Embed Size (px)
Citation preview

Histochemie 2, 435--441 (1962)
Aus dem Pathologischen Institut der Universit~t Tiibingen (Direktor: Prof. Dr. reed. E. LETTERER)
H I S T O C t I E M I S C H E R N A C H W E I S VON C H O L I N D E H Y D I ~ 0 G E N A S E I N T I E R I S C H E N G E W E B E N
Von
PETER GLOGNER* und WOLFGA~G G6SSNEn
Mit 3 Textabbildungen
(Eingegangen am 10. Januar 1962)
Cholinma.ngeldi~t ff ihrt bei verschiedenen Tie ra r ten zur Verfe t tung der Leber, diese wiederum zu diffuser F ib rose bzw. Cirrhose (BEST und HV~TSMAN 1934, CgAIKOFF und CONDOR 1940). Zum S tud ium der Pa thogenese dieser Leberver- ~nderungen erscheint es wfinsehenswert , Enzyme nachzuweisen, die ]i~r den Sto//weehsel des Cholins, einem essentiel len Nahrungs fak to r mi t Vi tamin- eharak te r , ve ran twor t l i ch sind.
Seit der E n t d e c k u n g der Chol inoxydat ion in t ier isehen Organen durch BEn~- HEI.~ und BEn~HEIM (1933), haben sieh zahlreiehe Unte r sucher um die Auf- kl / trung dieses Meeh~nismus bemiiht . Die Ergebnisse seien kurz zusammen- gefa[tt.
Cholin wird in einem ersten Stoffwechselschritt dureh Leber und Nierengewebe verschie- dener Tier~rten zu BetMnMdehyd oxydiert. Das diese Reaktion katalysierende Enzym Cholindehydrogenase (Cholinoxydase) ist an Mitochondrien gebunden und benStigt wahr- scheinlich keine Cofzktoren, m6glicherweise jedoch Mg++-Ionen. Seine Empfindlichkeit gegeniiber Fluorid und Cyanid ist gering. Als SH-Enzym ist es durch p-Chloromerkuri- benzoa$ hemmbar. ~rber den. Einflul~ yon Monojodacet~t sehw~nken die Angaben. Weitere Hemmstoffe sind Amine, Stiekstofflost, .~thylenimine und Aureomycin. In einem weiteren Reaktionssehritt wird entst~ndener BetMnMdehyd zu Betain oxydiert. Diese Dehydrogenase ist ebenfalls ein Mitochondrienenzym, benStigt jedoeh DPN Ms Cofaktor (lVIANX und QVASTEL 1937, KENSLER und L~GEMA~ 1951, WILLI~'vIS, LITWAC~: und ELVE~JE~ 1951, KLEL~ und H~NDL~ 1942, ROTHSCHILD und Bt~ON 1954, ROTHSCHILD, COt~I und B~nRON 1954, BARRON, BARTLETT und MILLER 1948, MILLER 1954, ZIMS~ERMANN und HUMOLLER 1953).
DUBNOFF (1949) wies auf die Bedeutung der Cholinoxydation hin: Zur Synthese der essentiellen Aminosaure Methionin kSnnen a.naerob nur die Methylgruppen des Betain, nicht aber die des Cholin verwertet werden.
I n der folgenden Arbe i t wird eine Methode zum histochemisehen Nachweis der Cholindehydrogenase mitgete i l t . Die Ver te i lung des Enzyms in t ier ischen 0 r g a n e n und seine Eigenschaf ten werden d iskut ie r t .
Bere i ts CH~N~ und H o ~ (1960) e rwahnen die h is tochemisehe Dars te l lung der Chol indehydrogenase ohne genaue Angabe der Methodik.
Material und Methoden a) Schnittherstellung. Als Versuchstiere dienten 150--200 g schwere weibliche Albino-
ratten und Albinomi~use einer h~useigenen Zucht. Die Tiere wurden durch Dekul0itieren get6tet und die Gewebsstiicke sofort nach der Ent-
nahme auf einem Gefriertisch aufgefroren. Schnitte yon 10# Dicke wurden bei - -20 o C im Kryosta~en hergestellt. Die Schnitte wurden auf Objekttr~ger aufgenommen, angetaut und
* Der Deutschen Forschungsgemeinschaft d~nke ich ~iir die Gew~hrung eines Stipendiums.

