Embed Size (px)
Citation preview

(Aus der Genetischen Abteilung des Kaiser Wilhelm-Instituts fiir Hirnforsehung, Berlin-Bueh.)
ZUR G E N E T I K D E R DROSOPHILA FUNEBI~IS. I. GESCHLECHTSGEBUNDENE VERERBUNG.
Von
N. W. TI~OF~EFF-R~ssovsKY.
Mit 8 Textabbildungen.
( Eingegangen am 8. Dezember 1930.)
Inhalt. seite 1. Einleitung . . . . . . . . . . . . . . . . . . . . . . . . . . . I54 2. Entstehung und Beschreibung Yon elf geschlechtsgebundenen Genoyaria-
tionen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 155 3. Lo](alisation der Gene N ~, ev, Va', l], m, lz un4 bb . . . . . . . . . . 164 4. Geschlechtsgebundene Letalfak~oren, Gynander und das Niehttrennen der
Geschleehtschromosomen . . . . . . . . . . . . . . . . . . . . . 167 5. Schlul~bemerkungen . . . . . . . . . . . . . " . . . . . . . . . . 170 6. Zusammenfassung . . . . . . . . . . . . . . . . . . . . . . . . 172 7. Literatur . . . . . . . . . . . . . . . . . . . . . . . . . . . . 173 8. Anhang: Tabetlen zu den Lokalisationsversuchen ". . . . . . . . . . 174
1. Einle i tung.
Drosphila/unebris ist viel weniger ausftihrlich genetisch untersucht als die anderen Drosophila-Arten, mit denen genetisch gearbeitet wird. Vor allem ist bei Drosophila ]unebris noch keine Grundlage ~iir die gene- tische Chromosomentopographie geschaffen worden, die die weitere gene- tische Analyse bedeutend erleichtern wfirde. I n dem MORGANScben La- borator ium wurden seinerzeit einige Genovariat ionen bei dieser Ar t ge- funden (MogR and STURTEVA~T 1919, MORGAn, BRIDGES and STURTE- VAST 1925, STURTEVANT 1918). Drei yon diesen Genovariat ionen (Notch, /orlced und bubble) waren geschlechtsgebunden; mit diesen drei Fak toren wurden Lokalisationsversuche durchgeffihrt, die gezeigt haben, dab /o r - ked etwa 28 und bubble etwa 70 Morganiden (units, Crossovereinheiten) yon Notch entfernt liegen. Die Arbei t wurde aber nicht weitergeffihrt und die Drosophila/unebris-Kulturen wurden aufgegeben. I m Laufe der le tz ten 5 - - 6 Jahre wurde sowohl yon uns wie auch yon anderen Autoren eine Reihe yon Genovariat ionen bei Drosophila /unebris gefunden und zum Tell besehrieben (ROMASC~OFF U. BALKASC~I~A 1929, SIlEnCeR 1928, TIMOF~EFF-RESSOVSKY, H. A, 1928, 1930, TIMOFI~EFF-REssovsKY, N. W.

Zur Genetik der Drosophila funebris. I. 155
1925, 1926, 1928, 1929). Die genauere Analyse und besonders die Lokali- sation dieser Genovariationen ist aber durch manche Umstande erschwert. Erstens haben fast alle bei Drosophila/unebris auftretenden Genovaria- tionen eine unvollkommene, oft sehr schwache Penetranz und schwan- kende Expressivitat (inkonstante lohanotypische Manifestierung). Zwei- tens werden yon den meisten dieser Genovariationen ausschlieBlich Flfigel- und Borstenmerkmale, oft in einander ahnlicher Weise betroffen, was die Identifikation in polyhybriden Kreuzungen erschwert. Diese bei- den Umstande erschweren betrachtlich auch die Lokalisationsarbeit.
Da wir aber jetzt, besonders nach der Durchffihrung yon R6ntgen- bestrahlungsversuchen, schon fiber eine ganze Reihe yon Genovariationen bei Drosophila/unebris verffigen, so scheint es doch angebracht, sie in systematischer Form zu beschreiben und zu versuchen, wenigstens ein vorlaufiges Schema der Lokalisationsverhaltnisse auszuarbeiten.
In der folgenden Mitteilung werden die yon uns untersuchten ge- schlechtsgebundenen Genovariationen beschrieben. Einige yon diesen Genovariationen konnten auch ffir Lokalisationsversuche verwendet werden. Neben der Beschreibung der geschlechtsgebundenen Genovaria- tionen und der Lokalisationsversuche wird auch kurz das wenige er- wahnt, was bisher fiber geschlechtsgebundene Letalfaktoren, Gynander und das Nichttrennen der Geschlechtschromosome bei Drosophila/unebris yon uns beobachtet wurde.
Die Zuchttechnik fiir Drosophila /unebris ist dieselbe wie ffir Droso- phila melanogaster. Man kann die Fliegen sowohl auf Maismehl-Syrup- Agar als auch auf Kartoffel-Rosinen-Agar (das Fut ter wird mit verdfinn- ter Here befeuchtet) ziichten. Die optimale Temperatur liegt ffir Droso- phila /unebris etwas tiefer als ffir Dsosophila melanogaster: optimal ist 20 25 o C. Schon bei 28--30 o C werden viele Fliegen steril. Bei 25 o C betragt die durchschnittliche Entwicklungsdauer etwa 16--20 Tage (also langer als bei Drosophila melanogaster).
2. Ents tehung und Beschreibung yon elf geschlechtsgebundenen Genovariationen.
1. Notch (N'), Abb. 1. Notch ist dominant und tuf t Ausschnitte am distalen Fliigelrande hervor; N' ist rezessiv-letal: Mannchen, die den Faktor N ' erhalten, sind nicht lebensfahig. In unseren Kulturen ist N ' zweimal entstanden: einmal in einer eversae-~C[assenkultur und das andere Mal im R6ntgenexperiment (It. A. T~OF~E~F-REssovsKY 1930). In beiden Fallen handelte es sich leider um sehr ,,schwache" Allele: die Penetranz und Expressivit~t war so schwach, dal~ das Merkmal flit ge- nauere Analysen fast unbrauchbar war (die Penetranz schwankte yon 15 40%). DaI~ es sich in beiden Fallen wahrscheinlich um homologe Genovariationen handelte, geht daraus hervor, dab beide Notchs mit

156 N. W. Timofdcff-Rcssovsky:
e v e r s a e etwa 8% Austausch ergaben (Tal)ellc l ) ; da abet keine genauere Lokalisation mit anderen Fak toren vorgenommen wurde, so kann es
Tabelle 1. Keppelungen zwischen eversae und den zwei unabh~ngig entstandenen ~Voteh- F a k t o r e n .
A~t der P-Kreuzung
~u + / \ + eV ~ - j ~ - 3 . . . .
N ~ ev + ev I ? T ~ - • . . . .
~aktorenaustausch"
Zahl der F1-Fliegen
281
317
~9
+ ~Yf t ev N P~v
39 52 87 o
109 4 I 50 51
8 ~9
94 9
.NI - - ev = 8,8% (5 N ' ev 9: allen N'~). A ~1 - - ev = 8,2% (8 + ~ : alle~l ~). 2 r ev = 7,3% (4 ~ ' ~ : allen N'~). N 2 - ev = 8,7% ( 9 e v G: allen ~).
Zahlenverhiiltni~ der Oeschlechter
1 : 0,52
1 : 0,48
Abb. 1. Zwei _7~))tch-Fliigel.
selbstverst~ndlich auch sein, dag es sich um zwei verschiedene, rechts und links yon e v e r s a e ]okalisierte Fak toren handelt . Die letztere Annahme scheint abet weniger wahrscheinlich zu sein. Die Kul tu r beider N'-Geno- variat ionen wurde nach vorl~ufiger Analyse aufgegeben.
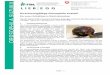
2. e v e r s a e ( ev ) , Abb. 2. e v e r s a e ist rezessiv und ist dem von H. NACHTS- ~ i ~ bei D r o s o p h i l a m e l a n o g a s t e r beschriebenen Fak to r w a v y phgnoty- pisch ~hnlich (NACHTSHEIlg 1928). Bei den ev-Fliegen ha t der ~ugere Flfigelrand an der Fltigelbasis eine oder zwei wellenartige Biegungen; deshalb sind auch meistens die Flfigelspitzen nach oben und augen ge- zogen, was eine dachart ige Ha l tung and eigenartige Form der l~lfigel
Zur Gcnetik dcr Drosophib~ flmcbris, i. 157
zttr Folge hat. l)iese Genov~riation wurde yon E. BA~K~SCmNA in eincr curved-Kultur cntdeckt (RO~ASCHOF~ U. BALKASCHINA 1929). Die ev-
Abb. 2. Ein eversae-Weibchen.
Fliegen haben normale Fertilit~t, Lebensf~higkeit und Entwicklungs- dauer. Die Expressivit~t des ev-Merkmals schwankt ziemlieh stark, die Penetranz ist aber in den meisten Y/tllen absolut. Dureh minusselektion ist es aber m6glich, inmanchen Kulturen die ]?enetranz bis auf etwa 80% abzuschw/i-
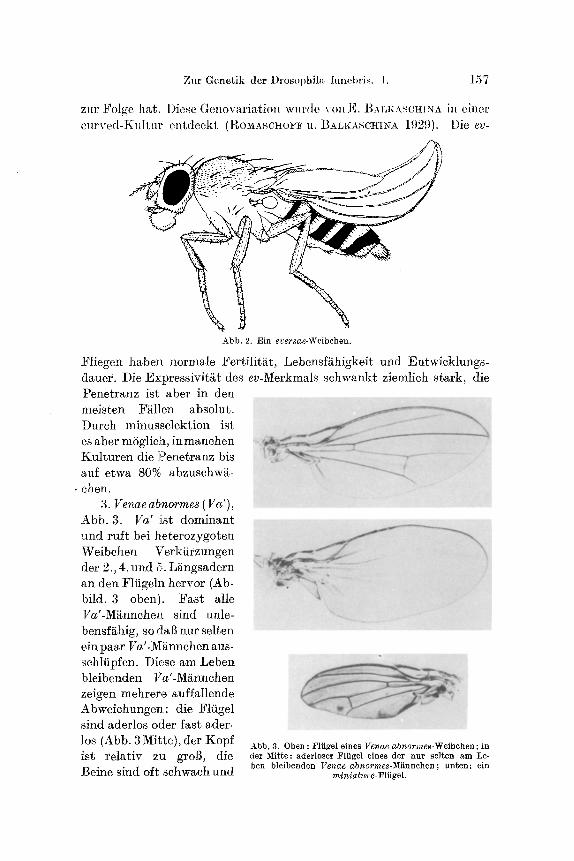
�9 ehen. 3. Venaeabnormes (Va'),
Abb. 3. Va' ist dominant und ruft bei heterozygoten Weibchen Verkiirzungen der 2., 4. und 5. L~ngsadern an den Flfigeln hervor (Ab- bild. 3 oben). :Fast alle Va'-Mgnnche~ sind unle- bensf~hig, so daft nur selten ein paar Va'.Mi~nnchen aus- sehliipfen. Diese am Leben bleibenden Va'-M~nnchen zeigen mehrere auffallende Abweichungen: die Flfigel sind aderlos oder fast ader-
los (Abb. 3 Mitre), der Kopf Abb. 3. Oben : ]~liigel eines Venae abnormes-Weibchen ; in i s t r e l a~s iv z l / g r o ~ , d i e der Mitte: aderloser Flfigel eines d e r n u r selten am Le- Beine sind oft schwach und ben bleibenden Venae abnormes-M~nnchen; unten: ein
miniature- Fliigel.

