Embed Size (px)
Citation preview

Aus dem Anatomischen Institut der Tierärztlichen Hochschule Hannover
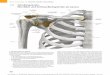
Zum Bandapparat des Ellbogengelenkes des Hundes insbesondere zu Verlauf und Struktur der
Kollateralbänder
INAUGURAL-DISSERTATION
Zur Erlangung des Grades einer Doktorin der Veterinärmedizin
(Dr. med. vet.) durch die Tierärztliche Hochschule Hannover
Vorgelegt von Miriam-Jasmin Hemmes
aus Leer
Hannover 2004

Aus dem Anatomischen Institut der Tierärztlichen Hochschule Hannover
Zum Bandapparat des Ellbogengelenkes des Hundes insbesondere zu Verlauf und Struktur der
Kollateralbänder
INAUGURAL-DISSERTATION
Zur Erlangung des Grades einer Doktorin der Veterinärmedizin
(Dr. med. vet.) durch die Tierärztliche Hochschule Hannover
Vorgelegt von Miriam-Jasmin Hemmes
aus Leer
Hannover 2004

Wissenschaftliche Betreuung: Univ.-Prof. Dr. med vet. H. Waibl 1. Gutachter: Univ.-Prof. Dr. med vet. H. Waibl 2. Gutachter: Univ.-Prof. Dr. med vet. M. Fehr Tag der mündlichen Prüfung: 27.05.2004

Meiner Familie und Heiko in Liebe und Dankbarkeit gewidmet
und in Erinnerung an meinen Opa Hemme Hemmes


Inhaltsverzeichnis
Abkürzungen
1. Einleitung
2. Schrifttum 2.1 Bau und Funktion synovialer Gelenke
2.2 Spezielle Anatomie des Ellbogengelenks (Articulatio cubiti)
2.2.1 Gelenktyp und Gelenkbewegung
2.2.2 Knöcherne Grundlage und Artikulationsflächen
2.2.3 Gelenkkapsel
2.2.4 Bandapparat
2.2.4.1 Allgemeines zum Aufbau von Gelenkbändern
2.2.4.2 Bänder des Ellbogengelenks
2.2.5 Muskeln
2.2.6 Schleimbeutel
2.2.7 Sesamoide Strukturen
3. Material und Methode
3.1 Makroskopische Untersuchung
3.1.1 Präparationstechnik
3.1.2 Längen- und Breitenmessung an den Gelenkbändern
Schematische Darstellung der Vermessungsart
3.1.3 Vorversuche
3.1.4 Dokumentation
3.1.5 Statistische Auswertung
3.2 Mikroskopische Untersuchung
3.2.1 Untersuchungsgut
3.2.2 Entnahmetechnik
3.2.3 Probenaufbereitung für die mikroskopische Untersuchung
3.2.4 Färbungen

3.2.5 Untersuchung mit dem Zeiss Photomikroskop III mit Polarisations-
ausrüstung
3.2.6 Dokumentation Mikroskopie
4. Ergebnisse 4.1 Makroskopische Untersuchung und deskriptive Statistik
4.1.1 Beschreibung des Bandapparates der Art. cubiti und der Art. radioulnaris
proximalis
4.1.1.1 Im Hinblick auf Ursprung, Verlauf und Ansatz
4.1.1.1.1 Ligamentum collaterale laterale
4.1.1.1.2 Ligamentum collaterale mediale
4.1.1.1.3 Ligamentum anulare radii
4.1.1.1.4 Ligamentum obliquum
4.1.1.1.5 Ligamentum olecrani
4.1.1.2 Statistik: Längen- und Breitenmessung des Bandapparates
4.2 Mikroskopische Untersuchung
4.2.1 Auswertung mit dem Lichtmikroskop
4.2.2 Auswertung im polarisierten Licht
5. Diskussion 5.1 Makroskopische Untersuchung
5.2 Messergebnisse
5.3 Mikroskopische Untersuchung
6. Zusammenfassung 7. Summary
8. Literaturverzeichnis 9. Anhang: Tabellen

Abkürzungen Art., Artt. Articulatio, Articulationes
Abb. Abbildung
coll. collateral(e)
cm Zentimeter
DSH Deutscher Schäferhund
DSH-Mix Deutscher Schäferhund-Mischling
ext. extensor
FPC Fragmentierter Processus coronoideus
lat. lateral
Lig., Ligg. Ligamentum, Ligamenta
M., Mm. Musculus, Musculi
med. medial
mm Millimeter
Proc., Procc. Processus, Processus
Tab. Tabelle

1 1. Einleitung In der Literatur gibt es ausführliche Beschreibungen zu Anatomie und Funktion des
Ellbogengelenkes beim Hund. Dennoch bestehen Unstimmigkeiten zu Ursprung,
Verlauf und Ansatz des Bandapparates der Articulatio cubiti und der Articulatio
radioulnaris proximalis. Die Kollateralbänder (Ligamentum collaterale laterale und
Ligamentum collaterale mediale) bilden für die Bewegung des Gelenkes eine
funktionelle Einheit. Auffällig ist zudem, dass im lateralen Kollateralband Knorpel
oder sogar Sesambeine gefunden werden.
Weitere Aussagen über bauliche Unterschiede zwischen dem lateralen und medialen
Seitenband fehlen der Literatur. Ebenso sind kaum Messwerte zum gesamten
Bandapparat bisher bekannt. Zur Schließung dieser Lücken werden alle Bänder des
Ellbogenglenkes vermessen (Längen und Breiten) sowie ihre Funktion besprochen.

2 2. Schrifttum
2.1 Bau und Funktion synovialer Gelenke
Das Ellbogengelenk ist ein synoviales Gelenk, Articulatio synovialis. Diese
Gelenke treten überall dort auf, wo eine große Beweglichkeit gefordert ist. Sie
werden auch echte Gelenke (EVANS 1993a) genannt und zeichnen sich durch einen
Gelenkspalt, Cavum articulare, mit einer Gelenkkapsel, Capsula articularis, aus.
Die Gelenkhöhle bzw. der Gelenkspalt ist von Gelenkflüssigkeit, Synovia, gefüllt. An
den Artikulationsflächen befindet sich Gelenkknorpel, Cartilago articularis, welcher
die Knochenenden überzieht. Zu den Merkmalen eines echten Gelenkes zählen auch
die für die Bewegungsführung wichtigen Gelenkbänder, Ligg. articularia (NICKEL et
al. 1992).
Die Capsula articularis besteht aus einem äußeren faserreichen Stratum fibrosum
und einem inneren lockeren Stratum synoviale (NICKEL et al. 1992). Das Stratum
fibrosum entspringt am Humerus, räumlich isoliert vom Stratum synoviale (STASZYK
u. GASSE 2001b).
Die Gelenkkapsel kann Ausbuchtungen (Recessus) bilden, die eine größere
Bewegungsfreiheit ermöglichen. Sie befinden sich dann an der Gelenkkehle und am
Gelenkscheitel. Leicht erreichbare Recessus eignen sich zur Gelenkinjektion
(EVANS 1993a; VOLLMERHAUS et al. 1994b). Kleinere Recessus können als
Gleitlager für Sehnen dienen, die nach SAJONSKI (1969) auch Kapselschleimbeutel
genannt werden.
Die Gelenkschmiere, Synovia, ist ein Ultrafiltrat des Blutplasmas. Sie überzieht die
Gelenkflächen. Die Synovia ist dünnflüssig bis fadenziehend, farblos bis leicht
gelblich (VOLLMERHAUS et al. 1994b). Sie wird vom Stratum synoviale der
Gelenkkapsel sezerniert und auch von dieser Schicht resorbiert. Die stoffliche
Zusammensetzung und ihre physikalischen Eigenschaften dienen der Ernährung des
bradytrophen Gelenkknorpels wie der Gewährleistung eines reibungsarmen Gleitens
der Gelenkflächen aufeinander (BANKS 1993).
Die Ligg. articularia sind als Führungs- und Hemmungsbänder aus kollagenem
Bindegewebe aufgebaut. Meistens liegen sie außerhalb der Kapsel und werden

3 deshalb auch Ligamenta extracapsularia genannt. Sie können auch
unterschiedliche Mengen elastischer Fasern enthalten. Neben den Ligamenta
extracapsularia gibt es auch die Ligg. capsularia. Als Verstärkungszüge sie sind in
die Gelenkkapsel eingelagert und werden so vor dem Einklemmen geschützt.
Schließlich gibt es auch Gelenkbänder, die durch den Gelenkspalt verlaufen, also
innerhalb der Gelenkkapsel liegen: Ligamenta intracapsularia (NICKEL et al. 1992;
VOLLMERHAUS et al.1994b).

4 2.2. Spezielle Anatomie des Ellbogengelenks
2.2.1 Gelenktyp und Gelenkbewegung
Gelenktyp
Zwischen dem distalen Endstück des Humerus und den proximalen Endstücken von
Radius und Ulna sind drei Teilgelenke ausgebildet: die Art. humeroradialis, die Art. humeroulnaris und die Art. radioulnaris proximalis, die in einer gemeinsamen
Gelenkkapsel eingefasst sind (NICKEL et al. 1992).
Das Ellbogengelenk ist im Standwinkel leicht gebeugt. Der Standwinkel beträgt beim
lebenden Tier: Deutscher Schäferhund 137°, beim Dobermann 138°, bei der
Deutschen Dogge 159° und beim Teckel 127-124° (MAI 1995).
Während nach der derzeitig gültigen Nomenklatur der Nomina Anatomica Veterinaria
(NAV 1994) nur die Art. humeroulnaris und die Art. humeroradialis zur Art. cubiti, Ellbogengelenk, gehören, zählt die humanmedizinische Nomina Anatomica (1985)
auch die Art. radioulnaris proximalis hinzu. Da die Art. radioulnaris proximalis beim
Hund mit den anderen Teilgelenken des Ellbogengelenkes kommuniziert, wird sie
auch von EVANS (1993a) diesem zugerechnet. Andere Autoren (siehe z. B. NICKEL
et al. 1992; KÖNIG u. LIEBICH 1999) schließen sich der Auflistung der NAV (1994)
an, was zur unterschiedlichen Beurteilung der Funktion des „Ellbogengelenkes“ führt.
So betrachten zum Beispiel KÖNIG u. LIEBICH (1999) die Art. cubiti (Art.
humeroulnaris und Art humeroradialis) des Hundes als Walzengelenk, Art. condylaris. Durch die Führungskämme und –rinnen der Gelenkwalze, Trochlea
humeri, sowie das tiefe Einsinken des Olecranon in die Fossa olecrani des Humerus
ist allein ein Strecken und Beugen des so definierten Ellbogengelenkes möglich. Das
Ellbogengelenk des Dackels hingegen ist ein Schraubengelenk, Art. cochlearis.
Durch die schräggestellte Gelenkwalze des Humerus wird der Unterarm bei der
Beugung gleichzeitig um 15° auswärtsgeführt (VOLLMERHAUS et al. 1994b). Bei

5 allen Rassen werden seitliche Bewegungsabläufe fast vollständig verhindert.
Rotationsbewegungen sind nur zwischen Radius und Ulna möglich.
Die Rotation nach außen ist bei den von MAI (1995) untersuchten Rassen (DSH,
Dobermann, Deutschen Dogge) größer, als die Rotation nach innen. Eine Ausnahme
stellt hier der Rauhhaarteckel dar.
Im Gegensatz dazu beschreiben VAN HERPEN (1988) und EVANS (1993a) die drei
Teilgelenke des Ellbogengelenks funktionell unabhängig voneinander: die Art.
humeroradialis trägt den Großteil des Gewichtes; die Art. humeroulnaris bewirkt die
strikte Wechselwirkung und die Art. radioulnaris proximalis erlaubt die Rotation des
Unterarmes.
Auch LIPPERT (1990) teilt das Ellbogengelenk des Menschen funktionell in seine
drei Teilgelenke ein:
Die Art. humeroulnaris ist ein einachsiges Scharniergelenk. Es führt Beugung und
Streckung aus.
Die Art. radioulnaris proximalis ist ebenfalls ein einachsiges Drehgelenk. Es führt
Radbewegungen aus: Einwärts- und Auswärtsdrehungen.
Die Art. humeroradialis ist ein zweiachsiges Gelenk. Es werden Scharnier- und
Drehbewegungen ausgeführt.
Die Befunde nach selektiver Durchtrennung der Kollateralbänder bestätigen diese
Sichtweise, da ein Großteil der Bewegung bei Pronation und Supination neben der
Art. radioulnaris proximalis im Humeroradialgelenk stattfindet. Das Humeroulnarge-
lenk wird hingegen nur wenig rotiert (MONTAVON u. SALVODELLI 1995).
Die Seitenbänder des Gelenkes, das Ligamentum collaterale laterale und das
Ligamentum collaterale mediale, sind stark entwickelt und tragen wesentlich zur
strikten Wechselwirkung der Art. humeroulnaris bei (EVANS 1993a).
Exzentrische Insertionsstellen der Seitenbänder über der Drehachse lassen die Art.
cubiti als ein Schnappgelenk wirken. Bei Streckung des Gelenkes wird also ein

6 Widerstand überwunden. In der Mittelstellung des Gelenkes sind die Seitenbänder
maximal angespannt, und der Processus anconeus schnappt regelrecht in die Fossa
olecrani ein (NICKEL et al. 1992; KÖNIG u. LIEBICH 1999).
Die Schnappwirkung beruht allerdings nicht auf einer elastischen Verkürzung des
Bandes, da es sich hier um ein straffes Band handelt, sondern auf einer
Komprimierung des Gelenkknorpels. In der Schnappstellung wird das Gelenk unter
einen hohen Druck gesetzt, die Bänder geben hierbei nicht nach, sondern der
Knorpel wird komprimiert. Da dieser durch die Synovia glatt und schlüpfrig an seiner
Oberfläche ist, genügt ein geringer Anstoß in eine Richtung und das Gelenk schnellt
über die Stelle der maximalen Spannung hinüber; es schnappt in die gewünschte
Position (AICHEL 1925). Die Stellung der maximalen Spannung wird in der Literatur
von FICK (1910) als kritische Stellung bezeichnet.
Gelenkbewegung
Die Rotationsbewegung des Unterarms erfolgt in der Weise, dass der Radius
gegenüber der feststehenden Ulna um seine Längsachse rotiert, was beim
Menschen in viel größerem Maße möglich ist als bei Hund und Katze. In der
Pronationsstellung kreuzen sich Ulna und Radius. Dagegen verlaufen in der
Supinationsstellung Ulna und Radius parallel zueinander (ZIETZSCHMANN 1943).
Der Radius muß in der Hangbeinphase bei seiner Drehung die Pfote mitnehmen, da
diese fester mit dem Radius und lockerer mit der Ulna verbunden ist. Folglich wird
der Radius in der Stützbeinphase von der Pfote als Fixpunkt benutzt. Der Körper
dreht sich mit der Ulna um diese Stütze (ROOS et al. 1992).
Diese Bewegungen werden durch das Lig. anulare radii, die Membrana interossea antebrachii und das Lig. radioulnare distale stabilisiert (KADLETZ 1932; VAN
HERPEN 1988; VOLLMERHAUS et al. 1994b).

7 Bewegungsumfang bei Rotation: Während bei der Katze eine aktive Supination
von bis zu 100° möglich ist (KÖNIG u. LIEBICH 1999), sind beim Hund nur passive
Supinationsbewegungen von ca. 50° und eine Einwärtsrotation von etwa 20° möglich
(ROOS et al. 1992). Nach EVANS (1993a) entspricht die Norm für die Pronation 40°-
50° und für die Supination 90° unter der Berücksichtigung, dass das Karpalgelenk
mitwirkt. Allerdings gibt es hierbei rassespezifische Unterschiede. Die Rotation findet
im Radioulnargelenk statt.
Der Bewegungsumfang bei Beugung und Streckung beträgt beim Dackel 100°
(ZIETZSCHMANN 1943; DOBBERSTEIN u. HOFFMANN 1961). Vermessungen bei
anderen Hunderassen ergaben höhere Werte wie zum Beispiel beim Basset 110°,
beim DSH 125° und beim Pudel 140° (VOLLMERHAUS et al. 1994b). Die Flexion
variiert von 60° bis 70°, während die Extension sich zwischen 65° bis 75° bewegt.
(VAN HERPEN 1988; EVANS 1993a).
Die Streckung wird durch drei Faktoren bremsend beeinflusst: durch das Anschlagen
der Processus anconeus in die Fossa olecrani, durch die Anspannung der vorderen
Kapselwand und durch den passiven Dehnungswiderstand der Beugemuskeln.
Die bremsenden Faktoren der Beugung sind: bei aktiver Beugung der Kontakt der
kontrahierten Muskeln kranial des Gelenkes, und bei passiver Beugung durch eine
Kraft, die von außen wirkt, sowie das Anschlagen des Radiuskopfes in der Fossa
radialis (FICK 1910; KAPANDJI 1984).
Die Kongruenz der Gelenkflächen ist nur gegeben, wenn die Ossa antebrachii ein
synchrones Längenwachstum aufweisen. Dies wiederum wird durch das Lig.
interosseum antebrachii (VOLLMERHAUS u. ROOS 1980) gewährleistet, sowie
durch das Lig. radioulnare, welches die distalen Enden der Ossa antebrachii
miteinander verbindet. Hier kommt es im Verlauf des Längenwachstums zum
sogenannten Mitnahmeeffekt. Falls dieser aussetzt, entwickeln sich die Kollateral-
bänder unterschiedlich und dadurch entsteht ein inkongruentes Gelenk (HENSCHEL
1977).

8 2.2.2 Knöcherne Grundlage und Artikulationsflächen
Die knöcherne Grundlage des Ellbogengelenks besteht aus dem distalen Anteil des
Humerus, dem Condylus humeri, und den proximalen Anteilen der beiden Ossa
antebrachii (NICKEL et al. 1992; HAHN VON DORSCHE u. SASSE 1994;
VOLLMERHAUS et al. 1994a; KÖNIG u. LIEBICH 1999).
Der Humerus besitzt bei den verschiedenen Hunderassen eine unterschiedliche
Länge und erscheint beim Dachshund und anderen chondrodystrophen Rassen
verdreht und gedrungen.
Der Humerusschaft, Corpus humeri, ist im Mittelteil drehrund und nur durch die
Crista humeri und die Crista supracondylaris, die den Sulcus musculi brachialis
begrenzen, strukturiert. Zudem ist das Corpus in der Längsachse leicht verdreht, so
daß die Querachse des Humeruskopfes und die Querachse durch den Condylus
humeri einen nach lateral offenen Winkel von ca. 15° bilden (VOLLMERHAUS et al.
1994a).
Das distale Ende des Humerus ist gegenüber dem Humerusschaft stark verbreitert
und wird, wie oben beschrieben, als Condylus humeri bezeichnet. Der Condylus trägt
zwei Gelenkkörper: Die Rolle, Trochlea, an der medialen Seite und das Köpfchen,
Capitulum humeri, an der lateralen Seite. Das Capitulum artikuliert mit der lateralen
und die Trochlea mit der medialen Hälfte der Fovea capitis radii. Die Trochlea dient
als Widerlager für die Incisura trochlearis ulnae (EVANS 1993b).
Lateral und medial befinden sich am Condylus seichte Bandgruben und kräftige
Bandhöcker. Proximal ist die Trochlea auf der kranialen Seite durch die Fossa
radialis und auf der kaudalen Seite durch die Fossa olecrani eingebuchtet. Die Fossa
olecrani nimmt in der Streckstellung des Gelenkes den Processus anconeus, den
Hakenfortsatz der Ulna, auf (DOBBERSTEIN u. HOFFMANN 1961; NICKEL et al.
1992; HAHN VON DORSCHE u. SASSE 1994; VOLLMERHAUS et al.1994a; KÖNIG
u. LIEBICH 1999).

9 Bei den meisten Hunderassen wird die Fossa radialis durch das Foramen
supratrochleare mit der Fossa olecrani verbunden. Die Häufigkeit des Auftretens bei
den einzelnen Rassen wurde noch nicht genau untersucht. Es ist lediglich
nachgewiesen, dass es bei kleineren Hunderassen nicht so häufig vorkommt. Es
fehlt meistens dem Dachshund und dem Bullterrier (MARTIN 1923; VOLLMERHAUS
et al. 1994a). Auch DETELS (1980) findet in seinen Untersuchungen, dass das
Auftreten des Foramen supratrochleare beim Dachshund die Ausnahme darstellt:
von 26 Dachshunden konnte es nur bei einem Langhaarteckel einseitig
nachgewiesen werden.
Die Rolle dieser Öffnung ist im Hinblick auf das Gelenk bzw. die Statik noch nicht
geklärt. Es könnte für die Mechanik notwendig sein. Laut SCHLEDERER (1999) ist
das Foramen supratrochleare durch ein Fettpolster ausgefüllt, in welches der
Processus anconeus bei maximaler Streckung eintaucht. Hierbei wird das Fettpolster
kranial vorgelagert. Damit kommt es zu einem Druckausgleich in der Fossa olecrani.
Fehlt dieser, kann die Traumatisierung des umgebenden Gewebes die Folge sein.
Laut STASZYK u. GASSE (2001a) ist das Foramen hingegen durch eine
bindegewebige Membran verschlossen, an der sich drei Schichten unterscheiden
lassen.
Kaudal der Trochlea erheben sich die Muskelknorren: kaudolateral der schwächere
Streckknorren, Epicondylus lateralis, und kaudomedial der stärkere Beugeknorren,
Epicondylus medialis. Am lateralen Epicondylus entspringen die Streckmuskeln des
Karpalgelenkes und der Zehengelenke am medialen die Beugemuskeln (NICKEL et
al. 1992; HAHN VON DORSCHE u. SASSE 1994). An beiden Muskelknorren heften
sich zusätzlich zur Muskulatur die Gelenkbänder, Ligg. collateralia, des
Ellbogengelenks an (KÖNIG u. LIEBICH 1999).
Der Epicondylus medialis humeri, der groß und eckig ist, verhindert, das sich die
Ulna nach medial verlagert. Dagegen ist der Epicondylus lateralis humeri abgerundet

10 und ermöglicht es dem Processus anconeus die Crista supracondylaris bei einer
Beugung über 90° zu überspringen.
Luxationen des Ellbogengelenkes stehen, nach der Luxation des Hüftgelenkes, an
zweiter Stelle der Luxationshäufigkeit (CAMPBELL 1969; STOYAK 1981;
BIDLINGMAIER 1988). Dabei ist die Luxatio antebrachii lateralis speziell in der
gebeugten Position aufgrund der anatomischen und mechanischen Gegebenheiten –
die Trochlea humeri ist sehr ausgeprägt und ragt medial weit heraus – am häufigsten
zu beobachten (HOURRIGAN 1940; CAMPBELL 1969; BUTLER 1974;
EISENMENGER 1981). In einer Untersuchung konnten bei 37 Hunden jeweils eine
Luxatio antebrachii festgestellt werden und davon hatten 91,9% eine Luxation nach
lateral (FEHR u. MEYER-LINDENBERG 1992).
Die Ossa antebrachii sind beim Hund vollständig ausgebildet und geringgradig
gegeneinander beweglich. Beide Knochen, Radius und Ulna, verbindet eine
Membrana interossea antebrachii, die das sehr enge Spatium interosseum
antebrachii überbrückt. Sowohl Radius als auch Ulna zeigen eine leichte Biegung
nach kranial, ihre proximalen Enden liegen sagittal hintereinander (MARTIN 1923).
Der Radius liegt bei der Pronationsstellung leicht schräg vor der Ulna und verläuft
von proximolateral nach distomedial. Der Kopf des Radius, Caput radii, trägt eine
querovale Gelenkfläche, Fovea capitis, und ist durch ein deutliches Collum radii vom
Schaft getrennt (VOLLMERHAUS et al. 1994a). Am Kaudalrand des Caput radii ist
eine konvexe Gelenkfläche, Circumferentia articularis radii, angeschliffen. Sie
artikuliert mit der Incisura radialis ulnae.
Vom Caput radii richtet sich ein Knochenvorsprung nach kraniolateral, der als sog.
„Caputprotuberanz“ (BERGER 1992) bzw. als „Protuberantia lateralis radii“
bezeichnet wird (HULTKRANTZ 1897). „Tuber globosum laevigatum“ ist eine weitere
Bezeichnung für diese Erhabenheit am Radius (KADLETZ 1932). Medial wird die
Protuberanz durch einen individuell unterschiedlich tiefen Sulcus in der kranialen
Kontur des Caput radii begrenzt. Die Funktion beider Strukturen liegt offensichtlich in

11 der Führung von Teilen des Kapselbandapparates. So wird unterstellt, dass die
Ursprungssehne des M. supinator durch den Sulcus im Caput radii an einer Stelle
gehalten wird (BERGER 1992).
Distal des Collum radii sind lateral ein kräftiger Bandhöcker und medial eine
schwächere Rauhigkeit, Tuberositas radii, ausgebildet. Der Bandhöcker bietet dem
lateralen Kollateralband und die Tuberositas radii dem M. biceps brachii und dem M.
brachialis Ansatz. Der Radiusschaft, Corpus radii, ist im Querschnitt oval und mit
einem lateralen bzw. einem medialen Rand ausgestattet. Die kaudale Fläche ist
aufgerauht und besitzt eine Kontaktfläche zur Ulna (VOLLMERHAUS et al. 1994a).
Die Ulna wird proximal vom vierkantigen Olecranon beherrscht. Dessen freies Ende,
Tuber olecrani, zeigt einen stumpfen kaudalen Muskelhöcker und zwei scharfe
kraniale Muskelleisten. Die Basis des Olecranon ist als halbkreisförmige Incisura
trochlearis zur Artikulation mit der Trochlea humeri eingeschnitten. Diese wird
proximal vom Processus anconeus und distal von den beiden ungleich großen
Processus coronoidei medialis bzw. lateralis begrenzt. Der Processus coronoideus
lateralis ist der schwächere der beiden Processus. Der mediale Processus bietet
dem M. brachialis Ansatz und ist deutlich stärker ausgebildet (VOLLMERHAUS et al.
1994a). Die beiden Processus coronoidei vergrößern die mit dem Humerus
artikulierende Gelenkfläche der Ulna und tragen 20 - 25% der vom Humerus
übertragenen Körperbelastung (FOX et al. 1983).
Zwischen den beiden Kronfortsätzen ist die Incisura radialis ulnae ausgespart. Sie
artikuliert mit der Circumferentia articularis radii, indem sie diese als 3 bis 5 mm hohe
Facette umfasst.
Der Ulnaschaft, Corpus ulnae, geht aus dem abgeplatteten Olecranon als
dreikantiger Knochen hervor. Er verjüngt sich beim Hund in distaler Richtung. Die
kraniale Fläche der Ulna ist beim Hund aufgerauht (VOLLMERHAUS et al.1994a).

12 2.2.3 Gelenkkapsel
Die Capsula articularis umfasst die Artt. humeroradialis, humeroulnaris und
radioulnaris proximalis in einer gemeinsamen Gelenkhöhle (NICKEL et al. 1992).
Nach proximal erstreckt sich ihr Ansatz soweit, dass sie das Foramen supra-
trochleare und den Processus anconeus in das Cavum articulare einschließt (VAN
HERPEN 1988). Auf der Beuge- und Streckseite ist die Kapsel weit und dehnbar, auf
der lateralen und medialen Seite liegt sie straff und eng seitlich an; so ist eine starke
Beugung des Ellbogengelenks möglich. Die straffen seitlichen Kapselabschnitte
überspannen den sehr schmalen Bereich zwischen dem distalen Rand der
Epicondyli humeri und der Incisura trochlearis ulnae bzw. des Processus
coronoideus medialis und lateralis ulnae. Diese schmalen Gelenkkapselstreifen
werden zum Teil durch die extrakapsulären Kollateralbänder überspannt und
dadurch eng an die Kontur des Gelenkes gepresst. Der Abschnitt, der die Art.
radioulnaris einschließt, ist entsprechend der geringradigen Bewegungsfreiheit
dieses Gelenkes, sehr eng anliegend (EVANS 1993a). Auf der Beugeseite wird die
Kapsel durch das schräg verlaufende Lig. obliquum verstärkt (ZIETZSCHMANN
1943).
Die Gelenkkapsel besteht aus dem Stratum fibrosum und dem Stratum synoviale.
Ersteres entspringt am Humerus räumlich isoliert vom Stratum synoviale. Das
Stratum synoviale folgt in seiner Ursprungslinie den Rändern des Gelenkknorpels,
während das Stratum fibrosum peripher der Fossa radialis bzw. der Fossa olecrani
entspringt (STASZYK u. GASSE 2001a). Je nach mechanischer Anforderung können
die Kollagenfasern aus dem Netzwerk der Fasern des Periosts hervorgehen, direkt
an den Osteonen des Knochens inserieren oder über eine Faserknorpelschicht
befestigt sein (STASZYK u. GASSE 2001b). An der lateralen Seite inseriert die
Gelenkkapsel 3 bis 8 mm proximal des distalen Trochlearandes und an der medialen
Seite 6 bis 10 mm SÜPPEL (1921).

13 Neben den für die Bewegungsfreiheit benötigten kranialen und kaudalen
Aussackungen werden von verschiedenen Autoren eine unterschiedliche Anzahl von
kleineren distalen Ausbuchtungen als Schutzeinrichtungen für Bänder und Muskeln
beschrieben. Eine Übersicht geben die Tabellen 1a und 1b.
NICKEL et al. (1992), VOLLMERHAUS et al. (1994b) und KÖNIG u. LIEBICH (1999)
sprechen an dieser Stelle von drei, KASER (1998) von vier Aussackungen, da er die
kaudale Ausbuchtung aufgrund histologischer Ergebnisse noch einmal unterteilt.
Diese Einteilung in kraniolateral, kraniomedial, kaudolateral und kaudomedial wurde
sinngemäß auf die Aussagen aller Autoren übertragen:
Die kraniolaterale Aussackung stülpt sich nur unter den M. extensor digitorum
communis (NICKEL et al. 1992; VOLLMERHAUS et al. 1994b; KÖNIG u. LIEBICH
1999), oder unter den Ursprungsbereich des M. extensor digitorum communis und
den des M. extensor carpi radialis (EVANS 1993b) oder nach der Auffassung von
SÜPPEL (1921) und BAUM u. ZIETZSCHMANN (1936) zusätzlich unter das laterale
Seitenband.
Dieser Recessus liegt distal des Lig. obliquum, wird durch das Lig. anulare radii
unterteilt und kaudal durch das Lig. collaterale cubiti laterale begrenzt (KASER
1998).
Kraniomedial schiebt sich die Kapsel unter den M. biceps brachii (SÜPPEL 1921;
BAUM u. ZIETZSCHMANN 1936; NICKEL et al. 1992; EVANS 1993b;
VOLLMERHAUS et al.1994b; KÖNIG u. LIEBICH 1999). Dieser Recessus wölbt sich
zwischen dem Lig. obliquum und dem Lig. anulare radii hervor. Seine kaudale
Begrenzung wird durch das Lig. collaterale cubiti mediale gebildet (KASER 1998).

14 Tab. 1a: Literaturangaben zu den kranialen Aussackungen der Gelenkkapsel am
Ellbogengelenk des Hundes.
Ausrichtung am Gelenk
Anzahl Lokalisation Autoren
M. extensor digitorum communis
NICKEL et al. (1992); VOLLMERHAUS et al. (1994b); KÖNIG u. LIEBICH (1999)
M. extensor digitorum communis und M. extensor carpi radialis
EVANS (1993b);
M. extensor digitorum communis und M. extensor carpi radialis und laterales Seitenband
SÜPPEL (1921); BAUM u. ZIETZSCHMANN (1936)
Kraniolateral 1
distal des Lig. obliquum, durch das Lig. anulare halbiert
KASER (1998)
M. biceps brachii SÜPPEL (1921); BAUM u. ZIETZSCHMANN (1936); EVANS (1993b); VOLLMERHAUS (1994b); KÖNIG und LIEBICH (1999)
1
proximal der Ligg. obliquum und anulare radii
KASER (1998)
Kraniomedial -distal
-proximal
2
zwischen M. pronator teres und M. flexor carpi radialis Ende des M. brachialis
SÜPPEL (1921) SÜPPEL (1921)
SÜPPEL (1921) beschreibt eine zusätzliche Bucht: die Kapsel springt vom
proximalen Teil der medialen Seite der Trochlea humeri taschenartig vor. Sie bildet
dadurch an dieser Stelle funktionell einen Schleimbeutel für das distale Ende des M.
brachialis.
Nach kaudolateral reicht die Gelenkkapsel bis zum proximalen Rand der Fossa
olecrani zwischen den Epicondylus lateralis humeri und dem Olecranon hindurch.
Diese Stelle wird für die Injektion durch den M. anconeus in das gebeugte Gelenk

15 benutzt. Geeigneter ist beim Hund allerdings der Einstich auf der Beugeseite von
kraniolateral in kaudodistaler Richtung (NICKEL et al. 1992; VOLLMERHAUS et al.
1994b; KÖNIG u. LIEBICH 1999). Die kraniale Begrenzung des Recessus wird durch
das Ligamentum collaterale cubiti laterale gebildet (KASER 1998).
Die vierte Aussackung richtet sich nach kaudomedial. Sie reicht vom Olecranon bis
hin zum Epicondylus medialis humeri (NICKEL et al. 1992; KASER 1998). SÜPPEL
(1921) und BAUM u. ZIETZSCHMANN (1936) unterteilen diesen kaudomedial
gerichteten Recessus noch in zwei Aussackungen: eine reicht medial vom Processus
anconeus proximal bis zum vorderen medialen Höcker des Olecranon. Die andere
Aussackung reicht (distal des Epicondylus medialis humeri) unter den M. flexor carpi
radialis und das Caput humerale des M. flexor digitorum profundus (EVANS 1993b).
Distal bildet dieser Recessus gewissermaßen einen Schleimbeutel zwischen dem M.
pronator teres und dem M. flexor carpi radialis. Hierdurch erscheint es so, als würde
letzterer frei an der Gelenkkapsel entspringen (SÜPPEL 1921).
Proximal der Fossa olecrani gehen beide kaudalen Recessus ineinander über. Diese
Fusion wird von KASER (1998) als Recessus caudoproximalis bezeichnet.
Tabelle 1b: Literaturangaben zu den kaudalen Aussackungen der Gelenkkapsel am
Ellbogengelenk des Hundes.
Ausrichtung am Gelenk
Anzahl Lokalisation Autoren
2 medial des Proc. anconeus bis zum vorderen Höcker des Olecranon unter Mm. flexor carpi radialis und caput humerale des flexor digitorum profundus
SÜPPEL (1921) BAUM u. ZIETZSCHMANN (1936) SÜPPEL (1921) BAUM u. ZIETZSCHMANN (1936) EVANS (1993b)
Kaudomedial
1 vom Olecranon bis zum Epicondylus medialis
NICKEL et al. (1992) KASER (1998)
Kaudolateral 1 vom Olecranon bis zum Epicondylus lateralis
VOLLMERHAUS (1994b); KASER (1998); KÖNIG u. LIEBICH (1999)

16 Die Bänder und einige Ursprungssehnen der Muskeln sind mit der Kapsel
verwachsen, so daß ihr Einklemmen verhindert wird (VOLLMERHAUS et al. 1994b).
Die Kapsel verschmilzt eng mit dem lateralen und dem Ursprung des medialen
Seitenbandes, ebenso mit den Enden des Lig. anulare radii (SÜPPEL 1921; EVANS
1993a).

17 2.2.4 Bandapparat
2.2.4.1 Allgemeines zum Bau von Gelenkbändern
Parallelfaseriges straffes Bindegewebe ist dort anzutreffen, wo sich Zugspannungen
nur in eine Vorzugsrichtung auswirken können, also in Sehnen und Bändern
(BUCHER u. WARTENBERG 1997). Gelenkbänder bestehen aus straffem,
kollagenfaserigem Bindegewebe mit einem unterschiedlich hohen Anteil an
elastischen Fasern. Eine Beimischung elastischer Fasern zu straffen Bändern äußert
sich funktionell als verzögernd wirkende Bremsvorrichtung (AICHEL 1925). Weil sie
sehr reich an Kollagen in der Grundsubstanz sind, sehen sie weißlich glänzend aus
(HEES 1990; BUCHER u. WARTENBERG 1997).
Im ausgereiften Zustand stellt die Interzellularsubstanz (Matrix) den Hauptbestandteil
des zellarmen Gewebes dar. Die wesentlichen Komponenten dieser intrazellulären
Matrix sind die fibrillären Strukturproteine und ein hydratisiertes Gel aus
Glykosaminoglykanen (STEVENS u. LOWE 1997). Die biochemische Zusammen-setzung der Sehnen und Bänder stellt sich daher wie folgt dar: 65-75 % Wasser
(AKESON et al. 1984), Trockensubstanz zu 70-80 % aus Kollagen (NEUBERGER u.
SLACK 1953).
Die lichtmikroskopisch sichtbaren kollagenen Fasern bestehen bei ultrastruktureller
Analyse aus Kollagenfibrillen. Diese formen zunächst Faserbündel und endlich
Faszikel (BENJAMIN u. RALPHS 1995), welche von einer Bindegewebsschicht
umhüllt werden, die als „Epiligament“ bezeichnet wird (CHOWDHURY et al. 1991).
Lichtmikroskopisch sind die Kollagenfasern in Längsschnitten auffällig „gewellt“.
Allgemein wird angenommen, das diese Kräuselung den Sehnen und Bändern als
Puffer dient, der es ihnen erlaubt, sich ohne Schaden zu verlängern (AMIEL et al.
1995). Im Gegensatz zu den elastischen Fasern sind die kollagenen Fasern fast
undehnbar. Eine gewisse Dehnbarkeit basiert auf dem Scherengitterprinzip der
Gesamtkonstruktion.

18 Ohne histologische Färbungen sind die kollagenen Fasern mehr oder weniger
farblos. Eine wichtige optische Eigenschaft ist die Doppelbrechung im polarisierten
Licht (BUCHER u. WARTENBERG 1997).
Die Fibrogenese beginnt mit der Synthese von Tropokollagen-Filamenten im rauen
endoplasmatischen Retikulum der Fibroblasten. Das ausgeschleuste Tropokollagen
wird an der Oberfläche der Zellen zu Mikrofibrillen kondensiert. Unter Einwirkung der
Zugspannung auf das Gewebe erfolgt die Zusammenlagerung der Mikrofibrillen zu
Fibrillen und Fasern (BUCHER u. WARTENBERG 1997).