436 P E ~ GLOGNER undWoLFGANG G6SS~E~:
entweder sofort, oder naeh Lafttrocknung weiterverarbeitet. Vor der Inkubation wurden die Sehnitte im a llgemeinen 5 rain in kalte 0,85%ige NaC1-LSsung eingestellt.
b) Inkubationsmedium. Das optimale Inkubationsmedium hat folgende Zusammen- setzung:
Cholinehlorid (100 mg/ml) . . . . . 0,3 ml 0,2 m Phosphatpuffer pH 7,5 . . . . 2,1 m] Neutrales Nitro-BT (5,0 mg/ml) . . . 0,3 ml Aqua dest . . . . . . . . . . . . . 0,3 ml
Die InkubationslSsung wird auf die Schnitte aufgetropft und diese werden in einer feuchten Kammer bei 370 C 45--60 min inkubiert. Naeh der Inkubation Abspfilen in physiologiseher NaCI-LSsung und Nachfixieren in 4%igem Formalin-Calcium (nach BAKEI~) 10 rain. Danach Eindecken in Karion-Gelatine oder nach Entw~sserung in der Alkoholreihe in Caedax.
Die Versuehe, dureh welehe dieses Inkubationsmedium bestimmt wurde, umfal~ten Variation der Konzentration yon Substrat, Puffer und Tetrazoliumsalz, Verwendung ver- sehiedener Puffer (Tris, Phosphat) und Elektronenakzeptoren (Nitro-BT, MTT, Tetrazolblau und Tetrazolpurpur).
An Cofaktoren wurden DPN und TPN (0,5 mg/ml GesamtlSsung) sowie CoClo, MgC] 2 und MnC12 (m/1000--m/200) getestet.
Als zus~tzliche prim~re Elektronenakzeptoren wurden Menadion (W~TTENBEaG und LEONG ]960) in versehiedenen Konzentrationen sowie Methylenblau (0,15 mg/ml Gesamt- 15sung naeh FARBER und LOUVIERE 1956) verwendet.
c) Sehnitt]ixierung. Zur Anwendung kamen 4%iges Formalin-Calcium (naeh BAKER), Aeeton und -~thano]. Die am Objekttr~ger aufgezogenen feuchten Kryostatsehnit~e wurden mit den Fixierungsmitteln 5 min bei Eissehranktemperatur (3--5 °C) behandelt.
d) Hemmsto]]e. Hemmstoffversuehe wurden mit Natriumeyanid, Monojodessigs~ure (JES), Natriumfluorid, Queeksilber(II)ehlorid, Stiekstofflost (Mitomen®) und Aureomycin durehgefiihrt.
Folgende Versuehsanordnungen wurden vorgenommen: 1. Vorinkubation der Schnitte mit gepufferter HemmstofflSsung 30 rain, Auswasehen in
physiologiseher NaC1-LSsung 10 rain; danaeh Einstellen in die Inkubationsl5sung. 2. Zusatz des Hemmstoffes zur InkubationslSsung. 3. Kombination von 1. und 2. Da bei den Versuehen mit HgC12 in Gegenwart yon Nitro-BT floekige gelbliehe Nieder-
sehl~ge auftraten, wurde dabei nur wie unter 1. besehrieben vorgegangen. Zusatzlieh wurde der Einflul~ yon Cystein (m/100) auf die Queeksilberehloridhemmung gepr[ift.
Ergebnisse Kontrollen. Nach Erh i tzen der Schni t te auf 100 ° C, sowie nach I n k u b a t i o n in
das S t a n d a r d m e d i u m ohne Subs t r a t ]ieB sich keine l~e~ktion beobachten . Zus~tz yon m/100 Semicarbaz id zur Inkuba t ions]Ssung zum Abfangen en t s t andenen Be ta ina ldehyds ergab p rak t i sch keine ~ n d e r u n g der Reakt ionss t£rke .
