Embed Size (px)
Citation preview

Research ArticleComparative Proteomic Analysis of DifferentialProteins in Response to Aqueous Extract of Quercus infectoriaGall in Methicillin-Resistant Staphylococcus aureus
Radhiah Khairon1 Noraziah Mohamad Zin1
Mariati Abdul Rahman2 and Dayang Fredalina Basri1
1School of Diagnostic amp Applied Health Sciences Faculty of Health Sciences Universiti Kebangsaan MalaysiaJalan Raja Muda Abdul Aziz 50300 Kuala Lumpur Malaysia2Department of Clinical Oral Biology Faculty of Dentistry Universiti Kebangsaan Malaysia Jalan Raja Muda Abdul Aziz50300 Kuala Lumpur Malaysia
Correspondence should be addressed to Dayang Fredalina Basri dayangukmedumy
Received 1 May 2016 Revised 27 July 2016 Accepted 3 August 2016
Academic Editor Djuro Josic
Copyright copy 2016 Radhiah Khairon et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
The aim of this study is to analyze the differential proteins in MRSA ATCC 33591 treated with aqueous extract from Q infectoriagall Protein extracts were obtained from MRSA cells by sonication and were separated by 2D polyacrylamide gels Protein spotsof interest were extracted from the gels and identified using LC-ESI-QTOF MS The concentration of Q infectoria extract usedfor 2D-gel electrophoresis was subinhibitory concentration Minimum inhibitory concentration (MIC) value of the extract againstMRSA was 1950 120583gmL with bacteriostatic action at 1x MIC from time-kill assay However the extract exhibited dose-dependentmanner and was bactericidal at 4x MIC with more than 3 log
10CFUmL reduction at 4 h 2D-GE map showed that 18 protein
spots were upregulated and another six were downregulated more than twofold (119901 lt 005) after treatment with subinhibitoryconcentration Out of six proteins being downregulated four proteins were identified as ferritin and catalase branched-chainalpha-keto acid dehydrogenase subunit E2 and succinyl-CoA ligase [ADP-forming] subunit beta Seven upregulated proteinswhich have been successfully identified were 3-hydroxyacyl-CoA dehydrogenase NAD binding domain protein formate C-acetyltransferase 3-hydroxyacyl-[acyl-carrier-protein] dehydratase FabZ NAD dependent epimerasedehydratase family proteinand phosphopantothenoyl cysteine decarboxylase It is postulated that the main mechanism of aqueous extract from gall of Qinfectoria was most likely involved in energy metabolism and protein stress
1 Introduction
Increasing prevalence and development of resistance toexisting antibacterial agent by the bacteria have become amajor threat to human health for centuries Staphylococcusaureus is a gram-positive pathogen which rapidly developedinto methicillin-resistant Staphylococcus aureus (MRSA) notlong after antibiotic was introduced [1] MRSA caused bothnosocomial acquired (HA) MRSA and community acquired(CA)MRSA [2] Glycopeptide derivative such as vancomycinis an agent of last resort for the treatment of this pathogendespite recent resistance of MRSA towards this drug [3]
Combating bacterial infection never ends and along with thedevelopment of resistance to current antibiotic an alternativephytotherapeutic agent is needed to overcome these problemswith MRSA infection
Phytomedicine has been widely used since ancient timeand became important source of pharmaceutical agent Inrecent year the world market in herbal industry is gettinghuge and increasing in demand The World Bank recordedan increase in trade for herbal medicine botanical medicineproduct and raw materials with yearly growth rate between5 and 15 [4] Most of natural plant compounds weredeveloped as both antimicrobial [5] and anticancer [6]
Hindawi Publishing CorporationInternational Journal of ProteomicsVolume 2016 Article ID 4029172 9 pageshttpdxdoiorg10115520164029172
2 International Journal of Proteomics
Natural products gain interest among many researchers dueto their chemotherapeutical properties exerted by their activemetabolite against various infections [7]
Quercus infectoria or ldquomanjakanirdquo is small oak tree indig-enous to Asia minor Malaysian women commonly amongthe Malays use the herb traditionally for postpartum careThe aqueous extract of galls from Q infectoria has skinwhitening effect and antioxidant property by inhibitingthe superoxide and 11-diphenyl-2-picrylhydrazyl (DPPH)radical scavenging activities and tyrosine activities [8] TheQ infectoria aqueous extract was reported to have highhydrolysable tannin content which inhibits the lethality of theNajakaouthia (Thai cobra) venom [9] The extract was alsohighly capable as an antimicrobial agent against Escherichiacoli O157H7 [10]
Proteomics is the study of the structure and function ofproteins in biological system to give a better understandingof the complex nature of the organism Nowadays the studyof proteomics is also applied as a tool to study adaptionregulation protein or global response of the bacteria to theenvironment including against antibiotic stress [11ndash13] Afew studies employed proteomic approach to elucidate theeffectiveness of natural product as antibacterial agent [14ndash17] However there is no study with regard to the proteinexpression profile ofMRSAon exposure to aqueous extract ofQ infectoria gall despite extensive studies on the anti-MRSAactivity of this plant Proteomics provide useful informationthat can be used to analyze differences in protein expressionbetween untreated bacterial cells and those treated withinhibitory concentrations of aqueous extract of Q infectoriagall Thus investigation of the proteins expressed in MRSAexposed to the aqueous extract of Q infectoria galls couldpotentially help to elucidate the response of MRSA in thepresence of the tested agent and perhaps can unravel itsmechanism of anti-MRSA through identification of the pro-tein markers
2 Materials and Methods
21 Preparation of Q infectoria Gall Extract The galls werecrushed to small pieces using pestle and mortar and pow-dered in an electric grinder Then the powdered specimenwas dissolved in distilled water for 24 hr at 45∘C and cen-trifuged at 3000timesg at 4∘C The supernatant was then filteredand the whole process was repeated using the remainingresidue with 300mL distilled water The filtrates were com-bined and freeze-dried at minus50∘C under vacuum for 12 hr toproduce a fine crystal-like crude aqueous extractThe extractswere stored in air-tight jars at 4∘C until further use
22 Bacterial Strains Only one bacterial strain was used inthis study namely MRSA ATCC 33591 The bacterial isolatewasmaintained and grown inMueller-Hinton Broth (Merck)for 18ndash20 hr at 37∘C
23 Determination of Minimal Inhibitory Concentration(MIC) The MIC values of the extract were determined by
microbroth dilution method according to Clinical Labo-ratory Standardization Institute [18] guideline with slightmodifications The aqueous extract was dissolved in steriledistilled water to a final concentration of 20mgmL andfiltered through a 045120583mmembrane filter (WhatmanUSA)Twofold serial dilution of the extract was prepared in a 96-well microtiter plate An exponential phase of MRSA growthwas diluted to obtain 108 CFUmL in Mueller-Hinton broth(MHB) which originally contained 5 times 105 CFUmL bacterialculture incubated at 37∘C for 16ndash20 hr The MIC value wasdefined as the lowest concentration of the extract that inhibitsthe bacterial growth
24 Time-Kill Assay An overnight culture of MRSA wasinoculated to 10mL of MHB each containing an aqueousextract at concentrations 05x 1x 2x and 4x MIC to a finalcell density of approximately 5 times 105 CFUmL The bacterialculture in the presence of vancomycin and the bacteria inMHB only were respectively used as positive control andgrowth control A 10 120583L aliquot of each dilution was platedout on Mueller-Hinton agar at 0 2 4 6 8 and 24 hr Afterincubating for 24 hr at 37∘C viable colonies of MRSA werecounted and a graph of log CFUmL against time was plotted
25 SDS-PAGE The protein samples of 100 120583g were sus-pended in sample buffer which comprised 7M urea 2Mtiourea 2 CHAPS and 05 IPG buffer and 005 (wv)bromophenol blueTheprotein sampleswere applied on acry-lamide gels and separated by gel electrophoresis apparatus(BioRad) Proteins were separated based on their molecularweight by electrophoresis at 100V through a stacking gel(4 acrylamide) for 10min and a separating gel (125acrylamide) at 150V for 1 hr The gels were then stainedby overnight incubation with gentle agitation in stainingsolution (03 (wv) Coomassie Brilliant Blue R-250 in 45(vv) methanol 40 (vv) acetic acid) Prestained proteinladder (Invitrogen) was used as SDS-PAGEmolecular weightmarker
26 Preparation of 2-DE Gel Samples MRSA 33591 culturewas grown overnight with constant shaking in MHB inthe presence or absence of the extract at subinhibitoryconcentration of 975120583gmL with three biological triplicatesThe suspension was centrifuged at 4000timesg for 30min at 4∘Cand was then washed by resuspending in 10mL phosphatebuffer
The bacteria pellet was then resuspended in cell lysisbuffer (7M urea 2M thiourea 4 CHAPS 2 IPGbuffer 40mM dithiothreitol and DTT) containing proteaseinhibitor (GE Healthcare) The mixtures were sonicated in atip-probe sonicator (Fisher Scientific) on ice for 1min Theprocedure was repeated five times with 1min interval onice The lysate was then centrifuged at 10000timesg at 4∘C for10min After precipitation with 100 ethanol the proteinwas centrifuged at 10000timesg for 15min at 4∘C The proteinsample was then dissolved in buffer solution (7M urea 2Mthiourea 4 CHAPS 2 IPG buffer and 40mM DTT) The
International Journal of Proteomics 3
concentration of protein sample was quantified by Bradfordprotein assay kit (Bio-Rad US)
27 Two-Dimensional Gel Electrophoresis (2-DE) Protocolsof 2-DE were performed according to previously describedmethod [19] with three technical triplicatesThe protein sam-ples were separated on immobilized pH gradient (IPG) stripsof 24 cmwith a pH range 4ndash7The IPG strips were rehydratedwith 450120583L IPG rehydration bufferThe sampleswere appliedonto the acidic region of the slot in the strip Isoelectric focus-ing (IEF) was performedwith the Ettan IPGPhor System (GEHealthcare) according to the manufacturerrsquos guideline at atotal voltage of 60 kVh IPG strips were equilibrated in 10mLof SDS-equilibrium buffer (6M urea 75mM Tris-HCl 30glycerol 2 SDS and 0002 bromophenol blue) containing50mg DTT for 15min at room temperature for the reductionreaction The strip was alkylated by immersing in 10mL ofequilibration buffer containing 250mg iodoacetamide (IAA)for 15min and placed on 125 SDS-polyacrylamide gel
The second dimension was run using EttanDalt Six (GEHealthcare) at 15∘C 10mAgelminus1 for 1 hr the current was thenincreased to 40mAgelminus1 and the runwas continued until thebromophenol blue dye had moved towards the end of the gelAfter the 2-GE run the gels were fixed in a mixture (04ethanol (vv) and 01 glacial acetic acid (vv)) and furtherstained using the silver staining with somemodification [20]
28 Image Analysis The gels were analyzed using ImageS-canner III Lab Scan (GEHealthcare) and Progenesis software(Nonlinear Dynamics) The statistic used in this study foranalysis was Anova in the Progenesis software The softwarewas used to detect the differences in the image of profileexpression protein between the images of proteins fromtreated aqueous extract cell with the untreated cell proteinsThe protein changes as well as similar protein patternmaps inthe gels were analyzed from three independent experiments(biological replicate) and three technical replicates Changesin protein expression spots were considered significant (119901 lt005) by Anova analysis only if the intensity of protein spotswas more than twofold higher
29 In-Gel Digestion and LC-ESI-QTOF MS Protein spotswere excised from silver stained 2-DE gels Protein spots weredestained using fresh washing solution (100mM ammoniumbicarbonate 15mM potassium ferrocyanide and 100mMsodium thiosulphate for 15min on orbital shaker) The gelpieces were reduced with 10mM DTT in 10mM ammoniumbicarbonate at 60∘C for 30min For alkylation process 55mMIAA in 100mM ammonium bicarbonate was added to thegel plugs and incubated at room temperature for 20min inthe dark The gel pieces were rinsed three times in 50acetonitrile (ACN) in 10mM ammonium bicarbonate for20min on orbital shaker Then 100 ACN was added untilthe gel plugs becomemilky white after 15min To perform in-gel digestion of proteins 25120583L of trypsin solution (7 ng120583Ltrypsin in 1 ACN40mM ammonium bicarbonate) wasadded to the gels and incubated at 37∘C overnight Afterincubation 25120583L of 30 ACN was added to keep the gels
immersed throughout the digestion processThe supernatantwhich was the extract peptides was collected and dried for1 hr Peptides were desalted using ZipTip C18 (Millipore US)after equilibration in 50 ACN in 01 formic acid Theextracted peptides were collected pooled dried by vacuumcentrifuge and kept at minus20∘C for further mass spectrometricanalysis The peptides were then eluted and injected into LC-ESI-QTOFMS (model G6550 Agilent US) with electrosprayat flow rate of 160 nLmin to a nanocolumn (G4240-62010analytical 75 120583m times 150mm Zorbax 300SB-C18 5 120583m) Asolvent gradient (solvent A 50 ACN in water solvent B70 ACN solvent C 70 ACN solvent D 5 acetonitrile)was run for 25min
210 Bioinformatics After all the distinct peptides have un-dergone validation process the mass spectra and sequenceswere analyzed using a public database (UniProtKB andNational Center for Biotechnology Information) with morethan two distinct peptides Protein-protein interactions werepredicted using Search Tool for the Retrieval of Interact-ing GenesProteins (STRING) database v90 (httpwwwstring-dborg)The Swiss-Prot identifier for the Staphylococ-cus aureus genes in ldquoprotein moderdquo was used to search usingthe STRING database Network analysis was set at mediumstringency (STRING score = 04) Proteins were linked basedon seven criteria neighbourhood gene fusion cooccurrencecoexpression experimental evidences existing databasesand text mining
3 Results
31 Antibacterial Activity The aqueous extract showinginhibitory activity against MRSA 33591 was 1975120583gmLwhich was ten times higher than the MIC value for van-comycin The result for the time-kill studies was presentedin Figure 1 whereby the MRSA treated with subinhibitoryconcentration showed colony count of viable cell lower thanthe growth control after 24 hr incubationMeanwhile bacteriagrowth was stable when treated with 1x and 2x MIC aqueousextract of Q infectoria gall after 6 hr incubationThe numberof viable cells after exposure to 4x MIC of extract decreasedby more than 3 log
10CFUmL faster at the 3rd hr compared
to vancomycin after 13 hr treatment
32 Protein Variation between Cell Cultures Preliminarystudy whether aqueous extract Q infectoria gall affectedstaphylococcal cell protein patterns was performed by SDS-PAGE MRSA treated with aqueous extract Q infectoria gallexhibited different protein expression pattern compared tountreated MRSA in view of the level of intensity of the bands(Figure 2) 2D-GE combined with LC-ESI-QTOF MS wasperformed to identify the differential proteins which wereaffected in response to exposure with the tested extract
Figure 3 showed the protein mapping of treated anduntreated cells observed in acidic range in which mostof alterations between the two proteomes were displayedMore than 1000 individual protein spots were resolved andvisualized on silver-staining The optimum concentration
4 International Journal of Proteomics
0
2
4
6
8
10
12
14
0 2 4 6 8 10 12 14 16 18 20 22 24 26Time (hours)
Time-kill of MRSA treated with aqueousQ infectoria at different concentrations
Growth controlVancomycin
log10
(CFU
mL)
(12)x MIC extract
1x MIC extract 2x MIC extract 4x MIC extract
Figure 1 Time-kill curve of aqueous extract from Q infectoriaagainst MRSA 33591 Viability of cells growth was counted atspecified hr by serial dilution Each point represents the mean oflog10plusmn SEM (119899 = 3)
1 2MW
10
15
2030
4050607085
120
Figure 2 SDS-PAGE map of MRSA 33591 after being treated withaqueous extract of Q infectoria MW protein ladder lane number1 untreated MRSA lane 2 extract-treated MRSA
of MRSA protein for 2D-GE analysis was 100120583g Imageanalysis identified 24 protein spots that exhibited significantdifferences after treatment with the extract There are 17protein spots that were shown to be upregulated and anotherfive were found to be downregulated with more than twofoldincrease (119901 lt 005) (data not shown) However only 12 ofthe protein spots were successfully identified four protein
spots showed downregulation whereas eight protein spotswere upregulated
33 Protein Network Analysis Predicted and identified func-tionally linked proteins and the determination of the poten-tial biological process involved after treatment with aqueousextract from Q infectoria gall were analyzed by STRINGnetwork analysis as shown in Figure 4 The network waspresented under confident view whereby stronger associa-tions were represented by thicker lines or edges and proteinswere represented as nodes The interaction between the 11proteins was tabulated (Table 1) in order to summarize thedifferentially expressed proteins Four proteins were found tobe linked either directly or indirectly through one or moreinteracting proteins suggesting the existence of reportedfunctional linkagesHowever another group of three proteinswere linked together without linkage to other groups
4 Discussion
Methicillin-resistant S aureus (MRSA) has become a com-mon pathogen particularly in the long-term care setting[21] Thus the treatment of MRSA infection using currentantibiotic has become limited due to the emergence ofmultidrug-resistant strains [22] Herbal medicine is one ofthe strategies to combat resistance from this infection in aneffort to overcome mortality and morbidity patient problemin healthcare Recent studies [23 24] reported that naturalproduct has high antimicrobial potential against MRSA Theaqueous extract from Quercus infectoria gall in this studyshowed strong anti-MRSA activity and this is in agreementwith [25] that this extract showed promising capacity as newphytotherapeutic candidate against MRSA
Time-kill assay indicated that the aqueous extract fromQ infectoria gall exhibited significant bactericidal activity at4x MIC in dose-dependent manner Bactericidal activity wasdefined as a reduction ofmore than 3 log
10CFUmL at certain
time compared to at 0 hr [26] However at lower dose theextract was bacteriostatic and subinhibitory concentration((12)x MIC) of 975120583gmL was chosen to run 2D-GE toensure that enough protein can be obtained in order toanalyze the protein expression profile ofMRSAThe presenceof crude aqueous extract of Q infectoria gall against MRSAremarkably interfered with the general metabolic pathwayOne of the affected proteins lost was the branched-chain120572-keto acid dehydrogenase subunit E2 It is one importantcomponent of the protein complex which is the dihydrolipoyltransacylase that catalyzes transacylation of the acyl groupfrom E2 to coenzyme A [27] Branched-chain 120572-keto aciddehydrogenase is an enzyme complex that catalyzes the earlystages of branched-chain fatty acid production [28] and playsa key role in catabolism of branched-chain amino acids inbacteria [29] This enzyme is important as adherence of Saureus to the host eukaryotic cells for its survival in a mousehost due to its function in the membrane fluidity of S aureus
Another protein which was downregulated was succinyl-coenzymeA (CoA) ligase [ADP-forming] subunit beta whichis involved in citric acid cycle for energy metabolism [30]
International Journal of Proteomics 5
pH 4 pH 7
2
4
7
9 1718
1921
2223
24
(a)
pH 7pH 4
2
4
7
9 1718
212223
24
(b)
Figure 3 Two 2D-GEmaps of the cellular MRSA proteome (a) Protein expression profile of untreated MRSA (b) Protein expression profileof extract treated-MRSA
ftnA
kat
SAV1089
fadB
sle1
fabZ
E25
SAV1211
SAV0128
sucC
pflB
Name of gene Identified protein
SAV0128 NAD-dependent epimerase dehydratasefamily protein
pflB Formate acetyltransferase
fadB 3-Hydroxyacyl-CoA dehydrogenaseSle 1 N-Acetylmuramoyl-L-alanine amidase
peptidoglycan hydrolaseSAV1089 Hypothetical protein (RNase J)E25 Branched-chain alpha-keto acid
dehydrogenase subunit E2SAV1211 Pantothenate metabolism flavoproteinsucC Succinyl-CoA synthetase subunit betakat CatalaseftnA Ferritin iron storage proteinFabZ The 3-hydroxyacyl-CoA dehydrogenase
NAD binding domain protein
Figure 4 STRING interaction network analysis showed the association between differentially expressed proteins The protein names andgene symbols used in this network are listed in the table Higher amount of lines indicates stronger interaction between the proteins
This enzyme reversibly catalyzes ATP-dependent ligationfrom succinate andCoA to form succinyl-CoA in either aero-bic or anaerobic condition During aerobic growth succinyl-CoA ligase catalyzes the hydrolysis of succinyl-CoA usingADP and inorganic orthophosphate (119875
119894) to form succinate
and ATP However during anaerobic condition succinyl-CoA ligase can freely interconvert ATP and succinate to formsuccinyl-CoA ADP and inorganic 119875
119894for anabolic reaction as
well [31]Exposure of MRSA culture to crude extract of Q infec-
toria gall suppressed the production of proteins that wereinvolved in oxidative stress These proteins were ferritinand catalase Ferritins are the major iron storage proteinsin organism and provide protection against metal toxicityand oxidative stress [32] Ferritin was downregulated after
treatment with Q infectoria gall possibly due to its effectin limiting the iron environment hence it interfered bylowering the expression of ferritin protein Tannin contentin Q infectoria extract is believed to retard the growthof bacteria by inhibiting the activity of metalloenzymes incomplex tannins and iron [33] Meanwhile catalase catalyzeshydrogen peroxide to form oxygen and water Catalase is alsoknown as one of the virulent factors and when its productionwas reduced the bacteria cells become more susceptible tobe killed by neutrophils in the host cells [34] The proposedstress-related protein obviously reflected an adaption of thebacteria to survival under stress condition
Cell wall in gram-positive bacteria is very importantfor its survival Bacterial cell wall is composed of thecross-linked polymer peptidoglycan also known as murein
6 International Journal of Proteomics
Table1Th
eprotein
andgenesidentified
intheS
TRIN
GanalysisnetworkTh
elist
ofproteins
identifi
edisdifferentially
expressedproteins
follo
wingtreatmentw
ithQinfectoria
gallextract
Spot
number
Accession
number
Proteins
identifi
edMascot
score
Nom
inal
mass(kD
a)Sequ
ence
coverage
()
Num
bero
fpeptides
matched
Foldsc
hanges
(untreated
totre
ated)
2FT
NSA
AR
Ferritin
9419590
345
minus24
4D6SHZ8
Branched-chain
alph
a-keto
acid
dehydrogenases
ubun
itE2
1317
846
4516
234
8minus27
7D6SHJ5
Succinyl-C
oAligase[ADP-form
ing]
subu
nitb
eta
6543
422837
131
5minus22
9D6SDX7
Form
ateC
-acetyltransfe
rase
14051
855525
157
1026
10D6SH75
Catalase
6678
587233
102
5minus24
17amp18
D6SBS
5N-Acetylm
uram
oyl-L
-alanine
amidase
2834
692891
33
253
19D6SDX0
3-Hydroxyacyl-C
oAdehydrogenaseNADbind
ingdo
mainprotein
5144
846729
63
426
21D6SHG5
Ribo
nucle
aseJ
7412
646507
925
24
22D6SHN0
Phosph
opantothenoylcysteined
ecarbo
xylasepho
spho
pantothenate-cysteineligase
3195
4419
6562
221
23D6SDL7
NADdepend
entepimerasedehydratasefam
ilyprotein
279
378017
762
21
24D6SEM
63-Hydroxyacyl-[acyl-carrie
r-protein]
dehydrataseF
abZ
2731
163096
123
222
International Journal of Proteomics 7
Peptidoglycan architecture consists of a disaccharide back-bone built of alternating 120573-1-4-N-acetylglucosamines andN-acetylmuramic acids [35] Bacterial cell produces severalhydrolases that specifically cleave various covalent bonds inthe peptidoglycan including N-acetylmuramoyl-L-alanineamidases N-acetylglucosaminidases N-acetylmuramidasesendopeptidases and transglycosylases N-acetylmuramoyl-L-alanine amidase plays important roles in the separationseptum of daughter cells during cell division [36] HenceN-acetylmuramoyl-L-alanine amidase binds to both alphaand beta human fibrinogen and fibronectin which play arole during colonization and pathogenicity in the host tissueduring infection Previous study [37] reported that thisenzyme was upregulated to activate and increase its ownproduction in the presence of antibiotic to the bacteria andeventually cause inhibition of the bacterial growth or celldeath
