Embed Size (px)
Citation preview

Heft 3 ] 279 1976 J
J. Orn. 117, 1976: s. 279-290
Populations- und syn6kologische Modelle in der Ornithologie
Von Udo Halbach
Modelle als Methoden der Erkenntnisgewinnung haben in der Biologie in den letzten Jahren zunehmend an Bedeutung gewonnen. Dies ist einerseits auf die Ent- wi&lung yon GroBrechenanlagen zurii&zufiihren, die komplizierte Berechnungen und Simulationen erm/Sglichen, andererseits aber auch auf einen Wandel in der heu- ristischen Einstellung vieler WissenschaPder (vgl. HALBACH 1974). Modelle sind for- male Analoga natiirlicher Gegebenheiten. Als solche miissen sie nicht notwendigerweise mathematische Abstraktionen sein. Gerade in der Ornithologie kennt man seit langem sehr handfeste Modelle, n~imlich Nachbildungen yon Tieren, mit denen man be- stimmte mechanische Funktionen iiberpriifen kann. Ich verweise auf den beriihmten kiinstlichen Enterich des barocken Automatenbauers VAUCANSON (vgl. ZrMANEK 196t, YON CIJB~ 1967) oder die mechanischen Flugmodelle YON HorsTs (1957) bzw. HER- ZOOs (1968), mit denen aerodynamische Experimente durchgefiihrt werden k~Snnen.
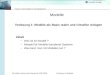
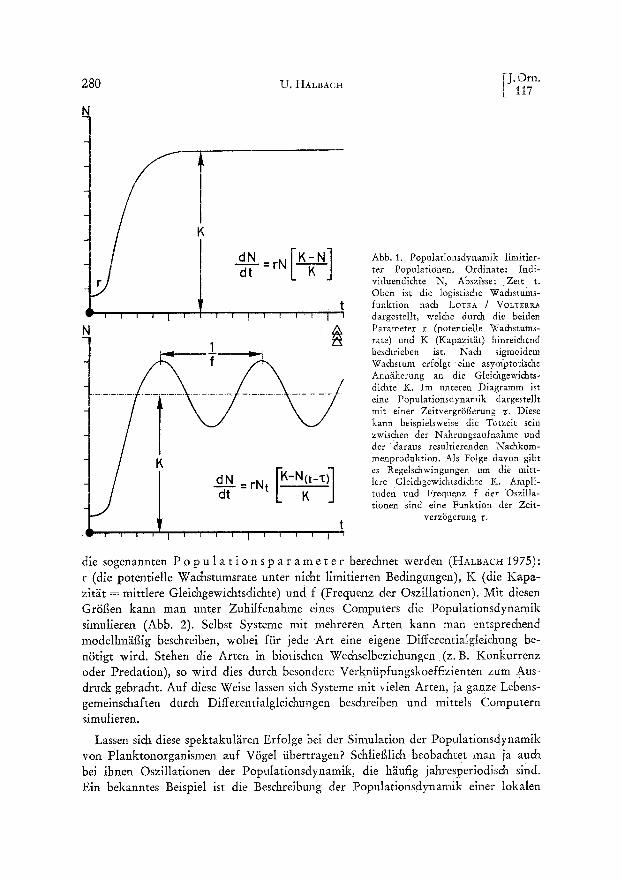
Von welt grSf~erer Bedeutung sind in der Ornithologie inzwischen mathematische und graphische Modelle der Populationsdynamik und interspezifischer Beziehungen. So wurden deterministische Modelle aufgrund theoretischer Ans~itze entwickelt (DuBLZN & LOTK~ 1925, VOLTERRA 1926), um nahrungs- oder areal-limitiertes Popu- lationswachstum mit Hilfe der sogenannten L o g i s t i s c h e n W a c h s t u m s - f u n k t i o n zu beschreihen (Abb. 1). Durch Einfiihrung einer Zeitverz~Sgerung (HuTcHINSON 1948, WANGrRSKY & CUNNINGHA~ 1957, HALBACr~ 1972) lassen sich auch regelm~iflige Schwankungen der Populationsdichte darstellen, wie sie in natiir- lichen Populationen h~iufig zu beobachten sin& Eine solche Zeitverz~Sgerung kann bei- spielsweise zwischen der Nahrungsaufnahme und der daraus resultierenden Nachkom- menproduktion liegen, wie dies fiir R~idertiere nachgewiesen wurde (Abb. 2). Es ist inzwischen sogar gelungen, aus Daten yon in der GefangenschafL gehaltenen isolierten Tieren (z. B. Lebensdauer, Zahl der Nachkommen, Biomasse) Voraussagen fiber die Dichteschwankungen ganzer Populationen zu machen. Diese lief~en sich experimentell sehr exakt verifizieren. Solche genauen Voraussagen des Populationsverlaufes sind nur m~Sglich mit Hilfe numerischer L~Ssungsmethoden, welche durch moderne Computer- techniken bew~iltigt werden k~Snnen, denn analytisch lassen sich diese komplizierten DifferentialgMchungen mit unserem heutigen mathematischen Rilstzeug nicht 15sen. Bei den erwiihnten Testorganismen, den R~idertieren, konnte auf diese Weise der Einflug eines ausgew~ihlten ~Skologischen Faktors, der Temperatur, auf die Popula- tionsschwankungen sehr exakt prognostiziert werden (Abb. 2). Die Kausalkette sieht dabei folgendermaf~en aus: iJ k o 1 o g i s c h e F a k t o r e n (wie Temperatur oder Lichtintensit~it ) beeinflussen p h y s i o 1 o g i s c h e E i g e n s c h a f t e n (wie Freg-, Assimilations- und Respirationsraten), diese wiederum die L e b e n s d a t e n (wie altersspezifische Geburts- und Sterberate, K~Srpergewicht). Aus diesen k~Snnen dann
280
N
U. HALBACH J. Orn. 117
N
J t i i t t J
dN =rN~IK--~ l d t t
i' ' ~ ' ~ 1 ' " ' I ' ~ ' ' m 'i
a t - ' [ K J
t
Abb. 1. Populationsdynamik limitler- ter Populationen. Ordinate: Indi- viduendichte N, Abszisse: Zeit t. Oben ist die logistis&e Wa&stums- funktion nach LOT~A / VOL'r~R~.~ dargestellt, welche dutch die beiden Parameter r (potentielle Wachstums- rate) und K (Kapazit~it) hinreichend beschrieben ist. Nach sigmoidem Wachstum erfolgt eine asymptotisch¢ Ann~iherung an die Gleichgewichts- dichte K. Im unteren Diagramm ist eine Populationsdynamlk dargestelh mit einer ZeitvergrL;flerung 7. Diese kann beispielsweise die Totzeit seir zwischen der Nahrungsaufnahme nnd der daraus resultierenden Nachkom- menproduktion Als Fotge davon gibt es Regelschwingungen um die mitt- lere Gleichgewichtsdichte K. Ampli- tuden und Frequenz f der Oszilla- tionen sind eine Funktion tier Zeit-
verzSgerung ~.
die sogenannten P o p u 1 a t i o n s p a r a m e t e r berechnet werden (HALBACH 1975) : r (die potentielle Wachstumsrate unter nicht l imit ierten Bedingungen), K (die Kapa - zit~it = mitt lere Gleichgewichtsdichte) und f (Frequenz der Oszit lat ionen). Mit diesen GrGt]en kann m an unter Zuhi l fenahme eines Computers die Popula t ionsdynamik simulieren (Abb. 2). Selbst Systeme mit mehreren Arten kann man entsprechend modellm[igig beschreiben, wobei fiir jede Ar t eine eigene DifferentiatgMchung be- nGtigt wird. Stehen die Ar ten in biotischen Wechselbeziehungen (z. B. Konkur renz oder Predation), so wi rd dies durch besondere Verkniipfungskoeffizienten zum Aus- druck gebracht. Auf diese Weise lassen sich Systeme rnit vielen Arten, ja ganze Lebens- gemeinsehaften durch Differentialgleichungen beschreiben und mittets Compute rn
simulieren.
