Embed Size (px)
Citation preview
The role of neutrophils and Porphyromonas gingivalis
Rheumatoid Arthritis & Periodontitis
Biomedical Sciences Internship Report
05-01-2015 / 28-08-2015
Written by Jarnick Hulzebos

University of Groningen
Biomedical Sciences Internship Report 28-08-2015
Rheumatology and Clinical Immunology department,
University Medical Centre Groningen, the Netherlands
Written by: J. Hulzebos, BSc
Supervisors: Dr. J. Westra
K.M.J. Janssen, MSc
Rheumatoid Arthritis &
Periodontitis
The role of neutrophils and Porphyromonas gingivalis

1 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Abstract
Introduction: Rheumatoid Arthritis (RA) is a chronic systemic autoimmune disease characterized by synovial inflammation resulting in damage to articular cartilage and the underlying bone. The formation of anti-citrullinated proteins (ACPAs) is a hallmark of RA. ACPAs target proteins which have been citrullinated by Peptidyl-Arginine Deiminase (PAD), such as PAD4 in neutrophils. Periodontitis (PD) is caused by periodontal pathogens, such as Porphyromonas gingivalis (PG). PG
is the only known bacterium to possess its own PAD enzyme; PPAD. Furthermore, neutrophils attracted to the inflamed periodontium can perform a process named Neutrophil Extracellular Trap formation; (NET)osis. During NETosis, neutrophils eject intracellular compartments, which are citrullinated by their own PAD. It is hypothesized that PPAD in the periodontium is able to hypercitrullinate these excreted extracellular components. Due to this hypercitrullination, susceptible individuals initiate a loss of tolerance to citrullinated proteins and begin to generate ACPAs. This ACPA formation can result in the induction of RA. The aim of this study was to assess the role of neutrophils when they come into contact with PG and PPAD in terms of inducing NETosis and citrullinating NET-constituents.
Methods: NETosis of stimulated or unstimulated healthy donor neutrophils was quantified using a SYTOX assay which measures extracellular DNA. PAD4 protein production has been visualized using the Western Blot method. Immunofluorescence Microscopy has been performed on coverslips to visualize NETosis and citrullination of histone H3. After optimization of these methods, co-cultures of neutrophils with several PG strains (PG-ATCC, PG-W83) and their PPAD
mutants (ΔPG-ATCC, ΔPG-W83) have been performed. An in-house made PAD-assay was used
to measure PAD4 activity. During these experiments, the effects of Cl-Amidine (CLA), a pan-PAD inhibitor, have been assessed.
Results: Neutrophil PAD4 protein production is upregulated after stimulation. CLA inhibits this effect and is also able to inhibit direct PAD-enzyme activity, but not NETosis itself. When PG-
ATCC or ΔPG-ATCC is co-cultured with neutrophils, it gets phagocytosed. However, when
neutrophils were pre-stimulated with PMA to form NETs and subsequently addition of PG-ATCC
or ΔPG-ATCC occurred, PG-ATCC – but not ΔPG-ATCC – would hypercitrullinate histone H3
within NETs. PG-W83 and ΔPG-W83 induced NETosis – and possibly hypercitrullination – when
co-cultured with neutrophils.
Discussion & Conclusion: A clear difference in interaction between different PG strains and neutrophils is observed in vitro. PG-ATCC and its mutant were both phagocytosed by neutrophils,
whereas both PG-W83 strains induced NETosis and supposedly hypercitrullination of histone H3. After pre-stimulation of neutrophils, even clearer hypercitrullination of histone H3 was observed. However, a larger diversity of PG strains should be used in these experiments to determine more differences in virulence. Moreover, if NETs – and preferably hypercitrullination – could be shown in periodontal tissue of PD/RA patients, the hypothesis would become more plausible. Even though no clear conclusion could be drawn due to inconsistency of the data, it can be stated that PG is probably able to citrullinate human proteins and is highly likely to be capable of hypercitrullinating NETs and could therefore very well be the causative pathogen of the pathogenesis of RA.

1 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Contents Abstract ................................................... 1
Introduction ............................................. 1
Rheumatoid arthritis ............................. 1
Neutrophils and NETosis ...................... 1
Periodontitis and P. gingivalis ................. 1
Rheumatoid arthritis and Periodontitis ... 2
Inhibiting (P)PAD................................. 2
Aim of the study ................................... 3
Methods ................................................... 3
Neutrophil Isolation, Inhibition and Stimulation ........................................... 3
P. gingivalis cultures ............................... 3
P. gingivalis growth and strains ............ 3
P. gingivalis co-culturing ..................... 4
SYTOX green ASSAY .......................... 4
SYTOX green ASSAY....................... 4
Western Blot ......................................... 4
Protein concentration ........................ 4
Immunoblotting ................................ 5
Antibodies ........................................ 5
Immunofluorescence Microscopy .......... 5
Immunofluorescent labelling .............. 5
PAD-assay............................................ 5
Illustrations .......................................... 6
Results ...................................................... 7
Quantifying NETosis ............................. 7
Inhibiting PAD4 protein production ....... 7
Inhibiting (P)PAD activity ..................... 8
Immunofluorescence Microscopy Optimization ......................................... 8
Immunofluorescence Microscopy PG/Neutrophil co-cultures .................... 9
PG-ATCC strain co-culture ................ 9
PG-W83 strain co-culture ................. 10
Effect of Cl-Amidine ........................ 12
Discussion & Conclusion......................... 12
Discussion ........................................... 12
Immunofluorescence Microscopy ..... 12
Induction and inhibition of NETosis . 13
Bacterial PAD activity remains unclear ....................................................... 14
General points of discussion within internship ........................................ 14
Carbamylation of human proteins ..... 14
In Conclusion ...................................... 15
Acknowledgements ................................. 15
References .............................................. 16
Personal Acknowledgements ................... 20
List of abbreviations of importance:
ACPA Anti-citrullinated protein antibodies HLA Human Leukocyte Antigen PD Periodontitis
APC Antigen Presenting Cell ION Calcium Ionophore PG P. gingivalis
CarP Carbamylated Protein(s) MPO Myeloperoxidase PPAD PG-PAD
CIA Collagen-Induced Arthritis NE Neutrophil Elastase PMA Phorbol 12-myristate
CitH3 Citrullinated Histone H3 NET Neutrophil Extracellular Trap 13-acetate
CLA Cl-Amidine PAD Peptidyl-Arginine Deiminase RA Rheumatoid Arthritis

