Embed Size (px)
Citation preview

Untersuchungen zur Kontrolle der
VDJ-Rekombination am IgH-Lokus
Den Naturwissenschaftlichen Fakultäten der
Friedrich-Alexander-Universität Erlangen-Nürnberg
zur
Erlangung des Doktorgrades
vorgelegt von
Johannes Lutz
aus Wiesbaden
2006


Als Dissertation genehmigt von den Naturwissenschaftlichen Fakultäten der
Friedrich-Alexander-Universität Erlangen-Nürnberg
Tag der mündlichen Prüfung: 21.12.2006
Vorsitzender der Promotionskommission: Prof. Dr. E. Bänsch
Erstberichterstatter: Prof. Dr. H.-M. Jäck
Zweitberichterstatter: Prof. Dr. T. Winkler


Inhaltsverzeichnis
1 Zusammenfassung ......................................................................................3
2 Summary.......................................................................................................5
3 Einleitung......................................................................................................7
3.1 B-Zellentwicklung und V(D)J-Rekombination ......................................................................... 7
3.2 Sekundäre Ig-Gen-Rearrangements..................................................................................... 12
3.3 Allelausschluss...................................................................................................................... 14
3.4 Nonsensense-vermittelter mRNA Abbau in Lymphozyten.................................................... 21
4 Aufgabenstellung.......................................................................................23
5 Ergebnisse..................................................................................................25
5.1 VH-Replacement bei Pro-B-Zellen mit zwei nicht-produktiven VDJ-Allelen.......................... 25
5.1.1 Etablierung einer transgenen Maus mit zwei nicht-produktiven VDJ–-Allelen ............ 27 5.1.2 B-Zellentwicklung in Knochenmark und Milz der IgHVDJ–/VDJ–-Mäuse ......................... 27 5.1.3 Sequenzanalyse produktiver µH-Ketten-Gene ........................................................... 30 5.1.4 Zeitpunkt des VH-Replacements ................................................................................. 32 5.1.5 B-Zellentwicklung in der Gegenwart langer CDR-3H-Regionen................................. 35 5.1.6 Konkurrenz zwischen B-Zellen mit einem VH-ersetzten und B-Zellen mit einem regulär
rekombinierten VDJ-Exon in heterozygoten IgHVDJ–/wt-Mäusen .................................. 37
5.2 Kontrolle der VDJ-Rekombination am IgH-Lokus durch eine nicht-produktive µ-mRNA...... 39
5.2.1 Etablierung einer transgenen Maus mit einer stabilisierten Nonsense-µ-mRNA........ 42 5.2.2 Bestimmung der Kopienzahl und des Integrationsortes des Transgens .................... 42 5.2.3 Expressions-Niveau der stabilisierten Ter3-Nonsense-µ-mRNA................................ 47 5.2.4 B-Zellentwicklung in Gegenwart der Ter3-µ-mRNA.................................................... 50 5.2.5 Komplementation der B-Zellentwicklung durch ein produktives VDJ-Exon ................ 54 5.2.6 Abwesenheit von µH-Ketten-Signalen in rekombinations-defizienten Ter3-transgenen
Pro-B-Zellen ................................................................................................................ 56 5.2.7 Expression der endogenen IgH-Keimbahn-Transkripte.............................................. 58 5.2.8 VH-Replacement in Gegenwart der Ter3-µ-mRNA...................................................... 60 5.2.9 VH-Repertoire in Ter3-transgenen Tieren ................................................................... 62 5.2.10 Anteil der B-Zellen mit VDJ+/VDJ–-Konfiguration ........................................................ 63
6 Diskussion..................................................................................................65
6.1 VH-Replacement in Pro-B-Zellen mit zwei nicht-produktiven VDJ-Allelen ............................ 65
6.2 Entwicklung von B-Zellen mit langen CDR3-H-Regionen..................................................... 68
6.3 Hemmung der VDJ-Rekombination am IgH-Lokus durch µH-Ketten-Transkripte................ 70

Inhaltsverzeichnis
6.3.1 Effekte von Ter3-µ-Transkripten auf die B-Zellentwicklung und VDJ-Rekombination 71 6.3.2 RNA-vermittelte Mechanismen.................................................................................... 72 6.3.3 Alternative Mechanismen ............................................................................................ 76 6.3.4 Erweitertes Feedback-Modell zur Allelausschluss am IgH-Lokus............................... 77 6.3.5 Zusammenfassung ...................................................................................................... 80
7 Material und Methoden ............................................................................. 81
7.1 Material.................................................................................................................................. 81
7.1.1 Antikörper .................................................................................................................... 81 7.1.2 Oligonukleotide............................................................................................................ 81 7.1.3 Mäusestämme ............................................................................................................. 83
7.2 Molekularbiologische Methoden............................................................................................ 83
7.2.1 Präparation von Plasmid-DNA und Restiktionsverdaus.............................................. 83 7.2.2 Herstellung und Transformation elektrokompetenter Bakterien.................................. 83 7.2.3 DNA für Oozyten-Injektion........................................................................................... 84 7.2.4 DNA für ES-Zell-Transfektion ...................................................................................... 84 7.2.5 Southern-Blot-Analyse................................................................................................. 85 7.2.6 PCR ............................................................................................................................. 86 7.2.7 RT-PCR ....................................................................................................................... 86 7.2.8 Site-directed Mutagenese............................................................................................ 86 7.2.9 DNA-Sequenzierungen................................................................................................ 87 7.2.10 Bestimmung des Transgen-Insertionspunkts per Zirkel-PCR ..................................... 87 7.2.11 Einzelzell-PCR............................................................................................................. 88 7.2.12 Genotypisierung transgener Mäuse ............................................................................ 89 7.2.13 Durchflusszytometrie (FACS) ...................................................................................... 89 7.2.14 Durchflusszyometrische (FACS) und magnetische (MACS) Zellsortierungen............ 90 7.2.15 Nachweis der Ig-Isotypen im Serum ........................................................................... 91
8 Literaturverzeichnis .................................................................................. 93
9 Anhang..................................................................................................... 105
9.1 Veröffentlichungen .............................................................................................................. 105
9.2 Lebenslauf ........................................................................................................................... 107

1. Zusammenfassung
3
1 Zusammenfassung Um den Organismus vor Pathogenen mit sich verändernden, unbekannten Strukturen zu schützen, produziert das erworbene Immunsystem eine gewaltige Anzahl von B-Zellen mit unterschiedlichen B-Zellrezeptoren (BZRs). Die Vielfalt der BZRs wird hierbei durch die VDJ-Rekombination erzeugt, bei der die Gene für die schwere und leichte Immunglobulin-(Ig-) Kette in den sich entwickelnden B-Zellen aus verschiedenen Gensegmenten zusammengestellt werden. Diese Promotionsarbeit beschäftigt sich mit zwei Aspekten der VDJ-Rekombination am Immunglobulin-Schwerketten-(IgH-) Lokus, die in Progenitor-(Pro-) B-Zellen stattfindet. Im ersten Teil werden die Folgen eines nicht-produktiven Rearrangements untersucht. Da die VDJ-Rekombination ein ungenauer Prozess ist, tragen rund Zwei-Drittel der erzeugten Ig-Gene eine Verschiebung im Leseraster, was zu einer großen Anzahl von Pro-B-Zellen mit zwei nicht-produktiv rearrangierten IgH-Allelen führt. Solche Zellen werden durch Apoptose eliminiert, da ihnen essentielle, durch die Expression einer schweren µH-Kette auf der Oberfläche ausgelöste Überlebenssignale fehlen. Allerdings wurden sekundäre Rearrangements von stromaufwärts gelegenen VH-Gensegmenten in rearrangierte VDJ-Exons in Mäusen beschrieben, die eine transgene produktive µH-Kette tragen. Falls dieses VH-Replacement unabhängig von µH-Ketten-Signalen verläuft, könnte es auch nicht-produktive VDJ-Exons verändern und so Pro-B-Zellen mit zwei nicht-produktiven Allelen retten. Um diese Hypothese zu testen, wurden die JH-Gensegmente einer Maus durch ein nicht-produktives VDJ–-Exon ersetzt. Überraschenderweise war die B-Zellentwicklung in IgHVDJ–/VDJ–-Mäusen nur geringfügig gestört, da durch ein RAG-abhängiges VH-Replacement eine beachtliche Anzahl an IgM-positiven B-Zellen produziert wurde. Eine weitere einmalige Eigenschaft der transgenen Mäuse war die Anwesenheit von IgH-Ketten mit ungewöhnlich langen Complementarity-Determining Regions 3 (CDR3-Hs). Diese IgH-Ketten waren trotzdem funktional und wurden nur geringfügig gegenselektioniert, was gegen strikte Längenbeschränkungen für die CDR3-H-Regionen spricht. Der zweite Teil der Promotionsarbeit beschäftigt sich mit dem Allelausschluss am IgH-Lokus. Obwohl beide IgH-Allele rekombiniert werden können, darf jede B-Zelle nur eine IgH-Kette produzieren, um die Monospezifität des BZRs zu wahren. Daher muss die Rekombination des zweiten IgH-Allels nach einem produktiven Rearrangement auf dem ersten Lokus gestoppt werden. Um zu testen, ob bereits die stabile µ-mRNA als direkte Folge eines produktiven Rearrangements die VDJ-Rekombination auf dem zweiten Lokus dämpfen kann, wurde eine transgene Maus etabliert, die eine stabile Nonsense-µ-mRNA mit einem Stopkodon im Leader-Peptid exprimiert. In diesen Tieren war die B-Zellentwicklung beim Übergang vom Pro-B- zum Prä-B-Zellstadium gestört und der Anteil der µH-Ketten-negativen Pro-B-Zellen erhöht. Da einerseits die Störung im Pro-B-Zellstadium durch ein produktives µH-Ketten-Transgen komplementiert werden konnte und andererseits das VH-Replacement in den transgenen B-Zellen ebenfalls reduziert ist, scheint die transgene µ-mRNA spezifisch die VDJ-Rekombination am IgH-Lokus in Pro-B-Zellen zu reduzieren. Aufgrund dieser Beobachtungen schlagen wir ein erweitertes Feedback-Modell zum Allelausschluss vor, bei dem die µ-mRNA einen kurzfristigen dämpfenden Effekt auf die Rekombination am zweiten Allel ausübt, bis Prä-BZR-Signale von der Oberfläche die Rekombination endgültig terminieren


2. Summary
5
2 Summary The adaptive immune system creates an enormous number of B cells with different B cell receptors
(BCRs) to protect the organism against pathogens with changing and unknown structures. The
diversity of the BCRs is created by VDJ recombination, which assembles the genes for the heavy and
light immunoglobulin (Ig) chains from different gene segments. This thesis focuses on two aspects of
the VDJ recombination at the Ig heavy (IgH) chain locus, which occurs in Progenitor-(Pro-) B cells.
In the first part, the consequences of a nonproductive rearrangement are investigated. Inaccurate VDJ
rearrangements generate a large number of pro-B cells with two nonproductive IgH alleles. Such cells
lack essential survival signals mediated by surface IgM heavy chain (µH chain) expression and are
normally eliminated. However, secondary rearrangements of upstream VH gene segments into
assembled VDJ exons have been described in mice transgenic for productive µH chains, a process
known as VH replacement. If VH replacement was independent of µH chain signals, it could also
modify nonproductive VDJ exons and thus rescue pro-B cells with unsuccessful rearrangements on
both alleles. To test this hypothesis, we homologously replaced the JH cluster of a mouse with a
nonproductive VDJ– exon. Surprisingly, B cell development in IgHVDJ–/VDJ– mice was only slightly
impaired and significant numbers of IgM-positive B cells were produced. DNA sequencing confirmed
that all VDJ sequences from µH chain-positive B lymphoid cells were generated by VH replacement in
a RAG-dependent manner. Another unique feature of our transgenic mice was the presence of IgH
chains with unusually long Complementarity-Determining Regions 3 regions (CDR3-Hs). Such IgH
chains were nevertheless functional and only modestly counter-selected, arguing against strict length
constraints for CDR3-H regions. In conclusion, VH replacement can occur in the absence of a µH chain
signal and provides a potential rescue mechanism for pro-B cells with two nonproductive IgH alleles.
The second part of this thesis deals with the establishment of allelic exclusion at the IgH locus. Even
though both IgH alleles can be rearranged each B cell may only express one IgH chain to maintain
monospecificity. Therefore, rearrangement of the second allele must be stopped once a productive
rearrangement has been performed on the first allele. To test the hypothesis, that already a stable µH
chain (µ-) mRNA as primary result of a productive rearrangement can reduce recombination of the
second IgH allele, a transgenic mouse was generated, which expresses a stable nonsense µ-mRNA
with a stop codon in the leader peptide. In these animals B cell development was impaired at the pro-
B to pre-B cell transition with an increased number of µH chain negative pro-B cells and a reduced VH
replacement. The effect of the transgenic µ-mRNA seems to specifically reduce VDJ recombination at
the IgH locus because this impairment could be complemented by a productive µH chain transgene.
Based on these observations we propose an extended feedback model for allelic exclusion at the IgH
locus, in which the µ-mRNA exerts a short-term reducing effect on the recombination of the second
allele until recombination is terminated by surface pre-BCR signals.


3. Einleitung
7
3 Einleitung
3.1 B-Zellentwicklung und V(D)J-Rekombination
Die B-Zellentwicklung, die anfangs in der fötalen Leber und später im adulten
Knochenmark stattfindet, beginnt mit der Differenzierung pluripotenter
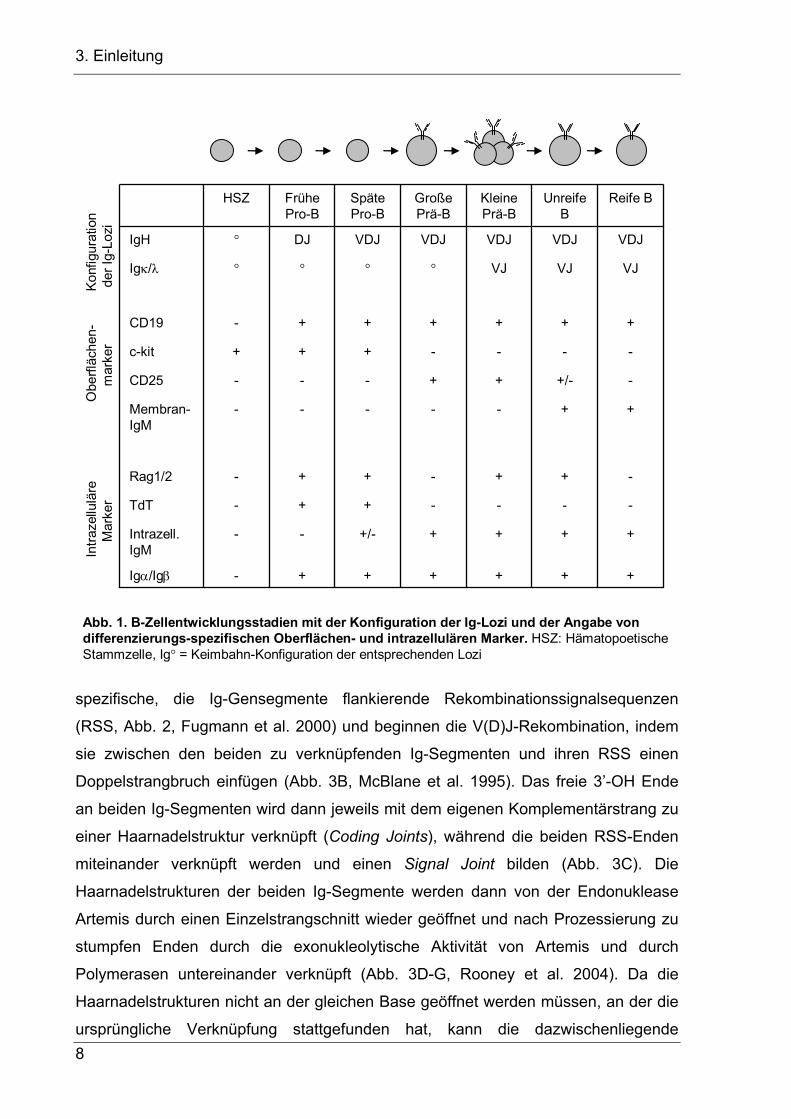
hämatopoetischer Stammzellen (HSZ) in Progenitor-(Pro-) B-Zellen (Abb. 1). Pro-B-
Zellen sind durch die Expression des Transkriptionsfaktors Pax5 auf die B-
Zellentwicklung festgelegt und tragen die Oberflächenmarker c-kit und, wie alle B-
Zellfolgestadien außer der Plasmazelle, CD19. Molekular sind Pro-B-Zellen durch die
somatische DNA-Rekombination am Immunglobulin-Schwerketten-(IgH-) Lokus
gekennzeichnet, bei der durch die Verknüpfung von Variable-(V-), Diversity-(D-) und
Joining-(J-) Gensegmenten ein VDJ-Exon erstellt wird. Das VDJ-Exon kodiert für die
V-Region der schweren Immunglobulin-Kette und bildet zusammen mit der V-Region
der leichten Ig-(IgL-) Kette die Antigenbindungstelle eines membrangebundenen B-
Zellrezeptors beziehungsweise eines sezernierten Antikörpers (zusammengefasst in
Jung et al. 2006). Die VDJ-Rekombination erzeugt eine große Anzahl
unterschiedlicher V-Regionen und ist damit die Grundlage für die immense
Antikörpervielfalt und das Funktionieren des erworbenen Immunsystems.
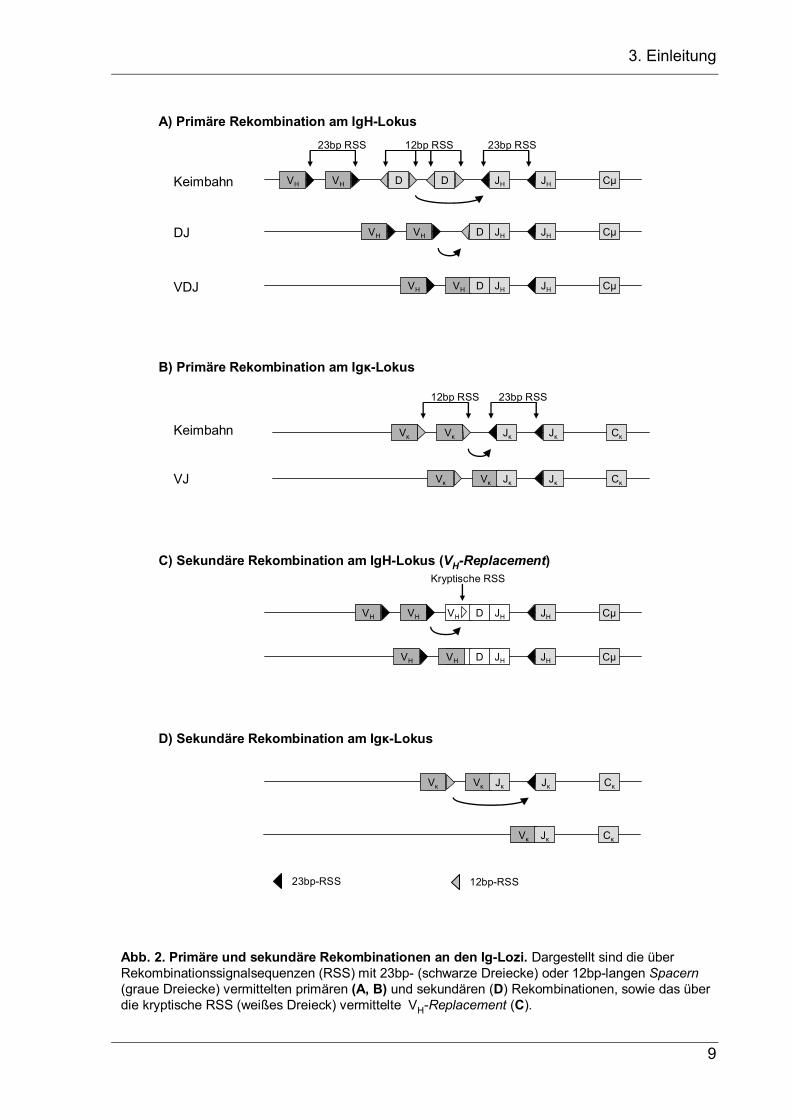
Am IgH-Lokus findet die VDJ-Rekombination in zwei Schritten statt, für die der Lokus
durch Chromatinmodifikationen schrittweise geöffnet wird (zusammengefasst in
Chowdhury and Sen 2004; Jung et al. 2006). In der frühen Pro-B-Zelle werden
zunächst auf beiden Allelen jeweils ein D- und ein JH-Gensegment durch nicht-
homologe Rekombination verknüpft (Abb. 2a). Anschließend werden in der späten
Pro-B-Zelle die stromaufwärts von den D- und JH-Segmenten liegenden VH-
Segmente geöffnet, der Lokus wird kontrahiert (Fuxa et al. 2004), und ein VH-
Segment wird mit einem DJ-Rearrangement verknüpft.
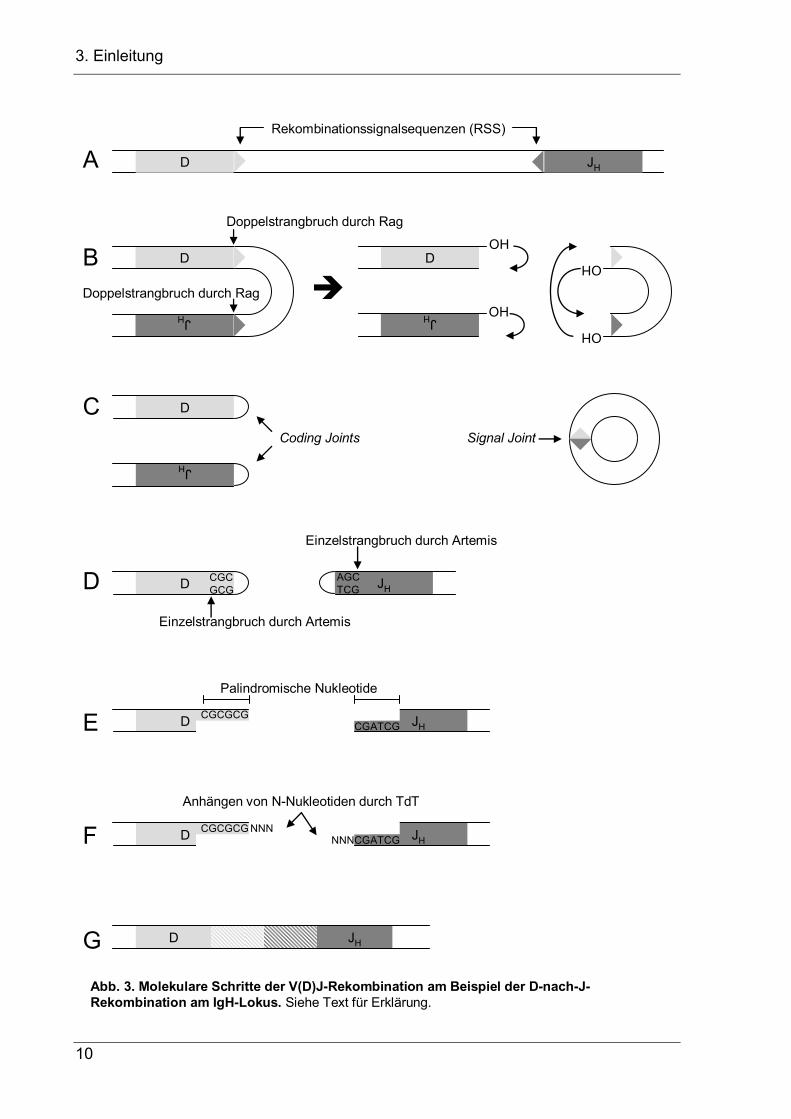
Molekular wird die Rekombination von den Lymphozyten-spezifischen Enzymen
RAG1 und RAG2 (Recombination-activating Genes 1 und 2) und den ubiquitär
vorkommenden DNA-Doppelstrangbruch-Reparaturenzymen Ku70, Ku80, XRCC4,
DNA-Ligase IV, DNA-abhänigen Proteinkinase (DNA-PK) und Artemis vermittelt
(Abb. 3, zusammengefasst in Bassing et al. 2002). Die RAG-Enzyme erkennen
3. Einleitung
++++++-Igα/Igβ
+++++/---Intrazell. IgM
----++-TdT
-++-++-Rag1/2
++-----Membran-IgM
-+/-++---CD25
----+++c-kit
++++++-CD19
VJVJVJ°°°°Igκ/λ
VDJVDJVDJVDJVDJDJ°IgH
Reife BUnreife B
Kleine Prä-B
Große Prä-B
Späte Pro-B
Frühe Pro-B
HSZ
Konf
igur
atio
nde
r Ig-
Lozi
O
berfl
äche
n-m
arke
rIn
traze
llulä
reM
arke
r
Abb. 1. B-Zellentwicklungsstadien mit der Konfiguration der Ig-Lozi und der Angabe von differenzierungs-spezifischen Oberflächen- und intrazellulären Marker. HSZ: Hämatopoetische Stammzelle, Ig° = Keimbahn-Konfiguration der entsprechenden Lozi
spezifische, die Ig-Gensegmente flankierende Rekombinationssignalsequenzen
(RSS, Abb. 2, Fugmann et al. 2000) und beginnen die V(D)J-Rekombination, indem
sie zwischen den beiden zu verknüpfenden Ig-Segmenten und ihren RSS einen
Doppelstrangbruch einfügen (Abb. 3B, McBlane et al. 1995). Das freie 3’-OH Ende
an beiden Ig-Segmenten wird dann jeweils mit dem eigenen Komplementärstrang zu
einer Haarnadelstruktur verknüpft (Coding Joints), während die beiden RSS-Enden
miteinander verknüpft werden und einen Signal Joint bilden (Abb. 3C). Die
Haarnadelstrukturen der beiden Ig-Segmente werden dann von der Endonuklease
Artemis durch einen Einzelstrangschnitt wieder geöffnet und nach Prozessierung zu
stumpfen Enden durch die exonukleolytische Aktivität von Artemis und durch
Polymerasen untereinander verknüpft (Abb. 3D-G, Rooney et al. 2004). Da die
Haarnadelstrukturen nicht an der gleichen Base geöffnet werden müssen, an der die
ursprüngliche Verknüpfung stattgefunden hat, kann die dazwischenliegende 8
3. Einleitung
9
23bp-RSS 12bp-RSS
Keimbahn
DJ
VDJ
23bp RSS
VH DDVH JHJH Cµ
VH DVH JHJH Cµ
VH DVH JHJH Cµ
12bp RSS 23bp RSS
A) Primäre Rekombination am IgH-Lokus
Keimbahn
VJ
VκVκ JκJκ Cκ
VκVκ JκJκ Cκ
12bp RSS 23bp RSS
B) Primäre Rekombination am Igκ-Lokus
Kryptische RSS
VH DVH JHJH CµVH
VH DVH JHJH CµVH
C) Sekundäre Rekombination am IgH-Lokus (VH-Replacement)
D) Sekundäre Rekombination am Igκ-Lokus
VκVκ JκJκ Cκ
Vκ Jκ Cκ
Abb. 2. Primäre und sekundäre Rekombinationen an den Ig-Lozi. Dargestellt sind die über Rekombinationssignalsequenzen (RSS) mit 23bp- (schwarze Dreiecke) oder 12bp-langen Spacern(graue Dreiecke) vermittelten primären (A, B) und sekundären (D) Rekombinationen, sowie das über die kryptische RSS (weißes Dreieck) vermittelte VH-Replacement (C).
3. Einleitung
D JH
D
JH
D
JH JH OH
DOH
D JH
OH
OH
D JH
D
CGCGCG
CGCGCG
AGCTCG
JHCGATCG
D CGCGCG NNN JHCGATCGNNN
A
B
C
D
E
F
G
Doppelstrangbruch durch Rag
Rekombinationssignalsequenzen (RSS)
Anhängen von N-Nukleotiden durch TdT
Coding Joints Signal Joint
Doppelstrangbruch durch Rag
Einzelstrangbruch durch Artemis
Einzelstrangbruch durch Artemis
Palindromische Nukleotide
AReko
bb. 3. Molekulare Schritte der V(D)J-Rekombination am Beispiel der D-nach-J-mbination am IgH-Lokus. Siehe Text für Erklärung.
10

3. Einleitung
Sequenz gespiegelt werden, was zu palindromischen (P-) Nukleotiden führt (Abb.
3D, E). Die Verknüpfungen werden durch die P-Nukleotide und die an der offenen
Bruchstelle durch die terminale Desoxynulkeotidyltransferase (TdT) zufällig
angefügten nontemplated-(N-) Nukleotide (Abb. 3F) diversifiziert, was als
Verknüpfungsdiversiät zusammen mit der rekombinatorischen Diversität das
Antikörperrepertoire erhöht. Um sicherzustellen, dass jeweils nur V- und D-
beziehungsweise D- und J-Segmente miteinander rekombiniert werden,
unterscheiden sich die RSS der beteiligten Gensegmente. Zwischen den relativ
konservierten Heptamer- und Nonamer-Sequenzen CACAGTG und ACAAAAACC,
die in allen RSS vorkommen, liegt eine Spacer-Sequenz, die bei den RSS von V-
und J-Gensegmenten 23bp und bei den RSS auf beiden Seiten des D-Segments
12bp lang ist (Abb. 2). Da die V(D)J-Rekombinationen fast ausschließlich zwischen
RSS mit unterschiedlich langen Spacern stattfinden (12/23-Regel), wird so eine
direkte Verknüpfung der V- und J-Segmente am IgH-Lokus verhindert (Early et al.
1980).
Nach einem produktiven Rearrangement wird die IgM-Schwere-(µH-) Kette
synthetisiert. Falls es sich um eine paarungsfähige µH-Kette handelt, bildet diese
zusammen mit der aus den Proteinen VpreB und λ5 bestehenden surrogaten
leichten (SL-) Kette und den Signalmolekülen Igα und Igβ den Prä-B-
Zellrezeptorkomplex (zusammengefasst in Bradl et al. 2005; Vettermann et al. 2006).
Dieser wird auf die Oberfläche transportiert, von wo er Signale induziert, die die
Diffrenzierung in frühe Prä-B-Zellen und deren klonale Expansion einleitet sowie die
Rekombination am IgH-Lokus terminiert. Falls bei dem VDJ-Rearrangement jedoch
das Leseraster durch das Einfügen von P- und N-Nukeotiden oder das Entfernen von
Nukleotiden verschoben wurde, das VDJ-Exon also nicht-produktiv ist, wird die
Rekombination auf dem zweiten Allel fortgesetzt. Ist auch das zweite Rearrangement
nicht-produktiv, so fehlen der Pro-B-Zelle essentielle Prä-BZR-vermittelte
Überlebenssignale, und sie geht durch Apoptose zugrunde.
Für Pro-B-Zellen mit einem produktiv rearrangierten VDJ-Exon und einer
paarungsfähigen µH-Kette bedeutet die Expression des Prä-BZRs den Übergang
vom Pro-B- in das in das frühe Precursor-(Prä-) B-Zellstadium, der mit einem Verlust
der c-kit- und der TdT-Expression einhergeht. Auch RAG1 und RAG2 werden
11

3. Einleitung
12
vorübergehend herunterreguliert (Grawunder et al. 1995). Stattdessen exprimieren
die frühen Prä-B-Zellen nun CD25 und durchlaufen eine kurze Proliferationsphase
mit 4-6 Zellteilungen, was zur selektiven Vervielfältigung von Prä-B-Zellen mit einer
funktionellen, mit IgL-Ketten paarungsfähigen IgH-Kette führt. Diese klonale
Expansion funktionaler Prä-B-Zellen trägt entscheidend zur Vergrößerung des Ig-
Repertoirs bei, da jeder daraus resultierende B-Zellvorläufer eine andere L-Kette und
somit einen Ig-Rezeptor mit einer anderen Spezifität produziert (Kawano et al. 2006,
Herrmann et al. unpublizierte Beobachtung). Der klonalen Expansion der frühen Prä-
B-Zellen folgt die Differenzierung in kleine, ruhende Prä-B-Zellen, die dann analog
zum Rearrangement am IgH-Lokus die κ- und λ-Leichtketten-Lozi umlagern (Abb.
2b). Nach erfolgreichem Rearrangement eines IgL-Gens wird eine IgL-Kette
produziert, die dann mit der IgH-Kette den für den Übergang ins unreife B-
Zellstadium notwendigen B-Zellrezeptor (BZR) bildet.
3.2 Sekundäre Ig-Gen-Rearrangements
Nach der Expression eines kompletten BZRs an der Zelloberfläche durchlaufen
unreife B-Zellen eine negative Selektion, bei der ihr BZR auf eine mögliche
Autoreativität überprüft wird. B-Zellen, die während dieser Phase Antigen binden,
werden je nach Signalstärke entweder durch Apoptose eliminiert oder verlieren ihre
Fähigkeit, auf Antigenstimuli zu reagieren, d.h., sie werden anergisch. Unreife B-
Zellen mit einem autoreaktiven BZR haben jedoch noch die Möglichkeit, durch
sekundäre Rearrangements der Ig-Lozi ihren BZR und damit ihre Antigen-Spezifität
zu ändern (Radic et al. 1993; Tiegs et al. 1993, zusammengefasst in Radic and
Zouali 1996; Nussenzweig 1998). Diese durch Antigenkontakt ausgelöste sekundäre
Umlagerung, die in diesem Fall als Rezeptor-Editing bezeichnet wird, findet
hauptsächlich durch Umlagerungen am IgL-Lokus statt, da die Keimbahn-VL- und -JL-
Segmente noch über die für die VJ-Rekombination benötigten RSS verfügen (Abb.
2c). Analog zum primären Rearrangement wird die RSS eines Keimbahn-VL-
Segments, das stromaufwärts vom rearrangierten VJ-Exon liegt, mit der RSS eines
stromabwärts gelegenen Keimbahn-JL-Segments verknüpft. Dieser Mechanismus ist
so effektiv, dass Mäuse, die im IgH- und Igκ-Lokus autoreaktive VDJ-Exons als
Transgene tragen, durch Rezeptorreditierung normale B-Zellzahlen erreichen (Chen
et al. 1997; Pelanda et al. 1997).

