Embed Size (px)
Citation preview

Aus dem Institut für Tierzucht
der Bundesforschungsanstalt für Landwirtschaft (FAL) in Mariensee
Untersuchungen zur Florfenicolresistenz bei
grampositiven und gramnegativen bakteriellen
Infektionserregern
Habilitationsschrift zur Erlangung der
VENIA LEGENDI
an der Tierärztlichen Hochschule Hannover
Vorgelegt von
Dr. Corinna Kehrenberg, Ph.D.
aus Lüdenscheid
Hannover 2007

für meine Familie für meine Familie für meine Familie für meine Familie und Frankund Frankund Frankund Frank

A journey of a thousand miles must begin with a single stepA journey of a thousand miles must begin with a single stepA journey of a thousand miles must begin with a single stepA journey of a thousand miles must begin with a single step
(Lao Tsu)(Lao Tsu)(Lao Tsu)(Lao Tsu)

1 Liste der Publikationen, die Bestandteil der Ha bilitationsschrift sind ............. 1
2 Einführung in die Thematik ................................................................................. 3
2.1 Florfenicol - Eingruppierung des Wirkstoffes und Molekülstruktur.................. 3 2.2 Chemische und pharmakologische Eigenschaften von Florfenicol ............... 5 2.3 Gründe für die Entwicklung und den Einsatz von Florfenicol ........................ 6 2.4 Zulassung und Anwendung des Wirkstoffes Florfenicol................................. 8 2.5 Wirkungsweise und Wirkungsspektrum von Florfenicol sowie der
Grundsubstanz Chloramphenicol ................................................................. 10 2.6 Resistenzentwicklung gegenüber Florfenicol im Unterschied zu
Chloramphenicol .......................................................................................... 12
2.6.1 Spezifische Chloramphenicol-Resistenzgene......................................... 13
2.6.2 Resistenzgene, die kombinierte Florfenicol- und Chloramphenicolresistenz vermitteln ...................................................... 16
2.7 Regulation der Expression von Chloramphenicol- und Florfenicol-
Resistenzgenen ........................................................................................... 19
2.8 Empfindlichkeitslage bakterieller Infektionserreger gegenüber Florfenicol ..... 22
2.8.1 Empfindlichkeitslage von Zielorganismen............................................... 23
2.8.2 Empfindlichkeitslage von Kommensalen oder Indikatorbakterien ........... 27
2.9 Florfenicol als Modellsubstanz ..................................................................... 34
3 Zielstellung der Arbeit ........................................................................................ 37
4 Einordnung der Resultate und zusammenfassende D iskussion ................... 39
4.1 Teil I: Atemwegsinfektionserreger von Rind u nd Schwein ...................... 39
4.1.1 Untersuchungen zur Resistenzlage bei Zielorganismen für Florfenicol .. 39
4.1.2 Auftreten und Analyse erster florfenicolresistenter Zielorganismen aus dem Vereinigten Königreich (UK)..................................................... 45
4.1.3 Auftreten und Analyse weiterer florfenicolresistenter Zielorganismen aus Frankreich ........................................................................................ 48
4.1.4 Auftreten erster florfenicolresistenter Zielbakterien in Deutschland ........ 51
4.1.5 Untersuchungen zur Florfenicol- und Chloramphenicolresistenz bei Bordetella bronchiseptica........................................................................ 52

4.2 Teil II: Kommensalen (oder Indikatorbakterie n) ........................................ 57
4.2.1 A) Escherichia coli ............................................................................ 57
4.2.1.1 Nachweis von floR-Genen bei Escherichia coli ................................. 57
4.2.1.2 Identifizierung des floR-tragenden Transposons TnfloR.................... 59
4.2.2 B) Staphylococcus spp. .................................................................... 61
4.2.2.1 Analyse des Plasmidtyps pSCFS2 mit Identifizierung des neuen Florfenicol-Resistenzgens fexA ......................................................... 64
4.2.2.2 Nachweis der Mobilität des fexA-Gens: Identifizierung eines neuen Transposons...................................................................................... 66
4.2.2.3 Analyse des Plasmidtyps pSCFS1 mit Identifizierung des neuen Resistenzgens cfr .............................................................................. 69
4.2.2.4 Lokalisation des Gens cfr auf einem Multiresistenzplasmid: Identifizierung des neuen Gens erm(33) ........................................... 71
4.2.2.5 Komplette Sequenzanalyse und Organisation des Plasmids pSCFS1 mit Identifizierung eines neuen ABC-Transporters.............. 73
4.2.2.6 Nachweis des durch das Gen cfr vermittelten Resistenz- mechanismus .................................................................................... 76
4.2.2.7 Multiresistenz durch den cfr-vermittelten Resistenzphänotyp PhLOPSA........................................................................................... 85
4.2.2.8 Verteilung der Resistenzgene cfr und fexA bei chloramphenicol-resistenten Staphylokokken............................................................... 92
4.2.2.9 Die Mobilität des cfr-Gens wird durch IS21-558-Elemente vermittelt............................................................................................ 97
4.2.2.10 Identifizierung des cfr-Gens bei einem MRSA-Isolat vom Menschen........................................................................................ 102
5 Zusammenfassung ........................................................................................... 105
6 Literaturverzeichnis .......................................................................................... 108
7 Darstellung des eigenen Anteils an den Publikat ionen ................................129
8 Publikationen .................................................................................................... 133

PUBLIKATIONSLISTE
1
1 Liste der Publikationen, die Bestandteil der Habili tations-
schrift sind
1. Schwarz S., C. Kehrenberg, B. Doublet and A. Cloeck aert:
Molecular basis of bacterial resistance to chloramphenicol and florfenicol.
FEMS Microbiol. Rev. (2004), 28, 519-542.
2. Kehrenberg C., J. Mumme, J. Wallmann, J. Verspohl, R. Tegeler, T. Kuhn and S. Schwarz:
Monitoring of florfenicol susceptibility among bovine and porcine respiratory tract pathogens collected in Germany during the years 2002 and 2003.
J. Antimicrob. Chemother. (2004), 54, 572-574.
3. Kehrenberg C. and S. Schwarz:
Plasmid-borne florfenicol resistance in Pasteurella multocida.
J. Antimicrob. Chemother. (2005), 55, 773-775.
4. Kehrenberg C., D. Meunier, H. Targant, A. Cloeckaer t, S. Schwarz and J.-Y. Madec:
Plasmid-mediated florfenicol resistance in Pasteurella trehalosi.
J. Antimicrob. Chemother. (2006), 58, 13-17.
5. Kadlec K., C. Kehrenberg and S. Schwarz:
Efflux-mediated resistance to florfenicol and/or chloramphenicol in Bordetella bronchiseptica: identification of a novel chloramphenicol exporter.
J. Antimicrob. Chemother. (2007), 59, 191-196.
6. Cloeckaert A., S. Baucheron, G. Flaujac, S. Schwarz , C. Kehrenberg, J.-L. Martel and E. Chaslus-Dancla:
Plasmid-mediated florfenicol resistance encoded by the floR gene in Escherichia coli isolated from cattle.
Antimicrob. Agents Chemother. (2000), 44, 2858-2860.
7. Doublet B., S. Schwarz, C. Kehrenberg and A. Cloeck aert:
Florfenicol resistance gene floR is part of a novel transposon.
Antimicrob. Agents Chemother. (2005), 49, 2106-2108.
8. Kehrenberg C. and S.Schwarz:
fexA, a novel Staphylococcus lentus gene encoding resistance to florfenicol and chloramphenicol.
Antimicrob. Agents Chemother. (2004), 48, 615-618.

PUBLIKATIONSLISTE
2
9. Kehrenberg C. and S. Schwarz:
Florfenicol-chloramphenicol exporter gene fexA is part of the novel transposon Tn558.
Antimicrob. Agents Chemother. (2005), 49, 813-815.
10. Schwarz S., C. Werckenthin and C. Kehrenberg:
Identification of a plasmid-borne chloramphenicol-florfenicol resistance gene in Staphylococcus sciuri.
Antimicrob. Agents Chemother. (2000), 44, 2530-2533.
11. Schwarz S., C. Kehrenberg and K.K. Ojo:
Staphylococcus sciuri gene erm(33), encoding inducible resistance to macrolides, lincosamides, and streptogramin B antibiotics, is a product of recombination between erm(C) and erm(A).
Antimicrob. Agents Chemother. (2002), 46, 3621-3623.
12. Kehrenberg C., K.K. Ojo and S. Schwarz:
Nucleotide sequence and organization of the multiresistance plasmid pSCFS1 from Staphylococcus sciuri.
J. Antimicrob. Chemother. (2004), 54, 936-939.
13. Kehrenberg C., S. Schwarz, L. Jacobsen, L.H. Hansen and B. Vester:
A new mechanism for chloramphenicol, florfenicol and clindamycin resistance: methylation of 23S ribosomal RNA at A2503.
Mol. Microbiol. (2005), 57, 1064-1073.
14. Long K., J. Poehlsgaard, C. Kehrenberg, S. Schwarz and B. Vester:
The Cfr rRNA methyltransferase confers resistance to Phenicols, Lincosamides, Oxazolidinones, Pleuromutilins, and Streptogramin A antibiotics.
Antimicrob. Agents Chemother. (2006), 50, 2500-2505.
15. Kehrenberg C. and S. Schwarz:
Distribution of florfenicol resistance genes fexA and cfr among chloramphenicol-resistant Staphylococcus isolates.
Antimicrob. Agents Chemother. (2006), 50, 1156-1163.
16. Kehrenberg C., F. Aarestrup and S. Schwarz:
IS21-558 insertion sequences are involved in the mobility of the multiresistance gene cfr.
Antimicrob. Agents Chemother. (2007) 51, 483-487. Auf diese Publikationen wird im Text Bezug genommen. Ihre Auflistung erfolgte
entsprechend der inhaltlichen Einteilung.

EINFÜHRUNG IN DIE THEMATIK
3
2 Einführung in die Thematik
2.1 Florfenicol - Eingruppierung des Wirkstoffes un d Molekülstruktur
Der Wirkstoff Florfenicol ist ein Antiinfektivum, welches in die Klasse der Phenicole
eingruppiert wird. In dieser Substanzklasse, die im weiteren durch Chloramphenicol,
Thiamphenicol oder Azidamphenicol vertreten wird, sind Phenylalanin-Derivate zu
finden. Diese Substanzen zeigen keine strukturelle Verwandtschaft zu anderen
Antibiotika und wurden zum Teil schon in den frühen Jahren der
Antibiotikaentwicklung entdeckt. Chloramphenicol konnte bereits 1947 aus dem
natürlichen Produzenten Streptomyces venezuelae isoliert werden und aufgrund der
recht einfachen Molekülstruktur gelang schon 1950 die chemisch-synthetische
Herstellung des Wirkstoffes.
Aufbauend auf diesem Molekül fanden Substitutionen an unterschiedlichen Resten
statt, die zur Entwicklung der weiteren Vertreter aus der Substanzklasse geführt
haben. Die Grundsubstanz Chloramphenicol sowie die vorgenommenen
Substitutionen bei den Wirkstoffen Florfenicol, Thiamphenicol und Azidamphenicol
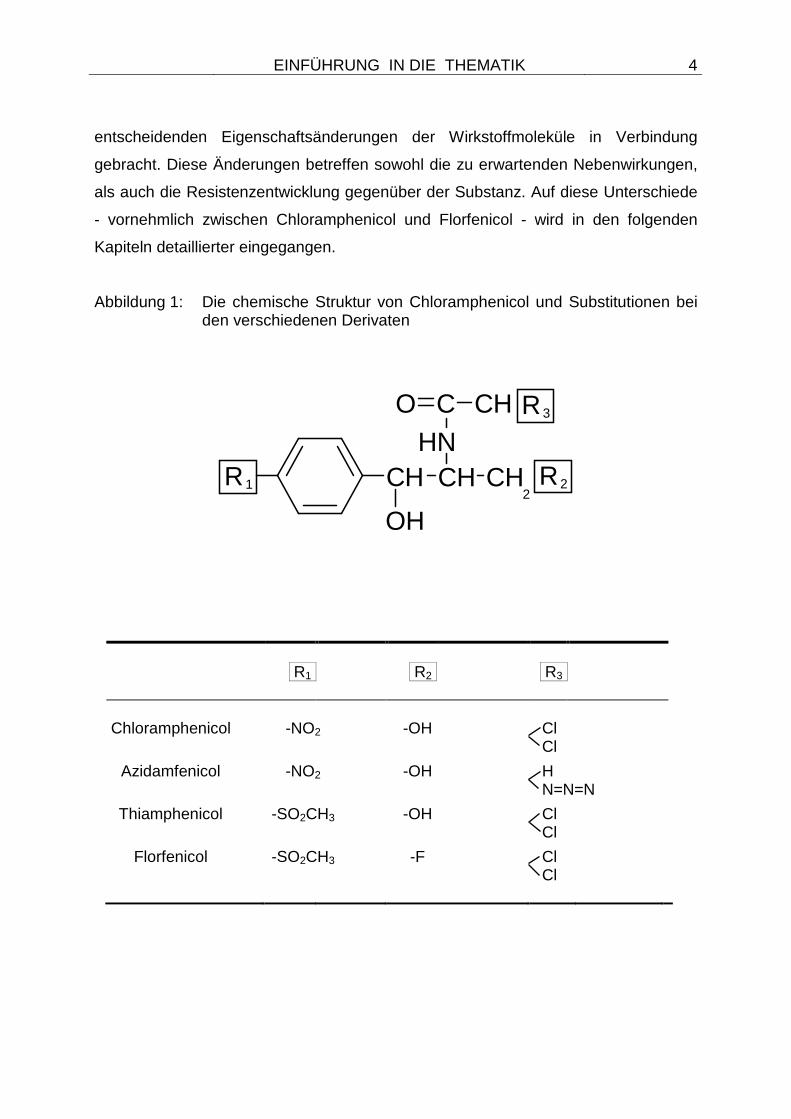
sind in Abbildung 1 dargestellt. Bei der Substanz Florfenicol handelt es sich um ein
fluoriertes Derivat von Chloramphenicol (WHITE et al. 2000), welches sich durch
zwei wesentliche strukturelle Änderungen von seiner Grundsubstanz unterscheidet.
So wurde die p-Nitrogruppe des Chloramphenicols beim Florfenicol durch eine
p-Methyl-Sulfonyl-Gruppe ersetzt sowie die Hydroxylgruppe in der terminalen
Alkoholfunktion durch ein Fluoratom substituiert (EMEA - THE EUROPEAN AGENCY
FOR THE EVALUATION OF MEDICINAL PRODUCTS 1995 - summary report 1).
Die gleiche Substitution der Nitrogruppe wurde auch bei dem Wirkstoff
Thiamphenicol vorgenommen, die Hydroxylgruppe blieb aber unverändert.
Azidamphenicol unterscheidet sich dagegen durch eine Dehalogenierung von der
Grundsubstanz Chloramphenicol (Abbildung 1). Obgleich für alle Substanzen aus der
Klasse der Phenicole ähnliche physiko-chemische Eigenschaften angegeben werden
(ROSIN 1992; THE UNITED STATES PHARMACOPEIAL CONVENTION 2007;
Publikation 1), werden die beschriebenen Substitutionen in der Molekülstruktur mit
EINFÜHRUNG IN DIE THEMATIK
4
entscheidenden Eigenschaftsänderungen der Wirkstoffmoleküle in Verbindung
gebracht. Diese Änderungen betreffen sowohl die zu erwartenden Nebenwirkungen,
als auch die Resistenzentwicklung gegenüber der Substanz. Auf diese Unterschiede
- vornehmlich zwischen Chloramphenicol und Florfenicol - wird in den folgenden
Kapiteln detaillierter eingegangen.
Abbildung 1: Die chemische Struktur von Chloramphenicol und Substitutionen bei den verschiedenen Derivaten
C CHO
CH
OH
CH CH2
R2
NR3
HR1
R1 R2 R3 Chloramphenicol -NO2 -OH Cl
Cl
Azidamfenicol -NO2 -OH H N=N=N
Thiamphenicol -SO2CH3 -OH Cl Cl
Florfenicol -SO2CH3 -F Cl Cl

EINFÜHRUNG IN DIE THEMATIK
5
2.2 Chemische und pharmakologische Eigenschaften v on Florfenicol
Im Gegensatz zu der Grundsubstanz Chloramphenicol, die ein Stoffwechselprodukt
einiger Actinomyceten ist und ursprünglich aus Streptomyces venezuelae isoliert
wurde (VINING u. STUTTARD 1995), handelt es sich bei dem Wirkstoff Florfenicol
um eine rein synthetische Verbindung. Diese Verbindung trägt den chemischen
Namen d-threo-3-fluoro-2-dichloracetamido-1-(4-methylsulfonylphenyl)-1-propanol
und zeichnet sich durch ein Molekulargewicht von 358,21 und einen Schmelzpunkt
zwischen 153 und 154°C aus (USP DICTIONARY OF USAN AND INTERNATIONAL
DRUG NAMES 2002; THE UNITED STATES PHARMACOPEIAL CONVENTION
2007; BUDAVARI 1996). Bei einem pH-Wert von 3 bis 9 liegt Florfenicol als nicht
ionisiertes Molekül vor (SAMS 1994). Ähnlich dem Chloramphenicol weist auch beim
Florfenicol nur das Isomer mit der D(-)-threo-Konfiguration antibakterielle Aktivität
auf. Florfenicol zeigt in organischen Lösungsmitteln (SAMS 1994) eine gute
Löslichkeit, aber auch in wässrigen Lösungen (BUDAVARI 1996; THE UNITED
STATES PHARMACOPEIAL CONVENTION 2007; SAMS 1994) sowie in alkalischen
Medien ist die Substanz löslich. Die Eigenschaft der Lipophilie stellt die Grundlage
für eine gute Penetration von Florfenicol in Gewebe dar. Im tierischen Organismus
wird die Substanz zu unterschiedlichen Metaboliten verstoffwechselt. Dazu zählen
das Florfenicol-Amin, Florfenicol-Alkohol, Florfenicol-Oxaminsäure, Monochlor-
Florfenicol und ein Glukuronsäure-Konjugat des Florfenicol-Amins. (SCHERING-
PLOUGH ANIMAL HEALTH 1998). Dabei bildet das Monochlor-Florfenicol mit
ungefähr 30 % die Hauptkomponente der faekalen Ausscheidung, weitere faekal
nachweisbare Komponenten sind Florfenicol-Oxaminsäure sowie Florfenicol.
Hauptbestandteil der Harnkomponente ist dagegen Florfenicol mit ungefähr 64 %, als
weitere Harnmetaboliten werden die Florfenicol-Oxaminsäure (ca. 12 %), Florfenicol-
Amin, (ca. 8 %), Florfenicol-Alkohol (ca. 7 %), und Monochlor-Florfenicol (< 2 %)
gefunden (SCHERING-PLOUGH ANIMAL HEALTH 1998; PRIEBE 2003). Von allen
Metaboliten erweist sich das nicht mehr antibiotisch aktive Florfenicol-Amin im
Lebergewebe des Rindes als langlebigster Metabolit. Daher stellt das Florfenicol-
Amin auch einen geeigneten Marker für Rückstandsuntersuchungen dar
(SCHERING-PLOUGH ANIMAL HEALTH 1998; PRIEBE 2003).

EINFÜHRUNG IN DIE THEMATIK
6
2.3 Gründe für die Entwicklung und den Einsatz von Flor fenicol
Vertreter aus der Gruppe der Phenicole, wie Chloramphenicol, Thiamphenicol oder
Azidamphenicol, befinden sich bereits über Jahre in der humanmedizinischen
Nutzung. Doch die anfängliche Hoffnung bei der Entdeckung von Chloramphenicol,
eine vielversprechende Substanz mit breitem antibakteriellen Wirkungsspektrum für
therapeutische Zwecke zur Verfügung zu haben (SHAW 1983), wurde bereits Mitte
der 60er Jahre getrübt. Zu diesem Zeitpunkt wurden schwere, unerwünschte
Wirkungen mit der Anwendung von Chloramphenicol in Verbindung gebracht
(MARTELO et al. 1964). Die schwerste dieser unerwünschten Wirkungen ist eine
dosisunabhängige, irreversible aplastische Anämie, für deren Auftreten je nach
Studie eine Frequenz von 1:10.000-1:40.000 (SIMON u. STILLE 2000) oder
1:20.000-1:600.000 (VINING u. STUTTART 1995) angegeben wird. Der ursächliche
Mechanismus für diese Reaktion ist unbekannt. Neben einer genetischen
Prädisposition des Patienten wird die Reaktion eines der Reduktionsprodukte der
Nitrogruppe des Wirkstoffmoleküls mit der DNA diskutiert, wobei Destabilisierungen
der Doppelhelix oder Strangbrüche auftreten können (ROSIN 1992). Als weitere
unerwünschte Wirkungen wurden eine dosisabhängige reversible
Knochenmarksuppression, das Gray-Syndrom bei Neugeborenen, Hypersensitivität
und anaphylaktische Reaktionen beschrieben (SHAW 1983; YAO u. MOELLERING
1999). All diese unerwünschten Wirkungen sowie die Möglichkeit, auf weniger
toxische Substanzen zurückgreifen zu können, haben zu einer starken Reduktion
des Einsatzes von Chloramphenicol beim Menschen geführt. Heute wird die
Substanz nur noch bei wenigen, lebensbedrohlichen Infektionen eingesetzt (SHAW
1983; YAO u. MOELLERING 1999), bei denen keine anderen Wirkstoffe (z.B. durch
Unverträglichkeiten) zur Verfügung stehen. Durch die Eigenschaft des Wirkmoleküls
die Blut-Hirn-Schranke rasch passieren zu können, zählen dazu die Behandlung von
Meningitiden, hervorgerufen durch empfindliche Stämme von Haemophilus
influenzae, Neisseria meningitidis oder Streptococcus pneumoniae bei Patienten mit
einer Penicillin-Allergie (MASCARETTI 2003) oder lebensbedrohliche intraokuläre
Infektionen, bei denen andere Antibiotika unwirksam oder kontraindiziert sind
(SIMON u. STILLE 2000). Auch das wasserlösliche Azidamphenicol, welches

EINFÜHRUNG IN DIE THEMATIK
7
ebenfalls die umstrittene Nitrogruppe enthält, ist noch für therapeutische Zwecke
beim Menschen verfügbar. Die Anwendung ist aber auf den Einsatz als Bestandteil
von Augentropfen limitiert (SIMON u. STILLE 2000).
Aufgrund der erheblichen unerwünschten Wirkungen von Chloramphenicol wurde der
Wirkstoff bereits 1994 für die Anwendung bei lebensmittelliefernden Tieren verboten.
Dabei stand der Schutz des Verbrauchers vor Chloramphenicol-Rückständen im
Fleisch von Schlachttieren im Vordergrund der Entscheidung. Da die beim Menschen
auftretende irreversible aplastische Anämie unabhängig von der verabreichten Dosis
ist, konnte kein NOEL (non-observed effect level) für Chloramphenicol festgelegt
werden. Der NOEL-Wert gibt dabei die Dosis eines Wirkstoffes in toxikologischen
Studien an, unterhalb der unerwünschte Wirkungen nicht auftreten (SCHWARZ u.
CHASLUS-DANCLA 2001). Da dieser Wert aber die Basis zur Berechnung des MRL
(maximum residue level) darstellt, welcher als maximale Konzentration eines
antimikrobiellen Wirkstoffes in Schlachtkörpern definiert ist, bei der keine
unerwünschten Wirkungen beim Menschen auftreten, konnte für Chloramphenicol
ebenfalls kein MRL-Wert festgelegt werden. Dies führte 1994 zu einem Verbot des
Einsatzes von Chloramphenicol bei lebensmittelliefernden Tieren nach
Rechtsprechung der Europäischen Union (EU). Nicht unter dieses Verbot fällt
allerdings der veterinärmedizinische Einsatz von Chloramphenicol bei Haus- und
Heimtieren, die nicht der Lebensmittelgewinnung dienen.
Da die schwerste unerwünschte Wirkung von Chloramphenicol, die
dosisunabhängige, irreversible aplastische Anämie, nicht bei Tieren zu beobachten
war, fand eine Weiterentwicklung dieses potenten Wirkstoffes für die
veterinärmedizinische Anwendung statt. Dabei wurden die zuvor beschriebenen
Substitutionen durchgeführt, von denen vor allem der Austausch der umstrittenen
Nitrogruppe für die Rückstandsproblematik in tierischen Lebensmitteln von
wesentlicher Bedeutung war. Nach Einschätzung der EMEA (European Agency for
the Evaluation of Medicinal Products), die eine zusammenfassende Bewertung
(„summary report“) von Arzneimittel-Wirkstoffen vornimmt, reicht diese Substitution
nicht für einen eindeutigen Beweis aus, dass Florfenicol kein Potential für Störungen
der Haematopoese besitzt. Trotzdem wurde es von der EMEA-Arbeitsgruppe als

EINFÜHRUNG IN DIE THEMATIK
8
höchst unwahrscheinlich eingestuft, dass Rückstände von Florfenicol in tierischen
Lebensmitteln zu ernsthaften Dyskrasien beim Menschen führen können (EMEA -
THE EUROPEAN AGENCY FOR THE EVALUATION OF MEDICINAL PRODUCTS
1995 - summary report 1).
2.4 Zulassung und Anwendung des Wirkstoffes Florfen icol
Der Wirkstoff Florfenicol wurde ab 1995, also genau ein Jahr nach dem Verbot von
Chloramphenicol, in der EU zugelassen. Die Zulassung war zunächst allerdings auf
den Einsatz bei Rindern und das Anwendungsgebiet der therapeutischen
Behandlung von respiratorischen Erkrankungen aufgrund von Infektionen mit
Mannheimia haemolytica, Pasteurella multocida und Histophilus somni (ehemals
Haemophilus somnus) beschränkt (Publikation 1). Dabei wird der Wirkstoff alleinig
als Lösung zur intramuskulären oder subkutanen Injektion angeboten. Die Dosierung
beträgt 20 mg/kg Körpergewicht bei intramuskulärer (i.m.) sowie 40 mg/kg
Körpergewicht bei subkutaner (s.c.) Injektion. Je nach Injektionsart ist eine Wartezeit
von 30 Tagen (i.m.) bzw. 44 Tagen (s.c.) einzuhalten. Florfenicol darf weder bei
Zuchtbullen noch bei trächtigen oder laktierenden Rindern angewendet werden
(ESSEX TIERARZNEI FACHINFORMATION 2006).
Im Jahr 2000 wurde die Zulassung auch für die Bekämpfung respiratorischer
Erkrankungen beim Schwein, verursacht durch Pasteurella multocida und
Actinobacillus pleuropneumoniae, erweitert. Auch für den Einsatz beim Schwein ist
der Wirkstoff nur als Injektionslösung erhältlich und kann in einer Dosierung von 15
mg/kg Körpergewicht intramuskulär verabreicht werden. Die entsprechende
Wartezeit ist mit 18 Tagen festgelegt worden. Eine Erweiterung der Zulassung auch
für die Behandlung von Infektionen mit Streptococcus suis beim Schwein wird
seitens des Herstellers angestrebt (Prof. Dr. Stefan Schwarz, persönliche Mitteilung)
Der Wirkstoff wird in Deutschland von Essex Tierarznei, München, unter dem
Handelsnamen Nuflor für die Anwendung beim Rind und unter dem Handelsnamen
Nuflor Schwein für den Einsatz bei Schweinen vertrieben.

EINFÜHRUNG IN DIE THEMATIK
9
Während die Zulassung von Florfenicol in der EU bis heute auf diese wenigen
Anwendungsgebiete beschränkt blieb, fand in anderen Ländern ein darüber
hinausgehender Einsatz des Wirkstoffes statt. So ist Florfenicol in den USA zur
Behandlung der infektiösen Pododermatitis (Interdigitalphlegmone) beim Rind,
verursacht durch empfindliche Bakterien der Spezies Fusobacterium necrophorum
und Bacteroides melaninogenicus, zugelassen (THE UNITED STATES
PHARMACOPEIAL CONVENTION 2007). In Kanada ist es zudem möglich, eine bei
Rindern auftretende infektiöse bovine Keratoconjunktivitis, verursacht durch
Moraxella bovis, mit dem Wirkstoff zu behandeln (DUEGER et al. 1999; ANGELOS
et al. 2000). Weiterhin ist Florfenicol in Ländern wie den USA, Kanada, Japan,
Korea, Chile oder Norwegen zur Behandlung der Furunkulose bei Lachsen,
hervorgerufen durch Aeromonas salmonicida, zugelassen. Für diese Anwendung auf
kommerziellen Fischfarmen steht ein Prämix mit dem Handelsnamen Aquaflor für
eine Wassermedikation zur Verfügung (AQUAFLOR® PRODUCT LABELING
SCHERING-PLOUGH - CANADA 2006). In Kanada ist es zudem möglich, Florfenicol
als orale Lösung beim Schwein einzusetzen, um damit Infektionen, verursacht durch
die Erreger Salmonella Choleraesuis oder Streptococcus suis Typ 2, einzudämmen.
Zusätzlich kann der Wirkstoff in Kanada beim Wels eingesetzt werden, um die
Mortalität einer enterischen Septikämie, verursacht durch Edwardsiella ictaluri,
einzudämmen (THE UNITED STATES PHARMACOPEIAL CONVENTION 2007). In
Asien wird sogar von einem Einsatz des Wirkstoffes in Aquakulturen seit 1980
berichtet (KEYES et al. 2000; FUKUI et al. 1987).
Mittlerweile sind von der EMEA-Arbeitsgruppe MRL-Werte für Florfenicol für die
Tierarten Rind, Schwein, Geflügel und Fische akzeptiert worden (EMEA - THE
EUROPEAN AGENCY FOR THE EVALUATION OF MEDICINAL PRODUCTS 1995,
1997, 1999, 1999a, 2000, 2002 - summary reports 1 - 6). Florfenicol wurde somit in
den Anhang I der VO 2377/90 (EWG) für alle zur Lebensmittelerzeugung genutzten
Arten sowie Fische und Geflügel aufgenommen. Daher ist zukünftig mit einer
erweiterten Zulassung der Substanz in der EU zu rechnen. Empfehlungen für
Dosierungen bei weiteren Tierarten wie Hund, Katze, Schaf, Ziege und Fische sind
bereits unter www.vetidata.de unter dem Stichwort Florfenicol zu finden.

EINFÜHRUNG IN DIE THEMATIK
10
Im Gegensatz zu Chloramphenicol war das fluorierte Derivat Florfenicol zu keiner
Zeit für eine Anwendung beim Menschen zugelassen.
2.5 Wirkungsweise und Wirkungsspektrum von Florfeni col sowie der
Grundsubstanz Chloramphenicol
Die beiden Wirksubstanzen Chloramphenicol und das fluorierte Derivat Florfenicol
haben eine recht ähnliche Wirkungsweise. Beide Substanzen sind spezifische und
potente Inhibitoren der bakteriellen Proteinsynthese und binden mit einer hohen
Affinität an das Peptidyltransferasezentrum der 50S Untereinheit der Ribosomen.
Dies führt zu einer effektiven Hemmung der Verlängerung der Polypeptidkette.
(SCHWARZ 1995; SCHLÜNZEN et al. 2001). Dabei fungieren lediglich 70S
Ribosomen als Zielstrukturen, die 80S Ribosomen von eukaryotischen Zellen werden
nicht beeinflusst. Allerdings wird vermutet, dass es auch zu einer Interaktion von
Chloramphenicol mit mitochondrialen Ribosomen, die den 70S Ribosomen der
Bakterien recht ähnlich sind, kommt. Diese Interaktion könnte die mitochondriale
Funktion von Stammzellen im Knochenmark beeinflussen und ursächlich sein für die
unter Anwendung von Chloramphencol auftretende Suppression der
Knochenmarksfunktion (MARTELO et al. 1964; FRANKLIN u. SNOW 1998).
Obgleich beide Substanzen als primär bakteriostatisch eingestuft werden, sind
bakterizide Effekte in höheren Konzentrationen oder bei hochempfindlichen
Bakterien nachgewiesen worden (THE UNITED STATES PHARMACOPEIAL
CONVENTION 2007; HAAS et al. 2002). Bei Erregern boviner Atemwegsinfektionen
wurde Florfenicol als bakterizid wirksam gegenüber Isolaten der Spezies
Mannheimia haemolytica, Pasteurella multocida, Actinobacillus pleuropneumoniae
und Histophilus somni eingestuft, wenn Konzentrationen im Bereich des minimalen
Hemmkonzentrationswertes (MHK-Wertes) der Erreger oder eine Verdünnungsstufe
darüber erreicht wurden (VARMA 1994; HAAS et al. 2002). Die minimale bakterizide
Konzentration (MBC) und der MHK-Wert für Florfenicol liegen also sehr dicht
beieinander (HAAS et al. 2002). Für den bakteriziden Effekt scheint eine gewisse
Abhängigkeit von der eingesetzten Konzentration vorzuliegen, die bei Bakterien wie

EINFÜHRUNG IN DIE THEMATIK
11
Haemophilus somni stärker zu beobachten ist als für Actinobacillus
pleuropneumoniae (HAAS et al. 2002). Trotz dieser festgestellten
Konzentrationsabhängigkeit ist Florfenicol als zeitabhängiges antimikrobielles
Chemotherapeutikum einzustufen. Die erreichte Verstärkung des Effektes in höheren
Konzentrationen ist nicht so ausgeprägt wie bei konzentrationsabhängigen
Antibiotika (HAAS et al. 2002).
Der genaue Transportweg von Florfenicol in die Bakterienzelle ist nicht bekannt, es
wird aber ein dem Chloramphenicol ähnlicher Mechanismus vermutet.
Chloramphenicol passiert mittels passiver Diffusion durch transmembranale Porine
die äußere Membran der Bakterienzelle und wird anschließend aktiv durch die
Zytoplasmamembran geschleust. An diesem Transport sind wahrscheinlich zwei
noch nicht näher identifizierte Transportsysteme beteiligt (RUSSEL u. CHOPRA
1996).
Alle Phenicole, so auch Florfenicol, haben ein ähnliches und breites
Wirkungsspektrum, welches sowohl grampositive als auch gramnegative aerobe und
anaerobe Bakterien umfasst (YAO u. MOELLERING 1999). Auch eine Wirksamkeit
gegenüber Chlamydien, Mykoplasmen und Rickettsien ist zu beobachten (YAO u.
MOELLERING 1999; ALEXANDER et al. 1995). Aufgrund der bislang sehr
eingeschränkten Zulassung von Florfenicol sind aber nur wenige, anerkannte
Grenzwerte vorhanden, die eine Einstufung der Bakterien in die Kategorien
empfindlich, intermediär oder resistent erlauben. Derartige valide Grenzwerte sind
von dem Clinical and Laboratory Standards Institute (CLSI - ehemals NCCLS) im
Jahr 2002 veröffentlicht worden und liegen für Isolate der Spezies Pasteurella
multocida, Mannheimia haemolytica und Haemophilus somni von
Atemwegserkrankungen vom Rind sowie für Isolate der Spezies Pasteurella
multocida, Actinobacillus pleuropneumoniae, Bordetella bronchiseptica und
Streptococcus suis von Atemwegserkrankungen von Schweinen vor (NATIONAL
COMMITTEE FOR CLINICAL LABORATORY STANDARDS - NCCLS 2002). Zudem
wurden Grenzwerte für Salmonella enterica serovar Choleraesuis, isoliert von
Infektionen des Schweines, erarbeitet und in den Richtlinien der CLSI publiziert
(NATIONAL COMMITTEE FOR CLINICAL LABORATORY STANDARDS - NCCLS

EINFÜHRUNG IN DIE THEMATIK
12
2004). Die Erreger von Atemwegsinfektionen des Rindes und des Schweines werden
als empfindlich eingestuft, wenn ihr MHK-Wert bei ≤ 2 µg/ml liegt, eine Einstufung als
resistent erfolgt bei einem MHK-Wert von ≥ 8 µg/ml. Lediglich für Salmonella
Choleraesuis wurde ein geringgradig höherer Grenzwert von ≤ 4 µg/ml für die
Einstufung als empfindlich festgelegt (NATIONAL COMMITTEE FOR CLINICAL
LABORATORY STANDARDS - NCCLS 2004).
2.6 Resistenzentwicklung gegenüber Florfenicol im Unter schied zu
Chloramphenicol
Obgleich Vertreter der verschiedenen bakteriellen Spezies und Genera durchaus
Unterschiede in ihrer basalen Empfindlichkeit gegenüber Florfenicol und
Chloramphenicol zeigen, wurden bislang keine intrinsisch resistenten Bakterien
beobachtet (YAO u. MOELLERING 1999; PRIEBE u. SCHWARZ 2003). Trotzdem
haben Bakterien im Laufe des mehrjährigen Einsatzes Mechanismen entwickelt, sich
vor den inhibitorischen Einflüssen der Phenicole zu schützen. Dabei differiert die
bakterielle Resistenzentwicklung gegenüber Chloramphenicol grundlegend von der
gegenüber Florfenicol. Die Publikation 1 fasst die molekularen Mechanismen der
Resistenzentwicklung gegenüber diesen Wirkstoffen zusammen und bietet eine
detaillierte und strukturierte Zusammenstellung aller zu diesem Zeitpunkt bekannten
Resistenzgene und Resistenzmechanismen.
Mit zunehmender Anzahl der bei Bakterien nachgewiesenen Phenicol-Resistenzgene
sind Inkonsequenzen in der Nomenklatur aufgetreten. So wurden sowohl gleiche
Bezeichnungen für unterschiedliche Gene gewählt, als auch gleiche Gene mit
unterschiedlichen Bezeichnungen versehen. Daher war es ebenfalls Ziel der
Publikation 1, eine sinnvolle und überschaubare Einteilung der Gene vorzunehmen.
Eine solche Einteilung erfolgte in Gruppen auf der Basis von funktionellen und
strukturellen Eigenschaften der Genprodukte sowie aufgrund ihrer
Aminosäuresequenz. Dabei wurde - analog zur Nomenklatur für Tetracyclin- oder
Makrolid-Resistenzgene - eine Grenze von 80% Aminosäureidentität für die
Zuordnung zur gleichen Gruppe gewählt. In der Publikation wurden alle bislang

EINFÜHRUNG IN DIE THEMATIK
13
bekannten Chloramphenicol- oder Florfenicol-Resistenzgene für diese
Gruppierungen berücksichtigt, für die eine Nukleotid- bzw. Aminosäuresequenz
vorliegt und bei denen eine funktionelle Aktivität des Genproduktes nachgewiesen
wurde.
2.6.1 Spezifische Chloramphenicol-Resistenzgene
Bei der Resistenz gegenüber Chloramphenicol war der zuerst beschriebene und
immer noch am häufigsten beobachtete Mechanismus der einer enzymatischen
Inaktivierung des Wirkstoffes durch Chloramphenicol-Acetyltransferasen (CATs)
(Publikation 1; MURRAY u. SHAW 1997). Diese CAT-Enzyme sind ebenfalls in der
Lage, die Derivate Thiamphenicol und Azidamphenicol zu inaktivieren. Dabei erfolgt
eine Übertragung einer Acetylgruppe auf das C3-Atom des Chloramphenicol-
Moleküls. Anschließend erfolgt ein nichtenzymatisches, intramolekulares
Rearrangement der Acetylgruppe an die C1-Position. Dieses Rearrangement wird
von einer erneuten Acetylierung des Chloramphenicol-Moleküls an der C3-Position
gefolgt. Diese zweite Acetylierung ist für eine Resistenzentstehung nicht zwingend
erforderlich, denn bereits durch eine C3-Acetylierung verliert das Wirkstoffmolekül
seine antibiotische Aktivität (SHAW u. LESLIE 1991). Da bei dem Wirkstoff
Florfenicol die C3-Position fluoriert vorliegt, kann kein Transfer einer Acetylgruppe an
diese Position stattfinden. Als Folge sind alle Stämme, deren
Chloramphenicolresistenz ausschließlich auf die Aktivität von CATs zurückzuführen
ist, florfenicolempfindlich (CANNON et al. 1990). Die zuvor beschriebene Substitution
der Hydroxylgruppe bei Florfenicol durch ein Fluoratom hat daher erheblichen
Einfluss auf die Resistenzentwicklung und führte diesbezüglich zu einer deutlichen
Verbesserung des Wirkstoffmoleküls.
Die CAT-Enzyme werden aufgrund struktureller Unterschiede in zwei Typen, A und
B, eingeteilt. CATs vom Typ A wurden bei einer Vielzahl von grampositiven und
gramnegativen Bakterien nachgewiesen (SHAW 1983; MURRAY u. SHAW 1997;
SCHWARZ u. WHITE 2005) und zeigen einige Gemeinsamkeiten, obgleich sie zum
Teil Variationen in ihren Aminosäuresequenzen aufweisen (Publikation 1). Die CAT

EINFÜHRUNG IN DIE THEMATIK
14
A-Varianten sind trimere Strukturen, die gewöhnlich aus drei identischen
Untereinheiten zusammengesetzt sind (SHAW u. LESLIE 1991). Jede dieser
Untereinheiten hat eine Größe zwischen 207 und 238 Aminosäuren (SCHWARZ u.
CHASLUS-DANCLA 2001). Bei allen CAT A-Typen scheinen einige Aminosäuren
konserviert vorzuliegen, die in bestimmte Funktionen wie Substratbindung,
katalytische Aktivitäten, Faltung des Monomers oder Zusammensetzung des Trimers
involviert sind (MURRAY u. SHAW 1997). Die einzelnen Monomere werden durch
cat-Gene kodiert. In seltenen Fällen kommen Heterodimere vor, wenn
unterschiedliche, aber ähnliche CATs in einem Bakterium vorhanden sind (MURRAY
u. SHAW 1997).
CATs vom Typ B wurden ursprünglich auch als xenobiotische Acetyltransferasen
bezeichnet und unterscheiden sich in ihrer Struktur von Typ A CATs. Sie zeigen
größere Verwandtschaft mit anderen acetylierenden Enzymen, wie Vat(D), Vat(E),
Vat(A) oder Vat(B), die bei Staphylokokken oder Enterokokken vorkommen und in
der Lage sind, Streptogramin A-Antibiotika zu inaktivieren (RENDE-FOURNIER et al.
1993; WERNER u. WITTE 1999; ALLIGNET u. EL SOLH 1995; ALLIGNET et al.
1993). Auch die Typ B CATs sind Homotrimere, bei denen das Monomer aus 209 bis
212 Aminosäuren besteht (MURRAY u. SHAW 1997). Im Gegensatz zu Typ A CATs,
bei denen aufgrund von Unterschieden in ihren Aminosäuresequenzen schon 2004
eine Einteilung in 16 Gruppen vorgenommen werden konnte, wurden CAT-Varianten
vom Typ B seltener beschrieben und ließen sich in nur 5 Gruppen einteilen
(Publikation 1). CATs beider Typen wurden als Bestandteile mobiler genetischer
Elemente, wie Plasmide, Transposons oder Genkassetten beschrieben.
Auch spezifische Transporter oder Multidrug-Transporter wurden identifiziert, die
Chloramphenicol aus der Bakterienzelle ausschleusen können. Dabei zeichnen sich
spezifische Transporter durch ein enges Substratspektrum aus, sie haben keine
physiologische Funktion in der Bakterienzelle und vermitteln höhere Resistenzlevel
im Vergleich zu Multidrug-Transportern. Ein spezifischer Chloramphenicol-Exporter
wurde zuerst 1997 in Pseudomonas aeruginosa identifiziert (RUBENS et al. 1979)
und später als Bestandteil des Transposons Tn1696 nachgewiesen. Dieser
Transporter wurde durch das Gen cmlA kodiert. Das abgeleitete Protein besteht aus

EINFÜHRUNG IN DIE THEMATIK
15
419 Aminosäuren und zeigt 12 transmembranale Domänen, die typisch sind für
Proteine aus der „Major facilitator superfamily“ von transmembranalen
Transportproteinen (GEORGE u. HALL 2002). In den folgenden Jahren wurden
identische oder eng verwandte cmlA-Gene - auch als Bestandteile von Genkassetten
- bei einer Vielzahl gramnegativer Bakterien nachgewiesen (POIREL et al. 1999;
CARATTOLI et al. 2002; POIREL et al. 2000; POIREL et al. 2003; PARTRIDGE et al.
2001; AUBERT et al. 2001; RICCIO et al. 2003). Ein nur 84 % identisches cmlA2-
Gen wurde 1998 bei Enterobacter aerogenes identifiziert (PLOY et al. 1998).
Während die Gene cmlA und cmlA2 bei human- oder tierpathogenen
Bakterienspezies nachgewiesen wurden, sind weitere spezifische Chloramphenicol-
Transporter bei Bodenbakterien oder Umweltkeimen beschrieben worden. Dazu
zählen die Gene cmr (DESOMER et al. 1992) und cmrA (NAGY et al. 1997),
nachgewiesen bei Rhodococcus spp., die Gene cmx (TAUCH et al. 1998) und cmr
(Genbank Zugangsnummer U85507) von Corynebacterium spp. oder die Gene cml
(DITTRICH et al. 1991) und cmlv (MOSHER et al. 1995) von Streptomyces spp..
Im Gegensatz zu den spezifischen Transportern umfasst das Substratspektrum von
Multidrug-Transportern eine weite Spannbreite unverwandter Substanzen
(SCHWARZ u. WHITE 2005). Ihre Funktion liegt in der Ausschleusung toxischer
Substanzen aus der Bakterienzelle, gegenüber antibiotisch wirksamen Substanzen
vermitteln sie nur niedrige Resistenzlevel. Multidrug-Transporter, deren
Substratspektrum Chloramphenicol einschließt, sind MdfA von Escherichia coli
(EDGAR u. BIBI 1997), die aus mehreren Proteinen zusammengesetzten Systeme
MexAB/OprM, MexCD/OprJ oder MexFE/OprN von Pseudomonas aeruginosa
(PAULSEN et al. 1996; POOLE 2002) oder CeoAB-OpcM von Burkholderia cepacia
(POOLE 2002; SCHWARZ u. WHITE 2005). Überexpression dieser
Transportsysteme führt zu einem Anstieg der MHK-Werte für Chloramphenicol,
funktionelle Deletionen der Systeme haben einen empfindlicheren Phänotyp zur
Folge (SULAVIK et al. 2001). Auch bei grampositiven Bakterien wurde von Multidrug-
Transportern berichtet, die Chloramphenicol ausschleusen können. Dazu zählen die
Transportsysteme NorA (YOSHIDA et al. 1990) und Blt (AHMED et al. 1995) von
Staphylococcus aureus bzw. Bacillus subtilis.

EINFÜHRUNG IN DIE THEMATIK
16
In dem Antibiotika-Produzenten Streptomyces venezuelae wurden zwei weitere
Chloramphenicol-Resistenzmechanismen entdeckt. Diese umfassen eine
O-Phosphorylierung (MOSHER et al. 1995) sowie eine hydrolytische Degradierung
von Chloramphenicol zu p-Nitrophenylserinol (MOSHER et al. 1990). Beide
Mechanismen haben die Funktion, den natürlichen Produzenten vor den
inhibitorischen Einflüssen der eigenen Stoffwechselprodukte zu schützen.
Auch von Permeabilitätsbarrieren wurde berichtet, die zu einem verminderten Influx
der Wirksubstanz in die Bakterienzelle führen. Dazu zählt der Verlust eines Proteins
der äußeren Zellmembran, OmpF, bei Salmonella Typhi, welches mit erhöhten MHK-
Werten für Chloramphenicol einhergeht (TORO et al. 1990). Ähnliche Barrieren, die
auf einer verminderten Permeabilität der äußeren Zellmembran beruhen, werden für
Haemophilus influenzae (BURNS et al. 1985) oder Burkholderia cepacia (BURNS et
al. 1989) angenommen. Zu einer Reduktion der ompF-Translation und damit zu einer
verminderten OmpF-Porinbildung führt über verschiedene Zwischenschritte auch die
Aktivierung von MarA, einem Transkriptionsaktivator bei Escherichia coli und
anderen Enterobacteriaceae (QUINTILIANI et al. 1999). Mutationen oder
Modifikationen der Angriffstelle für Chloramphenicol werden dagegen sehr selten
beobachtet. Eine mögliche Erklärung liegt darin, dass derartige Modifikationen im
Peptidyltransferasezentrum nicht mit einer zufriedenstellenden Funktion der
Ribosomen einhergehen (MURRAY 2000). Trotzdem wurden Mutationen in dem
ribosomalen Protein-Gencluster bei Escherichia coli (BAUGHMAN u. FAHNESTOCK
1979) und Bacillus subtilis (ANDERSON et al. 1984) sowie in der 23S rRNA bei
Escherichia coli (ETTAYEBI et al. 1985) beschrieben, die mit einer verminderten
Chloramphenicolempfindlichkeit einhergehen.
2.6.2 Resistenzgene, die kombinierte Florfenicol- und Chloramphenicolresistenz
vermitteln
Von übertragbarer Florfenicolresistenz wurde zuerst im Jahr 1993 berichtet (KIM et
al. 1993). In der entsprechenden Studie konnten acht Isolate des fischpathogenen
Bakteriums Photobacterium damselae subsp. piscicida (ehemals Pasteurella

EINFÜHRUNG IN DIE THEMATIK
17
piscicida) identifiziert werden, die erhöhte MHK-Werte gegenüber Florfenicol
aufwiesen. Die Autoren erläuterten, dass der Wirkstoff bereits seit 1990 zur
Behandlung der Pseudotuberkulose auf Fischfarmen in Japan eingesetzt wurde. In
den Untersuchungen zeigte sich, dass die Resistenzeigenschaft bei vier der Isolate
auf Plasmiden lokalisiert war. Diese Plasmide waren konjugativ und vermittelten den
entsprechenden Escherichia coli-Empfängerstämmen Resistenz gegenüber
Florfenicol, Chloramphenicol, Kanamycin, Sulfonamiden und Tetracyclinen. Die
Analyse von Plasmid-DNA mit Restriktionsendonukleasen ließ auf einen
gleichartigen Plasmidtyp bei allen vier Isolaten schließen (KIM et al. 1993). Aber erst
3 Jahre später konnte das Florfenicol-Resistenzgen identifiziert werden (KIM u. AOKI
1996). Dieses aus 1122 Nukleotiden bestehende Gen wurde mit pp-flo bezeichnet
und kodiert für ein Exporter-Protein aus der „Major Facilitator Superfamily“. Es ist in
der Lage, neben Florfenicol auch Chloramphenicol aus der Bakterienzelle
auszuschleusen (KIM u. AOKI 1996). Das abgeleitete Protein von Photobacterium
damselae würde allerdings aus nur 347 Aminosäuren bestehen und eine
Membrantopologie von 10 transmembranalen Segmenten aufweisen.
Später wurden weitere Florfenicol-Resistenzgene von Enterobacteriaceae und Vibrio
cholerae bekannt. Diese Gene erhielten unterschiedliche Bezeichnungen wie cmlA-
like, floSt, flo oder floR, sind aber untereinander eng verwandt und weisen eine 96 -
100%ige Identität in ihren Nukleotidsequenzen und eine 88 - 100%ige Identität in
ihren Aminosäuresequenzen auf. Die abgeleiteten Proteine sind einheitlich aus 404
Aminosäuren zusammengesetzt und zeigen eine Membrantopologie bestehend aus
12 transmembranalen Segmenten. Daher ist anzunehmen, dass die ursprüngliche
pp-flo Sequenz inkomplett ist oder Sequenzierfehler enthält (PAULSEN et al. 1996).
Heute hat sich die Bezeichnung floR für das Florfenicol-Chloramphenicol
Effluxprotein durchgesetzt. Es wurde in der Zwischenzeit auch als Bestandteil eines
chromosomalen Multiresistenzgenclusters bei Salmonella Typhimurium DT104
identifiziert (BRIGGS u. FRATAMICO 1999). Dieses Resistenzgencluster umfasst ca.
13 Kilobasen (kb) und ist Teil einer chromosomalen genomischen Insel, die als
„Salmonella Genomic Island 1 (SGI1)“ bezeichnet wird (BOYD et al. 2001). Dieses
43 kb SGI1 ist am 3´ Ende des thdF-Gens im Chromosom von Salmonella
Typhimurium DT104 lokalisiert. Das SGI1 oder Varianten davon wurden mittlerweile

EINFÜHRUNG IN DIE THEMATIK
18
auch bei anderen Phagentypen, wie beispielsweise DT120, oder bei anderen
Serovaren wie Salmonella Agona, Paratyphi B, Albany oder Newport nachgewiesen,
so dass ein horizontaler Transfer des SGI1 vermutet wird (BOYD et al. 2002;
CLOECKAERT et al. 2000; DOUBLET et al. 2003; DOUBLET et al. 2004; MEUNIER
et al. 2002). In einer dieser SGI1 Varianten ist das floR-Gen durch die Insertion eines
IS6100-Elementes unterbrochen. Andere Transpositionsvorgänge von IS6100-
Elementen innerhalb des SGI1 führten zu Inversionen von Teilen dieses
chromosomalen Genclusters. Eine vergleichende Darstellung der genetischen
Organisation der Resistenzgene innerhalb des SGI1 bei unterschiedlichen
Salmonella Serovaren ist in Abbildung 2 dargestellt. Weitere chromosomale
Lokalisationen des floR-Gens wurden in den Jahren 2001 und 2002 von HOCHHUT
et al. sowie BEABER et al. beschrieben. Dabei identifizierten die Autoren das floR-
Gen als Bestandteil des SXT-Elementes von Vibrio cholerae (HOCHHUT et al. 2001;
BEABER et al. 2002). Weiterhin sind plasmidäre Lokalisationen von floR auf dem
Plasmid R55 von Klebsiella pneumoniae beschreiben (CLOECKAERT et al. 2001)
und ein strukturell verwandtes Plasmid wurde 2003 auch bei Salmonella Newport
nachgewiesen (MEUNIER et al. 2003). Weitere Resistenzgene, die auch
Florfenicolresistenz vermitteln, waren nicht bekannt.
Bei Salmonella enterica Serovar Typhimurium DT104 konnte zudem nachgewiesen
werden, dass der Multidrug-Transporter AcrAB-TolC neben einer Vielzahl
antimikrobieller Substanzen auch Chloramphenicol und Florfenicol in sein
Substratspektum einschliesst und somit verminderte Empfindlichkeit gegenüber
diesen Wirksubstanzen vermittelt (BAUCHERON et al. 2004; SCHWARZ u. WHITE
2005).

EINFÜHRUNG IN DIE THEMATIK
19
Abbildung 2: Schematische Darstellung der genetischen Organisation des
Resistenzgenclusters innerhalb des Salmonella Genomic Island 1 (SGI1) bei verschiedenen Salmonella Serovaren. DR-L und DR-R bezeichnen die linken bzw. rechten „direct repeats“, die das Resistenzgencluster flankieren. Das floR-Gen sowie weitere Resistenzgene sind als Pfeile dargestellt. Diese Resistenzgene des SGI1 vermitteln Resistenz gegenüber Ampicillin (blaPSE-1), Gentamicin (aac(3)-Id), Streptomycin und Spectinomycin (aadA2, aadA7), Sulfonamiden (sul1), Tetracyclinen (tet(G)), und Trimethoprim (dfrA1, dfrA10).
2.7 Regulation der Expression von Chloramphenicol- und Florfenicol-
Resistenzgenen
Chloramphenicol-Resistenzgene vom Typ catA folgen bei gramnegativen Bakterien
einem konstitutiven Expressionstyp. Im Gegensatz dazu werden die catA-Gene bei
grampositiven Bakterien, wie Staphylococcus aureus, induzierbar exprimiert. Dabei
dient Chloramphenicol selber als Inducer eines Regulationssystems, dessen
SGI1-A (S. Agona)
SGI1 (S. Typhimurium DT104)
SGI1-H (S. Newport)
SGI1-F (S. Albany)
SGI1-E (S. Typhimurium DT104)
1 kb
tnpR intI1 aadA2qacE∆1
sul1∆ floR tetR tet(G) orf1 orf2groEL/intI1
pse-1qacE∆1
sul1orf5orf6IS6100S044
DR-L
yidY
DR-R
sul1qacE∆1
dfrA10orf513thdF int
tnpR intI1 aadA2qacE∆1
sul1∆ floR tetR tet(G) orf1 orf2groEL/intI1
pse-1qacE∆1
sul1orf5orf6IS6100S044
DR-L
yidY
DR-R
thdF int
tetRtet(G)orf1orf2groEL/intI1
pse-1qacE∆1
sul1orf5
orf6 IS6100S044 yidY
DR-R
tnpR intI1 aadA2qacE∆1
sul1∆ floR5’∆3’
DR-L
thdF intIS6100 floR3’∆5’
tnpR intI1qacE∆1
sul1∆ floR tetR tet(G) orf1 orf2groEL/intI1
pse-1qacE∆1
sul1orf5orf6IS6100S044
DR-L
yidY
DR-R
thdF int dfrA1orf
tnpR intI1qacE∆1
sul1∆ floR tetR tet(G) orf1 orf2groEL/intI1
pse-1qacE∆1
sul1orf5orf6IS6100S044
DR-L
yidY
DR-R
thdF int aadA7aac(3)-Id

EINFÜHRUNG IN DIE THEMATIK
20
Mechanismus als „attenuierte Translation“ bezeichnet wird (LOVETT 1990). Die
unterschiedlichen Strukturen dieses Regulationssystems sind in Abbildung 3
wiedergegeben. Bei dem Mechanismus der attenuierten Translation befinden sich
stromaufwärts des cat-Strukturgens zwei oder mehr Paare umgekehrt
komplementärer Sequenzwiederholungen. Diese als „inverted repeats, (IR)“
bezeichneten Sequenzen sind in der Lage, auf der Ebene der mRNA stabile
Sekundärstrukturen auszubilden. Diese Strukturen werden als Haarnadel-
Anordnungen oder „stem loops“ bezeichnet. Ebenfalls dem Strukturgen vorgeschaltet
findet sich ein Leserahmen für ein kleines Peptid von 6 bis 9 Aminosäuren. Innerhalb
dieses kleinen Peptids sind die Kodons 2 bis 5 hoch konserviert und schließen eine
Sequenz ein, die komplementär zu Sequenzen am 3´-Terminus der 16S rRNA sind.
Dieser Sequenzbereich wird im anglo-amerikanischen Sprachgebrauch als
„ribosome stall sequence“ bezeichnet. Wird ein Ribosom in diesem Bereich der
„ribosome stall sequence“ festgefahren, so überlappt es zum Teil die IR1 Sequenz
und verhindert das Ausbilden der Sekundärstruktur auf mRNA-Ebene. Durch den
Wegfall dieser mRNA-Sekundärstruktur wird eine Ribosomenbindungsstelle (RBS)
im IR2 für ein weiteres Ribosom zugänglich und damit auch die Translation des cat-
Strukturgens möglich. Dieser Regulationsmechanismus ist nur dann möglich, wenn
es durch die Anwesenheit von Chloramphenicol und die Bindung der Wirksubstanz
an ein Ribosom zu einer Konformationsänderung des Ribosoms kommt. Dabei
könnte die Bindung von Chloramphenicol eine strukturelle Veränderung bewirken,
die den relevanten Bereich der 16S rRNA in eine exponierte Position bringt und die
damit ein Festfahren des Ribosoms an der komplementären Sequenz der „ribosome
stall sequence“ erlaubt (LOVETT 1990). Ist kein Chloramphenicol anwesend, so wird
auch kein Ribosom an der „ribosome stall sequence“ festgefahren, die
Sekundärstruktur der mRNA bildet sich aus und die Translation des Strukturgens
wird damit verhindert.

EINFÜHRUNG IN DIE THEMATIK
21
Abbildung 3: Modell der Regulatorregion eines induzierbar exprimierten cat-Gens
nach dem Modell der attenuierten Translation gemäß Lovett 1990. Die inverted repeats 1 und 2 (IR1, IR2) sind als Pfeile dargestellt und bilden eine stabile Sekundärstruktur auf mRNA-Ebene. Die Ribosomenbindungsstellen (RBS), die vor dem Strukturgen und dem kleinen regulatorischen Peptid lokalisiert sind, umschließen als Kästchen die entsprechenden Sequenzen. Die Startkodons vor den Leserahmen sind unterstrichen, die Leserahmen selber sind in fettgedruckten Buchstaben dargestellt.
Während bei den cat-Genen von Staphylokokken, die dem Typ A zugerechnet
werden, ein Paar umgekehrt komplementärer Sequenzen (IR1 und IR2) dem
Strukturgen vorgeschaltet sind (Abbildung 3), so finden sich komplexere mRNA-
Sekundärstrukturen bei cat-Genen vom Typ B. Stromaufwärts des Gens catB1 von
Agrobacterium tumefaciens wurden vier unterschiedliche Paare von inverted repeats
nachgewiesen (RODGERS et al. 2002), von denen angenommen wird, dass sie eine
komplexere Struktur ausbilden. Viele cat-Gene vom Typ B sind zudem Teil von
C A U A U A U A A C A A A
U - AA - U
A - U A - U U - A C - G C - G U - A C - G C - G U - A U - A A - U UAC GAGGUGA UAUAACGUAUGAAAAAAUCAGAGGAUU UUAUAUAUGACUUUU
∆ G = -82.8 kJ/mol
start cat
RBS
RBS
start ORF9
stallsequence
IR1 IR2

EINFÜHRUNG IN DIE THEMATIK
22
Genkassetten und werden damit von einem Promotor aus transkribiert, der in dem
jeweiligen Empfängerintegron lokalisiert ist. Bei solchen Integrons werden diejenigen
Genkassetten, die näher am Promotor liegen, höher exprimiert als solche, die weiter
distal lokalisiert sind (ROWE-MAGNUS et al. 2002). Damit ist eine Beeinflussung der
Expression von Genen durch ihre Lokalisation im Empfängerintegron möglich. Diese
veränderte Expression je nach Lage im Integron ließ sich von Rowe-Magnus et al.
2002 auch in den jeweiligen MHK-Werten wiederspiegeln. War die catB9 Kassette
von Vibrio cholerae in den ersten vier Positionen eines Integrons positioniert, so
zeigte der Stamm einen MHK-Wert für Chloramphenicol von ≥ 25 µg/ml, war die
Kassette weiter distal an siebter Position lokalisiert, verringerte sich der MHK-Wert
auf < 1 µg/ml (ROWE-MAGNUS et al. 2002; HEIDELBERG et al. 2000). Eine
Ausnahme von dieser Feststellung stellen cmlA-Gene dar, die für Chloramphenicol-
Effluxproteine kodieren. Obgleich cmlA-Gene auch Bestandteile von Genkassetten
sind, so haben sie ihre eigene Promotorstruktur und ihre Expression wird über einen
Mechanismus ähnlich der attenuierten Translation von catA-Genen bei
Staphylokokken reguliert (STOKES u. HALL 1991). Stromaufwärts solcher cmlA-
Gene wurden Translationsattenuator-ähnliche Stukturen beschrieben (STOKES u.
HALL 1991). Für das einzig bekannte Gen, welches kombinierte Florfenicol- und
Chloramphenicolresistenz vermittelt, floR, ist kein Regulationsmechanismus bekannt.
Daher wird von einer konstitutiven Expression dieses Gens ausgegangen.
2.8 Empfindlichkeitslage bakterieller Infektionserr eger gegenüber
Florfenicol
Zu Beginn der eigenen Untersuchungen im Jahr 2000 lagen nur wenige Daten über
die Empfindlichkeitslage bakterieller Infektionserreger gegenüber Florfenicol vor
(HÖRMANSDORFER u. BAUER 1996; HÖRMANSDORFER u. BAUER 1998; KIM u.
AOKI 1993; KÜHN u. GOOSENS 1998). Gründe dafür mögen zum einen darin
liegen, dass der Wirkstoff erst wenige Jahre zuvor in die klinische Nutzung eingeführt
wurde und die Zulassung auf wenige Tierarten und Anwendungsgebiete beschränkt
war. Zum anderen könnte die Ursache darin liegen, dass weder der Wirkstoff zur

EINFÜHRUNG IN DIE THEMATIK
23
Herstellung der entsprechenden Medien für die MHK-Bestimmung, noch Plättchen
für eine Empfindlichkeitsbestimmung mittels Agardiffusion kommerziell erhältlich
waren (PRIEBE 2003). Ein Testen der Erregerempfindlichkeit wurde außerdem
erschwert, da Vergleichsdaten für die Bestimmung der minimalen
Hemmkonzentration von Florfenicol vom Clinical Laboratory Standards Institute
(CLSI - ehemals National Committee for Clinical Laboratory Standards, NCCLS) erst
1996 veröffentlicht wurden (MARSHALL et al. 1996). Die Ergebnisse verschiedener
Studien lassen sich zudem nur schwerlich vergleichen, da in diesen Studien zum Teil
unterschiedliche Methodiken, die auf unterschiedlichen Durchführungsvorschriften
basieren, zugrunde gelegt wurden. Auch die Klassifizierung der Isolate als
empfindlich oder resistent erfolgte in früheren Studien meist auf der Basis von
Herstellerinformationen (HÖRMANSDORFER u. BAUER 1996; HÖRMANSDORFER
u. BAUER 1998). Andere Studien verwendeten für die Klassifizierung eigene
Grenzwerte, die auf einer MHK-Wertverteilung aller getesteten Isolate aus früheren
Untersuchungen basierte (KIM u. AOKI 1993). Heute werden meist die anerkannten
Grenzwerte gemäß CLSI für eine Klassifizierung der Isolate in die Kategorien
empfindlich oder resistent herangezogen (SHIN et al. 2005; PRIEBE u. SCHWARZ
2003).
Generell gab es aber zu Beginn der eigenen Untersuchungen nur wenige
Veröffentlichungen, die über florfenicolresistente Bakterien berichtet haben (KIM et
al. 1993).
2.8.1 Empfindlichkeitslage von Zielorganismen
Zwei Studien von HÖRMANSDORFER und BAUER aus den Jahren 1996 und 1998
haben sich mit der Empfindlichkeit von Zielorganismen gegenüber Florfenicol
beschäftigt. Dabei wurden in der Studie von 1996 die Anzahl von 215 Pasteurella
multocida- und 160 „Pasteurella“ haemolytica-Isolaten getestet. Durch taxonomische
Änderungen nach ANGEN et al. 1999 werden die ehemals als trehalose-negativer
„Pasteurella haemolytica“-Komplex zusammengefassten Bakterien heute mit dem
Genusnamen Mannheimia bezeichnet. Die Isolate in dieser Studie wurden mit zwei

EINFÜHRUNG IN DIE THEMATIK
24
unterschiedlichen Methoden, der Agardiffusion und der Mikrodilution, auf ihre
Florfenicolempfindlichkeit geprüft. Alle Isolate waren bovinen Ursprungs und
stammten zu 80,53 % aus Bayern, zu 11,2 % aus Baden-Würtemberg, zu 4,27 %
aus den neuen Bundesländern und bei 4 % der Isolate war die Herkunft nicht zu
bestimmen. Die Autoren ermittelten Hemmhofdurchmesser zwischen 25 und 53 mm
und MHK-Werte für Florfenicol, die in einer Spannbreite von 0.25 µg/ml bis 1,0 µg/ml
lagen. Dabei zeigte die Mehrzahl der Isolate (57,9 %) einen MHK-Wert von 0,5
µg/ml, 33,3 % der Isolate hatten einen MHK-Wert von 0,25 µg/ml und 8,8 % der
Testisolate wiesen einen MHK-Wert von 1,0 µg/ml auf. Die Autoren stuften alle
getesteten Isolate als empfindlich gegenüber Florfenicol ein (HÖRMANSDORFER u.
BAUER 1996). Zwei Jahre später führten die Autoren (HÖRMANSDORFER u.
BAUER 1998) eine Folgeuntersuchung durch, bei der 60 Pasteurella multocida- und
29 „Pasteurella“ haemolytica- Isolate boviner Herkunft sowie 119 Pasteurella
multocida- und 13 „Pasteurella“ haemolytica- Isolate porciner Herkunft untersucht
wurden. Die Isolate vom Rind stammten zu 29,2 % aus Niedersachsen und zu
70,8 % aus Bayern, die Isolate vom Schwein waren zu 93,2 % niedersächsischer
Herkunft, nur 6,8 % stammten aus Bayern. Die Hemmhofdurchmesser aller
getesteten Isolate reichten von 23 bis 54,5 mm, MHK-Werte wurden in einem
Bereich von 0,25 µg/ml bis 1,0 µg/ml ermittelt. Dabei kam der MHK-Wert von 0,5
µg/ml sowohl bei den Pasteurella multocida (82,7 %) als auch bei den „Pasteurella“
haemolytica-Isolaten (35,7 %) am häufigsten vor. Einige „Pasteurella“ haemolytica-
Isolate (11 Isolate) ließen keine MHK-Wertbestimmung zu. Auch in dieser
Folgestudie von HÖRMANSDORFER und BAUER (1998) wurde kein
florfenicolresistentes Isolat ermittelt.
Weiterhin sind zwei Studien bekannt, die an der TU-München sowie am FZMB-Erfurt
durchgeführt wurden und die im Rahmen einer kontinuierlichen
Empfindlichkeitsprüfung von 1997 - 1999 insgesamt 305 Pasteurella multocida- und
135 „Pasteurella“ haemolytica- Isolate gegenüber Florfenicol getestet haben
(GOOSENS 1999; PRIEBE 2003). Die Untersuchung fand ebenfalls vergleichend
mittels Agardiffusionstests und Mikrodilutionsverfahren statt. Dabei wurden ja nach
Studie mittlere Hemmhofdurchmesser von 38,1 mm (TU-München) und 36,3 mm
(FZMB-Erfurt) für die Pasteurella multocida-Isolate und von 30,2 mm (TU-München)

EINFÜHRUNG IN DIE THEMATIK
25
bzw. 31,9 mm (FZMB-Erfurt) für die „Pasteurella“ haemolytica- Isolate gemessen. Die
MHK-Werte der Pasteurella multocida-Isolate zeigten einen mittleren Wert von 0,47
µg/ml (TU-München) und 0,79 µg/ml (FZMB-Erfurt), bei den „Pasteurella“
haemolytica- Isolaten lag der mittlere MHK-Wert geringfügig höher mit 0,68 µg/ml
(TU-München) bzw. 1,56 µg/ml (FZMB-Erfurt) (GOOSENS 1999; PRIEBE 2003).
Auch in diesen Studien wurde kein Isolat als resistent eingestuft.
Weitere Untersuchungen beschäftigten sich mit der Florfenicolempfindlichkeit von
Isolaten aus anderen Ländern. So wurden von MEVIUS und HARTMAN im Jahr
2000 Ergebnisse einer Monitoringstudie publiziert, deren Ziel es war, die Aktivität von
12 antimikrobiellen Chemotherapeutika gegenüber Pasteurella multocida- und
Mannheimia haemolytica-Isolaten von Kälbern aus den Niederlanden zu bestimmen
(MEVIUS u. HARTMAN 2000). Auch hier lagen die ermittelten MHK-Werte, bei
denen 50 bzw. 90 % der Isolate in ihrem Wachstum gehemmt oder abgetötet wurden
(MHK50- und MHK90-Werte) für Florfenicol mit maximal einer Verdünnungsstufe
Unterschied nah an dem von HÖRMANSDORFER und BAUER 1996 und 1998
publizierten Bereich.
In einer Untersuchung aus Japan wurde die In-vitro-Empfindlichkeit von 92
Actinobacillus pleuropneumoniae-Isolaten aus Schweinelungen gegenüber
Florfenicol festgestellt (UEDA u. SUENAGA 1995). Die Autoren ermittelten MHK-
Werte zwischen 0,2 µg/ml und 1,56 µg/ml, die für sämtliche Isolate eine Einstufung
als empfindlich zuließen.
In einer weiteren Studie an 68 japanischen Actinobacillus pleuropneumoniae-Isolaten
(YOSHIMURA et al. 2002) wurden MHK-Werte gegenüber Florfenicol zwischen 0,1
und 0,78 µg/ml festgestellt, die MHK50- und MHK90-Werte waren mit je 0,39 µg/ml in
einem ähnlichen Bereich wie die 1995 von UEDA u. SUENAGA ermittelten Werte
von 0,39 µg/ml (MHK50) und 0,78 µg/ml (MHK90). Die Ergebnisse dieser publizierten
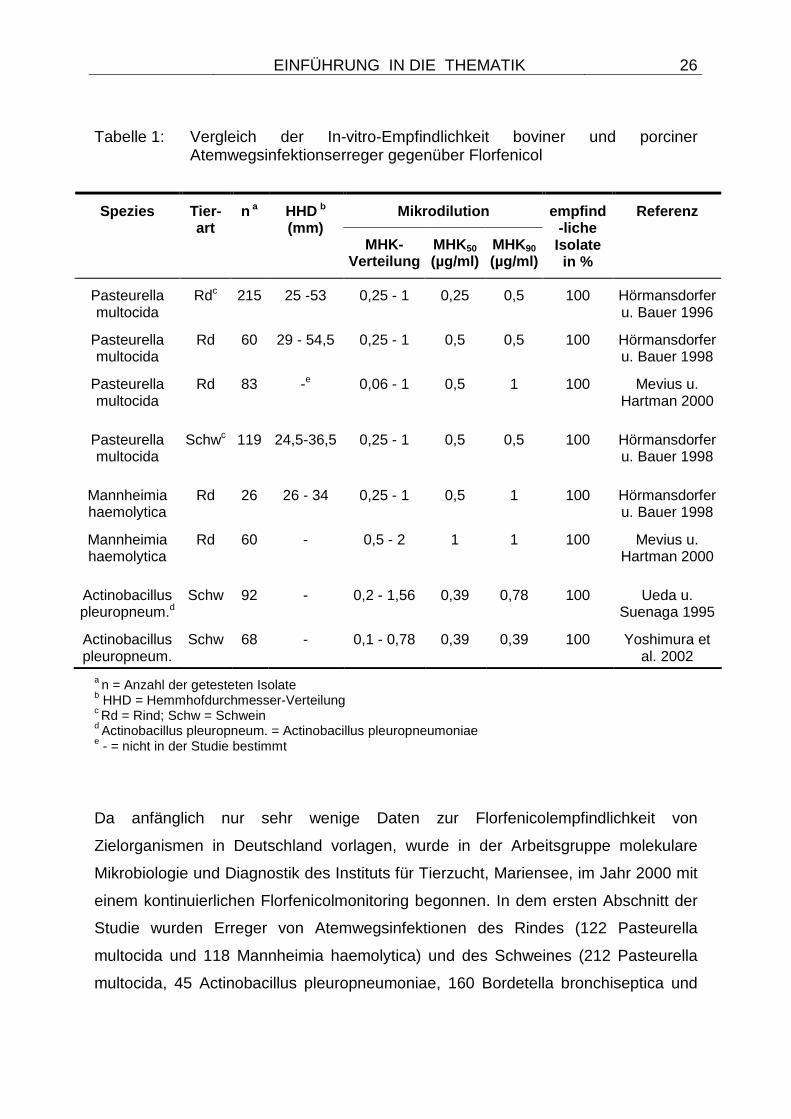
Studien sind in Tabelle 1 dargestellt.
EINFÜHRUNG IN DIE THEMATIK
26
Tabelle 1: Vergleich der In-vitro-Empfindlichkeit boviner und porciner Atemwegsinfektionserreger gegenüber Florfenicol
Mikrodilution Spezies Tier-art
n a HHD b
(mm) MHK-
Verteilung MHK50 (µg/ml)
MHK90
(µg/ml)
empfind-liche Isolate in %
Referenz
Pasteurella multocida
Rdc 215 25 -53 0,25 - 1 0,25 0,5 100 Hörmansdorfer u. Bauer 1996
Pasteurella multocida
Rd 60 29 - 54,5 0,25 - 1 0,5 0,5 100 Hörmansdorfer u. Bauer 1998
Pasteurella multocida
Rd 83 -e 0,06 - 1 0,5 1 100 Mevius u. Hartman 2000
Pasteurella multocida
Schwc 119 24,5-36,5 0,25 - 1 0,5 0,5 100 Hörmansdorfer u. Bauer 1998
Mannheimia haemolytica
Rd 26 26 - 34 0,25 - 1 0,5 1 100 Hörmansdorfer u. Bauer 1998
Mannheimia haemolytica
Rd 60 - 0,5 - 2 1 1 100 Mevius u. Hartman 2000
Actinobacillus pleuropneum.d
Schw 92 - 0,2 - 1,56 0,39 0,78 100 Ueda u. Suenaga 1995
Actinobacillus pleuropneum.
Schw 68 - 0,1 - 0,78 0,39 0,39 100 Yoshimura et al. 2002
a n = Anzahl der getesteten Isolate b HHD = Hemmhofdurchmesser-Verteilung c Rd = Rind; Schw = Schwein d Actinobacillus pleuropneum. = Actinobacillus pleuropneumoniae e - = nicht in der Studie bestimmt
Da anfänglich nur sehr wenige Daten zur Florfenicolempfindlichkeit von
Zielorganismen in Deutschland vorlagen, wurde in der Arbeitsgruppe molekulare
Mikrobiologie und Diagnostik des Instituts für Tierzucht, Mariensee, im Jahr 2000 mit
einem kontinuierlichen Florfenicolmonitoring begonnen. In dem ersten Abschnitt der
Studie wurden Erreger von Atemwegsinfektionen des Rindes (122 Pasteurella
multocida und 118 Mannheimia haemolytica) und des Schweines (212 Pasteurella
multocida, 45 Actinobacillus pleuropneumoniae, 160 Bordetella bronchiseptica und

EINFÜHRUNG IN DIE THEMATIK
27
99 Streptococcus suis) aus den Jahren 2000 und 2001 gesammelt und vergleichend
mittels den zwei Methoden Agardiffusion und Mikrodilution auf ihre In-vitro-
Empfindlichkeit gegenüber Florfenicol getestet (PRIEBE u. SCHWARZ 2003). Alle
Isolate wurden vor der Testung einer Speziesdifferenzierung unterzogen und
Referenzstämme der American Type Culture Collection (ATCC) dienten zur
Qualitätssicherung während der Testungen.
Parallel zu diesem spezifischen Florfenicol-Monitoringprogramm wurde im Jahr 2001
in Deutschland ein neues nationales Resistenzmonitoringprogramm für
veterinärpathogene Keime vom Bundesamt für Verbraucherschutz und
Lebensmittelsicherheit (BVL) unter der Leitung von Dr. J. Wallmann ins Leben
gerufen. In diesem GERM-Vet Programm, in dem man Fehler alter
Resistenzmonitoringprogramme aus Deutschland zu umgehen versuchte, wurden
neben weiteren Keimen und Indikationen Erreger respiratorischer Erkrankungen und
Bakterien vom Milchrind auf ihre Florfenicolempfindlichkeit getestet. Für diese
Untersuchungen wurden fortlaufend Isolate aus den Veterinäruntersuchungsämtern
der Länder nach einem statistischen Schlüssel an das BVL gesendet, dort wurde ihre
Spezieszugehörigkeit überprüft und eine zentrale Empfindlichkeitstestung
vorgenommen. Als Methodik für die Tests wurde die Mikrodilution gemäß den
Richtlinien des CLSI eingeführt. Alle Isolate werden zentral im BVL als Dauerkulturen
gelagert und stehen somit für nachträgliche Untersuchungen zur Verfügung. Sowohl
die untersuchten Wirkstoffe als auch die in das Monitoring einbezogenen Erreger und
Indikationen wurden kontinuierlich angepasst und erweitert.
Erste Ergebnisse aus diesen beiden deutschen Monitoringprogrammen standen im
Jahr 2003 zur Verfügung (WALLMANN et al. 2003; PRIEBE u. SCHWARZ 2003) und
werden mit den Daten aus den Folgejahren zusammen diskutiert (Kapitel 4.1.1).
2.8.2 Empfindlichkeitslage von Kommensalen oder Indikatorbakterien
Weitere Studien haben sich mit der Empfindlichkeitsprüfung von Bakterien
beschäftigt, die nicht als Zielorganismen für den Wirkstoff Florfenicol eingestuft

EINFÜHRUNG IN DIE THEMATIK
28
waren (SYRIOPOULOU et al. 1981; GRAHAM et al. 1988; CANNON et al. 1990;
ORDEN et al. 2000; SALMON u. WATTS 2000; HO et al. 2000; YOSHIMURA et al.
2000). Diese Untersuchungen verfolgten unterschiedliche Zielstellungen und dienten
zum einen dazu, die Aktivität von Florfenicol gegenüber weiteren bakteriellen Genera
und Spezies zu bestimmen, um ein Bild über die Wirkpotenz der Substanz zu
erhalten (SYRIOPOULOU et al. 1981; GRAHAM et al. 1988; CANNON et al. 1990).
Zum anderen wurden Studien initiiert, die gezielt nach resistenten Bakterien gesucht
haben, die als Kommensalen oder auch als Pathogene beim Tier vorkommen, die
aber nicht als Zielorganismen für den Wirkstoff definiert sind (KEYES et al. 2000;
ORDEN et al. 2000; ARCANGIOLI et al. 1999; BOLTON et al. 1999; SALMON u.
WATTS 2000). Diese Bakterien können ebenfalls Florfenicolresistenz entwickeln
oder erwerben und dienen unter anderem als Indikatorbakterien, um Änderungen in
der Empfindlichkeitssituation von bakteriellen Populationen zu erfassen oder um
einen möglichen Transfer von Resistenzgenen zu beobachten.
Eine der ersten Studien, die sich mit der Wirksamkeit von Florfenicol gegenüber
Enterobacteriaceae und Haemophilus influenzae beschäftigte, war die Publikation
aus dem Jahr 1981 von SYRIOPOULOU et al.. Dabei wurden mittels
Agardilutionsmethode die MHK-Werte von 18 Shigella dysenteriae-, 21 Salmonella
Typhi-, 6 Escherichia coli- und 7 Klebsiella pneumoniae-Isolaten gegenüber
Chloramphenicol, Thiamphenicol und drei fluorierten Derivaten ermittelt. Alle Isolate
waren bekanntlich chloramphenicolresistent, über ihre Herkunft lagen allerdings
keine Informationen vor. Zusätzlich wurden in der Studie 53 ampicillinresistente
Haemophilus influenzae-Isolate getestet (SYRIOPOULOU et al. 1981). Ein Ziel der
Untersuchung war es, die Wirkpotenz der fluorierten Derivate zu ermitteln, von denen
sich zu dem Zeitpunkt noch keines in der klinischen Nutzung befand. Unter diesen
Derivaten war auch die Substanz Sch25298, die später unter dem Wirkstoffnamen
Florfenicol bekannt wurde. Die in der Studie ermittelten MHK-Werte für diese nicht
als Zielorganismen definierten Isolate sind in Tabelle 2 dargestellt. Eine
Klassifizierung der Isolate in die Kategorien empfindlich oder resistent ist nicht
möglich, da keine anerkannten Grenzwerte für diese Bakterien verfügbar sind.

EINFÜHRUNG IN DIE THEMATIK
29
Tabelle 2: Vergleich der In-vitro-Empfindlichkeit von Enterobacteriaceae und Haemophilus influenzae gegenüber Florfenicol nach SYRIOPOULOU et al. (1981)
MHK-Werte c (µg/ml) Spezies n a Besonderheit der Isolate b
0,5 1 5 10 50 ≥≥≥≥100
Shigella dysenteriae
18 ausgewählt CHL-resistent
- 18
Salmonella Typhi
21 ausgewählt CHL-resistent
- 1 20
Escherichia coli
6 ausgewählt CHL-resistent
- 1 4 1
Klebsielle pneumonieae
7 ausgewählt CHL-resistent
- 3 3 1
Haemophilus influenzae
53 ausgewählt AMP-resistent
53 - - -
a n = Anzahl der getesteten Isolate b CHL = Chloramphenicol; AMP = Ampicillin c Anzahl der Isolate mit einem MHK-Wert von... - = nicht in der Studie bestimmt
Eine ähnliche Zielstellung verfolgte auch die Studie von GRAHAM et al. 1988. Die
Autoren verwendeten eine Agardilutionsmethode und testeten vergleichend die In-
vitro-Empfindlichkeit von klinischen Isolaten gegenüber Chloramphenicol und
Florfenicol. Auch wenn die Studie durch die Einbeziehung von 18 unterschiedlichen
bakteriellen Spezies recht interessant ist, so müssen die Ergebnisse aufgrund der
niedrigen Isolatzahlen von häufig ≤ 10 / Spezies dennoch vorsichtig bewertet werden
(GRAHAM et al. 1988). Eine Aussage über die MHK-Wertverteilung ist bei diesen
Spezies dementsprechend nicht möglich.
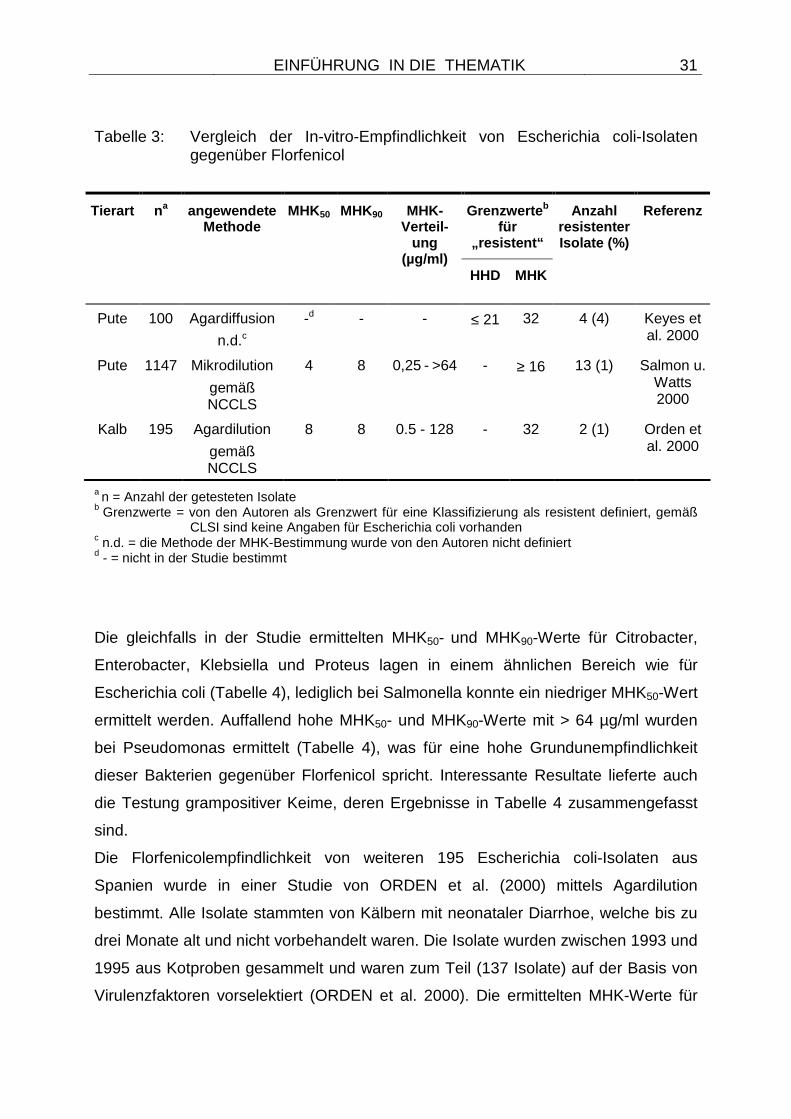
Eine spätere Studie (KEYES et al. 2000) beschäftigte sich mit der Empfindlichkeit
von 100 Escherichia coli- Isolaten gegenüber Florfenicol (Tabelle 3). Alle Isolate
stammten von erkrankten Puten aus den USA und vier der getesteten Isolate (4 %)
wurden von den Autoren als florfenicolresistent eingestuft. Diese Isolate zeigten in

EINFÜHRUNG IN DIE THEMATIK
30
der Agardiffusion einen Hemmhofdurchmesser von ≤ 21 mm beim Gebrauch von 30
µg Florfenicol Testplättchen und MHK-Werte von 32 µg/ml (2 Isolate) oder 64 µg/ml
(2 Isolate). Weitere sieben Isolate, die als chloramphenicolresistent eingestuft
wurden, zeigten deutlich niedrigere MHK-Werte für Florfenicol, die in einem Bereich
zwischen 4 und 8 µg/ml lagen. Erstaunlich war in der Studie, dass die
Florfenicolresistenz in allen Fällen auf der Präsenz des Gens floR basierte, die
Substanz Florfenicol aber in den USA zu keiner Zeit für die Anwendung beim
Geflügel zugelassen war und ein illegaler Kontakt der Tiere mit der Substanz als
äußerst unwahrscheinlich einzustufen war (KEYES et al. 2000). Auch wenn das Gen
floR zusätzlich Resistenz gegenüber Chloramphenicol vermittelt, so konnte auch Co-
Selektion durch eine Anwendung von Chloramphenicol keine mögliche Erklärung für
die Präsenz des floR-Gens liefern, da Chloramphenicol bereits 1988 in den USA für
lebensmittelliefernde Tiere verboten wurde. Daher spekulierten die Autoren, dass in
Escherichia coli Florfenicol-Resistenzgene präexistent sind und diese Gene einen
zukünftigen Einsatz der Substanz in weiteren Spezies limitieren könnten.
Auch von SALMON und WATTS wurden im Jahr 2000 aviäre Escherichia coli-Isolate
untersucht, wobei eine sehr große Anzahl von 1147 Isolaten auf ihre
Florfenicolempfindlichkeit getestet wurde (Tabelle 3). Die ermittelten MHK50- und
MHK90-Werte lagen bei 4 bzw. 8 µg/ml (SALMON u. WATTS 2000), was zumindest
einer weiteren Verbreitung von präexistenten floR-Genen bei Escherichia coli von
Puten (KEYES et al. 2000) entgegenspricht. Dennoch wurden 13 Isolate mit einem
MHK-Wert von ≥ 16 µg/ml identifiziert (1 %). Auch hier konnte die Ursache der hohen
MHK-Werte nicht erklärt werden, denn alle Isolate stammten, ähnlich den Isolaten
aus der Studie von KEYES et al. 2000, aus den USA oder Kanada (SALMON u.
WATTS 2000).
EINFÜHRUNG IN DIE THEMATIK
31
Tabelle 3: Vergleich der In-vitro-Empfindlichkeit von Escherichia coli-Isolaten gegenüber Florfenicol
Grenzwerte b für
„resistent“
Tierart na angewendete Methode
MHK50 MHK90 MHK-Verteil-
ung (µg/ml)
HHD MHK
Anzahl resistenter Isolate (%)
Referenz
Pute 100 Agardiffusion
n.d.c
-d - - ≤ 21 32 4 (4) Keyes et al. 2000
Pute 1147 Mikrodilution
gemäß NCCLS
4 8 0,25 - >64 - ≥ 16 13 (1) Salmon u. Watts 2000
Kalb 195 Agardilution
gemäß NCCLS
8 8 0.5 - 128 - 32 2 (1) Orden et al. 2000
a n = Anzahl der getesteten Isolate b Grenzwerte = von den Autoren als Grenzwert für eine Klassifizierung als resistent definiert, gemäß
CLSI sind keine Angaben für Escherichia coli vorhanden c n.d. = die Methode der MHK-Bestimmung wurde von den Autoren nicht definiert d - = nicht in der Studie bestimmt
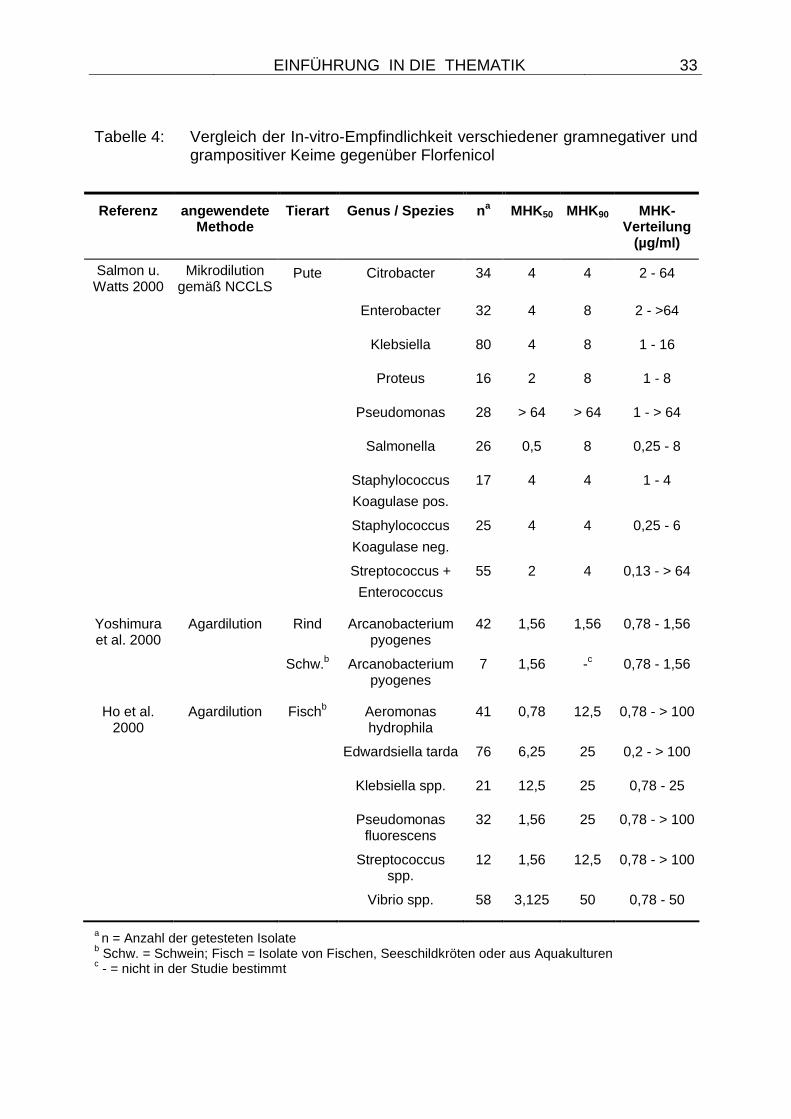
Die gleichfalls in der Studie ermittelten MHK50- und MHK90-Werte für Citrobacter,
Enterobacter, Klebsiella und Proteus lagen in einem ähnlichen Bereich wie für
Escherichia coli (Tabelle 4), lediglich bei Salmonella konnte ein niedriger MHK50-Wert
ermittelt werden. Auffallend hohe MHK50- und MHK90-Werte mit > 64 µg/ml wurden
bei Pseudomonas ermittelt (Tabelle 4), was für eine hohe Grundunempfindlichkeit
dieser Bakterien gegenüber Florfenicol spricht. Interessante Resultate lieferte auch
die Testung grampositiver Keime, deren Ergebnisse in Tabelle 4 zusammengefasst
sind.
Die Florfenicolempfindlichkeit von weiteren 195 Escherichia coli-Isolaten aus
Spanien wurde in einer Studie von ORDEN et al. (2000) mittels Agardilution
bestimmt. Alle Isolate stammten von Kälbern mit neonataler Diarrhoe, welche bis zu
drei Monate alt und nicht vorbehandelt waren. Die Isolate wurden zwischen 1993 und
1995 aus Kotproben gesammelt und waren zum Teil (137 Isolate) auf der Basis von
Virulenzfaktoren vorselektiert (ORDEN et al. 2000). Die ermittelten MHK-Werte für

EINFÜHRUNG IN DIE THEMATIK
32
Florfenicol lagen in einer Streubreite von 0.5 - 128 µg/ml, wobei 50 % bzw. 90 % der
Isolate mit einer Florfenicolkonzentration von 8 µg/ml zu hemmen waren. Nur zwei
Isolate (1 %) wurden als florfenicolresistent eingestuft, wobei die Autoren den
Breakpoint bei ≥ 32 µg/ml gesetzt haben (ORDEN et al. 2000). In der Tabelle 3
wurden die im Jahr 2000 ermittelten Daten zur Florfenicolempfindlichkeit bei
Escherichia coli-Isolaten und anderen Bakterien vergleichend gegenübergestellt.
Dabei ist zu beachten, dass unterschiedliche Methoden verwendet wurden und die
Breakpoints für eine Einstufung der Isolate als resistent von den Autoren festgesetzt
wurden.
Weitere Studien haben sich mit der Florfenicolempfindlichkeit recht spezifischer
Keime befasst. So bestimmten YOSHIMURA et al. (2000) MHK-Werte von 42
bovinen und 7 porcinen Arcanobacterium pyogenes-Isolaten aus Japan, während in
einer Studie aus Taiwan von HO et al. (2000) Keime von Fischen, Seeschildkröten
oder aus Wasserkulturen untersucht wurden. Zu beachten ist dabei, dass eine
Anwendung von Florfenicol für Wassermedikationen in einigen Ländern stattfindet,
die Substanz aber in Deutschland nicht für diese Zwecke zugelassen ist. Die
Ergebnisse dieser Testungen sind in Tabelle 4 zusammengefasst und zeigen, dass
bei einigen bakteriellen Genera und Spezies wie Enterobacter, Citrobacter,
Edwardsiella tarda oder Aeromonas hydrophila Isolate mit auffällig hohen MHK-
Werten vorkommen. Bei diesen Isolaten können spezifische Resistenzgene oder
andere Resistenzmechanismen (z.B. Mutationen) vermutet werden, eine
weiterführende Untersuchung der Isolate wurde aber nicht durchgeführt. Generell
lagen die MHK50- und MHK90-Werte der Indikatorbakterien aber deutlich über denen
der Zielorganismen, was auf eine höhere Grundempfindlichkeit dieser Bakterien
hindeutet.
EINFÜHRUNG IN DIE THEMATIK
33
Tabelle 4: Vergleich der In-vitro-Empfindlichkeit verschiedener gramnegativer und grampositiver Keime gegenüber Florfenicol
Referenz angewendete Methode
Tierart Genus / Spezies n a MHK50 MHK90 MHK-
Verteilung (µg/ml)
Salmon u. Watts 2000
Mikrodilution gemäß NCCLS
Pute Citrobacter 34 4 4 2 - 64
Enterobacter 32 4 8 2 - >64
Klebsiella 80 4 8 1 - 16
Proteus 16 2 8 1 - 8
Pseudomonas 28 > 64 > 64 1 - > 64
Salmonella 26 0,5 8 0,25 - 8
Staphylococcus
Koagulase pos.
17 4 4 1 - 4
Staphylococcus
Koagulase neg.
25 4 4 0,25 - 6
Streptococcus +
Enterococcus
55 2 4 0,13 - > 64
Yoshimura et al. 2000
Agardilution Rind Arcanobacterium pyogenes
42 1,56 1,56 0,78 - 1,56
Schw.b Arcanobacterium pyogenes
7 1,56 -c 0,78 - 1,56
Ho et al. 2000
Agardilution Fischb Aeromonas hydrophila
41 0,78 12,5 0,78 - > 100
Edwardsiella tarda 76 6,25 25 0,2 - > 100
Klebsiella spp. 21 12,5 25 0,78 - 25
Pseudomonas fluorescens
32 1,56 25 0,78 - > 100
Streptococcus spp.
12 1,56 12,5 0,78 - > 100
Vibrio spp. 58 3,125 50 0,78 - 50
a n = Anzahl der getesteten Isolate b Schw. = Schwein; Fisch = Isolate von Fischen, Seeschildkröten oder aus Aquakulturen c - = nicht in der Studie bestimmt

EINFÜHRUNG IN DIE THEMATIK
34
2.9 Florfenicol als Modellsubstanz
In der Literatur gibt es nur wenige Beispiele für antimikrobilelle Wirkstoffe, die
ausschließlich für eine Anwendung beim Tier zugelassen sind. Die Ursache hierfür
liegt darin, dass die meisten Altsubstanzen sowohl human- als auch tiermedizinisch
angewendet werden und neue Wirkstoffentwicklungen in der Regel dem
humanmedizinischen Gebrauch vorbehalten bleiben. Um aber einen Transfer von
Resistenzgenen vom Tier zum Menschen nachzuweisen, darf kein Selektionsdruck
durch die Anwendung des Wirkstoffes beim Menschen vorliegen. Zudem dürfen die
zu untersuchenden Gene keine Kreuzresistenzen zu anderen Wirkstoffen oder
Wirkstoffklassen vermitteln und sie sollten nicht vergesellschaftet mit anderen
Resistenzgenen auf mobilen genetischen Elementen lokalisiert sein. Eine solche
Lokalisation würde ihren Transfer und ihre Ausbreitung durch Co-Selektion
ermöglichen und Rückschlüsse auf einen Resistenzgentransfer durch einen
anwendungsbedingten Selektionsdruck des Wirkstoffes erschweren. Anhand einer
solchen Modellsubstanz können zudem Veränderungen der Empfindlichkeitslage auf
die Anwendung des Wirkstoffes zurückgeführt werden sowie die Verbreitung von
entsprechenden Resistenzgenen beobachtet werden.
Durch seine eingeschränkte Zulassung konnte Florfenicol als Modellbeispiel für
einen nur beim Tier eingesetzten Wirkstoff dienen. Da die bekannten und weit
verbreiteten Chloramphenicol-Resistenzgene (catA, catB, cmlA) keine
Kreuzresistenz zu Florfenicol bewirken (Publikation 1), sollten Veränderungen in der
Empfindlichkeitslage von Erregern direkt auf einen Einsatz der Wirksubstanz beim
Tier zurückzuführen sein. Besondere Bedeutung hat somit die Identifizierung
spezifischer Florfenicol-Resistenzgene und die Beobachtung ihrer Ausbreitung,
gerade wenn ein möglicher Transfer dieser Resistenzgene vom Tier zum Menschen
beobachtet werden soll.
Für solche Transferwege, die ihren Ursprung in einem beim Tier aufgebauten
Selektionsdruck finden, gibt es nur wenige Beispiele in der Literatur. Das
Streptothricin-Antibiotikum Nurseothricin war in der Deutschen Demokratischen
Republik (DDR) ab 1983 anstelle von Tetracyclinen als Leitungsförderer

EINFÜHRUNG IN DIE THEMATIK
35
ausschließlich beim Tier zugelassen. Obgleich anfänglich keine
Streptothricinresistenzen bei Enterobacteriaceae von Tieren und Menschen zu finden
waren (WITTE 2000), wurde das Nurseothricin-Resistenzgen sat im Jahr 1985, also
zwei Jahre nach der Zulassung des Wirkstoffs in der DDR, bei Escherichia coli-
Isolaten vom Schwein identifiziert (WITTE 1998). Das sat-Gen kam auf einem
Transposon lokalisiert vor und kodiert für eine Streptothricin-Acetyltransferase
(WITTE 2000). Bis zu dem Stopp der Anwendung von Nurseothricin im Zuge der
Deutschen Wiedervereinigung 1990 wurde das sat-Gen auch bei Escherichia coli-
Isolaten aus der Darmflora von Farmpersonal, von Familienangehörigen des
Farmpersonals, von städtischer Bevölkerung sowie bei Escherichia coli-Isolaten
humaner Harnwegsinfektionen identifiziert. Später gelang der Nachweis des
Resistenzgens auch bei Salmonella- und Shigella-Isolaten vom Menschen
(HUMMEL et al. 1986; TSCHÄPE 1994). Offensichtlich hatte eine Verbreitung dieses
Resistenzgens auch ohne direkten Selektionsdruck stattgefunden (WITTE 1997;
WITTE 1998) und ein Transfer des Gens vom Tier zum Menschen erschien
nachgewiesen. Allerdings wurde das sat-Gen mittlerweile auch als Bestandteil von
Mulitresistenz-Integrons identifiziert (TSAKRIS et al. 2007; SUNDSTRÖM et al.
1991), so dass eine Verbreitung über Co-Selektion ebenfalls in Betracht gezogen
werden muss.
Eine grundlegend andere Situation liegt für weitere Substanzen wie die zu den
Pleuromutilinen gehörenden Wirkstoffe Tiamulin oder Valnemulin vor, für die
ebenfalls nur eine veterinärmedizinische Zulassung besteht. Im Gegensatz zu
Nurseothricin, wurden für diese Substanzen lediglich Resistenzen nachgewiesen, die
auf Mutationen ribosomaler Proteine basieren (BÖCK et al. 1982). Von BØSLING et
al. wurde 2003 eine Tiamulinresistenz-vermittelnde Mutation im ribosomalen Protein
L3 bei einer Escherichia coli-Mutante beschrieben. Diese Mutation zog einen
Aminosäureaustausch an Position 149 von Asn zu Asp nach sich (BØSLING et al.
2003) und zeigte sich für den Resistenzphänotyp verantwortlich. Mutationen im L3
Protein wurden ebenfalls bei Brachyspira spp. identifiziert und gingen gleichfalls mit
verminderter Tiamulinempfindlichkeit einher (PRINGLE et al. 2004). In der gleichen
Studie konnten weitere Mutationen in der 23S rRNA von Brachyspira spp.
nachgewiesen werden, die eine verminderte Bindung von Tiamulin an das Ribsosom

EINFÜHRUNG IN DIE THEMATIK
36
bewirkten (PRINGLE et al. 2004). Transferable Pleuromutlin-Resistenzgene wurden
bislang allerdings nicht beschrieben. Somit konnten für die Modellsubstanzen
Tiamulin und Valnemulin weder spezifische Resistenzgene noch ein Gentransfer auf
humane Isolate beobachtet werden.
Wie zuvor erwähnt, findet bei den meisten älteren antimikrobiellen
Chemotherapeutika ein sowohl human- als auch veterinärmedizinischer Einsatz statt,
der nur eine eingeschränkte Aussage über die Herkunft der Resistenzen erlaubt. Ist
ein Resistenzgen erst einmal in verschiedenen Ökosystemen weit verbreitet, so ist
es stets schwierig, die Herkunft des Gens zu verfolgen (WITTE 2000). Da heute die
meisten neueren Substanzklassen, wie beispielsweise Glykopeptide, Carbapeneme,
Monobactame oder Oxazolidinone, eine ausschließlich humanmedizinische
Zulassung erhalten, ist auch in Kürze nicht mit vergleichbaren Modellsubstanzen wie
Florfenicol zur rechnen. Dies unterstreicht die besondere Bedeutung, die dem
Nachweis und der Verbreitung möglicher spezifischer Florfenicol-Resistenzgene
zukommt.

ZIELSTELLUNG DER ARBEIT
37
3 Zielstellung der Arbeit
Da nur sehr wenig Daten zur Florfenicolresistenz bekannt waren, wurde die
Zielstellung der Studie auf drei Bereiche ausgedehnt und wie folgt unterteilt:
• Bereich 1: Zielorganismen
In dem ersten Arbeitsschwerpunkt sollte die Empfindlichkeitslage von
Zielorganismen für Florfenicol in Deutschland über ein Monitoring erfasst
werden, um so mögliche Veränderungen der Resistenzlage zeitnah
festzustellen. Beim Auftreten von ersten resistenten Zielorganismen sollte der
genetische Hintergrund der Resistenz untersucht werden und Hinweise auf
eine mögliche Mobilität der Gene erarbeitet werden.
• Bereich 2: Kommensalen Escherichia coli
Da zu Beginn der Studie keine resistenten Zielorganismen beschrieben
waren, sollten die Untersuchungen auf „Indikatorbakterien“ ausgedehnt
werden, um Hinweise auf Veränderungen der Resistenzsituation zu erlangen.
Dafür wurden Escherichia coli-Isolate ausgewählt, die sowohl als
Kommensalen als auch als Pathogene beim Rind vorkommen, einer Tierart,
für die seit 1995 eine Zulassung von Florfenicol in Deutschland besteht. Auch
hier war es Ziel der Studie, die entsprechenden Resistenzgene zu
identifizieren und Transfermöglichkeiten der Gene aufzudecken.
• Bereich 3: Kommensalen Staphylokokken
Als „Indikatorbakterien“ für das Vorkommen und die Verbreitung von
Florfenicol-Resistenzgenen bei grampositiven Keimen wurden
Staphylokokken ausgewählt. Diese Bakterien sind - neben weiteren
Lokalisationen - als Kommensalen oder Pathogene im Respirationstrakt von
Tieren zu finden und könnten unter entsprechendem Selektionsdruck
gleichfalls Florfenicolresistenz ausbilden oder von anderen Bakterien des

ZIELSTELLUNG DER ARBEIT
38
gleichen Habitats erwerben. Da bislang keine Informationen zur
Florfenicolresistenz bei grampositiven Bakterien in Deutschland vorlagen,
sollten zunächst Staphylococcus spp. von Tieren auf ihren Resistenzphänotyp
untersucht werden. Ziel dieses Arbeitsschwerpunktes war es, mögliche
Resistenzgene zu identifizieren und diese Gene mit den Resistenzgenen
gramnegativer Bakterien zu vergleichen. Dabei sollten mobile genetische
Elemente, die Hinweise auf Transfermöglichkeiten der Gene geben, ermittelt
werden und ihre Funktionalität in gramnegativen Keimen (Escherichia coli und
Zielorganismen) getestet werden. Bei einer Identifizierung entsprechender
Gene sollte auch ihr Vorkommen und ihre Verbreitung bei humanen Isolaten
untersucht werden.
Ein weiterer Aspekt in allen Arbeitsbereichen war es, Hinweise auf eine mögliche Co-
Selektion von Florfenicol-Resistenzgenen unter Anwendung weiterer antimikrobieller
Chemotherapeutika zu erlangen und die genetischen Mechanismen zu identifizieren.