Lassen sich diese spektakul~iren Erfolge bei der Simulat ion der Popula t ionsdynamik von P lanktonorganismen auf V6gel iibertragen? Schliet~Iich beobachtet man ja auch bei ihnen Oszi l lat ionen der Popula t ionsdynamik , die h~iufig jahresperiodis& sind. Ein bekanntes Beispiel ist die Beschreibung der Popula t ionsdynamik einer lokalen
Heft 3 ] 1976 ] Modelle in der Ornithologie 281
100_
100
10 20 30 40 E 1oo _ . . _ . - . ,
.i2
-~ 100_ A
"O 10 20 30 40
150
1 0 0
5O
• 10 20 30 Tage
Abb. 2. Populationsdynamik des R~dertiers Brachionus calyciflorus bei drei verschiedenen Tempera- turen: 15°C (Oben) 20°C (Mitre), 25°C (Unten). Die drei oberen Kurven mit Punkten stellen jeweils empirische Experimentalpopulationen dar, die unteren Kurven aus den Lebensdaten erre&nete Com- puter-Simulationen. Die Ubereinstimmung ist augenf~illig: Mit zunehmender Temperatur nehmen
Frequenz und Amplitude der Oszillationen zu.
Population yon Wachteln durch EML~ (1940). Trotz der ~iut~eren Khnlichkeit der Kurven ist es jedoch bisher nicht gelungen, die Populationsdynamik der VSgel mittels so einfacher Modelle zu beschreiben, wie dies bel den Planktonorganismen mSglich war. Bei ihnen ist derzeit die Kausalkette yon den 5kologischen Faktoren bis zur Populationsdynamik nicht mit derselben Biindigkeit nachzuverfolgen. Die geschilder- ten Differentialgleichungsmodelle gehen n~imlich stillschweigend alle v o n d e r An- nahme aus, dai~ die Organismen entweder homogen oder zufallsgem~if~ verteilt sind, was bei den Planktonorganismen auch ann~ihernd der Fall ist, wenngleich auch hier ausgesprochene Heterogenit~iten (z.B. ,,Planktonwolken") vorkommen (HAL~ACH 1975). Bei VSgeln findet man eine ann~ihernd homogene Verteilung nut lokal bei extrem territofialen Arten. Aut~erdem setzen die geschilderten mathematischen

[J.Orn. 282 u. HALB•CH k 117
Modelle voraus, dag die Individuen einer Population alle glei& sind bzw. sich h6ch-. stens in ihrem Lebensalter unterscheiden. Tatsiichlich findet man bei WSgeln jedoch zwei Geschlechter sowie verschiedene Geno- und Phiinotypen. Man denke beispiels- weise nur an den Polymorphismus der Ku&u&sweibchen. Solche individuellen Varia- biliditen miissen in Populationsmodelte fiir V6gel inkorporiert werden, sollen sie einigermagen realistisch sein. Die als sehr wi&tig erkannte r~iumliche Heterogenit~it yon Populationen kann man mittels partieller Differentialgleichungen oder Diffu- sionsgleichungen simulieren. Erste Ans~itze zur Erarbeitung entsprechender mathema- tischer Methoden sind in Angriff genommen (vgl. HALI3ACI-I 1975). Mit sol&en pard- ellen Differentialgleichungen bzw. Diffusionsgleichungen wird sich vielleicht das Raum-Zeit-Muster der Individuendichte yon Populationen in absehbarer Zeit be- schreiben lassen. Die Individualidit der Organismen l~igt sich jedoch nicht auf diese Weise inkorporieren; hierzu mug die Digitalisierung in Organismen beibehalten werden (vgl. HALBACI-I 1975), wobei zwischen diesen Organismen biotische Bezie- hungen auftreten, die meist eine ethologische Grundlage haben. So k6nnen in der Verteilung der V6gel Abstand und Winkel der Einzeltiere zueinander yon der Rang- ordnung abh{ingen. McBRIDE, JAMES & SI~IOFFNER (1963) haben dies bei der Ver- teilung der Htihner auf einem Hiihnerhof gefunden (Abb. 3). Sie haben die Schar
Rb
j j JSb
/ / /
/ . . . ' / / .t /
L S "" -., / / / /
. ? . q / ....1 / - . . . / -
#s = f (db, Sb, R b / R )
/3 = ,~' - ~
Abb. 3. Stellung und Kopfhaltung der Hiihner auf einem Hiihnerhof zueinander. Die Bli&richtungs- winkel cc und fl sind Funktionen der Distanzen der Na&barn (da, db) , deren Bli&winkel (C~a, c~b) ,
sowie deren Rangordnung (Ra, Rb).
mehrfach yon oben fotografiert und die Verteilung und Anordnung geometrisch ana- lysiert. Dabei ergab sich: Die Tiere sind homogener verteilt als es dem Zufall ent- spricht. Auch die Kopfhaltung ist ni&t zufallsgem~ig, sondern sie weichen sich mit den Bli&en aus, wohl weil das direkte Anschauen als Aggression empfunden wlrd

Heft 2 ] 1976 Modelle in der Ornithologie 283
(vgl. Menschen im Aufzug!). Die Bli&winkel sind nicht zufiillig; sie sind die Resul- tante einer Vielzahl von Faktoren, wobei der Abstand der n~ichstbenachbarten Tiere ebenso eine Rolle spielt wie deren Bli&winkel und ihre Rangordnung. Dieses Modell ist natOrlich nur ein erster Ansatz, welcher die Individualit~it in einer Soziet~it berii&- si&tigt. Natalit~it, Mortalit~it und Migration lassen si& jedo& ohne grunds~tzliche S&wierigkeiten in das Modell inkorporieren. Es ist daher zu hoffen, dag auf diese Weise in Zukunfl: realistische Beschreibungen und Erkl~irungen der Populations- dynamik yon VSgeln m/Sgli& werden.
