Embed Size (px)
Citation preview

z.B. Lignin, Gerbstoffe (Phlobaphene) und Mineralsalze. Auf-
lagerungen auf die Zellwand (Adkrusten) sind Cutin und Sub-
erin. Pflanzliche Zellwande bestehen aus Grundsubstanzen
und Gerustsubstanzen. Wichtigste Gerustsubstanz der Zelle
hoherer Pflanzen ist Cellulose (Grundbaustein b-D-Glucose).
Grundsubstanzen sind Pektine (Grundbaustein Galacturon-
saure) und Hemicellulosen.
Die Zellwande von Pilzen konnen Cellulose oder Chitin(Grundbaustein N-Acetylglucosamin) als Gerustsubstanz ent-halten.
Tierische Zellen besitzen keine Zellwand, besitzen aber inder Regel eine komplexe extrazellulare Matrix aus fibrillarenProteinen und Glykoproteinen.
1.3 Biomembranen
1.3.1 Chemie und Aufbau
Membranen sind wesentliche Strukturelemente derZelle, fur deren Funktionen sie eine zentrale Rolle spie-len. Die Plasmamembran, bei pflanzlichen Zellen auchPlasmalemma genannt, grenzt den Protoplasten nachaußen ab. Bei Pflanzen setzt sich diese außere Plasma-membran uber die Plasmodesmen in den Membranender Nachbarzellen fort. Hier begrenzen also die Plasma-membranen eines Gewebes oder auch des gesamtenOrganismus eine Einheit, einen Symplasten. Membra-nen umschließen Vakuolen im Inneren der Zelle, z.B.die Tonoplastenmembran die große Zentralvakuole beiPflanzenzellen. Weiter werden wichtige Zellorganellen,wie Mitochondrien, Chloroplasten, Dictyosomen, derZellkern usw. von Membranen umgeben. Das Mem-bransystem des Endoplasmatischen Retikulums bildet
in der Zelle ein ausgedehntes System von Kanalen, de-ren Lage sich standig verandert. Durch Membranenwird die Zelle der Eukaryonten in zahlreiche Reaktions-raume, so genannte Kompartimente, gegliedert, die be-sondere Stoffwechsel-, Transport- und Speicherfunk-tionen ubernehmen. Etwa 60 bis 90% der Zellmassesind Membranen. Der geordnete Verlauf von Lebens-prozessen hangt wesentlich davon ab, dass bestimmteStoffe durch die Membran hindurchtransportiert, an-dere wiederum zuruckgehalten werden.
Membranen trennen zwar Zellkompartimente von-einander, erlauben jedoch einen spezifischen Stoff-transport zwischen den Kompartimenten. Sie sindalso im strengen Sinne stets selektiv permeabel und re-geln den spezifischen Ein- und Austritt von Molekulenund Ionen in die und aus der Zelle, resp. in die verschie-denen Kompartimente innerhalb des Protoplasten.
Biologische Membranen sind also keineswegs nurHullen. Es sind vielmehr hochspezifische Vermittlerzwischen Innen und Außen, also zwischen Zelle undUmgebung oder zwischen Organell und Cytosol. Dieunterschiedlichen Funktionen verschiedener Zellenund Organellen bedingen die Konstruktion der jewei-ligen Membran und die Eigenschaften der darin einge-lagerten Proteine.
Biomembranen dienen einerseits als Diffusionsbar-rieren, andererseits ermoglichen sie einen selektivenStoffaustausch. Sie erfullen somit Trenn- und Verbin-dungsfunktionen. Biomembranen bilden die struktu-relle Basis von Enzymen und konnen damit spezielleStoffwechselfunktionen erfullen. So werden z.B. zahl-reiche Energietransformationen im Zuge der Photo-synthese oder der Atmung durch membrangebundeneEnzyme katalysiert und laufen an Membranen ab.Schließlich sind Biomembranen beteiligt an Reizauf-nahme, Erregungsbildung, Reizleitung und chemischerInformationsubertragung (Tab. 1.13).
1.3.1.1 Stoffliche Zusammensetzung
Organisation und Funktion der Biomembranen beru-hen auf ihrer stofflichen Zusammensetzung: Lipide,Proteine, Kohlenhydrate. Lipide sind fur die Integritatder Membranen verantwortlich, Proteine regulierenden Stofftransport und dienen als Signalempfanger,Kohlenhydrate sind an Lipide oder Proteine (Glykoli-pide, Glykoproteine) gebunden und an Zell-Zellerken-nung oder der spezifischen Erkennung bestimmterMo-lekule beteiligt. Die Dicke der Biomembranen betragtdurchschnittlich 7 bis 8 nm. Biomembranen konnensich auch zu Doppel- oder Mehrfach-Membranen pa-rallel anordnen, wie etwa bei Mitochondrien und Zell-kern oder der Myelinscheide von Nervenzellen.
Tab. 1.13 Membranfunktionen
Abgrenzung und Kompartimentierung innerer Milieus
Diffusionsbarriere
Osmotische Regulation
Stoffaustausch (passiver, aktiver und Massen-Transport)
Energietransformation (Photosynthese, Atmung)
Elektronentransport
Drusenfunktion
Sensorische Erregungsbildung
Reizleitung in Nerven
Trager von Enzymen
Chemische Informationsubertragung
Stoffwechselvorgange
1.3.1 Chemie und Aufbau 27
1Cytolog
ie

Lipide (s. Kap. 4.4) bilden die Grundsubstanz, die Ma-trix der Membranstruktur. Auf Grund ihrer hydropho-ben Eigenschaft bilden sie die Phasengrenze zwischenzwei wassrigen Kompartimenten.
Fur die Stabilitat der Membranen sind ferner neu-trale Lipide, Steroide wie Cholesterol wesentlich. Cho-lesterol kommt v. a. in den Membranen tierischerZellen vor. Cholesterol lagert sich in die Zwischen-raume von benachbarten Phospholipidmolekulen ein,versteift damit die Lipiddoppelschicht und vermindertihre Fluiditat und Permeabilitat.
Die Proteine (s. Kap. 4.3) der Membranen konnenStrukturproteine oder Enzyme sein. Die Ausstattungmit Enzymen variiert stark, je nach den speziellenFunktionen einer Membran. Eine Gruppe von Enzy-men, die Adenosintriphosphatasen (ATPasen), scheintjedoch in allen Membranen vorzukommen. Diese En-zyme spalten ATP und setzen so die Energie frei, die furden aktiven Transport von Stoffen durch die Membrannotwendig ist.
Manche Biomembranen enthalten betrachtlicheMengen an Kohlenhydraten (s. Kap. 4.2). Diese be-finden sich an der Außenseite der Membran undsind kovalent an Lipide oder Proteine gebunden. DerKohlenhydratanteil von Glykolipiden kann sich veran-dern, z.B. wenn eine Zelle zur Tumorzelle entartet.
Außerdem sind ein- oder mehrwertige Kationen,insbesondere Ca2+ und Mg2+, Bestandteile der Mem-branen. Sie sind fur deren Stabilitat sehr wesentlich.
Diese grundsatzliche chemische Zusammensetzungist allen bisher untersuchten Zellmembranen gemein-sam, wohl kann sich aber der chemische Charakterder Lipide, ihr Mengenverhaltnis und die speziellenEigenschaften der Proteinschichten mit dem Zelltypund der Funktion der Zelle andern.
1.3.1.2 Struktur von Membranen
Biologische Membranen, also die Plasmamembran, derTonoplast und die Biomembranen von Zellorganellenim Inneren der Zelle, haben die gleiche Grundstruktur.Sie lassen sich im Elektronenmikroskop nach entspre-chender Kontrastierung als Doppellinien darstellen undbestehen aus Lipid- und Proteinmolekulen.
Biomembranen sind veranderliche, fließende Struk-turen. Die meisten der Lipid- und Proteinmolekule sindin der Membranebene beweglich.
Die Lipidmolekule sind in einer zusammenhangen-den Doppelschicht angeordnet. Diese bildet die Grund-struktur einer Biomembran. Die Lipiddoppelschichtdient als Diffusionsbarriere fur viele wasserlosliche(hydrophile) Molekule.
Die Proteinmolekule sind in die Lipiddoppelschicht in-tegriert oder an eine ihrer Oberflachen assoziiert(Abb. 1.27). Integrale Proteine bilden die Basis furdie Transportleistungen der Zelle, z.B. als Ionen-pumpen, oder Carrierproteine, sowie fur Signaltrans-duktion. Assoziierte Proteine sind reversibel an dieOberflache von Membranen gebunden.
Die Membranproteine vermitteln die meisten, spe-zifischen Funktionen einer Biomembran, z.B. alsTransportproteine, Enzyme, Rezeptoren oder Bin-dungsproteine zum Cytoskelett. Biomembranen sindasymmetrisch. Die innere und die außere Oberflacheunterscheiden sich in der Zusammensetzung ihrer Li-pide und Proteine. Dies spiegelt unterschiedliche Funk-tionen der verschiedenen Bereiche einer Biomembranwider.
Viele Proteine konnen frei in der Membran wan-dern, manche scheinen auch in spezifischen Membran-regionen verankert zu sein. Die freie Beweglichkeiteines Proteins in einer Membran kann dadurch behin-dert sein, dass es mit einer cytoplasmatischen Domanean das Cytoskelett gebunden ist oder dadurch, dass essich zusammenmit anderen Proteinen auf einem Lipid-floß (lipid raft) befindet. Die Lipide, aus denen dieseFloße aufgebaut sind, haben eine andere Zusammenset-zung als die umgebenden Phospholipide und konnenProteine festhalten.
Eine Biomembran besitzt daher eine Mosaikstruk-tur, die veranderlich ist und damit unterschiedlicheDomanen mit unterschiedlichen Funktionen bildenund verandern kann. Man spricht deshalb von einerflussigen Mosaikstruktur oder beschreibt die Mem-branen nach dem „Fluid mosaic“-Modell.
Biomembranen sind nicht fest und starr. Die Mem-branen unterschiedlicher Zellorgane konnen ineinan-der ubergehen und dabei ihre Funktionen wechseln.
Assoziiertes Membranprotein
Lipid-doppel-schicht
Transmembranprotein(Integrales Membranprotein)
Hydrophiles Endeder Lipidmoleküle
Lipophiles Endeder Lipidmoleküle
Abb. 1.27 Zellmembran. Schematische, dreidimensionale Abbil-dung eines kleinen Ausschnitts
1.3 Biomembranen28

Die Grundstruktur einer biologischen Membran besteht auseiner Doppelschicht von Lipidmolekulen.
1.3.1.3 Membranlipide
Die Lipidschicht biologischer Membranen ist ein Flus-sigkeitsfilm, dessen Molekule sich in seitlicher Richtungbewegen konnen. Dabei sind die Lipidmolekule in derMembran so angeordnet, dass ihr hydrophiles Endenach außen, ihr lipophiles Ende nach innen gerichtetist (Abb. 1.28). Die Lipidmolekule sind gewohnlichin standiger thermischer Bewegung und konnen sichinnerhalb der Membranebene frei bewegen. Trotzdemist die Doppelschicht stabil, da die Lipidmolekule in ihrdie gunstigste Orientierung haben.
Ein wichtiger Faktor fur die Fließeigenschaften derBiomembranen von tierischen Zellen ist das Choleste-rol. Daruber hinaus beeinflusst es die Durchlassigkeitfur kleinere wasserlosliche Molekule und erhoht diemechanische Festigkeit der Lipiddoppelschicht.
Wichtig fur die Fluiditat von Biomembranen ist auchder Bau der Fettsauremolekule in denMembranlipiden.
In der Regel liegt in einem Molekul eine ungesattigteund eine gesattigte Fettsaurekette unterschiedlicherLange vor. Dies verhindert Phasentrennungen in derLipiddoppelschicht.
Membranlipide haben selten eine spezifische bio-logische Funktion. Da sie aber die Grundsubstanzeiner Biomembran darstellen, bestimmen sie auch imWesentlichen deren physikochemischen Eigenschaften,vor allem die Flexibilitat und Fluiditat. Membran-lipide bestehen aus einer polaren (hydrophilen) Kopf-gruppe und einem unpolaren (hydrophoben) Schwanz-teil. Es sind also amphipathische (amphiphile) Ver-bindungen.
Den lipophilen, unpolaren Bereich bilden die Acyl-reste von langkettigen, gesattigten (Palmitinsaure,Stearinsaure) oder ungesattigten (Olsaure, Linolsaure,Linolensaure, Arachidonsaure, Myristinsaure) Fett-sauren. Die Fluiditat einer Biomembran wird durchihre chemische Zusammensetzung bedingt. Im Ver-gleich zur Palmitinsaure fuhrt ein hoherer Anteil vonMyristinsaure, Olsaure oder Linolsaure zu einer er-hohten Fluiditat einer Biomembran.
Bei den Glycerolipiden sind die Fettsauren uberEsterbindungen mit Glycerol verbunden.
Die polare Kopfgruppe der Membranlipide bestehtaus Phosphosaureestern oder Zuckerresten. Diesekonnen elektrisch neutral oder positiv bzw. negativ ge-laden sein.