RESULTATE UND DISKUSSION
39
4 Einordnung der Resultate und zusammenfassende
Diskussion
4.1 Teil I Atemwegsinfektionserreger von Rind und S chwein
4.1.1 Untersuchungen zur Resistenzlage bei Zielorga nismen für Florfenicol
Da nur sehr wenige Daten zur Empfindlichkeitslage boviner und porciner Erreger von
Atemwegsinfektionen in Deutschland verfügbar waren (HÖRMANSDORFER u.
BAUER 1996; HÖRMANSDORFER u. BAUER 1998; GOOSENS 1999) sollte in den
eigenen Untersuchungen die aktuelle Empfindlichkeitssituation gegenüber Florfenicol
anhand von Isolaten aus den Jahren 2002 und 2003 erfasst werden. Die Resultate
dieses spezifischen Monitorings wurden in Publikation 2 zusammengefasst und mit
Ergebnissen früherer Studien vergleichend analysiert. Dieser Vergleich diente dazu,
mögliche Veränderungen der Empfindlichkeitssituation einige Jahre nach Einführung
des Wirkstoffes in die veterinärmedizinische Nutzung in Deutschland erkennen zu
können.
Als Erreger respiratorischer Erkrankungen beim Rind wurden in diesem Zeitraum 95
Pasteurella multocida- und 98 Mannheimia haemolytica-Isolate gesammelt, vom
Schwein wurden 103 Pasteurella multocida- und 63 Actinobacillus
pleuropneumoniae-Isolate als Zielorganismen in die Testungen einbezogen
(Publikation 2). Zusätzlich wurden 131 Bordetella bronchiseptica- und 105
Streptococcus suis-Isolate getestet, da diese Erreger häufig bei Mischinfektionen
vergesellschaftet mit den Zielorganismen vorkommen und eine Erweiterung der
Zulassung von der Herstellerfirma auch für diese Erreger angestrebt wird. Zudem
liegen Grenzwerte nach den Richtlinien der CLSI für die Einstufung dieser Erreger
vor (NCCLS 2004). Die Durchführung dieses spezifischen Monitorings wurde so
angelegt, dass die Anforderungen an Empfindlichkeitsprüfungen im Rahmen von
Monitoringprogrammen erfüllt wurden. Dazu zählten die methodisch korrekte

RESULTATE UND DISKUSSION
40
Durchführung und Auswertung der Empfindlichkeitsprüfung gemäß einer
international anerkannten Durchführungsrichtlinie (NCCLS: M31-A2 2002), eine
einheitliche Speziesdifferenzierung der zu untersuchenden Bakterien auf der Basis
des aktuellen Kenntnisstandes sowie die Untersuchung einer ausreichend hohen
Anzahl epidemiologisch unverwandter Isolate der zu testenden Bakterien. Zudem
wurde eine ausführliche Dokumentation angefertigt, die neben dem angestrebten
Probenumfang auch Angaben über die Herkunftsbetriebe, die Tierart, die Probenart,
die Anamnese sowie eine eventuelle Prämedikation der Tiere enthielt. Auch der
Einbezug von Qualitätskontrollstämmen (Escherichia coli ATCC 25922,
Staphylococcus aureus ATCC 29213 und ATCC 25923, Actinobacillus
pleureopneumoniae ATCC 27090) gehörte zur Durchführung der
Empfindlichkeitsprüfung.
Auch wenn diese Grundvoraussetzungen bei jeder Empfindlichkeitsprüfung im
Rahmen von Monitoringprogrammen erfüllt werden sollten, so ist die praktische
Umsetzung dennoch häufig mit Problemen behaftet und erschwert eine
Vergleichbarkeit der Resultate. In älteren oder auch einigen aktuellen Studien zur
Florfenicolempfindlichkeit stand beispielsweise nur Untersuchungsmaterial aus
wenigen geographischen Regionen zur Verfügung (HÖRMANSDORFER u. BAUER
1996; HÖRMANSDORFER u. BAUER 1998; GUTIÉRREZ-MARTĺN et al. 2006). In
weiteren Studien wurde entweder eine abweichende Methodik gewählt (SHIN et al.
2005) oder es stand nur eine eingeschränkte Probenzahl zur Verfügung (MARTĺN
DE LA FUENTE et al. 2006). Wieder andere Studien legten eine andere Zielstellung
an, so dass zwar eine umfangreiche Anzahl von Isolaten untersucht wurde, aber
keine MHK50-; MHK90-Werte oder MHK-Bereiche errechnet wurden (MARIE et al.
2002). Daher war es Ziel der Publikation 2, Resultate zu erzielen, die eine gute
Vergleichbarkeit mit anderen nationalen oder internationalen
Empfindlichkeitsprüfungen ermöglichen.
Von besonderem Interesse in Publikation 2 waren die Ergebnisse für die bovinen
Atemwegsinfektionserreger, da Florfenicol bereits seit 8 Jahren für diese Indikation
eingesetzt wurde. Hierbei zeigte ein Vergleich der MHK50- und MHK90-Werte von

RESULTATE UND DISKUSSION
41
Isolaten aus unterschiedlichen Jahren (Tabelle 1 in Publikation 2), dass beide
Kennwerte über die Jahre stabil geblieben sind (HÖRMANSDORFER u. BAUER
1996; HÖRMANSDORFER u. BAUER 1998; PRIEBE u. SCHWARZ 2003). Die
Werte von 2002 / 2003 lagen auf einem Niveau, welches bereits vor der Zulassung
des Wirkstoffes bestanden hat. Somit war keine Resistenzentwicklung bei bovinen
Isolaten aus Deutschland gegenüber Florfenicol festzustellen.
Eine ähnlich günstige Empfindlichkeitslage war auch für die Isolate porciner Herkunft
zu ermitteln. Die MHK50- und MHK90-Werte für Pasteurella multocida und
Actinobacillus pleuropneumoniae lagen ebenfalls auf einem Niveau, welches vor
oder zum Zeitpunkt der Zulassung von Florfenicol für das Schwein bestanden hat
(HÖRMANSDORFER u. BAUER 1998; PRIEBE u. SCHWARZ 2003). Die Werte
waren zudem in guter Übereinstimmung mit den Ergebnissen für Pasteurella
multocida-Isolate aus dem Jahr 2001 des nationalen Resistenzmonitoring-
Programms GERM Vet (WALLMANN et al. 2003).
Für Streptococcus suis wurden zwar geringfügig höhere MHK50- und MHK90-Werte
ermittelt als für andere Zielorganismen, aber auch diese Werte zeigten sich über die
Testzeiträume stabil. Zusammenfassend waren alle getesteten Isolate der Spezies
Pasteurella multocida, Mannheimia haemolytica, Actinobacillus pleuropneumoniae
und Streptococcus suis als empfindlich gegenüber Florfenicol einzustufen und ließen
keine Resistenzentwicklung über die Testzeiträume erkennen. Auch eine Fortführung
des Resistenzmonitorings mit Isolaten aus den Jahren 2004 - 2006 (SCHWARZ u.
KEHRENBERG, nicht publizierte Daten) lieferte Ergebnisse, die maximal eine
Verdünnungsstufe Abweichung zu den Werten von 2002 / 2003 erkennen ließen.
Eine besondere Situation wurde nur bei Bordetella bronchiseptica-Isolaten ermittelt.
Für diese Spezies wurde in einer vorhergehenden Studie (PRIEBE u. SCHWARZ
2003) ein relativ hoher Prozentanteil (25,6 %) resistenter Isolate aus den Jahren
2000 / 2001 errechnet. Dabei blieben die Ursachen für die hohen Resistenzraten
ungeklärt (PRIEBE u. SCHWARZ 2003). Die Ergebnisse des Folgezeitraums
2002 / 2003 (Publikation 2) ließen einen Rückgang der Resistenzrate auf 0,8 %
erkennen und auch die MHK50- und MHK90-Werte verringerten sich um eine bzw.
zwei Verdünnungsstufen.

RESULTATE UND DISKUSSION
42
Werden die in Publikation 2 dargestellten Ergebnisse mit den Resultaten von
Monitoringstudien aus anderen Ländern verglichen (Tabelle 5), so zeigt sich, dass
die MHK-Werte in vergleichbaren Bereichen liegen. Die von MEVIUS u. HARTMAN
2000 publizierten MHK50- und MHK90-Werte boviner Pasteurella multocida-Isolate
aus den Niederlanden lagen nur eine Verdünnungsstufe über den in Deutschland
ermittelten Werten und auch für Mannheimia haemolytica zeigten sich kaum
abweichende Kennwerte (Tabelle 5). Genau wie bei MEVIUS u. HARTMAN 2000
wurden auch bei SHIN et al. 2005 alle mittels Agardiffusion getesteten Mannheimia
haemolytica-Isolate aus Korea als empfindlich gegenüber Florfenicol eingestuft.
Auch bei Isolaten von Schweinen wurden vergleichbare MHK50- und MHK90-
Kennwerte in Monitoringstudien anderer Länder ermittelt (UEDA u. SUENAGA 1995;
YOSHIMURA 2002; GUTIÉRREZ-MARTĺN et al. 2006; MARTĺN DE LA FUENTE et
al. 2006; WISSELINK et al. 2006; VERA LIZARAZO et al. 2006). Interessant war ein
Vergleich der Empfindlichkeitsentwicklung von Pasteurella multocida-Isolaten aus
Spanien aus den Testzeiträumen 1987 / 1988 und 2003 / 2004 (VERA LIZARAZO et
al. 2006). Hier wurde nach einem Zeitraum von 16 Jahren lediglich eine Abweichung
des MHK90-Wertes um eine Verdünnungsstufe nach oben festgestellt.
Für Actinobacillus pleuropneumoniae zeigte sich eine besonders gute
Übereinstimmung der MHK50- und MHK90-Ergebnisse mit den Studien aus Japan
(UEDA u. SUENAGA 1995; YOSHIMURA et al. 2002) und Spanien (GUTIÉRREZ-
MARTĺN et al. 2006). Auch für Streptococcus suis wurden ähnliche Kenndaten bei
Isolaten mit einer Herkunft aus Belgien, Frankreich, Italien, Deutschland, Spanien,
den Niederlanden und dem Vereinigten Königreich errechnet (WISSELINK et al.
2006). MARIE et al. 2002 stuften ebenfalls alle Streptococcus suis-Isolate vom
Schwein als empfindlich gegenüber Florfenicol ein. Isolate der Spezies Haemophilus
parasuis mit einer Herkunft aus dem Vereinigten Königreich oder Spanien wurden
von MARTĺN DE LA FUENTE et al. 2006 untersucht und zeigten gleichfalls niedrige
MHK50- und MHK90-Werte. Lediglich ein Bordetella bronchiseptica-Isolat aus Korea
wurde - wenn auch mit der abweichenden Methodik der Agardiffusion - als resistent
gegenüber Florfenicol getestet (SHIN et al. 2005).

RESULTATE UND DISKUSSION
43
Bei den in Tabelle 5 gegenübergestellten Resultaten aus den unterschiedlichen
Studien ist zu allerdings zu beachten, dass der Isolierungszeitpunkt der Bakterien
teilweise erheblich vom Publikationsjahr abweicht und zum Teil Isolate aus mehreren
Ländern (WISSELINK et al. 2006) oder aus unterschiedlichen Isolierungsjahren
(GUTIÉRREZ-MARTĺN et al. 2006; VERA LIZARAZO et al. 2006) in die
Untersuchungen einbezogen wurden.
Zusammenfassend lässt sich eine recht gute Übereinstimmung der Resultate aus
den verschiedenen Studien feststellen. Sowohl die ermittelten MHK50- und MHK90-
Werte als auch die MHK-Wertverteilung liegen in ähnlichen Konzentrationsbereichen
(Tabelle 5). Mit Ausnahme der Spezies Bordetella bronchiseptica wurden alle
getesteten Isolate als empfindlich gegenüber dem Wirkstoff eingestuft und eine
Resistenzentwicklung war - sofern Isolate aus unterschiedlichen Isolierungsjahren
einbezogen wurden - nicht zu erkennen.
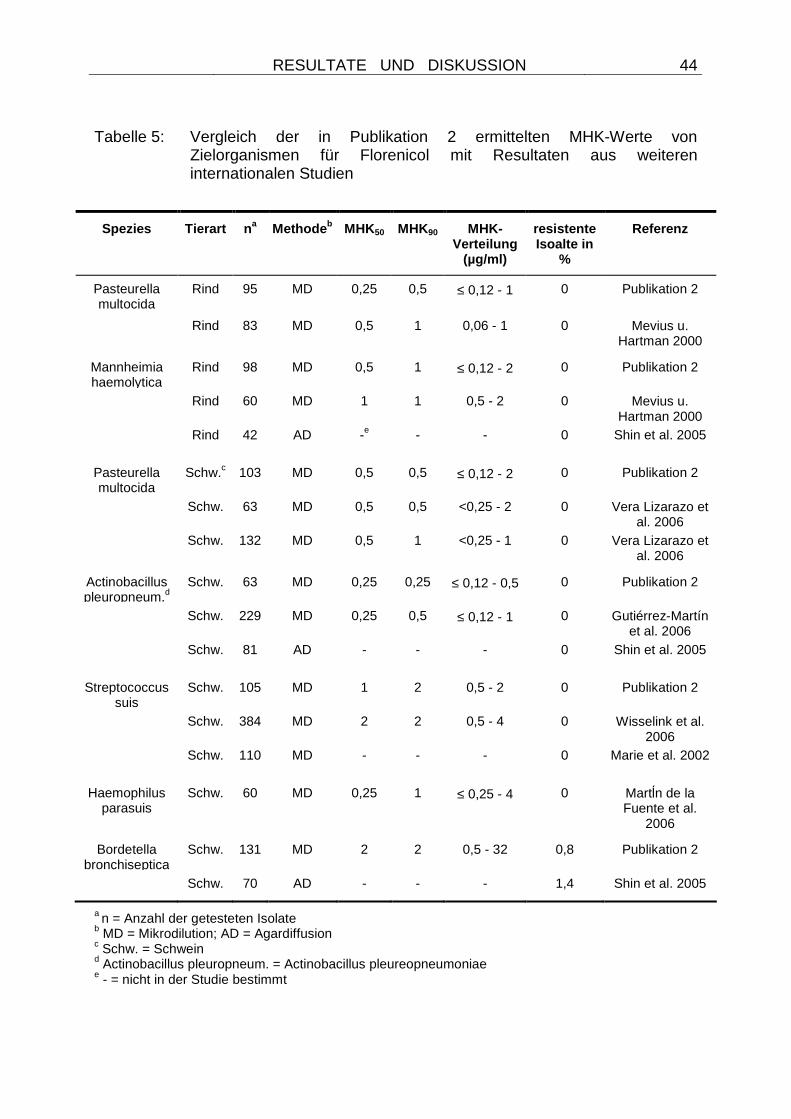
RESULTATE UND DISKUSSION
44
Tabelle 5: Vergleich der in Publikation 2 ermittelten MHK-Werte von Zielorganismen für Florenicol mit Resultaten aus weiteren internationalen Studien
Spezies Tierart na Methode b MHK50 MHK90 MHK-Verteilung
(µg/ml)
resistente Isoalte in
%
Referenz
Pasteurella multocida
Rind 95 MD 0,25 0,5 ≤ 0,12 - 1 0 Publikation 2
Rind 83 MD 0,5 1 0,06 - 1 0 Mevius u. Hartman 2000
Mannheimia haemolytica
Rind 98 MD 0,5 1 ≤ 0,12 - 2 0 Publikation 2
Rind 60 MD 1 1 0,5 - 2 0 Mevius u. Hartman 2000
Rind 42 AD -e - - 0 Shin et al. 2005
Pasteurella multocida
Schw.c 103 MD 0,5 0,5 ≤ 0,12 - 2 0 Publikation 2
Schw. 63 MD 0,5 0,5 <0,25 - 2 0 Vera Lizarazo et al. 2006
Schw. 132 MD 0,5 1 <0,25 - 1 0 Vera Lizarazo et al. 2006
Actinobacillus pleuropneum.d
Schw. 63 MD 0,25 0,25 ≤ 0,12 - 0,5 0 Publikation 2
Schw. 229 MD 0,25 0,5 ≤ 0,12 - 1 0 Gutiérrez-Martín et al. 2006
Schw. 81 AD - - - 0 Shin et al. 2005
Streptococcus suis
Schw. 105 MD 1 2 0,5 - 2 0 Publikation 2
Schw. 384 MD 2 2 0,5 - 4 0 Wisselink et al. 2006
Schw. 110 MD - - - 0 Marie et al. 2002
Haemophilus parasuis
Schw. 60 MD 0,25 1 ≤ 0,25 - 4 0 Martĺn de la Fuente et al.
2006 Bordetella
bronchiseptica Schw. 131 MD 2 2 0,5 - 32 0,8 Publikation 2
Schw. 70 AD - - - 1,4 Shin et al. 2005
a n = Anzahl der getesteten Isolate b MD = Mikrodilution; AD = Agardiffusion c Schw. = Schwein d Actinobacillus pleuropneum. = Actinobacillus pleureopneumoniae e - = nicht in der Studie bestimmt

RESULTATE UND DISKUSSION
45
4.1.2 Auftreten und Analyse erster florfenicolresis tenter Zielorganismen
aus dem Vereinigten Königreich (UK)
Auch wenn über viele Jahre der veterinärmedizinischen Anwendung von Florfenicol
keine resistenten Zielorganismen in Deutschland zu ermitteln waren, so wurde ein
erstes resistentes Pasteurella multocida-Isolat im Jahr 2003 von der Veterinary
Laboratories Agency, Thirsk Regional Laboratory, North Yorkshire, im Vereinigten
Königreich (UK) isoliert. Dieses Isolat stammte von einem 4-Monate alten und mit
Florfenicol, Oxytetracyclin und einem nicht weiter definiertem Kortikosteroid
vorbehandelten Kalb, welches an einer schweren Pneumonie erkrankt und nach 3
Tagen verstorben war. Parallel zu diesem Pasteurella multocida-Isolat wurde von
demselben Kalb post mortem ein florfenicolresistentes Salmonella enterica subsp.
enterica Serovar Dublin-Isolat identifiziert. Beide Isolate wurden von Jon Rogers und
Paul Todd, Veterinary Laboratories Agency, UK zur weiteren Untersuchung zur
Verfügung gestellt. Dadurch wurde die erste Identifizierung eines Florfenicol-
Resistenzgens bei einem Zielbakterium ermöglicht. Es zeigte sich, dass die
kombinierte Florfenicol- und Chloramphenicolresistenz bei Pasteurella multocida auf
der Präsenz des Effluxgens floR basierte. Dieses Gen war auf einem Plasmid
lokalisiert, welches die Bezeichnung pCCK381 erhielt. Eine genaue Analyse des
Resistenzgens sowie der flankierenden Bereiche ist in Publikation 3 dargestellt.
Durch diese Publikation konnten Spekulationen beendet werden, ob das von
Photobacterium damselae (KIM u. AOKI 1996) oder verschiedenen Salmonella
Serovaren (BOYD et al. 2002; CLOECKAERT et al. 2000) bekannte floR-
Resistenzgen nicht bei den Zielbakterien für Florfenicol vorkommt oder in diesen
Wirten nicht funktionell aktiv ist. Da eine Identität der Nukleotidsequenz von 97,2 -
99,7 % mit anderen floR-Genen (ARCANGIOLI et al. 1999, BOYD et al. 2001,
Publikation 1) festgestellt wurde, schienen umfangreiche Mutationen des Gens für
eine funktionelle Aktivität in Pasteurella nicht erforderlich zu sein. Allerdings wurden
relativ niedrige MHK-Werte für Florfenicol und Chloramphenicol von 16 und 32 µg/ml
sowohl bei dem Ausgangsstamm, als auch bei den Escherichia coli
JM101::pCCK381-Transformanden beobachtet (Publikation 3). Im Vergleich zeigten
Escherichia coli-Empfängerstämme, in die große floR-tragende originär aus

RESULTATE UND DISKUSSION
46
Escherichia coli stammende Plasmide konjugiert wurden, häufig höhere MHK-Werte
von 32 - 256 µg/ml für Florfenicol (Publikation 6). Ursächlich für diese Variabilität
könnte eine eingeschränkte Funktionalität des floR-Gens oder eine verminderte
Expression bei pCCK381 sein. Durch die hohe Nukleotidsequenzidentität zu anderen
floR-Genen scheint es aber wahrscheinlicher, dass das niedrige Resistenzniveau auf
eine abweichende Expression zurückzuführen ist.
Abbildung 4: Schematische Präsentation von Plasmid pCCK381. Die Leserahmen (repA, repB, repC: Plasmidreplikation; mob: Mobilisation, floR: Florfenicol- / Chloramphenicolresistenz) sind als Pfeile dargestellt, wobei die Pfeilspitze in die Transkriptionsrichtung zeigt. Eine Größenskala in kb ist unterhalb der Zeichnung angefertigt.
Das Plasmid pCCK381 (Abbildung 4) unterschied sich durch zwei wesentliche
Punkte von floR-tragenden Plasmiden, wie sie bei Escherichia coli gefunden wurden.
Der erste Unterscheidungspunkt war die Plasmidgröße von nur 11 kb bei Pasteurella
im Gegensatz zu 110 - 125 kb (Publikation 6) oder sogar 225 kb (WHITE et al. 2000)
bei Escherichia coli. Der zweite Unterscheidungspunkt war in dem
plasmidvermittelten Resistenzphänotyp zu finden. Während pCCK381 lediglich
Florfenicol- und Chloramphenicolresistenz vermittelte, wurden bei den Plasmiden
von Escherichia coli zusätzliche Resistenzmarker wie Sulfonamid-, Trimethoprim-,
Streptomycin- oder Ampicillinresistenz gefunden (Publikation 6). Aufgrund dieser
strukturellen Unterschiede wurde eine genaue Sequenzanalyse des gesamten
Pasteurella-Plasmids pCCK381 vorgenommen. Diese Analyse offenbarte, dass das
pCCK381
0 1 2 3 4 5 6 7 8 9 10
Ev CHp K Pv B DPE KHp P Hp
repA repC ∆tnp mobACB
0 1 2 3 4 5 6 7 8 9 10
floR
Ev CHp K Pv B DPE KHp P Hp
repArepC ∆tnp mobrepBACB repB pCCK381
0 1 2 3 4 5 6 7 8 9 10
Ev CHp K Pv B DPE KHp P Hp
repA repC ∆tnp mobACB
0 1 2 3 4 5 6 7 8 9 10
floR
Ev CHp K Pv B DPE KHp P Hp
repArepC ∆tnp mobrepBACB repB

RESULTATE UND DISKUSSION
47
Plasmid aus mehreren Segmenten anderer Plasmide zusammengesetzt ist
(Publikation 3). Diese in hohem Maße zu anderen Plasmiden identischen Regionen
waren zuvor bei Escherichia coli-Plasmiden von Rindern und Schweinen
beschrieben worden und schlossen die floR-Resistenzgenregion ein, oder die
Segmente wurden bereits bei fisch- (Vibrio salmonicida) oder rinderpathogenen
Bakterien (Dichelobacter nodosus) nachgewiesen. Die Analyse des Replikations-
Genbereichs zeigte nur wenige Aminosäureaustausche zu Plasmid pDN1
(Datenbank Zugangsnummer Y19120) von Dichelobacter nodosus (WHITTLE et al.
2000). Da keine Empfängerstämme dieser Spezies erhältlich sind, konnte die
Replikationsfähigkeit in diesen Wirten allerdings nicht geprüft werden. Zu bedenken
ist aber, dass Florfenicol zu der Zeit in vielen Ländern für die Bekämpfung dieser
Bakterien zugelassen war. Auch wenn die Entstehung des Pasteurella-Plasmids
retrospektiv nicht zu klären ist, so ist aufgrund der zusammengesetzten Struktur eine
interplasmidäre Rekombination wahrscheinlich.
Der gleiche Plasmidtyp wurde auch bei dem Salmonella Dublin-Isolat identifiziert. Er
ließ sich in Escherichia coli und Pasteurella multocida transformieren und replizierte
sich in beiden Wirten problemlos. Daher ist davon auszugehen, dass eine weitere
Verbreitung des Resistenzplasmids in andere Spezies oder Genera möglich ist.
Aufgrund der phänotypischen Resistenztestung von Transformanden sowie der
kompletten Sequenzanalyse des Plasmids (Abbildung 4) kann das auf pCCK381
lokalisierte floR-Resistenzgen allerdings nicht im Zuge von Co-
Selektionsmechanismen verbreitet werden. Derartige Co-Selektionsmechanismen
wurden in der Vergangenheit häufig als Ursache für die Persistenz und Verbreitung
von Resistenzgenen beschrieben (AARESTRUP 2000) und scheinen gerade bei
Pasteurella und Mannheimia eine wichtige Bedeutung zu haben (KEHRENBERG u.
SCHWARZ 2001; KEHRENBERG u. SCHWARZ 2002). So wurden bei diesen
Genera Co-Selektionsmechanismen für die Persistenz des catA3-Gens auch Jahre
nach dem Anwendungsverbot von Chloramphenicol für lebensmittelliefernde Tiere
verantwortlich gemacht (KEHRENBERG u. SCHWARZ 2001; KEHRENBERG u.
SCHWARZ 2002). Ursächlich war eine Organisation des Gens in einem sul2,-catA3-
strA Resistenzgencluster.

RESULTATE UND DISKUSSION
48
In Publikation 3 konnte gezeigt werden, dass das erste bekannte Florfenicol-
Resistenzplasmid aus einem Zielbakterium zwar in verschiedenen Wirten funktionell
aktiv ist (Pasteurella, Escherichia coli, Salmonella Dublin) und über Mobilisierung
transferiert werden kann, aber eine Verbreitung dieses Plasmid über Co-
Selektionsmechanismen nicht möglich ist.
Weitere Untersuchungen von Kälbern der gleichen Herkunftsfarm gaben keine
weiteren florfenicolresistenten Isolate (ROGERS u. TODD, persönliche Mitteilung).
Somit schien Florfenicolresistenz bei Zielorganismen zu der Zeit immer noch ein sehr
seltenes Ereignis zu sein.
4.1.3 Auftreten und Analyse weiterer florfenicolres istenter Zielorganismen
aus Frankreich
Im Jahr 2005 wurde in Frankreich im Rahmen des RESPATH network ein weiteres
florfenicolresistentes Isolat aus der Familie der Pasteurellaceae ermittelt. Auch
dieses Isolat wurde für eine Untersuchung der genetischen Basis des
Resistenzphänotyps zur Verfügung gestellt. Es stammte von der Lunge eines Kalbes
und konnte der Spezies Pasteurella trehalosi zugeordnet werden. Diese nur selten
bei Rindern auftretende Spezies wurde früher als „Pasteurella haemolytica Biotyp T“
bezeichnet. Aufgrund der seltenen Isolierung aus dem bovinen Respirationstrakt
wurde die Speziesdifferenzierung per 16S rDNA-Analyse bestätigt (Deutsche
Stammsammlung für Mikroorganismen und Zellkulturen DSMZ, Braunschweig).
Bislang waren noch keine Resistenzgene von Pasteurella trehalosi bekannt. Auch
bei dieser Spezies bestätigten Transformationsexperimente eine Plasmidlokalisation
des Florfenicol-Resistenzgens. Es war daher von Interesse zu ermitteln, ob das bei
dem Pasteurella multocida-Isolat aus dem Vereinigten Königreich identifizierte
Plasmid pCCK381 (Publikation 3) auch bei einem Isolat aus Frankreich zu finden ist.
Die in Publikation 4 dargestellte Plasmidanalyse identifizierte aber einen neuen
Plasmidtyp (pCCK13698) bei dem französischen Isolat.

RESULTATE UND DISKUSSION
49
Abbildung 5: Schemazeichnung von Plasmid pCCK13698 aus dem französischen Pasteurella trehalosi-Isolat. Eine Größenskala in kb ist in der Mitte eingetragen, die Leserahmen sind detaillierter ober- und unterhalb der Größenskala eingezeichnet. Die Trankskriptionsrichtung wird durch Pfeile dargestellt. Folgende Abkürzungen für die Leserahmen wurden verwendet: repA, repB, repC: Plasmidreplikation; tnp: Transposition; mobC: Mobilisation; rec: Rekombinationfunktion; int: Integrations-funktion; parA: DNA-Auftrennung; orfX: unbekannte Funktion; sul2: Sulfonamidresistenz; catA3: Chloramphenicolresistenz; floR: Florfenicol- / Chloramphenicolresistenz; strA: Streptomycinresistenz; L: lysR-ähnlicher Transkriptionsregulator; ∆: deletierter Leserahmen. Die Schnittstellen für Restriktionsendonukleasen wurden wie folgt abgekürzt: E (EcoRI), H (HpaI), Pv (PvuII), S (SspI) und X (XbaI).
Dieser neue Plasmidtyp (Abbildung 5) zeigte einige Gemeinsamkeiten, aber auch
Unterschiede zu pCCK381. Die Plasmidgrößen von ca. 15 kb (pCCK13698) und 11
kb (pCCK381) lagen in einem ähnlichen Bereich und waren damit deutlich kleiner als
floR-tragende Plasmide von Escherichia coli (Publikation 6; WHITE et al. 2000).
Beide Pasteurellaceae-Plasmide besitzen Gene, die ihren Transfer mittels
Mobilisierung ermöglichen. Ein konjugativer Plasmidtransfer ist aber
ausgeschlossen, da auf beiden Plasmiden keinen Gene lokalisiert sind, die einen
Transferkomplex kodieren. Der französische Plasmidtyp replizierte sich außerdem
0
mobC
0 5 10
1 2 3 4 5 6 7
parA rec orfX repB int
8
10 11 12
sul2 catA3
9
∆repA repC∆strA
13 14
floR L tnp
tnp
Pv S H S S S S S SS SPv EHX
IS1592
IS260
mobC
0 5 10
1 2 3 4 5 6 7
parA rec orfX repB int
8
10 11 12
sul2 catA3
9
∆repA repC∆strA
13 14
floR L tnp
tnp
Pv S H S S S S S SS SPv EHX
IS1592
IS26

RESULTATE UND DISKUSSION
50
nicht wie pCCK381 in kommerziellen Escherichia coli-Empfängerstämmen und
musste in einen Pasteurella multocida-Rezipienten transformiert werden. Die
Unfähigkeit zur Replikation in Escherichia coli oder anderen Empfängerstämmen
wurde bereits von weiteren Pasteurella-Plasmiden beschrieben (KEHRENBERG u.
SCHWARZ 2001) und kann Ursache für eine lediglich genus- oder
speziesbeschränkte Verbreitung eines Plasmidtyps sein. Zwei intakte
Replikationsgene, repC und repB, die zuvor auf Plasmiden von Bakterien der Genera
Mannheimia und Haemophilus identifiziert wurden, waren auf dem französischen
Plasmidtyp lokalisiert. Somit liegen Hinweise für eine genusübergreifende
Replikationsfähigkeit des französischen Plasmidtyps vor, ein experimenteller
Nachweis war aber mangels kommerziell erhältlicher Haemophilus-
Empfängerstämme nicht möglich. Ein weiterer Unterschied zwischen den Plasmiden
war in den von ihnen kodierten Resistenzmarkern zu finden. Während auf pCCK381
nur das floR-Resistenzgen lokalisiert war, wurden auf dem Pasteurella trehalosi-
Plasmid zwei weitere Resistenzgene (catA3 und sul2) identifiziert, die
Chloramphenicol- bzw. Sulfonamidresistenz vermitteln. Die funktionelle Aktivität der
beiden Gene wurde experimentell aufgezeigt (Publikation 4). Trotz der Anwesenheit
von zwei Chloramphenicolresistenzgenen (floR und catA3) auf dem gleichen Plasmid
wurden recht niedrige MHK-Werte von 16 bzw. 32 µg/ml für Florfenicol und
Chloramphenicol bei dem Ausgangsstamm und den Transformanden gemessen. Die
Gründe für das niedrige Resistenzniveau blieben aufgrund der funktionellen Aktivität
beider Gene allerdings unklar. Da Sulfonamide - im Gegensatz zu Chloramphenicol -
für therapeutische Zwecke bei Rindern zugelassen sind (www.vetidata.de) und in der
Fachliteratur auch empfohlen werden (RADOSTITS et al. 2000), ist aufgrund der
strukturellen Gegebenheiten bei pCCK13698 eine Verbreitung des floR-Gens im
Zuge von Co-Selektion möglich.
Die Analyse der Gesamtsequenzen beider Plasmide offenbarte eine
Zusammensetzung aus Segmenten unterschiedlicher bekannter Plasmide bei beiden
Typen. Während bei pCCK381 eine interplasmidäre Rekombination bei der
Plasmidentwicklung nur angenommen werden konnte (Publikation 3), wurden bei
dem französischen Plasmidtyp Rekombinationsstellen ermittelt und
Insertionselemente vom Typ IS1592 (KEHRENBERG et al. 1998) oder IS26

RESULTATE UND DISKUSSION
51
(MOLLET et al. 1983) nachgewiesen, die wahrscheinlich bei der Plasmidformation
beteiligt gewesen sind (Publikation 4).
Zusammenfassend ließen sich bei den florfenicolresistenten Isolaten aus Frankreich
und dem Vereinigten Königreich (UK) zwar in beiden Fällen floR-Gene nachweisen,
die plasmidäre Lokalisation unterschied sich aber voneinander. Offensichtlich kamen
in Frankreich und dem Vereinigten Königreich unterschiedliche Plasmide bei den
Zielbakterien vor, die Florfenicolresistenz vermitteln. Des weiteren wurden in
Publikation 4 erstmalig Resistenzgene bei Pasteurella trehalosi beschreiben.
4.1.4 Auftreten erster florfenicolresistenter Zielb akterien in Deutschland
Trotz günstiger Resistenzlage wurden 2005 einige florfenicolresistente Pasteurella
und Mannheimia-Isolate im Rahmen des deutschen Resistenzmonitoringprogramms
GERM Vet identifiziert. Eine weiterführende Analyse dieser Isolate (KEHRENBERG
u. SCHWARZ, nicht publizierte Daten) zeigte, dass auch hier ein Resistenzplasmid
vom Typ pCCK381 den kombinierten Florfenicol- und Chloramphenicol-
Resistenzphänotyp vermittelte. Damit zeigte sich erstmals ein grenzüberschreitendes
Vorkommen dieses Plasmidtyps. Aufgrund des zunehmenden Selektionsdrucks
durch eine erweiterte Zulassung von Florfenicol in Deutschland muss allerdings
zukünftig mit einer weiter ansteigenden Verbreitung gerechnet werden. Somit wird
die Wichtigkeit eines kontinuierlichen Monitorings in Deutschland unterstrichen.

RESULTATE UND DISKUSSION
52
4.1.5 Untersuchungen zur Florfenicol- und Chloramph enicolresistenz bei
Bordetella bronchiseptica
Im Gegensatz zu allen anderen Zielorganismen wurden relativ hohe Prozentanteile
(25,6 %) florfenicolresistenter Bordetella bronchiseptica-Isolate in Deutschland
bereits in den Jahren 2000 / 2001 ermittelt (PRIEBE u. SCHWARZ 2003). Daher
sollte auch bei dieser Spezies die genetische Grundlage dieser Resistenzeigenschaft
untersucht werden. Dabei war zunächst zu klären, ob in den Jahren 2000 / 2001 eine
temporäre Ausbreitung eines resistenten Klons in verschiedenen
Schweinebeständen stattgefunden hat oder es zu einem gehäuften Auftreten
resistenter Isolate in diesem Zeitraum gekommen ist. Die zur Verfügung gestellten
Isolate der entsprechenden Studie (PRIEBE u. SCHWARZ 2003) wurden einer
wiederholten Empfindlichkeitstestung unterzogen und ihre genetische
Verwandtschaft mittels Makrorestriktionsanalysen und anschließender Pulsfeld-
gelelektrophoretischer Auftrennung analysiert. Eine Darstellung der betreffenden
Bordetella bronchiseptica-Isolate nach Auftrennung der XbaI-Makrorestriktionsmuster
mit Hilfe der Pulsfeld-Gelelektrophorese ist in Abbildung 6 dargestellt. Dabei zeigte
sich, dass der Prozentwert resistenter Isolate auf 6,3 % korrigiert werden muss und
die Isolate zum Teil der gleichen klonalen Gruppe zuzuordnen sind (KEHRENBERG
u. SCHWARZ, nicht publizierte Daten). Somit ist es wahrscheinlich, dass in diesen
Jahren eine temporäre Ausbreitung klonaler Isolate stattgefunden hat.

RESULTATE UND DISKUSSION
53
M1 1 2 3 4 5 M2 M1 6 7 8 9 10 M2
Abbildung 6: Auftrennung von XbaI-Makrorestriktionsmustern der 10 florfenicolresistenen Bordetella bronchiseptica-Isolate aus den Jahren 2000 / 2001 (Spuren 1 - 5 und 6 - 10) mittels Pulsfeld-Gelelektropho-rese. Die Spuren M1 bzw. M2 enthalten SmaI-Fragmente von Staphylococcus aureus NCTC 8325 (M1) sowie XbaI-Fragmente von Salmonella Typhimurium LT2 (M2) als Größenstandards.
In Publikation 5 sind die Ergebnisse der molekularen Analyse der Florfenicolresistenz
dieser und weiterer Isolate dargestellt. In der Studie wurden insgesamt 496
Bordetella bronchiseptica-Isolate unterschiedlicher tierartlicher Herkunft auf ihre
Florfenicolempfindlichkeit getestet. Die 18 nach CLSI-Kriterien (NCCLS 2002) als
resistent bezeichneten Isolate wurden zusammen mit 2 nur
chloramphenicolresistenten- und 3 Kontrollisolaten für weitere Untersuchungen
ausgewählt. Von diesen Isolaten konnte bei 9 Bordetellen das Effluxgen floR
chromosomal lokalisiert nachgewiesen werden. Diese Isolate zeigten entweder nicht
oder kaum unterscheidbare XbaI-Makrorestriktionsmuster, weshalb sie der gleichen
klonalen Gruppe zugeordnet wurden und die Resistenzgenregion nur eines der
Isolate untersucht wurde. In dieser sequenzierten Region von 1638 Basenpaaren

RESULTATE UND DISKUSSION
54
(bp) wurde eine deutliche Übereinstimmung zu floR-Resistenzgenregionen von
Escherichia coli (Publikation 7) oder Pasteurella multocida (Publikation 5) festgestellt.
Zwar werden bei Bordetella häufiger Resistenzgene von Enterobacteriaceae
nachgewiesen (KADLEC et al. 2005; KADLEC et al. 2006), bislang sind aber keine
von Pasteurellen bekannten Resistenzgene bei diesem Genus identifiziert worden.
Auffällig war allerdings die unterschiedliche Lokalisation des floR-Gens in der
chromosomalen DNA (bei Bordetella) bzw. auf Plasmiden (bei Pasteurella). Trotz
Vergesellschaftung beider Erreger im Zuge von Mischinfektionen im
Respirationstrakt von Schweinen scheint somit ein Resistenzgentransfer zwischen
Bordetella und Pasteurella nicht oder nur selten vorzukommen. Die MHK-Werte der
floR-tragenden Isolate lagen in einem Bereich von 8 - 32 µg/ml für Florfenicol und
zeigten somit ähnlich niedrige Resistenzlevel wie floR-tragenden Pasteurellen
(Publikationen 4 und 5). Im Vergleich dazu lagen die MHK-Werte für
Chloramphenicol mit 128 - 256 µg/ml in einem deutlich höheren Bereich und
überstiegen den von florfenicolresistenten Pasteurellen bekannten Wert um 2 - 3
Verdünnungsstufen. Diese hohen MHK-Werte für Chloramphenicol könnten auf der
Präsenz weiterer Chloramphenicol-Resistenzgene bei den Bordetella-Isolaten
beruhen, wurden allerdings auch bei floR-tragenden Escherichia coli-Isolaten
beschrieben (BISCHOFF et al. 2002).
Bei den 9 weiteren Isolaten mit einem MHK-Wert für Florfenicol von ≥ 8 µg/ml waren
keine bekannten Florfenicol-Resistenzgene nachzuweisen. Daher wurde der Einfluss
von Effluxsystemen bei diesen Isolaten untersucht. Hierbei kam ein indirekter Assay
mit Phe-Arg-β-Naphthylamid (PAβN), einer Substanz, die spezifische Effluxpumpen
des „resistance nodulation cell division (RND)“-Typs hemmt (LOMOVSKAYA et al.
2001) zur Anwendung (Publikation 5). Unter dem Einfluss dieser Substanz fielen die
MHK-Werte der getesteten Isolate um 3 - 7 Verdünnungsstufen für Florfenicol und
3 - 5 Verdünnungsstufen für Chloramphenicol (Tabelle 1 in Publikation 5). Im
Gegensatz dazu wurde bei floR-tragenden oder florfenicolempfindlichen
Kontrollisolaten eine Reduktion um lediglich 1 - 2 Verdünnungsstufen erzielt.
Ähnliche Effekte unter dem Einfluss von PAβN konnten auch auf die MHK-Werte für
Nalidixinsäure, einem weiteren Substrat für RND-Effluxpumpen wie AcrAB-TolC
(BAUCHERON et al. 2004), beobachtet werden. Daher scheint ein noch nicht näher

RESULTATE UND DISKUSSION
55
charakterisiertes „Multidrug“-Effluxsystem vom RND-Typ an der Florfenicolresistenz
bei Bordetella beteiligt zu sein. Sequenzhomologien nicht näher charakterisierter
Effluxproteine (Genbank Zugangsnummern CAE34793 - CAE34795) aus der
Genomsequenz von Bordetella bronchiseptica (PARKHILL et al. 2003) mit AcrAB-
TolC-kodierenden Genen von Escherichia coli unterstreichen die Wahrscheinlichkeit,
dass ein AcrAB-TolC-ähnlicher „Multidrug-Transporter“ auch bei Bordetella
bronchiseptica vorkommt.
Im Zuge der Studie wurde ein neues Chloramphenicol-Resistenzgen, cmlB1, auf
einem ca. 50 kb großen Plasmid identifiziert. Dieses abgeleitete Protein zeigt nur
73,3 - 76,5 % Identität zu CmlA-Proteinen und repräsentiert damit eine neue Klasse
von Chloramphenicol-Exportproteinen. Sequenzanalysen der stromaufwärts
gelegenen Bereiche lassen eine Induzierbarkeit des Gens durch attenuierte
Translation vermuten. Eine genaue Analyse des Gens und seiner flankierenden
Regionen befindet sich in Publikation 5. Die aufgrund von Sequenzanalysen
vermutete induzierte Expression des Gens ließ sich experimentell nach Anzüchtung
in subinhibitorischen Konzentrationen von Florfenicol oder Chloramphenicol und
anschließender MHK-Wertbestimmung bestätigen. Dabei wurden Anstiege der MHK-
Werte von Ausgangsstamm und Transformanden um 2 - 4 Verdünnungsstufen
erzielt. Interessanterweise ließ sich das Regulationssystem auch durch Florfenicol
induzieren, obgleich dem Gen ein Attenuator vorgeschaltet war, wie er bei dem
cmlA1-Gen von Transposons Tn1696 zu finden war und für diesen Attenuator
bislang nur eine Induktion durch Chloramphenicol beschrieben war (STOKES u.
HALL 1991). Eine mögliche Erklärung für diese wechselseitige Induktion könnte in
der engen strukturellen Verwandtschaft beider Substanzen liegen. Betrachtet man
hingegen die MHK-Werte für Florfenicol der cmlB1-tragenden Transformanden, so
liegen sie ohne Induktion durch eine der beiden Substanzen im gleichen Bereich wie
die des Ausgangsstammes oder maximal eine Verdünnungsstufe höher. Somit
scheint Florfenicol ohne Induktion des Systems kein Substrat für das von cmlB1
kodierte Effluxprotein zu sein. Eine ähnliche Beobachtung wurde 1982 von DORMAN
u. FOSTER auch bei cmlA-Genen gemacht. Daraus schlussfolgerten die Autoren,
dass cmlA-Gene keine Florfenicolresistenz vermitteln. Trotzdem wurden immer

RESULTATE UND DISKUSSION
56
wieder cmlA tragende Escherichia coli-Isolate gefunden, die hohe MHK-Werte für
Florfenicol von bis zu 64 µg/ml aufwiesen, bei denen aber keine Florfenicol-
Resistenzgene nachgewiesen wurden (WHITE et al. 2000; BISCHOFF et al. 2002;
SINGER et al. 2004). Es bleibt daher zu klären, ob weitere Mechanismen bei diesen
Bakterien Florfenicolresistenz vermitteln oder eine durch cmlA-Gene vermittelte
Unempfindlichkeit vorliegt.

RESULTATE UND DISKUSSION
57
4.2 Teil II Kommensalen (oder Indikatorbakterien)
4.2.1 A) Escherichia coli
4.2.1.1 Nachweis von floR-Genen bei Escherichia coli
Bis zum Jahr 2003 wurden keine florfenicolresistenten Zielorganismen identifiziert,
wie sie in der Zulassung für Deutschland benannt waren. Daher wurden die
Untersuchungen zunächst mit Escherichia coli-Isolaten als Indikatorkeime begonnen.
Verschiedene In-vitro-Empfindlichkeitsprüfungen hatten bereits erste resistente
Isolate dieser bakteriellen Spezies ermittelt. In die Untersuchungen wurden
insgesamt 42 Isolate boviner Herkunft einbezogen, denn Florfenicol war zu dieser
Zeit in Deutschland und in der EU ausschließlich für diese Tierart zugelassen.
Außerdem wurden je ein Isolat von einem Schaf und einer Katze hinzugenommen,
die aber von derselben Farm wie ein erkranktes Rind stammten. Dreizehn der Isolate
waren deutscher und 31 Isolate französischer Herkunft. Ziel dieser anfänglichen
Untersuchungen war die Abklärung, ob das von Photobacterium damselae (KIM u.
AOKI 1996) bekannte und kürzlich bei Salmonella enterica serovar Typhimurium
beschriebene (ARCANGIOLI et al. 1999) Resistenzgen floR auch bei Escherichia coli
vorkommt.
Wie in Publikation 6 dargestellt, konnte bei allen Isolaten das floR-Gen
nachgewiesen werden, welches sich nur durch wenige Basenaustausche von den
bekannten floR-Genen unterschied. Das Gen war bei allen Isolaten auf großen
Plasmiden von 110 - 125 kb lokalisiert, die sich durch Konjugation auf entsprechende
Empfängerstämme übertragen ließen. Es konnte zwischen vier französischen und
einem deutschen Plasmidtyp unterschieden werden. Eine Anlayse der flankierenden
Bereiche zeigte zum einen Homologien mit stromauf- und stromabwärts von floR
oder sul2 gelegenen Plasmidregionen von Photobacterium damselae subsp.
piscicida (KIM u. AOKI 1996; KIM u. AOKI 1996a). Zum anderen wurden Homologien

RESULTATE UND DISKUSSION
58
zu dem floR-Genbereich von Salmonella enterica serovar Typhimurium dargestellt
(ARCANGIOLI et al. 1999; BRIGGS u. FRATAMICO 1999), die sich aber auf ein
kleineres Segment um das floR-Gen beschränkten. Aufgrund dieser Resultate aus
Publikation 6 wurde gefolgert, dass floR-Gene bei Escherichia coli wahrscheinlich
von Photobacterium damselae subsp. piscicida-Plasmiden stammen und nicht von
dem bei Salmonella Serovaren identifizierten chromosomalen Resistenzgencluster.
Alle Plasmidtypen, die im Rahmen der Studie identifiziert wurden, trugen 1 - 4
weitere Resistenzmarker. Von diesen kam Sulfonamidresistenz am häufigsten vor,
gefolgt von Trimethoprim-, Ampicillin- und Streptomycinresistenz bei je zwei
Plasmidtypen. Eine solche Lokalisation von floR-Genen auf konjugativen Plasmiden
mit mehreren Resistenzmarkern ist besonders in Hinblick auf ihre Verbreitung von
Interesse, denn auch unter der Anwendung anderer Substanzen, wie Sulfonamiden,
Trimethoprim, Ampicillin oder Streptomycin können nun die floR-Gene co-selektiert
werden. Eine Lokalisation auf Multiresistenzplasmiden kann damit auch das
Vorkommen von floR bei unterschiedlichen bakteriellen Spezies oder bei Isolaten
unterschiedlicher tierartlicher Herkunft erklären, auch wenn Florfenicol für diese
Tierarten nicht zugelassen war oder ist. So bestätigte eine im gleichen Jahr
publizierte Studie von KEYES et al. die Lokalisation des floR-Gens bei 3 von 4
florfenicolresistenten Escherichia coli-Isolaten vom Geflügel auf 186 und 204 kb
großen Plasmiden (KEYES et al. 2000). Leider lagen keine Daten über eine
Transferierbarkeit der Plasmide oder über mögliche weitere Resistenzmarker auf
ihnen vor. So spekulierten die Autoren zwar über die Präexistenz von Florfenicol-
Resistenzgenen in aviären Escherichia coli-Isolaten, wiesen aber nicht nach, ob Co-
Selektionsmechanismen ursächlich dafür in Frage kommen (KEYES et al. 2000).
Auch WHITE et al. 2000 identifizierten floR-Gene von bovinen Escherichia coli-
Isolaten aus den USA auf unterschiedlichen, ca. 225 kb großen Plasmiden, führten
aber ebenfalls keine Konjugations- oder Transformationsstudien zur Identifizierung
weiterer plasmidgebundener Resistenzmarker durch.
In der Studie von WHITE et al. vermuteten die Autoren erstmals auch eine
chromosomale Lokalisation von floR bei Escherichia coli-Isolaten (WHITE et al.
2000). Eine solche Lokalisation wurde dann 2002 bei deutschen und französischen

RESULTATE UND DISKUSSION
59
Escherichia coli-Isolaten nachgewiesen (DOUBLET et al. 2002). Durch die
unterschiedliche Lokalisation von floR auf Plasmiden oder im Chromosom stellte sich
die Frage nach der genetischen Mobilität des floR-Gens (DOUBLET et al. 2002;
WHITE et al. 2000). Erste Hinweise auf eine mögliche Mobilität lieferten die
Sequenzanalysen einer 6522 bp Region um den floR-Lokus bei einem bovinen
Escherichia coli-Plasmid, welche in Publikation 6 dargestellt sind. Hier konnte
aufgezeigt werden, dass das floR-Gen von zwei offenen Leserahmen (ORFs - open
reading frames) eingerahmt war, die Homologien zu ORFs für Transposaseproteine
(Genbank Zugangsnummern AB020531 und AF028594) zeigten. Aufgrund dieser
Struktur wurde bereits zu diesem Zeitpunkt vermutet, dass das floR-Gen Teil eines
transposablen Elementes ist. Der Nachweis gelang aber erst in Jahr 2005 durch die
Ergebnisse der Publikation 7.
4.2.1.2 Identifizierung des floR-tragenden Transposons Tn floR
In der Zwischenzeit wurde das floR-Gen bei einer Vielzahl gramnegativer Bakterien,
wie Salmonella enterica subsp. enterica, Vibrio chlolerae, Escherichia coli, Klebsiella
pneumoniae oder Photobacterium damselae, auf Plasmiden oder in der
chromosomalen DNA identifiziert (Publikation 7). Daher erhärtete sich der Verdacht,
dass das Gen Bestandteil eines transposablen Elementes ist.
In Publikation 7 konnten mit Hilfe von inversen Polymersase-Kettenreaktionen
(PCRs) zirkuläre Zwischenformen nachgewiesen werden, wie sie bei einigen
Transposons nach Exzision aus dem Donormolekül und vor Integration in die
Rezipienten-DNA vorkommen (MURPHY 1989; HAROCHE et al. 2002). Durch
Analyse der PCR-Amplifikate (Abbildung 7a) zeigte sich, dass das floR-Gen
Bestandteil eines Transposons ist, welches aus 4284 bp besteht und neben dem
floR-Gen einen ORF für ein Transposaseprotein von 497 aa trägt (Abbildung 7b).
Das Transposon wurde mit TnfloR bezeichnet. Es ist nicht konjugativ und trägt auch
keine umgekehrt komplementären („inverted repeats“) oder direkten
Sequenzwiederholungen („direkt repeats“) an seinen Enden. Es konnte eine
Insertionsstelle von 7 bp (TATACGT) identifiziert werden, die Teil des

RESULTATE UND DISKUSSION
60
Phosphoglucosamin-Mutasegens glmM ist und die bereits als Integrationsstelle für
andere Resistenzgene, wie dfrA20, beschrieben wurde (KEHRENBERG u.
SCHWARZ 2005).
(a) (b)
Abbildung 7: Transposon TnfloR. Die PCR-Amplifikate der zirkulären Zwischenformen (a) führten zur Identifikation der genetischen Organisation von TnfloR (b). Die Positionen und die Orientierung der Gene sind als Pfeile wiedergegeben. Die 7 bp Sequenz, die eine Integrationsstelle für TnfloR darstellt und die nun die Transposongrenzen aufzeigt, sind in Kästchen eingezeichnet. Die Lokalisation der PCR-Primer ist durch die Positionen von floRcirc1 und 2 dargestellt.
Die zirkulären Zwischenformen von TnfloR, die als funktionell aktives Element
angenommen werden, wurden nur bei einem von sechs untersuchten Stämmen
nachgewiesen (Publikation 7). Aber auch nur bei diesem Stamm wurde die komplette
Sequenz von TnfloR identifiziert. Das Vorkommen von weiteren deletierten Varianten
bei anderen Isolaten spricht für einen Verlust von Transposonsequenzen durch
Rekombinationen oder Deletionen im Anschluss an die Integration in Rezipienten-
DNA. Eine ähnliche Situation wurde für das Tetracyclin-Resistenztransposon Tn5706
beschreiben. Auch hier wurde bislang nur ein Isolat identifiziert, bei dem das
komplette tet(H)-tragende Element nachgewiesen wurde (KEHRENBERG et al.
3528 bp
1 2
∆∆∆∆ tnpA floR tnpA glmM
TATACGT TATACGT
TnfloR: 4284 bp
floRcirc1 floRcirc2
lysR
3528 bp
1 2
3528 bp
1 2
∆∆∆∆ tnpA floR tnpA glmM
TATACGT TATACGT
TnfloR: 4284 bp
floRcirc1 floRcirc2
lysR

RESULTATE UND DISKUSSION
61
1998). Zahlreiche deletierte Varianten sprechen aber für eine Mobilität von Tn5706,
die von Sequenzverlusten durch Rekombinationsprozesse gefolgt wird
(KEHRENBERG u. SCHWARZ 2000; KEHRENBERG et al. 2001).
Nach neueren Untersuchungen können für das Auftreten der zirkulären
Zwischenformen auch homologe Rekombinationsprozesse in Frage kommen
(TOLEMAN et al. 2006). Diese Prozesse könnten zwischen dem ORF für das
Transposaseprotein und seiner stromaufwärts gelegenen deletierten Variante
ablaufen und ebenfalls zu zirkulären, über PCR-Assays nachweisbaren Strukturen
führen (TOLEMAN et al. 2006). Eine endgültige Klärung über
rekombinationsdefiziente Escherichia coli-Empfängerstämme (recA-Mutanten) wird
daher angestrebt.
4.2.2 B) Staphylococcus spp.
Im Gegensatz zu gramnegativen Bakterien lagen für grampositive Bakterien zu
Beginn der Studien keine Informationen zur Florfenicolresistenz vor. Auch wenn
vereinzelt von Isolaten mit hohen MHK-Werten für Florfenicol berichtet wurde (HO et
al. 2000; SALMON u. WATTS 2000), so fehlten Untersuchungen zu deren
genetischer Grundlage. Da Staphylokokken genau wie die Zielbakterien im
Respirationstrakt von Tieren zu finden sind und unter entsprechendem
Selektionsdruck Resistenz entwickeln oder erwerben können, wurden Bakterien
dieses Genus für die weiteren Studien ausgewählt.
Für die Untersuchungen wurden insgesamt 302 chloramphenicolresistente
Staphylokokken auf ihre Florfenicolempfindlichkeit geprüft. Es wurden
chloramphenicolresistente Isolate ausgewählt, da alle bislang bekannten Florfenicol-
Resistenzgene kombinierte Phenicolresistenz vermitteln. Von diesen Isolaten
stammten 188 von Tieren, die zwischen den Jahren 1988 und 2005 von
Diagnostiklaboren in Deutschland gesammelt und zur Verfügung gestellt wurden und
die nunmehr ein Teil der Stammsammlung des Institutes für Tierzucht sind

RESULTATE UND DISKUSSION
62
(Tabelle 6). Weitere 114 Isolate waren humanen Ursprungs und wurden in ganz
Europa zwischen 1997 und 1999 im Zuge der SENTRY-Studie (FLUIT et al. 2001)
gesammelt (Tabelle 6). Von diesen 302 Isolaten zeigten nur 12 einen MHK-Wert für
Florfenicol von ≥ 16 µg/ml. Diese Isolate wurden als vermutlich florfenicolresistent
eingestuft. Eine vorbehaltliche Klassifizierung erfolgte, da kein anerkannter CLSI-
Grenzwert verfügbar war, eine Konzentration von ≥ 16 µg/ml im Tier aber nicht mehr
mit therapeutischen Dosierungen zu erreichen ist.
Tabelle 6: Auf Florfenicolresistenz getestete Staphylococcus spp. mit ihrer Herkunft von Tieren oder Menschen
Gesamtzahl animaler
oder humaner Isolate
Anzahl davon
Herkunft Zahl der als florfenicol-resistent
eingestuften Isolate a
Staphylococcus-Spezies der resistenten Isolate (n b)
188 45 Pferd 1 S. aureus
42 Hund -c
33 Geflügel -
22 Rind 8 S. lentus (3), S. simulans (3),
S. chromogenes, S. sciuri
15 Schwein 2 S. aureus, S. lentus
14 Nerz -
8 Katze -
7 Kaninchen -
2 Meerschweinchen -
114 114 Mensch 1 S. aureus
a = Isolate mit einem MHK-Wert für Florfenicol von ≥ 16 µg/ml wurden als vermutet resistent eingestuft b n = Anzahl der Isolate, wenn > 1 c - = kein Isolat wurde mit einem MHK-Wert ≥ 16 µg/ml getestet
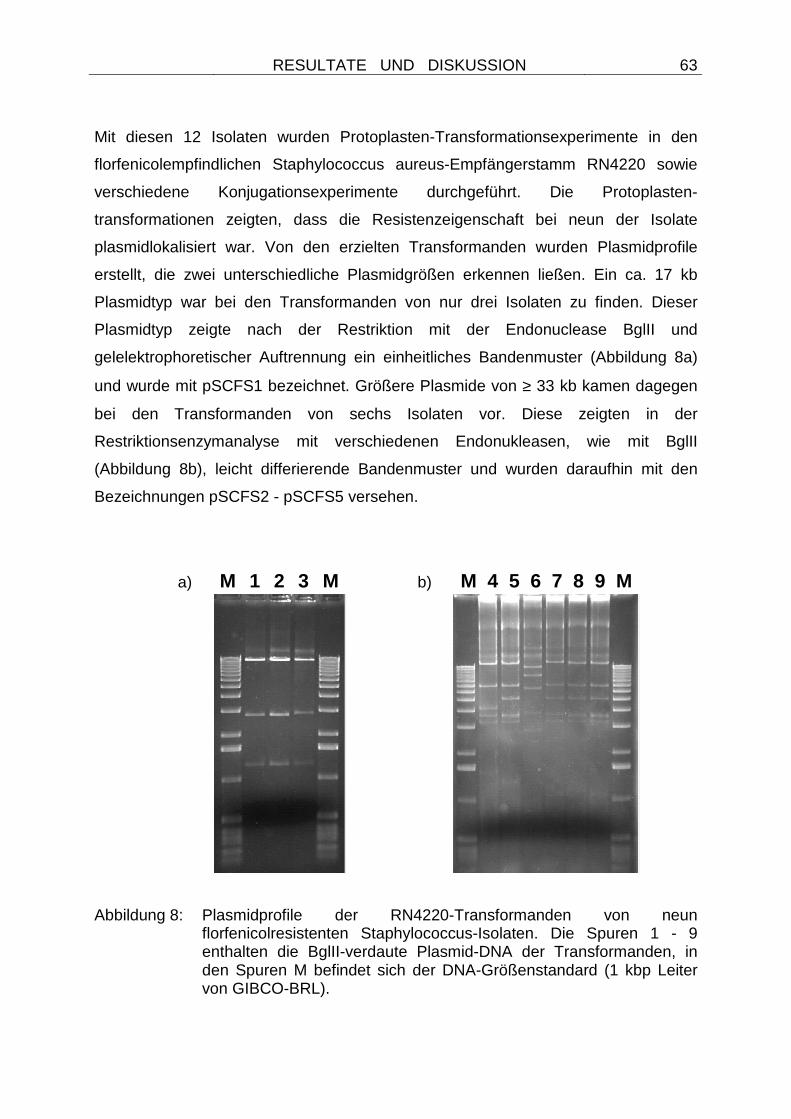
RESULTATE UND DISKUSSION
63
Mit diesen 12 Isolaten wurden Protoplasten-Transformationsexperimente in den
florfenicolempfindlichen Staphylococcus aureus-Empfängerstamm RN4220 sowie
verschiedene Konjugationsexperimente durchgeführt. Die Protoplasten-
transformationen zeigten, dass die Resistenzeigenschaft bei neun der Isolate
plasmidlokalisiert war. Von den erzielten Transformanden wurden Plasmidprofile
erstellt, die zwei unterschiedliche Plasmidgrößen erkennen ließen. Ein ca. 17 kb
Plasmidtyp war bei den Transformanden von nur drei Isolaten zu finden. Dieser
Plasmidtyp zeigte nach der Restriktion mit der Endonuclease BglII und
gelelektrophoretischer Auftrennung ein einheitliches Bandenmuster (Abbildung 8a)
und wurde mit pSCFS1 bezeichnet. Größere Plasmide von ≥ 33 kb kamen dagegen
bei den Transformanden von sechs Isolaten vor. Diese zeigten in der
Restriktionsenzymanalyse mit verschiedenen Endonukleasen, wie mit BglII
(Abbildung 8b), leicht differierende Bandenmuster und wurden daraufhin mit den
Bezeichnungen pSCFS2 - pSCFS5 versehen.
a) M 1 2 3 M b) M 4 5 6 7 8 9 M
Abbildung 8: Plasmidprofile der RN4220-Transformanden von neun florfenicolresistenten Staphylococcus-Isolaten. Die Spuren 1 - 9 enthalten die BglII-verdaute Plasmid-DNA der Transformanden, in den Spuren M befindet sich der DNA-Größenstandard (1 kbp Leiter von GIBCO-BRL).

RESULTATE UND DISKUSSION
64
4.2.2.1 Analyse des Plasmidtyps pSCFS2 mit Identifi zierung des neuen
Florfenicol-Resistenzgens fexA
Stellvertretend für die größeren Florfenicol-Resistenzplasmide wurde der Typ
pSCFS2 untersucht. Dieser Plasmidtyp wurde bei einem Staphylococcus lentus-
Isolat vom Kalb nachgewiesen. Er vermittelte ausschließlich kombinierte Florfenicol- /
Chloramphenicolresistenz und konnte in Publikation 8 als Träger eines neuen
Resistenzgens, fexA, identifiziert werden. Dieses Gen kodiert für ein 475 aa Protein,
dem ein kleines regulatorisches Peptid von 9 aa vorgeschaltet ist.
Die stromaufwärts gelegene Region um das regulatorische Peptid zeigte deutliche
Homologien zu Translationsattenuatoren, wie sie bei der induzierbaren Expression
von cat- oder cmlA-Genen beschrieben sind (LOVETT 1990). Es konnten typische
Strukturelemente, wie 11 bp umgekehrt komplementäre Sequenzwiederholungen
(inverted repeated sequences, IR1 und IR2) aufgezeigt werden, die auf mRNA-
Ebene eine stabile Sekundärstruktur bilden und die eine fexA assoziierte
Ribosomenbindungsstelle enthalten. Ferner wurde eine 12 bp „ribosome stall
sequence“, eine Sequenz, die komplementäre Bereiche zur 16S rRNA von
Staphylococcus aureus aufweist und die zum Festfahren der Ribosomen dient, in
dem regulatorischen Bereich ermittelt werden (Publikation 8). Ein Modell dieser
regulatorischen Region von Plasmid pSCFS2 ist in Abbildung 1a der Publikation 8,
dargestellt. Die auf diesem Modell basierte induzierte Expression konnte
experimentell über einen vierfachen Anstieg der MHK-Werte für Florfenicol und
Chloramphenicol nach Induktion des Systems bestätigt werden. Erstaunlicherweise
waren sowohl Florfenicol als auch Chloramphenicol als Induktoren geeignet. Daher
stellt sich die Frage, ob die durch attenuierte Translation regulierte Expression von
cat- oder cmlA-Genen (LOVETT 1990; HORINOUCHI u. WEISBLUM 1982;
PROJAHN et al. 1985; STOKES u. HALL 1991) ebenfalls durch Florfenicol zu
induzieren ist.
Das assoziierte fexA-Strukturgen zeigte bei der Analyse der Nukleotidsequenz keine
signifikanten Homologien zu Einträgen in den Datenbanken. Um den von fexA-
vermittelten Resistenzmechanismus klären zu können, wurde mit Hilfe des Tmpred-
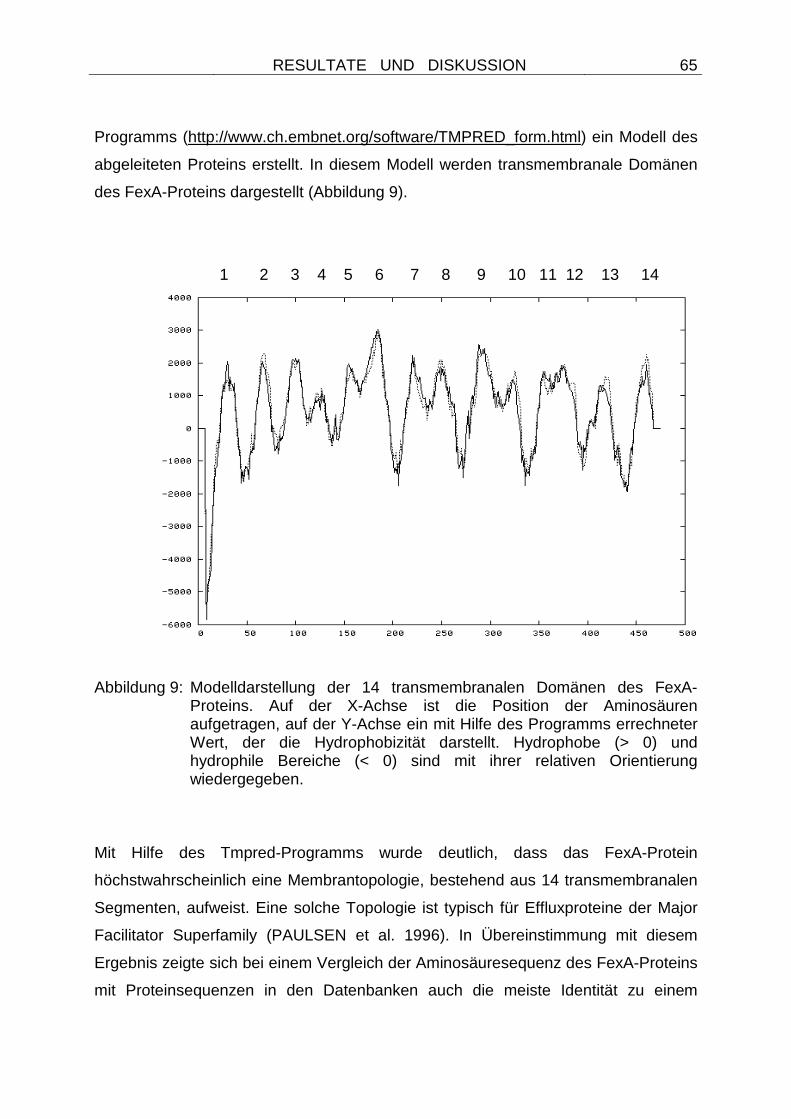
RESULTATE UND DISKUSSION
65
Programms (http://www.ch.embnet.org/software/TMPRED_form.html) ein Modell des
abgeleiteten Proteins erstellt. In diesem Modell werden transmembranale Domänen
des FexA-Proteins dargestellt (Abbildung 9).
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Abbildung 9: Modelldarstellung der 14 transmembranalen Domänen des FexA-Proteins. Auf der X-Achse ist die Position der Aminosäuren aufgetragen, auf der Y-Achse ein mit Hilfe des Programms errechneter Wert, der die Hydrophobizität darstellt. Hydrophobe (> 0) und hydrophile Bereiche (< 0) sind mit ihrer relativen Orientierung wiedergegeben.
Mit Hilfe des Tmpred-Programms wurde deutlich, dass das FexA-Protein
höchstwahrscheinlich eine Membrantopologie, bestehend aus 14 transmembranalen
Segmenten, aufweist. Eine solche Topologie ist typisch für Effluxproteine der Major
Facilitator Superfamily (PAULSEN et al. 1996). In Übereinstimmung mit diesem
Ergebnis zeigte sich bei einem Vergleich der Aminosäuresequenz des FexA-Proteins
mit Proteinsequenzen in den Datenbanken auch die meiste Identität zu einem

RESULTATE UND DISKUSSION
66
Effluxprotein von Lactobacillus plantarum oder einem Metall-Tetracyclin/H+-Antiporter
von Bacillus halodurans. Die Identität zu den Proteinen von Lactobacillus und
Bacillus war aber mit 28 bzw. 29 % vergleichsweise gering. Ein erstellter
Homologiebaum (Abbildung 2 in Publikation 8,) bestätigte, dass FexA offensichtlich
einen neuen Typ eines Chloramphenicol- / Florfenicol-Exporterproteins darstellt.
Durch den Nachweis des fexA-Gens bei einem Staphylococcus lentus-Isolat wurde
erstmals ein Phenicol-Exportergen bei Staphylokokken beschrieben. Auch wenn das
Isolat aus dem Respirationstrakt eines Kalbes stammt, so wurde nicht das von vielen
gramnegativen Bakterien bekannte floR-Gen (KIM u. Aoki 1996; BOYD et al. 2002;
CLOECKAERT et al. 2000, Publikationen 3 - 6) identifiziert. Obgleich Pasteurellen
oftmals funktionell aktive Resistenzgene tragen, die Verwandtschaft zu Genen
grampositiver Bakterien zeigen (KEHRENBERG u. SCHWARZ 2005), und eine
Funktionalität des fexA-Genproduktes auch in dem gramnegativen Bakterium
Escherichia coli nachgewiesen werden konnte, so scheint im Falle der
Florfenicolresistenz kein Gentransfer zwischen diesen grampositiven und
gramnegativen Genera vorzukommen. Dies bestätigte auch eine Analyse aller zur
Verfügung stehenden gramnegativen Isolate mit erhöhten MHK-Werten für
Florfenicol. Es konnte bei keinem dieser Isolate die Präsenz des fexA-Gens
nachgewiesen werden (KEHRENBERG u. SCHWARZ, nicht publizierte Daten).
4.2.2.2 Nachweis der Mobilität des fexA-Gens: Identifizierung eines neuen
Transposons
Das Gen fexA wurde in Publikation 8 plasmidlokalisiert nachgewiesen, wodurch eine
Verbreitung des Gens über horizontalen Gentransfer möglich ist. Eine Analyse der
fexA-flankierenden Regionen sollte weiterhin Hinweise auf eine mögliche Mobilität
des Resistenzgens innerhalb des bakteriellen Genoms geben. Dazu wurden
sämtliche BglII-Fragmente des Plasmids pSCFS2 in den Vektor pBluescriptIISK+
(Stratagene, Amsterdam, Niederlande) kloniert und Sequenzanalysen unterzogen.
Die Ergebnisse dieser Analysen sind in Publikation 9 dargestellt und zeigen, dass

RESULTATE UND DISKUSSION
67
das Gen fexA Bestandteil eines transposablen Elementes ist. Dieses Element
umfasst 6644 bp und zeigt in seiner Organisation Ähnlichkeit zu anderen
Transposons von Staphylokokken, wie Tn554 (MURPHY 1989; BASTOS u.
MURPHY 1988) oder Tn5406 (HAROCHE et al. 2002). Dabei konnte eine typische
Anordnung der Leserahmen für die Transposaseproteine tnpA, tnpB und tnpC
(Abbildung 10) aufgezeigt werden, wie sie bei dem Prototyp-Transposon Tn554 und
dem weiteren Vertreter dieser Transposonfamilie zu finden ist. Trotzdem zeigten die
entsprechenden Genprodukte TnpA, TnpB und TnpC des neuen Transposons nur
57,6 - 73,0 % Identität zu den von Tn554 oder Tn5406 bekannten Proteinen
(Publikation 9).
Abbildung 10: Organisation des bei Staphylococcus lentus identifizierten Transposons Tn558 mit Lokalisation der Primer zum Nachweis von zirkulären Zwischenformen
Unterschiede zu den beiden publizierten Vertretern Tn554 und Tn5406 dieser
Transposonfamilie (HAROCHE et al. 2002; Murphy 1990) konnten dagegen in dem
Resistenzgenbereich aufgezeigt werden. Während das Makrolid-Lincosamid-
Streptogramin B-Resistenzgen erm(A) sowie das Spectinomycin-Resistenzgen spc
Bestandteil von Tn554 sind (MURPHY 1989) und das Transposon Tn5406 eine
Variante des Streptogramin A-Resistenzgens vga(A) trägt (HAROCHE et al. 2002),
konnten auf dem neuen transposablen Element das Gen fexA sowie ein Leserahmen
für eine vermutete NAD(P)H-Oxidoreduktase (ORF138) identifiziert werden
(Publikation 9). Ein Vergleich der Organisation der drei bekannten Transposons aus
der Tn554-Familie ist in der Abbildung 1a in Publikation 9, zu finden. Neben den
EPv
0 1 2 3 4tnpA tnpB tnpC orf138
5 6fexA
circ-fw circ-rev
B EC Bg C C B GATCCAGATGTA EPv
0 1 2 3 4tnpA tnpB tnpC orf138
5 6fexA
circ-fw circ-rev
B EC Bg C C B GATCCAGATGTA

RESULTATE UND DISKUSSION
68
strukturellen Ähnlichkeiten konnten weitere Gemeinsamkeiten, die im wesentlichen
Merkmale des speziellen Transpositionsvorganges sind, festgestellt werden. So
fehlen dem neuen Transposon, welches die Bezeichnung Tn558 erhielt, umgekehrt
komplementäre Sequenzwiederholungen („inverted repeats“) an den Enden und an
der Integrationsstelle wird keine direkte Sequenzwiederholung („direct repeats“)
generiert. Dazu konnte eine typische „Kernsequenz“ („core“ sequence) mit der
Basenabfolge 5´-GATGTA-3´ am linken terminalen Ende aufgezeigt werden. Diese 6
Basenpaare stellen die typische Integrationssequenz von Tn554 im Chromosom von
Staphylococcus aureus dar (MURPHY 1990). Bei Transposons aus der Tn554-
Familie findet die Integration vorwiegend in einen Leserahmen mit Ähnlichkeit zu
DNA-Reparaturproteinen statt. Diese als att554 bezeichnete Region konnte in
ähnlicher Form auch auf dem Plasmid pSCFS2 identifiziert werden und wurde hier
mit att558 benannt. Da trotz der eindeutigen strukturellen Merkmale des neuen
Transposons Tn558 kein Nachweis seiner Funktionalität vorlag, wurden inverse
PCR-Assays generiert, die zirkuläre Zwischenformen des Transposons nachweisen
können. Solche zirkulären Zwischenformen bilden bei Tn554-ähnlichen Transposons
einen Bestandteil des Transpositionsvorganges (HAROCHE et al. 2002; MURPHY
1989; MURPHY 1990) und gehen einer Integration in eine neue Zielsequenz voraus.
Die Lokalisation der hierfür verwendeten Primer (circ-fw und circ-rv) ist in Abbildung
10 eingezeichnet. Da bei den inversen PCRs entsprechende Amplifikate (Abbildung
11) erzielt und über Sequenzanalysen bestätigt wurden, gelang mit der Publikation 9
auch der Nachweis, dass das fexA-tragende Transposon Tn558 funktionell aktiv ist.

RESULTATE UND DISKUSSION
69
1 M
871 bp
Abbildung 11: PCR-Nachweis einer zirkulären Zwischenform des neuen Transposons Tn558. Die Spur 1 enthält das PCR-Amplifikat des Staphylococcus aureus RN4220::pSCFS2-Transformanden, in der Spur M ist der Größenstandard (1 kbp Leiter von GIBCO-BRL) aufgetragen.
Somit konnte aufgezeigt werden, dass das fexA-Gen Bestandteil eines Transposons
aus der Tn554-Familie ist.
4.2.2.3 Analyse des Plasmidtyps pSCFS1 mit Identifi zierung des neuen
Resistenzgens cfr
Auch der Resistenzgenbereich des kleineren, ca. 17 kb großen Plasmidtyps
(pSCFS1) wurde, wie in Publikation 10 dargestellt, einer genauen Analyse
unterzogen. Dazu wurde Plasmid-DNA aus einem Staphylococcus aureus
RN4220::pSCFS1-Transformanden mit den Restriktionsenzymen EcoRI und BclI-
BamHI geschnitten. Die entstandenen pSCFS1-Fragmente wurden in Schnittstellen
des Vektors pBluescriptIISK+ (Stratagene, Amsterdam, Niederlande) kloniert.
Nachfolgende phänotypische Untersuchungen der Klone ermöglichten eine
Eingrenzung des Chloramphenicol / Florfenicol-Resistenzgenbereichs auf ein 3,8 kb

RESULTATE UND DISKUSSION
70
großes EcoRI-Fragment von pSCFS1. Verschiedene Subklonierungen dieses EcoRI-
Fragments folgten und engten die gesuchte Region weiter auf ein kleineres ClaI-
XbaI-Fragment ein. Die Analysen der unterschiedlichen Subklone sind in Publikation
10 detailliert beschreiben und zeigen, dass auf dem Plasmidtyp pSCFS1 drei
Leserahmen (ORFs) für die phänotypische Ausbildung der Florfenicolresistenz von
Bedeutung sind. Zwei kleine ORFs (Abbildung 12, ORF1 und ORF2) kodieren für
Proteine von 59 und 44 aa und scheinen für die Expression des nachgeschalteten
Gens essentiell zu sein. Wird einer dieser kleinen ORFs mit Hilfe eines der
Restriktionsenzyme HpaI oder MspI zerstört, so führt dies zum Verlust der dem
Empfängerstamm vermittelten kombinierten Resistenz. In diesem dem Strukturgen
vorgeschalteten Bereich konnten Strukturelemente identifiziert werden, die an
Translationsattenuatoren induzierbarer cat-Gene von Staphylococcus und Bacillus
erinnern (LOVETT 1990; STOKES u. HALL 1991). Somit scheint die induzierbare
Chloramphenicol- und Florfenicolresistenz auf Plasmid pSCFS1 auf dem
Mechanismus der attenuierten Translation zu basieren (Publikation 10). Eine
Induktion des Systems ließ sich, genau wie bei dem Gen fexA (Publikation 8), über
die beiden Substanzen Chloramphenicol und Florfenicol nachweisen.
Abbildung 12: Schematischer Aufbau des Chloramphenicol- / Florfenicol-
Resistenzgenbereichs von Plasmid pSCFS1 mit eingezeichneten Restriktionsschnittstellen für die Enzyme EcoRI (E), ClaI (C), HpaI (Hp), MspI (Ms), BamHI (Ba), PstI (Ps), PvuII (P) uns XbaI (X). Eine Größenskala in kb ist unter der Karte eingezeichnet.
0 1 2 3
EPs P XBaCCE
cfr
Hp Ms
1 2
0 1 2 3
EPs P XBaCCE
cfr
Hp Ms
1 2

RESULTATE UND DISKUSSION
71
Diesem regulatorischen Bereich war ein Strukturgen nachgeschaltet, welches für ein
349 aa Protein kodiert. Unerbrechung des Leserahmens durch Nutzung der BamHI-
Restriktionsschnittstelle führte zu einem Verlust der Resistenzeigenschaft. Daher
konnte angenommen werden, dass dieses Gen, welches die Bezeichnung cfr erhielt,
die kombinierte Chloramphenicol- und Florfenicolresistenz vermittelt. Der
Resistenzmechanismus blieb allerdings zu diesem Zeitpunkt ungeklärt (Publikation
10). Weder Vergleiche mit Sequenzen in den Datenbanken, noch Bioassays, die eine
enzymatische Inaktivierung oder eine Acetylierung von Chloramphenicol nachweisen
können (SCHWARZ et al. 1989), gaben Hinweise auf die Funktion des abgeleiteten
Cfr-Proteins. Es konnten auch keine Homologien zu Acetyltransferasen oder zu
Effluxproteinen (MURRAY u. SHAW 1997; KIM u. AOKI 1996) aufgezeigt werden.
Die Analyse der Proteinstruktur zeigte weder ATP-bindende Domänen noch mit Hilfe
der Tmpred-Software (http://www.ch.embnet.org/software/TMPRED_form.html) eine
typische Membrantopologie. Somit konnte in Publikation 10 zwar ein neues
Chloramphenicol- und Florfenicol-Resistenzgen identifiziert werden, es konnten aber
keine Hinweise auf den durch das Gen vermittelten Resistenzmechanismus
erarbeitet werden. Die Lokalisation von cfr auf einem Plasmid und die Funktionalität
des Gens in grampositiven (Staphylokokken) und gramnegativen Keimen
(Escherichia coli) ließ aber zukünftig eine weitere Verbreitung dieses Gens
befürchten.
4.2.2.4 Lokalisation des Gens cfr auf einem Multiresistenzplasmid:
Identifizierung des neuen Gens erm(33)
Zur Analyse des Resistenzgenbereichs von Plasmid pSCFS1 (Publikation 10) wurde
Plasmid-DNA aus einem Staphylococcus sciuri-Isolat gewonnen. Da der
Ausgangsstamm zusätzliche Resistenzen gegenüber Erythromycin, Kanamycin,
Spectinomycin und Tetracyclinen zeigte, sollte in weiteren Untersuchungen geklärt
werden, ob sich neben cfr weitere Resistenzgene auf pSCFS1 lokalisiert befinden.
Diese Resistenzgene könnten durch Co-Selektionsmechanismen zur Verbreitung
des neuen Resistenzgens cfr führen. Eine phänotypische Untersuchung der

RESULTATE UND DISKUSSION
72
Staphylococcus aureus RN4220::pSCFS1-Transformanden zeigte, dass durch den
Plasmidtyp pSCFS1 auch Resistenz gegenüber Makroliden, Lincosamiden und
Streptogramin B (MLSB)-Antibiotika vermittelt wird. Da PCR-Assays ein erm(C)-Gen
vermuten ließen, wurde das kleinste EcoRI-PstI-Fragment, welches mit einer erm(C)-
Gensonde hybridisierte, in den Vektor pBluescriptIISK+ kloniert und
Sequenzanalysen unterzogen. Die Ergebnisse dieser Sequenzanalysen sind in
Publikation 11 genauer beschrieben. Es wurde ein insgesamt 2196 bp Fragment
sequenziert und dabei ein Leserahmen für ein 243 aa Protein identifiziert. Das
abgeleitete Protein zeigte nur 71 - 79 % Identität zu unterschiedlichen Erm(A)- und
73 - 76 % Identität zu verschiedenen Erm(C)-Proteinen. Daher erhielt es vom
Nomenklaturcenter für MLSB-Gene (http://faculty.washington.edu/marilynr/) die neue
Bezeichnung Erm(33). Bei der genauen Untersuchung der Nukleotidsequenz des
Gens wurde deutlich, dass erm(33) ein Rekombinationsprodukt aus einem erm(C)
und einem erm(A)-Gen ist. Die 284 bp des 5´-Ende waren - abgesehen von 12 bp-
Austauschen - identisch mit einem erm(C)-Gen von Staphylococcus hyicus
(SCHWARZ et al. 1998), die terminalen 403 bp ließen sich nicht von dem erm(A)-
Gen des Transposons Tn554 (MURPHY 1985) unterscheiden. Eine genaue
Darstellung der Nukleotidsequenzidentität von erm(33) mit verwandten erm(C) und
erm(A)-Genen sowie ihrer abgeleiteten Proteinsequenzen ist in Publikation 11,
Tabelle 1 zu finden. Wird der Regulationsbereich des Gens betrachtet, so zeigt er
Ähnlichkeit mit Translationsattenuatoren von induzierbar exprimierten erm(C)-Genen.
Lediglich vier Basenaustausche und drei Deletionen in der nicht-kodierenden Region
unterscheiden ihn von stromaufwärts von erm(C)-Genen (HORINOUCHI u.
WEISBLUM 1982) gelegen Regulationsbereichen. Über die Entstehung eines
solchen Rekombinationsproduktes wie erm(33) lässt sich retrospektiv nur
spekulieren. Da aber weder eine Veränderung des vermittelten Resistenzspektrums
noch eine Veränderung der MHK-Werte im Vergleich zu erm(C)- und erm(A)-Genen
zu beobachten war, ist von einer zufälligen aufgetretenden Rekombination ohne
Selektionsvorteil auszugehen. Die wahrscheinliche Rekombinationsstelle von 45 bp,
die 93,3 bzw. 95,6 % Homologie zwischen einer Nukleotidsequenz von erm(C) und
erm(A) zeigt, konnte in Publikation 11 ausfindig gemacht werden.

RESULTATE UND DISKUSSION
73
Durch den Nachweis eines zweiten Resistenzgens wie erm(33) auf dem Plasmid
pSCFS1 ist es möglich, dass die von cfr vermittelte kombinierte Florfenicol- /
Chloramphenicol-Resistenzeigenschaft auch über Co-Selektion verbreitet wird.
Sowohl Makrolide als auch Lincosamide werden veterinärmedizinisch in größeren
Mengen eingesetzt als Florfenicol (SCHWARZ u. CHASLUS-DANCLA 2001), so
dass eine Verbreitung von cfr über diesen Mechanismus wahrscheinlicher ist, als
durch den Selektionsdruck, der durch die Anwendung des Wirkstoffes selber
entsteht.
4.2.2.5 Komplette Sequenzanalyse und Organisation d es Plasmids pSCFS1
mit Identifizierung eines neuen ABC-Transporters
Um weitere Informationen über das erste Chloramphenicol- / Florfenicol-
Resistenzplasmid von Staphylococcus sciuri zu erhalten, wurde eine komplette
Sequenzanalyse von Plasmid pSCFS1 vorgenommen. Diese Analyse sollte Hinweise
auf die Struktur des Plasmids und die Organisation der Resistenzgene geben. Dazu
wurden Fragmente, die durch Restriktionsverdaus mit unterschiedlichen
Endonukleasen erzielt wurden, in die Vektoren pBluescriptIISK+ oder pCR BluntII
TOPO (Invitrogen, Karlsruhe) kloniert und überlappend sequenziert. Die genaue
Strukturanalyse des Plasmids ist in Publikation 12 wiedergegeben. Es zeigte eine
Gesamtgröße von 17108 bp und trug 14 Leserahmen, die für Proteine über 100 aa
kodieren. Die Organisation dieser Leserahmen von pSCFS1 ist in Abbildung 13
dargestellt.

RESULTATE UND DISKUSSION
74
Abbildung 13: Strukturelle Organisation des kompletten Plamids pSCFS1. Die Leserahmen (ORFs) sind durch Pfeile dargestellt, wobei die Pfeilspitze das 3´-Ende des Gens angibt. Die ORFs, die in antimikrobielle Resistenz involviert sind, wurden farbig dargestellt.
Die 14 Leserahmen (ORFs) von pSCFS1 ließen sich unterschiedlichen Gruppen
zuordnen, wobei fünf ORFs Ähnlichkeit zu Rekombinasegenen zeigten (ORF 1, ORF
2, ORF 5, ORF 12, ORF 13), zwei ORFs zeigten Homologie zu
Plasmidrekombinationsgenen (ORF 11, ORF 14) und vier ORFs schienen in
antimikrobielle Resistenz involviert zu sein. Dabei kodierte ORF4 für das
Chloramphenicol- / Florfenicol-Resistenzgen cfr, ORF 7 für das MLSB-Resistenzgen
erm(33) und ORF 8 für das Spectinomycin-Resistenzgen spc, welches Bestandteil
von Transposon Tn554 (MURPHY 1989) ist. Ein weiterer kompletter und ein
deletierter Leserahmen, die für Transposaseproteine von Tn554 kodierten, wurden
mit ORF 9 und ORF 10 identifiziert. Einem Leserahmen, ORF 6, konnte keine
Funktion zugeordnet werden. Eine genaue Darstellung der Leserahmen mit ihrer
ORF 3 – „low level“Clindamycinresistenz
pSCFS1
ORF 1
ORF 2
ORF 4 -Chloramphenicol- / Florfenicolresistenz
ORF 5ORF 6
ORF 7 – MLSB- Resistenz
ORF 9
ORF 10
ORF 11
ORF 12
ORF 13
ORF 14
17108 bp
ORF 8 – Spectinomycin-resistenz
ORF 3 – „low level“Clindamycinresistenz
pSCFS1
ORF 1
ORF 2
ORF 4 -Chloramphenicol- / Florfenicolresistenz
ORF 5ORF 6
ORF 7 – MLSB- Resistenz
ORF 9
ORF 10
ORF 11
ORF 12
ORF 13
ORF 14
17108 bp
ORF 8 – Spectinomycin-resistenz

RESULTATE UND DISKUSSION
75
Größenangabe und Position, sowie der Identität zu anderen Proteinen, ist in
Publikation 12 tabellarisch aufgeführt.
Von besonderem Interesse dieser unterschiedlichen Leserahmen war der vierte, in
antimikrobielle Resistenz involvierte Leserahmen, ORF 3. Dieser Leserahmen kodiert
für einen ABC-Transporter von 492 aa und und zeigte die typischen Walker Motive A
und B in der abgeleiteten Proteinsequenz, die eine ATP-bindende Domäne
charakterisieren. Dem Gen vorgeschaltet befand sich ein Regulationsbereich, wie er
von der induzierbaren Expression antimikrobieller Resistenzgene grampositiver
Bakterien bekannt ist (ROBERTS 1996). Auch wenn die größte Identität des
abgeleiteten ABC-Transporterproteins mit 82 % zu einem ABC-Transporter
unbekannter Funktion von Bacillus anthracis bestand, so konnte immerhin noch 41 %
Identität zu dem ABC-Transporter Lsa (SINGH et al. 2002) beobachtet werden, der
eine intrinische Resistenz von Enterococcus faecalis gegenüber Clindamycin und
Streptogramin A-Antibiotika bewirkt. Um die genaue Funktion des neuen ABC-
Transporters bestimmen zu können, wurde das Gen samt seines
Regulationsbereiches in einen Plasmidvektor pT181MCS kloniert, der die Fähigkeit
zur Replikation in grampositiven Bakterien besitzt (AUGUSTIN et al. 1992). Das
rekombinante Plasmid wurde anschließend in den Empfängerstamm Staphylococcus
aureus RN4220 transformiert und mit den entsprechenden Staphylococcus aureus
RN4220::pT181MCS-Transformanden MHK-Wertbestimmungen durchgeführt. Ein
Vergleich der MHK-Werte von Stämmen mit und ohne einkloniertem ABC-
Transporter ließ darauf schließen, dass das Transporterprotein an der
Clindamycinresistenz beteiligt ist. Auch wenn der Transporter keine klinische
Resistenz vermittelte, konnte ein immerhin 16-facher Anstieg der MHK-Werte
beobachtet werden. Bei keinem der anderen getesteten Wirkstoffe konnte eine
Erhöhung der MHK-Werte > 1 Verdünnungsstufe festgestellt werden. Somit schien
der neue ABC-Transporter lediglich eine „low level“-Resistenz gegenüber
Clindamycin zu vermitteln. Er erhielt vom Nomenklaturcenter für MLSB-Gene
(http://faculty.washington.edu/marilynr/) die Bezeichnung Lsa(B). Um einen Hinweis
auf die Herkunft des ABC-Transporters zu erlangen, wurde der G + C-Gehalt

RESULTATE UND DISKUSSION
76
bestimmt. Mit 34,3 % lag er in dem von Staphylokokken-Genomen bekannten
Bereich, was auf eine ursprüngliche Herkunft von Staphylococcus spp. deutet.
Die auf dem Plasmid pSCFS1 nachgewiesenen Replikationsgene (rep) zeigten
Ähnlichkeit zu rep-Genen von Treponema denticola und Mannheimia varigena
(Publikation 12). Daher wurden weitere Untersuchungen vorgenommen, die eine
Replikationsfähigkeit von pSCFS1 in Escherichia coli- sowie in Pasteuella- und
Mannheimia-Empfängerstämmen testen sollten (KEHRENBERG u. SCHWARZ, nicht
publizierte Daten). Die Ergebnisse wiesen darauf hin, dass eine Replikation in diesen
Wirten nicht möglich ist. Das Vorkommen und die Replikationsfähigkeit dieses
Plasmidtyps war demnach auf Staphylokokken beschränkt.
Auf dem Plasmid wurden insgesamt drei neue Resistenzgene (lsa(B), erm(33) und
cfr) identifiziert. Somit scheint Staphylococcus sciuri ein potentielles Reservoir für
neue Resistenzgene zu sein. Das Vorkommen des Plasmidtyps auch bei anderen
Staphylococcus-Isolaten lässt weiterhin auf horizontale Gentransferprozesse
schließen. Diese können unter dem Selektionsdruck durch eine Anwendung von
Makroliden, Lincosamiden, Streptogramin B-Antibiotika, Spectinomycin,
Chloramphenicol oder Florfenicol gefördert werden und zu einer gleichzeitigen
Verbreitung aller auf pSCFS1 lokalisierten Gene führen.
4.2.2.6 Nachweis des durch das Gen cfr vermittelten Resistenzmechanismus
Durch PCR-Assays und Hybridisierungsexperimente konnte nachgewiesen werden,
dass cfr-Gene auch bei einigen der großen Plasmidtypen, wie pSCFS3, zu finden
sind. Auf diesem Plasmidtyp ließen sich die beiden neuen Florfenicol-Resistenzgene,
cfr und fexA, nachweisen. Transformationsexperimente von pSCFS3 in
Staphylococcus aureus RN4220 zeigten, dass das Plasmid neben der
Phenicolresistenz auch Resistenz gegenüber Clindamycin vermittelt. Da aber mittels
PCR-Assays kein bekanntes Clindamycin-Resistenzgen amplifiziert werden konnte,
wurden alle BglII-Fragmente von pSCFS3 in den Vektor pBluescriptIISK(+) kloniert.
Nach der Transformation in einen clindamycinempfindlichen Escherichia coli-

RESULTATE UND DISKUSSION
77
Empfängerstamm (AS19) wurden die Klone In-vitro-Empfindlichkeitstestungen
unterzogen. Es zeigte sich, dass lediglich Klone mit einem 3,6 kb BglII Insert dem
Empfängerstamm höhere MHK-Werte für Clindamycin vermittelten. Auf diesem 3,6
kb Fragment befanden sich neben zwei inkompletten Leserahmen ein kompletter
Leserahmen für ein cfr-Gen. Das abgeleitete Protein wich nur durch einen
Aminosäureaustausch von dem bekannten Cfr-Protein ab. Somit schien durch das
Gen cfr auch Resistenz gegenüber einem Lincosamid (Clindamycin) vermittelt zu
werden. Der cfr-vermittelte Resistenzphänotyp blieb bei der Analyse des Plasmides
pSCFS1 zunächst unentdeckt, da auf diesem Plasmidtyp auch die Makrolid-,
Lincosamid- und Streptogramin B-Resistenzgene erm(33) und lsa(B) lokalisiert
waren.
Da die Frage nach dem Cfr-Resistenzmechanismus noch ungeklärt war, aber
Chloramphenicol und Clindamycin bekanntlich die Proteinbiosynthese von Bakterien
durch eine Bindung an die 50S Untereinheit der Ribosomen hemmen, wurde die
Theorie aufgestellt, dass Cfr-Proteine zu einer Modifikation der ribosomalen
Bindungsstelle dieser Antibiotika führen. Zur Abklärung dieser Theorie wurde
zunächst ein Modell erstellt, welches die Bindungsstellen dieser Substanzen in der
50S Untereinheit der Ribosomen darstellt (Abbildung 14). Aus diesem Modell,
welches auf einer 50S ribosomalen Untereinheit von Deinococcus radiodurans
basiert (HARMS et al. 2001), geht hervor, dass die Bindungsstellen von
Chloramphenicol und Clindamycin sehr dicht beieinander im
Peptidyltransferasezentrum der 50S Untereinheit liegen. Daher ist es denkbar, dass
eine Modifikation in diesem Bereich zu einer verminderten Bindung der Substanzen
an das Ribosom führt.

RESULTATE UND DISKUSSION
78
Abbildung 14: Bindungsstellen von Chloramphenicol (CHL) und Clindamycin (CLI)
im Modell einer 50S ribosomalen Untereinheit von Deinococcus radiodurans nach HARMS et al. 2001. Die ribosomalen Proteine sind in blau, die RNA in grau dargestellt. Die Chloramphenicol-Bindungsstelle ist vergrößert und in grün gezeichnet, die von Clindamycin in dunkelblau.
Um zu untersuchen, ob Cfr eine Modifikation der Ribosomen durchführt, wurden
mehrere experimentelle Ansätze gewählt. Diese sind ausführlich in Publikation 13
beschrieben.
Eine ribosomale Modifikation sollte zu einer reduzierten Bindung von
Chloramphenicol, Florfenicol und Clindamycin an die 50S Untereinheit der
Ribosomen führen. Daher wurden „Footprint“-Analysen durchgeführt, um eine solche
reduzierte Bindung nachzuweisen. Ribosomen wurden dafür mit steigenden
Konzentrationen der Wirkstoffe Chloramphenicol, Florfenicol und Clindamycin in
Anwesenheit der Substanz CMCT (Cyclohexyl-Morpholinoethyl-Carbodiimid metho-
p-Toluensulphonat) inkubiert. Anschließend wurden mit der isolierten RNA und einer
reversen Transkriptase Primer-Extension-Analysen durchgeführt. Da die Substanz
CMCT bekanntlich zu Modifikationen der RNA an bestimmten Positionen, wie U2506,
führt (MOAZED u. NOLLER 1987), sollten diese Modifikationen in den Footprint-

RESULTATE UND DISKUSSION
79
Analysen erkennbar sein. Durch die Bindung verschiedener Antibiotika an das
Ribosom wird jedoch in der Nähe ihrer Bindungsstelle ein Schutzeffekt gegenüber
den CMCT-Modifikationen erzielt. Dieser konzentrationsabhängige Effekt kann
ebenfalls über die Footprints dargestellt werden und ermöglicht somit Rückschlüsse
auf die Stärke der Wirkstoffbindung an das Ribosom (DOUTHWAITE 1992).
Daher wurden vergleichende Analysen von CMCT-Modifikationen mit Ribosomen
aus Cfr-tragenden (Cfr+) und nicht tragenden (Cfr-) Escherichia coli-Zellen
durchgeführt. Da die CMCT-Modifikation an U2506 nicht stark ist, bestand eine
Variabilität zwischen den Experimenten. Die Ergebnisse sind daher als Spannbreite
des gemessenen Schutzeffektes bei 2 - 3 experimentellen Ansätzen wiedergegeben.
Es zeigte sich, dass die Wirkstoffe Chloramphenicol, Florfenicol und Clindamycin in
Cfr- Zellen einen deutlichen Schutzeffekt an der Position U2506 der 23S rRNA
gegenüber den CMCT-Modifikationen ausüben (Tabelle 7), dieser Schutzeffekt aber
in Cfr+ Zellen deutlich redziert ist (Tabelle 7). Daher konnte von einer verminderten
Bindung der drei Wirkstoffe an Ribosomen aus Cfr+ Zellen ausgegangen werden.
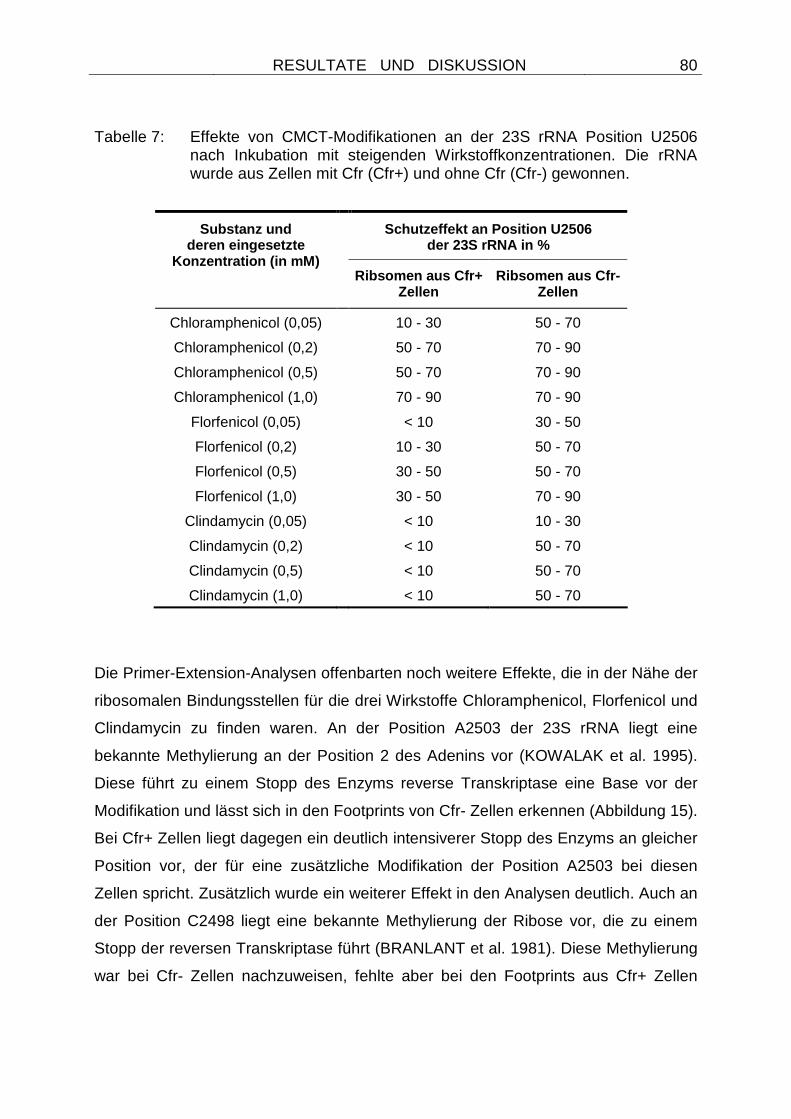
RESULTATE UND DISKUSSION
80
Tabelle 7: Effekte von CMCT-Modifikationen an der 23S rRNA Position U2506 nach Inkubation mit steigenden Wirkstoffkonzentrationen. Die rRNA wurde aus Zellen mit Cfr (Cfr+) und ohne Cfr (Cfr-) gewonnen.
Schutzeffekt an Position U2506 der 23S rRNA in %
Substanz und deren eingesetzte
Konzentration (in mM)
Ribsomen aus Cfr+ Zellen
Ribsomen aus Cfr- Zellen
Chloramphenicol (0,05) 10 - 30 50 - 70
Chloramphenicol (0,2) 50 - 70 70 - 90
Chloramphenicol (0,5) 50 - 70 70 - 90
Chloramphenicol (1,0) 70 - 90 70 - 90
Florfenicol (0,05) < 10 30 - 50
Florfenicol (0,2) 10 - 30 50 - 70
Florfenicol (0,5) 30 - 50 50 - 70
Florfenicol (1,0) 30 - 50 70 - 90
Clindamycin (0,05) < 10 10 - 30
Clindamycin (0,2) < 10 50 - 70
Clindamycin (0,5) < 10 50 - 70
Clindamycin (1,0) < 10 50 - 70
Die Primer-Extension-Analysen offenbarten noch weitere Effekte, die in der Nähe der
ribosomalen Bindungsstellen für die drei Wirkstoffe Chloramphenicol, Florfenicol und
Clindamycin zu finden waren. An der Position A2503 der 23S rRNA liegt eine
bekannte Methylierung an der Position 2 des Adenins vor (KOWALAK et al. 1995).
Diese führt zu einem Stopp des Enzyms reverse Transkriptase eine Base vor der
Modifikation und lässt sich in den Footprints von Cfr- Zellen erkennen (Abbildung 15).
Bei Cfr+ Zellen liegt dagegen ein deutlich intensiverer Stopp des Enzyms an gleicher
Position vor, der für eine zusätzliche Modifikation der Position A2503 bei diesen
Zellen spricht. Zusätzlich wurde ein weiterer Effekt in den Analysen deutlich. Auch an
der Position C2498 liegt eine bekannte Methylierung der Ribose vor, die zu einem
Stopp der reversen Transkriptase führt (BRANLANT et al. 1981). Diese Methylierung
war bei Cfr- Zellen nachzuweisen, fehlte aber bei den Footprints aus Cfr+ Zellen
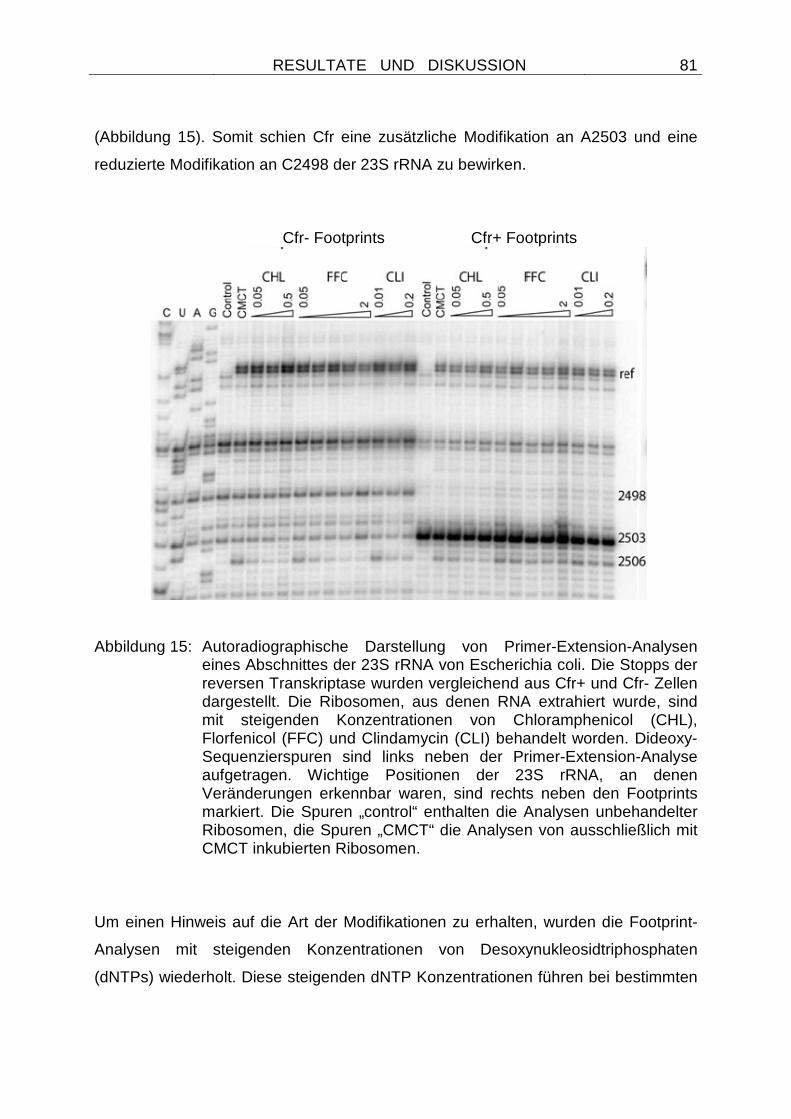
RESULTATE UND DISKUSSION
81
(Abbildung 15). Somit schien Cfr eine zusätzliche Modifikation an A2503 und eine
reduzierte Modifikation an C2498 der 23S rRNA zu bewirken.
Cfr- Footprints Cfr+ Footprints
Abbildung 15: Autoradiographische Darstellung von Primer-Extension-Analysen
eines Abschnittes der 23S rRNA von Escherichia coli. Die Stopps der reversen Transkriptase wurden vergleichend aus Cfr+ und Cfr- Zellen dargestellt. Die Ribosomen, aus denen RNA extrahiert wurde, sind mit steigenden Konzentrationen von Chloramphenicol (CHL), Florfenicol (FFC) und Clindamycin (CLI) behandelt worden. Dideoxy-Sequenzierspuren sind links neben der Primer-Extension-Analyse aufgetragen. Wichtige Positionen der 23S rRNA, an denen Veränderungen erkennbar waren, sind rechts neben den Footprints markiert. Die Spuren „control“ enthalten die Analysen unbehandelter Ribosomen, die Spuren „CMCT“ die Analysen von ausschließlich mit CMCT inkubierten Ribosomen.
Um einen Hinweis auf die Art der Modifikationen zu erhalten, wurden die Footprint-
Analysen mit steigenden Konzentrationen von Desoxynukleosidtriphosphaten
(dNTPs) wiederholt. Diese steigenden dNTP Konzentrationen führen bei bestimmten
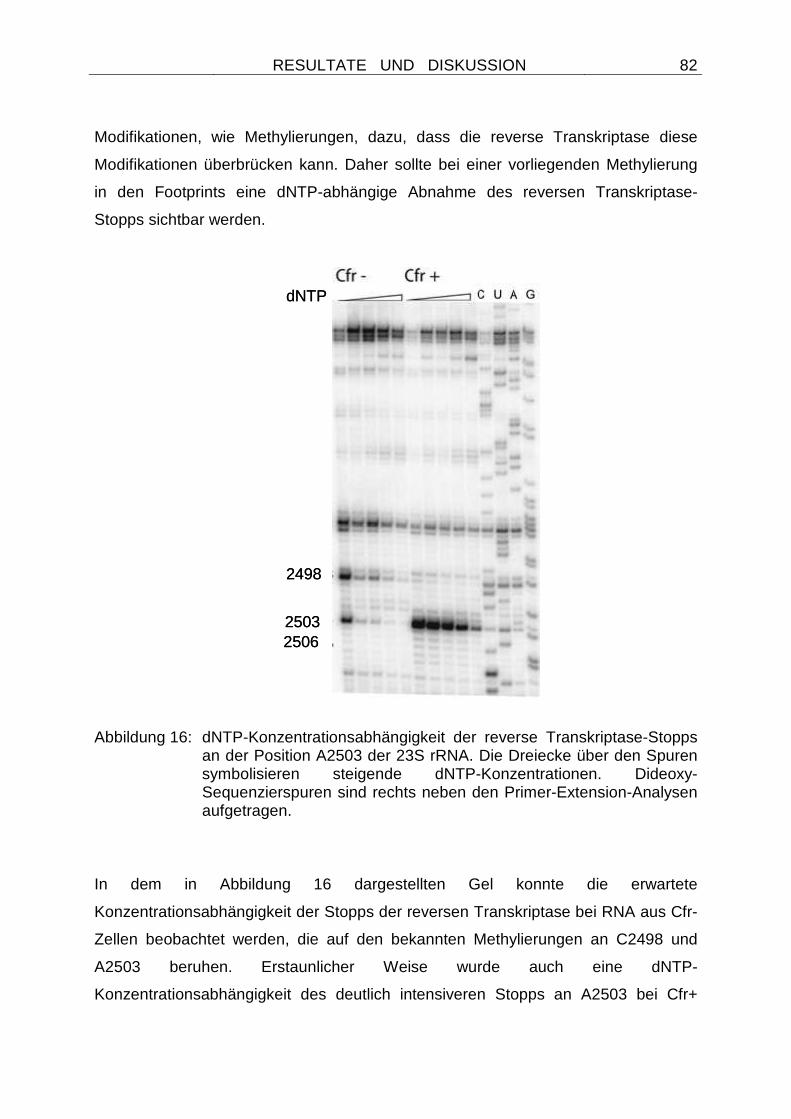
RESULTATE UND DISKUSSION
82
Modifikationen, wie Methylierungen, dazu, dass die reverse Transkriptase diese
Modifikationen überbrücken kann. Daher sollte bei einer vorliegenden Methylierung
in den Footprints eine dNTP-abhängige Abnahme des reversen Transkriptase-
Stopps sichtbar werden.
Abbildung 16: dNTP-Konzentrationsabhängigkeit der reverse Transkriptase-Stopps an der Position A2503 der 23S rRNA. Die Dreiecke über den Spuren symbolisieren steigende dNTP-Konzentrationen. Dideoxy-Sequenzierspuren sind rechts neben den Primer-Extension-Analysen aufgetragen.
In dem in Abbildung 16 dargestellten Gel konnte die erwartete
Konzentrationsabhängigkeit der Stopps der reversen Transkriptase bei RNA aus Cfr-
Zellen beobachtet werden, die auf den bekannten Methylierungen an C2498 und
A2503 beruhen. Erstaunlicher Weise wurde auch eine dNTP-
Konzentrationsabhängigkeit des deutlich intensiveren Stopps an A2503 bei Cfr+
2498
25032506
dNTP
2498
25032506
dNTP

RESULTATE UND DISKUSSION
83
Zellen festgestellt. Diese Veränderung deutet darauf hin, dass die zusätzliche
Modifikation an der Position A2503 wahrscheinlich auf einer weiteren Methylierung
der Ribose beruht.
Um die Art der Modifikation genauer festlegen zu können, wurden MALDI-TOF
(matrix assisted laser desorption / ionization time of flight) massenspektrometrische
Analysen durchgeführt. Dazu wurde ein 45 Nukleotidbereich (45n), der die fraglichen
Modifikationen an C2498 und A2503 einschließt, mit einem komplementären DNA-
Oligonukleotid hybridisiert. Nach der Behandlung mit einer Mung-Bohnen-Nuklease
zum Verdau der RNA-Einzelstränge wurde der isolierte 45n Bereich mit einer G-
spezifischen RNase T1 behandelt. Die dadurch entstandenden Fragmente wurden in
die MALDI-TOF-Analysen eingesetzt. Die erhaltenen Signale (Abbildung 17) wurden
dann mit theoretischen Werten, die aus der rRNA-Nukleotidsequenz berechnet
wurden, verglichen (Publikation 13).
Abbildung 17: MALDI-TOF Massenspektren aus den RNase T1 verdauten Fragmenten eines 45 Nukleotidbereiches der ribosomalen RNA. Die Spektren sind vergleichend aus Zellen mit (Cfr+) und ohne cfr-Gen (Cfr-) dargestellt. Die Nummern über den Signalen stehen für die erhaltenen Massen. Auf der Y-Achse ist die relative Intensität, auf der X-Achse die Masse / Ladung aufgetragen.