3. Einleitung
13
Ein Sonderfall der sekundären Rearrangements ist das VH-Replacement am IgH-
Lokus, bei dem das VH-Segment eines rearrangierten VDJ-Exons durch ein
stromaufwärts gelegenes Keimbahn-VH-Segment ersetzt wird (Abb. 2d, Kleinfield et
al. 1986; Reth et al. 1986; Chen et al. 1995, zusammengefasst in Zhang et al. 2004).
Mechanistisch unterscheidet sich das VH-Replacement von den primären
Rearrangements und dem Rezeptor-Editing am IgL-Lokus durch das Fehlen einer
regulären RSS im VDJ-Exon, mit der die RSS eines stromaufwärts gelegenen VH-
Segments verknüpft werden könnte. Stattdessen wird das VDJ-Exon an einer
kryptischen RSS intern am 3’-Ende des rekombinierten VH-Segments geöffnet. Die
kryptische RSS, die in fast allen VH-Gensegmenten vorhanden ist, besteht nur aus
einem Heptamer mit der Konsensus-Sequenz TACTGTG (Covey et al. 1990). Die
physiologische Funktion des VH-Replacements und die Häufigkeit, mit der es in
reifenden B-lymphoiden Zellen benützt wird, ist noch unklar. Ursprünglich wurden VH-
Replacements in Zelllinien entdeckt, die so ihre VDJ-Exons änderten (Kleinfield et al.
1986; Reth et al. 1986). Später wurden sie auch in transgenen Mausmodellen
beobachtet, bei denen über homologe Rekombination ein produktives VDJ-Exon mit
einer kryptischen RSS in den IgH-Lokus eingefügt worden war (Chen et al. 1995;
Taki et al. 1995; Cascalho et al. 1996). Besonders bei Transgenen, die für
autoreaktive BZRs kodierten, wurde eine hohe Anzahl an B-Zellen gefunden, die ihre
autoreaktive Spezifität durch VH-Replacements verloren hatten, weshalb es auch als
Toleranzmechanismus beschrieben wurde (Chen et al. 1995). Allerdings findet man
VH-Replacement auch in einer transgenen Maus mit einer nicht-autoreaktiven VH-
Kette (Cascalho et al. 1996), was darauf hindeutet, dass VH-Replacement auch in
nicht-autoreaktiven B-Zellvorläufer im Knochenmark stattfinden kann. Bei allen
bisherigen Untersuchungen blieb jedoch unklar, ob die beobachteten VH-
Replacements in vivo durch Antigenkontakt ausgelöst worden waren, oder ob VH-
Replacements auch in Abwesenheit von Ig-Signalen ablaufen können und nur
deshalb in transgenen Mäusen mit ursprünglich autoreaktiven BZRs so prominent
sind, weil die B-Zellen mit unveränderten BZRs durch Selektionsprozesse entfernt
werden.
Falls VH-Replacement in vivo auch in der Abwesenheit von Ig-Signalen stattfindet,
könnte dieses sekundäre Rearrangement auch wichtig sein, um Pro-B-Zellen vor

3. Einleitung
14
Apoptose zu retten, die entweder auf beiden Allelen ein nicht-produktives VDJ-
Rearrangement besitzen oder eine dysfunktionale, mit IgL paarungsunfähige µH-
Kette produzieren (Keyna et al. 1995; Kline et al. 1998). Diesen Zellen, die nicht in
der Lage sind einen Prä-BZR an der Oberfläche zu präsentieren und deshalb durch
Apoptose deletiert werden, könnte das VH-Replacement eine zweite Chance geben,
doch noch eine funktionelle µH-Kette zu rearrangieren und so die Entwicklung
fortzusetzen. Um diese Frage zu untersuchen, sollte im ersten Teil dieser Arbeit eine
transgene Maus etabliert werden, bei der alle Pro-B-Zellen ein nicht-produktives
VDJ-Exon tragen. Falls unsere Hypothese und VH-Replacements auch in
Abwesenheit von Ig-Rezeptorsignalen stattfinden können, sollte man in dieser Maus
reife B-Zellen mit modifizierten VDJ-Exons finden.
3.3 Allelausschluss
Potentiell kann eine B-Zelle am IgH-, Igκ- und Igλ-Lokus jeweils auf beiden Allelen
ein produktives V-Exon erzeugen. Die resultierenden zwei IgH- und vier IgL-Ketten
würden in verschiedenen Kombinationen zu einer Vielfalt an BZRs und Antikörpern
mit unterschiedlichen Antigen-Spezifitäten in einer B-Zelle beziehungsweise
Plasmazelle führen. Damit eine B-Zelle jedoch anhand ihres BZRs für oder gegen
eine Antigen-Spezifität selektioniert werden kann, ist es wichtig, dass jede B-Zelle
nur einen BZR produziert. Die Monospezifität der B-Zelle bildet die Grundlage von
Burnets klonaler Selektionstheorie (Clonal-Selection-Theory, Burnet 1960) und wird
molekular dadurch erreicht, dass jeweils nur ein IgH- und ein Igκ- oder Igλ-Allel
produktiv rearrangiert werden, und die anderen Allele von der Expression
ausgeschlossen werden. Experimentell wurde das Phänomen des Allelausschlusses
an den Ig-Lozi zuerst in Kaninchen mit spezifischen Antiseren gegen allotyp-
spezifische Epitope in der VH-Region und der konstanten Region des schweren µH-
Kettengens (CH) nachgewiesen. Diese Analyse ergab, dass mehr als 99% der B-
Zellen nur eine IgH exprimieren (Pernis et al. 1965; Cebra et al. 1966). Kurz darauf
wurde dieses Phänomen auch am IgH-Lokus der Maus mit Antikörper gegen allotyp-
spezische Epitope in der CH-region beobachtet (Weiler 1965), bei denen ein Bruch
des Allelausschluss’ ebenfalls sehr selten ist. Nur 0.01% der Milz-B-Zellen
exprimieren zwei verschiedene µH-Ketten, und oft handelt es sich bei einer der
beiden Ketten um eine dysfunktionelle Kette, die nicht an die Oberfläche gelangen

3. Einleitung
15
kann, so dass das Phänomen des Allelausschluss für den an der Oberfläche
exprimierten BZR gewahrt bleibt (Barreto and Cumano 2000).
Eine frühe Hypothese zum Allelausschluss schlug eine Steuerung auf Protein-Niveau
vor, bei der die von zwei Allelen produzierte IgH-Menge zur Deletion von Zellen mit
zwei µH-Ketten führen sollte (H-Chain-Toxicity-Theorie von Georges Köhler, Kohler
1980; Wabl and Steinberg 1982; Haas and Wabl 1984). Da diese Situation jedoch in
transgenen Tieren ohne Auswirkungen auf die B-Zellentwicklung blieb (Sonoda et al.
1997), wird heute hauptsächlich davon ausgegangen, dass der Allelausschluss
bereits auf genomischer Ebene etabliert wird.
Da alle Ig- und T-Zellrezeptor (TZR-) Gene durch die gleiche Rekombinations-
Maschinerie gebildet werden, muss das V(D)J-Rearrangement in B- und T-Zellen auf
der DNA-Ebene durch Chromatinstruktur, trans-aktivierende Faktoren und Lokus-
Zugänglichkeit reguliert werden (Zusammengefasst in Krangel 2003; Wilson and
Merkenschlager 2006). Grundlegend ist die Unterscheidung zwischen B- und T-
Zellen (Lineage-Specificity), da BZR-Gensegmente nur in B- und TZR-Gensegmente
nur in T-lymphoiden Vorläufern umgelagert werden dürfen. Aber auch innerhalb einer
Zelldifferenzierungsreihe wird die Zugänglichkeit der jeweils zu rekombinierenden
Lozi durch eine Vielzahl an Mechanismen schrittweise gesteuert, deren Abfolge und
jeweiliger Beitrag an der Rekombination experimentell noch nicht vollständig geklärt
sind.
In der B-Zelldifferenzierungsreihe beginnt die Rekombination am IgH-Lokus und lässt
sich im zeitlichen Ablauf folgendermaßen zusammenfassen (nach Corcoran 2005).
Zunächst werden beide IgH-Allele aus der repressiven nukleären Peripherie in das
Zentrum des Kerns gebracht (Kosak et al. 2002). Dort wird die DNA der JH-Lozi an
CpG-Sequenzen demethyliert und die RSS von Nukleosomen getrennt (Cherry and
Baltimore 1999), um ein Schneiden der DNA durch die RAG-Enzyme zu
ermöglichen. Gleichzeitig wird die Öffnung des Chromatins durch Histon-Azetylierung
und weitere Histon-Modifikationen verbeigeführt. Um die Rekombination eines D- mit
einem J-Gensegment zu erleichtert, werden in frühen Pro-B-Zellen zuerst die Histone
in der Keimbahn-D-J-Cµ-Region hyperazetyliert. Interessanterweise scheinen die
Umlagerungen von D- und J-Gensegmneten nicht reguliert zu sein, da sie auf beiden

3. Einleitung
16
Allelen stattfinden. Mit der Öffnung des Chromatins und der Aktivierung Zelltyp- und
Entwicklungsstadien-spezifischer Enhancer und Promotoren werden von beiden IgH-
Allelen sterile Keimbahntranskripten der konstanten Region produziert, deren
Transkription am DQ52-Promotor (µ°-Transkript) oder am intronischen µ-Enhacer
(Iµ-Transkripte) beginnen (Alt et al. 1982; Lennon and Perry 1985; Alessandrini and
Desiderio 1991; Kottmann et al. 1994). Zusätzlich wird nach dem DJ-Rearrangement
die Transkription von Keimbahn-VH-Gensegmenten induziert, die in der Synthese von
Sense-Transkripte einzelner VH-Gensegmente und Antisense-Transkripte über
größerer VH-Bereiche resultiert (Yancopoulos and Alt 1985; Bolland et al. 2004). Es
ist allerdings nicht geklärt, ob diese VH-Keimbahntranskripte nur eine Folge des
geöffneten Chromatins sind oder auch selber zur Öffnung des VH-Clusters beitragen.
Die Histone der VH-Segmente werden nun ebenfalls hyperazetylierten, und VH-
Segmente werden durch eine Kontraktion beider IgH-Allele (Fuxa et al. 2004) in die
Nähe des bereits rearrangierten DJ-Exons gebracht, um so eine Verknüpfung eines
Keimbahn-VH-Segments mit dem bereits rearrangierten DJ-Segments zu erleichtern.
Sobald ein produktives VDJ-Rearrangement stattgefunden hat und ein Prä-BZR
exprimiert wird, treten gegenläufige Prozesse ein. Der Lokus entspannt sich wieder
(Skok et al. 2001; Roldan et al. 2005) und die Sense- und Antisense-Transkription
am VH-Lokus wird, mit Ausnahme der auch in späterer B-Zellstadien vorhandenen
biallelischen Iµ-Transkription, beendet. Ein IgH-Allel, wahrscheinlich das nicht-
produktiv oder unvollständig arrangierte, assoziiert mit dem Heterochromatin, und die
entstandene große Prä-B-Zelle durchläuft die klonale Expansion (Vettermann et al.
2006).
In der kleinen Prä-B-Zelle werden anschließend die IgL-Kettengene rearrangiert, bei
denen es zusätzlich zum Allelausschluss auch zum Isotypen-Ausschluss kommt, so
dass nur eine produktive Kette vom κ- oder λ-Isotyp produziert wird (Hieter et al.
1981; Korsmeyer et al. 1982). Zunächst wird der Igκ-Lokus rearrangiert, der dazu
ebenfalls ins Kernzentrum gebracht und an den Histonen azetyliert wird. Wie beim
IgH-Lokus kommt es zu einer basalen Transkription von Igκ-Keimbahntranskripten
von beiden Allelen und einer Kontraktion beider Lozi (Singh et al. 2003). Allerdings
werden die Igκ-Allele im Gegensatz zum IgH-Lokus im nächsten Schritt
unterschiedlich prozessiert: Ein Allel wird bevorzugt DNA-demethyliert und scheint
häufiger zu rekombinieren (Mostoslavsky et al. 1998; Goldmit et al. 2002), während

3. Einleitung
Allelausschluss,Differenzierung
Öffnung und Rearrangement des zweiten Allels
V D J Cµ
V D J Cµ
V D J Cµ
V D J Cµ
mRNA
VDJ Cµ
V D J Cµ
Nicht-produktiv (2/3)
Produktiv (1/3)
Prä-BZR
Igα/Igβ
Abb. 4. Allelausschluss am IgH-Lokus durch Prä-BZR-Signale (µH-Ketten-Feedback-Modell nach Alt). Nach einem produktiven Rearrangement auf dem ersten IgH-Allel wird die µH-Kette in das endoplasmatische Retikulum (ER) synthetisiert und zusammen mit der SL-Kette und den Signalmolekülen Igα/Igβ als Prä-BZR an die Oberfläche transportiert. Die von dort initiierten Prä-BZR-Signale bewirken Differenzierung und Allelausschluss des zweiten Allels durch temporäre Terminierung der Rag-Expression und der anschließenden Schließung des zweiten IgH-Allels. Ist das erste Rearrangement jedoch nicht-produktiv, so wird das Rearrangement auf dem zweiten Allel fortgesetzt. Modell nach Alt et al. 1984.
das zweite Allel ins Heterochromatin rekrutiert wird und methyliert bleibt (Goldmit et
al. 2005). Nach einem produktiven Rearrangement wird der vollständige BZR an der
Oberfläche exprimiert und das primäre Rearrangement durch Terminierung der RAG-
Expression beendet (Grawunder et al. 1996).
Vor dem Hintergrund dieser die Rekombination begleitenden Ereignisse stellt sich
die Frage, wie es zum Allelausschluss am IgH-Lokus kommt. Hierfür werden
hauptsächlich zwei Modelle diskutiert. Das regulierte Feedback-Modell (Alt et al.
1980; Alt et al. 1984) geht davon aus, dass die Rekombination auf beiden Ig-Lozi
nacheinander stattfinden kann, die Umlagerung des zweiten Allels aber nach einem
produktiven Rearrangement des ersten Ig-Lokus' durch ein Signal der gebildeten Ig-
unterdrückt wird. Im Falle des IgH-Lokus' würde der Allelausschluss durch das Signal
einer µH-Kette beziehungsweise eines Prä-BZR kontrolliert, um so die Expression
zweier funktioneller IgH-Ketten zu verhindern. Das unregulierte, stochastische Modell
hingegen geht von einem ineffizienten Rearrangement der beiden IgH-Lozi aus, bei
17

3. Einleitung
18
dem es unwahrscheinlich ist, dass eine Zelle zwei produktive IgH-Rearrangements
erzeugt (Coleclough et al. 1981; Coleclough 1983; Liang et al. 2004). In beiden
Modellen wird das weitere Rearrangement des zweiten IgH-Lokus durch die mit der
Differenzierung einhergehenden Chromatin-Änderungen verhindert. Diese sind
besonders am IgH-Lokus zu erwarten, da die Prä-B-Zelle nach der VDJ-
Rekombination eine proliferative Phase mit 4-6 Replikationen durchläuft (Decker et
al. 1991; Karasuyama et al. 1994; Rolink et al. 1994).
Als wichtiger Beleg für das Feedback-Modell gilt die Beobachtung, dass Signal-
kompetente Ig-Transgene die Rekombination der endogenen Ig-Lozi unterdrücken
(Ritchie et al. 1984; Rusconi and Kohler 1985; Weaver et al. 1985; Nussenzweig et
al. 1987), während Signal-inkompetente Ig-Ketten, z.B. eine µH-Kette ohne
Transmembranregion (µMT, Kitamura and Rajewsky 1992), die Umlagerungen
endogener Lozi nicht verhindern. Auch ist der Allelausschluss in B-Zellen gestört, bei
denen die Signalleitung des Prä-BZRs oder BZRs durch Deletion oder Mutation der
Komponenten Igα (Minegishi et al. 1999) oder Igβ (Gong and Nussenzweig 1996)
oder durch eine µH-Kette, die nicht mit Igα/Igβ assoziieren kann (Papavasiliou et al.
1995), unterbrochen wurde. Diese Experimente weisen alle darauf hin, dass Signale
durch einen unreifen Prä-BZR oder reifen BZR wichtig sind, um die Umlagerung des
zweiten Ig-Allels zu verhindern und somit den Allelausschluss zu erreichen.
Allerdings muss hierbei beachtet werden, dass der zeitliche Verlauf der B-
Zellentwicklung und der Turnover der B-lymphoider Vorläufer durch diese Eingriffe
erheblich gestört wird. Produktive Ig-Transgene sorgen für eine Verkürzung des
Rekombinationsstadiums, was dadurch belegt wird, dass IgH-transgen-positive B-
Zellen im Gegensatz zu Wildtyp-B-Zellen z.T. unvollständige DJ-Rearrangements auf
den endogenen IgH-Lozi durchführen (Weaver et al. 1985). Signal-Mutanten
hingegen blockieren die Differenzierung B-lymphoider Vorläufer und verlängern so
das VDJ-rekombinationsaktive Pro-B-Zellstadium, so dass zeitliche Mechanismen
unterlaufen werden. Doch gerade der zeitliche Verlauf spielt eine große Rolle und
stellt für das Feedback-Modell am IgH-Lokus ein Problem dar. Da nach einem
produktiven Rearrangement die µH-Ketten erst neu synthetisiert und als Prä-BZR an
die Oberfläche transportiert werden müssen, können etwaige µH-Ketten-vermittelte
Feedback-Signale erst verzögert initiiert werden. Damit während dieser Zeit das
zweite Allel nicht arrangiert, muss davon ausgegangen werden, dass die Allele

3. Einleitung
19
nacheinander rearrangieren. Für den Igκ-Lokus wurde mit der unterschiedlichen
Replikation der beiden Igκ-Allele ein Mechanismus beschrieben, der für
unterschiedliche Chromatinstrukturen der beiden Allele sorgt und die Grundlage für
die benötigte Asynchronität des Rearrangements sein könnte. Hierbei repliziert eines
der beiden Igκ-Allele früh in der S-Phase, während das zweite Allel erst später
repliziert. Die Auswahl des früh-replizierenden Allels findet in einem frühen B-
Zellentwicklungstadium vor Öffnung des IgL-Lokuses zufällig statt und wird klonal
vererbt (Mostoslavsky et al. 1998). Beim IgL-Rearrangement ist es dann meistens
das früh-replizierende Allel, das zuerst demethyliert (Mostoslavsky et al. 1998),
Histon-azetyliert (Goldmit et al. 2005) und rearrangiert wird, während das zweite Allel
aus dem Zentrum des Kerns ins Heterochomatin rekrutiert wird. Diese
Beobachtungen am Igκ-Lokus scheinen jedoch nicht auf den IgH-Lokus übertragbar
zu sein. So findet am IgH-Lokus eine biallelische DNA-Demethylierung statt, und
beide kontrahierten Allele bleiben nach der DJ- und während der V-nach-DJ-
Umlagerung im Zentrum des Kerns (Skok et al. 2001). Erst nach einem Prä-BZR-
Signal findet man ein Allel in der Prä-B-Zelle wieder mit dem Heterochromatin
assoziiert (Kosak et al. 2002). Dies spricht für eine gleichzeitige Öffnung beider IgH-
Allele während der VDJ-Umlagerung und eine Stilllegung des nicht-komplett oder
nicht-produktiv rearrangierten Allels erst nach einer produktiven VDJ-Umlagerung.
Daher ist es unklar, wie nach einem produktiven Rearrangement des ersten IgH-
Allels die Rekombination auf dem zweiten Allel bis zur Initiierung von Prä-BZR-
Signalen von der Zelloberfläche unterdrückt wird.
Das stochastische Modell umgeht das zeitliche Problem durch die Annahme, dass
die VDJ-Rekombination so ineffizient ist, dass zwei produktive Rearrangements in
einer Zelle nur selten vorkommen können (Coleclough 1983; Schlissel 2003). Da die
DJ-Umlagerung jedoch in allen B-Zellen auf beiden IgH-Allelen stattfindet, kann die
ineffiziente Umlagerung erst bei der Verknüpfung eines Keimbahn-VH-Gensegments
mit einem bereits rearrangierten DJ zum Tragen kommen, was durch die
Beobachtung gestützt wird, dass die RSS von VH-Gensegmenten ein schlechtes
RAG-Substrat darstellen (Liang et al. 2002). Nach einem nicht-produktiven VDJ-
Rearrangement auf dem ersten Allel wird jedoch auf das zweite Allel gewechselt, und
das so effektiv, dass 40-50% der reifen B-Zellen ein produktives und ein nicht-
produktiv rearrangiertes IgH-Allel besitzen (Alt et al. 1984; Mostoslavsky et al. 2004).

3. Einleitung
20
Dies steht im Widerspruch zu der Annahme, dass das die Umlagerungen eines VH-
Gensegments mit einem DJ-Rearrangement einen ineffizienten Schritt darstellt
(Mostoslavsky et al. 2004). Das Verhältnis spiegelt eher eine Situation wider, bei der
1/3 der Zellen nach einem erfolgreichen VDJ-Rearrangement des ersten Allels das
zweite Allel unterdrücken, während die 2/3 der Zellen mit einem nicht-produktiv-
rearrangiertem ersten Allel mit der gleichen Wahrscheinlichkeit zügig das zweite Allel
rearrangieren. In Zahlen ausgedrückt sind 3/9 Zellen VDJ+/DJ und 2/9 VDJ–/VDJ+,
was ungefähr dem in reifen B-Zellen beobachteten Verhältnis von 60-zu-40
entspricht, auch wenn diese Berechnung weitere Faktoren wie z.B. die Leseraster
der D-Segmente außer acht lässt (Rajewsky 1996).
Ein neuer Gedankenansatz, wie der Allelausschluss am IgH-Lokus durch einen
Feedback-Mechanismus kontrolliert werden kann ohne eine Asynchronität in der
Rekombination der IgH-Allele vorauszusetzen, ergab sich aus Vorarbeiten in
unserem Labor mit einer transgenen Maus, die eine mit einer SL- und IgL-Kette
paarungsunfähige und somit Signal-inkompetente (dysfunktionelle) µH-Kette
produziert (Kline et al. 1998). Silke Meister konnte in ihrer Promotionsarbeit zeigen,
dass die dysfunktionale µH-Kette das VDJ-Rearrangement der endogenen IgH-Lozi
auch in Abwesenheit eines Prä-BZR-Signals reduziert jedoch nicht inhibiert (Kline et
al. 1998; Meister 2004). Dies legte den Schluss nahe, dass von einem produktiv
arrangierten IgH-Lokus neben dem µH-Ketten-Protein weitere Signale ausgehen
können, die das VDJ-Rearrangement beeinflussen. Ein idealer Kandidat hierfür ist
das µ-Transkript, das sofort nach einem produktiven VDJ-Rearrangement im Kern
zur Verfügung steht. Es könnte als elegante Erweiterung des Feedback-Modells ein
sofortiges Feedback erlaubten und damit die zeitliche Trennung der Rekombination
und einen Unterschied zwischen den IgH-Allelen überflüssig machen. Die wichtige
Unterscheidung zwischen einem produktiven und nicht-produktiven IgH-
Rearrangement, die im klassischen Feedback-Modell durch An- oder Abwesenheit
der µH-Kette getroffen wird, wird in unserem erweiterten Feedback-Modell durch die
mRNA-Stabilität erzielt, da eine von einem nicht-produktiv rearrangierten IgH-Allel
kodierte mRNA aufgrund von vorzeitigen Stopkodons (Nonsense-Kodons) im
Gegensatz zu einer produktiven mRNA durch den Nonsense-mediated mRNA-Decay
abgebaut wird (Baumann et al. 1985; Jack et al. 1989).

3. Einleitung
21
Um unser erweitertes Feedback-Modell zu überprüfen, sollte daher im zweiten Teil
dieser Arbeit untersucht werden, ob bereits eine stabile µ-mRNA als primäres
Resultat eines produktiven VDJ-Rearrangement die VDJ-Rekombination des zweiten
Allels reduzieren kann und so eine weitere Komponente des
Allelausschlussmechanismuses darstellt.
3.4 Nonsensense-vermittelter mRNA Abbau in Lymphozyten
Fehlerhafte mRNAs mit einem vorzeitiges Stopkodon (Premature Termination-
Codon, PTC) können aus viele Gründen entstehen, z.B. durch fehlerhaftes oder
alternatives Spleißen von Prä-mRNAs oder durch Mutationen, Deletionen oder
Insertionen im Genom (Maquat 2005). Würden diese Nonsense-mRNAs translatiert,
käme es zur Synthese verkürzter Proteine mit eventuell veränderten funktionellen
Domänen. Möglich wären zum Beispiel dominant-aktive oder -negative Rezeptoren.
Um dieser potentiellen Gefahr vorzubeugen, wird die mRNA-Synthese durch den
Nonsense-mediated mRNA-Decay (NMD) überwacht, der mRNAs mit PTCs erkennt
und schnell abbaut (zusammengefasst in Frischmeyer and Dietz 1999; Neu-Yilik et
al. 2004; Maquat 2005). Dieses könnte besonders in Lymphozyten notwendig sein,
da V(D)J-Umlagerungen in der Mehrheit der B- und T-Zellen nicht-produktive V(D)J-
Gene erzeugt, die für Nonsense-Ig-mRNAs kodieren (zusammengefasst in Li and
Wilkinson 1998). Rund 40% der reifen B-Zellen tragen einen nicht-produktiv
rearrangierten IgH-Lokus; hinzukommen die nicht-produktiven Rearrangements der
Leichtketten-Lozi. Interessanterweise ist der NMD in Lymphozyten besonders
effektiv, und Nonsense-µ-mRNA hat eine 50- bis 100-fach geringere Stabilität als
Sense-µ-mRNA (Baumann et al. 1985; Jack et al. 1989; Buhler et al. 2004).
Der NMD ist ein translationsabhängiger Prozess, der bisher in allen eukaryotischen
Organismen gefunden wurde, sich aber in seinem Mechanismus zwischen den
Spezies unterscheidet. In Säugern werden PTCs auf mRNAs nur unter bestimmten
Voraussetzungen entdeckt. Die beiden wichtigsten Voraussetzungen sind, dass die
mRNA gespleißt werden muss, und dass das PTC nicht im letzten Exon liegen darf.
Diese Bedingungen zeigen, dass der NMD in Säugern Exon-Exon-Grenzen als
Kriterium zur Erkennung fehlerhafter mRNAs heranzieht. Wird ein Gen, dessen
gespleißte, PTC-haltige mRNA vom NMD erkannt und abgebaut wird, als intronlose

3. Einleitung
22
cDNA exprimiert, so versagt der NMD, und das Transkript ist bei gleicher Sequenz
stabil (Zhang et al. 1998). Die molekularen Mechanismen der NMD in Säugerzellen
sind noch nicht vollständig geklärt. Ein allgemein akzeptiertes Modell geht davon
aus, dass nach dem Spleißen ein Teil der beteiligten Proteine mit der Exon-Exon-
Verküpfung assoziiert bleiben und einen Exon-Exon-Junction-Komplex (EJC) bilden
(Zusammengefasst in Lejeune and Maquat 2005). Dieser Komplex besteht unter
anderem aus Proteinen, die am NMD (Upf1, Upf2 und Upf3), dem Spleißen von Prä-
mRNAs (RNPS1, UAP56, SRm160 und Pnn/DRS) und dem mRNA-Export (REF/Aly,
Y14 und Magoh) beteiligt sind. Kommt es nun zur Translation der reifen mRNA, so
gleitet das Ribosom die mRNA entlang und streift die Exon-Exon-Junction-Komplexe
ab, bis es zu einem Stopkodon kommt und dort zerfällt. Ein postulierter
Überwachungskomplex setzt jedoch die Fahrt fort und überprüft die restliche mRNA.
Trifft dieser Überwachungskomplex auf einen weiteren Exon-Exon-Junction-
Komplex, so ist klar, dass die Translation bei einem vorzeitigen Stopkodon terminiert
wurde. Die betreffende mRNA wird als fehlerhaft deklariert. Als Konsequenz leitet der
Überwachungskomplex den schnellen Abbau der mRNA in 5’-3’- und 3’-5’-Richtung
ein (zusammengefasst in Frischmeyer and Dietz 1999; Neu-Yilik et al. 2004; Maquat
2005).
Die bisher bekannte Rolle des NMD in Lymphozyten beschränkt sich auf den Abbau
PTC-haltiger mRNAs von nicht-produktiv arrangierten Ig-Genen, um die Expression
verkürzter Ig-Rezeptoren zu verhindern (Baumann et al. 1985; Jack et al. 1989).
Sollte jedoch die Hypothese zutreffen, dass µ-mRNAs die VDJ-Rekombinationrate
des IgH-Lokus' reduzieren können, so wäre der NMD zusätzlich auch an der
Kontrolle der VDJ-Rekombination des IgH-Lokus beteiligt, da er zwischen Sense-
und Nonsense-µ-mRNAs unterscheidet und nur die Ansammlung von Sense-µ-
Transkripten zulässt, die dann die Rate der VDJ-Rekombination am zweiten IgH-
Lokus solange reduzieren, bis ein Prä-BZR-Signal weitere Rekombinationen am
zweiten Lokus endgültig verhindert.

4. Aufgabenstellung
23
4 Aufgabenstellung Bei der VDJ-Rekombination am IgH-Lokus werden Variable-(V-), Diversity-(D-) und Joining-(J-) Gensegmente zu einem VDJ-Exon zusammengesetzt, das die variable Region der schweren Ig-Kette kodiert. Dieser Vorgang ist von zentraler Bedeutung für die B-Zellentwicklung, da er die Grundlage der Antikörpervielfalt und der erworbenen B-Zell-vermittelten Immunität darstellt. Für die sich entwickelnde B-Zelle ist er jedoch ein zweischneidiges Schwert, das die Zelle vor zwei Probleme stellt, die in dieser Arbeit näher untersucht werden sollen. 1.) Die VDJ-Rekombination wird durch Rekombinationssignalsequenzen (RSS) vermittelt, welche die Segmente flankieren und bei der Rekombination entfernt werden. Kommt es bei der Genumlagerung, bei der Nukleotide an den Verknüpfungen hinzugefügt oder entfernt werden, zur einer Verschiebung im Leseraster, so kann das entstandene nicht-produktive VDJ-Exon nicht durch eine weitere VDJ-Rekombination ersetzt werden, da hierfür die RSS fehlen. Stattdessen wird die Rekombination auf dem zweiten Allel fortgesetzt. Da allerdings die meisten VDJ-Rekombinationen nicht-produktiv sind, entsteht eine große Anzahl an Pro-B-Zellen mit zwei nicht-produktiven VDJ–-Allelen, die keine µH-Ketten exprimieren können. Diesen Zellen fehlen essentielle Überlebenssignale, so dass sie durch Apoptose zugrunde gehen. In der vorliegenden Arbeit sollte nun untersucht werden, ob Pro-B-Zellen mit zwei nicht-produktiven VDJ–-Allelen nicht doch in der Lage sind, das VH-Segment des fehlerhaften VDJ-Exons durch ein stromaufwärts gelegenes VH-Segment zu ersetzten, und durch dieses VH-Replacement ein produktives IgH-Gen zu erzeugen, das eine weitere Entwicklung der Pro-B-Zelle zulässt. Hierfür sollten transgene Mäuse etabliert werden, bei denen der JH-Lokus durch ein nicht-produktives VDJ–-Exon ersetzt wird, um in ihnen dann die B-Zell-Entwicklung zu untersuchten. 2.) Während es nach einem nicht-produktiven Rearrangement auf dem ersten Allel vorteilhaft ist, dass die VDJ-Rekombination auf dem zweiten Allel fortgesetzt wird, muss nach einem produktiven Rearrangement auf dem ersten Allel das Rearrangement auf dem zweiten Lokus unterdrückt werden, damit die B-Zelle nur eine produktive IgH-Kette exprimiert. Obwohl dieser Allelausschluss die Voraussetzung für monospezifische B-Zellen bildet, ist der genaue Mechanismus noch unbekannt. Als zweites Ziel dieser Promotion sollte deshalb die Hypothese überprüft werden, dass eine stabile µ-mRNA, die als erste Konsequenz eines produktiven Rearrangements gebildet wird, einen dämpfenden Einfluss auf die V-nach-DJ-Rekombination haben und zum Allelausschluss beitragen könnte. Daher sollten transgene Mäusen hergestellt werden, die eine stabilisierte, nicht-produktive µ-mRNA exprimieren, um in ihnen dann die B-Zellentwicklung zu untersuchen.


5. Ergebnisse
5 Ergebnisse
5.1 VH-Replacement bei Pro-B-Zellen mit zwei nicht-produktiven VDJ-Allelen
Die Genumlagerungen am IgH-Lokus finden im Pro-B-Zellstadium statt. Ist das
gebildete VDJ-Exon produktiv, so wird eine µH-Kette synthetisiert, die mit der
surrogaten leichten (SL-) Kette und den Signalkomponenten Igα und Igβ assoziiert
und einen Prä-B-Zellerezeptor bildet (Vettermann et al. 2006). Durch das Hinzufügen
oder Entfernen von Nukleotiden während der Rekombination haben jedoch die
meisten VDJ-Exons eine Verschiebung im Leseraster, weshalb viele Pro-B-Zellen
nicht-produktive Rearrangements auf beiden Allelen enthalten (Rajewsky 1996).
Zusätzlich kodieren einige der produktiven VDJ-Rearrangements für dysfunktionale
µH-Ketten, die nicht mit der SL-Kette oder konventionellen leichten Ketten paaren
können (Keyna et al. 1995; ten Boekel et al. 1997). In beiden Fällen fehlen den Pro-
B-Zellen essentielle Differenzierungssignale, was zur Apoptose führt. Die meisten
rearrangierten VDJ-Exons stellen jedoch noch ein Substrat für RAG1/2 dar, da die
fast alle VH-Segmente an ihrem 3’-Ende eine nur aus dem konservierten Heptamer
TACTGTG bestehende kryptische RSS enthalten (Chen et al. 1995). Daher kann die
VH-Sequenz des VDJ-Exons durch ein stromaufwärts liegendes VH-Segment ersetzt
werden (Zhang et al. 2003). Dieses VH-Replacement wurde zuerst in murinen und
humanen Pro-B-Zelllinien beschrieben (Kleinfield et al. 1986; Reth et al. 1986; Zhang
et al. 2003) und später auch in transgenen Mäusen gefunden (Chen et al. 1995; Taki
et al. 1995; Cascalho et al. 1996; Zhang et al. 2004), bei denen der JH-Lokus durch
ein produktives VDJ-Exon ersetzt worden war. Während VH-Replacements in µH-
Ketten-negativen Pro-B-Tumorzelllinien nachgewiesen werden konnten, wurden VH-
Replacements in vivo nur in Mäusen mit einer transgenen funktionellen µH-Kette,
d.h. in Anwesenheit eines funktionalen Prä-BZR- und BZR-Signalwegs gefunden.
Daher war es zu Beginn der Arbeit nicht klar, ob VH-Replacements in vivo ein µH-
Ketten-Signal benötigen, oder ob sie auch in Abwesenheit eines Prä-BZR-Signals
vorkommen und so einen Rettungsmechanismus für Pro-B-Zellen mit zwei nicht-
produktiv rearrangierten IgH-Allelen darstellen können.
25
5. Ergebnisse
Wildtyp-IgH-Lokus
Targeting-Vektor
Veränderter IgH-Lokus (nach Deletion der neor-Kassette)
AseI
DQ52 Cµ1
HindIII
AseI
L VDJH4
AseI
4.6kb
7.4kb
neor
BamHI
SpeI
AseI
L VDJH4
DQ52-Sonde
SacI
I
JH4revVHQMfwdVDJ-
loxP-Stellen
DQ52
AseI
DQ52 JH1-4 Cµ1
HindIII
AseI
JH2revJH1fwdJH
1.2kb
C
7.4 (wt)
4.6 (VDJ-)
AseI-Verdau mit DQ52-Sonde
kb
5
3
6
IgH
wt/w
t
IgH
VDJ-
/wt
IgH
VDJ-
/VDJ
-
8
B
kb
5
3
6
8
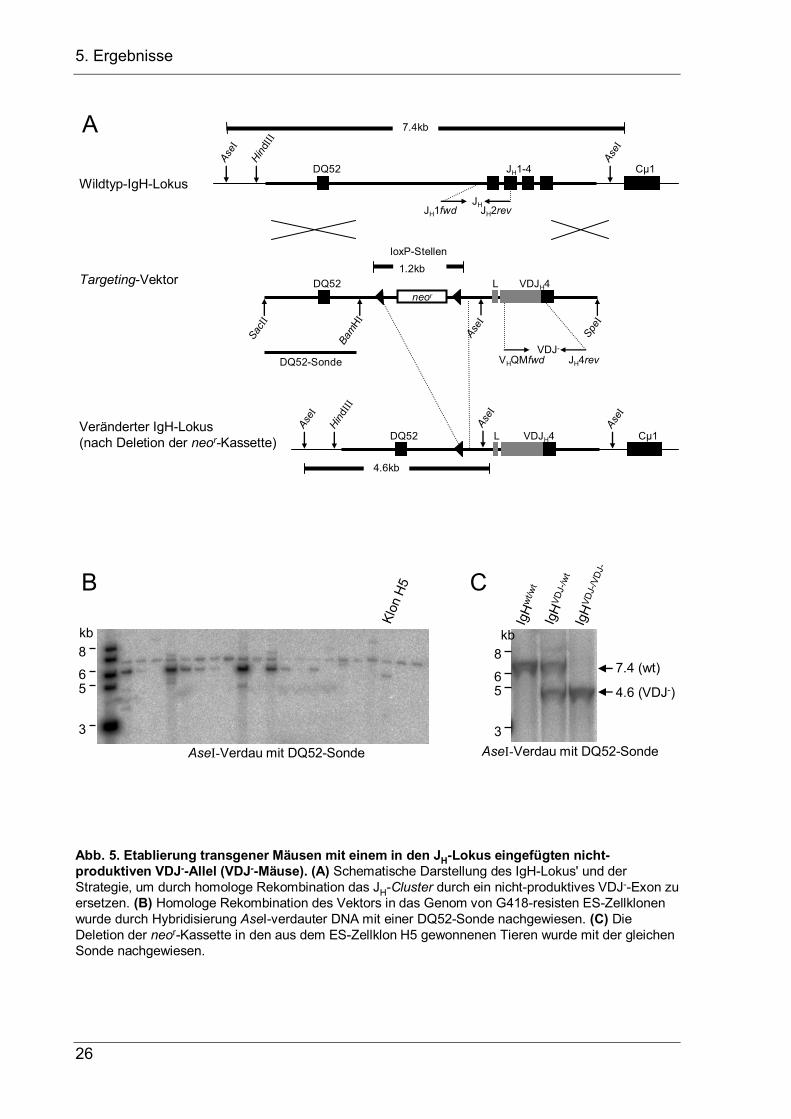
Abb. 5. Etablierung transgener Mäusen mit einem in den JH-Lokus eingefügten nicht-produktiven VDJ--Allel (VDJ--Mäuse). (A) Schematische Darstellung des IgH-Lokus' und der Strategie, um durch homologe Rekombination das JH-Cluster durch ein nicht-produktives VDJ--Exon zu ersetzen. (B) Homologe Rekombination des Vektors in das Genom von G418-resisten ES-Zellklonenwurde durch Hybridisierung AseI-verdauter DNA mit einer DQ52-Sonde nachgewiesen. (C) Die Deletion der neor-Kassette in den aus dem ES-Zellklon H5 gewonnenen Tieren wurde mit der gleichen Sonde nachgewiesen.
Klon
H5
AseI-Verdau mit DQ52-Sonde
A
26