19 2.2.4.2 Bänder des Ellbogengelenks
Der Bandapparat des Ellbogengelenks setzt sich gemäß NAV (1994) aus folgenden
Bändern zusammen:
Dem Ligamentum collaterale laterale und dem Ligamentum collaterale mediale.
Die Kollateralbänder haben die Aufgabe, die Gelenkenden der Knochen seitlich
miteinander zu verbinden und sie in der Wechselbewegung zu führen. Dabei
schränken sie Seitwärtsbewegungen der Articulatio cubiti ein (NICKEL et al.1992).
Sie sind an der lateralen Seite mit den Extensoren und an der medialen Seite mit den
Flexoren sehr eng verbunden (BRINKER et al. 1993).
Die klinische Prüfung der Stabilität der Kollateralbänder eines reponierten Gelenkes
wird von CAMPBELL (1971) beschrieben. Bei intakten Kollateralbändern sind beim
Hund in 90% Flexionsstellung eine Pronation der Pfote von 40°-50° und eine
Supination von 60°-70° möglich (MONTAVON u. SALVODELLI 1995).
Das mediale und laterale Kollateralband zeigen beim Längenwachstum der Knochen
eine Verlängerungstendenz. Sie verbinden die Epiphysen von proximal und distal der
am Gelenk beteiligten Knochen. Die Epiphysen vergrößern sich durch enchondrale
Ossi-fikation und Knorpelzellproliferation. Folglich müssen die Kollateralbänder sich
beim Wachstum verlängern, da sich ihre Ansatz- und Ursprungsstellen voneinander
entfernen. Die Kollateralbänder sind also genetisch auf Längenwachstum eingestellt
(HENSCHEL 1977).
Ligamentum collaterale laterale (Abb. 7, blau)
Das gesamte laterale Seitenband besteht aus äußerst straffen Bündeln kollagener
Fasern, zwischen denen flache Fibrozyten eingelagert sind (BERGER 1992).
Es ist 3,5 bis 6 mm breit, bis zu 2,5 mm dick (SÜPPEL 1921) und in Höhe des Ge-
lenkspalts mit der Kapsel verschmolzen (SÜPPEL 1921; BAUM u. ZIETZSCHMANN

20 1936). Dagegen beschreibt KASER (1998), daß es von Fettgewebe umgeben ist,
welches als Verschiebeschicht dient.
Ursprung
In der Literatur herrscht eine gewisse Uneinigkeit über Ursprung des Lig. collaterale
laterale am Ellbogengelenk. Es entspringt am „Epicondylus lateralis humeri“
zwischen dem Ursprung des M. extensor digitorum communis und des M. extensor
carpi ulnaris (EVANS 1993b; KÖNIG u. LIEBICH 1999); hier weist BERGER (1992)
auf zwei Anteile mit gemeinsamen Ursprung am Epicondylus lateralis humeri hin.
Von SÜPPEL (1921) und BAUM u. ZIETZSCHMANN (1936) wird der Ursprung in der
Bandgrube des Humerus beschrieben. KOCH u. BERG (1992) und NICKEL et al.
(1992) beschreiben ihn hingegen in der Bandgrube und am Bandhöcker des
Humerus.
Verlauf
Der Ursprungsschenkel überquert die kappenartige Vorwölbung der Supinatorsehne
und durchkreuzt sie oberflächlich. Er dient zusammen mit dem Lig. anulare radii dem
M. supinator als Ursprung und ist mit diesem untrennbar verwachsen (BERGER
1992). Distal in Höhe des Caput radii gehen aus dem kräftigen Ursprungsschenkel
zwei Schenkel hervor. Der stärkere, plattrundliche, kraniale oder auch „radiale“
Schenkel des lateralen Kollateralbandes ist 2 bis 2,5 mm breit und 1,5 mm dick
(SÜPPEL 1921). Der schwächere, platte, kaudale oder auch „ulnare“ Schenkel
nimmt kurz distal des Gelenkspaltes Fasern vom Lig. anulare radii auf (EVANS
1993a). Er misst in seiner Breite 1,5 bis 2 mm und in seiner Dicke 1 mm (SÜPPEL
1921).
Ansatz
a) Der kraniale Schenkel hat seinen Ansatz proximal am Radius (KOCH u. BERG
1992; KÖNIG u. LIEBICH 1999). Er wird von den einzelnen Autoren unterschiedlich
beschrieben (s. auch Tabelle 2). BAUM u. ZIETZSCHMANN (1936) sehen ihn wie
auch NICKEL et al. (1992) und BERGER (1992) am lateralen Bandhöcker. Wobei
BAUM u. ZIETZSCHMANN (1936) zusätzlich „distal am Collum radii“ als Ansatz
angeben. MARTIN (1923) und EVANS (1993a) sprechen hier von einer Erhabenheit
distal des Radiushalses. VOLLMERHAUS (1994b) beschränkt sich auf die

21 Bezeichnung „distal am Collum humeri“. SÜPPEL (1921) fügt dem Bandhöcker den
lateralen Rand des Radius als Ansatzpunkt hinzu.
b) Der kaudale Schenkel zieht zur Ulna (SÜPPEL 1921; MARTIN 1923; BAUM u.
ZIETZSCHMANN 1936; KOCH u. BERG 1992; NICKEL et al. 1992; EVANS 1993a;
KÖNIG u. LIEBICH 1999). Hier inseriert er mit einer breitgefächerten Ansatzfläche
distal des Processus coronoideus lateralis ulnae (VOLLMERHAUS et al. 1994b) und
geht in die Membrana interossea antebrachii über (BERGER 1992).

22 Tab. 2: Literaturangaben zu Ursprung und Ansatz des Ligamentum collaterale
laterale des Ellbogengelenkes beim Hund.
Ansatz Ursprung
Kranialer Schenkel Kaudaler Schenkel
Autor
proximal am Radius KÖNIG u. LIEBICH (1999)
an der Erhabenheit distal am Collum radii
an der Ulna
EVANS (1993a)
am Epicondylus lateralis humeri
lateraler am Bandhöcker Radius
fächerförmig distal des Processus coronoideus und an der Membrana interossea
BERGER (1992)
lateraler am Bandhöcker distal am Collum radii
BAUM u. ZIETZSCHMANN (1936)
in der Bandgrube des Humerus
lateraler am Bandhöcker distal am Collum radii und am lateralen Rand des Radius
SÜPPEL (1921)
am Radius KOCH u. BERG (1992)
in der Bandgrube und am Bandhöcker Humerus
am lateralen Bandhöcker des Radius
an der Ulna
NICKEL et al. (1992)
in der Bandgrube des Epicondylus lateralis humeri
distal des Collum radii
breitgefächert am Processus coronoideus lateralis ulnae
VOLLMERHAUS et al. (1994b)
an der lateralen Erhabenheit distal am Collum radii
an der Ulna MARTIN (1923)
Ligamentum collaterale mediale (Abb. 8, rot)
Das mediale Kollateralband ist etwas schwächer als das laterale. Es ist 2,5 bis 4 mm
breit, bis zu 2 mm dick (SÜPPEL 1921).

23 Ursprung
Es entspringt in der medialen Bandgrube des Humerus (SÜPPEL 1921; BAUM u.
ZIETZSCHMANN 1936; KOCH u. BERG 1992; NICKEL et al. 1992). Vergleichbar
wird von KÖNIG u. LIEBICH (1999) und EVANS (1993a) der Epicondylus medialis
humeri bzw. die Bandgrube am Epicondylus medialis humeri als Ursprungsstelle
(VOLLMERHAUS et al. 1994b) angegeben.
Verlauf
Das Lig. collaterale mediale verbindet sich mit der Gelenkkapsel und kreuzt das Lig.
anulare radii, mit dem es häufig an dieser Stelle verschmilzt. Es teilt sich distal davon
ebenfalls in zwei Schenkel (SÜPPEL 1921; VAN HERPEN 1988). Der schwächere,
kraniale Schenkel ist 1,5 bis 2 mm breit und 0,5 mm bis 1 mm dick. Der stärkere,
kaudale Schenkel misst 2,5 bis 3 mm in der Breite und 1 bis 1,5 mm in der Dicke
(SÜPPEL 1921).
Ansatz
a) Der kraniale Schenkel setzt proximal an der Tuberositas radii an (BAUM u.
ZIETZSCHMANN 1936; HLADIK 1986; EVANS 1993a). Diese Stelle ist meist als
rundliche Vertiefung nachzuweisen (SÜPPEL 1921). KÖNIG u. LIEBICH (1999)
sowie NICKEL et. al (1992) beschreiben den Ansatz am medialen Bandhöcker.
MARTIN (1923) erwähnt als eine teilweise Verschmelzung des kranialen Schenkels
im Ansatzbereich mit dem Ansatzschenkel des M. biceps brachii.
b) Der kaudale Schenkel verschwindet tief im Spatium interosseum, wo er zum
größten Teil an der Ulna, aber mit einigen Fasern auch am Radius inseriert (MARTIN
1923; BAUM u. ZIETZSCHMANN 1936; CAMPBELL 1969; VAN HERPEN 1988;
EVANS 1993a). Diese Insertionsstelle am Radius wird von SÜPPEL (1921) als
Grube oder gelegentlich auch als rinnenartige Vertiefung beschrieben, die sich
medial vom „lateralen Bandhöcker“ befindet. An der Ulna endet der kaudale
Schenkel an einer kleinen, länglichen Vertiefung. KÖNIG u. LIEBICH (1999) sowie
NICKEL et al. (1992) beschreiben den Ansatz des kaudalen Schenkels nur an der
Ulna.

24 Tab. 3: Literaturangaben zu Ursprung und Ansatz des Ligamentum collaterale
mediale des Ellbogengelenkes beim Hund.
Ursprung Ansatz
Autor
Kranialer Schenkel Kaudaler Schenkel im Spatium interosseum an Ulna und z.T. an den Radius
BAUM u. ZIETZSCHMANN (1936)
an der Tuberositas radii
an der Ulna SÜPPEL (1921) --- --- KOCH u. BERG (1992)
in der Bandgrube am Humerus
am medialen Bandhöcker des Radius
an der Ulna NICKEL et al. (1992)
in der Bandgrube am Epicondylus medialis humeri
--- --- VOLLMERHAUS et al. (1994b)
am medialer Bandhöcker des Radius
an der Ulna KÖNIG u. LIEBICH (1999)
am Epicondylus medialis humeri im Spatium
interosseum an Ulna und Radius
EVANS (1993a)
---
an der Tuberositas radii
an Ulna und Radius HLADIK (1986) --- verbunden mit der
Sehne des M. biceps brachii
im Spatium interosseum an Ulna und Radius
MARTIN (1923)
Zu den Bänder der Articulatio radioulnaris proximalis zählt die NAV (1994) das
Ligamentum anulare radii und das Ligamentum olecrani.
Ligamentum anulare radii (Abb. 7 und 9, grün)
Das Lig. anulare radii ist ein glänzendes Band, das an der Beugeseite des
Ellbogengelenkes ringförmig das Caput radii umfasst. Es ist nach SÜPPEL (1921) 2
bis 5 mm breit und meist sehr dünn.
Durch den Verlauf des Lig. anulare radii kranial um den Radius, wird dieser an die
Ulna gehalten. Es sorgt für die Stabilisierung der Supinations- und

25 Pronationsbewegungen im Ellbogengelenk (VOLLMERHAUS et al. 1994b). Das Lig.
anulare radii ist somit als besonderes Band der Art. radioulnaris proximalis
aufzufassen (SÜPPEL 1921), da es die Führung dieses Zapfengelenkes (MAI 1995)
übernimmt. Sollte es zu einer Entwicklungsstörung am Lig. anulare radii kommen,
wie zum Beispiel einer Hypoplasie, kann dies zur Instabilität in der Art. cubiti führen
mit eventueller (Sub-) Luxation des Caput radii als Folge (BINGEL u. RISER 1977).
Ursprung und Ansatz
Das Lig. anulare radii hat seinen Ursprung und Ansatz an den Ecken der Incisura
radialis der Ulna (BAUM u. ZIETZSCHMANN 1936; EVANS 1993a) bzw. am
lateralen und medialen Processus coronoideus ulnae (KOCH u. BERG 1992;
NICKEL et al. 1992). BERZON u. QUICK (1980) erweitern den Ursprung vom
lateralen Kollateralband bis zur Basis des Processus coronoideus lateralis ulnae. Der
Ansatz liegt am Processus coronoideus medialis ulnae.
MARTIN (1923) definiert den Ursprung am lateralen Bandhöcker des Radius und den
Ansatz am medialen Rand des Radius und am vorderen und medialen Rand der
Ulna, wobei an dieser Stelle eine Verschmelzung mit den Endsehnen der Mm. biceps
brachii und brachialis besteht.
Die genannten Autoren legen den Ursprung an die laterale und den Ansatz an die
mediale Seite. Die nun folgenden Autoren definieren Ursprung und Ansatz
entgegengesetzt: SÜPPEL (1921) stimmt in der Beschreibung der Knochenpunkte
mit BAUM u. ZIETZSCHMANN (1936) und EVANS (1993a) überein.
VOLLMERHAUS et al. (1994b) sieht den Ursprung ebenfalls medial. Er wird hier am
Processus coronoideus medialis und der Ansatz an der lateralen Seite jedoch am
lateralen Kollateralband beschrieben.
Verlauf
Eine detaillierte Beschreibung zum Verlauf des Lig. anulare radii stammt von
BERGER (1992). Das Band verläuft von der kranialen Spitze des Processus
coronoideus medialis ulnae bis zur Kreuzung mit der Ursprungssehne des M.
supinator zunächst intrakapsulär. An dieser Stelle zieht die Gelenkkapsel über das
Band hinweg und inseriert hier distal am Collum radii. In diesem Bereich liegt das
Lig. anulare radii distal des Gelenkspaltes, dem Collum radii somit kraniomedial

26 direkt an. Vom Kreuzungspunkt mit der Ursprungssehne des M. supinator bis zum
lateralen Ende eines Knorpelfalzes (siehe Kap. 2.2.7) ist es mit der Gelenkkapsel
verschmolzen und beteiligt sich am Aufbau dieser Falzstruktur. Danach nimmt das
Lig. anulare radii nach lateral einen extrakapsulären Verlauf. Es kommt zu einer
Teilung des Bandes in zwei Schenkel. Einer inseriert am „Epicondylus lateralis
humeri“ und der andere etwas proximal an der Spitze des schwach ausgeprägten
Processus coronoideus lateralis der Ulna. Dieses Ergebnis wird auch histologisch
bestätigt (BERGER 1992).
Tab. 4: Literaturangaben zu Ursprung und Ansatz des Lig. anulare radii der
Articulatio radioulnaris proximalis beim Hund.
Ursprung Ansatz
Autor
an den Ecken der Inc. radialis ulnae bzw. am Processus coronoideus lateralis
BAUM u. ZIETZSCHMANN (1936); EVANS (1993); KOCH u. BERG (1992); NICKEL et al. (1992); SÜPPEL (1921)*
am laterales Seitenband
VOLLMERHAUS et al. (1994b) *
am lateralen Seitenband und am Processus coronoideus lateralis
BERZON u. QUICK (1980)
1.) am Epicondylus lateralis humeri und 2.) am Processus coronoideus lateralis
am Processus coronoideus medialis
BERGER (1992) *
am lateralen Bandhöcker am Radius
am medialen Rand der Ulna, verschmilzt mit den Mm. biceps und brachialis humeri
MARTIN (1923)
∗ Der Ursprung ist bei diesen Autoren an der medialen Seite definiert und der Ansatz an der lateralen Seite der Art. cubiti.

27
Ligamentum olecrani (Abb. 10, türkis)
Im kaudomedialen Bereich der Art. cubiti verläuft das Lig. olecrani zwischen
Humerus und Olecranon. Es entspringt an der axialen Seite des Epicondylus
medialis humeri, nimmt seinen Verlauf über die Gelenkkapsel in kaudodistaler
Richtung und inseriert an der Erhabenheit zwischen medialem vorderen Höcker und
dem Processus anconeus des Olecranon (NICKEL et al. 1992). Seine Breite beträgt
2 bis 3 mm (SÜPPEL 1921).
Es handelt sich hier um ein elastisches Band (SÜPPEL 1921; ZIETZSCHMANN
1943; EVANS 1993a), welches eng mit der Gelenkkapsel verbunden ist (BAUM u.
ZIETZSCHMANN 1936). Diese Band sorgt für die Straffung des kaudalen
Kapselbereiches (KÖNIG u. LIEBICH 1999).
Ligamentum obliquum (Abb. 7, Abb. 8 und Abb. 9, orange)
Das Lig. obliquum findet in der NAV (1994) keine Erwähnung. Es liegt als kleines
schräges, aber festes Band an der Beugeseite der Kapsel und entspringt vom
dorsalen Rand des Foramen supratrochleare. Sein Verlauf ist extrakapsulär, schräg
mediodistal. Es ist fest mit der Gelenkkapsel verbunden. Durch seinen Verlauf dellt
es kranial die Gelenkhöhle in einen medialen und lateralen Bereich ein (STASZYK u.
GASSE 2001a). In Höhe der Endsehnen des M. biceps brachii und des M. brachialis
direkt neben dem Lig. anulare teilt es sich in zwei Schenkel. Der kürzere verschmilzt
mit dem kranialen Schenkel des Lig. collaterale mediale und der längere endet am
medialen Rand des Radius nach dem Umschlingen der Sehnen der eben genannten
Muskeln (SÜPPEL 1921; BAUM und ZIETZSCHMANN 1936; EVANS 1993a).
Nach KASER (1998) entspringt das Ligamentum obliquum lateroproximal des
Condylus humeri und zieht nach mediodistal. In seinem Verlauf teilt es sich in einen
kranialen und kaudalen Schenkel. Diese umfassen die Ansatzsehnen der Mm.
biceps brachii und brachialis. Der kaudale Schenkel mündet in den kranialen
Schenkel des medialen Seitenbandes und der kraniale Schenkel zieht an das
proximale Drittel des Radius.

28 Seine Funktion liegt in der Straffung des kranialen Abschnittes der Gelenkkapsel,
womit deren Überstreckung (KASER 1998) gemeinsam mit dem Anschlagen der
Olecranonspitze in der Fossa olecrani und dem passiven Widerstand der Beuge-
muskeln verhindert wird (KAPANDJI 1984).
Zudem hat die Gelenkkapsel an der Beugeseite nach BAUM u. ZIETZSCHMANN
(1936) zwei deutlich nach innen vorspringende Verstärkungszüge.

29 2.2.5 Muskeln
Die dem Ellbogengelenk anliegende Muskulatur ist aufgrund ihrer Wirkungsweise in
die Muskeln des Ellbogengelenkes und des Radioulnargelenkes zu unterteilen.
Zudem ist die Muskulatur zu beachten, die distal als Karpal- bzw. Zehenbeuger und
Zehenstrecker wirken. Im folgenden Kapitel wird auf die hier auftretenden Schleim-
beutel eingegangen, um deren Funktion oder mögliche Kommunikation mit der
Gelenkhöhle zu untersuchen.
Zu diesen Muskeln zählen die Beuger und Strecker des Ellbogengelenkes.
Der M. brachialis entspringt kaudal am Collum humeri. Er windet sich im Sulcus
brachialis von der lateralen auf die kraniale Fläche des Humerus und zieht dann
lateral vom M. biceps brachii über die Beugefläche der Art. cubiti. Seinen Ansatz
findet er mit zwei Sehnen: mit der Hauptsehne, die zwischen die beiden Endschenkel
des M. biceps brachii hindurchtritt, inseriert er am Processus coronoideus medialis
ulnae und mit der schwächeren Sehne zusammen mit einer Bizepsendsehne an die
Tuberositas radii. Der M. brachialis ist ein Beuger des Ellbogengelenkes (SEIFERLE
u. FREWEIN 1992).
Der M. biceps brachii entspringt mit einer Sehne am Tuberculum supraglenoidale
scapulae und zieht medial über die Streckseite des Schultergelenkes. Der
Muskelbauch liegt zuerst medial am Humerus, zieht dann nach kranial und
schließlich über die Beugeseite des Ellbogengelenkes hinweg. Hier geht er in zwei
Endsehnen über. Die Hauptsehne des M. biceps brachii setzt am Processus
coronoideus medialis ulnae an und die schwächere Sehne an der Tuberositas radii.
Der zweischenkelige y-förmige Ansatz des M. biceps brachii und die dazwischen in-
serierende Endsehne des M. brachialis scheinen mediokranial zur Stabilisierung des
Gelenkes beizutragen (BERGER 1992). Beim Hund ist zudem eine Struktur
ausgebildet, die dem Lacertus fibrosus des Menschen entspricht: ein Sehnenzug
zieht zwischen dem M. extensor carpi radialis und dem M. pronator teres hindurch in
die Tiefe und strahlt in die Fascia antebrachii ein (FREWEIN 1983; HERMANSON u.
EVANS 1993).

30 Es handelt sich bei diesem Muskel ebenfalls um einen Beuger des Ellbogengelenkes
(BAUM u. ZIETZSCHMANN 1936).
Der M. triceps brachii ist ein Muskel mit vier Köpfen: er besteht aus dem Caput
longum, Caput laterale, Caput mediale und Caput accessorium.
Das Caput longum entspringt an den zwei distalen Dritteln des kaudalen Scapula-
randes und inseriert von einem Schleimbeutel unterlagert am kaudalen Teil des
Olecranon.
Das Caput laterale nimmt seinen Ursprung an der Linea musculi tricipitis humeri und
verbindet sich weiter distal zum Teil mit dem Caput longum. Ein schwächerer Teil
strahlt in die Fascia antebrachii ein.
Das Caput mediale entspringt zwischen den Ansätzen des M. teres major und M.
coracobrachialis medial am Humerusschaft und inseriert medial am Olecranon.
Das Caput accessorium liegt tief zwischen den anderen Köpfen des Triceps. Es
entspringt an der Crista tuberculi minoris und lateral von dieser. Weiter distal
verbindet es sich mit dem Caput longum und Caput laterale.
Der M. triceps brachii ist der wichtigste Strecker des Ellbogengelenkes und das
Caput longum wirkt auch als Beuger des Schultergelenkes (FREWEIN 1983).
Der M. tensor fasciae antebrachii entspringt aus der lateralen Fläche des M.
latissimus dorsi. Sein Hauptteil strahlt in die Fascia antebrachii an. Er wirkt als
Strecker des Ellbogengelenkes (ADAMS 1986); kaudal und medial liegt er dem
Caput longum des Triceps eng an. Distal endet er teilweise mit diesem am
Olecranon (FREWEIN 1983).
Der M. anconeus hat seinen Ursprung an der kaudodistalen Seite des Humerus
(FREWEIN 1983). Er deckt die Fossa olecrani ab und liegt folglich zwischen den
Epicondyli humeri. Seinen Ansatz findet er an der lateralen Fläche des Olecranon,
und ist überwiegend vom M. triceps brachii bedeckt. Der proximalen Fläche des
Recessus caudalis der Ellbogengelenkkapsel liegt er direkt auf. Seine Funktion ist
die Streckung des Ellbogengelenkes und Spannung der Gelenkkapsel
(HERMANSON u. EVANS 1993).

31 Die Muskeln des Radioulnargelenkes sind für das Rotationsvermögen des Unterarms
verantwortlich.
Der M. brachioradialis ist ein länglicher, schmaler Muskel, der beim Hund häufig
auch fehlen kann. WAKURI u. KANO (1966) fanden ihn bei 35 von insgesamt 90
untersuchten Hunden. Ist er vorhanden, so entspringt er an der Crista
supracondylaris lateralis und zieht über die Beugeseite des Ellbogengelenkes auf
den M. extensor carpi radialis distal (FREWEIN 1983). Seinen Ansatz hat dieser
Muskel am Processus styloideus radii; er wirkt als Supinator des Unterarms und der
Pfote (SEIFERLE u. FREWEIN 1992).
Der M. supinator ist ein platter Muskel, welcher der Art. cubiti an seiner Beugefläche
lateral direkt aufliegt. Seinen Ursprung nimmt er am „Bandhöcker des Epicondylus
lateralis humeri“, dem lateralen Seitenband des Ellbogengelenkes und dem Lig.
anulare radii. Die Ursprungssehne des Muskels hat zwei Anteile. Der starke
Hauptsehnenanteil zieht vom Epicondylus lateralis schräg mediodistal und setzt mit
einem Muskeldreieck am Radiusschaft und dem Lig. collaterale mediale an.
Während dieses Verlaufs tritt der muskuläre Sehnenschenkel durch das Lig.
collaterale laterale hindurch, bevor sich dieses in seine zwei Schenkel aufteilt. Das
Band bildet somit eine Art Tunnel für die Sehne (BERGER 1992). Die Sehne des M.
supinator umgreift die sog. „Caputprotuberanz“ (HULTKRANTZ 1897) des Radius-
kopfes (siehe Kap. 2.2.2) und verflicht sich mit dem darunter liegenden Lig. anulare
radii und dem darüber hinwegziehenden Lig. collaterale laterale oberflächlich.
Dadurch kommt es an dieser Stelle zu einer kappenartigen Verstärkung der
Gelenkkapsel auf der Protuberanz des Caput radii. Der wesentlich schwächere
Sehnenschenkel des M. supinator entspringt in der Höhe des Processus coronoideus
lateralis der Ulna im Bereich des ulnaren Schenkels des Lig. collaterale laterale. Er
beteiligt sich an der Bildung der Kappe und setzt gemeinsam mit dem
Hauptsehnenanteil an. Es wird unterstellt, dass die kappenartige Verstärkung in der
Gelenkkapsel für eine feste Lagerung der Supinatorsehne sorgt und somit das
Gelenk ergänzend seitlich stabilisiert (BERGER 1992).
In der Ursprungssehne des M. supinator befindet sich ein fakultatives Sesambein,
welches aufgrund histologischer Untersuchungen als primär zur Sehne des M.

32 supinator zugehörig betrachtet werden muß. Seine Beziehungen zum lateralen
Kapselbereich sind sekundärer Natur (BERGER 1992). HERMANSON u. EVANS
(1993) weisen ebenfalls auf ein Sesambein in der Ursprungssehne des Muskels hin.
Nähere Ausführungen hierzu befinden sich im Kapitel 2.2.7.
Der Hauptsehnenanteil zieht schräg mediodistal und endet am proximalen Viertel
des medialen Radiusrandes (FREWEIN 1983). Er wirkt als Supinator der Pfote und
des Vorderfußes (SEIFERLE u. FREWEIN 1992).
Der M. pronator teres entspringt am Bandhöcker des Epicondylus medialis humeri,
zieht schräg distal an die dorsomediale Seite des Radius und liegt diesem im
proximalen Drittel auf. In seinem proximalen Verlauf wird er von der Arteria mediana
und dem Nervus medianus durchdrungen. Sein Ansatz befindet sich distal vom M.
supinator (ZIMMERMANN 1958; HERMANSON u. EVANS 1993). Der M. pronator
teres ist beim Hund ein kräftiger Muskel. Er hält den Unterarm und die Pfote in
Pronationsstellung (ROOS et al. 1992).
Der M. pronator quadratus liegt der Membrana interossea antebrachii auf und
erstreckt sich über die ganze Länge des Zwischenknochenspaltes. Die Muskelfasern
verlaufen senkrecht zur Längsachse des Unterarms. Sie beginnen an der medialen
Fläche der Ulna und ziehen an die kaudale Fläche und mediale Kante des Radius.
Dieser Muskel bewirkt ebenfalls die Pronation (FREWEIN 1983; SEIFERLE u.
FREWEIN 1992).
Der M. extensor carpi radialis wird den Muskeln des Karpalgelenkes zugeordnet.
Es handelt sich hier um einen ursprünglich zweiteiligen Muskel, wie man bei der
Katze noch erkennen kann (FREWEIN 1983). Er entspringt an der Crista
supracondylaris lateralis, beim Hund ist die Ursprungssehne noch zweiteilig
(SEIFERLE u. FREWEIN 1992) und verläuft über eine kurze Distanz gemeinsam mit
dem M. extensor digitorum communis. Diese beiden Muskeln werden durch ein
Muskelseptum, Septum intermusculare, fest verbunden (BAUM u. ZIETZSCHMANN
1936; HERMANSON u. EVANS 1993). Im weiteren Verlauf liegt er proximal der
medialen Fläche des M. extensor digitorum communis an (BAUM u.
ZIETZSCHMANN 1936). Der M. extensor carpi radialis zieht über die kraniale Fläche
der Art. cubiti nach distal. Über dem Gelenk wird er durch eine Ausbuchtung der

33 Gelenkkapsel unterlagert. Im Verlauf des distalen Drittels des Radius geht er in zwei
Endsehnen über (FREWEIN 1983). Sie inserieren proximal an der Dorsalfläche des
2. bzw. 3. Metakarpalknochens (ADAMS 1986). Dieser Muskel wirkt als Strecker des
Karpalgelenkes.
Der M. extensor digitorum communis entspringt am Epicondylus lateralis humeri
kranioproximal vom Ursprung des Lig. collaterale laterale. An seinem Ursprung ist er
über eine kurze Strecke in der Tiefe mit dem M. extensor carpi radialis durch einen
Sehnenspiegel verbunden, der schließlich für die einzelnen Muskeln getrennt
ausläuft (BAUM u. ZIETZSCHMANN 1936; SEIFERLE u. FREWEIN 1992;
HERMANSON u. EVANS 1993). Im Ursprungsbereich ist der Muskel von einem
Kapselschleimbeutel unterlagert (BAUM u. ZIETZSCHMANN 1936; SEIFERLE u.
FREWEIN 1992). Er liegt der kraniolateralen Fläche des Radius zwischen dem M.
extensor carpi radialis und dem M. extensor digitorum lateralis auf. Am Metakarpus
gehen die vorher eng aneinanderliegenden Sehnen fächerartig in einzelne über,
ziehen über die Streckseiten der entsprechenden Mittelhandknochen (II-V) und
zugehörigen Digiti hinweg und setzen am Krallenfalz des jeweiligen Os unguiculare
an. Er wirkt als Strecker der Zehengelenke und des Karpalgelenks (FREWEIN 1983).
Der M. extensor digitorum lateralis gehört ebenfalls zur Gruppe der langen
Zehenmuskel. Für die Betrachtung des Ellbogengelenkes ist er aufgrund seines
Ursprungs interessant. Beim Hund besitzt er einen einheitlichen Ursprung am
kranialen Rand des lateralen Seitenbandes des Ellbogengelenkes sowie am
lateralen Bandhöcker des Radius. Ein Sehnenspiegel zieht vom Lig. collaterale
laterale unter den Muskel, teilt sich in seiner distalen Hälfte in zwei Äste, von denen
jeder zu einer eigenständigen Sehne wird, die in einen Muskelbauch übergeht. Die
Muskelbäuche befinden sich lateral am Unterarm. Diese Endsehnen liegen eng
zusammen und sind am Karpalgelenk in einer gemeinsamen Sehnenscheide
eingeschlossen (BAUM u. ZIETZSCHMANN 1936; HERMANSON u. EVANS 1993).
Der M. extensor carpi ulnaris ist ein platter, sehnig durchsetzter Muskel. Er
entspringt kaudal des Lig. collaterale laterale cubiti am Epicondylus lateralis humeri.
In seinem distalen Verlauf zieht er an der lateralen Karpusseite entlang. Seinen
Ansatz findet er lateral am Proximalende des Mc IV. Eine schwache Abspaltung der

34 Endsehne verbindet sich mit dem Retinaculum extensorum und eine weitere
Abspaltung von einem Schleimbeutel unterlagert, mit dem Retinaculum flexorum
(SEIFERLE u. FREWEIN 1992). Der M. flexor carpi radialis entspringt kaudal des Lig. collaterale mediale cubiti am
Epicondylus medialis humeri und wird an seinem Ursprung von einer Aussackung
der Gelenkkapsel, einem Kapselschleimbeutel, unterlagert (SEIFERLE u. FREWEIN
1992). Er verläuft direkt unter der Haut und der Fascia antebrachii medial am
Unterarm zwischen dem M. pronator teres und dem M. flexor digitorum superficialis.
In der Mitte des Unterarms geht er in eine platte Sehne über (FREWEIN 1983).
Diese spaltet sich in zwei kurze Schenkel: ein Schenkel inseriert palmar am
Proximalende des Mc 2, der andere Schenkel setzt am Mc 3 an (ADAMS 1986).
Damit ist er ein Beuger des Karpalgelenkes (FREWEIN 1983).
Der M. flexor digitorum superficialis beginnt mit einer kurzen, aber starken Sehne
am Epicondylus medialis humeri zwischen den Humerusköpfen des tiefen
Zehenbeugers und dem M. flexor carpi ulnaris. Sein Ursprung ist von einem
Schleimbeutel unterlagert (SEIFERLE u. FREWEIN 1992). Er liegt oberflächlich
mediokaudal am Unterarm und bedeckt den M. flexor digitorum profundus sowie das
Caput humerale des M. flexor carpi ulnaris. Dicht proximal des Karpus geht er in die
oberflächliche Beugesehne über, die medial des Os carpi accessorium über die
Karpalbeuge zieht. Hier wird sie durch das Retinaculum flexorium von der tiefen
Beugesehne getrennt. Im proximalen Drittel des Metakarpus teilt sich die Sehne in
vier Teile, die divergierend zur 2. bis 5. Zehe verlaufen (HERMANSON u. EVANS
1993).
Dieser Muskel wirkt als Beuger des Zehengrundgelenkes, des Zehenmittelgelenkes
der Stützzehen und des Karpalgelenkes (BAUM u. ZIETZSCHMANN 1936;
FREWEIN 1983).
Der M. flexor digitorum profundus besteht aus drei Muskelbäuchen: dem Caput
humerale, dem Caput ulnare und dem Caput radiale.
Das Caput humerale entspringt mit einer kurzen, kräftigen Sehne, von einem
Kapselschleimbeutel unterlagert, vom medialen Epicondylus humeri unmittelbar
kaudal des Ursprungs vom M. flexor carpi radialis. Der M. flexor digitorum

35 superficialis und das Caput humerale des M. flexor carpi ulnaris verdecken ihn. Der
Muskelbauch liegt der kaudomedialen Seite des Unterarms an. An der Grenze
zwischen Karpus und Antebrachium laufen die Sehnen der drei Muskelbäuche zu
einer zusammen (HERMANSON u. EVANS 1993).
Das Caput radiale, der schwächste der drei Muskelbäuche, entspringt am medialen
Radiusrand und proximal an dessen Kaudalfläche. Es liegt dem Unterarm
kaudomedial an. Die dünne Endsehne fusioniert nahe dem Karpus mit der
Hauptsehne, trennt sich proximal des Metakarpus wieder von ihr und zieht dann an
die 1. Zehe (FREWEIN 1983).
Das stärkere Caput ulnare hat seinen Ursprung am Kaudalrand der Ulna vom
Olecranon bis zum distalen Ulnaviertel. Es liegt in seinem Verlauf zwischen dem M.
extensor und flexor carpi ulnaris, wobei es das Caput humerale lateral verdeckt.
Seine Endsehne strahlt unmittelbar proximal des Karpus in die des Caput humerale
ein. Im Anschluss an diese Vereinigung ist die tiefe Beugesehne breit und
rinnenförmig eingedellt (FREWEIN 1983). Ihren Ansatz findet sie am Tuberculum
flexorium des Krallenbeins (BAUM u. ZIETZSCHMANN 1936; SEIFERLE u.
FREWEIN 1992; HERMANSON u. EVANS 1993).
Der M. flexor carpi ulnaris, besteht aus zwei Muskelbäuchen, dem flachen Caput
ulnare und dem stärkeren Caput humerale.
Das Caput ulnare entspringt kaudomedial am Olecranon. Hier ist es von der
Ansatzsehne des Caput mediale des M. triceps brachii bedeckt. Proximal der
Unterarmmitte geht es in eine platte Sehne über, tritt unmittelbar danach unter das
Caput humerale und inseriert schließlich selbständig am Os carpi accessorium
(HERMANSON u. EVANS 1993).
Das Caput humerale ist wesentlich stärker ausgeprägt. Es entspringt am Epicondylus
medialis humeri mit einer kurzen, kräftigen Sehne nahe der Ursprungssehne des M.
flexor digitorum superficialis. Hier ist es von einem Schleimbeutel unterlagert, der mit
der Bursa des M. flexor digitorum superficialis kommuniziert. Der Muskelbauch wird
proximal am Unterarm medial vom M. flexor digitorum superficialis bedeckt. Die
Endsehne des Caput humerale findet ihren Ansatz ebenfalls am Os carpi
accessorium und beugt somit des Karpalgelenk (FREWEIN 1983).

