Embed Size (px)
Citation preview

------------------------------~~--¥v~Ochratoxin A und B: Ein dreidimensionalesmolekulares Modell zur mechanistischenErklarung ihrer Toxizitat
Angelo Vedani' und Arend Bruinink?1 Biografik-Labor, Schweiz. Institut fur Altemativen zu Tierversuchen (SlAT), CH-Basel2 Institut fur Toxikologie der ETH und Universitat Zurich, CH-Schwerzenbach ZH
ZusammenfassungOchratoxine sind Giftstoffe, die van Aspergillus aluta-ceus und anderen Schimmelpilzen produziert undfreigesetzt werden. Bisher wurde allgemein angenom-men, dajJ die Hemmung des Enzyms Phenylalanin-t-RNA-Synthetase for die Toxizitat von Ochratoxin Averantwortlich war. Neueste in vitro Untersuchungenweisen aber auf eine nicht-kompetitive Kinetik bezuglicbPhenylalanin und damit auf die Prasenr eines eigentli-chen Ochratoxinrezeptors hin. Basierend auf denStrukturen van Ochratoxin A und B, Mellein und Phenyl-alanin haben wir mittels Pseudorezeptormodeling eindreidimensionales molekulares Modell fur die Bindungs-stelle des vermuteten Ochratoxinrezeptors generiert. DasModell besteht aus insgesamt zwolf Aminosauren. Diehohe Affinitdt (und damit die rezeptor-gekoppelteToxizitat) van Ochratoxin A und B wird in diesemModell durch ein Wasserstof/briickennetzwerk erklart=ein Netzwerk, das weder mit Mellein noch mit Phenyl-alan in als Substrat moglicn ist. Diese Substanzen rufenbei vergleichbaren Konzentrationen denn auch keinetoxischen Effekte hervor. Die Relevanz des Modells wirddurch die quantitative Voraussage der Bindungsaffinitii-ten von drei Testsubstanzen gestutzt.
1 Einleitung
Ochratoxin A und B (Abbildung la,lb) sind Giftstoffe, die yon Aspergil-lus alutaceus (fruher: Aspergillusochraceusi und anderen Schimmel-pilzen produziert werden. Ochrato-xin A (7 -carboxyl-5-chloro-8-hydro-xyl-3,4-dihydro-3R-methylisocuma-rin-7-L-~-phenylalanin) wurde alsLebensmittelverumeinigung nachge-wiesen (Baumann und Zimmerli,1988). Relativ hohe Konzentrationen
124
Summary: Ochratoxin A and B: A three-dimensionalmolecular modelfor a mechanistic explanation of theirtoxicity.Ochratoxins are toxic substances produced and releasedby Aspergillus alutaceus and other molds. So far, it wasgenerally believed that the inhibition of the enzymephenylalanine-t-Rbl/v-synthetase was responsible for thetoxicity of ochratoxin A. Most recent in vitro results,however, suggest a non-competitive mechanism withrespect to phenylalanine and, consequently, the existenceof a proper ochratoxin receptor. Based on the structuresochratoxin A and B, melle in and phenylalanine, we havegenerated a three-dimensional molecular model for thebinding site of a putative ochratoxin receptor by meansof pseudo receptor modeling. The model consists oftwelve amino-acid residues. In this model, the highaffinity (and, hence, the receptor-mediated toxicity) ofochratoxin A and B is explained by a hydrogen-bondnetwork - a network that is not possible with bothmellein and phenylalanine as substrates. Consequently,these compounds do not exhibit toxic effects at compara-ble concentrations. The relevance of the model is sup-ported by the quantitative prediction of the bindingaffinity of three test compounds.
Keywords: pseudoreceptor modeling, 3D-QSAR, ochra-toxin receptor surrogate, receptor-mediated toxicity.
