Embed Size (px)
Citation preview

Z Lebensm Unters Forsch (1984) 178:376-381 Zeitschrift fLir
Lebensmittel- Untersuchung
und-Forschung © Springer-Verlag 1984
Lipoamiddehydrogenase, Citratsynthase und/ -Hydroxyacyl-CoA-dehydrogenase des Skelettmuskels IlL Aktivit~it und subzeilul~ire Verteilung in heHer und dunkler Muskulatur von Rind, Schwein und Gefliigel*
Reiner Hamm und Peter Gottesmann Institut fiir Chemie und Physik, Bundesanstalt fiir Fleischforschung, E.-C.-Baumann-Strage 20, D-8650 Kulmbach, Bundesrepublik Deutschland
Lipoamide Dehydrogenase, Citrate Synthase and /I-Hydroxyacyl-CoA-Dehydrogenase of Skeletal Muscle. III. Activity and Subcellular Distribution in Light and Dark Bovine, Porcine, and Poultry Muscles
Summary. The extractable total activities oflipoamide dehydrogenase (LIPDH), citrate synthase (CS), and /%hydroxyacyl-CoA-dehydrogenase (HADH) were determined in different muscles (longissirnus dorsi, semimembranosus, diaphragma) from cattle and pigs, and in the breast and leg muscles from chicken and ducks. The subcellular distribution of these enzymes was elucidated by determination of the enzyme ac- tivities in the pressjuice of the intact muscle tissue. In the muscles of the different species positive correla- tions between myoglobin content and the activities of the three enzymes were found, which were closer for pigs and chicken than for cattle and ducks. At least 90 percent of the total activity of LIPDH, CS and HADH was located in the mitochondria.
Zusammenfassung. In verschiedenen Muskeln (lon- gissimus dorsi, semimembranosus, diaphragma) yon Rind und Schwein sowie in Brust- und Schenkelmus- keln von Huhn und Ente wurde die extrahierbare ge- samte Enzymaktivit/it von Lipoamiddehydrogenase (LIPDH), Citratsynthase (CS) und /~-Hydroxyacyl- CoA-dehydrogenase (HADH) bestimmt. Die subcel- lulfire Verteilung dieser Enzyme wurde durch Bestim- mung der Enzymaktivit/iten im Muskelprel3saft ermit- telt. In den Muskeln der verschiedenen Tierarten wur- de eine positive Korrelation zwischen Myoglobinge- halt und der Aktivit/it der drei Enzyme gefunden; die-
* Diese Arbeit ist Tell der Dissertation yon P. Gottesmann (Technische Universit/it Miinchen, 1982); sie wurde durch Mittel des Bundesministeriums ffir Jugend, Familie und Gesundheit unterstiitzt
Offprint requests to: R. Hamm
se war beim Schwein enger als beim Rind, beim Huhn enger als bei der Ente. Mindestens 90 Prozent der ge- samten Aktivit/it von LIPDH, CS und HADH im Muskelgewebe sind an Mitochondrien gebunden.
Einleitung
Ffir systematische Untersuchungen, die zur Entwick- lung eines einfachen Gefrierfleischtests ffihren sollten [1], war es zun/ichst notwendig, die Aktivit/it und sub- cellul/ire Verteilung der Lipoamiddehydrogenase (LIPDH), der Citratsynthase (CS) und der /~-Hy- droxyacyl-CoA-dehydrogenase (HADH) in verschie- denen Muskeln einiger Tierarten zu ermitteln, da man weiB, dab die Aktivit/it von Mitochondrien-Enzymen sowohl vonder Muskelart als auch vonder Tierart ab- hfingt. Es ist schon lange bekannt, dab die Aktivitfit von Mitochondrien-Enzymen in roten Muskeln (ho- her Myoglobingehalt) aufgrund ihrer stfirkeren aero- ben Stoffwechselaktivit/it h6her ist als in hellen Mus- keln (niedriger Myoglobingehalt) [2]. Unter der sub- cellul/iren Verteilung eines Enzyms wird verstanden, welcher Anteil der gesamten Aktivitfit in den Zellorga- nellen (hier: Mitochondrien) und welcher Anteil im Sarkoplasma (MuskelpreBsaft) lokalisiert ist [2]. Am einfachsten 1/il3t sich die subcellul/ire Verteilung der Enzyme dadurch bestimmen, dab man die Enzymakti- vit/it im Prel3saft des unzerkleinerten Muskelgewebes mil3t und zu der extrahierbaren Gesamtaktivit/it in Be- ziehung setzt. Man nimmt dabei an, dab die Enzymak- tivitfit im Pregsaft mit der Aktivit/it des Enzyms im Sarkoplasma der unbesch/idigten Muskelzelle gleich- zusetzen ist [2].
