Embed Size (px)
Citation preview

J. SeHORMi3LLER und E. L ~ A ~ N : Beitr~ge zur Biochemie der Kasereifung. XVI 211
Nit ttilfe dieser Nethode, bei der die gleitenden Durchschnitte sowohl der Temperaturen Ms auch der Restaktivit~iten zueinander in Beziehung gesetzt werden, gelang es, die k]eineren Schwankun- gen weitgehend auszugleichen und die Kurve zu gl~tten. In unserem Fall wurde jeweils das arithmetisehe Mittel yon 5 aufeinanderfolgenden Temperaturwerten dem Nittel der dazugeh6rigen Werte ffir die Restaktivitiiten gegenfibergestellt. Diese gleitenden Durchschnitte ergaben dann die Trendkurve, die auf Seite 200 wiedergegeben ist.
Unter den Bedingungen der ttocherhitzung ffihrten wir Versuche im Bereich zwischen 80 und 85 ° C dureh, deren Ergebnisse in Tab. 3 niedergelegt sind. Die Zeit yon 12 see ffir Aufheizung entspricht etwa den Verh~ltnissen im I-Iocherhitzer der Praxis, da yon den 20 sec, die ffir die Erw~irmung der Milch yon 10 auf 85 ° C ben5tigt werden, nur etwa t0 see auf den inaktivierenden Bereich fiber 60 ° C entfallen. Die Zeit yon 3 bis 5 see ffir HeiBhaltung entspricht der Verweil- dauer der Milch in der Rohrleitung yon Erhitzerabteilung zum gfickkiihler. Aktivit~t der uner- hitzten Milehproben 15,15 bzw. 18,20.
Versuche mit Rahm Um den geeigneten Ansatz ffir die Bestimmungen yon Xanthindehydrase in l~ahm zu er-
mitteln, wurde ~hnlich wie bei Milch die Xanthin- und Methylenblaukonzentration variiert. Die ullgemeinen Versuchsbedingungen und die Arbeitsweise war dieselbe wie bei den Versuehen mit Milch. Deutliche Hemmung der Enzymreaktion konnten wir feststellen, wenn die Xanthin- menge fiber 10 -6 mol gesteigert wurde. Weft die Entf~rbungszeiten mit 2 ml Rahm zu kurz aus- fielen, so dab die Genauigkeit der Versuche leiden muBte, gingen wir dazu fiber, nur 1 ml Rahm je Ansatz zu verwenden. Dieser Ansatz wurde dann in allen Rahmversuchen angewendet, wobei die Xanthinmenge nie fiber 10 -6 tool gesteigert wurde.
Zusammen]assung
1. Mit Hilfe einer etwas ver~nder ten 5~ethode der T~uNBERG-Technik wurde die Hi tze inak t iv ie rung der Xan th indehydrase untersucht .
2. Die Kurzzei te rh i tzung (74 ° C) und Dauerpas teur is ierung sch~digen das E n z y m zwar in verschiedenem Ausma~% kSnnen aber auch bei l~ngerer He i~ha l tung keine vSllige In~kt iv ie rung bewirken.
3. Un te r den Bedingungen der Hocherhi tzung wird die Xan th indehydr~se bei Tempera tu ren fiber 85 ° C in~ktivier t .
4. Die In~k t iv ie rung des Enzyms bei der Rahmerh i t zung verl~uft in ~hnlicher Weise wie bei der 5~ilcherhitzung.
5. Ffir die Erh i t zung yon Milch und R a h m im Labora tor iumsversuch wurde eine n3ue Erh i tzungse ia r ich tung entwickelt , die sich aus Labora to r iumsmi t t e l a zusammen- stellen l i~ t .
Beitr~ige zur Biochemie der K~isereifung X VI. Mitteilung
Die Phosphatasensysteme des Sauermilchk~ises und ihre Entwicklung w~ihrend der Reifung
Von Z. SC|]ORMULLER und E. LAHMANN
Mitteilung aus dem Institut /i~r Lebensmittelchemie und Lebensmitteltechnologie der Technischen Universit~it J~erliu-Charlottenburg*
Nit 6 Textabbildungen
(Eingegangen am 25. August 1955)
I n einer fffiheren Arbei t 1 war fiber die E ignung verschiedener Methoden ffir die Bes t immung yon Phospha tasen in reifendem Sauermilchk~se ber ichtet worden. :Die
* Die vorliegende Arbeit wurde dureh Gew~hrung eines BATT~LLE-Stipendiums gefSrdert; hierffir sei auch an dieser Stelle verbindlichst gedankt.
i SC~O~ffLLE~, J., u. E. LA~MANN: Diese Z. 100, 114 (1955). 15"

212 J. SC]~OR~i~LLE~ und E. LA~IANN:
vorliegende Untersuchung beschaft igt sich mi t dem Studium der in der genannten Ki~seart auf t re tenden Phosphatasen-Systeme. Sie priift die Enzymentwick lung in verschiedenen, un ter wechselnden Bedingungen reifenden Ki~seproben, die Ferment- akt ivierung und Inakt iv ie rung durch chemische und physikalische Fak to ren sowie die enzymat ische Akt ivi t~t gegenfiber spezifischen Subst ra ten und versucht, die beiden hier festgestellten Fermentgruppen, die saure und alkalische Phosphatase, bes t immten Phospha tasen typen zuzuordnen.
Phosphatasen gelangen zuniichst aus der Milch in die K~isemasse, dariiber hinaus entstehen sie vor allem zus~tzlieh bei Stoffwechselprozessen vieler an der K~sereifung beteiligten Mikro- organismen 1. Was die yon ihnen katalysierten Prozesse betrifft, so wirken sie zuni~chst hydro- lysierend (dephosphorylierend) bei Casein, Phosphopeptonen, Phosphopeptiden und phosphorylier- ten Aminos~uren, daneben zweifellos aber auch phosphatatisch ~phosphorylierend) und trans- phosphorylierend. Und schlieBlich sind noch Unterschiede hinsichtlich des p,-Optimums dieser ttydrolasen zu erwarten, da ja K~se im Verlaufe der Reifung vom sauren zum alkalischen Milieu wechselt.
Die weite Verbreitung und wesentliche Bedeutung yon Phosphors~ureestern im gesamt~n Tier-, Pflanzcn- und Mikroorganismenreich brachte es mit sich, dab die Biochemie der Phos- ph~tasen zu einem wichtigen Gebiet der Fermentforschung wurde.
Je nach ihrer Substratspezffitat kSnnen wir die Phosphatasen in folgende Gruppen unter- teilen ~:
1. Phospho.monoesterasen, die Monoester der Orthophosphorsi~ure in Phosphorsiiure und alkoholischen oder phenolisehen Rest spalten;
2. Phospho.diesterasen, welehe Diester der Orthophosphors~ure in Phosphors~uremonoester und Alkohole zcrlegen;
3. Pyrophosphatasen, die aus Salzen und Diestern der Pyrophosphors~ure Salze bzw. Ester der Orthophosphorsfiure liefern;
4. Polyphosphatasen, die Salze der Polyphosphors~uren in solche der Orthophosphors~ure spalten;
5. Metaphosphatasen, die Metaphosphate zu Orthophosphaten hydrolysieren; 6. Phosphoamidasen, die N-substituierte Amidophosphorsi~uren in Amine und Phosphorsi~ure
zerlegen. 5[ach dieser Einteilung ist die Spezffitat der Phosph~tasen vorwiegend im Sgure~nteil des
Substr~tes begrtindet. Sie wird darfiber hinaus erweitert durch die Vielzahl jener Fermente, deren Wirkung yon der l~atur des alkoholischen Substratpartners abh~ngig ist: Adenosintriphosphatase, Apyrase, Phytase, Cholinphosphatase, Lezithase, 5-I~ucleotidase und Phosphoproteidphosphatase sind derartige Enzyme.
Die wichtigste Gruppe - - und sie soll im folgenden eingehender untersucht werden - - ist die der Phospho-monoesterasen, die allgemein kurz als Phospha tasen bezeichnet werden.
Wir unterscheiden gegenw~rtig 4 isodyname Enzymtypen, die sich durch ihr p~-Optimum, ihre BeeinfluBbarkeit dureh Mg ++ und ihre Spezffit~t gegeniiber ~- bzw. fl-Glycerophosphat (Tab. 1) abgrenzen lassen.
Tabelle 1. Einteilung der Phospho-monoesterasen
• Typ Akt iv ierbarkei t Spaltungsgrad ffir ~- bzw: fl- nach FOLLEY Typ nach AI~BERS a p ~ - 0 p t i m u m dutch Mg ++ Glycerophosphat und KAY ~
fi-I AI~ Am Air
Nierenphosphatase Erythroeytenphosphatase Milzphosphatase Oberhefenphosphatase
um 9 5--6
um 5 3--4
+ ÷
Hemmung
a > f l f l > a
i DoM~E~, G.: Landwirtseh. Jb. Schweiz 68, 196 (1954). 2 FOZLEY, S. J., u. H. D. K~: : Erg. Enzymforseh. 5, 159 (1936). 3 AL~ERS, It . : In F. F. No~D u. ~. W~IDE~rL~GE~: ttandbuch der Enzymologie. Bd. I
S. 408. Leipzig: Akad. Verlagsges. 1940.

Beitr~ge zur Bioehemie der K~isereifung. XVI 213
Wie die Tab. 1 zeigt, streuen die pH-Optima fiber einen weiten Bcreich. Man hat dement- spreohend die Begriffe der ,,alkalischen" und ,,sauren", nach BERSIN 1 auch den der ,,ncutralen" (ffir Typ A]I in Tab. 1) Phosphatasen gepr~gt, ohne damit die Wirkungsart der einzelnen Ferment- typen spezifisch umrei~en zu wollen.
I . P h o s p h a t a s e n im R a h m e n der I(~isereifung
W i e e r w £ h n t , s ind P h o s p h a t a s e n in d e r l e b e n d e n N a t u r sehr w e l t verbre i~e t . U n s i n t e r e s s i e r t ira R a h r a e n de r v o r l i e g e n d e n A r b e i t ]edigl ieh das V o r k o m m e n solcher F e r m e n t e in d e r Milch, den M i l c h p r o d u k t e n u n d in d e n f l i t d ie K ~ s e r e i f u n g in B e t r a c h t zu z i e h e n d e n Mikroorgan i s raen .
Die erste Mitteilung fiber das Vorkommen einer Phosphatase in Milch mit einem p~-Optimum um 8 s tammt yon DEMUTtt ~ aus dem Jahre 1925. Sp~itere Autoren geben hShere Werte an, die bei 9,03, 9,44, 9,75, 9,85--]0,26 odor 9--9,5 ~ liegen. Offenbar ist die milieubedingte Enzym- aktivit~t weitgehend abhiingig yon der Art des verwendeten Substrats, der Substratkonzentration und der Zusammensetzung der Pufferl6sungen. Die alkalische Milchphosphatase wird durch Mg ++ aktiviert und liil~t sich in die Gruppe AI der Klassifizierung nach KAY und FOLLEY (S. Tab. 1) einordnen. MULLEI~ s sowie JACQUET und S~NGT 4 beschreiben eine saute Phosphatase mit dem Wirkungsoptimum bei PH 4, die sieh gegenfiber dem alkalischen Enzym durch grSl~ere Hitze- stabilitiit auszeichnet 9, auBerdem wird ein weiteres Ferment dieses Typus erw~ihnt, dessen PH- Optimum bei 5,5--5,8 liegt 1°. K A ~ A ~ und Bxzv ~ sowie JA~EK~ ~2 konntcn das Vorkommen einer sauren Phosphatase in Kuhmilch nicht best~tigen.
Die Phosphataseprobe als Test auf stattgefundenc Pastcurisation yon Milch ~3 hat auch hin- slehtlich der Anwendung auf Milcherzeugnisse eingehende Bearbeitung gefunden. Das Ferment ist an die Fettkiigelchen bzw. deren Hfillsubstanz gebunden und gelangt daher beim Separieren vorwiegend in den Rahm. So konnte SJSsw~6~ z4 aus rohem Rahm durch ~thanol-Extrakt ion ein hochaktives Trockenpulver gewinnen. Bei der Verbutterung des Rahmes t r i t t der Haupttei l der Phosphatase in die Buttermilch fiber. Der in der Magermilch verbleibende Enzymanteil reiehert sich bei der Vcrk~sung im Quark bzw. Bruch an ~5.
Eine Fehlerquelle, die den Phosphatasentest bei der Milchpriifung zu stSren vermag, besitzt auch Bedeutung im Rahmen der Kasereifung. Es ist dies die Phosphatasenbildung im Mikro- organismenstoffweehsell% Die hier entstehenden Enzyme lassen sich zwar dureh ihre grSl]ere I-Ii~zestabilitiit gegenfiber Milchphosphatase abgrenzen~7; an gereiftem Kiise ist indessen die
1 BERS:~, T~. : Kurzes Lehrbuch der Enzymologie. 3. AufL, S. 68. Leipzig: Akad. Verlagsges. 1951.
Dr, MyTh, F.: Bioehcm. Z. 159, 415 (1925); 166, 162 (1925). 3 KAY, H. D., u. W. R. GRAm~M: J. Dairy Res. ~, 64 (1933); 6, 191 (1935).
JACQUET, J., u. O. SAI~GT: C. r. Soc. Biol. (Paris) 146, 1515 (1952). JANEKE, H., u. W. DIEMAIR: Z. an~lyt. Chem. 130, 56 (1949). Sx~D~s , G. P. : 12. Int. Milchwirtsch. Kongr. Stockholm ~, 757 (1949).
v ScHw~z , G., u. O. FISCHER: Milchwiss. 3, 41 (1948). s MULLEN, J. E. C.: J. Dairy Res. 17, 288 (1950). - - K~Y, H. D., R. ASGHAFFENBURG U.
J. E. C. MUL~E~: 12. Int. Milchwirtseh. Kongr. Stockholm ~, 743 (1949). MVLLE~, J . E. C. : Zit. diese S., Anm. 8 . - - H ~ K ~ s s o ~ , E. B., u. G. S JSS~6~ : Sv.Mejeritidn.
44, 15 (1952); ref. in Chem. Abstr. 46, 10238 {1952). ~0 J ~ c q u ~ , J., u. O. S ~ I ~ T : Zit. diese S., Anm. 4 . - - M V L L ~ , J. E. C. : Zit. diese S., Anm. 8. n K A ~ A ~ , A., u. K. K. B~zu: Indian J . Dairy Sci. ~, 51 (1949). ~ JANEKE, H.: Dtsch. Lebensmittcl-Rdsch. 46, 202 (1950). ~ Zusammenfassung bei H. D. KAY, R. ASCH~FF]~BUa~, U. J. E. C. MULL~: Zit. diese S.,
Anm. 8. - - Vgl. auch G. ScHw~az u. W. LANGE: Molkerei-Ztg. (Hildesheim) 195~, Nr. 43. - - Sc~c~, A.: Bull. Inst. agronom. Star. Rech. Gembloux ~ , 313 ( 1 9 5 2 ) . - RI~ER, W.: Sehweiz. Milchztg. 1953, Nr. 83. - - SA~DE~S, G. P., 0. S. SAVIOR U. J. A. HUPFE~: J . Dairy Sci. 37, 698 (1954). - - Mom~z~B, M.: Mi]chwirtseh. Forseh. Ber. Kiel Nr. 6 (1953); Nr. 1 (1954). - - ScHw~z , G., u. J . LuDwig: Dtseh. Molkerei-Ztg. ~5, 566 (1954). - - D~vIs, J. G.: Dairy Ind. 19, 383 (1954).
~ SJSSTaS~, G.: 12. Int . ~iehwirtsch. Kongr. Stockholm ~, 750 (1949). ~ RITUal, W.: Milchwiss. 7, 301 (1952). - - H ~ T a l K , H. G., u. P. I-I. T ~ c Y : J. Dairy Sci. ~1,
867 (1948). ~s NE~V~, F. K. : J . Dairy Res. 10, 475 (1939). - - BUCK, T. C. : Amer: J . Publ. Health Nation's
Health 3~, (1942). - - I ~ L ~ , E.: J . Assoc. Off. Agrie. Chemists 30, 422 (1947). ~ SX~D~.~S, G. P., u. O. S. S x ~ : J. Milk Food Technol. 11, 67 (1948).