wurden in Blut und Brustmilch desMenschen gefunden (Hald, 1991 ;Breitholz-EmanueIsson et aI., 1993).Bis heute wurden aber keine toxi-schen Effekte yon Ochratoxin A imMenschen nachgewiesen. Aufgrundvon in vivo Daten an Ratten undMausen wird jedoch vcrmutet, daBOchratoxin A im Menschen nephro-toxische und tcratogene Wi rkungenhervorruft. ZieIgewebe in der prana-tal en Phase sind hauptsachlich Au-gen und Nervensystem (Shirai, 1991;
Miki et aI., 1994). Der Wirkungsme-chanismus yon Ochratoxin A ist bisheute unbekannt; im allgemeinenwird aber angenommen, daB dieHemmung des Enzyms Phenylala-nin-t-RNA-Synthetase fur die Toxi-zitat von Ochratoxin A verantwort-lich ist (Creppy et al., 1983). Ktirz-lich wurde nachgewiesen (Studer eraI., 1995), daB sich 3R-Ochratoxin Abel erhohter Temperatur - wie siebeispielsweise beim Kochen oderRosten yon Nahrungsmitteln auftritt
ALTEX 13,3/96
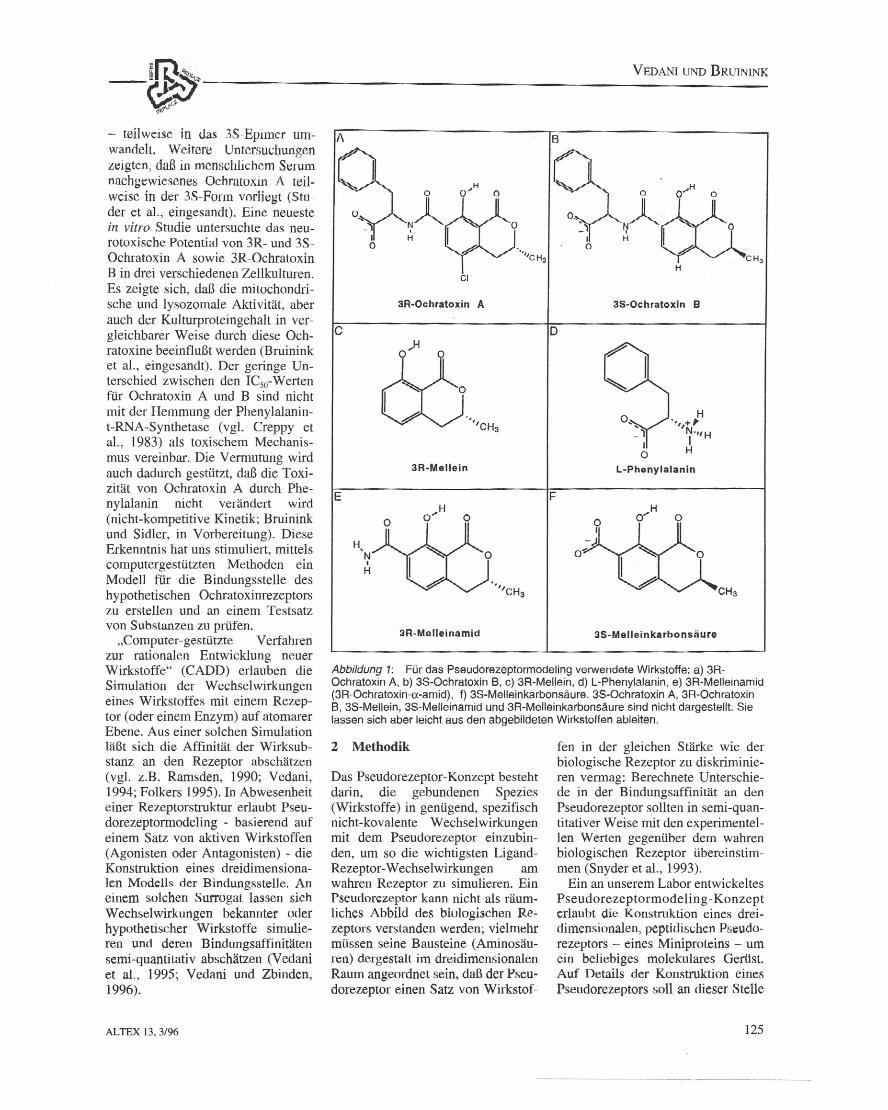
IITt~---~~----------------------------------------- teilweise in das 3S-Epimer um-wandelt, Weirere Untersuchungenzeigten, daB in mcnschlichcm Serumnachgewiesenes Ochratoxin A teil-weise in der 3S-Fonl1 vorliegt (Stu-der et al., eingesandt). Eine neuestein vitro Studie untersuchte das neu-rotoxische Potential von 3R- und 3S-Ochratoxin A sowie 3R-OchratoxinB in drei verschiedenen Zellkulturen.Es zeigte sich, daB die mitochondri-sche und lysozomale Aktivitat, aberauch der Ku1turproteingehalt in ver-gleichbarer Weise durch diese Och-ratoxine beeinfluBt werden (Bruininket a1., eingesandt). Der geringe Un-terschied zwischen den ICso-Wertenfur Ochratoxin A und B sind nichtmit der Hemmung der Phenylalanin-t-RNA-Synthetase (vg1. Creppy etal., 1983) als toxischem Mechanis-mus vereinbar. Die Vermutung wirdauch dadurch gestiitzt, daf die Toxi-zitat yon Ochratoxin A durch Phe-nylalanin nicht verandert wird(nicht-kompetitive Kinetik; Bruininkund Sidler, in Vorbereitung). DieseErkenntnis hat uns stirnuliert, mittelscomputergestiitzten Methoden einModell fur die Bindungsstelle deshypothetischen Ochratoxinrezeptorszu erstellen und an einem Testsatzvon Substanzen zu prufen,"Computer-gestiitzte Verfahren
zur rationalen Entwicklung neuerWirkstoffe" (CADD) erlauben dieSimulation der Wechselwirkungeneines Wirkstoffes mit einem Rezep-tor (oder einem Enzym) auf atomarerEbene. Aus einer solchen SimulationHH3tsich die Affinitat der Wirksub-stanz an den Rezeptor abschatzen(vgl. z.B. Ramsden, 1990; Vedani,1994; Folkers 1995). In Abwesenheiteiner Rezeptorstruktur erlaubt Pseu-dorezeptormodeling - basierend aufeinem Satz von aktiven Wirkstoffen(Agonisten oder Antagonisten) - dieKonstruktion eines dreidimensiona-len Modells der Bindungsstelle. Aneinem so1chen Surrogat lassen sichWechselwirkungen bekannter oderhypothetiseher Wirkstoffe simulie-ren und deren Bindungsaffinitatensemi-quantitativ abschatzen (Vedaniet al., 1995; Vedani und Zbinden,1996).
ALTEX 13. 3/96
VEDANI UND BRUININK
~~rY9~"CHS
CI
3R-Ochratoxin A
B
3S-0chratoxin B
C D
3R-Mellein L-Phenylalanin
E F..•.Ho 0
H,NI
H
3R-Melleinamid 3S-Melleinkarbonsiiure
CH3
Abbildung 1: FOrdas Pseudorezeptormodeling verwendete Wirkstoffe: a) 3R-Ochratoxin A. b) 3S-0chratoxin B, c) 3R-Mellein, d) L-Phenylalanin. e) 3R-Melleinamid(3R-Ochratoxin-a-amid), f) 3S-Melleinkarbonsaure. 3S-0chratoxin A, 3R-OchratoxinB. 3S-Mellein, 3S-Melleinamid und 3R-Melleinkarbonsaure sind nicht dargestellt. Sielassen sich aber leicht aus den abgebildeten Wirkstoffen ableilen.