158 N.W. Timofbeif-Ressovsky:
krumm und es fehlen bei diesen Ms viele yon den Kopf- und Thoraxborsten. Die iiberlebenden Va'-Ms sind steril und haben eine sebr verkfirzte Lebensdauer. Die phttno~ypisehe Manifestierung yon Va' wird durch Temperatur beeinflul~t: Unter hbherer Temperatur (25~28 ~ C) ist das Merkmal bei heterozygoten Weibehen absolut pene- t rant und stark expressiv (verkfirzt sind fast immer die 2., 4. und 5, L~ngs- adern), nnd Va'-M~nnehen sind nieht lebensf~hig und kommen nur als ganz seltene Ausnahmen vor. Bei tieferer Temperatur (15--180 C) ist die Penetranz bei heterozygoten Weibehen nicht absolnt und sinkt propor- tional der Temperaturherabsetzung; die Expressivit~t des Merkmals ist auch sehw~eher. Mit Herabsetzung der Temperatur erhbht sich aueti der Prozentsatz der am Leben bleibenden Va'-~t~nnehen, so dal~ in manchen Kreuzungen fiber 50% der Va'-M~nnchen aussehliipfen. Die heterozy- goten Va'-Weibehen haben eine normale Fertilitt~t und Entwieklungs- dauer, aber eine etwas herabgesetzte Lebensft~higkeit, so daI~, besonders in ,,fibervblkerten" Kreuzungen, weniger Va'-~. ~_ aussehlfipfen als zu erwarten ist. Diese Genovariation wurde yon W. R ~ m in einer nor- malen Kultur entdeekt (R~i~m 1928).
4. lf. Ein Letalfaktor, der in homozygotem Zustande die Weibchen tbtet. Zwei verwandte Kreuzungen yon miniature-Fliegen (die aus Rbntgenversuehen stammen) haben zweimal soviel M~nnehen als Weib- ehen ergeben. Diese Kreuzungen sind auf Tabelle 2 angeffihrt. Unter
Tabelle 2. Zwei verwandte miniature-Kreuzungen, die doppelt soviel Mt~nnchen wie Weibchen ergeben.
If m . If m
Kreuzungsnummer Zahl der ~ c:~ Zahlenverhtiltnis der Fliegen m m. Geschlechter
9 2 2 2 3 71 152 1 : 2 , 14
111 186 64 122 1 : 1,90
vielen versehiedenen Kreuzungen, ffir die miniature-Fliegen aus dem- selben Stamme vewendet wurden, haben zwei Kreuzungnn der Kon-
stitution ~ m • ~ + m ~ieder dasselbe Geschlechtsverh~ltnis ge- ev
zeigt. Diese Kreuzungen sind auf Tabelle 3 angeffibrt. Da fast alle minia- ture- ~ ~, die zu erwarten waren, feblen, so mu~ man annehmen, dub das miniature tragende X-Chromosom der Eltern, sowohl des ~ wie aueh des ~, den Letalfaktor enth~lt. Die wenigen miniature-Weibchen k6nnen dureh Faktorenaustausch zwischen miniature und dem Letalfaktor ent- stehen. Der Prozentsatz des Austausehes zwisches m u n d l], berechnet nach der Zahl der miniature-Weibchen ist in der ersten Kreuzung der