36
2.2.6 Schleimbeutel Das lockere Bindegewebe bildet Bursae mucosae, Schleimbeutel, als Hilfsein-
richtung aus, die den Reibungsweg der Sehnen zum Gleitweg gestalten (PREUß u.
WÜNSCHE 1974). Sie sind als wasserkissenartige Druckschutz- und
Gleitvorrichtungen (SEIFERLE u. FREWEIN 1992) zwischen hervorragenden
Knochenteilen und den verschieblich darüber hinwegziehenden Strukturen
eingelagert (SAJONSKI 1969). Bei kleineren Bewegungen können sie von lockerem
Bindegewebe ersetzt werden (HULTKRANTZ 1897).
Je nach Lage werden sie in subligamentöse, subtendinöse und subkutane
Schleimbeutel unterteilt (SAJONSKI 1969). Es handelt es sich bei Schleimbeuteln
mit Ausnahme der subkutanen, um embryonal angelegte Strukturen, deren
Vorkommen und Lage meistens konstant sind (SAJONSKI 1969; SEIFERLE u.
FREWEIN 1992).
Eine Kommunikation subligamentöser und subtendinöser Schleimbeutel mit der
Gelenkhöhle ist möglich; sie werden in diesem Fall als Kapselschleimbeutel
(SAJONSKI 1969) bezeichnet, wobei ihre fibröse Hülle den gleichen Wandaufbau
wie die der Gelenkhöhle aufweist (KRÖLLING u. GRAU 1960).
Die kollagenen Fasern der Bindegewebsschicht sind variierend in der Richtung
angeordnet. Abhängig ist diese Ordnung von den herrschenden Spannungs-
verhältnissen (HÄGGQVIST 1931). Elastische Fasern können reichlich vorhanden
sein (LUNGWITZ 1906). Sie befinden sich in der Membrana propria.
An den Gleitflächen oder den druckbelasteten Stellen durch die Unterlage oder die
darüber hinwegziehende Struktur, kann anstatt einer Membrana synovialis eine
Ansammlung von Knorpelzellen eingelagert sein (SAJONSKI 1969; SEIFERLE u.
FREWEIN 1992). Die Innenauskleidung der Schleimbeutel besteht aus einer
einschichtigen Lage abgeplatteter Bindegewebszellen (HÄGGQVIST 1931). Sie
bedecken nicht die gesamte Innenfläche des Schleimbeutels, sondern lassen

37 unregelmäßige Interzellularräume zwischen sich frei. Falls faserknorpelige
Einlagerungen in die Schleimbeutelwand eingelagert sind, sind diesen an ihrer
Innenseite keine Bindegewebszellen aufgelagert (LUNGWITZ 1906).
An der lateralen Seite der Art. cubiti ist aufgrund der Anheftung der Gelenkkapsel
(siehe Kapitel 2.2.3) ein Kapselschleimbeutel unter dem lateralen Seitenband und ein
weiterer unter dem Ursprungsteil der Mm. extensores carpi radialis und digitorum
communis zu finden (WALTHER 1908; SÜPPEL 1921: BAUM u. ZIETZSCHMANN
1936; FREWEIN 1983; SEIFERLE u. FREWEIN 1992; EVANS 1993a).
Zudem kommt bei älteren Hunden konstant ein Schleimbeutel unter der
Ursprungssehne des M. extensor carpi ulnaris vor (WALTHER 1908; BAUM u.
ZIETZSCHMANN 1936; EVANS 1993a). Dieser Schleimbeutel liegt unmittelbar
kaudal des lateralen Kollateralbandes. Er ist im Schnitt 1 bis 2 cm lang. Proximal
stößt er direkt an die Gelenkkapsel und distal verbindet er sich mit der Sehne des
Muskels. Es handelt sich hierbei um einen selbständigen Schleimbeutel, der nicht mit
der Gelenkkapsel in Verbindung steht (WALTHER 1908).
Von BERGER (1992) wird an der lateralen Seite des Ellbogengelenkes zwischen der
„Caputprotuberanz“ und der darüber befindlichen kappenartigen Verstärkung der
Gelenkkapsel ein Fettpolster erwähnt, welches Bestandteil des Stratum synoviale ist.

38 Tab. 5: Literaturangaben zu Kapselschleimbeutel und Schleimbeutel lateral am
Ellbogengelenk des Hundes.
Verbindung zur Gelenkhöhle
lateral an der Art. cubiti
Autor
laterales Seitenband
ja Kapselschleimbeutel WALTHER (1908); SÜPPEL (1921); BAUM u. ZIETZSCHMANN (1936); FREWEIN (1983); SEIFERLE u. FREWEIN (1992); EVANS (1993a)
M. extensor carpi radialis M. extensor digitorum communis
ja
ja
Kapselschleimbeutel unter der Ursprungssehne
WALTHER (1908); SÜPPEL (1921); BAUM u. ZIETZSCHMANN (1936); FREWEIN (1983); SEIFERLE u. FREWEIN (1992); EVANS (1993a)
M. extensor carpi ulnaris
nein Schleimbeutel unter der Ursprungssehne konstant bei älteren Hunden
WALTHER (1908); BAUM u. ZIETZSCHMANN (1936); EVANS (1993a)
An der medialen Seite bildet die Gelenkkapsel zwei Aussackungen, die einen
Schleimbeutel zwischen dem M. pronator teres (HULTKRANTZ 1897; SÜPPEL
1921) und dem M. flexor carpi radialis bilden, wie in Kapitel 2.2.3 beschrieben
(SÜPPEL 19921; BAUM u. ZIETZSCHMANN 1936; FREWEIN 1983; SEIFERLE u.
FREWEIN 1992; EVANS 1993a). Die zweite Aussackung bildet funktionell einen
Kapselschleimbeutel unter der Endsehne des M. brachialis (SÜPPEL 1921).
Ebenfalls an der medialen Seite befindet sich unter der Ursprungsehne des M. flexor
carpi ulnaris ein Schleimbeutel (BAUM u. ZIETZSCHMANN 1936; FREWEIN 1983;
SEIFERLE u. FREWEIN 1992; EVANS 1993a). Dieser kommuniziert i. d. R mit
einem zweiten unter der Ursprungssehne des Caput humerale des M. flexor digi-
torum superficialis befindlichen Schleimbeutel. Er kann auch mit diesem breitflächig
zusammenfließen (WALTHER 1908; BAUM u. ZIETZSCHMANN 1936).

39 Von WALTHER (1908) wird beschrieben, dass der Schleimbeutel direkt am Ursprung
des oberflächlichen Zehenbeugers beginnt. Er liegt genau zwischen dem
Ursprungsteil des oberflächlichen und tiefen Zehenbeugers. In seiner kaudalen
Wand befindet sich eine linsengroße Öffnung, die in einen zweiten Schleimbeutel
unterhalb des bereits oben erwähnten M. flexor carpi ulnaris mündet. Diese
Schleimbeutel stehen nicht mit der Gelenkkapsel in Verbindung.
Der Schleimbeutel unter der Sehne des oberflächlichen Zehenbeugers (BAUM u.
ZIETZSCHMANN 1936; FREWEIN 1983; SEIFERLE u. FREWEIN 1992; EVANS
1993a) wurde von BAUM u. ZIETZSCHMANN (1936) und EVANS (1993a)
vermessen und hierbei werden für große Hunde Durchmesser von 2 bis 2,5 cm
angegeben. Er kann bisweilen mit dem Schleimbeutel kommunizieren, der unter dem
Caput humerale des M. flexor carpi ulnaris liegt (BAUM u. ZIETZSCHMANN 1936).
Von WALTHER (1908) werden die Verhältnisse an der medialen Seite als
Ausbuchtungen beschrieben: die Gelenkkapsel buchtet sich sowohl nach proximal
wie auch nach distal aus. Proximal dient sie als Kapselschleimbeutel für das mediale
Kollateralband. Die distale Ausbuchtung formt einen Kapselschleimbeutel für die
End- bzw. Ursprungssehnen des M. biceps brachii, des M. pronator teres und des M.
flexor digitorum profundus.
Die Ausbuchtung unterlagert die Ursprungssehne des Caput humerale des M. flexor
digitorum profundus in Form eines Kapselschleimbeutels (SÜPPEL 1921; BAUM u.
ZIETZSCHMANN 1936; FREWEIN 1983; SEIFERLE u. FREWEIN 1992; EVANS
1993a). Dieser entsteht dadurch, dass sich die Gelenkkapsel distal des Processus
coronoideus medialis mit einer Entfernung von bis zu 12 mm am Knochen befestigt,
so kommt es an dieser Stelle zur Bildung einer geräumigen Ausbuchtung, die mit
Synovia gefüllt ist. Sie fungiert als Schleimbeutel für zwei Muskelursprungssehnen:
jene des M. flexor carpi radialis (SÜPPEL 1921; BAUM u. ZIETZSCHMANN 1936;
EVANS 1993a) und das Caput humerale des M. flexor digitorum profundus (SÜPPEL
1921; BAUM u. ZIETZSCHMANN 1936; FREWEIN 1983; SEIFERLE u. FREWEIN

40 1992; EVANS 1993a). WALTHER (1908) weist daraufhin, dass er an dieser Stelle
keinen Schleimbeutel beobachten konnte.
Da sich die Gelenkkapsel am Radius in unterschiedlicher Höhe dorsal, medial und
lateral befestigt, entstehen schleimbeutelartige Aussackungen der Gelenkkapsel für
das Ende des M. biceps brachii auf der medialen Seite (SÜPPEL 1921; BAUM u.
ZIETZSCHMANN 1936; FREWEIN 1983; SEIFERLE u. FREWEIN 1992; EVANS
1993a).

41 Tab. 6: Literaturangaben zu Schleimbeutel und Kapselschleimbeutel medial am Ell-
bogengelenk des Hundes.
Kapselschleim-beutel
Lokalisation Autor
M. biceps brachii
ja Kapselschleimbeutel unter den Ansatzsehnen
WALTHER (1908); SÜPPEL (1921); BAUM u. ZIETZ-SCHMANN (1936); FREWEIN (1983); SEIFERLE u. FREWEIN (1992); EVANS (1993a)
M. pronator teres
ja Kapselschleimbeutel unter der Ursprungssehne
WALTHER (1908); HULTKRANTZ (1897) ; SÜPPEL (1921)
M. flexor carpi radialis
ja Kapselschleimbeutel unter der Ursprungssehne kaudal des Seitenbandes
SÜPPEL (1921); BAUM u. ZIETZ-SCHMANN (1936); FREWEIN (1983); SEIFERLE u. FREWEIN (1992); EVANS (1993a)
M. flexor carpi ulnaris
nein Schleimbeutel unter der Ursprungssehne
BAUM u. ZIETZ-SCHMANN (1936); FREWEIN (1983); SEIFERLE u. FREWEIN (1992); EVANS (1993a)
M. flexor digitorum superficialis
nein Schleimbeutel unter der Ursprungssehne
BAUM u. ZIETZ-SCHMANN (1936); FREWEIN (1983); SEIFERLE u. FREWEIN (1992); EVANS (1993a)
M. flexor digitorum profundus, Caput humerale
ja Kapselschleimbeutel unter der Ursprungssehne
WALTHER (1908); SÜPPEL (1921); BAUM u. ZIETZ-SCHMANN (1936); FREWEIN (1983); SEIFERLE u. -FREWEIN (1992); EVANS (1993a)
mediales Seitenband
ja Kapselschleimbeutel unter dem Seitenband
WALTHER (1908)

42 An der kaudalen Seite des Gelenkes sind folgende Bursae synoviales angegeben:
Ein Schleimbeutel liegt unter den Trizepsansatzsehnen des Caput longum und Caput
mediale am Olecranon (WALTHER 1908; BAUM u. ZIETZSCHMANN 1936;
FREWEIN 1983). EVANS (1993a) beschreibt diesen Schleimbeutel mit einem
Durchmesser von 1 cm. Seine ventrale Wand ist fest mit dem Tuber olecrani
verbunden. Nach kranial zieht er aber über dieses hinaus, und dient somit auch dem
M. anconeus als Schleimbeutel (WALTHER 1908).
Ausnahmsweise befindet sich ein Schleimbeutel zwischen der Ansatzsehne des M.
tensor fasciae antebrachii und der medialen Fläche des Olecranon (BAUM u.
ZIETZSCHMANN 1936; FREWEIN 1983; SEIFERLE u. FREWEIN 1992; EVANS
1993). Dieser Schleimbeutel wird ebenfalls von WALTHER (1908) beschrieben, der
ihn bei 2 von 30 untersuchten Hunden gefunden hat. Es handelt sich hierbei um
einen Kapselschleimbeutel.
Tab. 7: Literaturangaben zu Schleimbeutel und Kapselschleimbeutel kaudal am
Ellbogengelenk des Hundes.
Kapselschleim-beutel
Lokalisation Autor
M. triceps brachii, hum. Anteil
nein Schleimbeutel unter der Ansatzsehne am Olecranon
HULTKRANTZ (1897); BAUM u. ZIETZSCHMANN (1936); EVANS (1993a)
M. tensor fasciae antebrachii
nein zwischen Ansatzsehne u. Olecranon
WALTHER (1908); BAUM u. ZIETZSCHMANN (1936); FREWEIN (1983); SEIFERLE u. FREWEIN (1992); EVANS (1993a)
M. triceps brachii, Caput mediale und Caput longum
ja zwischen Ansatzsehne und Olecranon
WALTHER (1908)

43
In der Literatur sind somit Angaben zu finden, dass sich Kapselschleimbeutel unter
folgenden Muskeln befinden: unter der Endsehen des M. biceps brachii, dem Ende
des M. brachialis, dem Ursprungsteil des M. extensor carpi radialis, M. extensor
digitorum communis und M. pronator teres und dem lateralen und medialen
Seitenband.
Selbständige Schleimbeutel besitzen an der Art. cubiti der M. extensor carpi ulnaris,
der M. flexor carpi ulnaris und der M. flexor digitorum superficialis. Letzterer kann mit
dem Schleimbeutel des M. flexor carpi ulnaris in Verbindung stehen.

44 2.2.7 Sesamoide Strukturen: Sesambeine und fibro-kartilaginöse Sonderbildungen
In Gelenkkapseln und -bändern bzw. in den Ursprungs- oder Ansatzsehnen
benachbarter Muskeln können fibröse bzw.fibrokartilaginöse Sonderbildungen (auch
mit Kalkeinlagerungen, diese sind von pathologischen Verkalkungen abzugrenzen)
oder Sesambeine eingelagert sein.
Entsprechend des histologischen Aufbaus verwenden PREUß u. WÜNSCHE (1974)
in ihrer Arbeit über die Schweinezehen auftretenden Strukturen folgende
Bezeichnungen: Corpora capsularia s. sesamoidea fibrosa, bei umschriebener
Verstärkung des kollagenen Materials, Corpora capsularia s. sesamoidea cartilaginea bei Einlagerung von Knorpelzellen und Corpora capsularia s. sesamoidea ossea bei Verknöcherung. Es handelt sich bei den sogenannten
Sesambeinen oder Sesamkörpern an den Schweinezehen, um primär reaktive
Verstärkungen der Gelenkkapsel, so dass für diese unterschiedlichen Einlagerungen
der Überbegriff „sesamoide Strukturen“ gewählt wurde.
Im Bereich des Ellbogengelenkes des Hundes werden sowohl medial als auch lateral
sesamoide Strukturen beschrieben. Eine Übersicht hierzu gibt die Tabelle 8, S. 51.
Sesamoide Struktur an der lateralen Seite Diese werden im lateralen Gelenkkapselbereich nahezu konstant, jedoch in unter-
schiedlicher Zuordnung zu den anliegenden Bändern (Lig. collaterale lat. und Lig.
anulare radii) oder in der Ursprungssehne des M. supinator beschrieben.
Grundsätzliche Unterschiede zu Lage und Bau werden folgendermaßen dargestellt:
a) fakultatives Sesambein
In Höhe des Gelenkspaltes ist das laterale Kollateralband höckerartig verdickt; es
enthält hier häufig ein Sesambein und geht an dieser Stelle in das Ligamentum
anulare radii über (BAUM u. ZIETZSCHMANN 1936; EVANS 1993a). Die Lage des
Sesambeins geben VAN HERPEN (1988) im kranialen Schenkel des lateralen

45 Seitenbandes an. KASER (1998) bestimmt die Lage im lateralen Kollateralband nicht
näher. HITZ (1976) sieht das Sesambein kraniolateral in Höhe des Caput radii in
oder unter dem Lig. anulare radii, jedoch nur mit dem Lig. collaterale laterale und der
Gelenkkapsel verbunden.
Dagegen ordnen WOOD et al. (1985) dieses Sesamoid einzig der Ursprungssehne
des M. supinator zu. Bei ihrer Untersuchung des Sesambeins aus der Ursprungs-
sehne dieses Muskels stellte sich heraus, daß die gelenknahe Seite mit hyalinem
Knorpel überzogen ist und die gelenkferne mit Faserknorpel ausgestattet ist. Die
Lage dieses Sesambeines wird von BERZON u. QUICK (1980) sowie von BERGER
(1992) sowohl der Ursprungssehne des M. supinator als auch Teilen des Lig. anulare
radii zugerechnet, während es von WEBBON u. CLAYTON-JONES (1976) nur dem
Lig. anulare radii zugeordnet wird. In einer Untersuchung von 112 Hunden
verschiedener Rassen werden Röntgenbilder angefertigt (MAYRHOFER 1980).
Hierbei wird festgestellt, dass der Zeitpunkt des Auftretens eines Sesambeins
vorwiegend in der Altersklasse der sechs bis sieben Monate alten Hunden liegt. Der
früheste röntgenologische Nachweis ist mit vier Monaten möglich. Im Alter von einem
Jahr ist die Bildung abgeschlossen. Sollte dieses fakultative Sesambein bis dahin
nicht aufgetreten sein, ist es nicht mehr zu erwarten.
Aufgrund der anatomischen Präparation beschreibt MAYRHOFER (1980) die
Beteiligung des Sesambeins an der Gelenkbildung. Es handelt sich um ein
fakultatives Sesambein im Sinne einer Ergänzung des Caput radii, dessen
Lokalisation im Lig. anulare radii an der lateralen Seite des Gelenkes unter dem M.
supinator angegeben wird. Bei der Präparation dieser Struktur stellen sich die
Bänder der Art. cubiti so straff dar, daß das Sesambein der proximalen Kontur des
Radius fest angefügt ist und somit jegliche „Einklemmphänomene“ und eine daraus
resultierende Lahmheit ausgeschlossen werden können. Bei arthrotischen
Erkrankungen des Ellbogengelenkes jedoch können sich an den Randpartien dieses
Sesambeins Exostosen bilden, die dann zu Funktionsstörungen wie Lahmheiten
beitragen können, aber nicht das Sesambein selbst. Dieses wird damit als eine
normale anatomische Bildung angesehen (MAYRHOFER 1980).

46 Gelegentlich wird dieses laterale Sesambein mit dem Auftreten von Lahmheiten in
Verbindung gebracht (VÄÄNÄNEN u. SKUTNABB 1978). Die Größe variiert von 2 x
2 bis 5 x 5 mm im Durchmesser. Es befindet sich in Höhe des Caput radii und kann
kranial des Lig. collaterale laterale palpiert werden. Bei der Entnahme wird sowohl
eine Verbindung zur Gelenkkapsel als auch zum Seitenband festgestellt. Eine
histologische Untersuchung ergibt Folgendes: Die Gelenkseite des Sesambeins ist
mit Knorpel überzogen, die gegenüberliegende Seite von Bindegewebe bedeckt.
Dieses bindegewebige Material hat seinen Ursprung in der Gelenkkapsel und im
Seitenband (VÄÄNÄNEN u. SKUTNABB 1978).
In drei Fällen von Lahmheiten bei Rottweilern werden im lateralen Kapselbereich
röntgenologisch knochendichte Verschattungen festgestellt (ZENZES et al. 1989).
Hier wird diese Erscheinung als ein „akzessorischer Knochenkern“ des Radiuskopfes
gedeutet, der nicht mit dem Caput radii verschmolzen ist.
b. Knorpel
Einige Autoren erkennen kein Sesambein an der lateralen Seite des
Ellbogengelenkes beim Hund, sondern lediglich Faserknorpel. So finden MARTIN
(1923) und VOLLMERHAUS et al. (1994b) an der Stelle, an der das Lig. anulare radii
in das laterale Seitenband übergeht, einen Knorpel, dessen Lage HULTKRANTZ
schon 1897 im „radialen Schenkel“ des Bandes dargestellt hat. Die Kreuzung und
intensive Verflechtung der vom Bandhöcker und der lateralen „Bandgrube des
Epicondylus lateralis humeri“ ausgehenden Fasern - das Lig. collaterale laterale mit
seinen zwei Schenkeln und der M. supinator - sind eine mögliche Ursache für die
Bildung eines Knorpels an dieser Stelle (HULTKRANTZ 1897). Knorpeliges Gewebe
kann unter Krafteinwirkung in Knochen umgewandelt werden. Bei älteren und
schwereren Hunden werden diese Krafteinwirkungen größer sein. Damit kommt es
besonders bei diesen mit größerer Wahrscheinlichkeit zur Ausbildung eines
fakultativen Sesambeins (BERGER 1992).

47 c. (meniskenartiger) Knorpel
In einer vergleichenden Untersuchung der Ellenbogengelenke verschiedener
Haussäugetiere – so auch beim Hund - wird an der inneren Oberfläche des Lig.
collaterale laterale eine halbmondförmige Falte beschrieben, die sich wie eine Lippe
zwischen Humerus und Radius des Hundes einschiebt. Hier vermutet CORNER
(1896) ein Homologon zur „internen Fibrocartilago“ des Kniegelenkes. Diese, auf
dem Querschnitt meniskusähnliche, annähernd dreieckige Erhöhung, passt sich dem
Gelenkspalt an, überragt kranial das Seitenband und bildet so einen Höcker, der oft
ein kleines Sesambein enthält und in das Lig. anulare radii übergeht (SÜPPEL 1921).
Dieser Interpretation der fibrokartilaginösen Sonderbildung als rudimentärer
Meniskus, der sich als wulstartige Verdickung des kranialen Schenkels des Lig.
collaterale laterale in den humeroantebrachialen Gelenkspalt einschiebt, schließt sich
BARONE (1968) an. Von KADLETZ (1932) wird diese Struktur ebenfalls erwähnt: Er
ordnet sie jedoch der Supinatorsehne, dem Lig. collaterale laterale und dem Lig.
anulare radii als „Fibrocartilago sesamoidea ligamenti lateralis“ zu.
d. meniskenartiger Knorpel mit Knochen- und Knorpelarealen
Eine histologische Untersuchung (BERGER 1992) des gesamten lateralen
Kapselbereiches und benachbarter Strukturen weist nach, dass der Knorpelfalz nur
aus zwei anatomisch voneinander unabhängigen Strukturen – dem Lig. anulare radii
und der sich medial anschließenden Ursprungssehne des M. supinator - gebildet
wird, aber dennoch den gleichen histologischen Aufbau besitzt. Dies kann als
Hinweis auf eine einheitliche funktionelle Belastung gedeutet werden.
Die Gewebegrundlage für diesen Falz und deren beider Kontaktflächen bilden
Faserknorpel, in dem Sehnenfibrozyten, chondroide Zellen und reife Fibrozyten
auftreten. Es sind mehrere fibröse Anteile an diesem meniskenartigen Knorpel von
innen nach aussen beteiligt. Die Ursprungssehne des M. supinator wird lateral durch
das Lig. collaterale laterale überlagert. Hierbei kommt es proximal des
meniskusähnlichen Falzes der Gelenkkapsel zu einem starken Faseraustausch
zwischen Seitenband und Ursprungssehne des Muskels. Im distalen Falzbereich
liegt das Lig. anulare radii dem Seitenband von innen an (BERGER 1992).

48 Der M. supinator entspringt mit zwei Sehnen im lateralen Kapselbereich und bildet
dadurch dessen Grundlage (siehe Kap. 2.2.5). Die Sehne des M. supinator umgreift
die sog. „Caputprotuberanz“ (HULTKRANTZ 1897) des Radiuskopfes (siehe Kap.
2.2.2) und verflicht sich mit dem innen liegenden Lig. anulare radii und dem außen
hinwegziehenden Lig. collaterale laterale oberflächlich. Dadurch kommt es an dieser
Stelle zu einer kappenartigen Verstärkung der Kapsel auf der Protuberanz des Caput
radii. Diese Verstärkung der Gelenkkapsel liegt distal des Falzes. Der inneren
Sehnenoberfläche ist das Lig. anulare radii angelagert, das von straffen kollagenen
Fasern durchsetzt und an der Innenseite überlagert wird. Diese Fasern stammen aus
der Ursprungssehne des M. supinator und bilden die Falzkante. So wird der
meniskenähnliche Falz in Substanz und Höhe ergänzt. Das Lig. anulare radii wird
zusätzlich von Fasern durchsetzt, die aus der Peripherie der Kapsel kommen. Sie
ziehen gegen die gelenkseitige Oberfläche der Kapsel. Ein Teil dieser Fasern endet
in der Falzkante. Dieses Doppelsystem der Fasern baut zwei Kontaktflächen zur
Artikulation um das Lig. anulare radii auf: Die proximale Seite artikuliert mit dem
Capitulum humeri und die distale Seite mit der sogenannten Caputprotuberanz
(siehe Kap. 2.2.2). Der meniskusähnliche Falz und die beiden Artikulationsflächen
tragen kein Stratum synoviale.
Weiter lateral rückt das Lig. anulare radii zentral vor, so dass es den Falz bis auf
einen schmalen Saum vollständig ausfüllt. Der laterale Bereich des Lig. anulare radii
weist sternförmige Fibrozyten auf, die zwischen Faserbündeln liegen. Das innere
Areal des Bandes enthält Chondrone, die in Reihen zwischen straffen Faserbündeln
angeordnet sind. Die Gestalt der Chondrone ist ellipsoid, wobei die längere Achse
parallel zu den Faserbündeln steht. Die Lakune der Zellen wird von einer intensiv
astralblau positiven Kapsel umgeben. Höfe sind nicht immer zu erkennen, zeichnen
sich aber stellenweise durch Kappenbildung aus. Das Gewebe zeigt folglich die
Charakteristika eines Faserknorpels. Die Färbung der Präparate wurde hier in der
Schnittfolge alternierend mit Hämalaun-Erythrosin und Astrablau (pH 3,2)-
Kernechtrot durchgeführt (BERGER 1992).
An der Grenze zwischen der Ursprungssehne des M. supinator und des
Seitenbandes vergrößern sich die beiden gelenkbildenden Flächen nach lateral

49 fortschreitend. Vor allem die Anlagerungsfläche für die „Caputprotuberanz“
vergrößert sich erheblich (BERGER 1992). Dieser Falz gleicht die im Ellbogengelenk
vorhandene Inkongruenz aus.
Röntgenologisch und histologisch konnte nachgewiesen werden, dass bei Tieren, bei
denen eine Verschattung in Form einer Spongiosierung in diesem Bereich
festgestellt wurde, es proximal zu einer Vergrößerung der Falzfläche kommt. In
diesem Fall wird der laterale Bereich des Capitulum humeri durch den Knorpelfalz
mitgetragen. Somit erfährt die Facies articularis des Radiuskopfes eine
Vergrößerung durch den seitlichen Falz (BERGER 1992).
Weiterhin finden sich neben Knochengewebe auch hyalinisierte Knorpelbereiche mit
oder ohne Mineralisation. Alle diese Strukturen bleiben auf die Ursprungssehne des
M. supinator im Bereich der distalen Falzkante und der distalen Anlagerungsfläche
beschränkt (BERGER 1992).
BERGER (1992) kommt aufgrund histologischer und röntgenologischer Unter-
suchungen zu dem Schluss, dass die als Sesambein bezeichnete Struktur sowohl
der Ursprungssehne des M. supinator als auch dem zentralen Bereich des Lig.
anulare radii zuzuordnen ist. Die Lage dieses Sesambeins befindet sich in der
Knorpelfalz distal des Lig. anulare radii und somit proximal der distalen
Artikulationsfläche. Folgerichtig spricht BERGER (1992) von einem knochen-
verstärkten Falz.
Eine enzymhistologische Untersuchung der Ursprungssehne des M. supinator zeigt
das Fehlen von „Stratum synoviale typischen“ Zellen in diesem Bereich. Ebenso
konnte histologisch kein Stratum fibrosum nachgewiesen werden. Dieses wird durch
zentralwärts ausscherende Fasern der Supinatorsehne vertreten. Damit fehlen
Verbindungen des Sesambeins zur Gelenkkapsel, und die Verbindung zur Sehne
wird dargelegt (BERGER 1992).

50 Laut BERGER (1992) handelt es sich bei der untersuchten Knorpelfalz um ein
genuines Bauelement, das konstant nachgewiesen werden konnte und bei der
knochendichten Verschattung um eine akzessorische Bildung bzw. seine Vorstufen.
Die untersuchten Tieren werden in verschiedene Gewichtsklassen (bis 10 kg, bis 20
kg, bis 30 kg und bis 40 kg) eingeteilt und die genannte Struktur in der Knorpelfalz
röntgenologisch dargestellt. Dabei ergibt sich folgendes Ergebnis: Die Verkalkung als
einfachste Form der Verschattung findet man in allen Gewichtsklassen, sie stellt sich
mit unscharfer Silhouette dar. Eine beginnende Spongiosierung fehlt der Gruppe bis
10 kg völlig. Die nächste Stufe ist die Spongiosierung; sie hat als Endstufe ihr
Maximum in der höchsten Gewichtsklasse. Bei der Entstehung eines Sesambeins
wird also eine Beziehung zwischen der Größe bzw. dem Gewicht des Probanden
hergestellt (BERGER 1992).

51 Tab. 8: Literaturangaben zur Lage sesamoider Strukturen an der lateralen Seite des
Ellbogengelenkes.
Art Lokalisation Autor Lig. coll. lat. und Lig. anulare radii
BAUM u. ZIETZSCHMANN (1936); EVANS (1993a)
Lig. coll. lat., kranialer Schenkel
VAN HERPEN (1988)
Lig. coll. lat. und Lig. anulare radii und Gelenkkapsel
HITZ (1976)
Lig. coll. lat. und Gelenkkapsel
VÄÄNÄNEN u. SKUTNABB (1978)
Gelenkkapsel ZENZES et al. (1989) Lig. anulare radii WEBBON u. CLAYTON –
JONES (1976); MAYRHOFER (1980);
Ursprungssehne des M. supinator und Lig. anulare radii
BERZON u. QUICK (1980) BERGER (1992)
a. fakultatives Sesambein
Ursprungssehne des M. supinator
WOOD et al. (1985)
Lig. coll. lat. und Lig. anulare radii
MARTIN (1923); VOLLMERHAUS et al. (1994b)
b. Knorpel
Lig. coll. lat., kranialer Schenkel
(HULTKRANTZ 1897)
Lig. coll. lat. SÜPPEL (1921) CORNER (1896)
Lig. coll. lat., kranialer Schenkel
BARONE (1968)
c. meniskenartiger Knorpel
Lig. coll. lat., Lig. anulare radii, Ursprungssehne des M. supinator
KADLETZ (1932)
d. meniskenartiger Knorpelfalz mit Knochen- und Knorpelarealen
Teile des Lig. anulare radii,Ursprungssehne des M. supinator
BERGER (1992)

52 Sesamoide Struktur an der medialen Seite
Im sehnigen Ursprungsbereich des humeralen Anteils des M. flexor digitorum
profundus wird zuweilen ein „Sesambein“ vorwiegend bei großen Hunderassen mit
Lahmheit gefunden (GRØNDALEN u. BRAUT 1976; PRICE u. KING 1977;
MAYRHOFER 1980; MEYER-LINDENBERG 2001). Eine ähnlich sesamoide Struktur
wird auch bei großen Hunden mit Lahmheit in der Ursprungssehne des M. flexor
digitorum superficialis (MEYER-LINDENBERG 2001), des M. flexor carpi ulnaris
(ZONTINE et al. 1989), des M. flexor carpi radialis (WALKER 1998) und des M.
pronator teres (MEYER-LINDENBERG 2001) gefunden.
Es gibt verschiedene Theorien über die Entstehung dieser Strukturen: Die fehlende
Verschmelzung des Verknöcherungskerns des medialen Epicondylus humeri mit der
distalen Epiphyse des Humerus (MAYRHOFR 1980) bzw. die Entstehung aus einem
vorgebildeten Ossifikationszentrum, also wie ein Sesambein (GRØNDALEN u.
BRAUT 1976; FEHR u. MEYER-LINDENBERG 1992), eine Abrissfraktur des M.
pronator teres, Corpora libera oder andere pathologische Veränderungen
(MAYRHOFER 1980) bzw. eine sekundäre dystrophische Kalzifizierung durch eine
chronische Arthropathia deformans (HLADIK 1986). Eine weitere Theorie dazu ist die
Metaplasie eines kollagenen Gewebes (GRØNDALEN u. BRAUT 1976; WALKER
1998; MEYER-LINDENBERG 2001). Warum hier eine Metaplasie vorliegt, konnte
noch nicht ausreichend geklärt werden. Auffällig ist die Beobachtung von MEYER-
LINDENBERG (2001), dass die Metaplasie bei 15 von insgesamt 26 Gelenken mit
einer anderen Erkrankung des Ellbogengelenkes einhergeht. Hierbei liegt am
häufigsten ein fragmentierter Processus coronoideus vor.

53 3. Material und Methoden 3.1. Makroskopische Untersuchung Für die Messungen an den Seitenbändern der Ellbogengelenke stehen die
Gliedmaßen von 22 aus medizinischen Gründen euthanasierten Schäferhunden und
Schäferhund-Mischlingen zur Verfügung. Insgesamt werden 40 Ellbogengelenke
präpariert.
Die Tierkörper werden bis zur Präparation tiefgefroren gelagert und für die
Untersuchungen aufgetaut. Die Vorderextremitäten werden mit dem Schulterblatt
vom Rumpf abgetrennt. Für die Untersuchung werden nur Gelenke verwendet, die
keinerlei pathologische Veränderungen aufweisen.
In der Tabelle 9 sind Rasse, Alter, Gewicht des Hundes, Geschlecht sowie teilweise
die Radiuslänge dokumentiert. Falls das Alter nicht bekannt ist, wird es nach
Zahnalter und Exterieur geschätzt.
Das Alter wird in drei Kategorien eingeteilt:
junger Hund bis 3 Jahre (n = 1)
mittelalter Hund 4 bis 6 Jahre (n = 12)
alter Hund 7 bis 14 Jahre (n = 9)
Das Körpergewicht in den einzelnen Altersklassen beträgt im Mittel:
„junger Hund“ = 20 kg (n = 1)
„mittelalter Hund“ = 28,5 kg (n = 11)
„alter Hund“ = 35,1 kg (n = 10)

54 Tab. 9: Übersicht zu Rasse, Alter, Gewicht und Geschlecht der untersuchten Hunde.
Tiernummer Präparat Nr.
Rasse Alter Gewicht in kg
Geschlecht
1 1/2 DSH alt 46 w 2 3/4 DSH mittelalt 30 m 3 5 DSH-Mix mittelalt 28 w 4 6 DSH-Mix alt 30 w 5 7/8 DSH-Mix mittelalt 26 w 6 9/10 DSH alt 38 m 7 11/12 DSH mittelalt 30 m 8 13/14 DSH alt 30,5 m 9 15/16 DSH-Mix alt 33 w 10 17/18 DSH-Mix mittelalt 25 w 11 19 DSH-Mix mittelalt 29,5 w 12 20/21 DSH alt 29 w 13 22/23 DSH mittelalt 23,5 m 14 24/25 DSH 10 J. 38 m 15 26/27 DSH mittelalt 38 m 16 28/29 DSH mittelalt 32 w 17 30/31 DSH-Mix 3 J. 20 m 18 32 DSH-Mix alt 35 w 19 33/34 DSH-Mix mittelalt 25 m 20 35/36 DSH-Mix alt 34 m 21 37/38 DSH mittelalt 26,5 w 22 39/40 DSH alt 37,5 m
DSH = Deutscher Schäferhund
DSH-Mix = Deutscher Schäferhund Mischling

55 3.1.1. Präparationstechnik Die Vordergliedmaßen werden vom Rumpf abgesetzt. Anschließend wird die Haut
abgezogen und die für das Ellbogengelenk relevanten Muskeln dargestellt (siehe
Kapitel 2.2.5.).
Die Muskeln werden einschließlich ihres Ansatzes bzw. Ursprungs entfernt, um den
Bandapparat der Art. cubiti isoliert darstellen zu können. Der Ursprungsbereich des
M. supinator und die Ursprungssehne des M. extensor digitorum communis, ebenso
die kaudale Ansatzsehnen der Mm. biceps brachii und brachialis verbleiben am
Präparat, da sie in engem Zusammenhang mit dem Bandapparat stehen. Nachdem
die Kollateralbänder in Ursprung, Ansatz und Verlauf freipräpariert sind, werden ihre
Längen- und Breitenmaße dokumentiert. Die Werte werden mit Hilfe eines
Stechzirkels abgenommen und auf ein Maßband übertragen.
Die weitere Lagerung der präparierten und vermessenen Beine erfolgt in Peterscher
Lösung.
3.1.2 Längen- und Breitenmessung an den Gelenkbändern a) Die Längen- und Breitenmessstrecken der Ligg. collateralia an der Art. cubiti des
Hundes sind den Schemazeichnungen, Abb. 1a bis Abb. 2b und ihren Legenden zu
entnehmen.
Die einzelnen Messstrecken werden jeweils für die Kollateralbänder der Art. cubiti bei
einer Gelenkwinkelung von 160° und von 30° in cm bestimmt. Das Lig. obliquum und
das Lig. anulare radii wird aus messtechnischen Gründen in der Gelenkstellung von
160° untersucht, denn in der Beugestellung des Ellbogengelenkes sind diese Bänder
vedeckt und somit nicht messbar. Das Lig. olecrani wird in einer Winkelung von 30°
gemessen, da es in der Streckstellung des Gelenkes einen s-förmigen Verlauf hat
und so die Gesamtlänge nicht zu bestimmen ist.