Cholesterol und andere Sterole besitzen als polareGruppe eine Hydroxylgruppe. Die starre, planareSteroidstruktur, sowie eine unpolare Kohlenwasser-stoff-Schwanzgruppe sind zwischen die Fettsaureresteeingelagert. Sie haben einen verfestigenden, stabilisie-renden Effekt auf die benachbarten Acylkettenbereiche.
1.3.1.4 Membranproteine
Spezifische Aufgaben biologischer Membranen werdenuberwiegend von Membranproteinen erfullt. VieleMembranproteine, so genannte Transmembranpro-teine, erstrecken sich durch die Lipiddoppelschichthindurch. Sie besitzen lipophile Bereiche, welche mitden Lipidmolekulen im Inneren der Doppelschichtin Wechselwirkung treten. Die hydrophilen Abschnitteder Transmembranproteine (= Tunnelproteine) ragenauf beiden Seiten aus der Lipiddoppelschicht heraus.Die Transmembranproteine sind meist glykosyliert.Ihre Oligosaccharidketten liegen stets auf der extra-zellularen Seite der Membran. Andere Proteine, diemit Membranen assoziiert sind, sind nur an eine derbeiden Membranaußenseiten gebunden (Abb. 1.29).
CH3
N+
CH3H3C
O
P O–O
O
O OO O
H3C CH3
Cholin
Phosphorsäure
Glycerin
Fettsäuren
Lecithin
HydrophilerMolekülteil
LipophilerMolekülteil
Abb. 1.28 Aufbau eines Phospholipids
1.3.1 Chemie und Aufbau 29
1Cytolog
ie

Funktionen der Membranproteine
Proteine sind an den selektiven aktiven und passivenTransportvorgangen durch Biomembranen beteiligt.Sie bieten die Grundlage fur die hochselektive Permea-bilitat der Membran. Andere Proteine dienen als spezi-fische Rezeptoren fur Hormone, Neurotransmittersub-stanzen, Antigene und Viren. Auch einige Enzyme sindin der Biomembran verankert. Damit ist sichergestellt,
dass bestimmte biochemische Reaktionen ortlich fest-gelegt ablaufen. Die Membranproteine sind spezifischfur jeden Membrantyp einer Zelle. Das Endoplasmati-sche Retikulum besitzt andere Membranproteine alsz.B. die Plasmamembran oder die Mitochondrien.Dies ist Ausdruck der unterschiedlichen Funktionenverschiedener Biomembranen. Mit einem Funktions-wechsel der Biomembran ist der Austausch von Mem-branproteinen verbunden. Beispiele hierfur bieten dieAusschleusung von Viren aus der Zelle (Kap. 6.2.2) so-wie Funktionswechsel von Biomembranen beim Mem-branfluss zwischen verschiedenen, membranumschlos-senen Organellen der Zelle.
1.3.1.5 Membrankohlenhydrate
Auf der Außenseite von Plasmamembranen von be-stimmten Eukaryontenzellen (bei Saugetieren) findensich Kohlenhydrate, die in der Regel als Oligosaccharidean Membranproteine (Glykoproteine) oder seltenerMembranlipide (Glykolipide) gebunden sind.
Ein einziges Glykoprotein kann viele Oligosaccha-ride tragen. Die kohlenhydratreiche Zone auf der Au-ßenseite solcher Plasmamembranen wird als cell coatoder Glykocalyx bezeichnet. Ihr kommt eine Funktionbei Zell-Zell-Erkennungsvorgangen zu (Abb. 1.30).
Glykoproteine besitzen meist kurze Oligosaccharid-ketten (bis 15 Monosaccharid-Einheiten). Spezifischausgeformte Oligosaccharide konnen an komplementarausgebildete Strukturen auf Nachbarzellen binden. AusMembranen abgespaltene Kohlenhydrate konnen Sig-nalfunktion haben, sie dienen z.B. als „Elicitoren“ einerAbwehrreaktion bei Pflanzen.
ExtrazellulärerRaum
Lipid-doppel-schicht
Cytoplasma
COOH
PP
NH2
1 2
3
4
5
Abb. 1.29 Verknupfungsarten von Membranproteinen mit der Lipiddoppelschicht. Transmembranproteine durchziehen die Lipid-doppelschicht als einzelne a-Helix (1) oder mit mehreren a-Helices (2). Andere Membranproteine sind nur uber ein kovalent gebundenesLipid mit der Doppelschicht verbunden (3). Auf der Außenseite der Membran konnen Oligosaccharide an der Bindung beteiligt sein (4).Viele Proteine sind auch durch nicht kovalente Wechselwirkungen mit anderen Membranproteinen an die Membran gebunden (5).
Glyko-calyx(cellcoat)
Lipid-doppel-schicht
Cytoplasma
adsorbiertesGlykoprotein
Transmembran-glykoprotein
Glykolipid
Transmembran-proteoglykan
= Zuckerrest
Abb. 1.30 Schematische Darstellung der Glykocalyx (cell coat). Siebesteht aus den Oligosaccharid-Seitenketten der Glykolipide undder integralen Membranproteine sowie aus den Polysaccharid-ketten der integralen Proteoglykane. Bei manchen Zellen gehorenzur Glykocalyx auch von außen adsorbierte Glykoproteine undProteoglykane (hier nicht dargestellt). Man beachte, dass alleZuckerreste sich ausschließlich auf der Außenseite der Membranbefinden.
1.3 Biomembranen30

1.3.2 Endozytose, Exozytose,Membranfluss
Makromolekule, wie Proteine, Nukleinsauren undPolysaccharide, konnen nicht von Transportproteinendurch Biomembranen transportiert werden. Ebensokann die Aufnahme von großen Partikeln, z.B. Bakte-rien und Viren, nicht durch Vermittlung von Trans-portproteinen erfolgen. Hierzu dienen die Mechanis-men der Endozytose oder der Exozytose. Hierbei er-folgt die Aufnahme oder Ausscheidung uber die Bil-dung und Fusion membranumhullter Vesikel(Abb. 1.31). Diese Transportvorgange sind also miteinem Verschmelzen von Biomembranen verbunden.
1.3.2.1 Endozytose
Durch Einstulpen von begrenzten Bereichen derPlasmamembran ins Innere der Zelle und Abschnurenkleiner Vakuolen konnen durch Pinozytose oderPhagozytose Bestandteile des Außenmediums in dieZelle aufgenommen werden.
Pinozytose und Phagozytose sind vor allem bei derAufnahme von sehr großen Molekulen oder Teilchen,die nicht durch die Membran hindurch diffundierenoder aktiv transportiert werden konnen, von Bedeu-tung. Der Großenbereich der pino- oder phagozytiertenPartikel reicht von Kolloiden bis zu Bakterien.
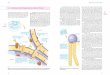
Einen Sonderfall stellt die rezeptorvermittelte Endo-zytose dar. Bei Tieren wird sie genutzt, um ganz be-stimmte Makromolekule zu erkennen und aufzuneh-men. Rezeptorgekoppelte Endozytose ist wichtig furdie Funktion der Immunantwort. PolymorphkernigeGranulozyten und Makrophagen phagozytieren inden Organismus eingedrungene Krankheitserreger,wenn diese vorher mit Antikorper oder Komponentendes Komplementsystems reagiert haben, also Signal-strukturen fur die zellgebundenen Rezeptoren tragen.Als integrale Membranproteine binden sie Substanzenspezifisch an bestimmten Orten der Plasmamembran.Diese Orte nennt man Coated Pits (uberzogene Gru-ben), die im elektronenmikroskopischen Bild als Ver-tiefungen in der Plasmamembran erkennbar sind. DieInnenseite der Coated Pits ist von Proteinen wieClathrin uberzogen (Abb. 1.32). Die Coated Pits stul-pen sich nach innen und schnuren sich als CoatedVesicles („Stachelsaumvesikel“) ab. Durch Abstoßender Clathrinhulle wandeln sie sich in Endosomenum. Das Endosom fusioniert mit primaren Lysosomenzu einem sekundaren Lysosom, in dem das endozytierteMaterial abgebaut wird. Die Abbauprodukte, z.B.Zucker oder Aminosauren, werden in das Cytoplasmatransportiert.
Die Bewegungsvorgange der Vesikel werden offensicht-lich durch das Cytoskelett der Zellen kontrolliert. EineStorung dieses Systems verhindert die Endozytose. BeiZellen von Saugetieren kann die Endozytose daher jenach Zelltyp uber die Nahrungsaufnahme hinaussehr spezielle Funktionen ubernehmen. Z.B. wirdCholesterol im Menschen durch rezeptorvermittelteEndozytose aufgenommen. Um das Korpergewebemit Lipiden zu versorgen, werden diese zusammenmit Cholesterol in Partikel verpackt. LDL-Partikel(low density lipoprotein) mussen von den Leberzellenzum Recycling aufgenommen werden. Die Aufnahmebeginnt mit der spezifischen Erkennung von LDL durchRezeptoren. Familiare Hypercholesterinamie ist eineerbliche Stoffwechselerkrankung, bei der ein LDL-Re-zeptor geschadigt ist. Als Folge davon ist der Choleste-rol-Wert erhoht.
Zellen des Immunsystems, so genannte Phagozyten(Makrophagen, und polymorphkernige Granulozyten),haben die Fahigkeit z.B. Bakterien oder Molekule auf-
EndoplasmatischesRetikulum
Endozytose
Exozytose
Phagozytose
Pinozytose
Pinosomen
Phagosomen
Cytoplasma-membran
Golgi-Vesikel
Dictyosom
Vakuole
Tonoplast
A
B
C
Abb. 1.31 Membranfluss. Endozytose: Aufnahme von Partikeln(Phagozytose), oder Flussigkeit (Pinozytose) durch Abschnurungvon Vesikeln an der Cytoplasmamembran. Exozytose: Ausschei-dung aus der Zelle z.B. des Inhalts von Golgi-Vesikeln. A. Uber-gang von Vesikeln des Endoplasmatischen Retikulums zum Dic-tyosom, B. Ubergang von Vesikeln des Endoplasmatischen Reti-kulums in eine Vakuole, C. Bildung einer Vakuole durch Vergro-ßerung der Zisterne des Endoplasmatischen Retikulums
1.3.2 Endozytose, Exozytose, Membranfluss 31
1Cytolog
ie
zunehmen und zu eliminieren. Die Endozytose verlauftbei Saugetierzellen sehr spezifisch uber die Bindung deraufzunehmenden Zellen und Strukturen an membran-standige Rezeptoren. So tragen die Phagozyten des Im-munsystems u. a. so genannte Fc-Rezeptoren. An diesekann der Fc-Teil eines Antikorpers binden, wenn derAntikorper seinerseits an ein Antigen gebunden ist.Bakterienzellen oder Viren, die an die Antikorper ge-bunden haben, konnen uber diese Fc-Rezeptoren ge-bunden und von dem Phagozyten endozytiert (= pha-gozytiert) werden (Abb. 1.31).
Viren und intrazellular sich vermehrende Bakterienund Parasiten bedienen sich des Endozytosewegs, umin Korperzellen des Wirts einzudringen. Sie bindenoft sehr spezifisch an Rezeptoren bestimmter Zielzellen.Solche intrazellular lebende Bakterien sind z.B. Rickett-sien und Chlamydien. Auch die Sporozoen von Toxo-plasma gondii und Plasmodien (Malaria-Erreger) ge-langen auf dem Wege der Endozytose in ihre Ziel-zellen.
Manche Bakterien vermogen nach Endozytose sogarin den Phagozyten (Makrophagen) zu uberleben, undsich in diesen Zellen zu vermehren. Beispiel hierfursind Legionella pneumophila, Tuberkelbazillen undMycobacterium leprae, der Erreger der Lepra.
1.3.2.2 Exozytose
Durch Exozytose konnen Stoffe aus der Zelle ausge-schleust werden (Abb. 1.31). Diese Moglichkeit istfur Sekretion und Exkretion von Bedeutung. DieMembranen von Vakuolen im Innern der Zelle, welchedie auszuscheidenden Stoffe enthalten, z.B. Golgi-Ve-sikel, verschmelzenmit der Plasmamembran, der Inhaltwird nach außen entleert. Danach geht die Vakuolen-membran in der Plasmamembran auf. Bei Exozytoseund Endozytose liegen die Makromolekule abgetrenntin membranumschlossenen Vesikeln. Sie vermischensich zunachst nicht mit anderen Makromolekulender Zelle. Die Vesikel verschmelzen nur mit ganz be-
Abb. 1.32 Rezeptorgekoppelte Endo-zytose. Die rezeptorgekoppelte Endo-zytose ermoglicht die gezielte Auf-nahme großer Molekule oder Partikel,z.B. Viren und Bakterien. Die Rezep-toren sind diffus uber die Zelloberfla-che verteilt. Sie sammeln sich, wennsie ein Partikel gebunden haben, in„Coated Pits“ (Stachelsaum-Grub-chen). Diese stulpen sich nach innenund schnuren sich als „Coated Vesic-les“ (Stachelsaum-Vesikel) ab. DieClathrinhulle wird abgestoßen, die soentstandenen Endosomen fusionierenmit Lysosomen, deren Enzyme dasaufgenommene Partikel abbauen.