Elelctronenalczeptoren. Ncben dem von NAC~LAS u. Mitarb . (1957) in die His to- chemie eingefi ihr ten Ni t ro -BT (2 ,2 ' -d i -p-n i t rophenyl 5 ,5 ' -d i -phenyl 3,3'-(3,3'-di- methoxy-4 ,4 ' -b iphenylen) -d i te t razo l iumchlor id ) wurden das yon PEARSE (1957) zuers t verwende te MTT[3-(4 ,5-dimethyl th iazolyl -2) 2 ,5-d iphenyl te t razol ium-bro- mid] , Te t r azo lpu rpu r [2 ,2 ' - (p-diphenylen)-bis - (3 ,5-diphenyl te t razol iumchlor id)] und Te t razo lb lau (Methoxyder iva t des Te t razo lpurpur ) vergle ichend untersucht .
MTT und Te t r azo lpu rpur wurden un te r den gew~hlten Bedingungen kaum, Te t razo lb lau n ich t reduzier t .
Durch Einff ihren zus~tzl icher E lek t ronenakzep to ren (Menadion, Methylen- blau) in das S t a n d a r d m e d i u m ergab sich, dal~ Menadion keinen EinfluB auf die F o r m a z a n b i l d u n g hut te , w/~hrend in Gegenwar t yon Methy lenb lau keiner le i Formazanniedersch l~ge beobach te t wurden. Es ist ~nzunehmen, dab dieser Redox-

Nachweis von Cholindebydrogenase 437
farbstoff infolge seines gegenfiber Nitro-BT giinstigeren gedoxpotentials ( + 0,011/ - -0 ,05 V) bevorzugt reduziert und anter aerol0en Bedingungen autoxycliert wird. Untersuchungen im anaeroben Milieu wurden nicht vorgenommen.
Pu//er uncl pmAbhitngigkeit . Bei vergleiehenden Untersuehungen ergab sich bei Verwendung yon Tris- an Stelle yon Phosphatpuffer stets eine schw~ehere Enzymreaktion.
Tabclle 1. p~-Optimum der Cholindehydrogenase im histochemischen Nachweis
Reaktionsstgrke.. + + + + + + + + + + +
Das pH-Optimum lag im histochemischen Test zwischen pK 7,3 und 7.8 (Tabelle 1).
Co/aktoren. Ein zus~tzlicher Bedarf an Cofaktoren konnte im histochemischen Test nicht gezeigt werden. Mit DPN, TPN sowie COC12, MgC12 und MnC12 ergab sich keine Verst~trkung der Reaktion. Jedoch kann ein durch die Vorbehandlung nicht entfernter Rest m6glicher Cofaktoren ange- nommen werden.
Fixierung. Durch verschiedene Fixierungsflfis- sigkeiten wird die Enzymaktiviti~t gehemmt, wie aus Tabelle 2 ersichtlich ist.
Hemmsto//e. Entsprechend den Untersuehungen yon BARRON (1948) mit Stickstofflost und von ZIM- MERMAN und HUMOLLEg (1953) mit Aureomycin, konnten wir auch beim histochemischen Naehweis eine deutliche Hemmwirkung dieser Substanzen beobachten.
Tabelle 2. Ein/lufl yon Fixie- rungs/li~ssig/ceiten an/die A lctivi-
t~it der Cholindehydrogenase
Re a k t ions - L6sung stf irke
Kontrolle . . . . 4% Formalin-Ca Aeeton . . . . . Athanol . . . .
+ + + + 0 0
Die erforderlichen Hemmstoffkonzentrationen lagen im Falle des Stickstofflost wesentlich h5her als im empfindlicheren bioehemischen Test. In (Jbereinstimmung mit Befunden anderer Autoren lieB sich auch die Unempfindliehkeit der Cholin- hydrogenase gegeniiber Cyanid und Jod- essigsaure zeigen (Tabelle 3).
Die grSBte Hemmwirkung wird durch Hg++-Ionen erreicht. Sehon Konzentra- tionen yon 10-4m verhindern die Cho- linoxydation im histochemischen Test vollstgndig. Offenbar handelt es sich hierbei um eine irreversible Blockierung reaktionsf~higer SH-Gruppen am Enzym- protein: Eine ttemmwirkung tritt ngm- lich nicht auf, wenn Cystein gleichzeitig dcr gepufferten HgC12-L6sung zugesetzt wird. Eine einmal eingetretene Hem- mung durch Hg++-Ionen kann dagegen durch Cystein nicht mehr aufgehoben werden (Tabelle 4).
