Embed Size (px)
Citation preview

This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution4.0 International License.
Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschungin Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung derWissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht:Creative Commons Namensnennung 4.0 Lizenz.
1 3 0 2 E. STEUDLE UND U.ZIMMERMANN
leukämie haben einen mehr malignen Verlauf als andere Myelom-Formen 5 4 ' 5 6 . Fälle mit Liehen myx-ödematosus verlaufen in der Regel gutartig.
Herrn Prof. H. E. BOCK möchte ich für die mir ge-währte Unterstützung bei der Durchführung der Arbeiten, Herrn Prof. W . SEYFFERT für statistische Konsultation und den Herrn Drs. HESS und HILSCH-MANN für die strukturelle Analyse unseres Bence-Jones-Proteins St danken, die unsere (antigenische) Klassifizierung desselben in die V;i-Gruppe bestätigte. Wir sind außerdem Drs. N. HILSCHMANN, C. MILSTEIN und F . W . PUTMAN für die Gabe komplett sequenzier-
56 W. PRUZANSKI and I. ROTHER, Canad. med. Assoc. J. 102, 1061 [1970].
ter Bence-Jones-Proteine bzw. A-Ketten der Prototyp-Sequenzen (Proteine New und Ha), VAIII (Bau, Kern und X), V n v (Vil und Bo) und V^v (Sh) dank-bar. Partiell sequenzierte Bence-Jones-Proteine bzw. A-Ketten der Prototypsequenzen V;u (HB]7, HB] 11, Daw) und der Prototypsequenz VAU (HBJ 15) erhielten wir dankenswerterweise von Drs. L. HOOD und E. M. PRESS, andere Bence-Jones-Proteine von Drs. C. BAGLIONI, D. BERGSAGEL, H. G. VAN EYK, H. H. FUDENBERG, J. HARTJE, F. HECKNER, G. MEYER- HOF-MANN, H. OHTA, W . PRUZANSKI und A. RUFFILLI. Für die Übermittlung der Daten der partiellen Amino-säurensequenz der A-Kette Daw (Ph. D. thesis London 1 9 7 0 — Nancy M . HOGG) danken wir Dr. E. M. PRESS. Diese Arbeit wurde zum Teil durch die Deutsche For-schungsgemeinschaft unterstützt.
Hydraulische Leitfähigkeit von Valonia utricularis Hydraulic Conductivity of Valonia utricularis
E. STEUDLE und U . ZIMMERMANN
Institut für Physikalische Chemie der Kernforschungsanlage Jülich GmbH
(Z. Naturforsch. 26 b, 1302—1311 [1971] ; eingegangen am 19. August 1971)
A method is described for the simultaneous determination of rapid changes of the cell turgor pressure (hydrostatic pressure) in algal cells (cell size must be at least 3 mm in diameter), and of the net volume flow across the cell membrane arising after a change of the cell turgor pressure or of the osmotic pressure in the outside medium. On the basis of the equations of irreversible thermodynamics it is possible to calculate the hydraulic conductivity of the cell membrane from these measurements, as it is theoretically shown.
The hydraulic conductivities of the marine alga Valonia utricularis determined in two indepen-dent ways (by osmotic and hydrostatic experiments) are equal. For exosmosis, Lpex (hydrostatic) and Lpex (osmotic) amounted to (9,6 ±1 ,0 ) '10~7 and (9,8 ±1 ,9 ) '10"7 respectively cm • sec"1 • atm-1, and for endomosis, Lpen (hydrostatic) was (9,4 ± 1,1)-10"7 cm-sec"1-atm"1.
A polarity in the water movement across the cell membranes as discussed in the literature could not be found for Valonia utricularis.
Der Volumenfluß, der bei einzelligen Algen zwi-schen Vakuole und Außenmedium als Folge einer hydrostatischen und/oder osmotischen Druckdiffe-renz im nichtstationären Zustand zu beobachten ist, wird nach den linearen Ansätzen der Thermodyna-mik irreversibler Prozesse durch Gl. (1 ) beschrie-ben:
Jv = = LP [Apz (t) - An-, (t) - os • Ajis (t) ] (1)
wobei den einzelnen Symbolen folgende Bedeutung zukomment:
Jy = das in der Zeiteinheit durch 1 cm2 der Membranoberfläche F hindurchtretende Volumen in cm3 .
Sonderdrudeanforderungen an Dr. U. ZIMMERMANN und Dipl.-Chem. E. STEUDLE, Institut für Physikalische Che-mie, KFA Jülich, D-5170 Jülich, Postfach 365.
^ P z W = P 1 ( 0 ~ P ° ( 0 = hydrostatische Drudedifferenz zwischen dem Zellinnern und Außenmedium.
Aji[(t) =jT i 1 — = die entsprechende osmotische Druckdifferenz der imper-meablen Teilchen.
Ang (2) = Jig1 — = die entsprechende osmotische Druckdifferenz der permeablen Teilchen.
(Die Indizes im Exponenten i und o beziehen sich auf das Zellinnere bzw. das Außenmedium.)
L p = Drudefiltrationskoeffizient oder hydraulische Leitfähig-keit ( c m - s e c - 1 - a t m - 1 ) ,
a s = Reflexionskoeffizient.

HYDRAULISCHE LEITFÄHIGKEIT VON VALONIA UTRICULARIS 1 3 0 3
Die Integration der Gl. ( 1 ) kann durchgeführt werden, wenn die Funktionen Apz = g(t), An\ — h(t) und A-a = k{t) bekannt sind. Aus der inte-grierten Gleichung lassen sich L p und os bestimmen, falls die Funktion V = f(t) experimentell ermittelt werden kann. Sind andererseits Lv und os aus ande-ren Messungen bekannt, so läßt sich das zu bestimm-ten Zeiten geflossene Volumen V, das in erster Nähe-rung dem Wasservolumen Vw entspricht, berech-nen.
Bei der Bestimmung der hydraulischen Leitfähig-keit und der Reflexionskoeffizienten an Pflanzen-zellen muß berücksichtigt werden, daß im Gegensatz zu tierischen Zellen der Wasser- und Teilchentrans-port durch mehrere, in Serie geschaltete Barrieren (Zellwand, Plasmalemma, Protoplasma und Tono-
plast) erfolgt. Daher lassen sich abgesehen von sehr wenigen Ausnahmen, bei denen die Eigenschaften der einzelnen Barrieren getrennt ermittelt werden konnten 1 , nur Lp- und o-Werte für die gesamte Barriere messen.
