
Arch. exper. Path. u. Pharmakol., Bd. 228, S. 288~301 (1956)
Aus dem Pharmakologisehen Institut der Universitit Erlangen (Direktor: Prof. Dr. F. HEIM)
Nachweis und Bestimmung mehrerer Coenzym-A-Verbindungen in der Leber
yon Meerschweinchen
Yon FRED LEUSCHNER
M-it 3 Textabbildungen
(Eingegangen am 2. Dezember 1955)
I n den vergangenen Jah ren wurde dem Coenzym A 1 wegen seiner Betei l igung an f u n d a m e n t a l e n biologischen Prozessen besondere Auf- merksamkei$ gewidmet. Dieses Interesse k a n n auf LIPMiNN zurfickge- f i ihr t werden, der CoA als notwendige Komponen¢e f ib die Acetytierung von Sulfani lamid in der Leber erkannte .
Fast zur gleiehen Zeit beobachteten NACH~NSOHN und B~RMA~, FF.LDBERG und MAm~ und LIPTON und B~RO~, dal3 CoA ebenfalls als Aktivator f'dr die Syn- these yon Aeetyleholin benStigt wird. In den folgenden Jahren warde die funktio- nelle Bedeutung des CoA f'dr die ~,~Tbertragung.von Carbonsiuren im Citronensiure- zyklus, bei der Fettsiuresynthese und -oxydation, bei der Synthese yon Steroiden, Fetten und Porphyrinen, sowie der Aeylierung yon Arylaminen, Histamin, Amino- siuren, Glueosamin u. a. erschlossen. CoA wirkt bei diesen Umsetzungen als Kata- lysator, indem sich seine funktioneUe Gruppe mit den zu iibertragenden Carbon- s~uren zu einer energiereichen Verbindung zusammenschlieBt. Aus dieser Bindung werden die Carbonsiuren dann durch speziilsehe Fermente auf Akzeptoren (z. B. Cholin) iiberf'tihrt. Was den ehemischen Aufbau des in der belebten Natur weir verbreiteten CoA betrifft, so haben Untersuchungen der Arbeitskreise um I~1~1~1~ und SNELL ergeben, dali es sich um ein Dinucleotid handelt, das neben Adenosin und Phosphorsfturen Pantethein enth~ilt, das einen aus Pantothens~ure und Thio~thanolamin aufgebauten Wuchsstoff f'tir Mikroorganismen (Lactobacillus bulgarieus-Faktor) darstellt.
NOYELLI u n d L I P ~ N ~ en tdeck ten die Abh~ngigkei~ der Acetatver- b r e n n u n g vom CoA-Gehalt der Zelle. LY~EN u / M i t a r b , e rg inz t en diese Beobachtung, indem sie umgekehr t die Abhing igke i t des CoA-Gehaltes yon der Ace ta tve rb rennung auffanden. Zur Erkl~rung der zwischen Ace ta tve rb rennung u n d CoA-Gehalt bes tehenden Beziehung muB yon der A n n a h m e ausgegangen werden, da~ die wirksame CoA-Form - -
i Die folgenden Abktirzungen wurden benutzt: Coenzym A ~ CoA und CoASH, Coenzym-A-Derivate mit Disulfidbindung ~ CoASSR, Acetyl- bzw. Aeyl-Coen- zym A ~ CoASCOCH~ bzw. CoASAc.

Nachweis und Bestimmung mehrerer Coenzym-A-Verbindungen in der Leber 289
CoASAe - - sich in der lebenden Zelle im Gleichgewicht mit ihren Vor- und Abbaustufen befindet. In diesem GleichgewichtsprozeI3 lguft die SpMtung des Cofermentes als hydrolytiseher ProzeB unter allen Um- st~nden ab, w~hrend die Synthese bzw. Resynthese der Spaltprodukte an die energieliefernden Dissimilationsprozesse (z. B. Acetatverbrennung) in der Zelle geknfipft ist. Andererseits ist ein Mindestgehalt an CoA Vor- aussetzung ffir den Ablauf normaler dissimilatorischer Prozesse mit mehreren CoA-induzierten Reaktionsstufen. Diese Koppelung ist be- sonders fiir gesch~digte Zellen folgenreieh, indem eine verminderte AtmungsgrSBe zur Anh~ufung vornehmlieh inaktiver CoA-Verbindungen neben wenig akt ivem CoA und dieser resultierende CoA-Mangel zur StSrung im Stoffweehsel fiihren muB.
Zur Aufkliixung dieser ffir den pathologischen Zellstoffwechsel be- deutenden Zusammenh~nge sind nicht nur qualitative und quanti ta t ive Kenntnisse fiber den gesamten CoA-Gehalt, sondern auch fiber die mit dem aktiven CoA im Gleichgewieht stehenden CoA-AbkSmmlinge yon Interesse.
Verschiedene Untersucher haben multiple Formen des CoA in hochgereinigten CoA-Pr~parationen durch papierchromatographische Trennung nachgewiesen. Doch ist es nieht bekannt, welche CoA-Formen im biologischen Ausgangsmaterial vor- liegen.
Die folgenden Untersuchungen haben deshalb zum Ziel: 1. Die schonende, ffir papierehromatographische Zweeke genfigende
Anreicherung (Acetonfraktionierung) der CoA-Verbindungen, die yon der Zelle zur enzymatischen Aeetylierung ausgenutzt werden kSnnen,
2. die papierchromatographische Trennung und Charakterisierung der Verbindungen und
3. die Best immung der mengenm~l~igen Beziehungen dieser Kom- ponenten zueinander.
Methodiseher Teil
l~fir alle Untersuchungen wurden ausgewachsene Meerschweinchen (Gewicht etwa 300 g) verwendet. Die Nahrung der Tiere bestand aus einem kalorien- lind vitaminreichen Misckfutter.
1. Heratellung des Kochaaftes aua Leberyewebe. Die TStung der Tiere erfolgte durch Dekapitation nach vorausgegangenem Nackenschlag. Die Leber gelangte etwa 30 see nach der Dekapitation in einen MSrser mit fliissiger Luft. l~ein zerrieben wurden etwa 5 g Leber in einen mit 10 ml Wasser beschickten, vorgewogenen V2A-Stahl- becher gef'fillt. Bei der Riickw~gung ergab sich das Gewicht des zu analysierenden Lebergewebes. ])as Analysengut wurde unter st~ndigem Riihren bis zum Sieden er- hitzt, darauftfin unter flieBendem Wasser abgekfihlt und in ein vorbereit~tes Eisbad gebracht. Dieser ErhitzungsprozeB benStigte etwa 60sec. Eine unnStige Ausdehnung soUte vermieden werden. Alle weiteren Arbeitsg~nge wurden bei Temperaturen zwischen --10 ° und +8 ° ausgefiihrt. Nun wurde 2 mln homogenisiert (etwa 20000 U. p. rain) und das denaturierte EiweiB abzentrifugiert (15 rain, 18000 × g).