Methodik
Material
Innerhalb 1 Std nach dem Schlachten den Schlachtk6rpern von 3 Rindern und 3 Schweinen jeweils den M. diaphragma (,,dunkler"
R. Harem u. P. Got tesmann: Lipoamiddehydrogenase, Citratsynthase und fl-Hydroxyacyl-CoA-dehydrogenase des Skelettmuskels III. 377
50
E 4C
--~ 3C
-~ 2C
<~
SCHWEIN
Ld. Sem. Dia.
RIND
Ld. Sem. Dia.
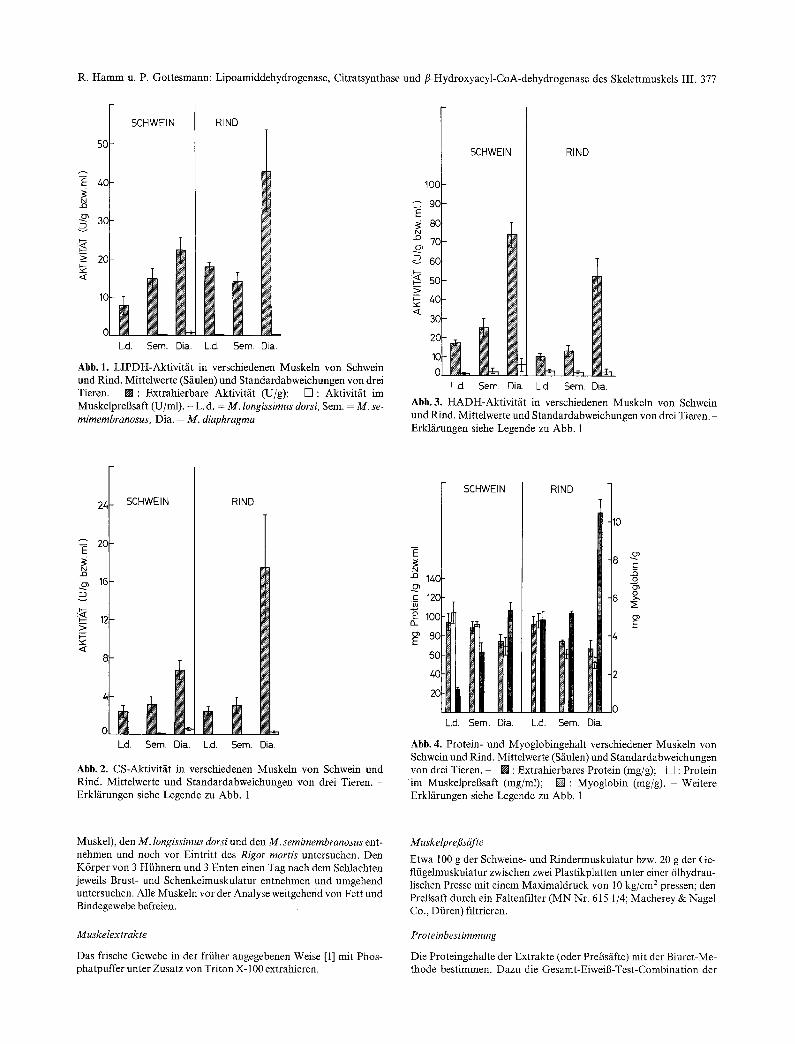
Abb. 1. LIPDH-Aktivitfi t in verschiedenen Muskeln yon Schwein und Rind. Mittelwerte (Sfiulen) und Standardabweichungen von drei Tieren. [] : Extrahierbare Aktivit/it (U/g); [] : Aktivitfit im MuskelpreBsaft (U/ml). - L.d. = M. longissimus dorsi, Sere. = M. se-
mimembranosus, Dia. = M. diaphragma
10C
9¢ --g
"' '7C
6C .~ • ~ 5C
4C
SCHWEIN RIND
Ld Sere Dia. L.d. Sere Dia.
Abb. 3. H A D H - A k t i v i t ~ t in verschiedenen Muskeln yon Schwein und Rind. Mittelwerte und Standardabweichungen von drei Tieren. - Erkl/irungen siehe Legende zu Abb. 1
24
20 E
N .13 .~ 16
8
SCHWEIN
L.d. Sem. Dia. L.d.
RIND
Sem. Dia.
Abb. 2. CS-Aktivitfit in verschiedenen Muskeln von Schwein und Rind. Mittelwerte und Standardabweichungen von drei Tieren. - Erklfirungen siehe Legende zu Abb. 1
'-' 1
c 1
n
E
SCHWEI N RIND
10
r -
25 o
£.
O) E
L.d. Sem. Dia. L.d. Sem. Dia.