214 J. SC]~O~i~L~ER und E. L A m ~ :
Durehffihrung des Phosphatasetestes zur Prfifung auf p~steurisierte Milch als Ausgangsmaterial der Ki~sebereitung kaum mLglieh, da im Verlaufe der Reifung erhebliche, mikrobiell bedingte Fermentmengen auftrcten. C ~ v s und ALIFASE ~ stellten bei Weiehk~se eine hohe Phosphatasen- entwicklung in der Rinde lest, die allerdings im Verlaufe der normalen Rcifung nicht his zum Kern durchdringt. Bei GruySre-K~se war jedoeh auch im Inncrn eine deutliche Aktivit~t mel]bar, die bei liingerer Lagerung des Kases abnahm. Die Autoren ftihren diese Abnahme auf proteolyti- schen Abbau der Phosphatase zurtick. CAMvs * gelang ferner der Naehweis, dal~ die an der Flora des Camembert-K~ises betefligten Schimmelpilze Penicilliura glaucum und candidum zur Bildung von Phosphatasen befahigt sind. Eingehendere Arbeiten ehemischer oder bakteriologiseher Art fiber die Natur und Entwicklung der in Ki~se vorkommenden Phosphat~sen sind bisher wenige bekannt geworden. I-IA~ER und OLso~ a untersuchten eine gr61~ere Zahl molkereiteehnisch wertvoller wie schadlicher Bakterien auf ihre Phospt~atasenbildungsf~higkeit. Zusammenfassend werten sie ihre Befunde dahingehend, dal~ phosphatasehaltige Keime wegen ihrer Empfindlichkeit gegenfiber S~ure im Kase sich nicht entwickeln. D O M ~ ~ hat vor kurzem eingehend u. ~. das alkalische und saure Phosphatasensystem von Tilsiterk~ise studiert. Die dort vorherrschenden st~ibehenfLrmigen Bakterien bilden keine der beiden Phosphatasen. In der gesamten Schmiere des Tilsiterk~ses hingegen, die zu 77 % aus peptonisierenden Befen der Gattung Endomycopsis bcsteht, wird sehr viel alkalische und saure Phosphatase gebildet. Auch dieser Autor weist auf die Schwierigkeiten hin, die dem Phosphatasetest in der Ki~seanalytik dureh Diffusion der Phosphatasen aus der Rindenpartie ins K~seinnere erwachs~n und h~lt weitere Untersuchungen ffir notwendig.
Was die im Rahmen unserer Problemstcllung besonders interessierende Frage zur Angreifbar- keit des Phosphoproteidcs Casein durch Phosphatasen betrifft, so sind hierfiber die Ansichten geteflt. Wiederholt wurde Resistenz des Caseins gegenfiber Phosphatasen beobachtet ~. P E R L ~ ~ fand in letzter Zeit, dal~ die einzelnen Fraktionen des Caseins auf Prostataphosphatase ver- schiedenartig reagieren. W~hi'end ~-Casein bei p . 5,6--6,6 leicht dephosphoryliert wird (bis zu 42 % des Phosphorgehaltes), wirkt sich ein Anteil yon fiber 30% des nicht spaltbaren fl-Caseins im Substrat hemmend aus. Ein endgttltiger Entscheid der Frage, ob Phosphorproteide und Phosphopeptone durch bestimmte Fermenttypcn dephosphoryliert werden, war bisher schwer zu treffen, da keine spezifische Phosphatase bekannt war, die solche Substrate aUein hydrolysiert. Kfirzlich beriehtete FOOTE ~ fiber eine hochaktive und spezifische Phosphoprotein-Phosphatase, die er als Acetontrockenpulver aus Hfihnerembryo gewinnen konnte. Ihr p~-Optimum liegt bei 5,8, sie ist aktiv gegen nicht fraktioniertes Casein, ~- und fl-Casein, Phosvitin und ein durch tryptische Verdauung aus Casein gewonnenes Phosphopepton. Einfaehe Phosphors~ureester werden nicht gespalten, die oben genannten Phosphorsubstrate jedoch vollkommen dephos- phoryliert. Im AnschhiB an sparer beschriebene Versuche soil auf diese Fragestellung dort (vgl. S. 237) naher eingegangen werden.
II . Phosphatasen in reifendem Sauermilchk~ise
1. ~Da~ Untersuchung~material
Die folgenden Versuche an Reffungsreihen wurden durchgeffihrt mit: a) Sauermilchquark, der ohne Kochsalz- und geffungssalzzusatz gleich normalen Reifungs-
ffihrungen fiber l~ingere Zcit gehalten wurde: Rei]ungsreihe I I (November 1953) und Rei/ungsreihe 1V (Mai 1954). b) Sauermilchkase aus Quark mit dem fabrikationsfiblichen Zusatz yon Kochsalz und
Reffungssalzen: Rei]ungsreihe I (November 1953; 3,6 % NaC1 und 1% NaI-ICO3) und ]~ei/ungsreihe 111 (Mai 1954; 3,6 % NaC1 und 1,25 % Rcifungssalz, bestehend aus NaHCO3 und
CaCOs).
z CAMUS, A., u. R. ALIFAS~: Lair 31, 367 (1951). 2 C ~ v s , A. : 12. Int. Milchwirtsch. Kongr. Stockholm 2, 114 (1949). a HAMM~, B. W., u. I-I. C. OLSO~: J. Milk Technol. 4, 83 (1941).
DoM~E~, G. :Zit. S. 212, Anm. 1. 5 RIMIN~TO~, C., u. It. D. KAY: Biochemic. J. 29, 777 (1926). - - SC~MIDT, G., u. S. J. Tm~NN-
~VSER: J. of Biol. Chem. 149, 369 (1943). - - A~AG~OSTOPOULOS, C., M. PAcer, E. BOURLA~D U. P. Ge~BA~: Bull. Soc. Chim. biol. 33, 699 (1951). - - Vgl. auch B. GLASS: Symposium on Phosphorus Metabolism. Bd. II, S. 846. Baltimore: The John ttopkins Press 1952.
6 PE~LMAN, G. E.: J. Amcr. Chem. Soe. 74, 3191 (1952). FOOTE, M. W. : Diss. Abstr., Univ. Microfilms Ann. Arbor, Michigan 15, 25 (1955).

Beitri~ge zur Bioehemie der K~sereifung. XVI 215
Einzelheiten fiber die Eeifungsfiihrung wurden mehrfaeh in fi'fiheren Arbeiten gebr~cht 1. Das Ausgangsm~terial der l%eifungsreihen, yon denen Mle 3--4 Tage eine Untersuohungsprobe gezogen wurde, ist aueh hier als O-Quark bezeiehnet.
Fiir die ~ehrzahl der nioht auf Reifungsreihen aufgebauten Un~ersuohungen diente duroh- gereifter, etwa 10---12 Tage alter Kgse; er zeigt erfahrungsgemgB hohe Phosphat~senaktivitii~ und liefert otme Schwierigkeit homogene Suspensionen.
2. Bestimmungsmethodil~ Die in einer vorhergehenden Arbeit 2 entwiokelte Methode zur Phosphat~senbestimmung in
Sauermilohk~tse mit Dinatriumphenylphospha~ als Subs~rat wurde auoh hier benu~zt. Zur Untersuohung der Substratspezifltgt yon Kgsephosphatasen war es in einigen Fgllen
notwendig, die Menge des abgespaltenen Phosphats zu ermitteln. Aueh hier wurde naoh einem frfiher mitgeteilten Verfahren gearbeitet. Im einzelnen ist folgendes zur Analytik nachzugragen:
a) Ve~uend~ng w n 1)ina#iu~phenylpt~osphat ala Substrat zur Bestimmung des enzymatisch abgespaltenen Phenols
20% ige K~sesuspensionen wurden aus dem Ausgangsm~terial dureh Verreiben mit toluol- ges/~ttigtem Wasser im M5rser und ansehliel3ende ttomogenisierung im ,,Multimix" gewonnen. Schaumbfldung 1/il]t sieh durch Zugabe yon 2--3 Tropfen OetylMkohol verhindern. Die Wahl des Pullers riehtet sich nach dem gewtinschten PH der Ansatzmisehung und dem p , des zu unter- suchenden K/ises. Ftir Ans~tze bis zum p~ 9 wurde der Veronal-Natriumacetat-Salzs~urepuffer nach Mm~_~]~LIS a verwendeL Fiir die Mehrzahl der Versuche, die an durchgereiftem K/ise beim pg-Optimum der sauren Phosphatase durchgefiihrt wurden, diente eine Puffermisehung yon PH 4,93. Zur Aufstellung yon p~-Aktivit~tskurven und zur Untersuehung yon K~sen jtingerer l~eifungsstufen gelangten die entsprechenden Puffermischungen hSheren oder niedrigeren p~- Wertes nach den Angaben yon MIC~AELIS zur Anwendung. Geringe p~-Korrekturen wurden, sofern erforderlich, mit wenigen Tropfen Salzs/iure oder Natronlauge entsprechender Konzen- tration vorgenommen. Ffir Ansiitze mit einem PH fiber 9 wurde der Natriumborat-Natronlauge- Puffer nach SS~E~SE~ ~ verwendet. Die Ans~t~ze beim p,-Optimum der alkalischen Phosphatase wurden, wenn nicht anders angegeben, in Puffermischung yon p~ 10,2 bebrfitet.
Die Bebriitungszeit der alkalischen Ansiitze betrug ~/2--1 Std., die der sauren im allgemeinen 3 Std. Vom Filtrat der Ans/itze wurden je nach Phenolgehalt aliquote Teile des Gesamtvolumens (8 ml) zur Bestimmung herangezogen. Die Einstellung des zur Messung erforderliehen p~ (9--9,5) und Volumens (12 ml) gesehieht auf folgende Weise:
Entnommenes Triehloressigsiiure-Filtrat . . . . . ml 4,0 2,0 • 1,0 0,5 0,2 Borat-NaOH-Puffer p~ 9,3 . . . . . . . . . . . ml 5,5 8,25 9,7 10 ,5 10,8
2,5%igeNaOH . . . . . . . . . . . . . . . . ml 1,5 0,75 0,3 - - - - Dibromehinonchlorimid-Reagens . . . . . . . . ml 1,0 1,0 1,0 1,0 1,0
Bei der Messung wurden friihere Beobach tungen best/~tigt, wonach a) die In tens i t / i t der F/~rbung recht lange kons t an t bleibt. Bei einer ~eg re ihe
mi t Ablesungen im Zei t raum yon 20 rain bis zu 2 Std. nach der l~eagenszugabe zeigten sich keine Ver/~nderungen der E x t i n k t i o n ;
b) keine Untersehiede feststellbar sind, wenn die Pheno]bes t immung einmal am Trichloressigs/~ure-Filtrat unmi t t e l ba r nach der Eiweif3f~llung und zum anderen nach zweit/itigem Stehen des Fi l t ra tes durchgefi ihrt wurde. Beide Befunde erleiehtern die Bew/~Itigung gr5i3erer t~eihenversuche, wo eine sofortige Ana]yse zeithch n icht mSg- lich ist.
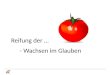
Ein/lu[3 der Substratkonzentration au/ die Fermenta~tivitiit : Wie Abb. 1 zeigt, erwie- sen sich 2 g D ina t r iumpheny lphospha t pro 1 1 Puffer als optimale Konzen t r a t i on fiir die Bes t immung der sauren wie der alkalischen Phosphatase. Vortci lhaft wirkt sich hierbei aus, dag einerseits eine hohe absolute Phenolabspa l tung zur Messung gelangt,
Vgl. J. Sc~o~/dnL~,R, I-I. WIESKE U. I-L W~NTE~: Diese Z. 99, 437 (1954). Se~O~iiLLE~, J., u. E. Ln~A~]~: Zit. S. 211, Anm. 1.
a ~m~ELrS, L.: Biochem. Z. 234, 139 (1931); J. of Biol. Chem. 87, 33 (1930). Ki2STE~, F. W., A. T~EL U. F. F~SC~BECK: Logarithmische gechentafeln. Berlin: W. de
Gruyter 1947,
216 J. SCHORI~I#3LLER und E. L ~ v ~ :
anderersei ts der Subs t ra t -Spa l tungsgrad selbs~ bei hoher F e r m e n t a k t i v i t ~ t un te r 10 % bleibt. Als molare Konzen t r a t i on an Subs t ra t ergibt sich ffir den Ansatz ein Wer t yon e twa 0,009 m.
Die besehriebene Methode ha t sich in den Fe rmen tve r suchen dieser Arbei tsre ihe gu t bew~hrt . Ffir jede E n z y m b e s t i m m u n g wurden 3 Paral le lans£tze durchgeffihrt . Abweichungen der E inze lwer te v o m Mit te lwer t lagen n ieh t fiber 3 %.
600 80
~abJr~#'onzemtr~h'on /m Pul~er Abb. 1. Einflu~ der Substratkonzentration auf die enzymatisehe Spaltung yon Dinatriumphen~lphosphat durch Phosphatasen ge- reiften Barzer K~ses. I = ( 9 - - O alkalische, II = (9---O saute
Phosphatase
/oo
'80
b) Die Bestimmung enzy. matisch abgespaltenen Phosphates
Auf Grund fr[iherer Angaben 1 gestaltet sieh die Ermittlung der Fermentaktivit~t folgendermal~en:
L6sungen: 1. 20 % ige K~sesuspension. Ihre
tIerstellung wurde oben besehrieben; 2. Puffergemische. Veronal-
natrium - Acetat- tIC1- Puffer naeh lV[ICHAELIS und Natriumborat- NaOH-Puffer naeh S6RENSEN. Ffir die Wahl des Puffers gelten dieselben Grunds~tze, die bei der unter a) be- sehriebeneu Fermentbestimmungs- methode erSrtert wurden. Bei den hier herrsehenden Verhaltnissen ist noeh besonders die Pufferungskapa- zit~t des hSheren K~seanteiles im Ansatz zu ber~ieksiehtigen.
3. Puffer-Substrat-Gemische. Die zur Untersuehung herangezogenen Substrate wurden zu 0,02 molar im Puffer gelSst. Dadurch ergibt sieh ffir die im folgenden geschilderten Ansatze eine Substratkonzentration yon 0,005 molar.
4. 10%ige Kieselwolframsaure in n-H2S04; 5. 10 n-I-I2S04; 6. 2,5% ige Ammonmolybdatl6sung in Wasser; 7. Reduktionsmittel. 0,5 g umkristallisiertes Eikonogen (1,2,4-Aminonaphtholsulfons~iure)
werden in 195 ml 15% iger NatriumbisulfitlSsung und 5 ml 20% iger Natriumsulfitl6sung gelSst. Das Reagens wird in einer braunen Glasstopfenflasehe auPoew~hrt und ist etwa 14 Tage haltbar.
Ansatz Test ml
1,5 3,0 1,5
K~sesuspension . . Puffer . . . . . . Puffer-Substrat . . Wasser . . . . . .
1. ]31indwert 12. ]31indwert ml ml
1,5 4,5 3,O - - ] , 5
- - 1 , 5
Nach dem Vorwarmen, Vermi- schen und Bebrfiten wird mit 2 ml Kieselwolframsaure sistiert, umge- sehfittelt und nach einiger Zeit fil- triert. Vom Filtrat werden aliquote Teile des Gesam~volumens zur Phos- phatb~stimmung abpipsttiert. Es folgt dann Zugabe yon 0,4ml 10 n-H2S04 (der aus dem Kieselwolfram- saure-i%eagens stammende H2SO~- Gehalt kann im gef~llten Ansatz bei
Ffltratanteilen bis zu 1 ml vernachl~ssigt werden), 0,8 ml Ammoam)[ybd~tl6sung, 1,0ml Eikonogenreagens und Aufffillen mit Wasser auf I0 ml. Naeh sofortigem kr~ftigem Umschfitteln wird nach 60 min die Extinktion der entwickelten Blaufarbung im PuLF~Ic~-Photometer (Filter S 66) abgelesen. Wie frfiher eingehend erSrtert, ist also zur Phosphorbestimmung im Kieselwolframs~urefiltrat der untersuchten Ans~tze gegentiber der Phosphat-Eichkurve in w~l~riger L5sung eine ErhShung der Eikonogenzugabe und eine Verl~ngerung der l~edaktionszeit erforderlieh.
Da der Kase yon vornherein betr~ehtliche Mengen anorganisehen Phosphates enthalt und zudem der Gehalt an freiem Phosphat sowie die Hydrolyse der verschiedenen benutzten Substrate unter den Versuchsbedingungen zu beriicksichtigen sind, mfissen ffir jeden Ansatz 3 Messungen durehgeffihrt werden:
1 SCEOR~ffLL~R, J., u. E. LA~A~N: Zit. S. 211, A_am. 1.

Beitr~ge zur Biochemie der K~sereifung. XVI 217
1. P-Bestimrnung im Test gegen Wasser; 2. P-Bestirnmung irn 1. Blindwert gegen Wasser; 3. P-Bestimmung irn 2. Blindwert gegen den Reagentienleerwert. Die Sumrne yon 2. und 3. wird vom Ergebnis der ersten Bestimmung subtrahiert.
Daraus ergibt sich die Menge des enzymatisch abgespaltenen Phosphors. Ffir jede Fermentbestimmung wurden 3 Parallelansi~tze durchgeffihrt. Die
Streuung der Einzelwerte vorn Mittelwert lag fast stets unter 3 %.
c) Vergleieh der Bestimmungsmethoden /i~r Phenol und Phosphat
Urn die Leistungsf~higkeit der beiden hier beschriebenen Verfahren kritisch zu fiberprfifen, wurden bei einem Ferrnentversuch an reifem Kiise (Dinatriurnphenyl- phosphat als Substrat) die Analysenergebnisse ffir beide Kornponenten des Substrates (Phenol und Phosphat) vergleichend ermittelt. Entsprechend den ~[olekularverh~lt- nissen (P --~ 30,98, Phenol ~-- 94,108) ist gegeniiber Phosphor die 3,04faehe Menge an Phenol zu erwarten. Die Versuchsffihrung entsprach der zuletzt geschilderten Arbeitsvorschrift. Es wurde 6 Std. b e i p . 5,5, dem Optimum der sauren Phosphatase, bebrfitet.
Die MeBergebnisse ffir Phosphor und Phenol gibt Tab. 2 wieder; sie zeigen sehr gute Ubereinstimrnung. Aus der Tabelle geht auch die grSl~ere Genauigkeit der Phenolbestirnrnung hervor. Bei ihr bewirkt im vorliegenden Fall ein Ablesefehler irn Gang der Colorimetrie yon 1% auch nut 1% Fehler irn Endergebnis gegenfiber 5 °0 bei der Phosphorbestimrnung, da hier der enzyrnatisch bedingte Phosphatgehalt im Vergleich zur gesamten Phosphorrnenge des Ansatzes relativ gering ist. Ffir die letztgenannte Bestimrnung bietet die dreifache Messung (irn hier beschriebenen Fall nut eine zweffache, weft die Eigenspaltung des Substrates auch in die Phenolbestim- rnung eingeht) neben der Urnst~ndlichkeit des Verfahrens eine weitere Fehlerquelle.
Tabelle 2. Vergleich der Phenol- und Phosphatbestimmung nach enzymatischer Spaltung von Dinatriumphenylphosphat (Konzentration im Ansatz 0,005 molar) dutch die saure Phosphatase eines
gerei/ten Harzer Kiises Ermitteltes Spaltprodukt Phenol Phosphor
Untersuchter Filtratanteil . . . . . . . . . . . . . . . . ml Spaltprodukt, ermittelt nach der Eichkurve . . . . . . . . #g Spaltprodukt, berechnet auf den Gesamtansatz . . . . . . #g Spaltungsgrad des Substrates . . . . . . . . . . . . . . %
0,5 19
304 t0,8
0,5 6,4
102,4 ll,0
d) p,-Messung Die Messung der p~-Werte erfolgte an der Glaselektrode (Metrohm-Gerat, t~Ierisuu). Di¢~'
p~-Kontrolle vor der Pneao|hestimmung wurde mit Spezi~l-Indicatorpapier (Merck) durchgeffihrt.
3. Die Abhdngigkeit der Phosphatasenaktiviti~t rei/enden Sauermilchkigses vom PII
Wie eingangs erw~hnt, unterscheidet man bei der hier irn Mittelpunkt der Unter- suchungen stchenden Phospho-monoesterase vier isodynarne Systeme, deren wesent- lichste Merkmale nach dem Stand unserer heutigen Kenntnis vor allem stark unter- schiedliche p,-Optima sind. Aus diesem Grunde erschien es uns zur Kennzeichnung des im Sauerrnilchk~se vorkomrnenden Phosphatasensystems zun~chst yon grund- s~tzlicher Bedeutung, entsprechende p,-Aktivit~tskurven zu erarbeiten. •icht zuletzt vor allem auch deshalb, well -- wie frfiher erSrtert -- fiber die pH-Optima der Milchphosphatasen einander widersprechende Befunde vorliegen.
218 j . SeHo~i)LLEa und E. L A H ~ N :
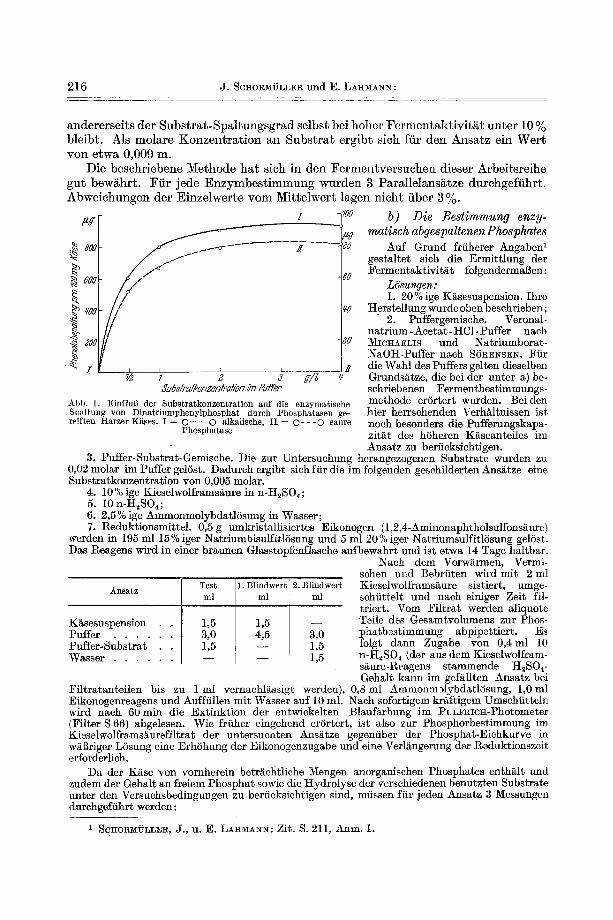
Wir haben deshalb versehiedene Proben durehgereiften Harzer K~ses, deren phosphatatische Aktivit~t durch Vorversuehe erwiesen war, yon dem genannten Gesichtspunkt her fiberpriift. Dis Versuchstechnik ist oben eingehend gesehildert; bei extrem sauren und alkalischen Werten war vor der Phenolbestimmung sine zus~tz- liehe Einstellung des zur Indophenolbildung erforderliehen p~ (9--9,5) notwendig. Die Abb. 2 und 3 bringen eine Zusammenfassung der Ergebnisse. Aus ihnen ist
/t:
.•30
% ,
// ! /
////
f ~ ]
I 5 7 8
~7oo
:5O f
I !
I l
/ 700 /
i
/ Abb. 2. p~-Optimum der ,,sauren'" Phosphatase yon hochreifem bzw. gereiftem Sauermilchkase, I = 10 Tage bzw. I1 = 14 Tage alt (Veronalnatrium-Acetat-ttC1- Puffer). i : 4 Std. ( O - - O ) und I I : 5 Std. ( O - - - O )
Bebrfitungszeit
r / / / \\
_~N ': ~ \,,xx
z 8,o 9,~ ~o,o N~ go p// gs
Abb. 3. p~-Optimum der ,,alkalischen" Phosphatase yon hochreifem bzw. gereiftem Sauermflchk~ise, I = 10 Tage bzw. I I = 1 4 Tage alt (Natriumborat-NaOtt-Puffer). I :
1]~ Std. ( O - - O ) und [I: 1 Std. (O " ' - O) Bebr~itungszeit
ersichtlich, dab im Sauermilchk~se zwei p : O p t i m a der Phosphatasenaktiviti~t auf- treten. Dementsprechend werden sich die folgenden Arbeitsreihen auf die Unter- suchung der durch derartige p,-Unterschiede gekennzeichneten ,,sauren" und,,alkali- schen" Phosphatase richten. Der Spaltungsgrad des Substrates bei den zwei p . - Optima war ffir die verschiedenen untersuchten Ki~seproben je nach ihrem geifungs- grad unterschiedlich.
Die s a u t e Phosphatase zeigt bei den beiden untersuchten Ki~seproben ein p . - Optimum zwischen 5,2 und 5,7, entsprechend den in der Einleitung angefiihrten Befunden verschiedener Autoren ffir saure Phosphatase aus Kuhmilch. Zur Erfas- sung dieses Fermentes im K~se ist mehrstfindige Bebrfitung (3--6 Std.) erforderlich.
Die all~aIische Phosphatase zeichnet sich gegenfiber der sauren durch eine bedeu- tend h5here Aktivit~t aus. Dis Fermentwirksamkeit ist demgemi~i3 hier bereits nach 1/2- 1 Std. gut meBbar. Das Opt imum liegt um P . 10 (Streuung zwischen 9,95 und 10,2), also im selben Bereich, den SA~I)nns I unter Verwendung yon Bariumborat- NaOI-I-Puffer fiir die alkalische Milchphosphatase land. Die ffir beide untersuchten Kiiseproben unterschiedliche Abnahme der Substratspaltung im stark alkalischen Gebiet dfirfte durch proteolytischen Abbau des Phosphatasen-Apofermentes mit- bedingt sein.
Zum Vergleich der Aktivitikt beider Fermenticypen daft der Ein/Iu[3 der verwendeten Puller nicht auBar acht gel~ssen werden 2. Um diese Frage zu kl~ren, wurden an hoohreifem K~se Fermentbastimmangen durchgeffihrt, bei denen der eine Ansatz mit Veronalnatrium-Acetat-HC1 und der ~ndere mit Natriumborat-NaOK gepuffert wurde. Die Bebrfitung erfolgte bei p~ 9, wo
SANDERS, G. P.: Zit. S. 213, Anm. 6. - - Vgl. auch G. DOM~EN: Zit. S. 212, Anm. 1. GRYDER, 1~. M., J. S. FI~IEDENWALD U. C. CARLSON: Arch. ofBiochem, a. Biophysics ~4,281
(1955).