Zur Genetik der Drosophila funebris. I. 159
Tabelle 3. Kreuzungen, die die Lokalisation des Faktors 1] im miniature-tragendm~ X-Chromosom und den Faktorenaustausch zwischert l[ und m zeigen.
P:9 9
Kreuzungsnummer Zahl der 1 Fliegen / +
118 119 132 73
Faktorenaustauseh: 1[-- m = g / - - m ==
+ I f m • + l fm. e v + +
18 63 101 7 84 69
13,1% 'Krz. Nr. 118 8,7% 'Krz. Nr. 132
.... Zahlenverhfiltnis m der Geschlechter
120 1 : 2,07 51 1 : 1,92
18 m 9: allen 9). 7 m ~: allen 9)*
Tabel le 3 gleich 13,1% und in der zwei ten = 8,7%. Es is t ferner gelungen, durch Anse tzen yon vielen Kreuzungen (da das Gen l / k e i n e s ich tbare morphologische Manifes t lerung hat) zwei Kreuzungen yon der Kons t i -
t u t i on ~Q Va' X ~ + If m zu erzielen. Diese Kreuzungen sind in If m
Tabel le 4 summie r t und zeigen, dab l / zwi schen Venae abnorm, und minia-
Tabelle 4. Lokalisationsversueh mit Venae abn. (Va'), l /und miniature (m). Va' + + + If m
P : 9 ~ + If m •
Kreuzungs- Zahl der 99 ~3 Zahlenverhgltnis nummer Fliegen + Va' m V a ' m + Va' [ m V a ' m der Geschlechter
401+422 11 143 1 19 2 7 t 6 /1152] 1 1:1,07 Faktorenaustausch:
V a ' - - m = 18,4% (11 + 9 + 19 V a ' m 9 + 2 x l i n g : a l l e n 9 9 ) . V a ' - - m = 15,1% (27 + 3 :27 + ~ + 152m 3). lZa'-- l] = 6,9% (11 + 2 + l i n g : a l l e n 9 9 ) .
1] - - m = 11,5% ( i r a 9 + t9 Va" m g: allen 99).
ture l iegt, m i t Va' etwa 7% und m i t m e twa 11,5% Aus tausch ergibt . Die geschlechtsbegrenzte le ta le W i r k u n g des Gens l / k a n n man en tweder phy- siologisch zu exklgren versuchen oder m a n muB annehmen, dab es sich um e inen / ihn l ichen Fa l l hande l t , wie bobbed bei Drosophila melanogaster. Der F a k t o r bobbed mani fes t i e r t sich bekannt l ich nur bei Weibchen. C. ST~R~ ha t bewiesen, dab im u der Drosophila melano- gaster das norrnale Allel yon bobbed (BB) vorhanden ist, wodurch sich die geschlechtsbegrenzte W i r k u n g e rk lgr t (STERN 1927). Eine ghnl iche Er- k lgrung k6nn te auch ffir 1/ zutreffen. Man hg t t e ferner d a r a n denken k6nnen, dab 1/ein le tales Allel yon bobbed ist. Bei Drosophila/unebris i s t ein bobbed-Faktor, d e r d e m bb bei Drosophila melanogaster v611ig ident isch is t , vorhanden . E r is t aber welt von l / a n einer ganz anderen Stelle des X-Chromosoms lokal is ier t , so dab diese le tz tere Ve rmu tung k a u m zu- t reffen k a n n . . D i r e k t e Kreuzungen yon bobbed mi t l / k o n n t e n leider n icht durchgeff ihr t werden, da inzwischen die l / -Ku l tu r wegen Schwier igkei t
N. W. Timof~eff-Ressovsky: 160
ihrer Weiterfiihrung verloren gegangen ist. Es ist sehr wahrseheinlieh, dag der Faktor l[ unter Einwirkung der I{Sntgenbestrahlung ent- standen ist.
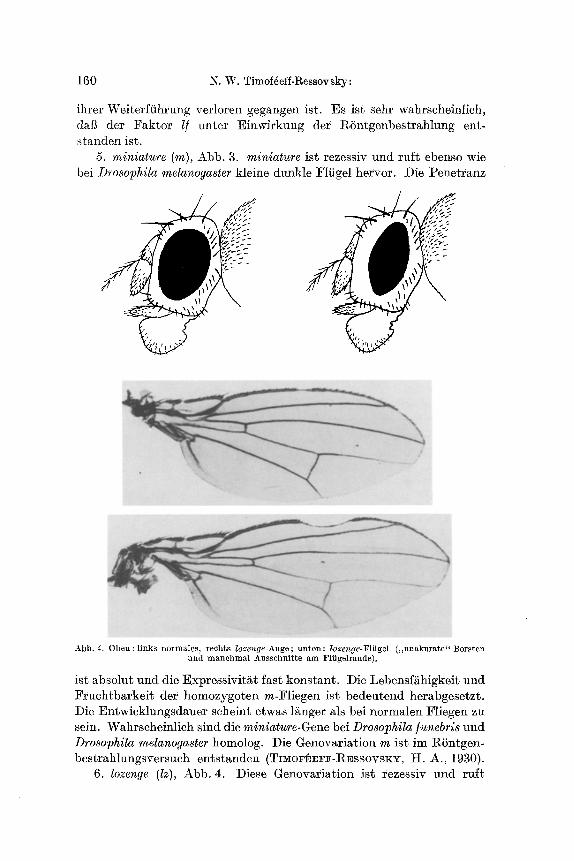
5. miniature (m), Abb. 3. miniature ist rezessiv und tuft ebenso wie bei Drosophila melanogaster kleine dunkle Flfigel hervor. Die Penetranz
y/~ 1/
Abb. 4. Oben: l inks normales~ reehts lozenge Auge; un ten : lozenge-Fliigel ( . unaku ra t e " Borsten und manchmal Ausschnit te am Fliigelrande).
ist absolut und die Expressivit~t fast konstant. Die Lebensfiihigkeit und Fruchtbarkeit der homozygoten m-t~liegen ist bedeutend herabgesetzt. I)ie Entwicklungsdauer scheint etwas l~nger als bei normalen Fliegen zu sein. Wahrscheinlich sind die miniature-Gene bei Drosophila/unebris und Drosophila melanogaster homolog. Die Genovariation m i s t im RSntgen- bestrahlungsversuch entstanden (TI~o~v, rF-RESSOVSKY, H. A., 1930).
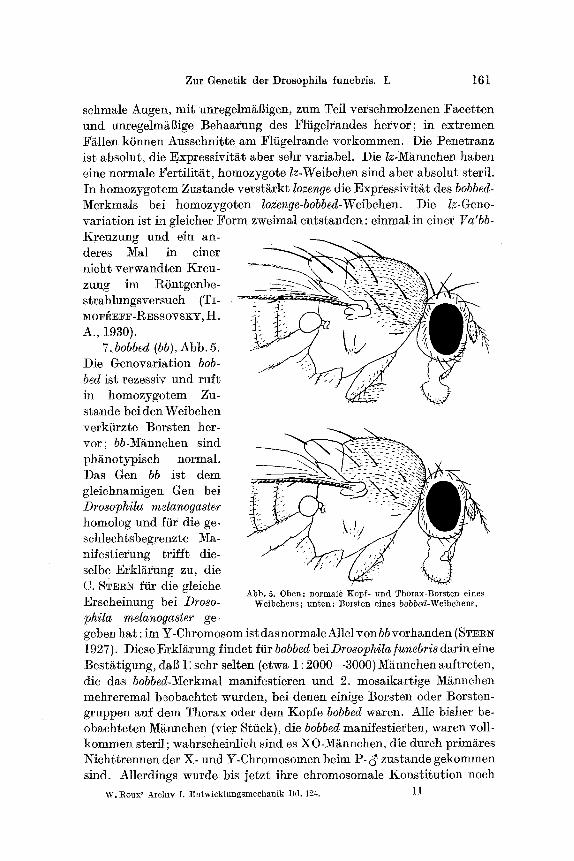
6. lozenge (Iz), Abb. 4. Diese Genovariation ist rezessiv und ruff
Zur Genetik der Drosophila tunebris. I. 161
schmale Augen, mit unregelm/~Bigen, zum Tefl versch~olzenen Faeetten und unfegelm~Sige Behaarung des Fliigelrandes horror; in extremen •/s k6nnen Aussehnitte am Fliigelrande vorkommen. Die Penetranz ist absolut, die Expressivit/~t abet sehr variabel. Die lz-M~nnchen haben eine normale ~'ertilit/~t, homozygote lz-Weibehen sind abet absolut steril. In homozygof.em Zustande verst~rkt lozenge die Expressivit/~t des bobbed- Merkmals bei homozygoten lozenge-bobbed-Weibchen. Die lz-Geno- variation ist in gleicher Form zweimal entstanden: einmal in einer Va'bb- Kreuzung und ein an- dares Mal in einer ~ _ ~ - ~ - ~ . ~ = ~ . ~ nicht verwandten Kreu- zung im RSntgenbe- strahlungsversueh (TI- M O F I ~ E F F - R E s S O V S K u I - I . _, , .7.__.~z.::
A., 1930). 7.bobbed (bb ), Abb. 5.
Die Genovariation bob- bed ist rezessiv und ruft in homozygotem Zu- stande bei den Weibchen verkiirzte Borsten her- vor; bb-Mgnnehen sind ph~notypiseh normal. Das Gen bb ist dem gleichnamigen Gen bei Drosophila melanogaster homolog und :[tir die ge- schlechtsbegrenzte Ma- nifestierung trifft die- selbe Erkl/~rung zu, die
C. STERN ftir die gleiche Abb. 5. Oben : normale Kopf= und Thorax-Bors ten eines
Erscheinung bei Droso- Weibchens ; u n t e n : t~orsten eines bobbed-Weibchens.
phila melanogaster ge- geben hat :im Y-Chromosom ist das normale Allel yon bb vorhanden (STaRer 1927). DieseErkl/~rung findet ffirbobbedbeiDrosophila]unebrisdarineine Best/~tigung, dal] 1: sehr selten (etwa 1 : 2000--3000) Mgnnchen auftreten, die das bobbed-Merkmal manifestieren und 2. mosaikartige M~nnehen mehreremal beobachtet wurden, bei denen einige Borsten odor Borsten- gruppen auf dam Thorax oder dam Kopfe bobbed waren. Alle bisher be- obachteten 1V[/~nnchen (vier Stiick), die bobbed manifestierten, waren voll- kommen steril; wahrscheinlich sind es XO-M~nnehen, die dureh primiires Niehttrennen der X- und u beim P- c~ zustande gekommen sind. Allerdings wurde his jetzt ihre chromosomale Konstitution noah
W. l toux ' Archiv f. E n t w i c k h m g s m e c h a n i k Bd. 124. 11
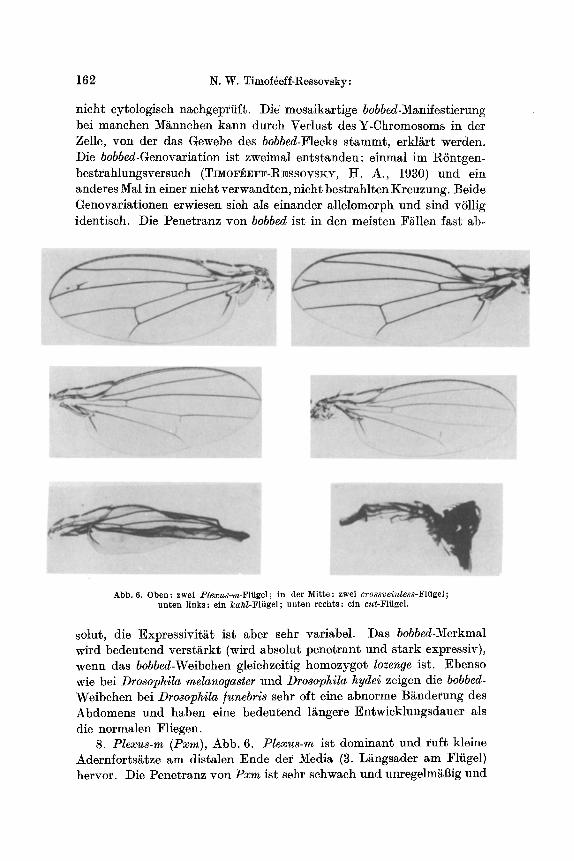
162 N. W. Timof6eff-Ressovsky:
nicht cytologisch n~chgeprfift. Die" mosaikartige bobbed-Manifestierung bei manchen M/innchen kann dureh Verlust des Y-Chromosoms in der Zelle, v0n der das Gewebe des bobbed-Flecks stammt, erkl/~rt werden. Die bobbed-Genovariation ist zweimal entstanden: einmal im RSntgen- bestrahlungsversueh (TnwoF~FF-R~ssovsxr, H. A., 1930) und ein anderes Mal in einer nicht verwandten, nicht bestrahlten Kreuzung. Beide Genovariationen erwiesen sieh als einander allelomorph und sind vSllig identisch. Die Penetranz yon bobbed ist in den meisten F/~llen fast ab-
Abb. 6. Oben : zwei _Plexus-m-Fliigel ; in der Mit te : zwei crossveinless-Fliigel; unten l inks : ein kah/ -F l i ige l ; un ten rechts : ein cut-Flfigel.
solut, die Expressivit/~t ist aber sehr variabel. Das bobbed-Merkmal wird bedeutend verst~rkt (wird absolut penetrant und stark expressiv), wenn das bobbed-Weibehen gleiehzeitig homozygot lozenge ist. Ebenso wie bei Drosophila melanogaster und Drosophila hydei zeigen die bobbed- Weibchen bei Drosophila ]unebris sehr oft eine abnorme B~nderung des Abdomens und haben eine bedeutend 1/~ngere Entwicklungsdauer als die normalen Fliegen.
8. Plexus-m (Pxm), Abb. 6. Plexus-m ist dominant und ruff kleine Adernforts/~tze am distalen Ende der Media (3. Ls am Fliigel) hervor. Die Penetranz von Pxm ist sehr sehwaeh und unregelm~Big und

Zur Genetik der Drosophila funebris. I. 163
die Expressivit~t variabel. Diese Genovariation ist im RSntgenbestrah- lungsversueh entstanden (TrMoF~F-R~ssovsKY, H.A., 1930).
9. crossveinless (clv), Abb. 6. Diese Genovariation ist ebenfalls naeh RSntgenbestrahlung entstanden (TIMOF~I~rF-REssovsKY, H.A., 1930), ist rezessiv und hat eine sehr schwaehe und sehwankende Penetranz und Expressivit~t. In homozygoten Kultnren sind etwa 40-50% der Fliegen phgnotypiseh normal, bei den fibrigen ist die 2. Querader unterbroehen. In extremen F/~llen kann die 2. Querader oder sogar beide Queradern ganz fehlen. Die cvl-Kultur wurde nach vorl~ufiger Analyse und Test- stellung der Gesehleehtsgebundenheit nieht weiter gefiihrt.
10. ~ahl (/~), Abb. 6. In P2 yon einer Kreuzung, deren PI- ~ bestrahlt wurde, sind M~nnchen aufgetreten, die mehrere auffallende Merkmale zeigten: die Fl~!gel waren in typiseher Weise ,,zerknittert" (Abb. 6 unten links), der Koiff war ,,kahl", da fast alle Borsten und Haare fehlten, mehrere yon den Thoraxborsten fehlten und die B/~nderung des Ab- domens war abnorm. I)iese Mgnnehen waren vollkommen steril. Aber in den n~iehsten Generationen yon manchen ihrer Sehwestern traten wie- der solehe Mgnnehen in etwa 40% auf. Dann wurde das Merkmal wegen Sehwierigkeit der Weiterzucht verloren. Allem Anschein nach handelte es sieh um einen rezessiven geschlechtsgebundenen Faktor (Tnuo]rgEF~- t~ESSOVSKY, H. A., 1930).
i1. cut (ct), Abb. 6. Ein rezessiver Faktor, der bei Mgnnchen vesti- gialghnliehe Fliigel (Abb. 6 unten reehts) und absolute Steriliti~t hervor- ruft. Weibehen waren immer normal, da wegen Steriliti~t der M/~nnehen keine homozygoten Weibchen erhalten werden kSnnen. Diese Geno- variation ist ebenfalls im RSntgenbestrahlungsversuch aufgetreten (TIMo~EF~'-R~ssovsKr, It. A., 1930). Die cut-Kultur wurde wegen Schwierigkeit der Weiterzucht und sehr schwacher Lebensf~higkeit der cut-Mgnnehen aufgegeben.
AuBer den soeben beschriebenen elf geschleehtsgebundenen Geno- variationen wurden zu verschiedener Zeit noch einige andere Merkmale beobachtet, die wahrseheinlieh durch Genovariationen im X-Chromosom entstanden waren. Sie wurden aber nicht n~her analysiert, meistens well sie eine ganz scl~wache und unregelm~Bige ph~notypische Manifestierung hatten oder weil zur Zeit noeh keine anderen ,,besseren" gesehlechts- gebundenen i~aktoren vorhanden waren, mit deren Hilfe man die Ver- erbung und Lokalisation dieser ,,schlechten" Genovariationen exakter analysieren konnte. Kurz erwi~hnt seien folgende: 1. Ein gesehlechts- gebundener Faktor, der in homozygotem Zustande die Expressiviti~t des rezessiven autosomalen Gens radius incompletus (Tr~or~]rF-RESSOVSKu 1925) bedeutend verstiirkt; 2. ein anscheinend rezessiver gesehlechts- gebundener Faktor, der die zweite Querader unterbricht (fihnlich wie der oben besehriebene Faktor crossveinless), aber sehr schwach penetrant ge-
ll*