5. Ergebnisse
5.1.1 Etablierung einer transgenen Maus mit zwei nicht-produktiven VDJ–-Allelen
Um zu untersuchen, ob VH-Replacement in vivo auch in der Abwesenheit von Ig-
Rezeptorsignalen vorkommen kann, wurde einen transgene Maus etabliert, bei der
alle Pro-B-Zellen zwei nicht-produktive VDJ–-Allele tragen. Dazu wurde in der
embryonalen Stammzelllinie IDG3.2 (freundlicherweise zur Verfügung gestellt von
Ralf Kühn, GSF München) der komplette JH-Lokus durch homologe Rekombination
gegen ein rearrangiertes VDJ–-Exon mit einem Stop-Kodon an Aminosäure-Position
+3 des Leader-Peptids ausgetauscht. Das daraus resultierende IgH-Gen wird zwar
transkribiert ist aber nicht-produktiv, da das vorzeitige Stop-Kodon nur die
Translation eines Dipeptids erlaubt. Die Targeting-Strategie ist in Abb. 5 dargestellt.
Nach der Transfektion wurden die ES-Zellen mit dem Antibiotikum G418 selektioniert
und DNA von 147 G418-resistenten Klonen auf homologe Integration des Transgens
geprüft. Dazu wurde die ES-Zell-DNA mit AseI verdaut und mit einer DQ52-Sonde
hybridisiert (Abb. 5B). Nur Klon H5 aus der Mikrotiterplatte Ter3#2-WM zeigte das für
eine homologe Rekombination diagnostische 5.8kb-Fragment und wurde zur
Blastozysteninjektion benutzt. Die chimären Mäuse wurden dann mit Cre-Deleter-
Mäusen (freundlicherweise zur Verfügung gestellt von Michael Wegner, Universität
Erlangen) verpaart, welche die Cre-Rekombinase unter dem ubiquitär aktiven EIIa-
Promotor exprimieren (Lakso et al. 1996). Die Deletion der von LoxP-Sequenzen
flankierten Neomyzin-Resistenz-Kassette (neor) wurde erfolgreich im Southern-Blot
über den Nachweis eines um 1.2kb auf 4.6kb verkürzten Fragments nachgewiesen
(Abb. 5C).
5.1.2 B-Zellentwicklung in Knochenmark und Milz der IgHVDJ–/VDJ–-Mäuse
Falls Pro-B-Zellen mit zwei nicht-produktiven VDJ–-Allelen sekundäre
Genumlagerungen am IgH-Lokus durchführen, würden wir eine B-Zellreifung über
das Pro-B-Zellstadium hinaus erwarten. Konsequenz hiervon wären IgM-positive B-
Zellen im Knochenmark und in der Milz von homozygoten Mäusen mit dem VDJ–-
Transgen. Tatsächlich gelang es, B-lymphoide Zellen jenseits des CD19/ckit-
doppelpositiven Pro-B-Zellstadiums im Knochenmark (Abb. 6A) und der Milz (Abb.
27
5. Ergebnisse
A
B
IgHwt/wt IgHVDJ-/wt IgHVDJ-/VDJ-
FI (CD19)
FI (c
-kit) 3.5 4.5 5.4
48.9 38.9 26.9
FI (CD19)
FI (I
gM)
28.8 20.5 15.4
23.5 22.8 16.7
IgHwt/wt IgHVDJ-/VDJ-
37.142.5
FI (I
gM)
FI (CD19)
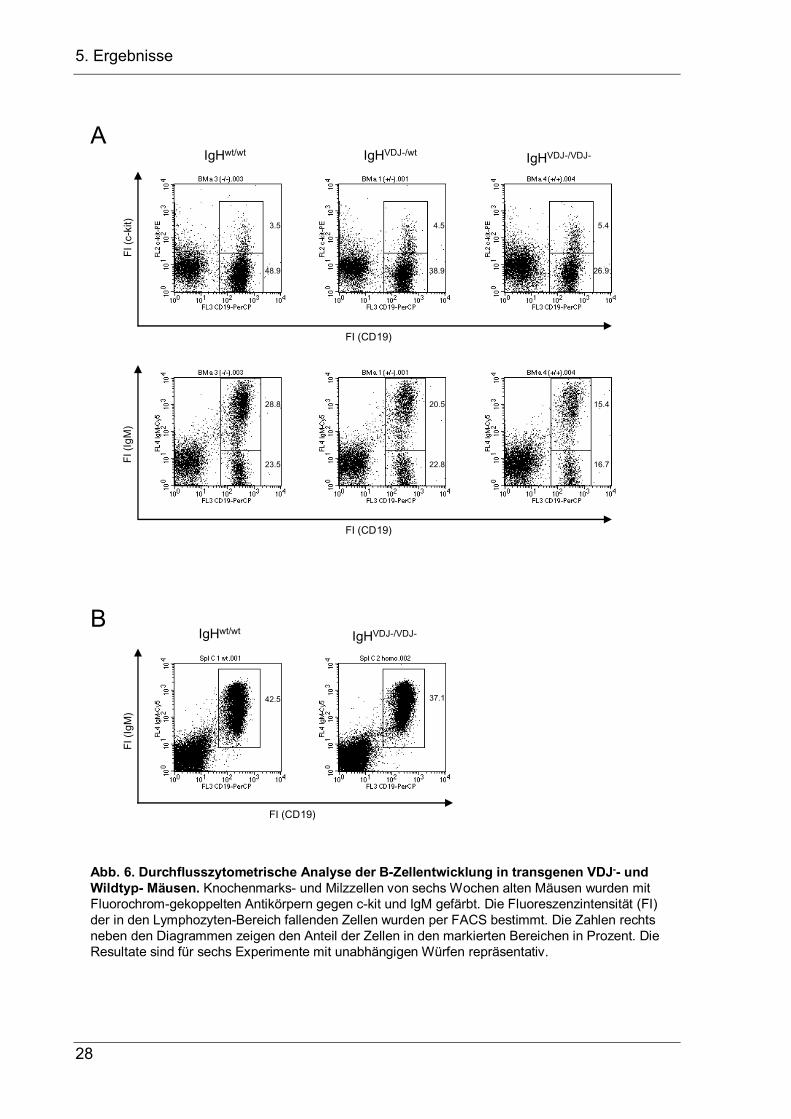
Abb. 6. Durchflusszytometrische Analyse der B-Zellentwicklung in transgenen VDJ-- und Wildtyp- Mäusen. Knochenmarks- und Milzzellen von sechs Wochen alten Mäusen wurden mit Fluorochrom-gekoppelten Antikörpern gegen c-kit und IgM gefärbt. Die Fluoreszenzintensität (FI) der in den Lymphozyten-Bereich fallenden Zellen wurden per FACS bestimmt. Die Zahlen rechts neben den Diagrammen zeigen den Anteil der Zellen in den markierten Bereichen in Prozent. Die Resultate sind für sechs Experimente mit unabhängigen Würfen repräsentativ.
28
5. Ergebnisse
IgHwt/wt, RAG2-/-IgHVDJ-/wt, RAG2-/-
11.5
1.3
18.2
9.3
0.052.6 0.0
FI (I
ntra
zellu
läre
s Ig
M)
SSC
IgHwt/wt
FI (CD19)
FI (c
-kit)
3.5
57.0
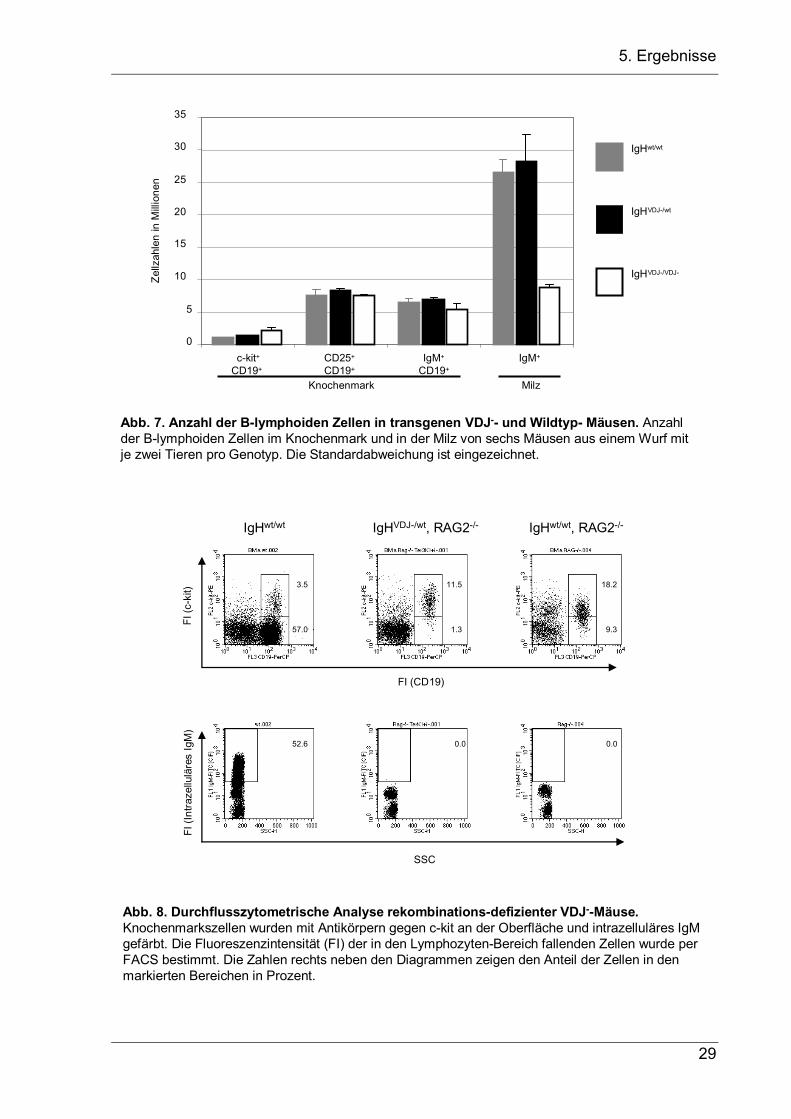
Abb. 8. Durchflusszytometrische Analyse rekombinations-defizienter VDJ--Mäuse.Knochenmarkszellen wurden mit Antikörpern gegen c-kit an der Oberfläche und intrazelluläres Igefärbt. Die Fluoreszenzintensität (FI) der in den Lymphozyten-Bereich fallenden Zellen wurde perFACS bestimmt. Die Zahlen rechts neben den Diagrammen zeigen den Anteil der Zellen in den markierten Bereichen in Prozent.
c-kit+CD19+
CD25+
CD19+IgM+
CD19+
gM
IgHVDJ-/wt
Zellz
ahle
n in
Milli
onen
Knochenmark Milz
0
5
10
15
20
25
30
35
IgM+
IgHVDJ-/VDJ-
IgHwt/wt
Abb. 7. Anzahl der B-lymphoiden Zellen in transgenen VDJ-- und Wildtyp- Mäusen. Anzahlder B-lymphoiden Zellen im Knochenmark und in der Milz von sechs Mäusen aus einem Wurf mje zwei Tieren pro Genotyp. Die Standardabweichung ist eingezeichnet.
it
29

5. Ergebnisse
6B) solcher Tiere nachzuweisen. In homozygoten Mäusen ist im Vergleich zu
Wildtyp-Tieren aus dem gleichen Wurf die Zahl der Pro-B-Zellen ungefähr auf das
Doppelte angestiegen, während die Zahl der CD19/CD25-doppelpositiven Prä-B- und
unreifen B-Zellen gleich blieb, und die Zahl der reifen Oberflächen-IgM-positiven
Zellen im Knochenmark leicht reduziert war (Abb. 7). Im Gegensatz zu diesem
leichten Phänotyp im Knochenmark war die Gesamtzellzahl und die Anzahl der
Oberflächen-IgM-positiven Zellen in der Milz deutlich reduziert auf 41%
beziehungsweise 33% der in Wildtyp-Tieren beobachteten Werte (Abb. 7). Bei der
Analyse der Marginalzonen und follikulären B-Zellen in der Milz und der peritonealen
B1-Population wurden keine reproduzierbaren Unterschiede gemessen (Daten nicht
gezeigt).
Um zu zeigen, dass das nicht-produktive VDJ–-Transgen nicht doch für eine
möglicherweise verkürzte µH-Kette kodiert, wurde das Transgen in einen
rekombinations-defizienten Rag2–/–-Hintergrund gekreuzt (Abb. 8). Das vollständige
Fehlen reifer B-Zellen und einer Differenzierung jenseits des Pro-B-Zellstadiums
sowie die Abwesenheit intrazelluläre µH-Ketten zeigen eindeutig, dass die
Produktion von µH-Ketten-positiven Zellen in RAG-exprimierenden homozygoten
IgHVDJ–/VDJ– Mäusen auf einer RAG-vermittelten Modifikation des Transgens beruht,
wahrscheinlich durch VH-Replacement. Gleichzeitig schließen diese Daten RAG-
unabhängige Mechanismen als Quelle produktiver µH-Ketten-Gene aus und belegen,
dass es nicht zu einer Reinitiation der Translation und der Expression einer
verkürzten µH-Kette vom Transgen kommt.
5.1.3 Sequenzanalyse produktiver µH-Ketten-Gene
Um zu überprüfen, ob die produktiven VDJ-Rearrangements in µH-Ketten-positiven
Zellen in homozygoten IgHVDJ–/VDJ–-Mäusen durch VH-Replacement des Transgens
erzeugt wurden, wurden B-Zellen unterschiedlicher Entwicklungsstufen sortiert und
deren Gesamt-RNA mit oligo-dT-Primern und dem SuperScriptIII Kit (Invitrogen) in
cDNA umgeschrieben. Dann wurden VH-Sequenzen mit einem Vorwärtsprimer, der
entweder an Sequenzen in der D-proximalen VH7183-Familie oder der distalen
VHJ558-Familie bindet, und einem Rückwärtsprimer im Cµ1-Exon amplifiziert. Alle
30

5. Ergebnisse
Akryptisches Heptamer Rekombinationssignalsequenzen
TACTATGCTATGGACTACTGGGGTCCTACTACAGGTAC AAGTATGGTATACTGTGCAAGACAA7183-1.06
TransgenN/PDN/PVH#
TACTATGCTATGGACTACTGGGGTCCTTAGGTAC GCCGACTGAGCCTACTGTGCAAGGGA7183-1.04TACTATGCTATGGACTACTGGGGTCCTACTATAGGTACGAGACTACGGTAGAGGGGTACTGTGCAAGAC7183-1.05
GGGA
GAGCTT
GGTTACGAC
CTACGGTAGTAGTATGGCAACTAC TACTATGCTATGGACTACTGGGGTCCTTACTATAGGTACTAGACA TACTGTGCAAG7183-1.02
TACTATGCTATGGACTACTGGGGTCCTTACTATAGGTACCTACTGTGCAAGA7183-1.03
TACTATGCTATGGACTACTGGGGTCCTTACTATAGGTACGAGTACTGTGCAAGJ558-5.13
JH4NDSP2.3VHTACTATGCTATGGACTACTGGGGTCCTTACTATAGGTACCAACACAGCCTACCTGCAGCTCAGCAGCCTGACATCTGAGGACACTGCCGTCTATTACTGTGCTAGAVDJ-
TACTATGCTATGGACTACTGGGGTCCTGTACAAAGCTCAACAGCCTGACATCTGAGGATTCTGCAGTGTATTTCTGTAJ558-5.14
TransgenN/PVH#
TACTATGCTATGGACTACTGGGGTCCTACTATAGGTAC ATGATGAGCAGTCTGAGGTCTGAGGACACAGCCTTGTATTACTGTGCAAGAC7183-3.04TACTATGCTATGGACTACTGGGGTCCTTACTCATGAGCAGTCTGAGGTCTGAGGACACGGCCATGTATTACTGTGCAAGAC7183-4.65
GGGG
CATGCCCC TACTATGCTATGGACTACTGGGGTCCTGGTAC ATGAGCAGTCTGAGGTCTGAGGACACAGCCATGTACTACTGTGCAAG7183-1.01
TACTATGCTATGGACTACTGGGGTCCTTACTATAGGTAC ATGAGCAGTCTGAAGTCTGAGGACACAGCCATGTATTACTGTGCAAGA7183-1.29
GCTATGGACTACTGGGGTATGAGCAGTCTGAAGTCTGAGGACACAGCCATGTATTACTGTGCAAGACG7183-4.19
a
b
Schnittstellekryptisches Heptamer
DSP2.3 JH4VH
DSP2.3 JH4VH DVH D VH DSP2.3 JH4
VH D VH DSP2.3 JH4
Direktes V-nach-VDJ-Replacement
Zweistufiges VD-nach-VDJ-Replacement
B
CDR3-H
a
b
Abb. 9. VH-Replacement auf dem VDJ--Allel. (A) Das inserierte VDJ--Transgen wird 3' von dem kryptischen Heptamer TACTGTG (fett in B) geschnitten und über Rekombinationssignalsequenzen (RSS) entweder (a) direkt mit einem stromaufwärts gelegenen VH-Segment (dunkelgrau) oder (b)in zwei Schritten mit einem D (hellgrau) und einem VH-Segment verknüpft. (B) Vergleich des unveränderten VDJ--Allels mit produktiven VH-Sequenzen aus IgHVDJ-/VDJ--Tieren. VDJ-Rearrangements wurden mittels RT-PCR mit einem für VH7183- oder VHJ558-Familienmitglieder spezifischen Primer amplifiziert. (a) Bei sechs von 114 Sequenzen wurde die transgene VH-Regiondurch ein stromaufwärts gelegenes VH-Segment ersetzt, ein Vorgang, der für das VH-Replacementin Wildtyp-Mäusen prototypisch ist. (b) Allerdings wurde in der Mehrheit der Sequenzen (108 von 114 Sequenzen) die transgene VH-Region durch ein stromaufwärts gelegenes VH- und D-Segment ersetzt. Sechs repräsentative Sequenzen werden gezeigt. In den meisten Rearrangements wurden N- und P-Nukleotide zu den Bruchstellen hinzugefügt und Basen 3' von der Rekombinationsstelleentfernt. Die CDR3-H-Region zwischen dem TGT im VH- und dem TGG im JH-Segment ist durch gestrichelte Linien angezeigt.
31

5. Ergebnisse
114 analysierten Sequenzen hatten das VH-Segment des transgenen VDJ–-Exons
durch ein stromaufwärts liegendes Ig-Segment ersetzt. Für das VH-Replacement
wurden die kryptische RSS TACTGTG im transgenen VDJ–-Exon und die reguläre
RSS des stromaufwärts liegenden Keimbahn-D- oder -VH-Segments benutzt (Abb.
9). Die Verknüpfungen waren zusätzlich durch das Hinzufügen von nontemplated (N-
) und palindromischen (P-) Nukleotiden, sowie durch das Entfernen von Nukleotiden
am 5’-Ende des verbleibenden VDJ–-Exons verändert worden.
Die produktiven VDJ-Sequenzen konnten in zwei Gruppen eingeteilt werden. Die
kleinere Gruppe (6 von 114 Sequenzen) hatte das VH-Segment des transgenen VDJ-
Exons durch ein Keimbahn-VH-Segment ersetzt und stellt den Prototyp für ein VH-
Replacement in Wildtyp-Mäusen dar (Abb. 9B,a). Die größere Gruppe (108 von 114
Sequenzen) hatte hingegen das VH-Segment durch ein VH- und ein D-Segment
ersetzt (Abb. 9B,b). Letzteres ist möglich, da in dieser Maus die D-Segmente noch
stromaufwärts vom VDJ–-Exon vorliegen und zur Rekombination benutzt werden
können, während sie beim regulären VDJ-Rearrangement deletiert werden.
5.1.4 Zeitpunkt des VH-Replacements
In Wildtyp-B-Zellen kann ein VH-Replacement erst in späten Pro-B-Zellen stattfinden,
nachdem die VH-Gensegmente geöffnet und das primäre VDJ-Exon erzeugt wurde.
In der VDJ–-transgenen Maus hingegen könnte bereits im frühen Pro-B-Zellstadium,
in dem normalerweise die D-nach-J-Umlagerung an beiden IgH-Lozi vollzogen wird,
ein unvollständiges D-nach-VDJ-Replacement stattfindet, dem später ein normales
V-nach-DJ-Rearrangement folgt. Alternativ dazu könnte das VDJ–-Allel auch erst im
späten Pro-B-Zellstadium parallel zu einer V-nach-DJ-Umlagerung am Wildtyp-Lokus
ersetzt werden. Da durch den Wechsel zwischen den Allelen nach einem nicht-
produktiven Rearrangement identische genetische Konfigurationen auf
unterschiedlichen Wegen erzeugt werden können, kann nur die vereinfachte Frage
gestellt werden, ob VH-Replacements in heterozygoten IgHVDJ–/wt B-Zellen bereits vor
der V-nach-DJ-Rekombination des Wildtyp-Lokus stattfinden, oder ob sie erst in
Zellen mit zwei nicht-produktiv rearrangierten Allelen stattfinden, wie man es in
IgHwt/wt B-Zellen erwarten würde.
32

5. Ergebnisse
Abb. 10. Einzelzell-PCR zur Bestimmung der Konfiguration der IgH-Lozi. (A) Nachweis einer Sequenz 5‘ vom ersten D-Lokus (DFL16.1), die nur vorhanden ist, wenn (a) das Wildtyp-Allel noch kein V-nach-DJ Rearrangement vollzogen hat oder (b) das VH-Segment im VDJ--Allel nicht durch ein Keimbahn-VH-Segment ersetzt wurde. Die Primerpaare sind als Pfeile dargestellt. (c) Eine repräsentative PCR von DNA aus sortierten IgMa- bzw. IgMb-positiven Milz-B-Zellen ist gezeigt. Ausgewertet wurden je drei einzelne IgMa+- oder IgMb+-Zellen. Die erste Spur enthält je 10 IgMa- und 10 IgMb-Zellen als Positivkontrolle, die letzte Spur ist eine DNA-Isolierung aus einer Mikrotitervertiefung ohne Zelle und dient als Negativkontrolle. (B) Nachweis des unveränderten VDJ-
-Allels (a) Lage der verwendeten PCR-Primer. (b) IgMa- und IgMb-positive Milz-B-Zellen wurden wie in (Ac) sortiert und das nicht-ersetzte VDJ--Allel per PCR nachgewiesen. (C) Tabellarische Auswertung von je 18 IgMa+- und IgMb+-Zellen. Konfigurationen, die nicht möglich sind, wurden mit einem Strich gekennzeichnet.
-
-
+
+
VDJ--PCR
-
-
+
+
VDJ--PCR Konfiguration der IgH-Lozi
Zellen5' of DFL-PCR
Konfiguration der IgH-Lozi
Zellen5' of DFL-PCR
-0--0-
IgMa: VDDJ-
IgMb: VDJ+
IgMa: DDJIgMb: VDJ+
IgMa: VDJ-
IgMb: VDJ+
IgMa: VDDJ+
IgMb: VDJ-
IgMa: VDDJ+
IgMb: DJ
-
1-10-
2+8+
15+0+
µb+-Zellenµa+-Zellen
A
B
C
a
b Nachweis eines nicht-ersetzten VDJ--Allels
Nachweis eines nicht V-DJ-rearrangierten Wildtyp-Allels
Nachweis eines nicht-ersetzten VDJ--Allels
c
ba
1 1 1 1 1 0120µa+ µb+
µa+µb+
Zellen
VH D JH CµbDFL
16.1
1 1 1 1 1 0120µa+ µb+
µa+µb+
Zellen
5’ of DFL16.1-PCR
VH D VDJ- Cµa
5’ of DFL16.1-PCR
DFL
16.1
VDJ--PCR
VH D VDJ- CµaDFL
16.1
33

5. Ergebnisse
Um diese Frage zu beantworten, wurde mit der Einzelzell-PCR-Technik der Anteil
der Zellen bestimmt, die ein produktives VH-Replacement des Transgens vor einem
V-nach-DJ-Rearrangement am Wildtyp-Lokus durchgeführt haben. Hierzu wurden
IgM-positive B-Zellen aus einer heterozygoten IgHVDJ–/wt Maus benutzt, bei denen die
beiden IgH-Allele durch ihren Allotyp in der Cµ-Region unterschieden werden können
(Abb. 10). In diesen Tieren haben die IgM+-B-Zellen entweder ein VH-Replacement
auf dem transgenen IgMa-Allel durchgeführt oder das Wildtyp-IgMb-Allel rearrangiert.
Für die Analyse wurden Milzzellen mit Antikörpern gegen IgMa und IgMb gefärbt, und
einzelne Zellen mit einem FACS-Sorter in PCR-Platten sortiert. Die Zellen wurden
dann mit Proteinase K verdaut und per Einzelzell-PCR die Anwesenheit einer
Keimbahnsequenz direkt stromaufwärts vom D-Lokus (DFL16.1-Gensegment)
nachgewiesen (Abb. 10A). Diese Sequenz wird bei jedem V-nach-DJ-
Rearrangement entfernt und ist deshalb ein Indikator für ein unrearrangiertes oder
DJ-rearrangiertes Allel. Zusätzlich wurde das nicht-ersetzte VDJ–-Exon amplifiziert
(Abb. 10B). Die Effizienz und Spezifität der Einzelzell-PCR wurde in einem
Vorversuch mit einer Zelllinie überprüft und in jedem Experiment folgendermaßen
kontrolliert: Erstens mussten alle IgMa-positiven Zellen negativ für das VDJ–-Signal
sein, da diese Sequenz durch das VH-Replacement entfernt wird; und zweitens
mussten alle Zellen mit einem VDJ–-Signal auch ein Signal für die Sequenz
stromaufwärts vom DLF16.1-Gensegment haben, da diese Sequenz auch oberhalb
des unveränderten VDJ–-Lokus’ liegt. Diese Bedingungen wurden bei allen PCRs
erfüllt.
Die Einzelzell-PCR-Analyse (Abb. 10C) ergab, dass acht von 18 IgMa-postiven Zellen
auf dem Wildtyp-Lokus noch kein V-nach-DJ-Rearrangement durchgeführt hatten. In
diesen Zellen hatte das VH-Replacement des Transgens also schon vor der V-nach-
DJ-Rekombination des Wildtyp-Allels stattgefunden, möglicherweise im frühen Pro-B-
Zellstadium während der D-nach-J-Rekombination. Bei einem VH-Replacement
während der D-nach-J-Rekombination würden aus Zellen mit einem VDJ–- und einem
Keimbahn-Allel erst Zellen mit einem D-nach-VDJ-ersetzten DDJ-Allel und einem
Wildtyp-DJ-Allel entstehen, die dann auf einem der beiden Allele V-nach-DJ-
rekombinieren können. In Einklang mit dieser Interpretation steht, dass zwei von 18
IgMb-positiven Zellen ein unvollständiges D-nach-VDJ-Replacement auf dem IgMa-
Lokus hatten. Allerdings sind die Daten nur schwer zu interpretieren, da das VDJ–-
34

5. Ergebnisse
Exon einen gravierenden Einfluss auf die B-Zellentwicklung haben könnte. So könnte
es mit seiner Promotor/Enhancer-Aktivität den Lokus öffnen und gegebenenfalls für
eine höhere Rekombinationsfrequenz des veränderten Allels in einem früheren B-
Zellstadium sorgen.
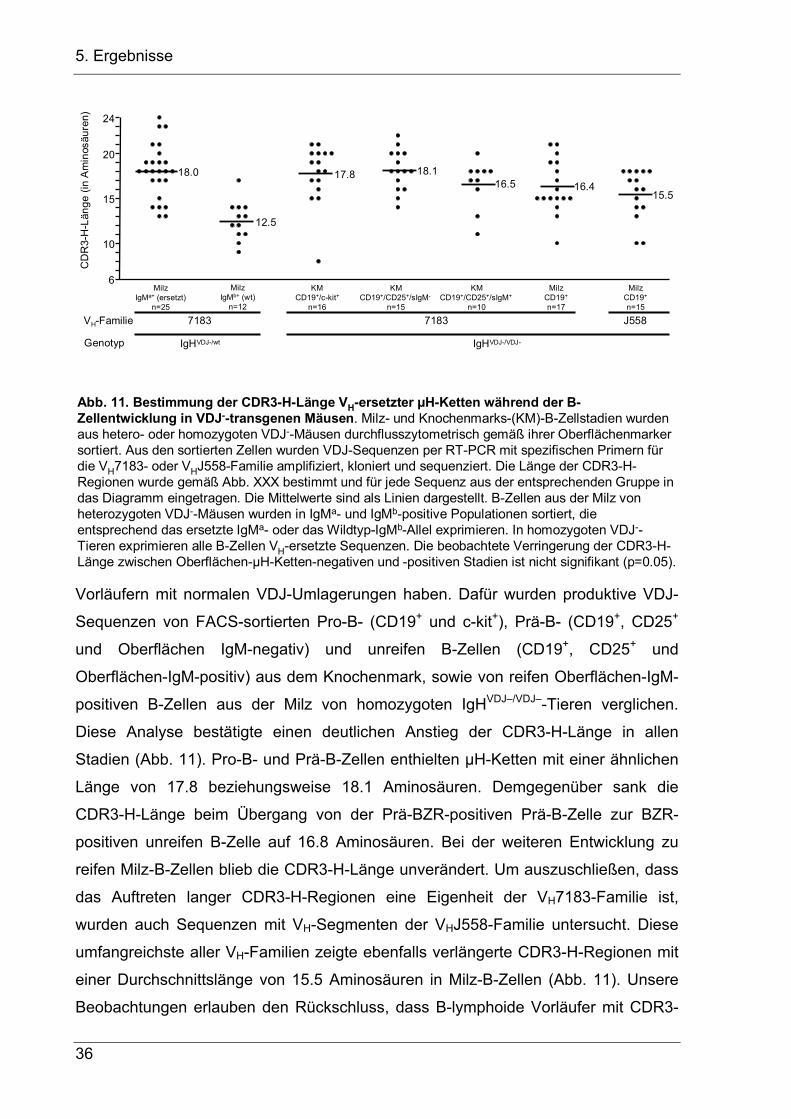
5.1.5 B-Zellentwicklung in der Gegenwart langer CDR-3H-Regionen
Da die meisten VH-Replacements auf dem VDJ–-Allel ein zweites D-Segment
enthalten, sollten sie für µH-Ketten kodieren, die im Vergleich zu µH-Ketten aus
Wildtyp-Mäusen deutlich längeren CDR3-H-Regionen besitzen. Um diese Frage zu
klären, wurden in produktiven VDJ-Sequenzen aus Milz-B-Zellen von heterozygoten
IgHVDJ–/wt-Mäusen die Länge der CDR3-H-Regionen bestimmt. Dazu wurden B-Zellen
im FACS-Sorter anhand der exprimierten µH-Ketten in eine IgMa-positive Population
mit VH-Replacement und in eine IgMb-positive Population mit normal rekombinierten
Wildtyp-VDJ-Rearrangements getrennt. Aus der Gesamt-RNA der Zellen wurden
dann VH-Sequenzen mit RT-PCR amplifiziert, kloniert und sequenziert. Da die
Änderungen in der Länge der CDR3-H-Region bei Sequenzen der VH7183-Familie
während der B-Zellentwicklung bereits ausgiebig untersucht worden sind (Ivanov et
al. 2005), wurde ein Vorwärtsprimer für diese D-proximale VH-Familie benutzt. VDJ-
Sequenzen mit dem VH7183-Familienmitglied VH81x wurden von der Analyse
ausgeschlossen, da IgH-Ketten mit VH81x-Sequenzen oft nicht mit der SL-Kette und
konventionellen leichten Ketten paaren können (Keyna et al. 1995; Kline et al. 1998).
Da alle VDJ-Sequenzen mit VH-Replacement das JH4-Segment des Transgens
enthalten, wurde als Rückwärtsprimer ein JH4-Primer benutzt, so dass alle
verglichenen Sequenzen das gleiche JH-Segment benutzen. Die analysierten
Sequenzen von IgMa-positiven B-Zellen mit einem VH-ersetzten V-Exon enthielten
CDR3-H-Regionen mit einer durchschnittlichen Länge von 18.0 Aminosäuren.
Demgegenüber hatten Sequenzen von IgMb-positiven B-Zellen mit einem normalen
VDJ-Wildtyp-Rearrangement CDR3-H-Regionen mit einer durchschnittlichen Länge
von 12.5 Aminosäuren (Abb. 11), ein Wert, der mit publizierten Befunde für VH7183-
Sequenzen in Wildtyp-Mäusen sehr gut übereinstimmt (Ivanov et al. 2005).
Als nächstes wurde die Frage untersucht, ob während der B-Zellreifung B-lymphoide
Vorläufer mit langen CDR3-H-Regionen einen Differenzierungsnachteil gegenüber
35
5. Ergebnisse
C
DR
3-H
-Län
ge (i
n A
min
osäu
ren)
7183 J5587183
IgHVDJ-/VDJ-IgHVDJ-/wt
VH-Familie
Genotyp
KMCD19+/c-kit+
n=16
KMCD19+/CD25+/sIgM-
n=15
KMCD19+/CD25+/sIgM+
n=10
MilzCD19+
n=17
MilzCD19+
n=15
MilzIgMa+ (ersetzt)
n=25
MilzIgMb+ (wt)
n=12
17.8 18.116.5 16.4
15.5
18.0
12.5
6
10
15
20
24
Abb. 11. Bestimmung der CDR3-H-Länge VH-ersetzter µH-Ketten während der B-Zellentwicklung in VDJ--transgenen Mäusen. Milz- und Knochenmarks-(KM)-B-Zellstadien wurden aus hetero- oder homozygoten VDJ--Mäusen durchflusszytometrisch gemäß ihrer Oberflächenmarker sortiert. Aus den sortierten Zellen wurden VDJ-Sequenzen per RT-PCR mit spezifischen Primern für die VH7183- oder VHJ558-Familie amplifiziert, kloniert und sequenziert. Die Länge der CDR3-H-Regionen wurde gemäß Abb. XXX bestimmt und für jede Sequenz aus der entsprechenden Gruppe in das Diagramm eingetragen. Die Mittelwerte sind als Linien dargestellt. B-Zellen aus der Milz von heterozygoten VDJ--Mäusen wurden in IgMa- und IgMb-positive Populationen sortiert, die entsprechend das ersetzte IgMa- oder das Wildtyp-IgMb-Allel exprimieren. In homozygoten VDJ--Tieren exprimieren alle B-Zellen VH-ersetzte Sequenzen. Die beobachtete Verringerung der CDR3-H-Länge zwischen Oberflächen-µH-Ketten-negativen und -positiven Stadien ist nicht signifikant (p=0.05).
Vorläufern mit normalen VDJ-Umlagerungen haben. Dafür wurden produktive VDJ-
Sequenzen von FACS-sortierten Pro-B- (CD19+ und c-kit+), Prä-B- (CD19+, CD25+
und Oberflächen IgM-negativ) und unreifen B-Zellen (CD19+, CD25+ und
Oberflächen-IgM-positiv) aus dem Knochenmark, sowie von reifen Oberflächen-IgM-
positiven B-Zellen aus der Milz von homozygoten IgHVDJ–/VDJ–-Tieren verglichen.
Diese Analyse bestätigte einen deutlichen Anstieg der CDR3-H-Länge in allen
Stadien (Abb. 11). Pro-B- und Prä-B-Zellen enthielten µH-Ketten mit einer ähnlichen
Länge von 17.8 beziehungsweise 18.1 Aminosäuren. Demgegenüber sank die
CDR3-H-Länge beim Übergang von der Prä-BZR-positiven Prä-B-Zelle zur BZR-
positiven unreifen B-Zelle auf 16.8 Aminosäuren. Bei der weiteren Entwicklung zu
reifen Milz-B-Zellen blieb die CDR3-H-Länge unverändert. Um auszuschließen, dass
das Auftreten langer CDR3-H-Regionen eine Eigenheit der VH7183-Familie ist,
wurden auch Sequenzen mit VH-Segmenten der VHJ558-Familie untersucht. Diese
umfangreichste aller VH-Familien zeigte ebenfalls verlängerte CDR3-H-Regionen mit
einer Durchschnittslänge von 15.5 Aminosäuren in Milz-B-Zellen (Abb. 11). Unsere
Beobachtungen erlauben den Rückschluss, dass B-lymphoide Vorläufer mit CDR3-
36

5. Ergebnisse
A B
IgM
a+al
s %
der
IgM
+
10
0
5
15
KM MilzFI
(IgM
b , W
ildty
p-Al
lel)
FI (IgMa, ersetztes Allel)
2.8 5.0
48.017.7
Knochenmark Milz
Abb. 12. Durchflusszytometrische Bestimmung der Frequenzen von IgM-positiven B-Zellen mit VH-ersetzten und Wildtyp-VDJ-Exons in heterozygoten VDJ--Mäusen. (A) Knochenmarks-und Milz-B-Zellen wurden mit Fluoreszenz-markierten Antikörpern gegen Oberflächen-IgM gefärbt, die zwischen VH-ersetzten µH-Ketten mit dem Allotyp des VDJ--Allels (IgMa) und Wildtyp-µH-Ketten mit dem Allotyp des Wildtyp-Allels (IgMb) unterscheiden können. (B) Frequenzen der IgMa-positivenZellen in sechs heterozygoten VDJ--Mäuse aus zwei unabhängigen Würfen. Die Mittelwerte sind als Striche eingetragen.
H-Regionen mit einer Länge von mehr als 16 Aminosäuren sich in reife B-Zellen
entwickeln können und nur eine geringfügige Gegenselektion beim Übergang vom
Prä-B- zum unreifen B-Zellstadium, möglicherweise über die Paarung mit IgL-Ketten,
erfahren.
5.1.6 Konkurrenz zwischen B-Zellen mit einem VH-ersetzten und B-Zellen mit einem regulär rekombinierten VDJ-Exon in heterozygoten IgHVDJ–/wt-Mäusen
Um die Differenzierung von B-Zellen mit langen CDR3-H-Regionen und B-Zellen mit
CDR3-H-Regionen mit normaler Länge direkt zu vergleichen, wurden wieder
heterozygote IgHVDJ–/wt-Mäuse untersucht. Wie oben beschrieben, können Zellen, die
ein VH-ersetztes transgenes IgMa-Allel mit einer langen CDR3-H-Region exprimieren,
durch ihren Allotyp in der Cµ-Region von Zellen mit einem Wildtyp-IgMb-Allel mit
normaler CDR3-H-Länge unterschieden werden. Falls B-Zellen mit einem BZR mit
einer langen CDR3-H-Region einen Nachteil haben, sollte die Frequenz der Zellen,
die das transgene IgMa-Allel exprimieren, in der IgM+-Population beim Übergang vom
unreifen zum reifen B-Zellstadium stark abnehmen. Jedoch sank der Anteil der
IgMa+-Zellen an den IgM+-Zellen beim Übergang vom Knochenmark in die Milz nur
um ein Drittel von 14% auf 9.2% (Abb. 12).
Die Beobachtung, dass B-Zellen mit deutlich längeren CDR3-H-Regionen als den im
Wildtyp-Repertoire gemessenen 11.5 Aminosäuren sich auch in Konkurrenz zu
37
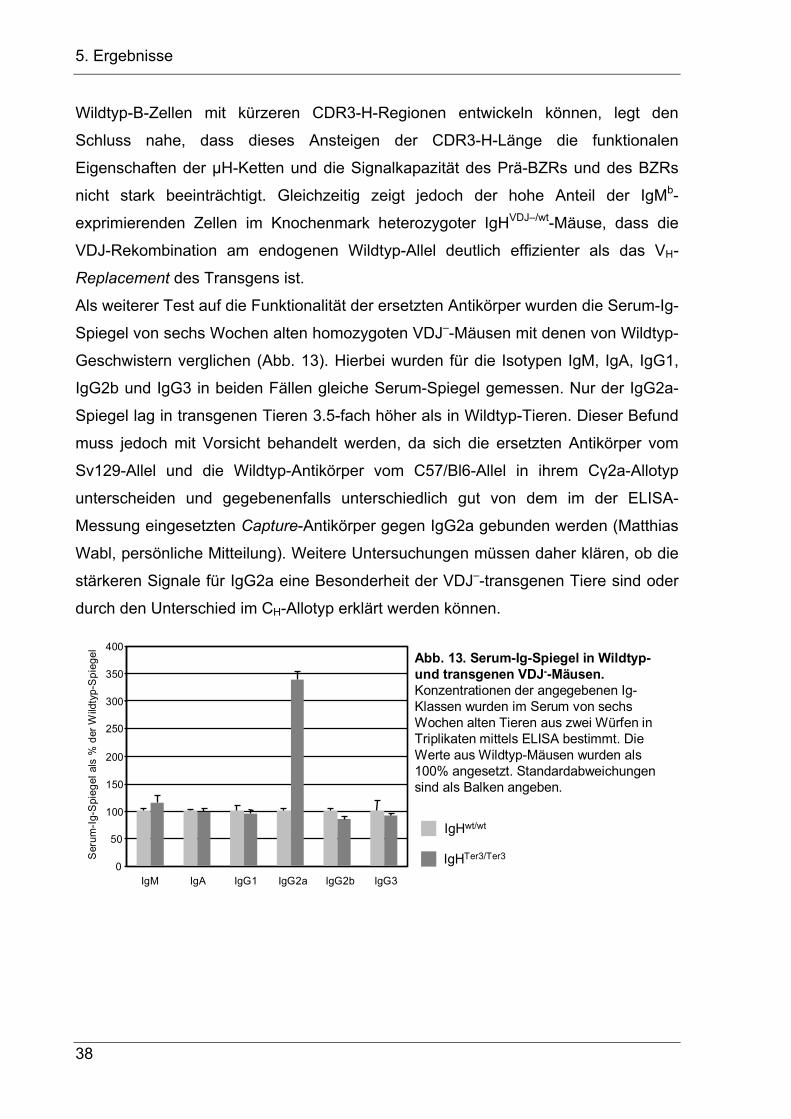
5. Ergebnisse
Wildtyp-B-Zellen mit kürzeren CDR3-H-Regionen entwickeln können, legt den
Schluss nahe, dass dieses Ansteigen der CDR3-H-Länge die funktionalen
Eigenschaften der µH-Ketten und die Signalkapazität des Prä-BZRs und des BZRs
nicht stark beeinträchtigt. Gleichzeitig zeigt jedoch der hohe Anteil der IgMb-
exprimierenden Zellen im Knochenmark heterozygoter IgHVDJ–/wt-Mäuse, dass die
VDJ-Rekombination am endogenen Wildtyp-Allel deutlich effizienter als das VH-
Replacement des Transgens ist.
Als weiterer Test auf die Funktionalität der ersetzten Antikörper wurden die Serum-Ig-
Spiegel von sechs Wochen alten homozygoten VDJ–-Mäusen mit denen von Wildtyp-
Geschwistern verglichen (Abb. 13). Hierbei wurden für die Isotypen IgM, IgA, IgG1,
IgG2b und IgG3 in beiden Fällen gleiche Serum-Spiegel gemessen. Nur der IgG2a-
Spiegel lag in transgenen Tieren 3.5-fach höher als in Wildtyp-Tieren. Dieser Befund
muss jedoch mit Vorsicht behandelt werden, da sich die ersetzten Antikörper vom
Sv129-Allel und die Wildtyp-Antikörper vom C57/Bl6-Allel in ihrem Cγ2a-Allotyp
unterscheiden und gegebenenfalls unterschiedlich gut von dem im der ELISA-
Messung eingesetzten Capture-Antikörper gegen IgG2a gebunden werden (Matthias
Wabl, persönliche Mitteilung). Weitere Untersuchungen müssen daher klären, ob die
stärkeren Signale für IgG2a eine Besonderheit der VDJ–-transgenen Tiere sind oder
durch den Unterschied im CH-Allotyp erklärt werden können.
Abb. 13. Serum-Ig-Spiegel in Wildtyp-und transgenen VDJ--Mäusen.Konzentrationen der angegebenen Ig-Klassen wurden im Serum von sechsWochen alten Tieren aus zwei Würfen in Triplikaten mittels ELISA bestimmt. Die Werte aus Wildtyp-Mäusen wurden als 100% angesetzt. Standardabweichungen sind als Balken angeben.
IgM IgA IgG1 IgG2a IgG2b IgG30
50
100
150
200
250
300
350
400
IgHwt/wt
IgHTer3/Ter3Ser
um-I
g-S
pieg
elal
s %
der
Wild
typ-
Spi
egel
38

