Embed Size (px)
Citation preview

Aus der Medizinischen Tierklinik
der Veterinärmedizinischen Fakultät
der Universität Leipzig
Mykotoxinscreening (Deoxynivalenol, Zearalenon) in Futter, Blut,
Milch und Galle bei Kühen mit Labmagenverlagerung ( LMV) und bei
gesunden Kühe n
Inaugural-Dissertation
zur Erlangung des Grades eines
Doctor medicinae veterinariae (Dr. med. vet.)
durch die Veterinärmedizinische Fakultät
der Universität Leipzig
eingereicht von
Ahmad Alkaassem
aus Hama (Syrien)
Leipzig, 2009
gefördert vom Sächsischen Staatsministerium für Umwelt und Landwirtschaft

Mit Genehmigung der Veterinärmedizinischen Fakultät der Universität Leipzig
Dekan: Prof. Dr. Arwid Daugschies
Betreuer: apl. Prof. Dr. habil. Manfred Fürll
Gutachter: apl. Prof. Dr. habil. Manfred Fürll,
Medizinische Tierklinik,
Veterinärmedizinische Fakultät, Universität Leipzig
Prof. Dr. habil. Sven Dänicke,
Institut für Tierernährung,
Bundesforschungsinstitut für Tiergesundheit (FLI),
Braunschweig
Prof. Dr. habil. em. Ute Schnurrbusch,
Schkeuditz
Tag der Verteidigung: 03.03.2009

Meinen Eltern und meiner Familie

Inhaltsverzeichnis
I
Inhaltsverzeichnis
1 Einleitung .............................................................................................................. 1
2 Literaturübersicht .................................................................................................. 3
2.1 Historische Betrachtungen............................................................................... 3
2.2 Mykotoxine....................................................................................................... 3
2.3 Mykotoxinbildung ............................................................................................. 4
2.4 Effekte von Mykotoxinen auf Tiergesundheit und Leistung beim Wiederkäuer..................................................................................................... 5
2.5 Mykotoxine der Fusariumspezies .................................................................... 6
2.5.1 Vorkommen der Fusariumspezies ........................................................... 6
2.5.2 Entwicklungsbedingungen ....................................................................... 7
2.5.3 Fusarientoxine ......................................................................................... 7
2.5.3.1 DON (Vomitoxin)............................................................................ 9
2.5.3.1.1 Physikalisch-chemische Eigenschaften von DON ................... 9
2.5.3.2 Zearalenon................................................................................... 10
2.5.3.2.1 Physikalisch-chemische Eigenschaften von Zearalenon ....... 11
2.5.4 Metabolismus von DON und ZON beim Wiederkäuer ........................... 11
2.5.5 Biologische Eigenschaften und Auswirkungen von DON bei
Nutztieren .............................................................................................. 13
2.5.6 Biologische Eigenschaften und Auswirkungen von ZON bei
Nutztieren .............................................................................................. 17
2.5.6.1 Wirkungsmechanismus von ZON................................................. 17
2.5.6.2 Klinische Auswirkungen von Zearalenon ..................................... 18
2.6 Labmagenverlagerung beim Rind.................................................................. 20
3 Tiere, Material und Methoden ............................................................................. 23
3.1 Tiere und Versuchsanordnung....................................................................... 23
3.2 Methoden....................................................................................................... 23
3.2.1 Klinische Untersuchung und Diagnosestellung...................................... 23
3.2.2 Entnahme von Futter-, Blut-, Milch- und Gallensaft- Proben.................. 24
3.2.3 Klinisch-chemische Analysen ................................................................ 25
3.2.4 Hämatologische Untersuchung.............................................................. 25
3.2.5 Trolox Equivalent Antioxidative Capacity (TEAC) .................................. 27
3.2.6 Bestimmung der Mykotoxine (DON, ZON, DOM-1, α-ZOL, ß-ZOL)
mittels HPLC.......................................................................................... 27

Inhaltsverzeichnis
II
3.2.6.1 Bestimmung von Deoxynivalenol und de-epoxy-Deoxynivalenol in
Futtermitteln, Blutserum, Milch und Gallensaft............................. 27
3.2.6.2 Zearalenon- (α- und ß-Zearalenol) Bestimmung mittels HPLC in
Futter, Blutserum, Milch und Gallensaft (mod. Zea-Lufa-Methode
IAC, HPLC) .................................................................................. 28
3.2.7 Statistische Auswertung......................................................................... 29
4 Ergebnisse .......................................................................................................... 30
4.1 Ergebnisse der Mykotoxinanalysen ............................................................... 30
4.1.1 Patienten (n=61) .................................................................................... 30
4.1.2 Gesunde Kühe (n=13) ........................................................................... 32
4.2 Klinische Untersuchungsbefunde .................................................................. 32
4.3 Ergebnisse der klinisch-chemischen Blutuntersuchung................................. 36
4.3.1 Gesamteiweiß........................................................................................ 36
4.3.2 Bilirubin .................................................................................................. 37
4.3.3 Kreatinin................................................................................................. 38
4.3.4 Glucose.................................................................................................. 39
4.3.5 Cholesterol............................................................................................. 40
4.3.6 Beta-Hydroxybutyrat .............................................................................. 41
4.3.7 Freie Fettsäuren .................................................................................... 42
4.3.8 Aspartat-Aminotransferase .................................................................... 43
4.3.9 Glutamat-Dehydrogenase...................................................................... 44
4.3.10 Gamma-Glutamyltransferase................................................................. 45
4.3.11 Alkalische Phosphatase......................................................................... 46
4.3.12 Creatinkinase......................................................................................... 47
4.3.13 Magnesium ............................................................................................ 48
4.3.14 Eisen...................................................................................................... 49
4.3.15 Anorg. Phosphat (Pi).............................................................................. 50
4.3.16 Trolox Equivalent Antioxidative Capacity ............................................... 51
4.3.17 Calcium.................................................................................................. 52
4.3.18 Kalium.................................................................................................... 53
4.3.19 Natrium .................................................................................................. 54
4.3.20 Chlorid ................................................................................................... 55
4.3.21 Säure-Basen-Haushalt........................................................................... 56
4.3.21.1 pH-Wert........................................................................................ 56

Inhaltsverzeichnis
III
4.3.21.2 Base Excess (BE) ........................................................................ 57
4.3.22 Ergebnisse der hämatologischen Untersuchungen ............................... 58
5 Diskussion........................................................................................................... 60
5.1 Ergebnisse der Mykotoxinanalysen ............................................................... 60
5.1.1 Patienten................................................................................................ 60
5.1.2 Gesunde Kühe....................................................................................... 62
5.2 Klinische Untersuchungsbefunde und Krankheitsausgang............................ 63
5.3 Klinisch-chemische Parameter ...................................................................... 67
5.3.1 Energiestoffwechsel............................................................................... 67
5.3.1.1 Beta-Hydroxybutyrat-Konzentrationen ......................................... 67
5.3.1.2 Glucosekonzentrationen .............................................................. 68
5.3.1.3 Freie Fettsäure-Konzentrationen ................................................. 68
5.3.2 Leberstoffwechsel.................................................................................. 69
5.3.2.1 Bilirubinkonzentrationen im Serum .............................................. 69
5.3.2.2 Aspartat-Aminotransferase-Aktivitäten......................................... 69
5.3.2.3 Glutamat-Dehydrogenase-Aktivitäten .......................................... 70
5.3.2.4 Gamma-Glutamyl-Transferase-Aktivitäten................................... 71
5.3.3 Eiweißstoffwechsel ................................................................................ 71
5.3.3.1 Gesamtproteinkonzentrationen .................................................... 71
5.3.3.2 Kreatininkonzentrationen ............................................................. 72
5.3.4 Muskelstoffwechsel................................................................................ 72
5.3.4.1 Creatinkinase-Aktivitäten ............................................................. 72
5.3.5 Mineralstoffwechsel ............................................................................... 73
5.3.5.1 Gesamtcalcium-Konzentrationen ................................................. 73
5.3.5.2 Anorg. Phosphat .......................................................................... 74
5.3.5.3 Magnesium-Konzentrationen ....................................................... 74
5.3.5.4 Natrium-Konzentrationen ............................................................. 75
5.3.5.5 Kalium-Konzentrationen............................................................... 75
5.3.5.6 Chlorid-Konzentrationen .............................................................. 76
5.3.6 Cholesterolkonzentrationen ................................................................... 76
5.3.7 Hämatologie........................................................................................... 77
5.3.7.1 Differentialblutbild ........................................................................ 77
5.3.8 Säure-Basen-Haushalt........................................................................... 78
5.3.9 Trolox Equivalent Antioxidative Capacity ............................................... 78

Inhaltsverzeichnis
IV
5.3.10 Eisen-Konzentrationen........................................................................... 78
5.3.11 Alkalische Phosphathase-Aktivitäten ..................................................... 79
6 Zusammenfassung.............................................................................................. 80
7 Summary............................................................................................................. 82
8 Literaturverzeichnis ............................................................................................. 84
9 Danksagung...................................................................................................... 109

Abkürzungsverzeichnis
V
Abkürzungen Abb. Abbildung
a. p. ante partum
AP Alkalische Phosphatase
AST Aspartat-Aminotransferase
BCS body condition scoring
BHB Beta-Hydroxybutyrat
CK Creatinkinase
DGKC Deutsche Gesellschaft für klinische Chemie
DON Deoxynivalenol
DOM-1 de-epoxy-Deoxynivalenol
EDTA Ethylendiamintetraacetat
EU Europäischen Union
F. Fusarien
Fa. Firma
FA Futteraufnahme
Fab antigen binding fragment
FAL Bundesforschungsanstalt für Landwirtschaft
FFS Freie Fettsäuren
ggr. geringgradig
GGT Gamma Glutamyltransferase
GLDH Glutamat-Dehydrogenase
hgr. hochgradig
HPLC High Performance Liquid Chromatography
ICAM interzelluläres Adhäsionsmolekül
IU international unit
i.v. intravenös
N, n Anzahl der Fälle
o. b. B. ohne besonderen Befund
PBW Pansenbewegung
p. p. post partum
S Serum
BE Base Excess
sog. sogenannt

Abkürzungsverzeichnis
VI
Tab. Tabelle
TEAC Trolox equivalent antioxidative capacity
TS Trockensubstanz
V Vollblut
v. a. vor allem
z. T. zum Teil
ZOL Zearalenol
ZON Zearalenon

Einleitung
1
1 Einleitung
Mykotoxinhaltige Futtermittel können die Gesundheit und Leistungsfähigkeit von
Nutztieren gefährden, aber auch der Mensch kann durch Aufnahme von mykotoxin-
belasteten Nahrungsmitteln tierischer und pflanzlicher Herkunft gesundheitlich beein-
trächtigt werden (JECFA 2001). Obwohl Wiederkäuer prinzipiell in der Lage sind, ei-
ne Reihe von Mykotoxinen im Pansen soweit zu metabolisieren und zu detoxifizieren,
so dass von ihnen keine oder nur geringe toxische Effekte ausgehen, können sie
dennoch negative Auswirkungen haben, wenn beispielsweise ein gestörtes Pansen-
milieu oder andere gesundheitlich relevante Probleme gleichzeitig auftreten. Insofern
sind die von der EU empfohlenen Richtwerte für kritische Konzentrationen von Deo-
xynivalenol (DON) und Zearalenon (ZON) in der Gesamtration von Wiederkäuern nur
unter optimalen Haltungs- und Fütterungsbedingungen ausreichend. Es wird empfoh-
len, bei Rindern (außer Kälbern) 5 mg DON und 0,5 mg ZON je kg der täglichen Ra-
tion (Referenztrockensubstanzgehalt 88 %) nicht zu überschreiten (EU 2006).
Mykotoxine sind toxische Sekundärstoffwechselprodukte von Schimmelpilzen, deren
chemische Strukturen sehr unterschiedlich sind (SCHUH 1989; BISCHOFF 1998).
Bisher wurden ca. 400 Mykotoxine nachgewiesen, die bei Säugtieren und Vögeln
dosisabhängig zu Vergiftungssymptomen führen können (BAUER u. GAREIS 1987;
REIß 1998; FINK-GREMMELS 1999; GAREIS u. WOLFF 2000; HOCHSTEINER et
al. 2000; ATROSHI et al. 2002; VÖLKL et al. 2004). Die Anfälligkeit gegenüber My-
kotoxinen ist abhängig von der jeweiligen Tierart, dem Alter und dem spezifischen
Giftstoff (WILKENS 2005). Die einzelnen Fusarientoxine haben unterschiedliche
Zielorgane, wie Nervensystem, blutbildende Organe, Haut, Verdauungs- und Genital-
trakt (LEIBETSEDER 1981).
Zu den häufig vorkommenden Mykotoxinen zählen unter anderem Deoxynivalenol
und Zearalenon. Deoxynivalenol (DON, Vomitoxin, 4-Deoxynivalenol, Rd-Toxin, De-
hydronivalenol) gehört zu den Trichothecenen, der wichtigsten von Fusarien gebilde-
ten Mykotoxingruppe. Es wird als sekundärer Stoffwechselmetabolit vorwiegend von
Fusarium graminearum und Fusarium culmorum gebildet und ist, wie andere Tri-
chothecene auch, chemisch und physikalisch sehr stabil. Die durch DON-
kontaminierte Futtermittel ausgelösten akuten Mykotoxikosen äußern sich bei Tieren
durch Nahrungsverweigerung und Erbrechen (Monogastrien) (D´MELLO et al. 1999).

Einleitung
2
In hohen Konzentrationen mit dem Futtermittel aufgenommen, können DON und an-
dere Trichothecene Leistungsdepressionen und vielfältige klinische Symptome bei
allen Nutztieren verursachen, wobei Rinder auf Grund der entgiftenden Aktivität des
Pansens nicht so anfällig sind wie andere Tierarten (DÄNICKE u. VALENTA 2004).
ZON ist weltweit eines der am häufigsten in Futtermitteln nachweisbaren Fusariento-
xine. Insbesondere in Jahren mit ungünstigen klimatischen Bedingungen für die Ge-
treideproduktion ist mit einer starken Kontamination zu rechnen. So konnte 1998 in
Abhängigkeit von der Ernteregion Zearalenon in bis zu 74 % der in Deutschland un-
tersuchten Weizenproben und in bis zu 95 % der getesteten Maisproben nachgewie-
sen werden (JANZ 1999; OLDENBURG et al. 2000). ZON und seine Derivate besit-
zen keine besonders hohe akute Toxizität. Es handelt sich vielmehr um nichtsteroi-
dale Makrolide, die aufgrund ihrer dem Östrogen ähnlichen räumlichen Struktur eine
kompetitive Bindung mit Östrogenrezeptoren eingehen können (MITTERBAUER et
al. 2005). Chronische Intoxikationen können bei Nutztiere so zu erheblichen Frucht-
barkeitsstörungen führen (BAUER u. GAREIS 1987).
Ziel diese Arbeit war es zu untersuchen, ob die genannten Mykotoxine (DON, ZON)
sowie deren Metaboliten an der Entstehung der häufigsten nichtinfektiösen Rinder-
krankheiten beteiligt sind. Deshalb wurden folgende Fragestellungen verfolgt:
- Kommen Mykotoxine bei Kühen mit Labmagenverlagerung und anderen Erkran-
kungen vor?
- Spielen Mykotoxine eine Rolle bei der Entstehung der Labmagenverlagerung?
- Bestehen Beziehungen zwischen dem Vorkommen von Mykotoxinen und klini-
schen Störungen?
- Eignet sich Gallensaft zum ZON-/ZOL-Nachweis bei Kühen?
- Gibt es sinnvolle Korrelationen zwischen Blutinhaltsstoffen und Mykotoxinbelas-
tungen?

Literatur
3
2 Literaturübersicht
2.1 Historische Betrachtungen
Vergiftungen durch Schimmelpilztoxine sind seit langem bekannt und in großem
Ausmaß über die Jahrhunderte aufgetreten. Historisch betrachtet lassen sich heute
Probleme und Phänomene der Geschichte, wie z.B. der Untergang mancher Hoch-
kulturen, eventuell auf Vergiftungen mit Mykotoxinen zurückführen. Auch die Ge-
heimnisse um den Tod der Entdecker der Pharaonengräber sollen auf akute Vergif-
tungen mit Mykotoxinen zurückzuführen sein (REIß 1998; SABATER et al. 2004).
Um 1945 wurden Berichte aus Russland bekannt, denen zu folge es durch den Ver-
zehr von stark schimmelpilzebelastetem Getreide zu schweren Gesundheitsschädi-
gungen beim Menschen gekommen war (BISCHOFF 1998). Anfang der 1960er Jah-
re beunruhigten Berichte über ein Massensterben von Truthühnern in England die
Öffentlichkeit. Die als "Turkey X" bezeichnete Krankheit führte zum Tod von 100000
jungen Truthühnern, wobei als auffälligste pathologische Veränderung Leberkarzi-
nome diagnostiziert wurden (BLOUNT 1961). In Folge erkrankten auch Entenküken,
Fasane und Forellen in außerordentlich hoher Anzahl sowie viele andere Nutztierar-
ten. Als Ursache wurden schließlich Aflatoxine, Stoffwechselprodukte des Schimmel-
pilzes Aspergillus flavus, im Futter, bei welchem es sich um brasilianisches Erd-
nussmehl handelte, nachgewiesen. Aufgrund der großen ökonomischen Auswirkun-
gen begann mit dem Auftreten der Turkey X-Disease die intensive wissenschaftliche
Aktivität der Mykotoxinforschung in vielen Ländern (BISCHOFF 1998).
2.2 Mykotoxine
Mykotoxine sind strukturell unterschiedliche, sekundäre giftige Stoffwechselprodukte
verschiedener Schimmelpilze (SCHUH 1989; LACEY u. MAGAN 1991; LEPSCHY
1992; OSWALD et al. 2003; SABATER et al. 2004; BAUER 2005). Sie stellen sozu-
sagen die „chemischen Waffen“ der Schimmelpilze dar, mit denen sie ihren Lebens-
raum gegenüber Nahrungskonkurrenten und Fressfeinden sichern können. Bisher
sind mehr als 400 verschiedene Mykotoxine von mehr als 250 Schimmelpilzarten
bekannt (GAREIS u. WOLFF 2000). Während einige Mykotoxine nur von einer be-
grenzten Zahl von Arten synthetisiert werden, werden andere wiederum von vielen

Literatur
4
Arten aus unterschiedlichen Pilzgattungen gebildet. Viele werden nur unter Laborbe-
dingungen in relevanten Mengen gebildet und nur eine relativ geringe Zahl kommt
häufig und in höheren Konzentrationen natürlich auf Getreide vor und ist damit für die
Lebensmittelsicherheit von Bedeutung (ENGELHARDT 2000; GAREIS u. WOLFF
2000). Mykotoxin-Produzenten sind vor allem Aspergillus-, Penicillium-, Fusarium-,
Claviceps- (Mutterkorn) und Rhizopus-Arten. Bekannte Mykotoxine sind Aflatoxine
(die stärksten natürlichen oral wirkenden Carcinogene), Citrinin, Byssochlamsäure,
Patulin, Ochratoxin, Sterigmatocystin, Moniliformin, Ergot-Alkaloide, Ergochrome,
Cytochalasane, Penicillinsäure, Zearalenon, Rubratoxine und Trichothecene (BI-
SCHOFF 1998; ENGELHARDT 2000; WHITLOW u. HAGLER 2005).
Man unterscheidet Feldpilze, deren Hauptvertreter die Gattungen Fusarium, Cla-
dosporium, Helminthosporium und Alternaria sind, und Lagerpilze, wie z.B. die Gat-
tungen Penicillium und Aspergillus. Bei Feldpilzen erfolgt der Befall der Pflanzen vor
der Ernte und die giftigen Metaboliten werden bereits auf dem Feld gebildet. Die
meisten Fusariumarten zeigen bei einer Feuchtigkeit von 22-24 % ein optimales
Wachstum (LACEY u. MAGAN 1991). Wenn der Wassergehalt des Getreides nach
der Ernte sinkt, vermehren sich hingegen die Lagerpilze. Ihr Wachstum kann bei ei-
nem Feuchtigkeitsgehalt von 13-15 % beobachtet werden. Die Bildung der toxischen
Stoffwechselprodukte beginnt während der Lagerung des Getreides (GEDEK 1980;
SCHUH 1981; REIß 1998). In Futtermitteln können mehrere Mykotoxine gleichzeitig
auftreten. Es handelt sich bei den in der Praxis auftretenden Mykotoxikosen i.d.R.
somit um ein multitoxisches Geschehen (PIER et al. 1980). Mykotoxine sind meist
niedermolekulare Substanzen. Dadurch wird ihre Stabilität gegenüber Umweltein-
flüssen erhöht und gleichzeitig verhindert, dass das Immunsystem eines Organismus
Antikörper gegen sie bilden kann (PIER et al. 1980; BISCHOFF 1998).
2.3 Mykotoxinbildung
Mykotoxine werden von Schimmelpilzen während des Wachstums gebildet. Die Pilze
wachsen nicht nur an der Oberfläche, sondern dringen tief in das Ernteprodukt oder
die Nahrung ein. Mykotoxine werden entweder kontinuierlich in das Substrat, auf
dem die Pilze wachsen, abgegeben oder erst dann freigesetzt, wenn das Pilzmyzel
(Zellverbund der Pilzfäden) auseinanderbricht. Auch die Sporen der Schimmelpilze
können Mykotoxine enthalten (MILCZEWSKI et al. 1981; STEYN 1998). Die Produk-
tionsrate von Toxinen und neuer Zellmasse verhält sich nicht proportional. Es liegt

Literatur
5
kein direkter Zusammenhang zwischen Myzelwachstum und Toxinbildung vor, so
dass eine starke Ausbreitung des Schimmelpilzes nicht automatisch eine hohe Myko-
toxinausschüttung bedeutet (REIß 1998).
Mykotoxine können über drei Wege in Futter- und Nahrungsmittel gelangen:
1. Primärkontamination
Die Ausgangsstoffe für das Futter- oder Nahrungsmittel sind von Toxinbildnern
befallen.
2. Sekundärkontamination
Das fertige Futter- oder Nahrungsmittel wird von Toxinbildnern kontaminiert.
3. Carry Over
Nutztiere, die toxinhaltige Futtermittel konsumieren, können einzelne
Mykotoxine auf Grund ihrer Stabilität in Organen oder in tierischen Produkten
anreichern (ENGELHARDT 2000; SCHNERR 2002).
2.4 Effekte von Mykotoxinen auf Tiergesundheit und Leistung beim Wiederkäuer
Die Auswirkungen von Mykotoxinen auf die Gesundheit und Leistung der Wieder-
käuer sind häufig schwer zu quantifizieren, weil Mykotoxine durch die Pansenflora
metabolisiert und damit inaktiviert werden können. Dies erklärt die relative Unemp-
findlichkeit dieser Spezies gegenüber Mykotoxinen. Die entgiftende Kapazität der
Pansenflora ist jedoch nicht unbegrenzt und abhängig von der Rationszusammen-
setzung und dem Pansenmilieu (FINK-GREMMELS 2008b). Die Anfälligkeit gegen-
über Mykotoxinen ist neben der jeweiligen Tierart auch abhängig vom Alter der Tiere,
der Expositionsdauer und dem jeweiligen Toxin. Junge Tiere sind gegenüber be-
stimmten Mykotoxinen empfindlicher als adulte Tiere (HOCHSTEINER et al. 2000).
HADLEY et al. (2006) und WENZ et al. (2007) konnten bei Milchkühen nach einer
Periode der Verfütterung von schimmelpilzkontaminierter Silage (schlechter Silage-
qualität) eine reduzierte Pansenmotorik, schlechtere Futterverwertung und Durchfall
beobachten. Die letztgenannten Symptome waren begleitet von einem Milchleis-
tungsabfall (Verluste >15 %) und einer erhöhten Inzidenz von subklinischen Mastiti-
den. Ein weiterer wichtiger ökonomischer und verbraucherschützerischer Aspekt ist
der potentiellen Übertritt der Mykotoxine in die Milch (FINK-GREMMELS 2005).
Man unterscheidet 3 Formen von Mykotoxikosen (PIER et al. 1980; FINK-
GREMMELS 1999; WILKENS 2005):

Literatur
6
1- Akute primäre Mykotoxikosen:
Diese treten dann auf, wenn hohe oder mittlere Konzentrationen des Toxins vom tie-
rischen Organismus aufgenommen werden. Sie gehen meist mit Hepatitis, Hä-
morrhagien, Nephritis, Nekrosen der Schleimhäute und des Verdauungstraktes und
schlussendlich dem Tod der Tiere einher.
2- Chronische primäre Mykotoxikosen:
Sie werden durch die Aufnahme von mittleren bis niedrigen Toxindosen über eine
längere Zeit hervorgerufen. Verminderte Fruchtbarkeit und Produktivität durch lang-
samere Wachstumsraten, verschlechterte Marktqualität und verminderte Futterver-
wertung sind die Folge. Diese Formen gehen meist nicht mit offensichtlichen Anzei-
chen einer primären Mykotoxikose einher und sind von großer wirtschaftlicher Be-
deutung.
3- Sekundäre Mykotoxikosen:
Diese entstehen durch Aufnahme von Mykotoxinkonzentrationen, die keine klini-
schen Symptome, jedoch eine Prädisposition gegenüber bakteriellen, viralen und
parasitären Erkrankungen durch Herabsetzung der natürlichen Resistenzmechanis-
men verursachen (PIER et al. 1980; SCHUH 1989).
2.5 Mykotoxine der Fusariumspezies
2.5.1 Vorkommen der Fusariumspezies
Fusarien sind weltweit verbreitet, können in allen Regionen wie auch in jedem Boden
vorkommen. Sie sind die wichtigsten toxinbildenden Schimmelpilze der nördlichen
Hemisphäre. Auch wenn die Temperatur- und Feuchtigkeitsbedingungen stark ab-
weichen, sind sie noch in der Lage zu überdauern, so dass sie selbst in extrem hei-
ßen und trockenen Regionen nachgewiesen werden können (PALTI 1978). Fusarie-
narten können z.B. in Abwesenheit einer Wirtspflanze jahrelang in einer Tiefe von
40-80 cm im Boden überleben (LEIBETSEDER 1982). F. graminearum, F. sporotri-
chioides und F. verticillioides sind die weltweit bedeutendsten Fusarienarten, die auf
Getreide (Körnermais) wachsen. In Europa sind insbesondere F. culmorum, F.
subglutinans und F. culmorum, F. graminearum und F. verticillioides präsent. Auch in
Deutschland treten am häufigsten F. graminearum, F. culmorum, F. avenaceum, F.
sporotrichioides und F. poae auf (CHELKOWSKI 1998).
Literatur
7
2.5.2 Entwicklungsbedingungen
Für ein optimales Wachstum benötigen die meisten Fusarienarten ein Temperaturop-
timum von 20-24°C, eine Substratfeuchte von 22-23 % und einen pH-Wert zwischen
5,5 und 6,5 (PALTI 1978; BÖHM 1989; LACEY u. MAGAN 1991). Auch ein Wachs-
tum im sauren Milieu (Silage) ist möglich (REIß 1998). Aufgrund des geringen Anteils
an stickstoffhaltigen Verbindungen und des hohen Kohlenhydratgehaltes ist Getreide
ein ideales Substrat für das Fusarienwachstum und damit verbundener Mykotoxinbil-
dung. Insbesondere in Weizen, Hafer, Gerste und Mais können Zearalenon und Tri-
chothecene nachgewiesen werden (THALMANN 1990; OLDENBURG et al. 2000).
2.5.3 Fusarientoxine
Man zählt zur Gruppe der Fusarium-Toxine diejenigen Mykotoxine, die von Fusarium
spp. gebildet werden: Trichothecene, Fumonisine, Moniliformin und Zearalenon, die
Butenolide, sowie die zur Stoffklasse der zyklischen Oligopeptide zählenden Enniati-
ne, Beauvericine und Fusaroproliferine (MARASAS et al. 1984; BOTTALICO 1998;
NIESSEN u. VOGEL 1998). Die beiden für die landwirtschaftliche Produktion und
Nutztierhaltung wichtigsten Vertreter dieser Toxingruppe sind das Zearalenon (ZON)
und das Deoxynivalenon (DON) (REIß 1998).
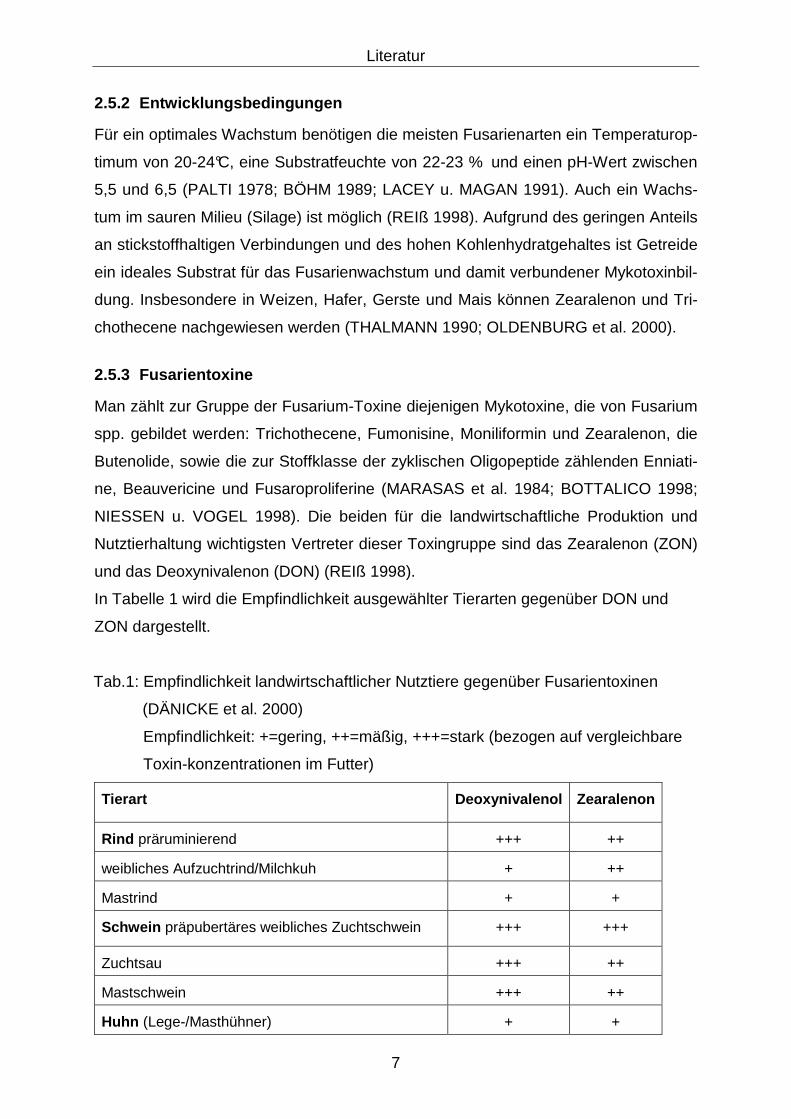
In Tabelle 1 wird die Empfindlichkeit ausgewählter Tierarten gegenüber DON und
ZON dargestellt.
Tab.1: Empfindlichkeit landwirtschaftlicher Nutztiere gegenüber Fusarientoxinen
(DÄNICKE et al. 2000)
Empfindlichkeit: +=gering, ++=mäßig, +++=stark (bezogen auf vergleichbare
Toxin-konzentrationen im Futter)
Tierart Deoxynivalenol Zearalenon
Rind präruminierend +++ ++
weibliches Aufzuchtrind/Milchkuh + ++
Mastrind + +
Schwein präpubertäres weibliches Zuchtschwein +++ +++
Zuchtsau +++ ++
Mastschwein +++ ++
Huhn (Lege-/Masthühner) + +
Literatur
8
Um einen Überblick über den Kontaminationsgrad von Futtermitteln mit Mykotoxinen
zu erhalten, werden weltweit immer wieder flächendeckende Untersuchungen durch-
geführt. Danach ist das Vorkommen von Fusarientoxinen großen lokalen Schwan-
kungen unterlegen und vom Erntejahr abhängig. Das Risiko einer Zearalenonkonta-
mination ist beim Mais am höchsten (JANZ 1999). So waren insbesondere Mais und
auch Weizen mit Zearalenongehalten von bis zu 26 mg/kg Mais (Kolben/Körner)
bzw. 8 mg/kg Weizen belastet (OLDENBURG et al. 2000).
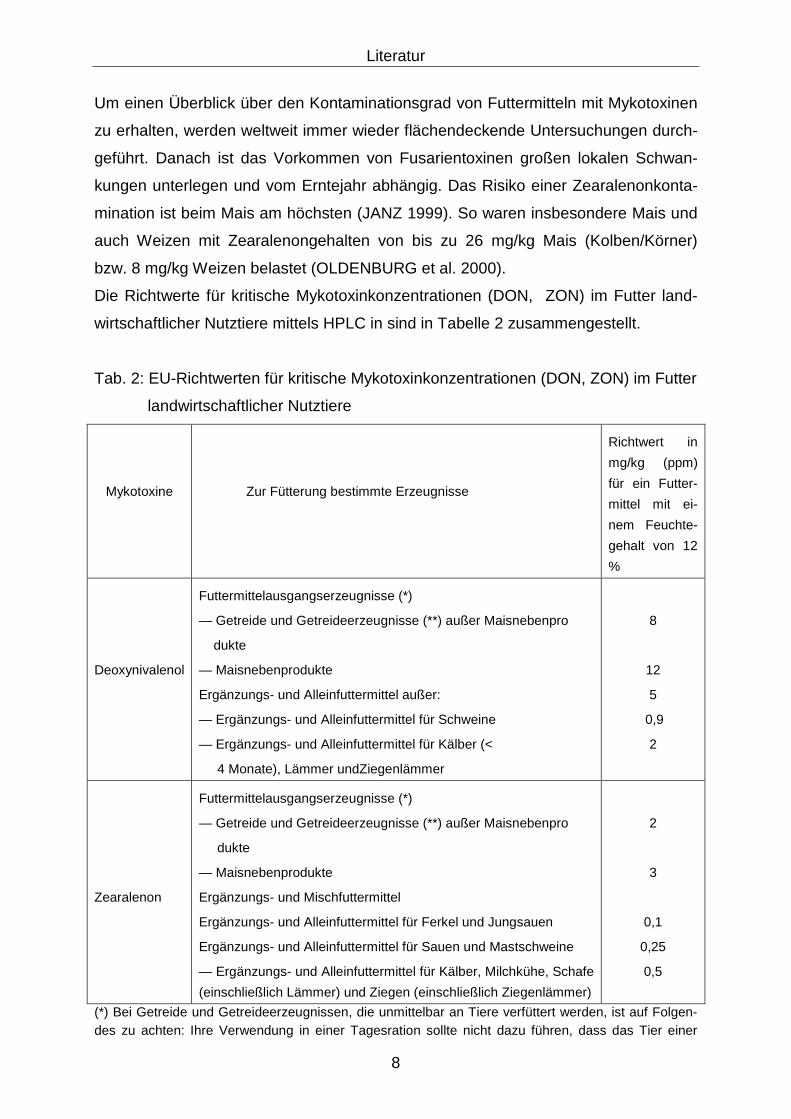
Die Richtwerte für kritische Mykotoxinkonzentrationen (DON, ZON) im Futter land-
wirtschaftlicher Nutztiere mittels HPLC in sind in Tabelle 2 zusammengestellt.
Tab. 2: EU-Richtwerten für kritische Mykotoxinkonzentrationen (DON, ZON) im Futter
landwirtschaftlicher Nutztiere
Mykotoxine
Zur Fütterung bestimmte Erzeugnisse
Richtwert in
mg/kg (ppm)
für ein Futter-
mittel mit ei-
nem Feuchte-
gehalt von 12
%
Deoxynivalenol
Futtermittelausgangserzeugnisse (*)
— Getreide und Getreideerzeugnisse (**) außer Maisnebenpro
dukte
— Maisnebenprodukte
Ergänzungs- und Alleinfuttermittel außer:
— Ergänzungs- und Alleinfuttermittel für Schweine
— Ergänzungs- und Alleinfuttermittel für Kälber (<
4 Monate), Lämmer undZiegenlämmer
8
12
5
0,9
2
Zearalenon
Futtermittelausgangserzeugnisse (*)
— Getreide und Getreideerzeugnisse (**) außer Maisnebenpro
dukte
— Maisnebenprodukte
Ergänzungs- und Mischfuttermittel
Ergänzungs- und Alleinfuttermittel für Ferkel und Jungsauen
Ergänzungs- und Alleinfuttermittel für Sauen und Mastschweine
— Ergänzungs- und Alleinfuttermittel für Kälber, Milchkühe, Schafe
(einschließlich Lämmer) und Ziegen (einschließlich Ziegenlämmer)
2
3
0,1
0,25
0,5
(*) Bei Getreide und Getreideerzeugnissen, die unmittelbar an Tiere verfüttert werden, ist auf Folgen-des zu achten: Ihre Verwendung in einer Tagesration sollte nicht dazu führen, dass das Tier einer

Literatur
9
höheren Menge an diesen Mykotoxinen ausgesetzt ist als bei einer entsprechenden Exposition, wenn in einer Tagesration nur die Alleinfuttermittel verwendet werden. (**) Der Begriff „Getreide und Getreideerzeugnisse“ umfasst nicht nur die unter der Überschrift 1 „Ge-treidekörner, deren Erzeugnisse und Nebenerzeugnisse“ des nicht ausschließlichen Verzeichnisses der wichtigsten Futtermittel-Ausgangserzeugnisse in Teil B des Anhangs zur Richtlinie 96/25/EG des Rates vom 29. April 1996 über den Verkehr mit Futtermittelausgangserzeugnissen (ABl. L 125 vom 3.5.1996, S. 35) aufgeführten Futtermittelausgangserzeugnisse, sondern auch andere aus Getreide gewonnene Futtermittelausgangserzeugnisse, vor allem Getreidegrünfutter und -raufutter.
2.5.3.1 DON (Vomitoxin)
Das Trichothecen DON ist erstmals in Japan isoliert worden (MOROOKA et al.
1972). Es wird vorwiegend von Fusarium graminearum und Fusarium culmorum ge-
bildet. DON ist durch seine allgemeine Zytotoxizität und Immunsuppressivität ge-
kennzeichnet (ROTTER et al. 1996) und ist nahezu weltweit im Getreide nachweis-
bar (SCF/CS/CNTM/MYC 1999; FUCHS et al. 2002; BENNET u. KLICH 2003;
VÖLKL et al. 2004). Aufgrund seines emetischen Effekts bei Schweinen wurde es als
„Vomitoxin“ bezeichnet.
2.5.3.1.1 Physikalisch-chemische Eigenschaften von DON
Deoxynivalenol (3",7",15-trihydroxy-12,13-epoxytrichothec-9-en-8-on) ist ein physika-
lisch und chemisch stabiles Molekül, welches während Lagerung, Erhitzen, Mahlpro-
zessen und anderen Produktionsschritten von Lebens- und Futtermitteln nicht zer-
stört wird (SCF/CS/CNTM/MYC 1999). Die Epoxidgruppe wird für die Toxizität der
Verbindungen verantwortlich gemacht. Zurzeit sind ungefähr 170 unterschiedliche
Trichothecene bekannt, die auf der Grund ihrer charakteristischen Eigenschaften in
vier Gruppen eingeteilt werden (Typ A - D), wobei das natürliche Vorkommen von
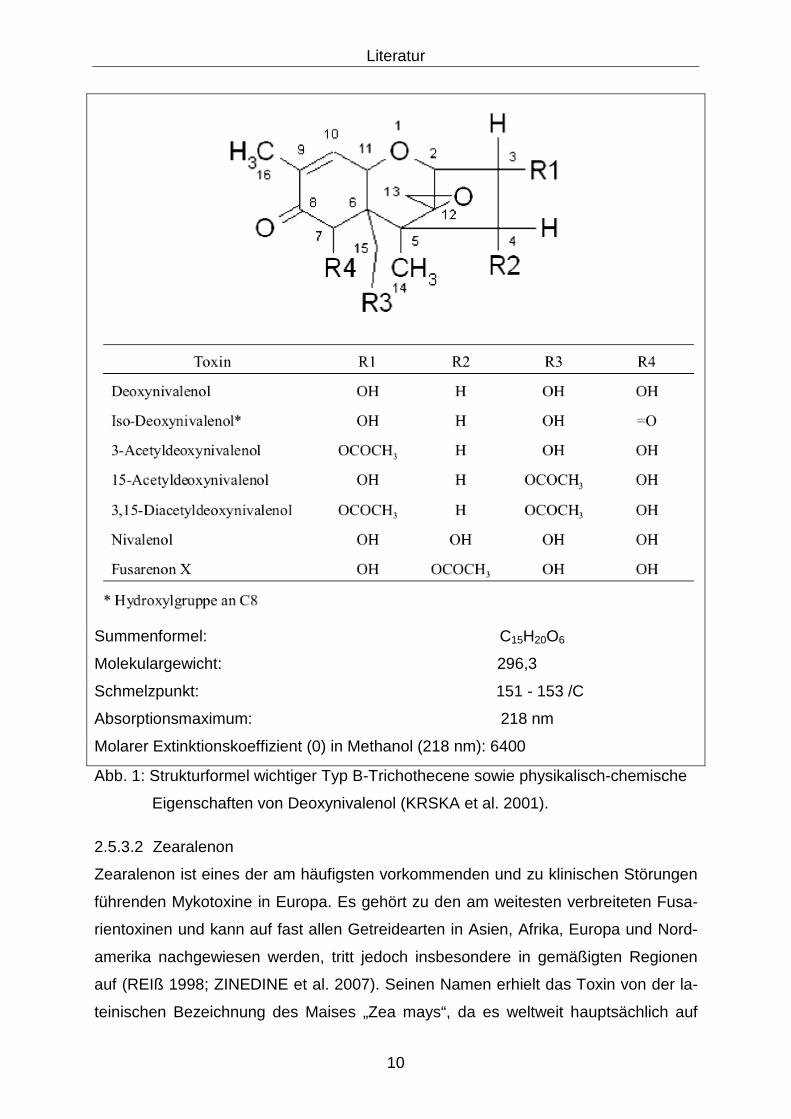
Typ C und D zu vernachlässigen ist (KRSKA et al. 2001). Typ B-Trichothecene, zu
denen auch Deoxynivalenol gehört, besitzen an C-8 eine Carbonylgruppe. Iso-
Deoxynivalenol, ein Isomer von DON, weist hingegen an C-8 eine Hydroxylgruppe
auf und de-epoxy-Deoxynivalenol (DOM-1) stellt ein mikrobielles Abbauprodukt des
Toxins dar. Einen Überblick über die Substituenten einiger Typen gibt die folgende
Abbildung 1.
Literatur
10
Summenformel: C15H20O6
Molekulargewicht: 296,3
Schmelzpunkt: 151 - 153 /C
Absorptionsmaximum: 218 nm
Molarer Extinktionskoeffizient (0) in Methanol (218 nm): 6400
Abb. 1: Strukturformel wichtiger Typ B-Trichothecene sowie physikalisch-chemische
Eigenschaften von Deoxynivalenol (KRSKA et al. 2001).
2.5.3.2 Zearalenon
Zearalenon ist eines der am häufigsten vorkommenden und zu klinischen Störungen
führenden Mykotoxine in Europa. Es gehört zu den am weitesten verbreiteten Fusa-
rientoxinen und kann auf fast allen Getreidearten in Asien, Afrika, Europa und Nord-
amerika nachgewiesen werden, tritt jedoch insbesondere in gemäßigten Regionen
auf (REIß 1998; ZINEDINE et al. 2007). Seinen Namen erhielt das Toxin von der la-
teinischen Bezeichnung des Maises „Zea mays“, da es weltweit hauptsächlich auf