Histocbemie. Bd. 2
Tabelle 3. Ein/lufl verschiedener Hemm- stof/e an/die Aktivit~it der Cholindehydro-
genase
H e m m s t o f f Konzen- t r a t i on
l:lem~nung in To*
NaCN . . . . . 10 -2 m 0
JES . . . . . . 10 -2 m 0
NaF . . . . . . 10 -2 m 0
HgC12 . . . . . l0 -8 m 100 10 -4 m 100 1 0 - 5 m 0
Stickstofflost . . 10 2 m 100 10 -2 m 50--75
Aureomycin . . . 10 -2 m 75 10 -8 m 25--50
* Geschatzt auf Grund der Farb- intensit~t.
,SOb
438 PETER GLOGNEI~ u n d WOLFGANO G6SSNEI~:
Tabelle 4. Hemmung der Cholindehydro- genase durch Hg++-Ionen
l~eaktions - Zus~tze st~rke
Keine . . . . . . . . . . .
I0 -~ In I-IgCl 2 . . . . . . . .
l0 -4 In HgCl~ + l0 -2 in Cystein
gleiehzeitig . . . . . . . .
successive . . . . . . . . .
+ + + 0
+ + + 0
Enzymlo]calisation in verschiedenen Geweben. Naeh Inkubation im Standard- medium tritt an Orten yon Enzymakti- vit/~t naeh ]0 min die fiir das reduzierte
Tabelle 5. Cholindehydrogenase in Organen yon Ratle und Maus
Organ Barge Maus
N i e r e . . . . . Leber . . . .
Pankreas . . . G1. subinandib. G1. subl ing . . . Magen . . . . Duodenum . . D iekda r in . . . I]brige 0rgane
+ + + + + + + +
bis + + + +
0 ( + ) b i s +
0
0 0 0
(@) sehwaeh bzw. inkonstant.
+ + + + + + bis + +
0 bis @ (+) bis +
(+) (+) (+) (+)
0
Nitro-BT typische Blaufarbung mi~ teils diffuser, tefls feink6rniger Formazan- ablagerung auf. Entsprechend den bisherigen Befunden biochemischer Unter-
suchungen lieB sich nur in Niere und Leber die Cholindehydrogenase in starkerer Aktivit~tt nachweisen (Tabelle 5).
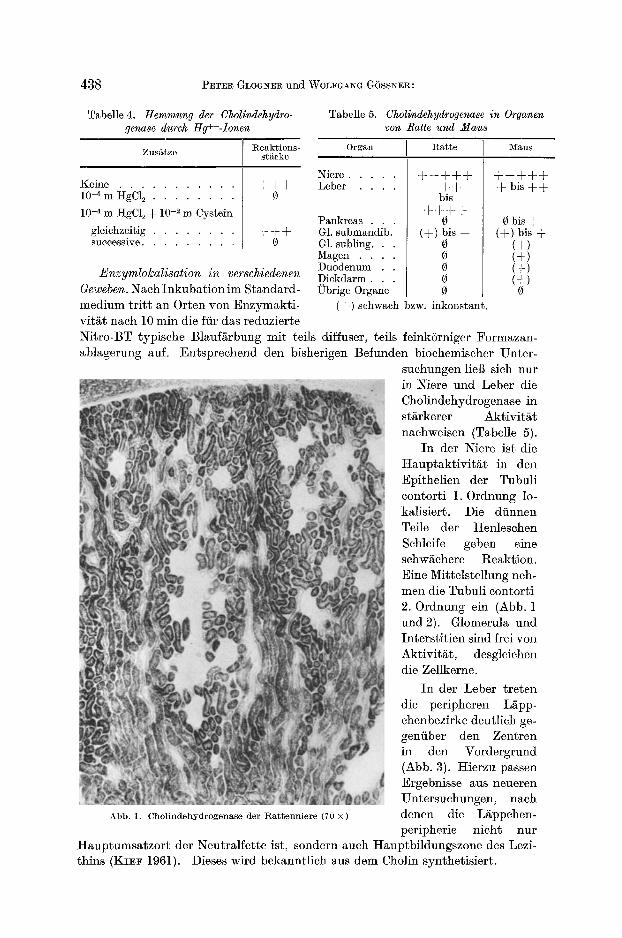
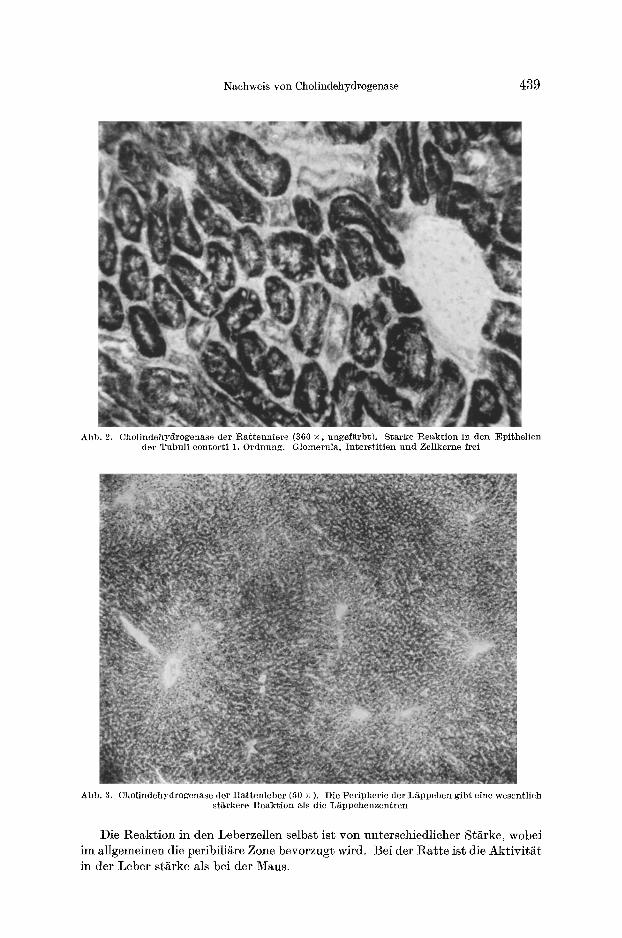