Abb. 4. Protein- und Myoglobingehalt verschiedener Muskeln von Schwein und Rind. Mittelwerte (Sfiulen) und Standardabweichungen von drei Tieren. - [] : Extrahierbares Protein (mg/g); [] : Protein
i m Muskelpregsaft (mg/ml); [ ] : Myoglobin (rag/g). - Weitere Erkl/irungen siehe Legende zu Abb. 1
Muskel), den M. longissimus dorsi und den M. semimembr anosus ent- nehmen und noch vor Eintritt des Rigor mortis untersuchen. Den K6rper von 3 Hiihnern und 3 Enten einen Tag nach dem Schlachten jeweils Brust- und Schenkelmuskulatur entnehmen und umgehend untersuchen. Alle Muskeln vor der Analyse weitgehend von Fett und Bindegewebe befreien.
Muske lex t rak t e
Das frische Gewebe in der frfiher angegebenen Weise [1] mit Phos- phatpuffer unter Zusatz yon Triton X-100 extrahieren.
Muskelpreflsafte
Etwa 100 g der Schweine- und Rindermuskulatur bzw. 20 g der Ge- flfigelmuskulatur zwischen zwei Plastikplatten unter einer 61hydrau- lischen Presse mit einem Maximaldruck von 10 kg/cm z pressen; den Pret3saft durch ein Faltenfilter (MN Nr. 615 1/4; Macherey & Nagel Co., Dtiren) filtrieren.
Proteinbest immung
Die Proteingehalte der Extrakte (oder PreBsfifte) mit der Biuret-Me- thode bestimmen. Dazu die Gesamt-EiweiB-Test-Combination der
378 R. Hamm u. P. Gottesmann: Lipoamiddehydrogenase, Citratsynthase und fl-Hydroxyacyl-CoA-dehydrogenase des Skelettmuskels III.
Fa. Boehringer, Mannheim, verwenden. Ansatz: 0,I mI Probe + 5 ml Biuret-Reagens. Nach 30 min die Extinktion der Probe gegen
das Biuret-Reagens als Leerwert bei 540 nm messen. Da die Proben z.T. selbst stark geffirbt sind, einen Probenleerwert aus 0,1 ml Probe und 5 ml Leerwert-Reagens ansetzen und gegen dest. Wasser mes- sen. Die hierbei erhaltene Extinktion yon der Extinktion der Probe abziehen. Fiir jede Test-Combination einmal die Extinktion des bei- gefiigten Eiweig-Standards ermitteln.
Myoglobinbestimmung
Da der Gehalt an Myoglobin in den untersuchten Muskeln demjeni- gen an HfimogIobin welt iiberwiegt, lediglich eine Gesamthfimin-Be- stimmung nach Hornsey [3, 4] durchffihren; Umrechnung in Myo- globingehalt nach Hornsey.
Bestimmung der Enzymaktivitdten
In Muskelextrakten und MuskelpreBsfiften die Aktivitfiten der Enzy- me LIPDH, CS und HADH nach den Angaben von Gottesmann u. Hamm [1] bestimmen. Die Aktivitfiten in der dort angegebenen Wei- se in internationalen Einheiten, bezogen auf Friscbgewicht Gewebe (U/g) bzw. aufVolumeneinheit Pregsaft (U/ml) angeben.
Ergebnisse
In den A b b i l d u n g e n 1-3 s ind die abso lu t en Akt iv i t / i - ten der L I P D H , CS u n d H A D H im E x t r a k t und PreB- sa l t von hel ler und dunk le r M u s k u l a t u r yon R i n d und Schwein dargeste l l t . W o die S/iulen ffir Muske lp reB- saft fehlen, lag im PreBsaft keine meBbare E n z y m a k t i - vit/it vor.
A b b i l d u n g 4 zeigt die Pro te in - u n d M y o g l o b i n g e - hal te im E x t r a k t u n d PreBsaft von hel ler und dunk le r M u s k u l a t u r yon R ind und Schwein.
In Tabe l le 1 ist angegeben , wieviel P rozen t der ex- t r a h i e r b a r e n Gesamtak t iv i t f i t im Muske lp reBsa f t vor- h a n d e n sind. Die Berechnung erfolgte nach fo lgender F o r m e l [2]:
Akt iv i t / i t im PreBsaft in P rozen t de r Gesamtak t iv i t f i t
Akt iv i t / i t im PreBsaf t /ml x 75
Akt iv i t / i t im E x t r a k t / g G e w e b e '
wobe i d a v o n ausgegangen wurde , d a b der durch- schni t t l iche W a s s e r g e h a l t des Muske lgewebes 75% be t rug und die E n z y m k o n z e n t r a t i o n im Muske lpreB- saft e twa die gleiche ist wie im nicht ausgepreBten Teil des S a r k o p l a s m a s . Die Tabe l le gibt somi t AufschluB fiber die subcellulfire Ver te i lung der L I P D H - , CS- und H A D H - A k t i v i t / i t .
In den Tabe l len 2 -5 s ind die Pro te ingeha l te , M y o - g lob ingeha l t e und Enzymak t iv i t / i t en von Ex t r ak t en und PreBs/iften des Brust - und Schenke lmuske lgewe- bes von H u h n und Ente aufgeffihrt .