Der Bestimmung von Reflexionskoeffizienten an Pflanzenzellen ist bisher nur wenig Bedeutung bei-gemessen worden. Für L p -Werte liegt dagegen eine wesentlich größere Zahl von Messungen vor. Die meisten Methoden sind aber — abgesehen von sehr
wenigen Verfahren 2> 3 — mit großen systematischen Fehlern behaftet. Für eine kritische Diskussion die-ser Meßverfahren sei auf WILKINS 4 verwiesen.
Im folgenden beschreiben wir eine Methode, die rasch und genau Messungen von Lp-Koeffizienten an Pflanzenzellen mit einem Mindestdurchmesser von 3 m m zuläßt. Das Verfahren beruht auf einer Messung der hydrostatischen Druckänderung in der Zelle als Folge eines Volumenflusses bzw. auf einer direkten Messung dieses Volumenflusses mit einer schon an anderer Stelle beschriebenen 5 , modifizier-ten Druckmeßapparatur. Diese Apparatur wurde be-reits zur Ermittlung von Reflexionskoeffizienten ver-schiedener Nichtelektrolyte an Valonia utricularis nach einer zero-flow-Methode verwandt6 .
E x p e r i m e n t e l l e r Tei l
Der Aufbau der Volumenfluß- bzw. Druckmeßappa-tur ist der Abb. 1 zu entnehmen.
Die Apparatur besteht aus einer Plexiglaskammer vom Volumen 0,1 cm3, in die eine Glaskapillare (Spitzendurchmesser von 60 p) und ein Metallstab (Durchmesser 0,5 mm) über Dichtungen eingeführt werden. Der Stab kann zur Veränderung des Kammer-volumens mit einer Mikrometerschraube bewegt wer-den, auf der 1/ioomm abgelesen werden können. In die
Voltmeter
Abb. 1. Druck- und Volumenflußmeßapparatur.
1 J. DAINTY U. B. Z. GINZBURG, Biochim. biophysica Acta [ A m s t e r d a m ] 7 9 , 1 1 2 [ 1 9 6 4 ] .
2 J. DAINTY U. B. Z. GINZBURG, Biochim. biophysica Acta [Amsterdam] 79, 102 [1964 b].
3 J. GUTKNECHT, Biochim. biophysica Acta [Amsterdam] 1 6 3 , 2 0 [ 1 9 6 8 ] .
4 M. B. WILKINS, The physiology of plant growth and deve-lopment, p. 421 ff., MacGraw-Hill, London 1969.
5 U . ZIMMERMANN, H . RAEDE U. E. STEUDLE, Naturwissen-schaften 5 6 , 6 3 4 [ 1 9 6 9 ] .
6 U . ZIMMERMANN U. E . STEUDLE, Z . Naturforsch. 2 5 b , 5 0 0 [ 1 9 7 0 ] .

1 3 0 4 E. STEUDLE UND U.ZIMMERMANN
Kammer, die einschließlich der Glaskapillare mit Sili-konöl gefüllt ist, ist ein Miniaturdruckwandler CPS 125 — 2 0 0 der Firma Kulite Semiconductor Pro-ducts, Ridgefield (New Jersey/USA) eingebaut, der den Druck in der Kammer proportional in elektrische Span-nung umformen kann. Die Funktionsweise des Druck-wandlers wurde bereits an anderer Stelle5 beschrie-ben. Die elektrische Spannung wird auf ein Voltmeter übertragen, an dem nach Eichung mit bekannten Druk-ken der Druck in der Kammer direkt abgelesen wer-den kann.
Bei schnellen Druckänderungen in der Zelle, wie sie nach Störung des stationären Zustandes auftreten, wird die elektrische Spannung auf einem mit dem Voltmeter verbundenen Schreiber aufgezeichnet. Zur Druck-messung wird die bis zur Spitze mit Öl gefüllte Kapil-lare mit Hilfe eines Mikromanipulators unter dem Mikroskop in die Zelle eingeführt. Beim Einstechen der Glaskapillare wird die Zellmembran kurzzeitig verletzt. Infolge des Verlustes an Zellwasser sinkt der hydro-statische Druck in der Vakuole zunächst ab, erreicht aber in den meisten Fällen nach 10 Min. wieder den ursprünglichen stationären Wert. Der Druck, der sich in der Kammer des Druckmeßgerätes aufbaut, ent-spricht dem hydrostatischen Drude in der Vakuole, da das gesamte System starr und das ö l in guter Nähe-rung inkompressibel ist.
Mit Hilfe des Metallstabes läßt sich durch Verschie-ben der Grenzschicht Öl/Wasser das Zellvolumen ver-größern oder verkleinern. Die Volumenänderung ergibt sich aus dem Vorschub des Metallstabes. Gleichzeitig kann die mit der Volumenänderung verbundene Druck-änderung gemessen werden. Die Verwendung eines Metallstabes zur Veränderung des Kammervolumens der Druckmeßapparatur an Stelle der bei früheren Messun-gen von Reflexionskoeffizienten eingesetzten elastischen Kunststoffmembran bringt außerdem den Vorteil, daß ein kontinuierliches Nachregulieren der Grenzschicht Öl/Zellsaft an der Kapillarspitze bei Druckänderungen in der Zelle unterbleiben kann.
Prinzipiell gibt es zwei Möglichkeiten, mit dieser Ap-paratur L p zu bestimmen:
Das erste Verfahren läuft auf eine direkte Bestim-mung der Funktion Vw = f(t) hinaus:
Nach Einstellung eines stationären hydrostatischen Druckes wird mit Hilfe des Meßstabes der Drude in der Kammer leicht erhöht, so daß sich an der Kapilla-renspitze ein Öltröpfchen bildet. Die Größe dieses Öl-tröpfchens, das bis zu einem Durchmesser von ca. 0,1 mm an der Kapillarspitze haften bleibt, hängt vom Vorschub des Meßstabes ab. Da durch diesen zusätz-lichen Druck das Gleichgewicht zwischen hydrostatischer und osmotischer Druckdifferenz gestört wird, tritt ein Volumenfluß aus der Zelle auf, der zu einer entspre-chenden Änderung des Zellvolumens führen würde. Hält man aber den Druck in der Zelle und damit das Zellvolumen durch kontinuierliches Nachregulieren kon-
7 U . Z I M M E R M A N N U. E . S T E U D L E , i n V o r b e r e i t u n g .
stant, so berechnet sich das zu bestimmten Zeiten ge-flossene Volumen aus dem Vorschub und dem Durdi-messer des Meßstabes.
Die Integration der Gl. (1) ist in diesem Fall be-sonders einfach, da Apz(t) = konstant ist7 . Experi-mentell können aber Sdiwierigkeiten auftreten:
Geringste Undichtigkeiten in der Apparatur stören die Messungen und können größere Werte für Lp vor-täuschen.
