Embed Size (px)
Citation preview

1
University of Bremen
International Max Planck Research School for Marine Microbiology
GENOMIC ANALYSIS OF THE ENDOSYMBIOTIC COMMUNITY OF A
GUTLESS MARINE WORM (OLAVIUS ALGARVENSIS)
by
Amelia Elena Rotaru
Supervised
by
Nicole Dubilier
A thesis submitted in partial fulfillment of the
requirements for the degree of
Master of Science in Marine Microbiology
Max Planck Institute for Marine Microbiology,
28359 Bremen, Germany
April, 2005

2
Erklärung
Statement
Hiermit versichere ich, dass ich diese Arbeit selbständig verfasst und keine anderen als die angegebenen Quellen und Hilfsmittel verwendet habe. I herewith confirm that I have written this thesis unaided and that I used no other resources than those mentioned. _________________________ _________________________________ (Ort und Datum / Place and Date) (Unterschrift / Signature )

3
Table of content
Summary
Chapter 1: Introduction
1.1. General introduction to marine oligochaetes symbioses
1.2. Molecular identification and phylogeny of O. algarvensis endosymbionts
1.3. Function of O. algarvensis symbionts
1.4. Metagenomics: microbial community genome analyses
Chapter 2: Materials and methods
2.1. Symbionts DNA isolation and BAC library construction
2.2. Plasmid preparation
2.3. BAC insert size determination via pulsed field gel electrophoresis
2.4. BAC sequencing and primer walking
2.5. Primer design
2.6. Sequence analysis
Chapter 3: Results
3.1. Symbionts DNA isolation and BAC library construction
3.2. Sequence analysis and primer walking
Chapter 4: Discussions
4.1. Carbon metabolism
4.2. Sulfur metabolism
4.3. Nitrogen metabolism
4.4. Transport proteins
4.5. Pathogenic / symbiotic associated traits
Annexes
Acknowledgements

4
Summary
The gutless marine worm, Olavius algarvensis, harbors a microbial community of five co-
occurring symbionts belonging to sulfide-oxidizers, sulfate-reducers, and spirochetes
(Rühland et al., In Prep.). In order to get access to the genetic capabilities of these symbionts
a metagenomic approach was used. Previously performed clone insert end-sequencing of the
entire BAC library had revealed sequences that are considered to be relevant for the
symbiotic community. In this study clones carrying interesting genes were further analyzed
using primer walking, in order to proceed further into the insert. For the obtained contiguous
sequences similarities were searched against the non-redundant GenBank protein database
applying the BLASTx algorithm.
Several sequences attracted special attention because their deduced amino acid sequences
exhibited a strong similarity to proteins involved in both oxidative and reductive sulfur
metabolisms. This indicates that the analyzed genomic fragments are derived from sulfur-
oxidizing and sulfate-reducing symbionts. Some of the genes offer insights into the
symbionts metabolic pathways that are presently not known. Particularly the degradation of
urea, the respiration of nitrate, one carbon metabolism, and the synthesis of cyanophycin
demand attention in the near future. Genes putatively encoding virulence associated proteins
can provide information on how the association between bacteria and oligochaetes has
developed and how bacteria adapt to host dependent lifestyle.
Although it has to be taken into consideration that the assigned functions by similarity search
is potential and that the actual role can be only validated experimentally, the sequence
information of the bacterial symbionts from O. algarvensis, obtained in this work, offers the
opportunity to examine the metabolic pathways and genetic interactions within the symbiotic
bacterial consortium in more detail, expanding the understanding of this symbiosis.

5
1. Chapter 1: Introduction
1.1. General introduction to marine oligochaete symbioses
Symbiosis, an interdependent relationship between two species is an important driver of
evolutionary novelty and ecological diversity (Margulis and Chapman, 1998; Wernegreen,
2004). The ties between symbiotic partners range from rather loose and occasional
associations over regular ectosymbiosis to obligatory incorporation into the host body. While
in ectosymbiosis the bacteria occur externally on the host surface, in endosymbiosis the
microbial partner lives within the host either extra- or intracellularly. Symbioses are diverse
and ubiquitous in aquatic and terrestrial habitats, and are particularly prominent in marine
ecosystems, such as coral reefs, hydrothermal vents or shallow waters. The first symbiotic
association between chemoautotrophic sulfur-oxidizing bacteria and marine invertebrates
was proposed for the tube worm Riftia pachyptila, inhabitant of the hydrothermal vent fauna
(Cavanaugh et al., 1981; Felbeck, 1981).
Within the annelid class Oligochaeta two genera of marine Tubificidae, Inanidrilus and
Olavius, are characterized by obligate endosymbiosis with extracellular bacteria. Gutless
marine oligochaetes are abundant in marine sediments through the tropics and subtropics of
the world and approximately 80 species have been described (Erseus, 1979; Felbeck, 1981;
Giere et al., 1998; Erseus, 2003). These marine worms are lacking completely both their
digestive and excretory systems having no gut, mouth or nephridia. Given the complete
reduction of digestive and excretory system an obligate association with autotrophic bacteria
is clearly obligate for the oligochaete partner (Giere et al., 1995; Dubilier et al., 2001).
Olavius algarvensis was first found in the subtidal sands of Algarve, Portugal and described
as a small tubificid worm (0.1 mm x 15-25 mm) found in subsurface layers at depths of 10-
15 cm (Giere et al., 1998). Later on, O. algarvensis was also found off the coast of Elba, Italy
in coarse-grained sands surrounding sea grass (Dubilier et al., 2001). O. algarvensis
endosymbiotic community was under focus, particularly the bacterial mutualistic
relationships in addition to their symbiotic relation to the host ((Dubilier et al., 2001).
Ultrastructural studies were performed on O. algarvensis and was observed a similar

6
arrangement of the symbionts as in other gutless species, occurring in a multicellular layer in
the subcuticular space between the cuticle and epidermis, called the symbiotic region (Giere
et al., 1998; Dubilier et al., 2001). Transmission electron micrography (TEM) revealed two
morphotypes: i) a large oval morphotype of 3-5 µm diameter with numerous intracellular
inclusions, ii) a smaller rod-shaped morphotype of 0.5-1 µm diameter without any inclusions
(Giere et al., 1995; Dubilier et al., 2001).
1.2 Molecular identification and phylogeny of O. algarvensis endosymbionts
In order to study the phylogeny of the bacterial symbionts the 16S ribosomal RNA gene was
used as a phylogenetic anchor. The full cycle rRNA approach (Amann et al., 1995) has been
used to correlate the morphotype of the symbionts with their phylotype. This approach
combines comparative rRNA sequence analysis with fluorescence in situ hybridization
(FISH).
The full cycle rRNA approach revealed five endosymbiotic phylotypes in O. algarvensis.
The large bacterial morphotype, with sulfur globules and polyhydroxybutyrate (PHB)
vesicles has been identified as a Gamma 1 phylotype and is closely related to the
endosymbionts of other marine oligochaetes (e.g. O. crassitunicatus, O. ilvae, I.
leukodermatus). These symbionts are distributed throughout the entire symbiotic region
positioned below the thin cuticle of the worm. The second gamma proteobacterial symbiont
is closely related to clone sequences from cold-seep communities form the Japan Trench (Li
et al., 1999c) and FISH analyses showed that corresponded to the smaller symbiotic
morphotype (Rühland et al., In Prep.; Dubilier et al., In press).
Two delta proteobacterial symbionts were found in O. algarvensis, both belonging to
Desulfococcus/Desulfosarcina/Desulfonema subgroup of sulfate-reducing bacteria. In situ
hybridization studies showed that these symbionts are small and cocci shaped and they are
distributed through the entire symbiotic region. The delta proteobacterial symbionts are in
close contact with the gamma proteobacterial symbionts suggesting a dependency on each
others metabolites (Rühland et al., In Prep.; Dubilier et al., In press).
The spirochete symbiont falls on a neighboring branch with free living marine spirochetes,
like Spirochaeta isovalerica (Harwood & Canale-Parola, 1983) and S. litoralis (Hespel &

7
Canale-Parola, 1973). TEM analyses of O. algarvensis showed no spirochete morphotype;
instead FISH with spirochete specific probes provided evidence for their presence (Rühland
et al., In Prep.).
1.3 Function of O. algarvensis symbionts
1.3.1. Physiological diversity
For long time was considered that symbiosis between bacteria and invertebrates is fueled by
symbionts chemoautotrophy. Chemoautotrophic bacteria derive metabolically useful energy
from the oxidation of inorganic compounds such as hydrogen, carbon monoxide, inorganic
reduced sulfur and nitrogen compounds and from divalent ions (Lengeler et al., 1999).
Thioautotrophs obtain energy by oxidation of reduced sulfur while their carbon is
incorporated from carbon dioxide via Calvin cycle.
The gamma proteobacterial symbionts from O. algarvensis were determined as being
evolutionary related to sulfur-oxidizing bacteria like A. vinosum. TEM analysis showed the
presence of sulfur globules in Gamma 1 symbiont and elemental sulfur is considered a good
indicator for chemoautotrophic symbioses (Fisher, 1990). Furthermore combined molecular
and immunocytochemical studies brought the final proof for the thioautotrophic nature of
Gamma 1 symbiont. Immunocytochemical analyses with an antiserum directed against the
form I of ribulose bisphosphate carboxilase consistently labeled this symbiont (Dubilier et
al., 2001). 16S rRNA FISH with probes designed for Gamma 1 phylotype together with TEM
and the immunocytochemical studies linked the phylogeny and morphology with the
autotrophic function of Gamma 1 in the endosymbiotic community.
The physiologic nature of Gamma 2 is presently unknown while their evolutionary
relationship with cold seeps bacteria suggest that they may also participate in chemosynthetic
pathways. Currently immunofluorescence studies with antiserum against the form I and II of
RubisCO are being used for a greater understanding of their metabolism (Rühland et al., In
Prep.; Dubilier et al., In press). The lack of any intracellular sulfur inclusions is not a proof
against thiotrophy (Fisher, 1990). It is well known that the oxidation of reduced sulfur
compounds is not performed using a single unifying enzymatic pathway and that many

8
sulfur-oxidizing bacteria sulfur-oxidizing bacteria do not store elemental sulfur (Robertson
and Kuenen, 1992; Kelly et al., 1997; Kappler and Dahl, 2001).
The delta proteobacterial symbionts, Delta 1 and Delta 2, are sulfate reducing bacteria based
on their close phylogenetic relationship to free living sulfate reducers, the presence of the
enzyme that catalyze the reduction of sulfite to sulfide, dissimilatory sulfite reductase, and
the detection of sulfate reduction rates at comparable levels with those of free-living sulfate
reducers (Dubilier et al., 2001).
Sulfate reducing bacteria (SRB) respire sulfate to sulfide and oxidize their substrates either
partially to acetate or completely to CO2 ((Widdel and Hansen, 1992a). Some sulfate reducers
(e.g Desulfovibrio) are non acetate oxidizers which utilize lactate, ethanol, short fatty acids
(succinate, fumarate, malate) and pyruvate, as electron donors. The resulting metabolic end
product is acetate. Other sulfate reducers are acetate oxidizers (e.g. Desulfobacter,
Desulfosarcina) and they are able to use acetate and other substrates like lactate, higher fatty
acids and phenyl-substituted organic acids. For these acetate oxidizers the end product is
CO2. Some acetate and non-acetate oxidizerst are able to use molecular hydrogen as an
electron donor. (Widdel and Hansen, 1992a)
Both O. algarvensis delta symbionts were classified by molecular analyses as part of the
Desulfosarcina/Desulfococcus/Desulfonema group. SRB are metabolically diverse in
particular within the Desulfosarcina group, where both chemoorganotrophy and
chemoautotrophy occurs and dissolved organic carbon and hydrogen are possible sources of
reducing power. Hypothesizing that endosymbiotic sulfate reducers take up fermentation
products (succinate, acetate, propionate) from a host that would otherwise excrete them
(Dubilier, 2004) and knowing that the free living relatives of the symbionts produce CO2 the
following questions arise. Is CO2 used by the chemoautotrophic symbiont so that there is no
need anymore for an external CO2 source? Can the delta proteobacterial symbionts use H2,
given that many SRB can utilize molecular hydrogen as an electron donor (facultative
chemolithotrophs)? During this mode of energy conservation cell material may be
synthesized from acetate and CO2 (chemolithoheterotrophs) or only from CO2 (autotrophs).
Perhaps one phylotype is using the fermentation products of the host and the other phylotype
uses molecular hydrogen. As the Desulfosarcina group is so diverse metabolically it is

9
possible for the two different delta endosymbionts to perform different functions in the
community (Dubilier et al., In press).
The spirochete symbiont is closely related to free living spirochetes like Spirochaeta litoralis
and S. isovalerica isolated from marine sediments and salt marshes. Spirochetes are
extremely diverse physiologically ranging from aerobes to facultative and obligate
anaerobes. Within these groupings there is a large diversity of nutritional requirements and
energy yielding mechanisms. Some spirochetes derive their energy exclusively from the
fermentation of plant polymers; others ferment a wide variety of sugars and amino acids,
while some dissimilate only certain long-chain fatty acids. Spirochaeta litoralis and S.
isovalerica are free living, strictly anaerobic spirochetes which are able to ferment glucose
mainly to acetate, ethanol, CO2 and H2 (Hespell and Canale-Parola, 1973; Harwood and
Canale-Parola, 1983, 1984).A spirochete symbiont utilizing glucose would be
disadvantageous for the worm. Recent research has showed that the metabolic possibilities of
spirochete symbionts are even broader than fermentation. Termite symbiotic spirochetes
were discovered as being chemoautotrophs, with the ability of H2-CO2 acetogenesis
(Leadbetter et al., 1999) and dinitrogen fixation (Lilburn et al., 2001). This type of
metabolism would be beneficial for the oligochaete host, providing an important carbon and
energy source beside the source of nitrogen (Dubilier et al., In press).
1.3.2. Mutualistic relationships: syntrophic sulfur cycle
The discovery of multiple symbionts was controversial and contrasting theories arouse about
whether competition or cooperation occurs between the symbionts. Recent advances in the
molecular characterization of uncultivable organisms, which many symbionts are, revealed a
large diversity of mutualistic relationships. Gutless oligochaetes are a good example for the
successful evolution of mutualistic associations with multiple symbionts. The sulfide-
oxidizing and sulfate-reducing symbionts do not appear to compete for host derived
resources, but cooperate instead in the use of resources from each other and the environment
(Dubilier, 2004).
The coexistence of sulfate reducing and sulfide oxidizing bacteria as endosymbionts in O.
algarvensis indicates that these are engaged in a syntrophic sulfur cycle in which oxidized

10
and reduced sulfur compounds are recycled between the symbionts (Fig. 1) (Dubilier et al.,
2001). The SRB are producing reduced sulfur compounds using as reducing power either
organic carbon or hydrogen. The reduced sulfur compounds are used by the sulfide oxidizer
as electron donors for autotrophic CO2 fixation via Calvin cycle. This syntrophic sulfur cycle
resembles a “perpetuum mobile”, and it is necessary for both host and symbionts to take up
form the environment energy sources like dissolved organic compounds (Dubilier et al.,
2001; Dubilier, 2004; Dubilier et al., In press). The role of the spirochete in this syntrophy is
unclear.
1.3.3. Host-symbiont relations: possible interactions
The existence of a sulfide oxidizer, the Gamma 1 symbiont, that donates
chemoautotrophically synthesized organic carbon to its host, is clearly benefic for the host,
nourishing the worm. While anaerobic metabolites like succinate, acetate, and propionate
produced by the worm when oxygen is limited might be taken up by the endosymbiotic
sulfate reducers. The absence of excretory and digestive systems for these oligochaetes made
them dependent on their symbionts The cycling of metabolic products between symbionts
could increase their energy yields, which in turn would benefit the host. SRB supply also the
sulfide oxidizing bacteria with internal produced sulfide, creating independence of
environmental sulfide sources. This increases the ability of the worm to colonize new
habitats with low sulfide concentration. Another advantage of having genetically diverse
symbionts would be the host versatility in adapting different environments (Dubilier, 2004).
Fig 1.Model of the endosymbiotic sulfur cycle in O.
algarvensis showing syntrophic cycling of oxidized
and reduced sulfur compounds between SRB and
sulfide-oxidizing symbionts (Dubilier et al., 2001 )

11
1.4. Metagenomics: microbial community genome analyses
The pure culture approach to the study of the microbial world seriously constrained the view
of microbial diversity because most microbes defy cultivation by standard methods (Pace,
1997)The study of oligochaete symbiotic associations remains limited by the fact that
bacterial symbionts are uncultivable despite numerous attempts. Nowadays, new tools are
available for investigating these associations without separating the partners; one of them, the
metagenomic approach, is defined as culture independent genomic analyses of microbial
communities (Schloss and Handelsman, 2003; Riesenfeld et al., 2004)
Two approaches functional driven analyses and sequence driven analyses have emerged to
extract information from metagenomic libraries. Functional analyses are based on
identification of clones that express a desired trait, followed by characterization of the active
clones by sequence and biochemical analysis. The limitations of this approach are that it
requires expression of the function of interest in the host cells and clustering of all of the
genes required for the function. It also depends on the availability of an assay for the function
of interest that can be performed efficiently on vast clone libraries, knowing that the
frequency of active clones is quite low (Schloss and Handelsman, 2003; Riesenfeld et al.,
2004). Sequence based approach relies on the use of conserved DNA sequences to design
hybridization probes or primers in order to screen metagenomic libraries for clones
containing genes of interest. This has proved effective for identification of clones carrying
phylogenetic anchors and genes encoding enzymes with highly conserved domains. (Piel,
2002) Significant discoveries resulted have also resulted from random sequencing of
metagenomic clones (Beja, 2004). There are contradictory opinions on the utility of random
sequencing of metagenomic clones, the method being considered too indirect to yield
biological understanding, but it is stressed that there is so little known about some divisions
of bacteria that any genomic sequence is helpful in guiding the design of new experiments to
reveal their biology (Schloss and Handelsman, 2003; Riesenfeld et al., 2004). Taking in
consideration advantages and disadvantages of both approaches can be emphasized that
function driven approach has the potential to identify genes that would not be recognizable
based on their sequences, but sequence based screening can identify sequences that are not

12
possible to express in the host species carrying the library. A combination of sequence-based
methods and functional screening is critical in order to define the full diversity of gene
function in the libraries. (Riesenfeld et al., 2004)
Metagenomic technology has been successful at all scales-it has been used to study single
genes e.g. for celullases (Healy et al., 1995), pathways e.g. antibiotic synthesis (Rondon et
al., 2000), organisms e.g. Archaea (Stein et al., 1996) and communities e.g. acid mine
drainage biofilm (Tyson et al., 2004). The ultimate goal of any genomic analyses is to use the
information for a better understanding of the biology of the organism, in the present study -
the biology of O algarvensis endosymbionts.
The objective of the study of O. algarvensis symbionts using the metagenomic approach is to
obtain information on the interactions between the endosymbionts with their host and their
environment. Therefore important issues are under question. How has the bacterial genome
been modified in the context of the demands of symbioses? What pathways are used for the
syntrophic interactions between the bacterial symbionts? Do the bacteria compete with each
other for substrates from the host or the environment? Which metabolic pathways are used by
the different symbionts to sequester inorganic and organic carbon, sulfur, and nitrogen from
the environment? (Dubilier et al., Proposal for the Community Sequencing Programm of the
Joint Genome Institute, USA).
The present survey is based on sequence-driven metagenomic analysis of a BAC (Bacterial
Artificial Chromosome) library, using vector primed insert sequencing and continuing with
primer walking. The obtained contiguous sequences were subjected to similarity searches
using BLASTx which aligns their inferred translation products against protein sequences
from protein databases. This project is focused on providing row data in order to become able
to discern interaction patterns in between symbiotic partners and also with their oligochaete
host. The goal of the project is to assemble as many fragments as possible to gain a better
understanding of the endosymbionts genetic potential and metabolic diversity.

