Embed Size (px)
Citation preview
202 | Pharmazie in unserer Zeit | 30. Jahrgang 2001 | Nr. 3
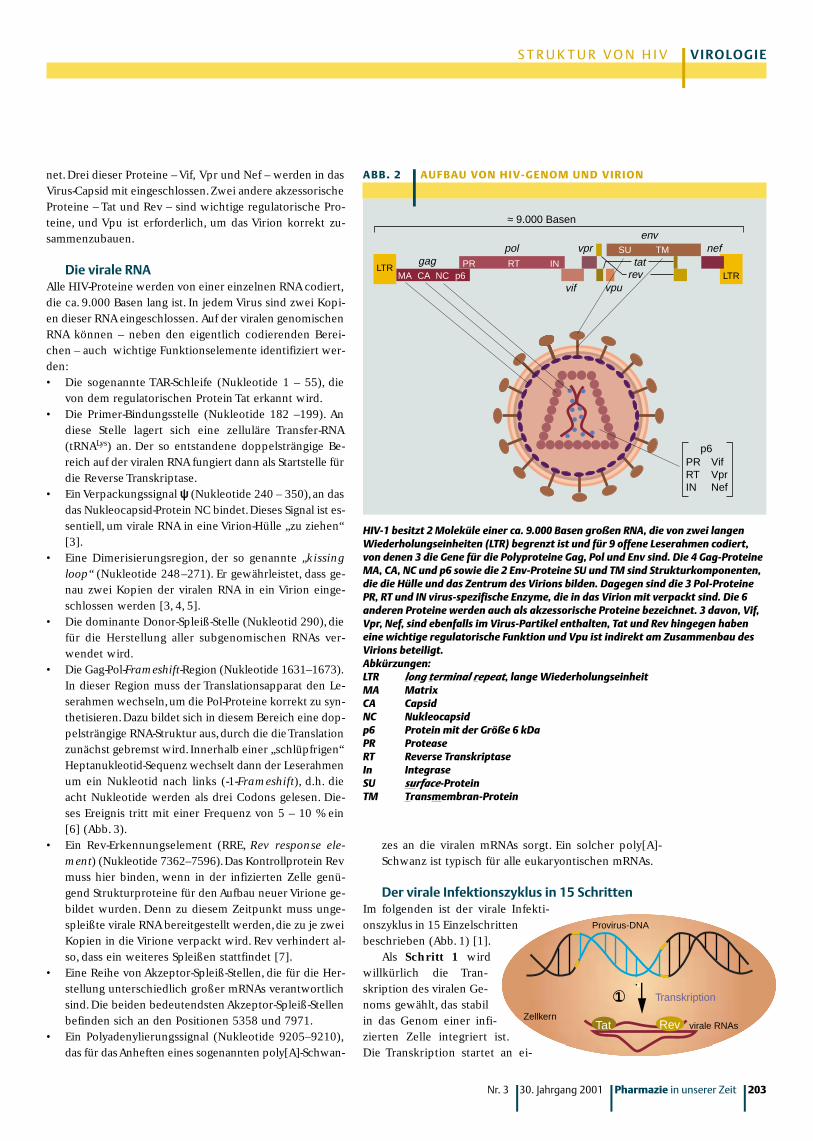
Die HIV-GeneDas HIV-Genom codiert neun offene Leserahmen, von de-nen zwei (tat und rev) durch Spleißen auf der Stufe der mRNA zusammengeführt werden (Abb. 2) [1]. Drei offeneLeserahmen tragen die genetische Information für die typi-schen retroviralen Polyproteine Gag (gruppenspezifischeAntigene), Pol (Reverse Transkriptase) und Env (Hüllpro-teine). Alle drei Polyproteine werden durch Proteasen inweitere Funktionseinheiten zerlegt.• Aus dem Gag-Polyprotein werden vier Proteine heraus-
geschnitten (Abb.2).Dies sind das Matrix-Protein (MA),das Capsid-Protein (CA),das Nukleocapsid-Protein (NC)und p6, eine kleines Protein mit einem Molekularge-wicht von 6 kDa.
• Aus dem Pol-Protein werden drei Enzyme hydrolytischfreigesetzt: Die HIV-Protease (PR), die Reverse Tran-skriptase (RT) und die Integrase (IN).
• Auf dem env-Leserahmen sind die beiden Proteine SU(surface-Protein) und TM (Transmembran-Protein) co-diert. Beide Proteine fungieren als wichtige Struktur-elemente, ohne deren korrekte Ausbildung ein Virionandere Zellen nicht infizieren kann. Als Synonym für SU wird auch gp120 verwendet. TM wird auch als gp41 bezeichnet. Bei diesen beiden Proteinen handeltes sich also um Glykoproteine mit Molekulargewichtenvon 120 bzw. 41 kDa.