164 N.W. Timof6eff-Ressovsky:
wesen ist, so dab nur etw~ 10--20% der Fliegen das Merkmal phano- typisch m~nifestierten; 3. ein anscheinend dominanter, geschlechts- gebundener (nicht letaler) Faktor, der bei etwa 5--20% der Fliegen das l~ehlen einer der vorderen dorsozentralen Borsten bewirkte und 4. ein rezessiver geschlechtsgebundener Faktor, der etwas kfirzere und breitere Flfigel hervorrief, sehr unregelmai~ig und schwach penetrant, so dab die Erkennung des betreffenden Merkmals sehr schwierig gewesen ist.
Nur die Gene ev, Va' m, Iz und bb sind ffir genauere AnMyse der Koppelungserscheinungen im X-Chromosom brauchbar. Deshalb wurden in weiteren Versuchen, die die Aufgabe hatten, ein Gerfist der ,,geneti- schen Kar te" des X-Chromosoms bei Drosophi la/unebris auszuarbeiten, nur diese ffinf Faktoren benutzt und die iibrigen Kultur~,l aufge- geben.
3. Lokal isat ion der Gene N', ev, Va', lf, m, lz and bb.
Die Ergebnisse aller Lok~lisationsversuche mit den Faktoren ev, Va', m, lz und bb sind auf den Tabellen im Anhang am Schlusse der Arbeit an- gegeben. In diesen Lokalisationsversuchen wurden etwa 18500 l~liegen durchgesehen und registriert. Im ganzen umfassen alle Lokalisations- versuche etwa 21000 Fliegen. Obwohl diese l~liegenzahl nicht zu gering ist, mfissen die gewonnenen Ergebnisse als vorlaufig und noch nicht ganz sicher bewertet werden, und zwar aus Iolgenden Grfinden. Die Variabili- ta t der Austauschwerte in verschiedenen Kreuzungen ist sehr stark ; viel- leicht erklart sich dieses zum Tell dadurch, dal3 fast an allen Kreuzungen Fliegen aus Kulturen, die friiher in R6ntgenbestrahlungsversuchen ent- standen sind, teilnahmen: es ist m6glich, dab unter der Bestrah'ung auch manche, den crossing-over modifizierende Faktoren aufgetreten sind. Als zweiter st6render Faktor kommt die Unm6glichkeit hinzu, manche yon den Merkmalen, besonders in Kombinationen, immer genau zu klassi- fizieren: eversae und bobbed ergeben oft viele phanotypisch fast normale Fliegen; Va' ist nicht immer mit Sicherheit in miniature-Fliegen zu klassi- fizieren, da m-Fliegen oft auch ahnliche Adernst6rungen zeigen; und die Kombination ev m ist iiberhaupt nicht yon m m i t Sicherheit Zu unter- scheiden, so dal3 der Austausch zwischen ev und m nur auf Grund des Ver- haltnisses zwischen normalen und eversae-Klassen berechnet werden kann. Schlieftlich haben verschiedene Kombinationen der in Frage kom- menden Genovariationen sehr verschiedene Lebensfahigkeiten, was auch die Zahlenverh/~ltnisse beeinfluf3t.
Der Mechanismus des crossing-overs (Variabilitat, doppelter und drei- facher crossingover, Koinzidenz), bei Drosophi la /unebr is wird in einer anderen Arbeit auf Grund exakterer Versuche mitgeteilt werden. Die je tz t vorliegenden Kreuzungsversuche sollen nur die l~eihenfolge und ungefahre Lage der bisher analysierten Gene im X-Chromosom angeben
Zur Genetik der Drosophila funebris. L 165
und damit eine Basis 1fir den weiteren Ausbau der ,,genetischen Kar te" des X-Chromosoms yon D r o s o p h i l a ] u n e b r i s bilden.
Au~ Tabelle 5 sind alle Austauschwerte aus den einzelnen Lokali- sationskreuzungen angeffihrt. Auf der rechten Seite der Tabelle sind die
Tabelle 5. Zusammenfassung aller Kreuzungsergebnisse fiber Faktorenkoppehngen ira X-Chromosom yon D r o s o p h i l a ] u n e b r i s .
Direkt aus Kreuzungsversuchen gewonnene Austauschwerte
Aus verschiedenen Kreuzungs- Gene versuchen gewonnene Faktoren-
austauschwerte
~ v _ e v
e v - - V a v
ev - lz
ev - bb
V a ' - I f
V a ' - lz
V a ' - bb
l f - m
m - b b
lz - bb
8,8; 8,2; 7,3; 8,7 28,8; 30,9; 26,5; 34,5; 39,8; 32,0; 35,9; 33,2; 38,0; 25,0;
30,7 ; 28,7; 32,8 39,2; 40,8; 44,8; 42,0
48,8; 47,3
53,1 ; 48,6 6,9
21,9; 19,2; 14,3; 15,6; 26,1; 27,7; 19,8; 19,0; 16,1; 18,4; 13,8; 23,5; 20,7; 22,6; 20,7;
22,7 ; 18,1 26,0; 22,6; 23,0; 26,3; 20,3;
25,4 31,2; 32,0
11,5; 13,1; 8,7
2,6; 4,7 36~0; 23,6; 33,8; 20,5
12,5; 11,2
Dutch Addition ktirzerer Strecken gewonnene Entfernungen
Ent- Gene fernungen in
Morganiden
52,0 ~V - - m
(ev - V a ' -- m )
V ~ r - I z
( V a ' - m - lz)
m - b b
( m - l z - bb)
e V - - 1Z
( e v - V a ' - m - lz)
V a ' - bb
( V a ' - m - lz - bb)
ev - bb
(ev - V a ' - m - lz - bb)
ev - bb
(ev - V a ' - m - bb)
23,6
15,6
55,6
35,6
67,6
80,5
Entfernungen zwischen einzelnen Faktoren angegeben, die auf Grund yon SummiernngeiL kfirzerer Strecken gewonnen wurden.
N o t c h ' . N o t c h wurde nur m i t e v gekreuzt; diese Kreuzungen wurden schon auf Tabelle 1 (im Text) angeffihrt. N ' - e v ergeben etwa 8% Arts- tausch, so dab N'-8 Morganiden nach rechts oder links yon e v liegen kann.
e v e r s a e . Dieses Gen wurde mit V a ' , m , l z und b b gekreuzt. Die Ta- bellen 1--4 des Anhanges zeigen verschiedene Kreuzungen, die e v und F a '
enth~lten. Der Austauschwert zwischen e v und V a ' variiert in einzelnen Kreuzungen yon 25--40% und ist im Durchschnitt etwa 32%. Auf Ta- bellen 5- -7 sind Kreuzungen zur Best immung des Austausches zwischen e v und m angeftihrt. D~ die doppelt rezessive Kombinat ion e v m bei fast

