Embed Size (px)
Citation preview

Short Communication
Zusammensetzung der Epikutikularwachse von Halimium halimifolium ( Cistaceae)
Composition of Epicuticular Waxes from Halimium halimifolium ( Cistaceae)
PAUL-GERHARD GOLZ, MONIKA ROSINSKI und CLAUDIA EICH
Botanisches Institut der Universitat Koln, Gyrhofstr. 15, D-5000 Koln 41, F.R.G.
Eingegangen am 1. Juli 1981· Angenommen am 16. Juni 1982
Summary
The epicuticular waxes from leaves and petals of Halimium halimifolium sampled at their natural habitat in Sardinia as well as from plants cultivated in Cologne were studied. From the two plant series of different habitats the composition of alkanes, free alcohols, free acids and wax esters including their constituent alcohols and fatty acids was analyzed. The patterns of homologous series found in the different wax components were identical in plants grown either in Sardinia or Cologne. Therefore, soil and climate differences between Cologne (humid climate) and Sardinia (mediterranean semiarid climate) may not influence the composition of epicuticular waxes from Halimium halimifolium. Only the patterns of free fatty acids showed primary tendency for environmental influence.
Key words: Cistaceae, Halimium, leaves, petals, epicuticular waxes, environmental influence.
Einleitung
Halimium halimi/olium (Cistaceae) ist vor allem im westlichen Mittelmeergebiet beheimatet (Proctor and Heywood, 1968). Blatter und Blliten dieser Pflanze wurden daher sowohl in ihrem natlirlichen Yerbreitungsgebiet, im vorliegenden Fall auf Sardinien, geerntet als auch von in Kaln kultivierten Pflanzen. Yon diesen Pflanzenorganen wurden die epikutikularen Wachse isoliert und analysiert und der Einflu8 der unterschiedlichen Herklinfte auf ihre Zusammensetzung hin diskutiert.
Material und Methoden
1. H halimifolium wurde aus Samen (Herkunft: Estremadura und Mallorca) gezogen und im Gewachshaus in Koln kultiviert. Ernte der Jahrestriebe: September 1977.
2. Blatter und Bliiten von H halimifolium wurden im natiirlichen Verbreitungsgebiet geerntet. Fundstelle: Mazzaccara, Sardinien, 20 m ii. NN. Erntezeit: jeweils im Mai 1977, 1978, 1979.
Z. PJlanzenphysiol. Ed. 107. S. 281-287. 1982.
Short Communication
Zusammensetzung der Epikutikularwachse von Halimium halimifolium ( Cistaceae)
Composition of Epicuticular Waxes from Halimium halimifolium ( Cistaceae)
PAUL-GERHARD GOLZ, MONIKA ROSINSKI und CLAUDIA EICH
Botanisches Institut der Universitat Koln, Gyrhofstr. 15, D-5000 Koln 41, F.R.G.
Eingegangen am 1. Juli 1981· Angenommen am 16. Juni 1982
Summary
The epicuticular waxes from leaves and petals of Halimium halimifolium sampled at their natural habitat in Sardinia as well as from plants cultivated in Cologne were studied. From the two plant series of different habitats the composition of alkanes, free alcohols, free acids and wax esters including their constituent alcohols and fatty acids was analyzed. The patterns of homologous series found in the different wax components were identical in plants grown either in Sardinia or Cologne. Therefore, soil and climate differences between Cologne (humid climate) and Sardinia (mediterranean semiarid climate) may not influence the composition of epicuticular waxes from Halimium halimifolium. Only the patterns of free fatty acids showed primary tendency for environmental influence.
Key words: Cistaceae, Halimium, leaves, petals, epicuticular waxes, environmental influence.
Einleitung
Halimium halimi/olium (Cistaceae) ist vor allem im westlichen Mittelmeergebiet beheimatet (Proctor and Heywood, 1968). Blatter und Blliten dieser Pflanze wurden daher sowohl in ihrem natlirlichen Yerbreitungsgebiet, im vorliegenden Fall auf Sardinien, geerntet als auch von in Kaln kultivierten Pflanzen. Yon diesen Pflanzenorganen wurden die epikutikularen Wachse isoliert und analysiert und der Einflu8 der unterschiedlichen Herklinfte auf ihre Zusammensetzung hin diskutiert.
Material und Methoden
1. H halimifolium wurde aus Samen (Herkunft: Estremadura und Mallorca) gezogen und im Gewachshaus in Koln kultiviert. Ernte der Jahrestriebe: September 1977.
2. Blatter und Bliiten von H halimifolium wurden im natiirlichen Verbreitungsgebiet geerntet. Fundstelle: Mazzaccara, Sardinien, 20 m ii. NN. Erntezeit: jeweils im Mai 1977, 1978, 1979.
Z. PJlanzenphysiol. Ed. 107. S. 281-287. 1982.
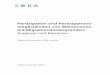
282 PAUL-GERHARD GULZ, MONIKA ROSINSKI und CLAUDIA EICH
Die Extraktion der kutikularen Wachse erfolgte sofort nach der Ernt~ mit Chloroform. Die extrahierten Wachse wurden uber Kieselgel-60-Saulen (Merck) in drei Fraktionen aufgetrennt (Konigs und Gulz, 1974). Mit Pentan wurden die Kohlenwasserstoffe eluiert, mit 2-Chlorpropan die Ester und mit Methanol freie Alkohole und Sauren. Die einzelnen Fraktionen wurden gaschrom~tographisch analysiert.