RESULTATE UND DISKUSSION
84
Die Analysen der MALDI-TOF Signale zeigten zwei wesentliche Veränderungen bei
der rRNA aus Cfr+ Zellen. Zum einen war das Signal 2233, welches der Sequenz
2496-CACmCUCG-2502 entspricht, reduziert und dafür ein Signal bei 2219 sichtbar.
Die Verschiebung des Signals entspricht dabei genau dem Verlust von 14 Dalton und
somit einer Methylgruppe. Damit war in den Cfr+ Proben die Sequenz 2496-
CACCUCG-2502 ohne Methylierung an C2498 nachgewiesen. Zum anderen waren
zwei zusätzliche Signale bei 3227 und 3241 bei den Cfr+ Proben sichtbar. Diese
entsprechen der Sequenz 2496-CACCUCGAUG-2505 mit zwei oder drei
Methylgruppen. Diese nicht bei Cfr- Proben zu findenden Signale lassen sich durch
einen unvollständigen Verdau durch die RNase T1 erklären. Ein derartiger
unvollständiger Verdau an G2502 beruht hächstwahrscheinlich auf einer zusätzlichen
Methylierung der nachfolgenden Position, A2503, da alle übrigen Fragmente des 45n
Bereiches komplett verdaut zu seien schienen. Um diese Schlussfolgerungen
abzusichern, wurden ergänzende Tandem-massenspektrometrische Analysen
durchgeführt. Die dabei erzielten Spektren der Cfr+ Proben stimmten mit der Präsenz
einer Dimethylierung an A2503 überein (Publikation 13). Somit konnte gezeigt
werden, dass das cfr-Gen eine Methylase kodiert, die eine zusätzliche Methylierung
der 23S rRNA an der Position A2503 einführt und eine verminderte Methylierung an
C2498 bewirkt.

RESULTATE UND DISKUSSION
85
Abbildung 18: Modell der in die Cfr-vermittelte antimikrobielle Resistenz involvierten
Region der 23S rRNA. Die Positionen A2503 und C2498 sind in rot bzw. rosa wiedergegeben. Der geringste Abstand des Nukleotids A2503 zur Bindungsstelle von Chloramphenicol und Clindamycin ist durch eine gepunktete Linie dargestellt.
Die Position der zusätzlichen Methylierung an A2503 ist in Abbildung 18 dargestellt.
Es wird deutlich, dass die Methylgruppe nur 5 Å von der Bindungsstelle für
Chloramphenicol und Clindamycin entfernt liegt und somit vermutlich die korrekte
Positionierung der Antibiotika an ihrer Bindungsstelle verhindert wird. Somit konnte
der durch das Gen cfr vermittelte Chloramphenicol-, Florfenicol- und Clindamycin-
Resistenzmechanismus aufgeklärt werden.
4.2.2.7 Multiresistenz durch den cfr-vermittelten Resistenzphänotyp
PhLOPS A
In Publikation 13 konnte nachgewiesen werden, dass das cfr-Gen eine Methylase
kodiert, die die Position A2503 der 23S rRNA methyliert und somit Resistenz
gegenüber Phenicolen und dem Lincosamid Clindamycin vermittelt. Da aber noch

RESULTATE UND DISKUSSION
86
weitere Klassen antimikrobieller Wirkstoffe an die 50S Untereinheit der Ribosomen
im Peptidyltransferase-Zentrum in der Nähe von A2503 binden, stellte sich die Frage,
ob die A2503 Methylierung auch die Bindung weiterer Wirkstoffe an das Ribosom
verringern oder verhindern kann. Für eine cfr-vermittelte Beeinflussung der
Wirkstoffbindung kamen drei weitere Substanzklassen in Frage: Pleuromutiline,
Oxazolidinone und Streptogramin A-Antibiotika.
Zunächst wurde geprüft, ob durch das cfr-Gen eine Beeinflussung des
Empfindlichkeitsstatus von Escherichia coli gegenüber diesen drei Wirkstoffklassen
vermittelt wird. Dazu wurde das cfr-Gen in den Vektor pBluescriptIISK(+) kloniert und
in den hochempfindlichen Escherichia coli-Empfängerstamm AS19 transformiert. Mit
diesen Escherichia coli AS19::pBglII-Transformanden wurden In-vitro-
Empfindlichkeitstestungen durchgeführt und die Werte mit dem Empfängerstamm
Escherichia coli AS19, einem Escherichia coli AS19-Transformanden mit leerem
Vektor pBluescriptIISK(+) (AS19::pBIISK+) und einem Escherichia coli AS19-
Transformanden, der den rekombinanten Vektor pBluescriptIISK(+) mit deletiertem
cfr-Gen enthält (AS19::pBamHI), verglichen. Die wichtigsten Ergebnisse dieser
Empfindlichkeitstests sind in Tabelle 8 dargestellt. Eine ausführliche Aufstellung aller
Testergebnisse befindet sich in Publikation 14.
Für die Empfindlichkeitstests wurden Vertreter der Wirkstoffklassen der
Pleuromutiline (Tiamulin, Valnemulin), der Streptogramine - entweder als reine A-
Komponente (Virginiamycin M1) oder als Kombination von A- und B-Komponente
(Virginiamycin, Quinupristin-Dalfopristin) - und das einzige auf dem Markt befindliche
Oxazolidinon (Linezolid) ausgewählt. Als Negativkontrollen dienten die Wirkstoffe
Erythromycin und Acetylisovaleryltylosin, die beide an andere Positionen der 50S
Untereinheit der Ribosomen binden und deren MHK-Werte somit nicht durch die Cfr-
Methyltransferase beeinflusst werden sollten.
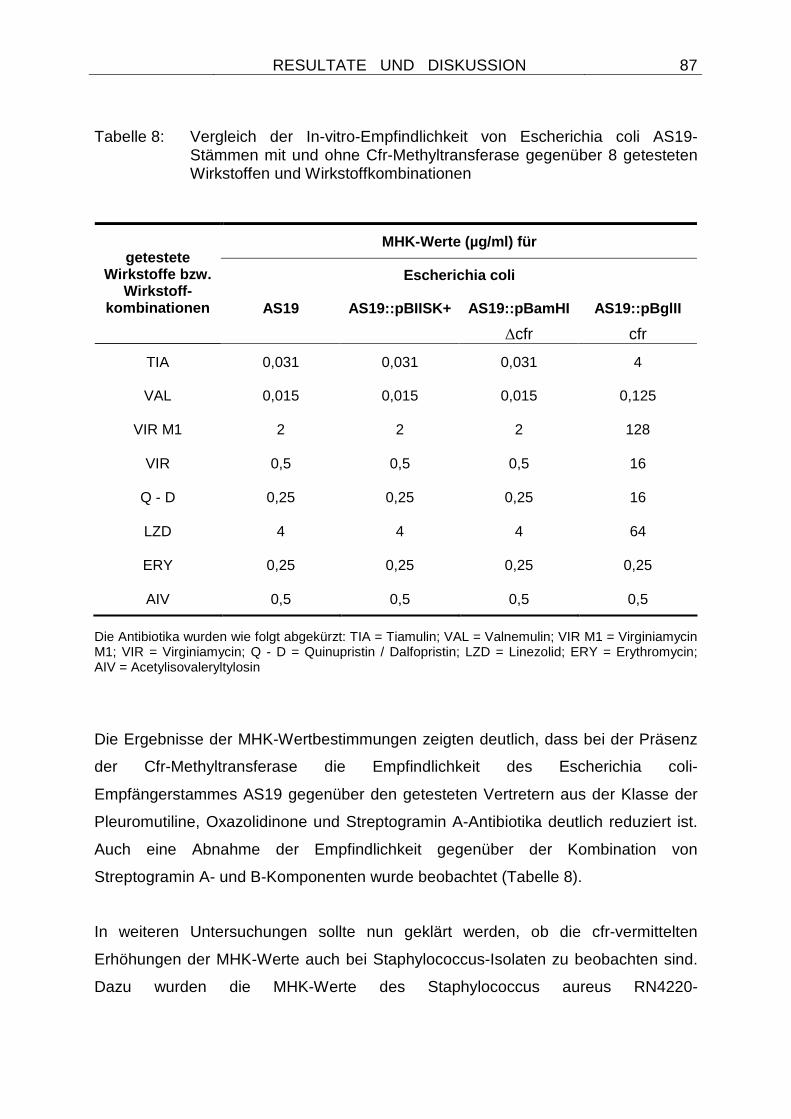
RESULTATE UND DISKUSSION
87
Tabelle 8: Vergleich der In-vitro-Empfindlichkeit von Escherichia coli AS19-Stämmen mit und ohne Cfr-Methyltransferase gegenüber 8 getesteten Wirkstoffen und Wirkstoffkombinationen
Die Antibiotika wurden wie folgt abgekürzt: TIA = Tiamulin; VAL = Valnemulin; VIR M1 = Virginiamycin M1; VIR = Virginiamycin; Q - D = Quinupristin / Dalfopristin; LZD = Linezolid; ERY = Erythromycin; AIV = Acetylisovaleryltylosin
Die Ergebnisse der MHK-Wertbestimmungen zeigten deutlich, dass bei der Präsenz
der Cfr-Methyltransferase die Empfindlichkeit des Escherichia coli-
Empfängerstammes AS19 gegenüber den getesteten Vertretern aus der Klasse der
Pleuromutiline, Oxazolidinone und Streptogramin A-Antibiotika deutlich reduziert ist.
Auch eine Abnahme der Empfindlichkeit gegenüber der Kombination von
Streptogramin A- und B-Komponenten wurde beobachtet (Tabelle 8).
In weiteren Untersuchungen sollte nun geklärt werden, ob die cfr-vermittelten
Erhöhungen der MHK-Werte auch bei Staphylococcus-Isolaten zu beobachten sind.
Dazu wurden die MHK-Werte des Staphylococcus aureus RN4220-
MHK-Werte (µg/ml) für
Escherichia coli getestete
Wirkstoffe bzw. Wirkstoff-
kombinationen AS19 AS19::pBIISK+ AS19::pBamHI
∆cfr
AS19::pBglII
cfr
TIA 0,031 0,031 0,031 4
VAL 0,015 0,015 0,015 0,125
VIR M1 2 2 2 128
VIR 0,5 0,5 0,5 16
Q - D 0,25 0,25 0,25 16
LZD 4 4 4 64
ERY 0,25 0,25 0,25 0,25
AIV 0,5 0,5 0,5 0,5
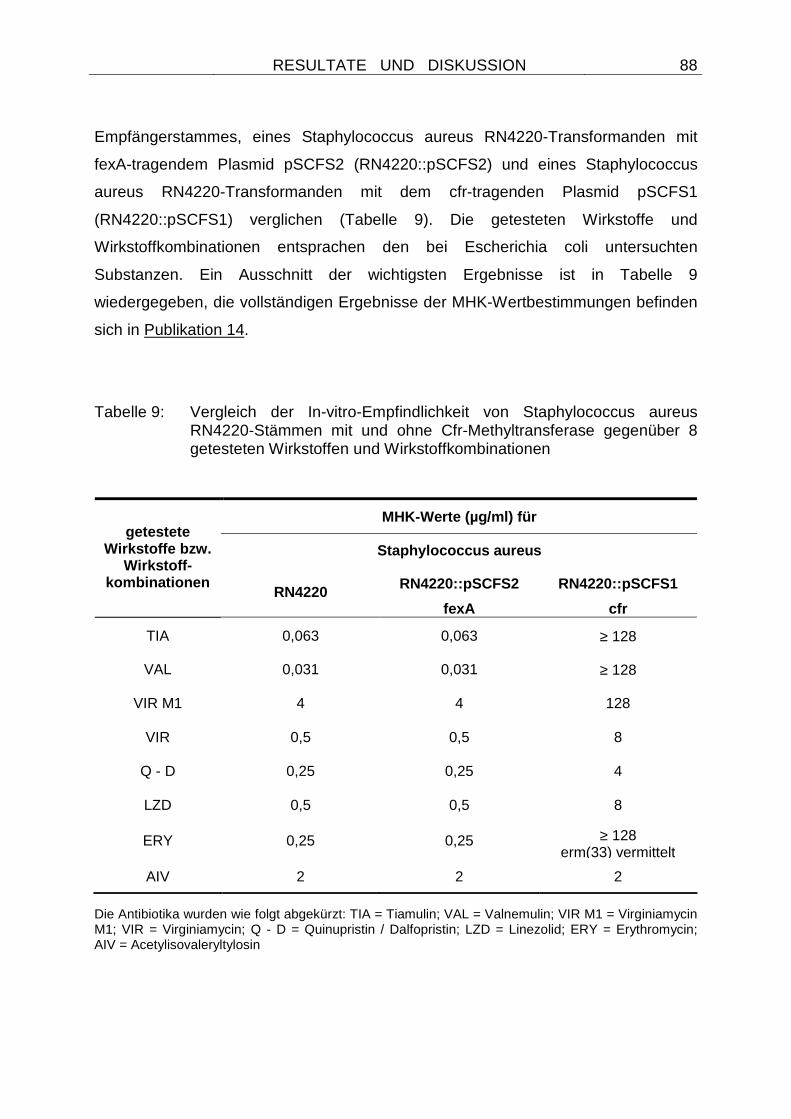
RESULTATE UND DISKUSSION
88
Empfängerstammes, eines Staphylococcus aureus RN4220-Transformanden mit
fexA-tragendem Plasmid pSCFS2 (RN4220::pSCFS2) und eines Staphylococcus
aureus RN4220-Transformanden mit dem cfr-tragenden Plasmid pSCFS1
(RN4220::pSCFS1) verglichen (Tabelle 9). Die getesteten Wirkstoffe und
Wirkstoffkombinationen entsprachen den bei Escherichia coli untersuchten
Substanzen. Ein Ausschnitt der wichtigsten Ergebnisse ist in Tabelle 9
wiedergegeben, die vollständigen Ergebnisse der MHK-Wertbestimmungen befinden
sich in Publikation 14.
Tabelle 9: Vergleich der In-vitro-Empfindlichkeit von Staphylococcus aureus RN4220-Stämmen mit und ohne Cfr-Methyltransferase gegenüber 8 getesteten Wirkstoffen und Wirkstoffkombinationen
Die Antibiotika wurden wie folgt abgekürzt: TIA = Tiamulin; VAL = Valnemulin; VIR M1 = Virginiamycin M1; VIR = Virginiamycin; Q - D = Quinupristin / Dalfopristin; LZD = Linezolid; ERY = Erythromycin; AIV = Acetylisovaleryltylosin
MHK-Werte (µg/ml) für
Staphylococcus aureus getestete
Wirkstoffe bzw. Wirkstoff-
kombinationen RN4220 RN4220::pSCFS2
fexA
RN4220::pSCFS1
cfr
TIA 0,063 0,063 ≥ 128
VAL 0,031 0,031 ≥ 128
VIR M1 4 4 128
VIR 0,5 0,5 8
Q - D 0,25 0,25 4
LZD 0,5 0,5 8
ERY 0,25 0,25 ≥ 128 erm(33) vermittelt
AIV 2 2 2

RESULTATE UND DISKUSSION
89
Auch bei diesen Testungen wurde deutlich, dass die Cfr-Methyltransferase die
antimikrobielle Empfindlichkeit von Staphylokokken gegenüber Pleuromutilinen,
Oxazolidinonen und Streptogramin A-Antibiotika beeinflusst. Es wurden MHK-
Wertdifferenzen festgestellt, die Erhöhungen der Werte um das 4 bis ≥ 4096-fache
bei Cfr-tragenden Stämmen bewirkten. Wurden die verfügbaren, anerkannten
Grenzwerte für die Beurteilung der Staphylococcus aureus RN4220::pSCFS1-
Transformanden zugrunde gelegt, so wurden sie als resistent gegenüber
Chloramphenicol, Linezolid, Quinupristin-Dalfopristin und Clindamycin eingestuft.
Wie durch die unterschiedliche Bindungsstelle von Makroliden zu erwarten, wurden
die MHK-Werte gegenüber Erythromycin und Acetylisovaleryltylosin nicht durch das
Cfr-Protein beeinflusst. Eine verminderte Empfindlichkeit des RN4220::pSCFS1-
Transformanden gegenüber Erythromycin konnte durch die Präsenz des MLSB-
Resistenzgens erm(33) auf dem Plasmid pSCFS1 erklärt werden.
Um die räumliche Anordnung der Bindungsstellen der unterschiedlichen Wirkstoffe
zu erfassen, wurde ein Modell erstellt, welches die Bindung von einem Phenicol,
Lincosamid, Pleuromutilin und Streptogramin A-Antibiotikum im
Peptidyltransferasezentrum einer 50S Untereinheit des Ribosoms zeigt (Abbildung
19).

RESULTATE UND DISKUSSION
90
Chloramphenicol Clindamycin Tiamulin Dalfopristin
Abbildung 19: Struktur der 50S Untereinheit eines Ribosoms, durch die ein Schnitt
geführt wurde (A), so dass ein Blick in das Peptidyltransferasezentrum ermöglicht wird (B). Das Target für die Cfr-Methyltransferase, das Nukleotid A2503, ist als rote Oberfläche gekennzeichnet. Die gebundenen Antibiotika Chloramphenicol, Clindamycin, Tiamulin und Dalfopristin sind als farbige Strukturen erkennbar. Die Strukturformeln der Wirkstoffe sind unterhalb der Abbildung dargestellt, wobei die Hintergrundfarbe der Farbe der Wirkstoffmoleküle im Abbildungsteil B entspricht.
Die Betrachtung der vergleichenden MHK-Werte und der Bindungsstellen der
Wirkstoffe ließ auf ihre reduzierte Bindung im Peptidyltransferasezentrum bei Cfr+
Stämmen schließen. Daher wurden chemische Footprint-Analysen durchgeführt, die
CMCT vermittelte Modifikationen an den Positionen U2506, U2584 und U2585 der
23S rRNA bei Stämmen mit und ohne Cfr-Methyltransferase betrachten sollten
(Publikation 14). Da Oxazolidinone keine Footprints bei leeren Ribosomen
verursachen, wurden die Wirkstoffe Tiamulin, Valnemulin und Virginiamycin M1 für
den experimentellen Ansatz ausgewählt. Die Ergebnisse sind in Abbildung 20
dargestellt und zeigen, dass der in Cfr- Zellen beobachtete Schutzeffekt durch die
drei Wirkstoffe vor CMCT-Modifikationen der rRNA in Cfr+ Stämmen deutlich
reduziert ist. Dies spricht für eine verminderte Bindung auch dieser Wirkstoffe an

RESULTATE UND DISKUSSION
91
Ribosomen von Stämmen, die eine Cfr-Methyltransferase bilden. Weiterhin konnte in
den Footprints eine intensive Bande in Cfr+ Stämmen gezeigt werden (Abbildung
20), die durch den Stopp der reversen Transkriptase aufgrund der nachgewiesenen
zusätzlichen Methylierung von Position A2503 verursacht wird.
Abbildung 20: Autoradiographische Darstellung von Primer-Extension-Analysen eines 23S rRNA-Abschnittes von Escherichia coli. Reverse Transkriptase-Stopps wurden vergleichend aus Cfr+ und Cfr- Zellen dargestellt. Die Ribosomen, aus denen RNA extrahiert wurde, sind in Abschnitt A mit steigenden Konzentrationen von Tiamulin (TIA) und Valnemulin (VAL) behandelt worden, in Abschnitt B mit der Streptogramin A Komponente Virginiamycin M1 (VIR M1). Dideoxy-Sequenzierspuren sind links neben den Primer-Extension-Analysen aufgetragen. Wichtige Positionen der 23S rRNA, an denen CMCT-Modifikationen zu erwarten waren, sind rechts neben den Footprints markiert. Die Spuren „control“ enthalten die Analysen unbehandelter Ribosomen, die Spuren „CMCT“ die Analysen von ausschließlich mit CMCT inkubierten Ribosomen.
Da die Wirkstoffkombinationen Virginiamycin und Quinupristin-Dalfopristin aus einer
A- und B-Streptograminkomponente zusammengesetzt sind, die an
unterschiedlichen Stellen des Ribosoms binden, konnte ihre relative Bindung nicht
über Footprint-Analysen festgestellt werden. Die beobachteten MHK-Wertanstiege
bei Cfr+ Stämmen gegenüber diesen Kombinationen werden aber

RESULTATE UND DISKUSSION
92
höchstwahrscheinlich durch die reduzierte Bindung der A-Komponente erreicht. Ein
ähnlicher Mechanismus der reduzierten Wirkstoffbindung kann auch bei der cfr-
vermittelten Linezolidresistenz vermutet werden. Denn es bestehen deutliche
Hinweise darauf, dass Oxazolidinone auch im Peptidyltransferasezentrum binden.
Alle bislang bekannten Linezolid-Resistenzmutationen liegen nämlich in einer nahe
am Peptidyltransferasezentrum gelegenen und sehr umschriebenen Region (MEKA
u. GOLD 2004).
In Publikation 14 konnte somit der Nachweis erbracht werden, dass die Cfr-
Methyltransferase zu einer verminderten Empfindlichkeit von Bakterien gegenüber
fünf Klassen unterschiedlicher antimikrobieller Wirkstoffe führt. Diese
Wirkstoffklassen sind von großer therapeutischer Bedeutung für die Human- und
Veterinärmedizin. Der Cfr-vermittelte Phänotyp wurde mit PhLOPSA bezeichnet, einer
Abkürzung, die für Resistenz gegenüber Phenicolen, Lincosamiden, Oxazolidinonen,
Pleuromutilinen und Streptogramin A-Antibiotika steht.
Zudem wurde in Publikation 14 erstmalig eine transferable Resistenz gegenüber
Pleuromutilinen und Oxazolidinonen beschrieben. Eine transferable Resistenz
gegenüber diesen Wirkstoffen kann eine wichtige Rolle bei der Verbreitung dieser
Resistenzeigenschaften führen.
4.2.2.8 Verteilung der Resistenzgene cfr und fexA bei chloramphenicol-
resistenten Staphylokokken
Wie zu Beginn des Kapitels 4.2.2 Staphylococcus spp. dargestellt, wurden insgesamt
302 chloramphenicolresistente Staphylokokken von Tieren und Menschen auf ihre
Florfenicolempfindlichkeit geprüft. Von diesen Isolaten zeigten 12 MHK-Werte von
≥ 16 µg/ml und wurden als resistent gegenüber Florfenicol eingestuft. Darunter
befanden sich 11 Isolate mit einer Herkunft von Tieren und ein Isolat vom Menschen.
Die Isolate wurden mit Hilfe des kommerziell erhältlichen Systems ID32 Staph
(bioMérieux, Marcy l´Etoile, Frankreich) auf ihre Spezieszugehörigkeit überprüft.
Nach ihren biochemischen Charakteristika ließen sich die Staphylokokken den

RESULTATE UND DISKUSSION
93
Spezies Staphylococcus lentus (4 Isolate), Staphylococcus aureus (3 Isolate),
Staphylococcus simulans (3 Isolate), Staphylococcus sciuri (1 Isolat) und
Staphylococcus chromogenes (1 Isolat) zuordnen. Von allen Isolaten wurden
Resistenzmuster erstellt, wobei die Empfindlichkeit gegenüber 11 Wirkstoffen
getestet wurde. Die Isolate erwiesen sich als multiresistent und zeigten Resistenzen
gegenüber 6 bis 11 der getesteten Substanzen.
Ziel der Publikation 15 war es nunmehr, Informationen über die Verteilung des
Phenicolresistenzgens fexA sowie des Multiresistenzgens cfr zu erlangen. Dazu
wurden PCR-Assays zum Nachweis der Gene entwickelt und die Ergebnisse mit
Hybridisierungsexperimenten bestätigt (Tabelle 10). Es wurde Gesamtzell-DNA
sowie Plasmid-DNA der Originalstämme und der Transformanden eingesetzt, um die
Lokalisation der Gene auf Plasmiden oder in der chromosomalen DNA zu
bestimmen. Da das fexA-Gen als Bestandteil eines funktionell aktiven Transposons
Tn558 identifiziert wurde (Publikation 9), sollten weitere PCR-Assays Bestandteile
dieses Transposons, das nahezu komplette Element sowie seine zirkulären
Zwischenformen nachweisen. Die hierfür entwickelten Primer, die Größe der
erzielten Amplifikate und ihre genaue Lokalisation sind in Publikation 15 aufgelistet.
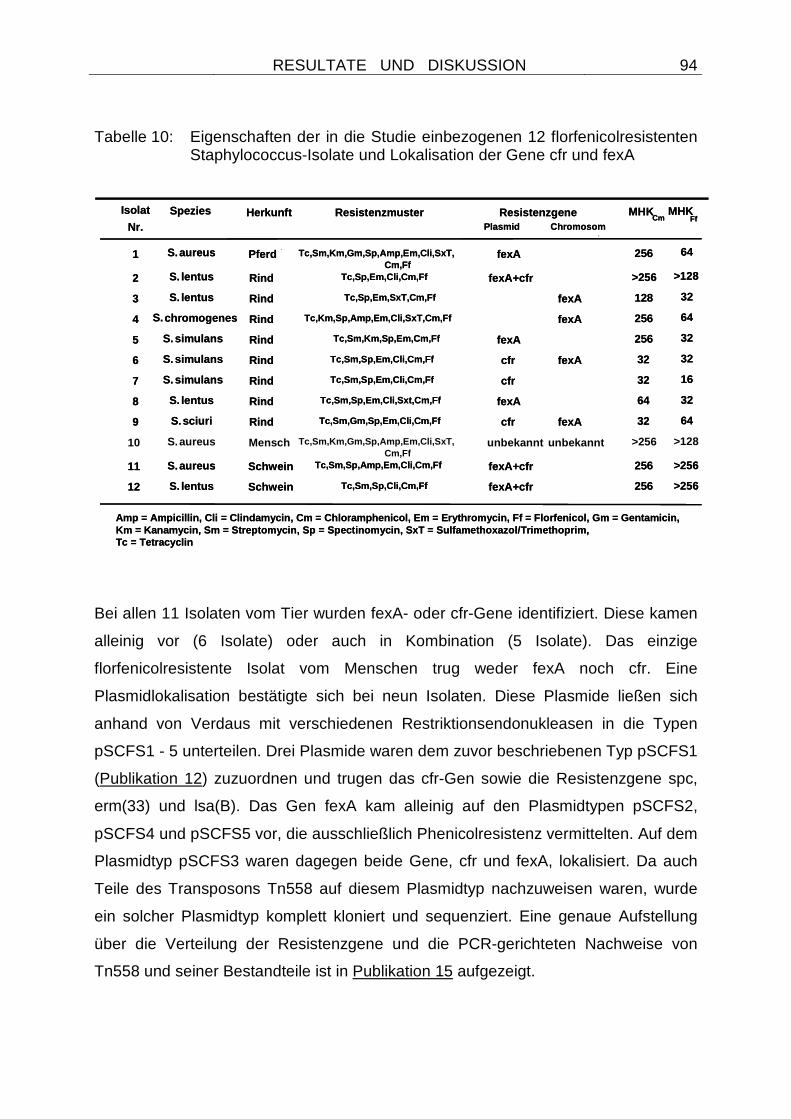
RESULTATE UND DISKUSSION
94
Tabelle 10: Eigenschaften der in die Studie einbezogenen 12 florfenicolresistenten Staphylococcus-Isolate und Lokalisation der Gene cfr und fexA
Bei allen 11 Isolaten vom Tier wurden fexA- oder cfr-Gene identifiziert. Diese kamen
alleinig vor (6 Isolate) oder auch in Kombination (5 Isolate). Das einzige
florfenicolresistente Isolat vom Menschen trug weder fexA noch cfr. Eine
Plasmidlokalisation bestätigte sich bei neun Isolaten. Diese Plasmide ließen sich
anhand von Verdaus mit verschiedenen Restriktionsendonukleasen in die Typen
pSCFS1 - 5 unterteilen. Drei Plasmide waren dem zuvor beschriebenen Typ pSCFS1
(Publikation 12) zuzuordnen und trugen das cfr-Gen sowie die Resistenzgene spc,
erm(33) und lsa(B). Das Gen fexA kam alleinig auf den Plasmidtypen pSCFS2,
pSCFS4 und pSCFS5 vor, die ausschließlich Phenicolresistenz vermittelten. Auf dem
Plasmidtyp pSCFS3 waren dagegen beide Gene, cfr und fexA, lokalisiert. Da auch
Teile des Transposons Tn558 auf diesem Plasmidtyp nachzuweisen waren, wurde
ein solcher Plasmidtyp komplett kloniert und sequenziert. Eine genaue Aufstellung
über die Verteilung der Resistenzgene und die PCR-gerichteten Nachweise von
Tn558 und seiner Bestandteile ist in Publikation 15 aufgezeigt.
Amp = Ampicillin, Cli = Clindamycin, Cm = Chloramphen icol, Em = Erythromycin, Ff = Florfenicol, Gm = Gent amicin, Km = Kanamycin, Sm = Streptomycin, Sp = Spectinomyci n, SxT = Sulfamethoxazol/Trimethoprim, Tc = Tetracyclin
Isolat
Nr.Spezies Herkunft Resistenzmuster Resistenzgene
Plasmid Chromosom
MHKCm
1
2
3
4
5
6
7
8
9
11
12
chromogenes
S. aureus
S. lentus
S. lentus
S.
S. simulans
S. simulans
S. simulans
S. lentus
S. sciuri
S. aureus
S. lentus
Pferd
Rind
Rind
Rind
Rind
Rind
Rind
Rind
Rind
Schwein
Schwein
Tc,Sm,Km,Gm,Sp,Amp,Em,Cli,SxT,Cm,Ff
Tc,Sp,Em,Cli,Cm,Ff
Tc,Sp,Em,SxT,Cm,Ff
Tc,Km,Sp,Amp,Em,Cli,SxT,Cm,Ff
Tc,Sm,Km,Sp,Em,Cm,Ff
Tc,Sm,Sp,Em,Cli,Cm,Ff
Tc,Sm,Sp,Em,Cli,Cm,Ff
Tc,Sm,Sp,Em,Cli,Sxt,Cm,Ff
Tc,Sm,Gm,Sp,Em,Cli,Cm,Ff
Tc,Sm,Sp,Amp,Em,Cli,Cm,Ff
Tc,Sm,Sp,Cli,Cm,Ff
fexA
fexA+cfr
fexA
fexA
fexA
cfr fexA
cfr
fexA
cfr fexA
fexA+cfr
fexA+cfr
256
>256
128
256
256
32
32
64
32
256
256
10 S. aureus Mensch Tc,Sm,Km,Gm,Sp,Amp,Em,Cli,SxT, unbekannt unbekannt >256
MHKFf
64
>128
32
64
32
32
16
32
64
>256
>256
>128Cm,Ff
Amp = Ampicillin, Cli = Clindamycin, Cm = Chloramphen icol, Em = Erythromycin, Ff = Florfenicol, Gm = Gent amicin, Km = Kanamycin, Sm = Streptomycin, Sp = Spectinomyci n, SxT = Sulfamethoxazol/Trimethoprim, Tc = Tetracyclin
Isolat
Nr.Spezies Herkunft Resistenzmuster Resistenzgene
Plasmid Chromosom
MHKCm
1
2
3
4
5
6
7
8
9
11
12
chromogenes
S. aureus
S. lentus
S. lentus
S.
S. simulans
S. simulans
S. simulans
S. lentus
S. sciuri
S. aureus
S. lentus
Pferd
Rind
Rind
Rind
Rind
Rind
Rind
Rind
Rind
Schwein
Schwein
Tc,Sm,Km,Gm,Sp,Amp,Em,Cli,SxT,Cm,Ff
Tc,Sp,Em,Cli,Cm,Ff
Tc,Sp,Em,SxT,Cm,Ff
Tc,Km,Sp,Amp,Em,Cli,SxT,Cm,Ff
Tc,Sm,Km,Sp,Em,Cm,Ff
Tc,Sm,Sp,Em,Cli,Cm,Ff
Tc,Sm,Sp,Em,Cli,Cm,Ff
Tc,Sm,Sp,Em,Cli,Sxt,Cm,Ff
Tc,Sm,Gm,Sp,Em,Cli,Cm,Ff
Tc,Sm,Sp,Amp,Em,Cli,Cm,Ff
Tc,Sm,Sp,Cli,Cm,Ff
fexA
fexA+cfr
fexA
fexA
fexA
cfr fexA
cfr
fexA
cfr fexA
fexA+cfr
fexA+cfr
256
>256
128
256
256
32
32
64
32
256
256
10 S. aureus Mensch Tc,Sm,Km,Gm,Sp,Amp,Em,Cli,SxT, unbekannt unbekannt >256
MHKFf
64
>128
32
64
32
32
16
32
64
>256
>256
>128Cm,Ff

RESULTATE UND DISKUSSION
95
Die Sequenzanalysen des Plasmidtyps pSCFS3 zeigten, dass ein aus 9491 bp
bestehendes Element auf dem Plasmid lokalisiert war. Dieses zeigte in den ersten
575 bp sowie in den terminalen 4242 bp Homologien zu Tn558. Die Tn558-
homologen Regionen wurden durch die Insertion eines 4674 bp Bereiches
(Abbildung 21) unterbrochen, wobei es zum Verlust des 3´-Endes des tnpA
Leserahmens sowie des 5´-Endes von tnpB gekommen war. Der inserierte Bereich
bestand aus einem Bereich, der das cfr-Gen einschloss sowie einem IS21-ähnlichen
Element. Dieses Element erhielt die Bezeichnung IS21-558.
Abbildung 21: Schematische Zeichnung der Tn558-Variante von Plasmid pSCFS3. Die Leserahmen sind als Pfeile dargestellt, wobei die Pfeilspitze die Transkriptionsrichtung anzeigt. Die Kästchen oberhalb der Zeichnung zeigen die Bereiche, in denen Homologien zu bekannten Strukturen aufgezeigt wurden.
Das abgeleitete Cfr-Protein von Plasmid pSCFS3 unterschied sich nur durch einen
Aminosäureaustausch von dem bei pSCFS1 beschriebenen Protein. Allerdings war
einer der dem Gen vorgeschalteten und scheinbar in die induzierbare Expression
des Gens involvierten Leserahmen (Publikation 10) deletiert. Durch diese Deletion
wurden aber die cfr-vermittelten Resistenzeigenschaften nicht beeinflusst. Eine
genaue Analyse auch der weiteren Leserahmen auf dem 9491 bp Element von
pSCFS3 (∆tnpA, ∆tnpB, tnpC, ORF138, fexA) befindet sich in Publikation 15.
0 1 2 3 4∆tnpA ∆tnpB tnpC orf138
5 6fexA
7 8 9istAS istBS cfr
IS21-558 homology to pSCFS1
homology to Tn558
homology to Tn558
no homology to any database entries
0 1 2 3 4∆tnpA ∆tnpB tnpC orf138
5 6fexA
7 8 9istAS istBS cfr
IS21-558 Homologie zupSCFS1
Homologie zu Tn558
Homologie zu Tn558
keine Homologie zu Datenbankeinträgen
0 1 2 3 4∆tnpA ∆tnpB tnpC orf138
5 6fexA
7 8 9istAS istBS cfr
IS21-558 homology to pSCFS1
homology to Tn558
homology to Tn558
no homology to any database entries
0 1 2 3 4∆tnpA ∆tnpB tnpC orf138
5 6fexA
7 8 9istAS istBS cfr
IS21-558 Homologie zupSCFS1
Homologie zu Tn558
Homologie zu Tn558
keine Homologie zu Datenbankeinträgen

RESULTATE UND DISKUSSION
96
Das neue IS21-ähnliche Element von pSCFS3 bestand aus 2446 bp und enthielt
zwei offene Leserahmen, die als istAS und istBS bezeichnet wurden. Diese beiden
überlappenden Leserahmen kodierten für Proteine von 445 und 250 aa und zeigten
weniger als 50 % bzw. 70 % Identität zu Transposaseproteinen der IS21-Familie
(MAHILLON u. CHANDLER 1998). Die abgeleitete IstAS Aminosäuresequenz zeigte
das typische DDE-Motiv, welches als aktiver Part von Transposaseproteinen
beschrieben wurde (MAHILLON u. CHANDLER 1998). Zwei imperfekte umgekehrt
komplementäre Sequenzwiederholungen („inverted repeats“) rahmten das neue
IS21-558-Element ein. Direkte Sequenzwiederholungen („direct repeats“) waren nicht
an der Integrationsstelle zu finden.
Abbildung 22: Schematische Darstellung der neuartigen Insertionssequenz IS21-
558. Die umgekehrt komplementären Sequenzwiederholungen „inverted repeats“ sind in dem Kästchen oberhalb der Schemazeichnung eingezeichnet. Die Leserahmen für die Transposaseproteine IstAS und IstBS sind als Pfeile dargestellt.
0 0.4 2.00.8 1.2 1.6 2.4
istAS istB
1 - TGTATATGTTAACTTAAAAGTGAGCCACCAACGCAATTGAAAAGTGAGCCACTTCTG - 57||||| | |||| |||| ||||||| ||||||| || |||||||||| ||
2446 - ACATAAAGCGTTGACTTTTAACTCGGT-AAAGCGTTAAATTAAAACTCGGTGAATAC - 2391
IS21-558
0 0.4 2.00.8 1.2 1.6 2.4
istAS istB
1 - TGTATATGTTAACTTAAAAGTGAGCCACCAACGCAATTGAAAAGTGAGCCACTTCTG - 57||||| | |||| |||| ||||||| ||||||| || |||||||||| ||
2446 - ACATAAAGCGTTGACTTTTAACTCGGT - 2391
0 0.4 2.00.8 1.2 1.6 2.4
istAS istB
0 0.4 2.00.8 1.2 1.6 2.4
istAS istBS
1 - TGTATATGTTAACTTAAAAGTGAGCCACCAACGCAATTGAAAAGTGAGCCACTTCTG - 57||||| | |||| |||| ||||||| ||||||| || |||||||||| ||
2446 - ACATAAAGCGTTGACTTTTAACTCGGT - 2391
imperfekte „inverted repeats“
IS21-558
0 0.4 2.00.8 1.2 1.6 2.4
istAS istB
0 0.4 2.00.8 1.2 1.6 2.4
istAS istB
1 - TGTATATGTTAACTTAAAAGTGAGCCACCAACGCAATTGAAAAGTGAGCCACTTCTG - 57||||| | |||| |||| ||||||| ||||||| || |||||||||| ||
2446 - ACATAAAGCGTTGACTTTTAACTCGGT-AAAGCGTTAAATTAAAACTCGGTGAATAC - 2391
IS21-558
0 0.4 2.00.8 1.2 1.6 2.4
istAS istB
0 0.4 2.00.8 1.2 1.6 2.4
istAS istB
1 - TGTATATGTTAACTTAAAAGTGAGCCACCAACGCAATTGAAAAGTGAGCCACTTCTG - 57||||| | |||| |||| ||||||| ||||||| || |||||||||| ||
2446 - ACATAAAGCGTTGACTTTTAACTCGGT - 2391
0 0.4 2.00.8 1.2 1.6 2.4
istAS istB
0 0.4 2.00.8 1.2 1.6 2.4
istAS istBS
1 - TGTATATGTTAACTTAAAAGTGAGCCACCAACGCAATTGAAAAGTGAGCCACTTCTG - 57||||| | |||| |||| ||||||| ||||||| || |||||||||| ||
2446 - ACATAAAGCGTTGACTTTTAACTCGGT - 2391
imperfekte „inverted repeats“
IS21-558

RESULTATE UND DISKUSSION
97
4.2.2.9 Die Mobilität des cfr-Gens wird durch IS 21-558-Elemente vermittelt
Zwar konnte in Publikation 15 ein IS21-558-Element in unmittelbarer Umgebung des
cfr-Gens nachgewiesen werden, Hinweise auf eine Beteiligung der
Insertionssequenz an der Mobilität des cfr-Gens lagen aber nicht vor. Da im Jahr
2005 auf zwei Schweinefarmen in Dänemark 23 weitere florfenicolresistente
Staphylokokken isoliert wurden, sollten auch diese Isolate auf die Präsenz der Gene
cfr und fexA untersucht werden. Beide Farmen befanden sich im Eigentum des
gleichen Besitzers und die Tiere waren bekanntlich mit Florfenicol und weiteren
Wirkstoffen vorbehandelt. Die biochemische Spezieszugehörigkeit mittels ID32
Staph-System identifizierte 14 Isolate als Staphylococcus hyicus, 8 als
Staphylococcus warneri und ein Isolat als Staphylococcus simulans. Da die Isolate
von zwei benachbarten Schweinefarmen stammten, wurden
Makrorestriktionsanalysen durchgeführt, die eine klonale Verwandtschaft der Isolate
aufdecken sollten. Basierend auf der Klonalität der Isolate (Publikation 16) wurden
zwei Staphylococcus warneri, zwei Staphylococcus hyicus und das Staphylococcus
simulans-Isolat für die weiteren Untersuchungen ausgewählt. Die Resistenzmuster
dieser Staphylokokken zeigten, dass es sich um multiresistente Isolate handelte
(Publikation 16). Durch Protoplastentransformationen wurde deutlich, dass die
Eigenschaft der Florfenicolresistenz bei beiden Staphylococcus warneri-Isolaten
sowie bei dem Staphylococcus simulans-Isolat auf einem großen Plasmid von ca. 43
kb lokalisiert war. Da es sich um einen neuen und anhand von
Restriktionsenzymanalysen nicht unterscheidbaren Plasmidtyp handelte, wurde er
mit pSCFS6 bezeichnet. PCR-Assays und Hybridisierungsexperimente zeigten, dass
sowohl das cfr- als auch das fexA-Gen auf pSCFS6 lokalisiert waren. Zudem
konnten die Gene tnpA, tnpB sowie die Leserahmen für die Transposasen istAS und
istBS nachgewiesen werden. Da bislang wenig Informationen über die genetische
Umgebung des Multiresistenzgens cfr vorlagen, wurde der Plasmidtyp pSCFS6
ebenfalls komplett kloniert und die Resistenzgenregion Sequenzanalysen
unterzogen. Bei den Staphylococcus hyicus-Isolaten konnte dagegen eine
chromosomale Lokalisation von cfr, fexA, istAS und istBS identifiziert werden. Dies

RESULTATE UND DISKUSSION
98
war mit der Publikation 16 auch der erste Bericht eines chromosomal lokalisierten
cfr-Gens.
Die Sequenzanalyse einer 22010 bp Region von pSCFS6 offenbarte eine neue
Variante des Transposons Tn558. Diese Variante bestand aus 16253 bp und war
zusammengesetzt aus einem 6659 bp Bereich mit Homologie zu Tn558 und einer
9594 bp Region, die zwischen den Tn558-homologen Bereichen inseriert war
(Abbildung 23). In dem Tn558-homologen Part waren nur wenige bp Austausche zu
dem von Plasmid pSCFS2 bekannten fexA-tragenden Transposon Tn558 zu finden.
Diese Austausche sind detailliert in Publikation 16 aufgezeigt. Auch die „core“
Sequenz an dem rechten terminalen Ende dieser Tn558 Variante differierte leicht
von den bislang bekannten „core“ Sequenzen und ließ sich durch eine variierende
ehemalige Integrationsstelle erklären.
Abbildung 23: Organisation der neuen Tn558 Variante von Plasmid pSCFS6, die im
Vergleich zu dem fexA-tragenden Transposon Tn558 dargestellt ist. Homologe Bereiche sind durch gelb unterlegte Balken hervorgehoben. Die Leserahmen sind durch Pfeile dargestellt, wobei die Pfeilspitze die Transkriptionsrichtung anzeigt. Die 6 bp direkten Sequenzwiederholungen an der Integrationsstelle des IS21-558-Elementes sind in Kästchen eingetragen. Eine Größenskala in kb ist unterhalb der Schemazeichnungen angebracht.
Tn558
tnpA tnpB tnpC orf138 fexA
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
0 1 2 3 4 5 6
tnpA tnpB lsa(B) orf138 fexAcfr???? tnpC IS21-558 ???? tnpCIS21-558
GACGTA GACGTA
16
Tn558
tnpA tnpB tnpC orf138 fexA
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
0 1 2 3 4 5 6
tnpA tnpB lsa(B) orf138 fexAcfrtnpC IS21-558 tnpCIS21-558
GACGTA GACGTA
16
Tn558
tnpA tnpB tnpC orf138 fexA
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
0 1 2 3 4 5 6
tnpA tnpB lsa(B) orf138 fexAcfr???? tnpC IS21-558 ???? tnpCIS21-558
GACGTA GACGTA
16
Tn558
tnpA tnpB tnpC orf138 fexA
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
0 1 2 3 4 5 6
tnpA tnpB lsa(B) orf138 fexAcfrtnpC IS21-558 tnpCIS21-558
GACGTA GACGTA
16

RESULTATE UND DISKUSSION
99
Von besonderem Interesse war die Analyse der 9594 bp Region, die sich zwischen
den Tn558-homologen Bereichen befand. Diese war inmitten des tnpC-Leserahmens
von Tn558 inseriert und führte zu einer Duplikation der Integrationsstelle 5´-
GACGTA-3´ und damit zu einer 6 bp direkten Sequenzwiederholung („direct
repeats“). An beiden Enden befanden sich Insertionssequenzen vom Typ IS21-558,
die in gleicher Orientierung angeordnet waren. Diese terminalen IS21-558-Elemente
unterschieden sich nur durch 4 bp Austausche voneinander und zeigten 99 %
Identität zu dem 6 bp kürzeren Element von pSCFS3. Der zentrale Part des
Elementes glich einem auf Plasmid pSCFS1 identifizierten Bereich und trug das Gen
lsa(B), welchen den in Publikation 11 nachgewiesenen „low-level“
Clindamycinexporter kodiert sowie das Multiresistenzgen cfr. Zwischen dem lsa(B)
und dem cfr-Part lag ein 94 bp Bereich ohne Homologie zu Datenbankeinträgen.
Durch die typische Anordnung der IS21-ähnlichen Elemente auf pSCFS6 konnte ein
Modell für die Mobilität von cfr - IS21-558 erstellt werden. Dieses basiert auf
Mobilitätswegen, die für Insertionssequenzen aus der IS21-Familie nachgewiesen
sind (BERGER u. HAAS 2001). Hierbei ist für das Genprodukt von istAS eine
Doppelfunktion als Transposase und Cointegrase beschrieben (BERGER u. HAAS
2001; SCHMID et al. 1998). Die jeweilige Funktion hängt dabei von dem Gebrauch
des Startkodons ab. Neben dem normalen ATG-Startkodon befindet sich ein
alternatives Startkodon einige Tripletts vor dem ATG, so dass bei Benutzung dieses
Startkodons das IstA-Genprodukt um 3 - 11 aa verlängert wird. Ein solches
alternatives GTG Startkodon wurde auch bei dem IS21-558-Element von pSCFS6
nachgewiesen (Publikation 16). Während das verlängerte IstA-Protein vornehmlich
Transpositionen unterstützt, werden durch das verkürzte IstA-Protein
Cointegrationen vermittelt. Die IS21-Transpositionen laufen durch einen einfachen
„cut and paste“ Mechanismus einer einzelnen IS21-Kopie ab und schließen eine
Zirkularisation des IS-Elementes ein (Abbildung 24a). Die Replikonfusionen laufen
dagegen bevorzugt zwischen zwei IS21-Elementen in einer Tandemformation und
einem zweiten Zielreplikon ab (Abbildung 24b) (BERGER u. HAAS 2001; MAHILLON
u. CHANDLER 1998; REIMANN u. HAAS 1990). Bei diesem Fusionsvorgang können
DNA-Fragmente von dem Donorreplikon zu einem Targetreplikon übertragen werden

RESULTATE UND DISKUSSION
100
(BERGER u. HAAS 2001; REIMANN u. HAAS 1990). Die Tandemformation der IS-
Elemente ist dabei für die Formation eines starken Promotors von Bedeutung, der
zur Ablesung des kürzeren istA-Leserahmens führt (BERGER u. HAAS 2001). Das
IstB-Protein fungiert während der Mobilitätsvorgänge als Helferprotein (SCHMID et
al. 1999). Als Resultat der Replikonfusion entsteht ein Cointegrat, welches zwei
IS21-Elemente in gleicher Orientierung aufweist. Ein solches Element konnte auf
dem Plasmid pSCFS6 identifiziert werden. Somit ist eine IS21-558 vermittelte
Mobilität des cfr-Gens als wahrscheinlich anzunehmen. Auch wenn keine IS21-558
Tandemformationen nachweisbar waren, so konnten doch die typische Struktur des
Elementes und die charakteristischen Sequenzwiederholungen an den Enden
aufgezeigt werden. Da nach IS21-558 vermittelten Replikonfusionen häufig
Deletionen von DNA-Bereichen durch Rekombinationsprozesse folgen, kann auch
das Auftreten des einzelnen IS21-558-Elementes auf Plasmid pSCFS3 (Publikation
15) durch diesen speziellen Mobilitätsweg erklärt werden. Zudem konnten bei den
cfr-tragenden Staphylococcus hyicus-Isolaten in Publikation 16 kreisförmige
Elemente („mini circles“) nachgewiesen werden, die typisch für IS21-vermittelte
Transpositionsvorgänge sind und die somit die funktionelle Aktivität der IS21-558
Insertionssequenzen nachweisen.
Zusammenfassend konnte in Publikation 16 gezeigt werden, dass IS21-558-
Elemente vermutlich die Mobilität des Multiresistenzgens cfr vermitteln. Zudem
konnte erstmalig eine Verbreitung des Gens auch bei Staphylokokken von
Schweinen aus Dänemark nachgewiesen werden. Somit scheint eine weitere
Verbreitung des cfr-Gens vorzuliegen, als ursprünglich vermutet wurde.
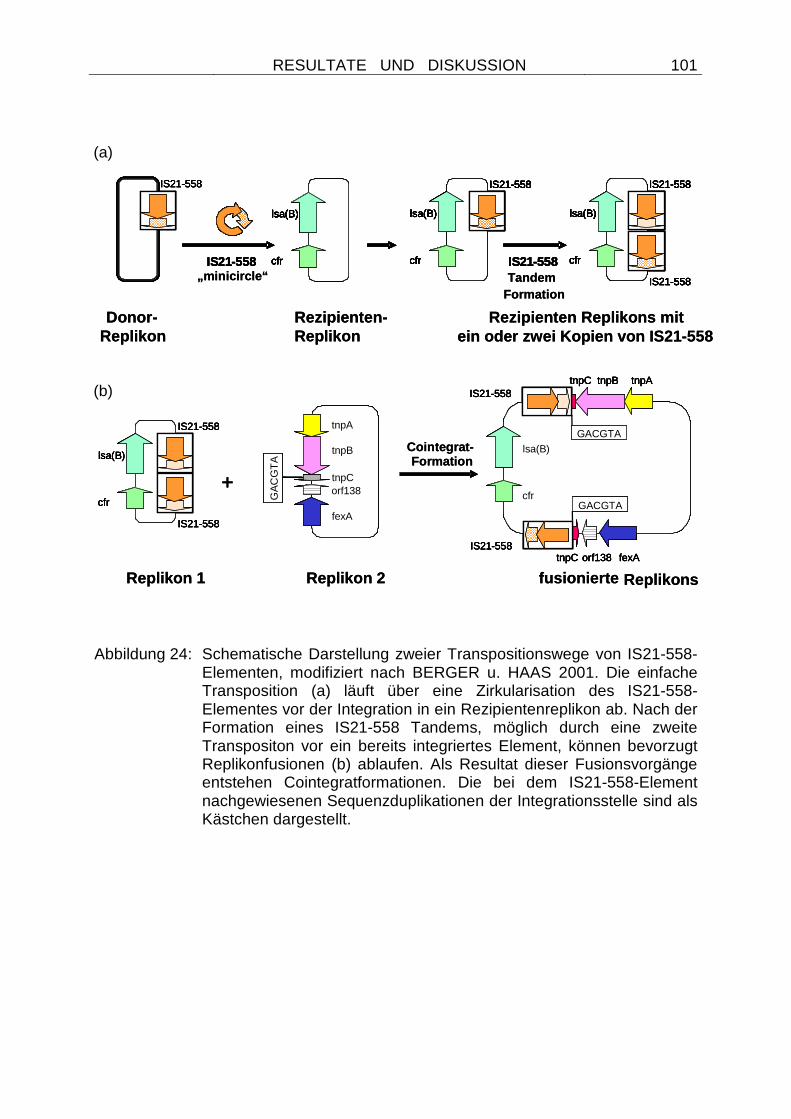
RESULTATE UND DISKUSSION
101
Abbildung 24: Schematische Darstellung zweier Transpositionswege von IS21-558-
Elementen, modifiziert nach BERGER u. HAAS 2001. Die einfache Transposition (a) läuft über eine Zirkularisation des IS21-558-Elementes vor der Integration in ein Rezipientenreplikon ab. Nach der Formation eines IS21-558 Tandems, möglich durch eine zweite Transpositon vor ein bereits integriertes Element, können bevorzugt Replikonfusionen (b) ablaufen. Als Resultat dieser Fusionsvorgänge entstehen Cointegratformationen. Die bei dem IS21-558-Element nachgewiesenen Sequenzduplikationen der Integrationsstelle sind als Kästchen dargestellt.
GACGTA
GACGTA
GA
CG
TA
tnpA
tnpB
tnpCorf138
fexAcfr
lsa(B)
IS21-558
IS21-558
+
Replikon 1 Replikon 2
Cointegrat-Formation
fusionierte Replikons
cfr
lsa(B)
IS21-558
IS21-558
cfr
lsa(B)
IS21-558
cfr
lsa(B)
IS21-558
(a)
(b)
IS21-558„minicircle“
IS21-558
IS21-558
cfr
lsa(B)
IS21-558
tnpAtnpBtnpC
orf138 fexAtnpC
GACGTA
GACGTA
GA
CG
TA
tnpA
tnpB
tnpCorf138
fexAcfr
lsa(B)
IS21-558
IS21-558
+
cfr
lsa(B)
IS21-558
IS21-558
cfr
lsa(B)
IS21-558
IS21-558
cfr
lsa(B)
IS21-558
cfr
lsa(B)
IS21-558
cfr
lsa(B)
cfr
lsa(B)
IS21-558
(a)
(b)
Donor-Replikon
Rezipienten-Replikon
IS21-558 IS21-558Tandem
Formation
Rezipienten Replikons mitein oder zwei Kopien von IS 21-558
IS21-558
cfr
lsa(B)
IS21-558
tnpAtnpBtnpC
orf138 fexAtnpC
GACGTA
GACGTA
GA
CG
TA
tnpA
tnpB
tnpCorf138
fexAcfr
lsa(B)
IS21-558
IS21-558
+
Replikon 1 Replikon 2
Cointegrat-Formation
fusionierte Replikons
cfr
lsa(B)
IS21-558
IS21-558
cfr
lsa(B)
IS21-558
IS21-558
cfr
lsa(B)
IS21-558
cfr
lsa(B)
IS21-558
cfr
lsa(B)
cfr
lsa(B)
IS21-558
(a)
(b)
IS21-558„minicircle“
IS21-558
IS21-558
cfr
lsa(B)
IS21-558
tnpAtnpBtnpC
orf138 fexAtnpC
GACGTA
GACGTA
GA
CG
TA
tnpA
tnpB
tnpCorf138
fexAcfr
lsa(B)
IS21-558
IS21-558
+
cfr
lsa(B)
IS21-558
IS21-558
cfr
lsa(B)
IS21-558
IS21-558
cfr
lsa(B)
IS21-558
cfr
lsa(B)
IS21-558
cfr
lsa(B)
cfr
lsa(B)
IS21-558
(a)
(b)
Donor-Replikon
Rezipienten-Replikon
IS21-558 IS21-558Tandem
Formation
Rezipienten Replikons mitein oder zwei Kopien von IS 21-558
IS21-558
cfr
lsa(B)
IS21-558
tnpAtnpBtnpC
orf138 fexAtnpC

RESULTATE UND DISKUSSION
102
4.2.2.10 Identifizierung des cfr-Gens bei einem MRSA-Isolat vom Menschen
In den letzten Jahren nahm die Bedeutung von grampositiven Bakterien - besonders
von methicillinresistenten Staphylokokken und auch Enterokokken - als Erreger
humaner nosokomialer Infektionen deutlich zu (BODMANN 2004). Mittlerweile wird
davon ausgegangen, dass mehr als die Hälfte aller schweren Infektionen durch
grampositive Bakterien verursacht werden, wobei solche Infektionen mit einem
deutlich erhöhten Mortalitätsrisiko einhergehen (BODMANN 2004). Durch die
zunehmende Verbreitung resistenter Stämme haben viele Antibiotika ihre gute
Wirksamkeit eingebüßt. Bei Staphylokokken kann der Erwerb eines Gens (mecA)
das ursprünglich β-Laktam-empfindliche Penicillin-bindende Protein (PBP) ersetzen
und so zur Methicillinresistenz führen (KEHRENBERG u. SCHWARZ 2005a).
Methicillinresistente Staphylokokken gelten als resistent gegenüber allen β-Laktam-
Antibiotika. Sind Infektionen durch solche methicillinresistenten Staphylococcus
aureus-Isolate (MRSA) verursacht, so waren Glykopeptide häufig die einzige
Therapiemöglichkeit. Heute stellt das Oxazolidinon Linezolid eine weitere
therapeutische Option dar (BODMANN 2004). Bei MRSA-Isolaten trat
Linezolidresistenz bislang recht selten auf und basierte auf Mutationen der 23S rRNA
(BECKER 2004). Somit war kein horizontaler Transfer dieser Resistenzeigenschaft
auf andere Bakterien möglich.
Erstmalig wurden von ARIAS et al. 2006 auf der 46th Interscience Conference on
Antimicrobial Agents and Chemotherapie (ICAAC) Ergebnisse vorgestellt, die ein
linezolidresistentes MRSA-Isolat zeigten, bei dem keine Mutation der 23S rRNA
vorlag (ARIAS et al. 2006). Das Isolat stammte von einem kolumbianischen
Patienten und hörte dem häufigsten klonalen Cluster an, welches zur Zeit in
Kolumbien zirkuliert und auch als „Chilean clone“ bezeichnet wird. Die genaue
Analyse dieses Isolates zeigte, dass die Linezolidresistenz auf der Präsenz eines
plasmidlokalisierten cfr-Gens basierte (MANKIN et al. 2006). Auf einem kleinen
sequenzierten Abschnitt des Plasmids wurde auch ein IS21-558-Element
nachgewiesen (MANKIN et al. 2006). Leider lagen keine Informationen darüber vor,
ob es sich bei dem Plasmid des humanen MRSA-Isolates um einen der bekannten

RESULTATE UND DISKUSSION
103
cfr-tragenden Plasmidtypen pSCFS1, pSCFS3 oder pSCFS6 handelt (Publikationen
12, 15, 16).
Trotzdem war dies ein sehr bedeutendes Ergebnis, da erstmalig der Nachweis eines
cfr-Multiresistenzgens bei einem Isolat vom Menschen geführt wurde. Auch wenn
primär an einen Transfer des Gens vom Tier zum Menschen gedacht werden muss,
kann ebenso eine Selektion beim Menschen durch die Anwendung von Linezolid
oder einer Streptogramin-Kombination stattgefunden haben. Immerhin wurden in
Deutschland innerhalb von 17 Jahren nur sechs cfr-tragende Isolate animaler
Herkunft isoliert (Publikation 15), was zumindest für eine geringe Prävalenz des
Gens bei animalen Staphylokokken spricht. Diese geringe Prävalenz mag ihre
Ursache in dem vergleichsweise geringen Selektionsdruck durch PhLOPSA-
Wirkstoffe haben. Von den relevanten Substanzen sind lediglich Lincosamide für
mehrere Indikationen bei Haus- und Heimtieren zugelassen. Florfenicol und
Pleuromutiline sind dagegen nur für wenige Anwendungen beim Rind und / oder
Schwein zugelassen und die Anwendung von Chloramphenicol ist bereits seit 1994
für lebensmittelliefernde Tiere verboten. Auch wenn ehemals eine Zulassung von
Virginiamycin oder Streptograminen als Leistungsförder vorlag, so sind diese
Wirkstoffe genau wie Oxazolidinone in der Europäischen Union mittlerweile dem
humanmedizinischen Gebrauch vorbehalten. Daher kann retrospektiv über die
Herkunft des cfr-Gens beim Menschen nur spekuliert werden. Leider lagen auch
keine Informationen darüber vor, ob der kolumbianische Patient Kontakt mit Tieren
hatte oder nicht. Ein eindeutiger Nachweis, der die Herkunft des Gens vom Tier
belegt, ist für dieses Multiresistenzgen nicht möglich.
Aufgrund des breiten cfr-vermittelten Resistenzphänotyps kann eine auf diesem Gen
basierende Florfenicolresistenz auch nicht als Modellbeispiel für den Transfer eines
Resistenzgens vom Tier zum Menschen dienen. Ein solches Modellbeispiel würde
voraussetzen, dass der auf die Bakterien ausgeübte Selektionsdruck zur
Wirkstoffresistenz ausschließlich beim Tier aufgebaut wird. Diese Möglichkeit
bestand bei der ausschließlich beim Tier eingesetzten Substanz Nurseothricin
(WITTE 1997), ist aber für den PhLOPSA- Phänotyp nicht gegeben.

RESULTATE UND DISKUSSION
104
Im Gegensatz dazu vermittelt das zweite neue Resistenzgen, fexA, lediglich
kombinierte Phenicolresistenz. Da Florfenicol nie für humanmedizinische
Anwendungen zur Verfügung stand und Chloramphenicol eine äußerst
eingeschränkte humanmedizinische Anwendung erfährt, wäre das Auftreten dieses
Gens beim Menschen eindeutiger auf eine Selektion beim Tier zurückzuführen.
Bislang konnte das Gen aber nicht bei humanen Isolaten nachgewiesen werden und
das einzige florfenicolresistente Isolat vom Menschen aus Deutschland trug weder
cfr noch fexA (Publikation 15).
Da nur sehr wenige neue Klassen antimikrobieller Wirkstoffe in den letzten Jahren
auf den Markt gekommen sind und die Cfr-Methylase Resistenz gegenüber fünf
unterschiedlichen Wirkstoffklassen vermittelt, sollte dem Auftreten und der
Verbreitung des entsprechenden Gens zukünftig größere Beachtung geschenkt
werden. Immerhin wurden bei Staphylokokken MHK-Werte Cfr-tragender Stämme
festgestellt, die - sofern Grenzwerte für die Wirkstoffe verfügbar sind - eine
Einstufung als resistent oder gerade noch empfindlich („borderline susceptible“)
zuließen (CLSI 2007). Zudem ist die Cfr-Methylase das einzige nachgewiesene
Protein, welches Oxazolidinon- und Pleuromutilinresistenz vermittelt. Sollte eine
Verbreitung ähnlich den Erm-Methyltransferasen stattfinden
(http://faculty.washington.edu/marilynr/), die lediglich Resistenz gegenüber
Makroliden, Lincosamiden und Streptogramin B-Antibiotika vermitteln, so würde sich
eine beängstigende Vorstellung ergeben. Die Verbreitung des cfr-Gens wird durch
die Lokalisation auf Plasmiden und auf einem mobilen genetischen Element
begünstigt. Da das Gen mittlerweile auch bei Isolaten aus Dänemark (Publikation 16)
und Kolumbien (MANKIN et al. 2006) aufgetreten ist, muß zukünftig mit einer
weiteren Verbreitung gerechnet werden. Verdächtige Isolate sollten daher stets auf
die Präsenz von cfr untersucht werden.

ZUSAMMENFASSUNG
105
5 Zusammenfassung In der vorliegenden Studie wurden die genetischen Grundlagen der
Florfenicolresistenz bei grampositiven und gramnegativen Bakterien erarbeitet. Da
bislang nur sehr wenige Daten zur Florfenicolresistenz bekannt waren, wurden die
Arbeitsschwerpunkte in drei Bereiche eingeteilt:
• In dem ersten Arbeitsbereich wurden bovine und porcine Erreger von
Atemwegsinfektionen, die als Zielorganismen für den Wirkstoff definiert sind, auf
ihre Empfindlichkeitslage gegenüber Florfenicol im Rahmen eines
Monitoringprogramms untersucht. Dabei zeigte sich, dass seit der Zulassung
des Wirkstoffes die MHK50- und MHK90-Werte bei Isolaten aus Deutschland
nahezu stabil geblieben sind und keine Resistenzentwicklung zu beobachten
war. Alle getesteten Bakterien der Spezies Pasteurella multocida, Mannheimia
haemolytica, Actinobacillus pleuropneumoniae und Streptococcus suis konnten
als empfindlich gegenüber dem Wirkstoff eingestuft werden. Lediglich bei
Bordetella bronchiseptica wurde ein geringer Anteil (0,8 %) resistenter Isolate
ermittelt. Da in anderen europäischen Ländern erste florfenicolresistente
Pasteurella multocida-Isolate auftraten, wurde die genetische Grundlage dieser
Resistenzeigenschaft untersucht. Sie basierte bei diesen Isolaten auf der
Präsenz des Exportergens floR, welches auf unterschiedlichen Plasmiden vom
Typ pCCK381 und pCCK13698 nachgewiesen wurde. Während auf dem
Plasmidtyp pCCK381 keine weiteren Resistenzgene lokalisiert waren, wurden
auf dem zweiten Plasmidtyp sul2- und catA3-Gene identifiziert, so dass eine
Verbreitung der Florfenicolresistenz auch im Zuge von Co-Selektion möglich ist.
Plasmide vom Typ pCCK381 waren später auch bei florfenicolresistenten
Pasteurella multocida-Isolaten aus Deutschland nachzuweisen. Auch bei
Bordetella bronchiseptica vermittelten floR-Gene den untersuchten
Resistenzphänotyp, allerdings lagen sie chromosomal lokalisiert vor. Bei
Bordetella scheint außerdem ein noch nicht näher charakterisiertes Multidrug-
Effluxsystem an dieser Resistenzeigenschaft beteiligt zu sein. Zudem wurde ein

ZUSAMMENFASSUNG
106
neues Chloramphenicol-Resistenzgen, cmlB1, bei Bordetella bronchiseptica
identifiziert.
• Da bis zum Jahr 2003 keine florfenicolresistenten Zielorganismen aufgetreten
waren, befasste sich der zweite Arbeitsbereich mit Untersuchungen bei
Escherichia coli-Isolaten, die als gramnegative Indikatorkeime ausgewählt
wurden. Auch bei diesen Studien wurden floR-Effluxgene nachgewiesen, die auf
110 - 125 kb großen, konjugativen Multiresistenzplasmiden unterschiedlicher
Struktur lokalisiert waren. So können floR-Gene auch unter der Anwendung
anderer Substanzen wie Sulfonamiden, Trimethoprim, Ampicillin oder
Streptomycin co-selektiert werden. Das Vorkommen des Gens floR in
unterschiedlichen Lokalisationen und bei verschiedenen bakteriellen Spezies
konnte durch die Identifizierung des Gens als Bestandteil eines neuartigen,
funktionell aktiven Transposons, TnfloR, erklärt werden.
• In dem dritten Arbeitsbereich wurden Studien zur Florfenicolresistenz bei
Staphylococcus spp. durchgeführt. Für grampositive Bakterien lagen zu Beginn
der Studien keine Informationen zur genetischen Basis der Florfenicolresistenz
vor. Insgesamt wurden 302 Staphylokokken von Menschen und Tieren auf ihre
Florfenicolempfindlichkeit überprüft und resistente Isolate von Diagnostiklaboren
in Deutschland gesammelt. Von diesen Isolaten zeigten 12 einen MHK-Wert für
Florfenicol von ≥ 16 µg/ml, wobei die Resistenzeigenschaft bei neun der Isolate
plasmidlokalisiert vorlag. Unterschiedliche Plasmidtypen wurden identifiziert und
näher charakterisiert. Dabei gelang der Nachweis der beiden ersten und bislang
einzigen Florfenicol-Resistenzgene, fexA und cfr, bei grampositiven Bakterien.
Das Gen fexA kodiert ein Effluxprotein aus der Major-Facilitator-Superfamily,
welches sich mittels attenuierter Translation durch Chloramphenicol und
Florfenicol induzierbar exprimieren ließ. Das abgeleitete Protein zeigte kaum
Homologien zu bislang bekannten Effluxproteinen und stellt einen neuen Typ
eines Chloramphenicol- / Florfenicol-Exporterproteins dar. Es konnte
nachgewiesen werden, dass das fexA-Gen Bestandteil eines neuen

ZUSAMMENFASSUNG
107
Transposons aus der Tn554-Familie ist und die funktionelle Aktivität dieses
Transposons konnte experimentell aufgezeigt werden.
Das Gen cfr kommt auf einem Multiresistenzplasmid, pSCFS1, vor. Auf diesem
Plasmid wurden das bekannte Spectinomycin-Resistenzgen spc sowie weitere
neue Gene lsa(B) und erm(33) identifiziert, die eine „low-level“
Clindamycinresistenz bzw. Resistenz gegenüber Makroliden, Lincosamiden und
Streptogramin B-Antibiotika vermitteln. Das Gen cfr kodiert eine Methylase, die
eine zusätzliche Methylierung an der Position A2503 der 23S rRNA vornimmt. In
weiteren Untersuchungen zeigte sich, dass durch die Cfr-Methylase
Multiresistenz gegenüber fünf human- und tiermedizinisch wichtigen Klassen
antimikrobieller Wirkstoffe sowohl bei Staphylokokken als auch bei Escherichia
coli-Empfängerstämmen vermittelt wird. Dieser Resistenzphänotyp wurde mit
PhLOPSA bezeichnet und schließt reduzierte Empfindlichkeit gegenüber
Phenicolen, Lincosamiden, Oxazolidinonen, Pleuromutilinen und Streptogramin
A-Antibiotika ein. Damit gelang auch erstmalig der Nachweis von transferabler
Resistenz gegenüber Pleuromutilinen und Oxazolidinonen. Die Verteilung der
neuen Resistenzgene cfr und fexA wurde bei den untersuchten Staphylokokken
ermittelt und ihre Lokalisation im Chromosom oder auf Plasmiden identifiziert.
Bei diesen Untersuchungen gelang der Nachweis einer neuen
Insertionssequenz, IS21-558, die auf einem Plasmidtyp stromaufwärts des cfr-
Gens zu finden war. Später konnte anhand weiterer Isolate aus Dänemark
gezeigt werden, dass die Mobilität des cfr-Gens höchstwahrscheinlich durch
IS21-558-Elemente vermittelt wird. Ein Modell, welches zwei mögliche
Transpositionswege dieser IS21-558-Elemente zeigt, wurde erstellt und die
Funktionalität der Insertionssequenzen konnte aufgezeigt werden.
Eine Verbreitung des Multiresistenzgens cfr wird durch eine Lokalisation auf
Plasmiden und auf einem mobilen genetischen Element begünstigt. Das
Auftreten des Gens bei verschiedenen Staphylokokkenspezies von
unterschiedlichen Tierarten und aus verschiedenen Ländern lassen eine weitere
Verbreitung des Gens befürchten und unterstreichen die Wichtigkeit einer
molekularer Analyse von Resistenzgenen und ihrer Verbreitung.

LITERATURVERZEICHNIS
108
6 Literaturverzeichnis
AARESTRUP, F. M. (2000): Characterization of glycopeptide-resistant Enterococcus faecium (GRE) from broilers and pigs in Denmark: Genetic evidence that persistence of GRE in pig herds is associated with coselection by resistance to macrolides. J. Clin. Microbiol. 38, 2774-2777 AHMED, M., L. LYASS, P.N. MARKHAM, S. S. TAYLOR, N. VAZQUEZ-LASLOP u. A. A. NEYFAKH (1995): Two highly similar multidrug transporters of Bacillus subtilis whose expression is differentially regulated. J. Bacteriol. 177, 3904-3910 ALEXANDER, M., C.-J. ESTLER u. F. LEGLER (1995): Antibiotika und Chemotherapeutika. 2. Aufl. Wissenschaftliche Verlagsgemeinschaft, Stuttgart ALLIGNET, J. u. N. EL SOLH (1995): Diversity among the grampositive acetyltransferases inactivating streptogramin A and structurally related compounds and characterization of a new staphylococcal determinant vatB. Antimicrob. Agents Chemother. 39, 2027-2036 ALLIGNET, J., V. LONCLE, C.SIMENEL, M. DELPIERRE u. N. EL SOLH (1993): Sequence of a staphylococcal gene, vat, encoding an acetyltransferase inactivating the A-type compounds of virginiamycin-like antibiotics. Gene 130, 91-98 ANDERSON, L. M., T. M. HENKIN, G. H. CHAMBLISS u. K . F. BOTT (1984): New chloramphenicol resistance locus in Bacillus subtilis. J. Bacteriol. 158, 386-388 ANGELOS, J. A., E. L. DUEGER, L. W. GEORGE; T. K. C ARRIER, J. E. MIHALYI, S. B. COSGROVE u. J. C. JOHNSON (2000): Efficacy of florfenicol for treatment of naturally occurring infectious bovine keratoconjunctivitis. J. Am. Vet. Med. Assoc. 216, 62-64

LITERATURVERZEICHNIS
109
ANGEN, Ø., R. MUTTERS, D. A. CAUGANT, J. E. OLSEN u . M. BISGAARD (1999): Taxonomic relationships of the [Pasteurella] haemolytica complex as evaluated by DNA-DNA hybridizations and 16S rRNA sequencing with proposal of Mannheimia haemolytica gen. nov., comb. nov., Mannheimia granulomatis comb. nov., Mannheimia glucosida sp. nov., Mannheimia ruminalis sp. nov. and Mannheimia varigena sp. nov. . Int. J. Syst. Bacteriol. 49, 67-86 AQUAFLOR® PRODUCT LABELING SCHERING-PLOUGH - CANADA (2006): Zu erhalten bei: www.scheringploughanimalhealth.ca vom 18.09.2006 ARCANGIOLI, M. A., S. LEROY-SETRIN, J. L. MARTEL u. E. CHASLUS-DANCLA (1999): A new chloramphenicol and florfenicol resistance gene linked to an integron structure in Salmonella typhimurium DT104. FEMS Microbiol. Lett. 174, 327-332 ARIAS, C. A., M. V. VILLEGAS, J. REYES, J. MORENO, L. XIONG, M. VALLEJO, K. LOLANS, A. MANKIN, E. CASTANEDA u. J. P. QUINN ( 2006): Linezolid resistance in the absence of 23S rRNA mutations in methicillin-resistant Staphylococcus aureus. Abstracts of the 46th Interscience Conference on Antimicrobial Agents and Chemotherapy (ICAAC), San Francisco/USA, C2-1147, S. 125 AUBERT, D., L. POIREL, J. CHEVALIER, S. LEOTARD, J. M. PAGES u. P. NORDMANN (2001): Oxacillinase-mediated resistance to cefepime and susceptibility to ceftazidime in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 45, 1615-1620
AUGUSTIN, J, R. ROSENSTEIN, B. WIELAND, U. SCHNEIDE R, N. SCHNELL, G. ENGELKE, K. D. ENTIAN u. F. GOTZ (1992): Genetic analysis of epidermin biosynthetic genes and epidermin-negative mutants of Staphylococcus epidermidis. Eur. J. Biochem. 204, 1149-1154 BASTOS, M. C. u. E. MURPHY (1988): Transposon Tn554 encodes three products required for transposition. EMBO J. 7, 2935-2941 BAUCHERON, S., H. IMBRECHTS, E. CHASLUS-DANCLA u. A . CLOECKAERT (2004): The AcrB multidrug transporter plays a major role in high-level fluoroquinolone resistance in Salmonella enterica serovar Typhimurium phage type DT204. Microb. Drug Resist. 8, 281-289

LITERATURVERZEICHNIS
110
BAUCHERON, S., S. TYLER, D. BOYD, M. R. MULVEY, E. CHASLUS-DANCLA u. A. CLOECKAERT (2004): AcrAB-TolC directs efflux-mediated multidrug resistance in Salmonella enterica serovar typhimurium DT104. Antimicrob Agents Chemother. 48, 3729-3735 BAUGHMAN, G. A. u. S. R. FAHNESTOCK (1979): Chloramphenicol resistance mutation in Escherichia coli which maps in the major ribosomal protein gene cluster. J. Bacteriol. 137, 1315-1323 BEABER, J. W., B. HOCHHUT u. M. K. WALDOR (2002): Genomic and functional analyses of SXT an integrating antibiotic resistance gene transfer element derived from Vibrio cholerae. J. Bacteriol. 184, 4259-4269 BECKER K. (2004): Oxazolidinon-Resistenz. In: VON EIFF, C. u. K. BECKER (Hrsg.): Linezolid: Neue Daten nach Einführung in die Klinik. 1. Aufl. Socio-medico, Wessobrunn, S. 71-92 BERGER, B. u. D. HAAS (2001): Transposase and cointegrase: specialized transposition proteins of the bacterial insertion sequence IS21 and related elements. Cell Mol Life Sci. 58, 403-419 BISCHOFF K. M., D. G. WHITE, P. F. McDERMOTT, S. ZH AO, S. GAINES, J. J. MAURER u. D. J. NISBET (2002): Characterization of Chloramphenicol resistance in beta-haemolytic Escherichia coli associated with diarrhea in neonatal swine. J. Clin. Microbiol. 40, 389-394 BODMANN, K.-F. (2004): Antibiotikaeinsatz bei MRSA-Infektionen: Die Rolle von Linezolid. In: VON EIFF, C. u. K. BECKER (Hrsg.): Linezolid: Neue Daten nach Einführung in die Klinik. 1. Aufl. Socio-medio, Wessobrunn, S. 145-160 BÖCK, A., F. TURNOWSKY u. G. HÖGENAUER (1982): Tiamulin resistance mutations in Escherichia coli. J. Bacteriol. 151, 1253-1260 BOLTON, L. F., L. C. KELLEY, M. D. LEE, P. J. FEDOR KA-CRAY u. J. J. MAURER (1999): Detection of Salmonella typhimurium DT104 based on the gene St-flo which confers resistance to florfenicol and chloramphenicol. J. Clin. Microbiol. 37, 1348-1351

LITERATURVERZEICHNIS
111
BØSLING, S. M. POULSEN, B. VESTER u. K. S. LONG (20 03): Resistance to the peptidyl transferase inhibitor tiamulin caused by mutation of ribosomal protein L3. Antimicrob. Agents Chemother. 47, 2892-2896 BOYD, D., A. CLOECKAERT, E. CHASLUS-DANCLA u. M. R. MULVEY (2002): Characterization of variant Salmonella genomic island 1 multidrug resistance regions from serovars Typhimurium DT104 and Agona. Antimicrob. Agents Chemother. 46, 1714-1722 BOYD, D., G. A. PETERS, A. CLOECKAERT, K. SIDI BOUM EDINE, E. CHASLUS-DANCLA, H. IMBERECHTS u. M. R. MULVEY (2001): Complete nucleotide sequence of a 43-kilobase genomic island associated with the multidrug resistance region of Salmonella enterica serovar Typhimurium DT104 and its identification in phage type DT120 and serovar Agona. J. Bacteriol. 183, 5725-5732 BRANLANT, C., A. KROL, M. A. MACHATT, J. POUYET, J. P. EBEL, K. EDWARDS u. H. KOSSEL (1981): Primary and secondary structures of Escherichia coli MRE 600 23S ribosomal RNA. Comparison with models of secondary structure for maize chloroplast 23S rRNA and for large portions of mouse and human 16S mitochondrial rRNAs. Nucleic Acids Res. 9, 4303-4324 BRIGGS, C. E. u. P. M. FRATAMICO (1999): Molecular characterization of an antibiotic resistance gene cluster of Salmonella typhimurium DT104. Antimicrob. Agents Chemother. 43, 846-849 BUDAVARI, S. (1996): In: The Merck Index. An encyclopedia of chemicals, drugs, and biologicals. 12. Aufl. Whitehouse Station, NJ: Merck Research Laboratories, S. 4146 BURNS, J. L., L. A. HEDI u. D. M. LIEN (1989): Chloramphenicol resistance in Pseudomonas cepacia because of decreased permeability. Antimicrob. Agents Chemother. 33, 136-141 BURNS, JL., P. M. MENDELMAN, J. LEVY, T. L. STULL u . A. L. SMITH (1985): A permeability barrier as a mechanism of chloramphenicol resistance in Haemophilus influenzae. Antimicrob. Agents Chemother. 27, 46-54 CANNON, M., S. HARTFORD u. J. DAVIES (1990): A comparative study on the inhibitory actions of chloramphenicol, thiamphenicol, and some fluorinated derivatives. J. Antimicrob. Chemother. 26, 307-317

LITERATURVERZEICHNIS
112
CARATTOLI, A., F. TOSINI, W. P. GILES, M. E. RUPP, S. H. HINRICHS, F. J. ANGULO, T. J. BARRETT u. P. D. FEY (2002): Characterization of plasmids carrying CMY-2 from expanded-spectrum cephalosporin-resistant Salmonella strains isolated in the United States between 1996 and 1998. Antimicrob. Agents Chemother. 46, 1269-1272 CLOECKAERT, A., S. BAUCHERON u. E. CHASLUS-DANCLA ( 2001): Nonenzymatic chloramphenicol resistance mediated by IncC plasmid R55 is encoded by a floR gene variant. Antimicrob. Agents Chemother. 45, 2381-2382 CLOECKAERT, A., K. SIDI BOUMEDINE, G. FLAUJAC, H. I MBERECHTS, I. D’HOOGHE u. E. CHASLUS-DANCLA (2000): Occurrence of a Salmonella enterica serovar Typhimurium DT104-like antibiotic resistance gene cluster including the floR gene in S. enterica serovar Agona isolates. Antimicrob. Agents Chemother. 44, 1359-1361 CLINICAL AND LABORATORY STANDARDS INSTITUTE (CLSI) (2007): Performance standards for antimicrobial susceptibility testing; seventeenth informational supplement. CLSI document M100-S17. CLSI, Wayne, PA, USA, 2007. DESOMER, J., D. VEREECKE, M. CRESPI u. M. VAN MONTA GU (1992): The plasmid-encoded chloramphenicol-resistance protein of Rhodococcus fascians is homologous to the transmembrane tetracycline efflux proteins. Mol. Microbiol. 6, 2377-2385 DITTRICH, W., M. BETZLER u. H. SCHREMPF (1991): An amplifiable and deletable chloramphenicol-resistance determinant of Streptomyces lividans 1326 encodes a putative transmembrane protein. Mol. Microbiol. 5, 2789-2799 DORMAN, C. J. u. T. J. FOSTER (1982): Nonenzymatic chloramphenicol resistance determinants specified by plasmids R26 and R55-1 in Escherichia coli K-12 do not confer high-level resistance to fluorinated analogs. Antimicrob. Agents Chemother. 22, 912-914 DOUBLET, B., R. LAILLER, D. MEUNIER, A. BRISABOIS, D. BOYD, M. R. MULVEY, E. CHASLUS-DANCLA u. A. CLOECKAERT (2003): Variant Salmonella genomic island 1 antibiotic resistance gene cluster in Salmonella enterica serovar Albany. Emerg. Infect. Dis. 9, 585-591

LITERATURVERZEICHNIS
113
DOUBLET, B., S. SCHWARZ, E. NUßBECK, S. BAUCHERON, J.-L. MARTEL, E. CHASLUS-DANCLA u. A. CLOECKAERT (2002): Molecular analysis of chromosomally florfenicol-resistant Escherichia coli isolates from France and Germany. J. Antimicrob. Chemother. 49, 49-54 DOUBLET, B., F. X. WEILL, L. FABRE, E. CHASLUS-DANC LA u. A. CLOECKAERT (2004): Variant Salmonella genomic Island 1 antibiotic resistance gene cluster containing a novel 3'-N-aminoglycoside acetyltransferase gene cassette, aac(3)-Id, in Salmonella enterica Serovar Newport Antimicrob. Agents Chemother. 48, 3806-3812 DOUTHWAITE, S. (1992): Interaction of the antibiotic clindamycin and lincomycin with Escherichia coli 23S ribosomal RNA. Nucleic Acids Res. 20, 4717-4720 DUEGER E. L., J. A. ANGELOS, S. COSGROVE, J. JOHNSO N u. L. W. GEORGE (1999): Efficacy of florfenicol in the treatment of experimentally induced infectiousbovine keratoconjunctivitis. Am. J. of Vet. Res. 60; 960-964 EDGAR, R. u. E. BIBI (1997): MdfA, an Escherichia coli multidrug resistance protein with an extraordinarily broad spectrum of drug recognition. J. Bacteriol. 179, 2274-2280 EMEA - THE EUROPEAN AGENCY FOR THE EVALUATION OF ME DICINAL PRODUCTS (1995): Committee for veterinary medicinal products: Florfenicol - Summary report (1) (http://www.emea.europa.eu/pdfs/vet/mrls/florfenicol.pdf) 7 Westferry circus, Canary Wharf, London, E14 4HB, UK EMEA - THE EUROPEAN AGENCY FOR THE EVALUATION OF ME DICINAL PRODUCTS (1997): Committee for veterinary medicinal products: Florfenicol (Extension to fish) - Summary report (2) (http://www.emea.europa.eu/pdfs/vet/mrls/025197en.pdf) 7 Westferry circus, Canary Wharf, London, E14 4HB, UK EMEA - THE EUROPEAN AGENCY FOR THE EVALUATION OF ME DICINAL PRODUCTS (1999): Committee for veterinary medicinal products: Florfenicol (Extension to chicken) - Summary report (3) (http://www.emea.europa.eu/pdfs/vet/mrls/058999en.pdf) 7 Westferry circus, Canary Wharf, London, E14 4HB, UK

LITERATURVERZEICHNIS
114
EMEA - THE EUROPEAN AGENCY FOR THE EVALUATION OF ME DICINAL PRODUCTS (1999a): Committee for veterinary medicinal products: Florfenicol (Extension to pigs) - Summary report (4) (http://www.emea.europa.eu/pdfs/vet/mrls/059199en.pdf) 7 Westferry circus, Canary Wharf, London, E14 4HB, UK EMEA - THE EUROPEAN AGENCY FOR THE EVALUATION OF ME DICINAL PRODUCTS (2000): Committee for veterinary medicinal products: Florfenicol (Extension to fish) - Summary report (5) (http://www.emea.europa.eu/pdfs/vet/mrls/076000en.pdf) 7 Westferry circus, Canary Wharf, London, E14 4HB, UK EMEA - THE EUROPEAN AGENCY FOR THE EVALUATION OF ME DICINAL PRODUCTS (2002): Committee for veterinary medicinal products: Florfenicol (Extension to all food producing species) - Summary report (6) (http://www.emea.europa.eu/pdfs/vet/mrls/082202en.pdf) 7 Westferry circus, Canary Wharf, London, E14 4HB, UK ETTAYEBI, M., S. M. PRASAD u. E. A. MORGAN (1985): Chloramphenicol-erythromycin resistance mutations in a 23S rRNA gene of Escherichia coli. J. Bacteriol. 162, 551-557 ESSEX TIERARZNEI: FACHINFORMATION (2006): Nuflor® Lösung zur Injektion. Für Tiere (Rinder). Ndl. der essex pharma GmbH, Thomas-Dehler-Strasse 27, 81737 München FLUIT, A. C., C. L. C. WIELDERS, J. VERHOEF u. F.-J . SCHMITZ (2001): Epidemiology and susceptibility of 3,051 Staphylococcus aureus isolates from 25 university hospitals participating in the European SENTRY study. J. Clin. Microbiol. 39, 3727-3732 FRANKLIN, T. J. u. G. A. SNOW (1998): Biochemistry and molecular biology of antimicrobial action. 5. Aufl. Kluwer Academic Publishers, Dordrecht, Boston, London FUKUI, H., Y. FUJIHARA u. T. KANO (1987): In vitro and in vivo antibacterial activities of florfenicol, a new fluorinated analog of thiamphenicol, against fish pathogens. Fish Pathol. 22, 201-207 GEORGE, A. M. u. R. M. HALL (2002): Efflux of chloramphenicol by the CmlA1 protein. FEMS Microbiol. Lett. 209, 209-213

LITERATURVERZEICHNIS
115
GOOSENS L. (1999): Fünf Jahre Nuflor® (Florfenicol), Rückblick, Ausblick. Abstractband des Essex Tierarznei Symposiums zum Themenkomplex Rinderproduktion/Rinderkrankheiten. München, 7. Mai 1999 GRAHAM, R., D. PALMER, B. C. PRATT u. C. A. HART (1 988): In vitro activity of florpheniol. Eur. J. Clin. Micorbiol. Infect. Dis. 7, 691-694 GUTIÉRREZ-MARTĺN, C. B., N. GARCIA DEL BLANCO, M. BLANCO, J. NAVAS u. E. F. RODRĺGUEZ-FERRI (2006): Changes in antimicrobial susceptibility of Actinobacillus pleuropneumoniae isolated from pigs in Spain durino the last decade. Vet. Micro. 115, 218-222 HAAS DE, V., M. BONNIER, M. GICQUEL, F. ETORE u. D. SHUSTER (2002): Florfenicol: a time- or concentration-dependent antibiotic? In: Schering-Plough animal health: New advances in calf disease management Proceedings of a symposium held in conjunction with the XXII World Buiatrics Congress, Hannover HARMS, J., F. SCHLUENZEN, R. ZARIVACH, A. BASHAN, S . GAT, I. AGMON, H. BARTELS, F. FRANCESCHI u. A. YONATH (2001): High resolution structure of the large ribosomal subunit from a mesophilic eubacterium. Cell 107, 679-688 HAROCHE, J., J. ALLIGNET u. N. EL SOLH (2002): Tn5406, a new staphylococcal transposon conferring resistance to streptogramin A and related compounds including dalfopristin. Antimicrob. Agents Chemother. 46, 2337-2343 HEIDELBERG, J. F., J. A. EISEN, W. C.NELSON, R. A.C LAYTON, M. L. GWINN, R. J. DODSON, D. H. HAFT, E. K. HICKEY, J. D. PETER SON, L. UMAYAM, S. R. GILL, K. E. NELSON, T. D. READ, H., TETTELIN, D. RI CHARDSON, M. D. ERMOLAEVA, J. VAMATHEVAN, S. BASS, H. QIN, , I. DRA GOI, P. SELLERS, L. McDONALD, T. UTTERBACK, R. D. FLEISHMANN, W. C. NIE RMAN u. O. WHITE (2000): DNA sequence of both chromosomes of the cholera pathogen Vibrio cholerae. Nature 40, 6477-6483 HO S.-P., T.-Y. HSU, M.-H. CHEN u. W.-S. WANG (2000): Antibacterial effect of chloramphenicol, thiamphenicol and florfenicol against aquatic animal bacteria. J. Vet. Med. Sci. 62, 479-485

LITERATURVERZEICHNIS
116
HOCHHUT, B., Y. LOTFI, D. MAZEL, S. M. FARUQUE, R. WOODGATE u. M. K. WALDOR (2001): Molecular analysis of antibiotic resistance gene clusters in Vibrio cholerae O139 and O1 SXT constins. Antimicrob. Agents Chemother. 45, 2991-3000 HÖRMANSDORFER, S. u. J. BAUER (1996): Zur Resistenz boviner und porciner Pasteurellen gegenüber Florfenicol und anderen Antibiotika. Berl. Münch. Tierärztl. Wschr. 111, 422-426 HÖRMANSDORFER, S. u. J. BAUER (1996): Zur Resistenzsituation boviner Pasteurellen. Berl. Münch. Tierärztl. Wschr. 109, 168-171 HORINOUCHI, S. u. B. WEISBLUM (1982): Nucleotide sequence and functional map of pC194, a plasmid that specifies inducible chloramphenicol resistance. J. Bacteriol. 150, 815-825 HUMMEL, R., H. TSCHÄPE u. W. WITTE (1986): Spread of plasmid mediated nourseothricin resistance due to antibiotic use in animal husbandry. J. Basic Microbiol. 26, 461-466 KADLEC, C., C. KEHRENBERG u. S. SCHWARZ (2005): Molecular basis of resistance to trimethoprim, chloramphenicol and sulphonamides in Bordetella bronchiseptica. J. Antimicrob. Chemother. 56, 485-490 KADLEC, C., C. KEHRENBERG u. S. SCHWARZ (2006): tet(A)-mediated tetracycline resistance in porcine Bordetella bronchiseptica isolates is based on plasmid-borne Tn1721 relics. J. Antimicrob. Chemother. 58, 225-227 KEHRENBERG, C., S. A. SALMON, J. L. WATTS u. S. SCH WARZ (2001): Tetracycline resistance genes in isolates of Pasteurella multocida, Mannheimia haemolytica, Mannheimia glucosida and Mannheimia varigena from bovine and swine respiratory disease: intergeneric spread of the tet(H) plasmid pMHT1. J. Antimicrob. Chemother. 48, 631-640 KEHRENBERG, C. u. S. SCHWARZ (2000): Identification of a truncated, but functionally active tet(H) tetracycline resistance gene in Pasteurella aerogenes and Pasteurella multocida. FEMS Microbiol. Lett. 188, 191-195

LITERATURVERZEICHNIS
117
KEHRENBERG, C. u. S. SCHWARZ (2001): Ocurrence and linkage of genes coding for resistance to sulfonamides, streptomycin and chloramphenicol in bacteria of the genera Pasteurella and Mannheimia. FEMS Microbiol. Lett. 205, 283-290 KEHRENBERG, C. u. S. SCHWARZ (2002): Nucleotide sequence and organization of plasmid pMVSCS1 from Mannheimia varigena: identification of a multiresistance gene cluster. J. Antimicrob. Chemother. 49, 383-386 KEHRENBERG, C. u. S. SCHWARZ (2005): dfrA20, a novel trimethoprim resistance gene from Pasteurella multocida. Antimicrob. Agents Chemother. 49, 414-417 KEHRENBERG, C. u. S. SCHWARZ (2005a): Antibiotika und Resistenzentstehung. In: EIKMANN, T., B. CHRISTIANSEN, M. EXNER, C. HERR u. A. KRÄMER (Hrsg.): Hygiene in Krankenhaus und Praxis. 3. Aufl. Ecomed Medizin, Landsberg, Kap. 4.4.2, S. 1-32 KEHRENBERG, C., C. WERCKENTHIN u. S. SCHWARZ (1998) : Tn5706, a transposon-like element from Pasteurella multocida mediating tetracycline resistance. Antimicrob. Agents Chemother. 42, 2116-2118 KEYES, K., C. HUDSON, J. J. MAURER, S. THAYER, D. G . WHITE u. M. D. LEE (2000): Detection of florfenicol resistance genes in Escherichia coli isolated from sick chickens. Antimicrob. Agents Chemother. 44, 421-424 KIM, E. u. T. AOKI (1993): Drug resistance and broad geographical distribution of identical R plasmids of Pasteurella piscicida isolated from cultured yellowtail in Japan. Microbiol. Immunol. 37, 103-109 KIM, E. u. T. AOKI (1996): Sequence analysis of the florfenicol resistance gene encoded in the transferable R plasmid from a fish pathogenPasteurella piscicida. Microbiol. Immunol. 40, 665-669 KIM, E. u. T. AOKI (1996a): Sulfonamide resistance gene in a transferable R plasmid of Pasteurella piscicida. Microbiol. Immunol. 40, 379-399

LITERATURVERZEICHNIS
118
KIM, E. H., T. YOSHIDA u. T. AOKI (1993): Detection of R plasmid encoded with resistance to florfenicol in Pasteurella piscicida. Fish Pathol. 28, 165-170 KOWALAK, J. A., E. BRUENGER u. J. A. McCLOSKEY (199 5): Posttranscriptional modifications of the central loop of domain V in Escherichia coli 23S ribosomal RNA. J. Biol. Chem. 270, 17758-17764 KÜHN, T. u. L. GOOSENS (1998): Resistenzsituation boviner Pasteurella-Isolate gegen Florfenicol. Abstractband des 15. Jenaer Symposiums “Respiratorische Erkrankungen des Kalbes und Zoonosen der Wiederkäuer”. Jena, 26.-27. Mai 1998 LOMOVSKAYA O., M. S. WARREN, A. LEE, J. GALAZZO, R. FRONKO, M. LEE, J. BLAIS, D. CHO, S. CHAMBERLAND, T. RENAU, R. LEGE R, S. HECKER, W. WATKINS,K. HOSHINO, H. ISHIDA u. V. J. LEE (2001): Identification and characterization of inhibitors of multidrug resistance efflux pumps in Pseudomonas aeruginosa: novel agents for combination therapy. Antimicrob. Agents Chemother. 45, 105-116 LOVETT, P. S. (1990): Translational attenuation as the regulator of inducible cat genes. J. Bacteriol. 172, 1-6 MAHILLON, J. u. M. CHANDLER (1998): Insertion sequences. Microbiol. Mol. Biol. Rev. 62, 725-774 MANKIN, A. S., S. M. TOH, L. XIONG, K. LOLANS u. J. P. QUINN (2006): New mechanism of linezolid resistance in MRSA based on posttranscriptional modification of ribosomal RNA. Abstracts of the 46th Interscience Conference on Antimicrobial Agents and Chemotherapy (ICAAC), San Francisco/USA, C1-942, S. 80 MARIE, J., H. MORVAN, F. BERTHEELOT-HÉRAULT, P. SAN DERS, I. KEMPF, A. V. GAUTIER-BOUCHARDON, E. JOUY u. M. KOBISCH (2002) : Antimicrobial susceptibility of Streptococcus suis isolated from swine in France and from humans in different countries between 1996 and 2000. J. Antimicrob. Chemother. 50, 201-209

LITERATURVERZEICHNIS
119
MARSHALL, S. A., R. N. JONES, A. WANGER, J. A. WASH INGTON, G. V. DOERN, A. L. LEBER u. T. H. HAUGEN (1996): Proposed MIC quality control guidelines for National Committee for Clinical Laboratory Standards susceptibility tests using seven veterinary antimicrobial agents: ceftiofur, enrofloxacin, florfenicol, penicillin G-novobiocin, pirlimycin, premafloxacin, and spectinomycin. J. Clin. Microbiol. 34, 2027-2029 MARTELO, O. J., D. R. MANYAN, U. S. SMITH u. A. A. YUNIS (1964): Chloramphenicol and bone marrow mitochondria. J. Lab. Clin. Med. 74, 927-942 MARTĺN DE LA FUENTE, A. J., A. W. TUCKER, J. NAVAS, M. B LANCO, S. J. MORRIS u. C. B. GUTIÉRREZ-MART ĺN (2006): Antimicrobial susceptibility patterns of Haemophilus parasuis from pigs in the United Kingdom and Spain. Vet. Micro. 120, 184-191 MASCARETTI, O. A. (2003): Bacteria versus antimicrobial agents, an integrated approach. ASM Press, Washington DC, USA MEKA, V. G. u. H. S. GOLD (2004): Antimicrobial resistance to linezolid. Clin. Infect. Dis. 39, 1010-1015
MEUNIER, D., S. BAUCHERON, E. CHASLUS-DANCLA, J. L. MARTEL u. A. CLOECKAERT (2003): Florfenicol resistance in Salmonella enterica serovar Newport mediated by a plasmid related to R55 from Klebsiella pneumoniae. J. Antimicrob. Chemother. 51, 1007-1009 MEUNIER, D., D. BOYD, M. R. MULVEY, S. BAUCHERON, C . MAMMINA, A. NASTASI, E. CHASLUS-DANCLA u. A. CLOECKAERT (2002): Salmonella enterica serotype Typhimurium DT104 antibiotic resistance genomic island I in serotype Paratyphi B. Emerg. Infect. Dis. 8, 430-433 MEVIUS, D. J. u. E. G. HARTMAN (2000): In vitro activiteit van 12 veterinair gebruikte antibacteriële midedelen tegen Mannheimia haemolytica en Pasteurella multocida geïsoleerd uit kalveren in Nederland (In vitro activity of twelve antibacterial drugs against Mannheimia haemolytica and Pasteurella multocida isolated from Dutch calves). Tijdschr. Diergeneeskd. 125, 147-152

LITERATURVERZEICHNIS
120
MOAZED D. u. H. F. NOLLER (1987): Chloramphenicol, erythromycin, carbomycin, and vernamycin B protect overlapping sites in the peptidyl transferase region of 23S rRNA. Biochimie 69, 879-884 MOSHER, R. H., D. J. CAMP, K. YANG, M. P. BROWN, W. V. SHAW u. L. C. VINING (1995): Inactivation of chloramphenicol by O-phosphorylation. J. Biol. Chem. 27, 27000-27006 MOLLET, B., S. IIDA, J. SHEPHERD u. W. ARDER (1983) : Nucleotide sequence of IS26, a new prokaryotic mobile genetic element. Nucleic Acids Res. 11, 6319-6330 MOSHER, R. H., N. P. RANADE, H. SCHREMPF u. L. C. V INING (1990): Chloramphenicol resistance in Streptomyces, cloning and characterization of a chloramphenicol hydrolase gene from Streptomyces venezuelae. J. Gen. Microbiol. 136, 293-301 MURPHY, E. (1985): Nucleotide sequence of ermA, a macrolide-lincosamide-streptogramin B determinant in Staphylococcus aureus. J. Bacteriol. 162, 633-640 MURPHY, E. (1989): Transposable elements in gram-positive bacteria. In: BERG, D. E. u. M. M. HOWE (Hrsg.): Mobile DNA. ASM Press, Washington D.C.; S. 269-288
MURPHY, E. (1990): Properties of the site-specific transposable element Tn554. In: NOVICK, R. P. (Hrsg.): Molecular biology of the staphylococci. VCH Publishers, New York, N.Y. S. 123-135 MURRAY, I. A. (2000): Chloramphenicol resistance. in: FISHETTI, V. A., R. P. NOVICK, J. J. FERRETTI, D. A. PORTNOY, J. I. ROOD (Hrsg.): Gram-positive pathogens. ASM Press, Washington, D.C. S. 678-684 MURRAY, I. A. u. W. V. SHAW (1997): O-Acetyltransferases for chloramphenicol and other natural products. Antimicrob. Agents Chemother. 41, 1-6 NAGY, I., G. SCHOOFS, J. VANDERLEYDEN u. R. DE MOT (1997): Transposition of the IS21-related element IS1415 in Rhodococcus erythropolis. J. Bacteriol. 179, 4635-4638

LITERATURVERZEICHNIS
121
NATIONAL COMMITTEE FOR CLINICAL LABORATORY STANDARD S (NCCLS) (2002): Performance standards for antimicrobial disk and dilution susceptibility tests for bacteria isolated from animals; Approved standard - 2nd edition. NCCLS document M31-A2. NCCLS, 940 West Valley Road, Suite 1400, Wayne, PA 19087, U.S.A. NATIONAL COMMITTEE FOR CLINICAL LABORATORY STANDARD S (NCCLS) (2004): Performance standards for antimicrobial disk and dilution susceptibility tests for bacteria isolated from animals; Informational Supplement, NCCLS document M31-S1. NCCLS, 940 West Valley Road, Suite 1400, Wayne, PA 19087, U.S.A. ORDEN, J. A., J. A. RUIZ-SANTA-QUITERIA, S. GARCIA, D. CID u. R. DE LA FUENTE (2000): In vitro susceptibility of Escherichia coli strains isolated from diarrhoeic dairy calves to 15 antimicrobial agents. J. Vet. Med. B 47, 329-335 PARKHILL, J., M. SEBAIHIA, A. PRESTON, L. D. MURPHY , N. THOMSON, D. E. HARRIS, M. T. HOLDEN, C. M. CHURCHER, S. D. BENTLEY , K. L. MUNGALL, A. M. CEREDENO-TARRAGA, L. TEMPLE, K. JAMES, B. HARRIS , M. A. QUAIL, M. ACHTMAN, R. ATKIN, S. BAKER, D. BASHIAM, N. BASON, I. CHEREVACH, T. CHILLINGWORTH, M. COLLINS, A. CRONIN, P. DAVIS, J. DOGGETT, T. FELTWELL, A. GOBLE, N. HAMLIN, H. HAUSER, S. HOLROY D, K. JAGELS, S. LEATHER, S. MOULE, H. NORBERCZAK, S. O´NEIL, D. ORM OND, C. PRICE, E. RABBINOWITSCH, S. RUTTER, M. SANDERS, D. SAUNDERS, K. SEEGER, S. SHARP, M. SIMMONDS, J. SKELTON, R. SQUARES, S. SQUA RES, K. STEVENS, L. UNWIN, S. WHITEHEAD, B. G. BARRELL u. D. J. MASK ELL (2003): Comparative analysis of the genome sequence of Bordetella pertussis, Bordetella parapertussis and Bordetella bronchiseptica. Nat. Genet. 35, 32-40 PARTRIDGE, S. R., G. D. RECCHIA, H. W. STOKES u. R. M. HALL (2001): Family of class 1 integrons related to In4 from Tn1696. Antimicrob. Agents Chemother. 45, 3014-3020 PAULSEN, I. T., M. H. BROWN u. R. A. SKURRAY (1996) : Proton-dependent multidrug efflux systems. Microbiol. Rev. 60, 575-608 PLOY, M.-C., P. COURVALIN u. R. LAMBERT (1998): Characterization of In40 of Enterobacter aerogenes BM2688a class 1 integron with two new gene cassettes, cmlA2 and qacF. Antimicrob. Agents Chemother. 42, 2557-2563

LITERATURVERZEICHNIS
122
POIREL, L., J. W. DECOUSSER u. P. NORDMANN (2003): Insertion sequence ISEcp1B is involved in expression and mobilization of a bla (CTX-M) beta-lactamase gene. Antimicrob. Agents Chemother. 47, 2938-2945 POIREL, L., I. LE THOMAS, T. NAAS, A. KARIM u. P. N ORDMANN (2000): Biochemical sequence analyses of GES-1a novel class A extended-spectrum beta-lactamase and the class 1 integron In52 from Klebsiella pneumoniae. Antimicrob. Agents Chemother. 44, 622-632 POIREL, L., T. NAAS, M. GUIBERT, E. B. CHAIBI, R. L ABIA u. P. NORDMANN (1999): Molecular and biochemical characterization of VEB-1a novel class A extended-spectrum beta-lactamase encoded by an Escherichia coli integron gene. Antimicrob. Agents Chemother. 43, 573-581 POOLE, K. (2002): Multidrug efflux pumps and antimicrobial resistance in Pseudomonas aeruginosa and related organisms. in: PAULSEN, I. T. u. K. LEWIS (Hrsg.): Microbial multidrug efflux. Horizon Scientific Press, Wymondham, UK. S. 273-298 PRIEBE, S. (2003): Untersuchungen zur in vitro Empfindlichkeit boviner und porciner Erreger von Infektionen des Respirationstraktes gegenüber Florfenicol. Hannover, Tierärztl. Hochsch., Diss. PRIEBE, S. u. S. SCHWARZ (2003): In vitro activities of florfenicol against bovine and porcine respiratory tract pathogens. Antimicrob. Agents Chemother. 47, 2703-2705 PRINGLE, M., J. POEHLSGAARD, B. VESTER u. K. S. LON G (2004): Mutations in ribosomal protein L3 and 23S ribosomal RNA at the peptidyl transferase centre are associated with reduced susceptibility to tiamulin in Brachyspira spp. Isolates. Mol. Microbiol. 54, 1295-1306 PROJAN, S. J., J. KORNBLUM, S. L. MOGHAZEH, I. EDEL MAN, M. L. GENNARO u. R. P. NOVICK (1985) Comparative sequence and functional analysis of pT181 and pC221, cognate plasmid replicons from Staphylococcus aureus. Mol. Gen. Genet. 199, 452-464 QUINTILIANI JR., R., D. F. SAHM u. P. COURVALIN (19 99): Mechanisms of resistance to antimicrobial agents. in: MURRAY, P. R., E. J. BARON, M. A. PFALLER, F. C. TENOVER, R. H. YOLKEN (Hrsg.): Manual of Clinical Microbiology. 7. Aufl. ASM Press, Washington D.C. S. 1505-1525

