Embed Size (px)
Citation preview

Zeitsehr. f. Hygiene 146, 26--47 (1959)
Aus dem Institut fiir Hygiene und Medizinisehe Mikrobiologie der Freien Universit~t Berlin
(Direktor: Professor Dr. med. BEmC~e~RD SC~--~gDT)
Myxovirus-It~imagglutinationsversuehe mit unterschiedlichen Erythrocytenarten
Vergleiehende quantitative Untersuehungen unter versehiedenen Bedingungen
Von ]~ERNHARD SCHMIDT und KLAUS GROSSGEBAUER
l~Iit 1 Textabbildung
(J~ingegangen am 22. Mai 1959)
A. Einleitung Myxoviron besitzen ein gegen verschiedene Kal t - und Warmbli i ter-
o ry th rocy ten geriohtetes Agglutinin. Eine Anzahl yon in vi t ro-Testen zur Diagnost ik, Difforenziorung und Typisierung basioren au f diesom Hi~magglutinationsph/inomen 13.. Es ist bekannt , dab zahlreiche Fak- boron den Titer des I-I/imagglutinationstestes beeinflusscn: z .B. der pH-Wer~, der Eloktrolytgohal t des Verdiinnungsmittels, die Konzen- t ra t ion und das Alter der Blu~kSrperehon, clio Reak t ions tempera tu r und die Zei~, naeh dor die Reak t ion abgelesen wirdg,56, 63.
Deshalb war es erforderlich, bei Vergleichsuntersuchungen diese Variabeln konstan~ zu halten, d .h . die routinemitBig dttrchgeffihr~o Virus- h/ imagglut inat ion zu standardisieren%
Viele Fak to ren beeinflussen die einzelnen agglutinationsf/ihlgen Ery- t la 'oeytenar ten in unterschiedlicher Weise, so dab die opt imalen Bedin- gungen fiir die jeweiligen Ery th rocy tensys t eme ungleieh sind2L
Aus den lYntersuehungsergebnissen, die yon l~Iyxovirus-H~magglutinationsver- suchen mit unterschiedlichen Erythrocytenarten vorliegen**, sei folgendes zu- sammenfassend hervorgehoben:
1. Die einzelnen St/~mme verm6gen eine untersehiedliehe Anzahl yon Erythro- cytenarten zu agglutinieren, d. h. sie besitzen versehiedene,,Agglutinationsspektren".
2. Variable yon seiten des Virus (Virusaktivit~t, Anpassungsphase, Virus- provenienz *** u.a.) sowie die den H~magglutinationstiter beeinfluBenden Faktoren yon seiten der Ery~hrocyCen und des Verdfinmmgsmittels erschweren einen exakten Vergleich der ,,Agglutina$ionsspektren" dutch einen anderen Untersueher. Daher sind die Angaben fiber diese Spektren oftmals wechselnd und widersprechend.
* SeRe 397. ** Siehe n.a , &,5,7,9,21,22,24,27,28,36,40,51,55-59,63,75,81,88,89,92.
*** Allantoisfliissigkeiten yon Enteneiern - - mi~ hfihnereiadaptierten Myxoviren infiziert -- zeigen im Vergleich zu solchen yon Hfihnereiern ein praktisch fiberein- stimmendes AgglutinationsvermSgen (eigene unverSffentlichte Untersuehungen).

Myxovirus-Hi~magglutinationsversuche 27
3. Es bestehen grSBtenteils keine Beziehungen zwischen der Agglutinierbarkeit der Erythroeyten unterschiedlicher Tierarten und der Empf~nglichkeit dieser Tiere fiir ein bestimmtes Virus.
4. ~Ian finder ein abweichendes Verhalten der Erythroeyten verschiedenster Species cinem bestimmten Infiuenzavirusstamm gegeniiber sowie umgekehrt der Ery~hrocyten einer bes~immten Art verschiedenen St~mmen gegeniiber. ])iese Diffe- renzen kSnnen sich u.a. im H~magglutinationsvermSgen, in der Virusbindungs- festigkeit und in der Absorptions- und Elutionsgesehwindigkei~ zeigen.
5. Die BlutkSrperchen verschiedener Tiere der gleichen Art kSnnen ein unter- schiedliches AgglutinationsvermSgen besitzen.
6. Dem Stamme B Lee werden viele, den A-Stammen mal~ig viele und dem Schweineinfluenzavirus wenig agglutinationsfahige Erythroeytenarten zuge- sehrieben.
"Wenn auch die speziellen Ergebnisse wei~gehend vom Material und der Technik des betreffenden Untersuchers abh~ngen, so sin4 doch diese reproduzierbaren aHgemeingfiltigen Regeln yon theoretischem und prak- tischem Werte erarbeitet wordcn.
Vergleichende Hiimagglutinationsversuche sind oftmals nut qualitativ, d.h. bei einer fixierten Virusverdfinnung oder quanti tat iv lediglich an einem kleineren Virusmaterial durchgcffilu't worden.
Dagegen sind gerade quantitative Untersuchungen, Versuche zur Ermittlung des unterschiedlichen tt/tmagglutinationsvermSgens, der Bindungsfestigkeit und -kapazit/it und des damit verbundenen Elu- tionsvermSgens schon deshalb yon Interesse, well sic u.a. Aufschlfissa fiber den Influenzavirusreceptor verschiedenartiger Erythrocyten zu geben versprechen.
M6glicherweise kann eine durch solche Untersuchungen aufgedeckte selektive Agglutinabilit~ zu diagnostischen Zwecken verwcndet werden. So sollen sich z. ]~. Erythrocyten vom Ziesel (Citellus eitellus) zu einer T}Tenunterscheidung innerhulb der Myxovirusgruppe eignen. Es hat sich aber im Verlaufe unserer Untersuchungen herausgestellt, dab dcr A/Asia57-St~mm bereits eine Ausnahme macht* (siehe D: Besprechung der Ergebnisse).
N~euerdings ist gezeigt worden aa, dab durch eine spezielle Formalinisierung be- stimmter Erythrocyteu eine vorher nieht vorhandene Agglutinierbarkeit durch Myxoviren erreicht werden kann.
Um zu derar~igen Fragen einen Bei~rag leisten zu kSnnen, haben wir des Verhalten verschiedenartiger Erythrocyten gegen/iber ]~'[yxoviron tm~ersucht. Dabei achteton wir zun/iehst auf 3 Punkte, die zur niihoren Charaktorisierung bzw. aueh als Hiffsmittel zur Diagnostik yon Myxo- viron dienen k6nnten, n/~mlich auf
1. die Gr6l~e der einzelnen ,,Agglu~inationsspektren", 2. eine m6glicherweiso bestehende selektive Agglutinabilit/it und 3. das Verhalten der Erythrocyten nach dor ~ormalinisierung.
* Inzwisehen dureh eine persSnliche Mitteilung besti~tigt (L. BORECK~:5).

28 ]~ERI~I-IARD SCltMIDT 11114 ~LAUS ~ROSSGEB&UER:"
Insbesondere schien uns der letzte 1)unkt von Interesse zu sein, da wit bereits bei Bakter ien eine erst nach der Formal in i s ie rung der Ery- th rocy ten einsetzende Aggluti~fierbarkeit beobachten k o n n t e n 75.
Wesent l ich schien die 1)riifung au f unterschiedliches Verha l ten hin- sichtlich folgender l~unkte zu sein:
1. des HSmagglu t ina t ionsverm6gens 2. des E lu t ionsverm6gcns ui~d 3. des Vert~altens yon Vi rusery throcytengemischen in Blutsenkungs-
pipet ten. Wi r un te r such ten da rau fh in :
a) die E r y t h r o e y t e n na t i v u n d formalinis ier t b) das Virusmater ia l ak t iv u n d h i tze inak t iv ie r t u n d e) die Viruserythroeytengemisehe bei unterschiedl ichen Tempera-
turen . Auf zwei Befunde, n~mlich das Zonen- u n d Panh~magglu t ina t ions -
phgnomen, sol] am Sehlufi e ingegangen werdcn.
B. Material und Mcthoden
1. Virusstdmme. Dic Virusstiimme ~urden der Stammsammlung unseres ~nstitu- tes entnommen. Es handelt sieh um Myxoviren: Schweine-Influenzavirus (,,Shope"), Infiuenza-A/WS, -A/PlY8, -A/Melbourne (,,Mel~ -A/WeiB, -A1/FM1, -A1/Eng- land/I/51 und 53 (,,A1]] 51" und ,,ALE 53"), -A~/Germany/1/53 (,,A1G"), -A55 , -A/Asia 57, -B Lee, -B195~, Mumps (E~DERS), Newcastle Disease-Virus (,,NDV'~).
Die Sti~mme waren durch zahlreiehe Eipassagen gegangen. :Die A'-Stgmme lagen in der D-l~hase vor.
2. Erythrocyten. IDle Blutk6rperchen wurden zum Teil dutch die (iblichen Punk- tionen gewonnen, zum Tell bezogen wit sie in Form yon Citratblut vom Schlachthof. Die Herstellung yon formalinisierten Erythroeyten ist nach FLICK at vorgenommen worden und an anderer Stelle besehriebenTL
3. Virus-Erythrocytenffemische. Zur Testung der nativen, formalinisierten und virusvorbehandelten Blutk6rperchen sind wlr yon einer 6--7~ Erythrocyten- suspension ausgegangen. Als Verdfinnungsmittel wurde (bis auf die im Text erwghn- ten Ausnahmen) eine phosphatgepufferte isotonische Kochsalzl6sung verwendet. 1 ml der Erythrocytensuspension wurde jewcils mit 1 ml der fraglichen infizicrten Allantoisflfissigkeit oder Normalserum (bei der Panhgmagglutination) vcrmischt und sofort in eine gew6hnliche Blutsenkungspipette aufgezogen 7*-
d. Hitzeinaktivierung des Virusmaterials. Die Inaktivierung dcr infizierten Allantoisflfissigkeiten erfo]gte im ~Vasserbad (30 rain bei 56~ unter Kontrolle eines geeichten Thermometers.
5. Absorptions-Elutlons-Schema. 10 ml der infizierten frisch geernteten Allantois- flfissigkeit mischten wir mit 1 ml des 50~ Erythroeytensedimentes und hielten das Gemisch 1 Std bei -k4~ Dan~ch wurde abzentrifugiert and der ~berstand auf seine verbliebene H~imagglutinationskraft nach SALK geprfift. ])as Erythro- cytenscdiment wurde zur 1. Elutio11 in 20 ml einer gepufferten KochsalzlSsung suspendiel~ und raehrere Stunden bei 37~ gehalten. Jetzt zentrifugierten wit" erneut ab, prfiften den ~berstand und eluierten weitere Male (zum Teil fiber Nacht), bis die Uberstiinde negativ waren.