Clathrin
Antikörper bindenan die Oberfläche
eines fremden Partikels
Stachelsaum-Grübchen
Rezeptor
Cyto-plasma-
membran
CURL
Endosom
Lysosom
Hydrolasen
Der Fc-Anteil desAntikörpers bindet an diezellständigen Rezeptoren
Aufnahme insCytoplasma
Fusion der Endosomenmit Lysosomen
Abbau der fremdenPartikel
Stachelsaum-Vesikel
vesikulärer Teil
tubulärerTeil
Fusion
1.3 Biomembranen32

stimmten Membranen. Hieraus resultiert ein gerichte-ter Stofftransport zwischen Zellumgebung und Zellin-nerem, aber auch ein gerichteter Transport zwischenmembranumschlossenen Organellen im Zellinneren.
Auch fur den intrazellularen Stoffaustausch spielensolche Vorgange eine wichtige Rolle. So konnen z.B.membranumschlossene Partikel vom Endoplasmati-schen Retikulum abgeschnurt werden und zu Golgi-Zisternen verschmelzen. Hierdurch werden Proteinevom Endoplasmatischen Retikulum zu den Dictyoso-men transportiert. Vesikel des Endoplasmatischen Re-tikulums konnen auch mit der Tonoplastenmembranverschmelzen und ihren Inhalt in die Vakuole ent-leeren. Des Weiteren konnen Stoffwechselreaktionendurch Verschmelzen verschiedener Vakuolen in Ganggesetzt werden. So werden etwa die abbauenden Enzy-me der Lysosomen unter Verschmelzung der Membra-nen beider Vakuolen in die Pinosomen entleert. Zumanderen kann sich auch die Pinosomenmembran auf-losen und mit dem Vakuoleninhalt im Grundplasmader Zelle aufgehen.
1.3.2.3 Membranfluss
Endozytose, Exozytose und intrazellularer Stoffaus-tausch uber membranumschlossene Vesikel ist alsomit einem Austausch von Membranstucken verbun-den. Teile der Plasmamembran, der Tonoplastenmem-bran, des Endoplasmatischen Retikulums, der Dictyo-somen und der Lysosomen konnen gegenseitig mitei-nander verschmelzen. Diese membranumschlossenenZellorganellen konnen sich auch gegenseitig aufbauen,z.B. das Endoplasmatische Retikulum die Kernmem-bran und die Dictyosomen. Die Plasmamembran ent-steht nach der Teilung pflanzlicher Zellen durch Zu-sammenfließen von Golgi-Vesikeln, also aus Dictyoso-menmembranen (vgl. Abb. 1.19).
Innerhalb der Zelle findet ein Austausch von Membranen, einMembranfluss statt. Ausgenommen hiervon sind die hoch-spezialisierten Membranen der Mitochondrien und Plastiden.
1.3.3 Semipermeabilitat, Osmose,Membranpotential
Die Lipiddoppelschicht stellt eine nichtwassrige Barrie-re zwischen zwei wassrigen Kompartimenten dar. DerAustausch von wasserloslichen Molekulen und Ionenzwischen diesen Kompartimenten ist daher stark einge-schrankt. Je lipophiler ein Molekul ist, desto besser dif-fundiert es durch eine Biomembran. Wasser und sehrkleine Molekule (z.B. Glycerol, Ethanol) bilden eine
Ausnahme: Sie passieren Biomembranen schneller alsman es von ihrer Lipidloslichkeit erwarten sollte.(Abb. 1.33).
1.3.3.1 Semipermeabilitat
Die Eigenschaft von Biomembranen, kleine hydrophileMolekule frei passieren zu lassen, großere jedoch nicht,wird als Semipermeabilitat bezeichnet. Sie ist dieGrundlage fur alle osmotischen Vorgange.
Molekule mit hydrophoben Eigenschaften konnendagegen in den lipophilen Bereich der Biomembraneindringen oder sich durch die Membran „hindurch-losen“. Zu dieser Gruppe von Molekulen gehorenz.B. die Steroidhormone.
Biomembranen sind also semipermeabel, oder bes-ser, selektiv permeabel. Sie sind gut durchlassig furWasser, jedoch weniger gut oder gar nicht fur inWassergeloste organische oder anorganische Stoffe. Ungela-dene, lipidlosliche Substanzen konnen recht gut durchBiomembranen permeieren. Dagegen sind Biomem-branen fur Ionen und organische polare Stoffe, wieGlucose oder Aminosauren, kaum oder gar nicht per-meabel. Die Moglichkeit einer Permeation (Diffusion)von Ionen durch Biomembranen nimmt mit steigenderLadungszahl und Ionengroße ab. Hierdurch halt z.B.die Plasmamembran ein osmotisches Gleichgewichtund ein Konzentrationsgefalle mit der Umgebungder Zelle aufrecht. Dadurch wird ein bestimmtes, furden Stoffwechsel unbedingt notwendiges inneres Mi-lieu gegenuber sehr unterschiedlich zusammengesetz-ten Außenlosungen aufrecht erhalten und verhindert,
Lipiddoppelschicht
O2
CO2
N2
Benzol
kleine hydrophobeMoleküle
H2OGlycerolEthanol
kleine ungeladenepolare Moleküle
GlucoseAminosäuren
Nukleotide
größere ungeladenepolare Moleküle
H+, Na+
HCO3–, K+
Ca2+, Cl–
Mg2+
Ionen
Abb. 1.33 Permeabilitatseigenschaften einer kunstlichen Lipid-doppelschicht fur unterschiedliche Molekulklassen
1.3.3 Semipermeabilitat, Osmose, Membranpotential 33
1Cytolog
ie

dass fur die Zellfunktion notwendige Stoffe aus der Zel-le diffundieren. Auch innerhalb der Zelle bilden die ver-schiedenen Membransysteme Barrieren gegen einenfreien Stoffaustausch. Funktionell unterschiedliche,membranumschlossene Kompartimente der Zelle un-terscheiden sich auch durch einen unterschiedlichenStoffbestand.
Die Funktionen der Membranen als Diffusionsbarrieren sindeng mit der Lebensfahigkeit der Zelle verbunden. Ein Er-loschen dieser Barrierenfunktion ist ein sicheres Zeichen furden Zelltod.
Die geringe Durchlassigkeit der Membran fur Ionen istfur die Resorption von Arzneimitteln von großer Be-deutung. Viele Arzneimittel dissoziieren in wassrigerLosung in positiv und negativ geladene Ionen. Da dieionisierte Form eines Arzneimittels biologische Mem-branen fast nicht oder sehr viel schlechter zu passierenvermag als die nichtionisierte, elektrisch neutrale, mog-licherweise auch lipidlosliche Substanz, spielt derDissoziationsgrad von Stoffen, z.B. Arzneimitteln inwassriger Losung, fur die Resorption und den Stoff-transport im Organismus eine wesentliche Rolle.
Schwache Sauren, wie Penicilline, werden besser ausdem Magen resorbiert, da sie im dort herrschendensauren Milieu nicht dissoziiert sind. Schwache Basenwie Phenazon konnen ebenfalls bereits im Magen auf-genommen werden, da sie trotz des sauren Milieus nurteilweise dissoziiert vorliegen. Starkere Basen werdenerst im Dunndarm aus dem alkalischen Speisebreiresorbiert. Quartare Ammoniumverbindungen, z.B.Curare, werden auf diesem Wege nur sehr langsamund in geringem Umfang aufgenommen.
1.3.3.2 Osmose
Semipermeable Membranen sind die Voraussetzungfur die Osmose. Unter Osmose versteht man Diffusionvon Wasser durch Membranen.
Die Aktivitat des Wassers zwischen zwei Losungenunterschiedlicher Konzentration, die durch eine semi-permeable, nur fur Wasser durchlassige, Membran ge-trennt sind, ist dem Verhaltnis der Molaritat des Was-sers in beiden Kompartimenten proportional.
Wasser diffundiert mit außerordentlicher Geschwin-digkeit durch biologische Membranen. Selbst wenn dieKonzentration desWassers in beiden Kompartimenten,d.h. zu beiden Seiten der Membran gleich ist, werdenstandig Wassermolekule durch die Membran hindurchausgetauscht. Die treibende Kraft hierbei ist die thermi-sche Energie der Wassermolekule.
Befinden sich zu beiden Seiten einer semipermea-blen Membran Losungen unterschiedlicher Wasser-konzentration, so stromt Wasser vom Kompartimentmit der hoheren in das mit der niedrigeren Wasser-konzentration. Die Wasserkonzentration, die Molaritatdes Wassers, wird durch darin geloste Stoffe verringert.Die Molaritat des Wassers in der Zelle ist um so gerin-ger, je hoher die Molaritat darin geloster Stoffe ist.
Es lasst sich also auch formulieren, dass Wasser ausdem Kompartiment mit der niedrigeren Konzentrationgeloster Stoffe in das mit der hoheren Konzentrationgeloster Stoffe fließt.
Durch die gelosten Stoffe wird die Beweglichkeit desWassers behindert. Hierdurch entsteht ein Druckgradi-ent in Richtung auf die konzentriertere Losung. Manspricht auch von einer Potentialdifferenz des Wasserszwischen Kompartimenten unterschiedlicher Konzen-tration. Diese Potentialdifferenz ist die Triebkraft dereinseitig gerichteten Diffusion von Wassermolekulendurch eine semipermeable Membran, also die Trieb-kraft der Osmose.
Die Hohe dieser Saugkraft bestimmt den osmoti-schen Wert des Kompartiments, resp. der Zelle. Der os-motische Wert eines Kompartiments bzw. einer Zellekann als osmotischer Druck gemessen werden. Kom-partimente, zwischen denen keine osmotische Druck-differenz besteht, werden als isoosmotisch bezeichnet.Dies trifft in den allermeisten Fallen fur die Zellen vonTieren und die sie umgebenden Korperflussigkeiten zu.Osmotisch wirksame Substanzen in der Zelle sind vorallem Elektrolyte und polare Nichtelektrolyte. Zu denElektrolyten zahlen anorganische und organischeIonen, zu den polaren Nichtelektrolyten Zucker, Alko-hole, Purine und Pyrimidine. Makromolekule, wieNukleinsauren, Proteine oder Polysaccharide sindwegen ihrer geringen Molaritat osmotisch praktischunwirksam.
Pflanzliche Zellen entwickeln immer einen hohenosmotischen Druck. Sie benotigen deshalb eine festeZellwand. Da die pflanzliche Zellwand keine rasche Vo-lumenanderung zulasst, baut sich ein Innendruck auf,den man als Turgor bezeichnet. Dieser halt krautigePflanzen aufrecht und ist Triebkraft fur die Vergroße-rung von Pflanzenzellen und damit auch fur das Pflan-zenwachstum.
Der Aufbau von Konzentrationsgradienten zwischendem inneren und außerenMilieu von Zellen ist ein wich-tiger Mechanismus, mit dessen Hilfe beispielsweise Ex-kretions- und Sekretionszellen einen passiven Wasser-transport ermoglichen. Absorptionsgewebe von Pflan-zen, z.B. Rhizodermiszellen, halten immer einen Kon-zentrationsgradienten mit dem Bodenwasser aufrechtund konnen so Wasser aus dem Boden aufnehmen.