13
2. Chapter 2: Materials and Methods
2.1. Symbionts DNA isolation and BAC library construction
DNA isolation and BAC library construction was performed at Texas A&M University,
Texas, USA, by Hong-Bin Zhang, Chengcang Wu, and Zhanyou Xu in 2004. High molecular
weight DNA was extracted from a pooled sample of approximately 500 Olavius algarvensis
individuals. The extracted DNA was used for the BAC library construction. Enzymatic
restriction was carried out and restriction fragments were ligated into the cloning vector
pECBAC1. Competent E. coli cells, strain ElectroMAX DH10B were transformed by
electroporation.
2.2. Plasmid preparation
For BAC DNA extraction the precultured, transformed Escherichia coli cells were grown 18
to 20 hours at 37°C in 5 ml Luria Bertani medium, containing 17 mg/l chloramphenicol (see
below). Addition of chloramphenicol assures that only transformed E. coli cells are growing.
The following BAC extraction protocol represents the standard alkaline lysis method
(Sambrook et al., 1989) and contains the next important steps: removal of Luria Broth, lysis
of the cells, DNA precipitation and purification.
Transformed E. coli cells from the overnight culture were precipitated by centrifuging on an
Eppendorf 5810 R centrifuge (Eppendorf) at 3000 rpm for 10 min. The supernatant was
removed to eliminate the Luria Broth. The remaining pellet was transferred into a 2 ml
microcentrifuge tube and resuspended by adding 200 µl alkaline lysis solution I (glucose 50
mM / EDTA 10 mM / TrisHCl 25 mM). The suspension was mixed by inverting the tube and
incubated on ice for 5 minutes. The bacterial cells were lysed in 400 µl solution II (SDS 1 %
/ NaOH 0.2 N) and the mixture was gently shaken prior to ice incubation for another 5
minutes. Cell debris and chromosomal DNA were precipitated by adding 300 µl solution III
(potassium acetate 3 M / acetic acid 1.15 M). The mixture was gently shaken, incubated on
ice for 15 min and centrifuged at 5600 rpm, 15 min, on an Eppendorf 5415 R microcentrifuge
(Eppendorf). The supernatant was transferred to a 1.5 ml microcentrifuge tube. The next

14
steps are important for precipitating the BAC DNA and removing RNA. Using 450 µl
isopropanol, plasmidial DNA was precipitated by centrifuging for 5 minutes at 11400 rpm on
an Eppendorf 5415 R microcentrifuge (Eppendorf). The supernatant was discarded from the
tube and the pellet was cleaned of salts and other small molecules by adding 450 µl of
ethanol 70 % and centrifuging at 11400 rpm for 2 minutes on an Eppendorf 5415 R
microcentrifuge (Eppendorf). The ethanol was removed, the nucleic acid pellet was air dried
and resuspended in 40 µl TE buffer (TrisCl 100 mM / EDTA 10 mM) prior to incubation at
65°C for to dissolve the BAC DNA and inhibit DNAses.
The presence of the BAC, after extraction, was verified by agarose gel electrophoresis using
1 % [w/v] SeaKem LE Agarose (BioZym) gels and run in TAE buffer 1x (Tris acetate 45mM
/ EDTA 1mM) at 75 mV, 20 minutes. Lambda digested with Hind III was used as ladder. Gel
pockets were loaded with 5 µl of BAC extract and 3µl loading buffer (0.4 % [w/v]
bromphenol blue solution) Agarose gels were stained with ethidium bromide (5 µg/ml), and
photographed.
2.2.1. Liquid Luria Bertani medium
Peptone from casein tryptic digest (Fluka Chemie GmbH) 10 g
Yeast extract (Fluka Chemie GmbH) 5 g
NaCl 5 g
Sterile water to1000 ml
The pH was adjusted to 7.00 with NaOH 5 N and was autoclaved
25 min; chloramphenicol was added to 17 mg/l final concentration;
2.3. BAC insert size determination via pulsed field gel electrophoresis
For BAC insert size determination pulse field gel electrophoresis in contour-clamped
homogenous electric field (CHEF) has been performed. This device comprises a hexagonal
array of electrodes in a voltage divider circuit and produces homogenous fields (oriented at
120°) approximating those of infinitely long parallel electrodes (Sambrook et al., 1989).

15
To obtain fragments of the desired size (2 kb to 100 kb) the BAC DNA was digested with a
rare cutter restriction enzyme, Not I, which has GC↓GGCCGC as recognition site. The size
of the restriction fragments depends on the GC content of the BAC insert since a higher GC
content raises the probability that Not I finds more restriction sites.
Analysis of inserted DNA was performed by digestion of each BAC with Not I at 37°C for 3
hours. The restriction reaction was performed with 40 mM spermidine, 4 µl Not I buffer 10x,
0.075 U/µl Not I (final concentration), 10 µl BAC DNA and adjusted to 40 µl with sterile
water. Loading dye 10x was added to the restriction reaction in a proportion of 1 to 10 [v/v]
and the restriction reaction was stopped by heating at 65° C for 10 min. Digests were
analyzed with PFGE using 1 % SeaKem LE agarose gel in TBE buffer 0.5x (AppliChem).
Gel pockets were loaded with 5 µl of restricted BAC and 1 µl loading buffer. PFGE was
performed on a BioRad CHEF-DR III system using the following parameters: linear pulse
time ramp from 5 to 15 sec, 10 h run time, 120° field angle and 6 V/cm field strength. The
CHEF gels were stained in an ethidium bromide bath (5 µg/ml) for 30 min. The size of the
restricted fragments was evaluated by comparison to high molecular weight markers in the
form of agarose plugs (Lambda Ladder PFG Marker, Low Range PFG Marker (New
England, BioLabs). For smaller fragments Lambda digested with Hind III (Invitrogen) was
used.
2.4. BAC sequencing and primer walking
Primer walking is a sequencing method used to obtain contiguous sequence information
(contigs). The initial sequences from each end of the BAC insert were obtained using
standard vector primers, T7 and SP6 (Biomers) (Table 1). To sequence further into the insert,
new primers were designed and synthesized (Biomers) (Table 2).
Sequencing on vectors that were earlier identified as containing cbb L gene, have been done
during this study with specific cbb L primers (Table 1).
Sequencing reactions were performed in 15 µl final volume using Big Dye reaction mix
(Applied Biosystems) or ET reaction mix (Amersham) with 0.67 µM primer (final
concentration) and 5 µl BAC DNA template. The sequencing reaction was carried out in an
Eppendorf Master Cycler (Eppendorf) starting with the denaturizing step at 96°C for 30 sec,

16
primer annealing at primer dependent temperatures (see Table 2) for 30 sec, and elongation
for 4 min at 60°C, repeated for 98 cycles. The products of the sequencing reactions were
purified on Sephadex G-50 Superfine (Amersham) columns according to the manufacturer’s
manual and run on an Applied Biosystems 3130 xl Genetic Analyzer (Applied Biosystems).
Vector primers Cbb L primers
Name Sequence Tm Name Sequence Tm
T7 5’-taa tac gac tca cta tag gg-3’ 56°C Cbb L 1b 5’-cag ctg gac cac vgt btg g-3’ 48°C
SP6 5’-tta ggt gac act ata gaa tac-3’ 50°C Cbb L 2c 5’-cgg tgy atg tgc agc agc att ccg-3’ 48°C
2.5. Primer design
Primers were designed manually starting with aligning and comparing the end-sequences
using the BioEdit program (www.mbio.ncsu.edu/BioEdit/bioedit.html). The priming site was
chosen with 100 nucleotides upstream the firs region of poor quality (areas beyond single
peak resolution of a chromatogram). During primer design the following issues were
considered: i) melting temperature (Tm) in the range of 52°C to 65°C; ii) absence of
dimerization capability; iii) absence of significant hairpin formation; iv) low specific binding
at the 3' end (lower GC content to avoid mispriming); v) high specific binding at the 5’ end;
vi) G+C to A+T ratio should be close to 1;.
All primers were designed accordingly, having from 20 to 24 bases in length, a GC content
of 40 to 60 % and a Tm range from 50° to 66°C (see Table 2).
2.6. Sequence analysis
Sequences were analyzed using BLASTx software which compares a nucleotide query
sequence translated in all reading frames against a protein sequence database
(http://www.ncbi.nlm.nih.gov/blast). The BLASTx software evaluates the quality of a
pairwise sequence alignment using the substitution matrix BLOSUM 62 with the word size 3.
The sequences were assembled into contiguous units with SEQUENCER® Contig Assembly
Table 1 List of primers used for end-sequencing (T7 and SP6) and Cbb L primers for sequencing the gene which codifies for RubisCO

17
Program (Gene Codes Corporation). These contiguous sequences were again compared
against protein sequences from the non-redundant protein data base
(http://www.ncbi.nlm.nih.gov/blast) using BLASTx. Known genes and putative functions
were assigned for each individual sequence by inspection of the search output. Information of
protein functions and their metabolic roles was searched using enzyme and protein data bases
easily accessible from DBGET (http://www.genome.jp/dbget/) which is an integrated
database retrieval system for a diverse range of molecular biology databases. In order to
assess the metabolic functions most of the retrieved sequence information was searched
against KEGG Pathway, release 33 (http://www.genome.jp/kegg/pathway.html). Diverse
roles in cellular functions, pathogenesis or symbiotic interactions were assessed by seeking
against protein databases like PIR release 79 (http://www.genome.jp/dbget-
bin/www_bfind?pir).
18
Primer I Primer II Primer III No BAC Size (kb)
Vector Primer Name Sequence Tm Name Sequence Tm Name Sequence Tm
1 1H4 71kb T7 drsF_H4_199 5’-atg agt tcg gaa acc cga gc-3’ 62°C dsrE-1H4_250 5’-cga aca tca ggc gta
ctt cc-3’ 62°C dsrE_1H4_611(p3) 5’-ttg gac gca ttg aag ctt gg-3’ 60°C
2 1B4 90kb T7 Dh_B4_119 5’-agg tta agg cga tgg tta cc-3’ 60°C FeSpr_1B4_475 5’-agg ata tct gta tcg
gct gc-3’ 60°C H2ase_1B4355(p3) 5’-ttg cct gga atc gat aca gg-3’ 60°C
3 2D12 67kb SP6 napA_1267 5’-tgt gcc ctg gtt cga cga gg-3’ 62°C napG_2D12_341 5’-ctt cga tac cct cga
tct gg-3’ 62°C napF_2D12 341 (p3)
5’-gcg aaa agg ttt gtc cga cg-3’ 62°C
4 2F5 48kb SP6 apr_F5_260 5’-tat gtc atg ggt tcg cac gc-3’ 62°C apr_2F5_254 5’-caa ggc cga gat ctt
caa gc-3’ 62°C Apr³_2F5_472 (p3) 5’-ttg gtg atg acg agc aat cc-3’ 60°C
5 2D4 45kb SP6 ITP_B_272 5’-cca cat atg gat caa cag gc-3’ 60°C FeoB_2D4_472 5’-ccc cgt tga gca gta
aat cc-3’ 62°C FeoB_2D4_363(p3) 5’-gga tgt cgt cgt ttt caa gc-3’ 60°C
6 3B6 39kb SP6 metr_B6_259 5’-cca agg ttt gca gca ttt cc-3’ 60°C metr_3B6_467(p2) 5’-aca tgg gga tat cga
gct gg-3’ 62°C - -
7 3E11 67kb SP6 soxA_ E11,6_361
5’-cac gat cga act gtt cga cg-3’ 62°C soxA_3E11_373 5’-gtc atg aat ggg ctg
caa cg-3’ 62°C - -
8 4H3 63kb SP6 SR_H3_329 5’-tga tcg acg ttc aag tca cc-3’ 60°C PcrfA_4H3_564 5’-atg tca aca agg tat
cct cg-3’ 58°C Hypth_4H3_571(p3) 5’-tta ccc att gct ggt tat cg-3’ 60°C
9 5C6 13kb SP6 COdh_C6_72 5’-acg atc tcg gtg aag aag gc-3’ 62°C - - - -
10 8G3 29kb T7 Octdh_G3_65 5’-gtc gct act acc cta atc gg-3’ 62°C OpoxB_8G3_378 5’-taa tga ccc ccg acg
gta ttc c-3’ 68°C op-ox_8G3_499(p3) 5’-tta ccc att gct ggt tat cg-3’ 58°C
11 8E6 41kb T7 soxA_ E11,6_361
5’-cac gat cga act gtt cga cg-3’ 62°C soxA_8E6_222 5’-cga tcc gga tca cta
cga cc-3’ 64°C soxA_8E6_402(p3) 5’-ttc ggc aac aat gac aaa cc-3’ 58°C
12 8D1 41kb T7 ureF_D1_250 5’-acg aaa tgt cgc aga cga cg-3’ 62°C UreG_8D1_385 5’-cta caa cga cct cga
tgt cg-3’ 62°C ureG_8D1_514(p3) 5’-tcc taa act caa tat cgc cg-3’ 58°C
13 9H6 64kb SP6 Mep_H6_151 5’-tat cga tca ttg ctg tcg gg-3’ 60°C Mep_9H6_517 5’-tac gcg gat aac atc
att cg-3’ 58°C - -
14 9A11 35kb T7 metr_A11_320 5’-ggc tgt cga aga aat cat gg-3’ 60°C metr_9A11_625(p2) 5’-tcg cga tac cga aga
aat gg-3’ 60°C - -
15 9D3 46kb T7 ureC_9D3_ 128 (p1)
5’-tga tgg gtg atg ggt gat gg-3’ 62°C - - - -
16 9C10 38kb T7 SR_C10_267 5’-gta aaa cct tcc ttc cca cc-3’ 60°C pSR_9C10_322 5’-cca tat gta taa gac
cat gg-3’ 56°C pSR_9C10_381(p3) 5’-ttc gga tac gat ccc cta cg-3’ 62°C
17 9B5 - SP6 COdh_B5_278 5’-gca tgc ccg gaa aat gat cg-3’ 62°C COdh_9B5_570(p2) 5’-gat cac gca tga gaa
gaa cc-3’ 60°C - -
18 10H7 - SP6 vapB_H7_357 5’-cga ggt aat cat caa gca cc-3’ 60°C vapC_10H7_422 5’-gct cga acg atc tcc
tga tgc-3’ 66°C vapC_10H7_ 538(p3)
5’-ctt gat acg cag ttg att gg-3’ 58°C
19 10H10 44kb T7 cya_ F5,H10_396
5’-cgg tca tat cgc ctt cac gg-3’ 64°C Cya-10H10_380 5’-atc ggt taa atc gat
tgc gg-3’ 58°C cya_10H10_523(p3) 5’-tat gcg tac cct cct tgt cg-3’ 62°C
20 10F5 46kb T7 cya_ F5,H10_396
5’-cgg tca tat cgc ctt cac gg-3’ 64°C Cya_10F5_414 5’-taa att tgc ggt cga
acg cag g-3’ 66°C - -
Table 2 List of primers designed and used during this study; the determined dimensions of the BACs, in
kilo bases (kb), are also listed below. Tm stands for the melting temperatures of the primers.