Formate C-acetyltransferase or pyruvate formate-lyasewas upregulated in MRSA treated with aqueous extract fromQ infectoria gall This is a cytoplasmic protein involved inenergy fermentationmetabolism pathway in themicroorgan-ism This enzyme was induced in anaerobic growth and lim-ited nitrate [38 39] Formate C-acetyltransferase functions tocatalyze pyruvate to acetyl-CoA and formate which is subse-quently used for biosynthesis of protein DNA and RNA [40]and also as source for citric acid cycle through acetyl-CoA[41] Formate C-acetyltransferase NAD-dependent formatedehydrogenase and formyltetrahydrofolate synthetase wereupregulated during transcription at proteome level for thesynthesis of S aureus biofilm growth [42] This enzyme wasupregulated as one of the survival strategies of bacteria understress [40]
Phosphopantothenoyl cysteine decarboxylasephospho-pantothenate-cysteine ligase is also known as acid pan-tothenic [43] This is the last enzyme involved in the biosyn-thesis of CoAwhich is a crucial substance in the biogenesis ofbacterial cell membrane and functions as important cofactorin mitosis and stability of DNA in microorganism [44]This protein was upregulated in the study probably due tocompensation of other pathways to increase in the synthesisof CoA for adaptation when exposed to subinhibitory con-centration of the Q infectoria gall extract
NAD-dependent epimerasedehydratase protein is anew protein that has high similarity with UDP-glucose-4-epimerase involved in Leloir pathway [45] The other proteinthat was shown to be upregulated after treatment was RNaseJ which is involved in maturation of RNA and regulationof posttranscription until degradation of RNA in familyFirmicute of bacteria
Other proteins that were upregulated during fatty acidmetabolism are 3-hydroxyacyl-CoA dehydrogenase NAD-binding domain and 3-hydroxyacyl-[acyl-carrier-protein]dehydratase FabZ The 3-hydroxyacyl-CoA dehydrogenaseNAD binding domain protein was involved in variousmetabolic pathways such as beta oxidation fatty acid bycatalyzing the oxidation of 3-hydroxyl coenzymeA to oxoacylCoA and production of butanol in bacteria gram-positiveorganism [46] Upregulation of this protein may be due tocompensation of low level of the enzyme to tricarboxylic acid
cycle [47] In addition another protein that was involved inthe synthesis of fatty acid was 3-hydroxyacyl-[acyl-carrier-protein] dehydratase FabZ The latter was reported to par-ticipate in the phospholipid membrane biosynthesis pathway[48]
5 Conclusions
In conclusion all the data reported in this studymay elucidateand give better understanding on antibacterial effect ofaqueous extract of Q infectoria towards MRSA throughinterference of various metabolism and functions of the bac-terial proteome Furthermore recent studies suggested thataqueous extract ofQ infectoria galls has no toxicity effect andmortality effect in animal [49 50] However due to its hightannin content which has a profound digestibility-reducingeffect in human it is ideal to be administered topically since ithas the advantage of possessing the anti-inflammatory effectandwoundhealing activityThis information haswidened thechances for alternative medicine as therapeutic agent to treatMRSA infection
Competing Interests
The authors declare that they have no competing interestswith regard to publication of this paper
Authorsrsquo Contributions
Dayang Fredalina Basri designed analyzed and supervisedthe study Noraziah Mohamad Zin provided lab facilitiesRadhiah Khairon performed all the experiments data anal-ysis and drafted the manuscript All authors read andapproved the final manuscript
Acknowledgments
This study was funded by the Ministry of Higher EducationGovernment of Malaysia under the research code GrantFRGS22014SG05UKM023
References
[1] N F Crum R U Lee S A Thornton et al ldquoFifteen-yearstudy of the changing epidemiology of methicillin-resistantStaphylococcus aureusrdquo The American Journal of Medicine vol119 no 11 pp 943ndash951 2006
[2] A Asensio R Canton J Vaque et al ldquoNosocomial andcommunity-acquiredmeticillin-resistant Staphylococcus aureusinfections in hospitalized patients (Spain 1993ndash2003)rdquo Journalof Hospital Infection vol 63 no 4 pp 465ndash471 2006
[3] K Hiramatsu H Hanaki T Ino K Yabuta T Oguri and FC Tenover ldquoMethicillin-resistant Staphylococcus aureus clini-cal strain with reduced vancomycin susceptibilityrdquo Journal ofAntimicrobial Chemotherapy vol 40 no 1 pp 135ndash136 1997
[4] WHO WHO Traditional Medicine Strategy 2002ndash2005World Health Organization Geneva Switzerland 2002WHO EDM TRM 20021 engpdf
8 International Journal of Proteomics
[5] D G Brown T Lister and T L May-Dracka ldquoNew naturalproducts as new leads for antibacterial drug discoveryrdquo Bioor-ganic amp Medicinal Chemistry Letters vol 24 no 2 pp 413ndash4182014
[6] G M Cragg P G Grothaus and D J Newman ldquoImpactof natural products on developing new anti-cancer agentsrdquoChemical Reviews vol 109 no 7 pp 3012ndash3043 2009
[7] D A Dias S Urban and U Roessner ldquoA historical overview ofnatural products in drug discoveryrdquo Metabolites vol 2 no 4pp 303ndash336 2012
[8] F Sharififar G Dehghan-Nudeh Z Raeiat B AmirheidariM Moshrefi and A Purhemati ldquoTyrosinase inhibitory activityof major fractions of Quercus infectoria gallsrdquo PharmacognosyCommunications vol 3 no 1 pp 21ndash26 2013
[9] P Pithayanukul P Ruenraroengsak R Bavovada N PakmaneeR Suttisri and S Saen-Oon ldquoInhibition of Naja kaouthiavenom activities by plant polyphenolsrdquo Journal of Ethnophar-macology vol 97 no 3 pp 527ndash533 2005
[10] S Voravuthikunchai A Lortheeranuwat W Jeeju T SririrakS Phongpaichit and T Supawita ldquoEffective medicinal plantsagainst enterohaemorrhagic Escherichia coli O157H7rdquo Journalof Ethnopharmacology vol 94 no 1 pp 49ndash54 2004
[11] S J Cordwell M R Larsen R T Cole and B J Walsh ldquoCom-parative proteomics of Staphylococcus aureus and the responseof methicillin-resistant and methicillin-sensitive strains to Tri-ton X-100rdquoMicrobiology vol 148 no 9 pp 2765ndash2781 2002
[12] C Kohler S Wolff D Albrecht et al ldquoProteome analysesof Staphylococcus aureus in growing and non-growing cellsa physiological approachrdquo International Journal of MedicalMicrobiology vol 295 no 8 pp 547ndash565 2005
[13] L Van Oudenhove and B Devreese ldquoA review on recent devel-opments in mass spectrometry instrumentation and quantita-tive tools advancing bacterial proteomicsrdquoAppliedMicrobiologyand Biotechnology vol 97 no 11 pp 4749ndash4762 2013
[14] R Dosselli R Millioni L Puricelli et al ldquoMolecular targets ofantimicrobial photodynamic therapy identified by a proteomicapproachrdquo Journal of Proteomics vol 77 pp 329ndash343 2012
[15] R Jenkins N Burton and R Cooper ldquoManuka honey inhibitscell division in methicillin-resistant Staphylococcus aureusrdquoJournal of Antimicrobial Chemotherapy vol 66 no 11 pp 2536ndash2542 2011
[16] J M Packer J Irish B R Herbert et al ldquoSpecific non-peroxideantibacterial effect of manuka honey on the Staphylococcusaureus proteomerdquo International Journal of Antimicrobial Agentsvol 40 no 1 pp 43ndash50 2012
[17] W Sianglum P Srimanote W Wonglumsom K Kittiniyomand S P Voravuthikunchai ldquoProteome analyses of cellularproteins in methicillin-resistant Staphylococcus aureus treatedwith rhodomyrtone a novel antibiotic candidaterdquo PLoS ONEvol 6 no 2 Article ID e16628 pp 1ndash10 2011
[18] Clinical and Laboratory Standards Institute ldquoPerformancestandards for antimicrobial susceptibility testingrdquo FifteenthInformational Supplement M100-S15 2005
[19] D F Basri L S Aik R Khairon and M A Rahman ldquo2-Dgel electrophoresis map of methicillinresistant Staphylococcusaureus treated with Quercus infectoria gall extractrdquo AmericanJournal of Biochemistry and Biotechnology vol 9 no 1 pp 19ndash26 2013
[20] M Chevallet S Luche and T Rabilloud ldquoSilver staining ofproteins in polyacrylamide gelsrdquo Nature Protocols vol 1 no 4pp 1852ndash1858 2006
[21] J Davies and D Davies ldquoOrigins and evolution of antibioticresistancerdquoMicrobiology andMolecular Biology Reviews vol 74no 3 pp 417ndash433 2010
[22] D M Livermore ldquoAntibiotic resistance in staphylococcirdquo Inter-national Journal of Antimicrobial Agents vol 16 supplement 1pp S3ndashS10 2000
[23] L Yu H Xiang J Fan et al ldquoGlobal transcriptional response ofStaphylococcus aureus toRhein a natural plant productrdquo Journalof Biotechnology vol 135 no 3 pp 304ndash308 2008
[24] M Doyle E-A Feuerbaum K R Fox J Hinds D EThurstonand P W Taylor ldquoResponse of Staphylococcus aureus to subin-hibitory concentrations of a sequence-selective DNA minorgroove cross-linking pyrrolobenzodiazepine dimerrdquo Journal ofAntimicrobial Chemotherapy vol 64 no 5 pp 949ndash959 2009
[25] D F Basri and S H Fan ldquoThe potential of aqueous and acetoneextracts of galls of Quercus infectoria as antibacterial agentsrdquoIndian Journal of Pharmacology vol 37 no 1 pp 26ndash29 2005
[26] J N Eloff ldquoA sensitive and quick microplate method to deter-mine the minimal inhibitory concentration of plant extracts forbacteriardquo Planta Medica vol 64 no 8 pp 711ndash713 1998
[27] L Li P Thipyapong D C Breeden and J C SteffensldquoOverexpression of a bacterial branched-chain 120572-keto aciddehydrogenase complex in Arabidopsis results in accumulationof branched-chain acyl-CoAs and alteration of free amino acidcomposition in seedsrdquo Plant Science vol 165 no 6 pp 1213ndash1219 2003
[28] V K Singh D S Hattangady E S Giotis et al ldquoInsertionalinactivation of branched-chain 120572-keto acid dehydrogenase inStaphylococcus aureus leads to decreased branched-chainmem-brane fatty acid content and increased susceptibility to certainstressesrdquo Applied and Environmental Microbiology vol 74 no19 pp 5882ndash5890 2008
[29] P N Lowe J A Hodgson and R N Perham ldquoDual role of asingle multienzyme complex in the oxidative decarboxylationof pyruvate and branched-chain 2-oxo acids in Bacillus subtilisrdquoBiochemical Journal vol 215 no 1 pp 133ndash140 1983
[30] V Kapatral X Bina and A M Chakrabarty ldquoSuccinyl coen-zyme A synthetase of Pseudomonas aeruginosa with a broadspecificity for nucleoside triphosphate (NTP) synthesis modu-lates specificity for NTP synthesis by the 12-kilodalton form ofnucleoside diphosphate kinaserdquo Journal of Bacteriology vol 182no 5 pp 1333ndash1339 2000
[31] S-J Park G Chao and R P Gunsalus ldquoAerobic regulationof the sucABCD genes of Escherichia coli which encode 120572-ketoglutarate dehydrogenase and succinyl coenzyme A syn-thetase roles of ArcA Fnr and the upstream sdhCDAB pro-moterrdquo Journal of Bacteriology vol 179 no 13 pp 4138ndash41421997
[32] S C Andrews ldquoIron storage in bacteriardquo Advances in MicrobialPhysiology vol 40 pp 281ndash351 1998
[33] K-T Chung Z Lu andMW Chou ldquoMechanism of inhibitionof tannic acid and related compounds on the growth ofintestinal bacteriardquo Food and Chemical Toxicology vol 36 no12 pp 1053ndash1060 1998
[34] G L Mandell ldquoCatalase superoxide dismutase and virulenceof Staphylococcus aureus In vitro and in vivo studies withemphasis on staphylococcal-leukocyte interactionrdquoThe Journalof Clinical Investigation vol 55 no 3 pp 561ndash566 1975
[35] M P Chapot-Chartier and S Kulakauskas ldquoCell structure andfunction in lactic acid in bacteriardquoMicrobial Cell Factories vol13 pp S1ndashS9 2014
International Journal of Proteomics 9
[36] V A Fischetti ldquoBacteriophage lytic enzymes novel anti-infectivesrdquo Trends in Microbiology vol 13 no 10 pp 491ndash4962005
[37] C Heidrich M F Templin A Ursinus et al ldquoInvolvementof N-acetylmuramyl-l-alanine amidases in cell separationand antibiotic-induced autolysis of Escherichia colirdquo MolecularMicrobiology vol 41 no 1 pp 167ndash178 2001
[38] S Fuchs J Pane-Farre C KohlerMHecker and S EngelmannldquoAnaerobic gene expression in Staphylococcus aureusrdquo Journal ofBacteriology vol 189 no 11 pp 4275ndash4289 2007
[39] L J Rasmussen P L Moller and T Atlung ldquoCarbonmetabolism regulates expression of the pfl (pyruvate formate-lyase) gene in Escherichia colirdquo Journal of Bacteriology vol 173no 20 pp 6390ndash6397 1991
[40] M Leibig M Liebeke D Mader M Lalk A Peschel andF Gotz ldquoPyruvate formate lyase acts as a formate supplierfor metabolic processes during anaerobiosis in Staphylococcusaureusrdquo Journal of Bacteriology vol 193 no 4 pp 952ndash962 2011
[41] W Buckel and B T Golding ldquoRadical enzymes in anaerobesrdquoAnnual Review of Microbiology vol 60 pp 27ndash49 2006
[42] A Resch S Leicht M Saric et al ldquoComparative proteomeanalysis of Staphylococcus aureus biofilm and planktonic cellsand correlation with transcriptome profilingrdquo Proteomics vol6 no 6 pp 1867ndash1877 2006
[43] T Kupke ldquoMolecular characterization of the 41015840-phosphopan-tothenoylcysteine synthetase domain of bacterial Dfp flavopro-teinsrdquo The Journal of Biological Chemistry vol 277 no 39 pp36137ndash36145 2002
[44] T Nakamura T Pluskal Y Nakaseko and M YanagidaldquoImpaired coenzyme A synthesis in fission yeast causes defec-tive mitosis quiescence-exit failure histone hypoacetylationand fragile DNArdquo Open Biology vol 2 no 9 Article ID 1201172012
[45] D D Simeonova I Susnea A Moise B Schnik and MPrzybylski ldquolsquoUnknown genomersquo proteomics a new NAD(P)-dependent epimerasedehydratase revealed by N-terminal se-quencing inverted PCR and high resolution mass spectrome-tryrdquo Molecular amp Cellular Proteomics vol 8 no 1 pp 122ndash1312009
[46] D P Clark and J E Cronan ldquoTwo carbon compounds andfatty acids as carbon sourcesrdquo inEscherichia coli and SalmonellaCellular and Molecular Biology ASM Press Washington DCUSA 1996
[47] Z L Boynton G N Bennett and F B Rudolph ldquoCloningsequencing and expression of clustered genes encoding betahydroxybutyryl-coenzyme A (CoA) dehydrogenase crotonaseand butyryl-CoA dehydrogenase from Clostridium aceto-butylicum ATCC 824rdquo Journal of Bacteriology vol 178 no 11pp 3015ndash3024 1996
[48] J Schiebel A Chang H Lu M V Baxter P J Tonge andC Kisker ldquoStaphylococcus aureus FabI inhibition substraterecognition and potential implications for in vivo essentialityrdquoStructure vol 20 no 5 pp 802ndash813 2012
[49] M Iminjan N Amat X-H Li H Upur D Ahmat and B HeldquoInvestigation into the toxicity of traditional uyghur medicinequercus infectoria galls water extractrdquo PLoS ONE vol 9 no 3Article ID e90756 2014
[50] A Hasmah Z Nurazila C Y Chow R Rina and M Rafiquz-zaman ldquoCytotoxic effects ofQuercus infectoria extracts towardscervical (Hela) and Ovarian (Caov-3) cancer cell linesrdquo Healthand the Environment Journal vol 1 no 2 pp 17ndash23 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 International Journal of Proteomics
Natural products gain interest among many researchers dueto their chemotherapeutical properties exerted by their activemetabolite against various infections [7]
Quercus infectoria or ldquomanjakanirdquo is small oak tree indig-enous to Asia minor Malaysian women commonly amongthe Malays use the herb traditionally for postpartum careThe aqueous extract of galls from Q infectoria has skinwhitening effect and antioxidant property by inhibitingthe superoxide and 11-diphenyl-2-picrylhydrazyl (DPPH)radical scavenging activities and tyrosine activities [8] TheQ infectoria aqueous extract was reported to have highhydrolysable tannin content which inhibits the lethality of theNajakaouthia (Thai cobra) venom [9] The extract was alsohighly capable as an antimicrobial agent against Escherichiacoli O157H7 [10]
Proteomics is the study of the structure and function ofproteins in biological system to give a better understandingof the complex nature of the organism Nowadays the studyof proteomics is also applied as a tool to study adaptionregulation protein or global response of the bacteria to theenvironment including against antibiotic stress [11ndash13] Afew studies employed proteomic approach to elucidate theeffectiveness of natural product as antibacterial agent [14ndash17] However there is no study with regard to the proteinexpression profile ofMRSAon exposure to aqueous extract ofQ infectoria gall despite extensive studies on the anti-MRSAactivity of this plant Proteomics provide useful informationthat can be used to analyze differences in protein expressionbetween untreated bacterial cells and those treated withinhibitory concentrations of aqueous extract of Q infectoriagall Thus investigation of the proteins expressed in MRSAexposed to the aqueous extract of Q infectoria galls couldpotentially help to elucidate the response of MRSA in thepresence of the tested agent and perhaps can unravel itsmechanism of anti-MRSA through identification of the pro-tein markers
2 Materials and Methods
21 Preparation of Q infectoria Gall Extract The galls werecrushed to small pieces using pestle and mortar and pow-dered in an electric grinder Then the powdered specimenwas dissolved in distilled water for 24 hr at 45∘C and cen-trifuged at 3000timesg at 4∘C The supernatant was then filteredand the whole process was repeated using the remainingresidue with 300mL distilled water The filtrates were com-bined and freeze-dried at minus50∘C under vacuum for 12 hr toproduce a fine crystal-like crude aqueous extractThe extractswere stored in air-tight jars at 4∘C until further use
22 Bacterial Strains Only one bacterial strain was used inthis study namely MRSA ATCC 33591 The bacterial isolatewasmaintained and grown inMueller-Hinton Broth (Merck)for 18ndash20 hr at 37∘C
23 Determination of Minimal Inhibitory Concentration(MIC) The MIC values of the extract were determined by
microbroth dilution method according to Clinical Labo-ratory Standardization Institute [18] guideline with slightmodifications The aqueous extract was dissolved in steriledistilled water to a final concentration of 20mgmL andfiltered through a 045120583mmembrane filter (WhatmanUSA)Twofold serial dilution of the extract was prepared in a 96-well microtiter plate An exponential phase of MRSA growthwas diluted to obtain 108 CFUmL in Mueller-Hinton broth(MHB) which originally contained 5 times 105 CFUmL bacterialculture incubated at 37∘C for 16ndash20 hr The MIC value wasdefined as the lowest concentration of the extract that inhibitsthe bacterial growth
24 Time-Kill Assay An overnight culture of MRSA wasinoculated to 10mL of MHB each containing an aqueousextract at concentrations 05x 1x 2x and 4x MIC to a finalcell density of approximately 5 times 105 CFUmL The bacterialculture in the presence of vancomycin and the bacteria inMHB only were respectively used as positive control andgrowth control A 10 120583L aliquot of each dilution was platedout on Mueller-Hinton agar at 0 2 4 6 8 and 24 hr Afterincubating for 24 hr at 37∘C viable colonies of MRSA werecounted and a graph of log CFUmL against time was plotted
25 SDS-PAGE The protein samples of 100 120583g were sus-pended in sample buffer which comprised 7M urea 2Mtiourea 2 CHAPS and 05 IPG buffer and 005 (wv)bromophenol blueTheprotein sampleswere applied on acry-lamide gels and separated by gel electrophoresis apparatus(BioRad) Proteins were separated based on their molecularweight by electrophoresis at 100V through a stacking gel(4 acrylamide) for 10min and a separating gel (125acrylamide) at 150V for 1 hr The gels were then stainedby overnight incubation with gentle agitation in stainingsolution (03 (wv) Coomassie Brilliant Blue R-250 in 45(vv) methanol 40 (vv) acetic acid) Prestained proteinladder (Invitrogen) was used as SDS-PAGEmolecular weightmarker
26 Preparation of 2-DE Gel Samples MRSA 33591 culturewas grown overnight with constant shaking in MHB inthe presence or absence of the extract at subinhibitoryconcentration of 975120583gmL with three biological triplicatesThe suspension was centrifuged at 4000timesg for 30min at 4∘Cand was then washed by resuspending in 10mL phosphatebuffer
The bacteria pellet was then resuspended in cell lysisbuffer (7M urea 2M thiourea 4 CHAPS 2 IPGbuffer 40mM dithiothreitol and DTT) containing proteaseinhibitor (GE Healthcare) The mixtures were sonicated in atip-probe sonicator (Fisher Scientific) on ice for 1min Theprocedure was repeated five times with 1min interval onice The lysate was then centrifuged at 10000timesg at 4∘C for10min After precipitation with 100 ethanol the proteinwas centrifuged at 10000timesg for 15min at 4∘C The proteinsample was then dissolved in buffer solution (7M urea 2Mthiourea 4 CHAPS 2 IPG buffer and 40mM DTT) The
International Journal of Proteomics 3
concentration of protein sample was quantified by Bradfordprotein assay kit (Bio-Rad US)
27 Two-Dimensional Gel Electrophoresis (2-DE) Protocolsof 2-DE were performed according to previously describedmethod [19] with three technical triplicatesThe protein sam-ples were separated on immobilized pH gradient (IPG) stripsof 24 cmwith a pH range 4ndash7The IPG strips were rehydratedwith 450120583L IPG rehydration bufferThe sampleswere appliedonto the acidic region of the slot in the strip Isoelectric focus-ing (IEF) was performedwith the Ettan IPGPhor System (GEHealthcare) according to the manufacturerrsquos guideline at atotal voltage of 60 kVh IPG strips were equilibrated in 10mLof SDS-equilibrium buffer (6M urea 75mM Tris-HCl 30glycerol 2 SDS and 0002 bromophenol blue) containing50mg DTT for 15min at room temperature for the reductionreaction The strip was alkylated by immersing in 10mL ofequilibration buffer containing 250mg iodoacetamide (IAA)for 15min and placed on 125 SDS-polyacrylamide gel
The second dimension was run using EttanDalt Six (GEHealthcare) at 15∘C 10mAgelminus1 for 1 hr the current was thenincreased to 40mAgelminus1 and the runwas continued until thebromophenol blue dye had moved towards the end of the gelAfter the 2-GE run the gels were fixed in a mixture (04ethanol (vv) and 01 glacial acetic acid (vv)) and furtherstained using the silver staining with somemodification [20]
28 Image Analysis The gels were analyzed using ImageS-canner III Lab Scan (GEHealthcare) and Progenesis software(Nonlinear Dynamics) The statistic used in this study foranalysis was Anova in the Progenesis software The softwarewas used to detect the differences in the image of profileexpression protein between the images of proteins fromtreated aqueous extract cell with the untreated cell proteinsThe protein changes as well as similar protein patternmaps inthe gels were analyzed from three independent experiments(biological replicate) and three technical replicates Changesin protein expression spots were considered significant (119901 lt005) by Anova analysis only if the intensity of protein spotswas more than twofold higher
29 In-Gel Digestion and LC-ESI-QTOF MS Protein spotswere excised from silver stained 2-DE gels Protein spots weredestained using fresh washing solution (100mM ammoniumbicarbonate 15mM potassium ferrocyanide and 100mMsodium thiosulphate for 15min on orbital shaker) The gelpieces were reduced with 10mM DTT in 10mM ammoniumbicarbonate at 60∘C for 30min For alkylation process 55mMIAA in 100mM ammonium bicarbonate was added to thegel plugs and incubated at room temperature for 20min inthe dark The gel pieces were rinsed three times in 50acetonitrile (ACN) in 10mM ammonium bicarbonate for20min on orbital shaker Then 100 ACN was added untilthe gel plugs becomemilky white after 15min To perform in-gel digestion of proteins 25120583L of trypsin solution (7 ng120583Ltrypsin in 1 ACN40mM ammonium bicarbonate) wasadded to the gels and incubated at 37∘C overnight Afterincubation 25120583L of 30 ACN was added to keep the gels
immersed throughout the digestion processThe supernatantwhich was the extract peptides was collected and dried for1 hr Peptides were desalted using ZipTip C18 (Millipore US)after equilibration in 50 ACN in 01 formic acid Theextracted peptides were collected pooled dried by vacuumcentrifuge and kept at minus20∘C for further mass spectrometricanalysis The peptides were then eluted and injected into LC-ESI-QTOFMS (model G6550 Agilent US) with electrosprayat flow rate of 160 nLmin to a nanocolumn (G4240-62010analytical 75 120583m times 150mm Zorbax 300SB-C18 5 120583m) Asolvent gradient (solvent A 50 ACN in water solvent B70 ACN solvent C 70 ACN solvent D 5 acetonitrile)was run for 25min