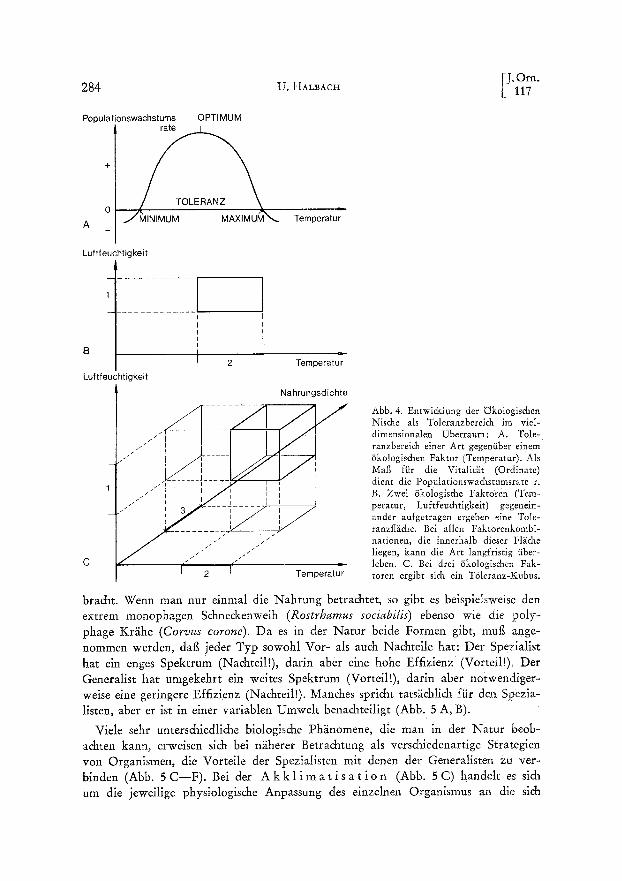
Ein anderer, auf dem fruchtbaren Konzept der/Skologis&en Nis&e fugender Typ yon Modellen hat inzwischen Bedeutung erlangt. Die Vorstellung yon der tikologi- schen Nis&e basierte urspriinglich auf der etwas vagen Idee vom ,,Beruf" einer Organismenart in einer Lebensgemeins&att. Inzwischen ist es gelungen, diesen Begriff zu quantifizieren und damit wissens&afHi& verwendbar zu ma&en. Be~cra&ten wir zun~i&st den Toleranzberei& einer Organismenart in Abh~ingigkeit yon einem iso- lierten 6kologis&en Faktor (Abb. 4 A). Hier ist die Populationswachstumsrate einer Art in Abh~ingigkeit yon einem iSkologischen Faktor (als BeispM die Temperatur) aufgetragen: In einem bestimmten Temperaturbereich ist sie positiv und hat ein Opti- mum; unterhalb einer Minimaltemperatur ist sie negativ, desglei&en oberhalb einer Maximaltemperatur. Im Temperaturbereich zwischen diesen beiden Kardinalpunkten kann die Population langfristig iiberleben. Dies ist der Toleranzbereich gegeniiber dem/5kologischen Faktor. Er daft nicht verwechselt werden mit dem engeren Pr~ife- renzbereich (der Vorzugstemperatur) und dem weiteren Resistenzbereich (f3berleben von Dauerstadien). Man kann entsprechend auch die Kurven fiir andere/3kologische Faktoren auftragen, zum Beispiel fiir die Luftfeuchtigkeit. Tr~igt man den Toleranz- bereich der LufLfeuchtigkeit gegeniiber dem der Temperatur auf, so erh~ilt man eine zweidimensionale Toleranzfl~iche (Abb. 4 B). Bei allen Faktorenkombinationen inner- halb dieser Fl~iche kann die Population langfristig iiberleben. Man kann das System um einen dritten 6kologischen Faktor erweitern, etwa die Nahrungsdichte, und erh~ilt dann einen Toleranzkubus innerhalb eines dreidimensionalen Koordinatensystems (Abb. 4 C). Dieses System l~igt sich um beliebig viele 8kologische Faktoren erweitern, wobei man ein vMdimensionales Koordinatensystem erh~ilt, das man zwar nicht mehr graphisch darstellen, das man sich aber sehr wohl vorstellen und vor allem mathema- tisch behandeln kann. Der Toleranzbereich in diesem vieldimensionalen Koordinaten- system ist die quantitative Definition fiir die ~Skologische Nische. Allerdings sind - - wie sp~iter dargelegt werden soll - - einige der tSkologischen Faktoren bei den V6geln yon iiberragender Bedeutung, w~ihrend man andere praktisch vernachl~issigen kann (vgl. Abb. 6).
Die sorgf~iltige Betrachtung der Natur zeigt, datt es VSgel mit einer groi~en und andere mit einer kleinen tikologischen Nische gibt. Welches ist fiir eine Vogelart im Rahmen der Evolution die bessere Strategie: eine groge 6kologische Nische zu be- kommen (d.h. ein Generalist zu werden) oder eine kleine tikologische Nische zu erreichen (d. h. ein Spezialist zu sein). Eine allgemeine Antwort hierauf gibt es nicht, denn die Evolution als Optimierungsprozef~ hat immerhin beide Typen hervorge-
284 U. HALBACH [J. ore. [ 117
Populationswachstums OPTIMUM
o Temperatur
Luftfeuchtigkeit
ZIIZI Temperatu~
Luftfeuchtigkeit Nahrungsdichte
( ( 2 Temperat2r
Abb. 4. Entwi&lung der ~kotogischen Nische als Toleranzbereich im viel- dimenslonalen Uberraum: A. Tole- ranzberelch elner Art gegeniiber einem /Skologischen Faktor (Temperatur). AIs Marl fi~r die Vitalit~.t (Ordinate) dient die Populationswachstumsrate r. B. Zwei ~Skologische Faktoren (Tem- peratur, Luftfeuchtigkeit) gegenein- ander aufgetragen ergeben eine Toie- ranzfl~iche. Bei allen Faktorenkombi- nationen, die innerhalb dieser Fl~iche liegen, kann die Art langfristig iiber- leben. C. Bel drel /Skologischen Fak- toren ergibt rich ein Toleranz-Kubus.
bracht. Wenn man nur einmal die Nahrung betrachtet, so gibt es beispielsweise den extrem monophagen Schneckenweih (Rostrhamus sociabilis) ebenso wie die poly- phage Kr~ihe (Corvus corone). Da es in der Natur beide Formen gibt, muf~ ange- nommen werden, dag jeder Typ sowohl Vor- als auch Nachteile hat: Der Spezialist hat ein enges Spektrum (Nachteit!), darin aber eine hohe Effizienz (Vorteil!). Der Generalist hat umgekehrt ein weites Spektrum (Vorteil!), darin aber notwendiger- weise eine geringere Effizienz (Nachteil!). Manches spricht tats~ichtich fi.ir den Spezia- listen, aber er ist in einer variablen Umwelt benachteiligt (Abb. 5 A, B).
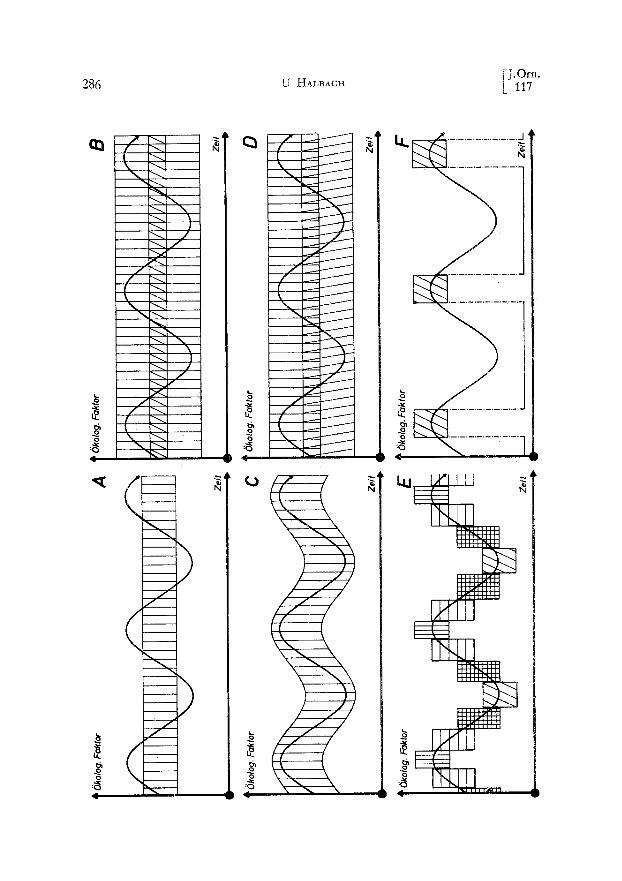
Viele sehr unterschiedliche biologische Ph:,inomene, die man in der Natur beob- achten kann, erweisen sich bei n~iherer Betrachtung als verschiedenartige Strategien yon Organismen, die Vorteile der Spezialisten mit denen der Generalisten zu ver- binden (Abb. 5 C--F). Bei der A k k 1 i m a t i s a t i o n (Abb. 5 C) handelt es rich um die jeweilige physiologische Anpassung des einzelnen Organismus an die sich

Heft 3] ~1976 J Modelie in der Ornithologie 285
{indernden Umweltbedingungen, wozu man beispielsweise auch einen jahresperiodi- s&en We&sel der Gefiederf~irbung re&nen kann. Beim P o 1 y m o r p h i s m u s (Abb. 5 D) setzt si& die Gesamtpopulation aus Spezialisten zusammen, yon denen jeder einen etwas anderen Toleranzberei& hat. Die einzelnen Morphen haben zwar eine genetis&e Grundlage, denno& handelt es si& ni&t um Rassen, da sich die Mor- phen untereinander paaren, ohne dag es intermedi~ire Formen gibt. Die betroffenen Merkmalskomplexe werden in Form eines Supergens als eine Einheit vererbt. Der Eipolymorphismus des Ku&u&s, die polymorphe Mimikry yon Schmetterlingen, aber auch der Ges&le&tsdimorphismus sind hierfiir augenf~illige Beispiele. Durch den Polymorphismus wird eine breite Ausnutzung der verfiigbaren Ressourcen durch die Population bei hoher Effizienz der einzelnen Individuen gew~ihrleistet. Ein nicht- genetisches Pendant zum Polymorphismus ist die konditionierte Nahrungsspezialisa- tion yon Individuen einer Population, wie sie beispielsweise bei MSwen zu beob- achten ist (VAuK, miindliche Mitteilung).