]~citrage zur Biochemie der K~sereifung. XVI 219
sich der 0bere Bereich des Veronalpuffers mit dem unteren des Boratpuffers trifft; letzterer besitzt hier allerdings eine erheblich hShere Pufferungskapazit~t. Die Bebrtitungszeit betrug 1 Std. Auf 100 nag Kase berechnet ergaben sich folgende Phenolabspaltungen aus Dinatrinmphenyl- phosphat:
ffir Veronalpuffer: 246#g, ftir Boratpuffer: 214 ttg. Die unter Verwendung yon Veronalpuffer erzielten Werte liegen also um 15 % hSher, was
auf einer wiederholt in der Literatur erwahnten kompetitiven Hemmung der Phosphatase durch Borat ~ beruhen dtirfte. Diese ist indessen nicht so stark, dab yore Boratpuffer fiir derartige vecgleichende Untersuchungen abzuraten ware. Seine Vorteile liegcn in der guten Pufferungs- kapazit~t und der Nichtbeeinflussung der Indophenolfarbreaktion.
4. Die Entwicklung der Phosphatasenaktiviti~t wii, hrend der Kiiserei/ung
a) Bestimmung der ,,alkalischen", ,,sauren" und ,,e//elctiven" Phosphatase in Rei/ungsreihen zunehmenden Alters
Anhand verschiedener, oben gekennzeichneter Reifungsreihen wurde in den fo]genden Untersuchungen die Aktivi t i i tsentwicklung der , ,sauren" wie der ,,alkali- schen" 1)hosphatase ermittelt . Auiterdem wurde die phosphatat ische Akt ivi t~t des K~ses beim p , der jeweils un tersuchten Reifungsstufe best immt. Es soll dami t die Frage der tats~tchlichen Enzymw-irksamkeit im K~se beantwor te t und auf die von GALn ~ entwickelten Gesichtspunkte fiber den Einflul~, den das PE des Mediums auf die Fermentb i ldung ausfibt, eingegangen werden.
GALn unterscheidet zwischen ,,potentieller" (helm pn-Optimum in Erschcinung tretender) und ,,e/#~tiver" (bci p~ des ~edinms, hier also des reifenden K~ses mel~barer) Aktivitat. Er stcllte lest, dal~ bei der Bildung muncher Enzymsysteme ein "protective mechanism" zu bcobach- ten ist. Da jedes Enzym ein definiertes pH-Optimum besitzt, wird seine Wirksamkeit gcringer, wenn das PH des Mediums yon diesem optimalen Milieu abweicht. GALE fand, dal~ lV[ikroorganis- men in solchen F~llen bef~higt sind, diesc Aktivitatsminderung durch crhi~hte Enzymproduktion zu komp~nsieren. Die ,,effektive Aktivitat" (Anzahl der Enzymeinheiten real Aktivitat jeder Einhcit beim herrschenden PH) bleibt hier also trotz des pH-Wcchsels im Medium annahernd konstant. Enzyme, deren Bildung im er5rterten Sinne zum PH des Kulturmediums in Beziehung steht, sind z. B. Urease, Katalase, Formico-dehydrase und Alkohol-dchydrogenasc.
Die beim p , der K~seproben wirksame Phosphatase wird im folgenden Ms ,,effek- t i r e " Phosphatase bezeichnet, ohne dal~ dami t zun~chst ein spezifischer Ferment- typus umrissen werden soll.
Die ansehliel~end beschriebenen Untersuchungen wurden wie tiblich mit Dinatriumphenyl- phosphat als Substrat durchgefiihrt. Bebrfitet wurde beim ,,alkalischen" Enzym 1 Std. (pH 10,0), bei der ,,sauren" Phosphatase 3 Std. (PH 5,6). Ffir die ,,effektive" Phosphatase betrug die Reak- tionsdauer 3 Std., und zwar beim p~ der jeweils untersuchten K~iseprobe (Veronal-Puffer).
Uber das Untersuchungsmaterial (t~eifungsreihen ohne und mit Salz- bzw. l~eifungssalzzusatz) wurde frfiher eingehend berichtet. Die als Mal3 der Fermentaktivitat herangezogene Phenol- abspaltung aus dem Substrat wurde auf asche/reie KSsetrockensubstanz bezogen.
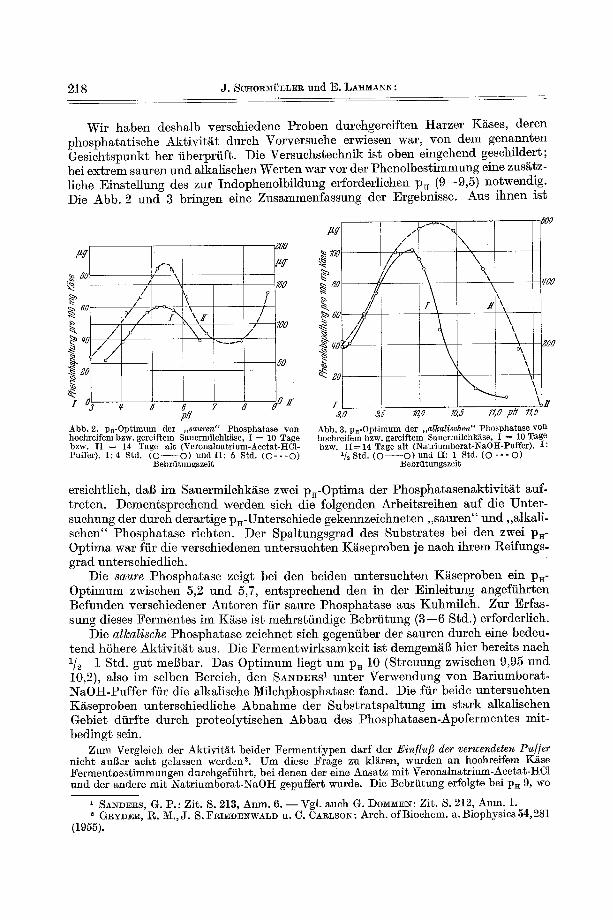
Untersuchungsergebnisse derart iger Arbeitsreihen sind in den Abb. 4 und 5 dar- gestellt, wobei die dor t (vgl. S. 220) benutz te Bezeichnung (l~eihen I mit IV) bei- behal ten wird. Zum Vergleich sind aul3erdem in den Tab. 3 und 4 die p . - W e r t e der einze]nen Reifungsffihrungen und Reifungsstadien angegeben.
Die Untersuchungen zeigten zun~chst, dal~ all~alische Phosphatase im Ausgangs- material ( , ,O-Quark") nicht vorhanden ist; sie bildet sich erst im Verlaufe der Rei- lung, und zwar bei K~seans~tzen mit Reffungs- und KochsMz in bedeutend hSherem Ausmal~ a]s in salzfreien Reifungsffihrungen. Dieser Befund zeigt, dab die hier
Vgl. z. B. C. A. ZI~LE u. E. S. D~nLA Mo~IeA: Arch. of Biochem. 26, 112 (1950). GALE, E. F. : The chemical activities of bacteria. 3. Ausg., S. 70--75. London: Univ. Tutorial
Press Ltd. 1951.
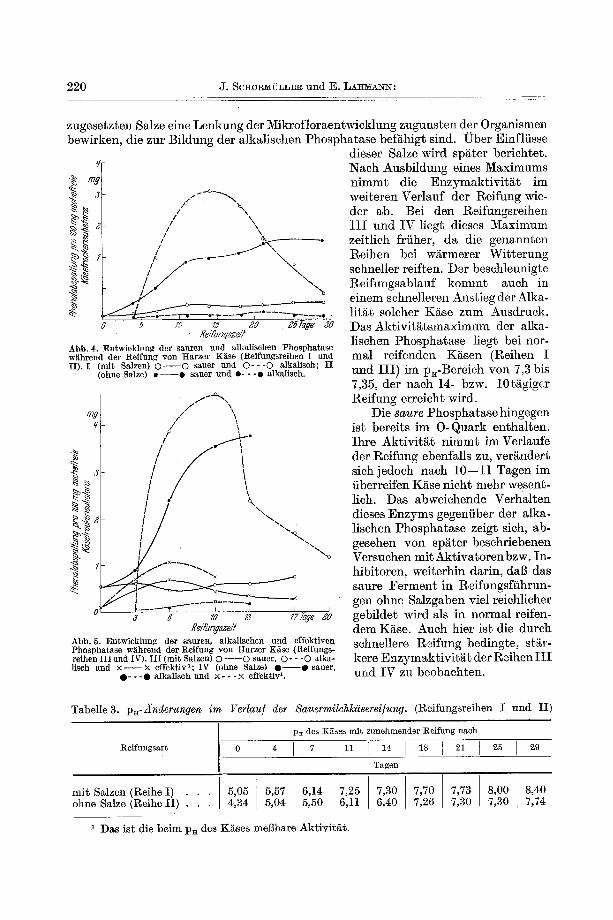
220 J. SC~O~ffLL~ und E. L ~ m a ~ :
zugese tz ten Salze eine Lenkung der Mikrof lo raen twick lung zuguns ten der Organismen bewirken, die zur Bi ldung der a lkal i schen Phospha t a se befi~higt sind. ~ b e r Einfl t isse
dieser Salze wird spi~ter ber ichte t .
~- ~ I / . ~ " , ,
, o
- - - , - ~ - - - ' " - ~ . . . . . "," . . . . . r g ~ 7o 1~ zo Zd~LTe 30
~e/ fun#sze// Abb. 4. Entwicklun~ der sauren und alkalischen Phospha&ase w{ihrend der Reiflmg yon l:l'arzer Kiise (Reifungsreihen I und II). [ (mi$ Salzen) O----O sauer and O- --O alkatisch; II
(ohne Salze) . - - $ sauer und ¢- -- • alkalisch.
O:
/ / / \ \ , \ /
/ \ °
! ! / / !
!
/ / -.... / / - . . I / "%-.\ I /
7o 17 jo zo l~e/f'ungszeil
Abb. 5. Entwicklung der sauren, alkalischen und effektiven Phosphatase wfihrend der Reifung von l:Iarzer K'hse (P~eifungs- reihen 111 und IV). [ i I (fair Salzen) O - - O sauer, O - " -O aika- lisch und x - - × eifektiv~; IV (otme Salze) $ - - e sauer,
$ - - - O alkalisch und × - ~- × effektiv~,
Nach Ausb i ldung eines ~ a x i m u m s n i m m t die E n z y m a k t i v i t ~ t im wei te ren Ver lauf de r l~eifung wie- der ab. Bei den l~eifungsreihen I I I und I V ]iegt dieses Max imum zeit l ich friiher, da die genannten Re ihen bei w~rmerer W i t t e r u n g schneller reif ten. Der beschleunigte Re i fungsab lau f k o m m t auch in e inem schnel]eren Anst ieg der Alka- l i t e r solcher K~se zum Ausdruck . Das A k t i v i t ~ t s m a x i m u m der alka- l ischen Phospha ta se l iegt bei nor- mal re i feuden K~sen (Reihea I und I I I ) im pa-Bere ich yon 7,3 bis 7,35, der nach 14- bzw. 10ti~gigcr Reffung er re icht wird.
Die s a u t e Phospha ta se hingegen is t bere i ts im 0 - Q u a r k en tha l ten . Ih re A k t i v i t ~ t n i m m t im Verlaufe der Rei fung ebenfal ls zu, ver~nder t s i ch j edoch nach 10--11 Tagen im fiberreifen K~se n ich t mehr wesent- lich. Das abweichendo Verha l t en dieses E n z y m s gegenfiber der a lka- ]ischen Phospha ta se zeigt sich, ab- gesehen yon spi i ter beschr iebenen Versuchen mi t A k t i v a t o r e n bzw. In- h ibi toren, wei terhin dar in , dag das saure F e r m e n t in Reifungsf i ihrun- gen ohne Sa lzgaben viel reichlicher gebi lde t wird als in normal reifen- dem Kiise. Auch hier is t die durch schuellere R~ifung bedingte , sti~r- kere E n z y m ~ k t i v i t ~ t der R~ihen I I I und IV zu beobachten .
Tabel[e 3. pa-it'nderungen im Verlau] der Sauermilch#gserei/ung. (Reifungsreihen I und II)
Reifungsart 0
mi~ S~lzen (ReiheI) . . . 5,05 ohne Salze (Reihe II) . . . 4,34
1 Das ist die beim p~ des K&ses me,bare Aktiviti~t.
p~ des K~ses mi t zunehmender Reifung nach
4 ] 7 I 11 14 18 21 25 29
Tagen I
5,57 6,14 7,25 7,30 7,70 7,73 5,04 5,50 6,11 6,40 7,26 7,30
8,00 7,30
8,40 7,74