Myxovirus-Hamagglutinationsversuche 29
6. HSmagglutinationstest. Unsere Auswertungsmethode lehnt sich weitgehend an das yon SALK 71 angegebene Verfahren an. Um Titerstreuungen klein zu halten, richten wir uns nach der yon BRAND 9 erstrebten Standardisierung.
7. Infektion und Bebriitunff der Eier sowie die Ernte der Allantoisflfissigkeiten erfolgten in fiblieher Weise und sind an anderer Stelle beschrieben 7.
C. E x p e r i m e n t e l l e r Tei l
1. VirusMimagglut inat ionsversuche m i t a k t i v e m Virusmater ia l
Die Tab. 1 zeigt die Ergebnisse der Untersuchungen. 2~ative Erythroeyten (,,NE") sind auf ihre Agglutinierbarkeit durch Virus auf der Salk-1)latte und in gewShnlichen Blutsenkungspipetten bei Zimmertemperatur u n d + 4 ~ gepriift wor- den. ~ormalinisierte Erythroeyten (, ,FE") haben wir lediglich in Blutsenkungs- pipetten getestet. U m einen Titervergleieh mit 2~E vom Huhn zu ermSgliehen, ist stets ein Virusstamm mit allen 7 Erythrocytenarten auf einer Plexiplatte ange- setzt worden.
Aus der Tab. 1 ist ersichtlieh, daft die verwendeten Virusst~mme (mit Ausnahme des 3[urnpsstammes) mit NE veto Huhn Himagglut inat ionst l ter yon 1:640--1280 ergeben haben. Damit lagen gute VergleichsmOglichkeiten mit Titern anderer Blut- arten vor. Die ,,Senkungszeit" betrug bei diesem Titer 1--3 rain, d.h. innerhalb dieser Zeit begann die mit unbewaffnetem Auge ablesbare Agglutination in der Sen- kungspipette. Bei geringercn Titern verl ingerten sich die ,,Senkungszeiten". Auch die FE veto Huhn lieSen die Beziehungen zwischen Agglutinationsbeginn und Himagglutinationsti ter erkennen. Lediglich der Mumpsstamm reagier te etwas schneller.
Die iibrigen :Erythrocytenarten wichen mehr oder weniger veto Verhalten der Huhnerythrocyten ab :
Von allen Virusst immen -- yon den meisten im Gegensatz zu den noeh aus- stehenden Erythrocytenarten sogar relativ hoehtitrig -- wurden 2~E veto Meer- schweinchert agglutiniert. Mit niedrigem Titer agglutinierten nur die Sti~mme A/PRS, A~5, Mumps und die beiden B-Stamme. Auffallend war, daft der NDV-Stamm mit einem Titer yon 1:320 keinc ,,Senkungswertc" mchr ergab und dal] die FE bei einigen St~mmen empfindlicher, d.h. sehneller reagierten sis die entspreehenden NE. Andere St~mme zeigten dagegen eine geringe ReaktionsverzSgerung mit FE. ,NE veto Hammel wurden ebenfalls von allen 15 Virussti~mmen - - im wesentliehen jedoch niedrigMtriger als 2~'E vom Huhn und Meerschweinehen -- auf der Salk-Platte agglutiniert. Die Reaktionsbereitschaft in der Senkungspipette war etwas verzOgert. Der A/PRS-Stamm und die beiden B-Stamme, die gegeniiber Erythrocyten veto Meerschweinchen nur einen geringen Httmagglutinationstiter anzeigten, wurden mit Hammelblutk6rpcrchen wicdcr hShcrtitrig agglutiniert. W~hrend ~NE veto Huhn, Meerschweinchen und Pferd keine wesentliehen Differenzenzwisehen den bei Zimmer- temperatur und + 4 ~ abgelesenen Plattenti~ern erkennen lieSen, fanden wit solche bei Verwendung yon NE vom Hammel, Rind und yon der Katze. Diese Erythro- cytenarten agglutinierten oft lmr bei + 4 ~ C.
Mit Ausnahme veto Shope-Stamm agglutinierten alle iibrigen such die N E veto Rind, besonders bei + 4 ~ Einen auffallend geringen Titer fanden wit bci den St~mmen AI/FM 1 AlE 51, A1G und NDV, einen praktisch unverandert hohen nur beim Stamme A/Asi~ 57. F E dieser Tierart reagierten bei den B-St~mmen und beim Mumps-Stamm schneller.
Von weniger Virusst~mmen wurden die NE und FE veto Pferd agglutiniert, namlieh veto A/Asia 57 und B195s regelmM3ig in einem hSheren, yon den A-Stammen ~tlteren Datums sowie vom B Lee und Mumps-Stamm in einem niedrigen Titer.

30 B~aa-~tnan Sc~r)~tn~ und K r ~ v s G ~ o s s a ~ v ~ , a :
8t~imme
A/WS
A]PR8
A/MEL
A/Weil3
A~/FM~
A~E51
A~E53
A~G
A/Asia 57
B Lee
Shope
Mumps
NDV
Tabelle 1. Ergebnisse der Hiimagglutinationsversuche mit as Virusmaterial
Meerschw. Hammel !l ! l=[uhn Rind , Pferd Schwein Katze
nat. form. nat. form. nat. form. nat. form. }nat.. form. L nat. form. nat.
+ + + + + + + + + + + : + + + + + + i ~ ;5 ~ ~ 1280++ 1280++ ,320++ 320+ I 8 0 + + n.n.
i + + + + ~ + I(+~ + ~6 ;5 / o + + ~ z: ~ 640+ 2 0 + + i 320 0 + + n .a .
+ + + + + + + + + ++] + + + (+) (+) ~ ~ 75 640 + 320 + + I 320 + 320 4 + + u .a . ;5
[ + + + + + + + + + + (+) (-:) ~ ;3 ;5 ~ + + + 1280 + + 1280 + 640 + + 160 + 40 -' ~ ~zf
+ + + + + + + + + J- +(4) 2 ~ ;5 ~ 75 ~ Y5 ~Z 640 + + 640 + + i60 + 20 ~ + ~ n .a .
I + + + + + + + + + + + + + + + i + (+) 75 ~ ~ ;-~ 1280++ 1280 ~-+ 40-~+ 4 0 b+ ~ Z
+ + + + + + + + + + * - ' + + + (+) ~ ~ !75 ~ 640 + + 640 + ~40 + + ~ 320 ~- ;5 I ~ .~
+ + + + + + + + + + I + + ;5 ~ ~ ~ ~ yj ;3 6 4 0 + + 640+ I 160+ 20+ ~ n .a .
I + + + + + + + + + l + + + 4 �9 75 ~ ~ ~ ~" ;zf 1280 + 80 + 320 + + 80 + 75 n . a .
+ + + + + 4 + + + + + + + + + + + + ( : ) 4 ~- + + (+) 75 1280++ 6 4 0 + + 6 4 0 + + . 1280 6 4 0 + + n.a .
-I
6 4 0 + + ' . . . . . 3 2 0 + + 3 2 0 + + + ~ 40+____ __ n .a . , 4 0 + +
+ + + + + + ~5 + + + + + 1 + + + + V~ + + + + + + + ] + 1280++ 1 4 0 + + 6 4 0 + + ]!640++ 6 4 0 + + n.a . ,] 160+
] + + + + + + " + + + + + + ~ ;5 ~ ;5 ~ ~ 75 6 4 0 + + ] 6 4 0 + + 3 2 0 + + 75 ,~ n . a .
. . ; ~ + + 75 + + + G ;5 (+) + ~ ~ + 160+ 4 0 4 + 8 0 + + 8 0 + + 8 0 + + n .a .
+ + + + + + ~ ~ ;5 + ~ Z ~ ~ U~ ~ a + + + i 1280++ 3 2 0 + + 80+ ] 40 i + ] 75 n . .
, ,Meer." u n d , ,Ha rem." : ~'Ieerschweinchen- u n d H~mmele ry th roey ten . , ,na t . " und , , form." : na t ive nnd formalinisierte E ry th r oey t e n . Fi ir jedert Virus- s ta ture wurde in die erste Reihe yon l inks nach reehts dns t t~maggla t ina t ions - ve rm6gen in Senkungsp ipe t ten , in die zweite l~eihe yon links nach rechts der ]:I~m- agglut in~t ionst i ter nach SM, K eingetragen.
Die Vi rusery throcy tengemisehe sind in B lu t senkungsp ipe t t en 30 rain bei Zim- m e r t e m p e r a t u r und bei + 4 ~ beobachte t worden: , , + + + " : mi t unbewaf fne tem Auge s ieh tbarer Agglu t ina t ionsbeginn innerhalb y o n 2 min, , , + + " innerha lb y o n 5 rain u n d ,, + " in mehr als 5 min. ,,(-}-)": Agglu t ina t ion n u r bei + 4 ~ C.
Der t I~magg lu t ina t ions t i t e r naeh S)~K (Znhlenwert) ist naeh zweist i indigem Aufen tha l t der Plexiplat~en bei + 4 ~ abgelesen worden: , ,n .u . " : n ieht ablesbar . ,, 75 " : keine H~magglu t ina t ion nach 30 rain (Pipette) bzw. 2 S td (Plexiplatte).
+ + 320++
;-j 20 + +