LITERATURVERZEICHNIS
123
RADOSTITS, O. M., C. C. GAY, D. C. BLOOD u. K. W. H INCHCLIFF (2000): A textbook of the diseases of cattle, sheep, pigs, goats, and horses. 9. Aufl. Saunders, Philadelphia, London, S. 829-867 REIMMANN, C. u. D. HAAS (1990): The istA gene of insertion sequence IS21 is essential for cleavage at the inner 3' ends of tandemly repeated IS21 elements in vitro. EMBO J. 9, 4055-4063 RENDE-FOURNIER, R., R. LECLERCQ, M. GALIMANT, J. DU VAL u. P. COURVALIN (1993): Identification of the satA gene encoding a streptogramin A acetyltransferase in Enterococcus faecium BM4145. Antimicrob. Agents Chemother. 37, 1896-1903 RICCIO, M. L., J. D. DOCQUIER, E. DELL'AMICO, F. LU ZZARO, G. AMICOSANTE u. G. M. ROSSOLINI (2003): Novel 3-N-aminoglycoside acetyltransferase gene, aac(3)-Ic, from a Pseudomonas aeruginosa integron. Antimicrob. Agents Chemother. 47, 1746-1748 ROBERTS, M. (1996): Tetracycline resistance determinants: mechanisms of action, regulation of expression, genetic mobility, and distribution. FEMS Microbiology Reviews 22, 1-20 RODGERS, E. J., M. S. RAHMAN, R. T. HILL u. P. S. L OVETT (2002): The chloramphenicol-inducible catB gene in Agrobacterium tumefaciens is regulated by translational attenuation. J. Bacteriol. 184, 4296-4300 ROSIN, H. (1992): Antibiotika und Chemoterapeutika. Antiinfektiöse Therapie. in: FORTH, W., D. HENSCHLER, W. RUMMEL, K. STARKE (Hrsg.): Allgemeine und spezielle Pharmakologie und Toxikologie. 6. Aufl. BI Wissenschaftsverlag, Mannheim, Leipzig, S. 613-721 ROWE-MAGNUS, D. A., A. M GUEROUT u. D. MAZEL (2002) : Bacterial resistance evolution by recruitment of super-integron gene cassettes. Mol. Microbiol. 43, 1657-1669 RUBENS, C.E., W. F. McNEILL u. W. E. FARRAR JR. (19 79): Transposable plasmid deoxyribonucleic acid sequence in Pseudomonas aeruginosa which mediates resistance to gentamicin and four other antimicrobial agents. J. Bacteriol. 139, 877-882

LITERATURVERZEICHNIS
124
RUSSEL, A. D. u. I. CHOPRA (1996): Understanding antibacterial action and resistance. 2. Aufl. Horwood, London, New York, S. 83-89 SALMON S. A. u. J. L. WATTS (2000): Minimum inhibitory concentration determinations for various antimicrobial agents against 1570 bacterial isolates from turkey poults. Avian Dis. 44, 85-98 SAMS, R. A. (1994): Florfenicol: chemistry and metabolism of a novel broad-spectrum antibiotic. in: Proceedings of the XVIII World Buiatrics Congress. Bologna, Italy. S. 13-17 SCHLÜNZEN, F., R. ZARIVACH, J. HARMS, A. BASHAN, A. TOCILJ, R. ALBRECHT, A. YONATH u. F. FRANCESCHI (2001): Structural basis for the interaction of antibiotics with the peptidyl transferase centre in eubacteria. Nature 413, 814-821 SCHMID, S., B. BERGER u. D. HAAS (1999): Target joining of duplicated insertion sequence IS21 is assisted by IstB protein in vitro. J. Bacteriol. 181, 2286-2289 SCHMID, S., T. SEITZ u. D. HAAS (1998): Cointegrase, a naturally occurring, truncated form of IS21 transposase, catalyzes replicon fusion rather than simple insertion of IS21. J. Mol. Biol. 282, 571-583 SCHWARZ, S. (1995): Genetische Grundlagen der Resistenz gegenüber Chloramphenicol und Tetracyclinen bei Staphylokokken animaler Herkunft. Hannover, Tierärztl. Hochsch., Habil. SCHWARZ, S., M. CARDOSO u. H. BLOBEL (1989): Plasmid-mediated chloramphenicol resistance in Staphylococcus hyicus. J Gen Microbiol.135, 3329-3336 SCHWARZ, S. u. E. CHASLUS-DANCLA (2001): Use of antimicrobials in veterinary medicine and mechanisms of resistance. Vet. Res. 32, 201-225 SCHWARZ, S., C. LANGE u. C. WERCKENTHIN (1998): Molecular analysis of the macrolide-lincosamide resistance gene region of a novel plasmid from Staphylococcus hyicus. J. Med. Microbiol. 47, 63-70

LITERATURVERZEICHNIS
125
SCHWARZ; S. u. D. G. WHITE (2005): Phenicol resistance. In: WHITE, D. G., M. N. ALEKSHUN U. P. F. McDERMOTT (Hrsg.): Frontiers in antimicrobial resistance - a tribute to Stuart B. Levy. ASM Press, Washington D.C. S. 124-147 SHAW, W. V. (1983): Chloramphenicol acetyltransferase, enzymology and molecular biology. Crit. Rev. Biochem. 14, 1-46 SHAW, W. V. u. A. G. W. LESLIE (1991): Chloramphencol acetytransferase. Annu. Rev. Biophys. Chem. 20, 363-386 SCHERING-PLOUGH ANIMAL HEALTH (1998): Nuflor®, New Therapeutic Applications. Proceedings of a symposium held in conjunction with the XX World Buiatrics Congress, Sydney, Australia SHIN, S. J., S. G. KANG, R. NABIN, M. L. KANG u. H. S. YOO (2005): Evaluation of the antimicrobial activity of florfenicol against bacteria isolated from bovine and porcine respiratory disease. Vet. Micro. 106, 73-77 SIMON, C. u. W. STILLE (2000): Chloramphenicol. in: SIMON, C. u. W. STILLE (Hrsg.): Antibiotika-Therapie in Klinik und Praxis. 10. Aufl. Schattauer, Stuttgart, New York, S. 172-175 SINGER, R. S., S. K. PATTERSON, A. E. MEIER, J. K. GIBSON, H. L. LEE u. C. W. MADDOX (2004): Relationship between phenotypic and genotypic florfenicol resistance in Escherichia coli. Antimicrob. Agents Chemother. 48, 4047-4049 SINGH, K. V., G. M. WEINSTOCK u. B. E. MURRAY (2002 ): An Enterococcus faecalis ABC homologue (Lsa) is required for the resistance of this species to clindamycin and quinupristin-dalfopristin. Antimicrob Agents Chemother. 46, 1845-1850 STOKES, H. W. u. R. M. HALL (1991): Sequence analysis of the inducible chloramphenicol resistance determinant in the Tn1696 integron suggests regulation by translational attenuation. Plasmid 26, 10-19

LITERATURVERZEICHNIS
126
SULAVIK, M.C., C. HOUSEWEART, C. CRAMER, N. JIWANI, N. MURGOLO, J. GREENE, B. DIDOMENICO, K. J., SHAW, G. H. MILLER, R . HARE u. G. SHIMER (2001): Antibiotic susceptibility profiles of Escherichia coli strains lacking multidrug efflux pump genes. Antimicrob. Agents Chemother. 45, 1126-1136 SUNDSTÖM, L., P. H. ROY u. O. SKÖLD (1991): Site-specific insertion of three structural gene cassettes in transposon Tn7. J. Bacteriol. 173, 3025-3028 SYRIOPOULOU V. P., A. L. HARDING, D. A. GOLDMANN u. A. L. SMITH (1981): In vitro antibacterial activity of fluorinated analogs of chloramphenicol and thiamphenicol. Antimicrob. Agents Chemother. 19, 294-297 TAUCH, A., Z. ZHENG, A. PUHLER u. J. KALINOWSKI (19 98): Corynebacterium striatum chloramphenicol resistance transposon Tn5564, genetic organization and transposition in Corynebacterium glutamicum. Plasmid 40, 126-139 THE UNITED STATES PHARMACOPEIAL CONVENTION (2007): USP Veterinary Pharmaceutical Information Monographs Florfenicol - Veterinary-systemic (http://www.usp.org/pdf/EN/veterinary/florfenicol.pdf) 12601 Twinbrook ParkwayRockville, Maryland 20852-1790, USA TOLEMAN M. A., P. M. BENNETT u. T. R. WALSH (2006): ISCR elements: novel gene-capturing systems of the 21st century? Microbiol. Mol. Biol. Rev. 70; 296-316 TORO, C. S., S. R. LOBOS, I. CALDERON, M. RODRIGUEZ u. G. C. MORA (1990): Clinical isolate of a porinless Salmonella typhi resistant to high levels of chloramphenicol. Antimicrob. Agents Chemother. 34, 1715-1719. TSAKRIS, A., A. IKONOMIDIS, N. SPANAKIS, A. POULOU u. S. POURNARAS (2007): Characterization of In3Mor, a new integron carrying VIM-1 metallo-{beta}-lactamase and sat1 gene, from Morganella morganii. J. Antimicob. Chemother. 59, 739-741 TSCHÄPE, H. (1994): The spread of plasmids as a function of bacterial adaptability. FEMS Microbiol. Lett. 15, 23-32

LITERATURVERZEICHNIS
127
UEDA, Y. u. I. SUENAGA (1995): In vitro antibacterial activity of florfenicol against Actinobacillus pleuropneumoniae. J. Vet. Med. Sci. 57, 363-364 USP DICTIONARY OF USAN AND INTERNATIONAL DRUG NAMES (2002): 2002 ed., Rockville, MD: The United States Pharmacopeial Convention, Inc, VARMA, K. J. (1994): Microbiology, pharmacokinetic disposition and safety of florfenicol in cattle. in: Proceedings of the XVIII World Buiatrics Congress. Bologna, Italy. S. 18-24 VERA LIZARAZO, Y. A., E. F. RODR ĺGUEZ FERRI, A. J. MART ĺN DE LA FUENTE u. C. B. GUTIÉRREZ-MART ĺN (2006): Evaluation of changes in antimicrobial susceptibility patterns of Pasteurella multocida subsp. multocida isolates from pigs in Spain in 1987-1988 and 2003-2004. Am. J. Vet. Res. 67, 663-668 VINING, L. C. u. C. STUTTARD (1995): Chloramphenicol. Biotechnol. 28, 505-530 WALLMANN, J., K. SCHRÖTER, L. H. WIELER u. R. KROKE R (2003): Antibiotikaempfindlichkeit ausgewählter pathogener Bakterien von erkrankten Lebensmittel liefernden Tieren in Deutschland. Tierärztl. Prax. 31, 122-131 WERNER, G. u. W. WITTE (1999): Characterization of a new enterococcal gene, satG, encoding a putative acetyltransferase conferring resistance to streptogramin A compounds. Antimicrob. Agents Chemother. 43, 1813-1814 WHITE, D. G., C. HUDSON, J. J. MAURER, S. AYERS, S. ZHAO, M. D. LEE, L. BOLTON, T. FOLEY u. J. SHERWOOD (2000): Characterization of chloramphenicol and florfenicol resistance in Escherichia coli associated with bovine diarrhea. J. Clin. Microbiol. 38, 4593-4598 WHITTLE; G., M. E. KATZ, E. H. CLAYTON u. B. F. CHE ETHAM (2000): Identification and characterization of a native Dichelobacter nodosus plasmid, pDN1. Plasmid 43, 230-234 WISSELINK, H. J., K. T. VELDMAN, C. VAN DE EEDE, S. A. SALMON u. D. J. MEVIUS (2006): Quantitative susceptibility of Streptococcus suis strains isolated from diseased pigs in seven European countries to antimicrobial agents licensed in veterinary medicine. Vet. Microbiol. 113, 73-82

LITERATURVERZEICHNIS
128
WITTE, W. (1997): Impact of antibiotic use in animal feeding on resistance of bacterial pathogens in humans. In: CHADWICK, D. J. u. J. GOODE (Hrsg.): Antibiotic resistance: origins, evolution, selection and spread - Ciba Foundation Symposium 207. Wiley, Chichester, New York, Weinheim S. 61-70 WITTE, W. (1998): Medical consequences of antibiotic use in agriculture. Science 279, 996-997 WITTE, W. (2000): Selective pressure by antibiotic use in livestock. Int. J. Antimicrob. Agents 16, Suppl.1, 19-24 YAO, J. D. C. u. R. C. MOELLERING Jr. (1999): Antibacterial agents. in: MURRAY, P. R., E. J. BARON, M. A. PFALLER, F. C. TENOVER u. R. H. YOLKEN (Hrsg.): Manual of Clinical Microbiology. 7. Aufl. ASM Press, Washington, D.C. S. 1474-1505 YOSHIDA, H., M. BOGAKI, S. NAKAMURA, K. UBUKATA u. M. KONNO (1990): Nucleotide sequence and characterization of the Staphylococcus aureus norA gene which confers resistance to quinolones. J. Bacteriol. 172, 6942-6949 YOSHIMURA H., A. KOJIMA u. M. ISHIMARU (2000): Antimicrobial susceptibility of Arcanobacterium pyogenes isolated from cattle and pigs. J. Vet. Med. B 47, 139-143 YOSHIMURA H., M. TAKAGI, M. ISHIMARU u. Y. S. ENDOH (2002): Comparative in vitro activity of 16 antimicrobial agnats against Actinobacillus pleuropneumoniae. Vet. Res. Commun. 26, 11-19

DARSTELLUNG DES EIGENEN ANTEILS
129
7 Darstellung des eigenen Anteils an den Publikatio nen
Publikation 1:
Schwarz S., C. Kehrenberg, B. Doublet and A. Cloeck aert: Molecular basis of bacterial resistance to chloramphenicol and florfenicol.
FEMS Microbiol. Rev. (2004), 28, 519-542.
Idee und Konzeption: Schwarz, Kehrenberg, Cloeckaert Experimentelle Umsetzung: Schwarz, Kehrenberg, Doublet, Cloeckaert Auswertung und Diskussion der Ergebnisse: Schwarz, Kehrenberg, Cloeckaert Verfassen des Manuskriptes: Schwarz, Kehrenberg, Cloeckaert
Publikation 2:
Kehrenberg C., J. Mumme, J. Wallmann, J. Verspohl, R. Tegeler, T. Kuhn and S. Schwarz: Monitoring of florfenicol susceptibility among bovine and porcine respiratory tract pathogens collected in Germany during the years 2002 and 2003.
J. Antimicrob. Chemother. (2004), 54, 572-574.
Idee und Konzeption: Schwarz, Kehrenberg Experimentelle Umsetzung: Kehrenberg, Schwarz, Mumme,
Wallmann, Verspohl. Tegeler, Kuhn Auswertung und Diskussion der Ergebnisse: Kehrenberg, Schwarz Verfassen des Manuskriptes: Schwarz, Kehrenberg
Publikation 3:
Kehrenberg C. and S. Schwarz: Plasmid-borne florfenicol resistance in Pasteurella multocida. J. Antimicrob. Chemother. (2005), 55, 773-775.
Idee und Konzeption: Kehrenberg Experimentelle Umsetzung: Kehrenberg Auswertung und Diskussion der Ergebnisse: Kehrenberg, Schwarz Verfassen des Manuskriptes: Kehrenberg, Schwarz
Publikation 4:
Kehrenberg C., D. Meunier, H. Targant, A. Cloeckaer t, S. Schwarz and J.Y. Madec: Plasmid-mediated florfenicol resistance in Pasteurella trehalosi. J. Antimicrob. Chemother. (2006), 58, 13-17. Idee und Konzeption: Kehrenberg, Schwarz, Meunier,
Targant, Cloeckaert, Madec Experimentelle Umsetzung: Kehrenberg Auswertung und Diskussion der Ergebnisse: Kehrenberg Verfassen des Manuskriptes: Schwarz, Kehrenberg

DARSTELLUNG DES EIGENEN ANTEILS
130
Publikation 5:
Kadlec K., C. Kehrenberg and S. Schwarz: Efflux-mediated resistance to florfenicol and/or chloramphenicol in Bordetella bronchiseptica: identification of a novel chloramphenicol exporter. J. Antimicrob. Chemother. (2007), 59, 191-196.
Idee und Konzeption: Schwarz, Kadlec, Kehrenberg Experimentelle Umsetzung: Kadlec, Kehrenberg Auswertung und Diskussion der Ergebnisse: Kadlec, Schwarz Verfassen des Manuskriptes: Schwarz, Kadlec, Kehrenberg
Publikation 6:
Cloeckaert A., S. Baucheron, G. Flaujac, S. Schwarz , C. Kehrenberg, J.L. Martel and E. Chaslus-Dancla: Plasmid-mediated florfenicol resistance encoded by the floR gene in Escherichia coli isolated from cattle. Antimicrob. Agents Chemother. (2000), 44, 2858-2860.
Idee und Konzeption: Cloeckaert, Schwarz, Kehrenberg, Chaslus-Dancla, Martel
Experimentelle Umsetzung: Baucheron, Kehrenberg, Flaujac Auswertung und Diskussion der Ergebnisse: Baucheron, Cloeckaert, Kehrenberg, Schwarz Verfassen des Manuskriptes: Cloeckaert, Schwarz
Publikation 7:
Doublet B., S. Schwarz, C. Kehrenberg and A. Cloeck aert: Florfenicol resistance gene floR is part of a novel transposon. Antimicrob. Agents Chemother. (2005), 49, 2106-2108.
Idee und Konzeption: Cloeckaert, Doublet Experimentelle Umsetzung: Doublet, Kehrenberg, Cloeckaert Auswertung und Diskussion der Ergebnisse: Schwarz, Kehrenberg, Cloeckaert Verfassen des Manuskriptes: Cloeckaert, Doublet, Schwarz, Kehrenberg
Publikation 8:
Kehrenberg C. and S. Schwarz: fexA, a novel Staphylococcus lentus gene encoding resistance to florfenicol and chloramphenicol. Antimicrob. Agents Chemother. (2004), 48, 615-618.
Idee und Konzeption: Kehrenberg Experimentelle Umsetzung: Kehrenberg Auswertung und Diskussion der Ergebnisse: Kehrenberg Verfassen des Manuskriptes: Kehrenberg, Schwarz

DARSTELLUNG DES EIGENEN ANTEILS
131
Publikation 9:
Kehrenberg C. and S. Schwarz: Florfenicol-chloramphenicol exporter gene fexA is part of the novel transposon Tn558.
Antimicrob. Agents Chemother. (2005), 49, 813-815.
Idee und Konzeption: Kehrenberg Experimentelle Umsetzung: Kehrenberg Auswertung und Diskussion der Ergebnisse: Kehrenberg, Schwarz Verfassen des Manuskriptes: Kehrenberg, Schwarz
Publikation 10:
Schwarz S., C. Werckenthin and C. Kehrenberg: Identification of a plasmid-borne chloramphenicol-florfenicol resistance gene in Staphylococcus sciuri. Antimicrob. Agents Chemother. (2000), 44, 2530-2533.
Idee und Konzeption: Schwarz, Werckenthin, Kehrenberg Experimentelle Umsetzung: Werckenthin, Kehrenberg Auswertung und Diskussion der Ergebnisse: Schwarz, Werckenthin, Kehrenberg Verfassen des Manuskriptes: Schwarz, Kehrenberg
Publikation 11:
Schwarz S., C. Kehrenberg and K.K. Ojo: Staphylococcus sciuri gene erm(33), encoding inducible resistance to macrolides, lincosamides, and streptogramin B antibiotics, is a product of recombination between erm(C) and erm(A). Antimicrob. Agents Chemother. (2002), 46, 3621-3623.
Idee und Konzeption: Schwarz, Kehrenberg Experimentelle Umsetzung: Kehrenberg, Ojo Auswertung und Diskussion der Ergebnisse: Schwarz, Kehrenberg, Ojo Verfassen des Manuskriptes: Schwarz, Kehrenberg, Ojo
Publikation 12:
Kehrenberg C., K.K. Ojo and S. Schwarz: Nucleotide sequence and organization of the multiresistance plasmid pSCFS1 from Staphylococcus sciuri. J. Antimicrob. Chemother. (2004), 54, 936-939.
Idee und Konzeption: Kehrenberg, Schwarz Experimentelle Umsetzung: Kehrenberg, Ojo Auswertung und Diskussion der Ergebnisse: Schwarz, Kehrenberg, Ojo Verfassen des Manuskriptes: Schwarz, Kehrenberg

DARSTELLUNG DES EIGENEN ANTEILS
132
Publikation 13:
Kehrenberg C., S. Schwarz, L. Jacobsen, L.H. Hansen and B. Vester: A new mechanism for chloramphenicol, florfenicol and clindamycin resistance: methylation of 23S ribosomal RNA at A2503. Mol. Microbiol. (2005), 57, 1064-1073.
Idee und Konzeption: Kehrenberg, Schwarz, Vester Experimentelle Umsetzung: Kehrenberg, Schwarz Vester, Jacobsen, Hansen Auswertung und Diskussion der Ergebnisse: Kehrenberg, Schwarz, Vester Verfassen des Manuskriptes: Vester, Schwarz, Kehrenberg
Publikation 14:
Long K., J. Poehlsgaard, C. Kehrenberg, S. Schwarz and B. Vester: The Cfr rRNA methyltransferase confers resistance to Phenicols, Lincosamides, Oxazolidinones, Pleuromutilins, and Streptogramin A antibiotics. Antimicrob. Agents Chemother. (2006), 50, 2500-2505.
Idee und Konzeption: Long, Vester, Schwarz, Kehrenberg Experimentelle Umsetzung: Long, Poehlsgaard, Kehrenberg, Schwarz, Auswertung und Diskussion der Ergebnisse: Schwarz, Kehrenberg, Long, Vester, Poehlsgaard Verfassen des Manuskriptes: Vester, Schwarz, Kehrenberg, Long
Publikation 15:
Kehrenberg C. and S. Schwarz: Distribution of florfenicol resistance genes fexA and cfr among chloramphenicol-resistant Staphylococcus isolates. Antimicrob. Agents Chemother. (2006), 50, 1156-1163.
Idee und Konzeption: Kehrenberg Experimentelle Umsetzung: Kehrenberg Auswertung und Diskussion der Ergebnisse: Kehrenberg Verfassen des Manuskriptes: Kehrenberg, Schwarz
Publikation 16:
Kehrenberg C., F. Aarestrup and S. Schwarz: IS21-558 insertion sequences are involved in the mobility of the multiresistance gene cfr. Antimicrob. Agents Chemother. (2007) 51, 483-487.
Idee und Konzeption: Kehrenberg, Aarestrup, Schwarz Experimentelle Umsetzung: Kehrenberg Auswertung und Diskussion der Ergebnisse: Kehrenberg Verfassen des Manuskriptes: Kehrenberg, Schwarz

PUBLIKATIONEN
133
Publikationen

PUBLIKATION 1
Schwarz S., C. Kehrenberg, B. Doublet and A. Cloeck aert:
Molecular basis of bacterial resistance to chloramphenicol and florfenicol.
FEMS Microbiol. Rev. (2004), 28, 519-542.

FEMS Microbiology Reviews 28 (2004) 519–542
www.fems-microbiology.org
Molecular basis of bacterial resistance tochloramphenicol and florfenicol
Stefan Schwarz a,*, Corinna Kehrenberg a, Benoıt Doublet b, Axel Cloeckaert b
a Institut f€ur Tierzucht, Bundesforschungsanstalt f€ur Landwirtschaft (FAL), H€oltystrasse 10, 31535 Neustadt-Mariensee, Germanyb Unit�e BioAgresseurs, Sant�e, Environnement, Institut National de la Recherche Agronomique (INRA), 37380 Nouzilly, France
Received 11 February 2004; received in revised form 5 April 2004; accepted 17 April 2004
First published online 10 May 2004
Abstract
Chloramphenicol (Cm) and its fluorinated derivative florfenicol (Ff) represent highly potent inhibitors of bacterial protein
biosynthesis. As a consequence of the use of Cm in human and veterinary medicine, bacterial pathogens of various species and
genera have developed and/or acquired Cm resistance. Ff is solely used in veterinary medicine and has been introduced into clinical
use in the mid-1990s. Of the Cm resistance genes known to date, only a small number also mediates resistance to Ff. In this review,
we present an overview of the different mechanisms responsible for resistance to Cm and Ff with particular focus on the two different
types of chloramphenicol acetyltransferases (CATs), specific exporters and multidrug transporters. Phylogenetic trees of the different
CAT proteins and exporter proteins were constructed on the basis of a multisequence alignment. Moreover, information is provided
on the mobile genetic elements carrying Cm or Cm/Ff resistance genes to provide a basis for the understanding of the distribution
and the spread of Cm resistance – even in the absence of a selective pressure imposed by the use of Cm or Ff.
� 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Chloramphenicol acetyltransferase; Cat and cml gene nomenclature; Enzymatic inactivation; Florfenicol resistance; Specific exporter;
Multidrug transporter; Co-selection
Contents
* Correspon
fax: +49-5034
E-mail add
0168-6445/$22
doi:10.1016/j.
1. Chemical structures and properties of chloramphenicol and florfenicol . . . . . . . . . . . . . . . . 520
2. Use in human and veterinary medicine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 520
3. Mode of action and spectrum of activity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 521
4. Bacterial resistance to chloramphenicol and florfenicol . . . . . . . . . . . . . . . . . . . . . . . . . . . 521
ding
-871-
ress:
.00 �femsr
4.1. Chloramphenicol acetyltransferases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 523
4.2. Chloramphenicol exporters. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 528
4.3. Other resistance mechanisms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 532
5. Dissemination, co-selection and persistence of chloramphenicol resistance genes . . . . . . . . . 532
6. Summary and conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 534
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 535
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 535
author. Tel.: +49-5034-871-241;
246.
[email protected] (S. Schwarz).
2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
e.2004.04.001

520 S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542
1. Chemical structures and properties of chloramphenicol
and florfenicol
Chloramphenicol (Cm), originally referred to as
chloromycetin, was isolated from Streptomyces venezu-
elae in 1947 [1] and shown to be a broad spectrum
antibiotic with a novel structure (Fig. 1), remarkable
both for a p-nitrophenyl group (at C-1) and an N -di-
chloroacetyl substituent (at C-2) attached to a 1,3-pro-
panediol with two chiral centers (C-1 and C-2) [2]. Cm
was the first naturally occurring substance described to
contain a nitro group. The relative simplicity of Cm
made it the first antibiotic to be marketed as the productof chemical synthesis and Cm has been produced ex-
clusively this way since 1950. Only one (DD-threo) of the
four possible diastereoisomers possesses antibiotic ac-
tivity. The C-3 primary hydroxyl group, initially
thought to be essential for inhibition of protein synthesis
through its affinity for the peptidyltransferase of 50S
ribosomes, can be replaced with fluorine [2]. Besides the
fluoro substitution at C-3 (in florfenicol), very few othersubstitutions are tolerated without adverse effects on
antimicrobial activity [2]. Among them, the substitution
of the nitro group (–NO2), which was considered to be
responsible for the dose-unrelated aplastic anemia, by a
sulfomethyl group (–SO2CH3) at the para position of
the 1-phenyl moiety became effective in thiamphenicol
and florfenicol (Fig. 1).
Cm is a highly stable antibiotic which can be storedfor prolonged times at room temperature. It is amphi-
philic and unionized at physiological pH [2]. Cm can
pass biological membranes to reach intracellular bacte-
O
CH
OH
R1
R1
Chloramphenicol – NO2
Azidamfenicol – NO2
Thiamphenicol – SO2CH3
Florfenicol – SO2CH3
Fig. 1. Structure of chloramphen
ria and is able to readily traverse the blood–brain barrier
[2–4]. The fluorinated Cm derivative florfenicol (Ff) is a
synthetic drug, of which also only the DD-threo stereo-
isomer is antibiotically active. Ff is unionized in a pH
range from 3 to 9 [5] and also poorly soluble in aqueoussolutions. Due to its lipophilicity Ff shows a good tissue
penetration.
2. Use in human and veterinary medicine
Cm and some derivatives, such as thiamphenicol and
azidamfenicol (Fig. 1), have been used over the years inhuman medicine. Certain esters of Cm, such as Cm
palmitate or Cm succinate, have been produced for
therapeutic applications. They do not exhibit antimi-
crobial activity until Cm is released after hydrolysis by
esterases [3]. Cm succinate shows a good solubility in
water and therefore is used for parenteral applications.
The water soluble azidamphenicol is only used in eye
drops [3]. In the early years after its introduction intoclinical use, Cm was considered as a promising broad
spectrum antibiotic [2]. However, a number of adverse
effects have been observed since the mid-1960s in con-
nection with the application of Cm [6]. These side-effects
include a dose-unrelated irreversible aplastic anemia
which occurs at frequencies of 1:10,000–1:40,000 [3] or
1:20,000–1:600,000 [7], a dose-related reversible bone-
marrow suppression, or the Gray syndrome in neonatesand infants [2,4]. Occasionally, hypersensitivity to Cm
ranging from skin rashes to anaphylaxis has been ob-
served, too [4]. Based on these adverse effects and on the
C CH
CH CH2
R2
NR3
H
R2 R3
– OH = Cl2
– OH
N N NH
– OH = Cl2
– F = Cl2
icol and related substances.

S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542 521
availability of less toxic antimicrobial agents with a
similar spectrum of activity, the use of Cm in humans is
nowadays limited to the therapy of a small number of
life threatening infections [2,4]. Since Cm readily crosses
the blood–brain barrier, it remains an alternative ther-apeutic agent for the treatment of meningitis caused by
susceptible strains of Haemophilus influenzae, Neisseria
meningitidis or Streptococcus pneumoniae when no other
antimicrobial agents can be used, e.g. in penicillin-
allergic patients [8].
The use of Cm in veterinary medicine in the Euro-
pean Union (EU) is currently limited to pets and non-
food-producing animals. It was banned in 1994 fromuse in any food-producing animals in the EU. The
main reason for this ban was protection of the con-
sumer from potential adverse effects arising from Cm
residues in carcasses of food animals. Because of the
dose-independent irreversible aplastic anemia in hu-
mans, the ‘‘non-observed effect level’’ (NOEL) could
not be determined for Cm. In toxicological studies,
NOEL represents the dose at and below which adverseeffects do not occur [9]. The calculation of the ‘‘maxi-
mum residue level’’ (MRL), which represents the
maximum level of antibiotic residues acceptable in
carcasses at slaughter without any adverse effect on
public health, is based on the NOEL and, therefore,
could not be determined. As a consequence, EU leg-
islation banned Cm along with several antibiotics, in-
cluding nitroimidazoles and furazolidinones, from usein food-producing animals.
Since adverse side-effects, in particular the dose-in-
dependent irreversible aplastic anemia, have not been
observed in animals, the fluorinated Cm derivative Ff
has been licensed for the control of bacterial respiratory
tract infections in cattle and pigs. Ff was approved in the
EU for the use in cattle and in pigs in 1995 and in 2000,
respectively. Moreover, Ff is also approved for thetreatment of infectious pododermatitis (interdigital
phlegmon) in cattle due to Fusobacterium necrophorum
and Bacteroides melaninogenicus in the USA. In com-
mercial salmon farming, a Ff premix is used for the
treatment of furunculosis in salmons caused by Aero-
monas salmonicida. Fluorinated Cm derivatives are
currently not used in human medicine.
3. Mode of action and spectrum of activity
In procaryotes, Cm is a highly specific and potent
inhibitor of protein biosynthesis. Cm-dependent inhibi-
tion of bacterial protein biosynthesis is mainly due to
the prevention of peptide chain elongation. Its bacte-
riostatic activity is based on a reversible binding to thepeptidyltransferase centre at the 50S ribosomal subunit
of 70S ribosomes [10]. 80S ribosomes of eucaryotic cells
are not targets of Cm and its derivatives. However, it
has been assumed that Cm may interact with mito-
chondrial ribosomes which are similar in their structure
to 70S ribosomes rather than to 80S ribosomes. As a
possible consequence, the mitochondrial function of
stem cells in bone marrow may be impaired resulting ina suppression of the bone marrow function [6,11].
The substrate spectrum of Cm includes Gram-posi-
tive and Gram-negative, aerobic and anaerobic bacteria,
but also chlamydiae, mycoplasmas, and rickettsiae [2,4].
Cm analogs including the fluorinated derivative Ff have
a similar spectrum of antimicrobial activity as Cm [4].
Intrinsic resistance to Cm and Ff has not been observed
although members of different bacterial species andgenera may differ in their basic levels of susceptibility to
both drugs as confirmed by the determination of mini-
mum inhibitory concentrations (MICs) [4,12].
The currently valid MIC breakpoints approved by
the National Committee for Clinical Laboratory Stan-
dards (NCCLS) [13] indicate that streptococci, including
S. pneumoniae, are considered as susceptible to Cm
when they exhibit MICs of 6 4 mg l�1 whereas thebreakpoint for susceptibility is at 8 mg l�1 in bacteria
other than streptococci. Approved breakpoints for Ff
are currently available for Pasteurella multocida,
Mannheimia haemolytica and H. somnus from respira-
tory diseases in cattle as well as for P. multocida, Acti-
nobacillus pleuropneumoniae, Bordetella bronchiseptica
and S. suis from respiratory diseases in pigs and Sal-
monella enterica serovar Choleraesuis from infections inpigs. Isolates of all aforementioned bacterial species,
except S. Choleraesuis, are considered as susceptible
when their MIC of Ff is 6 2 mg l�1. For S. Cholerae-
suis, the Ff-specific breakpoint for susceptibility is
4 mg l�1.
4. Bacterial resistance to chloramphenicol and florfenicol
Over the years, bacteria have developed a number of
mechanisms which enable them to circumvent the in-
hibitory effects of Cm. The first and still most frequently
encountered mechanism of bacterial resistance to Cm is
enzymatic inactivation by acetylation of the drug via
different types of chloramphenicol acetyltransferases
(CATs) [14]. However, there are also reports on othermechanisms of Cm resistance, such as efflux systems,
inactivation by phosphotransferases, mutations of the
target site and permeability barriers [2,14]. As the
number of genes associated with resistance to Cm and
related drugs increases, inconsistencies of their nomen-
clature were observed. These included the assignment of
identical designations for different Cm resistance genes
and that of different designations for virtually the sameresistance gene (Tables 1–3). As previously shown for
the nomenclature of tetracycline resistance [15] and
macrolide–lincosamide–streptogramin resistance genes
Table 1
Type A chloramphenicol acetyltransferases
Group Gene
designation(s)
Bacterial source(s) Plasmid/transposon/
chromosome% Identity Database
accession no(s).
References
DNA AAa
A-1 catI Escherichia coli Tn9,R429 98.3–100 97.3–100 V00622 [27]
catI Acinetobacter baumannii Chromosome (Tn2670) M62822 [28]
cat Acinetobacter calcoaceticus Tn2670-like M37690 [28]
pp-cat Photobacterium damselae
subsp. piscicida
pSP9351 D16171 [29]
– Pseudomonas putida Unknown E02706 [30]
cat Salmonella Typhi pHCM1 AL513383,
NC_003384
[31]
cat Serratia marcescens R478 NC_005211,
BX664015
[32]
cat Shigella flexneri Chromosome AF326777 [33]
A-2 catII Haemophilus influenzae pRI234, pMR375 89.3–99.7 92.0–99.5 X53797 [24]
catII Agrobacterium tumefaciens,
Escherichia coli
pSa X53796 [24]
cat Photobacterium damselae
subsp. piscicida
Plasmidb AB082569 [35]
A-3 catIII Shigella flexneri R387 99.8–100 99.5–100 X07848 [36]
catA3 Mannheimia taxon 10 pMHSCS1 AJ249249 [38]
catA3 Mannheimia varigena pMVSCS1 AJ319822 [39]
cat Uncultured eubacterium pIE1130 NC_004973,
AJ271879
[40]
A-4 cat Proteus mirabilis Chromosome M11587 [41]
A-5 cat Streptomyces acrimycini Chromosome P20074 [42]
A-6 cat86 Bacillus pumilus Chromosome K00544 [43]
A-7 cat(pC221),
catC
Staphylococcus aureus pC221 96.6–100 95.3–100 X02529 [45,46]
cat Staphylococcus aureus pKH7 U38429 [53]
cat Staphylococcus aureus pUB112 X02872 [47]
cat Staphylococcus intermedius pSCS1 M64281 [48]
cat Staphylococcus aureus pSCS6 X60827 [52]
cat Bacillus subtilis pTZ12 M16192 [63]
cat Streptococcus agalactiae pGB354 U83488 [58]
cat Streptococcus agalactiae pIP501 X65462 [57]
cat Enterococcus faecalis pRE25 X92945 [61]
A-8 cat(pC223) Staphylococcus aureus pC223 89.2–100 85.2–100 NC_005243,
AY355285
[54]
cat Staphylococcus aureus pSCS7 M58516 [50]
cat Listeria monocytogenes pWDB100 X68412 [64]
cat Staphylococcus aureus pSBK203 M90091 [51]
cat Lactococcus lactis pK214 X92946 [67]
cat Staphylococcus haemolyticus pSCS5 M58515 [49]
cat Enterococcus faecium pRUM NC_005000,
AF507977
[62]
A-9 cat(pC194) Staphylococcus aureus pC194 93.9–99.8 87.0–94.9 V01277 [44]
cat-TC Lactobacillus reuteri pTC82 U75299 [65]
cat Streptococcus suis TnSs1 AB080798 [60]
cat Staphylococcus aureus pMC524-MBM AJ312056 [55]
A-10 cat Bacillus clausii Chromosome AY238971 [74]
A-11 catP Clostridium perfringens pIP401:Tn4451 100 100 U15027 [75]
catP Neisseria meningitidis Chromosome AF031037 [81]
catD Clostridium difficile Chromosome (Tn4453) X15100,
AF226276
[83]
A-12 catS Streptococcus pyogenes Chromosome X74948 [84]
A-13 cat Campylobacter coli pNR9589 M35190 [85]
522 S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542
Table 1 (continued)
Group Gene
designation(s)
Bacterial source(s) Plasmid/transposon/
chromosome% Identity Database
accession no(s).
References
DNA AAa
A-14 cat Listonella anguillarum pJA7324 S48276 [86]
A-15 catB Clostridum butyricum Chromosome M93113 [78]
A-16 catQ Clostridium perfringens Chromosome M55620 [76]aAmino acid.bNo plasmid designation available.
Table 2
Type B chloramphenicol acetyltransferases
Group Gene
designation(s)
Bacterial source(s) Plasmid/transposon/
chromosome% Identity Database
accession no(s).
References
DNA AA
B-1 cat, catB1 Agrobacterium tumefaciens Chromosome 100 100 M58472 [98]
cat Agrobacterium tumefaciens Chromosome NC_003063 [99]
B-2 catB2 Escherichia coli pNR79:Tn2424 99.5–99.8 99.0–100 AF047479 [100]
catB2 Salmonella Enteritidis Plasmid AJ487034 [101]
catB2 Uncultured eubacterium pSp39 AY139601 [103]
catB2 Pasteurella multocida pJR1 NC_004771,
AY232670
[102]
B-3 catB3 Salmonella Typhimurium pWBH301 84.3–100 84.8–100 AJ009818 [105]
catB3 Salmonella Typhimurium IncF1 plasmid AJ310778 [106]
catB3 Acinetobacter baumannii Chromosome AF445082 [107]
catB3 Escherichia coli pHSH2 AY259086 [108]
catB4 Enterobacter aerogenes pWBH301 U13880 [105]
catB4 Klebsiella pneumoniae pEKP0787-1 AF322577 [109]
cat, catB5 Morganella morganii Tn840 X82455 [104]
catB6 Pseudomonas aeruginosa pPAM-101 AJ223604 [112]
catB8 Klebsiella pneumoniae pKB42 AF227506 [110]
catB8 Salmonella Typhi pST2301 AY123251 [111]
catB8 Pseudomonas aeruginosa Unknown AF418284 [113]
B-4 catB7 Pseudomonas aeruginosa Chromosome 100 100 AF036933 [114]
catB7 Pseudomonas aeruginosa Chromosome AE004506 [115]
B-5 catB9 Vibrio cholerae Chromosome 100 100 AF462019 [116]
catB9 Vibrio cholerae Chromosome NC_002506 [117]
S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542 523
[16], there is also an urgent need for a generally accepted
and unified nomenclature for genes involved in resis-
tance to Cm and Ff.
4.1. Chloramphenicol acetyltransferases
Cm acetyltransferases (CATs) are able to inactivate
Cm as well as thiamphenicol and azidamfenicol. Due to
the replacement of the hydroxyl group at C-3 by a fluor
residue, the acceptor site for acetyl groups was struc-
turally altered in Ff. This modification rendered Ff re-
sistant to inactivation by CAT enzymes, andconsequently, Cm-resistant strains, in which resistance is
exclusively based on the activity of CAT, are susceptible
to Ff [17]. There are two defined types of CATs which
distinctly differ in their structure: the classical CATs,
referred to in this review as type A CATs and the novel
CATs, also known as xenobiotic CATs [14], but referred
to in this review as type B CATs. In addition, annota-
tions of cat genes were found in the whole genome se-
quences of Rhodobacter capsulatus [18], Mesorhizobium
loti [19], S. agalactiae strain 2306 [20], Bacillus cereus
[21], and Brucella melitensis [22]. The potential CAT
variants encoded by these five presumed cat genes do
not exhibit structural features that allow their assign-
ment to either type A or type B. As long as functional
activity has not been confirmed, assignment of these
CAT-like proteins to further novel subtypes has to be
postponed.
4.1.1. Type A chloramphenicol acetyltransferases
Type A CATs have been detected in a wide variety of
bacteria [2,14]. Despite the differences in their amino
acid sequences, the type A CATs share some common
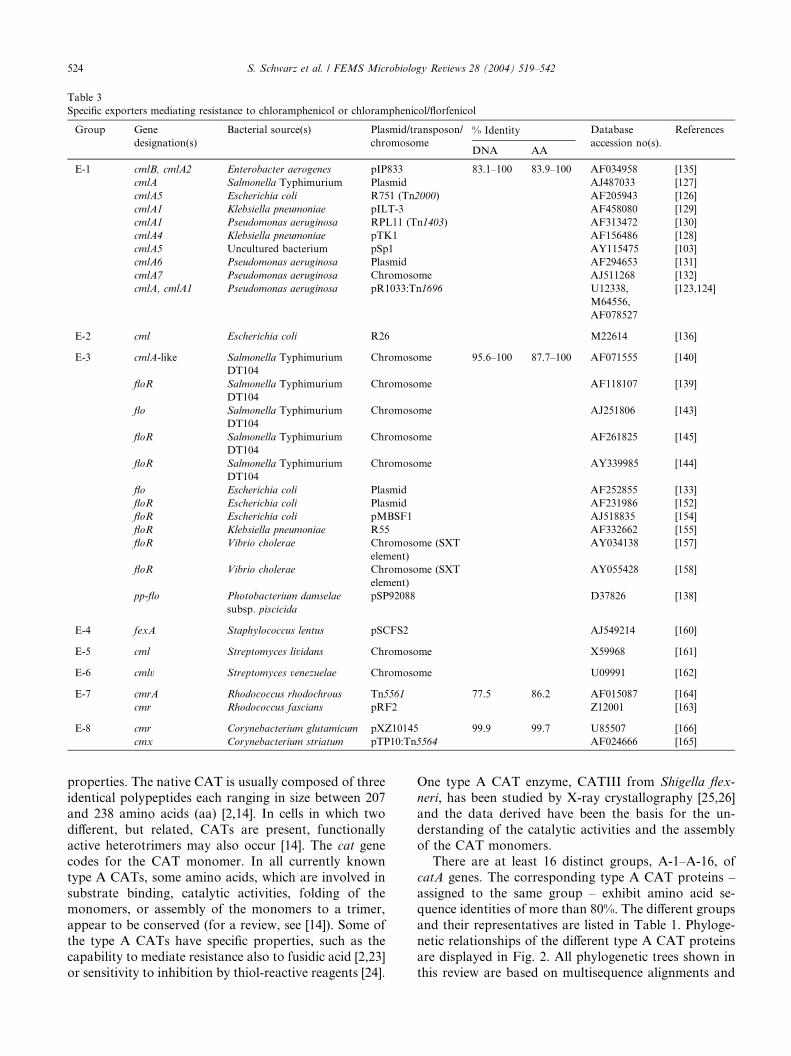
Table 3
Specific exporters mediating resistance to chloramphenicol or chloramphenicol/florfenicol
Group Gene
designation(s)
Bacterial source(s) Plasmid/transposon/
chromosome% Identity Database
accession no(s).
References
DNA AA
E-1 cmlB, cmlA2 Enterobacter aerogenes pIP833 83.1–100 83.9–100 AF034958 [135]
cmlA Salmonella Typhimurium Plasmid AJ487033 [127]
cmlA5 Escherichia coli R751 (Tn2000) AF205943 [126]
cmlA1 Klebsiella pneumoniae pILT-3 AF458080 [129]
cmlA1 Pseudomonas aeruginosa RPL11 (Tn1403) AF313472 [130]
cmlA4 Klebsiella pneumoniae pTK1 AF156486 [128]
cmlA5 Uncultured bacterium pSp1 AY115475 [103]
cmlA6 Pseudomonas aeruginosa Plasmid AF294653 [131]
cmlA7 Pseudomonas aeruginosa Chromosome AJ511268 [132]
cmlA, cmlA1 Pseudomonas aeruginosa pR1033:Tn1696 U12338,
M64556,
AF078527
[123,124]
E-2 cml Escherichia coli R26 M22614 [136]
E-3 cmlA-like Salmonella Typhimurium
DT104
Chromosome 95.6–100 87.7–100 AF071555 [140]
floR Salmonella Typhimurium
DT104
Chromosome AF118107 [139]
flo Salmonella Typhimurium
DT104
Chromosome AJ251806 [143]
floR Salmonella Typhimurium
DT104
Chromosome AF261825 [145]
floR Salmonella Typhimurium
DT104
Chromosome AY339985 [144]
flo Escherichia coli Plasmid AF252855 [133]
floR Escherichia coli Plasmid AF231986 [152]
floR Escherichia coli pMBSF1 AJ518835 [154]
floR Klebsiella pneumoniae R55 AF332662 [155]
floR Vibrio cholerae Chromosome (SXT
element)
AY034138 [157]
floR Vibrio cholerae Chromosome (SXT
element)
AY055428 [158]
pp-flo Photobacterium damselae
subsp. piscicida
pSP92088 D37826 [138]
E-4 fexA Staphylococcus lentus pSCFS2 AJ549214 [160]
E-5 cml Streptomyces lividans Chromosome X59968 [161]
E-6 cmlv Streptomyces venezuelae Chromosome U09991 [162]
E-7 cmrA Rhodococcus rhodochrous Tn5561 77.5 86.2 AF015087 [164]
cmr Rhodococcus fascians pRF2 Z12001 [163]
E-8 cmr Corynebacterium glutamicum pXZ10145 99.9 99.7 U85507 [166]
cmx Corynebacterium striatum pTP10:Tn5564 AF024666 [165]
524 S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542
properties. The native CAT is usually composed of three
identical polypeptides each ranging in size between 207
and 238 amino acids (aa) [2,14]. In cells in which two
different, but related, CATs are present, functionally
active heterotrimers may also occur [14]. The cat gene
codes for the CAT monomer. In all currently known
type A CATs, some amino acids, which are involved in
substrate binding, catalytic activities, folding of themonomers, or assembly of the monomers to a trimer,
appear to be conserved (for a review, see [14]). Some of
the type A CATs have specific properties, such as the
capability to mediate resistance also to fusidic acid [2,23]
or sensitivity to inhibition by thiol-reactive reagents [24].
One type A CAT enzyme, CATIII from Shigella flex-
neri, has been studied by X-ray crystallography [25,26]
and the data derived have been the basis for the un-
derstanding of the catalytic activities and the assembly
of the CAT monomers.
There are at least 16 distinct groups, A-1–A-16, of
catA genes. The corresponding type A CAT proteins –
assigned to the same group – exhibit amino acid se-quence identities of more than 80%. The different groups
and their representatives are listed in Table 1. Phyloge-
netic relationships of the different type A CAT proteins
are displayed in Fig. 2. All phylogenetic trees shown in
this review are based on multisequence alignments and

S. aureus U38429S. aureus X02872S. intermedius M64281
S. aureus X60827S. aureus X02529
E. faecalis X92945S. agalactiae X65462S. agalactiae U83488
S. aureus M58516
E. faecium NC_005000S. aureus NC_005243
S. aureus M90091L. monocytogenes X68412
S. haemolyticus M58515S. aureus V01277
B. clausii AY238971S. suis AB080798
L. reuteri U75299S. aureus AJ312056
100 N. meningitidis AF031037
C. difficile AF226276C. perfringens U15027
C. butyricum M93113C. coli M35190L. anguillarum S48276
C. perfringens M55620
S. pyogenes X74948
A.
A.
calcoaceticus M37690E. coli V00622
Typhi NC_003384baumannii M62822
P. putida E02706P
P
.
.
damselae D16171S. marcescens NC_005211S.
S.
flexneri AF326777 mirabilis M11587
H. influenzae X53797P. damselae AB082569E. coli X53796
uncultured eubacterium NC_004973M. varigena AJ319822Mannheimia taxon 10 AJ249249
S. flexneri X07848S. acrimycini P20074
B. pumilus K00544
A-3
A-2
A-4
A-1
A-5A-6
A-12
A-11
A-14A-13A-15A-16A-10
A-9
A-8
A-7B. subtilis M16192
100
100L. lactis X68412
99
99
10068
100
97
99
100100
53
82
97
96
62
100
100
100
100100
0.05
72
Fig. 2. Phylogenetic tree of the class A CATs. Branch lengths are scaled according to amino acid exchanges observed in a multisequence alignment.
The numbers at the major branch points refer to the percentage of times that a particular node was found in 1000 bootstrap replications. The
bacterial source and the database accession number are given for each CAT protein. Moreover, the groups according to Table 1 are indicated on the
right-hand side.
S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542 525
were produced using the DNAMAN software (Lynnon-
BioSoft, Ont., Canada).The prototype cat gene of group A-1, catI, was
originally identified as part of transposon Tn9 [27] in
Escherichia coli and has been detected on a variety of
resistance plasmids of Gram-negative bacteria, such as
Acinetobacter spp. [28], Photobacterium damselae
subsp. piscicida, formerly known as P. piscicida [29],
and Pseudomonas putida [30]. More recently cat genes
of this group have also been identified in S. Typhi [31],Serratia marcescens [32] and S. flexneri [33]. Enzymes
resembling that encoded by catI have also been de-
tected in B. ochraceus [2]. The cat genes of group A-2
are mainly found on plasmids of H. influenzae which
usually carry at least one more resistance gene [24,34].Genes closely related to catII have also been detected
in E. coli and Agrobacterium tumefaciens [24] and P.
damselae subsp. piscicida [35]. An CAT enzyme similar
to that encoded by catII was also reported to be
present in B. fragilis [2]. The members of group A-3 are
commonly found on plasmids in Enterobacteriaceae
[36] and Pasteurellaceae [37–39], but a cat gene of this
group has also been detected on a plasmid from anuncultured bacterium [40]. Most of these plasmids
carry one or more additional resistance genes. The
groups A-4–A-6 are represented by unique cat genes,

C A U A UA UA A C A
A AU - AA - UA - UA - UU - AC - GC - GU - AC - GC - GU - AU - AA - U
UAC GAGGUGA UAUAACGUAUGAAAAAAUCAGAGGAUU UUAUAUAUGACUUUU
∆ G = -82.8 kJ/mol
start cat
RBS
RBS
start ORF9
stallsequence
IR1 IR2
Fig. 3. Presentation of the regulatory region of the catA gene located
on the S. intermedius plasmid pSCS1 [48]. The ORF9- and catA-as-
sociated ribosome binding sites (RBS) are boxed. The start codons of
ORF9 and catA are underlined and the corresponding coding se-
quences are displayed in bold type letters. The inverted repeated se-
quences IR1 and IR2 are marked by arrows and a stable mRNA
secondary structure formed by these IR sequences is shown.
526 S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542
all of which are located in the chromosomal DNA of
either Proteus mirabilis [41], Streptomyces acrimycini
[42], or B. pumilus [43].
The cat genes commonly found in staphylococci
[44–55], streptococci [56–60] and enterococci [61,62],but also in B. subtilis [63], Listeria monocytogenes [64],
Lactobacillus reuteri [65] and Lactococcus lactis [66,67]
represent the three groups of cat genes A-7–A-9. The
prototype plasmids from which sequences of the re-
spective cat genes had first been deposited in the da-
tabases are pC221 [45,46], pSCS7 [50], and pC194 [44].
These three groups of cat genes are commonly located
on small multicopy plasmids which carry either the cat
gene alone or in combination with a streptomycin re-
sistance [68–70] or a macrolide resistance gene [57]. In
rare cases, such genes were found to be part of mul-
tiresistance plasmids [67,71,72] or conjugative trans-
posons [73]. The only representative of group A-10
was found in the chromosomal DNA of the probiotic
B. clausii [74]. Group A-11 comprises two cat genes, so
far reported in the literature as catP and catD. Thegene catP from Clostridium perfringens was identified
as part of the Cm resistance transposon Tn4451 [75]
which is able to integrate into plasmids as well as into
the chromosomal DNA. The gene catP has been de-
tected not only in clostridia [75–80], but also in N.
meningitidis [81,82]. An identical gene, designated
catD, was identified in C. difficile [83]. The gene catS
from S. pyogenes represents group A-12 and has beendetected in single streptococcal isolates of serogroups
A, B and G [84]. Even though only part of this gene is
deposited in the database, analysis of the amino acid
sequence deduced from this internal segment shows
approximately 77% identity to the catP and catD gene
products.
The cat gene of group A-13 was located on plasmid
pNR9589 of Campylobacter coli which also harbouredan aphA-3 gene for kanamycin resistance [85] whereas
that of group A-14 was found as part of a multiresis-
tance plasmid which also conferred resistance to tetra-
cycline, sulfonamides and streptomycin from Vibrio
anguillarum, meanwhile reclassified as Listonella an-
guillarum [86]. The representatives of the remaining
groups of cat genes A-15 and A-16, catB from C. bu-
tyricum [78] and catQ from C. perfringens [76] are lo-cated in the chromosomal DNA and have so far been
identified only in these clostridial hosts.
Database searches also revealed the presence of pu-
tative catA genes in Bacillus anthracis strain Ames [87],
Deinococcus radiodurans R1 [88], C. acetobutylicum
ATCC824 [89], C. tetani E88 [90], Zymomonas mobilis
[91], and B. thetaiomicron VPI-5482 [92]. These genes
were detected by whole genome sequencing. If CATactivity of their gene products is confirmed, they may be
considered as the representatives of another six indi-
vidual groups of catA genes.
Of the known catA genes, only cat86 and the cat
genes similar to those located on plasmids pC221,
pC223/pSCS7, or pC194 are inducibly expressed via
translational attenuation, with Cm itself acting as an
inducer [93]. The translational attenuators which arelocated immediately upstream of the cat structural genes
consist of a single pair of inverted repeated sequences
IR1 and IR2 as well as a reading frame for a short
peptide of 6–9 aa. Since a common mRNA transcript is
produced from the cat gene and its regulatory region,
IR1 and IR2 are able to form a stable stem-loop struc-
ture with the cat-associated ribosome binding site lo-
cated within IR2 (Fig. 3). The codons 2–5 of the shortpeptide are highly conserved and include a ribosome
stall sequence which is complementary to sequences at
the 30 terminus of the 16S rRNA. A ribosome stalled at
this position overlaps at least in part the IR1 sequence,
thereby preventing formation of a mRNA secondary
structure and rendering IR2 accessible to a second ri-
bosome. It is believed that binding of Cm to ribosomes
triggers conformational changes that eventually put therelevant area of the 16S rRNA in an exposed position
which then allows base pairing with the complementary
part of the cat transcripts. Inducible catA genes mediate
high level resistance to Cm with MICs of P 128 mg l�1.Expression of the catA gene found in P. mirabilis strain
PM13 was obviously based on a flip-flop control
mechanism involving an invertible promoter and a
trans-acting product [41]. As far as information isavailable, the remaining catA genes are expressed con-
stitutively and their MICs of Cm differ with regard to
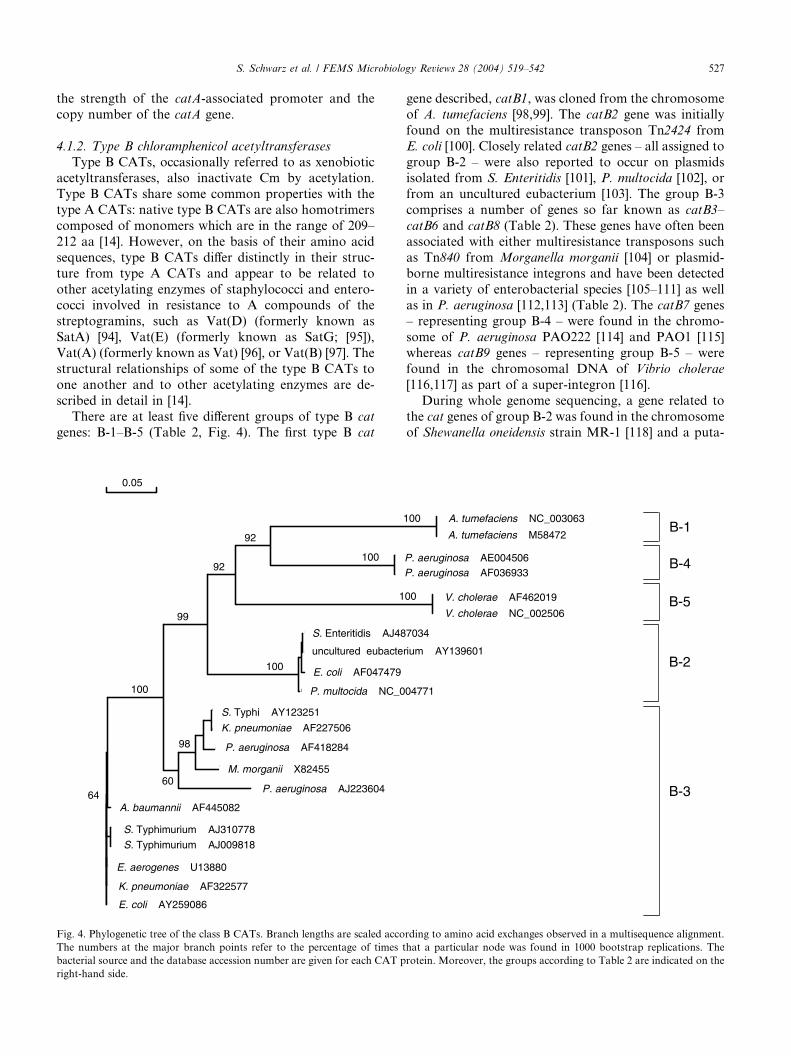
S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542 527
the strength of the catA-associated promoter and the
copy number of the catA gene.
4.1.2. Type B chloramphenicol acetyltransferases
Type B CATs, occasionally referred to as xenobioticacetyltransferases, also inactivate Cm by acetylation.
Type B CATs share some common properties with the
type A CATs: native type B CATs are also homotrimers
composed of monomers which are in the range of 209–
212 aa [14]. However, on the basis of their amino acid
sequences, type B CATs differ distinctly in their struc-
ture from type A CATs and appear to be related to
other acetylating enzymes of staphylococci and entero-cocci involved in resistance to A compounds of the
streptogramins, such as Vat(D) (formerly known as
SatA) [94], Vat(E) (formerly known as SatG; [95]),
Vat(A) (formerly known as Vat) [96], or Vat(B) [97]. The
structural relationships of some of the type B CATs to
one another and to other acetylating enzymes are de-
scribed in detail in [14].
There are at least five different groups of type B cat
genes: B-1–B-5 (Table 2, Fig. 4). The first type B cat
C
c a
C A T _ B2 _ S a l m _ E n t
C A T _ B2 _ u n c
c a t B 2
C A T _ B2 _ P a s
C A T _ B8 _ K l e b s i e l l
C A T _ B8 _ S a l
C A T _ B8 _ P s e
T n 8 4 0
ca t B 6
C A T _ B3 _ A c i
C A T _ B3 _ S a l m _ T y p h
ca t B 3
c a t B 4
C A T _ B4 _ K l e b s i e l l
C A T _ B3 _ E c o l i
S. Enteritidis AJ48
uncultured eubacte
E. coli AF047479
P. multocida NC_0
S. Typhi AY123251
K. pneumoniae AF227506
P. aeruginosa AF418284
M. morganii X82455
P. aeruginosa AJ223604
S. Typhimurium AJ310778
S. Typhimurium AJ009818
A. baumannii AF445082
E. aerogenes U13880
K. pneumoniae AF322577
E. coli AY259086
92
100 92
1
99
100
100
98
6064
0.05
Fig. 4. Phylogenetic tree of the class B CATs. Branch lengths are scaled acco
The numbers at the major branch points refer to the percentage of times t
bacterial source and the database accession number are given for each CAT p
right-hand side.
gene described, catB1, was cloned from the chromosome
of A. tumefaciens [98,99]. The catB2 gene was initially
found on the multiresistance transposon Tn2424 from
E. coli [100]. Closely related catB2 genes – all assigned to
group B-2 – were also reported to occur on plasmidsisolated from S. Enteritidis [101], P. multocida [102], or
from an uncultured eubacterium [103]. The group B-3
comprises a number of genes so far known as catB3–
catB6 and catB8 (Table 2). These genes have often been
associated with either multiresistance transposons such
as Tn840 from Morganella morganii [104] or plasmid-
borne multiresistance integrons and have been detected
in a variety of enterobacterial species [105–111] as wellas in P. aeruginosa [112,113] (Table 2). The catB7 genes
– representing group B-4 – were found in the chromo-
some of P. aeruginosa PAO222 [114] and PAO1 [115]
whereas catB9 genes – representing group B-5 – were
found in the chromosomal DNA of Vibrio cholerae
[116,117] as part of a super-integron [116].
During whole genome sequencing, a gene related to
the cat genes of group B-2 was found in the chromosomeof Shewanella oneidensis strain MR-1 [118] and a puta-
A T _ B _ P s e udo m o na
t B 7
e
V. cholerae AF462019
V. cholerae NC_002506B-5
A. tumefaciens NC_003063
A. tumefaciens M58472B-1
P. aeruginosa AE004506P. aeruginosa AF036933
B-4
7034
rium AY139601
04771
B-2
B-3
100
00
rding to amino acid exchanges observed in a multisequence alignment.
hat a particular node was found in 1000 bootstrap replications. The
rotein. Moreover, the groups according to Table 2 are indicated on the

528 S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542
tive catB gene was detected in the chromosome of V.
parahaemolyticus [119]. CAT activity of the gene prod-
ucts of these two genes, however, has not yet been
confirmed. In addition, the incomplete amino acid se-
quences of another four type B CAT proteins from S.
marcescens, P. aeruginosa, B. sphaericus and Staphylo-
coccus aureus have been reported [14]. These observa-
tions suggest a wider distribution of type B cat genes
among Gram-negative and Gram-positive bacteria than
initially assumed.
Translational attenuation has also been proposed as
the regulatory mechanism for the Cm-inducible catB1
gene from A. tumefaciens [120]. In contrast to the singlepair of inverted repeated sequences seen in the regula-
tory regions of the staphylococcal catA genes, four dif-
ferent pairs of inverted repeats were detected
immediately upstream of catB1 resulting in a more
complex mRNA secondary structure [120]. This gene
has been reported to confer only low level Cm resistance
with 5 to <20 mg l�1 [14]. Several catB genes are part of
gene cassettes and thus are transcribed from a promoterlocated in the integron. Cassette-borne genes located
closest to the promoter are more highly expressed than
distal cassettes [116]. Rowe-Magnus et al. investigated
the level of Cm resistance mediated by the cassette-
borne gene catB9 in relation to its position within a
multiresistance integron from V. cholerae consisting of
seven gene cassettes. When placed in the most distal
seventh position, the MIC of Cm was <1 mg l�1 whichcorresponded to a Cm susceptible phenotype. However,
when the catB9 cassette was placed in the first four
positions, Cm resistance at levels of P 25 mg l�1 were
observed [116].
4.2. Chloramphenicol exporters
The export of Cm or Ff from the bacterial cell can bemediated by either specific transporters and/or multi-
drug transporters. Specific transporters have a substrate
spectrum which is commonly limited to a small number
of structurally closely related compounds whereas that
of the multidrug transporters often includes a wide
range of unrelated substances. Specific transporters
commonly mediate distinctly higher levels of resistance
as compared to those of multidrug transporters. Whilespecific transporters involved in the export of Cm or Ff
have no known function in the physiological cell me-
tabolism, multidrug transporters play an important role
in the excretion of toxic compounds, occasionally also
including specific antimicrobial agents such as Cm and
Ff, from the bacterial cell.
4.2.1. Specific exporters
Genes associated with the export of Cm or Cm/Ff are
found in a wide variety of clinically relevant and envi-
ronmental bacteria. A short description of mobile genes
coding for specific efflux proteins, including those me-
diating the export of Cm or Cm/Ff from the bacterial
cell, was recently published by Butaye et al. [121].
At least eight different groups of specific exporters, E-1–
E-8, are currently known (Table 3). Their phylogeneticrelationships are shown in Fig. 5. Only the exporters
assigned to groups E-3 and E-4 have been reported to
mediate resistance to both, Cm and Ff.
Resistance to Cm not due to enzymatic inactivation
was first detected in 1979 in P. aeruginosa [122] and later
on shown to be based on the presence of the transposon
Tn1696. Sequence analysis of the Cm resistance gene of
Tn1696, cmlA, revealed that the corresponding proteinof 419 aa had 12 transmembrane domains and thus re-
sembled closely other transmembrane transport proteins
of the major facilitator superfamily [123–125]. The cmlA
gene proved to be part of a gene cassette. However, in
contrast to other cassette-borne resistance genes, the
cmlA gene had its own promoter and regulation of cmlA
expression was inducibly regulated via translational at-
tenuation. An attenuator-like structure – similar to thatof inducibly expressed catA genes – was detected up-
stream of the cmlA gene [123]. During the last decade, a
number of genes closely related to or indistinguishable
from cmlA – and all assigned to group E-1 – have been
identified in a wide variety of Gram-negative bacteria,
including E. coli [126], S. Typhimurium [127], Klebsiella
pneumoniae [128,129], and P. aeruginosa [130–132], but
also from an uncultured eubacterium [103] (Table 3).E. coli strains carrying the gene cmlA were reported to
exhibit MICs of Cm of P 32 to >256 mg l�1 and MICs
of Ff of 6 8 to 64 mg l�1 [133,134].
As compared to the other members of group E-1, the
gene cmlA2 (also referred to as cmlB) from Enterobacter
aerogenes [135] showed only 84% nucleotide sequence
identity and 85% identity in the amino acid sequence.
Database search also identified a Cm resistance gene,designated cml, which was located on plasmid R26 [136].
The Cml protein, which does not mediate resistance to
fluorinated Cm analogs [137], consists of 302 amino
acids and exhibits only five transmembrane segments. It
is in part similar to the distinctly larger CmlA protein of
Tn1696 and represents group E-2.
A number of genes, referred to in the published
literature as pp-flo, cmlA-like, floSt, flo, or floR, me-diate combined resistance to Cm and Ff and were
grouped together in group E-3. Despite their varying
designations, these genes are closely related and show
96–100% identity in their nucleotide sequences and 88–
100% identity in the amino acid sequences of their
products. The first member of this group, pp-flo, was
detected on plasmids in the fish pathogen P. damselae
subsp. piscicida in 1996 [138]. More recently, genes ofgroup E-3 were identified in a chromosomal multire-
sistance gene cluster of S. Typhimurium DT104 [139–
144]. This antibiotic resistance gene cluster of about 13
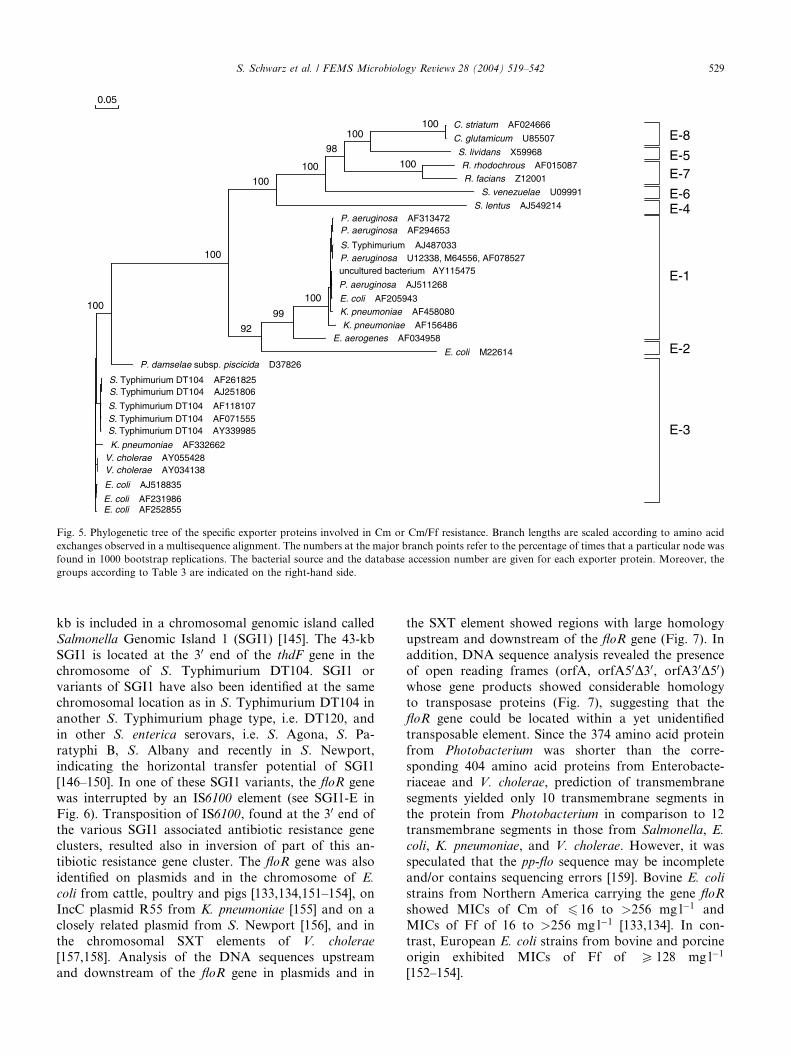
T n 5564
p XZ 10142
cm l
cm rA
rfas cia n s
cm lv
Fe xA
CmlA 1_ P_ ae rugi no
Cm lA 6_ Pae r
CmlA _ Sa lm_ T y phi m
tn 1696
CmlA 5_ u n c u ltu re d
Cm lA 7_ Pae r
Cm lA 5_ Eco l
CmlA 1_ K pne
Cm lA 4_ Kp n e
Cm lA 2E
pR 26
pp -flo
Flo S4
Flo S3
Flo S2
Flo S1
Flo S5
Flo K1
Flo V2
Flo V1
Flo E3
Flo E1
C. striatum AF024666
C. glutamicum U85507
S. lividans X59968R. rhodochrous AF015087R. facians Z12001
S. venezuelae U09991
S. lentus AJ549214P. aeruginosa AF313472P. aeruginosa AF294653
S. Typhimurium AJ487033P. aeruginosa U12338, M64556, AF078527uncultured bacterium AY115475
P. aeruginosa AJ511268
E. coli AF205943K. pneumoniae AF458080
K. pneumoniae AF156486E. aerogenes AF034958
E. coli M22614P. damselae subsp. piscicida D37826
S. Typhimurium DT104 AF261825S. Typhimurium DT104 AJ251806
S. Typhimurium DT104 AF118107S. Typhimurium DT104 AF071555S. Typhimurium DT104 AY339985
K. pneumoniae AF332662V. cholerae AY055428V. cholerae AY034138
E. coli AJ518835
E. coli AF231986E. coli AF252855
100
100
92
99
100
100
100
100
100
100
98
0.05
E-1
E-2
E-3
E-4E-6
E-5
E-8
E-7
Fig. 5. Phylogenetic tree of the specific exporter proteins involved in Cm or Cm/Ff resistance. Branch lengths are scaled according to amino acid
exchanges observed in a multisequence alignment. The numbers at the major branch points refer to the percentage of times that a particular node was
found in 1000 bootstrap replications. The bacterial source and the database accession number are given for each exporter protein. Moreover, the
groups according to Table 3 are indicated on the right-hand side.
S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542 529
kb is included in a chromosomal genomic island called
Salmonella Genomic Island 1 (SGI1) [145]. The 43-kb
SGI1 is located at the 30 end of the thdF gene in the
chromosome of S. Typhimurium DT104. SGI1 or
variants of SGI1 have also been identified at the samechromosomal location as in S. Typhimurium DT104 in
another S. Typhimurium phage type, i.e. DT120, and
in other S. enterica serovars, i.e. S. Agona, S. Pa-
ratyphi B, S. Albany and recently in S. Newport,
indicating the horizontal transfer potential of SGI1
[146–150]. In one of these SGI1 variants, the floR gene
was interrupted by an IS6100 element (see SGI1-E in
Fig. 6). Transposition of IS6100, found at the 30 end ofthe various SGI1 associated antibiotic resistance gene
clusters, resulted also in inversion of part of this an-
tibiotic resistance gene cluster. The floR gene was also
identified on plasmids and in the chromosome of E.
coli from cattle, poultry and pigs [133,134,151–154], on
IncC plasmid R55 from K. pneumoniae [155] and on a
closely related plasmid from S. Newport [156], and in
the chromosomal SXT elements of V. cholerae
[157,158]. Analysis of the DNA sequences upstream
and downstream of the floR gene in plasmids and in
the SXT element showed regions with large homology
upstream and downstream of the floR gene (Fig. 7). In
addition, DNA sequence analysis revealed the presence
of open reading frames (orfA, orfA50D30, orfA30D50)whose gene products showed considerable homologyto transposase proteins (Fig. 7), suggesting that the
floR gene could be located within a yet unidentified
transposable element. Since the 374 amino acid protein
from Photobacterium was shorter than the corre-
sponding 404 amino acid proteins from Enterobacte-
riaceae and V. cholerae, prediction of transmembrane
segments yielded only 10 transmembrane segments in
the protein from Photobacterium in comparison to 12transmembrane segments in those from Salmonella, E.
coli, K. pneumoniae, and V. cholerae. However, it was
speculated that the pp-flo sequence may be incomplete
and/or contains sequencing errors [159]. Bovine E. coli
strains from Northern America carrying the gene floR
showed MICs of Cm of 6 16 to >256 mg l�1 and
MICs of Ff of 16 to >256 mg l�1 [133,134]. In con-
trast, European E. coli strains from bovine and porcineorigin exhibited MICs of Ff of P 128 mg l�1
[152–154].

SGI1-A (S. Agona)
SGI1 (S. Typhimurium DT104)
SGI1-H (S. Newport)
SGI1-F (S. Albany)
SGI1-E (S. Typhimurium DT104)
1 kb
tnpR intI1 aadA2qacE∆1
sul1 floR tetR tet(G) orf1 orf2groEL/intI1
pse-1qacE∆1
sul1orf5orf6 IS6100S044
DR-L
yidY
DR-R
sul1qacE∆1
dfrA10orf513thdF int
tnpR intI1 aadA2qacE∆1
sul1 floR tetR tet(G) orf1 orf2groEL/intI1
pse-1qacE∆1
sul1orf5orf6 IS6100S044
DR-L
yidY
DR-R
thdF int
tetRtet(G)orf1orf2groEL/intI1
pse-1qacE∆1
sul1orf5
orf6 IS6100S044 yidY
DR-R
tnpR intI1 aadA2qacE∆1
sul1 floR5’∆3’
DR-L
thdF intIS6100 floR3’∆5’
tnpR intI1qacE∆1
sul1 floR tetR tet(G) orf1 orf2groEL/intI1
pse-1qacE∆1
sul1orf5orf6 IS6100S044
DR-L
yidY
DR-R
thdF int dfrA1orf
tnpR intI1qacE∆1
sul1 floR tetR tet(G) orf1 orf2groEL/intI1
pse-1qacE∆1
sul1orf5orf6 IS6100S044
DR-L
yidY
DR-R
thdF int aadA7aac(3)-Id
Fig. 6. Genetic organization of the antibiotic resistance gene clusters containing the floR gene of Salmonella Genomic Island 1 (SGI1) in different
Salmonella enterica serovars. SGI1, SGI1-A, SGI1-E, SGI1-F, and SGI1-H are schematized. SGI1 and variants are always located between the thdF
and yidY chromosomal genes. DR-L and DR-R are the left and right direct repeats, respectively, bracketing SGI1. The first SGI1 gene int codes for a
putative integrase probably involved in site-directed integration of SGI1 at the 30 end of thdF. The floR gene and other antibiotic resistance genes are
represented as black and grey arrows, respectively. Besides floR conferring cross-resistance to Cm and Ff, the other antibiotic resistance genes
mediate resistance to ampicillin (pse-1), gentamicin (aac(3)-Id), streptomycin and spectinomycin (aadA2, aadA7), sulfonamides (sul1), tetracyclines
(tet(G)), and trimethoprim (dfrA1, dfrA10).
530 S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542
A novel type of exporter gene, designated fexA, rep-
resents group E-4. This gene has recently been identified
on the 34-kb plasmid pSCFS2 from S. lentus [160]. TheFexA protein consists of 475 amino acids and exhibits
14 transmembrane domains. It mediates resistance to
Cm and Ff. Inducible expression of fexA is obviously
due to translational attenuation. An attenuator-like
structure was identified immediately upstream of the
fexA gene [160]. The S. lentus isolate carrying fexA
showed basic MICs of Cm of 64 mg l�1 and Ff of 32
mg l�1 which after induction with either Cm or Ff wereincreased to 128 mg l�1. This gene is the first exporter
gene identified in Gram-positive cocci which mediates
combined resistance to Cm and Ff. The gene fexA shows
no similarity to any other genes involved in the export of
Cm or Ff.
Several Cm exporters – distinctly different from the
aforementioned ones found in clinically relevant bac-
teria from humans and animals – were identified insoil and environmental bacteria of the genera Strep-
tomyces (groups E-6 and E-7), Rhodococcus (group
E-8) or Corynebacterium (group E-9). The cml gene
identified in the chromosome of S. lividans 1326 [161]
encodes a protein with 12 transmembrane segmentswhile the chromosomal gene cmlv of S. venezuelae
ISP5230, a chloramphenicol producer, encodes a
protein of 10 transmembrane segments assumed to be
involved in the export of Cm [162]. Cm exporter
proteins of 12 transmembrane domains were encoded
by the genes cmr and cmrA from Rhodococcus spp.
The gene cmr was found on the conjugative plasmid
pRF2 from Rhodococcus fascians [163] while the genecmrA appeared to be associated with transposon
Tn5561 in R. rhodochrous [164]. As part of the
transposon Tn5564 the gene cmx was detected on a
multiresistance plasmid in Corynebacterium striatum
[165]. A transposon-like element closely related to
Tn5564 and carrying a Cm resistance gene, designated
cmr, was found on plasmid pXZ10145 in C. glutami-
cum [166]. The Cmx and Cmr proteins from Coryne-
bacterium spp. appear to have 10 transmembrane
segments.

floR orfAorfA3’∆ 5’
E. coli plasmid 10660-1
floR orf1 orf2/orfA3’∆ 5’K. pneumoniae plasmid R55
strA strB orfA3’∆ 5’ floR tnpE. coli plasmid pMBSF1
V. cholerae SXT element
tnpA orfA dfr18 dcd orfA3’∆ 5’ floR orfA5'∆ 3’strB strA sul2 tnpA'
pp-flo orfAP. damselae plasmid pSP92088
1 kb
Fig. 7. Organization of the floR gene loci of different plasmids from E. coli, K. pneumoniae, P. damselae subsp. piscicida and of the chromosomal SXT
element from V. cholerae. The floR (or pp-flo) gene is shown as a black arrow while the putative transposase genes, orfA or parts of orfA, are
displayed as grey arrows. The regions of homology with the floR locus of the plasmid from E. coli 10660-1 are indicated by thick black bars. The
genes strA–strB, dfr18 and sul2 genes confer resistance to streptomycin, trimethoprim, and sulfonamides, respectively.
S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542 531
As previously seen with the catA and catB genes,
annotations of florfenicol resistance genes were alsodetected in whole genome sequences of C. tetani strain
E88 [90], B. melitensis [22], Yersinia pestis [167], F.
nucleatum subsp. nucleatum [168], Legionella pneumo-
phila [169], Heliobacillus mobilis [170], and the photo-
synthetic green-sulfur bacterium Chlorobium tepidum
TLS [171]. The functional activity of the corresponding
proteins in florfenicol resistance, however, has not been
confirmed yet. Moreover, they only exhibit low levelsof amino acid identity to the proteins FloR (8.5–
29.2%) and FexA (9.1–21.9%) known to export Ff
from the bacterial cell. In addition, whole genome se-
quencing revealed the presence of two genes coding for
a 399 amino acid protein from Sinorhizobium meliloti
[172] and a 385 amino acid protein from Mesorhizo-
bium sp. strain BNC1 [173] which exhibited identities
of 60% and 63%, respectively, and homologies of 75%to the FloR protein. But also for these two proteins, it
is unknown whether they are involved in the efflux of
Ff and Cm.
4.2.2. Multidrug transporters
In addition to specific exporters, a number of multi-
drug transporter systems have been identified whose
substrate spectrum includes Cm and/or Ff. In general,the levels of Cm and Ff resistance mediated by
multidrug transporters are lower than those mediated by
specific exporters.The AcrAB-TolC multidrug efflux system is able to
export Cm and Ff at low levels (MICs of Cm and Ff of
4 mg l�1). Overproduction of this system, due to muta-
tions at regulator loci, however, leads to clinical levels
of resistance to Cm (MIC 16–32 mg l�1), Ff (MIC
32 mg l�1) and other antimicrobials by active efflux
[174–177]. Another multidrug transporter, MdfA, which
also exports Cm has been identified in E. coli [178]. Itshows 96% amino acid identity to the E. coli protein
Cmr, a 12 TMS protein of 411 aa which specifies a Cm
efflux pump [179]. Multidrug transporters whose sub-
strate spectrum include Cm have also been described in
P. aeruginosa. Similarly to the AcrAB–TolC system in
E. coli, these multidrug transporters are also composed
of three components, a protein of the resistance/nodu-
lation/cell division family (MexB, MexD or MexF), amembrane fusion protein (MexA, MexC or MexE) and
an outer membrane protein (OmpM, OmpJ or OprN),
which interact cooperatively to enable export of the
drugs [159,180]. Multidrug transporters of a similar
structure which can also export Cm have been identified
in Burkholderia cepacia (CeoAB–OpcM) and P. putida
(ArpAB–ArpC; TtgAB–TtgC) [180]. Overexpression of
most of these multidrug transporter systems led to adistinct increase of the MIC of Cm [174,177,181]

532 S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542
whereas functional deletion resulted in a distinctly more
susceptible phenotype [182]. It should be noted that
several types of multidrug transporters may be present
in the same bacterial strain and that specific transporters
may occur side-by-side with multidrug transporters. Leeet al. investigated the effects of simultaneous expression
of several efflux pumps, including specific exporters such
as CmlA and multidrug transporters such as MdfA,
AcrAB–TolC or MexAB–OprM, and observed additive
as well as multiplicative effects on Cm resistance of
E. coli and P. aeruginosa [177].
Some multidrug transporters from Gram-positive
bacteria, such as NorA from S. aureus [183] or Blt fromB. subtilis [184] were reported to be able to export Cm.
However, studies on strains expressing the gene norA at
elevated levels showed that their MICs of Cm and Ff
were in the same low range as those of strains not car-
rying the gene norA [Schwarz, S. and Kehrenberg, C.,
unpublished data]. This observation suggested that
carriage of the gene norA is most probably not a relevant
factor in Cm and Ff resistance in staphylococci.
4.3. Other resistance mechanisms
Besides inactivation by acetylation, there are other
ways to inactivate Cm. Some of which, such as deha-
logination, glucuronidation, and reduction of the nitro
group, are usually seen during biotransformation in he-
patocytes of humans and animals [4], but have not beenidentified in bacteria. Other mechanisms such as O-
phosphorylation [162] and hydrolytic degradation of Cm
to p-nitrophenylserinol [185] are seen in the Cm producer
S. venezuelae ISP5230. These latter mechanisms seem to
have a self-defense function in the antibiotic producer.
Recently, the 3-O-phosphotransferase was crystallized
and its X-ray structure was determined [186].
Non-enzymatic Cm resistance mechanisms based onpermeability barriers have been described in various
bacteria. The loss of an outer membrane protein was
considered to play a role in Cm resistance of H. influ-
enzae strains which did not exhibit CAT activity [187].
Cm resistance due to decreased permeability of the outer
membrane was also observed in B. cepacia [188]. The
absence of a 50 kDa porin in Tn1696-carrying strains
may also enhance Cm resistance [189]. In S. Typhi, thelack of the OmpF protein, which is required for the
entry of Cm into the bacterial cell was found to result in
high level resistance to Cm [190]. The mar locus which is
present in bacteria of many enterobacterial genera, has
also been reported to contribute to Cm resistance of
E. coli. The transcriptional activator MarA is able to
activate the gene micF which produces an antisense
RNA that effectively inhibits ompF translation [191].Mutations in the major ribosomal protein gene clus-
ter of E. coli [192] and B. subtilis [193] as well as in the
23S rRNA gene of E. coli [194] are known to confer
resistance to Cm. However, in contrast to resistance to
other protein biosynthesis inhibitors, e.g. macrolide–
lincosamide–streptogramin antibiotics [195], Cm resis-
tance as a consequence of target site mutation/modifi-
cation is rarely seen. One plausible suggestion for thisobservation is that structural changes at the peptidyl
transferase center that might prevent Cm binding are
incompatible with satisfactory ribosome function [196].
Finally, a novel gene, cfr, which mediates resistance
to Cm and Ff by a yet unidentified mechanism has re-
cently been detected on plasmid pSCFS1 from S. sciuri
[197]. The corresponding gene product shows no
homology to any of the so far known Cm resistanceproteins, does not inactivate Cm or Ff, and also does
not display transmembrane topology. Structural com-
parisons revealed a certain degree of similarity with Fe–
S-binding oxidoreductases of the MoaA/NifB/PqqE
family. Two domains were detectable: the N-terminal
domain represented a putative Cys-rich Fe-S binding
sequence (CISSQCGCNFGCKFC), whereas the C-ter-
minal domain might contain a NAD-binding Rossmanfold. However, the target site of Cfr as well as the Cfr-
mediated mechanism of resistance remain to be deter-
mined. The MICs of Cm and Ff of the S. sciuri strain
carrying pSCFS1 were 32 and 64 mg l�1 and could be
increased after induction to 64 and 512 mg l�1, respec-tively. A potential regulatory region which resembled a
translational attenuator was detected immediately up-
stream of the cfr reading frame [197].
5. Dissemination, co-selection and persistence of chlor-
amphenicol resistance genes
There are few studies which describe the distribution
of specific cat or exporter genes within bacterial popu-
lations. A study on 28 Cm resistant E. coli isolates fromcattle, swine and poultry identified not further specified
catA genes in 68% and cmlA1-like genes in 36% of the
isolates whereas floR genes were not detected at all [198].
A study on the prevalence of floR and cmlA genes among
48 bovine E. coli isolates showed that floR genes were
present alone in 71%, cmlA genes alone in 5% and a
combination of floR and cmlA in 10% of the isolates
[133]. In contrast, 89.6% of E. coli strains from swinecarried the gene cmlA alone and another 8.3% in com-
bination with a cat gene of group A-2, while the gene floR
was detected only in a single isolate (2.1%) [134]. Another
study identified the cassette-borne Cm resistance gene
catB2 in 31 (10%) of 313 motile aeromonads from a fish
farming environment. In all cases the catB2 cassette was
located in a class 1 integron that also contained cassettes
for resistance to streptomycin and trimethoprim [199]. AcatA gene similar to that of plasmid pC194 was detected
in 44 of 100 erythromycin resistant S. pneumoniae iso-
lates [200], whereas this gene could only be detected in
single streptococcal isolates of serogroups A or B as well

S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542 533
as in a single Enterococcus faecium isolate [84]. In con-
trast, catA genes of group A-7 were detected in strepto-
cocci of serogroups A, B or G and in E. faecium, catA
genes of group A-8 in a single serogroup B Streptococcus
and in four E. faecalis isolates and catP as well as catQgenes in single serogroup B streptococci [84]. Such a
heterogeneity of cat genes has not been detected in
staphylococci. Plasmid-borne catA genes of groups A-7
and A-8 have been detected at almost similar frequencies
among S. aureus isolates from bovine mastitis [201],
S. lentus isolates frommink [202] and staphylococci from
pigeons [203], whereas catA genes of group A-7 genes
were the dominant genes in S. hyicus from pigs [204] andS. intermedius from dogs [205,206].
Many of the genes coding for any of the two types of
cat genes or specific exporters are located on mobile
genetic elements, such as plasmids, transposons or gene
cassettes (Tables 1–3). The location of the resistance
gene on a mobile element is an important prerequisite
for fast and efficient distribution among bacteria of the
same or different genera and species. Other requirementsare an appropriate selective pressure imposed by the use
of antimicrobial agents as well as the close contact be-
tween donor and recipient bacteria which is commonly
available in the mixed populations on the skin or the
mucosal surfaces of the intestinal, respiratory or genito-
urinary tract of humans and animals. In this regard, the
selective pressure does not have to be based only on the
use of Cm or Ff, but may also arise from the applicationof various other antimicrobials. The reason for this is
that plasmids carrying cat genes of type A (e.g. those
assigned to groups A-1–A-3) frequently also carry sev-
eral additional genes which code for resistance not only
to other antimicrobial agents [31,32,34,38–40], but also
to heavy metals such as mercury [31] or tellurium [32]. In
several plasmids from Pasteurella and Mannheimia
strains [38,39] and an uncultured bacterium [40] thecatA3 gene was found to be integrated between the
sulfonamide resistance gene sul2 and the streptomycin
resistance gene strA to form a multiresistance gene
cluster. Based on sequence comparisons, this integration
event was considered as an illegitimate recombination
between an R387-like plasmid carrying catA3 and an
RSF1010-like plasmid carrying sul2 and strA [39]. Since
there were no promoters upstream of catA3 and strA, itwas assumed that all three genes were transcribed from a
common promoter upstream of sul2 [38,39]. Persistence
of a catA3 gene in a multiresistance gene cluster repre-
sents a plausible explanation for the persistence of a Cm
resistance gene for more than eight years after the ban of
Cm use in cattle.
Some of the aforementioned plasmids are conjugative
[31,32] and thus can arrange their self-transfer to newhostcells. Smaller plasmids like pMHSCS1 and pMVSCS1
from Mannheimia spp. [38,39] and pIE1130 from uncul-
tured eubacterium [40] are mobilizable. Transformation
experiments also showed that the Mannheimia plasmids
pMHSCS1 and pMVSCS1 can replicate and express their
resistance properties even in distantly related hosts such
as E. coli [38,39]. When plasmids mediating resistance to
Cm or Cm/Ff are transferred from one bacterial host toanother, they are not always able to replicate in the new
host. Besides a general inability to replicate in the new
host, restriction-modification systems present in the
newhost, but also incompatibilities with plasmids already
resident in the new host may prevent efficient replication
of the new plasmid. Recombination between the new
plasmid and the plasmids resident in the new host is an
effective way to circumvent these replication problems.Such recombinations may lead to the formation of novel
resistance plasmids which carry the resistance genes of
both parental plasmids and are well adapted with regard
to their replication in the respective host. Such recombi-
nation events have not only been observed in the plasmids
from Mannheimia spp., but are also considered to play a
role in the formation of small staphylococcal plasmids
that carried a pC221-like catA gene and a pS194-likestreptomycin resistance gene [68–70,207]. Thewidespread
occurrence of pC221-like catA genes in large conjugative
multiresistance plasmids such as pIP501 [57] is most
probably also based on recombinational events. The fact
that broad host range plasmids such as pIP501 can rep-
licate in a wide variety of Gram-positive bacteria [71]
furthers the spread of the respective catA gene. The host
range of pIP501 was reported to include bacteria of thegenera Streptococcus, Enterococcus, Staphylococcus,
Clostridium, Listeria, and Pediococcus [71]. The spread of
Tn4451 – or Tn4453-associated catP genes from Clos-
tridium spp. toN.meningitidis [81–83] represents a serious
health threat since Cm is still the standard therapy for
meningococcal meningitis in developing countries [81].
This observation illustrates that transposon-borne cat
genes can also be spread across species and genus borders.Since catB genes have often been identified to be part
of gene cassettes which are located on chromosomal or
plasmidic multiresistance integrons, the co-selection is-
sue is also of major importance. The same is true for
cmlA genes which are also cassette-borne genes and are
most frequently found within plasmidic multiresistance
integrons. In this regard, a large number of multiresis-
tance integrons have been identified in a variety of Gram-negative bacteria. The observation that virtually the
same cat gene (e.g. those of groups B-2 or B-3) or ex-
porter gene (e.g. those of group E-1) have been identified
in different bacteria such as Salmonella, Escherichia,
Klebsiella and Pseudomonas underlines the efficient
transfer of these cassette-borne genes. Other exporter
genes, e.g. those of group E-3, have only been identified
either on plasmids [150,152,154,155] or in the chromo-somal DNA where they represented part of a multire-
sistance gene cluster of SGI1 in S. enterica serovars
[145–149] and part of the SXT element in V. cholerae

534 S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542
[158,159]. The SXT element of V. cholerae is a conjuga-
tive self-transmissible chromosomally integrating ele-
ment of about 100 kb, and has been related to integrative
conjugative elements (ICEs) [208,209]. Thus, SXT might
contribute to the spread of the Cm/Ff resistance genefloR by conjugal transfer to a variety of Gram-negative
bacteria. The identification of SGI1 or variants of SGI1
carrying the floR gene reported for S. enterica serovars
Typhimurium DT104 and DT120, Agona, Paratyphi B,
Albany and Newport (Fig. 6) indicates the horizontal
transfer potential of SGI1 [145–150]. Moreover, the
ant(40)-IIb gene of P. aeruginosa strain BM4492 has
been identified recently as part of a chromosomal mul-tiresistance gene cluster homologous to that of SGI1
which comprises the resistance genes floR and tetR-
tet(G) [210]. The horizontal transfer of SGI1 remains to
be experimentally confirmed, but could probably occur
in a similar way as the conjugal transfer of SXT. Thus,
the floR gene spreads together with other antibiotic re-
sistance genes by means of large mobile genomic islands
such as SXT and SGI1 that integrate into the chromo-some and may remain stable afterwards. A transposable
element has also been assumed to be involved in the
spread of floR since the floR genes detected on plasmids
of E. coli and K. pneumoniae, but also on the SXT ele-
ment, were flanked by transposase-like reading frames
[152,155] and/or transposon relics [154,158,159] (Fig. 7).
Again, all these floR-carrying plasmids and chromo-
somal elements also carried at least one additional re-sistance gene. Recently, conjugative plasmids carrying a
floR gene and the extended spectrum cephalosporin re-
sistance gene blaCMY�2 were detected in canine E. coli
strains [211], S. Typhimurium and S. Newport strains
from humans and retail foods [212] and S. Newport from
various animal and human sources [213].
In summary, the spread of cat genes or exporter genes
mainly depends on the genetic element on which therespective gene is located. Based on the transfer abilities
of this element, a distribution of the Cm or Cm/Ff re-
sistance gene across species and genus borders is medi-
ated by either conjugation, mobilization, transduction
or transformation. Another important aspect is the
presence of additional resistance genes on the same
mobile genetic element. Since other resistance genes are
commonly found on plasmids, transposons or integronsharbouring cat genes or exporter genes, co-selection of
Cm or Cm/Ff resistance genes needs to be taken into
account when discussing the spread and the persistence
of cat genes or genes coding for specific exporters.
6. Summary and conclusions
Even though the use of Cm has been reduced widely
in human medicine in the industrial countries, it is ex-
tensively used in developing countries throughout the
world for the treatment of severe diarrhea and pneu-
monia [214]. This practice is continued since Cm is in-
expensive, still effective against a wide range of bacteria
and does not require specific conditions for storage
[214]. In developing countries, antimicrobials, includingCm, are commonly available to consumers by over-the-
counter sale in pharmacies without prescription by
medical doctors. Self-medication and misuse of these
drugs favours the development of resistance [215]. Thus,
the major force to select for or maintain Cm resistance
in bacteria from humans, namely the widespread use of
Cm, is still present in those countries. Another impor-
tant aspect, which, however, is not only restricted todeveloping countries, is the use of Cm for prophylaxis or
therapy in food-producing animals including aquacul-
ture. The regulation of Cm application to food animals
in other parts of the world – if available at all – may
differ distinctly from the strict limitations of the use of
Cm in food-producing animals in the EU or Northern
America [216]. Import controls, done on the basis of
random samples, usually refer to Cm residues stillpresent in carcasses, but not to the presence of Cm re-
sistant bacteria. Thus, travel abroad, global trade or
open markets may lead to the ‘‘import’’ of Cm resistant
bacteria resident in or on humans, animals or food of
animal origin. A recent study from Denmark [217]
showed striking differences in the percentages of Cm
resistance in S. Typhimurium isolated in Danish pork
(22%) versus imported pork (61%), but also from humancases acquired domestically (14%) versus associated with
travel abroad (42%). This observation underlined the
role of imported food and travelling abroad as relevant
factors in the spread of Cm resistant strains. A third
aspect is the use of Cm in pets and other non-food-
producing animals which live in close contact to humans
in most industrial countries. Pet and companion animals
may represent a reservoir of resistant strains. Trans-mission of Staphylococcus strains between pets and hu-
mans has been demonstrated [218,219] and in one case
indistinguishable catA-carrying plasmids have been
identified from S. epidermidis strains of human and ca-
nine origins suggesting the exchange of such plasmids
[220]. Since many of the Cm resistance determinants
reside on mobile genetic elements (Tables 1–3), which
most often carry additional resistance genes, co-selec-tion and persistence of Cm or Ff resistance genes may
occur, even if there is no selective pressure imposed by
the use of Cm or Ff. Furthermore, the linkage of resis-
tance genes with virulence genes may also occur and
explain why resistance genes persist in the absence of
selection pressure [221].
New Cm derivatives, which are active against bac-
terial strains that harbour any of the known resistancemechanisms, are not currently under development, ei-
ther for use in human or in veterinary medicine. Ff is
one of the most recent antimicrobial agents exclusively

S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542 535
licensed for use in animals. For use in cattle and pigs,
Ff is only approved as an injectible drug for individual
animal therapy. This minimizes the risk of underdosing
– as compared with medication via feed and water –
and thus eliminates a major factor that contributes tothe resistance development in the veterinary field.
However, the use of Ff also resulted in the detection of
new resistance genes, such as floR, cfr or fexA. Since
these genes also mediate Cm resistance, it is most likely
that they have been developed prior to the introduction
of Ff into veterinary use. Nevertheless, increasing vet-
erinary use of Ff bears the possibility that all those
determinants which also mediate resistance to fluori-nated Cm analogs will spread or may give rise to new
Cm/Ff or Ff resistance determinants possibly specifying
new mechanisms. Therefore, prudent use of highly
potent antimicrobials, such as Ff, in veterinary medi-
cine is strongly required to pertain the efficacy of Ff for
the future.
All these aspects show that the problem of bacterial
resistance to Cm and Ff is multifaceted in its originsand manifestations. Knowledge of (i) the resistance
genes present in the different bacteria, (ii) the mobile
elements on which these resistance genes are located,
(iii) the transfer capacities and host range of these
mobile elements and (iv) the organisation of the Cm
and Cm/Ff resistance genes in multiresistance integ-
rons and gene clusters as summarized in this review
will provide a substantial database for the under-standing of the spread and the persistence of Cm and
Cm/Ff resistance genes, but also the co-selection of
genes conferring resistance to other classes of antimi-
crobial agents, such as extended spectrum cephalo-
sporins, which are even more relevant than Cm or Ff
for therapeutic interventions in human and veterinary
medicine.
Acknowledgements
S.S. and C.K. were supported by grants of the
Deutsche Forschungsgemeinschaft (SCHW 382/6-1,
SCHW 382/6-2). The authors would like to thank Wil-
liam V. Shaw for helpful discussions.
References
[1] Ehrlich, J.Q., Bartz, Q.R., Smith, R.M., Joslyn, D.A. and
Burkholder, P.R. (1947) Chloromycetin a new antibiotic from a
soil actinomycete. Science 106, 417.
[2] Shaw, W.V. (1983) Chloramphenicol acetyltransferase, enzymol-
ogy and molecular biology. Crit. Rev. Biochem. 14, 1–46.
[3] Simon, C. and Stille, W. (2000) Chloramphenicol. In: Antibio-
tika-Therapie in Klinik und Praxis (Simon, C. and Stille, W.,
Eds.), 10th. edn, pp. 172–175. Schattauer, Stuttgart, New York.
[4] Yao, J.D.C. and Moellering Jr., R.C. (1999) Manual of Clinical
Microbiology. In: (Murray, P.R., Baron, E.J., Pfaller, M.A.,
Tenover, F.C. and Yolken, R.H.,, pp. 1474–1505 ASM Press,
Washington, D.C..
[5] Sams, R.A. (1995) Florfenicol: Chemistry and metabolism of a
novel broad-spectrum antibiotic. Tier€arzt. Umschau 50, 703–707.
[6] Martelo, O.J., Manyan, D.R., Smith, U.S. and Yunis, A.A.
(1964) Chloramphenicol and bone marrow mitochondria. J. Lab.
Clin. Med. 74, 927–942.
[7] Vinning, L.C. and Stuttard, C. (1995) Chloramphenicol. Bio-
technology 28, 505–530.
[8] Mascaretti, O.A. (2003) Bacteria versus Antimicrobial Agents,
an Integrated Approach. ASM Press, Washington DC, USA.
[9] Schwarz, S. and Chaslus-Dancla, E. (2001) Use of antimicrobials
in veterinary medicine and mechanisms of resistance. Vet. Res.
32, 201–225.
[10] Schl€unzen, F., Zarivach, R., Harms, J., Bashan, A., Tocilj, A.,
Albrecht, R., Yonath, A. and Franceschi, F. (2001) Structural
basis for the interaction of antibiotics with the peptidyl trans-
ferase centre in eubacteria. Nature 413, 814–821.
[11] Franklin, T.J. and Snow, G.A. (1998) Biochemistry and Molec-
ular Biology of Antimicrobial Action, 5th edn. Kluwer Academic
Publishers, Dordrecht, Boston, London.
[12] Priebe, S. and Schwarz, S. (2003) In vitro activities of florfenicol
against bovine and porcine respiratory tract pathogens. Anti-
microb. Agents Chemother. 47, 2703–2705.
[13] National Committee for Clinical Laboratory Standards
(NCCLS) (2002) Performance standards for antimicrobial disk
and dilution susceptibility tests for bacteria isolated from
animals; Approved standard – 2nd ed. NCCLS document
M31-A2. NCCLS, 940 West Valley Road, Suite 1400, Wayne,
PA 19087, USA.
[14] Murray, I.A. and Shaw, W.V. (1997) O-acetyltransferases for
chloramphenicol and other natural products. Antimicrob.
Agents Chemother. 41, 1–6.
[15] Levy, S.B., McMurray, L.M., Barbosa, T.M., Burdett, V.,
Courvalin, P., Hillen, W., Roberts, M.C., Rood, J.I. and Taylor,
D.E. (1999) Nomenclature for new tetracycline resistance deter-
minants. Antimicrob. Agents Chemother. 43, 1523–1524.
[16] Roberts, M.C., Sutcliffe, J., Courvalin, P., Jensen, L.B., Rood, J.
and Seppala, H. (1999) Nomenclature for macrolide and mac-
rolide-lincosamide-streptogramin B resistance determinants. An-
timicrob. Agents Chemother. 43, 2823–2830.
[17] Cannon, M., Hartford, S. and Davies, J. (1990) A comparative
study on the inhibitory actions of chloramphenicol, thiamphen-
icol, and some fluorinated derivatives. J. Antimicrob. Chemo-
ther. 26, 307–317.
[18] Vlcek, C., Paces, V., Maltsev, N., Paces, J., Haselkorn, R. and
Fonstein, M. (1997) Sequence of a 189-kb segment of the
chromosome of Rhodobacter capsulatus SB1003. Proc. Natl.
Acad. Sci. USA 94, 9384–9388.
[19] Kaneko, T., Nakamura, Y., Sato, S., Asamizu, E., Kato, T.,
Sasamoto, S., Watanabe, A., Idesawa, K., Ishikawa, A., Kawa-
shima, K., Kimura, T., Kishida, Y., Kiyokawa, C., Kohara, M.,
Matsumoto, M., Matsuno, A., Mochizuki, Y., Nakayama, S.,
Nakazaki, N., Shimpo, S., Sugimoto, M., Takeuchi, C., Yamada,
M. and Tabata, S. (2000) Complete genome structure of the
nitrogen-fixing symbiotic bacterium Mesorhizobium loti. DNA
Res. 7, 331–338.
[20] Tettelin, H., Masignani, V., Cieslewicz, M.J., Eisen, J.A.,
Peterson, S., Wessels, M.R., Paulsen, I.T., Nelson, K.E.,
Margarit, I., Read, T.D., Madoff, L.C., Wolf, A.M., Beanan,
M.J., Brinkac, L.M., Daugherty, S.C., DeBoy, R.T., Durkin, S.,
Kolonay, J.F., Umayam, L.A., Madupu, R., Lewis, M.R.,
Radune, D., Fedorova, N.B., Scanlan, D., Khouri, H., Mulligan,
S., Carty, H.A., Cline, R.T., Gill, J., Scarselli, M., Mora, M.,
Iacobini, E.T., Brettoni, C., Galli, G., Mariani, M., Vegni, F.,

536 S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542
Maione, D., Rinaudo, D., Rappuoli, R., Telford, J.L., Kasper,
D.L., Grandi, G. and Fraser, C.M. (2002) Complete genome
sequence and comparative genomic analysis of an emerging
human pathogen serotype V Streptococcus agalactiae. Proc. Natl.
Acad. Sci. USA 99, 12391–12396.
[21] Ivanova, N., Sorokin, A., Anderson, I., Galleron, N.,
Candelon, B., Kapatral, V., Bhattacharyya, A., Reznik, G.,
Mikhailova, N., Lapidus, A., Chu, L., Mazur, M., Goltsman,
E., Larsen, N., D’Souza, M., Walunas, T., Grechkin, Y.,
Pusch, G., Haselkorn, R., Fonstein, M., Ehrlich, D.S.D.,
Overbeek, R. and Kyrpides, N. (2003) Genome sequence of
Bacillus cereus and comparative analysis with Bacillus anthra-
cis. Nature 423, 87–91.
[22] DelVecchio, V.G., Kapatral, V., Redkar, R.J., Patra, G., Mujer,
C., Los, T., Ivanova, N., Anderson, I., Bhattacharyya, A.,
Lykidis, A., Reznik, G., Jablonski, L., Larsen, N., D’Souza, M.,
Bernal, A., Mazur, M., Goltsman, E., Selkov, E., Elzer, P.H.,
Hagius, S., O’Callaghan, D., Letesson, J.-J., Haselkorn, R.,
Kyrpides, N. and Overbeek, R. (2002) The genome sequence of
the facultative intracellular pathogen Brucella melitensis. Proc.
Natl. Acad. Sci. USA 99, 443–448.
[23] V€olker, T.A., Iida, S. and Bickle, T.A. (1982) A single gene
coding for resistance to both fusidic acid and chloramphenicol. J.
Mol. Biol. 154, 417–425.
[24] Murray, I.A., Martinez-Suarez, J.V., Close, T.J. and Shaw, W.V.
(1990) Nucleotide sequences of genes encoding the type II
chloramphenicol acetyltransferases of Escherichia coli and Hae-
mophilus influenzae, which are sensitive to inhibition by thiol-
reactive reagents. Biochem. J. 272, 505–510.
[25] Leslie, A.G.W. (1990) Refined crystal structure of chloramphen-
icol acetyltransferase at 1.75 A resolution. J. Mol. Biol. 213, 167–
186.
[26] Leslie, A.G.W, Moody, P.C.E. and Shaw, W.V. (1988) Structure
of chloramphenicol acetyltransferase at 1.75 A resolution. Proc.
Natl. Acad. Sci. USA 85, 4133–4137.
[27] Alton, N.K. and Vapnek, D. (1979) Nucleotide sequence analysis
of the chloramphenicol resistance transposon Tn9. Nature 282,
864–869.
[28] Elisha, B.G. and Steyn, L.M. (1991) Identification of an
Acinetobacter baumannii gene region with sequence and organi-
zational similarity to Tn2670. Plasmid 25, 96–104.
[29] Kim, E. and Aoki, T. (1993) The structure of the chloramphenicol
resistance gene on a transferable R plasmid from the fish
pathogen Pasteurella piscicida. Microbiol. Immunol. 37, 705–712.
[30] Nomoto, A. and Yamagishi, K. (1991) DDBJ database accession
no. E02706.
[31] Parkhill, J., Dougan, G., James, K.D., Thomson, N.R., Pickard,
D., Wain, J., Churcher, C., Mungall, K.L., Bentley, S.D.,
Holden, M.T.G., Sebaihia, M., Baker, S., Basham, D., Brooks,
K., Chillingworth, T., Connerton, P., Cronin,, A., Davis, P.,
Davies, R.M., Dowd, L., White, N., Farrar, J., Feltwell, T.,
Hamlin, N., Haque, A., Hien, T.T., Holroyd, S., Jagels, K.,
Krogh, A., Larsen, T.S., Leather, S., Moule, S., O’Gaora, P.,
Parry, C., Quail, M., Rutherford, K., Simmonds, M., Skelton, J.,
Stevens, K., Whitehead, S. and Barrell, B.G. (2001) Complete
genome sequence of a multiple drug resistant Salmonella enterica
serovar Typhi CT18. Nature 413, 848–852.
[32] Gilmour, M.W., Thomson, N.R., Saunders, M., Parkhill, J. and
Taylor, D.E. (2003) GenBank accession no. NC_005211.
[33] Luck, S.N., Turner, S.A., Rajakumar, K., Sakellaris, H. and
Adler, B. (2001) Ferric dicitrate transport system (Fec) of
Shigella flexneri 2a YSH6000 is encoded on a novel pathogenicity
island carrying multiple antibiotic resistance genes. Infect.
Immun. 69, 6012–6021.
[34] Roberts, M.C., Corne, A. and Shaw, W.V. (1982) Molecular
characterization of three chloramphenicol acetyltransferases
isolated from Haemophilus influenzae. J. Bacteriol. 151, 737–741.
[35] Morii, H., Hayashi, N. and Uramoto, K. (2003) Cloning and
nucleotide sequence analysis of the chloramphenicol resistance
gene on conjugative R plasmids from the fish pathogen Photo-
bacterium damselae subsp. piscicida. Dis. Aquat. Organ. 53, 107–
113.
[36] Murray, I.A., Hawkins, J., Keyte, J.W. and Shaw, W.V. (1988)
Nucleotide sequence analysis and overexpression of the gene
encoding a type III chloramphenicol acetyltransferase. Biochem.
J. 252, 173–179.
[37] Vassort-Bruneau, C., Lesage-Descauses, M.-C., Martel, J.-L.,
Lafontand, J.-P. and Chaslus-Dancla, E. (1996) CAT III
chloramphenicol resistance in Pasteurella haemolytica and Pas-
teurella multocida isolated from calves. J. Antimicrob. Chemo-
ther. 38, 205–213.
[38] Kehrenberg, C. and Schwarz, S. (2001) Occurrence and linkage
of genes coding for resistance to sulfonamides, streptomycin and
chloramphenicol in bacteria of the genera Pasteurella and
Mannheimia. FEMS Microbiol. Lett. 205, 283–290.
[39] Kehrenberg, C. and Schwarz, S. (2002) Nucleotide sequence and
organization of plasmid pMVSCS1 from Mannheimia varigena,
identification of a multiresistance gene cluster. J. Antimicrob.
Chemother. 49, 383–386.
[40] Tietze, E. and Smalla, K. (2000) GenBank accession no.
NC_004973.
[41] Charles, I.G., Keyte, J.W. and Shaw, W.V. (1985) Nucleotide
sequence analysis of the cat gene of Proteus mirabilis,
comparison with the type I (Tn9) cat gene. J. Bacteriol. 164,
123–129.
[42] Murray, I.A., Gil, J.A., Hopwood, D.A. and Shaw, W.V. (1989)
Nucleotide sequence of the chloramphenicol acetyltransferase
gene of Streptomyces acrimycini. Gene 85, 283–291.
[43] Harwood, C.R, Williams, D.M. and Lovett, P.S. (1983) Nucle-
otide sequence of Bacillus pumilus gene specifying chloramphen-
icol acetyltransferase. Gene 24, 163–169.
[44] Horinouchi, S. and Weisblum, B. (1982) Nucleotide sequence
and functional map of pC194, a plasmid that specifies
inducible chloramphenicol resistance. J. Bacteriol. 150, 815–
825.
[45] Brenner, D.G. and Shaw, W.V. (1985) The use of synthetic
oligonucleotides with the universal templates for rapid DNA
sequencing, results with staphylococcal replicon pC221. EMBO
J. 4, 561–568.
[46] Projan, S.J, Kornblum, J., Moghazeh, S.L., Edelman, I.,
Gennaro, M.L. and Novick, R.P. (1985) Comparative sequence
and functional analysis of pT181 and pC221, cognate plasmid
replicons from Staphylococcus aureus. Mol. Gen. Genet. 199,
452–464.
[47] Br€uckner, R. and Matzura, H. (1985) Regulation of the inducible
chloramphenicol acetyltransferase gene of the Staphylococcus
aureus plasmid pUB112. EMBO J. 4, 2295–2300.
[48] Schwarz, S., Spies, U. and Cardoso, M. (1991) Cloning and
sequence analysis of a plasmid-encoded chloramphenicol acetyl-
transferase from Staphylococcus intermedius. J. Gen. Microbiol.
137, 977–981.
[49] Schwarz, S. and Cardoso, M. (1991) Molecular cloning, purifi-
cation and properties of a plasmid-encoded chloramphenicol
acetyltransferase from Staphylococcus haemolyticus. Antimicrob.
Agents Chemother. 35, 1277–1283.
[50] Schwarz, S. and Cardoso, M. (1991) Nucleotide sequence and
phylogeny of a chloramphenicol acetyltransferase encoded by the
plasmid pSCS7 from Staphylococcus aureus. Antimicrob. Agents
Chemother. 35, 1551–1556.
[51] Byeon, W.-H. (1992) GenBank accession no. M90091.
[52] Cardoso, M. and Schwarz, S. (1992) Nucleotide sequence and
structural relationships of a chloramphenicol acetyltransferase
encoded by the plasmid pSCS6 from Staphylococcus aureus. J.
Appl. Bacteriol. 72, 289–293.