Eine weitere Fehlerquelle, die man zunächst vermu-ten sollte und die auch bei dem zweiten Verfahren auftreten kann, läßt sich dagegen ausschließen:
Innerhalb eines Flüssigkeitströpfchens erzeugt die Oberflächenspannung, die die Grenzfläche zu verklei-nern sucht, einen ins Innere gerichteten Druck, den so-genannten Kapillardrude PK , der um so größer ist je kleiner der Radius r des Tröpfchens ist8 . Bei einer Grenzflächenspannung zwischen Öl und Wasser von etwa 50 dyn/cm und einem Durchmesser des Öltröpf-chens von r 3 0 / 4 berechnet sich der maximal mög-liche Kapillardruck PK zu 3 - 1 0 - 2 atm. Dieser Wert liegt in der Größenordnung der Meßgenauigkeit der Druckmeßapparatur und ist demnach zu vernachlässi-gen. Übereinstimmend mit diesem Ergebnis konnte auch bei Kontrollversuchen in Seewasser ein derartiger Kapillardruck mit der Druckmeßapparatur nicht nach-gewiesen werden.
Nach dem zweiten Verfahren, über das in dieser Arbeit berichtet werden soll, wird Lp aus einer Druck-messung ermittelt:
Nach Störung des stationären Zustandes durch Än-derung der Osmolalität des Außenmediums oder durch Änderung des hydrostatischen Druckes in der Zelle wird der hydrostatische Druck in der Zelle als Funk-tion der Zeit gemessen.
Der Durchmesser des Öltröpfchens an der Kapilla-renspitze wird hierbei nach der Störung des stationä-ren Zustands mit Hilfe eines Okularmikrometers kon-stant gehalten. Eventuelle Undiditigkeiten in der Appa-ratur gehen nicht in die Messung ein, da die Größe des Öltröpfchens mit Hilfe der Mikrometerschraube nachreguliert werden kann.
Aus der gemessenen Funktion /fpz = g(i) lassen sich der Volumenfluß und die hydraulische Leitfähigkeit be-rechnen, wenn das Zellvolumen im untersuchten Druck-bereich eine eindeutige Funktion des hydrostatischen Druckes im Zellinnern ist. Die Funktion Vz = l(Apz) muß experimentell bestimmt werden.
Im folgenden soll zum Verständnis der durchgeführ-ten Experimente die Berechnung der hydraulischen Leitfähigkeit aus der zeitlichen Änderung des hydro-statischen Drudees in der Zelle nach Störung des sta-tionären Gleichgewichtes gezeigt werden.
Eine vollständige Abhandlung über die Beziehungen zwischen Zellvolumen, hydrostatischem Druck und Volu-menfluß einschließlich der Bestimmung der Lp - und ö-Koeffizienten, wird in Kürze publiziert 7 .
8 G. KORTÜM, Einführung in die Thermodynamik, Verlag Chemie, Weinheim 1960.

HYDRAULISCHE LEITFÄHIGKEIT VON VALONIA UTRICULARIS 1305
Theoretischer Teil
Die Versuchsbedingungen seien der Einfachheit halber zunächst so gewählt, daß = 0 und
und na°(t) =^s(jv=o) sind. na ( jv=o) ent-spricht einer Konzentration cs° des betreffenden permeablen Stoffes im Außenmedium, bei der kein Volumenfluß auftritt. Diese Konzentration kann z. B. der Abb. 3 der bereits mehrfach erwähnten Publi-kation 6 entnommen werden.
Unter diesen Bedingungen ( / v = 0) muß nach Gl. (1)
A po — JijA — os-7r?(j,=o) (2)
sein, wobei JZJA der osmotische Anfangsdruck der impermeablen Teilchen in der Zelle ist und Ap0 der hydrostatischen Druckdifferenz im stationären Zu-stand entspricht. Gl. (2) besagt nichts anderes, als daß dann die osmotische Druckdifferenz gleich der hydrostatischen Druckdifferenz sein muß.
Bei Störung dieses stationären Zustandes durch Erhöhung des Druckes in der Zelle in der oben be-schriebenen Weise (Ausbildung eines Öltröpfchens an der Kapillarspitze) müssen zum Zeitpunkt t = 0 folgende Gleichungen gelten:
VZA = VA + AVA (3) und
ApA = Ap0 + Ap(t = 0), (4)
wobei Vza de m Anfangsvolumen der Zelle ent-spricht, das sich aus dem Anfangsvolumen der Wasserphase in der Vakuole V\ und der Volumen-vergrößerung der Zelle AV\, die dem Volumen des Öltröpfchens entspricht, zusammensetzt.
Entsprechend ist Ap\ die Anfangsdruckdifferenz zwischen Zelle und Außenmedium und Ap der zu-sätzlich zum hydrostatischen Gleichgewichtsdruck durch das Öltröpfchen erzeugte Druck in der Zelle.
Die zeitliche Änderung des osmotischen Druckes der impermeablen Teilchen die infolge des einsetzenden Wasserflusses aus der Zelle auftritt, kann durch Gl. (5) beschrieben werden
Tl i
n?(t) Va
(1 -f Vw/VA) (I + VW/VA).
Setzt man Gl. (2) und Gl. (6) in Gl. (1) ein, so er-hält man nach entsprechender Umformung:
m + "t"vL»'F Vw = Lp• F[Apz(t) - Apo]. (7)
Nach PHILIP 9 besteht zwischen dem Zellvolu-men und dem Druck in der Zelle folgender Zusam-menhang :
Apz(t) -APo=e(Vz(t)/VA-l).
bzw., da Apz(t) =Ap0 + Ap(t)
sein muß (Abb. 2 ) ,
Ap(t)=e(Vz(t)/VA-l) sowie
d(Jpz) dt
d(Ap) dt
s V A
dVz dt
(8)
( 9 )
(10)
(11)
£ ist eine Konstante, die die Eigenschaft eines Volumenelastizitätsmoduls hat. Sie muß experimen-tell ermittelt werden (s. w. u.) . Gl. (8 ) , die sich zur Gleichung von PHILIP nur in der Wahl des Be-zugssystems unterscheidet7, gilt unter der Bedin-gung, daß sich die Zelle wie ein klassisches Osmo-meter verhält, d. h. daß Volumen und Elastizitäts-eigenschaften des Protoplasten vernachlässigt wer-den können und nur die Zellwand elastische Eigen-schaften besitzt.
Diese Voraussetzung wurde auch stillschweigend bei der Gl. (5) bzw. Gl. (6) zugrundegelegt.
