Embed Size (px)
Citation preview

Biochem. Physiol. Pflanzen 176, 744-752 (1981)
Regulation der Aktivitat granulumgebundener Starkesynthetase durch einwertige Kationen
ERHARD PREUSSER, FA WZY ANDRA WISS KHALIL und HORST GORING Humboldt-Universitat zu Berlin, Sektion Biologie, Bereiche Biochemie und Pflanzenphysiologie
Berlin, DDR
Regulation of Activtiy of the Granule-bound Starch Synthetase by Monovalent Cations
Key Term Index: granule-bound starch synthetase, monovalent cations; Zea mays, Solanum tuberosum, Yicia (aba.
Summary
Granule-bound starch synthetase was isolated from Zea mays seeds (mild-waxy stage), Solanum tuberosum tubers, and Vicia faba leaves. The activity of the enzyme system was studied in vitro under the influence of various cations applied over a large concentration range. Tlje activity of the granulebound starch synthetase is considerably increased by relatively high K+ or Rb+ concentrations (50 to 100 mM). Other monovalent cations have a slightly activating effect at concentrations up to 20 mM only. Their effect is an additive one at both suboptimal K + concentrations and mutually low concentrations of the other univalent cations concerned. At higher concentrations the K+ effect is substantually abolished. The results are discussed in relation to regulatory problems.
Einleitung
\Viederholt wurden Vorstellungell entwickelt, nach denen primare Licht- und Hormonwirkungen eine Veranderung von Kationenbalancen in pflanzlichen Zellen bzw. Zellkompartimenten induzieren sollen (GORING und MARDANOV 1976; HAUPT 1980). 1m gleichen Zusammenhang ist die regulierende Wirkung von Kationen auf verschiedene Enzymsysteme beschrieben worden (EVANS und WILDE 1971; EVANS und SORGER 1966). Zu diesen kationenaktivierbaren Enzyrnen gehort die starkekorngebundene Starkesynthetase (ADP-Glucose: 1,4-tX-D-glucose-4-tX-glucosyltransferase, EC 2.4.1.b) der Chloroplasten ebenso, wie die Glycosyltransferase (EC 2.4.1.21), die vorwiegend in den Amyloplasten der Speichergewebe lokalisiert ist(MEISEL 1974; NITSOS und EVANS 1969; HAWKER et al. 1974; MURATA und AKAZAWA 1969).
Die granulumgebundene Starke synthetase ist fest in das Starkekorn integriert und relativ stabil. Bezuglich ihrer Aktivitat ist sie im starken MaI3e von der Wechselwirkung mit Kationen und einigen anderen Verbindungen abhangig.
Die starkekorngebundene Starke synthetase sollte in den vorliegenden Untersuchungen als Modell fur ein kationenaktivierbares Enzymsystem genutzt werden, urn den Einflu13 verschiedener monovalenter Kationen uber einen grofieren Konzentrationsbereich zu analysieren.

Regulation der Aktivitat von Starkesynthetase 745
Material und Methoden
Zur Durchfiihrung der nachfolgenden Versuche wurde Starke aus Blattern von Vicia taba minor (nach Belichtung), aus Knollen von Solanum tuberosum (Handelsware) und aus Karyopsen von Zea mays im Stadium der Milch-Wachs-Reife isoliert. Das Pflanzenmaterial wurde bei 0 bis 4 °C im TrisAcetat-Puffer (0,1 M, pH 8,4) unter Zusatz von Dithioerythrol und Natriumthiosulfat als Stabilisatoren sorgfaltig homogenisiert. Das Homogenat wurde unter Druck schnell durch Perlongewebe filtriert. Das Filtrat wurde dann sofort in vorgekuhlten Zentrifugenbechern 15 min bei etwa 1500 x g zentrifugiert. Der resultierende Starkeniederschlag ist noch stark mit Ohloroplasten und anderen Zellbruchstucken verunreinigt. Nach Entfernung der oberen gelblichen Schicht wurde die verbleibende Starke im gieichen Puffer bei 0 bis 4 °0 wiederholt zentrifugiert. AbschlieBend wurde die einheitliche, weiBliehe Starkegranulafraktion bei -15 °0 3 x mit Aceton gewaschen. Die Acetonbehandlung dient nicht nur der Entfernung von Membranbruchstucken (Lipide), sondern auch der Inaktivierung der Phosphorylasen.
Die auf diese Weise gewonnene Starke wurde bei -18 00 zum Teil uber einen langeren Zeitraum aufbewahrt. Dabei kommt es zu einem langsamen Aktivitatsverlust, der unter unseren Bedingungen bei 40-50 % im Verla ufe einer ein j ahrigen Lagerung lag.
Fur die Inkubationsversuche wurden 40 mg des Starkepraparats mit 4 mi Tris-Acetat-Puffer (0,1 M, pH 8,4) versetzt. Dem Ansatz wurden O,l,ttM Dithioerythrol und zuletzt 20,tt1 einer ADPGLosung (Glucose uniform HO-markiert, Gesamtradioaktivitat 2 x 105 bzw. 2 X 106 Imp./min) zugesetzt. Nach sorgfaltigem Durchmischen bei 0 °0 wurden je 0,2 mi dieses Ansatzes in kleine Glaschen gegeben, die bereits 0,2 ml der Ionenlosung enthielten (siehe Legende der Abbildungen). Diese Ansatze wurden bei 34°C 45 min inkubiert. Durch Uberfuhren in ein kochendes Wasserbad (3 min) und durch Zusatz von 0,4 ml Xthanol wurde der Versuch abgebrochen. In diesen Ansatzen wurde die Starke durch Zentrifugation (1500 x g, 5 min) abgetrennt und durch wiederholtes Waschen mit Xthanol so lange gereinigt, bis der Uberstand frei von Radioaktivitat war.