In der Niere ist die Hauptaktivitat in den Epithelien der Tubuli contorti 1. Ordnung lo- kalisier~. Die dfinnen Teile der ttenleschen Sehleife geben eine sehw~ehere Reaktion. Eine Mittelstellung neh- men die Tubuli eontorti 2. Ordnung ein (Abb. 1 und 2). Glomerula und Interstitien sind frei yon Aktivit~t, desgleiehen die Zellkerne.
In der Leber treten die peripheren Lapp- ehenbezirke deutlieh ge- geniiber den Zentren in den Vordergrund (Abb. 3). Itierzu passen Ergebnisse aus neueren Untersuehungen, naeh
Abb. 1. Chol indehydrogenase der t t a t t enn ie re (70 x ) denen die L&ppehen- peripherie nicht nur
Hauptumsatzort der Neutralfette ist, sondern auch Hauptbildungszone des Lezi- thins (KIEF 1961). Dieses wird bekanntlich aus dem Cholin synthetisiert.
Nachweis yon Cholindehydrogenase 439
Abb. 2. Cholindehydrogenase der Ra t tenn ie re (360 ×, ungef~rbt) . Sta~'ke Reak t ion in den Epithelien der Tubul i contor t i 1. Ordnung. Glomerula, In te rs t i t i en und Zellkerne irei
Abb. 3. Chol indehydrogenase der Ra t t en lebe r (50 × ). Die Per ipher ie der L~ppchen gibt eine wesentl ich st~rkere Reak t ion als die L~ppchenzentren
Die Reaktion in den Leberzellen selbst ist yon unterschiedlicher St/~rke, wobei im allgemeinen die peribili/~re Zone bevorzugt wird. Bei der Ratte ist die Aktivit~t in der Leber st~rke als bei der Maus.

440 PETER GLOONER undWoLFGANG G6SSN~R:
Eine schwache, aber konstante Reaktion geben die Drfisenendstficke und Gangepithelien der Glandu]a submandibularis. Im exkretorischen Parenchym des Pankreas und der Schleimhaut yon Magen, Duodenum und Dickdarm der Maus finder sich ebenfalls eine schwache Aktiviti~t. Neben den in der Tabelle aufgeffihrten Organen wurde yon uns untersucht: Milz, Gro]hirn, Kleinhirn, Thymus, Nebenniere, Hypophyse, Ovar, glatte und quergestreifte Muskulatur und Lymphknoten.