Aus Gl. (7) und Gl. (11) folgt, wenn man berück-sichtigt, daß
dVz dFw dt dt >
sein muß,
VA d (Apz) ffrU-Lp-F + tt —Vw = Lv-F[Apz(t) -Ap0]. dt Va
( 5 ) ^ ( 0 = 1 y j ^ - R - T .
Die Gleichung geht, falls V\ Vw angenommen werden darf, in Gl. (6) über
nrR-T
(6)
9 J. R. PHILIP, Plant Physiol. 33, 264 [1958].
(13)
Zum Zeitpunkt t = 0 ist Vw = 0. Gl. (13) geht des-halb in Gl. (14) über:
/ d(Apz) \ S-Lp-F [ dt L o ( ^ P A - ^ P O ) . ( 1 4 )
Andererseits wurde bereits in früheren Unter-suchungen bei Störung des stationären Gleichgewich-tes zwischen Vakuole und Außenmedium durch Er-niedrigung der Osmolalität des Außenmediums ex-perimentell gefunden6, daß sich der Druck in der Zelle mit der Zeit exponentiell ändert. Da sich unter den hier diskutierten Versuchsbedingungen (Er-höhung des Druckes in der Zelle durch ein Öltröpf-chen) diese Gesetzmäßigkeit ebenfalls bestätigte

1306 E. STEUDLE UND U.ZIMMERMANN
Abb. 2. Zeitliche Änderung des Zellturgors (Apz) einer Valoniazelle (Zelle Nr. 13) nach einer Erhöhung bzw. Erniedrigung des Zellturgors (Versuch I und II) bzw. nach einer Störung des stationären Zustandes durch Änderung des osmotischen Druk-kes des Außenmediums (Versuch III und IV). I) Lpex (hydrostatisch) = 9 ,8 - IO - 7 , II) Lpen (hydrostatisch) = 12 ,8 -10 - 7 ,
III) Lpex (osmotisch) = 14 ,4 - IO - 2 und IV) Lpen (osmotisch) = 1 4 , 5 - 1 0 - 7 cm-sec _ 1 -atm _ : t .
(s. w. u . ) , muß für / d(Jpz) auch folgender Zu-V d« /«= o
sammenhang gelten, der sich aus den Gin. ( 1 5 ) bis ( 1 8 ) herleitet: Es gilt:
Jpz(t) = (ApA-ApE)-e-kt + /lpE (15)
oder unter Berücksichtigung der Gl. ( 9 )
Ap(t) = (ApA - ApE) • e " k t + (ApE - Apo) (16) wobei & ( m i n - 1 ) die sog. Druckabfallkonstante und ApE der Enddruck ist, der sich bei Erreichen eines neuen stationären Zustandes einstellt. Aus der Druckabfallkonstanten k läßt sich die Halbwertszeit für die Einstellung des neuen stationären Zustandes berechnen ( T y t = In 2/k).
Durch Differenzieren von Gl. ( 1 5 ) erhält man:
d (Apz) dt
Bei Erniedrigung der Osmolalität im Außenme-dium, läßt sich für k eine analoge Gleichung ab-leiten7 , in der statt der „Druckglieder" im Zähler „osmotische Größen" und der Reflexionskoeffizient der permeablen Teilchen auftreten:
£-Lp-F-oa(jts° — Ji°e(j -o)) k =
bzw. für Lr
VA(AP0-Ape)
k-VA- (Ap0—Ape)
(21)
(22)
= -k(Ap\ — Ape) -e kt (17)
und
= -k(ApA-ApE). (18) (d (Apz) \ d t /<=0
Nach Einsetzen von Gl. ( 1 8 ) in Gl. ( 1 4 ) erhält man für k folgenden Zusammenhang:
E-Lp-F(ApA-Ap0) k =
bzw. für Lr
Va- (Apa-Ape)
L D = U-Va-(Apa-Ape)
e-F-(Apa-AP0)
(19)
(20)
£ - F - ( J T S ° - ^ V - 0 , ) - O S '
Ergebnisse
Die Gl. ( 2 0 ) stellt die Meßvorschrift für die Be-stimmung der hydraulischen Leitfähigkeit bei Än-derung der hydrostatischen Druckdifferenz dar. Da die Gleichung keine „osmotischen Glieder" enthält, beschreibt sie auch — in Erweiterung der bei der Ableitung dieser Gleichung gemachten Bedingun-gen — Experimente in Seewasser, das dieselben Ionen wie die Zellvakuole (nur in anderer Zusam-mensetzung) enthält.
Gl. ( 2 2 ) stellt die Meßvorschrift für die Bestim-mung von L p aus osmotischen Versuchen dar. Bei der Bestimmung von Reflexionskoeffizienten6 wur-den die Algen in Lösungen inkubiert, die verschie-

HYDRAULISCHE LEITFÄHIGKEIT VON VALONIA UTRICULARIS 1 3 0 7
dene Konzentrationen an permeabler Substanz, wie z. B. Glucose, enthielten.
Bei den vorliegenden Untersuchungen wurde von diesen Versuchsbedingungen, die der Ableitung die-ser Gleichung zunächst zugrunde gelegt wurden, ebenfalls abgewichen, um die auf osmotischem W e g e gemessenen L p -Werte mit denen aus den hy-drostatischen Experimenten gewonnenen L p -Wer -ten vergleichen zu können.
Die Inkubation der Algen in Seewasser, dem ver-schiedene Konzentrationen an Glucose zugesetzt werden, bietet außerdem den Vorteil, daß osmoti-sche und hydrostatische Experimente an einer Zelle durchgeführt werden können, da die Algen über mehrere Stdn. funktionstüchtig bleiben. Die Diffe-renz n6° — 7is(Jv=o) in der Gl. ( 2 2 ) entspricht unter diesen Versuchsbedingungen der zugesetzten Kon-zentration an Glucose und wird im folgenden als An%\n bezeichnet. Bei der Berechnung von AnGIu ist der aus früheren Messungen 6 bekannte Reflexions-koeffizient von Glucose (a = 0 , 9 5 ) berücksichtigt. Abb . 2 gibt den typischen Verlauf eines Experimentes bei Änderung der hydrostatischen und osmotischen Druckdifferenzen an einer Zelle wieder.
Für die Versuche wurden Valonia-ZeWen mit einer geometrischen Form ausgesucht, die sich unter dem Mikroskop zur Bestimmung des Zellvolumens und der geometrischen Zelloberfläche gut ausmessen ließen.
Die Zellen wurden in einer kleinen Kammer be-festigt, die mit Lösung durchströmt wurde. Das Kammervolumen wurde etwa fünfmal in der Min. ausgetauscht, um ausgeprägte ungerührte Schichten auf der Zelloberfläche zu vermeiden.