Darüber hinaus codiert das HIV-Genom für sechs weitereProteine, die man auch als akzessorische Proteine bezeich-
Innerhalb von eineinhalb Jahrzehnten ist es gelungen, ein tief-greifendes Verständnis zu Struktur und Funktion des huma-nen Immundefizienz-Virus zu erlangen. Dass es gelang, einsolch detailliertes Bild eines so komplexen Virus zu erarbeiten,zeugt von der effektiven Kooperation der beteiligten Wissen-schaftler. Es belegt aber auch, dass eine außergewöhnlicheFörderung der wissenschaftlichen Arbeit zu außergewöhnli-chen Resultaten führen kann. Detaillierte Struktur- und Funk-tionskenntnisse sind die Voraussetzung dafür, rationale Wirk-stoffe entwickeln zu können, die in der Lage sind, effektiv mitder Biologie dieses todbringenden Virus zu interferieren.
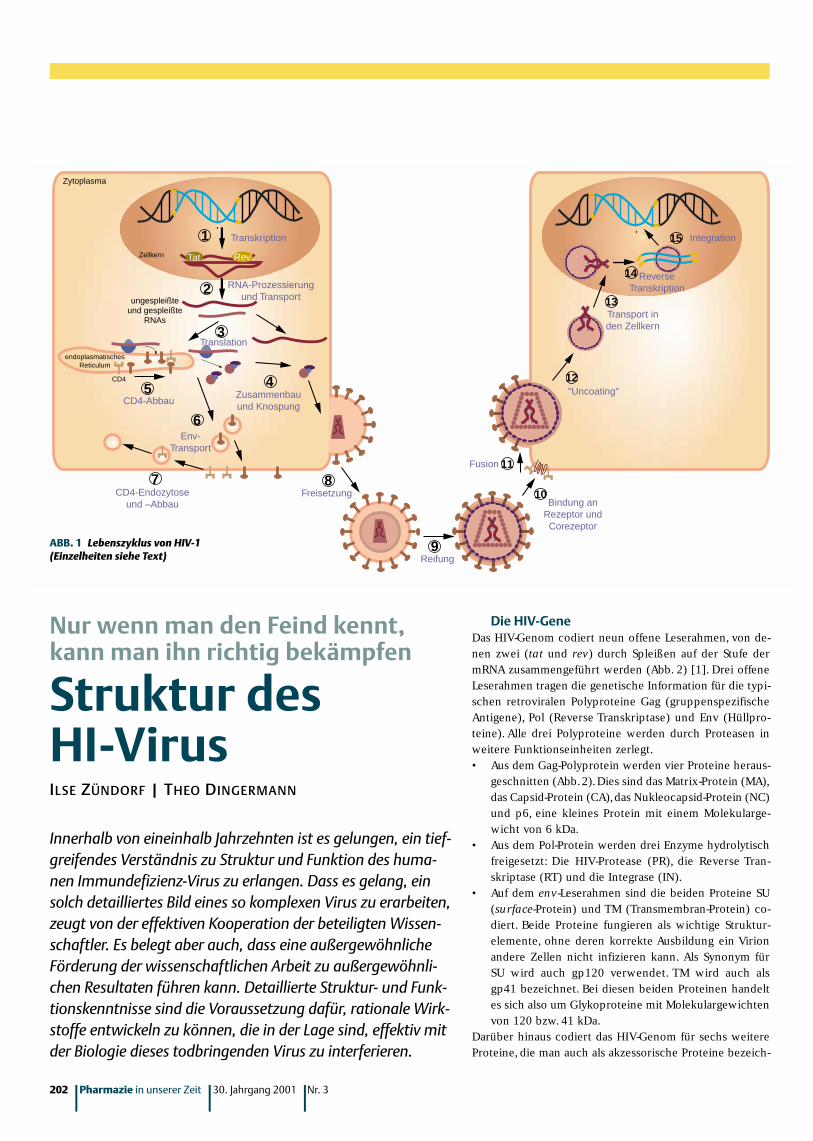
Zellkern
Transkription
Zytoplasma
CD4
Reifung
1Tat
ungespleißteund gespleißte
RNAs
Rev
2 RNA-Prozessierungund Transport
3Translation
endoplasmatischesReticulum
4Zusammenbauund Knospung
5CD4-Abbau
6Env-
Transport
7CD4-Endozytose
und –AbbauFreisetzung
8
9
Fusion 11
Bindung anRezeptor undCorezeptor
10
"Uncoating"12
ReverseTranskription
Transport inden Zellkern
13
14
Integration15
ABB. 1 Lebenszyklus von HIV-1(Einzelheiten siehe Text)
Nur wenn man den Feind kennt,kann man ihn richtig bekämpfen
Struktur des HI-VirusILSE ZÜNDORF | THEO DINGERMANN
Nr. 3 | 30. Jahrgang 2001 | Pharmazie in unserer Zeit | 203
S T R U K T U R V O N H I V | V I RO LO G I E
net.Drei dieser Proteine – Vif, Vpr und Nef – werden in dasVirus-Capsid mit eingeschlossen.Zwei andere akzessorischeProteine – Tat und Rev – sind wichtige regulatorische Pro-teine, und Vpu ist erforderlich, um das Virion korrekt zu-sammenzubauen.