210 Bioinformatics After all the distinct peptides have un-dergone validation process the mass spectra and sequenceswere analyzed using a public database (UniProtKB andNational Center for Biotechnology Information) with morethan two distinct peptides Protein-protein interactions werepredicted using Search Tool for the Retrieval of Interact-ing GenesProteins (STRING) database v90 (httpwwwstring-dborg)The Swiss-Prot identifier for the Staphylococ-cus aureus genes in ldquoprotein moderdquo was used to search usingthe STRING database Network analysis was set at mediumstringency (STRING score = 04) Proteins were linked basedon seven criteria neighbourhood gene fusion cooccurrencecoexpression experimental evidences existing databasesand text mining
3 Results
31 Antibacterial Activity The aqueous extract showinginhibitory activity against MRSA 33591 was 1975120583gmLwhich was ten times higher than the MIC value for van-comycin The result for the time-kill studies was presentedin Figure 1 whereby the MRSA treated with subinhibitoryconcentration showed colony count of viable cell lower thanthe growth control after 24 hr incubationMeanwhile bacteriagrowth was stable when treated with 1x and 2x MIC aqueousextract of Q infectoria gall after 6 hr incubationThe numberof viable cells after exposure to 4x MIC of extract decreasedby more than 3 log
10CFUmL faster at the 3rd hr compared
to vancomycin after 13 hr treatment
32 Protein Variation between Cell Cultures Preliminarystudy whether aqueous extract Q infectoria gall affectedstaphylococcal cell protein patterns was performed by SDS-PAGE MRSA treated with aqueous extract Q infectoria gallexhibited different protein expression pattern compared tountreated MRSA in view of the level of intensity of the bands(Figure 2) 2D-GE combined with LC-ESI-QTOF MS wasperformed to identify the differential proteins which wereaffected in response to exposure with the tested extract
Figure 3 showed the protein mapping of treated anduntreated cells observed in acidic range in which mostof alterations between the two proteomes were displayedMore than 1000 individual protein spots were resolved andvisualized on silver-staining The optimum concentration
4 International Journal of Proteomics
0
2
4
6
8
10
12
14
0 2 4 6 8 10 12 14 16 18 20 22 24 26Time (hours)
Time-kill of MRSA treated with aqueousQ infectoria at different concentrations
Growth controlVancomycin
log10
(CFU
mL)
(12)x MIC extract
1x MIC extract 2x MIC extract 4x MIC extract
Figure 1 Time-kill curve of aqueous extract from Q infectoriaagainst MRSA 33591 Viability of cells growth was counted atspecified hr by serial dilution Each point represents the mean oflog10plusmn SEM (119899 = 3)
1 2MW
10
15
2030
4050607085
120
Figure 2 SDS-PAGE map of MRSA 33591 after being treated withaqueous extract of Q infectoria MW protein ladder lane number1 untreated MRSA lane 2 extract-treated MRSA
of MRSA protein for 2D-GE analysis was 100120583g Imageanalysis identified 24 protein spots that exhibited significantdifferences after treatment with the extract There are 17protein spots that were shown to be upregulated and anotherfive were found to be downregulated with more than twofoldincrease (119901 lt 005) (data not shown) However only 12 ofthe protein spots were successfully identified four protein
spots showed downregulation whereas eight protein spotswere upregulated
33 Protein Network Analysis Predicted and identified func-tionally linked proteins and the determination of the poten-tial biological process involved after treatment with aqueousextract from Q infectoria gall were analyzed by STRINGnetwork analysis as shown in Figure 4 The network waspresented under confident view whereby stronger associa-tions were represented by thicker lines or edges and proteinswere represented as nodes The interaction between the 11proteins was tabulated (Table 1) in order to summarize thedifferentially expressed proteins Four proteins were found tobe linked either directly or indirectly through one or moreinteracting proteins suggesting the existence of reportedfunctional linkagesHowever another group of three proteinswere linked together without linkage to other groups
4 Discussion
Methicillin-resistant S aureus (MRSA) has become a com-mon pathogen particularly in the long-term care setting[21] Thus the treatment of MRSA infection using currentantibiotic has become limited due to the emergence ofmultidrug-resistant strains [22] Herbal medicine is one ofthe strategies to combat resistance from this infection in aneffort to overcome mortality and morbidity patient problemin healthcare Recent studies [23 24] reported that naturalproduct has high antimicrobial potential against MRSA Theaqueous extract from Quercus infectoria gall in this studyshowed strong anti-MRSA activity and this is in agreementwith [25] that this extract showed promising capacity as newphytotherapeutic candidate against MRSA
Time-kill assay indicated that the aqueous extract fromQ infectoria gall exhibited significant bactericidal activity at4x MIC in dose-dependent manner Bactericidal activity wasdefined as a reduction ofmore than 3 log
10CFUmL at certain
time compared to at 0 hr [26] However at lower dose theextract was bacteriostatic and subinhibitory concentration((12)x MIC) of 975120583gmL was chosen to run 2D-GE toensure that enough protein can be obtained in order toanalyze the protein expression profile ofMRSAThe presenceof crude aqueous extract of Q infectoria gall against MRSAremarkably interfered with the general metabolic pathwayOne of the affected proteins lost was the branched-chain120572-keto acid dehydrogenase subunit E2 It is one importantcomponent of the protein complex which is the dihydrolipoyltransacylase that catalyzes transacylation of the acyl groupfrom E2 to coenzyme A [27] Branched-chain 120572-keto aciddehydrogenase is an enzyme complex that catalyzes the earlystages of branched-chain fatty acid production [28] and playsa key role in catabolism of branched-chain amino acids inbacteria [29] This enzyme is important as adherence of Saureus to the host eukaryotic cells for its survival in a mousehost due to its function in the membrane fluidity of S aureus
Another protein which was downregulated was succinyl-coenzymeA (CoA) ligase [ADP-forming] subunit beta whichis involved in citric acid cycle for energy metabolism [30]
International Journal of Proteomics 5
pH 4 pH 7
2
4
7
9 1718
1921
2223
24
(a)
pH 7pH 4
2
4
7
9 1718
212223
24
(b)
Figure 3 Two 2D-GEmaps of the cellular MRSA proteome (a) Protein expression profile of untreated MRSA (b) Protein expression profileof extract treated-MRSA
ftnA
kat
SAV1089
fadB
sle1
fabZ
E25
SAV1211
SAV0128
sucC
pflB
Name of gene Identified protein
SAV0128 NAD-dependent epimerase dehydratasefamily protein
pflB Formate acetyltransferase
fadB 3-Hydroxyacyl-CoA dehydrogenaseSle 1 N-Acetylmuramoyl-L-alanine amidase
peptidoglycan hydrolaseSAV1089 Hypothetical protein (RNase J)E25 Branched-chain alpha-keto acid
dehydrogenase subunit E2SAV1211 Pantothenate metabolism flavoproteinsucC Succinyl-CoA synthetase subunit betakat CatalaseftnA Ferritin iron storage proteinFabZ The 3-hydroxyacyl-CoA dehydrogenase
NAD binding domain protein
Figure 4 STRING interaction network analysis showed the association between differentially expressed proteins The protein names andgene symbols used in this network are listed in the table Higher amount of lines indicates stronger interaction between the proteins
This enzyme reversibly catalyzes ATP-dependent ligationfrom succinate andCoA to form succinyl-CoA in either aero-bic or anaerobic condition During aerobic growth succinyl-CoA ligase catalyzes the hydrolysis of succinyl-CoA usingADP and inorganic orthophosphate (119875
119894) to form succinate
and ATP However during anaerobic condition succinyl-CoA ligase can freely interconvert ATP and succinate to formsuccinyl-CoA ADP and inorganic 119875
119894for anabolic reaction as
well [31]Exposure of MRSA culture to crude extract of Q infec-
toria gall suppressed the production of proteins that wereinvolved in oxidative stress These proteins were ferritinand catalase Ferritins are the major iron storage proteinsin organism and provide protection against metal toxicityand oxidative stress [32] Ferritin was downregulated after
treatment with Q infectoria gall possibly due to its effectin limiting the iron environment hence it interfered bylowering the expression of ferritin protein Tannin contentin Q infectoria extract is believed to retard the growthof bacteria by inhibiting the activity of metalloenzymes incomplex tannins and iron [33] Meanwhile catalase catalyzeshydrogen peroxide to form oxygen and water Catalase is alsoknown as one of the virulent factors and when its productionwas reduced the bacteria cells become more susceptible tobe killed by neutrophils in the host cells [34] The proposedstress-related protein obviously reflected an adaption of thebacteria to survival under stress condition
Cell wall in gram-positive bacteria is very importantfor its survival Bacterial cell wall is composed of thecross-linked polymer peptidoglycan also known as murein
6 International Journal of Proteomics
Table1Th
eprotein
andgenesidentified
intheS
TRIN
GanalysisnetworkTh
elist
ofproteins
identifi
edisdifferentially
expressedproteins
follo
wingtreatmentw
ithQinfectoria
gallextract
Spot
number
Accession
number
Proteins
identifi
edMascot
score
Nom
inal
mass(kD
a)Sequ
ence
coverage
()
Num
bero
fpeptides
matched
Foldsc
hanges
(untreated
totre
ated)
2FT
NSA
AR
Ferritin
9419590
345
minus24
4D6SHZ8
Branched-chain
alph
a-keto
acid
dehydrogenases
ubun
itE2
1317
846
4516
234
8minus27
7D6SHJ5
Succinyl-C
oAligase[ADP-form
ing]
subu
nitb
eta
6543
422837
131
5minus22
9D6SDX7
Form
ateC
-acetyltransfe
rase
14051
855525
157
1026
10D6SH75
Catalase
6678
587233
102
5minus24
17amp18
D6SBS
5N-Acetylm
uram
oyl-L
-alanine
amidase
2834
692891
33
253
19D6SDX0
3-Hydroxyacyl-C
oAdehydrogenaseNADbind
ingdo
mainprotein
5144
846729
63
426
21D6SHG5
Ribo
nucle
aseJ
7412
646507
925
24
22D6SHN0
Phosph
opantothenoylcysteined
ecarbo
xylasepho
spho
pantothenate-cysteineligase
3195
4419
6562
221
23D6SDL7
NADdepend
entepimerasedehydratasefam
ilyprotein
279
378017
762
21
24D6SEM
63-Hydroxyacyl-[acyl-carrie
r-protein]
dehydrataseF
abZ
2731
163096
123
222
International Journal of Proteomics 7
Peptidoglycan architecture consists of a disaccharide back-bone built of alternating 120573-1-4-N-acetylglucosamines andN-acetylmuramic acids [35] Bacterial cell produces severalhydrolases that specifically cleave various covalent bonds inthe peptidoglycan including N-acetylmuramoyl-L-alanineamidases N-acetylglucosaminidases N-acetylmuramidasesendopeptidases and transglycosylases N-acetylmuramoyl-L-alanine amidase plays important roles in the separationseptum of daughter cells during cell division [36] HenceN-acetylmuramoyl-L-alanine amidase binds to both alphaand beta human fibrinogen and fibronectin which play arole during colonization and pathogenicity in the host tissueduring infection Previous study [37] reported that thisenzyme was upregulated to activate and increase its ownproduction in the presence of antibiotic to the bacteria andeventually cause inhibition of the bacterial growth or celldeath
Formate C-acetyltransferase or pyruvate formate-lyasewas upregulated in MRSA treated with aqueous extract fromQ infectoria gall This is a cytoplasmic protein involved inenergy fermentationmetabolism pathway in themicroorgan-ism This enzyme was induced in anaerobic growth and lim-ited nitrate [38 39] Formate C-acetyltransferase functions tocatalyze pyruvate to acetyl-CoA and formate which is subse-quently used for biosynthesis of protein DNA and RNA [40]and also as source for citric acid cycle through acetyl-CoA[41] Formate C-acetyltransferase NAD-dependent formatedehydrogenase and formyltetrahydrofolate synthetase wereupregulated during transcription at proteome level for thesynthesis of S aureus biofilm growth [42] This enzyme wasupregulated as one of the survival strategies of bacteria understress [40]
Phosphopantothenoyl cysteine decarboxylasephospho-pantothenate-cysteine ligase is also known as acid pan-tothenic [43] This is the last enzyme involved in the biosyn-thesis of CoAwhich is a crucial substance in the biogenesis ofbacterial cell membrane and functions as important cofactorin mitosis and stability of DNA in microorganism [44]This protein was upregulated in the study probably due tocompensation of other pathways to increase in the synthesisof CoA for adaptation when exposed to subinhibitory con-centration of the Q infectoria gall extract
NAD-dependent epimerasedehydratase protein is anew protein that has high similarity with UDP-glucose-4-epimerase involved in Leloir pathway [45] The other proteinthat was shown to be upregulated after treatment was RNaseJ which is involved in maturation of RNA and regulationof posttranscription until degradation of RNA in familyFirmicute of bacteria
Other proteins that were upregulated during fatty acidmetabolism are 3-hydroxyacyl-CoA dehydrogenase NAD-binding domain and 3-hydroxyacyl-[acyl-carrier-protein]dehydratase FabZ The 3-hydroxyacyl-CoA dehydrogenaseNAD binding domain protein was involved in variousmetabolic pathways such as beta oxidation fatty acid bycatalyzing the oxidation of 3-hydroxyl coenzymeA to oxoacylCoA and production of butanol in bacteria gram-positiveorganism [46] Upregulation of this protein may be due tocompensation of low level of the enzyme to tricarboxylic acid
cycle [47] In addition another protein that was involved inthe synthesis of fatty acid was 3-hydroxyacyl-[acyl-carrier-protein] dehydratase FabZ The latter was reported to par-ticipate in the phospholipid membrane biosynthesis pathway[48]
5 Conclusions
In conclusion all the data reported in this studymay elucidateand give better understanding on antibacterial effect ofaqueous extract of Q infectoria towards MRSA throughinterference of various metabolism and functions of the bac-terial proteome Furthermore recent studies suggested thataqueous extract ofQ infectoria galls has no toxicity effect andmortality effect in animal [49 50] However due to its hightannin content which has a profound digestibility-reducingeffect in human it is ideal to be administered topically since ithas the advantage of possessing the anti-inflammatory effectandwoundhealing activityThis information haswidened thechances for alternative medicine as therapeutic agent to treatMRSA infection
Competing Interests
The authors declare that they have no competing interestswith regard to publication of this paper
Authorsrsquo Contributions
Dayang Fredalina Basri designed analyzed and supervisedthe study Noraziah Mohamad Zin provided lab facilitiesRadhiah Khairon performed all the experiments data anal-ysis and drafted the manuscript All authors read andapproved the final manuscript
Acknowledgments
This study was funded by the Ministry of Higher EducationGovernment of Malaysia under the research code GrantFRGS22014SG05UKM023
References
[1] N F Crum R U Lee S A Thornton et al ldquoFifteen-yearstudy of the changing epidemiology of methicillin-resistantStaphylococcus aureusrdquo The American Journal of Medicine vol119 no 11 pp 943ndash951 2006
[2] A Asensio R Canton J Vaque et al ldquoNosocomial andcommunity-acquiredmeticillin-resistant Staphylococcus aureusinfections in hospitalized patients (Spain 1993ndash2003)rdquo Journalof Hospital Infection vol 63 no 4 pp 465ndash471 2006
[3] K Hiramatsu H Hanaki T Ino K Yabuta T Oguri and FC Tenover ldquoMethicillin-resistant Staphylococcus aureus clini-cal strain with reduced vancomycin susceptibilityrdquo Journal ofAntimicrobial Chemotherapy vol 40 no 1 pp 135ndash136 1997
[4] WHO WHO Traditional Medicine Strategy 2002ndash2005World Health Organization Geneva Switzerland 2002WHO EDM TRM 20021 engpdf
8 International Journal of Proteomics
[5] D G Brown T Lister and T L May-Dracka ldquoNew naturalproducts as new leads for antibacterial drug discoveryrdquo Bioor-ganic amp Medicinal Chemistry Letters vol 24 no 2 pp 413ndash4182014
[6] G M Cragg P G Grothaus and D J Newman ldquoImpactof natural products on developing new anti-cancer agentsrdquoChemical Reviews vol 109 no 7 pp 3012ndash3043 2009
[7] D A Dias S Urban and U Roessner ldquoA historical overview ofnatural products in drug discoveryrdquo Metabolites vol 2 no 4pp 303ndash336 2012
[8] F Sharififar G Dehghan-Nudeh Z Raeiat B AmirheidariM Moshrefi and A Purhemati ldquoTyrosinase inhibitory activityof major fractions of Quercus infectoria gallsrdquo PharmacognosyCommunications vol 3 no 1 pp 21ndash26 2013
[9] P Pithayanukul P Ruenraroengsak R Bavovada N PakmaneeR Suttisri and S Saen-Oon ldquoInhibition of Naja kaouthiavenom activities by plant polyphenolsrdquo Journal of Ethnophar-macology vol 97 no 3 pp 527ndash533 2005
[10] S Voravuthikunchai A Lortheeranuwat W Jeeju T SririrakS Phongpaichit and T Supawita ldquoEffective medicinal plantsagainst enterohaemorrhagic Escherichia coli O157H7rdquo Journalof Ethnopharmacology vol 94 no 1 pp 49ndash54 2004
[11] S J Cordwell M R Larsen R T Cole and B J Walsh ldquoCom-parative proteomics of Staphylococcus aureus and the responseof methicillin-resistant and methicillin-sensitive strains to Tri-ton X-100rdquoMicrobiology vol 148 no 9 pp 2765ndash2781 2002
[12] C Kohler S Wolff D Albrecht et al ldquoProteome analysesof Staphylococcus aureus in growing and non-growing cellsa physiological approachrdquo International Journal of MedicalMicrobiology vol 295 no 8 pp 547ndash565 2005
[13] L Van Oudenhove and B Devreese ldquoA review on recent devel-opments in mass spectrometry instrumentation and quantita-tive tools advancing bacterial proteomicsrdquoAppliedMicrobiologyand Biotechnology vol 97 no 11 pp 4749ndash4762 2013
[14] R Dosselli R Millioni L Puricelli et al ldquoMolecular targets ofantimicrobial photodynamic therapy identified by a proteomicapproachrdquo Journal of Proteomics vol 77 pp 329ndash343 2012
[15] R Jenkins N Burton and R Cooper ldquoManuka honey inhibitscell division in methicillin-resistant Staphylococcus aureusrdquoJournal of Antimicrobial Chemotherapy vol 66 no 11 pp 2536ndash2542 2011
[16] J M Packer J Irish B R Herbert et al ldquoSpecific non-peroxideantibacterial effect of manuka honey on the Staphylococcusaureus proteomerdquo International Journal of Antimicrobial Agentsvol 40 no 1 pp 43ndash50 2012
[17] W Sianglum P Srimanote W Wonglumsom K Kittiniyomand S P Voravuthikunchai ldquoProteome analyses of cellularproteins in methicillin-resistant Staphylococcus aureus treatedwith rhodomyrtone a novel antibiotic candidaterdquo PLoS ONEvol 6 no 2 Article ID e16628 pp 1ndash10 2011
[18] Clinical and Laboratory Standards Institute ldquoPerformancestandards for antimicrobial susceptibility testingrdquo FifteenthInformational Supplement M100-S15 2005
[19] D F Basri L S Aik R Khairon and M A Rahman ldquo2-Dgel electrophoresis map of methicillinresistant Staphylococcusaureus treated with Quercus infectoria gall extractrdquo AmericanJournal of Biochemistry and Biotechnology vol 9 no 1 pp 19ndash26 2013
[20] M Chevallet S Luche and T Rabilloud ldquoSilver staining ofproteins in polyacrylamide gelsrdquo Nature Protocols vol 1 no 4pp 1852ndash1858 2006
[21] J Davies and D Davies ldquoOrigins and evolution of antibioticresistancerdquoMicrobiology andMolecular Biology Reviews vol 74no 3 pp 417ndash433 2010
[22] D M Livermore ldquoAntibiotic resistance in staphylococcirdquo Inter-national Journal of Antimicrobial Agents vol 16 supplement 1pp S3ndashS10 2000
[23] L Yu H Xiang J Fan et al ldquoGlobal transcriptional response ofStaphylococcus aureus toRhein a natural plant productrdquo Journalof Biotechnology vol 135 no 3 pp 304ndash308 2008
[24] M Doyle E-A Feuerbaum K R Fox J Hinds D EThurstonand P W Taylor ldquoResponse of Staphylococcus aureus to subin-hibitory concentrations of a sequence-selective DNA minorgroove cross-linking pyrrolobenzodiazepine dimerrdquo Journal ofAntimicrobial Chemotherapy vol 64 no 5 pp 949ndash959 2009
[25] D F Basri and S H Fan ldquoThe potential of aqueous and acetoneextracts of galls of Quercus infectoria as antibacterial agentsrdquoIndian Journal of Pharmacology vol 37 no 1 pp 26ndash29 2005
[26] J N Eloff ldquoA sensitive and quick microplate method to deter-mine the minimal inhibitory concentration of plant extracts forbacteriardquo Planta Medica vol 64 no 8 pp 711ndash713 1998
[27] L Li P Thipyapong D C Breeden and J C SteffensldquoOverexpression of a bacterial branched-chain 120572-keto aciddehydrogenase complex in Arabidopsis results in accumulationof branched-chain acyl-CoAs and alteration of free amino acidcomposition in seedsrdquo Plant Science vol 165 no 6 pp 1213ndash1219 2003
[28] V K Singh D S Hattangady E S Giotis et al ldquoInsertionalinactivation of branched-chain 120572-keto acid dehydrogenase inStaphylococcus aureus leads to decreased branched-chainmem-brane fatty acid content and increased susceptibility to certainstressesrdquo Applied and Environmental Microbiology vol 74 no19 pp 5882ndash5890 2008
[29] P N Lowe J A Hodgson and R N Perham ldquoDual role of asingle multienzyme complex in the oxidative decarboxylationof pyruvate and branched-chain 2-oxo acids in Bacillus subtilisrdquoBiochemical Journal vol 215 no 1 pp 133ndash140 1983
[30] V Kapatral X Bina and A M Chakrabarty ldquoSuccinyl coen-zyme A synthetase of Pseudomonas aeruginosa with a broadspecificity for nucleoside triphosphate (NTP) synthesis modu-lates specificity for NTP synthesis by the 12-kilodalton form ofnucleoside diphosphate kinaserdquo Journal of Bacteriology vol 182no 5 pp 1333ndash1339 2000
[31] S-J Park G Chao and R P Gunsalus ldquoAerobic regulationof the sucABCD genes of Escherichia coli which encode 120572-ketoglutarate dehydrogenase and succinyl coenzyme A syn-thetase roles of ArcA Fnr and the upstream sdhCDAB pro-moterrdquo Journal of Bacteriology vol 179 no 13 pp 4138ndash41421997
[32] S C Andrews ldquoIron storage in bacteriardquo Advances in MicrobialPhysiology vol 40 pp 281ndash351 1998
[33] K-T Chung Z Lu andMW Chou ldquoMechanism of inhibitionof tannic acid and related compounds on the growth ofintestinal bacteriardquo Food and Chemical Toxicology vol 36 no12 pp 1053ndash1060 1998
[34] G L Mandell ldquoCatalase superoxide dismutase and virulenceof Staphylococcus aureus In vitro and in vivo studies withemphasis on staphylococcal-leukocyte interactionrdquoThe Journalof Clinical Investigation vol 55 no 3 pp 561ndash566 1975
[35] M P Chapot-Chartier and S Kulakauskas ldquoCell structure andfunction in lactic acid in bacteriardquoMicrobial Cell Factories vol13 pp S1ndashS9 2014
International Journal of Proteomics 9
[36] V A Fischetti ldquoBacteriophage lytic enzymes novel anti-infectivesrdquo Trends in Microbiology vol 13 no 10 pp 491ndash4962005
[37] C Heidrich M F Templin A Ursinus et al ldquoInvolvementof N-acetylmuramyl-l-alanine amidases in cell separationand antibiotic-induced autolysis of Escherichia colirdquo MolecularMicrobiology vol 41 no 1 pp 167ndash178 2001
[38] S Fuchs J Pane-Farre C KohlerMHecker and S EngelmannldquoAnaerobic gene expression in Staphylococcus aureusrdquo Journal ofBacteriology vol 189 no 11 pp 4275ndash4289 2007
[39] L J Rasmussen P L Moller and T Atlung ldquoCarbonmetabolism regulates expression of the pfl (pyruvate formate-lyase) gene in Escherichia colirdquo Journal of Bacteriology vol 173no 20 pp 6390ndash6397 1991
[40] M Leibig M Liebeke D Mader M Lalk A Peschel andF Gotz ldquoPyruvate formate lyase acts as a formate supplierfor metabolic processes during anaerobiosis in Staphylococcusaureusrdquo Journal of Bacteriology vol 193 no 4 pp 952ndash962 2011
[41] W Buckel and B T Golding ldquoRadical enzymes in anaerobesrdquoAnnual Review of Microbiology vol 60 pp 27ndash49 2006
[42] A Resch S Leicht M Saric et al ldquoComparative proteomeanalysis of Staphylococcus aureus biofilm and planktonic cellsand correlation with transcriptome profilingrdquo Proteomics vol6 no 6 pp 1867ndash1877 2006