I Protein- undMyoglobingehalt
Die D i a p h r a g m a - M u s k e l n von R ind und Schwein ent- hie l ten weniger ex t r ah ie rba res P ro t e in als die be iden ande ren Muske l a r t en . A n s o n s t e n be s t and kein Ein-
TabeUe 1. Enzymaktivitfiten im MuskelpreBsaft von Rind und Schwein in Prozent der extrahierbaren Gesamtaktivit/it (Mittelwert und Standardabweichungen yon je 3 Tieren)
Tier Muskelart LIPDH CS HADH
Rind Longissimusdorsi 2,4_+0,40 0 19,0_+2,80 Semimembranosus 0,6+0,35 0 9,1_+2,35 Diaphragma 0,5_+0,32 0,9_+0,25 3,0__+0,31
Schwein Longissimus dorsi 0 0 4,7_+ 3,81 Semimembranosus 1,6_+1,40 3,3__.0,85 6,5-+4,61 Diaphragma 2,9_+ 1,85 5,9 _+ 2,27 6,1 _+ 3,62
Tabelle 2. Myoglobin- und Proteingehalt sowie Enzymaktivit/iten in Extrakt und PreBsaft des Brustmuskels von Huhn. - In Klammern: Angaben in Prozent der extrahierbaren Gesamtaktivit/it bzw. des extrahierbaren Proteingehaltes
Tier I Tier II Tier III
Extrakt Myoglobingebalt (mg/g) 0,70 0,51 0,53 Proteingehalt (rag/g) 106,00 124,80 121,00 LIPDH (U/g) 2,00 9,50 4,80 CS (U/g) 1,80 2,10 3,50 HADH (U/g) 15,00 20,00 28,20
Pregsaft Proteingehalt (mg/ml) 97,2 (72,5) 111,2 (81,0) 114,2 (91,2) LIPDH (U/ml) 0 0 0 CS (U/ml) 0 0 0 HADH (U/ml) 0 0 0
Tabelle 3. Myoglobin- und Proteingehalt sowie Enzymaktivitfiten in Extrakt und PreBsaft des Schenkelmuskels vom Huhn. - In Klam- mern: Angaben in Prozent der extrahierbaren Gesamtaktivitfit bzw. des extrahierbaren Proteingehaltes
Tier I Tier II Tier III
Extrakt Myoglobingehalt (rag/g) 3,11 3,38 4,10 Proteingehalt (rag/g) 55,70 67,40 50,20 LIPDH (U/g) 3,10 17,80 10,70 CS (U/g) 8,90 9,40 9,20 HADH (U/g) 40,20 48,40 64,90
PreBsaft Proteingehalt (mg/ml) 56,4 (75,9) 52,8 (58,8) 37,0 (55,2) LIPDH (U/ml) 0,1 (2,9) 0,6 (2,5) 1,9 (13,3) CS (U/ml) 0,3 (2,5) 0,4 (3,2) 1,2 (9,8) HADH (U/ml) 1,8 (3,4) 2,7 (4,2) 8,8 (10,1)
f lug der Tier- und M u s k e l a r t a u f den Pro te ingeha l t der M u s k e l e x t r a k t e und PreBs/ifte (Abb. 4).
De r M y o g l o b i n g e h a l t im M. diaphragma von R i n d und Schwein war bedeu t end h6he r als in den be iden a nde re n Muske ln . D e r Un te r sch i ed zwischen M. lon- gissimus dorsi und M.semimembranosus war be im Schwein ausgepr / ig te r als be im R i n d (Abb . 4).