5. Ergebnisse
5.2 Kontrolle der VDJ-Rekombination am IgH-Lokus durch eine nicht-produktive µ-mRNA
Ein zentraler Aspekt der Rekombination eines Keimbahn-VH-Gensegments mit einem
bereits umgelagerten DJ-Segment (V-nach-DJ-Rekombination) am IgH-Lokus ist der
Allelausschluss, durch den sichergestellt wird, dass nur eines der beiden IgH-Allele
eine produktive µH-Kette kodiert und die B-Zelle monospezifisch ist. Daher muss das
Rearrangement des zweiten Allels sofort nach einer produktiven Rekombination auf
dem ersten Allel unterdrückt werden. Ein Mechanismus hierfür ist die
Herunterregulierung der RAG-Rekombinations-Maschinerie durch Oberflächen-µH-
Ketten-Signale (Grawunder et al. 1995) und die Schließung der IgH-Lozi, bevor die
Rekombinationsenzyme zur Umlagerung der Leichtketten-Lozi erneut re-exprimiert
werden. Die Notwendigkeit einer Signal-kompetenten µH-Kette für die permanente
Schließung des IgH-Lokus’ für weitere V-nach-DJ-Rekombinationen belegen
Experimenten mit Mäusen, in denen die Prä-BZR-Signalleitung durch Mutationen in
der Membranregion der µH-Kette (µMT, Kitamura and Rajewsky 1992) oder durch
die Deletion oder Mutation der Prä-BZR-Signalkomponenten Igα und Igβ verhindert
wird (Papavasiliou et al. 1995; Gong and Nussenzweig 1996; Minegishi et al. 1999).
In diesen transgenen Zellen ist der Allelausschluss unvollständig, und es können
Zellen mit zwei µH-Ketten detektiert werden. Gleichzeitig sind die Zellen jedoch auch
in ihrer Entwicklung im Pro-B-Zellstadium arretiert und haben dadurch eine
verlängerte Zeitspanne, in der sie V-nach-DJ-Rearrangements durchführen können.
Dieser Entwicklungsblock könnte dazu geführt haben, dass schwächere, kurzfristige
Signale übersehen wurden, die in Wildtyp-Mäusen nach einem produktiven
Rearrangement auf einem Allel die V-nach-DJ-Rekombination auf dem zweiten Allel
solange verhindern, bis das µH-Ketten-Signal einsetzt und die Zelle ins Prä-B-
Zellstadium differenziert.
Hinweise für einen Mechanismus, der unabhängig von einem µH-Ketten-Signal die
VDJ-Rekombination und damit die Differenzierung früher Pro-B-Zellen kontrolliert,
ergaben sich aus Experimenten mit einer transgenen Maus, die eine transport-
inkompetente µH-Kette produziert, die weder das Endoplasmatische Retikulum (ER)
39

5. Ergebnisse
verlassen noch Differenzierungssignale auslösen kann (Kline et al. 1998; Mielenz et
al. 2003). In diesen Tieren war trotz der Gegenwart endogener Wildtyp-IgH-Lozi die
B-Zellentwicklung am Übergang vom Pro-B- zum Prä-B-Zellstadium beeinträchtigt,
was zu einer erhöhten Anzahl an Pro-B-Zellen und weniger Zellen in den B-
lymphoiden Folgestadien führte. Negative Effekte durch die Ansammlung
ungefalteter µH-Ketten und die Induktion der Unfolded-Protein-Response (UPR)
konnten ausgeschlossen werden, da die B-Zellentwicklung durch das Einbringen
einer funktionalen µH-Kette wiederhergestellt werden konnte (Meister 2004). Der
beobachtete Phänotyp ist vergleichbar mit einem partiellen Allelausschluss, bei dem
die VDJ-Rekombination in Pro-B-Zellen reduziert wird, es am Ende aber doch zu
einer Rekombination kommt. Da die transgene µH-Kette jedoch nicht signalisieren
kann, muss ein zweites, Transgen-abhängiges Signal beteiligt sein.
Daher sollte im zweiten Teil der Arbeit untersucht werden, ob schon die
Akkumulierung einer µ-mRNA alleine, d.h. in Abwesenheit eines µH-Ketten-Signals,
die Rate der VDJ-Rekombination am IgH-Lokus reduzieren kann. Die Menge an µ-
mRNA wäre ein idealer Indikator für eine produktive VDJ-Rekombination: Ist ein VDJ-
Rearrangement produktiv, so ist die transkribierte µ-mRNA stabil, liegt deshalb in
hohen Mengen vor und wird translatiert. Ist das VDJ-Rearrangement jedoch nicht-
produktiv, so enthält die transkribierte µ-mRNA ein vorzeitiges Translations-
Stopkodon (Nonsense- beziehungsweise Premature-Termination-Codon, PTC)
stromaufwärts von Exon-Exon-Grenzen in der Cµ-Region. Solche PTC-haltigen
Nonsense-mRNAs werden von der Nonsense-Mediated-mRNA-Decay (NMD)-
Maschinerie erkannt und rasch abgebaut, was zu einer 50- bis 100-fachen
Reduzierung in der Menge an Nonsense-µ-mRNA führt (Baumann et al. 1985; Jack
et al. 1989). Falls die µ-mRNA an der Kontrolle der VDJ-Rekombination am IgH-
Lokus beteiligt wäre, könnte sie die Zeitlücke zwischen dem Entstehen eines
produktiven VDJ-Rearrangement und der Induktion eines Prä-BZR-Signals von der
Oberfläche überbrücken.
40
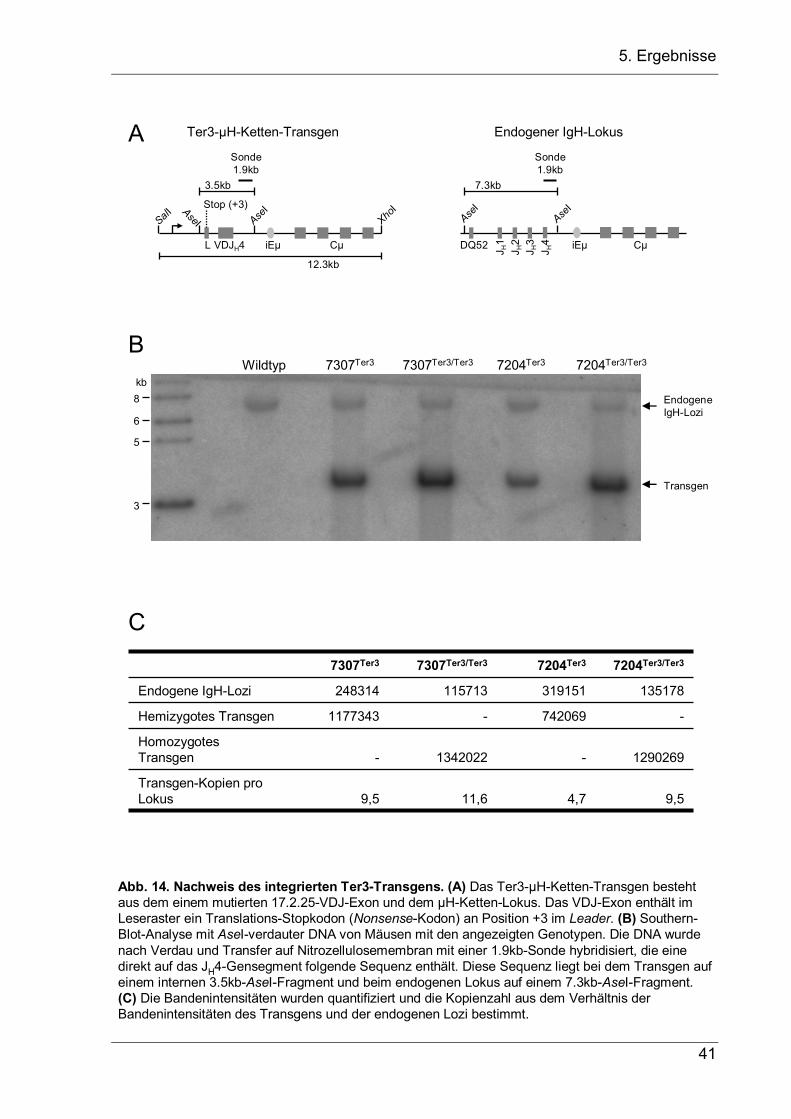
5. Ergebnisse
Ter3-µH-Ketten-TransgenA Endogener IgH-Lokus
B
12.3kb
3.5kb
Sonde1.9kb
L VDJH4
SalI XhoIAseI Ase
I
Cµ
Stop (+3)
iEµ
7.3kb
Sonde1.9kb
DQ52 J H4
AseI
AseI
CµJ H3
J H2
J H1 iEµ
Abb. 14. Nachweis des integrierten Ter3-Transgens. (A) Das Ter3-μH-Ketten-Transgen besteht aus dem einem mutierten 17.2.25-VDJ-Exon und dem µH-Ketten-Lokus. Das VDJ-Exon enthält im Leseraster ein Translations-Stopkodon (Nonsense-Kodon) an Position +3 im Leader. (B) Southern-Blot-Analyse mit AseI-verdauter DNA von Mäusen mit den angezeigten Genotypen. Die DNA wurde nach Verdau und Transfer auf Nitrozellulosemembran mit einer 1.9kb-Sonde hybridisiert, die eine direkt auf das JH4-Gensegment folgende Sequenz enthält. Diese Sequenz liegt bei dem Transgen auf einem internen 3.5kb-AseI-Fragment und beim endogenen Lokus auf einem 7.3kb-AseI-Fragment. (C) Die Bandenintensitäten wurden quantifiziert und die Kopienzahl aus dem Verhältnis der Bandenintensitäten des Transgens und der endogenen Lozi bestimmt.
kb
5
3
6
8
Wildtyp 7307Ter3 7307Ter3/Ter3 7204Ter3 7204Ter3/Ter3
Endogene IgH-Lozi
Transgen
9,54,711,69,5Transgen-Kopien pro Lokus
1290269-1342022-Homozygotes Transgen
-742069-1177343Hemizygotes Transgen
135178319151115713248314Endogene IgH-Lozi
7204Ter3/Ter37204Ter37307Ter3/Ter37307Ter3
C
41

5. Ergebnisse
5.2.1 Etablierung einer transgenen Maus mit einer stabilisierten Nonsense-µ-mRNA
Um die Hypothese zu überprüfen, dass eine stabile µ-mRNA unabhängig von der
Anwesenheit einer µH-Kette weitere Umlagerungen am IgH-Lokus beeinflussen
kann, wurde eine transgene Maus etabliert, die eine stabilisierte Nonsense-µ-mRNA
exprimiert. Ausgehend vom Expressionsvektor pµPGT wurde dazu der transgene
Expressionsvektor pµTer3 konstruiert, in dem das dritte Kodon (+3) im Leader-Peptid
in ein Stopkodon umgewandelt wurde, da ein Stopkodon an dieser Position die
Stabilität der Nonsense-µ-mRNA in Plasmazellen nur um 50% reduziert (Buzina and
Shulman 1999; Buhler et al. 2004). Um eine Re-Initiation der Translation und die
Synthese einer verkürzten µH-Kette zu verhindern, wurden zusätzlich im pµTer3-
Vektor alle im Leseraster vorhandenen ATG-Kodons des VDJ-Exons in Ala-Kodons
umgewandelt. Anschließend wurde das 12.3kb SalI/XhoI-Fragment (Ter3-Transgen,
Abb. 14A), das den VH-Promotor und das mutierte VDJ-Exon des Hybridomas
17.2.25 sowie die genomische Cµ-Region aus Sv129-Mäusen enthält, aus dem
Vektor pµTer3 herausgeschnitten, gelelektrophoretisch isoliert und in den Pronukleus
von C57/Bl6-Oozyten injiziert. Durch Verpaaren der Founder-Mäuse 7204, 7302 und
7307 mit C57/Bl6-Tieren wurden drei unabhängige Ter3-transgene Linien etabliert,
die den gleichen Knochenmarks-Phänotyp zeigten. Von den weitergeführten Linien
7204 und 7307 wurde für die meisten Untersuchungen die 7307-Linie benutzt.
5.2.2 Bestimmung der Kopienzahl und des Integrationsortes des Transgens
Zuerst wurde die Anzahl der integrierten Transgenkopien durch Southern-Blot-
Analyse bestimmt. Dazu wurde DNA aus Schwanzbiopsien aufgereinigt, mit AseI
verdaut, gelelektrophoretisch aufgetrennt, auf eine Membran transferiert und mit
einer 1.9kb langen NaeI/SpeI-Sonde aus dem Vektor pµGPT hybridisiert (Abb. 14A).
Mit dieser Sonde konnte nach dem Verdau mit AseI ein transgenes 3.5kb-Fragment
und ein endogenes 7.3kb-Fragment nachgewiesen werden. Die Anzahl der
integrierten Transgenkopien wurde dann aus dem Verhältnis der Bandenintensitäten
42

5. Ergebnisse
5'-Ende des Transgens
IgMTer113 revMseI MseI (419)
IgMTer386 fwd
IgMTer344 rev
Bruchpunkt
FlankierendeSequenz
MseI
IgMTer113rev
IgMTer344 rev
IgMTer386fwd
FlankierendeSequenz
5'-Ende des Transgens
A B
Abb. 15. Bestimmung des Ter3-Transgen-Integrationsorts durch Zirkel-PCR.(A) Schema eines Insertionspunktes mit den Positionen der für die Zirkel-PCR verwendeten Primer. (B) Konfiguration eines nach MseI-Verdau von Schwanz-DNA und anschließender Ligierungentstandenen DNA-Zirkels mit Bruchpunktsequenz, der neben dem 5'-Ende des Transgens auch die flankierende endogene Sequenz bis zur nächsten MseI-Schnittstelle enthält. Die flankierende Sequenz wurde in einer semi-nested PCR mit den als Pfeilen eingezeichneten Primern amplifiziert, kloniert und sequenziert.
für das transgene 3.5kb-Fragment und für das endogene 7.3kb-Fragment
abgeschätzt (Abb. 14B, C) und ergab für die 7204-Linie 5-10 Transgenkopien und für
die 7307-Linie ca. 10 Transgenkopien.
Um den Einfluss des Integrationsortes auf die VDJ-Rekombination beziehungsweise
auf ein für die B-Zellentwicklung wichtiges Gen auszuschließen, wurden die
Integrationsorte der Transgene in beiden transgenen Mauslinien bestimmt. Dazu
wurde die aufgereinigte Schwanz-DNA mit dem 4bp-erkennenden Enzym MseI
verdaut, und die Fragmente nach Verdünnung zu Zirkeln ligiert (Abb. 15). Da MseI
an einer unbekannten Stelle im Genom stromaufwärts vom inserierten Transgen und
an Position 419 des Transgens schneidet, enthielten einige der entstandenen Zirkel
das 5’-Ende des Transgens und die flankierende genomische Sequenz bis zur
nächsten MseI-Schnittstelle. Diese Bruchpunktsregion wurde mit Primern in der
Transgen-Sequenz (IgMTer386 fwd und IgMTer344 rev oder IgMTer113 rev) in einer
semi-nested PCR amplifiziert, kloniert und sequenziert (Abb. 16). Die genomische
Position der Integrationsorte wurde mit dem Maus-BLAT-Programm der Genome-
Bioinformatics-Group an der UC Santa Cruz (http://genome.ucsc.edu) bestimmt und
basiert auf der "essentially complete" Zusammenstellung des Mus musculus-Genoms
von NCBI und dem Mouse-Genome-Sequencing-Consortium vom Februar 2006
(mm8, Build 36). Die beiden Integrationsorte werden im folgenden als 7204- und
7307-Lokus bezeichnet, und Hemizygotie (Ter3) oder Homozygotie (Ter3/Ter3) der
Transgene entsprechend angezeigt.
43

5. Ergebnisse
A
B
5'-Ende des Transgens
IgMTer 113 rev 3' of 7204 rev5' of 7204 fwd
Sequenzierter Bruchpunkt
MseI (41) MseI (148) MseI (225)
IgMTer 113 rev 3' of Ter7307 rev35' of Ter7307 fwd2
MseI (62) MseI (351)
5'-Ende des Transgens
Sequenzierter Bruchpunkt
C
D
Abb. 16. Bruchpunktsequenzen der inserierten Ter3-Transgene. Der Bruchpunkt in der transgenen Linie 7204 als Schema (A) und Sequenz (B) und in der Linie 7307 als Schema (C) und Sequenz (D). Die zur Genotypisierung verwendeten Primer sind als Pfeile eingezeichnet.
44

5. Ergebnisse
A B Abb.
17.
Anal
yse
der d
as T
rans
gen
7307
flan
kier
ende
n ge
nom
isch
enSe
quen
zen
auf f
unkt
ione
lle G
ene.
(A) I
nteg
ratio
nsor
tdes
Tra
nsge
ns 7
307
auf C
hrom
osom
5 a
n Po
sitio
n 10
.473
.198
. (B
) Pos
ition
en v
on G
enen
, mR
NAs
und
miR
NA
sin
ein
em B
erei
ch v
on 2
38kb
stro
mau
f-un
d ab
wär
ts z
um
Inte
grat
ions
orts
. Der
ver
größ
erte
Ber
eich
ist a
ls ro
ter K
aste
n in
(A)m
arki
ert.
Seq
uenz
anal
yse
und
Dar
stel
lung
erfo
lgte
mit
BLA
T S
earc
hG
enom
e
Inte
grat
ions
ort d
es T
rans
gens
730
7
(http
://ge
nom
e.uc
sc.e
du).
45
5. Ergebnisse
A B Ab
b. 1
8. A
naly
se d
er d
as T
rans
gen
7204
flan
kier
ende
n ge
nom
isch
enSe
quen
zen
auf f
unkt
ione
lle G
ene.
(A) I
nteg
ratio
nsor
tdes
Tra
nsge
ns 7
204
auf C
hrom
osom
5 a
n Po
sitio
n 10
9.97
5.63
3. (B
)Pos
ition
en v
on G
enen
, mR
NA
s un
d m
iRN
Asin
ein
em B
erei
ch v
on 2
38kb
stro
mau
f-un
d ab
wär
ts z
um
Inte
grat
ions
orts
. Der
ver
größ
erte
Ber
eich
ist a
ls ro
ter K
aste
n in
(A) m
arki
ert.
Seq
uenz
anal
yse
und
Dar
stel
lung
erfo
lgte
mit
BLA
T S
earc
hG
enom
e(h
ttp://
geno
me.
ucsc
.edu
).
Inte
grat
ions
ort d
es T
rans
gens
720
4
46

5. Ergebnisse
In der Linie 7307 ist das Ter3-Transgen auf Chromosom 5 an Position 10.473.198
integriert, wo zu beiden Seiten in einem Bereich von 238kb keine Gene oder miRNAs
in Santa-Cruz-Browser angezeigt wurden (Abb. 17). Die Region ist zwar homolog
zum Gen Anapc5 (Anaphase-promoting-complex-subunit 5) im Krallenfrosch
(Xenopus), Zebrafisch (Danio) und Huhn (Gallus), doch liegt der Anapc5-Lokus in
der Maus an einem anderen Ort.
In der Linie 7204 ist das Ter3-Transgen ebenfalls auf Chromosom 5 integriert (Abb.
18). In der Nähe des Integrationsortes an Position 109.975.633 befinden sich zwei
Gene. In direkter Nachbarschaft liegt das uncharakterisierte Gen BC065392 (Similar
to Kruppel-Related-Zinc-Finger-Protein F80-L). Das zweite Gen, Tpte2 (CRLM2,
Tslpr), liegt 175kb stromaufwärts vom Integrationsort und kodiert die Common-γ-
Like-Chain (Fujio et al. 2000), die ähnlich zur Common-γ-Chain des Interleukin-7-
Rezeptors ist. Die Common-γ-Like-Chain bildet als Heterodimer mit der Interleukin-7-
Rezeptor-α-Kette den Thymic-Stromal-Lymphopoietin-Receptor (TSLP-R). Dieser
Rezeptor erkennt das Thymic-Stromal-Lymphopoietin, das ursprünglich aus einer
murinen Thymusstroma-Zelllinie isoliert wurde und die Entwicklung unreifer B-Zellen
in vitro unterstützt (Friend et al. 1994; Levin et al. 1999). Trotz des Abstands von
175kb lässt sich ein Einfluss des Transgens auf die Expression des Tpte2-Lokus’
nicht komplett ausschließen. Allerdings sollte eine verringerte TSLP-R-Expression
keine größeren Auswirkungen auf die B-Zellentwicklung haben, da Mäuse mit einer
Deletion des TSLP-R-Lokus’ eine normale B-Zellentwicklung mit unverändertem Pro-
B-Zellstadium durchlaufen (Carpino et al. 2004). In nächster Nachbarschaft des
7204-Transgens befindet sich zusätzlich eine Region, die homolog zu dem Gen
Golga7 (Golgi-Autoantigen, Golgin Subfamily A, 7) in Ratte, Mensch und anderen
Spezies ist. In der Maus liegt der Golga7-Lokus jedoch an einer anderen Position.
5.2.3 Expressions-Niveau der stabilisierten Ter3-Nonsense-µ-mRNA
Um zu überprüfen, ob die durch das Stopkodon an Position +3 stabilisierte Ter3-
Nonsense-µ-mRNA als Modell für eine endogene Sense-µ-mRNA dienen kann und
in ähnlichen Mengen wie Sense-µ-mRNA vorliegt, wurde zunächst in der transgene
Mauslinie 7307 das Expressionsniveau der Ter3-µ-mRNA mit dem einer endogenen,
47
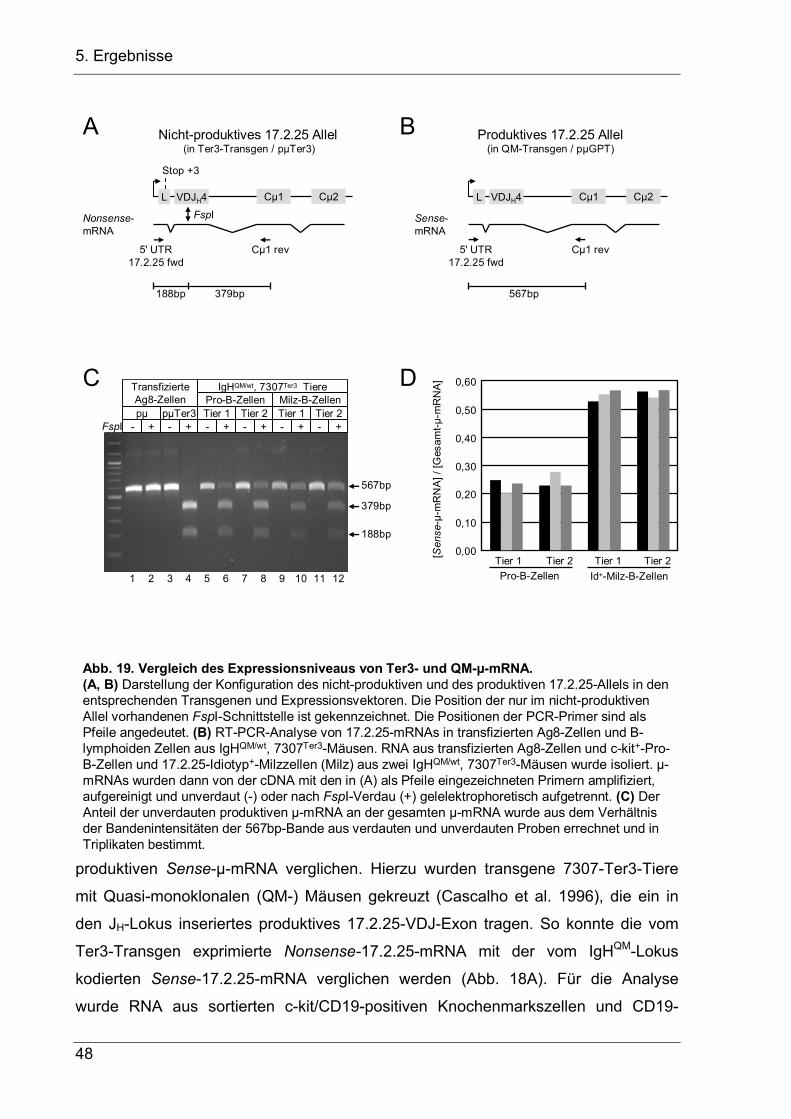
5. Ergebnisse
Produktives 17.2.25 Allel(in QM-Transgen / pµGPT)
Nicht-produktives 17.2.25 Allel(in Ter3-Transgen / pµTer3)
A
C D
Abb. 19. Vergleich des Expressionsniveaus von Ter3- und QM-µ-mRNA.(A, B) Darstellung der Konfiguration des nicht-produktiven und des produktiven 17.2.25-Allels in den entsprechenden Transgenen und Expressionsvektoren. Die Position der nur im nicht-produktiven Allel vorhandenen FspI-Schnittstelle ist gekennzeichnet. Die Positionen der PCR-Primer sind als Pfeile angedeutet. (B) RT-PCR-Analyse von 17.2.25-mRNAs in transfizierten Ag8-Zellen und B-lymphoiden Zellen aus IgHQM/wt, 7307Ter3-Mäusen. RNA aus transfizierten Ag8-Zellen und c-kit+-Pro-B-Zellen und 17.2.25-Idiotyp+-Milzzellen (Milz) aus zwei IgHQM/wt, 7307Ter3-Mäusen wurde isoliert. µ-mRNAs wurden dann von der cDNA mit den in (A) als Pfeile eingezeichneten Primern amplifiziert, aufgereinigt und unverdaut (-) oder nach FspI-Verdau (+) gelelektrophoretisch aufgetrennt. (C) Der Anteil der unverdauten produktiven µ-mRNA an der gesamten µ-mRNA wurde aus dem Verhältnis der Bandenintensitäten der 567bp-Bande aus verdauten und unverdauten Proben errechnet und in Triplikaten bestimmt.
Sense-mRNA
567bp
Cµ1 rev5' UTR17.2.25 fwd
Nonsense-mRNA
0,00
0,10
0,20
0,30
0,40
0,50
0,60
Tier 1 Tier 2 Tier 1 Tier 2Pro-B-Zellen Id+-Milz-B-Zellen
FspI
379bp188bp
Cµ1 rev5' UTR17.2.25 fwd
VDJH4 Cµ2Cµ1L
Stop +3
VDJH4 Cµ2Cµ1L
B
[Sen
se-µ
-mR
NA
] / [G
esam
t-µ-m
RN
A]FspI
Milz-B-ZellenIgHQM/wt, 7307Ter3 Tiere
+-+-+-+-+-+-Tier 2Tier 1Tier 2Tier 1pµTer3pµ
Pro-B-ZellenTransfizierteAg8-Zellen
567bp
379bp
188bp
121110987654321
produktiven Sense-µ-mRNA verglichen. Hierzu wurden transgene 7307-Ter3-Tiere
mit Quasi-monoklonalen (QM-) Mäusen gekreuzt (Cascalho et al. 1996), die ein in
den JH-Lokus inseriertes produktives 17.2.25-VDJ-Exon tragen. So konnte die vom
Ter3-Transgen exprimierte Nonsense-17.2.25-mRNA mit der vom IgHQM-Lokus
kodierten Sense-17.2.25-mRNA verglichen werden (Abb. 18A). Für die Analyse
wurde RNA aus sortierten c-kit/CD19-positiven Knochenmarkszellen und CD19-
48

5. Ergebnisse
/17.2.25-Idiotyp-positiven Milz-B-Zellen von IgHQM/wt, 7307Ter3-Tieren gewonnen und
mit Oligo-dT-Primern in cDNA umgeschrieben. Sense- und Nonsense-Transkripte
wurden dann im gleichen PCR-Ansatz mit Primern amplifiziert, die an die 5’-UTR des
17.2.25-Exons und an die Cµ1-Region binden. Die PCR-Produkte wurden
gelelektrophoretisch aufgereinigt, und die Hälfte der Probe wurde mit dem
Restriktionsenzym FspI verdaut. Da FspI nur in den Nonsense-Amplifikaten
schneidet (Abb. 19A), konnte so der relative Anteil der Nonsense-µ-mRNA an der
Gesamt-µ-mRNA aus dem Verhältnis zwischen den Bandintensitäten der
unverdauten Proben und den Intensitäten der Sense-Amplifikate (567bp in Abb. 19C)
in verdauten Proben errechnet werden (Abb. 19D). Die Validität des diagnostischen
FspI-Verdaus wurde mit cDNA aus Ag8-Hybridomalinien überprüft, die entweder mit
dem Sense-pµGPT- oder dem Nonsense-pµTer3-Plasmid stabil transfiziert worden
waren (Abb. 19C, Spuren 1-4).
Die Analyse ergab etwa dreimal mehr Ter3-Nonsense- als Sense-Transkripte in c-kit-
positiven Pro-B-Zellen und etwa gleiche Mengen beider Transkripte in 17.2.25-
Idiotyp-positiven Milz-B-Zellen. Allerdings ist die scheinbar höhere Expression der
Ter3-Nonsense-µ-mRNA in Pro-B-Zellen wahrscheinlich eine Überschätzung, da ein
signifikanter Anteil an Pro-B-Zellen in IgHQM-Mäusen das QM-Allel durch VH-
Replacement verliert und die Sense-17.2.25-mRNA somit nicht mehr exprimiert
(siehe unten). Bei der Analyse der RNA aus sortierten 17.2.25-Idiotyp-positiven
Milzzellen, die alle das Sense-Transkript exprimieren, sollte dieses Problem nicht
auftreten, weshalb die 17.2.25-Idiotyp-positiven Milzzellen besser zur Abschätzung
der relativen Sense- und Nonsense-µ-mRNA-Mengen geeignet sind. Aus diesen
Ergebnissen kann geschlossen werden, dass in B-lymphoiden Zellen der Ter3-
transgenen 7307-Linie die Menge der Ter3-Nonsense-µ-mRNA in etwa vergleichbar
mit der Menge an endogener Sense-µ-mRNA ist. Die Ter3-transgenen Mäuse sollten
deshalb ein ideales Modellsystem sein, um den Einfluss einer µ-mRNA auf die VDJ-
Rekombination am IgH-Lokus in Abwesenheit einer µH-Kette zu untersuchen.
49