19
3. Chapter 3: Results
Vector primed end sequencing on the 500 clones from O. algarvensis BAC library
revealed functional genes of potential importance for the metabolism of the symbionts.
Twenty from these clones were previously chosen for primer walking based on the reason
that they might bear genes that encode for proteins involved in carbon, sulfur, nitrogen
metabolisms, and symbiotic/pathogenic related traits.
3.1. BAC insert size determination
Dimension of the chosen BACs was determined using pulsed field gel electrophoresis.
The NotI restricted fragments of the BAC inserts were compared to markers which
ranged from 125 bp (Lambda digest of Hind III) to 1000 kb (Lambda Ladder PFG
Marker). For each clone the size of its insert was assessed by summing up the size of the
restriction fragments. The average insert size was determined as being 50 kb with the
smallest value 13 kb and the largest 90 kb (Table 2).
3.2. Sequence analysis and primer walking
The sequences of the functional genes discovered during the first screening of the library
by Anna Blazejack were confirmed during this study and subjected to primer walking.
Numerous contiguous sequences were obtained and their translated consensus sequences
showed similarity to proteins involved in carbon, sulfur and nitrogen metabolism,
transport processes and also other relevant pathogenic or symbiotic functions. Assigning
a function based on sequence similarity is a matter of prediction; proofs for function can
only be brought using functional tests.
Similarity search against protein databases of the obtained contiguous sequences was
done using BLASTx and similarities with E-values less than 1E-10 were considered
relevant (Annex 1). Similarities higher than 40 % for identical amino acid residues and 60
% for positive amino acids together with similarity scores less than 1E-10 were
considered significant for maintaining the protein function and were taken into account.

20
The encoded protein information was classified into five categories: carbon metabolism,
sulfur metabolism, nitrogen metabolism, transport proteins and pathogenic/symbiotic
interactions.
3.2.1 Carbon metabolism
Autotrophy requires the ability to synthesize cell material from inorganic carbon. The key
enzyme for autotrophy is RubisCO which catalyzes the fixation of CO2. cbbL is the gene
that encodes for the large subunit of RubisCO. In this study were sequenced five cbbL
genes, from five BAC clones, which were previously identified by Anna Blazejack as
presenting this gene.
In this project were sequenced, from the vector, other genes encoding for enzymes which
might be involved in one carbon metabolism like CO dehydrogenase, formate
dehydrogenase, trimethylamine methyltransferase, sarcosine oxidase and opine oxidase.
3.2.1.1 RubisCO key enzyme of Calvin cycle
The gene encoding for the large subunit of RubisCO form I (cbbL) was identified by PCR
screening with specific cbbL primers on five clones in the BAC library of O. algarvensis
(by Anna Blazejak).
In this study sequencing of the cbbL gene on these five BAC clones was performed using
as primer cbbL 1b. The five sequences obtained were aligned with BioEdit and showed to
be identical (see Annex 2). The consensus sequence was analyzed with BLASTx and the
inferred translation was found similar to the large chain of RubisCO from the
gammaproteobacterial sulfur oxidizer, Allochromatium vinosum (92 % identical and 97 %
positive amino acids).
3.2.1.2 CO dehydrogenases
Two BAC clones were identified as having genes putatively encoding for CO
dehydrogenase. Using BioEdit no similarity between the two sequences has been found.

21
Primer walking on both clones ended with two contiguous sequences. One contig was
only 545 nucleotides long (Fig. 2) and the BLASTx analysis showed that the inferred
translation was similar to anaerobic CO dehydrogenase form I from the
alphaproteobacterial marine phototroph Rugeria sp.. BLASTx showed a shift in the
translation and both fragments were assigned to neighboring regions in CO
dehydrogenase from Rugeria sp. (Annex 1) The percentages of identical and positive
amino acids was calculated by making the average of identical and positive residues from
the two fragments which had the same E-value (85 % identical and 89 % positive amino
acids).
The second contig was of 937 nucleotides (Fig. 2) and the first part of its translated
consensus sequence showed similarity to the end of CO dehydrogenase from the
methanogen Methanosarcina mazei (36 % identical and 55 % positive amino acids).
Contig clone 9B5
in O. alg. symbionts
CODH1 in M. mazei
SP6
p2
p1
CoxL in Rugeria sp.
Contig clone 5C6
CoxL in O. alg. symbionts
SP6
p1
Fig. 2 Graphic representation of two contigs which contain regions encoding for CO dehydrogenase; the arrows shows the correspondence between nucleotide sequence and amino acid sequences found similar to CO dehydrogenases from other organisms; The red stripe represent the cut in the translation.

22
The second part of its deduced translation product was similar to protein 3 from the
ComE operon of the deltaproteobacterial sulfate reducer Desulfotaela psychrophila (32 %
identical and 48 % positive amino acids). The end of this translated contig was related to
a sensory transduction histidine kinase from an uncultured archaeon (52 % identitical and
85 % positive amino acids).
3.2.1.3 Formate dehydrogenase
Primer walking on a clone, which was previously identified as containing a gene
encoding for formate dehydrogenase, provided a contiguous sequence of 1485
nucleotides which was analyzed with BLASTx (Fig. 3). The first part of the translated
consensus sequence was similar to the end of formate dehydrogenase α subunit from the
alphaproteobacterial marine isolate Silicibacter pomeroyi (59 % identical and 70 %
positive amino acids). The second part of the translated consensus sequence was similar
to Fe-S containing formate dehydrogenase from S. pomeroyi. This region presented a cut
in its conceptual translation and the fragments were assigned by BLASTx to neighboring
parts form the iron sulfur subunit of formate dehydrogenase. The percentage (63 %
identical and 74 % positive amino acids) represents the average of the identical and
positive amino acids from the two fragments. The end of the translated consensus
sequence had no similarity to any protein in the protein data bases.
T7
p1
p2
p3
FDH O. alg. symbionts α Fe/S
FDH genes S. pomeroyi
αβ // β γ αFe/S
Fig. 3 Graphic representation of one contig which contain regions encoding for formate dehydrogenase; the arrows shows the correspondence between nucleotide sequence and amino acid sequences found similar to formate dehydrogenases from other organisms; α, β, γ and Fe/S represent the α, β, γ and Fe/S subunits of FDH. The red stripe represents the cut in the translation. “//” represents a non scaled region.
23
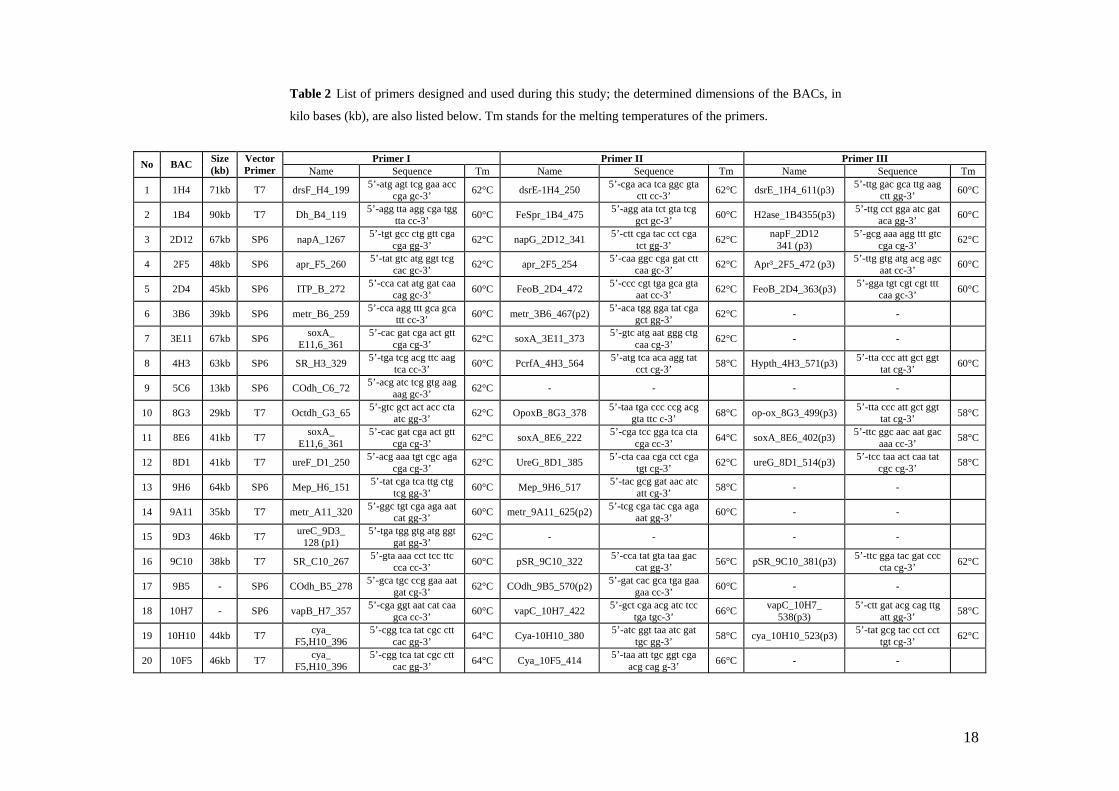
3.2.1.4 Trimethylamine methyltransferase
Two BACs in the library were identified by end sequencing as containing trimethylamine
methyltransferase genes. The two end sequences showed no similarity to each other.
Therefore primer walking was initiated on both clones and two contiguous sequences
were obtained. The BLASTx analysis of a 1512 nucleotides contig (Fig. 4) showed that
the inferred translation product was similar to trimethylamine methyltransferase (MttB)
from the alphaproteobacterial plant symbiont Mesorhizobium loti (58 % identical, 73 %
positive amino acids). The end of the conceptual translation of this contig had no
similarity to any protein stored in the protein data bases.
The second contig of 1516 nucleotides long (Fig. 4) have been identified as encoding in
the first part a protein similar to MttB1 form Methanosarcina acetivorans (31 % identical
and 51 % positive amino acids). A cut in the translation resulted in two fragments
MttB
MttB
In O. alg. symbionts
In M. loti
SP6 p1
p2
T7
p1 p2
In O. algarvensis ABC
Mtt
Mtt
In M. acetivorans
ABC
Contig on clone 9A11
In F. nucleatus
Contig clone 3B6
Fig. 4 Graphic representation of two contigs which contain regions encoding for trimethylamine methyltransferase (MttB); the arrows shows the correspondence between nucleotide sequence and amino acid sequences found similar to MttB from other organisms; The red stripe represent the cut in the translation.

24
assigned to neighboring regions from MttB1. The average similarity percentage was
calculated for the entire region that putatively encodes for a trimethylamine
methyltransferase. The second part of the conceptual translation had no similarity to any
proteic sequence from the available data bases, while the third region was similar to
spermidine/putresceine ABC transporter from the anaerobic pathogen Fussobacterium
nucleatum (35 % identical and 57 % positive amino acids). This region presented two
shifts in its translation and the three fragments obtained were related to three near regions
in the amino acid sequence of the spermidine/putresceine transporter F. nucleatum. The
average similarity percentage was calculated and the E-value was the same for all three
fragments (Annex 1).
3.2.1.5 Sarcosine oxidase
End sequencing on two clones showed that these BACs have sarcosine oxidase genes and
BioEdit alignment of these two sequences proved that they are identical. Primer walking
on one clone resulted in a contiguous sequence of 1592 nucleotides (Fig. x) whose
inferred translation showed similarity to sarcosine oxidase, subunit A from S. pomeroyi
(identical 45 %, positive 59 % amino acids). The conceptual translation presented a cut in
its sequence and the two resulted fragments were similar to adjacent regions from SoxA
of S. pomeroyi. The similarity of the entire inferred translation was considered by making
an average of the percentages of identical and positive amino acids, and knowing that the
E-value was identical for both fragments (Annex 1).
T7
T7 p1
p2
SoxA
Sox B D Sox G Sox A
In O. alg. symbionts
In S. pomeroyi
Fig. 5 Graphic representation of the contigs which encodes for subunit A of sarcosine oxidase; SoxA, B, D and G represents the subunits of sarcosine oxidase ; the arrows shows the correspondence between nucleotide sequence and amino acid sequences; The red stripe represent the cut in the translation.

25
3.2.1.6 Opine oxidase
The contig of 1894 nucleotides (Fig. 6) was the result of primer walking on a clone that
was previously identified as possible bearing one gene for opine oxidase. This contig was
subjected to BLASTx analysis and the inferred translation of the first part resembled
opine oxidase subunit B from the pSymA plasmid of Sinorhizobium meliloti (43 %
identical and 54 % positive amino acids).
The next part from the translated contig was related to an uncharacterized NAD / FAD
dependent dehydrogenase from the betaproteobacteria Burkholderia fungorum (48 %
identical and 60 % positive amino acids). The final part of this translated contig was
similar to opine oxidase subunit A from the alphaproteobacterial plant symbiont
Bradyrhizobium japonicum (38 % identical and 55 % positive amino acids).
3.2.2 Sulfur metabolism
Four contiguous sequences obtained by primer walking were putatively encoding for
enzymes involved in assimilatory and dissimilatory sulfur metabolism.
T7 T7
p1 p2
OoxA
OoxA
OoxB
OoxB SoxA SoxB
OoxB OoxA In O. alg. symbionts
In S. meliloti
In B. japonicum
Fig. 6 Representation of the contig which encodes for subunit A and B of opine oxidase; OoxA, OoxB, represents the subunits of opine oxidase SoxD and SoxB represents subunits of sarcosine oxidase; the arrows shows the correspondence between nucleotide sequence and amino acid sequences; The red stripe represent the cut in the translation.

26
BLASTx analysis of one contig revealed the existence of an encoded beta subunit of
sulfite reductase which is involved in assimilatory sulfur metabolism.
Two enzymes which appear to be involved in both oxidative and reductive sulfur
metabolism APS reductase and sirohaem sulfite reductase might be encoded by
sequences from contigs obtained on two different clones from the BAC library. Another
contig restrained a possible polysulfide reductase encoding region.
3.2.2.1 Sulfite reductase
A clone possibly bearing a cysI gene was identified and primer walking gave a
contiguous sequence of 1923 nucleotides. The inferred translation product of the first
region of this contig was similar to sulfite reductase from Chromobacterium violaceum,
form the Betaproteobacteria class. BLASTx assigned the three fragments that were the
result of two cuts in the translation to adjacent regions in the amino acid sequence of
sulfite reductase. The average percentage of identical and positive amino acids was
calculated (75 % identity and 89 % positive amino acids).
The next region from the translated consensus of this contig shows similarity to a
hypothetical protein from the alphaproteobacterial plant symbiont Bradyrhizobium
japonicum (32 % identical and 46 % positive amino acids). Next region resembled
protein chain releasing factor A from the gammaproteobacterial marine isolate
Microbulbifer degradans (71 % identical and 83 % positive). The rest of the inferred
translation showed no similarity to any protein in the available data bases.
3.2.2.2 Adenosine-5’-phosphosulfate (APS) reductase
The α subunit of adenosine-5’-phophosulfate reductase was previously identified as
being encoded on a clone from O. algarvensis BAC library and using primer walking on
a contig of 1956 nucleotides, have been established (Fig. 8). The BLASTx analysis
showed that the first region from the conceptual translation was similar to α subunit of
APS reductase from the gammaproteobacterial sulfur oxidizer A. vinosum (54 % identical
and 60 % positive amino acids).The middle part was found not similar to known proteins