Die Ex t r ak t e und PreBs~ifte des Brus tmuske l s von H u h n und Ente wiesen h6here P ro te ingeha l t e a u f als
R. Hamm u. P. Gottesmann: Lipoamiddehydrogenase, Citratsynthase und fl-Hydroxyacyl-CoA-dehydrogenase des Skelettmuskels III. 379
Tabelle 4. Myoglobin- und Proteingehalt sowie Enzymaktivitiiten in Extrakt und PreBsaft des Brustmuskels der Ente. - In Klammern: Angaben in Prozent der extrahierbaren Gesamtaktivit/it bzw. des extrahierbaren Proteingehaltes
Tier I Tier II Tier III
Extrakt Myoglobingehalt (mg/g) 5,13 7,27 4,73 Proteingehalt (rag/g) 100,60 102,90 93,90 LIPDH (U/g) 51,90 31,50 22,50 CS (U/g) 14,70 13,50 11,30 HADH (U/g) 78,40 82,80 42,60
Pregsaft Proteingehalt (mg/ml) 94,5 (70,5) 89,6 (65,3) 80,0 (63,9) LIPDH (U/ml) 3,1 (4,5) 2,6 (6,2) 1,9 (6,3) CS (U/ml) 2,4 (12,2) 1,6 (8,9) 1,2 (9,5) HADH (U/ml) 7,9 (7,6) 7,1 (6,5) 4,5 (7,9)
Tabelle 5. Myoglobin- und Proteingehalt sowie Enzymaktivitfiten in Extrakt und PreBsaft des Schenkelmuskels der Ente. - In Klammern: Angaben in Prozent der extrahierbaren GesamtaktivitS.t bzw. des extrahierbaren Proteingehaltes
Tier I Tier II Tier III
Extrakt Myoglobingehalt (rag/g) 3,54 4,63 4,60 Proteingehalt (rag/g) 70,60 67,20 56,00 LIPDH (u/g) 22,10 24,30 16,80 CS (U/g) 10,70 13,20 12,90 HADH (U/g) 48,90 68,00 57,20
PreBsaft Proteingehalt (mg/ml) 54,3 (57,7) 54,9 (61,3) 46,0 (61,6) LIPDH (U/ml) 2,2 (7,5) 1,4 (4,3) 1,5 (6,7) CS (U/ml) 2,2 (15,4) 2,0 (11,4) 2,6 (15,1) HADH (U/ml) 7,9 (12,1) 7,5 (8,3) 9,8 (12,8)
der betreffende Schenkelmuskel. Der Myoglobinge- halt des Brustmuskels des Huhns war niedriger als derjenige des Schenkelmuskels. Bei der Ente wurde in der Brustmuskulatur ein etwas h6herer Gehalt an ex- trahierbarem Myoglobin als in der Schenkelmuskula- tur gemessen (Tabellen 2-5).
2 Lipoamiddehydrogenase (LIPDH)
Die gesamte extrahierbare LIPDH-Aktivitilt war beim Diaphragma-Muskel des Rindes am h6chsten, w/ihrend sie im M. longissimus dorsi etwas h6her war als im M. semimembranosus. Weiterhin war die Aktivi- t/it im M. longissimus dorsi des Rindes bedeutend h6- her als im gleichen Muskel des Schweins. Beim Schwein waren die Unterschiede in der Gesamtaktivi- tilt zwischen den drei Muskelarten deutlicher ausge- pr/igt als beim Rind (Abb. 1). Zwischen der gesamten LIPDH-Aktivit/it im Muskel und dem Myoglobinge- halt bestand bei beiden Tierarten eine hochsignifikan- te Korrelation (r= +0,973 beim Rind; r= +0,877 beim Schwein).
Die Aktivitilt im Prel3saft des Rindermuskelgewe- bes war verMltnismilgig gering (Abb. 1 und Tabel- le 1). Beim Schwein war der Anteil an der Gesamtak- tivit/it ftir M. diaphragma (2,9%) und M. semimembra- nosus (1,6%) etwas h6her. Im Sarkoplasma yon M. longissimus dorsi konnte keine LIPDH-Aktivitilt festgestellt werden.
Die extrahierbare Gesamtaktivit/it der LIPDH war in der Brustmuskulatur des Huhns niedriger als in der Schenkelmuskulatur (Tabellen 2 und 3). Die bei 3 Enten ermittelte LIPDH-Gesamtaktivit/it lag im Brustmuskel ebenfalts h6her als im Schenkelmuskel (Tabellen 4 und 5). Demnach wiesen, wie bei den S/iu- getiermuskeln, die dunkleren (myoglobinreicheren) Muskeln des Huhns und der Ente eine h6here LIPDH-Gesamtaktivit/it auf als die helleren (myoglo- binarmen) Muskeln. Zwischen der extrahierbaren LIPDH-Aktivit/it im Muskelgewebe und dem Myo- globingehalt beider Tierarten bestand eine signifikant positive Korrelation (r= +0,491 beim Huhn; r = + 0,348 bei der Ente).
Im Pregsaft des Brustmuskels des Huhns wurde keine LIPDH-Aktivit/it festgestellt, w/ihrend im Schenkelmuskel eine gewisse absolute LIPDH-Aktivi- tilt gemessen wurde (Tabellen 2 und 3). Das Sarko- plasma des Brustmuskels der Ente besal3 eine etwas h6here LIPDH-Aktivitfit als das des Schenkelmuskels (Tabellen 4 und 5).
3 Citratsynthase (CS)
Der Unterschied in der extrahierbaren CS-Aktivitfit zwischen M. longissimus dorsi und M. semimembra- nosus von Rind und Schwein war gering, wfihrend bei beiden Tierarten der M. diaphragma eine bedeu- tend h6here CS-Aktivit/it aufwies. Die CS-Gesamtak- tivit/it im M. longissimus dorsi und M. semimembra- nosus des Rindes war der im betreffenden Muskel des Schweins/ihnlich (Abb. 2). Auch bier bestand zwi- schen der CS-Aktivit/it und dem Myoglobingehalt der drei Muskelarten ffir Rind und Schwein eine signifi- kante positive Korrelation (Schwein: r= +0,93; Rind: r = +0,918).