Bei Arten mit mehreren bis vMen Generationen im Jahr kSnnen die aufeinander- folgenden Generationen vers&iedene - - der jeweiligen Jahreszeit angepaflte - - Toleranzbereiche haben. Eine solche Erscheinung ist yon den Planktonorganismen unserer Teiche und Seen bekannt. Dieses als Z y k 1 o m o r p h o s e (Abb. 5 E) be- zeichnete Ph~inomen hat der biologischen Forschung lange Zeit grof~e R~itsel aufge- geben. Heute weig man, datg es sich um Modifikationen handelt, also um dutch die Umwelt setbst gesteuerte ph~inotypische Ver~inderungen (JAcoBs 196i, HALBACH 1970, 1971). Eine andere Strategie, mit einer variablen Umwelt fertig zu werden, ist die D i a p a u s e (Abb. 5 F). Hier zieht sich der Organismus w~ihrend ungiinstiger Perioden in der Form anabiotischer Zust~inde (Dauereier, Samen oder Winterschlaf) voriibergehend aus dem aktiven Leben zurii&. Der V o g e 1 z u g ist eine andere Form derselben Strategie; hier wird eine Region w~ihrend unwirtlicher Zeiten g~inz- lich verlassen.
Die 5kotogische Nische ist demnach ein einfaches konzeptionelles ModeI1 (vgl. HAL~ACH 1974), mit dem eine generalisierende Gesamtschau ~iut~erlich sehr unter- schiedlicher Ph~inomene wie Polymorphismus, Vogelzug, Diapause, Zyklomorphose, Winterschlaf und Akklimatisation nach einem einheitlichen Aspekt mSglich ist. Es sind verschiedene Strategien, die das gleiche ZM haben: die Erhaltung einer hohen Effizienz in einem variablen Milieu.
Aus dem Konzept der 5kologischen Nische ist ein zweites, noch wichtigeres er- wachsen" das K o n k u r r e n z - A u s s c h l u i ~ - P r i n z i p . Arten mit denselben oder sich iiberlappenden 5kologischen Nis&en sollen nach diesem Konzept auf Dauer nicht nebeneinander in demselben Biotop koexistieren kSnnen. Es kommt entweder auf evolutivem Wege zu einer M e r k m a 1 s v e r s c h i e b u n g, wofiir die diver- gierenden Schnabell~ingen der KMber Sitta tephronota und Sitta neumeyer im Uber- lappungsgebiet des Hochlandes von Iran ein klassisches BeispM darstelten. Inzwischen sind viele weitere Beispiele in der Vogelwelt entdeckt worden. Generell beobachtet man beim Vorkommen zweier verwandter und ~/hnlicher Arten am selben Standort
[3.Om. 286 U. HaLBACU ~_ 117
o
:8 4
o
:8 t
f l
J \
" x
f J
\
(
.
~ ' X . I
"-, J l
"" .,..
" , / I
" i l
\
J
,,,, !
, /
Z
%----
o~ :8
- - 7 N ZZI j
:. - j
m ~ j
. . . . . . . . . / - - -
~ J J j - - - - ~ j
"- j
- x -.----- j ~ J /
1
t /
v w ~ j J .~=:::: J
: j
: / j
J

Heft ] 1976 Modelle in der Ornithologie 287
Unterschiede in der Lebensweise. Xormoran (Phalacrocorax carbo) und Kr~ihen- scharbe (Pha!acrocorax aristotelis) beispielsweise zeigen bei gemeinsamem Vorkom- men unterschiedli&e Brutpl~ttze, Brutzeiten und Jagdareale.
Eine an&re miSgliche Folge des Konkurrenz-Ausschlut~-Prinzips ist die g e o - g r a p h i s c h e S e p a r i e r u n g oder auch die Disaggregation entlang eines HShen- gradienten, welche wir sp~iter noch an einem Beispiel kennen lernen werden (Abb. 12). Weiterhin beobachtet man nicht selten eine vertikale oder horizontale Aufteilung der Areale in einem Habitat. Ein klassisches Beispiel hierfiir sind 5 yon MAcARTHUtt (1958) untersuchte Baumwalds~inger-Arten (Dendroica tigrina, D. castanea, D. fusca, D. virens, D. coronata), welche zwar in denselben B~iumen leben, sich zur Nahrungs- suche aber bevorzugt in verschiedenen Bezirken der Baumkrone aufhalten.
Die Richtung der r~iumlichen Segregation oder der Merkmalsverschiebung ist nicht pr~idestiniert. Sie kann in vers&iedenen geographischen Regionen unterschiedlich oder sogar gegensinnig verlaufen. Bei den Sperlingen Sardiniens leben Passer hispanicus und P. montanus bevorzugt in den St~idten und P. domesticus auf dem Land, w~ih- rend dies auf dem Kontinent umgekehrt ist (MAcARTHUR 1972).
Sind nun bei V6geln, die in einem Habitat leben, die ~Skologischen Nischen stets sauber getrennt - - wie es das Konkurrenz-Ausschlug-Prinzip fordert - - oder gibt es fdberlappungen? Bildhaflc kann man sich s~imtliche Ressourcen eines Biotopes auf eine Stre&e projiziert vorstellen. Die Teilstre&en, die yon den einzelnen Vogelarten be- setzt gehalten werden, sollten dann nicht iiberlappen. Sie sollten sich l~ingenm~it~ig verteilen wie bei einem Glasstab, der auf den Boden geworfen in viele verschieden grofle Stiicke zerspringt. MAcARTHUR hat fiir dieses yon ibm erfundene ,,Modell des zerbrochenen Stabes" eine mathematische Formel flit das Verteilungsmuster ent- wi&elt. Ausgehend yon der Annahme, daf~ die Individuenzahl einer Art in einem Habitat die Breite der iSkologischen Nische dieser Art widerspiegelt, hat er die Zu- sammensetzung yon Vogel-Lebensgemeinschaften in bestimmten Lebensr~iumen unter- sucht und Diskrepanzen zwischen den empirischen Daten und den Erwartungen (din Nullhypothese) gefunden und daraus auf teilweises Uberlappen der Nischen ge schlossen. Dieses vieldiskutierte Modell ist insbesondere wegen seiner Annahmen nicht
4 Abb. 5. Verschiedene Strategien der Organismen, innerhalb einer variablen Umwelt mlt miSglichst hoher Effizienz zu existieren. A. Der Spezialist mit einem engen iSkologischen Spektrum ist in einer variablen Umwelt nicht zu allen Zeiten existenzf~ihig. B. Der Generalist mit breitem 8kologischen Spektrum kann auch bei variabler Umwelt dauernd existieren, allerdings mit geringer Effizienz. C. Bei der Akklimatisation passen si& die Organismen physiologisch den jeweiligen Umweltbedlngungen an. D. Beim Polymorphismus existieren verschiedene Morphen mit engen aber unterschiedlichen Toleranz- bereichen. Die Population besteht aus verschiedenartigen Spezialisten und hat dadurch insgesamt eine breite 6kologische Valenz. E. Bei der Zyklomorphose folgen Generationen mit unterschiedlichen Tole- ranzbereichen aufeinander, wobei die Anpassung an die jeweiligen Milieubedingungen modifikatorisch determiniert ist. F. Bei der Diapause ziehen rich die Organismen w~ihrend ungtinstiger Perioden in der Form anabiotischer Zust~inde ans dem aktiven Leben zurti& (z. B. Dauereier, Winterschlaf). Der Vogel-
zug ist eine ,ihnliche Strategie.