27
while the end of the translated contig resembled the beginning of FKBP type peptydil-
prolyl cis-trans isomerase 2 from T. denitrificans (72 % identical and 84 % positive
amino acids).
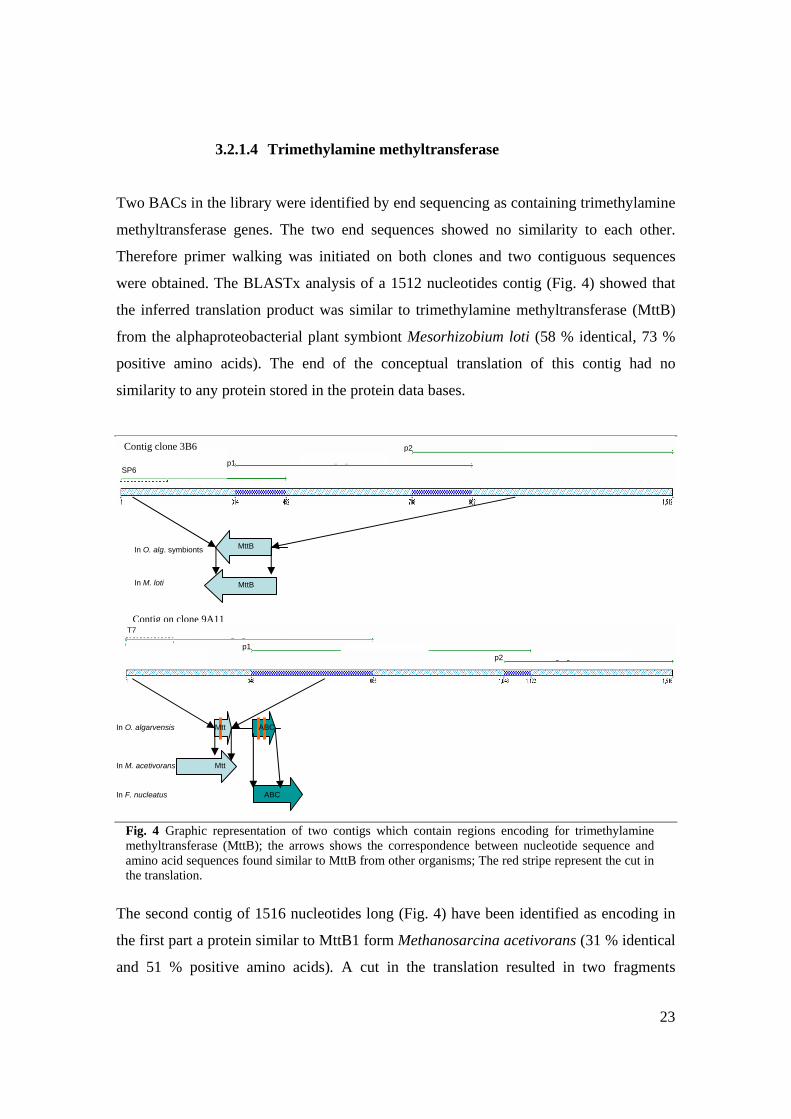
3.2.2.3 Dissimilatory sulfite reductase
Four BACs were identified as containing genes which potentially encode for different
proteins involved in dissimilatory sulfite reduction from A. vinosum. Primer walking was
performed on one clone and a 1232 nucleotides contig was assembled (Fig. 8). BLASTx
analysis of the consensus sequence showed that the translation product of the first region
resembled DsrF from A. vinosum (71 % identical and 80 % positive amino acids). The
next region form the translated consensus resembled DsrE also from A. vinosum (90 %
identical and 95 % positive amino acids). The end of the translated contig showed
similarity with the end of dissimilatory sirohaem sulfite reductase beta subunit from the
same A. vinosum (58 % identical and 69 % positive amino acids).
Using BioEdit the consensus sequence of this contig was aligned against the dsr gene
cluster (see Annex 2) of an entirely sequenced BAC (Anna Blazejack, unpublished data).
For a better perception of this difference these two sequences were subjected to the
bl2seq which gave a nucleotide similarity of 75 % for dsrE and dsrF genes and 88% for
the end of dsrB gene.
SP6
p1
p2
p3
Sat AprM
In O. alg. symbionts
AprB AprA
AprA
In A. vinosum
Fig. 7 Representation of the contig which encodes for AprA which is one of the three subunits of APS reductase, the other two are AprM and AprB; Sat is sulfate adenylyl transferase; the arrows shows the correspondence between nucleotide sequence and amino acid sequences;
28
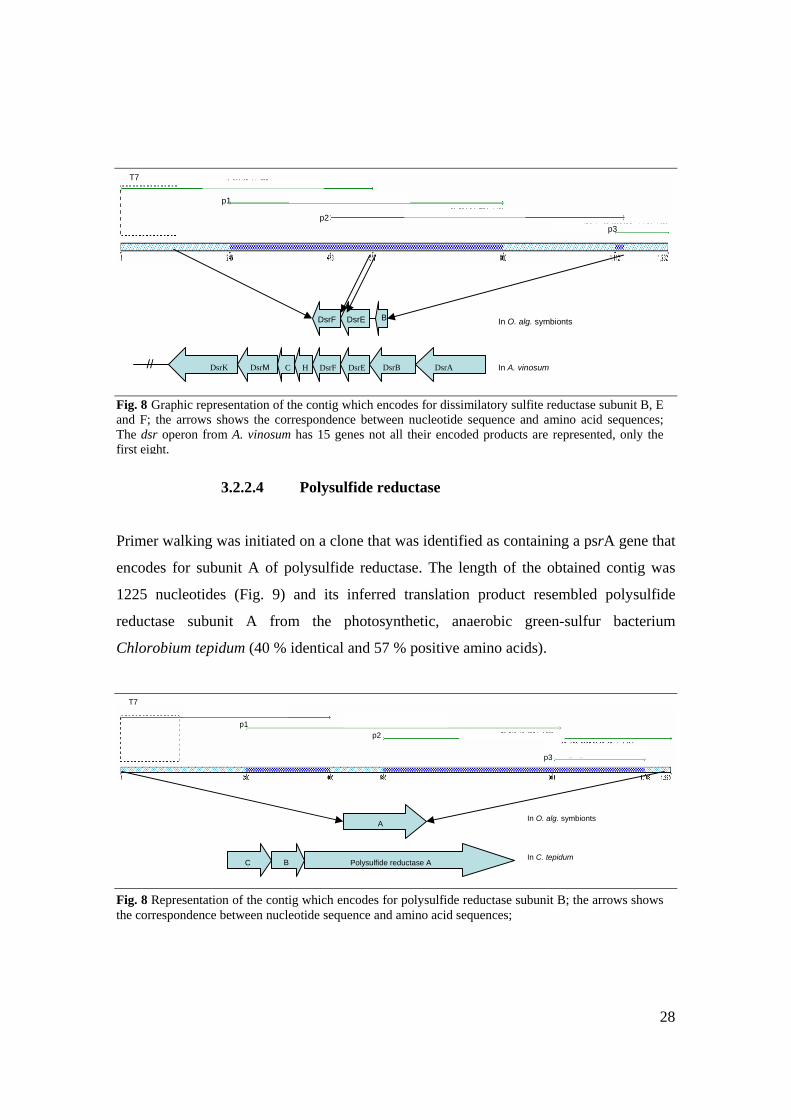
3.2.2.4 Polysulfide reductase
Primer walking was initiated on a clone that was identified as containing a psrA gene that
encodes for subunit A of polysulfide reductase. The length of the obtained contig was
1225 nucleotides (Fig. 9) and its inferred translation product resembled polysulfide
reductase subunit A from the photosynthetic, anaerobic green-sulfur bacterium
Chlorobium tepidum (40 % identical and 57 % positive amino acids).
DsrF DsrE B
// DsrE DsrF DsrB DsrA H C DsrM DsrK
In O. alg. symbionts
In A. vinosum
T7
p1
p2 p3
T7
p1 p2
p3
C
A
B Polysulfide reductase A
In O. alg. symbionts
In C. tepidum
Fig. 8 Graphic representation of the contig which encodes for dissimilatory sulfite reductase subunit B, E and F; the arrows shows the correspondence between nucleotide sequence and amino acid sequences; The dsr operon from A. vinosum has 15 genes not all their encoded products are represented, only the first eight.
Fig. 8 Representation of the contig which encodes for polysulfide reductase subunit B; the arrows shows the correspondence between nucleotide sequence and amino acid sequences;

29
3.2.3 Nitrogen metabolism
During this survey several regions were discovered that codify for proteins which could
play a role in assimilatory, dissimilatory nitrogen metabolism and nitrogen storage for the
symbionts of O. algarvensis. These included genes which encode for urease,
periplasmatic nitrate reductase involved in the dissimilatory nitrate respiration and
cyanophycin synthetase involved in nitrogen storage.
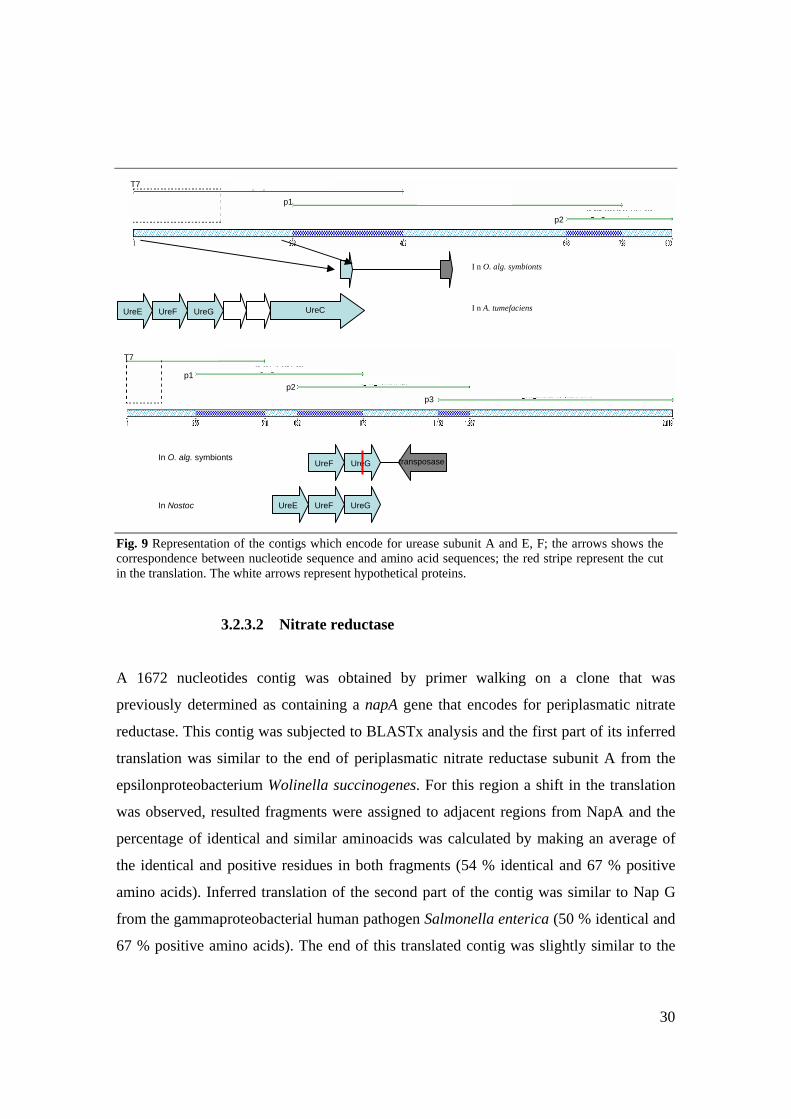
3.2.3.1 Urease
Two BAC clones were previously identified as bearing genes encoding for different
subunits of urease. Therefore primer walking was initiated on both BAC clones and two
contiguous sequences were assembled on which BLASTx analyses were performed. One
contig of 803 nucleotides (Fig. 9) in the first part of its inferred translation was similar to
the end of urease alpha subunit from the alphaproteobacterial plant pathogen
Agrobacterium tumefaciens (81 % identical and 84 % positives). No similarity was found
for the middle part of the contig while the end of its translation was vaguely similar to a
region from anthranilate synthase component II of Corynebacterium efficiens (37 %
identical and 48 % positive residues).
The second contig of 2019 nucleotides presents in the first part a region whose inferred
translation resembled UreF from the gammaproteobacterial pathogen Vibrio
parahaemolyticus (40 % identical and 56 % positive amino acids).
The next region from the translated contig was similar to UreG from cyanobacterium
Nostoc sp. (72 % identical and 89 % positive amino acids). This region presents a shift in
its conceptual translation and both fragments are aligned to neighboring segments in the
amino acid sequence of UreG. The average content of identical and positive amino acids
was calculated. The end of the translated consensus sequence was similar to a transposase
from the betaproteobacterium Polaromonas sp. (37 % identical and 56 % positive amino
acids)
30
3.2.3.2 Nitrate reductase
A 1672 nucleotides contig was obtained by primer walking on a clone that was
previously determined as containing a napA gene that encodes for periplasmatic nitrate
reductase. This contig was subjected to BLASTx analysis and the first part of its inferred
translation was similar to the end of periplasmatic nitrate reductase subunit A from the
epsilonproteobacterium Wolinella succinogenes. For this region a shift in the translation
was observed, resulted fragments were assigned to adjacent regions from NapA and the
percentage of identical and similar aminoacids was calculated by making an average of
the identical and positive residues in both fragments (54 % identical and 67 % positive
amino acids). Inferred translation of the second part of the contig was similar to Nap G
from the gammaproteobacterial human pathogen Salmonella enterica (50 % identical and
67 % positive amino acids). The end of this translated contig was slightly similar to the
T7
p1
p3 p2
UreF UreG transposase
UreF UreG UreE
In O. alg. symbionts
In Nostoc
T7
I n O. alg. symbionts
p1
p2
UreG UreF UreE UreC I n A. tumefaciens
Fig. 9 Representation of the contigs which encode for urease subunit A and E, F; the arrows shows the correspondence between nucleotide sequence and amino acid sequences; the red stripe represent the cut in the translation. The white arrows represent hypothetical proteins.

31
beginning of NapH from W. succinogenes. BLASTx output revealed, for this part of the
inferred translation, a translation cut that resulted in two fragments similar to near regions
from NapH. For these two fragments an average percentage of identical and positive
amino acids was calculated and assigned to the entire region (43 % identical and 56 %
positive amino acids).
3.2.3.3 Cyanophycin synthetase
Two BACs were determined as having genes which putatively codify for cyanophycin
synthetase. BioEdit alignment showed that these two sequences were identical so that
only one clone was sequenced. The result of the primer walking was a contig of 1587
nucleotides which was analyzed using BLASTx. Three cleavages occur in the translation
and the fragments were all assigned to neighboring regions in the amino acid sequence of
cyanophycin synthetase from the gammaproteobacterial intracellular pathogen
Francisella tulariensis. The average of identical and similar aminoacids for all these three
fragments was calculated (74 % identical and 88 % positive aminoacids).
3.2.4 Transport proteins
Transport systems allow the uptake of essential nutrients and ions, excretion of end
products, metabolites and deleterious substances but also facilitate the communication
between cells and the environment. These facts make the ‘transporter protein’ category,
appropriate for studying the interactions of the symbiotic community from O.
algarvensis.
During this study three BAC clones were analyzed and contiguous sequences were
assembled. The translation products were related to proteins like ferrous iron transport
protein, natrium driven multidrug efflux pump and spermidine/putresceine transporter.
The last one was analyzed previously as part of a contig described in the carbon
metabolism section.

32
3.2.4.1 Ferrous iron transport protein (FeoB)
Primer walking was performed on a BAC which previously was identified that might
encode FeoB. All sequences obtained with the designed primers were assembled into a
contig of 1345 nucleotides. Similarity searches were performed using BLASTx and the
inferred translation product of this contig was similar to ferrous iron transport protein B
from the pathogen Porphyromonas gingivalis member of the Bacteroides. BLASTx
showed that the translated contig had three regions of similarity to neighboring segments
from FeoB. The E-value was identical for all three fragments whether the ratio of
identical and positive aminoacid was different. For addressing the entire region assigned
to FeoB an average of the percentages was calculated (47 % identical and 65 % positive
amino acids).
3.2.4.2 Na+ driven multidrug efflux pump (NorM)
A BAC clone previously identified as possible bearing a gene encoding for a multidrug
efflux pump was subjected to primer walking and the assembled contig of 831
nucleotides was analyzed using BLASTx. The translated consensus was related to a
natrium driven multidrug efflux pump, NorM from the alphaproteobacterium
Rhodopseudomonas palustris (35 % identical and 52 % positive amino acids). BLASTx
showed a cut in the conceptual translation and the fragments were similar to near
segments from the proteic sequence of NorM. For a general idea of the similarity to
NorM of the translated contig an average value of the identical and positive amino acids
from both fragments together.
3.2.5 Pathogenic/symbiotic associated traits
Traits that are generally associated with pathogenic or symbiotic lifestyle were found
encoded in the genome of O. algarvensis symbionts: urease, ferrous iron transporter, and
virulence associated proteins. Urease contigs that were analyzed are described in the
‘nitrogen metabolism’ section whether the ferrous iron transporter is explained in the

33
‘transporter proteins’ section. In this section only the virulence associated proteins are
detailed.
3.2.5.1 Virulence associated proteins (VapB and VapC)
A contig of 1482 nucleotides was obtained on a clone which has been determined as
possibly containing a gene for a protein associated with virulence. BLASTx analyses of
this contig showed that the first part of the translated consensus sequence was similar to
virulence associated protein B form Leptospira interogans. BLASTx output showed a
shift in the translation and the resulted fragments were assigned to neighboring regions in
virulence associated protein B. The E-value is identical for both fragments and the
average of their identical and positive amino acid content was calculated (52 % identical
and 77 % positive). The next part from the translated contig was found similar to
virulence associated protein C from the deltaproteobacterium Geobacter sulfurreducens
and also in this region a modification was detected and two slightly separated fragments
were similar to neighboring regions that cover the entire VapC, and the percentages of
identical and positive amino acids assigned to the entire region was calculated taking in
consideration the values for both fragments (47 % identical and 68 % positive amino
acids).The next region in the conceptual translation showed no similarity to any known
protein.

