
Immunsystem
110/08
Alfons Meyer
Grundlagen der ImmunologieEine kurzweilige Einführung
Was oft übersehen wird: Das Immunsystem gehört mit einer Masse von zwei bis dreiKilogramm zu den großen Organen des menschlichen Organismus, denn Immunzel-len und lymphatisches Gewebe sind im gesamten Körper verteilt. Neben seiner Kom-plexität besitzt das Immunsystem eine beeindruckende Dynamik hinsichtlich Zelltei-lung und Zelltod, Umbau der Organe durch Ein- und Auswanderung von Zellen. Ver-änderungen durch Differenzierung ist hier die Regel, nicht die Ausnahme. MeinerMeinung nach ist das Immunsystem das wichtigste Organ zur Integrität, Individuali-tät und Gesunderhaltung des Organismus. Das Immunorgan gliedert sich in zwei Haupt-funktionsgruppen: Das angeborene und das adaptive Immunsystem.
Die Zellen des angeborenen(unspezifischen)Immunsystems
Die Zellen des angeborenen Immunsystemssind u. a. die Makrophagen resp. Monozyten.In den Geweben haben sie geringfügig unter-schiedliche Eigenschaften. Da sich die Ma-krophagen in allen Geweben befinden, ge-hören sie meistens zu den ersten, die einge-drungene Infektionserreger erkennen und pha-gozytieren. Gleichzeitig werden von ihnen dieZellen des Organismus gewarnt und durch dieSekretion von Zytokinen, den Botenstoffendes Immunsystems, die Kaskade der spezifi-schen Abwehr in Gang gesetzt.
Weiterhin gehören zu den Zellen des ange-borenen Immunsystems die polymorphkerni-gen Granulozyten. Die neutrophilen, basophi-len und eosinophilen Granulozyten sind seltenim Gewebe anzutreffen, sie befinden sich imperipheren Blut. Bei Infektionen werden sie ver-mehrt im Knochenmark gebildet und dann in
größerer Anzahl zum Entzündungsherd ge-bracht, um ihre spezifischen Aufgaben zu er-füllen.
Weiterhin findet man nur im Gewebe (beson-ders in der Haut und in den Schleimhäuten)die Mastzellen. Sofern sie Infektionserregererkennen oder spezielle Aktivierungsreize (z. B. IgE) erhalten, setzen sie in Sekunden-schnelle toxische Inhaltsstoffe frei. Durch Zy-tokin-Sekretionen sind die Mastzellen auch inder Lage, weitere Zellen anzulocken und Ent-züngungsreaktionen zu erwirken.
Die interdigitierenden dendritischen Zellen,kurz dendritische Zellen (dendritic cells, DCs)genannt, wandern als unreife Zellen aus demBlut in die Gewebe ein und bilden dort zahl-reiche zarte Verästelungen aus. Als Langer-hans-Zellen flechten sie in der Haut mit ihrenFortsätzen ein dichtes Netz. Erfolgt eine Ak-tivierung durch Infektionserreger, hören die-se Zellen auf zu pinozytieren und wandern mitdem Lymphstrom in die Lymphknoten undpräsentieren den Lymphozyten diejenigen An-
tigene, die sie soeben im Gewebe aufge-nommen haben. Eine andere Form sind die fol-likulären dendritischen Zellen (follicular den-dritic cells, FDCs). Sie bilden ein Netzwerk inden B-Zell-Follikeln der sekundären lymphati-schen Organe. Sie sind darauf spezialisiert,antigene Substanzen, die sie mit dem Lymph-strom erreichen, aufzunehmen und den B-Lymphozyten zu präsentieren.
Die Natürlichen Killer-(NK-) Zellen sind Lym-phozyten, welche darauf spezialisiert sind, in-fizierte Zellen, Tumorzellen und durch Anti-körper „markierte“ Zellen zu vernichten.