[J.Orn. 288 u. HALBACH [ 117
unwidersprochen geblieben. Es tauchte daher das Problem der direkten Messung mSg- liner Oberlappungen auf, wobei dies bei Konzentration auf zuniichst einen Faktor keine Probleme bereitet (Abb. 6 A). Mitgt man diese rdberlappung fiir 3 als bei VSgeln
A
R e s s o u r c e . , _ _ .
. . [ u i
I n d i v i d u e n d i c h t e
a o ( . = - -
G
0-<o(.-<1
B C D
HHS H orizontale Habitat S elektion
I - - - I
A I ,
el B
~ _ _ - J
VHS FS V ertikale Habitat F utter S elektion S elektion
~ A
. . . . . . . . . . . . J
Abb. 6. Messung der Nischen-r3berlappung o~ an Hand einzelner 6koiogis&er Faktoren. A. Prinzip der (a = ii/G). B. Oberlappung yon Territorien (HHS). C. Oberlappung des Vorkommens entlang eines
H6hengradienten (VHS). D. Oberlappung der Beute-Gr6ge (FS).
besonders wichtig erachtete Parameter (die Horizontale Habitat-Selektion HHS - - Abb. 6 B -- , die Vertikale Habitat-Selektion VHS - - Abb. 6 C - - und die Futter-- Selektion FS - - Abb. 6 D --), dann zeigt sich, datg ihre Summe in konkreten F~illen konstant bleibt, was naturgem~it~ auch fiir das Gegenstiick, die Separierung, gilt (Abb. 7). Oberlappungen auf einem Sektor k6nnen also durch Separierung auf einem anderen kompensiert werden.
Modelle sind fiir das Verfahren deduktiver Erkenntnisgewinnung pr~idestiniert (HAL~ACH 1974): Aus logischen Annahmen und Voraussetzungen werden Modelle als Hypothesen konstruiert, aus diesen spezielle Folgerungen abgeleitet, welche an Hand natiirlicher Verh~iltnisse oder auch yon Experimenten iiberpriil~ werden kSnnen.
Zum Abschlut~ soll eine Modell-Untersuchung yon Come (1974) etwas ausfiihrlicher behandelt werden, um exemplarisch die praktische Arbeit mit einem Modell zu illu- strieren. Hierbei geht es um die Aufteilung yon Ressourcen-Gradienten zwischen 2 verwandten Arten. COPY untersuchte dies am Beispiel zweier Baumwalds~inger, die an den Bergh~ingen der Arizona-Mountains leben: Grace's Baumwalds~inger (Den- droica graciae) und der Schwarzkehlige Graue Baumwalds~inger (Dendroica nigres- cens). Beide Arten suchen als Nahrung Insekten in B~iumen. D. nigrescens ist jedo& vorwiegend auf Eichen spezialisiert, die insbesondere am Futge der Berge vorkommen, D. graciae dagegen auf Kiefern, die an den hSheren Hanglagen verbreitet sin& Auf

Heft 3 ] 1976 ] Modelle in der Ornithologie 289
~IoVHS Abb. 7. Die Summe der Separierungen yon Vertikaler Habitat-Selektion (VHS), Horizontaler Habitat- Selektion (HHS), und Futter-Selektion (FS) sind konstant. Im 3dimensionalen Koordinatensystem Iiegen die Punkte ann~ihernd auf einer (dreieckigen) Fl~iche, deren Koordinatenabschnitte nicht signifi- kant divergierem Dies bedeutet, dat~ Uberlappungen in einem Faktor durch Separierung in anderen ausgeglichen werden ktinnen. Kreise beziehen sich auf nordamerikanische Vogel-Lebensgemeinschaften,
Dreie&e auf siidamerikanis&e.
mittleren H6hen gibt es Mischw~ilder. CODY stellte sich die Frage: Wie ist die opti- male Strategie der Nahrungssuche f~ir die beiden Arten in dem Oberlappungsgebiet der beiden Waldtypen? Dazu konstruierte er ein Modell, das auf 3 einfachen An- nahmenbas ie r t : 1. die P r o p o r t i o n d e r E i c h e n p ver~indert s ich im ~ber - lappungsgebiet linear (p sinkt yon 1 nach 0). Das ist nattirlich eine Vereinfachung des konkreten Falles, die jedoch im Durchschnitt bei Betrachtung vieler Hanglagen stimmt. Solche Vereinfachungen mtissen in Modellen gemacht werden, um tiberhaupt zu generellen Aussagen zu kommen. 2. W~ihrend der Nahrungssuche teilen sich die Aktivit~iten des Vogels auf in a) F 1 i e g e n v o n B a u m z u B a u m u n d b) N a 11-- r u n g s s u ch e i m B a um. Das Fliegen nimmt den Anteil der insgesamt zur Ver- £tigung stehenden Zeit k in Anspruch, das Fressen demgem~if~ den Anteil ( l - -k) . k ist dabei umgekehrt proport ional der Baumdichte. Je di&ter die B~iume stehen, desto weniger Zeit mug f ib das Fliegen aufgewendet werden. Im Mis&wald muff daher D. nigrescens, wenn er nur Eichen aufsucht, die Zeit k /p f ib das Fliegen aufbringen. 3. Der Vogel kann zwei S t r a t e g i e n der Nahrungsaufnahme verfolgen: Er kann entweder als S p e z i a 1 i s t nut Eichen absuchen, oder er kann als G e n e r a I i s t Eichen und Kiefern absuchen. Es ist anzunehmen, daft in letzterem Fall seine Effi-

290 U. HALBACn [J. Ore. L l l7
zienz der Nahrungsaufnahme nicht in beidea Baumarten gleich grof~ ist. In Kiefern ist sie eine Fraktion q derjenigen in Eichen, wobei q < 1.
Tab. 1 gibt die Fitness-Werte W ftir den S p e z i a 1 i s t e n und den G e n e r a - 1 i s t e n. Fitness bedeutet in diesem Fall die Effizienz der Nahrungsaufnahme oder Netto-Energie-Gewinn. Dies ist die Energie der aufgenommenen Nahrung nach Ab- zug der aufgewendeten Energien fiir Fliegen, Suchen und Fressen. Die Fitness- Formeln der Tab. 1 enthalten ausschlief~lich folgende Parameter: die Fitness in reinem
Dendro ica n i q r e s c e n s
Funk t ionen der F i t n e s s - W e r t e (W)
S__pezialist •
G e n e r a l i s t :
W ~. W(1 - k l p)
= W I' -~p ( q W ) l ' - " ~ ' - ~
Die Parameter: W = Fitness im reinen Eichenwaid
p = Proportion der Eichenb~.ume q = Proportion der Fitness
k = Proportion der Flugzeit
Tab. 1. Formeln zur Berechnung der Fitness-Werte des Spezialisten und des Generalisten.