Literatur
11
diesem gebildet wird (GEDEK 1985). Die Fähigkeit zur Synthese von Zearalenon und
seinen Derivaten konnte nur für die Gattung Fusarium nachgewiesen werden (LE-
POM et al. 1990). Insbesondere die Arten F. culmorum und F. graminearum werden
für die Zearalenonbildung verantwortlich gemacht (RANFFT et al. 1990), aber auch
F. acuminatum, F.avenaceum, F. moniliforme, F. oxysporum und F. sambucinum
sind zur Produktion des Toxines fähig (GEDEK 1980; GEDEK 1985).
2.5.3.2.1 Physikalisch-chemische Eigenschaften von Zearalenon
Die chemische Bezeichnung lautet 6-(10-Hydroxy-6-oxo-trans-1-undeceny1)-ß-
Resorcylsäurelakton mit der Summenformel C18H22O5 (Abbildung 2) (SCHWADORF
1995). Zearalenon ist eine weiße, kristalline Substanz, erweist sich gegenüber Hitze
als sehr beständig (MÜLLER 1982; ENDERS 1984) und zeichnet sich durch eine
gute UV-Absorption mit Absorptionsmaxima bei 236, 274 und 316 nm aus, wobei der
Schmelzpunkt bei 164-165°C liegt (YOSHIZAWA 1983; S CHWADORF 1995). Die
Löslichkeit von Zearalenon in Wasser beträgt nur 20 ng/ml, d.h. das Toxin ist in
Wasser nahezu unlöslich. In Aceton, Alkohol, Chloroform, Ether und wässrigen Alka-
lien liegt dagegen eine gute Löslichkeit vor (URRY et al. 1966; MIROCHA et al.
1977).
Abb. 2: Chemische Strukturformel von Zearalenon (SCHWADORF 1995)
2.5.4 Metabolismus von DON und ZON beim Wiederkäuer
Im Pansen werden die genannten Mykotoxine vor allem durch die Infusorien zu we-
niger toxischen Stoffwechselprodukten umgebaut (HÖLTERSHINKEN et al. 1996;
BAUER 2005; DÄNICKE et al. 2005; SORENSEN u. ELBAEK 2005; FINK-
GREMMELS 2008a, 2008b). DON wird zum de-epoxy-deoxynivalenol (KING et al.
1984; COTE et al. 1986; SWANSON et al. 1987; ROTTER et al. 1996; FUCHS et al.
2002; SEELING et al. 2006b) und ZON zu α und ß Zearalenol umgebaut
(KIESSLING et al. 1984; MILES et al. 1996; VALENTA u. VEMMER 1996;

Literatur
12
KENNEDY et al. 1998; VÖLKL et al. 2004; ZINEDINE et al. 2007). Dadurch, dass
DON sehr schnell als de-epoxy-DON über den Harn ausgeschieden wird und nicht
über die Milch, ist davon auszugehen, dass DON fast vollständig durch die Pansen-
flora umgebaut wird (PING et al. 1992; DÄNICKE et al. 2005; SEELING et al. 2006b;
FINK-GREMMELS 2008b). Aufgrund des Abbaus im Pansen können die Stoffwech-
selmetaboliten von DON weder in der Muskulatur noch in der Leber nachgewiesen
werden (BÖHM 1992). In vitro wurden nach 96 Stunden Inkubation von Pansensaft
mit 1 mg DON ca. 35 % davon abgebaut (PING et al. 1992). Zudem wurde DON bei
der Inkubation mit Pansensaft teilweise zu de-epoxy-DON abgebaut (SWANSON et
al. 1987). KIESSLING et al. (1984) konnten hingegen in vitro keinen Effekt der Mik-
roorganismen des Pansensaftes auf den DON-Abbau feststellen.
ZON wird hingegen nicht vollständig im Pansen umgebaut. In Untersuchungen von
SEELING et al. (2005a) sowie DÄNICKE et al. (2005) wurden ca. 43 % im Dünndarm
nachgewiesen. ZON wurde bei der Inkubation mit Pansensaft in vitro nur teilweise
abgebaut. Nach 24 Stunden waren noch 50 % der Ausgangskonzentration nach-
weisbar (VALENTA u. VEMMER 1996). Andere Publikationen sprechen ZON, nach
Inkubation im Pansensaft in vitro, eine fast vollständige Umsetzung von Zearalenon
zu α- und ß-ZOL zu (KIESSLING et al. 1984; LERCH 1990; DUARTE 2003). Rück-
stände von ZON und seinen Derivaten konnten beim Milchrind in Gallensaftproben
nachgewiesen werden (BAUER 2002; DÄNICKE et al. 2002; KLEINOVA et al. 2002)
(vgl. Abbildung 3).

Literatur
13
ÜbergangsrateDON+ZON
(Futter)
ZON
ZON
α-ZOL ß-ZOL
α-ZAL ß-ZAL
ZAL
ZON
Absorption??
Pansen
DON DON
DOM-1DON ??Absorption
Magen- Darm- Trakt
Leber
Gallensaft
Blut
?
Absorption
Kot
MilchHarnGewebe
Fettgewebe?
Stressfaktoren
Geburt
Haltung
Wetter
?
Abb. 3: DON und ZON Metabolisierung bei Rindern (modifiziert nach KIESSLING et
al. 1984; LERCH 1990; DUARTE 2003; SEELING 2005)
2.5.5 Biologische Eigenschaften und Auswirkungen vo n DON bei Nutztieren
Durch DON-kontaminierte Futtermittel hervorgerufene akute Mykotoxikosen äußern
sich bei Tieren durch Nahrungsverweigerung und Erbrechen (D'MELLO et al. 1999).
Die Aufnahme von DON ab 0,05-2 mg/kg Futter führt bei Schweinen zu Abmage-
rung, Erbrechen und Anorexie. Akute Vergiftungen wurden auch bei Menschen be-
schrieben. Nach Aufnahme hoher Dosen kam es zu Übelkeit, Verdauungsstörungen,
Schwindelgefühl und Kopfschmerzen (JECFA 2001). Nach oraler Aufnahme und Re-
sorption erreicht Deoxynivalenol die Area postrema des Gehirns, wo es über Interak-
tion mit Dopaminrezeptoren einen Brechreiz auslöst (PESTKA et al. 1987; PRE-
LUSKY 1996).
Deoxynivalenol kann wie andere Trichothecene die Proteinsynthese im Organismus
hemmen. Zellen des Immunsystems, die sich durch eine hohe Zellteilungsrate aus-
zeichnen, können hiervon betroffen sein. Dabei hängt das Ausmaß der Beeinträchti-
gung von der chemischen Struktur der einzelnen Toxine und von der Tierart ab
(SCF/CS/CNTM/MYC 1999; SZKUDELSKA et al. 2002). Bei Milchrindern konnten
bei Dosierungen mit 66 mg DON/kg TS Futter über 5 Tage (COTE et al. 1986), 6,4
mg DON/kg TS Futter (TRENHOLM et al. 1985), 5,2 und 8,1 mg DON/100 kg Kör-
permasse (DÄNICKE et al. 2005), 12,6 mg DON/kg TS Futter (BOLAND et al. 1994),
21 mg DON/kg TS Futter (DICONSTANZO et al. 1995; WINDELS et al. 1995) bzw.
?

Literatur
14
mit 36,8 mg DON/kg TS Futter über 8 Tage während der Trächtigkeit und 12 Tage
während der Laktation (ANDERSON et al. 1995) sowie mit 1,5 bzw. 6,4 mg/kg TS
Futter (BAUER 2005) keine negativen Einflüsse auf den Gesundheitszustand der
Tiere festgestellt werden. Auch bei Mastbullen konnten nach Aufnahme von fusa-
rienkontaminiertem Futter (2,2 mg DON und 0,1 mg ZON/kg TS Futter) keine negati-
ven Auswirkungen nachgewiesen werden (DÄNICKE et al. 2002). Bei der Untersu-
chung der Effekte von DON auf die Leistung von Bullen und Färsen bei definierter
Futterration (1mg DON/kg TS Futter/d) während einer Periode von 142 Tagen, konn-
ten bezüglich der täglichen Körpermassezunahmen, der Energieaufnahme und der
Leistungsfähigkeit keine negativen Wirkungen nachgewiesen werden (DEHANN et
al. 1984; DICOSTANZO et al. 1996). Auf der anderen Seite wurden bei höheren My-
kotoxinkonzentrationen von mehr als 5 mg DON/kg TS Futter eine verminderte Fut-
teraufnahme, langes, struppiges Haarkleid und schlechtere Fleischqualität von Mast-
bullen beobachtet (SCHUH 1996). Bei einer Dosis von 0,4 bis 0,6 mg DON/kg Kör-
permasse wurden bei Rindern ein Rückgang der Milchleistung, Aborte und blutige
Diarrhöe beschrieben (GEDEK 1980). Die Fütterung von gemischten Rationen mit 50
% Kraftfutteranteil und einer Belastung von 5,3 mg DON/kg TS über einen Zeitraum
von 11 Wochen bei Milchkühen führte zu Veränderungen des ruminalen Gärmusters
sowie zu einem erniedrigten pH-Wert des Pansensaftes in der Woche 4 und 8. Ähnli-
ches wurde bei einer Erhöhung des Kraftfutteranteils auf 60 % und einer DON-
Konzentration von 4,6 mg/kg TS sowie bei 30 % Kraftfutteranteil in der Ration mit
einer Konzentration von 4,4 mg/kg TS festgestellt (KEESE et al. 2008b). In Fütte-
rungversuchen wurde festgestellt, dass keine negativen Effekte auf Tiergesundheit
und Leistung gefunden wurden, wenn Rationen mit DON-Gehalten zwischen 4.4 und
5.3 mg/kg TS und Konzentratanteilen zwischen 30 und 60 % über einen Gesamtzeit-
raum von 29 Wochen eingesetzt wurden. In Serum, Galle und Milch wurde haupt-
sächlich DOM-1 nachgewiesen. Nicht-metabolisiertes DON trat im Serum nur in sehr
niedrigen Konzentrationen auf (KEESE 2008c). In beiden Perioden zeigten die Tiere
in den Gruppen mit Fusarium-Toxin Kontamination eine höhere TS-Aufnahme (in
Gruppe Myko-30), die vermutlich verursacht wurde durch die stimulierenden Effekte
des Fusarium-Toxins kontaminierten Triticale auf die Ingesta-Passagerate. Auch ein
erhöhter Konzentratanteil von 60 % stimulierte die TS-Aufnahme.
In Bezug auf die Milchleistung konnten KEESE et al. (2008a) bei Fütterung verschie-
dener Rationen mit unterschiedlichen Kraftfutteranteilen und DON-Konzentrationen

Literatur
15
beobachten, dass die Milchleistung unter stärkeren Mykotoxinbelastungen höher war
als unter Fütterung schwach kontaminierter Rationen. Klinische Daten von 300
Milchvieh-Herden zeigen, dass DON mit einem Rückgang in der Milchleistung in
Verbindung zu bringen ist, können aber keinen direkten Zusammenhang zwischen
Ursache und Wirkung herstellen (WHITLOW et al. 1994). DÄNICKE et al. (2005)
konnte bei einer Dosierung von 3,1 und 3,5 mg DON/kg tägliche Ration (88 % TS)
keine reduzierte Futteraufnahme oder verringerte Milchleistung beobachten. In einem
Versuch mit natürlichem kontaminiertem Futter mit einer DON-Konzentration von ca.
3,6 µg/g TS über 56 Tage wurde festgestellt, dass das Gesamt-Serum-Protein und
die Globulin-Konzentration nach 42 d angestiegen waren. Es gab jedoch keine Aus-
wirkungen auf Leistungsfähigkeit, hämatologische Blutparameter, Körpergewicht,
Milchproduktion und Milchzusammensetzung (KOROSTELEVA et al. 2007). Auch die
in einem weiterem Versuch beschriebene Aufnahme von DON-Gehalt bis zu 190,8
mg/d über 135 Tage (DICOSTANZO et al. 1995) zeigte keinen Einfluss auf die tägli-
chen Zunahme, die Leistungsfähigkeit, klinisch-chemische und hämatologische Pa-
rameter von Ochsen. In einer anderen Arbeit konnte nach Fütterung von 50 % Kraft-
futter mit einer Belastung von 5,3 mg DON/kg TS über einen Zeitraum von 11 Wo-
chen nur de-epoxy DON (≤ 3,2 µg/kg) in der Milch nachgewiesen werden. Ebenso
war dies bei einer Fütterung von 30 % Kraftfutteranteil mit 4,4 mg DON/kg TS und
bei einer Ration mit 60 % Kraftfutteranteil und 4,6 mg DON/kg TS der Fall. Der Über-
gang von DON als DOM-1 in die Milch war unbedeutend (zwischen 0,0001 und
0,0011). DOM-1 wurde in Blut, Zwölffingerdarm, Urin und Kot in höheren Konzentra-
tionen als DON festgestellt (KEESE et al. 2008).
Bei einer einmaligen Verabreichung von 920 mg DON pro Tier (PRELUSKY et al.
1984) an Milchkühe wurde die freie und konjugierte DON-Konzentration in der Milch
mit unter 4 ng/ml im Nachweis angegeben. In einem weiteren Versuch mit einer Do-
sierung von 66 mg/kg TS Futter DON/Tag über 5 Tage (COTE et al. 1986) blieb die
Milchleistung unbeeinflusst, allerdings wurden 26 ng/ml DOM-1 in der Milch nachge-
wiesen. In einer anderen Studie wurde bei 27 Kühen (drei Gruppen T, K, M) der Ef-
fekt von natürlich mit DON und ZON kontaminiertem Futtermittel auf labormedizini-
sche und produktionsbezogene Parameter untersucht. Die Konzentration von DON
lag bei 12,4 mg (Gruppe T), 14,1 mg/kg TS (Gruppe K), 14,3 mg/kg TS (Gruppe M)
und die von ZON bei 0,94 mg/kg TS (T), 0,67 mg/kg TS (K) und 0,68 mg/kg TS (M).
Lediglich die Mittelwerte der Aspartat-Aminotransferase (AST)- und Glutamat-

Literatur
16
Dehydrogenase (GLDH)-Aktivitäten aller Gruppen waren geringgradig erhöht, alle
weiteren Parameter lagen im physiologischen Bereich. Bezüglich der Milchleistung
und Milchbestandteile konnte keine nennenswerter Unterschied zwischen den ver-
schiedenen Versuchsgruppen beobachtet werden (HOCHSTEINER et al. 2000). In
zwei Fütterungsversuchen mit folgenden Dosierungen: 8,21 mg DON/kg und 0,08 mg
ZON/kg TS Futter (MATTHÄUSE et al. 2004) sowie 5 mg DON/kg Futter (88 % TS)
(SEELING et al. 2005a u. 2005b) blieben die Aktivitäten von AST, GLDH, GGT und
die Konzentration von Kreatinin bei den untersuchten Milchkühen unbeeinflusst. In
einer Untersuchung an Kühen nach Gabe von 2,3 und 10 mg DON/kg TS Futter wur-
den keine klinische Veränderungen des histologisch untersuchte Gewebes, der Leu-
kozytenzahl und -differenzierung oder der Konzentrationen von Glucose, Harnstoff,
Kreatinin, Calcium, Phosphat, Natrium und Kalium im Blut sowie keine reduzierte
Trockensubstanzaufnahme festgestellt (NELSON 1984; WHITLOW et al. 1986). In
einem weiteren Fütterungsversuch mit 10 mg DON/kg TS Futter und 0,76 mg
ZON/kg TS Futter an Jungbullen waren die Serumaktivitäten der AST und GLDH
auch unbeeinflusst (DÄNICKE et al. 2002). Dies stimmt mit der Beschreibung von
SEELING et al. (2006 a) überein.
In einem Versuch an Ferkeln, die mit DON-kontaminierten Triticale gefütterte wur-
den, konnten nur geringe Auswirkungen auf die Leberfunktion, klinisch-chemischen
Parameter und die Organ-Histopathologie festgestellt werden (DÄNICKE et al. 2008).
Wiederkäuer gelten als relativ unempfindlich gegenüber DON durch die Fähigkeit
ihrer Mikroorganismen im Pansen, DON zu dem fast ungiftigen de-epoxy-DON
(DOM-1) zu metabolisieren. Es gibt Untersuchungen, die bestätigen, dass keine ne-
gativen Einflüsse von DON sowie bei Nachweisen von de-epoxy-DON auf die Ge-
sundheit und Leistung bestehen (HOCHSTEINER et al. 2000; BAUER 2005; DÄNI-
CKE et al. 2005; KEESE 2008c). Allerdings gibt es bislang keine Studien über Lang-
zeiteffekte moderater DON-Konzentrationen bei Milchkühen. In einem weiteren Füt-
terungsversuch mit 8,21 mg DON/kg TS Futter und 0,08 mg ZON/kg TS Futter waren
die Serumaktivitäten der AST, GLDH und GGT unverändert trotz Nachweis von de-
epoxy-DON im Zwölffingerdarm von 94 % bis 99 % des ursprünglichen DON (SEE-
LING et al. 2005). Dies sind Hinweise auf eine Detoxifizierung von DON im Pansen.
Ähnlich ist das Ergebnis von DÄNICKE et al. (2004) mit 89 % de-epoxy-DON vom
ursprünglichen DON im Zwölffingerdarm.

Literatur
17
2.5.6 Biologische Eigenschaften und Auswirkungen vo n ZON bei Nutztieren
Die Intoxikation mit Zearalenon, das Zearalenon-Syndrom, wird in der Literatur teil-
weise unter anderen Bezeichnungen geführt, so z.B. Hyperöstrogenismus, Östroge-
nismus, Vulvovaginitis, Vulva-Ödem, Östrogen-Syndrom, Pseudobrunst oder F-2 To-
xikose (PALYUSIK 1977; GEDEK 1980). Am empfindlichsten gegenüber Zearalenon
zeigt sich das Schwein, aber auch beim Rind, Geflügel, Pferd und bei Labortieren
konnte durch Zearalenon ein Hyperöstrogenismus ausgelöst werden (DROCHNER
1990; DÄNICKE et al. 2000; ZINEDINE et al. 2007).
2.5.6.1 Wirkungsmechanismus von ZON
Zearalenon und seine Derivate besitzen keine besonders hohe akute Toxizität. Es
handelt sich vielmehr um nichtsteroidale Makrolide, die aufgrund ihrer dem Östrogen
ähnlichen räumlichen Struktur eine kompetitive Bindung mit Östrogenrezeptoren ein-
gehen können (KIANG et al. 1978; KATZENELLENBOGEN et al. 1979), wodurch
das sexualzyklische Geschehen der Tiere beeinflusst wird (KARLSON 1979; KAWA-
BATA et al. 1982; KOLB 1989; COULOMBE 1993; ENGELHARDT 2000; BAUER
2005). Es gelang der Nachweis, dass Zearalenon, α-Zearalenol und ß-Zearalenol auf
diesem Weg spezifisch zur Synthese entsprechender Proteine, führen können (Ab-
bildung 4).
Abb. 4: Schematische Darstellung des Wirkungsmechanismus von Zearalenon (mo-
difiziert nach OLDENBURG et al. 2000).

Literatur
18
Eine unterschiedliche Östrogenrezeptoraffinität wurde für Zearalenon und seine De-
rivate festgestellt. Die relative Bindungsaffinität von α-Zearalenol war 10-20mal stär-
ker als die von Zearalenon, und 100mal stärker als die von ß-Zearalenol. Zearalenon
wiederum hat eine 2-3mal höhere Rezeptoraffinität am Uterus als 17ß-Estradiol
(BLANKENSHIP et al. 1982). Die Effekte auf den Uterus, wie z.B. eine Gewichtszu-
nahme, sind beim Zearalenon etwa 2000fach geringer als bei einer entsprechenden
Estradiolmenge (KIANG et al. 1978).
2.5.6.2 Klinische Auswirkungen von Zearalenon
Die Symptome treten in der Regel fünf bis sieben Tage nach Aufnahme des toxinhal-
tigen Futters auf (BAUER u. GEDEK 1978; BAUER 1982). Beim Rind wurden neben
Futterverweigerung und erheblichem Rückgang der Milchleistung vermehrt Ödeme
des Euters und der Vulva, Vaginitis und zunächst klarer, später trüber und eitriger
Scheidenausfluss, Ovarialzysten, verlängerte Brunst und ein schlechter Kontrakti-
onszustand des Uterus als Symptome beschrieben (VANYI et al. 1974; PALYUSIK
1977; MÜLLER 1978; SCHUH 1981; SCHUH u. BAUMGARTNER 1988; MANSFELD
et al. 1989; KAMPHUES 1991; SCHUH 1996). Bereits eine Zearalenonkonzentration
von 17,3 mg/kg TS Futter konnte bei Rindern verschlechterte Konzeptionsraten, Un-
fruchtbarkeit, ödematisierte Vulven, zystöse Entartungen der Ovarien bzw. Dys-
zyklien und schlaffe Uteri bei Kalbinnen hervorrufen (SCHUH 1981). Fruchtbarkeits-
störungen beim Milchrind und ein Abfall der Milchleistung konnten zudem bei folgen-
den Dosierungen beschrieben werden: Dosis von 1,6 mg ZON/kg TS Futter/d (BAU-
ER et al. 1989), 1 mg ZON/kg TS Futter (SCHWEIGHARDT 1980), 5 und 75 mg/kg
TS Futter (VANYI u. SZAILER 1974) sowie 14 mg ZON/kg TS Futter (KURTZ et al.
1969). Auch eine Gabe von 1,5 mg ZON/kg TS Futter führte zu vorzeitiger Entwick-
lung und Vergrößerung des Euters bei Kalbinnen, Sterilitäten, Vaginitiden, Brunster-
scheinungen, welche nicht mit dem ovariellen Zyklus korrelierten, zu brünstigem
Verhalten im 3. Trächtigkeitsdrittel und zu verlängerten Brunstzeiten (2-5 Tage) bei
einer Kuh- bzw. Kalbinnenherde (COPPOCK et al. 1990). DROCHNER (1990) konn-
te bei Milchkühen, die mit mehr als 100 µg ZON/kg TS Futter gefüttert wurden, eine
Vulva- und Euterhypertrophie und vermehrten Abgang von Brunstschleim feststellen.
Geringe Fusarientoxinkonzentrationen (DON und ZON) im Futter zeigten beim Milch-
rind keine Wirkung auf Leistungsparameter (HOCHSTEINER et al. 2000). Bei höhe-
ren Konzentrationen wurden jedoch negative Auswirkungen auf den Organismus
vermutet und weiterführende Untersuchungen konnten ZON (10 mg/kg TS Heu) für

Literatur
19
Frühaborte bzw. Fruchtresorption verantwortlich machen (KALLELA u. VASENIUS
1982). Die Aufnahme von kontaminiertem Weizen (10 mg DON und 0,76 mg ZON/kg
TS Futter) beeinflusste die Leistung von wachsenden Stieren nicht nachteilig (DÄNI-
CKE et al. 2002). In einem anderen Versuch wurde eine Tiergruppe mit zearalenon-
kontaminiertem Haferschrot (1,25 mg ZON/kg Futter) in einer Menge von 2 kg Fut-
ter/Tag/8 Wochen gefüttert. Die 2.Tiergruppe wurden mit fast ZON-freiem (0,079 mg
ZON/kg Futter) in derselben Menge gefüttert. Tiere der Gruppe 3 bekamen zusätzlich
ein östrogenwirksames Präparat in Form von zwei Applikationen zu je 25 mg Zeranol
im Abstand von 6 Wochen. Die Tiere zeigten sich klinisch unauffällig und wiesen kei-
ne Abweichungen im Brunstzyklus auf. Das Blutbild, die Enzyme und Stoffwechsel-
produkte zeigten in allen Gruppen keine Abweichungen, zudem bestanden keine pa-
thologisch-histologischen Veränderungen am Genitaltrakt (MOESER 2001).
In einer anderen Studie wurden mit 8,21 mg DON/kg TS und 0,09 mg ZON/kg TS
sowie als Kontrolle Weizen mit 0,25 mg DON/kg TS und 51 µg ZON/kg TS keine be-
deutenden Veränderungen festgestellt werden (SEELING et al. 2006a). Die relative
Bindungsaffinität von α-Zearalenol ist 10-20mal stärker als die von Zearalenon und
100mal stärker als die von ß-Zearalenol und hat eine große Bedeutung beim
Schwein (BLANKENSHIP et al. 1982). Bei Färsen, die mit 2 kg Zeralenon-
kontaminiertem Hafer (1370 µg/kg) über einen Gesamtzeitraum von 84 Tagen gefüt-
tert wurden als auch bei Färsen, die mit 25 mg Zeranol-Pellets (α-Zearalanol, ein in
der EU streng verbotener hormoneller Leistungsförderer) gefüttert wurde, konnten
keine klinischen, pathologischen und histologischen Veränderungen am Genitaltrakt
und bei der Milchleistung festgestellt werden. Es zeigte sich, dass etwa 80 % von
ZON im Urin, Muskel- und Lebergewebe als ß-ZOL und der restliche Anteil als α-ZOL
oder andere Metaboliten nachweisbar waren. Im Gegensatz dazu steht das Schwein,
bei dem mehr α-ZOL als ß-ZOL gebildet wird. Diese Ergebnisse ergeben keinen
Hinweis auf einen negativen Einfluss von Zearalenon, α-ZOL, ß-ZOL oder Zeranol
auf die Reproduktion oder die Milchproduktion (KLEINOVA et al. 2002). Der Umbau
zu α-ZOL steigert die Toxizität, da α-ZOL eine höhere Affinität zum Östrogenrezeptor
aufweist als ZON. Demgegenüber scheint ß-ZOL eine geringere oder vergleichbare
Toxizität wie ZON zu besitzen. Ähnlich wird von DÄNICKE et al. (2005) und MEYER
et al. (2002) berichtet, dass ZON mehr zu ß-ZOL als zu α-ZOL bei Wiederkäuer me-
tabolisiert wird, ohne das auffällige Auswirkungen durch ZON, β-ZOL oder α-ZOL
auftreten. In einer Studie von BAUER (2002) wurde festegestellt, dass ZON und sei-

Literatur
20
ne Derivate (α- und ß-ZOL) keine Ursache für auftretende Probleme bei Milchkühen
darstellen. Auch 10 mg DON und 0,76 mg ZON/kg TS Futter beeinflussten die Leis-
tung von wachsenden Mastbullen nicht nachteilig (DÄNICKE et al. 2002), trotz
Nachweis von 24 % ZON, 8 % α-ZOL und 68 % ß-ZOL im Gallensaft. In einem weite-
ren Fütterungsversuch mit 8,21 mg DON/kg TS Futter und 0,08 mg ZON/kg TS Futter
an Kühe waren die Serumaktivitäten der AST, GLDH und GGT unbeeinflusst, trotz
des Nachweises von ZON, α-ZOL und ß-ZOL im Zwölffingerdarm. Zusammenfas-
send lässt sich sagen, dass eine schnellere ruminale Passage von Futter und/oder
ZON als Folge einer erhöhten Futteraufnahme die Metabolisierung von ZON zu ß-
ZOL beeinträchtigen kann. Infolgedessen gelangt ein erhöhter Anteil an ZON und ein
niedriger Anteil der Metaboliten ß-ZOL und α-ZOL in den Zwölffingerdarm (SEELING
et al. 2005), die einen geringen Umfang des Abbaus im Pansen bzw. eine ZON-
Metabolit aus dem entero-hepatischen-Kreislauf vermuten lassen (DÄNICKE et al.
2005). Allerdings gibt es bislang keine Studien über Langzeiteffekte moderater ZON-
Konzentrationen bei Milchkühen.
2.6 Labmagenverlagerung beim Rind
Die Labmagenverlagerung kommt weltweit vor. Vor allem in Regionen mit intensiver
Rinderhaltung tritt diese Erkrankung gehäuft auf. Bei Kühen treten 85-96 % der Fälle
von Labmagenverlagerung linksseitig und 4-15 % rechtsseitig auf (CONSTABLE et
al. 1992; WOLF et al. 2001). Die Labmagenverlagerung ist als multikausale Fakto-
renkrankheit aufzufassen (STAUFENBIEL 1998).
Als mögliche Ursachen gelten die Fütterung, Stress, Stoffwechselstörungen und
Krankheiten anderer Organe. Die Labmagenverlagerung kommt vorwiegend bei Kü-
hen mit hoher individueller Milchleistung im Alter von 4 bis 7 Jahren in den ersten
vier Wochen nach der Abkalbung vor (CONSTABLE et al. 1992; FÜRLL et al. 1996;
WOLF et al. 2001; WITTEK et al. 2005). Stoffwechselstörungen wie die Ketose,
Puerperalstörungen, Metritis, Mastitis und Klauenleiden wurden als Primär- oder Be-
gleiterkrankungen der Labmagenverlagerung festgestellt. Diese können vor der
Labmagenverlagerung, zur gleichen Zeit oder im Anschluss an diese auftreten
(CONSTABLE et al. 1992; FÜRLL u. KRÜGER 1999a; ROHRBACH et al. 1999). Die
Labmagenverlagerung wurde auch im Zusammenhang mit dem Fettmobilisations-
syndrom beobachtet (FÜRLL et al. 2000). Eine besondere Rolle spielt der Verlauf der
Kalbung. Zwillingsträchtigkeiten sowie Schwer- und Totgeburten werden vorbericht-

Literatur
21
lich oft erwähnt (FÜRLL u. KRÜGER 1999a; WOLF et al. 2001). Zudem wird vermu-
tet, dass Endotoxine das Auftreten von Labmagenverlagerungen fördern können, da
Endotoxine die Entleerung des Labmagens beeinträchtigen (VLAMINCK et al. 1985;
FÜRLL u. KRÜGER 1999b; FÜRLL et al. 2000).
Wichtigster Einflußfaktor für die Entstehung einer Labmagenverlagerung ist die Füt-
terung. Besonders gefährdet sind Kühe, die eine Verfettung ante partum aufweisen
und im peripartalen Zeitraum in ein Energiedefizit geraten. Hinzu kommen starker
Geburtsstress und eine gesteigerte Lipolyse, die zur Leberverfettung führt. Dies wie-
derum bewirkt eine reduzierte Neutralisation und Ausscheidung der Endotoxine. In
diesem Zusammenhang läuft weiterhin eine massive Akute-Phase-Reaktion ab.
Letztendlich kommt es zur Entspannung der glatten Muskulatur der Hohlorgane, was
auch am Labmagen zu Motilitätsstörungen führt. Der Labmagen füllt sich mit Gas,
und es kommt zur Labmagenverlagerung. Die Prädispositionsfaktoren bei Kühen
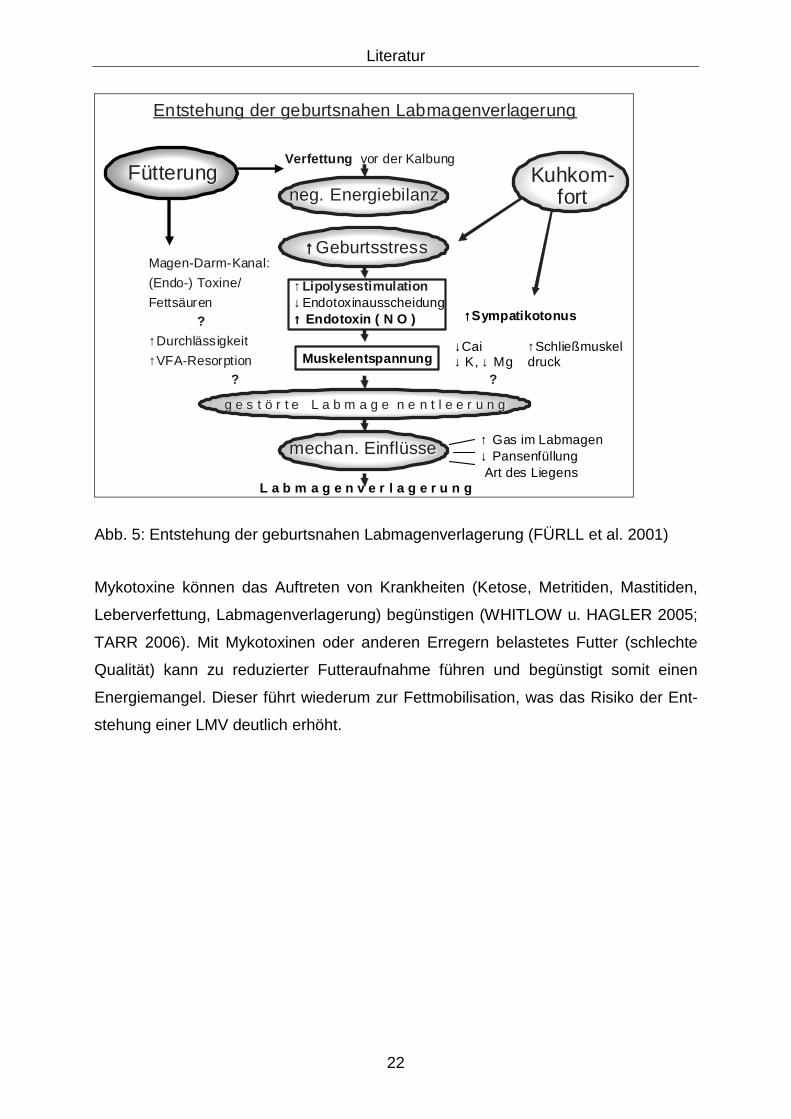
vom Trockenstellen bis zur Erkrankung an LMV sind in der Tabelle 3 und Entstehung
der geburtsnahen LMV in der Abbildung 5 dargestellt (FÜRLL et al. 2001).
Tab. 3: Prädispositionsfaktoren bei Kühen vom Trockenstellen bis zur Erkrankung
an LMV (FÜRLL et al. 2001)
Trockenstehperiode um die Geburt Frühlaktation
• häufig Überkonditio- nierung
bzw.Verfettung • Energieunterversor- gung am Ende der Hochträchtigkeit (↑Ketonkörper, ↑Fettsäuren u.a.) • häufig Zwillings- trächtigkeiten mit höherem Energiebe- darf und subklinischer Ketose
• häufig Wehenschwä- che und Schwerge- burten
• häufig schwere,
männliche Kälber so-wie Zwillinge
• → starker physischer und Stoffwechselstress während der Geburt (starke partusinduzier- te Akute-Phase- Reaktion)
• stärkere postpartale Stressreaktion mit gesteigerter Lipolyse (↑Ketonkörper, ↑Fettsäuren, ↑Carni-
tin, ↓Lipoproteine, ↓Cholesterol, ↓Albumin,
↓2+Kationen) • Leberverfettung und Fettstoffwechselstö- rungen = Fettmobilisations- syndrom
Literatur
22
FFüütterungtterungFFüütterungtterung KuhkomKuhkom--fortfort
↑↑↑↑↑↑↑↑GeburtsstressGeburtsstress
mechanmechan. . EinflEinflüüssesse ↑↑ Gas Gas imim LabmagenLabmagen↓↓ PansenfPansenfüüllungllungArt des Art des LiegensLiegens
L a b m a g e n v e r l a g e r u n gL a b m a g e n v e r l a g e r u n g
g e s t g e s t öö r t e L a b m a g e n e n t l e e r u n gr t e L a b m a g e n e n t l e e r u n g
VerfettungVerfettung vorvor derder KalbungKalbung
MagenMagen--DarmDarm--Kanal:Kanal:
((EndoEndo--) Toxine/) Toxine/
FettsFettsääuren uren ??
↑↑DurchlDurchläässigkeitssigkeit
↑↑VFAVFA--ResorptionResorption
??
EntstehungEntstehung derder geburtsnahengeburtsnahen LabmagenverlagerungLabmagenverlagerung
↑↑LipolysestimulationLipolysestimulation↓Endotoxinausscheidung↑↑↑↑↑↑↑↑ Endotoxin ( N O )Endotoxin ( N O )
MuskelentspannungMuskelentspannung
neg. neg. EnergiebilanzEnergiebilanz
↑↑↑↑↑↑↑↑SympatikotonusSympatikotonus
↓Cai ↑Schließmuskel↓ K, ↓ Mg druck
??
Abb. 5: Entstehung der geburtsnahen Labmagenverlagerung (FÜRLL et al. 2001)
Mykotoxine können das Auftreten von Krankheiten (Ketose, Metritiden, Mastitiden,
Leberverfettung, Labmagenverlagerung) begünstigen (WHITLOW u. HAGLER 2005;
TARR 2006). Mit Mykotoxinen oder anderen Erregern belastetes Futter (schlechte
Qualität) kann zu reduzierter Futteraufnahme führen und begünstigt somit einen
Energiemangel. Dieser führt wiederum zur Fettmobilisation, was das Risiko der Ent-
stehung einer LMV deutlich erhöht.
Material und Methoden
23
3 Tiere, Material und Methoden
3.1 Tiere und Versuchsanordnung
Für die Untersuchungen wurden 61 Milchkühe (1 bis 6 Wochen p.p.) aus dem Pati-
entengut der Medizinischen Tierklinik der Universität Leipzig genutzt. Die Tiere ge-
hörten der Rasse Schwarzbunte (SB) an. Das Alter der Tiere lag zwischen zwei und
acht Jahren. Die Patientenkühe stammten aus 39 Milchviehbetrieben aus dem Ein-
zugsgebiet der Klinik und wurden ausschließlich wegen einer Labmagenverlagerung
(Dislocatio abomasi) mit unterschiedlichen Begleiterkrankungen (Endometritis, Masti-
tis, Enteritis, Peritonitis, Laminitis u.a.) eingewiesen.
Die gesunden Kontrollkühe (n = 13, 2 bis 6 Wochen p.p.) stammten aus zwei Betrie-
ben im Leipziger Umland. Die Kontrollkühe wiesen innerhalb der letzten drei Monate
keine Erkrankungen auf. Die zurückliegende Kalbung verlief komplikationslos und
ohne Nachgeburtsverhaltung. Die Tiere hatten einen Ernährungszustand mit einem
BCS von 2,0 bis 4,0 (EDMONSON et al. 1989). Allen Kühe wurden nach Einlieferung
in die Klinik ein zentraler Venenkatheter (Cavafix Certo Splittocan 338, B. Braun Mel-
sungen AG, Melsungen) in die V. jugularis externa eingesetzt. Für die Mykotoxin-
bestimmungen wurden von 20 Patienten- bzw. 13 Kontrollkühen Blut, Milch, Galle
und Futter sowie von 41 Patientenkühen Blut und Galle entnommen (Tabelle 4).
Tab.4: Verteilung der Patienten- und Kontrollkühe auf die einzelnen untersuchten
Substrate
Tiere Serum Galle Milch Futter
n Patienten 61(41+20) 61(41+20) 20 20
Kontrolle 13 13 13 2
3.2 Methoden
3.2.1 Klinische Untersuchung und Diagnosestellung
Die klinische Untersuchung der Kühe erfolgte nach dem in der Medizinischen Tierkli-
nik der Universität Leipzig etablierten allgemeinen Untersuchungsgang. Die Labma-

Material und Methoden
24
genverlagerung wurde mittels Perkussions- und Schwingauskultation diagnostiziert
(BAUMGARTNER 2002). Die Bestimmung des Säure-Basen-Status wurde ebenfalls
zur Diagnosesicherung routinemäßig herangezogen. Der Zugang zur Bauchhöhle
erfolgte bei allen an Labmagenverlagerung erkrankten Kühen zur Reposition im Be-
reich der rechten Hungergrube, eine handbreit kaudal der letzten Rippe und eine
handbreit ventral der Processus transversi der Lendenwirbel. Das weitere operative
Vorgehen erfolgte entsprechend der von GABEL u. HEATH (1969) beschriebenen
Methode. Dabei wurden die Organe der Bauch- und Beckenhöhle explorativ unter-
sucht (insbesondere Pansen, Blättermagen, Labmagen, Leber, Gallenblase, Niere,
Uterus), wobei pathologisch veränderte Organbefunde dokumentiert wurden. Die Di-
agnosestellung der Endometritis erfolgte durch Palpation des Uterus und der Ovarien
während der Laparatomie. Dabei wurde auf die Größe, den Inhalt, den Tonus und die
Kontraktilität des Uterus geachtet. Weiterhin wurde vorhandener Vaginalausfluss
nach Menge, Farbe und Geruch beurteilt. Die Untersuchung des Euters bestand in
der Adspektion und Palpation des Euters sowie der grobsinnlichen Untersuchung
des Milchsekrets. Bei jedem erkrankten Tier wurde zudem ein Schalmtest (SCHALM
1960) durchgeführt.
3.2.2 Entnahme von Futter-, Blut-, Milch- und Galle nsaft- Proben
Das Futter aus den Herkunftsbetrieben (totale Mischration je 2 kg) wurde mit den
Patienten geliefert und bei -70°C tiefgefroren. Die Entnahme von Blut erfolgte unter
sterilen Kautelen mit einer Kanüle der Stärke 18 G (Neolus 1½”, 1,2 x 40 mm, TE-
RUMO®, Leuven, Belgien). Zur Blutentnahme wurde die Vena jugularis externa ge-
nutzt. Die Blutzentrifugation erfolgte bei 3800 g über 10 min. Das Serum wurde bei -
20°C tiefgefroren. Für die Milchproben wurden die K ühe ausgemolken und 0,5 l Milch
je Kuh bei -20°C eingefroren. Die Entnahme von Gall ensaft erfolgte bei den Patien-
ten durch Punktion der Gallenblase während der operativen Labmagenreposition.
Hierzu wurde mit einer an einem Schlauch befestigten Kanüle von dorsal in die Gal-
lenblase eingestochen und Gallensaft (5 ml) abgesaugt. Bei den gesunden Kühen
erfolgte die Entnahme durch transkutane Punktion der Gallenblasen unter so-
nographischer Kontrolle 5cm dorsal bis 5cm ventral der Buggelenkslinie im 9. bis 11.
Interkostalraum an der Stelle, wo die Gallenblase sonographisch am besten darge-
stellt werden konnte. Die Kanüle wurde in einem Winkel von 45° in den oben be-
schriebenen Bereich eingeführt.