Diskussion Unsere im histoehemischen Test erhobenen Befunde fiber Eigenschaften und
Organlokalisation des Enzyms stimmen weitgehend mit denen biochcmischer Untersuchungen fiberein. Die quantitativen Unterschiede in Hemmstoff- und Substratkonzentrationen dfirften mit den besonderen Gegebenheiten des Enzym- nachweises im Gewebsschnitt zusammenhi~ngen. Es ist anzunehmen, daI~ in unserem System nur die Oxydation yon Cholin zu Betainaldehyd errant wird. Zuni~chst liegt das pmOptimum der BetMnaldehyddehydrogenase, die in einem zweiten Reaktionsschritt BetainMdehyd zu Betain oxydiert, ira alkalischen Be- reich (zwischen pH 8,8 und 9,2) (ROTttSCHILD, CORI und BAnno~ 1951). Zum anderen konnte in orientierenden Versuchen, bei denen Semicarbazid zum Abfangen des Betainaldehyd der Inkubationsl6sung zugeffigt war, keine signifikante Ab- schwi~chung der Reaktion beobachtet werden. Ware der Anteil der BetMnMde- hyddehydrogenase in unserem System groB, mfiBte eine solehe erwartet werden. Zudem ist erfahrungsgem~l~ beim histochemisehen Nachweis DPN-abh~ngiger Dehydrogenasen stets ein DPN-Zusatz zur Inkubationsl6sung erforderlich.
Bemerkenswert ist die Verteilung der Cholindehydrogenase im Si~ugetier- organismus, vor allem im Vergleieh mit anderen histochemisch nachweisbaren Dehydrogenasen. Sie ist in deutlieh fM~barer Form nur in Leber und Niere vor- handen. In allen anderen untersuchten Organen findet man nur eine ganz geringe bzw. keine Aktivitat. Es ist jedoch m6glich, dM~ eine etwa vorhandene Aktivitat unterhalb der Erfahrungsgrenze der histochemisehen Methode liegt.
Auffallend ist die unterschiedlieh starke Aktivit~t in versehiedenen Absehnitten des Nephron und in verschiedenen Zonen des Leberl~ppchens. In diesem Zu- sammenhang scheint der st£rkere EnzymgehMt in der Peripherie des Leber- l~ppchens bemerkenswert, wie sie bereits ffir andere Enzyme festgestellt wurde (G6ssN]~ ]958, NOVlKOFF 1959, KITZlNG und SCHUMACH]~R 1961).
Die Erfassung der Muster verschiedener Enzyme innerhalb des Leberli~ppehens ergibt einen neuen Aspekt ffir die Beurteflung yon Stoffwechselvorgi~ngen und -st6rungen in der Leber.
lJber die Zuordnung der histoehemisch erfaBbaren Cholindehydrogenase zu bestimmten intrazelluliiren Strukturen m6chten wir auf Grund der bisher vor- liegenden Befunde noch keine Stellung nehmen, vor allem im Hinbliek auf die in letzter Zeit yon verschiedener Seite festgestellten Schwierigkeiten bei der Inter- pretation intrazellul~rer Formazanablagerungen.
Zusammenfassung Eine Methode zum histochemischen Nachweis der Cholindehydrogen~se wird
mitgeteilt. In diesem System werden keine zus£tzlichen Cofaktoren ben6tigt. Das Enzym wird durch Hg++-Ionen, Stickstofflost und Aureomycin gehemmt.

Nachweis yon Cholindehydrogenase 441
Es ist resistent gegenfiber Cyanid, Fluor id und Jodessigs~ure. Sein p~-Op t imum liegt zwischen p~ 7,3 und 7,8.
I m wesentl ichen ist das E n z y m auf Leber und Niere der un te r such ten Nager beschr~nkt.
L i t e r a t u r
BARI~ON, E. S. G., G. L. BARTLETT and Z. BAKER MILLER." The effect of nitrogen mustard on enzymes and tissue metabolism. J. exp. Med. 87, 489--501 (1948).
BERNHEI~, F., and 2K. C. L. BE~N~EIM: Oxydation of Aeetylcholine by tissues. Amer. J. Physiol. 104, 4 3 8 ~ 4 0 (1933).