[43] T Kupke ldquoMolecular characterization of the 41015840-phosphopan-tothenoylcysteine synthetase domain of bacterial Dfp flavopro-teinsrdquo The Journal of Biological Chemistry vol 277 no 39 pp36137ndash36145 2002
[44] T Nakamura T Pluskal Y Nakaseko and M YanagidaldquoImpaired coenzyme A synthesis in fission yeast causes defec-tive mitosis quiescence-exit failure histone hypoacetylationand fragile DNArdquo Open Biology vol 2 no 9 Article ID 1201172012
[45] D D Simeonova I Susnea A Moise B Schnik and MPrzybylski ldquolsquoUnknown genomersquo proteomics a new NAD(P)-dependent epimerasedehydratase revealed by N-terminal se-quencing inverted PCR and high resolution mass spectrome-tryrdquo Molecular amp Cellular Proteomics vol 8 no 1 pp 122ndash1312009
[46] D P Clark and J E Cronan ldquoTwo carbon compounds andfatty acids as carbon sourcesrdquo inEscherichia coli and SalmonellaCellular and Molecular Biology ASM Press Washington DCUSA 1996
[47] Z L Boynton G N Bennett and F B Rudolph ldquoCloningsequencing and expression of clustered genes encoding betahydroxybutyryl-coenzyme A (CoA) dehydrogenase crotonaseand butyryl-CoA dehydrogenase from Clostridium aceto-butylicum ATCC 824rdquo Journal of Bacteriology vol 178 no 11pp 3015ndash3024 1996
[48] J Schiebel A Chang H Lu M V Baxter P J Tonge andC Kisker ldquoStaphylococcus aureus FabI inhibition substraterecognition and potential implications for in vivo essentialityrdquoStructure vol 20 no 5 pp 802ndash813 2012
[49] M Iminjan N Amat X-H Li H Upur D Ahmat and B HeldquoInvestigation into the toxicity of traditional uyghur medicinequercus infectoria galls water extractrdquo PLoS ONE vol 9 no 3Article ID e90756 2014
[50] A Hasmah Z Nurazila C Y Chow R Rina and M Rafiquz-zaman ldquoCytotoxic effects ofQuercus infectoria extracts towardscervical (Hela) and Ovarian (Caov-3) cancer cell linesrdquo Healthand the Environment Journal vol 1 no 2 pp 17ndash23 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Proteomics 3
concentration of protein sample was quantified by Bradfordprotein assay kit (Bio-Rad US)
27 Two-Dimensional Gel Electrophoresis (2-DE) Protocolsof 2-DE were performed according to previously describedmethod [19] with three technical triplicatesThe protein sam-ples were separated on immobilized pH gradient (IPG) stripsof 24 cmwith a pH range 4ndash7The IPG strips were rehydratedwith 450120583L IPG rehydration bufferThe sampleswere appliedonto the acidic region of the slot in the strip Isoelectric focus-ing (IEF) was performedwith the Ettan IPGPhor System (GEHealthcare) according to the manufacturerrsquos guideline at atotal voltage of 60 kVh IPG strips were equilibrated in 10mLof SDS-equilibrium buffer (6M urea 75mM Tris-HCl 30glycerol 2 SDS and 0002 bromophenol blue) containing50mg DTT for 15min at room temperature for the reductionreaction The strip was alkylated by immersing in 10mL ofequilibration buffer containing 250mg iodoacetamide (IAA)for 15min and placed on 125 SDS-polyacrylamide gel
The second dimension was run using EttanDalt Six (GEHealthcare) at 15∘C 10mAgelminus1 for 1 hr the current was thenincreased to 40mAgelminus1 and the runwas continued until thebromophenol blue dye had moved towards the end of the gelAfter the 2-GE run the gels were fixed in a mixture (04ethanol (vv) and 01 glacial acetic acid (vv)) and furtherstained using the silver staining with somemodification [20]
28 Image Analysis The gels were analyzed using ImageS-canner III Lab Scan (GEHealthcare) and Progenesis software(Nonlinear Dynamics) The statistic used in this study foranalysis was Anova in the Progenesis software The softwarewas used to detect the differences in the image of profileexpression protein between the images of proteins fromtreated aqueous extract cell with the untreated cell proteinsThe protein changes as well as similar protein patternmaps inthe gels were analyzed from three independent experiments(biological replicate) and three technical replicates Changesin protein expression spots were considered significant (119901 lt005) by Anova analysis only if the intensity of protein spotswas more than twofold higher
29 In-Gel Digestion and LC-ESI-QTOF MS Protein spotswere excised from silver stained 2-DE gels Protein spots weredestained using fresh washing solution (100mM ammoniumbicarbonate 15mM potassium ferrocyanide and 100mMsodium thiosulphate for 15min on orbital shaker) The gelpieces were reduced with 10mM DTT in 10mM ammoniumbicarbonate at 60∘C for 30min For alkylation process 55mMIAA in 100mM ammonium bicarbonate was added to thegel plugs and incubated at room temperature for 20min inthe dark The gel pieces were rinsed three times in 50acetonitrile (ACN) in 10mM ammonium bicarbonate for20min on orbital shaker Then 100 ACN was added untilthe gel plugs becomemilky white after 15min To perform in-gel digestion of proteins 25120583L of trypsin solution (7 ng120583Ltrypsin in 1 ACN40mM ammonium bicarbonate) wasadded to the gels and incubated at 37∘C overnight Afterincubation 25120583L of 30 ACN was added to keep the gels
immersed throughout the digestion processThe supernatantwhich was the extract peptides was collected and dried for1 hr Peptides were desalted using ZipTip C18 (Millipore US)after equilibration in 50 ACN in 01 formic acid Theextracted peptides were collected pooled dried by vacuumcentrifuge and kept at minus20∘C for further mass spectrometricanalysis The peptides were then eluted and injected into LC-ESI-QTOFMS (model G6550 Agilent US) with electrosprayat flow rate of 160 nLmin to a nanocolumn (G4240-62010analytical 75 120583m times 150mm Zorbax 300SB-C18 5 120583m) Asolvent gradient (solvent A 50 ACN in water solvent B70 ACN solvent C 70 ACN solvent D 5 acetonitrile)was run for 25min
210 Bioinformatics After all the distinct peptides have un-dergone validation process the mass spectra and sequenceswere analyzed using a public database (UniProtKB andNational Center for Biotechnology Information) with morethan two distinct peptides Protein-protein interactions werepredicted using Search Tool for the Retrieval of Interact-ing GenesProteins (STRING) database v90 (httpwwwstring-dborg)The Swiss-Prot identifier for the Staphylococ-cus aureus genes in ldquoprotein moderdquo was used to search usingthe STRING database Network analysis was set at mediumstringency (STRING score = 04) Proteins were linked basedon seven criteria neighbourhood gene fusion cooccurrencecoexpression experimental evidences existing databasesand text mining
3 Results
31 Antibacterial Activity The aqueous extract showinginhibitory activity against MRSA 33591 was 1975120583gmLwhich was ten times higher than the MIC value for van-comycin The result for the time-kill studies was presentedin Figure 1 whereby the MRSA treated with subinhibitoryconcentration showed colony count of viable cell lower thanthe growth control after 24 hr incubationMeanwhile bacteriagrowth was stable when treated with 1x and 2x MIC aqueousextract of Q infectoria gall after 6 hr incubationThe numberof viable cells after exposure to 4x MIC of extract decreasedby more than 3 log
10CFUmL faster at the 3rd hr compared
to vancomycin after 13 hr treatment
32 Protein Variation between Cell Cultures Preliminarystudy whether aqueous extract Q infectoria gall affectedstaphylococcal cell protein patterns was performed by SDS-PAGE MRSA treated with aqueous extract Q infectoria gallexhibited different protein expression pattern compared tountreated MRSA in view of the level of intensity of the bands(Figure 2) 2D-GE combined with LC-ESI-QTOF MS wasperformed to identify the differential proteins which wereaffected in response to exposure with the tested extract
Figure 3 showed the protein mapping of treated anduntreated cells observed in acidic range in which mostof alterations between the two proteomes were displayedMore than 1000 individual protein spots were resolved andvisualized on silver-staining The optimum concentration
4 International Journal of Proteomics
0
2
4
6
8
10
12
14
0 2 4 6 8 10 12 14 16 18 20 22 24 26Time (hours)
Time-kill of MRSA treated with aqueousQ infectoria at different concentrations
Growth controlVancomycin
log10
(CFU
mL)
(12)x MIC extract
1x MIC extract 2x MIC extract 4x MIC extract
Figure 1 Time-kill curve of aqueous extract from Q infectoriaagainst MRSA 33591 Viability of cells growth was counted atspecified hr by serial dilution Each point represents the mean oflog10plusmn SEM (119899 = 3)
1 2MW
10
15
2030
4050607085
120
Figure 2 SDS-PAGE map of MRSA 33591 after being treated withaqueous extract of Q infectoria MW protein ladder lane number1 untreated MRSA lane 2 extract-treated MRSA
of MRSA protein for 2D-GE analysis was 100120583g Imageanalysis identified 24 protein spots that exhibited significantdifferences after treatment with the extract There are 17protein spots that were shown to be upregulated and anotherfive were found to be downregulated with more than twofoldincrease (119901 lt 005) (data not shown) However only 12 ofthe protein spots were successfully identified four protein
spots showed downregulation whereas eight protein spotswere upregulated
33 Protein Network Analysis Predicted and identified func-tionally linked proteins and the determination of the poten-tial biological process involved after treatment with aqueousextract from Q infectoria gall were analyzed by STRINGnetwork analysis as shown in Figure 4 The network waspresented under confident view whereby stronger associa-tions were represented by thicker lines or edges and proteinswere represented as nodes The interaction between the 11proteins was tabulated (Table 1) in order to summarize thedifferentially expressed proteins Four proteins were found tobe linked either directly or indirectly through one or moreinteracting proteins suggesting the existence of reportedfunctional linkagesHowever another group of three proteinswere linked together without linkage to other groups
4 Discussion
Methicillin-resistant S aureus (MRSA) has become a com-mon pathogen particularly in the long-term care setting[21] Thus the treatment of MRSA infection using currentantibiotic has become limited due to the emergence ofmultidrug-resistant strains [22] Herbal medicine is one ofthe strategies to combat resistance from this infection in aneffort to overcome mortality and morbidity patient problemin healthcare Recent studies [23 24] reported that naturalproduct has high antimicrobial potential against MRSA Theaqueous extract from Quercus infectoria gall in this studyshowed strong anti-MRSA activity and this is in agreementwith [25] that this extract showed promising capacity as newphytotherapeutic candidate against MRSA
Time-kill assay indicated that the aqueous extract fromQ infectoria gall exhibited significant bactericidal activity at4x MIC in dose-dependent manner Bactericidal activity wasdefined as a reduction ofmore than 3 log
10CFUmL at certain
time compared to at 0 hr [26] However at lower dose theextract was bacteriostatic and subinhibitory concentration((12)x MIC) of 975120583gmL was chosen to run 2D-GE toensure that enough protein can be obtained in order toanalyze the protein expression profile ofMRSAThe presenceof crude aqueous extract of Q infectoria gall against MRSAremarkably interfered with the general metabolic pathwayOne of the affected proteins lost was the branched-chain120572-keto acid dehydrogenase subunit E2 It is one importantcomponent of the protein complex which is the dihydrolipoyltransacylase that catalyzes transacylation of the acyl groupfrom E2 to coenzyme A [27] Branched-chain 120572-keto aciddehydrogenase is an enzyme complex that catalyzes the earlystages of branched-chain fatty acid production [28] and playsa key role in catabolism of branched-chain amino acids inbacteria [29] This enzyme is important as adherence of Saureus to the host eukaryotic cells for its survival in a mousehost due to its function in the membrane fluidity of S aureus
Another protein which was downregulated was succinyl-coenzymeA (CoA) ligase [ADP-forming] subunit beta whichis involved in citric acid cycle for energy metabolism [30]
International Journal of Proteomics 5
pH 4 pH 7
2
4
7
9 1718
1921
2223
24
(a)
pH 7pH 4
2
4
7
9 1718
212223
24
(b)
Figure 3 Two 2D-GEmaps of the cellular MRSA proteome (a) Protein expression profile of untreated MRSA (b) Protein expression profileof extract treated-MRSA
ftnA
kat
SAV1089
fadB
sle1
fabZ
E25
SAV1211
SAV0128
sucC
pflB
Name of gene Identified protein
SAV0128 NAD-dependent epimerase dehydratasefamily protein
pflB Formate acetyltransferase
fadB 3-Hydroxyacyl-CoA dehydrogenaseSle 1 N-Acetylmuramoyl-L-alanine amidase
peptidoglycan hydrolaseSAV1089 Hypothetical protein (RNase J)E25 Branched-chain alpha-keto acid
dehydrogenase subunit E2SAV1211 Pantothenate metabolism flavoproteinsucC Succinyl-CoA synthetase subunit betakat CatalaseftnA Ferritin iron storage proteinFabZ The 3-hydroxyacyl-CoA dehydrogenase
NAD binding domain protein
Figure 4 STRING interaction network analysis showed the association between differentially expressed proteins The protein names andgene symbols used in this network are listed in the table Higher amount of lines indicates stronger interaction between the proteins
This enzyme reversibly catalyzes ATP-dependent ligationfrom succinate andCoA to form succinyl-CoA in either aero-bic or anaerobic condition During aerobic growth succinyl-CoA ligase catalyzes the hydrolysis of succinyl-CoA usingADP and inorganic orthophosphate (119875
119894) to form succinate
and ATP However during anaerobic condition succinyl-CoA ligase can freely interconvert ATP and succinate to formsuccinyl-CoA ADP and inorganic 119875
119894for anabolic reaction as
well [31]Exposure of MRSA culture to crude extract of Q infec-
toria gall suppressed the production of proteins that wereinvolved in oxidative stress These proteins were ferritinand catalase Ferritins are the major iron storage proteinsin organism and provide protection against metal toxicityand oxidative stress [32] Ferritin was downregulated after
treatment with Q infectoria gall possibly due to its effectin limiting the iron environment hence it interfered bylowering the expression of ferritin protein Tannin contentin Q infectoria extract is believed to retard the growthof bacteria by inhibiting the activity of metalloenzymes incomplex tannins and iron [33] Meanwhile catalase catalyzeshydrogen peroxide to form oxygen and water Catalase is alsoknown as one of the virulent factors and when its productionwas reduced the bacteria cells become more susceptible tobe killed by neutrophils in the host cells [34] The proposedstress-related protein obviously reflected an adaption of thebacteria to survival under stress condition
Cell wall in gram-positive bacteria is very importantfor its survival Bacterial cell wall is composed of thecross-linked polymer peptidoglycan also known as murein
6 International Journal of Proteomics
Table1Th
eprotein
andgenesidentified
intheS
TRIN
GanalysisnetworkTh
elist
ofproteins
identifi
edisdifferentially
expressedproteins
follo
wingtreatmentw
ithQinfectoria
gallextract
Spot
number
Accession
number
Proteins
identifi
edMascot
score
Nom
inal
mass(kD
a)Sequ
ence
coverage
()
Num
bero
fpeptides
matched
Foldsc
hanges
(untreated
totre
ated)
2FT
NSA
AR
Ferritin
9419590
345
minus24
4D6SHZ8
Branched-chain
alph
a-keto
acid
dehydrogenases
ubun
itE2
1317
846
4516
234
8minus27
7D6SHJ5
Succinyl-C
oAligase[ADP-form
ing]
subu
nitb
eta
6543
422837
131
5minus22
9D6SDX7
Form
ateC
-acetyltransfe
rase
14051
855525
157
1026
10D6SH75
Catalase
6678
587233
102
5minus24
17amp18
D6SBS
5N-Acetylm
uram
oyl-L
-alanine
amidase
2834
692891
33
253
19D6SDX0
3-Hydroxyacyl-C
oAdehydrogenaseNADbind
ingdo
mainprotein
5144
846729
63
426
21D6SHG5
Ribo
nucle
aseJ
7412
646507
925
24
22D6SHN0
Phosph
opantothenoylcysteined
ecarbo
xylasepho
spho
pantothenate-cysteineligase
3195
4419
6562
221
23D6SDL7
NADdepend
entepimerasedehydratasefam
ilyprotein
279
378017
762
21
24D6SEM
63-Hydroxyacyl-[acyl-carrie
r-protein]
dehydrataseF
abZ
2731
163096
123
222
International Journal of Proteomics 7
Peptidoglycan architecture consists of a disaccharide back-bone built of alternating 120573-1-4-N-acetylglucosamines andN-acetylmuramic acids [35] Bacterial cell produces severalhydrolases that specifically cleave various covalent bonds inthe peptidoglycan including N-acetylmuramoyl-L-alanineamidases N-acetylglucosaminidases N-acetylmuramidasesendopeptidases and transglycosylases N-acetylmuramoyl-L-alanine amidase plays important roles in the separationseptum of daughter cells during cell division [36] HenceN-acetylmuramoyl-L-alanine amidase binds to both alphaand beta human fibrinogen and fibronectin which play arole during colonization and pathogenicity in the host tissueduring infection Previous study [37] reported that thisenzyme was upregulated to activate and increase its ownproduction in the presence of antibiotic to the bacteria andeventually cause inhibition of the bacterial growth or celldeath
Formate C-acetyltransferase or pyruvate formate-lyasewas upregulated in MRSA treated with aqueous extract fromQ infectoria gall This is a cytoplasmic protein involved inenergy fermentationmetabolism pathway in themicroorgan-ism This enzyme was induced in anaerobic growth and lim-ited nitrate [38 39] Formate C-acetyltransferase functions tocatalyze pyruvate to acetyl-CoA and formate which is subse-quently used for biosynthesis of protein DNA and RNA [40]and also as source for citric acid cycle through acetyl-CoA[41] Formate C-acetyltransferase NAD-dependent formatedehydrogenase and formyltetrahydrofolate synthetase wereupregulated during transcription at proteome level for thesynthesis of S aureus biofilm growth [42] This enzyme wasupregulated as one of the survival strategies of bacteria understress [40]
Phosphopantothenoyl cysteine decarboxylasephospho-pantothenate-cysteine ligase is also known as acid pan-tothenic [43] This is the last enzyme involved in the biosyn-thesis of CoAwhich is a crucial substance in the biogenesis ofbacterial cell membrane and functions as important cofactorin mitosis and stability of DNA in microorganism [44]This protein was upregulated in the study probably due tocompensation of other pathways to increase in the synthesisof CoA for adaptation when exposed to subinhibitory con-centration of the Q infectoria gall extract
NAD-dependent epimerasedehydratase protein is anew protein that has high similarity with UDP-glucose-4-epimerase involved in Leloir pathway [45] The other proteinthat was shown to be upregulated after treatment was RNaseJ which is involved in maturation of RNA and regulationof posttranscription until degradation of RNA in familyFirmicute of bacteria
Other proteins that were upregulated during fatty acidmetabolism are 3-hydroxyacyl-CoA dehydrogenase NAD-binding domain and 3-hydroxyacyl-[acyl-carrier-protein]dehydratase FabZ The 3-hydroxyacyl-CoA dehydrogenaseNAD binding domain protein was involved in variousmetabolic pathways such as beta oxidation fatty acid bycatalyzing the oxidation of 3-hydroxyl coenzymeA to oxoacylCoA and production of butanol in bacteria gram-positiveorganism [46] Upregulation of this protein may be due tocompensation of low level of the enzyme to tricarboxylic acid
cycle [47] In addition another protein that was involved inthe synthesis of fatty acid was 3-hydroxyacyl-[acyl-carrier-protein] dehydratase FabZ The latter was reported to par-ticipate in the phospholipid membrane biosynthesis pathway[48]
5 Conclusions
In conclusion all the data reported in this studymay elucidateand give better understanding on antibacterial effect ofaqueous extract of Q infectoria towards MRSA throughinterference of various metabolism and functions of the bac-terial proteome Furthermore recent studies suggested thataqueous extract ofQ infectoria galls has no toxicity effect andmortality effect in animal [49 50] However due to its hightannin content which has a profound digestibility-reducingeffect in human it is ideal to be administered topically since ithas the advantage of possessing the anti-inflammatory effectandwoundhealing activityThis information haswidened thechances for alternative medicine as therapeutic agent to treatMRSA infection
Competing Interests
The authors declare that they have no competing interestswith regard to publication of this paper
Authorsrsquo Contributions
Dayang Fredalina Basri designed analyzed and supervisedthe study Noraziah Mohamad Zin provided lab facilitiesRadhiah Khairon performed all the experiments data anal-ysis and drafted the manuscript All authors read andapproved the final manuscript
Acknowledgments
This study was funded by the Ministry of Higher EducationGovernment of Malaysia under the research code GrantFRGS22014SG05UKM023
References
[1] N F Crum R U Lee S A Thornton et al ldquoFifteen-yearstudy of the changing epidemiology of methicillin-resistantStaphylococcus aureusrdquo The American Journal of Medicine vol119 no 11 pp 943ndash951 2006
[2] A Asensio R Canton J Vaque et al ldquoNosocomial andcommunity-acquiredmeticillin-resistant Staphylococcus aureusinfections in hospitalized patients (Spain 1993ndash2003)rdquo Journalof Hospital Infection vol 63 no 4 pp 465ndash471 2006
[3] K Hiramatsu H Hanaki T Ino K Yabuta T Oguri and FC Tenover ldquoMethicillin-resistant Staphylococcus aureus clini-cal strain with reduced vancomycin susceptibilityrdquo Journal ofAntimicrobial Chemotherapy vol 40 no 1 pp 135ndash136 1997
[4] WHO WHO Traditional Medicine Strategy 2002ndash2005World Health Organization Geneva Switzerland 2002WHO EDM TRM 20021 engpdf
8 International Journal of Proteomics
[5] D G Brown T Lister and T L May-Dracka ldquoNew naturalproducts as new leads for antibacterial drug discoveryrdquo Bioor-ganic amp Medicinal Chemistry Letters vol 24 no 2 pp 413ndash4182014
[6] G M Cragg P G Grothaus and D J Newman ldquoImpactof natural products on developing new anti-cancer agentsrdquoChemical Reviews vol 109 no 7 pp 3012ndash3043 2009
[7] D A Dias S Urban and U Roessner ldquoA historical overview ofnatural products in drug discoveryrdquo Metabolites vol 2 no 4pp 303ndash336 2012
[8] F Sharififar G Dehghan-Nudeh Z Raeiat B AmirheidariM Moshrefi and A Purhemati ldquoTyrosinase inhibitory activityof major fractions of Quercus infectoria gallsrdquo PharmacognosyCommunications vol 3 no 1 pp 21ndash26 2013
[9] P Pithayanukul P Ruenraroengsak R Bavovada N PakmaneeR Suttisri and S Saen-Oon ldquoInhibition of Naja kaouthiavenom activities by plant polyphenolsrdquo Journal of Ethnophar-macology vol 97 no 3 pp 527ndash533 2005
[10] S Voravuthikunchai A Lortheeranuwat W Jeeju T SririrakS Phongpaichit and T Supawita ldquoEffective medicinal plantsagainst enterohaemorrhagic Escherichia coli O157H7rdquo Journalof Ethnopharmacology vol 94 no 1 pp 49ndash54 2004
[11] S J Cordwell M R Larsen R T Cole and B J Walsh ldquoCom-parative proteomics of Staphylococcus aureus and the responseof methicillin-resistant and methicillin-sensitive strains to Tri-ton X-100rdquoMicrobiology vol 148 no 9 pp 2765ndash2781 2002
[12] C Kohler S Wolff D Albrecht et al ldquoProteome analysesof Staphylococcus aureus in growing and non-growing cellsa physiological approachrdquo International Journal of MedicalMicrobiology vol 295 no 8 pp 547ndash565 2005
[13] L Van Oudenhove and B Devreese ldquoA review on recent devel-opments in mass spectrometry instrumentation and quantita-tive tools advancing bacterial proteomicsrdquoAppliedMicrobiologyand Biotechnology vol 97 no 11 pp 4749ndash4762 2013
[14] R Dosselli R Millioni L Puricelli et al ldquoMolecular targets ofantimicrobial photodynamic therapy identified by a proteomicapproachrdquo Journal of Proteomics vol 77 pp 329ndash343 2012
[15] R Jenkins N Burton and R Cooper ldquoManuka honey inhibitscell division in methicillin-resistant Staphylococcus aureusrdquoJournal of Antimicrobial Chemotherapy vol 66 no 11 pp 2536ndash2542 2011
[16] J M Packer J Irish B R Herbert et al ldquoSpecific non-peroxideantibacterial effect of manuka honey on the Staphylococcusaureus proteomerdquo International Journal of Antimicrobial Agentsvol 40 no 1 pp 43ndash50 2012