2 Methodik
Das Pseudorezeptor-Konzept bestehtdarin, die gebundenen Spezies(Wirkstoffe) in genugend, spezifischnicht-kovalente Wechselwirkungenmit dem Pseudorezeptor einzubin-den, urn so die wichtigsten Ligand-Rezeptor-Wechselwirkungen amwahren Rezeptor zu simulieren. EinPseudorezeptor kann nieht als raum-liches Abbild des biologischen Re-zeptors verstanden werden; vielmehrmussen seine Bausteine (Aminosau-ren) dergestalt irn dreidimensionalenRaum angeordnet sein, daB der Pseu-dorezeptor einen Satz von Wirkstof-
fen in der gleichen Starke wie derbiologisehe Rezeptor zu diskriminie-ren vermag: Berechnete Unterschie-de in der Bindungsaffinitat an denPseudorezeptor soUten in semi-quan-titativer Weise mit den experimentel-len Werten gegeniiber dem wahrenbiologischen Rezeptor ubereinstim-men (Snyder et al., 1993).Ein an unserem Labor entwickeltes
Pseudorezeptormodeling -Konzeptcrlaubt die Konstruktion eines drei-dimensionalen, pepridischen Pseudo-rezeptors - eines Miniproteins - umein beliebiges mo1ekulares Geriist.Auf Details der Konstruktion einesPseudorezeptors soll an dieser Stelle
125

VEDANI UND BRUJN1NK ~r;')"------------~J~ ..-~~~.
nicht eingegangen worden (vgl. hie-zu Vedani et al., 1995; Vedani undZbinden, 19%); hingegen soll dieAbschatzung der Bindungsalfinitataus einem solchen dreidimensiona-len, molekularen Modell hergeleitetwerden.Zwischen einer molekularen
Struktur und ihrer Energie bestehteine quantitative Beziehung. Furkleine Molekule kann diese mitquantenchemischen Methoden sehrgenau berechnet werden. Fur grosse-re Systeme wie Proleine und Rezep-toren ubersteigt die Komplexitat sol-cher Rechnungen die heute zur Ver-fugung stehende Computerleistungbei weitem. Hier wird auf empirischeVerfahren (z.B. Kraftfeldrnethoden)zuruckgegriffen. Der in unseremKonzept verwendete Algorithmusberechnet mit Hilfe eines solchenKraftfeldes (Vedani, 1994; Vedaniund Huhta, 1990) fur jeden Wirk-stoff eine Wechselwirkungsenergie,~, mit dem Pseudorezeptor. Urnaus der relativen Wechselwirkungs-energie eines einzelnen Wirkstoffs,ME, (beziiglich eines Referenzwirk-stoffs) die Bindungsaffinitat, k, ge-geniiber dem wahren biologischenRezeptor abzuleiten, benutzen wireine Naherung von Blaney und Mit-arbeitern (1984):
~(~GQb<J '" t.(~Eber) - ~(Tt.SBindung)- t.(~GSOlvatation, Wirkstuff)
SOWle:k == e-LiGoIRTbzw.k/k2 == e-MG'IRT
Die freie Solvatationsenergie derWirkstoffe, t.GSolvatation. Wirksrofrs berech-nen wir nach einem semi-analyti-schen Ansatz (Still et al., 1990);Unterschiede in der Bindungsentro-pie, T~S8indUng,konnen aufgrund derTorsionsfreiheitsgrade der einzelnenWirkstoffe abgeschatzt werden (Se-arle and Williams, ] 992).Quantitativ gesehen, diskrirniniert
ein Pseudurezeptor nicht immer imgleichen AusmaB -wie der entspre-chen de biologische Rezeptor. Urnsemi-quantitative Voraussagen zumachen, konnen berechnete Werteanhand einer linearen Regression
126
(Steigung a, Achsenuhschnitt b) mitden Datenpaaren des Trainingssatzcskorrigiert werden:
Um wahrend der Strukturvcrfeine-rung die Korrelation zu maximieren,doh. ein aussagekraftiges Modell zuerhalten, wird im Kraftfeld-Energie-ausdruck ein zusatzlicher Kopp-lungsterm eingefuhrt (Vedani et al.,eingcsandt):
Dieser Ansatz erlaubt es, fur einengegebenen Trainingssatz an Wirk-stoffen ein Modell mit hoher Korre-lation (>0,95) zu finden. Fur dieGiite des Surrogates ist eine solchenotwendig, aber nicht hinreichend,Vielmehr muB das RezeptonnodellanschlieBend an einem Satz von Test-wirkstoffen validiert werden, d.h. anSubstanzen, die nicht zur Erstellungdes Modelles verwendet wurden.Um die gegenseitige Anpassung
von Wirkstoff und Rezeptor zu simu-lieren, haben wir den Algorithmus soerweitert, daf zunachst der Rezeptormit Korrelationskopplung optimiertwird. Dies fuhrt zu einem Modell mithoher Korrelation (>0,95) fur dieWirkstoffe des Trainingssatzes. Ineinem zweiten Schritt werden dieWirkstoffe frei (d.h. ohne Korrelati-onskopplung) relaxiert. Dies fuhrtzwangsweise zu einem Modell mitgeringerer Korrelation. Danach wirdwiederum der Rezeptor mit Korrela-tionskopplung optimiert, und dieKorrelation wird wieder hoch. Imnachsten Schritt (Relaxation derWirkstoffe) wird die Korrelationabermals abnehmen, aber nicht mehrso stark wie beirn ersten Mal, da sichdas Rezeptormodell im zweitenSchritt den relaxierten Wirkstoffenbereits angepaBt hat. In einem iterati-yen Verfahren werden nun diesebeiden Schritte so lange ausgefuhrt,bis das relaxierte Modell ebenfallseine hohe Korrelation aufweist. Manspricht dann von einem "equilibrier-ten System".