1.3 Biomembranen34
Plasmolyse
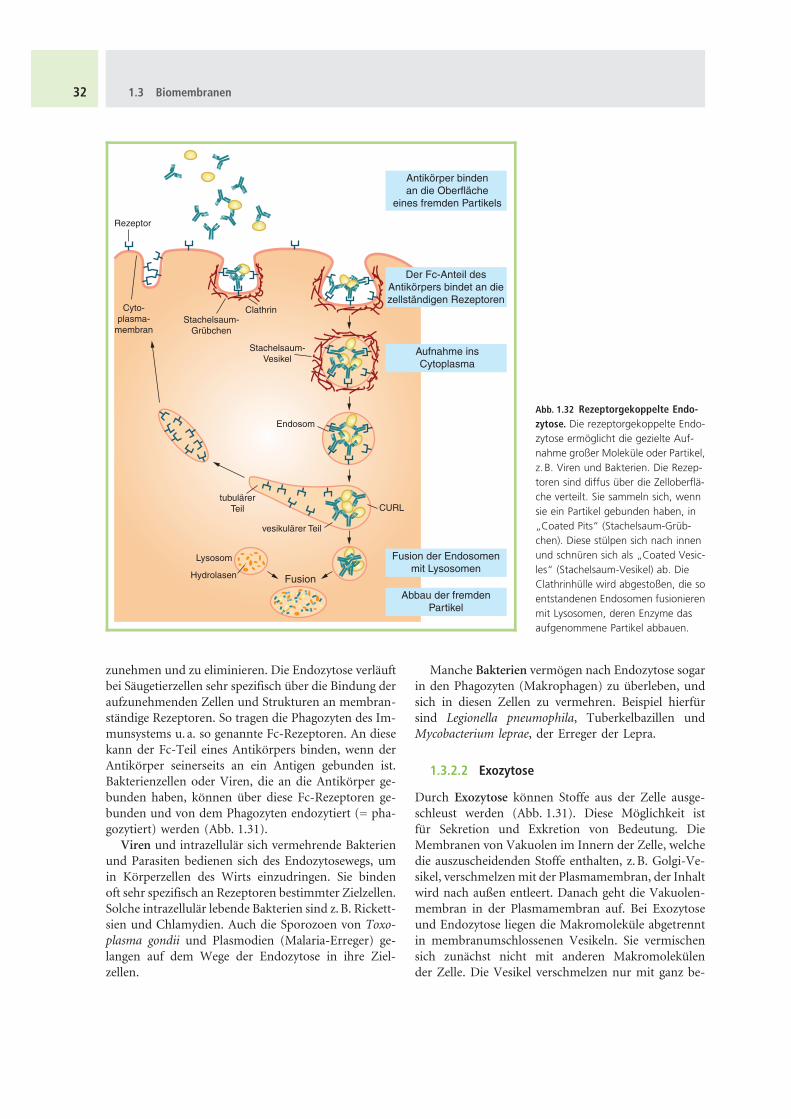
Der osmotische Druck einer pflanzlichen Zelle kannu. a. durch Plasmolyse gemessen werden. Bringt manZellen, z.B. Epidermiszellen, in eine hypertonischeLosung, d. h. eine Losung mit hoherer Konzentrationgeloster, osmotisch wirksamer Substanzen als in derZentralvakuole, so wird der Zelle Wasser entzogen.Der Plasmaschlauch lost sich von der Zellwand. Bringtman die plasmolysierte Zelle wieder inWasser (= hypo-tonische Losung), so nimmt die Zelle umgekehrt wiederWasser auf. Der Plasmaschlauch druckt sich wieder festan die Zellwand.
Bringt man die Zelle in eine Losung mit gleicherKonzentration geloster Stoffe wie in der Zentralva-kuole, also in eine aquimolare (isotonische, isoosmoti-sche) Losung, so verliert die Zelle ihren Turgor, derPlasmaschlauch lost sich gerade etwas von der Zellwandab. Dieser Zustand wird als Grenzplasmolyse bezeich-net (Abb. 1.34). Aus dem bekannten Druck der Außen-losung ergibt sich der osmotische Druck der Zelle. Er istgleich dem der Außenlosung. Der osmotische Druckeiner Losung lasst sich in einem Osmometer messen.
1.3.3.3 Membranpotential
Ionen konnen keine Biomembran durchqueren, wennsie dabei nicht durch Kanale und Transportproteine
unterstutzt werden (s. unten). Protonenpumpen fuh-ren z.B. dazu, dass der Zellinnenraum einer Pflanzen-zelle im Vergleich zur Umgebung stark negativ wird.Eine derartige Ladungsdifferenz uber eine Membranbezeichnet man alsMembranpotential. Das Membran-potential kann mitMikroelektroden gemessen werden.Die meisten Pflanzenzellen halten ein Membranpoten-tial von mindestens -120 mV aufrecht.
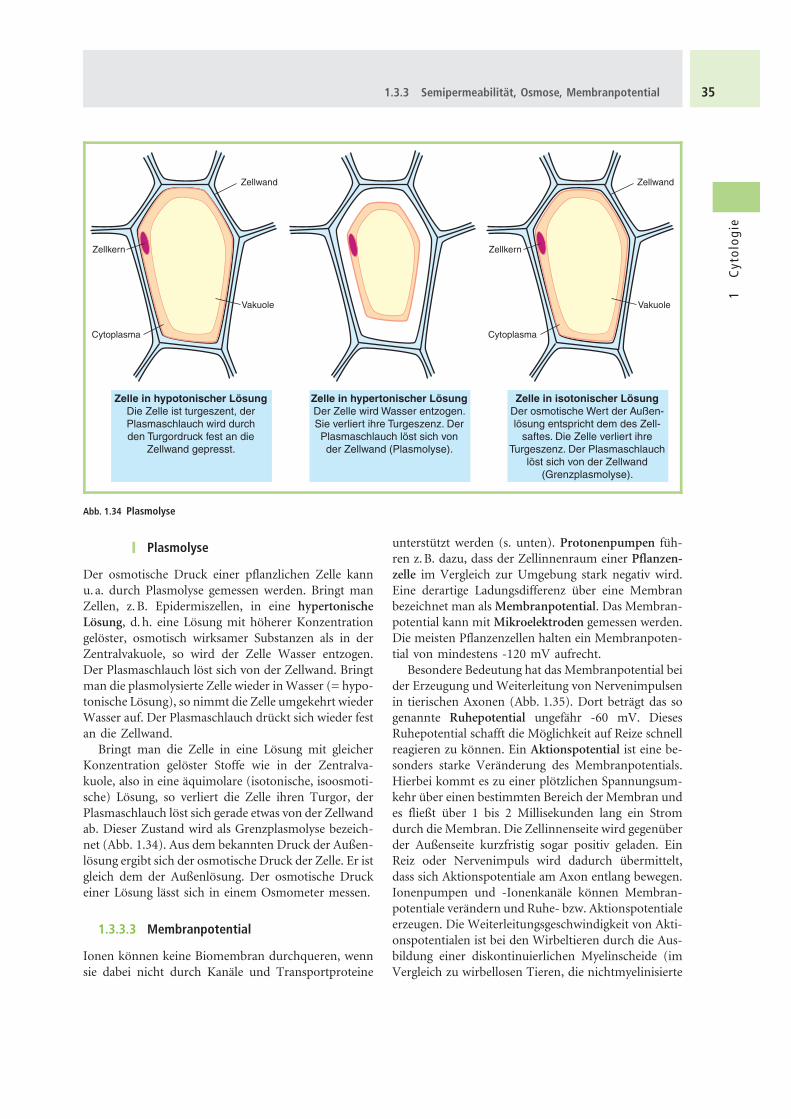
Besondere Bedeutung hat das Membranpotential beider Erzeugung und Weiterleitung von Nervenimpulsenin tierischen Axonen (Abb. 1.35). Dort betragt das sogenannte Ruhepotential ungefahr -60 mV. DiesesRuhepotential schafft die Moglichkeit auf Reize schnellreagieren zu konnen. Ein Aktionspotential ist eine be-sonders starke Veranderung des Membranpotentials.Hierbei kommt es zu einer plotzlichen Spannungsum-kehr uber einen bestimmten Bereich der Membran undes fließt uber 1 bis 2 Millisekunden lang ein Stromdurch die Membran. Die Zellinnenseite wird gegenuberder Außenseite kurzfristig sogar positiv geladen. EinReiz oder Nervenimpuls wird dadurch ubermittelt,dass sich Aktionspotentiale am Axon entlang bewegen.Ionenpumpen und -Ionenkanale konnen Membran-potentiale verandern und Ruhe- bzw. Aktionspotentialeerzeugen. Die Weiterleitungsgeschwindigkeit von Akti-onspotentialen ist bei den Wirbeltieren durch die Aus-bildung einer diskontinuierlichen Myelinscheide (imVergleich zu wirbellosen Tieren, die nichtmyelinisierte
Zelle in hypotonischer LösungDie Zelle ist turgeszent, derPlasmaschlauch wird durchden Turgordruck fest an die
Zellwand gepresst.
Zellwand
Zellkern
Vakuole
Cytoplasma
Zelle in hypertonischer LösungDer Zelle wird Wasser entzogen.Sie verliert ihre Turgeszenz. DerPlasmaschlauch löst sich vonder Zellwand (Plasmolyse).
Zelle in isotonischer LösungDer osmotische Wert der Außen-lösung entspricht dem des Zell-
saftes. Die Zelle verliert ihreTurgeszenz. Der Plasmaschlauch
löst sich von der Zellwand(Grenzplasmolyse).
Zellwand
Zellkern
Vakuole
Cytoplasma
Abb. 1.34 Plasmolyse
1.3.3 Semipermeabilitat, Osmose, Membranpotential 35
1Cytolog
ie
Axone besitzen) dramatisch erhoht. Die Aktionspoten-tiale „springen“ in myelinisierten Axonen von einemRanvier-Schnurring zum nachsten. An diesen Stellen,die dicht mit Ionenkanalen besetzt sind, lasst die elek-trisch isolierende Myelinscheide die Axonmembranfrei. Ein ausgelostes Aktionspotential kann aber nichtuber die von Myelin umhullte und daher isolierteMembran selbst weitergeleitet werden. Der Ionenstromerfolgt daher durch das Cytoplasma. Dieser Strom kanndie Membran am nachsten Schnurring depolarisierenund ein Aktionspotential auslosen. Diese Art der Im-pulsverarbeitung nennt man saltatorische Erregungs-leitung.
1.3.4 Zellkontakte
Zellen, die miteinander verbunden sind oder verbun-den werden sollen, bilden bestimmte Strukturen aus,die entweder der Haftung oder dauerhaften Bindungoder Kommunikation dienen. Diese spezialisiertenZell-Zell-Verbindungen nennt man bei tierischen Zel-len auch „Junctions“. Drei wichtige Typen von Verbin-
dungen sind die Tight Junctions, Desmosomen undGap Junctions
Tight Junctions sind spezielle Strukturen der Plas-mamembran, die benachbarte Epithelzellen miteinan-der verbinden. Es handelt sich hierbei um fest mitei-nander verknupfte Membranproteine. Sie versiegelnGewebe, speziell Epithelien, und verhindern den freienTransport geloster Stoffe durch die Zellzwischenraume.So mussen alle Stoffe bestimmte Zellen passieren undkonnen zielgerichtet in den einen oder anderen Teil desKorpers geleitet werden.
Desmosomen sind mit der Plasmamembran verbun-dene Strukturen, die Zellen druckknopfartig miteinan-der verbinden, aber die Bewegung von Stoffen kaumbehindern. Auf der cytoplasmatischen Seite besitzt je-des Desmosom einen so genannten desmosalen Plaque,an den spezielle Zelladhasionsproteine der Plasma-membran angeheftet sind. Uber diese Proteine erfolgtdie Bindung an die Nachbarzelle. Außerdem sind dieDesmosomen mit Intermediarfilament-Proteinen desCytoskeletts verbunden (s. Kap. 1.4.12). Diese ziehensich quer durch die Zellen und verleihen z.B. den epi-thelialen Geweben ihre hohe mechanische Stabilitat.
Die Basalmembran, eine spezielle Form der extra-zellularen Matrix tierischer Zellen (s. Kap. 1.2.3) istuber so genannte Hemidesmosomen mit den Epithel-zellen verknupft.
Wahrend die Tight Junctions und Desmosomenmechanische Aufgaben erfullen, dienen die Gap Junc-tions der Zell-Zell-Kommunikation. Sie konnen bis zu25% der Plasmamembran tierischer Zellen ausmachenund hunderte kleine Kanale bilden. Im Bereich solcherZell-Zell-Verbindungen treffen so genannte Conne-xone aufeinander, die miteinander verknupft werden.Ein Connexon kann als Halbkanal aufgefasst werden,der sich mit einem Connexon der Nachbarzellen zueinem funktionellen Kanal verbindet. Uber diese Kana-le konnen kleine Signalmolekule, Metaboliten oderIonen von einer Zelle in die andere gelangen, siesind jedoch zu eng, um Proteine durchzulassen. Con-nexone sind aus sechs identischen Connexin-Unterein-heiten aufgebaut. Connexin ist ein Protein. Die GapJunctions ermoglichen den miteinander verknupftenZellen eine Kooperation im Bereich Energie- und Bau-steinstoffwechsel sowie der Signalubermittlung.
Die Plasmodesmen (Plasmodesmata) der Pflanzen-zellen (s. Kap. 1.2.2.1) entsprechen funktionell den GapJunctions der tierischen Zellen. Eine typische Pflanzen-zelle besitzt viele Tausend Plasmodesmen. Im Gegen-satz zu den Gap Junctions sind bei den Plasmodesmennicht nur Kanalproteine miteinander verbunden, son-dern es handelt sich um von der Plasmamembran aus-gekleidete Zell-Zell-Verbindungen, das heißt in diesen
Myelinscheide
Dendrit
Zellkern
Axon
Ranvier-Schnürring
Abb. 1.35 Nervenzelle (Neuron). Myelinisiertes Axon mit Ranvier-Schnurringen
1.3 Biomembranen36

Bereichen wurden die Plasmamembranen benachbarterZellen miteinander fusioniert. Die so gebildeten Kanalebesitzen einen etwa viermal großeren Durchmesser alsdie Connexone der Gap Junctions. Die Stoffpassagewird allerdings durch die Einlagerung eines kompaktenZylinders, dem so genannten Desmotubulus erschwert,so dass auch zwischen den Pflanzenzellen normalerwei-se nur kleine Molekule transportiert werden konnen.Allerdings konnen auch großere Molekule uber Plas-modesmen transportiert werden. Pflanzen produzierenhierfur eigene Bewegungsproteine (movement pro-teins), die die Poren vorubergehend erweitern.