1 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Introduction
Rheumatoid arthritis
heumatoid arthritis (RA) is the most common chronic systemic autoimmune disease in high-income countries, with a prevalence of 0.5% to 1% in adults [1,2]. Even though the first descriptions of RA date back to 400 BCE, classification criteria have only been
developed 55 years ago. As of today, there is still no curative treatment available for this disease. RA can best be described as a clinical syndrome which spans several disease subsets. These several subsets will then come together in a final common pathway, which exists of persistent synovial inflammation and damage to underlying bone and articular cartilage [1].
In one of these subsets, generation of anti-citrullinated protein antibodies (ACPA) occurs, one of the hallmarks of RA [3]. These antibodies target citrullinated proteins, which are generated by conversion of arginine amino acid residues into citrulline amino acid residues by the calcium-dependent Peptidyl-arginine Deiminase (PAD) enzyme. In humans, there are several subtypes of PAD: 1, 2, 3, 4 and 6 [4]. However, citrullination also occurs in regular – healthy – homeostasis
[5]. ACPAs are disease-specific and are not present in healthy individuals. ACPAs can be found in blood years before symptoms of RA occur [6]. Therefore a loss of tolerance to citrullinated proteins must have occurred, which possibly triggers the development of RA [7].
The cause of RA remains debatable still. The discovery of the reason of the loss of tolerance to citrullinated proteins has yet to be made. One of the hypothesis is that inflamed periodontium initiates ACPA production, which subsequently triggers the pathogenesis of RA [8].
Neutrophils and NETosis
Neutrophils play a major role during bacterial infection. Neutrophils respond to inflammatory stimuli and migrate to infected tissues from the circulating blood. They can then bind, engulf and kill bacteria through several processes. For instance, neutrophils are able to release antimicrobial factors into their extracellular matrix (degranulation) and kill bacteria in that fashion. Neutrophils are also capable of phagocytosis, which kills the internalized bacteria with proteolytic enzymes, reactive oxygen species and antimicrobial proteins [9,10].
Up and until 2004, it was generally believed that neutrophils were only capable of killing bacteria in these ways. After Brinkmann et al. published their literature about Neutrophil Extracellular Traps (NETs), another mode of bacterial clearance was proposed. Neutrophils are capable of generating extracellular fibers which exists of several nuclear and granular constituents, such as Neutrophil Elastase (NE), Myeloperoxidase (MPO), histones and DNA. These molecules are also capable of binding and killing extracellular bacteria [10]. This process, named NETosis, is not only another way of bacterial clearance, but it is also another form of cell death next to apoptosis and necrosis [11,12].
In human granulocytes, PAD4 is expressed, which is essential for the formation of NETs
[13]. During NETosis, calcium is mobilized by PLCγ activation, reactive oxygen species are
generated and nuclear delobulation occurs – which involves enzymatic activity of MPO and NE [14] – and PAD4 citrullinates intracellular constituents such as histones [15], which are then excreted from the cell. PAD4 is also able to citrullinate other proteins which are present in the surrounding extracellular matrix such as fibrinogen [16] and vimentin [17]. Previously our group showed that citrullinated histone H3 (CitH3) is a target of ACPAs. It can thus be hypothesized that
this PAD4 activity could play a big role in ACPA generation and thus in the pathogenesis of RA [18].
Periodontitis and P. gingivalis
Periodontitis (PD) is a chronic inflammatory disease, characterized by damage of the tooth-supporting tissues, the periodontium. The periodontium exists of the gingiva, alveolar bone and periodontal ligament. This oral inflammation is caused by several oral anaerobic bacteria such as Treponema denticola, Tannerella forsythia but most importantly; Porphyromonas gingivalis (PG) [19,20].
R
2 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
In the past few years, PD has been linked to several chronic and inflammatory diseases such as atherosclerosis, Crohn’s disease and diabetes mellitus. The genome of the patients is very often the reason of this linkage [21-23]. In the past decade, PD susceptibility has often been linked to the expression of HLA-DR1 antigen, as has the development of RA. It is therefore not surprising that a link between RA and PD has been established as well [3,24-26].
PG is a gram-negative, rod-shaped bacterium which possesses several virulence factors, such as gingipains (proteases) [27]. PG is also capable of inhibiting apoptosis when phagocytosed
[28]. It is also the only currently known bacterium which possesses its own, calcium-independent, PAD enzyme called P. gingivalis PAD (PPAD) [29]. This PPAD is able to citrullinate C-terminal
residues of proteins or peptides [30]. PG is therefore thought to be able to (hyper)citrullinate human proteins. In 2012, Nesse et al., were the first to show that citrullinated proteins are present in PD affected periodontium [31].
Rheumatoid arthritis and Periodontitis
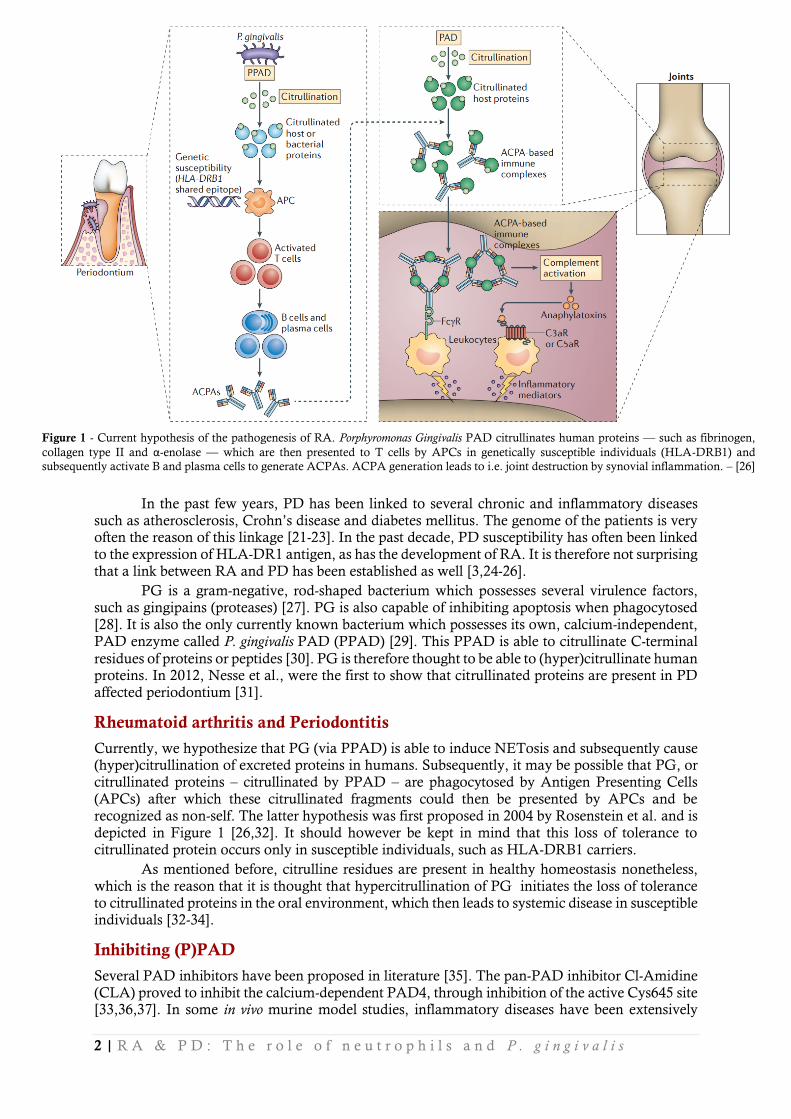
Currently, we hypothesize that PG (via PPAD) is able to induce NETosis and subsequently cause (hyper)citrullination of excreted proteins in humans. Subsequently, it may be possible that PG, or citrullinated proteins – citrullinated by PPAD – are phagocytosed by Antigen Presenting Cells (APCs) after which these citrullinated fragments could then be presented by APCs and be recognized as non-self. The latter hypothesis was first proposed in 2004 by Rosenstein et al. and is depicted in Figure 1 [26,32]. It should however be kept in mind that this loss of tolerance to citrullinated protein occurs only in susceptible individuals, such as HLA-DRB1 carriers.
As mentioned before, citrulline residues are present in healthy homeostasis nonetheless, which is the reason that it is thought that hypercitrullination of PG initiates the loss of tolerance to citrullinated proteins in the oral environment, which then leads to systemic disease in susceptible individuals [32-34].
Inhibiting (P)PAD
Several PAD inhibitors have been proposed in literature [35]. The pan-PAD inhibitor Cl-Amidine (CLA) proved to inhibit the calcium-dependent PAD4, through inhibition of the active Cys645 site [33,36,37]. In some in vivo murine model studies, inflammatory diseases have been extensively
Figure 1 - Current hypothesis of the pathogenesis of RA. Porphyromonas Gingivalis PAD citrullinates human proteins –– such as fibrinogen,
collagen type II and α-enolase –– which are then presented to T cells by APCs in genetically susceptible individuals (HLA-DRB1) and
subsequently activate B and plasma cells to generate ACPAs. ACPA generation leads to i.e. joint destruction by synovial inflammation. – [26]

3 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
reduced in severity of disease by intake of CLA, such as collagen-induced arthritis [38] and colitis
[39]. These studies have shown that PAD-inhibiting treatment is potentially a better treatment than currently used treatments. Therefore, PAD inhibition should be further assessed.
CLA was expected not to inhibit PPAD, because PPAD has a completely different protein structure than human PADs [40].
Aim of the study
In this study, the interaction between P. gingivalis and neutrophils was studied in an in vitro
environment to attempt to discover the missing link between PD and RA. The current hypothesis is that PG is responsible for the induction of NETosis and hypercitrullinating excreted proteins, leading to loss of tolerance against citrullinated proteins and thus pathogenesis of RA (Figure 1).
To do so, citrullination of histone H3 has been assessed with the use of immuno-fluorescence microscopy. Histone H3 has been specifically chosen because of previous data about ACPAs being able to target CitH3. Quantification of NETosis has been measured after stimulation of neutrophils with either chemical substances or by co-culturing neutrophils with PG.
Also, an Enzyme-Linked Immuno Sorbent Assay (ELISA)-based assay has been established in order to quantify citrullination by (P)PAD directly on human fibrinogen. If PPAD is able to citrullinate human fibrinogen, it might be able to citrullinate other human proteins such as histones.
During the course of these experiments, the pan-PAD inhibitor CLA has been used occasionally to disable the human PAD, whilst bacterial PPAD was expected to preserve its functionality.
Methods
Neutrophil Isolation, Inhibition and Stimulation
Human neutrophils were obtained from whole blood samples, collected from healthy donors. Whole blood was collected in EDTA tubes and subsequently diluted 1:1 with Phosphate Buffered Saline (PBS). This diluted blood was carefully brought upon a layer of lymphoprep (Axis Shield, LYS 3773) in a 2:1 fashion and centrifuged for 20 minutes, 2500RPM without brake. After centrifugation, three layers of suspension appear, after which the upper two are removed. The lower segment contains erythrocytes and neutrophils, which was washed on ice with NH4Cl twice, to lyse the erythrocytes. Both wash steps were centrifuged for 3 minutes, 2500RPM without brake. The remaining pallet was solved in 1ml Hank’s Balanced Salt Solution (HBSS) without calcium and magnesium (Gibco, 14175-095). Cells where counted and diluted further, depending on the following experiment, with HBSS with calcium and magnesium (Gibco, 14025-050). 0.1% BSA (Sigma Life Science, A9647-1KG) was added to this solution. After isolation, cells were incubated
for 30 minutes in a 37⁰C, 5% CO2 incubator to adhere to the plates.
Stimulation of neutrophils either occurred with 20nM phorbol 12-myristate 13-acetate (PMA) which is the standard for inducing NETosis, or various concentrations of Calcium Ionophore (ION) ranging from 1.9µM to 11.4µM. Stimulations occurred in varying time intervals,
in a 37⁰C, 5% CO2 incubator. PMA and ION were on stock in-house.
In some cases, CLA was added to the neutrophils to inhibit intrinsic NETosis and
citrullination. After plating, neutrophils were pre-incubated with 200µM CLA for 1 hour at 37⁰C,
5% CO2. CLA was obtained from Calbiochem (506282-10MG) and dissolved in either H2O or DMSO.
P. gingivalis cultures
P. gingivalis growth and strains PG wild type (WT) strains were used for this project, to wit: PG-ATCC 33277 (PG-ATCC) and
PG-W83. Mutants of these strains (ΔPG-ATCC and ΔPG-W83), lacking the PPAD enzyme, were provided by the research group of K. Nguyen (Institute of Dental Research, University of Sydney,