Beitr~ge zur Bioehemie der K/~sereifung. XVI 221
Tabelle4. p,-A'nderungen im Verlau] der Sauermilchkiiserei/ung. (geifungsreihen llI und IV)
tteifungsart
mit Salzen (Reihe III) . . . . . . . . ohne Salze (Reihe IV) . . . . . . . .
p. des K~ses mit zunehmender Reifung nach
0 3 6 I 10 13 17 20
Tagen
3,7 5,6 7,0 4,1 5,7
7,35 6,4
7,70 7,75 7,05
7,90
Die Akt iv i tg t der e//elctiver~ Phosphatase zeigt trotz der betrgcht l ichen pH-~nde- rungen in beiden F/~llen, der normalen wie der anomalen Reifung, ausgesprochen flachen Verlauf, so dag hier der yon GALE erTrterte und oben besproehene ,,protective mechanism" festzustellen ist.
b) Die Wirkung von Kochsalz und Rei/ungssalzen au] die Phosphatasenaktivit~it
Die betr~cht l ichen Unterschiede der Phosphatasenaktivi t~t t salzhaltiger u n d salz- freier Reifungsff ihrungen legen die Frage nahe, ob die bei der Hers te l lung yon K~se zugesetzten Salzmengen das Phosphatasensys tem hemmend oder akt iv ierend beein- f]ussen kTnnen. Die im folgenden beschriebenen Versuche wurden an einer hoch- reifen, ohne Kochsalz und i~eifungssalze aufbewahr ten Quarkmasse durchgefiihrt .
Im Puffer-Substratgemisch wurden derartige Mengen an NaC1 (pro anal.) und NattCO~ (reinst) gelds~, dM] naeh Zagabe yon 6 ml soleher LSsungsgemische zu o,5 mlKasesuspension in den einzelnen Ansatzen folgende Salzkonzentrationen vorlagen:
1. 3,6 % NaC1, 4. 1% NattCOa, 2. 0,72% NaC1, 5. 0,2% NaHC08, 3. 0,0554% NaC1, 6. 0,0154% NaI-IC08. Die Versuehsreihen 1. und 4. enthalten pro ml Ansatz jeweils dieselben Salzkonzentrationen,
wie sie 1 g Kase normaler tterstellung aufweist; die geihen 2. und 5. haben pro ml Ansatz den- selben SalzgehMt wie 1 ml der 20% igen Suspension normalen K~ises; in 1 ml AnsatzlSsung der Versuche 3. und 6. schlieBlich liegt dieselbe Salzkonzentration vor wie in I ml Fermentsatz bei Verwendung normalen Kases (entsprechend der oben unter der Bezeichnung,,t~eifungsreihe I" gefiihrten Fertigung). Emem mitgemessenen Vergleichsansatz wurden keinerlei S~lze zugegeben. Die ,,sauren" Ansatze wurden 3 Std. bei Pn 5,6, die ,,alkalisehen" 1 Std. bei PH 1%0 bebrfitet. Bei der Pufferwahl ist die puffernde Wirkung des NaHCO3 zu berfieksiehtigen; aul3erdem ist bei Gegenwart dieses Salzes eine ErhShung der zugegebenen Triehloressigs/~uremengen bei der Eiweigf~tllung notwendig. Ergebnisse soleher Untersuchungen bringt die ']Jab. 5.
Tabelle 5. Ein/lu[3 von NaCl und NaHC08 au/ die Aktivit~it der sauren und der alkalischen Phosphatase einer hochrei]en, ohne Salz und Rei/ungssalz au/bewahrten Quarkmasse
0 3,6 0,72 0,0554
1 0,2 0,0154
8alzkonzentration im Ansatz
%
NaCI ...........
NaHC03
Phenolabspaltung, bet. auf 100 mg aschefreie K~isetrockensubstanz
sauer a~aliseh ~g #g
2664 94 1959 63 2399 88 2576 96
2091 48 2576 83 2664 87
Hemmung
sauer alkaliseh % %
26 33 10 6 3 0
21 51 3 12 0 7
Aus den Versuchen geht hervor, dal3 bei den im normalen Bebr i i tungsansatz vor- l iegenden Salzkonzent ra t ionen (Versuche 3. u n d 6.) kein nennenswer te r EinfluB der

222 J. SC~ORMULLER und E. LAHMA~N:
Salze zu beobaehten ist. Lediglich fibernormale Salzgaben (Versuchsreihen 1. und 4.) bewirken deutliche Hemmung beider Phosphatasentypen.
Es zeigt sieh also, dat~ die untersehiedliche Phosphatasenaktivit~t yon Ki~sen mit und ohne Salzgehalt nicht einfach auf einer primi~ren Wirkung der zugesetzten Salze beruht; w~re dies der Fall, dann mfil~ten, wie aus den unter a) beschriebenen Ergeb- nissen (Seite 2]9) an Reifungsreihen hervorgeht, die in Rede stehenden Salze eine Hemmung der sauren Phosphatase einerseits, eine Aktivierung des alkalisehen Fer- ment typus andererseits bewirken. Tats~chlieh wird yon h6heren Salzkonzentra- tionen aueh das letztgenannte Enzym gehemmt. Diese t t emmung finder keine befriedigende Erkli~rung dureh die Tatsache, dab bei den vor]iegenden Versuchen insofern dem teehnologisch fiblichen Gebrauch nicht genfigt wird, als die Salze erst dem fertigen Fermentansatz (einem flfissigen System) und nicht bereits der Kiise- masse zugesetzt wurden. Es bleibt also die Bereehtigung zur Annahme, da~ die Reifungssalze ein Milieu sehaffen, das ffir alkalische Phosphatasen produzierende Mikroorganismen besonders gfinstig ist. F fir saure Phosphatase ist das Gegenteil anzunehmen; hier kann au[~erdem eine Hemmung dutch die geprfiften Salze eintreten.
c) Die Phosphatasenalctivit~t in , ,Kern" und , ,Rinde" des Kgses
Frfihere Untersuchungen, so bei Aminos~urebilanzen ffir Tyrosin ~ und Trypto- phan ~ oder bei Untersuchungen fiber den Prolinasegehalt des KiSses a hat ten gezeigt, dal~ , ,Kern" und ,,Rinde" recht verschiedenartige Verteilung der betreffenden Amino- s~uren bzw. des Fermentes aufweisen. Es lag nahe, aueh hier die Phosphatasen- aktivit~t cler, ,Rinde" mit ihrem hohen Gehalt an fermentliefernden Mikroorgani ~men gegenfiber der Enzymakt iv i t~t des , ,Kernes" abzugrenzen. Bei der beim Harzer K~se sehnell von au~en nach innen erfolgenden Reifung ist eine einwandfreie Trennung dieser beiden Frakt ionen in ausreichenden Mengen nur w~hrend einer relativ kurzen Periode mSglieh. Tab. 6 bringt die nach iiblicher Methodik erhal~enen Ergebnisse.
Tabelle 6. Alctivi~tsverteilung der alkalischen, sauren und e//e]ctiven Phosphatase in verschiedenen Partien von Kdseproben, die ohne sowie mit Salzeu gerei/t wareu
(Werte bezogen auf 100 nag aschefreie Trockensubstanz)
Probe
a) alkal. Phosphatase
b) saute Phosphatase
c) effektive Phosphatase
Gesamtk~se Reifungs-
dauer Phenol- p~ abspalt, p~
Tage ~ g
1. K~isemasse ohne Salze (Reihe IV)
i 57 I 1 6,4 i 249 7,35 5,7 i 2445 6,9
10 6,4 3637 7,35 6 - - - - 6,9
l0 7,35
Rinde Kern
Phenol- abspalt.
~g
146 399
3547 8806 1096 754
1)K
3,9 4,17 3,9 4,17 3,9 4,17
Phenol- abspalt.
/~g
0 15
489 551 104 256
2. Kasemasse mit Rei]uugssalzen (~eihe III) a) alkal. Phosphatase [ 6 I 7,0 3904 7,35 7219
saure Phospha%ase I 7,0 427 7,35 825 bl effektive Phosphatase I 6 - - - - 7 , 3 5 692
5,9 5,9 5,9
79 315 229
1 SCKORMULLER, J., K. J. Br~N u. I-I. WINTEr: Diese Z. 98, 420 (1954). 2 SCHO~Mi~LLE~, J., K. J. BI~N u. H. WINTER: Diese Z. 99, 106 (1954). 3 SCKORMi)LLE~, J., u. H. Mi~LLE~: Diese Z. 100, 391 (1955).
Beitr~ge zur Bioohemie der Kgsereifung. XVI 223
Die Versuchsreihe zeigt, dab die Bildung der alkalischen sowie die Aktivit~ts- erhShung der sauren Phosphatase yon der Rinde her erfolgt ~. In den Kern dringt nur wenig Ferment ein. Bei der effektiven Phosphatase sind keine derart krassen Unter- schiede festzustellen. Die bier beobaahtete RTivellierung dfirfte in erster Linie darauf zuriickzufiihren sein, dal7 die mit der Reifung eintretende pn-Anderung bei der Rinde sich vom p~-Optimum der sauren Phosphatase entfernt, bairn Kern jedoch diesem Optimum zustrebt. Eine Bestimmung der effektiven Phosphatase im Gesamtk~tse wurde nicht durchgefiihrt, da deren Ermitt lung in diesem Zusammenhang definitions- gemA{~ nur untar Berficksichtigung des im Medium herrsehenden p~-Wertes Sinn hat. Am Beispiel der 6 Tage ohne Salzzusatz gereiften K~semasse (Tab. 6) l~tl~t sieh ein- ]euehtend zeigen, dal~ der pa-Wert des Gesamtk~ses (5,7) aus dem ps der Rinde (6,9) und dam p~ des Kernes (3,9) resultiert. Da bei p~ 5,7 das Optimum der sauren Phos- phatasa liegt, wfirde der Gesamtk~tse scheinbar einen viel h5heren Gehalt an ,,effek- river" Phosphatase aufwaisen als Rinde und Kern.
d) Die Abhi~ngigkeit der Phosphatasenalctiviti~t yon der Bebri~tungsdauer
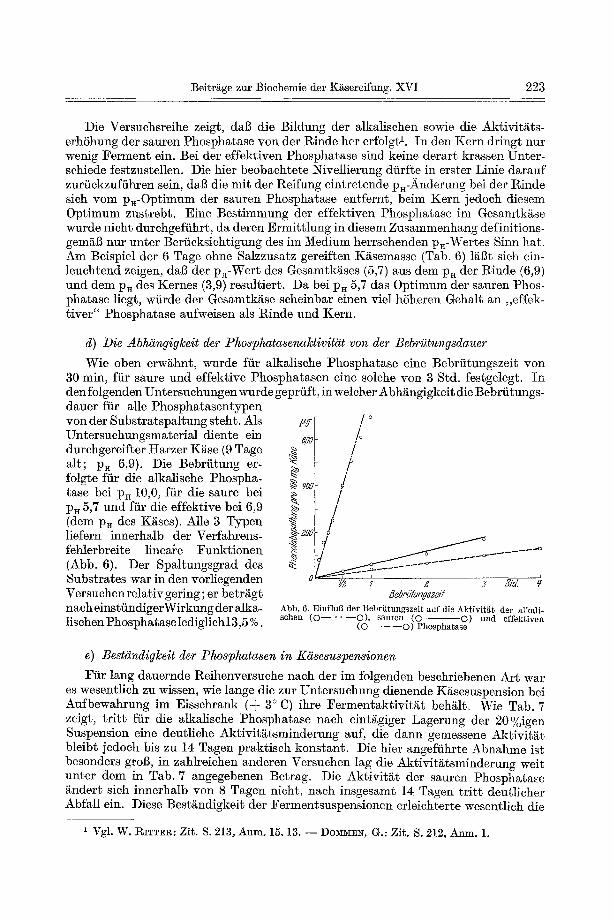
Wie oben erw~hnt, wurde fiir alka]ische Phosphatase eine Bebriitungszeit yon 30 rain, fiir saure und effektive Phosphatasen eine solehe yon 3 Std. festgelegt. In den folgenden Untersuchungen wurde geprfift, in welaher Abh~ngigkeit die Bebriitungs- dauer fiir alle Phosphatasentypen yon der Substratspa!tung steht. Als Untersuahungsmaterial diente ein durehgeraifter t tarzer K~se (9 Tage alt; p~ 6,9). Die Bebriitung er- folgte fiir die alkalische Phospha- tase bei p~ 10,0, ffir die saure bei p~ 5,7 und fiir die effektive bei 6,9 (dem pa des K~sas). Alle 3 Typen liefern innerhalb der Verfahrens- fehlerbreite lineare Funktionen (Abb. 6). Der Spaltungsgrad des Substrates war in den vorliegenden Versuchen relativ gering; er betr~g~ nach einstiindigerWirkung der alka- lisahen Phosphatase lediglichl3,5 %.
/~
/
o N 7 x 3 2k/ f 8ehrUlung3zel7
Abb. 6. Einflul~ der Bobrfitungszei~ aLif die Aktivit~t der alkali- schen (O . . . . O), sauren (O ©) und effektiven
( O - - - - - O ) Phosphatase
e) Besti~ndiglceit der Phosphatasen in Kiisesuspensionen
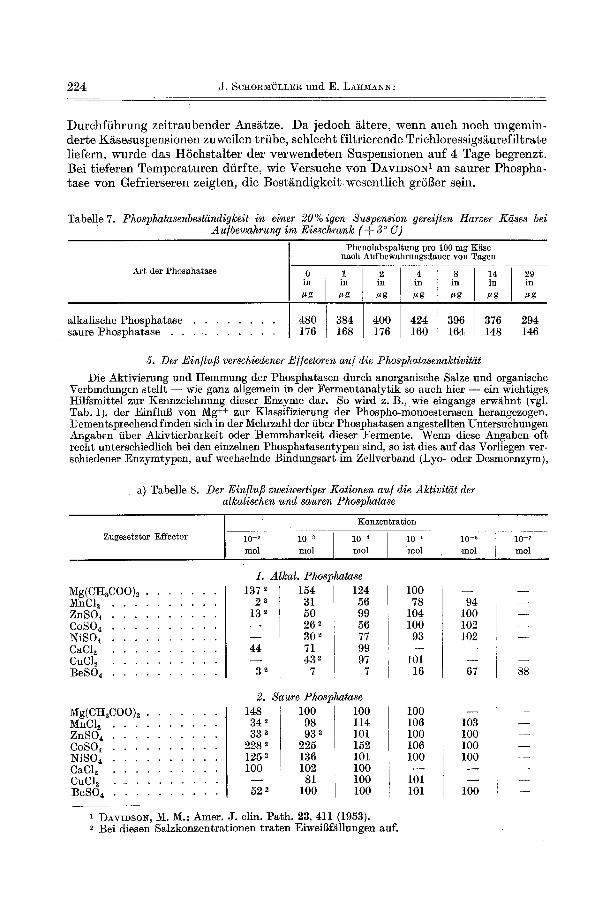
Fiir lang dauernde Reihenversuehe nach der im folgenden beschriabenen Art war es wesentliah zu wissen, wie lunge die zur Untersuchung dienende K~sesuspansion bei Aufbewahrung im Eisschrank (-t- 3 ° C) ihre Fermentaktivit~t beh~lt. Wie Tab. 7 zeigt, t r i t t fiir die alkalische Phosphatase naah eint~giger Lagerung der 20%igen Suspension eine deutliche Aktivit~tsminderung auf, die dann gemessene Aktivit~t bleibt jedoeh bis zu 14 Tagen praktisch konstant. Die hier angefiihrte Abnahme ist besonders grol], in zahlreichen anderen Versuchan lag die Aktivit~tsminderung weir unter dam in Tab. 7 angegebenen Betrag. Die Aktivit~t der sauren Phosphatase gndert siah innerhalb yon 8 Tagen nicht, nach insgesamt 14 Tagen t r i t t deutlicher Abfall ein. Diese Best~ndigkeit der Fermentsuspensionen erleichterte wesentlieh die
1 Vgl. W. I~IWTER: Zit. S. 213, Anm. 15, 13. - - DOM~E~, G." Zit. S. 212, Anm. 1.
224 J. SCHORMiYLLER und E. LAHhIA1N'h':
D u r C h f t i h r u n g z e i t r a u b e n d e r A n s ~ t z e . D a j e d o c h El tere , w e n n a u c h n o c h u n g e m i n - d e r t e K / i s e s u s p e n s i o n e n z u w e i l e n t r i i be , s c h l e c h t f f l t r i e r e n d e T r i c h l o r e s s i g s ~ u r e f i l t r a t e l i e fe rn , w u r d e d a s H S c h s t a l t e r d e r v e r w e n d e t e n S u s p e n s i o n e n a u f 4 T a g e b e g r e n z t . B e i t i e f e r e n T e m p e r a t u r e n d f i r f t e , wie V e r s u c h e y o n I)AVIDSOI~ 1 a n s a u r e r P h o s p h a - t a s e y o n G e f r i e r s e r e n z e i g t e n , d ie B e s t i ~ n d i g k e i t w e s e n t l i c h g rSBer se in .
Tabelle 7. Phosphatasenbesttindigkeit in einer 20% igen Suspension gerei/ten Harzer Kgses bei Au/bewahrung im Eissehrank (-~ 3 ° U)
Art der Phosphatase
alkalisehe Phosphatase . . . . . . . . saure Phosphatase . . . . . . . . . .
P h e n o l a b s p a l t u n g p r o I 0 0 m g K ~ s e n a c h A u f b e w a h r u n g s d a u e r y o n T a g e n
0 I in in /~g # g
480 384 176 168
2 4 in in /zg /~g
400 424 176 160
8 14 in in /~g /~g
396 376 164 148
29 in ~ g
294 146
5. Der Ein/Iufi versehiedener E/]eetoren au/ die Phosphatasenaktivigit
Die Aktivierung und Hemmung der Phosphatasen du tch anorganische Salze und organische Verbmdungen stell t - - wie ganz allgemein in der Fermentana ly t ik so auch hier - - ein wichtiges Hiffsmittel zur Kennzeichnung dieser Enzyme dar. So wird z. B . , wie eingangs erwtihnt (vgl. Tab. 1), der Einflul~ yon Mg ++ zur Klassffizierung der Phospho-monoesterasen herangezogen. ]~ementsvechend l inden sich in der Mehrzahl der fiber Phosphatasen angestellten Untersuchungen Angaben fiber Akivt ierbarkei t oder ~ e m m b a r k e i t dieser ]fermente. Wean diese Angaben oft recht unterschiedlich bei den einzelnen Phosphatasentypen sind, so ist dies auf das Vortiegen ver- schiedener Enzymtypen, auf wechselnde Bmdungsar t im Zellverband (Lyo- oder Desmoenzym),
Z u g e s e t z t e r E f f e c t o r
a) Tabelle 8. Der Einflufi zweiwertiger Kationen au/ die Aktivittit der alkalischen und sauren Phosphatase
Konzen tration
1 0 - : 10 - a 10 -4 10 -~ 10 -6
tool mol tool tool mol
Mg(CH3COOh . . . . . . . M_nCI~ . . . . . . . . . . ZnSO~ . . . . . . . . . . COS04 . . . . . . . . . . NiS04 . . . . . . . . . . CaCI~ . . . . . . . . . .
CuCI~ . . . . . . . . . .
BeS04 . . . . . . . . . .
Mg(Ctt3COOh . . . . . . . MnCI~ . . . . . . . . . . ZnS04 . . . . . . . . . . CoSO~ . . . . . . . . . . :NiSO~ . . . . . . . . . . CaC12 . . . . . . . . . . CuCI~ . . . . . . . . . . . BeSOa . . . . . . . . . .
1. AIkal. Phos ohatase 137 2
2 3 13 ~
44
154 31 50 26 ~ 30 3 71 432
7
124 56 99 56 77 99 97
7
100 78
104 100
93
101 16
100 102 102
67
i0-~ tool
J
88
2. Saure Phos 148 100
34 ~ 98 33 ~ 93 ~
228 e 225 125 ~ 136 100 i 102
I 81 5 2 ~ ! 100
ohatase 100 114 101 152 101 100 100 100
100 106 100 106 100
1 0 1 101
103 100 100 100
1 0 0
m
1 DAWDSON, M. 1YI.: Amer. J . d in . Pa th . 23, 411 (1953). 2 Bei diesen Salzkonzentrat ionen t r a t en EiweiBf/illungen auf.