290 F. LEusCHNER:
Es k6nnen etwa 2/s des Ausgangsvolumens als Kochsaft gewonnen werden. 2. Anreicherung der CoA-Verbindun~jen durch Aceton/raktionierung aua Koch.
,a/t. Die Anreicherung der Verbindungen wurde mittels des Aeetylierungstestes yon KAPLA~ und LrP~t~¢~ kontroUiert.
Zur Priifung der Acetonf~llbarkeit der CoA-Derivate wurden 10 ml Koehsaft mit den aus Abb. 1 zu entnehmenden Volumen gekiihlten Aceton stufenweise versetzt und sofort zentrifugiert (15 min, 18000 × g). Bis zur 4. Fraktion wurde der Boden- kSrper einmal mit 2 ml des entsprechenden Aceton-Wasser-Gemisches gewaschen. Bet so gewonnene BodenkSrper wurde jeweils mit Wasser auf 0,5 g aufgef'fillt, gelSst und das Uberstehende der n~chsten Fraktionierungsstufe unterzogen (Aceton- zusatz unter Beriicksichtigung der Wasehfliissigkeit). Nach der 12. Fraktionierung (12 ml Aeeton/ml w~Briger Phase) wurde das ~berstehende mit 5 ml 10°/oiger Queeksilber-acetat-L(isung versetzt, wobei ein l~iederscblag ausfiel. Nach Abzentri- fugieren wurde der BodenkSrper in 1 ml Wasser suspendiert (A), alles Quecksilber dureh Einleiten yon H2S als Sulfid ausgef~llt und erneut abzentrffugiert. Um das Fliissigkeitsvolumen nicht unnStig zu vergriiBern, wurde auf das Waschen des Bodenki~rpers verzichtet. A wurde zur Bestimmung des CoA-Gehaltes mit festem NaHCO a neutralisiert.
Das Fraktionierverfahren fiir papierchromatographische Zwecke wurde inso- welt vereinfacht, als die Fraktion I (Fraktionsbezeiehnungen vgl. Abb. 1) durch 1,5 ml Aceton/ml Kochsaft abgetrennt und mit 5 ml 60°/oigem w/~Brigen Aceton (V/V) gewasehen wurde. Die Fraktionen l I und I I I wurden aus den vereinigten Fliissigkeiten mit 10,5 ml Aceton/ml w~Briger Phase ausgef~llt. Die Fraktion I be- stand aus einem r6tlichen voluminSsen BodenkSrper, Fraktion I I / I I I war eine dureh Flavinderivate gelbbraun gef/~rbte 5hge Fliissigkeit yon etwa 10 mg/ml Kochsaft. Sie wurde mit Wasser auf 0,5 g aufgef'ffllt (zu diesem Zwecke erfolgte die Fraktionie- rung in vorgewogenen V2A-Stahlbeehern). Diese LSsung wurde papierchromato- graphisch und im enzymatischen Acetylierungstest naeh K~LA~ u. LIrMA~¢ und LY~EN u. Mitarb. welter untersucht. In der Regel wurde nut die auf diesem Wege erzielte 7--10fache CoA-Anreicherung ftir die folgenden Untersuchungen ver- wendet, gelegentlich eine 20fache.
3. Enzymatische Methoden. Quantitative CoA-Bestimmung nach KArLA~ U. LIPMA~: Die quantitative Erfassung yon CoA-Verbindungen und einigen Vor- stufen (Pantethein, Pantetheinphosphat, Dephospho-CoA) gelingt durch Acetylie- rung yon Sulfanilamid in Gegenwart yon Adenosintriphosphat und einem Tauben- leberenzymsystem. Zur Materialersparnis wurde die Modifikation yon L¥~E~ ver- wandt.
Ansatz in ZentrffugenglKsern:
0,10 ml der auf CoA zu priifenden Substanz, (bzw. 0,1 - - x ml Wasser), 0,04 ml m/10 Glutathion-LOsung,
0,02 ml einer frisch bereitef~n m NaHCOa-Lfsung,
0,10 ml Reaktionsgemiseh (10 Teile 0,006 in Sulfanilamid-Li~sung, 2,5- Teile m Na-acetat-LOsung, 10 Tefle 0,05 m Na4-Adenosintriphos- phat-LSsung und 10 Teile 0,2 m Na-eitrat-L~sung)
0,06 ml gealtertes Taubenleberfermentsystem (Darstellung nach EAr-
Nach 2stiindiger Inkubation bei 37 ° wurde die Reaktion dutch Zugabe yon 1 ml 5~oiger Trichloressigs~ure abgestoppt, denaturiertes Eiweill abzentrifugiert und an- schlieBend in 0,6 ml der klaren LSsung Sulfanilamid nach BtcA~O~¢ u. MAI~HALL kolorimetrisch bestimmt. Die Standardisierung erfolgte mit einem 75°/oigen CoA- Pr~parat. Das verwendbare MeBbereieh betrug 0,5--4 #g CoA.

Nachweis und Bestimmung mehrerer Coenzym-A-Verbindungen in der Leber 291
Quantitative Bestimmung yon CoASCOCH a nach LYNEN u. MiC~rb.: Dieser Bestimmungsmethode liegt die stSehiometrische Transacetylierung yon CoASCOCH 3 auf Sulfanilamid zugrunde. In Abwesenheit yon Adenosintriphosphat wird lediglieh CoASCOCH 8 zur Transacetylierung ausgenutzt.
Ansatz in Zentrifugengl~sern:
0,20 ml der auf CoASCOCH 3 zu priifenden LSsung, bzw. 0 , 2 - × ml Wasser).
0,05 ml m/5 Pyrophosphatpuffer (8,92 g Na4P207 . 10 H~O mit 8 ml n HC1 und Wasser auf 100 ml),
0,02 ml 0,4 m Glutathion-Liisung, 0,032 ml 0,006 m Sulfanilamid-LSsung, 0,06 ml EnzymlSsung.
Die Inkubation betrug 60 rain bei 37% Die Sulfanilamidbestimmung erfolg¢~ nach BRATTON U. MARSHALL. ~ EnzymlSsung diente eine nach LYI~EN u. Mitarb. gereinigte ProteinlSsung aus Taubenleber. Das zur Priifung der Methode erforder- liehe CoASCOCH 8 wurde naeh STAI)TMA~ arts CoA und Essigs~ureanhydrid dar- gestellt.
d. Papierchromatographisches Ver]ahren. Schleieher & Sehiill-Papier 2043b 20×50 em wurde 1 Std in eine 0,5%ige w~Brige LSsung yon ~thylendiamintetra- essigsaurem Natrium (PH 8,5) eingelegt und ansehlieBend mehrfach in Wasser ge- wasehen. Durch diese Behandiung wurde der Gehalt an Ionen, die Naehweismethoden stSren odor in ihrer Empfindiichkeit beeintr~ehtigen k(innen, sehr reduzier~.