4 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Australia). Methods are described by Wegner et al., [41]. PCR validation of the presence of the
PPAD enzyme has been performed by M. du Teil. All used PG strains were cultured on Blood
Agar Base No.2 plates (BA2), anaerobic for 5 days at 37⁰C. Next, PG were inoculated into open
serum bottles containing Brain Heart Infusion broth (BHI) supplemented with 5% L-cysteine, 5mg/L hemin and 1mg/L menadione. PG was grown until the early stationary phase was reached and the corresponding OD600 values were measured during that culture. Now the bacteria were quantified. FITC staining was performed as described by Pathirana et al. [42].
P. gingivalis co-culturing P. gingivalis was co-cultured with neutrophils with a Multiplicity of infection (MOI) of 100. MOI
stands for the amount of bacteria used to infect one single cell. Co-culture duration lasted 90
minutes, incubated at 37⁰C, 5% CO2. Fluorescein isothiocyanate (FITC)-stained bacteria (green
fluorescent label) were only used during immunofluorescence microscopy.
SYTOX green ASSAY
To quantify the amount of extracellular DNA from supernatants of cultured neutrophils, SYTOX Green Assays were performed.
To prepare supernatants for SYTOX assays, neutrophils were seeded in 24-wells plates (Corning, 3524) with a concentration of 2x106 neutrophils/ml, 1ml in every well. After stimulation was finished, 250mU MNase (Worthington, LS004798) was added to cut off the NETs and
incubated for 10 minutes at 37⁰C, 5% CO2. The MNase reaction was blocked by adding 2.5mM
EGTA, 3 minutes on a shaker. Subsequently, medium was collected and centrifuged for 5 minutes,
3000RPM. Supernatants were collected in 1.5ml tubes and stored at -20⁰C until the SYTOX assay
took place. If co-cultured with PG, metronidazole was added to kill the bacteria before storage.
SYTOX green ASSAY Samples were thawed on ice and 50µl of each sample and 5µM SYTOX Green (Invitrogen, MP 07020) was added in duplo to a dark 96-wells plate (Fluotrac 200). A standard sample was used to form a calibration curve, expressed in arbitrary units (aU) in duplo.
The plate was incubated for 15 minutes at Room Temperature (RT) on a shaker. Next, air bubbles were removed and the plate was read in a Victor3 1420 multilabel counter (Perkin Elmer Precisely) at ex487 em535 (1s) TOP in triplo, with the use of the Wallac 1420 manager software. Triplicates were averaged, subsequently the duplicates of this data was used for analysis.
Western Blot
To visualize the protein expression of PAD4 after stimulation and/or co-culture with PG, the Western Blot method was used.
To prepare lysates for Western Blotting, neutrophils were seeded in 6-wells plates (Corning, 3516) with a concentration of 2x106 neutrophils/ml, 2ml in every well. Cells were adhered to plates
after 30 minutes of incubation at 37⁰C, 5% CO2. After the inhibition and stimulation was finished,
wells were washed with PBS. Neutrophils were suspended in 200 µl lysisbuffer (Cell Signaling) supplemented with protease inhibitors (Sigma-Aldrich), plates were incubated on ice for 5 minutes. Cells and residue were loosened from the wells using a cell scraper and homogenized with a syringe
and 25G needle. The suspension was centrifuged for 10 minutes in a cold (4⁰C) microfuge.
Supernatant was stored at -20⁰C. If co-cultured with PG, metronidazole was added to kill the
bacteria before storage.
Protein concentration The protein concentrations of the previously made lysates were measured using a PierceTM BCA protein assay kit (Thermo Scientific, 23225).

5 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Immunoblotting Lysates were diluted to obtain uniform concentrations with non-reducing SDS buffer. Samples were brought upon a pre-cast CriterionTM 12.5% Tris-HCl gel (Bio-Rad, 345-0015). Gel-electrophoresis was performed and gels were transferred to PVDF membranes to visualize blots using Bio-Rad CriterionTM cells. Membranes were blocked with Odyssey® blocking buffer (PBS) (Li-Cor, 927-40000).
Antibodies To visualize PAD4, mouse anti-PAD4 was used (Abcam, ab128086; 1:2000). Mouse anti-actin was used to visualize baseline actin expression (UMCG; 1:2000). Blots were incubated overnight on an
orbital shaker at 4⁰C.
Secondary Odyssey goat anti-mouse IRDye 800cw (Li-Cor, 925-68020; 1:5000) was used to visualize primary antibodies with the Odyssey CLx infra-red imaging scanner (Li-Cor).
Immunofluorescence Microscopy
Fluorescence microscopy was performed to visualize NETosis in general and citrullination of histone H3 after inhibition and/or stimulation of neutrophils, or after co-culturing neutrophils with PG.
Before neutrophils were plated, Ø12mm coverslips (Menzel, CB00120RA1) were placed into 24-wells plates (Corning, 3524). Neutrophils were added in a 0,25x106 neutrophils/ml fashion,
1ml per well. Plates were incubated for 30 to 60 minutes at 37⁰C, 5% CO2 to let the cells adhere to
the coverslips. Next, neutrophils were inhibited, stimulated or left untreated for a total of 1 to 3 hours. Cells were either co-cultured with PG, or incubated without PG for 90 minutes. Afterwards,
cells were fixated by the addition of 4% formalin. Plates were stored at 4⁰C in the dark for
microscopy the day after, or in PBS at -20⁰C for long term storage.
Immunofluorescent labelling To visualize NETs, CitH3 and PG, coverslips were blocked with 2% BSA/PBS for 1 hour. NETs
were stained with rabbit anti-MPO (DAKO, A0398; 1:200) and/or mouse Anti-NE (DAKO, M0752; 1:200). CitH3 was stained with Rabbit anti-CitH3 (Abcam, ab5103; 1:250) instead of MPO. Primary antibodies were incubated for 1 hour at RT in PBS supplemented with 0.05% Tween-20, 0.5% BSA (PBS+), 100µl/coverslip. Coverslips were washed with PBS before adding secondary antibodies.
Secondary Alexa 488 goat anti-mouse (Invitrogen, A11001; 1:400) and Alexa Fluor 568 goat anti-rabbit (Invitrogen, A11011; 1:400) were used to visualize the primary antibodies. Secondary antibodies were added in PBS+ with DAPI (Roche, 10 236 276 001; 1:5000) and incubated for 30 minutes before mounting the coverslips upside-down in citifluor (Citifluor Ltd., AF1). Slides were analyzed using the Leica DFC450 C fluorescence microscope with the Leica Application Suite Version 4.2.0 software. The magnification used in these pictures is always 160x.
PAD-assay
Quantification of in vitro citrullination of fibrinogen by (P)PAD enzyme activity was assessed using
an in-house made ELISA-based PAD-assay. This PAD-assay was based on Zendman’s ABAP-assay and Damgaard’s use of (whole) fibrinogen, but improved for and optimized to our experiments [43,44]. A 96 wells plate (Costar, 9018) was coated overnight with 1µg/ml fibrinogen (Calbiochem, 341578) in 30mM Na2CO3, 70mM NaHCO3 pH=9.6 buffer 100µl/well. Plates were washed 5 times with washing buffer (0.15M NaCl, 0.25M Tris-HCl, and 0.05% Tween-20 (Sigma, P2287) in demineralized water (dH2O)). For the calibration curve, a 10x step-wise decreasing range of recombinant human PAD4 (rhPAD4) (Modiquest, MQ16.203) was used, starting from 2 mU until 0.0002 mU in 100µl deimination buffer (40mM Tris-HCl pH7.5, 5mM CaCl2, 1mM DTT in