Die virale RNAAlle HIV-Proteine werden von einer einzelnen RNA codiert,die ca. 9.000 Basen lang ist. In jedem Virus sind zwei Kopi-en dieser RNA eingeschlossen. Auf der viralen genomischenRNA können – neben den eigentlich codierenden Berei-chen – auch wichtige Funktionselemente identifiziert wer-den:• Die sogenannte TAR-Schleife (Nukleotide 1 – 55), die
von dem regulatorischen Protein Tat erkannt wird.• Die Primer-Bindungsstelle (Nukleotide 182 –199). An
diese Stelle lagert sich eine zelluläre Transfer-RNA (tRNALys) an. Der so entstandene doppelsträngige Be-reich auf der viralen RNA fungiert dann als Startstelle fürdie Reverse Transkriptase.
• Ein Verpackungssignal ψψ (Nukleotide 240 – 350),an dasdas Nukleocapsid-Protein NC bindet.Dieses Signal ist es-sentiell, um virale RNA in eine Virion-Hülle „zu ziehen“[3].
• Eine Dimerisierungsregion, der so genannte „kissingloop“ (Nukleotide 248–271). Er gewährleistet, dass ge-nau zwei Kopien der viralen RNA in ein Virion einge-schlossen werden [3, 4, 5].
• Die dominante Donor-Spleiß-Stelle (Nukleotid 290),diefür die Herstellung aller subgenomischen RNAs ver-wendet wird.
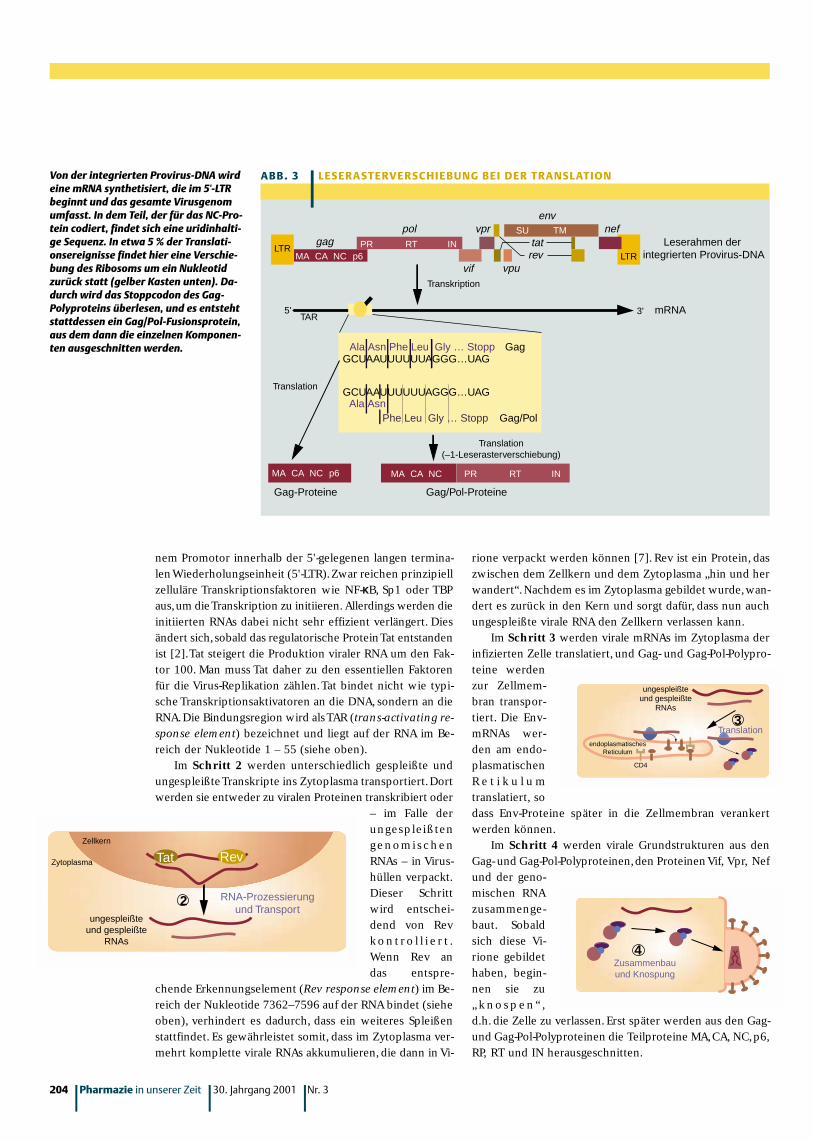
• Die Gag-Pol-Frameshift-Region (Nukleotide 1631–1673).In dieser Region muss der Translationsapparat den Le-serahmen wechseln,um die Pol-Proteine korrekt zu syn-thetisieren.Dazu bildet sich in diesem Bereich eine dop-pelsträngige RNA-Struktur aus,durch die die Translationzunächst gebremst wird. Innerhalb einer „schlüpfrigen“Heptanukleotid-Sequenz wechselt dann der Leserahmenum ein Nukleotid nach links (-1-Frameshift), d.h. dieacht Nukleotide werden als drei Codons gelesen. Die-ses Ereignis tritt mit einer Frequenz von 5 – 10 % ein[6] (Abb. 3).