Table 1: Yields of epicuticular wax fractions from H. halimifolium.
leaves petals Cologne Sardinia Sardinia
fresh weight (g) 616 251 628 dry weight (g) 228 93 113 % dry weight 37 37 18 wax
mmg 1106 463 453 wax
in % of dry weight 0.5 0.5 0.4 pentane fraction
inmg 90 51 135 in % of dry weight 0.04 0.05 0.12
2-~hloropropane fraction 488 212 114 mmg
in % of dry weight 0.22 0.23 0.10 methanol fraction
mmg 408 136 130 in % of dry weight 0.18 0.15 0.12
Die Ausbeuten der Fraktionierung sind in Tabelle 1 zusammengefafk Die Auftrennung von Alkanen und Alkenen geschah auf Kieselgelplatten, die mit AgN03 (0,6 g in 10 ml Acetonitril) impragniert worden waren. Laufmittel: Isooctan (Gulz et al., 1978). Die Hydrierung der Alkene wurde mit Palladium als Katalysator durchgefiihrt. Die Verseifung der Wachsester erfolgte mit athanolischer HCI (6%) (Tulloch und Weenink, 1969). Die freien Alkohole und Fettsauren der Methanolfraktion wurden uber pdparative Kieselgelplatten getrennt; Laufmittel: Benzol. Zu diesem Zweck wurden die freien Fettsauren mit methanolischer H2S04 (5 %) in die Methylester uberfuhrt.
IR-Spektren: Perkin Elmer, Modell Infracord 137, KBr-PreBlinge. Gaschromatographie: Gerat: Hewlett-Packard Modell 5750 und 5830 mit Flammenionisationsdetektor. Saulen: Kohlenwasserstoffe: 25 m Glaskapillarsaule OV 101
Temperaturprogramm 1500 -220 oC, 4°C/Min. Wachsester: 25 m Glaskapillarsaule OV 101 Temperaturprogramm 1800 -300 oC, 4°C/Min. Fettsauremethylester, Fettsaureathylester, Alkohole: 10 m und 25 m Glaskapillarsaule FF AP Temperaturprogramm 1500 -220 oC, 4°C/Min. Gas-Vordruck 1,5kglcm2 N2
Die Mittelwerte it und ihre Standardabweichungen s resultieren 1. aus drei parallelen Aufarbeitungen der in Koln kultivierten Pflanzen und 2. aus den drei Aufarbeitungen auf Sardinien in den Jahren 1977, 1978, 1979.
z. Pjlanzenphysiol. Bd. 107. S. 281-287. 1982.
282 PAUL-GERHARD GULZ, MONIKA ROSINSKI und CLAUDIA EICH
Die Extraktion der kutikularen Wachse erfolgte sofort nach der Ernt~ mit Chloroform. Die extrahierten Wachse wurden uber Kieselgel-60-Saulen (Merck) in drei Fraktionen aufgetrennt (Konigs und Gulz, 1974). Mit Pentan wurden die Kohlenwasserstoffe eluiert, mit 2-Chlorpropan die Ester und mit Methanol freie Alkohole und Sauren. Die einzelnen Fraktionen wurden gaschrom~tographisch analysiert.
Table 1: Yields of epicuticular wax fractions from H. halimifolium.
leaves petals Cologne Sardinia Sardinia
fresh weight (g) 616 251 628 dry weight (g) 228 93 113 % dry weight 37 37 18 wax
mmg 1106 463 453 wax
in % of dry weight 0.5 0.5 0.4 pentane fraction
inmg 90 51 135 in % of dry weight 0.04 0.05 0.12
2-~hloropropane fraction 488 212 114 mmg
in % of dry weight 0.22 0.23 0.10 methanol fraction
mmg 408 136 130 in % of dry weight 0.18 0.15 0.12
Die Ausbeuten der Fraktionierung sind in Tabelle 1 zusammengefafk Die Auftrennung von Alkanen und Alkenen geschah auf Kieselgelplatten, die mit AgN03 (0,6 g in 10 ml Acetonitril) impragniert worden waren. Laufmittel: Isooctan (Gulz et al., 1978). Die Hydrierung der Alkene wurde mit Palladium als Katalysator durchgefiihrt. Die Verseifung der Wachsester erfolgte mit athanolischer HCI (6%) (Tulloch und Weenink, 1969). Die freien Alkohole und Fettsauren der Methanolfraktion wurden uber pdparative Kieselgelplatten getrennt; Laufmittel: Benzol. Zu diesem Zweck wurden die freien Fettsauren mit methanolischer H2S04 (5 %) in die Methylester uberfuhrt.
IR-Spektren: Perkin Elmer, Modell Infracord 137, KBr-PreBlinge. Gaschromatographie: Gerat: Hewlett-Packard Modell 5750 und 5830 mit Flammenionisationsdetektor. Saulen: Kohlenwasserstoffe: 25 m Glaskapillarsaule OV 101
Temperaturprogramm 1500 -220 oC, 4°C/Min. Wachsester: 25 m Glaskapillarsaule OV 101 Temperaturprogramm 1800 -300 oC, 4°C/Min. Fettsauremethylester, Fettsaureathylester, Alkohole: 10 m und 25 m Glaskapillarsaule FF AP Temperaturprogramm 1500 -220 oC, 4°C/Min. Gas-Vordruck 1,5kglcm2 N2
Die Mittelwerte it und ihre Standardabweichungen s resultieren 1. aus drei parallelen Aufarbeitungen der in Koln kultivierten Pflanzen und 2. aus den drei Aufarbeitungen auf Sardinien in den Jahren 1977, 1978, 1979.
z. Pjlanzenphysiol. Bd. 107. S. 281-287. 1982.