Anschlieliend werden die Wirk-stoffe des Tcstsatzes zugefugt undohne Korrelationskopplung relaxiert(denn sonst wurden sie lediglicheinern erweiterten Trainingssatz ent-sprechen) Das Rezeptormodell unddie Wirkstoffe des Trainingssatzeswerden dabei nicht mehr verandert.Die Wirkstoffe des Testsatzes kon-nen sich so - wahrend der Simulati-on - individuell an die Rezeptortopo-logie anpassen, was ja auch ambiologischen Rezeptor der Fall ist.AuBerdem werden so die Wirkstoffedes Testsatzes gleich wie diejenigendes Trainingssatzes behandelt.
3 Resultate und Diskussion
Unser Modell fur den vermutetenOchratoxinrezeptor wurde basierendauf einem Trainingssatz von vierWirkstoffen (3R-Ochratoxin A, 3R-Ochratoxin B, 3S-0chratoxin B, 3S-Mellein) erstellt und anschlieBend aneinem Testsatz von drei Wirkstoffen(3S-0chratoxin A, 3R-Mellein, L-Phenylalanin) gepruft. Die ICso-Wer-te von 3R-Ochratoxin A, 3S-0chra-toxin A, 3R-Ochratoxin B, ±Melleinund L-Phenylalanin stammen ausExperimenten, in welchen die Effek-te dieser Substanzen auf den Protein-gehalt von Hirn- und Hirnhautzellendes Hiihnerembryos in Flachkulturenbestimmt wurden (Bruinink et al.,eingesandt). Die ICso-Werte von 3R-Ochratoxin A und 3S-0chratoxin Aunterscheiden sich im Experimenturn einen Faktor 12. Derselbe Faktorwurde verwendet, um die ICso-Wertefur 3S-0chratoxin B und 3S-Melleinzu bestimrnen. Dabei wurde ange-nommen, daf sich die Bindungsaffi-nitaten der 3R und 3S-Formen vonOchratoxin B und Mellein um den-selben Faktor 12 unterscheiden wiediejenigen des Ochratoxin A. Zu-satzlich wurde angenomrnen, daBalle Testsubstanzen gleich stabil wie3R-Ochratoxin A, 3S-0chratoxin A,3R-Ochratoxin B (Bruinink et a1.,eingesandt) sind.Die dreidimensionalen Strukturen
der uruersuchten Wirkstoffe wurdenmit MacroModel 5.0 (Mohamadi etal., 1990) erzeugt und unter Beruck-
ALTEX 13,3/96
~rl\..~ VEOANI UNO BRUININK--~~~-------------------------------~.sicluigung von Solvntatiouseftcktenoptimiert. Die Sauregruppe der Och-ratoxine ist bei physiologischem pHnicht protoniert (Ahb. I a, lb); L-Pheuylalanin Iiegt in wassrigcr Lo-sung als Zwitterion vor (Abb. 1d).ESP (electrostatic potential fitted)-Ladungen wurden mit MOPAC 6.0(QCPE, 1992) bestimmt, die Solvat-ationsenergien der freien Wirkstoffenach einem serni-analytischen An-satz berechnet (Still, 1990). Das Mo-dell fur den Ochratoxinrezeptor wur-de mit der an unserem Labor entwik-kelten Pseudorezeptormodelingsoft-ware Yak 4.1 (Vedani et aI., 1995;Vedani und Zbinden, 1996, Vedani etaI., eingesandt) erstellt und optimiert.In diesem Modell betragt der Kor-