Der Anfangsdruck Ap0j wird durch die Ausbil-dung eines Öltröpfchens in der Vakuole auf A p A j erhöht. Die Drudeabnahme in der Vakuole infolge des Wasseraustrittes aus der Zelle (Exosmose) er-folgt exponentiell. Die osmotische Konzentration in der Vakuole steigt entsprechend an. Der stationäre Zustand wird nach etwa 1 2 Min. erreicht, wenn hy-drostatische und osmotische Druckdifferenz ent-gegengesetzt gleich sind. Dieser stationäre End-druck Apvi ist größer als der Ausgangsdrude, wie es zu erwarten ist, wenn kein Teilchenfluß stattfindet. Stellt man nun durch entsprechend rasches Drehen des Metallstabes die Grenzschicht Öl/Zellsaft auf die
Kapillarspitze ein, so erfolgt der Wasserfluß in umgekehrter Richtung vom Außenmedium in die Zelle (Endosmose) . Der Druck fällt zunächst von A p 0 u auf Ap^ii und steigt dann exponentiell an. Der ursprüngliche Ausgangsdrude wird ebenfalls in etwa 12 Min. erreicht. Die Störung des stationären Zustandes durch Zugabe von Glucose führt zu einer exp onentieilen Abnahme des hydrostatischen Druk-kes. Austausch dieses Mediums gegen natürliches Seewasser zeigt, daß auch dieser Vorgang reversibel ist.
A u s den Drudeabfall- bzw. -anstiegskurven läßt sich k bzw. die Halbwertszeit für die Exosmose und Endosmose entnehmen. Zur Berechnung der ent-sprechenden L p e x ( osmotisch und hydrostatisch)-und L p en (hydrostatisch)-Werte muß der Volumen-elastizitätsmodul £ bestimmt werden.
Nach Gl. ( 1 0 ) läßt sich £ aus der differentiellen Volumenänderung der Zelle, die bei einer differen-tiellen Druckänderung auftritt, ermitteln:
VA
bzw.
/ dVz \ \ &Ap )jv=o
(Avl\ \ Ap /Jv=o
VA
( 2 3 )
( 2 4 )
Experimentell muß so verfahren werden, daß die Druck- und Volumenänderungen so rasch erfolgen und gemessen werden, daß Volumenverluste durch den nach Störung des stationären Zustandes ein-setzenden Wasserfluß vermieden werden.
KAMIYA et a l . 1 0 beobachteten bei Elastizitäts-untersuchungen an der Zellwand von Nitella, daß nach Erhöhung des hydrostatischen Druckes die Ex-pansion der Zelle erst nach 2 0 Min. abgeschlossen ist. Bei analogen Untersuchungen an Chaetomorpha linum11 fanden wir, daß über 9 0 % der Volumen-ausdehnung bereits nach wenigen Sekn. erreicht wer-den. Auch bei Valonia ist die Volumenausdehnung wenige Sekn. nach der Druckerhöhung abgeschlos-sen.
D a die Halbwertszeiten für die Einstellung des neuen stationären Zustands dagegen zwischen 3 und 1 0 Min. liegen (Tabn. 1 — 3 ) , wird die Bestimmung von L p hierdurch nicht beeinflußt.
Die Drudeänderung wird, wie beschrieben, durch Hineindrücken eines Öltröpfchens in die Zelle be-wirkt. Aus dem Vorschub und dem Durchmesser
1 0 N . K A M I Y A , M . T A Z A W A U. T . T A K A T A , P r o t o p l a s m a 5 7 , 5 0 1 [ 1 9 6 3 ] .
1 1 U . Z I M M E R M A N N U. E . S T E U D L E , M a r i n e B i o l o g y 1 1 , 1 3 2 [ 1 9 7 1 ] .
1 3 0 8 E. STEUDLE UND U.ZIMMERMANN
Zelle Nr.
FA-102
[cm3]
F
[cm2]
A po
[atm]
Apx — Ap0 ApE
[atm] [atm]
e
[atm]
k-103
[sec - 1]
T\
[min]
Lpex * 107
[cm • sec - 1 • atm - 1 ]
Lpex-107
[cm • sec - 1 • atm - 1 ]
1 3,24 0,598 1,01 0,51 1,14 89,0 2,57 4,49 11,1 11,9 ± 0 , 7 1,13 0,51 1,26 89,0 2,81 4,11 11,9 1,17 1,03 1,47 89,0 2,82 4,10 12,0 1,26 1,03 1,48 89,0 2,60 4,44 12,7
2 1,63 0,561 1,40 0,51 1,48 71,3 2,31 5,00 8,1 6,2 ± 1,3 1,27 0,51 1,37 74,5 2,20 5,25 6,2 1,17 1,03 1,36 71,2 1,52 7,60 5,1 1,08 1,03 1,27 69,8 1,60 7,22 5,4
3 5,48 1,02 0,93 0,41 0,99 82,9 2,31 5,00 12,9 10,9 ± 2,8 0,82 0,41 0,93 82,9 1,70 6,79 8,9
4 2,79 0,339 1,21 0,51 1,27 120,3 1,60 7,22 10,7 10,7 5 2,53 0,485 0,48 0,51 0,52 84,9 1,25 9,24 7,3 6,9 ± 0,7