Die Messung der Radioaktivitat erfolgte im Packard-Tricarb-liquid-Scintillationsspektrometer. In allen Versuchen wurden zwei Kontrollen mitgefuhrt: eine "Kochprobe", die so fort nach Zugabe der markierten Verbindung in der beschriebenen Weise ina.ktiviert wurde und eine 0-Variante, die anstelle der TestlOsung 0,2 ml Tris-Acetat-Puffer enthielt.
In Vorbereitung <Luf die Proteinbestimmung wurde die Starkeprobe im kochenden Wasserbad 15 min mit 0,1 M KOH behandelt. Die anschlief3ende Proteinbestimmung erfolgte nach LOWRY et aJ. (1951). Der Na+-, K +- und Oa++~Gehalt wurde flammenphotometrisch nach feuchter Veras chung der Starkekorner bestimmt. Die Phosphatbestimmung erfolgte kolorimetrisch nach Mineralisierung der Starke (BARTLETT 1959). Zur Lipidbestimmung wurde die Starke nach einer Vorbehandlung mit Amyloglucosidase und Amylase 3 Std. mit Athanol gekoeht und anschlieBend eingeengt. Die Extraktion der "Roh"-Lipide erfolgte mit Ather.
Ergebnisse
Allgemeine Charakterisierung der granulumgebundenen Stiirkesynthetase
Starkegranula enthalten neben Amylopektin und Amylose immer geringe Mengen anderer Stoffe (Tabelle 1). Es ist ersichtlich, daB sich in quantitativer Hinsicht bei diesen "Starkebegleitstoffen" erhebliche Schwankungen abzeichnen. Diese unterschiedlichen Befunde sind im wesentlichen mit dem Objekt bzw. mit dem Reifegrad des Starkekorns und weniger mit der Isolierung und Reinigung der Starke selbst verbunden. Fur den Glucosetransfer von ADPG auf die Starkegranula wurde ein pH-Optimum von 8,2 bis 8,4 und ein Temperaturoptimum von 34°C ermittelt.
746 E. PREUSSER U. a.
Tabelle 1. Zusammensetzung enzymatisch aktiver Stiirkegranula (Angabeu in % vom Trockengewicht, n = 4-5)
Objekt
Protein Lipid Kalium Natrium Calcium Phosphat
Zea mays (Karyopsen, Milch-W achs-Reife)
0,35-0,72 0,82-0,92 0,037-0,043 0,12-0,18 0,1:23 0,16
Solanum tuberosum (Knollen)
° "') ,0 ...
0,60 0,026 0,10 0,02 0,10
Vicia faba (Blatter)
1,45-1,75 1,08-1,15 0,052 0,22 0,01 0,42
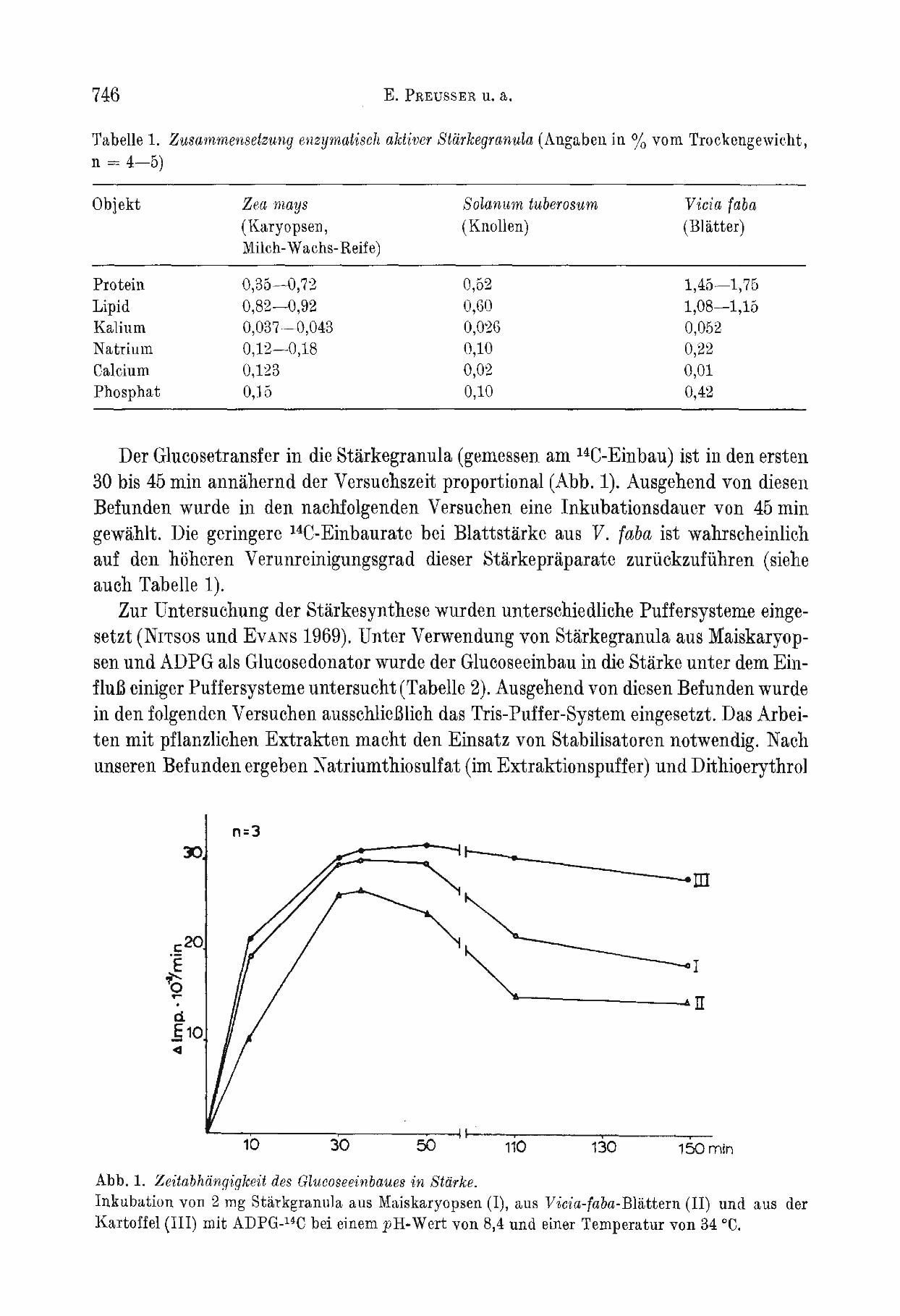