Das eisgekiihlte Analysengut (50--150 ~ul) wurde mit Mikropipetten in Por- tionen yon 10/~1 auf der Startlinie m emer Breite yon 12--15 mm aufgetragen und im kalten Luftstrom getrocknet.
Fiir die 1. Dimension wurde folgendes LSsungsmittel benutzt:
40 Teile m/5 Acetatpuffer p~ 3,6 naeh WALPOLE (LSsung A: 12,0 ml Eisessig und Wasser auf 1000 ml, LSsung B: 27,2 g Iqa-acetat • 3It20 und Wasser auf 1000 ml, 18,5 Teile A und 1,5 Tei]e B)
10 Teile Isoamylalkohol 75 Tefle Aceton.
Die in der absteigenden Durchlaufteehnik vorgenommene Chromatographie dauerte 24--30 Std bei Zimmertemperatur.
Naeh Trocknung der Papiere wurde das 2. LSsungsmittel zur Anwendung ge- bracht:
50 Teile l~Iethylalkohol 50 Teile Aeetatpuffer (LSsung A: 12 ml Eisessig und Wasser auf 1000 ml
LSsung B: 13,6 g Na-acetat. 3H~0 und Wasser auf 1000 ml, 1 Tell A nnd 1 Teil B).
Zur Entwieklung wurden die Papiere in Streffen gesehnitten, an 40 × 50 cm grol3e PapierbSgen sehmalkantig angen~ht, zu einem Zylinder geroUt nnd in das LSsungsmittelgemisch gestellt. Entwicklungszeit 14 Std bei Zimmertemperatur.
Lokalisation der bei 260 m/~ absorbierenden Flecke im UV-Lieht: Zur Anwen- dung gelangte eine Queeksilberniederdruekri~hre mit maximaler Strahlung bei 254 m/~. Fiir die Photographie wurde Agfa-Copex-Papier benutzt.
Entwicklung der P-haltigen Flecke nach HANES U. ISHERWOOD." Die Papiere wurden mit Molybdat-Reagens (25 ml 4%iger AmmoniummolybdatlSsung, 5 ml 60%iger Perchlors~ure, 10 ml n HCI und Wasser auf 100 ml) bespriiht und infeuehtem ZusC~nd 7 min einer Temperatur yon 85 ° zur Spaltung der Ester ausgesetzt. Naeh kurzer Behandiung mit Wasserdampf und H2S ersehienen P-haltige SteUeu als blaue Fleeke.

292 F. LEUSCttNER :
l~achweis des Sulfid-Schwefels (ira wesentliehen naoh TOENNIES und KOLB): Reagens I : 1,5 g Na-Nitroprussid gelSst in 5 ml 2 n H2SO 4 ~ 95 ml Methanol ~- 10 ml 25~oiges Ammoniak, filtrieren und in der KMte aufbewahren. Reagens I I : 2 g KCN gelSst in 5 ml Wasser -~ 95 ml Methanol. Reagens 1TI: 3 g NaOH gelSst in 10 ml Wasser Jr 50 ml Methanol. Sulfhydrylgruppen wurden durch Eintauchen in das Reagens I nachgewiesen. Es entwickelten sieh im Laufe yon Sekunden rStliehe Flecke. CN-spaltbare Sehwefelverbindungen wurden nacheinander mit dem Rea- gens I and LI behandelt. Abhi~ngig yon der Leichtigkeit der Spaltung trat die F~r- bung sofort oder bis zu Stunden verzSgert auf. Die F~ixbung der Sulfhydrylgruppen wurde durch Reagens I I verst~ixkt. Dureh das Reagens I I I warden Acylmercaptane gespalten.
Naehweis unsubstitutierter Glykolgruppiertmgen nach BUCHA~A-~, DEKKER und LonG: Das Chromatogramm warde 5 rain bei 80 ° getrocknet, anschliel3end mit l~oigem w~Brigen NaJO 4 besprtiht und 10 min lung bei Zimmertemperatur gehalten. Sodann wurde das Papier SO s (Na~SO a und H2SOa) ausgesetzt, bis das freigewordene Jod verbraucht war. Schliel31ich wurde das Papier mit SeHiF~s-Reagens bespriiht (1 g Rosanilin in 50 ml Wasser mit SO s entf~rbt und auf 1000 ml aufgefiillt) und 10 rain auf 85 ° erhitzt. Die violetten Flecke wurden naeh 10---20 Std deutlich.
Ergebnisse und Bespreehung Anreicherung der CoA-Verbindungen aus Meerschweinchenleber. Ffir
die A b t r e n n u n g CoA-ak t ive r F r a k t i o n e n aus Leber war eine quan t i t a t i ve B e s t i m m u n g s m e t h o d e erforderl ich, deren Spezif i t~t sich au f CoA, sowie die yon der Leber le ieht ausnu tzba ren Vors tufen ers t reekt . Zu diesem Zwecke wurde die enzymat i sche Aee ty l i e rung yon Suffani lamid naeh K A P L ~ und LIPMA~N verwendet . Dabe i d i en t eine acetongef~l l te und gea l te r te P ro t e in f r ak t i on aus Taubenleber , Adenos in t r iphospha t , Glu- t a t h i o n oder Cyste in und Sul fan i lamid als Enzymansa t z . Mit te ls dieser Tes tme thode war es mSglieh, eine CoA-angere ieher te Lebe r f r ak t ion ab- zu t rennen, die sieh ffir eine pap ie reh romatograph i sche Unte r suehung eignete.
Als Ausgangsma te r i a l fi ir die Anre ieherung wurde ein Koehsa f t aus Lebergewebe benu tz t . Wie der Ver l au f der Unte r suehungen zeigte, t r i t t 4ureh das kurzf r i s t ige E rh i t zen des Gewebes bei der Hers te l lung des Koehsa f t e s keine wesent l iehe Ver~nderung der CoA-Zusammense tzung ein.
Als Anre ieherungsver fahren aus Koehsa f t eignen sich ffir diese Zwecke k a u m Sehwermetal lsa lzf~l lungen. Mehrere Gri inde spreehen dagegen. Nach a l len b isher igen Er fah rungen l iegt ein Tell des CoA als Aeylmer - c a p t a n vor. Diese Verb indungsklasse zeigt jedoeh eine groBe Empf ind- l iehkei t gegen das in F r a g e k o m m e n d e Quecksilber. Wie SACHs gefunden h a t und L Y ~ . ~ u. Mi tarb . an einer wei teren Anzah l A c y l m e r e a p t a n e be- s t~ t igen konnte , zerfal len diese Verb indungen be im Versetzen m i t e iner L6sung yon Queeks i lber -Aee ta t in die Carbons~ure und das Mereap tan , das sieh als Queeksi lbersalz niedersehl~gt . Verwendet m a n andererse i t s B a r i u m - A c e t a t oder S i lbe r -Ni t r a t - - bei en t spreehendem PH - - zur
ii~il
i
?