6 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
dH2O) per well. As a positive control, 3mL 100µg/ml native fibrinogen plus approximately 75mU
rhPAD4 in deimination buffer was incubated at 37⁰C, 5% CO2 overnight. Recombinant PPAD
was manufactured at the medical microbiology department (UMCG) and used in several dilutions in the PAD-assay in pH9.5 buffer (50mM Tris-HCl, pH9.5). After adding the samples, the plate
was put on a shaker for 15 minutes, and incubated for at least 1 hour and 45 minutes at 37⁰C, 5%
CO2. If used, CLA was pre-incubated for 30 minutes with enzyme samples, on ice, before addition
to the fibrinogen-coated plate. After incubation, the plate was washed 5 times. Antibodies were diluted in 1% BSA, 0.05% Tween-20 in PBS. Mouse anti-citrullinated fibrinogen (Modiquest, 20B2; 1:1000) was added, 100µl/well and incubated for 1 hour on a shaker at RT. Washing steps were performed once more after this. Secondary goat anti-mouse (Jackson ImmunoResearch, 115-035-146; 1:2000) was added 100µl/well and incubated for 1 hour on a shaker at RT. The plate was washed 5 more times after which 100µl 3,3’,5,5’-Tetramethylbenzidine (TMB) diluted in acetate buffer with 0.0006% H2O2 was added, to start a color-reaction with the secondary HRP-label, which was incubated for 5 to 15 minutes on a shaker at RT. The reaction was stopped with 100µl 1M HsSO4 and light emission was read in a microplate reader (Tunable versa max, Molecular Devices) at 450nm. Plate analysis was performed using the SoftMax Pro 5.4 software.
Illustrations
Graphs and error bars were made using the GraphPad Prism 5 (version 5.03) software. Pictures were edited in Microsoft® PowerPoint® 2013 (15.0.4737.1003).
7 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Results
Quantifying NETosis
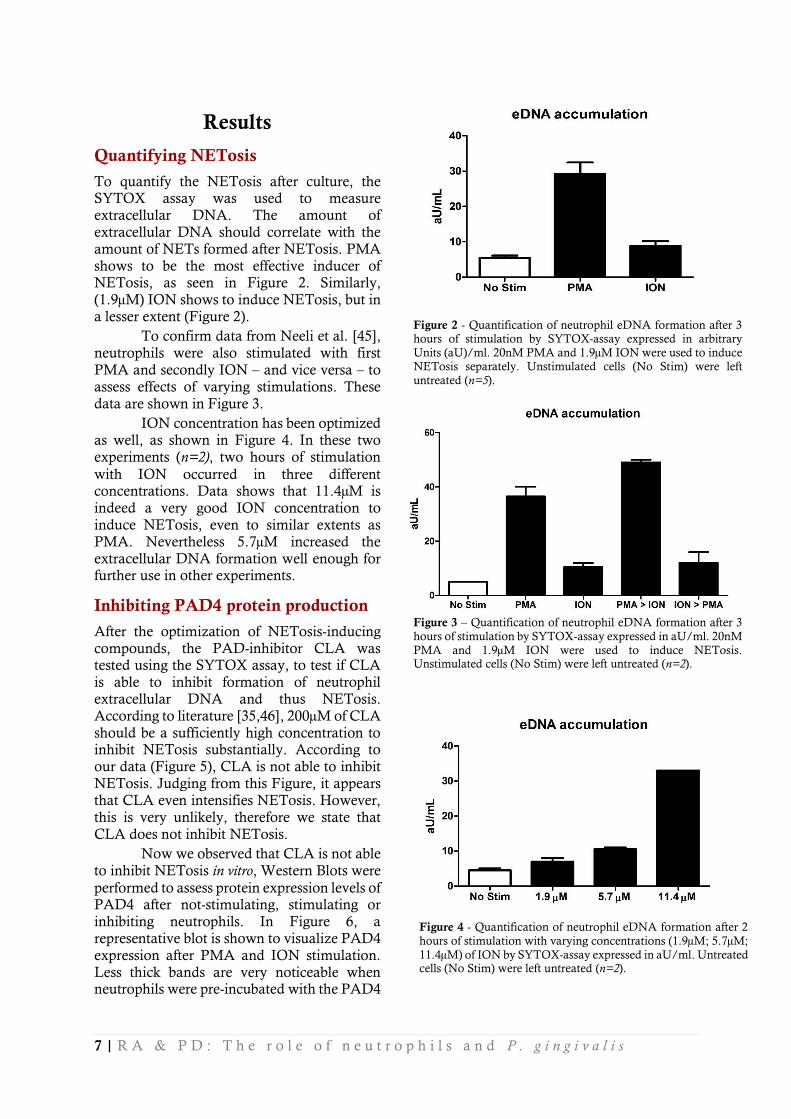
To quantify the NETosis after culture, the SYTOX assay was used to measure extracellular DNA. The amount of extracellular DNA should correlate with the amount of NETs formed after NETosis. PMA shows to be the most effective inducer of NETosis, as seen in Figure 2. Similarly, (1.9µM) ION shows to induce NETosis, but in a lesser extent (Figure 2).
To confirm data from Neeli et al. [45], neutrophils were also stimulated with first
PMA and secondly ION – and vice versa – to assess effects of varying stimulations. These data are shown in Figure 3.
ION concentration has been optimized as well, as shown in Figure 4. In these two experiments (n=2), two hours of stimulation
with ION occurred in three different concentrations. Data shows that 11.4µM is indeed a very good ION concentration to induce NETosis, even to similar extents as PMA. Nevertheless 5.7µM increased the extracellular DNA formation well enough for further use in other experiments.
Inhibiting PAD4 protein production
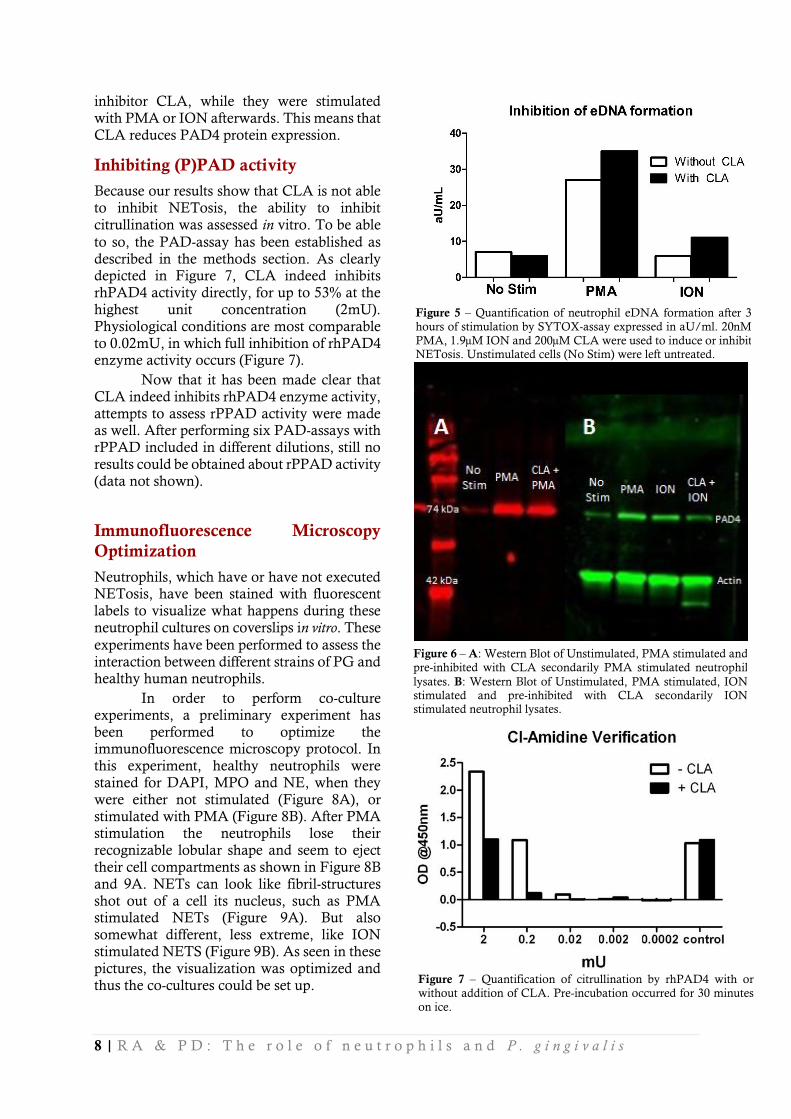
After the optimization of NETosis-inducing compounds, the PAD-inhibitor CLA was tested using the SYTOX assay, to test if CLA is able to inhibit formation of neutrophil extracellular DNA and thus NETosis. According to literature [35,46], 200µM of CLA should be a sufficiently high concentration to inhibit NETosis substantially. According to our data (Figure 5), CLA is not able to inhibit NETosis. Judging from this Figure, it appears that CLA even intensifies NETosis. However, this is very unlikely, therefore we state that CLA does not inhibit NETosis.
Now we observed that CLA is not able
to inhibit NETosis in vitro, Western Blots were
performed to assess protein expression levels of PAD4 after not-stimulating, stimulating or inhibiting neutrophils. In Figure 6, a representative blot is shown to visualize PAD4 expression after PMA and ION stimulation. Less thick bands are very noticeable when neutrophils were pre-incubated with the PAD4
Figure 2 - Quantification of neutrophil eDNA formation after 3 hours of stimulation by SYTOX-assay expressed in arbitrary
Units (aU)/ml. 20nM PMA and 1.9µM ION were used to induce NETosis separately. Unstimulated cells (No Stim) were left
untreated (n=5).
Figure 4 - Quantification of neutrophil eDNA formation after 2
hours of stimulation with varying concentrations (1.9µM; 5.7µM;
11.4µM) of ION by SYTOX-assay expressed in aU/ml. Untreated cells (No Stim) were left untreated (n=2).
Figure 3 – Quantification of neutrophil eDNA formation after 3
hours of stimulation by SYTOX-assay expressed in aU/ml. 20nM
PMA and 1.9µM ION were used to induce NETosis. Unstimulated cells (No Stim) were left untreated (n=2).
8 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
inhibitor CLA, while they were stimulated
with PMA or ION afterwards. This means that CLA reduces PAD4 protein expression.
Inhibiting (P)PAD activity
Because our results show that CLA is not able to inhibit NETosis, the ability to inhibit citrullination was assessed in vitro. To be able
to so, the PAD-assay has been established as described in the methods section. As clearly depicted in Figure 7, CLA indeed inhibits rhPAD4 activity directly, for up to 53% at the highest unit concentration (2mU). Physiological conditions are most comparable to 0.02mU, in which full inhibition of rhPAD4 enzyme activity occurs (Figure 7).
Now that it has been made clear that CLA indeed inhibits rhPAD4 enzyme activity, attempts to assess rPPAD activity were made as well. After performing six PAD-assays with rPPAD included in different dilutions, still no results could be obtained about rPPAD activity (data not shown).
Immunofluorescence Microscopy
Optimization
Neutrophils, which have or have not executed NETosis, have been stained with fluorescent labels to visualize what happens during these
neutrophil cultures on coverslips in vitro. These
experiments have been performed to assess the interaction between different strains of PG and healthy human neutrophils.
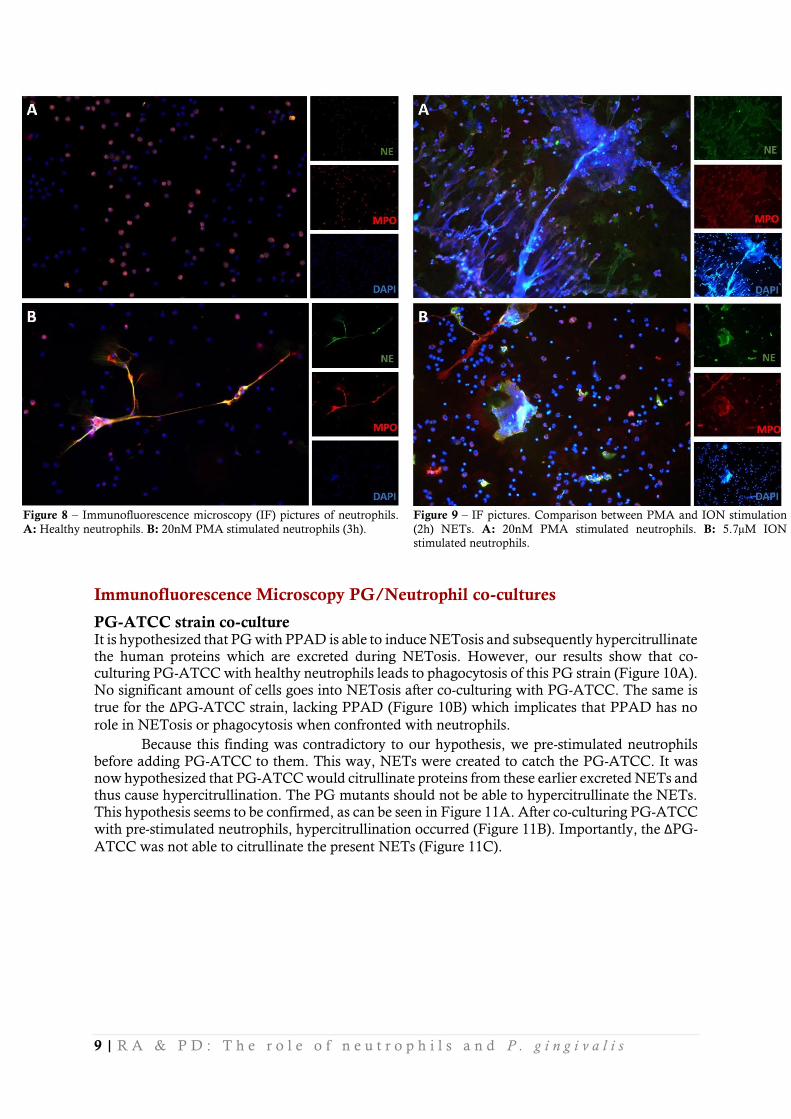
In order to perform co-culture experiments, a preliminary experiment has been performed to optimize the immunofluorescence microscopy protocol. In this experiment, healthy neutrophils were stained for DAPI, MPO and NE, when they were either not stimulated (Figure 8A), or stimulated with PMA (Figure 8B). After PMA stimulation the neutrophils lose their recognizable lobular shape and seem to eject
their cell compartments as shown in Figure 8B and 9A. NETs can look like fibril-structures shot out of a cell its nucleus, such as PMA stimulated NETs (Figure 9A). But also somewhat different, less extreme, like ION stimulated NETS (Figure 9B). As seen in these pictures, the visualization was optimized and thus the co-cultures could be set up.
Figure 5 – Quantification of neutrophil eDNA formation after 3
hours of stimulation by SYTOX-assay expressed in aU/ml. 20nM
PMA, 1.9µM ION and 200µM CLA were used to induce or inhibit NETosis. Unstimulated cells (No Stim) were left untreated.
Figure 6 – A: Western Blot of Unstimulated, PMA stimulated and
pre-inhibited with CLA secondarily PMA stimulated neutrophil
lysates. B: Western Blot of Unstimulated, PMA stimulated, ION stimulated and pre-inhibited with CLA secondarily ION stimulated neutrophil lysates.
Figure 7 – Quantification of citrullination by rhPAD4 with or
without addition of CLA. Pre-incubation occurred for 30 minutes
on ice.
9 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Immunofluorescence Microscopy PG/Neutrophil co-cultures
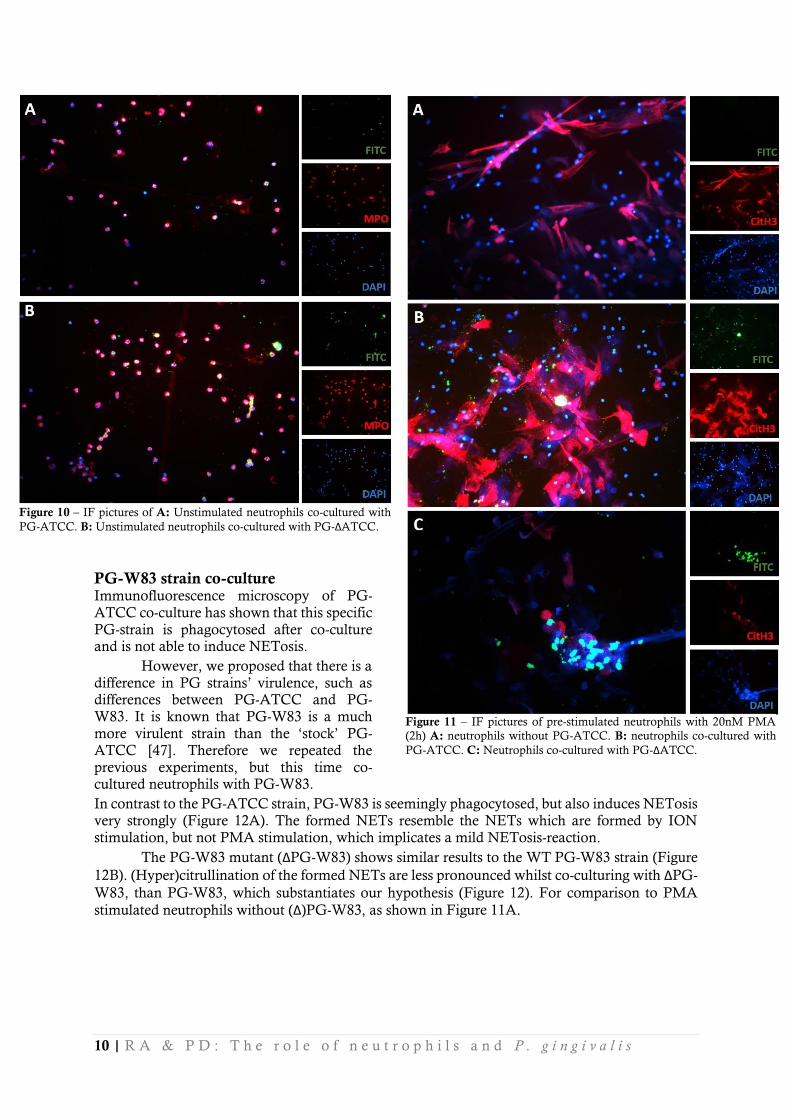
PG-ATCC strain co-culture It is hypothesized that PG with PPAD is able to induce NETosis and subsequently hypercitrullinate the human proteins which are excreted during NETosis. However, our results show that co-culturing PG-ATCC with healthy neutrophils leads to phagocytosis of this PG strain (Figure 10A). No significant amount of cells goes into NETosis after co-culturing with PG-ATCC. The same is
true for the ΔPG-ATCC strain, lacking PPAD (Figure 10B) which implicates that PPAD has no
role in NETosis or phagocytosis when confronted with neutrophils.
Because this finding was contradictory to our hypothesis, we pre-stimulated neutrophils before adding PG-ATCC to them. This way, NETs were created to catch the PG-ATCC. It was now hypothesized that PG-ATCC would citrullinate proteins from these earlier excreted NETs and thus cause hypercitrullination. The PG mutants should not be able to hypercitrullinate the NETs. This hypothesis seems to be confirmed, as can be seen in Figure 11A. After co-culturing PG-ATCC
with pre-stimulated neutrophils, hypercitrullination occurred (Figure 11B). Importantly, the ΔPG-
ATCC was not able to citrullinate the present NETs (Figure 11C).
Figure 8 – Immunofluorescence microscopy (IF) pictures of neutrophils.
A: Healthy neutrophils. B: 20nM PMA stimulated neutrophils (3h).
Figure 9 – IF pictures. Comparison between PMA and ION stimulation
(2h) NETs. A: 20nM PMA stimulated neutrophils. B: 5.7µM ION
stimulated neutrophils.
10 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
PG-W83 strain co-culture Immunofluorescence microscopy of PG-ATCC co-culture has shown that this specific
PG-strain is phagocytosed after co-culture and is not able to induce NETosis.
However, we proposed that there is a difference in PG strains’ virulence, such as differences between PG-ATCC and PG-W83. It is known that PG-W83 is a much more virulent strain than the ‘stock’ PG-ATCC [47]. Therefore we repeated the previous experiments, but this time co-cultured neutrophils with PG-W83.
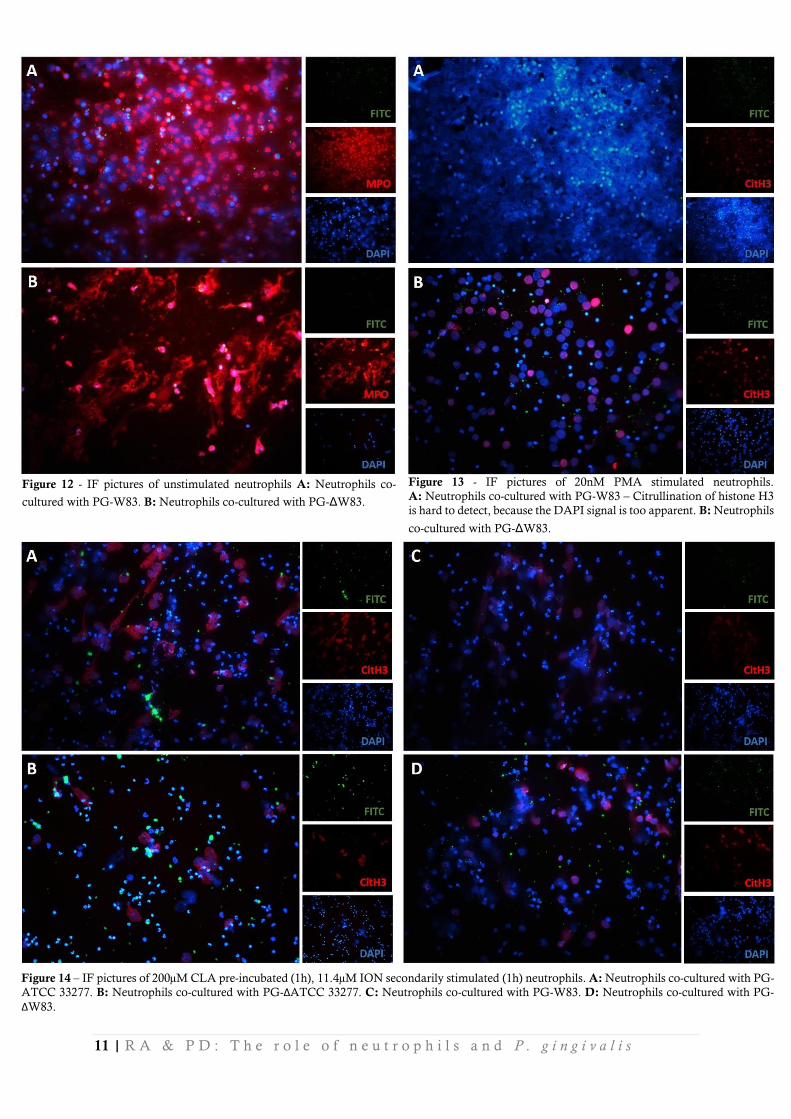
In contrast to the PG-ATCC strain, PG-W83 is seemingly phagocytosed, but also induces NETosis very strongly (Figure 12A). The formed NETs resemble the NETs which are formed by ION stimulation, but not PMA stimulation, which implicates a mild NETosis-reaction.
The PG-W83 mutant (ΔPG-W83) shows similar results to the WT PG-W83 strain (Figure
12B). (Hyper)citrullination of the formed NETs are less pronounced whilst co-culturing with ΔPG-
W83, than PG-W83, which substantiates our hypothesis (Figure 12). For comparison to PMA
stimulated neutrophils without (Δ)PG-W83, as shown in Figure 11A.
Figure 11 – IF pictures of pre-stimulated neutrophils with 20nM PMA
(2h) A: neutrophils without PG-ATCC. B: neutrophils co-cultured with
PG-ATCC. C: Neutrophils co-cultured with PG-ΔATCC.
Figure 10 – IF pictures of A: Unstimulated neutrophils co-cultured with
PG-ATCC. B: Unstimulated neutrophils co-cultured with PG-ΔATCC.
11 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Figure 14 – IF pictures of 200µM CLA pre-incubated (1h), 11.4µM ION secondarily stimulated (1h) neutrophils. A: Neutrophils co-cultured with PG-
ATCC 33277. B: Neutrophils co-cultured with PG-ΔATCC 33277. C: Neutrophils co-cultured with PG-W83. D: Neutrophils co-cultured with PG-
ΔW83.
Figure 12 - IF pictures of unstimulated neutrophils A: Neutrophils co-
cultured with PG-W83. B: Neutrophils co-cultured with PG-ΔW83.
Figure 13 - IF pictures of 20nM PMA stimulated neutrophils.
A: Neutrophils co-cultured with PG-W83 – Citrullination of histone H3
is hard to detect, because the DAPI signal is too apparent. B: Neutrophils
co-cultured with PG-ΔW83.