Im PreBsaft des Rindermuskelgewebes wurde nur beim M. diaphragma eine geringe CS-Aktivit/it (0,9%) gefunden. Beim Schweinemuskel enthielten der M. diaphragma (5,9%) und der M.semimembranosus (3,3%) einen gewissen CS-Aktivit/itsanteil im Sarko- plasma (Abb. 2 und Tabelle 1).
Die CS-Gesamtaktivitilt im Brustmuskel des Huhns war geringer als im Schenkelmuskel, w/ihrend fiir Muskelgewebe der Ente kein Unterschied festzu- stellen war (Tabellen 2-5). Zwischen der CS-Aktivit/it und dem Myoglobingehalt der beiden Muskelarten ffir Huhn und Ente bestand wiederum eine signifikant

380 R. Hamm u. P. Gottesmann: Lipoamiddehydrogenase, Citratsynthase und/~-Hydroxyacyl-CoA-dehydrogenase des Skelettmuskels III.
positive Korrelation (Huhn: r = + 0,967; Ente: r = + 0,559).
Der Prel3saft des Brustmuskels des Huhns enthielt keine CS-Aktivit/it; der Schenkelmuskel wies jedoch aufgrund der h6heren CS-Gesamtaktiviffit eine gewis- se CS-Aktivit/it im Sarkoplasma auf (Tabellen 2 und 3).
4 [l-Hydroxyacyl-CoA-dehydrogenase (HADH)
Bei allen drei Muskelarten des Schweins war eine be- deutend h6here extrahierbare HADH-Aktivit/it fest- zustellen als bei den gleichen Muskeln des Rindes. Der Unterschied in den Aktivit/iten zwischen dem M. longissimus dorsi und M. semimembranosus des Schweins war deutlicher ausgepr/igt als der Unter- schied zwischen den entsprechenden Muskeln des Rindes (Abb. 3). Zwischen der extrahierbaren HADI-I-Gesamtaktivit/it und dem Myoglobingehalt bestand f/Jr Rind und Schwein eine signifikant posi- tive Korrelation (Schwein: r = +0,992; Rind: r = +0,965).
Im Prel3saft aller Muskelgewebe beider Tierarten war eine mehr oder weniger deutliche HADH-Aktivi- t/it zu beobachten (Abb. 3 und Tabelle 1). Am h6ch- sten war sie im M. diaphragma des Rindes.
Der Unterschied in der extrahierbaren HADH- Aktivit/it zwischen Brust- und Schenkelmuskel des Huhns war wiederum auffallend (Tabellen 2 und 3). Die HADH-Gesamtaktivit/it war in der Brustmusku- latur der Ente etwas h6her als im Schenkelmuskel (Ta- bellen 4 und 5). Zwischen der gesamten HADH-Akti- vit/it im Muskelgewebe und dem Myoglobingehalt beider Tierarten bestand eine signifikant positive Kor- relation (r= +0,936 beim Huhn; r = +0,735 bei der Ente). Im Prel3saft des Brustmuskels der drei unter- suchten Hiihner wurde keine HADH-Aktivit/it gefun- den, w/ihrend im Prel3saft des Schenkelmuskels teil- weise eine verhfiltnism/il3ig hohe HADH-Aktivit/it ge- messen wurde (Tabellen 2 und 3). Bei der Ente waren die absoluten Aktivit/iten der HADH im PreBsaft des Schenkelmuskels etwas h6her als beim Brustrr/uskel, obwohl die HADH-Gesamtaktivitfit im Extrakt des Schenkelmuskels niedriger war als im Extrakt des Brustmuskels (Tabellen 4 und 5).
Diskussion
Es wurde eine - teilweise sehr enge - positive Korrela- tion zwischen Myoglobingehalt und Aktivit/it der am Citrat-Cyclus (CS, LIPDH) und am Fetts/iureabbau (HADH) beteiligten Enzyme in den Muskeln der un- tersuchten Tierarten gefunden. Die dunklen Muskeln yon Rind, Schwein, Huhn und Ente, die einen haupt-
s/ichlich aerob ausgerichteten Stoffwechsel besitzen (viele Mitochondrien), wiesen eine h6here Aktivit/it aller drei untersuchten Enzyme auf als die hellen, mehr anaerob orientierten Muskeln (wenige Mitochondri- en). Mit zunehmender Beanspruchung der Muskeln steigen die Aktivit/iten derjenigen Enzyme an, die in Hauptwegen der aeroben Substrat-Endoxidation wir- ken [2, 5]. Ein weiteres Beispiel daffir ist die hohe CS- Aktivit/it im Herzmuskel [6]. Die gemachten Beobach- tungen best/itigen Ergebnisse anderer Autoren. So fanden Bficher et al. [7] im ,,roten" Soleus-Muskel des Kaninchens h6here CS- und HADH-Aktivit/iten als im ,,hellen" Psoas-Muskel. Bass et al. machten diesel- be Feststellung in dunklen und hellen Muskeltypen yon Ktiken [8] und Kaninchen [9]. Eine signifikante Korrelation (r = 0,66) zwischen CS-Aktivit/it und dem H/imgehalt von 34 verschiedenen Muskeln des Schweins wiesen Laborde et al. [10] nach.