Der Glucosetransfer in die Starkegranula (gemessen am 14C-Einbau) ist in den ersten 30 bis 45 min annahernd der Versuchszeit proportional (Abb. 1). Ausgehend von diesen Befunden wurde in den nachfolgenden Versuchen eine Inkubationsdauer von 45 min gewahlt. Die geringere 14C-Einbaurate bei Blattstarke aus V. (aha ist wahrscheinlich auf den hoheren Verunreinigungsgrad dieser Starkepraparate zuruckzufiihren (siehe auch Tabelle 1).
Zur Untersuchung der Starkesynthese wurden unterschiedliche Puffersysteme eingesetzt (NITSOS und EVANS 1969). Unter Verwendung von Starkegranula aus Maiskaryopsen und ADPG als Glucosedonator wurde der Glucoseeinbau in die Starke unter dem EinfluB einiger Puffersysteme untersucht (Tabelle 2). Ausgehend von diesen Befunden wurde in den folgenden Versuchen ausschlieBlich das Tris-Puffer-System eingesetzt. Das Arbeiten mit pflanzlichen Extrakten macht den Einsatz von Stabilisatoren notwendig. Nach unseren Befunden ergeben Natriumthiosulfat (im Extraktionspuffer) und Dithioerythrol
n=3 30 r--------_____
m
c20 E I ~ .,...
II • d .510 oCI
10 30 50 110 ,
130 ,
150 min
Abb. 1. Zeitabhiingigkeit des Glucoseeinbaues in Starke. Inkubation von 2 mg Starkgranula aus Maiskaryopsen (I), aus Vicia-faba-Blattern (II) und aus der Kartoffel (III) mit ADPG-14C bei einem pH. Wert von 8,4 und einer Temperatur von 34 °C.
Regulation der Aktivitat von Starkesynthetase 747
Tabelle 2. Ghwoseeinbau in Starke mit ADPG als Donator wIter Verwendung unterschiedlicher Puffer. systeme (Inkubationsbedingungen: 2 mg Starke, ADPG [Glucose uniform He, 2 x 105 Imp.jminl, 45 min bei 34 °e, n = 3)
Puffersystem
Tris-Acetat (0,1 M) pH 8,4 pH 7,6
Glycylglycin (0,1 M) pH 8,4 pH 7,6
~a-Phosphat (0,1 M) pH 8,4 pH 7,6
HEPES (0,1 M) pH 8,4 pH 7,6
Radioaktivitat der Starke (Imp./min)
3200 (2775) 1861 (1742)
3282 (3625) 2441 (2992)
1785 1543
3116 2603
In Klammern Werte, die unter Verwendung eines Starkepraparates aus jiingeren Maiskaryopsen (frfthe Milcbreife) erzielt wurden.
0--0 K"; l.m. ...-... Rb:Z.m. _K~Vf.
50 o---D K~ S.t.