5. Ergebnisse
5.2.4 B-Zellentwicklung in Gegenwart der Ter3-µ-mRNA
Falls die transgene Ter3-µ-mRNA tatsächlich die VDJ-Umlagerung am endogenen
IgH-Lokus negativ beeinflusst, sollte sich in Ter3-transgenen Tieren die Anzahl der
Pro-B-Zellen erhöhen und die Anzahl der Zellen in den darauf folgenden B-
Zellstadien erniedrigen. Zur Überprüfung dieser Vorhersage wurden in den beiden
Ter3-Linien 7307 und 7204 B-lymphoide Differenzierungsstadien im Knochenmark
durchflusszytometrisch analysiert (Abb. 20A, B und 21A, B). Hierbei zeigten Ter3-
transgene Tiere in beiden Linien im Vergleich zu Wildtyp-Mäusen aus dem gleichen
Wurf im Knochenmark eine vergrößerte Pro-B-Zellpopulation und eine Reduktion in
den Folgestadien. In homozygoten Ter3-Mäusen war die Frequenz (Abb. 20, 21) und
die absolute Zellzahl (Tab. 1) der Pro-B-Zellpopulation verdoppelt, während in
heterozygoten Ter3-Mäuse einen intermediären Phänotyp beobachtet wurde.
Gleichzeitig waren die CD25-positiven Prä-B- und IgM-positiven unreifen B-Zellen im
Knochenmark in Frequenz und absoluter Zellzahl in beiden transgenen Ter3-Linien
erniedrigt. In der Milz waren hingegen die IgM-positiven Populationen nicht verändert
(Abb. 22). Ähnliche Ergebnisse wurden auch in der dritten Ter3-transgenen Linie
7302 beobachtet (Daten nicht gezeigt).
Um zu untersuchen, ob die Pro-B-Zellen vor oder nach dem VDJ-Rearrangement in
ihrer Differenzierung beeinträchtigt sind, wurden CD19-/c-kit-postive Pro-B-Zellen
fixiert und für intrazelluläre µH-Ketten angefärbt (Abb. 20C, D, 21C, D). In beiden
transgenen Ter3-Linien war der Anteil der µH-Ketten-positiven Zellen in der Pro-B-
Zellpopulation in homozygoten transgenen Tieren im Vergleich zu Wildtyp-
Geschwistern auf die Hälfte reduziert. Diese Erhöhung des Anteils an µH-Ketten-
negativen Pro-B-Zellen in transgenen Ter3-Mäusen deutet auf eine Störung der VDJ-
Rekombination hin.
Um einen unspezifischen negativen Einfluss des Transgens auf RAG-vermittelte
Rekombinationen auszuschließen, wurde die T-Zellentwicklung und die Umlagerung
der T-Zellrezeptor-(TZR-) Gene im Thymus von Ter3-transgenen und Wildtyp-
Mäusen analysiert (Abb. 23). Diese Analyse ergab keinen signifikanten Unterschied
zwischen Ter3-transgenen und nicht-transgenen Geschwistern in der Frequenz an
doppelt-negativen (DN) Zellen, die den TZR-β-Lokus umlagern und dann den Prä-
50
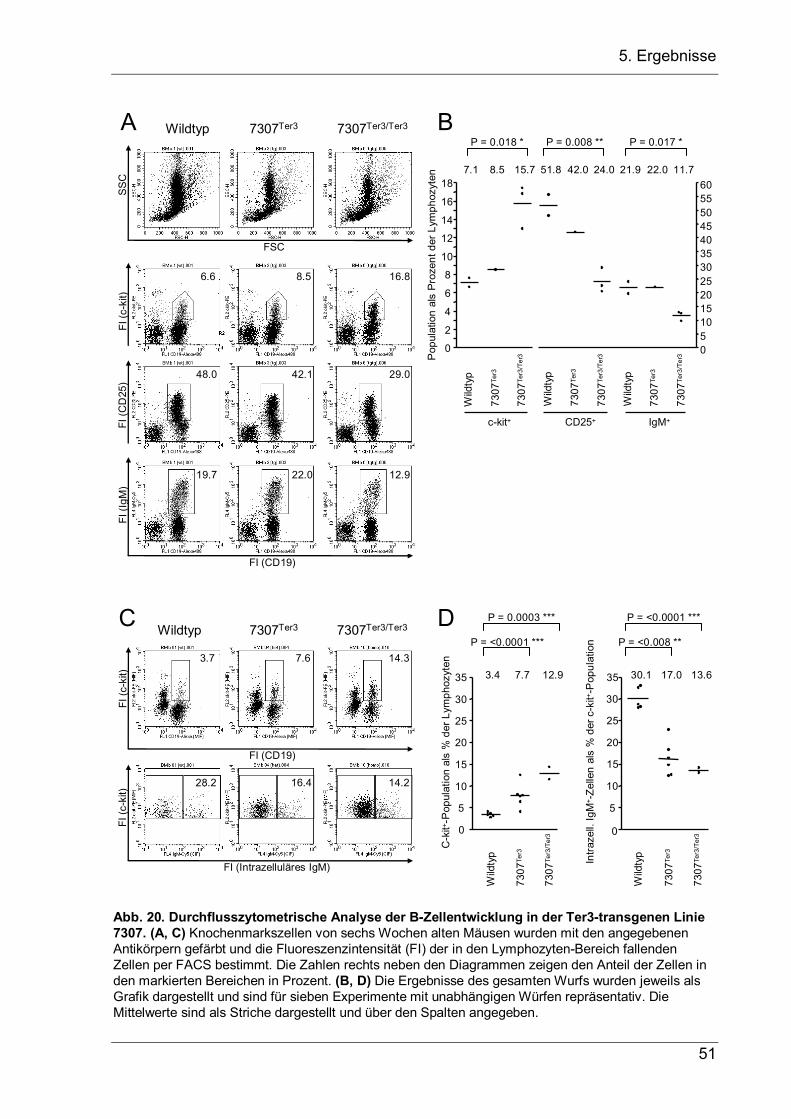
5. Ergebnisse
FI (c
-kit)
FI (CD19)
FI (C
D25
)FI
(IgM
)7307Ter3A Wildtyp 7307Ter3/Ter3
SSC
FSC
FI (c
-kit)
FI (CD19)
6.6 8.5 16.8
19.7 22.0 12.9
48.0 42.1 29.0
7307Ter3C Wildtyp 7307Ter3/Ter3
FI (c
-kit)
FI (Intrazelluläres IgM)
28.2 16.4 14.2
3.7 7.6 14.3
B
D
Abb. 20. Durchflusszytometrische Analyse der B-Zellentwicklung in der Ter3-transgenen Linie 7307. (A, C) Knochenmarkszellen von sechs Wochen alten Mäusen wurden mit den angegebenen Antikörpern gefärbt und die Fluoreszenzintensität (FI) der in den Lymphozyten-Bereich fallenden Zellen per FACS bestimmt. Die Zahlen rechts neben den Diagrammen zeigen den Anteil der Zellen in den markierten Bereichen in Prozent. (B, D) Die Ergebnisse des gesamten Wurfs wurden jeweils als Grafik dargestellt und sind für sieben Experimente mit unabhängigen Würfen repräsentativ. Die Mittelwerte sind als Striche dargestellt und über den Spalten angegeben.
P = 0.018 * P = 0.008 ** P = 0.017 *
0
2
4
6
8
10
12
14
16
18
051015202530354045505560
c-kit+ CD25+ IgM+
7.1 8.5 15.7 51.8 42.0 24.0 21.9 22.0 11.7
7307
Ter3
/Ter
3
7307
Ter3
Wild
typ
7307
Ter3
/Ter
3
7307
Ter3
Wild
typ
7307
Ter3
/Ter
3
7307
Ter3
Wild
typ
Pop
ulat
ion
als
Proz
ent d
er L
ymph
ozyt
en
P = 0.0003 *** P = <0.0001 ***
0
5
10
15
20
25
30
35 3.4 7.7 12.9
7307
Ter3
/Ter
3
7307
Ter3
Wild
typ
C-k
it+-P
opul
atio
nal
s %
der
Lym
phoz
yten
Intra
zell.
IgM
+ -Ze
llen
als
% d
er c
-kit+
-Pop
ulat
ion
7307
Ter3
/Ter
3
7307
Ter3
Wild
typ
P = <0.0001 *** P = <0.008 **
30.1 17.0 13.635
30
25
20
15
10
5
0
51
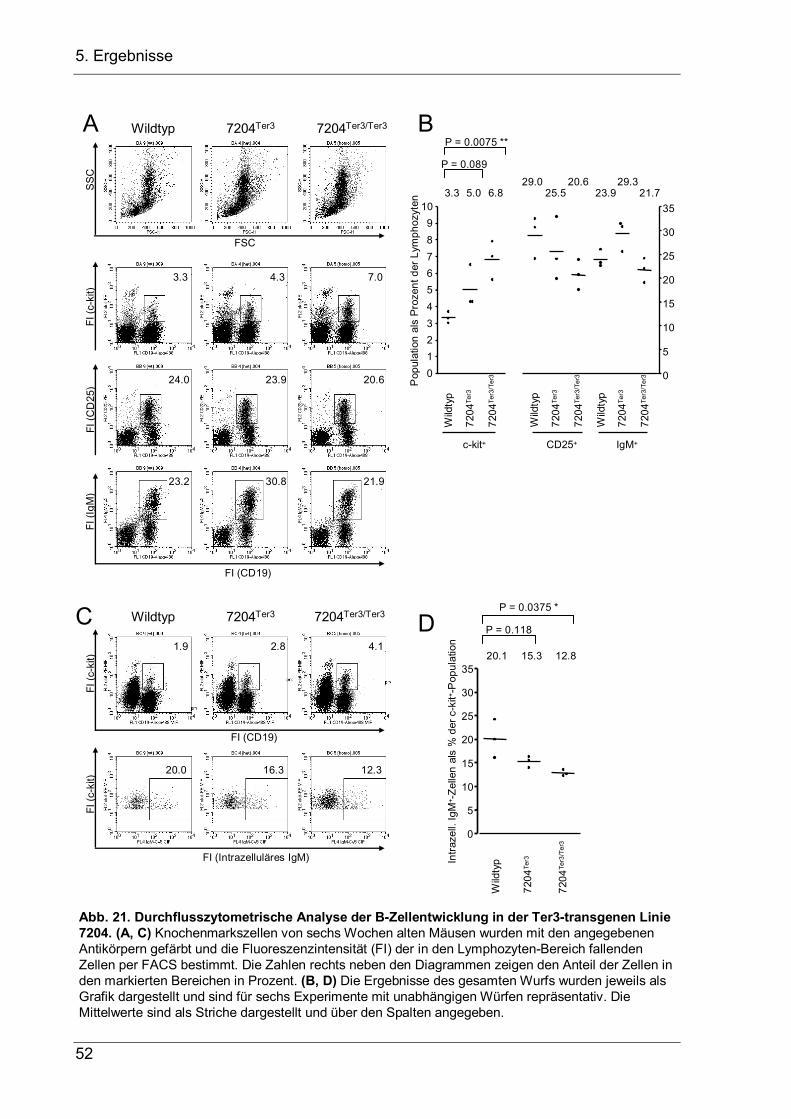
5. Ergebnisse
FI
(c-k
it)
FI (CD19)
FI (C
D25
)FI
(IgM
)
A 7204Ter3Wildtyp 7204Ter3/Ter3
SSC
FSC
23.2 30.8 21.9
24.0 23.9 20.6
3.3 4.3 7.0
FI (Intrazelluläres IgM)
FI (c
-kit)
C
B
FI (CD19)
FI (c
-kit)
Abb.7204.AntikZellenden mGrafiMittel
D7204Ter3Wildtyp 7204Ter3/Ter3
21. Durchflusszytometrische Analyse der B-Zellentwicklung in der Ter3-transgenen Linie (A, C) Knochenmarkszellen von sechs Wochen alten Mäusen wurden mit den angegebenen
örpern gefärbt und die Fluoreszenzintensität (FI) der in den Lymphozyten-Bereich fallenden per FACS bestimmt. Die Zahlen rechts neben den Diagrammen zeigen den Anteil der Zellen in arkierten Bereichen in Prozent. (B, D) Die Ergebnisse des gesamten Wurfs wurden jeweils als
k dargestellt und sind für sechs Experimente mit unabhängigen Würfen repräsentativ. Die werte sind als Striche dargestellt und über den Spalten angegeben.
0123456789
10
0
5
10
15
20
25
30
35
c-kit+ CD25+ IgM+
3.3 5.0 6.829.0
25.520.6
23.929.3
21.7
Pop
ulat
ion
als
Pro
zent
der
Lym
phoz
yten
7204
Ter3
/Ter
3
7204
Ter3
Wild
typ
7204
Ter3
/Ter
3
7204
Ter3
Wild
typ
7204
Ter3
/Ter
3
7204
Ter3
Wild
typ
0
5
10
15
20
25
30
3520.1 15.3 12.8
Intra
zell.
IgM
+ -Ze
llen
als
% d
er c
-kit+
-Pop
ulat
ion
7204
Ter3
/Ter
3
7204
Ter3
Wild
typ
1.9 2.8 4.1
20.0 16.3 12.3
P = 0.0075 **
P = 0.089
P = 0.0375 *
P = 0.118
52
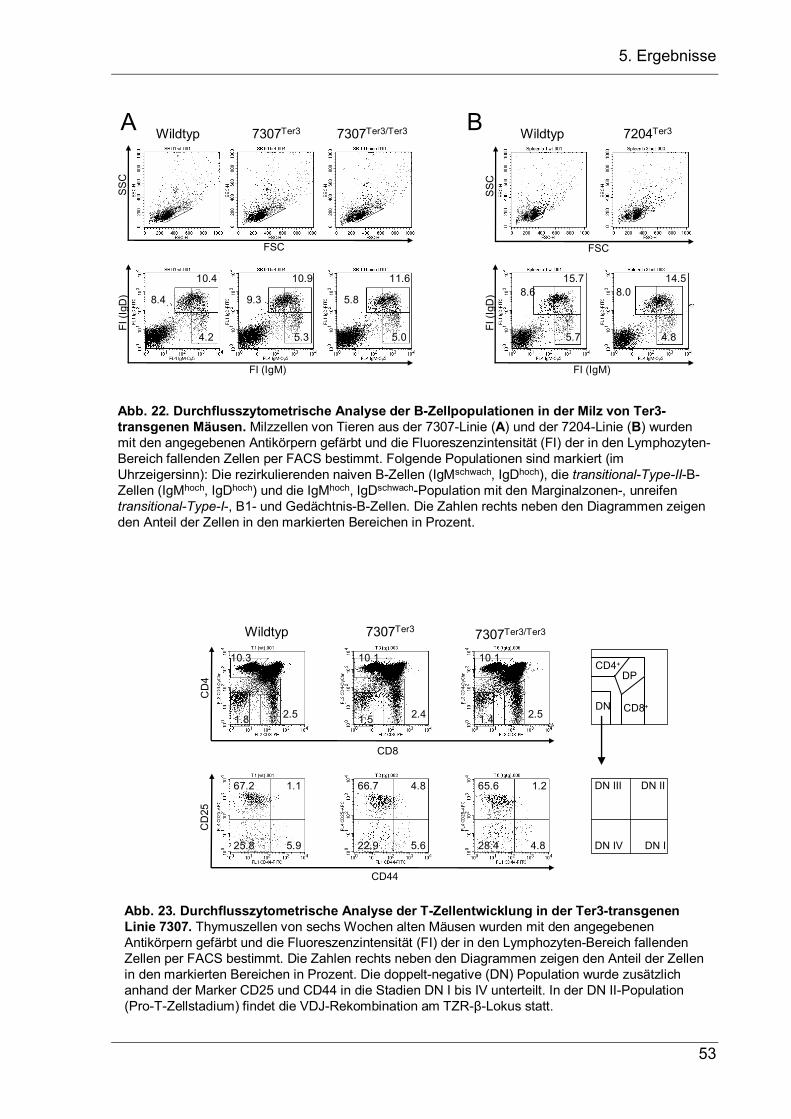
5. Ergebnisse
7307Ter3A Wildtyp 7307Ter3/Ter3S
SC
FSC
FI (IgM)
FI (I
gD)
10.4 10.9 11.6
B
Abb. 22. Durchflusszytometrische Analyse der B-Zellpopulationen in der Milz von Ter3-transgenen Mäusen. Milzzellen von Tieren aus der 7307-Linie (A) und der 7204-Linie (B) wurmit den angegebenen Antikörpern gefärbt und die Fluoreszenzintensität (FI) der in den LymphozBereich fallenden Zellen per FACS bestimmt. Folgende Populationen sind markiert (im Uhrzeigersinn): Die rezirkulierenden naiven B-Zellen (IgMschwach, IgDhoch), die transitional-Type-IIZellen (IgMhoch, IgDhoch) und die IgMhoch, IgDschwach-Population mit den Marginalzonen-, unreifen transitional-Type-I-, B1- und Gedächtnis-B-Zellen. Die Zahlen rechts neben den Diagrammen zeiden Anteil der Zellen in den markierten Bereichen in Prozent.
den yten-
-B-
gen
FI (IgM)
FI (I
gD)
15.7 14.5
SSC
FSC
7204Ter3Wildtyp
5.7 4.8
8.6 8.0
4.2 5.3 5.0
8.4 9.3 5.8
7307Ter3/Ter37307Ter3Wildtyp
Abb. 23. Durchflusszytometrische Analyse der T-Zellentwicklung in der Ter3-transgenen Linie 7307. Thymuszellen von sechs Wochen alten Mäusen wurden mit den angegebenen Antikörpern gefärbt und die Fluoreszenzintensität (FI) der in den Lymphozyten-Bereich fallendenZellen per FACS bestimmt. Die Zahlen rechts neben den Diagrammen zeigen den Anteil der Zein den markierten Bereichen in Prozent. Die doppelt-negative (DN) Population wurde zusätzlich anhand der Marker CD25 und CD44 in die Stadien DN I bis IV unterteilt. In der DN II-Population(Pro-T-Zellstadium) findet die VDJ-Rekombination am TZR-β-Lokus statt.
llen
CD44
CD
25
66.7 4.8
22.9 5.6
67.2 1.1
25.8 5.9
65.6 1.2
28.4 4.8
DN III DN II
DN IV DN I
CD8
CD
4
10.1
1.4 2.5
10.1
1.5 2.4
10.3
1.8 2.5
CD4+
DN CD8+
DP
53

5. Ergebnisse
TZR exprimieren, an doppelt-positiven (DP) Zellen, die den TZR-α-Lokus umlagern
und dann den TZR exprimieren, und an den darauf folgenden CD4- und CD8-einzel-
positiven Zellen. Um die T-Zellstadien, in denen die dem IgH-Lokus entsprechende
Rekombination am TZR-β-Lokus stattfindet, genauer zu untersuchen, wurden die
DN-Stadien durch das Anfärben spezifischer Oberflächenmarker weiter aufgetrennt.
Auch hierbei war keine Änderung in der Frequenz von Zellen am Übergang vom Pro-
T-Zellstadium (DN II), in dem die TZR-β-Rekombination stattfindet, zum Prä-TZR-
positiven Prä-T-Zellstadium (DN III) nachweisbar. Diese Daten zeigen, dass das
Ter3-Transgen nur die Entwicklung von Pro-B-Zellen beeinträchtigt. Ob damit auch
ein Einfluss der Ter3-µ-mRNA auf die T-Zellentwicklung ausgeschlossen werden
kann, ist noch nicht klar. Zwar werden in µH-Ketten-transgenen Mäusen vom Ig-
Promoter/Enhancer getriebene Transgene auch in T-Zellen exprimiert (Grosschedl et
al. 1984), allerdings wurde das Expressionsniveau des Ter3-Transgens in T-Zellen
noch nicht untersucht.
5.2.5 Komplementation der B-Zellentwicklung durch ein produktives VDJ-Exon
Falls die erhöhte Anzahl an µH-Ketten-negativen Pro-B-Zellen in Ter3-transgenen
Mäusen auf einer Störung der VDJ-Rekombination am IgH-Lokus beruht, sollte die
B-Zellentwicklung durch eine transgene funktionelle QM-µH-Kette (siehe oben)
wieder in den in Wildtyp-Mäusen beobachten Phänotyp revertiert werden. Daher
wurde die B-Zellentwicklung in heterozygoten QM-Mäusen mit und ohne 7307-Ter3-
Transgen verglichen. Durchflusszytometrische Analysen mit Knochenmark aus
entsprechenden transgenen Mäusen ergaben vergleichbare Frequenzen an c-kit-
positiven Pro-B-Zellen in IgHQM/wt, 7307Ter3- und IgHQM/wt-Mäusen (Abb. 24). Dies
zeigt, dass die in Ter3-transgenen Mäusen beobachtete Beeinträchtigung der
Differenzierung von Pro-B-Zellen (Abb. 20, 21) durch eine funktionelle µH-Kette
vollständig aufgehoben werden kann (Abb 24). Der geringe Anstieg der c-kit-
positiven Pro-B-Zellpopulation in Ter3-transgenen QM-Mäusen lässt sich
wahrscheinlich durch Pro-B-Zellen erklären, die ihr produktives QM-Allel durch VH-
Replacement verloren haben und in Gegenwart der Ter3-Transkripte den endogenen
IgH-Lokus schlechter rearrangieren können.
54
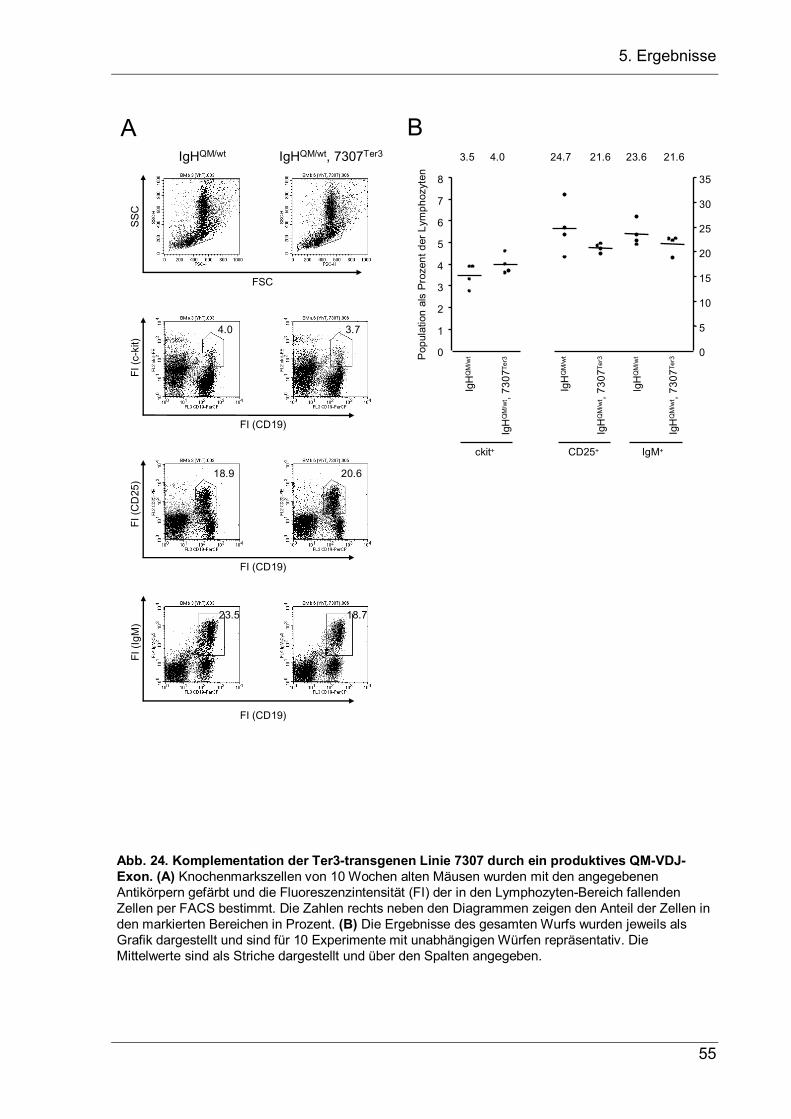
5. Ergebnisse
A BFI
(c-k
it)FI
(CD
25)
IgHQM/wt IgHQM/wt, 7307Ter3
SSC
FSC
FI (I
gM)
FI (CD19)
FI (CD19)
FI (CD19)
4.0 3.7
18.9 20.6
23.5 18.7
Abb. 24. Komplementation der Ter3-transgenen Linie 7307 durch ein produktives QM-VDJ-Exon. (A) Knochenmarkszellen von 10 Wochen alten Mäusen wurden mit den angegebenen Antikörpern gefärbt und die Fluoreszenzintensität (FI) der in den Lymphozyten-Bereich fallenden Zellen per FACS bestimmt. Die Zahlen rechts neben den Diagrammen zeigen den Anteil der Zellen in den markierten Bereichen in Prozent. (B) Die Ergebnisse des gesamten Wurfs wurden jeweils als Grafik dargestellt und sind für 10 Experimente mit unabhängigen Würfen repräsentativ. Die Mittelwerte sind als Striche dargestellt und über den Spalten angegeben.
3.5 4.0 24.7 21.6 23.6 21.6
8
CD25+ckit+ IgM+
7 30
6 255
204
153
102
51
0 0
35
Pop
ulat
ion
als
Proz
ent d
er L
ymph
ozyt
en
IgH
QM
/wt
IgH
QM
/wt ,
7307
Ter3
IgH
QM
/wt
IgH
QM
/wt ,
7307
Ter3
IgH
QM
/wt
IgH
QM
/wt ,
7307
Ter3
55

5. Ergebnisse
Da QM-Mäuse wie Wildtyp-Mäuse im Prä-B-Zellstadium eine endogene leichte Kette
rearrangieren müssen, um in das IgM-positive Stadium zu differenzieren, ließ sich in
ihnen auch der Einfluss der Ter3-µ-Transkripte auf die VJ-Rekombination an den
Leichtketten-Lozi untersuchen. Hierbei stellte sich heraus, dass die CD25-positiven
Prä-B- und die IgM-positiven B-Zellpopulationen in IgHQM/wt, 7307Ter3-Mäusen durch
das Ter3-Transgen nicht verändert wurden und mit IgHQM/wt-Mäusen vergleichbar
waren (Abb. 24).
Die erfolgreiche Reversion der in Ter3-transgenen Mäusen beobachteten
Beeinträchtigung der B-Zellentwicklung durch eine produktive µH-Kette und die
unveränderten Prä-B-Zellpopulationen deuten also darauf hin, dass die in Ter3-
transgenen Mäusen beobachte Störung im Pro-B-Zellstadium durch eine
Beeinträchtigung der VDJ-Rekombination am IgH-Lokus ausgelöst wird, während die
Rekombination der Leichtketten-Lozi nicht behindert wird.
5.2.6 Abwesenheit von µH-Ketten-Signalen in rekombinations-defizienten Ter3-transgenen Pro-B-Zellen
Um auszuschließen, dass von dem Ter3-Transgen durch Reinitiierung der
Translation an einem Leseraster-ATG der Cµ-Region verkürzte µH-Ketten exprimiert
werden, die dann Differenzierungssignale auslösen könnten, wurde das Ter3-
Transgen in einen rekombinations-defizienten Mausstamm gekreuzt, der durch die
homozygote Deletion des Rag2-Lokus’ keine endogenen µH-Ketten umlagern und
exprimieren kann. In diesen Rag2-defizienten Ter3-transgenen Mäusen wurden
keine Zellen mit intrazellulären µH-Ketten nachgewiesen (Abb. 25A). Das Transgen
wurde zusätzlich in einen ebenfalls rekombinations-defizienten Mausstamm mit einer
Deletion des JH-Lokus’ gekreuzt. Da die B-Zellentwicklung in beiden rekombinations-
defizienten Hintergründen auf dem c-kit-positiven Pro-B-Zellstadium arretiert war
(Abb. 25B, C), können vom Transgen ausgehende Differenzierungssignale
ausgeschlossen werden. Interessanterweise ist jedoch in beiden Hintergründen die
Pro-B-Zellpopulation in Anwesenheit des Transgens vergrößert (Abb. 25B-D).
56
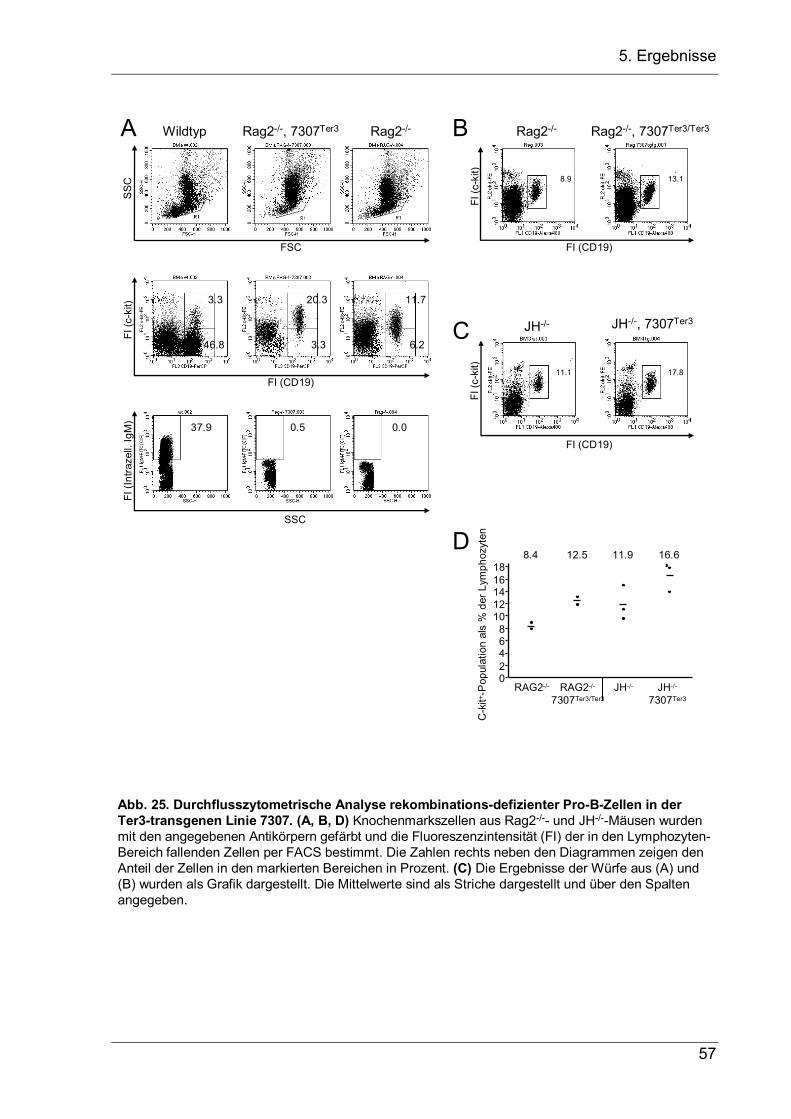
5. Ergebnisse
D
B
C
FI (CD19)
JH-/- JH-/-, 7307Ter3
FI (c
-kit) 11.1 17.8
Rag2-/- Rag2-/-, 7307Ter3/Ter3
FI (c
-kit)
FI (CD19)
8.9 13.1
Abb. 25. Durchflusszytometrische Analyse rekombinations-defizienter Pro-B-Zellen in der Ter3-transgenen Linie 7307. (A, B, D) Knochenmarkszellen aus Rag2-/-- und JH-/--Mäusen wurden mit den angegebenen Antikörpern gefärbt und die Fluoreszenzintensität (FI) der in den Lymphozyten-Bereich fallenden Zellen per FACS bestimmt. Die Zahlen rechts neben den Diagrammen zeigen den Anteil der Zellen in den markierten Bereichen in Prozent. (C) Die Ergebnisse der Würfe aus (A) und (B) wurden als Grafik dargestellt. Die Mittelwerte sind als Striche dargestellt und über den Spalten angegeben.
A
RAG2-/- RAG2-/-
7307Ter3/Ter3JH-/- JH-/-
7307Ter3
02468
1012141618
C-k
it+-P
opul
atio
nal
s %
der
Lym
phoz
yten
8.4 12.5 11.9 16.6
Rag2-/-, 7307Ter3 Rag2-/-WildtypS
SC
FSC
FI (I
ntra
zell.
IgM
)
SSC
0.00.537.9
FI (CD19)
FI (c
-kit) 3.3 20.3 11.7
46.8 3.3 6.2
57

5. Ergebnisse
5.2.7 Expression der endogenen IgH-Keimbahn-Transkripte
Falls die transgenen Ter3-µ-Transkripte die VDJ-Rekombination durch einen
Mechanismus reduzieren, bei dem die Zugänglichkeit der endogenen IgH-Allele
verringert wird, dann sollte auch die Transkription der IgH-Lozi reduziert sein. Um
dies zu untersuchen, wurden die Expressionsniveaus des sterilen Keimbahn-µ°-
Transkripts gemessen, das während der VDJ-Rekombination in Pro-B-Zellen von
einem Promotor vor dem DQ52-Segment exprimiert und ausschließlich vom
Keimbahn-IgH-Lokus kodiert wird (Abb. 26A,a). Hierzu wurde RNA aus FACS-
sortierten CD19-positiven Zellen aus rekombinations-defizienten Rag2–/–-Mäusen mit
und ohne Ter3-Transgen isoliert und in cDNA umgeschrieben. In semiquantiativen
RT-PCRs mit in 1:3-Schritten verdünnten cDNA-Mengen wurden dann die
Expressionsniveaus der µ°-Keimbahntranskripte abgeschätzt (Abb. 26B).
Interessanterweise ist das µ°-Expressionsniveau in transgenen Tieren leicht
reduziert, was auf eine geringere Zugänglichkeit des IgH-Lokus’ in Ter3-Pro-B-Zellen
hinweist.
Die in den transgenen Tieren exprimierte Ter3-µ-mRNA besteht hauptsächlich aus
der Sequenz der Cµ-Region. Doch auch in unrearrangierten Pro-B-Zellen werden
bereits Cµ-haltige Transkripte exprimiert, nämlich das µ°-Transkript und das Iµ-
Transkript, das am intronischen µ-Enhancer (iEµ) initiiert (Abb. 26A). Beide
Transkripte enthalten die Cµ-Region und unterscheiden sich nur in ihren 5’-
Bereichen von der Ter3-µ-mRNA. Falls die Ter3-µ-mRNA ihren Effekt durch die Cµ-
Sequenz ausübt, müssten sich die Expressionsniveaus der Ter3-µ-mRNA und der
Keimbahntranskripte deutlich unterscheiden. Daher wurde die Gesamtmenge der
Cµ-haltigen Transkripte in Pro-B-Zellen aus Rag-defizienten Mäusen mit und ohne
Ter3-Transgen mit einem Cµ-spezifischen PCR (Primer Cµ1 fwd und Cµ1/2 rev in
Abb. 26A) vergleichen, die in den Rag-2-defizienten Mäusen ohne Ter3-Transgen die
Keimbahntranskripte (Abb. 26A,a) und in den Ter3-transgenen Mäusen zusätzlich
die transgene µ-mRNA amplifiziert (Abb. 26A,b). Überraschenderweise war die
Gesamtmenge Cµ-haltiger Transkripte in transgenen Tieren nur auf das dreifache
erhöht (Abb. 26B).
58
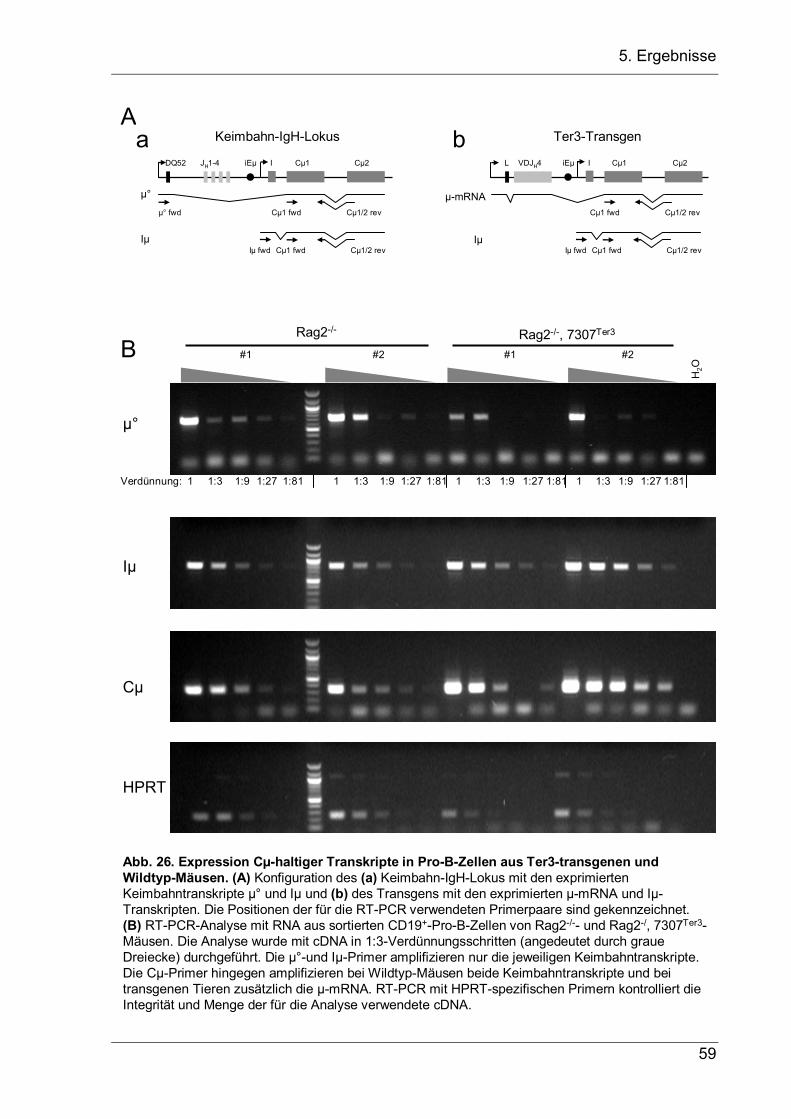
5. Ergebnisse
Abb. 26. Expression Cµ-haltiger Transkripte in Pro-B-Zellen aus Ter3-transgenen und Wildtyp-Mäusen. (A) Konfiguration des (a) Keimbahn-IgH-Lokus mit den exprimierten Keimbahntranskripte µ° und Iµ und (b) des Transgens mit den exprimierten µ-mRNA und Iµ-Transkripten. Die Positionen der für die RT-PCR verwendeten Primerpaare sind gekennzeichnet. (B) RT-PCR-Analyse mit RNA aus sortierten CD19+-Pro-B-Zellen von Rag2-/-- und Rag2-/, 7307Ter3-Mäusen. Die Analyse wurde mit cDNA in 1:3-Verdünnungsschritten (angedeutet durch graue Dreiecke) durchgeführt. Die µ°-und Iµ-Primer amplifizieren nur die jeweiligen Keimbahntranskripte. Die Cµ-Primer hingegen amplifizieren bei Wildtyp-Mäusen beide Keimbahntranskripte und bei transgenen Tieren zusätzlich die µ-mRNA. RT-PCR mit HPRT-spezifischen Primern kontrolliert die Integrität und Menge der für die Analyse verwendete cDNA.
B
Cµ
Iµ
HPRT
A
µ°
#1 #2 #1 #2
Rag2-/-, 7307Ter3Rag2-/-
1 1:3 1:9 1:27
H2O
1:81 1 1:3 1:9 1:27 1:81 1 1:3 1:9 1:27 1:81 1 1:3 1:9 1:27 1:81Verdünnung:
a b
µ°
Iµ
Keimbahn-IgH-Lokus
JH1-4DQ52 iEµ I Cµ1 Cµ2
Cµ1/2 revCµ1 fwd
Cµ1/2 revCµ1 fwdIµ fwd
µ° fwd
µ-mRNA
Ter3-Transgen
VDJH4L iEµ I Cµ1 Cµ2
Cµ1/2 revCµ1 fwd
Cµ1/2 revCµ1 fwdIµ fwdIµ
59