Beitrgge zur Biochemie der K/isereifung. XVI 225
auf die Natur des Sub- strafes 1 oder auf die Art des Effectors zurtickzuffih- ren. So berichten CAClOPPO und Mitarbciter ~, dab ffir gereinigte Phosphatase aus Rinderhirn bei Anwesenheit yon Mg ++ das pH-Optimum eine Verschiebung yon 9,0 auf 7,3 erf/ihrt. SchlieBlich ist die Wirkung derartiger Zusgtze aueh davon abhgn- gig, ob sie im fermenthal- tigen Rohprodukt oder im hochgereinigten Prgparag geprfift wird 3.
In den folgenden Ver- suchen wurden den An- sgtzen verschiedene Sub- stanzen zugegeben, yon
b) Tabelle 9. Der Ein/lu# versehiedener Anionen au/ die Aktivitiit der sauren und alkalischen Phosphatase
Zugesetzter Effector
Konzentrat ion
10-2 10-3 10-~ 10-~ mol mol rnol tool
1. Alkal. Phosphatase l~Ta ~ I-IP O 4 . . . . [ 55 10 o NaCN . . . . . . - - u NaF . . . . . . [ - - 83 Na-eitrat . . . . 55 9i
2. Saure Phosphatase KH2P04 . . . . . ] 57 89 NaCN . . . . . . [ - - 86 N~F . . . . . . - - 22 Na-citrat . . . . 93 91
100 53 100
100 99
97 98 93 95
denen eine aktivit~tsbeeinflussende Wirkung zu erwarten war. Die im Mkalischen und sauren Bereich wirksamen Phosphatasen wurden in getrennten Versuchsffihrungen fiberprtift; diese waren folgendermaBen zusammengesetzt:
0,5 ml K/~sesuspension fiblicher Herstellung aus gereiftem Kgse; 5,35 ml Puffer-Substratgemisch; 0,65 ml 10-x tool w~Brige SubstanzlSsung. Ftir den zu untersuchenden Effector ergibt sieh damit die Konzentration im Ansatz zu
10-(x+l) mol. Die Blindwerte enthalten Puffer start Puffer-Substratgemisch, die Yergleichswerte Wasser s ta t t SubstanzlSsung. Versuehsfiihrung und Phenolbestimmung erfolgten wie fiblich. Zur Ermit t lung der sanren Phosphatase wurde 3 Std. bci Pn 5,4--5,7, der alkalischen Phosphatase ~/2 Std. bei p~ 9,85--10 bebrtitet. Der Einflug der zugesetzten Substanzen wird auf den jeweils mitlaufendenVergleiehsansatz ohne Effeetorbeigabe bezogen, dessen AktivitKt zu 100 gesetztwird. Angaben unter 100 drficken demnach die prozentuale Hemmung, solche fiber 100 die entspre- chende Aktivierung aus.
I n d e n v o r s t e h e n d e n Tab . 8 u n d 9 s ind E rgebn i s s e d e r a r t i g e r V e r s u e h e zu- s a m m e n g e f a B t . I h r e E r S r t e r u n g e r fo lg t e i n g e h e n d a m SehluB de r Arbe i t .
c) Der Ein/lu[3 o~vanischer •//ectoren au/ die Aktivitiit der sauren und alkalischen Phosphatase
l~lber die W i r k u n g y o n Aminos~turen a u f P h o s p h a t a s e n s ind in de r L i t e r a t u r w i d e r s p r e c h e n d e A n g a b e n zu f i n d e n 4. W i r p r f i f t en Cystein u n d Histidin, d e r e n u n t e r - seh ied l iches V e r h a l t e n gegenf iher P h o s p h a t a s e n v e r s c h i e d e n e r H e r k u n f t y o n I~oc~E u n d vA~¢ TItOAI in de r a n g e z o g e n e n l J b e r s i e h t e i n g e h e n d r e f e r i e r t wird . N a c h unse r en U n t e r s u e h u n g s e r g e b n i s s e n i n a k t i v i e r t Cys t e in (10 -2 tool) die a lka] ische P h o s p h a t a s e m e r k l i c h (10 -8 tool zu e t w a 15 %), a u f die sau re i s t es in a l len K o n z e n t r a t i o n s b e r e i c h e n 0hne E in f luB . H i s t i d i n h e m m t das a lka l i sche E n z y m bei 10 -a tool zu 22 %, das sau re bei 10 -~ too l zu 5 1 % 5.
Mono]odessigsiiure a]s t y p i s e h e r D e h y d r a s e n i n h i b i t o r wirk~ a u f a lka l i sche Phos- p h a t a s e n ieh t , a u f s au t e P h o s p h a t a s e d e u t l i c h h e m m e n d (22% bei 10 -2 mo]). Jodoso- benzoesiiure u n d Phenylmercuriborat s ind als spezff i sehe I t e m m s t o f f e (,,Thioprivine")
1 RoegE, J. , u. S. BOUCHILLOUX: Arch. Sci. physiol. 2, 283 (1948). CAelO~rO, F., E. QUAGLIAI~ELLO, ~V[. COLTOII, I U. G. D. PIETRA: Boll. Soc. ital. Biol. sperim.
28, 1609 (1952). BA~ANN, E . , E . I~IEDEL U. K. DIEDERICI-IS: Hoppe-Seylers Z. 230, 175 (1934). Zusammenfassung bei J . ROC~rE u. N. van T~OAI: Advances in Enzymol, 10, 100 (1950),
V g l . a u c h CIt , A . ZITTLE U. E . S. DELLA MONICA: Arch.of Biochem. a. Biophysics 26, 135 (1950). 5 Vgl. O. BODANSK:g: J . of Biol. Chem. 174, 465 (1948).
Z. Lebensmittel-Unters. u. -Forsch., Band 103 16

226 J. SC~O~iiL~E~ und E. L ~ w ~ :
ffir E n z y m e bekann t , die funkt ionel le S H - G r u p p e n en tha l ten , also sog. , ,Thiol- f e rmen te" dars te l len 1. 1NTach fr i iheren Un te r suchungen unseres Arbei t skre ises 2 ent - fa l ten sic am Transaminasensys t em des Sauermilehk~ses sehr s t a rke Hemmwirkung . Die P h o s p h a t a s e n des Sauermilehk~ses werden durch derar t ige Thio l inhib i toren n icht nennenswer t gehemmt , so dai3 weder die saure noch die a lkal ische Sauermilchk~se- P h o s p h a t a s e ak t i ve S H - G r u p p e n zur W i r k s a m k e i t ben5t ig t .
I n wei te ren Versuchen wurde die W i r k u n g von Bradoso l ( /~-Phenoxy~thyl- d i me thy ] -dodecy l - ammon iumbromid ) und Na-desoxycho la t un te rsuch t . Bradosol a ls Beispiel e iner qua te rn~ren Ammoniumbase , wie sie in der Milchindust r ie mi t zu- nehmender Bedeu tung Verwendung f inden, ak t i v i e r t die s~ure Phospha ta se gering- ffigig (weniger als 10 %), au f die a lkal ische is t es ohne Einfluf]. 0 f f e n b a r t r i t t hier d ie yon K c ~ N und BInL~G a e rSr te r te R e a k t i o n solcher , , Inver t se i fen" mi t P ro t e inan ionen oder S y m p l e x e n n ich t s tSrend in Erscheinung. Die f e rme n the mme nde W i r k u n g y o n quatern~tren A m m o n i u m b a s e n wird vie l faeh in der L i t e r a t u r erw~hnO. Dai~ diese Stoffe a u c h - unseren Un te r suchungen an saurer Kgsephosph~tase zufo]ge - - ak t i - v ie rend wirken kSnnen, zeigen Beobach tungen yon MEIS~S~ und Mi ta rbe i t e rn s bei Glu tamins~ure -decarboxy lase , yon HUGHES s bei Bak te r i endeca rboxy la sen u n d Glu taminase .
Das Verha l t en von Na -desoxycho l a t gegenfiber den yon uns un te r such ten K~tse- p h o s p h a t a s e n wurde deshalb fiberprfift , weft sich gezeigt ha t te , dal~ Salze yon Gallen- s~turen die ~lkalische Geh i rnphospha tase ~ sowie auch andere Phospha t a sen s zu h e m m e n vermSgen, wi~hrend Geh i rnpyrophosph~tase s sowie P o l y m e t a p h o s p h a t a s e aus Aspergillus niger und Saccharomyces cerevisiae ~° durch Tauroeho la t oder Cholat ke ine A k t i v i t ~ t s m i n d e r u n g erfahren. Unseren Versuchen zufolge spreehen die h ier geprf i f ten Phospha t~sen in be iden R ich tungen an : das a lkal ische F e r m e n t wird dureh
10 -~ mol Na -desoxycho la t zu immerh in 31%, Tabelle 10. Ein/lufi von Fluorid und das saure E n z y m nicht merkl ich gehemmt , Cyanid au/ die ,,e/]e]~tive' Phosphatase so da~ auch bier ein wesent l ich untersch ied-
eines gerei/ten Kggses (p~6,8) ]iches Verha l t en be ider P h o s p h a t a s e n t y p e n fes tzus te l len ist. Phenolab-
Zugese tz ter spa l tung durch l t e m m u n g Effec tor 100 m g K~se
10 -~ tool t tg % d) Einflufi spezi]ischer Hemmsto]]e au/ die ,,eHektive " Phosphatase
- - 94 0 Mit Hilfe der spezif ischen I n h i b i t o r e n CN- 90 4 F luo r id (fiir saure) und Cyan id (fiir a lkal i - F - 51,5 45
sche Phospha tase ) wurde geprfift , welchem der be iden E n z y m t y p e n d i e , , e f f ek t i ve" Phos-
pha t a se auf G r u n d ihres VerhMtens gegen die genann ten Hemmsto f f e zugeordnet werden kann . Das p~ des un t e r such ten K~ses lag bei 6,8. Demen t sp rechend wurden
1 BARROW, E. S. G.: Adv. in Enzymol. 11, 235 (1951). 2 SCHORMi~LLE~, J., u. W. GET.L~Ie~: Diese Z. 100, 216 (1955). a Kurd , R., u. H. J. BIELI¢: Ber. dtsch, chem. Ges. 73, 1080 (1940).
Vgl. z; B. F. BV, RG~ANN U. A. S~I~oNI: Biochem. et biophysica Acta (Amsterd.) 9, 473 (1952). - - C~Oe~:E~ C. H.: J. Milk Food Teehnol. 14, 138 (1951). - - W~LLS, E.D. : Biochemic. J. 53, 20 (1953).
5 MEISTE~, A., H. A. SOBER U. S. V. TICE: J. of Biol. Chem. 189, 591 (1951). 6 I-IvG]t]~S, D. E. : Int. Congr. Biochem., Abstracts of Communs. 1. KongreB. S. 324. Cam-
bridge 1949. FLEISe~i~KEI~, H. H. : J. Mental Sci. 84, 947 (1938).
s URAXI, Z.: J. Biochem. (Japan) 14, 123 (1931). Go~DoN, J . J . : Biochemic. J. 46, 96 (1950).
10 MAL~GlCE~, I-I.: Acta chem. scand. 3, 1331 (1949).

Beitr~ge zur Biochemic der K~isereifung. XVI 227
A n s a t z e m i t 10 -3 ree l F - , 10 -a m o l CI~- sowie solche o h n e S a l z z u s a t z 3 S td . bei d ieser pH-Stufe bebr f i t e t . Tab . 10 zeigt , daI3 die , , e f f e k t i v e " P h o s p h a t a s e de r e r w a h n t e n R e i f u n g s s t u f e in i h r e m V e r h a l t e n w e i t g e h e n d d e m s a u r e n F e r m e n t e n t s p r i c h t .
e) Der Ein/lu[3 yon Penicillin und Streptomycin au/ die Phosphatasen-Aktivit~it Die Behandlung euterkranker Kiihe mit Penicillin sowic die Verwendung anderer Antibiotica
in der Tierheilkunde wirft das Problem der Kiisereitauglichkeit der yen solchcn Tieren gewonne- nen Milch auf. Da das Antibioticum in die Milch fibergeht und die zur Kfisereifung notwendigen Mikroorganismen teilwcise abtLtct 1, wird die Herstcllung eines cinwandfreien K~ses aus der- artiger Milch empfindlich gest(irt 2. Zusatz des als Penicillinase (Pcnasc) bezcichneten Fermcntcs zur Ki~sereimilch wurde als eine MLgtichkeit zur Bescitigung des Pcnicillins erLrtert ~.
I n de r v o r l i e g e n d e n A r b e i t w u r d e n V e r s u c h e f iber d e n Einflul~ y o n Pen ic i l l i n G (Grf inen tha l ) sowie y o n S t r e p t o m y c i n s u l f a t ( B a y e r - L e v e r k u s e n ) a u f d ie i m Sauer - milchk~ise v o r l i e g e n d e n P h o s p h a t a s e n d u r c h g e f i i h r t . V o n d e n be iden A n t i b i o t i c a sche in t Pen i c i l l i n vo rzugswe i se a m Eiweil~- u n d a m N u c l e o t i d s t o f f w e c h s e l anzu- g re i fen 4.
Die Gcwebsatmung wird in thcrapcutisch wirksamen Doscn nicht bccinflufit. Da die genann- ten Stoffwechselprozcsse durch viele SH-haltige Fermenttypen katalysiert werden, ist dcr Befund verschicdcncr Autoren nicht vcrwunderlich 5, wonach Penicillin und andcre Antibiotica essentielle SI-I-Gruppen blockieren. Streptomycin hemmt nach UMBREIT ~ insbesondere die enzymatische Kondensation der Oxalessigsiiure mit Brenztraubens~iure und inhibiert verschicdcnc oxydativc Stoffwechselprozesse 7. Uber die Wirkung beider Antibiotica auf phosphatatische Fermente ist bisher wenig bckannt gewordcn. MmABEAU CRUZ s beobachtcte bei Penicillin Aktivit~itssteigerung fiir eine Glycerophosphatase, FRO~MEL und Mitarbeiter 9 Hemmung der Knochenphosphatase. Streptomycin hemmt ticrische Glycerophosphatase ~°. Alkalische Phosphatase aus Rattcnniere wird durch Penicillin G nicht merklich, durch Streptomycin im p.-Bercich 6--8 zu 20~35 % gehe.mmt 11.
Uber das Verhalten dcr Milchphosphatasc gegenfibcr antibiotisch wirkcnden Stoffen liegen widersprechende Mitteilungen vor. STOLTZ und HA~:I~SO~ ~2 beschreiben ein Verfahren, das auf Grund dcr Sch/~digung der Rohmilchphosphatasc die Fcststellung hLheren Antibioticagehaltes (Penicillin, Streptomycin, Aureomycin, Thyrothricin) crmLglichen solh C~URC~ILL und Mit- arbeitcr 13 hingegen konntcn keinen Einflul~ antibiotischer Substanzen auf die Milchphesphatase feststellen.
WILKOWSKE, It . tI., u. W. A. KaIE~KE: J . Dairy Sci. 34, 1030 (1951). 2 Vgl. z. B. M. SEELEMA~ U. F. TI~M: Milchwiss. 10, 113 (1955). - - J o ~ s , C .K. : J. Dairy
Sei. ~6, 1241 (1953). ~- JACQUET, J. : 13. Int . Milchwirtsch. Kongr. (s'Gravenhage)3,1143 (1953) . - TI~ROT~, I .V.: Sv. Mc]critidn. Nr. 2--3, 6 (1953). - - CLAYB~UGlZ, G.A. , u. F. E. NELSON: J . Milk. Food Technol. 14, 5 (1951). - - JEPSE~, A., u. A. J . OVERB¥: Netherl. Milk Dairy J . ~, 157 (1951).
W~SS~ER, P.: 13. Int . Milchwirtscl~. Kongr. (s'Gravcnhage) ~, l l21 (1953). GALE, E . F . : Zit. S. 219, Anm. 2, daselbst S. 198, 199. - - M_~CH~OE~, M., u. F. GRos:
2. Kongr. int. Biochemie. Symposium sur le mode d'aetion des antibiotiques. S. 5; 101. Paris: 1952; zit nach K. P. JACOBSOI~ U. M. D. DE AZEVEDO: Exper. Medicine Surgery 11, 149 (1953); C. r. Soc. Biol. (Paris) 148, 199 (1954); dort weitere Literatur.
SAVELS~ER¢, W., u. J . STRLI)~R: Schweiz. mud. Wschr. 19~2, 140. U~rBR~,IT, W.W." Zit. bci K. P. JACOBSOI~ u. M. D. DE AZEVEDO" Zit. diese S., Anm. 4. - -
GALE, E . F . : Zit. S. 219, Anm. 2, dasclbst S. 199. 7 PAINE JR., TH. F., u .L . ST. CLARK" Science (Washington) 118, 73 (1953). - - Antibiotica
Chemotherapy 4, 262 (1953). - - ROSENBLUM, E. D., u. V. B~YsoN: Antibiotica Chemotherapy 8, 957 (1953). - - GERATgI~I, S., u. A. G. LEONARDI: At t i Soc. Lombarda Scienze reed. Biol. 8, 289 (1953). - - BAm~lZ~is, I. I . : J. Bactero 6~, 337 (1953).
s MIRABEAU CRUZ, J . : Arch. portug. Sei. biol., Suppl. 9, 23 (1946); 9, 97 (1947). FROlg~EL, :E., A. GOLDFIEDER U. J . PIQUET: Acta pharm, rex. (Kopenhagen) 2, 207 (1946).
10 JAco~soI~, P .K . , u. J. MIRA~EAU CRUZ: Arch. portug. Sci. biol. Suppl. 10, 29 (1948). 1~ G~ATAK, S., u. C. R. K a I S ~ A MUI~TI: J . sei. ind. Res. IPB, 160 (1953). ~ STOLTZ, E. I., u. D. J . IqA~KI~SO~: Amer. Milk Rev., Dez. 1950, S. 60. 1~ C~IUI~C~ILL, E. S., A. R. D R u ~ , B. D. LEWIS, C. L. FRANK U. C. S. BRYAn: Milk P lan t
Monthly 40, 28 (1951).
16"