56 a) Längenmessungen an den Kollateralbändern:
Abb. 1a: Längenmessstrecken am Lig. collaterale laterale des Ellbogengelenkes
des Hundes, Gelenkwinkelung = 160°
1 Länge gesamt
2 Länge Ursprungsbereich
3 Länge freier Teil
4 Länge Ansatzbereich
5 Länge Humerusabschnitt
6 Länge freier Teil
Humerusabschnitt
7 Länge Unterarmabschnitt
a) kranialer Schenkel
b) kaudaler Schenkel
8 Länge freier Teil
Unterarmabschnitt
9 Länge vom Ursprung bis
zur Aufspaltung

57
Abb. 2a: Längenmessstrecken am Lig. collaterale
mediale des Ellbogengelenkes des Hundes,
Gelenkwinkelung = 160°
1 Gesamtlänge
Das Band wird proximal vom Ursprung bis distal zum Ansatz für jeden Schenkel
einzeln gemessen (Abb. 1a und Abb. 2a). Beim kaudalen Schenkel des Lig.
collaterale laterale entspricht dies der Länge des Unterarmabschnitts (siehe Abb
1b/7b).
Da der kaudale Schenkel des lateralen Kollateralbandes fächerförmig inseriert, wird
proximal von seinem Hervortreten am kranialen Schenkel bis zum distalen Ansatz-
rand gemessen, so dass die Messstrecke hier nicht parallel zu den Fasern verläuft.
1 Länge gesamt
a) kranialer Schenkel
b) kaudaler Schenkel
2 Länge Ursprungsbereich
3 Länge freier Teil
4 Länge Ansatzbereich
a) kranialer Schenkel
b) kaudaler Schenkel
5 Länge Humerusabschnitt
6 Länge freier Teil
Humerusabschnitt
7 Länge Unterarmabschnitt
a) kranialer Schenkel
b) kaudaler Schenkel
8 Länge freier Teil
Unterarmabschnitt
a) kranialer Schenkel
b) kaudaler Schenkel
9 Länge vom Ursprung bis
zur Aufspaltung

58
2 Länge des Ursprungsbereiches
Dies bezeichnet die Strecke vom proximalen Rand, an dem die Bandfasern mit dem
Periost verwachsen sind, bis zu der Stelle, an der man das Kollateralband wieder
von seiner Unterlage abheben kann (Abb. 1a/2 und Abb. 2a/2). Die distale Grenze
wird durch das Unterschieben einer Knopfkanüle bestimmt.
3 Länge des „freien Teils“
Die Länge des „freien Teils“ wird ebenfalls durch das Unterschieben einer Knopf-
kanüle abgegrenzt. Dies ist die Strecke, die sich vom Periost abheben läßt und somit
nicht zum Ursprungs- oder Ansatzbereich gehört (Abb. 1a/3 und Abb. 2a/3). Für den
kaudalen Schenkel des Lig. collaterale laterale wird dieser Wert nicht vermessen, da
dies der breitgefächerte Ansatz nicht zulässt.
4 Länge des Ansatzbereiches
Dieser Abschnitt schließt sich distal an den „freien Teil“ an. Hier sind die Bandfasern
wieder mit der Knochenhaut verwachsen. Er wird für den kranialen Schenkel des
lateralen Kollateralbandes (Abb. 1a/4), für den kranialen Schenkel des medialen
Kollateralbandes (Abb. 2a/4a) und für den kaudalen Schenkel des medialen
Kollateralbandes (Abb. 2a/4b) bestimmt. Für den kaudalen Schenkel des Lig.
collaterale laterale wird dieser Wert nicht bestimmt (siehe 3).
5 Länge des Humerusabschnittes
Mit diesem Wert wird die gesamte Länge des Bandteiles bezeichnet, der dem
Humerus aufliegt. Der Wert bezeichnet die Strecke vom Proximalrand des Ursprungs
bis zur Höhe des Gelenkspalts der Art. cubiti (Abb. 1a/5 und Abb. 2a/5).
6 Länge des „freien Teils“ des Humerusabschnittes
Hierbei handelt es sich um den auf dem Humerus gelegenen Teil, der mit der
Knopfkanüle abhebbar ist, also nicht zum Ursprungsbereich gehört (Abb. 1a und
Abb. 2a/6).

59 7 Länge des Unterarmabschnitts
Die gesamte Länge des Kollateralbandes, die auf Radius bzw. Ulna liegt inklusive
des Ansatzbereiches. Dies wird sowohl für den kranialen Schenkel des lateralen
(Abb. 1a/7a) und medialen Kollateralbandes (Abb. 2a/7a) als auch den kaudalen
Schenkel beider Bänder (Abb. 1a/7b und 2a/7b) vermessen.
8 Länge des „freien Teils“ des Unterarmabschnitts
Dies bezeichnet den Bereich des Kollateralbandes, der dem Unterarm anliegt und
mit der Knopfkanüle abhebbar ist, also exklusive des Ansatzbereiches.
Vom Lig. collaterale laterale wird nur der kraniale Schenkel (Abb. 1a/8) vermessen
(siehe 2), jedoch vom Lig. collaterale mediale sowohl der kraniale (Abb. 2a/8a) als
auch der kaudale Schenkel (Abb. 2a/8b) bestimmt.
9 Länge vom Ursprung des lateralen und medialen Kollateralbandes bis zur
Aufspaltung in zwei Schenkel (Abb. 1b/9 und Abb. 2b/9).

60 b) Breitenmessungen an den Kollateralbändern
Abb. 1b: Breitenmessstrecken des Lig. collaterale laterale und des Lig. anulare radii
am Ellbogengelenk des Hundes, Gelenkwinkelung = 160°
10 Breite Ursprung Lig.
coll. laterale
a) ungeteiltes Band
b) kaudaler Schenkel
11 Breite Gelenkspalt Lig.
coll. laterale
12 Breite Ansatzbereich
Lig. coll. laterale
a) kranialer Schenkel
b) kaudaler Schenkel

61
Abb. 2b: Breitenmessstrecken am Lig. collaterale mediale des Ellbogengelenkes des
Hundes, Gelenkwinkelung = 160°
10 Breite im Ursprungsbereich
In diesem Fall wird der unter Punkt 2 beschriebene Ursprungsbereich in seiner Breite
bestimmt (Abb. 1b/10a und Abb. 2b/10). Am Lig. collaterale laterale wird diese Breite
zusätzlich für den kaudalen Schenkel an der Stelle, an der er aus dem ungeteilten
Band hervorgeht, bestimmt (Abb. 1b/10b).
11 Breite im Gelenkspaltbereich
Die Breite des jeweiligen Bandes wird auf der Höhe des Gelenkspaltes gemessen
(Abb. 1b und Abb. 2b). Für den kaudalen Schenkel des Lig. collaterale laterale ist
dies nicht möglich, da er sich erst unterhalb des Gelenkspaltes aus dem kranialen
Schenkel löst.
10 Breite Ursprung
11 Breite Gelenkspalt
12 Breite Ansatzbereich
a) kranialer Schenkel
b) kaudaler Schenkel

62 12 Breite im Ansatzbereich
Die kranialen (Abb. 1b/12a und Abb. 2b/12a) und kaudalen Schenkel (Abb. 1b/ 12b
und Abb. 2b/12b) der Kollateralbänder werden in ihrer Ansatzbreite vermessen.
c) Messungen am Lig. obliquum und an den Bänder der Art. radioulnaris proximalis Die Messungen am Lig. obliquum, Lig. anulare radii und Lig. olecrani sind in den
Schemazeichnungen Abb. 3a, Abb. 3b und Abb. 4 dargestellt.
Abb. 3a: Längenmessstrecken am Lig. obliquum der Art. radioulnaris proximalis des
Hundes, Gelenkwinkelung = 160°
1 Länge gesamt
a) bis zum
Ansatz an der
Bizepssehne
b) bis zum
Ansatz am
medialen
Seitenband
2 Länge bis zur
Aufteilung in zwei
Schenkel

63
Abb. 3b: Breitenmessstrecken am Lig. anulare radii und Lig. obliquum der Art.
radioulnaris proximalis des Hundes, Gelenkwinkelung = 160°
a) Längen (Abb. 3a und Abb. 3b)
1 Gesamtlänge
Für die Gesamtlänge des Lig. obliquum müssen zwei Messungen durchgeführt
werden: vom Ursprung bis zum Ansatz am M. biceps brachii (Abb. 3a/1a) und vom
Ursprung bis zum Ansatz am medialen Seitenband (Abb. 3a/1b). Diese Werte
konnten jedoch nicht bei allen Präparaten bestimmt werden (siehe Tab. 37 im
Anhang), da eine präparatorische Trennung nicht immer eindeutig möglich war.
Das Lig. anulare radii wird in seiner gesamten Länge durch das Anlegen eines
Fadens von Processus coronoideus zu Processus coronoideus bestimmt. Die Länge
des Fadens wird schließlich an einem Maßband abgelesen. Diese Messstrecke ist
schematisch in Abb. 3b/1 dargestellt.
1 Gesamtlänge des Lig.
anulare radii
3a Breite Ursprungsbereich
Lig. anulare radii
3b Breite Ursprungsbereich
Lig. obliquum
4a Breite Lig. anulare radii
4b Breite Gelenkspaltsbe-
reich Lig. obliquum
5a Breite Ansatzbereich
Lig. anulare radii
5b Breite Ansatzbereich
M. biceps brachii
5c Breite Ansatzbereich
mediales Seitenband

64 2 Länge vom Ursprung des Lig. obliquum bis zur Aufspaltung in zwei Schenkel (Abb.
3a/2).
b) Breiten (Abb. 3b)
3 Breite im Ursprungsbereich
Am Lig. anulare radii erfolgt die Bestimmung dieses Wertes an der lateralen Seite
(Abb. 3b/3a).
Es wird die Bandbreite des Lig. obliquum im Ursprungsbereich bestimmt (Abb3b/3b).
4 Breite im Gelenkspaltsbereich
Hier wird die Messung der Breite des Bandabschnittes unterhalb des höchsten
Punktes des Caput radii für das Lig. anulare radii (3b/4a) bestimmt bzw. die Breite
jenes Abschnitts des Lig. obliquum am Gelenkspalt vermessen (Abb. 3b/4b).
5 Breite im Ansatzbereich
Beim Lig. anulare radii bezeichnet dieser Wert die Breite an der medialen Seite des
Gelenkes. Der Ansaz ist durch das Lig. collaterale mediale verdeckt (Abb. 3b/5a).
Für das Lig. obliquum werden zwei Werte bestimmt: die Breite am Ansatz an der
Bizepssehne (Abb. 3b/5b) und die Breite zum Ansatz am medialen Seitenband (Abb.
3b/5c).

65
Abb 4: Längen- und Breitenmesslinien am Lig. olecrani der Art. radioulnaris
proximalis des Hundes , Gelenkwinkelung = 30°
a) Längen (Abb. 4)
1 Gesamtlänge
Das Lig. olecrani wird bei maximal gebeugtem Gelenk gemessen, da in der
Streckstellung keine exakte Messung durch den s-förmigen Verlauf des Bandes
möglich ist. Die Gesamtlänge wird vom Ursprung bis zum Ansatz bestimmt (Abb.
4/1).
1 Länge gesamt
3 Breite im
Ursprungsbereich
4 Breite im
Gelekspaltbereich
5 Breite im
Ansatzbereich

66 b) Breiten (Abb. 4)
3 Breite im Ursprungsbereich an der axialen Seite des Epicondylus medialis humeri
(Abb 4/3).
4 Breite in Höhe des Gelenkspaltes in der Beugestellung, dies entspricht etwa der
Mitte der Gesamtlänge (Abb. 4/4).
5 Breite im Ansatzbereich am Olecranon (Abb. 4/5).
3.1.3 Vorversuche Eine erste Messreihe wurde (bei fünf Hunden bzw. acht Vordergliedmaßen) mit den
oben definierten Strecken durchgeführt. Hier wurden jeweils Gelenkstellungen von
160°, 140°, 90° und 30° im Ellbogengelenk gewählt. Da sich die vier Messwerte in
den unterschiedlichen Winkelstellungen unwesentlich voneinander unterscheiden,
wurde im Weiteren nur in der Winkelung 160° und 30° gemessen (als extreme
Streck- bzw. Beugestellung).
3.1.4 Dokumentation der Messergebnisse Die einzelnen Werte der Messergebnisse wurden in Excel-Tabellen festgehalten und
für jedes Band und jede Gelenkstellung dokumentiert (siehe Tab. 21-40 im Anhang).
Etwaige Besonderheiten, die bei der Messung bzw. bei der Präparation auffällig
waren, werden in einem Protokollbogen dokumentiert und im Kapitel 4 erwähnt.
Ausgewählte Präparate werden fotografiert, um die Verhältnisse am Bandapparat
bzw. deren Sondereinrichtungen darzustellen.
3.1.5 Statistische Auswertung Die statistische Auswertung erfolgte mit Hilfe des Institutes für Biometrie,
Epidemiologie und Informationsverarbeitung der Tierärztlichen Hochschule
Hannover.
Alle in Excel angefertigten Dateien wurden mit dem „Statistical Analysis Programm“
(SAS) bearbeitet.

67 3.2 Mikroskopische Untersuchung
3.2.1 Untersuchungsgut Die Proben stammen von 5 euthanasierten DSH-Mischlingen, die aus medizinischen
Gründen getötet wurden. Der Bandapparat wird bis maximal zehn Stunden post
mortem entnommen.
Die Tabelle 10 gibt einen Überblick über den Materialumfang, Rasse, Alter, Gewicht,
Geschlecht und den Zeitpunkt der Probenentnahme.
Tab. 10: Übersicht zu Rasse, Alter, Gewicht, Geschlecht und Probenentnahme post
mortem der verwendeten Schäferhund-Mischlinge.
Alle verwendeten Schäferhund-Mischlinge waren adulte Tiere mit einem
Körpergewicht zwischen 32 kg und 50 kg. Es wurden drei weibliche Tiere und zwei
männliche Tiere verwendet. Die Entnahme des Probenmaterials post mortem variiert
zwischen einer und zehn Stunden.
Tiernummer Bein Nr. Rasse Alter in
Jahren
Gewicht
in kg
Geschlecht Probeentnahme
post mortem in
h
H1 1+2 DSH-
Mix
9 45 w 10
H2 3+4 DSH-
Mix
5 35 w 4
H3 5+6 DSH-
Mix
14 32 m 8
H4 7+8 DSH-
Mix
13 50 m
2
H5 9+10 DSH
9 35 w 1

68 3.2.2 Entnahme der Bänder Die Kollateralbänder wurden nach dem Entfernen der Muskulatur freipräpariert,
schließlich in ihrer gesamten Länge vom Ursprung bis zum Ansatz von den
jeweiligen Knochenpunkten entnommen und entsprechend auf einer Korkplatte mit
Seide vernäht. Die Lage des Seitenbandes auf der Korkplatte wird anhand einer
Skizzenzeichung protokolliert. Die Entnahme des lateralen Kollateralbandes erfolgt
so, dass der Ursprung des M. supinator und ein Stumpf des Lig. anulare radii am
Seitenband belassen werden.
3.2.3 Probenaufbereitung für die mikroskopische Untersuchung Alle Proben werden in BOUINscher-Lösung (BÖCK 1989) gebracht und dort für
mindestens 48 Stunden belassen. Lediglich die Probe H 4 Nr. 8 wird in 4%iger
Formol-Kalzium-Lösung nach LILLIE u. FULLMER (1976) fixiert, um Gefrierschnitte
anzufertigen. Hierzu wird das Gefriermikrotom Bright Typ 5030 verwendet. Die
Anfertigung der Gefrierschnitte dient zum Nachweis elastischer Fasern.
Bevor die Proben weiterbehandelt werden, werden sie in kleine Blöcke von ca. 4 mm
Höhe zugeschnitten. Die Breite variiert je nach Bandbreite. Um bei der Auswertung
einen genauen Vergleich zwischen dem lateralen und medialen Kollateralband zu
erhalten, wird jeweils ein Block aus der gleichen Höhe geschnitten (Abb.5 und Abb.
6). Hierzu wird der „freie Teil“ des Bandes proximal des Gelenkspaltes gewählt. In
diesem Teil sind keine Fasern des Lig. anulare radii bzw. des M. supinator enthalten.

69
Abb. 5 und Abb. 6: Lokalisation der Probenblöcke im lateralen und medialen Kollate-
ralband.
Entkalkung
Da das Material knorpelige Areale aufweisen kann, die zum Teil mineralisiert sind,
werden die zugeschnittenen Proben für 21 Tage in einer EDTA-Lösung (Ansatz: 500
g EDTA, 1900 ml Aqua dest., 100 ml 40%ige Natronlauge, pH-Wert 7,4) entkalkt, die
jeden zweiten Tag erneuert wird.
Wässerung
Nach der Entkalkung werden die Proben für 24 Stunden in fließendem Leitungs-
wasser gewässert.
Entwässerung und Paraffineinbettung
Für die Einbettung wird das in wässriger Fixationslösung liegende Gewebe über eine
aufsteigende Alkoholreihe entwässert. Hierfür wird es nach dem Zuschneiden für 24
Entnahme des Proben-
blocks aus dem medialen
Kollateralband
Entnahme des Proben-
blocks aus dem lateralen
Kollateralband

70 Stunden in 70%igen Alkohol und anschließend für 24 Stunden in 80%igen Alkohol
überführt. Schließlich werden die Proben für jeweils 90 Minuten in 100%igem
Alkohol, Isopropanol, Xylol I und Xylol II belassen. Nach der Entwässerung folgt das
Umfüllen der Proben für 90 Minuten in Parafin I sowie jeweils 2 Stunden in Paraffin II
und Paraffin III. Anschließend werden die Proben in flüssigem, 60° Celsius heißen
Paraffin eingebettet und bei Raumtemperatur ausgehärtet.
Schneiden
Von den eingebetteten Proben des Bandapparates werden mit Hilfe eines
Rotationsmikrotoms (Jung) 4 µm dicke Schnitte angefertigt und auf einen
Objektträger gebracht. Anschließend trocknen sie im Wärmeschrank bei 37° C an.
Es werden pro Block sowohl drei Längs- als auch drei Querschnitte in Folge
angefertigt.
Behandlung vor dem Färben
Bevor die Proben gefärbt werden können, müssen sie nach der Entparaffinierung
über eine absteigende Alkoholreihe wieder gewässert werden: zweimal für jeweils 10
Minuten in Xylol, anschließend je 3 Minuten Isopropanol, 96%igen Alkohol, 80%igen
Alkohol, 70%igen Alkohol und schließlich spülen in Aqua dest..
3.2.4 Färbungen Es werden für die Darstellung des Faserverlaufs zwei Färbungen verwendet, um
einmal, wie methodisch üblich, in Pikrosiriusrot (JUNQUEIRA et al. 1979), je nach
Abhängigkeit von der Farbintensität, den Faserverlauf und Richtungsänderungen der
Fasern deutlich hervorzuheben bzw. den gleichen Ausschnitt in H.E. mit seinen
unterschiedlichen Kernformen der Fibroblasten/Fibrozyten in Bezug auf den
Faserverlauf zu vergleichen.
Darstellung der Zellkerne
Als histologische Übersichtsfärbung wird die Hämatoxylin-Eosin-Färbung (H-E.) nach
DELAFIELD (BÖCK 1989) verwendet.
Darstellung der Kollagenfasern
Kollagenanfärbung erfolgt mit Pikrosiriusrot nach JUNQUEIRA et al. (1979) durch-
geführt: Die Färbung wird an entparaffinierten Schnitten mit 0,1%igem Pikrosiriusrot

71 durchgeführt (Ansatz: 400 ml wässrige und gesättigte Pikrinsäure, 0,6 g
Säurefuchsin [Rubin S, Merck], 0,1 g Siriusrot F3BA [Chroma] und 1 ml 37%ige HCl)
in der Mikrowelle (500 Watt für 2,5 Minuten). Die so gefärbten Präparate werden mit
fließendem Leitungswasser und Aqua dest. gespült, mit dem Fön getrocknet und mit
EUKITT eingedeckt.
Darstellung der elastischen Fasern
Die Anfärbung der elastischen Fasern erfolgt mit der Orcein-Färbung nach
TAENZER in der Modifikation nach SHELLOW u. KLIGMAN (1967). Der Farbstoff ist
eine alkoholsaure 1%ige Orcein-Lösung (Ansatz: 1 g Orcein auf 100 ml 70%igen
Alkohol und 0,6 ml konz. HCl). Entparaffinierte Schnitte werden für 1 Stunde in die
Farblösung gestellt, mit 70%igem Alkohol differenziert, entwässert auf die übliche Art
und eingedeckt mit EUKITT.
Elastische Fasern stellen sich nun im Lichtmikroskop als braun-schwarze Strukturen
dar.
3.2.5 Untersuchung mit dem Zeiss Photomikroskop III mit Polarisationsaus-rüstung Die Anwendung des polarisierten Lichtes ermöglicht es, Strukturunterschiede, also
unterschiedliche Faserverläufe, in dreidimensionaler Form darzustellen. Bei den
Kollateralbändern handelt es sich um doppeltbrechendes Material (GERLACH 1976):
Es leuchtet bei einer 90° Drehung des zentrierten Drehtisches hell auf und wird in
den Zwischenstellungen wieder dunkel. Hierzu sei angefügt, dass geordnete
Molekülverbände, die in verschiedenen Richtungen unterschiedliche optische
Eigenschaften aufweisen – Anisotropie – doppelbrechend sind. Das durchtretende
Licht wird in zweilinear polarisierte Strahlen zerlegt. Die Schwingungsebenen dieser
Strahlen stehen senkrecht aufeinander. Die kollagenen Fasern sind positiv einachsig
doppelbrechend; die optische Achse entspricht der Längsachse der Fibrillen. Sie
werden im polarisierten Licht am Stärksten aufleuchten, wenn sie – längsgetroffen –
in die Diagonale der senkrecht aufeinander stehenden Schwingungsebenen der
beiden Polarisationsfilter gedreht werden (BUCHER u. WARTENBERG 1997).

72 3.2.6 Dokumentation Mikroskopie Fotographien von identischen Ausschnitt in der H.E. und Pikrosiriusrot Färbung,
werden in verschiedenen Vergrößerungen mit und ohne Polarisation angefertigt, um
das laterale und das mediale Kollateralband zu vergleichen. Die fotographische
Dokumentation wird beispielhaft an den Bänder des Hundes Nr. 1 durchgeführt
(siehe Tab. 10, Kap. 3.2.1).

73 4. Ergebnisse
4.1 Makroskopische Untersuchung und deskriptive Statistik
4.1.1 Es folgt eine Darstellung der Bänder der Art. cubiti und der Art. radioulnaris
proximalis im Hinblick auf Ursprung, Verlauf, Ansatz, der Lokalisation der
vorkommenden Schleimbeutel und der Messergebnisse (siehe auch Anhang, Tab.
21-40).
4.1.1.1 Ligamentum collaterale laterale (Abb. 7, blau)
Ursprung
Der Ursprung des lateralen Seitenbandes liegt mit einigen Fasern in der
laterodistalen Bandgrube und dem benachbarten Bandhöcker des Condylus humeri.
Verlauf
Als kräftiges Band verläuft es einheitlich bis zur Höhe des Caput radii und teilt sich 1
bis 2 mm distal des Gelenkspaltes in zwei Schenkel auf. Die Fasern, des im
Querschnitt querovalen kranialen Schenkels, verlaufen in distaler Richtung, während
jene des kaudalen Schenkels schräg kaudodistal zur Ulna ziehen. Der kaudale
Schenkel ist im Querschnitt platt und hat eine annähernd dreieckige Form.
Ansatz
Der Ansatz des kranialen Schenkels liegt proximal an der lateralen Seite des Radius
auf einer kleinen Erhabenheit. Der kaudale Schenkel setzt mit einer breiten Basis
distal des Processus coronoideus lateralis ulnae an und verschmilzt teilweise mit den
proximalen Fasern der Membrana interossea antebrachii.
Schleimbeutel
Eine kleine Kapselausbuchtung tritt von proximal unter das laterale Seitenband.
Zusätzlich ist ein Fettkörper von 2 bis 4 mm Durchmesser ist an der Stelle zu finden,
an der das Lig. collaterale laterale, der M. supinator und das Lig. anulare radii
teilweise fusionieren bzw. sich kreuzen. Er befindet sich zwischen dem Bandgeflecht
und einer kleinen knöchernen Erhabenheit (Abb. 7a/A) lateral am Caput radii kurz
distal des Gelenkspaltes an der laterale Seite der Art. cubiti. Kraniomedial dieser

74 Knochenerhabenheit befindet sich eine Rinne (Abb. 7a/B), die lateral vom medialen
Muskelbauchrand des M. supinator begrenzt wird.
Sesamoide Strukturen
Das Bandgeflecht, bestehend aus dem Lig. anulare radii, dem Lig. collaterale laterale
und der Ursprungssehne des M. supinator, wird an der knöchernen Erhabenheit des
Caput radii von einem Fettkörper unterlagert und besitzt hier eine knorpelharte
Struktur von 4 bis 5 mm Durchmesser, die zu palpieren ist (Abb. 7/B). Diese ist in
den Tieren der Altersklasse „junger Hund“ (n = 1) geringradig, in der Altersklasse
„mittelalter Hund“ (n = 12) mittelgradig und in der Altersklasse „alter Hund“ (n = 9)
stark ausgeprägt.
Wird der laterale Bandapparat vom Ellbogengelenk abpräpariert (Abb. 7b), so ist an
der, dem Gelenk zu gewandten Seite, ein knorpelharter Falz (Abb 7b/Pfeile) in Höhe
des Gelenkspaltes zu erkennen. Das Lig. anulare radii zieht in diesen Falz hinein.

75
Abb. 7: Lig. collaterale laterale (blau), Lig. anulare radii (grün) und Lig. obliquum
(orange) der Art. cubiti des Hundes; laterale Ansicht, Gelenkwinkel = 160°
U = Ursprung
A1 = Ansatz des kranialen Schenkels Lig. collaterale laterale
A2 = Ansatz des kaudalen Schenkels des lateralen Kollateralbandes
C = knorpelharte Struktur
E = Reste des M. extensor digitorum communis
S = Reste des M. supinator
H = Humerus
R = Radius
Ul = Ulna

76
Abb. 7a: Die knöcherne Erhabenheit („Caputprotuberanz“) am Caput radii (A) und die
kraniomedial davon befindliche Rinne (B) an der Art. cubiti des Hundes, laterale
Ansicht, Gelenkstellung = 160°
A = Erhabenheit am Caput radii
B = Knochenrinne
C = Unterarmabschnitt des Lig. collaterale laterale
D = Muskelbauch des M. supinator
H = Humerus
R = Radius
U = Ulna

77
Abb. 7b: Darstellung des Knorpelfalzes am lateralen Bandapparat (Lig. coll laterale
mit kranialem und kaudalem Schenkel, Lig. anulare radii) der Art. cubiti und des M.
supinator; im Zusammenhang entfernt; Ansicht von medial
A1 = Lig. collaterale laterale (Humerusabschnitt)
A2 = Kranialer Schenkel des Lig. collaterale laterale
A3 = Kaudaler Schenkel des Lig. collaterale laterale
B1 = Lig. anulare radii (Ursprungsbereich)
B2 = Lig. anulare radii (Bereich, der kraniomedial dem Radius anliegt)
C = M. supinator
Pfeile weisen auf den Knorpelfalz an der dem Gelenk zugewandten Seite des
Bandapparates hin.

78 Messergebnisse des Lig. collaterale laterale
(siehe Material und Methoden, Kap. 3, Abb 1a und 1b)
a) Längen
Das laterale Kollateralband (= einschließlich kranialer Schenkel) hat im Mittel eine
Gesamtlänge von 4,22 cm bei einer Gelenkwinkelung von 160°. Der kaudale
Schenkel tritt erst in Gelenkspalthöhe aus dem Band hervor, deshalb ist seine Länge
dem Unterarmabschnitt gleichzusetzen. Sein Längenmaß beträgt im Mittel 2,75 cm.
Die Länge das kranialen Schenkels beträgt in der Winkelung von 30° im Mittel 4,12
cm und die des kaudalen Schenkels 2,7 cm.
Die Länge des gemeinsamen Ursprungsbereiches ist mit durchschnittlich 0,76 cm
(sowohl bei 160° als auch bei 30°) anzugeben.
Die Länge des freien Teils misst 2,55 cm (160°) bzw. 2,45 cm (30°).
Für die Länge des Ansatzbereiches ergibt sich am kranialen Schenkel des lateralen
Kollateralbandes ein Wert von 0,91 cm (160° und 30°). Die Länge des Humerus-
abschnittes hat für das noch ungeteilte Band eine Länge von 1,82 cm (160°) und
1,61 cm (30°) und entspricht der Bandlänge bis zur Aufteilung in zwei Schenkel.
Für den freien Teil des Humerusabschnittes ergeben sich Werte von 1,07 cm (160°)
und 8,6 cm (30°). Die Länge des Unterarmabschnittes beträgt 2,4 cm (160°) und 2,52
cm (30°) für den kranialen Schenkel. Beim kaudalen Schenkel entspricht dieser Wert
der Gesamtlänge (s.o.). Der freie Teil des Unterarmabschnittes beträgt am kranialen
Schenkel 1,44 cm (160°) und 1,61 cm (30°). Die Strecke vom Ursprung des unge-
teilten Bandes bis zur Aufteilung in zwei Schenkel misst 1,88 cm (160°) bzw. 1,92 cm
(30°).
b) Breiten
Im Ursprungsbereich hat das laterale Kollateralband eine Breite von 0,61 cm (160°)
und 0,6 cm (30°). Die Breite im Gelenkspalt beträgt für das an dieser Stelle noch
ungeteilte Band 0,72 cm (160°) und 0,66 cm (30°). Die Breite des kranialen
Schenkels im Ansatzbereich hat einen Wert von 0,67 cm (160° und 30°), die des
kaudalen Schenkels beträgt 2,59 cm (160° und 30°), wobei er sich mit einer Breite
von 1,43 cm (160° und 30°) aus der Teilungsstelle löst.

79 4.1.1.2 Ligamentum collaterale mediale (Abb. 8, rot) Ursprung
Nahezu alle Bandfasern entspringen am medialen Bandhöcker des Humerus; ein
kleiner Anteil der Fasern nimmt seinen Ursprung an der benachbarten Bandgrube.
Verlauf
Die Fasern des medialen Kollateralbandes verlaufen proximodistal parallel zur
Knochenoberfläche. Das mediale Band verschmilzt wenige Millimeter proximal des
Gelenkspaltes mit der Gelenkkapsel und ist schwer von dieser zu trennen.
Distal des Gelenkspaltes teilt sich das Lig. collaterale mediale in zwei Schenkel.
Diese Aufteilung in den kranialen und kaudalen Schenkel ist in allen Fällen distal der
Höhe der Aufteilung des Lig. collaterale laterale.
Der kraniale Schenkel inseriert proximomedial an der Tuberositas radii. Die
Darstellung des Ansatzes ist erst nach dem Entfernen der kaudalen Ansatzsehne
des M. biceps brachii und des M. brachialis möglich, da diese beiden Strukturen hier
miteinander verschmolzen sind.
Ansatz
Der kaudale Schenkel zieht tief in das Spatium interosseum. Hier setzt er mit einem
großen Teil seiner Fasern an der proximokranialen Seite der Ulna im Spatium
interosseum, mit wenigen Fasern am kraniolateralen Rand der Ulna, aber auch mit
einigen Fasern kaudal am Radius an.
Schleimbeutel
Zwischen der kaudalen Ansatzsehne des M. biceps brachii bzw. des M. brachialis
und dem medialen Seitenband ist ein selbstständiger Schleimbeutel eingelagert, der
eine durchschnittliche Größe von 4 mm im Durchmesser besitzt. Unter dem medialen
Seitenband verläuft die Gelenkkapsel, die hier als Kapselschleimbeutel dient.
Sesamoide Strukturen
Einige Millimeter proximal des Gelenkspaltes, verbindet sich der kürzere Schenkel
des Lig. obliquum mit dem kranialen Schenkel des medialen Seitenbandes (Abb.
9/A1). An der Stelle, an der die Bandverbindung über den mediokranialen Rand der
Trochlea zieht, ist eine knorpelharte Struktur von 3 bis 4 mm Durchmesser zu sehen
(Abb. 8a/B). Es lässt sich ein Unterschied in den verschiedenen Altersklassen

80 feststellen; bei den Hunden in der Altersklasse „alter Hund“ ist diese knorpelharte
Einlagerung deutlicher ausgeprägt als bei den Hunden geringeren Alters.
Abb. 8: Lig. collaterale mediale (rot), Lig. anulare radii (grün) und Lig. obliquum
(orange) der Art. cubiti des Hundes; mediale Ansicht, Gelenkwinkel = 160°
U = Ursprung
A1 = Ansatz des kranialen Schenkels des medialen Kollateralbandes
A2 = Ansatz des kaudalen Schenkels des medialen Kollateralbandes
H = Humerus
R = Radius
Ul = Ulna

81
Abb. 8a: Knorpelharte Struktur an der medialen Seite des Ellbogengelenkes;
im Übergang des kürzeren Schenkels des Lig. obliquum in den kranialen Schenkel
des Lig. collaterale mediale; mediale Ansicht der Art. cubiti, Gelenkwinkelung = 160°.
A1 = Humerusabschnitt des Lig. collaterale laterale
A2 = Kaudaler Schenkel des Lig. collaterale laterale
A3 = Kranialer Schenkel des Lig. collaterale mediale
R = Radius
B = Knorpelharte Struktur
H = Humerus
U = Ulna

82 Messergebnisse des Lig. collaterale mediale
(siehe Material und Methoden, Kap. 3, Abb. 2a und Abb. 2b)
a) Längen
Der kraniale Schenkel des medialen Kollateralbandes hat im Mittel eine Gesamt-
länge von 4,24 cm (160°) bzw. 4,15 cm (30°), der kaudale Schenkel misst hier 4,68
cm (160°) bzw. 4,66 cm (30°). Die Strecke vom Ursprung des ungeteilten Bandes bis
zur Aufteilung in zwei Schenkel beträgt 2,24 cm (160°) bzw. von 2,22 cm (30°).
Die Länge des Ursprungsbereiches misst 0,42 cm (160°) und 0,44 cm (30°); jene des
freien Teils beträgt 3,42 cm (160°) und 3,36 cm (30°) für den kranialen Schenkel und
3,55 cm (160°) und 3,48 cm (30°) für den kaudalen.
Am kranialen Schenkel des medialen Kollateralbandes ergibt sich für die Länge des
Ansatzbereiches ein Wert von 0,37 cm (160° und 30°). Für den kaudalen Schenkel
lässt sich ein Wert von 0,71 cm in beiden Gelenkstellungen messen.
Die Länge des Humerusabschnittes beträgt für das noch ungeteilte Band 1,96 cm
(160°) und 1,93 cm (30°). Für den freien Teil des Humerusabschnittes ergibt sich ein
Wert von 1,52 cm (160°) bzw. ein Wert von 1,49 cm (30°). Die Länge des
Unterarmabschnittes beträgt 2,3 cm (160°) bzw. 2,22 cm (30°) für den kranialen
Schenkel. Für den kaudalen Schenkel ergeben sich für diesen Teilabschnitt 2,7 cm
(bei 160° und 30°). Der freie Teil des Unterarmabschnittes besitzt für den kranialen
Schenkel eine Länge von 1,93 cm (160°) und 1,85 cm (30°), für den kaudalen
Schenkel in den genannten Winkelungen jeweils 1,99 cm.
b) Breiten
Für die Breite im Ursprungsbereich lässt sich ein Wert von 0,53 cm (160° und 30°)
für das ungeteilte Band bestimmen. Die Breite in Höhe des Gelenkspaltes beträgt
0,72 cm (160°) und 0,67 cm (30°). Der Ansatzbereich des kranialen Schenkels hat
eine Breite von 0,29 cm (160° und 30°); demgegenüber lässt sich für den kaudalen
Schenkel ein Wert von 0,37 cm (160° und 30°) bestimmen.

83 4.1.1.3 Ligamentum anulare radii (Abb. 7 und Abb. 9, grün) Ursprung
Das Lig. anulare radii hat seinen Ursprung an allen untersuchten Präparaten an der
Spitze des Processus coronoideus lateralis ulnae.
Verlauf
In seinem Verlauf umfasst es das Caput radii wie ein Halbring. Dabei unterkreuzt es
von kaudolateral nach medial das laterale Kollateralband, die Ursprungssehne des
M. supinator und das mediale Kollateralband. Hierbei gibt es an der dem
Kollateralband zugewandten Seite einige Fasern an den kranialen Schenkel des
medialen Kollateralbandes ab. Durch diese Verbindung mit dem Lig. anulare radii
liegt der kraniale Schenkel in der Beugestellung des Gelenkes leicht s-förmig der
Knochenoberfläche auf.
Von seinem Ursprung an der lateralen Seite bis zum Hervortreten unter dem M.
supinator ist der Verlauf des Bandes extrakapsulär, anschließend verläuft es bis zu
seinem Ansatz an der medialen Seite innerhalb der Kapsel.
Ansatz
Der Ansatz liegt ca. 1 bis 3 mm distal des Processus coronoideus medialis ulnae.

84
Abb. 9: Lig. collaterale laterale (blau), Lig. coll mediale (rot), Lig. anulare radii (grün)
und Lig. obliquum (orange) der Art. cubiti des Hundes; kraniale Ansicht,
Gelenkwinkel = 160°
U1 = Ursprung
A1 = Ansatz des Lig. obliquum am medialen Seitenband
A2 = Ansatz des Lig. obliquum an der kaudalen Ansatzsehne des M. biceps brachii
U2 = Ursprung Lig. anulare radii
A3 = Ansatz Lig. anulare radii
H = Humerus
R = Radius
Ul = Ulna

85 Messergebnisse (siehe Material und Methoden, Kap. 3, Abb. 3b)
Bei einer Gelenkwinkelung von 160° beträgt die Gesamtlänge des Bandes im Mittel
2,2 cm. Die Breite misst am Ursprung, im Gelenkspalt und am Ansatz jeweils 0,3 cm.
4.1.1.4 Ligamentum obliquum (Abb. 7, Abb. 8 und Abb. 9, orange)
Ursprung
Das Band entspringt als breitgefächerte Verstärkung zusammen mit der
Gelenkkapsel am dorsolateralen Rand des Foramen supratrochleare.
Verlauf
Distal des Ursprungs des M. extensor digitorum communis (Abb. 7/E) verläuft es
schmaler schräg distomedial in der Gelenkkapsel über die Beugeseite des Gelenkes
auf das Lig. collaterale mediale bzw. die Ansatzsehne des M. biceps brachii zu. Dort
liegt es extrakapsulär, ist allerdings fest mit der Gelenkkapsel verbunden. Kurz bevor
es am medialen Seitenband bzw. am Radius ansetzt, teilt es sich in zwei Schenkel.
Ansatz
Mit einem kürzeren Schenkel setzt das Lig. obliquum am kranialen Schenkel des Lig.
collaterale mediale an. Mit einem etwas längeren Schenkel inseriert es proximal an
der medialen Seite des Radius, nachdem es die Endsehnen des M. biceps brachii
und des M. brachialis umschlungen hat.
Sesamoide Struktur
Kurz vor dem Ansatz des Bandes an dem kranialen Schenkel des medialen
Kollateralbandes zeigt sich bei allen Hunden eine im Durchmesser ca. 3 bis 4 mm
große, mehr oder weniger knorpelharte Einlagerung (siehe Kap. 4.1.1.2; Abb. 8a/B).