B~ST, C. It., and M. E. ttUNTS~AN: The effect of choline on the liver fat of rats in various states of nutrition. J. Physiol. (Lond.) 83, 255--274 (1934).
CI~AIKOFE, I. L., and C. L. CONNOR: Production of cirrhosis of the liver of the normal dog by high-fat diet. Proe. Soc. exp. Biol. (N.Y.) 43, 638--641 (1940).
CHANG, J .P. , and S. K. HoRI: 1 st Internat. Congr. Histoehem. Cytoehem., Paris, 1960. DUENOEF, J. W. : The role of eholinoxidase in labilizing choline methyl. Arch. Bioehem. 24,
251--262 (1949). FARBE~, E., and C. D. LOUVIEEE: Histochemieal loeMisation of specific oxidative enzymes.
IV. Soluble oxidation-reduction dyes as aids in the histochemieal locMisation of oxidative enzymes with tetrazolium salts. J. Histochem. Cytoehem. 4, 347--356 (1956).
G/)SSNER, W. : Histoehemischer Nachweis hydrolytiseher Enzyme mit Hilfe der Azofarbstoff- methode. Histochemie l, 48--96 (1958).
KENSLEt~, C. J., and H. LANGEMANN: The distribution of choline oxidase activity in rat liver. J. biol. Chem. 192, 551--559 (1951).
KIEF, It. : Habilitationsschrift. Tiibingen 1961. KITZING, W., u. H. H. SCHU~AC~ER: Ontogenese und topographische Histoehemie der Bern-
steinsi~uredehydrogenase in der Lebcr einiger Nager. Z. Zellforsch. 54, 443--467 (1961). KLEIN, R.~ and P. HANDLER: Effects of diphosphopyridine nueleotide on the rate of oxidation
of betMne aldehyde. J. biol. Chem. 144, 537--539 (1942). MANN, P. C. G., and J. It. QUASTEL: The oxydution of choline by rat liver. Biochem. J.
31, 869--878 (1937). MILLE~, Z. : Influence of the ethyleniminc structure on the oxidation of choline, formaldehyde
and sarcosine. Arch. Biochem. 47, 137--147 (1953). NAC~LAS, M. M., K. C. Tsov, E. ])E SOVZA, C. S. CHE~G and A. M. S~LmMAN: CytochemicM
demonstration of succinic dehydrogenase by the use of p-nitrophenyl substituted ditetra- zole. J. tIistoehem. Cytochem. 5, 420--436 (1957).
NOVIt~oFF, A. B. : Cell heterogenity within the hepatic lobnle of the rat (staining reactions). J. Histoehem. Cytochem. 7, 240--244 (1959).
PEARSE, A . G. E . : Intracellular locMisation of dehydrogenase systems using monotetrazolium salts and metal chelation of their formazans. J. histochem. Cytochem. 5, 515--527 (1957).
ROTHSCmLD, H.A., and E. S. G. BARRON: The oxidation of betMne aldehyde by betaine Mdehyde dehydrogenase. J. biol. Chem. 209, 511--523 (1954).
- - O. CoRI and E. S. G. BARRON : The components of choline oxidase and aerobic phosphory- lation coupled with choline oxidation. J. biol. Chem. 208, 41--53 (1954).
WATT]gNBEI%G, L. W., and J. L. LEONG: Effects of coenzyme Q10 and Menadione on succinic dehydrogenase activity as measured by tetrazolium salt reduction. J. tIistochem. Cyto- chem. 8, 296--303 (1960).
WILLIAMS, J. N. jr., G. LITWACK and C. A. ELV]~HJE~: Studies on rat liver choline oxidase: An assay method. J. biol. Chem. 192, 73--80 (1951).
ZIMMERMAN~, I-I. J., and 1~. L. HUMOLL~R: Effect of aureomycin on choline oxidase and other enzymes of rat liver. Amer. j . Physiol. 175, 468--472 (1953).
Dr. P. GLOGNER und Priv.-Doz. Dr. W. G6SSN~R, Pathologisches Institut der Universitiit,
Tiibingen, Liebermeisterstr. 8





