[17] W Sianglum P Srimanote W Wonglumsom K Kittiniyomand S P Voravuthikunchai ldquoProteome analyses of cellularproteins in methicillin-resistant Staphylococcus aureus treatedwith rhodomyrtone a novel antibiotic candidaterdquo PLoS ONEvol 6 no 2 Article ID e16628 pp 1ndash10 2011
[18] Clinical and Laboratory Standards Institute ldquoPerformancestandards for antimicrobial susceptibility testingrdquo FifteenthInformational Supplement M100-S15 2005
[19] D F Basri L S Aik R Khairon and M A Rahman ldquo2-Dgel electrophoresis map of methicillinresistant Staphylococcusaureus treated with Quercus infectoria gall extractrdquo AmericanJournal of Biochemistry and Biotechnology vol 9 no 1 pp 19ndash26 2013
[20] M Chevallet S Luche and T Rabilloud ldquoSilver staining ofproteins in polyacrylamide gelsrdquo Nature Protocols vol 1 no 4pp 1852ndash1858 2006
[21] J Davies and D Davies ldquoOrigins and evolution of antibioticresistancerdquoMicrobiology andMolecular Biology Reviews vol 74no 3 pp 417ndash433 2010
[22] D M Livermore ldquoAntibiotic resistance in staphylococcirdquo Inter-national Journal of Antimicrobial Agents vol 16 supplement 1pp S3ndashS10 2000
[23] L Yu H Xiang J Fan et al ldquoGlobal transcriptional response ofStaphylococcus aureus toRhein a natural plant productrdquo Journalof Biotechnology vol 135 no 3 pp 304ndash308 2008
[24] M Doyle E-A Feuerbaum K R Fox J Hinds D EThurstonand P W Taylor ldquoResponse of Staphylococcus aureus to subin-hibitory concentrations of a sequence-selective DNA minorgroove cross-linking pyrrolobenzodiazepine dimerrdquo Journal ofAntimicrobial Chemotherapy vol 64 no 5 pp 949ndash959 2009
[25] D F Basri and S H Fan ldquoThe potential of aqueous and acetoneextracts of galls of Quercus infectoria as antibacterial agentsrdquoIndian Journal of Pharmacology vol 37 no 1 pp 26ndash29 2005
[26] J N Eloff ldquoA sensitive and quick microplate method to deter-mine the minimal inhibitory concentration of plant extracts forbacteriardquo Planta Medica vol 64 no 8 pp 711ndash713 1998
[27] L Li P Thipyapong D C Breeden and J C SteffensldquoOverexpression of a bacterial branched-chain 120572-keto aciddehydrogenase complex in Arabidopsis results in accumulationof branched-chain acyl-CoAs and alteration of free amino acidcomposition in seedsrdquo Plant Science vol 165 no 6 pp 1213ndash1219 2003
[28] V K Singh D S Hattangady E S Giotis et al ldquoInsertionalinactivation of branched-chain 120572-keto acid dehydrogenase inStaphylococcus aureus leads to decreased branched-chainmem-brane fatty acid content and increased susceptibility to certainstressesrdquo Applied and Environmental Microbiology vol 74 no19 pp 5882ndash5890 2008
[29] P N Lowe J A Hodgson and R N Perham ldquoDual role of asingle multienzyme complex in the oxidative decarboxylationof pyruvate and branched-chain 2-oxo acids in Bacillus subtilisrdquoBiochemical Journal vol 215 no 1 pp 133ndash140 1983
[30] V Kapatral X Bina and A M Chakrabarty ldquoSuccinyl coen-zyme A synthetase of Pseudomonas aeruginosa with a broadspecificity for nucleoside triphosphate (NTP) synthesis modu-lates specificity for NTP synthesis by the 12-kilodalton form ofnucleoside diphosphate kinaserdquo Journal of Bacteriology vol 182no 5 pp 1333ndash1339 2000
[31] S-J Park G Chao and R P Gunsalus ldquoAerobic regulationof the sucABCD genes of Escherichia coli which encode 120572-ketoglutarate dehydrogenase and succinyl coenzyme A syn-thetase roles of ArcA Fnr and the upstream sdhCDAB pro-moterrdquo Journal of Bacteriology vol 179 no 13 pp 4138ndash41421997
[32] S C Andrews ldquoIron storage in bacteriardquo Advances in MicrobialPhysiology vol 40 pp 281ndash351 1998
[33] K-T Chung Z Lu andMW Chou ldquoMechanism of inhibitionof tannic acid and related compounds on the growth ofintestinal bacteriardquo Food and Chemical Toxicology vol 36 no12 pp 1053ndash1060 1998
[34] G L Mandell ldquoCatalase superoxide dismutase and virulenceof Staphylococcus aureus In vitro and in vivo studies withemphasis on staphylococcal-leukocyte interactionrdquoThe Journalof Clinical Investigation vol 55 no 3 pp 561ndash566 1975
[35] M P Chapot-Chartier and S Kulakauskas ldquoCell structure andfunction in lactic acid in bacteriardquoMicrobial Cell Factories vol13 pp S1ndashS9 2014
International Journal of Proteomics 9
[36] V A Fischetti ldquoBacteriophage lytic enzymes novel anti-infectivesrdquo Trends in Microbiology vol 13 no 10 pp 491ndash4962005
[37] C Heidrich M F Templin A Ursinus et al ldquoInvolvementof N-acetylmuramyl-l-alanine amidases in cell separationand antibiotic-induced autolysis of Escherichia colirdquo MolecularMicrobiology vol 41 no 1 pp 167ndash178 2001
[38] S Fuchs J Pane-Farre C KohlerMHecker and S EngelmannldquoAnaerobic gene expression in Staphylococcus aureusrdquo Journal ofBacteriology vol 189 no 11 pp 4275ndash4289 2007
[39] L J Rasmussen P L Moller and T Atlung ldquoCarbonmetabolism regulates expression of the pfl (pyruvate formate-lyase) gene in Escherichia colirdquo Journal of Bacteriology vol 173no 20 pp 6390ndash6397 1991
[40] M Leibig M Liebeke D Mader M Lalk A Peschel andF Gotz ldquoPyruvate formate lyase acts as a formate supplierfor metabolic processes during anaerobiosis in Staphylococcusaureusrdquo Journal of Bacteriology vol 193 no 4 pp 952ndash962 2011
[41] W Buckel and B T Golding ldquoRadical enzymes in anaerobesrdquoAnnual Review of Microbiology vol 60 pp 27ndash49 2006
[42] A Resch S Leicht M Saric et al ldquoComparative proteomeanalysis of Staphylococcus aureus biofilm and planktonic cellsand correlation with transcriptome profilingrdquo Proteomics vol6 no 6 pp 1867ndash1877 2006
[43] T Kupke ldquoMolecular characterization of the 41015840-phosphopan-tothenoylcysteine synthetase domain of bacterial Dfp flavopro-teinsrdquo The Journal of Biological Chemistry vol 277 no 39 pp36137ndash36145 2002
[44] T Nakamura T Pluskal Y Nakaseko and M YanagidaldquoImpaired coenzyme A synthesis in fission yeast causes defec-tive mitosis quiescence-exit failure histone hypoacetylationand fragile DNArdquo Open Biology vol 2 no 9 Article ID 1201172012
[45] D D Simeonova I Susnea A Moise B Schnik and MPrzybylski ldquolsquoUnknown genomersquo proteomics a new NAD(P)-dependent epimerasedehydratase revealed by N-terminal se-quencing inverted PCR and high resolution mass spectrome-tryrdquo Molecular amp Cellular Proteomics vol 8 no 1 pp 122ndash1312009
[46] D P Clark and J E Cronan ldquoTwo carbon compounds andfatty acids as carbon sourcesrdquo inEscherichia coli and SalmonellaCellular and Molecular Biology ASM Press Washington DCUSA 1996
[47] Z L Boynton G N Bennett and F B Rudolph ldquoCloningsequencing and expression of clustered genes encoding betahydroxybutyryl-coenzyme A (CoA) dehydrogenase crotonaseand butyryl-CoA dehydrogenase from Clostridium aceto-butylicum ATCC 824rdquo Journal of Bacteriology vol 178 no 11pp 3015ndash3024 1996
[48] J Schiebel A Chang H Lu M V Baxter P J Tonge andC Kisker ldquoStaphylococcus aureus FabI inhibition substraterecognition and potential implications for in vivo essentialityrdquoStructure vol 20 no 5 pp 802ndash813 2012
[49] M Iminjan N Amat X-H Li H Upur D Ahmat and B HeldquoInvestigation into the toxicity of traditional uyghur medicinequercus infectoria galls water extractrdquo PLoS ONE vol 9 no 3Article ID e90756 2014
[50] A Hasmah Z Nurazila C Y Chow R Rina and M Rafiquz-zaman ldquoCytotoxic effects ofQuercus infectoria extracts towardscervical (Hela) and Ovarian (Caov-3) cancer cell linesrdquo Healthand the Environment Journal vol 1 no 2 pp 17ndash23 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
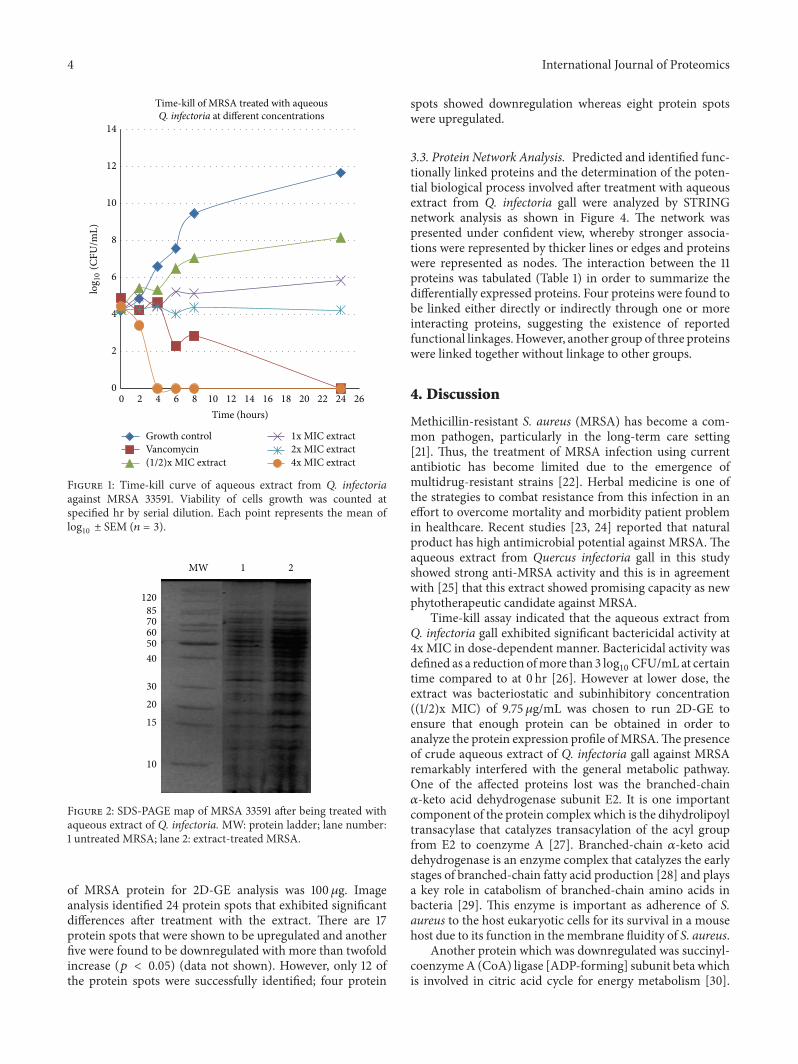
4 International Journal of Proteomics
0
2
4
6
8
10
12
14
0 2 4 6 8 10 12 14 16 18 20 22 24 26Time (hours)
Time-kill of MRSA treated with aqueousQ infectoria at different concentrations
Growth controlVancomycin
log10
(CFU
mL)
(12)x MIC extract
1x MIC extract 2x MIC extract 4x MIC extract
Figure 1 Time-kill curve of aqueous extract from Q infectoriaagainst MRSA 33591 Viability of cells growth was counted atspecified hr by serial dilution Each point represents the mean oflog10plusmn SEM (119899 = 3)
1 2MW
10
15
2030
4050607085
120
Figure 2 SDS-PAGE map of MRSA 33591 after being treated withaqueous extract of Q infectoria MW protein ladder lane number1 untreated MRSA lane 2 extract-treated MRSA
of MRSA protein for 2D-GE analysis was 100120583g Imageanalysis identified 24 protein spots that exhibited significantdifferences after treatment with the extract There are 17protein spots that were shown to be upregulated and anotherfive were found to be downregulated with more than twofoldincrease (119901 lt 005) (data not shown) However only 12 ofthe protein spots were successfully identified four protein
spots showed downregulation whereas eight protein spotswere upregulated
33 Protein Network Analysis Predicted and identified func-tionally linked proteins and the determination of the poten-tial biological process involved after treatment with aqueousextract from Q infectoria gall were analyzed by STRINGnetwork analysis as shown in Figure 4 The network waspresented under confident view whereby stronger associa-tions were represented by thicker lines or edges and proteinswere represented as nodes The interaction between the 11proteins was tabulated (Table 1) in order to summarize thedifferentially expressed proteins Four proteins were found tobe linked either directly or indirectly through one or moreinteracting proteins suggesting the existence of reportedfunctional linkagesHowever another group of three proteinswere linked together without linkage to other groups
4 Discussion
Methicillin-resistant S aureus (MRSA) has become a com-mon pathogen particularly in the long-term care setting[21] Thus the treatment of MRSA infection using currentantibiotic has become limited due to the emergence ofmultidrug-resistant strains [22] Herbal medicine is one ofthe strategies to combat resistance from this infection in aneffort to overcome mortality and morbidity patient problemin healthcare Recent studies [23 24] reported that naturalproduct has high antimicrobial potential against MRSA Theaqueous extract from Quercus infectoria gall in this studyshowed strong anti-MRSA activity and this is in agreementwith [25] that this extract showed promising capacity as newphytotherapeutic candidate against MRSA
Time-kill assay indicated that the aqueous extract fromQ infectoria gall exhibited significant bactericidal activity at4x MIC in dose-dependent manner Bactericidal activity wasdefined as a reduction ofmore than 3 log
10CFUmL at certain
time compared to at 0 hr [26] However at lower dose theextract was bacteriostatic and subinhibitory concentration((12)x MIC) of 975120583gmL was chosen to run 2D-GE toensure that enough protein can be obtained in order toanalyze the protein expression profile ofMRSAThe presenceof crude aqueous extract of Q infectoria gall against MRSAremarkably interfered with the general metabolic pathwayOne of the affected proteins lost was the branched-chain120572-keto acid dehydrogenase subunit E2 It is one importantcomponent of the protein complex which is the dihydrolipoyltransacylase that catalyzes transacylation of the acyl groupfrom E2 to coenzyme A [27] Branched-chain 120572-keto aciddehydrogenase is an enzyme complex that catalyzes the earlystages of branched-chain fatty acid production [28] and playsa key role in catabolism of branched-chain amino acids inbacteria [29] This enzyme is important as adherence of Saureus to the host eukaryotic cells for its survival in a mousehost due to its function in the membrane fluidity of S aureus
Another protein which was downregulated was succinyl-coenzymeA (CoA) ligase [ADP-forming] subunit beta whichis involved in citric acid cycle for energy metabolism [30]
International Journal of Proteomics 5
pH 4 pH 7
2
4
7
9 1718
1921
2223
24
(a)
pH 7pH 4
2
4
7
9 1718
212223
24
(b)
Figure 3 Two 2D-GEmaps of the cellular MRSA proteome (a) Protein expression profile of untreated MRSA (b) Protein expression profileof extract treated-MRSA
ftnA
kat
SAV1089
fadB
sle1
fabZ
E25
SAV1211
SAV0128
sucC
pflB
Name of gene Identified protein
SAV0128 NAD-dependent epimerase dehydratasefamily protein
pflB Formate acetyltransferase
fadB 3-Hydroxyacyl-CoA dehydrogenaseSle 1 N-Acetylmuramoyl-L-alanine amidase
peptidoglycan hydrolaseSAV1089 Hypothetical protein (RNase J)E25 Branched-chain alpha-keto acid
dehydrogenase subunit E2SAV1211 Pantothenate metabolism flavoproteinsucC Succinyl-CoA synthetase subunit betakat CatalaseftnA Ferritin iron storage proteinFabZ The 3-hydroxyacyl-CoA dehydrogenase
NAD binding domain protein
Figure 4 STRING interaction network analysis showed the association between differentially expressed proteins The protein names andgene symbols used in this network are listed in the table Higher amount of lines indicates stronger interaction between the proteins
This enzyme reversibly catalyzes ATP-dependent ligationfrom succinate andCoA to form succinyl-CoA in either aero-bic or anaerobic condition During aerobic growth succinyl-CoA ligase catalyzes the hydrolysis of succinyl-CoA usingADP and inorganic orthophosphate (119875
119894) to form succinate
and ATP However during anaerobic condition succinyl-CoA ligase can freely interconvert ATP and succinate to formsuccinyl-CoA ADP and inorganic 119875
119894for anabolic reaction as
well [31]Exposure of MRSA culture to crude extract of Q infec-
toria gall suppressed the production of proteins that wereinvolved in oxidative stress These proteins were ferritinand catalase Ferritins are the major iron storage proteinsin organism and provide protection against metal toxicityand oxidative stress [32] Ferritin was downregulated after
treatment with Q infectoria gall possibly due to its effectin limiting the iron environment hence it interfered bylowering the expression of ferritin protein Tannin contentin Q infectoria extract is believed to retard the growthof bacteria by inhibiting the activity of metalloenzymes incomplex tannins and iron [33] Meanwhile catalase catalyzeshydrogen peroxide to form oxygen and water Catalase is alsoknown as one of the virulent factors and when its productionwas reduced the bacteria cells become more susceptible tobe killed by neutrophils in the host cells [34] The proposedstress-related protein obviously reflected an adaption of thebacteria to survival under stress condition
Cell wall in gram-positive bacteria is very importantfor its survival Bacterial cell wall is composed of thecross-linked polymer peptidoglycan also known as murein
6 International Journal of Proteomics
Table1Th
eprotein
andgenesidentified
intheS
TRIN
GanalysisnetworkTh
elist
ofproteins
identifi
edisdifferentially
expressedproteins
follo
wingtreatmentw
ithQinfectoria
gallextract
Spot
number
Accession
number
Proteins
identifi
edMascot
score
Nom
inal
mass(kD
a)Sequ
ence
coverage
()
Num
bero
fpeptides
matched
Foldsc
hanges
(untreated
totre
ated)
2FT
NSA
AR
Ferritin
9419590
345
minus24
4D6SHZ8
Branched-chain
alph
a-keto
acid
dehydrogenases
ubun
itE2
1317
846
4516
234
8minus27
7D6SHJ5
Succinyl-C
oAligase[ADP-form
ing]
subu
nitb
eta
6543
422837
131
5minus22
9D6SDX7
Form
ateC
-acetyltransfe
rase
14051
855525
157
1026
10D6SH75
Catalase
6678
587233
102
5minus24
17amp18
D6SBS
5N-Acetylm
uram
oyl-L
-alanine
amidase
2834
692891
33
253
19D6SDX0
3-Hydroxyacyl-C
oAdehydrogenaseNADbind
ingdo
mainprotein
5144
846729
63
426
21D6SHG5
Ribo
nucle
aseJ
7412
646507
925
24
22D6SHN0
Phosph
opantothenoylcysteined
ecarbo
xylasepho
spho
pantothenate-cysteineligase
3195
4419
6562
221
23D6SDL7
NADdepend
entepimerasedehydratasefam
ilyprotein
279
378017
762
21
24D6SEM
63-Hydroxyacyl-[acyl-carrie
r-protein]
dehydrataseF
abZ
2731
163096
123
222
International Journal of Proteomics 7
Peptidoglycan architecture consists of a disaccharide back-bone built of alternating 120573-1-4-N-acetylglucosamines andN-acetylmuramic acids [35] Bacterial cell produces severalhydrolases that specifically cleave various covalent bonds inthe peptidoglycan including N-acetylmuramoyl-L-alanineamidases N-acetylglucosaminidases N-acetylmuramidasesendopeptidases and transglycosylases N-acetylmuramoyl-L-alanine amidase plays important roles in the separationseptum of daughter cells during cell division [36] HenceN-acetylmuramoyl-L-alanine amidase binds to both alphaand beta human fibrinogen and fibronectin which play arole during colonization and pathogenicity in the host tissueduring infection Previous study [37] reported that thisenzyme was upregulated to activate and increase its ownproduction in the presence of antibiotic to the bacteria andeventually cause inhibition of the bacterial growth or celldeath
Formate C-acetyltransferase or pyruvate formate-lyasewas upregulated in MRSA treated with aqueous extract fromQ infectoria gall This is a cytoplasmic protein involved inenergy fermentationmetabolism pathway in themicroorgan-ism This enzyme was induced in anaerobic growth and lim-ited nitrate [38 39] Formate C-acetyltransferase functions tocatalyze pyruvate to acetyl-CoA and formate which is subse-quently used for biosynthesis of protein DNA and RNA [40]and also as source for citric acid cycle through acetyl-CoA[41] Formate C-acetyltransferase NAD-dependent formatedehydrogenase and formyltetrahydrofolate synthetase wereupregulated during transcription at proteome level for thesynthesis of S aureus biofilm growth [42] This enzyme wasupregulated as one of the survival strategies of bacteria understress [40]
Phosphopantothenoyl cysteine decarboxylasephospho-pantothenate-cysteine ligase is also known as acid pan-tothenic [43] This is the last enzyme involved in the biosyn-thesis of CoAwhich is a crucial substance in the biogenesis ofbacterial cell membrane and functions as important cofactorin mitosis and stability of DNA in microorganism [44]This protein was upregulated in the study probably due tocompensation of other pathways to increase in the synthesisof CoA for adaptation when exposed to subinhibitory con-centration of the Q infectoria gall extract
NAD-dependent epimerasedehydratase protein is anew protein that has high similarity with UDP-glucose-4-epimerase involved in Leloir pathway [45] The other proteinthat was shown to be upregulated after treatment was RNaseJ which is involved in maturation of RNA and regulationof posttranscription until degradation of RNA in familyFirmicute of bacteria
Other proteins that were upregulated during fatty acidmetabolism are 3-hydroxyacyl-CoA dehydrogenase NAD-binding domain and 3-hydroxyacyl-[acyl-carrier-protein]dehydratase FabZ The 3-hydroxyacyl-CoA dehydrogenaseNAD binding domain protein was involved in variousmetabolic pathways such as beta oxidation fatty acid bycatalyzing the oxidation of 3-hydroxyl coenzymeA to oxoacylCoA and production of butanol in bacteria gram-positiveorganism [46] Upregulation of this protein may be due tocompensation of low level of the enzyme to tricarboxylic acid
cycle [47] In addition another protein that was involved inthe synthesis of fatty acid was 3-hydroxyacyl-[acyl-carrier-protein] dehydratase FabZ The latter was reported to par-ticipate in the phospholipid membrane biosynthesis pathway[48]
5 Conclusions
In conclusion all the data reported in this studymay elucidateand give better understanding on antibacterial effect ofaqueous extract of Q infectoria towards MRSA throughinterference of various metabolism and functions of the bac-terial proteome Furthermore recent studies suggested thataqueous extract ofQ infectoria galls has no toxicity effect andmortality effect in animal [49 50] However due to its hightannin content which has a profound digestibility-reducingeffect in human it is ideal to be administered topically since ithas the advantage of possessing the anti-inflammatory effectandwoundhealing activityThis information haswidened thechances for alternative medicine as therapeutic agent to treatMRSA infection
Competing Interests
The authors declare that they have no competing interestswith regard to publication of this paper
Authorsrsquo Contributions
Dayang Fredalina Basri designed analyzed and supervisedthe study Noraziah Mohamad Zin provided lab facilitiesRadhiah Khairon performed all the experiments data anal-ysis and drafted the manuscript All authors read andapproved the final manuscript
Acknowledgments
This study was funded by the Ministry of Higher EducationGovernment of Malaysia under the research code GrantFRGS22014SG05UKM023
References
[1] N F Crum R U Lee S A Thornton et al ldquoFifteen-yearstudy of the changing epidemiology of methicillin-resistantStaphylococcus aureusrdquo The American Journal of Medicine vol119 no 11 pp 943ndash951 2006
[2] A Asensio R Canton J Vaque et al ldquoNosocomial andcommunity-acquiredmeticillin-resistant Staphylococcus aureusinfections in hospitalized patients (Spain 1993ndash2003)rdquo Journalof Hospital Infection vol 63 no 4 pp 465ndash471 2006
[3] K Hiramatsu H Hanaki T Ino K Yabuta T Oguri and FC Tenover ldquoMethicillin-resistant Staphylococcus aureus clini-cal strain with reduced vancomycin susceptibilityrdquo Journal ofAntimicrobial Chemotherapy vol 40 no 1 pp 135ndash136 1997
[4] WHO WHO Traditional Medicine Strategy 2002ndash2005World Health Organization Geneva Switzerland 2002WHO EDM TRM 20021 engpdf
8 International Journal of Proteomics
[5] D G Brown T Lister and T L May-Dracka ldquoNew naturalproducts as new leads for antibacterial drug discoveryrdquo Bioor-ganic amp Medicinal Chemistry Letters vol 24 no 2 pp 413ndash4182014
[6] G M Cragg P G Grothaus and D J Newman ldquoImpactof natural products on developing new anti-cancer agentsrdquoChemical Reviews vol 109 no 7 pp 3012ndash3043 2009
[7] D A Dias S Urban and U Roessner ldquoA historical overview ofnatural products in drug discoveryrdquo Metabolites vol 2 no 4pp 303ndash336 2012
[8] F Sharififar G Dehghan-Nudeh Z Raeiat B AmirheidariM Moshrefi and A Purhemati ldquoTyrosinase inhibitory activityof major fractions of Quercus infectoria gallsrdquo PharmacognosyCommunications vol 3 no 1 pp 21ndash26 2013
[9] P Pithayanukul P Ruenraroengsak R Bavovada N PakmaneeR Suttisri and S Saen-Oon ldquoInhibition of Naja kaouthiavenom activities by plant polyphenolsrdquo Journal of Ethnophar-macology vol 97 no 3 pp 527ndash533 2005
[10] S Voravuthikunchai A Lortheeranuwat W Jeeju T SririrakS Phongpaichit and T Supawita ldquoEffective medicinal plantsagainst enterohaemorrhagic Escherichia coli O157H7rdquo Journalof Ethnopharmacology vol 94 no 1 pp 49ndash54 2004
[11] S J Cordwell M R Larsen R T Cole and B J Walsh ldquoCom-parative proteomics of Staphylococcus aureus and the responseof methicillin-resistant and methicillin-sensitive strains to Tri-ton X-100rdquoMicrobiology vol 148 no 9 pp 2765ndash2781 2002
[12] C Kohler S Wolff D Albrecht et al ldquoProteome analysesof Staphylococcus aureus in growing and non-growing cellsa physiological approachrdquo International Journal of MedicalMicrobiology vol 295 no 8 pp 547ndash565 2005
[13] L Van Oudenhove and B Devreese ldquoA review on recent devel-opments in mass spectrometry instrumentation and quantita-tive tools advancing bacterial proteomicsrdquoAppliedMicrobiologyand Biotechnology vol 97 no 11 pp 4749ndash4762 2013
[14] R Dosselli R Millioni L Puricelli et al ldquoMolecular targets ofantimicrobial photodynamic therapy identified by a proteomicapproachrdquo Journal of Proteomics vol 77 pp 329ndash343 2012
[15] R Jenkins N Burton and R Cooper ldquoManuka honey inhibitscell division in methicillin-resistant Staphylococcus aureusrdquoJournal of Antimicrobial Chemotherapy vol 66 no 11 pp 2536ndash2542 2011
[16] J M Packer J Irish B R Herbert et al ldquoSpecific non-peroxideantibacterial effect of manuka honey on the Staphylococcusaureus proteomerdquo International Journal of Antimicrobial Agentsvol 40 no 1 pp 43ndash50 2012
[17] W Sianglum P Srimanote W Wonglumsom K Kittiniyomand S P Voravuthikunchai ldquoProteome analyses of cellularproteins in methicillin-resistant Staphylococcus aureus treatedwith rhodomyrtone a novel antibiotic candidaterdquo PLoS ONEvol 6 no 2 Article ID e16628 pp 1ndash10 2011
[18] Clinical and Laboratory Standards Institute ldquoPerformancestandards for antimicrobial susceptibility testingrdquo FifteenthInformational Supplement M100-S15 2005