86 Messergebnisse des Lig. obliquum
(siehe Material und Methoden, Kap. 3, Abb. 3a und Abb. 3b)
a) Länge
Das Band hat bei einer Gelenkstellung von 160° vom Ursprung bis zum Ansatz am
medialen Kollateralband eine Gesamtlänge von im Mittel 3,24 cm und eine Länge
vom Ursprung bis zum Ansatz an der medialen Seite des Radius von im Mittel 3,4
cm.
b) Breite
Die Breite im Ursprungsbereich beträgt unter diesen Bedingungen 0,33 cm, in Höhe
des Gelenkspaltbereichs 0,24 cm und im Ansatzbereich 0,2 cm für den Ansatz des
längeren Schenkels medial am Radius sowie einen Wert von 0,31 cm für den Ansatz
des kürzeren Schenkels am medialen Seitenband.
4.1.1.5 Ligamentum olecrani (Abb. 10, türkis)
Ursprung
Dieses Band entspringt an der axialen Seite des Epicondylus medialis humeri mit
breitgefächerten Ursprungsfasern parallel zum sagittalen Rand des Epicondylus
medialis humeri.
Verlauf
Es verläuft schräg distokaudal zwischen Humerus und Olecranon und liegt dabei
verbunden der kaudomedialen Aussackung der Gelenkkapsel an. Bei den Hunden
der Altersklasse „alter Hund“ ist das Band deutlicher in seinem Verlauf von der
Gelenkkapsel trennbar.
Ansatz
Das Lig. obliquum hat seinen Ansatz an einer flachen Erhabenheit zwischen
medialem vorderen Höcker und dem Processus anconeus des Olecranon.

87 Messergebnisse des Lig. olecrani
(siehe Material und Methoden, Kap. 3, Abb. 4)
a) Länge
Die Gesamtlänge des Bandes beträgt bei maximal gebeugtem Gelenk im Mittel 3,2
cm.
b) Breite
Die Breite im Ursprungsbereich misst 0,9 cm, in Höhe des Gelenkspaltes 0,4 cm und
im Ansatzbereich 0,8 cm.

88
Abb. 10: Lig. olecrani (türkis) der linken Art. cubiti des Hundes; kaudale Ansicht,
Gelenkwinkel = 160°
U = Ursprung
A = Ansatz
H = Humerus
O = Olecranon

89 4.1.1.2 Statistische Untersuchung
Eine Übersicht der Tabellen 21 bis 40 mit den Messergebnissen an den Bänder der
Art. cubiti des Hundes befindet sich im Anhang. Sie sind die Grundlage der hier
beschriebenen statistischen Untersuchungen.
Es werden die Bänder der Ellbogengelenke von 22 Hunden vermessen und
statistisch untersucht. Von den Hunden mit der Nr. 3, 4, 11 und 18 steht jeweils nur
eine Gliedmaße zur Verfügung, von den übrigen Tieren werden beide
Vorderextremitäten untersucht (siehe Kap. 3 Material und Methoden).
Vergleich der Messergebnisse an der rechten und linken Extremität Mit dem gepaarten Students-t-Test wird untersucht, ob es einen Unterschied
zwischen den Werten des rechten Beines zu den gleichen Parametern des linken
Beines gibt.
Für alle errechneten Variablen ergeben sich keine signifikanten Unterschiede
zwischen rechtem und linkem Ellbogengelenk. Bei den statistischen Tests entsprach
ein p-Wert von<0,05 einem signifikanten Ergebnis. Alle p-Werte der getesteten
Individuen lagen oberhalb dieses Wertes. Die Hypothese, dass die Differenz der
beiden gegenübergestellten Variablen gleich Null ist, wird angenommen.
Folglich können für den weiteren Verlauf der Untersuchung die gemittelten Werte aus
den rechten und linken Gliedmaßen (_rl) zur weiteren statistischen Auswertung
herangezogen werden.
Test auf Normalverteilung der Residuen Um festzustellen, ob die Residuen einer Normalverteilung unterliegen, wird der
Shapiro-Wilk-Test (Test einer Stichprobe auf Zugehörigkeit zur Normalverteilung)
durchgeführt. Ergebnis: Die Werte sind normalverteilt.

90 Vergleich der Messergebnisse des lateralen und medialen Kollateralbandes Aufgrund der Normalverteilung der Residuen wird mit dem gepaarten Students t-Test
(parametrischer Test zum Vergleich der Mittelwerte zweier unabhängiger
Stichproben aus normal verteilten Grundgesamtheiten) fortgefahren. Es erfolgt der
Vergleich des medialen mit dem lateralen Kollateralband für alle relevanten
Parameter (Kap. 3.1.2) in den Winkelungen 160° und 30°. Ein p-Wert von<0,05
entspricht einem signifikanten Unterschied zwischen den zu vergleichenden
Parametern, bei einem p-Wert>0,05 unterscheiden sich die Variablen nicht
signifikant. Mittelwert und Standardabweichung der Differenzen werden bestimmt.
Einzelbetrachtung der Messwert-Differenzen zwischen lateralem und medialem Kollateralband a) Vergleich der Gesamtlängen der kranialen Schenkel der lateralen und medialen
Kollateralbänder (Tab. 11).
Ein Mittelwert der Differenzen von –0,03 bzw. -0,02 ist mathematisch zu
vernachlässigen, da er annähernd Null ist. Die Standardabweichung ist bei einer
Gelenkstellung von 30° um 0,09 größer als bei einem Winkel von 160°.
Durch den t-Test wird das Ergebnis des Mittelwertes untermauert: Es besteht kein
signifikanter Unterschied in der Gesamtlänge zwischen dem lateralen und dem
medialen Kollateralband bezüglich ihrer kranialen Schenkel.
Die Hypothese, dass der laterale Wert minus dem medialen Wert gleich Null ist, wird
angenommen.
Tab. 11: Statistische Auswertung der Differenzen der Gesamtlängen (LG Diff.) der
kranialen Schenkel der lateralen und medialen Kollateralbänder.
Strecke Var. Diff. Mittelwert Standard-
abweichung
gepaarter
t-Test
LG (Diff.) dif_llm_skr160_rl 7 -0,03 0,24 >0,05
LG (Diff.) dif_llm_skr30_rl 8 -0,02 0,33 >0,05 7dif_llm_skd160_rl = Lig. coll. lat. - Lig. coll. med. (kranialer Schenkel, Winkel 160°, gemittelt re/li Bein) 8dif_llm_skd30_rl = Lig. coll. lat. - Lig. coll. med. (kranialer Schenkel, Winkel 30°, gemittelt re/li Bein

91 b) Vergleich der Gesamtlängen der kaudalen Schenkel der lateralen und medialen
Kollateralbänder (Tab. 12).
Der Mittelwert der Differenzen zeigt, dass die Gesamtlängen der kaudalen Schenkel
der medialen Kollateralbänder im Mittel 1,93 cm (160°) bzw. 1,96 cm (30°) länger
sind als die der kaudalen Schenkel der lateralen Kollateralbänder. Ein Unterschied
sowohl im Mittelwert als auch in der Standardabweichung ist zwischen den beiden
Gelenkwinkelungen von 160° und 30° auszumachen.
Der t-Test beschreibt hier ebenfalls einen signifikanten Unterschied (p<0,05)
zwischen den Gesamtlängen der kaudalen Schenkel der medialen und lateralen
Kollateralbänder.
Tab. 12: Statistische Auswertung der Differenzen der Gesamtlängen (LG Diff.) der
kaudalen Schenkel der lateralen und medialen Kollateralbänder.
Strecke Var. Diff. Mittelwert Standard-
abweichung
gepaarter
t-Test
LG (Diff.) dif_llm_skd160_rl 7 -1,93 0,49 <0,05
LG (Diff.) dif_llm_skd30_rl 8 -1,96 0,45 <0,05 7dif_llm_skd160_rl = Lig. coll. lat. - Lig. coll. med. (kaudaler Schenkel, Winkel 160°, gemittelt re/li Bein) 8dif_llm_skd30_rl = Lig. coll. lat. - Lig. coll. med. (kaudaler Schenkel, Winkel 30°, gemittelt re/li Bein)
c) Vergleich der Differenzen der Ursprungslängen der lateralen und medialen
Kollateralbänder (Tab.13).
Der Mittelwert der Differenzen besagt, dass der Ursprung der lateralen
Kollateralbänder in seiner Länge im Mittel 0,32 cm bei 160° und 0,33 cm bei 30°
größer ist, als der Ursprung der medialen Kollateralbänder. Sowohl Standard-
abweichung als auch Mittelwert zeigen hier keinen Unterschied in den verschiedenen
Winkelstellungen des Gelenkes.
Ein signifikanter Unterschied zwischen dem lateralen und medialen Kollateralband im
Hinblick auf die Länge des Ursprungsbereiches ist zu verzeichnen.

92 Tab. 13: Statistische Auswertung der Differenzen der Ursprungslängen (LU Diff.) der
kranialen Schenkel der lateralen und medialen Kollateralbänder.
Strecke Var. Diff. Mittelwert Standard-
abweichung
gepaarter
t-Test
LU (Diff.) dif_llm_s160_rl 9 0,32 0,17 < 0,05
LU (Diff.) dif_llm_s30_rl 10 0,33 0,17 < 0,05 9dif_llm_s160_rl = Lig. coll. lat. - Lig. coll. med. (ungeteiltes Band, Winkel 160°, gemittelt re/li Bein) 10dif_llm_s30_rl = Lig. coll. lat. - Lig. coll. med. (ungeteiltes Band, Winkel 30°, gemittelt re/li Bein
d) Vergleich der Differenzen der Längen der freien Teile der kranialen Schenkel der
lateralen und medialen Kollateralbänder (Tab. 14).
Ein negativer Mittelwert von –0,81 bei 160° bzw. –0,91 bei 30° zeigt, dass der freie
Teil des medialen Kollateralbandes um 0,81 cm bzw. um 0,91 cm länger ist als der
des lateralen Kollateralbandes. Zwischen den Standardabweichungen in den beiden
Gelenkstellungen gibt es einen deutlichen Unterschied, denn das Band verlängert
sich medial bei 30° Gelenkwinkelung um 0,1 cm.
Es besteht laut t-Test ein signifikanter Unterschied zwischen den Längen der freien
Teile an der lateralen und medialen Seitenbänder.
Tab. 14: Statistische Auswertung der Differenzen der Längen der freien Teile (LfT
Diff.) der kranialen Schenkel der lateralen und medialen Kollateralbänder.
Strecke Var. Diff. Mittelwert Standard-
abweichung
gepaarter
t-Test
LfT (Diff.) dif_llm_skr160_rl 11 -0,81 0,38 < 0,05
LfT (Diff.) dif_llm_skr30_rl 12 -0,91 0,26 < 0,05 11dif_llm_skr160_rl = Lig. coll. lat. - Lig. coll. med. (kranialer Schenkel, Winkel 160°, gemittelt re/li Bein) 12dif_llm_skr30_rl =Lig. coll. lat. - Lig. coll. med. (kranialer Schenkel, Winkel 30°, gemittelt re/li Bein)

93 e.) Vergleich der Differenzen der Ansatzlängen der kranialen Schenkel der lateralen
und medialen Kollateralbänder (Tab. 15).
Diese Streckenabschnittdifferenzen sind am lateralen Kollateralband im Mittel um
0,54 cm größer als am medialen Kollateralband. Es gibt keinen Unterschied in
Mittelwert und Standardabweichung zwischen den beiden Winkelungen des
Gelenkes.
Die Hypothese, dass die Länge des medialen Ansatzbereiches und die Länge des
lateralen Ansatzbereiches gleich sind, wird durch den t-Test abgelehnt: Es liegt ein
signifikanter Unterschied vor.
Tab.15: Statistische Auswertung der Differenzen der Längen der Ansatzbereiche (LA
Diff.) der kranialen Schenkel der lateralen und medialen Kollateralbänder.
Strecke Var. Diff. Mittelwert Standard-
abweichung
gepaarter
t-Test
LA (Diff.) dif_llm_skr160_rl 13 0,54 0,26 < 0,05
LA (Diff.) dif_llm_skr30_rl 14 0,54 0,26 < 0,05 13dif_llm_skr160_rl = Lig. coll. lat. - Lig. coll. med. (kranialer Schenkel, Winkel 160°, gemittelt re/li Bein) 14dif_llm_skr30_rl = Lig. coll. lat. - Lig. coll. med. (kranialer Schenkel, Winkel 30°, gemittelt re/li Bein)
f) Vergleich der Differenzen der freien Teile der Humerusabschnitte der kranialen
Schenkel der lateralen und medialen Kollateralbänder (Tab. 16).
Der negative Mittelwert der Differenzen beweist hier, dass der freie Teil des
Humerusabschnittes des Seitenbandes an der medialen Seite größer ist als an der
lateralen Seite: Es herrscht ein Größenunterschied von 0,46 cm (160°) bzw. von 0,63
cm (30°) des medialen zum lateralen Band vor. Die Standardabweichung der
Messwerte ist bei 160° annähernd doppelt so groß wie bei 30°. Es besteht ein
deutlicher Unterschied zwischen den beiden Gelenkstellungen.
Der t-Test beschreibt einen signifikanten Unterschied zwischen der Länge des freien
Teiles am Humerusabschnitt des medialen und des lateralen Kollateralbandes.

94 Tab. 16: Statistische Auswertung der Differenzen der Längen der freien Teile der
Humerusabschnitte (LfTH Diff.) der kranialen Schenkel der lateralen und medialen
Kollateralbänder.
Strecke Var. Diff. Mittelwert Standard-
abweichung
gepaarter
t-Test
LfTH (Diff.) dif_llm_s160_rl 15 -0,46 0,28 < 0,05
LfTH (Diff.) dif_llm_s30_rl 16 -0,63 0,15 < 0,05 15dif_llm_s160_rl = Lig. coll. lat. - Lig. coll. med. (ungeteiltes Band, Winkel 160°, gemittelt re/li Bein) 16dif_llm_s30_rl =Lig. coll. lat. - Lig. coll. med. (ungeteiltes Band, Winkel 30°, gemittelt re/li Bein)
g) Vergleich der Differenzen der Längen der freien Teile der Unterarmabschnitte der
kranialen Schenkel der lateralen und medialen Kollateralbänder (Tab. 17).
Das negative Vorzeichen des Mittelwertes weist auf den größeren freien Teil des
Unterarmabschnitts am medialen Kollateralband hin. Es gibt einen deutlichen
Unterschied zwischen den beiden Gelenkwinkelungen. Das mediale Band ist im
Mittel 0,49 cm (160°) bzw. 0,25 cm (30°) länger als das laterale Band. Zwischen den
Standardabweichungen gibt es einen Unterschied zwischen 160° und 30°.
Der gepaarte t-Test zeigt einen signifikanten Unterschied zwischen der Länge des
freien Teils am Unterarmabschnitt des medialen Bandes zu jener Länge des
lateralen Bandes.
Tab. 17: Statistische Auswertung der Differenz der Längen der freien Teile der
Unterarmabschnitte LfTU (Diff.) der kranialen Schenkel der lateralen und medialen
Kollateralbänder.
Strecke Var. Diff. Mittelwert Standard-
abweichung
gepaarter
t-Test
LfTU (Diff.) dif_llm_skr160_rl 17 -0,49 0,35 < 0,05
LfTU (Diff.) dif_llm_skr30_rl 18 -0,25 0,30 < 0,05 17dif_llm_skr160_rl = Lig. coll. lat. - Lig. coll. med. (kranialer Schenkel, Winkel 160°, gemittelt re/li Bein) 18dif_llm_skr30_rl = Lig. coll. lat. - Lig. coll. med. (kranialer Schenkel, Winkel 30°, gemittelt re/li Bein)

95 h) Vergleich Ursprungsbreiten der Differenzen der lateralen und medialen Kollateral-
bänder (Tab. 18).
Im Mittel ist das laterale Band in seinem Ursprungsbereich um 0,07 bzw. 0,08 cm
breiter als das mediale Band. Dies lässt sich aus dem positiven Vorzeichen des
Mittelwertes folgern. Zwischen 160° und 30° lässt sich im Mittelwert und in der
Standardabweichung kein nennenswerter Unterschied feststellen.
Der t-Test besagt, dass die Breiten des medialen und die des lateralen Bandes sich
signifikant unterscheiden.
Tab. 18: Statistische Auswertung der Differenz der Ursprungsbreiten (BU Diff.) der
kranialen Schenkel der lateralen und medialen Kollateralbänder.
Strecke Var. Diff. Mittelwert Standard-
abweichung
gepaarter
t-Test
BU (Diff.) dif_llm_skr160_rl 1 0,07 0,12 < 0,05
BU (Diff.) dif_llm_skr30_rl 2 0,08 0,13 < 0,05 1dif_llm_skr160_rl = Lig. coll. lat. - Lig. coll. med. (kraniale Schenkel, Winkel 160°, gemittelt re/li Bein) 2dif_llm_skr30_rl = Lig. coll. lat. - Lig. coll. med. (kraniale Schenkel, Winkel 30°, gemittelt re/li Bein)
i) Vergleich der Differenzen der Breiten in Höhe des Gelenkspaltes der lateralen und
medialen Kollateralbänder (Tab. 19).
Der Mittelwert der Differenzen beträgt hier –0,05 bei 160° Gelenkwinkelung und -0,01
bei 30°, d. h. das mediale Band ist im Mittel 0,05 cm bzw. 0,01 cm breiter als das
laterale Band. Mathematisch können diese geringen Differenzen vernachlässigt
werden. Die Standardabweichung zeigt ebenfalls keinen nennenswerten
Unterschied. Ein Unterschied zwischen den beiden Gelenkstellungen ist vorhanden.
Wird der t-Test berücksichtigt, so lässt sich feststellen, dass es bei dieser
Breitenmessung zwischen dem lateralen und medialen Kollateralband keinen
signifikanten Unterschied gibt.

96 Tab. 19: Statistische Auswertung der Differenzen der Breiten (BGs Diff.) der noch
ungeteilten Kollateralbänder in Höhe des Gelenkspaltes.
Strecke Var. Diff. Mittelwert Standard-
abweichung
gepaarter
t-Test
BGs (Diff.) dif_llm_s160_rl 3 -0,05 0,13 >0,05
BGs (Diff.) dif_llm_s30_rl 4 -0,01 0,16 >0,05 3dif_llm_s160_rl = Lig. coll. lat. - Lig. coll. med. (ungeteiltes Band, Winkel 160°, gemittelt re/li Bein) 4dif_llm_s30_rl = Lig. coll. lat. - Lig. coll. med. (ungeteiltes Band, Winkel 30°, gemittelt re/li Bein)
j) Vergleich der Differenzen der Ansatzbreiten der kaudalen Schenkel der lateralen
und medialen Kollateralbänder (Tab. 20).
Der Mittelwert der Differenzen beträgt 2,26 bei 160°, d.h. dass der kaudale Schenkel
an der lateralen Seite im Ansatzbereich im Mittel 2,26 cm breiter ist als der kaudale
Schenkel an der medialen Seite. Bei einer Winkelung des Gelenkes von 30° ist der
kaudale Schenkel an der lateralen Seite 2,22 cm breiter. Es besteht ein Unterschied
zwischen den Mittelwerten von 0,04 cm bei unterschiedlichen Gelenkstellungen. Die
Standardabweichung bei 160° ist größer als die Standardabweichung bei 30°, d.h.,
dass die Streuung um den Mittelwert bei 160° größer ist.
Der t-Test zeigt einen signifikanten Unterschied zwischen den Ansatzbreiten des
lateralen und medialen Kollateralbandes.
Tab. 20: Statistische Auswertung der Differenzen der Ansatzbreiten (BA Diff.) der
kaudalen Schenkel der lateralen und medialen Kollateralbänder.
Strecke Var. Diff. Mittelwert Standard-
abweichung
gepaarter
t-Test
BA (Diff.) dif_llm_skd160_rl 5 2,26 0,41 <0,05
BA (Diff.) dif_llm_skd30_rl 6 2,22 0,37 <0,05 5dif_llm_skd160_rl = Lig. coll. lat. - Lig. coll. med. (kaudale Schenkel, Winkel 160°, gemittelt re/li Bein) 6dif_llm_skd30_rl = Lig. coll. lat. - Lig. coll. med. (kaudale Schenkel, Winkel 30°, gemittelt re/li Bein)

97 Zusammenfassung der statistischen Auswertung. 1. In der Gesamtlänge besteht zwischen den kranialen Schenkeln der Ligg.
collaterale laterale und mediale kein signifikanter Unterschied.
2. Die Gesamtlängen der kaudalen Schenkel der Bänder unterscheiden sich
signifikant voneinander. Die kaudalen Schenkel der medialen Seitenbänder sind
länger als die der lateralen Kollateralbänder. Ein minimaler Unterschied zwischen
den beiden Gelenkstellungen ist zu vernachlässigen.
3. Die Differenzen der Ursprungslängen der beiden Bänder unterscheiden sich
signifikant voneinander. Das mediale Band hat einen kürzeren Ursprungsbereich
als das laterale Band.
4. Die Differenzen der Längen der freien Teile der medialen und lateralen Kollateral-
bänder unterscheiden sich ebenfalls signifikant. Die freien Teile der medialen
Kollateralbänder sind länger als die der lateralen Kollateralbänder. Zwischen den
beiden Gelenkwinkelungen gibt es einen Unterschied.
5. Die Differenzen der Längen der Ansatzbereiche der kranialen Schenkel der Ligg.
collaterale laterale und mediale weisen einen signifikanten Unterschied auf. Die
Ansatzlängen der kranialen Schenkel der lateralen Kollateralbänder sind größer
als die der medialen Bänder.
6. Die Längen der freien Teile der Humerusabschnitte sind zwischen den lateralen
und medialen Kollateralbändern signifikant unterschiedlich. An den medialen
Bändern sind die freien Teile länger als an den lateralen Kollateralbändern. Es
gibt einen deutlichen Unterschied zwischen den Gelenkstellungen 160° und 30°.
Die Standardabweichung ist bei 160° fast doppelt so groß wie bei 30°.
7. Die Längen der freien Teile der Unterarmabschnitte zeigen einen signifikanten
Unterschied zwischen den lateralen Kollateralbändern und den medialen
Kollateralbändern. Diese Messstrecken sind ebenfalls an den medialen
Seitenbändern länger als an den lateralen. Ein Unterschied zwischen den
Gelenkwinkelungen ist auch hier vorhanden.
8. Die Differenzen der Ursprungsbreiten zeigen einen signifikanten Unterschied
zwischen den medialen und lateralen Kollateralbändern. Im Mittel sind die
lateralen Kollateralbänder hier breiter als die medialen.

98 9. Die Differenzen der Breiten der beiden Kollateralbänder in Gelenkspalthöhe
weisen keinen signifikanten Unterschied.
10. Die Ansatzbreiten der Kollateralbänder sind in ihren Differenzen signifikant
unterschiedlich. Die Breiten der lateralen Kollateralbänder sind größer als die der
medialen Kollateralbänder. Es besteht auch ein Unterschied zwischen den beiden
Winkelungen des Gelenkes.
Die Hypothese, dass die Bänder medial und lateral gleich sind, ist folglich durch den
Students t-Test mit hoher Signifikanz in acht von zehn Fällen abzulehnen.
Angenommen wird diese Hypothese für die Parameter „Breite in der Höhe des
Gelenkspaltes“ und „Gesamtlänge des Bandes“ im Hinblick auf den kranialen
Schenkel sowohl bei 160° als auch bei 30°.
Alle anderen Messstrecken (2.-8. und 10.) weisen zwischen dem lateralen und
medialen Kollateralband einen signifikanten Unterschied auf.

99 4.2 Mikroskopische Untersuchung (Abb. 11 bis Abb. 22)
Diese Untersuchung erfolgt an H.E., Orcein und Pikrosiriusrot gefärbten Schnitten:
Bei der Gegenüberstellung des lateralen mit dem medialen Kollateralband werden an
beiden Schnittrichtungen (längs zum Faserverlauf; quer zum Faserverlauf) die
unterschiedliche Faserarchitektur und die unterschiedliche Form der Zellkerne deut-
lich.
Die Entnahmestelle der Probenblöcke ist im Kapitel Material und Methoden be-
schrieben (siehe Kap. 3.2.3, Abb. 5 und Abb. 6).
4.2.1 Auswertung mit dem Lichtmikroskop H.E. Färbung (Vergrößerung 2,5x) Allgemein ist am lateralen und medialen Kollateralband eine Bindegewebshülle gut
zu erkennen. An der Gelenkseite ist sie ebenso dicker und die Vaskularisation
stärker ausgeprägt als an der vom Gelenk abgewandten Seite.
Als Bindegewebszellen sind Fibroblasten und Fibrozyten zu unterscheiden. Die Zell-
kerne der Fibroblasten sind groß und abgerundet, die der Fibrozyten schmal und
dicht.
Die Kollagenfasern sind in ihrem Verlauf gut zu beobachten. Sie stellen sich dabei
nicht gewellt dar, da sie während der Fixation straff auf die Korkplatten aufgenäht
worden sind. Diese Aussagen treffen sowohl auf das laterale als auch auf das
mediale Band zu.
An der Gelenkseite des medialen Kollateralband unter dem „freien Teil“ des Bandes
finden sich regelmäßig Fettzellen.
4.2.1.1 Längsschnitt H.E. Färbung (Abb. 11/Abb. 10, Vergrößerung 40x)
Ein deutlicher Unterschied zwischen dem lateralen (Abb. 11) und dem medialen
(Abb. 12) Band besteht darin, dass im lateralen Band hauptsächlich runde bis breit-
ovale Zellkerne zu finden sind, und im medialen Band nahezu nur längliche,

100 spindelförmige Zellkerne. Die Anordnung der Kerne im lateralen Band lässt kein
einheitliches System erkennen, im Gegensatz zu den Kernen im medialen Band. Hier
liegen die spindelförmigen Zellen in Reihen angeordnet zwischen den parallel
ausgerichteten Fasern. Das mediale Band vermittelt den Eindruck eines straff
organisierten Verbandes von Fasern und Zellen.
4.2.1.2 Querschnitt H.E. Färbung (Abb. 13, Vergrößerung 40x/Abb. 14, Vergrößerung 20x)
Auch in diesem Anschnitt wird deutlich, dass die Kerne des lateralen Bandes (Abb.
13) vorwiegend eine rundliche Form haben. Die Kerne des medialen Bandes (Abb.
14) sind im Durchmesser flach. Eine unterschiedliche Faseranordnung der beiden
Kollateralbänder wird im Querschnitt deutlich.
Orcein Färbung Diese Schnitte werden hergestellt, um elastische Fasern nachzuweisen: Es sind
keine bzw. nur wenige elastische Fasern zu beobachten. Dies liegt nicht an der
mangelnden Qualität der Färbung, denn die Gefäßwände haben farblich gut reagiert,
d.h. die Tunica media ist mit ihren elastischen Anteilen deutlich zu erkennen.
Um dieses Ergebnis abzusichern, wird ein vorher in Formol fixierter Gefrierschnitt
angefertigt. Es werden auch hierbei keine bzw. nur sehr wenige elastische Fasern
beobachtet.

101
Abb. 11: Laterales Kollateralband, Längsschnitt, H.E. Färbung, 40x Auffällig ist die runde Form der Zellkerne und deren „zufällige“ Anordnung.
Abb. 12: Mediales Kollateralband, Längsschnitt, H.E. Färbung, 40x Zellkerne sind langgestreckt und zwischen den straffen Kollagenfasern angeordnet.
Abb. 13: Laterales Kollateralband, Querschnitt, H.E. Färbung, 40x Die runde Form der Zellkerne und der lockere Verband der Kollagenfasern wird deutlich.
Abb. 14: Mediales Kollateralband, Querschnitt, H.E. Färbung, 20x Die spindelförmige, langgestreckte Form der Fibrozyten und deren Fortsätze sind zu erkennen.

102
4.2.2 Auswertung im polarisierten Licht 4.2.2.1 Längsschnitt Pikrosiriusrot Färbung (Abb. 15 und Abb. 16, Vergrößerung 2,5x)
In dieser Färbung werden das laterale und das mediale Kollateralband im
polarisierten Licht betrachtet. Es fällt eine starke Anisotropie der Kollagenfasern auf.
In der Übersicht wird die scherengitterartige Anordnung der Fasern des lateralen
Kollateralbandes (Abb. 15) deutlich. Es ist keine einheitliche Verlaufsrichtung der
Fasern auszumachen. Die Fasern des medialen Kollateralbandes (Abb. 16) ziehen
parallel zur Schnittrichtung in proximodistaler Richtung und liegen geordnet
nebeneinander.
H.E. Färbung (Abb. 17 und Abb. 18, Vergrößerung 10x)
Im lateralen Band ist keine deutliche Zugrichtung der Fasern auszumachen. Hier sind
mehr dunkle Bereiche als im medialen Band zu sehen. Die straffe Organisation der
Fasern im medialen Kollateralband (Abb. 18) mit ihrer proximodistalen Verlaufs-
richtung, wie auch die Flechtstruktur des lateralen Kollateralbandes (Abb. 17) werden
hier sehr deutlich.

103 4.2.2.2 Querschnitt Pikrosiriusrot Färbung (Abb.19 und Abb. 20, Vergrößerung 2,5x)
In diesem Anschnitt werden im lateralen Band (Abb. 19) verschiedene Verlaufs-
formen sichtbar. Es sind sowohl Längsfasern als auch Querfasern zu beobachten. Im
medialen Band (Abb. 20) verlaufen die Fasern einheitlich vertikal zur Schnittrichtung.
Dreht man den Objekttisch, erscheint nach einer Drehung um 90° das gesamte Band
durch Lichtbrechung hell aufleuchtend. Bei weiterer 90° Drehung bleibt das Band
einheitlich schwarz, d.h. die Fasern werden so getroffen, dass das Licht sich nicht an
ihnen brechen kann. Im lateralen Band lassen sich dagegen nur wenige solcher
dunklen Areale finden.
H.E Färbung (Abb. 21 und Abb. 22, Vergrößerung 10x)
Im Lig. collaterale laterale (Abb. 21) sind sowohl Längs- als auch Querschnitte der
Fasern zu sehen; hier wechseln sich sehr helle mit sehr dunklen Bereichen ab. Die
parallele Anordnung der Fasern im medialen Kollateralband (Abb. 22) ist deutlich zu
erkennen und zeigt die gleichmäßige Verteilung der dunklen Zonen.

104 xcfvshdjfhweur
Abb. 15: Laterales Kollateralband Längsschnitt, Pikrosiriusrot, 2,5x Scherengitterartige Anordnung der Fasern
Abb. 16: Mediales Kollateralband Längsschnitt, Pikrosiriusrot, 2,5x Proximodistaler Faserverlauf, die Fasern liegen streng nebeneinander.
Abb. 17: Laterales Kollateralband Längsschnitt, H.E. Färbung, 10x Verwobene Faserstruktur, keine eindeutige Faserzugrichtung
Abb. 18: Mediales Kollateralband Längsschnitt, H.E. Färbung, 10x Paralleler Faserverlauf noch deutlicher. Es ist eine straffe Organisation der Fasern zu sehen

105
fhgjhj
Abb. 19: Laterales Kollateralband Querschnitt, Pikrosiriusrot, 2,5x Auch im Querschnitt werden sowohl Quer- als auch Längsfasern getroffen.
Abb. 20: Mediales Kollateralband Querschnitt, Pikrosiriusrot, 2,5x Es sind nur Längsfasern zu sehen, die in ihrem Verlauf quer angeschnitten sind.
Abb. 21: Laterales Kollateralband Querschnitt, H.E. Färbung, 10x Es sind Queranschnitte und Längsanschnitte der Fasern zu sehen. Helle und dunkle Bereiche wechseln sich miteinander ab.
Abb. 22: Mediales Kollateralband Querschnitt, H.E. Färbung, 10x Die Farbverteilung, provoziert durch das polarisierte Licht, ist homogen. Die dunklen Areale dominieren.

106 5. Diskussion
Es werden die Kollateralbänder am Ellbogengelenk des Hundes untersucht. Dies
wird sowohl makroskopisch als auch mikroskopisch durchgeführt. Für die
makroskopische Untersuchung werden 40 Vordergliedmaßen von 22 Deutschen
Schäferhunden oder Schäferhund-Mischlingen präpariert und vermessen. Die
mikroskopische Untersuchung wird an zehn Vordergliedmaßen von fünf frisch toten
Schäferhund-Mischlingen durchgeführt.
5.1 Makroskopische Untersuchung Nach der Darstellung der Kollateralbänder der Articulatio cubiti, der Bänder der
Articulatio radioulnaris proximalis und des Ligamentum obliquum, wurden Ursprung,
Verlauf, Ansatz und Längen und Breiten der Bänder bestimmt und etwaige
Sondereinrichtungen des Bandapparates dokumentiert.
Ligamentum collaterale laterale (Abb. 7, blau)
Maßangaben zum lateralen Kollateralband des Ellbogengelenks beim Hund werden
in der zugänglichen Literatur nur von SÜPPEL (1921) gemacht. Er erkennt Breiten
des Lig. collaterale laterale von 3,6 bis 6,0 mm. Allerdings lassen sich in seiner Arbeit
weder Angaben über die Lokalisation der Breitenmessungen am Band, noch die
Stellung des Gelenkes während der Messung bzw. Angaben über Alter, Rasse oder
Gewicht der verwendeten Hunde finden.
Bei dem hier untersuchten Material lässt sich für die Breite im Ursprungsbereich im
Mittel ein Wert von 0,61 cm (bei 160°) und ein Wert von 0,6 cm (bei 30°) für den noch
ungeteilten Schenkel bestimmen. Für den kaudalen Schenkel an jener Stelle, an der
er aus dem noch ungeteilten Band hervortritt, liegt die Breite sogar bei 1,43 cm (160°
und 30° Gelenkstellung). Die Breite im Gelenkspalt liegt für das an der Stelle noch
ungeteilte Band bei 0,72 cm (160°) und bei 0,66 cm (30°). Im Ansatzbereich hat die
Breite einen Wert von 0,67 cm (160°/30°) für den kranialen Schenkel. Für den
kaudalen Schenkel lässt sich ein Wert von 2,59 cm (160°/30°) bestimmen.