40
. ci. 20 E <I
10
10 30
ill
50 75 100 mM
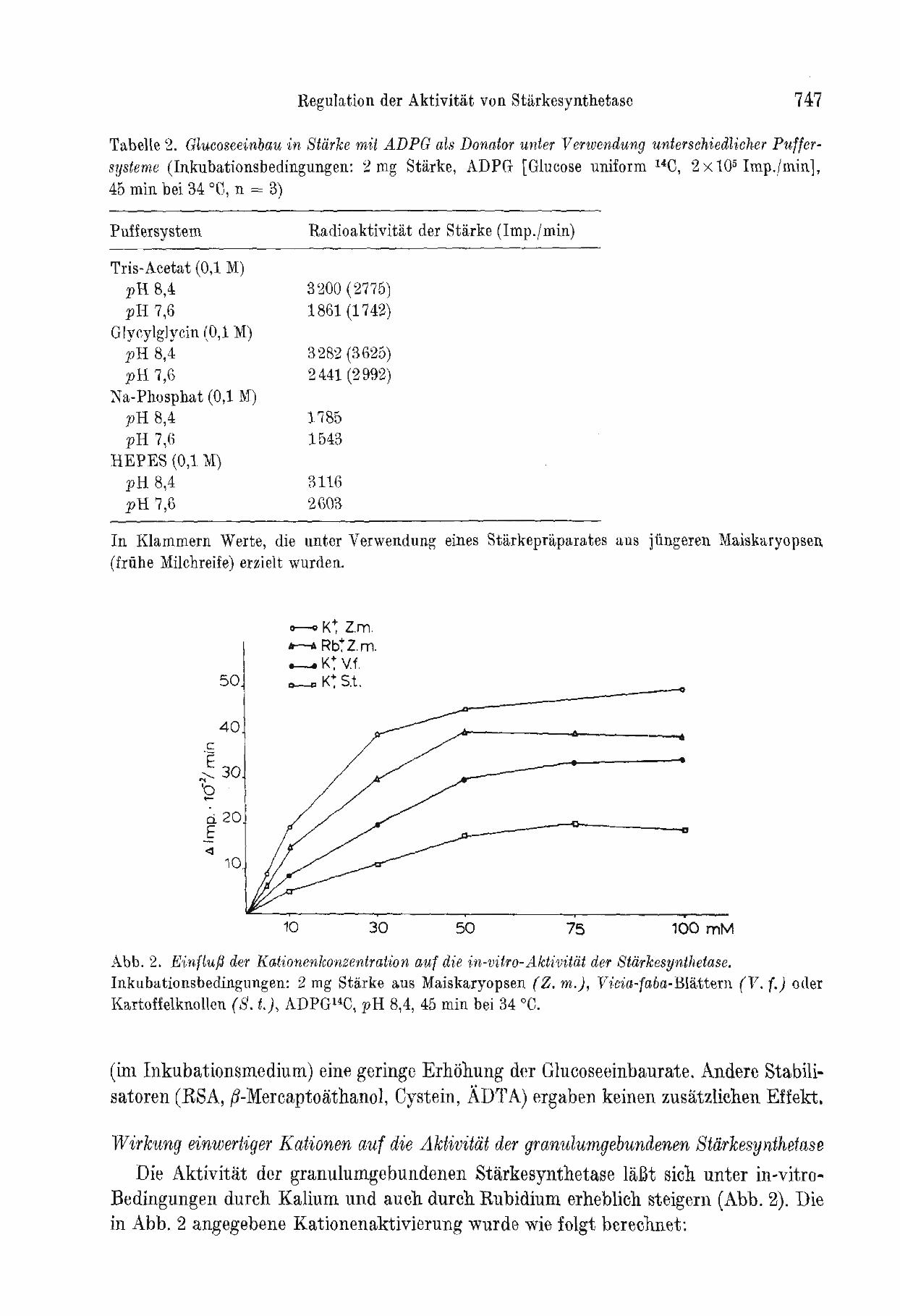
Abb. 2. EinflufJ der Kationenkonzentration auf die in-vitro-Aktivitiit der Starkesynthetase. Inkubationsbedingungen: 2 mg Starke aus Maiskaryopsen (Z. m.j, V1:cia-faba-Blattem (V. (.) oder KartoffelknoHen (S. t.), ADPG14C, pH 8,4, 45 min bei 34 °e.
(inl Inkubationsmedium) eine geringe Erhohung der Glucoseeinbaurate. Andere Stabilisatoren (RSA, p-Mercaptoathanol, Cystein, ADTA) ergaben keinen zusatzlichen Effekt.
Wirkung einwertiger Kationen auf die Aktivitiit der granulumgebundenen Stiirkesynthetase Die Aktivitat def granulumgebundenen Starkesynthetase laJ3t sich unter in-vitro ..
Bedingungen durch Kalium und auch durch Rubidium erheblich steigern (Abb. 2). Die in Abb. 2 angegebene Kationenaktivierung wurde wie folgt berechnet:
748
Kationenaktivierung (Imp.jmin)