34
4. Chapter IV Discussions
The symbiotic community of O. algarvensis consists of five phylotypes: two
gammaproteobacterial, two deltaproteobacterial and one spirochete (Rühland et al., In
Prep.). To address the genomic encoded function of these uncultured symbionts the
sequence driven metagenomic approach was used. End-sequencing was previously
performed on the 500 clones of the BAC library and sequences that have been considered
relevant for the symbiotic community were confirmed during this study, prior to primer
walking for proceeding further into the inserts. The outcome was represented by longer
contiguous sequences that were analyzed with BLASTx.
In the present survey BLASTx assignments were performed based on the similarity of the
inferred translation products to aminoacid sequences of proteins stored in the GenBank
protein database. Assigning a function based on sequence similarity is a matter of
prediction; proofs for function can only be brought using functional tests, e.g. expression
tests (Riesenfeld et al., 2004). BLASTx gave in some cases results in different frames for
fragments which were assigned to neighboring regions in the same protein and it might
occur due to sequencing errors, knowing that deletion or misreading of only one
nucleotide from the chromatogram can induce a modification in the inferred translation.
The existence of five phylotypes in the endosymbiotic community of O. algarvensis
makes difficult the link between the functional information encoded in the community
genome to any of the symbionts without having a 16 S rRNA phylogenetic anchor in the
same clone.
The similarities are given taking in consideration both E-values (see Annex 1) and
percentages of identical and positive aminoacids. The positive amino acids were
considered since it is known that there are groups of aminoacids with comparable
biochemical traits and these amino acids are considered as producing a positive change,
maintaining the folding of the protein hereby keeping its function. In fact the function of
a protein can be preserved also if only functional domains are conserved, which would
allow a lot of sequence variation in unconserved parts of the protein.
Classification of the potentially encoded protein information was done into five
categories: carbon metabolism, sulfur metabolism, nitrogen metabolism, transport

35
proteins and pathogenic/symbiotic relevant traits. This classification is maybe helpful for
this study since we did not deal with such a high amount of sequence information like in
a complete genome sequencing project were more categories are considered (e.g. Riley.
1993, Kaneko et al. 2000).
4.1. Carbon metabolism
The ability of O. algarvensis symbionts to grow with CO2 was demonstrated earlier
(Dubilier et al., 2001) by showing the occurrence of ribulose bisphosphate carboxylase,
the key enzyme of the CO2 fixation pathway. Immunocytochemical analyses showed the
presence of this enzyme in the large morphotype, now considered as being the Gamma 1
phylotype (Rühland et al., In Prep.). Previously five genes encoding for RubisCO form I
were determined by PCR screening and in the present study they were sequenced and
showed sequence equivalence (Annex 2) facilitating the assumption that they might
belong to the same symbiont, namely the Gamma 1. Autotrophy of the symbionts via
Calvin cycle is nourishing the worm with reduced carbon substrates.
Using the sequence driven metagenomic approach were discovered encoded proteins
which might be involved in one carbon metabolism: CO dehydrogenase, formate
dehydrogenase, trimethylamine methyltransferase, opine oxidase and sarcosine oxidase.
It seem that O. algarvensis symbiotic community might use one carbon compounds like
CO, formic acid but also compounds that are known as methyl-group-donors like
trimethylamines, sarcosine (N-methyl-alanine) or opines. The reductive-acetyl-CoA
pathway for autotrophic CO2 fixation can be used also for assimilation of various one
carbon compounds (Lengeler et al., 1999). Methylotrophs use CO, formate,
formaldehyde, methanol, methylamine, methylmercaptane, or methane as sole sources of
carbon. Some methylotrophs are able to use one-carbon units linked via a hetero-atom (N,
S, O) to the rest of an organic molecule(Lengeler et al., 1999). Methylotrophs are known
as symbionts for some marine invertebrates (Fisher, 1990) This kind of metabolism with
synthesis of macromolecules starting from one carbon compounds, is certainly beneficial
for a host which has no ability to use them but is incorporating the symbiotically
synthesized macromolecules. The finding of encoded CO dehydrogenase and formate

36
dehydrogenase show that substrates like CO and formate might also support symbiotic
growth in O. algarvensis.
The host also can nourish its symbionts with products of its metabolism like
trimethylamine, opines, sarcosine or urea. A proof for such way of cycling carbon
compounds is the finding of potentially encoded trimethylamine methyltransferase, opine
oxidase and sarcosine oxidase.
4.1.1. CO dehydrogenase
CO dehydrogenases are present in physiologically and phylogenetically diverse microbes
where the enzyme functions to either oxidize CO, synthesize acetyl-CoA, or cleave
acetyl-CoA. Aerobic microbes utilize Mo-Fe-flavin CO dehydrogenases to oxidize CO in
respiratory pathways. Phototrophic anaerobes converts CO and water to CO2 and H2,
process catalyzed by Ni-(Fe-S) CO dehydrogenase. Acetate-producing anaerobes employ
a Ni-(Fe-S) CO dehydrogenase to synthesize acetyl-CoA from a methyl group, CO and
CoA. A similar enzyme is responsible for the cleavage of acetyl-CoA by methanogens
that obtain energy by fermenting acetate to CH4 and CO2. Acetotrophic sulfate reducers
also utilize CO dehydrogenase to cleave acetyl-CoA yielding methyl and carbonyl groups
and they obtain energy for growth via a respiratory pathway in which the methyl and
carbonyl groups are oxidized to CO2 and sulfate is reduced to sulfide (Ferry, 1995).
Autotrophic sulfate reducers fix CO2 into their cell carbon through the non cyclic
reductive acetyl-CoA pathway which has as key enzyme the same CO
dehydrogenase/acetyl-CoA synthase. The reductive acetyl-CoA pathway for autotrophic
CO2 fixation can be used also for assimilation of various one carbon compounds
(Lengeler et al., 1999)
The conceptual translation products of two contigs from the studied BAC library were
found related to CO dehydrogenases from the methanogen Methanosarcina mazei and
from the anaerobic phototroph Rugeria sp.
Knowing in which kind of organisms the CO dehydrogenases could occur and relating
with the members of O. algarvensis symbiotic community, could be speculated that any
of the sulfate-reducing delta symbionts might utilize the CO dehydrogenase/acetyl-CoA

37
synthase pathway, either just for gaining energy by complete oxidation to CO2 being a
complete oxidizing sulfate-reducer but might be that they perform CO2 fixation being
autotophs, by using the reductive CO dehydrogenase pathway.
4.1.2. Formate dehydrogenase
Formate dehydrogenase oxidizes formate to carbon dioxide with the release of a proton
and two electrons. In some anaerobic bacteria formate is a fermentation product, and
formate dehydrogenase is a component of anaerobic formate hydrogen lyase complex.
The ability to grow with formate has been observed in most genera of sulfate reducing
eubacteria. Furthermore, spirilloid sulfur reducers and Archaeoglobus fulgidus are known
to grow with formate. Formate dehydrogenase has been found in Desulfovibrio and in
acetate oxidizing sulfate reducers, except Desulfobacter. Formate dehydrogenase in
acetate oxidizers is part of the carbon monoxide pathway for acetyl CoA oxidation. In
autotrophic sulfate reducers formate dehydrogenase is the first carbon fixing enzyme of
the reductive acetyl-CoA pathway. Formate dehydrogenase in the sulfur reducer,
Wolinella succinogenes, is membrane bound and involved in the oxidation of formate to
CO2 resulting in electron transport to sulfur reductase which is in close contact to formate
dehydrogenase. (Widdel and Hansen, 1992a; Lengeler et al., 1999).
In this study the highest similarity of the potential encoded formate dehydrogenase is to
the alpha and iron-sulfur subunits of formate dehydrogenase form the marine isolate
Silicibacter pomeroyi. The genes for these two subunits in Silicibacter are placed in the
following context: fdhβ, fdhα, followed by a large genomic sequence and than fdhγ, fdh
for Fe-S subunit and fdhα are encountered (NCBI complete genome for S. pomeroyi,
CP000031). S pomeroyi seems to rely upon a litoheterotrophic strategy using inorganic
compounds (CO and sulfide) to supplement heterotrophy (Moran et al., 2004).
Taking in consideration the types of symbionts that O. algarvensis bears could be
therefore suggested that formate dehydrogenase might belong to a sulfate reducing
member from the endosymbiotic community, given that in acetate oxidizers and
autotrophic SRB this enzyme occurs. Another possibility is that the spirochete ferments

38
carbohydrates to formate with formate dehydrogenase as part of the formate hydrogen
lyase complex.
4.1.3. Trimethylamine methyltransferase
Trimethylamine is a degradation product of trimethylammonium compounds like choline,
carnitine, or lecithine which are present in all animal tissues. Trimethylamine in
eukaryotes is oxidized to trimethylamine oxide which is an osmoregulators in muscle
tissues of marine organisms protecting proteins by counteracting destabilizing forces like
urea, ammonia, hydrostatic pressure, temperature stress, and salt-stress. (Lengeler et al.,
1999; Seibel and Walsh, 2001).
Trimethylamine methyltransferase is catalyzing the transfer of methyl groups either to a
corrinoid protein in methanogens or to tetrahydrofolate in methylotrophs (Lengeler et al.,
1999).
Similarities of the conceptual translations of two contigs to trimethylamine
methyltransferase from Silicibacter pomeroyi, and Methanosarcina mazei represent an
indicator that O. algarvensis symbionts could utilize trimethylamines.
Trimethylamine represents a precursor for worm’s osmolytes e.g. trimethylamine oxide,
but in the same time trimethylamines could represent a new carbon source for its
symbionts. Trimethylamines are known substrates for some sulfate reducers which have
the ability to degrade N-methylated compounds (Widdel and Hansen, 1992a). In Olavius
algarvensis one possibility would be that the sulfate reducing symbionts take use of the
break down of the worm osmoregulators, being able to use trimethylamines as methyl
group donors for the reductive acetyl-CoA pathway. Symbiotic utilization of the
precursors of worm’s osmoregulators might not be valuable from the host point of view.
4.1.4. Sarcosine oxidase
Sarcosine or N-methylglycine is a eukaryotic metabolite found in eukaryotic tissues
which can be used as sole carbon and energy source by microorganisms. Sarcosine
oxidase is a flavoprotein that catalyzes the oxidative demethylation of sarcosine yielding

39
glycine, H2O2, 5,10-methylenetetrahydrofolate in a reaction requiring tetrahydrofolate
and oxygen. In the absence of tetrahydrofolate, the methyl group from sarcosine is
released as formaldehyde Bacterial sarcosine oxidases have been isolated from different
organisms and fall into two classes: heterotetrameric (α β γ and δ subunits) and
monomeric the last ones being similar to β subunit (SoxB) from the heterotetrameric
enzymes. (Chlumsky et al., 1995)
One contig was found during this study as potentially encoding for a protein similar to
sarcosine oxidase subunit α from S. pomeroyi. In S. pomeroyi the gene cluster that
encodes for sarcosine oxidase shows a similar organization like the described operon
which encodes for the heterotetrameric enzyme in Corynebacterium sp (Chlumsky et al.,
1995) which is soxBDAG with soxA encoding for subunit α, soxB for subunit β, soxC for
γ whether soxD encodes for the δ subunit (Moran et al., 2004).
Sarcosine which results from the host metabolism could be used by the endosymbionts
most probably by the sulfate reducers which are known for their ability to use methyl
groups bounded to a heteroatom like nitrogen, in the case of sarcosine.
4.1.5. Opine oxidase
Opines are the products of the NAD(P)H- dependent reductive condensation between α-
keto acid and the α- or ω-NH2 group of an amino acid. This kind of compounds have
been isolated from eukaryotic cells, plant tumors, bacteria and marine invertebrates
(Britton et al., 1998). In marine invertebrates, muscular metabolism has its own
characteristics with glycolytic end products different than lactate. In invertebrates opines
are formed instead of lactate and they represent condensation products of pyruvate with
amino acids (e.g. alanine for alanopine, glycine for strombine, arginine for octopine).
(Portner, 2002). Opine oxidase catalyze the degradation of opines and this enzyme is met
in syntrophic or pathogenic bacteria that can use this kind of metabolites from their hosts,
like the plant pathogen A. tumefaciens or the leguminous plant symbiont, S. meliloti
which is presumed that can parasitize plant tumors produced by the first one (Barnett et
al., 2001).

40
The translated contiguous sequences of one contig was considered similar in the first part
to subunit B from S. meliloti and the neighboring region seemed similar to subunit A of
opine oxidase from B. japonicum. Might be possible that O. algarvensis has an advantage
if it has endosymbionts which might consume the acidifying end products of its anaerobic
glycolysis.
4.2. Sulfur metabolism
The existence of both sulfur oxidizing and sulfate reducing bacteria in O. algarvensis
makes possible the cycling of reduced and oxidized sulfur species between symbionts.
The idea of this kind of syntrophic sulfur cycle was supported initially by phylogenetic
analysis, showing the relations of O. algarvensis symbionts to other sulfur oxidizing and
sulfate reducing bacteria (Dubilier et al., 2001). Evidences for the reductive side of this
syntrophic sulfur cycle beside 16S rRNA phylogeny were the determination of sulfate
reduction rates at levels similar to free living sulfate reducers and the PCR amplification
of dissimilatory sulfite reductase (Dubilier et al., 2001). Evidences for the oxidative side
of this syntrophic sulfur cycle beside 16S rRNA phylogenetic analyses was the
occurrence of intracytoplasmatic sulfur globules and immunohistochemical analyses with
an antiserum directed against RubisCO (Dubilier et al., 2001). For these endosymbionts
the enzymology of oxidative sulfur metabolism and reductive sulfur metabolism is
unknown. Furthermore it is important to establish which kind of sulfur substrates are
used, and how they are metabolized, which could be possible only by looking for specific
enzymes.
The present study found similarities for inferred translation products to proteins involved
in sulfur assimilation, dissimilatory sulfur reduction and oxidation but also in polysulfide
respiration. During this survey was found encoded a sulfite reductase which is involved
in assimilation of sulfur into cysteine. Encoded APS reductase and dissimilatory sulfite
reductase involved in both sulfur reduction and oxidation were also revealed. The genes
encoding for these two enzymes are considered good “clocks” for the evolution of the
dissimilatory sulfur metabolism for both SRB and sulfur oxidizing bacteria (Hipp et al.,

41
1997). Another finding of this study was an encoded polysulfide reductase which shows
the potential of O. algarvensis symbionts to utilize polysulfide as final electron acceptor.
4.2.1. Sulfite reductase
Reductive sulfate assimilation is a pathway widely occurring in plants, algae, fungi and
bacteria. The reduction of sulfite to sulfide is catalyzed by sirohaem sulfite reductases
and is one of the final steps in cysteine biosynthesis. The electron donor for this the
reduction of sulfite is NAD(P)H. Sulfide is immediately incorporated into O-acetyl-L-
serine yielding cysteine (Lengeler et al. 1999, Neumann et al., 2000).
The inferred translation of one contig was similar to sulfite reductase hemoprotein beta
subunit, from Chromobacterium violaceum. This assimilatory sulfite reductase could
belong to any member from O. algarvensis endosymbiotic community, given that
reductive sulfur assimilation occurs in any bacterial group.
4.2.2. APS reductase
APS reductase is found in sulfate-reducing bacteria and archaea, and in some
chemoautotrophic and phototrophic sulfur oxidizing bacteria (Hipp et al. 1997). The
reduction of APS and its conversion to sulfite and AMP is catalyzed by APS reductase in
sulfate reducers, whether the same enzyme in sulfur oxidizers catalyzes the reverse
reaction. In sulfur oxidizing bacteria two pathways for sulfite oxidation were described:
direct oxidation via a sulfite acceptor-oxidoreductase and indirect AMP-dependent
oxidation known as the APS pathway. The direct pathway for sulfite oxidation is
considered to be far more widespread between sulfur oxidizers (Robertson & Kuenen
1992, Kappler & Dahl 2001) but for the sulfur-oxidizing endosymbionts of invertebrates
so far the APS reductase pathway has been demonstrated (Nelson & Hagen, 1995).
The inferred translation of one contig was found similar to ATP sulfurylase from the
phototrophic sulfur oxidizer A. vinosum. In A. vinosum an operon is formed by the gene
for ATP sulfurylase (sat) and APS reductase (aprMBA) with aprA and aprB encoding for
APS reductase α and β subunits and aprM encoding a putative membrane anchor. The

42
two gamma endosymbiotic phylotypes are also closely related to A. vinosum at the 16S
rRNA gene level (Rühland et al. In Prep), and corroborating with the fact that APS
reductase is used as an evolutionary marker for dissimilatory sulfur metabolism (Hipp et
al., 1997) seems reasonably to assign APS reductase to the gamma sulfur oxidizing
symbionts.
4.2.3. Dissimilatory sulfite reductase
Dissimilatory sulfite reductase (DSR) in sulfate reducing bacteria catalyzes the reduction
of sulfite to sulfide involving the transfer of six electrons whether in sulfur oxidizing
bacteria the enzyme catalyze the reverse reaction.
The similarity between translation product of one contig to DsrF and DsrE and the end of
the β subunit of DSR form A. vinosum corroborated to the 16S rRNA relationships to A.
vinosum for the gamma symbionts from O. algarvensis ensures that this sequence might
only belong to the sulfur oxidizing symbionts. In A. vinosum the dsr gene cluster has 15
genes dsrABEFHCMKLJOPNSR, and with interposon mutagenesis in dsrB and dsrL the
was demonstrated the essential role of the Dsr proteins in the oxidation of intracellular
sulfur. dsrA and dsrB encode the α and β subunits of dissimilatory sulfite reductase.
DsrEFH together with DsrK, DsrAB, DsrC,DsrO and DsrJ are considered as being
associated in a super complex involved in the oxidation of intracellular sulfur. The
formation of all Dsr gene products is enhanced by sulfide (Dahl et al., 2005). Hereby
inside O. algarvensis sulfide is currently produced by the sulfate reducers and might
enhance the growth rates for the sulfur oxidizers.
The comparison of the nucleotide sequence of this contig against the sequence of a dsr
locus obtained by sequencing the entire clone from the same BAC library (unpublished
data Anna Blazejack) revealed a high sequence difference (Annex 3). This might be due
to the fact that the sequences belong to a different gamma phylotype from the two known
in O. algarvensis.

43
4.2.4. Polysulfide reductase
Polysulfides are formed abiotically form sulfur and sulfide and its reduction to sulfide is
catalyzed by polysulfide reductase. This process is typical for the catabolism of extremely
thermophilic archebacteria, of the thermophilic eubacterium Thermotoga, and of certain
proteobacteria including Desulfuromonas, Sulfurospirillum deleyianum and Wolinella
succinogenes. The function of polysulfide reductase is determined in W. succinogenes
where this enzyme is a component of the phosphorylative transport system with
polysulfide as terminal electron acceptor (Widdel and Hansen, 1992a; Krafft et al., 1995).
By similarity search the inferred translation of one contig was related to polysulfide
reductase from the green sulfur bacterium C. tepidum. The genome sequence of this
organism revealed a psrA gene followed by psrB and psrC. The encoded function was not
yet determined experimentally but it is assumed that the products of these genes might
function in sulfide oxidation to form polysulfides (Eisen et al., 2002).
In O. algarvensis which is like a “closed system” accumulation of sulfur and sulfide
could occur as a result of fast growth rates of bacterial cells, resulting in enough
substrates for the chemical formation of polysulfides. It is possible that the sulfate-
reducing endosymbionts could reduce polysulfides to sulfide but their energy yield would
be rather low.
4.3. Nitrogen metabolism
Enzymes involved in ammonia release from urea (urease), synthesis of storage
nitrogenous compounds (cyanophycin synthetase) or energy metabolism (dissimilatory
nitrate reductase) were found encoded on clones from the BAC library of O. algarvensis
symbionts. This shows the potential of these symbionts to use ammonia detoxification
products, like urea released from the host metabolism but also the ability to synthesize
storage compounds like cyanophycin, which is not a common trait among bacteria. The
ability to respire nitrate for gaining energy seems also possible. Some other compounds
like opines or sarcosine are also considered potential nitrogen sources besides their role
as carbon sources for bacteria, and both are discussed in the carbon metabolism section.