Die Zellen des adaptiven (erworbenen, spezifischen)Immunsystems
Die Zellen des adaptiven Immunsystems sindLymphozyten mit einem großen, fast rundenKern und einem schmalen Zytoplasmasaum.Es werden B-Lymphozyten und T-Lymphozy-ten unterschieden. Diese Zellen zeichnen sichdurch klonal verteilte Rezeptoren für Antige-ne aus. Ein Klon ist die aus einer Zelle her-vorgegangene Nachkommenschaft. Das be-deutet, dass sich die Antigenrezeptoren indi-vidueller B- und T-Lymphozyten voneinanderunterscheiden. Die klonal verteilten Antigen-rezeptoren bilden die molekulare Grundlageder außerordentlichen Unterscheidungsfähig-keit des adaptiven Immunsystems. Die Zellendes adaptiven Immunsystems sind die B-Lym-phozyten und T-Lymphozyten. Die B-Lympho-zyten reifen beim Menschen im Knochenmark(bone marrow) heran. Die Antigenrezeptorenheißen B-Zell-Rezeptoren. Es sind membran-verankerte Immunglobuline oder Antikörper.Wenn B-Lymphozyten aktiviert werden, diffe-renzieren sie sich zu Plasmazellen, die daraufspezialisiert sind, große Mengen Immunglo-buline zu synthetisieren und in löslicher Formzu sezernieren. Eine Plasmazelle kann bis zu2000 Antikörpermolekülen pro Sekunde pro-duzieren. Die T-Lymphozyten werden aus demKnochenmark ausgeschleust und im Thymuszu T-Helfer-Zellen und zytotoxische T-Lym-phozyten (CTLs) geprägt. Ihre Antigene er-kennen die T-Lymphozyten durch klonal ver-teilte T-Zell-Rezeptoren (TCRs).
T-Helfer-Zellen entscheiden die Immunantwort.
Abb. 1: Die Zellen des angeborenen (unspezifischen) Immunsystems
Der
nac
hfol
gend
e A
rtik
el is
t m
it fr
eund
liche
r G
eneh
mig
ung
entn
omm
en a
us
Aus
gabe
10/
08. F
orde
rn S
ie Ih
r Pr
obeh
eft
an!
Tel
.: 0
61 4
6 - 9
0 74
- 0
• Fa
x: 0
61
46 -
90 7
4-44
• w
ww
.com
edve
rlag
.de

Immunsystem
2 10/08
gene an Peptidfragmenten, die an Proteine desHaupthistokompatibilitäts-Komplexes (MHC)gebunden sind. Durch das angeborene Kom-plementsystem werden Krankheitserregerauch humoral mittels Proteasen opsonisiert,um sie für die Zerstörung durch Phagozytenzu markieren und auf diese Art und Weise dasImmunsystem zu unterstützen, zu „komple-mentieren“. Monozyten sind stets im Gewebevorhanden und werden in der Entzündungsre-aktion zu phagozytierenden Makrophagen um-gewandelt. Die Reaktion vom Makrophagen aufbakterielle Lipopolysaccharide (LPS) ist ab-hängig von der Bindung des CD14 (LPS-Rezeptorkomplex) an den Toll-like-Rezeptor 4(TLR-4), welcher anschließend die Produktionvon entzündungsfördernden Cytokinen undChemokinen, sowie die Expression co-stimu-lierender Moleküle bewirkt.
Chemokine locken weitere Abwehrzellen zumInfektionsherd. Neutrophile Granulozyten wer-den u. a. durch Interleukin-8 (IL-8) und Tumor-Nekrosefaktor- α (TNF-α) aktiviert. Sie durch-queren als erste die Blutgefäßwände und drin-gen ins Entzündungsgebiet ein. Hier erzeugensie über den „respiratory burst“ Sauerstoffra-dikale und Stickstoffmonoxid (NO) als Waffegegen Eindringlinge. Um Wasserstoffperoxidund Superoxid-Anionen bereit zu stellen, besitzen sie NADPH (Nicotinamid-Adenin-
Sie können z. B. B-Lymphozyten zur Antikör-perproduktion anregen und den Makrophagenhelfen, aufgenommene Mikroorganismen ab-zutöten. Regulatorische T-Helfer-Zellen (Treg)wirken entgegengesetzt und supprimieren dieImmunantwort. Sie haben die lebenswichtigeFunktion, den Organismus selbst vor Angriffendes Immunsystems zu schützen, sowie die Im-munreaktionen und damit den Schaden durchaggressive Effektormechanismen des Im-munsystems zu begrenzen. Die CTLs sind da-rauf spezialisiert, infizierte Zellen abzutöten.