S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542 537
[53] Moon, K.H., Lee, B.R., Yoon, S.J., Koh, C.H., Jeong, H.D. and
Lee, D.S. (1995) Nucleotide sequences of the REP and CAT
proteins encoded by the chloramphenicol resistance plasmid
pKH7. Yakhak Hoeji 39, 676–680.
[54] Smith, M.C.A. and Thomas, C.D. (2003) GenBank accession no.
NC_005243.
[55] Bhakta, M. and Bal, M. (2003) Identification and characteriza-
tion of a shuttle plasmid with antibiotic resistance gene from
Staphylococcus aureus. Curr. Microbiol. 46, 413–417.
[56] Pepper, K., de Cespedes, G. and Horaud, T. (1988) Heteroge-
neity of chromosomal genes encoding chloramphenicol resistance
in streptococci. Plasmid 19, 71–74.
[57] Trieu-Cuot, P., de Cespedes, G. and Horaud, T. (1992) Nucle-
otide sequence of the chloramphenicol resistance determinant of
the streptococcal plasmid pIP501. Plasmid 28, 272–276.
[58] Lee, M.H., Nittayajarn, A. and Rubens, C.E. (1997) GenBank
accession no. U83488.
[59] Widdowson, C.A., Adrian, P.V. and Klugman, K.P. (2000)
Acquisition of chloramphenicol resistance by the linearization
and integration of the entire staphylococcal plasmid pC194 into
the chromosome of Streptococcus pneumoniae. Antimicrob.
Agents Chemother. 44, 393–395.
[60] Takamatsu, D., Osaki, M. and Sekizaki, T. (2003) Chloram-
phenicol resistance transposable element TnSs1 of Streptococcus
suis, a transposon flanked by IS6-family elements. Plasmid 49,
143–151.
[61] Schwarz, F.V., Perreten, V. and Teuber, M. (2001) Sequence of
the 50-kb conjugative multiresistance plasmid pRE25 from
Enterococcus faecalis RE25. Plasmid 46, 170–187.
[62] Grady, R.E. and Hayes, F. (2003) Axe-Txe, a broad-spectrum
proteic toxin-antitoxin system specified by a multidrug-resistant
clinical isolate of Enterococcus faecium. Mol. Microbiol. 47,
1419–1432.
[63] Aoki, T., Noguchi, N., Sasatsu, M. and Kono, M. (1987)
Complete nucleotide sequence of pTZ12, a chloramphenicol-
resistance plasmid of Bacillus subtilis. Gene 51, 107–111.
[64] Hadorn, K.F., Kayser, F.H. and H€achler, H. (1994) Miniplasmid
derived from Listeria monocytogenes multiresistance plasmid
pWDB100 upon conjugal transfer into Staphylococcus epidermi-
dis carries chloramphenicol resistance gene identical with staph-
ylococcal gene. System. Appl. Microbiol. 17, 492–500.
[65] Lin, C.F, Fung, Z.F., Wu, C.L. and Chung, T.C. (1996)
Molecular characterization of a plasmid-borne (pTC82) chlor-
amphenicol resistance determinant (cat-TC) from Lactobacillus
reuteri G4. Plasmid 36, 116–124.
[66] Perreten, V., Moschetti, G. and Teuber, M. (1995) Genbank
accession no. X92945.
[67] Perreten, V., Schwarz, F., Boeglin, M., Cresta, L., Dasen, G. and
Teuber, M. (1998) Antibiotic resistance spread in food. Nature
391, 801–802.
[68] Gillespie, M.T. and Skurray, R.A. (1988) Structural relationships
among chloramphenicol resistance plasmids of Staphylococcus
aureus. FEMS Microbiol. Lett. 51, 205–210.
[69] Schwarz, S. and Gr€olz-Krug, S. (1991) A chloramphenicol-
streptomycin resistance plasmid from a clinical strain of Staphy-
lococcus sciuri and its structural relationships to other staphylo-
coccal resistance plasmids. FEMSMicrobiol. Lett. 66, 1551–1556.
[70] Schwarz, S. and Noble, W.C. (1994) Structure and putative
origin of a plasmid from Staphylococcus hyicus that mediates
chloramphenicol and streptomycin resistance. Lett. Appl. Mi-
crobiol. 18, 281–284.
[71] Macrina, F.L. and Archer, G.L. (1993) Conjugation and broad
host range plasmids in streptococci and staphylococci. In:
Bacterial Conjugation (Clewell, D.B., Ed.), pp. 313–329. Plenum
Press, New York.
[72] Werckenthin, C., Cardoso, M., Martel, J.L. and Schwarz, S.
(2001) Antimicrobial resistance in staphylococci from animals
with particular reference to bovine Staphylococcus aureus,
porcine Staphylococcus hyicus, and canine Staphylococcus inter-
medius. Vet. Res. 32, 341–362.
[73] Ayoubi, P., Kilic, O. and Vijayakumar, M.N. (1991) Tn5253, the
pneumococcal X (cat tet) BM6001 element is a composite
structure of two conjugative transposons, Tn5251 and Tn5252.
J. Bacteriol. 173, 1617–1622.
[74] Galopin, S. and Leclercq, R. (2003) GenBank accession no.
AY238971.
[75] Bannam, T.L., Crellin, P.K. and Rood, J.I. (1995) Molecular
genetics of the chloramphenicol-resistance transposon Tn4451
from Clostridium perfringens, the TnpX site-specific recombinase
excises a circular transposon molecule. Mol. Microbiol. 16, 535–
551.
[76] Bannam, T.L. and Rood, J.I. (1991) The relationship between the
Clostridium perfringens catQ gene product and chloramphenicol
acetyltransferases from other bacteria. Antimicrob. Agents
Chemother. 35, 471–476.
[77] Bannam, T.L. and Rood, J.I. (1993) Clostridium perfringens–
Escherichia coli shuttle vectors that carry single antibiotic
resistance determinants. Plasmid 29, 233–235.
[78] Huggins, A.S., Bannam, T.L. and Rood, J.I. (1992) Comparative
sequence analysis of the catB gene from Clostridium butyricum.
Antimicrob. Agents Chemother. 36, 2548–2551.
[79] Steffen, C. and Matzura, H. (1989) Nucleotide sequence analysis
and expression studies of a chloramphenicol-acetyltransferase-
coding gene from Clostridium perfringens. Gene 75, 349–354.
[80] Wren, B., Mullany, P., Clayton, C. and Tabaqchali, S. (1989)
Nucleotide sequence of a chloramphenicol acetyltransferase gene
from Clostridium difficile. Nucl. Acids Res. 17, 4877.
[81] Galimand, M., Gerbaud, G., Guibourdenche, M., Riou, J.Y. and
Courvalin, P. (1998) High-level chloramphenicol resistance in
Neisseria meningitidis. N. Engl. J. Med. 339, 868–874.
[82] Shultz, T.R., Tapsall, J.W., White, P.A., Ryan, C., Slyras, D.,
Rood, J.I., Binotto, E. and Richardson, C.J. (2003) Chloram-
phenicol-resistant Neisseria meningitidis containing catP isolated
in Australia. J. Antimicrob. Chemother. 52, 856–859.
[83] Lyras, D. and Rood, J.I. (2000) Transposition of Tn4451 and
Tn4453 involves a circular intermediate that forms a promoter
for the large resolvase TnpX. Mol Microbiol. 38, 588–601.
[84] Trieu-Cuot, P., de Cespedes, G., Bentorcha, F., Delbos, F.,
Gasparand, E. and Horaud, T. (1993) Study of heterogeneity of
chloramphenicol acetyltransferase (CAT) genes in streptococci
and enterococci by polymerase chain reaction, characterization
of a new CAT determinant. Antimicrob. Agents Chemother. 37,
2593–2598.
[85] Wang, Y. and Taylor, D.E. (1990) Chloramphenicol resistance in
Campylobacter coli, nucleotide sequence, expression, and cloning
vector construction. Gene 94, 23–28.
[86] Zhao, J. and Aoki, T. (1992) Cloning and nucleotide sequence
analysis of a chloramphenicol acetyltransferase gene from Vibrio
anguillarum. Microbiol. Immunol. 36, 695–705.
[87] Read, T., Peterson, S., Tourasse, N., Baillie, L., Paulsen, I.,
Nelson, K., Tettelin, H., Fouts, D., Eisen, J., Gill, S., Holtzapple,
E., Okstad, O., Helgason, E., Rilstone, J., Wu, M., Kolonay, J.,
Beanan, M., Dodson, R., Brinkac, L., Gwinn, M., DeBoy, R.,
Madupu, R., Daugherty, S., Durkin, A., Haft, D., Nelson, W.,
Peterson, J., Pop, M., Khouri, H., Radune, D., Benton, J.,
Mahamoud, Y., Jiang, L., Hance, I., Weidman, J., Berry, K.,
Plaut, R., Wolf, A., Watkins, K., Nierman, W., Hazen, A., Cline,
R., Redmond, C., Thwaite, J., White, O., Salzberg, S., Thomason,
B., Friedlander, A., Koehler, T., Hanna, P., Kolsto, A.-B. and
Fraser, C. (2003) The genome sequence ofBacillus anthracisAmes
and comparison to closely related bacteria. Nature 42, 381–386.
[88] White, O., Eisen, J.A., Heidelberg, J.F., Hickey, E.K., Peterson,
J.D., Dodson, R.J., Haft, D.H., Gwinn, M.L., Nelson, W.C.,
Richardson, D.L., Moffat, K.S., Qin, H., Jiang, L., Pamphile,

538 S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542
W., Crosby, M., Shen, M., Vamathevan, J.J., Lam, P., McDon-
ald, L., Utterback, T., Zalewski, C., MakarovaL, K.S., Aravind,
L., Daly, M.J., Minton, K.W., Fleischmann, R.D., Ketchum,
K.E., Nelson, K.E., Salzberg, S., Smith, H.O., Venter, J.C. and
Fraser, C.M. (1999) Genome sequence of the radioresistant
bacterium Deinococcus radiodurans R1. Science 286, 1571–1577.
[89] Nolling, J., Breton, G., Omelchenko, M.V., Markarova, K.S.,
Zeng, Q., Gibson, R., Lee, H.M., Dubois, J., Qiu, D., Hitti, J.,
Wolf, Y.I., Tatusov, R.L., Sabathe, F., Doucette-Stamm, L.,
Soucaille, P., Daly, M.J., Bennett, G.N., Koonin, E.V. and
Smith, D.R. (2001) Genome sequence and comparative analysis
of the solvent-producing bacterium Clostridium acetobutylicum.
J. Bacteriol. 183, 4823–4838.
[90] Bruggemann, H., Baumer, S., Fricke, W.F., Wiezer, A., Liese-
gang, H., Decker, I., Herzberg, C., Martinez-Arias, R., Merkl,
R., Henne, A. and Gottschalk, G. (2003) The genome sequence
of Clostridium tetani, the causative agent of tetanus disease. Proc.
Natl. Acad. Sci. USA 100, 1316–1321.
[91] Lee, H.J. and Kang, H.S. (1999) Genbank accession no.
AF124757.
[92] Xu, J., Bjursell, M.K., Himrod, J., Deng, S., Carmichael, L.K.,
Chiang, H.C., Hooper, L.V. and Gordon, J.I.A. (2003) Genomic
view of the human–Bacteroides thetaiotaomicron symbiosis.
Science 299, 2074–2076.
[93] Lovett, P.S. (1990) Translational attenuation as the regulator of
inducible cat genes. J. Bacteriol. 172, 1–6.
[94] Rende-Fournier, R., Leclercq, R., Galimant, M., Duval, J. and
Courvalin, P. (1993) Identification of the satA gene encoding a
streptogramin A acetyltransferase in Enterococcus faecium
BM4145. Antimicrob. Agents Chemother. 37, 1896–1903.
[95] Werner, G. and Witte, W. (1999) Characterization of a new
enterococcal gene, satG, encoding a putative acetyltransferase
conferring resistance to streptogramin A compounds. Antimic-
rob. Agents Chemother. 43, 1813–1814.
[96] Allignet, J. and El Solh, N. (1995) Diversity among the
grampositive acetyltransferases inactivating streptogramin A
and structurally related compounds and characterization of a
new staphylococcal determinant vatB. Antimicrob. Agents Che-
mother. 39, 2027–2036.
[97] Allignet, J., Loncle, V., Simenel, C., Delpierre, M. and El Solh,
N. (1993) Sequence of a staphylococcal gene, vat, encoding an
acetyltransferase inactivating the A-type compounds of virginia-
mycin-like antibiotics. Gene 130, 91–98.
[98] Tennigkeit, J. and Matzura, H. (1991) Nucleotide sequence
analysis of a chloramphenicol-resistance determinant from Ag-
robacterium tumefaciens and identification of its gene product.
Gene 98, 113–116.
[99] Goodner, B., Hinkle, G., Gattung, S., Miller, N., Blanchard, M.,
Qurollo, B., Goldman, B.S., Cao, Y., Askenazi, M., Halling, C.,
Mullin, L., Houmiel, K., Gordon, J., Vaudin, M., Iartchouk, O.,
Epp, A., Liu, F., Wollam, C., Allinger, M., Doughty, D., Scott,
C., Lappas, C., Markelz, B., Flanagan, C., Crowell, C., Gurson,
J., Lomo, C., Sear, C., Strub, G., Cielo, C. and Slater, S. (2001)
Genome sequence of the plant pathogen and biotechnology agent
Agrobacterium tumefaciens C58. Science 294, 2323–2328.
[100] Parent, R. and Roy, P.H. (1992) The chloramphenicol acetyl-
transferase gene of Tn2424: a new breed of cat. J. Bacteriol. 174,
2891–2897.
[101] Villa, L., Mammina, C., Miriagou, V., Tzouvelekis, L.S.,
Tassios, P.T., Nastasi, A. and Carattoli, A. (2002) Multidrug
and broad-spectrum cephalosporin resistance among Salmonella
enterica serotype Enteritidis clinical isolates in Southern Italy. J.
Clin. Microbiol. 40, 2662–2665.
[102] Wu, J., Shieh, H.K., Shien, J., Gong, S. and Chang, P. (2003)
GenBank accession no. NC_004771.
[103] Tennstedt, T., Szczepanowski, R., Braun, S., Puehler, A. and
Schlueter, A. (2003) Occurrence of integron-associated resistance
gene cassettes located on antibiotic resistance plasmids isolated
from a wastewater treatment plant. FEMS Microbiol. Ecol. 45,
239–252.
[104] Kupzig, S. and Bennett, P.M. (1994) Genbank accession no.
X82455.
[105] Bunny, K.L., Hall, R.M. and Stokes, H.W. (1995) New mobile
gene cassettes containing an aminoglycoside resistance gene
aacA7 and a chloramphenicol resistance gene, catB3, in an
integron in pBWH301. Antimicrob. Agents Chemother. 39, 686–
693.
[106] Tosini, F., Visca, P., Luzzi, I., Dionisi, A.M., Pezzella, C.,
Petrucca, A. and Carattoli, A. (1998) Class 1 integron-borne
multiple-antibiotic resistance carried by IncFI and IncL/M
plasmids in Salmonella enterica serotype Typhimurium. Anti-
microb. Agents Chemother. 42, 3053–3058.
[107] Houang, E.T., Chu, Y.W., Lo, W.S., Chu, K.Y. and Cheng, A.F.
(2003) Epidemiology of rifampin ADP-ribosyltransferase (arr-2)
and metallo-beta-lactamase (bla (IMP-4)) gene cassettes in class 1
integrons in Acinetobacter strains isolated from blood cultures in
1997–2000. Antimicrob. Agents Chemother. 47, 1382–1390.
[108] Wang, M., Tran, J.H., Jacoby, G.A., Zhang, Y., Wang, F. and
Hooper, D.C. (2003) Plasmid-mediated quinolone resistance in
clinical isolates of Escherichia coli from Shanghai, China.
Antimicrob. Agents Chemother. 47, 2242–2248.
[109] Yan, J.J., Ko, W.C. and Wu, J.J. (2001) Identification of a
plasmid encoding SHV-12, TEM-1, and a variant of IMP-2
metallo-beta-lactamase, IMP-8, from a clinical isolate of Klebsi-
ella pneumoniae. Antimicrob. Agents Chemother. 45, 2368–2371.
[110] Kim, J. (2003) GenBank accession no. AF227506.
[111] Pai, H., Byeon, J.-H., Yu, S,, Lee, B.K. and Kim, S. (2003)
Salmonella enterica serovar Typhi strains isolated in Korea
containing a multidrug resistance class 1 integron. Antimicrob.
Agents Chemother. 47, 2006–2008.
[112] Laraki, N., Galleni, M., Thamm, I., Riccio, L., Amicosante, G.,
Frere, J.M. and Rossolini, G.M. (1999) Structure of In31, a
blaIMP -containing Pseudomonas aeruginosa integron phyletically
related to In5, which carries an unusual array of gene cassettes.
Antimicrob. Agents Chemother. 43, 890–901.
[113] Lee, K., Yum, J.H., Yong, D., Roh, K.H., Chong, Y. and
Rossolini, G.M. (2001) GenBank accession no. AF418284.
[114] White, P.A., Bunny, K.L., Hall, R.M. and Stokes, H.W. (1999)
Characterization of a chloramphenicol acetyltransferase deter-
minant found in the chromosome of Pseudomonas aeruginosa.
FEMS Microbiol. Lett. 175, 27–35.
[115] Stover, C.K., Pham, X.-Q.T., Erwin, A.L., Mizoguchi, S.D.,
Warrener, P., Hickey, M.J., Brinkman, F.S.L., Hufnagle, W.O.,
Kowalik, D.J., Lagrou, M., Garber, R.L., Goltry, L., Tolentino,
E., Westbrock-Wadman, S., Yuan, Y., Brody, L.L., Coulter,
S.N., Folger, K.R., Kas, A., Larbig, K., Lim, R.M., Smith, K.A.,
Spencer, D.H., Wong, G.K.-S., Wu, Z. and Paulsen, I.T. (2000)
Complete genome sequence of Pseudomonas aeruginosa PAO1 an
opportunistic pathogen. Nature 40, 6959–6964.
[116] Rowe-Magnus, D.A., Guerout, A.M. and Mazel, D. (2002)
Bacterial resistance evolution by recruitment of super-integron
gene cassettes. Mol. Microbiol. 43, 1657–1669.
[117] Heidelberg, J.F., Eisen, J.A., Nelson, W.C., Clayton, R.A.,
Gwinn, M.L., Dodson, R.J., Haft, D.H., Hickey, E.K., Peterson,
J.D., Umayam, L., Gill, S.R., Nelson, K.E., Read, T.D., Tettelin,
H., Richardson, D., Ermolaeva, M.D., Vamathevan, J., Bass, S.,
Qin, H., Dragoi, I., Sellers, P., McDonald, L., Utterback, T.,
Fleishmann, R.D., Nierman, W.C. and White, O. (2000) DNA
sequence of both chromosomes of the cholera pathogen Vibrio
cholerae. Nature 40, 6477–6483.
[118] Heidelberg, J., Paulsen, I., Nelson, K., Gaidos, E., Nelson, W.,
Read, T., Eisen, J., Seshadri, R., Ward, N., Methe, B., Clayton,
R., Meyer, T., Tsapin, A., Scott, J., Beanan, M., Brinkac, L.,
Daugherty, S., DeBoy, R., Dodson, R., Durkin, A., Haft, D.,

S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542 539
Kolonay, J., Madupu, R., Peterson, J., Umayam, L., White, O.,
Wolf, A., Vamathevan, J., Weidman, J., Impraim, M., Lee, K.,
Berry, K., Lee, C., Mueller, J., Khouri, H., Gill, J., Utterback,
T., McDonald, L., Feldblyum, T., Smith, H., Venter, J., Nealson,
K. and Fraser, C. (2002) Genome sequence of the dissimilatory
metal ion-reducing bacterium Shewanella oneidensis. Nat. Bio-
technol. 20, 1118–1123.
[119] Makino, K., Oshima, K., Kurokawa, K., Yokoyama, K., Uda,
T., Tagomori, K., Iijima, Y., Najima, M., Nakano, M.,
Yamashita, A., Kubota, Y., Kimura, S., Yasunaga, T., Honda,
T., Shinagawa, H., Hattori, M. and Iida, T. (2003) Genome
sequence of Vibrio parahaemolyticus, a pathogenic mechanism
distinct from that of V. cholerae. Lancet 361, 743–749.
[120] Rodgers, E.J., Rahman, M.S., Hill, R.T. and Lovett, P.S. (2002)
The chloramphenicol-inducible catB gene in Agrobacterium
tume-faciens is regulated by translational attenuation. J. Bacte-
riol. 184, 4296–4300.
[121] Butaye, P., Cloeckaert, A. and Schwarz, S. (2003) Mobile genes
coding for efflux-mediated antimicrobial resistance in Gram-
positive and Gram-negative bacteria. Int. J. Antimicrob. Agents
22, 205–210.
[122] Rubens, C.E., McNeill, W.F. and Farrar Jr., W.E. (1979)
Transposable plasmid deoxyribonucleic acid sequence in Pseu-
domonas aeruginosa which mediates resistance to gentamicin and
four other antimicrobial agents. J. Bacteriol. 139, 877–882.
[123] Stokes, H.W. and Hall, R.M. (1991) Sequence analysis of the
inducible chloramphenicol resistance determinant in the Tn1696
integron suggests regulation by translational attenuation. Plas-
mid 26, 10–19.
[124] Bissonnette, L., Champetier, S., Buisson, J.P. and Roy, P.H.
(1991) Characterization of the nonenzymatic chloramphenicol
resistance (cmlA) gene of the In4 integron of Tn1696, similarity
of the product to transmembrane transport proteins. J. Bacteriol.
173, 4493–4502.
[125] George, A.M. and Hall, R.M. (2002) Efflux of chloramphenicol
by the CmlA1 protein. FEMS Microbiol. Lett. 209, 209–
213.
[126] Poirel, L., Naas, T., Guibert, M., Chaibi, E.B., Labia, R. and
Nordmann, P. (1999) Molecular and biochemical characteriza-
tion of VEB-1a novel class A extended-spectrum beta-lactamase
encoded by an Escherichia coli integron gene. Antimicrob.
Agents Chemother. 43, 573–581.
[127] Carattoli, A., Tosini, F., Giles, W.P., Rupp, M.E., Hinrichs,
S.H., Angulo, F.J., Barrett, T.J. and Fey, P.D. (2002) Charac-
terization of plasmids carrying CMY-2 from expanded-spectrum
cephalosporin-resistant Salmonella strains isolated in the United
States between 1996 and 1998. Antimicrob. Agents Chemother.
46, 1269–1272.
[128] Poirel, L., Le Thomas, I., Naas, T., Karim, A. and Nordmann, P.
(2000) Biochemical sequence analyses of GES-1, a novel class A
extended-spectrum beta-lactamase and the class 1 integron In52
from Klebsiella pneumoniae. Antimicrob. Agents Chemother. 44,
622–632.
[129] Poirel, L., Decousser, J.W. and Nordmann, P. (2003) Insertion
sequence ISEcp1B is involved in expression and mobilization of a
bla (CTX-M) beta-lactamase gene. Antimicrob. Agents Chemo-
ther. 47, 2938–2945.
[130] Partridge, S.R., Recchia, G.D., Stokes, H.W. and Hall, R.M.
(2001) Family of class 1 integrons related to In4 from Tn1696.
Antimicrob. Agents Chemother. 45, 3014–3020.
[131] Aubert, D., Poirel, L., Chevalier, J., Leotard, S., Pages, J.M. and
Nordmann, P. (2001) Oxacillinase-mediated resistance to cefe-
pime and susceptibility to ceftazidime in Pseudomonas aerugin-
osa. Antimicrob. Agents Chemother. 45, 1615–1620.
[132] Riccio, M.L., Docquier, J.D., Dell’Amico, E., Luzzaro, F.,
Amicosante, G. and Rossolini, G.M. (2003) Novel 3-N-amino-
glycoside acetyltransferase gene, aac(3)-Ic, from a Pseudomonas
aeruginosa integron. Antimicrob. Agents Chemother. 47, 1746–
1748.
[133] White, D.G., Hudson, C., Maurer, J.J., Ayers, S., Zhao, S., Lee,
M.D., Bolton, L., Foley, T. and Sherwood, J. (2000) Character-
ization of chloramphenicol and florfenicol resistance in Esche-
richia coli associated with bovine diarrhea. J. Clin. Microbiol. 38,
4593–4598.
[134] Bischoff, K.M., White, D.G., McDermott, P.F., Zhao, S.,
Gaines, S., Maurer, J.J. and Nisbet, D.J. (2002) Characterization
of chloramphenicol resistance in beta-hemolytic Escherichia coli
associated with diarrhea in neonatal swine. J. Clin. Microbiol. 40,
389–394.
[135] Ploy, M.-C., Courvalin, P. and Lambert, R. (1998) Character-
ization of In40 of Enterobacter aerogenes BM2688, a class 1
integron with two new gene cassettes, cmlA2 and qacF.
Antimicrob. Agents Chemother. 42, 2557–2563.
[136] Dorman, C.J., Foster, T.J. and Shaw, W.V. (1986) Nucleotide
sequence of the R26 chloramphenicol resistance determinant and
identification of its gene product. Gene 41, 349–353.
[137] Dorman, C.J. and Foster, T.J. (1982) Nonenzymatic chloram-
phenicol resistance determinants specified by plasmids R26 and
R55-1 in Escherichia coli K-12 do not confer high-level resistance
to fluorinated analogs. Antimicrob. Agents Chemother. 22, 912–
914.
[138] Kim, E. and Aoki, T. (1996) Sequence analysis of the florfenicol
resistance gene encoded in the transferable R plasmid from a fish
pathogen Pasteurella piscicida. Microbiol. Immunol. 40, 665–
669.
[139] Arcangioli, M.A., Leroy-Setrin, S., Martel, J.L. and Chaslus-
Dancla, E. (1999) A new chloramphenicol and florfenicol
resistance gene linked to an integron structure in Salmonella
typhimurium DT104. FEMS Microbiol. Lett. 174, 327–332.
[140] Briggs, C.E. and Fratamico, P.M. (1999) Molecular characteriza-
tion of an antibiotic resistance gene cluster of Salmonella typhimu-
riumDT104. Antimicrob. Agents Chemother. 43, 846–849.
[141] Bolton, L.F., Kelley, L.C., Lee, M.D., Fedorka-Cray, P.J. and
Maurer, J.J. (1999) Detection of Salmonella typhimurium DT104
based on the gene St-flo which confers resistance to florfenicol
and chloramphenicol. J. Clin. Microbiol. 37, 1348–1351.
[142] Cloeckaert, A. and Schwarz, S. (2001) Molecular characteriza-
tion, spread and evolution of multidrug resistance in Salmonella
enterica Typhimurium DT104. Vet. Res. 32, 301–310.
[143] Frech, G., Kehrenberg, C. and Schwarz, S. (2003) Resistance
phenotypes and genotypes of multiresistant Salmonella enterica
subsp. enterica serovar Typhimurium var. Copenhagen isolates
from animal sources. J. Antimicrob. Chemother. 51, 180–182.
[144] Kim, S. (2003) GenBank accession no. AY339985.
[145] Boyd, D., Peters, G.A., Cloeckaert, A., Sidi Boumedine, K.,
Chaslus-Dancla, E., Imberechts, H. and Mulvey, M.R. (2001)
Complete nucleotide sequence of a 43-kilobase genomic island
associated with the multidrug resistance region of Salmonella
enterica serovar Typhimurium DT104 and its identification in
phage type DT120 and serovar Agona. J. Bacteriol. 183, 5725–
5732.
[146] Boyd, D., Cloeckaert, A., Chaslus-Dancla, E. and Mulvey, M.R.
(2002) Characterization of variant Salmonella genomic island 1
multidrug resistance regions from serovars Typhimurium DT104
and Agona. Antimicrob. Agents Chemother. 46, 1714–1722.
[147] Cloeckaert, A., Sidi Boumedine, K., Flaujac, G., Imberechts, H.,
D’Hooghe, I. and Chaslus-Dancla, E. (2000) Occurrence of a
Salmonella enterica serovar Typhimurium DT104-like antibiotic
resistance gene cluster including the floR gene in S. enterica
serovar Agona isolates. Antimicrob. Agents Chemother. 44,
1359–1361.
[148] Doublet, B., Lailler, R., Meunier, D., Brisabois, A., Boyd, D.,
Mulvey, M.R., Chaslus-Dancla, E. and Cloeckaert, A. (2003)
Variant Salmonella genomic island 1 antibiotic resistance gene

540 S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542
cluster in Salmonella enterica serovar Albany. Emerg. Infect. Dis.
9, 585–591.
[149] Doublet, B., Weill, F.X., Fabre, L., Chaslus-Dancla, E. and
Cloeckaert, A. (2003) GenBank accession no. AY458224.
[150] Meunier, D., Boyd, D., Mulvey, M.R., Baucheron, S., Mam-
mina, C., Nastasi, A., Chaslus-Dancla, E. and Cloeckaert, A.
(2002) Salmonella enterica serotype Typhimurium DT104 anti-
biotic resistance genomic island I in serotype Paratyphi B.
Emerg. Infect. Dis. 8, 430–433.
[151] Keyes, K., Hudson, C., Maurer, J.J., Thaye, S., White, D.G. and
Lee, M.D. (2000) Detection of florfenicol resistance genes in
Escherichia coli from sick chickens. Antimicrob. Agents Chemo-
ther. 44, 421–424.
[152] Cloeckaert, A., Baucheron, S., Flaujac, G., Schwarz, S.,
Kehrenberg, C., Martel, J.L. and Chaslus-Dancla, E. (2000)
Plasmid-mediated florfenicol resistance by the floR gene in
Escherichia coli isolated from cattle. Antimicrob Agents Chemo-
ther. 44, 2858–2860.
[153] Doublet, B., Schwarz, S., Nußbeck, E., Baucheron, S., Martel,
J.L., Chaslus-Dancla, E. and Cloeckaert, A. (2002) Molecular
analysis of chromosomally florfenicol resistant Escherichia coli
isolates from France and Germany. J. Antimicrob. Chemother.
49, 49–54.
[154] Blickwede, M. and Schwarz, S. (2004) Molecular analysis of
florfenicol-resistant Escherichia coli isolates from pigs. J. Anti-
microb. Chemother. 53, 58–64.
[155] Cloeckaert, A., Baucheron, S. and Chaslus-Dancla, E. (2001)
Nonenzymatic chloramphenicol resistance mediated by IncC
plasmid R55 is encoded by a floR gene variant. Antimicrob.
Agents Chemother. 45, 2381–2382.
[156] Meunier, D., Baucheron, S., Chaslus-Dancla, E., Martel, J.L. and
Cloeckaert, A. (2003) Florfenicol resistance in Salmonella enterica
serovar Newport mediated by a plasmid related to R55 from
Klebsiella pneumoniae. J. Antimicrob. Chemother. 51, 1007–1009.
[157] Hochhut, B., Lotfi, Y., Mazel, D., Faruque, S.M., Woodgate, R.
and Waldor, M.K. (2001) Molecular analysis of antibiotic
resistance gene clusters in Vibrio cholerae O139 and O1 SXT
constins. Antimicrob. Agents Chemother. 45, 2991–3000.
[158] Beaber, J.W., Hochhut, B. and Waldor, M.K. (2002) Genomic
and functional analyses of SXT, an integrating antibiotic
resistance gene transfer element derived from Vibrio cholerae. J.
Bacteriol. 184, 4259–4269.
[159] Paulsen, I.T., Brown, M.H. and Skurray, R.A. (1996) Proton-
dependent multidrug efflux systems. Microbiol. Rev. 60, 575–608.
[160] Kehrenberg, C. and Schwarz, S. (2004) fexA, a novel Staphylo-
coccus lentus gene encoding resistance to florfenicol and chlor-
amphenicol. Antimicrob. Agents Chemother 48, 615–618.
[161] Dittrich, W., Betzler, M. and Schrempf, H. (1991) An amplifiable
and deletable chloramphenicol-resistance determinant of Strep-
tomyces lividans 1326 encodes a putative transmembrane protein.
Mol. Microbiol. 5, 2789–2799.
[162] Mosher, R.H., Camp, D.J., Yang, K., Brown, M.P., Shaw, W.V.
and Vining, L.C. (1995) Inactivation of chloramphenicol by O-
phosphorylation. J. Biol. Chem. 27, 27000–27006.
[163] Desomer, J., Vereecke, D., Crespi, M. and van Montagu, M.
(1992) The plasmid-encoded chloramphenicol-resistance protein
of Rhodococcus fascians is homologous to the transmembrane
tetracycline efflux proteins. Mol. Microbiol. 6, 2377–2385.
[164] Nagy, I., Schoofs, G., Vanderleyden, J. and de Mot, R. (1997)
Transposition of the IS21-related element IS1415 in Rhodococcus
erythropolis. J. Bacteriol. 179, 4635–4638.
[165] Tauch, A., Zheng, Z., Puhler, A. and Kalinowski, J. (1998)
Corynebacterium striatum chloramphenicol resistance transposon
Tn5564, genetic organization and transposition in Corynebacte-
rium glutamicum. Plasmid 40, 126–139.
[166] Zheng, Z., Shi., S., Jiang, X., Wang, Z. and Caro, L. (1997)
GenBank accession no. U85507.
[167] Parkhill, J., Wren, B.W., Thomson, N.R., Titball, R.W., Holden,
M.T.G., Prentice, M.B., Sebaihia, M., James, K.D., Churcher,
C., Mungall, K.L., Baker, S., Basham, D., Bentley, S.D., Brooks,
K., Cerdeno-Tarraga, A.M., Chillingworth, T., Cronin, A.,
Davies, R.M., Davis, P., Dougan, G., Feltwell, T., Hamlin, N.,
Holroyd, S., Jagels, K., Leather, S., Karlyshev, A.V., Moule, S.,
Oyston, P.C.F., Quail, M., Rutherford, K., Simmonds, M.,
Skelton, J., Stevens, K., Whitehead, S. and Barrell, B.G. (2001)
Genome sequence of Yersinia pestis, the causative agent of
plague. Nature 413, 523–527.
[168] Kapatral, V., Anderson, I., Ivanova, N., Reznik, G., Los, T.,
Lykidis, A., Bhattacharyya, A., Bartman, A., Gardner, W.,
Grechkin, G., Zhu, L., Vasieva, O., Chu, L., Kogan, Y., Chaga,
O., Goltsman, E., Bernal, A., Larsen, N., D’Souza, M., Walunas,
T., Pusch, G., Haselkorn, R., Fonstein, M., Kyrpides, N. and
Overbeek, R. (2002) Genome sequence and analysis of the oral
bacterium Fusobacterium nucleatum strain ATCC 25586. J.
Bacteriol. 184, 2005–2018.
[169] Rankin, S., Li, Z. and Isberg, R.R. (2002) Macrophage-induced
genes of Legionella pneumophila: Protection from reactive inter-
mediates and solute imbalance during intracellular growth.
Infect. Immun. 70, 3637–3648.
[170] Raymond, J., Zhaxybayeva, O., Gogarten, J.P., Gerdes, S.Y. and
Blankenship, R.E. (2002) Whole-genome analysis of photosyn-
thetic prokaryotes. Science 298, 1616–1620.
[171] Eisen, J.A., Nelson, K.E., Paulsen, I.T., Heidelberg, J.F., Wu,
M., Dodson, R.J., Deboy, R., Gwinn, M.L., Nelson, W.C., Haft,
D.H., Hickey, E.K., Peterson, J.D., Durkin, A.S., Kolonay, J.L.,
Yang, F., Holt, I., Umayam, L.A., Mason, T., Brenner, M.,
Shea, T.P., Parksey, D., Nierman, W.C., Feldblyum, T.V.,
Hansen, C.L., Craven, M.B., Radune, D., Vamathevan, J.,
Khouri, H., White, O., Gruber, T.M., Ketchum, K.A., Venter,
J.C., Tettelin, H., Bryant, D.A. and Fraser, C.M. (2002) The
complete genome sequence of Chlorobium tepidum TLS, a
photosynthetic, anaerobic, green-sulfur bacterium. Proc. Natl.
Acad. Sci. USA 99, 9509–9514.
[172] Capela, D., Barloy-Hubler, F., Gouzy, J., Bothe, G., Ampe, F.,
Batut, J., Boistard, P., Becker, A., Boutry, M., Cadieu, E.,
Dreano, S., Gloux, S., Godrie, T., Goffeau, A., Kahn, D., Kiss,
E., Lelaure, V., Masuy, D., Pohl, T., Portetelle, D., Puehler, A.,
Purnelle, B., Ramsperger, U., Renard, C., Thebault, P., Van-
denbol, M., Weidner, S. and Galibert, F. (2001) Analysis of the
chromosome sequence of the legume symbiont Sinorhizobium
meliloti strain 1021. Proc. Natl. Acad. Sci. USA 98, 9877–
9882.
[173] NCBI Microbial Genomes Annotation Project (2004) GenBank
accession no. NZ_AAED01000007.
[174] McMurray, L.M., George, A.M. and Levy, S.B. (1994) Active
efflux of chloramphenicol in susceptible Escherichia coli strains
and in multiple-antibiotic-resistant (Mar) mutants. Antimicrob.
Agents Chemother. 38, 542–546.
[175] Okusu, H., Ma, D. and Nikaido, H. (1996) AcrAB efflux pump
plays a major role in the antibiotic resistance phenotype of
Escherichia coli multiple-antibiotic-resistance (Mar) mutants. J.
Bacteriol. 178, 306–308.
[176] Baucheron, S., Imberechts, H., Chaslus-Dancla, E. and Cloe-
ckaert, A. (2002) The AcrB multidrug transporter plays a major
role in high-level fluoroquinolone resistance in Salmonella
enterica serovar Typhimurium phage type DT204. Microb. Drug
Resist. 8, 281–289.
[177] Lee, A., Mao, W., Warren, M.S., Mistry, A., Hoshino, K.,
Okumura, R., Ishida, H. and Lomovskaya, O. (2000) Interplay
between efflux pumps may provide either additive or multiplica-
tive effects on drug resistance. J. Bacteriol. 182, 3142–3150.
[178] Edgar, R. and Bibi, E. (1997) MdfA, an Escherichia coli
multidrug resistance protein with an extraordinarily broad
spectrum of drug recognition. J. Bacteriol. 179, 2274–2280.

S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542 541
[179] Nilsen, I.W., Bakke, I., Vader, A., Olsvik, O. and El-Gewely,
M.R. (1996) Isolation of cmr, a novel chloramphenicol resistance
gene encoding a putative efflux pump. J. Bacteriol. 178, 3188–
3193.
[180] Poole, K. (2002) Multidrug efflux pumps and antimicrobial
resistance in Pseudomonas aeruginosa and related organisms. In:
Microbial Multidrug Efflux (Paulsen, I.T. and Lewis, K., Eds.),
pp. 273–298. Horizon Scientific Press, Wymondham, UK.
[181] Li, X.-Z., Livermore, D.M. and Nikaido, H. (1994) Role of efflux
pump(s) in intrinsic resistance of Pseudomonas aeruginosa:
resistance to tetracycline, chloramphenicol, and norfloxacin.
Antimicrob. Agents Chemother. 38, 1732–1741.
[182] Sulavik, M.C., Houseweart, C., Cramer, C., Jiwani, N., Murg-
olo, N., Greene, J., DiDomenico, B., Shaw, K.J., Miller, G.H.,
Hare, R. and Shimer, G. (2001) Antibiotic susceptibility profiles
of Escherichia coli strains lacking multidrug efflux pump genes.
Antimicrob. Agents Chemother. 45, 1126–1136.
[183] Yoshida, H., Bogaki, M., Nakamura, S., Ubukata, K. and
Konno, M. (1990) Nucleotide sequence and characterization of
the Staphylococcus aureus norA gene which confers resistance to
quinolones. J. Bacteriol. 172, 6942–6949.
[184] Ahmed, M., Lyass, L., Markham, P.N., Taylor, S.S., Vazquez-
Laslop, N. and Neyfakh, A.A. (1995) Two highly similar
multidrug transporters of Bacillus subtilis whose expression is
differentially regulated. J. Bacteriol. 177, 3904–3910.
[185] Mosher, R.H., Ranade, N.P., Schrempf, H. and Vining, L.C.
(1990) Chloramphenicol resistance in Streptomyces, cloning and
characterization of a chloramphenicol hydrolase gene from
Streptomyces venezuelae. J. Gen. Microbiol. 136, 293–301.
[186] Izard, T. and Ellis, J. (2000) The crystal structures of chloram-
phenicol phosphotransferase reveal a novel inactivation mecha-
nism. EMBO J. 19, 2690–2700.
[187] Burns, J.L., Mendelman, P.M., Levy, J., Stull, T.L. and Smith,
A.L. (1985) A permeability barrier as a mechanism of chloram-
phenicol resistance in Haemophilus influenzae. Antimicrob.
Agents Chemother. 27, 46–54.
[188] Burns, J.L., Hedi, L.A. and Lien, D.M. (1989) Chloramphenicol
resistance in Pseudomonas cepacia because of decreased perme-
ability. Antimicrob. Agents Chemother. 33, 136–141.
[189] Burns, J.L., Rubens, C.E., Mendelman, P.M. and Smith, A.L.
(1986) Cloning and expression in Escherichia coli of a gene
encoding nonenzymatic chloramphenicol resistance from Pseu-
domonas aeruginosa. Antimicrob. Agents Chemother. 29, 445–
450.
[190] Toro, C.S., Lobos, S.R., Calderon, I., Rodriguez, M. and Mora,
G.C. (1990) Clinical isolate of a porinless Salmonella typhi
resistant to high levels of chloramphenicol. Antimicrob. Agents
Chemother. 34, 1715–1719.
[191] Quintiliani Jr., R., Sahm, D.F. and Courvalin, P. (1999)
Mechanisms of resistance to antimicrobial agents. In: Manual
of Clinical Microbiology (Murray, P.R., Baron, E.J., Pfaller,
M.A., Tenover, F.C. and Yolken, R.H., Eds.), 7th edn, pp. 1505–
1525. ASM Press, Washington D.C..
[192] Baughman, G.A. and Fahnestock, S.R. (1979) Chloramphenicol
resistance mutation in Escherichia coli which maps in the major
ribosomal protein gene cluster. J. Bacteriol. 137, 1315–1323.
[193] Anderson, L.M., Henkin, T.M., Chambliss, G.H. and Bott, K.F.
(1984) New chloramphenicol resistance locus in Bacillus subtilis.
J. Bacteriol. 158, 386–388.
[194] Ettayebi, M., Prasad, S.M. and Morgan, E.A. (1985) Chloram-
phenicol-erythromycin resistance mutations in a 23S rRNA gene
of Escherichia coli. J. Bacteriol. 162, 551–557.
[195] Vester, B. and Douthwaite, S. (2001) Macrolide resistance
conferred by base substitutions in 23S rRNA. Antimicrob.
Agents Chemother. 45, 1–12.
[196] Murray, I.A. (2000) Chloramphenicol resistance. In: Gram-
positive Pathogens (Fishetti, V.A., Novick, R.P., Ferretti, J.J.,
Portnoy, D.A. and Rood, J.I., Eds.), pp. 678–684. ASM Press,
Washington, D.C..
[197] Schwarz, S., Werckenthin, C. and Kehrenberg, C. (2000)
Identification of a plasmid-borne chloramphenicol/florfenicol
resistance gene in Staphylococcus sciuri. Antimicrob. Agents
Chemother. 44, 2530–2533.
[198] Guerra, B., Junker, E., Schroeter, A., Malorny, B., Lehmann, S.
and Helmuth, R. (2003) Phenotypic and genotypic characteriza-
tion of antimicrobial resistance in German Escherichia coli
isolates from cattle, swine and poultry. J. Antimicrob. Chemo-
ther. 52, 489–492.
[199] Schmidt, A.S., Bruun, M.S., Dalsgaard, I. and Larsen, J.B.
(2001) Incidence, distribution, and spread of tetracycline resis-
tance determinants and integron-associated antibiotic resistance
genes among motile aeromonads from fish farming environment.
Appl. Environ. Microbiol. 67, 5675–5682.
[200] Seral, C., Castillo, F.J., Rubio-Calvo, M.C., Fenoll, A., Garcia,
C. and Gomez-Luis, R. (2001) Distribution of resistance genes
tet(M), aph 30-III, catpC194 and the integrase gene of Tn1545 in
clinical Streptococcus pneumoniae harbouring erm(B) and mef(A)
genes in Spain. J. Antimicrob. Chemother. 47, 863–866.
[201] Cardoso, M. and Schwarz, S. (1992) Chloramphenicol resistance
plasmids in Staphylococcus aureus isolated from bovine subclin-
ical mastitis. Vet. Microbiol. 30, 223–232.
[202] Schwarz, S. (1994) Emerging chloramphenicol resistance in
Staphylococcus lentus from mink following chloramphenicol
treatment: characterisation of the resistance genes. Vet. Micro-
biol. 41, 51–61.
[203] Schwarz, S. and Werckenthin, C. (1994) Antibiotic resistance in
staphylococci from pigeons. Vet. Dermatol. 5, 9–12.
[204] Schwarz, S., Cardoso, M. and Blobel, H. (1989) Plasmid-
mediated chloramphenicol resistance in Staphylococcus hyicus.
J. Gen. Microbiol. 135, 3329–3336.
[205] Greene, R.T. and Schwarz, S. (1992) Small antibiotic resistance
plasmids in Staphylococcus intermedius. Zentralbl. Bakteriol. 276,
380–389.
[206] Schwarz, S., Werckenthin, C., Pinter, L., Kent, L.E. and Noble,
W.C. (1995) Chloramphenicol resistance in Staphylococcus
intermedius from a single veterinary centre: evidence for plasmid
and chromosomal location of the resistance genes. Vet. Micro-
biol. 43, 151–159.
[207] Lyon, B.R. and Skurray, R. (1987) Antimicrobial resistance
in Staphylococcus aureus: genetic basis. Microbiol. Rev. 51, 88–
137.
[208] Burrus, V. and Waldor, M.K. (2003) Control of SXT integration
and excision. J. Bacteriol. 185, 5045–5054.
[209] Burrus, V., Pavlovic, G., Decaris, B. and Guedon, G. (2002)
Conjugative transposons: the tip of the iceberg. Mol. Microbiol.
46, 601–610.
[210] Sabtcheva, S., Galimand, M., Courvalin, P. and Lambert, T.
(2003) The ant(40)-IIb gene of Pseudomonas aeruginosa BM4492
is part of a multiple antibiotic gene cluster homologous to the
genomic island of Salmonella enterica serovar Typhimurium
DT104. In: Abstracts of the 43rd Interscience Conference on
Antimicrobial Agents and Chemotherapy (ICAAC), C1-1181,
85, ASM Press Washington D.C.
[211] Sanchez, S., McCrackin Stevenson, M.A., Hudson, C.R., Maier,
M., Buffington, T., Dam, Q. and Maurer, J.J. (2002) Character-
ization of multidrug-resistant Escherichia coli isolates associated
with nosocomial infections in dogs. J. Clin. Microbiol. 40, 3586–
3595.
[212] Doublet, B., Carattoli, A., Whichard, J.M., White, D.G.,
Baucheron, S., Chaslus-Dancla, E. and Cloeckaert, A. (2004)
Plasmid-mediated florfenicol and ceftriaxone resistance encoded
by the floR and blaCMY�2 genes in Salmonella enterica serovars
Typhimurium and Newport isolated in the United States. FEMS
Microbiol. Lett. 233, 301–305.

542 S. Schwarz et al. / FEMS Microbiology Reviews 28 (2004) 519–542
[213] Berge, A.C.B., Adaska, J.M. and Sischo, W.M. (2004) Use of
antibiotic susceptibility patterns and pulsed-field gel electropho-
resis to compare historic and contemporary isolates of multi-
drug resistant Salmonella enterica subsp. enterica serovar New-
port. Appl. Environm. Microbiol. 70, 318–323.
[214] Levy, S.B. (1992) The Antibiotic Paradox: How Miracle Drugs
are Destroying the Miracle. Plenum Press, New York.
[215] Fabricant, S.J. and Hirschhorn, N. (1987) Deranged distribution,
perverse prescription, unprotected use: the irrationality of
pharmaceuticals in the developing world. Health Policy Planning
2, 204–213.
[216] Gilmore, A. (1996) Chloramphenicol and the politics of health.
Can. Med. Assoc. J. 134, 423–435.
[217] DANMAP (2002) Use of antimicrobial agents and occurrence of
antimicrobial resistance in bacteria from food animals, foods and
humans in Denmark. ISSN 1600-2032.
[218] Simoons-Smit, A.M., Savelkoul, P.H.M., Stoof, J., Starink, T.M.
and Vanderbroucke-Grauls, C.M.J. (2000) Transmission of
Staphylococcus aureus between humans and domestic animals
in a household. Eur. J. Clin Microbiol. Infect. Dis. 19, 150–152.
[219] Tanner, M.A., Everett, C.L. and Youvan, D.C. (2000) Molecular
phylogenetic evidence for noninvasive zoonotic transmission of
Staphylococcus intermedius from a canine pet to a human. J. Clin.
Microbiol. 38, 1628–1631.
[220] Schwarz, S., Cardoso, M., Gr€olz-Krug, S. and Blobel, H. (1990)
Common antibiotic resistance plasmids in Staphylococcus aureus
and Staphylococcus epidermidis from human and canine infec-
tions. Zentralbl. Bakteriol 273, 369–377.
[221] Harnett, N.M. and Gyles, C.L. (1985) Linkage of genes for heat-
stable enterotoxin, drug resistance, K99 antigen, and colicin in
bovine and porcine strains of enterotoxigenic Escherichia coli.
Am. J. Vet. Res. 46, 428–433.

PUBLIKATION 2
Kehrenberg C., J. Mumme, J. Wallmann, J. Verspohl, R. Tegeler, T. Kuhn and S. Schwarz:
Monitoring of florfenicol susceptibility among bovine and porcine respiratory tract pathogens collected in Germany during the years 2002 and 2003.
J. Antimicrob. Chemother. (2004), 54, 572-574.

Journal of Antimicrobial Chemotherapy
DOI: 10.1093/jac/dkh371
Advance Access publication 14 July 2004
Monitoring of florfenicol susceptibility among bovineand porcine respiratory tract pathogens collectedin Germany during the years 2002 and 2003
Corinna Kehrenberg1, Jurgen Mumme2, Jurgen Wallmann3,
Jutta Verspohl4, Regina Tegeler5, Tilman Kuhn6 and
Stefan Schwarz1*
1Institut fur Tierzucht, Bundesforschungsanstalt furLandwirtschaft (FAL), Holtystr. 10, 31535 Neustadt-Mariensee; 2Institut fur Tiergesundheit, LUFA Nord-West,Hannover; 3Bundesamt fur Verbraucherschutz undLebensmittelsicherheit (BVL), Berlin; 4Institut furMikrobiologie, Zentrum fur Infektionsmedizin, StiftungTierarztliche Hochschule Hannover, Hannover; 5Aubenstellefur Epidemiologie, Stiftung Tierarztliche HochschuleHannover, Bakum; 6Thuringer Landesamt furLebensmittelsicherheit und Verbraucherschutz, Jena,Germany
Keywords: susceptibility testing, Pasteurella multocida,Mannheimia haemolytica, Streptococcus suis, Actinobacilluspleuropneumoniae
*Corresponding author. Tel: +49-5034-871-241;Fax: +49-5034-871-246; E-mail: [email protected]
Sir,The fluorinated chloramphenicol derivative florfenicol is exclu-sively used since veterinary medicine. In Europe, it has beenlicensed since 1995 for the control of bacterial respiratory tractinfections in cattle due to Pasteurella multocida, Mannheimiahaemolytica, or Haemophilus somnus and since 2000 for thetherapy of respiratory tract infections in pigs due to P. multocidaor Actinobacillus pleuropneumoniae. The aim of this study wasto investigate whether the most recently collected bovine andporcine respiratory tract pathogens from Germany have changedin their florfenicol susceptibility patterns since the introductionof florfenicol into clinical veterinary use.
For this, respiratory tract pathogens were collected on thebasis of one isolate per herd in 2002 and 2003, respectively. Allisolates were obtained from nasal swabs or lung tissue samplesof animals suffering from respiratory tract infections. As far asinformation on antimicrobial pre-treatment was available, theanimals from which samples had been taken had not been pre-treated with any antimicrobial agents within the weeks beforesample collection. A total of 595 isolates was included in thisstudy: 95 bovine P. multocida, 98 M. haemolytica, 103 porcineP. multocida, 63 A. pleuropneumoniae, 131 Bordetella bronchi-septica, and 105 Streptococcus suis isolates. All isolates werebiochemically identified to species level1 and comparativelytested for their in vitro susceptibilities to florfenicol by twodifferent methods, agar disc diffusion and broth microdilution.Discs charged with 30mg florfenicol (Becton Dickinson,
Heidelberg, Germany) were used for disc diffusion, and microti-tre plates (Sensititre, Westlake, OH, USA) containing florfenicolconcentrations of 0.12–128 mg/L in serial two-fold dilutions forthe broth microdilution method. Both susceptibility tests werecarried out and evaluated according to the recommendationsgiven in the National Committee for Clinical Laboratory Stand-ards (NCCLS) document M31-A2.2 The four reference strainsEscherichia coli ATCC 25922, Staphylococcus aureus ATCC29213, S. aureus ATCC 25923 as well as A. pleuropneumoniaeATCC 27090 were used for quality control purposes. The flor-fenicol-specific breakpoints presented in the NCCLS documentM31-A22 are valid for bovine P. multocida, M. haemolytica andH. somnus isolates. These breakpoints have recently also beenaccepted by the Antimicrobial Susceptibility Testing Subcom-mittee of the NCCLS not only for porcine P. multocida andA. pleuropneumoniae isolates, but also for B. bronchiseptica andS. suis isolates from porcine respiratory tract infections. For allbacteria included in this study, the approved florfenicol-specificbreakpoints for susceptibility are zone diameters of >_19 mm andMIC values of <_2 mg/L. Those for classification as intermediateare 15–18 mm and 4 mg/L, and those for resistance are <_14 mmand >_8 mg/L.2 MIC50 and MIC90 values were calculated as theflorfenicol concentrations that inhibited growth of 50% and 90%of the isolates, respectively.
Table 1 shows the results of in vitro susceptibility testing ofthe isolates from 2002/2003 in comparison to the results fromearlier studies conducted in Germany.3 – 6 All bovine P. multocidaand M. haemolytica isolates collected during 2002/2003 werefound to be susceptible to florfenicol by both methods. A com-parison of the data from 2002/2003 with data from previousyears revealed that the most recent bovine P. multocida andM. haemolytica isolates had the same MIC50 and MIC90 valuesas the isolates collected in the pre-approval phase in 1993/19944
(Table 1). A comparison between the data collected at the timeof approval of florfenicol for use in cattle5 and those from2002/2003 also revealed no increases in the MIC50 and MIC90
values of bovine P. multocida and M. haemolytica isolates(Table 1). This comparison showed that after 8 years of veterin-ary use, bovine P. multocida and M. haemolytica isolates fromrespiratory tract infections in Germany are still susceptible toflorfenicol and that so far no resistance development has beendetected in these target bacteria.
A similar situation was seen among the porcine pathogens(Table 1). All porcine P. multocida, A. pleuropneumoniae, andS. suis isolates collected in 2002 and 2003 were susceptible toflorfenicol. The S. suis isolates exhibited the same MIC50 andMIC90 values as those isolates collected at the time of introduc-tion of florfenicol into use in pigs or shortly thereafter.3 Changesof not more than one dilution step in the MIC50 or MIC90 valueswere recorded when comparing the porcine P. multocida andA. pleuropneumoniae isolates collected in 2002 and 2003 withthose collected either in the pre-approval period or at the time offlorfenicol approval for pigs.3,5,6 A surprising development wasseen among the B. bronchiseptica isolates. During the years2000–2003, the number of resistant isolates decreased from25.6% in 2000/2001 to 0.8% in 2002/2003. Moreover, the per-centage of B. bronchiseptica isolates classified as ‘intermediate’dropped from 40.0% in 2000/2001 to 6.9% in 2002/2003. Thereasons for this development towards increased susceptibility areunknown. However, it is likely that the temporal occurrence anddisappearance of B. bronchiseptica isolates that exhibit varying
Correspondence
572
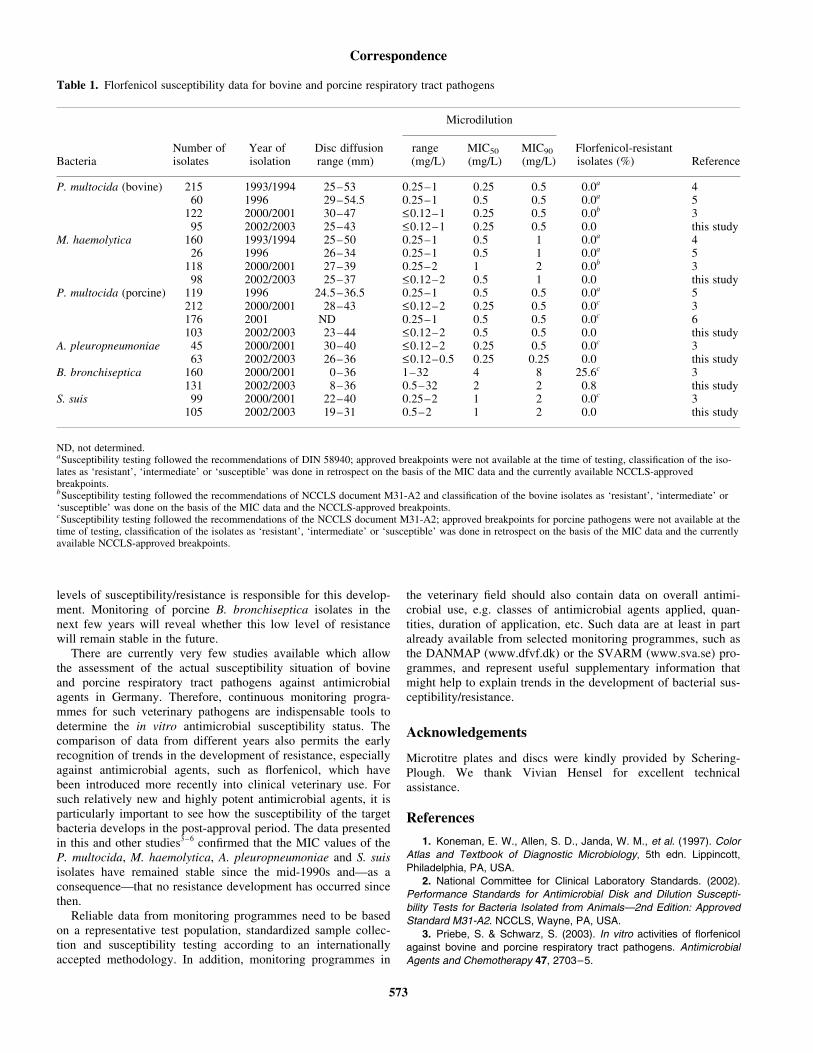
levels of susceptibility/resistance is responsible for this develop-ment. Monitoring of porcine B. bronchiseptica isolates in thenext few years will reveal whether this low level of resistancewill remain stable in the future.
There are currently very few studies available which allowthe assessment of the actual susceptibility situation of bovineand porcine respiratory tract pathogens against antimicrobialagents in Germany. Therefore, continuous monitoring progra-mmes for such veterinary pathogens are indispensable tools todetermine the in vitro antimicrobial susceptibility status. Thecomparison of data from different years also permits the earlyrecognition of trends in the development of resistance, especiallyagainst antimicrobial agents, such as florfenicol, which havebeen introduced more recently into clinical veterinary use. Forsuch relatively new and highly potent antimicrobial agents, it isparticularly important to see how the susceptibility of the targetbacteria develops in the post-approval period. The data presentedin this and other studies3 – 6 confirmed that the MIC values of theP. multocida, M. haemolytica, A. pleuropneumoniae and S. suisisolates have remained stable since the mid-1990s and—as aconsequence—that no resistance development has occurred sincethen.
Reliable data from monitoring programmes need to be basedon a representative test population, standardized sample collec-tion and susceptibility testing according to an internationallyaccepted methodology. In addition, monitoring programmes in
the veterinary field should also contain data on overall antimi-crobial use, e.g. classes of antimicrobial agents applied, quan-tities, duration of application, etc. Such data are at least in partalready available from selected monitoring programmes, such asthe DANMAP (www.dfvf.dk) or the SVARM (www.sva.se) pro-grammes, and represent useful supplementary information thatmight help to explain trends in the development of bacterial sus-ceptibility/resistance.
Acknowledgements
Microtitre plates and discs were kindly provided by Schering-Plough. We thank Vivian Hensel for excellent technicalassistance.
References
1. Koneman, E. W., Allen, S. D., Janda, W. M., et al. (1997). Color
Atlas and Textbook of Diagnostic Microbiology, 5th edn. Lippincott,
Philadelphia, PA, USA.
2. National Committee for Clinical Laboratory Standards. (2002).
Performance Standards for Antimicrobial Disk and Dilution Suscepti-
bility Tests for Bacteria Isolated from Animals—2nd Edition: Approved
Standard M31-A2. NCCLS, Wayne, PA, USA.
3. Priebe, S. & Schwarz, S. (2003). In vitro activities of florfenicol
against bovine and porcine respiratory tract pathogens. Antimicrobial
Agents and Chemotherapy 47, 2703–5.
Table 1. Florfenicol susceptibility data for bovine and porcine respiratory tract pathogens
Microdilution
BacteriaNumber ofisolates
Year ofisolation
Disc diffusionrange (mm)
range(mg/L)
MIC50
(mg/L)MIC90
(mg/L)Florfenicol-resistantisolates (%) Reference
P. multocida (bovine) 215 1993/1994 25–53 0.25–1 0.25 0.5 0.0a 460 1996 29–54.5 0.25–1 0.5 0.5 0.0a 5
122 2000/2001 30–47 <_0.12–1 0.25 0.5 0.0b 395 2002/2003 25–43 <_0.12–1 0.25 0.5 0.0 this study
M. haemolytica 160 1993/1994 25–50 0.25–1 0.5 1 0.0a 426 1996 26–34 0.25–1 0.5 1 0.0a 5
118 2000/2001 27–39 0.25–2 1 2 0.0b 398 2002/2003 25–37 <_0.12–2 0.5 1 0.0 this study
P. multocida (porcine) 119 1996 24.5–36.5 0.25–1 0.5 0.5 0.0a 5212 2000/2001 28–43 <_0.12–2 0.25 0.5 0.0c 3176 2001 ND 0.25–1 0.5 0.5 0.0c 6103 2002/2003 23–44 <_0.12–2 0.5 0.5 0.0 this study
A. pleuropneumoniae 45 2000/2001 30–40 <_0.12–2 0.25 0.5 0.0c 363 2002/2003 26–36 <_0.12–0.5 0.25 0.25 0.0 this study
B. bronchiseptica 160 2000/2001 0–36 1–32 4 8 25.6c 3131 2002/2003 8–36 0.5–32 2 2 0.8 this study
S. suis 99 2000/2001 22–40 0.25–2 1 2 0.0c 3105 2002/2003 19–31 0.5–2 1 2 0.0 this study
ND, not determined.aSusceptibility testing followed the recommendations of DIN 58940; approved breakpoints were not available at the time of testing, classification of the iso-lates as ‘resistant’, ‘intermediate’ or ‘susceptible’ was done in retrospect on the basis of the MIC data and the currently available NCCLS-approvedbreakpoints.bSusceptibility testing followed the recommendations of NCCLS document M31-A2 and classification of the bovine isolates as ‘resistant’, ‘intermediate’ or‘susceptible’ was done on the basis of the MIC data and the NCCLS-approved breakpoints.cSusceptibility testing followed the recommendations of the NCCLS document M31-A2; approved breakpoints for porcine pathogens were not available at thetime of testing, classification of the isolates as ‘resistant’, ‘intermediate’ or ‘susceptible’ was done in retrospect on the basis of the MIC data and the currentlyavailable NCCLS-approved breakpoints.
Correspondence
573

4. Hormansdorfer, S. & Bauer, J. (1996). Zur Resistenzsituation
boviner Pasteurellen [Resistance pattern of bovine pasteurellae].
Berliner Munchener Tierarztliche Wochenschrift 109, 168–71.
5. Hormansdorfer, S. & Bauer, J. (1998). Zur Resistenz boviner
und porciner Pasteurellen gegenuber Florfenicol und anderen Anti-
biotika [Resistance of bovine and porcine pasteurellae against
florfenicol and other antibiotics]. Berliner Munchener Tierarztliche
Wochenschrift 111, 422–6.
6. Wallmann, J., Schroter, K., Wieler, L. H. et al. (2003).
Antiibiotikaempfindlichkeit ausgewahlter pathogener Bakterien von
erkrankten Lebensmittel liefernden Tieren in Deutschland: Ergebnisse
aus der Modellstudie 2001 des nationalen Resistenzmonitorings
[Antibiotic sensitivity in selected veterinary pathogens from sick food
animals in Germany: Results 2001 from the study model of the national
antibiotic resistance monitoring]. Tierarztliche Praxis 31, 122–31.
Journal of Antimicrobial Chemotherapy
DOI: 10.1093/jac/dkh323
Advance Access publication 23 June 2004
CTX-M enzymes are the most commonextended-spectrum b-lactamases amongEscherichia coli in a tertiary Greek hospital
Spyros Pournaras1, Alexandros Ikonomidis1, Ioulia Kristo1,
Athanassios Tsakris2* and Antonios N. Maniatis1
1Department of Medical Microbiology, University ofThessalia, Mezourlo, Larissa; 2Department of Microbiology,Faculty of Nursing, School of Health Sciences, University ofAthens, 123 Papadiamantopoulou Street, 11527 Athens,Greece
Keywords: ESBLs, hospital-acquired infections, CTX-Mb-lactamases, Etest, PFGE, PCR, Greece
*Corresponding author. Tel: +30-210-7461483;Fax: +30-210-7461489; E-mail: [email protected]
Sir,Extended-spectrum b-lactamases (ESBLs) have emerged amongGram-negative bacteria; predominantly Klebsiella pneumoniaeand, to a lesser extent, Escherichia coli and other species. MostESBLs are mutants of the classical TEM and SHV enzymes, butsince 1995 a rapid increase in the number of CTX-M variantshas been reported among enterobacterial isolates from hospital-ized patients.1 CTX-M enzymes hydrolyse and confer resistanceto cefotaxime preferentially over ceftazidime and exhibit ahigher susceptibility to tazobactam than to clavulanic acid. Cur-rently, the CTX-M family includes almost 40 variants, dividedbetween five major amino acid sequence subtypes (see www.la-hey.org/studies/other.asp#table1).1 They are most prevalent inSouth America and the Far East, but have also been dissemi-nated in several European countries.1 – 3 Recently, CTX-M-typeb-lactamases were reported among E. coli isolates from Greece.4
During February 2003–February 2004, 426 E. coli isolateswere recovered consecutively from hospital-acquired infectionsof separate patients hospitalized at the University Hospital ofLarissa, Thessalia, Greece. Twenty-three (5.4%) of the isolateswere confirmed as ESBL producers by the Etest ESBL screeningmethod (AB Biodisk, Solna, Sweden) using cefotaxime and cef-tazidime plus clavulanate. MICs of b-lactams against these iso-lates were determined using Etest, whereas susceptibility testingagainst other antimicrobials was performed by the disc diffusionmethod on Mueller–Hinton agar.5 Against all but four of theseisolates, the cefotaxime MIC was at least eight-fold higher thanthat of ceftazidime. Third-generation cephalosporins and aztreo-nam exhibited a notably variable efficacy against the ESBL pro-ducers (Table 1).
To type the b-lactamase genes carried by the ESBL-produ-cing isolates, PCR was performed using primers specific forblaTEM, blaSHV, blaCTX-M and blaIBC.6 Genes encoding CTX-M-type determinants were detected in 20 of the ESBL-positive iso-lates with two others being positive for both blaSHV and blaTEM
and one was negative for all bla genes tested. Fourteen of theCTX-M-positive isolates produced an additional b-lactamase ofthe TEM-type. Sequencing of the 873 bp blaCTX-M amplicons onboth strands revealed that six encode CTX-M-1 and 14 encodeCTX-M-3. Genotyping using pulsed-field gel electrophoresis(PFGE) of Xba I-digested genomic DNA7 showed that the CTX-M-producers belonged to 13 distinct groups. CTX-M-producingisolates were mated in nutrient agar with E. coli 26R793 (lac �
rif R) and transconjugants were selected on Muller–Hinton agarcontaining rifampicin 100 mg/L and cefotaxime 2 mg/L. Transferof cefotaxime resistance was obtained in all but four cases alongwith other antibiotic-resistance determinants, including chloram-phenicol, gentamicin, tetracycline and trimethoprim/sulfa-methoxazole (Table 1). Transfer frequencies varied from3.8� 10�5–4.1� 10�3 per donor cell. Following plasmid iso-lation by an alkaline lysis method,8 transconjugants containedone to three plasmids that varied in size from 8–130 kb. Thefour isolates with non-transferable resistance contained plasmidsthat ranged from 30–150 kb and were different in size fromthose with transferable resistance.
The CTX-M-positive isolates were recovered from seven chil-dren and 13 adults who were hospitalized in seven differentunits of the hospital. Four of them had been treated with third-generation cephalosporins prior to the isolation. Sixteen of thepatients had a severe urinary tract infection whereas the remain-ing four exhibited purulent infections.
This report documents the predominance of blaCTX-M genesamong ESBL-positive E. coli recovered from clinical infectionsin a tertiary Greek hospital. The isolates exhibited plasmid-mediated resistance that affected the antimicrobial activity ofpenicillins and cephalosporins but also to several alternativeantibiotics used to treat E. coli infections. The observation thatdifferent CTX-M subtypes, encoded by plasmids of varying size,are being carried by distinct strains of E. coli implies that thegenes, and not simply the organisms carrying the genes, arespreading within our hospital. Our findings support the hypoth-esis that CTX-M enzymes will become the dominant ESBLsamong E. coli worldwide. Since ESBL detection procedures arenot always sensitive, the predominance of CTX-M enzymessuggests that it is important for our laboratories to perform
Correspondence
574

PUBLIKATION 3
Kehrenberg C. and S. Schwarz:
Plasmid-borne florfenicol resistance in Pasteurella multocida.
J. Antimicrob. Chemother. (2005), 55, 773-775.

Plasmid-borne florfenicol resistance in Pasteurella multocida
Corinna Kehrenberg and Stefan Schwarz*
Institut fur Tierzucht, Bundesforschungsanstalt fur Landwirtschaft (FAL), Holtystr. 10,
31535 Neustadt-Mariensee, Germany
Received 24 January 2005; returned 20 February 2005; revised 21 February 2005; accepted 22 February 2005
Objectives: A florfenicol-resistant Pasteurella multocida isolate from a calf was investigated for thegenetic basis of florfenicol resistance and the location of the resistance gene.
Methods: The P. multocida isolate 381 was investigated for its in vitro susceptibility to antimicrobialagents and its plasmid content. A 10.8 kb florfenicol–chloramphenicol resistance plasmid, designatedpCCK381, was identified by transformation into Escherichia coli. The plasmid was mapped with restric-tion endonucleases, cloned and sequenced completely.
Results: Of the antimicrobials tested, plasmid pCCK381 conferred resistance only to chloramphenicoland florfenicol. It showed extended similarity to the 5.1 kb plasmid pDN1 from Dichelobacter nodosusin the part carrying the mobilization and replication genes. An adjacent 3.2 kb segment was highlyhomologous to the florfenicol resistance gene region of plasmid pMBSF1 from E. coli. In pCCK381,combined resistance to chloramphenicol and florfenicol was based on the presence of a floR gene thatshowed 97.2–99.7% identity to so far known floR genes.
Conclusions: The results of this study showed that a plasmid-borne floR gene was responsible forchloramphenicol and florfenicol resistance in the bovine respiratory tract pathogen P. multocida. Thisis, to the best of our knowledge, the first report of a florfenicol resistance gene in a target bacterium.
Keywords: floR gene, respiratory tract pathogens, antimicrobial resistance, gene transfer
Introduction
Florfenicol, a fluorinated chloramphenicol derivative, is one ofthe few antimicrobial agents that are exclusively licensed for usein veterinary medicine.1 In Europe, it has been licensed in 1995and 2000 for the control of respiratory pathogens from cattle andpigs, respectively, whereas in the USA, florfenicol is alsoapproved for the treatment of infectious pododermatitis ( = inter-digital phlegmon) in cattle. In addition, a florfenicol premix islicensed in countries such as the USA, Canada, Chile, Norway,Korea and Japan for the treatment of various fish diseases.1
Since the introduction of florfenicol into clinical veterinaryuse, continuous monitoring programmes have been conducted todetermine MICs of florfenicol of bovine and porcine respiratorytract pathogens. The results of these monitoring programmesindicated that virtually all target bacteria obtained from cattle(Pasteurella multocida, Mannheimia haemolytica and Histophi-lus somni) and pigs (P. multocida and Actinobacilluspleuropneumoniae) were florfenicol-susceptible and that theirMIC50 and MIC90 values had remained stable over the last
decade.2,3 To date, no florfenicol resistance genes have beendetected in any of these target bacteria. In contrast to the situ-ation in the target bacteria, florfenicol resistance in variousGram-negative enteric bacteria has been detected and related tothe gene floR.1 This gene codes for a membrane-associatedexporter protein that promotes the efflux of chloramphenicol andflorfenicol from the bacterial cell. Closely related floR geneshave been detected so far on plasmids of Escherichia coli,Klebsiella pneumoniae and Salmonella enterica subsp. entericaserovar Newport, but also in the chromosomal DNA of E. coli,various Salmonella serovars and Vibrio cholerae.1
In the present study, we analysed a florfenicol-resistantbovine P. multocida isolate for the genetic basis of florfenicolresistance.
Material and methods
The P. multocida isolate 381 was obtained from the lung of a4-month-old calf submitted to the Veterinary Laboratories Agency,Thirsk Regional Laboratory, North Yorkshire, UK. The clinical
..........................................................................................................................................................................................................................................................................................................................................................................................................................
*Corresponding author. Tel: +49-5034-871-241; Fax: +49-5034-871-246; E-mail: [email protected]..........................................................................................................................................................................................................................................................................................................................................................................................................................
Journal of Antimicrobial Chemotherapy (2005) 55, 773–775
doi:10.1093/jac/dki102
Advance Access publication 6 April 2005JAC
773q The Author 2005. Published by Oxford University Press on behalf of the British Society for Antimicrobial Chemotherapy. All rights reserved.
For Permissions, please e-mail: [email protected]

signs were reported as pneumonia, with a 3 day duration prior todeath. The treatment history of this calf included the application of
florfenicol and a corticosteroid (not further specified) 2 days prior todeath and oxytetracycline 1 day prior to death. The species assign-
ment was confirmed biochemically and by a species-specific PCR.4,5
In addition, the capsular type was also determined by PCR.6 Inanother lung sample from the same calf, an S. enterica subsp. enter-
ica serovar Dublin isolate (no. 336) was identified.4 Both isolateswere initially screened for florfenicol resistance by the Veterinary
Laboratories Agency, Regional Laboratory Rougham Hill, Bury StEdmunds, Suffolk, UK and then sent to our laboratory. In vitro
susceptibility testing was performed by disc diffusion with discscontaining ampicillin (10mg), chloramphenicol (30mg), florfenicol(30mg), gentamicin (10mg), kanamycin (30mg), streptomycin
(10mg), sulfamethoxazole (300mg), tetracycline (30mg) or tri-methoprim (5mg).7 MICs of florfenicol were determined by broth
macrodilution according to the NCCLS document M31-A2.7 PCRanalyses for the florfenicol–chloramphenicol resistance gene floR
followed a previously described protocol.8 Plasmid preparation byalkaline lysis and transformation experiments into E. coli recipientstrains HB101 and JM101 were conducted as previously described.8
Plasmid DNA obtained from the transformants was subjected torestriction mapping with 18 restriction endonucleases. Overlapping
Cla I–Pst I fragments of � 5.3 and 2.5 kb, Hpa I fragments of 5.5and 1.1 kb, as well as a Hpa I–Dra I fragment of 1.5 kb of the plas-
mid from P. multocida 381 were cloned into either pBluescript IISK+ (Stratagene, Amsterdam, The Netherlands) or pCR-Bluntw II-TOPO (Invitrogen, Groningen, The Netherlands) and transformed
into E. coli recipient strains JM109 or TOP10, respectively.Sequence analyses were started with the M13 reverse and forward
primers and completed with primers derived from sequencesobtained with the aforementioned standard primers. A final segment
of 2.5 kb was sequenced by primer walking using DNA from the
E. coli JM101:pCCK381 transformant (MWG, Ebersberg, Germany).
Sequence comparisons were performed with the Blast programs
blastn and blastp (http://www.ncbi.nlm.nih.gov/BLAST/; last
accessed 14 January 2005) and with the ORF finder program (http://
www.ncbi.nlm.nih.gov/gorf/gorf.html; last accessed 14 January
2005). The nucleotide sequence of plasmid pCCK381 has been
deposited in the European Molecular Biology Laboratory database
under accession number AJ871969.
Results and discussion
P. multocida 381 proved to be of capsular type A and was resist-ant to chloramphenicol, florfenicol and sulphonamides, whereasS. Dublin 336 showed the same resistance phenotype in additionto kanamycin resistance. The MICs of florfenicol and chloram-phenicol were 16 and 32 mg/L, respectively, for P. multocida381 and S. Dublin 336. In both cases, plasmid profiling revealedthe presence of a single plasmid of � 11 kb. E. coli HB101 andJM101 transformants were obtained on Luria–Bertani agarsupplemented with either 15 mg/L chloramphenicol or 10 mg/Lflorfenicol. Subsequent screening of the transformants for theirplasmid content and their in vitro susceptibility revealed thattransformants from both donor isolates 381 and 336 carried onlythe 11 kb plasmid and that this plasmid mediated only resistanceto chloramphenicol and florfenicol.
PCR analysis of the transformants indicated the presence ofthe chloramphenicol–florfenicol resistance gene floR. Since thesmallest floR-carrying plasmid known to date was � 35 kb insize,8 the detection of this gene on relatively small plasmids of11 kb was an interesting new observation. For a better character-ization, the 11 kb plasmids from P. multocida 381 and S. Dublin
Figure 1. Schematic presentation of plasmid pCCK381 (accession no. AJ871969) in comparison with plasmid pDN1 (accession no. Y19120) as well as parts
of plasmids pMBSF1 (accession no. AJ581835) and pRVS1 (accession no. AJ289135). The reading frames are shown as arrows with the arrowhead indicating
the direction of transcription (repA, repB, repC: plasmid replication; tnp: transposition; mob: mobilization; sul2: sulphonamide resistance; floR: chlorampheni-
col-florfenicol resistance; strA, strB: streptomycin resistance). The reading frames marked as A, B and C in the maps of pCCK381 and pDN1 indicate in part
overlapping mobilization genes mobA, mobB and mobC. The D symbol indicates a truncated gene. A distance scale in kb is shown below each map. The
grey-shaded areas mark the areas of >_ 99% nucleotide sequence identity between pCCK381 and the other three plasmids. Restriction sites are abbreviated as
follows: B (Bam HI), C (Cla I), D (Dra I), E (Eco RI), EV (Eco RV), Hp (Hpa I), K (Kpn I), P (Pst I) and Pv (Pvu II).
Kehrenberg and Schwarz
774

336 were subjected to extended restriction analysis, and restric-tion mapping revealed identical results for both plasmids. Basedon this observation, the results of PCR analysis and MIC testing,as well as the finding that both strains were from the lung of thesame calf, further analysis focused on the floR-carrying plasmidfrom P. multocida 381. A map of this plasmid, designatedpCCK381, is shown in Figure 1. Sequence analysis revealed atotal plasmid size of 10874 bp.
The initial 5144 bp of plasmid pCCK381 correspondedclosely to the entire 5112 bp plasmid pDN1 from Dichelobacternodosus.9 This part contained three—in places overlapping—reading frames for the 99 amino acid (aa) protein MobC, the136 aa protein MobB and the 703 aa protein MobA, all involvedin plasmid mobilization. While MobC and MobB from plasmidpCCK381 were identical to the corresponding proteins fromplasmid pDN1, MobA from pCCK381 revealed three amino acidexchanges in comparison with MobA from pDN1.9 The 30 endof the mobC reading frame has been reported to constitute a sep-arate reading frame for a 325 aa RepB protein involved in thereplication of plasmid pDN1. Such a potential reading framewas also available in pCCK381, and the corresponding RepBprotein differed by two amino acids from that of pDN1. Furtherdownstream of mobC and repB in pCCK381, another two read-ing frames for replication proteins were detected. The gene repAcoded for a 279 aa protein that was indistinguishable from RepAof pDN1.9 The repC reading frame overlapped the repA readingframe by 112 bp and coded for a 316 aa protein that differedfrom the corresponding protein of pDN1 by two amino acids.These comparisons showed that the entire part comprising thegenes involved in plasmid replication and mobilization is highlyconserved between pCCK381 and pDN1 (Figure 1). Previousstudies on pDN1 suggested that its replication and mobilizationgenes are responsible for a broad host range and that derivativesof pDN1 also replicate in E. coli.9 Our finding, that plasmidpCCK381—which may also be considered as a pDN1 deriva-tive—replicates in Pasteurella, Salmonella, and E. coli hosts,confirms this observation. Moreover, the fact that derivatives ofpDN1 were mobilized by plasmid RP4 in E. coli has led to thesuggestion that the mobilization determinants of pDN1 are func-tionally active.9 Since the entire mob–rep gene area of plasmidpCCK381 is virtually identical to that of pDN1, we assume thatplasmid pCCK381 may also be mobilizable.
Adjacent to the pDN1-homologous region, an � 3.2 kb regionwas found to exhibit 99% sequence identity to the floR generegions of the 35 kb plasmid pMBSF1 from porcine E. coli8
(Figure 1) and a 110 kb plasmid from bovine E. coli 10660.10
This region included a truncated transposase gene (Dtnp) and thefloR gene coding for an exporter protein of the Major facilitatorsuperfamily that specifically exports phenicol antibiotics. ThefloR gene of plasmid pCCK381 revealed 97.2–99.7% nucleotidesequence identity to the so far known floR genes.1 The initial0.8 kb of this floR-homologous region, including part of Dtnpand its downstream region, also showed 99% identity to a partof the Vibrio salmonicida plasmid pRVS1 (accession no.AY171244) (Figure 1). Further downstream of the floR-homolo-gous region, an � 1.6 kb region of pCCK381 exhibited again99% identity to another part of plasmid pRVS1 (Figure 1).Similarity to pRVS1 ended within a reading frame for a putativeMob-like protein of 269 amino acids whose N-terminal 180amino acids closely resembled the N-terminal 180 amino acidsof the 333 aa Mob protein from Bartonella grahamii (accession
no. NP_696963) and the 329 aa Mob protein from Bordetellabronchiseptica (accession no. CAA47269). The C-terminal partbetween amino acids 144 and 269 was virtually identical to theN-terminal 126 amino acids of the 165 aa Mob protein fromplasmid pRVS1.
The structural analysis of plasmid pCCK381, the first florfeni-col resistance plasmid of P. multocida, revealed that this plasmidis composed of several segments previously found on other plas-mids. All these other plasmids have been found either in bacteriasuch as E. coli from cattle and pigs, which have previously beenshown to carry the floR genes, or in bacteria that cause diseasesin fish and ruminants, such as coldwater vibriosis (V. salmoni-cida) or infectious pododermatitis (D. nodosus), for the controlof which florfenicol is used. Although it is not possible to deter-mine in retrospect where and when this plasmid has evolved, thestructural analysis suggested that plasmid pCCK381 is mostlikely the result of interplasmid recombination. The presence ofthe pDN1-analogous rep–mob gene region bears the danger of afurther dissemination of this plasmid and its floR gene.
Acknowledgements
We thank Vera Noding for excellent technical assistance as wellas Jon Rogers and Paul Todd of the Veterinary LaboratoryAgency, Rougham Hill, Bury St Edmunds, Suffolk, UK forproviding the two florfenicol-resistant strains and helpfuldiscussions.
References
1. Schwarz, S., Kehrenberg, C., Doublet, B. et al. (2004).
Molecular basis of bacterial resistance to chloramphenicol and
florfenicol. FEMS Microbiol Rev 28, 519–42.
2. Priebe, S. & Schwarz, S. (2003). In vitro activities of florfenicol
against bovine and porcine respiratory tract pathogens. Antimicrob
Agents Chemother 47, 2703–5.
3. Kehrenberg, C., Mumme, J., Wallmann, J. et al. (2004).
Monitoring of florfenicol susceptibility among bovine and porcine
respiratory tract pathogens collected in Germany during the years
2002 and 2003. J Antimicrob Chemother 54, 572–4.
4. Koneman, E. W., Allen, S. D., Janda, W. M., et al. (1997). Color
Atlas and Textbook of Diagnostic Microbiology, 5th edn. Lippincott,
Philadelphia.
5. Townsend, K. M., Frost, A. J., Chiang, W. L. et al. (1998).
Development of PCR assays for species- and type-specific identifi-
cation of Pasteurella multocida isolates. J Clin Microbiol 36, 1096–100.
6. Townsend, K. M., Boyce, J. D., Chung, J. Y. et al. (2001).
Genetic organization of Pasteurella multocida cap loci and develop-
ment of a multiplex capsular PCR typing system. J Clin Microbiol 39,
924–9.
7. National Committee for Clinical Laboratory Standards. (2002).
Performance Standards for Antimicrobial Disk and Dilution Suscepti-
bility Tests for Bacteria Isolated From Animals—Second Edition:
Approved Standard M31-A2. NCCLS, Wayne, PA, USA.
8. Blickwede, M. & Schwarz, S. (2004). Molecular analysis of
florfenicol-resistant Escherichia coli from pigs. J Antimicrob Chemother
53, 58–64.
9. Whittle, G., Katz, M. E., Clayton, E. H. et al. (2000).
Identification and characterization of a native Dichelobacter nodosus
plasmid, pDN1. Plasmid 43, 230–4.
10. Cloeckaert, A., Baucheron, S., Flaujac, G. et al. (2000).
Plasmid-mediated florfenicol resistance encoded by the floR gene in
Escherichia coli isolated from cattle. Antimicrob Agents Chemother 44,
2858–60.
Florfenicol-resistant P. multocida
775

PUBLIKATION 4
Kehrenberg C., D. Meunier, H. Targant, A. Cloeckaer t, S. Schwarz and J.-Y. Madec:
Plasmid-mediated florfenicol resistance in Pasteurella trehalosi.
J. Antimicrob. Chemother. (2006), 58, 13-17.

Journal of Antimicrobial Chemotherapy (2006) 58, 13–17
doi:10.1093/jac/dkl174
Advance Access publication 2 May 2006
Plasmid-mediated florfenicol resistance in Pasteurella trehalosi
Corinna Kehrenberg1†, Daniele Meunier2†, Hayette Targant2, Axel Cloeckaert3,
Stefan Schwarz1* and Jean-Yves Madec2
1Institut fur Tierzucht, Bundesforschungsanstalt fur Landwirtschaft (FAL), Holtystr. 10,
31535 Neustadt-Mariensee, Germany; 2Agence Francaise de Securite Sanitaire des Aliments (AFSSA),
69007 Lyon, France; 3Institut National de la Recherche Agronomique (INRA), Plasticite Genomique,
Biodiversite, Antibioresistance UR1282, 37380 Nouzilly, France
Received 7 February 2006; returned 22 March 2006; revised 4 April 2006; accepted 7 April 2006
Objectives: A florfenicol-resistant Pasteurella trehalosi isolate from a calf was investigated for thepresence and the location of the gene floR.
Methods: The P. trehalosi isolate 13698 was investigated for its in vitro susceptibility to antimicrobialagentsand itsplasmidcontent.A14.9kbplasmid,designatedpCCK13698,was identifiedby transformationinto Pasteurella multocida to mediate resistance to florfenicol, chloramphenicol and sulphonamides. Theplasmid was sequenced completely and analysed for its structure and organization.
Results:PlasmidpCCK13698exhibitedextendedsimilarity toplasmidpHS-RecfromHaemophilusparasuisincluding the region carrying theparA, repB, rec and intgenes.Moreover, it revealed similarities to plasmidRSF1010 in the parts covering the mobC and repA-repC genes and to plasmid pMVSCS1 in the partscovering thesul2-catA3-strAgenecluster.Moreover, the floRgeneareacorresponded to thatof transposonTnfloR. In addition, two complete insertion sequences were detected that were highly similar to IS1593from Mannheimia haemolytica and IS26 from Enterobacteriaceae. Several potential recombination siteswere identified that might explain the development of plasmid pCCK13698 by recombination events.
Conclusions: The results of this study showed that in the bovine pathogen P. trehalosi, floR-mediatedresistance to chloramphenicol and florfenicol was associated with a plasmid, which also carried function-ally active genes for resistance to sulphonamides (sul2) and chloramphenicol (catA3). This is to the best ofour knowledge the first report of resistancegenes inP. trehalosi andonly the second report of thepresenceof a florfenicol-resistance gene in target bacteria of the family Pasteurellaceae.
Keywords: floR gene, respiratory tract pathogens, antimicrobial resistance, gene transfer, recombination, insertionsequences
Introduction
Relatively few novel antimicrobial agents have been approved forveterinary use during recent years. One of them, the fluorinatedchloramphenicol derivative florfenicol, was licensed in 1995 and2000 for the control of respiratory pathogens from cattle and pigs,respectively.1 A drug-specific monitoring programme in Ger-many that aimed at determining MICs of florfenicol amongbovine (Pasteurella multocida, Mannheimia haemolytica) andporcine respiratory tract pathogens (P. multocida andActinobacillus pleuropneumoniae) revealed that virtually all
target bacteria were florfenicol-susceptible and that theirMIC50 and MIC90 values have remained stable over the pastdecade.2 Recently, the first florfenicol-resistant bovine P. multo-cida isolate from the UK carrying the florfenicol resistance genefloR on a plasmid has been described.3
The gene floR codes for a membrane-associated efflux proteinof the major facilitator superfamily and specifically exports pheni-cols from the bacterial cell.4 This gene was previously detected aspart of the Salmonella genomic island 1 associated multiresistancegene cluster, but was also detected in various Gram-negativeenteric bacteria on plasmids or in the chromosomal DNA.1
.............................................................................................................................................................................................................................................................................................................................................................................................................................
*Corresponding author. Tel: +49-5034-871-241; Fax: +49-5034-871-246; E-mail: [email protected]†These authors contributed equally to this work.
.............................................................................................................................................................................................................................................................................................................................................................................................................................
13� The Author 2006. Published by Oxford University Press on behalf of the British Society for Antimicrobial Chemotherapy. All rights reserved.
For Permissions, please e-mail: [email protected]

In 2005, the floR gene was identified to be part of the small non-conjugative transposon TnfloR.5
In contrast to P. multocida, little is known about antimicrobialresistance in other Pasteurella species.6 This applies in particularto Pasteurella trehalosi. Originally described as biotype T ofPasteurella haemolytica, P. trehalosi was recognized as a sepa-rate species in 1990.7 P. trehalosi is mainly a pathogen of sheepwhere it causes septicaemia in older lambs.6 It has also beenisolated from other ruminants including goats,8 bisons9 and cat-tle.10 In the present study, we identified the first florfenicol-resistant bovine P. trehalosi isolate and investigated the geneticbasis of this resistance trait and the localization of the corre-sponding resistance gene.
Materials and methods
Bacterial strain and species identification
The P. trehalosi isolate 13698 was obtained from the lung of a calf.The clinical signs were reported as respiratory disease. The strainwas collected in France through the RESAPATH network. For spe-cies assignment, biochemical tests11 were backed up by 16S rDNAsequencing. The latter approach was performed at the DSMZ,the German National Resource Centre for Biological Material inBraunschweig, Germany.
Antimicrobial susceptibility testing
In vitro susceptibility testing was performed by disc diffusion withdiscs containing ampicillin (10 mg), chloramphenicol (30 mg),enrofloxacin (5 mg), florfenicol (30 mg), gentamicin (10 mg), kana-mycin (30 mg), neomycin (30 mg), spectinomycin (100 mg), strepto-mycin (10 mg), sulfamethoxazole (300 mg), tetracycline (30 mg) ortrimethoprim (5 mg). MICs of florfenicol, chloramphenicol and sul-phonamides were determined by broth macrodilution according tothe NCCLS document M31-A2. The transformant P. multocidaP4000::pCCK13698 was investigated for chloramphenicol acetyl-transferase (CAT) activity by a colorimetric CAT assay.12 Thechloramphenicol-susceptible recipient strain P. multocida P4000served as a negative control.
DNA techniques
PCR detection of florfenicol, chloramphenicol and sulphonamideresistance genes followed previously described protocols and pri-mers.4,13 Plasmid preparation by alkaline lysis and transformationexperiments (heat-shock transformation into CaCl2-treated competentcells or electrotransformation) into Escherichia coli recipient strainsJM109, JM110, JM101, HB101 and C600 as well as P. multocidastrain P4000 were conducted as described previously.13,14 Transfor-mants were selected on Luria–Bertani agar or sheep blood agar sup-plemented with either chloramphenicol (15 mg/L) or florfenicol(10 mg/L). Plasmid DNA obtained from the P. multocida transformantswas subjected to restriction mapping; SspI fragments were cloned intopCR-Blunt� II-TOPO (Invitrogen, Groningen, The Netherlands) andtransformed into E. coli recipient strain TOP10. Sequence analysis bythe dideoxy chain termination method (MWG Biotech, Martinsried,Germany) was started with the M13 reverse and forward primers andcompleted with primers derived from sequences obtained with theaforementioned standard primers. Sequence comparisons were per-formed with the BLAST programs blastn and blastp (http://www.ncbi.nlm.nih.gov/BLAST/; last accessed 29 January 2006) and withthe ORF finder program (http://www.ncbi.nlm.nih.gov/gorf/gorf.html;last accessed 29 January 2006).15 The nucleotide sequence of plasmid
pCCK13698 has been deposited in the European Molecular BiologyLaboratory (EMBL) database under accession number AM183225.
Results and discussion
Identification of plasmid pCCK13698
P. trehalosi 13698 carried a plasmid of �15 kb, designatedpCCK13698, which mediated resistance to chloramphenicol, flor-fenicol and sulphonamides when transferred into the P. multocidarecipient strain P4000. No transformants were obtained inrepeated transformation experiments with any of the E. colirecipient strains used, suggesting that this plasmid does not repli-cate in E. coli, but in Pasteurella hosts. The MICs for the originalP. trehalosi strain and the P. multocida P4000::pCCK13698transformant of florfenicol, chloramphenicol and sulphonamideswere 16, 32 and 512 mg/L, respectively. PCR analysis of thetransformant indicated the presence of not only the chlorampheni-col-florfenicol resistance gene floR, but also the chloramphenicolacetyltransferase gene catA3 and the sulphonamide resistancegene sul2. Because of the comparatively low MIC of chloram-phenicol, we performed a CAT assay that showed a >20-foldhigher deacetylation rate of acetyl-CoA in the cell-free lysatesof P. multocida P4000::pCCK13698 as compared with the plas-mid-free recipient strain P. multocida P4000 and hence con-firmed pCCK13698-based CAT activity. The reason for thelow MIC of chloramphenicol in the presence of two functionallyactive chloramphenicol resistance genes, catA3 and floR, remainsto be answered.
For a better characterization of the florfenicol resistance plas-mid from P. trehalosi and a structural comparison with thefloR-carrying plasmid pCCK381, previously identified inP. multocida,3 plasmid pCCK13698 was sequenced completely.Sequence analysis revealed a total plasmid size of 14 969 bp. Amap of plasmid pCCK13698 is shown in Figure 1.
Structure and organization of plasmid pCCK13698
The initial 635 bp of plasmid pCCK13698 as presented in Figure 1consists of small segments that exhibited similarity to plasmidpCCK3813 (positions 1–225) and to the mobC gene region of plas-mid RSF101016 (positions 169–635). The RSF1010-like part endedat a 96 bp sequence that was identical to an internal segment of thetransposase reading frames of IS1592 and IS1593, the latter ofwhich is an insertion sequence fromM. haemolytica (EMBL data-base accession no. AJ439064). A large part of plasmidpCCK13698, comprising 6770 bp (positions 731–7500), corre-sponded closely (99.5% identity) to plasmid pHS-Rec from Hae-mophilus parasuis.17 This segment included the genes parA codingfor a 207-amino-acid partition protein, rec coding for a 255-amino-acid recombinase protein, orfX coding for a 168-amino-acid hypo-thetical protein of not further specified function, repB coding for a372-amino-acid plasmid replication protein and int coding for a261-amino-acid integrase protein (Figure 1). Immediately after thepHS-Rec homologous part, a complete copy of an IS1593-likeinsertion sequence was detected (positions 7497–8527). This inser-tion sequence, designated IS1592, was 1031 bp in size, had perfect20 bp inverted repeated sequences at its termini and coded for atransposase protein of 294 amino acids that differed by three aminoacid exchanges from that of IS1593 from M. haemolytica. Furtherdownstream of the IS1592 element, a 2812 bp RSF1010-like
Kehrenberg et al.
14

segment (positions 8526–11 337) was detected. It included a trun-cated DrepA gene and a complete repC gene, which represent partof the rep gene area of the broad-host-range plasmid RSF1010, andan RSF1010-associated sul2 gene.16 The sul2 gene, which codes fora 272-amino-acid sulphonamide-resistant dihydropteroate syn-thase, was followed by a catA3 gene coding for a 213-amino-acidCAT and a largely truncated, functionally inactive strA gene codingfor a streptomycin phosphotransferase. Only the initial 73 codons ofthe strA reading frame were present in plasmid pCCK13698. The1831 bp segment (positions 10 475–12 305), comprising the genessul2, catA3 andDstrA, showed 99.7% identity to themultiresistancegene cluster previously identified on plasmid pMVSCS1 fromMannheimia varigena.18 The DstrA gene was followed by a1801 bp segment (positions 12 306–14 149) that exhibited 99.7%identity to the sequence of TnfloR.5 It contained the complete genefloR coding for a 404-amino-acid exporter protein, the readingframe for a 101-amino-acid LysR-like transcriptional regulatorand the initial 51 bp of the TnfloR-associated transposase readingframe. Immediately thereafter, a complete copy of the insertionsequence IS26 was detected (positions 14 150–14 969). This IS26element was 820 bp in size, had perfect 14 bp terminal invertedrepeats and coded for a transposase protein of 234 amino acids. Itwas identical to that associated with the kanamycin resistancetransposon Tn268019 and to a large number of IS26 sequences
from various enterobacterial species deposited in the databases,such as E. coli (AF550679), Salmonella enterica (AY333434),Klebsiella pneumoniae (AY123253), Citrobacter freundii(AF550415), Enterobacter cloacae (AY532647), Serratia marces-cens (BX664015) and Proteus vulgaris (AP004237). It should alsobenoted thatan IS26-flankedregionencompassing thegenesDrepA,repC, sul2, strA and strB has been described recently on a plasmidfrom S. enterica subsp. enterica serovar Enteritidis.20
Integration and recombination sites in plasmid
pCCK13698
The presence of a 96 bp IS1592 relic, which was in the oppositeorientation to the complete IS1592 sequence, suggested thatoriginally a not further specifiable part of pHS-Rec or of apHS-Rec-related plasmid was flanked by two inverted copiesof IS1592. While the right-hand copy remained complete, theleft-hand copy was found to be largely truncated. A comparisonof the pCCK13698 sequence with the corresponding sequences ofthe two plasmids RSF1010 and pHS-Rec revealed the presence oftwo potential recombination sites (see Figure 2). Recombinationsite 1 is 17 bp in size with 16/17 bases matching the IS sequenceand 12/17 bases matching the RSF1010 sequence. Recombinationsite 2 consists of 15 bp with 15/15 bases matching the IS sequence
0
mobC
0 5 10
1 2 3 4 5 6 7
rec repB int
8
Homology to pHS-Rec IS1592
10 11 12
sul2 catA3
9
∆repA repC∆strA
13 14
floR L tnp
Homology to RSF1010 IS26Homology topMVSCS1
Homologyto TnfloR
pCCK13698
Pv S H S S S S S S S SPv EHX
1
parA orfX
3
2
tnp
Figure 1. Schematic presentation of plasmid pCCK13698 (accession no. AM183225). A distance scale in kb is shown below the restriction map in the middle. The
reading frames are presented as arrows in more detail either above or below the map with the arrowhead indicating the direction of transcription (repA, repB, repC:
plasmid replication; tnp: transposition;mobC: mobilization; rec: recombination functions; int: integration functions; parA: DNA partition; orfX: unknown function;
sul2: sulphonamide resistance; catA3: chloramphenicol resistance; floR: chloramphenicol-florfenicol resistance; strA: streptomycin resistance). The reading frame
marked as L indicates the lysR-like transcriptional regulator, and the black box downstream of this reading frame indicates the 50 end of the TnfloR-associated
transposase reading frame. The D symbol indicates a truncated gene. Restriction sites are abbreviated as follows: E (EcoRI), H (HpaI), Pv (PvuII), S (SspI) and
X (XbaI).Areas of>99%sequence identity to plasmidspHS-Rec,RSF1010andpMVSCS1; to insertion sequences IS1592/IS1593 and IS26; and to transposonTnfloR
are indicated. The black circles with numbers 1–3 refer to the potential recombination sites 1–3 shown in Figure 2.
Florfenicol-resistant P. trehalosi
15

and 9/15 bases matching the pHS-Rec sequence. Recombinationinvolving these two sites might explain the deletion of the left-hand IS1592 copy. A third recombination site was detected in thestrA sequence and might have been used for recombination withpart of the floR upstream sequence of TnfloR. This recombinationsite 3 comprised 16 bp with 12/16 bases matching the strAsequence of pMVSCS1 and 14/16 bases matching the TnfloRsequence.
A closer look at the sequences flanking the two completeinsertion sequences revealed no direct repeats. For IS1592, itis not known whether it produces directly repeated sequencesat the integration site. However, the Tn5706-associated elementsIS1596 and IS1597, which represent close derivatives of IS1592,are known to produce a 7 bp direct repeat at the integration site.21
For IS26, it is known that it generates an 8 bp direct repeat.12 Thelack of direct repeats might suggest that besides the integration ofthese IS elements, further processes have occurred by which thedirectly repeated sequences at one or both ends were deleted ormodified. The observation that the sequences up- and down-stream of both IS elements were different might support thisassumption.
In conclusion, this is the first report of antimicrobial resistancegenes in P. trehalosi. The structural analysis of plasmidpCCK13698 revealed that it is almost exclusively composed ofsegments previously associated with other plasmids, transposonsor insertion sequences and that interplasmid recombination pro-cesses might have played a role in the formation of this plasmid.Most of these plasmid and transposon segments have already beenfound in members of the family Pasteurellaceae. However, the
occurrence of IS26—an insertion sequence widely distributedamong Enterobacteriaceae—in the genus Pasteurella is a novelobservation that supports the hypothesis of a gene flow betweenPasteurellaceae and Enterobacteriaceae.
Acknowledgements
We thank Vera Noding for excellent technical assistance. Weacknowledge the veterinary laboratories participating in RESAP-ATH. Part of this work was supported by a grant of the FrenchMinistry of Agriculture (Direction Generale de l’Alimentation).
Transparency declarations
None to declare.
References
1. Schwarz S, Kehrenberg C, Doublet B et al. Molecular basis of
bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol
Rev 2004; 28: 519–42.
2. KehrenbergC,MummeJ,Wallmann J et al. Monitoring of florfenicol
susceptibility among bovine and porcine respiratory tract pathogens
collected in Germany during the years 2002 and 2003. J Antimicrob
Chemother 2004; 54: 572–4.
3. Kehrenberg C, Schwarz S. Plasmid-borne florfenicol resistance in
Pasteurella multocida. J Antimicrob Chemother 2005; 55: 773–5.
4. Braibant M, Chevalier J, Chaslus-Dancla E et al. Structural and
functional study of the phenicol-specific efflux pump FloR belonging to
458 - AGGGGAG GGCGCACTT-ACCGGTTT CTCTTC - 488 - RSF1010 ||||||| ||| || || || ||| | 626 - AGGGGAG GGCACA-TTGACGACTTT TACCAA - 655 - pCCK13698
| |||| ||||||||||| |||||| 120 - TTATACT GCCACA-TTGACGACTTT TACCAA - 149 - IS1593
207 - AAGAGCA TGCA-AGATAAGTGTA CCAGAG – 230 - IS1593 ||||||| |||| ||||||||||| | 709 - AAGAGCA TGCA-AGATAAGTGTA TTTCAT - 736 - pCCK13698 | |||| || | | | |||||| 9135 - TCAATAT TGCATGGAGATGGAAA TTTCAT – 9106 - pHS-Rec
3577 - TCAACTG GCA-GGAGGAACAGGAG GGTGCA - 3605 - pMVSCS1 ||||||| ||| |||| | || || 12292 - TCAACTG GCA-GGAGAAGCAAAAG ATAATC – 12320 - pCCK13698 || ||| | |||||||||| |||||| 3098 CCATGCC GCATTGCGAAGCAAAAG ATAATC - 3127 -
1
2
3
plasmid 10507-1 (TnfloR)
Figure 2. Potential recombination sites in the pCCK13698 sequence. Recombination site 1 is located at the junction between RSF1010-homologous sequences and
the left-hand IS1592 relic; recombination site 2 is located at the junction between the left-hand IS1592 relic and pHS-Rec-homologous sequences; and recombination
site 3 is located within the strA reading frame at the junction with the TnfloR sequence. The numbers refer to the database entries of RSF1010 (X04830), IS1593
(AJ439064), pHS-Rec (AY862436), pMVSCS1 (AJ319822)andTnfloR (AF231986).Vertical bars indicate identical bases as compared to thepCCK13698sequence.
The recombination sites, where cross-over is believed to have occurred, are boxed.
Kehrenberg et al.
16

the major facilitator superfamily. Antimicrob Agents Chemother 2005; 49:2965–71.
5. Doublet B, Schwarz S, Kehrenberg C et al. Florfenicol resistance
gene floR is part of a novel transposon. Antimicrob Agents Chemother
2005; 49: 2106–8.
6. Kehrenberg C,Walker RD,WuCC et al. Antimicrobial resistance in
members of the family Pasteurellaceae. In: Aarestrup FM, ed. Antimicro-
bial Resistance inBacteria of AnimalOrigin.Washington,DC:ASMPress,
2005; 167–86.
7. Sneath PH, StevensM.Actinobacillus rossi sp. nov.,Actinobacillus
seminis sp. nov., nom. rev., Pasteurella bettii sp. nov., Pasteurella
lymphangitidis sp. nov., Pasteurella mairi sp. nov., and Pasteurella
trehalosi sp. nov. Int J Syst Bacteriol 1990; 40: 148–53.
8. Ward AC, Weiser GC, DeLong WJ et al. Characterization of
Pasteurellaspp. isolated fromhealthydomesticpackgoatsandevaluation
of the effects of a commercial Pasteurella vaccine. Am J Vet Res 2002;
63: 119–23.
9. Dyer NW, Ward AC, Weiser GC et al. Seasonal incidence and
antibiotic susceptibility patterns of Pasteurellaceae from American
bison (Bison bison). Can J Vet Res 2001; 65: 7–14.
10. CatryB,BaeleM,OpsomerGet al. tRNA-intergenic spacerPCR for
the identification of Pasteurella andMannheimia spp. Vet Microbiol 2004;
98: 251–60.
11. KonemanEW, Allen SD, JandaWM et al.Color Atlas and Textbook
of Diagnostic Microbiology, 5th edn. Philadelphia, New York: Lippincott,
1997; 416–23.
12. Azemun P, Stull T, Roberts M et al. Rapid detection of chloram-
phenicol resistance in Haemophilus influenzae. Antimicrob Agents
Chemother 1981; 20: 168–70.
13. KehrenbergC,SchwarzS.Occurrence and linkageof genescoding
for resistance to sulfonamides, streptomycin and chloramphenicol in bac-
teria of the genera Pasteurella and Mannheimia. FEMS Microbiol Lett
2001; 205: 283–90.
14. Kehrenberg C, Schwarz S. Molecular analysis of tetracycline resis-
tance in Pasteurella aerogenes. Antimicrob Agents Chemother 2001; 45:2885–90.
15. AltschulSF,MaddenTL,SchafferAAetal.GappedBLASTandPSI-
BLAST: a new generation of protein database search programs. Nucleic
Acids Res 1997; 25: 3389–402.
16. Scholz P, Haring V, Wittmann-Liebold B et al. Complete nucleotide
sequence and gene organization of the broad-host-range plasmid
RSF1010. Gene 1989; 75: 271–88.
17. Lancashire JF, Terry TD, Blackall PJ et al. Plasmid-encoded Tet B
tetracycline resistance in Haemophilus parasuis. Antimicrob Agents
Chemother 2005; 49: 1927–31.
18. Kehrenberg C, Schwarz S. Nucleotide sequence and organization
of plasmid pMVSCS1 fromMannheimia varigena: identification of a multi-
resistance gene cluster. J Antimicrob Chemother 2002; 49: 383–6.
19. Mollet B, Iida S, Shepherd J et al. Nucleotide sequence of IS26, a
new prokaryotic mobile genetic element. Nucleic Acids Res 1983; 11:6319–30.
20. DalyM,Villa L,PezzellaCetal.Comparisonofmultiresistancegene
regions between two geographically unrelated Salmonella serotypes.
J Antimicrob Chemother 2005; 55: 558–61.
21. Kehrenberg C, Werckenthin C, Schwarz S. Tn5706, a transposon-
like element fromPasteurellamultocidamediating tetracycline resistance.
Antimicrob Agents Chemother 1998; 42: 2116–8.
Florfenicol-resistant P. trehalosi
17

PUBLIKATION 5
Kadlec K., C. Kehrenberg and S. Schwarz:
Efflux-mediated resistance to florfenicol and/or chloramphenicol in Bordetella bronchiseptica: identification of a novel chloramphenicol exporter.
J. Antimicrob. Chemother. (2007), 59, 191-196.