Myxovirus-Hiimagglutinationsversuche 31
]:)as Agglutinationsph~nomen war sowohl bei Zimmertemperatur als aueh bei -[-4~ ablesbar. I~E veto Pferd (wie auch solche vein Sehwein und yon der Katze) neigten zu Spontanagglutinationen und eignen sich zumindes~ in dieser Form wenig zur Testung yon Virusmaterial.
NE veto Schwein wurden nur in der Senkungspipette, und auch da lediglich yon wenigen St~mmen (regelm~Big stark vom B1955-Stamm und schwach yon den Stem- men B Lee und A/Asia 57), agglutiniert. Die entspreehenden Werte auf der Plexi- platte waren oftmals nicht klar ablesbar.
Ery~hrocyten yon der Katze haben wit nur im nativen Zustand auf ihre Agglu- ~inabilit~t untersucht. Die St~mme A/Asia 57, ]3 Lee, B105a und ~Iumps gabon ein reproduzierbares positives ResultaC.
Zusammenfassend l~Bt sich folgendos aussagen:
1. Nur 4 yon 15 Myxovirusst~mmen, n~mlieh A/Asia 57, Mumps und die beiden B - S ~ m m e , haben mi t allen 7 E r y t h r o c y ~ n a r t e n das Hiim- agglutinationsph/inoInen gezeigt.
2. Von diesen 4 agglutinierten lediglich die St/imme A/Asia 57 und B195~ regelm~Big in einom h6heren Titer, unterschieden sich aber in auf- fallender Weise ira Vorhalten gegeniiber Ery throcyten vom Schwein:
Der B~gss-St~mm war als einziger S tamm in der Lage, bei -[-4~ Zimmertemperatur und ~ 3 7 ~ (!) N E veto Schwein in der Senkungs- pipette rasch zu agglutinieren. Das muB als eino Stammesoigontfimlich- keit herausgestellt werden.
3. NE vom Huhn, ~eerschweinchon und Hammel wurden yon allen 15, NE veto Rind yon 14 St/immen agglu~iniert.
4. N E yon der Katze zeigten nur mi t den Sti~mmen A/Asia 57 (regel- m/iBig in einem h6heren Titer), B Lee (nur bei ~-4~ einen Titer fiber 1:20), B~gs5 und ~ u m p s das tI~magglutinationsph~nomen.
5. Die A-St~mme/~lteren Datums agglutinierten N E veto Ps Die fibrigen A-St/imme neueren Datums (mi~ Ausnahme des A/Asia 57) er- gabon mi t dieser Blu tar t ein negatives Agglutinationsergebnis.
6. Ery throcyten vom Hammel , Sehwein, Rind und yon der Katze zeigten Differenzen zwisehen den bei Zimmer~empera~ur und H-4~ abgelosenen ttitmagglutinationsergebnissen.
7. FE verhielten sieh in der Senkungspipette im Hinblick auf ihre Reaktionsfreudigkoit untersehiedlich. Einige Virusstitmme waren erst nach der Formalinisierung der BlutkSrperchen in der Lage, diese zu agglutinieren.
8. F E veto Pferd, Sehwein und yon der Katze neigten zu Spontan- agglutina~ionen.
2. VirusMimagglutinationsversuche mlt hitzelnaktiviertem Virusmaterial
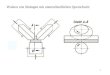
Die Abb. i zeigt die Ergebnisse der Versuche. Die Inaktivierung der infizierten AUantoisfliissigkeiten erfolgte ira Wasserb~d (30 rain bei 56~ unter Kontrolle eines geeiehten Thermometers.

32 BERI~Hal~D SCH~nDm und KLAVS GROSSGEBAUER:
Die Ergebnisse lassen sich folgendermal~en zusammenfassen:
1. Die Hitzeinaktivierung war ira allgemeinen nur yon geringem Ein- flus auf die HShc dos tI~magglutinationstit~rs.
2. Die einzelnen Erythrocytenarten (I-Iuhn, l~'[cerschweinchen, Ham- reel und Rind) verhielten sich bis auf wenige Ausnahmen (s. Punkt 3) gegeniiber den inaktivierten Virusst~mmen praktisch gleich.
/iT zsoo F
~0 ~
1 2 3 r A/WS
'IH ,'gO
80
20
A/Asia 57
AIPs A/NEL A/Wez~7 AJF~
8 Lee B lgSS
A~EsI AIES3 A~" A~;~-
S'hope Humps Nz~V
//tuhn 2/'/ezrschweziTchen 3 ttarnrne/ 4, ,~/nd
Abb. 1. Die Ordinatcnwertc geben den auf der Salk-Platte erhaltencn H~imagglutinationstitcr ( , , t IT") an, der nach 2 Std Aufenthalt bei 4 + ~ abgelesen wurde, Alle 15 ~iyxovirusstfimme Driiften wir akt iv und hitzeinaktiviert (30 rain bei 56~G im Wasserbad) mit nat iven Erythrocyten: 1 Huhn-, 2 5Ieerschweinchen-, 3 t Iammel- und 4 Rindcrerythrocyien. Der EinfluB der Inaktivierung wurde folgendermaiJen kenntlich gemacht: [ ] t~bereinstimmender l iT yon akt ivem und inaktiviertem Virusmaterial. [ ] Abfan des l IT nach der hmkt ivierung um einen best immten Betrag. [ ] Zunahme des HT nach der Inakt ivierung um einen best immten Betrag. [ ] Ausbleiben der t t~magglutination nach der Inaktivierung. Die SKulen geben den mit akt ivem Virusmaterial erhalteaen 14T an. Die in der
Legcnde angegebenen Siiulen entsprechen in Abb.1 einzeln numericrten ganzen S~iulcn
3. Der Stamm A/PR8 und dio boiden B-St~mmo zoigten mi~ Ery- throcyten vom Rind einen gr6Beren Titorverlust.
4. Der AlE 51-St~mm hatte als einziger Influenzavirusstamm (:lurch den Inak~ivierungsprozeB seinen (geringen) Hgmagglutinationstiter gegeniiber den Erythrocyten yore Hammel und Rind verloren.
5. Die St/imme A/WeiB und A1/FM 11leiden wiederholt nach der Inakti- vicrung einen etwas hSheren H~magglutinutionstiter erkennen.
6. Die St4imme Mumps und NDV verloren durch den Inaktivierungs- proze8 ihro h/imagglutinierende Aktivititt.

5Iyxovirus.H~magglutinationsversuche 33
3. Absorptions- und Elutionsversuche mit aktiven und hitzeinaktivierten Myxoviren
Die Tab. 2 zeigt die Ergebnisse der /)riifungen auf H~tmagglutinations- und Elutionsfiihigkeit auf Salk-Plexiplatten unter Verwendung yon aktivem u n d - dureh 30 Inin Aufenthalt im Wasserbad bei 56~ -- inaktiviertem Virusmaterial. Dic H~magglutinations~iter, die sich nach zweistiindigem Stehen bei +4~ ergaben (s. Tab. 1 u. Abb. 1), wurden vermerk~ und je eine Platte bei Zimmertemperatur und +4~ belassen. Nach insgesamt 3 und 24 S~d lasen wir erncut die I-I~magglutina- tionstiter ab. Die sich ergebenden Zonenph~nomene sind in Tab. 2 nicht beriick- sichtigt worden (s. Absohnitt: Versuche zur Deutung der beobachteten Zonen- phiLnomene).
Die Ergebnisse k6nnen folgendermal3on zusammengefaBt werden :
1. Die Virusst~mme zeigten abh~ngig yon der Temperatur, bei der dio Reaktion abl~uft, der VJrusaktivit~t und der verwondeten Erythroeyton- art oftmals ein unterschiodliehes H~magglutinations- und Elutionsver- m6gen.
2. Erythroey~en vom Huhn und yore Meersehweinehen neigten im allgemeinon weniger zur Elution auf der Platte als solehe vom Hammel und Rind.
3. Bei Verwendung yon Er3r~hroeyten vom Hammel und Rind ergaben sieh folgendo Besonderheiten:
a) Hammelerythrocyten. Einigo Myxovirusst~mmo (A/WeiB, A1/FM 1 und A1G ) vermochten bei Zimmertemperatur nur im inalctivierten Zustand eine tt~magglutination herbeizuffihren. I n der K~ilte bei + 4 ~ agglu- tinierten sio dagegen dio Erythrocyten dieser Blntart auch in der aktiven Form.
b) Rindererythrocyten. Die Mehrzahl der Virusstiimme agglutinierte die Rindererythrocyten nur in der Kdlte bei +d ~ C. Lediglich dio St~mmo mit cincm grSI3eren ,,Agglutinationsspoktrum" (A/Asia 57 und clio beiden ]3- Sti~mmo) vermochten aktiv und inaktivier~ sowohl bei Zimmortempera- tur als auch bei + 4 ~ clio Blutk5rperchen dieser Tierart zu h~magglu- tinicrcn.
4. Der AlE 51-Stamm hatte als einziger Influenzavirusstamm (wie bereits erwi~hnt) dutch dell InaktivierungsprozeB seinen geringen (!) Hamagglutinationstiter gegenfibor den BlutkOrperchen vom Hammel und Rind verloren.
5. Die S~s Mumps und NDV bfiBten durch den Inaktivierungs- prozoB ihre Hiimagglutinationsfiihigkoit ein.
6. Allgemein ist zu sagen, dab das Elutionsphiinomen nach 24 Std vollsti~ndiger war und besondors mit aktivem Virusmaterial bei Zimmer- temperatur in Erscheinung trat.
Diese zum Tell sehr unterschiedlichen Agglutinationsergebnisse lassen sich zwanglos durch verschiedeno Elutionsgeschwindigkeiten erldgren. Es koImte gezeigt werden, dab ein Virusstamm yon verschiedenartigen Erythrocyten unterschiedlich
Z. t Iyg. Infekt . -Kr. , Bd. 146 3

O00
O
OO
O0
OO
O0
0 O
@ 6
~
0()0
@
00@
@
OO
oo
00
~6
~
OQ
O0
OO
OO
QO
OQ
O0
o~
o~
@O
@O
OO
O0
t4
t~
1@
t~
00@
0
0000
oo,
001~
i
00@
@
0000
:ooo
OO
O
00
~0
0000
O0
oO
0
00
0
00
00
O
OO
0
0000
0000
QO
QO
0000
OO
OO
OO
OO
@@
00
0000
OQ
O0
0000
00@
0'00
00
O0~
OlO
000
o@
oo
00~
0 ~ @
IL
@@
0 ~ C
~o
o
o0o0
oe
I
I
I I0
0 ~
Oo@
I
r
�9
�9
I I
~00
0
~@
@0
000
OO
oO
I /I
I I
I
I @
0~
I
I O
Oo
O
[
Illl
II
II
IIO
OG
Oo@
I111
~
1oo
oO
oQ
0
00
�9
~ �9
0000
O
000
OO
OO
O
000
OO
OO
O
0@@
OO
OO
00
@@
0006
) 00
o~
OO
OO
00
~0
I I
I I
~[
I I
O0
@
O
Oo
0
T.a
�9
�9