Das angeborene underworbene immun -regulatorische Netzwerk der AntigenpräsentationDie Zellen der unspezifischen Abwehr sind es-sentiell für die Funktion der T-Helfer-Zellen. DieAktivierung antigenpräsentierender Zellen(APC), wie Makrophagen, dendritische Zellenund antigenpräsentierende B-Zellen sind ein ers-ter notwendiger Schritt zur Auslösung eineradaptiven Immunantwort. T-Lymphozyten sinderforderlich für die Kontrolle intrazellulärerKrankheitserreger und die Aktivierung von B-Lymphozyten gegen die meisten Antigene.Dabei erkennen T-Lymphozyten Fremd-Anti-
Abb. 2: Entzündungsreaktionen im Gewebe
Abb. 3: Die Zellen des erworbenen (adaptiven, spezifischen) Immunsystems
Dinucleotid-Phosphat)-Oxidasen in ihren Lyso-somen. Abgestorbene neutrophile Granulozy-ten mit phagozytierten Erregerresten bildendementsprechend den Hauptbestandteil desEiters. Damit sich die Entzündung nicht ausbreitet, wird von aktivierten MakrophagenTNF-α lokal freigesetzt, was zur Blutgerinnungin umgebenen Blutgefäßen führt. Kommt es jedoch zur systemischen Freisetzung vonTNF-α in Folge einer Sepsis, so entwickelt sicheine Verbrauchskoagulopathie mit Multior-ganversagen. Die von den Phagozyten freige-setzten Cytokine (IL-1, IL-6) aktivieren die Aku-te-Phasen-Proteine, wie C-reaktives Protein(CRP). In Folge steigt die Körpertemperatur an.Gleichzeitig wird neben der angeborenen un-spezifischen Immunreaktion auch die spezifi-sche eingeleitet, indem einige Makrophagenund vorwiegend dendritische Zellen im Ent-zündungsbereich Erreger und deren Antigeneaufnehmen und zu den regionalen Lymphkno-ten transportieren, um sie dort den T-Lym-phozyten zu präsentieren. Diese veranlassendaraufhin die Ausschüttung von Interleukinenzur adaptiven Immunantwort. Natürliche Kil-lerzellen (NK-Zellen) werden u. a. durch frei-gesetztes Interferon-γ (IFN-γ) und Cytokine (In-terleukin-12) aktiviert. Gleichzeitig verstärkenIFN-γ und IL-12 die Expression von MHC-I Mo-lekülen auf körpereigenen T-Zellen zum Schutzvor einem Angriff aktivierter NK-Zellen, sowiedie Expression von viralen Peptidfragmentenim Komplex mit MHC-I auf infizierten Zellen, umzytotoxische T-Lymphozyten (CD8-CTL) anzu-regen.
Gleichzeitig verhindern andere T-Zellen durchunmittelbare Gegenregulationen eine Überre-aktion der Abwehrfunktionen. Solche T-regu-latorische Zellen (Treg) sezernieren beispiels-weise bei Überstimulation von IL-2, als CD4-CD25-Treg markiert, IL-I0, IL-4 und TGF-β undbei Überexpression von CD8-CTL, als CD8-CD28-Treg, IL-10 im Sinne einer Suppression.TR-I-Zellen verhindern bereits bei der Bildungvon Th1- bzw. Th2- Zellen jeweils eine unkon-trollierte Überexpression.