Efflux-mediated resistance to florfenicol and/or chloramphenicolin Bordetella bronchiseptica: identification of a novel
chloramphenicol exporter
Kristina Kadlec, Corinna Kehrenberg and Stefan Schwarz*
Institut fur Tierzucht, Bundesforschungsanstalt fur Landwirtschaft (FAL), Holtystr.
10, 31535 Neustadt-Mariensee, Germany
Received 25 August 2006; returned 14 November 2006; revised 14 November 2006; accepted 14 November 2006
Objectives: Twenty florfenicol- and/or chloramphenicol-resistant Bordetella bronchiseptica isolates ofporcine and feline origin were investigated for the presence of floR and cml genes and their locationon plasmids.
Methods: The B. bronchiseptica isolates were investigated for their susceptibility to antimicrobialagents by broth micro- or macrodilution and for their plasmid content. Hybridization experiments andPCR assays were conducted to identify resistance genes. Transformation and conjugation studieswere performed to show their transferability. Representatives of both types of genes includingtheir flanking regions were sequenced. Moreover, inhibitor studies with the efflux pump inhibitor Phe-Arg-b-naphthylamide (PAbN) were performed.
Results: The gene floR was found in the chromosomal DNA of 9 of the 18 florfenicol/chloramphenicol-resistant isolates. Sequence analysis revealed that the deduced FloR protein sequence differed by asingle amino acid exchange from FloR of Vibrio cholerae. A chloramphenicol-resistant, but florfenicol-susceptible isolate carried a novel plasmid-borne cml gene, designated cmlB1. The CmlB1 proteinrevealed only 73.8–76.5% identity to known CmlA proteins. The gene cmlB1 was not part of a gene cass-ette. The results of inhibitor studies with PAbN suggested that a so-far unidentified efflux system mightplay a role in phenicol resistance of the remaining florfenicol- and/or chloramphenicol-resistant isolates.
Conclusions: This is to the best of our knowledge the first report of a floR gene in B. bronchiseptica iso-lates. The identification of the first member of a new subclass of cml genes, cmlB1 fromB. bronchiseptica, extends our knowledge on specific chloramphenicol exporters.
Keywords: cmlB1 gene, floR gene, efflux pump inhibitors, respiratory tract pathogen
Introduction
Bordetella bronchiseptica is frequently involved in respiratorytract infections of food-producing animals and companionanimals.1 Antimicrobial agents are commonly used to treatthese infections in animals. Initial studies of antimicrobialresistance in B. bronchiseptica from pigs revealed a decreasedsusceptibility to most of the antimicrobial agents currentlyapproved for the treatment of respiratory tract infections, suchas tilmicosin and ceftiofur, with MIC90 values of 16 mg/L and32 mg/L, respectively.2 For other antimicrobial agents, such asflorfenicol, a fluorinated chloramphenicol derivative, the corre-sponding MIC values were distinctly lower.2 Following theEuropean Union’s ban of chloramphenicol use in food-producing animals in 1994, florfenicol has been approved for
the treatment of respiratory tract infections in cattle (1995)and in pigs (2000). By contrast, chloramphenicol is stillapproved for use in dogs, cats and other non food-producinganimals, and based on its favourable susceptibility situation itis used for the control of a wide variety of infections in theseanimals.
Although florfenicol-resistant B. bronchiseptica isolates fromrespiratory tract infections in pigs have been detected in recentyears,3,4 the genetic basis for this resistance in B. bronchisepticahad not been elucidated. In the present study, we analysed iso-lates classified as florfenicol/chloramphenicol-resistant or onlychloramphenicol-resistant for the presence of known florfenicoland/or chloramphenicol resistance genes. In addition, an effluxpump inhibitor was used to assess whether efflux may play arole in phenicol resistance of B. bronchiseptica.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
*Corresponding author. Tel: þ49-5034-871-241; Fax: þ49-5034-871-246; E-mail: [email protected]
Journal of Antimicrobial Chemotherapy (2007) 59, 191–196
doi:10.1093/jac/dkl498
Advance Access publication 15 January 2007
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
191
# The Author 2007. Published by Oxford University Press on behalf of the British Society for Antimicrobial Chemotherapy. All rights reserved.
For Permissions, please e-mail: [email protected]

Materials and methods
Bacterial isolates and susceptibility testing
A total of 496 B. bronchiseptica isolates from animals sufferingfrom respiratory tract infections in Germany, including 349 iso-lates from pigs collected between 2000 and 2003,2 as well as 105isolates from pigs, 8 isolates from cats and 34 from dogs, all col-lected between 2004 and 2006, were investigated for their suscep-tibility to florfenicol and chloramphenicol. Biochemical speciesidentification was confirmed by genus- and species-specific PCRanalysis.5 Macrorestriction analysis with XbaI was performed asdescribed previously.6 MIC determination by broth micro- ormacrodilution followed the recommendations of the CLSI as laiddown in documents M31-A2 and M31-S1.7,8 The reference strainsEscherichia coli ATCC 25922 and/or Staphylococcus aureusATCC 25923 were included for quality control purposes. Toinduce phenicol resistance gene expression, the strains weregrown overnight either on MH-agar plates containing chloram-phenicol (0.5 mg/L) or florfenicol (0.5 mg/L). MIC determinationswere performed at least twice on independent occasions.
Detection of phenicol resistance genes
PCR assays for the genes conferring combined resistance tochloramphenicol and florfenicol, namely floR, fexA and cfr, butalso for the most common chloramphenicol resistance genescatA1, catA2, and catA3 were performed according to previouslydescribed protocols.6,9 – 11 For the detection of the chlorampheni-col resistance gene cmlA, previously described primers were usedat an annealing temperature of 608C.12 In addition, all isolateswere investigated for class 1 integrons and their associated catBgene cassettes.6 For the gene floR, additional new primers weredesigned to amplify the entire floR gene and used with theannealing temperature of 508C. The forward primer(50-AGGGTTGATTCGTCATGACCA-30) contained the startcodon and the reverse primer (50-CGGTTAGACGACTGGCGACT-30) the stop codon of the floR gene. To detectcircular forms of the floR-carrying transposon TnfloR, theprimers floRcirc1 and floRcirc2 were used.13 In addition, PCRassays were conducted for the oqxAB operon14 which hasrecently been described to mediate the efflux of chloramphenicolin addition to that of olaquindox.
Plasmid profiling, transfer experiments
and Southern blot hybridization
Plasmid profiles were prepared by alkaline lysis as described.6
Conjugation into E. coli HK225 was performed.6
Electrotransformation into B. bronchiseptica B543 and intoE. coli HB101 was carried out as described previously forPasteurella15 with the Gene Pulser II electroporation system(Bio-Rad, Munich, Germany). Transfer was confirmed by MICdetermination and by plasmid isolation with subsequentrestriction analysis and PCR assays. Southern blot hybridizationwas performed with a PCR-generated floR gene probe16 usingeither EcoRI- or SacI-digested whole cell DNA or uncut plasmidprofiles as target DNA. Probe labelling was achieved withthe DIG-High Prime DNA labelling and detectionsystem. Hybridization and signal detection followed the rec-ommendations given by the manufacturer (Roche DiagnosticsGmbH, Mannheim, Germany).
Sequencing of resistance gene regions
For sequence analysis, the floR and cmlA PCR amplicons werecloned into the vector pCR-blunt (Invitrogen, Groningen, TheNetherlands) as described previously.6 To sequence the flankingregions of floR, chromosomal DNA was first digested with therestriction enzyme AgeI and the fragments re-ligated with T4DNA ligase. Subsequently, inverse PCR with the floRcirc primerswas conducted. The resulting amplicon was cloned intopCR-blunt and sequenced. To sequence the flanking regions ofthe plasmid-borne cmlA-like gene, the plasmid was digested withEcoRI and KpnI and the fragments cloned into pBluescript IISK+ (Stratagene, Amsterdam, The Netherlands). The clones wereconfirmed by plasmid profiling, restriction analysis, PCR-directeddetection of the cmlA-like gene, and expression of chlorampheni-col resistance. Sequences were deposited in the EMBL databaseunder accession nos. AM296480 (floR) and AM296481 (cmlB1).
Inhibition of efflux mechanisms
The efflux pump inhibitor Phe-Arg-b-naphthylamide (PAbN)was used for inhibition studies.17,18 First, the strain-specific sus-ceptibility to the efflux inhibitor was determined by brothmacrodilution. Then, MICs of chloramphenicol and florfenicol,but also nalidixic acid, were determined in parallel in the pre-sence and absence of the inhibitor. An inhibitor concentration of80 mg/L was used, representing 1/4 of the MIC of PAbN.18
Results and discussion
Susceptibility testing
Of all the isolates analysed, only 18 B. bronchiseptica isolates, 17from pigs and one from a cat, were classified as florfenicol-resistant by CLSI criteria with MICs of florfenicol of �8 mg/L.7
All florfenicol-resistant isolates (nos. 1–18) exhibited MICs ofchloramphenicol of �16 mg/L (Table 1). In addition, two isolates(nos. 19 and 20) were chloramphenicol-resistant, but notflorfenicol-resistant, and had MICs for chloramphenicol of128 mg/L and 32 mg/L, respectively. The high MIC for chloram-phenicol of 128 mg/L for isolate no. 19 suggested the presence ofa specific chloramphenicol resistance gene. The remaining isolates(nos. 21–23) were used for control purposes and showed low MICvalues of 4 mg/L for chloramphenicol and florfenicol (Table 1).
FloR-mediated florfenicol/chloramphenicol resistance
In isolates 3–11 (Table 1), the gene floR was detected by PCR.Hybridization studies confirmed that this gene was located in thechromosomal DNA in all nine cases. Plasmid profiling revealedthat the floR-carrying strains were plasmid-free. Since all theseB. bronchiseptica isolates shared indistinguishable or closelyrelated XbaI macrorestriction patterns, one of these isolates, no. 5,was chosen for further analysis. Analysis of a 1638 bp regionincluding the floR gene and 66 bp in its upstream and 356 bp inits downstream flanking regions revealed a single base-pairexchange as compared with the corresponding sequence ofVibrio cholerae (accession no. AY822603). This base-pairexchange resulted in an amino acid exchange of His-202 inB. bronchiseptica versus Arg-202 in V. cholerae. Compared withFloR proteins so far found in other respiratory tract pathogens,
Kadlec et al.
192
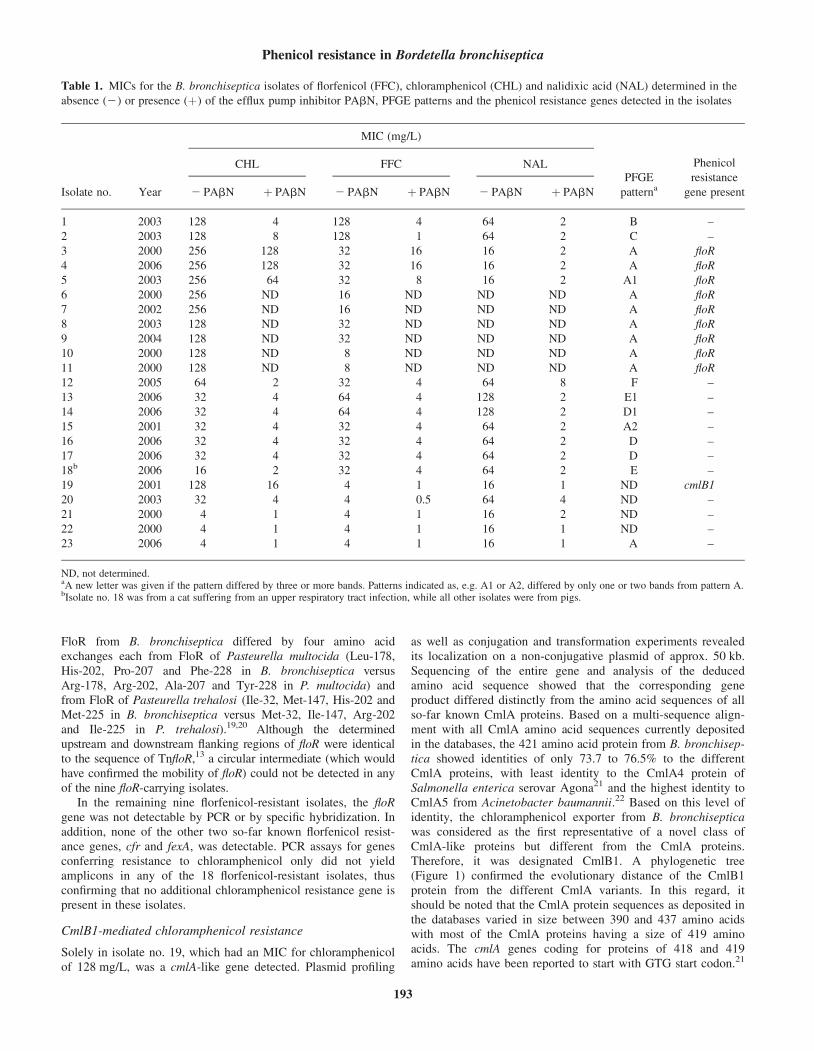
FloR from B. bronchiseptica differed by four amino acidexchanges each from FloR of Pasteurella multocida (Leu-178,His-202, Pro-207 and Phe-228 in B. bronchiseptica versusArg-178, Arg-202, Ala-207 and Tyr-228 in P. multocida) andfrom FloR of Pasteurella trehalosi (Ile-32, Met-147, His-202 andMet-225 in B. bronchiseptica versus Met-32, Ile-147, Arg-202and Ile-225 in P. trehalosi).19,20 Although the determinedupstream and downstream flanking regions of floR were identicalto the sequence of TnfloR,13 a circular intermediate (which wouldhave confirmed the mobility of floR) could not be detected in anyof the nine floR-carrying isolates.
In the remaining nine florfenicol-resistant isolates, the floRgene was not detectable by PCR or by specific hybridization. Inaddition, none of the other two so-far known florfenicol resist-ance genes, cfr and fexA, was detectable. PCR assays for genesconferring resistance to chloramphenicol only did not yieldamplicons in any of the 18 florfenicol-resistant isolates, thusconfirming that no additional chloramphenicol resistance gene ispresent in these isolates.
CmlB1-mediated chloramphenicol resistance
Solely in isolate no. 19, which had an MIC for chloramphenicolof 128 mg/L, was a cmlA-like gene detected. Plasmid profiling
as well as conjugation and transformation experiments revealedits localization on a non-conjugative plasmid of approx. 50 kb.Sequencing of the entire gene and analysis of the deducedamino acid sequence showed that the corresponding geneproduct differed distinctly from the amino acid sequences of allso-far known CmlA proteins. Based on a multi-sequence align-ment with all CmlA amino acid sequences currently depositedin the databases, the 421 amino acid protein from B. bronchisep-tica showed identities of only 73.7 to 76.5% to the differentCmlA proteins, with least identity to the CmlA4 protein ofSalmonella enterica serovar Agona21 and the highest identity toCmlA5 from Acinetobacter baumannii.22 Based on this level ofidentity, the chloramphenicol exporter from B. bronchisepticawas considered as the first representative of a novel class ofCmlA-like proteins but different from the CmlA proteins.Therefore, it was designated CmlB1. A phylogenetic tree(Figure 1) confirmed the evolutionary distance of the CmlB1protein from the different CmlA variants. In this regard, itshould be noted that the CmlA protein sequences as deposited inthe databases varied in size between 390 and 437 amino acidswith most of the CmlA proteins having a size of 419 aminoacids. The cmlA genes coding for proteins of 418 and 419amino acids have been reported to start with GTG start codon.21
Table 1. MICs for the B. bronchiseptica isolates of florfenicol (FFC), chloramphenicol (CHL) and nalidixic acid (NAL) determined in the
absence (2) or presence (þ) of the efflux pump inhibitor PAbN, PFGE patterns and the phenicol resistance genes detected in the isolates
Isolate no. Year
MIC (mg/L)
PFGE
patterna
Phenicol
resistance
gene present
CHL FFC NAL
2 PAbN þ PAbN 2 PAbN þ PAbN 2 PAbN þ PAbN
1 2003 128 4 128 4 64 2 B –
2 2003 128 8 128 1 64 2 C –
3 2000 256 128 32 16 16 2 A floR
4 2006 256 128 32 16 16 2 A floR
5 2003 256 64 32 8 16 2 A1 floR
6 2000 256 ND 16 ND ND ND A floR
7 2002 256 ND 16 ND ND ND A floR
8 2003 128 ND 32 ND ND ND A floR
9 2004 128 ND 32 ND ND ND A floR
10 2000 128 ND 8 ND ND ND A floR
11 2000 128 ND 8 ND ND ND A floR
12 2005 64 2 32 4 64 8 F –
13 2006 32 4 64 4 128 2 E1 –
14 2006 32 4 64 4 128 2 D1 –
15 2001 32 4 32 4 64 2 A2 –
16 2006 32 4 32 4 64 2 D –
17 2006 32 4 32 4 64 2 D –
18b 2006 16 2 32 4 64 2 E –
19 2001 128 16 4 1 16 1 ND cmlB1
20 2003 32 4 4 0.5 64 4 ND –
21 2000 4 1 4 1 16 2 ND –
22 2000 4 1 4 1 16 1 ND –
23 2006 4 1 4 1 16 1 A –
ND, not determined.aA new letter was given if the pattern differed by three or more bands. Patterns indicated as, e.g. A1 or A2, differed by only one or two bands from pattern A.bIsolate no. 18 was from a cat suffering from an upper respiratory tract infection, while all other isolates were from pigs.
Phenicol resistance in Bordetella bronchiseptica
193

This unusual start codon has also been identified in the novelcmlB1 gene. A closer look at the reading frames for the390 amino acid proteins (accession nos. AAY43147, AAY43150,ABH07981, ABB71444, CAD31707) strongly suggested that thesecmlA genes also have the GTG start codon rather than the proposedATG start codon and thus code for a protein of 419 amino acids aswell. A wrong annotation of the start codon (ATC at positions43 516–43 518) in the nucleotide sequence of the cmlA5 gene ofA. baumannii (CT025832) resulted in the uncommon size of 437amino acids of the respective gene product (CAJ77046). Mostlikely, the cmlA5 gene of A. baumannii also starts with GTG (atpositions 43 570–43 572) and codes for a 419 amino acid protein.
The analysis of a 2291 bp region encompassing the cmlB1gene revealed a 582 bp region in the upstream part which dif-fered only by 1 bp from the respective part in the whole genomesequence of B. bronchiseptica strain RB50 (BX640441).23
Immediately downstream of the cmlB1 gene, an incompletereading frame was detected which resembled the N-terminusof a transposase from Marinobacter aquaeolei VT8(ZP_00818190). Although many cmlA genes are part of genecassettes located in class 1 integrons,24 no structures resemblingthe 50- and 30-conserved segments of integrons were detectableupstream and downstream of the cmlB1 gene. Moreover, no
59-base element was detectable downstream of the translationaltermination codon of cmlB1.
In the area immediately upstream of the cmlB1 gene, a puta-tive regulatory region comprising a small reading frame for a9 amino acid peptide and two pairs of imperfect invertedrepeated (IR) sequences of 12 and 10 bp, respectively, weredetected. Such an arrangement has also been described for thecmlA1 gene of Tn1696 and is assumed to play a role in thechloramphenicol-inducible expression of the cmlA1 gene by atte-nuated translation.25 The IR1 sequence in the cmlB1 upstreamregion was detected immediately after the translational stopcodon of the small reading frame, whereas the IR4 sequencecomprised the start of the cmlB1 gene. Calculation of themRNA stabilities suggested that IR1 : IR2 (DG ¼ 90.3 kJ/mol)and IR3 : IR4 (DG ¼ 79.4 kJ/mol), but also IR2 : IR3(DG ¼ 74.4 kJ/mol) may be able to form stable mRNA second-ary structures.26 In addition, the small reading frame also con-tained a ribosome stall sequence 50-AAGAAAGCAGAC-30
which was indistinguishable from that in the small readingframe upstream of the inducibly expressed chloramphenicolresistance gene of the staphylococcal plasmid pC194.27 Allthese sequence features may support the assumption that cmlB1expression is also regulated by translational attenuation. MIC
CmlA2 E. aerogenes AAD22144
CmlA4 K. pneumoniae AAF27726
CmlA4 S. Agona CAI29522
CmlA5 uncultured bacterium AAM77075
CmlA5 E. coli AAG45719
CmlA7 P. aeruginosa CAD53576
CmlA5 A. baumannii CAJ77046
CmlA1 P. aeruginosa AAK50387
CmlA6 P. aeruginosa AAK52606
CmlA S. Choleraesuis AAS76336
CmlA1 P. aeruginosa AAA26057, AAB60004
CmlA E. coli AAY43147
CmlA E. coli AAY43150
CmlA6 E. coli ABH07981
CmlA P. aeruginosa ABB71444
CmlA S. Typhimurium CAD31707
CmlA1 K. pneumoniae AAO15535
CmlA1 E. coli BAD98312
CmlB1 B. bronchiseptica CAL30186
100
73
0.05
Figure 1. Phylogenetic tree of the CmlA amino acid sequences deposited in the databases. Branch lengths are scaled according to amino acid exchanges
observed in a multi-sequence alignment produced with the DNAMAN software (Lynnon-BioSoft, Ontario, Canada). The numbers at the major branch points
refer to the percentage of times that a particular node was found in 10 000 bootstrap replications. The bacterial source and the database accession number are
given for each CmlA protein.
Kadlec et al.
194
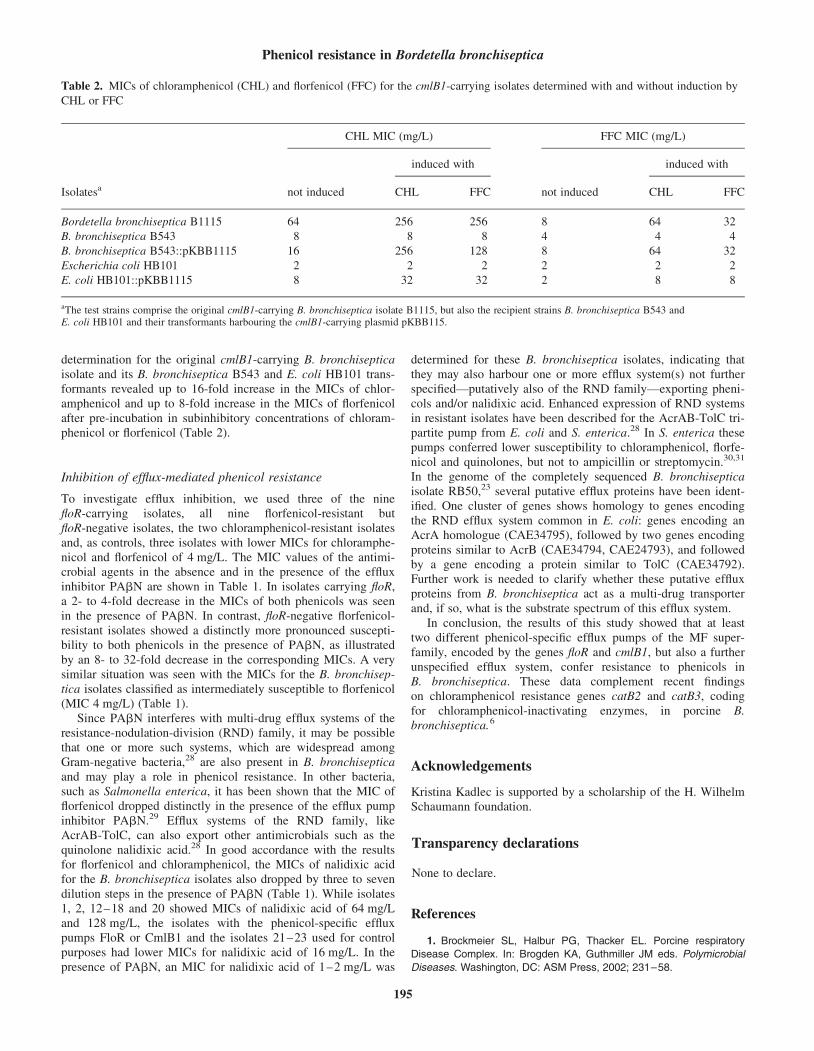
determination for the original cmlB1-carrying B. bronchisepticaisolate and its B. bronchiseptica B543 and E. coli HB101 trans-formants revealed up to 16-fold increase in the MICs of chlor-amphenicol and up to 8-fold increase in the MICs of florfenicolafter pre-incubation in subinhibitory concentrations of chloram-phenicol or florfenicol (Table 2).
Inhibition of efflux-mediated phenicol resistance
To investigate efflux inhibition, we used three of the ninefloR-carrying isolates, all nine florfenicol-resistant butfloR-negative isolates, the two chloramphenicol-resistant isolatesand, as controls, three isolates with lower MICs for chloramphe-nicol and florfenicol of 4 mg/L. The MIC values of the antimi-crobial agents in the absence and in the presence of the effluxinhibitor PAbN are shown in Table 1. In isolates carrying floR,a 2- to 4-fold decrease in the MICs of both phenicols was seenin the presence of PAbN. In contrast, floR-negative florfenicol-resistant isolates showed a distinctly more pronounced suscepti-bility to both phenicols in the presence of PAbN, as illustratedby an 8- to 32-fold decrease in the corresponding MICs. A verysimilar situation was seen with the MICs for the B. bronchisep-tica isolates classified as intermediately susceptible to florfenicol(MIC 4 mg/L) (Table 1).
Since PAbN interferes with multi-drug efflux systems of theresistance-nodulation-division (RND) family, it may be possiblethat one or more such systems, which are widespread amongGram-negative bacteria,28 are also present in B. bronchisepticaand may play a role in phenicol resistance. In other bacteria,such as Salmonella enterica, it has been shown that the MIC offlorfenicol dropped distinctly in the presence of the efflux pumpinhibitor PAbN.29 Efflux systems of the RND family, likeAcrAB-TolC, can also export other antimicrobials such as thequinolone nalidixic acid.28 In good accordance with the resultsfor florfenicol and chloramphenicol, the MICs of nalidixic acidfor the B. bronchiseptica isolates also dropped by three to sevendilution steps in the presence of PAbN (Table 1). While isolates1, 2, 12–18 and 20 showed MICs of nalidixic acid of 64 mg/Land 128 mg/L, the isolates with the phenicol-specific effluxpumps FloR or CmlB1 and the isolates 21–23 used for controlpurposes had lower MICs for nalidixic acid of 16 mg/L. In thepresence of PAbN, an MIC for nalidixic acid of 1–2 mg/L was
determined for these B. bronchiseptica isolates, indicating thatthey may also harbour one or more efflux system(s) not furtherspecified—putatively also of the RND family—exporting pheni-cols and/or nalidixic acid. Enhanced expression of RND systemsin resistant isolates have been described for the AcrAB-TolC tri-partite pump from E. coli and S. enterica.28 In S. enterica thesepumps conferred lower susceptibility to chloramphenicol, florfe-nicol and quinolones, but not to ampicillin or streptomycin.30,31
In the genome of the completely sequenced B. bronchisepticaisolate RB50,23 several putative efflux proteins have been ident-ified. One cluster of genes shows homology to genes encodingthe RND efflux system common in E. coli: genes encoding anAcrA homologue (CAE34795), followed by two genes encodingproteins similar to AcrB (CAE34794, CAE24793), and followedby a gene encoding a protein similar to TolC (CAE34792).Further work is needed to clarify whether these putative effluxproteins from B. bronchiseptica act as a multi-drug transporterand, if so, what is the substrate spectrum of this efflux system.
In conclusion, the results of this study showed that at leasttwo different phenicol-specific efflux pumps of the MF super-family, encoded by the genes floR and cmlB1, but also a furtherunspecified efflux system, confer resistance to phenicols inB. bronchiseptica. These data complement recent findingson chloramphenicol resistance genes catB2 and catB3, codingfor chloramphenicol-inactivating enzymes, in porcine B.bronchiseptica.6
Acknowledgements
Kristina Kadlec is supported by a scholarship of the H. WilhelmSchaumann foundation.
Transparency declarations
None to declare.
References
1. Brockmeier SL, Halbur PG, Thacker EL. Porcine respiratory
Disease Complex. In: Brogden KA, Guthmiller JM eds. Polymicrobial
Diseases. Washington, DC: ASM Press, 2002; 231–58.
Table 2. MICs of chloramphenicol (CHL) and florfenicol (FFC) for the cmlB1-carrying isolates determined with and without induction by
CHL or FFC
Isolatesa
CHL MIC (mg/L) FFC MIC (mg/L)
not induced
induced with
not induced
induced with
CHL FFC CHL FFC
Bordetella bronchiseptica B1115 64 256 256 8 64 32
B. bronchiseptica B543 8 8 8 4 4 4
B. bronchiseptica B543::pKBB1115 16 256 128 8 64 32
Escherichia coli HB101 2 2 2 2 2 2
E. coli HB101::pKBB1115 8 32 32 2 8 8
aThe test strains comprise the original cmlB1-carrying B. bronchiseptica isolate B1115, but also the recipient strains B. bronchiseptica B543 andE. coli HB101 and their transformants harbouring the cmlB1-carrying plasmid pKBB115.
Phenicol resistance in Bordetella bronchiseptica
195

2. Kadlec K, Kehrenberg C, Wallmann J et al. Antimicrobial sus-
ceptibility of Bordetella bronchiseptica isolates from porcine respiratory
tract infections. Antimicrob Agents Chemother 2004; 48: 4903–6.
3. Priebe S, Schwarz S. In vitro activities of florfenicol against
bovine and porcine respiratory tract pathogens. Antimicrob Agents
Chemother 2003; 47: 2703–5.
4. Kehrenberg C, Mumme J, Wallmann J et al. Monitoring of florfe-
nicol susceptibility among bovine and porcine respiratory tract patho-
gens collected in Germany during the years 2002 and 2003.
J Antimicrob Chemother 2004; 54: 572–4.
5. Hozbor D, Fouque F, Guiso N. Detection of Bordetella bronchi-
septica by the polymerase chain reaction. Res Microbiol 1999; 150:
333–41.
6. Kadlec K, Kehrenberg C, Schwarz S. Molecular basis of resist-
ance to trimethoprim, chloramphenicol and sulphonamides in
Bordetella bronchiseptica. J Antimicrob Chemother 2005; 56: 485–90.
7. Clinical and Laboratory Standards Institute. Performance
Standards for Antimicrobial Disk and Dilution Susceptibility Tests for
Bacteria Isolated from Animals – Second edition: Approved standard
M31-A2. CLSI, Wayne, PA, USA, 2002.
8. Clinical and Laboratory Standards Institute. Performance
Standards for Antimicrobial Disk and Dilution Susceptibility Tests for
Bacteria Isolated from Animals; Informational Supplement: M31-S31.
CLSI, Wayne, PA, USA, 2002.
9. Kehrenberg C, Schwarz S, Jacobsen L et al. A new mechanism
for chloramphenicol, florfenicol and clindamycin resistance: methylation
of 23S ribosomal RNA at A2503. Mol Microbiol 2005; 57: 1064–73.
10. Kehrenberg C, Schwarz S. fexA, a novel Staphylococcus lentus
gene encoding resistance to florfenicol and chloramphenicol.
Antimicrob Agents Chemother 2004; 48: 615–8.
11. Blickwede M, Schwarz S. Molecular analysis of florfenicol-
resistant Escherichia coli isolates from pigs. J Antimicrob Chemother
2004; 53: 58–64.
12. Keyes K, Hudson C, Maurer JJ et al. Detection of florfenicol
resistance genes in Escherichia coli isolated from sick chickens.
Antimicrob Agents Chemother 2000; 44: 421–4.
13. Doublet B, Schwarz S, Kehrenberg C et al. Florfenicol resist-
ance gene floR is part of a novel transposon. Antimicrob Agents
Chemother 2005; 49: 2106–8.
14. Hansen LH, Johannesen E, Burmolle M et al. Plasmid-encoded
multidrug efflux pump conferring resistance to olaquindox in
Escherichia coli. Antimicrob Agents Chemother 2004; 48: 3332–7.
15. Kehrenberg C, Schwarz S. Molecular analysis of tetracycline
resistance in Pasteurella aerogenes. Antimicrob Agents Chemother
2001; 45: 2885–90.
16. Doublet B, Schwarz S, Nussbeck E et al. Molecular analysis of
chromosomally florfenicol-resistant Escherichia coli isolates from
France and Germany. J Antimicrob Chemother 2002; 49: 49–54.
17. Li XZ, Nikaido H. Efflux-mediated drug resistance in bacteria.
Drugs 2004; 64: 159–204.
18. Preisler A, Mraheil MA, Heisig P. Role of novel gyrA mutations in
the suppression of the fluoroquinolone resistance genotype of vaccine
strain Salmonella Typhimurium vacT (gyrA D87G). J Antimicrob
Chemother 2006; 57: 430–6.
19. Kehrenberg C, Meunier D, Targant H et al. Plasmid-mediated
florfenicol resistance in Pasteurella trehalosi. J Antimicrob Chemother
2006; 58: 13–17.
20. Kehrenberg C, Schwarz S. Plasmid-borne florfenicol resistance
in Pasteurella multocida. J Antimicrob Chemother 2005; 55: 773–5.
21. Michael GB, Cardoso M, Schwarz S. Class 1 integron-associated
gene cassettes in Salmonella enterica subsp. enterica serovar Agona
isolated from pig carcasses in Brazil. J Antimicrob Chemother 2005; 55:
776–9.
22. Fournier PE, Vallenet D, Barbe V et al. Comparative genomics
of multidrug resistance in Acinetobacter baumannii. PLoS Genet
2006; 2: e7.
23. Parkhill J, Sebaihia M, Preston A et al. Comparative analysis of
the genome sequences of Bordetella pertussis, Bordetella parapertus-
sis and Bordetella bronchiseptica. Nat Genet 2003; 35: 32–40.
24. Schwarz S, Kehrenberg C, Doublet B et al. Molecular basis of
bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol
Rev 2004; 28: 519–42.
25. Stokes HW, Hall RM. Sequence analysis of the inducible chlor-
amphenicol resistance determinant in the Tn1696 integron suggests
regulation by translational attenuation. Plasmid 1991; 26: 10–9.
26. Tinoco I, Borer P, Dengler B. et al. Improved estimation of sec-
ondary structure in ribonucleic acid. Nature New Biol 1973; 246:
171–2.
27. Horinouchi S, Weisblum B. Nucleotide sequence and functional
map of pC194, a plasmid that specifies inducible chloramphenicol
resistance. J Bacteriol 1982; 150: 815–25.
28. Poole K. Efflux-mediated antimicrobial resistance. J Antimicrob
Chemother 2005; 56: 20–51.
29. Baucheron S, Imberechts H, Chaslus-Dancla E et al. The AcrB
multidrug transporter plays a major role in high-level fluoroquinolone
resistance in Salmonella enterica serovar Typhimurium phage type
DT204. Microb Drug Resist 2002; 8: 281–9.
30. Baucheron S, Tyler S, Boyd D et al. AcrAB-TolC directs efflux-
mediated multidrug resistance in Salmonella enterica serovar
Typhimurium DT104. Antimicrob Agents Chemother 2004; 48:
3729–35.
31. Quinn T, O’Mahony R, Baird AW et al. Multi-drug resistance in
Salmonella enterica: efflux mechanisms and their relationships with the
development of chromosomal resistance gene clusters. Curr Drug
Targets 2006; 7: 849–60.
Kadlec et al.
196

PUBLIKATION 6
Cloeckaert A., S. Baucheron, G. Flaujac, S. Schwarz , C. Kehrenberg, J.-L. Martel and E. Chaslus-Dancla:
Plasmid-mediated florfenicol resistance encoded by the floR gene in Escherichia coli isolated from cattle.
Antimicrob. Agents Chemother. (2000), 44, 2858-2860.

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY,0066-4804/00/$04.0010
Oct. 2000, p. 2858–2860 Vol. 44, No. 10
Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Plasmid-Mediated Florfenicol Resistance Encoded by the floRGene in Escherichia coli Isolated from Cattle
AXEL CLOECKAERT,1* SYLVIE BAUCHERON,1 GERALDINE FLAUJAC,1 STEFAN SCHWARZ,2
CORINNA KEHRENBERG,2 JEAN-LOUIS MARTEL,3 AND ELISABETH CHASLUS-DANCLA1
Station de Pathologie Aviaire et Parasitologie, Institut National de la Recherche Agronomique, 37380 Nouzilly,1 andAgence Francaise de Securite Sanitaire des Aliments, 69007 Lyon,3 France and Institut fur Tierzucht und Tierverhalten,
Bundesforschungsanstalt fur Landwirtschaft Braunschweig (FAL), 29223 Celle, Germany2
Received 24 February 2000/Returned for modification 30 May 2000/Accepted 5 July 2000
A florfenicol resistance gene almost identical to floR of Salmonella enterica serovar Typhimurium DT104 wasdetected on 110- to 125-kb plasmids in Escherichia coli isolates of animal origin. Analysis of the floR geneflanking regions of one of the plasmids showed that they were different from those encountered in S. entericaserovar Typhimurium DT104.
Florfenicol (FFC) is a fluorinated derivative of chloram-phenicol (CM) approved for use in cattle in Europe since 1995.Ffc is active against CM-resistant isolates coding for either CMacetyltransferases (6) or CM exporters, such as CmlA (3). Thefirst Ffc resistance gene, pp-flo, was detected in 1996 on aplasmid in the fish pathogen Photobacterium damselae subsp.piscicida (formerly known as Pasteurella piscicida) (9). Ffc re-sistance has gained interest with its description in the multi-drug-resistant Salmonella enterica serovar Typhimurium phage-type DT104 worldwide epidemic strains (1, 2, 4, 5). SerovarTyphimurium DT104 strains harbor a chromosomal locus inwhich the floR gene (1) and the tetracycline resistance operontetR-tet(G) are bracketed by two integron structures. This an-tibiotic resistance gene cluster including the floR gene hasrecently been found in serovar Agona isolates from poultry,suggesting the mobility of this locus (7). In this study, weinvestigated FFC-resistant Escherichia coli isolates from
France and Germany for the presence of the floR gene and itstransferability. Analysis of the floR flanking areas was per-formed to obtain information on whether the floR gene area inE. coli might have originated from Photobacterium or Salmo-nella.
All 31 French E. coli isolates included in this study werefrom cattle, while the 13 German isolates included 11 iso-lates from cattle and single isolates from a sheep and a catliving on the same farm as one of the diseased cattle. Allisolates were multidrug resistant, and the FFC MICs variedbetween 64 and 256 mg/ml. Conjugation experiments withthe E. coli strain BM14 (pro met azi) as the recipient con-firmed the FFC resistance genes to be located on conjuga-tive plasmids. The plasmids of the original E. coli isolates aswell as those of all FFC-resistant transconjugants were sub-jected to PCR with internal primers of the floR gene (1, 2,7). The floR-specific internal 496-bp amplicon was seen in all
TABLE 1. Characteristics of the E. coli strains used in this study
Straina Originb Serotype Antibiotic resistanceprofilec
MIC(mg/ml)of FFC
floRPCR
floRhybridization
Plasmidsize(kb)
EcoRI plasmidrestriction
profile
floR hybridizationEcoRI fragment
size (kb)
BN10337 F K991 Ap Cm Ffc Sm Su Tc Tp 256 1 1BN10337-1 F K991 Cm Ffc Su 64 1 1 120 A 5.5
BN10507 F K992 Ap Cm Ffc Sm Su Tc Tp 128 1 1BN10507-1 F K992 Cm Ffc Su 32 1 1 115 B 9
BN10660 F K992 Ap Cm Ffc Sm Su Tc Tp 128 1 1BN10660-1 F K992 Cm Ffc Su Tp 64 1 1 110 C 5.7
BN10870 F K992 Ap Cm Ffc Sm Su Tc Tp 256 1 1BN10870-1 F K992 Ap Cm Ffc Sm Su Tp 64 1 1 120 D 9
EC-23-99 G K992 Ap Cm Ffc Sm Su Tc Tp 256 1 1EC-23-99-1 G K992 Ap Cm Ffc Sm Su Tc 256 1 1 125 E 12.4
a For each strain the transconjugant is indicated by the suffix “1” at the end of the strain number.b F, France; G, Germany.c Antibiotics; Ap, ampicillin; Cm, chloramphencol; Ffc, florfenicol; Sm, streptomycin; Su, sulphonamides; Tc, tetracyclines; Tp, trimethoprim.
* Corresponding author. Mailing address: Station de PathologieAviaire et Parasitologie, Institut National de la RechercheAgronomique, 37380 Nouzilly, France. Phone: (33) 2 47427750. Fax:(33) 2 47427774. E-mail: [email protected].
2858
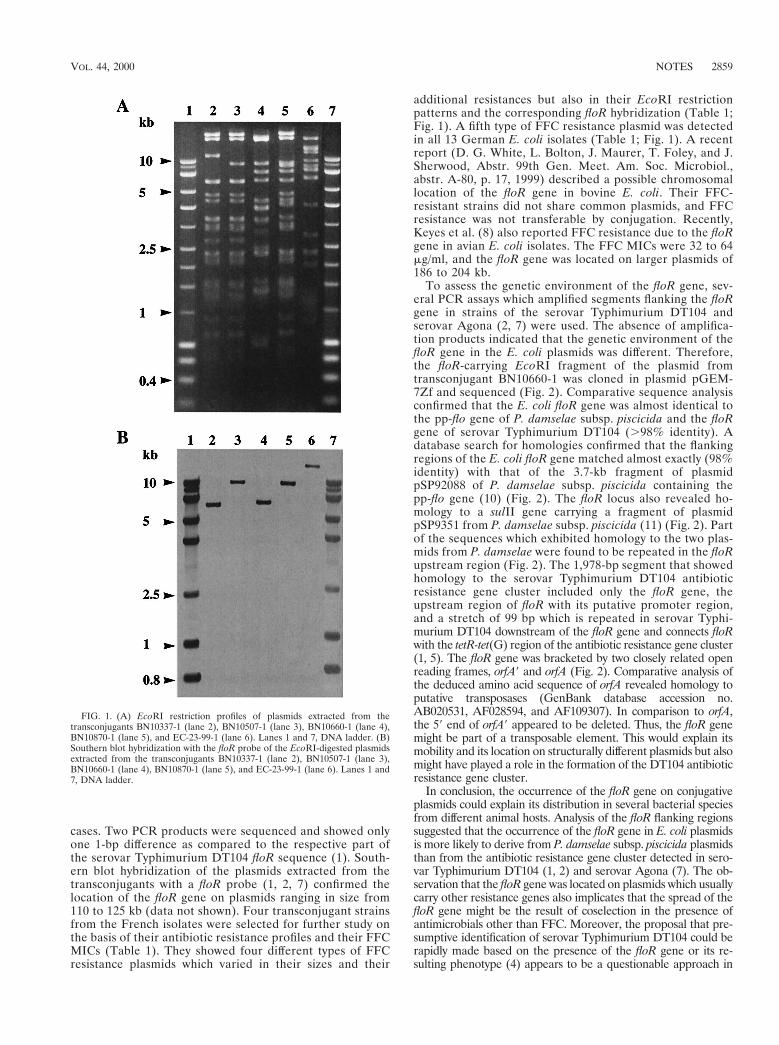
cases. Two PCR products were sequenced and showed onlyone 1-bp difference as compared to the respective part ofthe serovar Typhimurium DT104 floR sequence (1). South-ern blot hybridization of the plasmids extracted from thetransconjugants with a floR probe (1, 2, 7) confirmed thelocation of the floR gene on plasmids ranging in size from110 to 125 kb (data not shown). Four transconjugant strainsfrom the French isolates were selected for further study onthe basis of their antibiotic resistance profiles and their FFCMICs (Table 1). They showed four different types of FFCresistance plasmids which varied in their sizes and their
additional resistances but also in their EcoRI restrictionpatterns and the corresponding floR hybridization (Table 1;Fig. 1). A fifth type of FFC resistance plasmid was detectedin all 13 German E. coli isolates (Table 1; Fig. 1). A recentreport (D. G. White, L. Bolton, J. Maurer, T. Foley, and J.Sherwood, Abstr. 99th Gen. Meet. Am. Soc. Microbiol.,abstr. A-80, p. 17, 1999) described a possible chromosomallocation of the floR gene in bovine E. coli. Their FFC-resistant strains did not share common plasmids, and FFCresistance was not transferable by conjugation. Recently,Keyes et al. (8) also reported FFC resistance due to the floRgene in avian E. coli isolates. The FFC MICs were 32 to 64mg/ml, and the floR gene was located on larger plasmids of186 to 204 kb.
To assess the genetic environment of the floR gene, sev-eral PCR assays which amplified segments flanking the floRgene in strains of the serovar Typhimurium DT104 andserovar Agona (2, 7) were used. The absence of amplifica-tion products indicated that the genetic environment of thefloR gene in the E. coli plasmids was different. Therefore,the floR-carrying EcoRI fragment of the plasmid fromtransconjugant BN10660-1 was cloned in plasmid pGEM-7Zf and sequenced (Fig. 2). Comparative sequence analysisconfirmed that the E. coli floR gene was almost identical tothe pp-flo gene of P. damselae subsp. piscicida and the floRgene of serovar Typhimurium DT104 (.98% identity). Adatabase search for homologies confirmed that the flankingregions of the E. coli floR gene matched almost exactly (98%identity) with that of the 3.7-kb fragment of plasmidpSP92088 of P. damselae subsp. piscicida containing thepp-flo gene (10) (Fig. 2). The floR locus also revealed ho-mology to a sulII gene carrying a fragment of plasmidpSP9351 from P. damselae subsp. piscicida (11) (Fig. 2). Partof the sequences which exhibited homology to the two plas-mids from P. damselae were found to be repeated in the floRupstream region (Fig. 2). The 1,978-bp segment that showedhomology to the serovar Typhimurium DT104 antibioticresistance gene cluster included only the floR gene, theupstream region of floR with its putative promoter region,and a stretch of 99 bp which is repeated in serovar Typhi-murium DT104 downstream of the floR gene and connects floRwith the tetR-tet(G) region of the antibiotic resistance gene cluster(1, 5). The floR gene was bracketed by two closely related openreading frames, orfA9 and orfA (Fig. 2). Comparative analysis ofthe deduced amino acid sequence of orfA revealed homology toputative transposases (GenBank database accession no.AB020531, AF028594, and AF109307). In comparison to orfA,the 59 end of orfA9 appeared to be deleted. Thus, the floR genemight be part of a transposable element. This would explain itsmobility and its location on structurally different plasmids but alsomight have played a role in the formation of the DT104 antibioticresistance gene cluster.
In conclusion, the occurrence of the floR gene on conjugativeplasmids could explain its distribution in several bacterial speciesfrom different animal hosts. Analysis of the floR flanking regionssuggested that the occurrence of the floR gene in E. coli plasmidsis more likely to derive from P. damselae subsp. piscicida plasmidsthan from the antibiotic resistance gene cluster detected in sero-var Typhimurium DT104 (1, 2) and serovar Agona (7). The ob-servation that the floR gene was located on plasmids which usuallycarry other resistance genes also implicates that the spread of thefloR gene might be the result of coselection in the presence ofantimicrobials other than FFC. Moreover, the proposal that pre-sumptive identification of serovar Typhimurium DT104 could berapidly made based on the presence of the floR gene or its re-sulting phenotype (4) appears to be a questionable approach in
FIG. 1. (A) EcoRI restriction profiles of plasmids extracted from thetransconjugants BN10337-1 (lane 2), BN10507-1 (lane 3), BN10660-1 (lane 4),BN10870-1 (lane 5), and EC-23-99-1 (lane 6). Lanes 1 and 7, DNA ladder. (B)Southern blot hybridization with the floR probe of the EcoRI-digested plasmidsextracted from the transconjugants BN10337-1 (lane 2), BN10507-1 (lane 3),BN10660-1 (lane 4), BN10870-1 (lane 5), and EC-23-99-1 (lane 6). Lanes 1 and7, DNA ladder.
VOL. 44, 2000 NOTES 2859

light of the occurrence of floR genes in bacteria other than serovarTyphimurium DT104.
Nucleotide sequence accession number. The sequence of theEcoRI floR containing fragment from plasmid of transconju-gant BN10660-1 has been deposited in GenBank under acces-sion no. AF231986.
We thank C. Mouline and V. Noding for expert technical assistance.
REFERENCES1. Arcangioli, M. A., S. Leroy-Setrin, J. L. Martel, and E. Chaslus-Dancla.
1999. A new chloramphenicol and florfenicol resistance gene flanked by twointegron structures in Salmonella typhimurium DT104. FEMS Microbiol.Lett. 174:327–332.
2. Arcangioli, M. A., S. Leroy-Setrin, J. L. Martel, and E. Chaslus-Dancla.2000. Evolution of chloramphenicol resistance, with emergence of cross-resistance to florfenicol, in bovine Salmonella Typhimurium strains impli-cates definitive phage type (DT) 104. J. Med. Microbiol. 49:103–110.
3. Bissonnette, L., S. Champetier, J. P. Buisson, and P. H. Roy. 1991. Charac-terization of the nonenzymatic chloramphenicol resistance (cmlA) gene ofthe In4 integron of Tn1696: similarity of the product to transmembranetransport proteins. J. Bacteriol. 173:4493–4502.
4. Bolton, L. F., L. C. Kelley, M. D. Lee, P. J. Fedorka-Cray, and J. J.Maurer. 1999. Detection of multidrug-resistant Salmonella enterica sero-type typhimurium DT104 based on a gene which confers cross-resistanceto florfenicol and chloramphenicol. J. Clin. Microbiol. 37:1348–1351.
5. Briggs, C. E., and P. M. Fratamico. 1999. Molecular characterization of anantibiotic resistance gene cluster of Salmonella typhimurium DT104. Anti-microb. Agents Chemother. 43:846–849.
6. Cannon, M., S. Harford, and J. Davies. 1990. A comparative study on theinhibitory actions of chloramphenicol, thiamphenicol and some fluorinatedderivatives. J. Antimicrob. Chemother. 26:307–317.
7. Cloeckaert, A., K. Sidi Boumedine, G. Flaujac, H. Imberechts, I. D’Hooghe,and E. Chaslus-Dancla. 2000. Occurrence of a Salmonella enterica serovarTyphimurium DT104-like antibiotic resistance gene cluster including thefloR gene in S. enterica serovar Agona. Antimicrob. Agents Chemother.44:1359–1361.
8. Keyes, K., C. Hudson, J. J. Maurer, S. Thayer, D. G. White, and M. D. Lee.2000. Detection of florfenicol resistance genes in Escherichia coli isolatedfrom sick chickens. Antimicrob. Agents Chemother. 44:421–424.
9. Kim, E. H., and T. Aoki. 1996. Sequence analysis of the florfenicol resistancegene encoded in the transferable R-plasmid of a fish pathogen, Pasteurellapiscicida. Microbiol. Immunol. 40:665–669.
10. Kim, E. H., and T. Aoki. 1996. Sulfonamide resistance gene in a trans-ferable R plasmid of Pasteurella piscicida. Microbiol. Immunol. 40:397–399.
FIG. 2. Structural organization of the 6,522-bp floR locus of the plasmid from E. coli isolate BN10660. Regions which exhibit homology to the pp-flo-carryingplasmid pSP92088 (GenBank accession no. D37826) and the sulII-carrying plasmid pSP9351 (GenBank accession no. D37825), both from P. damselae subsp. piscicida,but also to the serovar Typhimurium DT104 antibiotic resistance gene cluster, to plasmid ColIb-P9 from Shigella sonnei, and to the enterobacterial plasmid pKM101are indicated. The numbers of the homologous segments refer to their position within the sequence of the floR locus. The extent and the direction of transcription ofthe floR, orfA, and orfA9 reading frames are marked by arrows. The black box upstream of floR indicates the 99-bp direct repeat. Restriction sites are abbreviated asfollows: Ba (BamHI), D (DraI), EV (EcoRV), H (HpaI), K (KpnI), P (PstI), S (SmaI), and X (XhoI). Distances (in kilobases) are given for constructs shown belowthe map of the floR locus.
2860 NOTES ANTIMICROB. AGENTS CHEMOTHER.

PUBLIKATION 7
Doublet B., S. Schwarz, C. Kehrenberg and A. Cloeck aert:
Florfenicol resistance gene floR is part of a novel transposon.
Antimicrob. Agents Chemother. (2005), 49, 2106-2108.

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, May 2005, p. 2106–2108 Vol. 49, No. 50066-4804/05/$08.00�0 doi:10.1128/AAC.49.5.2106–2108.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Florfenicol Resistance Gene floR Is Part of a Novel TransposonBenoıt Doublet,1 Stefan Schwarz,2 Corinna Kehrenberg,2 and Axel Cloeckaert1*
Unite BioAgresseurs, Sante, Environnement, Institut National de la Recherche Agronomique, 37380 Nouzilly,France,1 and Institut fur Tierzucht, Bundesforschungsanstalt fur Landwirtschaft (FAL), Holtystr.
10, 31535 Neustadt-Mariensee, Germany2
Received 9 November 2004/Accepted 21 January 2005
The florfenicol/chloramphenicol resistance gene floR was found to be part of the novel 4,284-bp transposonTnfloR from Escherichia coli. TnfloR consists of the gene floR, a putative regulatory gene, and the transposasegene tnpA. A circular form of TnfloR was detected and suggested the potential mobility of this transposon.
Florfenicol is a fluorinated structural analogue of thiam-phenicol and chloramphenicol approved exclusively for veter-inary use (23). So far, three florfenicol resistance genes, cfr,fexA, and floR, which also mediate resistance to chloramphen-icol, have been described (23). While cfr (24) and fexA (16)have been found exclusively in staphylococci, floR has beenreported to occur in various gram-negative bacteria, e.g., inseveral Salmonella enterica subsp. enterica serovars as part ofthe Salmonella genomic island 1 (SGI1) (1, 3–5, 8, 10, 12, 13,20, 21), in the SXT element from Vibrio cholerae (15), in thechromosomal DNA of Escherichia coli (2, 11), and on conju-gative and nonconjugative plasmids in E. coli (2, 7), Klebsiellapneumoniae (6), Photobacterium damselae subsp. piscicida (19),S. enterica serovar Typhimurium, and S. enterica serovar New-port (9). The different locations of the gene floR suggest theinvolvement of a yet unidentified transposable element. A re-cent study showed that the florfenicol/chloramphenicol resis-tance gene fexA is part of a nonconjugative transposon whoseintermediate circular forms were identified by inverse PCR(18). A similar approach was used in this study to identify thetransposable element carrying floR.
During recent years, the floR gene areas present on thechromosome or on different-sized plasmids in a number ofunrelated Salmonella and E. coli isolates had been character-ized by hybridization studies and/or sequence analysis (1–4,7–12). From this collection, we selected six isolates which re-vealed striking differences in the floR flanking regions. Theseisolates included one E. coli isolate with two chromosomal floRgene copies (11), one E. coli isolate with floR on a nonconju-gative plasmid (2), an E. coli transconjugant with floR on aconjugative plasmid (7), and transconjugants of two S. entericaserovar Typhimurium isolates and one S. enterica serovar New-port isolate in which floR is located on conjugative plasmids(9). SGI1-carrying Salmonella isolates were not included, sincesequence analysis of the floR flanking regions within the SGI1-associated multiresistance gene cluster gave no hints toward atransposable element. Whole-cell DNA of these six isolateswas prepared (11) and subjected to inverse PCR to detectcircular intermediate forms which may occur after resolution
of the transposon from the donor molecule and precede inte-gration into the new recipient molecule. For this, the twoprimers floRcirc1 (5�-ATCATAGCGGGCGTCGTGTT-3�)and floRcirc2 (5�-GCCGTCATTCCTCACCTTCA-3�) (an-nealing temperature of 62°C) and Taq DNA polymerase (Pro-mega, Charbonnieres, France) were used. Only one of the sixstrains, namely the E. coli BN10660-1 transconjugant (7),yielded a PCR product and thus was considered as a suitablecandidate to detect the transposable element carrying floR. Ina previous study, the sequence of a 6,522-bp EcoRI-BamHIfragment of this conjugative plasmid of E. coli BN10660—including the floR gene—had already been reported (7). Anal-ysis of the sequences upstream and downstream of the floRgene revealed the presence of two open reading frames, �tnpAand tnpA (formerly referred to as orfA� and orfA [7]), whoseproducts show considerable homology to transposase proteins.The sequence downstream of tnpA, missing in the former study(7), was determined by sequencing the terminus of the cloned7-kb EcoRI fragment of the conjugative plasmid of E. coliBN10660 using the standard M13 reverse primer (MWG Bio-tech, Ebersberg, Germany). Thus, another 513 bp was added,and database entry AF231986 was updated accordingly.
Sequence analysis of the complete 7,035-bp EcoRI-fragmentrevealed the presence of a transposon-like element of 4,284 bp.This novel element, designated TnfloR, comprised the resis-tance gene floR, which codes for a membrane-associated pro-tein of 404 amino acids (aa) that exports florfenicol and chlor-amphenicol from the bacterial cell. Further components ofTnfloR are an open reading frame coding for a putative LysR-like transcriptional regulator of 101 aa and the gene tnpAcoding for a putative transposase of 497 aa (Fig. 1a, b). TnfloRstarts with a 7-bp sequence (TATACGT) located 903 bp up-stream of the start codon of the floR gene, whereas 228 bpdownstream of tnpA, the same sequence is found again andconstitutes most likely the right-end junction of the transpo-son. TnfloR does not exhibit inverted or direct repeats at itstermini. Inverse PCR using primers complementary to se-quences in the 5� and the 3� end of the floR gene yielded afragment of 3,528 bp (Fig. 1c). Sequence analysis of this am-plicon revealed the presence of the 3� end of the floR gene, theputative transcriptional regulator gene, the complete tnpAgene, and its downstream region, the 7-bp sequence (TATACGT), the upstream region of floR, and the 5� end of the floRgene. This observation confirmed that the transposon-like el-
* Corresponding author: Mailing address: Unite BioAgresseurs,Sante, Environnement, Institut National de la Recherche Agrono-mique (INRA), 37380 Nouzilly, France. Phone: (33) 2 47 42 77 50. Fax:(33) 2 47 42 77 74. E-mail: [email protected].
2106

ement identified by sequence analysis is in fact a functionallyactive transposon which produces circular intermediate formsand that the 7-bp sequence is part of TnfloR. In this regard,TnfloR closely resembles several site-specific integrating trans-posons, such as Tn554 (22), Tn5406 (14), and Tn558 (18),which are known to produce circular intermediate forms, donot exhibit inverted repeats at their termini, and have 6- to7-bp sequences at their left- and right-hand junctions, one ofwhich is part of the transposon while the other represents thetarget sequence. The TnfloR-associated 7-bp target sequence islocated within the phosphoglucosamine mutase gene glmMand has also been detected flanking the dfrA20 trimethoprimresistance gene area in the recently described Pasteurella mul-tocida plasmid pCCK154 (17).
Homology searches revealed that the complete TnfloR is notpresent in any other sequence deposited in the databases.However, there are several sequences where part of tnpA andthe adjacent integration site are present (for a recent review,see reference 23). This might suggest either that there areother transposons that carry the same transposase gene andintegrate at the same target site or that a TnfloR element wassubject to recombination or deletion events after its integrationinto a new target sequence resulting in such largely truncatedforms. Although TnfloR is a nonconjugative transposon, itslocation on a large conjugative plasmid—as in the presentcase—facilitates the spread of this transposon across speciesand genus borders. The observation that the TnfloR-carrying
plasmid from E. coli BN10660 also mediates resistance to sul-fonamides and trimethoprim (7), both of which are used inveterinary medicine at much higher quantities than florfenicol,indicate the potential of coselection of the floR gene by the useof other antimicrobial agents.
This work was supported by a grant from the French Institut Na-tional de la Recherche Agronomique (INRA, Action Transversalite2001–2003) and grants of the Deutsche Forschungsgemeinschaft(SCHW 382/6-1, SCHW 382/6-2).
REFERENCES
1. Arcangioli, M. A., S. Leroy-Setrin, J. L. Martel, and E. Chaslus-Dancla.1999. A new chloramphenicol and florfenicol resistance gene linked to anintegron structure in Salmonella typhimurium DT104. FEMS Microbiol. Lett.174:327–332.
2. Blickwede, M., and S. Schwarz. 2004. Molecular analysis of florfenicol-resistant Escherichia coli isolates from pigs. J. Antimicrob. Chemother. 53:58–64.
3. Boyd, D., A. Cloeckaert, E. Chaslus-Dancla, and M. R. Mulvey. 2002. Char-acterization of variant Salmonella genomic island 1 multidrug resistanceregions from serovars Typhimurium DT104 and Agona. Antimicrob. AgentsChemother. 46:1714–1722.
4. Boyd, D., G. A. Peters, A. Cloeckaert, K. Sidi Boumedine, E. Chaslus-Dancla, H. Imberechts, and M. R. Mulvey. 2001. Complete nucleotide se-quence of a 43-kilobase genomic island associated with the multidrug resis-tance region of Salmonella enterica serovar Typhimurium DT104 and itsidentification in phage type DT120 and serovar Agona. J. Bacteriol. 183:5725–5732.
5. Briggs, C. E., and P. M. Fratamico. 1999. Molecular characterization of anantibiotic resistance gene cluster of Salmonella typhimurium DT104. Anti-microb. Agents Chemother. 43:846–849.
6. Cloeckaert, A., S. Baucheron, and E. Chaslus-Dancla. 2001. Nonenzymatic
FIG. 1. (a) Genetic organization of the floR transposon on plasmid from E. coli strain BN10660-1. The position and orientation of the genesare indicated by arrows. The 7-bp nucleotide sequences at the transposon junctions are shown in boxes. The positions of primers floRcirc1 andfloRcirc2 used for the detection of a circular TnfloR intermediate are indicated by arrows. A distance scale in bp is given above the map. (b)Schematic view of the 4,284-bp circular form of the floR transposon. The integration site of TnfloR is shown in a box. (c) PCR product of thecircular form of TnfloR (3,528 bp) obtained by using primers floRcirc1 and floRcirc2. Lane 1, Smart Ladder (Eurogentec, Angers, France); lane2, E. coli BN10660-1 transconjugant.
VOL. 49, 2005 NOTES 2107

chloramphenicol resistance mediated by IncC plasmid R55 is encoded by afloR gene variant. Antimicrob. Agents Chemother. 45:2381–2382.
7. Cloeckaert, A., S. Baucheron, G. Flaujac, S. Schwarz, C. Kehrenberg, J. L.Martel, and E. Chaslus-Dancla. 2000. Plasmid-mediated florfenicol resis-tance encoded by the floR gene in Escherichia coli isolated from cattle.Antimicrob. Agents Chemother. 44:2858–2860.
8. Doublet, B., P. Butaye, H. Imberechts, D. Boyd, M. R. Mulvey, E. Chaslus-Dancla, and A. Cloeckaert. 2004. Salmonella genomic island 1 multidrugresistance gene clusters in Salmonella enterica serovar Agona isolated inBelgium in 1992 to 2002. Antimicrob. Agents Chemother. 48:2510–2517.
9. Doublet, B., A. Carattoli, J. M. Whichard, D. G. White, S. Baucheron, E.Chaslus-Dancla, and A. Cloeckaert. 2004. Plasmid-mediated florfenicol andceftriaxone resistance encoded by the floR and blaCMY-2 genes in Salmonellaenterica serovars Typhimurium and Newport isolated in the United States.FEMS Microbiol. Lett. 233:301–305.
10. Doublet, B., R. Lailler, D. Meunier, A. Brisabois, D. Boyd, M. R. Mulvey, E.Chaslus-Dancla, and A. Cloeckaert. 2003. Variant Salmonella genomic is-land 1 antibiotic resistance gene cluster in Salmonella enterica serovar Al-bany. Emerg. Infect. Dis. 9:585–591.
11. Doublet, B., S. Schwarz, E. Nu�beck, S. Baucheron, J. L. Martel, E.Chaslus-Dancla, and A. Cloeckaert. 2002. Molecular analysis of chromo-somally florfenicol resistant Escherichia coli isolates from France and Ger-many. J. Antimicrob. Chemother. 49:49–54.
12. Doublet, B., F.-X. Weill, L. Fabre, E. Chaslus-Dancla, and A. Cloeckaert.2004. Variant Salmonella genomic island 1 antibiotic resistance gene clustercontaining a novel 3�-N-aminoglycoside acetyltransferase gene cassette,aac(3)-Id, in Salmonella enterica serovar Newport. Antimicrob. Agents Che-mother. 48:3806–3812.
13. Ebner, P., K. Garner, and A. Mathew. 2004. Class 1 integrons in variousSalmonella enterica serovars isolated from animals and identification ofgenomic island SGI1 in Salmonella enterica var. Meleagridis. J. Antimicrob.Chemother. 53:1004–1009.
14. Haroche, J., J. Allignet, and N. El Solh. 2002. Tn5406, a new staphylococcal
transposon conferring resistance to streptogramin A and related compoundsincluding dalfopristin. Antimicrob. Agents Chemother. 46:2337–2343.
15. Hochhut, B., Y. Lotfi, D. Mazel, S. M. Faruque, R. Woodgate, and M. K.Waldor. 2001. Molecular analysis of antibiotic resistance gene clusters inVibrio cholerae O139 and O1 SXT constins. Antimicrob. Agents Chemother.45:2991–3000.
16. Kehrenberg, C., and S. Schwarz. 2004. fexA, a novel Staphylococcus lentusgene encoding resistance to florfenicol and chloramphenicol. Antimicrob.Agents Chemother. 48:615–618.
17. Kehrenberg, C., and S. Schwarz. 2005. dfrA20, a novel trimethoprim resis-tance gene from Pasteurella multocida. Antimicrob. Agents Chemother. 49:414–417.
18. Kehrenberg, C., and S. Schwarz. 2005. The florfenicol-chloramphenicol ex-porter gene fexA is part of the novel transposon Tn558. Antimicrob. AgentsChemother. 49:813–815.
19. Kim, E., and T. Aoki. 1996. Sequence analysis of the florfenicol resistancegene encoded in the transferable R-plasmid of a fish pathogen, Pasteurellapiscicida. Microbiol. Immunol. 40:665–669.
20. Meunier, D., D. Boyd, M. R. Mulvey, S. Baucheron, C. Mammina, A. Nas-tasi, E. Chaslus-Dancla, and A. Cloeckaert. 2002. Salmonella enterica sero-type Typhimurium DT104 antibiotic resistance genomic island I in serotypeParatyphi B. Emerg. Infect. Dis. 8:430–433.
21. Mulvey, M. R., D. Boyd, A. Cloeckaert, R. Ahmed, L. K. Ng, and the Pro-vincial Public Health Laboratories. 2004. Emergence of multidrug-resistantSalmonella Paratyphi B dT�, Canada. Emerg. Infect. Dis. 10:1307–1310.
22. Murphy, E. 1989. Transposable elements in gram-positive bacteria, p. 269–288. In D. E. Berg and M. M. Howe (ed.), Mobile DNA. ASM Press,Washington, D.C.
23. Schwarz, S., C. Kehrenberg, B. Doublet, and A. Cloeckaert. 2004. Molecularbasis of bacterial resistance to chloramphenicol and florfenicol. FEMS Mi-crobiol. Rev. 28:519–542.
24. Schwarz, S., C. Werckenthin, and C. Kehrenberg. 2000. Identification of aplasmid-borne chloramphenicol-florfenicol resistance gene in Staphylococ-cus sciuri. Antimicrob. Agents Chemother. 44:2530–2533.
2108 NOTES ANTIMICROB. AGENTS CHEMOTHER.

PUBLIKATION 8
Kehrenberg C. and S.Schwarz:
fexA, a novel Staphylococcus lentus gene encoding resistance to florfenicol and chloramphenicol.
Antimicrob. Agents Chemother. (2004), 48, 615-618.

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Feb. 2004, p. 615–618 Vol. 48, No. 20066-4804/04/$08.00�0 DOI: 10.1128/AAC.48.2.615–618.2004Copyright © 2004, American Society for Microbiology. All Rights Reserved.
fexA, a Novel Staphylococcus lentus Gene EncodingResistance to Florfenicol and
ChloramphenicolCorinna Kehrenberg and Stefan Schwarz*
Institut fur Tierzucht, Bundesforschungsanstalt fur Landwirtschaft,31535 Neustadt-Mariensee, Germany
Received 23 April 2003/Returned for modification 21 September 2003/Accepted 19 October 2003
The Staphylococcus lentus plasmid pSCFS2 carries a novel florfenicol-chloramphenicol resistance gene,designated fexA, encoding a protein of 475 amino acids with 14 transmembrane domains. The FexA proteindiffers from all previously known proteins involved in the efflux of chloramphenicol and florfenicol. Inductionof fexA expression by chloramphenicol and florfenicol occurs via translational attenuation.
Florfenicol is a fluorinated derivative of chloramphenicolwhich was licensed in Germany for the control of bacterialrespiratory tract infections in 1995 for cattle and in 2000 forswine. In contrast to chloramphenicol, florfenicol is exclusivelyused in veterinary medicine (11), and the known chloramphen-icol acetyltransferases are unable to inactivate florfenicol (6).While chloramphenicol acetyltransferase genes (cat genes)have been detected in a wide variety of gram-positive bacteria,including staphylococci of human and animal origin (5, 16),only a single gene, cfr, from bovine Staphylococcus sciuri, hasbeen observed to mediate combined resistance to florfenicoland chloramphenicol by a yet-unknown mechanism (13). Thecfr gene is located on the 17-kb plasmid pSCFS1, which alsoconfers resistance to macrolide-lincosamide-streptogramin Bantibiotics and spectinomycin (12).
In this study, the bovine Staphylococcus lentus isolate no. 8,obtained from the nasal swab of a calf suffering from a respi-ratory tract infection, was shown to be resistant to chloram-phenicol, clindamycin, erythromycin, florfenicol, streptomycin,and tetracycline by agar disk diffusion (7) and was shown tocarry six plasmids of ca. 2 to 34 kb. After transformation intoprotoplasts of Staphylococcus aureus RN4220, a 34-kb plasmid,designated pSCFS2, mediated only resistance to florfenicoland chloramphenicol. MICs for S. aureus RN4220:pSCFS2were 32 �g of florfenicol/ml and 64 �g of chloramphenicol/ml(Table 1) as determined by the microdilution broth method(7). Preincubation in the presence of either 0.5 �g of chlor-amphenicol/ml or 0.5 �g of florfenicol/ml led to a fourfoldincrease of both MICs (Table 1). Since PCR assays and hy-bridization experiments did not reveal the presence of the cfrgene, cloning experiments using BglII-digested pSCFS2 DNAand BamHI-digested vector pBluescript II SK(�) (Stratagene,Amsterdam, The Netherlands) were performed. After trans-formation into Escherichia coli JM109, only transformants that
carried a 7-kb BglII insert grew on Luria-Bertani agar platessupplemented with 10 �g of florfenicol/ml. They exhibitedMICs of 16 �g of florfenicol/ml and 64 �g of chloramphenicol/ml, which could be increased to 64 �g of florfenicol/ml and 128�g of chloramphenicol/ml, respectively, by preincubation inthe presence of either 0.5 �g of chloramphenicol/ml or 0.5 �gof florfenicol/ml (Table 1). These observations suggested thatthe resistance gene in question is expressed inducibly in gram-positive and gram-negative hosts and that both antimicrobialagents, florfenicol and chloramphenicol, are effective as induc-ers.
Within the 7-kb BglII fragment, a single EcoRI site wasdetected. Subclones which carried EcoRI/BglII inserts of 5.2and 1.8 kb proved to be susceptible to florfenicol and chlor-amphenicol, suggesting that this EcoRI site is located eitherwithin the gene in question or in its regulatory region. Se-quence analysis was performed on both strands by primerwalking, starting at the EcoRI sites of both subclones and usingthe M13 universal and reverse primers. The nucleotide se-quence of a 1,674-bp fragment of plasmid pSCFS2 was deter-mined. Analysis of this region confirmed the presence of twoopen reading frames (ORFs), one of them coding for a proteinof 475 amino acids (aa) (position 177 to 1604), the othercoding for a small peptide of 9 aa (position 118 to 147) pre-ceding the aforementioned ORF.
The reading frame for the 475-aa protein was designatedfexA (for florfenicol exporter), and the EcoRI site was locatedwithin this reading frame at position 1334. Analysis of the fexAupstream region revealed striking homologies to the transla-tional attenuators upstream of chloramphenicol-induciblestaphylococcal cat genes (4) and also to that of the chloram-phenicol-inducible gene cmlA from Tn1696 (14). In addition tothe reading frame for the 9-aa peptide, a pair of invertedrepeated sequences (IR1 and IR2) of 11 bp was detected.These inverted repeats might be able to form a stable mRNAstem-loop structure (�G � �74.4 kJ/mol) (15) with the fexA-associated ribosome binding site (5�-AGGAGG-3�; position164 to 169) located within IR2) (Fig. 1a). The sequence of thecodons 2 to 5 of the 9-aa peptide (5�-GTGAAAGCAGTG-3�;
* Corresponding author. Mailing address: Institut fur Tierzucht,Bundesforschungsanstalt fur Landwirtschaft, Holtystr. 10, 31535 Neu-stadt-Mariensee, Germany. Phone: 49-5034-871242. Fax: 49-5034-871246. E-mail: [email protected].
615

position 121 to 132) demonstrated significant homology to theribosome stall sequences previously identified in the regulatoryregions of cat57 and cat86 from Bacillus spp. but also to thoseof staphylococcal cat genes (2, 4, 9, 10) (Fig. 1b). Stalling of aribosome in the reading frame of the 9-aa peptide might pre-vent the formation of an mRNA secondary structure betweenIR1 and IR2 and thus allow translation of the fexA transcriptby a second ribosome.
Comparisons of the fexA nucleotide sequence using the Na-tional Center for Biotechnology Information standard nucleo-tide BLAST program (http://www.ncbi.nlm.nih.gov/BLAST/)revealed no significant homology to known sequences. TheTmpred program (http://www.ch.embnet.org/software/TMPRED_form.html) predicts that the FexA protein has 14 transmem-brane helices. This feature is seen in a wide variety of effluxproteins of the major facilitator superfamily (8), including acluster of efflux proteins from gram-positive bacteria whichconfer resistance to antimicrobial agents such as tetracyclinesand lincomycin. The deduced FexA amino acid sequence re-veals only minor homologies to other proteins deposited in thedatabases. Closest similarities of 28 or 29% amino acid se-quence identity were observed between FexA and a multidrugefflux protein from Lactobacillus plantarum (accession no.NP_84696.1) as well as a metal-tetracycline/H� antiporterfrom Bacillus halodurans (NP_242832.1). Comparisons of theamino acid sequences revealed no more than 19% amino acididentity between FexA and known chloramphenicol or flor-fenicol/chloramphenicol exporter proteins (Fig. 2). Thus, FexArepresents a novel type of florfenicol/chloramphenicol effluxsystem which is distinctly different from those of the FloRsubgroup and the CmlA subgroup, both found in clinicallyimportant gram-negative bacteria, as well as from the chlor-amphenicol exporters so far detected in soil and environmentalbacteria of the genera Streptomyces, Corynebacterium, andRhodococcus (Fig. 2).
The detection of the florfenicol-chloramphenicol resistancegene fexA in staphylococci is to the best of our knowledge thefirst report of a gene coding for a florfenicol efflux protein instaphylococci. The location of this gene on a plasmid and theobservation that it is functionally active even in E. coli suggesta potential transfer of fexA-mediated resistance between mem-bers of different bacterial species and genera.
Nucleotide sequence accession number. The sequence of thefexA gene has been deposited with the EMBL database underaccession number AJ549214.
FIG. 1. (a) Presentation of the fexA regulatory region. The ORF9-and fexA-associated ribosome binding sites are boxed. The start codonsof ORF9 and fexA are underlined, and the corresponding coding se-quences are displayed in bold letters. The inverted repeated sequencesIR1 and IR2 are marked by arrows, and a stable mRNA secondarystructure formed by these IR sequences is shown. Calculation of thestability of this stem-loop structure followed the specifications given byTinoco et al. (15). (b) Comparison of the potential ribosome stallsequence in the fexA regulatory region with those of chloramphenicol-inducible cat genes from Bacillus (4) and Staphylococcus (2, 9, 10). The16S rRNA sequence was taken from the whole genome sequence of S.aureus N315 (3) (accession no. NC_002745) and is identical for all fiverRNA operons detected in this strain. The “G” at position 1300 is a“U,” and the “U” at position 1301 is a “C” in the B. subtilis 16S rRNA(1). Since pairing of U:G and G:U has no negative impact of thestability of the binding (15), such pairing was not considered a mis-match. Matching bases in the stall sequences with regard to the S.aureus 16S rRNA are displayed in bold capital letters, whereas mis-matches are displayed as lowercase letters.
TABLE 1. MICs of florfenicol and chloramphenicol for the strains used in this studya
StrainMIC of florfenicol (�g/ml) MIC of chloramphenicol (�g/ml)
Noninduced Ff induced Cm induced Noninduced Ff induced Cm induced
S. lentus 8 32 128 128 64 128 128S. aureus RN4220 2 2 2 2 2 2S. aureus RN4220:pSCFS2 32 128 128 64 256 256E. coli JM109 8 8 8 4 4 4E. coli JM109 Bgl30b 16 64 64 64 128 128
a Ff, florfenicol; Cm, chloramphenicol.b E. coli JM109 carrying pBluescript II SK(�) with the 7-kb BglII fragment of pSCFS2 in its BamHI site.
616 NOTES ANTIMICROB. AGENTS CHEMOTHER.

This study was supported by grants of the Deutsche Forschungsge-meinschaft (SCHW 382/6-1 and SCHW 382/6-2).
We thank Stefan Hormansdorfer for providing S. lentus isolate 8 andVera Noding for excellent technical assistance.
REFERENCES
1. Green, C. J., G. C. Stewart, M. A. Hillis, B. S. Vold, and K. F. Bott. 1985.Nucleotide sequence of the Bacillus subtilis ribosomal RNA operon rrnB.Gene 37:261–266.
2. Horinouchi, S., and B. Weisblum. 1982. Nucleotide sequence and functionalmap of pC194, a plasmid that specifies inducible chloramphenicol resistance.J. Bacteriol. 150:815–825.
3. Kuroda, M., T. Ohta, I. Uchiyama, T. Baba, H. Yuzawa, I. Kobayashi, L. Cui,A. Oguchi, K. Aoki, Y. Nagai, J. Lian, T. Ito, M. Kanamori, H. Matsumaru,A. Maruyama, H. Murakami, A. Hosoyama, Y. Mizutani-Ui, N. K. Taka-hashi, T. Sawano, R. Inoue, C. Kaito, K. Sekimizu, H. Hirakawa, S. Kuhara,S. Goto, J. Yabuzaki, M. Kanehisa, A. Yamashita, K. Oshima, K. Furuya, C.Yoshino, T. Shiba, M. Hattori, N. Ogasawara, H. Hayashi, and K. Hira-matsu. 2001. Whole genome sequencing of methicillin-resistant Staphylococ-cus aureus. Lancet 357:1225–1240.
4. Lovett, P. S. 1990. Translational attenuation as the regulator of inducible catgenes. J. Bacteriol. 172:1–6.
5. Lyon, B. R., and R. Skurray. 1987. Antimicrobial resistance of Staphylococ-cus aureus: genetic basis. Microbiol. Rev. 51:88–134.
FIG. 2. Homology tree of the so far known chloramphenicol exporter proteins. For the different exporter proteins, information on bacterialhosts, database accession numbers, and designations (as given in the database entries) are provided. The tree was constructed using the DNAMANsoftware (Lynnon Biosoft, Vaudreuil, Quebec, Canada).
VOL. 48, 2004 NOTES 617

6. Murray, I. A., and W. V. Shaw. 1997. O-Acetyltransferases for chloramphen-icol and other natural products. Antimicrob. Agents Chemother. 41:1–6.
7. National Committee for Clinical Laboratory Standards. 2002. Performancestandards for antimicrobial disk and dilution susceptibility tests for bacteriaisolated from animals. Approved standard. NCCLS document M31-A2. Na-tional Committee for Clinical Laboratory Standards, Wayne, Pa.
8. Paulsen, I. T., M. H. Brown, and R. A. Skurray. 1996. Proton-dependentmultidrug efflux systems. Microbiol. Rev. 60:575–608.
9. Projan, S. J., J. Kornblum, S. L. Moghazeh, I. Edelman, M. L. Gennaro, andR. P. Novick. 1985. Comparative sequence and functional analysis of pT181and pC221, cognate plasmid replicons from Staphylococcus aureus. Mol.Gen. Genet. 199:452–464.
10. Schwarz, S., and M. Cardoso. 1991. Nucleotide sequence and phylogeny ofa chloramphenicol acetyltransferase encoded by the plasmid pSCS7 fromStaphylococcus aureus. Antimicrob. Agents Chemother. 35:1551–1556.
11. Schwarz, S., and E. Chaslus-Dancla. 2001. Use of antimicrobials in veteri-nary medicine and mechanisms of resistance. Vet. Res. 32:201–225.
12. Schwarz, S., C. Kehrenberg, and K. K. Ojo. 2002. Staphylococcus sciuri gene,erm(33), encoding inducible resistance to macrolides, lincosamides, andstreptogramin B antibiotics is a product of recombination between erm(C)and erm(A). Antimicrob. Agents Chemother. 46:3621–3623.
13. Schwarz, S., C. Werckenthin, and C. Kehrenberg. 2000. Identification of aplasmid-borne chloramphenicol-florfenicol resistance gene in Staphylococ-cus sciuri. Antimicrob. Agents Chemother. 44:2530–2533.
14. Stokes, H. W., and R. M. Hall. 1991. Sequence analysis of the induciblechloramphenicol resistance determinant in the Tn1696 integron suggestsregulation by translational attenuation. Plasmid 26:10–19.
15. Tinoco, I., P. Borer, B. Dengler, M. Levine, O. Uhlenbeck, D. Crothers, andJ. Gralla. 1973. Improved estimation of secondary structure in ribonucleicacid. Nat. New Biol. 246:171–172.
16. Werckenthin, C., M. Cardoso, J.-L. Martel, and S. Schwarz. 2001. Antimi-crobial resistance in staphylococci from animals with particular reference tobovine Staphylococcus aureus, porcine Staphylococcus hyicus and canineStaphylococcus intermedius. Vet. Res. 32:341–362.
618 NOTES ANTIMICROB. AGENTS CHEMOTHER.

PUBLIKATION 9
Kehrenberg C. and S. Schwarz:
Florfenicol-chloramphenicol exporter gene fexA is part of the novel transposon Tn558.
Antimicrob. Agents Chemother. (2005), 49, 813-815.

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Feb. 2005, p. 813–815 Vol. 49, No. 20066-4804/05/$08.00�0 doi:10.1128/AAC.49.2.813–815.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Florfenicol-Chloramphenicol Exporter Gene fexA Is Part of the NovelTransposon Tn558
Corinna Kehrenberg and Stefan Schwarz*Institut fur Tierzucht, Bundesforschungsanstalt fur Landwirtschaft (FAL), Neustadt-Mariensee, Germany
Received 6 July 2004/Returned for modification 16 September 2004/Accepted 17 October 2004
The florfenicol-chloramphenicol exporter gene fexA is part of the novel transposon Tn558 from Staphylococ-cus lentus. Similarities between Tn558 and Tn554 from Staphylococcus aureus included the arrangement of thetransposase genes tnpA to -C and an att554-like target sequence. Circular forms of Tn558 were detected andsuggest the functional activity of this transposon.
Recently the first staphylococcal florfenicol-chlorampheni-col efflux gene, fexA, was detected on plasmid pSCFS2 ofStaphylococcus lentus (4). To investigate whether fexA is part ofa transposable element, the fexA flanking sequences in pSCFS2were analyzed with regard to similarities to other staphylococ-cal transposons.
Plasmid pSCFS2 was digested with BglII, and the six resultingfragments of sizes between 1.2 and 14.0 kb were cloned separatelyinto the BamHI-digested vector pBluescript II SK(�) (Strat-agene, Amsterdam, The Netherlands). Confirmation of the fexAgene on the 7.1-kb BglII fragment was done by PCR and byhybridization experiments (data not shown). The PCR primersfexA-fw (5�-GTACTTGTAGGTGCAATTACGGCTGA-3�) andfexA-rev (5�-CGCATCTGAGTAGGACATAGCGTC-3�) (am-plicon size, 1,272 bp; annealing temperature, 57°C) were usedalong with Pwo polymerase (Peqlab, Erlangen, Germany). Todetermine a sufficiently long sequence up- and downstream offexA, sequence analysis included parts of the 7.1-kb BglII frag-ment and the adjacent 14.0-kb BglII fragment. The nucleotidesequence of the fexA flanking regions was determined by primerwalking on both strands starting from the terminal parts of thefexA gene (MWG-Biotech, Ebersberg, Germany).
Analysis of a 7,718-bp region revealed the presence of atransposon-like element of 6,644 bp, designated Tn558. Thiselement consisted of five reading frames of more than 120amino acids (aa), which accounted for 78.5% of the Tn558sequence (Fig. 1a). Three of these reading frames exhibitedsimilarities to the genes tnpA, tnpB, and tnpC, whose productsare involved in the transposition of the Staphylococcus aureustransposons Tn554 (1, 5–8) and Tn5406 (3) (Fig. 1a). It isnoteworthy that despite the same arrangement and the similarsizes, the tnpA, tnpB, and tnpC genes of the three transposonsTn554, Tn5406, and Tn558 differed considerably in their nu-cleotide sequences and in the deduced amino acid sequencesof their gene products (Table 1). The remaining two openreading frames of Tn558 were orientated in the opposite di-rection and code for the 475-aa florfenicol-chloramphenicolexporter protein FexA and a putative oxidoreductase of 138 aa,
respectively (Fig. 1a). The deduced amino acid sequence of thelatter open reading frame exhibited 45 to 47% identity and76% similarity to the NAD(P)H oxidoreductases of Fusobac-terium nucleatum subsp. nucleatum (NP_604130) or Fusobac-terium nucleatum subsp. vincentii (ZP_00143945).
As previously reported for Tn554 and Tn5406 (3, 9), Tn558also did not contain inverted repeats at its ends and failed togenerate a duplication of the target sequence at the integrationsite. Tn558 exhibited the hexanucleotide sequence 5�-GATGTA-3� at the left-end junction and a similar sequence, 5�-GATCCA-3�, at the right-end junction. The sequence 5�-GATGTA-3� haspreviously been described as the “core” sequence of Tn554 andTn5406 in the primary target site att554 in the S. aureus chromo-some (Fig. 1b) (2, 3, 11–13). Studies on serial transposition ofTn554 into primary and secondary target sites revealed that thesequences at the junctions of Tn554 varied with respect to thetarget sites: with each new transposition event, the sequence orig-inally present in the target site is found at the left end of Tn554,whereas the former left-end junction is now found at the right endand the former right-end junction is lost (5, 10). A similar processis assumed to be responsible for the altered sequence found at theright-end junction of Tn558 in plasmid pSCFS2. Analysis of theregions flanking the Tn558 insertion in plasmid pSCSF2 identi-fied a sequence similar to that of att554. A comparison of thispSCFS2 region, designated att558, with the sequences up- anddownstream of insertion sites of Tn554 and Tn5406 is shown inFig. 1b. The att554 sites of Tn554 and Tn5406 are located withinreading frames for proteins of 222 aa which show similarity toDNA repair proteins (3). The reading frame including the att558site codes for a putative protein of 140 aa which showed 51%identity and 68% similarity to a 147-aa DNA repair protein fromListeria monocytogenes (ZP_00231288).
Since transposition of Tn554 and Tn5406 includes the forma-tion of circular forms which precede the integration of the trans-poson into a new target sequence (3, 5), inverse PCR assays wereconducted to detect these circular intermediates. For this, thePwo polymerase (Peqlab) and the two primers circ-fw (5�-CGGTGCCTAATCATTCGTATGC-3�) and circ-rev (5�-CGCTTAACCGGTTCTATCACTTCA-3�) (amplicon size, 871 bp; anneal-ing temperature, 62°C) were used; the primers positions areshown in Fig. 1a. Amplicons of the expected size were obtained inrepeated experiments from several different S. aureus RN4220:pSCFS2 transformants (data not shown). The sequence of such
* Corresponding author. Mailing address: Institut fur Tierzucht,Bundesforschungsanstalt fur Landwirtschaft (FAL), Holtystr. 10,31535 Neustadt-Mariensee, Germany. Phone: 49-5034-871-241. Fax:49-5034-871-246. E-mail: [email protected].
813

814
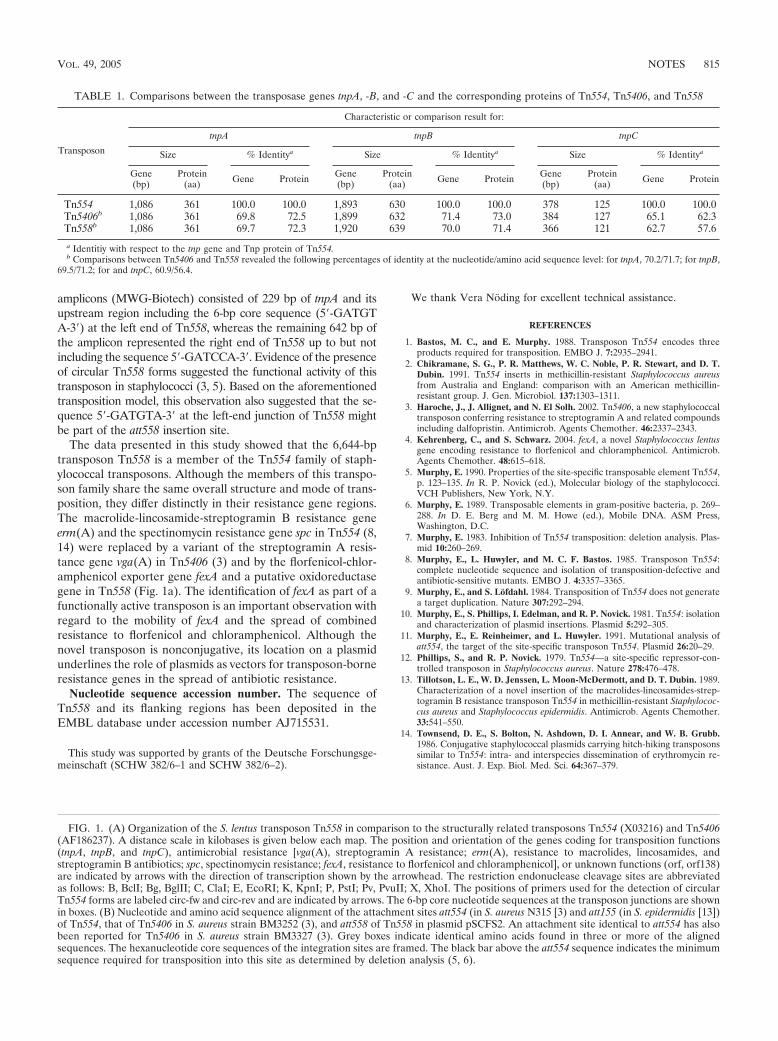
amplicons (MWG-Biotech) consisted of 229 bp of tnpA and itsupstream region including the 6-bp core sequence (5�-GATGTA-3�) at the left end of Tn558, whereas the remaining 642 bp ofthe amplicon represented the right end of Tn558 up to but notincluding the sequence 5�-GATCCA-3�. Evidence of the presenceof circular Tn558 forms suggested the functional activity of thistransposon in staphylococci (3, 5). Based on the aforementionedtransposition model, this observation also suggested that the se-quence 5�-GATGTA-3� at the left-end junction of Tn558 mightbe part of the att558 insertion site.
The data presented in this study showed that the 6,644-bptransposon Tn558 is a member of the Tn554 family of staph-ylococcal transposons. Although the members of this transpo-son family share the same overall structure and mode of trans-position, they differ distinctly in their resistance gene regions.The macrolide-lincosamide-streptogramin B resistance geneerm(A) and the spectinomycin resistance gene spc in Tn554 (8,14) were replaced by a variant of the streptogramin A resis-tance gene vga(A) in Tn5406 (3) and by the florfenicol-chlor-amphenicol exporter gene fexA and a putative oxidoreductasegene in Tn558 (Fig. 1a). The identification of fexA as part of afunctionally active transposon is an important observation withregard to the mobility of fexA and the spread of combinedresistance to florfenicol and chloramphenicol. Although thenovel transposon is nonconjugative, its location on a plasmidunderlines the role of plasmids as vectors for transposon-borneresistance genes in the spread of antibiotic resistance.
Nucleotide sequence accession number. The sequence ofTn558 and its flanking regions has been deposited in theEMBL database under accession number AJ715531.
This study was supported by grants of the Deutsche Forschungsge-meinschaft (SCHW 382/6–1 and SCHW 382/6–2).
We thank Vera Noding for excellent technical assistance.
REFERENCES
1. Bastos, M. C., and E. Murphy. 1988. Transposon Tn554 encodes threeproducts required for transposition. EMBO J. 7:2935–2941.
2. Chikramane, S. G., P. R. Matthews, W. C. Noble, P. R. Stewart, and D. T.Dubin. 1991. Tn554 inserts in methicillin-resistant Staphylococcus aureusfrom Australia and England: comparison with an American methicillin-resistant group. J. Gen. Microbiol. 137:1303–1311.
3. Haroche, J., J. Allignet, and N. El Solh. 2002. Tn5406, a new staphylococcaltransposon conferring resistance to streptogramin A and related compoundsincluding dalfopristin. Antimicrob. Agents Chemother. 46:2337–2343.
4. Kehrenberg, C., and S. Schwarz. 2004. fexA, a novel Staphylococcus lentusgene encoding resistance to florfenicol and chloramphenicol. Antimicrob.Agents Chemother. 48:615–618.
5. Murphy, E. 1990. Properties of the site-specific transposable element Tn554,p. 123–135. In R. P. Novick (ed.), Molecular biology of the staphylococci.VCH Publishers, New York, N.Y.
6. Murphy, E. 1989. Transposable elements in gram-positive bacteria, p. 269–288. In D. E. Berg and M. M. Howe (ed.), Mobile DNA. ASM Press,Washington, D.C.
7. Murphy, E. 1983. Inhibition of Tn554 transposition: deletion analysis. Plas-mid 10:260–269.
8. Murphy, E., L. Huwyler, and M. C. F. Bastos. 1985. Transposon Tn554:complete nucleotide sequence and isolation of transposition-defective andantibiotic-sensitive mutants. EMBO J. 4:3357–3365.
9. Murphy, E., and S. Lofdahl. 1984. Transposition of Tn554 does not generatea target duplication. Nature 307:292–294.
10. Murphy, E., S. Phillips, I. Edelman, and R. P. Novick. 1981. Tn554: isolationand characterization of plasmid insertions. Plasmid 5:292–305.
11. Murphy, E., E. Reinheimer, and L. Huwyler. 1991. Mutational analysis ofatt554, the target of the site-specific transposon Tn554. Plasmid 26:20–29.
12. Phillips, S., and R. P. Novick. 1979. Tn554—a site-specific repressor-con-trolled transposon in Staphylococcus aureus. Nature 278:476–478.
13. Tillotson, L. E., W. D. Jenssen, L. Moon-McDermott, and D. T. Dubin. 1989.Characterization of a novel insertion of the macrolides-lincosamides-strep-togramin B resistance transposon Tn554 in methicillin-resistant Staphylococ-cus aureus and Staphylococcus epidermidis. Antimicrob. Agents Chemother.33:541–550.
14. Townsend, D. E., S. Bolton, N. Ashdown, D. I. Annear, and W. B. Grubb.1986. Conjugative staphylococcal plasmids carrying hitch-hiking transposonssimilar to Tn554: intra- and interspecies dissemination of erythromycin re-sistance. Aust. J. Exp. Biol. Med. Sci. 64:367–379.
TABLE 1. Comparisons between the transposase genes tnpA, -B, and -C and the corresponding proteins of Tn554, Tn5406, and Tn558
Transposon
Characteristic or comparison result for:
tnpA tnpB tnpC
Size % Identitya Size % Identitya Size % Identitya
Gene(bp)
Protein(aa) Gene Protein Gene
(bp)Protein
(aa) Gene Protein Gene(bp)
Protein(aa) Gene Protein
Tn554 1,086 361 100.0 100.0 1,893 630 100.0 100.0 378 125 100.0 100.0Tn5406b 1,086 361 69.8 72.5 1,899 632 71.4 73.0 384 127 65.1 62.3Tn558b 1,086 361 69.7 72.3 1,920 639 70.0 71.4 366 121 62.7 57.6
a Identitiy with respect to the tnp gene and Tnp protein of Tn554.b Comparisons between Tn5406 and Tn558 revealed the following percentages of identity at the nucleotide/amino acid sequence level: for tnpA, 70.2/71.7; for tnpB,
69.5/71.2; for and tnpC, 60.9/56.4.
FIG. 1. (A) Organization of the S. lentus transposon Tn558 in comparison to the structurally related transposons Tn554 (X03216) and Tn5406(AF186237). A distance scale in kilobases is given below each map. The position and orientation of the genes coding for transposition functions(tnpA, tnpB, and tnpC), antimicrobial resistance [vga(A), streptogramin A resistance; erm(A), resistance to macrolides, lincosamides, andstreptogramin B antibiotics; spc, spectinomycin resistance; fexA, resistance to florfenicol and chloramphenicol], or unknown functions (orf, orf138)are indicated by arrows with the direction of transcription shown by the arrowhead. The restriction endonuclease cleavage sites are abbreviatedas follows: B, BclI; Bg, BglII; C, ClaI; E, EcoRI; K, KpnI; P, PstI; Pv, PvuII; X, XhoI. The positions of primers used for the detection of circularTn554 forms are labeled circ-fw and circ-rev and are indicated by arrows. The 6-bp core nucleotide sequences at the transposon junctions are shownin boxes. (B) Nucleotide and amino acid sequence alignment of the attachment sites att554 (in S. aureus N315 [3) and att155 (in S. epidermidis [13])of Tn554, that of Tn5406 in S. aureus strain BM3252 (3), and att558 of Tn558 in plasmid pSCFS2. An attachment site identical to att554 has alsobeen reported for Tn5406 in S. aureus strain BM3327 (3). Grey boxes indicate identical amino acids found in three or more of the alignedsequences. The hexanucleotide core sequences of the integration sites are framed. The black bar above the att554 sequence indicates the minimumsequence required for transposition into this site as determined by deletion analysis (5, 6).
VOL. 49, 2005 NOTES 815

PUBLIKATION 10
Schwarz S., C. Werckenthin and C. Kehrenberg:
Identification of a plasmid-borne chloramphenicol-florfenicol resistance gene in Staphylococcus sciuri.
Antimicrob. Agents Chemother. (2000), 44, 2530-2533.

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY,0066-4804/00/$04.0010
Sept. 2000, p. 2530–2533 Vol. 44, No. 9
Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Identification of a Plasmid-Borne Chloramphenicol-FlorfenicolResistance Gene in Staphylococcus sciuri
STEFAN SCHWARZ,1* CHRISTIANE WERCKENTHIN,1,2
AND CORINNA KEHRENBERG1
Institut fur Tierzucht und Tierverhalten der Bundesforschungsanstalt fur Landwirtschaft Braunschweig (FAL),29223 Celle,1 and Institut fur Medizinische Mikrobiologie, Infektions- und Seuchenmedizin,
Tierarztliche Fakultat der Ludwig-Maximilians-Universitat Munchen,80539 Munich,2 Germany
Received 18 January 2000/Returned for modification 23 March 2000/Accepted 14 June 2000
The 16.5-kbp plasmid pSCFS1 from Staphylococcus sciuri mediated combined resistance to chloramphenicoland florfenicol. The gene responsible for this resistance property, cfr, was cloned and sequenced. The aminoacid sequence of the Cfr protein revealed no homology to known acetyltransferases or efflux proteins involvedin chloramphenicol and/or florfenicol resistance or to other proteins whose functions are known.
Staphylococcus sciuri is a common inhabitant of the physio-logical skin flora of most rodents, ungulates, carnivora, andmarsupials. Although classified as rarely pathogenic (6), S.sciuri isolates have been obtained occasionally from cases ofmastitis in goats (10) and bronchopneumonia in cattle (13).Antimicrobial resistance is common among S. sciuri isolates,and a number of plasmids carrying one or more resistancegenes have been identified (11, 13, 14). Resistance to chlor-amphenicol (CM) in staphylococci has usually been associatedwith plasmid-borne cat genes (11, 13), whose gene productsinactivate CM by diacetylation. CM acetyltransferases, how-ever, are unable to inactivate florfenicol (FF), a fluorinatedCM derivative which was licensed in Germany in 1995 as atherapeutic agent to control bacterial respiratory infections incattle. Genes whose gene products mediate combined resis-tance to CM and FF by efflux of both drugs have been identi-fied in gram-negative bacteria, such as Salmonella enterica se-rovar Typhimurium (2) and Photobacterium damselae subsp.piscicida, formerly known as Pasteurella piscicidae (5). In staph-ylococci and related organisms, FF resistance genes have notbeen described yet.
An S. sciuri isolate obtained from the nasal swab of a calfsuffering from an infection of the respiratory tract proved to beresistant to tetracycline, erythromycin, kanamycin, CM, andFF. Plasmid analysis revealed the presence of six plasmids inthe size range between 1.5 and 16.5 kbp. Experiments involvingtransformation into protoplasts of Staphylococcus aureusRN4220 (12) and subsequent selection of the transformants onregeneration media containing 20 mg of FF/ml (Essex, Munich,Germany) identified only the 16.5-kbp plasmid, designatedpSCFS1, as the mediator of resistance to CM and FF. Thisplasmid also mediated resistance to erythromycin by an induc-ibly expressed ermC gene as confirmed by PCR analysis (7).Cloning experiments revealed that the ermC gene was locatedon a 2.5-kbp PstI fragment of pSCFS1 (data not shown). Theoriginal S. sciuri isolate and S. aureus RN4220:pSCFS1 showedFF MICs of 64 mg/ml and CM MICs of 32 mg/ml. Preincuba-tion of these isolates in the presence of either 0.5 mg of FF or
0.5 mg of CM increased the FF MICs to 512 mg/ml and the CMMICs to 64 mg/ml, suggesting that pSCSF1-mediated resis-tance to FF and CM in both staphylococcal hosts is inducibleby FF as well as CM. Plasmid pSCFS1 was mapped (Fig. 1) andsubjected to cloning experiments. Restriction fragments ofpSCFS1 generated by the enzymes EcoRI and BclI-BamHIwere cloned into pBluescript SKII1. The recombinant plas-mids were transformed into the recipient strain Escherichia coliHB101 and plated on Luria-Bertani (LB) agar supplementedwith 20 mg of FF/ml. Only E. coli HB101 clones which carrieda 3.8-kbp EcoRI fragment of pSCFS1 (Fig. 1) grew on theseselective plates. Subclones of this EcoRI fragment were pro-duced and tested for their ability to grow on LB agar supple-mented with 20 mg of FF/ml (Fig. 1). Subclones which carrieda 3-kbp ClaI-EcoRI fragment, a 2.9-kbp EcoRI-XbaI fragment,or a 2-kbp ClaI-XbaI fragment grew on this selective mediumand also on LB agar supplemented with 15 mg of CM/ml. TheMICs of FF and CM for these subclones were 32 mg/ml; pre-incubation in the presence of subinhibitory concentrations ofFF or CM increased the FF MICs to 64 mg/ml but had no effecton the CM MICs. A lack of increase in CM MICs has also beenobserved when inducible cat genes from Staphylococcus spp.were expressed in E. coli hosts (15). All subclones generated byBamHI digestion, e.g., those carrying 1.1-kbp ClaI-BamHI and0.95-kbp BamHI-XbaI fragments (Fig. 1), failed to exhibit re-sistance to FF and CM.
The sequence of the smallest restriction fragment that con-ferred resistance to FF and CM, the 2,037-bp ClaI-XbaI frag-ment, was determined on both strands. Three open readingframes (ORFs) were detected. The BamHI site was locatedwithin an ORF for a peptide of 349 amino acids (aa) (positions570 to 1619). This reading frame, designated cfr (CM and FFresistance) was followed by a pair of inverted repeated se-quences of 13 bp, which may represent the transcriptionalterminator. The cfr reading frame was preceded by a potentialpromoter structure (235: TTTACA, positions 168 to 173; 210:TTACAG, positions 190 to 195; A, position 204) and twooverlapping reading frames, ORF1 (positions 237 to 416) andORF2 (positions 371 to 505), coding for putative peptides of 59and 44 aa, respectively. The amino acid sequences encoded byboth small ORFs did not exhibit significant homology to pro-tein sequences deposited in the databases. Deletion of the cfrgene upstream region as shown in the HpaI-XbaI and theMspI-XbaI subclones (Fig. 1) resulted in sensitivity to FF and
* Corresponding author. Mailing address: Institut fur Tierzucht undTierverhalten der Bundesforschungsanstalt fur Landwirtschaft Braun-schweig (FAL), Dornbergstr. 25-27, 29223 Celle, Germany. Phone:49-5141-384673 or -384675. Fax: 49-5141-381849. E-mail: [email protected].
2530

CM, suggesting that this region is essential for the expressionof combined resistance to FF and CM. Further analysis of theupstream region revealed similarities to the upstream regionsof inducible cat genes from Staphylococcus and Bacillus spp. (8,16). The region between the stop codon of ORF2 and the startcodon of cfr comprised a pair of inverted repeated sequences(IR1: positions 515 to 527; IR2: positions 549 to 563) whichmight be able to form a stable mRNA secondary structure(DG 5 260.3 kJ/mol). The cfr-associated ribosome binding sitewas located within the IR2 sequence. Moreover, the terminalpart of ORF2 (59-GTGCAAAAAGAAATTGATTCT-39)showed considerable homology to previously identified ribo-some stall sequences in the reading frames of the regulatorypeptides involved in inducible CM resistance (8, 16). A ribo-some stalled in the terminal part of ORF2 will overlap the IR1sequence and abolish mRNA secondary structure formation,thus rendering the cfr-associated ribosome binding site acces-sible to ribosomes and allowing translation of the cfr tran-scripts. Assuming that inducible expression of cfr occurs via atranslational attenuation-like process (8, 16), deletion of theupstream region which comprises relevant elements for such aregulatory system may explain the loss of resistance to FF andCM.
Comparison of the Cfr amino acid sequence as deducedfrom the nucleotide sequence revealed no homology to acetyl-transferases or efflux proteins (2, 5, 9) so far known to beassociated with resistance to FF and/or CM. However, homol-ogy to a number of proteins from a wide variety of bacteria,including Mycobacterium tuberculosis H37RV (accession no.Q10806), Treponema pallidum (accession no. AAC65061),Haemophilus influenzae Rd (accession no. P44665), Pseudomo-nas aeruginosa PAO1 (accession no. Q51385), E. coli K12 (ac-cession no. P36979), Bacillus subtilis 16 (accession no.CAA74265), the soil bacterium Streptomyces coelicolor A3(2)(accession no. CAA19907), the cyanobacterium Synechocystissp. strain PCC6803 (accession no. Q55880), and the archaeo-bacterium Thermotoga maritima MSB8 (accession no.AAD36781) was detected (Fig. 2). The reading frames encod-ing most of these proteins were identified during whole-ge-
nome sequencing of the respective organisms. These proteinshave some properties in common: they exhibit similar sizes of340 to 390 aa, have no known functions, and do not exhibit anyspecific features such as ATP binding domains which mightpoint to their possible functions. Recently, the terminal 133 aaof a protein from S. aureus (accession no. CAB60749) whichshows 53% homology to the Cfr protein have been reported(3). This protein was assumed to be an auxiliary protein whichmight play a role in the expression of methicillin resistance (3).Analysis of the Cfr protein sequence confirmed the lack ofATP binding domains (1). Use of the TMpred program (http://www.ch.embnet.org/software/TMPRED_form.html) did notresult in the detection of any topology typical for transmem-brane proteins. This observation suggested that the Cfr proteinis unlikely to be secreted or anchored to the membrane (4).Moreover, the negative results of a CM acetyltransferase assayand a bioassay to demonstrate the enzymatic inactivation of FFand CM (12) confirmed that neither the original S. sciuri northe S. aureus RN4220:pSCFS1 transformant was resistant toFF and CM by enzymatic inactivation of the drugs. Eventhough the mechanism of Cfr-mediated FF and CM resistanceremains to be elucidated, these observations indicate that thecfr gene represents a novel type of transferable CM-FF resis-tance gene, the product of which confers resistance to bothdrugs not only in staphylococci but also in E. coli and obviouslyis not associated with any of the so far known mechanisms ofFF and CM resistance.
Nucleotide sequence accession number. The nucleotide se-quence of the cfr gene and its adjacent regions has been sub-mitted to the EMBL database and was assigned accession no.AJ249217.
C.K. received a scholarship from the Gesellschaft der Freunde derFAL (GdF). This study was supported by a grant from the DeutscheForschungsgemeinschaft (SCHW 382/6-1).
We thank Georg Wolf for providing the S. sciuri isolate, Keith G. H.Dyke for helpful discussions, and B. Otto for help with sequenceanalysis.
FIG. 1. Restriction map of plasmid pSCFS1 from S. sciuri and subcloning strategy for obtaining restriction fragments which mediate (or do not mediate) resistanceto CM and FF. Restriction enzyme abbreviations: B, BclI; Ba, BamHI; Bs, BstEII; C, ClaI; E, EcoRI; EV, EcoRV; Hp, HpaI; Ms, MspI; P, PvuII; Ps, PstI; S, SacI; X,XbaI. A distance scale in kilobase pairs is given below each map. Arrows, locations of the cfr reading frames, ORF1 and ORF2, and their directions of transcription.
VOL. 44, 2000 NOTES 2531

FIG. 2. Amino acid alignment of the Cfr protein from S. sciuri with similar proteins from M. tuberculosis H37RV, S. coelicolor A3(2), E. coli K12, H. influenzae Rd,P. aeruginosa PAO1, Synechocystis sp. strain PCC6803, T. maritima MSB8, B. subtilis 16, and T. pallidum produced with the DNAMAN sequence analysis software(Lynnon BioSoft, Vaudreuil, Quebec, Canada). Black boxes, identical amino acids; gray boxes, homologous amino acids which are present in at least 40% of the alignedsequences.
2532 NOTES ANTIMICROB. AGENTS CHEMOTHER.