• Ein Rev-Erkennungselement (RRE, Rev response ele-ment) (Nukleotide 7362–7596).Das Kontrollprotein Revmuss hier binden, wenn in der infizierten Zelle genü-gend Strukturproteine für den Aufbau neuer Virione ge-bildet wurden. Denn zu diesem Zeitpunkt muss unge-spleißte virale RNA bereitgestellt werden,die zu je zweiKopien in die Virione verpackt wird. Rev verhindert al-so, dass ein weiteres Spleißen stattfindet [7].
• Eine Reihe von Akzeptor-Spleiß-Stellen, die für die Her-stellung unterschiedlich großer mRNAs verantwortlichsind. Die beiden bedeutendsten Akzeptor-Spleiß-Stellenbefinden sich an den Positionen 5358 und 7971.
• Ein Polyadenylierungssignal (Nukleotide 9205–9210),das für das Anheften eines sogenannten poly[A]-Schwan-
zes an die viralen mRNAs sorgt. Ein solcher poly[A]-Schwanz ist typisch für alle eukaryontischen mRNAs.
Der virale Infektionszyklus in 15 SchrittenIm folgenden ist der virale Infekti-onszyklus in 15 Einzelschrittenbeschrieben (Abb. 1) [1].
Als Schritt 1 wirdwillkürlich die Tran-skription des viralen Ge-noms gewählt, das stabilin das Genom einer infi-zierten Zelle integriert ist.Die Transkription startet an ei-
MA CA NC p6
gagLTR PR RT IN
pol
vif
vpr SU TM
vpu
tat
env
rev
nef
LTR
p6PR VifRT VprIN Nef
≈ 9.000 Basen
A B B . 2 | AU F BAU VO N H I V- G E N O M U N D V I R I O N
HIV-1 besitzt 2 Moleküle einer ca. 9.000 Basen großen RNA, die von zwei langenWiederholungseinheiten (LTR) begrenzt ist und für 9 offene Leserahmen codiert,von denen 3 die Gene für die Polyproteine Gag, Pol und Env sind. Die 4 Gag-ProteineMA, CA, NC und p6 sowie die 2 Env-Proteine SU und TM sind Strukturkomponenten,die die Hülle und das Zentrum des Virions bilden. Dagegen sind die 3 Pol-ProteinePR, RT und IN virus-spezifische Enzyme, die in das Virion mit verpackt sind. Die 6anderen Proteine werden auch als akzessorische Proteine bezeichnet. 3 davon, Vif,Vpr, Nef, sind ebenfalls im Virus-Partikel enthalten, Tat und Rev hingegen haben eine wichtige regulatorische Funktion und Vpu ist indirekt am Zusammenbau desVirions beteiligt.Abkürzungen:LTR long terminal repeat, lange WiederholungseinheitMA MatrixCA CapsidNC Nukleocapsidp6 Protein mit der Größe 6 kDaPR ProteaseRT Reverse TranskriptaseIn IntegraseSU surface-Protein TM Transmembran-Protein
Zellkern
Transkription1
Tat Rev
Provirus-DNA
virale RNAs
204 | Pharmazie in unserer Zeit | 30. Jahrgang 2001 | Nr. 3
nem Promotor innerhalb der 5'-gelegenen langen termina-len Wiederholungseinheit (5'-LTR).Zwar reichen prinzipiellzelluläre Transkriptionsfaktoren wie NF-κκB, Sp1 oder TBPaus,um die Transkription zu initiieren. Allerdings werden dieinitiierten RNAs dabei nicht sehr effizient verlängert. Diesändert sich,sobald das regulatorische Protein Tat entstandenist [2].Tat steigert die Produktion viraler RNA um den Fak-tor 100. Man muss Tat daher zu den essentiellen Faktorenfür die Virus-Replikation zählen.Tat bindet nicht wie typi-sche Transkriptionsaktivatoren an die DNA, sondern an dieRNA.Die Bindungsregion wird als TAR (trans-activating re-sponse element) bezeichnet und liegt auf der RNA im Be-reich der Nukleotide 1 – 55 (siehe oben).
Im Schritt 2 werden unterschiedlich gespleißte undungespleißte Transkripte ins Zytoplasma transportiert.Dortwerden sie entweder zu viralen Proteinen transkribiert oder
– im Falle derungespleißtengenomischenRNAs – in Virus-hüllen verpackt.Dieser Schrittwird entschei-dend von Revk o n t r o l l i e r t .Wenn Rev andas entspre-
chende Erkennungselement (Rev response element) im Be-reich der Nukleotide 7362–7596 auf der RNA bindet (sieheoben), verhindert es dadurch, dass ein weiteres Spleißenstattfindet. Es gewährleistet somit, dass im Zytoplasma ver-mehrt komplette virale RNAs akkumulieren, die dann in Vi-
rione verpackt werden können [7]. Rev ist ein Protein, daszwischen dem Zellkern und dem Zytoplasma „hin und herwandert“.Nachdem es im Zytoplasma gebildet wurde,wan-dert es zurück in den Kern und sorgt dafür, dass nun auchungespleißte virale RNA den Zellkern verlassen kann.