Myxovirus-H~magglutinationsversuche 35
B195s
Shope
Mumps
NDV
StSmme
3 a. 24
3 i. 24
3 a. 24
3 i. 24
3 a. 24
3 i. 24
3 a. 24
3 i. 24
Tabelle 2
Huhn
W K
O 0 0 0 0 0 0 0
0 0 �9
0 0 �9 O
0 0 0 0
0
Fortsetzung)
Meerschw.
W K
0 0 0 0 0 0 0 0
0 0 0 O 0 0 0 0
O O 0 0
0 0 �9 @
tIammel W K
@ @ �9 (p
0 0 0 0
-- 0
--
-- ()
--
0 0 O 0
- - O - - @
Rind
W K
@ @ �9 (ll
p
0 O 0 0
Es wurden die nativen Erythrocyten yore Huhn, l~[eerschweinchen, Hammel und Rind verwendet. Den H~magglutinationstiter ermittelten wir auf der Salk- Plexiplatte. Die Inaktivierung des Virusmaterials erfolgte 30 min bei 56~ im Wasserbad (,,a.": aktives, ,,i.": inaktiviertes Virusmaterial). Die Priifung auf E[u- tion wurde nach 3 und 24 Std sowohl in der W~rme (, ,W": Zimmertemperatur) als auch in der KMte ( , ,K": + 4 ~ vorgenommen:
O : Der H~magglutinationstiter hat sich praktisch nicht ver~tndert
(~ @ @ �9 : Verlust yon 25~ 50~ 75~ und Verlust aller urspriinglich vor- handenen Hamagglutinationseinheiten.
, , - - " : Keine H~magglutination
schncll eluiert. Im Falle des Huhnerythrocyten verl~uft die Elution relativ langsam, so dab selbst bei Zimmertemperatur eine H~tmagglutination beobachtct werden kann. Bei anderen Erythrocytenartcn (in unscren Untcrsuchungen z. B. Blutk6rper- chen yore Hammel, Rind und yon dcr Katze) kann die Absorption des Virus bci Zimmertemperatur Hand in Hand mit der Elut ion gehen. Erst durch bestimmte ,,Kunstgriffe", die die Elutionsphase verzSgern bzw. ausschalten (Ansetzen der Reaktion bei + 4 ~ oder Inaktivierung des Virusmaterials), kann es zur H~magglu- tination kommen2L Das ist auch ein Grund ffir die Empfehlung, den Hi~magglu- tinationstest grunds~tzlich bei ~ 4 ~ durchzufiihren%
Die in Tab . 2 v c r m e r k t e n E l u t i o n s e r g e b n i s s e s ind a u f d e r S a l k - P l e x i -
p l a t t e abge l e sen worden . E s w a r n u n y o n I n t e r e s s o zu pr i i fen , i n w i e w e i t
die a u f d e r P l a t t e s i c h t b a r c E l u t i o n (d. h. zun i i chs t n u r das Z u s a m m e n -
s in t e rn des A g g l u t i n a t e s zu e i n e m k n S p f c h e n f S r m i g e n S e d i m e n t ) y o n
a k t i v e m u n d auch hitzeinaktiviertem V i r u s m a t e r i a l m i t e i n e m A b b a u de r
v i r u s g e r i c h t e t e n R e c e p t o r e n a u f de r E r y t h r o c y t e n m e m b r a n v e r b u n d e n ist .
3 "

36 ~ER~HARD SCH~HDT undKLAUS GROSSCEBAUER:
Zu diesem Zweeke prfiften wir einige Virusst~mme (u.a. AaE 51, A/Asia 57 und Shope) unter Itinzuziehung der Ery~hroeytenarten, mit denen ~uf der l~latte aueh mit hitzein~ktiviertem Virusmaterial eine Elution abgelesen wurde, auf die F~higkeit der Absorption, Elution, ~Viederagglutination (mit dem Ausgangsvirusmatcrial) und Panh~magglu- tination. Erythrocyten vom ttuhn wurden zum Vergleiehe mit heran- gezogen.
Die h~magglutinierende Aktivitiit der ~berstfinde naeh der jeweiligen Absorption bzw. Elution gab uns dabei des AusmaB der Beladung bzw. Absprengung, der Hitmagglutinationstiter des Wiederagglutinationsver- suches das 5'IaB ffir die Receptorzerst6rung am Erythrocyten an. Die Technik der Durchfiihrung dieser Versuche, yon denen ]edig]ich einer herausgegriffen wird, siehe unter ,,Material und Methoden".
Die Tab. 3 zeigt einen derartigen Versueh. Zun~chst zeigte es sich wieder, dab der InaktivierungsprozeB kaum
einen EinfluB auf die h~magglutinierende Aktivit~t des Virus ausiibte. Sowohl in der Wfirme als auch in der Ki~lte konnte auf der Salk-Platte ein gleichhoher Titer (~- +-4- : etwa 1280 H'/imagglutinationseinheiten) abgelesen werden. Die Absorptionsversuehe bestStigen die bekannte Tat- saehe, dab bei -4-4 ~ mehr Hs als bei Zimmer- temperatur absorbiert und entsprechend wieder eluiert werden. Unter den ]tier gegebenen Bedingungen konnte eine Elution des hitzeinaktivierten Virusmateri~ls nachgewiesen werden. Ein untcrschiedliches Verhalten ergaben abet diejenigen Untersuchungen, die die Priifung des Zustandes der vh'usgerichteten Receptoren auf den Erythrocyten zum Ziele hatten. Im Falle des tIuhnerythrocyten konnte nach praktisch vollst~ndiger Elu- tion des Virusmaterials keine Wiederagglutination mehr erreicht werden. Die Panhiimagglutination fiel in der Senkungspipette positiv aus, d.h. es trat sofort nach dem Misehen des Serums mit den ,,virusmodifizierten" Erythroeyten eine mit blol3em Auge deutlich sichtbare It~magglutination tin. Das Phdnomen der Panhiimagglutination lr auch mit inalctiviertem Virusmaterial ausgeldst werden. ])as serologische Verhalten dcr ,virus- modifizierten" Ituhnerythrocyten l~Bt den SchluB zu, dab durch enzy- matisehe Prozesse, die aber Iceineswegs durch die hier angewandte Inaktivie- rungstemperatur ausgeschaltet werden, yon seiten des Virus ein Abbau der Reeeptoren am Erythrocyten stattgefunden hat.
Anders verhielten sich die B]utk6rperchen veto l~ind. Trotz eines hohen Hs und einer nachweisbaren Absorptions- und Elutionsrate traten auf der Salk-P]atte hochtitrige Wiederagg]utina- tionen auf. Das Panh~magglutinationsphi~nomen war eindeutig negativ. Dieses Verhalten kann durch die Unversehrtheit der Influenzavirus- receptoren erkl~rt werden, die m6glicherweise am I~indererythrocyten anders gelagert bzw. auch andersartig vorliegen.

hiyxovirus-Hamagglutinationsversuehe 37
Es muB bo ton t werden, dab die in Tab. 3 ve rmerk ten Ergebnisse n u r fiir den A/Asia 57-Stature zutreffen. Bei Verwendung anderer Virus- s t amme u n d E r y t h r o c y t e n a r t e n ergaben sich oftmals sehr ungerschiedliche Ergebnisse.
Tabelle 3
E r y t h r . H u h n /~ind
Virus-M. a k t i v i n a k t i v a k t i v inak t iv Temper . W :K W K W K W K
1. HA 2. Abs.
a. EL. b. PHA e. WA
§ + + +
+ + + +
+ + §
+ §
+ + + + + +
+ + + +
+ + + + +
+ +
+ + + + +
+ + §
§ §
++ (+)
§ + + +
(+) ++
§ +§
++ (+)
Es wurde der A/Asia 57-Stamm mit nativen Erythrocyten vom Huhn und Rind getestet. Das Hamagglutinations- und AbsorptionsvermSgen (,,HA" und ,,Abs.") priiften wit bei Zimmertemperatur (,,W") und +4~ (,,K"). Die beiden virusbela- denen Erythrooytensedimente, die sich naeh der Absorption bei Zimmertemperatur und +4~ ergaben, wurden zur Ermittlung der Viruselution weiterverwandt. Mit beiden nach der ]~lution praktiseh virusfreien Erythroeytenfraktionen wurde eine Panh~magglutination (,,PHA") mit normalemPferdeserum und eine Wiederagglu- tination (,,~VA") mi~ dem Ausgangsvirusmaterial durchgeffihrt.
4. Bemerkungen zur Verwendung von /ormalinisierten Erythrocyteu ( , ,FE")
FE sind zur Beantwortung bestimmter Fragen in der Virologie und Bakterio- logie herangezogen worden. So z.B. bei den Arbeiten fiber das Verhalten der Influ- enzaviren in elektrolytfreiem Milieu und Fragen der Absorption-Elution und bei der Anreicherung yon Influenzaviren aus RachenspiilflfissigkeitS,s4,ss. 53. )Teuerdings wird fiber die Verwendbarkeit formalingehi~rteter Erythrocyten als Antigentrager beriehtet 2s,$2,~,~7,s2,93,94. Wir haben Mikroorganismen (Bakterien, Bacillen und Viren) beschrieben, die FE zu agglutinieren vermochten TM. Dabei ergab sich weiterhin, da~ die unterschiedIiehsten Bakteriensuspensionen, die allein FE nieht zur Agglu- tination braehten, zusammen mit bestimmten Hormalseren FE agglutinierten. Inwieweit dieses Hamagglutinationsphanomen in das Gebiet der Panagglutina- tionen gehSrt, ist noeh Gegenstand yon Untersuchungen.
Hinsichtlich des Verhaltens der FE gegeniiber Myxoviren ist hervorzuheben, dal~ FE yore Menschen nach FLICK $4 dieselbe Bindungskapazit~t fiir Influenzaviren besitzen wie nutive Blutkbrperchen. Eine dennoch mSgliche unspezifische, nicht receptorgebundene Adsorption-Elution wird durch BRA~D U. LIPPELT S ausgeschlos- sen. Bis auf eine verminderte l~eaktionsbereitschaft der formalinisierten Receptor- substanz und eine dadurch bedingte Verzbgerung der Absorptions-Elutionsabl~ufe fanden sie die virusgeriehteten Receptoren funktionstfichtig und im Sinne des Receptorgradienten spezifiseh.
Eine Vereinfachung ware es gewesen, derartige Flieksche FE statt der nativen Erythrocyten zurBestimmung desVirushamagglutinationstiters auf der Salk-Platte heranzuziehen, da sie sich fiber lange Zeitraume unbeschadet konservieren lassen, beim Zentrifugieren schneller absetzen und ein kompakteres Sediment geben, was wiederum eine grSl3ere Genauigkeit bei der Herstellung yon Erythrocytensuspen- sionen mit sich bringt. Weiterhin ware die Gewinnung yon Blutzellen nur in groi~en