166 N. W. Timof~eff-Ressovsky:
allen solchen Fliegen yon m allein nicht zu unterscheiden ist, so kann der Austauschwert nur auf Grund des Verhgltnisses zwischen den Klassen der normalen und eversae-Fliegen berechnet werden. Der durchschnitt- liche Austauschwert zwischen ev und mis t etwa 42% gleich. Auf Tabelle 8
ev lz sind Kreuzungen yon ~ ~ 9 mit ev c~ (~ summiert, die einen Aus-
tauschwert yon 48% Iiir die Entfernung zwischen ev und lz ergeben. Die Faktoren ev und bb (Tabelle 9--10) zeigen praktiseh eine freie Spaltung.
Venae abnormes. Am ni~chsten zu Va' liegt der Faktor l[, dessen Lo- kalisation oben schon beschrieben wurde (Tabelle 4 im Text). Die Kreu- zungen der Tabellen 11--15 geben den Austauschwert zwischen Va' und m an, der yon etwa 13% bis etwa 28% in einzelnen Kreuzungen variiert; im Durchschnitt betr~gt er 20%. Schon daraus geht hervor, dab Va'
zwisehen ev und m liegen mul3 ; dieses wird direkt best/~tigt durch Kreu- zungen der Tabellen 13 und 15, die aul~er m u n d Va' aueh ev enthalten. Mit lz ergibt Va' etwa 24% Austausch (Tabellen 16, 25 und 26); diese Kreuzungen enthalten auch den Faktor ev und zeigen, dab Iz reehts yon Va' lokalisiert ist (dab also Va' zwisehen ev und lz liegt). Auf den Ta- bellen 17--18 sind Kreuzungen summiert, die den Austausch zwisehen Va' und bb zeigen, der etwa 32% betri~gt ; die Kreuzungen der Tabelle 18 enthalten ev und zeigen, daB. Va' zwischen ev und bb liegt (dab also bb
rechts yon Va' liegt). miniature . Die Tabellen 19--20 zeigen den Austauseh zwischen m und
lz, tier im Durchschnitt etwa 3,6% betriigt; die Kreuzungen der Tabelle 20 zeigen auBerdem, dab lz rechts yon m lokalisiert ist, dab also m zwisehen Va' und Iz liegt. Dieses s t immt auch gut fiberein mit den friiher erwiihn- ten Austausehwerten Va ' -m (etwa 20% ) und Va'-lz (etwa 24% ). Die Kreu- zungen der Tabellen 21--24 geben den Austausehwert zwischen m u n d bb an, der im Durchschnitt gleieh etwa 28% ist. Aus den Tabellen 23---24 ist auBerdem zu ersehen, dab m zwisehen Va' und bb liegt.
Auf den Tabellen 25--26 sind sehliel31ich Vierfaktorenkreuzungen an- geffihrt (ev, Va' , Iz, bb). Diese Kreuzungen zeigen wieder, dal~ die l~eihen- folge dieser Fektoren im X-Chromosom ev-Va'-Iz-bb ist. Die Austausch- werte zwisehen lz und bb, die aus diesen Kreuzungen gewonnen wurden, sind aber bedeutend geringer als zu erwarten wgre. Der Austausch zwischen m und bb betrug etwa 28% und der zwischen m u n d lz etwa 4%. Man miiBte also ennehmen, dab der Austausch zwischen lz und bb etwa 24% ausmacht. Die tats~chlich erhaltenen Resultate ergeben aber etwa 12% Austauseh. Es ist vorli~ufig noeh unm6glich, zu entscheiden, weleher yon diesen zwei Austausehprozentsi~tzen der riehtigere ist.
Alle Lokalisationsversuehe kann man folgendermaBen zusammen- fassen. Die tgeihenfolge der Faktoren ist: ev - - Va' ~ l / - - m - - lz ~ bb.
Die Entfernungen dieser Gene voneinander betragen: ev - - -Va ' = 32 ; Va'

Zur Genetik der Drosophila funebris. I. 167
- - l / ~ - 7 , I / - - m = 1 3 ; m - - l z - ~ 4 , l z - - b b ~ 2 4 (oder 12). Wenn man nun annimmt, daf/ N ' links yon ev liegt, so wird die vorl~ufige Kar te des X-Chromosoms so aussehen wie sie auf Abb. 7 dargestetlt ist. Auf Abb. 7 sind auSer den schon beschriebenen Faktoren noch drei geschlechts-
0 8 32 40 r 60 64 7Q g5 H [ I [ [ Z P I I #' e# z~ vi zr ~4 ~ bb Z~
Abb. 7. Vorl'~ufige Lokalisa~ion der Gene Notch (N'), eversae (ev)~ Venae-abnor~. (Va% Weib- chen-letal (lf), miniatzo'e (m), lozenge (lz) und bobbed (bb) und der Le ta l fak to ren I b 1.~ und l~ im
X-Chromosom yon .Drosol)hila funebris.
gebundene Le:~algenovariationen lokalisiert, die im n~chsten Paragraphen beschrieben werden.
4. Geschlechtsgebundene Letalfaktoren, Gynander und das ]Nichttrennen der X-Chromosomen.
Letal/aktoren. Aui~er den sichtbaren, morphologisch sich manifestie- renden Genovariationen wurden bei Drosophila/unebris ebenso wie bei Drosophila melanogaster, auch geschlechtsgebundene Letalfaktoren ge- funden, die die sie enthaltenden M~nnchen t6ten und dadurch das Ge- schlechtsverhitltnis 2 : 1 (zugunsten der Weibchen) hervorrufen.
In l~6ntgenbestrahlungsversuchen sind viele letale Genovariationen entstanden und es hat sich gezeigt, dal~ ebenso wie bei Drosophila melano- gaster sie viel h~ufiger auftreten als die ,,sichtbaren" Genovariationen (TIMoF~nFF-RESSOVSKY, H. A., 1930). Abet auch in nicht bestrahl tem Material wurde ihr Auftreten mehrere Male beobachtet.
Da zur Ze~s des Auftretens der meistens Letalfaktoren die Kar te des X-Chromosoms bei Drosophila/unebris noch nicht ausgearbeitet war, so wurden diese Genovariationen nicht welter analysiert. Nur drei letale Genovariationen, die in fiir die Lokalisation gfinstigen Kreuzungskombi- nationen auftraten, wurden lokalisiert. Sie wurden als l~, 12 und 13 be- zeichnet und sind auf der Kar t e des X-Chromosoms auf Abb. 7 angefiihrt.
Tabelle 6. Kreuzung mit dem geschlechtsgebundenen rezessiven Letalfaktor Iz. + Va' + + + m
P : $ . - - • c ~ - - " l ~ + m
Zahl der 99 ~cP Zahlenver- h~ltnis d. Ge-
Fliegen + Va' ~ I Va' u~ + I Va' m Va%~ schlechter
154 13 61 53 -- 5 6 ! 1:0,08 Faktorer austausch: Va" - - m = 18,1% (13 + ~ und 15 Va' m 9: allen ~)
11 - - Va' = 8,6% (6 m c~: 70 ~, die zu erwarten w~ren).
Auf Tabelle 6 ist die 11 enthaltende Kreuzung angeffihrt. Normaler- weise mill]ten in dieser Kreuzung auBer den wenigen fiberlebenden Va'- und Va'm-M~nnchen, noch etwa 70 M~nnchen schltipfen, yon denen der

16 8 N.W. Timof6eff-Ressovsky:
grSl~te Tell minia ture und die fibrigen (etwa 20%) normal sein miii~ten. Es sind ~ber nut 6 minia ture und keine normalen M/s geschlfipft. Dieses erklirt sich durch die Annahme, dal~ das m4ragende X-Chromo- sore einen Letalfaktor enth~lt. Dieser Faktor muI~ links yon Va' liegen (da dutch Austauseh 6 m-Minnchen und keine normalen entstanden sind) und ergibt mit Va" etwa 8% Austausch. 3 yon den 53 m-Weibchen dieser Kreuzung wurden welter gezfichtet und ergaben alle ein 2 : 1-Geschlechts- verh~ltnis. Dadurch wird die Annahme best~tigt, dal3 das Fehlen der m-Minnchen nicht auf Zufall beruhte, sondern durch einen Letalfaktor im m-tragenden X-Chromosom hervorgerufen wurde.
Auf Tabelle 7 ist die l~ enthaltende Kreuzung angeffihrt. Das ev-
tragende Chromosom des P- ~ enth/~lt den Letalfaktor, der rechts yon m liegt und m i t m etwa 9% Austausch ergibt. ])as eine normale M~nnchen mu[3 (lurch doppelten Austausch ents~anden sein.
T~belle 7. Kreuzung mit dem rezessiven geschlechtsgebundenen Letalfaktor le. ev § l ~ ev § §
P : 9 + m + •
Zahl der Fliegen Zahlenverhiltnis der Geschlechter
278 1 : 0,53 Faktorenaustausch: m - - lu = 9,3% (1 + ~ und 8 ev ~: allen ~).
Tabelle 8. zeigt die 18 enthaltende Kreuzung. Dieser Letalfaktor ist links yon ev lokalisiert und ergibt mit ev etwa 8--9% Austausch.
Tabelle 8 Kreuzung mi~ dem rezessiven geschlechtsgebundenen Letalfaktor l:~.
l 3 e v + • + e v + P : - § m
Zahl der Fliegen Zahlenverhiltnis der Geschlechter
191 1 : 0,56 Faktorenaustausch: ev - - m = 42,0% (29 + ~: allen ~)
l e - - e v = 8,7% (6ev 3:allen ~).
Gynander. Ein Gynander bei Drosophi la [unebris wurde zuerst yon W. P. Spv,~ccw~ beschrieben (1927b). Nachdem wurden noeh einige Gy- nander bei Drosophi la /unebris gefunden und beschrieben (Tnwo~kE~F- RESSOVSKY, H. A., 1928). Im Laufe der weiteren Arbeit mit Drosophi la
/unebris wurden noch mehrere Gynander gefunden. Es soll hier keine ausfiihrliche Beschreibung aller Gyn~nder gegeben werden. Es sei nut erw/~hnt, dab mit NiNe der jetzt bekannten gescMechtsgebundenen Gene es m6glich ist, nicht nut die extremen Gynandertypen, sondern auch kleinere ,,mgnnliehe Flecke" bei heterozygoten Weibchen zu identifi-

Zur Genetik der Drosophila funebris. I. 169
zieren. So warden z. B. Weibchen gefunden, die heterozygot in Bezug auf bobbed waren und bei denen ein Teil der Thorax- oder Kopfborsten
bobbed manifestierten. Bei einem Weibchen der Konsti tut ion m b__~b war + +
der linke Flfigel miniature und gleichzeitig waren einige der Borsten an der linken Thoraxseite und fast alle Kopfborsten bobbed.
Das N~cht~rennen der Geschleehtsehromosomen. Es wurden in einer Kreuzung zwischen einem normalen ~ und eversae-~ unter 104 F I - ~ , 3 ev- ~ ~ gefunden. Anscheinend hat te das P- 9 die Konstitution X X Y. Dieser Fall wurde aber nicht weiter verfolgt.
ev + + In einer Kreuzung ~ ~ Va' • ~ m wurde in F1 ein Ausnahme-
weibchen gefunden, das eversae war. Dieses ~ wurde mit einem minia- ture-~ gekreuzt; diese Kreuzung ist auf Tabelle9 unter Nr. 213 an- gegeben. I)as Kreuzungsergebnis zeigt ein typisches Bild des sekundBren Nichttrennens und beweist damit, dab das P - ~ die Konsti tution X X Y hatte.
Tabelle 9. Sekund~res Nichttrennen in der Nachkommenschaft eines eversae- Weibchens, das durch prim/~res, aquationelles Nichttrennen der X-Chromosomen
entstanden ist. ev +
P : ? e--~-~- x ,~ - - -
Nr. der Zah lde r ~9 Kreuzmagen Yliegen + e v i
213 187 89 9 319 149 68 8 322 173 83 3
e v m !
83 6 7O 3 82 5
Total
% der Ausnahme-
9,2 10,5 3,5
7,7 i
% der Ausnahme-
6,7 4,1 5,7
% der Ausnahmen (?~+~)
8,0 7,4 4,6
6,6
2 yon den 9 ev- ~ der Kreuzung Nr. 213 wurden wieder mit miniature- ~ gekreuzt (Kreuzungen Nr. 319 und 322 der Tabelle 9) und haben auch sekund~res Nichttrennen ergeben. Man mut~ also annehmen, dab das urspriingliche eversae-Ausnahmeweibchen durch/~quationelles Nicht- t rennen zustande gekommen ist.
Das gesam~e Material der Tabelle 9, das allerdings sehr gering ist, er- gibt 6,6% sekund/ire Ausnahmen; diesel- Prozentsatz ist etwas hSher als bei Drosophila melanogaster (4,3), es ist aber vorl~ufig noeh unmSglich zu sagen, ob der Unterschied nicht rein zuf~llig ist.
Unter etwa 10000--12000 untersuchten bobbed- ~ ~ (die ph/s normal in Bezug auf bobbed sind) wurden 4 1Vf/~nnchen gefunden, die auch phBnotypisch das bobbed-Merkmal manifestierten. Diese 4 M~nnchen waxen alle absolut steril. Als Erkl~rung kann die Annahme dienen, dab diese M~nnchen XO sind und durch Befruchtung no~maler (bobbed ent-