E. PREUSSER U. a.
Radioaktivitat der Probe (Imp.jmin)
Radioaktivitat der O-Variante (Imp.jmin)
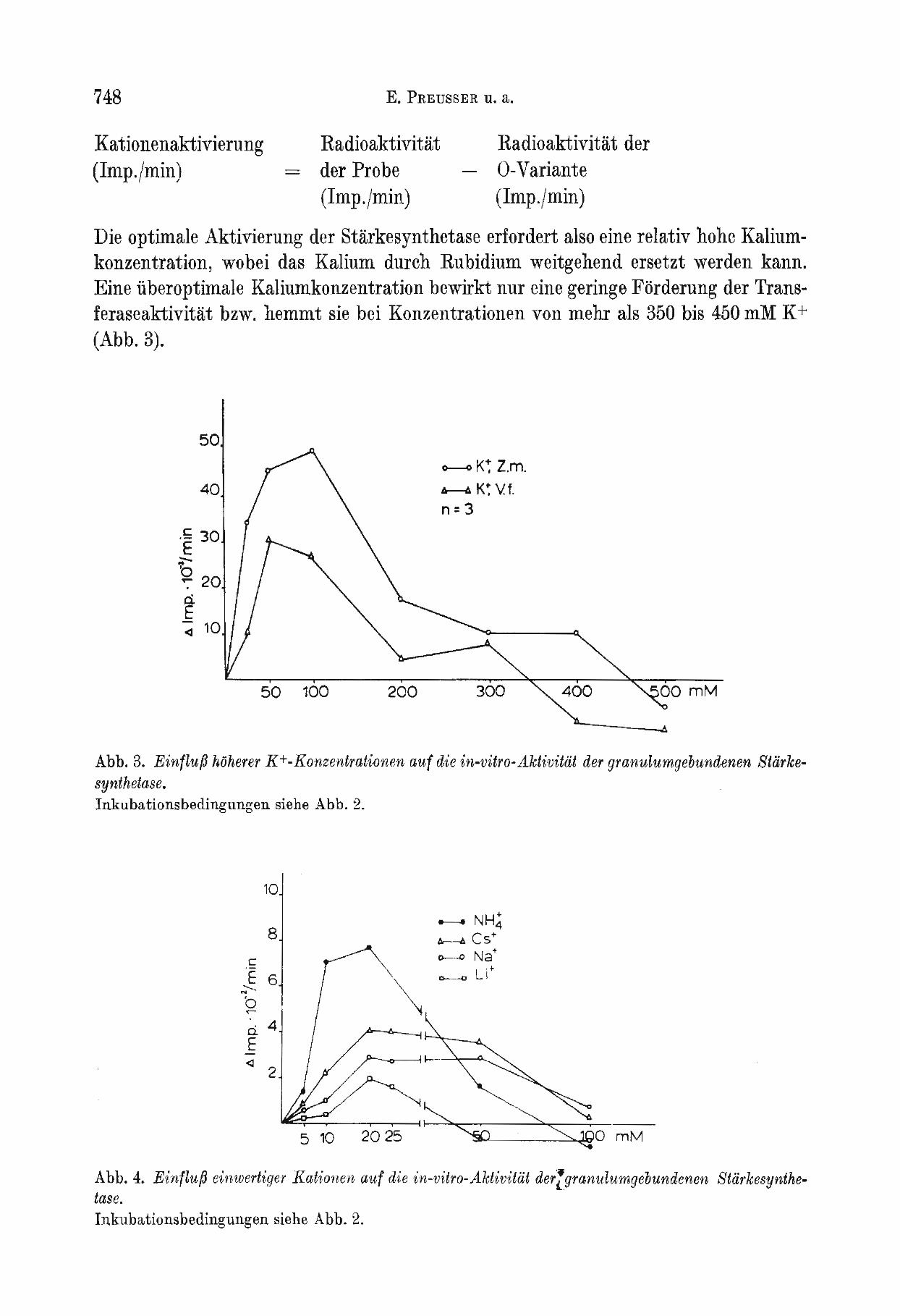
Die optimale Aktivierung der Starkesynthetase erfordert also eine relativ hohe Kaliumkonzentration, wobei das Kalium durch Rubidium weitgehend ersetzt werden kann. Eine iiberoptimale Kaliumkonzentration bewirkt nur eine geringe Forderung der Transferaseaktivitat bzw. hemmt sie bei Konzentrationen von mehr als 350 bis 450 mM K + (Abb.3).
50
40
.£ 30 E
;;-
'~ 20 • ci E ~ 10
50 100 200
0--0 K; Z.m. 4---6 K~ '1.1. n=3
300
Abb.3. EinflufJ hOherer K+-Konzenirationen auf d'ie in-vitro-Aktivitiit der granulumgebundenen Starkesynthetase. Inkubationsbedingungen siehe Abb. 2.
10
8 ..- NH~ f>---.6 Cs·
+
c ().--...C Na
E L '+ 6
\ G--~ I
;;-.. b .... ti. 4
-i E <l
2
5 10 2025 .......... _~ __ ....,:,.",~o mM
Abb.4. EinflufJ einweriiger Kaiionen auf die in-v'iiro-Aktivitiit der'(granulumgebundenen Stiirkesynthelase. Inkubationsbedingungen siehe Abb. 2.
Regulation der Aktivitat von Starkesynthetase 749
Die Aktivitat der granulumgebundenen Starkesynthetase wird unter in-vitro-Bedingungen auch durch andere einwertige Kationen beeinfluJ3t. Aktivierende Effekte ergeben sich hier allerdings in wesentlich niedrigeren Konzentrationsbereichen, wobei die fordernde Wirkung jedoch betrachtlich unter derjenigen der K +-Aktivierung bleibt (Abb.4).