relationskoeffizient berechneter undexperimenteller Bindungsaffinitatenfur die vier Wirkstoffe des Trai-ningssatzes 1,00 (Tabelle). Die Stei-gung der Regressionsgeraden errech-net sich zu 0,94, was zeigt, daB derPseudorezeptor die gepriiften Wirk-stoffe fast gleich wie der biologischeRezeptor diskriminiert. Der mittlereFehler in der Voraussage der Bin-dungsaffinitat ftir einen Testsatz vondrei Wirkstoffen betragt 0,182 kcal/mol, entsprechend einem Faktor 1,4im ICso-Wert. Ein Vertrauensbereichmit 99,7% Wahrscheinlichkeit (3cr)entspricht 0,669 kcallmol bzw. ei-nem Faktor 3,2 im ICso-Wert. Diesist bei einem Affinitatsbereich von105 und einem mittleren Fehler in derexperimentellen Bestimmung derICso-Werte yon einem Faktor 1,5(Bruinink et al., eingesandt) hinrei-chend klein, um cine sichere Klassie-rung der gepruften Substanzen vor-zunehmen.Das Surrogat fur die Bindungsstel-
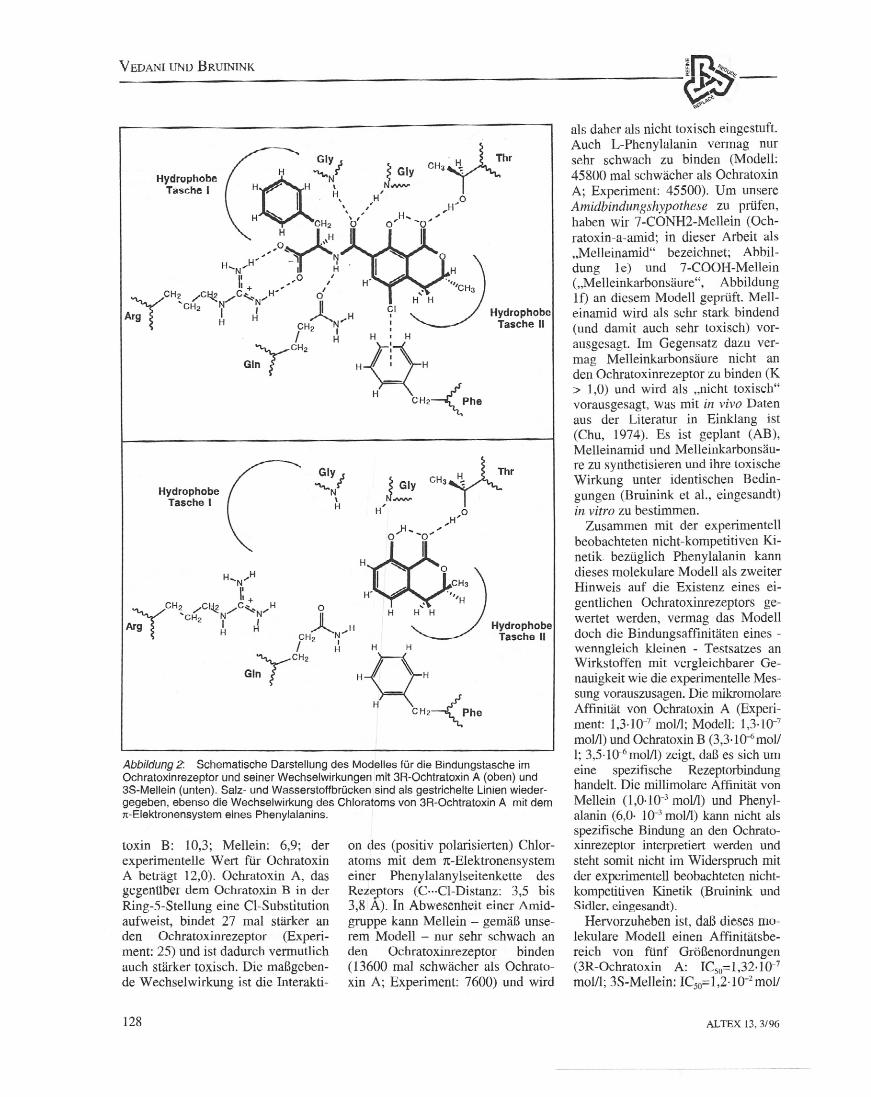
le des Ochratoxinrezeptors (Abbil-dung 2) besteht aus insgesamt zwolfAminosauren (Arg, Gly, Gly, Phe,Tyr, GIn, Trp, Phe, He, Thr, Val,Ala). Die hohe Affinitat von Ochra-toxin A und B wird in dies emModell durch vier Wechselwirkun-gen erklart, die weder bei Melleinnoch bei Phenylalanin als Substratmoglich sind: Eine Salzbrucke' zurGuanidiniumgruppe eines Argininsund drei Wasserstoftbriicken ZlI zwei
ALTEX 13,3/96
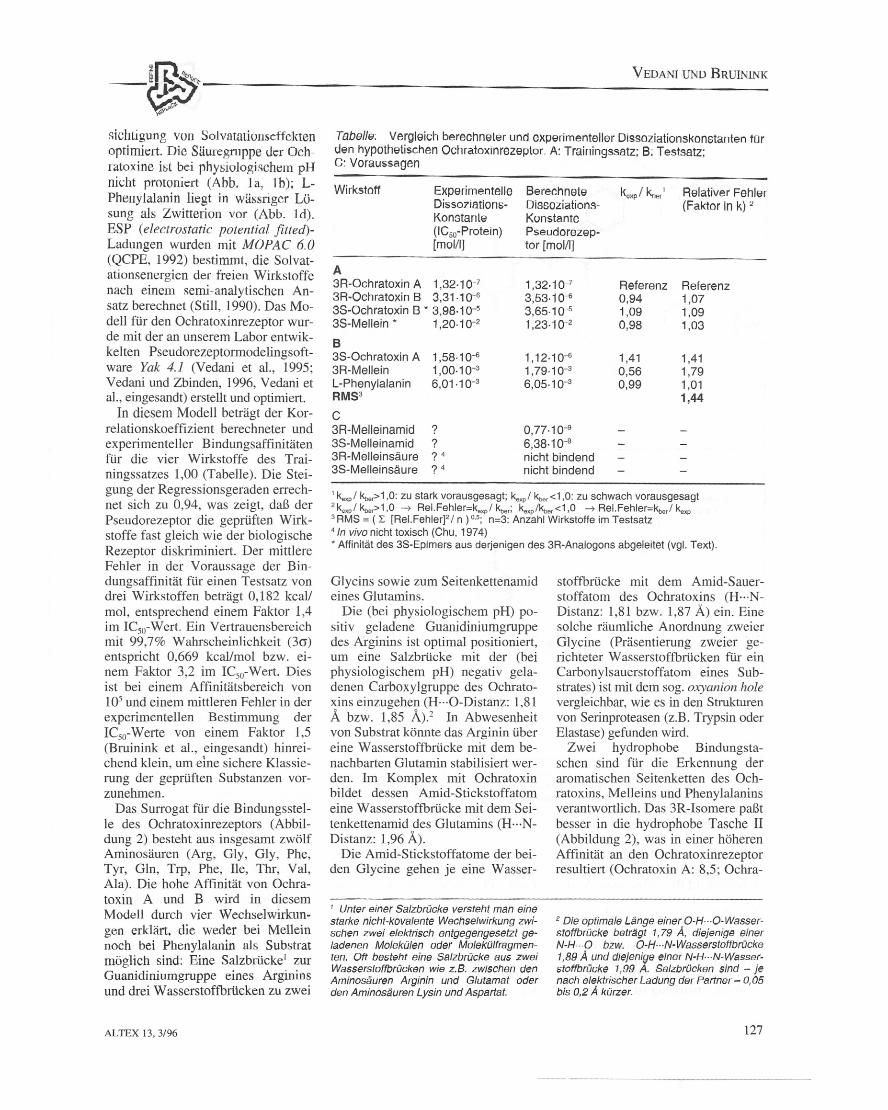
Tsbolle: Vergleich berechneler und experimenteller Dissoziationskonstanten fOrden hypothetischen Ochratcxinrezeptor. A: Trainingssatz; B; Testsatz;c. Voraussagen
Wirkstoff Experimentelle Berechnete kcxp / kner' Relativer FehlerDissoziatlons- Dissoziations- (Faktor in k) 2
Konstanle Konstante(ICso-Protein) Pseudorezep-[mol/l] tor [molll]
A3R-Ochratoxin A 1,32.10-7 1,32.10-7 Referenz Referenz3R-Ochratoxin B 3,31·10-6 3,53·10-6 0,94 1,073S-0chratoxin 8 • 3,98.10-5 3,65.10-5 1,09 1,093S-Mellein * 1,20.10-2 1,23.10-2 0,98 1,03
B3S-0chratoxin A 1,58·10-6 1,12·10-6 1,41 1,413R-Mellein 1,00.10-3 1,79.10-3 0,56 1,79L-Phenylalanin 6,01.10-3 6,05.10-3 0,99 1,01RMS3 1,44C3R-Melleinamid ? 0,77.10-93S-Melleinamid ? 6,38.10-93R-Melleinsaure ? 4 nicht bindend3S-Melleinsaure ? 4 nicht bindend
1 kexpl kber>1,0: zu stark vorausgesagt; kexpl kber<1,0: zu schwach vorausgesagt2 kexpl kber>1,0 --7 Rel.Fehler=kexpl kber; kexp/kber<1,0 --7 ReI. Fehler=kbe/ kexp3 RMS = (L [Rel.Fehler]" In) 0.5; n=3: Anzahl Wirkstoffe im Testsatz41n vivo nicht toxisch (Chu, 1974)* Aftlnitat des 3S-Epimers aus derjenigen des 3R-Analogons abgeleitet (vgl. Text).