170 N. W. Timof6eff-Ressovsky:
haltender) Eier mit Spermien ohne X- und ohne u ent- standen sind. Solche Spermien kSnnen dureh prim~res Niehttrennen w~hrend der Spermatogenese entstehen, wie es bei Drosophila melano- gaster der Fall ist.
Unter den M~nnehen, die genotypisch bobbed sind, wurden in 9 F~llen solche gefunden, die attf dem Thorax oder Kopf einen ph~notypisehen ,,bobbed-Fleck" hatten: eine lokal begrenzte Gruppe von Borsten war bobbed. Diese FMle deuten in dieselbe Riehtung, wie die oben angeffihrten sterilen bobbed.M~nnehen: im XO-Gewebe kann sich das Merkmal bobbed aueh bei M~nnchen manifestieren.
Eine eytologische Analyse des Niehttrennens der Gesehlechtsehromo- somen wurde bei Drosophila/unebris noch nieht durehgeffihrt.
5. Schluflbemerkungen.
Das bisher untersuchte Material fiber gesehleehtsgebundene Ver- erbung yon Drosophila ]unebris ist noch verhMtnism~l]ig sehr gering. Man kann aber daraus doch schon manehe, wenn auch nur vorl~ufige Schlul]folgerungen ziehen,
Es hat sieh vor allem gezeigt, dab die gesehleehtsgebundene Ver- erbung yon Drosophila /unebris im allgemeinen einen weitgehenden Parallelismus zu tier yon Drosophila melanogaster aufweist: neben ,,sieht- baren" Genovariationen entstehen auch Letalfaktoren; die letzteren treten ebenso wie bei Drosophila melanogaster bedeutend h~ufiger auf als die ,,siehtbaren"; yon Zeit zu Zeit werden Gynander gebildet; das pri- m~re Nichttrennen der Gesehlechtsehromosomen kann sowohl beim Weibehen als auch beim M~nnchen stattfinden; die XO-M~nnehen sind, wie bei Drosophila melanogaster, sterfl.
Einige yon den ,,siehtbaren" Genovariationen bei Drosophila/unebris und Drosophila melanogaster sind parallel: die Faktoren Notch, eversae, miniature, lozenge und bobbed yon Drosophila ]unebris sind den Faktoren Notch, wavy, miniature, lozenge und bobbed von Drosophila melanogaster ph~notypiseh i~uBerst ~hnlieh. Die Gene N', mund lz k5nnen mit grol]er Wahrseheinlichkeit und das Gen bb mit Sieherheit bei den beiden Arten als homolog bezeiehnet werden. Besonders interessant ist in dieser Hin- sieht bobbed.
bobber wurde bisher bei vier Arten gefunden: bei Drosophila melano- gaster, Drosophila simulans, Drosophila hydei und Drosophila /unebris. Bei allen vier Arten weist bobbed denselben typischen ~r auf: kurze Borsten, abnorme Abdomenb~nderung, verl~ngerte Entwick- lungsdauer und geschlechtsbegrenzte ~ani/estierung. Diese letztere ver- h~lt sieh in ganz gleieher Weise bei Drosophila melanogaster, Drosophila hydei und Drosophila/unebris: die bobbed-M~nnchen sind ph~notypiseh normal. Bei Drosophila simulans dagegen manifestiert sieh bobbed bei

Zur Genetik der Drosophila funebris. I. 171
Mannchen starker als bei Weibchen (STURTEVANT 1929). Fiir Droso- phila melanogaster wurde der Beweis erbracht, dab im u der Mannchen ein Faktor liegt, der die Manffestierung yon bobbed unter- drfickt; wahrscheinlich ist es das normale Allel yon bobbed (STE~ 1928). Dieselbe Erkl,~rung muB auch ffir bobbed bei Drosophila hydei und Droso- phila/unebris zutreffen. Drosophila simulans enth~lt aber im Y-Chro- mosom anscheinend einen , ,mutierten" bobbed-Faktor und dieser ist ein ,,extremeres" Allel als die bobbed.Genovariation im X-Chromosom.
Auf Abb. 8 sind die Geschlechtschromosomenkarten yon Drosophila melanogazter und Drosophila/unebris gezeichnet, auf denen die ,,paral- lelen" Genovariationen angegeben sind. Wenn man diese Genovariatio- nen bei den beiden Arten ffir homolog h~lt, so ergibt sich, dab die lz-m-wy. Strecke des X-Chromosoms yon Drosophila melanogaster bei Drosophila ]unebris in Form einer Inversion vorhanden ist: die entspreehenden Fak- toren liegen in entgegengesetzter Reiher~olge. Es muB aber betont wer- den, dab ohne die )r direkter Kreuzung, nur auf Grund der
,v' k, I I
I I I 1 IV' eu ~ l z Z~
Abb. 8. u der Lokalisation yon .parallelen '~ Genovariationen in den i-Chromosomen yon Drosophi~ me~anogaster (oben) und Drosophila fu~br i s (unten).
ph~notypischen (und zum Teil,,loka|isatorisehen") J~hnliehkeit, man nut mit einer gewissen Wahrseheinlichkeit yon der Homologie yon Geno- variationen bei verschiedenen Arten sprechen kann.
Nachdem wit kurz die Parallelismen in der geschlechtsgebundenen Vererbung bei Drosophila/unebris und Drosophila melanogaster erw~hnt haben, muB auf die Unterschiede in der Mutabilit~t dieser zwei Arten hingewiesen werden.
Es f~llt vor allem auf, dab die am h~ufigsten bei Drosophila melano. gaster auftretenden geschlechtsgebundenen Genovariationen yellow, white und vermilion bei Drosophila/unebris noch nie beobaehtet wurden; bei Drosophila melanogaster wtirde man sie bei der gleichen Fliegenzahl be- st immt finden.
AuBerdem i~uBert sich in der geschlechtsgebundenen Vererbung eine Eigenschaft, die im allgemeinen ffir die gesamte Genovariabflit~t de r Drosophila/unebris gilt und allen, die mit dieser Art gearbeitet haben, aufgefallen ist : fast alle Genovariationen zeigen eine unvollkommene und sehwankende Manifestierung, die durch Selektion und durch auBere Be- dingungen beeinfluBt werden kann. Von den gesehleehtsgebundenen

172 N. W. Timof~eff-Ressovsky:
Genovariationen hat nur miniature eine konstante und alternative Mani'- festierung; bei den fibrigen sehwankt die Expressivit~t und die Pene- tranz ist in manchen F~llen sehr. schwaeh.
In diesem Zusammenhange ist auch noch folgendes interessant. Die Eigenschaften des ,,normalen" wilden Typus sind bei Drosophila melano- gaster anscheinend viel konstanter als bei Drosophila /unebr~s. Augen- farbe, KSrperf~rbung und -grSJ]e, die Zahl, Lage und L~nge der Borsten zeigen bei Drosophila ]unebris aueh innerhalb einer lokalen Population eine bedeutende Variabflit~t, die zum Teil wenigstens nicht erblich ist.
Uber die Ursachen der oben aufgez~hlten Untersehiede zwisehen Drosophila melanogaster und Drosophila/unebris kann vorl~ufig noeh nichts bestimmtes ausgesagt werden.
Das Fehlen mancher Genovariationen bei Drosophila/unebris, die bei Drosophila melanogaster besonders h~ufig auftreten, kSnnte zweierlei Er- kl~rungen linden: entweder sind die entsprechenden Gene bei Drosophila /unebris dutch stabilere Allele vertreten oder sind bei Drosophila/unebris Gene vorhanden, dis die Wirkung der entspreehenden Genovariationen unterdriicken. Solehe ,,Unterdriieker" (Supressors) wurden bei Droso- phila melanogaster in einigen F~llen tats~chlich beobaehtet (MORGA); 1929, PLOVG~ 1928). Eine ~hnliche ,Unterdriickung" kann auch bei Verdoppehmgen (dureh Translokation, Polyploidie) entspreehender nor- maler Allele vorkommen.
Die variable Manifestierung der meisten Genovariationen und der erb- liche Anteil an der st~rkeren ,,normalen Variabilit~t" der Drosophila/u- nebris kSnnte vielleicht zum Teil au~ einer im Prinzip ~hnlichen Ersehei- hung beruhen: auf grSflerer Zahl analog wirkender, zum Tefl identiseher Gene. Drosophila/unebris hat eine hShere Chromosomenzahl als Droso- phila melanogaster und vielleicht ist ein Teil ihres Chromosomenkom- plexes phylogenetisch dutch Verdoppelungen yon Chromosomenteilen einer einfacheren urspriinglichen Chromosomengarnitur entstanden.
Die oben angedeuteten und auch andere m5gliche Erkl~rungsversuche bleiben vorl~ufig unbeweisbare Vermutungen. Begrtindete Einsichten in die Ursaehen der verschiedenen Gestaltung der erbliehen Variabilit/~t werden nur nach weiterem Ausbau der vergleichenden Genetik versehie- dener Drosophila-Arten m6glich sein.
6. Zusammenfassung. 1. Es wurden elf geschleehtsgebundene Genovariationen yon Droso-
phila ]unebris beschrieben, darunter ein rezessiver Letalfaktor, der in homozygotem Zust~nde die Weibchen tStet; die l~/~nnehen werden nieht betroffen, wahrscheinlich wegen Anwesenheit des normalen Alleles dieses Gens im Y-Chromosom.
2. Seehs yon den ,siehtbaren" Genovariationen, der die Weibchen