228 J. ScHoRMffLLER und E. LAHMANN:
Unsere Versuche zeigten, dal~ weder Penici l l in G (5 und 25 I E / m l Ansatz) noch S t r ep tomyc in su l f a t (0,1, 1 u n d 5 mg/ml Ansatz) die sau te und alkal ische P h o s p h a t a s e beeinfluBten. Al lerdings is t dabe i zu berf icksichtigen, da{3 die Zugabe dieser Subs tan- zen ers t zum eigent l iehen F e r m e n t a n s a t z , also u n t e r Ausscha l tung des vorhe rgehenden mikrobie l len Stoffweehsels de r Ki~seflora erfolgte. Nach GALE 1 ist der Wi rkungs - meehanismus des Peniei l l ins versch ieden je nachdem, ob er an ruhenden oder wach- senden Ze l lku l tu ren s tud ie r t wird.
6. Der Ein/lufl physilcalischer Faktoren au] die Phosphatasenaktiviti~t a) Die Hitze-lnaktivierung A u f die B e d e u t u n g der H i t ze inak t i v i e rung der a lka] ischen Mi lchphospha tase als
Tes tg rund lage zum Naehweis s t a t t ge fundene r Pas teur i s i e rung wurde bere i ts ein- l e i tend hingewiesen. Die folgenden Versuche wurden vor al lem m i t der Abs ich t
durchgeff ihr t , die Hi tzes tab i l i - Tabelle 11. Die Hitze-Inaktivierung der alkalischen und
sauren Phosphatase gerei/ten Harzer KiSses
]~rhitzungs- temperatur Erbitzungs-
dauer °C
i ° . Phenol- Inaktlvmrung abspaltung gegenfiber dean
durch nicht erhitzten 100 mg tt:~se Vergleichsansatz
gg %
0 50
62
72
85 100
0 50
62
72
85 100
1. Alkal. Phosphatase 0 min 2384
10 min 2352 30 rain 2352 5 min 2032
10 min 1600 30 rain 744
5 sec 648 40 sec 306 10 rain 31,5 5 sec 27,5
t5 rain ' 0
2. Saure Phosphatase 0 rain 188
l0 rain l l 0 30 rain 94
5 min 67 10 min 55 30 min 24
5 sec 23 40 sec 19 10 min 9 5 sec 9
15 rain 0
ti~t yon im Sauermilchki~se nachgewiesenen P h o s p h a t a s e n mi t der yon Mi lchphospha tase zu vergleichen. Als Erh i tzungs- t e m p e r a t u r e n und -zei ten wur- den deshalb vorwiegend die zur Mi lchpas teur i s ie rung v0r-
0 geschr iebenen Bedingungen ge- l wi~hlt (Dauererh i tzung 30 min 1 auf 62 ° C; Kurzze i t e rh i t zung
15 40 sec au f 72 ° C; Moment- 33 erhi tzung auf 85 ° C), die zu 69 73 96- - 100 %iger I n a k t i v i e r u n g 87 dieses F e r m e n t e s ffihren. Die 99 fiber 15 rain ausgedehnte Er- 99 h i tzung auf 1 0 0 ° C l ieferte
100 einen fiir die sparer beschrie- benen Dia lyseversuche not-
0 wendigen, i nak t i ven , ,Koch- 41 sa l t " . 50 Die H i t zebehand lung er- 64 71 folgte an der 20%igen K~tse- 87 suspension gereif ten Ha rze r 88 K~ses (p~ 7,2) im H6PPLE~- 90 Thermos ta t . A]s Erh i tzungs- 95 95 beginn wurde der Z e i t p u n k t
100 gewiihlt, bei dem die in sehma- len geagenzg l~e rn bef indl iche
Suspension die geforder te T e m p e r a t u r angenommen hat te .Wi~hrend des Versuches wurde mehrma l s gerfihrt . Die alka] ischen Ansi~tze wurden 1 Std. bei PH 9,9, die sauren 4 Std. bei p~ 5,7 wie fiblich bebr f i te t und ana lys ie r t .
Die in de r vors tehenden Tabel le geb rach ten Ergebnisse zeigen, dal~ die a lkal ische Phospha t a se h i tze res i s ten te r als das sau te F e r m e n t ist. Beide E n z y m e haben nach d e m Erh i t zen au f 85 ° C ihre A k t i v i t ~ t p r ak t i s ch v511ig ver loren. Vergl iehen mi t dem a lka l i s ehen .Fe rmen t der l~ohmilch, is t die H i t ze s t ab i l i t £ t der a]kal ischen P h o s p h a t a s e
GALE, E . F . : Zit. nach Angew. Chem. 66, 422 (1954).

Beitrage zur Biochemie der K/~sereifung. XVI 229
des Harzer K/~ses betr~Lchtlich hSher; diese Tatsache mach t sich besonders bemerkbar beim halbs t t indigen Erh i tzen auf 62 ° C, wo lediglich eine 70 %ige Sch~digung e int r i t t . Der , ,Kochsaft" zeigte keine phosphata t ische Akt ivi t / i t Inehr. E ine an mehreren der~rt erhi tz ten Proben nach 7 t~giger Aufbewahrung der LSsung (im Eisschrank u n d bei Z immer tempera tur ) durchgeftihrte Nachun te r suchung erg~b keine Reakt iv ie rung der Phosphatasen.
b) Der Ein/lufi yon Ultraschall Angaben fiber die Wirkung yon Ultraschall auf Mikroorganismen zeigen, dal~ diese sich recht
verschieden verhalten ~. Entsprechend unterschiedlich sind auch die Mitteilungen fiber den Ein- flu] you Ultraschall auf Fermente. I-Iyaluronidase ~, Blutkatalase 3, 0xydasen ~, Pepsin (vor allem in verdfinnter LSsung) ~ sowie alkalische Phosphatase ~ werden zerstSrt oder inaktiviert, Carb- oxyl~se wird nicht ver/~ndert ~. Angaben fiber Milchfermente sind nur sp/~rlich zu finden. ~NTELSO~ und WIl~nm~ s geben an, dab Ultraschall die K/isereifung beschleunige, ohne den hier ablaufenden Meehanismus n/~her zu erSrtern. Milchkatalase wird aktiviert ~. Xanthindehydrase, vor allem im gereinig~en Zustand, schnell zerst5rt '0, Phosphatase nicht beeinflul]t ~. Die Best~indigkeit vieler Bak~erienfermente gegen Ultraschall schuf die M6gliehkeit, dutch derartige Behandlung aus Mikroorganismen zellfreie FermentlSsungen zu gewinnen, wie z. B. solebe der ,,Racemase" aus Streptococcus/aecalis 1~ oder der Desulfurase aus Proteus morgani ~.
Zu unsereu Versuchen benutzten wit die iibliche 20% ige Suspension eines gereiften ttarzer K~ses, die in einem Siemens-Ultraschallger/~t bei 430 kHz und 6,5 W/cm ~ behandelt wurde ~. Dureh Vorkfihlen der Suspension auf -~5 ° C und zus~tzliche Ktihlung des Reaktionsgefi~l~es wurde die Temperatur w/£hrend der Ultraschall-Einwirkung auf maximal 29 ° C gehalten.
Die Versuche zeigten, dal~ die alkalische Phosphatase, f ibere ins t immend mi~ An- gaben yon TOBLER n fiber das Verhal ten der Milchphosphatase, t rotz der hohen Schall- intensit/~t auch nach 12 min keine Akt iv i t~ t sminderung erfahren hat te . Die saure Phosphatase ist empfindlicher. Sic erleidet un t e r denselben Bedingungen nach 4 rain dauernder Behand lung eine Inak t iv i e rung um 23%; 1/£ngere Behand lung erhSht diesen Betrag n u t unwesentl ich. Die hohe Best~ndigkei t der K/tsephosphat~sen zeig~, dal] diese Fe rmen tg ruppe weder durch Desaggregierung des Apoproteins, wie dies N~w~oN u n d K i s s E n ~ amTabakmosa ikv i rus zeigen konnten , noch durch Oxydat ions- prozesse, hervorgerufen infolge des Auf t re tens freier OH-Radikale bei der Schall- beh~ndlung l~, gesch~digt wird. Auch der E inf lu~ etwa ents tehender toxischer Sub- stanzen, den ANDERSON 1~ bei der Beschal lung yon Saccharomyces cerevisiae erSrterte, ist auszuschliel~en.
1 Zusammenfassende Arbeiten: It. I-IO~PESClt: Zbl. Bakter. I. Orig. 159, 358 (1953). - - WAI~-WECKE, B. : Z. I-Iyg. 188,17 (1953).-- VELTIVIAlq, G. : Z. exper. Med. llS, 55 (1951).-- BEI~O~rA~¢~¢, G.: Der Ultraschall. 5. Aufl., S. 663. Stuttgart: I-Iirze11949.-- I-InlN, H.: Z. Hyg. 13"2, 519 (1951).
LEvi, F.A.: Enzymologia 15, 285 (1952). s A~IcI, A.: Boll. Soc. ital. Biol. sper. 28, 1930 (1952). 4 GRABAR, P., L. VOINOVlTCH U. 1~. O. PRUDI~O~E : Biochem. et Biophysiea Acta (Amsterd.)
3, 412 (1949). NAI~ARK, G. M., u. W. A. 1V[OSEER: J. Acoust. Soc. America 2~, 289 (1953). CozzAl~I, L. V., u. F. A. LEVI: Boll. Soc. itah Biol. sper. 29, 1 (1953).
7 EULEI~, H. v., u. B. SKARZlNSKY: Naturwiss. 31, 389 (1943). s NELSON, J. H., u. W. C. WIICD~,I~: J. Dairy Sci. 87, 638 (1954).
BUI~O~R, M., u. W. C. WnCD~R: J. Dairy Sci. 37, 645 (1954). ~0 RonEa% L., u. N. NOLLA: Bull. Soc. Chim. biol. 85, 1363 (1953). ~1 TOBLm~, F .R. : 13. Int. Milchwirtsch. Kongre~ (s'Gravenhage) 2, 703 (1953). 12 Woon, W. A., u. I. C. GUlVSALUs: J. of Bioh Chem. 190, 403 (1951). ~3 KALLIO, R. E.: J. of Biol. Chem. 192, 371 (1951). ~4 Ffir die tJberlassung des Ger/~tes sind wir den Siemens-Schuekert-Werken AG sehr zu Dank
verbunden. (GHO 03a ffir 350 kHz, 300 W Ultraschall-Leist.) 1~ £NEWTOI% N., U. J. W. KISSEL: Arch. Biochem. Biophysics 47, 424 (1953). 16 GR-4BA1R, P., u. R. 0. PRUDHOMME: Der Ultraschall in der Medizin. Kongr. ber. Er]anger
Ultraschall-Tagung 1949, 114 u. 118. ~7 A~I)EI~SO~, J. M. : Biochem. et Biophysica Acta 11, (Amsterd.) 122 (1953).

230 J. SCHORMi~LLER und E. LAttMANI~:
c) Das Verhalten der Phosphatasen bei der Dialyse Die im folgenden besehriebenen Dialyseversuohe sollten zur Kl~rung der Frage beitragen, ob
durch eine derartige Behandlung wasserl6sliehe Stoffe organischer oder anorganischer Art ent- fernt und dabei die Kasephosphatasen aktivierend oder hemmend beeinfluBt wfirden. AuBer an die Beseitigung soleher Aktivatoren oder Inhibitoren ist aueh an eine Abtrenfiung yon Coferment- ffaktionen zu denken, die yon der Bindungsart des Fermentes an die Zellsubstanz (Lye-, Endo- oder Desmoenzym), daneben auch yon der p.-bedingten Dissoziation abhangig ist. AL~.RS ~ gelang die Abtrennung des Coenzyms dureh Dialyse und die Reaktivierung durch Oberhefen- coferment bei alkalischer Nierenphosphatase. EK, ECLER und HAH~ 2 berichten fiber ~hnliehe Inaktivierungsm6glichkeiten bei der Dialyse anderer tierischer Phosphatasen. Die weitere, oben er6rterte MSgliehkeit einer Aktivit~tsbeeinflussung durch Dialyse, nfimlich die Beseitigung yon Inhibitoren, geht aus Untersuehungen y o n :DELoRY u n d HETI=IERINGTON 3 hervor, wonaeh unter derartigen Bedingungen die saute Harnphosphatase eine Aktivitatssteigerung erfahrt.
In den vorliegenden Versuchen wurden je 100 ml Kfisesuspension 24 und 48 Std. in Cellophan- sehlfiuchen gegen dest. Wasser dialysiert. Zum Ausgleich der ¥ olumzunahme wurde jeweils auf200 ml erg~inzt. Als Vergleich diente die 10% ige, nicht dialysierte Suspension derselben KfisepIobe. Die Phenolabspaltung wurde auf 1 ml 10% ige K~sesuspension (dialysiert bzw. nicht dia]ysiert), ent- spreehend 100 mg Kase, bezogen. Durch Kochsaft- und Mg++. Zus~tz Zu den Dia]ysatcn wulde auf eine AktivierungsmSglichkeit gepriift. Der Kochsaft wurde aus der nicht dialysierten 10 % igen K~sesuspension durch Erhitzen fiber 15 rain auf 100 ° C gewonnen und in einer Menge yon 0,5 ml pro Ansa~z zugegeben.
Den Versuchsergebnissen zufo]ge wurde die alkalische Phosphatase bei der Dialyse nach 24 Std. u m 27 % inakt iv ier t , nach 48 Std. war wieder ein leichter Akt ivi t~ts-
anstieg zu beobachten. Tabelle 12. ~t'nderung der A]ctivi~t von al]calischer und saurer K~isephosphatase nach .Dialyse und Kochsa/t- bzw. Mg++-Zugabe
Ansatz
Vergleiehsansatz (nicht dialysiert) . . . . Dialyse fiber 24 Std. (Ansatz I) . . . . Dialyse fiber 48 Std. (Ansatz II) . . . . Ansatz I u. Kochs~ft . . . . . . . . . Ansatz I I u. Kochsaft . . . . . . . . Ansatz' I u. Mg++ * . . . . . . . . . . Ansatz I I u. Mg ++ ~ . . . . . . . . . .
Dutch 1 ml Suspension (100 mg Kase) enzymatisch
abgespaltenes Phenol
alkalische I saure Phosphatase Phosphatase
/~g /~g
2360 372 1720 368 1840 368 2520 372 2700 356 2640 412 284o 404
durch Mg ++ aktiviert . Wie weitere Versuche zeigten,
Nach Kochsaft- sowie nach Mg++-Zusatz ]iegt die Wirksamkei t der dialysier ten Ferment - 15sung fiber der der n icht dia lysier ten Vergleichs- suspension. Die saure Phosphatase wird durch Dialyse nicht beein- f lug t ; dami t ist eine M6g- lichkei~ zur weiteren l~einigung des sauren Fermentes gegeben. Die dialysierten Suspensio- ne n werden gleiehfalls
fibertrifft aueh in diesen Ansi i tzen die akt ivierende Wirkung yon Co ++ die des Mg ++ betri~chtlich. Zusammen- fassend ergibt sich, dag bei K~sephosphatasen alle oben a n h a n d der L i te ra tu r er- 6r te r ten MSglichkeiten einer Fe rmen tak t iv i e rung bzw - inakt iv ierung zu rea- lisieren sind.
d) Fermentverteilung im [liissigen ~: nd ]esten Anteil der KSse~uspension ; Versuche zur Extral~tion der Phosphatasen
Zu den bisher beschriebenen Versuchen diente eine 20 % ige Kiisesuspension, die sich fiir alle Arbeitsg~nge gut bew~hrte. Sie ist bei der Gewinnung aus genfigend durchgereiftem Kiise derart
1 ALBERS, H., E. BEYER, A. BOHNENKA~P U. G. MULLER: Ber. dtsch, chem. Ges. 71, 1913 (1938).
s EK, B., H. v. EvLE~ u. L. H A ~ : Ark. Kemi 1, 117 (1949). 3 DELO~Y, G. E., u. M. HE~E~I~GTO~: Canad. J. Med. Sci. 80, 1 (1952).
Konzentration im alkalisehen Ansatz: 10 -s tool, im sanren Ansatz: l0 -smo].

Beitr~tge zur Biocheraie der K~sereifung. XVI 231
horaogen, da] dureh einfaehes Zentrifugieren (30 rain bei 3000 U/rain) keine nennenswerte Fraktio- nierung erreicht werden kann. Der bei dieser Prozedur anfallende ]~odensatz umfal~t nur etwa 2 % des Gesamtvolumens; in ihm tritt zwar starke Anreicherung der alkalischen Phosphatase ein, sic kann jedoeh wegen der geringen Niederschlagsraenge und des erhebliehen Fermentgehaltes im fiberstehenden Zentrifugat praktisch vernachlassi~t werden.
So wurden bei einem Ansatz durch gleiche Volumina Fermentl5sung aus Dinatrium-phenyl- phosphat folgende Phenolmengcn in Freiheit gesetzt (1 Std. Bebrfitung bei p~9,8; NaOH- Boratpuffer) :
1. Durch 1 ral Gesamt-Suspension (20%ige Suspension fiblicher I-Ierstellung aus durch- gereiftem K~se) . . . . . . . . . . . . . . . . . . . . . . . . . . . . 328 #g Phenol
2. Dureh 1 ml Zentri]ugat (iiberstehende Flfissigkeit naeh Zentrifugieren fiber 30 rain bei 3000 U/rain; Anteil betr~igt etwa 98 Vol.- % der Gesamtsuspension) . . . . . . 256 #g Phenol
3. Dureh 1 ml Boclensatz (Niedersehlag yon 2; Voluraen betragt deragemal~ etwa 2 % der Gesamtsuspension) . . . . . . . . . . . . . . . . . . . . . . . . . . . 1800 pg Phenol
Diese wie die folgenden Ans~tze wurden nur im alkalischen Gebiet bebrfitet, da die hier statt- findende starke Substratspaltung eine besonders klare Ubersicht fiber die herrsehenden Verh~lt- nisse liefert.
Im weiteren Verlauf dieser Untersuehungen wurde geprfift, ob nach dem yon MoR~os ~ be- schriebenen Verfahren der Butanol-Extraktion die Gewinnung klarer, w~l~riger Phosphatasen- 15sungen aus Sauermilehk~ise mSglich ist. Bei Nachprfifung der Angaben yon MORTOS zeigte sieh jedoeh, dal~ Zentrifugat wie Bodensatz der n-Butanolbehandlung gleieherma~en reich an Ferment sind. So lieferte das Zentrifugat eine Phenolabspaltung yon 1896/~g pro ral, der Niederschlag eine solche yon 2352 #g. Trotz Variation der Versuehsbedingungen und des Ausgsngsmaterials konnte das Zie] einer Anreicherung der Phosphatase in wal]riger LSsung durch Butanolextraktion naeh MO~TO~ in unseren Untersuchungen nicht erreieht werden.
7. Die Substratspezi[itiit der K~isephosphatasen
a) Enzymatische ItydroIyse ve~chiedener Phosphorsi~ureester bei den p~-Optima der al]calischen und sauren Phosphatase
I m fo]genden sol]te un te r such t werden, wieweit die phosphata t i schen E n z y m e des Sauermilchk~ses bef~higt sind, eine Reihe biochemisch bedeutsamer Verb indungen zu dephosphorylieren. Es w~r im l~ahmen dieser Arbei t n ieht mSglich, al]e Gesichts- p u n k t e zu berficksichtigen, die eine erseh5pfende Kennze ichnung der ffir die einzelnen Subs t ra te wesentl ichen Verh~]tnisse erlauben.
Wir haben deshalb einheitlich ffir alle Ans~tze eine Substratkonzentration yon 0,005 molar ge- w~hlt sowie die Bebrfitung bei den p~-0ptima der im Sauerrailehkase vorkoramenden Phosphatasen (p~ 5,5 und 9,9) durehgef~hrt. Hierbei wurde in Kauf genommen, dal~ die Spaltung besonders der Verbindungen, ffir deren Dephosphorylierung ein spezifisehes Ferment anzunehraen ist (Pyro- phosphat, Adenosintriphosphat) nieht immer unter optiraalen Bedingungen, jedenfalls aber ira Wirkungsbereich der K~sephosphatasen verl~uft.
Als Beispiel eines yon den gew~hlten pE-Stufen im Optimum divergierenden Phosphatasen- systems sei die Pyrophosphatase des Sauermilehkases genannt, deren optimale Wirkung naeh sparer beschriebenen Versuchen bei p~ 6,3 liegt. Im fibrigen jedoch dfirften die bier geprfiften Substrate weitgehend ira Rahraen der yon uns gew~hlten Bedingungen liegen. So zeigen mikro- bielle Phosphomonoesterasen nach DoM~I]~ 2 maxiraale Aktivitat bei p~ 10 und zwisehen p~ 4--6. :Die Dephosphorylierung des Thiarainpyrophosphates verl~uft optimal bei p~ 9 und 58, die des Adenosintriphosphates vorzugsweise bei p~ 9, daneben im Neutralgebiet sowie ira sauren Medium bei p~ 4--5% Casein bzw. Phosphopeptone werden bei p~ 5,85, fl-Glycerophosphat bei p~ 9,2 6 gespalten.
1 Mo~To~, R. K." Nature (London) 166, 1092 (1950). D o ~ 1 ~ , G. : Zit. S. 212, Anm. 1. ER£N~:5, O., u. J. I~AsA~." Aeta path. raierobiol, scand. (Copenh.) ]5, 563 (1954). Vgl. z. B. J. GERmanY: J. of Biol. Chem. ~00, 543 (1953). - - HO~FMAN~--Os~OF, 0., U.
W. W ~ m ~ : Mh. Chem. 83, 1180 (1952). Foozle, M. W.: Zit. S. 214, Anra. 7;--FOOTE, W. M., u. C. A. KIND: Arch. Bioehera. Bio-
physics 46, 255 (1953). - - RocHe, J., N. v~N T~OAI u. P. PI~-: C.r. Soc. Biol. (Paris) 147, 409 ~(1953). - - No~m~R~, B. : Acta chem. scand. (Copenh.) 4, 1206 (1950).
GRYDER, ]:~. M., J. S. FRIEDENWALD U. C. CARLSON: Zit. S. 218, Anm. 2.