Glycins sowie zum Seitenkettenamideines Glutamins.Die (bei physiologischern pH) po-
sitiv geladene Guanidiniumgruppedes Arginins ist optimal positioniert,um eine Salzbrucke mit der (beiphysiologischem pH) negativ gel a-denen Carboxylgruppe des Ochrato-xins einzugehen (H···O-Distanz: 1,81A bzw. 1,85 A).2 In Abwesenheitvon Substrat konnte das Arginin ubereine Wasserstoffbrucke mit dem be-nachbarten Glutamin stabilisiert wer-den. Im Komplex mit Ochratoxinbildet dessen Amid-Stickstoffatomeine Wasserstoffbrucke mit dem Sei-tenkettenamid des Glutamins (H···N-Distanz: 1,96 A).Die Arnid-Stickstoffatome der bei-
den Glycine gehen je eine Wasser-
stoffbrucke mit dem Amid-Sauer-stoffatom des Ochratoxins (H···N-Distanz: 1,81 bzw. 1,87 A) ein. Einesolche raumliche Anordnung zweierGlycine (Prasentierung zweier ge-richteter Wasserstoffbrticken fur einCarbonylsauerstoffatom eines Sub-strates) ist mit dem sog. oxyanion holevergleichbar, wie es in den Strukturenyon Serinproteasen (z.B. Trypsin oderElastase) gefunden wird.Zwei hydrophobe Bindungsta-
schen sind fur die Erkennung deraromatischen Seitenketten des Och-ratoxins, Melleins und Phenylalaninsverantwortlich. Das 3R-Isomere pabtbesser in die hydrophobe Tasche II(Abbildung 2), was in einer hoherenAffinitat an den Ochratoxinrezeptorresultiert (Ochratoxin A: 8,5; Ochra-
1 Unter einer Selzbtucke versteht man einestarke nichl-kovalente Wechselwirkung zwi-schen zwei elektriscn entgegengesetzt ge-ladenen Molekillen oder Molekulfragmen-ten. Oft bestebt eine Salzbrucke eus zweiWasserstoffbrucken wie z.B. zwischen denAmlnosiiuren Arginin und Glutamat oderden Aminoseuren Lysin und Aspartat.
2 Die optimale Lange einer O-H···O-Wasser-stoffbrucke betragt 1,79 A, diejenige einerN-H·.Q bzw. O-H-··N-WasserstoffbrOcke1,89 A und diejeniQe einer N-H···N-Wasser·sfoffbrucke 1,99 A. Selzbtucken sind - jenach etekittscher Ladung der Partner- 0,05bis 0,2 A kurzer.
127
_V_E_D_A_N_I_U_N_U_B_R_u_I_N_m_K ~~~ __ --
~~
HydrophobeTasche I
HydrophobeTasche II
HydrophobeTasche I
Abbildung 2: Schematische Darstellung des Modelles fUr die Bindungstasche imOchratoxinrezeplor und seiner Wechselwirkungen mil 3R-Ochlratoxin A (oben) und3S-Mellein (unten). Salz- und Wasserstoffbrlicken sind als gestrichelte Linien wieder-gegeben, ebenso die Wechselwirkung des Chloratoms von 3R-Ochlraloxin A mit demn-Elektronensystem eines Phenylalanins.