Material und Methoden
25
3.2.3 Klinisch-chemische Analysen
Die Bestimmung der klinisch-chemischen Parameter erfolgte im Labor der Medizini-
schen Tierklinik der Universität Leipzig. Für die anschließend dargestellten klinisch-
chemischen Analysen wurden folgende Analysengeräte, Testkits und Material ver-
wendet:
Analysengeräte
-1- = Hitachi 912 Automatic Analyzer, Fa. Roche Diagnostics GmbH, Mannheim
-2- = UNICAM 929 Atomabsorptionsspektrometer (AAS), Fa. ATI UNICAM,
Cambridge, Großbritannien
-3- = Hämatologieautomat Technicon H1, Fa. Bayer Vital GmbH, Fernwald
bzw.Cellcounter Typ Alvet, Fa. möLAB GmbH, Langenfeld
-4- = Analysenautomat ABL 555 Radiometer Copenhagen, Fa. Radiometer GmbH,
Willich
Testkits
(A) = Fa. Roche Diagnostics GmbH, Mannheim
(B) = Fa. Randox Laboratories GmbH, Krefeld
Material
S = Serum V = Vollblut
VK S % = Präzisionskontrollen in der Serie in % (n = 10)
VK T % = Präzisionskontrollen von Tag zu Tag in % (n > 30)
DGKC = Deutsche Gesellschaft für Klinische Chemie
3.2.4 Hämatologische Untersuchung
Die Analyse des Blutbildes erfolgte am Technicon H1 im Labor der Medizinischen
Tierklinik der Universität Leipzig mittels Vollblut.
In Tabelle 5 sind die erfassten Parameter, angewandten Methoden und Variations-
koeffizienten in der Serie von Tag zu Tag zusammengestellt:
Material und Methoden
26
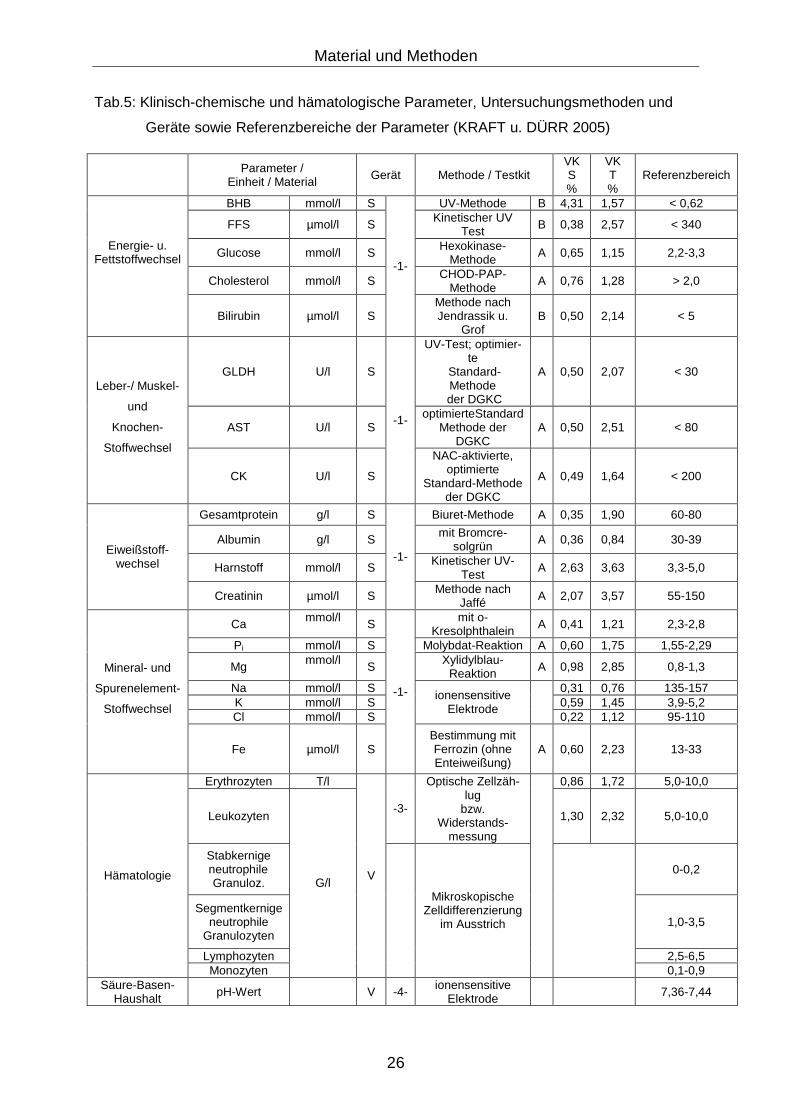
Tab.5: Klinisch-chemische und hämatologische Parameter, Untersuchungsmethoden und
Geräte sowie Referenzbereiche der Parameter (KRAFT u. DÜRR 2005)
Parameter / Einheit / Material Gerät Methode / Testkit
VK S %
VK T %
Referenzbereich
BHB mmol/l S UV-Methode B 4,31 1,57 < 0,62
FFS µmol/l S Kinetischer UV Test B 0,38 2,57 < 340
Glucose mmol/l S Hexokinase- Methode A 0,65 1,15 2,2-3,3
Cholesterol mmol/l S CHOD-PAP- Methode
A 0,76 1,28 > 2,0
Energie- u. Fettstoffwechsel
Bilirubin µmol/l S
-1-
Methode nach Jendrassik u.
Grof B 0,50 2,14 < 5
GLDH U/l S
UV-Test; optimier-te
Standard-Methode
der DGKC
A 0,50 2,07 < 30
AST U/l S optimierteStandard
Methode der DGKC
A 0,50 2,51 < 80
Leber-/ Muskel-
und
Knochen-
Stoffwechsel
CK U/l S
-1-
NAC-aktivierte, optimierte
Standard-Methode der DGKC
A 0,49 1,64 < 200
Gesamtprotein g/l S Biuret-Methode A 0,35 1,90 60-80
Albumin g/l S mit Bromcre-solgrün A 0,36 0,84 30-39
Harnstoff mmol/l S Kinetischer UV- Test
A 2,63 3,63 3,3-5,0
Eiweißstoff- wechsel
Creatinin µmol/l S
-1-
Methode nach Jaffé A 2,07 3,57 55-150
Ca mmol/l S mit o-Kresolphthalein A 0,41 1,21 2,3-2,8
Pi mmol/l S Molybdat-Reaktion A 0,60 1,75 1,55-2,29
Mg mmol/l S Xylidylblau- Reaktion A 0,98 2,85 0,8-1,3
Na mmol/l S 0,31 0,76 135-157 K mmol/l S 0,59 1,45 3,9-5,2 Cl mmol/l S
ionensensitive Elektrode
0,22 1,12 95-110
Mineral- und
Spurenelement-
Stoffwechsel
Fe µmol/l S
-1-
Bestimmung mit Ferrozin (ohne Enteiweißung)
A 0,60 2,23 13-33
Erythrozyten T/l 0,86 1,72 5,0-10,0
Leukozyten -3-
Optische Zellzäh-lug
bzw. Widerstands-
messung
1,30 2,32 5,0-10,0
Stabkernige neutrophile
Granuloz. 0-0,2
Segmentkernige neutrophile
Granulozyten 1,0-3,5
Lymphozyten 2,5-6,5
Hämatologie
Monozyten
G/l V
Mikroskopische
Zelldifferenzierung im Ausstrich
0,1-0,9 Säure-Basen-
Haushalt pH-Wert V -4- ionensensitive Elektrode 7,36-7,44

Material und Methoden
27
3.2.5 Trolox Equivalent Antioxidative Capacity (TEA C)
Die Herstellung und Verwendung der Reagenzien sowie die Durchführung der Ana-
lysen erfolgte nach der Methode von MILLER et al. (1996). Die TEAC wurde durch
Messung bei 734 nm am UV/VIS Spektralphotometer DU 640B der Fa. Beckman
Coulter GmbH, Krefeld, ermittelt. Die Methode der Bestimmung der TEAC beruht auf
der Reaktion von Antioxidantien mit dem stabilen grüngefärbten 2,2´-Azino-bis(3-
ethylbenzothiazolin-6-sulfonsäure)-Radikalkation (ABTS�+). Die Antioxidantien bewir-
ken dabei eine Entfärbung der Lösung, die durch photometrische Messung verfolgt
werden kann. In Anwesenheit von Antioxidantien resultiert folglich eine Extinktion-
serniedrigung. Als Standardsubstanz wurde Trolox (6-Hydroxy-2, 5, 7, 8-
tetramethylchroman-2-carbon-säure), ein hydrophiles Vitamin E-Analogon genutzt,
das ebenfalls eine Entfärbung der ABTS�+-Lösung bewirkt. Nach Erstellung einer
Kalibrierkurve durch Messung verschiedener geeigneter Troloxkonzentrationen kann
die durch die Probe verursachte Extinktionserniedrigung der entsprechenden Trolox-
konzentration zugeordnet und das Ergebnis als TEAC angegeben werden.
3.2.6 Bestimmung der Mykotoxine (DON, ZON, DOM-1, α-ZOL, ß-ZOL) mittels
HPLC
Die HPLC (High Performance Liquid Chromatography, Hochleistungs-Säulen-
Flüssig-Chromatographie) ist eine Methode, welche Substanzen trennen und diese
über Standards identifizieren und quantifizieren kann. Die Funktionsweise beruht auf
einem chromatographischen Trennverfahren, bei dem die zu untersuchende Sub-
stanz zusammen mit einem Laufmittel, der „mobilen Phase“ (“Eluent“), durch eine so
genannte Trennsäule quantifiziert wird.
Eine Übersicht der HPLC Bedingungen ist Tabelle 6 zu entnehmen.
3.2.6.1 Bestimmung von Deoxynivalenol und de-epoxy-Deoxynivalenol in Futtermit-
teln, Blutserum, Milch und Gallensaft
Die Bestimmung von Deoxynivalenol und DOM-1 wurde in Futtermitteln, Blutserum,
Milch und Gallensaft nach der Methode von VALENTA et al. (2003) bestimmt. Diese
Methode wird im Folgenden kurz beschrieben. Zum Nachweis von DON und DOM-1
wurden alle Proben, außer den Futtermittelproben, mit ß-Glucuronidase behandelt.
Danach wurden die Proben in einer Immunoaffinitätssäule (IAC) (DONprep, r-
Biopharm) aufgereinigt. Die so entstandenen Eluate wurden mit HPLC bei einer Wel-
lenlänge von 218 nm analysiert. Die HPLC- Analyse wurde auf einer C18-RP-Säule
Material und Methoden
28
(Phenomenex), mit einem Acetonitril-Wassergemisch (13+87 oder 12,5+87,5) durch-
geführt. Aufgrund der Vorreinigung der Proben mit der Immunoaffinitätssäule konn-
ten gute Nachweisgrenzen erreicht werden. Für Futter lag die Nachweisgrenze bei
0,03 mg/kg, für Serum bei 0,002 µg/ml, für Milch, bezogen auf die Trockenmasse, bei
0,004 µg/g und für Galle bei 0,004 µg/ml.
3.2.6.2 Zearalenon- (α- und ß-Zearalenol) Bestimmung mittels HPLC in Futter, Blut-
serum, Milch und Gallensaft (mod. Zea-Lufa-Methode IAC, HPLC)
Zearalenon, α- und ß- Zearalenol wurden in Futter, Blutserum, Milch und Gallensaft
mit der modifizierten Zea-Lufa-Methode nach UEBERSCHÄR (1999) und DÄNICKE
et al. (2001) bestimmt. Dazu wurden die Futterproben mit ß-Glucuronidase inkubiert.
Serum, Milch und Galle wurde ebenfalls mit ß-Glucuronidase inkubiert und mit der
IAC Easi-Extract™ Zearalenon (Firma r- Biopharm) aufgereinigt. Alle Proben wurden
mit HPLC bei einer Wellenlänge von 238 nm analysiert. Die HPLC- Analyse wurde
ebenfalls auf einer C18-RP-Säule (Phenomenex), mit einem Acetonitril-
Wassergemisch, durchgeführt. Die Nachweisgrenzen lagen für Futter bei 0,3 µg/kg,
für Serum, Milch und Gallensaft bei 2 ng ZON/g, 2 ng α-ZOL/g sowie 5 ng ß-ZON/g.
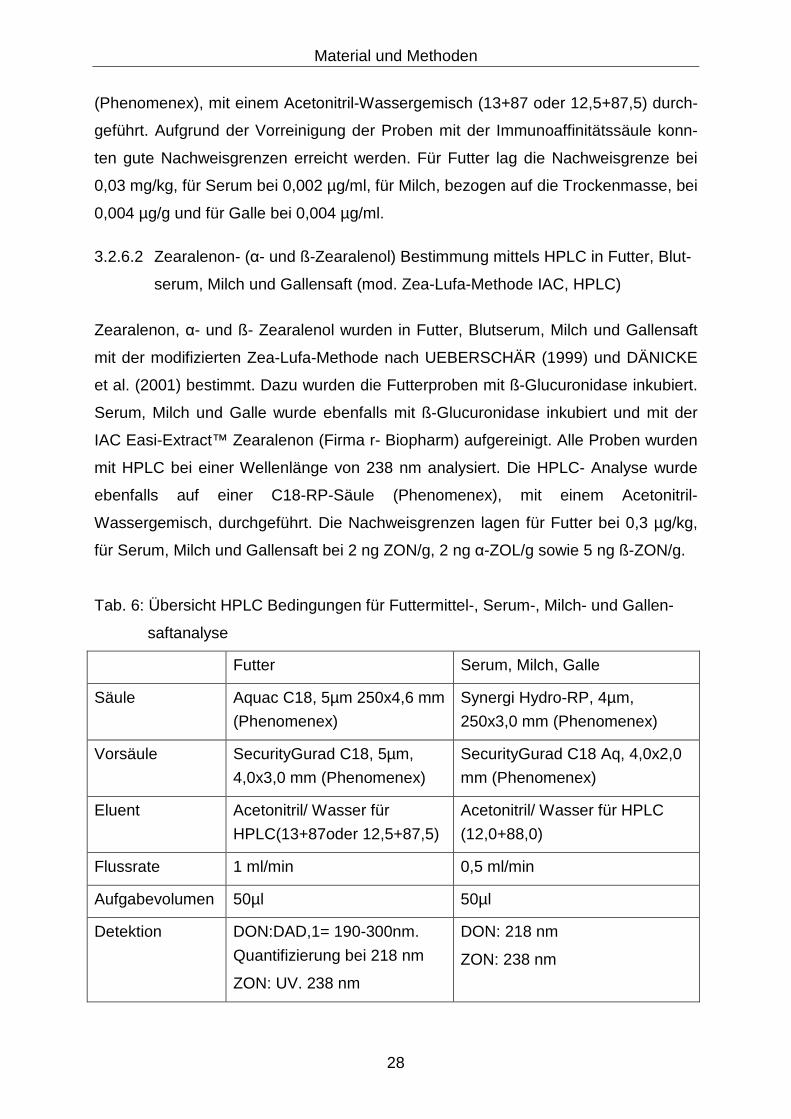
Tab. 6: Übersicht HPLC Bedingungen für Futtermittel-, Serum-, Milch- und Gallen-
saftanalyse
Futter Serum, Milch, Galle
Säule Aquac C18, 5µm 250x4,6 mm
(Phenomenex)
Synergi Hydro-RP, 4µm,
250x3,0 mm (Phenomenex)
Vorsäule SecurityGurad C18, 5µm,
4,0x3,0 mm (Phenomenex)
SecurityGurad C18 Aq, 4,0x2,0
mm (Phenomenex)
Eluent Acetonitril/ Wasser für
HPLC(13+87oder 12,5+87,5)
Acetonitril/ Wasser für HPLC
(12,0+88,0)
Flussrate 1 ml/min 0,5 ml/min
Aufgabevolumen 50µl 50µl
Detektion DON:DAD,1= 190-300nm.
Quantifizierung bei 218 nm
ZON: UV. 238 nm
DON: 218 nm
ZON: 238 nm

Material und Methoden
29
3.2.7 Statistische Auswertung
Die statistische Auswertung der Daten erfolgte mit dem Programm SPSS 11.5. Die
Prüfung auf Normalverteilung erfolgte mit dem Kolmogorov-Smirnov-Test. Es wur-
den die Medianwerte und 1. bis 3. Quartile berechnet.
Die Prüfung der Daten auf statistisch signifikante Unterschiede wurde auf Grund der
signifikanten Abweichungen von der Normalverteilung mit dem nicht parametrischen
H-Test nach Kruskal-Wallis durchgeführt. Bei Signifikantem H-Test wurde die Signifi-
kanzprüfung zwischen jeweils 2 Gruppen mit dem Test nach Mann-Whitney durchge-
führt (p≤0,05).
Die grafische Darstellung im Ergebnisteil erfolgte zum überwiegenden Teil mit Box-
Plots. Die untere und obere Grenze der Box repräsentieren das 1. und 3. Quartil. Die
Länge der Box entspricht dem Interquartilbereich, so dass eine Box die mittleren 50
% der Werte einer Gruppe enthält. Die Linie in der Box gibt die Lage des Medians
wieder. Die von der Box weggehenden Linien (Whiskers) reichen jeweils bis zum
letzten Wert, der weniger als einen Interquartilbereich außerhalb der Box liegt. Dabei
ist das 1. Quartil (auch 25. Perzentil) wie folgt bestimmt: 25 % der beobachteten Fäl-
le sind kleiner oder gleich diesem Wert und 75 % (auch 75. Perzentil) der beobachte-
ten Fälle sind größer oder gleich diesem Wert.
Ergebnisse
30
4 Ergebnisse
4.1 Ergebnisse der Mykotoxinanalysen
4.1.1 Patienten (n=61)
Mykotoxine konnten regelmäßig in den Futterproben aus den Herkunftsbetrieben der
Patienten nachgewiesen werden (Tabelle 7), ohne dass grobsinnlichen Veränderun-
gen feststellbar waren.
Tab.7: Mykotoxinkonzentrationen in den Futterproben aus den Herkunftsbetrieben
der Patienten mit LMV (n=20)
DON (mg/kg) ZON (mg/kg)
Grenzwert <5,000 <0,050
Median 0,161 0,00635
1. Quartil 0,086 0,00488
2.Quartil 0,191 0,00785
Die Ergebnisse des Mykotoxinnachweises (DON, DOM-1, ZON, α-ZOL, ß-ZOL) bei
den Patienten mit LMV werden in den Tabellen 8 und 9 präsentiert. In der Milch wa-
ren keine Mykotoxine nachweisbar.
Tab. 8: Anteiliger Mykotoxinnachweis (%) bei Kühen mit LMV in Serum, Gallensaft
und Milch
Proben Tiere positiv Mykotoxinnachweis
Serum 61 5 (8 %)
Gallensaft 61 23 (38 %)
Milch 20 0 (0 %)
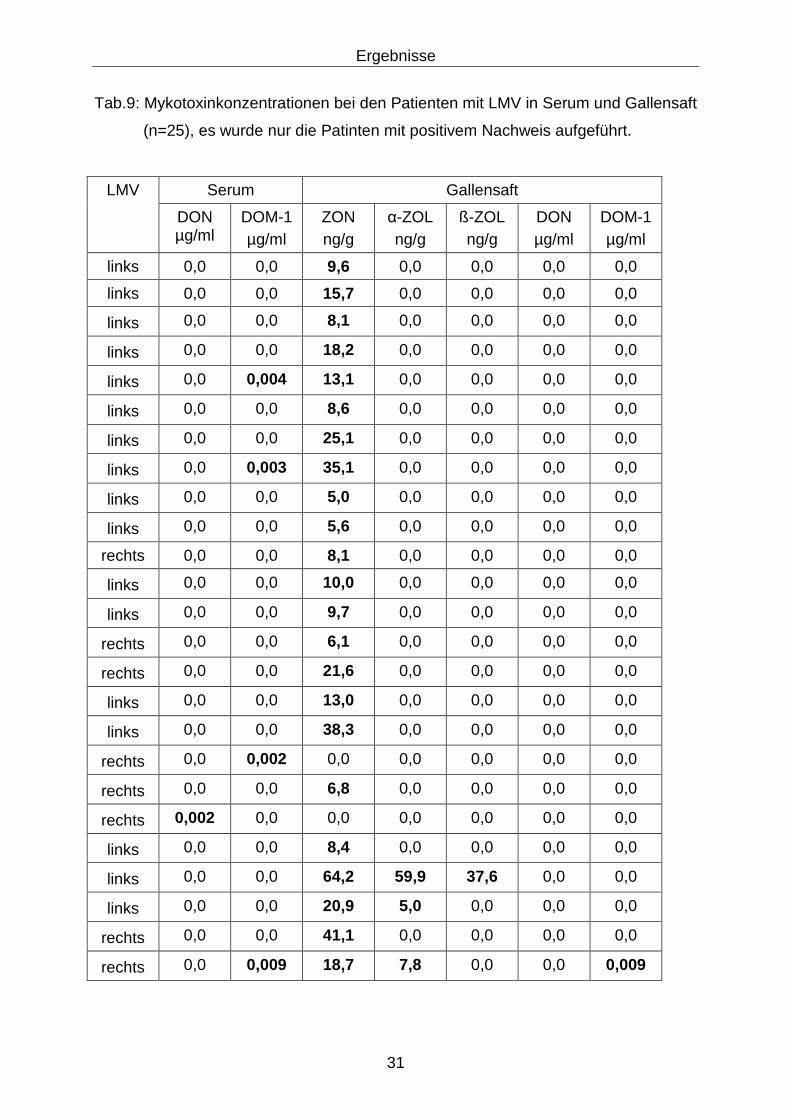
Ergebnisse
31
Tab.9: Mykotoxinkonzentrationen bei den Patienten mit LMV in Serum und Gallensaft
(n=25), es wurde nur die Patinten mit positivem Nachweis aufgeführt.
Serum Gallensaft LMV
DON µg/ml
DOM-1 µg/ml
ZON ng/g
α-ZOL ng/g
ß-ZOL ng/g
DON µg/ml
DOM-1 µg/ml
links 0,0 0,0 9,6 0,0 0,0 0,0 0,0
links 0,0 0,0 15,7 0,0 0,0 0,0 0,0
links 0,0 0,0 8,1 0,0 0,0 0,0 0,0
links 0,0 0,0 18,2 0,0 0,0 0,0 0,0
links 0,0 0,004 13,1 0,0 0,0 0,0 0,0
links 0,0 0,0 8,6 0,0 0,0 0,0 0,0
links 0,0 0,0 25,1 0,0 0,0 0,0 0,0
links 0,0 0,003 35,1 0,0 0,0 0,0 0,0
links 0,0 0,0 5,0 0,0 0,0 0,0 0,0
links 0,0 0,0 5,6 0,0 0,0 0,0 0,0
rechts 0,0 0,0 8,1 0,0 0,0 0,0 0,0
links 0,0 0,0 10,0 0,0 0,0 0,0 0,0
links 0,0 0,0 9,7 0,0 0,0 0,0 0,0
rechts 0,0 0,0 6,1 0,0 0,0 0,0 0,0
rechts 0,0 0,0 21,6 0,0 0,0 0,0 0,0
links 0,0 0,0 13,0 0,0 0,0 0,0 0,0
links 0,0 0,0 38,3 0,0 0,0 0,0 0,0
rechts 0,0 0,002 0,0 0,0 0,0 0,0 0,0
rechts 0,0 0,0 6,8 0,0 0,0 0,0 0,0
rechts 0,002 0,0 0,0 0,0 0,0 0,0 0,0
links 0,0 0,0 8,4 0,0 0,0 0,0 0,0
links 0,0 0,0 64,2 59,9 37,6 0,0 0,0
links 0,0 0,0 20,9 5,0 0,0 0,0 0,0
rechts 0,0 0,0 41,1 0,0 0,0 0,0 0,0
rechts 0,0 0,009 18,7 7,8 0,0 0,0 0,009