3~ ]~ERI~HARD SCttMIDT und KL~US GROSSGEBALrER:
Zeitabst/~nden notwendig und eine Beurteilung von Titeri~nderungen fiber diese lange Zeit hinweg mit denselben Erythrocytenans/~tzen erfal~bar. Es wurde aber schon yon FLint; a4 festgestellt, dab Spontanagglutinationen dcr FE vom Mensehen auf der Salk-Platte auftreten. Cox 2~ suehte deshalb mit I-Iilfe yon besonders pr~- parierten FE vom Mensehen diese Vorteile der FE zu wahren. Wir haben fest- gestellt, dait sich die verschiedenartigen Erythrocyten nicht gleich gut zur For- malinisierung eignen. FE yon der Katze, yore Rind und vom Sehwein neigen besonders stark zu Spontanagglutinationen, wenn die Formalinhgrtung nach FLicX durchgeffihrt wird. Abet auch FE vom Pferd und vom Menschen waren bei weitem nicht so stabil wie z. B. diejenigen vom Huhn, Kaninchen, ttammel und Meerschwein- chen.
Interessant ist eine Beobachtung, nach der durch eine spezielle Formalinisierung yon Mauserythrocyten ein Ansteigen der Agglutinabilit/~t gegentiber ~iyxoviren erreicht werden konnte aa. Nach Ansicht der Verfasser kSnnten sieh derartige Ery- throeyten bei der Suche nach weiteren h~magglutinationsf/ihigen Viren bew~hren. Eine erst nach der Formalinisierung der Erythrocyten einsetze~e Agglutinierbarkeit haben wir unter Verwendung yon .Bakterien beobachtet 7~. Die Erythroeyten wurden dabei nach FLICK formalinisiert. Unserc jetzigen Versuche mit FE haben ergeben, dab einige Virusst/imme tats/~chlich erst nach der Formalinisierung bestimmter BlutkSrperchen in der Lage waren, diese zu agglutinieren (s. auch ,,Besprechung der Ergebnisse"). Es wird yon Fall zu Fall zu prfifen sein, inwieweit bei derartigen I-I/~magglutinationen ein echtes Reeeptorgeschehen vorliegt.
Um eine qualitative und anni~herungsweise quantitative Aussage iiber den H~m- agglutiningehalt einer myxovirusinfizierten Allantoisflfissigkeit zu maehen, haben wir fiber die Brauehbarkeit yon nach FLICl~ behandelten FE vom I-Iuhn berichtet 7a. Unter Ber/icksiehtigung des Agglutinationsbeginnes, d.h. der Zeit, die yon der Bereitung des Virus-Erythrocytengemisches (das sofort in eine gewShnliehe Blut- senkungspipette* aufgezogen wird) bis zum Auftreten einer mit bloSem Auge sichtbaren Hiimagglutination vergeht, kann man sich ein orientierendes Bild fiber den H/~magglutiningehalt verschaffen. Bemerkenswert ist, dab Kochsalzfehler oder andere Fehler der Verdtirmungsfliissigkeiten sowie Schwankungen der Erythrocyten- konzentrationen, die sieh bei der Salk-Methode durch Titeranderungen bemerkbar machen, hierbei wenig in Erseheinung treten. Hoehtitrige Allantoisflfissigkeiten ffihren -- i~hnlich der Objekttr/tgermethode 6r -- schneller zu einer makroskopiseh sichtbaren It~magglutination als niedrigtitrige.
Zusammenfassend kann gesagt werden, dag FE in neuerer Zeit in der Serologie Verwendung finden. Ihr praktiseher Wert wird sieh aber noch erweisen miissen.
5. Versuche zur Deutung der beobachteteu Zonenphi~nomene
Beim Ablesen der Elut ionsergebnisse au f der Salk-Ploxiplat~o ze ig ten sich bei einigen Vi rus s t~mmen sogenanute Zonenph'~nomeno 2~,a6. ~Nach
2, 6 oder m a n c h m a l auch ers t nach 24 Std waren nur noch in den K u p p e n
der mi t t l e r en Vi rusverd i innungen ~ypische Agglu t ina te zu sehen, w~hrend die Agglu t ina te m i t den sti~rksten Vi ruskonzen t r a t ionen vol ls t i indig
e luier t waren.
Die Tab .4 zeig~ die jenigen Virusst~imme (8 yon 15 un te r such ten ) , die
m i t den versch iedonar t igs ten E r y t h r o c y t e n dieses Ph~nomen e rgeben
haben .
* Ahnlich der Biutgruppenbestimmung in B[utsenkungspipetten 73.

Myxovirus- H~imagglutinationsversuche 39
Zun~chst f~llt das Verhalten des A/Asia 57-Stammes auf, der aktiv und hitze- inaktiviert (!) mit allen 4Erythrocytenarten das ZonenphAnomcn auslSste. Weiter- hin ist bemerkenswert, dab im ganzen 4 A-St~mme neueren Datums (die Mehrzahl der yon uns gepriiften) mit Huhnerythrocyten dieses Ph~nomen zeigten. Daran kSnnte man etwas Gemelnsames im Elutionsgeschehen erblicken. Die in Tab. 4 auf- gefiihrten Zonenphi~nomene bildeten sich nur bei Zimmertemperatur, nicht bei +4~
Das mit dem NVD-S tamm und I iuhnerythroeyten ausgelSste Zonen- ph~nomen wurde in Zusammenhang gebracht mit einem elutionsfSrdern- den Fak tor in der infizierten Allantoisflfissigkeit 21. Gereinigtes Newcastle- Disease-Virus eluierte we- sentlieh langsamer yon Huhnerythrocyten. "Wir haben nun die mit A/Asia 57 infizierto Allantoisfliis- sigkeit durch hoeh~ouriges Zentrifugieren (3 Std bei 20000 Upm) sowie dureh Absorption-Elution und an- schliei]endes hochtouriges Zentrifugieren weitgehend yon der Allantoisflfissigkeib befreit, in eino gleich grol~e Menge isotonische KoehsalzlSsung aufgenom- men und danach mit
S('~mme
Mel A,E 51 AlE 53 A55 A/Asia 57 Shope NDV Mumps
Tabe]le 4. Erythrocyten
Huhn
+
+
+ +*
+
Meerschw. IIammel
+
+
+
+* +*
+
+
:Rind
+0
+*
+ :Zonenphiinomen mit aktivem Virus- material.
+*: mit aktivem und inaktiviertem Virus- material.
+o : nur mit inaktiviertem Virusnmterial.
guhn- , Hammcl- und 5[eerschweinehenerythrocyten auf die Ausbildung des Zonenph/inomens untersueht. Dabei zeigte es sich, dai~ praktisch keine Untersehiede zwischcn den gereinigten Fr~ktionen und einer als Kontrolle verwendeten A/Asia infizierten Allantoisfliissigkeit bestanden.
6. Das Phiinomen der PanMimagglutination ( , ,PHA")
Menschlicbe Erythrocyten, die einer Myxoviruseinwirkung ausgesetz~ waren, erhielten best immte neue serologische Eigenschaften ~. Unter an- derem konnten sie gegenfiber ~inigenNormalserenpanagglutinabel gemacht werden. Dieses Ph~nomen hat groi]e ~ n l i c h k e i t mi t der von T~OMSEN beschriebenen PHA, hervorgerufen durch best immte Bakterien bzw. auch keimfreie Kulturfiltratel%ls3L
Fiir uns war in diesem Zusammenhang die Frage yon Intoresse, wie sich Erythrocyten anderer Species im virusvorbehandeltem Zustand zu- sammen mi t einigen Normalseren verhalten.
Aus diesem Grunde prfiften wir alle 15 Myxovirusst';imme, die wit jeweils auf Ery throcytcn yore t tuhn, I tammel , Meerschweinehen, Pferd
3,14--16,19,25,27,30,37,40,4{,82--86

40 BERNItARD SCttMIDT und KLAUS GROSSGEBAUER:
und R ind einwirken lieBen, mi t Normalseren vom Pferd, Kaninchen, Meerschweinchen und Menschen au f die Fahigkoit , zu panhamagglut inie- ren. Einige infizierte Allantoisfl/issigkeiten wurden auch hitzeinaktiviert , E ry th roey t en ve to H u h n auch formalinisiert verwendet .
Zu je 15 ml einer hochtitrigen Allantoisflfissigkeit gaben wir 2 ml des etwa 50~ Erythrocytensedimentes, lieBen das Gemiseh 2 Std bei +4~ stehen und eluierten danaeh d~s Virus fiber h~aeht bei 37~ Eine etw~ 6~ Erythrocyten- suspension wurde 1 : 1 mit dem jeweiligen Normalserum gemischt und zur Beobach- tung der PHA in eine gew6hrdiehe Blutsenkungspipette aufgezogen. DiG Kontrolle (start Serum nahmen wir isotonisehe Kochsalzl6sung) war stets negativ.
A u f eine tabellarische Wiedergabe der Ergebnisse sol aus Grfinden der Raumersparnis verziehtet . Wi r m6ehten sie folgendermaBen zusammen- fassen :
1. Normalserum ve to Menschen (ein Gemisch versehiedener blut- gruppenuntersehiedlicher Seren) agglutinierte keine der virusvorbehan- delten Ery th roey tenar ton .
2. I )urch Normalseren ve to Pferd und Kaninehen konnto mi t allen 15 Frak t ionen der nat iven modifizierten E r y t h r o e y t e n vom H u h n inner- halb yon 3 rain (oft sehon naeh wenigen Sekundon) das Panhamagglu- t ina t ionsphanomen hervorgorufen werden.
3. Die mi t einigen hi tzeinakt ivier ten Viren vorbehandel ten nat iven Blutk6rperchen ve to H u h n agglutinierten gleichfalls zusammen mit Pferdeseren.
4. Virusveriinderte formalinisierte H u h n e r y t h r o e y t e n verhielten sieh untersehiedlich. I m allgemeinen zeigte sich ein - - im Gegensatz zu den verander ten nat iven Blutk6rperchen - - verz6gerter E i n t r i t t der P H A und auch nnr mi t Pferdeserum. B Leo vorbehandel te F E agglut inier tcn nicht, Shope vorbehandel te z.B. besonders stark.
5. ,,Virusmodifizier~e" nat ive Meersehweinchenerythroeyten wurden zwar yon Pferdenormalserum prompt agglutiniert , dagegen nicht durch ihr homologes Serum.
6. E r y t h r o e y t e n vom R i n d zeigten in keinem Falle eine P H A , dic vom Pferd und H a m m e l nur be iVorbehandlung von einigen wenigen S tammen; aber aueh d a n n handel te es sich m n schwache, erst spat einsetzende Agglut inat ionen.
Bei den hier durchgefiihrten Serumagglutinationen war zu beachten, dab einige Seren in der Lage waren, bereits normalerweise nichtvorbehandelte Erythrocyten zu agglutinieren. Das wurde durch Kontrollen ausgesehlossen.
Da uns mit mensehlichem Normalserum eine PHA in keinem Falle gelang, versuchten wir dureh Trypsin-, Virus-, RDE- und Hitzevorbehandlung des Serums einen m6glieherweise vorliegenden Hemmfaktor2, ~2 zu beseitigen. Da die Versuehc negativ verliefen, sell hier nieht n/~her darauf eingegangen werden.