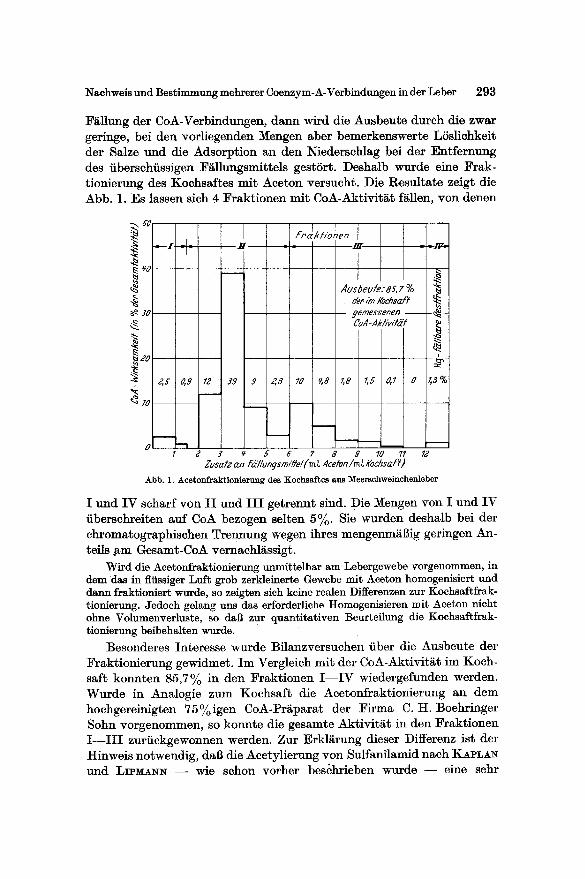
Nachweis und Bestimmung mehrerer Coenzym-A-Verb'mdungen in der Leber 293
Fgllung der CoA-Verbindungen, dann wird die Ausbeute durch die zwar geringe, bei den vorliegenden Mengen aber bemerkenswerte LSslichkeit der Salze und die Adsorption an den Niedersehlag bei der Entfernung des iibersehiissigen Fgllungsmittels gestSrt. Deshalb wurde eine Frak- tionierung des Kochsaftes mit Aceton versucht. Die Resultate zeigt die Abb. 1. Es lassen sigh 4 l%aktionen mit CoA-Aktivitgt fgllen, yon denen
fPo;/r//onen I 1
Ausbeu/e." SS, r % .~ o'er ,'co Koc&aPt
$0 ]emes~enen ~= <~ CoA - Ak//vtY@l
~20
g,4 49 12 $8 $ z,8 7a ~,8 ~, o 1,5 o,1 a 7,3%
~21o
1 g $ ~ 5 6 1 8 9 la 11 Zus~zlz con Fdc)'lungsmiltel(~t Acetan/ml, l(ochs~zf'l)
Abb. 1. Acetonfrakt ionierung des Kochsaftes aus Meerschweinchenleber
I und IV scharf yon I I und I I I getrennt sind. Die Mengen yon I u n d IV iibersehreiten auf CoA bezogen selten 5%. Sie wurden deshalb bei der chromatographischen Trennung wegen ihres mengenmgBig geringen An- tells ~am Gesamt-CoA vernachlgssigt.
Wird die Acetonfraktionierung unmittelbar am Lebergewebe vorgenommen, in dem das in fliissiger Luft grob zerkleinerte Gewebe mit Aceton homogenisiert und dann fraktioniert wurde, so zeigten sich keine realen Differenzen zur Kochsaftfrak- tionierung. Jedoch gelang uns das erforderliche Homogenisieren mit Aceton nicht ohne Volumenverluste, so dab zur quantitativen Beurteilung die Kochsaftfrak- tionierung beibehalten wurde.
Besonderes Interesse wurde Bflanzversuchen fiber die Ausbeute der Fraktionierung gewidmet. Im Vergleieh mit der CoA-Aktivitgt im Koch- saft konnten 85,7% in den Fraktionen I - - I V wiedergefunden werden. Wurde in Analogie zum Kochsaft die Acetonfraktionierung an dem hoehgereinigten 75%igen CoA-Pr~parat der Firma C. H. Boehringer Sohn vorgenommen, so konnte die gesamte Aktivit~t in den Fraktionen I - - - I I I zuriiekgewonnen werden. Zur Erkl~rung dieser Differenz ist der Hinweis notwendig, dab die Acetylierung von Sulfanilamid naeh KAPLAN und LIPM~_~N - - wie schon vorher bes~hrieben wurde - - eine sehr
2 9 4 F . L E U S C H N E R :
komplexe Reaktion ist und nicht nur CoA selbst, sondern auch Teilstruk- turen zu erfassen vermag (soweit sie durch das Taubenleberfermentsystem zu aktivem CoA umgewandelt werden k~innen). W~hrend die in hoch- gereinigten CoA-Pr~parationen vorliegenden CoA-Verbindungen (CoA SH, CoASSR) sieh durch Aeeton quantitativ ausf~llen lassen und unver- i~ndert aktiv sind, werden bei der Acetonfraktionierung des Koehsaftes Substanzen abgetrennt, die entweder 1. den Ausfall des Acetylierungs- testes fSrdern oder 2. nur mit Teflstrukturen in den Fraktionen I - - I V zu CoA-aktiven Verbindungen aufgebaut werden kSnnen. Das Vorliegen gr51~erer Mengen CoA-Verbindungen mit guter LSslichkeit in Aceton konnte durch Zusatz yon Queeksilber-Aeetat zum Uberstehenden der I I I . Fraktionierung ausgeschlossen werden. Die so gewonnene Fraktion IV besaB eine Restaktivit~t yon nur wenigen Prozenten (Fraktion IV, Abb. 1).