12 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Effect of Cl-Amidine During these experiments, there were several coverslips prepared on which the neutrophils were pre-incubated with CLA and stimulated afterwards with ION to assess the effects of this pan-PAD inhibitor on the citrullinating effects of the PG strains. No differences could be distinguished between the PAD mutants and wild types in terms of histone 3 citrullination. Still, after CLA incubation, NETosis occurred and citrullination of histone H3 took place (Figure 14).
Discussion & Conclusion After looking into the results thoroughly, several concluding remarks can be made. Most importantly, it can be stated that there is a clear difference in interaction between different PG
strains and neutrophils when co-culturing them in vitro. In our case, PG-ATCC and PG-ΔATCC
were both phagocytosed by neutrophils, whereas PG-W83 and PG-ΔW83 both induced NETosis and supposedly hypercitrullination of histone H3. After pre-stimulating neutrophils with PMA and subsequently co-culturing them with PG-ATCC, even clearer hypercitrullination of histone H3
could be observed.
When neutrophils are stimulated to induce NETosis in vitro, PAD4 protein production is
enhanced. The pan-PAD inhibitor CLA seems to inhibit this production. It has also been established that CLA is able to inhibit rhPAD4 activity and probably also PAD4 itself, but it cannot inhibit NETosis. Moreover, no distinguishable effects could be observed when neutrophils were pre-incubated with CLA and subsequently co-cultured with PG.
Discussion
After several experiments had been performed to optimize several NETosis inducing compounds such as PMA and ION, the optimal concentrations have been determined. PMA stimulation is optimal after 4 hours at a concentration of 20nM. In this research, PMA stimulation often occurred for 3 hours, or sometimes even 2 hours. It might have been that NETosis was not fully induced or finished in this shorter timeframe. This is one of the reasons that ION was included as a secondary
stimulant.
According to literature [14,48], 1.9µM ION would have been sufficient to induce NETosis to some extent. In this research, the same result could not always be achieved. Hence, ION concentrations were optimized to a higher concentration, to be certain that NETosis would be induced (Figure 4). The conclusion that higher ION concentrations would induce not only more NETosis, but also faster NETosis has been supported by Barrientos et al. They have found that high concentrations of ION may even be as effective as PMA-induced NETosis [48].
Immunofluorescence Microscopy During immunofluorescence microscopy with PG-W83 and PG-ATCC strains, differences in interaction with neutrophils were observed. The more virulent strain, PG-W83, induced NETosis
in healthy neutrophils. ΔPG-W83 showed similar results as PG-W83 itself: induction of NETosis
(Figure 12). Because of this, the role of PG-W83 PPAD could not clearly be distinguished. PG-
ATCC and ΔPG-ATCC were phagocytosed, therefore we can conclude that PPAD does not play
a distinct role in internalization of PG-ATCC (Figure 10). These findings can be compared to other findings in literature [49]. For instance, Baek et al., found degradation of inflammatory cytokines by PG – which is another capability of PG – is not as important as its invasive capabilities. This invasive capability can differ between strains greatly [50]. That study also used PG-ATCC 33277 – but not PG-W83 – and found that the invasive ability of clinical isolates can be up to 300x higher than PG-ATCC-strains. This implicates that in our experiments, perhaps a larger diversity of PG strains should be used for co-culturing. The currently used PG-ATCC might not be virulent enough
to gather enough results about realistic in vivo situations.

