Form der enzootischen Leukose des Rindes nachgewiesen werden. In Neutralisationsversuchen an Mausen konnte das Agens mit Blutplasma und Vollblut einer leukosefreien
35*

538 E. MITSCHERLICH
Kuh nicht neutralisiert werden. Seine Neutralisation gelang auch nicht rnit Blutplasma und ebenso nicht mit Blutzellen einer leukoseinfizierten Kuh, die an einer prognostisch giinstig zu beurteilenden persistierenden Lymphozytose erkrankt war. Die Neutralisation gelang jedoch sowohl mit Vollblut als auch mit ausgewaschenen Blutzellen dieser Kuh, die bis zu ihrem zuvor natiirlicher- weise im Tier gegebenen Blutzellen-Plasma-Verhaltnis in Blutplasma resuspen- diert worden waren, das zu 2 O/o der fertigen Suspension aus Plasma derselben leukoseinfizierten Kuh und zum Rest aus Plasma der leukosefreien Kuh bestand.
Die Resistenz des Agens in der Umwelt ist gering. Es halt sich in der Gefriertruhe bei - 70’ C in einer 20 “/oigen Mauselebersuspension in Earle- Losung 5-7 Monate und biii3t bei Zimmertemperatur in einer 0,05 O/oigen Lebersuspension in Earle-Losung in 1 Stunde seine Wirksamkeit ein.
Die Versuchsergebnisse fiihren zu der Annahme, dai3 das den Leberstoff- wechsel der Versuchsmause stor.ende Agens mit dem Erreger der Rinderleukose in direktem Zusammenhang steht oder mit ihm identisch ist, und dai3 im Blute von Rindern, die das leukamische Stadium der Rinderleukose iiberstehen, Abwehrkrafte wirksam sind, zu denen Blutzellen und bestimmte Plasmafrak- tionen gehoren.
Summary Studies on the transmission to white mice of an agent present in bovine
leucotic tumours Tumor material of bovine leukosis was experimentally transmitted to
male NMRI-mice. In 3 out of 9 experiments a haemolytic substance was found in the livers of the mice some days after the transmission of the material. The substance reacted chemically as lysolecithin. The agent which caused this disturbance of liver metabolism was transmitted in 3-4 passages from mouse to mouse. Liver suspensions of mice containing the haemolytic sub- stance were inoculated into 13 calves. Three of them showed a persistent lymphocytosis 13 months after inoculation. Histologically in one of these animals the tumorous form of enzootic bovine leucosis could be found. In a limited number of experiments it was found that the agent which caused the synthesis of a haemolysin in the livers of mice was neutralized by blood cells plus plasma of a cow which was apparently recovering from bovine leucosis 4 years after experimental infection. N o neutralizing effect was observed when blood cells alone or plasma alone of this cow were employed. Whole blood of a non-infected healthy cow did not neutralize the agent. The experi- ments lead to the assumption that the agent which causes the production of the haemolytic substance in the livers of mice is closely related to or identical with, the agent of bovine leukosis.
RCsumC Recherches sur la transmissibiliti sur la souris blanche d’un agen trouvC
dam des prklbvements de tumeurs leucotiques bovines On rapporte des experiences cosistant h transmettre des prklbvements
de tumeurs leucotiques bovines sur des souris blanches miles de la souche NMRI. Dans 3 de ces expkriences de transmission, on observe la formation d’une substance hkmolysante, trbs proche ou identique ?i la lysolkcithine, qui fait son apparition dans le foie des souris dans un laps de temps de quel- ques jours, commensant environ 3 jours p. i. L’agent qui provoque ce trouble du mktabolisme a pu &re maintenu en culture pendant 3 A 4 passages. On a retransmis des suspensions de foies de souris contenant de l’hkmolysine, provenant de ces passages, sur 13 veaux d’expkrience pris dans des exploita- tions exemptes de leucose. Trois d’entre eux sont atteints a p r h environ 13

Untersuchungen uber die Ubertragbarkeit eines Agens auf weiBe Mause 539
mois p. i. d’une lymphocytose persistante de degrk moyen, dans un des tumeurs leucotiques se sont dkveloppks. On ne parvient pas A neutraliser l’agent par le plasma sanguin et le sang complet d’une vache exempte de leucose dans des expkriences de neutralisation. On ne parvient pas davantage A sa neutralisation par le plasma sanguin ou par des globules sanguins d’une vache leucotique, atteinte d’une lymphocytose persistante, au pronostic favorable. Mais la neutralisation rkussit pourtant avec du sang complet, de mkme qu’avec des globules sanguins de cette vache lavks et remis en suspension, dans des proprotions globules sanguins - plasma naturelles, dans un plasma sanguin constituk de 2 O/o de cette suspension de plasma de la vache leucotique et complktk avec le plasma de la vache aleucotique.
La rksistance de l’agent est faible dans le milieu ambiant. I1 se conserve 5-7 mois dans le congklateur A -70’ C dans une suspension A 20 O/o de foie de souris en solution Earle et perd son activitk en 1 heure A la tempkrature ambiante dans une suspension A 0,05 O/o en solution Earle.