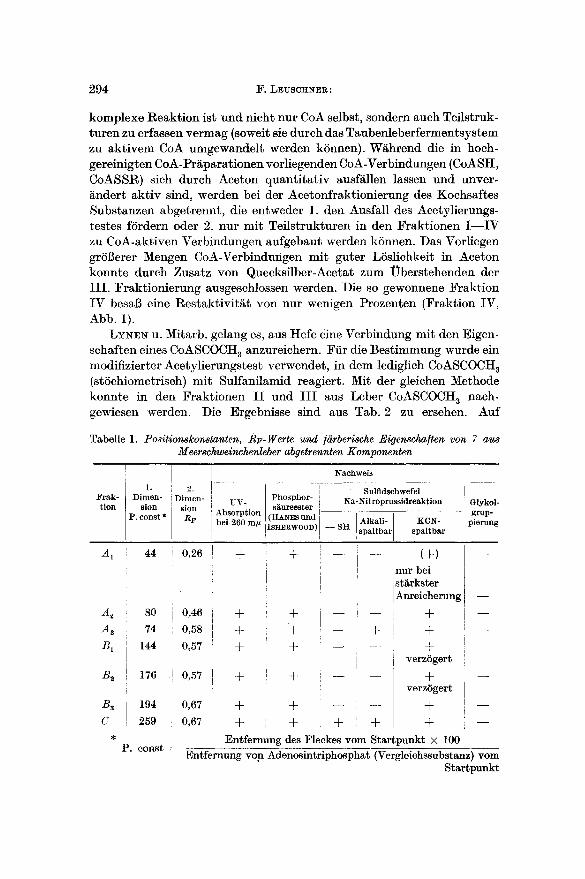
L Y ~ N u. Mitarb. gelang es, aus Hefe eine Verbindung mit den Eigen- schaften eines CoASCOCHa anzureichern. Ffir die Bestimmung wurde ein modifizierter Acetylierungstest verwendet, in dem lediglich CoASCOCH 3 (stSehiometriseh) mit Suffanilamid reagiert. Mit der gleichen Methode konnte in den Fraktionen I I und I I I aus Leber CoASCOCH 3 nach- gewiesen werden. Die Ergebnisse sind aus Tab. 2 zu ersehen. Auf
Tabelle 1. Positionskonstanten, RF-Werte und /iirberische Eigenscha/ten yon 7 au~ Meerschweinchenleber abgetrennten Komponenten
Frak - t ion
AI
A~
A 8
B,
B e
Ba C
1° 2. Dimen- I Dimen-
sion sion P. const * R F
44
80 74
144
176
194 259
P. c o n s t
Phosphor- i UV- siiureester i
Absorption (tIANES und bei 260 mg IISHERWOOD) - -
0,26 + -~
0,46 + +
0,58 ] ÷ ÷ 0,57 1
i
0,57! +
÷
÷
÷ ÷
Nachweis
0,67 ~- 0,67 ~-
I , Sulfldschwefel
Na-Ni t ro ~russidreaktion
÷
SH KCN- spal tbar
- - (+)
nur bei st~rkster Anreicherung
+ ÷ + - - +
verzbgert - - +
verz0gert - - +
+ ÷
Entfernung des Fleckes vom Startpunkt × 100
Glykol- grup-
pierung
Entfernung yon Adenosintriphosphat (Vergleichssubstanz) vom Startpunkt
Nachweis und Bestimmung mehrerer Coenzym-A-Verbindungen in der Leber 295
Frischgewebe bereehnet konnte pro Gramm 0,5--1,2 #g ,,aktive Essig- si~ure" - - entsprechend 9--24 /~g CoASCOCtt S - - gefunden werden. Dieser hohe CoASCOCH3-Gehalt konnte bei der folgenden Identifi- zierung der chromatographisch getrennten Komponenten besti£tigt werden.
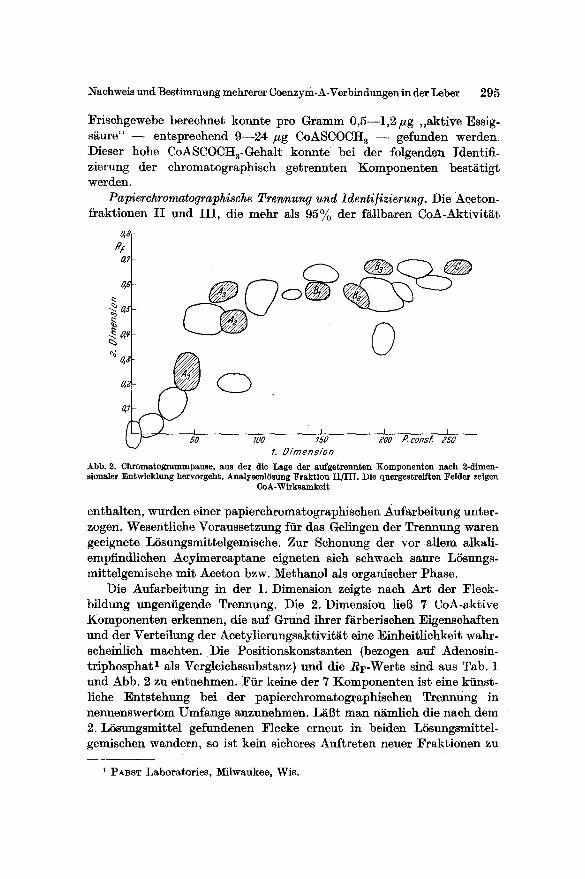
Papierchromatographische Trennung und Identi]izierung. Die Aeeton- fraktionen II und III, die mehr als 95% der f/~llbaren CoA-Aktivit~t
RE
".~ 0,
9 so
<ZZ)
I I I I 700 ;ZSO 200 .~. cons~ 2S0
7. O/mens/on
Abb. 2. Chromatogrammpause, aus der die Lage der aufgetrennten Komponenten nach 2-dimen- sionaler Entwicklung hervorgeht. Analysenl~sung Fraktion II/III . Die quergestreiften Felder zeigen
CoA-Wirksamkeit
enthalten, warden einer papierehromatographisehen Aufarbeitung unter- zogen. Wesentliehe Voraussetzung ffir das Gelingen der Trennung waren geeignete LSsungsmittelgemisehe. Zur Sehonung der vor ahem alkali- empfindliehen Aeylmereaptane eigneten sieh schwach saute LSsungs- mittelgemische mit Aceton bzw. Methanol als organischer Phase.
Die Aufarbeitung in der 1. Dimension zeigte naeh Art der Fleck- bildung ungeniigende Trennung. Die 2. Dimension lieB 7 CoA-aktive Komponenten erkennen, die auf Grund ihrer farberisehen Eigensehaften und der Verteilung der Aeetylierungsaktivit~t eine Einheitliehkeit wahr- sehei~ich maehten. Die Positionskonstanten (bezogen auf Adenosin- triphosphat 1 als Vergleichssubstanz) und die Rr-Werte sind aus Tab. 1 und Abb. 2 zu entnehmen. Fiir keine der 7 Komponenten ist eine kiinst- liche Entstehung bei der papierehromatographischen Trennung in nennenswertem Umfange anzunehmen. L~13t man n~mlieh die naeh dem 2. LSsungsmittel gefundenen Flecke erneut in beiden LSsungsmittel- gemischen wandern, so ist kein sicheres Auftreten neuer Fraktionen zu
1 PABST Laboratories, Milwaukee, Wis.

2 9 6 F. LEUSCHNER :
beobaehten. Ebenso waren keine wesentlich anderen Ergebnisse zu er- zielen, wenn die Trocknung nach Auftropfen der AnalysenlSsung und die Wanderung in einer Stickstoff-Kohlens~ure-Atmosph~re (95~o/5~o) vorgenommen wurde (Tab. 3, Nr. 11).