0,52 0,51 0,55 84,9 1,13 10,2 6,5 6 3,24 0,598 0,72 0,41 0,92 63,1 1,41 8,19 5,9 6,6 ± 0,9
0,77 0,82 1,01 69,3 1,65 7,00 7,2 7 2,71 0,471 1,34 0,51 1,54 91,2 2,51 4,60 10,1 10,6 ± 0,6
1,35 1,03 1,74 90,6 2,83 4,08 11,0 8 12,9 1,44 1,40 0,51 1,50 144 2,11 5,47 10,7 9,8 ± 0,8
1,45 0,31 1,50 142 1,77 6,53 9,4 1,44 0,82 1,63 140 1,99 5,80 9,2
9 4,48 0,697 1,75 0,51 1,84 141 1,77 6,53 6,9 6,9 10 3,50 0,595 1,55 0,51 1,66 87,3 3,89 2,97 (17,8) 12,4
1,54 0,31 1,62 87,3 2,54 4,55 12,4 11 8,50 1,14 1,64 0,51 1,75 161 3,03 3,81 11,3 11,0 ± 0 , 4
1,61 0,51 1,70 161 2,69 4,29 10,7 12 4,86 0,707 1,35 0,51 1,47 123 3,00 3,85 13,8 10,6 ± 2,5
1,30 0,51 1,38 123 2,27 5,09 11,1 1,27 0,51 1,38 123 1,82 6,35 8,6 1,17 0,51 1,28 123 1,97 5,86 8,7
13 3,22 0,526 1,55 0,41 1,74 148 3,73 3,10 9,7 10,8 ± 1,8 1,66 0,62 1,75 159 3,01 3,84 9,8 1,75 0,31 1,77 159 3,56 3,24 12,8
14 12,5 1,46 1,36 0,31 1,49 96 1,67 6,92 9,4 9,6 ± 0,2 1,39 0,31 1,44 96 1,37 8,43 9,7
15 14,1 1,41 1,05 0,31 1,10 115 1,50 7,70 9,5 9,5
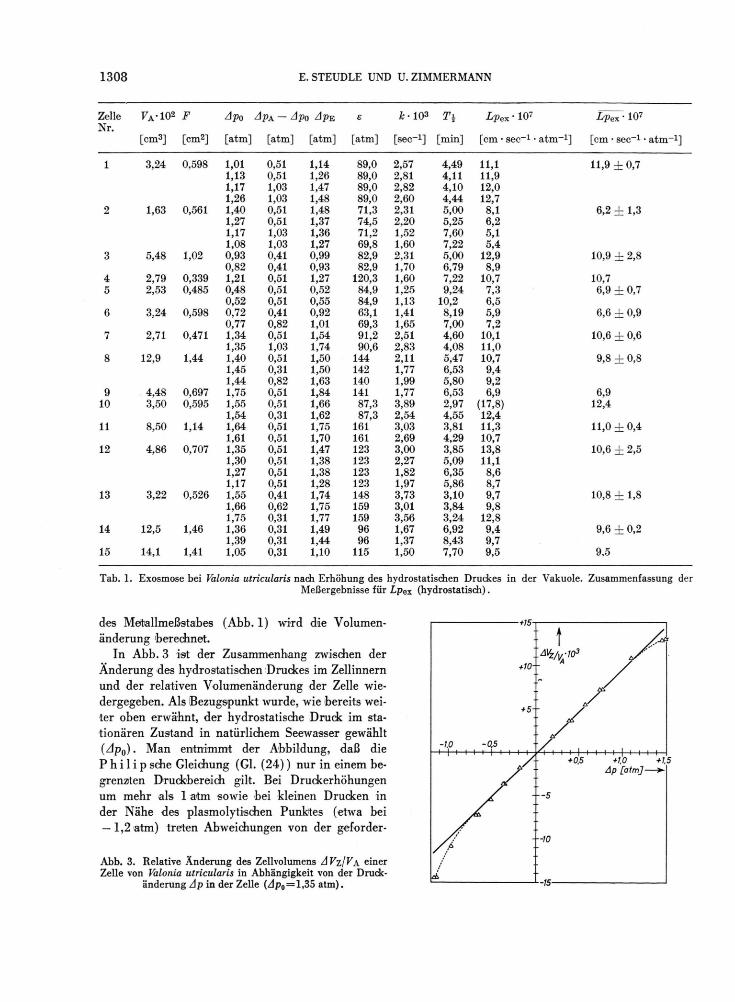
Tab. 1. Exosmose bei Valonia utricularis nach Erhöhung des hydrostatischen Druckes in der Vakuole. Zusammenfassung der Meßergebnisse für Lpex (hydrostatisch).
des Metallmeßstabes (Abb . 1) wird die Volumen-änderung berechnet.
In A b b . 3 ist der Zusammenhang zwischen der Änderung des hydrostatischen Druckes im Zellinnern und der relativen Volumenänderung der Zelle wie-dergegeben. Als Bezugspunkt wurde, wie bereits wei-ter oben erwähnt, der hydrostatische Druck im sta-tionären Zustand in natürlichem Seewasser gewählt ( /dp 0 ) . M a n entnimmt der Abbildung, daß die P h i l i p sehe Gleichung (Gl. ( 2 4 ) ) nur in einem be-grenzten Druckbereich gilt. Bei Druckerhöhungen um mehr als 1 atm sowie bei kleinen Drucken in der Nähe des plasmolytischen Punktes (etwa bei — 1 ,2 atm) treten Abweichungen von der geforder-
Abb. 3. Relative Änderung des Zellvolumens AVz/V\ einer Zelle von Valonia utricularis in Abhängigkeit von der Druck-
änderung zip in der Zelle ( J p 0 = l , 3 5 atm).
+0.5 +1.0 +1.5 Ap [atm]—
- - - 5
--10
-15-

HYDRAULISCHE LEITFÄHIGKEIT VON VALONIA UTRICULARIS 1 3 0 9
Zelle Nr.
FA-102 F
[cm3] [cm2]
Apo Ap0 — Apa ApE
[atm] [atm] [atm]
e
[atm]
k • 103
[sec -1]
Ti
[min]
Lpen • 107
[cm • sec -1 • a tm - 1 ]
Lpen • 10'
[cm • sec - 1 • atm - 1 ]
1 3,24 0,508 1,14 0,43 1,13 81,1 1,01 6,05 12,5 10,0 ± 1,1 1,27 0,47 1,17 86,0 2,18 5,30 10,6 1,48 1,11 1,26 67,3 1,50 7,26 10,4 1,48 1,07 1,27 67,3 1,57 7,35 10,1
2 1,63 0,561 1,47 0,56 1,27 68,8 2,00 5,78 5,8 5,1 ± 0,7 1,37 0,52 1,17 67,4 1,70 6,70 4,4 1,36 0,80 1,08 54,3 1,27 0,10 4,7 1,27 0,88 0,00 54,3 1,48 7,80 5,5
3 5,48 1,02 0,00 0,46 0,82 77,4 2,22 5,20 0,0 0,0 6 3,24 0,508 0,03 0,41 0,77 57,3 1,13 10,2 6,4 6,5 ± 0,1
1,12 0,60 0,60 54,5 1,63 7,00 6,5 7 2,71 0,471 1,54 0,60 1,35 87,3 2,21 5,23 10,2 10,2 8 12,0 1,44 1,50 0,52 1,43 142 1,38 8,37 7,6 7,5 ± 0,1
1,50 0,30 1,43 142 1,43 8,08 7,4 0 4,48 0,607 1,84 0,55 1,62 00,0 2,00 5,53 0,6 0,6
10 3,50 0,505 1,66 0,58 1,44 00,0 2,46 4,70 13,3 12,8 ± 0,8 1,62 0,33 1,58 00,0 2,18 5,30 12,2
11 8,50 1,14 1,75 0,63 1,61 140 2,00 5,53 8,3 0,0 ± 1,0 1,70 0,62 1,57 140 2,55 4,53 0,7
12 4,86 0,707 1,47 0,56 1,30 114 2,20 5,04 10,4 8,5 ± 1,6 1,37 0,41 1,27 114 1,04 5,05 0,1 1,37 0,52 1,10 114 1,02 6,02 7,0 1,28 0,45 1,14 114 1,60 7,22 6,7
13 3,22 0,526 1,74 0,40 1,65 141 3,16 3,66 11,2 12,0 ± 1,1 1,75 0,75 1,61 141 3,67 3,15 12,8
14 12,5 1,46 1,44 0,31 1,31 102 1,76 6,56 0,8 0,8 15 14,1 1,41 1,11 0,31 1 02 110 1,66 6,06 10,0 10,0
Tab. 2. Endosmose bei Valonia utricularis nach Erniedrigung des hydrostatischen Druckes in der Vakuole. Zusammenfassung der Meßergebnisse für Lpen (hydrostatisch).