Les rksultats de l’expkrience nous amhen t A l’hypothkse que l’agent perturbant le mktabolisme du foie des souris d’expkrience se trouve en rela- tion directe avec l’agent de la leucose bovine ou est identique avec lui et que, dans le sang des bovins qui surmontent le stade leuckmique, agit un mkcanisme de dkfense, auquel participent les globules sanguins et des fractions dkfinies du plasma.
Resumen Estudios sobre la transmisibilidad a1 rat6n blanco de un agente que aparece
en el material tumoral leuc6tico de 10s bovinos Se inforina sobre ensayos, en 10s que se transmiti6 material tumoral
leucbtico de reses vacunas a ratones blancos, machos, de la estirpe NMRI. En 3 de tales ensayos de transmisidn se form6 en el higado de 10s ratones en un espacio de tiempo que abarcaba pocos dias, comenzando hacia 10s 3 dias p. i., una substancia hemolizante, que tenia un parentesco muy estrecho o era idkn- tica a la lisolecitina. El agente que desencadenaba este trastorno metab6lico se podia seguir cultivando durante 3 6 4 pases por ratones. Suspensiones de higados murinos, que contenian hemolisina, de estos pases se retrotransmitie- ron a 13 terneras de ensayo procedentes de explotaciones libres de leucosis. Tres de las mismas enfermaron unos 13 meses p. i. de una linfocitosis persis- tente en la gradacibn ligeramente aumentada. En ensayos de neutralizaci6n en ratones no se pudo neutralizar el agente con plasma sanguiiieo y sangre completa de una vaca libre de leucosis. Su neutralizaci6n tampoco se logr6 con plasma sanguineo ni tampoco con cklulas hemiticas de una vaca infectada de leucosis, que habia enfermado de una linfocitosis persistente de pron6stico favorable. Sin embargo, la neutralizacidn se lo& tanto con sangre completa como con cklulas lavadas de esta vaca, que habian sido resuspendidas en plasma sanguineo hasta la relaci6n natural cklulas hemiticas-plasma dada antes en el animal, consistente en un 2 O/o de la suspensi6n preparada de plasma de la misma vaca infectada de leucosis y hasta completar en plasma de la vaca libre de leucosis.
La resistencia del agente en el medio ambiente es escasa. Se mantiene eficaz en el congelador a - 7OoC en una suspensi6n de higado murino a1 20 O/o en soluci6n Earle durante 5-7 meses, perdiendo su actividad a la tem- peratura de habitaci6n en una suspensi6n de higado a1 0,05 O/o en soluci6n Earle a1 cab0 de 1 hora.
Los resultados obtenidos hacen suponer que el agente que altera el inetabolismo hepitico de 10s ratones de experimentacibn se halla en relaci6n

540 E. MITSCHERLICH
intima con el agente etiolbgico de la leucosis bovina o es idkntico a kl, y que en la sangre de reses vacunas que se recuperan del estadio leuckmico de la leucosis bovina intervienen fuerzas de defensa, a las que pertenecen las cklulas hemiticas y ciertas fracciones plasmiticas.
Literaturverzeichnis 1 . BEDERKE, G., und A. TOLLE, 1964: Zur Ubertragbarkeit der Rinderleukose durch
das Blut und den Kontakt mit experimentell behandelten Tieren. Zbl. Vet. Med. B, 11, 433-442. 2. BEDERKE, G., A. TOLLE und H. LOPPNOW, 1967: Ubertragungsversuche mit leukotischem Tumormaterial auf das Rind. Zbl. Vet. Med. B, 14, 32-48. 3. BERGENHEM, B., und R. FAHRAEUS, 1936: Ober spontane Hamolysinbildung im Blut unter besonderer Berucksichtigung der Physiologie der Milz. Z. exper. Med. 97, 555-687. 4. DAWSON, R.M.C., 1956: The phospholipase B of liver. Biochem. J. 64, 192-196. 5. DEUEL, H . H . jr., 1951 : The lipids. Their chemistry and biochemistry. Band I. Interscience publisher, New York. 6. Derselbe, 1955: The lipids. Band 111. Interscience publisher, New York. 7. EVANS, H., and ST. ROGERS, 1967: Relative depletion of an arginine-rich deoxyribonucleo- histone component during tumor induction by the Shope papilloma virus. Exper. a. molec. Pathol. 7, 105-115. 8. GROSS, L., 1947: Hemolytic action of mouse mammary carcinoma filtrate on mouse erythrocytes in vitro. Proc. SOC. exp. Biol. a. Med. 65, 292-293. 