Ergebnisse
32
4.1.2 Gesunde Kühe (n=13)
Die makroskopische Untersuchung der Futterproben ergab keine grobsinnlichen
Veränderungen. Mykotoxine (DON, ZON) konnten regelmäßig in den Futterproben
aus den Herkunftsbetrieben der gesunden Kühe, nicht jedoch im Serum, in der Milch
oder den Gallensaftproben nachgewiesen werden (Tabelle 10).
Tab. 10: Mykotoxingehalt in den untersuchten Futtermitteln aus den Herkunftsbetrie-
ben der gesunden Kühe
Futter DON µg/g ZON ng/g
Betrieb 1 0,272 11,1
Betrieb 2 0,0 7,3
4.2 Klinische Untersuchungsbefunde
Von den in die Studie aufgenommenen Patienten der Medizinischen Tierklinik, Uni-
versität Leipzig, wurden bei 43 Kühen eine LMV nach links und bei 18 Kühen eine
LMV nach rechts diagnostiziert. Am Tag der Aufnahmeuntersuchung zeigten alle Kü-
he (mit oder ohne positiven Mykotoxinnachweis) ein geringgradig bis mittelgradig
gestörtes Allgemeinbefinden, mittelgradige Abmagerung, gering- bis mittelgradig re-
duzierte Futteraufnahme sowie gering- bis mittelgradig reduzierte Pansenbewegun-
gen. Die Atemfrequenz lag bei 24 ± 12/min. Die Klauen waren bei 52 % der Tiere
entzündlich verändert und wiesen gerötete Kronsäume auf. Die Pulsfrequenzen der
Tiere lagen am Tag der Klinikeinweisung im Referenzbereich von 60-80/Minute. Die
durchschnittliche innere Körpertemperatur lag bei allen untersuchten Kühen zu Be-
ginn der Untersuchung im Referenzbereich von 38,3 bis 38,8°C. Die Rektaluntersu-
chungen zeigten, dass die Ovarien palpatorisch ohne besonderen Befund waren. Die
Untersuchung des Euters erfolgte durch Adspektion und Palpation desselben sowie
durch die grobsinnliche Beurteilung des Milchsekrets. Bei jedem Tier wurde ein
Schalmtest durchgeführt, wobei sich folgende Ergebnisse ergaben: 1-3 Plus (subkli-
nisch positiv). Der Kot der Tiere war olivgrün, dünnbreiig bis breiig und hatte den für
einen ruminierenden Wiederkäuer typischen Geruch. Die Ergebnisse der klinischen
Befunde sind in Abbildung 6 dargestellt.
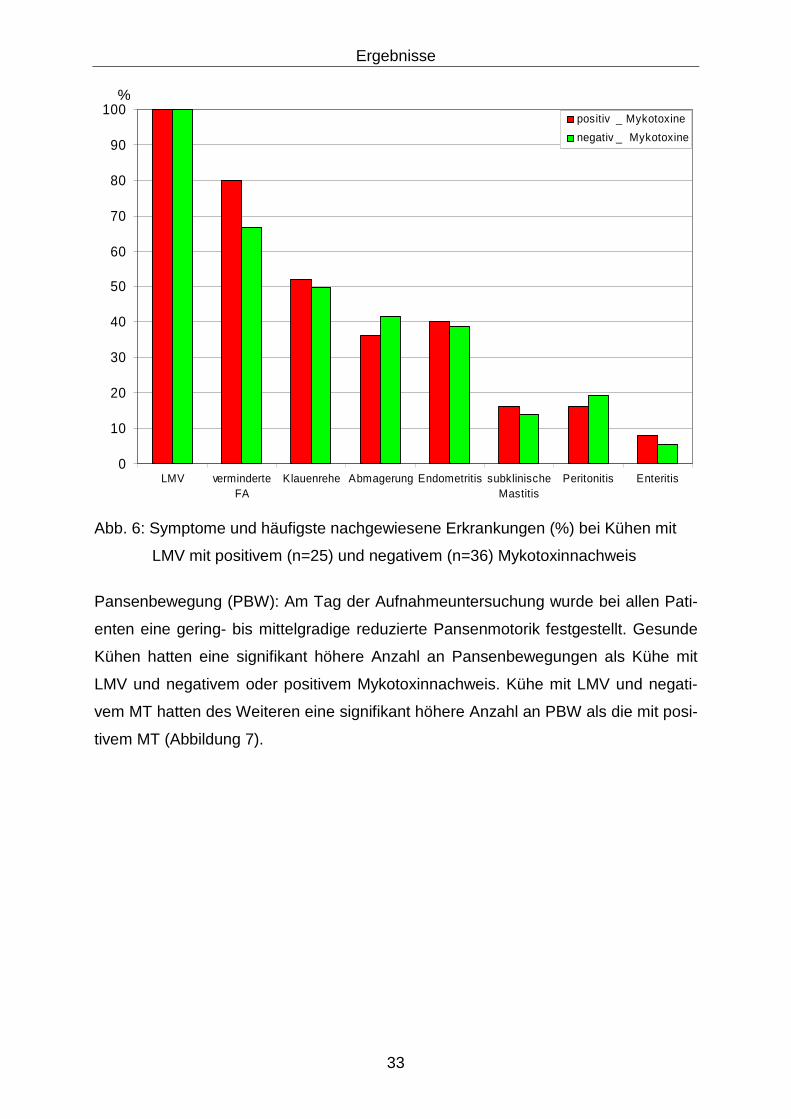
Ergebnisse
33
0
10
20
30
40
50
60
70
80
90
100
LMV verminderteFA
Klauenrehe Abmagerung Endometritis subklinischeMastitis
Peritonitis Enteritis
positiv _ Mykotoxine
negativ _ Mykotoxine
Abb. 6: Symptome und häufigste nachgewiesene Erkrankungen (%) bei Kühen mit
LMV mit positivem (n=25) und negativem (n=36) Mykotoxinnachweis
Pansenbewegung (PBW): Am Tag der Aufnahmeuntersuchung wurde bei allen Pati-
enten eine gering- bis mittelgradige reduzierte Pansenmotorik festgestellt. Gesunde
Kühen hatten eine signifikant höhere Anzahl an Pansenbewegungen als Kühe mit
LMV und negativem oder positivem Mykotoxinnachweis. Kühe mit LMV und negati-
vem MT hatten des Weiteren eine signifikant höhere Anzahl an PBW als die mit posi-
tivem MT (Abbildung 7).
%
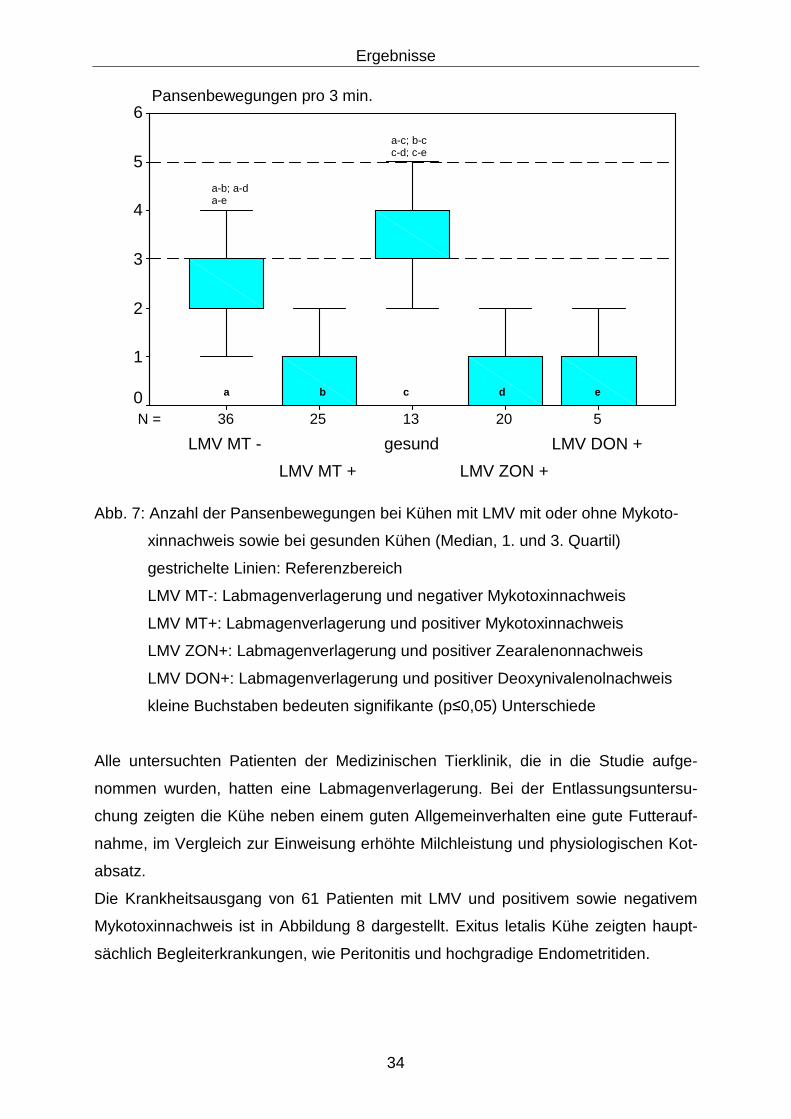
Ergebnisse
34
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
6
5
4
3
2
1
0
Abb. 7: Anzahl der Pansenbewegungen bei Kühen mit LMV mit oder ohne Mykoto-
xinnachweis sowie bei gesunden Kühen (Median, 1. und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0,05) Unterschiede
Alle untersuchten Patienten der Medizinischen Tierklinik, die in die Studie aufge-
nommen wurden, hatten eine Labmagenverlagerung. Bei der Entlassungsuntersu-
chung zeigten die Kühe neben einem guten Allgemeinverhalten eine gute Futterauf-
nahme, im Vergleich zur Einweisung erhöhte Milchleistung und physiologischen Kot-
absatz.
Die Krankheitsausgang von 61 Patienten mit LMV und positivem sowie negativem
Mykotoxinnachweis ist in Abbildung 8 dargestellt. Exitus letalis Kühe zeigten haupt-
sächlich Begleiterkrankungen, wie Peritonitis und hochgradige Endometritiden.
Pansenbewegungen pro 3 min.
a-c; b-c c-d; c-e
a b c d e
a-b; a-d a-e
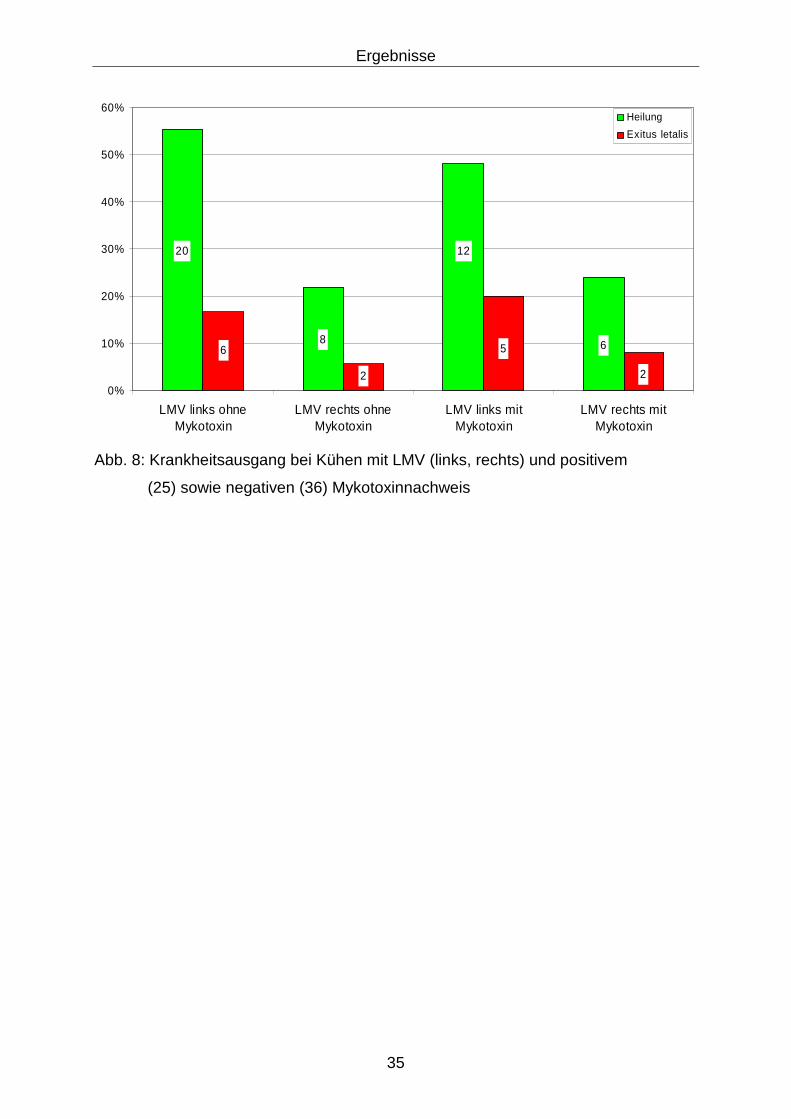
Ergebnisse
35
68
1220
22
56
0%
10%
20%
30%
40%
50%
60%
LMV links ohneMykotoxin
LMV rechts ohneMykotoxin
LMV links mitMykotoxin
LMV rechts mitMykotoxin
Heilung
Exitus letalis
Abb. 8: Krankheitsausgang bei Kühen mit LMV (links, rechts) und positivem
(25) sowie negativen (36) Mykotoxinnachweis
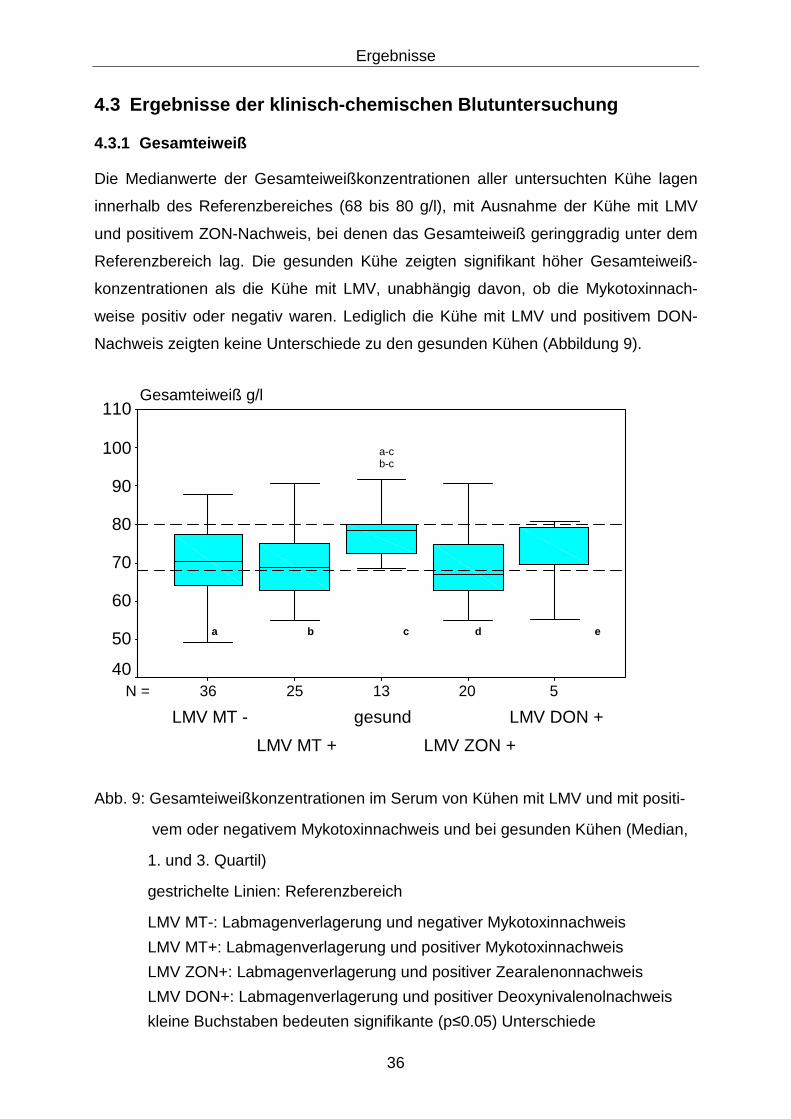
Ergebnisse
36
4.3 Ergebnisse der klinisch-chemischen Blutuntersuc hung
4.3.1 Gesamteiweiß
Die Medianwerte der Gesamteiweißkonzentrationen aller untersuchten Kühe lagen
innerhalb des Referenzbereiches (68 bis 80 g/l), mit Ausnahme der Kühe mit LMV
und positivem ZON-Nachweis, bei denen das Gesamteiweiß geringgradig unter dem
Referenzbereich lag. Die gesunden Kühe zeigten signifikant höher Gesamteiweiß-
konzentrationen als die Kühe mit LMV, unabhängig davon, ob die Mykotoxinnach-
weise positiv oder negativ waren. Lediglich die Kühe mit LMV und positivem DON-
Nachweis zeigten keine Unterschiede zu den gesunden Kühen (Abbildung 9).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
110
100
90
80
70
60
50
40
Abb. 9: Gesamteiweißkonzentrationen im Serum von Kühen mit LMV und mit positi-
vem oder negativem Mykotoxinnachweis und bei gesunden Kühen (Median,
1. und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
a c b d e
a-c b-c
Gesamteiweiß g/l
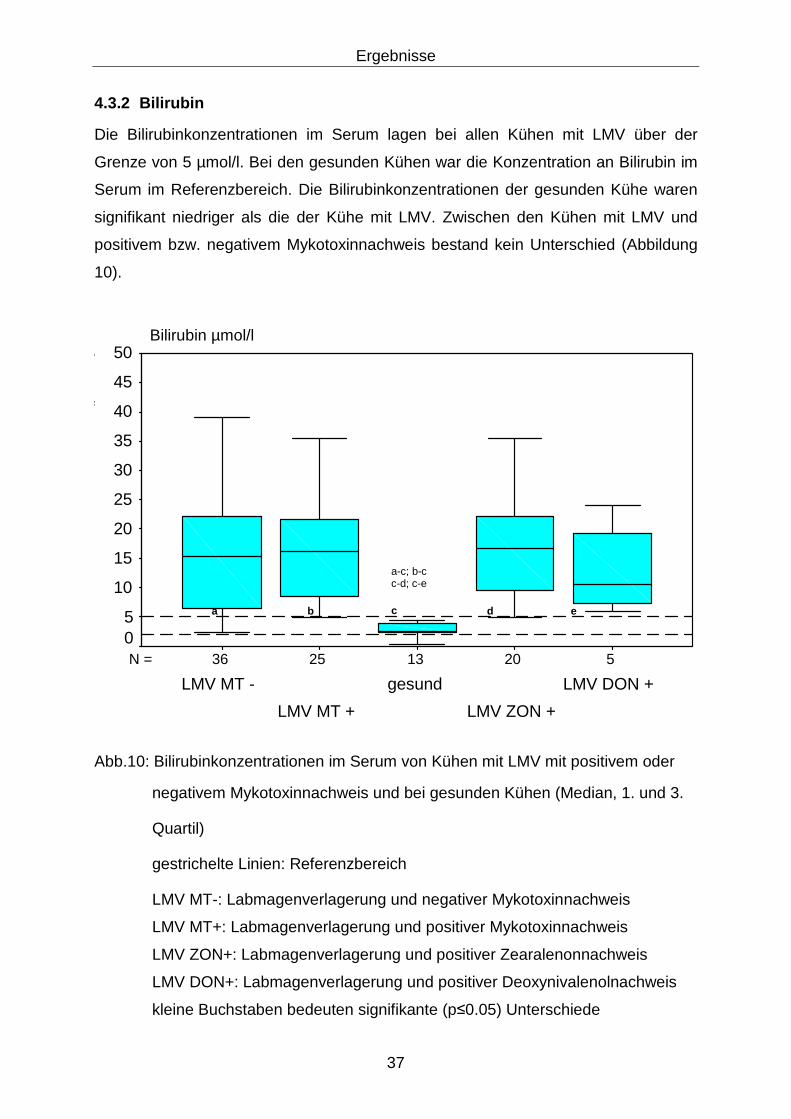
Ergebnisse
37
4.3.2 Bilirubin
Die Bilirubinkonzentrationen im Serum lagen bei allen Kühen mit LMV über der
Grenze von 5 µmol/l. Bei den gesunden Kühen war die Konzentration an Bilirubin im
Serum im Referenzbereich. Die Bilirubinkonzentrationen der gesunden Kühe waren
signifikant niedriger als die der Kühe mit LMV. Zwischen den Kühen mit LMV und
positivem bzw. negativem Mykotoxinnachweis bestand kein Unterschied (Abbildung
10).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
Bili
rubi
n (µ
mol
/l) 50
45
40
35
30
25
20
15
10
50
Abb.10: Bilirubinkonzentrationen im Serum von Kühen mit LMV mit positivem oder
negativem Mykotoxinnachweis und bei gesunden Kühen (Median, 1. und 3.
Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
Bilirubin µmol/l
a b c
d e
a-c; b-c c-d; c-e
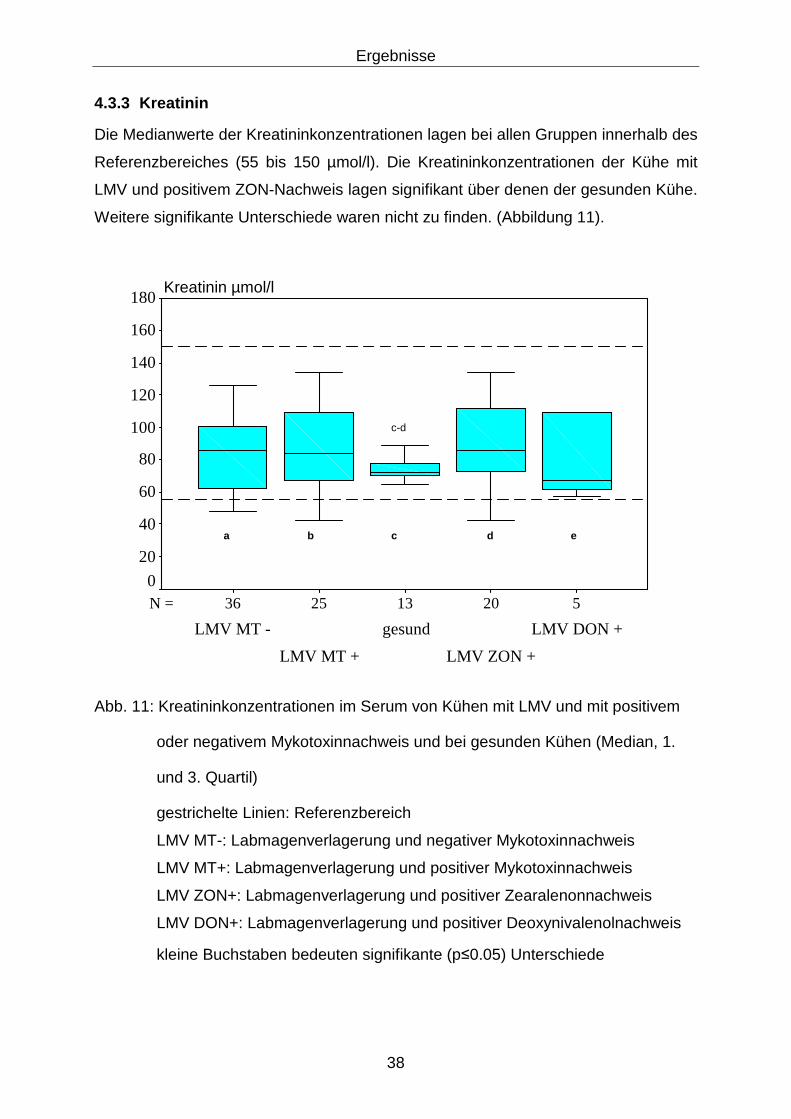
Ergebnisse
38
4.3.3 Kreatinin
Die Medianwerte der Kreatininkonzentrationen lagen bei allen Gruppen innerhalb des
Referenzbereiches (55 bis 150 µmol/l). Die Kreatininkonzentrationen der Kühe mit
LMV und positivem ZON-Nachweis lagen signifikant über denen der gesunden Kühe.
Weitere signifikante Unterschiede waren nicht zu finden. (Abbildung 11).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
180
160
140
120
100
80
60
40
20
0
Abb. 11: Kreatininkonzentrationen im Serum von Kühen mit LMV und mit positivem
oder negativem Mykotoxinnachweis und bei gesunden Kühen (Median, 1.
und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
Kreatinin µmol/l
c-d
a b c d e
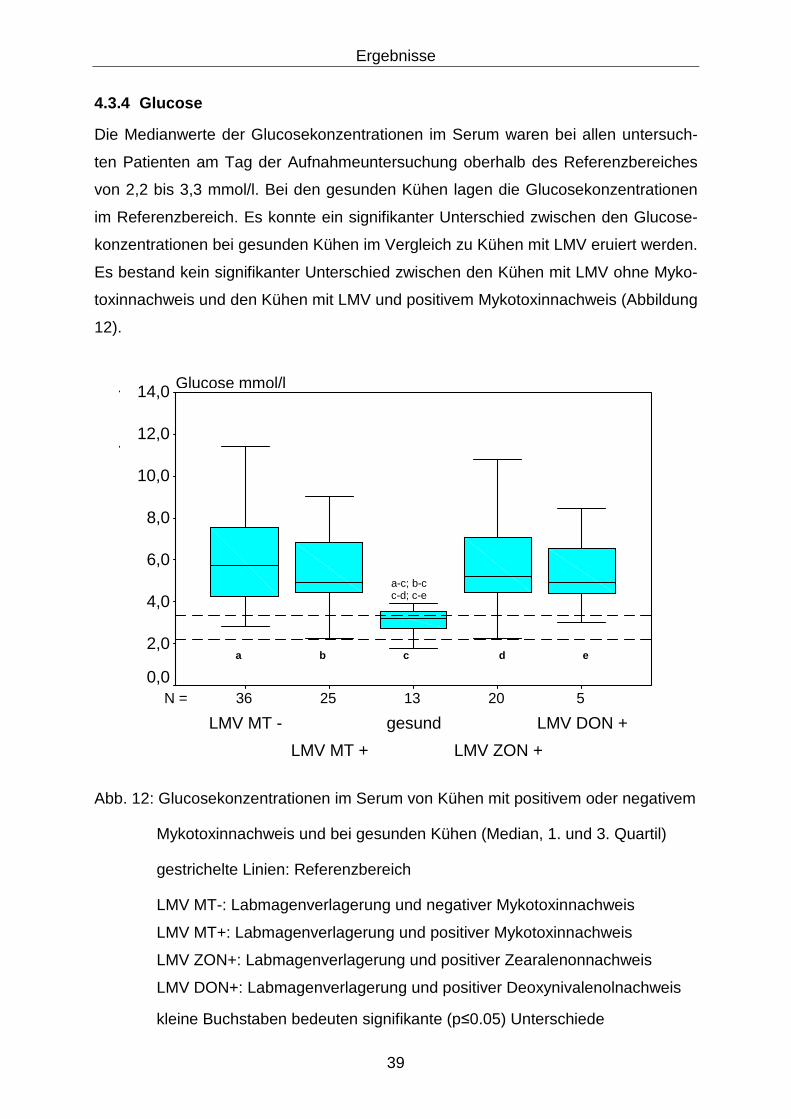
Ergebnisse
39
4.3.4 Glucose
Die Medianwerte der Glucosekonzentrationen im Serum waren bei allen untersuch-
ten Patienten am Tag der Aufnahmeuntersuchung oberhalb des Referenzbereiches
von 2,2 bis 3,3 mmol/l. Bei den gesunden Kühen lagen die Glucosekonzentrationen
im Referenzbereich. Es konnte ein signifikanter Unterschied zwischen den Glucose-
konzentrationen bei gesunden Kühen im Vergleich zu Kühen mit LMV eruiert werden.
Es bestand kein signifikanter Unterschied zwischen den Kühen mit LMV ohne Myko-
toxinnachweis und den Kühen mit LMV und positivem Mykotoxinnachweis (Abbildung
12).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
Glu
cose
(m
mol
/l) 14,0
12,0
10,0
8,0
6,0
4,0
2,0
0,0
Abb. 12: Glucosekonzentrationen im Serum von Kühen mit positivem oder negativem
Mykotoxinnachweis und bei gesunden Kühen (Median, 1. und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
Glucose mmol/l
a-c; b-c c-d; c-e
a b c d e
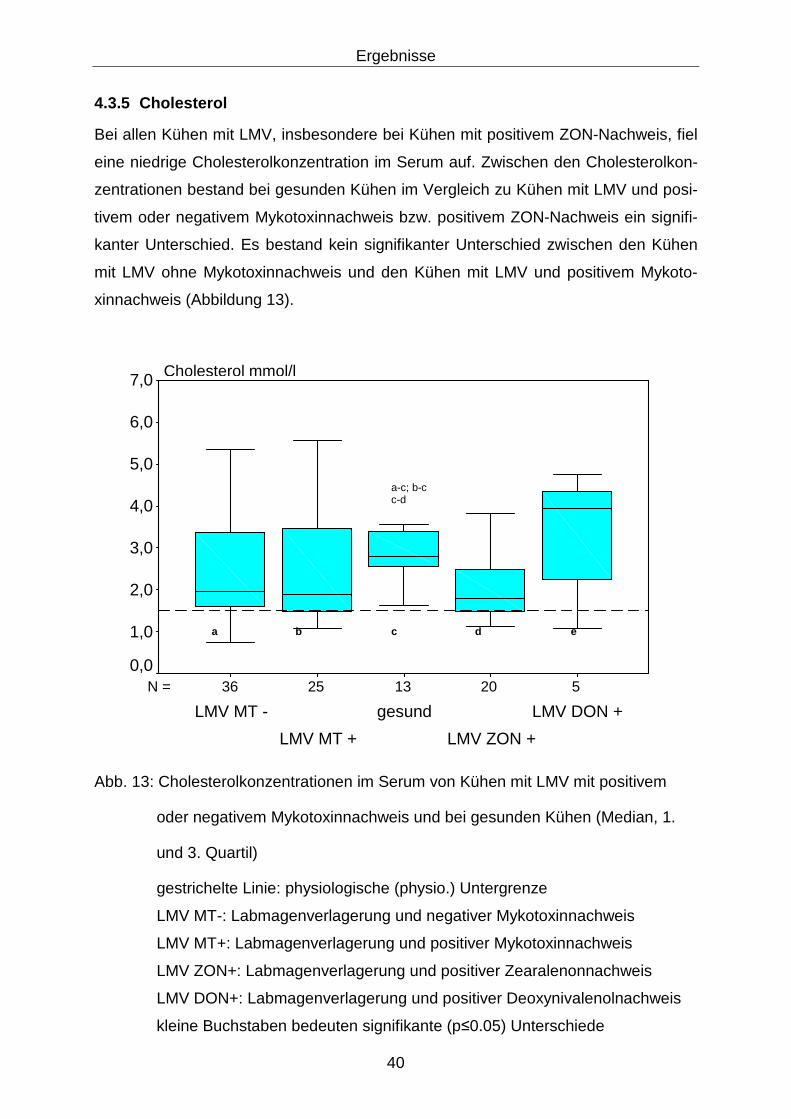
Ergebnisse
40
4.3.5 Cholesterol
Bei allen Kühen mit LMV, insbesondere bei Kühen mit positivem ZON-Nachweis, fiel
eine niedrige Cholesterolkonzentration im Serum auf. Zwischen den Cholesterolkon-
zentrationen bestand bei gesunden Kühen im Vergleich zu Kühen mit LMV und posi-
tivem oder negativem Mykotoxinnachweis bzw. positivem ZON-Nachweis ein signifi-
kanter Unterschied. Es bestand kein signifikanter Unterschied zwischen den Kühen
mit LMV ohne Mykotoxinnachweis und den Kühen mit LMV und positivem Mykoto-
xinnachweis (Abbildung 13).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
7,0
6,0
5,0
4,0
3,0
2,0
1,0
0,0
Abb. 13: Cholesterolkonzentrationen im Serum von Kühen mit LMV mit positivem
oder negativem Mykotoxinnachweis und bei gesunden Kühen (Median, 1.
und 3. Quartil)
gestrichelte Linie: physiologische (physio.) Untergrenze
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
Cholesterol mmol/l
a-c; b-c c-d
a b c d e
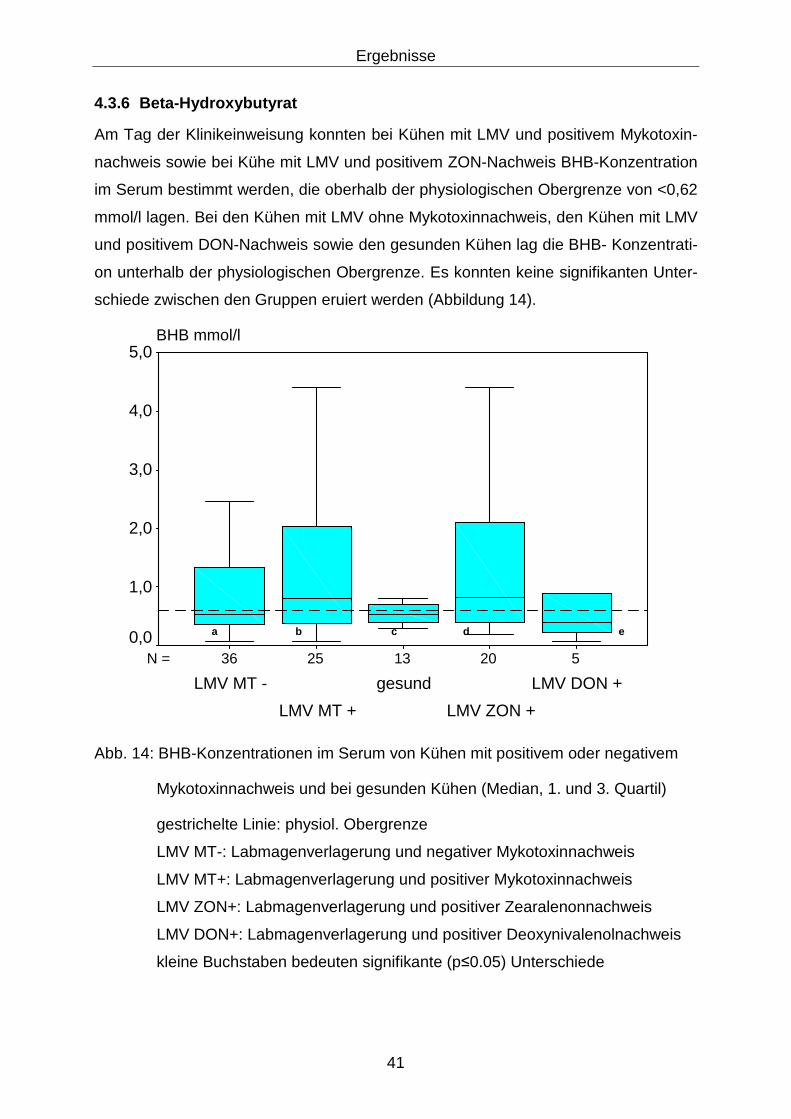
Ergebnisse
41
4.3.6 Beta-Hydroxybutyrat
Am Tag der Klinikeinweisung konnten bei Kühen mit LMV und positivem Mykotoxin-
nachweis sowie bei Kühe mit LMV und positivem ZON-Nachweis BHB-Konzentration
im Serum bestimmt werden, die oberhalb der physiologischen Obergrenze von <0,62
mmol/l lagen. Bei den Kühen mit LMV ohne Mykotoxinnachweis, den Kühen mit LMV
und positivem DON-Nachweis sowie den gesunden Kühen lag die BHB- Konzentrati-
on unterhalb der physiologischen Obergrenze. Es konnten keine signifikanten Unter-
schiede zwischen den Gruppen eruiert werden (Abbildung 14).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
5,0
4,0
3,0
2,0
1,0
0,0
Abb. 14: BHB-Konzentrationen im Serum von Kühen mit positivem oder negativem
Mykotoxinnachweis und bei gesunden Kühen (Median, 1. und 3. Quartil)
gestrichelte Linie: physiol. Obergrenze
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
BHB mmol/l
a b c d e
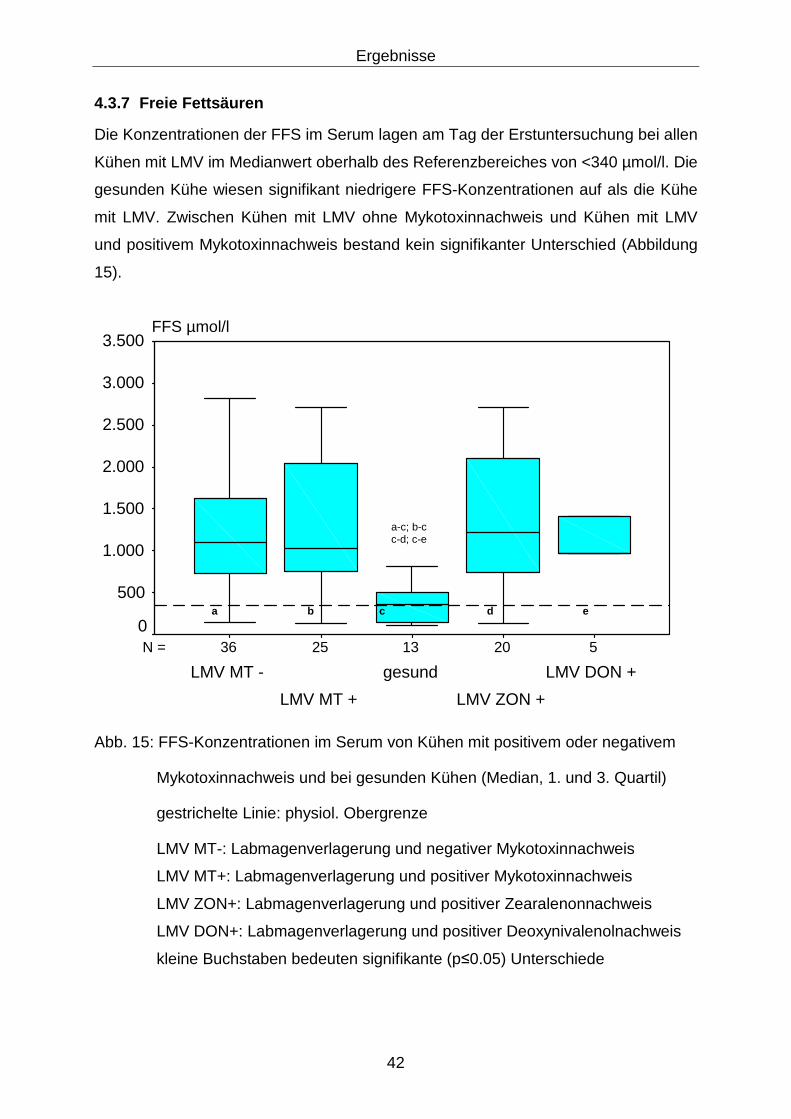
Ergebnisse
42
4.3.7 Freie Fettsäuren
Die Konzentrationen der FFS im Serum lagen am Tag der Erstuntersuchung bei allen
Kühen mit LMV im Medianwert oberhalb des Referenzbereiches von <340 µmol/l. Die
gesunden Kühe wiesen signifikant niedrigere FFS-Konzentrationen auf als die Kühe
mit LMV. Zwischen Kühen mit LMV ohne Mykotoxinnachweis und Kühen mit LMV
und positivem Mykotoxinnachweis bestand kein signifikanter Unterschied (Abbildung
15).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
3.500
3.000
2.500
2.000
1.500
1.000
500
0
Abb. 15: FFS-Konzentrationen im Serum von Kühen mit positivem oder negativem
Mykotoxinnachweis und bei gesunden Kühen (Median, 1. und 3. Quartil)
gestrichelte Linie: physiol. Obergrenze
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
FFS µmol/l
a-c; b-c c-d; c-e
a b c d e
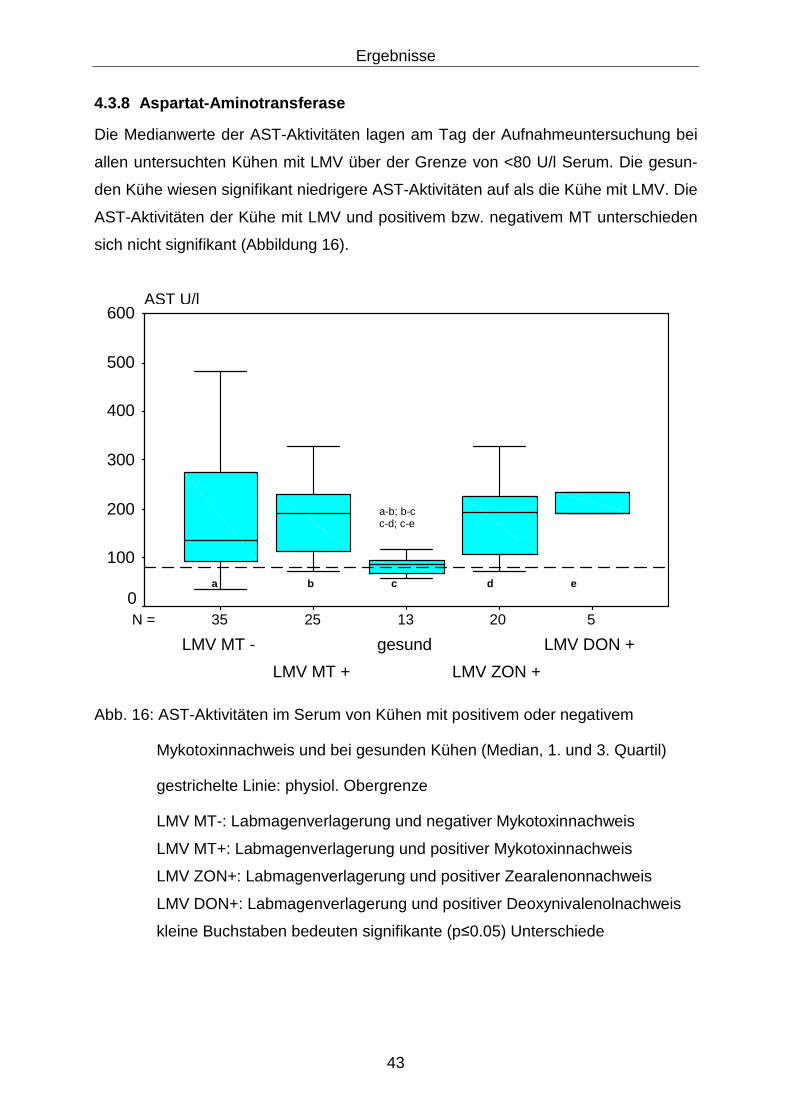
Ergebnisse
43
4.3.8 Aspartat-Aminotransferase
Die Medianwerte der AST-Aktivitäten lagen am Tag der Aufnahmeuntersuchung bei
allen untersuchten Kühen mit LMV über der Grenze von <80 U/l Serum. Die gesun-
den Kühe wiesen signifikant niedrigere AST-Aktivitäten auf als die Kühe mit LMV. Die
AST-Aktivitäten der Kühe mit LMV und positivem bzw. negativem MT unterschieden
sich nicht signifikant (Abbildung 16).
520132535N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
600
500
400
300
200
100
0
Abb. 16: AST-Aktivitäten im Serum von Kühen mit positivem oder negativem
Mykotoxinnachweis und bei gesunden Kühen (Median, 1. und 3. Quartil)
gestrichelte Linie: physiol. Obergrenze
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
AST U/l
a-b; b-c c-d; c-e
a b c d e
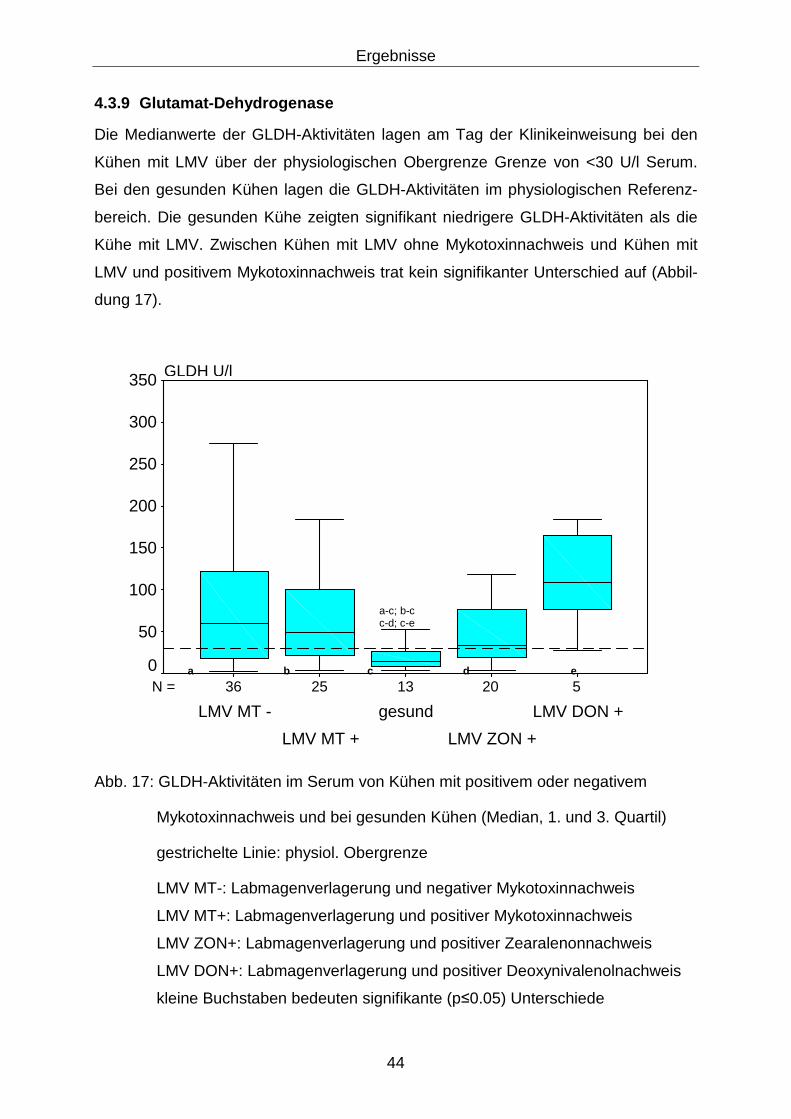
Ergebnisse
44
4.3.9 Glutamat-Dehydrogenase
Die Medianwerte der GLDH-Aktivitäten lagen am Tag der Klinikeinweisung bei den
Kühen mit LMV über der physiologischen Obergrenze Grenze von <30 U/l Serum.
Bei den gesunden Kühen lagen die GLDH-Aktivitäten im physiologischen Referenz-
bereich. Die gesunden Kühe zeigten signifikant niedrigere GLDH-Aktivitäten als die
Kühe mit LMV. Zwischen Kühen mit LMV ohne Mykotoxinnachweis und Kühen mit
LMV und positivem Mykotoxinnachweis trat kein signifikanter Unterschied auf (Abbil-
dung 17).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
350
300
250
200
150
100
50
0
Abb. 17: GLDH-Aktivitäten im Serum von Kühen mit positivem oder negativem
Mykotoxinnachweis und bei gesunden Kühen (Median, 1. und 3. Quartil)
gestrichelte Linie: physiol. Obergrenze
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
GLDH U/l
a-c; b-c c-d; c-e
a b c d e
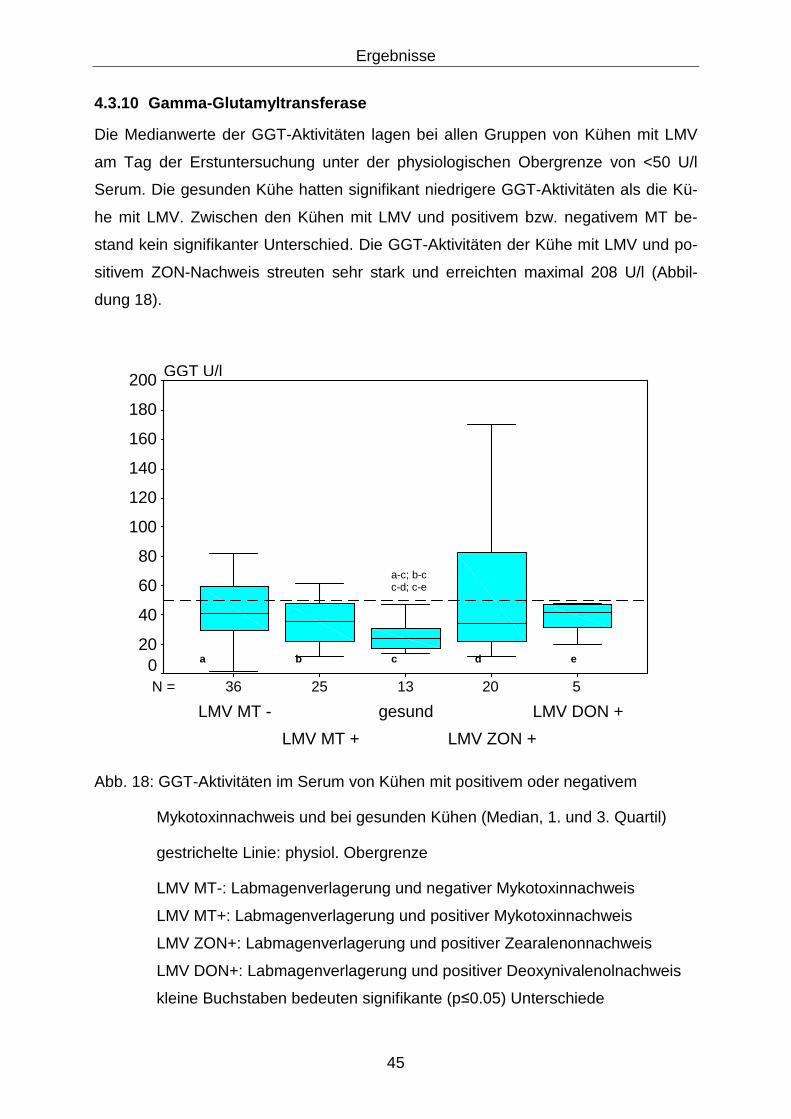
Ergebnisse
45
4.3.10 Gamma-Glutamyltransferase
Die Medianwerte der GGT-Aktivitäten lagen bei allen Gruppen von Kühen mit LMV
am Tag der Erstuntersuchung unter der physiologischen Obergrenze von <50 U/l
Serum. Die gesunden Kühe hatten signifikant niedrigere GGT-Aktivitäten als die Kü-
he mit LMV. Zwischen den Kühen mit LMV und positivem bzw. negativem MT be-
stand kein signifikanter Unterschied. Die GGT-Aktivitäten der Kühe mit LMV und po-
sitivem ZON-Nachweis streuten sehr stark und erreichten maximal 208 U/l (Abbil-
dung 18).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
200
180
160
140
120
100
80
60
40
200
Abb. 18: GGT-Aktivitäten im Serum von Kühen mit positivem oder negativem
Mykotoxinnachweis und bei gesunden Kühen (Median, 1. und 3. Quartil)
gestrichelte Linie: physiol. Obergrenze
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
GGT U/l
a-c; b-c c-d; c-e
a b c d e
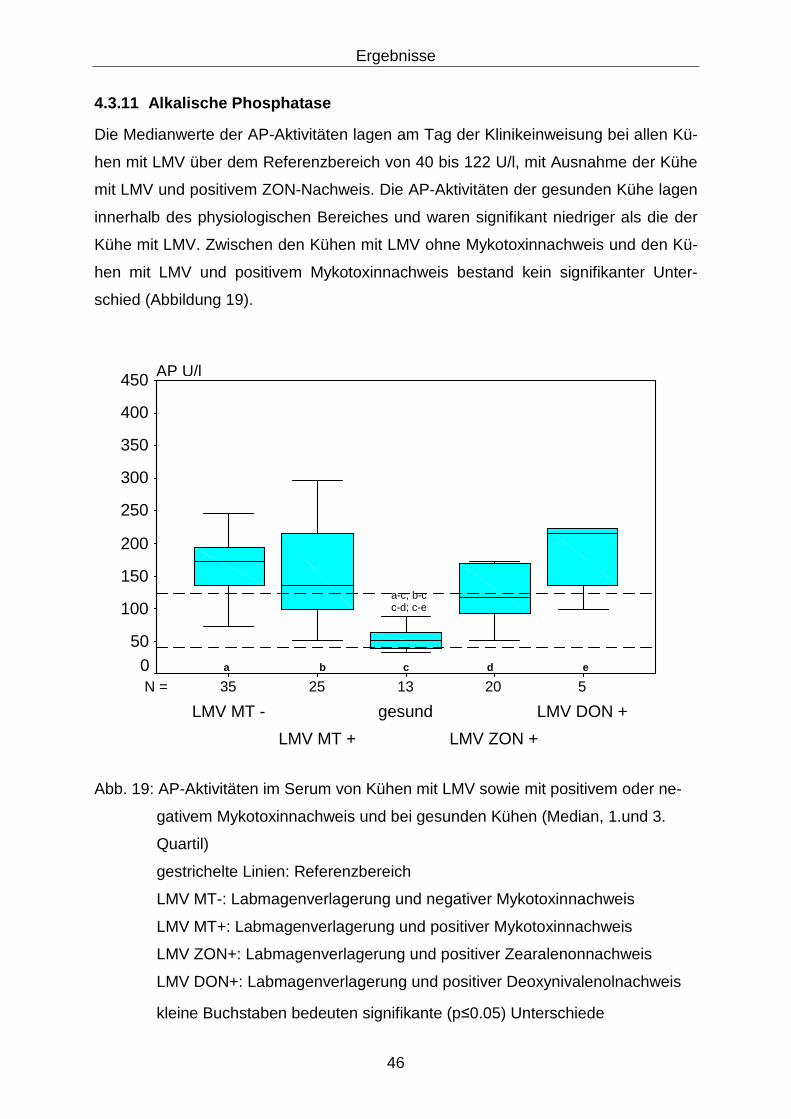
Ergebnisse
46
4.3.11 Alkalische Phosphatase
Die Medianwerte der AP-Aktivitäten lagen am Tag der Klinikeinweisung bei allen Kü-
hen mit LMV über dem Referenzbereich von 40 bis 122 U/l, mit Ausnahme der Kühe
mit LMV und positivem ZON-Nachweis. Die AP-Aktivitäten der gesunden Kühe lagen
innerhalb des physiologischen Bereiches und waren signifikant niedriger als die der
Kühe mit LMV. Zwischen den Kühen mit LMV ohne Mykotoxinnachweis und den Kü-
hen mit LMV und positivem Mykotoxinnachweis bestand kein signifikanter Unter-
schied (Abbildung 19).
520132535N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
450
400
350
300
250
200
150
100
500
Abb. 19: AP-Aktivitäten im Serum von Kühen mit LMV sowie mit positivem oder ne-
gativem Mykotoxinnachweis und bei gesunden Kühen (Median, 1.und 3.
Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
AP U/l
a-c; b-c c-d; c-e
a b c d e
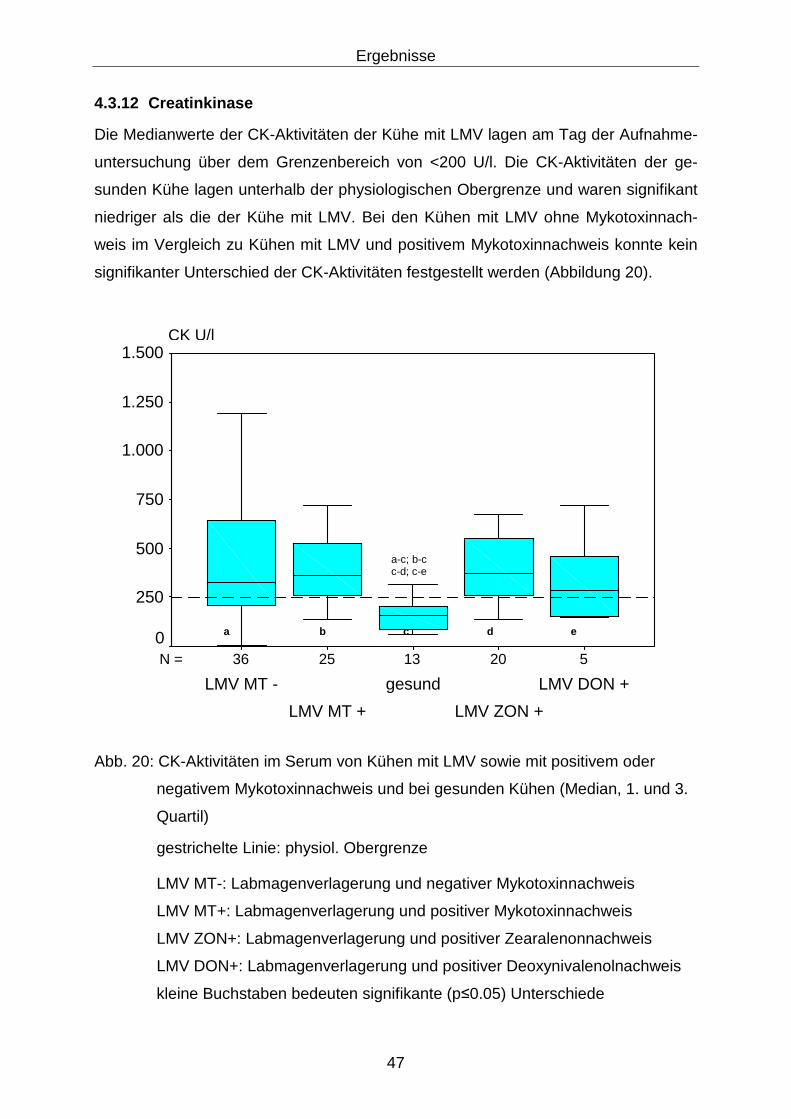
Ergebnisse
47
4.3.12 Creatinkinase
Die Medianwerte der CK-Aktivitäten der Kühe mit LMV lagen am Tag der Aufnahme-
untersuchung über dem Grenzenbereich von <200 U/l. Die CK-Aktivitäten der ge-
sunden Kühe lagen unterhalb der physiologischen Obergrenze und waren signifikant
niedriger als die der Kühe mit LMV. Bei den Kühen mit LMV ohne Mykotoxinnach-
weis im Vergleich zu Kühen mit LMV und positivem Mykotoxinnachweis konnte kein
signifikanter Unterschied der CK-Aktivitäten festgestellt werden (Abbildung 20).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
1.500
1.250
1.000
750
500
250
0
Abb. 20: CK-Aktivitäten im Serum von Kühen mit LMV sowie mit positivem oder
negativem Mykotoxinnachweis und bei gesunden Kühen (Median, 1. und 3.
Quartil)
gestrichelte Linie: physiol. Obergrenze
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
CK U/l
a-c; b-c c-d; c-e
a b c d e
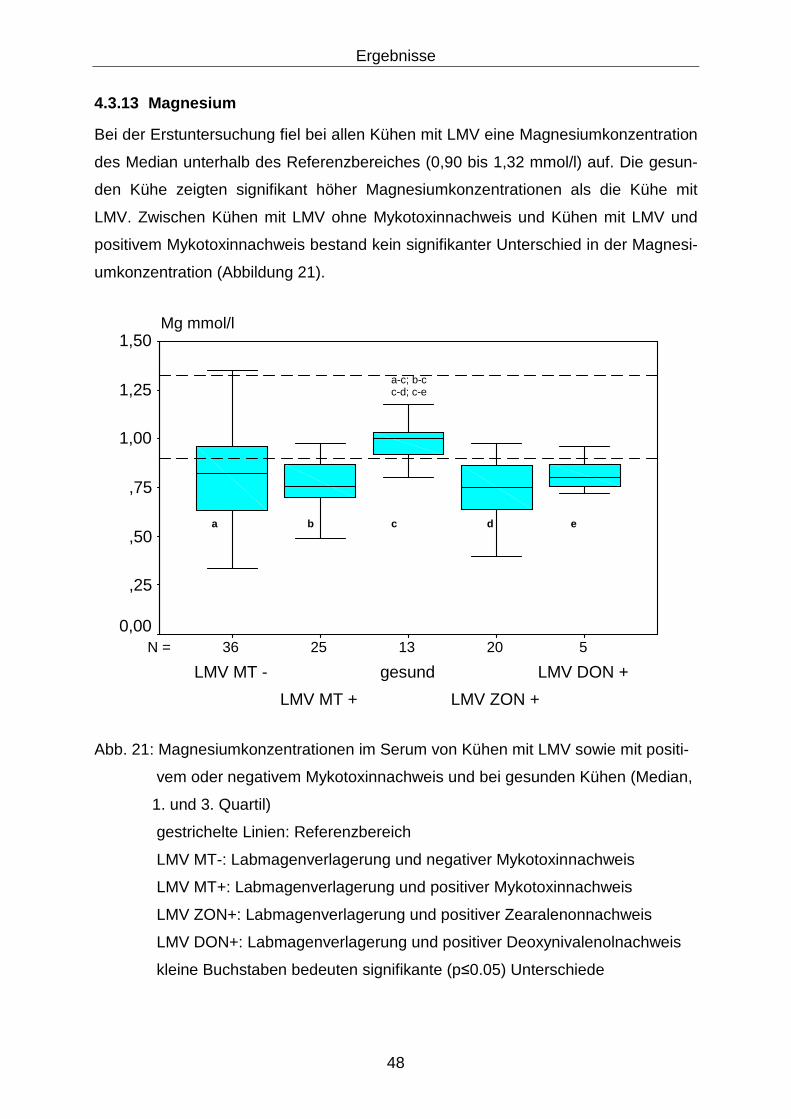
Ergebnisse
48
4.3.13 Magnesium
Bei der Erstuntersuchung fiel bei allen Kühen mit LMV eine Magnesiumkonzentration
des Median unterhalb des Referenzbereiches (0,90 bis 1,32 mmol/l) auf. Die gesun-
den Kühe zeigten signifikant höher Magnesiumkonzentrationen als die Kühe mit
LMV. Zwischen Kühen mit LMV ohne Mykotoxinnachweis und Kühen mit LMV und
positivem Mykotoxinnachweis bestand kein signifikanter Unterschied in der Magnesi-
umkonzentration (Abbildung 21).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
1,50
1,25
1,00
,75
,50
,25
0,00
Abb. 21: Magnesiumkonzentrationen im Serum von Kühen mit LMV sowie mit positi-
vem oder negativem Mykotoxinnachweis und bei gesunden Kühen (Median,
1. und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
Mg mmol/l
a-c; b-c c-d; c-e
a b c d e
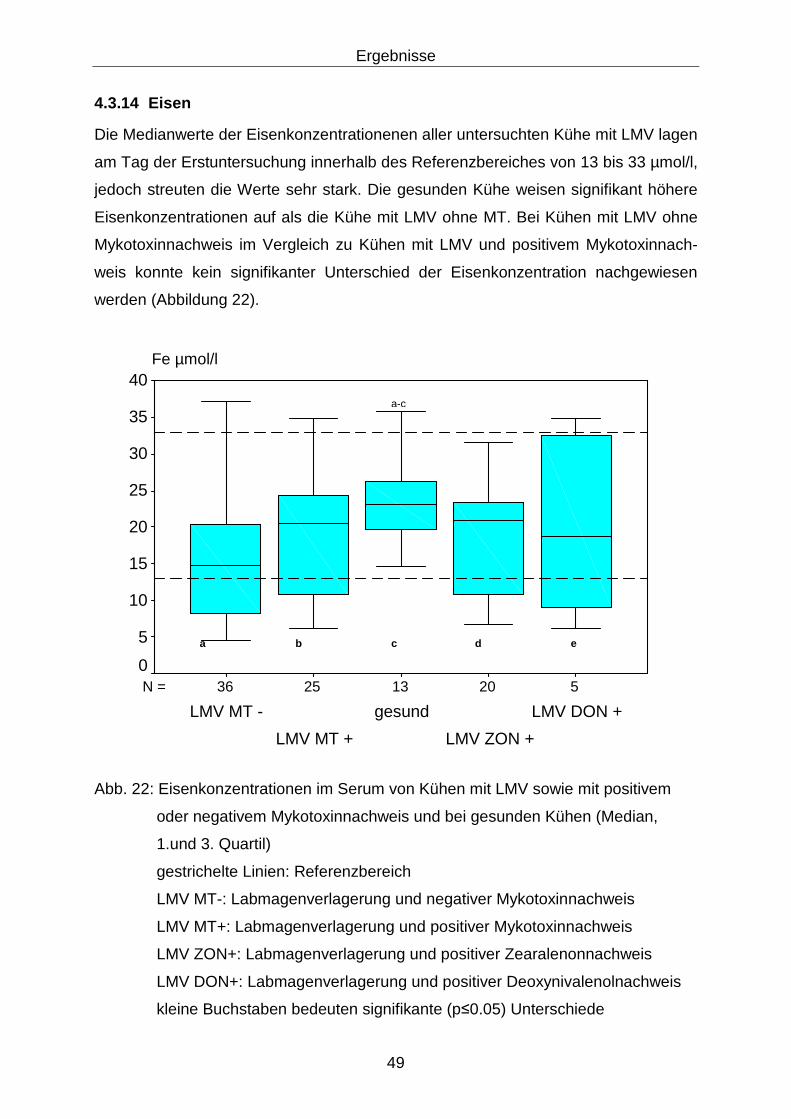
Ergebnisse
49
4.3.14 Eisen
Die Medianwerte der Eisenkonzentrationenen aller untersuchten Kühe mit LMV lagen
am Tag der Erstuntersuchung innerhalb des Referenzbereiches von 13 bis 33 µmol/l,
jedoch streuten die Werte sehr stark. Die gesunden Kühe weisen signifikant höhere
Eisenkonzentrationen auf als die Kühe mit LMV ohne MT. Bei Kühen mit LMV ohne
Mykotoxinnachweis im Vergleich zu Kühen mit LMV und positivem Mykotoxinnach-
weis konnte kein signifikanter Unterschied der Eisenkonzentration nachgewiesen
werden (Abbildung 22).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
40
35
30
25
20
15
10
5
0
Abb. 22: Eisenkonzentrationen im Serum von Kühen mit LMV sowie mit positivem
oder negativem Mykotoxinnachweis und bei gesunden Kühen (Median,
1.und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
Fe µmol/l
a b c d e
a-c
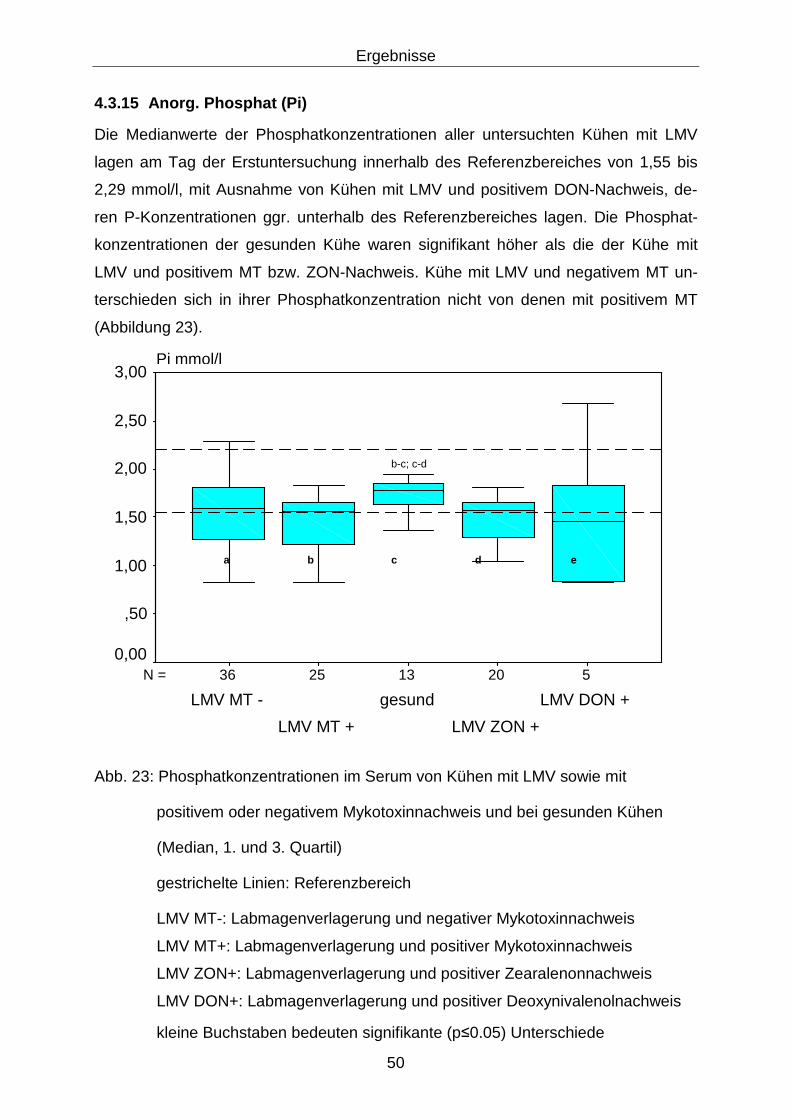
Ergebnisse
50
4.3.15 Anorg. Phosphat (Pi)
Die Medianwerte der Phosphatkonzentrationen aller untersuchten Kühen mit LMV
lagen am Tag der Erstuntersuchung innerhalb des Referenzbereiches von 1,55 bis
2,29 mmol/l, mit Ausnahme von Kühen mit LMV und positivem DON-Nachweis, de-
ren P-Konzentrationen ggr. unterhalb des Referenzbereiches lagen. Die Phosphat-
konzentrationen der gesunden Kühe waren signifikant höher als die der Kühe mit
LMV und positivem MT bzw. ZON-Nachweis. Kühe mit LMV und negativem MT un-
terschieden sich in ihrer Phosphatkonzentration nicht von denen mit positivem MT
(Abbildung 23).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
3,00
2,50
2,00
1,50
1,00
,50
0,00
Abb. 23: Phosphatkonzentrationen im Serum von Kühen mit LMV sowie mit
positivem oder negativem Mykotoxinnachweis und bei gesunden Kühen
(Median, 1. und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
Pi mmol/l
b-c; c-d
a b c d e
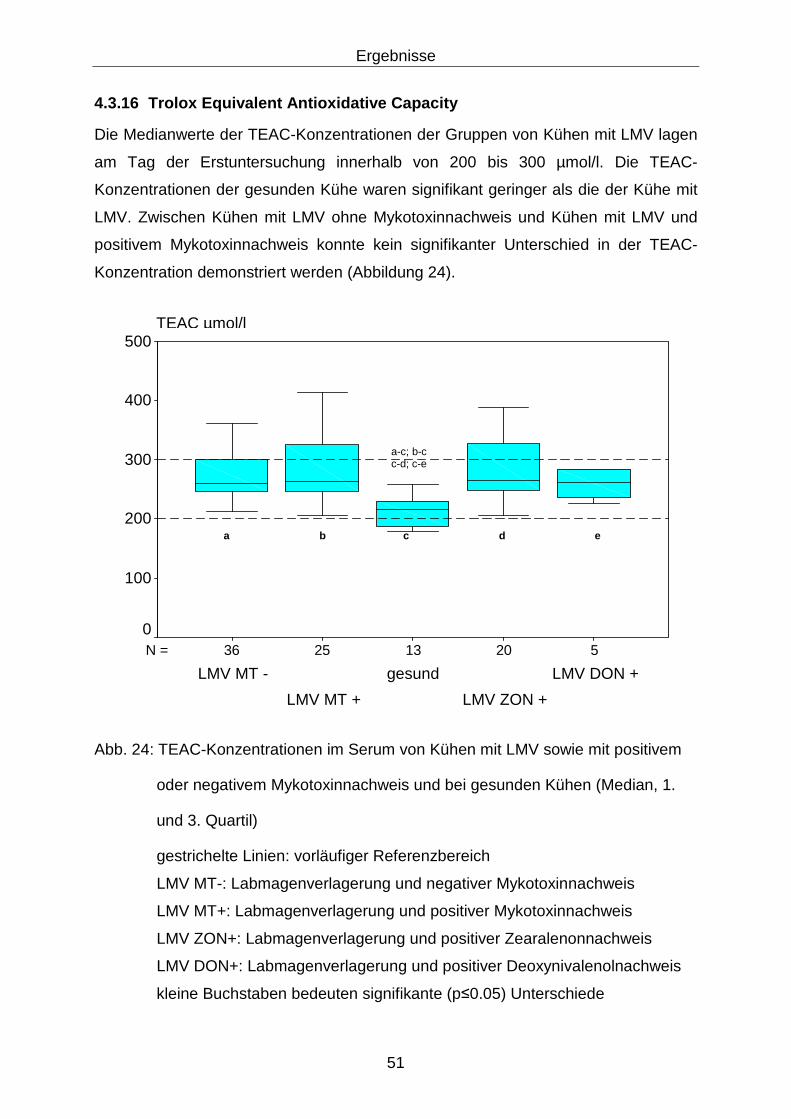
Ergebnisse
51
4.3.16 Trolox Equivalent Antioxidative Capacity
Die Medianwerte der TEAC-Konzentrationen der Gruppen von Kühen mit LMV lagen
am Tag der Erstuntersuchung innerhalb von 200 bis 300 µmol/l. Die TEAC-
Konzentrationen der gesunden Kühe waren signifikant geringer als die der Kühe mit
LMV. Zwischen Kühen mit LMV ohne Mykotoxinnachweis und Kühen mit LMV und
positivem Mykotoxinnachweis konnte kein signifikanter Unterschied in der TEAC-
Konzentration demonstriert werden (Abbildung 24).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
500
400
300
200
100
0
Abb. 24: TEAC-Konzentrationen im Serum von Kühen mit LMV sowie mit positivem
oder negativem Mykotoxinnachweis und bei gesunden Kühen (Median, 1.
und 3. Quartil)
gestrichelte Linien: vorläufiger Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
TEAC µmol/l
a-c; b-c c-d; c-e
a b c d e
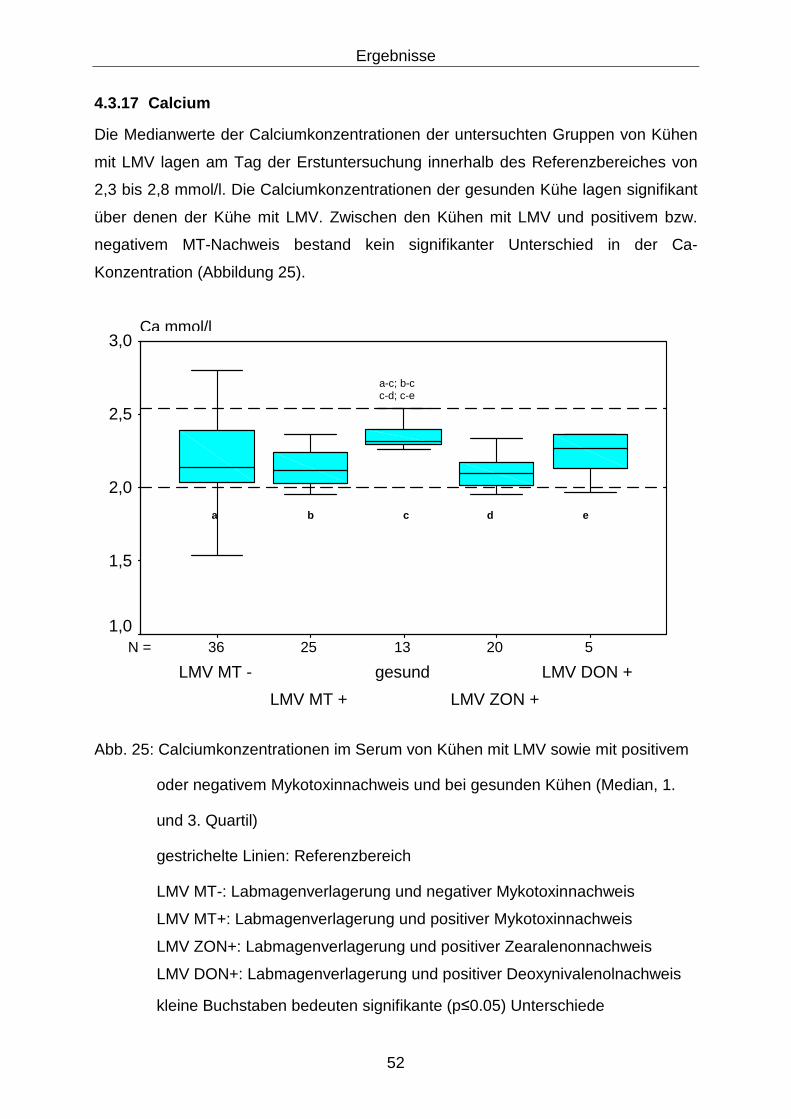
Ergebnisse
52
4.3.17 Calcium
Die Medianwerte der Calciumkonzentrationen der untersuchten Gruppen von Kühen
mit LMV lagen am Tag der Erstuntersuchung innerhalb des Referenzbereiches von
2,3 bis 2,8 mmol/l. Die Calciumkonzentrationen der gesunden Kühe lagen signifikant
über denen der Kühe mit LMV. Zwischen den Kühen mit LMV und positivem bzw.
negativem MT-Nachweis bestand kein signifikanter Unterschied in der Ca-
Konzentration (Abbildung 25).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
3,0
2,5
2,0
1,5
1,0
Abb. 25: Calciumkonzentrationen im Serum von Kühen mit LMV sowie mit positivem
oder negativem Mykotoxinnachweis und bei gesunden Kühen (Median, 1.
und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
Ca mmol/l
a-c; b-c c-d; c-e
a b c d e
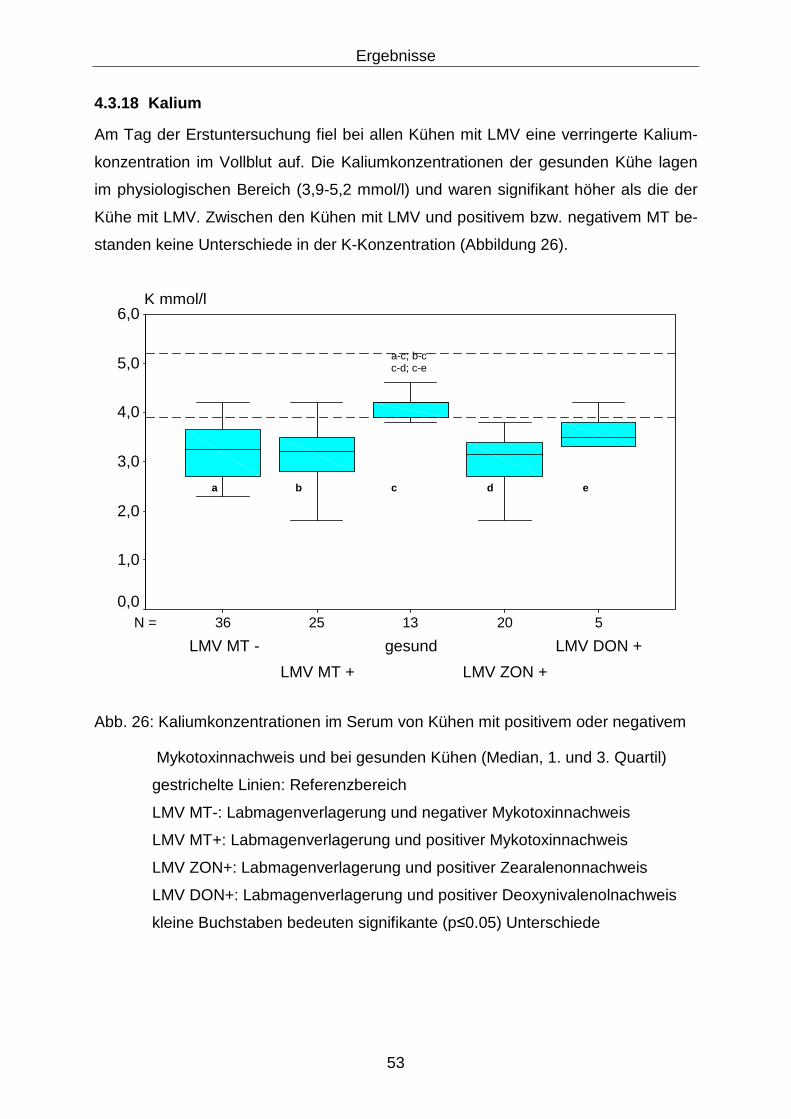
Ergebnisse
53
4.3.18 Kalium
Am Tag der Erstuntersuchung fiel bei allen Kühen mit LMV eine verringerte Kalium-
konzentration im Vollblut auf. Die Kaliumkonzentrationen der gesunden Kühe lagen
im physiologischen Bereich (3,9-5,2 mmol/l) und waren signifikant höher als die der
Kühe mit LMV. Zwischen den Kühen mit LMV und positivem bzw. negativem MT be-
standen keine Unterschiede in der K-Konzentration (Abbildung 26).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
6,0
5,0
4,0
3,0
2,0
1,0
0,0
Abb. 26: Kaliumkonzentrationen im Serum von Kühen mit positivem oder negativem
Mykotoxinnachweis und bei gesunden Kühen (Median, 1. und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
K mmol/l
a-c; b-c c-d; c-e
a b c d e
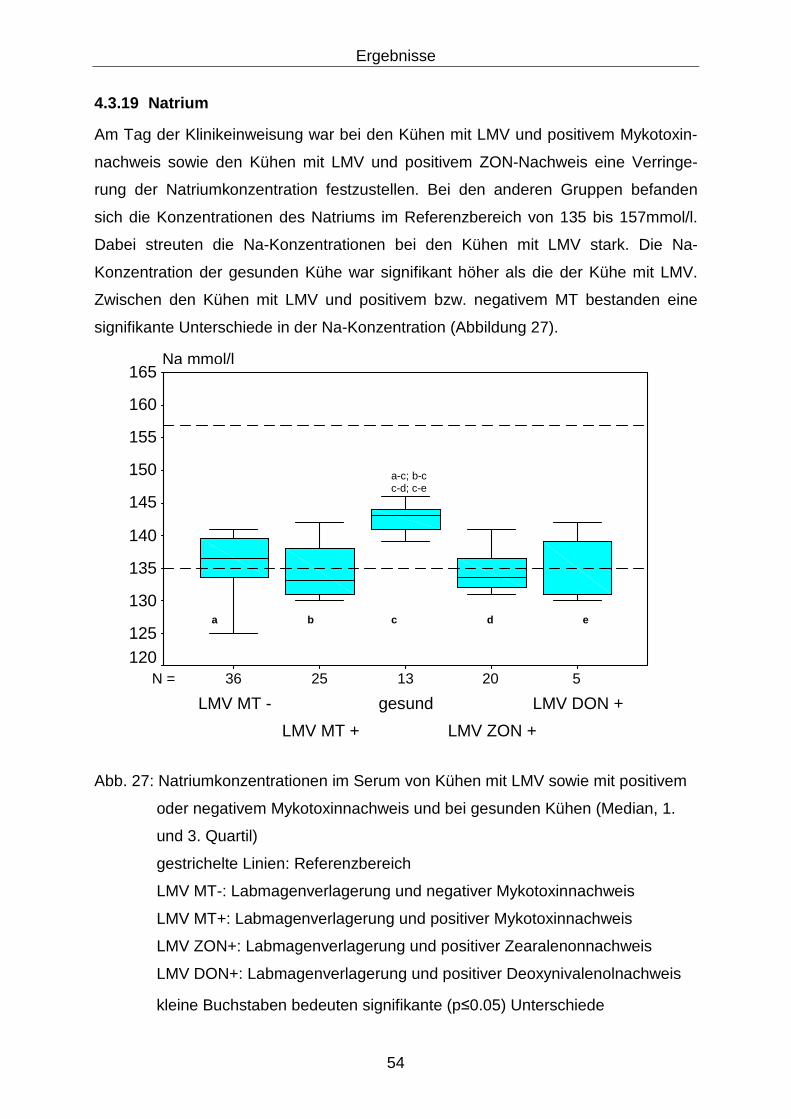
Ergebnisse
54
4.3.19 Natrium
Am Tag der Klinikeinweisung war bei den Kühen mit LMV und positivem Mykotoxin-
nachweis sowie den Kühen mit LMV und positivem ZON-Nachweis eine Verringe-
rung der Natriumkonzentration festzustellen. Bei den anderen Gruppen befanden
sich die Konzentrationen des Natriums im Referenzbereich von 135 bis 157mmol/l.
Dabei streuten die Na-Konzentrationen bei den Kühen mit LMV stark. Die Na-
Konzentration der gesunden Kühe war signifikant höher als die der Kühe mit LMV.
Zwischen den Kühen mit LMV und positivem bzw. negativem MT bestanden eine
signifikante Unterschiede in der Na-Konzentration (Abbildung 27).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
165
160
155
150
145
140
135
130
125120
Abb. 27: Natriumkonzentrationen im Serum von Kühen mit LMV sowie mit positivem
oder negativem Mykotoxinnachweis und bei gesunden Kühen (Median, 1.
und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
Na mmol/l
a-c; b-c c-d; c-e
a b c d e
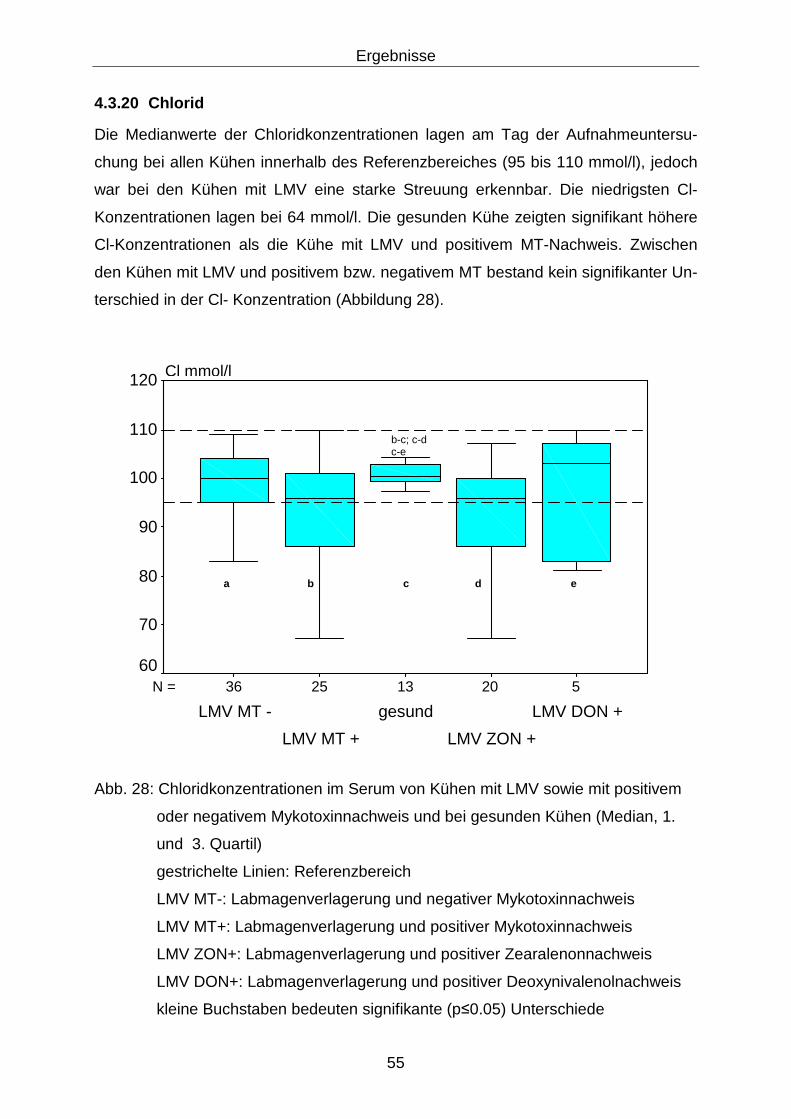
Ergebnisse
55
4.3.20 Chlorid
Die Medianwerte der Chloridkonzentrationen lagen am Tag der Aufnahmeuntersu-
chung bei allen Kühen innerhalb des Referenzbereiches (95 bis 110 mmol/l), jedoch
war bei den Kühen mit LMV eine starke Streuung erkennbar. Die niedrigsten Cl-
Konzentrationen lagen bei 64 mmol/l. Die gesunden Kühe zeigten signifikant höhere
Cl-Konzentrationen als die Kühe mit LMV und positivem MT-Nachweis. Zwischen
den Kühen mit LMV und positivem bzw. negativem MT bestand kein signifikanter Un-
terschied in der Cl- Konzentration (Abbildung 28).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
120
110
100
90
80
70
60
Abb. 28: Chloridkonzentrationen im Serum von Kühen mit LMV sowie mit positivem
oder negativem Mykotoxinnachweis und bei gesunden Kühen (Median, 1.
und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
Cl mmol/l
b-c; c-d c-e
a b c d e
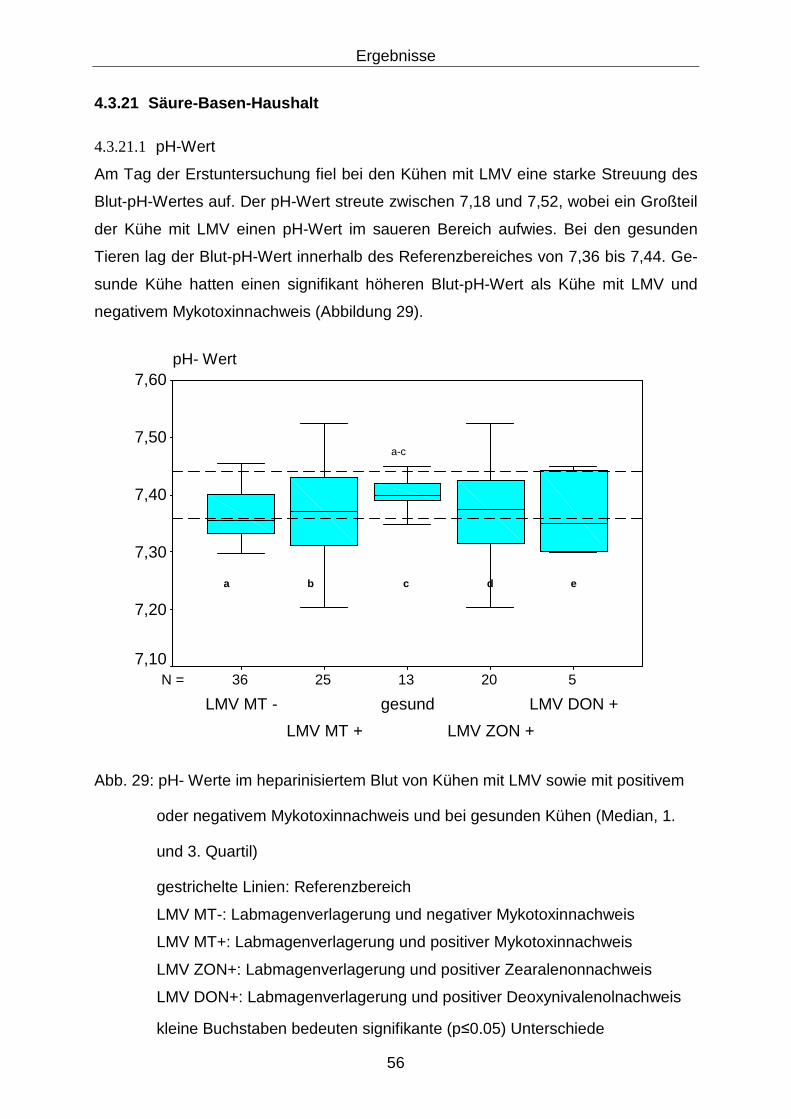
Ergebnisse
56
4.3.21 Säure-Basen-Haushalt
4.3.21.1 pH-Wert
Am Tag der Erstuntersuchung fiel bei den Kühen mit LMV eine starke Streuung des
Blut-pH-Wertes auf. Der pH-Wert streute zwischen 7,18 und 7,52, wobei ein Großteil
der Kühe mit LMV einen pH-Wert im saueren Bereich aufwies. Bei den gesunden
Tieren lag der Blut-pH-Wert innerhalb des Referenzbereiches von 7,36 bis 7,44. Ge-
sunde Kühe hatten einen signifikant höheren Blut-pH-Wert als Kühe mit LMV und
negativem Mykotoxinnachweis (Abbildung 29).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
7,60
7,50
7,40
7,30
7,20
7,10
Abb. 29: pH- Werte im heparinisiertem Blut von Kühen mit LMV sowie mit positivem
oder negativem Mykotoxinnachweis und bei gesunden Kühen (Median, 1.
und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
pH- Wert
a-c
a b c d e
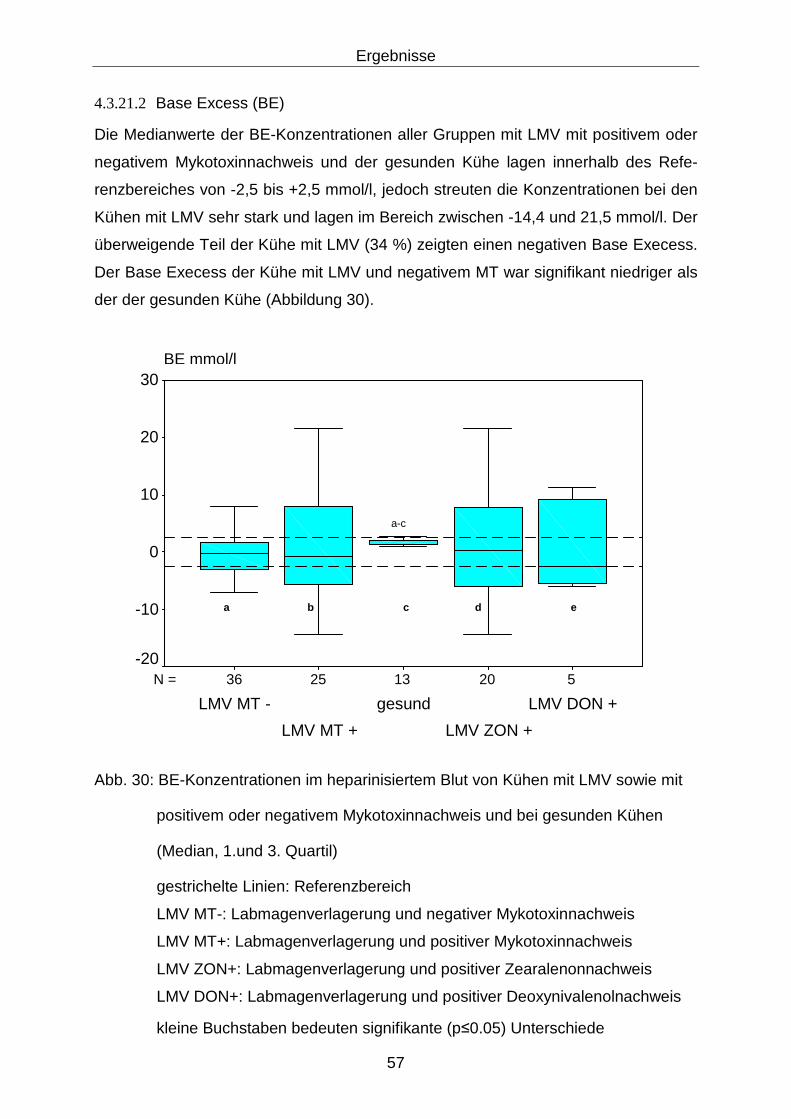
Ergebnisse
57
4.3.21.2 Base Excess (BE)
Die Medianwerte der BE-Konzentrationen aller Gruppen mit LMV mit positivem oder
negativem Mykotoxinnachweis und der gesunden Kühe lagen innerhalb des Refe-
renzbereiches von -2,5 bis +2,5 mmol/l, jedoch streuten die Konzentrationen bei den
Kühen mit LMV sehr stark und lagen im Bereich zwischen -14,4 und 21,5 mmol/l. Der
überweigende Teil der Kühe mit LMV (34 %) zeigten einen negativen Base Execess.
Der Base Execess der Kühe mit LMV und negativem MT war signifikant niedriger als
der der gesunden Kühe (Abbildung 30).
520132536N =
LMV DON +
LMV ZON +
gesund
LMV MT +
LMV MT -
30
20
10
0
-10
-20
Abb. 30: BE-Konzentrationen im heparinisiertem Blut von Kühen mit LMV sowie mit
positivem oder negativem Mykotoxinnachweis und bei gesunden Kühen
(Median, 1.und 3. Quartil)
gestrichelte Linien: Referenzbereich
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
kleine Buchstaben bedeuten signifikante (p≤0.05) Unterschiede
BE mmol/l
a-c
a b c d e