5. Ergebnisse
Sterile Transkripte vom Igκ-Lokus (κ°), die ein Hinweis für das Öffnen des IgL-Lokus
sind und vor der VJ-Rekombination in Prä-B-Zellen initiiert werden, waren in Zellen
aus beiden Genotypen nicht messbar (Daten nicht gezeigt).
5.2.8 VH-Replacement in Gegenwart der Ter3-µ-mRNA
Falls Ter3-µ-Transkripte die V-nach-DJ-Umlagerung am IgH-Lokus hemmen, sollte man erwarten, dass sie auch das VH-Replacement an einem bereits rearrangierten VDJ-Exon beeinträchtigen. Um diese Vorhersage zu überprüfen und so den hemmenden Einfluss des Ter3-Transgens auf das Rearrangement in einer unabhängigen Analyse zu bestätigen, wurde das VH-Replacement in heterozygoten QM-Mäusen (IgHQM/wt) mit und ohne Ter3-Transgen verglichen (Abb. 27). In QM-Mäusen können mittels idio- und allotypischer Marker drei „Arten“ von B-Zellen unterschieden werden (Abb. 27A): (1) Zellen mit einem unveränderten QM-Transgen exprimieren die transgene µH-Kette vom IgMa-Allotyp mit dem VH17.2.25-Idiotyp. (2) Wurde das QM-17.2.25-VDJ mit seiner kryptischen RSS jedoch durch ein VH-Replacement verändert, so exprimieren die Zellen bei einem produktiven Replacement eine µH-Kette vom IgMa-Allotyp ohne VH17.2.25-Idiotyp. (3) Schließlich rearrangieren die Pro-B-Zellen bei einem nicht-produktiven Replacement am QM-Allel das Wildtyp-C57/BL6-Allel und produzieren eine µH-Kette vom IgMb-Allotyp. Anhand der idio- und allotypischen Marker konnte nun in transgenen IgHQM/wt-Mäusen mit und ohne Ter3-Allel der Anteil der B-Zellen mit einen ersetzten QM-Allel durchflusszytometrisch bestimmt werden. Diese Analyse bestätigte unsere Vorhersage, da der Anteil der Zellen mit einem ersetzten VH-Exon (d.h. idiotyp-negativ) in der IgMa-positiven Population im Knochenmark von IgHQM/wt-Mäusen mit Ter3-Transgen im Vergleich zu IgHQM/wt-Mäusen ohne Ter3-Allel auf die Hälfte reduziert war (Abb. 27Ba, Ca). Das gleiche Bild ergabt sich in QM-Mäusen für den Anteil der Zellen in der Gesamt-IgM-positiven B-Zellpopulation, die das endogene Wildtyp-Allel vom IgMb-Allotyp rearrangiert haben (Abb. 27Bb, Cb). Auch hier war die IgMb-positive Population in Anwesenheit des Ter3-Allels auf die Hälfte reduziert. Diese Ergebnisse zeigen, dass in Anwesenheit von Ter3-Transkripten nicht nur die klassische VDJ-Rekombination an den endogenen Wildtyp-IgH-Lozi sondern auch das nicht-klassische VH-Replacement behindert ist.
60
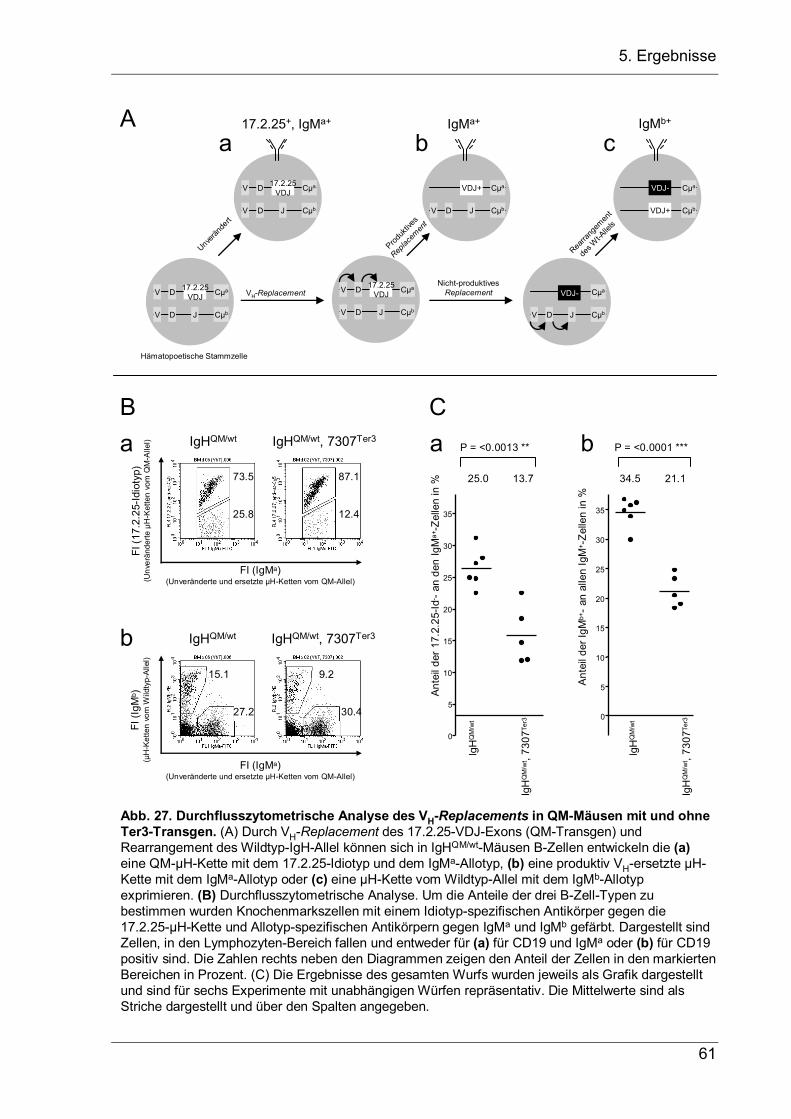
5. Ergebnisse
B C
Abb. 27. Durchflusszytometrische Analyse des VH-Replacements in QM-Mäusen mit und ohne Ter3-Transgen. (A) Durch VH-Replacement des 17.2.25-VDJ-Exons (QM-Transgen) und Rearrangement des Wildtyp-IgH-Allel können sich in IgHQM/wt-Mäusen B-Zellen entwickeln die (a)eine QM-µH-Kette mit dem 17.2.25-Idiotyp und dem IgMa-Allotyp, (b) eine produktiv VH-ersetzte µH-Kette mit dem IgMa-Allotyp oder (c) eine µH-Kette vom Wildtyp-Allel mit dem IgMb-Allotypexprimieren. (B) Durchflusszytometrische Analyse. Um die Anteile der drei B-Zell-Typen zu bestimmen wurden Knochenmarkszellen mit einem Idiotyp-spezifischen Antikörper gegen die 17.2.25-µH-Kette und Allotyp-spezifischen Antikörpern gegen IgMa und IgMb gefärbt. Dargestellt sind Zellen, in den Lymphozyten-Bereich fallen und entweder für (a) für CD19 und IgMa oder (b) für CD19 positiv sind. Die Zahlen rechts neben den Diagrammen zeigen den Anteil der Zellen in den markierten Bereichen in Prozent. (C) Die Ergebnisse des gesamten Wurfs wurden jeweils als Grafik dargestellt und sind für sechs Experimente mit unabhängigen Würfen repräsentativ. Die Mittelwerte sind als Striche dargestellt und über den Spalten angegeben.
a
b
a b
FI (I
gMb )
(µH
-Ket
ten
vom
Wild
typ-
Alle
l)
FI (IgMa)(Unveränderte und ersetzte µH-Ketten vom QM-Allel)
IgHQM/wt IgHQM/wt, 7307Ter3
27.2
15.1
30.4
9.2
0
5
10
15
20
25
30
35
FI (1
7.2.
25-Id
ioty
p)(U
nver
ände
rte µ
H-K
ette
n vo
m Q
M-A
llel)
FI (IgMa)(Unveränderte und ersetzte µH-Ketten vom QM-Allel)
12.4
87.1
25.8
73.5
IgHQM/wt IgHQM/wt, 7307Ter3
A
V D 17.2.25VDJ Cµa
V D J Cµb
V D 17.2.25VDJ Cµa
V D J Cµb
Cµa
V D J Cµb
VDJ-
V D 17.2.25VDJ Cµa
V D J Cµb
17.2.25+, IgMa+
VDJ+ Cµa
V D J Cµb
IgMa+
VDJ- Cµa
VDJ+ Cµb
IgMb+
Unverän
dert
Produk
tives
Replac
emen
t
Rearra
ngem
ent
des W
t-Alle
ls
VH-ReplacementNicht-produktives
Replacement
a b c
Hämatopoetische Stammzelle
Ant
eil d
er Ig
Mb+
-an
alle
n Ig
M+ -
Zelle
nin
%
0
5
10
15
20
25
30
35
IgH
QM
/wt
IgH
QM
/wt ,
7307
Ter3
34.5 21.1
P = <0.0001 ***An
teil
der 1
7.2.
25-Id
- -an
den
IgM
a+-Z
elle
nin
%
IgH
QM
/wt
IgH
QM
/wt ,
7307
Ter3
25.0 13.7
P = <0.0013 **
61

5. Ergebnisse
Homozygot #1
Homozygot #2
Wildtyp #1
Homozygot #1
Homozygot #2
Wildtyp #1
Wildtyp #2
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
2.0
2.2
2.4
2.6
2.8
7204
7307
Abstand zwischen dem verwendeten VH- und dem DFL16.1 D-Segment in Mb
DFL
16.1
VH-Segmente D-Seg. JH-Seg. Cµ
Abb. 28. Chromosomale Position der verwendeten VH-Gensegmente in produktiven VDJ-Sequenzen aus homozygoten Ter3-transgenen Tieren. RNA aus IgM+-Milzzellen von 7204Ter3/Ter3- und 7307Ter3/Ter3-Mäusen, sowie den jeweiligen Wildtyp-Geschwistern, wurde revers transkribiert, und produktive µH-Sequenzen mit einem degenerierten VH- und einem das Ter3-Transgen nicht erkennenden JH3-Primer amplifiziert, kloniert und sequenziert. Für jede produktive VDJ-Sequenz wurde der Abstand zwischen dem verwendeten VH-Segment und dem ersten D-Segment (DFL16.1) bestimmt und als Punkt aufgetragen. Der Mittelwert ist für jede Gruppe als Linie eingezeichnet
IgH-Lokus
5.2.9 VH-Repertoire in Ter3-transgenen Tieren
Um die Auswirkungen der Ter3-µ-Transkripte auf die Öffnung und Zugänglichkeit der
VH-Lozi und deren Verwendung bei der VDJ-Rekombination zu untersuchen, wurden
produktive VDJ-Rearrangements auf den endogenen Wildtyp-Allelen in homozygoten
7204- und 7307-Ter3-Mäusen analysiert. Dazu wurde RNA aus Milz-B-Zellen isoliert,
revers transkribiert und VDJ-Sequenzen mit einem universellen VH-Primer und einem
JH3-Primer, der nicht an Transgensequenzen bindet, amplifiziert, kloniert und
sequenziert. Die chromosomale Position der VH-Gensegmente wurde mit Mouse-
BLAT-Search bestimmt, und der Abstand in Mb zum ersten D-Segment (DFL16.1) in
Abb. 28 dargestellt. Die Verteilung der benutzen VH-Segmente zeigt keinen
Unterschied zwischen homozygoten Ter3- und nicht-transgenen Wildtyp-Mäusen.
Die Beobachtung, dass auch in transgenen Tieren distale VH-Segmente benutzt
62
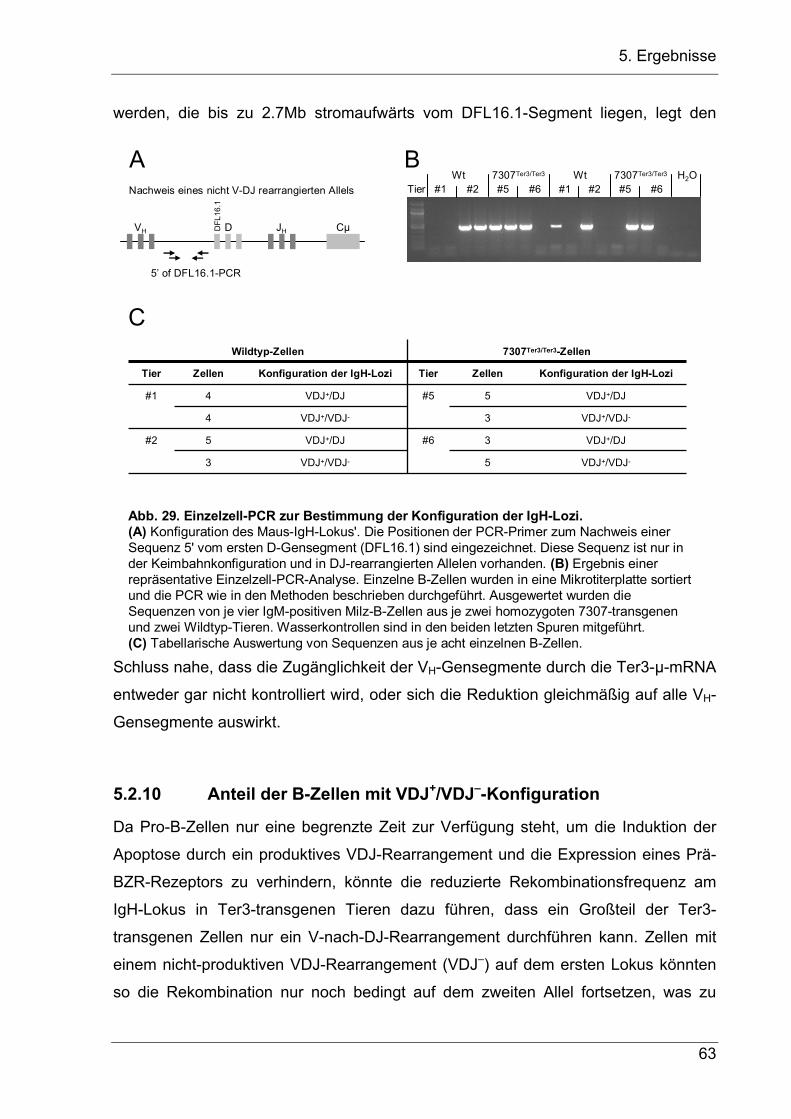
5. Ergebnisse
werden, die bis zu 2.7Mb stromaufwärts vom DFL16.1-Segment liegen, legt den
Abb. 29. Einzelzell-PCR zur Bestimmung der Konfiguration der IgH-Lozi.(A) Konfiguration des Maus-IgH-Lokus'. Die Positionen der PCR-Primer zum Nachweis einer Sequenz 5' vom ersten D-Gensegment (DFL16.1) sind eingezeichnet. Diese Sequenz ist nur in der Keimbahnkonfiguration und in DJ-rearrangierten Allelen vorhanden. (B) Ergebnis einer repräsentative Einzelzell-PCR-Analyse. Einzelne B-Zellen wurden in eine Mikrotiterplatte sortiert und die PCR wie in den Methoden beschrieben durchgeführt. Ausgewertet wurden die Sequenzen von je vier IgM-positiven Milz-B-Zellen aus je zwei homozygoten 7307-transgenen und zwei Wildtyp-Tieren. Wasserkontrollen sind in den beiden letzten Spuren mitgeführt. (C) Tabellarische Auswertung von Sequenzen aus je acht einzelnen B-Zellen.
Konfiguration der IgH-LoziZellenTierKonfiguration der IgH-LoziZellenTier
VDJ+/VDJ-3VDJ+/VDJ-4
VDJ+/VDJ-
VDJ+/DJ
VDJ+/DJ
VDJ+/VDJ-
VDJ+/DJ
VDJ+/DJ
53
3#65#2
5#54#1
7307Ter3/Ter3-ZellenWildtyp-Zellen
A
C
Nachweis eines nicht V-DJ rearrangierten Allels
BWt 7307Ter3/Ter3 H2OWt
#1 #2 #5 #6 #1 #2 #5 #67307Ter3/Ter3
Tier
VH D JH Cµ
5’ of DFL16.1-PCR
DFL
16.1
Schluss nahe, dass die Zugänglichkeit der VH-Gensegmente durch die Ter3-µ-mRNA
entweder gar nicht kontrolliert wird, oder sich die Reduktion gleichmäßig auf alle VH-
Gensegmente auswirkt.
5.2.10 Anteil der B-Zellen mit VDJ+/VDJ–-Konfiguration
Da Pro-B-Zellen nur eine begrenzte Zeit zur Verfügung steht, um die Induktion der
Apoptose durch ein produktives VDJ-Rearrangement und die Expression eines Prä-
BZR-Rezeptors zu verhindern, könnte die reduzierte Rekombinationsfrequenz am
IgH-Lokus in Ter3-transgenen Tieren dazu führen, dass ein Großteil der Ter3-
transgenen Zellen nur ein V-nach-DJ-Rearrangement durchführen kann. Zellen mit
einem nicht-produktiven VDJ-Rearrangement (VDJ–) auf dem ersten Lokus könnten
so die Rekombination nur noch bedingt auf dem zweiten Allel fortsetzen, was zu
63

5. Ergebnisse
einer erhöhten Frequenz von Zellen in der Peripherie führen sollte, die ein
produktives VDJ+-Allel und ein unvollständig rearrangiertes DJ-Allel tragen. Um diese
Hypothese zu testen wurde die Konfiguration der IgH-Lozi in einzelnen IgM+-Milz-B-
Zellen aus homozygoten 7307Ter3/Ter3- und Wildtyp-Mäusen per Einzelzell-PCR
miteinander verglichen (Abb. 29). Hierzu wurde mit Einzelzell-PCR ein DNA-
Fragment amplifiziert, das stromaufwärts vom DFL16.1-Gensegment liegt und bei
jedem V-nach-DJ-Rearrangement entfernt wird (Abb. 29A). IgM-positive Zellen, bei
denen dieses Fragment nicht nachgewiesen werden kann, besitzen also neben
ihrem VDJ+-Allel noch ein nicht-produktives VDJ–-Allel, während in Zellen, in denen
diese Sequenz noch detektierbar ist, am zweite Allel nur ein DJ-Rearrangement
vorliegt. Die Einzelzell-PCR-Analyse ergab, dass acht von 16 IgM-positiven B-Zellen
aus 7307-Ter3-Mäusen und sieben von 16 IgM-positiven Zellen aus Wildtyp-Mäusen
ein nicht-produktiv rearrangiertes IgH-Allel besaßen (Abb. 29C). Die resultierenden
Verhältnisse von Zellen mit VDJ+/DJ-Konfiguration zu Zellen mit VDJ+/VDJ–-
Konfiguration von 50-zu-50 für transgene beziehungsweise 56-zu-44 für Wildtyp-
Tiere entsprechen dem in der Literatur dokumentiertem Wert von ungefähr 60-zu-40
(Alt et al. 1984). Dies zeigt, dass die transgenen Pro-B-Zellen nach einem nicht-
produktiven Rearrangement die Rekombination auf dem zweiten Allel fortsetzen
können.
64

6. Diskussion
6 Diskussion
6.1 VH-Replacement in Pro-B-Zellen mit zwei nicht-produktiven VDJ-Allelen
Angesichts des fehlerbehafteten VDJ-Rekombinationsmechanismus' könnte das VH-
Replacement ein wichtiger Mechanismus sein, der nicht nur B-Zellen mit einem
autoreaktiven BZR rettet, sondern auch Pro-B-Zellen mit entweder nicht-produktiven
VDJ-Rearrangements oder produktiven Rearrangements, die für Transport-
inkompetente dysfunktionale µH-Ketten kodieren (Zusammengefasst in Zhang et al.
2004). Dieser Mechanismus würde allerdings voraussetzten, dass VH-Replacement
in Pro-B-Zellen in der Abwesenheit von Prä-BZR-Signalen stattfinden kann.
Fortwährendes VH-Replacement wurde in verschiedenen murinen und humanen Pro-
B- und Prä-B-Zelllinien entdeckt (Kleinfield et al. 1986; Reth et al. 1986; Zhang et al.
2003), was bedeutet, dass in transformierten B-Zelllinen VH-Replacement in der
Abwesenheit eines µH-Ketten-Signals stattfinden kann. Weitere Indizien für ein µH-
Ketten unabhängiges VH-Replacement kommen von Mäusen, die transgen für
funktionale µH-Ketten sind. In diesen Mäusen wurden VH-Replacements per PCR
bereits in großen Pro-B-Zellen (Hardy Fraction C) detektiert, die normalerweise keine
µH-Ketten exprimieren (Taki et al. 1995). Außerdem enthalten VH-Replacements oft
N-Nukleotide, die von der nur in Pro-B-Zellen exprimierten Terminalen
Deoxynucleotidyltransferase (TdT) eingefügt werden. Allerdings beginnen Vorläufer-
B-Zellen in diesen transgenen Mauslinien bereits mit einem produktiv rearrangierten
VDJ-Exon, so dass die Expression der transgenen µH-Kette früher beginnen könnte
als die Expression normaler µH-Ketten und sich mit der Expression von Pro-B-
Zellmarkern überschneiden könnte.
Um eindeutig zu untersuchen, ob VH-Replacements von nicht-produktiven VDJ–-
Exons in der Abwesenheit von µH-Ketten-Signalen stattfinden, wurde eine Knock-In-
Maus etabliert, bei der die JH-Gensegmente durch ein nicht-produktives VDJ-Exon
ersetzt wurden. Diese Maus gleicht der Quasi-Monoklonalen (QM-) Maus (Cascalho
et al. 1996), in der VH-Replacement ausgiebig untersucht worden ist (Cascalho et al.
65

6. Diskussion
1996; Cascalho et al. 1997; Bertrand et al. 1998; Cascalho et al. 1999; Madan et al.
2000; Golub et al. 2001), allerdings mit dem Unterschied, dass das VDJ-Exon ein
Translations-Stopkodon (Nonsense-Kodon) im Leseraster der µH-Kette an Position
+3 enthält. Die Analyse homozygoter IgHVDJ–/VDJ–-Mäuse zeigte eindeutig, dass Pro-
B-Zellen mit zwei nicht-produktiven VDJ–-Allelen in vivo VH-Replacement benutzen
können, um produktive VDJ-Exons zu erstellen und so in reife IgM-positiven B-Zellen
zu differenzieren. Zusätzlich konnten VH-Replacements nur in RAG-exprimierenden
homozygoten IgHVDJ–/VDJ–-Mäusen detektiert werden, was sich mit früheren Daten
deckt (Cascalho et al. 1999) und die Notwendigkeit von RAG beim VH-Replacement
unterstreicht. Andere DNA-Rekombinationsmechanismen, wie die durch die
Activation-induced-Cytidine-Deaminase (AID) vermittelte Genkonversion von Ig-
Exons in einigen Vögeln und Säugern (Knight and Barrington 1998; Arakawa et al.
2002), können daher für murine Pro-B-Zellen ausgeschlossen werden.
Die Sequenzanalyse von 114 produktiven VDJ-Exons zeigte, dass alle VH-
Replacements die kryptische RSS im VH-Segment des VDJ–-Exons benutzt hatten.
Ähnlich zu den VH-Replacements in anderen transgenen Mauslinien (Chen et al.
1995; Taki et al. 1995; Cascalho et al. 1996) konnten die VH-Replacements in zwei
Gruppen unterteilt werden. In sechs Sequenzen wurde die VH-Region im transgenen
VDJ-Exon durch ein stromaufwärts gelegenes VH-Gensegment ersetzt, ein Vorgang,
der prototypisch für ein VH-Replacement in Wildtyp-Mäusen ist. In den restlichen 108
Sequenzen wurden jedoch durch das VH-Replacement sowohl ein VH- als auch ein
D-Gensegment eingefügt. Dies ist möglich, da in IgHVDJ–/VDJ-Knock-in-Mäusen die D-
Gensegmente noch stromaufwärts vom transgenen VDJ–-Exon liegen. Ein Grund für
die bevorzugte Rekombination der kryptischen RSS mit stromaufwärts positionierten
D-Gensegmenten, die in Wildtyp-Mäusen bei der primären VDJ-Rekombination
entfernt werden, könnten der Unterschied in den Abständen (Spacer) zwischen dem
Nonamer und dem Heptamer in den 3’-RSS der D- und der VH-Segmente sein, die
12bp beziehungsweise 23bp lang sind. Generell findet VDJ-Rekombination
bevorzugt zwischen einer RSS mit einem 12bp- und einer RSS mit einem 23bp-
Spacer statt (Tonegawa 1983). Allerdings ist bei der kryptischen RSS im V-
Gensegment des transgenen VDJ-Exon unklar, ob das Heptamer mit einem
Nonamer assoziiert ist. Chen et al. (Chen et al. 1995) haben ein potentielles
66

6. Diskussion
Nonamer-Motiv 12bp stromaufwärts des Heptamers beschrieben. Aber in diesem
Falle würde die 12/23-Regel eine bessere Kompatibilität des putativen 12bp-Spacers
mit dem 23bp-Spacers der VH-RSS als mit dem 12bp-Spacer des bevorzugt
benutzten D-RSS vorhersagen. Daher ist es wahrscheinlicher, dass VH-
Replacements mit einem zusätzlichen D-Segment hauptsächlich ihre Ursache im
kürzeren Abstand zwischen dem transgenen VDJ-Exon und den D-Gensegmenten
und der geordneten Öffnung des Chromatins am IgH Lokus haben (Alt et al. 1984;
Chowdhury and Sen 2004). Aufgrund von letzterem können die D-Gensegmente
bereits im frühen Pro-B-Zellstadium rekombiniert werden, wo normalerweise die D-
nach-J-Rekombination stattfindet; die VH-Gensegmente werden hingegen erst in dem
darauf folgenden späten Pro-B-Zellstadium zugänglich, in dem die V-nach-DJ-
Rekombination stattfindet. VH-Replacements des bereits rearrangierten transgenen
VDJ–-Exons, die D-Gensegmente benutzen, könnten daher bereits im frühen Prä-B-
Zellstadium erfolgt sein. Jedoch zeigen die analysierten Sequenzen mit direkten V-
nach-VDJ-Replacements in unserer Maus, dass VH-Replacements auch nach
Öffnung des VH-Clusters in der späten Pro-B-Zelle erfolgen, und so möglicherweise
Pro-B-Zellen mit zwei nicht-produktiven IgH-Allelen in Wildtyp-Mäusen retten können.
Diese Beobachtung wurde mit einem ähnlichen Knock-In-Mausmodell in einer
kürzlich erschienenen Studie bestätigt (Koralov et al. 2006).
Die Idee, dass VH-Replacement in der B-Zellentwicklung ein häufiger Vorgang sein
könnte, kommt von einer Computeranalyse humaner Ig-Sequenzen. Hierbei wurden
in 5-13% der Sequenzen Hinweise für ein VH-Replacement gefunden, das zu
verschiedenen Zeitpunkten der B-Zellentwicklung stattgefunden haben könnte
(Zhang et al. 2003). Diese Ergebnisse widersprechen jedoch sowohl einer Studie, die
nur eine geringe Rekombinationsfrequenz in Substraten mit kryptischen RSS
gefunden hat (Nadel et al. 1998), als auch der Beobachtung, dass in TdT-defizienten
Mäusen, in denen VH-Replacement eindeutig nachgewiesen werden kann, VH-
Replacements nur selten gefunden wurden (Watson et al. 2006). Daher können der
Umfang des VH-Replacements in Pro-B-Zellen von nicht-transgenen Tieren und sein
quantitativer Beitrag zum Schicksal von Pro-B-Zellen mit zwei nicht-produktiven VDJ–
-Exons erst in weiteren Studien geklärt werden.
67

6. Diskussion
6.2 Entwicklung von B-Zellen mit langen CDR3-H-Regionen
Interessanterweise haben µH-Ketten, die in IgHVDJ–/VDJ–-transgenen Tieren durch VH-
Replacement erzeugt werden, im Durchschnitt erheblich längere CDR3-H-Regionen
als µH-Ketten von Wildtyp-B-Zellen. Dies bietet die Möglichkeit, den Einfluss von
langen CDR3-H-Regionen auf die B-Zellentwicklung und die Funktionalität des Prä-
BZRs und BZRs zu untersuchen.
Die CDR3-H-Region ist hauptsächlich für die Antigenspezifität des Antikörpers
verantwortlich, und beinhaltet das D-Segment und die benachbarten Enden des VH-
und des JH-Segments. In Wildtyp-Mäusen erzeugt die VDJ-Rekombination CDR3-H-
Regionen unterschiedlicher Länge, die mit dem Beginn der TdT-Aktivität in adulten
B-Zellen länger werden (Bangs et al. 1991). Trotzdem folgt die Längenverteilung der
CDR3-H-Regionen im peripheren Repertoire einer engen Gaußschen Verteilung
(Zemlin et al. 2003). Diese enge Verteilung mit einem Mittelwert von 11.5
Aminosäuren in Wildtyp-Mäusen (Zemlin et al. 2003) könnte zwei verschiedene
Ursachen haben. Einerseits könnte sie einfach intrinsische Eigenschaften des VDJ-
Rekombinationsmechanismuses in Mäusen reflektieren, wie die Länge der D- und
JH-Segmente oder die TdT-Aktivität. Andererseits könnte sie auch das Ergebnis einer
Selektion für B-lymphoide Zellen mit µH-Ketten mit bevorzugten strukturellen und
funktionalen Eigenschaften sein. Letzteres wird durch das Ansteigen der
durchschnittlichen Länge der CDR3-H während der B-Zellentwicklung in adulten
Wildtyp-Mäusen unterstützt (Ivanov et al. 2005; Schroeder 2006). Frühere Studien
haben gezeigt, dass die Struktur der CDR3-H-Region die Bindung der µH-Kette mit
der surrogaten leichten (SL-) und einer konventionellen leichten Kette, und damit den
Zusammenbau eines signalkompetenten Prä-BZRs und BZRs, beeinflussen kann
(Keyna et al. 1995; Kline et al. 1998; Kawano et al. 2005). Außerdem kann die
Struktur der CDR3-H-Region auch die Menge eines Prä-BZRs und BZRs an der
Zelloberfläche, und damit die Signalstärke, durch ihre Affinität zu dem im
endoplasmatischen Retikulum (ER) vorkommenden Chaperon BiP regulieren
(Herrmann, Vettermann, Jäck, unpubliziert). Schließlich kann die CDR3-H-Region
auch zu autoreaktiven Spezifitäten beitragen.
68

6. Diskussion
In VDJ–-transgenen Tieren enthalten die meisten VH-Replacements auf dem VDJ–-
Allel ein zweites D-Segment und erzeugen VDDJ-Exons mit einer CDR3-H-Länge
von bis zu 22 Aminosäuren. Gleichzeitig entstehen aber auch CDR3-H-Regionen von
Wildtyp-Länge durch direkte V-nach-VDJ-Replacements oder die Entfernung von
Basen stromabwärts der geöffneten kryptischen RSS. Falls die in reifen B-Zellen von
Wildtyp-Mäusen beobachtete Durchschnittslänge für CDR3-H-Regionen von 11.5
Aminosäuren (Zemlin et al. 2003) das Resultat einer Selektion für strukturelle
Eigenschaften der µH-Kette sind, sollte sich die CDR3-H-Längenverteilung in
homozygoten IgHVDJ–/VDJ–-Mäusen während der Entwicklung B-lymphoider Zellen von
frühen zu späten Entwicklungsstadien von längeren zu kürzeren CDR3-H-Regionen
verschieben. Ist die durchschnittliche Länge von CDR3-H-Regionen jedoch durch
den VDJ-Prozess und seine Eigenschaften bestimmt und sind µH-Ketten mit
längeren CDR3-H-Regionen bezüglich ihrer Assoziation mit SL- und IgL-Ketten
genauso funktional wie µH-Ketten mit kürzeren CDR3-H-Regionen, so sollten sich
die CDR3-H-Längenverteilungen in Pro-B- und reifen B-Zellen ähneln.
Um diese Vorhersagen zu überprüfen, wurden VDJ-Exons mit VH7183-
Gensegmenten untersucht, da die Änderungen in der CDR3-H-Länge in dieser VH-
Familie ausgiebig untersucht worden sind (Ivanov et al. 2005). In Wildtyp-Mäusen
liegt die Durchschnittslänge der CDR3-H-Regionen in Pro-B-Zellen bei 11.4
Aminosäuren und in reifen B-Zellen bei 12.5 Aminosäuren (Ivanov et al. 2005), Werte
die auch in dieser Studie für Milzzellen mit Wildtyp-Sequenzen aus heterozygoten
IgHVDJ–/wt-Mäuse bestätigt wurden. Im Gegensatz dazu lag die durchschnittliche
CDR3-H-Länge in homozygoten VDJ–-Tieren bei 17.8 Aminosäuren in unselektierten
Pro-B-Zellen und bei 16.5 Aminosäuren in reifen Milz-B-Zellen. Bezeichnenderweise
lag die durchschnittliche CDR3-H-Länge von VH-ersetzten VDJ–-Exons in Milz-B-
Zellen, die sich in heterozygoten (also in Konkurrenz von B-Zellen mit kürzeren
CDR3-H-Regionen) VDJ–-Mäusen entwickelten, ebenfalls bei etwa 17 Aminosäuren,
was darauf hinweist, dass B-Zellen mit längeren CDR3-H-Sequenzen keinen
Wachstumsnachteil gegenüber B-Zellen mit kürzeren CDR3H-Regionen haben.
Gleichzeitig sank in heterozygoten Mäusen während des Übergangs vom
Knochenmark in die Milz die Frequenz der B-Zellen mit einem VH-ersetzten BZR nur
um ein Drittel. B-Zellen mit längeren CDR3-H-Regionen waren auch funktional im
69

6. Diskussion
Hinblick auf ihre Möglichkeit, in Antikörper-produzierende Plasmazellen zu
differenzieren, da die IgM-, IgA- und IgG1-Titer im Serum vom IgHVDJ–/VDJ–-Mäusen
sich nicht von denen in Wildtyp-Tieren aus dem gleichen Wurf unterschieden. Auch
die Toleranz wurde nicht beeinflusst, da in sechs Monate alten homozygoten IgHVDJ–
/VDJ–-Weibchen keine Antikörper gegen doppelsträngige DNA im ELISA
nachgewiesen werden konnten (Daten nicht gezeigt), obwohl lange CDR3-H-
Regionen oft mit antinukleären Spezifitäten und Autoreaktivität assoziiert sind (Bangs
et al. 1991; Eilat and Fischel 1991; Klonowski et al. 1999). Diese Ergebnisse zeigen,
dass die in Wildtyp-Mäusen beobachtete geringe Frequenz von B-Zellen, die eine
µH-Kette mit einer CDR3-H-Länge von mehr als 16 Aminosäuren exprimieren, eher
auf den VDJ-Rekombinationsmechanismus zurückzuführen ist, als auf eine starke
Selektion gegen B-Zellen mit längeren CDR3-H-Regionen.
Zusammengefasst konnte in diesem Abschnitt der Promotionsarbeit gezeigt werden,
dass VH-Replacement in Pro-B-Zellen in der Abwesenheit eines Ig-Rezeptor-Signals
stattfinden kann. Dies deutet stark darauf hin, dass VH-Replacement ein potentieller
Rettungsmechanismus sein könnte, um Pro-B-Zellen mit zwei nicht-produktiv
rearrangierten Allelen durch Editieren eines der nicht-funktionellen VDJ-Exons vor
Apoptose zu retten. Zusätzlich wurde gezeigt, dass die enge CDR-3-H-
Längenverteilung in Wildtyp-Mäusen nicht ausschließlich das Ergebnis negativer
Selektion gegen längere CDR3-H-Regionen ist, sondern eher auf mechanistische
Eigenschaften der VDJ-Rekombination zurückzuführen ist.
6.3 Hemmung der VDJ-Rekombination am IgH-Lokus durch µH-Ketten-Transkripte
In späten Pro-B-Zellen können auf beiden IgH-Lozi V-nach-DJ-Rekombinationen
stattfinden. Um jedoch die Expression zweier IgH-Ketten in einer Zelle zu vermeiden,
muss nach einer produktiven VDJ-Rekombination auf dem ersten Allel die V-nach-
DJ-Rekombination des zweiten Allels verhindert werden. In dieser Arbeit wurde
untersucht, ob bereits die Expression eines produktiven µH-Ketten-Transkriptes (µ-
Transkriptes) vor der Initiierung eines Prä-BZR-Signals von der Oberfläche einen
70

6. Diskussion
dämpfenden Einfluss auf die V-nach-DJ-Rekombination am zweiten Allel haben
kann.
6.3.1 Effekte von Ter3-µ-Transkripten auf die B-Zellentwicklung und VDJ-Rekombination
Normalerweise korreliert die Stabilität einer µ-mRNA direkt mit ihrer Produktivität, da
nicht-produktive Nonsense-mRNAs schnell durch den Nonsense-Mediated mRNA-
Decay (NMD) abgebaut werden (Jack et al. 1989). Ein produktives und ein nicht-
produktives VDJ-Rearrangement am IgH-Lokus unterscheiden sich daher nicht nur in
der An- oder Abwesenheit eines µH-Ketten-Proteins, sondern auch in der An- oder
der fast kompletten Abwesenheit eines µ-Transkriptes. Um den Effekt einer
produktiven µ-mRNA auf die VDJ-Rekombination in Abwesenheit der kodierten µH-
Kette zu studieren, wurde eine transgene Maus etabliert, die eine Nonsense-µ-
mRNA exprimiert, deren Stabilität der einer Sense-µ-mRNA ähnelt. Während
physiologische Nonsense-µ-mRNAs ein vorzeitiges Stopkodon (Premature
Termination Codon, PTC) in oder nach der CDR3-H-Region haben und durch den
NMD 50-100-fach herunterreguliert werden (Jack et al. 1989; Buhler et al. 2004),
wurde im Ter3-Transgen das Stopkodon an Position +3 im Leader-Peptid platziert.
Dies führte in den Ter3-transgenen Mäusen zu einer Akkumulation der transgenen
Nonsense-Ter3-µ-mRNA, die mit dem Niveau einer produktiven Sense-µ-mRNA
vergleichbar ist.
Die Ter3-transgenen Mäuse zeigten eine gestörte B-Zellentwicklung mit einer
vergrößerten Pro-B-Zellpopulation und einer Minderung der Zellzahlen in den B-
lymphoiden Folgestadien. Diese Störung wurde in drei unabhängigen Linien
beobachtet und war in homozygoten Tieren am stärksten ausgeprägt. Mehrere
Beobachtungen deuten darauf hin, dass dem Phänotyp einen Defekt in der VDJ-
Rekombination am IgH-Lokus zugrunde liegt. So war im Vergleich zu Wildtyp-
Mäusen in den transgenen Tieren hauptsächlich die intrazellulär µH-Ketten-negative
Fraktion der Pro-B-Zellpopulation erhöht, in der die VDJ-Rekombination stattfindet.
Auch konnte die gestörte B-Zellentwicklung in Ter3-transgenen Mäusen durch das
produktiv rearrangierte µH-Gen der QM-Maus komplementiert werden, was andere,
71