Entscheidender Hinweis ffir die ZugehSrigkeit der Komponenten zur CoA-Klasse war der Acetylierungstest nach KAPLAN und LreMANN.
Auf Grund der geringen CoA-Konzentrationen, die in den Papierflecken zu er- warten waren, wurden in der Regel die ausgeschnittenen Flecke unmittelbar dem Reaktionsansatz hinzugefiigt. Klein gew~hlte Stiicke, die g~nzlich yon der Fliissig- keit des Ansatzes bedeckt waren, gew~hrleisteten die analytischen Bedingungen.
Tabelle 2. Gesamt-CoA- und CoASCOCHa-Gehalt in Meerschweinchenleber
Gesamtmenge der CoASCOCH3 ** CoA-wirksamen Verbindungen*
VersuchS-~r. i im Kochsaft I in Fraktion I I / I I I in Fraktion I I / I I I in :Komponente As
in eMol in ~hIol (bezogen auf I Frischgewebe) (bezogen auf I Frischgewebe)
1. 2. 3. 4. 5. 6. 7. 8. 9.
10. 11. 12.
0,107 0,147 0,173 0,186 0,131 0,125 0,183 0,105 0,210 0,186 0,155 0,095
0,101 0,126 0,124 0,160 0,120 0,128 0,148 0,071 0,168 0,121 0,105 0,082
0,017 0,011 0,025 0,012 0,012 0,030
Mittelwert 0,150 0,121 0,018
* Bestimmt nach KAPLA~ und Ln~MA~N, berechnet auf CoASH. ** Bestimmt naeh LY~E~ u. Mitarb.
w
0,013 0,006 0,025 0 0,013 0,015
0,012
Der Aeetylierungstest l~Bt die chemische Natur der Verbindungen weitgehend often. Es war deshalb yon Interesse, die ~bereinstimmung unserer Chromatogrammflecke mit CoASH oder CoA-verwandten Ver- bindungen zu priifen.
Die funktionelle Wirkung des CoA ist an die SH-Gruppe gebunden. Ihr Nachweis gelingt mit der Na-nitroprussid-Reaktion yon TOEN~IES und KOLB. Beim positiven Ausfall zeigen sich rote Fleeke auf dem Chro- matogramm. CoASAe als Aeylmercaptan gibt keine positive Na-nitro- prussid-Reaktion fiir SH-Gruppen. Doch werden solche Verbindungen leicht in stark alkalischem Milieu hydrolysiert und reagieren daraufhin mit diesem Reagens. Aueh Disulfidbhldungen sind mit I~a-nitroprussid erst nach Spaltung mit KCN nachweisbar.
Nachweis und Bestimmung mehrerer Coenzym-A-Verbindungen in der Leber 297
Die Beurteilung der Chromato- gramme auf ihren Schwefel-Gehalt und dessen Bindungsart aus dem Er- gebnis der Reaktion von TOENNIES und KOLB bietet einige Schwierigkeiten durch die im Verhaltnis zum Ace- tylierungstest geringe Empfindlich- keit. Dazu wird die Rotf/ixbung des positiven SH-Nachweises durch KCN (disulfidspaltendes Reagens) ver- st~rkt. Liegt nun die CoAStt-Konzen- tration etwas unterhalb der Nach- .~ weisgrenze, dann f/tllt die einfache Na-nitroprussid-Reaktion negativ aus. Die verst/irkende Wirkung des KCN kann in diesem Fall das Vorliegen des Schwefels in der Disulfidform vor- t/~uschen. Eine Identifizierung der Schwefelbindung im CoA auf dem Chromatogramm gelingt nur in ver- h/~ltnism/~Big stark angereicherten Fraktionen.
Die Ergebnisse des Sehwefel- nachweises s ind aus Tab. 1 zu ersehen. Ledigl ieh die sehwache K o m p o n e n t e C zeichnet sich dureh freie S H - G r u p p e n aus. Die -~ F r a k t i o n Aa reag ie r t In der A r t "~ von A c y l m e r c a p t a n e n . Alle. ubri- ~ gen F lecke zeigen eine Reak t ions - ~ weise, die den Disulf iden ent- spr icht . Bemerkenswer t ist, dab die K o m p o n e n t e n B 1 und B~ sich ~- n a c h / q a - n i t r o p r u s s i d - und KCN- "~ Behand lung nur sehr verzSger t ro t f/~rben.
CoA tr/~gt in 3'- S te l lung einen Phosphors~ureres t . E r wird als l e tz te r Schr i t t der Biosynthese ¢~®
= des CoA durch Adenos in t r iphos- p h a t in 3 ' -Dephospho-CoA ein- geftigt. Nueleoside und Nucleo- t ide, die in Pos i t ion 2' und 3' unsubs t i t u i e r t sind, werden nun du tch P e r j o d a t zu D ia ldehyden oxydier t , was auf dem Chroma- t o g r a m m durch Spr i ihen mi t
o~ e~
o~
~zv.9
298 F. LEVSCH~ER:
SCHIFFs-Reagens nachgewiesen werden kann. CoA mit seinem Phosphor- si~urerest in 3'-Stellung gibt keine Farbreaktion. Bei Anwendung dieses Verfahrens auf die 7 Komponenten aus Meerschweinchenleber 1/~13t sich nicht sicher eine CoA-aktive Verbindung nachweisen, die in 2'- und 3'-Stellung unsubstituiert ist (Tab. 1).