[19] D F Basri L S Aik R Khairon and M A Rahman ldquo2-Dgel electrophoresis map of methicillinresistant Staphylococcusaureus treated with Quercus infectoria gall extractrdquo AmericanJournal of Biochemistry and Biotechnology vol 9 no 1 pp 19ndash26 2013
[20] M Chevallet S Luche and T Rabilloud ldquoSilver staining ofproteins in polyacrylamide gelsrdquo Nature Protocols vol 1 no 4pp 1852ndash1858 2006
[21] J Davies and D Davies ldquoOrigins and evolution of antibioticresistancerdquoMicrobiology andMolecular Biology Reviews vol 74no 3 pp 417ndash433 2010
[22] D M Livermore ldquoAntibiotic resistance in staphylococcirdquo Inter-national Journal of Antimicrobial Agents vol 16 supplement 1pp S3ndashS10 2000
[23] L Yu H Xiang J Fan et al ldquoGlobal transcriptional response ofStaphylococcus aureus toRhein a natural plant productrdquo Journalof Biotechnology vol 135 no 3 pp 304ndash308 2008
[24] M Doyle E-A Feuerbaum K R Fox J Hinds D EThurstonand P W Taylor ldquoResponse of Staphylococcus aureus to subin-hibitory concentrations of a sequence-selective DNA minorgroove cross-linking pyrrolobenzodiazepine dimerrdquo Journal ofAntimicrobial Chemotherapy vol 64 no 5 pp 949ndash959 2009
[25] D F Basri and S H Fan ldquoThe potential of aqueous and acetoneextracts of galls of Quercus infectoria as antibacterial agentsrdquoIndian Journal of Pharmacology vol 37 no 1 pp 26ndash29 2005
[26] J N Eloff ldquoA sensitive and quick microplate method to deter-mine the minimal inhibitory concentration of plant extracts forbacteriardquo Planta Medica vol 64 no 8 pp 711ndash713 1998
[27] L Li P Thipyapong D C Breeden and J C SteffensldquoOverexpression of a bacterial branched-chain 120572-keto aciddehydrogenase complex in Arabidopsis results in accumulationof branched-chain acyl-CoAs and alteration of free amino acidcomposition in seedsrdquo Plant Science vol 165 no 6 pp 1213ndash1219 2003
[28] V K Singh D S Hattangady E S Giotis et al ldquoInsertionalinactivation of branched-chain 120572-keto acid dehydrogenase inStaphylococcus aureus leads to decreased branched-chainmem-brane fatty acid content and increased susceptibility to certainstressesrdquo Applied and Environmental Microbiology vol 74 no19 pp 5882ndash5890 2008
[29] P N Lowe J A Hodgson and R N Perham ldquoDual role of asingle multienzyme complex in the oxidative decarboxylationof pyruvate and branched-chain 2-oxo acids in Bacillus subtilisrdquoBiochemical Journal vol 215 no 1 pp 133ndash140 1983
[30] V Kapatral X Bina and A M Chakrabarty ldquoSuccinyl coen-zyme A synthetase of Pseudomonas aeruginosa with a broadspecificity for nucleoside triphosphate (NTP) synthesis modu-lates specificity for NTP synthesis by the 12-kilodalton form ofnucleoside diphosphate kinaserdquo Journal of Bacteriology vol 182no 5 pp 1333ndash1339 2000
[31] S-J Park G Chao and R P Gunsalus ldquoAerobic regulationof the sucABCD genes of Escherichia coli which encode 120572-ketoglutarate dehydrogenase and succinyl coenzyme A syn-thetase roles of ArcA Fnr and the upstream sdhCDAB pro-moterrdquo Journal of Bacteriology vol 179 no 13 pp 4138ndash41421997
[32] S C Andrews ldquoIron storage in bacteriardquo Advances in MicrobialPhysiology vol 40 pp 281ndash351 1998
[33] K-T Chung Z Lu andMW Chou ldquoMechanism of inhibitionof tannic acid and related compounds on the growth ofintestinal bacteriardquo Food and Chemical Toxicology vol 36 no12 pp 1053ndash1060 1998
[34] G L Mandell ldquoCatalase superoxide dismutase and virulenceof Staphylococcus aureus In vitro and in vivo studies withemphasis on staphylococcal-leukocyte interactionrdquoThe Journalof Clinical Investigation vol 55 no 3 pp 561ndash566 1975
[35] M P Chapot-Chartier and S Kulakauskas ldquoCell structure andfunction in lactic acid in bacteriardquoMicrobial Cell Factories vol13 pp S1ndashS9 2014
International Journal of Proteomics 9
[36] V A Fischetti ldquoBacteriophage lytic enzymes novel anti-infectivesrdquo Trends in Microbiology vol 13 no 10 pp 491ndash4962005
[37] C Heidrich M F Templin A Ursinus et al ldquoInvolvementof N-acetylmuramyl-l-alanine amidases in cell separationand antibiotic-induced autolysis of Escherichia colirdquo MolecularMicrobiology vol 41 no 1 pp 167ndash178 2001
[38] S Fuchs J Pane-Farre C KohlerMHecker and S EngelmannldquoAnaerobic gene expression in Staphylococcus aureusrdquo Journal ofBacteriology vol 189 no 11 pp 4275ndash4289 2007
[39] L J Rasmussen P L Moller and T Atlung ldquoCarbonmetabolism regulates expression of the pfl (pyruvate formate-lyase) gene in Escherichia colirdquo Journal of Bacteriology vol 173no 20 pp 6390ndash6397 1991
[40] M Leibig M Liebeke D Mader M Lalk A Peschel andF Gotz ldquoPyruvate formate lyase acts as a formate supplierfor metabolic processes during anaerobiosis in Staphylococcusaureusrdquo Journal of Bacteriology vol 193 no 4 pp 952ndash962 2011
[41] W Buckel and B T Golding ldquoRadical enzymes in anaerobesrdquoAnnual Review of Microbiology vol 60 pp 27ndash49 2006
[42] A Resch S Leicht M Saric et al ldquoComparative proteomeanalysis of Staphylococcus aureus biofilm and planktonic cellsand correlation with transcriptome profilingrdquo Proteomics vol6 no 6 pp 1867ndash1877 2006
[43] T Kupke ldquoMolecular characterization of the 41015840-phosphopan-tothenoylcysteine synthetase domain of bacterial Dfp flavopro-teinsrdquo The Journal of Biological Chemistry vol 277 no 39 pp36137ndash36145 2002
[44] T Nakamura T Pluskal Y Nakaseko and M YanagidaldquoImpaired coenzyme A synthesis in fission yeast causes defec-tive mitosis quiescence-exit failure histone hypoacetylationand fragile DNArdquo Open Biology vol 2 no 9 Article ID 1201172012
[45] D D Simeonova I Susnea A Moise B Schnik and MPrzybylski ldquolsquoUnknown genomersquo proteomics a new NAD(P)-dependent epimerasedehydratase revealed by N-terminal se-quencing inverted PCR and high resolution mass spectrome-tryrdquo Molecular amp Cellular Proteomics vol 8 no 1 pp 122ndash1312009
[46] D P Clark and J E Cronan ldquoTwo carbon compounds andfatty acids as carbon sourcesrdquo inEscherichia coli and SalmonellaCellular and Molecular Biology ASM Press Washington DCUSA 1996
[47] Z L Boynton G N Bennett and F B Rudolph ldquoCloningsequencing and expression of clustered genes encoding betahydroxybutyryl-coenzyme A (CoA) dehydrogenase crotonaseand butyryl-CoA dehydrogenase from Clostridium aceto-butylicum ATCC 824rdquo Journal of Bacteriology vol 178 no 11pp 3015ndash3024 1996
[48] J Schiebel A Chang H Lu M V Baxter P J Tonge andC Kisker ldquoStaphylococcus aureus FabI inhibition substraterecognition and potential implications for in vivo essentialityrdquoStructure vol 20 no 5 pp 802ndash813 2012
[49] M Iminjan N Amat X-H Li H Upur D Ahmat and B HeldquoInvestigation into the toxicity of traditional uyghur medicinequercus infectoria galls water extractrdquo PLoS ONE vol 9 no 3Article ID e90756 2014
[50] A Hasmah Z Nurazila C Y Chow R Rina and M Rafiquz-zaman ldquoCytotoxic effects ofQuercus infectoria extracts towardscervical (Hela) and Ovarian (Caov-3) cancer cell linesrdquo Healthand the Environment Journal vol 1 no 2 pp 17ndash23 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
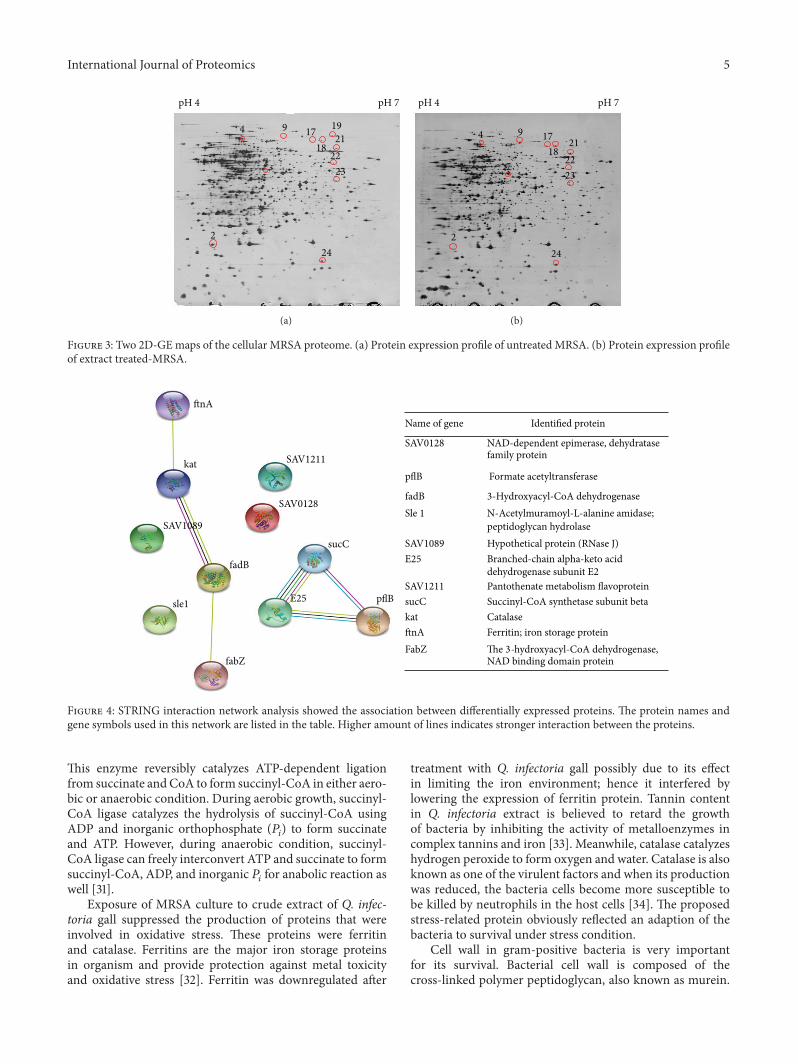
International Journal of Proteomics 5
pH 4 pH 7
2
4
7
9 1718
1921
2223
24
(a)
pH 7pH 4
2
4
7
9 1718
212223
24
(b)
Figure 3 Two 2D-GEmaps of the cellular MRSA proteome (a) Protein expression profile of untreated MRSA (b) Protein expression profileof extract treated-MRSA
ftnA
kat
SAV1089
fadB
sle1
fabZ
E25
SAV1211
SAV0128
sucC
pflB
Name of gene Identified protein
SAV0128 NAD-dependent epimerase dehydratasefamily protein
pflB Formate acetyltransferase
fadB 3-Hydroxyacyl-CoA dehydrogenaseSle 1 N-Acetylmuramoyl-L-alanine amidase
peptidoglycan hydrolaseSAV1089 Hypothetical protein (RNase J)E25 Branched-chain alpha-keto acid
dehydrogenase subunit E2SAV1211 Pantothenate metabolism flavoproteinsucC Succinyl-CoA synthetase subunit betakat CatalaseftnA Ferritin iron storage proteinFabZ The 3-hydroxyacyl-CoA dehydrogenase
NAD binding domain protein
Figure 4 STRING interaction network analysis showed the association between differentially expressed proteins The protein names andgene symbols used in this network are listed in the table Higher amount of lines indicates stronger interaction between the proteins
This enzyme reversibly catalyzes ATP-dependent ligationfrom succinate andCoA to form succinyl-CoA in either aero-bic or anaerobic condition During aerobic growth succinyl-CoA ligase catalyzes the hydrolysis of succinyl-CoA usingADP and inorganic orthophosphate (119875
119894) to form succinate
and ATP However during anaerobic condition succinyl-CoA ligase can freely interconvert ATP and succinate to formsuccinyl-CoA ADP and inorganic 119875
119894for anabolic reaction as
well [31]Exposure of MRSA culture to crude extract of Q infec-
toria gall suppressed the production of proteins that wereinvolved in oxidative stress These proteins were ferritinand catalase Ferritins are the major iron storage proteinsin organism and provide protection against metal toxicityand oxidative stress [32] Ferritin was downregulated after
treatment with Q infectoria gall possibly due to its effectin limiting the iron environment hence it interfered bylowering the expression of ferritin protein Tannin contentin Q infectoria extract is believed to retard the growthof bacteria by inhibiting the activity of metalloenzymes incomplex tannins and iron [33] Meanwhile catalase catalyzeshydrogen peroxide to form oxygen and water Catalase is alsoknown as one of the virulent factors and when its productionwas reduced the bacteria cells become more susceptible tobe killed by neutrophils in the host cells [34] The proposedstress-related protein obviously reflected an adaption of thebacteria to survival under stress condition
Cell wall in gram-positive bacteria is very importantfor its survival Bacterial cell wall is composed of thecross-linked polymer peptidoglycan also known as murein
6 International Journal of Proteomics
Table1Th
eprotein
andgenesidentified
intheS
TRIN
GanalysisnetworkTh
elist
ofproteins
identifi
edisdifferentially
expressedproteins
follo
wingtreatmentw
ithQinfectoria
gallextract
Spot
number
Accession
number
Proteins
identifi
edMascot
score
Nom
inal
mass(kD
a)Sequ
ence
coverage
()
Num
bero
fpeptides
matched
Foldsc
hanges
(untreated
totre
ated)
2FT
NSA
AR
Ferritin
9419590
345
minus24
4D6SHZ8
Branched-chain
alph
a-keto
acid
dehydrogenases
ubun
itE2
1317
846
4516
234
8minus27
7D6SHJ5
Succinyl-C
oAligase[ADP-form
ing]
subu
nitb
eta
6543
422837
131
5minus22
9D6SDX7
Form
ateC
-acetyltransfe
rase
14051
855525
157
1026
10D6SH75
Catalase
6678
587233
102
5minus24
17amp18
D6SBS
5N-Acetylm
uram
oyl-L
-alanine
amidase
2834
692891
33
253
19D6SDX0
3-Hydroxyacyl-C
oAdehydrogenaseNADbind
ingdo
mainprotein
5144
846729
63
426
21D6SHG5
Ribo
nucle
aseJ
7412
646507
925
24
22D6SHN0
Phosph
opantothenoylcysteined
ecarbo
xylasepho
spho
pantothenate-cysteineligase
3195
4419
6562
221
23D6SDL7
NADdepend
entepimerasedehydratasefam
ilyprotein
279
378017
762
21
24D6SEM
63-Hydroxyacyl-[acyl-carrie
r-protein]
dehydrataseF
abZ
2731
163096
123
222
International Journal of Proteomics 7
Peptidoglycan architecture consists of a disaccharide back-bone built of alternating 120573-1-4-N-acetylglucosamines andN-acetylmuramic acids [35] Bacterial cell produces severalhydrolases that specifically cleave various covalent bonds inthe peptidoglycan including N-acetylmuramoyl-L-alanineamidases N-acetylglucosaminidases N-acetylmuramidasesendopeptidases and transglycosylases N-acetylmuramoyl-L-alanine amidase plays important roles in the separationseptum of daughter cells during cell division [36] HenceN-acetylmuramoyl-L-alanine amidase binds to both alphaand beta human fibrinogen and fibronectin which play arole during colonization and pathogenicity in the host tissueduring infection Previous study [37] reported that thisenzyme was upregulated to activate and increase its ownproduction in the presence of antibiotic to the bacteria andeventually cause inhibition of the bacterial growth or celldeath
Formate C-acetyltransferase or pyruvate formate-lyasewas upregulated in MRSA treated with aqueous extract fromQ infectoria gall This is a cytoplasmic protein involved inenergy fermentationmetabolism pathway in themicroorgan-ism This enzyme was induced in anaerobic growth and lim-ited nitrate [38 39] Formate C-acetyltransferase functions tocatalyze pyruvate to acetyl-CoA and formate which is subse-quently used for biosynthesis of protein DNA and RNA [40]and also as source for citric acid cycle through acetyl-CoA[41] Formate C-acetyltransferase NAD-dependent formatedehydrogenase and formyltetrahydrofolate synthetase wereupregulated during transcription at proteome level for thesynthesis of S aureus biofilm growth [42] This enzyme wasupregulated as one of the survival strategies of bacteria understress [40]
Phosphopantothenoyl cysteine decarboxylasephospho-pantothenate-cysteine ligase is also known as acid pan-tothenic [43] This is the last enzyme involved in the biosyn-thesis of CoAwhich is a crucial substance in the biogenesis ofbacterial cell membrane and functions as important cofactorin mitosis and stability of DNA in microorganism [44]This protein was upregulated in the study probably due tocompensation of other pathways to increase in the synthesisof CoA for adaptation when exposed to subinhibitory con-centration of the Q infectoria gall extract
NAD-dependent epimerasedehydratase protein is anew protein that has high similarity with UDP-glucose-4-epimerase involved in Leloir pathway [45] The other proteinthat was shown to be upregulated after treatment was RNaseJ which is involved in maturation of RNA and regulationof posttranscription until degradation of RNA in familyFirmicute of bacteria
Other proteins that were upregulated during fatty acidmetabolism are 3-hydroxyacyl-CoA dehydrogenase NAD-binding domain and 3-hydroxyacyl-[acyl-carrier-protein]dehydratase FabZ The 3-hydroxyacyl-CoA dehydrogenaseNAD binding domain protein was involved in variousmetabolic pathways such as beta oxidation fatty acid bycatalyzing the oxidation of 3-hydroxyl coenzymeA to oxoacylCoA and production of butanol in bacteria gram-positiveorganism [46] Upregulation of this protein may be due tocompensation of low level of the enzyme to tricarboxylic acid
cycle [47] In addition another protein that was involved inthe synthesis of fatty acid was 3-hydroxyacyl-[acyl-carrier-protein] dehydratase FabZ The latter was reported to par-ticipate in the phospholipid membrane biosynthesis pathway[48]
5 Conclusions
In conclusion all the data reported in this studymay elucidateand give better understanding on antibacterial effect ofaqueous extract of Q infectoria towards MRSA throughinterference of various metabolism and functions of the bac-terial proteome Furthermore recent studies suggested thataqueous extract ofQ infectoria galls has no toxicity effect andmortality effect in animal [49 50] However due to its hightannin content which has a profound digestibility-reducingeffect in human it is ideal to be administered topically since ithas the advantage of possessing the anti-inflammatory effectandwoundhealing activityThis information haswidened thechances for alternative medicine as therapeutic agent to treatMRSA infection
Competing Interests
The authors declare that they have no competing interestswith regard to publication of this paper
Authorsrsquo Contributions
Dayang Fredalina Basri designed analyzed and supervisedthe study Noraziah Mohamad Zin provided lab facilitiesRadhiah Khairon performed all the experiments data anal-ysis and drafted the manuscript All authors read andapproved the final manuscript
Acknowledgments
This study was funded by the Ministry of Higher EducationGovernment of Malaysia under the research code GrantFRGS22014SG05UKM023
References
[1] N F Crum R U Lee S A Thornton et al ldquoFifteen-yearstudy of the changing epidemiology of methicillin-resistantStaphylococcus aureusrdquo The American Journal of Medicine vol119 no 11 pp 943ndash951 2006
[2] A Asensio R Canton J Vaque et al ldquoNosocomial andcommunity-acquiredmeticillin-resistant Staphylococcus aureusinfections in hospitalized patients (Spain 1993ndash2003)rdquo Journalof Hospital Infection vol 63 no 4 pp 465ndash471 2006
[3] K Hiramatsu H Hanaki T Ino K Yabuta T Oguri and FC Tenover ldquoMethicillin-resistant Staphylococcus aureus clini-cal strain with reduced vancomycin susceptibilityrdquo Journal ofAntimicrobial Chemotherapy vol 40 no 1 pp 135ndash136 1997
[4] WHO WHO Traditional Medicine Strategy 2002ndash2005World Health Organization Geneva Switzerland 2002WHO EDM TRM 20021 engpdf
8 International Journal of Proteomics
[5] D G Brown T Lister and T L May-Dracka ldquoNew naturalproducts as new leads for antibacterial drug discoveryrdquo Bioor-ganic amp Medicinal Chemistry Letters vol 24 no 2 pp 413ndash4182014
[6] G M Cragg P G Grothaus and D J Newman ldquoImpactof natural products on developing new anti-cancer agentsrdquoChemical Reviews vol 109 no 7 pp 3012ndash3043 2009
[7] D A Dias S Urban and U Roessner ldquoA historical overview ofnatural products in drug discoveryrdquo Metabolites vol 2 no 4pp 303ndash336 2012
[8] F Sharififar G Dehghan-Nudeh Z Raeiat B AmirheidariM Moshrefi and A Purhemati ldquoTyrosinase inhibitory activityof major fractions of Quercus infectoria gallsrdquo PharmacognosyCommunications vol 3 no 1 pp 21ndash26 2013
[9] P Pithayanukul P Ruenraroengsak R Bavovada N PakmaneeR Suttisri and S Saen-Oon ldquoInhibition of Naja kaouthiavenom activities by plant polyphenolsrdquo Journal of Ethnophar-macology vol 97 no 3 pp 527ndash533 2005
[10] S Voravuthikunchai A Lortheeranuwat W Jeeju T SririrakS Phongpaichit and T Supawita ldquoEffective medicinal plantsagainst enterohaemorrhagic Escherichia coli O157H7rdquo Journalof Ethnopharmacology vol 94 no 1 pp 49ndash54 2004
[11] S J Cordwell M R Larsen R T Cole and B J Walsh ldquoCom-parative proteomics of Staphylococcus aureus and the responseof methicillin-resistant and methicillin-sensitive strains to Tri-ton X-100rdquoMicrobiology vol 148 no 9 pp 2765ndash2781 2002
[12] C Kohler S Wolff D Albrecht et al ldquoProteome analysesof Staphylococcus aureus in growing and non-growing cellsa physiological approachrdquo International Journal of MedicalMicrobiology vol 295 no 8 pp 547ndash565 2005
[13] L Van Oudenhove and B Devreese ldquoA review on recent devel-opments in mass spectrometry instrumentation and quantita-tive tools advancing bacterial proteomicsrdquoAppliedMicrobiologyand Biotechnology vol 97 no 11 pp 4749ndash4762 2013
[14] R Dosselli R Millioni L Puricelli et al ldquoMolecular targets ofantimicrobial photodynamic therapy identified by a proteomicapproachrdquo Journal of Proteomics vol 77 pp 329ndash343 2012
[15] R Jenkins N Burton and R Cooper ldquoManuka honey inhibitscell division in methicillin-resistant Staphylococcus aureusrdquoJournal of Antimicrobial Chemotherapy vol 66 no 11 pp 2536ndash2542 2011
[16] J M Packer J Irish B R Herbert et al ldquoSpecific non-peroxideantibacterial effect of manuka honey on the Staphylococcusaureus proteomerdquo International Journal of Antimicrobial Agentsvol 40 no 1 pp 43ndash50 2012
[17] W Sianglum P Srimanote W Wonglumsom K Kittiniyomand S P Voravuthikunchai ldquoProteome analyses of cellularproteins in methicillin-resistant Staphylococcus aureus treatedwith rhodomyrtone a novel antibiotic candidaterdquo PLoS ONEvol 6 no 2 Article ID e16628 pp 1ndash10 2011
[18] Clinical and Laboratory Standards Institute ldquoPerformancestandards for antimicrobial susceptibility testingrdquo FifteenthInformational Supplement M100-S15 2005
[19] D F Basri L S Aik R Khairon and M A Rahman ldquo2-Dgel electrophoresis map of methicillinresistant Staphylococcusaureus treated with Quercus infectoria gall extractrdquo AmericanJournal of Biochemistry and Biotechnology vol 9 no 1 pp 19ndash26 2013
[20] M Chevallet S Luche and T Rabilloud ldquoSilver staining ofproteins in polyacrylamide gelsrdquo Nature Protocols vol 1 no 4pp 1852ndash1858 2006
[21] J Davies and D Davies ldquoOrigins and evolution of antibioticresistancerdquoMicrobiology andMolecular Biology Reviews vol 74no 3 pp 417ndash433 2010
[22] D M Livermore ldquoAntibiotic resistance in staphylococcirdquo Inter-national Journal of Antimicrobial Agents vol 16 supplement 1pp S3ndashS10 2000
[23] L Yu H Xiang J Fan et al ldquoGlobal transcriptional response ofStaphylococcus aureus toRhein a natural plant productrdquo Journalof Biotechnology vol 135 no 3 pp 304ndash308 2008
[24] M Doyle E-A Feuerbaum K R Fox J Hinds D EThurstonand P W Taylor ldquoResponse of Staphylococcus aureus to subin-hibitory concentrations of a sequence-selective DNA minorgroove cross-linking pyrrolobenzodiazepine dimerrdquo Journal ofAntimicrobial Chemotherapy vol 64 no 5 pp 949ndash959 2009
[25] D F Basri and S H Fan ldquoThe potential of aqueous and acetoneextracts of galls of Quercus infectoria as antibacterial agentsrdquoIndian Journal of Pharmacology vol 37 no 1 pp 26ndash29 2005
[26] J N Eloff ldquoA sensitive and quick microplate method to deter-mine the minimal inhibitory concentration of plant extracts forbacteriardquo Planta Medica vol 64 no 8 pp 711ndash713 1998
[27] L Li P Thipyapong D C Breeden and J C SteffensldquoOverexpression of a bacterial branched-chain 120572-keto aciddehydrogenase complex in Arabidopsis results in accumulationof branched-chain acyl-CoAs and alteration of free amino acidcomposition in seedsrdquo Plant Science vol 165 no 6 pp 1213ndash1219 2003
[28] V K Singh D S Hattangady E S Giotis et al ldquoInsertionalinactivation of branched-chain 120572-keto acid dehydrogenase inStaphylococcus aureus leads to decreased branched-chainmem-brane fatty acid content and increased susceptibility to certainstressesrdquo Applied and Environmental Microbiology vol 74 no19 pp 5882ndash5890 2008
[29] P N Lowe J A Hodgson and R N Perham ldquoDual role of asingle multienzyme complex in the oxidative decarboxylationof pyruvate and branched-chain 2-oxo acids in Bacillus subtilisrdquoBiochemical Journal vol 215 no 1 pp 133ndash140 1983
[30] V Kapatral X Bina and A M Chakrabarty ldquoSuccinyl coen-zyme A synthetase of Pseudomonas aeruginosa with a broadspecificity for nucleoside triphosphate (NTP) synthesis modu-lates specificity for NTP synthesis by the 12-kilodalton form ofnucleoside diphosphate kinaserdquo Journal of Bacteriology vol 182no 5 pp 1333ndash1339 2000
[31] S-J Park G Chao and R P Gunsalus ldquoAerobic regulationof the sucABCD genes of Escherichia coli which encode 120572-ketoglutarate dehydrogenase and succinyl coenzyme A syn-thetase roles of ArcA Fnr and the upstream sdhCDAB pro-moterrdquo Journal of Bacteriology vol 179 no 13 pp 4138ndash41421997
[32] S C Andrews ldquoIron storage in bacteriardquo Advances in MicrobialPhysiology vol 40 pp 281ndash351 1998
[33] K-T Chung Z Lu andMW Chou ldquoMechanism of inhibitionof tannic acid and related compounds on the growth ofintestinal bacteriardquo Food and Chemical Toxicology vol 36 no12 pp 1053ndash1060 1998
[34] G L Mandell ldquoCatalase superoxide dismutase and virulenceof Staphylococcus aureus In vitro and in vivo studies withemphasis on staphylococcal-leukocyte interactionrdquoThe Journalof Clinical Investigation vol 55 no 3 pp 561ndash566 1975
[35] M P Chapot-Chartier and S Kulakauskas ldquoCell structure andfunction in lactic acid in bacteriardquoMicrobial Cell Factories vol13 pp S1ndashS9 2014
International Journal of Proteomics 9
[36] V A Fischetti ldquoBacteriophage lytic enzymes novel anti-infectivesrdquo Trends in Microbiology vol 13 no 10 pp 491ndash4962005
[37] C Heidrich M F Templin A Ursinus et al ldquoInvolvementof N-acetylmuramyl-l-alanine amidases in cell separationand antibiotic-induced autolysis of Escherichia colirdquo MolecularMicrobiology vol 41 no 1 pp 167ndash178 2001
[38] S Fuchs J Pane-Farre C KohlerMHecker and S EngelmannldquoAnaerobic gene expression in Staphylococcus aureusrdquo Journal ofBacteriology vol 189 no 11 pp 4275ndash4289 2007
[39] L J Rasmussen P L Moller and T Atlung ldquoCarbonmetabolism regulates expression of the pfl (pyruvate formate-lyase) gene in Escherichia colirdquo Journal of Bacteriology vol 173no 20 pp 6390ndash6397 1991