Die besondere Rolle der NK-ZellenNK-Zellen erkennen Fremdantigene auch ohneMHC-Markierung, was sie so bedeutend imKampf gegen Karzinom-Zellen macht, dennNK-Zellen greifen insbesondere Zellen an, dieeine verzögerte bis keine MHC-I-Expression ander Zelloberfläche präsentieren. Krebszellenbilden IL-lO, TGF-β und PGE-2, um die Th1-Zel-len auf zellulärer Ebene zu schwächen. Er-höhte PGE-2-Spiegel unterdrücken das Im-munsystem und insbesondere die NK-Aktivität1.Darüber hinaus fördert PGE-2 die Neoangio-genese. Ähnliche immunologische Verhältnis-se finden sich in der Schwangerschaft. Auchhier sind GM-CSF (Granulozyten / Monozyten-Kolonie stimulierender Faktor) sowie IL-4 undIL-1O zur Unterdrückung der Th1-Reaktionen
1 Biesalski, Ernährungsmedizin, 323
Immunsystem
310/08
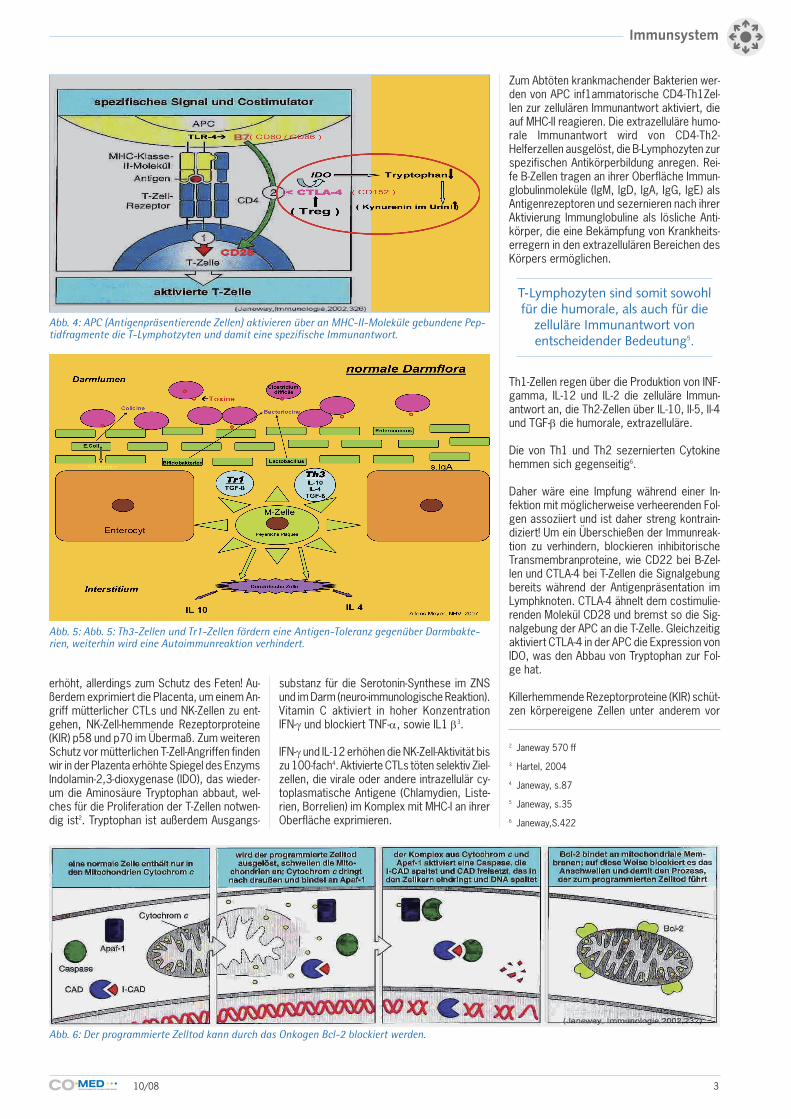
Abb. 4: APC (Antigenpräsentierende Zellen) aktivieren über an MHC-II-Moleküle gebundene Pep-tidfragmente die T-Lymphotzyten und damit eine spezifische Immunantwort.
Abb. 5: Abb. 5: Th3-Zellen und Tr1-Zellen fördern eine Antigen-Toleranz gegenüber Darmbakte-rien, weiterhin wird eine Autoimmunreaktion verhindert.
erhöht, allerdings zum Schutz des Feten! Au-ßerdem exprimiert die Placenta, um einem An-griff mütterlicher CTLs und NK-Zellen zu ent-gehen, NK-Zell-hemmende Rezeptorproteine(KIR) p58 und p70 im Übermaß. Zum weiterenSchutz vor mütterlichen T-Zell-Angriffen findenwir in der Plazenta erhöhte Spiegel des EnzymsIndolamin-2,3-dioxygenase (IDO), das wieder-um die Aminosäure Tryptophan abbaut, wel-ches für die Proliferation der T-Zellen notwen-dig ist2. Tryptophan ist außerdem Ausgangs-
substanz für die Serotonin-Synthese im ZNSund im Darm (neuro-immunologische Reaktion).Vitamin C aktiviert in hoher Konzentration IFN-γ und blockiert TNF-α, sowie IL1 β3.
IFN-γ und IL-12 erhöhen die NK-Zell-Aktivität biszu 100-fach4. Aktivierte CTLs töten selektiv Ziel-zellen, die virale oder andere intrazellulär cy-toplasmatische Antigene (Chlamydien, Liste-rien, Borrelien) im Komplex mit MHC-I an ihrerOberfläche exprimieren.
Zum Abtöten krankmachender Bakterien wer-den von APC inf1ammatorische CD4-Th1Zel-len zur zellulären Immunantwort aktiviert, dieauf MHC-II reagieren. Die extrazelluläre humo-rale Immunantwort wird von CD4-Th2-Helferzellen ausgelöst, die B-Lymphozyten zurspezifischen Antikörperbildung anregen. Rei-fe B-Zellen tragen an ihrer Oberfläche Immun-globulinmoleküle (IgM, IgD, IgA, IgG, IgE) alsAntigenrezeptoren und sezernieren nach ihrerAktivierung Immunglobuline als lösliche Anti-körper, die eine Bekämpfung von Krankheits-erregern in den extrazellulären Bereichen desKörpers ermöglichen.