Eichenwald W, die Proportion der Eichenb~ume p, die Proportion der Fitness q und die Proportion der Flugzeit k. Alle 4 Gr6t~en lassen sich grunds~itzlich bestimmen. Solange sie nicht bekannt sind, kann marl jedoch auch zun~ichst einmal fiktive Werte einsetzen um zu pr~ifen, wie die Fitness-Werte sich in dem Gradienten prinzipiell verhalten.
In der Abb. 8 A ist die berechnete Fitness W fiir D. nigrescens im Eichengradienten aufgetragen, u.z. ftir einen angenommenen k-Wert yon 0,1, was bedeutet, datl im reinen Eichenwald 10°/0 der Gesamtzeit ftir das Fliegen und 90 % ftir das Fressen aufgewendet wird. Die Fitness des Spezialisten (durchgezogene Linie) nimmt mit zu- nehmender BerghShe erst langsam und dann immer schneller ab und erreicht kurz vor dem reinen Kiefernwald den Weft Null. Die gestrichelte Kurve gibt die Fitness eines Generalisten an, der auch Kiefern besucht, dort aber nut 70 % der Effizienz hat im Vergleich zu den Eichen (q = 0,7). Die Kurven des Spezialisten und des Generalisten schneiden sich bei P. Rechts yon P hat der Spezialist, links yon P der Generalist eine gr6f~ere Fitness. Fiir D. nigrescens ist es dementsprechend aus energetischen GrLinden die beste Strategie, vom Tal bis P Spezialist und bei gr6f~eren H/Shen Generalist zu sein. Der P-Wert, bei dem der Vogel sein Verhalten tunlichst umschalten mike, tiegt fLir q = 0,7 bei p = 0,23 (23 % der B{iume Eichen, 77 0/0 Kiefern).

Heft 3 ] 1976 J Modelle in der Omithologie 291
2 0 - - ~ 2.0 - - 2.0 E S ~ 2.0 I E S ~ 2.0
1.0- 1.0 1.0 . . . . . . 1.0 - !.0
o . . . . I i ' " ! . . . . I - ° o o 0.5 1.0 0.5 1.0 0 5 1.0 0 0.5 1.0
A B C D
Abb. 8. Fitness-Werte W {iir D. nigrescens im Eichengradienten. A. k = 0,1; q = 0,7. B. k = 0,1; q = 0,3; C. k = 0,5; q = 0,3. D. Fitness yon D. nigrescens als Eichenspezialist (ES), als Eichen- generalist (EG) und als Eichengeneralist in Gegertwart des Kiefemspezialisten D. graciae. Der Um-
schlagspunkt yon SpeziaIist auf Generalist sinkt in letzterem Fall yon p = 0,31 auf p =: 0,12.
Betra&ten wir nun den Fall, dag die Kiefern sehr viel schlechter genutzt werden. Wenn die Effizienz in den Kiefern nut 30 % derjenigen in Ei&en ist ( q = 0,3), dann erfolgt der Umschlag bei p = 0,11 ( l l % Ei&en), wobei die Fitness sehr viel tiefer sinkt als bei der vorherigen Konstellation (Abb. 8 B). Man kann in dem Modell auch k veriindern, indem man die Dichte der Niume geringer macht. Dann miissen die VSgel mehr Zeit zum Fliegen aufwenden, z.B. 50 °/0 (k = 0,5). Dieser Fall ist in der Abb. 8 C dargestellt. Die Fitness ist dur&gehend geringer als bei der vorherigen Konstel!ation. Der Ums&lag yon Spezialist nach Generalist sollte in diesem Fall frtiher erfolgen, niimlich berelts bei p = 0,58 (58'% Ei&en). Hier wird der Vorteil solcher Modelle deutlich: Man kann Parameter willkiirlich veriindern und die daraus resultierenden Folgen prtifen. Auf diese Weise lassen si& Strategien dur&spielen, was im Experiment nicht mSglich ist, denn p, q und k lassen si& ni&t willktirlich in be- tiebigem Ausmag veHindern.
Die Aussagekra8 des Modelles ist jedo& noch nicht ers&Sp~. Bisher haben wit gezeigt, wie si& eine einzelne Art (D. nigrescens) am besten in einem Gradienten verh~ilt. Wir wollten jedo& priKen, welche Strategien zwei verwandte Arten beim Aufeinandertreffen in einem Mis&wald ausiiben sollten, um zu optimalen Effizien- zen zu kommen. Was ges&ieht also, wenn in dem Gradienten Grace's Baumwald- s~inger, der Kie£ernspezialist, hinzukommt? Man kann ein ganz allgemeines Resultat voraussehen, n~imli& dag fiir den Eichenspezialisten der Umschlag zum Generalisten verz~Sgert werden sollte, da der Konkurrent die Ausnutzbarkeit der in den Kiefern lebenden Insekten herabsetzt. Dies kann 3 Griinde haben: 1. Der Konkurrent {rigt die Insekten und reduziert sic damit zahlenm~igig. 2. Unter seinem Feinddru& ver- ste&en si& die Insekten m6glicherweise besser. 3. Der Kontrahent kann direkt atta&iert und vom Nahrungsbaum vertrieben werden. Durch den Konkurrenten kann also k und q gesenkt werden. Tabelle 2 gibt die Fitness-Werte •r D. nigrescens allein (u. z. als Spezialist und als Generalist) sowie in Mischpopulationen mit D. graciae als Kiefernspezialist und als Generalist. Es handett si& um einfa&e alge-

FJ.Orn. 292 U. HaL~aCH k 117
kann also k und q gesenktwerden. Tabelle 2 gibt die Fitness-Werte l a r D . nigrescens
Typ
ES
EG
ES
EG
ES
EG
Konkurrenz
- - W ( ; - k I p)
_ W e , - , l ' P ( q W ) o - , I ( I - .I
K S KEINE BEEINFLUSSUNG
qa~ - ,m - p ) w ( 1 - k)(2 -p) KS
KG
KG
Dendro ica nN.rescens
W [1 - k I (1 - p) ) . l . q (1 - k)(1 - p) W ( 1 - k)(1 - p)
W 2 1 1 - k / (1 - P ) I
W{1 -k I(I -p)l+qO k)(1 -p)w(1 -k)(1 -p)
KOMMT NICHT VOR
W " FITNESS IN REINEN EtCHENWALO
Funktionen der Fitness-Werte (0) p ~ PROPORTION DER EICHENB.AUME q • PRQPORTION DER FITNESS
k " PROPORTION DER FLUGZE[T
Tab. 2. Fimess-Werte fiir Dendroica nigrescens allein als Spezialist (ES) und als Generalist (EG) sowie mit D. graclae als Kiefern-Spezialist (KS) und als Generalist (KG).
braische Ausdrticke, die sich aus den eingangs gemachten vier Annahmen ableiten lassen. Als Parameter treten wiederum nur W, p, q und k auf.