[40] M Leibig M Liebeke D Mader M Lalk A Peschel andF Gotz ldquoPyruvate formate lyase acts as a formate supplierfor metabolic processes during anaerobiosis in Staphylococcusaureusrdquo Journal of Bacteriology vol 193 no 4 pp 952ndash962 2011
[41] W Buckel and B T Golding ldquoRadical enzymes in anaerobesrdquoAnnual Review of Microbiology vol 60 pp 27ndash49 2006
[42] A Resch S Leicht M Saric et al ldquoComparative proteomeanalysis of Staphylococcus aureus biofilm and planktonic cellsand correlation with transcriptome profilingrdquo Proteomics vol6 no 6 pp 1867ndash1877 2006
[43] T Kupke ldquoMolecular characterization of the 41015840-phosphopan-tothenoylcysteine synthetase domain of bacterial Dfp flavopro-teinsrdquo The Journal of Biological Chemistry vol 277 no 39 pp36137ndash36145 2002
[44] T Nakamura T Pluskal Y Nakaseko and M YanagidaldquoImpaired coenzyme A synthesis in fission yeast causes defec-tive mitosis quiescence-exit failure histone hypoacetylationand fragile DNArdquo Open Biology vol 2 no 9 Article ID 1201172012
[45] D D Simeonova I Susnea A Moise B Schnik and MPrzybylski ldquolsquoUnknown genomersquo proteomics a new NAD(P)-dependent epimerasedehydratase revealed by N-terminal se-quencing inverted PCR and high resolution mass spectrome-tryrdquo Molecular amp Cellular Proteomics vol 8 no 1 pp 122ndash1312009
[46] D P Clark and J E Cronan ldquoTwo carbon compounds andfatty acids as carbon sourcesrdquo inEscherichia coli and SalmonellaCellular and Molecular Biology ASM Press Washington DCUSA 1996
[47] Z L Boynton G N Bennett and F B Rudolph ldquoCloningsequencing and expression of clustered genes encoding betahydroxybutyryl-coenzyme A (CoA) dehydrogenase crotonaseand butyryl-CoA dehydrogenase from Clostridium aceto-butylicum ATCC 824rdquo Journal of Bacteriology vol 178 no 11pp 3015ndash3024 1996
[48] J Schiebel A Chang H Lu M V Baxter P J Tonge andC Kisker ldquoStaphylococcus aureus FabI inhibition substraterecognition and potential implications for in vivo essentialityrdquoStructure vol 20 no 5 pp 802ndash813 2012
[49] M Iminjan N Amat X-H Li H Upur D Ahmat and B HeldquoInvestigation into the toxicity of traditional uyghur medicinequercus infectoria galls water extractrdquo PLoS ONE vol 9 no 3Article ID e90756 2014
[50] A Hasmah Z Nurazila C Y Chow R Rina and M Rafiquz-zaman ldquoCytotoxic effects ofQuercus infectoria extracts towardscervical (Hela) and Ovarian (Caov-3) cancer cell linesrdquo Healthand the Environment Journal vol 1 no 2 pp 17ndash23 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
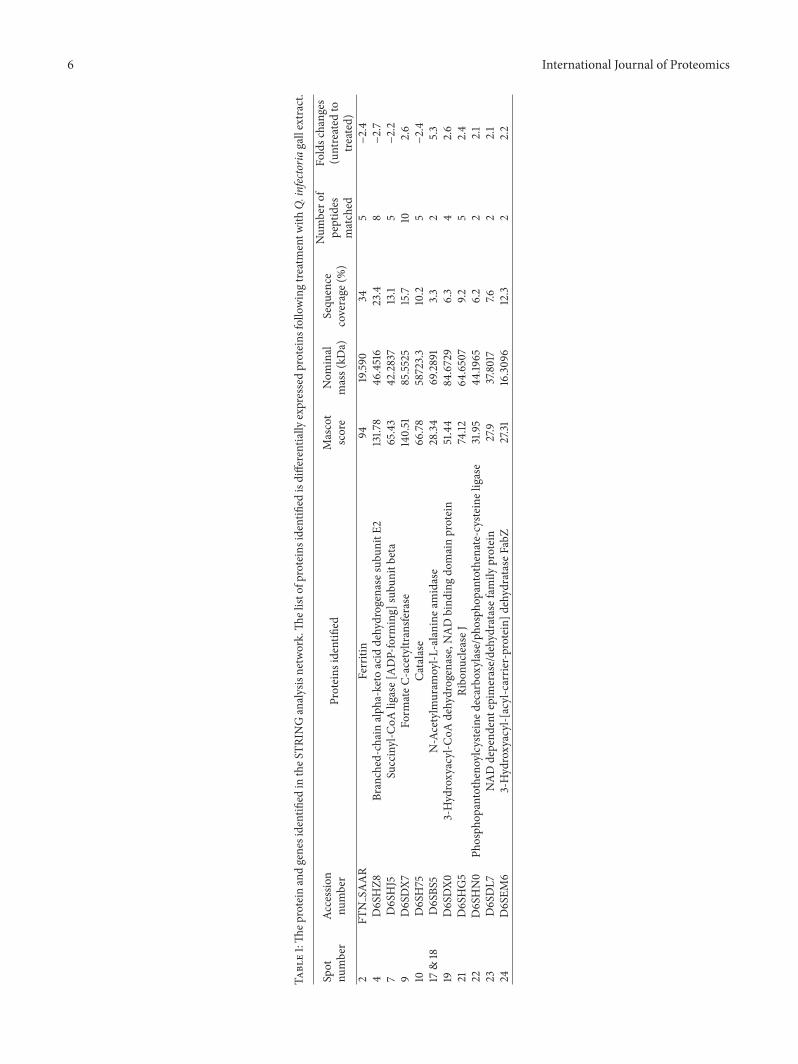
6 International Journal of Proteomics
Table1Th
eprotein
andgenesidentified
intheS
TRIN
GanalysisnetworkTh
elist
ofproteins
identifi
edisdifferentially
expressedproteins
follo
wingtreatmentw
ithQinfectoria
gallextract
Spot
number
Accession
number
Proteins
identifi
edMascot
score
Nom
inal
mass(kD
a)Sequ
ence
coverage
()
Num
bero
fpeptides
matched
Foldsc
hanges
(untreated
totre
ated)
2FT
NSA
AR
Ferritin
9419590
345
minus24
4D6SHZ8
Branched-chain
alph
a-keto
acid
dehydrogenases
ubun
itE2
1317
846
4516
234
8minus27
7D6SHJ5
Succinyl-C
oAligase[ADP-form
ing]
subu
nitb
eta
6543
422837
131
5minus22
9D6SDX7
Form
ateC
-acetyltransfe
rase
14051
855525
157
1026
10D6SH75
Catalase
6678
587233
102
5minus24
17amp18
D6SBS
5N-Acetylm
uram
oyl-L
-alanine
amidase
2834
692891
33
253
19D6SDX0
3-Hydroxyacyl-C
oAdehydrogenaseNADbind
ingdo
mainprotein
5144
846729
63
426
21D6SHG5
Ribo
nucle
aseJ
7412
646507
925
24
22D6SHN0
Phosph
opantothenoylcysteined
ecarbo
xylasepho
spho
pantothenate-cysteineligase
3195
4419
6562
221
23D6SDL7
NADdepend
entepimerasedehydratasefam
ilyprotein
279
378017
762
21
24D6SEM
63-Hydroxyacyl-[acyl-carrie
r-protein]
dehydrataseF
abZ
2731
163096
123
222
International Journal of Proteomics 7
Peptidoglycan architecture consists of a disaccharide back-bone built of alternating 120573-1-4-N-acetylglucosamines andN-acetylmuramic acids [35] Bacterial cell produces severalhydrolases that specifically cleave various covalent bonds inthe peptidoglycan including N-acetylmuramoyl-L-alanineamidases N-acetylglucosaminidases N-acetylmuramidasesendopeptidases and transglycosylases N-acetylmuramoyl-L-alanine amidase plays important roles in the separationseptum of daughter cells during cell division [36] HenceN-acetylmuramoyl-L-alanine amidase binds to both alphaand beta human fibrinogen and fibronectin which play arole during colonization and pathogenicity in the host tissueduring infection Previous study [37] reported that thisenzyme was upregulated to activate and increase its ownproduction in the presence of antibiotic to the bacteria andeventually cause inhibition of the bacterial growth or celldeath
Formate C-acetyltransferase or pyruvate formate-lyasewas upregulated in MRSA treated with aqueous extract fromQ infectoria gall This is a cytoplasmic protein involved inenergy fermentationmetabolism pathway in themicroorgan-ism This enzyme was induced in anaerobic growth and lim-ited nitrate [38 39] Formate C-acetyltransferase functions tocatalyze pyruvate to acetyl-CoA and formate which is subse-quently used for biosynthesis of protein DNA and RNA [40]and also as source for citric acid cycle through acetyl-CoA[41] Formate C-acetyltransferase NAD-dependent formatedehydrogenase and formyltetrahydrofolate synthetase wereupregulated during transcription at proteome level for thesynthesis of S aureus biofilm growth [42] This enzyme wasupregulated as one of the survival strategies of bacteria understress [40]
Phosphopantothenoyl cysteine decarboxylasephospho-pantothenate-cysteine ligase is also known as acid pan-tothenic [43] This is the last enzyme involved in the biosyn-thesis of CoAwhich is a crucial substance in the biogenesis ofbacterial cell membrane and functions as important cofactorin mitosis and stability of DNA in microorganism [44]This protein was upregulated in the study probably due tocompensation of other pathways to increase in the synthesisof CoA for adaptation when exposed to subinhibitory con-centration of the Q infectoria gall extract
NAD-dependent epimerasedehydratase protein is anew protein that has high similarity with UDP-glucose-4-epimerase involved in Leloir pathway [45] The other proteinthat was shown to be upregulated after treatment was RNaseJ which is involved in maturation of RNA and regulationof posttranscription until degradation of RNA in familyFirmicute of bacteria
Other proteins that were upregulated during fatty acidmetabolism are 3-hydroxyacyl-CoA dehydrogenase NAD-binding domain and 3-hydroxyacyl-[acyl-carrier-protein]dehydratase FabZ The 3-hydroxyacyl-CoA dehydrogenaseNAD binding domain protein was involved in variousmetabolic pathways such as beta oxidation fatty acid bycatalyzing the oxidation of 3-hydroxyl coenzymeA to oxoacylCoA and production of butanol in bacteria gram-positiveorganism [46] Upregulation of this protein may be due tocompensation of low level of the enzyme to tricarboxylic acid
cycle [47] In addition another protein that was involved inthe synthesis of fatty acid was 3-hydroxyacyl-[acyl-carrier-protein] dehydratase FabZ The latter was reported to par-ticipate in the phospholipid membrane biosynthesis pathway[48]
5 Conclusions
In conclusion all the data reported in this studymay elucidateand give better understanding on antibacterial effect ofaqueous extract of Q infectoria towards MRSA throughinterference of various metabolism and functions of the bac-terial proteome Furthermore recent studies suggested thataqueous extract ofQ infectoria galls has no toxicity effect andmortality effect in animal [49 50] However due to its hightannin content which has a profound digestibility-reducingeffect in human it is ideal to be administered topically since ithas the advantage of possessing the anti-inflammatory effectandwoundhealing activityThis information haswidened thechances for alternative medicine as therapeutic agent to treatMRSA infection
Competing Interests
The authors declare that they have no competing interestswith regard to publication of this paper
Authorsrsquo Contributions
Dayang Fredalina Basri designed analyzed and supervisedthe study Noraziah Mohamad Zin provided lab facilitiesRadhiah Khairon performed all the experiments data anal-ysis and drafted the manuscript All authors read andapproved the final manuscript
Acknowledgments
This study was funded by the Ministry of Higher EducationGovernment of Malaysia under the research code GrantFRGS22014SG05UKM023
References
[1] N F Crum R U Lee S A Thornton et al ldquoFifteen-yearstudy of the changing epidemiology of methicillin-resistantStaphylococcus aureusrdquo The American Journal of Medicine vol119 no 11 pp 943ndash951 2006
[2] A Asensio R Canton J Vaque et al ldquoNosocomial andcommunity-acquiredmeticillin-resistant Staphylococcus aureusinfections in hospitalized patients (Spain 1993ndash2003)rdquo Journalof Hospital Infection vol 63 no 4 pp 465ndash471 2006
[3] K Hiramatsu H Hanaki T Ino K Yabuta T Oguri and FC Tenover ldquoMethicillin-resistant Staphylococcus aureus clini-cal strain with reduced vancomycin susceptibilityrdquo Journal ofAntimicrobial Chemotherapy vol 40 no 1 pp 135ndash136 1997
[4] WHO WHO Traditional Medicine Strategy 2002ndash2005World Health Organization Geneva Switzerland 2002WHO EDM TRM 20021 engpdf
8 International Journal of Proteomics
[5] D G Brown T Lister and T L May-Dracka ldquoNew naturalproducts as new leads for antibacterial drug discoveryrdquo Bioor-ganic amp Medicinal Chemistry Letters vol 24 no 2 pp 413ndash4182014
[6] G M Cragg P G Grothaus and D J Newman ldquoImpactof natural products on developing new anti-cancer agentsrdquoChemical Reviews vol 109 no 7 pp 3012ndash3043 2009
[7] D A Dias S Urban and U Roessner ldquoA historical overview ofnatural products in drug discoveryrdquo Metabolites vol 2 no 4pp 303ndash336 2012
[8] F Sharififar G Dehghan-Nudeh Z Raeiat B AmirheidariM Moshrefi and A Purhemati ldquoTyrosinase inhibitory activityof major fractions of Quercus infectoria gallsrdquo PharmacognosyCommunications vol 3 no 1 pp 21ndash26 2013
[9] P Pithayanukul P Ruenraroengsak R Bavovada N PakmaneeR Suttisri and S Saen-Oon ldquoInhibition of Naja kaouthiavenom activities by plant polyphenolsrdquo Journal of Ethnophar-macology vol 97 no 3 pp 527ndash533 2005
[10] S Voravuthikunchai A Lortheeranuwat W Jeeju T SririrakS Phongpaichit and T Supawita ldquoEffective medicinal plantsagainst enterohaemorrhagic Escherichia coli O157H7rdquo Journalof Ethnopharmacology vol 94 no 1 pp 49ndash54 2004
[11] S J Cordwell M R Larsen R T Cole and B J Walsh ldquoCom-parative proteomics of Staphylococcus aureus and the responseof methicillin-resistant and methicillin-sensitive strains to Tri-ton X-100rdquoMicrobiology vol 148 no 9 pp 2765ndash2781 2002
[12] C Kohler S Wolff D Albrecht et al ldquoProteome analysesof Staphylococcus aureus in growing and non-growing cellsa physiological approachrdquo International Journal of MedicalMicrobiology vol 295 no 8 pp 547ndash565 2005
[13] L Van Oudenhove and B Devreese ldquoA review on recent devel-opments in mass spectrometry instrumentation and quantita-tive tools advancing bacterial proteomicsrdquoAppliedMicrobiologyand Biotechnology vol 97 no 11 pp 4749ndash4762 2013
[14] R Dosselli R Millioni L Puricelli et al ldquoMolecular targets ofantimicrobial photodynamic therapy identified by a proteomicapproachrdquo Journal of Proteomics vol 77 pp 329ndash343 2012
[15] R Jenkins N Burton and R Cooper ldquoManuka honey inhibitscell division in methicillin-resistant Staphylococcus aureusrdquoJournal of Antimicrobial Chemotherapy vol 66 no 11 pp 2536ndash2542 2011
[16] J M Packer J Irish B R Herbert et al ldquoSpecific non-peroxideantibacterial effect of manuka honey on the Staphylococcusaureus proteomerdquo International Journal of Antimicrobial Agentsvol 40 no 1 pp 43ndash50 2012
[17] W Sianglum P Srimanote W Wonglumsom K Kittiniyomand S P Voravuthikunchai ldquoProteome analyses of cellularproteins in methicillin-resistant Staphylococcus aureus treatedwith rhodomyrtone a novel antibiotic candidaterdquo PLoS ONEvol 6 no 2 Article ID e16628 pp 1ndash10 2011
[18] Clinical and Laboratory Standards Institute ldquoPerformancestandards for antimicrobial susceptibility testingrdquo FifteenthInformational Supplement M100-S15 2005
[19] D F Basri L S Aik R Khairon and M A Rahman ldquo2-Dgel electrophoresis map of methicillinresistant Staphylococcusaureus treated with Quercus infectoria gall extractrdquo AmericanJournal of Biochemistry and Biotechnology vol 9 no 1 pp 19ndash26 2013
[20] M Chevallet S Luche and T Rabilloud ldquoSilver staining ofproteins in polyacrylamide gelsrdquo Nature Protocols vol 1 no 4pp 1852ndash1858 2006
[21] J Davies and D Davies ldquoOrigins and evolution of antibioticresistancerdquoMicrobiology andMolecular Biology Reviews vol 74no 3 pp 417ndash433 2010
[22] D M Livermore ldquoAntibiotic resistance in staphylococcirdquo Inter-national Journal of Antimicrobial Agents vol 16 supplement 1pp S3ndashS10 2000
[23] L Yu H Xiang J Fan et al ldquoGlobal transcriptional response ofStaphylococcus aureus toRhein a natural plant productrdquo Journalof Biotechnology vol 135 no 3 pp 304ndash308 2008
[24] M Doyle E-A Feuerbaum K R Fox J Hinds D EThurstonand P W Taylor ldquoResponse of Staphylococcus aureus to subin-hibitory concentrations of a sequence-selective DNA minorgroove cross-linking pyrrolobenzodiazepine dimerrdquo Journal ofAntimicrobial Chemotherapy vol 64 no 5 pp 949ndash959 2009
[25] D F Basri and S H Fan ldquoThe potential of aqueous and acetoneextracts of galls of Quercus infectoria as antibacterial agentsrdquoIndian Journal of Pharmacology vol 37 no 1 pp 26ndash29 2005
[26] J N Eloff ldquoA sensitive and quick microplate method to deter-mine the minimal inhibitory concentration of plant extracts forbacteriardquo Planta Medica vol 64 no 8 pp 711ndash713 1998
[27] L Li P Thipyapong D C Breeden and J C SteffensldquoOverexpression of a bacterial branched-chain 120572-keto aciddehydrogenase complex in Arabidopsis results in accumulationof branched-chain acyl-CoAs and alteration of free amino acidcomposition in seedsrdquo Plant Science vol 165 no 6 pp 1213ndash1219 2003
[28] V K Singh D S Hattangady E S Giotis et al ldquoInsertionalinactivation of branched-chain 120572-keto acid dehydrogenase inStaphylococcus aureus leads to decreased branched-chainmem-brane fatty acid content and increased susceptibility to certainstressesrdquo Applied and Environmental Microbiology vol 74 no19 pp 5882ndash5890 2008
[29] P N Lowe J A Hodgson and R N Perham ldquoDual role of asingle multienzyme complex in the oxidative decarboxylationof pyruvate and branched-chain 2-oxo acids in Bacillus subtilisrdquoBiochemical Journal vol 215 no 1 pp 133ndash140 1983
[30] V Kapatral X Bina and A M Chakrabarty ldquoSuccinyl coen-zyme A synthetase of Pseudomonas aeruginosa with a broadspecificity for nucleoside triphosphate (NTP) synthesis modu-lates specificity for NTP synthesis by the 12-kilodalton form ofnucleoside diphosphate kinaserdquo Journal of Bacteriology vol 182no 5 pp 1333ndash1339 2000
[31] S-J Park G Chao and R P Gunsalus ldquoAerobic regulationof the sucABCD genes of Escherichia coli which encode 120572-ketoglutarate dehydrogenase and succinyl coenzyme A syn-thetase roles of ArcA Fnr and the upstream sdhCDAB pro-moterrdquo Journal of Bacteriology vol 179 no 13 pp 4138ndash41421997
[32] S C Andrews ldquoIron storage in bacteriardquo Advances in MicrobialPhysiology vol 40 pp 281ndash351 1998
[33] K-T Chung Z Lu andMW Chou ldquoMechanism of inhibitionof tannic acid and related compounds on the growth ofintestinal bacteriardquo Food and Chemical Toxicology vol 36 no12 pp 1053ndash1060 1998
[34] G L Mandell ldquoCatalase superoxide dismutase and virulenceof Staphylococcus aureus In vitro and in vivo studies withemphasis on staphylococcal-leukocyte interactionrdquoThe Journalof Clinical Investigation vol 55 no 3 pp 561ndash566 1975
[35] M P Chapot-Chartier and S Kulakauskas ldquoCell structure andfunction in lactic acid in bacteriardquoMicrobial Cell Factories vol13 pp S1ndashS9 2014
International Journal of Proteomics 9
[36] V A Fischetti ldquoBacteriophage lytic enzymes novel anti-infectivesrdquo Trends in Microbiology vol 13 no 10 pp 491ndash4962005
[37] C Heidrich M F Templin A Ursinus et al ldquoInvolvementof N-acetylmuramyl-l-alanine amidases in cell separationand antibiotic-induced autolysis of Escherichia colirdquo MolecularMicrobiology vol 41 no 1 pp 167ndash178 2001
[38] S Fuchs J Pane-Farre C KohlerMHecker and S EngelmannldquoAnaerobic gene expression in Staphylococcus aureusrdquo Journal ofBacteriology vol 189 no 11 pp 4275ndash4289 2007
[39] L J Rasmussen P L Moller and T Atlung ldquoCarbonmetabolism regulates expression of the pfl (pyruvate formate-lyase) gene in Escherichia colirdquo Journal of Bacteriology vol 173no 20 pp 6390ndash6397 1991
[40] M Leibig M Liebeke D Mader M Lalk A Peschel andF Gotz ldquoPyruvate formate lyase acts as a formate supplierfor metabolic processes during anaerobiosis in Staphylococcusaureusrdquo Journal of Bacteriology vol 193 no 4 pp 952ndash962 2011
[41] W Buckel and B T Golding ldquoRadical enzymes in anaerobesrdquoAnnual Review of Microbiology vol 60 pp 27ndash49 2006
[42] A Resch S Leicht M Saric et al ldquoComparative proteomeanalysis of Staphylococcus aureus biofilm and planktonic cellsand correlation with transcriptome profilingrdquo Proteomics vol6 no 6 pp 1867ndash1877 2006
[43] T Kupke ldquoMolecular characterization of the 41015840-phosphopan-tothenoylcysteine synthetase domain of bacterial Dfp flavopro-teinsrdquo The Journal of Biological Chemistry vol 277 no 39 pp36137ndash36145 2002
[44] T Nakamura T Pluskal Y Nakaseko and M YanagidaldquoImpaired coenzyme A synthesis in fission yeast causes defec-tive mitosis quiescence-exit failure histone hypoacetylationand fragile DNArdquo Open Biology vol 2 no 9 Article ID 1201172012
[45] D D Simeonova I Susnea A Moise B Schnik and MPrzybylski ldquolsquoUnknown genomersquo proteomics a new NAD(P)-dependent epimerasedehydratase revealed by N-terminal se-quencing inverted PCR and high resolution mass spectrome-tryrdquo Molecular amp Cellular Proteomics vol 8 no 1 pp 122ndash1312009
[46] D P Clark and J E Cronan ldquoTwo carbon compounds andfatty acids as carbon sourcesrdquo inEscherichia coli and SalmonellaCellular and Molecular Biology ASM Press Washington DCUSA 1996
[47] Z L Boynton G N Bennett and F B Rudolph ldquoCloningsequencing and expression of clustered genes encoding betahydroxybutyryl-coenzyme A (CoA) dehydrogenase crotonaseand butyryl-CoA dehydrogenase from Clostridium aceto-butylicum ATCC 824rdquo Journal of Bacteriology vol 178 no 11pp 3015ndash3024 1996
[48] J Schiebel A Chang H Lu M V Baxter P J Tonge andC Kisker ldquoStaphylococcus aureus FabI inhibition substraterecognition and potential implications for in vivo essentialityrdquoStructure vol 20 no 5 pp 802ndash813 2012
[49] M Iminjan N Amat X-H Li H Upur D Ahmat and B HeldquoInvestigation into the toxicity of traditional uyghur medicinequercus infectoria galls water extractrdquo PLoS ONE vol 9 no 3Article ID e90756 2014
[50] A Hasmah Z Nurazila C Y Chow R Rina and M Rafiquz-zaman ldquoCytotoxic effects ofQuercus infectoria extracts towardscervical (Hela) and Ovarian (Caov-3) cancer cell linesrdquo Healthand the Environment Journal vol 1 no 2 pp 17ndash23 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Proteomics 7
Peptidoglycan architecture consists of a disaccharide back-bone built of alternating 120573-1-4-N-acetylglucosamines andN-acetylmuramic acids [35] Bacterial cell produces severalhydrolases that specifically cleave various covalent bonds inthe peptidoglycan including N-acetylmuramoyl-L-alanineamidases N-acetylglucosaminidases N-acetylmuramidasesendopeptidases and transglycosylases N-acetylmuramoyl-L-alanine amidase plays important roles in the separationseptum of daughter cells during cell division [36] HenceN-acetylmuramoyl-L-alanine amidase binds to both alphaand beta human fibrinogen and fibronectin which play arole during colonization and pathogenicity in the host tissueduring infection Previous study [37] reported that thisenzyme was upregulated to activate and increase its ownproduction in the presence of antibiotic to the bacteria andeventually cause inhibition of the bacterial growth or celldeath
Formate C-acetyltransferase or pyruvate formate-lyasewas upregulated in MRSA treated with aqueous extract fromQ infectoria gall This is a cytoplasmic protein involved inenergy fermentationmetabolism pathway in themicroorgan-ism This enzyme was induced in anaerobic growth and lim-ited nitrate [38 39] Formate C-acetyltransferase functions tocatalyze pyruvate to acetyl-CoA and formate which is subse-quently used for biosynthesis of protein DNA and RNA [40]and also as source for citric acid cycle through acetyl-CoA[41] Formate C-acetyltransferase NAD-dependent formatedehydrogenase and formyltetrahydrofolate synthetase wereupregulated during transcription at proteome level for thesynthesis of S aureus biofilm growth [42] This enzyme wasupregulated as one of the survival strategies of bacteria understress [40]
Phosphopantothenoyl cysteine decarboxylasephospho-pantothenate-cysteine ligase is also known as acid pan-tothenic [43] This is the last enzyme involved in the biosyn-thesis of CoAwhich is a crucial substance in the biogenesis ofbacterial cell membrane and functions as important cofactorin mitosis and stability of DNA in microorganism [44]This protein was upregulated in the study probably due tocompensation of other pathways to increase in the synthesisof CoA for adaptation when exposed to subinhibitory con-centration of the Q infectoria gall extract
NAD-dependent epimerasedehydratase protein is anew protein that has high similarity with UDP-glucose-4-epimerase involved in Leloir pathway [45] The other proteinthat was shown to be upregulated after treatment was RNaseJ which is involved in maturation of RNA and regulationof posttranscription until degradation of RNA in familyFirmicute of bacteria
Other proteins that were upregulated during fatty acidmetabolism are 3-hydroxyacyl-CoA dehydrogenase NAD-binding domain and 3-hydroxyacyl-[acyl-carrier-protein]dehydratase FabZ The 3-hydroxyacyl-CoA dehydrogenaseNAD binding domain protein was involved in variousmetabolic pathways such as beta oxidation fatty acid bycatalyzing the oxidation of 3-hydroxyl coenzymeA to oxoacylCoA and production of butanol in bacteria gram-positiveorganism [46] Upregulation of this protein may be due tocompensation of low level of the enzyme to tricarboxylic acid
cycle [47] In addition another protein that was involved inthe synthesis of fatty acid was 3-hydroxyacyl-[acyl-carrier-protein] dehydratase FabZ The latter was reported to par-ticipate in the phospholipid membrane biosynthesis pathway[48]
5 Conclusions
In conclusion all the data reported in this studymay elucidateand give better understanding on antibacterial effect ofaqueous extract of Q infectoria towards MRSA throughinterference of various metabolism and functions of the bac-terial proteome Furthermore recent studies suggested thataqueous extract ofQ infectoria galls has no toxicity effect andmortality effect in animal [49 50] However due to its hightannin content which has a profound digestibility-reducingeffect in human it is ideal to be administered topically since ithas the advantage of possessing the anti-inflammatory effectandwoundhealing activityThis information haswidened thechances for alternative medicine as therapeutic agent to treatMRSA infection
Competing Interests
The authors declare that they have no competing interestswith regard to publication of this paper
Authorsrsquo Contributions
Dayang Fredalina Basri designed analyzed and supervisedthe study Noraziah Mohamad Zin provided lab facilitiesRadhiah Khairon performed all the experiments data anal-ysis and drafted the manuscript All authors read andapproved the final manuscript
Acknowledgments
This study was funded by the Ministry of Higher EducationGovernment of Malaysia under the research code GrantFRGS22014SG05UKM023
References
[1] N F Crum R U Lee S A Thornton et al ldquoFifteen-yearstudy of the changing epidemiology of methicillin-resistantStaphylococcus aureusrdquo The American Journal of Medicine vol119 no 11 pp 943ndash951 2006