Im Schritt 3 werden virale mRNAs im Zytoplasma derinfizierten Zelle translatiert, und Gag- und Gag-Pol-Polypro-teine werdenzur Zellmem-bran transpor-tiert. Die Env-mRNAs wer-den am endo-plasmatischenR e t i k u l u mtranslatiert, sodass Env-Proteine später in die Zellmembran verankert werden können.
Im Schritt 4 werden virale Grundstrukturen aus denGag- und Gag-Pol-Polyproteinen,den Proteinen Vif, Vpr, Nefund der geno-mischen RNAzusammenge-baut. Sobaldsich diese Vi-rione gebildethaben, begin-nen sie zu„ k n o s p e n “ ,d.h. die Zelle zu verlassen. Erst später werden aus den Gag-und Gag-Pol-Polyproteinen die Teilproteine MA,CA, NC,p6,RP, RT und IN herausgeschnitten.
MA CA NC p6
gagLTR PR RT IN
pol
vif
vpr SU TM
vpu
tat
env
rev
nef
LTR
Leserahmen der integrierten Provirus-DNA
3'5'TAR
mRNA
Transkription
Translation
GCUAAUUUUUUAGGG…UAGAla Asn Phe Leu Gly … Stopp Gag
GCUAAUUUUUUAGGG…UAGAla Asn
Phe Leu Gly … Stopp Gag/Pol
MA CA NC p6 MA CA NC PR RT IN
Gag-Proteine Gag/Pol-Proteine
Translation(–1-Leserasterverschiebung)
A B B . 3 | L E S E R A S T E RV E R S C H I E B U N G B E I D E R T R A N S L AT I O NVon der integrierten Provirus-DNA wirdeine mRNA synthetisiert, die im 5'-LTRbeginnt und das gesamte Virusgenomumfasst. In dem Teil, der für das NC-Pro-tein codiert, findet sich eine uridinhalti-ge Sequenz. In etwa 5 % der Translati-onsereignisse findet hier eine Verschie-bung des Ribosoms um ein Nukleotidzurück statt (gelber Kasten unten). Da-durch wird das Stoppcodon des Gag-Polyproteins überlesen, und es entstehtstattdessen ein Gag/Pol-Fusionsprotein,aus dem dann die einzelnen Komponen-ten ausgeschnitten werden.
Zellkern
Zytoplasma Tat
ungespleißteund gespleißte
RNAs
Rev
2 RNA-Prozessierungund Transport
CD4
ungespleißteund gespleißte
RNAs
3Translation
endoplasmatischesReticulum
4Zusammenbauund Knospung

Nr. 3 | 30. Jahrgang 2001 | Pharmazie in unserer Zeit | 205
S T R U K T U R V O N H I V | V I RO LO G I E
Vif ist ein Protein mit 192 Aminosäuren. Es scheint füreine hohe Infektiösität der Viren wichtig zu sein, denn Vi-ren, die im Vif-Gen mutiert sind, produzieren instabile Re-plikationsintermediate [8,9]. Auch die Morphologie der Vi-rus-Partikel ist verändert,so dass Vif auch für den viralen Zu-sammenbau und für die Reifung der Virione von Bedeutungsein könnte [8] (siehe auch Schritt 9). Man hat kalkuliert,dass 7 – 100 Vif-Proteine in ein Virion verpackt werden.Dies wiederum könnte bedeuten,dass Vif seine Funktion in-nerhalb der Viruspartikel ausübt.
Vpr wird ebenfalls in die Virushülle eingeschlossen.Die-ses Protein wird wichtig, wenn das Virus in einer anderenZelle wieder seine Hüllen ablegt (siehe Schritt 12).Das Pro-tein besteht aus 96 Aminosäuren. Ganz besonders wichtigscheint dieses Protein zu sein, um in ruhenden Zellen denNukleoprotein-Komplex in den Kern zu transportieren(Schritt 12). Ruhende, d.h. sich nicht teilende Zellen, sindvor allem die Makrophagen. Eine weitere wichtige Funkti-on von Vpr scheint darin zu bestehen, dass es ein Anhaltendes Zellzyklusses in der G2-Phase induzieren kann.Dadurchscheint bei T-Zellen Apoptose induziert zu werden [10].