REFERENCES
1. Allignet, J., V. Loncle, and N. El Solh. 1992. Sequence of a staphylococcalplasmid gene, vga, encoding a putative ATP-binding protein involved inresistance to virginiamycin A-like antibiotics. Gene 117:45–51.
2. Arcangioli, M. A., S. Leroy-Setrin, J. L. Martel, and E. Chaslus-Dancla.1999. A new chloramphenicol and florfenicol resistance gene linked to anintegron structure in Salmonella typhimurium DT104. FEMS Microbiol. Lett.174:327–332.
3. De Lencastre, H., S. W. Wu, M. G. Pinho, A. M. Ludovice, S. Filipe, S.Gardete, R. Sobral, S. Gill, M. Chung, and A. Tomasz. 1999. Antibioticresistance as a stress response: complete sequencing of a large number ofchromosomal loci in Staphylococcus aureus strain COL that impact on theexpression of resistance to methicillin. Microb. Drug Resist. 5:163–175.
4. Foulger, D., and J. Errington. 1998. A 28 kbp segment from the spoVMregion of Bacillus subtilis 168 genome. Microbiology 144:801–805.
5. Kim, E., and T. Aoki. 1996. Sequence analysis of the florfenicol resistancegene encoded in the transferable R plasmid from a fish pathogen, Pasteurellapiscicida. Microbiol. Immunol. 40:665–669.
6. Kloos, W. E., and K. H. Schleifer. 1986. Genus IV. Staphylococcus, p. 1013–1035. In P. H. A. Sneath, N. S. Mair, M. E. Sharpe, and J. G. Holt (ed.),Bergey’s manual of systematic bacteriology, vol. 2. Williams & Wilkins,Baltimore, Md.
7. Lodder, G., C. Werckenthin, S. Schwarz, and K. G. H. Dyke. 1997. Molecularanalysis of naturally occurring ermC-encoding plasmids in staphylococci iso-lated from animals with and without previous contact with macrolide/linco-
samide antibiotics. FEMS Immunol. Med. Microbiol. 18:7–15.8. Lovett, P. S. 1990. Translational attenuation as the regulator of inducible cat
genes. J. Bacteriol. 172:1–6.9. Murray, I. A., and W. V. Shaw. 1997. O-Acetyltransferases for chloramphen-
icol and other natural products. Antimicrob. Agents Chemother. 41:1–6.10. Poutrel, B. 1984. Udder infection of goats by coagulase-negative staphylo-
cocci. Vet. Microbiol. 9:131–137.11. Schwarz, S., M. Cardoso, and H. Blobel. 1990. Detection of a novel chlor-
amphenicol resistance plasmid from “equine” Staphylococcus sciuri. J. Vet.Med. B 37:674–679.
12. Schwarz, S., M. Cardoso, and H. Blobel. 1990. Plasmid-mediated chloram-phenicol resistance in Staphylococcus hyicus. J. Gen. Microbiol. 135:3329–3336.
13. Schwarz, S., and S. Grolz-Krug. 1991. A chloramphenicol/streptomycin-resistance plasmid from a clinical strain of Staphylococcus sciuri and itsstructural relationships to other staphylococcal resistance plasmids. FEMSMicrobiol. Lett. 82:319–322.
14. Schwarz, S., and W. C. Noble. 1994. Tetracycline resistance genes in staph-ylococci from the skin of pigs. J. Appl. Bacteriol. 76:320–326.
15. Schwarz, S., U. Spies, and M. Cardoso. 1991. Cloning and sequence analysisof a plasmid-encoded chloramphenicol acetyltransferase gene from Staphy-lococcus intermedius. J. Gen. Microbiol. 137:977–981.
16. Stokes, H. W., and R. M. Hall. 1991. Sequence analysis of the induciblechloramphenicol resistance determinant in the Tn1696 integron suggestsregulation by translational attenuation. Plasmid 26:10–19.
VOL. 44, 2000 NOTES 2533

PUBLIKATION 11
Schwarz S., C. Kehrenberg and K.K. Ojo:
Staphylococcus sciuri gene erm(33), encoding inducible resistance to macrolides, lincosamides, and streptogramin B antibiotics, is a product of recombination between erm(C) and erm(A).
Antimicrob. Agents Chemother. (2002), 46, 3621-3623.

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Nov. 2002, p. 3621–3623 Vol. 46, No. 110066-4804/02/$04.00�0 DOI: 10.1128/AAC.46.11.3621–3623.2002Copyright © 2002, American Society for Microbiology. All Rights Reserved.
Staphylococcus sciuri Gene erm(33), Encoding Inducible Resistance toMacrolides, Lincosamides, and Streptogramin B Antibiotics, Is a
Product of Recombination between erm(C) and erm(A)Stefan Schwarz,1* Corinna Kehrenberg,1 and Kayode K. Ojo1,2
Institut fur Tierzucht, Bundesforschungsanstalt fur Landwirtschaft (FAL), 31535 Neustadt-Mariensee, Germany,1
and Department of Pharmaceutical Microbiology and Clinical Pharmacy,College of Medicine, University of Ibadan, Nigeria2
Received 28 March 2002/Returned for modification 10 May 2002/Accepted 25 July 2002
A gene which mediates inducible resistance to macrolides, lincosamides, and streptogramin B antibiotics,designated erm(33), was detected on the Staphylococcus sciuri plasmid pSCFS1. Analysis of the erm(33) readingframe suggested that this gene was the product of a recombination between an erm(C) gene and an erm(A) gene.Such a recombination event is a novel observation for erm genes.
Staphylococcus sciuri, a common inhabitant of the skin ofrodents and other mammals, has been reported to carry anumber of resistance plasmids, such as the tetracycline resis-tance plasmid pSTS9 (12), the chloramphenicol resistanceplasmid pSCS13 (10), and also the chloramphenicol-strepto-mycin resistance plasmid pSCS12 (11), which differ in sizeand/or structure from the resistance plasmids commonly foundin staphylococci. More recently, the first and, to date, onlyknown staphylococcal chloramphenicol-florfenicol resistanceplasmid was isolated from a bovine S. sciuri isolate (13). Anal-ysis of this plasmid, designated pSCFS1, showed that it alsomediated inducible resistance to macrolides, lincosamides, andstreptogramin B antibiotics (MLSB antibiotics). Based on theresults of PCR analysis, the MLSB resistance gene of plasmidpSCFS1 was considered to be an erm gene of class C (13).Since erm(C) genes are commonly located on small 2.3- to4.3-kb plasmids (1–3, 16) and have very rarely been detectedon larger plasmids, we decided to clone and sequence the ermgene and its adjacent regions of the ca. 17-kb plasmid pSCFS1.This approach should provide information on how the erm(C)gene has become part of plasmid pSCFS1.
To localize the erm gene in plasmid pSCFS1, hybridizationstudies were conducted with a gene probe that consisted of the378-bp SacI-BclI fragment of plasmid pSES5 (3). This geneprobe comprised the entire erm(C) translational attenuatorand the first 219 bp of the erm(C) gene. The smallest hybrid-izing fragment was an EcoRI-PstI fragment of ca. 2.2 kb. Thisfragment was cloned into pBluescript II SK(�), and the re-combinant plasmid was transformed into Escherichia coliJM107. Sequence analysis on both strands was performed byprimer walking, starting with the M13 universal and reverseprimers.
The sequence of this EcoRI-PstI fragment consisted of 2,196bp. At the EcoRI end, the first 602 bp represented the 5� endof a reading frame whose product showed similarity to plas-
mid-borne recombination-mobilization proteins of gram-posi-tive bacteria (Fig. 1). The amino terminal 200 amino acids (aa)of this reading frame were most closely related to the corre-sponding parts of a recombination-mobilization protein of Lis-teria monocytogenes (U40997) with 82% amino acid identity,the recombination protein of the Bacillus plasmid pTB913(X15670; 81% amino acid identity), the mobilization proteinof Geobacillus stearothermophilus (M63891; 81% amino acididentity), and the recombination-mobilization protein ofStaphylococcus cohnii (AF015628; 79% amino acid identity).Sequence homology to the expected erm(C) region startedabout 340 bp from the start codon of the aforementionedreading frame. A 66-bp region that showed similarity to thepre-mob upstream region as well as to the erm(C) upstreamregion indicated the junction of both sequences and mighthave served as a putative recombination site. This site did notexhibit similarity to the staphylococcal recombination sitesRSA (on which Pre acts) or RSB (6, 7) previously found onsmall erm(C)-carrying plasmids (6, 7). The 8-bp sequence GTATTCTT within this region did not occur in the two partnersequences (Fig. 1). It represented an imperfect tandem dupli-cation which might have arisen during the recombination pro-cess.
Further downstream, a small reading frame for a peptide of19 aa and two pairs of inverted repeated sequences were foundto precede a reading frame for a 243-aa protein (Fig. 1). Thisregion corresponded closely to the translational attenuatorsseen upstream of the inducibly expressed erm(C) genes ofplasmids pT48 (1), pE194 (2) or pSES5 (3). The invertedrepeats appear to form different mRNA secondary structuresin the presence or absence of inducing macrolides, therebyallowing or preventing translation of the erm(C) transcripts(15). The regulatory region of the erm gene of plasmid pSCFS1differed only by four base pair exchanges and three base pairdeletions from the translational attenuators of inducibly ex-pressed erm(C) genes (1–3). All these sequence alterationswere located in the noncoding parts of the regulatory regionand also did not affect the inverted repeats. The 243-aa proteinrepresented a rRNA methylase that mediates MLSB resis-tance. A comparison of its amino acid sequence with those of
* Corresponding author. Mailing address: Institut fur Tierzuchtder Bundesforschungsanstalt fur Landwirtschaft (FAL), Holtystr. 10,31535 Neustadt-Mariensee, Germany. Phone: 49-5034-871-241. Fax:49-5034-871-246. E-mail: [email protected].
3621

other rRNA methylase proteins revealed highest overall iden-tity of 79% to Tn554-analogous Erm(A) proteins (A25101,AP003137, AP003363, and AB037671) (Table 1), but only 71%amino acid identity to the Erm(A) from Streptococcus pyogenes(AF002716), formerly known as Erm(TR) (14). Identity to theErm(C) proteins found on small plasmids from Bacillus andStaphylococcus varied between 73% [Erm(C) of the Staphylo-coccus hominis plasmid pSES5 (Y09001)] and 76% [Erm(C) ofthe Staphylococcus hyicus plasmid pSES21 (Y09003)] (Table
1). Identity of the Erm protein of plasmid pSCFS1 to Erm(T)of the Lactobacillus reuteri plasmid pGT633 (M64090) was only67% and was even lower at 65% to Erm(Y) of Staphylococcusaureus (AB014481) and Erm(G) of Bacillus sphaericus (M15332).Since this Erm protein exhibited �79% amino acid identity tothe next most closely related Erm proteins (8), it received thedesignation Erm(33) from the Nomenclature Center for MLSGenes (http://faculty.washington.edu/marilynr/; M. Roberts,personal communication). Further analysis of the erm(33) gene
FIG. 1. Restriction map and structural organization of the 2.2-kb EcoRI-PstI fragment of plasmid pSCFS1. The 5� end of a pre-mob-like geneis shown as a box. The white box indicates the small reading frame for the 19-aa regulatory peptide. The black triangles represent the two pairsof inverted repeated sequences detected in the erm(33) translational attenuator. The erm(33) gene and the 3� end of the spc reading frame aredisplayed as arrows, with the arrowhead showing the direction of transcription. The sequences of two potential recombination sites A and B(displayed as boxes) and their adjacent sequences are shown in detail below the map.
TABLE 1. Comparison of different parts of the erm(33) gene and Erm(33) protein sequences with the respective partsof the most closely related erm genes and Erm proteins
Sequence
% Identity
erm(33) gene Erm(33) protein
5� end(284 bp)
Recombinationarea (45 bp)
3� end(403 bp)
Entire gene(732 bp)
N terminus(95 aa)
Recombinationarea (15 aa)
C terminus(133 aa)
Entire protein(243 aa)
erm(A) Tn554 59.5 95.6 100.0 84.0 50.5 86.7 100.0 79.4erm(C) pSES21 99.3 88.9 58.9 76.5 97.9 86.7 59.4 76.1erm(C) pE194 95.4 91.1 60.4 76.0 94.7 86.7 58.3 74.5erm(C) pSES5 94.7 93.3 59.6 75.4 93.3 93.3 57.9 73.7
3622 NOTES ANTIMICROB. AGENTS CHEMOTHER.

showed that the first 284 bp at the 5� end of the reading framewere almost identical to those of erm(C) genes (12 base pairexchanges that caused five amino acid alterations), while the403 bp at the 3� end of erm(33) were indistinguishable fromthose of erm(A) from Tn554 (Table 1). At the junction oferm(C)-homologous and erm(A)-homologous sequences, astretch of 45 bp was seen which displayed sequence identitiesof 95.6% to erm(A) and 93.3% to erm(C) (Fig. 1; Table 1).Assuming that erm(33) resulted from a recombination betweenan erm(C) and an erm(A) gene, it is most probable that this45-bp sequence had served as the site for the recombination.The knowledge of the entire erm(33) sequence also explainsthe initial misidentification of the gene as an erm(C) gene (13)since the PCR primers and the erm(C) gene probe used (3)bound exclusively in the erm(C)-homologous part of erm(33).
The 125 bp immediately downstream of erm(33) corre-sponded exactly to the noncoding sequence downstream oferm(A) in Tn554 (5). Furthermore, the final 166 bp of theEcoRI-PstI fragment represented the 3� end of the spc gene ofTn554 coding for a spectinomycin adenyltransferase (5). Theobservation that additional Tn554-homologous sequenceswere detected in the sequenced part downstream of erm(33)confirmed the involvement of a Tn554-associated erm(A) gene(4) in the development of erm(33).
To the best of our knowledge, this is the first report of anatural recombination between two known erm genes resultingin the formation of a functionally active new erm gene. Bothgenes erm(A) and erm(C) which show ca. 62% identity in theirnucleotide sequences and 58% identity in their deduced aminoacid sequences are widespread among human and animalstaphylococci and occasionally have been detected in the sameisolates (9). Under such conditions, a recombination mighthave occurred by chance. Since Erm(33) did not differ in itssubstrate spectrum, nor in the MICs as determined for induc-ibly expressed Erm(A) or Erm(C) proteins (9), there is nobiological need for the bacteria to develop a recombinationproduct such as Erm(33). However, in PCR-directed studieson the occurrence of different erm genes in staphylococci,misidentification of erm(33) or failure to detect this gene byusing previously described PCR assays must be taken intoconsideration. Therefore, a suitable primer system that specif-ically recognizes this gene should be used for the correct iden-tification of erm(33).
Nucleotide sequence accession number. The sequence of athe erm(33) gene has been deposited with the EMBL databaseunder accession number AJ313523.
This study was supported by grants of the Deutsche Forschungsge-meinschaft (SCHW 382/6-1, SCHW 382/6-2). K. K. Ojo was financiallysupported by the Gesellschaft der Freunde der Kleintierforschung(GdFuF).
We thank Keith G. H. Dyke for helpful discussions.
REFERENCES
1. Catchpole, I., C. Thomas, A. Davies, and K. G. H. Dyke. 1988. The nucleo-tide sequence of Staphylococcus aureus plasmid pT48 conferring induciblemacrolide-lincosamide-streptogramin B resistance and comparison with sim-ilar plasmids expressing constitutive resistance. J. Gen. Microbiol. 134:697–709.
2. Horinouchi, S., and B. Weisblum. 1982. Nucleotide sequence and functionalmap of pE194, a plasmid that specifies inducible resistance to macrolide,lincosamide and streptogramin type B antibiotics. J. Bacteriol. 150:804–814.
3. Lodder, G., C. Werckenthin, S. Schwarz, and K. Dyke. 1997. Molecularanalysis of naturally occurring ermC-encoding plasmids in staphylococci iso-lated from animals with and without previous contact with macrolide/lincos-amide antibiotics. FEMS Immunol. Med. Microbiol. 18:7–15.
4. Murphy, E. 1985. Nucleotide sequence of ermA, a macrolide-lincosamide-streptogramin B determinant in Staphylococcus aureus. J. Bacteriol. 162:633–640.
5. Murphy, E., L. Huwyler, and M. C. F. Bastos. 1985. Transposon Tn554:complete nucleotide sequence and isolation of transposition-defective andantibiotic-sensitive mutants. EMBO J. 4:3357–3365.
6. Novick, R. P. 1989. Staphylococcal plasmids and their replication. Annu.Rev. Microbiol. 43:537–565.
7. Novick, R. P., S. J. Projan, W. Rosenblum, and I. Edelman. 1984. Staphy-lococcal plasmid cointegrates are formed by host- and phage-mediated gen-eral rec systems that act on short regions of homology. Mol. Gen. Genet.195:374–377.
8. Roberts, M. C., J. Sutcliffe, P. Courvalin, L. B. Jensen, J. Rood, and H.Seppala. 1999. Nomenclature for macrolide and macrolide-lincosamide-streptogramin B resistance determinants. Antimicrob. Agents Chemother.43:2823–2830.
9. Schmitz, F.-J., J. Petridou, D. Milatovic, J. Verhoef, A. C. Fluit, and S.Schwarz. 2002. In vitro activity of new ketolides against macrolide-suscepti-ble and -resistant Staphylococcus aureus isolates with defined resistance genestatus. J. Antimicrob. Chemother. 49:580–582.
10. Schwarz, S., and H. Blobel. 1993. Molecular analysis of chloramphenicolresistance in Staphylococcus sciuri. Mh. Vet.-Med. 48:123–127.
11. Schwarz, S., and S. Grolz-Krug. 1991. A chloramphenicol-streptomycin re-sistance plasmid from a clinical strain of Staphylococcus sciuri and its struc-tural relationships to other staphylococcal plasmids. FEMS Microbiol. Lett.82:319 to 322.
12. Schwarz, S., and W. C. Noble. 1994. Tetracycline resistance genes in staph-ylococci from the skin of pigs. J. Appl. Bacteriol. 76:320–326.
13. Schwarz, S., C. Werckenthin, and C. Kehrenberg. 2000. Identification of aplasmid-borne chloramphenicol-florfenicol resistance gene in Staphylococ-cus sciuri. Antimicrob. Agents Chemother. 44:2530–2533.
14. Seppala, H., M. Skurnik, H. Soini, M. C. Roberts, and P. Huovinen. 1998. Anovel erythromycin resistance methylase gene (ermTR) in Streptococcus pyo-genes. Antimicrob. Agents Chemother. 42:257–262.
15. Weisblum, B. 1995. Insights into erythromycin action from studies of itsactivity as inducer of resistance. Antimicrob. Agents Chemother. 39:797–805.
16. Werckenthin, C., S. Schwarz, and H. Westh. 1999. Structural alterations inthe translational attenuator of constitutively expressed ermC genes. Antimi-crob. Agents Chemother. 43:1681–1685.
VOL. 46, 2002 NOTES 3623

PUBLIKATION 12
Kehrenberg C., K.K. Ojo and S. Schwarz:
Nucleotide sequence and organization of the multiresistance plasmid pSCFS1 from Staphylococcus sciuri.
J. Antimicrob. Chemother. (2004), 54, 936-939.

Nucleotide sequence and organization of the multiresistanceplasmid pSCFS1 from Staphylococcus sciuri
Corinna Kehrenberg1, Kayode K. Ojo1,2 and Stefan Schwarz1*
1Institut fur Tierzucht, Bundesforschungsanstalt fur Landwirtschaft (FAL), Holtystrasse 10, 31535 Neustadt-
Mariensee, Germany; 2Department of Pathobiology, University of Washington, Seattle, WA 98195, USA
Received 16 July 2004; returned 2 September 2004; revised 7 September 2004; accepted 12 September 2004
Objectives: The multiresistance plasmid pSCFS1 from Staphylococcus sciuri was sequenced com-pletely and analysed with regard to its gene organization and the putative role of a novel ABCtransporter in antimicrobial resistance.
Methods: Plasmid pSCFS1 was transformed into Staphylococcus aureus RN4220, overlapping restric-tion fragments were cloned into Escherichia coli plasmid vectors and sequenced. For further analysisof the ABC transporter, a � 3 kb Eco RV–Hpa I fragment was cloned into the staphylococcal plasmidpT181MCS and the respective S. aureus RN4220 transformants were subjected to MIC determination.
Results: A total of 14 ORFs coding for proteins of >100 amino acids were detected within the 17 108 bpsequence of pSCFS1. Five of them showed similarity to recombination/mobilization genes whileanother two were similar to plasmid replication genes. In addition to the previously described genescfr for chloramphenicol/florfenicol resistance and erm(33) for inducible resistance to macrolide-lincosamide-streptogramin B resistance, a Tn554-like spectinomycin resistance gene and Tn554-relatedtransposase genes were identified. Moreover, a novel ABC transporter was detected and shown tomediate low-level lincosamide resistance.
Conclusion: Plasmid pSCFS1 is composed of various parts which show similarity to sequences knownto occur on plasmids or transposons of Gram-positive, but also Gram-negative bacteria. It is likely thatpSCFS1 represents the result of inter-plasmid recombination events also involving the truncation of aTn554-like transposon.
Keywords: macrolide resistance, ABC transporter, low-level clindamycin resistance, florfenicol resistance,spectinomycin resistance
Introduction
Members of the species Staphylococcus sciuri are commoninhabitants of the skin of rodents, insectivores, ruminants andungulates, and have rarely been associated with diseases in ani-mals or humans. The finding that S. sciuri strains harbour mecA-related sequences led to the assumption that the staphylococcalgene mecA, responsible for methicillin resistance, may originatefrom S. sciuri.1 Antimicrobial resistance is common amongS. sciuri isolates and several small resistance plasmids isolatedfrom S. sciuri have already been described.2 Plasmid pSCFS1 isthe first multiresistance plasmid detected in S. sciuri and hasbeen reported to mediate resistance to chloramphenicol and itsfluorinated analogue florfenicol, to macrolide, lincosamide andstreptogramin B (MLSB) antibiotics as well as to spectino-mycin.3,4 Two novel resistance genes, cfr for resistance to
chloramphenicol/florfenicol and erm(33) for resistance to MLSB
antibiotics, have already been detected on this plasmid.3,4 Sinceerm(33) in part closely resembled erm(A) of transposon Tn554,and Tn554 also carried a spectinomycin resistance gene in closeproximity to erm(A), the pSCFS1 sequence was analysed forsequences similar to that of Tn554.5
To gain detailed insight into the structure of plasmid pSCFS1,an approach was made to sequence this plasmid completely andto analyse the predicted reading frames for their similarities toother sequences deposited in the databases.
Materials and methods
The S. sciuri isolate harbouring plasmid pSCFS1 was obtained fromthe nasal swab of a calf suffering from a respiratory tract infection.Plasmid pSCFS1 was transformed into Staphylococcus aureus
..........................................................................................................................................................................................................................................................................................................................................................................................................................
*Corresponding author. Tel: +49-5034-871-241; Fax: +49-5034-871-246; E-mail: [email protected]..........................................................................................................................................................................................................................................................................................................................................................................................................................
Journal of Antimicrobial Chemotherapy (2004) 54, 936–939
DOI: 10.1093/jac/dkh457
Advance Access publication 7 October 2004JAC
936
JAC vol.54 no.5 q The British Society for Antimicrobial Chemotherapy 2004; all rights reserved.

RN4220 and the transformants were checked for their in vitro
susceptibility to antimicrobial agents.3,4 MICs were determined by
either the microdilution or the macrodilution broth method accord-
ing to NCCLS document M31-A2.6 Restriction mapping of plasmid
pSCFS1 with various restriction endonucleases and cloning of the
Eco RV, Pst I, Cla I and Bcl I fragments into either pBluescript II
SK+ (Stratagene, Amsterdam, The Netherlands) or pCR Blunt II
TOPO (Invitrogen, Groningen, The Netherlands) followed standard
protocols. Sequencing of these overlapping fragments was achieved
by primer walking on both strands starting with the M13 universal
and reverse primers. For separate analysis of ORF3, apart from the
other resistance genes, a 3 kb Eco RV–HpaI fragment containing
the entire ORF3 and its adjacent sequences was initially cloned into
pCR Blunt II TOPO and later cloned into pT181MCS.7 The recom-
binant plasmid, designated pT181MCS-ABC, was transformed into
S. aureus RN4220. Transformants were checked for their MICs
of chloramphenicol, florfenicol, spectinomycin, erythromycin, til-
micosin, clindamycin and quinupristin/dalfopristin. The complete
sequence of pSCFS1 has been deposited in the EMBL database
under accession number AJ579365. Potential open reading frames
were identified by using the ORF finder system (http://www.ncbi.
nlm.nih.gov/gorf/gorf.html; last accessed 6 July 2004) while com-
parisons of the nucleotide sequences or the deduced amino acid
sequences were obtained using the BLAST system (http://www.
ncbi.nlm.nih.gov/BLAST/; last accessed 6 July 2004).
Results and discussion
Organization of plasmid pSCFS1 and analysis
of the reading frames
Sequence analysis confirmed the size of plasmid pSCFS1 to be17 108 bp. A total of 14 reading frames potentially coding forproteins of >100 amino acids was detected (Figure 1, Table 1).Similarity searches allowed the grouping of 13 of the 14 readingframes into any of five categories: resistance genes, transposasegenes, recombinase/integrase/mobilization genes, plasmid repli-cation genes and hypothetical genes.
The resistance genes were represented by the two previouslydescribed genes cfr (ORF4), coding for a putative oxidoreduc-tase that mediates combined resistance to chloramphenicol andflorfenicol, and the gene erm(33) (ORF7), a novel MLSB resist-ance gene that represents an in vivo derived ‘in-frame’ recombi-nation product of the MLSB resistance genes erm(C) anderm(A).3,4 The third resistance gene is the gene spc (ORF8)which codes for a spectinomycin adenyltransferase identical tothat of transposon Tn554.5
Immediately downstream of spc, two more Tn554-relatedreading frames were found: the complete reading frame for the125 amino acid transposase C (ORF9) and a truncated readingframe (ORF10) whose deduced C-terminal 45 amino acids wereidentical to the C terminus of transposase B of Tn554. The2030 bp segment located from bases 10182 to 12211 showed99% identity to the corresponding part of Tn554 and includesthe 30 end of erm(33) and the genes spc, tnpC and DtnpB.
The putative products of the five ORFs 1, 2, 5, 12 and 13showed more or less extended similarities to DNA-bindingproteins which may play a role in integration, recombination ormobilization processes. The ORF1 product exhibits 44% identityand 64% similarity to an integrase protein of Enterococcusfaecium (accession no. ZP_00036382). The ORF2 protein shows29% identity and 51% similarity to a putative DNA-binding pro-tein of Bacillus subtilis (accession no. NP_053778). The productof ORF5 shows 52% identity and 66% similarity to the N termi-nus of a recombinase/mobilization protein from S. aureus (acces-sion no. AAF85649) whereas the product of ORF12 exhibits35% identity and 56% similarity to the C terminus of the samerecombinase/mobilization protein. The product of ORF13 shows71% identity and 83% similarity to an internal 153 amino acidsegment of the 361 amino acid recombinase/mobilizationproteins of Lactobacillus plantarum and Lactobacillus hilgardii(accession nos. AAA25252 and AAA98162).
Two reading frames were detected whose products showedsimilarity to plasmid replication proteins: that of ORF11 shows38% identity and 55% similarity to the Rep protein of plasmidpTS1 from the oral spirochaete Treponema denticola (accessionno. NP_073755) while that of ORF14 exhibits 29% identity and46% similarity to a 220 amino acid internal segment of theRep protein of the Mannheimia varigena plasmid pMVSCS1(accession no. NP_573540). Finally, the deduced amino acidsequence of ORF6, which in part overlaps ORF5 (Table 1),revealed low levels of similarity to small hypothetical proteinsdetected during whole-genome sequencing of S. aureus strainN315.
Analysis of the ABC transporter and its role
in antibiotic resistance
All ORFs described previously either played a defined role inresistance to chloramphenicol/florfenicol, MLSB antibiotics orspectinomycin, or were supposed to have other functions basedon the results of the structural comparisons. The only remainingreading frame, ORF3, was considered a candidate for a putativerole in antimicrobial resistance. ORF3 codes for an ABC trans-porter of 492 amino acids. The Walker A motif (G RNG RGKT)was detected at positions 38–45 and the Walker B motif(FLLID) at positions 137–142 in the deduced amino acidsequence. Both motifs characterize the ATP-hydrolysingdomains and are supposed to constitute a nucleotide-binding
Figure 1. Circular map of plasmid pSCFS1. The reading frames are num-
bered with reference to Table 1; grey-shaded reading frames are involved in
antimicrobial resistance.
Antibiotic multiresistance in S. sciuri
937
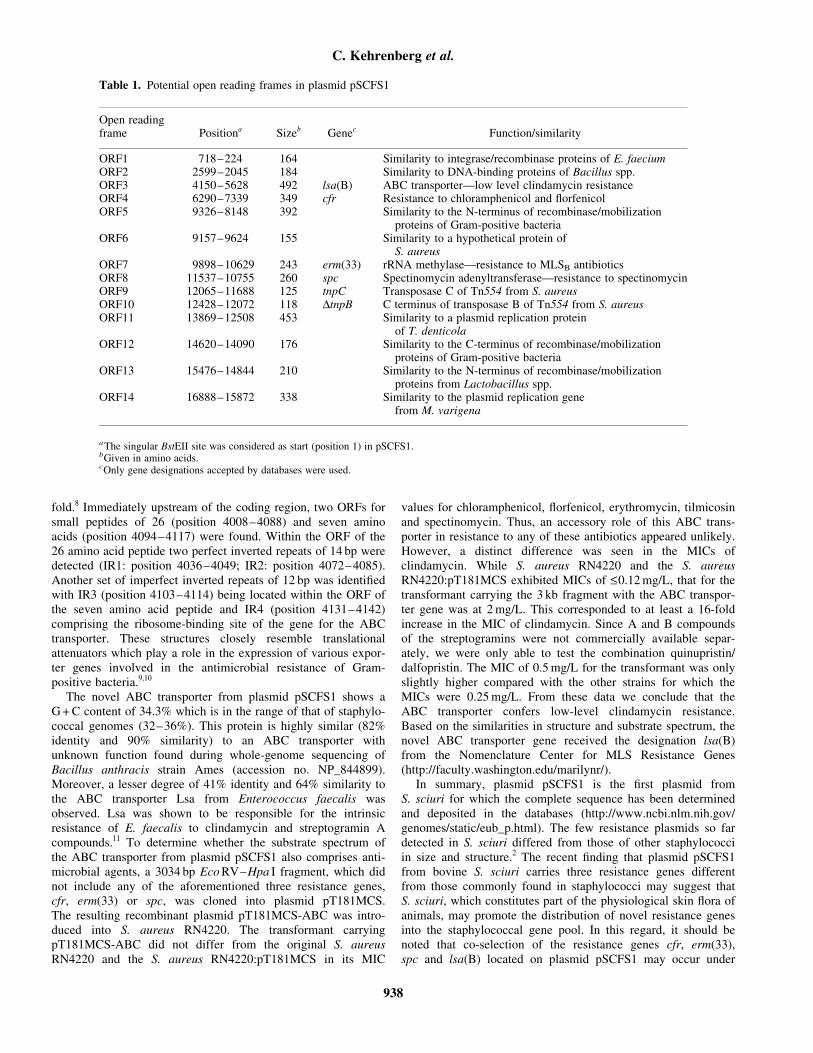
fold.8 Immediately upstream of the coding region, two ORFs forsmall peptides of 26 (position 4008–4088) and seven aminoacids (position 4094–4117) were found. Within the ORF of the26 amino acid peptide two perfect inverted repeats of 14 bp weredetected (IR1: position 4036–4049; IR2: position 4072–4085).Another set of imperfect inverted repeats of 12 bp was identifiedwith IR3 (position 4103–4114) being located within the ORF ofthe seven amino acid peptide and IR4 (position 4131–4142)comprising the ribosome-binding site of the gene for the ABCtransporter. These structures closely resemble translationalattenuators which play a role in the expression of various expor-ter genes involved in the antimicrobial resistance of Gram-positive bacteria.9,10
The novel ABC transporter from plasmid pSCFS1 shows aG + C content of 34.3% which is in the range of that of staphylo-coccal genomes (32–36%). This protein is highly similar (82%identity and 90% similarity) to an ABC transporter withunknown function found during whole-genome sequencing ofBacillus anthracis strain Ames (accession no. NP_844899).Moreover, a lesser degree of 41% identity and 64% similarity tothe ABC transporter Lsa from Enterococcus faecalis wasobserved. Lsa was shown to be responsible for the intrinsicresistance of E. faecalis to clindamycin and streptogramin Acompounds.11 To determine whether the substrate spectrum ofthe ABC transporter from plasmid pSCFS1 also comprises anti-microbial agents, a 3034 bp Eco RV–Hpa I fragment, which didnot include any of the aforementioned three resistance genes,cfr, erm(33) or spc, was cloned into plasmid pT181MCS.The resulting recombinant plasmid pT181MCS-ABC was intro-duced into S. aureus RN4220. The transformant carryingpT181MCS-ABC did not differ from the original S. aureusRN4220 and the S. aureus RN4220:pT181MCS in its MIC
values for chloramphenicol, florfenicol, erythromycin, tilmicosinand spectinomycin. Thus, an accessory role of this ABC trans-porter in resistance to any of these antibiotics appeared unlikely.However, a distinct difference was seen in the MICs ofclindamycin. While S. aureus RN4220 and the S. aureusRN4220:pT181MCS exhibited MICs of <_0.12 mg/L, that for thetransformant carrying the 3 kb fragment with the ABC transpor-ter gene was at 2 mg/L. This corresponded to at least a 16-foldincrease in the MIC of clindamycin. Since A and B compoundsof the streptogramins were not commercially available separ-ately, we were only able to test the combination quinupristin/dalfopristin. The MIC of 0.5 mg/L for the transformant was onlyslightly higher compared with the other strains for which theMICs were 0.25 mg/L. From these data we conclude that theABC transporter confers low-level clindamycin resistance.Based on the similarities in structure and substrate spectrum, thenovel ABC transporter gene received the designation lsa(B)from the Nomenclature Center for MLS Resistance Genes(http://faculty.washington.edu/marilynr/).
In summary, plasmid pSCFS1 is the first plasmid fromS. sciuri for which the complete sequence has been determinedand deposited in the databases (http://www.ncbi.nlm.nih.gov/genomes/static/eub_p.html). The few resistance plasmids so fardetected in S. sciuri differed from those of other staphylococciin size and structure.2 The recent finding that plasmid pSCFS1from bovine S. sciuri carries three resistance genes differentfrom those commonly found in staphylococci may suggest thatS. sciuri, which constitutes part of the physiological skin flora ofanimals, may promote the distribution of novel resistance genesinto the staphylococcal gene pool. In this regard, it should benoted that co-selection of the resistance genes cfr, erm(33),spc and lsa(B) located on plasmid pSCFS1 may occur under
Table 1. Potential open reading frames in plasmid pSCFS1
Open readingframe Positiona Sizeb Genec Function/similarity
ORF1 718–224 164 Similarity to integrase/recombinase proteins of E. faeciumORF2 2599–2045 184 Similarity to DNA-binding proteins of Bacillus spp.ORF3 4150–5628 492 lsa(B) ABC transporter—low level clindamycin resistanceORF4 6290–7339 349 cfr Resistance to chloramphenicol and florfenicolORF5 9326–8148 392 Similarity to the N-terminus of recombinase/mobilization
proteins of Gram-positive bacteriaORF6 9157–9624 155 Similarity to a hypothetical protein of
S. aureusORF7 9898–10629 243 erm(33) rRNA methylase—resistance to MLSB antibioticsORF8 11537–10755 260 spc Spectinomycin adenyltransferase—resistance to spectinomycinORF9 12065–11688 125 tnpC Transposase C of Tn554 from S. aureusORF10 12428–12072 118 DtnpB C terminus of transposase B of Tn554 from S. aureusORF11 13869–12508 453 Similarity to a plasmid replication protein
of T. denticolaORF12 14620–14090 176 Similarity to the C-terminus of recombinase/mobilization
proteins of Gram-positive bacteriaORF13 15476–14844 210 Similarity to the N-terminus of recombinase/mobilization
proteins from Lactobacillus spp.ORF14 16888–15872 338 Similarity to the plasmid replication gene
from M. varigena
aThe singular BstEII site was considered as start (position 1) in pSCFS1.bGiven in amino acids.cOnly gene designations accepted by databases were used.
C. Kehrenberg et al.
938

selective pressure imposed by the use of either macrolides,spectinomycin, chloramphenicol, florfenicol or lincosamides.Although plasmids such as pSCFS1 have been detected veryrarely to date, the occurrence of a plasmid that was indistin-guishable from pSCFS1 in its restriction map and resistance phe-notype/genotype, in a bovine Staphylococcus simulans isolateindicates the potential of this plasmid to be spread horizontallyto other staphylococcal species.
Acknowledgements
We wish to thank Vera Noding for expert technical assistance.This study was supported by grants from the DeutscheForschungsgemeinschaft (SCHW 382/6-1, SCHW 382/6-2).
References
1. Couto, I., de Lencastre, H., Severina, E. et al. (1996).
Ubiquitous presence of mecA homologue in natural isolates of
Staphylococcus sciuri. Microbial Drug Resistance 2, 377–91.
2. Werckenthin, C., Cardoso, M., Martel, J.-L. et al. (2001).
Antimicrobial resistance in staphylococci from animals with particular
reference to bovine Staphylococcus aureus, porcine Staphylococcus
hyicus and canine Staphylococcus intermedius. Veterinary Research
32, 341–62.
3. Schwarz, S., Kehrenberg, C. & Ojo, K. K. (2002). Staphylococ-
cus sciuri gene, erm(33), encoding inducible resistance to macrolides,
lincosamides and streptogramin B antibiotics is a product of
recombination between erm(C) and erm(A). Antimicrobial Agents and
Chemotherapy 46, 3621–3.
4. Schwarz, S., Werckenthin, C. & Kehrenberg, C. (2000).
Identification of a plasmid-borne chloramphenicol-florfenicol resistance
gene in Staphylococcus sciuri. Antimicrobial Agents and Chemotherapy
44, 2530–3.
5. Murphy, E., Huwyler, L. & Bastos, M. C. F. (1985).
Transposon Tn554: complete nucleotide sequence and isolation of
transposition-defective and antibiotic-sensitive mutants. EMBO Jour-
nal 4, 3357–65.
6. National Committee for Clinical Laboratory Standards. (2002).
Performance Standards for Antimicrobial Disk and Dilution Suscep-
tibility Tests for Bacteria Isolated from Animals:—2nd edition.
Approved Standard M31-A2. NCCLS, Wayne, PA, USA.
7. Augustin, J., Rosenstein, R., Wieland, B. et al. (1992). Genetic
analysis of epidermin biosynthetic genes and epidermin-negative
mutants of Staphylococcus epidermidis. European Journal of
Biochemistry 204, 1149–54.
8. Schneider, E. & Hunke, S. (1998). ATP-binding-cassette (ABC)
transport systems: functional and structural aspects of the
ATP-hydrolyzing subunits/domains. FEMS Microbiology Reviews 22,
1–20.
9. Roberts, M. C. (1996). Tetracycline resistance determinants:
mechanisms of action, regulation of expression, genetic mobility,
and distribution. FEMS Microbiology Reviews 19, 1–24.
10. Kehrenberg, C. & Schwarz, S. (2004). fexA, a novel
Staphylococcus lentus gene encoding resistance to florfenicol
and chloramphenicol. Antimicrobial Agents and Chemotherapy 48,
615–8.
11. Singh, K. V., Weinstock, G. M. & Murray, B. E. (2002).
An Enterococcus faecalis ABC homologue (Lsa) is required
for the resistance of this species to clindamycin and
quinupristin–dalfopristin. Antimicrobial Agents and Chemotherapy 46,
1845–50.
Antibiotic multiresistance in S. sciuri
939

PUBLIKATION 13
Kehrenberg C., S. Schwarz, L. Jacobsen, L.H. Hansen and B. Vester:
A new mechanism for chloramphenicol, florfenicol and clindamycin resistance: methylation of 23S ribosomal RNA at A2503.
Mol. Microbiol. (2005), 57, 1064-1073.

Molecular Microbiology (2005)
57
(4), 1064–1073 doi:10.1111/j.1365-2958.2005.04754.x
© 2005 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 2005
? 2005
57
410641073
Original Article
Drug resistance by methylation of A2503 in 23S rRNAC. Kehrenberg
et al.
Accepted 31 May, 2005. *For correspondence. [email protected]; Tel. (
+
45) 65 502 377; Fax (
+
45) 65 502 467.
A new mechanism for chloramphenicol, florfenicol and clindamycin resistance: methylation of 23S ribosomal RNA at A2503
Corinna Kehrenberg,
1
Stefan Schwarz,
1
Lene Jacobsen,
2
Lykke H. Hansen
2
and Birte Vester
2
*
1
Institut für Tierzucht, Bundesforschungsanstalt für Landwirtschaft (FAL), Höltystrasse 10, 31535 Neustadt-Mariensee, Germany.
2
Department of Biochemistry and Molecular Biology, University of Southern Denmark, DK-5230 Odense M, Denmark.
Summary
The gene product of
cfr
from
Staphylococcus sciuri
confers resistance to chloramphenicol, florfenicoland clindamycin in
Staphylococcus
spp. and
Escher-ichia coli
. Cfr is not similar to any other knownchloramphenicol resistance determinant. Compara-tive investigation of
E. coli
with and without aplasmid-encoded Cfr showed a decreased drug bind-ing to ribosomes in the presence of Cfr. As chloram-phenicol/florfenicol and clindamycin have partlyoverlapping drug binding sites on the ribosome, themost likely explanation is that Cfr modifies the RNAin the drug binding site. This hypothesis was sup-ported by drug footprinting data that showed both adecreased drug binding and an enhanced reversetranscriptase stop at position 2504, which corre-sponds to a modification at position A2503 at the drugbinding site. A 45 n long RNA fragment containing theappropriate region was isolated and MALDI-TOF massspectrometry in combination with tandem mass spec-trometry showed an additional methylation at positionA2503. Moreover, reduced methylation was detectedat nucleotide C2498. The results show that Cfr is anRNA methyltransferase that targets nucleotide A2503and inhibits ribose methylation at nucleotide C2498,thereby causing resistance to chloramphenicol, flor-fenicol and clindamycin.
Introduction
Phenicol antibiotics, such as chloramphenicol (CHL) andflorfenicol (FFC), and also lincosamide antibiotics like clin-
damycin (CLI), are potent inhibitors of bacterial proteinbiosynthesis. Members of both classes of antimicrobialagents have been used for several decades in human andveterinary medicine and resistance is common among awide variety of bacteria of medical or veterinary impor-tance. Recent reviews have shown that resistance tophenicols or lincosamides can be based on several differ-ent resistance mechanisms specified by a considerablenumber of genes or mutations (Roberts
et al
., 1999;Schwarz
et al
., 2004). In the case of combined resistanceto CHL and FFC, however, only three different resistancegenes have been detected. Two of these,
floR
(Arcangioli
et al
., 1999; Briggs and Fratamico, 1999) and
fexA
(Kehrenberg and Schwarz, 2004), code for membrane-associated efflux systems of the Major Facilitator Super-family that mediate the export of both drugs from thebacterial cell. The third gene,
cfr
from
Staphylococcussciuri
(Schwarz
et al
., 2000), was the first CHL–FFCresistance gene found in Gram-positive bacteria. Thisgene was initially detected on the 17 kb multiresistanceplasmid, pSCFS1, that also carried the macrolide–lincosamide–streptogramin B resistance gene
erm
(33)(Schwarz
et al
., 2002), the spectinomycin resistance gene
spc
, and the ABC transporter gene
lsa
(B) that mediateslow-level CLI resistance (Kehrenberg
et al
., 2004).Analysis of the amino acid sequence of the Cfr protein
revealed no homology to efflux proteins or enzymes, suchas acetyltransferases or phosphotransferases, involved inresistance to CHL (Schwarz
et al
., 2000). Furthermore, anacetyltransferase assay and a bioassay to demonstrateenzymatic inactivation of CHL and FFC gave negativeresults. In addition, the Cfr sequence apparently does notcontain any motifs typical for transmembrane proteinssuggesting that the Cfr protein is unlikely to be anchoredto the membrane. Moreover, no ATP-binding domainswere detectable. The Cfr sequence shows homology to anumber of proteins from a wide variety of bacteria(Schwarz
et al
., 2000), but no function has been assignedto these proteins. Database searches indicate that Cfr hassome homology to a protein superfamily called RadicalSAM that catalyses diverse reactions, including unusualmethylations, isomerization, sulphur insertion, ring forma-tion, anaerobic oxidation and protein radical formation(Sofia
et al
., 2001). The
cfr
gene thus represents a noveltype of transferable CHL–FFC resistance gene, the prod-

Drug resistance by methylation of A2503 in 23S rRNA
1065
© 2005 Blackwell Publishing Ltd,
Molecular Microbiology
,
57
, 1064–1073
uct of which confers resistance to both drugs not only instaphylococci, but also in
Escherichia coli
(Schwarz
et al
.,2000). However, the mechanism of Cfr-mediated CHL–FFC resistance is still unknown.
If Cfr neither modifies the drugs that it confers resis-tance to nor affects the efflux of the drugs, it is likely toaffect the drug target site. It is well known that resistanceto many protein biosynthesis inhibitors can be due tomutations in the genes for ribosomal RNA or ribosomalproteins, or may be caused by methylation of rRNA atspecific positions (Weisblum, 1995; Vester and Douth-waite, 2001). CHL and CLI bind to the 50S ribosomalsubunit and interact with the central loop of 23S rRNAdomain V in the peptidyl transferase centre (Vasquez,1979; Moazed and Noller, 1987; Douthwaite, 1992) wherethey inhibit peptidyl transferase activity. The detailed bind-ing of CHL to 50S subunits from
Deinococcus radiodurans
and
Haloarcula marismortui
(Schlünzen
et al
., 2001;Hansen
et al
., 2003) was reported in X-ray studies as wasthe binding of CLI to 50S subunits from
D. radiodurans
(Schlünzen
et al
., 2001). The binding properties of CHLand CLI in
D. radiodurans
are believed to resemble closelythe functional binding of these antimicrobial agents toribosomes from bacteria of medical and veterinary impor-tance. The CHL and CLI binding sites correlate well withfootprinting studies where protections against modifica-tion of specific nucleotides indicate the binding sites(Moazed and Noller, 1987; Douthwaite, 1992; Rodriguez-Fonseca
et al
., 1995). It is also in accordance with theresults of a number of mutagenesis studies in which muta-tions in the RNA at the binding site affected CHL bindingas shown by footprinting and cross-linking analyses (Longand Porse, 2003 and references herein). The CHL bindingsite, which is believed to be identical to the FFC bindingsite, and the CLI binding site are in the peptidyl trans-ferase region that is composed entirely of RNA (Nissen
et al
., 2000). Hence it is likely that an effect on the drugtarget site has to be found in this RNA region, which isdepicted in Fig. 1.
The aim of this study was to investigate whether Cfrmodifies the CHL/FFC/CLI binding site on the ribosomeand, if so, to determine the exact position and the type ofmodification using footprinting analysis, MALDI-TOF MSand tandem MS.
Results
Revised resistance phenotype mediated by Cfr
The
cfr
gene was initially reported to mediate resistanceonly to CHL and FFC (Schwarz
et al
., 2000). The first
cfr
-carrying plasmid, pSCFS1, also carried two genes,
erm
(33) and
lsa
(B), which mediate CLI resistance, andtherefore an association of
cfr
with CLI resistance was not
previously taken into account. Most
E. coli
strains, includ-ing those used as recipients in the cloning experiments,exhibited noticeably high levels of basic insensitivity toCLI. It was thus difficult to recognize a change in theCLI susceptibility patterns of subclones carrying the
cfr
gene as compared to those carrying other segments ofpSCFS1. Recently, a larger plasmid of approximately36 kb, designated pSCFS3, from a porcine
Staphylococ-cus aureus
strain that carried the two staphylococcalCHL–FFC resistance genes
cfr
and
fexA
was identified(C. Kehrenberg and S. Schwarz, unpublished). Transfor-mation studies into
S. aureus
RN4220 showed thatplasmid pSCFS3 mediated high-level CLI resistance(minimum inhibitory concentration, MIC: 256
m
g ml
-
1
) inaddition to CHL–FFC resistance (MICs for CHL and FFC:
≥
128
m
g ml
-
1
). Cloning of all BglII fragments of plasmidpSCFS3 and
in vitro
susceptibility testing of the respective
E. coli
clones (Table 1) showed that only transformantscarrying the cloning vector with the 3.6 kb BglII insert(pBglII) exhibited an MIC
CLI
of
≥
128
m
g ml
-
1
and anMIC
CHL,FFC
of 32
m
g ml
-
1
, whereas
E. coli
HB101 carryingthe empty cloning vector or other cloned fragments hadthe
E. coli
HB101 basic MIC
CLI
of 32
m
g ml
-
1
and MIC
CHL,FFC
of 2
m
g ml
-
1
(Table 1).The 3.6 kb BglII fragment was sequenced and found to
carry a
cfr
gene whose deduced amino acid sequencediffered from that of Cfr from pSCFS1 by a single aminoacid substitution (A254
Æ
D254 in Cfr from pSCFS3). Twoincomplete reading frames of 290 aa and 150 aa brack-eted the
cfr
gene. The larger one in the upstream partshowed similarities to transposases of the IS
21
familyfrom
Symbiobacterium thermophilum
(database acces-sion no. BAD41683) and the smaller one in the down-stream part closely resembled part of the transposase B
Table 1.
Correlation of MIC values for CHL, FFC and CLI and CMCTmodification effects at
E. coli
23S RNA position U2506 in Cfr
+
andCfr–
E. coli
HB101.
E. coli
HB101::pBglII (Cfr
+
)
E. coli
HB101::pBluescript (Cfr–)
MICCHL 32
m
g ml
-
1
2
m
g ml
-
1
FFC 32
m
g ml
-
1
2
m
g ml
-
1
CLI
≥
128
m
g ml
-
1
32
m
g ml
-
1
Drug protection of U2506CHL (0.05 mM) 10–30% 50–70%CHL (0.2 mM) 50–70% 70–90%CHL (0.5 mM) 50–70% 70–90%CHL (1.0 mM) 70–90% 70–90%FFC (0.05 mM)
<
10% 30–50%FFC (0.2 mM) 10–30% 50–70%FFC (0.5 mM) 30–50% 50–70%FFC (1 mM) 30–50% 70–90%CLI (0.05 mM)
<
10% 10–30%CLI (0.2 mM)
<
10% 50–70%CLI (0.5 mM)
<
10% 50–70%CLI (1 mM)
<
10% 50–70%
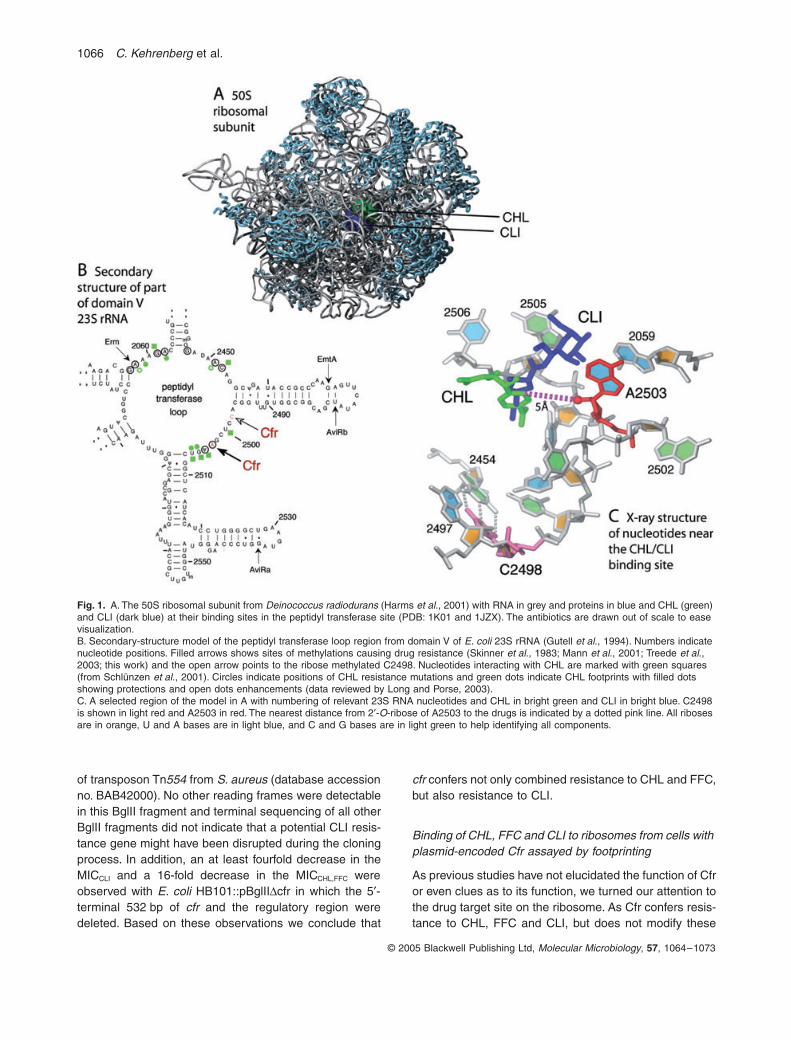
1066
C. Kehrenberg
et al.
© 2005 Blackwell Publishing Ltd,
Molecular Microbiology
,
57
, 1064–1073
of transposon Tn
554
from
S. aureus
(database accessionno. BAB42000). No other reading frames were detectablein this BglII fragment and terminal sequencing of all otherBglII fragments did not indicate that a potential CLI resis-tance gene might have been disrupted during the cloningprocess. In addition, an at least fourfold decrease in theMIC
CLI
and a 16-fold decrease in the MIC
CHL,FFC
wereobserved with
E. coli
HB101::pBglII
Dcfr in which the 5¢-terminal 532 bp of cfr and the regulatory region weredeleted. Based on these observations we conclude that
cfr confers not only combined resistance to CHL and FFC,but also resistance to CLI.
Binding of CHL, FFC and CLI to ribosomes from cells with plasmid-encoded Cfr assayed by footprinting
As previous studies have not elucidated the function of Cfror even clues as to its function, we turned our attention tothe drug target site on the ribosome. As Cfr confers resis-tance to CHL, FFC and CLI, but does not modify these
Fig. 1. A. The 50S ribosomal subunit from Deinococcus radiodurans (Harms et al., 2001) with RNA in grey and proteins in blue and CHL (green) and CLI (dark blue) at their binding sites in the peptidyl transferase site (PDB: 1K01 and 1JZX). The antibiotics are drawn out of scale to ease visualization.B. Secondary-structure model of the peptidyl transferase loop region from domain V of E. coli 23S rRNA (Gutell et al., 1994). Numbers indicate nucleotide positions. Filled arrows shows sites of methylations causing drug resistance (Skinner et al., 1983; Mann et al., 2001; Treede et al., 2003; this work) and the open arrow points to the ribose methylated C2498. Nucleotides interacting with CHL are marked with green squares (from Schlünzen et al., 2001). Circles indicate positions of CHL resistance mutations and green dots indicate CHL footprints with filled dots showing protections and open dots enhancements (data reviewed by Long and Porse, 2003).C. A selected region of the model in A with numbering of relevant 23S RNA nucleotides and CHL in bright green and CLI in bright blue. C2498 is shown in light red and A2503 in red. The nearest distance from 2¢-O-ribose of A2503 to the drugs is indicated by a dotted pink line. All riboses are in orange, U and A bases are in light blue, and C and G bases are in light green to help identifying all components.
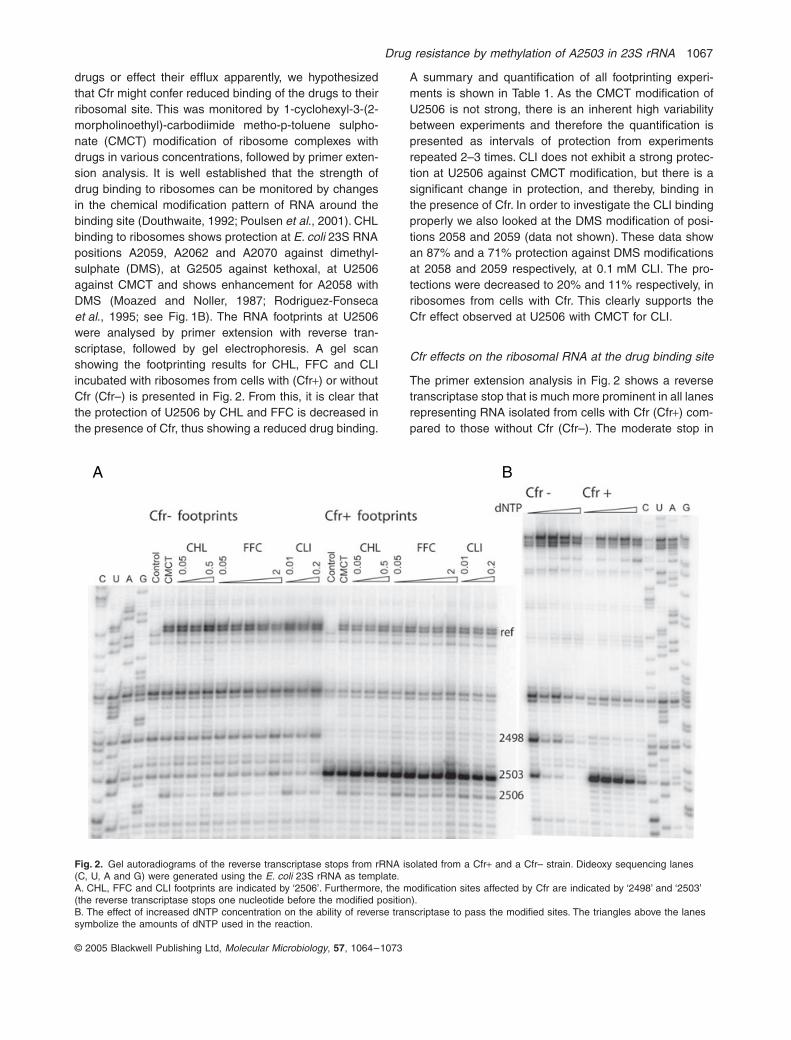
Drug resistance by methylation of A2503 in 23S rRNA 1067
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1064–1073
drugs or effect their efflux apparently, we hypothesizedthat Cfr might confer reduced binding of the drugs to theirribosomal site. This was monitored by 1-cyclohexyl-3-(2-morpholinoethyl)-carbodiimide metho-p-toluene sulpho-nate (CMCT) modification of ribosome complexes withdrugs in various concentrations, followed by primer exten-sion analysis. It is well established that the strength ofdrug binding to ribosomes can be monitored by changesin the chemical modification pattern of RNA around thebinding site (Douthwaite, 1992; Poulsen et al., 2001). CHLbinding to ribosomes shows protection at E. coli 23S RNApositions A2059, A2062 and A2070 against dimethyl-sulphate (DMS), at G2505 against kethoxal, at U2506against CMCT and shows enhancement for A2058 withDMS (Moazed and Noller, 1987; Rodriguez-Fonsecaet al., 1995; see Fig. 1B). The RNA footprints at U2506were analysed by primer extension with reverse tran-scriptase, followed by gel electrophoresis. A gel scanshowing the footprinting results for CHL, FFC and CLIincubated with ribosomes from cells with (Cfr+) or withoutCfr (Cfr–) is presented in Fig. 2. From this, it is clear thatthe protection of U2506 by CHL and FFC is decreased inthe presence of Cfr, thus showing a reduced drug binding.
A summary and quantification of all footprinting experi-ments is shown in Table 1. As the CMCT modification ofU2506 is not strong, there is an inherent high variabilitybetween experiments and therefore the quantification ispresented as intervals of protection from experimentsrepeated 2–3 times. CLI does not exhibit a strong protec-tion at U2506 against CMCT modification, but there is asignificant change in protection, and thereby, binding inthe presence of Cfr. In order to investigate the CLI bindingproperly we also looked at the DMS modification of posi-tions 2058 and 2059 (data not shown). These data showan 87% and a 71% protection against DMS modificationsat 2058 and 2059 respectively, at 0.1 mM CLI. The pro-tections were decreased to 20% and 11% respectively, inribosomes from cells with Cfr. This clearly supports theCfr effect observed at U2506 with CMCT for CLI.
Cfr effects on the ribosomal RNA at the drug binding site
The primer extension analysis in Fig. 2 shows a reversetranscriptase stop that is much more prominent in all lanesrepresenting RNA isolated from cells with Cfr (Cfr+) com-pared to those without Cfr (Cfr–). The moderate stop in
Fig. 2. Gel autoradiograms of the reverse transcriptase stops from rRNA isolated from a Cfr+ and a Cfr– strain. Dideoxy sequencing lanes(C, U, A and G) were generated using the E. coli 23S rRNA as template.A. CHL, FFC and CLI footprints are indicated by ‘2506’. Furthermore, the modification sites affected by Cfr are indicated by ‘2498’ and ‘2503’ (the reverse transcriptase stops one nucleotide before the modified position).B. The effect of increased dNTP concentration on the ability of reverse transcriptase to pass the modified sites. The triangles above the lanes symbolize the amounts of dNTP used in the reaction.
A B

1068 C. Kehrenberg et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1064–1073
the Cfr– samples is assigned to the methylation at position2 in the adenine at position 2503 (Kowalak et al., 1995).Primer extension stops with reverse transcriptase can becaused by structures in RNA or modifications, and bybreakage of RNA caused by enzymatic degradation orsimple water hydrolysis. An RNA modification that inter-feres with base pairing will stop or pause reverse tran-scriptase one nucleotide before the modified base. Aribose methylation will usually cause a pause of reversetranscriptase at the modified nucleotide or one nucleotidebefore or both, but the effect is typically most pronouncedwhen the dNTP concentration is limited (Maden et al.,1995). The strong stop observed in Fig. 2A in lanes rep-resenting Cfr+ samples indicates that different modifica-tions are present at A2503 compared to the wild-typem2A2503. The stop/pause patterns from non-ribose RNAmodifications have not been systematically investigated,but it is reported that m6A does not affect reverse tran-scriptase extension, whereas dimethylation at position 6at A causes a complete stop (Zalacain and Cundliffe,1989).
The reverse transcriptase stop at position C2498 in thewild-type samples is assumed to be caused by the 2¢-O-ribose methylation previously reported at this position(Branlant et al., 1981). This stop is decreased in samplesfrom cells with Cfr indicating that the presence of Cfrdecreases the amount of Cm2498 in ribosomes. Thus, thedata in Fig. 2 show that Cfr somehow modifies A2503 anddecreases modification at C2498.
To obtain additional information about the nature of theobserved modifications, the stops were investigated atvarious dNTP concentrations (Fig. 2B). The gel shows adNTP concentration dependence of the Cm2498 stopin the Cfr– RNA as expected. Surprisingly, the stop atm2A2503 is apparently also dNTP concentration-depen-dent similarly to what is observed for ribose methylations.The same pattern is obtained for RNA from cells with Cfr,but the transcription stop is much more prominent. Thisindicates that the additional modification on A2503 alsocauses a dNTP-dependent reverse transcriptase stop andthat it could be a 2¢-O-ribose methylation.
MALDI-TOF MS analysis of rRNA modified by Cfr
To investigate the effects observed on the gels and furtherdefine the modifications, we employed MALDI-MS analysison a 45n rRNA fragment encompassing positions 2483–2527 of 23S rRNA. The RNA fragment was isolated byspecific hybridization of a complementary DNA oligonu-cleotide, followed by single-strand RNA digestion and gelelectrophoresis as described by Andersen et al. (2004).The isolated 45n RNA was digested with the G-specificRNase T1 to obtain fragments suitable for detailed analy-sis. Some modifications, e.g. 2¢-O-ribose methylations,
inhibit RNase T1 digestions, whereas other modificationsdo not (Lankat-Buttgereit et al., 1987; Maden, 1990;Mengel-Jørgensen and Kirpekar, 2002). MALDI-TOF massspectrometry was carried out on the fragment mixtures asdescribed previously (Kirpekar et al., 2000; Andersenet al., 2004). The obtained signals were compared to the-oretical values derived from the gene sequence of therRNA. Mono- and dinucleotides are not considered in theMALDI-TOF mass spectra because their mass region isdominated by signals from the matrix or buffers. The rel-evant regions of the spectra are shown in Fig. 3 with thetheoretical mass values shown in the box. There is arelative decrease in the 2233 signal (2496-CACmCUCG-2502) when comparing the Cfr– and Cfr+ samples. Themagnification of the 2100–3300 m z-1 region in Fig. 3Balso shows the appearance of a 2219 signal (2496-CACCUCG-2502) when comparing the Cfr– and Cfr+ sam-ples. The loss of 14 Da corresponds to the loss of a methylgroup. The presence of Cfr thus decreases the originalribose methylation at C2498 and, as a consequence, thesignal intensity from the 2496-CACmCUCG-2502 frag-ment. The spectrum also shows the presence of a 3227.5signal and a 3241.5 signal in the Cfr+ sample that corre-spond to 2496-CACCUCGAUG-2505 with two and threemethyl groups respectively. This signal is not present inthe Cfr– sample. The fragment contains an internal G, butis not digested by RNase T1 although all other signalsindicate a complete digestion. This points to the presenceof a methylation interfering with RNase T1 cleavage ofG2502. The region around 1000 m z-1 (Fig. 3C) shows acorresponding change with the 2503-m2AYG-2505 signaldisplaced +14 Da in presence of Cfr. The 1027 signal isweak as to be expected from the decrease in RNase T1digestion at G2502. An inhibited RNase T1 cleavage wouldusually indicate the presence of a ribose methylation atthe G to be cleaved, but this contradicts the position ofthe primer extension stop and the presence of the 1027signal. Apparently, the additional methylation at the 2503-m2AYG-2505 strongly decreases the RNase T1 cleavageat G2502. An extra methylation at A2503 could possiblycause decreased RNase T1 digestion at the neighbouringG2502, and thereby explain the occurrence of the 3227.5and 3241.5 signals, the shift and decrease of the 1013signal, and the decrease in the 2219/2233 signals.
The 998 signal corresponds to an ACG fragment thatis not expected from the 45n sequence. It is probably aresult of incomplete RNA digestion with Mung beannuclease leaving an additional A2482 just outside thehybridized sequence. This phenomenon has beenobserved previously (T. Andersen, pers. comm.). The 980signal is the cyclic phosphate from the ACG fragment andis due to incomplete conversion of cyclic phosphate in theacid treatment. Similar -18 Da signals are also observedfor the other fragments.
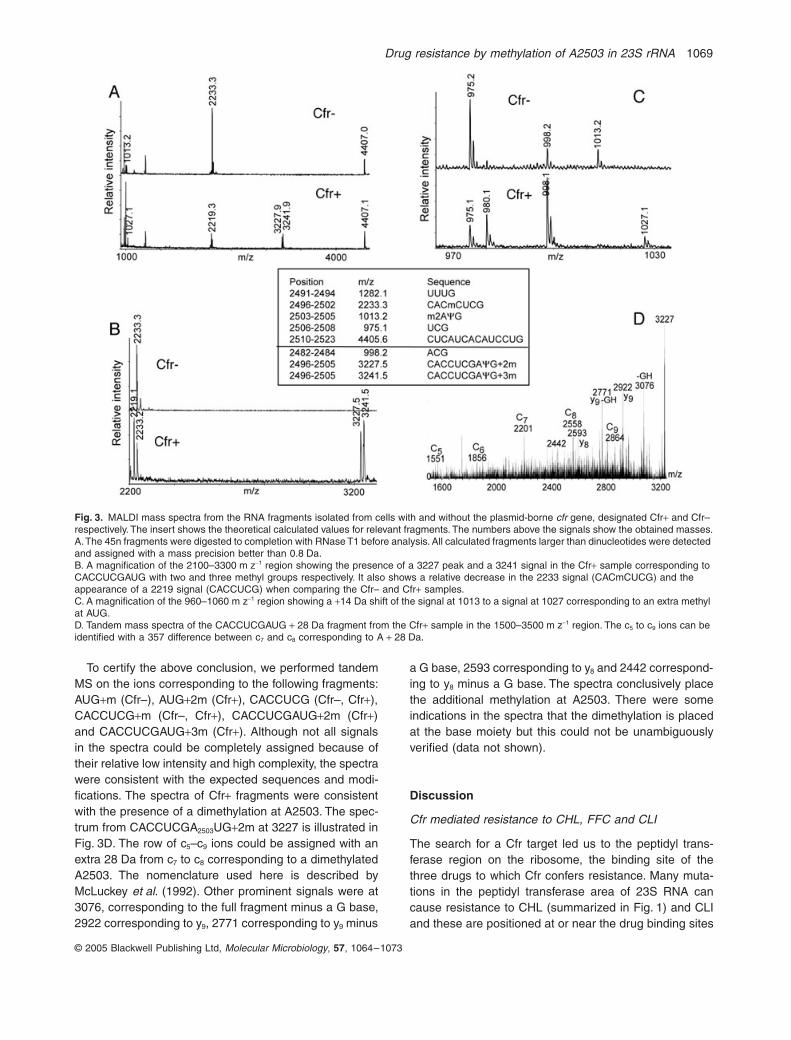
Drug resistance by methylation of A2503 in 23S rRNA 1069
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1064–1073
To certify the above conclusion, we performed tandemMS on the ions corresponding to the following fragments:AUG+m (Cfr–), AUG+2m (Cfr+), CACCUCG (Cfr–, Cfr+),CACCUCG+m (Cfr–, Cfr+), CACCUCGAUG+2m (Cfr+)and CACCUCGAUG+3m (Cfr+). Although not all signalsin the spectra could be completely assigned because oftheir relative low intensity and high complexity, the spectrawere consistent with the expected sequences and modi-fications. The spectra of Cfr+ fragments were consistentwith the presence of a dimethylation at A2503. The spec-trum from CACCUCGA2503UG+2m at 3227 is illustrated inFig. 3D. The row of c5–c9 ions could be assigned with anextra 28 Da from c7 to c8 corresponding to a dimethylatedA2503. The nomenclature used here is described byMcLuckey et al. (1992). Other prominent signals were at3076, corresponding to the full fragment minus a G base,2922 corresponding to y9, 2771 corresponding to y9 minus
a G base, 2593 corresponding to y8 and 2442 correspond-ing to y8 minus a G base. The spectra conclusively placethe additional methylation at A2503. There were someindications in the spectra that the dimethylation is placedat the base moiety but this could not be unambiguouslyverified (data not shown).
Discussion
Cfr mediated resistance to CHL, FFC and CLI
The search for a Cfr target led us to the peptidyl trans-ferase region on the ribosome, the binding site of thethree drugs to which Cfr confers resistance. Many muta-tions in the peptidyl transferase area of 23S RNA cancause resistance to CHL (summarized in Fig. 1) and CLIand these are positioned at or near the drug binding sites
Fig. 3. MALDI mass spectra from the RNA fragments isolated from cells with and without the plasmid-borne cfr gene, designated Cfr+ and Cfr– respectively. The insert shows the theoretical calculated values for relevant fragments. The numbers above the signals show the obtained masses.A. The 45n fragments were digested to completion with RNase T1 before analysis. All calculated fragments larger than dinucleotides were detected and assigned with a mass precision better than 0.8 Da.B. A magnification of the 2100–3300 m z-1 region showing the presence of a 3227 peak and a 3241 signal in the Cfr+ sample corresponding to CACCUCGAUG with two and three methyl groups respectively. It also shows a relative decrease in the 2233 signal (CACmCUCG) and the appearance of a 2219 signal (CACCUCG) when comparing the Cfr– and Cfr+ samples.C. A magnification of the 960–1060 m z-1 region showing a +14 Da shift of the signal at 1013 to a signal at 1027 corresponding to an extra methyl at AUG.D. Tandem mass spectra of the CACCUCGAUG + 28 Da fragment from the Cfr+ sample in the 1500–3500 m z-1 region. The c5 to c9 ions can be identified with a 357 difference between c7 and c8 corresponding to A + 28 Da.

1070 C. Kehrenberg et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1064–1073
as observed in X-ray structures of these ribosome com-plexes (Schlünzen et al., 2001) (Fig. 1). It is also knownfrom the literature that CHL binding results in partial pro-tection against CMCT modification of U2506 (Moazedand Noller, 1987; Rodriguez-Fonseca et al., 1995). Wetherefore decided to use U2506 protection to analyse fora potentially reduced CHL binding in ribosomes fromcells harbouring Cfr. Moreover, we expected that FFCwould have a similar effect as the two drugs are verysimilar in structure and are expected to bind to the samesite. As shown by the footprinting presented in Fig. 2,both CHL and FFC protected U2506 against CMCT mod-ification. We cannot conclude from this observation, thatCHL and FFC occupy the same site on the ribosomebecause that will require investigation of all the CHL foot-printing sites, and that is beyond the scope of this study.However, we can monitor the drug binding, and that theribosomes from cells with Cfr show a clear reduced bind-ing of both, CHL and FFC, as compared to ribosomesfrom cells not expressing Cfr. The reduced binding,observed as a reduced protection of U2506 againstCMCT (Fig. 2; Table 1), strongly indicates that the mech-anism of resistance is due to an altered ribosomal bind-ing site.
We tested CLI binding in the same manner. As men-tioned previously the CLI binding site on the ribosomepartly overlaps the CHL site according to X-ray studies ofribosome complexes (Schlünzen et al., 2001). This is inaccordance with its footprinting pattern, with protectionagainst DMS modification at A2058, A2059 and A2451,and with protection against kethoxal at G2505 and weaklyat G2061 (Douthwaite, 1992). As no CMCT modificationstudies of CLI binding have been reported so far, weincluded them in our CMCT study of the U2506 region.Cfr also reduced CLI binding although not as strongly asfor CHL and FFC (Fig. 2; Table 1). The effect was difficultto quantify because the CMCT modification of U2506 isnot strong and the effect of CLI binding on the modificationis relatively weak. This is probably because the drug isnot binding that close to U2506 (Fig. 2B). The strong DMSfootprint at A2058 was therefore used to corroborate theeffect observed at U2506 and showed a clearly reducedCLI protection, and thereby reduced binding, with ribo-somes from cells with Cfr.
Cfr mediated modifications at the RNA drug binding site
The primer extensions in Fig. 2A show an enhancedreverse transcriptase stop at position A2503 in the pres-ence of Cfr and a decrease in the stop at Cm2498.Figure 2B shows both stops to be dependent on the con-centration of dNTP available for reverse transcriptaseand this is observed both in the absence and presenceof Cfr.
To further define the changes affecting the A2503 andC2498 stops, this part of 23S RNA was investigated withmass spectrometry. As shown in Fig. 3, both a decreasein the signal encoding the Cm2498 and the appearanceof the corresponding signal without the methylation areobserved. Examination of the signal from the fragmentwith A2503 was complicated by an inhibition of the RNaseT1 digest at G2502 as mentioned in the results section,but a dimethylated fragment containing A2503 is never-theless observed. Furthermore, tandem mass spectrom-etry established the presence of two methyl groups atA2503. Several theoretical possibilities from known mod-ified nucleotides are suggested assuming that eitherthe m2 remains present plus another methylation or isreplaced by a dimethylation at one position: m2m6A2503,m6
2A2503, m1m2A2503 and m2Am2503. Of thesem6
2A2503 and m1m2A2503 can be excluded based on theprimer extension pattern.
C2498 is relatively close to, but not abutting, the CHLand CLI ribosomal binding sites, as illustrated in Fig. 1C,which shows nucleotides directly interacting with CHL(green) and some of the neighbouring positions (fromSchlünzen et al., 2001). Although it cannot be ruled outthat the absence of the C2498 ribose methylation has anindirect effect on the drug binding site, we believe this isa side-effect of the Cfr activity and not the main effectresponsible for decreased drug binding. It could be thatribose methylation at C2498 is inhibited by steric hin-drance or alteration of folding kinetics caused by the extramethylation at A2503. In contrast, A2503 is close to theCHL site and a mutation of A2503 to C can confer CHLresistance (Vester and Garrett, 1988). The ribose moietyof A2503 (shown in red in Fig. 1C) is pointed towards theCHL binding site with a distance of only 5 Å from the 2¢-O of the ribose to the benzyl ring in CHL. The CLI bindingsite (Schlünzen et al., 2001) is superimposed in blue inFig. 1C and shows that the overlapping part of the drug-binding sites is the part closest to the ribose moiety ofA2503. In addition, the other end of CLI are even closerto A2503 than CHL. A very recent X-ray structure of CLIbound to the 50S ribosomal subunit of the archaeaHaloarcula marismortui shows a similar occupationalthough not identical (Tu et al., 2005). A methyl group atA2503 could readily perturb the local structure or evendirectly interfere with positioning of the drug. Preliminaryinvestigations of the Cfr-modified A2503 performed by F.Kirpekar and S.S. Jensen using On-line Electrospray iontrap tandem MS strongly indicate that the two methyla-tions are present on the base (pers. comm.). Therefore,even though the most straightforward interpretationaccording to the X-ray structure is that Cfr is a ribosemethyltransferase, we suggest that Cfr methylatesm2A2503, resulting in m2m6A2503, and thereby perturbdrug binding.

Drug resistance by methylation of A2503 in 23S rRNA 1071
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1064–1073
Cfr and other rRNA methyltransferases causing drug resistance
Antibiotic resistance resulting from RNA methyltrans-ferases is not uncommon to the ribosome. The sites ofaction of four methyltransferases are shown in Fig. 1B:Erm methyltransferases (m6
2A2058) conferring macrolideresistance, EmtA (m1G2470), AviRa (m1G2470) and aviRb(Um2479) mediating avilamycin resistance. Other knownsites are m1G745 23S RNA (viomycin resistance),m1G748 23S RNA (tylosin resistance), Am1067 23S RNA(thiostrepton resistance) m1A964 16S RNA (pactamycinresistance), m7G1405 16S RNA (gentamicin resistance),m1A1408 16S RNA (neamine resistance) and the reversecase where the absence of RsmA causing m6
2A1518-916S RNA provides kasugamycin resistance (reviewed byDouthwaite et al., 2005). All these RNA positions are partof functional sites where the RNA structure plays a role inbinding factors or mediates conformational changes.
Cfr is currently the only methyltransferase providingdrug resistance by acting right in the peptidyl transferasecentre. The reason for this could be that very few struc-tural changes are allowed in this functionally importantregion.
Conclusions
This study shows that the presence of Cfr provides ribo-somes with a reduced binding affinity to CHL, FFC andCLI. Cfr causes a reduced ribose methylation at C2498and the addition of an extra methylation at A2503. Wehave thus identified Cfr as an rRNA methyltransferase thatcan methylate A2503 in E. coli 23S RNA and it is inferredthat the resistance mechanism for Cfr is this methylation.From primer extension, mass spectrometry and visualexamination of the X-ray structure of CHL bound to 50Sribosomal subunits, we found the methyltransferase Cfr toconfer resistance by targeting m2A2503.
Experimental procedures
Bacterial strains, plasmids, MIC determination,DNA manipulations
Staphylococcus aureus strain 405/2002 was obtained from apig during routine screening of animal staphylococci for FFCresistance. The plasmid profile was prepared by a previouslydescribed modification of the alkaline lysis procedure(Schwarz et al., 1989). Transformation of plasmids into therecipient strain S. aureus RN4220 was achieved by polyeth-ylene glycol-mediated protoplast transformation (Schwarzet al., 1989) and transformants were selected on regenera-tion plates supplemented with 15 mg ml-1 FFC. Transformantsthat grew on these plates and carried a single plasmid of c.36 kb, designated pSCFS3, were initially investigated for theirantimicrobial susceptibility patterns by disc diffusion. The
determination of the minimum inhibitory concentrations bybroth micro- or macrodilution was performed according to theNCCLS guideline M31-A2 (NCCLS, 2002) using S. aureusATCC29213 and E. coli ATCC25922 as quality controlstrains.
Plasmid pSCFS3 was digested with BglII and all fragmentswere cloned separately into the BamHI site of pBluescript IISK+ (Stratagene, Amsterdam, the Netherlands). Recombi-nant plasmids were transformed into E. coli recipient strainsJM109 and HB101 by the CaCl2 method (Schwarz et al.,1989). DNA from the pBluescript vector with a c. 3.6 kb BglIIinsert (designated pBglII) was prepared by alkaline lysis andsubsequently purified by affinity chromatography on QiagenMidi columns (Qiagen, Hilden, Germany). Confirmation ofcarriage of the gene cfr on plasmid pSCFS3 as well as onthe 3.6 kb BglII insert was done by polymerase chain reactionusing primers cfr-fw: 5¢-TGAAGTATAAAGCAGGTTGGGAGTCA-3¢ and cfr-rev: 5¢-ACCATATAATTGACCACAAGCAGC-3¢with annealing temperature: 57∞C.
A deletion variant of pBglII was constructed by BamHIdigestion and subsequent religation of the largest fragment.This variant, designated pBglIIDcfr, showed a 1664 bp dele-tion, which comprised the 5¢-terminal 532 bp of the cfr geneand its regulatory region. E. coli recipient strains carryingeither pBglII or pBglIIDcfr were comparatively investigated fortheir MICs to CHL, FFC and CLI. Sequence analyses wereconducted on both strands using the standard M13 forwardand reverse primers as well as primers derived from thecfr gene sequence (MWG-Biotech, Ebersbach, Germany).Sequence comparisons were performed with the BLAST pro-grams BLASTn and BLASTp (http://www.ncbi.nlm.nih.gov/BLAST/) and with the open reading frame finder program(http://www.ncbi.nlm.nih.gov/gorf/gorf.html). The nucleotidesequence of the 3594 bp BglII fragment has been depositedin the European Molecular Biology Laboratory (EMBL) data-base under accession number AJ879565.
Preparation of ribosomes for drug footprinting and isolation of rRNA fragment
Escherichia coli HB101/pBluescript and HB101/pBglII strainswere grown in liquid Luria–Bertani (LB) medium to an opticaldensity of 0.3 at 450 nm, and harvested by centrifugation.The cells were washed and resuspended in TMN buffer(50 mM Tris-HCl at pH 7.8, 10 mM MgCl2, 100 mM NH4Cl).Then the cells were lysed by sonication, and cell debris wasremoved by centrifugation (12 min twice at 6000 r.p.m.).Ribosomes were collected from the supernatants by centrif-ugation at 18 000 r.p.m. for 16 h at 4∞C in a Beckman Ti50rotor. The pellet was resuspended in TMN buffer and storedat 80∞C. Total rRNA was isolated by extractions with phenol,phenol/chloroform and chloroform followed by ethanolprecipitation.
Footprinting of drugs and identification of methylation sites by primer extension analysis
Five pmoles of ribosomes were incubated with various con-centrations of drugs in modification buffer (50 mM Hepes-OH, pH 8.0, 10 mM MgCl2, 100 mM KCl, 5 mM DTT) for

1072 C. Kehrenberg et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1064–1073
30 min at 37∞C. The complexes were modified with CMCT [1-cyclohexyl-3-(2-morpholinoethyl)-carbodiimide metho-p-tolu-ene sulfonate, 42 mg ml-1 in modification buffer] for 20 min at37∞C or with DMS [2 ml of a 1:6 dilution in 96% (w/v) ethanol]for 10 min at 37∞C and rRNA recovered essentially asdescribed by Poulsen et al. (2001). The modification patternin the chloramphenicol binding area and around positionA2058 in 23S RNA was monitored by primer extension anal-ysis with AMV reverse transcriptase (Life Sciences) (Poulsenet al., 2001 and references therein). The 5¢-[32P]-labelleddeoxyoligonucleotide primers (5¢-GGACAGCCATACCCTTG-3¢), complementary to nucleotides 2640–2656 of E. coli 23SrRNA, and 5¢-GCCTCCCACCTATCCTAC-3¢, complementaryto nucleotides 2112–2129, were used. The cDNA primerextension products were separated on 6% polyacrylamidesequencing gels. The positions of the stops were visualizedby autoradiography and identified by referencing to dideoxysequencing reactions on 23S rRNA that were electrophore-sed in parallel. The intensities of the modifications were quan-tified using a phosphorimager. The chemical footprintingexperiments were performed 2–3 times with reproducibleresults. The effect of increasing amounts of dNTP wereassayed at the following concentrations: 0.004 mM, 0.02 mM,0.04 mM, 0.1 mM and 0.5 mM.
MALDI-MS analysis of RNA
Isolation of a 45n rRNA fragment encompassing positions2483–2527 of 23S rRNA by specific hybridization and single-strand digestion with Mung bean nuclease was performedessentially as described by Andersen et al. (2004). Furtherfragmentation of the isolated RNA sequence by RNase T1(USB) also followed the specifications given by Andersenet al. (2004).
MALDI-TOF mass spectrometry was carried out asdescribed previously (Kirpekar et al., 2000; Andersen et al.,2004). Briefly, the RNase T1 digested sample, 0.7 ml of 0.5 M3-HPA matrix, and a small amount of suspended ammonium-loaded ion exchange beads were mixed on the target plate.The sample was allowed to dry and the beads were subse-quently removed. Spectra were recorded in reflector andpositive ion mode on a Bruker mass spectrometer andsmoothed using the software ‘m/z’ (Proteometrics).
The sample for MALDI tandem mass spectrometry wasprepared as for MALDI-TOF mass spectrometry. The tandemmass spectra were recorded in positive ion mode on a Micro-Mass MALDI Q-TOF Ultima mass spectrometer and all spec-tra were smoothed using the MassLynx software supplied bythe manufacturer.
Acknowledgements
The Danish National Research Foundation is gratefullyacknowledged for financial support by B.V. Finn Kirpekar andThomas E. Andersen have been very helpful with perfor-mance and interpretation of the MS experiments, StephenDouthwaite is thanked for discussion of experiments, JacobPoehlsgaard for help with Fig. 1 and Katherine Long arethanked for critical reading of the manuscript. This study wassupported by grants of the Deutsche Forschungsgemein-schaft (SCHW 382/6-1, SCHW 382/6-2).
References
Andersen, T.E., Porse, B.T., and Kirpekar, F. (2004) A novelpartial modification at C2501 in Escherichia coli 23S ribo-somal RNA. RNA 6: 907–913.
Arcangioli, M.A., Leroy-Setrin, S., Martel, J.L., and Chaslus-Dancla, E. (1999) A new chloramphenicol and florfenicolresistance gene linked to an integron structure in Salmo-nella typhimurium DT104. FEMS Microbiol Lett 174: 327–332.
Branlant, C., Krol, A., Machatt, M.A., Pouyet, J., Ebel, J.P.,Edwards, K., and Kossel, H. (1981) Primary and secondarystructures of Escherichia coli MRE 600, 23S ribosomalRNA. Comparison with models of secondary structure formaize chloroplast 23S rRNA and for large portions ofmouse and human 16S mitochondrial rRNAs. NucleicAcids Res 9: 4303–4324.
Briggs, C.E., and Fratamico, P.M. (1999) Molecular charac-terization of an antibiotic resistance gene cluster of Salmo-nella typhimurium DT104. Antimicrob Agents Chemother43: 846–849.
Douthwaite, S. (1992) Interaction of the antibiotic clindamycinand lincomycin with Escherichia coli 23S ribosomal RNA.Nucleic Acids Res 20: 4717–4720.
Douthwaite, S., Fourmy, D., and Yoshizawa, S. (2005) Nucle-otide methylations in rRNA that confer resistance toribosome-targeting antibiotics. In Topics in Current Genet-ics 12: Fine-tuning of RNA Functions by Modification andEditing. Grosjean, H. (ed.). New York: Springer Verlag, pp.287–309.
Gutell, R.R., Larsen, N., and Woese, C.R. (1994) Lessonsfrom an evolving rRNA: 16S and 23S rRNA structures froma comparative perspective. Microbiol Rev 58: 10–26.
Hansen, J., Moore, P.B., and Steitz, T.A. (2003) Structuresof five antibiotics bound at the peptidyl transferase centerof the large ribosomal subunit. J Mol Biol 289: 905–920.
Harms, J., Schluenzen, F., Zarivach, R., Bashan, A., Gat, S.,Agmon, I., et al. (2001) High resolution structure of thelarge ribosomal subunit from a mesophilic eubacterium.Cell 107: 679–688.
Kehrenberg, C., and Schwarz, S. (2004) Identification offexA, a novel gene from Staphylococcus lentus coding forresistance to florfenicol and chloramphenicol. AntimicrobAgents Chemother 48: 615–618.
Kehrenberg, C., Ojo, K.K., and Schwarz, S. (2004) Nucle-otide sequence and organisation of the multiresistanceplasmid pSCFS1 from Staphylococcus sciuri. J AntimicrobChemother 54: 936–939.
Kirpekar, F., Douthwaite, S., and Roepstorff, P. (2000) Map-ping posttranscriptional modifications in 5S ribosomal RNAby MALDI mass spectrometry. RNA 6: 296–306.
Kowalak, J.A., Bruenger, E., and McCloskey, J.A. (1995)Posttranscriptional modifications of the central loop ofdomain V in Escherichia coli 23 S ribosomal RNA. J BiolChem 270: 17758–17764.
Lankat-Buttgereit, B., Gross, H.J., and Krupp, G. (1987)Detection of modified nucleosides by rapid RNA sequenc-ing methods. Nucleic Acids Res 15: 7649.
Long, K.S., and Porse, B.T. (2003) A conserved chloram-phenicol binding site at the entrance to the ribosomal pep-tide exit tunnel. Nucleic Acid Res 31: 7208–7215.
Maden, B.E. (1990) The numerous modified nucleotides in

Drug resistance by methylation of A2503 in 23S rRNA 1073
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1064–1073
eukaryotic ribosomal RNA. Prog Nucleic Acid Res Mol Biol39: 241–303.
McLuckey, S.A., Van Berkel, G.J., and Glish, G.L. (1992)Tandem mass spectrometry of small multiply charged oli-gonucleotides. J Am Soc Mass Spectrom 3: 60–70.
Maden, B.E., Corbett, M.E., Heeney, P.A., Pugh, K., andAjuh, P.M. (1995) Classical and novel approaches to thedetection and localization of the numerous modifiednucleotides in eukaryotic ribosomal RNA. Biochimie 77:22–29.
Mann, P.A., Xiong, L., Mankin, A.S., Chau, A.S., Mendrick,C.A., Najarian, D.J., et al. (2001) EmtA, a rRNA methyl-transferase conferring high-level evernimicin resistance.Mol Microbiol 41: 1349–1356.
Mengel-Jørgensen, J., and Kirpekar, F. (2002) Detection ofpseudouridine and other modifications in tRNA by cyano-ethylation and MALDI mass spectrometry. Nucleic AcidsRes 30: e135.
Moazed, D., and Noller, H.F. (1987) Chloramphenicol, eryth-romycin, carbomycin and vernamycin B protect overlap-ping sites in the peptidyl transferase region of 23S rRNA.Biochimie 69: 879–884.
National Committee for Clinical Laboratory Standards(NCCLS) (2002) Performance standards for antimicrobialdisk and dilution susceptibility tests for bacteria isolatedfrom animals. Approved standard. NCCLS document M31-A2. NCCLS, 940 West Valley Road, Suite 1400, Wayne, PA19087, USA.
Nissen, P., Hansen, J., Ban, N., Moore, P.B., and Steitz, T.A.(2000) The structural basis of ribosome activity in peptidebond synthesis. Science 289: 920–930.
Poulsen, S.M., Karlsson, M., Johansson, L.B., and Vester, B.(2001) The pleuromutilin drugs tiamulin and valnemulinbind to the RNA at the peptidyl transferase centre on theribosome. Mol Microbiol 41: 1091–1100.
Roberts, M.C., Sutcliffe, J., Courvalin, P., Jensen, L.B.,Rood, J., and Seppala, H. (1999) Nomenclature formacrolide and macrolide–lincosamide–streptogramin Bresistance determinants. Antimicrob Agents Chemother43: 2823–2830.
Rodriguez-Fonseca, C., Amils, R., and Garrett, R.A. (1995)Fine structure of the peptidyl transferase centre on 23S-like rRNAs deduced from chemical probing of antibiotic-ribosome complexes. J Mol Biol 247: 224–235.
Schlünzen, F., Zarivach, R., Harms, J., Bashan, A., Tocilj, A.,Albrecht, R., et al. (2001) Structural basis for the interactionof antibiotics with the peptidyl transferase centre in eubac-teria. Nature 413: 814–821.
Schwarz, S., Cardoso, M., and Blobel, H. (1989) Plasmid-
mediated chloramphenicol resistance in Staphylococcushyicus. J Gen Microbiol 135: 3329–3336.
Schwarz, S., Werckenthin, C., and Kehrenberg, C. (2000)Identification of a plasmid-borne chloramphenicol/florfeni-col resistance gene in Staphylococcus sciuri. AntimicrobAgents Chemother 44: 2530–2533.
Schwarz, S., Kehrenberg, C., and Ojo, K.K. (2002) Staphy-lococcus sciuri gene erm(33), encoding inducible resis-tance to macrolides, lincosamides and streptogramin Bantibiotics, is a product of recombination between erm(C)and erm(A). Antimicrob Agents Chemother 46: 3621–3623.
Schwarz, S., Kehrenberg, C., Doublet, B., and Cloeckaert,A. (2004) Molecular basis of bacterial resistance tochloramphenicol and florfenicol. FEMS Microbiol Rev 28:519–542.
Skinner, R., Cundliffe, E., and Schmidt, F.J. (1983) Site ofaction of a ribosomal RNA methylase responsible for resis-tance to erythromycin and other antibiotics. J Biol Chem258: 12702–12706.
Sofia, H.J., Chen, G., Hetzler, B.G., Reyes-Spindola, J.F.,and Miller, N.E. (2001) Radical SAM, a novel protein super-family linking unresolved steps in familiar biosynthetic path-ways with radical mechanisms: functional characterizationusing new analysis and information visualization methods.Nucleic Acids Res 29: 1097–1106.
Treede, I., Jacobsen, L., Kirpekar, F., Vester, B., Weitnauer,G., Bechthold, A., and Douthwaite, S. (2003) The avilamy-cin resistance determinants AviRa and AviRb methylate23S rRNA at the guanosine 2535 base and the uridine2479 ribose. Mol Microbiol 49: 309–318.
Tu, D., Blaha, G., Moore, P.B., and Steitz, T.A. (2005)Structures of MLSBK antibiotics bound to mutated largeribosomal subunits provide a structural explanation forresistance. Cell 121: 257–270.
Vasquez, D. (1979) Inhibitors of Protein Synthesis. Berlin:Springer-Verlag.
Vester, B., and Douthwaite, S. (2001) Macrolide resistanceconferred by base substitutions in 23S rRNA. AntimicrobAgents Chemother 45: 1–12.
Vester, B., and Garrett, R.A. (1988) The importance of highlyconserved nucleotides in the binding region of chloram-phenicol at the peptidyl transferase centre of Escherichiacoli 23S ribosomal RNA. EMBO J 7: 3577–3587.
Weisblum, B. (1995) Erythromycin resistance by ribosomemodification. Antimicrob Agents Chemother 39: 577–585.
Zalacain, M., and Cundliffe, E. (1989) Methylation of 23SrRNA caused by tlrA (ermSF), a tylosin resistance deter-minant from Streptomyces fradiae. J Bacteriol 171:4254–4260.

PUBLIKATION 14
Long K., J. Poehlsgaard, C. Kehrenberg, S. Schwarz and B. Vester:
The Cfr rRNA methyltransferase confers resistance to Phenicols, Lincosamides, Oxazolidinones, Pleuromutilins, and Streptogramin A antibiotics.
Antimicrob. Agents Chemother. (2006), 50, 2500-2505.