Zur Genetik der Drosophila funebris. I. 173
tS tende Letal faktor und drei , ,gewShnliche" Leta l fak toren wurden lokall-
siert (siehe Abb. 7). 3. Das Vorkommen yon Letalfaktoren, Gynande r u n d des Nieht-
t r ennens der Geschlechtschromosomen bei Drosophila /unebris wurde erw~hnt .
4. Die gesehleehtsgebundene Vererbung der Drosophila /unebr~s wurde mi t der yon Drosophila melanogaster vergliehen. Wahrseheinl ich s ind die Genovar ia t ionen Notch, eversae, miniature, lozenge u n d bobbed der Drosophila/unebris den Genovar ia t ionen Notch, wavy, miniature, lozenge u n d bobbed der Drosophila melanogaster homolog. Anderersei ts wird der Untersehied in der erbl iehen Variabi l i t~t dieser zwei Ar t en betont , der dar in besteht , da~ : a) manehe bei Drosophila ~nelanogaster besonders h~u- fige Genovar ia t ionen bei Drosophila/unebris noeh nie beobachte t wurden und b) fast alle Genovar ia t ionen der Drosophila /une~r~ eine unvoll- kommene und variable ph~notypisehe Manifest ierung zeigen.
7. Literatur. 1. Balkasehina, E . I . : EinfluB des Genotyps auf die mannigfaltige Mani-
festierung der Genovariation alae curvatae bei Drosophila ]unebris. Z. ~ksper. Biol. (russ.) 2 (1926). - - 2. Mohr, 0. a. Sturtevant, A. H.: A semi-lethal in Drosophila funebris. Proc. Soc. exper. Biol. a. Med. 16 (1919). - - 3. Morgan, T. H.: Data relating to six mutants of Drosophila. Carn. Inst. of Wash. Publ. Nr 399 (1929). - - 4. Morgan, T. H., Bridges, C. B. a. Sturtevant, A. H.: The genetics of Drosophila. Bibliogr. Genet. 2 (1925). - - 5. Nachtsheim, H.: Bei- trag zur Topographie des X-Chromosoms yon Drosophila melanoffaster. Z. Ind. Abst. Vererb. 48 (1928). - - 6. Plough, H.: Black suppressor-a sex linked gene in Drosophila causing apparent anomalies in crossing over in the _second chromo- some. Verh. V. intern. Kongr. Vererb. 2 (1928). - - 7. Reinig, W. F.: ~ber das Manifestieren :~,weier Genovariationen bei Drosophila /unebris. Biol. Zbl. 48 (1928). - - 8. Romasehoff, D. D.: Die Mutation Alae divergentes bei Drosophila ]unebris. Z. ~ksper. Biol. (russ.) 1 (1925a). - - 9. ~ber die Variabilitgt in der Manifestierung eines erbliehen Merkmales bei Drosophila/unebris. J. Psychol. u. Neur. 31 (1925b). - - 10. Romasehoff, D. D. u. Balkasehina, E. I.: Materialien zur Genetik yon Drosophila funebris. Z. ~ksper. Biol. (russ.) 5 (1929). - - 11. Spen- cer, W. P.: The X-Chromosome of Drosophila hydel. J. of exper. Zool. 47 (1927a). - - 12. A gynandromorph in Drosophila funebris. Amer. Naturalist 61 (1927 b). - - 13. New mutations in Drosophila ]unebris. J. of exper. Zool. 51 (1928). - - 14. Stern, C.: Ein genetischer und zytologischer Beweis fiir Vererbung im X-Chromosom yon Drosophila ~nelanogaster. Z. Ind. Abst. Vererb. 44 (1927). - - 15. Sturtevant, A. H.: A parallel mutation in Drosophila /unebris. Science (N. u 48(1918) . - 16. The genetics of Drosophila simulans. Carn. Inst. of Wash. Publ. Nr 399 (1929). - - 17. Timof~eff-Ressovsky, H.A.: Gynandromorphen und Genitalienabnormit~ten bei Drosophila /unebris. Roux' Arch. 113 (1928). - - 18. Induced Gene-variations in Drosophila ]unebris. J. Hered. 2I (1930a). - - 19. RSntgenbestrahlungsversuche mit Drosophila [unebris. Naturwiss. 18 (1930b). - - 20. Timof~eIf-Ressovsky, N. W.: Studies on the phaen0typic manifestation of here- ditary factors. I. The genovariation radius incompletus in Drosophila funebris. Z. ~ksper. Biol. (russ.) 1 (1925a); dasselbe engliseh: Genetics 12 (1927). - - 21. A re-

174 N. W. Timof4eff-Ressovsky:
verse genovariation in Drosophi la lunebris . Z. ~ksper. Biol. (russ.) 1 (1925b); dasselbe englisoh: Genetics 12 (1927). - - 22. Timof~eff-Ressovsky, N. W. u. H. A.: Ober das ph~notypische Manifestieren des Genotyps. II . Ober idiosomatischo Variationsgruppen bei Drosophi la]unebr i s . Roux' Arch. 108 (1926a). - - 23. Timo- I6ell-Ressovsky, N. W. u. Vogt, 0.: Ober idiosomatische Variationsgruppen und ihre Bedeutung fiir die Klassifikation der Krankheitseinheiten. Naturwiss. 14: (1926b). - - 24. Timof~eff-Ressovsky, N. W.: Der EinfluB der Temperatur auf die Ausbildung der Queradern bei einer Genovariation yon Drosoph i la /unebr@. Z . ~ksper. Biol. (russ.) 4 (1928); dasselbe deutseh: & Psychol. u. Neur. 38 (1929). 25. The phaenotypic realization of the gene vtl in Drosophi la ]unebri8. Trudy Vsesoj. Sjesda po Genetike, Leningrad (russ.) (1929).
8. Anhang: Tabellen zu den Lokal i sa t ionsversuchen .
Tabelle 1.
+ Va' + + P: ? 9 eT-~- x d ~ - - .
Zahl der 99 ~4' Zahlenverh~ltnis der Ftiegen + Va' [ + ev iVa' I Geschlechter
F 1 643 I 220 205 57 141 20 I 1:0,51
Faktmrenaustausch: e v - - Va ' = 28,8% (57 + ~7c~: 57 + ~ + 141 e v ~ ) .
Tabelle 2. + Ira ' ev + p:? ,~xaa
Zahlen- verh~tnis der Geschlechter
1 : 0,61
:Faktorenaustausch: ev - - Va" = 30,9% (84 + 9 9 + 93 ev Y a ' 99 : allen 9~)- e v - - Va" ~ 2 6 , 5 % (79 + 0 0 : 7 9 + ~ + 2 1 9 e v ~ ) .
Tabelle 3.
ev Va' ev + P : 9 ~ x 3 ~ I "
ZahI der ~9 I ~6 Zahlen- verh~ltnis der Fliegen + I ev Va' evVa r + ev Va' Geschlechter
[ 872 Ii99 1111f 89 1181 r172 t114 [ 0 i 1:0,50 I
Faktorenaustausch: ev - - Va" =34,5% (111 ev ~.~. + 89 Va" 9~ : allen ~ ) . e v - Va" = 39,8% (114 e v ~ : 172 + ~ + l l 4 e v ~ ) .

Zur Genet ik der Drosophila funebris. I. 175
P : ~ g ~ - -
Tabel le 4. ev Va ' • + + m
ev + +
I Zahlen- Zahl der[ ~ ~ , verh~iltnis der
I m Va'm Gesehlechter
F1 73 11oo1162 47 I 8 I19 1.o,5o Faktorenaustauseh:
ev - - Va" = 32,0% (73 ev ~ + 100 Va" ! ~ : allen ~!~). Ira " _ m -- 2]L,9%(47 + ~ # + 8 e v ~ : 4 7 + ~c~ + 8 e v ~ + 196m ~c~).
Tabelle 5. e v -k ev -k
I Zahl der 99 4'@ [ Zahlenverh~ltnis tier Fliegen q_ ev q- [ ev [ m ] Geschleehter
/~, 285 la5 I 209 I 241 1:0,97 Fak to renaus tauseh : ev - - m = 39,2% (135 + @~: 135 + @~ + 209 e v # ~ ) .
Tabelle 6.
+ m + m
Zahl der ?$ J J Zahlenverh~iltnis der o- o ,o
FI 310 1 : 0,98 Fak to renaus tauseh : ev - - m = 40,8% (214 + ~ ~: 214 + ~ ~ + 310 ev ~ ~ ) .
Tabelle 7. ev m + §
Zahl der ~ 9 @ d Zahlenverh~lltnis der Fliegen + + ev [ m Geschlechter
F 1 215 1 : 0,97 F~ktorenaus tausch : ev - - m = 44,8% (56 e v ~ : 56 e v ~ + 69 + ~ ) .
Tabel le 8.
ev + ev l z P : ? ? - T ~ - x c? ~
Zahl der I ~ ] @c? Zahlen- 1 verhitltnis d.
Fliegen ~ I ev l z evlz + ev l z evlz Geschlechter
F ~ [ 617 I 86 1 88 74 69 82 93 6 5 60 1 :0 ,95 Fak to renaus t ausch : e v - l z = 48,8% (alle + und evlz ~ : a l l e n 9!~).
ev - - lz = 47,3% (alle + trod e v l z ~ : a l l e n , 3 ) .