6. Diskussion
rekombinations-unabhängige Effekte der Ter3-µ-Transkripte ausschließt. Der
stärkste Hinweis auf eine gestörte VDJ-Rekombination ist jedoch die reduzierte Rate
des VH-Replacements des QM-Transgens in Ter3-transgenen Tieren.
Übereinstimmend mit einer spezifischen Reduktion der Rekombination am IgH-Lokus
ist auch die Beobachtung, dass die V-nach-J-Rekombination der Leichtketten-Lozi im
Prä-B-Zellstadium nicht beeinträchtigt ist. Dies wird in IgHQM/wt-Mäusen deutlich, die
wegen der Anwesenheit des transgenen, produktiven QM-VDJ-Exons zwar kein IgH-
Kettengen im Pro-B-, aber ein IgL-Kettengen im Prä-B-Zellstadium rearrangieren
müssen. Trotzdem ist der Anteil der CD25-positiven Prä-B- und unreifen B-Zellen
und der Ausstoß an IgM-positiven B-Zellen in An- und Abwesenheit des Ter3-
Transgens unverändert.
6.3.2 RNA-vermittelte Mechanismen
Der molekulare Mechanismus, mit dem die transgenen Ter3-µ-Transkripte in Pro-B-
Zellen die VDJ-Rekombination am IgH-Lokus reduzieren, ist allerdings noch unklar.
Mehrere Möglichkeiten sind denkbar. So könnten die µ-Transkripte die VDJ-
Rekombination entweder durch eine Repression der an der VDJ-Rekombination
beteiligten Enzyme oder durch Chromatinmodifikationen an den IgH-Lozi reduzieren.
Im Folgenden werden mögliche Mechanismen diskutiert, wie die Akkumulierung von
µ-Transkripten ohne Ig-Rezeptorsignale die VDJ-Rekombination am IgH-Lokus
reduzieren könnten.
Die erste Möglichkeit ist ein posttranskriptionelles Gene-Silencing (PTGS), bei dem
die Rekombinations-Enzyme durch den Abbau oder die translationelle Inhibition ihrer
mRNA in ihrem Expressionsniveaus beeinflusst werden. Dieser Mechanismus wird
von microRNAs (miRNAs) benutzt, um Gene in ihrer Expression zu regulieren, und
basiert auf kurzen, polyadenylierten pri-miRNAs mit einem 5'-Cap, die sekundäre
Haarnadel-Strukturen ausbilden (Ambros et al. 2003). Die Haarnadel-Motive werden
im Kern von der RNase III Drosha abgeschnitten und als pre-miRNAs von 60-70
Nukleotiden ins Zytoplasma exportiert. Dort wird die Haarnadel-Schlaufe von der
RNase III Dicer entfernt, so dass 21-23 Nukleotid lange Duplexe entstehen.
Anschließend werden die beiden Stränge getrennt und die reife miRNA in den
72

6. Diskussion
miRNA-haltigen Ribonukleoprotein (miRNP) Komplex geladen, der an die Ziel-
mRNAs entweder ohne Basen-Fehlpaarungen bindet und sie schneidet, oder mit
Basen-Fehlpaarungen bindet und ihre Translation hemmt. Falls nun das stabile µ-
Transkript pri-miRNA-ähnliche sekundäre Haarnadel-Strukturen ausbilden würde, so
könnte es neben seiner Funktion als mRNA ebenfalls von der miRNA-Maschinerie
prozessiert werden. Dies könnte zur Bildung kurzer µ-miRNAs führen, die dann z.B.
Rekombinations-Enzyme posttranskriptionell unterdrücken könnten. Dieser
Mechanismus hat jedoch zwei gravierende Nachteile. Einerseits ist die Stadien-
Spezifität nicht gewährleistet, da die gleiche µ-mRNA auch bei der Rekombination
der Leichtketten-Lozi vorliegt, und zweitens wird die mRNA hierbei verbraucht und
kann nicht zur Proteinbiosynthese verwendet werden.
Eine Lokus-, Differenzierungs- und Linien-spezifische Regulation der VDJ-
Rekombination könnte durch die Bildung doppelsträngiger RNAs (dsRNA) erreicht
werden, bei der stabile µ-Transkripte mit Antisense-Transkripten von VH-
Gensegmenten hybridisieren. Diese Möglichkeit ist besonders attraktiv, da die
Expression der Antisense-Transkripte die VH-Exons und intergenische Bereiche des
VH-Clusters abdeckt und zeitlich genau auf die V-nach-DJ-Rekombination beschränkt
ist. Die aus µ-mRNA und VH-Antisense-Transkripten gebildeten dsRNAs könnten von
der RNA-Interferenz-(RNAi)-Maschinerie erkannt und in siRNA-ähnliche Strukturen
verarbeitet werden (Hamilton and Baulcombe 1999; Tuschl et al. 1999; Hammond et
al. 2000; Zamore et al. 2000). Hierbei schneidet Dicer die dsRNA im Zytosol in
Duplexe von small-interfering RNAs (siRNAs) mit ungefähr 21 Nukleotiden, die dann
in den RNA-induced Silencing-complex (RISC) geladen werden und den Abbau von
mRNAs mit homologen Sequenzen einleiten (Bernstein et al. 2001; Hammond 2005).
Zusätzlich zu diesem posttranskriptionellen Gene-Silencing können siRNAs aber
auch direkte Chromatinmodifikationen am Lokus bewirken, die zu
Heterochromatinbildung und transkriptionellem Gene-Silencing (TGS) führen
(zusammengefasst in Bernstein and Allis 2005). So können siRNAs in Säugerzellen
an Promotoren binden und durch Rekrutierung von entsprechenden Enzymen durch
DNA-Methylierung und H3K9-Methylierung von Histonen den Lokus deaktivieren
(Kawasaki and Taira 2004; Weinberg et al. 2006). In der Spalthefe
(Schizosaccharomyces pombe) sind siRNAs Teil des RNA-induced inititation of
73

6. Diskussion
Transcriprional Gene Silencing (RITS)-Komplexes, der zur Heterochromatin-Bildung
notwendig ist (Verdel and Moazed 2005) und vermutlich Histon-Methyltransferasen
enthält. Die siRNAs übernehmen hierbei die Rekrutierung von Chromatin-
modifizierenden Enzymen an das betreffende Zielgen. Da die VH-Antisense-
Transkription wahrscheinlich von schwächeren Promotoren getrieben wird als die
Transkription der rearrangierten µ-mRNA, könnte die µ-mRNA im Überschuss
vorliegen, so dass nicht alle Sense-Transkripte mit Antisense-Transkripten
hybridisieren. Die µ-mRNA könnte dann in zwei Formen unterschiedliche Funktionen
erfüllen. Die hybridisierten µ-mRNAs könnten als dsRNAs regulative Aufgaben in der
Modifikation des nicht komplett rearrangierten IgH-Allels übernehmen, während die
freien µ-mRNAs zur Proteinsynthese herangezogen würden. Da Dicer jedoch nur im
Zytosol vorliegt, in das die nicht-polyadenylierten sterilen VH-Transkripte alleine nicht
exportiert werden, müssten sich die dsRNAs bereits im Kern bilden und gemeinsam
exportiert werden.
µ-Transkripte könnten aber auch ohne Prozessierung als lange einzel- oder, in
Verbindung mit Antisense-Transkripten, doppelsträngige RNAs den IgH-Lokus
modifizieren. Ein Beispiel hierfür ist die Inaktivierung des zweiten X-Chromosoms in
weiblichen Säugern-Zellen durch lange, nichtkodierende Sense-Xist- und Antisense-
Tsix-RNAs, die zu kovalenten Histonmodifikationen führen (Avner and Heard 2001).
Ein vierter Mechanismus, bei dem µ-Transkripte abhängig von ihrer Menge die VDJ-
Rekombination beeinflussen könnten, beruht auf den potentiellen RNA-
Bindungsdomänen des RAG1-Enzyms (Spanopoulou et al. 1995). Dadurch könnten
RAG1 mit einem µ-mRNA/Antisense-VH-RNA-Hybrid interagieren und so in seiner
Aktivität reduziert werden. Auch hierbei würde die Stadien-Spezifität durch die auf
das Pro-B-Zellstadium beschränkte Expression der VH-Antisense-RNA erreicht
werden und so das IgL-Rearrangement trotz der Anwesenheit großer Mengen an µ-
mRNA nicht beeinflussen. Als mögliches Bindemotiv auf der RNA könnten die
kryptischen RSS dienen, die in fast allen VH-Gensegmenten vorkommen.
Bei allen oben diskutierten Mechanismen zum transkriptionellen und post-
transkriptionellen Gene-Silencing stellt sich allerdings die Frage nach den
74

6. Diskussion
Zielsequenzen auf dem IgH-Lokus beziehungsweise in den mRNAs, die für Faktoren
der Rekombinations-Maschinerie kodieren. Die Untersuchung des VH-Repertoires in
Ter3-transgenen Tieren hat keine Unterschiede in der Verwendung von distalen und
proximalen VH-Regionen bei der VDJ-Rekombination gezeigt und damit keinen
Hinweis auf eine reduzierte Zugänglichkeit der distalen Region des IgH-Lokus’
ergeben, wie sie z.B. bei IL-7Rα-defizienten Mäusen auftritt (Corcoran et al. 1998;
Chowdhury and Sen 2003). Daher ist nicht anzunehmen, dass die einzelnen VH-
Gensegmente durch die µ-mRNA modifiziert werden. Allerdings wäre es für einen
regulativen Mechanismus auch effektiver, anstelle des langen VH-Clusters die Region
zwischen den VH- und D-Segmenten zu modifizieren, die kürzer und vor allem nur
auf dem noch nicht vollständig rearrangierten DJ-Allel vorhanden ist, dessen
Rearrangement unterdrückt werden soll. Unsere Daten schließen auch einen
Einfluss der µ-mRNA auf die Lokus-Kontraktion und -Dekontraktion aus, da hierbei in
transgenen Tieren ebenfalls weniger VDJ-Exons mit distalen VH-Regionen erwarten
würden.
Verbunden mit der Frage nach der Zielsequenz ist die Frage nach der Effektor-
Sequenz in den µ-Transkripten. Hierbei muss beachtet werden, dass die
rearrangierten µ-mRNAs nicht die einzigen Sense-Transkripte sind, die während der
V-nach-DJ-Rekombination vom IgH-Lokus exprimiert werden. Zusätzlich zu sterilen,
gespleißten Transkripten der VH-Gensegmenten (Yancopoulos and Alt 1985; Duber
et al. 2003) werden auch vom intronische µ-Enhancer das sterile Iµ und von den
rearrangierten DJ-Segmenten DJCµ-Transkripte erzeugt, die beide gespleißte Cµ-
Regionen enthalten. Von diesen Transkripten unterscheidet sich die rearrangierte µ-
mRNA dadurch, dass sie sowohl eine VH- als auch eine Cµ-Region besitzt. Weitere
Untersuchungen müssen zeigen, ob die rearrangierte µ-mRNA ihren Effekt durch
Sequenzmerkmale ausübt, oder ob dieser einfach durch die höheren
Transkriptmengen ausgelöst wird. Letzteres ist zumindest für die Cµ-Region
unwahrscheinlich, da die semiquantitative PCR gezeigt hat, dass durch die
Expression der µ-mRNA nur eine 3-9fach höhere Mengen an Cµ-haltigen
Transkripten erzeugt wird. Man kann allerdings auch nicht ausschließen, dass die
sterilen Transkripte ähnlich wie unstabile Nonsense-Transkripte in speziellen
75

6. Diskussion
Bereichen in der Zelle, den so genannten P-Bodies, konzentriert sind und so für eine
Regulation des IgH-Lokus' nicht zur Verfügung stehen (Teixeira et al. 2005).
Die grundlegende Frage, ob die µ-mRNA spezifisch die Rekombination am IgH-
Lokus oder unspezifisch die Aktivität der gesamten Rekombinations-Maschinerie
reduziert, wird hoffentlich durch Experimente geklärt, die momentan durchgeführt
werden und bei denen die Rekombinationsaktivität in transgenen Pro-B-Zellen mit
transfizierten extrachromosomalen Substraten gemessen werden soll. Parallel zu
diesem funktionellen Ansatz sollen die Expressionsniveaus der RAG-mRNAs in den
transgenen Pro-B-Zellen mit und ohne Ter3-µ-mRNA gemessen werden, um
mögliche PTGS-Mechanismen zu erkennen. Im Rahmen dieser Studien, die durch
Probleme in der Zucht aufgehalten wurden, sollen außerdem die RNA-Mengen der
Keimbahn-Sense- und der Antisense-Transkripte im VH-Cluster und der Sense-
Transkripte in der Cµ-Region mit quantitativer PCR genauer bestimmt werden. Erste
Ergebnisse zeigen eine leicht reduzierte Expression des µ°-Keimbahntranskripts in
Ter3-transgenen Pro-B-Zellen, was auf eine reduzierte Zugänglichkeit des IgH-
Lokus' hindeuten könnte.
6.3.3 Alternative Mechanismen
Die Ansammlung von Pro-B-Zellen in Ter3-transgenen Tieren ließe sich auch durch
einen durch die Expression des Transgens ausgelösten Mangel an
Transkriptionsfaktoren erklären, der zu einer verringerten Expression der endogenen
µH-Kette und daher zu verringerten Prä-BZR-Rezeptorsignalen führt. Gegen diese
Hypothese spricht jedoch, dass die Expression von endogenen µH-Ketten und von
CD19, das ein Zielgen des die B-Zellentwicklung dominierenden
Transkriptionsfaktors Pax5 ist, in B-Zellen durch die Anwesenheit des Ter3-Transgen
nicht reduziert wird (Daten nicht gezeigt) und Transkriptionsfaktoren normalerweise
im starke Überschuss vorliegen (Mark Schlissel, persönliche Mitteilung). Auch könnte
ein Mangel an Transkriptionsfaktoren nicht durch ein produktives µH-Ketten-
Transgen komplementiert werden und widerspricht der Beobachtung, dass die µ°-
Expression in Ter3-transgenen Pro-B-Zellen nur leicht reduziert ist. Am wichtigsten
76

6. Diskussion
ist jedoch die Beobachtung, dass sich µH-Ketten-negative Pro-B-Zellen ansammeln,
die Störung also vor der Expression der µH-Kette und des Prä-BZRs liegt.
Es lässt sich auch nicht ausschließen, dass der Effekt auf die Differenzierung von
Pro-B-Zellen nicht von der transkribierten Ter3-µ-mRNA, sondern von dem in das
Genom integrierten Transgensequenz ausgelöst wird. Möglich wäre hier, dass durch
die Intergation Gene beeinträchtigt wurden, die für die B-Zellentwicklung notwendig
sind. Dies ist jedoch unwahrscheinlich, da der Phänotyp in drei Ter3-Linien mit
unterschiedlichen Integrationsorten auftrat, und zumindest bei der hauptsächlich
untersuchten 7307-Linie keine Gene in der Nähe des Transgens lagen. Eine weitere
Möglichkeit wäre die Bindung von RAG an die kryptische RSS im Transgen, so dass
weniger freies RAG für die VDJ-Rekombination zur Verfügung steht. In Anbetracht
der Tatsache, dass in C56/BL6 Mäusen pro Allel 110 funktionale VH-Segmente
während des V-nach-DJ-Rearrangements zur Verfügung stehen (Johnston et al.
2006), die je eine Konsensus-RSS und in 95% der Fälle auch eine kryptischen RSS
enthalten (Chen et al. 1995), scheint es unwahrscheinlich, dass die 10 Transgen-
Kopien in heterozygoten, beziehungsweise die 20 Transgen-Kopien in homozygoten
Ter3-Tieren mit je einer kryptischen RSS die verfügbare RAG-Menge in einem
Ausmaß reduzieren, das den Phänotyp erklären würde. Außerdem ist die Bindung
von RAG an kryptische RSS fünfmal weniger effizient als die an konventionelle RSS
(Zhang et al. 2003).
6.3.4 Erweitertes Feedback-Modell zur Allelausschluss am IgH-Lokus
Zusammengenommen lässt sich die Ansammlung µH-Ketten-negativer Pro-B-Zellen
und die reduzierte Rate des VH-Replacement in Ter3-tansgenen Mäusen am besten
durch einen dämpfenden Effekt der Ter3-µ-mRNA auf die VDJ-Rekombination der
endogenen IgH-Allele erklären. Wir schlagen daher für den Allelausschluss am IgH-
Lokus ein erweitertes Feedback-Modell vor (Abb. 30), das im Gegensatz zum
klassischen Feedback-Modell (Alt et al. 1980; Alt et al. 1984, Abb. 4) nicht nur das
µH-Ketten-Protein sondern auch die µ-mRNA berücksichtigt. Dieses Modell
unterscheidet zwischen einer kurzfristigen, unvollständigen Dämpfung der
Rekombination, die direkt nach einem produktiven Rearrangement auftritt und durch
77

6. Diskussion
Stabile mRNA dämpft Rekombination am zweiten Lokus
Oberflächen-µH-Signalbeendet Rekombination am
zweiten Lokus
Nicht-produktive mRNA wirdvom NMD abgebaut
V DJ CµV DJ Cµ
mRNA
VDJ CµV DJ Cµ
mRNA
VDJ CµV DJ Cµ
VDJ CµV DJ Cµ
mRNANMD
Allelausschluss,Differenzierung
Öffnung und Rearrangement deszweiten Allels
Abb. 30. Das erweiterte Feedback-Modell zum Allelausschluss am IgH-Lokus. Im Gegensatz zum klassischen Feedback-Modell (Abb. 4) berücksichtigt das erweiterte Feedback-Modell zum Allelausschluss nicht nur das µH-Ketten-Protein und die von ihm initiierten Prä-BZR-Signale, sondern auch die µ-mRNA und die von ihr ausgehenden Effekte. Direkt nach dem produktiven Rearrangement wird ein stabiles Sense-µ-Transkript synthetisiert, das die Rekombination auf dem zweiten Lokus solange dämpft bis die neu-synthetisierte µH-Ketten mit SL-Kette und Igα/Igβ als Prä-BZR an die Oberfläche gelangt und von dort Signale zur Modifizierung des zweiten IgH-Allels und zur Differenzierung initiiert. Bei einem nicht-produktiven VDJ-Rearrangement hingegen entsteht eine Nonsense-mRNA, die vom Nonsense-Mediated-mRNA-Decay (NMD) abgebaut wird und die Öffnung und das Rearrangement des zweiten Allels nicht unterdrückt.
die Menge an produktiver mRNA kontrolliert wird, und einer langfristigen,
vollständigen Beendigung der Rekombination, die durch Oberflächen-Prä-BZR-
Signale vermittelt wird. Wichtiger Bestandteil des erweiterten Feedback-Modells ist
der Nonsense-mediated mRNA-Decay (NMD), da er zwischen den produktiven
Sense- und den nicht-produktiven Nonsense-µ-mRNAs unterscheiden und durch
raschen Abbau der Nonsense-mRNAs gewährleisten muss, dass sich nur Sense-µ-
mRNAs ansammeln und die Rekombination dämpfen können.
Der große Vorteil des erweiterten Feedback-Modells besteht darin, dass durch die
sofortige Verfügbarkeit der µ-mRNA nach einem produktiven Rearrangement schnell
Signale unabhängig von der Produktion einer IgH-Kette erzeugt werden können, um
die Rekombination am zweiten Lokus solange zu verhindern, bis Prä-BZR-Signale
78

6. Diskussion
initiiert werden können. Dies schließt die Zeitlücke, die zwischen Rearrangement und
Oberflächensignal klafft und in der kein Rearrangement des zweiten Lokus
stattfinden darf. Versuche in Zelllinien haben gezeigt, dass zwischen der VDJ-
Rekombination und der Oberflächenexpression eines BZRs mindestens zwei
Stunden vergehen (Zhang et al. 1998). Im klassischen Feedback-Modell wird
deshalb ein asynchrones Rearrangement der IgH-Lozi postuliert, bei dem zwischen
den Rekombinationen genügend Zeit liegen muss, um ein mögliches Proteinsignal
abzuwarten. Eine Asynchronität der Allele, wie sie beim Igκ-Lokus durch
monoallelische Demethylierung und unterschiedliche nukleäre Positionierung der
Allele erreicht wird (Goldmit et al. 2005), ist im erweiterten Feedback-Modell für den
IgH-Lokus nicht mehr unbedingt notwendig. Das passt zu der Beobachtung, dass im
Gegensatz zu Igκ-Lokus bisher keine klaren Belege für Asynchronität am IgH-Lokus
gefunden wurden (Corcoran 2005).
Die über die µ-RNA-Menge vermittelte Dämpfung der VDJ-Rekombination ist ein
schwacher Effekt, der in unserem Mausmodell zwar zu einer Ansammlung von Pro-
B-Zellen führt, ihre Differenzierung jedoch nicht verhindert. Dies liegt allerdings auch
daran, dass unser Mausmodell nur den ersten Teil des erweiterten Feedback-
Modells abbildet, da die transgene Ter3-mRNA eben nicht für eine µH-Kette kodiert,
die in der Wildtyp-Situation nach der für Synthese und Transport der µH-Kette
benötigten Zeit ein starkes Oberflächensignal gibt. Die endogenen Lozi werden in
Abwesenheit dieses Prä-BZR-Signals nicht vollkommen geschlossen, so dass es
dann doch noch zu einer Rekombination kommt. Dies ist auch der Grund, wieso der
Effekt der µ-mRNA nicht in den Maus-Modellen beobachtet wurde, in denen kein
Prä-BZR-Signal erzeugt (Kitamura and Rajewsky 1992) oder weitergeleitet
(Papavasiliou et al. 1995; Gong and Nussenzweig 1996; Minegishi et al. 1999)
werden kann und die als Belege des klassischen Feedback-Modells gelten. In diesen
Tieren können die Pro-B-Zellen nach einem produktiven Rearrangement nicht
weiterdifferenzieren, sondern bleiben unphysiologisch lange im rekombinations-
aktiven Pro-B-Zellstadium. In dieser Situation kann der temporäre, dämpfende Effekt
der µ-mRNA die Rekombination nicht verhindern, und es kommt zum Bruch des
Allelausschluss'. Das erweiterte Feedback-Modell steht somit im Einklang mit den
bisherigen Beobachtungen.
79

6. Diskussion
Ohne Zweifel sind weitere Versuche sind notwendig, um die mechanistische
Grundlage des beobachteten Effekts zu klären und weitere Belege für das erweiterte
Feedback-Modell zu finden. Es wäre außerdem sehr interessant, die VDJ-
Rekombination in NMD-defizienten Mäusen zu studieren, in denen es nach einem
nicht-produktiven Rearrangement zur Expression stabiler Nonsense-µ-mRNAs
kommen sollte. Als Folge wäre in der Peripherie ein geringerer Anteil an B-Zellen zu
erwarten, die nach einem nicht-produktiven Rearrangement das zweite Allel
rearrangiert haben, da die stabilisierten Nonsense-µ-mRNAs ein Rearrangement am
zweiten Lokus dämpfen sollten. Während dieser Arbeit wurde versucht, hierzu zwei
NMD-defiziente Mausmodelle zu etablieren, bei denen jedoch die Aktivität des NMDs
nicht beeinträchtigt war.
6.3.5 Zusammenfassung
Im zweiten Teil dieser Arbeit wurde zum ersten Mal gezeigt, dass bereits eine stabile
µ-mRNA einen dämpfenden Einfluss auf die Pro-B-Zelldifferenzierung und die VDJ-
Rekombination am IgH-Lokus hat und so am Allelausschluss beteiligt sein kann. Wir
schlagen daher für den Allelausschluss am IgH-Lokus ein erweitertes Feedback-
Modell vor (Abb. 30), bei dem die stabile µ-mRNA als primäres Produkt einer
produktiven VDJ-Rekombination durch bisher unbekannte Mechanismen die
Rekombination am zweiten Allel kurzfristig dämpft, bis die Rekombination durch Prä-
BZR-Signale beendet wird.
80

7. Material und Methoden
7 Material und Methoden
7.1 Material
7.1.1 Antikörper
Folgende Antikörper wurden für die FACS-Analyse benutzt:
Spezifität Spezies Bezugsquelle Verdünn.
Maus-CD19-PerCP (1D3) Ratte BD, Heidelberg 1:200
Maus-CD25-PE (PC61) Ratte BD, Heidelberg 1:50
Maus-c-kit-PE (ack45) Ratte BD, Heidelberg 1:50
Maus-IgM (µH)-FITC Ziege BD, Heidelberg 1:200
Maus-IgM (µH) Ziege Southern Biotech., USA 1:200
Maus-IgM (µH)-Cy5 Ziege Southern Biotech., USA 1:200
Maus-Igλ-PE Ziege Southern Biotech., USA 1:200
Maus-Igκ-PE Ziege Southern Biotech., USA 1:200
Maus-IgMa-FITC (DS-1) Maus BD, Heidelberg 1:200
Maus-IgMb-PE (AF6-78) Maus BD, Heidelberg 1:200
7.1.2 Oligonukleotide
Alle Oligonukleotide wurden von Invitrogen synthetisiert und in einer Konzentration
von 100µM in H2O bei -70°C gelagert.
Name ER# Sequenz
IgMTer139 fwd 1650 GCACTGATCAAGGATAATCAAGGGAG
IgMTer113 rev 1649 CTGTCAAAGCTACTTGATGAGGATGC
IgMTer386 fwd 1652 CAGGGTATGGTCCTCATTTACAAAGC
IgMTer344 rev 1651 CACAGAATAAGGTTATTTTATGGTCTCAC
5' of JH1 fwd 997 GAACAGAGGCAGAACAGAGACTGTG
JH2 rev 998 ACCTGAGGAGACTGTGAGAGTGGTG
JH-/- Neo fwd 999 CTGAAGAGCTTGGCGGCGAAT
81
7. Material und Methoden
JH -/- rev 1000 GACTCCACCAACACCATCACACAGA
5’ of 7204 fwd 1646 CAAGACAACTCTCGACTACATGTGAG
3’ of 7204 rev 1645 CTGTCCTGAAACTCACCTTGTAGACC
5' of Ter7307 fwd2 1359 AACATCAAGTTTCCAAGTAGTGGTGG
3' of Ter7307 rev3 1360 GCTTCTACTAGATTCAGTGTATCTGG
RAG2-for 14892 GACGTTCATACATGCCTTCTACCC
RAG2-rev 14893 TGTCAAATTCATCGTCACCATCAA
Neo-rev 14894 GGCCACACGCGTCACCTTA
QM-VDJ-for 972 TGCTAGATACTATAGGTACCCTTACTATGC
JH4-rev 973 ATCGGATACTGTATAAATGCTGTCAC
Cre fwd 1077 CCTGGAAAAATGCTTCTGTCCG
Cre rev 1078 CAGGGTGGTATAAGCAATCCC
5’UTR of 17.2.25 fwd 1376 CTACAGACACTGAATCTCAAGGTCC
VH7183 fwd 1101 TGCGAGGTCGACCTGGTGGAGTCTGGG
J558 leader1 1012 ATG GGA TGG AGC TGG ATC TT
J558 leader3 1014 ATG GAA TGG AGC TGG GTC TT
mCµ1 rev 1377 GAAGGAAATGGTGCTGGGCAGG
JH4exon rev 1589 ACGGTGACTGAGGTTCCTTGACC
Cµ1 fwd 1633 AGTCAGTCCTTCCCAAATGTCTTCC
Cµ1/2 rev 1634 GGGGTTCATCTCTGCGACAGCTGG
Ter3KI fwd 1st 1325 GCCTGAGCTATGAGGAAGTAGAGAGGC
Ter3KI fwd 2nd 1326 GTGCAGAGAAGAACGAGTTACTGTGG
Ter3KI rev 1st 1327 GACGCTGGGCCCGACGTCGCATGCTCC
Ter3KI rev 2nd 1328 CGTCGCATGCTCCTCTAGACTCGAGG
5’ DFL16.1 fwd 1007 GAGTGCCATCAGACACCACAG
5’ DFL16.1 rev 1008 GTGTGGAAAGCTGTGTATCCCC
DFL16.1 2nd fwd 1643 CAGGAGAGATGAGTGTACCTAGTGC
DFL16.1 2nd rev 1644 GGTAGTGGAGATTCACAGCTTCCC
HPRT-for 415 GCTGGTGAAAAGGACCTC
HPRT-rev 416 CACAGGACTAGAAGACCTGC
82

7. Material und Methoden
7.1.3 Mäusestämme
Folgende Mäusestämme wurden in individuell belüfteten Käfigen im Tierstall des
Nikolaus-Fiebiger-Zentrums gemäß der Institutsrichtlinien gehalten:
• Rag2–/– (Shinkai et al. 1992)
• JH–/– (Chen et al. 1993)
• Quasi-monoklonale (QM) (Cascalho et al. 1996)
• EIIa-cre (Lakso et al. 1996)
• C57/BL6 (Charles River)
7.2 Molekularbiologische Methoden
Die meisten molekularbiologischen Methoden wurden gemäß der Protokolle von
Maniatis (Sambrook 1989), beziehungsweise gemäß der Herstellerprotokolle
durchgeführt. Daher erfolgt hier nur ein kurzer Überblick.
7.2.1 Präparation von Plasmid-DNA und Restiktionsverdaus
Plasmid-DNA wurde je nach benötigter Menge mit den Mini-, Midi-, oder Maxi-Kits
der Firma QIAgen (Hilden) aufgereinigt.
Restriktionsverdaus und PCR-Produkte wurden auf 0.8-1.5% Agarosegelen mit TAE
Puffer (50x TAE: 2 M Tris, 50 mM EDTA, 143 ml Eisessig für 2,5 L Puffer, um einen
pH-Wert von ca. 8,5 zu erhalten) aufgetrennt, mit Ethidiumbromid angefärbt, und die
Fragmente mit QIAquick Gel Extraction Kit (QIAgen) bei kleinen Fragmenten oder
QIAexII (Qiagen) bei Fragmenten über 8kb aus dem Gel extrahiert.
7.2.2 Herstellung und Transformation elektrokompetenter Bakterien
Für die Klonierungen wurden GeneHog-Bakterien (Invitrogen) benutzt, die selbst
elektrokompetent gemacht wurden.
Dafür wurde 1l LB-Medium mit 1/100 Volumen einer GeneHog-Übernachtkultur
angeimpft, und die Kultur bis zu einer OD600 von 0.5-0.6 inkubiert. Die Kultur wurde
dann 30min auf Eis gekühlt, die Bakterien 15min bei 3000 Upm auf 4°C in einem
JA14-Rotor abzentrifugiert. Der Überstand wurde vollständig entfernt, und das Pellet
83

7. Material und Methoden
dreimal mit kleiner werdenden Volumina (1l, 500ml, 20ml) eiskalter 10%
Glyzerinlösung gewaschen. Anschließend wurde das Pellet in 2ml eiskalter 10%
Glyzerinlösung aufgenommen und in 50µl-Aliquots bei -70°C eingelagert.
Zur Transformation wurden 50µl kompetente Bakterien mit 1-2µl DNA-Lösung
vermischt und auf Eis in eine Elektroporationsküvette (Spaltbreite 1mm) überführt.
Die Elektroporation wurde in einem GenePulser (BioRad Laboratories, München) mit
den Einstellungen 25µF, 1.66kV und 200Ω durchgeführt. Die Bakterien wurden nach
der Transformation in 1ml reinem LB-Medium aufgenommen und 45min bei 37°C
inkubiert. Anschließend wurde die Kultur auf LB-Agar-Platten mit den
entsprechenden Selektionsmitteln ausplattiert und über Nacht bei 37°C bebrütet.
7.2.3 DNA für Oozyten-Injektion
Der Transgenvektor pµTer3 wurde mit einem QIAgen Maxi-Kit aufgereinigt, mit SalI
und XhoI verdaut und auf einem Agarosegel aufgetrennt. Das 12.3kb SalI-XhoI-
Fragment wurde anschließend mit QiagenExII Glasmilch (Qiagen) extrahiert, über
Schleicher&Schüll EluTips (Whatman, Dassel) aufgereinigt und durch Zugabe von
1/10 Volumen 3 M NaAcetat pH 4.8 und 2.5 Volumenteilen 100% Ethanol gefällt. Der
Ansatz wurde dann 20min bei 4°C und 13000 rpm in einer Heraeus Biofuge Fresco
abzentrifugiert, und die präzipitierte DNA zweimal mit 70% Ethanol gewaschen. Von
der gefällten DNA wurden 2.8µg in der Transgene-Facility der University of Alabama
in Birmingham in den Pronukleus von C57Bl/6 Oozyten injiziert.
7.2.4 DNA für ES-Zell-Transfektion
Der Knock-In-Vektor pBS-DQNTe3 wurde mit dem Qiagen-Maxi-Kit-Endofree
aufgereinigt, mit SacII und SpeI verdaut und auf einem Agarosegel aufgetrennt. Das
7.4kb SacII-SpeI-Fragment, das keine bakteriellen Sequenzen enthielt, wurde
anschließend mit QiagenExII Glasmilch (Qiagen) extrahiert, über Schleicher&Schüll
EluTips aufgereinigt und mit Ethanol gefällt. In TE gelöst wurden 15µg DNA an
Werner Müller (GBF - Gesellschaft für Biotechnologische Forschung / Helmholtz-
Zentrum für Infektionsforschung, Braunschweig) geschickt und dort in die von Ralf
Kühn (GSF - Forschungszentrum für Umwelt und Gesundheit, München) aus
84

7. Material und Methoden
(C57BL/6J x 129S6/SvEvTac) F1-Mäusen etablierte embryonale Stammzelllinie
IDG3.2 (unpubliziert) transfiziert.
7.2.5 Southern-Blot-Analyse
Für die Southern-Blot-Analyse wurden Schwanz-Biopsien mit 10µl Proteinase K
(20mg/ml, PeqLab, Erlangen) in 500µl Lysepuffer (50mM Tris-HCl pH 8.3, 5mM
EDTA, 100mM NaCl, 0.5% SDS) unter starkem Schütteln bei 56°C über Nacht
verdaut, die DNA mit Phenol/Chloroform extrahiert und in Isopropanol gefällt. Dann
wurden 5µg DNA in einem 30µl Ansatz mit den entsprechenden Restriktionsenzymen
für 3h verdaut und auf einem Agarosegel bei 20V über Nacht aufgetrennt. Das Gel
wurde anschließend beschnitten und 2x 20min in Denaturierungspuffer (0.5M NaOH,
1.5M NaCl) denaturiert.
Die positiv geladene Nylon-Membran (GeneScreenPlus, Perkin Elmer, Rodgau-
Jügesheim) wurde im Denaturierungspuffer getränkt und auf drei Whatman-Papiere
gelegt, die wiederum auf einem Stapel von Papiertüchern lagen. Auf die Membran
wurde dann das Gel gelegt, dessen DNA per Kapillartransfer auf die Membran
übertragen wurde. Nach dem Transfer wurde die Membran kurz in H2O
abgewaschen und in Neutralisierungspuffer (0.5M Tris-HCl, pH 7.2, 1M NaCl)
neutralisiert.
Hybridisierungslösung Formamid 50%(10ml)
SSC 5x (5ml von 20x)
Denhardts 5x (2ml von 50x)
SDS 1.5% (1.5ml von 20%)
Lachssperma-DNA 5min auf 95°C, dann auf Eis;100µl/ml Endkonzentration
H2O ad 20ml
Die Membran wurde bei 42°C in einem rotierenden Ofen mit der
Hybridisierungslösung vorinkubiert. Währenddessen wurden 25ng Sonden-DNA mit
dem RadPrime Labeling Kit (Invitrogen) und 5µl α-32P dCTP [10mCi/ml, Amersham
Bioscience, GE Healthcare Europe, München] radioaktiv markiert. Die Sonde wurde
85

7. Material und Methoden
zur Hybridisierungslösung hinzugegeben und für mindestens 16h hybridisiert.
Anschließend wurde die Membran in drei verschiedenen Waschlösungen
gewaschen:
Waschlösung 1: 2x SSC, 0.5% SDS für 5min bei RT
Waschlösung 2: 2x SSC, 0.1% SDS für 15min bei RT
Waschlösung 3: 0.1x SSC, 0.1% SDS für 30min bei 65°C
Die Membran wurde dann in einer Plastiktüte für zwei Tage auf eine
Phosphoimagerplatte gelegt, die Phosphoimagerplatte anschließend eingelesen
(FLA-3000, Fujifilm, Düsseldorf) und das Bild mit BASReader 3.14 Software (Raytest,
Straubenhardt) ausgewertet.
7.2.6 PCR
PCR-Reaktionen wurden auf einer GeneAmp PCR System 9700 (Perkin Elmer) oder
einer Primus advanced 96 (Peqlab) durchgeführt. Das zur Amplifikation verwendete
Standard-PCR-Protokoll war: 95°C, 3 min; 35 Zyklen (95°C, 30 s / 60°C, 30 s / 72°C,
1 min); 72 °C, 5 min. Die Bindetemperatur und die Elongationszeit wurden je nach
Primer und PCR-Produkt angepasst. Ein PCR-Ansatz enthielt bei 20 µl
Gesamtvolumen je 5 pmol Primer (1 µl der 1:20 Verdünnung einer 100 µM
Stammlösung), 0.25 mM dNTPs, 0.2 U Taq-Polymerase (Genaxxon Bioscience,
Biberach) und 2 µl des 10x Reaktionspuffer mit MgCl2.
7.2.7 RT-PCR
RNA wurde mit dem RNeasy Kit (Qiagen) isoliert und dem Superscript II Kit
(Invitrogen) in cDNA umgeschrieben.
7.2.8 Site-directed Mutagenese
Die benötigten Mutationen wurden mit dem QuikChange-Site-Directed Mutagenesis
Kit (Stratagene, Cedar Creek, TX) in den Vektor pµGPT eingefügt. Dieses Kit benutzt
86

7. Material und Methoden
die PfuTurbo DNA-Polymerase, um den kompletten Vektor mit zwei Primern zu
amplifizieren, die gegenläufig an der gleichen Stelle binden und die gewünschte
Mutation in der Mitte tragen. Nach der Amplifikation wurden die methylierten,
unmutierten Vektoren mit DpnI verdaut. Die unmethylierten, mutierten PCR-
Amplifikate blieben hierbei unverdaut und wurden in GeneHog-Bakterien
transformiert.
7.2.9 DNA-Sequenzierungen
Zur Sequenzierung wurde das BigDye Terminator Cycle Sequencing Kit Version 1.1
(Applied Biosystems, Foster City, CA) benutzt. Abweichend vom Hersteller-Protokoll
wurde folgendes Protokoll angewandt.
6µl 200-300ng Plasmid-DNA in H2O (HPLC-Grade)
2µl BigDye Kit Ver. 1.1 Sequenzier-Mix (2.5x)
1µl 5x BigDye Sequenzierpuffer
1µl Primer (5pmol/µl)
PCR Programm: 94°C 3min, 25 Zyklen (96°C 20sec, 52°C 15sec, 60°C 4min)
Die Sequenzier-Reaktion wurde anschließend in Eppendorff-Gefäße überführt und
durch die Zugabe von 90µl H2O (HPLC-Grade), 1/10 Volumen 3M NaAc pH 5.8 und
2.5 Volumen 100% Ethanol in einer Zentrifuge 20min bei 13000rpm auf 4°C gefällt.
Der Überstand wurde verworfen und das Pellet zweimal mit 200µl 70% Ethanol
gewaschen und getrocknet. Anschließend wurde das Pellet in 15µl HighDye-
Formamid (Applied Biosystems) gelöst und auf einem ABI3130 Kapillarsequenzierer
(Applied Biosystems) aufgetrennt. Die Chromatogramme wurden mit der Software
SeqMan (DNASTAR, GATC Biotech, Konstanz) analysiert.
7.2.10 Bestimmung des Transgen-Insertionspunkts per Zirkel-PCR
Zur Bestimmung des Transgen-Insertionspunkts wurde DNA wurde aus Schwanz-
Biopsien wie für eine Southern Blot Analyse aufgereinigt. 5µg DNA wurden in einem
50µl-Ansatz für 10h mit MseI verdaut, und das Enzym anschließend 30min bei 65°C
87