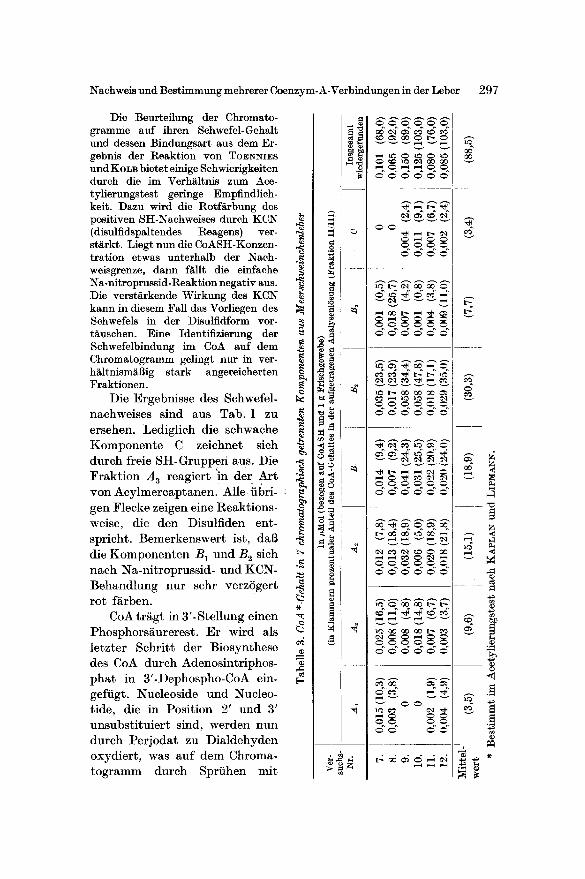
Das empfindliche Verfahren von H A ~ s und ISHERWOOD kenn- zeichnet den Phosphorgehalt der Fraktionen. Ihre Adeninkomponente
Nahtl inie --~
1. Dimension
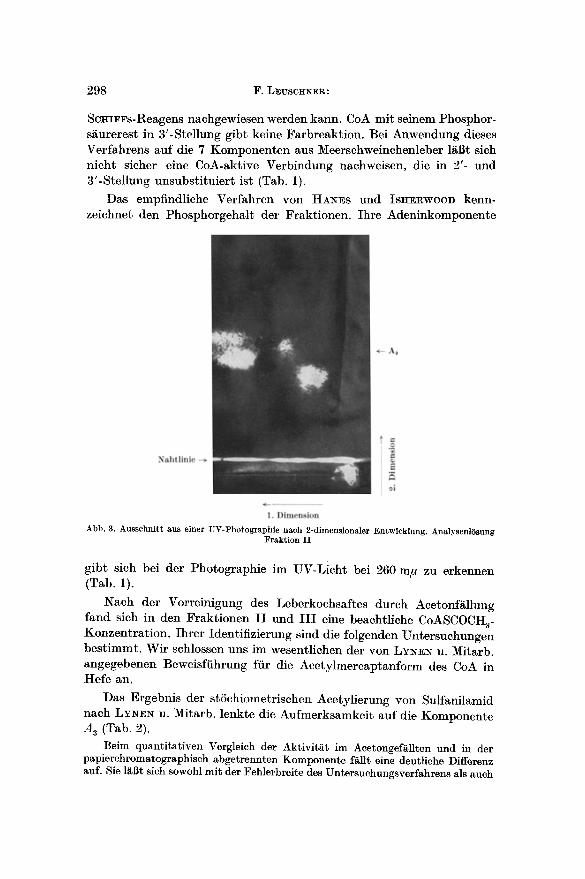
Abb. 3. Ausschni t t aus einer UV-Photographie nach 2-dimensionaler Entwicklung. AnalysenlSsung Frakt ion I I
gibt sich bei der Photographie im UV-Licht bei 260 m# zu erkennen (Tab. 1).
Nach der Vorreinigung des Leberkochsaftes durch Acetonfi~llung land sich in den Fraktionen I I und I I I eine beachtliche CoASCOCHa- Konzentration. Ihrer Identifizierung sind die folgenden Untersuchungen best immt. Wir schlossen uns im wesentlichen der yon LYNEN U. Mitarb. angegebenen Beweisfiihrung fiir die Acetylmercaptanform des CoA in Here an.
Das Ergebnis der st5chiometrischen Acetylierung von Sulfanilamid nach LYNEN u. Mitarb. lenkte die Aufmerksamkeit auf die Komponente A 3 (Tab. 2).
Beim quantitativen Vergleich der Aktivit~t im Acetongef~llten und in der papierchromatographisch abgetrennten Komponente fMlt eine deutliche Differenz auf. Sie l~]t sich sowohl mit der Fehlerbreite des Untersuchungsverfahrens als auch

Nachweis und Bestimmung mehrerer Coenzym-A-Verbindungen in der Leber 299
dem Vorliegen weiterer unter der Emptindlichkeitsgrenze liegender Acetylformen des CoA auf dem Papierchroma~gramm deuten. Wit mSchten uns beiden Griinden anschlie~en.
In guter ~bereins t immung mit dem st6chiometrischen Aeetylierungs- test ist die Na-nitroprussid-Reaktion. Erst nach Behandlung mit alko- holischer NaOH-LSsung zeigte sich Rotfgrbung. Als weitere charakte, ristische Eigenschaft der Acylmercaptane prfiften wir die Spaltbarkeit dureh Queeksflber. Wenn die Acetonfraktion I I / I I I vor dem Auftropfen mit Quecksilber-Acetat versetzt, zentrifugiert und das in Wasser aufge- nommene Pr~cipitat mit H2S behandelt wurde, so fehlte die Komponente A 3 auf dem Chromatogramm. Die ~bereinst immung mit CoASC OCH~ und Differenzierung yon CoASH l~l]t sich weiterhin durch die fehlende Oxydierbarkeit mi t Jod belegen. Wird die Aeetonfraktion I I / I I I mit alkoholischer JodlSsung versetzt, dann werden freie Mercaptane zu Di- sulfiden oxydiert. Naeh der papierehromatographischen Trennung zeigte sich, dab die Komponente A 3 durch Jod nieht ver~tndert worden war.
Fiir die Auftrennung der beschriebenen 7 Komponenten wurden die Aceton- fraktionen I I und I I I verwendet. Wurde lediglich die Fraktion I I chromatographiert, so konnte besonders gut der Fleck A 3 erhalten werden (Abb. 3). Allerdings war da- mit ein Aktivit~tsverlust verbunden.
Die wirksame Sulfhydxylform des CoA kann durch Oxydation in eine Disulfidbindung iiberffihrt werden. Dann ist das Coenzym katalyt isch unwirksam. Diese inaktive Form, deren Disulfidkomponente ver- schiedener Na tur ist, kann in der lebenden Zelle leicht wieder aktiviert werden. Wahrscheinlich i ibernimmt Glutathion oder ein spezifisches Enzym dort diese Rolle. Die Acetyfform des CoA, das funktionell wichtigste Aeylderivat des CoA, kann im tierischen Organismus unmittel- bar enzymatiseh aus Acetat und ATP, aus Brenztraubens~ure oder mSglicherweise durch Dehydrierung eines Halbmercaptals gebildet werden. CoASH, CoASCOCH 3 und CoASSR stehen in enger Beziehung, die dureh das folgende Schema gekennzeichnet werden kann:
CoASSR ~ CoASH ~ Co2;SCOCH~
3'-Dephospho-CoASH
Ihr Auftreten konnte mit dem beschriebenen Verfahren in der Leber yon ~¢[eerschweinchen beobachtet werden. Das ~vIengenverh~ltnis der Verbin- dungen zueinander betr~gt : 23 : 5:1 (CoASSR: CoASCOCH~ : CoASH). Das in diesem Schema ebenfalls eingetragene 3'-Dephospho-CoASH, das als letzte Stufe auf dem biosynthetischen Wege durehlaufen wird, konnte nicht naehgewiesen werden. Da die Verbindung im Acetylierungstest zu CoASH phosphoryliert und somit enzymatiseh akt iv wlrd, so kann sie lediglich mater 1% des gesamten CoA betragen. (Dieser Grenzwert ergibt sieh aus der Empfindlichkeit des Acetylierungstestes.)