Epikutikularwachse von H halimifolium 283
Die aus diesen zwei MeBreihen erhaltenen Mittelwerte wurden mittels des t-Testes miteinander verglichen und auf signifikante Unterschiede hin gepriift; Signifikanz-Niveau 5 %; t (95, n); n = NJ + N2-2 (Kaiser, 1969).
Ergebnisse
KohlenwasserstojJe
Die Kohlenwasserstoffe der Blattwachse von H halimi/olium bestehen aus einer homologen Reihe von n-Alkanen der Kettenlange C19-C37 mit einer eindeutigen Hauptkomponente n-Nonacosan (61 %) (Tabelle2). Die Standardabweichungen der Prozent-Mittelwerte betragen bei in Koln kultivierten Pflanzen bis zu 2 %, bei den Pflanzen von Sardinien bis zu 5 %. Die Verteilungsmuster beider Herkiinfte sind gleich. Die quantitativen Werte liegen innerhalb der Standardabweichungen.
Bei den Petalen von H halimi/olium ist ebenfalls eine homologe Reihe von Alkanen mit gleichen Kettenlangen zu beobachten, jedoch ist die Hauptkomponente zu kiirzerer Kettenlange hin verschoben, n-Heptacosan. Auch sind in diesen Kohlenwasserstoffen Alkene der Kettenlange C23-C35 nachzuweisen, die nach Hydrierung zu n-Alkanen iiberfiihrt werden konnen. Die IR-Spektren weisen auf eine cis-Konfiguration der Alkene hin. Hier diirften, wie bei Cistus (Giilz et al., 1979 b), vorwiegend isomere Monoene vorliegen, die bei den Petalen von H halimi/olium 12 % der Kohlenwasserstoffe ausmachen (Tabelle 3).
Wachsester
Die Blattwachse enthalten Wachsester in einer homologen Reihe mit Kettenlangen von C34-CS4 • Sie bilden flache Verteilungsmuster mit schwach ausgepragten Hauptkomponenten (C44, C46, C48) (Tabelle 2). Die Gaschromatographie der Wachsester bedarf extrem hoher Temperaturbedingungen. Die Analyse der Wachsester fuhrt bei den verwendeten OV-101-Saulen zur Auftrennung nach Kettenlangen und la6t Doppelbindungen in diesen Verbindungen nicht in Erscheinung treten. Doppelbindungen in Wachsestern sind auf diese Weise nur iiber ihre Verseifungsprodukte nachzuweisen. Die Standardabweichungen der einzelnen Mittelwerte betragen bei in Koln kultivierten Pflanzen bis zu ± 5 %, bei Pflanzen von Sardinien bis zu ± 7 %. Die Verteilungsmuster entsprechen einander, die quantitativen Werte liegen innerhalb der Schwankungsbreiten. Gewisse Differenzen sind bei'C4o- und C48-Estern zu erwahnen.
Bei den Petalen sind die gleichen Kettenlangen der Wachsester zu beobachten, allerdings ist der Anteil der kiirzerkettigen Verbindungen angestiegen und bei C38 und C40 ein weiteres Maximum neben C46 (Tabelle3). In der Esterfraktion der Petalen sind auch T riterpenolester anzutreffen. Z wei weitere, bisher nicht identifizierte, Verbindungen sind im Gaschromatogramm der Esterfraktion vor den Wachsestern zu beobachten.
Z. Pf/anzenphysiol. Bd. 107. S. 281-287. 1982.
Epikutikularwachse von H halimifolium 283
Die aus diesen zwei MeBreihen erhaltenen Mittelwerte wurden mittels des t-Testes miteinander verglichen und auf signifikante Unterschiede hin gepriift; Signifikanz-Niveau 5 %; t (95, n); n = NJ + N2-2 (Kaiser, 1969).
Ergebnisse
KohlenwasserstojJe
Die Kohlenwasserstoffe der Blattwachse von H halimi/olium bestehen aus einer homologen Reihe von n-Alkanen der Kettenlange C19-C37 mit einer eindeutigen Hauptkomponente n-Nonacosan (61 %) (Tabelle2). Die Standardabweichungen der Prozent-Mittelwerte betragen bei in Koln kultivierten Pflanzen bis zu 2 %, bei den Pflanzen von Sardinien bis zu 5 %. Die Verteilungsmuster beider Herkiinfte sind gleich. Die quantitativen Werte liegen innerhalb der Standardabweichungen.
Bei den Petalen von H halimi/olium ist ebenfalls eine homologe Reihe von Alkanen mit gleichen Kettenlangen zu beobachten, jedoch ist die Hauptkomponente zu kiirzerer Kettenlange hin verschoben, n-Heptacosan. Auch sind in diesen Kohlenwasserstoffen Alkene der Kettenlange C23-C35 nachzuweisen, die nach Hydrierung zu n-Alkanen iiberfiihrt werden konnen. Die IR-Spektren weisen auf eine cis-Konfiguration der Alkene hin. Hier diirften, wie bei Cistus (Giilz et al., 1979 b), vorwiegend isomere Monoene vorliegen, die bei den Petalen von H halimi/olium 12 % der Kohlenwasserstoffe ausmachen (Tabelle 3).