13 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Jayaprakash et al. [51], state that the difference in PG virulence factors, specifically
gingipains, are the reason of differences in induction of NETosis. When gingipains-mutants – lacking gingipains – were used, NETosis was induced. Gingipains are proteolytic enzymes which are able to cleave inflammatory signals, but also other human proteins such as fibrinogen. Jayaprakash used PG-ATCC and W50 PG strains [51]. However, in our research is shown that PG-W83 is less internalized by neutrophils, but also are trapped in NETs (Figure 12A). Therefore we can conclude that Jayaprakash’s study is incomplete and should have been expanded with several other PG strains (such as PG-W83, or even better, clinical isolates). Nevertheless, their finding that PG-ATCC PG strains are internalized are equal to our results.
In the study of Maresz et al., the PG-W83 PG strain caused enhancement of Collagen Induced Arthritis (CIA) in a murine model [34]. They had found that, compared to a CIA model without PG-W83, the addition of PG-W83 causes more citrullination of murine proteins. Additionally, production of antibodies against citrullinated proteins increased similarly. Could this murine model be equal to the human in vivo situation? In our results we have seen that PG-W83
indeed causes NETosis to healthy neutrophils (Figure 12). In this case, our in vitro data is indeed
somewhat comparable to Maresz’ in vivo murine data. We state ‘somewhat comparable’ because
Maresz’ study does not include all vital data and of course, in vivo data is (almost) never comparable
to in vitro data. In their article they do not state anything about NETosis, only about citrullinated
proteins. Therefore, our data cannot be compared to their literature. As mentioned before, the use of only one PG strain is not ideal as well.
This research has also shown that when neutrophils were pre-stimulated with PMA,
hypercitrullination of histone H3 occurs during co-culture with PG-ATCC (Figure 11B). ΔPG-
ATCC was indeed barely (or not) able to induce citrullination or cause citrullination of histone H3 itself after NETosis has occurred (Figure 11C). These experiments however, have only been performed twice so far and should therefore be reproduced. Moreover, the second time this experiment was performed, the results weren’t quite as convincing as the previous attempt.
Disappointingly, PG is almost never co-localized with the CitH3. It could thus be stated that PG does not account for hypercitrullination, but that the citrullination of histones is merely sporadic. This statement is possibly refuted by the fact that the anti-CitH3 antibody might not have been able to bind all the citrullinated histones, for they could have still been packed into chromatin
fibers of the DNA. PPAD is at the same time known to only citrullinate terminal arginine ends of proteins [40]. The used PG strains might not have been able to cut the histones free from the matrix, or are not able to do so at all. When hypercitrullination was observed, it might have not been a direct influence of PG, but only the indirect activation of PAD4 by PG that caused this phenomenon. It is a possibility that PG triggers intracellular mechanisms which possible upregulate and activate the cells’ PAD enzymes. This is not restricted to PAD4, but could also include PAD2. PAD2 for instance is able to citrullinate the same amount of proteins but required a lower concentration of PAD2 to be present [44]. Since upregulation of PAD4 occurs after stimulation of neutrophils (Figure 6) it might also be that PAD2 is also upregulated. If upregulation of PAD indeed occurs after stimulation with PG, it could be hypothesized that PG only accounts for indirect hypercitrullination of human proteins. This could possibly be verified by western blotting co-cultured neutrophils with PG and subsequently performing Real time-PCR on these samples.
Induction and inhibition of NETosis Stimulation by both PMA and ION proved to increase the PAD4 protein levels in healthy
neutrophils (Figure 6). This result has never been shown in literature before, to our knowledge. It
has however, been proposed often that the already available PAD4 is activated by the PKCζ
pathway after PLCγ activation [14]. We therefore propose another separate mechanism which also
induces transcription of PADI4 genes. Several molecular mechanisms have already been proposed such as the induction of Interleukin-8 transcription by PG [27]. However, to our knowledge, no mechanism has been proposed to activate transcription of PAD genes.
Judging from the Western Blot data, PAD4 protein level elevation is inhibited by CLA (Figure 6). It is known that CLA actively inhibits PAD4 through its active Cys645-site [33].