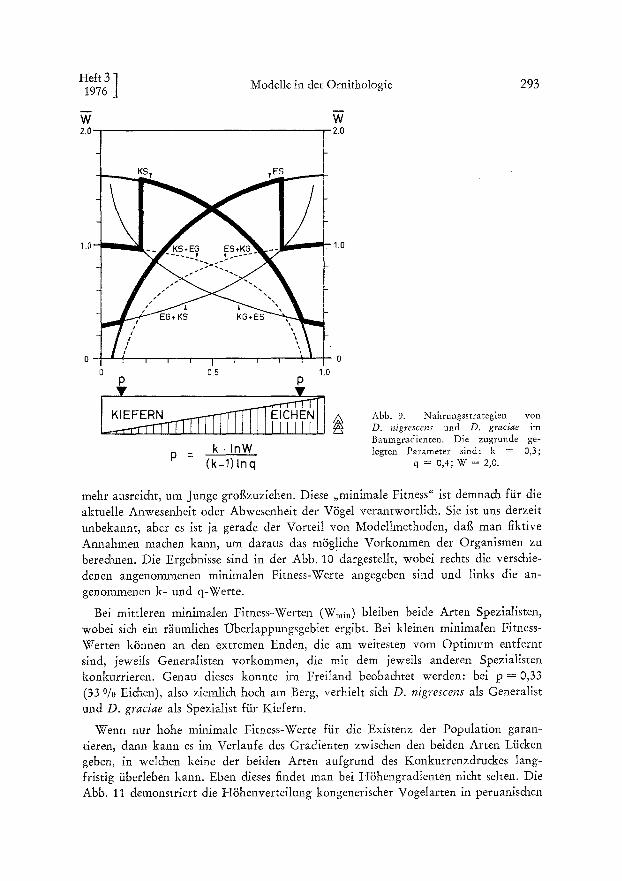
Die Abb. 8 D zeigt, dat~ D. nigrescens bei Abwesenheit des Konkurrenter. D. graciae bei p = 0,23 (23 o/0 Eichen) vom Spezialisten auf einen Generalisten um- s&ahen solhe. Bei Anwesenheit des Konkurrenten wird die Effizienz des D. nigres- cens-Generalisten drastisch gesenkt und damit der Umschlagpunkt bis auf p = 0,12 (12 0/o Ei&en) verzSgert. Fiir die konkurrierende Art gilt natiirlich splegelbildli& das Gleiche (Abb. 9). Die bier zugrunde gelegten Parameter sind k = 0,3, q = 0,4 unc W = 2,0. Neben den bereits bekannten Sa&verhahen tritt hier iiberraschend noch eine interessante Ers&einung auf: Nahe dem reinen Eichenwald soll die Fitness yon D. nigrescens erstaunlicherweise noch einmal drastis& absinken, weil hier n~imlich D. graciae yon dem £iir ihn unerhebli&en Kiefernspezialisten auf einen Generalisten umsteigt und damit plStzlich D. nigrescens als Konkurrent gegeniiber steht, wodurch die Fitness yon D. nigrescens auf die untere Kurve der Abb, 9 absinkt. In der Praxis beobachtet man natiirli& nicht einen so s&arfen Abfall der Fitness, aber doch immer- bin einen Rti&gang der Effizienz. Au& dies gilt spiegelbildli& £iir beide Arten.
Es ist demonstriert worden, dag die Fitness-Werte unter dem Einflug der Kon- kurrenz erhebli& sinken kSnnen. Dabei taucht die Frage auf, ob die P°puIati°nen bei den nledrigen Fimess-Werten no& langfristig f~berleben kSnnen. Es wird ja mit Sicherheit einen unteren Fimess-Wert geben, bei dem die gewonnene Energie nicht
Heft 3 ] 1976
w 2 . 0 ' ,
1.0"
Modelle in der Omithologie
W . 2 . ( 1
293
. . . . "1.0
i ~ ' ' ' I ~ r , 0
O 0.5 1.0 P P T T
k • l nW P = ( k - 1 ) l n q
Abb. 9. Nahrungss t ra teg ien yon D. nigrescens und D. graciae im Baumgradien ten . Die zugrunde ge- legten Pa rame te r sind: k = 0,3;
q = 0,4; W = 2,0.
mehr ausreicht, um Junge grot~zuziehen. Diese ,,minimale Fitness" ist demnach fiir die aktuelle Anwesenheit oder Abwesenheit der V6gel verantwortli&. Sie ist uns derzeit unbekannt, aber es ist ja gerade der Vorteil yon Modellmethoden, daf~ man fiktive Annahmen ma&en kann, um daraus das m/ig!iche Vorkommen der Organismen zu bere&nen. Die Ergebnisse sind in der Abb. 10 dargestellt, wobei re&ts die verschie- denen angenommenen minimalen Fimess-Werte angegeben sind und links die an- genommenen k- und q-Werte.
Bei mittleren minimalen Fimess-Werten (Wmin) bleiben beide Arten Spezialisten, wobei si& ein r~iumli&es 12Ibertappungsgebiet ergibt. Bei kleinen minimalen Fitness- Werten k/Snnen an den extremen Enden, die am weitesten vom Optimum entfernt sind, jeweils Generalisten vorkommen, die mit dem jeweils anderen Spezialisten konkurrieren. Genau dieses konnte im Freiland beoba&tet werden: bei p = 0,33 (33 % Ei&en), also ziemli& ho& am Berg, verhielt sich D. nigrescens als Generalist und D. graciae als Spezialist fiir Kiefern.
Wenn nur hohe minimale Fimess-Werte fiir die Existenz der Population garan- tieren, dann kann es im Verlaufe des Gradienten zwis&en den beiden Arten Lii&en geben, in welchen keine der beiden Arten aufgrund des Konkurrenzdru&es Iang- fristig iiberleben kann. Eben dieses findet man bei H6hengradienten nicht selten. Die Abb. 11 demonstriert die H6henverteilung kongeneris&er Vogelarten in peruanischen

294 U . H A L B A C H
I , EG ~ E$ k=o3 q = O ~ i i i i i i
' ' • ' i I i I i i I ', Ks I KG i ! ! i ! i ~ = : '
' l ; E g , = o ~ = o ~ i i i i i i i ! i
• i : i i I i i I i I ~s I KG ~ i i { i i i i i i F'EG I ES
: 1 1 ; ; ~ 1 ; 1 1 ! 1 , ~ i ; i ; !
k = 0 . 3 q = 0 . 4
I . . K ~ I . . , i i i ! i , ~ i i !, ,,
i i ' ' E S ' k=03 q = 0 . 4
Wmin = 0.85
Wmin = 0.B5
Wmin = 0.3
Wmin • 0.4
Wmin > 1.0
k = 0 5 q=0-4 i i i i K ; ~ i I [ ; Kd
{ I I~GI; ' ; ~s I ' I
~ = ° ~ = ° ~ b ~ : ~ ~ , i l ¢ ~ j i i i i i i ! i i ~ i ; : I I ; ; ] i i l i I i E*
, , i i i
k = 0 . 3 ' ~ i : i l i ] i °=°' i , i i : ; i PaOPONT ON DE N E CHE~ i i t , I | / I * i 0 0.5 1.0
W m i n > 0 . 3
W m i n > 0 . 4
J.Orn. 117
Abb. 10. Verteilung zweier Voge!arten wmin-l.0 endang eines H6hengradienten. EG
= Eichengeneralist, ES = Ei&en- spezialist, KG = Kieferngeneralist, KS = Kiefernspezialist. Bel hoherl
Wmin > 1.4 minimalen Fimess-Werten k6nnen Lticken im Verteilungsmuster auf-
t r e l : e n .
Bergen. Hier fallen die vorausgesagten Lticken im Verteilungsmuster auf. Sie weisen darauf hin, dat~ diese Arten einen hohen minimalen Fimess-Wert haben. Aus dem geschilderten Modell lassen sich noch weitere Folgerungen ziehen (CoDY 1974). Es ist gezeigt worden, daf~ das einfache Modell der optimaten Nahrungsstrategie in Oberlappungsgebieten der Vegetation auf der Basis sehr weniger logischer Annah- men, die wegen ihrer Einfachheit kaum anzufechten sind, deduktiv eine ganze Reihe yon Folgerungen zul~if~t, die aufgrund der logischen Konstruktion des Modeiles be- reits einen hohen Wahrscheinlichkeitsgrad haben, die man aber zudem im Freiland testen kann, was im Vorigen an zwei Beispielen exemplarisch demonstriert worden ist.