9. Derselbe, 1948 a : Increased hemolytic potency of mouse mammary carcinoma extracts following incubation with tumor cells. Proc. SOC. exp. Biol. a. Med. 67, 341-343. 10. Derselbe, 1948 b: Destructive action of mouse and rat tumor extracts on red blood cells in vitro. J. Immunol. 59, 173-179. 11. HOLMAN, R. T., W. 0. LUNDBERG and T. MALKIN, 1957: Progress in the chemistry of fats and other lipids. 4, 107, 142. Pergamon Press, Lon- don-New York-Paris. * 12. KELLER, R., 1965: Zur Bedeutung von Lysolecithin fur die Immunocytolyse von Mastzellen. Path. Microbiol. 28, 185-196. 13. KLIBANSKY, C I ~ . and A. DE VRIES, 1963: Quantitative study of erythrocytelysolecithin interaction. Biochini. biophys. Acta 70, 176-187. 14. LEHNERT, E., 1964: Zur Frage der Existenz leukosespe- zifischer Antikorper im Blutserum leukotischer Rinder. Berl. Munch. Tierarztl. Wschr. 77, 113-115. 15. MARINETTI, G. V., R. F. WITTER and E. STOTZ, 1957 a : The incorpora- tion in vivo of Pse - labelled orthophosphate into individual phosphatides of rat tissues 5. biol. Chem. 226, 475-483. 16. MARINETTI, G. V., J. KOCHEN, J. ERBLAND and E. STOTZ, 1957 b: The lipids composition of a purified cytohrome preparation of pig heart. 1. biol. Chem. 229, 1027-1035. 17. MARINETTI, G. V., J. ERBLAND, R. F. WITTER, J. PERIX and E. STOTZ, 1958: Metabolic pathways of lysolecithin in a soluble rat-liver system. Biochem. biophys. Acta 30, 223. 18. MARINETTI, G.V., M. ALBRECHT, T. FORD and E. STOTZ, 1959: Analysis of human plasma phosphatides by paper chromatography. Biochirn. biophys. Acta 36, 4-13. 19. MONTEMAGNO, F., 1967: Viral etiology of the bovine Ivmpholeucosis. 3rd. intern. Symp. comp. leucemia Res., Paris. 20. PHILLIPS, G. B., 1958: The isolation and quantitation of the principle phospholipids components of human serum using chromatography on silicic acid. Biochim. biophys. Acta 29, 594-602. 21. POLLARD, M., 1967: Germ-free animals in comparative medicine. Adv. vet. Sci. 11, 139-157. 22. ROGERS, ST., 1959: Induction of arginase in rabbit epithelium by the Shope rabbit papil- loma virus. Nature 183, 1815-1816. 23. Derselbe, 1966: Shope papilloma virus: A passen- ger in man and its significance to the potential control of the host genome. Nature 212, 1220-1222. 24. ROSENBERGER, G., 1961 : Ergebnisse zehnjahriger Leukoseuntersuchungen an der Rinderklinik Hannover. Tagungsbericht 49, Leukose des Rindes. Deutsche Akademie d. Landwirtschaftswissenschft. Berlin. 25. SCHMIDT, F.-W., 1968: Prufung des Latextestes zur serologischen Diagnose der Rinderleukose. Zbl. Vet. Med. B, 15, 174-184. 26. SCHREIER, CH., und I.-R. SCHIMMELPFENNIG, 1965: Histologische Untersuchungen an Primar- tumoren weiblicher NMRI-Mause. Arch. exp. Vet. Med. 19, 1037-1051. 27. SCHREIER, CH., A. TOLLE, F.-W. SCHMIDT und I.-R. SCHIMMELPFENNIG, 1967: Zur Tumorbildung bei Mausen des Insuchtstammes NMRI nach Injektion von Rinderleukosematerial und Versuche zur Ruckubertragung dieses Maustumors auf Kalber. Zbl. Vet. Med., B, 14, 585-597. 28. STRAUB, 0. C., 1965: The difficulty in interpreting results of mouse transmission studies of bovine leukosis. Proc. int. Wenner-Gren Symp. Stockholm, Comp. leukaeniia Res 245-249 (1966). 29. TITUS, E., H. WEISS and ST. HAYDU, 1956: Isolation of a cardiacactive principle from mammalian tissues. Science 124, 1205-1206. 30. TOLLE, A., 1966: Zur Obertragbarkeit der Rinderleukose. Zbl. Bakt. Abt. I. Orig. 198, 142-149. 31 . WITTMANN, W., 1963: Zu einigen Problemen der Leukoseforschung beim Rind. Mh. Vet. Med. 18, 843-849.
Anschrift des Verfassers: Prof. Dr. E. Mitscherlich, Tierarztliches Institut der Universi- tat Gottingen, 34 Gottingen, Groner LandstraBe 2.





















![Definition · 2019. 5. 16. · (HMP06/MAR.00155 Hamburger Morgenpost, 02.03.2006, S. 8\; Die Wilden Kerle 3 [\;Ausführliche Zitierung nicht verfügbar]\;) Agens: Beispiel:Immer wieder](https://img.pdfslide.org/doc/110x75/60c0dcbd4b7999558f05d7a1/definition-2019-5-16-hmp06mar00155-hamburger-morgenpost-02032006-s.jpg)