T-Lymphozyten sind somit sowohlfür die humorale, als auch für die
zelluläre Immunantwort von entscheidender Bedeutung5.
Th1-Zellen regen über die Produktion von INF-gamma, IL-12 und IL-2 die zelluläre Immun-antwort an, die Th2-Zellen über IL-10, Il-5, Il-4und TGF-β die humorale, extrazelluläre.
Die von Th1 und Th2 sezernierten Cytokinehemmen sich gegenseitig6.
Daher wäre eine Impfung während einer In-fektion mit möglicherweise verheerenden Fol-gen assoziiert und ist daher streng kontrain-diziert! Um ein Überschießen der Immunreak-tion zu verhindern, blockieren inhibitorischeTransmembranproteine, wie CD22 bei B-Zel-len und CTLA-4 bei T-Zellen die Signalgebungbereits während der Antigenpräsentation imLymphknoten. CTLA-4 ähnelt dem costimulie-renden Molekül CD28 und bremst so die Sig-nalgebung der APC an die T-Zelle. Gleichzeitigaktiviert CTLA-4 in der APC die Expression vonIDO, was den Abbau von Tryptophan zur Fol-ge hat.
Killerhemmende Rezeptorproteine (KIR) schüt-zen körpereigene Zellen unter anderem vor
2 Janeway 570 ff
3 HarteI, 2004
4 Janeway, s.87
5 Janeway, s.35
6 Janeway,S.422
Abb. 6: Der programmierte Zelltod kann durch das Onkogen Bcl-2 blockiert werden.

Immunsystem
4 10/08
dem Angriff der NK-Zellen. Th3-Zellen des Dar-mes sezernieren ähnlich den Th2-Zellen IL-4,IL-10 und TGF-ß zur mäßigen Hemmung derTh1-Zellen, um eine Antigen-Toleranz gegen-über Darmbakterien zu erreichen. Infolge wirdsekretorisches IgA als Schleimhaut assoziier-ter Antikörper gebildet. Daher reagiert das Im-munsystem der Schleimhaut ohne Entzün-dungsreiz auf Fremdantigene tolerant. Zudemverhindern T-regulatorischen Zellen (Treg-Zel-len) vor Ort eine Überreaktion von Th1-Zellenund beugen Autoimmunerkrankungen vor. Den-dritische Zellen exprimieren in PeyerschenPlaques IL-10 und IL-4, während sie in peri-pheren Lymphknoten INF-gamma und IL-12synthetisieren.
Um die Homöostase der Lymphozytenpopula-tion zu erhalten, werden nach einer durchge-machten Infektion die nicht mehr benötigten ak-tivierten Effektor-T-Zellen durch Wechselwir-kung mit dem Fas-Liganten zur Apoptose ver-anlasst. Der programmierte Zelltod kann durchdas Onkogen Bcl-2 blockiert werden. Bcl-2 ver-hindert das bei der Apoptose induzierte An-schwellen und somit das Austreten des Cyto-chrom c aus den Mitochondrien. Erst das Frei-setzen von Cytochrom c aus den Mitochondriender Zelle löst die Apoptose aus, weswegen Kar-zinomzellen unsterblich werden, da sie kaumMitochondrien besitzen und zudem ihre Hä-moxigenase zum Abbau des Cytochrom c starkexprimiert ist.
L i t e r a t u r h i n w e i s e1. Janeway, Travers, Walport, Shlomchik, Im-munologie, Spektrum-Verlag, 20022. Georg A. Holländer, Immunologie, Urban &Fischer-Verlag 20063. Biesalski, Ernährungsmedizin, Thieme-Ver-lag, 19994. Claudius Christopher Rauscher, Immunolo-gische Behandlungsmethoden, Haug-Ver-lag,19965. Kremer, Heinrich, Die stille Revolution derKrebs- und AIDS-Medizin,Ehlers Verlag,20046. Berg, Tymoczko, Stryer, Biochemie, Spek-trum Verlag, 20037. Vorlesungsskript Immunologie und Infek-tiologie Uniklinik Düsseldorf 20078. Beyer, Peters, Mikrobiologische Therapie,Forum Medizin-Verlag 2003
Alfons Meyerist Facharzt für Allge-meinmedizin, Naturheil-verfahren und Akupunkturmit den Schwerpunktenbiologische Krebsmedizinund Orthomolekularmedi-zin (Vitamintherapie).
Kon takt:Kaiser-Friedrich-Ring 77
D-65185 WiesbadenTel.: 0611 / 2055280, Fax: 0611 / 2055281
Recommended

