Ergebnisse
58
4.3.22 Ergebnisse der hämatologischen Untersuchunge n
Die Ergebnisse der hämatologischen Untersuchungen sind in Tabelle 11 dargestellt.
Die Zahlen für Leukozyten, Erythrozyten, Hämoglobin, Hämatokrit, Thrombozyten,
Stabkernige Neutrophile, Segmentkernige Neutrophile Granulozyten, Lymphozyten,
Monozyten, Eosinophile und Basophile sind im physiologischen Bereich.
Tab 11: Ergebnisse der Hämatologischen Untersuchung von Kühen mit LMV sowie
mit positivem oder negativem Mykotoxinnachweis und bei gesunden Kühen
(Median, 1. und 3. Quartil)
LMV MT-: Labmagenverlagerung und negativer Mykotoxinnachweis
LMV MT+: Labmagenverlagerung und positiver Mykotoxinnachweis
LMV ZON+: Labmagenverlagerung und positiver Zearalenonnachweis
LMV DON+: Labmagenverlagerung und positiver Deoxynivalenolnachweis
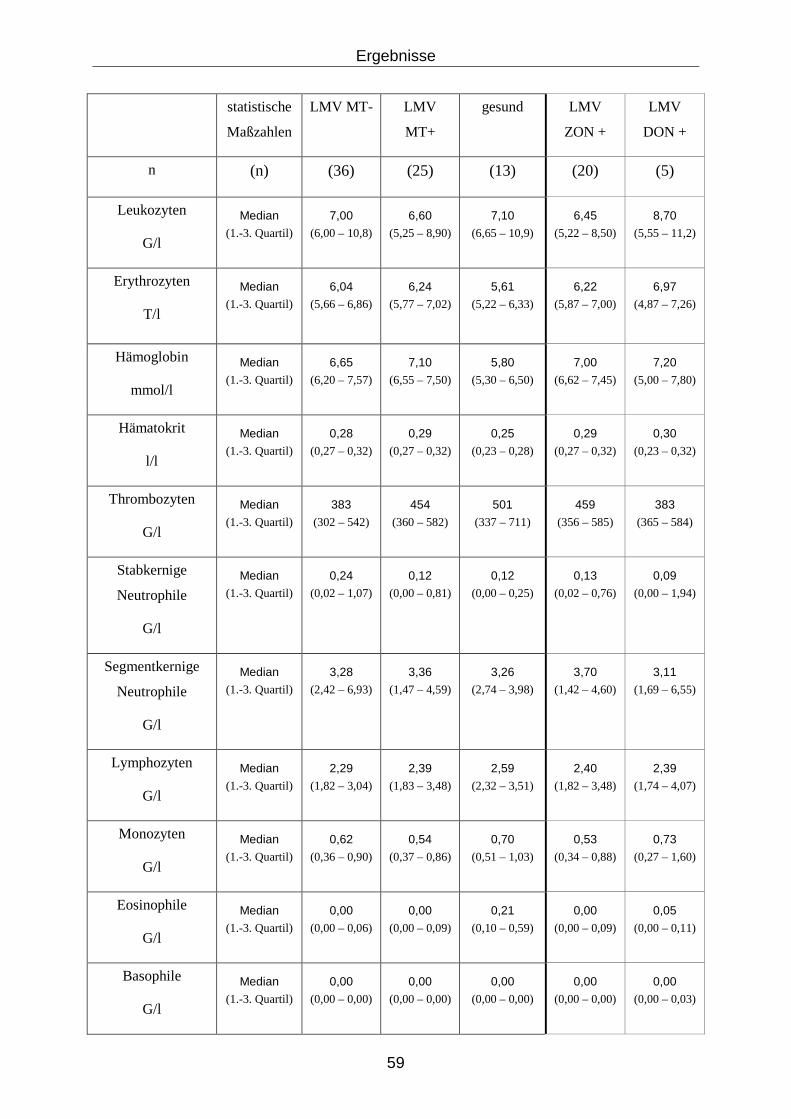
Ergebnisse
59
statistische
Maßzahlen
LMV MT- LMV
MT+
gesund LMV
ZON +
LMV
DON +
n (n) (36) (25) (13) (20) (5)
Leukozyten
G/l
Median
(1.-3. Quartil)
7,00
(6,00 – 10,8)
6,60
(5,25 – 8,90)
7,10
(6,65 – 10,9)
6,45
(5,22 – 8,50)
8,70
(5,55 – 11,2)
Erythrozyten
T/l
Median
(1.-3. Quartil)
6,04
(5,66 – 6,86)
6,24
(5,77 – 7,02)
5,61
(5,22 – 6,33)
6,22
(5,87 – 7,00)
6,97
(4,87 – 7,26)
Hämoglobin
mmol/l
Median
(1.-3. Quartil)
6,65
(6,20 – 7,57)
7,10
(6,55 – 7,50)
5,80
(5,30 – 6,50)
7,00
(6,62 – 7,45)
7,20
(5,00 – 7,80)
Hämatokrit
l/l
Median
(1.-3. Quartil)
0,28
(0,27 – 0,32)
0,29
(0,27 – 0,32)
0,25
(0,23 – 0,28)
0,29
(0,27 – 0,32)
0,30
(0,23 – 0,32)
Thrombozyten
G/l
Median
(1.-3. Quartil)
383
(302 – 542)
454
(360 – 582)
501
(337 – 711)
459
(356 – 585)
383
(365 – 584)
Stabkernige
Neutrophile
G/l
Median
(1.-3. Quartil)
0,24
(0,02 – 1,07)
0,12
(0,00 – 0,81)
0,12
(0,00 – 0,25)
0,13
(0,02 – 0,76)
0,09
(0,00 – 1,94)
Segmentkernige
Neutrophile
G/l
Median
(1.-3. Quartil)
3,28
(2,42 – 6,93)
3,36
(1,47 – 4,59)
3,26
(2,74 – 3,98)
3,70
(1,42 – 4,60)
3,11
(1,69 – 6,55)
Lymphozyten
G/l
Median
(1.-3. Quartil)
2,29
(1,82 – 3,04)
2,39
(1,83 – 3,48)
2,59
(2,32 – 3,51)
2,40
(1,82 – 3,48)
2,39
(1,74 – 4,07)
Monozyten
G/l
Median
(1.-3. Quartil)
0,62
(0,36 – 0,90)
0,54
(0,37 – 0,86)
0,70
(0,51 – 1,03)
0,53
(0,34 – 0,88)
0,73
(0,27 – 1,60)
Eosinophile
G/l
Median
(1.-3. Quartil)
0,00
(0,00 – 0,06)
0,00
(0,00 – 0,09)
0,21
(0,10 – 0,59)
0,00
(0,00 – 0,09)
0,05
(0,00 – 0,11)
Basophile
G/l
Median
(1.-3. Quartil)
0,00
(0,00 – 0,00)
0,00
(0,00 – 0,00)
0,00
(0,00 – 0,00)
0,00
(0,00 – 0,00)
0,00
(0,00 – 0,03)