[2] A Asensio R Canton J Vaque et al ldquoNosocomial andcommunity-acquiredmeticillin-resistant Staphylococcus aureusinfections in hospitalized patients (Spain 1993ndash2003)rdquo Journalof Hospital Infection vol 63 no 4 pp 465ndash471 2006
[3] K Hiramatsu H Hanaki T Ino K Yabuta T Oguri and FC Tenover ldquoMethicillin-resistant Staphylococcus aureus clini-cal strain with reduced vancomycin susceptibilityrdquo Journal ofAntimicrobial Chemotherapy vol 40 no 1 pp 135ndash136 1997
[4] WHO WHO Traditional Medicine Strategy 2002ndash2005World Health Organization Geneva Switzerland 2002WHO EDM TRM 20021 engpdf
8 International Journal of Proteomics
[5] D G Brown T Lister and T L May-Dracka ldquoNew naturalproducts as new leads for antibacterial drug discoveryrdquo Bioor-ganic amp Medicinal Chemistry Letters vol 24 no 2 pp 413ndash4182014
[6] G M Cragg P G Grothaus and D J Newman ldquoImpactof natural products on developing new anti-cancer agentsrdquoChemical Reviews vol 109 no 7 pp 3012ndash3043 2009
[7] D A Dias S Urban and U Roessner ldquoA historical overview ofnatural products in drug discoveryrdquo Metabolites vol 2 no 4pp 303ndash336 2012
[8] F Sharififar G Dehghan-Nudeh Z Raeiat B AmirheidariM Moshrefi and A Purhemati ldquoTyrosinase inhibitory activityof major fractions of Quercus infectoria gallsrdquo PharmacognosyCommunications vol 3 no 1 pp 21ndash26 2013
[9] P Pithayanukul P Ruenraroengsak R Bavovada N PakmaneeR Suttisri and S Saen-Oon ldquoInhibition of Naja kaouthiavenom activities by plant polyphenolsrdquo Journal of Ethnophar-macology vol 97 no 3 pp 527ndash533 2005
[10] S Voravuthikunchai A Lortheeranuwat W Jeeju T SririrakS Phongpaichit and T Supawita ldquoEffective medicinal plantsagainst enterohaemorrhagic Escherichia coli O157H7rdquo Journalof Ethnopharmacology vol 94 no 1 pp 49ndash54 2004
[11] S J Cordwell M R Larsen R T Cole and B J Walsh ldquoCom-parative proteomics of Staphylococcus aureus and the responseof methicillin-resistant and methicillin-sensitive strains to Tri-ton X-100rdquoMicrobiology vol 148 no 9 pp 2765ndash2781 2002
[12] C Kohler S Wolff D Albrecht et al ldquoProteome analysesof Staphylococcus aureus in growing and non-growing cellsa physiological approachrdquo International Journal of MedicalMicrobiology vol 295 no 8 pp 547ndash565 2005
[13] L Van Oudenhove and B Devreese ldquoA review on recent devel-opments in mass spectrometry instrumentation and quantita-tive tools advancing bacterial proteomicsrdquoAppliedMicrobiologyand Biotechnology vol 97 no 11 pp 4749ndash4762 2013
[14] R Dosselli R Millioni L Puricelli et al ldquoMolecular targets ofantimicrobial photodynamic therapy identified by a proteomicapproachrdquo Journal of Proteomics vol 77 pp 329ndash343 2012
[15] R Jenkins N Burton and R Cooper ldquoManuka honey inhibitscell division in methicillin-resistant Staphylococcus aureusrdquoJournal of Antimicrobial Chemotherapy vol 66 no 11 pp 2536ndash2542 2011
[16] J M Packer J Irish B R Herbert et al ldquoSpecific non-peroxideantibacterial effect of manuka honey on the Staphylococcusaureus proteomerdquo International Journal of Antimicrobial Agentsvol 40 no 1 pp 43ndash50 2012
[17] W Sianglum P Srimanote W Wonglumsom K Kittiniyomand S P Voravuthikunchai ldquoProteome analyses of cellularproteins in methicillin-resistant Staphylococcus aureus treatedwith rhodomyrtone a novel antibiotic candidaterdquo PLoS ONEvol 6 no 2 Article ID e16628 pp 1ndash10 2011
[18] Clinical and Laboratory Standards Institute ldquoPerformancestandards for antimicrobial susceptibility testingrdquo FifteenthInformational Supplement M100-S15 2005
[19] D F Basri L S Aik R Khairon and M A Rahman ldquo2-Dgel electrophoresis map of methicillinresistant Staphylococcusaureus treated with Quercus infectoria gall extractrdquo AmericanJournal of Biochemistry and Biotechnology vol 9 no 1 pp 19ndash26 2013
[20] M Chevallet S Luche and T Rabilloud ldquoSilver staining ofproteins in polyacrylamide gelsrdquo Nature Protocols vol 1 no 4pp 1852ndash1858 2006
[21] J Davies and D Davies ldquoOrigins and evolution of antibioticresistancerdquoMicrobiology andMolecular Biology Reviews vol 74no 3 pp 417ndash433 2010
[22] D M Livermore ldquoAntibiotic resistance in staphylococcirdquo Inter-national Journal of Antimicrobial Agents vol 16 supplement 1pp S3ndashS10 2000
[23] L Yu H Xiang J Fan et al ldquoGlobal transcriptional response ofStaphylococcus aureus toRhein a natural plant productrdquo Journalof Biotechnology vol 135 no 3 pp 304ndash308 2008
[24] M Doyle E-A Feuerbaum K R Fox J Hinds D EThurstonand P W Taylor ldquoResponse of Staphylococcus aureus to subin-hibitory concentrations of a sequence-selective DNA minorgroove cross-linking pyrrolobenzodiazepine dimerrdquo Journal ofAntimicrobial Chemotherapy vol 64 no 5 pp 949ndash959 2009
[25] D F Basri and S H Fan ldquoThe potential of aqueous and acetoneextracts of galls of Quercus infectoria as antibacterial agentsrdquoIndian Journal of Pharmacology vol 37 no 1 pp 26ndash29 2005
[26] J N Eloff ldquoA sensitive and quick microplate method to deter-mine the minimal inhibitory concentration of plant extracts forbacteriardquo Planta Medica vol 64 no 8 pp 711ndash713 1998
[27] L Li P Thipyapong D C Breeden and J C SteffensldquoOverexpression of a bacterial branched-chain 120572-keto aciddehydrogenase complex in Arabidopsis results in accumulationof branched-chain acyl-CoAs and alteration of free amino acidcomposition in seedsrdquo Plant Science vol 165 no 6 pp 1213ndash1219 2003
[28] V K Singh D S Hattangady E S Giotis et al ldquoInsertionalinactivation of branched-chain 120572-keto acid dehydrogenase inStaphylococcus aureus leads to decreased branched-chainmem-brane fatty acid content and increased susceptibility to certainstressesrdquo Applied and Environmental Microbiology vol 74 no19 pp 5882ndash5890 2008
[29] P N Lowe J A Hodgson and R N Perham ldquoDual role of asingle multienzyme complex in the oxidative decarboxylationof pyruvate and branched-chain 2-oxo acids in Bacillus subtilisrdquoBiochemical Journal vol 215 no 1 pp 133ndash140 1983
[30] V Kapatral X Bina and A M Chakrabarty ldquoSuccinyl coen-zyme A synthetase of Pseudomonas aeruginosa with a broadspecificity for nucleoside triphosphate (NTP) synthesis modu-lates specificity for NTP synthesis by the 12-kilodalton form ofnucleoside diphosphate kinaserdquo Journal of Bacteriology vol 182no 5 pp 1333ndash1339 2000
[31] S-J Park G Chao and R P Gunsalus ldquoAerobic regulationof the sucABCD genes of Escherichia coli which encode 120572-ketoglutarate dehydrogenase and succinyl coenzyme A syn-thetase roles of ArcA Fnr and the upstream sdhCDAB pro-moterrdquo Journal of Bacteriology vol 179 no 13 pp 4138ndash41421997
[32] S C Andrews ldquoIron storage in bacteriardquo Advances in MicrobialPhysiology vol 40 pp 281ndash351 1998
[33] K-T Chung Z Lu andMW Chou ldquoMechanism of inhibitionof tannic acid and related compounds on the growth ofintestinal bacteriardquo Food and Chemical Toxicology vol 36 no12 pp 1053ndash1060 1998
[34] G L Mandell ldquoCatalase superoxide dismutase and virulenceof Staphylococcus aureus In vitro and in vivo studies withemphasis on staphylococcal-leukocyte interactionrdquoThe Journalof Clinical Investigation vol 55 no 3 pp 561ndash566 1975
[35] M P Chapot-Chartier and S Kulakauskas ldquoCell structure andfunction in lactic acid in bacteriardquoMicrobial Cell Factories vol13 pp S1ndashS9 2014
International Journal of Proteomics 9
[36] V A Fischetti ldquoBacteriophage lytic enzymes novel anti-infectivesrdquo Trends in Microbiology vol 13 no 10 pp 491ndash4962005
[37] C Heidrich M F Templin A Ursinus et al ldquoInvolvementof N-acetylmuramyl-l-alanine amidases in cell separationand antibiotic-induced autolysis of Escherichia colirdquo MolecularMicrobiology vol 41 no 1 pp 167ndash178 2001
[38] S Fuchs J Pane-Farre C KohlerMHecker and S EngelmannldquoAnaerobic gene expression in Staphylococcus aureusrdquo Journal ofBacteriology vol 189 no 11 pp 4275ndash4289 2007
[39] L J Rasmussen P L Moller and T Atlung ldquoCarbonmetabolism regulates expression of the pfl (pyruvate formate-lyase) gene in Escherichia colirdquo Journal of Bacteriology vol 173no 20 pp 6390ndash6397 1991
[40] M Leibig M Liebeke D Mader M Lalk A Peschel andF Gotz ldquoPyruvate formate lyase acts as a formate supplierfor metabolic processes during anaerobiosis in Staphylococcusaureusrdquo Journal of Bacteriology vol 193 no 4 pp 952ndash962 2011
[41] W Buckel and B T Golding ldquoRadical enzymes in anaerobesrdquoAnnual Review of Microbiology vol 60 pp 27ndash49 2006
[42] A Resch S Leicht M Saric et al ldquoComparative proteomeanalysis of Staphylococcus aureus biofilm and planktonic cellsand correlation with transcriptome profilingrdquo Proteomics vol6 no 6 pp 1867ndash1877 2006
[43] T Kupke ldquoMolecular characterization of the 41015840-phosphopan-tothenoylcysteine synthetase domain of bacterial Dfp flavopro-teinsrdquo The Journal of Biological Chemistry vol 277 no 39 pp36137ndash36145 2002
[44] T Nakamura T Pluskal Y Nakaseko and M YanagidaldquoImpaired coenzyme A synthesis in fission yeast causes defec-tive mitosis quiescence-exit failure histone hypoacetylationand fragile DNArdquo Open Biology vol 2 no 9 Article ID 1201172012
[45] D D Simeonova I Susnea A Moise B Schnik and MPrzybylski ldquolsquoUnknown genomersquo proteomics a new NAD(P)-dependent epimerasedehydratase revealed by N-terminal se-quencing inverted PCR and high resolution mass spectrome-tryrdquo Molecular amp Cellular Proteomics vol 8 no 1 pp 122ndash1312009
[46] D P Clark and J E Cronan ldquoTwo carbon compounds andfatty acids as carbon sourcesrdquo inEscherichia coli and SalmonellaCellular and Molecular Biology ASM Press Washington DCUSA 1996
[47] Z L Boynton G N Bennett and F B Rudolph ldquoCloningsequencing and expression of clustered genes encoding betahydroxybutyryl-coenzyme A (CoA) dehydrogenase crotonaseand butyryl-CoA dehydrogenase from Clostridium aceto-butylicum ATCC 824rdquo Journal of Bacteriology vol 178 no 11pp 3015ndash3024 1996
[48] J Schiebel A Chang H Lu M V Baxter P J Tonge andC Kisker ldquoStaphylococcus aureus FabI inhibition substraterecognition and potential implications for in vivo essentialityrdquoStructure vol 20 no 5 pp 802ndash813 2012
[49] M Iminjan N Amat X-H Li H Upur D Ahmat and B HeldquoInvestigation into the toxicity of traditional uyghur medicinequercus infectoria galls water extractrdquo PLoS ONE vol 9 no 3Article ID e90756 2014
[50] A Hasmah Z Nurazila C Y Chow R Rina and M Rafiquz-zaman ldquoCytotoxic effects ofQuercus infectoria extracts towardscervical (Hela) and Ovarian (Caov-3) cancer cell linesrdquo Healthand the Environment Journal vol 1 no 2 pp 17ndash23 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 International Journal of Proteomics
[5] D G Brown T Lister and T L May-Dracka ldquoNew naturalproducts as new leads for antibacterial drug discoveryrdquo Bioor-ganic amp Medicinal Chemistry Letters vol 24 no 2 pp 413ndash4182014
[6] G M Cragg P G Grothaus and D J Newman ldquoImpactof natural products on developing new anti-cancer agentsrdquoChemical Reviews vol 109 no 7 pp 3012ndash3043 2009
[7] D A Dias S Urban and U Roessner ldquoA historical overview ofnatural products in drug discoveryrdquo Metabolites vol 2 no 4pp 303ndash336 2012
[8] F Sharififar G Dehghan-Nudeh Z Raeiat B AmirheidariM Moshrefi and A Purhemati ldquoTyrosinase inhibitory activityof major fractions of Quercus infectoria gallsrdquo PharmacognosyCommunications vol 3 no 1 pp 21ndash26 2013
[9] P Pithayanukul P Ruenraroengsak R Bavovada N PakmaneeR Suttisri and S Saen-Oon ldquoInhibition of Naja kaouthiavenom activities by plant polyphenolsrdquo Journal of Ethnophar-macology vol 97 no 3 pp 527ndash533 2005
[10] S Voravuthikunchai A Lortheeranuwat W Jeeju T SririrakS Phongpaichit and T Supawita ldquoEffective medicinal plantsagainst enterohaemorrhagic Escherichia coli O157H7rdquo Journalof Ethnopharmacology vol 94 no 1 pp 49ndash54 2004
[11] S J Cordwell M R Larsen R T Cole and B J Walsh ldquoCom-parative proteomics of Staphylococcus aureus and the responseof methicillin-resistant and methicillin-sensitive strains to Tri-ton X-100rdquoMicrobiology vol 148 no 9 pp 2765ndash2781 2002
[12] C Kohler S Wolff D Albrecht et al ldquoProteome analysesof Staphylococcus aureus in growing and non-growing cellsa physiological approachrdquo International Journal of MedicalMicrobiology vol 295 no 8 pp 547ndash565 2005
[13] L Van Oudenhove and B Devreese ldquoA review on recent devel-opments in mass spectrometry instrumentation and quantita-tive tools advancing bacterial proteomicsrdquoAppliedMicrobiologyand Biotechnology vol 97 no 11 pp 4749ndash4762 2013
[14] R Dosselli R Millioni L Puricelli et al ldquoMolecular targets ofantimicrobial photodynamic therapy identified by a proteomicapproachrdquo Journal of Proteomics vol 77 pp 329ndash343 2012
[15] R Jenkins N Burton and R Cooper ldquoManuka honey inhibitscell division in methicillin-resistant Staphylococcus aureusrdquoJournal of Antimicrobial Chemotherapy vol 66 no 11 pp 2536ndash2542 2011
[16] J M Packer J Irish B R Herbert et al ldquoSpecific non-peroxideantibacterial effect of manuka honey on the Staphylococcusaureus proteomerdquo International Journal of Antimicrobial Agentsvol 40 no 1 pp 43ndash50 2012
[17] W Sianglum P Srimanote W Wonglumsom K Kittiniyomand S P Voravuthikunchai ldquoProteome analyses of cellularproteins in methicillin-resistant Staphylococcus aureus treatedwith rhodomyrtone a novel antibiotic candidaterdquo PLoS ONEvol 6 no 2 Article ID e16628 pp 1ndash10 2011
[18] Clinical and Laboratory Standards Institute ldquoPerformancestandards for antimicrobial susceptibility testingrdquo FifteenthInformational Supplement M100-S15 2005
[19] D F Basri L S Aik R Khairon and M A Rahman ldquo2-Dgel electrophoresis map of methicillinresistant Staphylococcusaureus treated with Quercus infectoria gall extractrdquo AmericanJournal of Biochemistry and Biotechnology vol 9 no 1 pp 19ndash26 2013
[20] M Chevallet S Luche and T Rabilloud ldquoSilver staining ofproteins in polyacrylamide gelsrdquo Nature Protocols vol 1 no 4pp 1852ndash1858 2006
[21] J Davies and D Davies ldquoOrigins and evolution of antibioticresistancerdquoMicrobiology andMolecular Biology Reviews vol 74no 3 pp 417ndash433 2010
[22] D M Livermore ldquoAntibiotic resistance in staphylococcirdquo Inter-national Journal of Antimicrobial Agents vol 16 supplement 1pp S3ndashS10 2000
[23] L Yu H Xiang J Fan et al ldquoGlobal transcriptional response ofStaphylococcus aureus toRhein a natural plant productrdquo Journalof Biotechnology vol 135 no 3 pp 304ndash308 2008
[24] M Doyle E-A Feuerbaum K R Fox J Hinds D EThurstonand P W Taylor ldquoResponse of Staphylococcus aureus to subin-hibitory concentrations of a sequence-selective DNA minorgroove cross-linking pyrrolobenzodiazepine dimerrdquo Journal ofAntimicrobial Chemotherapy vol 64 no 5 pp 949ndash959 2009
[25] D F Basri and S H Fan ldquoThe potential of aqueous and acetoneextracts of galls of Quercus infectoria as antibacterial agentsrdquoIndian Journal of Pharmacology vol 37 no 1 pp 26ndash29 2005
[26] J N Eloff ldquoA sensitive and quick microplate method to deter-mine the minimal inhibitory concentration of plant extracts forbacteriardquo Planta Medica vol 64 no 8 pp 711ndash713 1998
[27] L Li P Thipyapong D C Breeden and J C SteffensldquoOverexpression of a bacterial branched-chain 120572-keto aciddehydrogenase complex in Arabidopsis results in accumulationof branched-chain acyl-CoAs and alteration of free amino acidcomposition in seedsrdquo Plant Science vol 165 no 6 pp 1213ndash1219 2003
[28] V K Singh D S Hattangady E S Giotis et al ldquoInsertionalinactivation of branched-chain 120572-keto acid dehydrogenase inStaphylococcus aureus leads to decreased branched-chainmem-brane fatty acid content and increased susceptibility to certainstressesrdquo Applied and Environmental Microbiology vol 74 no19 pp 5882ndash5890 2008
[29] P N Lowe J A Hodgson and R N Perham ldquoDual role of asingle multienzyme complex in the oxidative decarboxylationof pyruvate and branched-chain 2-oxo acids in Bacillus subtilisrdquoBiochemical Journal vol 215 no 1 pp 133ndash140 1983
[30] V Kapatral X Bina and A M Chakrabarty ldquoSuccinyl coen-zyme A synthetase of Pseudomonas aeruginosa with a broadspecificity for nucleoside triphosphate (NTP) synthesis modu-lates specificity for NTP synthesis by the 12-kilodalton form ofnucleoside diphosphate kinaserdquo Journal of Bacteriology vol 182no 5 pp 1333ndash1339 2000
[31] S-J Park G Chao and R P Gunsalus ldquoAerobic regulationof the sucABCD genes of Escherichia coli which encode 120572-ketoglutarate dehydrogenase and succinyl coenzyme A syn-thetase roles of ArcA Fnr and the upstream sdhCDAB pro-moterrdquo Journal of Bacteriology vol 179 no 13 pp 4138ndash41421997
[32] S C Andrews ldquoIron storage in bacteriardquo Advances in MicrobialPhysiology vol 40 pp 281ndash351 1998
[33] K-T Chung Z Lu andMW Chou ldquoMechanism of inhibitionof tannic acid and related compounds on the growth ofintestinal bacteriardquo Food and Chemical Toxicology vol 36 no12 pp 1053ndash1060 1998
[34] G L Mandell ldquoCatalase superoxide dismutase and virulenceof Staphylococcus aureus In vitro and in vivo studies withemphasis on staphylococcal-leukocyte interactionrdquoThe Journalof Clinical Investigation vol 55 no 3 pp 561ndash566 1975
[35] M P Chapot-Chartier and S Kulakauskas ldquoCell structure andfunction in lactic acid in bacteriardquoMicrobial Cell Factories vol13 pp S1ndashS9 2014
International Journal of Proteomics 9
[36] V A Fischetti ldquoBacteriophage lytic enzymes novel anti-infectivesrdquo Trends in Microbiology vol 13 no 10 pp 491ndash4962005
[37] C Heidrich M F Templin A Ursinus et al ldquoInvolvementof N-acetylmuramyl-l-alanine amidases in cell separationand antibiotic-induced autolysis of Escherichia colirdquo MolecularMicrobiology vol 41 no 1 pp 167ndash178 2001
[38] S Fuchs J Pane-Farre C KohlerMHecker and S EngelmannldquoAnaerobic gene expression in Staphylococcus aureusrdquo Journal ofBacteriology vol 189 no 11 pp 4275ndash4289 2007
[39] L J Rasmussen P L Moller and T Atlung ldquoCarbonmetabolism regulates expression of the pfl (pyruvate formate-lyase) gene in Escherichia colirdquo Journal of Bacteriology vol 173no 20 pp 6390ndash6397 1991
[40] M Leibig M Liebeke D Mader M Lalk A Peschel andF Gotz ldquoPyruvate formate lyase acts as a formate supplierfor metabolic processes during anaerobiosis in Staphylococcusaureusrdquo Journal of Bacteriology vol 193 no 4 pp 952ndash962 2011
[41] W Buckel and B T Golding ldquoRadical enzymes in anaerobesrdquoAnnual Review of Microbiology vol 60 pp 27ndash49 2006
[42] A Resch S Leicht M Saric et al ldquoComparative proteomeanalysis of Staphylococcus aureus biofilm and planktonic cellsand correlation with transcriptome profilingrdquo Proteomics vol6 no 6 pp 1867ndash1877 2006
[43] T Kupke ldquoMolecular characterization of the 41015840-phosphopan-tothenoylcysteine synthetase domain of bacterial Dfp flavopro-teinsrdquo The Journal of Biological Chemistry vol 277 no 39 pp36137ndash36145 2002
[44] T Nakamura T Pluskal Y Nakaseko and M YanagidaldquoImpaired coenzyme A synthesis in fission yeast causes defec-tive mitosis quiescence-exit failure histone hypoacetylationand fragile DNArdquo Open Biology vol 2 no 9 Article ID 1201172012
[45] D D Simeonova I Susnea A Moise B Schnik and MPrzybylski ldquolsquoUnknown genomersquo proteomics a new NAD(P)-dependent epimerasedehydratase revealed by N-terminal se-quencing inverted PCR and high resolution mass spectrome-tryrdquo Molecular amp Cellular Proteomics vol 8 no 1 pp 122ndash1312009
[46] D P Clark and J E Cronan ldquoTwo carbon compounds andfatty acids as carbon sourcesrdquo inEscherichia coli and SalmonellaCellular and Molecular Biology ASM Press Washington DCUSA 1996
[47] Z L Boynton G N Bennett and F B Rudolph ldquoCloningsequencing and expression of clustered genes encoding betahydroxybutyryl-coenzyme A (CoA) dehydrogenase crotonaseand butyryl-CoA dehydrogenase from Clostridium aceto-butylicum ATCC 824rdquo Journal of Bacteriology vol 178 no 11pp 3015ndash3024 1996
[48] J Schiebel A Chang H Lu M V Baxter P J Tonge andC Kisker ldquoStaphylococcus aureus FabI inhibition substraterecognition and potential implications for in vivo essentialityrdquoStructure vol 20 no 5 pp 802ndash813 2012
[49] M Iminjan N Amat X-H Li H Upur D Ahmat and B HeldquoInvestigation into the toxicity of traditional uyghur medicinequercus infectoria galls water extractrdquo PLoS ONE vol 9 no 3Article ID e90756 2014
[50] A Hasmah Z Nurazila C Y Chow R Rina and M Rafiquz-zaman ldquoCytotoxic effects ofQuercus infectoria extracts towardscervical (Hela) and Ovarian (Caov-3) cancer cell linesrdquo Healthand the Environment Journal vol 1 no 2 pp 17ndash23 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Proteomics 9
[36] V A Fischetti ldquoBacteriophage lytic enzymes novel anti-infectivesrdquo Trends in Microbiology vol 13 no 10 pp 491ndash4962005
[37] C Heidrich M F Templin A Ursinus et al ldquoInvolvementof N-acetylmuramyl-l-alanine amidases in cell separationand antibiotic-induced autolysis of Escherichia colirdquo MolecularMicrobiology vol 41 no 1 pp 167ndash178 2001
[38] S Fuchs J Pane-Farre C KohlerMHecker and S EngelmannldquoAnaerobic gene expression in Staphylococcus aureusrdquo Journal ofBacteriology vol 189 no 11 pp 4275ndash4289 2007
[39] L J Rasmussen P L Moller and T Atlung ldquoCarbonmetabolism regulates expression of the pfl (pyruvate formate-lyase) gene in Escherichia colirdquo Journal of Bacteriology vol 173no 20 pp 6390ndash6397 1991
[40] M Leibig M Liebeke D Mader M Lalk A Peschel andF Gotz ldquoPyruvate formate lyase acts as a formate supplierfor metabolic processes during anaerobiosis in Staphylococcusaureusrdquo Journal of Bacteriology vol 193 no 4 pp 952ndash962 2011
[41] W Buckel and B T Golding ldquoRadical enzymes in anaerobesrdquoAnnual Review of Microbiology vol 60 pp 27ndash49 2006
[42] A Resch S Leicht M Saric et al ldquoComparative proteomeanalysis of Staphylococcus aureus biofilm and planktonic cellsand correlation with transcriptome profilingrdquo Proteomics vol6 no 6 pp 1867ndash1877 2006
[43] T Kupke ldquoMolecular characterization of the 41015840-phosphopan-tothenoylcysteine synthetase domain of bacterial Dfp flavopro-teinsrdquo The Journal of Biological Chemistry vol 277 no 39 pp36137ndash36145 2002
[44] T Nakamura T Pluskal Y Nakaseko and M YanagidaldquoImpaired coenzyme A synthesis in fission yeast causes defec-tive mitosis quiescence-exit failure histone hypoacetylationand fragile DNArdquo Open Biology vol 2 no 9 Article ID 1201172012
[45] D D Simeonova I Susnea A Moise B Schnik and MPrzybylski ldquolsquoUnknown genomersquo proteomics a new NAD(P)-dependent epimerasedehydratase revealed by N-terminal se-quencing inverted PCR and high resolution mass spectrome-tryrdquo Molecular amp Cellular Proteomics vol 8 no 1 pp 122ndash1312009
[46] D P Clark and J E Cronan ldquoTwo carbon compounds andfatty acids as carbon sourcesrdquo inEscherichia coli and SalmonellaCellular and Molecular Biology ASM Press Washington DCUSA 1996
[47] Z L Boynton G N Bennett and F B Rudolph ldquoCloningsequencing and expression of clustered genes encoding betahydroxybutyryl-coenzyme A (CoA) dehydrogenase crotonaseand butyryl-CoA dehydrogenase from Clostridium aceto-butylicum ATCC 824rdquo Journal of Bacteriology vol 178 no 11pp 3015ndash3024 1996
[48] J Schiebel A Chang H Lu M V Baxter P J Tonge andC Kisker ldquoStaphylococcus aureus FabI inhibition substraterecognition and potential implications for in vivo essentialityrdquoStructure vol 20 no 5 pp 802ndash813 2012
[49] M Iminjan N Amat X-H Li H Upur D Ahmat and B HeldquoInvestigation into the toxicity of traditional uyghur medicinequercus infectoria galls water extractrdquo PLoS ONE vol 9 no 3Article ID e90756 2014
[50] A Hasmah Z Nurazila C Y Chow R Rina and M Rafiquz-zaman ldquoCytotoxic effects ofQuercus infectoria extracts towardscervical (Hela) and Ovarian (Caov-3) cancer cell linesrdquo Healthand the Environment Journal vol 1 no 2 pp 17ndash23 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology





