1.3.5 Spezifischer Stofftransportdurch Biomembranen
Neben ihrer Trennfunktion sind die BiomembranenOrganelle des Stoff- und Informationsaustausches inder Zelle. Wasser, Nahrstoffe, z.B. Glucose, Amino-sauren, Ionen sowie Nukleotide und zahlreiche Zell-metaboliten mussen von der Zelle aufgenommen, End-produkte des Stoffwechsels ausgeschieden werden unddabei die Plasmamembran passieren. Auch zwischenden einzelnen Reaktionsraumen in der Zelle mussein spezifischer, kontrollierter Stoffaustausch ermog-licht werden.
Fur den vielfaltigen Stoffaustausch zwischen denKompartimenten einer Zelle sowie der Zelle und ihrerUmgebung enthalten Biomembranen zahlreiche spezi-fische Translokatoren. Dies sind spezielle Membran-proteine, die man als Membrantransportproteine be-zeichnet.
Jedes dieser Proteine ist darauf spezialisiert, einebestimmte Klasse von Verbindungen oder nur ein be-stimmtes Molekul zu transportieren.
Alle bisher bekannten Membrantransportproteinesind Proteine, welche die Lipiddoppelschicht mit meh-reren a-Helices durchdringen (Multipass Transmem-branproteine, s. Abb. 1.29).
Es gibt zwei Hauptklassen von Transportproteinen:Carrierproteine und Kanalproteine. DieCarrierproteinebinden spezifisch die zu transportierenden Molekuleund transportieren diese mittels Konformationsande-rung auf die andere Seite der Biomembran.
Die Kanalproteine dagegen formen wassergefulltePoren durch die Lipiddoppelschicht. Wenn diese Porengeoffnet sind, erlauben sie z.B. anorganischen Ionen denDurchtritt durch die Membranen (Abb. 1.36, 1.37).
In beiden Fallen wird ein spezifischer Transport er-moglicht, einmal durch passiven Transport (= erleich-terte Diffusion), zum anderen durch aktive Transport-vorgange (Tab. 1.14).
Der spezifische Transport ist von der freien Diffusi-on durch folgende Kriterien zu unterscheiden:
ExtrazellulärerRaum
Lipid-doppel-schicht
Cytoplasma
EinfacheDiffusion
Kanal-vermittelteDiffusion
Kanalprotein
Transportierte Moleküle
Carrier-vermittelteDiffusion
Carrierprotein
Passiver Transport(erleichterte Diffusion)
Aktiver Transport
Energie
Elektro-chemischer
Gradient
Abb. 1.36 Schematische Darstellung des passiven Transports, der einem elektrochemischen Gradienten folgt, und des aktiven Transports,der in der entgegen gesetzten Richtung verlauft. Die einfache Diffusion und der von Membrantransportproteinen vermittelte passiveTransport (auch „erleichterte Diffusion“ genannt) laufen spontan ab; der aktive Transport dagegen erfordert die Zufuhr von Stoff-wechselenergie. Durch einfache Diffusion konnen nur nichtpolare und kleine, ungeladene, polare Molekule die Lipiddoppelschichtdurchqueren; andere polare Molekule werden mit nennenswerter Geschwindigkeit nur von Carrier- oder Kanalproteinen transportiert.
1.3.5 Spezifischer Stofftransport durch Biomembranen 37
1Cytolog
ie

& Er ist normalerweise schneller als die freie Diffusion& Er wird durch spezifische Translokatoren vermittelt& Er ist substratspezifisch& Er verlauft bis zu einer Sattigung& Er ist spezifisch hemmbar
Es werden grundsatzlich zwei Formen von spezifischemTransport unterschieden (Abb. 1.38, 1.39)
& Passiver Transport (katalysierte Diffusion)& Aktiver Transport
1.3.5.1 Passiver Transport (katalysierte,erleichterte Diffusion)
Die katalysierte Diffusion kann wie die freie Diffusionnur zu einem Konzentrationsausgleich zwischen zweiKompartimenten fuhren. Katalysierte Diffusion imStoffaustausch mit der Umwelt ist nicht bekannt. Of-fensichtlich sind hier selektive Anreicherungsvorgange
durch aktiven Transport unerlasslich. Bei Zellen imInneren vielzelliger Organismen, die von einer korper-eigenen Flussigkeit mit relativ konstanter molekularerZusammensetzung umgeben sind, konnen jedoch Ami-nosauren oder Glucose durch katalysierte Diffusionaufgenommen werden, z.B. aus dem Blut. Epithelzellenkonnen umgekehrt Molekule an das Blut abgeben, ohnedass hierfur Energie aufgewendet werden muss.
Ein gut untersuchtes Beispiel fur ein solches Trans-portsystem ist der Glucose-Translokator der Erythro-zytenmembran beim Menschen. Uber diesen kannGlucose um den Faktor 105 schneller aufgenommenwerden als durch freie Diffusion.
Es sind auch Translokator-Systeme nach dem Prin-zip der katalysierten Diffusion fur Ionen bekannt. Diessind die so genannten Ionenkanale in den Plasmamem-branen elektrisch erregbarer Zellen, Nervenzellen undMuskelzellen, fur Na+, K+, Ca2+ und Cl–.
Spezifische Transportsysteme beschleunigen dabeilediglich den Transport von Stoffen durch die Mem-
ExtrazellulärerRaum
Lipid-doppel-schicht
Cytoplasma
Gelöste Substanz
Carrierprotein Kanalprotein
WassergefülltePore
Abb. 1.37 Die beiden Klassen von Membrantransportproteinen in vereinfachter, schematischer Darstellung. Ein Carrierprotein kann zweiverschiedene Konformationen einnehmen und so die Bindungsstelle fur das zu transportierende Molekul zuerst auf der einen und dannauf der anderen Seite der Membran zuganglich machen. Ein Kanalprotein dagegen bildet eine wassergefullte Membranpore, durch diespezifische Ionen hindurchfließen konnen.
Tab. 1.14 Stofftransport durch Biomembranen (Ubersicht)
1. Freie DiffusionDiffusion kleiner hydrophiler bzw. lipophiler Molekule
2. Erleichterte DiffusionUber Translokatoren, substratspezifisch, sattigbar, hemmbar,schneller als freie Diffusion
Passiv, ohne Energieverbrauchnur mit Konzentrationsgradienten
3. Aktiver Transport
Primarer aktiver Transport
Sekundarer aktiver Transport
Gruppentranslokation
Polyprenolzyklus
Aktiv, unter Energieverbrauchgegen Konzentrationsgradienten,nur in einer Richtung
1.3 Biomembranen38
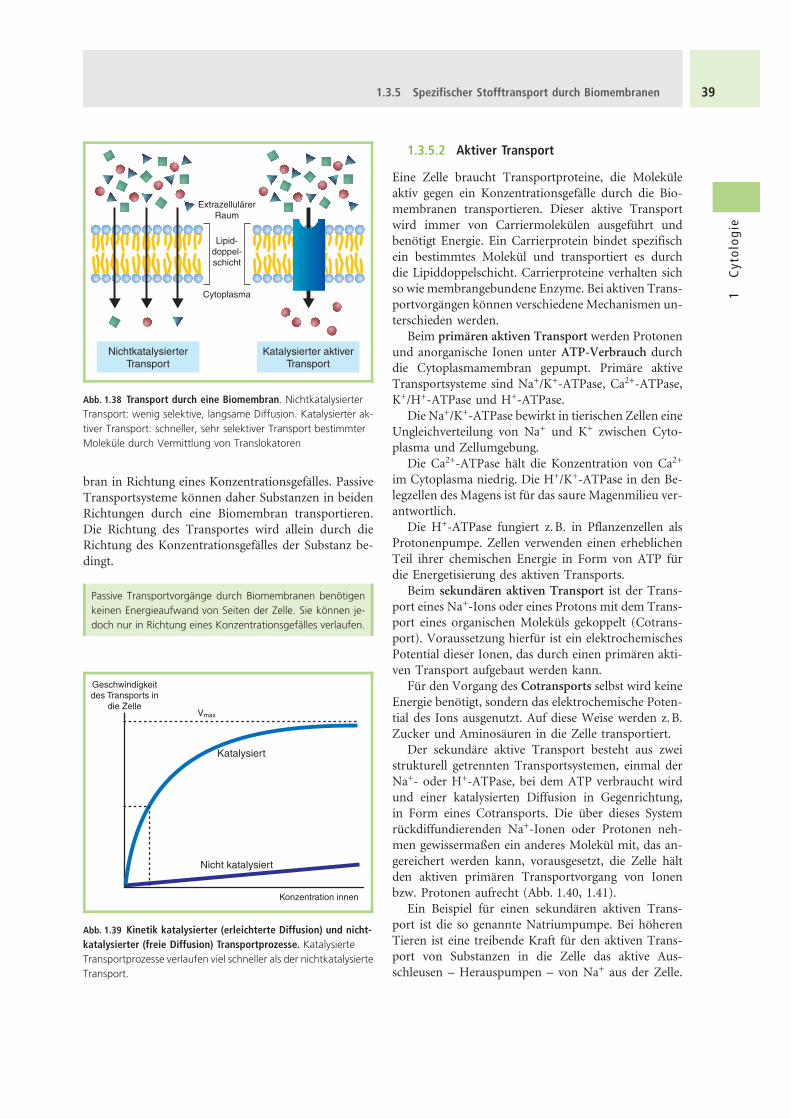
bran in Richtung eines Konzentrationsgefalles. PassiveTransportsysteme konnen daher Substanzen in beidenRichtungen durch eine Biomembran transportieren.Die Richtung des Transportes wird allein durch dieRichtung des Konzentrationsgefalles der Substanz be-dingt.
Passive Transportvorgange durch Biomembranen benotigenkeinen Energieaufwand von Seiten der Zelle. Sie konnen je-doch nur in Richtung eines Konzentrationsgefalles verlaufen.
1.3.5.2 Aktiver Transport
Eine Zelle braucht Transportproteine, die Molekuleaktiv gegen ein Konzentrationsgefalle durch die Bio-membranen transportieren. Dieser aktive Transportwird immer von Carriermolekulen ausgefuhrt undbenotigt Energie. Ein Carrierprotein bindet spezifischein bestimmtes Molekul und transportiert es durchdie Lipiddoppelschicht. Carrierproteine verhalten sichso wie membrangebundene Enzyme. Bei aktiven Trans-portvorgangen konnen verschiedene Mechanismen un-terschieden werden.
Beim primaren aktiven Transport werden Protonenund anorganische Ionen unter ATP-Verbrauch durchdie Cytoplasmamembran gepumpt. Primare aktiveTransportsysteme sind Na+/K+-ATPase, Ca2+-ATPase,K+/H+-ATPase und H+-ATPase.
Die Na+/K+-ATPase bewirkt in tierischen Zellen eineUngleichverteilung von Na+ und K+ zwischen Cyto-plasma und Zellumgebung.
Die Ca2+-ATPase halt die Konzentration von Ca2+
im Cytoplasma niedrig. Die H+/K+-ATPase in den Be-legzellen des Magens ist fur das saure Magenmilieu ver-antwortlich.
Die H+-ATPase fungiert z.B. in Pflanzenzellen alsProtonenpumpe. Zellen verwenden einen erheblichenTeil ihrer chemischen Energie in Form von ATP furdie Energetisierung des aktiven Transports.
Beim sekundaren aktiven Transport ist der Trans-port eines Na+-Ions oder eines Protons mit dem Trans-port eines organischen Molekuls gekoppelt (Cotrans-port). Voraussetzung hierfur ist ein elektrochemischesPotential dieser Ionen, das durch einen primaren akti-ven Transport aufgebaut werden kann.
Fur den Vorgang des Cotransports selbst wird keineEnergie benotigt, sondern das elektrochemische Poten-tial des Ions ausgenutzt. Auf diese Weise werden z.B.Zucker und Aminosauren in die Zelle transportiert.
Der sekundare aktive Transport besteht aus zweistrukturell getrennten Transportsystemen, einmal derNa+- oder H+-ATPase, bei dem ATP verbraucht wirdund einer katalysierten Diffusion in Gegenrichtung,in Form eines Cotransports. Die uber dieses Systemruckdiffundierenden Na+-Ionen oder Protonen neh-men gewissermaßen ein anderes Molekul mit, das an-gereichert werden kann, vorausgesetzt, die Zelle haltden aktiven primaren Transportvorgang von Ionenbzw. Protonen aufrecht (Abb. 1.40, 1.41).
Ein Beispiel fur einen sekundaren aktiven Trans-port ist die so genannte Natriumpumpe. Bei hoherenTieren ist eine treibende Kraft fur den aktiven Trans-port von Substanzen in die Zelle das aktive Aus-schleusen – Herauspumpen – von Na+ aus der Zelle.