Hamm et al. [2] untersuchten einige andere Enzy- me des Citrat-Cyclus in verschiedenen Muskeln von Rind und Schwein. Sie fanden eine hochsignifikante Korrelation zwischen Myoglobingehalt und den Akti- vit/iten von Aconitase, Fumarase und Succinat-Dehy- drogenase. Eine enge Beziehung ergab sich auch zwi- schen Myoglobingehalt von Rind- und Schweinemus- kel und den Aktivit/iten der Aspartat- und Alanin- Aminotransferase [11]. Entsprechendes ergab sich fiir die unterschiedlichen Muskeltypen von Huhn und Ente [12]. Diese Unterschiede in den ver- schiedenen Aktivit/iten der Enzyme im Brust- und Schenkelmuskel des Huhns dfirften darin begriin- det liegen, dab das Huhn haupts/ichlich 1/iuft und nur selten flattert. Bass et al. [8] stellten in der Brustmuskulatur des Huhns CS-Aktivit/iten lest, die den von uns gemessenen/ihnlich waren. Der Myo- globingehalt in der Brustmuskulatur der Ente war we- sentlich h6her als in der Brustmuskulatur des Huhns. Diese Tatsache scheint darauf hinzudeuten, dab die Ente h/iufiger fliegt als das Huhn. Masic et al. [12] fan- den die h6chste Glutamat-Oxalacetat-Transaminase- Aktivit/it im Brustmuskel der Taube, einem Tier, das haupts/ichlich fliegt. Der Unterschied im Myoglobin- gehalt zwischen Brust- und Schenkelmuskulatur der Ente war nicht, wie beim Huhn, deutlich ausgepr/igt. Dies kommt auch in den geringen Aktivit/itsunter- schieden der drei untersuchten Enzyme zum Aus- druck. Entsprechend den h6heren Myoglobingehalten waren auch die Gesamtaktivit/iten der LIPDH, CS und HADH in den Muskeln der Ente h6her als in der Hiihnermuskulatur.
Die oben dargestellten Beziehungen zwischen Myoglobingehalt und Aktivit/iten der LIPDH, CS und HADH sollten vermuten lassen, dab die Rinder- muskeln wegen ihres h6heren Myoglobingehaltes eine h6here Aktivit/it der drei Enzyme aufweisen als

R. Hamm u. P. Gottesmann: Lipoamiddehydrogenase, Citratsynthase und/~-Hydroxyacyl-CoA-dehydrogenase des Skelettmuskels III. 381
Schweinemuskeln. So zeigten zwar M. diaphragma und M. longissimus dorsi des Rindes eine h6here LIPDH- und CS-Aktivit/it als die entsprechenden Muskeln des Schweins, die Aktivitfiten der LIPDH und CS im M. semimembranosus beider Tierarten wa- ren jedoch/ihnlich. Die HADH-Aktivit/it in den drei verschiedenen Schweinemuskeln war deutlich ausge- pr/igter als in den Rindermuskeln; dies lfil3t auf einen stfirkeren Fetts/iureabbau schliel3en. Die durch den Myoglobingehalt zum Ausdruck kommende unter- schiedliche Stoffwechsellage ist also f/Jr die biochemi- schen Unterschiede zwischen Rinder- und Schweine- muskel nicht allein mal3gebend. Wahrscheinlich spie- len genetische Einflfisse eine ebenso grol3e Rolle. Dies zeigt auch ein Vergleich der Myoglobingehalte und der Enzymaktivitfiten zwischen Rind-Schwein und Huhn-Ente. So besitzt z.B. der Schenkelmuskel des Huhns einen niedrigeren Myoglobingehalt als der M. semimembranosus des Rindes, die Aktivit/iten der CS und HADH im Schenkelmuskel des Huhns sind aber bedeutend h6her.