Aktivitiit der granulumgebundenen Stiirkesynthetase bei gleichzeitiger Einwirkung von mehreren eiriwertigen Kationen
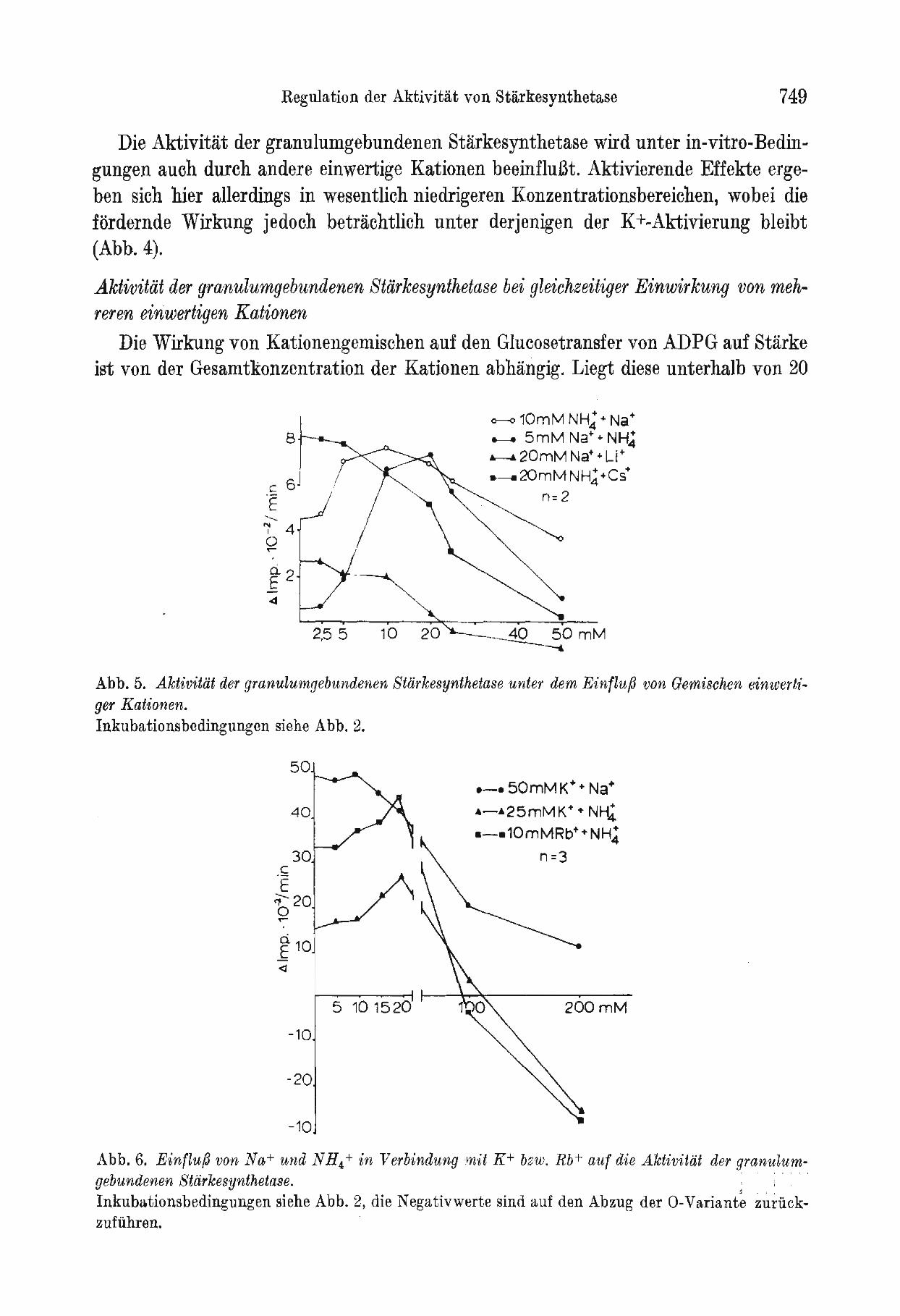
Die Wirkung von Kationengemischen auf den Glucosetransfer von ADPG auf Starke ist von der Gesamtkonzentration der Kationen abhangig. Liegt diese unterhalb von 20
8 0----<) 10mM NH;'" Na· --- 5mM Na ...... NH4 ~ 20mM Na· ... U'" ._ 20mM N H; +Cs'"
n=2 c 6 !
E -----.... 4 I
0 .-
0.2 E <I
. , 2,55 10 ~OmM
Abb. o. Aktivitiit der granulumgebundenen Stlirkesynthetase unter dem Einfluf3 von Gemischen einwertiger Kationen. Inkubationsbedingungen siehe Abb. 2.
50
40
30
~10 <I
5 101520
-10
-20
-10
.-e 50mM K·· Na+
A-A25mMK+· NH.;: _-_lOmMRb·+NH;
n=3
200mM
Abb.6. Einflu/3 von Na+ und NH4+ in Verbindung In'it K+ bzw. Rb+ auf die Aktivitdt der granulumgebundenen Stlirkesynthetase. : ..... Inkubationsbedingungen siehe Abb. 2, die Negativwerte sind auf den Abzug der 0-Variante zuriickzufiihren.

750 E. PREUSSER U. a.
mM, SO ergeben sich im wesentlichen additive Aktivierungseffekte. Ubersteigt die Gesamtkonzentration der einwertigen Kationen aber diesen Wert, so ergeben sich Hemmeffekte (Abb. 5). Prinzipiell die gleichen GesetzmaEigkeiten zeigen sich auch, wenn mehr als zwei unterschiedliche einwertige Kationen gleichzeitig im Inkubationsmedium vorliegen. Auf eine Darstellung wurde deshalb verzichtet.