ExtrazellulärerRaum
Cytoplasma
NichtkatalysierterTransport
Lipid-doppel-schicht
Katalysierter aktiverTransport
Abb. 1.38 Transport durch eine Biomembran. NichtkatalysierterTransport: wenig selektive, langsame Diffusion. Katalysierter ak-tiver Transport: schneller, sehr selektiver Transport bestimmterMolekule durch Vermittlung von Translokatoren
Konzentration innen
Geschwindigkeitdes Transports in
die ZelleVmax
Katalysiert
Nicht katalysiert
Abb. 1.39 Kinetik katalysierter (erleichterte Diffusion) und nicht-katalysierter (freie Diffusion) Transportprozesse. KatalysierteTransportprozesse verlaufen viel schneller als der nichtkatalysierteTransport.
1.3.5 Spezifischer Stofftransport durch Biomembranen 39
1Cytolog
ie
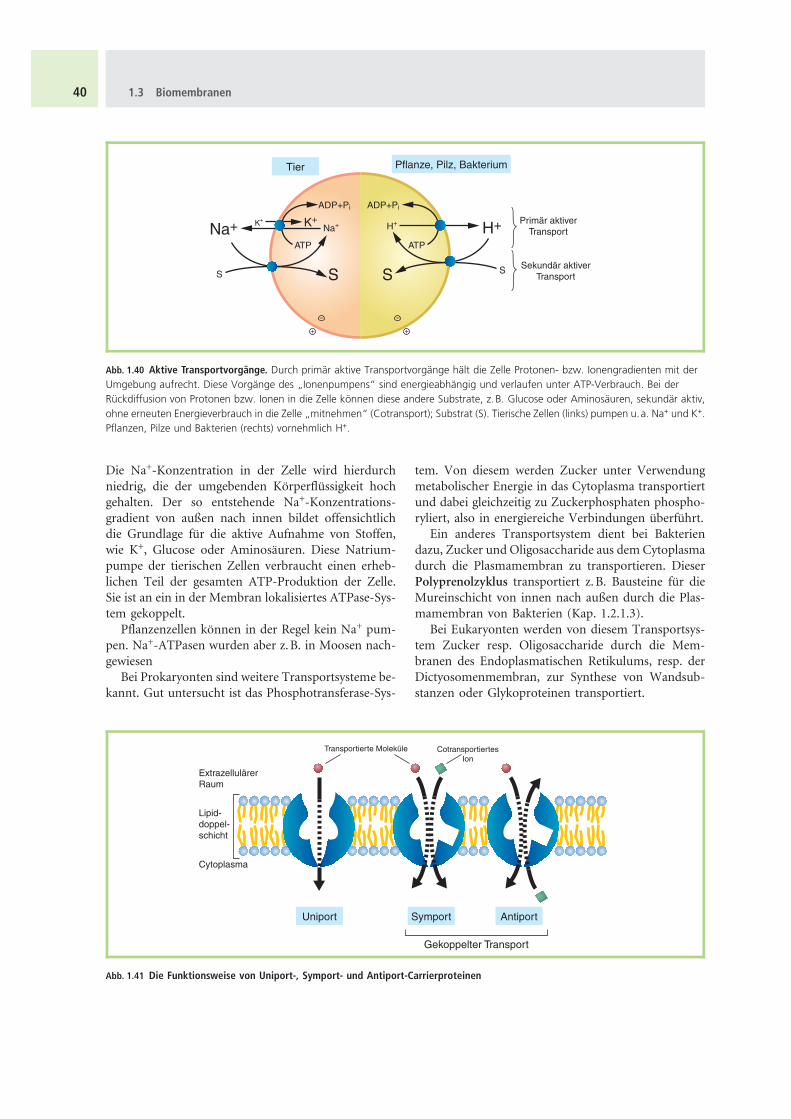
Die Na+-Konzentration in der Zelle wird hierdurchniedrig, die der umgebenden Korperflussigkeit hochgehalten. Der so entstehende Na+-Konzentrations-gradient von außen nach innen bildet offensichtlichdie Grundlage fur die aktive Aufnahme von Stoffen,wie K+, Glucose oder Aminosauren. Diese Natrium-pumpe der tierischen Zellen verbraucht einen erheb-lichen Teil der gesamten ATP-Produktion der Zelle.Sie ist an ein in der Membran lokalisiertes ATPase-Sys-tem gekoppelt.
Pflanzenzellen konnen in der Regel kein Na+ pum-pen. Na+-ATPasen wurden aber z.B. in Moosen nach-gewiesen
Bei Prokaryonten sind weitere Transportsysteme be-kannt. Gut untersucht ist das Phosphotransferase-Sys-
tem. Von diesem werden Zucker unter Verwendungmetabolischer Energie in das Cytoplasma transportiertund dabei gleichzeitig zu Zuckerphosphaten phospho-ryliert, also in energiereiche Verbindungen uberfuhrt.
Ein anderes Transportsystem dient bei Bakteriendazu, Zucker und Oligosaccharide aus dem Cytoplasmadurch die Plasmamembran zu transportieren. DieserPolyprenolzyklus transportiert z.B. Bausteine fur dieMureinschicht von innen nach außen durch die Plas-mamembran von Bakterien (Kap. 1.2.1.3).
Bei Eukaryonten werden von diesem Transportsys-tem Zucker resp. Oligosaccharide durch die Mem-branen des Endoplasmatischen Retikulums, resp. derDictyosomenmembran, zur Synthese von Wandsub-stanzen oder Glykoproteinen transportiert.
Tier Pflanze, Pilz, Bakterium
ADP+Pi
ATP
Na+K+K+
Na+
SS
+
–
+
–
S S
H+
ADP+Pi
ATP
H+ Primär aktiverTransport
Sekundär aktiverTransport
Abb. 1.40 Aktive Transportvorgange. Durch primar aktive Transportvorgange halt die Zelle Protonen- bzw. Ionengradienten mit derUmgebung aufrecht. Diese Vorgange des „Ionenpumpens“ sind energieabhangig und verlaufen unter ATP-Verbrauch. Bei derRuckdiffusion von Protonen bzw. Ionen in die Zelle konnen diese andere Substrate, z.B. Glucose oder Aminosauren, sekundar aktiv,ohne erneuten Energieverbrauch in die Zelle „mitnehmen“ (Cotransport); Substrat (S). Tierische Zellen (links) pumpen u.a. Na+ und K+.Pflanzen, Pilze und Bakterien (rechts) vornehmlich H+.
ExtrazellulärerRaum
Lipid-doppel-schicht
Cytoplasma
Transportierte Moleküle
Uniport
CotransportiertesIon
Symport Antiport
Gekoppelter Transport
Abb. 1.41 Die Funktionsweise von Uniport-, Symport- und Antiport-Carrierproteinen
1.3 Biomembranen40

Der aktive Transport ist fur den Zellstoffwechsel unentbehr-lich. Hiermit konnen Nahrstoffe aus der Umgebung spezifischaufgenommen und in der Zelle angereichert werden. DieIonenkonzentration innerhalb der Zelle oder bei Tieren auchin Korperflussigkeiten wird mit Hilfe aktiver Transportvorgan-ge reguliert.
In einigen Fallen ist die Biosynthese eines Transport-systems genetisch mit der Synthese spezifischer Enzymefur den Abbau der in die Zelle transportierten Substanzgekoppelt. Ein Beispiel dafur bietet das Transport-system fur b-Galactoside bei E. coli (Kap. 3.2.1.3 ff).
1.3.6 Signaltransduktion undInformationsverarbeitung
Die Plasmamembran muss die Beziehung der Zelle zuihrer Umgebung und zu anderen Zellen vermitteln.Plasmamembranen von Saugetierzellen enthalten Gly-koproteine und Glykolipide, deren Oligosaccharid-Sei-tenketten ausschließlich auf der Außenseite der Mem-bran lokalisiert sind (s. Abb. 1.30). Diese Glykoproteineund Glykolipide werden im Endoplasmatischen Reti-kulum und den Dictyosomen gebildet und gelangenvon den Membranen dieser Organellen durch Mem-branfluss in die Plasmamembran. Die Struktur der Oli-gosaccharidketten kann sehr vielfaltig sein. Vielen die-ser Oberflachenstrukturen kommen Rezeptorfunktio-nen zu. An sie binden z.B. Viren bei der „Adsorption“an die Zelloberflache. Zu diesen Strukturen zahlen auchdie zellstandigen Antigene. Die verschiedenen Zelltypen
eines Organismus unterscheiden sich in der Strukturund Zusammensetzung ihrer Glykocalyx und damitauch in ihren zellstandigen Antigenen. Auf solchenOberflachenstrukturen beruhen die Phanomene derZell-Zell-Erkennung. Die Bindung einer bestimmtenSubstanz kann als Signal dienen, um eine bestimmteZellfunktion zu regulieren. Gerade in der Medizinsind solche Erkennungsphanomene von weit reichen-der Bedeutung. Um die Glucose-Homoostase aufrechtzu erhalten, die durch Insulin und Glukagon gesteuertwird, mussen diese Hormone an bestimmte Rezeptorender Zielzellen binden, damit eine Zellantwort erfolgenkann.
Bei Bluttransfusionen und Organ- bzw. Gewebe-transplantationen konnen Zellen und Gewebe mit kor-perfremden Oberflachenstrukturen vom Immunsystemdes Empfangers angegriffen werden. Die Blutgruppen-antigene sind auf den Plasmamembranen der Erythro-zyten lokalisiert. Die antigenen Determinanten desAB0-Systems sind Zuckerreste (Abb. 1.42). Die wich-tigsten Rezeptoren fur die Erkennung korpereigenerZelloberflachen gehoren zum HLA-System (HumanesLymphozyten-Antigen), auch Transplantations- oderHistokompatibilitatssystem genannt. Es findet sich inden Plasmamembranen aller Zellen, mit Ausnahmeder Erythrozyten.
1.3.6.1 Signale und Signaltransduktion
Fur Zellen ist die Kommunikation mit der Umgebungvon essentieller Bedeutung, um Differenzierungspro-zesse oder Zellbewegung zu steuern. Diese Steuerungerfolgt z.B. durch Ionen, Neurotransmitter, Cytokineoder Hormone. Viele Hormone, z.B. Peptidhormone,Catecholamine wie Adrenalin und Noradrenalin undalle bekannten Neurotransmitter, konnen Plasmamem-branen nicht passieren. Sollen sie ihre Wirkung ent-falten, mussen sie an der Oberflache von Zellen von Re-zeptoren gebunden werden. Von dieser Bindung auskonnen sie in der Zelle Stoffwechselvorgange auslosen.Die Ubertragung eines Nervenimpulses z.B. geschiehtdurch Neurotransmittersubstanzen. In der postsynap-tischen Plasmamembran finden sich Rezeptoren zurBindung dieser Transmittermolekule. Durch derenBindung werden Ionenkanale in dieser Membran geoff-net. Der Impuls wird so von Zelle zu Zelle weiter ge-reicht.
Innerhalb eines vielzelligen Organismus erreichenchemische Signale ihre Ziele durch Diffusion oder Zir-kulation. Man unterscheidet (Abb. 1.43):
& Autokrine Signalstoffe. Sie beinflussen die Zellenvon denen sie produziert wurden
GlucoseGalactose
N-Acetyl-glucosamin Fucose
O-Antigen
A-AntigenN-Acetyl-
galactosamin
B-AntigenGalactose
Abb. 1.42 Determinanten des AB0-Blutgruppensystems als Bei-spiel fur einen Oberflachenrezeptor in einer Cytoplasmamembran.Erythrozyten der Blutgruppe 0 tragen nur die „Kern-Zucker“Glucose, Galactose, N-Acetylglucosamin und Fucose. Bei derBlutgruppe A kommt noch ein endstandiges N-Acetylglucosamin,bei der Blutgruppe B eine endstandige Galactose hinzu.
1.3.6 Signaltransduktion und Informationsverarbeitung 41
1Cytolog
ie
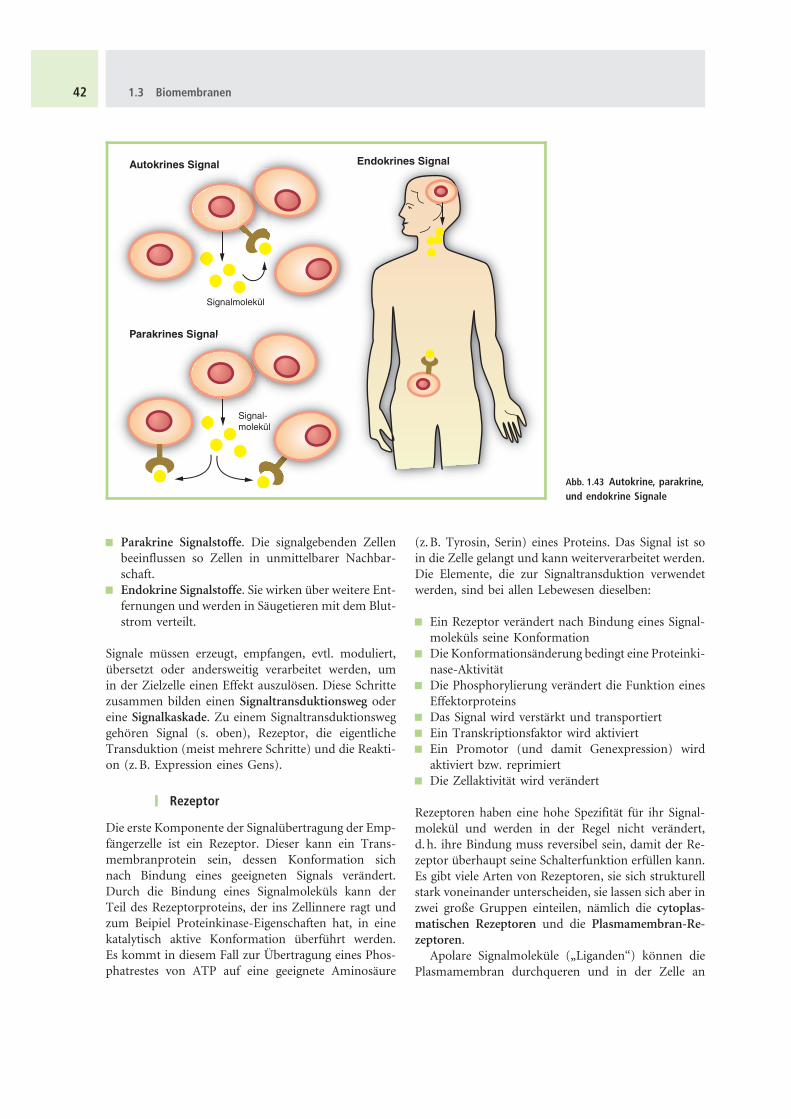
& Parakrine Signalstoffe. Die signalgebenden Zellenbeeinflussen so Zellen in unmittelbarer Nachbar-schaft.