Wachsester
Die Blattwachse enthalten Wachsester in einer homologen Reihe mit Kettenlangen von C34-CS4 • Sie bilden flache Verteilungsmuster mit schwach ausgepragten Hauptkomponenten (C44, C46, C48) (Tabelle 2). Die Gaschromatographie der Wachsester bedarf extrem hoher Temperaturbedingungen. Die Analyse der Wachsester fuhrt bei den verwendeten OV-101-Saulen zur Auftrennung nach Kettenlangen und la6t Doppelbindungen in diesen Verbindungen nicht in Erscheinung treten. Doppelbindungen in Wachsestern sind auf diese Weise nur iiber ihre Verseifungsprodukte nachzuweisen. Die Standardabweichungen der einzelnen Mittelwerte betragen bei in Koln kultivierten Pflanzen bis zu ± 5 %, bei Pflanzen von Sardinien bis zu ± 7 %. Die Verteilungsmuster entsprechen einander, die quantitativen Werte liegen innerhalb der Schwankungsbreiten. Gewisse Differenzen sind bei'C4o- und C48-Estern zu erwahnen.
Bei den Petalen sind die gleichen Kettenlangen der Wachsester zu beobachten, allerdings ist der Anteil der kiirzerkettigen Verbindungen angestiegen und bei C38 und C40 ein weiteres Maximum neben C46 (Tabelle3). In der Esterfraktion der Petalen sind auch T riterpenolester anzutreffen. Z wei weitere, bisher nicht identifizierte, Verbindungen sind im Gaschromatogramm der Esterfraktion vor den Wachsestern zu beobachten.
Z. Pf/anzenphysiol. Bd. 107. S. 281-287. 1982.
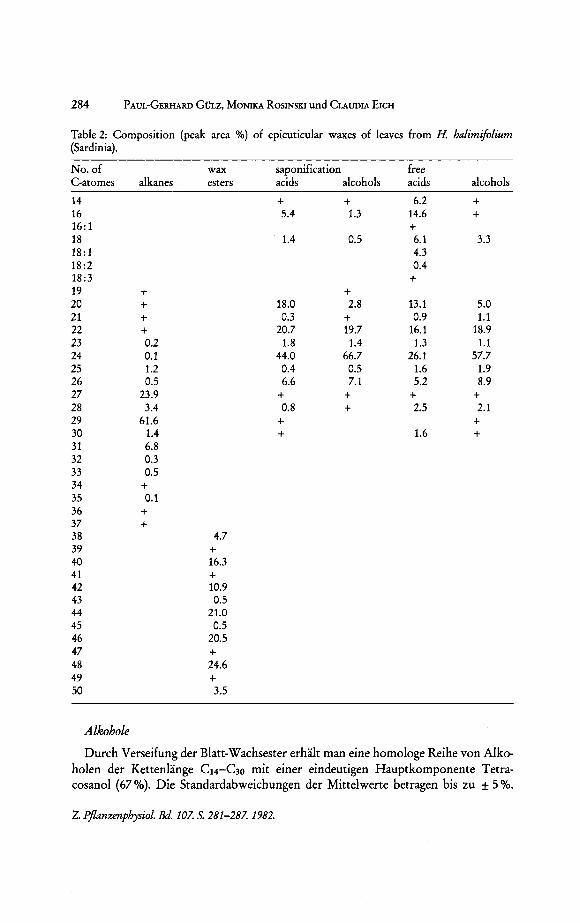
284 PAUL-GERHARD GOLZ, MONIKA ROSINSKI und CLAUDIA EICH
Table 2: Composition (peak area %) of epicuticular waxes of leaves from H. halimifolium (Sardinia).
No. of C-atomes
14 16 16:1 18 18: 1 18:2 18:3 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50
Alkohole
alkanes
+ + + + 0.2 0.1 1.2 0.5
23.9 3.4
61.6 1.4 6.8 0.3 0.5
+ 0.1
+ +
wax esters
4.7 + 16.3 + 10.9 0.5
21.0 0.5
20.5 + 24.6 + 3.5
saponification acids alcohols
+ 5.4
1.4
18.0 0.3
20.7 1.8
44.0 0.4 6.6
+ 0.8
+ +
+ 1.3
0.5
+ 2.8
+ 19.7 1.4
66.7 0.5 7.1
+ +
free acids
6.2 14.6 + 6.1 4.3 0.4
+
13.1 0.9
16.1 1.3
26.1 1.6 5.2
+ 2.5
1.6
alcohols
+ +
3.3
5.0 1.1
18.9 1.1
57.7 1.9 8.9
+ 2.1
+ +
Durch Verseifung der Blatt-Wachsester erhaIt man eine homologe Reihe von Alkoholen der Kettenlange C14-C30 mit einer eindeutigen Hauptkomponente TetracosanoI (67 %). Die Standardabweichungen der Mittelwerte betragen bis zu ± 5 %.
Z. Pjlanzenphysiol. Ed. 107. S. 281-287. 1982.
284 PAUL-GERHARD GOLZ, MONIKA ROSINSKI und CLAUDIA EICH
Table 2: Composition (peak area %) of epicuticular waxes of leaves from H. halimifolium (Sardinia).