toxin B: 10,3; Mellein: 6,9; derexperimentelle Wert fur OchratoxinA betragt 12,0). Ochratoxin A, dasgegenuber dem Ochratoxin B in derRing-5-Stellung eine Cl-Substitutionaufweist, bindet 27 mal starker anden Ochratoxinrezeptor (Experi-ment: 25) und ist dadurch vermutlichauch starker toxisch. Die maBgeben-de Wechselwirkung ist die Interakti-
on des (positiv polarisierten) ChI or-atoms mit dem n-Elektronensystemeiner Phenylalanylseitenkette desRezeytors (C··CI-Distanz: 3,5 bis3,8 A). In Abwesenheit einer Amid-gruppe kann Mellein - gemaf unse-rem Modell - nur sehr schwach anden Ochratoxinrezeptor binden(13600 mal schwacher als Ochrato-xin A; Experiment: 7600) und wird
128
als daher als nicht toxisch eingestuft.Auch L-Phenylalanin vermag nursehr schwach zu binden (Modell:45800 mal schwacher als OchratoxinA; Experiment: 45500). Urn unsereAmidbindungstiypothese zu prufen,haben wir 7-CONH2-Mellein (Och-ratoxin-a-amid; in dieser Arbeit als"Melleinamid" bezeichnet; Abbil-dung Ie) und 7-COOH-Mellein("Melleinkarbonsaure", AbbildungIf) an dies em Modell gepriift. Mell-einamid wird als sehr stark bindend(und damit aueh sehr toxisch) vor-ausgesagt. Irn Gegensatz dazu ver-mag Melleinkarbonsaure nicht anden Ochratoxinrezeptor zu binden (K> l,O) und wird als .nicht toxisch"vorausgesagt, was mit in vivo Datenaus der Literatur in Einklang ist(Chu, 1974). Es ist geplant CAB),Melleinamid und Melleinkarbonsau-re zu synthetisieren und ihre toxischeWirkung unter identischen Bedin-gungen (Bruinink et al., eingesandt)in vitro zu bestimmen.Zusammen mit der experimentell
beobachteten nicht-kompetitiven Ki-netik beziiglich Phenylalanin kanndieses molekulare Modell als zweiterHinweis auf die Existenz eines ei-gentlichen Ochratoxinrezeptors ge-wertet werden, vermag das Modelldoch die Bindungsaffinitaten eines -wenngleich kleinen - Testsatzes anWirkstoffen mit vergleichbarer Ge-nauigkeit wie die experimentelle Mes-sung vorauszusagen. Die rnikromolareAffinitat van Ochratoxin A (Experi-ment: 1,3.10-7 mol/l; Modell: 1,3.10-7molll) und Ochratoxin B (3,3·1~mollI; 3,5·10-6 mol/l) zeigt, daB es sich urneine spezifische Rezeptorbindunghande1t. Die millimolare Affinitat vonMellein (1,0.10-3 mol/l) und Phenyl-alan in (6,0· 10-3 mol/l) kann nicht alsspezifische Bindung an den Ochrato-xinrezeptor interpretiert werden undsteht somit nicht im Widerspruch mitder experimentell beobaehteten nicht-kompetitiven Kinetik (Bruinink undSidler, eingesandt).Hervorzuheben ist, daB dieses mo-
lekulare Modell einen Affinitatsbe-reich von funf Grotienordnungen(3R-Ochratoxin A: ICsu=I,32·1O-7rnol/l; 3S-Mellein: ICso=l ,2.10-2 moll
ALTEX 13,3/96

VEDANr UND BRUTNINK
I) zu crklaren vcrmag. Ebcnso erklartes die Bindung elCktriDCh unter-schiedlichst geladener Wirkstoffe:AlJe Ochratoxine und Melleinkar-bonsauren binden bei physiologi-schem pH als Anionen; L-Phenylala-nin ist ein Zwitterion, wahrend dieMelleine und Melleinamide neutraleSpezies darstellen. Was im Modellnicht simuliert worden ist, sind Pha-nomene der Metabolisierung, Elimi-nation, Aufnahme und Verteilungder Wirkstoffe im Gewebe. Es wurdevielmehr angenommen, daB die un-tersuchten Wirkstoffe in der obenbeschriebenen Form an den hypothe-tischen Rezeptor binden. Dies istjedoch mit den in vitro-Studien(Bruinink und Sidler, eingesandt) inEinklang.Dieses molekulare Modell liefert
nebst den semi-quantitativen Wertenfur die Affinitat einer Serie yonWirkstoffen an den hypothetischenOchratoxinrezeptor auch einen indi-rekten Hinweis fur ihre toxischeWirkung, indem es das Vorliegeneiner rezeptor-gekoppelten Toxizitatvermuten laBt. Ein Schwachpunktdes Modelles kann in der geringfugi-gen Anzahl an Ochratoxin-Deriva-ten, die zur Zeit zur Verfiigungstehen, urn es zu iiberpriifen, gesehenwerden.Der Sinn eines kombinierten Ein-
satzes yon in vitro und computer-gestiitzten Methoden kann natiirlichnicht darin gesehen werden, "alter-native Toxine" zu finden, sondernvielmehr - im Sinne einer 3R-For-schung - als Screeningmethode zurPrufung potentieller Toxine. 1m wei-teren konnen damit mechanistischeErklarungen fur (rezeptor-gekoppel-te) toxische Phanome gefunden wer-den. Basierend auf einem solchenmolekularen Modell konnten in ei-nem nachsten Schritt Substanzenentwickelt werden, welche die toxi-sche Wirkung von Ochratoxin Aantagonisieren. Wunschenswert er-scheint, daB in vitro Tests unterBerucksichtigung von pharmakoki-netischen und metabolischen Phano-menen moglichst fruhzeitig mit derModellierung verknupft werden(Folkers, 1995).
ALTEX 13,3/96
LiteraturverzeichnlsBaumann, U. und Zimmerli, B. (I YR8). Einfa-
chc Ochratoxin-A-Bt:stimmung in Lehens-rniueln. Min. Gebiete Lebensm. Hyg. 79,15H58.