Zelle FA-102 F Apo Ap~Ei e -103 T1/2 Lpex • 107 Lpex- 107
Nr. [cm3] [cm2] [atm] [atm] [atm] [atm] [sec -1] [min] [cm • sec - 1 • atm - 1 ] [cm • sec - 1
8 12,0 1,44 1,63 2,30 0,30 142 1,53 7,55 5,2 5,2 0 4,48 0,607 1,63 2,30 0,16 00 1,74 6,64 8,0 8,0
10 3,50 0,505 1,62 3,45 0,03 00 3,77 3,06 11,4 11,4 11 8,50 1,14 1,58 4,55 0,01 148 6,11 1,80 10,6 10,6 12 4,86 0,707 1,20 4,62 0,00 114 6,14 1,88 0,6 0,7 ± 0,1
1,13 2,32 0,00 104 3,28 3,52 0,7 13 3,22 0,526 1,65 1,16 0,65 134 3,71 3,11 14,6 14,5 ± 0,1 0,526
1,61 1,16 0,68 136 4,00 2,80 14,4 14 12,5 1,46 1,31 2,20 0,18 73,0 1,78 6,40 10,1 8,0 ± 1,8
1,10 1,15 0,67 73,0 1,44 8,02 7,6 15 14,1 1,41 1,02 3,45 0,02 112 4,12 2,80 10,6 0,7 ± 1,3
0,07 4,60 0,01 112 4,75 2,43 8,8
Tab. 3. Exosmose bei Valonia utricularis nach Erhöhung des osmotischen Druckes des Außenmediums durch Zusatz von Glu-cose (50, 100, 150 und 200 mMol/1) zum Seewasser (/JJTGIU )• Zusammenfassung der Meßergebnisse für Lpex (osmotisch).
ten Linearität auf. Der Druckbereich, in dem die P h i l i p sehe Gleichung erfüllt ist, kann sich von Zelle zu Zelle ändern. Ebenso variiert der Volumen-elastizitätsmodul £, der nach Gl. ( 2 4 ) aus der Stei-gung der Geraden entnommen wird, zwischen 5 0 atm und 1 6 0 atm (Tabn. 1 — 3 ) .
Die Tabellen zeigen weiterhin, daß £ für eine einzige Zelle, abgesehen von wenigen Ausnahmen,
bei variablen Versuchsbedingungen (Exosmose, Endosmose, Änderung des hydrostatischen bzw. des osmotischen Druckes) über längere Zeiten innerhalb von 1 0 % konstant bleibt.
Hysteresis-Phänomene, wie sie von KAMIYA an Nitella beobachtet wurden, treten bei Valonia an-scheinend im untersuchten Druckbereich nicht in meßbarer Größe auf.

1310 E. STEUDLE UND U.ZIMMERMANN
In den Tabn. 1 — 3 sind die gemessenen k-, Ty,-und Lp-Werte zusammengefaßt.
In der letzten Spalte der Tabn. 1 — 3 sind die Mittelwerte für die Lp-Werte für die einzelnen Zel-len mit der Standardabweichung angegeben. Im Mittel ergeben sich hieraus für die hydraulischen Leitfähigkeiten an Valonia utricularis (in 10 7 cm s e c - 1 a t m - 1 ) :
Lp ex (hydrostatisch) = 9,6 ± 1 , 0 ( 1 0 , 4 % ) ( 1 5 ) .
Lp en (hydrostatisch) = 9 , 4 + 1 , 1 ( 1 1 , 7 % ) ( 1 3 ) .
Lp ex (osmotisch) = 9,8 ± 1 , 9 (19 ,4%) ( 8 ) .
wobei der mittlere Fehler des Mittelwerts für ein 95-proz. Konfidenzintervall angegeben ist (in Klam-mern die Zahl der untersuchten Zellen).
Diskussion
In der vorliegenden Arbeit wurden die theoreti-schen Grundlagen und experimentellen Voraus-setzungen für eine neue Bestimmungsmethode von hydraulischen Leitfähigkeitskoeffizienten beschrie-ben. Die Methode ist bei allen Zellen anwendbar, so-weit eine Glaskapillare mit einem Spitzendurchmesser von etwa 60 ju ohne Schädigung der Zellmembran eingeführt werden kann und soweit die den Gin. (20) und (22) zugrundeliegende Forderung, daß sich die Zellen wie ideale Osmometer verhalten, erfüllt ist. Die beiden auf voneinander unabhängigen We-gen gefundenen Werte für die hydraulische Leit-fähigkeit Lp ex von Valonia utricularis stimmen mit 9 ,6 - 1 0 ~ 7 und 9 , 8 - 1 0 - 7 c m - s e c - 1 - a t m - 1 gut über-ein, was für die Leistungsfähigkeit der Methode spricht. Für die End- und Exosmose ergibt sich die gleiche hydraulische Leitfähigkeit. Nach den ersten Messungen von Lp e n( osmotisch) (vgl. Abb. 2) sind auch die Werte für Lp en (hydrostatisch) und L p e n
(osmotisch) gleich7.
Für eine Fehlerbetrachtung von Lp ersetzt man in Gl. (20) bzw. (22) zweckmäßigerweise e durch AP-VA/AV7J (Gl. ( 2 4 ) ) . Die Größe des Fehlers hängt von der Genauigkeit ab, mit der die Zellober-fläche und die hydrostatischen bzw. osmotischen Druckdifferenzen bestimmt werden können. Der Fehler bei der Bestimmung der anderen Meßgrößen ist vergleichsweise gering.
Die geforderte Meßgenauigkeit für die Bestim-mung der hydrostatischen Druckdifferenzen wird
von der Drudemeßapparatur erfüllt, die die Mes-sung von Druckunterschieden in der Größenordnung von 1/ioo atm erlaubt, so daß für die Bestimmung dieser Meßgrößen ein Fehler von 2 — 4 % resultiert. Da der Reflexionskoeffizient für Glucose mit einer Genauigkeit von 3 — 4 % ermittelt wurde 6 , liegt auch der Fehler in der osmotischen Druckdifferenz in der gleichen Größenordnung.