No. of C-atomes
14 16 16:1 18 18: 1 18:2 18:3 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50
Alkohole
alkanes
+ + + + 0.2 0.1 1.2 0.5
23.9 3.4
61.6 1.4 6.8 0.3 0.5
+ 0.1
+ +
wax esters
4.7 + 16.3 + 10.9 0.5
21.0 0.5
20.5 + 24.6 + 3.5
saponification acids alcohols
+ 5.4
1.4
18.0 0.3
20.7 1.8
44.0 0.4 6.6
+ 0.8
+ +
+ 1.3
0.5
+ 2.8
+ 19.7 1.4
66.7 0.5 7.1
+ +
free acids
6.2 14.6 + 6.1 4.3 0.4
+
13.1 0.9
16.1 1.3
26.1 1.6 5.2
+ 2.5
1.6
alcohols
+ +
3.3
5.0 1.1
18.9 1.1
57.7 1.9 8.9
+ 2.1
+ +
Durch Verseifung der Blatt-Wachsester erhaIt man eine homologe Reihe von Alkoholen der Kettenlange C14-C30 mit einer eindeutigen Hauptkomponente TetracosanoI (67 %). Die Standardabweichungen der Mittelwerte betragen bis zu ± 5 %.
Z. Pjlanzenphysiol. Ed. 107. S. 281-287. 1982.
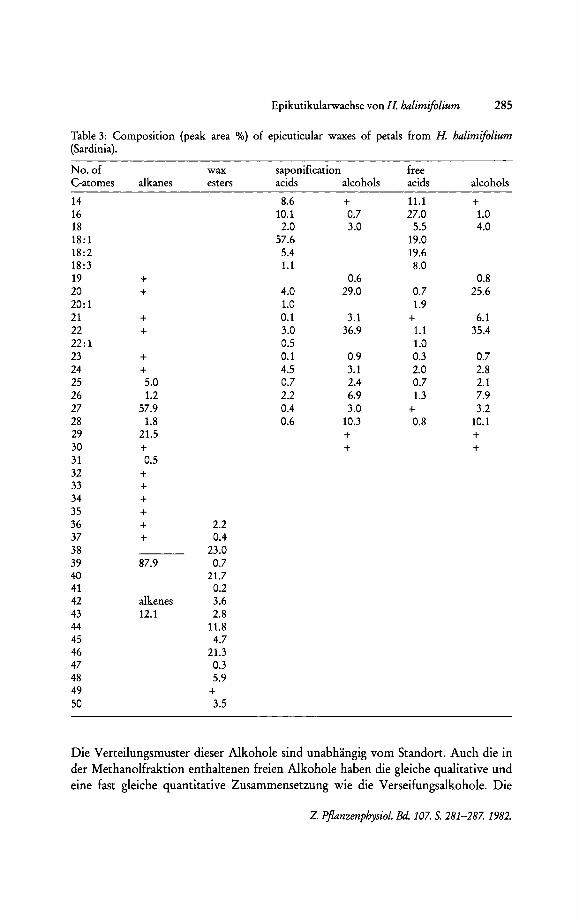
Epikutikularwachse von H. halimifolium 285
Table 3: Composition (peak area %) of epicuticular waxes of petals from H. halimifolium (Sardinia).
No. of wax saponification free C-atomes alkanes esters acids alcohols acids alcohols
14 8.6 + 11.1 + 16 10.1 0.7 27.0 1.0 18 2.0 3.0 5.5 4.0 18:1 57.6 19.0 18:2 5.4 19.6 18:3 1.1 8.0 19 + 0.6 0.8 20 + 4.0 29.0 0.7 25.6 20:1 1.0 1.9 21 + 0.1 3.1 + 6.1 22 + 3.0 36.9 1.1 35.4 22:1 0.5 1.0 23 + 0.1 0.9 0.3 0.7 24 + 4.5 3.1 2.0 2.8 25 5.0 0.7 2.4 0.7 2.1 26 1.2 2.2 6.9 1.3 7.9 27 57.9 0.4 3.0 + 3.2 28 1.8 0.6 10.3 0.8 10.1 29 21.5 + + 30 + + + 31 0.5 32 + 33 + 34 + 35 + 36 + 2.2 37 + 0.4 38 23.0 39 87.9 0.7 40 21.7 41 0.2 42 alkenes 3.6 43 12.1 2.8 44 11.8 45 4.7 46 21.3 47 0.3 48 5.9 49 + 50 3.5
Die Verteilungsmuster dieser Alkohole sind unabhangig vom Standort. Auch die in der Methanolfraktion enthaltenen freien Alkohole haben die gleiche qualitative und eine fast gleiche quantitative Zusammensetzung wie die Verseifungsalkohole. Die
Z. Pjlanzenphysiol. Ed. 107. S. 281-287. 1982.
Epikutikularwachse von H. halimifolium 285
Table 3: Composition (peak area %) of epicuticular waxes of petals from H. halimifolium (Sardinia).
No. of wax saponification free C-atomes alkanes esters acids alcohols acids alcohols
14 8.6 + 11.1 + 16 10.1 0.7 27.0 1.0 18 2.0 3.0 5.5 4.0 18:1 57.6 19.0 18:2 5.4 19.6 18:3 1.1 8.0 19 + 0.6 0.8 20 + 4.0 29.0 0.7 25.6 20:1 1.0 1.9 21 + 0.1 3.1 + 6.1 22 + 3.0 36.9 1.1 35.4 22:1 0.5 1.0 23 + 0.1 0.9 0.3 0.7 24 + 4.5 3.1 2.0 2.8 25 5.0 0.7 2.4 0.7 2.1 26 1.2 2.2 6.9 1.3 7.9 27 57.9 0.4 3.0 + 3.2 28 1.8 0.6 10.3 0.8 10.1 29 21.5 + + 30 + + + 31 0.5 32 + 33 + 34 + 35 + 36 + 2.2 37 + 0.4 38 23.0 39 87.9 0.7 40 21.7 41 0.2 42 alkenes 3.6 43 12.1 2.8 44 11.8 45 4.7 46 21.3 47 0.3 48 5.9 49 + 50 3.5
Die Verteilungsmuster dieser Alkohole sind unabhangig vom Standort. Auch die in der Methanolfraktion enthaltenen freien Alkohole haben die gleiche qualitative und eine fast gleiche quantitative Zusammensetzung wie die Verseifungsalkohole. Die