& Endokrine Signalstoffe. Sie wirken uber weitere Ent-fernungen und werden in Saugetieren mit dem Blut-strom verteilt.
Signale mussen erzeugt, empfangen, evtl. moduliert,ubersetzt oder andersweitig verarbeitet werden, umin der Zielzelle einen Effekt auszulosen. Diese Schrittezusammen bilden einen Signaltransduktionsweg odereine Signalkaskade. Zu einem Signaltransduktionsweggehoren Signal (s. oben), Rezeptor, die eigentlicheTransduktion (meist mehrere Schritte) und die Reakti-on (z.B. Expression eines Gens).
Rezeptor
Die erste Komponente der Signalubertragung der Emp-fangerzelle ist ein Rezeptor. Dieser kann ein Trans-membranprotein sein, dessen Konformation sichnach Bindung eines geeigneten Signals verandert.Durch die Bindung eines Signalmolekuls kann derTeil des Rezeptorproteins, der ins Zellinnere ragt undzum Beipiel Proteinkinase-Eigenschaften hat, in einekatalytisch aktive Konformation uberfuhrt werden.Es kommt in diesem Fall zur Ubertragung eines Phos-phatrestes von ATP auf eine geeignete Aminosaure
(z.B. Tyrosin, Serin) eines Proteins. Das Signal ist soin die Zelle gelangt und kann weiterverarbeitet werden.Die Elemente, die zur Signaltransduktion verwendetwerden, sind bei allen Lebewesen dieselben:
& Ein Rezeptor verandert nach Bindung eines Signal-molekuls seine Konformation
& Die Konformationsanderung bedingt eine Proteinki-nase-Aktivitat
& Die Phosphorylierung verandert die Funktion einesEffektorproteins
& Das Signal wird verstarkt und transportiert& Ein Transkriptionsfaktor wird aktiviert& Ein Promotor (und damit Genexpression) wird
aktiviert bzw. reprimiert& Die Zellaktivitat wird verandert
Rezeptoren haben eine hohe Spezifitat fur ihr Signal-molekul und werden in der Regel nicht verandert,d. h. ihre Bindung muss reversibel sein, damit der Re-zeptor uberhaupt seine Schalterfunktion erfullen kann.Es gibt viele Arten von Rezeptoren, sie sich strukturellstark voneinander unterscheiden, sie lassen sich aber inzwei große Gruppen einteilen, namlich die cytoplas-matischen Rezeptoren und die Plasmamembran-Re-zeptoren.
Apolare Signalmolekule („Liganden“) konnen diePlasmamembran durchqueren und in der Zelle an
Abb. 1.43 Autokrine, parakrine,und endokrine Signale
Autokrines Signal
Signalmolekül
Parakrines Signal
Signal-molekül
es Signal
Signaal-külal
Endokrines Signal
1.3 Biomembranen42
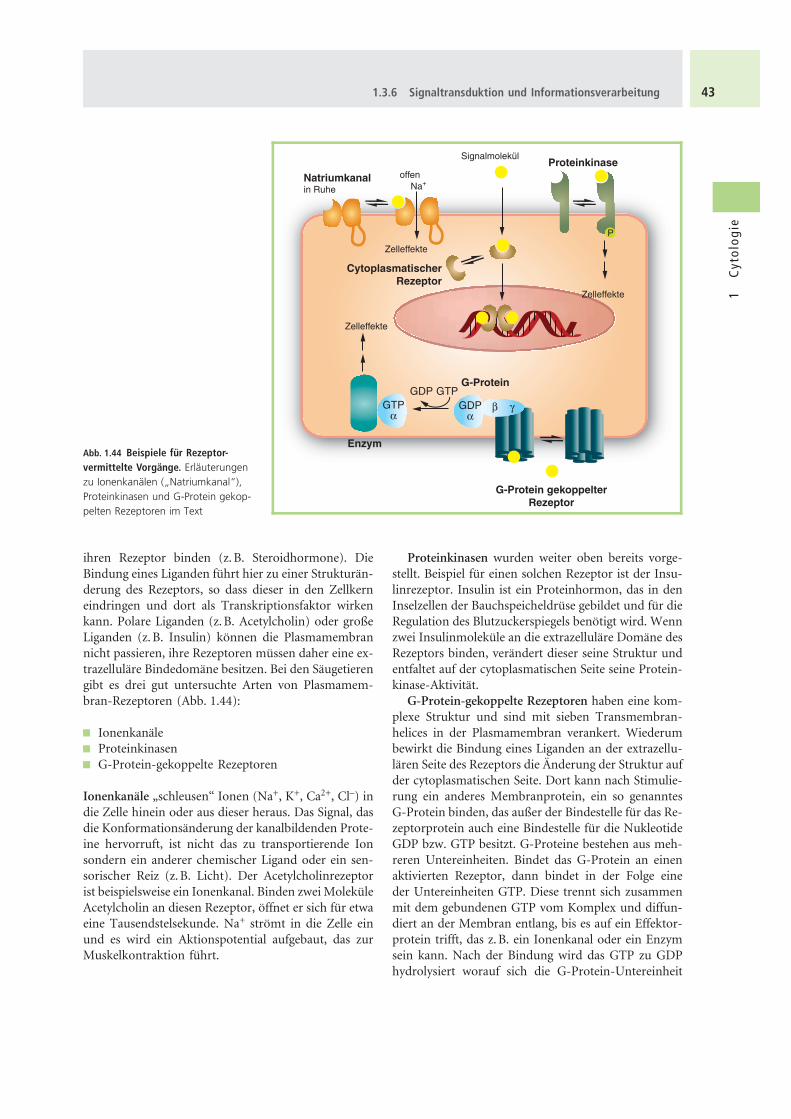
ihren Rezeptor binden (z.B. Steroidhormone). DieBindung eines Liganden fuhrt hier zu einer Strukturan-derung des Rezeptors, so dass dieser in den Zellkerneindringen und dort als Transkriptionsfaktor wirkenkann. Polare Liganden (z.B. Acetylcholin) oder großeLiganden (z.B. Insulin) konnen die Plasmamembrannicht passieren, ihre Rezeptoren mussen daher eine ex-trazellulare Bindedomane besitzen. Bei den Saugetierengibt es drei gut untersuchte Arten von Plasmamem-bran-Rezeptoren (Abb. 1.44):
& Ionenkanale& Proteinkinasen& G-Protein-gekoppelte Rezeptoren
Ionenkanale „schleusen“ Ionen (Na+, K+, Ca2+, Cl–) indie Zelle hinein oder aus dieser heraus. Das Signal, dasdie Konformationsanderung der kanalbildenden Prote-ine hervorruft, ist nicht das zu transportierende Ionsondern ein anderer chemischer Ligand oder ein sen-sorischer Reiz (z.B. Licht). Der Acetylcholinrezeptorist beispielsweise ein Ionenkanal. Binden zwei MolekuleAcetylcholin an diesen Rezeptor, offnet er sich fur etwaeine Tausendstelsekunde. Na+ stromt in die Zelle einund es wird ein Aktionspotential aufgebaut, das zurMuskelkontraktion fuhrt.
Proteinkinasen wurden weiter oben bereits vorge-stellt. Beispiel fur einen solchen Rezeptor ist der Insu-linrezeptor. Insulin ist ein Proteinhormon, das in denInselzellen der Bauchspeicheldruse gebildet und fur dieRegulation des Blutzuckerspiegels benotigt wird. Wennzwei Insulinmolekule an die extrazellulare Domane desRezeptors binden, verandert dieser seine Struktur undentfaltet auf der cytoplasmatischen Seite seine Protein-kinase-Aktivitat.
G-Protein-gekoppelte Rezeptoren haben eine kom-plexe Struktur und sind mit sieben Transmembran-helices in der Plasmamembran verankert. Wiederumbewirkt die Bindung eines Liganden an der extrazellu-laren Seite des Rezeptors die Anderung der Struktur aufder cytoplasmatischen Seite. Dort kann nach Stimulie-rung ein anderes Membranprotein, ein so genanntesG-Protein binden, das außer der Bindestelle fur das Re-zeptorprotein auch eine Bindestelle fur die NukleotideGDP bzw. GTP besitzt. G-Proteine bestehen aus meh-reren Untereinheiten. Bindet das G-Protein an einenaktivierten Rezeptor, dann bindet in der Folge eineder Untereinheiten GTP. Diese trennt sich zusammenmit dem gebundenen GTP vom Komplex und diffun-diert an der Membran entlang, bis es auf ein Effektor-protein trifft, das z.B. ein Ionenkanal oder ein Enzymsein kann. Nach der Bindung wird das GTP zu GDPhydrolysiert worauf sich die G-Protein-Untereinheit
Abb. 1.44 Beispiele fur Rezeptor-vermittelte Vorgange. Erlauterungenzu Ionenkanalen („Natriumkanal“),Proteinkinasen und G-Protein gekop-pelten Rezeptoren im Text
Signalmolekül
Natriumkanalin Ruhe
offenNa+
P
Proteinkinase
G-Protein gekoppelterRezeptor
GDP
G-Protein
αβ γ
GTPGDPGTP
α
Enzym
Zelleffekte
Zelleffekte
Zelleffekte
CytoplasmatischerRezeptor
1.3.6 Signaltransduktion und Informationsverarbeitung 43
1Cytolog
ie

vom Effektorprotein lost. Diese Untereinheit muss mitden anderen G-Protein-Untereinheiten einen Komplexbilden, um dann wieder an den G-Protein-gekoppeltenRezeptor binden zu konnen. Kommt die Bindung zu-stande, wird das noch anhaftende GDP gegen GTP aus-getauscht und das neue Signal kann verarbeitet werden.G-Proteine konnen ihre Effektoren entweder aktivieren(Adrenalin-Wirkung am Herzen uber die Bildung vonzyklischem Adenosinmonophosphat, s. unten) oderhemmen (Adrenalin-Wirkung an der glatten Muskula-tur fuhrt zur Muskelentspannung).
Signaltransduktion
Das vom Rezeptor empfangene Signal lost eine Kaskadevon biochemischen Reaktionen aus, wodurch das Sig-nal verstarkt und weiter getragen werden kann. DasSignal kann entweder direkt oder indirekt transduziertwerden. Die direkte Signaltransduktion ist eine Funk-tion des Rezeptors selbst, die indirekte Signaltransduk-tion benotigt zusatzliche Molekule, die das Signal in dieTiefe der Zelle ubermitteln konnen. Man bezeichnetsolche Botenstoffe als „second messenger“.
Reine Proteinkinasekaskaden benotigen keinensecond messenger. In diesem Fall wird eine Folgevon Proteinen der Reihe nach durch Kinasen phospho-ryliert und damit aktiviert. Eukaryontische Genomecodieren fur hunderte oder tausende von Kinasen.Nicht alle Kinasen sind in allen Zellen oder Gewebengleichzeitig aktiv. Am Ende einer Proteinkinasekaskadesteht immer ein Protein, das nach Aktivierung in den
Zellkern eindringt und dort die Transkription beein-flussen kann. Proteinkinasekaskaden kann man folgen-dermaßen charakterisieren:
& Bei jedem Schritt der Kaskade wird das Signal ver-starkt
& Die Vielzahl der Schritte erlaubt Spezifitat und Va-riation
Sekundare Botenstoffe
Sekundare Botenstoffe (second messenger) sind Ver-bindungen, die im Cytoplasma freigesetzt werden,nachdem ein erster Botenstoff, namlich das Signalmo-lekul, an seinen Rezeptor gebunden hat. Auch ubersekundare Botenstoffe kann ein Eingangssignal ver-starkt werden.