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, July 2006, p. 2500–2505 Vol. 50, No. 70066-4804/06/$08.00�0 doi:10.1128/AAC.00131-06Copyright © 2006, American Society for Microbiology. All Rights Reserved.
The Cfr rRNA Methyltransferase Confers Resistance to Phenicols,Lincosamides, Oxazolidinones, Pleuromutilins, and
Streptogramin A AntibioticsKatherine S. Long,1 Jacob Poehlsgaard,2 Corinna Kehrenberg,3 Stefan Schwarz,3 and Birte Vester2*
Institute of Molecular Biology and Physiology, University of Copenhagen, Sølvgade 83 H, DK-1307 Copenhagen K,1 and Department ofBiochemistry and Molecular Biology, University of Southern Denmark, DK-5230 Odense M,2 Denmark, and Institut fur Tierzucht,
Bundesforschungsanstalt fur Landwirtschaft (FAL), Holtystrasse 10, 31535 Neustadt-Mariensee, Germany3
Received 1 February 2006/Returned for modification 15 March 2006/Accepted 27 April 2006
A novel multidrug resistance phenotype mediated by the Cfr rRNA methyltransferase is observed inStaphylococcus aureus and Escherichia coli. The cfr gene has previously been identified as a phenicol andlincosamide resistance gene on plasmids isolated from Staphylococcus spp. of animal origin and recently shownto encode a methyltransferase that modifies 23S rRNA at A2503. Antimicrobial susceptibility testing shows thatS. aureus and E. coli strains expressing the cfr gene exhibit elevated MICs to a number of chemically unrelateddrugs. The phenotype is named PhLOPSA for resistance to the following drug classes: Phenicols, Lincosamides,Oxazolidinones, Pleuromutilins, and Streptogramin A antibiotics. Each of these five drug classes containsimportant antimicrobial agents that are currently used in human and/or veterinary medicine. We find thatbinding of the PhLOPSA drugs, which bind to overlapping sites at the peptidyl transferase center that abutnucleotide A2503, is perturbed upon Cfr-mediated methylation. Decreased drug binding to Cfr-methylatedribosomes has been confirmed by footprinting analysis. No other rRNA methyltransferase is known to conferresistance to five chemically distinct classes of antimicrobials. In addition, the findings described in this studyrepresent the first report of a gene conferring transferable resistance to pleuromutilins and oxazolidinones.
The bacterial ribosome is the site of protein synthesis andthe target for many chemically diverse classes of antimicrobialagents. The antimicrobial drugs target important functionalcenters of the ribosome and most often bind to rRNA. Re-cently, a new phenicol and clindamycin resistance phenotypewas found to be caused by an RNA methyltransferase desig-nated Cfr. A detailed analysis by drug footprinting studies andmatrix-assisted laser desorption–ionization time of flight/tan-dem mass spectrometry showed that Cfr adds an additionalmethyl group at position A2503 of 23S rRNA (9). Since A2503is located in close proximity to the overlapping ribosomal bind-ing sites of phenicols and clindamycin, it was concluded thatthe Cfr-mediated methylation confers resistance to these twoclasses of antimicrobial agents by interfering with the position-ing of the drugs (9).
The cfr gene was first discovered in 2000 during a surveil-lance study for florfenicol resistance among staphylococci fromanimals. It was initially detected on the 17.1-kb multiresistanceplasmid pSCFS1 from a bovine strain of Staphylococcus sciuri(24) and has also been found in bovine strains of Staphylococ-cus simulans (6). In addition to cfr, the pSCFS1 plasmid carriesthe rRNA methylase gene erm(33), the aminocyclitol phospho-transferase gene spc, and the ABC transporter gene lsa(B),which confer resistance to macrolide-lincosamide-streptograminB (MLSB) antibiotics, spectinomycin, and lincosamides, re-spectively. The cfr gene was recently detected on the 35.7-kbplasmid, pSCFS3, from a porcine Staphylococcus aureus strain,
together with the chloramphenicol/florfenicol exporter genefexA (8). Cloning of the cfr gene and expression in Escherichiacoli revealed that Cfr conferred resistance not only in theoriginal gram-positive hosts but also in gram-negative bacteria.Comparison with other protein sequences deposited in thedatabases showed that the Cfr protein is not related to otherknown resistance-conferring rRNA methyltransferases but ratherto the Radical SAM superfamily (9), which includes a widerange of enzymes from a diverse set of bacteria involved inprotein radical formation, isomerization, sulfur insertion, an-aerobic oxidation, and unusual methylations (26).
As the Cfr-mediated methylation of position A2503 of 23SrRNA confers resistance to chloramphenicol and florfenicol(phenicol drugs) and clindamycin (a lincosamide drug) (9), itmay also affect binding of other drugs to the ribosomal peptidyltransferase center. Therefore, we assayed strains harboring thecfr gene for decreased susceptibility to a number of importantantimicrobial drugs that are known to bind close to A2503 atthe peptidyl transferase center. These included pleuromutilins,oxazolidinones, and streptogramin A antibiotics. The effect ofcfr on drug susceptibility was investigated both in gram-nega-tive E. coli and gram-positive S. aureus strains with plasmidslacking and carrying the cfr gene. Moreover, drug binding toCfr-methylated ribosomes was investigated by footprintingstudies.
MATERIALS AND METHODS
Bacterial strains and plasmids and antimicrobial susceptibility testing. A3,594-bp BglII fragment carrying the cfr gene from plasmid pSCFS3 (EMBLdatabase accession number AJ879565) was inserted into the pBluescript IISK(�) cloning vector (Stratagene, Amsterdam, The Netherlands). The recom-binant plasmid, designated pBglII, was transformed into E. coli recipient strains
* Corresponding author. Mailing address: Department of Biochem-istry and Molecular Biology, University of Southern Denmark, Cam-pusvej 55, DK-5230 Odense M, Denmark. Phone: 45-6550-2377. Fax:45-6550-2467. E-mail: [email protected].
2500

HB101 or AS19 (25) by the CaCl2 method (23). To investigate the effects of cfr,a deletion variant of pBglII was constructed by BamHI digestion and subsequentreligation of the largest fragment. This variant, designated pBamHI, had a1,664-bp deletion which included the 5� terminal 532 bp of the cfr gene and itsregulatory region. Transformation of the staphylococcal plasmids pSCFS1 car-rying the cfr gene (24), pSCFS3 harboring the cfr plus fexA genes (7), or pSCFS2carrying only the fexA gene (8) into the recipient strain S. aureus RN4220 (14)was achieved by polyethylene glycol-mediated protoplast transformation (23).
The original E. coli recipient strain, AS19, E. coli AS19 transformants thatcarried either the empty cloning vector or the recombinant vector pBglII orpBamHI, as well as the original S. aureus recipient strain, RN4220, and S. aureusRN4220 transformants carrying plasmid pSCFS1, pSCFS2, or pSCFS3, werecomparatively investigated for their MICs to the antimicrobial agents listed inTable 1. The determination of MICs by broth macrodilution or broth microdi-lution was performed according to guideline M31-A2 of the Clinical and Labo-ratory Standards Institute (formerly NCCLS) (12) using S. aureus ATCC 29213and E. coli ATCC 25922 as quality control strains. All MIC determinations wereperformed at least three times. The MICs of the test strains and the qualitycontrol strains were validated according to the data presented in Clinical andLaboratory Standards Institute documents M31-S1 (13) and M100-S14 (1).
Growth of cells and preparation of ribosomes for drug footprinting. E. colistrains carrying an intact cfr gene or not (9) were grown in LB broth to an opticaldensity of 0.3 at 450 nm and harvested by centrifugation. The cells were washedand resuspended in TMN buffer (50 mM Tris-HCl at pH 7.8, 10 mM MgCl2, 100mM NH4Cl). Then, the cells were lysed by sonication, and cell debris wasremoved by centrifugation (twice for 12 min at 6,000 rpm). Ribosomes werecollected from the supernatants by centrifugation at 18,000 rpm for 16 h at 4°Cin a Beckman Ti50 rotor. The pellet was resuspended in TMN buffer and storedat 80°C.
Chemical modification and primer extension analysis. E. coli 70S ribosomes(2.5 pmol) were incubated with 0.5, 2, or 10 �M tiamulin (a gift from Novartis),0.2, 0.5, or 2 �M valnemulin (a gift from Novartis), 0.5, 5, or 50 �M virginiamycinM1 (Sigma-Aldrich), or with no antimicrobial agent in modification buffer (50mM HEPES-OH, pH 8.0, 10 mM MgCl2, 100 mM KCl, 5 mM dithiothreitol) for30 min at 37°C. The ribosome complexes (12.5 �l) were modified with 12.5 �lCMCT [1-cyclohexyl-3(2-morpholinoethyl)-carbodiimide metho-p-toluene sulfo-nate] (42 mg/ml in modification buffer) for 20 min at 37°C. The reactions wereterminated by precipitating the ribosomes with ethanol. The ribosomes wererecovered by centrifugation, resuspended in 0.3 M sodium acetate, and extractedwith phenol and chloroform. rRNA was precipitated with ethanol, resuspendedin water, and monitored by primer extension analysis with avian myeloblastosisvirus reverse transcriptase (Finnzymes). The 5�-32P-labeled deoxyoligonucleotideprimer Ec2654 (5�-TCCGGTCCTCTCGTACT-3�), complementary to nucleo-tides 2654 to 2670 of E. coli 23S rRNA, was used. The cDNA extension productswere separated on 8% polyacrylamide sequencing gels. The positions of the stopswere visualized by autoradiography and identified by referencing to dideoxysequencing reactions on 23S rRNA that were electrophoresed in parallel. Re-
verse transcriptase stops one nucleotide before the corresponding nucleotide inthe sequencing lanes.
Visualization of the ribosomal subunits and drug-ribosomal subunit com-plexes. The expanded view of the drug binding site (Fig. 1B) was created byaligning the coordinates of the four antibiotic-50S ribosomal subunit cocrystalstructures relative to the RNA surrounding the peptidyl transferase center in themolecular modeling package MolMol (10). The image showing the slice plane(Fig. 1A) was generated using VMD (5). A molecular surface of the RNAsurrounding the drug-binding cavity was generated and the surface area ofnucleotide A2503 colored red. The surface was cut by a slice plane to show theinternal components of the binding site. The image was postprocessed in adrawing package to emphasize the location of the slice plane.
RESULTS AND DISCUSSION
Decreased antimicrobial susceptibility in the presence ofCfr. The effect of Cfr on the antimicrobial susceptibility ofbacterial strains was investigated. The rationale for drug selec-tion was based on current knowledge of antibiotic binding siteson the ribosome derived from footprinting, mutational, cross-linking, and crystallographic data. The drugs are all of clinicalor veterinary importance and bind at the peptidyl transferasecenter in close proximity to 23S rRNA nucleotide A2503, theposition targeted by the Cfr methyltransferase. In addition tothe previously investigated phenicols (florfenicol and chloram-phenicol) and clindamycin (a member of the lincosamidegroup), we have investigated the drugs described below.Oxazolidinones (in this study represented by linezolid) andstreptogramins are clinically important antimicrobial agentsfor the control of infections in humans caused by gram-positivecocci. Both the natural origin and the clinical use of strepto-gramins are as drug mixtures composed of an A and B com-ponent that act synergistically. Only the A component (as, forexample, virginiamycin M1) interacts at the peptidyl trans-ferase center, while the B component targets the macrolidebinding site on the ribosome. The availability of these drugs asseparate compounds sets some limits on their investigation. Asthe streptogramin A compound dalfopristin is not available,the only commercially available streptogramin A, virginiamy-cin M1, was included in this study as a representative of thisclass. The pleuromutilin antibiotics tiamulin and valnemulin
TABLE 1. Comparison of antimicrobial susceptibilities to 10 drugs and 2 drug mixtures in the absenceor presence of the Cfr methyltransferase in E. coli AS19 and S. aureus RN4220 strains
Bacterium cfrcarriage
Additionalresistancegene(s)a
MIC (�g/ml)b,c
TIA VAL VIRM1 VIR Q-D LZD CLI CHL FFC ERY AIV TEL
E. coli AS19 0.031 0.015 2 0.5 0.25 4 8 4 0.5 0.25 0.5 0.25E. coli AS19::pBluescript
II SK(�)blaTEM-116 0.031 0.015 2 0.5 0.25 4 8 4 0.5 0.25 0.5 0.25
E. coli AS19::pBamHI �cfr blaTEM-116 0.031 0.015 2 0.5 0.25 4 8 4 0.5 0.25 0.5 0.25E. coli AS19::pBglII cfr blaTEM-116 4 0.125 128 16 16 64 �512 16 8 0.25 0.5 0.25S. aureus RN4220 0.063 0.031 4 0.5 0.25 0.5 0.125 4 2 0.25 2 �0.063S. aureus RN4220::pSCFS3 cfr fexA �128 1 128 4 1 4 256 256 128 0.25 2 �0.063S. aureus RN4220::pSCFS1 cfr erm(33), spc,
lsa(B)�128 �128 128 8 4 8 256 64 32 �128 2 �0.063
S. aureus RN4220::pSCFS2 fexA 0.063 0.031 4 0.5 0.25 0.5 0.125 64 32 0.25 2 �0.063
a Additional resistance genes located on the same plasmid/cloning vector: blaTEM-116 (resistance to ampicillin), fexA (resistance to phenicols), erm(33) (inducibleresistance to macrolide, lincosamide, and streptogramin B antibiotics), spc (resistance to spectinomycin), and lsa(B) (low-level lincosamide resistance).
b Antimicrobial agents are abbreviated as follows: TIA, tiamulin; VAL, valnemulin; VIR M1, virginamycin M1; VIR, virginiamycin; Q-D, quinupristin/dalfopristin;LZD, linezolid; CLI, clindamycin; CHL, chloramphenicol; FFC, florfenicol; ERY, erythromycin; AIV, acetylisovaleryltylosin; TEL, telithromycin.
c Breakpoints for susceptibilities of staphylococci (according to CLSI document M100-S14): quinupristin/dalfopristin, �1 mg/liter; linezolid, �4 mg/liter; chloram-phenicol, �8 mg/liter; clindamycin, �0.5 mg/liter. There are no approved CLSI breakpoints for staphylococci and tiamulin, valnemulin, virginiamycin M1, virginiamycin,or florfenicol.
VOL. 50, 2006 Cfr-MEDIATED MULTIPLE RESISTANCE 2501

are used in the treatment of economically important infectionsof the intestinal and respiratory tracts of pigs caused by Brach-yspira spp., Mycoplasma spp., and Actinobacillus pleuropneumo-niae, as well as respiratory tract infections in poultry due toMycoplasma spp. Additional pleuromutilin derivatives are cur-rently being developed for use in humans.
Antimicrobial susceptibility testing was conducted with bothCfr-positive (�Cfr) and Cfr-negative (�Cfr) S. aureus RN4220(14) and E. coli AS19 (25) to investigate whether differences inantimicrobial susceptibility were detectable in gram-positiveand gram-negative host bacteria. As E. coli has a relatively lowsusceptibility to many drugs, the hyperpermeable strain E. coliAS19 was used to emphasize the observed effects. All teststrains were exposed to 10 antimicrobial drugs and two drugmixtures at various concentrations, and the MIC results areshown in Table 1.
The presence of the Cfr methyltransferase substantially re-duces the susceptibilities to all antimicrobials known to bindclose to A2503. The relative MIC difference between strainslacking or harboring the Cfr methyltransferase varied from 4-to �4,096-fold (Table 1 and Fig. 1C). Although the overallpattern was the same for both E. coli and S. aureus (Table 1),the magnitude of the MIC increases for the pleuromutilinsdiffered distinctly between the two host organisms. For E. coli,the tiamulin and valnemulin MICs increased 128- and 8-fold,respectively, whereas the corresponding MIC increases for S.
aureus are �2,048- and �4,096-fold. According to the ap-proved clinical breakpoints for the antimicrobial agents thatare available, strains with the Cfr methyltransferase exhibitMICs that allow their classification as either resistant (e.g.,S. aureus RN4220::pSCFS1 for chloramphenicol, linezolid,quinupristin-dalfopristin, and clindamycin; S. aureus RN4220::pSCFS3 for chloramphenicol and clindamycin) or borderlinesusceptible (e.g., S. aureus RN4220::pSCFS3 for linezolid andquinupristin-dalfopristin) (see footnote c of Table 1).
As a control that the Cfr methyltransferase only affects sen-sitivity to a subset of antimicrobials, susceptibility testing wasalso performed with the macrolides erythromycin, acetyliso-valeryltylosin, and telithromycin. Macrolides bind in the pep-tide exit tunnel in a cleft adjacent to the peptidyl transferasecenter (15), but they are not in direct contact with nucleotideA2503 of 23S rRNA. As expected from the different ribosomalbinding site of macrolides, the Cfr-mediated methylation ofA2503 did not affect erythromycin, acetylisovaleryltylosin, andtelithromycin MICs for E. coli and S. aureus (Table 1). S.aureus RN4220::pSCFS1 is erythromycin resistant becausepSCFS1 also carries the inducibly expressed MLSB resistancegene erm(33), for which erythromycin is an excellent inducer.The same strain has low MICs for acetylisovaleryltylosin andtelithromycin, since 16-membered macrolides, such as tylosinderivatives, and ketolides are not efficient inducers. The highMICs for chloramphenicol and florfenicol observed for S. au-
FIG. 1. Binding of the phenicol, lincosamide, pleuromutilin, and streptogramin A classes of antimicrobials to overlapping sites at the ribosomalpeptidyl transferase center. (A) The structure of the bacterial 50S ribosomal subunit showing the slice plane used in panel B. (B) An expandedview showing the structures of four drugs bound at the peptidyl transferase center. The structural data can be found in reference 22 and referencestherein. The names and chemical structures of the four antimicrobial agents are shown at the bottom on background colors that correspond to thebound structures (depicted in stick representation). The target of the Cfr methyltransferase, nucleotide A2503, is shown in red. The surroundingRNA is shown in light gray. (C) The Cfr-mediated resistance patterns with S. aureus for chloramphenicol, clindamycin, tiamulin, and virginiamycinM1. The data are from Table 1. The MICs are depicted on a logarithmic scale with strains lacking Cfr shown in the left column of each pair ofbars (marked �), whereas those of strains containing Cfr are shown in the right column of each pair of bars (marked �). The numbers above the�Cfr columns are the n-fold differences in MICs between �Cfr and �Cfr strains. Details on the visualization of the 50S ribosomal subunit andantibiotic-50S subunit complexes are provided in Materials and Methods.
2502 LONG ET AL. ANTIMICROB. AGENTS CHEMOTHER.

reus RN4220::pSCFS3 are most likely due to a synergistic effectof the two different resistance mechanisms including target sitemodification via Cfr and phenicol efflux via FexA (7). Thefourfold-higher MIC for quinupristin-dalfopristin of S. aureusRN4220::pSCFS1 than that of S. aureus RN4220::pSCFS3 ispresumably the result of a synergistic effect of Cfr-mediatedstreptogramin A resistance and low-level expression of thepSCFS1-borne macrolide-lincosamide-streptogramin B resis-tance erm(33) gene in the absence of an inducer.
Antibiotic binding to Cfr-methylated ribosomes is reduced.A reduced level of chloramphenicol, florfenicol, and clindamy-cin binding to Cfr-modified ribosomes was established previ-ously using chemical footprinting (9). Here we show that it isalso true for the pleuromutilin drugs tiamulin and valnemulinand the streptogramin A drug virginiamycin M1 (Fig. 2). Areduction in drug binding to ribosomes can be monitored bychanges in the chemical modification pattern of rRNA at thedrug binding site. In the chemical footprints of tiamulin, val-nemulin, and virginiamycin M1, nucleotides U2506, U2584,and U2585 of 23S rRNA are protected from CMCT modifi-cation (16, 19). Binding of antimicrobials to ribosomes isolatedfrom E. coli strains lacking or harboring the Cfr methyltrans-ferase was assayed by CMCT modification of ribosome-drugcomplexes, followed by primer extension analysis with reversetranscriptase and gel electrophoresis. Autoradiograms of thetiamulin, valnemulin, and virginiamycin M1 footprints on ribo-somes are shown in Fig. 2. The protections observed at U2506and U2584/U2585 in the presence of the drugs are significantlydecreased in ribosomes from �Cfr cells compared to thosefrom �Cfr cells, indicating that drug binding is reduced in�Cfr cells. The prominent additional band present in the �Cfrsamples is from the reverse transcriptase stop caused by theCfr-mediated methylation at nucleotide A2503. The drug mix-tures virginiamycin (virginiamycin S and M1) and quinupristin-dalfopristin are composed of streptogramin B and A compo-nents, respectively. These bind synergistically to overlappingsites on the ribosome, and therefore an evaluation of theirrelative binding by footprinting is not feasible. Since the foot-printing data show that binding of virginiamycin M1 to �Cfrrelative to �Cfr ribosomes is reduced (Fig. 2B), it is believed
that the attenuated binding affinities of the streptogramin Acomponents of the mixtures are responsible for the observedMIC increases for virginiamycin and quinupristin-dalfopristinin strains expressing the cfr gene (Table 1). Taken togetherwith our previous data (9), the footprinting data demonstratethat the binding of phenicol, lincosamide, pleuromutilin, andstreptogramin A drugs to ribosomes modified by Cfr is dimin-ished.
Binding of drugs to the ribosomal peptidyl transferase cen-ter. The exact position of single representatives from the afore-mentioned four groups of antimicrobial agents bound to abacterial 50S ribosomal subunit is available from X-ray crys-tallography. The crystal structures of bound chloramphenicol,clindamycin, tiamulin, and dalfopristin (4, 20, 21) show thatthey bind to overlapping sites at the peptidyl transferase site atthe entrance of the ribosomal exit tunnel (Fig. 1A and B). Thecorresponding data for clindamycin and virginiamycin M1 withthe archaeon Haloarcula marismortui show the same bindingsites with only minor differences in orientation and interactionsand thus support the bacterial sites (27). The target of Cfrmethylation, nucleotide A2503, comprises a considerable partof the exposed surface area in the peptidyl transferase cavity(shown in red in Fig. 1B), and chloramphenicol, clindamycin,tiamulin, and dalfopristin bind in close proximity to this surfacearea. A simple model of resistance is suggested, in which themodification at A2503 interferes with drug binding. The meth-ylation either disturbs binding directly or indirectly by causinga shift in the position of A2503, which in turn leads to de-creased drug binding.
Various lines of evidence suggest that oxazolidinones alsotarget the peptidyl transferase center. Most of the 23S rRNAresistance mutations known to confer linezolid resistance arelocated near the site of peptide bond formation (11). In vivocross-linking data show that oxazolidinones cross-link to nu-cleotide A2602, a position that is also cross-linked to sparso-mycin in the presence of a P-site-bound tRNA (2). Like spar-somycin, oxazolidinones do not produce a footprint on emptyribosomes. However, a weak linezolid footprint has been re-ported on a ribosome-mRNA-tRNA complex (30). In addition,an X-ray structure of an oxazolidinone-50S complex shows that
FIG. 2. Gel autoradiograms comparing antibiotic binding to E. coli 70S ribosomes isolated from cells lacking (�Cfr) or harboring (�Cfr) theCfr methyltransferase. Footprints of the pleuromutilin drugs tiamulin and valnemulin (panel A) and the streptogramin A drug virginiamycin M1(panel B) are shown. Control lanes for each experiment contain unmodified 70S ribosomes. Lanes labeled CMCT denote 70S ribosomes modifiedwith CMCT in the absence of drug. Wedges are used to indicate the increase in tiamulin (TIA), valnemulin (VAL), and virginiamycin M1 (VIRM1)concentration in reactions of 70S subunits modified with CMCT in the presence of 0.5, 2, or 10 �M tiamulin, 0.2, 0.5, or 2 �M valnemulin, or 0.5,5, or 50 �M virginiamycin M1. CMCT modifications are detected through primer extension analysis. The nucleotide positions in domain V of 23SrRNA exhibiting altered CMCT reactivity in the presence of the drugs are indicated. The Cfr modification at nucleotide A2503 is labeled (Cfr md.).Lanes marked G, A, U, and C denote dideoxy sequencing reactions.
VOL. 50, 2006 Cfr-MEDIATED MULTIPLE RESISTANCE 2503

this drug binds in the A-site of the peptidyl transferase centerof the ribosome (J. Ippolito, Z. Kanyo, B. Wimberly, D. Wang,E. Skripkin, J. Devito, B. Freeborn, J. Sutcliffe, E. Duffy, andF. Franceschi, Abstr. 45th Intersci. Conf. Antimicrob. AgentsChemother., abstr. F-1254, 2005). The observed increases inlinezolid MICs (Table 1) in strains expressing the Cfr methyl-transferase can thereby be rationalized in the same manner asfor the drugs described above. It is thus expected that bacterialstrains expressing Cfr will show decreased susceptibility to alldrugs belonging to these five groups of antimicrobial agents.Therefore, we suggest naming the phenotype caused by theCfr methyltransferase PhLOPSA for Phenicol, Lincosamide,Oxazolidinone, Pleuromutilin, and Streptogramin A resistance.
Perspectives of Cfr dissemination. A frightening scenariocan be foreseen if the appearance and spread of the Cfr methyl-transferase parallels the situation observed with the Ermmethyltransferases and combined resistance to MLSB. TheErm family consists of approximately 40 different classes ofmethylases (http://faculty.washington.edu/marilynr/), all actingat position A2058 of 23S rRNA and causing MLSB resistance(18). Mutations in 23S rRNA can also cause the MLSB phe-notype (28), but these mutations are not transferable by hor-izontal gene transfer. In contrast, erm methylase genes havebeen identified in a wide range of gram-positive and gram-negative bacteria with the transposon-borne erm(B), erm(F),and erm(A) genes, as well as the plasmid-borne erm(C) genehaving the broadest host range (18). About 10 years ago, thenumber of reports of Erm-mediated MLSB resistance rose sud-denly and it was discovered that this type of resistance appearedworldwide in a number of different bacteria. The high incidenceof resistance was probably caused by the extensive use of mac-rolides for treatment of bacterial infections in humans andanimals and by their use as growth promoters in the farmingindustry. As the acquired resistance is not detrimental to bac-teria, it can persist for a long time, which in turn promotes itsspread.
The cfr gene has been identified on structurally related mul-tiresistance plasmids from animal staphylococci and can, inprinciple, be easily disseminated among staphylococci. How-ever, surveillance studies in Germany have identified only 6cfr-carrying staphylococcal strains during the past 17 years (7).The low prevalence of the cfr gene might result from a com-paratively low selective pressure imposed by the PhLOPSA
drugs on animal staphylococci. Of the relevant drug classes,only lincosamides are approved for several indications in foodand companion animals in the European Union. Florfenicoland pleuromutilins are approved only for selected infections incattle and/or swine. Moreover, the use of chloramphenicol hasbeen prohibited in food animals in the European Union since1994. Although previously used as growth promoter, neithervirginiamycin nor other streptogramin antibiotics are currentlyapproved in the European Union for use on animals, and theoxazolidinone drug class is licensed exclusively for human useworldwide.
The clinical and veterinary importance of Cfr-mediated re-sistance. As very few new antimicrobial agents appear on themarket, the fact that Cfr confers resistance to five differentclasses of relevant antimicrobial agents warrants attention. Forthe drugs with approved clinical breakpoints with Staphylococ-cus spp., the elevated MICs observed in strains expressing Cfr
classifies them as resistant or borderline susceptible (Table 1).In the case of the streptogramins, the Cfr-mediated decrease insusceptibility to the A component, in combination with otherresistance mechanisms, such as Erm-mediated resistance tothe B component as seen with the erm(33) gene present to-gether with cfr on plasmid pSCFS1, could seriously affect drugefficiency. The oxazolidinone linezolid is a last-resort antimi-crobial agent for the control of gram-positive bacterial patho-gens against which no other antimicrobials are effective any-more. Therefore, transferable resistance or largely decreasedsusceptibility to oxazolidinones is of particular concern.
The detection of the plasmid-borne resistance gene cfr is, toour knowledge, the first report of transferable resistance topleuromutilins and oxazolidinones. In Brachyspira isolates withreduced susceptibility to tiamulin, mutations in ribosomal pro-tein L3 and 23S rRNA genes have been identified (17). Thereports of linezolid resistance published to date have also de-scribed only mutations in 23S rRNA or ribosomal protein L4genes (11, 29). Thus, it was believed that the likelihood of trans-ferable resistance to either pleuromutilins or oxazolidinonesbased on these mechanisms was rather low, and it has beenclaimed that there are no mechanisms of cross-resistance to lin-ezolid (3). However, this assumption must be revised on the basisof the findings presented in this study.
We conclude that expression of the Cfr methyltransferaseconfers a PhLOPSA resistance phenotype. As the Cfr-modifiednucleotide A2503 abuts the overlapping binding sites ofPhLOPSA drugs and drug binding to Cfr-modified ribosomes isimpaired, we infer that the resistance is caused by perturbationof the drug-binding site on the ribosome.
ACKNOWLEDGMENTS
We thank L. H. Hansen and Vera Noding for excellent technicalassistance and Pfizer for providing linezolid.
This work was supported by The Danish National Research Foun-dation, the European Commission’s 5th Framework Program (grantQLK2-CT-2002-00892), and the Deutsche Forschungsgemeinschaft(SCHW 382/6-2 and SCHW 382/6-3).
REFERENCES
1. Clinical and Laboratory Standards Institute. 2005. Performance standardsfor antimicrobial susceptibility testing; 15th informational supplement.NCCLS document M100-S15. Clinical and Laboratory Standards Institute,Wayne, Pa.
2. Colca, J. R., W. G. McDonald, D. J. Waldon, L. M. Thomasco, R. C. Gad-wood, E. T. Lund, G. S. Cavey, W. R. Mathews, L. D. Adams, E. T. Cecil, J. D.Pearson, J. H. Bock, J. E. Mott, D. L. Shinabarger, L. Xiong, and A. S.Mankin. 2003. Cross-linking in the living cell locates the site of action ofoxazolidinone antibiotics. J. Biol. Chem. 278:21972–21979.
3. Fines, M., and R. Leclercq. 2000. Activity of linezolid against Gram-positivecocci possessing genes conferring resistance to protein synthesis inhibitors. J.Antimicrob. Chemother. 45:797–802.
4. Harms, J. M., F. Schlunzen, P. Fucini, H. Bartels, and A. Yonath. 1 April2004. Alterations at the peptidyl transferase centre of the ribosome inducedby the synergistic action of the streptogramins dalfopristin and quinupristin.BMC Biol. 2:4. doi:10.1186/1741-7007-2-4.
5. Humphrey, W., A. Dalke, and K. Schulten. 1996. VMD: visual moleculardynamics. J. Mol. Graph. 14:33–38, 27–28.
6. Kehrenberg, C., K. K. Ojo, and S. Schwarz. 2004. Nucleotide sequence andorganization of the multiresistance plasmid pSCFS1 from Staphylococcussciuri. J. Antimicrob. Chemother. 54:936–939.
7. Kehrenberg, C., and S. Schwarz. 2006. Distribution of florfenicol resistancegenes fexA and cfr among chloramphenicol-resistant Staphylococcus isolates.Antimicrob. Agents Chemother. 50:1156–1163.
8. Kehrenberg, C., and S. Schwarz. 2004. fexA, a novel Staphylococcus lentusgene encoding resistance to florfenicol and chloramphenicol. Antimicrob.Agents Chemother. 48:615–618.
9. Kehrenberg, C., S. Schwarz, L. Jacobsen, L. H. Hansen, and B. Vester. 2005.
2504 LONG ET AL. ANTIMICROB. AGENTS CHEMOTHER.

A new mechanism for chloramphenicol, florfenicol and clindamycin resis-tance: methylation of 23S ribosomal RNA at A2503. Mol. Microbiol. 57:1064–1073.
10. Koradi, R., M. Billeter, and K. Wuthrich. 1996. MOLMOL: a program fordisplay and analysis of macromolecular structures. J. Mol. Graph. 14:51–55,29–32.
11. Meka, V. G., and H. S. Gold. 2004. Antimicrobial resistance to linezolid.Clin. Infect. Dis. 39:1010–1015.
12. National Committee for Clinical Laboratory Standards. 2002. Performancestandards for antimicrobial disk and dilution susceptibility tests for bacteriaisolated from animals; approved standard, 2nd ed. NCCLS document M31-A2. National Committee for Clinical Laboratory Standards, 9Wayne, Pa.
13. National Committee for Clinical Laboratory Standards. 2004. Performancestandards for antimicrobial disk and dilution susceptibility tests for bacteriaisolated from animals; informational supplement. NCCLS document M31-S1. National Committee for Clinical Laboratory Standards, Wayne, Pa.
14. Novick, R. 1967. Properties of a cryptic high-frequency transducing phage inStaphylococcus aureus. Virology 33:155–166.
15. Poehlsgaard, J., and S. Douthwaite. 2005. The bacterial ribosome as a targetfor antibiotics. Nat. Rev. Microbiol. 3:870–881.
16. Poulsen, S. M., M. Karlsson, L. B. Johansson, and B. Vester. 2001. Thepleuromutilin drugs tiamulin and valnemulin bind to the RNA at the pep-tidyl transferase centre on the ribosome. Mol. Microbiol. 41:1091–1099.
17. Pringle, M., J. Poehlsgaard, B. Vester, and K. S. Long. 2004. Mutations inribosomal protein L3 and 23S ribosomal RNA at the peptidyl transferasecentre are associated with reduced susceptibility to tiamulin in Brachyspiraspp. isolates. Mol. Microbiol. 54:1295–1306.
18. Roberts, M. C., J. Sutcliffe, P. Courvalin, L. B. Jensen, J. Rood, and H.Seppala. 1999. Nomenclature for macrolide and macrolide-lincosamide-streptogramin B resistance determinants. Antimicrob. Agents Chemother.43:2823–2830.
19. Rodriguez-Fonseca, C., R. Amils, and R. A. Garrett. 1995. Fine structure ofthe peptidyl transferase centre on 23 S-like rRNAs deduced from chemicalprobing of antibiotic-ribosome complexes. J. Mol. Biol. 247:224–235.
20. Schlunzen, F., E. Pyetan, P. Fucini, A. Yonath, and J. M. Harms. 2004.Inhibition of peptide bond formation by pleuromutilins: the structure of the
50S ribosomal subunit from Deinococcus radiodurans in complex with tiamu-lin. Mol. Microbiol. 54:1287–1294.
21. Schlunzen, F., R. Zarivach, J. Harms, A. Bashan, A. Tocilj, R. Albrecht, A.Yonath, and F. Franceschi. 2001. Structural basis for the interaction ofantibiotics with the peptidyl transferase centre in eubacteria. Nature 413:814–821.
22. Schuwirth, B. S., M. A. Borovinskaya, C. W. Hau, W. Zhang, A. Vila-Sanjurjo, J. M. Holton, and J. H. Cate. 2005. Structures of the bacterialribosome at 3.5 A resolution. Science 310:827–834.
23. Schwarz, S., M. Cardoso, and H. Blobel. 1989. Plasmid-mediated chloram-phenicol resistance in Staphylococcus hyicus. J. Gen. Microbiol. 135:3329–3336.
24. Schwarz, S., C. Werckenthin, and C. Kehrenberg. 2000. Identification of aplasmid-borne chloramphenicol-florfenicol resistance gene in Staphylococ-cus sciuri. Antimicrob. Agents Chemother. 44:2530–2533.
25. Sekiguchi, M., and S. Iida. 1967. Mutants of Escherichia coli permeable toactinomycin. Proc. Natl. Acad. Sci. USA 58:2315–2320.
26. Sofia, H. J., G. Chen, B. G. Hetzler, J. F. Reyes-Spindola, and N. E. Miller.2001. Radical SAM, a novel protein superfamily linking unresolved steps infamiliar biosynthetic pathways with radical mechanisms: functional charac-terization using new analysis and information visualization methods. NucleicAcids Res. 29:1097–1106.
27. Tu, D., G. Blaha, P. B. Moore, and T. A. Steitz. 2005. Structures of MLSBKantibiotics bound to mutated large ribosomal subunits provide a structuralexplanation for resistance. Cell 121:257–270.
28. Vester, B., and S. Douthwaite. 2001. Macrolide resistance conferred by basesubstitutions in 23S rRNA. Antimicrob. Agents Chemother. 45:1–12.
29. Wolter, N., A. M. Smith, D. J. Farrell, W. Schaffner, M. Moore, C. G.Whitney, J. H. Jorgensen, and K. P. Klugman. 2005. Novel mechanism ofresistance to oxazolidinones, macrolides, and chloramphenicol in ribosomalprotein L4 of the pneumococcus. Antimicrob. Agents Chemother. 49:3554–3557.
30. Xu, J., A. Golshani, H. Aoki, J. Remme, J. Chosay, D. L. Shinabarger, andM. C. Ganoza. 2005. Protected nucleotide G2608 in 23S rRNA confersresistance to oxazolidinones in E. coli. Biochem. Biophys. Res. Commun.328:471–476.
VOL. 50, 2006 Cfr-MEDIATED MULTIPLE RESISTANCE 2505

PUBLIKATION 15
Kehrenberg C. and S. Schwarz:
Distribution of florfenicol resistance genes fexA and cfr among chloramphenicol-resistant Staphylococcus isolates.
Antimicrob. Agents Chemother. (2006), 50, 1156-1163.

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Apr. 2006, p. 1156–1163 Vol. 50, No. 40066-4804/06/$08.00�0 doi:10.1128/AAC.50.4.1156–1163.2006Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Distribution of Florfenicol Resistance Genes fexA and cfr amongChloramphenicol-Resistant Staphylococcus Isolates
Corinna Kehrenberg and Stefan Schwarz*Institut fur Tierzucht, Bundesforschungsanstalt fur Landwirtschaft, 31535 Neustadt-Mariensee, Germany
Received 17 October 2005/Returned for modification 13 December 2005/Accepted 12 January 2006
A total of 302 chloramphenicol-resistant Staphylococcus isolates were screened for the presence of theflorfenicol/chloramphenicol resistance genes fexA and cfr and their localization on mobile genetic elements. Ofthe 114 isolates from humans, only a single Staphylococcus aureus isolate showed an elevated MIC to florfenicol,but did not carry either of the known resistance genes, cfr or fexA. In contrast, 11 of the 188 staphylococci fromanimal sources were considered florfenicol resistant and carried either cfr (one isolate), fexA (five isolates), orboth resistance genes (five isolates). In nine cases we confirmed that these genes were carried on a plasmid.Five different types of plasmids could be differentiated on the basis of their sizes, restriction patterns, andresistance genes. The gene fexA, which has previously been shown to be part of the nonconjugative transposonTn558, was identified in 10 of the 11 resistant isolates from animals. PCR assays were developed to detectdifferent parts of this transposon as well as their physical linkage. Complete copies of Tn558 were found in fivedifferent isolates and shown by inverse PCR to be functionally active. Truncated copies of Tn558, in which thetnpA-tnpB area was in part deleted by the integration of a 4,674-bp segment including the gene cfr and a novel2,446-bp IS21-like insertion sequence, were seen in a plasmid present in three staphylococcal isolates.
Florfenicol[d-threo-3-fluoro-2-dichloroacetamido-1-(4-methyl-sulfonylphenyl)-1-propanol] is a synthetic, broad-spectrum fluor-inated analogue of thiamphenicol. Like chloramphenicol andthiamphenicol, it shows activity against many gram-positive andgram-negative bacteria. Bacterial resistance to chloramphenicoland thiamphenicol is most commonly mediated by mono- anddiacetylation via chloramphenicol acetyltransferase (CAT) en-zymes. Due to the replacement of the hydroxyl group at posi-tion C-3 with a fluorine residue, the acceptor site for acetylgroups was structurally altered in florfenicol. This modificationrendered florfenicol resistant to inactivation by CAT enzymes,and consequently, chloramphenicol-resistant strains, in whichresistance is solely based on CAT activity, are susceptible toflorfenicol (36, 39).
Florfenicol has been licensed exclusively for use in veter-inary medicine. In the European Union, it has been ap-proved for the treatment of respiratory tract infections incattle in 1995 and in swine in 2000. However, in severalnon-European Union countries florfenicol is also licensedfor the treatment of infectious pododermatitis in cattle andvarious bacterial diseases of commercially reared fish (36).The results of monitoring studies indicated that virtually alltarget bacteria isolated from respiratory tract infections ofcattle and pigs (Pasteurella multocida, Mannheimia haemo-lytica, Histophilus somni, and Actinobacillus pleuropneumo-niae) are susceptible to florfenicol (19, 34, 37), although afirst florfenicol-resistant P. multocida isolate carrying a plas-mid-borne floR gene, coding for a chloramphenicol/florfeni-col exporter, has been reported recently (22). The gene floRis located on a small nonconjugative transposon (13) and
has been identified in various gram-negative bacteria, in-cluding several Salmonella enterica serovars (3–6, 10–12,29), Escherichia coli (1, 2, 9, 14, 41), Klebsiella pneumoniae(8), Photobacterium damselae (27), and Vibrio cholerae (17).
In staphylococci, two different florfenicol resistance geneshave been identified so far. The gene cfr was initially foundon the 17.1-kb plasmid pSCFS1 from a Staphylococcus sciuriisolate (20, 38) and recently shown to code for an rRNAmethylase which mediates combined resistance to chloram-phenicol, florfenicol, and clindamycin by methylation of the23S rRNA at position A2503 (26). In contrast, the gene fexAencodes a protein of 475 amino acids with 14 transmem-brane domains which represents a novel type of efflux pro-tein within the major facilitator superfamily (24). Its sub-strate spectrum includes only florfenicol and chloramphenicol.The gene fexA was first identified on the 34-kb plasmid pSCFS2from Staphylococcus lentus and shown to be part of the Tn554-liketransposon Tn558 (23).
The present study was conducted to gain detailed informa-tion about the presence of the two resistance genes, cfr andfexA, among chloramphenicol-resistant Staphylococcus isolatesof animal and human origin which exhibited elevated MICs offlorfenicol. Besides the localization of these genes on plasmidsor in chromosomal DNA, particular attention was paid to thedetection of complete and truncated versions of Tn558 in fexA-positive isolates.
MATERIALS AND METHODS
Bacterial isolates and antimicrobial susceptibility testing. A total of 302chloramphenicol-resistant Staphylococcus isolates of animal (n � 188) and hu-man (n � 114) origin were screened for florfenicol resistance. The isolates ofanimal origin were collected during the years 1988 to 2005 from geographicallydistinct locations all over Germany and represent part of the strain collection ofthe Institut fur Tierzucht (Bundesforschungsanstalt fur Landwirtschaft). Most ofthese isolates were kindly provided by local and county veterinary diagnosticlaboratories in Germany and included isolates from horses (n � 45), dogs (n �
* Corresponding author. Mailing address: Institut fur Tierzucht,Bundesforschungsanstalt fur Landwirtschaft (FAL), Holtystr. 10,31535 Neustadt-Mariensee, Germany. Phone: 49-5034-871-241. Fax:49-5034-871-246. E-mail: [email protected].
1156
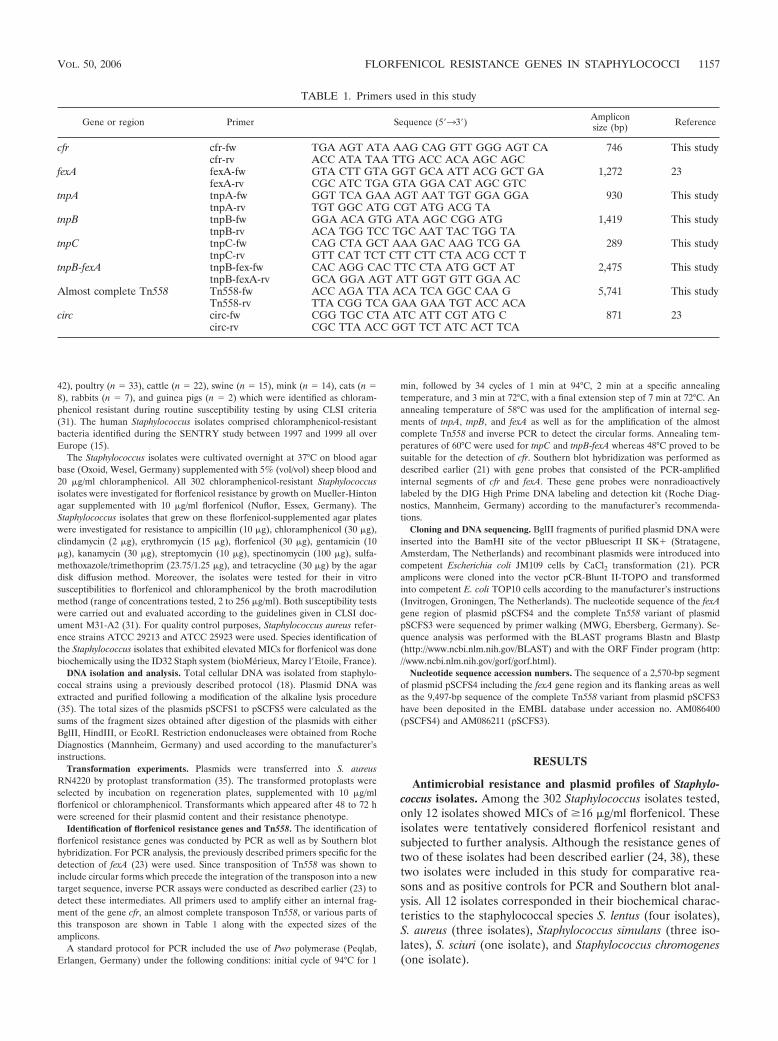
42), poultry (n � 33), cattle (n � 22), swine (n � 15), mink (n � 14), cats (n �8), rabbits (n � 7), and guinea pigs (n � 2) which were identified as chloram-phenicol resistant during routine susceptibility testing by using CLSI criteria(31). The human Staphylococcus isolates comprised chloramphenicol-resistantbacteria identified during the SENTRY study between 1997 and 1999 all overEurope (15).
The Staphylococcus isolates were cultivated overnight at 37°C on blood agarbase (Oxoid, Wesel, Germany) supplemented with 5% (vol/vol) sheep blood and20 �g/ml chloramphenicol. All 302 chloramphenicol-resistant Staphylococcusisolates were investigated for florfenicol resistance by growth on Mueller-Hintonagar supplemented with 10 �g/ml florfenicol (Nuflor, Essex, Germany). TheStaphylococcus isolates that grew on these florfenicol-supplemented agar plateswere investigated for resistance to ampicillin (10 �g), chloramphenicol (30 �g),clindamycin (2 �g), erythromycin (15 �g), florfenicol (30 �g), gentamicin (10�g), kanamycin (30 �g), streptomycin (10 �g), spectinomycin (100 �g), sulfa-methoxazole/trimethoprim (23.75/1.25 �g), and tetracycline (30 �g) by the agardisk diffusion method. Moreover, the isolates were tested for their in vitrosusceptibilities to florfenicol and chloramphenicol by the broth macrodilutionmethod (range of concentrations tested, 2 to 256 �g/ml). Both susceptibility testswere carried out and evaluated according to the guidelines given in CLSI doc-ument M31-A2 (31). For quality control purposes, Staphylococcus aureus refer-ence strains ATCC 29213 and ATCC 25923 were used. Species identification ofthe Staphylococcus isolates that exhibited elevated MICs for florfenicol was donebiochemically using the ID32 Staph system (bioMerieux, Marcy l�Etoile, France).
DNA isolation and analysis. Total cellular DNA was isolated from staphylo-coccal strains using a previously described protocol (18). Plasmid DNA wasextracted and purified following a modification of the alkaline lysis procedure(35). The total sizes of the plasmids pSCFS1 to pSCFS5 were calculated as thesums of the fragment sizes obtained after digestion of the plasmids with eitherBglII, HindIII, or EcoRI. Restriction endonucleases were obtained from RocheDiagnostics (Mannheim, Germany) and used according to the manufacturer’sinstructions.
Transformation experiments. Plasmids were transferred into S. aureusRN4220 by protoplast transformation (35). The transformed protoplasts wereselected by incubation on regeneration plates, supplemented with 10 �g/mlflorfenicol or chloramphenicol. Transformants which appeared after 48 to 72 hwere screened for their plasmid content and their resistance phenotype.
Identification of florfenicol resistance genes and Tn558. The identification offlorfenicol resistance genes was conducted by PCR as well as by Southern blothybridization. For PCR analysis, the previously described primers specific for thedetection of fexA (23) were used. Since transposition of Tn558 was shown toinclude circular forms which precede the integration of the transposon into a newtarget sequence, inverse PCR assays were conducted as described earlier (23) todetect these intermediates. All primers used to amplify either an internal frag-ment of the gene cfr, an almost complete transposon Tn558, or various parts ofthis transposon are shown in Table 1 along with the expected sizes of theamplicons.
A standard protocol for PCR included the use of Pwo polymerase (Peqlab,Erlangen, Germany) under the following conditions: initial cycle of 94°C for 1
min, followed by 34 cycles of 1 min at 94°C, 2 min at a specific annealingtemperature, and 3 min at 72°C, with a final extension step of 7 min at 72°C. Anannealing temperature of 58°C was used for the amplification of internal seg-ments of tnpA, tnpB, and fexA as well as for the amplification of the almostcomplete Tn558 and inverse PCR to detect the circular forms. Annealing tem-peratures of 60°C were used for tnpC and tnpB-fexA whereas 48°C proved to besuitable for the detection of cfr. Southern blot hybridization was performed asdescribed earlier (21) with gene probes that consisted of the PCR-amplifiedinternal segments of cfr and fexA. These gene probes were nonradioactivelylabeled by the DIG High Prime DNA labeling and detection kit (Roche Diag-nostics, Mannheim, Germany) according to the manufacturer’s recommenda-tions.
Cloning and DNA sequencing. BglII fragments of purified plasmid DNA wereinserted into the BamHI site of the vector pBluescript II SK� (Stratagene,Amsterdam, The Netherlands) and recombinant plasmids were introduced intocompetent Escherichia coli JM109 cells by CaCl2 transformation (21). PCRamplicons were cloned into the vector pCR-Blunt II-TOPO and transformedinto competent E. coli TOP10 cells according to the manufacturer’s instructions(Invitrogen, Groningen, The Netherlands). The nucleotide sequence of the fexAgene region of plasmid pSCFS4 and the complete Tn558 variant of plasmidpSCFS3 were sequenced by primer walking (MWG, Ebersberg, Germany). Se-quence analysis was performed with the BLAST programs Blastn and Blastp(http://www.ncbi.nlm.nih.gov/BLAST) and with the ORF Finder program (http://www.ncbi.nlm.nih.gov/gorf/gorf.html).
Nucleotide sequence accession numbers. The sequence of a 2,570-bp segmentof plasmid pSCFS4 including the fexA gene region and its flanking areas as wellas the 9,497-bp sequence of the complete Tn558 variant from plasmid pSCFS3have been deposited in the EMBL database under accession no. AM086400(pSCFS4) and AM086211 (pSCFS3).
RESULTS
Antimicrobial resistance and plasmid profiles of Staphylo-coccus isolates. Among the 302 Staphylococcus isolates tested,only 12 isolates showed MICs of �16 �g/ml florfenicol. Theseisolates were tentatively considered florfenicol resistant andsubjected to further analysis. Although the resistance genes oftwo of these isolates had been described earlier (24, 38), thesetwo isolates were included in this study for comparative rea-sons and as positive controls for PCR and Southern blot anal-ysis. All 12 isolates corresponded in their biochemical charac-teristics to the staphylococcal species S. lentus (four isolates),S. aureus (three isolates), Staphylococcus simulans (three iso-lates), S. sciuri (one isolate), and Staphylococcus chromogenes(one isolate).
TABLE 1. Primers used in this study
Gene or region Primer Sequence (5�33�) Ampliconsize (bp) Reference
cfr cfr-fw TGA AGT ATA AAG CAG GTT GGG AGT CA 746 This studycfr-rv ACC ATA TAA TTG ACC ACA AGC AGC
fexA fexA-fw GTA CTT GTA GGT GCA ATT ACG GCT GA 1,272 23fexA-rv CGC ATC TGA GTA GGA CAT AGC GTC
tnpA tnpA-fw GGT TCA GAA AGT AAT TGT GGA GGA 930 This studytnpA-rv TGT GGC ATG CGT ATG ACG TA
tnpB tnpB-fw GGA ACA GTG ATA AGC CGG ATG 1,419 This studytnpB-rv ACA TGG TCC TGC AAT TAC TGG TA
tnpC tnpC-fw CAG CTA GCT AAA GAC AAG TCG GA 289 This studytnpC-rv GTT CAT TCT CTT CTT CTA ACG CCT T
tnpB-fexA tnpB-fex-fw CAC AGG CAC TTC CTA ATG GCT AT 2,475 This studytnpB-fexA-rv GCA GGA AGT ATT GGT GTT GGA AC
Almost complete Tn558 Tn558-fw ACC AGA TTA ACA TCA GGC CAA G 5,741 This studyTn558-rv TTA CGG TCA GAA GAA TGT ACC ACA
circ circ-fw CGG TGC CTA ATC ATT CGT ATG C 871 23circ-rv CGC TTA ACC GGT TCT ATC ACT TCA
VOL. 50, 2006 FLORFENICOL RESISTANCE GENES IN STAPHYLOCOCCI 1157
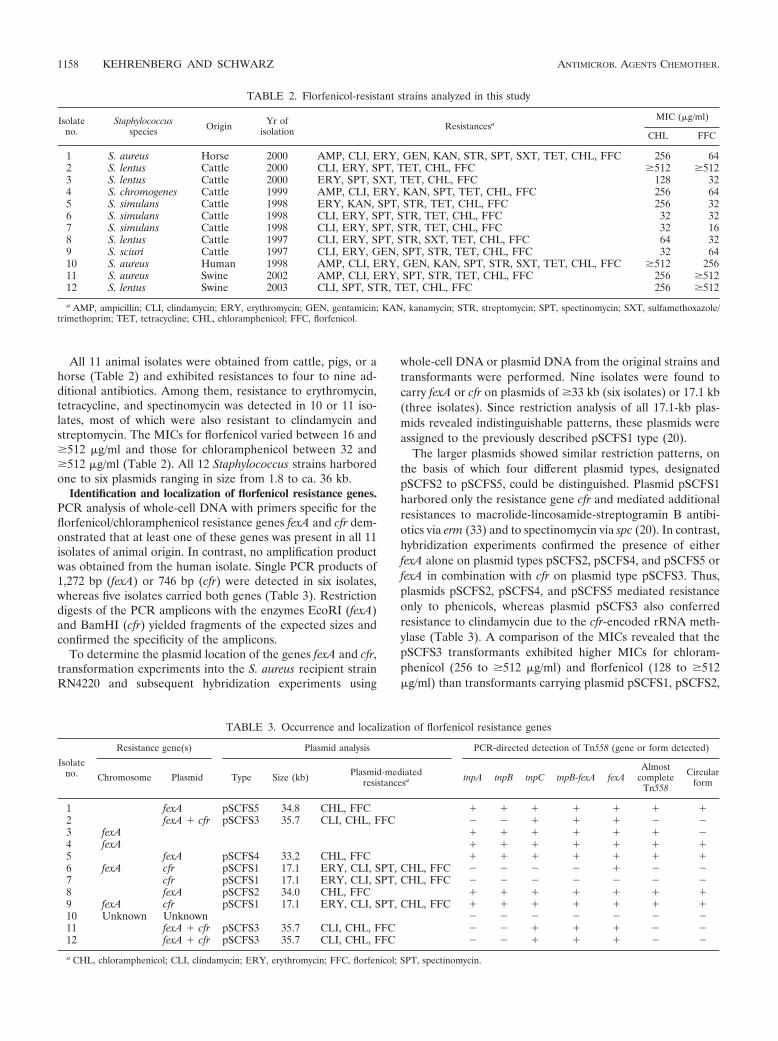
All 11 animal isolates were obtained from cattle, pigs, or ahorse (Table 2) and exhibited resistances to four to nine ad-ditional antibiotics. Among them, resistance to erythromycin,tetracycline, and spectinomycin was detected in 10 or 11 iso-lates, most of which were also resistant to clindamycin andstreptomycin. The MICs for florfenicol varied between 16 and�512 �g/ml and those for chloramphenicol between 32 and�512 �g/ml (Table 2). All 12 Staphylococcus strains harboredone to six plasmids ranging in size from 1.8 to ca. 36 kb.
Identification and localization of florfenicol resistance genes.PCR analysis of whole-cell DNA with primers specific for theflorfenicol/chloramphenicol resistance genes fexA and cfr dem-onstrated that at least one of these genes was present in all 11isolates of animal origin. In contrast, no amplification productwas obtained from the human isolate. Single PCR products of1,272 bp (fexA) or 746 bp (cfr) were detected in six isolates,whereas five isolates carried both genes (Table 3). Restrictiondigests of the PCR amplicons with the enzymes EcoRI (fexA)and BamHI (cfr) yielded fragments of the expected sizes andconfirmed the specificity of the amplicons.
To determine the plasmid location of the genes fexA and cfr,transformation experiments into the S. aureus recipient strainRN4220 and subsequent hybridization experiments using
whole-cell DNA or plasmid DNA from the original strains andtransformants were performed. Nine isolates were found tocarry fexA or cfr on plasmids of �33 kb (six isolates) or 17.1 kb(three isolates). Since restriction analysis of all 17.1-kb plas-mids revealed indistinguishable patterns, these plasmids wereassigned to the previously described pSCFS1 type (20).
The larger plasmids showed similar restriction patterns, onthe basis of which four different plasmid types, designatedpSCFS2 to pSCFS5, could be distinguished. Plasmid pSCFS1harbored only the resistance gene cfr and mediated additionalresistances to macrolide-lincosamide-streptogramin B antibi-otics via erm (33) and to spectinomycin via spc (20). In contrast,hybridization experiments confirmed the presence of eitherfexA alone on plasmid types pSCFS2, pSCFS4, and pSCFS5 orfexA in combination with cfr on plasmid type pSCFS3. Thus,plasmids pSCFS2, pSCFS4, and pSCFS5 mediated resistanceonly to phenicols, whereas plasmid pSCFS3 also conferredresistance to clindamycin due to the cfr-encoded rRNA meth-ylase (Table 3). A comparison of the MICs revealed that thepSCFS3 transformants exhibited higher MICs for chloram-phenicol (256 to �512 �g/ml) and florfenicol (128 to �512�g/ml) than transformants carrying plasmid pSCFS1, pSCFS2,
TABLE 2. Florfenicol-resistant strains analyzed in this study
Isolateno.
Staphylococcusspecies Origin Yr of
isolation ResistancesaMIC (�g/ml)
CHL FFC
1 S. aureus Horse 2000 AMP, CLI, ERY, GEN, KAN, STR, SPT, SXT, TET, CHL, FFC 256 642 S. lentus Cattle 2000 CLI, ERY, SPT, TET, CHL, FFC �512 �5123 S. lentus Cattle 2000 ERY, SPT, SXT, TET, CHL, FFC 128 324 S. chromogenes Cattle 1999 AMP, CLI, ERY, KAN, SPT, TET, CHL, FFC 256 645 S. simulans Cattle 1998 ERY, KAN, SPT, STR, TET, CHL, FFC 256 326 S. simulans Cattle 1998 CLI, ERY, SPT, STR, TET, CHL, FFC 32 327 S. simulans Cattle 1998 CLI, ERY, SPT, STR, TET, CHL, FFC 32 168 S. lentus Cattle 1997 CLI, ERY, SPT, STR, SXT, TET, CHL, FFC 64 329 S. sciuri Cattle 1997 CLI, ERY, GEN, SPT, STR, TET, CHL, FFC 32 6410 S. aureus Human 1998 AMP, CLI, ERY, GEN, KAN, SPT, STR, SXT, TET, CHL, FFC �512 25611 S. aureus Swine 2002 AMP, CLI, ERY, SPT, STR, TET, CHL, FFC 256 �51212 S. lentus Swine 2003 CLI, SPT, STR, TET, CHL, FFC 256 �512
a AMP, ampicillin; CLI, clindamycin; ERY, erythromycin; GEN, gentamicin; KAN, kanamycin; STR, streptomycin; SPT, spectinomycin; SXT, sulfamethoxazole/trimethoprim; TET, tetracycline; CHL, chloramphenicol; FFC, florfenicol.
TABLE 3. Occurrence and localization of florfenicol resistance genes
Isolateno.
Resistance gene(s) Plasmid analysis PCR-directed detection of Tn558 (gene or form detected)
Chromosome Plasmid Type Size (kb) Plasmid-mediatedresistancesa tnpA tnpB tnpC tnpB-fexA fexA
Almostcomplete
Tn558
Circularform
1 fexA pSCFS5 34.8 CHL, FFC � � � � � � �2 fexA � cfr pSCFS3 35.7 CLI, CHL, FFC � � � � � � �3 fexA � � � � � � �4 fexA � � � � � � �5 fexA pSCFS4 33.2 CHL, FFC � � � � � � �6 fexA cfr pSCFS1 17.1 ERY, CLI, SPT, CHL, FFC � � � � � � �7 cfr pSCFS1 17.1 ERY, CLI, SPT, CHL, FFC � � � � � � �8 fexA pSCFS2 34.0 CHL, FFC � � � � � � �9 fexA cfr pSCFS1 17.1 ERY, CLI, SPT, CHL, FFC � � � � � � �10 Unknown Unknown � � � � � � �11 fexA � cfr pSCFS3 35.7 CLI, CHL, FFC � � � � � � �12 fexA � cfr pSCFS3 35.7 CLI, CHL, FFC � � � � � � �
a CHL, chloramphenicol; CLI, clindamycin; ERY, erythromycin; FFC, florfenicol; SPT, spectinomycin.
1158 KEHRENBERG AND SCHWARZ ANTIMICROB. AGENTS CHEMOTHER.

pSCFS4, or pSCFS5, with MICs for chloramphenicol of 32 to128 �g/ml and MICs for florfenicol of 16 to 64 �g/ml.
Negative results for transformation and the lack of hybrid-ization to plasmid profiles suggested that the fexA genes mightbe located in the chromosomal DNA of four isolates (Table 3).Hybridization of DraI-digested whole-cell DNA with the fexAgene probe yielded single hybridizing bands of 1.9 kb in each ofthe four Staphylococcus isolates. Two of these isolates alsoharbored the cfr-carrying plasmid pSCFS1, whereas no secondplasmid-borne florfenicol resistance gene was detected in theremaining two isolates. The only S. aureus isolate of humanorigin did not hybridize with any of the gene probes tested(Table 3).
Identification of complete and truncated Tn558 elements.Since fexA was identified as part of the transposon Tn558 (23),PCR assays were developed to detect the different parts of thistransposon or circular forms that suggest transpositional activ-ity of this element (Fig. 1). Hence, four primer pairs werechosen that allow the amplification of internal segments of thetransposase genes tnpA, tnpB, and tnpC, as well as the resis-tance gene fexA (Table 1). In addition to the previously de-scribed primers for the circular intermediate form, another twoprimer pairs were used to confirm the linkage between tnpBand fexA as well as to amplify an internal ca. 5.7-kb segment ofTn558 (Table 1).
The results obtained with all these primer pairs suggestedthat complete and functionally active Tn558 elements arepresent in five isolates (Table 3), with the original bovine S.lentus isolate no. 8 (23), an equine S. aureus isolate (no. 1), aswell as a bovine S. simulans isolate (no. 5) carrying Tn558 onplasmids of types pSCFS2, pSCFS5, and pSCFS4, respectively.Chromosomal copies of Tn558 were detected in the singlebovine S. chromogenes (no. 4) and S. sciuri (no. 9) isolates. Itshould be noted that all PCR assays except that for the circularintermediate form were positive for another bovine S. lentusisolate (no. 3), whereas only an amplicon for fexA was obtained
from the chromosomal DNA of a pSCFS1-carrying bovine S.simulans isolate (no. 6).
Additional sequence analysis of the PCR amplicon of thecircular form obtained from S. simulans isolate no. 5, whichhad its Tn558 on plasmid pSCFS4, confirmed the expectedcomposition of 229 bp of tnpA and its upstream region and 642bp representing the right arm of Tn558. No base pair exchangewas observed in comparison to the published S. lentus circularform. Moreover, when whole-cell DNA from S. aureus RN4220transformants carrying either plasmid pSCFS2, pSCFS4, orpSCFS5 was used, positive results were again obtained in theseinverse PCR assays.
As previously seen with PCR assays and hybridization ex-periments using gene probes for fexA and cfr, human S. aureusisolate no. 10 did not yield amplicons for any of the Tn558-specific segments (Table 3).
Analysis of the Tn558 variant located on plasmid pSCFS3.Amplicons for the resistance gene fexA, the transposase genetnpC, and the tnpB-fexA segment, but neither for the transposasegenes tnpA and tnpB nor for the almost complete Tn558 and thecircular form, were obtained during analysis of plasmid pSCFS3(Table 3). These observations suggested that at least a part ofTn558 was present on this plasmid type. Sequence analysis ofthe entire Tn558 variant from plasmid pSCFS3 confirmed theseobservations. A 9,491-bp element which closely resembled the6,644-bp transposon Tn558 was detected. The Tn558-homolo-gous part comprised the initial 575 bp, including the upstreamnoncoding region and the 5� end of the tnpA gene, which corre-sponded exactly to the respective Tn558 sequence and the ter-minal 4,242 bp, including the 3� end of tnpB, tnpC, orf138, fexA,and the downstream noncoding region, which differed by 4 bpfrom the corresponding Tn558 sequence (Fig. 2a). The 3� end oftnpA, the 5� end of tnpB, and the short spacer between these twotransposase genes was replaced by a 4,674-bp segment whichconsisted of an insertion sequence (IS)-like structure and theresistance gene cfr (Fig. 2a).
FIG. 1. (a) PCR amplicons specific for the detection of transposase genes tnpA (lane 1), tnpB (lane 2), and tnpC (lane 3), the linkage betweentnpB-fexA (lane 4), an internal 5,741-bp fragment representing the almost complete Tn558 (lane 5), and a circular intermediate of Tn558 indicatingfunctional activity (lane 6). Lanes M contain the size standards (HindIII fragments of lambda DNA [Gibco-BRL]). (b) Schematic drawing of Tn558and illustration of the different fragments amplified by the PCR assays. The different reading frames are shown as arrows, with the arrowheadindicating the direction of transcription. A distance scale in kb is shown below the map.
VOL. 50, 2006 FLORFENICOL RESISTANCE GENES IN STAPHYLOCOCCI 1159

The IS-like element, designated IS21-558 (Fig. 2b), com-prised 2,446 bp and contained two reading frames. The firstreading frame, istAS, coded for a protein of 445 amino acidswhich revealed less than 30% identity and less than 50% sim-ilarity to a large number of transposase proteins similar to IstAof the IS21 family (28). The deduced IstAS amino acid se-quence contained the DDE motif which is believed to formpart of the active site of transposases with D79-D141-E187 asdescribed by Mahillon and Chandler (28). The second readingframe, designated istBS, overlapped the first one by 8 bp andcoded for a 250-aa protein which exhibited less than 50%identity and less than 70% similarity to a wide range of trans-posase-like proteins similar to IstB of the IS21 family (28). Twoimperfect inverted repeated (IR) sequences of 57 and 56 bpwere detected. The 57-bp IR sequence was located at thejunction between tnpA-homologous and tnpA-nonhomologoussegments about 250 bp upstream of the translational startcodon of istAS. The 56-bp IR sequence included the terminal5 bp of the istBS reading frame. No direct repeats were de-tected up- and downstream of this IS-like element.
Immediately downstream of the 56-bp IR sequence, a2,124-bp region, homologous to the cfr gene region of plasmidpSCFS1 (20), was detected. In the cfr upstream part, the pu-
tative promoter and the partly overlapping reading frames forthe 59-aa and 44-aa peptides (38) were present, while a 35-bpdeletion in the region between the stop codon of the readingframe for the 44-aa peptide and the start codon of cfr wasdetected. This deletion comprised one of the inverted repeatedsequences believed to play a role in the inducible expression ofcfr (38). The high clindamycin MIC of 256 �g/ml determinedfor the S. aureus RN4220::pSCFS3 transformants, however,suggested that this deletion had no effect on the cfr-mediatedresistance properties.
The cfr structural gene from plasmid pSCFS3 coded for aprotein of 349 aa, which differed by one amino acid from theCfr protein of S. sciuri plasmid pSCFS1 (D254 in Cfr frompSCFS3 versus A254 in Cfr from pSCFS1). The noncodingpSCFS1-homologous 584 bp downstream of cfr differed fromthe corresponding pSCFS1 sequence by four base pair ex-changes. The cfr-associated transcriptional terminator, locatedin this region, was not affected. Between this pSCFS1-homol-ogous region and the tnpB-homologous region, a stretch of 104bp which did not resemble any sequence deposited in thedatabases was detected (Fig. 2a).
The adjacent truncated tnpB reading frame correspondedexactly to codons 396 to 639 of TnpB from Tn558. Similarly,
FIG. 2. (a) Schematic drawing of the Tn558 variant detected on plasmid pSCFS3. The areas of homology to Tn558 in plasmid pSCFS2 (23) andcfr-carrying plasmid pSCFS1 (20) are indicated. (b) Detailed schematic drawing of the novel insertion sequence IS21-558 detected in the Tn558variant of plasmid pSCFS3. The imperfect terminal inverted repeats are displayed in a box above the IS element; vertical bars indicatecomplementary bases. The overlap area between the 3� end of istAS and the 5� end of istBS is shown in a box below the IS element. Amino acidsare presented in the single-letter code, the ribosome binding site (RBS) is underlined, and the translational stop codon is indicated by an asterisk.
1160 KEHRENBERG AND SCHWARZ ANTIMICROB. AGENTS CHEMOTHER.

the TnpC, Orf138, and FexA proteins showed no variations intheir amino acid sequences compared to the correspondingproteins encoded by Tn558. Moreover, analysis of 400-bp se-quences immediately up- and downstream of this Tn558 vari-ant revealed 97 and 99% sequence identity, respectively, to theatt558 sequence of plasmid pSCFS2 (23).
Sequence analysis of the fexA gene and its flanking regionsof plasmid pSCFS4. Plasmid pSCFS4 was detected in bovine S.simulans isolate no. 5 and showed major variations in its re-striction patterns compared to the other fexA-carrying plasmidtypes. The internal 2,550-bp sequenced segment from plasmidpSCFS4 differed by only 4 bp from the Tn558 sequence. Anal-ysis of the pSCFS4-associated fexA gene revealed that thecorresponding 475-aa protein differed from the FexA proteinsof pSCFS2 and pSCFS3 by only two amino acid exchanges,V131 and L288 in FexA from pSCFS4 versus I131 and F288 inFexA of pSCFS2 and pSCFS3. The reading frame for theputative oxidoreductase (orf138) was indistinguishable fromthose found on plasmids pSCFS2 and pSCFS3.
DISCUSSION
Since no information has been available about the occur-rence and distribution of the florfenicol resistance genes cfrand fexA in staphylococci, we screened 302 Staphylococcusisolates for the presence of these genes. The isolates wereselected as chloramphenicol resistant since all known florfeni-col resistance genes mediate combined resistance to chloram-phenicol and florfenicol (36, 39). The observation that wedetected only 11 florfenicol-resistant staphylococcal isolatesfrom animals and a single isolate of human origin might sug-gest that florfenicol resistance is still very rare among staphy-lococci. This assumption was supported by recent data fromthe GERM-Vet (41) monitoring program in Germany, whichrevealed that none of the chloramphenicol-resistant staphylo-cocci from cattle and swine (chloramphenicol resistance rate,�5%) showed elevated MICs to florfenicol. However, it shouldalso be noted that florfenicol is not approved for the control ofstaphylococcal infections in animals and hence staphylococcalisolates are not routinely tested for their susceptibility to flo-rfenicol. Moreover, human staphylococci are not at all checkedfor this resistance in routine diagnostics since florfenicol islicensed exclusively for use in animals. Based on these aspects,an unknown number of staphylococcal isolates with elevatedMICs for florfenicol might remain undetected.
Another problem is the tentative breakpoint of �8 �gflorfenicol/ml for susceptibility. During our studies, we noticedthat only isolates that exhibited an MIC of at least 16 �gflorfenicol/ml carried either cfr or fexA (Kehrenberg andSchwarz, unpublished observations). Although this tentativebreakpoint is in good accordance with the breakpoints forsusceptibility to chloramphenicol published by CLSI (31), theBritish Society for Antimicrobial Chemotherapy, the Comitede l’Antibiogramme de la Societe Francaise de Microbiologie,and the Deutsches Institut fur Normung (all �8 �g chloram-phenicol/ml), it should not be regarded as a valid breakpoint.It only served to select isolates suitable for further studies onthe presence of the resistance genes cfr and fexA.
As far as information on antibiotic pretreatment was avail-able, at least bovine isolates no. 8 and 9 were from animals
suffering from respiratory tract infections that had received flo-rfenicol treatment. Surprisingly, one florfenicol-resistant S. au-reus was isolated from a horse, although approved florfenicoluse is restricted to cattle and swine. Since fluorinated chlor-amphenicol derivatives are not used in human medicine, it wasinteresting to investigate a highly phenicol-resistant human S.aureus isolate for the presence of cfr and fexA. In contrast tothe findings of other studies, which indicated animal-to-humantransfer of resistance genes (43), neither fexA nor cfr could bedetected in human S. aureus isolate no. 10.
The highest levels of florfenicol resistance were seen withthe original strains and transformants harboring the cfr- andfexA-carrying plasmid pSCFS3. This might be due to the addi-tive effect of the cfr-encoded rRNA methylase, which inhibitsphenicol binding to the ribosome, and the fexA-encoded ex-porter, which mediates the active efflux of phenicols from thebacterial cell. Why this phenomenon could not be observed instrains carrying a plasmid-borne cfr gene in addition to a chro-mosomally located fexA remains unclear.
The presence of more than one phenicol resistance gene inthe same isolate has already been described in Salmonellaenterica and E. coli. In these cases, genes coding for differentphenicol resistance mechanisms were identified, such as catA1and floR in S. enterica serovar Typhimurium var. Copenhagen(16) or catA2 and cmlA in S. enterica serovar Choleraesuis (7)and E. coli (1). However, Chen et al. (6) also found the genescatA1 and catA2, both coding for different type A chloram-phenicol acetyltransferases, together in the same isolates of S.enterica serovar Derby and S. enterica serovar Typhimurium.White and coworkers (42) identified the genes floR and cmlA,both coding for phenicol efflux systems, in bovine E. coli iso-lates.
Like floR (13), fexA has also been identified as part of a smalltransposon. Until now, no information has been availableabout the presence of the fexA-carrying transposon Tn558 (23)in staphylococci. The finding that half of the fexA-carryingisolates in the present study harbored a complete Tn558 eitheron a plasmid or in the chromosomal DNA suggests that Tn558remains stable after integration into the att558 attachment site,independently of the genomic localization. This observation isin good accordance with the finding that the prototype trans-poson of this family, Tn554 (30), commonly is present as acomplete element in the genomic DNA of different staphylo-coccal species (40). In contrast, other transposons, such as thetetracycline resistance transposons Tn1721, Tn10, and Tn5607,were frequently found to be deleted after integration intoplasmids or chromosomal DNA (21, 25, 32, 33).
Using the PCR assays described in this study, we were ableto identify complete and functionally active forms of Tn558on different types of plasmids, namely pSCFS2, pSCFS4, andpSCFS5, and in chromosomal DNA. In addition, a Tn558variant was detected on plasmid pSCFS3 in which part of tnpAand tnpB was replaced by a 4,674-bp segment that contained anovel IS21-like element, IS21-558, and the resistance gene cfr.Since TnpA and TnpB are essential for transposition (30), partialdeletion of these genes resulted in the immobility of this Tn558variant, as confirmed by lack of detection of circular forms.
Insertion sequences of the IS21 family have been identifiedin a number of gram-negative and gram-positive bacterial gen-era (28). To the best of our knowledge, IS21-558 from plasmid
VOL. 50, 2006 FLORFENICOL RESISTANCE GENES IN STAPHYLOCOCCI 1161

pSCFS3 is the first member of this family identified in staph-ylococci. With a total size of 2,446 bp, this IS element is in thesize range of 1.9 to 2.5 kb known for members of the IS21family (28). Moreover, it exhibited the typical two readingframes and the conserved terminal base pairs 5�-TG-3� in theimperfect inverted repeats characteristic of the IS21 family(28). Only the 4-bp direct repeats commonly seen at the inte-gration site of IS21-like insertion sequences were missing inplasmid pSCFS3. The processes by which the IS21-558-cfr seg-ment had integrated into a Tn558 element remain speculative,but it is likely that the integration of IS21-558 together withinterplasmid recombination processes led to the formation ofthe structures detected on plasmid pSCFS3.
In conclusion, the results of this study showed that florfenicolresistance genes occur in different staphylococcal species fromdifferent animal sources. Their location on mobile genetic ele-ments which may carry additional resistance genes might facilitatetheir dissemination.
ACKNOWLEDGMENTS
We thank Vera Noding for expert technical assistance.This study was supported by grants from the Deutsche Forschungs-
gemeinschaft (SCHW 382/6-2 and SCHW 382/6-3).
REFERENCES
1. Bischoff, K. M., D. G. White, P. F. McDermott, S. Zhao, S. Gaines, J. J.Maurer, and D. J. Nisbet. 2002. Characterization of chloramphenicol resis-tance in beta-hemolytic Escherichia coli associated with diarrhea in neonatalswine. J. Clin. Microbiol. 40:389–394.
2. Blickwede, M., and S. Schwarz. 2004. Molecular analysis of florfenicol-resistant Escherichia coli isolates from pigs. J. Antimicrob. Chemother. 53:58–64.
3. Boyd, D., A. Cloeckaert, E. Chaslus-Dancla, and M. R. Mulvey. 2002. Char-acterization of variant Salmonella genomic island 1 multidrug resistanceregions from serovars Typhimurium DT104 and Agona. Antimicrob. AgentsChemother. 46:1714–1722.
4. Boyd, D., G. A. Peters, A. Cloeckaert, K. Sidi Boumedine, E. Chaslus-Dancla, H. Imberechts, and M. R. Mulvey. 2001. Complete nucleotide se-quence of a 43-kilobase genomic island associated with the multidrug resis-tance region of Salmonella enterica serovar Typhimurium DT104 and itsidentification in phage type DT120 and serovar Agona. J. Bacteriol. 183:5725–5732.
5. Briggs, C. E., and P. M. Fratamico. 1999. Molecular characterization of anantibiotic resistance gene cluster of Salmonella typhimurium DT104. Anti-microb. Agents Chemother. 43:846–849.
6. Chen, S., S. Zhao, D. G. White, C. M. Schroeder, R. Lu, H. Yang, P. F.McDermott, S. Ayers, and J. Meng. 2004. Characterization of multiple-antimicrobial-resistant Salmonella serovars isolated from retail meat. Appl.Environ. Microbiol. 70:1–7.
7. Chiu, C. H., P. Tang, C. Chu, S. Hu, Q. Bao, J. Yu, Y. Y. Chou, H. S. Wang, andY. S. Lee. 2005. The genome sequence of Salmonella enterica serovar Choler-aesuis, a highly invasive and resistant zoonotic pathogen. Nucleic Acids Res.33:1690–1698.
8. Cloeckaert, A., S. Baucheron, and E. Chaslus-Dancla. 2001. Nonenzymaticchloramphenicol resistance mediated by IncC plasmid R55 is encoded by afloR gene variant. Antimicrob. Agents Chemother. 45:2381–2382.
9. Cloeckaert, A., S. Baucheron, G. Flaujac, S. Schwarz, C. Kehrenberg, J. L.Martel, and E. Chaslus-Dancla. 2000. Plasmid-mediated florfenicol resis-tance encoded by the floR gene in Escherichia coli isolated from cattle.Antimicrob. Agents Chemother. 44:2858–2860.
10. Doublet, B., A. Carattoli, J. M. Whichard, D. G. White, S. Baucheron, E.Chaslus-Dancla, and A. Cloeckaert. 2004. Plasmid-mediated florfenicol andceftriaxone resistance encoded by the floR and blaCMY-2 genes in Salmonellaenterica serovars Typhimurium and Newport isolated in the United States.FEMS Microbiol. Lett. 233:301–305.
11. Doublet, B., D. Boyd, M. R. Mulvey, and A. Cloeckaert. 2005. The Salmonellagenomic island 1 is an integrative mobilizable element. Mol. Microbiol.55:1911–1924.
12. Doublet, B., R. Lailler, D. Meunier, A. Brisabois, D. Boyd, M. R. Mulvey, E.Chaslus-Dancla, and A. Cloeckaert. 2003. Variant Salmonella genomic is-land 1 antibiotic resistance gene cluster in Salmonella enterica serovar Al-bany. Emerg. Infect. Dis. 9:585–591.
13. Doublet, B., S. Schwarz, C. Kehrenberg, and A. Cloeckaert. 2005. Florfenicol
resistance gene floR is part of a novel transposon. Antimicrob. Agents Che-mother. 49:2106–2108.
14. Doublet, B., S. Schwarz, E. Nussbeck, S. Baucheron, J. L. Martel, E.Chaslus-Dancla, and A. Cloeckaert. 2002. Molecular analysis of chromo-somally florfenicol resistant Escherichia coli isolates from France and Ger-many. J. Antimicrob. Chemother. 49:49–54.
15. Fluit, A. C., C. L. C. Wielders, J. Verhoef, and F.-J. Schmitz. 2001. Epide-miology and susceptibility of 3,051 Staphylococcus aureus isolates from 25university hospitals participating in the European SENTRY study. J. Clin.Microbiol. 39:3727–3732.
16. Frech, G., C. Kehrenberg, and S. Schwarz. 2003. Resistance phenotypes andgenotypes of multiresistant Salmonella enterica subsp. enterica serovar Ty-phimurium var. Copenhagen isolates from animal sources. J. Antimicrob.Chemother. 51:180–182.
17. Hochhut, B., Y. Lotfi, D. Mazel, S. M. Faruque, R. Woodgate, and M. K.Waldor. 2001. Molecular analysis of antibiotic resistance gene clusters inVibrio cholerae O139 and O1 SXT constins. Antimicrob. Agents Chemother.45:2991–3000.
18. Jordens, J. Z., and L. M. C. Hall. 1988. Characterization of methicillin-resistant Staphylococcus aureus isolates by restriction endonuclease digestionof chromosomal DNA. J. Med. Microbiol. 27:117–123.
19. Kehrenberg, C., J. Mumme, J. Wallmann, J. Verspohl, R. Tegeler, T. Kuhn,and S. Schwarz. 2004. Monitoring of florfenicol susceptibility among bovineand porcine respiratory tract pathogens collected in Germany during theyears 2002 and 2003. J. Antimicrob. Chemother. 54:572–574.
20. Kehrenberg, C., K. K. Ojo, and S. Schwarz. 2004. Nucleotide sequence andorganisation of the multiresistance plasmid pSCFS1 from Staphylococcussciuri. J. Antimicrob. Chemother. 54:936–939.
21. Kehrenberg, C., S. A. Salmon, J. L. Watts, and S. Schwarz. 2001. Tetracy-cline resistance genes in isolates of Pasteurella multocida, Mannheimia hae-molytica, Mannheimia glucosida and Mannheimia varigena from bovine andswine respiratory disease: intergeneric spread of the tet(H) plasmid pMHT1.J. Antimicrob. Chemother. 48:631–640.
22. Kehrenberg, C., and S. Schwarz. 2005. Plasmid-borne florfenicol resistancein Pasteurella multocida. J. Antimicrob. Chemother. 55:773–775.
23. Kehrenberg, C., and S. Schwarz. 2005. Florfenicol-chloramphenicol exportergene fexA is part of the novel transposon Tn558. Antimicrob. Agents Che-mother. 49:813–815.
24. Kehrenberg, C., and S. Schwarz. 2004. fexA, a novel Staphylococcus lentusgene encoding resistance to florfenicol and chloramphenicol. Antimicrob.Agents Chemother. 48:615–618.
25. Kehrenberg, C., and S. Schwarz. 2001. Molecular analysis of tetracyclineresistance in Pasteurella aerogenes. Antimicrob. Agents Chemother. 45:2885–2890.
26. Kehrenberg, C., S. Schwarz, L. Jacobsen, L. H. Hansen, and B. Vester. 2005.A new mechanism for chloramphenicol, florfenicol and clindamycin resis-tance: methylation of 23 S ribosomal RNA at A2503. Mol. Microbiol. 57:1064–1073.
27. Kim, E., and T. Aoki. 1996. Sequence analysis of the florfenicol resistancegene encoded in the transferable R-plasmid of a fish pathogen, Pasteurellapiscicida. Microbiol. Immunol. 40:665–669.
28. Mahillon, J., and M. Chandler. 1998. Insertion sequences. Microbiol. Mol.Biol. Rev. 62:725–774.
29. Meunier, D., D. Boyd, M. R. Mulvey, S. Baucheron, C. Mammina, A. Nastasi,E. Chaslus-Dancla, and A. Cloeckaert. 2002. Salmonella enterica serotype Ty-phimurium DT104 antibiotic resistance genomic island I in serotype ParatyphiB. Emerg. Infect. Dis. 8:430–433.
30. Murphy, E., L. Huwyler, and M. C. F. Bastos. 1985. Transposon Tn554:complete nucleotide sequence and isolation of transposition-defective andantibiotic-sensitive mutants. EMBO J. 4:3357–3365.
31. National Committee for Clinical Laboratory Standards. 2002. Performancestandards for antimicrobial disk and dilution susceptibility tests for bacteriaisolated from animals. Approved standard, 2nd ed. NCCLS document M31-A2. National Committee for Clinical Laboratory Standards, Wayne, Pa.
32. Ojo, K. K., C. Kehrenberg, H. A. Odelola, and S. Schwarz. 2003. Structuralanalysis of the tetracycline resistance gene region of a small multiresistanceplasmid from uropathogenic Escherichia coli isolated in Nigeria. J. Antimi-crob. Chemother. 52:1043–1044.
33. Pasquali, F., C. Kehrenberg, G. Manfreda, and S. Schwarz. 2005. Physicallinkage of Tn3 and part of Tn1721 in a tetracycline and ampicillin resistanceplasmid from Salmonella Typhimurium. J. Antimicrob. Chemother. 55:562–565.
34. Priebe, S., and S. Schwarz. 2003. In vitro activities of florfenicol againstbovine and porcine respiratory tract pathogens. Antimicrob. Agents Che-mother. 47:2703–2705.
35. Schwarz, S., M. Cardoso, and H. Blobel. 1989. Plasmid-mediated chloram-phenicol resistance in Staphylococcus hyicus. J. Gen. Microbiol. 135:3329–3336.
36. Schwarz, S., C. Kehrenberg, B. Doublet, and A. Cloeckaert. 2004. Molecularbasis of bacterial resistance to chloramphenicol and florfenicol. FEMS Mi-crobiol. Rev. 28:519–542.
37. Schwarz, S., C. Kehrenberg, S. A. Salmon, and J. L. Watts. 2004. In vitro
1162 KEHRENBERG AND SCHWARZ ANTIMICROB. AGENTS CHEMOTHER.

activities of spectinomycin and comparator agents against Pasteurella multo-cida and Mannheimia haemolytica from respiratory tract infections of cattle.J. Antimicrob. Chemother. 53:379–382.
38. Schwarz, S., C. Werckenthin, and C. Kehrenberg. 2000. Identification of aplasmid-borne chloramphenicol-florfenicol resistance gene in Staphylococ-cus sciuri. Antimicrob. Agents Chemother. 44:2530–2533.
39. Schwarz, S., and D. G. White. 2005. Phenicol resistance, p. 124–147. In D. G.White, M. N. Alekshun, and P. F. McDermott (ed.), Frontiers in antimicro-bial resistance: a tribute to Stuart B. Levy. ASM Press, Washington, D.C.
40. Tillotson, L. E., W. D. Jenssen, L. Moon-McDermott, and D. T. Dubin. 1989.Characterization of a novel insertion of the macrolides-lincosamides-strep-togramin B resistance transposon Tn554 in methicillin-resistant Staphylococ-
cus aureus and Staphylococcus epidermidis. Antimicrob. Agents Chemother.33:541–550.
41. Wallmann, J., H. Kasper, and R. Kroker. 2004. Data on the prevalence ofantimicrobial susceptibility of veterinary pathogens from cattle and pigs:national antibiotic resistance monitoring 2002/2003 of the BVL. Berl.Munch. Tierarztl. Wschr. 117:480–492.
42. White, D. G., C. Hudson, J. J. Maurer, S. Ayers, S. Zhao, M. D. Lee, L.Bolton, T. Foley, and J. Sherwood. 2000. Characterization of chloramphen-icol and florfenicol resistance in Escherichia coli associated with bovinediarrhea. J. Clin. Microbiol. 38:4593–4598.
43. Witte, W. 1998. Medical consequences of antibiotic use in agriculture. Sci-ence 279:996–997.
VOL. 50, 2006 FLORFENICOL RESISTANCE GENES IN STAPHYLOCOCCI 1163

PUBLIKATION 16
Kehrenberg C., F. Aarestrup and S. Schwarz:
IS21-558 insertion sequences are involved in the mobility of the multiresistance gene cfr.
Antimicrob. Agents Chemother. (2007) 51, 483-487.

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Feb. 2007, p. 483–487 Vol. 51, No. 20066-4804/07/$08.00�0 doi:10.1128/AAC.01340-06Copyright © 2007, American Society for Microbiology. All Rights Reserved.
IS21-558 Insertion Sequences Are Involved in the Mobility of theMultiresistance Gene cfr�
Corinna Kehrenberg,1 Frank M. Aarestrup,2 and Stefan Schwarz1*Institut fur Tierzucht, Bundesforschungsanstalt fur Landwirtschaft (FAL), Holtystr. 10, 31535 Neustadt-Mariensee, Germany,1 and
Danish Institute for Food and Veterinary Research (DFVF), Bulowsvej 27, DK-1790 Copenhagen V, Denmark2
Received 26 October 2006/Returned for modification 13 November 2006/Accepted 24 November 2006
During a study of florfenicol-resistant porcine staphylococci from Denmark, the genes cfr and fexA weredetected in the chromosomal DNA or on plasmids of Staphylococcus hyicus, Staphylococcus warneri, andStaphylococcus simulans. A novel variant of the phenicol resistance transposon Tn558 was detected on the ca.43-kb plasmid pSCFS6 in S. warneri and S. simulans isolates. Sequence analysis of a 22,010-bp segmentrevealed that the new Tn558 variant harbored an additional resistance gene region integrated into the tnpCreading frame. This resistance gene region consisted of the clindamycin exporter gene lsa(B) and the gene cfrfor combined resistance to phenicols, lincosamides, oxazolidinones, pleuromutilins, and streptogramin Aantibiotics bracketed by IS21-558 insertion sequences orientated in the same direction. A 6-bp target siteduplication was detected at the integration site within tnpC. Transpositionally active forms of the IS21-558element, known as minicircles, were detected by PCR and suggest that this insertion sequence is involved inthe mobility of the multiresistance gene cfr. Based on the knowledge of the transposition pathways of IS21-likeinsertion sequences and the sequence features detected, the resistance gene region of plasmid pSCFS6 isbelieved to have developed via IS21-558-mediated cointegrate formation. The data obtained in this studyidentified the multiresistance gene cfr not only in three novel host species but also in a novel genetic contextwhose further analysis suggested that insertion sequences of the type IS21-558 are likely to be involved in thedissemination of cfr.
The gene cfr was initially discovered in a bovine Staphylo-coccus sciuri isolate during a florfenicol surveillance study ofstaphylococci from animals (19). In 2005, it was shown that theCfr protein represents a methyltransferase which modifies the23S rRNA (6). Detailed footprint analysis and mass spectro-metric studies showed that the Cfr enzyme adds an additionalmethyl group at position A2503 (6). This Cfr-mediated modi-fication affected not only the binding of phenicols but also thatof members of another four different drug classes to the bac-terial ribosome. The corresponding multidrug resistance phe-notype was characterized by elevated MICs to phenicols, lin-cosamides, oxazolidinones, pleuromutilins, and streptograminA (PhLOPSA) antibiotics, all of which are important anti-microbial agents for the treatment of bacterial infections inhumans and/or animals (7). The PhLOPSA phenotype wasexpressed not only in Staphylococcus aureus but also in Esch-erichia coli carrying the gene cfr (7).
So far, the gene cfr has been detected in six staphylococcalisolates of animal origin, located either on the 17.1-kb plasmidtype pSCFS1 or on the 35.7-kb plasmid type pSCFS3 (4).Recently, cfr has also been detected in a methicillin-resistant S.aureus isolate of human origin (8a). In the completely se-quenced plasmid pSCFS1, the gene cfr was located in closeproximity to other antibiotic resistance genes, such as lsa(B),erm(33), and spc, which mediated low-level clindamycin resis-tance; inducible resistance to macrolides, lincosamides, and
streptogramin B antibiotics; or spectinomycin resistance, re-spectively (5). When the cfr gene is located on this plasmidtype, coselection of the gene may occur under selective pres-sure imposed by the use of macrolides or spectinomycin. In thesecond cfr-carrying plasmid type, pSCFS3, the regions up- anddownstream of the gene cfr differ distinctly from the situationin plasmid pSCFS1. The cfr gene was found to be integratedinto a partially deleted variant of transposon Tn558 in plasmidpSCFS3 (4). This transposon carries the gene fexA, whichcodes for a phenicol-specific efflux protein of the major facil-itator superfamily (2, 3). In this Tn558 variant of plasmidpSCFS3, two of the three transposase genes, tnpA and tnpB,assumed to be involved in the mobility of Tn558 were trun-cated. The 3� end of tnpA and the 5� end of tnpB were replacedby a 4,674-bp segment consisting of the resistance gene cfr anda novel IS21-like insertion sequence, designated IS21-558 (4).This novel insertion sequence contained two overlapping read-ing frames, istAS and istBS, that coded for 445-amino-acid (aa)and 250-aa proteins, respectively. Although the novel insertionsequence was located close to cfr, its involvement in the mo-bility of the gene cfr remained to be clarified.
In the present study, we identified a novel plasmid-bornevariant of Tn558 with a large inserted segment. This segmentharbored the genes cfr and lsa(B) bracketed by copies of theinsertion sequence IS21-558. Based on the knowledge of themode of transfer of IS21-like insertion sequences and furtheranalysis, a model for the mobility of cfr involving the IS21-558sequences was developed.
MATERIALS AND METHODS
Bacterial isolates and antimicrobial susceptibility testing. In 2005, a total of23 florfenicol-resistant staphylococci were collected from two pig farms in Den-
* Corresponding author. Mailing address: Institut fur Tierzucht,Bundesforschungsanstalt fur Landwirtschaft (FAL), Holtystr. 10, 31535Neustadt-Mariensee, Germany. Phone: 49-5034-871-241. Fax: 49-5034-871-246. E-mail: [email protected].
� Published ahead of print on 4 December 2006.
483

mark. Both farms, owned by the same farmer, had a known history of florfenicolpretreatment of the animals and additional usage of other classes of antimicro-bials to treat and/or prevent respiratory tract infections in 2004 and 2005. Thedrugs used at these farms included penicillin, amoxicillin, florfenicol, gentamicin,lincomycin, spectinomycin, tetracycline, tiamulin, tylosin, and trimethoprim plussulfonamides. The staphylococcal isolates were cultivated overnight at 37°C onblood agar (Oxoid, Wesel, Germany) supplemented with 5% (vol/vol) sheepblood. The species identification was confirmed biochemically using the ID32Staph system (bioMerieux, Marcy l’Etoile, France). Antimicrobial susceptibilitytesting was conducted by agar disk diffusion with disks containing ampicillin(10 �g), chloramphenicol (30 �g), clindamycin (2 �g), erythromycin (15 �g),florfenicol (30 �g), gentamicin (10 �g), kanamycin (30 �g), streptomycin (10�g), spectinomycin (100 �g), sulfamethoxazole-trimethoprim (23.75/1.25 �g,respectively), or tetracycline (30 �g). MICs were determined for chloramphen-icol, florfenicol, and clindamycin by the broth macrodilution method (range ofconcentrations tested, 2 to 256 �g/ml). Staphylococcus aureus strains ATCC29213 and ATCC 25923 served as reference strains for quality control purposes.All susceptibility tests were performed and evaluated according to CLSI guide-lines M31-A2 and M31-S1 (12, 13).
Macrorestriction analysis. Since the bacterial strains were collected from onlytwo neighboring pig farms, macrorestriction analysis was performed to investi-gate the clonality of the isolates. DNA preparation and restriction analysis wereconducted as described previously (20). The SmaI (Roche Diagnostics, Mann-heim, Germany) fragments of staphylococcal whole-cell DNA were separated ina CHEF DR III system (Bio-Rad) at 15 V/cm with 0.5� Tris-borate-EDTAbuffer as running buffer. The pulse time was increased from 7 to 12 s for 12 h andfrom 20 to 65 s for another 13 h.
Isolation of DNA and transformation experiments. The preparation of whole-cell DNA followed a previously described protocol (4). Plasmids were preparedaccording to a modification of the alkaline lysis procedure (4). For the determi-nation of plasmid sizes, the sum of fragment sizes obtained after digestion of theplasmids with either BglII or EcoRI (Roche Diagnostics, Mannheim, Germany)was calculated. The use of the restriction endonucleases followed the manufac-turer’s recommendations. Comparisons of plasmids were performed on the basisof their fragment patterns. Transformation experiments into protoplasts of therecipient strain S. aureus RN4220 were used to identify phenicol resistanceplasmids (18). Transformants were picked after 48 to 72 h on regeneration platessupplemented with either 10 �g/ml florfenicol or 10 �g/ml chloramphenicol.
PCR assays and hybridization experiments. The presence of the resistancegenes cfr and fexA as well as of parts of the transposon Tn558 was confirmed byPCR assays and Southern blot hybridization experiments using previously de-scribed primers and probes (4). To detect the IS21-558 element and the associ-ated reading frames istAS and istBS, the following PCR primers were used:istAS-fw (5�-GAGATTGTGGAAAGGCTCAAGG-3�), istAS-rv (5�-TGTTCAAGCACTCCTTCATCGAC-3�), istBS-fw (5�-GAATTACGTCTTCCTGGCATCC-3�), and istBS-rv (5�-GTAAGATGGGCCGATCATATTCA-3�). A combina-tion of istAS-fw and istBS-rv was used to detect almost the complete IS21-558.The expected amplicon sizes were 1,322 bp (istAS), 672 bp (istBS), and 2,029 bp(almost complete IS21-558); the annealing temperature of 58°C was used for allthree PCR assays.
Transposition activities of IS21 insertion sequences and related elements arereported to occur via different transposition pathways. A single copy of IS21 isthought to generate simple transpositions via a cut-and-paste mechanism (1),and since this specific mechanism involves a circularization of IS21, inverse PCRassays were performed to detect this minicircle formation. For this, a combina-tion of the primers istBS-fw and istAS-rv and the Phusion polymerase (NewEngland Biolabs, Frankfurt, Germany) were used. The PCR conditions were thesame as those for the istAS amplification protocol. To determine whether tandemrepeats of IS21-558 are present, plasmid and chromosomal DNA were digestedwith either EcoRV or BglII (the single restriction sites for these enzymes inIS21-558 are located within istAS) and hybridized with the istBS amplicon ordigested with HindIII (the single restriction site for this enzyme in IS21-558 islocated within istBS) and hybridized with the istAS amplicon.
Cloning and sequencing of plasmid DNA. The BglII fragments of plasmidDNA obtained from the transformants were cloned into the BamHI site of vectorpBluescript II SK(�), and recombinant plasmids were introduced into E. coliJM109. In total, a segment of 22,010 bp, including the resistance gene regionsand their up- and downstream flanking parts, was sequenced on both strands byprimer walking (MWG-Biotech, Martinsried, Germany). Cloning of PCR am-plicons of the minicircles into vector pCR-Blunt II-TOPO and transformationinto E. coli TOP10 followed the manufacturer’s instructions (Invitrogen, Gro-ningen, The Netherlands). The minicircle amplicons were sequenced using stan-dard primers (MWG-Biotech).
Nucleotide sequence accession number. The 22,010-bp sequence of the novelTn558 variant of pSCFS6 including the up- and downstream flanking regions hasbeen deposited in the EMBL database under accession number AM408573.
RESULTS AND DISCUSSION
Species identification, antimicrobial resistance, and clonal-ity of the isolates. Among the 23 staphylococci included in thisstudy, 14 corresponded in their biochemical characteristics tothe species Staphylococcus hyicus, 8 were identified as Staphy-lococcus warneri, and a single isolate was classified as S. simu-lans. Macrorestriction analysis of the 14 S. hyicus isolates de-tected only slight differences in the SmaI patterns of twoisolates, while the fragment patterns of the remaining S. hyicusisolates were indistinguishable from each other (Fig. 1). Asimilar situation was seen with the S. warneri isolates, amongwhich only two very similar SmaI macrorestriction patternswere identified. As expected, the S. simulans isolate showed aunique SmaI restriction pattern. Therefore, all the S. hyicus aswell as the S. warneri isolates were considered closely related,a finding that was expected with regard to the origin of thesestrains from only two neighboring farms. Based on the clonalityof the isolates, two S. hyicus and two S. warneri isolates withslight differences in their macrorestriction profiles and thesingle S. simulans isolate were chosen for further analysis.
Agar disk diffusion (12, 13) revealed that all selected isolateswere resistant to florfenicol, chloramphenicol, clindamycin,erythromycin, streptomycin, and tetracyclines. In addition,both S. hyicus isolates were also resistant to gentamicin andkanamycin and the two S. warneri isolates showed no zone ofgrowth inhibition around the spectinomycin disk. The MICsfor chloramphenicol and florfenicol varied between 128 and�256 �g/ml; those for clindamycin were �256 �g/ml for all fiveisolates.
FIG. 1. SmaI macrorestriction patterns of 10 S. hyicus isolates(lanes 1 to 10). Lanes M contain the SmaI pattern of S. aureus 8325with the fragment sizes given in kilobases on the right-hand side.
484 KEHRENBERG ET AL. ANTIMICROB. AGENTS CHEMOTHER.

Localization of the resistance genes cfr and fexA. Protoplasttransformation experiments using plasmid DNA obtained fromthe two S. warneri isolates as well as from the S. simulansisolate identified a ca. 43-kb plasmid to confer phenicol andclindamycin resistance on the recipient strain S. aureusRN4220. Comparisons of the restriction patterns of the 43-kbplasmids from transformants originating from either S. warnerior S. simulans did not detect any differences (data not shown),suggesting that a single plasmid type was present among theseporcine staphylococci. This plasmid type was designatedpSCFS6. The MICs for the RN4220::pSCFS6 transformants asdetermined by the macrodilution broth method (12) were 256�g/ml for florfenicol as well as 256 �g/ml for chloramphenicoland increased to �256 �g/ml after preincubation in the pres-ence of 0.5 �g of the respective drug. The clindamycin MICsfor all transformants were �256 �g/ml.
PCR assays as well as hybridization experiments confirmedthe localization of the phenicol exporter gene fexA as well asthe multidrug resistance gene cfr on plasmid type pSCFS6.When pSCFS6 plasmid DNA was used as template, PCR am-plicons for the transposase genes tnpA and tnpB, but not fortnpC or the circular form of transposon Tn558, were obtained.Furthermore, PCR assays and Southern blot hybridizationidentified the open reading frames istAS and istBS as well astwo almost complete IS21-558 sequences on plasmid pSCFS6.When the primer combination istAS-fw and istBS-rv was used,only the expected amplicon size of 2,029 bp was obtained andno larger amplification products were identified which mightgive a hint towards a tandem formation of IS21-558 elements.
In contrast, repeated negative results for protoplast trans-formation experiments as well as hybridization of plasmid pro-files but positive hybridization results when whole-cell DNAwas used as template suggested that the genes fexA and cfrwere located in the chromosomal DNA of the S. hyicus iso-lates. The istAS and istBS reading frames and the almost com-plete IS21-558 sequence were also identified in the chromo-somal DNA of these isolates. While previous studies identifiedthe gene cfr exclusively on plasmids (4, 5, 19), this is the firstreport of a chromosomal location of cfr.
Sequence analysis of the cfr–IS21-558 resistance gene areaon plasmid pSCFS6. Since only a single localization of thenovel IS21-558 element has been described so far (4), theresistance gene area of plasmid pSCFS6 was cloned and se-quenced. Analysis of this sequenced part of 22,010 bp led tothe detection of a novel variant of transposon Tn558. Thisvariant comprised in total 16,253 bp and consisted of a Tn558-homologous part of 6,659 bp that was interrupted by the in-sertion of a 9,594-bp region. The Tn558-homologous part in-cluded the complete transposase genes tnpA and tnpB as wellas the phenicol exporter gene fexA and orf138. While orf138corresponded exactly to the respective open reading frame ofTn558 (accession no. AJ715531), single bp exchanges wereidentified in tnpA (4 bp), tnpB (2 bp), and fexA (1 bp) or in thenoncoding regions. Compared to all so-far-known Tn558 se-quences, the novel variant exhibited the typical 6-bp core se-quence 5�-GATGTA-3� at the left-end junction but differedslightly in the right-end junction (5�-GATCCG-3�). Serialtransposition experiments with Tn554 (9), the best-studied
FIG. 2. Organization of the novel variant of transposon Tn558 of plasmid pSCFS6 carrying an lsa(B)-cfr resistance gene region bracketed byIS21-558 elements. This Tn558 variant is shown in comparison to the original fexA-carrying transposon Tn558 located on plasmid pSCFS2(AJ715531) and to the other so-far-known Tn558 variant detected on plasmid pSCFS3 (AM086211). The positions and orientations of the genescoding for transposition functions (tnpA, tnpB, and tnpC), antimicrobial resistance [fexA, resistance to florfenicol and chloramphenicol; cfr,resistance to phenicols, lincosamides, oxazolidinones, pleuromutilins, and streptogramin A antibiotics; lsa(B), reduced susceptibility to lincos-amides], or unknown functions (orf138) are indicated by arrows with the direction of transcription shown by the arrowhead. A distance scale inkilobases is given below each map. The 6-bp direct repeats at the junctions of the IS21-558 elements in plasmid pSCFS6 are shown in boxes.
VOL. 51, 2007 TRANSFERABILITY OF THE GENE cfr 485

transposon of the family to which Tn558 belongs, revealed thatthe junctions varied with respect to the previous target sites(10). The observed altered core sequence at the right-endjunction in plasmid pSCFS6 is therefore likely to have devel-oped during transposition from a former target site. Analysis ofthe attachment site of the Tn558 variant on plasmid pSCFS6identified a sequence similar to att554 (11) and att558 onpSCFS3 (3). Circular intermediates of Tn558, previously de-scribed as a hint towards a functional activity of this transposon(3), were not detectable for the Tn558 variant of plasmidpSCFS6. The transposase gene tnpC of the novel Tn558 variantwas interrupted by the insertion of a 9,594-bp region, with 270bp of tnpC located upstream and 102 bp downstream of thisintegrated segment (Fig. 2). The presumable target sequencefor the inserted region, 5�-GACGTA-3�, was duplicated at theright- and left-end junctions of the novel region, forming 6-bpdirect repeats (Fig. 2).
Analysis of the inserted 9,594-bp region revealed a structureconsisting of two terminal IS21-558 insertion sequencespresent in the same orientation and a central part carrying themultidrug resistance gene cfr and the low-level clindamycinexporter gene lsa(B) (Fig. 2). Both IS21-558 elements were2,452 bp in size, differed by only 4-bp exchanges from each
other, and contained the two overlapping reading frames istASand istBS, coding for proteins of 445 aa and 250 aa, respec-tively. Both IS21-558 elements of pSCFS6 exhibited 99% nu-cleotide sequence identity to the 6-bp-shorter IS21-558 of plas-mid pSCFS3 (accession no. AM086211). Moreover, bothpSCFS6-associated insertion sequences exhibited imperfectterminal inverted repeats of 56 or 57 bp ending with the nu-cleotides CA which are characteristic for members of the IS21family. The multiple repeats, previously reported to be presentnear the termini in other members of the IS21 family (1, 8),could not be detected in these pSCFS6-associated IS21-558elements.
A 4,690-bp region with striking homology to the cfr-lsa(B)resistance gene area of plasmid pSCFS1 (accession no.AJ579365) was located between the two IS21-558 elements(Fig. 2). While cfr was indistinguishable from the gene de-scribed on plasmid pSCFS1 and only two base pair exchangeswere detected in the pSCFS1-homologous, noncoding regions,lsa(B) differed in 2 bp from lsa(B) of plasmid pSCFS1 andthree single-base-pair insertions and one 14-bp insertion weredetectable in the lsa(B) up- and downstream areas. A stretch of94 bp without homology to any database entries was identifiedbetween these cfr and lsa(B) parts (Fig. 2).
FIG. 3. Schematic presentation of the two transposition pathways of IS21-like elements, simple transposition and cointegrate formation(replicon fusion), according to reference 1. (a) Simple transposition is believed to involve minicircle formation of IS21 elements prior to integrationinto a new recipient replicon. Tandem formation may arise from transposition of an IS21-like element next to the terminus of an IS21-like elementalready present on a replicon or by recombination between an IS21-like element located on a replicon and an IS21-like minicircle (1). (b)Cointegrate formation involves an IS21-mediated fusion of two replicons and generates typical target site duplications. The 6-bp target duplicationin plasmid pSCFS6, GACGTA (which is part of the tnpC coding sequence), flanking the inserted segment (replicon 2), is shown in boxes.
486 KEHRENBERG ET AL. ANTIMICROB. AGENTS CHEMOTHER.

Analysis of potential mobility pathways for cfr–IS21-558.For the IS21 family of insertion sequences, a double func-tion of the istA gene product as a transposase or a cointe-grase was described, depending on the use of alternativestart codons that may enlarge the IstA protein by 3 to 11 aa(1, 17). For the pSCFS6-associated IS21-558 elements, suchan alternative start codon, GTG, was also detected 12 bpupstream of the ATG start codon. While the enlarged IstAprotein has been reported to promote mainly simple trans-position events, the shorter IstA protein primarily promotesreplicon fusions (17). The simple transposition event isthought to occur via a “cut-and-paste” mechanism of a sin-gle IS21 copy and involves circularization of the IS21 ele-ment (Fig. 3a), while the replicon fusion occurs preferen-tially between two IS21 elements arranged in a tandemformation and a second target replicon (Fig. 3b) (1, 8, 14,15). All these processes are assisted by the IstB helperprotein (16) and enable the translocation of DNA fragmentsfrom a donor site to a target site. As a result of this repliconfusion pathway, a cointegrate is formed which carries twoIS21 elements located in the same orientation at the formerreplicon junctions (1, 14). Such a cointegrate, flanked by twoIS21-558 elements, was detected on plasmid pSCFS6. Al-though a tandem formation of IS21 elements normally pre-cedes the cointegrate formation (1, 15), IS21-558 tandemscould not be detected by sequence analysis, PCR assays, orhybridization experiments in any of the strains investigatedin this study. Nevertheless, this specific transposition or“replicon fusion” process (17) seems to be the most likelyway by which the insertion of the resistance genes cfr andlsa(B) into Tn558 has occurred. This assumption is sup-ported not only by the characteristic arrangement of theIS21-558 insertion sequences but also by the 6-bp directrepeats at the junctions of the inserted element. Concerningtheir size, these duplications are in the typical range (4 to 7bp) of target site duplications described elsewhere for IS21-like elements (1). PCR assays and subsequent sequenceanalysis were conducted to detect the circular intermediatespromoted by the IstAS transposase. Amplicons were ob-tained from both S. hyicus isolates, but neither from the S.warneri or S. simulans isolates nor from pSCFS6-carrying S.aureus RN4220 transformants. Sequence analysis of the am-plicons confirmed the linkage between the 3� and 5� ends ofIS21-558, as expected from a circularization of the element,and both ends were separated by two additional base pairs,5�-CA-3�. Since the results of hybridization experiments didnot confirm the presence of an IS21-558 tandem formation,the amplicons obtained are considered indicative for thepresence of minicircles in S. hyicus and thus suggest a func-tional activity of the IS21-558 elements.
In contrast to the simple transposition pathway, IS21-medi-ated transposition events resulting in cointegrate formationare occasionally followed by recombination-derived deletionswhich may affect one insertion sequence element. In suchcases, flanking DNA can be deleted concomitantly (1). Recom-bination processes might explain the presence of a single IS21-558 element and the cfr gene on plasmid pSCFS3 (4) (Fig. 2).
In conclusion, the data described in this study show thepresence of the multidrug resistance gene cfr not only in sev-eral new staphylococcal host species but also in a new genetic
context. Its identification in pathogenic porcine staphylococci,including S. hyicus, underlines a wider distribution than origi-nally thought (4). Moreover, the detection of the entire cointe-grate structure on plasmid pSCFS6 might explain a potentialIS21-558-mediated transfer of the resistance genes cfr andlsa(B) and bears the danger of a further dissemination of themultidrug resistance gene cfr.
ACKNOWLEDGMENTS
This study was supported by grant SCHW382/6-3 of the DeutscheForschungsgemeinschaft and grant 274-05-0117 from the Danish Re-search Agency.
Many thanks to Vera Noding for excellent technical assistance.
REFERENCES
1. Berger, B., and D. Haas. 2001. Transposase and cointegrase: specializedtransposition proteins of the bacterial insertion sequence IS21 and relatedelements. Cell. Mol. Life Sci. 58:403–419.
2. Kehrenberg, C., and S. Schwarz. 2004. fexA, a novel Staphylococcus lentusgene encoding resistance to florfenicol and chloramphenicol. Antimicrob.Agents Chemother. 48:615–618.
3. Kehrenberg, C., and S. Schwarz. 2005. Florfenicol-chloramphenicol exportergene fexA is part of the novel transposon Tn558. Antimicrob. Agents Che-mother. 49:813–815.
4. Kehrenberg, C., and S. Schwarz. 2006. Distribution of florfenicol resistancegenes fexA and cfr among chloramphenicol-resistant Staphylococcus isolates.Antimicrob. Agents Chemother. 50:1156–1163.
5. Kehrenberg, C., K. K. Ojo, and S. Schwarz. 2004. Nucleotide sequence andorganization of the multiresistance plasmid pSCFS1 from Staphylococcussciuri. J. Antimicrob. Chemother. 54:936–939.
6. Kehrenberg, C., S. Schwarz, L. Jacobsen, L. H. Hansen, and B. Vester. 2005.A new mechanism for chloramphenicol, florfenicol and clindamycin resis-tance: methylation of 23S ribosomal RNA at A2503. Mol. Microbiol. 57:1064–1073.
7. Long, K. S., J. Poehlsgaard, C. Kehrenberg, S. Schwarz, and B. Vester. 2006.The Cfr rRNA methyltransferase confers resistance to phenicols, lincos-amides, oxazolidinones, pleuromutilins, and streptogramin A antibiotics.Antimicrob. Agents Chemother. 50:2500–2505.
8. Mahillon, J., and M. Chandler. 1998. Insertion sequences. Microbiol. Mol.Biol. Rev. 62:725–774.
8a.Mankin, A. S., S. M. Toh, L. Xiong, K. Lolans, and J. P. Quinn. 2006. Abstr.46th Intersci. Conf. Antimicrob. Agents Chemother., abstr. C1-942.
9. Murphy, E., L. Huwyler, and M. C. F. Bastos. 1985. Transposon Tn554:complete nucleotide sequence and isolation of transposition-defective andantibiotic-sensitive mutants. EMBO J. 4:3357–3365.
10. Murphy, E., and S. Lofdahl. 1984. Transposition of Tn554 does not generatea target duplication. Nature 307:292–294.
11. Murphy, E., E. Reinheimer, and L. Huwyler. 1991. Mutational analysis ofatt554, the target of the site-specific transposon Tn554. Plasmid 26:20–29.
12. NCCLS. 2002. Performance standards for antimicrobial disk and dilutionsusceptibility tests for bacteria isolated from animals; approved standard,2nd ed. NCCLS document M31-A2. NCCLS, Wayne, PA.
13. NCCLS. 2004. Performance standards for antimicrobial disk and dilutionsusceptibility tests for bacteria isolated from animals; informational supple-ment (May 2004). NCCLS document M31-S1. NCCLS, Wayne, PA.
14. Reimmann, C., and D. Haas. 1990. The istA gene of insertion sequence IS21is essential for cleavage at the inner 3� ends of tandemly repeated IS21elements in vitro. EMBO J. 9:4055–4063.
15. Reimmann, C., R. Moore, S. Little, A. Savioz, and N. Willetts. 1989. Geneticstructure, function and regulation of the transposable element IS21. Mol.Gen. Genet. 215:416–424.
16. Schmid, S., B. Berger, and D. Haas. 1999. Target joining of duplicatedinsertion sequence IS21 is assisted by IstB protein in vitro. J. Bacteriol.181:2286–2289.
17. Schmid, S., T. Seitz, and D. Haas. 1998. Cointegrase, a naturally occurring,truncated form of IS21 transposase, catalyzes replicon fusion rather thansimple insertion of IS21. J. Mol. Biol. 282:571–583.
18. Schwarz, S., M. Cardoso, and H. Blobel. 1989. Plasmid-mediated chloramphen-icol resistance in Staphylococcus hyicus. J. Gen. Microbiol. 135:3329–3336.
19. Schwarz, S., C. Werckenthin, and C. Kehrenberg. 2000. Identification of aplasmid-borne chloramphenicol-florfenicol resistance gene in Staphylococ-cus sciuri. Antimicrob. Agents Chemother. 44:2530–2533.
20. Strommenger, B., C. Kehrenberg, C. Kettlitz, C. Cuny, J. Verspohl, J. Witte,and S. Schwarz. 2006. Molecular characterization of methicillin-resistantStaphylococcus aureus strains from pet animals and their relationship tohuman isolates. J. Antimicrob. Chemother. 57:461–465.
VOL. 51, 2007 TRANSFERABILITY OF THE GENE cfr 487

Danksagung
Mein Dank gilt Herrn Prof. Dr. Stefan Schwarz für viele fachliche Anregungen und Diskussionen, für die Unterstützung sowie die stete Bereitschaft, Personal- und Sachmittel für die Projekte zur Verfügung zu stellen. Herrn Prof. Dr.sc.agr. Dr.habil. Dr.h.c. Ellendorff danke ich für die Möglichkeit, die Studien im Institut für Tierzucht in Mariensee und im ehemaligen Institutsteil Celle der Bundesforschungsanstalt für Landwirtschaft (FAL) durchgeführt haben zu können. Ich danke für die freundliche Unterstützung und Förderung. Bei der Deutschen Forschungsgemeinschaft (DFG) möchte ich mich für die langjährige finanzielle Förderung der Projekte bedanken. Ich danke Herrn Dr. Jürgen Wallmann für die Bereitstellung von Isolaten und für hilfreiche Erläuterungen. Bei Herrn Dr. Axel Cloeckaert und Benoît Doublet bedanke ich mich ebenfalls für wertvolle Ratschläge und die gute Zusammenarbeit. Auch allen anderen Kooperationspartnern - insbesondere Prof. Dr. Birte Vester und ihrem Team, Prof. Dr. Jean-Yves Madec und Mitarbeitern, Dr. Jon Rogers, Dr. Paul Todd und Prof. Dr. Frank Aarestrup - sei für ihr Vertrauen und ihre wertvolle Hilfe gedankt. Den ehemaligen Doktoranden und Kollegen/innen Kristina Kadlec, Petra Lüthje, Geovana Brenner Michael, Gideon Kikuvi und Maren Blickwede sei für viele fachliche Gespräche, gemeinsame Projekte und ihre Hilfsbereitschaft gedankt. Auch den Gästen in der Arbeitsgruppe Boudewijn Catry, Frédérique Pasquali, Mόnica Blanco, Nga T. Tham, Kayode K. Ojo, Diana Waturangi und Tomasz Hauschild möchte ich meinen besonderen Dank aussprechen. Durch sie wurde nicht nur eine erfolgreiche Zusammenarbeit möglich, sondern auch zahlreiche heitere Momente in einer stressigen Zeit. Für hervorragende technische Assistenz über lange Jahre und bei sämtlichen Projekten möchte ich mich bei Vera Nöding bedanken. Ihre natürliche, freundliche und energische Art hat sehr zu einem angenehmen Arbeitsklima beigetragen. Natürlich gilt mein Dank auch Roswitha Becker, Kerstin Meyer, Vivian Hensel, Regina Ronge und den ehemaligen Kolleginnen Erika Nußbeck, Heidrun Meyer und Giesela Niemann für die gute Zusammenarbeit und ihre stete Bereitschaft, sich für die Projekte zu engagieren. Frau Astrid Sass und Frau Monika Strack sei für die Behebung von PC-Problemen gedankt. Ebenso danke ich allen Mitarbeitern des Institutes für ihre Unterstützung. Außerdem bedanke ich mich bei meinen Freunden für viel Verständnis, zahlreiche Freundlichkeiten und ihre Hilfsbereitschaft. Dank an meine Familie für das Wissen, einen steten Rückhalt zu finden. Herzlichen Dank schulde ich Frank für die unbezahlbare Rolle, die er gespielt hat, während ich an den Studien und der Entstehung dieser Schrift gearbeitet habe.























![S2k-Leitlinie: Diagnostik bei Verdacht auf eine ...€¦ · von Oralcephalosporinen anstelle von Penicillin-derivaten werden Überempfindlichkeiten ange-führt [10]. Darüber hinaus](https://img.pdfslide.org/doc/110x75/6061763e77829b193251e501/s2k-leitlinie-diagnostik-bei-verdacht-auf-eine-von-oralcephalosporinen-anstelle.jpg)



![Menschen Präparate: Mundflora des - mgw.or.at · tetracycline, chloramphenicol, and trimethoprim -sulfamethoxazole.[1] Variable or pleomorphic in shape and similar to Actinomyces](https://img.pdfslide.org/doc/110x75/5e0accf09a10763f5578a699/menschen-prparate-mundflora-des-mgworat-tetracycline-chloramphenicol-and.jpg)

![Strukturoptimierung von Benzo[b]carbazol- Derivaten als](https://img.pdfslide.org/doc/110x75/61b0180d80860209e408b5f2/strukturoptimierung-von-benzobcarbazol-derivaten-als-.jpg)