176 N. W. Timof~eff-I~essovsky:
Tabel le 9.
Q ev + ev bb P : 9 .-+-g-fiX4`4`--.
Zahl der ~9 Jd' Zahlenverh~iltnis der Fliegen + ev I bb l evbb + l e v Geschlechter
F 1 78 66 I : 1,07 Fak~orenaustauseh: e v - - bb = 53,1% (78 + 9 + 65 evbb 9: al len !~).
Tabelle 10.
ev bb ev bb P : 9 %-%- x 4 ' - - .
Zahl der $9 $3 Zahlenverh~ltnis der Fliegen + t ev bb I evbb + ev Geschlechter
F1 48 ] 44 81 1 :0 ,95
Fak to renaus tausch : ev - - b b = 48,6% (44 ev 9 + 42 bb 9: al len 9)-
Tabelle 11.
p : ~ . _ Va ' + X4`4` + +
Zahl dot ~9 $c~ Zahlenverhaltnis der Yliegen - + Va' + Va' m Va'm Geschlechter
$'1 144 I 136 29 33 122 4 1 : 0,67 Fak to renaus tausch : V a ' - - m ~ 19,2% (29 + 4 ` ~ : 2 9 + 4`4` + 122 m4`4`).
Tabet le 12.
Va' + X .P: o 9 %---~- 4 ' 4 - - - + m
F1
Zahl der ~!~ $$ Zahlen- verh~ltnis d.
Fltegen + ' Va' m Va'm + t Va' m Va'm Geschleehter
40 ] 242 I 218 I 41 ~ 46 221 t 1 :0 ,59 Fak to renaus tausch : V a ~ - - m = 14,3% (40 + 99 + 37 V a ~ m 99 : al len 99)-
V a ' - - m -~ 15,6%(41 + 4 ` ~ : 4 1 + 4`4` + 221 m 4 ` ~ ) .
P : 9 9 - -
Tabel le 13.
ev Va ' + x 3 g , + + m + + m
Zahl del ~ d~ Zahlen- verh~ltnls d
Fllegen + Vat m I Va'm + I Fat m Va'm ev Gesehlechte~
F 1 39 I 1771 120 I 31 33 1 99 - - 1 :0 ,51 :Faktorenaustauseh: Ira' - - m = 26,1% (39 + ~9 + 31 Va" m 99 : al len 99)- V a ' - - m = 27,7% (33 + ~ + 5 e v ~ : 3 3 + ~ + 9 9 m ~ + 5 e v ~ ) .

Zur Genetik der Drosophila funebris. I.
P : 9 9 - -
Tabelle 14.
Va' m X#(~ + +
+ m
177
F1
I Zahlen- Zahl der! 97 88' verhiiltnis d Fliegen I + Var m Va'm + Var m Va'm Geschlechte:
57 50 194 225 -- 53 3 1 : 0,53
Fak to renaus tauseh : V a ' - - m = 19,8% (57 Va" 9 9 + 50 m 99 : al len 99). V a ' - - m = 19,0% ( 5 3 m o ~ : 5 3 m ~ 3 + 225 + ~ ) .
P : 9 5 - - -
Tabelle 15.
+ Va ' m x # # - -
ev + +
+ + m
Zahl der 99 ~ 8 Zahlen- verhgltnis d
Fliegen 4- I Va' I m I Va'm + Va' I m Va'm ev Oeschlechte:
FI 167 7 64 40 233 ] 1 :0 ,47 i
Fak to renaus t ausch : V a ' - - m = 16,1% (101 Va" 9 9 + 74 m 99 : al len 99). V a ' - - m = 13,8% ( 6 4 m ~ : 167 + ~ + 6 4 m ~ + 233 e v ~ ) .
e v - - V a ' = 35,9% (167 + ~ : 167 + ~ + 6 4 m ~ + 2 3 3 e v 6 ~ ) .
Tabelle 16.
ev Va ' + X d ~ - - . P : 9 T 7 Iz
+ + lz
F1
Zahl der 99 88 Fliegen + Va' lz Va'lz + ev lz evlz
24 60 57 17 18 1 45 20
Fak to renaus t ausch : Va" - - Iz = 26,0% (alle + und Va" lz 99 : al len 99). V a " - - l z = 22,6% (alle + und e v ~ : a l l e n 3 ~ ) .
ev - - Va" = 25,0% (alle ev und evlz ~ 3 : allen ~ ~).
Zahlen- verh~ltnis d Geschlechte
1 : 0,53
Tabelle 17.
Va' + + bb
Zahl der 99 88 Zahlenverh~ltnis der Fliegen + Va' bb Va'bb ]+82 Va' Geschlechter
2' 1 45 92 93 42 1 : 0,48
Fak to renaus tausch : V a ' - - b b = 32,0% (45 + 9 + 42 V a ' b b 9: al ien 9)-
w. Roux' Archiv f. Entwicklungsmechanik Bd. 124. 12

178 N. W. Timoi~eff-Ressovsky:
P : 9 9
Tabelle 18. ev V a ' +
x # # - - . + + bb
ev + bb
F1
79 Zahl del Fliegen + ev Va' bb evVa' evbb Far evVa '
bb bb + ev Va I
Faktorenaustausch: ev - - V a " = 33,2% (124 e v ( b b ) 9 + 101 V a ' ( b b ) 9 : allen 9). e v - V a " - - 38,0% (122 evc~: 199 + ~' + 122 ev~).
V a ' - - bb = 31,2% (115 + ( ev ) 9 + 97 V a " bb (ev ) 9: allen 9). e v - bb = 64,4% (alle c r . - ov . 9: allen 9).
Zahlen- verh~ltnis der Geschlechter
1 : 0,48
Tabelle 19. m+•
P : 9 9 7 ~ - 4 m §
Zahl der 99 6'g' Zahlenverhgltnis der Fliegen § I m § m l z m l z Geschlechter
F 1 5 132 166 3 I 1:0,94 Faktorenaustausch: m - lz = 2,6% (5 + ~ + 3 m l z ~ : a l l e n , ) .
P : 9 9
Tabelle 20. V a ' m §
x3~-- ++Iz
+ m §
F1
Zahl der 99 ~3 Zahlen- verhgltnis d
Fliegen 22§ Va' I m ] Va 'm + m l z m l z ~eschlechte,
10 51 172 1 ' 1:0,43 Faktorenaustauseh:
V a " - - m = 20,7% (alle V a " und m 99: allen 99). V a " - - m = 22,2% (alle m u n d m l z ~ (~ : a l l e n ~ ) .
m - - Iz = 4,7% (alle + und m l z ~ : a l l e n ~ ) .
V a ' - - Iz = 26,9% (alle cr. - - ov . ~ c~ : allen 3 3).
Tabelle 21. m + P : 9 9 ~ X ~ 4 - - " m bb
Zahl der 99 ~ Zahlenverhgltnis der Fliegen § m bb m b b + I m Geschlechter
-~1 32 71 18 90 I 64 1 :0 ,96 Faktorenaustausch: m - - bb = 36,0% (40 + 9 + 18 m b b 9: allen 9).

Zur Genetik der Drosophila funebris. I. 179
Tabelle 22.
m bb P : ? 9 -4---4- x 4 4 - - .
m bb
Zahl der ?9 ~ Zahlenverh~iltnis der Fliegen + 'm bb mbb -k ] m Geschlechter
/
F1 116 31 30 I 106 1 :0 ,95 f
Fak to renaus t ausch : m - - bb = 23,6% (31 m !~ + 30 bb 9: al len 9).
Tabelle 23.
Va ' m + + m bb P : ? ? + + b ~ •
FI
Zahl der Fliegen
?? ~ Zahlen- i verhiiltnis der
+ Va' m bb Va'm Va' mbb Va'm + m Varm Geschlechter bb bb
Faktorenaus~ausch: Va" - - m = 23,5% V a ' - - m - 20,7%
m - - b b = 33,8% Va" - - bb = 57,3%
P : ? 9 - -
(30 Va ' (bb ) ~. + 23 m ( b b ) ~: al len 9)- ( 2 4 m 4 : 9 2 + 4 + 2 4 m ~ ) . 46 + ( V a / ) 9 + 3 0 m b b ( V a ' ) 9: al len 9). (alle cr. - - ov. 9 : allen 9)-
Tabelle 24. Va ' + + + m bb
x 4 4 - - - + m b b
1 : 0,52
?g ~J Zahlen- Zahl der l verh~ltnis der Fliegen + ira ~ m bb Va' m Va' mbb Va' m + m I Ira' Oeschlechter
bb bb
1 : 0,37 Faktorenaus~ausch:
Va" - - m = 22,6% (44 + (bb) 9 + 30 V a ' m ( b b ) 9: allen 9). m - - b b = 20,5% (26 m ( V a ' ) 9 + 41 b b ( V a ' ) 9: al len 9).
V a ' - - bb = 43,i% (alle cr. - - ov. 9: al len 9).
Tabelle 25. ev Va ' + + + + lz bb
P : 9 ? + + Iz bb •
Zahl ~? d~ Zahlen- der verh~ltnis d,
Flie- Va' Va' lzbb Va' + ev lz evlz Gesehlechte~ gen + Ira r Iz bb lz bb lzbb
1 : 0,47 Fak~orenaus tausch:
V a ' - - lz = 23,0% (alle + , b b V a ' l z und Va ' l zbb 99 : al len g~9). V a ' - - l z = 26,3% (alle + und e v 3 ~ : a l l e n 4 ~ ) .
Iz - - bb = 12,5% (alle lz , bb, V a ' l z und V a ' b b ~9: al len 99). V a ' - - b b = 35,5% (alle c r . - ov. 99 : al len 99). ev - - Va" = 30,7% (alle ev und evlz 4 4 : al len 4 4).
12"
180 N.W. Timof6eff-Ressovsky: Zur Genetik der Drosophila funebris. I.
Tabelle 26. ev V a ' + + ev + lz bb
P : ~ • + + lz bb
Zahl ~? 33 der
Flie- ev ev ev Va' Va' lz ~ l ~a' evl~ Va'iz va'eV [+ ev lg ev gen + ev Ira ~ lz bb ira ~ lz bb lz bb bb lz bb bb bb ~zbb lz
208 Faktorenaustausch:
ev - - V a ' = 28,7% (nach ~!~ berechnet). ev - - Va" = 32,8% (nach ~ ~ berechnet).
V a " - - Iz = 20,3% (nach ~ berechnet). V a " - - lz = 25,4% (nach c~ ~ berechnet).
lz - - bb = 11,2% (nach i~ 9 berechnet). V a ' - - bb = 31,5%.
e v - - bb = 60,2%.
Zahlen- verhalt his d.Ge schlech
ter
1 : 0,45