Nef besteht aus 206 Aminosäuren. Am N-Terminus istNef mit einem Myristinsäure-Molekül modifiziert.Eine wich-tige Funktion von Nef besteht darin, dafür zu sorgen, dassnicht zu viele CD4-Moleküle in der infizierten Zelle syn-thetisiert werden (siehe Schritte 5 und 7). Wäre das derFall, würden sich die CD4-Moleküle und die sich bildendenViren gegenseitig daran hindern, zur Zellmembran zu ge-langen. Auch Nef wird in die Virushülle mit eingeschlossen.Man hat kalkuliert, dass ca. 70 Nef-Moleküle in einem Virusenthalten sind.
Im Schritt 5 muss dafür gesorgt werden,dass die am en-doplasmatischen Retikulum translatierten CD4-Moleküle ab-
gebaut werden.Diese interagie-ren nämlich mitden Env-Prote-inen, die hier –wie in Schritt 3beschrieben –ebenfalls transla-tiert werden.Die
Degradation von CD4-Molekülen wird durch Vpu unter-stützt.Vpu (81 Aminosäuren) ist ein phosphoryliertes Mem-branprotein. Es gleitet quasi durch das zelluläre Membran-netzwerk und spürt so CD4-Moleküle auf.
In Schritt 6 wird Env zunächst als Polyprotein in derZellmembran verankert. Erst später (Schritt 8) wird dieses
Polyprotein indie Teilproteinegp120 (SU) undgp41 (TM) zer-legt. Bei derKnospung um-hüllt sich das Vi-rion mit einer
Schicht aus der zellulären Membran.So gelangt dann auch Env in die äuße-re Membran des HI-Virus.
In Schritt 7 müssen wiederumCD4-Moleküle degradiert bzw. durchEndozytose von der Zellmembran ent-fernt werden,damit sich die gebildetenHI-Virionen auch von der Zelle ablö-sen können. Dabei leistet Nef (sieheoben) einen entscheidenden Beitrag.
Durch die Zerteilung von Env ingp120 und gp41 (Schritt 8) unddurch die Hydrolyse der Gag- und Gag-Pol-Polyproteine durch die HIV-Pro-tease (PR) werden erhebliche mor-phologische Änderungen induziert(Schritt 9). In diesem Stadium scheintauch Vif eine wichtige Rolle zu spielen.Allerdings kennt man die genaue Funk-tion von Vif bisher noch nicht.
CD4
endoplasmatischesReticulum
5CD4-Abbau
7CD4-Endozytose
und –Abbau
Freisetzung8
Reifung9
6Env-Transport
endoplasmatischesReticulum
TA B . 1 a PROT E I N E VO N H I V- 1 : S T R U K T U R PROT E I N E
1. MembranproteineVorläuferproteine gp 160 glykosyliertexternes Glykoprotein gp 120 glykosyliert, Adsorption, Bil-(EP, SU) dung neutralisierender Anti-
körper, Zelltropismustransmembranes gp 41 glykosyliert, Fusion, Bildung Glykoprotein neutralisierender Antikörper
2. gruppenspezifische Antigene (Gag)Gag-Vorläuferprotein pr 55 myristyliert, Bildung virus-
ähnlicher PartikelMatrixprotein (MA) p 17 myristyliert, phosphoryliert
mit der Hüllmembran assoziiert
Capsidprotein (CA) p 24 Strukturprotein des CapsidsNukleocapsidprotein (NC) p 7 Zinkfingermotiv, Assoziation
mit RNA-GenomLink-Protein p 6 Verbindung zwischen Capsid
und Membran
Protein Kurzbezeichnung/ MerkmaleGröße
TA B . 1 b PROT E I N E VO N H I V- 1 : R EG U L ATO R PROT E I N E
Transaktivator der p 9/14 bindet an TAR an den 5'-Enden Transkription (Tat) der RNA und ermöglicht Elon-
gationposttranskriptioneller p 19 bindet an RRE beziehungs-Transaktivator (Rev) weise RxRE und fördert den
Transport einfach gespleißterund ungespleißter mRNA ausdem Kern
Protein Kurzbezeichnung/ MerkmaleGröße

206 | Pharmazie in unserer Zeit | 30. Jahrgang 2001 | Nr. 3
Jedenfalls ist das gereifteVirus nun in der Lage, eineandere Zelle zu infizieren(Schritt 10). Dabei bindetdas aus 515 Aminosäuren be-stehende SU (gp 120) mithoher Affinität (Kd ca. 4 nM)an ein CD4-Molekül. Dies
reicht jedoch für eine effiziente Auf-nahme des Virus in die Zelle nochnicht aus. Co-Rezeptoren sind erfor-derlich.
Durch die Bindung von gp 120an das CD4-Molekül ändert sich dieräumliche Struktur von Env, so dass nun eine zweite Bin-dung mit einem Co-Rezeptor möglich wird [11]. Co-Re-zeptoren für HI-Viren sind Chemokin-Rezeptoren,G-Pro-tein-gekoppelte Rezeptoren, die intrazelluläres Calciummobilisieren und Leukozyten-Chemotaxis induzieren[12].