Z. Pjlanzenphysiol. Ed. 107. S. 281-287. 1982.

286 PAUL-GERHARD GtiLZ, MONIKA ROSINSKI und CLAUDIA EICH
Verseifungsalkohole der Petalen zeigen ein nicht SO steil ausgepragtes Verteilungsmuster mit einer Verschiebung der Hauptkomponente zu C22 (37%) und C20 (29%). Fast gleiche Profile sind von den freien Alkoholen aus Petalen zu erhalten (Tabelle 3).
Fettsauren
Die Verseifungsfettsauren werden in Form der Athylester analysiert. Sie liegen vor als homologe Reihe mit Kettenlangen von C I4-C30. Bei den Blattern bestehen sie liberwiegend aus gesattigten Fettsauren mit nur Spuren (2 %) an Monoenfettsauren. Ais Hauptkomponente beobachtet man Tetracosansaure (44 %). Die Standardabweichungen der Mittelwerte betragen bis zu ± 3 % bei Werten der in Kaln kultivierten Pflanzen und bis zu ± 6 % bei den auf Sardinien geernteten Pflanzen. Die Verteilungs muster sind gleich, unabhangig von der Herkunft. Die aus der Methanolfraktion isolierten freien Fettsauren haben ein ahnliches Verteilungsmuster wie die Verseifungsfettsauren. Abweichungen beobachtet man aber bei den auf Sardinien extrahierten Wachsen bei den Fettsauren C I6 und C24 (Tabelle2). Die Verseifungsfettsauren aus den Epikutikularwachsen der Petalen weisen einen hohen Anteil an ungesattigten Fettsauren der Kettenlange C I8 (64%) auf, und zwar vorwiegend Olsaure mit 57 %. Weitere ungesattigte Fettsauren in geringer Konzentration sind CI6:1, CI8:2,
C I8:3, C20:1, C22:1, C24:1•
Auch das Verteilungsmuster der freien Fettsauren aus Petalen zeigt ein ahnliches Profil. Deutlich ist aber ein Anstieg von C14- und CwFettsauren zu beobachten, und vor allem ist das Verhaltnis der ungesattigten C l8-Fettsauren zueinander stark verschoben; CI8:1, CI8:2, C18:3 verhalten sich nun wie 2: 2: 1 (Tabelle 3).
Annahernd gleiche Verteilungsmuster wie bei H halimi/olium erhalt man auch bei in Kaln kultivierten Pflanzen von H alyssoides. AIle typischen Charakteristika der Verteilungsmuster sind auch bei dieser Halimium-Art festzustellen. Nur einzelne geringe quantitative Unterschiede konnten beobachtet werden.
Diskussion
Die Zusammensetzung der Epikutikularwachse von H halimi/olium gleicht in sehr starkem Ma£e den Befunden, die bei Pflanzen der Gattung Cistus (Glilz et al., 1979 a, 1980) erhalten wurden, und sind ein Zeichen flir die nahe Verwandtschaft der beiden Cistaceae-Gattungen.
Die Mittelwerte aus drei parallelen Aufarbeitungen der in Kaln kultivierten Pflanzen haben natlirlich geringere Standardabweichungen zu verzeichnen, liberwiegend kleiner als ± 2 % bei Alkanen und kleiner als ± 5 % bei Wachsestern, wie die in drei aufeinanderfolgenden Jahren auf Sardinien geernteten Pflanzen (± 5 % bzw. ± 7 %).
Ein Vergleich der aus diesen zwei MeBreihen gefundenen Mittelwerte der einzelnen Wachskomponenten von Blattern wurde mit Hilfe des t-Testes durchgefiihrt und auf Ubereinstimmung oder signifikante Unterschiede hin gepriift. Bei einem SignifikanzNiveau von 5 % konnte eine Ubereinstimmung aller Mittelwerte der folgenden Ver-
Z. Pjlanzenphysiol. Bd. 107. S. 281-287. 1982.
286 PAUL-GERHARD GtiLZ, MONIKA ROSINSKI und CLAUDIA EICH
Verseifungsalkohole der Petalen zeigen ein nicht SO steil ausgepragtes Verteilungsmuster mit einer Verschiebung der Hauptkomponente zu C22 (37%) und C20 (29%). Fast gleiche Profile sind von den freien Alkoholen aus Petalen zu erhalten (Tabelle 3).