107 Somit ist nur eine vergleichende Betrachtung der Breitenmessungen im
Ursprungsbereich des lateralen Kollateralbandes möglich; die Messergebnisse von
SÜPPEL (1921) sind kleiner, aber liegen doch nahe den eigenen Werten; jedoch
fehlen der Literatur vergleichende Angaben zum Untersuchungsgut.
Die Beschreibungen des Ursprungs in der Literatur sind unterschiedlich. Die eigene
Untersuchung schließt sich der Beschreibung von KOCH u. BERG (1992) und
NICKEL et al. (1992) an, dass der Ursprung sich in der laterodistalen Bandgrube und
am benachbarten Bandhöcker des Humerus befindet. Dies lässt sich durch die
Präparation ausnahmslos bestätigen. BERGER (1992), EVANS (1993a) und KÖNIG
u. LIEBICH (1999) beschreiben ihn hingegen am Epicondylus lateralis humeri, wobei
BERGER (1992) auf einen geteilten Ursprung am Epicondylus hinweist.
VOLLMERHAUS et al. (1994b) geben hier die Bandgrube am Epicondylus lateralis
humeri an. Hier bedarf es zur einheitlichen Darstellung einer Klärung in der NAV
(1994): liegen die Bandgruben und der Bandhöcker am Epicondylus humeri oder am
Condylus humeri? Die von BERGER (1992) beschriebenen zwei Anteile mit
gemeinsamem Ursprung können weder makroskopisch noch histologisch an dem
untersuchten Material bestätigt werden.
Der beschriebene Verlauf stimmt mit den Aussagen der Literatur überein. Das Band
zieht einheitlich bis zum Caput radii und teilt sich 1 bis 2 mm distal des Gelenk-
spaltes in zwei Schenkel auf. Hierbei wird von BERGER (1992) eine enge Ver-
bindung das Lig. collaterale laterale mit dem Lig. anulare radii und dem M. supinator
histologisch nachgewiesen. Präparatorisch ist eine Trennung dieser drei Strukturen
nicht durchführbar.
Der Ansatz des kranialen Schenkels wird von MARTIN (1923) und EVANS (1993a)
definiert und bestätigt sich so ausnahmslos in den eigenen Untersuchungen. Er
befindet sich an einer knöchernen Erhabenheit, die distal des Collum radii liegt.
Diese Erhabenheit an der lateralen Seite des Radius findet in der aktuellen NAV
(1994) keine Erwähnung. MARTIN (1923) und auch SÜPPEL (1921) beschreiben sie
als Bandhöcker distal am Collum radii und EVANS (1993a) spricht von einer
Erhabenheit distal des Radiushalses. Die Beschreibungen der übrigen Autoren wie

108 proximal am Radius (KÖNIG u. LIEBICH 1999) bzw. distal des Collum radii
(VOLLMERHAUS et. al 1994b) sind mehr allgemein gehalten. Der Ansatz des
kaudalen Schenkels wird ähnlich allgemein an der Ulna beschrieben (MARTIN 1923;
SÜPPEL 1921; BAUM u. ZIETZSCHMANN 1936; KOCH u. BERG 1992; NICKEL et
al. 1992; EVANS 1993a; KÖNIG u. LIEBICH 1999). Nur VOLLMERHAUS et. al
(1994b) und BERGER (1992) treffen eine detailgenaue Beschreibung: Der Ansatz
befindet sich fächerförmig, distal des Processus coronoideus lateralis, mit einem
Übergang in die Membrana interossea. Diese letztgenannte Verbindung beschränkt
sich bei den von hier untersuchten Hunden auf wenige Fasern. Abgesehen davon ist
die Aussage der genannten Autoren bei den eigenen Präparaten zu bestätigen.
Das Vorkommen des, in der Literatur beschriebenen, Kapselschleimbeutels an der
lateralen Seite der Art. cubiti unter dem Lig. collaterale laterale stimmt mit den
eigenen Untersuchungen überein. Der Fettkörper, der die kappenartige Verstärkung
der Gelenkkapsel unterlagert, wird in der zugänglichen Literatur lediglich von
BERGER (1992) beschrieben.
Die oben erwähnte enge Verbindung des Lig. collaterale laterale mit dem Lig.
anulare radii und der Ursprungssehne des M. supinator bildet eine kappenartige
Verstärkung, die sich in ihrer Wölbung der Kontur des darunter liegenden
Knochenpunktes genau anpasst. Diese kappenartige Verstärkung wurde so auch
von BERGER (1992) erwähnt. Es wird unterstellt, dass diese für die Stabilität und die
feste Lagerung der Ursprungssehne des M. supinator sorgt und somit ergänzend das
Gelenk seitlich stabilisiert.
Der Knochenpunkt, über dem sich die kappenartige Verstärkung ausbildet, findet so
in der NAV (1994) keine Erwähnung, wohl aber in der Literatur. Er befindet sich am
Caput radii und richtet sich von diesem kraniolateral aus. Als sogenannte
„Caputprotuberanz“ wird er bei BERGER (1992) angegeben bzw. als „Protuberantia
lateralis radii“ von HULTKRANTZ (1897) oder „Tuber globosum laevigatum“ bei
(KADLETZ 1932). Medial wird die Protuberanz durch einen individuell unterschiedlich
tiefen Sulcus in der kranialen Kontur des Caput radii begrenzt. BERGER (1992) sieht
in diesen beiden Strukturen die Führung von Teilen des Kapselbandapparates,
insbesondere die Fixation der Ursprungssehne des M. supinator. In den eigenen

109 Untersuchungen wird diese „Caputprotuberanz“ mit angrenzendem Sulcus (Kap.
4.1.1.2., Abb 7a/A und Abb. 7a/B) ausnahmslos dokumentiert.
Die sesamoide Struktur (Abb. 7/C) im lateralen Bandapparat wird in der Literatur
hinsichtlich Lokalisation und Beschaffenheit unterschiedlich dargestellt. Nahezu
vollständig wird sie im lateralen Gelenkbereich erwähnt, hier jedoch
unterschiedlichen Anteilen des lateralen Bandapparates zugeordnet. Als fakultatives
Sesambein wird es von einigen Autoren dem Lig. collaterale laterale und dem Lig.
anulare radii zugeordnet (BAUM u. ZIETZSCHMANN 1936; EVANS 1993a). Auch
MARTIN (1923) und VOLLMERHAUS et al. (1994b) erkennen an dieser Stelle eine
Struktur, beschreiben sie allerdings als Knorpel. Ein Sesambein wird weiterhin im
Lig. collaterale laterale, dem Lig. anulare radii und der Gelenkkapsel (HITZ 1976)
bzw. nur dem Lig. anulare radii (WEBBON u. CLAYTON-JONES 1976;
MAYRHOFER 1980) gesehen. VAN HERPEN (1988) stellt eine Zugehörigkeit des
Sesambeins zum kranialen Schenkel des Lig. collaterale laterale dar, diese
Zugehörigkeit wurde bereits von HULTKRANTZ (1897) als knorpelige Struktur bzw.
von BARONE (1968) als meniskenartiger Knorpel erkannt. Von SÜPPEL (1921) wird
das Sesamoid im Lig. collaterale laterale erwähnt. Wohingegen VÄÄNÄNEN u.
SKUTNABB (1978) diese knöcherne fakultative Struktur dem Lig. collaterale laterale
und der Gelenkkapsel zuordnen. Nur zur Gelenkkapsel zugehörig wird das
Sesambein von ZENZES et al. (1989) beschrieben. Als ein Teil der Ursprungssehne
des M. supinator wird es von WOOD et al. (1985) gesehen. KADLETZ (1932) und
BERZON u. QUICK (1980) wie auch BERGER (1992) betonen eine Zugehörigkeit
zum Lig. collaterale laterale, dem Lig. anulare radii und der Ursprungssehne des M.
supinator. Hierbei wird es als meniskenartiger Knorpel (KADLETZ 1932), als
fakultatives Sesambein (BERZON u. QUICK 1980) oder als meniskenartige
Knorpelfalz mit Knochen- bzw. Knorpelarealen beobachtet (BERGER 1992).
Die gelenknahe Seite des Sesamoids ist mit hyalinem Knorpel überzogen
(VÄÄNÄNEN u. SKUTNABB 1978; WOOD et al. 1980) und an der Gelenkbildung
beteiligt. Dieser Beobachtung schließt MAYRHOFER (1980) sich an, deutet es als
Ergänzung des Caput radii und somit als normalanatomische Bildung. Ein möglicher
Grund für das Entstehen des Sesambeins an dieser Stelle stellt die intensive

110 Verflechtung der Band- und Muskelsehnenfasern dar, bzw. der Kräfte, die über die
genannten Strukturen wirken (HULTKRANTZ 1897). BERGER (1992) gibt eine
Erklärung für die von den einzelnen Autoren beobachtete unterschiedliche
Beschaffenheit des Sesamoids. Knorpeliges Gewebe kann unter Krafteinwirkung in
Knochen umgewandelt werden, deshalb ist die Struktur bei den Hunden der
Altersklasse „alter Hund“ und „mittelalter Hund“ deutlicher zu palpieren. Bei einem
jungen Hund wirken nicht so große Kräfte über einen kürzeren Zeitraum. BERGER
(1992) wie vorher schon SÜPPEL (1921) erkannte einen Knorpelfalz, der manchmal
ein Sesambein erhält. Die Gewebegrundlage für diesen Falz bildet Faserknorpel. Die
Lage des Sesambeins in dieser Falz befindet sich distal des Lig. anulare radii,
welches durch den Falz zieht. BERGER (1992) spricht somit von einem
knochenverstärkten Falz. Dieser konnte auch an den eigenen Präparaten
durchgängig nachgewiesen werden. Der Zusammenhang zwischen der deutlicheren
Ausprägung in der Altersklasse „alter Hund“ hängt mit dem höheren gemittelten
Körpergewicht (35,1 kg) in dieser Gruppe zusammen (siehe Kap. 3). So stellt auch
BERGER (1992) in ihren Untersuchungen eine Verbindung zwischen dem
Vorhandensein des Knorpelfalzes und dem Gewicht bzw. der Körpergröße der
Probanden her. Der Falz gleicht die im Ellbogen vorhandene Inkongruenz aus
(BERGER 1992).
Ligamentum collaterale mediale (Abb. 8, rot)
Die Breite des medialen Kollateralbandes wurde ebenfalls nur von SÜPPEL (1921)
bestimmt. Nach seinen Angaben ist es 2,5 bis 4 mm breit. Die eigenen Breiten-
messungen bestimmen einen gemittelten Wert von 0,53 cm (bei 160° und bei 30°)
Gelenkstellung im Ursprungsbereich, und einer Breite von 0,72 cm (bei 160°) bzw.
0,67 cm (bei 30°) in Höhe des Gelenkspaltes. Sie liegt also deutlich über den von
SÜPPEL (1921) angegebenen Werten.
Die zitierten Autoren geben als Ursprungspunkt die Bandgrube des Humerus
(SÜPPEL 1921; BAUM u. ZIETZSCHMANN 1936; KOCH u. BERG 1992; NICKEL et
al. 1992; VOLLMERHAUS et al. 1994b) bzw. den Epicondylus humeri (EVANS

111 1993a; KÖNIG u. LIEBICH 1999) an. Nach den eigenen Ergebnissen nehmen zudem
wenige Fasern am benachbarten Bandhöcker ihren Ursprung.
SÜPPEL (1921) und VAN HERPEN (1988) beschreiben im Verlauf des Bandes eine
Verschmelzung mit dem Lig. anulare radii. Hierbei handelt es sich jedoch nur um
wenige Fasern, die das Lig. anulare radii lediglich an den kranialen Schenkel des Lig.
collaterale mediale abgibt.
Die Aufteilung des medialen Kollateralbandes in ihre beiden Schenkel liegt im
Vergleich zur lateralen Seite in allen untersuchten Fällen etwas weiter distal. Beide
Bänder teilen sich ausnahmslos distal des Gelenkspaltes auf. Die Breiten der beiden
Schenkel wurden von SÜPPEL (1921) vermessen: Der schwächere, kraniale
Schenkel ist 1,5 bis 2 mm breit, der stärkere, kaudale Schenkel misst 2,5 bis 3 mm in
der Breite (SÜPPEL 1921). In den durchgeführten Messungen wird die Breite der
einzelnen Schenkel am Ansatz des Bandes bestimmt. Es ergibt sich ein Wert von
0,29 cm für den kranialen Schenkel bei 160° und bei 30° Gelenkstellung. Für den
kaudalen Schenkel lässt sich in beiden Winkelungen ein Wert von 0,37 cm
bestimmen. Auch hier liegen die Messungen mit ihren Werten deutlich über den von
SÜPPEL (1921) angegebenen Werten.
Der Ansatz des kranialen Schenkels wird in der Literatur weitestgehend einheitlich an
der Tuberositas radii gesehen (BAUM u. ZIETZSCHMANN 1936; HLADIK 1986;
EVANS 1993a). NICKEL et al. (1992) sowie KÖNIG u. LIEBICH (1999) beschreiben
diesen Knochenpunkt als medialen Bandhöcker, was so nicht der Auflistung der NAV
(1994) entspricht. Eine, wie von SÜPPEL (1921) beobachtete, Vertiefung an dieser
Stelle, kann durch diese Untersuchungen in keinem Fall bestätigt werden. Zur
Darstellung des Ansatzes des kranialen Schenkels des medialen Kollateralbandes ist
das Entfernen der kaudalen Ansatzsehnen der Mm. biceps brachii und brachialis
notwendig, da diese ebenfalls an der Tuberositas radii ihren Ansatz finden und somit
den des Kollateralbandes an dieser Stelle verdecken. Auf diese Verbindung am
Ansatzpunkt der beiden Strukturen, wies lediglich MARTIN (1923) in seiner Arbeit
hin. Die Hauptsehne des M. biceps brachii setzt am Processus coronoideus medialis
ulnae an. Der zweischenkelige y-förmige Ansatz des M. biceps brachii und des M.

112 brachialis scheinen mediokranial zur Stabilisierung des Gelenkes beizutragen
(BERGER 1992).
Der Ansatz des kaudalen Schenkels befindet sich nicht nur an der proximokranialen
Seite der Ulna (SÜPPEL 1921; NICKEL et al. 1992; KÖNIG u. LIEBICH 1999),
sondern auch mit wenigen Fasern kaudal am Radius tief im Spatium interosseum
(MARTIN 1923; BAUM u. ZIETZSCHMANN 1936; CAMPBELL 1969; VAN HERPEN
1988; EVANS 1993a) sowie an den eigenen Präparaten mit wenigen Fasern am
kraniolateralen Rand der Ulna (Abb. 8). Die von SÜPPEL (1921) beschriebenen
Vertiefungen im Knochen an den Ansatzstellen können nicht bestätigt werden. Dies
kann am unterschiedlichen Alter der untersuchten Hunde liegen, zudem bei SÜPPEL
(1921) keine Angaben zum Material zu finden sind.
In der Literatur wird ein Kapselschleimbeutel beschrieben, der die Ansatzsehne des
M. biceps brachii und des M. brachialis unterlagert (SÜPPEL 1921; BAUM u.
ZIETZSCHMANN 1936; FREWEIN 1983; SEIFERLE u. FREWEIN 1992; EVANS
1993a). Der von mir beschriebene Schleimbeutel, der ausnahmslos nachgewiesen
werden konnte, stellt jedoch einen selbstständigen Schleimbeutel dar, der nie
Verbindung zur Kapsel besitzt.
Als Gleitlager für das mediale Kollateralband dient die Gelenkkapsel, die im
Gegensatz zur lateralen Seite ihren Insertionsrand weiter proximal besitzt. Somit hat
das Lig. collaterale mediale einen größeren Gleitweg als das laterale Band. Wie
bereits von SÜPPEL (1921) durch eine Messung bestimmt wurde, nimmt die Kapsel
an der medialen Seite ihren Ansatz 6 bis 10 mm proximal des Trochlearandes und
an der lateralen Seite 3 bis 8 mm hiervon proximal.
Die knorpelharte Verstärkung am kranialen Rand des Lig. collaterale mediale findet
in der Literatur keine Erwähnung. Eine Einlagerung einer solchen Struktur an dieser
Stelle, hängt mit den Reibe- und Druckkräften in der Bewegung auf dem medialen
Trochlearand zusammen, so dass sie bei jungen Hunden schwächer als bei älteren
Hunden ausgebildet ist.

113 Ligamentum anulare radii ( Abb. 9, grün)
Von SÜPPEL (1921) wurde die Breite dieses Bandes vermessen und mit 2 bis 5 mm
angegeben. Die Breite misst am Ursprung, im Gelenkspalt und am Ansatz jeweils 0,3
cm im Mittel. Die Gesamtlänge dieses Bandes an den eigenen Präparaten beträgt im
Mittel 2,2 cm. Ein Vergleich ist aufgrund der fehlenden Materialangaben in der Arbeit
von SÜPPEL (1921) nicht möglich.
Das Lig. anulare radii verläuft ringförmig um den Radius und fixiert diesen dabei an
die Ulna, um die Rotationsbewegungen zu stabilisieren. Es ist laut NAV (1994) der
Art. radioulnaris proximalis zugehörig.
Eine Definition, an welcher Seite der Ursprung bzw. der Ansatz des Bandes zu
setzen ist, wurde bisher nicht festgelegt. MARTIN (1923) bestimmt den Ursprung
lateral am Bandhöcker des Radius und den Ansatz medial an der Ulna.
VOLLMERHAUS et al. (1994b) beschreiben den Ursprung am lateralen Seitenband
bzw. BERZON u. QUICK (1980) am lateralen Seitenband und dem Processus
coronoideus lateralis. Dies widerspricht meinen Untersuchungen und auch denen
von SÜPPEL (1921), BERGER (1992), BAUM u. ZIETZSCHMANN (1936), EVANS
(1993a) und NICKEL et al. (1992). Hier liegt der Ursprung bzw. der Ansatz jeweils
am Processus coronoideus lateralis bzw. medialis. VOLLMERHAUS et al. (1994b)
schreiben dem Lig. anulare radii eine stabilisierende Funktion im Hinblick auf die
Supinations- und Pronationsbewegungen im Ellbogengelenk zu. Das Band wäre
allerdings bei einem Ursprung am lateralen Kollateralband funktionell etwas
eingeschränkt für die Stabilisation während der Rotationsbewegungen der Art.
radioulnaris proximalis. In den eigenen Untersuchungen hat sich durchgehend
bestätigt, dass Ursprung und Ansatz jeweils an den Processus coronoidei lateralis
und medialis liegen. Wobei das Band an der lateralen Seite direkt an der Spitze des
Processus coronoideus entspringt und an der medialen Seite ca. 1 bis 3 mm unter-
halb des Processus ansetzt. Hingegen ist das Ansetzen einiger Fasern des Lig.
anulare radii am kranialen Schenkel des medialen Kollateralbandes, wie in den
eigenen Untersuchungen festgestellt wurde, so von keinem der genannten Autoren
beschrieben worden. Das Lig. anulare radii zieht vom Processus coronoideus
lateralis unter das Lig. collaterale laterale in den axial vorspringenden Knorpelfalz,

114 um schließlich unter der Ursprungssehne des M. supinator distal des Gelenkspaltes
wieder aufzutauchen und auf das Lig. collaterale mediale zu zulaufen. Die von
BERGER (1992) beschriebene Zweiteilung des Bandes an der lateralen Seite, an
den Epicondylus lateralis humeri und den Processus coronoideus lateralis ulnae, die
auch von BERZON u. QUICK (1980) als zweiteiliger Ursprung, allerdings an
divergierenden Punkten beobachtet wird, kann durch die hier untersuchten Präparate
nicht bestätigen werden.
Ligamentum obliquum (Abb. 10, türkis)
Dieses schräge Band findet in der aktuellen NAV (1994) keine Erwähnung. In der
Literatur (s.u.) wird es beobachtet und bei der Präparation aller untersuchten
Gelenke war es immer darzustellen. Es stellt sich als Verstärkungszug der Kapsel an
der Beugeseite des Gelenks dar. Hier stimmen die in der Literatur (SÜPPEL 1921;
BAUM u. ZIETZSCHMANN 1936; EVANS 1993a; STASZYK u. GASSE 2001a) mit
den eigenen Beobachtungen weitestgehend überein. Es verläuft extrakapsulär und
ist mit der Kapsel fest verbunden. Der Ursprung befindet sich breitgefächert
zusammen mit der Gelenkkapsel am dorsolateralen Rand des Foramen
supratrochleare. Von hier zieht es mediodistal und teilt sich einige Millimeter proximal
des Gelenkspaltes der Art. cubiti in zwei Schenkel. Ein kürzerer inseriert am
medialen Kollateralband und ein längerer Schenkel umschlingt die kaudalen
Ansatzsehnen der Mm. biceps brachii und brachialis und setzt schließlich am
medialen Rand im proximalen Drittel des Radius an.
Eine Vermessung des Lig. obliquum war in der gesichteten Literatur nicht zu finden.
Die von mir durchgeführten Messungen wurden im Winkel von 160° durchgeführt
(siehe Kap. 3). Das Lig. obliquum hat eine Länge von im Mittel 3,24 cm und eine
Länge vom Ursprung bis zum Ansatz an der medialen Seite des Radius von im Mittel
3,4 cm. Die Breite im Ursprungsbereich beträgt unter diesen Bedingungen 0,33 cm,
die Breite im Gelenkspaltbereich 0,24 cm und die Breite im Ansatzbereich 0,20 cm
für den Ansatz des längeren Schenkels medial am Radius und 0,31 cm für den
Ansatz des kürzeren Schenkels am medialen Seitenband. Die Messung des
kürzeren Schenkels, der an den kranialen Schenkel des medialen Kollateralbandes

115 zieht, wurde durch die Einlagerung der knorpelharten Struktur an der
Verbindungsstelle erschwert (Abb. 8a/B).
Als Funktion des Bandes wird die Straffung des kranialen Kapselabschnittes und
daraus ein Verhindern des Anschlagens des Processus anconeus in die Fossa
olecrani in Streckstellung diskutiert (KAPANDJI 1984; KASER 1998). Durch eigene
Beobachtungen wird eine mögliche Funktion des Lig. obliquum zusätzlich in einer
Zugwirkung auf die Gelenkkapsel durch die Verbindung mit den Mm. biceps brachii
und brachialis sowie das mediale Seitenband erwogen.
5.2 Messergebnisse Da eine Vermessung der Längen und Breiten der Kollateralbänder des Ellbogen-
gelenkes bzw. der Bänder der Art radioulnaris proximalis und des Lig. obliquum in
der Literatur nicht zu finden ist, können zu den eigenen Messergebnissen nur
Schlußfolgerungen angeboten werden.
Mit Hilfe des Statistikprogramms SAS und der darin verfügbaren Tests (t-Test,
Shapiro-Wilk Test, gepaarter Students t-Test, siehe Kap. 3) werden das laterale und
das mediale Kollateralband auf signifikante Unterschiede untersucht. Es ergeben
sich für zwei Messstrecken keine signifikanten Unterschiede und für acht Mess-
strecken signifikante Unterschiede.
Keine Signifikanzen ergeben sich für die Strecken der Gesamtlänge der kranialen
Schenkel der beiden Kollateralbänder. Ebenfalls nicht signifikant sind die Breiten der
Bänder in der Höhe des Gelenkspaltbereiches. Somit bieten das laterale und das
mediale Kollateralband an dieser Stelle die gleiche Bandbreite für die seitliche
Stabilisierung und Führung des Gelenkes.
Die Gesamtlänge der kaudalen Schenkel der Kollateralbänder ist signifikant unter-
schiedlich. Ein Unterschied der kaudalen Schenkel kann schon makroskopisch
erkannt werden, da hier die Faserverläufe und die Ansatzbreiten einen deutlichen
Unterschied aufweisen (siehe Kap. 4). Der kaudale Schenkel des lateralen
Kollateralbandes setzt breitbasig an der Ulna an und hat einen schräg kaudodistalen
Faserverlauf. Die Fasern des kaudalen Schenkels des medialen Kollateralbandes
verlaufen dagegen nach distal und setzen tief im Spatium interosseum an Radius

116 und Ulna an. Somit hat das laterale Band über den kaudalen Schenkel nur eine
Verbindung zu einem Unterarmknochen, während das mediale Seitenband über den
kaudalen Schenkel eine Verbindung zu beiden Unterarmknochen eingeht. Dies weist
auf eine unterschiedliche Funktion der beiden Kollateralbänder hin (siehe unten).
Die Längen im Ursprungs- und Ansatzbereich der Kollateralbänder weisen einen
signifikanten Unterschied auf. Sie sind im Mittel an der lateralen Seite länger als an
der medialen Seite. In den beiden Gelenkstellungen gibt es keinen erwähnenswerten
Unterschied. Der Mittelwert der Differenz bedeutet, dass das laterale Kollateralband
bei einer Ursprungslänge von 0,76 cm um 0,32 cm bzw. 0,33 cm länger ist (160°
sowie 30°) als der Ursprung an der medialen Seite. Dieses Verhältnis trifft ebenso für
die Länge des Ansatzes zu: hier ist der laterale Ansatz des kranialen Schenkels bei
einer Gesamtlänge von 0,91 cm (160° und 30°) im Mittel um 0,54 cm länger als an
der medialen Seite. Die Ansatzlängen der kaudalen Schenkel können nicht
miteinander verglichen werden, weil es aus messtechnischen Gründen nicht möglich
war, diese Strecke am kaudalen Schenkel des lateralen Kollateralbandes zu
bestimmen (siehe Kap. 3).
Für das Verhältnis der „freien Teile“ der beiden Bänder zueinander ergibt sich ein
umgekehrtes Verhältnis: Die Differenz der Mittelwerte ist am lateralen Band größer
als am medialen Band. Auch hier ist von einem signifikanten Unterschied zu
sprechen. Der gesamte freie Teil des kranialen Schenkels ergibt im Vergleich des
medialen zum lateralen Band eine Differenz von im Mittel 0,81 cm (160°) bzw. 0,91
cm (30°) und ist also im Schnitt medial einen knappen Zentimeter länger als lateral.
Für den freien Teil der beiden Humerusabschnitte zueinander lässt sich eine
Differenz von 0,46 cm (160°) bzw. 0,63 cm (30°) errechnen. Der „freie Teil“ des
Unterarmabschnittes ist im Mittel an der medialen Seite 0,49 cm (160°) bzw. 0,25 cm
(30°) länger als an der lateralen Seite.
Zusammenfassend lässt sich für die Ursprungs- und Ansatzlängen und die freien
Teile der beiden Kollateralbänder sagen, dass die Ursprungslängen der
Kollateralbänder an der lateralen Seite 1,8 mal länger sind als an der medialen Seite,
die Ansatzlängen der kranialen Schenkel 2,5 mal länger sind an der lateralen Seite.
Hingegen ist die Länge des gesamten freien Teils des kranialen Schenkels des

117 medialen Kollateralbandes 1,3 mal länger bei 160° und 1,4 mal länger bei 30° als die
des kranialen Schenkels des lateralen Kollateralbandes.
Damit ist bewiesen, dass die Strecke ohne Verbindung mit dem Periost (= freier Teil)
medial länger ist als lateral. Das Lig. collaterale mediale hat somit eine größere
Strecke, an der Kräfte in der Bewegung des Gelenkes wirken können. Dadurch
erklären sich die in der mikroskopischen Untersuchung gemachten Beobachtungen:
eine Struktur, an der die Kräfte auf eine größere Strecke wirken, ist straffer
organisiert (siehe Kap. 5.3).
Wird dies auf die Aussage von MARTIN (1923) bezogen, dass das mediale
Seitenband auch die Pronation des Unterarms beschränkt, wird deutlich, dass hier in
zwei Achsen Kräfte wirken (Beugung und Streckung; Pronation und Supination).
Auch von Hultkrantz (1897) wird in seinem Kapitel über die Caniden erwähnt, dass
das „Lig. epitrochleare radiale“ (= mediales Kollateralband) bei der Pronation
gespannt wird, wobei es den Unterarm etwas abduziert.
An einer Stabilisierung in diesen zwei Achsen ist auch das laterale Kollateralband
beteiligt. Darauf weist schon der hier viel breitere kaudale Schenkel hin, der eine
starke Verbindung von Radius und Ulna gewährleistet. Durch seinen breiten
Ursprung distal des Gelenkspaltes aus dem bis dahin ungeteilten Band, bewirkt der
kaudale Schenkel auch eine starke Bindung der beiden Unterarmknochen an den
Humerus. Die Ligg. collateralia tragen somit nicht nur zur strikten Wechselwirkung
des Gelenkes bei (Evans 1993a).
In klinischen Untersuchungen hat sich ergeben, dass die Luxation bzw. Subluxation
nach lateral am häufigsten vorkommt (EISENMENGER 1981; BIDLINGMAIER 1988).
Die Luxation nach medial ist aufgrund der schräg nach distal vorstehenden
Gelenkfläche des medialen „Epicondylus humeri“ (= Trochlea humeri) und des
kräftigen lateralen Seitenbandes selten (EISENMENGER 1981). Das laterale
Seitenband fungiert hier somit weniger als Führungsband (EVANS 1993a) sondern
als Halteband, wenn hierzu die Definition von FICK (1910) übertragen wird.
Allerdings sei hier angemerkt, dass an der lateralen Seite Muskulatur vorhanden ist,
um Krafteinwirkungen abzupuffern, diese jedoch an der medialen Seite fehlt.

118 Anhand der Ergebnisse einer Untersuchung zur Luxatio antebrachii (BIDLINGMAIER
1988) bei verschiedenen Hunderassen fällt auf, das der Anteil luxierter Gelenke bei
großen Rassen prozentual mehr beträgt als bei kleinwüchsigen. Das ist darauf
zurückzuführen, dass bei Hunden kleinwüchsiger Rassen die Kollateralbänder relativ
kräftiger sind als bei Hunden großwüchsiger Rassen, so dass es bei den kleinen
Rassen eher zu einer gelenknahen Fraktur als zu einer Luxation kommt.
Ähnliche Ergebnisse stammen von MEYER–LINDENBERG (1991) wobei nur 34,2%
kleinwüchsige, jedoch 42,1% großwüchsige Hunderassen hiervon betroffen waren.
Die Bedeutung der Kollateralbänder in diesem Zusammenhang muss demnach
genauer untersucht werden. Es wäre folglich sinnvoll, nicht nur Hunde einer Größe
und Rasse miteinander zu vergleichen, sondern auch kleine Rassen in solch eine
Vermessung der Gelenkbänder miteinzubeziehen.
5.3 Mikroskopische Untersuchung
Gelenkbänder bestehen aus straffem, kollagenfaserigem Bindegewebe. Dieses
Bindegewebe enthält einen unterschiedlich hohen Anteil an elastischen Fasern
(AICHEL 1925). Da diese Bänder sehr reich an Kollagen in der Grundsubstanz sind,
sehen sie weißlich glänzend aus (HEES 1990; BUCHER u. WARTENBERG 1997).
Wo sich Zugspannungen nur in eine Vorzugsrichtung auswirken können, liegt
parallelfaseriges straffes Bindegewebe vor.
Die lichtmikroskopisch darstellbaren kollagenen Fasern bestehen aus
Kollagenfibrillen. Diese formen zunächst Faserbündel und endlich Faszikel
(BENJAMIN u. RALPHS 1995), welche von einer Bindegewebsschicht, dem
„Epitendineum“ bzw. dem „Epiligament“ umhüllt werden (CHOWDHURY et al. 1991).
Um eine plastische Darstellung der Bandkonstruktionen zu erreichen, wurden sowohl
Längs- als auch Querschnitte der Probenblöcke angefertigt.
Lichtmikroskopisch sind die Kollagenfasern in Längsschnitten auffällig „gewellt“
(AMIEL et al. 1995). Diese Wellung ist in den angefertigten Abbildungen nicht zu
sehen, da die Bänder unter Spannung auf die Korkplatten aufgenäht wurde (siehe
Kap. 3). Im Gegensatz zu den elastischen Fasern sind die kollagenen Fasern fast
undehnbar. In der Literatur wird beschrieben, dass die Ligg. collateralia eine

119 unterschiedliche Menge an elastischen Fasern enthalten können (NICKEL et al.
1992; VOLLMERHAUS et. al 1994b). Es konnten mit der Orcein Färbung keine
elastischen Fasern beobachtet werden. Eine Dehnbarkeit der Bänder, basiert im
lateralen Kollateralband, wie durch die Untersuchung mit dem Polarisations-
mikroskop nachgewiesen werden konnte, auf der besonderen Gesamtkonstruktion
des Scherengitterprinzips der kollagenen Fasern (BUCHER u. WARTENBERG
1997). Dies wird sowohl im Längs- als auch im Querschnitt sehr deutlich, denn in
beiden Ebenen sieht man im lateralen Kollateralband sowohl Längs- als auch
Querfasern. Im medialen Kollateralband sind im Längsschnitt nur Längsfasern und
im Querschnitt nur quer angeschnittene Längsfasern zu sehen. Im lateralen Band
sind im Längsschnitt (Polarisation der H.E. Färbung) mehr dunkle Bereiche als im
medialen Band zu erkennen. Im Querschnitt hingegen erscheint das mediale Band
teilweise einheitlich dunkel: diese Dunkelfärbung tritt immer dann auf, wenn das Licht
parallel zur optischen Achse der Kollagenfasern einfällt.
Die Anordnung der Fasern nach dem Scherengitterprinzip bedeutet somit, da sich
die Kollagenfasern gemäß BUCHER u. WARTENBERG (1997) immer in Richtung
der Zugkräfte ausrichten, dass hier kein Zug vornehmlich in einer Richtung besteht.
Das laterale Band sorgt nicht nur für die Verbindung zwischen Humerus, Radius und
Ulna während der Beuge- und Streckbewegung. Es ist auch an der Pro- und
Supination des Unterarms unterstützend beteiligt, unterstützt in dieser Bewegung für
den Kontakt von Radius und Ulna zueinander und sichert die Drehbewegungen
gegen den Oberarm. Auch hier ist es richtig, wie schon von EVANS (1993a) und
LIPPERT (1990) angewandt, sich in der Listung des Ellbogenglenkes der
humanmedizinischen Nomina Anatomica (1985) anzuschließen und die einzelnen
Teilgelenke nach ihrer Funktion zu betrachten. Es erscheint jedoch für die Erklärung
der baulichen Struktur der Bänder und damit der Funktion sinnvoll zu sein, die Art.
radioulnaris proximalis in die Mechanik und Definition der Art. cubiti
miteinzubeziehen. Unterstützt wird diese Vermutung auch durch den makroskopisch
schon erkennbaren Faserverlauf des kaudalen Schenkels nach kaudodistal und den
fächerförmigen Ansatz an der Ulna.

120 Diese scherengitterartige Anordnung der Fasern ermöglicht dem lateralen Kollateral-
band folglich die notwendige Dehnbarkeit ohne Einbau elastischer Fasern.
Beim medialen Kollateralband wirken folglich vornehmlich die Kräfte in einer
Richtung. Hier verlaufen, wie durch die Untersuchungen mit dem polarisierten Licht
nachgewiesen wurde, die Fasern parallel zueinander; somit fehlt ein Dehnungs-
bereich durch die Anordnung der Fasern. Das mediale Band ist also nicht in der
Lage, sich zu verlängern.
In der H.E. Färbung wird die straffe Organisation im medialen Kollateralband im
Hinblick auf die Anordnung und Form der Zellkerne ebenso deutlich; die Fibrozyten
sind mit ihren länglichen, schmalen Zellkernen zwischen den Fasern in Reihen
angeordnet. Im lateralen Kollateralband hingegen sind vornehmlich runde Zellkerne
anzutreffen, die wie zufällig angeordnet zwischen den Fasern liegen.
Aufgrund des histologischen Bauplans der Bänder ist anzunehmen, dass das
mediale Band einer größeren Zugbelastung in einer Richtung ausgesetzt ist,
während am lateralen Band verschiedene Kräfte in unterschiedliche Richtungen
ziehen und somit die Faserkonstruktion eines netzartigen Scherengitters entstehen
lassen.
Auf die Funktion des medialen Kollateralbandes bezogen, kann hier gefolgert
werden, dass es durch seinen Ansatz an beiden Unterarmknochen zwar die
Pronation und Supination mitunterstützt, in erster Linie jedoch für den straffen
Zusammenhalt von Humerus und Unterarmknochen sorgt.
Wird hierzu der Bauplan des Humerusendes betrachtet, fällt auf, dass der mediale
Epicondylus humeri groß und eckig ist, und somit eine Verlagerung der Ulna nach
medial verhindert. Auf der lateralen Seite wird dies durch den kleinen Epicondylus
lateralis humeri nicht gewährleistet. Die Verlagerung muss auf der lateralen Seite
durch ein Kollateralband ausgeglichen werden. Diese Tatsache wurde bereits von
CAMPBELL (1969) und BRINKER et al. (1993) erwähnt.

121 6. Zusammenfassung Miriam-Jasmin Hemmes
Zum Bandapparat des Ellbogengelenkes des Hundes insbesondere zu Verlauf und Struktur der Kollateralbänder. In der vorliegenden Arbeit wurde der Bandapparat am Ellbogengelenk des Hundes,
insbesondere die Kollateralbänder, im Seitenvergleich makroskopisch hinsichtlich
Ursprung, Verlauf, Ansatz und Sondereinrichtungen (Schleimbeutel, sesamoide
Einlagerungen) untersucht. Hierzu wurden 40 Vordergliedmaßen von 22
verschiedenen Hunden (DSH und DSH-Mix) verwendet. An den medialen und den
lateralen Seitenbändern wurden definierte Strecken gemessen. Die erhaltenen Werte
wurden dokumentiert und mit Hilfe des Statistikprogramms SAS ausgewertet.
Die mikroskopische Untersuchung wurde an insgesamt zehn Vordergliedmaßen von
fünf Schäferhund-Mischlingen durchgeführt. Es wurden sowohl Längs- als auch
Querschnitte in den Färbungen Pikrosiriusrot, H.E. und Orcein angefertigt. Anhand
der H.E. gefärbten Pärparate wurde die Form der Zellkerne untersucht und mit den
Orcein gefärbten Präparaten wurde das Vorhandensein elastischer Fasern nicht
nachgewiesen. Der Faserverlauf in den Kollateralbändern wurde mit dem
Polarisationsmikroskop an Pikrosiriusrot und H.E. gefärbten Schnitten nachgeprüft.
Die Untersuchung zur Topographie des Bandapparates ergab nur teilweise
Übereinstimmungen mit der Literatur in der Definition des Ursprungs und des
Ansatzes. Das laterale und mediale Kollateralband als Verbindungsbänder der
Knochen des Ober- und Unterarms und Führungsbänder der Beuge- und
Streckbewegungen sowie unterstützend der Pronations- und Supinations-
bewegungen unterscheiden sich in folgenden Punkten voneinander:
Am Lig. collaterale laterale findet sich an jedem untersuchten Ellbogengelenk ein die
Inkongruenz des Gelenkes ausgleichender Knorpelfalz, der eine sesamoide Struktur
enthalten kann. Der M. supinator bildet über einem Knochenvorsprung am Caput
radii eine kappenartige Verstärkung, die die Ursprungssehne in der Bewegung führt.
Durch die statistische Auswertung wurde deutlich, dass das laterale Kollateralband

122 im Verhältnis zum medialen Kollateralband einen 1,8 mal längeren Ursprungs-
bereich und einen 2,5 mal längeren Ansatzbereich hat als das mediale Band.
Hingegen ist die Länge des gesamten „freien Teils“ im Lig. collaterale mediale 1,3
mal (Winkelung 160°) bzw. 1,4 mal (30°) länger als die des lateralen
Kollateralbandes. Folglich existiert am medialen Kollateralband eine größere
Teilstrecke, die nicht mit dem Periost verwachsen ist und somit eine längere Strecke
an der Kräfte wirken können.
Im lateralen Kollateralband sind die Kollagenfasern in einer scherengitterartigen
Struktur angeordnet und die rundlichen Zellkerne liegen zwischen den Fasern. Im
medialen Kollateralband sind die Fasern streng parallel nebeneinander angeordnet
und die länglich, schmalen Zellkerne liegen dazwischen. Es wirkt sehr straff
organisiert und somit höheren Kräften an der längeren freien Strecke entgegen.
Zu einer ganzheitlichen Betrachtung des Bandapparates des Ellbogengelenkes sind
die Daten des Lig. obliquum, Lig. olecrani und des Lig. anulare radii
miteinzubeziehen.

123 7. Summary Miriam-Jasmin Hemmes On the ligamental system of the canine elbow joint, with special regard to course and structure of the collateral ligaments. This study investigated the ligamental system of the canine elbow joint, regarding the
origin, course, insertion and specific structures (bursae synovialis, sesamoid
structures) of the collateral ligaments. For this purpose, 40 forelimbs of 22 dogs
(German shepherds and shepherd mongrels) were used.
Defined parts of the lateral and medial collateral ligaments were measured, and the
values received were evaluated statistically by SAS. The microscopic observations
were made on ten forelimbs of five German shepherd mongrels, using longitudinal
and transverse histological sections of the samples, which had been stained with H.
E., picrosirius red and orcein. In the H. E. staining, the form of the cell nuclei was
checked, and the orcein samples were analysed for elastic fibres. In the picrosirius
red and the H. E. sections, the course of the collagen fibre bundles was followed.
The results obtained on the topography of the ligamental systems were only in
accordance with the literature as far as the origin and insertion was concerned. The
lateral and medial collateral ligaments, defined as connecting ligaments of humerus,
ulna, and radius, and as guiding ligaments of extension and flexion movements and
pronation and supination, differed in following points: In the lateral collateral ligament
of each elbow joint, a cartilaginous fold could be found, which smoothed the
incongruity of the joint. This fold could contain a sesamoid structure. The supinator
muscle formed a cap-like thickening above a bony protuberance of the caput radii.
This thickening seemed to guide the muscle during movement. Along the side of the
medial collateral ligament, lateral movements were limited by a large medial
epicondyle.
Statistical evaluation made obvious that the origin of the lateral collateral ligament, in
proportion to that one of the medial collateral ligament, was 1.8 times longer, with an

124 insertion 2.5 times longer. The length of the “free part” of the medial collateral
ligament was 1.3 times (angle of 160°), resp. 1.4 times (angle of 30°) longer than that
of this part on the lateral side of the joint. According to these aspects, a longer part
on the medial side of the elbow joint exists, as not closely attached to the periost,
where forces could be active.
This structure defined the different constructions of the collateral ligaments, as
detectable by histological analysis. In the lateral collateral ligament, the collagen fibre
bundles were arranged in a folding trellis-like manner, and round cell nuclei were
lying between the fibre bundles. In the medial collateral ligaments, the fibre bundles
were arranged parallel to each other, with long and small cell nuclei in between. The
medial ligament seemed to be much more tight, and, thus, resistant to strong forces
at the "free part".
For a more complete functional analysis of the ligamental system of the elbow joint,
data of the Lig. obliquum, Lig. olecrani and Lig. anulare radii has to be included.