Modelle sind mathematisch ausformulierte Arbeitshypothesen, die einer experi- mentellen Nachprtifung bediirfen. Als solche haben sie - - insbesondere in amerikani- schen Forschungsgruppen - - fruchtbare Anregungen gegeben. So lieferten sie plausible Aussagen zu folgenden Problemen (MAcARTHUI~ 1972, CODY 1974): 1. Wie erkl~iren sich die Unterschiede zwischen Insel-Lebensgemeinschaften gegeniiber denen des Fest- landes? 2. Wie iassen sich die saisonellen Unterschiede yon Lebensgemeinschaften, ins- besondere die Eingliederung yon Winterg~isten, deuten? 3. Welche energetische

Heft 3 ] 1976 J Modelle in der Ornithologie 295
V- i 2 I
[ 2 1
I __~ ~ 4 I
I I~]
I i 2 II~
,,r-T--1
i i i r - - r -
I 2 ] i i i
t i | 2000 1000 Meter 3000
Coe[igeno. violifer (1) C. co[eligna (2)
Myloborus me[anocepha[us (1) M. miniatus (2)
Cacicus holosericeus (1) C. [eucorhamphus (2) C. uropygialis (3) C, cela (4)
Scytaiopus unicolor (1) S. femoraiis (2)
Grallaria rufula (1) G. erythroleuca (2) G. squamigera (3) G. guatema[ensis (4)
Thripadectes scrutator (1) Th. hoiostictus (2) Th, melanorhynchus (3)
Xiphorhynchus triangu[aris (1) X. oce[latus (2) X. guttatus (3)
Veniiiornis nigriceps (1) V. dignis (2) V. affinis (3)
Eubucco versicolor (1) E. richc~rdsoni (2)
Iridisornis reinhardti (1) t. analis (2)
Abb. 11. Die H6henverteilung kongenerischer Vogelarten in peruanischen Bergen.
Strategie liegt den gemischten Vogelschw~rmen im Winter zugrunde? 4. Wie erkl~iren sich die Unterschiede in den Lebensgemeinschaften gem~iffigter Zonen und der Tropen?
Es ist zu wtinschen, dall die auf~erordentlich anregenden Modellmethoden auch in der deutschen Ornithologie eine verbreitetere Anwendung linden*).
S u m m a r y
P o p u l a t i o n m o d e l s a n d s y n e c o l o g i c a l m o d e l s i n o r n i t h o l o g y
Mathematical models are frequently used in modern ecology and have become a necessary tool for studying the complex systems of populations and communitles. - - Here the logistic growth function which describes the sigmoid growth of the individual density of populations in a limited environment is presented as an example of a deterministic model. The incor- poration of time lags results in realistic descriptions of the population oscillations empirically found (Fig. 1). Thus it is possible to predict population dynamics using life table data measured from single individuals in laboratory cultures. Ecological factors such as tempera- ture etc., which modify the life table data, influence the frequency and amplitude of the population oscillations (Fig. 2). Yet the deterministic models based on differential equations do not consider the individual variability of the organisms. To incorporate it, stochastic models based on the computer language STMULA are used.
The very productive concept of the ecological niche (Fig. 4) has resulted in explaining the various evolutionary strategies by one common conception (Fig. 5): Specialists, generalists, different adaptations, polymorphlsm, cyclomorphosis, diapause, and annual migration are different strategies to survive in a variable environment. Based on the concept of the eeo-
* Fiir Diskussion, Durchsicht nnd Bearbeitung des Manuskriptes danke ich ROSWITHA und WOLF- GANG WILTSeHKO, ftir die Herstellung der Zeichnungen CLAUDIA NITSCHE und [-IEINZ-B. M/JLLER.

296 U. HALBACH r C~,,~[J.117..
logical niche, the following principles are established describing the effects of competition: competition-exclusion-principle, character displacement, geographical and spatial separation. These principles are closely related with each other, yet they may produce different or even opposite results.
To find evidence for possible overlaps of ecological niches in a bird community, MacArthur used the well known "broken stick model", which later was questioned because of its formal inadequacies. - - One way of measuring the overlap in isolated ecological parameters is shown in Fig. 6. The sum of overlaps in the three main factors - - vertical habitat selection, hori- zontal habitat selection and food selection - - is nearly constant (Fig. 7), which means that a large overlap in one factor can be compensated by a small overlap in another factor.
The explanatory value and the power of prediction of a model is demonstrated for the model based on a few simple assumptions concerning the foraging strategies of two closely related bird species (Dendroica graciae, D. nigrescens) in a food gradient, introduced by Cody: In specific situations the birds change from being specialists to being generalists (Fig. 8 and 9). Minimum fitness values cause the specific distributions of the bird species and their gaps and overlaps (Fig. 10 and 11).
The use of mathematical models as scientific tool is still debated. But in ecology, they have resulted in so many useful concepts that a further introduction into ornithology appears desirable.
L i te ra tur
Copy, M.L. (1974): Competition and the Structure of Bird Communities. Princeton (New Jersey). • CtmE, F. yon (1967): Was ist Kybernetik? Sch~inemann Verlag (Bremen). • DUZLIN, L. I., LOTXA, A.I. (1925): On the True Rate of Natural Increase. J. Amer. statist. Ass. 20, 305--339. ° E~LEN, J.T. (1940): Sex and Age Ratios in Survival of the California Quail. J. Wildlife Mint 4, 29 99. ° HALBACH, U. (1970): Die Ursachen der Temporal- variation yon Brachionus calyciflorus PALLAS (Rotatoria). Oecologia (Berl.) 4, 262 318. • Ders. (1971): Zum Adaptivwert der zyklomorphen Dornenbildung von Brachionus calyci- florus PALLAS (Rotatoria). I. R~iuber-Beute-Beziehung in Kurzzeit-Versuchen. Oecologia (Berl.) 6, 267 288. ° Ders. (1974): Modelle in der Biologic. Naturw. Rdsch. 27, 3--15. • Ders. (1975): Methoden in der Populations~Skologie. Verh. Ges. (Skologie, Erlangen 1974, I 24. • HALBACH. U. ~; BURKHARDT, H.-J. (1972): Sind einfache Zeitverzbgerungen die Ur- sachen fiir perlodische Populationsschwankungen? Vergleich experimenteller Untersuchungen an Brachionus calyciflorus PALLAS (Rotatoria) mit Computer-Simulationen. Oecologia (Berl.) 9, 215- -222 . . HEP, ZOG, K. (1968): Anatomie und Flugbiologie der VtSgel. Fischer Verlag (Stuttgart). • HOLST, E. yon (1957) : Wie flog Rhamphorhynchus? Natur und Volk 87, 81--87. • HUTCHINSON, G. E. (1948): Circular Causal Systems in Ecology. Amn. N. Y. Acad. Sci. 50, 221--246. • JACOBS, J. (1961): Cyclomorphosis in Daphnia galeata rnendotae BIRGE, a Case of Environmentally Controlled Allometry. Arch. Hydrobiol. 58, 7 71. ® MAcARTHtm, R. H. (1958): Population Ecology of Some Warblers of Northeastern Coniferous Forests. Ecology 39, 599--619. • Ders. (1972): Geographical Ecology. Patterns in the Distri- bution of Species. Harper & Row (London). ° VOLTERRA, V. (~926): Variazione e fluttuazioni del numero d'induvidii in specie animall conviventi. Mem. Accad Naz Lincei 2, 31--113. • WANGERSKY, P. J. ~ CUNNINGHAM, W. J. (1957): On Time Lags in Equations of Growth. Cold Spr. Harb. Symp. Quant. Biol. 22, 329--338. ° Z~AneII, H. (1961): Technische und Kyber- netische Modelle. In: H. MITTELSTAEDT (Hrsgb.): Regelungsvorglinge in Iebenden Wesen. Oldenbourg Verlag (Miinchen).
Anschriflc des Verfassers: Zoologisches Institut der Universifilt, 6000 Frankfurt/M, Sies- mayerstraf~e 70