Fettsauren
Die Verseifungsfettsauren werden in Form der Athylester analysiert. Sie liegen vor als homologe Reihe mit Kettenlangen von C I4-C30. Bei den Blattern bestehen sie liberwiegend aus gesattigten Fettsauren mit nur Spuren (2 %) an Monoenfettsauren. Ais Hauptkomponente beobachtet man Tetracosansaure (44 %). Die Standardabweichungen der Mittelwerte betragen bis zu ± 3 % bei Werten der in Kaln kultivierten Pflanzen und bis zu ± 6 % bei den auf Sardinien geernteten Pflanzen. Die Verteilungs muster sind gleich, unabhangig von der Herkunft. Die aus der Methanolfraktion isolierten freien Fettsauren haben ein ahnliches Verteilungsmuster wie die Verseifungsfettsauren. Abweichungen beobachtet man aber bei den auf Sardinien extrahierten Wachsen bei den Fettsauren C I6 und C24 (Tabelle2). Die Verseifungsfettsauren aus den Epikutikularwachsen der Petalen weisen einen hohen Anteil an ungesattigten Fettsauren der Kettenlange C I8 (64%) auf, und zwar vorwiegend Olsaure mit 57 %. Weitere ungesattigte Fettsauren in geringer Konzentration sind CI6:1, CI8:2,
C I8:3, C20:1, C22:1, C24:1•
Auch das Verteilungsmuster der freien Fettsauren aus Petalen zeigt ein ahnliches Profil. Deutlich ist aber ein Anstieg von C14- und CwFettsauren zu beobachten, und vor allem ist das Verhaltnis der ungesattigten C l8-Fettsauren zueinander stark verschoben; CI8:1, CI8:2, C18:3 verhalten sich nun wie 2: 2: 1 (Tabelle 3).
Annahernd gleiche Verteilungsmuster wie bei H halimi/olium erhalt man auch bei in Kaln kultivierten Pflanzen von H alyssoides. AIle typischen Charakteristika der Verteilungsmuster sind auch bei dieser Halimium-Art festzustellen. Nur einzelne geringe quantitative Unterschiede konnten beobachtet werden.
Diskussion
Die Zusammensetzung der Epikutikularwachse von H halimi/olium gleicht in sehr starkem Ma£e den Befunden, die bei Pflanzen der Gattung Cistus (Glilz et al., 1979 a, 1980) erhalten wurden, und sind ein Zeichen flir die nahe Verwandtschaft der beiden Cistaceae-Gattungen.
Die Mittelwerte aus drei parallelen Aufarbeitungen der in Kaln kultivierten Pflanzen haben natlirlich geringere Standardabweichungen zu verzeichnen, liberwiegend kleiner als ± 2 % bei Alkanen und kleiner als ± 5 % bei Wachsestern, wie die in drei aufeinanderfolgenden Jahren auf Sardinien geernteten Pflanzen (± 5 % bzw. ± 7 %).
Ein Vergleich der aus diesen zwei MeBreihen gefundenen Mittelwerte der einzelnen Wachskomponenten von Blattern wurde mit Hilfe des t-Testes durchgefiihrt und auf Ubereinstimmung oder signifikante Unterschiede hin gepriift. Bei einem SignifikanzNiveau von 5 % konnte eine Ubereinstimmung aller Mittelwerte der folgenden Ver-
Z. Pjlanzenphysiol. Bd. 107. S. 281-287. 1982.

Epikutikularwachse von H halimifolium 287
bindungsklassen abgesichert werden: Alkane, Wachsester, Verseifungsalkohole, Verseifungsfettsauren und freie Alkohole. Die freien Fettsauren weisen jedoch einige signifikante Unterschiede bei den Fettsauren C14, C16, C26 auf.
Trotz der groBen klimatischen Unterschiede zwischen dem semiariden Mittelmeerklima auf Sardinien und dem feucht gemaBigten Klima in Koln sind bei Aufarbeitungen von diesen verschiedenen Herklinften sehr groBe Ohereinstimmungen in der Zusammensetzung der Epikutikularwachse bei H halimi/alium zu beobachten.
Lediglich bei den F ettsauren dieser Wachse scheinen die U mwelteinfllisse zu einer geringen Modifikation ihrer quantitativen Zusammensetzung zu flihren. Die Erklarung dlirfte darin zu suchen sein, daB die Fettsauren die Primarprodukte der Biosynthese alIer Komponenten der Kutikularwachse darstelIen (Kolattukudy et al., 1976).
Von drei auf Sardinien vorkommenden Cistus-Arten, C. manspeliensis, C. salvi/alius und C. incanus wurden ebenfalIs die Blatter und Blliten geerntet und die Epikutikularwachse isoliert und analysiert. Auch bei diesen Cistus-Arten war, unabhangig von ihrer Herkunft, eine weitgehende Obereinstimmung in der Zusammensetzung der Wachskomponenten zu beobachten. TendenzielI zeichneten sich jedoch bei den sehr kurzlebigen Petalen groBere Schwankungen urn die Mittelwerte der in Koln kultivierten Pflanzen ab, als bei den Blattern, und primar sind es wieder die Fettsauren, die aufgrund der unterschiedlichen Umweltfaktoren je nach ihrer Herkunft modifizierte Verteilungsmuster aufweisen. Die Zusammensetzung der Kutikularwachse scheint bei Pflanzen der Gattung Cistus und Halimium somit im wesentlichen eng spezifisch determiniert und nicht standortabhangig variabel zu sein.
Literatur
GULZ, P.-G., P. FAXEL, G. B. FEIGE, B. SCHMITZ und H. EGGE: Z. Pflanzenphysiol. 89, 159-167 (1978).