Man unterscheidet zwei verschiedene HIV-1-Typen:Solche,die in erster Linie Makrophagen infizieren (M-tro-pe HI-Viren) und solche, die T-Zellen infizieren (T-trope
HI-Viren). Als Co-Rezeptor für T-trope Viren fungiert derCXCR4/Fusin-Rezeptor. M-trope Viren können diesen Co-
Rezeptor nicht nutzen. Sie binden vielmehr anden CCR5.
Hat das Virus gebunden, verändert TM(gp41) seine Konformation, wodurch die äuße-re Virus-Membran und die Membran der„attackierten“ Zelle fusionieren können(Schritt 11).
In Schritt 12 verliert das Virus auch das Core-ProteinCA,so dass nun ein viraler Nukleoprotein-Komplex aus MA,RT, IN,Vpr und den beiden RNA-Kopien freigelegt wird.
Dieser Komplex wird in Schritt 13 in den Zellkerntransportiert, wo die virale RNA durch die Reverse Tran-
skriptase (RT) in eine doppel-strängige, lineare DNA transkri-biert wird (Schritt 14).
Der virale Infektionszykluswird geschlossen, indem die In-tegrase (IN) die virale DNA indas Wirtsgenom integriert(Schritt 15).
R E V E R S E T R A N S K R I P T I O N|Die Reverse Transkription beginnt am 3’-Ende eines tRNA-Moleküls, das in der Nähe des 5’-Endes der Virus-RNA bindet.Am effektivsten startet die Reverse Transkription an einem tRNALys, das bereits beim Zusammenbau des Virions mit in-korporiert wird. Das Enzym Reverse Transkriptase katalysiertdann sowohl die RNA-abhängige als auch die DNA-abhängigeDNA-Polymerisation. Gleichzeitig hat das Enzym eine RNase-H-Domäne, die die RNA innerhalb des RNA-DNA-Hybrids zer-schneidet. Die eigentliche Reaktion findet wahrscheinlichnach dem „uncoating“ des Virus im Zytoplasma statt. Ab-geschlossen ist die Reverse Transkription im Zellkern, wo derDNA-Doppelstrang in das Genom der Wirtszelle intergriertwird. Darüber lässt sich auch erklären, weshalb einige Auto-ren die Reverse Transkription in ihren Abbildungen im Zyto-plasma ansiedeln (siehe Clement- und Schake-Artikel),während andere Quellen das Ende der Reaktion im Zellkernmarkieren (siehe Dingermann-Artikel).
Bindung anRezeptor undCorezeptor
10
Fusion 11
"Uncoating"12
Transport inden Zellkern
13
Die besondere Bedeutung der HIV-ProteaseEin HIV-Partikel wird erst dann infektiös, wenn Gag unddas Gag-Pol-Polyprotein durch die HIV-Protease (PR) pro-zessiert werden (Schritt 9). Der genaue zeitliche Verlaufdieser wichtigen Prozesse ist nicht bekannt.Klar ist jedoch,dass durch die Aktivität der HIV-Protease die oben be-schriebenen wichtige Produkte entstehen.
Die HIV-Protease funktioniert als ein Dimer und istzunächst Teil des Pol-Proteins. Durch Autokatalyse „schnei-det“ sie sich förmlich selber aus dem Polyprotein heraus.Bei diesem Prozess könnten die benachbart liegenden p6-Sequenzen eine regulatorische Rolle spielen [13]. Die un-terschiedlichen Substratstellen für die HIV-Protease werdenmit deutlich unterschiedlichen Effizienzen geschnitten.Wirddieser Prozess gestört – beispielsweise indem man die Pro-tease überexprimiert – nimmt die Infektiösität der Virendeutlich ab.
Die zentrale funktionelle Rolle der HIV-Protease bei derVirus-Reifung erklärt die besondere Stellung dieses Molekülsunter den Zielstrukturen für HIV-Virustatika.
TA B . 1 c PROT E I N E VO N H I V- 1 : E N Z Y M E
Gag/Pol-Vorläuferprotein pr 160Reverse Transkriptase (RT) p 51/66 RNA- und DNA-abhängige Po-
lymerase, RNase HProtease (PR) p 9 spaltet Gag- und Gag/Pol-Vor-
läuferproteine in die Einzel-komponenten
Integrase (IN) p 38 Endonuclease, Ligase, verant-wortlich für die Integrationdes des Virusgenoms in dieWirts-DNA
Protein Kurzbezeichnung/ MerkmaleGröße
Nr. 3 | 30. Jahrgang 2001 | Pharmazie in unserer Zeit | 207
S T R U K T U R V O N H I V | V I RO LO G I E
TA B . 1 d PROT E I N E VO N H I V- 1 : H I L F S PROT E I N E
Virion-Infektionsfaktor (Vif) p 23 beschleunigt die Infektion inperipheren Blutlymphozyten
Virion-assoziiertes p 11/p 15 Virion-assoziiert, Protein R (Vpr) Transaktivatorvirales Protein U (Vpu) p 14/16 phosphoryliert, mit der Mem-
bran des EndoplasmatischenReticulums assoziiert, verhin-dert die intrazelluläre Kom-plexbildung zwischen gp 160und CD4-Rezeptoren
Nef-Protein p 25/27 myristyliert, Ähnlichkeit mit G-Proteinen, teilweise mit derZytoplasmamembranassoziiert
Strukturproteine Kurzbezeichnung/ MerkmaleGröße
Zitierte Literatur:[1] Frankel, A.D., Young, J.A.: HIV-1: fifteen proteins and an RNA. Annu.