44
4.3.1. Urease
Animals can either excrete ammonia directly, or synthesize urea or uric acid. Hydrolysis
of urea to ammonia and carbon dioxide is catalyzed by the nickel containing enzyme,
urease. Many eukaryotes synthesize urease, but urease activity is widely observed among
prokaryotes, including many eubacteria and at least one archaebacterium,
Methanobacterium thermoautotrophicum. Microbial urease is associated in general with
pathogenesis that causes direct damage to the host due to ammonium or alkali production
(Mobley and Hausinger, 1989; Toffanin et al., 2002). Analysis of urease from Klebsiella
aerogenes, the model organism for this enzyme, has revealed that the three catalytic
subunits: α β and γ associate in a (αβγ)3 structure containing three bi-nickel catalytic
sites. Assembly of the bi nickel metallocenter requires accessory proteins, UreD, UreE,
UreF and UreG (Lee et al., 1992).
By similarity search the inferred translations of two contigs were related to α subunit of
urease from the alphaproteobacterial plant pathogen A. tumefaciens, and to the urease
accessory proteins UreF and UreG from the cyanobacterium Nostoc sp.(see Fig. 9).
A source of nitrogen for O. algarvensis endosymbionts could be the ammonia
detoxification products from the worm, like urea; The problem remains unsolved since it
is not possible to assign this enzyme to a symbiont in the community, but it shows that
the community adapted to use the resources from its host.
4.3.2. Periplasmic nitrate reductase
There are two pathways for nitrate respiration and many organisms are able to use nitrate
as electron acceptor, which is reduced to nitrite and further either via nitric oxide and
nitrous oxide to molecular nitrogen (denitrification) or more directly reduced with
NAD(P)H to ammonia (ammonification). The dissimilatory ammonification pathway
differs from the assimilatory pathway by having the enzymatic systems integrated in the
membranes or located in the periplasm (Lengeler et al., 1999). Hereby a periplasmic
located nitrate reductase is considered as being involved in the dissimilatory reduction of
nitrate to nitrite.

45
During this study was identified potential encoded periplasmic nitrate reductase with
similarity to NapA from the epsilonproteobacterium W. succinogenes, NapG from the
gammaproteobacterial pathogen Salmonella enterica and to NapH also from W.
succinogenes. In W. succinogenes the gene cluster encoding for periplasmic nitrate
reductase is napAGHBFLD (Simon et al., 2003) whether in S. enterica is napFAGHBC.
Studies on respiratory nitrate reductase were done on W. succinogenes and it was proved
with interposon mutation in napA that the only respiratory nitrate reductase is NapA.
NapG and NapF might be involved in electron transfer while NapH in menaquinol
oxidation (Simon et al., 2003)
This encoded enzyme shows the potential of O. algarvensis symbionts to reduce nitrate,
and could be suggested that it belongs to the sulfate reducing symbionts forasmuch
neither spirochete or sulfur reducing symbionts seem likely to perform nitrate respiration.
4.3.3. Cyanophycin synthetase
Cyanophycin is a branched, non-ribosomally synthesized polypeptide consisting of
aspartate in the backbone and a roughly equimolar amount of arginine in the side chain.
The capability for cyanophycin synthesis is not only restricted to cyanobacteria, as it was
initially supposed, but it occurs among other bacterial groups representing a temporary
nitrogen, energy and possibly carbon source (e.g. Accinetobacter, Clostridium,
Nitrosomonas ). Cyanophycin was described as accumulating under conditions of intense
light at high carbon dioxide concentrations, and phosphate or sulfur starvation but also a
high N/C ratio promotes cyanophycin biosynthesis. Cyanophycin synthetase is encoded
by cphA and is involved in synthesis of cyanophycin via an ATP dependent mechanism
(Krehenbrinck et al., 2002; Krehenbrinck and Steinbüchel, 2004).
The encoded cyanophycin synthetase from O. algarvensis symbionts was found similar to
its counterpart in the betaproteobacterial human pathogen Francisella tulariensis and it is
intricate to make an assumption on the symbiont that might present cyanophycin
synthetase capacity but it seems interesting to determine the reason for synthesizing
storage compounds. Could be that compounds that are synthesized by such a

46
metabolically diverse community accumulate and instead of eliminating them in the
environment they are stored by the microorganisms.
4.4. Transport proteins
Transport is an essential aspect of all life-endowing processes: metabolism,
communication, reproduction and both cooperative and antagonistic inter-organismal
behavior (Busch and Saier, 2004). In conclusion the transporters are appropriate for
studying the interactions of the symbiotic community from O. algarvensis.
The translation products for three sequences were related to proteins like ferrous iron
transport protein, natrium driven multidrug efflux pump and spermidine/putresceine
transporter.
4.4.1. Ferrous iron transport protein B
Most bacteria that live in oxic environments produce iron (III) chelators, called
siderophores, to satisfy their needs for iron, which forms highly insoluble ferric
hydroxides at neutral pH. The ferrous iron (II) uptake proteins was proved in E. coli to be
encoded by the feoAB (Kammler et al., 1993) and now in almost 50 % of the bacterial
genomes a feoB-like gene was found. The FeoB protein from E. coli was identified as
having GTPase activity that is necessary for ferrous iron (II) transport whether the uptake
is directly accomplished by the FeoB protein. The iron transport function of Feo appears
to be of particular importance during low oxygen conditions when ferrous iron (II)
predominates over ferric iron (III) (Andrews et al., 2003; Hantke, 2003). The ability to
utilize ferrous iron efficiently is considered to be a major factor in pathogenesis for those
microbes that try to colonize essentially anaerobic environments(Guerinot, 1994).
During the present study the determined translation of one contig had high similarity to
FeoB from the pathogen Porphyromonas gingivalis. This ferrous iron transporter shows
that the bacteria adapted to the oligochaete host, while living in conditions of low oxygen

47
concentration, inside the worm, are able to take up their essential micronutrients, iron
being one of them.
4.4.2. Na+ driven multidrug efflux pump
Drug efflux mechanism is one of the major mechanisms of drug resistance. Multidrug
efflux pumps are integral membrane transporters that have the ability to extrude a variety
of structural unrelated drugs from the cell. NorM is a multidrug efflux pump driven by an
electrochemical potential of Na+ instead of a proton motive force or ATP dependency
like in other categories of transporters (Brown et al., 1999). A Na+ driven multidrug
efflux pump was described for the marine bacterium Vibrio parahaemolyticus with a
homologue in E. coli (Morita et al., 1998; Morita et al., 2000) and in the causative agent
of cholera, Vibrio cholerae (Huda et al., 2003).
One clone from O. algarvensis BAC library after contig assembly was identified as
bearing the encoded information for NorM similar to its correspondent from
Rhodopseudomonas palustris, showing the possible ability of these symbionts to extrude
drugs.
4.4.3. Spermidine/putresceine transporter
Polyamines (putrescine, spermidine and spermine) are necessary for cell growth.
Polyamines are among the major polycations in cells together with Ca 2+ and Mg2+ and
their content in cells is mediated by biosynthesis, degradation and transport. The
putresceine-specific and the spermidine-preferential uptake systems are ABC
transporters. Polyamine transport systems were identified in most bacteria for which
whole genome were sequenced and it even exists in archaea. (Igarashi and Kashiwagi,
1999)
During this study was found a PotF encoded enzyme similar to PotF from
Fussobacterium nucleatum. PotF is the substrate binding protein and its presence shows
that polyamines might be used as metabolites by the symbionts of O. algarvensis.

48
4.5. Pathogenic/symbiotic relevant traits
Pathogenic associated traits correlate with the expression of disease related factors
that are present in pathogenic organisms and lack from the other non-pathogenic species.
Symbiotic associated traits are represented by various factors which can increase the
adaptability and versatility of the bacterium. Some pathogenic associated traits are also
met in symbiotic organisms, offering to the last ones a better fitness, and several of these
pathogenic associated traits were found in the genome on O. algarvensis symbionts:
ferrous iron transporter, urease and virulence associated proteins but also opine oxidase .
4.5.1. Virulence associated proteins BC
vapABCD genomic region was first characterized in the pathogen Dichelobacter nodosus
(Katz et al., 1992) and was associated with virulence. Novel studies on Leptospira
interogans associated the vapBC to physiological regulation and not to virulence, being
characterized as a toxin-antitoxin module (Zhang et al., 2004). The same study stated that
vapBC is widely present and expressed in both non pathogenic and pathogenic
leptospires. Therefore it was considered more related to physiological regulation than to
virulence. It was demonstrated in the same study that the product of vapC gene is toxic
for E.coli cells, while vapB gene product counteracts VapC toxicity.
A region potentially encoding for vapBC was found in this project by similarity search
and was related to VapB from Leptospira interrogans and VapC from Geobacter
sulfurreducens. It is possible that in O. algarvensis symbionts its presence is associated to
physiological regulation than to virulence, enhancing the fitness of the symbionts the
oligochaete host.
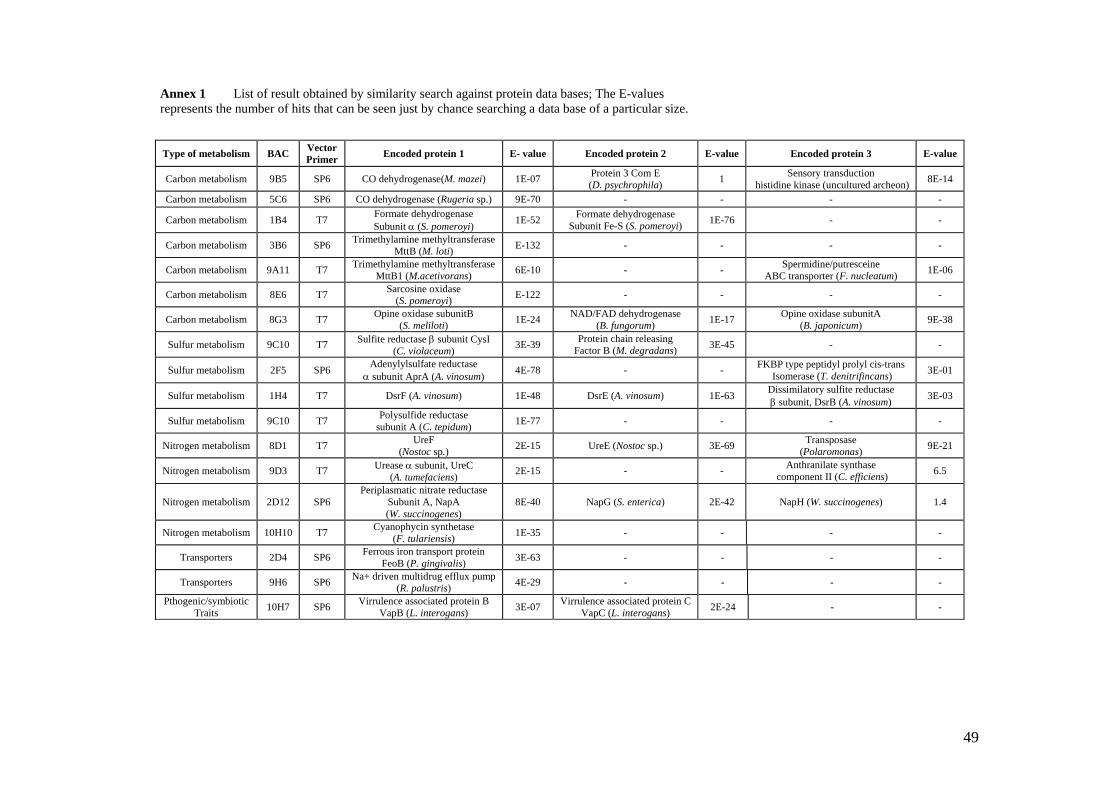
49
Type of metabolism BAC Vector Primer Encoded protein 1 E- value Encoded protein 2 E-value Encoded protein 3 E-value
Carbon metabolism 9B5 SP6 CO dehydrogenase(M. mazei) 1E-07 Protein 3 Com E (D. psychrophila) 1 Sensory transduction
histidine kinase (uncultured archeon) 8E-14
Carbon metabolism 5C6 SP6 CO dehydrogenase (Rugeria sp.) 9E-70 - - - -
Carbon metabolism 1B4 T7 Formate dehydrogenase Subunit α (S. pomeroyi) 1E-52 Formate dehydrogenase
Subunit Fe-S (S. pomeroyi) 1E-76 - -
Carbon metabolism 3B6 SP6 Trimethylamine methyltransferase MttB (M. loti) E-132 - - - -
Carbon metabolism 9A11 T7 Trimethylamine methyltransferase MttB1 (M.acetivorans) 6E-10 - - Spermidine/putresceine
ABC transporter (F. nucleatum) 1E-06
Carbon metabolism 8E6 T7 Sarcosine oxidase (S. pomeroyi) E-122 - - - -
Carbon metabolism 8G3 T7 Opine oxidase subunitB (S. meliloti) 1E-24 NAD/FAD dehydrogenase
(B. fungorum) 1E-17 Opine oxidase subunitA (B. japonicum) 9E-38
Sulfur metabolism 9C10 T7 Sulfite reductase β subunit CysI (C. violaceum) 3E-39 Protein chain releasing
Factor B (M. degradans) 3E-45 - -
Sulfur metabolism 2F5 SP6 Adenylylsulfate reductase α subunit AprA (A. vinosum) 4E-78 - - FKBP type peptidyl prolyl cis-trans
Isomerase (T. denitrifincans) 3E-01
Sulfur metabolism 1H4 T7 DsrF (A. vinosum) 1E-48 DsrE (A. vinosum) 1E-63 Dissimilatory sulfite reductase β subunit, DsrB (A. vinosum) 3E-03
Sulfur metabolism 9C10 T7 Polysulfide reductase subunit A (C. tepidum) 1E-77 - - - -
Nitrogen metabolism 8D1 T7 UreF (Nostoc sp.) 2E-15 UreE (Nostoc sp.) 3E-69 Transposase
(Polaromonas) 9E-21
Nitrogen metabolism 9D3 T7 Urease α subunit, UreC (A. tumefaciens) 2E-15 - - Anthranilate synthase
component II (C. efficiens) 6.5
Nitrogen metabolism 2D12 SP6 Periplasmatic nitrate reductase
Subunit A, NapA (W. succinogenes)
8E-40 NapG (S. enterica) 2E-42 NapH (W. succinogenes) 1.4
Nitrogen metabolism 10H10 T7 Cyanophycin synthetase (F. tulariensis) 1E-35 - - - -
Transporters 2D4 SP6 Ferrous iron transport protein FeoB (P. gingivalis) 3E-63 - - - -
Transporters 9H6 SP6 Na+ driven multidrug efflux pump (R. palustris) 4E-29 - - - -
Pthogenic/symbiotic Traits 10H7 SP6 Virrulence associated protein B
VapB (L. interogans) 3E-07 Virrulence associated protein C VapC (L. interogans) 2E-24 - -
Annex 1 List of result obtained by similarity search against protein data bases; The E-values represents the number of hits that can be seen just by chance searching a data base of a particular size.

50
Annex 2 BioEdit alignment of the five clones found as bearing a gene encoding for RubisCO. Boxed nucleotides represent the conserved regions whether unboxed parts shows the differences between the two DNA fragments.
1 0 2 0 3 0 4 0 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - G A C G C G C C T A C T C G A T 1- - - - - - - - - A C C T G A C C T C G A T C A C T A C A A T G G G G A C G C G C C T A C T C G A T 2- - - - - - - - G A C C T G A C C T C G A T C A C T A C A A T G G G G A C G C G C C T A C T C G A T 3- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 4C T T T G T T T G A C C T G A C C T C G A T C A C T A C A A T G G G G A C G C G C C T A C T C G A T 5
6 0 7 0 8 0 9 0 1 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C G A G G A C G T A C C C G G C G A C G A C A G C T G T T T T T A C G C T T T C A T C G C T T A C C 1C G A G G A C G T A C C C G G C G A C G A C A G C T G T T T T T A C G C T T T C A T C G C T T A C C 2C G A G G A C G T A C C C G G C G A C G A C A G C T G T T T T T A C G C T T T C A T C G C T T A C C 3- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 4C G A G G A C G T A C C C G G C G A C G A C A G C T G T T T T T A C G C T T T C A T C G C T T A C C 5
1 1 0 1 2 0 1 3 0 1 4 0 1 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C G A T C G A T C T G T T C G A G G A A G G C T C G G T A G T T A A T G T A T T T A C C T C G C T G 1C G A T C G A T C T G T T C G A G G A A G G C T C G G T A G T T A A T G T A T T T A C C T C G C T G 2C G A T C G A T C T G T T C G A G G A A G G C T C G G T A G T T A A T G T A T T T A C C T C G C T G 3- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 4C G A T C G A T C T G T T C G A G G A A G G C T C G G T A G T T A A T G T A T T T A C C T C G C T G 5
1 6 0 1 7 0 1 8 0 1 9 0 2 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |G T C G G T A A C G T A T T C G G T T T C A A G G C C G T T C G C G G G C T A C G C C T G G A A G A 1G T C G G T A A C G T A T T C G G T T T C A A G G C C G T T C G C G G G C T A C G C C T G G A A G A 2G T C G G T A A C G T A T T C G G T T T C A A G G C C G T T C G C G G G C T A C G C C T G G A A G A 3- - - - - - - - - - - - - - - - - - - - - - - - G C C G T T C G C G G G C T A C G C C T G G A A G A 4G T C G G T A A C G T A T T C G G T T T C A A G G C C G T T C G C G G G C T A C G C C T G G A A G A 5
2 1 0 2 2 0 2 3 0 2 4 0 2 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C G T G C G T T T C C C G A T T G C C T A C G T C A T G A C C T G T A A C G G C C C G C C C A A C G 1C G T G C G T T T C C C G A T T G C C T A C G T C A T G A C C T G T A A C G G C C C G C C C A A C G 2C G T G C G T T T C C C G A T T G C C T A C G T C A T G A C C T G T A A C G G C C C G C C C A A C G 3C G T G C G T A T C C C G A T T G C C T A C G T C A T G A C C T G T A A C G G C C C G C C C A A C G 4C G T G C G T T T C C C G A T T G C C T A C G T C A T G A C C T G T A A C G G C C C G C C C A A C G 5
2 6 0 2 7 0 2 8 0 2 9 0 3 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |G C A T C C A G G T C G A G C G C G A C A A A A T G A A C A A A T A C G G T C G A C C G C T G C T A 1G C A T C C A G G T C G A G C G C G A C A A A A T G A A C A A A T A C G G T C G A C C G C T G C T A 2G C A T C C A G G T C G A G C G C G A C A A A A T G A A C A A A T A C G G T C G A C C G C T G C T A 3G C A T C C A G G T C G A G C G C G A C A A A A T G A A C A A A T A C G G T C G A C C G C T G C T A 4G C A T C C A G G T C G A G C G C G A C A A A A T G A A C A A A T A C G G T C G A C C G C T G C T A 5
3 1 0 3 2 0 3 3 0 3 4 0 3 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |G G C T G C A C C A T C A A G C C C A A A C T G G G C T T G T C G G C G A A A A A T T A C G G T C G 1G G C T G C A C C A T C A A G C C C A A A C T G G G C T T G T C G G C G A A A A A T T A C G G T C G 2G G C T G C A C C A T C A A G C C C A A A C T G G G C T T G T C G G C G A A A A A T T A C G G T C G 3G G C T G C A C C A T C A A G C C C A A A C T G G G C T T G T C G G C G A A A A A T T A C G G T C G 4