Diskussion
60
5 Diskussion
5.1 Ergebnisse der Mykotoxinanalysen
5.1.1 Patienten
Ziel dieser Arbeit war es, von klinisch gesunden Kühen und Kühen mit Labmagenver-
lagerung Futter, Blut, Milch und Gallensaft auf die Rückstände von DON, ZON sowie
deren Metaboliten zu untersuchen und ihre Beziehungen zur Tiergesundheit zu
überprüfen.
Zur Untersuchung von Futtermitteln auf Mykotoxinbelastung stehen zum einen che-
misch-physikalische Methoden, wie die Dünnschicht-Chromatographie oder die
Hochleistungsflüssigkeitschromatographie (HPLC) sowie immunologische Methoden,
wie der Enzymimmunoessay (ELISA), zur Verfügung (BAUER et al. 2000). Der ELI-
SA ist vor allem im Rahmen von Screeninguntersuchungen bei stark kontaminierten
Einzelfuttermitteln sinnvoll und liefert ein qualitatives, jedoch nur eingeschränkt quan-
titatives Ergebnis (SATOR u. SOMMER 2003; SIEVERERDING 2003). Im Vergleich
zum ELISA ist die HPLC ein sehr kosten- und zeitaufwendiges Verfahren mit hohem
apparativem Aufwand, das jedoch eine höhere Sensitivität und Spezifität aufweist
(BAUER et al. 2000; PAPADOPOULOU-BOURAOUI et al. 2004). Mit dieser Methode
können DON wie auch ZON und deren Derivate bestimmt werden (KOCH 2004;
ZHENG et al. 2005; KRSKA et al. 2008). Überdies ermöglicht sie eine genaue Quan-
tifizierung des Mykotoxingehaltes einer untersuchten Probe. Die HPLC gilt als Refe-
renzverfahren. Proben, die im ELISA-Test positiv sind, sollten zur Ermittlung des ge-
nauen Toxingehaltes mittels HPLC-Verfahrens nachuntersucht werden (SATOR u.
SOMMER 2003). Für die vorliegende Untersuchung wurde daher die HPLC als Ana-
lysemethode gewählt.
Die in der vorliegenden Untersuchung ermittelten DON- und ZON-Konzentrationen in
einer Teilstichprobe des Futters von 20 Patienten (Tabelle 7) liegen unter den
Grenzwerten (EU 2006) und entsprechen einer durchschnittlichen Kontamination,
wie sie regelmäßig nachgewiesen werden kann. In der Regel liegen die Gehalte in
einer Größenordnung von Mikro- bis Milligramm pro Kilogramm (BAUER et al. 2000).
Daher kann anhand der Grenzwerte, die nach dem derzeitigen Kenntnisstand fest-

Diskussion
61
gesetzt wurden, nicht von einer übermäßigen Belastung der Tiere durch das Futter
ausgegangen werden.
Für die Gewinnung von Gallensaft am lebenden Tier bietet sich die Möglichkeit der
Probennahme durch Punktion der Gallenblase während einer Laparotomie mit Lab-
magenreposition an. Durch die transkutane Punktion unter Ultraschallkontrolle lässt
sich ebenfalls Gallensaft gewinnen, insbesondere wenn die Gallenblase leicht ge-
staut ist (BRAUN u. GERBER 1992). Bei gesunden Kühen ist das jedoch selten der
Fall, was die Punktion der Gallenblase mitunter erschwert. Bei kranken Kühen mit
Störungen im Gastrointestinaltrakt, wie der Labmagenverlagerung, hingegen ist sie
infolge der Gallenstauung bei reduzierter Verdauungstätigkeit meist umfangsver-
mehrt und somit gut darstellbar und punktierbar (DELLING 2000).
Wie in den Tabellen 8 und 9 dargestellt, gelang der Mykotoxinnachweis am häufigs-
ten in der Gallenflüssigkeit, während im Serum nur bei 8 % der Tiere Mykotoxine
nachweisbar waren. In der Milch wurden keine Mykotoxine nachgewiesen. Im Allge-
meinen ist die Ausscheidung der Mykotoxine über die Milch ohnehin gering, selbst
wenn Veränderungen in der Blut-Milch- Barriere durch systemische oder vor allem
lokale Infektionen (Mastitis) vorhanden sind (FINK-GREMMELS 2008a). COTE
(1986) berichtete, dass bei einer Dosis von 66 mg DOM/kg Futter über 5 Tage 26
ng/ml DOM-1 in der Milch nachgewiesen werden konnten. Dies stimmt mit der Be-
schreibung von KEESE et al. (2008) überein. KEESE et al. (2008) konnten mit einer
Belastung von 5,3 mg DON/kg TS über einen Zeitraum von 11 Wochen nur de-epoxy
DON (≤ 3,2 µg/kg) in der Milch nachweisen. Bei Dosierungen von 50 sowie 165 mg
ZON/kg Futter über 21 Tage konnten ZON und seine Derivate α- und ß-ZOL in der
Milch nachgewiesen werden (PRELUSKY et al. 1990; GAREIS u. WOLFF 2000).
Dies stimmt mit der Beschreibung von SEELING (2005) überein.
In den hier aufgeführten Quellen lagen die aufgenommenen Mykotoxinmengen um
ein Vielfaches über denen, die von den in der vorliegenden Untersuchung untersuch-
ten Rindern über das Futter hätten aufgenommen werden können. Als weiterer Fak-
tor kommt hinzu, dass Mykotoxine ungleichmäßig im Futter verteilt sein können und
somit die Aufnahme je nach verfütterter Partie noch geringer sein kann. Dies kann
von BAUER et al. (2000) bestätigt werden. Ähnliche Resultate ergaben die Ergebnis-
se der Arbeitsgruppen um TRENHOLM (1985), COTE (1986), CHARMLEY (1993),
DÄNICKE et al. (2002, 2005) sowie KOROSTELEVA et al. (2007). Überdies zeigten
die Patienten, wie aus den reduzierten Pansenbewegungen (Abbildung 7) geschlos-

Diskussion
62
sen werden kann, ohnehin eine reduzierte Futteraufnahme, bei der davon auszuge-
hen ist, dass sie auf Grund der Erkrankung bereits im Herkunftsbetrieb bestanden
hat.
Manche Mykotoxine, wie ZON, reichern sich im Körper und besonders auch im Gal-
lensaft an. Der Vorteil einer Gallensaftuntersuchung ist, dass man die Belastung über
einen längeren Zeitraum zurück feststellen kann. DON hingegen unterliegt im Körper
einer schnellen Umwandlung (PING et al. 1992; DÄNICKE et al. 2005; SEELING et
al. 2006b; FINK-GREMMELS 2008b) und ist deshalb für einen Nachweis in der Galle
weniger gut geeignet als ZON. Zu beachten ist, dass durch die Ein- und Auslagerung
von Mykotoxinen in Fettdepots und durch die Eindickung des Gallensaftes einige
Stunden nach der Futteraufnahme die Mykotoxingehalte in der Galle stark schwan-
ken (BAUER et al. 2000). Wie aus Tabelle 9 ersichtlich, war in der vorliegenden Un-
tersuchung ZON öfter als DON, DOM-1, α-ZOL sowie ß-ZOL nachweisbar, jedoch
lagen selbst hier die Konzentrationen an der Nachweisgrenze (Abschnitt 3.2.6.1 und
3.2.6.2) (UEBERSCHÄR 1999; DÄNICKE et al. 2001; VALENTA et al. 2003). Über-
dies ist die DON-Konzentration im Gallensaft von der DON-Aufnahme abhängig.
Dies stimmt mit den Beschreibungen anderer Autoren (SEELING et al. 2006b; KEE-
SE et al. 2008) überein.
In Serum war de-epoxy DON öfter als DON nachweisbar jedoch lagen die Konzent-
rationen wiederum an der Nachweisgrenze (Tabelle 9). Im Blut beeinflusst der Zeit-
abstand nach der letzten Futteraufnahme das Ergebnis, weil einige Mykotoxine im
Körper einer schnellen Umwandlung unterliegen (DÄNICKE et al. 2005; SEELING et
al. 2006b; FINK-GREMMELS 2008b).
Somit sind der seltene Mykotoxinnachweis in den untersuchten Medien und die nied-
rigen nachgewiesenen Konzentrationen in der vorliegenden Untersuchung auf die
durch verschiedene Faktoren begründete geringe Mykotoxinaufnahme zurückzufüh-
ren.
5.1.2 Gesunde Kühe
Auch in den beiden Betrieben, aus denen die Kontrolltiere stammten, wurden DON
und ZON im Futter nachgewiesen (Tabelle 10). Auch hier sind die Konzentrationen
sehr gering und liegen unter den Grenzwerten (EU 2006). Die Konzentrationen in
den Futtermitteln sind zwar geringer als der Median der Patientenbetriebe (Tabelle
7), dennoch kommen auch in diesen Betrieben Labmagenverlagerungen vor. Die

Diskussion
63
Tatsache, dass bei keiner der gesunden Tiere in den drei Körperflüssigkeiten die un-
tersuchten Mykotoxine nachgewiesen wurden, könnte als Hinweis darauf gewertet
werden, dass die Anreicherung von Mykotoxinen im Körper mit der Labmagenverla-
gerung in Verbindung steht. Jedoch sind in den Kontrollbetrieben die Tiere mit Lab-
magenverlagerung nicht untersucht worden, so dass eine solche Schlussfolgerung
nicht überprüft werden kann.
5.2 Klinische Untersuchungsbefunde und Krankheitsau sgang
Der Krankheitsausgang von 61 Patienten mit LMV und positivem sowie negativem
Mykotoxinnachweis ist in Abbildung 8 dargestellt. Sowohl bei den klinischen Sym-
ptomen (Abbildung 6), wie auch beim klinischen Ausgang gab es keine gesicherten
Unterschiede zwischen mykotoxinbelasteten und –unbelasteten Kühen. Die Kühe mit
Exitus letalis zeigten schwere Begleiterkrankungen, wie Peritonitis oder hochgradige
Endometritiden mit hochgradigen hämatologischen sowie klinisch-chemischen Ab-
weichungen im Blut, wie sie für Labmagenverlagerungen typisch sind (SCHREIBER
2006). Mykotoxine konnten somit in dieser Untersuchung weder für spezifische Sym-
ptome, noch für den Krankheitsausgang, d.h. Heilung oder Exitus letalis, verantwort-
lich gemacht werden. Dies wurde ebenfalls von PRELUSKY et al. (1990),
HOCHSTEINER et al. (2000), KOROSTELEVA et al. (2007) beobachtet. Für die Er-
klärung muss bedacht werden, dass die DON- und ZON-Belastungen im Futter ge-
ring waren (Tabelle 7) und die Aufnahme durch weitere Faktoren (Abschnitt 5.1.1)
vermindert wurde.
Die gering- bis mittelgradig reduzierte Futteraufnahme in den vorliegenden Ergebnis-
sen steht so wahrscheinlich in Zusammenhang mit der Allgemeinerkrankung und der
Verdauungsstörung in Form einer LMV. Eine reduzierte Futteraufnahme kann auch
eine Folge von Mykotoxinbelastung sein (PRELUSKY et al. 1990; SCHUH 1996;
DUARTE 2003). In den vorliegenden Ergebnissen ist dies jedoch nicht der Fall, weil
die Kühe, mit oder ohne positiven Mykotoxinnachweis, nach Reposition des verlager-
ten Labmagens eine verbesserte Futteraufnahme gezeigt haben (Abschnitt 4.2).
Es wurde bei 52 % der Kühe mit LMV und positivem Mykotoxinnachweis sowie 50 %
der Patienten mit LMV und negativem Mykotoxinnachweis eine Klauenrehe festge-
stellt.
Hauptursachen der Klauenrehe in der Milchviehhaltung sind Fütterungsfehler und
überhöhte mechanische Belastung. Infektionsbedingte Vergiftungserscheinungen

Diskussion
64
(Endotoxine) und Nachwirkungen von Stoffwechselstörungen sind weitere krank-
heitsauslösende Faktoren. Aus Sicht der Fütterung sind vor allem ein hoher Rations-
anteil leichtfermentierbarer Kohlenhydrate und eine damit verbundene azidotische
Stoffwechsellage problematisch. Hohe Säurebildung und fehlende Puffersubstanz
führen zu einem sinkenden pH-Wert im Pansen. Nachfolgend steigt die Milchsäure-
produktion an. Dadurch werden Pansenmikroben und Pansenwand angegriffen und
Endotoxine freigesetzt. Histamine und andere Gefäß verändernde Substanzen ge-
langen nachfolgend verstärkt in das Gefäßsystem und verursachen eine entzündli-
che Veränderung der Lederhaut (WEAVER 1997; KOFLER 2001). Ebenfalls Leber
schädigend wirkt sich eine hohe ruminale Stickstoffbilanz aus. Diese kann vor allem
in Betrieben mit hohem Gras- bzw. Leguminosenanteil in der Futterration als Ursa-
che der Klauenrehe ausgemacht werden. Infektionen wie z.B. Mastitis oder Metritis
können direkt durch die Freisetzung von Giftstoffen (KOFLER 2001) oder indirekt
durch eine mangelhafte Rauhfutteraufnahme und selektives Fressen der Kuh eine
Klauenrehe auslösen. Aus Sicht der Haltung sind vor allem eine zu geringe Durch-
blutung der Klauenlederhaut auf Grund von Bewegungsmangel und Mängel in der
Ausführung der Laufflächen als die Klauenrehe begünstigende Faktoren anzusehen
(NOCEK 1997). Verstärkte Lipolyse und Endotoxin-Freisetzung bzw. eine verminder-
te Endotoxin- Clearance können auch eine Ursache sein (FÜRLL 2000).
Einen weiteren Einfluss auf die Klauenrehe können auch indirekt Mykotoxine haben.
Diese bewirken eine Schädigung der Mikrobenflora des Pansens.
Am Tag der Aufnahmeuntersuchung wurde bei 40 % der Kühe mit LMV und positi-
vem Mykotoxinnachweis sowie bei 39 % der Kühe mit LMV und negativem Mykoto-
xinnachweis eine Endometritis beobachtet. Dieser relativ hohe Anteil an Kühen, die
eine Endometritis hatten, könnte durch den verringerten Rohfaser-Gehalt in der Fut-
terration bedingt sein, der aufgrund einer azidotischen Belastung zu einer verminder-
ten Abwehrleistung des Tieres führt (HOOPS 2007). In den vorliegenden Ergebnis-
sen wurde nach lokaler sowie systemischer antibiotischer Behandlung der Endo-
metritis eine deutliche Besserung der Gebärmutterentzündung beobachtet.
Bei der Aufnahmeuntersuchung wurde bei 16 % der Patienten mit LMV und positi-
vem Mykotoxinnachweis und bei 14 % der Patienten mit LMV und negativem Myko-
toxinnachweis eine Mastitis festgestellt. Der verminderte Anteil der strukturwirksa-
men Rohfaser und eine Erhöhung des Anteils an Kraftfutter in der Futterration im
Sommer führt zu einer verminderten Wiederkauaktivität und damit zu einer verringer-

Diskussion
65
ten Speichelproduktion. Durch die geringere Pufferkapazität im Pansen besteht das
Risiko einer subklinischen Pansenazidose (ENEMARK et al. 2002). Die azidotische
Belastung bedingt dann eine verminderte Futteraufnahme und eine ketotische Stoff-
wechsellage. Durch die azidotische Stoffwechsellage wird besonders die Phagozyto-
seaktivität der Leukozyten im Blut sowie der Alveolarmakrophagen im Euter vermin-
dert (LACHMANN 1981). Dies führt zu einer verminderten Abwehrleistung des Kör-
pers bzw. Eutergewebes, was wiederum die Entstehung einer Mastitis begünstigt.
Zusätzlich kann sich die Belastung der Kühe durch hohe Umgebungstemperaturen
im Sommer mit Werten bis zu 35 °C negativ auf die F utteraufnahme ausgewirkt ha-
ben (SHALIT et al. 1991; FÜRLL 2002a). LOTTHAMMER et al. (1988) beobachteten
auch einen ungünstigen Einfluss von azidotischen Zuständen auf die Eutergesund-
heit. In den eigenen Untersuchungen ist es jedoch nicht der Fall, dass die Mykotoxi-
ne eine Rolle spielen, weil die Kühe, sowohl mit als auch ohne Mykotoxinnachweis,
nach Behandlung des Euters eine Verbesserung gezeigt haben.
Bei 7 % der Patienten mit LMV und positivem Mykotoxinnachweis und 5 % der Pati-
enten mit LMV und negativem Mykotoxinnachweis wurde eine Diarrhoe festgestellt.
Die häufigste Ursache für eine Diarrhoe sind die Fehler in der Fütterung (kaltes oder
schwer verdauliches Futter, plötzlicher Futterwechsel, unregelmäßige Fütterung,
Überfütterung u.a.).
Weitere Ursachen sind Überanstrengung, Fremdkörper, bestimmte Giftstoffe, Auf-
nahme von speziell auf den Magendarmtrakt wirkende Krankheitserreger wie Darm-
parasiten, bestimmte Vieren, Darmbakterien bzw. Bakterientoxine und Pilze bzw.
Mykotoxine (BAUMGARTNER 2000).
Hier ist es jedoch eher unwahrscheinlich, dass die alle Kühe nach der Reposition und
Behandlung der Labmagenverlagerung physiologischen Kotabsatz zeigten.
Die Anzahl der Pansenbewegungen gilt als indirekter Marker für die Futteraufnahme
und die Verdauungsfunktion. Bei Kühen mit Labmagenverlagerung wird häufig eine
Pansenatonie diagnostiziert, wobei die Angaben zwischen 46 % (SATTLER u.
FÜRLL 2004), 25 % (MÜLLER 1992) und 40 % der Tiere mit LMV (DIRKSEN 1961)
variieren. Andere Quellen verweisen auch auf die puerperale Septikämie (FÜRLL u.
FÜRLL 2006) sowie Endotoxämie (EADES 1997), die zu Pansenatonie führen kön-
nen. In den vorliegenden Untersuchungen wurde bei 88 % der Patienten mit LMV
und positivem Mykotoxinnachweis und bei 67 % der Patienten mit LMV und negati-
vem Mykotoxinnachweis im Vergleich zu gesunden Kühen eine signifikant niedrigere

Diskussion
66
Anzahl an Pansenbewegungen festgestellt (Abbildung 7). Kühe mit LMV und negati-
vem Mykotoxinnachweis hatten des Weiteren eine signifikant höhere Anzahl an Pan-
senbewegungen als die mit positivem Mykotoxinnachweis (Abbildung 7). Ähnliche
Resultate zeigten Arbeiten von MANSFELD (1989) sowie SCHUH und BAUM-
GARTNER (1988). Auf der einen Seite erklärt sich der relativ hohe Anteil von Kühen
mit Pansenatonie durch die massiv ausgeprägten Energiestoffwechselstörungen mit
hepatischer Lipidose und zusätzlichen Puerperalstörungen in Form einer Endometri-
tis. Laut den Angaben von REHAGE et al. (1996) leiden etwa 50 % der an einer
Labmagenverlagerung erkrankten Kühe zusätzlich an einer Begleiterkrankung, wobei
die Endometritis an erster Stelle steht. Auf der anderen Seite wird berichtet, dass der
Abbau von Mykotoxinen eine Pansenatonie bedingen kann. Die Pansenflora stellt für
die Toxine der Schimmelpilze eine entgiftende Barriere dar (DÄNICKE et al. 2005;
FINK-GREMMELS 2008a, 2008b). Dabei werden die genannten Mykotoxine insbe-
sondere durch Infusorien zu weniger giftigen Stoffwechselprodukten umgebaut
(KING et al. 1984; SWANSON et al. 1987; BAUER 2005; WILKENS 2005). DON und
ZON können die Stoffwechselleistungen der Pansenbakterien und –protozoen
nachteilig beeinflussen und somit den Vormagenstoffwechsel der Kühe beeinträchti-
gen (DUARTE 2003, FINK-GREMMELS 2008b). In den vorliegenden Untersuchun-
gen erklärt sich die reduzierte Pansenbewegung durch Verdauungsstörungen in
Form von LMV und nicht durch die Mykotoxinbelastung, weil nach Reposition des
Labmagens alle Kühe eine gute Pansenmotorik entwickelten (Abschnitt 4.2).
Zearalenon und seine Derivate verursachen durch ihre östrogene Wirkung haupt-
sächlich Fruchtbarkeitsprobleme bei Zuchttieren. Bei Aufnahme von ZON wiesen
Milchkühe, die mit fusarienkontaminiertem Getreide gefüttert wurden, Vaginitis, ver-
längerte Brunst, Aborte und blutige Diarrhöe auf (ZEPAUER et al. 1985; MANSFELD
et al. 1989; DROCHNER 1990). In den vorliegenden Ergebnissen bei Patienten mit
positivem Nachweis von ZON und seinen Metaboliten α- und ß-ZOL konnten keine
entsprechenden Veränderungen am Genitaltrakt festgestellt werden (Abschnitt 4.2).
Dies spricht dafür, dass hier die DON- und ZON-Konzentrationen zu gering im Futter
waren. Diese Ergebnisse entsprechen denen von WEAVER et al. (1986b),
HOCHSTEINER et al. (2000), DÄNICKE et al. (2002) und KLEINOVA (2002).
Als Ursache für die geringen oder nicht nachweisbaren Beziehungen zwischen My-
kotoxinrückständen in den Futterproben und den hier erhobenen klinischen Befunden
ist einerseits zu berücksichtigen, dass die DON- und ZON-Konzentrationen in den

Diskussion
67
Futterproben sehr gering waren (Tabellen 7 und 10), andererseits, dass die Mykoto-
xine DON und ZON durch die Pansenmikroorganismen entgiftet werden (ROTTER et
al. 1996; VALENTA u. VEMMER 1996; DÄNICKE et al. 2005). Zur Darstellung von
Zusammenhängen eignet sich am besten Gallensaft für den Nachweis von ZON, α-
ZOL sowie ß-ZOL. Dies stimmt mit Untersuchungen von KLEINOVA et al. (2002),
BAUER (2002), DÄNICKE et al. (2002), MEYER et al. (2002) überein.
Mykotoxine können indirekt zu einer LMV bei Kühen beitragen. Aufgrund der
schlechten Futterqualität kann ein Energiedefizit entstehen. Durch eine hohe Milch-
einsatzleistung mit einem entsprechenden Energiedefizit mobilisieren die Kühe Kör-
perfett aus den Depots. Ein unzureichender Energiegehalt in der Ration verschärft
die Situation und fehlende Futterstruktur wirkt zusätzlich krankheitsauslösend für
LMV (DIRKSEN 1961; FÜRLL et al. 2000). Abschließend kann in diesen Untersu-
chungen in Übereinstimmung mit BÖHM (1992), SABATER et al. (2004), SEELING
et al. (2006b) sowie KEESE et al. (2008, 2008a, 2008b) bestätigt werden, dass die
DON- und ZON-Belastungen in geringen Konzentrationen beim Milchrind keine be-
sondere Gefahrenquelle darstellen, da diese Mykotoxine von der Pansenflora offen-
sichtlich metabolisiert werden, wenn die Haltungs- und Fütterungsbedingungen opti-
mal gestaltet werden.
5.3 Klinisch-chemische Parameter
In den vorliegenden Ergebnissen waren in den klinisch-chemischen Parametern kei-
ne signifikanten Unterschiede zwischen Kühen mit LMV mit oder ohne Mykotoxin-
nachweis festzustellen. Mögliche Erklärungen dafür sind die geringe Mykotoxinbe-
lastung in den Futterproben sowie die Metabolisierung der Mykotoxine im Pansen.
5.3.1 Energiestoffwechsel
5.3.1.1 Beta-Hydroxybutyrat-Konzentrationen
Die Medianwerte der BHB-Konzentrationen befanden sich in den Gruppen der Kühe
mit LMV und negativem Mykotoxinnachweis, der Kühe mit LMV und positivem DON-
Nachweis und der gesunden Kühe innerhalb des Grenzwertes von <0,62 mmol/l
(FÜRLL 2005b). Bei Kühen mit LMV und positivem Mykotoxinnachweis und bei Kü-
hen mit LMV und positivem ZON-Nachweis lagen sie oberhalb des Grenzwertes
(Abbildung 14). Zwischen allen Gruppen war kein signifikanter Unterschied der BHB-
Konzentrationen nachzuweisen. Die erhöhten BHB-Konzentrationen sind auf die

Diskussion
68
Energiemangelsituation infolge Geburtsbelastung und einsetzender Milchleistung
zurückzuführen (TEUFEL 1999). Auch CITIL (1999) beschreibt eine erhöhte BHB-
Konzentration bei Kühen mit LMV. Verringerte Futteraufnahme verschiedener Ätiolo-
gie führt zu einem Mangel an glukoplastischen Substanzen und zur Fettmobilisation,
die eine Ketose zur Folge haben kann. Somit ist generell auch eine Beteiligung der
Mykotoxine am Ketosegeschehen durch die negative Beeinflussung von Pansenflora
und Pansenmotorik denkbar. Durch diese wird die Futteraufnahme verringert, so
dass die Tiere in eine negative Energiebilanz geraten können (TEUFEL 1999; BU-
SATO et al. 2002).
5.3.1.2 Glucosekonzentrationen
Die Medianwerte der Glucosekonzentrationen lagen in den vorliegenden Untersu-
chungen bei allen Patienten (Kühe mit LMV und mit positivem und negativem Myko-
toxinnachweis), mit Ausnahme der gesunden Kühe, oberhalb des Referenzbereiches
von 2,22-3,30 mmol/l (FÜRLL 2005b). Es gab keinen signifikanten Unterschied zwi-
schen Kühen mit LMV und negativem Mykotoxinnachweis sowie Kühen mit LMV und
positivem Mykotoxinnachweis. Zwischen gesunden Kühen und Kühen mit LMV be-
stand ein signifikanter Unterschied (Abbildung 12). Dies stimmt mit den Beschrei-
bungen anderer Autoren überein (BOLAND et al. 1994; DICOSTANZO et al. 1996;
HOCHSTEINER et al. 2000; MOESER 2001; KLEINOVA et al. 2002). Auch sie konn-
ten keinen Einfluss von DON, ZON und deren Derivaten auf die Glucosekonzentrati-
on im Serum feststellen. Hohe Glucosekonzentrationen lassen sich zum einen durch
den Transportstress, zum anderen durch eine beginnende Endotoxämie erklären
(LING et al. 2005). Aufgrund der pathophysiologischen Zusammenhänge ist es be-
kannt, dass freies Endotoxin zu erhöhten Blutglucosekonzentrationen führt (FÜRLL
u. LEIDEL 2002).
5.3.1.3 Freie Fettsäure-Konzentrationen
Die Medianwerte der FFS-Konzentrationen befanden sich bei allen Patienten (Kühe
mit LMV und positivem oder negativem Mykotoxinnachweis) oberhalb des Referenz-
bereiches von <340 µmol/l (FÜRLL 2005b). Kühe mit LMV zeigten im Vergleich zu
gesunden Kühen eine signifikant höhere Konzentration (Abbildung 15). Kühe mit
LMV und negativem Mykotoxinnachweis zeigten jedoch im Vergleich zu Kühen mit
LMV und positivem Mykotoxinnachweis keinen signifikanten Unterschied. Die signifi-
kant erhöhten FFS-Konzentrationen p. p. weisen auf eine verstärkte Lipolyse hin.

Diskussion
69
Diese Erhöhung der FFS-Konzentrationen p. p. wurde in der Literatur schon häufiger
beschrieben (BUSATO et al. 2002; KOLLER et al. 2003; WILKEN 2004) und wird u.
a. durch den Fettansatz a. p., den Geburtstress und die Ausprägung des Energiede-
fizits p. p. mit besonderer regulatorischer Wirkung durch das Insulin beeinflusst
(FÜRLL et al. 1992).
5.3.2 Leberstoffwechsel
5.3.2.1 Bilirubinkonzentrationen im Serum
Die Medianwerte der Bilirubin-Konzentrationen waren bei Kühen mit LMV oberhalb
des physiologischen Bereiches von 3,30-5,30 µmol/l (FÜRLL 2005b). Kühe mit LMV
und negativem Mykotoxinnachweis zeigten keinen signifikanten Unterschied zu Kü-
hen mit LMV und positivem Mykotoxinnachweis. Ein signifikanter Unterschied konnte
nur im Vergleich zu den gesunden Kühen erhoben werden (Abbildung 10). Der An-
stieg der Bilirubinkonzentration ist eine Folge des Anstieges der FFS-
Konzentrationen im Sinne des Inanitionsikterus (FÜRLL et al. 1992). Die erhöhten
Bilirubinkonzentrationen, die alle Patienten am Tag der Erstuntersuchung aufwiesen,
können unter anderem Ausdruck der Cholestase oder Parenchymveränderungen
(Hepatosteatose) (REHAGE et al. 1996), aber auch der verringerten Futteraufnahme,
damit verbundener ungenügender Energieaufnahme und resultierender Verfettung
von Organen, besonders der Leber (FÜRLL 2002), sein. Der gleichzeitige Anstieg
des Bilirubins wird durch die Konkurrenz des Bilirubins mit den freien Fettsäuren um
dieselben Transportproteine in der Leber verursacht (FÜRLL 1989; BUSATO et al.
2002). Die erhöhten Bilirubinkonzentrationen zeigen sich in den Untersuchungen als
unabhängig von den Mykotoxinkonzentrationen (KLEINOVA et al. 2002). Dies stimmt
mit den Beschreibungen von SCHUH (1989) überein.
5.3.2.2 Aspartat-Aminotransferase-Aktivitäten
In den Untersuchungen lagen die Medianwerte der AST-Aktivitäten bei Kühen mit
LMV oberhalb des Referenzbereiches von <80,0 U/l (FÜRLL 2005b). Es konnte kein
signifikanter Unterschied zwischen Kühen mit LMV und negativem Mykotoxinnach-
weis mit Kühen mit LMV und positivem Mykotoxinnachweis festgestellt werden (Ab-
bildung 16). Die geringgradig erhöhten Medianwerte der AST-Aktivitäten bei Kühen
mit positivem Mykotoxinnachweis stimmen mit den Beschreibungen von
HOCHSTEINER et al. (2000) und MOESER (2001) überein. Diese konnten jedoch

Diskussion
70
keine bedeutenden Veränderungen der AST-Aktivitäten bei Kühen feststellen, die mit
mykotoxinbelasteten Futter gefüttert wurden (SEELING et al. 2006a). Aber es konnte
ein signifikanter Unterschied zwischen gesunden Kühen und Kühen mit LMV erho-
ben werden. Bei der AST handelt es sich um ein Enzym, das aus zwei Isoenzymen
besteht und vor allem in Leber, Skelett- und Herzmuskulatur vorkommt und im Zytop-
lasma als auch in den Mitochondrien aktiv ist (FRANK 1975; HOCHSTEINER et al.
2000). Untersuchungen von MATTHÄUSE et al. (2004) und SEELING et al. (2006a)
hatten ergeben, dass die erhöhte Enzymaktivität von AST nicht von der Mykotoxinbe-
lastung des Futters, sondern von einer erhöhten Aufnahme organischen Materials
abhängt. Die erhöhten AST-Aktivitäten können auch die Folge von Resorptions- bzw.
Involutionsvorgängen an der Gebärmutter sein (EULENBERGER 1984; WEMHEU-
ER 1987; SATTLER u. FÜRLL 2004). Diese Erhöhung wird von den Autoren als phy-
siologisch angesehen (KLEINOVA et al. 2002). Weiterhin beschrieben KARSAI und
SCHÄFER (1984) sowie FÜRLL (1989) erhöhte AST-Aktivitäten bei intensiver Lipo-
lyse mit nachfolgender Leberverfettung.
5.3.2.3 Glutamat-Dehydrogenase-Aktivitäten
Die mittleren GLDH-Aktivitäten bei Kühen mit LMV und negativem oder positivem
Mykotoxinnachweis lagen über dem Referenzbereich von 30 U/l Serum (FÜRLL
2005b). Zwischen gesunden Kühen und Kühen mit LMV und positivem oder negati-
vem Mykotoxinnachweis wurde ein signifikanter Unterschied erhoben. Keine signifi-
kanten Unterschiede gab es aber zwischen Kühen mit LMV und negativem Mykoto-
xinnachweis im Vergleich zu Kühen mit LMV und positivem Mykotoxinnachweis. Die
Medianwerte der GLDH-Aktivitäten waren bei Kühen mit LMV und positivem DON
bzw. DOM-1 geringgradig höher als bei Kühen mit LMV und negativem Mykotoxin-
nachweis (Abbildung 17). Diese Beobachtungen stimmen mit der Beschreibung an-
derer Studien (JAKSCH u. GLAWISCHNIG 1990; HOCHSTEINER et al. 2000; SA-
BATER et al. 2004) überein. Sie konnten zudem eine geringgradige Erhöhung der
GLDH-Aktivität bei Kühen, die mit mykotoxinbelastetem Futter gefüttert wurden, fest-
stellen. Hingegen konnten andere Autoren (MATTHÄUSE et al. 2004; SEELING et
al. 2006a) belegen, dass die erhöhte Enzymaktivität von GLDH nicht von der Myko-
toxinbelastung des Futters, sondern von der erhöhten Aufnahme organischen Mate-
rials abhängt. Da die GLDH als leberspezifisches Enzym angesprochen werden kann
und sie ausschließlich in den Mitochondrien der Leberzellen lokalisiert ist, sind Aktivi-
tätserhöhungen als Hinweise auf Schädigungen der Leberzellen anzusehen (HIEBL

Diskussion
71
2005). Die GLDH macht eine selbstständige, vom Leberfettgehalt relativ unabhängi-
ge Aussage über den Grad eingetretener Leberzellschäden, was zur Abgrenzung
(STAUFENBIEL et al. 1992) einer physiologischen von einer pathologischen Leber-
fetteinlagerung dienen kann. REHAGE (1996) bestätigte bei Kühen mit histologisch
nachweisbaren hochgradigen Leberverfettungen höhere AST-, GGT- und GLDH-
Aktivitäten als bei Kühen mit gering- bzw. mittelgradiger Leberverfettung. Erhöhte
Aktivitäten der leberspezifischen Enzyme und hohe Konzentrationen an freien Fett-
säuren, in Zusammenhang mit erhöhten Bilirubinkonzentrationen, deuten auf eine
Leberverfettung hin (FÜRLL 1989; REHAGE et al. 1996; FÜRLL u. JÄCKEL 2005).
Mit zunehmender Steigerung der Lipolyserate weist insbesondere der Anstieg der
GLDH auf eine starke Ausprägung von Leberzellschäden hin (STAUFENBIEL et al.
1992, HIEBL 2005).
5.3.2.4 Gamma-Glutamyl-Transferase-Aktivitäten
Die Medianwerte der GGT-Aktivitäten lagen bei allen Tieren innerhalb des Referenz-
bereiches von <50 U/l (FÜRLL 2005b). Signifikante Unterschiede zwischen Kühen
mit LMV und negativem Mykotoxinnachweis und Kühen mit LMV und positivem My-
kotoxinnachweis waren nicht ersichtlich (Abbildung 18). Dies deckt sich nicht mit den
Beschreibungen weiterer Untersuchungen (HOCHSTEINER et al. 2000; MOESER
2001; KLEINOVA et al. 2002; SEELING et al. 2006a). Diese konnten geringgradig
erhöhte GGT-Aktivitäten bei Kühen, die mit mykotoxin-kontaminiertem Futter ver-
sorgt worden waren, feststellen. Es bestand zudem ein relevanter signifikanter Un-
terschied zwischen gesunden Kühen und Kühen mit LMV. Kühe mit LMV und positi-
vem Zearalenonnachweis zeigten im Vergleich zu allen anderen Gruppen geringgra-
dig erhöhte Medianwerte. Die erhöhten Aktivitäten der leberspezifischen Enzyme und
die zu hohen Konzentrationen an freien Fettsäuren, im Zusammenhang mit erhöhten
Bilirubinkonzentrationen, deuten auf eine Leberverfettung hin (FÜRLL 1989; REHA-
GE et al. 1996; FÜRLL 2005; KRETZSCHMAR 2008).
5.3.3 Eiweißstoffwechsel
5.3.3.1 Gesamtproteinkonzentrationen
In den Untersuchungen lagen die Medianwerte der Proteinkonzentrationen in allen
Gruppen im Referenzbereich von 68-82 g/l (FÜRLL 2005b). Die gesunden Kühe