Arch. exper. Path. u. Pharmakol. , Bd. 228 20

300 F. LEUSem~ER:
Die multiplen Formen in hoehgereinigten, bis zu 80O/o enthaltenden CoA- Pr~parationen aus Hefe und Streptomyees fradiae (DE V~Es u. Mit~rb., GREGORY U. Mitarb., BEgiRT U. Mitarb. und BuYSXE u. Mitarb.), die dutch teflweise nicht indifferente Arbeitsg~nge gewonnen wurden (F~llung als Sehwermetallsalze, EIuierung mit Pyridin-Wasser, Reduktion u. a.), enthalten vornehmlieh CoA mit freier Sulfhydrylgruppe neben Disulfidformen. Naehweisbare Mengen CoASCOCH a erkannte L¥~EN in Here. Von BASFORD und HUENNEKENS wird auf Grund far- beriseher Eigenschaften bei der Papierchromatographie eine Thiazolin-Struktur des CoA diskutiert. Dieselben Untersucher fanden in einem hoehgereinigten CoA- Pr~parat 4 Komponenten mit Suffhydryl- und Disulfideigenschaften im Ver- h~ltnis 79 : 21 (bezogen auf die Aktivit~t im Acetylierungstest).
Unsere Untersuchungen lassen erkennen, daB die Verteilung in diesen Pr~parationen keinen Vergleich mit den Verh~ltnissen in der Meer- schweinehenleber zul~Bt.
Die Charakterisierung der sogenannten Disulfidverbindungen des CoA, die mengenm~Big bedeutenden Anteil besitzen und in unseren Ver- suchen in 5 Fraktionen (bei Vernachl~ssigung der Aceton-Fraktionen I und IV) anzutreffen sind, stiitzt sich bislang fast ausschlieBlich auf die KCN-Spaltbarkeit im Na-nitroprussid-Test. Jedoch lassen die recht unterschiedliehe Spaltungsdauer und die Erfahrungen yon BASFORD und H U E ~ K E N S an verwandten Fraktionen aus Here mit Indophenol ver- tauten, dab sich hinter den KC~q-spaltbaren Komponenten neben Di- sulfidformen andere schwerer spaltbare Schwefelverbindungen verbergen.
CoA, 75~oig, wurde freundliehcrweise yon der ~irma C. H. Boehringer Sohn, Ingelheim am Rhein, und Glutathion yon der Firma Deutsche Hoffmann-La Roche A.G., Grenzach (Laden), zur Verfiigung gestellt.
Zusammenfassung
1. Durch schonende Acetonfraktionierung und papierchromato- graphische Trennung lassen sich aus dem Kochsaft yon Meerschweinchen- ]eber 7 Komponenten darstellen, die Coenzym-A-Wirksamkeit zeigen.
2. Durch Analyse der Papierflecke lassen sich neben Coenzym A mit freien SH-Gruppen, Acetyl-Coenzym A und 5 Verbindungen mit KCN-spaltbaren Schwefelbindungen im Verh~ltnis 1 : 5 : 23 (Coenzym A-SIt : Coenzym A-SCOCH 3 : Coenzym A-SSR) naehweisen.
3. ])as auf dem biosynthetischen Wege des Coenzyms A als ]etzte Stufe auftretende 3'-Dephospho-Coenzym A - - nachweisbar fiber seine freie Glykolgruppierung - - konnte nieht geftmden werden. Seine Kon- zentration muB unter 1°/o des gesamten Coenzyms A liegen.
4. Die beschriebene Arbeitsweise bietet die MSgliehkeit, wesentliche Coenzym-A-])erivate in analytisehe Studien einzubeziehen.
Literatur
BASFORD, R. E., and F. M. HU~.~EKENS: J. Amer. Chem. Soe. 77, 3878 (1955). - - BEINERT, H., R. W. v. KORFF, D. E. GREEN, D. A. BUYSKE, R. E. HANDSCHU- MACHER, H. I-IIGGINS and F. M. STRONG: J. of Biol. Chem. 200, 385 (1953). - -

Nachweis und Best lmmung mehrerer Coenzym-A-Verbindungen in der Leber 301
BRATTOI~, A. C., and E. K. MARS~Ar.r~" J . of Biol. Chem. 127, 537 (1939). - - BU- CHANAN, J . G., C. A. DEKKER and A. G. LONG: J . Chem. Soc. (Lond.) 1950, 3162. - - BUYSKE, D. M., R. ]~. HANDSCHUMACHER, H. HIGGINS, T. E. K~NG, F. M. STRONG, V. I-I. CHELDELIN, L. J . TEPLY and G. C. MUELLER: J . of Biol. Chem. 198, 307 (1951). - - FELDBERG, W., and T. MAN~: J . of Physiol. 104, 411 (1946). - - GRE- GORY, J . D., G. D. NOVELLI and F. I ~ P M ~ : J . Amer. Chem. Soc. 74, 854 (1952). - - HA~ES, C. S., and F. A. ISHERWOOD: Nature (Lond.) 164, 1107 (1949). - - KAPLAN, N. 0. , and F. L I P ~ A ~ : J . of Biol. Chem. 174, 37 (1948). - - LIP~.A~, F. : J . of Biol. Chem. 160, 173 (1945). - - LIPMANI~, F., N. O. KAPLA~, G. D. NOVELLI, L. C. TUTTLE and B. M. GU~RARD : J . of Biol. Chem. 186, 235 (1950). - - LIPTOn, M. A., and E. S. G. BARRON: J . of Bioh Chem. 166, 367 (1946). - - LYNEN, F., E. REICHERT U. L. RU]~FF: Liebigs Ann. 574, 1 ( 1 9 5 1 ) . - NACHMX~SOH~, D., and M. BER~A~: J . of Biol. Chem. 165, 551 (1946). - - NOVELLI, G. I)., N. O. KAPLAN and F. LIP~A~N: Federat . Proc. 9, 209 (1950). --NOVELLI, G. D., and F. L I P ~ A ~ : J . of Biol. Chem. 182, 213 (1950). - - SACHs, G.: Ber. Dtsch. Chem. Ges. 54, 1849 (1921). - - SNELL, E. E. , G. M. BROWl~, V. J . PETERS, J . A. CRAIG, E. L. WITTLE, J . A. MOORE, V. ~/~. MCGLOHOI~" and C. D. BIBD: J . Amer. Chem. Soc. 72, 5349 (1950). - - STADTMA~, E. R., aus G. D. NOVELLI: Methods of Biochemical Analysis 2, 189 (1955). - - ToEnai l .s , G., and J . I. KOLB: Analyt . Chemistry 28, 823 (1951).
Dr. I~ED LEUSCHNER, Erlangen, Pharmakol . In s t i t u t d. Univ.
20*
Recommended

