51
G G C T G C A C C A T C A A G C C C A A A C T G G G C T T G T C G G C G A A A A A T T A C G G T C G 5
3 6 0 3 7 0 3 8 0 3 9 0 4 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C G C G G T T T A C G A A T G T C T G C G C G C G G A C T C G A C T T C A C C A A G G A C G A T G A 1C G C G G T T T A C G A A T G T C T G C G C G C G G A C T C G A C T T C A C C A A G G A C G A T G A 2C G C G G T T T A C G A A T G T C T G C G C G C G G A C T C G A C T T C A C C A A G G A C G A T G A 3C G C G G T T T A C G A A T G T C T G C G C G C G G A C T C G A C T T C A C C A A G G A C G A T G A 4C G C G G T T T A C G A A T G T C T G C G C G C G G A C T C G A C T T C A C C A A G G A C G A T G A 5
4 1 0 4 2 0 4 3 0 4 4 0 4 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |A A A C G T C A A C T C C C A G C C G T T C A T G C G C T G G A A T C A C C G T T T C G A T T T C G 1A A A C G T C A A C T C C C A G C C G T T C A T G C G C T G G A A T C A C C G T T T C G A T T T C G 2A A A C G T C A A C T C C C A G C C G T T C A T G C G C T G G A A T C A C C G T T T C G A T T T C G 3A A A C G T C A A C T C C C A G C C G T T C A T G C G C T G G A A T C A C C G T T T C G A T T T C G 4A A A C G T C A A C T C C C A G C C G T T C A T G C G C T G G A A T C A C C G T T T C G A T T T C G 5
4 6 0 4 7 0 4 8 0 4 9 0 5 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |T T A T G G A A G C G A T C C A T A A G G C C G A A C A G G A A A C C G G C G A A C G C A A G G G A 1T T A T G G A A G C G A T C C A T A A G G C C G A A C A G G A A A C C G G C G A A C G C A A G G G A 2T T A T G G A A G C G A T C C A T A A G G C C G A A C A G G A A A C C G G C G A A C G C A A G G G A 3T T A T G G A A G C G A T C C A T A A G G C C G A A C A G G A A A C C G G C G A A C G C A A G G G A 4T T A T G G A A G C G A T C C A T A A G G C C G A A C A G G A A A C C G G C G A A C G C A A G G G A 5
5 1 0 5 2 0 5 3 0 5 4 0 5 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C A C T A C C T C A A C G T T A C C G C G C C G A C G C C G G A A G A A A T G T A C A A G C G C G C 1C A C T A C C T C A A C G T T A C C G C G C C G A C G C C G G A A G A A A T G T A C A A G C G C G C 2C A C T A C C T C A A C G T T A C C G C G C C G A C G C C G G A A G A A A T G T A C A A G C G C G C 3C A C T A C C T C A A C G T T A C C G C G C C G A C G C C G G A A G A A A T G T A C A A G C G C G C 4C A C T A C C T C A A C G T T A C C G C G C C G A C G C C G G A A G A A A T G T A C A A G C G C G C 5
5 6 0 5 7 0 5 8 0 5 9 0 6 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C G A G T A C G C C A A G C A A C T G G G C G C A C C G A T C A T C A T G C A C G A C T A C C T G A 1C G A G T A C G C C A A G C A A C T G G G C G C A C C G A T C A T C A T G C A C G A C T A C C T G A 2C G A G T A C G C C A A G C A A C T G G G C G C A C C G A T C A T C A T G C A C G A C T A C C T G A 3C G A G T A C G C C A A G C A A C T G G G C G C A C C G A T C A T C A T G C A C - - - - - - - - - - 4C G A G T A C G C C A A G C A A C T G G G C G C A C C G A T C A T C A T G C A C G A C T A C C T G A 5
6 1 0 6 2 0 6 3 0 6 4 0 6 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C C G G C G G C T T C T G C G C C A A T A C C G G T C T C G C G C A A T G G T G C C G C G A C A A C 1C C G G C G G C T T C T G C G C C A A T A C C G G T C T C G C G C A A T G G T G C C G C G A C A A C 2C C G G C G G C T T C T G C G C C A A T A C C G G T C T C G C G C A A T G G T G C C G C G A C A A C 3- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 4C C G G C G G C T T C T G C G C C A A T A C C G G T C T C G C G C A A T G G T G C C G C G A C A A C 5
6 6 0 6 7 0. . . . | . . . . | . . . . | . . . . | . . .G G C A T C T G C T G C A C A T G C A C C G A 1G G C A T C T G C T G C A C A T - - - - - - - 2
Annex 2

52
5 4 1 0 5 4 2 0 5 4 3 0 5 4 4 0 5 4 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - C o n t i g d s r _ 1 H 4C C C G C C C T G C A C G C C G A T G C A A A T C A A C G A C C C C G A A C A T A C C A A G C T A G d s r _ B e l i n
5 4 6 0 5 4 7 0 5 4 8 0 5 4 9 0 5 5 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - G G T C T T T T T C T T C C T C o n t i g d s r _ 1 H 4C T A T C T G G G T C G G C G G T A A C C A C T C C A A C G C G C G C G G C A A G C C G A C T T T C d s r _ B e l i n
5 5 1 0 5 5 2 0 5 5 3 0 5 5 4 0 5 5 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C G G G G C C T C C T C T T T A A A G C G C C A G G G A T T G G T G C G G G T T T T T C G T G C G G C o n t i g d s r _ 1 H 4C A G A A A C T C G T C G C C G C C G G C A T T C C G A A C A A C C C G C C G C G C T G G C C C G A d s r _ B e l i n
5 5 6 0 5 5 7 0 5 5 8 0 5 5 9 0 5 6 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |G A - G C C T T T C A T C G G T G G C C T C G T T T C T C G - - C G G A G G T C G A A G C T G C C C C o n t i g d s r _ 1 H 4G G C G A C C G C G A T C G T T A A A A A G A T T T T A C A A A C C T A T A A G G A T A A C G C C C d s r _ B e l i n
5 6 1 0 5 6 2 0 5 6 3 0 5 6 4 0 5 6 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |T T C A C C A A G T A C C C A T A T C G A C A C C T G G C G C G A G C G C C C C G G G C C A G C T T C o n t i g d s r _ 1 H 4G C G A T T G G G A G C G C A T G - - A A C G A C T G G A T C G A A C G C A T C G G - C T G G C C G d s r _ B e l i n
5 6 6 0 5 6 7 0 5 6 8 0 5 6 9 0 5 7 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C A A T G C G T C C A A C T A T - - - A T C C G T T T C T G A G C G G A T A - - - - A T C G G G A A C o n t i g d s r _ 1 H 4C A A T T C T T C G A A T T G A C C G A A C T G C C G T T T A C C A A A T T C C A C G T C G A T A A d s r _ B e l i n
5 7 1 0 5 7 2 0 5 7 3 0 5 7 4 0 5 7 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C C C G T A C - - - C C G C T C C C C G C C C G G A G A G G G G G G A G C G G G G A G C A A C C A C C o n t i g d s r _ 1 H 4C T G G C G C G G A T C G C G G C A T T C G C T C A A C G C T T C A A C C C A T A T C C G C T T T T d s r _ B e l i n
5 7 6 0 5 7 7 0 5 7 8 0 5 7 9 0 5 8 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |A G C G A G G T C G C T A - - T G A A G T T C G C A A T C C A G A T C A A C G A G G G G C C C T A C C o n t i g d s r _ 1 H 4A A C G G G G T C T G C A G G T G A A A T T C G G A A T A C T A G T C A A C G A A G G G C C C T A C d s r _ B e l i n
5 8 1 0 5 8 2 0 5 8 3 0 5 8 4 0 5 8 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C A G C A C C A G G C G T C C G A C T C G G C G T A C T T C T T C A C G A A G G C C G T T C T G G A C o n t i g d s r _ 1 H 4A C G C A C C A G G C G T C C G A T T C G G C T T A C C A T T T C A C C G C C G C G G C G C T G C G d s r _ B e l i n
5 8 6 0 5 8 7 0 5 8 8 0 5 8 9 0 5 9 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |G A A G G G A C A C G A G G T A T T T C G G G T C T T C T T C T A C C A T G A T G G C G T C A A C A C o n t i g d s r _ 1 H 4T G C C G G G C A T G A A A T C G T G C G C G T G T T T T T T T A T C A T G A C G G C G T C A A C A d s r _ B e l i n
5 9 1 0 5 9 2 0 5 9 3 0 5 9 4 0 5 9 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |A C G G G A C C C G G C T G A C C A C G C C G C C C C A G G A C G A C C G T A A C G T C G T C A A C C o n t i g d s r _ 1 H 4A C G G T A C C C G C C T G A G C G T G C C G C C A C A G G A T G A C C G C A A C A T C A G C G A A d s r _ B e l i n
Annex 3 BioEdit alignment of the contig assembled on BAC clone 1H4 against the region that contains a dsr operon, from a completely sequenced BAC from O. algarvensis library. Boxed nucleotides represent the conserved regions whether unboxed parts shows the differences between the two DNA fragments.

53
5 9 6 0 5 9 7 0 5 9 8 0 5 9 9 0 6 0 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C G C T G G G G G G A G C T G G C C G A G C A G C A C G A A C T C G A C C T G G T G G T C T G C G T C o n t i g d s r _ 1 H 4C G C T G G T C G G C G C T G G C G G A A G C G C A C G A G G T C G A G T T G A T T C T A T G C G T d s r _ B e l i n
6 0 1 0 6 0 2 0 6 0 3 0 6 0 4 0 6 0 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C G C C G C G G C C C A G C G T C G C G G C G T C G T G G A C G A G G G C G A G G C C C A G C G T A C o n t i g d s r _ 1 H 4G G C G G C G G C C C A A C G C C G C G G C C T G C T G G A C G G G G A C G A A G C C A A A C T C G d s r _ B e l i n
6 0 6 0 6 0 7 0 6 0 8 0 6 0 9 0 6 1 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |A C G G C A A G G A C G C C A C C A A C A T C C A T C C G A A A T T C C G C A T C T C C G G T T T G C o n t i g d s r _ 1 H 4C C G G C A A G G A T A G C G A C A A C A T G G C G C C C G G G T T T C A G A T T T C C G G G C T G d s r _ B e l i n
6 1 1 0 6 1 2 0 6 1 3 0 6 1 4 0 6 1 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |G G G C A A T T G G T C G A G G C G G C C A T C C A G G C G G A T C G G C T C G T C G T G T T C G G C o n t i g d s r _ 1 H 4G G C C A G C T G A T C G A T G C C G G C A T T C A G G C G G A T C G T T T A A T C G T A T T C G G d s r _ B e l i n
6 1 6 0 6 1 7 0 6 1 8 0 6 1 9 0 6 2 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |T G A C T A A G G G G G T C C G C C A T G T C T G A A A C C A T C A A G A A A T T C T T G T A C C T C o n t i g d s r _ 1 H 4T G A T T G A A - - - - - - - - - - - - - - C T A T G G C G A C T A A A A A G T T T T T A T A C G T d s r _ B e l i n
6 2 1 0 6 2 2 0 6 2 3 0 6 2 4 0 6 2 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C A A T C G C A A G G C C C C C T A C G G C A C C A T C T A T G C C T G G G A A T C C C T G G A A G C o n t i g d s r _ 1 H 4A A A C C G C A A G G C G C C T C A T G G C A G C A T T T A T G C C C T G G A A T C G T T G G A A G d s r _ B e l i n
6 2 6 0 6 2 7 0 6 2 8 0 6 2 9 0 6 3 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |T G G T G C T C A T C G G C G C C G C C T T C G A T C A G G A A G T A C G C C T G A T G T T C G T C C o n t i g d s r _ 1 H 4T G G T A C T C A T A G C C G C G G C C T T C G A A C A G G A T G T C A G C C T C G C C T T C G T C d s r _ B e l i n
6 3 1 0 6 3 2 0 6 3 3 0 6 3 4 0 6 3 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |G A C G A T G G G G T C T A C C A G C T C A C C A A G G G C C A G G A T A C C G C C G G T A T C G G C o n t i g d s r _ 1 H 4G A C G A T G G G G T T T A C C A G C T G C T G C A A A A C C A G G A T A C C G G C A A G C T C G G d s r _ B e l i n
6 3 6 0 6 3 7 0 6 3 8 0 6 3 9 0 6 4 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C A T G A A A A A C T T C T C G C C G A C C T A C C G G A C C C T G G G T G A C T A C G A G G T A A C o n t i g d s r _ 1 H 4A T C G A A G A A T T T T T C G C C A A C C T A C A A G G C G C T G G G C G A C T A C G A C G T C A d s r _ B e l i n
6 4 1 0 6 4 2 0 6 4 3 0 6 4 4 0 6 4 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |A A A A C A T C T A C G T G G A C C A G G A C T C G C T T A A A A C C C G C G G A C T G A C C C A G C o n t i g d s r _ 1 H 4G C A A A A T A T A C G T C G A T C A G G A A T C G C T C G A A C A A A G A G G T C T C A A C A A G d s r _ B e l i n
6 4 6 0 6 4 7 0 6 4 8 0 6 4 9 0 6 5 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |G A C G A C C T G G T G G C G A T C G C C T G G G A G G A C T G G G A G A C C G A G G A A G A G A T C o n t i g d s r _ 1 H 4
Annex 3
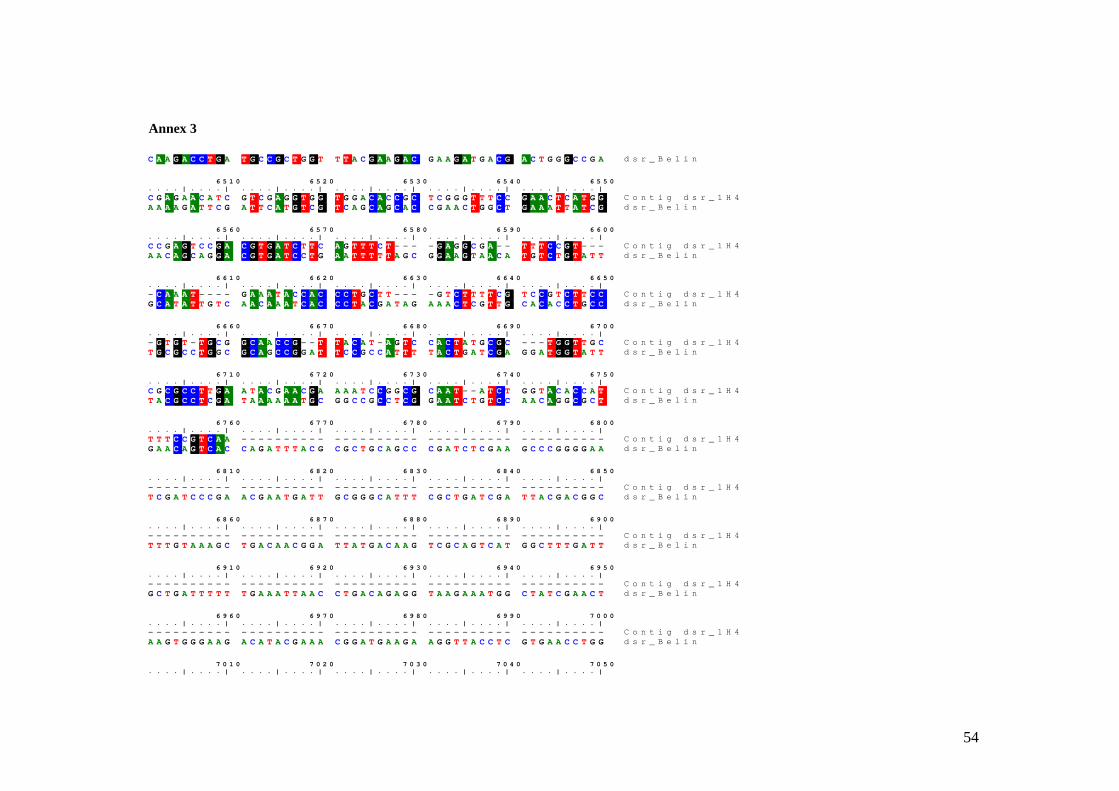
54
C A A G A C C T G A T G C C G C T G G T T T A C G A A G A C G A A G A T G A C G A C T G G G C C G A d s r _ B e l i n
6 5 1 0 6 5 2 0 6 5 3 0 6 5 4 0 6 5 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C G A G A A C A T C G T C G A G G T G G T G G A C A C C G C T C G G G T T T C C G A A C T C A T G G C o n t i g d s r _ 1 H 4A A A A G A T T C G A T T C A T G T C G T C A G C A G C A C C G A A C T G G C T G A A A T T A T C G d s r _ B e l i n
6 5 6 0 6 5 7 0 6 5 8 0 6 5 9 0 6 6 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C C G A G T C C G A C G T G A T C T T C A G T T T C T - - - - G A G G C G A - - T T T C C G T - - - C o n t i g d s r _ 1 H 4A A C A G C A G G A C G T G A T C C T G A A T T T T T A G C G G A A G T A A C A T G T C T G T A T T d s r _ B e l i n
6 6 1 0 6 6 2 0 6 6 3 0 6 6 4 0 6 6 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |- C A A A T - - - - G A A A T A C C A C C C T G C T T - - - - G T C T T T T C G T C C G T C T T C C C o n t i g d s r _ 1 H 4G C A T A T T G T C A A C A A A T C A C C C T A C G A T A G A A A C T C G T T G C A C A C C T G C C d s r _ B e l i n
6 6 6 0 6 6 7 0 6 6 8 0 6 6 9 0 6 7 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |- G T G T - T G C G G C A A C C G - - T T A C A T - A G T C C A C T A T G C G C - - - T G G T T G C C o n t i g d s r _ 1 H 4T G C G C C T G G C G C A G C C G G A T T C C G C C A T T T T A C T G A T C G A G G A T G G T A T T d s r _ B e l i n
6 7 1 0 6 7 2 0 6 7 3 0 6 7 4 0 6 7 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |C G C G C C T T G A A T A C G A A C G A A A A T C C G G C G C A A T - - A T C T G G T A C A C C A T C o n t i g d s r _ 1 H 4T A C G C C T C G A T A A A A A A T G C G G C C G C C T C G G A A T C T G T C C A A C A G G C G C T d s r _ B e l i n
6 7 6 0 6 7 7 0 6 7 8 0 6 7 9 0 6 8 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |T T T C C G T C A A - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - C o n t i g d s r _ 1 H 4G A A C A G T C A C C A G A T T T A C G C G C T G C A G C C C G A T C T C G A A G C C C G G G G A A d s r _ B e l i n
6 8 1 0 6 8 2 0 6 8 3 0 6 8 4 0 6 8 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - C o n t i g d s r _ 1 H 4T C G A T C C C G A A C G A A T G A T T G C G G G C A T T T C G C T G A T C G A T T A C G A C G G C d s r _ B e l i n
6 8 6 0 6 8 7 0 6 8 8 0 6 8 9 0 6 9 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - C o n t i g d s r _ 1 H 4T T T G T A A A G C T G A C A A C G G A T T A T G A C A A G T C G C A G T C A T G G C T T T G A T T d s r _ B e l i n
6 9 1 0 6 9 2 0 6 9 3 0 6 9 4 0 6 9 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - C o n t i g d s r _ 1 H 4G C T G A T T T T T T G A A A T T A A C C T G A C A G A G G T A A G A A A T G G C T A T C G A A C T d s r _ B e l i n
6 9 6 0 6 9 7 0 6 9 8 0 6 9 9 0 7 0 0 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - C o n t i g d s r _ 1 H 4A A G T G G G A A G A C A T A C G A A A C G G A T G A A G A A G G T T A C C T C G T G A A C C T G G d s r _ B e l i n
7 0 1 0 7 0 2 0 7 0 3 0 7 0 4 0 7 0 5 0. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Annex 3