In Verbindung mit K+ oder Rb+ heben Na+, NH4+, Cs+ und Li+ den aktivierenden Effekt dieser beiden Kationen wieder auf, sobald der optimale Konzentrationsbereich (etwa 20 mM) iiberschritten wird. Bei suboptimalen Kalium- oder Rubidiumkonzelltrationenfuhren diese Kationen im Konzentrationsbereich bis zu 20 mM zu einer geringen zusatzlichen Aktivierung der granulumgebundenen Starke synthetase (Abb. 6).
Diskussion
Das Starkekorn ist als ein dynamisches Gebilde aufzufassen, bei dem quantitative Veranderungen der Zusammensetzung neben dem Amylose-Amylopektinverhaltnis auch andere "Inhaltsstoffe" betreffen. Neben pflanzen- und oganspezifischen Unterschieden kommt z. B. dem Reifezustand bzw. der Endospermentwicklung groEe Bedeutung zu (BAUN et al. 1970). Assirnilationsstarke besitzt einen hoheren Proteingehalt als Reservestarke. Verunreinigungen durch Chloroplastenbruchstucke o. a. sind durch mikroskopische Kontrolle weitgehend auszuschlieEen (vgl. auch MURATA und AKAZAWA 1964). Auch der Proteingehalt von Reservestarken unterliegt erheblichen Schwankungen. So wurde fiir die Starke aus Kartoffelknollen nur ein Proteinwert von 0,08 % ermittelt (W ASHUTTL et al. 1966).
Der K +-Gehalt unterschiedlicher Starken differiert ebenfalls betrachtlich: in Sarnen von Vicia (aba 1,350/0' in Karyopsen von Oryza sativa etwa 0,3 % (MURATA und AKAZA WA 1969). Diese Werte liegen weit iiber den von uns ermittelten. Ais Ursache miissen auch unterschiedliche Reinigungsverfahren in Betracht gezogen werden. Diese "Restgehalte" an Kationen sind sicherlich von Bedeutung fiir die Aktivitat der starkekorngebUlldenen Synthetase, die ohne Zusatz von Kationen zum Inkubationsmedium gemessen werden kann (Abb. 1). Allerdings ist die Glucosetransferreaktion dann nur in den ersten 30 min linear mit der Zeit. Die darauf folgende Abnahme der 14C-Einbaurate diirfte auf hydrolytische Reaktionen (Amylasen?) zuriickzufiihren sein. K + (50 bis 100 mM) ver hindert diesen A bfall der 14C-Mar kierung (vgl. auch N ITSOS und EVANS 1969).
Die vorliegenden Ergebnisse bestatigen die Notwendigkeit des K+ fiir die Starkesynthese und die regulatorische Funktion dieses Kations. In Dbereinstimmung mit den Befunden von AKATSUKA und NELSON (1966); GREENBERG und PREISS (1965); MURATA und AKAZAWA (1968) sowie NITSOS und EVANS (1969) ist die aktivierende Wirkung des Kaliums auf mehrere Erscheinungen zuruckzufiihren. HAWKER et al. (1974) nehmen Konformationsanderungen im Enzymmolekiil bedingt durch Ionenanlagerung an. Damit wiirde sich eine Dbereinstirnrnung mit anderen Enzymen, 80 z. B. mit der Pyruvatkinase, ergeben (NOWAK 1976). Nicht auszuschlieEen ist auch die Annahme, daE die Kaliumaktivierung auf eine festere Bindung des Enzyms an die Starke und damit auf eine Enzymstabilisierung zuruckzufiihren sei. Der Schutzeffekt des Kaliums gegen Hitzedena-

Regulation der Aktivitiit von Starkesynthetase 751
turierung konnte cine solche Annahme stutzen. Da Kalium die Quellung der Starkekor" ncr erhoht, ist naturlich auch die Substratzuganglichkeit erhoht. In diesem Zusammenhang sind moglicherweise auch die Befunde mit iso-Propanol einzuordnen, das eine kaliumahnliche Aktivierung der granulumgebundenen Starke synthetase bewirkt (JUDEWICZ et al. 1972).
Andere einwertige Kationen beeinflussen die granulumgebundene Starke synthetase in unterschiedlichem Ma13e. Die von uns gefundene Reihenfolge der Aktivierung (Li+, Na+, Cs+, NH,+, Rb+, K+) ist mit geringen Abweichungen auch bei anderen Objekten gefunden worden (MURATA und AKAZAWA 1968; NITSOS und EVANS 1969). Unterschiede ergeben sich vor aHem in den optimal en Konzentrationen dieser Kationen. So haben nach NITSOS und EVANS (1969) auch hohere Konzentrationen dieser Kationen (bis zu 50 mM) noch cine stimulierende Wirkung.
Zur Wirkungsweise dieser einwertigen Kationen lassen sieh noch keine endgiiltigen Aussagen treffen. Sieher ist davon auszugehen, daI3 eine sekundare Wirkung - mogli~
cherweise uber eine Freisetzung von unspezifisch gebundenem Kalium - vorliegt. Die vorliegenden Ergebnisse machen deutlich, daI3 die Aktivitat der granulumgebun
denen Starkesynthetase in starkem MaI3e durch einwertige Kationen beeinflu13t wird. Fur eine Aktivierung des Enzyms unter in-vivo-Bedingungen diirfte jedoch nur K + relevant sein, dessen Konzentration in cytoplasmatischen Kompartimenten sich durchaus im Bereich einer maximalen Aktivierung des Enzyms bewegen diirfte (vgl. BESFORD und MAw 1976).