Myxovirus-H~magglutinationsversuehe 41
D. Besprechung der Ergebnisse
Es wurde das Verhalten verschiedenartiger Erythrocyten gegeniiber Myxoviron unter mSglichst vielseitigen Versuchsanordnungen verglei- chend untersucht. Die Ergebnisso, die zum groBen Tcil sinngem/iB mit den in der Literatur vorlicgendcn Resultaton fiberoinstimmen, zeigon eino Vielzahl feiner Unterschiedo im I-I~magglutinations-, Elutions- und ReceptorabbauvermSgen zwischen den einzelnen Virusst~mmen, sowohl einer bestimmten Erythrocytenar t als auch versehiedenartigen Blut- kSrperchen gegenfiber.
Zun~ehst wollen wir auf die l~unkte eingehen, doren Beantwortung yon praktischem Interesse zu sein semen:
Die GrSBe der ,,Agglutinationsspektren", die selektivo Agglutina- bilit/it und das VorhMten der formalinisierten Erythroeyten (,,FE").
Obwohl zwischen den gepr/iften 15 Myxovirusst/immen zum Tell charakteristische Unterschiede, die in Wiederholungsversuehen reprodu- zierbar waren, tfinsichtlich der GrSBo der Spektren, der ttShe dor I{fim- agglutinationstiter und der Reaktionsbereitschaft dor Virus-Erythro- cytengemisehe in der Blutsenkungspipette bcobaehtet werdcn konnten, m6chten wir ihren praktischen Wert nur als gering bemessen. Mit den hier verwendeten 7 Erythroeytenar ten kann keine Typendiagnoso be- triebon werden. Diese muB unseres Erachtens auch fiir andore Erythro- cyten in Frage gestellt werdon, da man die GrSBo des ,,Agglutinations. spoktrums" eines neu isolierten Virusstammes vorher nicht kennt. So konnten wir zeigen, dab aueh Erythroey~en veto Ziesel (Citollus ci~ellus *), die A- und A'-St~mme nich~ zu agglutinieren vormSgen und deshalb praktische Bedeutung ffir eine Typendifferenzierung besitzen sollen 5, vom A/Asia 57-Stamm doch agglutiniert werden. Das Agglutinationsspektrum scheint eben yon der immunologischen Spezi/itiit unbeeinlluflt zu bleiben.
Dagegen k6nnen einzelne St~mme (unabh/ingig yon ihrer Typon- zugehSrigkeit) u.a. auch durch ihre ,,Agglutinationsspek~ron" n~her charakterisiorb werdon13, 92. Aber selbs~ hier wisson wir noch nicht mit Sieherheit, ob bei allen gleichnamigen St~mmen mit demselbon Agglu- tina~ionsergebnis zu rechnen ist 79. Die allgemeine Labilits des Influenza- virus, Adaptationsvorg/~nge s9 und die in der Einleitung beschricbenen Sehwierigkeiten bei Vorgleichsuntorsuchungen kSnnton sich auoh in oinom differierenden I-I~m~gglutinationsvermSgen zeigen.
Der Versueh einer Typendifferenzierung ist auch mit Hilfe norrnalen Ratten- serums gemaeht worden s~.
Es wurde yon anderer Seite mitgeteilt, dal~ durch eine spezielle Formalinisierung yon Mauserythrocyten eine vordem nich~ vorhandene Aggluti~ierbarkeit dureh
:Fiir die freundliche ~berlassung zweier Ziesel mSchten wir auch an dieser Stelle Herrn Dr. L. BO~ECK~: (Bratislava, ~SAV-Virologisehe Abteilung) unseren verbindlichsten Dank sagen.

42 BElZl~HARD SCI{MIDT und KLAUS GROSSGEBAUEt{:
Myxoviren erreicht werden konnte an. Dieser Befund ist deshMb yon Interesse, well deraICcige Erythroeyten bei der Suehe nach weiteren, noch nicht bekannten ]l~Yn- agglutinationsf/ihigen Viren eingesetzt werden kSnnten.
Unsere Versuehe mit verschiedenartigen nach FLICK hergestellten formalinisierten Erythrocyten haben ergeben, dab einige Virus- st~mme -- in Ubereinstimmung mit den oben angegebenen Befunden -- erst naeh der Formalinisierung der BlutkSrperchen diese in der Senkungs- pipette zu agglutinieren vermochten. Es toni3 aber betont werden, dab gerade diese Erythrocyten zu Spontanagglutinationen neigten. W/ihrend die Viruserytt~'oeytengemische innerhalb von 1--3 rain zum It/~magglu- tinationsphi~nomen fiihrten, zeigte die Kochsalzkontrolle naeh etwa 20 min die Zeichen der Agglutination. Durch Variation des Formalini- sierungsprozesses 1/il~t sieh auck das tt~magglutinationsergebnis beein- flussen (eigene Versuche). M5glicherweise k6nnen bestimmte Erythro- cyten durch eine spezielle Formalinisierung oder auch Sublimath/irtung (eigene Versuche*) zum Naehweis h/imagglutinat, ionsf/~higer Partikeln verwendet werden.
Nebon diesen Fragen yon praktischem Interesse stehen woitere, die u.a. das Verhalten yon hitzeinaktiviertem Virus, den Influenzavirus- receptor verschiedenartiger Erythrocyten und das Problem der Panh~m- agglutination betreffen.
Es ist bekannf., dab das durch 30 rain Aufentllalt bei 56~ im Wasserbad inaktivierte Influenzavirus im al|gemeinen nur wenig yon sehler h~magglutinieren- den Aktivit~t einbiiBt42.43,:~,9~m u.a.). Es kann sogar eine ErhShung derselben ein- treten 21 (eigene Beobachtung : s. Tab. 2). Eine spontane Elution des erhitzten Virus soil im a]lgemeinen nieht mebr erfolgenX0,20m-45, 4s. Cn-g ~1 beobachtete eine Virus- elution (erhitztes B Lee-Virus) yon Rindererythroeyten, die nich~ mit einem l~eceptorabbau verbunden war.
Wir habcn die verschiedensten Myxovirusst/imme im hitzeinaktivier- ten Zustand auf Erythroeyten der unterschiedlichsten Species einwirken lassen und dabei festgestellt, dab es einzelne bei 56 ~ inaktivierte Virus- st/~mme gibt, die nicht nur die F/ihigkcit zur tt/~magglutination, sondern auch zur Elution, Panh/~magglutination, ReceptorzerstOrung und Ausbildung des sogenannten Zonenphs besitzen. Erst bei wenig hSheren Inaktivierungstemperaturen wurden die. onzymatischen Leistungen yon seiten dieser Virusst/~mme ausgeschaltet. Die Tatsache der Elution yon hitzeinaktiviertem Virusmaterial kSnnte mit einer geringen Bindungsfestigkcit des Virus am Erythroeyten zusammenh/ingen. Bei einer sehr schwaehen Virusbindung erfolgt eine Dissoziation (,,Disagglu- tination") sogar in Abwesenheit yon Enzymaktivit/i t ss. Wir haben in einigen F/fllen (mit Ausnahme der Rindererythroeyten) eine Recoptor- zerstSrung (Ausbleiben der Agglutination der virusvorbehandelten Blur-
Die Sublimathartung der Blutk5rperchen wurde in Anlehnung an eine Vor- schrift von M. MATTHES6~ 6s durchgefiihrt.

Myxovirus-H/~magglutinationsv:ersuche 43
k6rperehen mit dem Ausgangsvirusmaterial) und eine Panh/tmaggluti- nation erreichen k6nnen, so dab wir annehmen miissen, dab von diesen hitzeinaktivierten Viren noch Enzymaktivi t i i t ausgeht.
Von hitzeinaktivierten Viren, die die F/ihigkeit zu einer spontanon Elution und die der Reeeptorzerst6rung verloren haben, wissen wir, dab berei~s eine Bindung der Viren an die Blutk6rperehen deren Agglu- tination zur Folge haben kann. Die ZerstSrung der virusgerichteten Receptoren ist also nicht die notwendige Voraussetzung f/it den Eint r i t t der H/imagglutination. Umgekehrt ffihrt der Abbau dieser Receptoren night unbedingt zum I-I/imagglutinationsph/inomen30, 64 (RDE-Einwir- kung). Daraus kann geschlossen werden, dab der H/imagglutinationstiter kein zuverl/tssiges MaB fiir die receptorzerst6rende Virusaktivitiit sein kann. Um die ViImsaffinit/it auszudrfieken, wird yon anderer Seite das AusmaB der Virusadsorption angenommen s~ Es ist schwer zu priifen, ob die Enzymaktivi t / i t parallel mi t der Adsorptionskapazitit t verl~uft 11.
In unseren Panhgmagglutinationsversuchen konnten wir zeigen, dab - - unter den yon uns gew~ihlton Bediugungen - - keiner der 15 Myxo- virusstitmme in der Lage war, die verwendeten Ery throcyten so weir zu ver/~ndern, dab sie in der Senkungspipette mit einem menschliehen Nor- malserum zusammen agglutinierten. Gegen virusvorbehandelte Rinder- blutk6rperehen waren alle Seren wirkungslos ; beziiglieh ihres HSmagglu- tinationsverm6gens verhalten sic sich aber oftmals auflergew6hnlieh**. MSglicherweise k6unen die yon uns orhobenen Befundo zur Unterschei- dung der Thomsensehon und virusbedingten Panh~magglutinat ion bei~ragen 30.
Mannigfache Fragen, die die Verschiedenartigkeit der virusgerichteten Receptorenl,Ss,ss, ~ die Natur des Receptors al, den Mechanismus der Receptorzerst6rung (u.a.V0,:s), den Sitz des Virusenzyms3S, 39 sowie die Untersehiedliehkeit des Virusreceptors zum bakteriell gerichteten betref- fen 29, sind zum groBen Tell noch Gegenstand intensivon Studiums.
Zusammenfassung
1. It/ imagglutinationsversuche unter Verwendung yon 15 Myxovirus- st/immen und 7 verschiedcnartigen Erythrocytenspecics bestiitigen, dab es eine Vielzahl felner Unterschicde zwischcn den einzelnen Virusst~m- men gibt, sowohl einer Ery throcytenar t als auch verschiedenartigen Blut- k6rperchen gegeniiber.
2. Die , ,Agglutinationsspektren" scheinen yon der immunologisehen Spezifit/~t unbeeinfluBt zu bleiben.
* Zitiert nach Sinkovics, S. 37. ~ * 24j30,54,65,66~69~76,91 U . a ,