Blaney, J. M., Weincr, P. K., Dearing, A.,Kollman, P. A .. Jorgensen, E. C., Oatley.S. J., Bnrridge, 1. M. und Blake. J. F.(1982). Molecular mechanics simulation ofprotein-ligand interactions; Binding of thy-roid analogues to prealbumin. 1. Am.Chern. Soe. 104,6424-6434.
Brninink, A., Rasonyi, T. Sidler, C. undDietrich, D. R. (eingesandt). Effects ofochratoxins on embryonic chick neuralretina, brain and meningeal cell cultures.Toxicol. Appl. Pharmacal.
Brninink, A. und Sidler, C. (in Vorbereitung).The neurotoxic effects of ochratoxin A arecell-type specific, are rednced by proteinbinding but are not affected by L-phena-lalanine. Arch. Toxicol.
Breitholz-Emanuelsson, A., Olsen, M., Os-karsson, A., Palminger, 1. und Hult, K.(1993). Ochratoxin A and citrinin uponbiochemical function of procine kidney. 1.AOAC 76, 842-846.
Chu, F. S. (1974). Studies on ochratoxines.CRC Critical Reviews in Toxicology 2,499-523.
Creppy, E. E., Kern, D., Steyn, P. S. Vieggar,R., Roschenthaler, R. und Dirheimer, G.(1983). Comparative study of the effects ofochratoxin A analogues on yeast phenyl-alanyl-tRNA-synthetase and on the growthof protein synthesis of hepatoma cells. Tax.Leu. 19,217-224.
Folkers, G. (1995). Modelliernng von Prote-in-Ligand-Kornplexen alsBasis einer ratio-nalen Arzneistoffentwicklung. ALTEX 12,138-151.
Hald, B. (1991). Ochratoxin A in humanblood in European countries. IARC Sci.Publ. 115, 159-164.
Mohamadi, F., Richards, N. G. J., Gnida, W.c., Liskamp, R., Lipton, M., Caufield, C.,Chang, G., Hendrickson, T. und Still, W.C. (1990). MacroModel - An integratedsoftware system for modeling organic andbioorganic molecules using molecular me-chanics.l. Comput. Chem. 11,440-448.
Miki, T., Fukui, Y., Uemura, N. und Takegu-chi, Y. (1994). Regional difference in theneurotoxicity if ochratoxin A on the de-veloping cerebral cortex in mice. Dev!.Brain. Res. 82, 259-264.
QCPE (1994). Quantum Chemistry ProgramExchange. University of Indiana, Bloo-mington, Indiana, USA.
Ramsden, C. A. (Hrsg, 1990). Quantitativedrng design. Comprehensive MedicinalChemistry 4, Oxford: Pergamon Press.
Searle, M. S. und Williams, D. H. (1992). Thecost of conformational order; Entropychanges in molecular associations. 1. Am.Chern. Soc. 114, 10690-10697.
Shirai, S. (1991). Developmental mechanismof congenital eye abnormalities. NipponGanka Gakkai Zasshi 95, 1206--1237.
Snyder, J. P., Rao, S. N., Koehler. K. F. undVedani, A. (1993). Pseudoreceptors. In H.Kubinyi, (Hrsg.), 3D-QSAR in Drug De-sign (336--354). Leiden NL: ESCOM Sci-
encc Publishers B. V.Still, W. C, Tcmpczyk, A .. Hawley, R. C.
und Hendrickson, T. (1990). Semianalyti-cal ueatrnent uf solvation for molecularmecnanics and dynamics. 1. Am. Chern.Soc. 112,6127-6129.
Studcr-Rohr, L, Dietrich, D. R., Schlatter, 1.und Schlatter, Ch. (eingereicht). Intraindi-vidual variations in plasma levels andkinetic parameters of ochratoxin A inhumans. Toxicol. Appl. Pharmacal.
Studer-Rohr, 1.. Dietrich, D. R., Schlatter, J.und Schlatter, Ch. (1995). The occurenceof ochratoxin A in coffee. Food Chern.Toxicol. 33, 341-355.
Vedani, A., Dobler, M., Folkers, G. nndZbinden, P. (eingesandt). Pseudoreceptormodeling. A molecular model for thebinding site of the cannabinoid receptor. 1.Med. Chern.
Vedani, A. und Zbinden, P. (1996). Compu-termodelle im pharmakologischen und to-xikologischen Screening. In F. P. Gruberund H. Spielmann (Hrsg.), Alternativme-thoden zu Tierversuchen (146-163). Hei-delberg: Spektrum Akademischer Verlag.
Vedani, A., Zbinden, P., Snyder, J. P. undGreenidge, P. A. (1995). Pseudoreceptormodeling: The construction of three-di-mensional receptor surrogates. 1. Am.Chern. Soc. 117,4987-4994.
Vedani, A. (1994). Das Konzept des Pseudo-rezeptors fur das pharrnakologische Scree-ning. ALTEX 11. 11-21.
Vedani, A. und Huhta, D. W. (1990). A newforce field for modeling metalloproteins. 1.Am. Chern. Soc. 112, 4759-4767.
VerdankungDieses Projekt wurde durch die groB-zugige finanzielle Unterstutzung derMargaret und Francis Fleitmann-Stiftung (Luzern), der Stiftung Fondsfur versuchstierfreie Forschung(FFVFF, Zurich) und der StiftungSlAT (Zurich) ermoglicht.
ZusatzmaterialDie dreidimensionalen Koordinatendes in diesem Artikel diskutiertenModelles der Bindungsstelle des ver-muteten Ochratoxinrezeptors konnenvia E-Mail angefordert werden(siat@ dial.eunet.ch).
KorrespondenzdresseDr. Angelo VedaniSlAT Biografik-LaborMissionsstraBe 60CH-4055 BaselTel. +41-61-321-0533,Fax: +41-61-321-0540,E-Mail: [email protected].
129




