125
Literaturliste
AICHEL, O. (1925): Über die Wirkung der Ligamenta collateralia nebst Bemerkungen über die sogenannten Schnappgelenke. Z. Anat. Entwicklungsgesch. 76, 1–15 ADAMS, D. R. (1986): Canine anatomy. Iowa State University Press, Ames, 29-83 AKESON, W. H., S. L.-Y. WOO, D. AMIEL u. C. B. FRANK (1984): The biology of ligaments. In: FUNK, F. J. u. L. Y. HUNTER (Eds.): Rehabilitation of the injured knee. Mosby, St. Louis, 93-148 AMIEL, D., C. R. CHU u. J. LEE (1995) : Effect of loading on metabolism and repair of tendons and ligaments. In: GORDON, S. L., S. J. BLAIR u. L. J. FINE (Eds.): Am. Acad. Orthop. Surg. Rosemont, 217-230 BANKS, W. J. (1993): Applied veterinarian histology. 3. Aufl. Verlag Mosby Year Book, St. Louis, 96-106
BAUM, H. u. O. ZIETZSCHMANN (1936): Das Muskelsystem.
Bd. 1: Skelett- und Muskelsystem In: ZIETZSCHMANN, O. (Hrsg.): Handbuch der Anatomie des Hundes. 2. Aufl., Parey, Berlin
BARONE, R. R. (1968): Anatomie comparee des mammiferes domestique. Bd. 2 : Arthrologie et Myologie Laboratoires d Anatomie. Ecole Nationale Veterinaire, Lyon BENJAMIN, M. u. J. R. RALPHS (1995) : Functional and developmental anatomy of tendons and ligaments. In: GORDON S. L., S. J. BLAIR u. L. J. FINE (Eds.): Repetitive motion disorders of the upper extremity. Am. Acad. Orthop. Surg. Rosemont, 185-203

126 BERGER, C. (1992):
Röntgenologische, anatomische und histologische Untersuchungen zu einem fakultativ vorhandenen Sesambein der Articulatio cubiti des Hundes. Berlin, Univ., Veterinärmed. Fak., Diss.
BERZON, J. u. C. B. QUICK (1980): Fragmental coronoid process: anatomical, clinical and radiographic considerations with case analyses. J. Amer. Anim. Hosp. 16, 241-251 BIDLINGMAIER, B. (1988): Klinische Untersuchungen und Erhebungen zur Luxatio antebrachii und der Monteggia – Fraktur bei Hund und Katze in den Jahren 1978–87. München, Univ. Veterinärmed. Fak., Diss. BINGEL, S. A. u. W. H. RISER (1977): Congenital elbow luxation in the dog. J. Small Anim. Pract. 18, 445
BÖCK, P. (Hrsg.) (1989): Romeis mikroskopische Technik. 17. Aufl., Urban und Schwarzenberg, München
BRINKER, W. O., D. L. PIERMATTEI u. G. L. FLO (1993): Orthopädie und Frakturbehandlung beim Kleintier. Schattauer, Stuttgart, N. Y., 386-449
BUCHER, O. u. H. WARTENBERG (1997): Cytologie, Histologie und mikroskopische Anatomie des Menschen. 12. Aufl., Verlag Hans Huber, Bern
BUTLER, H. C. (1974): Tendon, muscle and fascia. In: ARCHIBALD, J. (Ed.) Canine Surgery. Amer. Vet. Publ., INC Drawer KK, Santa Barbara, California CAMPBELL, J. R. (1969): Nonfracture injuries to the canine elbow. J. Amer. Med. Vet. Assoc. 155, 735-744
CAMPBELL, J. R. (1971): Luxation and ligamentous injuries of the elbow of the dog. Vet. Clin. North Am. 1, 429-440

127
CORNER, M. (1896): Some structures in the elbow-joint. J. Anat. Physiol. 30, 369–376 CHOWDHURY, P., MATYAS, J. R. u. FRANK, C. B. (1991): The “epiligament” of the rabbit medial collateral ligament: a quantitative morphological study. Connect. Tissue Res. 27, 33-50 DETELS, A. (1980): Das Gliedmaßenskelett des Dackels (Dachshund, Teckels). München, Veterinärmed. Fak., Diss.
DOBBERSTEIN J. u. G. HOFFMANN (1961): Skelett der Gliedmaßen. In: Lehrbuch der vergleichenden Anatomie der Haustiere. Bd. 1: Bewegungsapparat. 2. Aufl., Hirzel Verlag, Leipzig, 26-42
EISENMENGER, E. (1981): Ellbogengelenksluxation – konservative Behandlung und Ergebnisse. Wien. Tierärztl. Mschr. 68, 156–158 EVANS, H. E. (1993 a): Arthrologie. In: Millers anatomy of the dog. 3rd Edition, W.B. Saunders Company, Philadelphia, London EVANS, H. E. (1993 b): The Skeleton. In: Millers Anatomy of the dog. 3rd Edition, W.B. Saunders Company, Philadelphia, London
FEHR, M. u. A. MEYER–LINDENBERG (1992): Über die Ellbogenerkrankungen beim Hund. Kleintierpraxis 37, 427–438
FICK, R. (1910): Anatomie der Gelenke unter Berücksichtigung der bewegenden Muskeln. 3. Spezielle Gelenk- und Muskelmechanik. In: VON BARDELEBEN, K.-H. (Hrsg.): Handbuch der Anatomie des Menschen. 2. Band, 1. Abteilung, Teil 3, Fischer Verlag, Jena FOX, S. M., S. BLOOMBERG u. R. M. BRIGHT (1983): Developmental anomalies of the canine elbow. J. Am. Anim. Hosp. Assoc. 19, 605-615

128
FREWEIN, J. (1983): Muskulatur. In: FREWEIN, J. u. B. VOLLMERHAUS (Hrsg.): Anatomie von Hund und Katze. Blackwell – Wissenschaft, Berlin GERLACH D. (1976): Das Lichtmikroskop. Thieme Verlag, Stuttgart GR∅NDALEN J. u. T. BRAUT (1976): Lameness in two young dogs caused by a calcified body in the joint capsule of the elbow. J. Small Anim. Pract. 17, 681-684 HÄGGQVIST, G. (1931): In: MÖLLENDORF, W. v. (Hrsg.): Handbuch der mikroskopischen Anatomie des Menschen. Bd. 2, Teil 3, Springer Verlag Berlin
HAHN von DORSCHE, H. u. D. SASSE (1994): Taschenbuch der Anatomie. Bd. 1: Histologie, Bewegungsapparat. Verlag Fischer, Stuttgart, Jena
HEES, H. (1990):
Binde- und Stützgewebe. In: MOSIMANN, W. u. T. KOHLER (Hrsg.): Zytologie, Histologie, und mikroskopische Anatomie der Haussäugetiere. Verlag Paul Parey, Berlin, Hamburg, 48-61
HENSCHEL, E. (1977): Asynchrones Längenwachstum der Ossa antebrachii und seine Wirkung auf die Art. cubiti beim Hund. Tierärztl. Praxis 7, 227–233
HERMANSON J. W. u. H. E. EVANS (1993): The muscular system. In: Millers anatomy of the dog. 3rd Edition W.B. Saunders Company, Philadelphia, London, 258-384 HITZ, D. (1976): Obligatorische Untersuchung der Ellbogengelenke auf Dysplasie beim Bassethound. Schw. Arch. Tierheilk. 118, 167-172

129
HLADIK, G. (1986): Röntgenologisch erfassbare Tendinosen im Schulter- und Ellbogengelenksbereich beim Hund: ihre Diagnose, Lokalisation, Klinik und Verlauf. Wien, Veterinärmed. Univ., Diss.
HOURRIGAN, J. L. (1940):
Medial luxation of the elbow joint in a collie. Vet. Med. 35, 367-368
HULTKRANTZ, W. (1897): Das Ellenbogengelenk und seine Mechanik. Verlag Fischer, Jena JUNQUEIRA L. C. U., G. BINOLAS u. R. R. BRETANI (1979): Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histoch. J. 11, 447-455 KADLETZ, M. (1932): Anatomischer Atlas der Extremitätengelenke von Pferd und Hund. Verlag Urban & Schwarzenberg, Berlin, Wien, 53-54 KAPANDJI, I. A. (1984): Funktionelle Anatomie der Gelenke. Bd. 1: Obere Extremität.
Enke, Stuttgart KASER, W. (1998): Untersuchung zur funktionellen Anatomie des Ellbogengelenkes (Art. cubiti) des Hundes. München, Univ., Veterinärmed. Fak., Diss. KÖNIG, H. E. u. H.-G. LIEBICH (1999): Anatomie der Haussäugetiere. Bd. 1: Bewegungsapparat, Schattauer, Stuttgart KOCH, T. u. R. BERG (1992): Lehrbuch der Veterinär–Anatomie. Bd. I: Bewegungsapparat. 5. Aufl. Fischer, Stuttgart, 221-222
KRÖLLING, O. u. H. GRAU (1960): Lehrbuch der Histologie und vergleichender mikroskopischer Anatomie der Haustiere. 10. Aufl.
Verlag Paul Parey, Berlin, Hamburg, 51

130
KÜMMEL, H. (1984): Ein Beitrag zur Distractio cubiti beim Teckel. Berlin, Freie Univ., Fachber. Vetmed., Diss.
LILLIE, R. D. u. H. M. FULLMER (1976): Histopathological technic and practical histochemistry. Mc Graw-Hill Book Company, New York, Toronto LIPPERT, H. (1990): Obere Extremität In: Lehrbuch der Anatomie. 2.Aufl. Urban & Schwarze, München, Wien, Baltimore LJUNGGREN, G., A. J. CAWLEY u. J. ARCHIBALD (1966): The elbow dysplasia in dog. J. Am. Vet. Med. Assoc. 148, 887-891 LUNGWITZ, M. (1906): In: ELLENBERGER, W. (Hrsg.): Handbuch der vergleichenden Anatomie der Haustiere. 11. Aufl., Paul Parey, Berlin MAI, M. (1995): Funktionell-anatomische Untersuchungen an den Articulationes membri thoracici von Deutschem Schäferhund, Dobermann, Deutscher Dogge, Rauhaar-und Langhaarteckel. München, Univ., Veterinärmed. Fak., Diss. MARTIN, P. (1923): Lehrbuch der Anatomie der Haustiere. 2. Aufl., Bd. 4 Schikhardt und Ebner, Stuttgart, 121-302 MAYRHOFER, E. (1980): Die Skelettentwicklung des Junghundes, entwicklungsbedingte Varianten und ihre Abgrenzung gegenüber pathologischen Prozessen im Röntgenbild. Wien, Vet. Med. Univ., Habil.-Schrift
MEYER–LINDENBERG, A. (1991): Angeborene und erworbene Erkrankungen des Ellbogengelenkes beim Hund: Häufigkeit, Diagnostik und Behandlungsergebnisse der Jahre 1985 bis 1989. Hannover, Tierärztl. Hochschule, Diss. MEYER-LINDENBERG, A. (2001): Zur Diagnostik und Therapie entwicklungsbedingter Ellbogengelenkserkrankungen des Hundes. Hannover, Tierärztl. Hochschule, Habil.-Schrift

131
MONTAVON, P. M. u. D. SAVOLDELLI (1995): Clinical and radiologic evaluation of the integrity of medial and lateral collateral ligaments of the elbow in dogs. Schw., Arch. f. Tierheilk. 137, 475–479 NEUBERGER, A. u. B. SLACK (1953): The metabolism of collagen from liver, bone, skin and tendon in the normal rat. Biochem. J. 53, 47-52
NICKEL, R., A. SCHUMMER, K. H. WILLE u. H. WILKENS (1992): Passiver Bewegungsapparat, Skelettsystem. In: NICKEL, R., A. SCHUMMER u. E. SEIFERLE: Lehrbuch der Anatomie der Haussäugetiere. 6. Aufl., Bd.1: Bewegungsapparat. Verlag Paul Parey, Berlin, Hamburg, 15-272 NOMINA ANATOMICA (1985): International Anatomical Nomenclature Committee International Congress of Anatomists 12, London 6. Aufl.Edinburgh, Livingstone
NOMINA ANATOMICA VETERINARIA (1994): In: Nomina Anatomica Veterinaria. 4. Aufl. zusammen mit Nomina Anatomica Histologica. 2. Aufl. International Commitee on Veterinary Gnoss Anatomical Nomenclature. Vet. Histol. nomenclature of the World Assoc. Vet. Anat. Zürich, Ithaka, New York
PREUß, F. u. A. WÜNSCHE (1974): Über einige neue Kapselkörper an den Schweinezehen und andere Gleitdruckstrukturen bei verschiedenen Spezies, zugleich ein Beitrag zur Sesambeinfrage und zur Interosseusfrage. Beiheft zum Zentralbl. Veterinärmed. 21 Verlag Paul Parey, Berlin Hamburg PRICE, C. J. u. S. C. KING (1977): Elbow lameness in a young dog caused by an ossified disc in the joint capsule. Vet. Rec. 100, 566 RAUSCHER, T. (1986): Metrische Untersuchungen an den Gelenken und Muskel der Schultergliedmaße bei zwei Hunderassen – ein Vergleich zwischen Dackel (Teckel und Dachshund) und dem Deutschen Schäferhund. München, Univ., Veterinärmed. Fak., Diss.

132
ROOS, H., S. BRUGGER u. T. RAUSCHER (1992): Über die biologische Wertigkeit der Bewegung in den Radioulnargelenken bei Katze und Hund. Anat. Histol. Embryol. 21, 199-205
SAJONSKI, H. (1969): Schleimbeutel. Normalanatomische Vorbemerkungen.
In: DOBBERSTEIN, J. (Hrsg.): Handbuch der speziellen pathologischen Anatomie der Haustiere. 3. Aufl., Bd. 1 Parey Berlin, Hamburg, 199-201 SCHLEDERER, J. (1999): Ellbogengelenkdysplasie beim Hund: Röntgenologische und Pathomorphologische Befunde im Vergleich. Wien, Veterinärmed. Univ., Diss. SEIFERLE, E. u. J. FREWEIN (1992): Aktiver Bewegungsapparat, Muskelsystem, Myologie. In: NICKEL, R., A. SCHUMMER u. E. SEIFERLE: Lehrbuch der Anatomie der Haussäugetiere. 6. Aufl., Bd.1: Bewegungsapparat. Verlag Paul Parey, Berlin, Hamburg, 273-569 SHELLOW W. V. R. u. A. M. KLIGMAN (1967): Three-dimensional visualisation of elastic fibres in thick skin sections. Arch. Derm. 95, 221-224
STASZYK, C. u. H. GASSE (2001a): Das Foramen supratrochleare humeri: Ein Passageweg in der Ellbogenarthroskopie beim Hund. Kleintierpraxis 46, 219-22
STASZYK, C. u. H. GASSE (2001b): The enthesis of the elbow-joint capsule of the dog humerus. Europ. J. morph. 39, 319-323
STEVENS, A. u. J. LOWE (1997): Bewegungsapparat. In. STEVENS, A. u. J. LOWE (Hrsg.): Histologie des Menschen. 2. Aufl. Verlag Chapman und Hall, London, Weinheim

133
STOYAK, J. M. (1981): Dislokation des Ellbogens. In: BOJRAB, M. J.: Praxis der Kleintierchirurgie. Ferdinand Enke Verlag, Stuttgart
SÜPPEL, R. (1921): Bänder des Schulter-, Ellbogen- und Karpalgelenks des Hundes. Leipzig, Veterinärmed. Fak., Diss.
VAN HERPEN, H. (1988): Anatomy and development of the elbow joint. Tijdschr. Dierengeneeskd. 113 (Suppl. 1), 38–41 VAN SICKLE, D. C., H. D. DELLMANN u. E. BROWN (1993): Connective and supportive tissues. in: DELLMANN H. D. (Hrsg.:) An Outline Text – Atlas. 4 th Ed., Lea & Felbiger, Philadelphia, 29-53
VÄÄNÄNEN, M. u. K. SKUTNABB (1978): Elbow lameness in the young dog caused by a sesamoidal fragment. J. Small Anim. Pract. 19, 363-371 VOLLMERHAUS, B. u. H. ROOS (1980): Postnatal development of the forearm bones in dogs during functional adaption. Tierärztl. Wschrift. 93, 150-155 VOLLMERHAUS, B., J. FREWEIN, H. ROOS u. C. KNOSPE (1994a): Skelett. In: FREWEIN, J. u. B. VOLLMERHAUS (Hrsg.): Anatomie von Hund und Katze. Blackwell – Wissenschaft, Berlin VOLLMERHAUS, B., H. WAIBL u. H. ROOS (1994b): Gelenke. In: FREWEIN, J. u. B. VOLLMERHAUS (Hrsg.): Anatomie von Hund und Katze. Blackwell – Wissenschaft, Berlin WALKER, T. M. (1998): A redefined type of elbow dysplasia in the dog – 2 cases. Can. Vet. J. 39, 573-575

134
WALTHER, C. (1908): Sehnenscheiden und Schleimbeutel des Hundes. Leipzig, Vetmed. Fak., Diss. WAKURI H. u. Y. KANO (1966): Anatomical studies on the brachioradial muscle in dogs. Kaibogaku Zasshi. 4, 222-231.
WEBBON, P. M. u. D. G. CLAYTON-JONES (1976): The elbow. J. Small Anim. Prac. 17, 395-401
WOOD, A. K. W., P. H. MC CARTHY u. C. R. HOWLETT (1985): Anatomic and radiographic appearance of a sesamoid bone in the tendon of origin of the supinator muscle in dogs. Amer. J. Vet. Res. 46, 259-272 ZENZES, W., SCHREIBER, M. u. W. KÜPPER (1989): Verknöcherungskerne am Radius als Lahmheitsursache bei Rottweilerhunden. Prakt. Tierarzt 12, 40-43
ZIETZSCHMANN, O. (1943): Das Skelettsystem. Der passive Bewegungsapparat. In: ELLENBERGER W. u. H. BAUM (Hrsg.) Handbuch der vergleichenden Anatomie der Haustiere. 18. Aufl., Springer Verlag, Berlin
ZIMMERMANN, A. (1958):
Über die Sesambeine. Acta. Vet. Hung. 8, 137-142 ZONTINE, W. J., R. A. WEITKAMP u. C. L. LIPPINCOTT (1989): Redefined type of elbow dysplasia involving calcified flexor tendons attached to the medial humeral epicondyle in three dogs. J. Am. Vet. Med. Assoc. 194, 1082-1085

135 9. Anhang Tabellarische Dokumentation der Messergebnisse der Bänder der Articulatio cubiti
(Lig. collaterale laterale und Lig. collaterale mediale), des Lig. obliquum, des Lig.
olecrani und des Lig. anulare radii.
Tab. 21 und 22: Messergebnisse der Vorversuche zu den Gesamtlängen der
kranialen und kaudalen Schenkel des Lig. collaterale laterale bzw. des Lig.
collaterale mediale von fünf Hunden bei einer Gelenkwinkelung von 160°, 140°, 90°
und 30°.
Tab. 23 bis 33: Messergebnisse zu den Längen und Teillängen der Ligg. collateralia
der Artt. cubiti von 22 Hunden bei einer Gelenkwinkelung von 160° und 30°.
Tab. 34 bis 40: Messergebnisse zu den Breiten der Gelenkbänder der Artt. cubiti von
22 Hunden in den Gelenkwinkelungen 160° und 30°.

136
9. A
nhan
g: T
abel
len
Tab.
21:
Mes
serg
ebni
sse
der V
orve
rsuc
he z
u de
n G
esam
tläng
en d
er k
rani
alen
und
kau
dale
n S
chen
kel d
es L
ig. c
oll.
late
rale
der
Art.
cubi
ti vo
n fü
nf H
unde
n be
i ein
er G
elen
kwin
kelu
ng v
on 1
60°,
140
°, 9
0° u
nd 3
0°, g
emes
sen
in c
m.
Lig.
col
l. la
tera
le
16
0°
140°
90
° 30
°
Tier
num
mer
kr
ania
ler
Sch
enke
l ka
udal
er
Sch
enke
l kr
ania
ler
Sch
enke
l ka
udal
er
Sch
enke
l kr
ania
ler
Sch
enke
l ka
udal
er
Sch
enke
l kr
ania
ler
Sch
enke
l ka
udal
er
Sch
enke
l 1r
4,
1 2,
9 4
2,7
4 2,
7 3,
8 2,
7 1l
4
2,6
4 2,
6 4
2,6
3,5
2,6
2r
4,9
3,1
4,8
3,1
4,8
3,1
4,6
3,1
2l
4,3
3,1
4,3
3,1
4,3
3,1
4,2
3 3r
4
3 4
3 4
3 3,
6 3
4r
4,1
2,8
4,1
2,7
4,1
2,7
3,9
2,8
5r
3,8
3 3,
8 2,
9 3,
8 2,
9 3,
6 3
5l
3,6
2,6
3,6
2,6
3,6
2,6
3,6
2,6
Mitt
elw
ert
4,1
2,9
4,1
2,8
4,1
2,8
3,9
2,9
Sta
bw
0,4
0,2
0,4
0,2
0,4
0,2
0,4
0,2
r = re
chte
Ext
rem
ität
l = li
nke
Ext
rem
ität
Sta
bw =
Sta
ndar
dabw
eich
ung

137
Tab.
22:
Mes
serg
ebni
sse
der V
orve
rsuc
he z
u de
n G
esam
tläng
en d
er k
rani
alen
und
kau
dale
n S
chen
kel d
er L
ig. c
oll.
med
iale
der
Art.
cubi
ti vo
n fü
nf H
unde
n be
i ein
er G
elen
kwin
kelu
ng v
on 1
60°,
140
°, 9
0° u
nd 3
0°, g
emes
sen
in c
m.
Li
g. c
oll.
med
iale
160°
14
0°
90°
30°
Tier
num
mer
kr
ania
ler
Sch
enke
l ka
udal
er
Sch
enke
l kr
ania
ler
Sch
enke
l ka
udal
er
Sch
enke
l kr
ania
ler
Sch
enke
l ka
udal
er
Sch
enke
l kr
ania
ler
Sch
enke
l ka
udal
er
Sch
enke
l 1r
4,
1 4,
8 3,
8 4,
5 3,
8 4,
5 3,
9 4,
5 1l
4
4,3
3,8
4,2
3,8
4,2
3,8
4,2
2r
4,3
5,2
4,3
5,2
4,3
5,2
4,5
5 2l
4,
6 4,
9 4,
6 4,
8 4,
6 4,
8 4,
2 4,
8 3r
4,
1 4,
1 4,
1 4,
1 4,
1 4,
1 4,
2 4,
6 4r
4,
2 4,
8 4,
2 4,
8 4,
2 4,
8 4,
1 4,
7 5r
3,
5 3,
9 3,
5 3,
9 3,
5 3,
9 3,
5 3,
9 5l
3,
6 3,
9 3,
6 3,
9 3,
6 3,
9 3,
6 4,
1 M
ittel
wer
t 4,
1 4,
5 4,
0 4,
4 4,
0 4,
4 4,
0 4,
5 S
tabw
0,
4 0,
5 0,
4 0,
5 0,
4 0,
5 0,
3 0,
4
r = re
chte
Ext
rem
ität
l = li
nke
Ext
rem
ität
Sta
bw =
Sta
ndar
dabw
eich
ung

138
Tab. 23: Messergebnisse zu den Gesamtlängen der kranialen und kaudalen
Schenkel des Lig. coll. laterale der Art. cubiti von 22 Hunden bei einer Gelenk-
winkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale 160° 30°
Tiernummer kranialer Schenkel
kaudaler Schenkel
kranialer Schenkel
kaudaler Schenkel
1r 4,1 2,9 3,8 2,7 1l 4 2,6 3,5 2,6 2r 4,9 3,1 4,6 3,1 2l 4,3 3,1 4,2 3 3r 4 3 3,6 3 4r 4,1 2,8 3,9 2,8 5r 3,8 3 3,6 3 5l 3,6 2,6 3,6 2,6 6r 4,4 3 4,3 3 6l 4,2 3 4,1 3 7r 5 3,5 4,7 3,5 7l 4,9 3,1 4,8 3,1 8r 4,3 2,5 3,9 2,5 8l 4,1 3,2 3,9 2,8 9r 4 3 4 2,5 9l 4,4 3 4 3
10r 4 2,5 4 2,4 10l 3,6 2,3 4,1 2,3 11r 3,9 2,6 4,2 2,6 12r 4,5 2,8 3,9 3 12l 4,6 2,6 4,5 2,5 13r 3,9 1,7 4 1,8 13l 4 2,7 3,6 1,6 14r 5 2,7 4,8 3 14l 5 2,7 4,8 2,9 15r 4,4 2,9 4,4 3,2 15l 4,3 2,8 4,6 2,8

139
Fortsetzung Tab. 23: Messergebnisse zu den Gesamtlängen des kranialen und
kaudalen Schenkels des Lig. coll. laterale der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale 160° 30°
Tiernummer kranialer Schenkel
kaudaler Schenkel
kranialer Schenkel
kaudaler Schenkel
16r 4,8 2,8 4,7 2,8 16l 5 2,9 4,8 3 17r 3,1 2,3 3,3 2,2 17l 3,3 2,3 2,6 2,1 18r 3,7 2,5 4,1 2,2 19r 4 2,8 4 2,7 19l 4,3 2,5 4,3 2,5 20r 3,7 2,5 4,1 2,2 20l 3,9 2,6 4,2 2,6 21r 4,5 2,8 3,9 3 21l 4,6 2,6 4,5 2,5 22r 4,2 2,9 4,4 3,2 22l 4,3 2,8 4,6 2,8
Mittelwert 4,22 2,75 4,12 2,7 Stabw 0,47 0,31 0,47 0,39
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

140
Tab. 24: Messergebnisse zu den Gesamtlängen des kranialen und kaudalen
Schenkels des Lig. coll. mediale der Art. cubiti von 22 Hunden bei einer Gelenk-
winkelung von 160° und 30°, gemessen in cm.
Lig. coll. mediale 160° 30°
Tiernummer kranialer Schenkel
kaudaler Schenkel
kranialer Schenkel
kaudaler Schenkel
1r 4,1 4,8 3,9 4,5 1l 4 4,3 3,8 4,2 2r 4,3 5,2 4,5 5 2l 4,6 4,9 4,2 4,8 3r 4,1 4,1 4,2 4,6 4r 4,2 4,8 4,1 4,7 5r 3,5 3,9 3,5 3,9 5l 3,6 3,9 3,6 4,1 6r 3,8 4,2 3,9 4,4 6l 4 4,6 3,8 4,6 7r 4,5 5,2 4,7 5,4 7l 5,5 5,7 5,2 5,6 8r 4,4 4,9 4,6 5 8l 4,1 4,7 4,4 5,2 9r 4 4,5 4,3 4,1 9l 4,5 4,6 4,3 4,6
10r 4,2 4,5 4,2 4,2 10l 4,6 4,4 4 4,6 11r 4 4,8 3,9 4,8 12r 4,6 5 4,3 5 12l 4,7 5 4,3 4,9 13r 3,7 4,2 4 4,3 13l 4 4,3 4,3 4,4 14r 4,7 5,3 4,7 5,3 14l 4,6 5 4,6 5,5 15r 4,8 5,2 4,6 5,1 15l 4,5 5,2 4,5 4,9

141
Fortsetzung Tab. 24: Messergebnisse zu den Gesamtlängen des kranialen und
kaudalen Schenkels des Lig. coll. mediale der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. mediale 160° 30°
Tiernummer kranialer Schenkel
kaudaler Schenkel
kranialer Schenkel
kaudaler Schenkel
16r 4,7 5,3 4,7 5,1 16l 4,5 4,8 4,1 4,8 17r 3,7 3,5 3,5 3,5 17l 3,3 3,4 3,3 3,4 18r 3,7 4,4 3,7 4,3 19r 4,2 4,7 3,8 4,4 19l 3,7 4,3 3,2 3,9 20r 3,6 4,4 3,7 4,2 20l 4 4,8 3,9 4,8 21r 4,6 5 4,3 5 21l 4,7 5 4,3 4,9 22r 4,7 5,2 4,5 5 22l 4,6 5,2 4,4 4,9
Mittelwert 4,24 4,68 4,15 4,65 Stabw 0,45 0,50 0,43 0,51
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

142
Tab. 25: Messergebnisse zu den Längen der Ursprungsbereiche des Lig. coll.
laterale und des Lig. coll. mediale der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale Tiernummer 160° 30° 160° 30°
1r 0,9 0,9 0,8 0,8 1l 0,7 0,7 0,5 0,5 2r 0,5 0,5 0,7 0,7 2l 0,8 0,8 0,4 0,4 3r 0,6 0,6 0,4 0,4 4r 0,8 0,8 0,4 0,4 5r 0,7 0,7 0,4 0,4 5l 0,6 0,6 0,3 0,3 6r 0,8 0,8 0,2 0,2 6l 0,5 0,5 0,3 0,3 7r 1 1 0,5 0,5 7l 1 1 0,5 0,5 8r 0,9 0,9 0,6 0,6 8l 0,8 0,8 0,5 0,5 9r 0,5 0,5 0,3 0,3 9l 0,5 0,5 0,7 0,7
10r 0,9 0,9 0,2 0,2 10l 0,5 0,5 0,2 0,2 11r 1 1 0,5 0,5 12r 1 1 0,5 0,5 12l 0,7 0,7 0,6 0,6 13r 0,7 0,7 0,5 0,5 13l 0,6 0,6 0,3 0,3 14r 0,9 0,9 0,6 0,6 14l 0,8 0,8 0,4 0,4 15r 0,7 0,7 0,5 0,5 15l 0,8 0,8 0,6 0,6

143
Fortsetzung Tab. 25: Messergebnisse zu den Längen der Ursprungsbereiche des
Lig. coll. laterale und des Lig. coll. mediale der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale Tiernummer 160° 30° 160° 30°
16r 0,9 0,9 0,5 0,5 16l 0,7 0,7 0,6 0,6 17r 0,4 0,4 0,2 0,2 17l 0,4 0,4 0,2 0,2 18r 0,7 0,7 0,2 0,2 19r 1 1 0,4 0,4 19l 1,2 1,2 0,4 0,4 20r 0,7 0,7 0,2 0,2 20l 0,8 0,8 0,2 0,2 21r 1 1 0,5 0,5 21l 0,7 0,7 0,6 0,6 22r 0,7 0,7 0,5 0,5 22l 0,8 0,8 0,6 0,6
Mittelwert 0,76 0,76 0,42 0,44 Stabw 0,19 0,19 0,18 0,16
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

144
Tab. 26: Messergebnisse zu den Gesamtlängen der freien Teile des kranialen
Schenkels des Lig. coll. laterale und des kranialen und kaudalen Schenkels des Lig.
coll. mediale der Art. cubiti von 22 Hunden bei einer Gelenkwinkelung von 160° und
30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale 160° 30° 160° 30°
Tiernummer kranialer Schenkel
kranialer Schenkel
kranialerSchenkel
kaudalerSchenkel
kranialer Schenkel
kaudaler Schenkel
1r 2,5 2,2 3,2 2,8 3 2,5 1l 2,2 1,7 2,7 3,6 2,5 3,5 2r 3,2 2,9 3,3 4,1 3,5 3,9 2l 2,7 2,6 3,7 4,4 3,3 4,3 3r 2,3 1,9 2,7 2,5 2,8 3 4r 2,8 2,6 3,6 4 3,5 3,9 5r 2,1 1,9 2,9 2,8 2,9 2,8 5l 2,3 2,3 3,1 3,1 3,1 3,3 6r 2,6 2,5 3,4 3,4 3,5 3,6 6l 2,7 2,6 3,6 3,3 3,4 3,3 7r 2,8 2,5 3,5 3,9 3,7 4,1 7l 2,7 2,6 4,5 4,5 4,2 4,4 8r 2,3 1,9 3,2 3,3 3,4 3,4 8l 2,2 2 3 2,9 3 3,4 9r 2,6 2,6 3,2 3,4 3,5 3 9l 2,9 2,5 3,3 3,2 3,2 3,2
10r 1,9 1,9 3,5 3,5 3,3 3,2 10l 2,3 2,8 3,9 3,4 3,3 3,6 11r 1,9 2,2 3,3 3,8 3,3 3,8 12r 2,5 1,9 3,9 3,5 3,7 3,5 12l 2,7 2,6 3,8 3,3 3,7 3,2 13r 2,3 2,4 2,7 3,2 2,8 3,3 13l 2,9 2,5 3,2 3,3 3,4 3,4 14r 3,4 3,2 3,6 3,8 3,7 3,8 14l 3,3 3,1 3,6 4 3,6 4,5 15r 3,1 3,1 3,9 3,8 3,7 3,7 15l 2,6 2,9 3,6 3,5 3,5 3,2

145
Fortsetzung Tab. 26: Messergebnisse zu den Gesamtlängen der freien Teile des
kranialen Schenkels des Lig. coll. laterale und des kranialen und kaudalen Schenkels
des Lig. coll. mediale der Art. cubiti von 22 Hunden bei einer Gelenkwinkelung von
160° und 30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale 160° 30° 160° 30°
Tiernummer kranialer Schenkel
kranialer Schenkel
kranialerSchenkel
kaudalerSchenkel
kranialer Schenkel
kaudaler Schenkel
16r 3 2,9 3,7 4,1 3,7 3,9 16l 3,2 3 3,7 3,8 3,6 3,8 17r 2,4 2,6 3 2,7 2,8 2,7 17l 2,5 1,8 3 2,8 3 2,8 18r 2 2,4 3,4 3,9 3,4 3,8 19r 2,1 2,1 3,5 3,9 3,1 3,6 19l 2,1 2,1 3 3,4 2,9 3 20r 2 2,2 3,4 3,9 3,4 3,7 20l 2 2,1 3,2 4 3,3 4,3 21r 2,5 2,4 3,9 3,5 3,7 3,5 21l 2,8 2,5 3,7 3,5 3,7 3,2 22r 2,9 3,3 3,8 3,7 3,7 3,6 22l 2,6 2,7 3,6 3,5 3,5 3,2
Mittelwert 2,55 2,45 3,42 3,53 3,36 3,5 Stabw 0,40 0,41 0,39 0,46 0,35 0,47
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

146
Tab. 27: Messergebnisse zu den Längen der Ansatzbereiche des kranialen
Schenkels des Lig. coll. laterale und des kranialen und kaudalen Schenkels des Lig.
coll. mediale der Art. cubiti von 22 Hunden bei einer Gelenkwinkelung von 160° und
30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale 160° 30° 160° 30°
Tiernummer kranialer Schenkel
kranialerSchenkel
kranialerSchenkel
kaudalerSchenkel
kranialer Schenkel
kaudalerSchenkel
1r 0,7 0,7 0,1 1,2 0,1 1,2 1l 1,1 1,1 0,8 0,2 0,8 0,2 2r 1,2 1,2 0,3 0,4 0,3 0,4 2l 0,8 0,8 0,5 0,1 0,5 0,1 3r 1,1 1,1 1 1,2 1 1,2 4r 0,5 0,5 0,2 0,4 0,2 0,4 5r 1 1 0,2 0,7 0,2 0,7 5l 0,7 0,7 0,2 0,5 0,2 0,5 6r 1 1 0,2 0,6 0,2 0,6 6l 1 1 0,1 1 0,1 1 7r 1,2 1,2 0,5 0,8 0,5 0,8 7l 1,2 1,2 0,5 0,7 0,5 0,7 8r 1,1 1,1 0,6 1 0,6 1 8l 1,1 1,1 0,6 1,3 0,6 1,3 9r 0,9 0,9 0,5 0,8 0,5 0,8 9l 1 1 0,5 0,7 0,5 0,7
10r 1,2 1,2 0,5 0,8 0,5 0,8 10l 0,8 0,8 0,5 0,8 0,5 0,8 11r 1 1 0,2 0,5 0,2 0,5 12r 1 1 0,2 1 0,2 1 12l 1,2 1,2 0,3 1,1 0,3 1,1 13r 0,9 0,9 0,5 0,5 0,5 0,5 13l 0,5 0,5 0,5 0,7 0,5 0,7 14r 0,7 0,7 0,5 0,9 0,5 0,9 14l 0,9 0,9 0,6 0,6 0,6 0,6 15r 0,6 0,6 0,4 0,9 0,4 0,9 15l 0,9 0,9 0,3 1,1 0,3 1,1

147
Fortsetzung Tab. 27: Messergebnisse zu den Längen der Ansatzbereiche des
kranialen Schenkels des Lig. coll. laterale und des kranialen und kaudalen Schenkels
des Lig. coll. mediale der Art. cubiti von 22 Hunden bei einer Gelenkwinkelung von
160° und 30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale 160° 30° 160° 30°
Tiernummer kranialer Schenkel
kranialerSchenkel
kranialerSchenkel
kaudalerSchenkel
kranialer Schenkel
kaudalerSchenkel
16r 0,9 0,9 0,5 0,7 0,5 0,7 16l 1,1 1,1 0,2 0,4 0,2 0,4 17r 0,3 0,3 0,5 0,6 0,5 0,6 17l 0,4 0,4 0,1 0,4 0,1 0,4 18r 1 1 0,1 0,3 0,1 0,3 19r 0,9 0,9 0,3 0,4 0,3 0,4 19l 1 1 0,3 0,5 0,3 0,5 20r 1 1 0,1 0,3 0,1 0,3 20l 0,9 0,9 0,1 0,3 0,1 0,3 21r 1 1 0,2 1 0,2 1 21l 1,2 1,2 0,3 1,1 0,3 1,1 22r 0,6 0,6 0,4 0,9 0,4 0,9 22l 0,8 0,8 0,3 1,1 0,3 1,1
Mittelwert 0,91 0,91 0,37 0,71 0,37 0,71 Stabw 0,23 0,23 0,21 0,31 0,21 0,31
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

148
Tab. 28: Messergebnisse zu den Längen der Humerusabschnitte des in diesem
Abschnitt noch ungeteilten Lig. coll. laterale und Lig. coll. mediale der Art. cubiti von
22 Hunden bei einer Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale Tiernummer 160° 30° 160° 30°
1r 1,9 1,1 2,1 2,2 1l 1,8 1,5 2,3 1,9 2r 2,2 1,5 2 2,2 2l 1,8 1,5 2,3 2 3r 1,1 1,5 1,7 1,8 4r 1,7 1,7 2 1,8 5r 1,5 1,3 1,6 1,6 5l 1,5 1,4 1,7 1,4 6r 1,7 1,4 1,6 1,6 6l 1,8 1,7 2 1,9 7r 2 1,8 2,2 2,2 7l 1,9 1,9 2,4 2,3 8r 1,7 1,5 2 2 8l 1,6 1,6 2 2,1 9r 1,6 1,5 2 2,1 9l 1,7 1,4 1,8 1,8
10r 1,6 1,6 2,5 1,6 10l 1,4 1,8 1,7 1,9 11r 2,1 1,7 1,9 2 12r 2,1 1,7 2 2,1 12l 2 1,7 2,2 2 13r 2 1,8 1,8 1,9 13l 1,7 1,4 1,8 1,9 14r 2,4 1,8 2 2,2 14l 2,1 1,7 2 2 15r 2,1 1,8 2,3 2,2 15l 2 2 2,1 2,1