232 J. SC~O~MULLER und E. LA~MANN:
I m einzelnen wurden folgende Substrate gepriift : 1. Dinatr iumphenylphosphat (Merck) ; 2. Dinatrium-fi-glycerophosphat (purum, Fluka AG) ; 3. Dinatrium-~-glycerophosphat (purum, Fluka AG); 4. Fructose-diphosphorsaures Natr ium (Sammlungspr~parat); 5. Pyridoxalphosphat (Ammonium-pyridoxal-phosphat, Reinheit e twa 62%.
California Foundation for Biochemical Research, Los Angeles). Die in Ansatz gebrachte Menge wurde dem Reinheitsgrad entsprechend erhSht.
6. Ein aus hochreifem Harzer K~se isoliertes Phosphopepton 1. Das Pr~parat enthielt 3,5 % t t20; 5,29 % P und 11,79 % N. Seine Konzentrat ion
im Ansatz wurde derart gew~hlt, dab eine an P 0,005m-LSsung resultierte. Wegen der starken Hydrolyse des Peptons im alka]isehen Bereich war hier keine Aktivit~ts- messung mSglich, l~ber die n~here Kennzeichnung derartiger Phosphopeptone wird demn~chst berichtet.
7. Natr iumpyrophosphat . Beim Vergleich der Phosphatabspal tung aus den verschiedenen Substraten ist zu
beriicksichtigen, dab die Hydrolyse eines Mol Pyrophosphat 2 Mol Orthophosphat liefert.
8. Adenosintriphosphorsaures Natr ium. In dem sparer beschriebenen Versuch wurde lediglich das freigesetzte Phosphat
best immt. Es bleibt also die Frage often, wieviel Phosphat-Bindungen im einzelncn Substratmolekiil gelSst werden. Das als Adenosintriphosphatase bezeichnete spe- zifische Ferment spaltet nur 1 mol Phosphors~ure (unter Bildung yon Adenosin- diphosphat), die , ,Apyrase" hingegen 2 mol Phosphors~ure ab.
9. Thiaminpyrophosphat (Cocarboxylase). Entsprechend der eingangs geschilderten Versuchsfiihrung wurde das enzymatisch abgespal-
tene Phosphat in den sauren Ans~tzen nach 6, in den alkalischen nach 4 Std. Bebriitung ermittelt. Die Versuchsergebnisse zeigen, dal~ die dephosphorylierende Aktivit~t der Phos-
phatasen in Sauermilchk~se wenig spezifisch, im Grade ihrer Wirksamkeit indessen stark p~-abhiingig ist.
Tabelle 13. Enzymatische Spaltung versehiedener Phosphorsiiure. ester dutch die 20 % ige Suspension eines gerei]ten Harzer Kiises. Bebriitungszeit 4 Std. bei Pn 9,9 (Borat-NaOH) bzw. 6 Std. bei
PH 5,5 (Veronal-Na-acetat) Dttrch 300 mg Kgse Spaltungsgrad
enzymatisch (hydrolysierter freigesetzter P P/Gesamt- Substrat-P)
Substrat alkalisch sauer alkalisch sauer
ttg /tg /~g t*g
Phenylphosphat 624 ~ 70 67 7,5 fl- Glycerophosphat 262 27 28 3 a- Glycerophosphat 147 32 16 3,4 Fructosediphosphat 400 105 22 5,6 Pyridoxalphosphat 93 432 10 46,5 Pyrophosphat 78 374 4 20,1 Adenosintriphosphat 576 384 21 13,8 Thiaminpyrophosph. 123 0 7 0 Phosphopepton - - 13 - - 1,4
1 SC~OnM~LLE~, J., u. It. HUT~: Diese Z. 99, 280 (1954). 2 Dieser Wert wurde mit einstfindiger Bebriitungszeit erh~lten; eine Umrechnung auf 4 Std.
ist nicht mSglich, da bereits nach 1 Std. weir mehr als die tt~lfte des Substrates hydroly- siert ist.
Bei p~ 9,9 ist die Spal- tung yon Phenylphos- phat weitaus am stark- stem Bemerkenswert gut wird Fructosediphos- phat gespalten, fi-Glyce- rophosphat ist leichter angreifbar als das ~-Iso- mere.Pyrophosphatester werden recht unter- schiedlich angegriffen ; hier liegt es nahe, an einen spezifischen Fer- ment typus zu denken, dessen Vorkommen in Sauermilchk~se durch

Beitr/~ge zur Bioehemie der K/~sereifung. XVI 233
spgter beschr iebene Versuehe wei tgehend erh•rtet werden konnte . Besonders auf- sehlugreieh sind die bei 1% 5,5 gewonnenen Ergebnisse, da dieser Mil ieubereieh vorzugsweise im Ver lauf der no rma len K~tsereifung du rehseh r i t t en wird. H ie r t r i t t de r Spa l tungsg rad des P h e n y l p h o s p h a t e s gegeni iber dem andere r Subs t r a t e s t a rk zuriiek. Beaeht l i eh wei tgehend werden P y r i d o x a l p h o s p h a t , Py rophospha t , Adenosin- t r i p h o s p h a t und aueh F r u e t o s e d i p h o s p h a t gespal ten. I m Gegensatz zum alkal isehen E n z y m hydro lys i e r t das saure F e r m e n t e - G l y e e r o p h o s p h a t besser als das ~-Isomere . Das un te r sueh te P h o s p h o p e p t o n wird k a u m angegndffen. Dies erseheint n ieht ver- wunder l ich , da ja die I so l ie rungsmSgl iehkei t yon P h o s p h o p e p t o n e n aus hoehrei fem K/~se zweifellos dem U m s t a n d zuzusehreiben ist, dag derar t ige Case inf rakt ionen gegeni iber phospha t a t i s eh wi rksamen F e r m e n t e n der K~isemasse besonders res is tent sind. I m Gegensatz zu dem als Coferment bedeu t samen P y r i d o x a l p h o s p h a t konn te fiir T h i a m i n i n p y r o p h o s p h a t keine Dephosphory l i e rung dureh die saure Phospha t a se festgeste!I t werden.
b) p~-Optimum der enzymatischen Pyrophosphat-Hydrolyse
Wie erw~hnt , i s t die Spa l tung des P y r o p h o s p h a t e s ve rmut l i ch der T~t igke i t eines besonderen F e r m e n t e s zuzuschreiben. Es erschien daher aufschlugreich, den Spal- t ungsg rad dieses Subs t r a t e s in Abh/~ngigkeit yon der t~eakt ionslage zu un te rsuchen .
Viele Nitteilungen der Literatur weisen darauf hin, dab auch bei der Pyrophosphatase ver- schiedene isodyname Systeme vorkommen, deren ~¥irkungsoptima sich fiber einen weiten p~-Be- reich erstrecken. Wit unterseheiden demgem~B ,,saure", ,,neutrale" und ,,alkalische" Pyro- phosphatasen, wobei mehrere dieser Typen in ein und demselben System vorkommen k5nnen. GUITONNEAU und Mitarbeiter 1 fanden in Milch zwei Pyrophosphatasen mit den p~-Optimas bei 4,2 bzw. 7,6--7,8, die sich durch ihre verschiedene Hitzeresistenz abgrenzen lassen und bei der Milchverarbeitung in der Buttermilch angereichert werden. NO,BERG 2 beschreibt mehrere iso- dyname Enzyme in Rattenleber, deren Aktiviti~tsoptima bei p~ 4,5, 5,8--7 (je nach Pufferwahl) und 8 liegen. Ein typisch ,,sautes" Enzym enth~lt ,,Ragi" und ,,Ragi-~Mz "a, wiihrend die Mehr- zahl der Literaturangaben ,,neutrale" Pyrophosphatasen erw~hnt, so in Rattenleber (PH 7--8) a und Rattengehirn (p~ 7,2 bzw. 6,5--7,4) 5, in Meersehweinchengewebe (PH 7,2--7,9) 6 in Erythro- eyten (p. 7,1) 7 und in B/~ckerhefc (PH 7,0 bzw. 7,2) s. So entdeekte K~ISH~A~ 9 in Aspergillus niger und PeniciUium chrysogenum zwei derartige Fermentsysteme, yon denen das eine bei p . 3--4 optimal wirksam ist und durch Mg ++ gehemmt wird, w/~hrend das andere ein pR-Optimnm im Neutralpunkt und eine Aktivierung durch Mg ++ zcigt.
Zur Untersuchung der Pyrophosphatase yon gereiftem I-[arzer K~se wurden Versuehe nach der fiblichen Methode (Substratkonzentration 0,005 molar; Bebrfitungsdauer in beiden pH-Bereichen 6 Std.) durchgeffihrt. Die Notwendigkeit eines Mg++-Zusatzes konnte bei den vorliegenden Ver- suchen nicht erwiesen werden. Ffir eine Konzentration yon 0,002 molar Mg ++ im Ansatz wurde die enzymatische Aktivit~t der alkalischen Phosphatase lediglieh um 30 % gesteigert.
Die Versuche (Tab. 14) zeigen, dal] im Un te r suchungsma te r i a l (durchgere i f te r K/~se, PH 7,3) ledigl ich ein W i r k u n g s o p t i m u m , und zwar bei p . 6,3 beobach te t werden konnte . Hie r be t r~g t der Spa l tungsg rad des Subs t r a t e s rund 3 0 0 , also wei t mehr als bei f r i iher beschr iebenen Versuchen zur a lkal i schen und sauren Bebr i i tung.
1 GUITONNEAU, G., R. CttEVALLIER U. H. JAROUSSE: C.r. Acad. Sci. (Paris) 218, 1006 (1944). NORBERG, B.: Acta chem. scand. (Copenh.) 4, 601 (1950); 5,.325 (1951).
3 C]:[ANDRASEKIIARA, ~V[. I{., U. M. SWA2CIINATI~AN: Bull. Central Food Technol. Res. Inst. 2, 212 (1953).
4 SwANsox, A.: J. of Biol. Chem. 194, 685 (1952). Ro~BI~S, E.A. , M. P. S~ULBEa~ U. P. D. BOYE~: Arch. of Bioehem. a. Biophysics 54, 215
1955). - - Go~Do~r, J. J . : Zit. S. 226, Anm. 9. s NAGANNA, B.: Current Sci. 20, 231 (1951). 7 BLOCH-FRANKENTttAL, L.: Biochemie. J. 57, 87 (1954). s H ~ ' ~ L , L.A., u. 1~. J. H~LMO~: J. of Biol. Chem. 192, 87 (1951). - - K c ~ z , M.: J. Gem
Physiol. 35, 423 (1952). KR~s~z~, P. S. : Arch. of Biochem. a. Biophysics 32, 230 (1951).

234 J. SCHOI~MULLE]~, und E. LAH_~A~:
im
Tabelle 14. pu-Optimum der Pyrophosphatase gerei]tenHarzerKiises. Substrat: ~a-pyrophosphat; Bebrtitung jeweils 6 Std. mit Borat-NaOtt bzw.
Veronal-lqa-aeetat-Puffer
Dami t dfirfte erwiesen sein, dab ffir die Dephosphoryl ierung des Pyrophosphates Sauermilehk~se ein besonderes Ferment vorkommt, das dem Typus der oben
erw~hnten ,,neutralen" Pyrophos- phatasen zuzurechnen ist.
Enzymatisch Enzymatisch p~ freigesetzter P p~ freigesetzter P
#g #g
4,0 4,6 4,95 5,35 5,8 6,3 6,8 7,15 7,5
7,8 8,2 8,6 8,8 9,0 9,25 9,5 9,7
10,0
240 248 224 216 216 200 192 152
88
Diskussion Die Spal tbarkei t yon Dinatr ium-
phenylphosphat durch w~Brige Sus- pensionen reifenden Harzer KiSses weist auf die Anwesenheit yon Phos- phomonoesterasen hin. Zwei isody- name Systeme dieses Phosphatasen- typus lieBen sieh insbesondere kennzeiehnen.
181 304 309 472 576 608 508 ~36 312
1. Die ,,allcalische" Phosphatase
bedingungen ein pa -Opt imum zwischen 9,95 und 10,2. Sie ist dureh Mg ++ aktivierbar n n d spa] te t /~-Glyeerophosphat besser als das ~-Isomere. Das Ferment zi~hlt demnach z u m Typ A I d e r Phosphomonoesterasen, entspreehend der Klassifikation yon FOLLEY und KAY. Die relat iv mi~Bige Aktivierbarkei t durch Mg ++ l~Bt vermuten, dab es sigh im vorliegenden Fall um Gin Desmoenzym handelt , da Lyofrakt ionen nach BAMANN 1 um 5000 % und mehr aktiviert werden. Die spezifische H e m m u n g durch Be ++ und CN -~ entspricht dem Verhal ten des in tierisehen Geweben vorkommenden alkalisehen Fer- mentes. Als ~ul~erst wirksamer Inhibi tor erwies sich in Ubere ins t immung mit Angaben der Li tera tur ~ insbesondere Be++; noch 10 -7 tool Be ++ zeigen deutliche Akt ivi t~tsminderung. Auffallend ist - - im Gegensatz zur sauren Phosphatase -- die H e m m u n g durch Mn ++, ZW +, Co ++ und Ni ++. Das alkalisehe Ferment des Sauer- milehk~ses zeigt gegeniiber versehiedenen Effectoren Gin ~hn]iches Verhalten wie die Yon HOFFMANN-OsTENI~OF ~ beschriebene alka]ische Phospho-monoesterase aus Ober- und Unterhefe : Akt ivierung durch Mg ++, H e m m u n g durch Mn ++, Zn ++, Co +e, Ni ++, Ca ++, CN- und Cystein sowie nur schwache H e m m u n g dureh F - und Jodace ta t . Die Eigensehaften der alkalischen Phosphatase yon Sauermilchk£se s t immen auBerdem weitgehend mit denen des entsprechenden gohmilc l~ermentes fiberein, so im ps- Opt imum, in der Mg++-Aktivierbarkeit, t t e m m u n g durch Zn ++, CN- und Cystein und Nichtbeeinflussung durch F , Antibiotica oder Ultrascha]l 5. Beide Fermente unter- sGheiden sich jedoch in ihrem Verhalten beim Erhitzen. Milchphosphatase ist ver- h~ltnismgBig hitzelabil, eine Eigenschaft , die ja zur Ausarbei tung eines Testes au f
1 BAMANN, E., u. K. MYRBXCK: Die Methoden der Fermentforsehung. Bd. 2, S. 1424. Leipzig: Thieme 1941.
2 ROCHE, J., u. N: VAN T~OAI: Zit. S. 231, Anm. 5. s KLEMPEI~E~, F . W . , J . M. MILLER U. C. J . I-JILL: J. of Biol. Chem. 180, 28I (1949). - -
KLEMPE~ER, F. W. : J. of Biol. Chem. 187, 189 (1950). - - CgEV~EMO~¢, M., u. It. FINEST : Nature (London) 107, 772 (1951). - - ALD~DGE, W.N.: Natm'e (London) 165, 772 (1950).-- VEE~KAMt', T~. A., u, G. SNrrs: Nature (London) 172, 589 (1953). - - SCgUBER% J., u. A. LI~D]~-BAU~: J. of Biol. Chem. 208, 359 (1954). - - D[: BOlS, K. P., K. W. COCm~AN u. IV[. MAzv~: Science (Lancaster, Pa.) 110, 420 (1949). - - BOCHE, J., N. VAN THOAI u. J. LoEwY: C.r . Soe. Biol. (Paris) 144, 638 (1950).
4 HOFFIVI&~-OSTE~]tOF, O., I-I. MOSER U. R . EHP~E2qI~EIG~[: ~V~h. C h e m . 82, 295 (1951). MdV[EEKIN, T. L. , u . B . D . POLIS: A d v . P r o t e i n C h e m . 5, 221 (1949). - - KANNA~, A. , u.
K. K. BAZ~: Indian J. D~iry Sci. 2, 51 (1949).
Die ,alkalisehe" Phosphatase zeigt unter den gew~hlten Versuehs-