Wie aus den verschiedenen Tabellen zu ersehen ist, findet man im Sarkoplasma (Prel3saft) der verschiede- nen Muskeln der untersuchten Tierarten nur eine ge- ringe LIPDH- und CS-Aktivit/it. Je h6her die Ge- samtaktivit/it des betreffenden Enzyms im Extrakt des jeweiligen Muskels war, um so h6her war die Aktivit/it des Enzyms im Sarkoplasma. Beim Rind war die Sar- koplasma-Aktivit/it der LIPDH im roten Muskel niedriger als in den helleren Muskeln. Im Schweine- muskelgewebe lagen die Verh/iltnisse umgekehrt. Hier war auch eine gewisse CS-Aktivit/it im Prel3saft der dunkleren Muskeln zu beobachten. Der auf die Ge- samtaktivitfit bezogene Anteil der Aktivit/it der LIPDH (ca. 5%) und der CS (ca. 10%) im Prel3saft von Hfihner- und Entenmuskelgewebe war etwas h6- her als im S/iugetiermuskel.
Aus den Ergebnissen lfiBt sich schliel3en, dab die LIPDH und CS in der Skelettmuskulatur von Rind, Schwein, Huhn und Ente fast ausschliel31ich in den Mitochondrien lokalisiert sind. Nach Beenakkers [13] handelt es sich bei der CS um ein rein mitochondriales Enzym (ca. 97% der Gesamtaktivit~it sind an die Mi- tochondrien gebunden). Pette [14] fand mit Hilfe der sogenannten Mehrfachextraktion in der Leber von Ratten erst nach dem letzten Schritt (Homogenisie-
rung des Gewebes) durch Zerst6rung der Mitochon- drien ca. 97 % der CS-Aktivitfit.
Im Sarkoplasma der Rindermuskeln wurden durchschnittlich 10%, im Sarkoplasma der Schweine- muskeln ca. 6%, im Prel3saft von Huhn- und Enten- muskelgewebe ca. 5% der HADH-Gesamtaktivit/it gemessen. Die HADH ist ebenfalls ein Enzym, das in der Skelettmuskulatur fast ausschliel31ich an Mito- chondrien gebunden ist. Nach Beenakkers [13] liegen im Flugmuskel der Wanderheuschrecke etwa 97% der HADH-Gesamtaktivit/it in den Mitochondrien vor. Im Lebergewebe von Ratten wurde in den Peroxiso- men ein Isoenzym der HADH entdeckt, das sich in ih- ren Eigenschaften von der mitochondrialen HADH unterscheidet [15-17]. Obwohl in der Muskelzelle Peroxisomen nicht vorkommen, ist es nicht auszu- schliegen, dab im Sarkoplasma der Muskelzelle ein Isoenzym der HADH existiert.
Literatur
I. Gottesmann P, Hamm R (1984) Z Lebensm Unters Forsch 178:366-370
2. E1-Badawi AA, Harem R (1972) Z Lebensm Unters Forsch 149:87
3. Hornsey HC (1956) J Sci Food Agric 8:534 4. Hornsey HC (1957) J Sci Food Agric 9:547 5. Pette D (1977) In: Jahnke K, Jahnke M, Reis HE (eds) Muskel-
stoffwechsel, k6rperliche Leistungsf/ihigkeit und Diabetes melli- tus. Schattauer Stuttgart New York, p 21
6. Pette D, D61ken G (1975) In: Weber G (ed) Advances in enzyme regulation. Pergamon Press Oxford New York, p 355
7. Biicher T, Pette D (1965) In: Verhandlungen der Deutschen Ge- sellschaft f/ir inhere Medizin. JF Bergmann, M/inchen, p 104
8. Bass A, Lusch G, Pette D (1970) Eur J Biochem 13:289 9. Bass A, Brdiczka D, Eyer P, Hofer S, Pette D (1969) Eur J
Biochem 10:198 10. Laborde D, Talmant A, Monin G (1983) Vortrag, R6union vian-
de et produits carries, Paris 1 I. Hamm R (1969) J Food Sci 34:449 12. Masic D, Hamm R, TetzlaffL (1971) Z Lebensm Unters Forsch
246:205 13. Beenakkers AM (1963) Biochem Z 337:436 14. Pette D (1968) In: Schmidt FW (ed) Praktische Enzymologie,
Hans Huber, Bern Stuttgart, p 15 15. Osumi T, Hashimoto T (1979) Arch Biochem Biophys Res Com-
mun 89:580 16. Osumi T, Hashimoto T (1979) Biochim Biophys Acta 574:258 17. Osumi T, Hashimoto T (1980) Arch Biochem Biophys 203:372
Eingegangen am 13. Dezember 1983. Angenommen am 11. Januar 1984.













![Kapitel 6) Ctitratcyklus+Atmungskette [Kompatibilitätsmodus]biochemietrainingscamp.de/stoff/ci/Ctitratcyklus.pdf · TPP Liponsäure NAD FAD CoA ( Vergleiche die Pyruvat-Dehydrogenase)](https://img.pdfslide.org/doc/110x75/5e1287ca50baaa02a71d88e6/kapitel-6-ctitratcyklusatmungskette-kompatibilittsmodusbi-tpp-liponsure.jpg)