149
Fortsetzung Tab. 28: Messergebnisse zu den Längen der Humerusabschnitte des
in diesem Abschnitt noch ungeteilten Lig. coll. laterale und Lig. coll. mediale der Art.
cubiti von 22 Hunden bei einer Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale Tiernummer 160° 30° 160° 30°
16r 2,1 1,7 2 2 16l 2,3 1,7 2 2 17r 1,3 1,1 1,5 1,6 17l 1,5 1,2 1,3 1,5 18r 1,6 1,6 1,7 1,7 19r 1,5 1,4 1,8 1,9 19l 2 1,8 1,7 1,7 20r 1,6 1,6 1,7 1,7 20l 1,5 1,8 1,9 1,9 21r 2,1 1,7 2 2,1 21l 2 1,7 2,2 2 22r 2,1 1,8 2,3 2,2 22l 2 2 2,1 2,1
Mittelwert 1,82 1,61 1,96 1,93 Stabw 0,29 0,22 0,26 0,22
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

150
Tab. 29: Messergebnisse zu den Längen der freien Teile der Humerusabschnitte
des in diesem Abschnitt noch ungeteilten Lig. coll. laterale und Lig. coll. mediale der
Art. cubiti von 22 Hunden bei einer Gelenkwinkelung von 160° und 30°, gemessen in
cm.
Lig. coll. laterale Lig. coll. mediale Tiernummer 160° 30° 160° 30°
1r 1 0,2 1,3 1,4 1l 1,1 0,8 1,8 1,4 2r 1,7 1 1,3 1,5 2l 1 0,7 1,9 1,6 3r 0,5 0,9 1,3 1,4 4r 0,9 0,9 1,6 1,4 5r 0,8 0,6 1,2 1,2 5l 0,9 0,8 1,4 1,1 6r 1,1 0,6 1,4 1,2 6l 1,3 1,2 1,7 1,6 7r 1 0,8 1,7 1,7 7l 0,9 0,9 1,9 1,8 8r 0,8 0,6 1,4 1,4 8l 0,8 0,8 1,5 1,6 9r 1,1 1 1,7 1,8 9l 1,2 0,9 1,1 1,1
10r 0,7 0,7 2,3 1,4 10l 0,9 1,3 1,5 1,7 11r 1,1 0,7 1,4 1,5 12r 1,1 0,7 1,5 1,6 12l 1,3 1 1,6 1,4 13r 1,3 1,1 1,3 1,4 13l 1,1 0,8 1,5 1,6 14r 1,5 0,9 1,4 1,6 14l 1,3 0,9 1,6 1,6 15r 1,4 1,1 1,8 1,7 15l 1,2 1,2 1,5 1,5

151
Fortsetzung Tab. 29: Messergebnisse zu den Längen der freien Teile der Humerusabschnitte des in diesem Abschnitt noch ungeteilten Lig. coll. laterale und
Lig. coll. mediale der Art. cubiti von 22 Hunden bei einer Gelenkwinkelung von 160°
und 30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale Tiernummer 160° 30° 160° 30°
16r 1,2 0,8 1,5 1,5 16l 1,6 1 1,4 1,4 17r 0,9 0,7 1,3 1,4 17l 1,1 0,8 1,1 1,3 18r 0,9 0,9 1,5 1,5 19r 0,5 0,4 1,4 1,5 19l 0,8 0,6 1,3 1,3 20r 0,9 0,9 1,5 1,5 20l 0,7 1 1,7 1,7 21r 1,1 0,7 1,5 1,6 21l 1,3 1 1,6 1,4 22r 1,6 1,1 1,8 1,7 22l 1,2 1,2 1,5 1,5
Mittelwert 1,07 0,85 1,52 1,49 Stabw 0,28 0,22 0,23 0,17
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

152
Tab. 30: Messergebnisse zu den Längen der Unterarmabschnitte des kranialen
und kaudalen Schenkels des Lig. coll. laterale der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale 160° 30°
Tiernummer kranialer Schenkel
kaudaler Schenkel
kranialer Schenkel
kaudaler Schenkel
1r 2,2 2,9 2,7 2,7 1l 2,2 2,6 2 2,6 2r 2,7 3,1 3,1 3,1 2l 2,5 3,1 2,7 3 3r 2,9 3 1,6 3 4r 2,4 2,8 2,2 2,8 5r 2,3 3 2,3 3 5l 1,6 2,6 2,2 2,6 6r 2,7 3 2,9 3 6l 2,6 3 2,4 3 7r 3 3,5 3,1 3,5 7l 3 3,1 3,1 3,1 8r 2,6 2,5 2,4 2,5 8l 2,5 3,2 2,3 2,8 9r 2,4 3 2,5 2,5 9l 2,7 3 2,6 3
10r 2,4 2,5 2,4 2,4 10l 2,2 2,3 2,3 2,3 11r 1,8 2,6 2,5 2,6 12r 2,4 2,8 2,2 3 12l 2,6 2,6 2,8 2,5 13r 1,9 1,7 2,2 1,8 13l 2,3 2,7 2,2 1,6 14r 2,6 2,7 3 3 14l 2,9 2,7 3,1 2,9 15r 2,3 2,9 2,6 3,2 15l 2,3 2,8 2,6 2,8

153
Fortsetzung Tab. 30: Messergebnisse zu den Längen der Unterarmabschnitte des
kranialen und kaudalen Schenkels des Lig. coll. laterale der Art. cubiti von 22
Hunden bei einer Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale 160° 30°
Tiernummer kranialer Schenkel
kaudaler Schenkel
kranialer Schenkel
kaudaler Schenkel
16r 2,7 2,8 3 2,8 16l 2,7 2,9 3,1 3 17r 1,8 2,3 2,2 2,2 17l 1,8 2,3 1,4 2,1 18r 2,1 2,5 2,5 2,2 19r 2,5 2,8 2,6 2,7 19l 2,3 2,5 2,5 2,5 20r 2,1 2,5 2,5 2,2 20l 2,4 2,6 2,8 2,6 21r 2,4 2,8 2,2 3 21l 2,6 2,6 2,8 2,5 22r 2,1 2,9 2,6 3,2 22l 2,3 2,8 2,6 2,8
Mittelwert 2,40 2,75 2,52 2,70 Stabw 0,33 0,31 0,39 0,39
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

154
Tab. 31: Messergebnisse zu den Längen der Unterarmabschnitte des kranialen
und kaudalen Schenkels des Lig. coll. mediale der Art. cubiti von 22 Hunden bei
einer Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. mediale 160° 30°
Tiernummer kranialer Schenkel
kaudaler Schenkel
kranialer Schenkel
kaudaler Schenkel
1r 2 2,7 1,7 2,3 1l 1,7 2 1,9 2,3 2r 2,3 3,2 2,3 2,8 2l 2,3 2,6 2,2 2,8 3r 2,6 2,4 2,4 2,8 4r 2,2 2,8 2,3 2,9 5r 1,9 2,3 1,9 2,3 5l 1,9 2,2 2,2 2,7 6r 2,2 2,6 2,3 2,8 6l 2 2,6 1,9 2,7 7r 2,3 3 2,5 3,2 7l 3,1 3,3 2,9 3,3 8r 2,4 2,9 2,6 3 8l 2,1 2,7 2,3 3,1 9r 2 2,5 2,2 2 9l 2,7 2,8 2,5 2,8
10r 1,7 2 2,6 2,6 10l 2,9 2,7 2,1 2,7 11r 2,1 2,9 1,9 2,8 12r 2,6 3 2,2 2,9 12l 2,5 2,8 2,3 2,9 13r 1,9 2,4 2,1 2,4 13l 2,2 2,5 2,4 2,5 14r 2,7 3,33 2,5 3,1 14l 2,6 2,9 2,6 2,5 15r 2,5 3,1 2,4 2,9 15l 2,4 3,2 2,4 2,8

155
Fortsetzung Tab. 31: Messergebnisse zu den Längen der Unterarmabschnitte des
kranialen und kaudalen Schenkels der Lig. coll. mediale der Art. cubiti von 22
Hunden bei einer Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. mediale 160° 30°
Tiernummer kranialer Schenkel
kaudaler Schenkel
kranialer Schenkel
kaudaler Schenkel
16r 2,7 2,8 2,7 3,1 16l 2,5 2 2,1 2,8 17r 2,2 2 1,9 1,9 17l 2 2,1 1,8 1,9 18r 2 2,6 2 2,6 19r 2,6 2,9 1,9 2,5 19l 2 2,6 1,5 2,2 20r 1,9 2,7 2 2,5 20l 2,1 2,9 2 2,9 21r 2,6 3 2,2 2,9 21l 2,5 2,8 2,3 2,9 22r 2,4 2,9 2,3 2,8 22l 2,5 3,1 2,3 2,8
Mittelwert 2,30 2,70 2,22 2,69 Stabw 0,33 0,36 0,29 0,33
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

156
Tab. 32: Messergebnisse zu den Längen der freien Teile der Unterarmabschnitte des kranialen Schenkels der Lig. coll. laterale und des kranialen und kaudalen
Schenkels der Lig. coll. mediale der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale 160° 30° 160° 30°
Tiernummer kranialer Schenkel
kranialerSchenkel
kranialerSchenkel
kaudalerSchenkel
kranialer Schenkel
kaudalerSchenkel
1r 1,5 2 1,9 1,5 1,6 1,1 1l 1,1 0,9 0,9 1,8 1,1 2,1 2r 1,5 1,9 2 2,8 2 2,4 2l 1,7 1,9 1,7 2,5 1,7 2,7 3r 1,8 0,4 1,6 1,2 1,4 1,6 4r 1,9 1,7 2 2,4 2,1 2,5 5r 1,3 1,3 1,7 1,5 1,7 1,6 5l 0,9 1,5 1,7 1,7 2 2,2 6r 1,7 1,9 2 2 2,1 2,2 6l 1,6 1,4 1,9 1,6 1,8 1,7 7r 1,8 1,9 1,8 2,2 2 2,4 7l 1,8 1,9 2,6 2,6 2,4 2,6 8r 0,5 1,3 1,8 1,9 2 2 8l 0,4 1,2 1,5 1,4 1,7 1,8 9r 1,5 1,6 1,5 1,7 1,7 1,2 9l 1,7 1,6 2,2 2,1 2 2,1
10r 1,2 1,2 1,2 1,2 2,1 1,8 10l 1,4 1,5 2,4 1,9 1,6 1,9 11r 0,8 1,5 1,9 2,4 1,7 2,3 12r 1,4 1,2 2,4 2 2 1,9 12l 1,4 1,6 2,2 1,7 2 1,8 13r 1 1,3 1,4 1,9 1,6 1,9 13l 1,8 1,7 1,7 1,8 1,9 1,8 14r 1,9 2,3 2,2 2,4 2 2,3 14l 2 2,2 2 2,3 2 1,9 15r 1,7 2 2,1 2,2 2 2 15l 1,4 1,7 2,1 2,1 2,1 1,7

157
Fortsetzung Tab. 32: Messergebnisse zu den Längen der freien Teile der Unterarmabschnitte des kranialen Schenkels des Lig. coll. laterale und der
kranialen und kaudalen Schenkel des Lig. coll. mediale der Art. cubiti von 22 Hunden
bei einer Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale 160° 30° 160° 30°
Tiernummer kranialer Schenkel
kranialerSchenkel
kranialerSchenkel
kaudalerSchenkel
kranialer Schenkel
kaudalerSchenkel
16r 1,8 2,1 2,2 2,1 2,2 2,4 16l 1,6 2 2,3 1,8 1,9 2,4 17r 1,5 1,9 1,7 1,4 1,4 1,3 17l 1,4 1 1,9 1,7 1,7 1,5 18r 1,1 1,5 1,9 2,3 1,9 2,3 19r 1,6 1,7 2,3 2,5 1,6 2,1 19l 1,3 1,5 1,7 2,1 1,2 1,7 20r 1,1 1,5 1,8 2,4 1,9 2,3 20l 1,5 1,9 2 2,6 1,9 2,6 21r 1,4 1,2 2,4 2 2 1,9 21l 1,4 1,6 2,2 1,7 2 1,8 22r 1,5 2 2 2 1,9 1,9 22l 1,5 1,8 2,2 2 2 1,7
Mittelwert 1,44 1,61 1,93 1,99 1,85 1,99 Stabw 0,36 0,38 0,34 0,39 0,27 0,38
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

158
Tab. 33: Messergebnisse zur Länge vom Ursprung des ungeteilten Bandes bis zur Aufspaltung in zwei Schenkel des Lig. coll. laterale und des Lig. coll. mediale
der Art. cubiti von 22 Hunden bei einer Gelenkwinkelung von 160° und 30°,
gemessen in cm.
Lig. coll. laterale Lig. coll. mediale Tiernummer 160° 30° 160° 30°
1r 1,6 1,6 2,6 2,5 1l 1,7 1,6 2,5 2,2 2r 2,2 1,5 2 2,2 2l 1,5 1,7 2,9 2,4 3r 1,3 1,7 2,6 2,8 4r 1,7 1,8 2,8 2,4 5r 1,5 1,5 1,6 1,6 5l 1,6 1,6 1,7 1,7 6r 1,2 1,4 1,9 2,1 6l 1,6 1,6 2 1,9 7r 1,7 2 2,5 2,5 7l 1,9 2,2 2,6 2,4 8r 2,2 2,1 2,3 2,5 8l 1,8 1,8 2,2 2,3 9r 2 1,9 2,3 2,3 9l 1,8 2,1 2,1 2,3
10r 1,9 1,9 2,1 1,8 10l 1,7 1,7 1,8 1,8 11r 2 2,3 2 1,9 12r 2,3 2,1 2 2,3 12l 2,2 2 2,7 2,5 13r 2 2,2 1,9 2 13l 1,7 2 2,3 2,3 14r 2,3 2,2 2 2,2 14l 2,4 2,3 2,3 2,8 15r 2,1 2,3 2,5 2,3 15l 2,3 2,1 2,5 2

159
Fortsetzung Tab. 33: Messergebnisse zur Länge vom Ursprung des ungeteilten Bandes bis zur Aufspaltung in zwei Schenkel des Lig. coll. laterale und des Lig.
coll. mediale der Art. cubiti von 22 Hunden bei einer Gelenkwinkelung von 160° und
30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale Tiernummer 160° 30° 160° 30°
16r 2 2 2,5 2,5 16l 2,2 2,1 2,5 2,3 17r 1,4 1,6 1,9 1,7 17l 1,5 1,6 2 2 18r 1,8 1,9 2,1 2,1 19r 2 2,1 2,5 2,4 19l 1,8 2,3 1,9 1,9 20r 1,8 1,9 2,1 2,1 20l 1,7 1,8 2 2,1 21r 2,3 2,1 2 2,3 21l 2,2 2 2,7 2,5 22r 2,1 2,1 1,9 2,3 22l 2,2 2 2,7 2,4
Mittelwert 1,88 1,92 2,24 2,22 Stabw 0,31 0,26 0,33 0,28
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

160
Tab. 34: Messergebnisse zu den Breiten im Ursprungsbereich des kranialen und
kaudalen Schenkels des Lig. coll. laterale und des Lig. coll. mediale der Art. cubiti
von 22 Hunden in den Gelenkwinkelungen 160° und 30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale 160° 30° 160° 30°
Tiernummer kranialer Schenkel
kaudalerSchenkel
kranialerSchenkel
kaudalerSchenkel
ungeteiltes Band
ungeteiltes Band
1r 0,8 1,5 0,8 1,5 0,8 0,8 1l 0,8 1,5 0,8 1,5 0,7 0,7 2r 0,5 1,8 0,5 1,8 0,8 0,8 2l 0,6 1,7 0,6 1,7 0,6 0,6 3r 0,7 1,8 0,7 1,8 0,6 0,6 4r 0,6 1,5 0,6 1,5 0,4 0,4 5r 0,6 1,5 0,6 1,5 0,5 0,5 5l 0,4 1,5 0,4 1,5 0,4 0,4 6r 0,6 1,7 0,6 1,7 0,5 0,5 6l 0,6 1,6 0,6 1,6 0,5 0,5 7r 0,9 2,3 0,9 2,3 0,5 0,5 7l 0,7 1,9 0,7 1,9 0,7 0,7 8r 0,6 1,8 0,6 1,8 0,4 0,4 8l 0,6 1,8 0,6 1,8 0,5 0,5 9r 0,5 1,4 0,4 1,4 0,4 0,4 9l 0,6 1,5 0,6 1,5 0,5 0,5
10r 0,4 1,4 0,4 1,4 0,4 0,4 10l 0,4 1 0,4 1 0,3 0,3 11r 0,7 1,2 0,7 1,2 0,5 0,5 12r 0,5 1,3 0,5 1,3 0,5 0,5 12l 0,7 1,2 0,5 1,2 0,6 0,6 13r 0,6 1 0,6 1 0,5 0,5 13l 0,7 1,3 0,7 1,3 0,5 0,5 14r 0,7 1,5 0,7 1,5 0,5 0,5 14l 0,8 1,3 0,8 1,3 0,4 0,4 15r 0,8 1,6 0,8 1,6 0,6 0,6 15l 0,7 1,5 0,7 1,5 0,6 0,6

161
Fortsetzung Tab. 34: Messergebnisse zu den Breiten im Ursprungsbereich des
kranialen und kaudalen Schenkels des Lig. coll. laterale und des Lig. coll. mediale
der Art. cubiti von 22 Hunden in den Gelenkwinkelungen 160° und 30°, gemessen in
cm.
Lig. coll laterale Lig. coll.mediale 160° 30° 160° 30°
Tiernummer kranialer
Schenkel kaudalerSchenkel
kranialerSchenkel
kaudalerSchenkel
ungeteiltes Band
ungeteiltes Band
16r 0,8 1,4 0,8 1,4 0,6 0,6 16l 0,6 1,2 0,6 1,2 0,4 0,4 17r 0,4 1 0,4 1 0,4 0,4 17l 0,5 0,9 0,5 0,9 0,4 0,4 18r 0,5 1,3 0,5 1,3 0,6 0,6 19r 0,5 1,2 0,5 1,2 0,5 0,5 19l 0,5 1,1 0,5 1,1 0,6 0,6 20r 0,5 1,3 0,5 1,3 0,6 0,6 20l 0,4 1,2 0,4 1,2 0,6 0,6 21r 0,5 1,3 0,5 1,3 0,5 0,5 21l 0,7 1,2 0,5 1,2 0,6 0,6 22r 0,8 1,6 0,8 1,6 0,5 0,5 22l 0,7 1,5 0,7 1,5 0,6 0,6
Mittelwert 0,61 1,43 0,60 1,43 0,53 0,53 Stabw 0,13 0,28 0,14 0,28 0,11 0,11
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

162
Tab. 35: Messergebnisse zu den Breiten des Lig. coll. laterale und des Lig. coll.
mediale in Höhe des Gelenkspaltes der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale Lig. coll. mediale Tiernummer 160° 30° 160° 30°
1r 1 0,8 0,8 0,7 1l 1,1 1 1,1 0,9 2r 0,7 0,7 0,9 0,8 2l 0,6 0,8 0,7 0,6 3r 0,7 0,8 0,6 0,5 4r 0,8 0,8 0,7 0,6 5r 0,6 0,6 0,8 0,8 5l 0,7 0,7 0,4 0,4 6r 0,8 0,8 0,7 0,6 6l 0,7 0,7 0,8 0,8 7r 0,9 0,7 1 0,9 7l 0,9 0,9 1 1 8r 0,7 0,6 0,7 0,7 8l 0,7 0,7 1 0,8 9r 0,7 0,6 0,7 0,6 9l 0,6 0,4 0,6 0,6
10r 0,6 0,8 0,8 0,7 10l 0,8 0,8 1 0,5 11r 0,8 0,6 1,2 0,8 12r 0,7 0,4 0,8 0,8 12l 0,8 0,8 0,9 0,9 13r 0,7 0,8 0,7 0,6 13l 0,6 0,5 0,7 0,7 14r 0,7 0,5 0,9 0,5 14l 0,8 0,8 0,6 0,5 15r 0,7 0,8 0,8 0,7 15l 0,8 0,8 0,7 0,6

163
Fortsetzung Tab. 35: Messergebnisse zu den Breiten des Lig. coll. laterale und des
Lig. coll. mediale in Höhe des Gelenkspaltes der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll laterale Lig. coll.mediale Tiernummer 160° 30° 160° 30°
16r 0,7 0,6 0,5 0,5 16l 0,8 0,6 0,9 0,8 17r 0,4 0,4 0,3 0,5 17l 0,5 0,5 0,5 0,4 18r 0,6 0,4 0,7 0,6 19r 0,5 0,5 0,8 0,7 19l 0,4 0,4 0,6 0,4 20r 0,6 0,4 0,7 0,6 20l 0,8 0,4 0,6 0,5 21r 0,7 0,4 0,8 0,8 21l 0,8 0,8 0,9 0,9 22r 0,7 0,8 0,8 0,7 22l 0,8 0,8 0,7 0,6
Mittelwert 0,71 0,66 0,76 0,67 Stabw 0,14 0,17 0,18 0,15
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

164
Tab. 36: Messergebnisse zu den Breiten im Ansatzbereich des kranialen und
kaudalen Schenkels des Lig. coll. laterale der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale Tiernummer 160° 30°
kranialer Schenkel
kaudaler Schenkel
kranialer Schenkel
kaudaler Schenkel
1r 0,9 2,8 0,9 2,8 1l 0,9 2,5 0,9 2,5 2r 0,5 2,9 0,5 2,9 2l 0,6 3 0,6 3 3r 0,6 3,3 0,6 3,3 4r 0,8 2,4 0,8 2,4 5r 0,5 2,3 0,5 2,3 5l 0,4 2,7 0,4 2,7 6r 0,7 3 0,7 3 6l 0,7 3 0,7 3 7r 0,9 3 0,9 3 7l 0,8 3,1 0,8 3,1 8r 0,7 3 0,7 3 8l 0,6 3,1 0,6 3,1 9r 0,6 2,5 0,6 2,5 9l 0,6 2,6 0,6 2,6
10r 0,5 2,8 0,5 2,8 10l 0,5 2 0,5 2 11r 0,7 2,1 0,7 2,1 12r 0,6 2,2 0,6 2,2 12l 0,7 2,3 0,7 2,3 13r 0,7 2,5 0,7 2,5 13l 0,6 2 0,6 2 14r 0,7 3,5 0,7 3,5 14l 0,7 2,7 0,7 2,7 15r 0,8 2,8 0,8 2,8 15l 0,8 2,5 0,8 2,5

165
Fortsetzung Tab. 36: Messergebnisse zu den Breiten im Ansatzbereich des
kranialen und kaudalen Schenkels der Lig. coll. laterale der Art. cubiti von 22 Hunden
bei einer Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. laterale Tiernummer 160° 30°
kranialer Schenkel
kaudaler Schenkel
kranialer Schenkel
kaudaler Schenkel
16r 0,8 2,5 0,8 2,5 16l 0,9 3,1 0,9 3,1 17r 0,5 2 0,5 2 17l 0,4 2 0,4 2 18r 0,6 2,2 0,6 2,2 19r 0,6 2,9 0,6 2,9 19l 0,6 2,3 0,6 2,3 20r 0,6 2 0,6 2 20l 0,5 2,2 0,5 2,2 21r 0,6 2,2 0,6 2,2 21l 0,7 2,3 0,7 2,3 22r 0,8 2,8 0,8 2,8 22l 0,8 2,5 0,8 2,5
Mittelwert 0,66 2,59 0,66 2,59 Stabw 0,14 0,40 0,14 0,40
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

166
Tab. 36a: Messergebnisse zu den Breiten im Ansatzbereich des kranialen und
kaudalen Schenkels des Lig. coll. mediale der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. mediale 160° 30°
Tiernummer kranialer Schenkel
kaudaler Schenkel
kranialer Schenkel
kaudaler Schenkel
1r 0,2 0,4 0,2 0,4 1l 0,3 0,4 0,5 0,5 2r 0,5 0,3 0,5 0,3 2l 0,4 0,5 0,4 0,5 3r 0,3 0,4 0,3 0,4 4r 0,2 0,4 0,2 0,4 5r 0,2 0,3 0,2 0,3 5l 0,2 0,4 0,2 0,4 6r 0,2 0,2 0,2 0,2 6l 0,1 0,3 0,1 0,3 7r 0,4 0,2 0,4 0,2 7l 0,3 0,3 0,3 0,3 8r 0,3 0,4 0,3 0,4 8l 0,6 0,4 0,6 0,4 9r 0,3 0,3 0,3 0,3 9l 0,2 0,3 0,2 0,3
10r 0,2 0,3 0,2 0,3 10l 0,3 0,3 0,3 0,3 11r 0,2 0,3 0,2 0,3 12r 0,2 0,3 0,2 0,3 12l 0,3 0,4 0,3 0,4 13r 0,5 0,4 0,5 0,4 13l 0,3 0,4 0,3 0,4 14r 0,3 0,4 0,3 0,4 14l 0,3 0,5 0,3 0,5 15r 0,4 0,5 0,4 0,5 15l 0,3 0,5 0,3 0,5

167
Fortsetzung Tab. 36a: Messergebnisse zu den Breiten im Ansatzbereich des
kranialen und kaudalen Schenkels des Lig. coll. mediale der Art. cubiti von 22
Hunden bei einer Gelenkwinkelung von 160° und 30°, gemessen in cm.
Lig. coll. mediale 160° 30°
Tiernummer kranialer Schenkel
kaudaler Schenkel
kranialer Schenkel
kaudaler Schenkel
16r 0,2 0,4 0,2 0,4 16l 0,2 0,3 0,2 0,3 17r 0,3 0,4 0,3 0,4 17l 0,2 0,3 0,2 0,3 18r 0,3 0,3 0,3 0,3 19r 0,3 0,3 0,3 0,3 19l 0,4 0,3 0,4 0,3 20r 0,3 0,3 0,3 0,3 20l 0,2 0,5 0,2 0,5 21r 0,2 0,3 0,2 0,3 21l 0,3 0,4 0,3 0,4 22r 0,4 0,5 0,4 0,5 22l 0,3 0,5 0,3 0,5
Mittelwert 0,29 0,37 0,30 0,37 Stabw 0,10 0,08 0,11 0,09
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

168
Tab. 37: Messergebnisse zu den Gesamtlängen des Lig. anulare radii
(Gelenkwinkelung 160°), Lig. olecrani (Gelenkwinkelung 30°) und des Lig. obliquum
(Gelenkwinkelung 160°) der Art. cubiti von 22 Hunden bei einer Gelenkwinkelung von
160°, gemessen in cm.
Lig. obliquum Schenkel zum
Tiernummer Lig. anulare radii Lig. olecrani M. biceps brachii Lig. coll. mediale 1r 2,8 3,2 3,0 3,0 1l 2,8 3,0 2,6 2,8 2r 3,0 3,8 2l 3,6 3,2 3r 2,3 3,6 3,4 4r 2,7 3,2 5r 2,2 2,8 2,3 2,6 5l 2,1 2,9 6r 2,7 3,4 6l 2,4 3,0 7r 2,4 3,7 3,7 3,7 7l 3,0 4,1 3,5 3,9 8r 2,8 3,3 3,5 3,7 8l 2,6 3,6 4,1 9r 2,2 3,2 3,9 3,9 9l 2,8 3,1 3,3 3,9
10r 2,9 2,9 3,4 3,5 10l 3,0 3,4 2,8 2,9 11r 2,7 3,6 3,0 3,3 12r 2,4 3,8 3,4 3,8 12l 2,7 3,8 3,6 3,8 13r 3,6 13l 2,5 3,4 14r 3,0 3,5 14l 2,4 3,9 15r 2,3 3,3 15l 2,7 3,6

169
Fortsetzung Tab. 37: Messergebnisse zu den Gesamtlängen des Lig. anulare radii
(Gelenkwinkelung 160°), Lig. olecrani (Gelenkwinkelung 30°) und des Lig. obliquum
(Gelenkwinkelung 160°) der Art. cubiti von 22 Hunden bei einer Gelenkwinkelung von
160°, gemessen in cm.
Lig. obliquum Schenkel zum
Tiernummer Lig. anulare radii Lig. olecrani M. biceps brachii Lig. coll. mediale 16r 2,8 3,5 3,6 3,7 16l 2,9 3,7 3,2 3,3 17r 1,4 1,9 17l 1,5 2,5 2,8 2,8 18r 2,0 2,9 3,4 3,3 19r 2,0 2,8 2,7 2,9 19l 2,3 2,6 2,7 2,5 20r 1,9 2,9 3,4 3,5 20l 2,0 2,8 3,2 3,6 21r 2,0 2,9 3,5 3,7 21l 2,0 2,8 22r 2,7 3,5 22l 2,3 3,3
Mittelwert 2,26 3,20 3,24 3,4 Stabw 0,45 0,48
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

170
Tab. 38: Messergebnisse zu den Breiten im Ursprungsbereich des Lig. anulare
radii (Gelenkwinkelung 160°), Lig. olecrani (Gelenkwinkelung 30°) und des Lig.
obliquum (Gelenkwinkelung 160°) der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160°, gemessen in cm.
Tiernummer Lig. anulare radii Lig. olecrani Lig. obliquum 1r 0,5 0,9 0,5 1l 0,4 0,9 0,4 2r 0,5 0,9 2l 0,4 1,1 3r 0,5 0,4 4r 0,3 1,0 5r 0,2 0,7 0,3 5l 0,2 0,6 6r 0,2 1,0 6l 0,3 0,9 7r 0,4 1,4 0,4 7l 0,4 1,0 0,5 8r 0,4 1,0 0,4 8l 0,4 0,9 0,5 9r 0,3 0,4 0,3 9l 0,4 1,0 0,5
10r 0,3 1,0 0,3 10l 0,2 1,3 0,2 11r 0,4 0,9 0,6 12r 0,2 1,0 0,3 12l 0,2 0,9 0,4 13r 0,3 1,0 0,3 13l 0,4 1,0 14r 0,5 0,7 14l 0,5 0,9 15r 0,3 0,9 15l 0,2 1,1

171
Fortsetzung Tab. 38: Messergebnisse zu den Breiten im Ursprungsbereich des
Lig. anulare radii (Gelenkwinkelung 160°), Lig. olecrani (Gelenkwinkelung 30°) und
des Lig. obliquum (Gelenkwinkelung 160°) der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160°, gemessen in cm.
Tiernummer Lig. anulare radii Lig. olecrani Lig. obliquum 16r 0,3 0,8 0,3 16l 0,3 1,0 0,3 17r 0,3 0,6 0,2 17l 0,3 0,4 0,3 18r 0,2 0,8 0,2 19r 0,3, 0,8 0,2 19l 0,4 0,8 0,3 20r 0,3 0,7 0,3 20l 0,4 0,8 0,1 21r 0,2 0,6 0,2 21l 0,2 0,9 22r 0,5 0,7 22l 0,3 1,0
Mittelwert 0,33 0,88 0,33 Stabw 0,1 0,21
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

172
Tab. 39: Messergebnisse zu den Breiten in Höhe des Gelenkspalts des Lig.
anulare radii (Gelenkwinkelung 160°), Lig. olecrani (Gelenkwinkelung 30°) und des
Lig. obliquum (Gelenkwinkelung 160°) der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160°, gemessen in cm.
Tiernummer Lig. anulare radii Lig. olecrani Lig. obliquum 1r 0,4 0,3 0,4 1l 0,5 0,4 0,25 2r 0,5 0,3 2l 0,3 0,3 3r 0,3 0,3 0,3 4r 0,2 0,5 5r 0,2 0,4 0,1 5l 0,2 0,2 6r 0,2 0,4 6l 0,3 0,5 7r 0,4 0,4 0,3 7l 0,4 0,5 0,3 8r 0,5 0,3 0,3 8l 0,4 0,3 0,3 9r 0,4 0,4 0,3 9l 0,4 0,5 0,2
10r 0,3 0,4 0,3 10l 0,3 0,4 0,2 11r 0,4 0,3 0,3 12r 0,4 0,4 0,3 12l 0,2 0,3 0,2 13r 0,3 0,5 0,2 13l 0,3 0,3 14r 0,4 0,2 14l 0,4 0,5 15r 0,2 0,3 15l 0,2 0,3

173
Fortsetzung Tab. 39: Messergebnisse zu den Breiten in Höhe des Gelenkspalts
des Lig. anulare radii (Gelenkwinkelung 160°), Lig. olecrani (Gelenkwinkelung 30°)
und des Lig. obliquum (Gelenkwinkelung 160°) der Art. cubiti von 22 Hunden bei
einer Gelenkwinkelung von 160°, gemessen in cm.
Tiernummer Lig. anulare radii Lig. olecrani Lig. obliquum 16r 0,4 0,4 0,2 16l 0,4 0,3 0,2 17r 0,3 0,3 0,2 17l 0,3 0,2 0,3 18r 0,3 0,3 0,2 18l 0,3 0,4 0,2 19r 0,3 0,4 0,2 19l 0,3 0,3 0,2 20r 0,2 0,3 0,1 20l 0,4 0,4 0,3 21r 0,2 0,3 0,1 21l 0,2 0,3 22r 0,2 0,3 22l 0,2 0,3
Mittelwert 0,32 0,35 0,24 Stabw 0,09 0,08
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

174
Tab. 40: Messergebnisse zu den Breiten des Ansatzbereiches des Lig. anulare
radii (Gelenkwinkelung 160°), Lig. olecrani (Gelenkwinkelung 30°) und des Lig.
obliquum (Gelenkwinkelung 160°) der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160°, gemessen in cm.
Lig. obliquum Schenkel zum
Tiernummer Lig. anulare radii Lig. olecrani M. biceps rachii Lig. coll. mediale 1r 0,5 0,7 0,2 0,3 1l 0,5 0,8 0,2 0,3 2r 0,3 0,9 2l 0,6 0,5 3r 0,3 0,9 0,4 0,5 4r 0,2 1 5r 0,2 0,8 0,1 0,3 5l 0,3 1 6r 0,2 1 6l 0,3 0,7 7r 0,4 0,8 0,3 0,4 7l 0,3 1,5 0,2 0,4 8r 0,6 0,8 0,3 0,4 8l 0,5 0,8 0,2 0,4 9r 0,3 0,7 0,2 0,3 9l 0,4 1,1 0,2 0,3
10r 0,3 1 0,2 0,4 10l 0,4 1 0,1 0,2 11r 0,3 1 0,3 0,3 12r 0,4 1 0,1 0,2 12l 0,4 0,7 0,2 0,3 13r 0,2 1 0,2 0,2 13l 0,4 0,7 14r 0,5 1 14l 0,3 1,1 15r 0,2 0,6 15l 0,3 0,6

175
Fortsetzung Tab. 40: Messergebnisse zu den Breiten des Ansatzbereiches des
Lig. anulare radii (Gelenkwinkelung 160°), Lig. olecrani (Gelenkwinkelung 30°) und
des Lig. obliquum (Gelenkwinkelung 160°) der Art. cubiti von 22 Hunden bei einer
Gelenkwinkelung von 160°, gemessen in cm.
Lig. obliquum Schenkel zum
Tiernummer Lig. anulare radii Lig. olecrani M. biceps brachii Lig. coll. mediale16r 0,4 0,7 0,2 0,2 16l 0,3 0,8 0,2 0,3 17r 0,4 0,6 0,2 0,2 17l 0,3 0,4 0,2 0,2 18r 0,2 0,8 0,2 0,4 19r 0,2 0,6 0,2 0,4 19l 0,2 0,5 0,1 0,2 20r 0,2 0,8 0,1 0,4 20l 0,3 0,7 0,2 0,3 21r 0,3 0,7 0,1 0,2 21l 0,3 0,8 22r 0,3 0,9 22l 0,2 0,6
Mittelwert 0,33 0,82 0,20 0,31 Stabw 0,11 0,21
r = rechte Extremität
l = linke Extremität
Stabw = Standardabweichung

176
Danksagung Herrn Prof. Dr. Helmut Waibl danke ich sehr herzlich für die Überlassung des
Themas und seine große Geduld und Hilfe.
Herrn Prof. Dr. Wilfried Meyer gilt mein herzlicher Dank für die allzeit gewährte und
freundliche Hilfe speziell im histologischen Teil der Arbeit und für so manches
persönliche Wort.
Herrn Dr. Rüdiger Koch möchte ich für die Unterstützung bei der Anfertigung dieser
Arbeit und das Erstellen der Fotos sehr danken.
Herrn Dr. Karl Rohn (Institut für Biometrie und Epidemiologie) möchte ich danken für
die freundliche Unterstützung bei der statistischen Auswertung der Ergebnisse.
Frau Dr. Elisabeth Engelke und Frau Dr. Anke Schnapper danke ich herzlich für die
stets hilfsbereite Unterstützung zu allen PC Fragen.
Frau Marion Gähle und Britta Schönnagel danke ich herzlich nicht nur für die
Unterstützung im histologischen Labor, sondern auch für das Korrektur lesen.
Frau Ines Blume danke ich herzlich für die Korrekturen und die ständige
Hilfsbereitschaft am PC.
Der akad. Zeichnerin Frau Caren-Imme von Stemm danke ich herzlich für die
Anfertigung der Zeichnungen.
Frau Marlies Bewarder, Herrn Stünkel, Herrn Stahlhut und Herrn Mörstedt danke ich
für die freundliche Unterstützung im Anatomischen Institut.
Mein weiterer Dank gilt allen Angehörigen des Institutes für ihre hilfsbereite und
freundliche Zusammenarbeit.
Meiner Mutter, meiner Großmutter und meinem Onkel möchte ich sehr herzlich dafür
danken, dass sie immer an mich geglaubt haben und dass ich allzeit auf ihre
Unterstützung zählen konnte.
Meinen Freunden möchte ich für die schöne Zeit und ihre Freundschaft danken –
ganz besonders Wiebke, die immer für mich da war.
Und schließlich möchte ich Heiko herzlich danken, der diese Arbeit überhaupt erst
möglich gemacht hat.