Beitr/~ge zur Biochemie der Kasereifung. XVI 235
stattgefundene Milchpasteurisierung diente. Die grSgere Hitzeresistenz der im Harzer Kase auftretenden alkalisehen Phosphatase stellt einen weiteren Beweis ffir ihren bakteriellen Ursprung dar, da auch andere Mikrobienphosphatasen diese Eigenschaft zeigen 1. Allerdings sind auch Ausnahmen bekannt geworden. So berichteten SANDERS und SAGER 2 fiber eine im Cheddarkase vorkommende Phosphatase mit einer im Ver- gleich zum Milchferment geringeren Hitzeresistenz. Auch yon der ,,reaktivierten" Milchphosphatase ist bekannt, dag sie hitzelabiler als das Enzym der Rohmilch ist 3.
Die alkalische Phosphomonoesterase des Sauermilchk~tses spaltet bei p~ 9,9 haupt- saeh]ieh Pheny]phosphat, andere Substrate werden in viel geringerem Ausmal~ dephosphoryliert. Das Ferment ist vorwiegend zellgebunden; der Versueh einer Butanolextraktion blieb erfolglos. Dialyse mindert die Aktivitat um etwa ein Vierte]. SH-Gruppen scheinen ohne Bedeutung ftir die Wirksamkeit des Fermentes zu sein, da ffir diesen Enzymtypus (Thiolfermente) spezifische Inhibitoren die enzymatische Aktivitgt nieht nennenswert beeinflussen. Andere Yhosphatasen indessen, wie z. B. die yon GOI~DON 4 beschriebene Gehirn-Phosphatase oder die saure Phosphatase der Erythrocyten a scheinen Thio]gruppen a]s wesent]iche Komponente des aktiven Enzyms zu enthalten.
,,O-Quark", das Ausgangsmaterial der hier untersuehten geifungsreihen, ist frei yon analytiseh nachweisbaren MeRgeR an alkalischer Phosphatase. Das Ferment bildet sich offenbar erst im Verlauf der Kasereifung; seine Aktivitat erreicht nach etwa 10--14 Tagen im reifenden Kase ein Maximum und nimmt dann wieder betrgchtlieh ab. Im normalen t~eifungsansatz (Zugabe yon Kochsalz und geifungs- salzen, Behandlung mit Rotsehmierebakterien) ist der Fermentgehalt bedeutend hSher als in salzfreien Reifungsffihrungen, obgleich die Salze NaC1 und NaHCO3 bei den in Frage kommenden Konzentrationen keinen nennenswerten Einflug auf die Aktivitat ausiiben. Die Salzwirkung erstreckt sich also nicht auf das Ferment an sieh, sondern in erster Linie auf die Phosphatasen liefernde Mikroflora. In allen F/~llen nimmt die Enzymbildung ihren Ausgang yon der Rinde her, ein weiterer Beweis dafiir, dab die Bildung der a]kalischen Phosphatase dutch die Mikroorganismenflora der Kasereifung hervorgerufen wird. Die Aktivitatsabnahme im fiberreifen Kase ist vermutlich mitbedingt durch enzymatisehe Proteolyse der Phosphatase. SCHOR- /¢I/)LLEg und Mitarbeiter 6 haben das Vorkommen yon Proteinasen und Peptidasen im reifenden Sauermilchkase nachgewiesen, CAMUS und ALIFASE 7 ste]Iten derartigen proteolytisehen Abbau der Phosphatase in Gruygre-Kase lest. Aueh die yon GALE s erSrterten Beziehungen zwischen Fermentaktivit~t und Alter der Mikroorganismen- kultur spieler hier zur Beurteilung des Abfailes der Phosphatasenaktivitgt mit eine t~olle. Zur K]arung der umstrittenen Frage, inwieweit der Phosphatasengehalt ein Kriterium ffir den Reifezustand yon Kase darstellt~ bedarf es einer Ausweitung sol- chef Untersuchungen, vor allem auch an anderen Kasesorten. Bei den yon uns geprfiften t~eifungsreihen entsprach die maximale Aktivit/~t des a]kalischen Fer- mentes einem PH yon 7,3 in der Kasemasse.
1 NEAVE, F. K. : Zit. S. 213, ARm. 16.--BUCK, T. C. : Zit. S. 213, ARm. 16. - - KAPLAN, E. : Zit. S. 213, Amn. 16. - - SANDERS, G. P., u. O. S. SAGER: Zit. S. 213, ARm. 17.
2 SANDEUS, G. P., u. 0. S. SAGER: J. Dairy Sci. 31, 845 (1948). 3 SIEGES-TKALEE, E.: Mitt. Lebensmittelunters. ttyg. 45, 84 (1954). - - FucHs, A. : Mitt.
Lebensmittelunters. tIyg. 44, 162 (1953). GORDON, J. J." Zit. S. 226, ARm. 9.; Biochemic. J. 55, 812 (1953).
5 TSUBOI, K. K., u. P. B. IguDso~: Arch. Biochem. Biophysics 55, 206 (1955). 6 SCttORIVIiJLLER, J., ~::~. WIESKE U. H. WINTER: Diese Z. 100, 35 (1955). 7 CAMus,.A., u. 1%. ALIFASE: Zit. S. 214, ARm. 1. s GALE, E.F.: Zit. S. 219, ARm. 2, daselbst S. 78.

236 J. ScHoR~ULLER und E. LA~MA~:
II. Die ,,saute" Phosphatase
Die im Sauermilchk~tse auftretende ,,saure" Phosphatase zeigt ein pH-Optimum bei 5,2--5,7. Auf Grund ihrer Aktivierbarkeit durch Mg ++ und der grSi~eren Spal- tungsgeschwindigkeit des ~-Glycerophosphates gegenfiber dem/3-Isomeren z~hlt sie zur Gruppe An der yon FOLLEY und KAY klassifizierten Phosphomonoesterasen. Das Ferment wird durch F - spezifisch gehemmt, auffallend stark durch Co ++ aktiviert. Die spezifische Hemmung der sauren Phosphatase durch Fluoride ist vielfach belegt 1, wobei die Fluoridwirkung stark substratabh~ngig 2 ist. W~hrend ffir das Substrat fl-Glycerophosphat im weiten Bereich vollstgndige Inaktivierung eintritt, ist dieser Effekt ffir Phenylphosphat erst bei geringer Substrat- und hoher F--Konzentrat ion zu erzielen.
Die saure Phosphatase des Harzer K~ses wird durch Dialyse nicht beeinflul3t. Ultraschall sch~digt das Ferment, wenn auch nicht betriichtlich. F fir SH--Gruppen spezifische Inhibitoren beeinflussen die Aktivit~t des Fermentes nur wenig, es scheint sich also auch hier nicht um ein typisches Thiolferment zu handeln. Die saure Phos- phatase ist bereits im , ,0-Quark" enthalten. Ihre Aktivit~tt steigt im Laufe der Rei- lung bis zu 10--11 Tagen an und ~ndert sich dann im hochreifen K~se nicht mehr wesentlich. Im GegenSatz zur alkalischen Phosphatase wird das saure Enzym bei anomaler Reifungsffihrung (also ohne jeden Salzzusatz) wesentlich reiehlicher als bei normaler K~seherstellung gebildet. Auch hier t r i t t die Fermentzunahme yon der Rinde her ein, ein Beweis ffir die bakteriell bedingte Aktivit~tssteigerung der sauren Phosphatase. Der geringere Fermentgehalt normal reifenden K~ses ist mitbedingt durch den in vitro naehgewiesenen Hemmungseinflul~ zugesetzter Salze. Wieweit hier aul~erdem ver~nderte Entwicklungsbedingungen ffir die phosphatasenliefernde Mikroflora eine Rolle spielen, bedarf weiterer Untersuchungen der Phosphatasen- aktiviti~t entsprechender Reinkulturen.
Da das p . -Optimum der sauren Phosphatase in einem Bereich liegt, den der K~se bei normal ablaufender Reifung durchschreitet, verspricht das Studium der Dephos- phory]ierung verschiedener Phosphors~ureester ni~here Aufschlfisse fiber die Bedeu- tung dieses Phosphatasentypus. Hier t r i t t die Spa]tung des Phenylphosphates gegen- fiber der yon Pyridoxalphosphat, Adenosintriphosphat und Pyrophosphat weitgehend zurfiek. Besonders auffallend ist auch die deutliche Spaltbarkeit des Fructose- diphosphates. Da Hexosephosphorsi~ureester Zwischenprodukte der wi~hrend der Vorreifung ablaufenden Milchs~ureg~rung darstellen, scheint deren Dephosphorylie- rung eine wesentliche Aufgabe der sauren Phosphatase zu sein. In diese Richtung weist eine Mitteflung yon ARNAUDI und FI~ANCIOLI 3, wonach Phosphatasen eine besehleunigende Wirkung auf die Milchsgureg~rung ausfiben. Wesentlich fiir Prozesse der K~tsereifung erscheint weiterhin die starke Spaltung des Pyridoxalphosphates. Dieser Phosphors~ureester spielt als Coferment verschiedener Enzymsysteme eine bedeutungsvolle RoUe. F fir Deearboxylasen 4 und Transaminasen 5 sowie ffir den Abbau yon Serin und Threonin ~ haben wir in vorhergehenden Arbeiten dies im Rahmen der Ki~sereifung eingehend studiert und er6rtert. Dort hatte sich gezeigt, da[~ die Wirkung der genannten Fermente einem Maximum zustrebt und bei fiberreifem K~se abnimmt. Es liegt nahe, den Grund ffir diese Abnahme in der enzymatischen
1 COURTOIS, J., u. C. ANAGNOSTOPOVLOS: Bull. Soc. Chim. biol. 81, 1494, 1504 (1949). 2 KUTSC]IER, W., u. H. WriST: Biochem. Z. ~10, 292 (1942). 3 AI~NAUDI, C., u. 1V[. FRANCIOLI: Biochem. Z. 250, 125 (1932). 4 SC]IORMifLLER, J . , u. L. LEICHTER: Diese Z. 102, 13 (1955). '~ SO]IORM~LLER, J . , u. W. GELLRICH: Diese Z. 100, 200 (1955). 6 SCItOP~MULLER, J., M. GLAT]IE 11. H. RUTH: Diese Z., im Druck.

Beigr~ge zur Bioehemie der K~sereifung. XVI 237
Spaltung des Pyridoxalphosphates zu suehen, das damit als Coferment der I)eearboxylasen und Transaminasen ausf~llt. Diese Annahme erf~hrt Best~tigung einmal dureh die friiher mitgeteilten Befunde, wonaeh Pyridoxalphosphatzusatz zu hoehreifem K~se eine betr~chtliehe Aktivit~tssteigerung der in Rede stehenden Fer- mente bewirkt. Sie wird weiterhin gestfitzt dureh Ergebnisse dieser Arbeit, denen zufolge Pyridoxalphosphat durch die saure Phosphatase des K~tses einen starken Abbau erf~hrt.
I)er geringe Spaltungsgrad eines yon uns aus hoehgereiftem Kgse isolierten Phosphopeptons bei Einwirkung der sauren Phosphatase ist verst~ndlieh. Die An- reieherung soleher Caseinabbauprodukte in gereiftem K/ise ist ja an sieh sehon als Beweis fiir deren Enzymresistenz anzusehen. Die Alkalilabilit~t der Phosphopeptone ist bekannt und aueh yon uns 1 bestgtigt worden. I)araus kann gesehlossen werden, dab der Abbau soleher Phosphorverbindungen im sp~teren Reifungsstadium des K~tses dureh die herrsehende Alkalit~t vorzugsweise auf ehemisehem, nieht aber auf enzymatisehem Wege vor sich geht. Untersuehungen von MATTENHEIMEU 2 zeigten allerdings, dab Casein und Phosphopeptone dureh Phosphorproteidphosphatasen ver- sehiedenen Ursprungs dephosphoryliert werden k5nnen. Neuerdings hat P~RL~A~ a die Bindungsart des Phosphors im e-Casein untersueht. P E ~ L ~ sehliel3t aus Ver- suehen mit Phospho-diesterase (p~ 8,2), Pyrophosphatase (p~ 7,2) und Darm- bzw. Prostata-phosphatasen (p~ 6,0), wobei der gesamte Phosphor des Substrates frei- gesetzt wird, dab im ~-Casein 40% des Phosphors als Mono-ester der Formel --O--P(O)(OH)~, 20% als Pyrophosphate des Typus - - O - - P - - O - - P - - O und 40% als Di-ester entspreehend der Formulierung - - N - - P - - O vorkomme. Eine endgtiltige Entseheidung der Frage, welehe Rolle die Phosphatasen des Sauermilehk/~ses bei der ])ephosphorylierung des Caseins und seiner phosphorhaltigen Abbauprodukte spielen, kann nur unter Verwendung hoehgereinigter Fermentpr~parate spezifischer Wirkung getroffen werden (vgl. S. 214). Die yon P E U L ~ fiir e-Casein aufgestellte Bindungs- bilanz des Proteidphosphors deutet darauf hin, dag bei der Freisetzung des Gesamt- phosphors aus dem Casein eine I~eihe versehiedener Phosphatasen mitwirken diirften.
I I I . Die e//elctive Pho~phatase Die beim p~ des jeweils gepriiften K~ses wirksame Fermentkomponente, also die
,,e[/elctive" Phosphatase, ist auf Grund ihres Verh~ltens gegeniiber spezifisehen Inhi- bitoren dem Typus der sauren Phosphatase zuzureehnen. Normaler K~se zeigt die -con GALE als ,,protective mechanism" gezeigte Erseheinung, der zufolge in reifendem K~se ein Ausgleieh zwisehen Fermentmenge und pH-Lage des Mediums stat tf indet mit dem Resultat, dab die tatsiiehlieh wirksame Fermentaktivit~t praktiseh gteieh- bleibt. Das fiir anomal gereiften Quark stark ausgebildete Aktivit~tsmaximum liegt bei friiheren Reifungsstadien als die HSehstwerte der alkalischen und sauren Phos- phatase. Der Aktivit~tsuntersehied zwisehen l~inde und Kern nimmt ftir die ,,effek- t i r e " Phosphatase mit zunehmender geifung stark ab.
IV. Die P yrophosphatase I)ie Pyrophosphatase des reifenden Sauermilehk~tses weist ein pu-Optimum von 6,3
au f und rangiert damit in der Gruppe der ,,neutralen" Vertreter dieses Phosphatasen- typus, den, wie erw~hnt, versehiedene Autoren aueh in anderem Material fanden. Sie wurde im Rahmen der vorliegenden Arbeit eingehender untersueht.
1 SCHOgMi~LL~g, J., u. H. I-IvT~: Zig. S. 232, Anm. 1. M~TTE~EI~ER, tI.: Hoppe-Seylers Z. 292, 276 (1953). PERLMA~, G.E.: Nature (London) 174, 273 (1954).

23 8 Kurznaehriehten
Die beachtliche hohe Sivaltung best immter Substrate, wie Pyridoxalivhosivhat , Adenosintriivhosivhat und Fruetosediivhosivhat , vor allem im sauren Gebiet, legt weiterhin die Annahme sivezifischer Phosivhatasen im Sinne der f/ir Phosivhatasen er6rterten Gesichtsivunkte nahe. Ihre genaue Kennzeichnung im System des reifen- den Sauermilchki~ses bedarf weiterer Untersuehungen. Dies gilt insbesondere fiir Pyridoxalivhosivhat als bedeutsames Coferment enzymkatalysierter Reifungsmecha- nismen.
V. Zusammen/assende Betrachtung Zusammenfassend ergibt sich, dab die ,,saure" wie die ,,alkalische" Phosphatase
in Sauermilehk~se durch ihr grunds~tzlich verschiedenes Verhalten gegeniiber sivezifi- schen Effectoren Mar gegeneinander abgrenzbar sind. Das ,,alka]isehe" Enzym wird insbesondere durch CN- und Be ++, aul~erdem durch Mn ++, Zn ++, Ca ++, Cystein und Citrat gehemmt, durch F - nicht wesentlich beeinflugt. Die ,,saure" Phosivhatase erfi~hrt sivezifische Hemmung durch F-, nieht aber oder nur geringffigig durch Ca ++, Citrat, CN-, Cystein und insbesondere nicht dureh Be ++. Besonders auffallend ist hier die starke Aktivierung durch Co ++ und Ni ++, auBerdem auch durch Mn ++. Vou den Anionen hemmt Phosivhat in der Konzentrat ion 10 -~ tool beide Ferment typen stark. Diese Wirkung ist ausgesprochen komivetitiv: entsivrechend dem allgemeinen l~eaktionsmechanismus der Fermente verl~uft die kata]ytisehe Substratsivaltung nur bis zum Gleiehgewicht; sie muB deshalb dureh den Ubersehug eines Sivaltivroduktes gehemmt werden.
Zusammen/assung In gereiftem Sauermilchk~se wurde eine ,,alkalische" Phosivhatase mit dem IVR-
Opt imum 9,95--10,2 und eine ,,saure" Phosivhatase mit dem Wirkungsoivtimum bei IV, 5,2--5,7 gefunden. Das alkalisehe Ferment ist bedeutend reich]icher vorhanden als das saure Enzym. Die Bildung beider Phosivhatasen im Verlauf der K~sereifung sowie die Aktivit~tsentwicklung yon der Rinde her, li~Bt auf ihren mikrobiellen Ursivrung schlieBen. Normale und anomale Reifungsfiihrungen zeigen grundsiitzlich versehiedene Enzymbildung, sowohl nach Art wie nach Menge beider Phosivhatasen- tyiven. Verhalten beim Erhitzen und bei der Dialyse sowie Wirkung yon Effectoren und Sivaltung versehiedener Phosivhosubstrate grenzen die hier untersuchten Phos- Ivhatasen seharf gegeneinander, auBerdem auch gegeniiber Rohmilchivhosphatase ab. Die, ,effekt ive" Phosivhatase zeigt den yon GALE diskutierten ,,pr0tective mechanism". Als Beispiel substratsivezifischer Phosivhatasenwirkung wurde die enzymatische Hydro- lyse des Na-ivyrophosphates nigher untersucht, die optimal bei iv. 6,3 verl/iuft. Die Bedeutung Ivhosivhatatischer Sivaltungen ftir Cofermente yon der Art des PyridoxM- phosphates wurde erSrtert.
Kurznachrichten Persiinliche Naehrichten
Dr. F. Egger "}- Unser Wunsch zu seinem 65. Geburtstag [vgl. diese Z. 95, 275 (1952)], unseren liebenDr. EGGER
noch recht lange Ms Mitarbeiter zu haben, elffiillte sich ]eider nicht. Schon Ende 1953 bat er uns, aus gesundheitlichen Griinden yon der Redaktion des Gesetzesteils entlastet zu werden. Nur sehweren tterzens kamen wir seinem Wunsche naeh, denn wir wugten keinen, der eine so lang ji~hrige Erfahrung fiber dieses fiir alle Fachgenossen so wichtige Gebiet besML In liebenswiirdiger und bereitwilliger Weise hat er seinen Nachfolger, Herrn Oberchemierat Dr. PATZSC~ (Duisburg) eingearbeitet. Seine Liebe zum Fach hat ihn, obwohl es ihm immer miihs~mer wurde, dennoch


![$XVI KUXQJVEHVWLPPXQJHQ %779 · 2020. 1. 10. · '77% :hwwvslhorugxqj $xvi kuxqjvehvwlppxqjhq %779 '77% :2 6wdqg yrp ]xohw]w ehduehlwhw )dvvxqj ghu $xvi kuxqjvehvwlppxqjhq $% ghv](https://img.pdfslide.org/doc/110x75/601f50bc083e116c8e1e9850/xvi-kuxqjvehvwlppxqjhq-779-2020-1-10-77-hwwvslhorugxqj-xvi-kuxqjvehvwlppxqjhq.jpg)