44 BERNIIARD SC~I~DT und KL~US GROSSGEBAUER:
3. Die optimalen Bedingungen ffir die jeweiligen Erythrocytensysteme sind ungleich; eine Inagglutinabiliti~t kann zum Teil unter anderen Um- st~inden a u f g e h o b e n we rden .
4. E i n z e l n e (30 m i n be t 86 ~ i m W a s s e r b a d ) i nuk t iv i e rbe M y x o v i r u s -
s t /~mme k 6 n n e n n e b e n d e r F ~ h i g k e i t zt tr H a m a g g l u t i n a t i o n a u c h d ie z u r E l u t i o n , P a n h / i m a g g l u t i n a t i o n , R e e e p t o r z e r s t 6 r u n g u n d A u s b i l d u n g des
s o g e n a n n t e n Z o n c n p h h n o m e n s bes i t zen . 5. E s w i r d e ine U b e r s i c h t f iber die e r h a l t e n e n Z o n e n p h a n o m e n e gegeben .
6. I m Z u s a m m e n h a n g m i t d e m P r o b l e m des I n f l u e n z a v i r u s r e c e p t o r s
v e r s e h i e d e n a r t i g e r E r y t h r o e y t e n w i r d e in B e i t r a g zu r F r a g e de r P a n h f i m -
a g g l u t i n a t i o n ge le i s t e t . 7. E i n e spez ic l le F o r m a l i n bzw. S u b l i m a t h i i r t u n g b e s t i m m t e r E r y t h r o -
c y t e n k a n n z u m N a e h w e i s w e i t e r e r h~ imagg lu t ina t ions f i ih ige r P a r t i k e l n
e m p f o h l e n w e r d e n .
Frau IlZMG.~.rCD B~AVN danken wir ffir ihre sorgfiiltige technische Mitarbeit.
Literatur 1 ADA, G. L., and J. D. STO~E: Electrophoretie studies of virus-red cell inter-
action: Mobility gradient of cells treated with viruses of the influenza group and the receptor-destroying enzyme of V. cholerae. Brit. J. exp. P~th. 81, 263 (1950). -- 2 ALLEN, •., ~ . _A. ~INKELSTEIN and S. E. S~LKIN: Viral inhibitors in normal animal sera. Tex. Rep. Biol. Med. 16,391 (1958). -- a ANDErtSON, S. G. : The reaction between red ceils and viruses of the influenz~ group. Studies with Newcastle Disease Virus. Austr. J . exp. Biol. reed. Set. 25, 163 (1947). -- 4 BIEL]NG, R., u. L. OEL- RICHS: Untersuchungen fiber die H~magglutination durch Grippevirus. Z. Hyg. Infekt.-Kr. 127, 535 (1948). -- ~ BOREeK~, L.: On the question of virus receptors. The selective agglutinabflity of erythrocytes of the ground squirrel (Citellus citellus). Folia biol. (Praha) II , 344 (1956). - - G BaAeD, G.: ~t)-ber ein Anreicherungsver- fahren bet der Ziichtung yon Influenza-Virusst~mmen aus Rachenspfilflfissigkeit. Zbl. Bakt., I. Abt. Orig. 164, 185 (1955). -- ~ BRA~D, G.: Zur Antigenstruktur des Influenzavirus. Habilitationsschrift, ~Vestberlin 1956. - - s B ~ N n , G., u. H. LrP- PENT: Untersuchungen fiber den Influenzavirus-Rezeptor formalinisierter Erythro- cyten. Arch. ges. Virusforsch. 6, 134 (1955). - - 9 BICAND, G., H. LIPPELT U. ~. MUL- LE~: Untersuchungen tiber den EinfiuB der Blutk6rpcrchcn auf dcn Hhmagglutin~- tionstest mit Influenza- und Mumps-Virus. Z. Hyg. Infekt.-Kr. 139, 293 (1954). -- xo BalODY, B. A. : Characterization of the , ,enzymic" action of influenza virus on human red cells. J . Imlnunol. 59, 115 (1948). -- 11 BUI~N~T, F. M.: Variation in influenza viruses. Handbuch d. Virusforschung, I I . Erg~nzungsband. Wien: Springer 1950, S. 47. - - ~2 BURNET, F. M. : Recent work on the intrinsic qualities of influenza virus. Aus: World Health Organisation: Influenza. A review of current research. Monograph series Nr. 20, Genf 1954. -- xa BTJItNET, F. M. : Principles of animal virology. New York 1955. -- 14 BURN-ET, F. M., J. F. I~ICCREA and J . D. STOle-E" Modification of human red cells by virus action. 1. The receptor gradicnt for virus action in human red ceils. Brit. g. exp. :Path. 27,228 (1946). - - t~ BURNET, F. M., and S. G. ANDm~SO~ : Modification of human red cells by virus action. 2. Agglutina- tion of modified human red cells by sera from cases of infectious mononucleosis. Brit. J. exp. Path. 27, 236 (1946). - - la BUlCNET, F. M.: Modification of human red cells by virus action. 3. A sensitive test for mumps antibody in human serum by

Myxovirus-Hitmagglutinationsversuche 45
the agglut inat ion of h u m a n red cells coated wi th a virus antigen. Brit . J . exp. Path. 27, 244 (1946). - - 1: CASELITZ, F. H., u. G. STEIN: Experimentel le Beitrage zum Problem der Hamagglu t ina t ion nach T~OMSE.~. (1. Mitteilung) Z. Immun. - Forsch. l lO, 165 (1953). - - is CASELITZ, F. H., u. K. )LxRTENS: Experimcntel le Beitrage zum Problem der Hamagglu t ina t ion nach THO.~ISEN. (2. Mitteilung) Z. Immun.-Forsch. 110, 206 (1953). - - 1~ CHv, C. M.: Enzymic act ion of viruses and bacterial products on h u m a n red cells. Na ture (Lond.) 161, 606 (1948). - - 20 Clzu, C. M. : Inac t iva t ion of hamagglu t in in and infect ivi ty of influenza and Newcastle Disease viruses by hea t and by formalin. J. Hyg. (Lond.) 46, 247 (1948). - - 21 CHu, C. M. : Agglut inat ion of red blood cells of different animal species by influenza and Newcastle Disease viruses. J . Hyg. (Lond.) 46, 239 (1948). - - ~.2 CLARK, E., and F. P. O. NAGr.E~: Ham-Agglu t ina t ion by viruses. The range of susceptible cells with special reference to agglut inat ion by vaccinia virus. Austr. J . exp. Biol. reed. Sci. 21, 103 (1943). - - ~3 CoLLE, L. 1~., and V. R. F.~RELr.: A method for coupling protein antigens to erythrocytes. J . exp. Med. 102, 631 (1955). - - :~ Coo.~n3s, 1~. Ir A., M. H. GLEESoN-Wn-ITE and J . G. HALL: Factors influencing the agglut inabi l i ty of red cells. Bri t . J . exp. Pa th . 32, 195 (1951). - - -,5 Cox, C. D., A. ZERSCItLI~CG and R. MELOY: Hemolysis of PR8-virus sensitized erythrocytes. Proc. Soc. exp. Biol. (N.Y.) 84, 693 (1953). - - 26 Cox, C. D., and E. C. PmTLV.: T i t ra t ion of influenza virus and viral antibodies by preserved h u m a n erythrocytes. Proc. Soc. exp. Biol. (N.Y.) 93, 373 (1956). - - 2~ DEIBEr., R. : Virushamagglut inat ion. Klin. Wschr. 85, 265 (1957). - - 2s D E ~ ' o~ , 1~., and P. FISET: Serological behaviour of influenza viruses. 1. The effect of different fowl erythrocytes on the results of serological tests. Bull. Wld H l t h Org. 11, 981 (1954). - - -~ DRIMMER-HErCRN~EIS~R, H., and A. L. OLLITZKI: A_lcaligenes hamagglu t inans : Factors determining the agglut inabil i ty of the red blood cells and the mechanism of the agglutination. Schweiz. Z. allg. Path. 21, 93 (1958). - - 30 EmLEIr M. : Veranderungen yon Ery th roey ten du tch den Vibrio cholerae und Trypsin. Z. I-Iyg. Infekt . -Kr. 133, 543 (1952). - - al FAILLARD, H. : (~ber die Abspal tung yon N-Acetyl-neuraminsaure aus Mucinen durch das ,,l~ecep- to t -Dest roying-Enzyme" aus Vibrio cholerae, t toppe Seyiers Z. physiol. Chem. 307, 62 (1957). - - ~2 F~tVCO~'~r~EIL B. : Ut i l isat ion des hLmaties hyperformol6es en virologie. I. Pr6para t ion et propri4t~s g6n6rales de ces h6maties. I I . Mise au poin t d 'un complexe virus-hLmaties de longue conservation, util isable pour le diagnostic des oreillons par une m6thode d ' inhib i t ion de l 'h6magglut inat ion simplifi6e. Ann. Inst. Pas teur T 95, 777 (1958); T 96, 110 (1958/59). - - aa FAtrCO.WNXF~R, B., and D. BARUA: Increase of viral haemagglut inabi l i ty of red cells of the mouse af ter t rea tment wi th formalin. Na tu re (Lend.) 183, 629 (1959). - - 3~ FLrCr:, g. A.: Use of formalin- treated red cells for the s tudy of influenza virus hcmagglu t ina t ing activity. Prec. Soc. exp. Biol. (N.Y.) 68, 448 (1948). - - aa Fr.icK, J . A., B. Sa~-rORD and S. MVDD: The effect of sal t concentra t ion on the in terac t ion of i~dtuenza A virus and erythroeytes. J . Immunol . 61, 65 (1949). - - ar Fr.om~r.t.w~, A. L. : Hem- agglutination wi th Newcastle Disease virus (NDV). Prec. Soc. exp. Biol. (N.Y.) 64, 458 (1947). -- ar FLOe~a~'~, A. L.: Some al terat ions in chicken erythrocytes wlfich follow t r ea tmen t wi th influenza and Newcastle Disease virus. J. Bact. 55, 183 (1948). - - as GOTTSCItALK, A., and BEVERLEY T. PEm~Y: Ovomucin: A sub- strate for the enzyme of the influenza virus (III) . Bri t . J . exp. Pa th . 32, 408 (1951). - - ~9 GOTTSC~AL]~, A. : Virus enzymes and virus templates. Physiol. Rev. 87, 66 (1957). - - a0 HALLAVEZ, C. : Die Hamagglu t ina t ion durch Virusarten. (Phanomen yon G. K. HIRST). H a n d b u c h d. Virusforschg., II . Erg.-band. XVien: Springer-VErlag 1950, S. 141. - - ~ HA.WIn, M. : Electrokinet ic change in h u m a n erythrocytes during adsorp- tion and elution of PlY8 influenza virus.Proc.Soc, exp. Biol. (N.Y.) 68,385 (1948). - - ~'~ HmST, G. K. : Adsorpt ion of influenza hemagglutinins and virus by red blood