55
Amann, R.I., Ludwig, W., and Schleifer, K.-H. (1995) Phylogenetic identification and in
situ detection of individual microbial cells without cultivation. Microbiol. Rev. 59: 143-169.
Andrews, S.C., Robinson, A.K., and Rodriguez-Quinones, F. (2003) Bacterial iron homeostasis. Fems Microbiology Reviews 27: 215-237.
Barnett, M.J., Hung, D.Y., Reisenauer, A., Shapiro, L., and Long, S.R. (2001) A homolog of the CtrA cell cycle regulator is present and essential in Sinorhizobium meliloti. Journal of Bacteriology 183: 3204-3210.
Beja, O. (2004) To BAC or not to BAC: marine ecogenomics. Current Opinion in Biotechnology: 187-190.
Britton, K.L., Asano, Y., and Rice, D.W. (1998) Crystal structure and active site location of N-(1-D-carboxylethyl)-L-norvaline dehydrogenase. Nature Structural Biology 5: 593-601.
Brown, M.H., Paulsen, I.T., and Skurray, R.A. (1999) The multidrug efflux protein NorM is a prototype of a new family of transporters. Molecular Microbiology 31: 394-395.
Busch, W., and Saier, M.H. (2004) The IUBMB-endorsed transporter classification system. Molecular Biotechnology 27: 253-262.
Cavanaugh, C.M., Gardiner, S.L., Jones, M.L., Jannasch, H.W., and Waterbury, J.B. (1981) Prokaryotic Cells in the Hydrothermal Vent Tube Worm Riftia-Pachyptila Jones - Possible Chemoautotrophic Symbionts. Science 213: 340-342.
Chlumsky, L.J., Zhang, L.N., and Jorns, M.S. (1995) Sequence-Analysis of Sarcosine Oxidase and Nearby Genes Reveals Homologies with Key Enzymes of Folate One-Carbon Metabolism. Journal of Biological Chemistry 270: 18252-18259.
Dubilier, N. (2004) Multiple bacterial endosymbionts in gutless marine worms: Competitors or Partners? In Nova Acta Leopoldina NF 88, pp. 107-112.
Dubilier, N., Blazejak, A., and Rühland, C. (In press) Symbioses between bacteria and gutless marine oligochaetes. In Molecular basis of symbiosis. J., O. (ed). Berlin: Springer Verlag.
Dubilier, N., Glöckner, F.O., and Amann, R. (Proposal for the Community Sequencing Programm of the Joint Genome Institute, USA) Comparative metagenomics of co-occuring bacterial symbionts in a gutless marine worm. In, p. 15.
Dubilier, N., Mulders, C., Ferdelman, T., de Beer, D., Pernthaler, A., Klein, M. et al. (2001) Endosymbiotic sulphate-reducing and sulphide-oxidizing bacteria in an oligochaete worm. Nature 411: 298-302.
Eisen, J.A., Nelson, K.E., Paulsen, I.T., Heidelberg, J.F., Wu, M., Dodson, R.J. et al. (2002) The complete genome sequence of Chlorobium tepidum TLS, a photosynthetic, anaerobic, green-sulfur bacterium. Proceedings of the National Academy of Sciences of the United States of America 99: 9509-9514.
Erseus, C. (1979) Taxonomic Revision of the Marine Genus Phallodrilus Pierantoni (Oligochaeta, Tubificidae), with Descriptions of 13 New Species. Zoologica Scripta 8: 187-208.
Erseus, C. (2003) The gutless Tubificidae (Annelida: Oligochaeta) of the Bahamas. In Meiofauna Marina. Pfeil, F. (ed). München: Verlag Dr. Friedrich Pfeil, pp. 59-84.
Felbeck, H. (1981) Chemoautotrophic Potential of the Hydrothermal Vent Tube Worm, Riftia-Pachyptila Jones (Vestimentifera). Science 213: 336-338.

56
Ferry, J.G. (1995) CO dehydrogenase. Annu. Rev. Microbiol. 49: 305-333. Fisher, C.R. (1990) Chemoautotrophic and methanotrophic symbioses in marine
invertebrates. Rev. Aquat. Sci. 2: 399-436. Giere, O., Erseus, C., and Stuhlmacher, F. (1998) A new species of Olavius (Tubificidae)
from the Algarve coast in Portugal, the first East Atlantic gutless oligochaete with symbiotic bacteria. Zoologischer Anzeiger 237: 209-214.
Giere, O., Nieser, C., Windoffer, R., and Erseus, C. (1995) A Comparative Structural Study on Bacterial Symbioses of Caribbean Gutless Tubificidae (Annelida, Oligochaeta). Acta Zoologica 76: 281-290.
Guerinot, M.L. (1994) Microbial Iron Transport. Annual Review of Microbiology 48: 743-772.
Hantke, K. (2003) Is the bacterial ferrous iron transporter FeoB a living fossil? Trends in Microbiology 11: 192-195.
Harwood, C.S., and Canale-Parola, E. (1983) Spirochaeta-Isovalerica Sp-Nov, a Marine Anaerobe That Forms Branched-Chain Fatty-Acids as Fermentation Products. International Journal of Systematic Bacteriology 33: 573-579.
Harwood, C.S., and Canale-Parola, E. (1984) Ecology of Spirochetes. Annual Review of Microbiology 38: 161-192.
Healy, F.G., Ray, R.M., Aldrich, H.C., Wilkie, A.C., Ingram, L.O., and Shanmugam, K.T. (1995) Direct Isolation of Functional Genes Encoding Cellulases from the Microbial Consortia in a Thermophilic, Anaerobic Digester Maintained on Lignocellulose. Applied Microbiology and Biotechnology 43: 667-674.
Hespell, R.B., and Canale-Parola, E. (1973) Glucose and Pyruvate Metabolism of Spirochaeta-Litoralis, an Anaerobic Marine Spirochete. Journal of Bacteriology 116: 931-937.
Huda, N., Chen, J., Morita, Y., Kuroda, T., Mizushima, T., and Tsuchiya, T. (2003) A Na+-coupled multidrug efflux pump, from Vibrio cholerae Non-O1. Microbiol. Immunol. 47: 419-427.
Igarashi, K., and Kashiwagi, K. (1999) Polyamine transport in bacteria and yeast. Biochemical Journal 344: 633-642.
Kammler, M., Schon, C., and Hantke, K. (1993) Characterization of the Ferrous Iron Uptake System of Escherichia-Coli. Journal of Bacteriology 175: 6212-6219.
Kappler, U., and Dahl, C. (2001) Enzymology and molecular biology of prokaryotic sulfite oxidation. Fems Microbiology Letters 203: 1-9.
Kelly, D.P., Shergill, J.K., Lu, W.P., and Wood, A.P. (1997) Oxidative metabolism of inorganic sulfur compounds by bacteria. Antonie Van Leeuwenhoek International Journal of General and Molecular Microbiology 71: 95-107.
Krafft, T., Gross, R., and Kroger, A. (1995) The function of Wolinella succinogenes psr genes in electron transport with polysulphide as the terminal electron acceptor. Eur J Biochem 230: 601-606.
Krehenbrinck, M., and Steinbüchel, A. (2004) Partial purification and characterization of a non-cyanobacterial cyanophycin synthetase from Acinetobacter calcoaceticus strain ADP1 with regards to substrate specificity, substrate affinity and binding to cyanophycin. Microbiology 150: 2599-2608.
Krehenbrinck, M., Oppermann-Sanio, F.-B., and Steinbüchel, A. (2002) Evaluation of non-cyanobacterial geneome sequences for occurence of genes encoding proteins

57
homologues to cyanophycin synthetase and cloning of an active cyanophycin synthetase from Acinetobacter sp. strain DSM 587. Archives of Microbiology 177: 371-380.
Leadbetter, J.R., Schmidt, T.M., Graber, J.R., and Breznak, J.A. (1999) Acetogenesis from H-2 plus CO2 by spirochetes from termite guts. Science 283: 686-689.
Lee, M.H., Mulrooney, S.B., Renner, M.J., Markowicz, Y., and Hausinger, R.P. (1992) Klebsiella-Aerogenes Urease Gene-Cluster - Sequence of Ured and Demonstration That 4 Accessory Genes (Ured, Uree, Uref, and Ureg) Are Involved in Nickel Metallocenter Biosynthesis. Journal of Bacteriology 174: 4324-4330.
Lengeler, J.W., Drews, G., and Schlegel, H.G. (eds) (1999) Biology of the prokaryotes. Stuttgart: G. Thieme Verlag.
Li, L., Kato, C., and Horikoshi, K. (1999c) Microbial diversity in sediments collected from the deepest cold-seep area, the Japan trench. Mar. Biotechnol. 1: 391-400.
Lilburn, T.C., Kim, K.S., Ostrom, N.E., Byzek, K.R., Leadbetter, J.R., and Breznak, J.A. (2001) Nitrogen fixation by symbiotic and free-living spirochetes. Science 292: 2495-2498.
Margulis, L., and Chapman, J. (1998) Endosymbioses:cyclical and permanent in evolution. Trends in microbiology 6: 342-345.
Mobley, H.L.T., and Hausinger, R.P. (1989) Microbial Ureases - Significance, Regulation, and Molecular Characterization. Microbiological Reviews 53: 85-108.
Moran, M.A., Buchan, A., Gonzalez, J.M., Heidelberg, J.F., Whitman, W.B., Kiene, R.P. et al. (2004) Genome sequence of Silicibacter pomeroyi reveals adaptations to the marine environment. Nature 432: 910-913.
Morita, Y., Kataoka, A., Shiota, S., Mizushima, T., and Tsuchiya, T. (2000) NorM of Vibrio parahaemolyticus is a Na+-driven multidrug efflux pump. Journal of Bacteriology 182: 6694-6697.
Morita, Y., Kodama, K., Shiota, S., Mine, T., Kataoka, A., Mizushima, T., and Tsuchiya, T. (1998) NorM, a putative multidrug efflux protein, of Vibrio parahaemolyticus and its homolog in Escherichia coli. Antimicrobial agents and chemotherapy 42: 1778-1782.
Pace, N.R. (1997) A molecular view of microbial diversity and the biosphere. Science 276: 734-740.
Piel, J. (2002) A polyketide synthase-peptide synthetase gene cluster from an uncultured bacterial symbiont of Paederus beetles. Proceedings of the National Academy of Sciences of the United States of America 99: 14002-14007.
Portner, H.O. (2002) Environmental and functional limits to muscular exercise and body size in marine invertebrate athletes. Comp Biochem Physiol A Mol Integr Physiol 133: 303-321.
Riesenfeld, C.S., Schloss, P.D., and Handelsman, J. (2004) Metagenomics: Genomic analysis of microbial communities. Annual Review of Genetics 38: 525-552.
Robertson, L.A., and Kuenen, J.G. (1992) The colorless sulfur bacteria. In The Prokaryotes. Balows, A., Trüper, H.G., Dworkin, M., Harder, W., and Schleifer, K.-H. (eds). New York: Springer-Verlag, pp. 385-413.
Rondon, M.R., August, P.R., Bettermann, A.D., Brady, S.F., Grossman, T.H., Liles, M.R. et al. (2000) Cloning the soil metagenome: a strategy for accessing the genetic and

58
functional diversity of uncultured microorganisms. Appl. Environ. Microbiol. 66: 2541-2547.
Rühland, C., Blazejak, A., Loy, A., Wagner, M., Erseus, C., Amann, R., and Dubilier, N. (In Prep.) Multiple symbioses with bacterial sulfide-oxidizers and sulfate-reducers in two co-occuring species of gutless worms (Oligochaeta) from Mediterranean.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecular cloning: a laboratory manual. cold spring harbor, N.Y.: Cold spring harbor laboratory press.
Schloss, P.D., and Handelsman, J. (2003) Biotechnological prospects from metagenomics. Current Opinion in Biotechnology 14: 303-310.
Seibel, B.A., and Walsh, P.J. (2001) Carbon cycle - Potential, impacts of CO2 injection on deep-sea biota. Science 294: 319-320.
Simon, J., Sanger, M., Schuster, S.C., and Gross, R. (2003) Electron transport to periplasmic nitrate reductase (NapA) of Wolinella succinogenes is independent of a NapC protein. Molecular Microbiology 49: 69-79.
Stein, J.L., Marsh, T.L., Wu, K.Y., Shizuya, H., and DeLong, E.F. (1996) Characterization of uncultivated prokaryotes- isolation and analysis of a 40-kilobase-pair genome fragment from a planktonic marine Archaeon. J. Bact. 178: 591-599.
Toffanin, A., Cadahia, E., Imperial, J., Ruiz-Argueso, T., and Palacios, J.M. (2002) Characterization of the urease gene cluster from Rhizobium leguminosarum bv. viciae. Archives of Microbiology 177: 290-298.
Tyson, G.W., Chapman, J., Hugenholtz, P., Allen, E.E., Ram, R.J., Richardson, P.M. et al. (2004) Community structure and metabolism through reconstruction of microbial genomes from the environment. Nature 428: 37-43.
Wernegreen, J.J. (2004) Endosymbiosis: lessons in conflict resolution. PLoS Biol 2: E68. Widdel, F., and Hansen, T. (1992a) The dissimilatory sulfate- and sulfur-reducing
bacteria. In The Prokaryotes. Balows, A., Trüper, H.G., Dworkin, M., Harder, W., and Schleifer, K.-H. (eds). New York: Springer-Verlag, pp. 583-624.
Zhang, Y.X., Guo, X.K., Wu, C., Bi, B., Ren, S.X., Wu, C.F., and Zhao, G.P. (2004) Characterization of a novel toxin-antitoxin module, VapBC, encoded by Leptospira interrogans chromosome. Cell Research 14: 208-216.

59
Aknowledgements
I want to thank to Nicole Dubilier for supervision, to Anna Blazejak for good advices
during my work in the Molecular Ecology Department, to Silke Wetzel, Anke
Meyerdierks and Michael Richter and not the last to Niculina Musat for her support and
advices.
I thank to the entire Molecular Ecology group for the great working atmosphere and
finally to Max Planck Geselshaft for the financial support.





