Diskussion
72
zeigten signifikant höher Gesamteiweißkonzentrationen als die Kühe mit LMV, unab-
hängig von Mykotoxinnachweis. Bei den gesunden Kühen lagen Gesamtproteinkon-
zentrationen im oberen, bei den Patientenkühen im unteren Drittel des Referenzbe-
reichs (Abbildung 9). Bei gesunden Kühen mit einer Milchleistung von über 8750
kg/Jahr wurde festgestellt, dass die Proteinkonzentrationen über den physiologi-
schen Bereich hinaus reichen (WILKEN 2004). Erhöhte Proteinkonzentrationen kön-
nen auch für chronische Entzündungen sprechen (PAYNE et al. 1974). Zwischen
Kühen mit LMV sowie negativem Mykotoxinnachweis und Kühen mit LMV sowie po-
sitivem Mykotoxinnachweis konnten keine signifikanten Unterschiede beobachtet
werden. Die Ergebnisse stimmen auch hier mit den bereits angegebenen Quellen
überein (HOCHSTEINER et al. 2000; MOESER 2001; KLEINOVA et al. 2002; SEE-
LING et al. 2006a). Zudem konnten keine Abweichungen der Proteinkonzentrationen
der untersuchten Kühe, die mykotoxinbelastetes Futter erhielten, nachgewiesen wer-
den.
5.3.3.2 Kreatininkonzentrationen
Die Medianwerte der Kreatininkonzentrationen lagen bei allen Kühen im Referenzbe-
reich von 55-150 µmol/l (FÜRLL 2005b). Es konnte kein signifikanter Unterschied
zwischen Kühen mit LMV sowie negativem Mykotoxinnachweis und Kühen mit LMV
sowie positivem Mykotoxinnachweis festgestellt werden (Abbildung 11). Auch hier
finden sich Übereinstimmungen mit den aufgeführten Autoren (HOCHSTEINER et al.
2000; MOESER 2001; KLEINOVA et al. 2002). Untersuchungen der Kreatininkon-
zentrationen nach Fütterung von mykotoxinbelastetem Futter ergaben keine Abwei-
chungen vom Referenzbereich. Zwischen den gesunden Kühen und Kühen mit LMV
sowie positivem ZON-Nachweis konnte ein relevanter signifikanter Unterschied er-
hoben werden (Abbildung 11).
5.3.4 Muskelstoffwechsel
5.3.4.1 Creatinkinase-Aktivitäten
Die CK-Aktivität lag bei allen Patienten deutlich über der oberen Grenze des Refe-
renzbereichs von 200 U/l (FÜRLL 2005b) (Abbildung 20). Es gab keine signifikanten
Unterschiede zwischen Kühen mit LMV und negativem Mykotoxinnachweis und Kü-
hen mit LMV und positivem Mykotoxinnachweis. Bestätigt wird dies in Arbeiten von
HOCHSTEINER (2000), MOESER (2001) und KLEINOVA (2002). Zwischen gesun-

Diskussion
73
den Kühen und Kühen mit LMV gab es signifikante Unterschiede. Dabei sind wohl
weniger die Mykotoxine für die Veränderung der CK-Aktivität verantwortlich als viel-
mehr die Primärerkrankung und deren Begleitkrankheiten. Die Ergebnisse bestätigen
die Untersuchungen von KLEISER und FÜRLL (1998), die bei Kühen im Vorfeld ei-
ner Labmagenverlagerung erhöhte CK-, als auch AST-Aktivitäten messen konnten,
denn auch die Medianwerte der AST-Aktivitäten lagen deutlich über der Toleranz-
obergrenze von 80 U/l. Eine Erklärung für die moderat erhöhten CK-Aktivitäten wird
in Entzündungen des Endometriums gesehen (SATTLER u. FÜRLL 2004), so dass
CK-Aktivitäten von über 200 U/l bei Kühen in der Frühlaktation auf Puerperalstörun-
gen in Form von Endometritiden hinweisen können. Dieser Sachverhalt konnte bes-
tätigt werden, da alle Kühe im Versuch an einer Endometritis erkrankt waren und CK-
Aktivitäten im Serum über der Toleranzobergrenze von 200 U/l (FÜRLL 2005b)
messbar waren. Es wurde beobachtet, dass bei Kühen mit Labmagenverlagerung die
CK-Aktivitäten bei zusätzlicher Erkrankung an mittel- bis hochgradigen Endometriti-
den signifikant anstiegen (SATTLER 2001). Eine weitere Erklärung wäre das gehäuf-
te Auftreten von Klauenerkrankungen, welche zu vermehrtem Liegen der Tiere und
damit verbundenen Muskelschädigungen führen (SHAFFER et al. 1981).
5.3.5 Mineralstoffwechsel
5.3.5.1 Gesamtcalcium-Konzentrationen
Die Medianwerte der Ca-Konzentrationen im Serum waren bei allen Kühen im Refe-
renzbereich von 2,3-2,8 mmol/l (FÜRLL 2005b). Es war kein Unterschied zwischen
Kühen mit LMV sowie negativem Mykotoxinnachweis und Kühen mit LMV sowie po-
sitivem Mykotoxinnachweis ersichtlich (Abbildung 25). Auch diese Ergebnisse de-
cken sich mit den Angaben mehrerer Autoren (HOCHSTEINER et al. 2000; MOE-
SER 2001; KLEINOVA et al. 2002). Abweichungen der Ca-Konzentration konnten bei
Kühen, die mit mykotoxin-kontaminiertem Futter versorgt worden sind, nicht eruiert
werden. Zwischen gesunden Kühen und Patiententieren konnte eine signifikant nied-
rigere Gesamtkalziumkonzentration ausgemacht werden. Ein Abfall an die Unter-
grenze von 2 mmol/l p.p. wurde bereits von BOSTEDT (1974) und FÜRLL (1994)
beschrieben. Mögliche Ursache könnte die Milchsynthese sein (DRACKLEY 2002).
Als Ursache für die niedrigen Ca-Konzentrationen an der Untergrenze des Referenz-
bereiches, könnte ein verringerter Rohfaser-Gehalt im Futter und die damit verbun-

Diskussion
74
dene reduzierte Futteraufnahme sein (HOOPS 2007). Diese Ergebnisse bestätigen
sich bei SCHUH (1989).
5.3.5.2 Anorg. Phosphat
In den Untersuchungen lagen die Medianwerte der Phosphatkonzentrationen aller
untersuchten Kühen mit LMV lagen am Tag der Erstuntersuchung innerhalb des Re-
ferenzbereiches von 1,55 bis 2,29 mmol/l (FÜRLL 2005b), mit Ausnahme von Kühen
mit LMV und positivem DON-Nachweis, deren P-Konzentrationen ggr. unterhalb des
Referenzbereiches lagen. Es zeigte sich kein signifikanter Unterschied zwischen Kü-
hen mit LMV sowie negativem Mykotoxinnachweis und Kühe mit LMV sowie positi-
vem Mykotoxinnachweis (Abbildung 23). Abweichungen der Phosphat-
Konzentrationen konnten bei Kühen, die mit mykotoxin-kontaminiertem Futter ver-
sorgt worden sind, nicht beobachtet werden (HOCHSTEINER et al. 2000; MOESER
2001; KLEINOVA et al. 2002). SCHUH (1989) untersuchte die Phosphat-
Konzentration bei Masttieren nach Fütterung mit kontaminiertem Futter, stellte dabei
aber keine Veränderungen zur Kontrollgruppe fest. Zwischen den gesunden Kühen
und den Kühen mit LMV und positivem Mykotoxinnachweis bestanden relevante sig-
nifikante Unterschiede in der Konzentration des anorganischen Phosphates im Blut
(Abbildung 23). Als Ursache dafür kommen verminderte Futteraufnahme a.p. und
p.p, aber auch endotoxisch-entzündliche Prozesse in Betracht (GOFF 2000; FÜRLL
et al. 2002). Auch der erhöhte Bedarf für die Milchsynthese (BOSTEDT 1974; HÖ-
RÜGEL u. FÜRLL 1998b) könnte ätiologisch beteiligt sein. Da anorganisches Phos-
phat ein fütterungsabhängiger Parameter ist (KLIMIENE et al. 2005), sind die niedri-
gen Konzentrationen entweder auf einen geringen Gehalt im Futter zurückführbar
oder auf eine verminderte Futteraufnahme, jedoch nicht auf Mykotoxinbelastung.
5.3.5.3 Magnesium-Konzentrationen
Die Medianwerte der Mg-Konzentration lagen bei allen Patienten (Kühe mit LMV und
negativem oder positivem Mykotoxinnachweis) in den vorliegenden Ergebnissen un-
terhalb des Referenzbereiches von 0,90-1,32 mmol/l (FÜRLL 2005b). Die gesunden
Kühe zeigten signifikant höher Magnesiumkonzentrationen als die Kühe mit LMV. Es
bestand kein signifikanter Unterschied zwischen Kühen mit LMV sowie negativem
Mykotoxinnachweis und Kühen mit LMV sowie positivem Mykotoxinnachweis (Abbil-
dung 21). Dies stimmt mit den Beschreibungen von (HOCHSTEINER et al. 2000;
MOESER 2001; KLEINOVA et al. 2002) überein. Die Tendenz der niedrigsten Mg-

Diskussion
75
Konzentrationen einige Tage p.p. entspricht den Resultaten aus der Literatur (GIB-
SON et al. 1987, FÜRLL et al. 1998b) und spiegelt die verminderte Futteraufnahme
um die Geburt (FÜRLL et al. 1998a) bzw. bei einer Labmagenverlagerung (STAU-
FENBIEL 2001) wider. KUME et al. (1998) berichteten über erniedrigte Magnesium-
konzentrationen infolge von Hitzestress, welcher wiederum eine verminderte Futter-
aufnahme nach sich zieht (SRIKANDAKUMAR u. JOHNSON 2004). SCHUH (1989)
konnte feststellen, dass auch nach Verfütterung von fusarientoxinbelastetem Futter
keine Veränderungen der Mg-Konzentration bei Masttieren auftraten. Da Magnesium
ein fütterungsabhängiger Parameter ist (KLIMIENE et al. 2005), ist die insgesamt
erniedrigte Magnesiumkonzentration hinweg auf eine zu geringe Konzentration im
Futter zurück zu führen. Die klassische Hypomagnesämie, die durch zu wenig Mag-
nesium im frischen Gras der Frühjahrsweide ausgelöst wird (McCOY 2004), kann
aber auch durch einen zu hohen Gehalt an Kalium oder freien Fettsäuren im Futter
bedingt sein.
5.3.5.4 Natrium-Konzentrationen
In den vorliegenden Untersuchungen lagen die Medianwerte der Na-Konzentrationen
bei Kühen mit LMV sowie negativem Mykotoxinnachweis im Normbereich und bei
Kühen mit LMV sowie positivem Mykotoxinnachweis geringgradig unterhalb des Re-
ferenzbereiches von 135-157 mmol/l (FÜRLL 2005b). Es bestand ein signifikanter
Unterschied zwischen den gesunden Kühen und den Kühen mit LMV. Ein signifikan-
ter Unterschied konnte auch zwischen Kühen mit LMV sowie negativem Mykotoxin-
nachweis und Kühen mit LMV sowie positivem Mykotoxinnachweis (ZON) eruiert
werden (Abbildung 27). Die Na-Konzentrationen im Serum steigen um die Kalbung
hin an (ROWLANDS et al. 1975, FÜRLL et al. 1994) bzw. verändern sich nicht signi-
fikant und fallen nach der Geburt bis in die Frühlaktation ab (SETZ 2000, AE-
BERHARD et al. 2001, KASTNER 2002). Die signifikant niedrigeren Na-
Konzentrationen p.p. sind durch das Einsetzen der Milchsekretion bzw. die steigende
Milchleistung begründet (Hoops 2007).
5.3.5.5 Kalium-Konzentrationen
Die Medianwerte der K-Konzentrationen lagen in den vorliegenden Untersuchungen
bei allen Patienten unterhalb des Referenzbereiches von 3,9-5,2 mmol/l (FÜRLL
2005b). Es wurden signifikante Unterschiede zwischen den gesunden Kühen und
den Kühen mit LMV festgestellt (Abbildung 26). Zwischen den Kühen mit LMV sowie

Diskussion
76
negativem Mykotoxinnachweis und den Kühen mit LMV sowie positivem Mykotoxin-
nachweis ergaben sich keine signifikanten Unterschiede (JAKSCH u. GLA-
WISCHNIG 1990; HOCHSTEINER et al. 2000; MOESER 2001; KLEINOVA et al.
2002). Nach Fütterung von mykotoxinbelastetem Futter konnten keine signifikanten
Veränderungen der K-Konzentrationen bestimmt werden. WITTEK und FÜRLL
(1998) konnten Kaliumkonzentrationen bei Kühen mit LMV bestimmen, die deutlich
unter den Angaben in der Literatur lagen. Dies liegt an der reduzierten Futterauf-
nahme bei LMV.
5.3.5.6 Chlorid-Konzentrationen
In den vorliegenden Untersuchungen lagen die Medianwerte der Cl-Konzentration
bei allen Kühen im Referenzbereich von 95-110 mmol/l (FÜRLL 2005b). Es gab ei-
nen signifikanten Unterschied zwischen den gesunden Kühen den Kühen mit LMV
und positivem Mykotoxinnachweis. Die Medianwerte der Cl-Konzentrationen lagen
im unteren Drittel des Referenzbereiches (Abbildung 28). Die Cl-Konzentrationen im
Serum verhielten sich um die Kalbung wie die Na-Konzentrationen im Serum. Die
abfallenden Cl-Konzentrationen stimmen mit den Ergebnissen von weiteren Autoren
(FÜRLL et al. 1998b; SETZ 2000; KASTNER 2002) überein. Bei beiden Labmagen-
verlagerungsformen können sich schwere Störungen des Säure-Basen-Haushaltes
einstellen. Sie sind ätiologisch u.a. durch Reflux (inneres Erbrechen) von Labmagen-
inhalt in den Pansen bedingt, wodurch Chloridionen in den Pansen gelangen, durch
dessen Schleimhaut sie nicht adäquat resorbiert werden können. Im Blut entstehen
ein Hypochlorämie (< 90 bis 95 mmol Cl/l) und eine leichte Hypokaliämie (3,2 bis 3,5
mmol K/l).
5.3.6 Cholesterolkonzentrationen
Cholesterol gilt als ein Stoffwechselparameter, der die Resorption aus dem Darm
widerspiegelt und sich in seiner Entwicklung parallel zum Futteraufnahmeverhalten
verhält (FÜRLL u. JÄCKEL 2005). In den vorliegenden Ergebnissen lagen die Medi-
anwerte der Cholesterolkonzentrationen bei allen Patienten oberhalb des unteren
Grenzwertes (1,5 mmol/l) (FÜRLL 2005b). Bei den gesunden Kühen und den Kühen
mit LMV sowie positivem DON-Nachweis lagen die Cholesterolkonzentrationen im
Referenzbereich (Abbildung 13). Hohe Cholesterolkonzentrationen 10 Tage a.p. bis
28 Tage p.p. entsprechen den Beobachtungen von SHAFFER et al. (1981) und
KAPPEL et al. (1984). Es bestanden signifikante Unterschiede zwischen den gesun-

Diskussion
77
den Kühen und den Kühen mit LMV und zwischen Kühen mit LMV und negativem
Mykotoxinnachweis im Vergleich zu Kühen mit LMV und positivem ZON-Nachweis
(Abbildung 13). Dies erklärt sich durch die zu geringe Futteraufnahme der Kühe be-
reits vor der Klinikeinweisung aufgrund des Bestehens einer Labmagenverlagerung.
REHAGE et al. (1996) erklärt sich die geringen Cholesterolkonzentrationen in einer
Leberfunktionseinschränkung. Im Zusammenhang mit den niedrigeren Cholesterol-
konzentrationen fällt eine dramatische Erniedrigung der strukturwirksamen Rohfaser
auf. Diese Verringerung führt zu einer azidotischen Belastung und reduziert die Fut-
teraufnahme, was sich in den erniedrigten Cholesterolkonzentrationen widerspiegelt
(WILKEN 2004).
5.3.7 Hämatologie
Die Leukozytenzahlen, Erythrozyten, Hämoglobin, Hämatokrit, MCV, MCH und die
Thrombozyten befanden sich bei allen Kühen im physiologischen Bereich (FÜRLL
2005a). Es konnten keine signifikanten Unterschiede zwischen Kühen mit LMV und
negativem Mykotoxinnachweis sowie Kühen mit LMV und positivem Mykotoxinnach-
weis beobachtet werden. Die Ergebnisse stimmen mit denen anderer Untersuchun-
gen überein (WEAVER et al. 1986a; WEAVER et al. 1986b; SCHUH u. BAUM-
GARTNER 1988; JAKSCH u. GLAWISCHNIG 1990; HOCHSTEINER et al. 2000;
MOESER 2001; BAUER 2002; KLEINOVA et al. 2002; KOROSTELEVA et al. 2007).
Auch die in einem Fütterungsversuch mit mykotoxinbelastetem Futter versorgten Tie-
re wiesen keine Veränderungen des Blutbildes auf (SEELING et al. 2006a).
5.3.7.1 Differentialblutbild
Die Medianwerte der Anzahl der eosinophilen Granulozyten lagen bei den Patienten
an der unteren Grenze des Referenzbereichs von 0,05-1,0 G/l, die Anzahl der stab-
kernigen Granulozyten lag bei allen Kühen im Referenzbereich von 0-0,3 G/l und
auch die Anzahl der segmentkernigen Granulozyten lag im Referenzbereich von 1,3-
4,5 G/l. Die Lymphozytenzahlen der gesunden Tiere lagen im Referenzbereich von
2,5-6,5 G/l (FÜRLL 2005a). Auch die Monozyten lagen im Referenzbereich von 0,1-
0,9 G/l (FÜRLL 2005a). Die Patiententiere hingegen zeigten bei positivem oder nega-
tivem Mykotoxinnachweis eine Erniedrigung der Medianwerte der Lymphozyten und
Monozyten unterhalb des Referenzbereichs (FÜRLL 2005a). Es konnten keine signi-
fikanten Unterschiede zwischen Kühen mit LMV ohne Mykotoxinnachweis und Kühen
mit LMV und Mykotoxinnachweis festgestellt werden Ähnliche Ergebnisse wurden

Diskussion
78
auch in der Literatur beschrieben (WEAVER et al. 1986a; WEAVER et al. 1986b;
SCHUH u. BAUMGARTNER 1988; JAKSCH u. GLAWISCHNIG 1990; HOCHSTEI-
NER et al. 2000; MOESER 2001; BAUER 2002; KLEINOVA et al. 2002; KOROSTE-
LEVA et al. 2007). Zusammenfassend konnten keine Veränderungen des Differenti-
alblutbildes von Kühen festgestellt werden, bei denen Mykotoxine in der Ration
nachweisbar waren.
5.3.8 Säure-Basen-Haushalt
Die Medianwerte der pH-Werte lagen bei allen Kühen im Referenzbereich von 7,36-
7,41. Die Medianwerte des pCO2 waren im Referenzbereich von 4,8-6,4 kPa. Die
Medianwerte der BE lagen ebenso im Referenzbereich von -2,5 bis +2,5 mmol/l
(FÜRLL 2005b). Es konnte kein signifikanter Unterschied zwischen Kühen mit LMV
und negativem Mykotoxinnachweis sowie Kühen mit LMV und positivem Mykotoxin-
nachweis beobachtet werden. Die Ergebnisse stimmen mit den Beschreibungen wei-
terer Autoren (HOCHSTEINER et al. 2000; MOESER 2001; KLEINOVA et al. 2002)
überein.
5.3.9 Trolox Equivalent Antioxidative Capacity
Die Trolox Aquivalent Antioxidative Capacity (TEAC) ist bisher vorwiegend als Me-
thode zur Bestimmung der antioxidativen Kapazität von pflanzlichen Nahrungsmitteln
und Einzelstoffen eingesetzt worden (MILLER et al. 1996). Bei den vorliegenden Un-
tersuchungen lagen die Medianwerte der TEAC-Konzentrationen im Blut bei den Kü-
hen innerhalb 200-300 µmol/l, was mit den Untersuchungen der TEAC bei Milchkü-
hen im peripartalen Zeitraum übereinstimmt. Die ermittelten Messdaten lagen zwi-
schen 200 und 300 µmol/l, wobei die Werte ante partum tendenziell am niedrigsten
waren und post partum langsam anstiegen (FÜRLL et al. 1998b). Es konnten keine
Unterschiede zwischen Kühen mit LMV und negativem Mykotoxinnachweis und Küh-
nen mit LMV und positivem Mykotoxinnachweis festgestellt werden.
5.3.10 Eisen-Konzentrationen
Die Medianwerte der Fe-Konzentrationen im Blut lagen innerhalb des Referenzbe-
reichs von 13-33 µmol/l (FÜRLL 2005b). Es gab keinen signifikanten Unterschied
zwischen Kühen mit LMV und negativem Mykotoxinnachweis sowie Kühen mit LMV
und positivem Mykotoxinnachweis. Ein relevanter signifikanter Unterschied wurde
zwischen gesunden Kühen und Kühen mit LMV mit negativem Mykotoxinnachweis

Diskussion
79
festgestellt (Abbildung 22). Dies liegt wahrscheinlich an der reduzierte Futteraufnah-
me oder niedrigen Fe-Konzentration im Futter. Die Ergebnisse stimmen mit den Be-
schreibungen anderer Autoren überein (MOESER 2001; KLEINOVA et al. 2002).
5.3.11 Alkalische Phosphathase-Aktivitäten
Die Medianwerte der Alkalischen Phosphathase lagen in den vorliegenden Untersu-
chungen bei Kühen mit LMV oberhalb des Referenzbereichs von 40-122 U/l (FÜRLL
2005b). Es bestand kein signifikanter Unterschied bezüglich der AP-Aktivitäten von
Kühen mit LMV und negativem Mykotoxinnachweis sowie Kühen mit LMV und positi-
vem Mykotoxinnachweis (Abbildung 19). Eine geringgradige Erhöhung kann auf eine
Galleabflussstörung innerhalb der Leber zurückgeführt werden. Kommt es zum
Rückstau der Galle, z.B. bei LMV, steigt die AP im Blut an. Differentialdiagnostisch
sind Leberentzündungen (THIMME et al. 2002) und Knochenerkrankungen (HAIMA
2006) etabliert.
Abschließend kann festgestellt werden, dass die DON- und ZON-Kontaminationen in
geringen Konzentrationen beim Milchrind keine besondere Gefahrenquelle darstel-
len, wenn die Haltungs- und Fütterungsbedingungen optimal gestaltet werden.

Zusammenfassung
80
6 Zusammenfassung
Ahmad Alkaassem
Mykotoxinscreening (Deoxinivalenol, Zearalenon) in Futter, Blut, Milch und Gal-
le bei Kühen mit Labmagenverlagerung (LMV) und bei gesunden Kühen
Medizinische Tierklinik, Veterinärmedizinische Fakultät der Universität Leipzig
Eingereicht im Dezember 2008
83 Seiten, 30 Abbildungen, 11 Tabellen, 231 Literaturangaben
Schlüsselworte: Kühe, Mykotoxine, DON, ZON, Labmagenverlagerung
Einleitung: Ziel der vorliegenden Arbeit war es, mögliche Zusammenhänge zwi-
schen einer Exposition von Milchkühen mit Mykotoxinen (DON, ZON und deren Me-
taboliten) und dem Entwickeln und Bestehen einer LMV sowie weiteren Primär- und
Sekundärerkrankungen zu prüfen. Es bestand die Frage nach einer möglichen Be-
ziehung zu klinischen Störungen, nach einer möglichen Korrelation von Veränderun-
gen der Blutparameter und nach der Eignung von Gallensaft zum Mykotoxinnach-
weis.
Material und Methoden: Für die Untersuchungen wurden 61 Kühe der Rasse
Schwarzbunte aus dem Patientengut der Medizinischen Tierklinik mit der Diagnose
LMV genutzt. Die gesunden Kontrollkühe (n = 13) stammten aus zwei Betrieben im
Leipziger Umland. Bei der Aufnahme in die Klinik bzw. der Probenentnahme wurden
alle Kühe klinisch untersucht. Für die Mykotoxinbestimmungen wurden von 20 Pati-
enten- und 13 Kontrollkühen Futter, Serum, Milch und Galle sowie von 41 Patienten-
kühen Serum und Galle entnommen. Es wurden klinisch-chemische Parameter im
Serum untersucht. Die Mykotoxine wurden mittels High-Performance-Liquid-
Chromatography (HPLC) analysiert.
Ergebnisse: Bei 36 von 61 Patienten konnten keine Mykotoxine in der Galle nach-
gewiesen werden. Bei den anderen 25 Tieren war ein positiver Mykotoxinnachweis
erfolgt. Von den 36 Kühen mit negativem Mykotoxinnachweis konnten 20 Kühe (56
%) mit linksseitiger LMV und 8 Kühe (22 %) mit rechtsseitiger LMV als geheilt entlas-
sen werden. Sechs Kühe (17 %) mit LMV nach links und negativem Mykotoxinnach-
weis und zwei Kühe (6 %) mit LMV nach rechts und negativem Mykotoxinnachweis
hatten einen Exitus letalis. Von 25 Kühen mit positivem Mykotoxinnachweis wurden
zwölf Kühe (48 %) mit der Diagnose LMV nach links und sechs Kühe (24 %) mit LMV

Zusammenfassung
81
nach rechts geheilt. Fünf Kühe (20 %) mit LMV nach links und zwei Kühe (8 %) mit
LMV nach rechts hatten einen Exitus letalis.
Die makroskopische Untersuchung der Futterproben ergab keine grobsinnlichen
Veränderungen. Mykotoxine konnten hingegen regelmäßig in den Futterproben aus
den Herkunftsbetrieben der Patienten und der gesunden Kühe nachgewiesen wer-
den. Der Median betrug für DON 0,161 mg/kg (0,086-0,191) und für ZON 6,35 µg/kg
(4,88-7,85). Im Serum enthielten eine von 61 Proben DON (0,002 µg/mL) und weite-
re vier Proben DOM-1 (0,002-0,003-0,004-0,009 µg/mL).
Im Gallensaft konnte bei 61 Proben nur einmal DOM-1 (37,6 µg/mL) und kein DON
nachgewiesen werden. Die untersuchten Galleproben waren zu 39 % mit ZON bzw.
dessen Metabolite belastet. Dabei lag ZON (ng/g) im Mittel bei 9,85 (8,10-16,33), α-
ZOL bei 59,9 (5-78) ng/g und β-ZOL bei 37,6 ng/g. Bei Patienten mit positivem
Nachweis von ZON und seinen Metaboliten α- und ß-ZOL konnten keine Verände-
rungen am Genitaltrakt festgestellt werden.
Bei den gesunden Kühen waren keine Mykotoxine in Serum, Milch oder Gallensaft
nachweisbar. In den eigenen Untersuchungen ließen sich klinisch sowie biochemisch
weder DON- noch ZON-spezifische Effekte nachweisen. Entweder waren die geprüf-
ten Parameter im physiologischen Bereich, wie z.B. Protein, Kreatinin, TEAC und im
Blutbild, oder die Veränderungen waren auch bei den Kühen ohne nachweisbare
Mykotoxine vorhanden, wie z.B. bei Bilirubin, Glucose, FFS, Cholesterol, BHB, CK,
GGT, Mg, Ca, Na, K, Cl, FFS und im Säure-Basen-Status. Die mittleren GLDH-
Aktivitäten aller Patienten mit DON-Nachweis waren geringgradig höher als bei Pati-
enten ohne Mykotoxinnachweis.
Schlussfolgerung: Mykotoxine konnten in dieser Untersuchung weder für spezifi-
sche Symptome oder hämatologische sowie klinisch-chemische Abweichungen im
Blut noch für den Krankheitsausgang, d.h. Heilung oder Exitus letalis, verantwortlich
gemacht werden. Eine Ausnahme stellt die Frequenz und Intensität der Pansenkon-
traktionen dar, die bei Patienten mit positivem Mykotoxinnachweis im Futter reduziert
waren. ZON- bzw. ZOL-assoziierte Veränderungen an den Ovarien und dem Uterus
wurden makroskopisch in keinem Fall festgestellt.
Als zusätzliche Erkenntnis stellte sich im Verlaufe dieser Arbeit unabhängig von der
eigentlichen Fragestellung heraus, dass die Gallensaftentnahme in der Praxis eine
leicht durchführbar, diagnostisch wertvolle Maßnahme darstellt.
Abschließend kann festgestellt werden, dass die DON- und ZON-Kontaminationen in
geringen Konzentrationen beim Milchrind keine besondere Gefahrenquelle darstel-
len, wenn die Haltungs- und Fütterungsbedingungen optimal gestaltet werden.

Summary
82
7 Summary
Ahmad Alkaassem
Mycotoxin screening (Deoxnivalenol, Zearalenon) in foodstuff, blood, milk and
bile of healthy dairy cows and cows with abomasal d isplacement
Large Animal Clinic for Internal Medicine, Faculty of Veterinary Medicine, University
of Leipzig
Submitted in December 2008
83 Pages, 30 figures, 11 tables, 231 references
Keywords: cows, mycotoxins, DON, ZON, abomasal displacement
Introduction: This study was aimed to examine exposure of mycotoxins: Deoxyniva-
lenol (DON), Zearalenon (ZON) and their metabolites; de-epoxy-DON (DOM-1), α-
Zearalenol (α-ZOL) and β-Zearalenol (β-ZOL) in dairy cows to investigate their role in
the abomasal displacement and its related primary and secondary diseases. Its clini-
cal effects, correlation with haematological changes and application of mycotoxin de-
tection in bile samples were also examined.
Materials and Methods: 61 Schwarzbunte dairy cattle with displaced abomasums,
hospitalized from September 2005- March 2006 in large animal clinic for internal
medicine, University of Leipzig, were examined. The healthy controls (n=13) were
from two different dairy farms of Leipzig. General clinical examinations were done in
all cows in admission and before sample collection. Foodstuff, serum, milk and bile
from 20 patients and 13 control cows and serum and bile from 41 patients were ex-
amined for mycotoxins and also for serum clinical chemistry. Mycotoxins were ana-
lysed with high performance liquid chromatography (HPLC).
Results: 36/61 cows were mycotoxins negative and 25/61 were mycotoxin positive in
bile. From 36 cows with negative mycotoxins, 20 cows (56 %) with LDA and 8 cows
(22 %) with RDA were completely recovered and discharged from the clinic. 6 cows
(17 %) with LDA and 2 cows (6 %) with RDA from this group were euthanized. From
25 cows with positive mycotoxins, 12 cows (48 %) with LDA and 6 cows (24 %) with
RDA were also completely recovered. But 5 cows (20 %) with LDA and 2 cows (8 %)
with RDA were euthanized. No macroscopic changes of foodstuff were found. My-
cotoxins were regularly detected in the foodstuff of the healthy cows and the patients
from their original area. But median values of 0.161 mg/kg for DON (0.086-0.191)
and 6.35 µg/kg (4.88-7.85) for ZON were measured in foodstuff of all patients and

Summary
83
healthy cows. 0.002 µg/ml 8 % of cows with DON (1/61) and (4/61) with DOM-1
(0.002, 0.003, 0.004 and 0.009 µg/ml) were measured in serum.
DOM-1 (37.6 µg/ml) was measured by 1/61 bile sample but DON was not found.
ZON or its metabolites were detected in 39 % of all examined bile samples. Median
values of ZON, α-ZOL and β-ZOL were 9.85 (8.10-16.33) ng/g, 59.9 (5-78) ng/g and
37.6 ng/g respectively. No genital tract changes were found in patients with detect-
able ZON and its metabolites (α- and ß-ZOL). No mycotoxins were detected in the
milk, serum and bile of healthy controls.
The haematological and other biochemical parameters such as protein, creatine and
TEAC of the patients with detectable DON and ZON were within physiological range.
Other parameters including bilirubin, glucose, FFS, cholesterol, BHB, CK, GGT, Mg,
Ca, Na, K, Cl and acid-base balance were also changed in mycotoxins negative
cows. GLDH activities were slightly higher in mycotoxins positive cows than those of
mycotoxins negative.
Conclusions: Mycotoxins didn’t play a role on the haematological parameters, se-
rum clinical chemistry and clinical outcome, i.e. complete recovery or euthanasia in
this study. But the frequency and intensity of ruminal contraction was reduced in pa-
tients with positive mycotoxins in foodstuff. Macroscopic changes of ovaries and
uterus due to ZON and ZOL were not found. As an additional fanding, regadless of
the orginal question, we found out, that obtaining bile fluid is easy to perform and is
af high diagnostic value for the practitioner. The present study can be concluded that
contamination of DON and ZON in low concentrations played no special role in dairy
cattle if feeding system and other dairy-management-programs were optimal.

Literaturverzeichnis
84
8 Literaturverzeichnis
Aeberhard K, Bruckmaier RM, Blum JW. Metabolic enzymatic and endocrine status
in high-yielding dairy cows. Part 2 J Vet Med A Physiol Pathol Clin Med.
2001;48:111-27.
Anderson VL, Boland EW, Caspet HH. The effects of vomitoxin (DON) from scab in-
fested barley fed to gestating and lactating heifers. Carrington Research Center Beef
Production Field Day 1995;18:10-2.
Atroshi F, Rizzo A, Westermarck T, Ali-Vehmas T. Antioxidant nutrients and mycotox-
ins. Toxicology 2002;15:151-67.
Bauer J, Gedek B. Mykotoxikosen beim Schwein. Prakt Tierarzt. 1978;59:780-5.
Bauer J, Gedek B. Fusariumtoxine als Ursache von Ursache von Futterverweigerung
und Fruchtbarkeitsstörungen beim Pferd. Tierärztl Umsch. 1980;35:600-3.
Bauer J. Mykotoxikosen in der tierischen Produktion-Bedeutung und Diagnose. Berl
Münch Tierarztl Wschr. 1982;95:301-7.
Bauer J, Gareis M. Veränderungen am Genitaltrakt des weiblichen Schweines nach
Verfütterung praxisrelevanter Zearalenonmengen. J Vet Med. 1987;34:613-23.
Bauer S, Schneidler D, Kroll J. A standardized test method for determination of afla-
toxin M1 in milk and milk products. Nahrung. 1989;33:217-9.
Bauer J, Eckstein B, Dänicke S, Gropp J, Meyer HHD, Obst A, et al. Mykotoxine:
Vermeiden statt Bekämpfen. In: DLG Mitteilungen 2000;8:13-7.
Bauer J. Mycotoxins in feedstoffs for ruminants: biochemical effects and clinical rele-
vance. In: Kaske M, Scholz H, Höltershinken M. (eds), Recent Developments and

Literaturverzeichnis
85
Perspectives in Bovine Medicine. - Lectures XXII World Buiatrics Congress 18-23
August. 2002:168-80.
Bauer J. Mykotoxine im Hochleistungsfutter von Milchkühen. Nutztierpraxis Aktuell,
Tagungsband zu den 4 Haupttagungen der Agrar- und Veterinär-Akademie (AVA).
2005:14-5.
Baumgartner W. Interne Medizin und klinische Seuchenlehre Rind, Schaf, Ziege Teil.
1, II Medizinische Universitätsklinik für Klauentiere Wien. 2000:82-118.
Baumgartner W. Klinische Propädeutik der inneren Krankheiten und Hautkrankheiten
der Haus- und Heimtiere. 5. aktualisierte Aufl. Berlin, Wien: Blackwell Wissenschafts-
Verlag; 2002.
Bennet JW, Klich M. Mycotoxins. Clinical Microbiology Reviews. 2003;16:497-16.
Bischoff M. Mykotoxine im Getreide unter Berücksichtigung der Ernte 1998:17-37.
<http://www.lufa-nord-west.de/lufa2/downloads/pdf/mykotoxine_bischoff.pdf>.
Blankenship LT, Dickey JF, Bodine AB. In vitro mycotoxin binding to bovine uterine
steroid hormone receptors. Theriogenology. 1982;17:325-31.
Blount WP. ‘Turkey’ X diseases’. Journal of the British Turkey Federation. 1961;9:52-
8.
BML (The German federal Ministry of Nutrition AaF). Orientation values for critical
concentrations of deoxynivalenol and zearalenon in diets for pigs, ruminants and gal-
linaceous poultry. VDM. 2000;27/00:2-3.
Böhm KH. Entwicklungsbedingungen für toxinbildende Pilze. Dtsch Tierärztl Wsch.
1989;96:339-41.
Böhm J. Über die Bedeutung der Mykotoxine Desoxynivalenol, Zearalenon und Och-
ratoxin A für landwirtschaftliche Nutztiere. Arch Anim Nutr. 1992;42:95-111.

Literaturverzeichnis
86
Boland EW, Anderson VL, Caspet HH, Berg PT, Dhuyvetter DV. The effects of in-
creasing vomitoxin (DON) levels from scab infested barley fed to feedlot steers. Beef
Prod. Field Day. Carrington Res. Ext. Ctr. Livestock Unit. North Dakota State Univer-
sity Ag Expt Sta. 1994;17:34.
Bostedt H. Untersuchungen über den Calcium-, anorganischen Phosphat- und Mag-
nesiumgehalt im Blutserum von Rindern in Abhängigkeit vom Schweregrad der Ge-
burt. Fortschritte der Veterinärmedizin. 10. Kongressbericht. 1974;20:172-8.
Bottalico A. Fusarium diseases of cereals: Species complex and related mycotoxin
profiles in Europe. J Plant Pathol. 1998;80:85-103.
Braun U, Gerber D. Percutaneous ultrasound-guided cholecystocentesis in cows. Am
J Vet Res. 1992;53:1079-84.
Busato A, Faissler D, Küpfer U, Blum JW. Body Condition Scores in Dairy Cows: As-
sociations with Metabolic and Endocrine Changes in Healthy Dairy Cows. J Vet Med
A. 2002;49:455-60.
Charmley E, Trenholm HL, Thompson BK, Vudathala D, Nicholson JW, Prelusky DB,
et al. Influence of level of deoxynivalenol in the diet of dairy cows on feed intake, milk
production, and its composition. J Dairy Sci. 1993;76:3580-7.
Chelkowski J. Mycotoxins in agriculture and food safety. In: Sinha KK, Bhatnager D.
editors. Mycopathologia. New York: Marcel Dekker Inc.; 1998.
Citil M. Untersuchungen zur Carnitinkonzentration und zu weiteren biochemischen
Parametern im Blutserum bei Milchkühen mit besonderer Berücksichtigung der Dis-
locatio abomasi und von Puerperalstörungen. [Dissertation med. vet]. Leipzig: Univ.
Leipzig; 1999.
Constable PD, Miller GY, Hoffsis BL, Rings DM. Risk factors for abomasal volvulus
and left abomasal displacement in cattle. Am J Vet Res. 1992;53:1184-92.

Literaturverzeichnis
87
Coppock RW, Mostrom MS, Sparling CG, Jacobsen B, Ross SC. Apparent
zearalenone intoxication in a dairy herd from feeding spoiled acid-treated corn. Vet
Hum Toxicol. 1990;32:246-8.
Cote LM, Dahlem AM, Yoshizawa T, Swanson SP, Buck WB. Excretion of deoxyniva-
lenol and its metabolite in milk, urine and feces of lactating dairy cows. J Dairy Sci.
1986;69:2416-23.
Coulombe RA Jr. Biological action of mycotoxins. J Dairy Sci. 1993;76:880-91.
Dänicke S, Valenta H, Ueberschär KH. Risikoabschätzung und Vermeidungsstrate-
gien bei der Fütterung. In: Risikofaktoren für die Fusariumtoxinbildung und Vermei-
dungsstrategien bei der Futtermittelerzeugung und Fütterung. Landbauforschung
Völkenrode, Sonderheft Nr. 216, Hrsg. S. Dänicke und E. Oldenburg 2000:35-138.
Dänicke S, Ueberschär KH, Halle I, Valenta H, Flachowsky G. Excretion kinetics and
metabolim of zearalenon in broilers in dependence on a detoxifying agent. Arch Anim
Nutr. 2001;55:299-313.
Dänicke S, Gadeken D, Ueberschär KH, Meyer U, Scholz H. Effects of Fusarium
toxin contaminated wheat and of a detoxifying agent on performance of growing
bulls, on nutrient digestibility in wethers and on the carry over of zearalenone. Arch
Tierernähr. 2002;56:245-61.
Dänicke S, Valenta H. Mykotoxine im Futter- Gefahr für landwirtschafliche Nutztiere.
Forschungsreport 2004:10-3.
Dänicke S, Matthaus K, Lebzien P, Valenta H, Stemme K, Ueberschär KH, et al. Ef-
fects of Fusarium toxin-contaminated wheat grain on nutrient turnover, microbial pro-
tein synthesis and metabolism of deoxynivalenol and zearalenone in the rumen of
dairy cows. J Anim Physiol Anim Nutr (Berl). 2005;89:303-15.
Dänicke S, Beineke A, Goyarts T, Valenta H, Beyer M, Humpf HU. Effects of a Fusa-
rium toxin-contaminated triticale, either untreated or treated with sodium metabisul-

Literaturverzeichnis
88
phite (Na2S2O5, SBS), on weaned piglets with a special focus on liver function as de-
termined by the 13C-methacetin breath test. Arch Anim Nutr. 2008 Aug;62:263-86.
DeHaan K, Stock R, Brink D, Klopfenstein T, Schneider N. Scabby wheat-influence
on performance. Nebraska Beef Cattle Report MP. 1984;47:6.
Delling U. Intraoperative Ultraschalluntersuchung der Leber und der Gallenblase des
Rindes. [Dissertation med. vet]. Leipzig: Univ. Leipzig; 2000.
Dicostanzo A, Meiske JC, Murphy M, Epley R, Felice L, Johnston L, et al. Effects of
Feeding vomitoxin-contaminated barley on performance of feedlot steers. Minnesota
Cattle Feeders Rep. 1995;B-418:6-8.
Dicostanzo A, Johanston L, Windels H, Murphy M. A Review of the Effects of Molds
and Mycotoxins in Ruminants. The Professional Animal Scientist. 1996;12:138-50.
Dirksen G. Die Erweiterung, Verlagerung und Drehung des Labmagens beim Rind.
Zentralbl Vet Med. 1961;8:101-3.
D´Mello JPF, Placinta CM, McDonald AMC. Fusarium mycotoxins: a review of global
implications for animal health, welfare and productivity. Anim Feed Sci Tech.
1999;80:183-205.
Drackley JK. Die Rolle von Ernährung und Management für die Prävention von
Stoffwechselerkrankungen der Milchkuh im peripartalen Zeitraum. In: Bundesver-
band Praktischer Tierärzte e. V. (BPT), Hrsg. Vortragszusammenfassungen: BPT-
Kongress; 2002 Nov 7-10; Nürnberg, Germany. 2002;46-60.
Drochner W. Aktuelle Aspekte zur Wirkung von Phytohomonen, Mykotoxinen und
ausgewählten schädlichen Pflanzeninhaltsstoffen auf die Fruchtbarkeit beim weibli-
chen Rind. Übers Tierernährung. 1990;18:177-96.
Duarte D. Das Geheimnis von Mykotoxinen: Neue Wege zu Lösung des Problems.
Alltechs 17. Europäische Vortragveranstaltung 2003:39-50.