Enzyme des Typs der granulumgebundenen Starkesynthetase konntcn somit durehaus durch Veranderungen der K+-Aktivitat im gegebenen Kompartiment reguliert wer" den. Die Relevanz eines solchen Regulationsmechanismus soIl weiteren Untersuchungen vorbehalten bleiben.
Literatur
AKATSUKA, T., and NELSON, O. E.: Starch granule-bound adenosine diphosphate glucose starch glucosyltransferase of maize seeds. J. BioI. Chem. 241, 2280-2286 (1966).
BARTLETT, G. R.: Phosphorus assay in column chromatography. J. BioI. Chem. 234, 466 (1959). R<\UN, C. C., PALMIANO, E. P., PEREZ, C. M., and JULIANO, B. D.: Enzymes of starch metabolism in
the developing rice grain. Plant Physiol. 48, 429-434 (1970). BESFORD, R. T., and MAW, G. A.: Effect of potassium nutrition on some enzymes of the tomato plant.
Ann. Bot. 40, 461-471 (1976). EVANS, H. J., and SORGER, G. J.: Role of mineral elements with empha.sis on the univalent cations.
Ann. Rev. Plant Physiol. 17, 47-76 (1966). ~ and WILDE, R. A.: Potassium and its role iu enzyme activation. 8th Colloq. Int. Potash Inst.
13-39 (1971), Uppsala, Int. Potash Inst. Berne. GORING, R., und l\L>\'RDANOV, A. A.: Beziehung zwischeu dem K+/Ca++-Verhiiltnis im Gewebe und
der Wirkung von Cytokininen in hoheren Pflanzen. Diol. Rundschau 14, 177-189 (1976). GREENBERG, E., und PREISS, J.: Biosynthes of bacterial glycogen. II. Purification and properties of
the adenosine diphosphoglucose-glycogen transglycosylase of Arthrobacter species NRRL B1973. J. BioI. Chem. 240. 2341-2348 (1965).
HAUPT, W.: Panel discussion. Sensory transduction and photobehaviors. Final considerations and imerging themes. In: ,. Photoreception and sensory transduction in aneural organisms" Ed.: LENCI, F., and COLOMBETTI, Plenum Press, New York-London 1980.
50 Biochem. Physio). Pflanzen, Bd. 176

752 E. PREUSSER u. a., Regulation der Aktivitat von Starkesynthetase
HAWKER, J. S., MARSCHNER, H., and DOWNTON, W. J. S.: Effects of sodium and potassium on starch synthesis in leaves. Aust. J. Plynt Physiol. 1, 491-501 (1974).
JUDEWICZ, N. D., LAVINTMANN, N., and CARDINI, C. E.: Effect of isopropanol on the activity of particulate starch synthetase. Phytochem. 11, 2213-2215 (1972).
LOWRY, O. H., ROSEBROUGH, N. J., FARR, A. L., and RANDALL, R. J.: Protein measurement with the folin phenol reagent. J. BioI. Chem. 193, 265-275 (1951).
MEISEL, P.: Die Biosynthese der Starke. In: Handbuch der Starke in Einzeldarstellungen. Bd. VI, 48-54. Berlin und Hamburg 1974.
MURATA, T., and AKAZAWA, T.: The role of adenosine diphosphate glucose in leaf starch formation. Biochem. Biophys. Res. Commun. 16, 6-11 (1964).
- - Enzymic mechanism of starch synthesis in sweet potato roots. Arch. Biochem. Biophys. 126, 873-879 (1968).
- - Stimulative effect of potassium ion on starch synthetase of different plant origins. Plant Cell Physiol. 10, 457-460 (1969).
N ITSOS, R. E., and EVANS, H. J.: Effects of univalent cations on the activity of particulate starch synthetase. Pla.nt Physiol. 44, 1260-1266 (1969).
NOWAK, T.: Conformational changes required for pyruvate kinase activity as modulated by monovalent cations. J. BioI. Chem. 201, 73-78 (1976).
WASHUTTL, J., HOLZL, J., und BANCHER, E.: Diinnschichtchromatographischer Nachweis von Phosphatiden und Aminosauren in Weizenstarke. Z. Pflanzenphysiol. 50, 20-30 (1966).
Eingegangen am 5. Ma·i 1981.
Anschrift der Verfasser: Dozent Dr. E. PREUSSER, FAWZY ANDRAWISS KHALIL und HORST GoRING, Sektion Biologie der Humboldt-Universitat Berlin, DDR - 1040 Berlin, InvalidenstraBe 43.

![Kristallstrukturen und Eigenschaften der Hochdruck ... · Struktur[24] vor. Die Kationen bestehen aber in dieser Betrachtungsweise formal aus heterokuban- Die Kationen bestehen aber](https://img.pdfslide.org/doc/110x75/5d4d2d6788c993c16f8b518c/kristallstrukturen-und-eigenschaften-der-hochdruck-struktur24-vor-die.jpg)



