Rev. Biochem. 67 (1998): 1 - 25[2] Jones, K.A., Peterlin, B.M.: Control of RNA initiation and elongation
at the HIV-1 promoter. Annu. Rev. Biochem. 63 (1994): 717-743[3] Clever, J.L, Parslow, T.G.: Mutant human immunodeficiency virus ty-
pe 1 genomes with defects in RNA dimerization or encapsidation. J.Virol. 71 (1997) 3407-3414
[4] Laughrea, M., Jette, L., Mak, J., Kleiman, L., Liang C., Wainberg,M.A.: Mutations in the kissing-loop hairpin of human immunodefi-ciency virus type 1 reduce viral infectivity as well as genomic RNApackaging and dimerization. J. Virol. 71 (1997) 3397-3406
[5] Paillart, J.C., Skripkin, E., Ehresmann, B., Ehresmann, C., Marquet,R.: The use of chemical modification interference and inverse PCRmutagenesis to identify the dimerization initiation site of HIV-1 ge-nomic RNA. Pharm Acta Helv 71 (1996) 21-8
[6] Cassan, M., Delaunay, N., Vaquero, C., Rouset, J.P.: Translational fra-meshifting at the gag-pol junction of human immunodeficiency vi-rus type 1 is not increased in infected T-lymphoid cells. J. Virol. 71(1994) 1501-1508
[7] Hope, T.J.: Viral RNA export. Chem. Biol. 4 (1997) 335-344.[8] Cohen, E.A., Subbramanian, R.A., Gottlinger, H.G.: Role of auxiliary
proteins in retroviral morphogenesis. Curr. Top. Microbial. Immun-ol. 214 (1996) 219-235.
[9] Simon, J.H., Malim, M.H.: The human immunodeficiency virus type1 Vif protein modulates the postpenetration stability of viralnucleoprotein complexes. J. Virol. 70 (1996) 5297-5305.
[10] Emerman, M.: HIV-1, Vpr and the cell cycle. Curr. Biol. 6 (1996)1096-1103.
[11] Clapham, P.R.: HIV and chemokines: ligands sharing cell-surface re-ceptors. Trends Cell Biol. 7 (1997) 264 - 268
[12] Clapham, P.R., Weiss, R.A.: Immunodeficiency viruses. Spoilt forchoice of co-receptors. Nature 388 (1997) 230 - 231
[13] Zybarth, G., Carter, C.: Domains upstream of the protease (PR) inhuman immunodeficiency virus type 1 Gag-Pol influence PR auto-processing. J. Virol. 69 (1995) 3878 - 3884
Zu den AutorenDr. Ilse Zündorf, geb. 1965. Studium der Biologie inErlangen. Promotion 1991–1995 in Frankfurt a.M.1994 Forschungsaufenthalt an der University ofKentucky in Lexington. Seit 1995 Akademische Rätinan der Johann Wolfgang Goethe-Universität inFrankfurt a.M. Forschungsschwerpunkt: Mono-klonale Antikörper gegen sekundäre Pflanzenin-haltsstoffe und Transmittersubstanzen, Herstellungund Modifikation rekombinanter Antikörper-fragmente.
Prof. Dr. Theodor Dingermann, geb. 1948. Studiumder Pharmazie in Erlangen. Promotion 1980 im FachBiochemie. Zweijähriger Postdoc-Aufenthalt an derYale-University USA. 1987 Habilitation in denFächern Biochemie und Molekularbiologie. 1990 Rufauf eine C4-Professur an die Johann WolfgangGoethe-Universität in Frankfurt a.M. Mitglied imwissenschaftlichen Beirat der Bundesapothekerkam-mer und im Vorstand der Akademie für Pharmazeu-tische Fortbildung der LAK Hessen. Vorsitzender derArbeitsgruppe „Biotechnologisch hergestellte Arz-neimittel“ und stellvertretender Vorsitzender desAusschusses „Pharmazeutische Biologie“ der Arznei-buchkommission. Mitglied der Deligiertenversamm-lung der LAK Hessen. 1998–2000 Vizepräsident derJohann Wolfgang Goethe-Universität Frankfurt. Von1996 bis 1999 Vizepräsident und seit 2000 Präsi-dent der Deutschen Pharmazeutischen Gesellschaft.
Anschrift:Dr. Ilse Zündorf und Prof. Theo Dingermann,Institut für Pharmazeutische Biologie,Johann Wolfgang Goethe-Universität,
Marie-Curie-Str. 9, 60439 Frankfurt a.M. E-mail: [email protected], [email protected]





