14 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Therefore we believed that CLA only inhibits the already available PAD4 and that upregulation of
the PAD4 protein might be an explanation of why CLA apparently does not inhibit NETosis in
vitro (Figure 5). It might also be that the sterically bound PAD4 by CLA cannot be labeled by the
anti-PAD4 antibody and thus cannot show up on the western blots due to steric hindrance. Another possibility is that CLA does not inhibit PAD4 activity of live neutrophil-cultures (Figure 5). CLA however does inhibit PAD4 in vitro (Figure 7).
In many published papers, CLA inhibits NETosis in vivo and in vitro, which could not be
reproduced during my internship. For instance, CLA functions as an immunomodulatory and vasculoprotective compound in mice against murine lupus [52]. Remarkably, it can even alleviate the severity of CIA in a murine model [38]. When neutrophils were stimulated with ION, after pre-inhibition with CLA for 1 hour, NETosis still occurred (Figure 14). First, attempts were made to dissolve CLA in H2O to inhibit NETosis, which did not work. A second batch was ordered and dissolved in DMSO, which also did not seem to inhibit NETosis (data not shown).
While CLA is not able to inhibit NETosis, the question arose if CLA was bioactive at all. After the Western Blots, it was apparent that CLA did have a certain effect, but the Western Blot data fell short of concluding anything. Therefore, a PAD-assay was performed to assess the
bioactivity of CLA on rhPAD4, which proved to inhibit rhPAD4 directly for up to 53% at the highest concentration (2mU) (Figure 7).
Bacterial PAD activity remains unclear The rPPAD has been tested in our PAD-assay to assess whether it is capable of citrullinating human fibrinogen, or to discover if it is bioactive at all. Sadly, no results could be gather to state that the rPPAD is bioactive or is able to citrullinate human proteins. The gathered data from rPPAD PAD-assays was too variously to make any concluding remarks regarding its (bio-)activity (data not shown).
Recently, we were informed that PPAD is inhibited by CLA (Gawron, PG conference London 2015). If this is confirmed, CLA can no longer be used in our experiments to test the influence of PPAD solely on human neutrophils.
General points of discussion within internship Healthy individuals were asked to give blood to isolate neutrophils from. According to many different publications, neutrophils differ greatly between individuals in terms of immunological reaction. Dwivedi et al. [14], have shown that neutrophils from patients with different diseases, such as Felty’s Syndrome or systemic lupus erythematosus, respond differently to NET-inducing compounds. Compared to healthy controls’ neutrophils, the neutrophils from diseased patients showed enhanced NETosis. More importantly, NETosis has shown to be enhanced in synovial fluid and peripheral blood of RA patients [53]. The same has been shown for RA patient sera, which even shows to be a possible diagnostic utility [54].
These findings implicate that the use of neutrophils from varying individuals might be an interesting follow-up. In this research only healthy individuals were asked to donate blood. It might be interesting to see how neutrophils from RA or PD patients will respond to PG co-culture. The interaction between PG and healthy neutrophils is of greatest importance to study the initiation of RA. Nevertheless, it would have been (or would be) interesting to have compared healthy controls’ neutrophils with neutrophils from RA or PD patients in terms of interaction with PG.
Carbamylation of human proteins Currently, the hypothesis states that citrullination of human proteins is the cause of pathogenesis of RA. Bright et al. [7], point out that there is another very important process which occurs within inflamed periodontal tissues: carbamylation. A similar mechanism as ACPA generation occurs: the generation of anti-carbamylated proteins (anti-CarP) in RA patients. PD is – probably – a risk factor for, or even the cause of ACPA formation. In their paper it is stated that this inflammatory-mediated carbamylation is a very potent drug target to eventually prevent RA. However, Janssen et al. [55], have shown that many PD patients do not possess anti-CarP antibodies (3.5%).

15 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Therefore, I state that carbamylation and PG are not necessarily connected directly. Moreover, our
group has shown that RA patients possess antibodies against CitH3 (ACPAs) but PD patients do not. These differences between antibody formation in PD and RA patients do not necessarily mean that both disease cannot be linked to one another.
In Conclusion
Realistically, the internship was too short to find the missing link between PD and RA. This is not surprising, because many different aspects of the story are still undiscovered or not fully understood.
Several of the performed experiments should be performed again. Western Blot data has to be confirmed by performing Real time-PCR. And still a promising target remains; to stain for NETs in periodontal tissue itself.
When all findings from this internship are combined, we could state that PG can indeed hypercitrullinate human proteins and might even induce the loss of tolerance that causes RA.
Altogether, there is need for more research on this topic. In the last decade, great progress
has been made in understanding the pathogenesis of RA and PD. (P)PAD inhibitors could become novel drug therapies for treating RA and/or PD in the coming decades.
Acknowledgements
Marines du Teil and Tim Stobernack are kindly acknowledged for their help with the co-cultures and provision of PG strains. The rPPAD was provided by Giorgio Gabarrini which is also kindly acknowledged. I am grateful for all the blood donors who have given blood once or several times.

16 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
References
[1] Scott DL, Wolfe F, Huizinga TW Rheumatoid arthritis. Lancet 2010; 376:1094-1108.
[2] Rudan I, Sidhu S, Papana A et al. Prevalence of rheumatoid arthritis in low- and middle-income countries: A systematic review and analysis. J Glob Health 2015; 5:010409.
[3] Pratesi F, Petit Teixeira E, Sidney J et al. HLA shared epitope and ACPA: just a marker or an active player? Autoimmun Rev 2013; 12:1182-1187.
[4] Wang S, Wang Y Peptidylarginine deiminases in citrullination, gene regulation, health and pathogenesis. Biochim Biophys Acta 2013; 1829:1126-1135.
[5] Wijnands KA, Meesters DM, van Barneveld KW et al. Citrulline Supplementation Improves Organ Perfusion and Arginine Availability under Conditions with Enhanced Arginase Activity.
Nutrients 2015; 7:5217-5238.
[6] Jilani AA, Mackworth-Young CG The role of citrullinated protein antibodies in predicting erosive disease in rheumatoid arthritis: a systematic literature review and meta-analysis. Int J Rheumatol 2015; 2015:728610.
[7] Bright R, Proudman SM, Rosenstein ED, Bartold PM Is there a link between carbamylation and citrullination in periodontal disease and rheumatoid arthritis? Med Hypotheses 2015; 84:570-576.
[8] de Smit MJ, Westra J, Brouwer E et al. Periodontitis and Rheumatoid Arthritis: What do we Know? J Periodontol 2015:1-10.
[9] Amanzada A, Malik IA, Nischwitz M et al. Myeloperoxidase and elastase are only expressed
by neutrophils in normal and in inflamed liver. Histochem Cell Biol 2011; 135:305-315.
[10] Brinkmann V, Reichard U, Goosmann C et al. Neutrophil extracellular traps kill bacteria. Science 2004; 303:1532-1535.
[11] Fuchs TA, Abed U, Goosmann C et al. Novel cell death program leads to neutrophil extracellular traps. J Cell Biol 2007; 176:231-241.
[12] Steinberg BE, Grinstein S Unconventional roles of the NADPH oxidase: signaling, ion homeostasis, and cell death. Sci STKE 2007; 2007:pe11.
[13] Leshner M, Wang S, Lewis C et al. PAD4 mediated histone hypercitrullination induces heterochromatin decondensation and chromatin unfolding to form neutrophil extracellular trap-like structures. Front Immunol 2012; 3:307.
[14] Dwivedi N, Radic M Citrullination of autoantigens implicates NETosis in the induction of autoimmunity. Ann Rheum Dis 2014; 73:483-491.
[15] Wang Y, Li M, Stadler S et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J Cell Biol 2009; 184:205-213.
[16] Sebbag M, Moinard N, Auger I et al. Epitopes of human fibrin recognized by the rheumatoid arthritis-specific autoantibodies to citrullinated proteins. Eur J Immunol 2006; 36:2250-2263.