7. Material und Methoden
hitzeinaktiviert. Vom Verdau wurden 5µl mit H2O und Ligationspuffer auf 500µl
verdünnt, 5µl T4 DNA-Ligase hinzugefügt und über Nacht bei 14°C inkubiert, um
DNA-Zirkel zu erhalten. Die Ligase wurde danach 15min bei 65°C inaktiviert.
Die Amplifikation der Bruchpunkte geschah in zwei PCR-Schritten. Zunächst wurden
1.5µl des Ligationsansatzes in einer 20µl PCR-Reaktion mit den Primern IgMTer113
rev und IgMTer139 fwd oder IgMTer344 rev und IgMTer386 fwd eingesetzt. Von der
ersten PCR-Reaktion wurde dann 1µl für die zweite PCR-Reaktion mit den Primern
IgMTer113 rev und IgMTer386 fwd benutzt. Das PCR-Programm war für beide
Amplifikationen gleich: 5min 94°C, 20 Zyklen (30sec 94°C, 30sec 65°C, 3min 72°C),
3min 72°C. Das zweite PCR-Produkt wurde auf einem Gel aufgetrennt, die Banden
extrahiert, mit dem TA Cloning Kit (Invitrogen) kloniert und mit dem Primer T7
sequenziert. Dabei wurden sowohl die Bruchpunkte, als auch der endogene Lokus
identifiziert.
7.2.11 Einzelzell-PCR
Für die Einzelzell-PCR wurden einzelne B-Zellen am FACS-Sorter (Coulter, Krefeld)
in PCR-Platten (PeqLab) sortiert, in die in jede Vertiefung 40µl H2O vorgelegt waren.
Die Platten wurden sofort auf Trockeneis eingefroren und bei -20°C gelagert. Zur
PCR wurden die Platten aufgetaut, 10µl Lysepuffer (6µ PCR Puffer mit 15mM Mg2+
und 4µl Proteinase K [0.1mg/ml]) zu jeder Vertiefung hinzugegeben und 1h bei 56°C
inkubiert. Anschließend wurde die Proteinase 15min bei 95°C inaktiviert, die Platten
auf Eis gekühlt und abzentrifugiert. Nun wurde in jede Vertiefung 10µl PCR-Mix (2µl
NTPs, 2µl von jedem Primer, 0.25µl Taq, H2O ad 10µl) gefüllt und die PCR mit
folgenden Bedingungen gestartet: 5min 94°C, 30 Zyklen (30sec 94°C, 30sec 65°C,
40sec 72°C), 3min 72°C.
In der ersten PCR wurden das äußere Primerpaar für die Sequenz 5’ von DFL16.1
(5’DFL16.1 fwd, 5’DFL16.1 rev) und das Transgen (Ter3KI fwd 1st, Ter3KI rev 1st)
gemeinsam eingesetzt. Je 1µl des PCR-Produkts der ersten Runde wurde dann mit
den gleichen Bedingungen ein zweites Mal in getrennten Ansätzen amplifiziert mit
den inneren Primerpaaren DFL16.1 2nd fwd und DFL16.1 2nd rev oder Ter3KI fwd
2nd und Ter3KI rev 2nd.
88

7. Material und Methoden
Zur Analyse der Konfiguration der IgH-Allele in den Splenozyten von heterozygoten
IgHVDJ–/wt-Mäusen wurden die Zellen mit Antikörpern gegen IgMa und IgMb angefärbt
und nach folgendem Schema sortiert:
1 2 3 4 5 6 7 8 9 10 11 12
A 20 20 20 20 20 20 20 20 20 20 20 20 Je 10 µa+ und µb+ Zellen
B 1 1 1 1 1 1 1 1 1 1 1 1 Eine µa+ Zelle
C 1 1 1 1 1 1 1 1 1 1 1 1 Eine µa+ Zelle
D 1 1 1 1 1 1 1 1 1 1 1 1 Eine µa+ Zelle
E 1 1 1 1 1 1 1 1 1 1 1 1 Eine µb+ Zelle
F 1 1 1 1 1 1 1 1 1 1 1 1 Eine µb+ Zelle
G 1 1 1 1 1 1 1 1 1 1 1 1 Eine µb+ Zelle
H 0 0 0 0 0 0 0 0 0 0 0 0 Keine Zelle
Bei der Analyse der Nonsense-transgenen Mäuse wurde hingegen in alle
Vertiefungen der Platten je eine IgM-positive Milzzelle aus 7307Ter3/Ter3- oder Wildtyp-
Tieren sortiert.
7.2.12 Genotypisierung transgener Mäuse
Zur Genotypisierung transgener Mäuse wurden Schwanzbiopsien entnommen und in
jeweils 300µl PBND (PCR-Buffer with nonionic-detergent)-Puffer (50mM KCl, 10mM
Tris-HCl pH 8.3, 2.5mM MgCl2, 0.1mg/mgl Gelatine, 0.45%v/v NP-40, 0.45% v/v
Tween 20) mit 2µl Proteinase K (20mg/ml, PeqLab) unter starkem Schütteln bei 56°C
über Nacht verdaut. Die Proteinase wurde anschließend 15min bei 95°C inaktiviert,
die Proben auf Eis gekühlt und die unverdauten Bestandteile abzentrifugiert. Für die
PCR wurde 1µl des Verdaus eingesetzt.
7.2.13 Durchflusszytometrie (FACS)
Primärzellen wurden je nach Gewebe unterschiedlich präpariert. Zur Isolation von
Knochenmarkszellen wurden Femur und Tibia präpariert, an den Enden
aufgeschnitten und mit FACS-PBS (PBS, 5% FCS, 0.01% Azid) gespült. Durch
89

7. Material und Methoden
erneutes Aufziehen in die Spritze wurde aus dem Knochenmark eine
Einzelzellsuspension erzeugt. Milzen wurden in RPMI (Invitrogen) aufgenommen und
mit einem Sieb (Cell Strainer, BD, Heidelberg, Maschenweite 70µm) homogenisiert,
und peritonealen B-Zellen durch Ausspülen der Peritoneums mit PBS gewonnen.
Die Einzelzellsuspensionen wurden dann 5min bei 1500 rpm auf 4°C in einer
Heraeus Megafuge abzentrifugiert, und in 0.15M NH4Cl, 20mM HEPES
aufgenommen, um die Erythrozyten zu lysieren. Nach einer 5minütigen Inkubation
bei RT wurde die Lyse durch Zugabe von PBS gestoppt, die Zellen abzentrifugiert
und das Pellet erneut in FACS-PBS aufgenommen.
Zur Oberflächenfärbung wurden die Zellen in Micronix-Röhrchen (Lelystad, NL)
abzentrifugiert, in 50µl FACS-PBS mit den entsprechenden Antikörpern
aufgenommen, und 45min auf Eis inkubiert. Abschließend wurden die Zellen zweimal
mit PBS gewaschen und auf einem FACS Calibur (BD) analysiert und mit CellQuest-
Software ausgewertet.
Für intrazelluäre Färbungen wurden die Zellen, ggf. nach vorheriger
Oberflächenfärbung, in Micronix-Röhrchen abzentifugiert, in 50µl Fixierlösung
(Solution A, Fix und Perm Kit (An der Grub Biosciences)) aufgenommen und 15min
bei RT fixiert. Die Zellen wurden danach zweimal mit PBS gewaschen, in 50µl
Permeabilisierungslösung (Solution B) mit den entsprechenden Antikörpern
aufgenommen und erneut 15min bei RT inkubiert. Abschließend wurden die Zellen
zweimal mit PBS gewaschen und analysiert.
7.2.14 Durchflusszyometrische (FACS) und magnetische (MACS) Zellsortierungen
Um definierte Zellpopulationen aus der Gesamtpopulation aufzureinigen, wurden die
gewünschten Zellen anhand ihrer Oberflächenmarker entweder mit
floureszenzgekoppelten Antikörpern im FACS-Sorter (Coulter) oder mit an
magnetische Kügelchen gekoppelten Antikörpern im AutoMACS (Miltenyi, Bergisch
Gladbach) sortiert. Die Vorbereitung der Zellen für den FACS-Sorter entspricht dem
FACS-Protokoll. Für die MACS-Sortierung wurden 1x107 Zellen nach
Erythrozytenlyse durch einen Nylonfilter (Pre-Separation Filter, Miltenyi) gefiltert, in
90

7. Material und Methoden
10µl CD19 Magnetic Beads (Milteny) und 90µl MACS-PBS (PBS, 2% FCS, 0.01%
Azid) aufgenommen, 20min auf Eis inkubiert, und zweimal in MACS-PBS
gewaschen. Die Zellen wurden dann im AutoMACS separiert.
7.2.15 Nachweis der Ig-Isotypen im Serum
Serum von sechs Wochen alten Tieren wurde durch einen ELISA mit dem
Clonotyping Sytem/HRP (Southern Biotechnology, Birmingham, AL) nach dem
Herstellerprotokoll nachgewiesen. Die Platten wurden mit Ziege-Anti-Maus-Ig-
Antikörpern beschichtet, mit dem Serum inkubiert und mit HRP-gekoppelten Isotyp-
spezifischen Antikörpern und dem entsprechenden HRP-Substrat entwickelt.
91

7. Material und Methoden
92

8. Literaturverzeichnis
8 Literaturverzeichnis
Alessandrini, A. and S. V. Desiderio (1991). "Coordination of immunoglobulin DJH transcription and D-to-JH rearrangement by promoter-enhancer approximation." Mol Cell Biol 11(4): 2096-107.
Alt, F. W., V. Enea, et al. (1980). "Activity of multiple light chain genes in murine myeloma cells producing a single, functional light chain." Cell 21(1): 1-12.
Alt, F. W., N. Rosenberg, et al. (1982). "Multiple immunoglobulin heavy-chain gene transcripts in Abelson murine leukemia virus-transformed lymphoid cell lines." Mol Cell Biol 2(4): 386-400.
Alt, F. W., G. D. Yancopoulos, et al. (1984). "Ordered rearrangement of immunoglobulin heavy chain variable region segments." Embo J 3(6): 1209-19.
Ambros, V., B. Bartel, et al. (2003). "A uniform system for microRNA annotation." Rna 9(3): 277-9.
Arakawa, H., J. Hauschild, et al. (2002). "Requirement of the activation-induced deaminase (AID) gene for immunoglobulin gene conversion." Science 295(5558): 1301-6.
Avner, P. and E. Heard (2001). "X-chromosome inactivation: counting, choice and initiation." Nat Rev Genet 2(1): 59-67.
Bangs, L. A., I. E. Sanz, et al. (1991). "Comparison of D, JH, and junctional diversity in the fetal, adult, and aged B cell repertoires." J Immunol 146(6): 1996-2004.
Barreto, V. and A. Cumano (2000). "Frequency and characterization of phenotypic Ig heavy chain allelically included IgM-expressing B cells in mice." J Immunol 164(2): 893-9.
Bassing, C. H., W. Swat, et al. (2002). "The mechanism and regulation of chromosomal V(D)J recombination." Cell 109 Suppl: S45-55.
Baumann, B., M. J. Potash, et al. (1985). "Consequences of frameshift mutations at the immunoglobulin heavy chain locus of the mouse." Embo J 4(2): 351-9.
93

8. Literaturverzeichnis
Bernstein, E. and C. D. Allis (2005). "RNA meets chromatin." Genes Dev 19(14): 1635-55.
Bernstein, E., A. A. Caudy, et al. (2001). "Role for a bidentate ribonuclease in the initiation step of RNA interference." Nature 409(6818): 363-6.
Bertrand, F. E., R. Golub, et al. (1998). "V(H) gene replacement occurs in the spleen and bone marrow of non-autoimmune quasi-monoclonal mice." Eur J Immunol 28(10): 3362-70.
Bolland, D. J., A. L. Wood, et al. (2004). "Antisense intergenic transcription in V(D)J recombination." Nat Immunol 5(6): 630-7.
Buhler, M., A. Paillusson, et al. (2004). "Efficient downregulation of immunoglobulin mu mRNA with premature translation-termination codons requires the 5'-half of the VDJ exon." Nucleic Acids Res 32(11): 3304-15.
Burnet, F. M. (1960). "Theories of immunity." Perspect Biol Med 3: 447-58.
Buzina, A. and M. J. Shulman (1999). "Infrequent translation of a nonsense codon is sufficient to decrease mRNA level." Mol Biol Cell 10(3): 515-24.
Carpino, N., W. E. Thierfelder, et al. (2004). "Absence of an essential role for thymic stromal lymphopoietin receptor in murine B-cell development." Mol Cell Biol 24(6): 2584-92.
Cascalho, M., A. Ma, et al. (1996). "A quasi-monoclonal mouse." Science 272(5268): 1649-52.
Cascalho, M., D. A. Martin, et al. (1999). "A mouse with a monoclonal primary immunoglobulin repertoire not further diversified by V-gene replacement." Dev Immunol 7(1): 43-50.
Cascalho, M., J. Wong, et al. (1997). "VH gene replacement in hyperselected B cells of the quasimonoclonal mouse." J Immunol 159(12): 5795-801.
Cebra, J. J., J. E. Colberg, et al. (1966). "Rabbit lymphoid cells differentiated with respect to alpha-, gamma-, and mu- heavy polypeptide chains and to allotypic markers Aa1 and Aa2." J Exp Med 123(3): 547-58.
94

8. Literaturverzeichnis
Chen, C., Z. Nagy, et al. (1995). "Immunoglobulin heavy chain gene replacement: a mechanism of receptor editing." Immunity 3(6): 747-55.
Chen, C., E. L. Prak, et al. (1997). "Editing disease-associated autoantibodies." Immunity 6(1): 97-105.
Chen, J., M. Trounstine, et al. (1993). "Immunoglobulin gene rearrangement in B cell deficient mice generated by targeted deletion of the JH locus." Int Immunol 5(6): 647-56.
Cherry, S. R. and D. Baltimore (1999). "Chromatin remodeling directly activates V(D)J recombination." Proc Natl Acad Sci U S A 96(19): 10788-93.
Chowdhury, D. and R. Sen (2003). "Transient IL-7/IL-7R signaling provides a mechanism for feedback inhibition of immunoglobulin heavy chain gene rearrangements." Immunity 18(2): 229-41.
Chowdhury, D. and R. Sen (2004). "Regulation of immunoglobulin heavy-chain gene rearrangements." Immunol Rev 200: 182-96.
Coleclough, C. (1983). "Chance, necessity and antibody gene dynamics." Nature 303(5912): 23-6.
Coleclough, C., R. P. Perry, et al. (1981). "Aberrant rearrangements contribute significantly to the allelic exclusion of immunoglobulin gene expression." Nature 290(5805): 372-8.
Corcoran, A. E. (2005). "Immunoglobulin locus silencing and allelic exclusion." Semin Immunol 17(2): 141-54.
Corcoran, A. E., A. Riddell, et al. (1998). "Impaired immunoglobulin gene rearrangement in mice lacking the IL-7 receptor." Nature 391(6670): 904-7.
Covey, L. R., P. Ferrier, et al. (1990). "VH to VHDJH rearrangement is mediated by the internal VH heptamer." Int Immunol 2(6): 579-83.
Decker, D. J., N. E. Boyle, et al. (1991). "The expression of the Ig H chain repertoire in developing bone marrow B lineage cells." J Immunol 146(1): 350-61.
95

8. Literaturverzeichnis
Duber, S., H. Engel, et al. (2003). "Germline transcripts of immunoglobulin light chain variable regions are structurally diverse and differentially expressed." Mol Immunol 40(8): 509-16.
Early, P., H. Huang, et al. (1980). "An immunoglobulin heavy chain variable region gene is generated from three segments of DNA: VH, D and JH." Cell 19(4): 981-92.
Eilat, D. and R. Fischel (1991). "Recurrent utilization of genetic elements in V regions of antinucleic acid antibodies from autoimmune mice." J Immunol 147(1): 361-8.
Friend, S. L., S. Hosier, et al. (1994). "A thymic stromal cell line supports in vitro development of surface IgM+ B cells and produces a novel growth factor affecting B and T lineage cells." Exp Hematol 22(3): 321-8.
Frischmeyer, P. A. and H. C. Dietz (1999). "Nonsense-mediated mRNA decay in health and disease." Hum Mol Genet 8(10): 1893-900.
Fugmann, S. D., A. I. Lee, et al. (2000). "The RAG proteins and V(D)J recombination: complexes, ends, and transposition." Annu Rev Immunol 18: 495-527.
Fujio, K., T. Nosaka, et al. (2000). "Molecular cloning of a novel type 1 cytokine receptor similar to the common gamma chain." Blood 95(7): 2204-10.
Fuxa, M., J. Skok, et al. (2004). "Pax5 induces V-to-DJ rearrangements and locus contraction of the immunoglobulin heavy-chain gene." Genes Dev 18(4): 411-22.
Goldmit, M., Y. Ji, et al. (2005). "Epigenetic ontogeny of the Igk locus during B cell development." Nat Immunol 6(2): 198-203.
Goldmit, M., M. Schlissel, et al. (2002). "Differential accessibility at the kappa chain locus plays a role in allelic exclusion." Embo J 21(19): 5255-61.
Golub, R., D. Martin, et al. (2001). "VH gene replacement in thymocytes." J Immunol 166(2): 855-60.
Gong, S. and M. C. Nussenzweig (1996). "Regulation of an early developmental checkpoint in the B cell pathway by Ig beta." Science 272(5260): 411-4.
96

8. Literaturverzeichnis
Grawunder, U., T. M. Leu, et al. (1995). "Down-regulation of RAG1 and RAG2 gene expression in preB cells after functional immunoglobulin heavy chain rearrangement." Immunity 3(5): 601-8.
Grawunder, U., T. H. Winkler, et al. (1996). "Regulation of recombination activating gene expression during lymphocyte development." Curr Top Microbiol Immunol 217: 31-43.
Grosschedl, R., D. Weaver, et al. (1984). "Introduction of a mu immunoglobulin gene into the mouse germ line: specific expression in lymphoid cells and synthesis of functional antibody." Cell 38(3): 647-58.
Haas, I. G. and M. R. Wabl (1984). "Immunoglobulin heavy chain toxicity in plasma cells is neutralized by fusion to pre-B cells." Proc Natl Acad Sci U S A 81(22): 7185-8.
Hamilton, A. J. and D. C. Baulcombe (1999). "A species of small antisense RNA in posttranscriptional gene silencing in plants." Science 286(5441): 950-2.
Hammond, S. M. (2005). "Dicing and slicing: the core machinery of the RNA interference pathway." FEBS Lett 579(26): 5822-9.
Hammond, S. M., E. Bernstein, et al. (2000). "An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells." Nature 404(6775): 293-6.
Hieter, P. A., S. J. Korsmeyer, et al. (1981). "Human immunoglobulin kappa light-chain genes are deleted or rearranged in lambda-producing B cells." Nature 290(5805): 368-72.
Ivanov, II, R. L. Schelonka, et al. (2005). "Development of the expressed Ig CDR-H3 repertoire is marked by focusing of constraints in length, amino acid use, and charge that are first established in early B cell progenitors." J Immunol 174(12): 7773-80.
Jack, H. M., J. Berg, et al. (1989). "Translation affects immunoglobulin mRNA stability." Eur J Immunol 19(5): 843-7.
Johnston, C. M., A. L. Wood, et al. (2006). "Complete sequence assembly and characterization of the C57BL/6 mouse Ig heavy chain V region." J Immunol 176(7): 4221-34.
97

8. Literaturverzeichnis
Jung, D., C. Giallourakis, et al. (2006). "Mechanism and control of V(D)J recombination at the immunoglobulin heavy chain locus." Annu Rev Immunol 24: 541-70.
Karasuyama, H., A. Rolink, et al. (1994). "The expression of Vpre-B/lambda 5 surrogate light chain in early bone marrow precursor B cells of normal and B cell-deficient mutant mice." Cell 77(1): 133-43.
Kawano, Y., S. Yoshikawa, et al. (2005). "Selection of stereotyped VH81X-microH chains via pre-B cell receptor early in ontogeny and their conservation in adults by marginal zone B cells." Int Immunol 17(7): 857-67.
Kawano, Y., S. Yoshikawa, et al. (2006). "Pre-B cell receptor assesses the quality of IgH chains and tunes the pre-B cell repertoire by delivering differential signals." J Immunol 177(4): 2242-9.
Kawasaki, H. and K. Taira (2004). "Induction of DNA methylation and gene silencing by short interfering RNAs in human cells." Nature 431(7005): 211-7.
Keyna, U., S. E. Applequist, et al. (1995). "Ig mu heavy chains with VH81X variable regions do not associate with lambda 5." Ann N Y Acad Sci 764: 39-42.
Kitamura, D. and K. Rajewsky (1992). "Targeted disruption of mu chain membrane exon causes loss of heavy-chain allelic exclusion." Nature 356(6365): 154-6.
Kleinfield, R., R. R. Hardy, et al. (1986). "Recombination between an expressed immunoglobulin heavy-chain gene and a germline variable gene segment in a Ly 1+ B-cell lymphoma." Nature 322(6082): 843-6.
Kline, G. H., L. Hartwell, et al. (1998). "Pre-B cell receptor-mediated selection of pre-B cells synthesizing functional mu heavy chains." J Immunol 161(4): 1608-18.
Klonowski, K. D., L. L. Primiano, et al. (1999). "Atypical VH-D-JH rearrangements in newborn autoimmune MRL mice." J Immunol 162(3): 1566-72.
Knight, K. L. and R. A. Barrington (1998). "Somatic diversification of IgH genes in rabbit." Immunol Rev 162: 37-47.
Kohler, G. (1980). "Immunoglobulin chain loss in hybridoma lines." Proc Natl Acad Sci U S A 77(4): 2197-9.
98

8. Literaturverzeichnis
Koralov, S. B., T. I. Novobrantseva, et al. (2006). "Antibody repertoires generated by VH replacement and direct VH to JH joining." Immunity 25(1): 43-53.
Korsmeyer, S. J., P. A. Hieter, et al. (1982). "Normal human B cells display ordered light chain gene rearrangements and deletions." J Exp Med 156(4): 975-85.
Kosak, S. T., J. A. Skok, et al. (2002). "Subnuclear compartmentalization of immunoglobulin loci during lymphocyte development." Science 296(5565): 158-62.
Kottmann, A. H., B. Zevnik, et al. (1994). "A second promoter and enhancer element within the immunoglobulin heavy chain locus." Eur J Immunol 24(4): 817-21.
Krangel, M. S. (2003). "Gene segment selection in V(D)J recombination: accessibility and beyond." Nat Immunol 4(7): 624-30.
Lakso, M., J. G. Pichel, et al. (1996). "Efficient in vivo manipulation of mouse genomic sequences at the zygote stage." Proc Natl Acad Sci U S A 93(12): 5860-5.
Lejeune, F. and L. E. Maquat (2005). "Mechanistic links between nonsense-mediated mRNA decay and pre-mRNA splicing in mammalian cells." Curr Opin Cell Biol 17(3): 309-15.
Lennon, G. G. and R. P. Perry (1985). "C mu-containing transcripts initiate heterogeneously within the IgH enhancer region and contain a novel 5'-nontranslatable exon." Nature 318(6045): 475-8.
Levin, S. D., R. M. Koelling, et al. (1999). "Thymic stromal lymphopoietin: a cytokine that promotes the development of IgM+ B cells in vitro and signals via a novel mechanism." J Immunol 162(2): 677-83.
Li, S. and M. F. Wilkinson (1998). "Nonsense surveillance in lymphocytes?" Immunity 8(2): 135-41.
Liang, H. E., L. Y. Hsu, et al. (2002). "The "dispensable" portion of RAG2 is necessary for efficient V-to-DJ rearrangement during B and T cell development." Immunity 17(5): 639-51.
Liang, H. E., L. Y. Hsu, et al. (2004). "Variegated transcriptional activation of the immunoglobulin kappa locus in pre-b cells contributes to the allelic exclusion of light-chain expression." Cell 118(1): 19-29.
99

8. Literaturverzeichnis
Madan, M. K., R. Golub, et al. (2000). "Specific antibody production by V(H)-gene replacement." Eur J Immunol 30(8): 2404-11.
Maquat, L. E. (2005). "Nonsense-mediated mRNA decay in mammals." J Cell Sci 118(Pt 9): 1773-6.
McBlane, J. F., D. C. van Gent, et al. (1995). "Cleavage at a V(D)J recombination signal requires only RAG1 and RAG2 proteins and occurs in two steps." Cell 83(3): 387-95.
Meister, S. (2004). "Untersuchungen zu Signalkompetenz paarungsunfähiger und paarungsfähiger µ-schwerer Immunglobulinketten in Abwesenheit von leichten Ketten." Promotionsarbeit.
Mielenz, D., A. Ruschel, et al. (2003). "Immunoglobulin mu heavy chains do not mediate tyrosine phosphorylation of Ig alpha from the ER-cis-Golgi." J Immunol 171(6): 3091-101.
Minegishi, Y., E. Coustan-Smith, et al. (1999). "Mutations in Igalpha (CD79a) result in a complete block in B-cell development." J Clin Invest 104(8): 1115-21.
Mostoslavsky, R., F. W. Alt, et al. (2004). "The lingering enigma of the allelic exclusion mechanism." Cell 118(5): 539-44.
Mostoslavsky, R., N. Singh, et al. (1998). "Kappa chain monoallelic demethylation and the establishment of allelic exclusion." Genes Dev 12(12): 1801-11.
Nadel, B., A. Tang, et al. (1998). "V(H) replacement is unlikely to contribute significantly to receptor editing due to an ineffectual embedded recombination signal sequence." Mol Immunol 35(4): 227-32.
Neu-Yilik, G., N. H. Gehring, et al. (2004). "Nonsense-mediated mRNA decay: from vacuum cleaner to Swiss army knife." Genome Biol 5(4): 218.
Nussenzweig, M. C. (1998). "Immune receptor editing: revise and select." Cell 95(7): 875-8.
Nussenzweig, M. C., A. C. Shaw, et al. (1987). "Allelic exclusion in transgenic mice that express the membrane form of immunoglobulin mu." Science 236(4803): 816-9.
100

8. Literaturverzeichnis
Papavasiliou, F., Z. Misulovin, et al. (1995). "The role of Ig beta in precursor B cell transition and allelic exclusion." Science 268(5209): 408-11.
Pelanda, R., S. Schwers, et al. (1997). "Receptor editing in a transgenic mouse model: site, efficiency, and role in B cell tolerance and antibody diversification." Immunity 7(6): 765-75.
Pernis, B., G. Chiappino, et al. (1965). "Cellular localization of immunoglobulins with different allotypic specificities in rabbit lymphoid tissues." J Exp Med 122(5): 853-76.
Radic, M. Z., J. Erikson, et al. (1993). "B lymphocytes may escape tolerance by revising their antigen receptors." J Exp Med 177(4): 1165-73.
Radic, M. Z. and M. Zouali (1996). "Receptor editing, immune diversification, and self-tolerance." Immunity 5(6): 505-11.
Rajewsky, K. (1996). "Clonal selection and learning in the antibody system." Nature 381(6585): 751-8.
Reth, M., P. Gehrmann, et al. (1986). "A novel VH to VHDJH joining mechanism in heavy-chain-negative (null) pre-B cells results in heavy-chain production." Nature 322(6082): 840-2.
Ritchie, K. A., R. L. Brinster, et al. (1984). "Allelic exclusion and control of endogenous immunoglobulin gene rearrangement in kappa transgenic mice." Nature 312(5994): 517-20.
Roldan, E., M. Fuxa, et al. (2005). "Locus 'decontraction' and centromeric recruitment contribute to allelic exclusion of the immunoglobulin heavy-chain gene." Nat Immunol 6(1): 31-41.
Rolink, A., U. Grawunder, et al. (1994). "IL-2 receptor alpha chain (CD25, TAC) expression defines a crucial stage in pre-B cell development." Int Immunol 6(8): 1257-64.
Rooney, S., J. Chaudhuri, et al. (2004). "The role of the non-homologous end-joining pathway in lymphocyte development." Immunol Rev 200: 115-31.
Rusconi, S. and G. Kohler (1985). "Transmission and expression of a specific pair of rearranged immunoglobulin mu and kappa genes in a transgenic mouse line." Nature 314(6009): 330-4.
101

8. Literaturverzeichnis
Schlissel, M. S. (2003). "Regulating antigen-receptor gene assembly." Nat Rev Immunol 3(11): 890-9.
Schroeder, H. W., Jr. (2006). "Similarity and divergence in the development and expression of the mouse and human antibody repertoires." Dev Comp Immunol 30(1-2): 119-35.
Shinkai, Y., G. Rathbun, et al. (1992). "RAG-2-deficient mice lack mature lymphocytes owing to inability to initiate V(D)J rearrangement." Cell 68(5): 855-67.
Singh, N., Y. Bergman, et al. (2003). "Biallelic germline transcription at the kappa immunoglobulin locus." J Exp Med 197(6): 743-50.
Skok, J. A., K. E. Brown, et al. (2001). "Nonequivalent nuclear location of immunoglobulin alleles in B lymphocytes." Nat Immunol 2(9): 848-54.
Sonoda, E., Y. Pewzner-Jung, et al. (1997). "B cell development under the condition of allelic inclusion." Immunity 6(3): 225-33.
Spanopoulou, E., P. Cortes, et al. (1995). "Localization, interaction, and RNA binding properties of the V(D)J recombination-activating proteins RAG1 and RAG2." Immunity 3(6): 715-26.
Taki, S., F. Schwenk, et al. (1995). "Rearrangement of upstream DH and VH genes to a rearranged immunoglobulin variable region gene inserted into the DQ52-JH region of the immunoglobulin heavy chain locus." Eur J Immunol 25(7): 1888-96.
Teixeira, D., U. Sheth, et al. (2005). "Processing bodies require RNA for assembly and contain nontranslating mRNAs." Rna 11(4): 371-82.
ten Boekel, E., F. Melchers, et al. (1997). "Changes in the V(H) gene repertoire of developing precursor B lymphocytes in mouse bone marrow mediated by the pre-B cell receptor." Immunity 7(3): 357-68.
Tiegs, S. L., D. M. Russell, et al. (1993). "Receptor editing in self-reactive bone marrow B cells." J Exp Med 177(4): 1009-20.
Tonegawa, S. (1983). "Somatic generation of antibody diversity." Nature 302(5909): 575-81.
102

8. Literaturverzeichnis
Tuschl, T., P. D. Zamore, et al. (1999). "Targeted mRNA degradation by double-stranded RNA in vitro." Genes Dev 13(24): 3191-7.
Verdel, A. and D. Moazed (2005). "RNAi-directed assembly of heterochromatin in fission yeast." FEBS Lett 579(26): 5872-8.
Vettermann, C., K. Herrmann, et al. (2006). "Powered by pairing: the surrogate light chain amplifies immunoglobulin heavy chain signaling and pre-selects the antibody repertoire." Semin Immunol 18(1): 44-55.
Wabl, M. and C. Steinberg (1982). "A theory of allelic and isotypic exclusion for immunoglobulin genes." Proc Natl Acad Sci U S A 79(22): 6976-8.
Watson, L. C., C. S. Moffatt-Blue, et al. (2006). "Paucity of V-D-D-J Rearrangements and VH Replacement Events in Lupus Prone and Nonautoimmune TdT-/- and TdT+/+ Mice." J Immunol 177(2): 1120-8.
Weaver, D., F. Costantini, et al. (1985). "A transgenic immunoglobulin mu gene prevents rearrangement of endogenous genes." Cell 42(1): 117-27.
Weiler, E. (1965). "Differential activity of allelic gamma-globulin genes in antibody-producing cells." Proc Natl Acad Sci U S A 54(6): 1765-72.
Weinberg, M. S., L. M. Villeneuve, et al. (2006). "The antisense strand of small interfering RNAs directs histone methylation and transcriptional gene silencing in human cells." Rna 12(2): 256-62.
Wilson, C. B. and M. Merkenschlager (2006). "Chromatin structure and gene regulation in T cell development and function." Curr Opin Immunol 18(2): 143-51.
Yancopoulos, G. D. and F. W. Alt (1985). "Developmentally controlled and tissue-specific expression of unrearranged VH gene segments." Cell 40(2): 271-81.
Zamore, P. D., T. Tuschl, et al. (2000). "RNAi: double-stranded RNA directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals." Cell 101(1): 25-33.
Zemlin, M., M. Klinger, et al. (2003). "Expressed murine and human CDR-H3 intervals of equal length exhibit distinct repertoires that differ in their amino acid composition and predicted range of structures." J Mol Biol 334(4): 733-49.
103

8. Literaturverzeichnis
Zhang, J., X. Sun, et al. (1998). "At least one intron is required for the nonsense-mediated decay of triosephosphate isomerase mRNA: a possible link between nuclear splicing and cytoplasmic translation." Mol Cell Biol 18(9): 5272-83.
Zhang, Y., J. Wienands, et al. (1998). "Induction of the antigen receptor expression on B lymphocytes results in rapid competence for signaling of SLP-65 and Syk." Embo J 17(24): 7304-10.
Zhang, Z., P. D. Burrows, et al. (2004). "The molecular basis and biological significance of VH replacement." Immunol Rev 197: 231-42.
Zhang, Z., Y. H. Wang, et al. (2003). "Molecular mechanism of serial VH gene replacement." Ann N Y Acad Sci 987: 270-3.
Zhang, Z., M. Zemlin, et al. (2003). "Contribution of Vh gene replacement to the primary B cell repertoire." Immunity 19(1): 21-31.
104

9. Anhang
9 Anhang
9.1 Veröffentlichungen
Paul E, Lutz J, Erikson J, Carroll MC
Germinal center checkpoints in B cell tolerance in 3H9 transgenic mice.
Int Immunol. 2004 Feb;16(2):377-84.
Lutz J, Muller W, Jack HM
VH Replacement Rescues Progenitor B Cells with Two Nonproductive VDJ Alleles
J. Immunol., Nov 2006; 177: 7007 - 7014.
Vettermann C, Lutz J, Selg M, Bosl M and Jack HM
Genomic suppression of murine B29/Ig-b promoter-driven transgenes
Eur J Immunol. 2006 Dec, in press
105

9. Anhang
106

9. Anhang
9.2 Lebenslauf
Persönliche Daten Name: Johannes Lutz
Geboren: 14.5.1975 in Wiesbaden
Familienstand: Ledig
Schulbildung Sept. 1981 bis Juli 1985 Grundschule, Wiesbaden.
Sept. 1985 bis Juli 1991 Leibniz-Gymnasium Wiesbaden.
Sept. 1991 bis Juli 1994 Carl-von-Ossietzky Oberstufen-Gymnasium Wiesbaden,
abgeschlossen mit Abitur.
Zivildienst Nov. 1994 bis Sept. 1995 Zivildienstleistender im Rettungsdienst, Wiesbaden.
Studium Okt. 1995 bis März 2001 Biologiestudium auf Diplom an der Universität Konstanz.
Okt. 1998 bis Jan. 1999 Auslandssemester an der University of Wales,
Aberystwyth, UK.
März 2000 bis Feb. 2001 Diplomarbeit am Center for Biomedical Research
(CBR), Harvard Medical School, Boston, USA. Thema
der Diplomarbeit: "Autoimmunity in Complement
Deficient Mice".
März 2001 Abschluss als Diplom-Biologe.
April 2001 bis April 2002 Fortführung des Diplom-Projektes als wissenschaftlicher
Angestellter am CBR.
Mai 2002 bis Dez. 2006 Promotion in der Abteilung für Molekulare Immunologie,
Medizinische Klinik III, Nikolaus-Fiebiger-Zentrum,
Universität Erlangen-Nürnberg. Thema der Promotion:
"Untersuchungen zur Kontrolle der VDJ-Rekombination
am IgH-Lokus".
107

9. Anhang
108

9. Anhang
9.3 Danksagung
Besonders danken möchte ich Herrn Prof. Dr. Hans-Martin Jäck für die Überlassung
des attraktiven Themas, die großzügige Unterstützung, die exzellenten
Arbeitsbedingungen und die große Freiheit bei der experimentellen Planung und
Umsetzung meines Projektes.
Herrn Prof. Dr. Thomas Winkler danke ich für die vielen wissenschaftlichen
Anregungen und die Übernahme des Zweitgutachtens.
Christian Vettermann danke ich für unzählige wissenschaftliche (und nicht-
wissenschaftliche) Diskussionen und kritische Anregungen, und Jürgen Wittmann
besonders für die Einführung in das RNA-Feld, den NMD und das Klonieren.
Edith Roth und Christine Albert danke ich für die Unterstützung bei der Durchführung
einiger Experimente, Prof. Dr. Werner Müller, Anke Samuels, Maria Ebel und Dr.
Martin Hafner für die ES-Zellkultur und die Herstellung Keimbahn-kompetenter
chimärer Mäuse und Dr. Chander Raman für die Hilfe bei der Herstellung der
transgenen Mäuse.
Bei allen Mitarbeitern der Molekularen Immunologie möchte ich mich für
immerwährende Hilfs- und Diskussionsbereitschaft und die schöne Zeit bedanken.
Ein besonderer Dank gilt meiner Freundin Christina, die alle Höhen und Tiefen
meines Forscher-Daseins mitbekam, sowie meinen Eltern und meiner Familie, die
meinen Weg nicht nur immer unterstützt haben, sondern mir diesen auch erst
ermöglicht haben.
109





