Zyklisches Adenosinmonophosphat (cAMP)(Abb. 1.45) ist ein universeller sekundarer Botenstoff.Es wird von einer Adenylat-Cyclase aus ATP gebildet.Das Enzym ist an der cytoplasmatischen Seite der Plas-mamembran lokalisiert. In der Regel wird es durchG-Proteine (siehe oben) aktiviert. cAMP kann als Co-faktor oder allosterischer Regulator von Zielenzymendienen. Zu diesen zahlen z.B. Ionenkanalproteineoder Proteinkinasen. So kann direkte und indirekteSignaltransduktion kombiniert und feinreguliert wer-den.
Andere wichtige sekundare Botenstoffe sind Dia-cylglycerol (DAG) und Inositoltrisphosphat (IP3)(Abb. 1.45), die beide nach Hydrolyse eines Membran-
N
NN
N
NH2
O
OHO
OP
O O–
zyklischesAdenosinmonophosphat
(cAMP)
OHOHHO OPO32–
OPO32–2–3OPO
Inositol-1,4,5-trisphosphat(IP3)
H3C O
O
O
OHO
H3C
Diacylglycerid(DAG)
NH
N
N
O
NH2NO
OHO
OP
O O–
zyklischesGuanosinmonophosphat
(cGMP)
Abb. 1.45 Beispiele fur sekundare Botenstoffe (Second Messenger)
1.3 Biomembranen44

lipids (Phosphatidylinositol-4,5-bisphosphat, PIP2)durch Phospholipasen – ein wichtiger Vertreter istdie Phospholipase C – entstehen. Die Initiation derSignaltransduktionskaskade uber diese sekundarenBotenstoffe kann wiederum uber G-Proteine (s. oben)erfolgen. Nach der Hydrolyse verbleibt das lipophileDAG in der Plasmamembran, wo es die ProteinkinaseC (PKC), ein membranstandiges Enzym, aktiviert. DiePKC ist Ca2+-abhangig. Ca2+-Ionen konnen selbst se-kundare Botenstoffe sein (s. unten). Jetzt kommt IP3,das zweite Produkt der Hydrolyse ins Spiel: Es diffun-diert zum glatten Endoplasmatischen Retikulum undmoduliert dort einen Ionenkanal so, dass Ca2+ ins Cy-toplasma entlassen wird. Dies fuhrt zusammen mitDAG zur Aktivierung der PKC, die dann verschiedeneProteine phosphorylieren kann.
Die Konzentration von Ca2+ im Cytosol ist in der Re-gel sehr gering. Transportproteine pumpen dieses Ionaus der Zelle hinaus oder in das Endoplasmatische Re-tikulum hinein. Dazu wird Energie in Form von ATPverbraucht. Die intrazelluare Ca2+-Konzentration wirddurch das Offnen und Schließen von Kanalen zusammenmit den membranstandigen Ionenpumpen reguliert.
Stickstoffmonoxid (NO) ist ein gasformiger sekun-darer Botenstoff, der in lebenden Geweben nur eineHalbwertszeit von funf Sekunden besitzt. NO wird inEndothelzellen durch die NO-Synthase aus Arginingebildet. Dieses Enzym ist durch Ca2+ aktivierbar. Inder glatten Muskulatur aktiviert NO die Guanylat-Cy-clase und zyklisches Guanosinmonophosphat (cGMP)wird gebildet. Diese Reaktionsfolge fuhrt schließlich zurMuskelerschlaffung, Blutgefaße werden erweitert, derBlutzufluss zum Herzen und in den Beckenbereichwird verstarkt. Dieser Mechanismus erklart die Wirk-samkeit von Nitroglyzerin bei coronaren Durchblu-tungsstorungen (Angina pectoris) und Aktivatorender NO-Synthese bei Erektionsstorungen.
Viele rezeptorvermittelte Signale werden uber so genanntesekundare Botenstoffe (second messenger) verstarkt undweitergegeben. Zu diesen Stoffen zahlen zyklisches Adenosin-monophosphat (cAMP), zyklisches Guanosinmonophosphat(cGMP), Diacylglycerol und Inositoltrisphosphat, Ca2+ undStickstoffmonoxid.
1.3.7 Plasmamembran der Bakterien
Einziges Biomembransystem der Bakterienzelle istdie Plasmamembran. Wie die Plasmamembran derZellen von Pflanzen und Tieren ist die der Bakterieneine Lipoproteinmembran. In ihrer chemischen Zu-sammensetzung weicht sie aber von entsprechenden
Membranen der Eukaryonten deutlich ab. Wie beider Eukaryontenzelle dient sie jedoch als osmotischeBarriere und Regulationsorganell des Stofftransportes.Sie regelt den Stoffaustausch der Bakterienzelle mit derUmgebung und besitzt Strukturen und Enzyme, die denpassiven und aktiven Stoffaustausch der Zelle mit derUmgebung ermoglichen und regulieren. Ein Erloschendieser Membranfunktion bedeutet den Zelltod. Anti-biotika, die diese Funktionen der Plasmamembran sto-ren, wirken daher primar bakterizid, d.h. zelltotendauch auf ruhende Keime. Zu solchen Antibiotika geho-ren z.B. Polymyxin, Colistin und Tyrothricin. Da derBau der Cytoplasmamembran der Zellen des Menschendemjenigen der Plasmamembran der Bakterien ahnelt,wirken solche Antibiotika nicht spezifisch. Sie sind da-her nur bei strenger Indikationsstellung, vornehmlichlokal anwendbar und zeigen starke Nebenwirkungen.
Bei den Prokaryonten hat die Plasmamembran da-ruber hinaus noch weitere Aufgaben als Ort wichtigerStoffwechselfunktionen (Tab. 1.15). Sie ist Sitz desElektronentransportsystems und zumindest einigerEnzyme des Citratzyklus. Weiterhin ist die Plasma-membran hier ein Ort aktiver Proteinsynthese; anihr sind Ribosomen assoziiert. Auch bei der Biosyn-these der Zellwand und der Kapselkomponenten spieltsie eine Rolle.
Bakterien besitzen keine Mitochondrien. DerenFunktion als Trager der Enzyme der Endoxidationwird von der Plasmamembran ubernommen.
Lamellenformige Einfaltungen der Plasmamembrantragen Photosynthesepigmente. Solche Thylakoide sindbei manchen photoautotrophen Bakterien ausgebildet.Sie entsprechen funktionell den Thylakoiden der Chlo-roplasten der Pflanzen.
1.3.8 Andere Aufgaben vonMembranen
Bisher wurden die Hauptfunktionen von Membranendargestellt: Kompartimentierung, Transport, Zell-Zell-Erkennung sowie Signalempfanger und -umsetzer.
Tab. 1.15 Funktionen der prokaryontischen Cytoplasmamembran
Diffusionsbarriere
Aktiver Transport
Proteinbiosynthese (+Ribosomen)
Energiestoffwechsel (evtl. Mesosomen)
Photosynthese (evtl. Thylakoide)
Angriffsort von Antibiotika
1.3.8 Andere Aufgaben von Membranen 45
1Cytolog
ie

Weitere wichtige Aufgaben erfullen sie bei der Infor-mationsverarbeitung, der Energieumwandlung undder Kopplung chemischer Reaktionen.
Membranen dienen als strukturelle Basis fur eineganze Reihe von Enzymen, die nur in Bindung anMembranen aktiv sind. Beispiele hierfur bieten dieMembranen der Mitochondrien und Chloroplasten.Membrangebundene Enzyme katalysieren hier sehrwichtige Stoffwechselvorgange, namlich Elektronen-transport (s. Kap. 4.5.7.1), oxidative Phosphorylierung(s. Kap. 4.5.7.3) und Photosynthese (s. Kap. 4.6.1).
Biosynthesen und andere Zellleistungen erforderneine definierte Abfolge biochemischer Reaktionen.Diese werden in der Regel durch Enzyme katalysiert,die jeweils nur eine chemische Reaktion durchfuhrenkonnen. Zwar gibt es auch sehr effiziente multifunktio-nelle Enzyme und Enzymkomplexe, die Spezialisierunghat aber auch ihre Vorteile, z.B. weil die monofunktio-nellen Enzyme vielfaltiger eingesetzt werden konnen.Da meist das Produkt einer bestimmten Enzymreaktiondas Substrat fur ein weiteres Enzym liefert, wurde eineBildung der Zielprodukte nur sehr langsam und unge-richtet verlaufen, wenn man nicht annimmt, dass dieEnzymkomponenten eines bestimmten Biosynthesewe-ges in einer geeigneten Reihenfolge z.B. an Membranengebunden vorliegen. Dadurch konnen in einem so ge-nannten Metabolite Channelling die Zwischenpro-dukte wie an einem Fließband von Enzym zu Enzymweiter gereicht werden; die Reaktionen laufen soschneller und effizienter ab.
*** Zusammenfassung
Biomembranen bestehen aus Lipiden und Proteinen. Sie ent-halten Aminoalkohole, Aminosauren, Fettsauren, Sterole undZucker. Sie gliedern die Zellen von Eukaryonten in zahlreicheReaktionsraume, Kompartimente. Biomembranen haben eineflussige Mosaikstruktur. Grundstruktur ist ein doppelter Lipid-film, in den Proteine eingelagert oder angelagert sind. Mem-branproteine verleihen der jeweiligen Membran ihre spezifi-sche Funktion. Sie vermitteln z.B. die Transportvorgangedurch die Membran. Andere sind Rezeptoren fur Hormoneoder Neurotransmitter. InMembranen sind zahlreiche Enzymeeingelagert.Biomembranen dienen als Diffusionsbarrieren. Sie sind semi-permeabel. Die Semipermeabilitat von Biomembranen ist dieGrundlage fur alle osmotischen Vorgange. Lipophile Stoffedagegen konnen sich durch die Lipidschicht hindurchlosen.Pflanzliche Zellen entwickeln immer einen hohen osmotischenDruck. Sie benotigen daher eine feste Zellwand. Osmotischwirksame Substanzen sind Ionen und polare Nichtelektrolyte,z.B. Zucker, Alkaloide. Makromolekule sind osmotisch un-wirksam. Die Lipidschichten von Biomembranen sind fur diemeisten wasserloslichen Molekule und Ionen undurchlassig.
Zum Transport derartiger Molekule dienen zahlreiche spezifi-
sche Transportproteine, die in die Biomembran integriert sind.
Es sind dies Carrier- und Kanalproteine.
Carrierproteine binden niedermolekulare Stoffe und transpor-tieren sie durch Konformationsanderung durch die Biomem-bran.
Dieser Transport kann ohne Energieaufwand als katalysierte,resp. erleichterte Diffusion entlang eines Konzentrationsgra-dienten erfolgen. Andere Carrier-vermittelte Transportvor-gange verlaufen aktiv unter Energieaufwand, meist ubereine Hydrolyse von ATP, und konnen das gebundene Molekulgegen einen Konzentrationsgradienten transportieren.
Kanalproteine bilden wassergefullte Poren, welche die Lipid-doppelmembran durchdringen. Sie ermoglichen anorgani-schen Ionen entsprechend ihrem Konzentrationsgradientenden Durchtritt durch die Biomembran. Solche Ionenkanale off-nen sich gewohnlich nur als Antwort auf spezifische Reize, diedie Membran treffen.
Durch Ein- bzw. Ausstulpungen und Bildung von Transportva-kuolen sind Biomembranen an den Vorgangen der Endozytoseund Exozytose beteiligt. Der Großenbereich der so transpor-tierten Partikel reicht von Kolloiden bis zu Bakterien, ein-schließlich Zellen und Zellbestandteilen.
Innerhalb der Zelle kann ein Austausch von Membranteilenzwischen verschiedenen Membranen stattfinden. An diesenVorgangen des Membranflusses sind die Plasmamembran,die Tonoplastenmembran, das Endoplasmatische Retikulum,die Dictyosomen, Lysosomen und andere Vakuolenmembra-nen beteiligt. Nicht am Membranfluss beteiligt sind die Mem-branen der Mitochondrien und Plastiden.
Biomembranen vermitteln Erregungsleitung und Erregungs-ubertragung.
Die direkte Kommunikation zwischen Zellen erfolgt uber GapJunctions (tierische Zellen) oder Plasmodesmen (pflanzlicheZellen). Desmosomen heften tierische Zellen fest aneinander,hemmen aber nicht den Stoffdurchtritt. Tight Junctions ver-hindern den Durchtritt von Molekulen durch den Interzellular-raum.
Zellen empfangen Signale von der Umwelt und anderenZellen. Die Signalubertragung umfasst drei Schritte: Aufnah-me des Signals (Rezeptor), die Ubertragung des Signals (Si-gnaltransduktion) und die zellulare Reaktion. Rezeptoren kon-nen Ionenkanale, Proteinkinasen und G-Protein-gekoppelteRezeptoren sein. Signaltransduktion kann direkt oder indirektuber sekundare Botenstoffe erfolgen.Wichtige sekundare Bo-tenstoffe sind cAMP, cGMP, Ca2+, NO, DAG und IP3.
1.3 Biomembranen46