17 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
[17] Vossenaar ER, Despres N, Lapointe E et al. Rheumatoid arthritis specific anti-Sa antibodies
target citrullinated vimentin. Arthritis Res Ther 2004; 6:R142-50.
[18] Wright HL, Moots RJ, Edwards SW The multifactorial role of neutrophils in rheumatoid arthritis. Nat Rev Rheumatol 2014; 10:593-601.
[19] Socransky SS, Haffajee AD Periodontal microbial ecology. Periodontol 2000 2005; 38:135-187.
[20] Olsen I, Potempa J Strategies for the inhibition of gingipains for the potential treatment of periodontitis and associated systemic diseases. J Oral Microbiol 2014; 6:10.3402/jom.v6.24800. eCollection 2014.
[21] Vavricka SR, Manser CN, Hediger S et al. Periodontitis and gingivitis in inflammatory bowel disease: a case-control study. Inflamm Bowel Dis 2013; 19:2768-2777.
[22] Kalsi DS, Chopra J, Sood A Association of lipid profile test values, type-2 diabetes mellitus, and periodontitis. Indian J Dent 2015; 6:81-84.
[23] Menegazzo L, Ciciliot S, Poncina N et al. NETosis is induced by high glucose and associated with type 2 diabetes. Acta Diabetol 2015; 52:497-503.
[24] Routsias JG, Goules JD, Goules A et al. Autopathogenic correlation of periodontitis and rheumatoid arthritis. Rheumatology (Oxford) 2011; 50:1189-1193.
[25] Kundu D, Bandyopadhyay P, Nair V et al. Aggressive periodontitis: A clinico-hematological appraisal. J Indian Soc Periodontol 2014; 18:166-171.
[26] Hajishengallis G Periodontitis: from microbial immune subversion to systemic inflammation. Nat Rev Immunol 2015; 15:30-44.
[27] Zenobia C, Hajishengallis G Porphyromonas gingivalis virulence factors involved in subversion of leukocytes and microbial dysbiosis. Virulence 2015; 6:236-243.
[28] Mao S, Park Y, Hasegawa Y et al. Intrinsic apoptotic pathways of gingival epithelial cells modulated by Porphyromonas gingivalis. Cell Microbiol 2007; 9:1997-2007.
[29] McGraw WT, Potempa J, Farley D, Travis J Purification, characterization, and sequence analysis of a potential virulence factor from Porphyromonas gingivalis, peptidylarginine deiminase. Infect Immun 1999; 67:3248-3256.
[30] Konig MF, Paracha AS, Moni M et al. Defining the role of Porphyromonas gingivalis peptidylarginine deiminase (PPAD) in rheumatoid arthritis through the study of PPAD biology. Ann Rheum Dis 2014.
[31] Nesse W, Westra J, van der Wal JE et al. The periodontium of periodontitis patients contains citrullinated proteins which may play a role in ACPA (anti-citrullinated protein antibody) formation. J Clin Periodontol 2012; 39:599-607.
[32] Rosenstein ED, Greenwald RA, Kushner LJ, Weissmann G Hypothesis: the humoral immune response to oral bacteria provides a stimulus for the development of rheumatoid arthritis. Inflammation 2004; 28:311-318.

18 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
[33] Rohrbach AS, Slade DJ, Thompson PR, Mowen KA Activation of PAD4 in NET
formation. Front Immunol 2012; 3:360.
[34] Maresz KJ, Hellvard A, Sroka A et al. Porphyromonas gingivalis facilitates the development and progression of destructive arthritis through its unique bacterial peptidylarginine deiminase (PAD). PLoS Pathog 2013; 9:e1003627.
[35] Luo Y, Arita K, Bhatia M et al. Inhibitors and inactivators of protein arginine deiminase 4: functional and structural characterization. Biochemistry 2006; 45:11727-11736.
[36] Causey CP, Jones JE, Slack JL et al. The development of N-alpha-(2-carboxyl)benzoyl-N(5)-(2-fluoro-1-iminoethyl)-l-ornithine amide (o-F-amidine) and N-alpha-(2-carboxyl)benzoyl-N(5)-(2-chloro-1-iminoethyl)-l-ornithine amide (o-Cl-amidine) as second generation protein arginine deiminase (PAD) inhibitors. J Med Chem 2011; 54:6919-6935.
[37] Jang B, Kim HW, Kim JS et al. Peptidylarginine deiminase inhibition impairs Toll-like
receptor agonist-induced functional maturation of dendritic cells, resulting in the loss of T cell-proliferative capacity: a partial mechanism with therapeutic potential in inflammatory settings. J Leukoc Biol 2015; 97:351-362.
[38] Willis VC, Gizinski AM, Banda NK et al. N-alpha-benzoyl-N5-(2-chloro-1-iminoethyl)-L-ornithine amide, a protein arginine deiminase inhibitor, reduces the severity of murine collagen-induced arthritis. J Immunol 2011; 186:4396-4404.
[39] Chumanevich AA, Causey CP, Knuckley BA et al. Suppression of colitis in mice by Cl-amidine: a novel peptidylarginine deiminase inhibitor. Am J Physiol Gastrointest Liver Physiol 2011; 300:G929-38.
[40] Goulas T, Mizgalska D, Garcia-Ferrer I et al. Structure and mechanism of a bacterial host-protein citrullinating virulence factor, Porphyromonas gingivalis peptidylarginine deiminase. Sci
Rep 2015; 5:11969.
[41] Wegner N, Wait R, Sroka A et al. Peptidylarginine deiminase from Porphyromonas gingivalis citrullinates human fibrinogen and alpha-enolase: implications for autoimmunity in rheumatoid arthritis. Arthritis Rheum 2010; 62:2662-2672.
[42] Pathirana RD, O'Brien-Simpson NM, Visvanathan K et al. Flow cytometric analysis of adherence of Porphyromonas gingivalis to oral epithelial cells. Infect Immun 2007; 75:2484-2492.
[43] Zendman AJ, Raijmakers R, Nijenhuis S et al. ABAP: antibody-based assay for peptidylarginine deiminase activity. Anal Biochem 2007; 369:232-240.
[44] Damgaard D, Senolt L, Nielsen MF et al. Demonstration of extracellular peptidylarginine deiminase (PAD) activity in synovial fluid of patients with rheumatoid arthritis using a novel assay for citrullination of fibrinogen. Arthritis Res Ther 2014; 16:498-014-0498-9.
[45] Neeli I, Radic M Opposition between PKC isoforms regulates histone deimination and neutrophil extracellular chromatin release. Front Immunol 2013; 4:38.
[46] Subramanian V, Knight JS, Parelkar S et al. Design, synthesis, and biological evaluation of tetrazole analogs of Cl-amidine as protein arginine deiminase inhibitors. J Med Chem 2015; 58:1337-1344.

19 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
[47] Laheij AM, van Loveren C, Deng D, de Soet JJ The impact of virulence factors of
Porphyromonas gingivalis on wound healing in vitro. J Oral Microbiol 2015; 7:27543.
[48] Barrientos L, Marin-Esteban V, de Chaisemartin L et al. An improved strategy to recover large fragments of functional human neutrophil extracellular traps. Front Immunol 2013; 4:166.
[49] Walter C, Zahlten J, Schmeck B et al. Porphyromonas gingivalis strain-dependent activation of human endothelial cells. Infect Immun 2004; 72:5910-5918.
[50] Baek KJ, Ji S, Kim YC, Choi Y Association of the invasion ability of Porphyromonas gingivalis with the severity of periodontitis. Virulence 2015; 6:274-281.
[51] Jayaprakash K, Demirel I, Khalaf H, Bengtsson T The role of phagocytosis, oxidative burst and neutrophil extracellular traps in the interaction between neutrophils and the periodontal pathogen Porphyromonas gingivalis. Mol Oral Microbiol 2015.
[52] Knight JS, Zhao W, Luo W et al. Peptidylarginine deiminase inhibition is immunomodulatory and vasculoprotective in murine lupus. J Clin Invest 2013; 123:2981-2993.
[53] Khandpur R, Carmona-Rivera C, Vivekanandan-Giri A et al. NETs are a source of citrullinated autoantigens and stimulate inflammatory responses in rheumatoid arthritis. Sci Transl Med 2013; 5:178ra40.
[54] Sur Chowdhury C, Giaglis S, Walker UA et al. Enhanced neutrophil extracellular trap generation in rheumatoid arthritis: analysis of underlying signal transduction pathways and potential diagnostic utility. Arthritis Res Ther 2014; 16:R122.
[55] Janssen KM, de Smit MJ, Brouwer E et al. Rheumatoid arthritis-associated autoantibodies in non-rheumatoid arthritis patients with mucosal inflammation: a case-control study. Arthritis Res Ther 2015; 17:174-015-0690-6.

20 | R A & P D : T h e r o l e o f n e u t r o p h i l s a n d P . g i n g i v a l i s
Personal Acknowledgements
First of all, I would like to thank Koen Janssen for his supervision, support and technical assistance and Hannie Westra for her supervision and support during my internship at the UMCG.
I also would like to thank Berber Doornbos-Van der Meer, Johan Bijzet, Gerda Horst, Fleur Schaper, Qi Wang, Lucas Lintermans, Aad Withaar, Lisa Schouten, Janneke Pietersma, Alicia Borneman, Nienke Sinnema, Rianne Lovers, Renate Akkerman, Tim Miedema and Gerjan Dekkema for either their technical support in the laboratory, or being so kind to donate blood for my experiments. Or of course, both.
I show equal gratitude to Liesbeth Brouwer, Arjan Vissink, Menke de Smit, Arie-Jan van Winkelhoff, Tim Stobernack and Giorgio Gabarrini and all other participants for our informative RA/PD-Friday meetings.
Special thanks to Marines du Teil and her supervisor Tim Stobernack for providing us the PG cultures and taking care of the co-cultures themselves. Also appreciation for Giorgio Gabarrini is in place for providing us with recombinant PPAD.