Literaturverzeichnis
89
Eades SC. Endotoxaemia in Dairy Cattle: Mechanism of Reticulorumen Stasis. The
Vet J. 1997;153:321-7.
Edmonson AJ, Lena IJ, Weaver LD, Farver T, Webster G. A body condition scoring
chart for Holstein dairy-cows. J Dairy Sci. 1989;72:68-78.
Enders C. Zur Analytik von Zearalenon, alpha- und beta-Zearalenol in Material
pflanzlichen und tierichen Ursprungs [Dissertation med. vet]. München: Univ. Mün-
chen; 1984.
Enemark JMD, Jorgensen RJ, Enemark PS. Rumen acidosis with special emphasis
on diagnostic aspects of subclinical rumen acidosis: a review Veterinarija ir Zootech-
nika. 2002;20: 16-27.
Engelhardt G. Mykotoxine - Giftige Stoffwechselprodukte von Schimmelpilzen 2000:
<http://www.lfe.bayern.de/lebensmittel/mykotox.html>. In Bayerische Landesanstalt
für Ernährung (Hrsg).
EU (2006): Empfehlung der Kommission vom 17. August 2006 betreffend das Vor-
handensein von Deoxynivalenol, Zearalenon, Ochratroxin A, T-2- und HT-2-Toxin
sowie von Fumonisinen in zur Verfütterung an Tiere bestimmten Erzeugnissen (Text
von Bedeutung für den EWR) (2006/576/EG) Amtsblatt der Europäischen Union L
229/7 (23.8.2006).
Eulenberger K. Physiologische und pathophysiologische Grundlagen für Maßnahmen
zur Steuerung des Puerperiums beim Rind. [Habilschr. Med. vet]. Leipzig: Univ Leip-
zig; 1984.
Fink-Gremmels J. Mycotoxins: their implications for human and animal health. The
Veterinary Quarterly. 1999;21:115-20.
Fink-Gremmels J. Mycotoxins in forages. In: Diaz DE. Ed. The Mycotoxin Blue Book.
Nottingham University Press, Nottingham, United Kingdom, 2005:249–68.

Literaturverzeichnis
90
Fink-Gremmels J. Mycotoxins in cattle feeds and carry-over to dairy milk: a review.
Food Addit Contam Part A Chem Anal Control Expo Risk Assess. 2008a Feb;25:172-
80.
Fink-Gremmels J. The role of mycotoxins in the health and performance of dairy
cows. Vet J. 2008b Apr;176:84-92.
Frank K. Enzymatisch Untersuchungen bei verschiedenen Erkrankungen der
Schweine in der tierärzlichen Praxis [Dissertation med. vet]. München: Univ.
München; 1975.
Fuchs E, Binder EM, Heidler D, Krska R. Structural characterization of metabolites
after the microbial degradation of type A trichothecenes by the bacterial strain BBSH
797. Food Addit Contam. 2002;19:379-86.
Fürll M. Vorkommen, Ätiologie, Pathogenese, Diagnostik und medikamentelle Beein-
flussung von Leberschäden beim Rind. [Habilschr. Med. vet]. Leipzig: Univ. Leipzig.
1989.
Fürll M, Schäfer M. Lipolyse und Hyperbilirubinämie- ein Beitrag zur Pathogenese
des Ikterus. Monatsh Veterinärmed. 1992;47:181-6.
Fürll M, Eckermann W, Amin M. Säure-Basen- und Mineralstoff-Haushalt bei gesun-
den Kühen im geburtsnahen Zeitraum. Tierärztl Umsch. 1994;49:107-15.
Fürll M, Schmidt E, Jäkel L, Dabbagh MN, Schwarzer U, Leidel T, et al. Zum Vor-
kommen der Dislocatio abomasi in Ostdeutschland. Tierärztl Umsch. 1996;51:211-5.
Fürll M, Kastner A, Fritsche D, Kleiser L, Jäkel L, Richter V, et al. Fettstoffwechsel-
Endotoxin- Interaktionen bei Kühen und deren mögliche pathogenetische für die Ent-
stehung der Dislocatio abomasi. In: Fürll M. Hrsg. Stoffwechselbelastung, -diagnostik
und -stabilisierung beim Rind, Leipziger Samstagsakademie, 1998 Apr 25; Leipzig:
Medizinische Tierklinik der Universität Leipzig; 1998a, S.105-16.

Literaturverzeichnis
91
Fürll M, Bialek N, Kirbach H, Gruys E, Jäkel L, Krüger M. Verhalten Akuter Phasen
Proteine (APP) sowie weiterer klinisch-chemischer Parameter im Vorfeld der Disloca-
tio abomasi (DA) bei Milchkühen. In: Fürll M. Hrsg. Stoffwechselbelastung, -
diagnostik und -stabilisierung beim Rind, Leipziger Samstagsakademie, 1998 Apr 25;
Leipzig: Medizinische Tierklinik der Universität Leipzig; 1998b, S.182-99.
Fürll M, Krüger M. Alternative Möglichkeiten zur Prophylaxe der Dislocatio abomasi
(DA) beim Rind. Prakt. Tierarzt, Colleg. Vet. XXIX 1999a;81-90.
Fürll M, Krüger M. Zur Entstehung der geburtsnahen Labmagenverlagerung bei Kü-
hen. Lohmann Information 1999b;4/99:19-26.
Fürll M, Bialek N, Dabbagh MN, JÄKEL L, Kastner A, Kleiser L, et al. Stertenbrinck
W. Zur Pathogenese der Dislocatio abomasi. Großtierpraxis 2000;1:6–17.
Fürll M. Dislocatio abomasi (DA) beim erwachsenen Rind. In: Wiesner E. Hrsg.
Handlexikon der Tierärztlichen Praxis, Enke Verlag Stuttgart, 2001:181 xa-181 xw
(23 S).
Fürll M, Leidel I. Untersuchungen zur Gesundheitsstabilisierung im peripartalen Zeit-
raum bei Milchkühen. Tierärztl Umsch. 2002;57:423-38.
Fürll M. Damit der Stoffwechsel auch in kritischen Phasen nicht verrückt spielt. In:
Fürll M. Hrgb. Stoffwechselstörungen bei Wiederkäuern: Erkennen - Behandeln-
Vorbeugen, Leipzig: Medizinische Tierklinik, Universität Leipzig; 2002a, S. 2-12.
Fürll M, Jäckel F. Glucocorticoideinflüsse auf Parameter des Fett- und Leberstoff-
wechsels, auf hämatologische Parameter sowie auf die Milchleistung bei Hochleis-
tungskühen in der Frühlaktation. Berl Münch Tierärztl Wschr. 2005;118:2-10.
Fürll M. Hämatologie. In: Kraft W, Dürr UM. Hrsg. Klinische Labordiagnostik in der
Tiermedizin, 6. Aufl. Stuttgart, New York: Schattauer; 2005a;S.49-86.

Literaturverzeichnis
92
Fürll M. Spezielle Untersuchungen beim Wiederkäuer. In: Kraft W, Dürr UM. Hrsg.
Klinische Labordiagnostik in der Tiermedizin, 6. Aufl. Stuttgart, New York: Schattau-
er, 2005b:444-71.
Fürll M, Fürll B. Puerperale Septikämie- Ursachen, Diagnostik, Therapie und Prophy-
laxe. Nutztierpraxis Aktuell. 2006:59-66.
Gabel AA, Heath RB. Correction and right-sided omentopexy in treatment of left-
sided displacement of the abomasum in dairy cattle. J Am Vet Med Ass.
1969;155:632-41.
Gareis M, Wolff J. Relevance of mycotoxin contaminated feed for farm animals and
carryover of mycotoxins to food of animal origin. Mycoses. 2000;43 Suppl 1:79-83.
Gedek B. Kompendium der medizinischen Mykologie. 1. Aufl. Verlag Paul Parey,
Berlin-Hamburg. 1980;S.324-33.
Gedek B. Fusarienbesatz und Mykotoxinbelastung der Grundfuttermittel. DLG-
Forschungsbericht über Tierernährung. 1985.
Gibson JP, Field AC, Wiener G. Concentrations of blood constituents in genetically
high and low milkproductionlines of British Friesian and Jersey cattle around calving
and in early lactation. Anim Prod. 1987;44:183-99.
Goff JP. Pathophysiology of calcium and phosphorus disorders. Vet Clin North Am
Food Anim Pract. 2000;16:319-37.
Hadley G, Wolf C, Harsh S. Dairy cattle culling patterns, explanations, and implica-
tions. J Dairy Sci. 2006;89:2286–96.
Haima P. Knochenspezifische alkalische Phosphatase (BAP): ein biochemischer
Marker für den Knochenaufbau. OSTEOmedical Group/TECOmedical AG, Switzer-
land. 2006;06:1-33.

Literaturverzeichnis
93
Hiebl B. Untersuchungen zur Bedeutung erhöhter Bilirubin-Konzentrationen und Glu-
tamatdehydrogenase-Aktivitäten im Blutserum klinisch kranker Kühe [Dissertation
med. vet]. Leipzig: Univ Leipzig; 2005.
Hochsteiner W, Schuh M, Luger K, Baumgartner W. Effect of mycotoxin contami-
nated feed on production parameters of dairy cows. Berl Münch Tierärztl Wschr.
2000;113:14-21.
Höltershinken M, Maiworm K, Scholz H. Mykotoxikosen beim Rind- Problem auch in
Norddeutschland. Prakt Tierarzt Colleg Vet. 1996;77:9-14.
Hoops M. Peripartaler Stoffwechsel und Morbidität bei Hochleistungskühen während
eines Jahres [Dissertation med. vet]. Leipzig: Univ. Leipzig; 2007.
Hörügel U, Fürll M. Untersuchungen zur Früherkennung der Gebärparesegefährdung
bei Kühen. Prakt Tierarzt Colleg Vet. 1998;XXVIII:86-92.
Jaksch W, Glawischnig E. Klinische Propädeutik der inneren Krankheiten und Haut-
krankheiten der Haus- und Heimtiere. 3. Aufl. Berlin, Hamburg: Verlag Paul Parey;
1990.
Janz U. Untersuchungen zum Vorkommen von Fusarien und deren Toxinen in Wei-
zenkorn und Weizenstroh nach Einsatz fungizider Wirkstoffe [Dissertation med. vet].
Hannover: Tierärztliche Hochschule Hannover; 1999.
JECFA. Joint FAO/WHO Expert committee on food additives. Fifty-sixth meeting.
Geneva, 6-15 February 2001. p.1-33.
<http://www.who.int/pcs/jecfa/JECFA_publications.htm>.
Kallela K, Vasenius l. The effects of rumen fluid on the content of zearalenone in
animal fodder. Nord Vet Med. 1982;34:336-9.
Kamphues J. Fütterungsbedingte Gesundheitsstörungen in Nutztierbeständen. Übers
Tierernährung. 1991;19:247-72.

Literaturverzeichnis
94
Kappel LC, Ingraham RH, Morgan EB, Zeringue L, Wilson D, Babcock DK. Relation-
ship between fertility and blood glucose and cholesterol concentrations in Holstein
cows. Am J Vet Res. 1984;45:2607-12.
Karlson P. Hormonrezeptoren und Hormonwirkung. Klin Wschr. 1979;57:607-12.
Karsai F, Schäfer M. Diagnostische Erfahrungen bei metabolisch bedingten Leber-
krankheiten der Milchkühe. Mh Vet Med. 1984;39:181-6.
Kastner A. Untersuchungen zum Fettstoffwechsel und Endotoxin-Metabolismus bei
Milchkühen vor dem Auftreten der Dislocatio abomasi [Dissertation med. vet]. Leip-
zig: Univ. Leipzig; 2002.
Katzenellenbogen BS, Katzenellenbogen JA, Mordecai D. Zearalenones: Characteri-
sation of the estrogenic potenties and receptor interactions of a series of fungal -
resorcylic acid lactones. Endocrinology. 1979;105:33-40.
Kawabata Y, Tashiro F, Ueno Y. Synthesis of a specific protein induced by
zearalenone and its derivatives in rat uterus. J Biochem. 1982;91:801-8.
Keese C, Meyer U, Valenta H, Schollenberger M, Starke A, Weber I, et al. No carry
over of unmetabolised deoxynivalenol in milk of dairy cows fed high concentrate pro-
portions. Mol Nutr Food Res. 2008;52:1514-29.
Keese C, Meyer U, Rehage J, Spilke J, Boguhn J, Breves G, et al. On the effects of
the concentrate proportion of dairy cow rations in the presence and absence of a
fusarium toxin-contaminated triticale on cow performance. Arch Anim Nutr. 2008a
Jun;62:241-62.
Keese C, Meyer U, Rehage J, Spilke J, Boguhn J, Breves G, et al. Ruminal fermen-
tation patterns and parameters of the acid base metabolism in the urine as influenced
by the proportion of concentrate in the ration of dairy cows with and without Fusarium
toxin-contaminated triticale. Arch Anim Nutr. 2008b Aug;62:287-302.

Literaturverzeichnis
95
Keese C. Investigations on the interactions between the concentrate proportion in the
ration of dairy cows and the presence or absence of a Fusarium toxin-contaminated
triticale on animal health, performance and deoxynivalenol residues in serum, bile
and milk [Dissertation med. vet]. Hannover: Tierärtzliche Hochschule Hannover;
2008c.
Kennedy DG, Hewitt SA, Mcevoy JD, Currie JW, Cannavan A, Blanchflower WJ et al.
Zeranol is formed from Fusarium spp. toxins in cattle in vivo. Food Additives and
Contaminants. 1998;15:393-400.
Kiang DT, Kennedy BJ, Pathre SV, Mirocha CJ. Binding characteristics of
zearalenone analogs to estrogen receptors. Cancer Res. 1978;38:3611-5.
King RR, McQueen RE, Levesque D, Greenhalgh R. transformation of deoxyniva-
lenol (vomitoxin) by rumen microorganisms. J Agric Food Chem. 1984;32:1181-3.
Kiessling KH, Petterson H, Sandholm K, Olsen M. Metabolism of aflatoxin, ochra-
toxin, zearalenone, and three trichothecenes by intact rumen fluid, rumen protozoa,
and rumen bacteria. Appl Environ Microbiol. 1984;47:1070-3.
Kleinova M, Zollner P, Kahlbacher H, Hochsteiner W, Lindner W. Metabolic profiles
of the mycotoxin zearalenone and of the growth promoter zeranol in urine, liver, and
muscle of heifers. J Agric Food Chem. 2002;50:4769-76.
Kleiser L, Fürll M. Screening zur Früherkennung einer Disposition für die Dislocatio
abomasi bei Kühen. In: Fürll M. Hrsg. Stoffwechselbelastung, -diagnostik, -
stabilisierung beim Rind. Leipzig. 1998.
Klimiene I, Spakauskas V, Matusevicius A. Correlation of different biochemical
parameters in blood sera of healthy and sick cows. Vet Res Commun. 2005;29:95-
102.
Koch P. State of the art of trichothecenes analysis. Toxicol Lett. 2004;153:109-12.

Literaturverzeichnis
96
Kofler J. Beziehungen zwischen Fütterung und Gliedmaßenerkrankungen bei Rin-
dern. in: 28. Viehwirtschaftliche Fachtagung Gumpenstein. 2001.
Kolb E. Lehrbuch der Physiologie der Haustiere. 5. Aufl. München: Verlag Urban &
Fischer; 1989.
Koller A, Reist M, Blum JW, Küpfer U. Time empty and ketone body status in the
early postpartum period of dairy cows. Reprod Domest Anim. 2003;38:41-9.
Korosteleva SN, Smith TK, Boermans HJ. Effects of Feedborne Fusarium Mycotox-
ins on the Performance, Metabolism, and Immunity of Dairy Cows. J. Dairy Sci. 2007;
90:3867–73.
Kraft W, Dürr U. Klinische Labordiagnostik in der Tiermedizin. 6. Aufl. Stuttgart, New
York: Schattauer; 2005.
Kretzschmar AM. Die Aktivität der g-Glutamyltransferase im Serum des Rindes- eine
retrospektive Studie [Dissertation med. vet]. Leipzig: Univ. Leipzig; 2008.
Krska R, Baumgartner S, Josephs R. The state-of-the-art in the analysis of type-A
and -B trichothecene mycotoxins in cereals. Fresenius J Anal Chem. 2001;371:285-
99.
Krska R, Schubert-Ullrich P, Molinelli A, Sulyok M, MacDonald S, Crews C. My-
cotoxin analysis: an update. Food Addit Contam Part A Chem Anal Control Expo
Risk Assess. 2008;25:152-63.
Kume S, Toharmat T, Bobayashi N. Effect of restricted feed intake of dams and heat-
stress on mineral status of newborn calves. J Dairy Sci. 1998;81:1581-90.
Kurtz HJ, Nairn GH, Nelson GH, Christensen CM, Mirocha CJ. Histologic changes in
the genital tracts of swine fed estrogenic mycotoxin. Am J Vet Res. 1969;30:551-6.

Literaturverzeichnis
97
Lacey J, Magan N. Fungi in cereal grains: Their occurence and water and tempera-
ture relationships. In: Chelowski J, editor. Cereal grain-Mycotoxins, fungi and quality
in drying and storage. Amsterdam: Elsevier Science B. V.; 1991. p. 77-118.
Lachmann G. Ätiologie, Pathogenese, Diagnostik, Therapie und Prophylaxe der ali-
mentären metabolischen Azidose des Rindes [Habilschr. med. vet]. Leipzig: Universi-
tät Leipzig; 1981.
Leibetseder J. Mykotoxikosen - eine kurze Einführung. Übers Tierernährung.
1981;9:1-10.
Leibetseder J. Mykotoxinprobleme in der Schweineproduktion. 11. Aufl. 1982:428-32.
Lepom P, Knabe O, Baath O. Vorkommen von Fusarium-Arten und ihren Mykotoxi-
nen auf Silomais. 6. Bildung von Zearalenon und Trichthecenen (TypA) durch ein-
heimische Fusarium-Isolate. Arch Anim Nutr. 1990;40:871-83.
Lepschy J. Fusarientoxine in Getreide - Ihre Entstehung und Vorbeugmaßnahmen.
Gesunden Pflanzen 1992;44:35-40.
Lerch C. Metabolismus von Ochratoxin A und Zearalenon in Pansenflüssigkeit von
Milchkühen in Vitro [Dissertation med. vet]. Stuttgart: Univ. Stuttgart-Hohenheim;
1990.
Ling PR, Smith RJ, Bistrian BR. Hyperglycemia enhances the cytokine production
and oxidative responses to a low but not high dose of endotoxin in rats. Crit Care
Med. 2005;33:1084-9.
Lotthammer KH, Boehnke HJ, Morawietz M. Beziehungen zwischen verschiedenen
Blutparametern als Kriterien für Stoffwechselstörungen und dem Milchzellgehalt bei
Milchrindern. Dtsch Tierärztl Wschr. 1988;95:379-84.
Mansfeld R, Grunter E, Kautni J. Mykotoxine als Bestandproblem bei Milchkühen -
ein Fallbericht. Mh Vet Med. 1989;44:409-12.

Literaturverzeichnis
98
Marasas WFO, Nelson PE, Toussoun TA. Toxigenic Fusarium species: Identity and
mycotoxicology. The Pennsylvania State University Press, University Park and Lon-
don.1984:328 pp.
Matthäuse K, Dänicke S, Lebzien P, Valenta H, Ueberschär KH, Flachowski G. On
the effects of a Fusarium-contaminated wheat and the feed intake level on ruminal
fermentation and toxin-turnover of cows. 26. Mykotoxin-Workshop Herrsching am
Ammersee 17.-19. Mai 2004 Tagungsband. 2004:38.
McCoy MA. Hypomagnesaemia and new data on vitreous humour magnesium con-
centration as a post-mortem marker in ruminants. Magnes Res. 2004; 17:137-45.
Meyer K, Usleber E, Dietrich R, Märtlbauer E, Bauer J. Zearalenone metabolites in
bovine bile. Archiv für Lebensmittelhygiene. 2002;53:115-7.
Milczewski KEV, Engel G, Teuber M. Übersicht über die wichtigen toxinbildenden
Schimmelpilze und ihre Toxine. In: Reiß J. Mykotoxine in Lebensmitteln. 1981:13-85.
Miles CO, Erasmuson AF, Wilkins AL, Towers NR, Smith BL, Garthwaite I et al.
Ovine Metabolism of Zearalenone to Zearalenol (Zeranol). J Agric Food Chem.
1996;44:3244-50.
Miller NJ, Sampson J, Candeisas LP, Rice-Evans CA. Antioxidant activities of caro-
tenes and xanthophylls. FEBS Lett. 1996;384:240-2.
Mirocha CJ, Pathere SV, Christensen CM. Chemistry of Fusarium and Stachybotrys
mycotoxins. 1977:365-420.
Mitterbauer R, Wondrasch K, Peruci M, Berthiller F, Schaumacher R, Kraska R et al.
Biosynthese und Metabolismus von Zearalenon. ALVA-Mitteilungen 2005;2:22-4.
Moeser S. Effects of zearalenone and zeranol in heifers [Dissertation med. vet].
Wien: Veterinärmedizinische Universität Wien; 2001.

Literaturverzeichnis
99
Morooka N, Uraguchi N, Yoshizzawa T, Yamamoto H. Studies on the toxic substan-
ces in barley infected with Fusarium spp. Jpn J food Hyg. 1972;13:368-75.
Müller HM. Zearalenon-Ein Östrogen Wirksames Mykotoxin. Übers Tierernärung.
1978;6:265-300.
Müller HM. Entgiftung von Mykotoxinen. 1. Physikalisches Verfahren. Übers Tierer-
nährung. 1982;10:95-122.
Müller U. Untersuchungen über das Vorkommen und die Diagnostik der Labmagen-
verlagerung beim Rind. [Dissertation med. vet]. Leipzig: Univ. Leipzig; 1992.
Nelson M, Schneider NR, Doster AR, Carlson MP, Klopfenstein T. Vomitoxinconta-
minated wheat-pathology, toxicity in cattle. Nebraska Beef Cattle Rep. MP
1984;47:3-12.
Niessen L, Vogel RF. Group specific PCR-detection of potential trichothecen- Pro-
ducing Fusarium-Species in pure cultures and cereal samples. System. Appl Micro-
biol. 1998;21:618-31.
Nocek JE. Bovine Acidosis: Implications on Laminitis. J Dairy Sci. 1997;80:1005-28.
Oldenburg E, Valenta H, Sator C. Risikoabschätzung und Vermeidungsstrategien bei
der Futtermittelerzeugung. Landbauforschung Völkenrode, Sonderheft. 2000;216:5-
34.
Oswald I, Bouhet S, Marin DE, Pinton P, Taranu I. Pilzgifte im Schweinefutter – Aus-
wirkungen von Mykotoxinen auf das Immunsystem von Schweinen. Großtierpraxis.
2003;4:5-10.
Palti J. Toxigenic Fusaria, their distribution and significance as causes of disease in
animal and man. Berlin und Hamburg: Verlag Paul Parey; 1978:49.
Palyusik M. Mykotoxikosen (2. Teil). Wien Tierärztl Mh. 1977;64:259-66.

Literaturverzeichnis
100
Papadopoulou-Bouraoui A, Vrabcheva T, Valzacchi S, Stroka J, Anklam E. Screen-
ing survey of deoxynivalenol in beer from the European market by an enzyme-linked
immunosorbent assay. Food Addit Contam. 2004;21:607-17.
Payne JM, Rowlands GJ, Manston R, Dew SM, Parker WH. A statistical appraisal of
the results of the metabolic profile tests on 191 herds in the B.V.A.-A.D.A.S Joint Ex-
ercise in Animal Health and Productivity. Br Vet J. 1974;130:34-44.
Pestka JJ, Lin WS, Miller ER. Emetic activity of the trichothecene 15-
acetyldeoxynivalenol in swine. Food Chem Toxicol. 1987;25:855-8.
Pier AC, Richard JL, Cysewski SJ. Implications of mycotoxins in animal diseases. J
Am Vet Med Ass. 1980;176:719-24.
Ping HE, Young LG, Forsberg C. Microbial Transformation of Deoxynivalenol (Vomi-
toxin). Appl Environ Microbiol. 1992;58:3857-63.
Prelusky DB, Trenholm HL, Lawrence GA, Scott PM. Nontransmission of deoxyniva-
lenol (vomitoxin) to milk following oral administration to dairy cows. J Environ Sci
Health B. 1984;19:593-609.
Prelusky DB, Warner RM, Trenholm HL. Sensitive analysis of the mycotoxin
zearalenone and its metabolites in biological fluids by high-performance liquid chro-
matography alpha. J Chromatogr. 1989;29:267-77.
Prelusky DB, Scott PM, Trenholm HL, Lawrence GA. Minimal transmission of
zearalenone to milk of dairy cows. J Environ Sci Health B25.1990;1:87-103.
Prelusky DB. A study on the effect of deoxynivalenol on serotonin receptor binding in
pig brain membranes. J Environ Sci Health B. 1996;31:1103-17.
Ranfft K, Gerst R, Mayer G. Determination and occurrence of zearalenone in cereals
and mixed feeds. European Food Research and Technology. 1990;191:449-53.

Literaturverzeichnis
101
Rehage J, Mertens M, Stockhofe-Zurwieden N, Kaske M, Scholz H. Post surgical
convalescence of dairy cows with left abomasal displacement in relation to fatty liver.
Schweiz Arch Tierheilk. 1996;138:361-8.
Reiß J. Schimmelpilze - Lebensweise, Nutzen, Schaden, Bekämpfung. 2. Aufl. Ber-
lin: Springer Verlag; 1998.
Rohrbach BW, Cannedy AL, Freeman K, Slenning BD. Risk factors for abomasal
displacement in dairy cows. J Am Vet Med Ass. 1999;214:1660-63.
Rotter BA, Prelusky DB, Pestka JJ. Toxicology of deoxynivalenol (vomitoxin). J Toxi-
col Environ Health. 1996;48:1-34.
Rowlands GJ, Little W, Manston R, Dew SM. The effect of season on the composi-
tion of the blood of lactating and non-lactating cows as revealed from repeated meta-
bolic profile tests on 24 dairy herds. J Agric Sci Camb. 1974; 83:27-35.
Sabater-Vilar M, Maas RFM, De Bosschere H, Ducatelle R, Fink-Gremmels J. Patulin
produced by an Aspergillus clavatus isolated from feed containing malting residues
associated with a lethal neurotoxicosis in cattle. Mycopathologia 2004;158:419-26.
Sator M, Sommer W. Werden Mykotoxine in der Schweinfütterung zum Problem?
2003.
<http://www.landwirtschaftskammer.de/fachangebot/tierproduktion/schweinehaltung/f
uetterung/mykotoxine.htm>.
Sattler T. Untersuchungen zum antioxidativen Status von Kühen mit Labmagenverla-
gerung [Dissertation med. vet]. Leipzig: Univ. Leipzig; 2001.
Sattler T, Fürll M. Creatine Kinase and Aspartate Aminotransferase in cows as indi-
cators for endometritis. J Vet Med Ass. 2004;51:132-7.

Literaturverzeichnis
102
SCF/CS/CNTM/MYC. Scientific committee on food of the European commission
(SCF) (1999): Opinion on Fusarium toxins. Part 1: Deoxynivalenol (DON). SCIEN-
TIFIC COMMITTEE ON FOOD. 1999.
Schalm O. Ein neuer Mastitis-Test. Tierärztl Umsch. 1960;15:151-4.
Schnerr H. Quantitativer Nachweis von Deoxynivalenol und Trichothecene-bildenden
Fusarium spp. mit Biosensor und PCR in Getreide. [Dissertation rer. nat]. München:
TU München; 2002.
Schreiber K. Der Xyloseresorptionstest bei gesunden und kranken Kühen. [Disserta-
tion med. vet]. Leipzig: Univ. Leipzig; 2006.
Schuh M. Klinische Auswirkungen der in Österreich vorkommenden Mykotoxine.
Wien Tierärztl Mh. 1981;68:308-12.
Schuh M, Baumgartner W. Mycrobiological and mycotoxicological feedstuffs as dis-
ease causing agents in cattle. Wien Tierärztl Mh. 1988;75:329-33.
Schuh M. The significance of mycotoxin assimilation for the productivity and health of
animals. Dtsch Tierärztl Wschr. 1989;96:353-5.
Schuh M. Schimmelpilzgifte - Probleme beim Rind. Der fortschrittliche Landwirt
(Verminderung von Futterverpilzungen). 1996;21:7-8.
Schwadorf K. Pilzbesatz und Mykotoxine in Futtermitteln [Dissertation med. vet].
Stuttgart: Univ. Stuttgart-Hohenheim; 1995.
Schweighardt H. Mikrobielle bedingte Ernährungsschäden beim Geflügel und deren
Nachweis. Wien Tierärztl Mh. 1980;67:316-8.
Seeling K. Investigations of the interactions between feed intake level (passage rate)
and Fusarium contaminated wheat on ruminal nutrient fermentation as well as on

Literaturverzeichnis
103
metabolism and carry over of deoxynivalenol and zearalenone into the milk [Disserta-
tion med. vet]. Halle-Wittenberg. Univ. Martin-Luther; 2005.
Seeling K, Dänicke S, Gädeken D, Lebzien P, Valenta H, Uberschär KH, et al. On
the effects of Fusarium-contaminated wheat and the feed intake level on ruminal
fermentation and toxin -turnover of cows. Mycotoxin Res. 2005a;21:132-35.
Seeling K, Danicke S, Ueberschar KH, Lebzien P, Flachowsky G. On the effects of
Fusarium toxin-contaminated wheat and the feed intake level on the metabolism and
carry over of zearalenone in dairy cows. Food Addit Contam. 2005b;22:847-55.
Seeling K, Lebzien P, Danicke S, Spilke J, Sudekum KH, Flachowsky G. Effects of
level of feed intake and Fusarium toxin-contaminated wheat on rumen fermentation
as well as on blood and milk parameters in cows. J Anim Physiol Anim Nutr (Berl).
2006a;90:103-15.
Seeling K, Dänicke S, Valenta H, Van Egmond HP, Schothorst RC, Jekel AA, et al.
Effects of Fusarium toxin-contaminated wheat and feed intake level on the biotrans-
formation and carry-over of deoxynivalenol in dairy cows. Food Addit Contam. 2006b
Oct;23:1008-20.
Setz M. Die Bedeutung der Lipoproteine [Dissertation med. vet]. Leipzig: Univ. Leip-
zig; 2000.
Shaffer L, Roussel JD, Koonce KL. Effects of age, temperature-season and breed on
blood characteristics of dairy cattle. J Dairy Sci. 1981;64:62-70.
Shalit U, Maltz N, Silanikove N, Berman A. Water, sodium, potassium, and chlorine
metabolism of dairy cows at the onset of lactation in hot weather. J Dairy Sci.
1991;74:1874-83.
Sieverding E. Nicht ganz so einfach wie man denkt. Mykotoxine in der Gallenflüssig-
keit. Nutztierpraxis Aktuell. Ausgabe 7. Dezember 2003.

Literaturverzeichnis
104
Sorensen LK, Elbaek TH. Determination of mycotoxins in bovine milk by liquid chro-
matography tandem mass spectrometry. J Chromatogr B Analyt Technol Biomed Life
Sci. 2005;820:183-96.
Srikandakumar A, Johnson EH. Effect of heat stress on milk production, rectaltem-
perature, respiratory rate and blood chemistry in Holstein, Jersey and Australian Milk-
ing Zebu cows. Trop Anim Health Prod. 2004; 36:685-92.
Staufenbiel R, Johannsen U, Dargel H, Rossow N. Experimentelle Untersuchung zur
Leberverfettung der Milchkuh bei Futterrestriktion – Versuchsmodell und klinisch-
chemische Parameter. Mh Vet Med. 1992;47:559-66.
Staufenbiel R. Ansätze zur Prophylaxe der Labmagenverlagerung. In: Fürll M. Ätiolo-
gie, Pathogenese, Diagnostik, Prognose, Therapie und Prophylaxe der Dislocatioa-
bomasi. Proceedings Internationaler Workshop, 14. Oktober 1998: 291-309.
Staufenbiel R. Labmagenverlagerung- Ansätze zur Prophylaxe. Grosstierpraxis 2.
2001;9:31-40.
Steyn PS. The biosynthesis of mycotoxins. Rev Med Vet. 1998;149:469-78.
Swanson SP, Nicoletti J, Rood HD Jr, Buck WB, Cote LM, Yoshizawa T. Metabolism
of three trichothecene mycotoxins, T-2 toxin, diacetoxyscirpenol and deoxynivalenol,
by bovine rumen microorganisms. J Chromatogr. 1987;414:335-42.
Szkudelska K, Szkudelski T, Nogowski L. Short-time deoxynivalenol treatment in-
duces metabolic disturbances in the rat. Toxicol Lett. 2002;36:25-31.
Tarr B. Moulds and Mycotoxins - Effects of Mouldy Feed and Mycotoxins on Cattle
2006. Ministry of Agriculture, Food and Rural Affairs.
< http://www.omafra.gov.on.ca/english/livestock/dairy/herd/food/mico2.htm>.
Thalmann A. Mykotoxinen in Getreide. Angew Botanik. 1990;64:167-73.

Literaturverzeichnis
105
Thimme R, Opitz OG, Kreisel W. Cholestatische Lebererkrankungen: Diagnostik und
Therapie der primär biliären Zirrhose. Dtsch Med Wschr. 2002;127:1823-6.
Teufel EM. Verlaufsuntersuchungen zu Serum-Amyloid A, L-Carnitin sowie ausge-
wählte Stoffwechselparameter bei hochleistenden Milchkühen im peripartalen Zeit-
raum. [Dissertation med. vet]. Leipzig: Univ. Leipzig; 1999.
Trenholm HL, Thompson BK, Hartin KE, Greenhalgh R, McAllister AJ. Ingestion of
vomitoxin (deoxynivalenol) - contaminated wheat by nonlactating dairy cows. J Dairy
Sci. 1985;68:1000-5.
Uberschär KH. Einfluß von Zearalenon auf Wachstum und Rückstände in den Ge-
weben von Mastkaninchen. VDLUFA-Schriftenreihe. 1999;52:425-8.
Urry WH, Wehrmeister HL, Hodge EB, Hidy PH. The structure of zearalenone. Te-
thrahedron. 1966;27:3109-14.
Valenta H, Goll M. Determination of zearalenon, α-zearalenol and ß-zearalenol in
plasma and milk by HPLC and GC/MS. Institut für Tierernährung –
Bundesforschungsanstalt für Landwirtschaft (FAL) - Braunschweig. 1995:168-71.
Valenta H, Vemmer H. In vitro-Untersuchungen zum Metabolismus von Zearalenon
bei Inkubation mit Pansensaft. Mycotoxin Res. 1996;12:185-91.
Valenta H, Dänicke S, Döll S. Methode zur Bestimmung von Deoxynivalenon (DON)
und Deepoxy-deoxynivalenol (DOM-1) in Blutserum, Milch und Galle mit HPLC, Rei-
nigung über Immunoaffinitätssäulen (IAC). Mycotoxin Res. 2003;19:51-55.
Vanyi A, Szailer E. Investigation of the cytotoxic effect of F-2 toxin (zearalenone) in
various monolayer cell cultures. Acta Vet Acad Sci Hung. 1974;24:407-12.
Vlaminck K, van Meirhaeghe H, van den Hende C, Oyaert W, Muylle E. Einfluss von
Endotoxinen auf die Labmagenentleerung beim Rind. Dtsch Tierärztl Wschr.
1985;92:392-5.

Literaturverzeichnis
106
Völkl A, Volger B, Schollenberger M, Kalrovsky P. Microbial detoxification of my-
cotoxin deoxynivalenol. Basic Microbiol. 2004;44:147-56.
Weaver GA, Kurtz HJ, Behrens JC, Robison TS, Seguim BE, Bates FY, et al. Effect
of zearalenone on the fertility of virgin dairy heifers. Am J Vet Res. 1986a;47:1395-7.
Weaver GA, Kurtz HJ, Behrens JC, Robison TS, Segium BE, Bates FY, et al. Effect
of zearalenone on dairy cows. Am J Vet Res. 1986b;47:1826-8.
Weaver D. Subclinical Laminitis. in: 10th International Symposium on Lameness in
Ruminants Luzern. 1997.
Wemheuer W. Auswertung von Blutparametern aus fruchtbarkeitsgestörten Milch-
viehbeständen. Tierärztl Praxis. 1987;15:353-60.
Wenz J, Jensen S, Lombard J, Wagner B, Dinsmore R. Herdmanagement practices
and their association with bulk tank somatic cellcount on United States dairy opera-
tions. J of Dairy Sci. 2007;90:3652–59.
Wilken H. Endotoxin-Status und Antioxidative Kapazität sowie ausgewählte Stoff-
wechselparameter bei gesunden Milch- und Mutterkühen [Dissertation med. vet].
Leipzig: Univ. Leipzig; 2004.
Wilkens C. Umgang mit Mykotoxinen und deren sekundären folgen beim Rind. Nutz-
tierpraxis Aktuell.Tagungsabend zur 4. Haupttagung der Agrar- und Vetrinär- Aka-
demie (AVA). 2005:31-4.
Whitlow LW, Nebel RF, Hagler WM, Brownie CFG. Mycotoxins in North Carolina
dairy feeds - urvey of 100 dairy farms. J Dairy Sci. 1986;69(Suppl.1):223.
Whitlow LW, Nebel RL, Hagler WM. The assiociation of deoxynivalenol in grain with
milk production loss in dairy cows. 1994;131-139. In: G.C. Llewellyn, W.V. Dashek
and C. E. O= Rear, (eds.), Biodeterioration Research 4. Plenum Press, New York.

Literaturverzeichnis
107
Whitlow LW and Hagler WM. Mycotoxins in Dairy Cattle: Occurrence, Toxicity, Pre-
vention and Treatment. Proc Southwest Nutr Conf. 2005:124-38.
Windels HP, Dicostanzo A, Goodrich D. Effect of Deoxynivalenol from barley on per-
formance and health of large frame crossbred steers. Minnesota Cattle Feeders Rep.
1995; B-417. St. Paul, MN.
Wittek T, Fürll M. Prognostic value of pre-operative clinical and clinical-chemical pa-
rameters in cows with abomasal dislocation. In: Fürll M. Hrsg. Stoffwechselbelastung,
- diagnostik und stabilisierung beim Rind; Leipziger Samstagsakademie; 1998 Apr
25; Leipzig, BRD. Leipzig: Medizinische Tierklinik der Universität Leipzig. Auflage
1998.
Wittek T, Schreiber K, Fürll M, Constable PD. Use of the D-Xylose Absorption Test to
Measure Abomasal Emptying Rate in Healthy Lactating Holstein-Friesian Cows and
in Cows with Left Displaced Abomasum or Abomasal Volvulus. J Vet Intern Med.
2005;19:905–13.
Wolf V, Hamann H, Scholz H, Distl O. Einflüsse auf das Auftreten von Labmagenver-
lagerungen bei Deutschen Holstein Kühen. Dtsch Tierärztl Wschr. 2001;108:403-08.
Yoshizawa T. Fusarium matabolites other than trichothececnes /in Ueno Y. (Edit):
Trichothecenes-Chemical, Biological and Toxicological Aspects. Development in food
science 4, Elsevier Science Publishers B. V., Amsterdam. 1983.
Zepezauer V, Gerisch V, Schacht KH. Beitrag zur Mykotoxikose beim Rind (Spei-
chelsyndrom). Mh Vet Med. 1985;40:622-4.
Zheng Z, Hanneken J, Houchins D, King RS, Lee P, Richard JL. Validation of an
ELISA test kit for the detection of ochratoxin A in several food commodities by com-
parison with HPLC. Mycopathologia. 2005;159:265-72.

Literaturverzeichnis
108
Zinedine A, Soriano JM, Molto JC, Manes J. Review on the toxicity, occurrence, me-
tabolism, detoxification, regulations and intake of zearalenone: an oestrogenic my-
cotoxin. Food and Chemical Toxicology 2007;45:1–18.

Danksagung
109
9 Danksagung
Zum Schluss dieser Arbeit möchte ich mich bei allen bedanken die mich unterstützt
haben.
Mein Dank gilt vor allem Prof. Dr. M. Fürll für die freundliche Unterstützung und die
ständige Beratung und Betreuung. Ohne seine Mithilfe wäre dies nicht möglich ge-
wesen.
Mein Dank gilt ebenfalls allen Mitarbeiterinnen des Labors der Medizinischen Tierkli-
nik, die mich bei der Probenaufbereitung und Messung der Stoffwechselparameter
unterstützt haben.
Herrn Andreas Richter möchte ich herzlich für die ausdauernde Hilfe seitens der
statistischen Auswertung danken.
Danken möchte auch Herr Prof. Dr. Dänicke und seinen Mitarbeiter für die Mykotoxi-
nanalysen.
Ich bedanke mich ganz herzlich bei allen Mitarbeitern der Medizinischen Tierklinik
der Universität Leipzig für die Unterstützung.
Weiterhin möchte ich mich bei allen Tierarzthelfern und Tierpfleger für die Zusam-
menarbeit in der Klinik bedanken.
Mein besonderer Dank gilt meiner Familie (Vater, Mutter, Kadug, Mohamad, Mah-
mod, Mahmod, Kasem, Nuha, Huda, Rana, Adel), Schwiegerbrüder, Schwiegereltern
und meine Frau Afaf Dreie für die moralische Unterstützung und ihr grenzenloses
Verständnis.




