Bei Valonia utricularis ist gegenüber Algen mit geometrisch einfacher Form wie Nitella und Valonia ventricosa der Fehler, der aus der Oberflächenbe-stimmung resultiert, relativ groß und liegt bei 15 bis 20 Prozent.
Damit hängt die Größe des Fehlers für die Einzel-bestimmung von Lp praktisch ausschließlich von der Genauigkeit ab, mit der die Zelloberfläche be-stimmt werden kann.
Eine weitere Fehlerquelle, die nach DAINTY 12
allgemein die Messungen von Lp und vor allem von Permeabilitätskoeffizienten verfälschen kann, tritt durch ungerührte Schichten auf der Zelloberfläche auch bei starker Durchmischung des gesamten Lö-sungsvolumens auf.
Nach DAINTY besteht zwischen der Konzentration c& in der ungerührten Schicht und der Konzentra-tion csi in der Lösung folgender Zusammenhang:
Jv'Ö
c8 = c s r e " i) ' (25)
wobei c5 die Dicke der ungerührten Schicht in cm und D der Diffusionskoeffizient des gelösten Stoffes im Außenmedium ist. Unter unseren Versuchsbedin-gungen ist Jv bei einer Druckdifferenz von 1 atm ca. 1 • 1 0 - 6 c m - s e c - 1 und <5 im ungünstigsten Fall 1 0 - 2 cm.
Der Hauptanteil der gelösten Substanz ist Na-triumchlorid (D = 1,5 • 1 0 - 5 c m 2 - s e c - 1 ) bzw. NaCl und Glucose (D = 5 • 1 0 - 6 cm2 - s e e - 1 ) . Nach Ein-setzen dieser Werte in Gl. (25) ergibt sich, daß praktisch cs = csi ist, d. h., daß Konzentrationsände-rungen durch ungerührte Schichten bei den gerin-gen Volumenflüssen, wie sie bei Valonia utricularis zu beobachten sind, vernachlässigt werden können.
Der Gl. (21) entnimmt man, daß bei einem kon-stanten Oberflächen/Volumenverhältnis (F/V\), einem konstanten e-Wert und hydrostatischen An-fangsdrude Ap0 k linear von der Teilchenkonzen-tration im Außenmedium abhängt, falls Ap^ ^ /Ipo ist. Dieser lineare Zusammenhang wurde bereits
12 J. DAINTY, Advan. Botan. Res. 1, 279 [1963] .

METABOLIC ENERGY, ION PUMP AND RECEPTOR POTENTIAL 1 3 1 1
früher gefunden und konnte auch unter den Ver-suchsbedingungen dieser Arbeit (s. Tab. 3 ) bestä-tigt werden.
Überraschend ist der experimentelle Befund — wenn man den komplexen Aufbau der Membran-barrieren bedenkt — , daß die hydraulische Leit-fähigkeit unter exosmotischen und endosmotischen Bedingungen innerhalb der angegebenen Fehler-grenzen gleich ist. Eine Polarität des Wasserflusses, d. h. eine Abhängigkeit der Flußgröße bei gleicher chemischer Potentialdifferenz von der Richtung durch die Membran, liegt demnach anscheinend bei Valo-nia utricularis nicht vor.
In der Literatur finden sich eine Reihe von ex-perimentellen Nachweisen dieses Phänomens (s. z. B. die Zusammenfassung bei DAINTY 1 2 ) . DAINTY hat gezeigt, daß eine Polarität bei hintereinander-
geschalteten Membranen, die sich in ihrem Hydrata-tionszustand, bzw. in ihrem Reflexionskoeffizienten unterscheiden, zu erwarten ist. Er hat aber auch gleichzeitig auf die Fehlermöglichkeiten, die vor allem durch die bereits diskutierten ungerührten Schichten auftreten können, hingewiesen. Deshalb bleibt es zur Zeit fraglich, wie weit dieses Phänomen tatsächlich in der Natur vorkommt. Für Valonia utricularis läßt sich auf jeden Fall aufgrund der ab-gehandelten Fehlerbetrachtung eine Polarität des Wasserflusses ausschließen.
Wir danken der Deutschen Forschungsgemeinschaft für die Unterstützung dieser Arbeit durch Personal-und Reisemittel sowie für die Bereitstellung von Arbeitsplätzen in der Biologischen Anstalt Helgoland auf Helgoland und in der Stazione Zoologica, Neapel.
The Significance of Metabolie Energy and the Ion Pump for the Receptor Potential of the Crayfish Photoreceptor Cell *
H. STIEVE, H . BOLLMANN-FISCHER, and B . BRAUN
Institut für Neurobiologie der KFA Jülich
(Z. Naturforsdi. 26 b, 1311—1321 [1971]; received August 3, 1971)
Isolated crayfish retinas were poisoned by dinitrophenol (DNP) or ouabain (OU). The changes of the extracellularly recorded receptor potential (ReP) evoked by short or long stimuli were measured.
The changes of the shape of the ReP after poisoning by OU are the same, except the transient increase, as appear after depolarizing the retina by high external potassium concentration. DNP has different effects on the ReP. It causes a prolongation of the amplitude hmax and an increase of the shape quotient hma,x/he (Fig. 9).
Expecially the rate of decrease of the excitability under different stimulus programs was measured (Tab. 5).
DNP experiments show that the inexcitability occurs the earlier the higher the incident light intensity is.
The time needed depends on the stimulating efficiency of the light rather than on the direct amount of energy.
The loss of excitability rather depends on the energy of the total light applied than on the stimulus pattern.
Concerning the inexcitability OU shows the same effect as DNP: it occurs the earlier the more the retina is excited.
These results contradict the hypothesis that the receptor potential is a consequence of a light induced change of the activity of an electrogenic pump mechanism. Fig. 10 shows the electrogenic pump mechanism (EPM) as opposed to the conductance increase mechanism (CIM).
Following former investigations about the effect of some metabolic inhibitors on the receptor poten-tial of the retina of the hermit crab we wanted to
Requests for reprints should be sent to Prof. Dr. H. STIEVE, Institut für Neurobiologie der KFA Jülich, D-5170 Jülich 1, Postfach 365.
study the influence of 2 , 4 dinitrophenol more thoroughly for short ( < latency) and long stimuli. Our main interest was attached to the influence of
* In remembrance of Prof. Dr. HEINZ STEINITZ (Jerusalem) with gratitude and friendship.





