46 :BERNHAED S c ~ m ) w und K~Aus G n o s s ~ B A v ~ :
cells. J . exp. Med. 76, 195 (1942). - - 43 HIP, ST, G. K. : The quant i ta t ive determinat ion of influenza virus and antibodies by means of red cell agglutination, g. exp. Med. 75, 49 (1942). - - 44 HmST, G. K. : The na ture of the virus receptors of red cells. (I). J . exp. Med. 87, 301, 315 (1948). - - ~ HIRST, G. K. : The na ture of tile virus receptors of red cells. (II). J . exp. Med. 89, 223, 233 (1949). - - 46 INCRAI~A~i, g. S. : The prepara t ion and use of formalinized erythrocytes wi th a t tached antigens or haptens to t i t r a te antibodies. Proe. Soc. exp. Biol. (N.Y.). 99, 452 (1958). - - 47 INGRA- ~A~I, J . S. : Effect of t r ea tmen t wi th formalin on agglut inat ion of red cells by ant iserum homologous to artificially a t t ached antigens. Fed. Proe. 17, 518 (1958). - - as IsnAcs, A. : React ivat ions of neut ra l mixtures of influenza virus and serum by virus inac t iva tcd by heat . Brit . J . exp. Pa th . 29, 529 (1948). -- ~9 JocgE.~i, E. M.: RSle des streptocoques dans la panh~magglut inat ion et la polyagglutinabili t~. (I). RSle de bacillus anthracis dans la panh~magglut inat ion. Ann. Inst . Pas teur T 95, 756, 760 (1958) (II). - - s0 JONES, M.: Adapt ion of influenza virus to heat . Proc. Soe. exp. Biol. (N.Y.). 58, 315 (1945). - - 51 K_Aw]~cKI, Z.: Dete rmina t ion of the difference in the properties of adsorpt ion of the influenza virus by various kinds of erythrocytes. Bull. Inst . mar. trop. Med. Gds 7, 76 (1956). - - ~2 KLOSE, A.: Ober die En t s t ehung der Hemmstoffe gegen Grippevirus im menschlichen Normal- serum. Z. Hyg. Infekt .-Kr. 181, 46 (1950). - - as LrerEUT, H., u. G. BRAND: Anreiche. rung des Influenza.Virus bei Erstisolierungen. Arch. ges. Virusforsch. 5, 277 (1954).-- 5a LIPPELT, H., u. J . ~OGALSKI: Die Hetero-H~magglut inat ion. (1. Mitteflung). Z. Hyg. Infekt . -Kr. 136, 177 (1953). - - ~s L6FFLER, }t. : Zur H~magglu t ina t ion durch Influenzavirus A-Asia. Schweiz. Z. allg. Pa th . 21, 1002 (1958). - - as LOWEr.L, F. C., and ~I. BUCKINGtIASf: A comparision of the effect of various salt concentrat ions on the agglut inat ion of red cells by influenza A virus and ant ibody. J. Immunol . 58, 229 (1948). - - 57 MAOILL, T. P. : The sorption of i~ffiuenzavirus by chicken erythro- eytes. J . exp. Med. 94, 31 (1951). - - ~s MAGILL, T. P., and J . Y. SucG: Physical- chemical factors in agglut inat ion of sheep erythrocytes by influenza virus. Proc. Soc. exp. Biol. (N.Y.) 66, 89 (1947). - - ~9 MAGILL, T. P., and J . Y. SvGo: The reversibi l i ty of the O - - D Type of influenza virus variat ion. J. exp. Med. 87, 535 (1948). - - 6~ M.: Experimentel ler Beitrag zur Frage der H~molyse. Miinch. med. Wschr. 1, 8 (1902). - - G1 McCttEA, J. F . : Modification of red-cell agglut inabi l i ty by C1. Welchii toxins. Austr. J . exp. Biol. reed. Sci. 25,127 (1947). - - 62 McKENlUA, J. M. : A stable prepara t ion of antigcn-sensitizcd erythroeytes. Proc. Soc. exp. Biol. (N.Y.) 95, 591 (1957). - - 63 MILLEn, G. L., and W. M. STANLEY: Quant i ta t ive aspects of the red blood cell agglut inat ion tes t for influenza virus. J. exp. Med. 79, 185 (1944). - - 6~ OVEm~IAN, J. 1~., and W. F. FRIEDEWALD: Cold hcmagglut inat ion reactions wi th influenza virus. J. Immunol . 62, 415 (1949). - - 6~ PoPe, L. : Uber die Verstarkung der H~tmagglutinationswirkung yon Tb-Immun- Seren durch Liquoid ,,Roche". Z. Immun.-Forseh. 114, 62 (1957). - - 6s P o r r , L.: ~ b e r die ~%utralisation yon TB-Ant ikSrpern in vivo. Z. Immun.-Forsch. 115, 56 (1958). - - ~ R~ETTIa, H. : Untersuchungen zur Immunit&t und Serologic der In- fluenza. ( I . - - iV. Mitteilung), Zbl. Bakt . , I. Abt. Orig. 152, 159, 186, 381 (1947). - - ~s SACHS, H. : Ober den Ans t r i t t des Hamoglobins aus sublimatgehi~rteten Blut- kOrperehen. Miinch. Med. Wschr. 5, 189 (1902). -- ~9 S~c~sE, I t . H. : Zur Frage der Bezichungcn zwischcn T-Ferment und Hamosensi t inen. Z. Hyg. Infekt . -Kr. 145,471 (1959). - - 70 SAGI~:, B., T. PUCK and SEY~mt~R LEvi tE : Quant i ta t ive aspects of the spontaneous elution of influenza virus from red cells. J . exp. Med. 99, 25 i (1954). -- 71 S ~ K , J. E. : A simplified procedure for t i t ra t ing hemagglut inat ing capacity of influenza-virus and the corresponding ant ibody. J . Immunol . 49, 87 (1944). - - ~ S~L~, J. E. : .Variation in influenza viruses. A s tudy of heat s tabi l i ty of the red ccll agglut inat ing factor. Proc. Soc. exp. Biol. (N u 63,134 (1946). - - ~a SA~WDER, F.,

Myxovirus-Hiimagglutinationsversuche 47
u. M. SANDER: Die Blutgruppenbestimmung als Blutsenkungsreaktion. Z. 2mmun.- Forsch. 95, 268 (1939). - - :t SCHMIDT, B., u. K. GROSSGE~AgER: Beobachtungcn an Viruserythrocytengemischen in Blutsenkungspipetten. Z. Hyg. Infekt.-Kr. 144, 549 (1958). - - 75 SCm~IDT, B., u. K. GROSSaEBAtTER: H~magglutinationsversuche mit verschiedcnartigcn nativen und formalinisierten Erythrocyten. Z. Hyg. Infckt.- Kr. 145, 276 (1958). - - 7~ SCm~IDT, G. : Ubcr den Einfluit yon Periston auf einige serologische Ph~nomene in vitro. Z. Immun.-Forsch. 107,341 (1950). - - 77 Scm~xDT, H. : Fortschri t te der Serologic. Darmstadt : Verlag Dr. Dietrich Steinkopff 1955. - - 78 S~EFFIELD, F. W., WmSOH SMIT~ and G. BELYAVIS: Purification of influenza virus by red-ceU adsorption and elution. Brit. J . exp. Path. 85, 214 (1954). -- 79 SIGEL, M. M. : Differences in the hemagglutinating and antigenic properties of strains of influenza virus isolated from one outbreak. J. Immunol. 62, 81 (1949). - - s0 SINKOVICS, J . : Die Grundlagen der Virusforschung. Verlag der ungarisehen Akademic der Wissenschaften 1956. - - sl SMADEL, J. E. : Serologic reactions in viral and rickettsial infections. Aus: T. M. RIV~a~s: Viral and rickettsial infections of man. Philadelphia 1948, S. 78. - - se SeRIN(~ER, G. F. : Wcitere Untcrsuchungcn tiber den Einflul] yon Influenzaviren auf Blutgruppenantigene. Klin. Wschr. 86, 442 (1958) - - 8a STANLEY, W. M. : Studies on listeria monocytogenes. 1. Isolation of a monocytosis producing agent. 2. R61e of listeria in infectious mononucleosis. Austr. J. exp. Biol. med. Sei. 27, 123, 133 (1949). - - s4 STONE, J . D.: Comparision of the action of V. cholerae enzyme and of viruses on the red cell surface. Austr. J. exp. Biol. med. Sci. 25, 137 (1947). - - 85 STONE, J . D., and G. L. AriA: Electro- phorctic studies of virus-red cell interaction: Additive effect of viruses of the influenza group and the receptor-destroying enzyme of V. cholerae. Brit. J . exp. Path. 82, 275 (1950). - - se STO~E, J. D., and G. L. AI)A: Eleetrophoretic studies of virus-red cell interaction: Relationship between agglutinability and electropbo- retie mobility. Brit. J . exp. Path. 83, 428 (1952). - - s7 STYK, B.: ~ o n specific inhibitors in normal rat serum for the influenza C type virus. Folia biol. (Praha) I, 207 (1955). - - ss TAMM, 2. : Influenza virus-erythrocyte interaction. 1. Reversible reaction between Lee virus and cat erythroeytes. J. Immunol. 73, 180 (1954). - - 8~ TAM.~, I. : Influenza virus-erythrocytc interaction. 2. Effect of egg passage on the reaction between a recently recovered influenza A virus and sheep erythrocytes. J. Immunol. 73, 190 (1954). - - ~0 TA)I.~I, I., and F. L. HORSFALL jr. : A mucoprotein derived from human urine which reacts with influenza, mumps, and Newcastle disease viruses. J. exp. Mcd. 95, 71 (1952). - - 91 "WALTnlZ, G., U. O. JAM~N: Untcr- suchungen tiber die Steigerung der Agglutinabilit~t roter BlutkSrperchen durch Fermente aus Virus und Bakterien-Kulturen. Z. Hyg. 2nfekt.-Kr. 181,407 (1950). - - 92 WANG, SAH-PIH, and T s u - Y A ~ L ~ : Laboratory studies on some biological properties of the far east influenza virus. J. infect. Dis. 103, 178 (1958). - - 9a WE~'- BnCH, R. : Die Verwendbarkeit formolbehandelter Erythrocytcn als Antigentr~iger in der indirekten H~magglutination. 1. Mitteilung. Schweiz. Z. allg. Path. 21, 1043 (1958). - - 9t WE~.~C~ , R. : Die Verwendbarkeit formolbehandelter Erythrocyten als Antigentr~ger in der indirekten tt~magglutination. Schweiz. Z. allg. Path. 22, 1 (1959). - - s~ WHITE, J. : The position of Taylor's 1233 strain of virus in the human red cell receptor gradient. Brit. J . exp. Path. 84, 668 (1953). - - 9~ W~LSO~ S.~IITII, and J. C. ~ . WESTWOOD: Influenza virus hacmagglutination. The mechanism of the Francis phenomenon. Brit. J. exp. Path. 31, 725 (1950). - - 9~ W~LSO.~ S.~HTH, and M. A. WESTWOOD: Factors involvcd in influenza h~magglutination reactions. Brit. J. exp. Path. 80, 48 (1949).
Prof. Dr. med. BERNHARD SCHMIDT und Dr. recd. ]~LAUS GROSSGEBAUER, Berlin ~ 65, FShrer Str. 14,
Inst i tut fiir Hygiene u. Medizinische Mikrobiologie der Freien Universit~t