GULZ, P.-G., P. PROKSCH und D. SCHWARZ: Z. Pflanzenphysiol. 92, 341-347 (1979 a). GULZ, P.-G., E. AMIGONI, B. SCHMITZ und H. EGGE: Z. Pflanzenphysiol. 94, 35-43 (1979 b). Gtkz, P.-G.: In: Biogenesis and function of plant lipids. P. MAZLlAK et al. (eds.). Elsevier/North-
Holland Biomedical Press, Amsterdam, pp. 275-279 (1980). KAISER, R.: Chromatographie in der Gasphase, Bd. IV, Quantitative Auswertung 1. Teil. Hoch
schultaschenbuch-Verlag, Mannheim, Zi.irich, 1969. KONIGS, R. und P.-G. GULZ: Z. Pflanzenphysiol. 72,237-248 (1974). KOLATTUKUDY, P. E., R. CROTEAU, and J. S. BUCKNER: Biochemistry of plant waxes. In: Chemis
try and biochemistry of natural waxes. P. E. KOLATTUKUDY (eds.). Elsevier, Amsterdam, pp. 289-347 (1976).
PROCTOR, M. C. F. and V. H. HEYWOOD: Halimium, In: Flora Europaea Vol. 2, 284 Cambridge (1968).
TULLOCH, A. P. and R. O. WEENINK: Can. J. Chern. 47, 3119-3126 (1969).
Z. Pjlanzenphysiol. Ed. 107. S. 281-287. 1982.
Epikutikularwachse von H halimifolium 287
bindungsklassen abgesichert werden: Alkane, Wachsester, Verseifungsalkohole, Verseifungsfettsauren und freie Alkohole. Die freien Fettsauren weisen jedoch einige signifikante Unterschiede bei den Fettsauren C14, C16, C26 auf.
Trotz der groBen klimatischen Unterschiede zwischen dem semiariden Mittelmeerklima auf Sardinien und dem feucht gemaBigten Klima in Koln sind bei Aufarbeitungen von diesen verschiedenen Herklinften sehr groBe Ohereinstimmungen in der Zusammensetzung der Epikutikularwachse bei H halimi/alium zu beobachten.
Lediglich bei den F ettsauren dieser Wachse scheinen die U mwelteinfllisse zu einer geringen Modifikation ihrer quantitativen Zusammensetzung zu flihren. Die Erklarung dlirfte darin zu suchen sein, daB die Fettsauren die Primarprodukte der Biosynthese alIer Komponenten der Kutikularwachse darstelIen (Kolattukudy et al., 1976).
Von drei auf Sardinien vorkommenden Cistus-Arten, C. manspeliensis, C. salvi/alius und C. incanus wurden ebenfalIs die Blatter und Blliten geerntet und die Epikutikularwachse isoliert und analysiert. Auch bei diesen Cistus-Arten war, unabhangig von ihrer Herkunft, eine weitgehende Obereinstimmung in der Zusammensetzung der Wachskomponenten zu beobachten. TendenzielI zeichneten sich jedoch bei den sehr kurzlebigen Petalen groBere Schwankungen urn die Mittelwerte der in Koln kultivierten Pflanzen ab, als bei den Blattern, und primar sind es wieder die Fettsauren, die aufgrund der unterschiedlichen Umweltfaktoren je nach ihrer Herkunft modifizierte Verteilungsmuster aufweisen. Die Zusammensetzung der Kutikularwachse scheint bei Pflanzen der Gattung Cistus und Halimium somit im wesentlichen eng spezifisch determiniert und nicht standortabhangig variabel zu sein.
Literatur
GULZ, P.-G., P. FAXEL, G. B. FEIGE, B. SCHMITZ und H. EGGE: Z. Pflanzenphysiol. 89, 159-167 (1978).
GULZ, P.-G., P. PROKSCH und D. SCHWARZ: Z. Pflanzenphysiol. 92, 341-347 (1979 a). GULZ, P.-G., E. AMIGONI, B. SCHMITZ und H. EGGE: Z. Pflanzenphysiol. 94, 35-43 (1979 b). Gtkz, P.-G.: In: Biogenesis and function of plant lipids. P. MAZLlAK et al. (eds.). Elsevier/North-
Holland Biomedical Press, Amsterdam, pp. 275-279 (1980). KAISER, R.: Chromatographie in der Gasphase, Bd. IV, Quantitative Auswertung 1. Teil. Hoch
schultaschenbuch-Verlag, Mannheim, Zi.irich, 1969. KONIGS, R. und P.-G. GULZ: Z. Pflanzenphysiol. 72,237-248 (1974). KOLATTUKUDY, P. E., R. CROTEAU, and J. S. BUCKNER: Biochemistry of plant waxes. In: Chemis
try and biochemistry of natural waxes. P. E. KOLATTUKUDY (eds.). Elsevier, Amsterdam, pp. 289-347 (1976).
PROCTOR, M. C. F. and V. H. HEYWOOD: Halimium, In: Flora Europaea Vol. 2, 284 Cambridge (1968).
TULLOCH, A. P. and R. O. WEENINK: Can. J. Chern. 47, 3119-3126 (1969).
Z. Pjlanzenphysiol. Ed. 107. S. 281-287. 1982.








![Tabelle 14: Zusammensetzung von In-Ceram Zirkoniawebdoc.sub.gwdg.de/ebook/diss/2003/fu-berlin/2002/80/kap8.pdf · 122 Tabelle 14: Zusammensetzung von In-Ceram Zirkonia [130] Gerüstkeramik](https://img.pdfslide.org/doc/110x75/5e1070452c90ef6c6e2e9ec2/tabelle-14-zusammensetzung-von-in-ceram-122-tabelle-14-zusammensetzung-von-in-ceram.jpg)