Embed Size (px)
Citation preview

ZYTOSKELETT
ZELLTEILUNG
ZELLZYKLUS

22
Krebs ist die Ursache der meisten Todesfälle weltweit: 7.4 Million Todesfälle/Jahr (ungefähr 13% aller Todesfälle).

3

4
Rb: RETINOBLASTOMA

5
CYTOSKELETT

Funktionen des Cytoskeletts:
1. Es verleiht der Zelle Form und Reissfestigkeit
2. Es ermöglicht verschiedene Arten zellulärer
Bewegung.
3. Separiert die Chromosomen bei der Zellteilung
4. Es liefert „Schienen” für Motorproteine, die an der
Bewegung von Zellbestandteilen beteiligt sind.
5. Wechselwirkt mit extracellulären Strukturen, um die
Zelle in Position zu halten

Heilung einer Wunde
DAS CYTOSKELETT

Wachstum eines Axons
DAS CYTOSKELETT

Die schnelle Bewegung der Kenatocyte ist duch die Koordination zwischen der Ausstülpung des Lamellipodiums und der Translokation der
Zellkörpers ermöglicht. Fluoreszente Partikel auf dem Substrat zeigen die rollende Bewegung der Zelle Während der Bewgung haften diese
Partikel an den Zelloberfläche. Anderson et al., 1996.
DAS CYTOSKELETT

Weiße Blutzelle folgt ein Bakterium
DAS CYTOSKELETT

11Zellteilung
DAS CYTOSKELETT

MIKROFILAMENTE MICROTUBULI INTERMEDIÄRFILAMENTE
A B C
DAS CYTOSKELETT

13

14
INTERMEDIÄR-
FILAMENTE

15
INTERMEDIÄRFILAMENTE

16
Fibrilläre Untereinheit
Intermediärfilamente
• Intermediärfilamente bestehen aus fibrilläre Proteinen
Stabilisieren die Form der Zelle und verleihen ihr Reissfestigkeit.
• Manche sind an Desmosomen verankert unterstützen so den Zusammenhalt von Nachbarzellen.
• Andere bilden die Kernlamina

17
Typen:
I., II, Keratine (Desmosomen, Haar, Nagel)
III., Vimentin (Anfang der Zelldifferenzierung) Desmin
(Muskeln).
IV., Neurofilamente (Axone von Neuronen);
V., Lamin (Innere Seite der Kernmembran);
Krankheiten:
epidermolysis bullosa simplex, (Keratin)
amyotrophic lateral sclerosis (ALS) (Neurofilament)
INTERMEDIÄRFILAMENTE

18
AKTIN-
FILAMENTE
(Microfilamente)
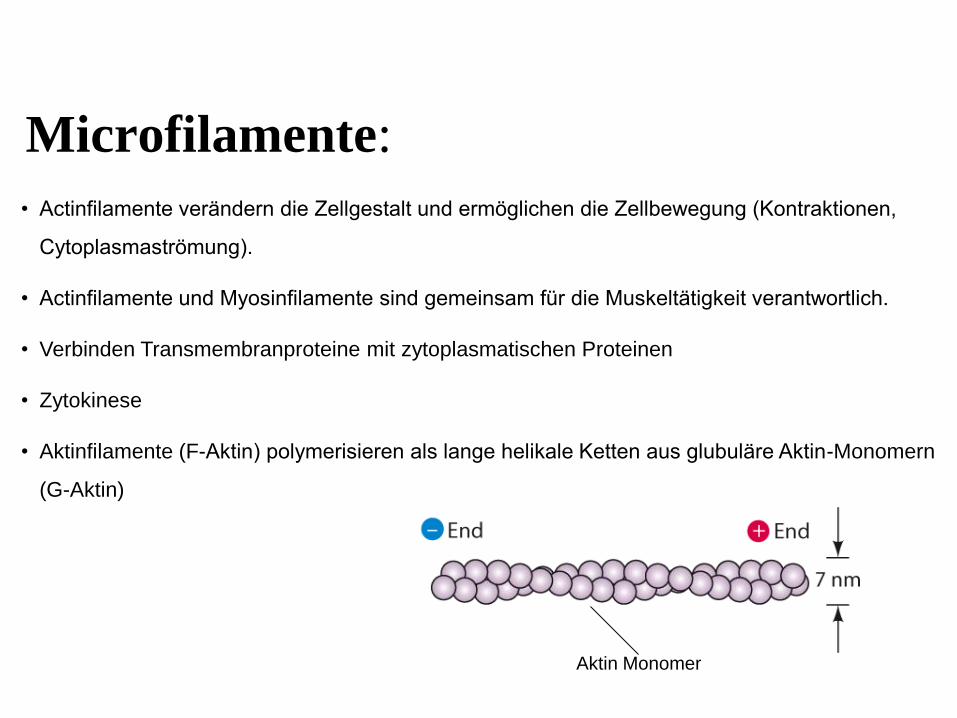
Microfilamente:
• Actinfilamente verändern die Zellgestalt und ermöglichen die Zellbewegung (Kontraktionen,
Cytoplasmaströmung).
• Actinfilamente und Myosinfilamente sind gemeinsam für die Muskeltätigkeit verantwortlich.
• Verbinden Transmembranproteine mit zytoplasmatischen Proteinen
• Zytokinese
• Aktinfilamente (F-Aktin) polymerisieren als lange helikale Ketten aus glubuläre Aktin-Monomern
(G-Aktin)
Aktin Monomer

20
ACTIN-BINDENDE PROTEINE
Quervernetzung: filamin
Depolimerisierung: KofilinMembranverbindung: spectrin, distrofin
Fragmentierung: Gelsolin
Actin Monomer
Actin Polymer
Monomer-
Sequstrierung:
Timozin
Capping: CapZ Bündelung: Fascin, Fimbrin

21
Viele Zellen bewegen sich durch Ausstülpungen an der
Zellfront (LAMELLIPODIUM) and ziehen danach das
Zellende zürück.
Zellbewegung - Lamellipodia

22
Zellbewegung - Filopodia
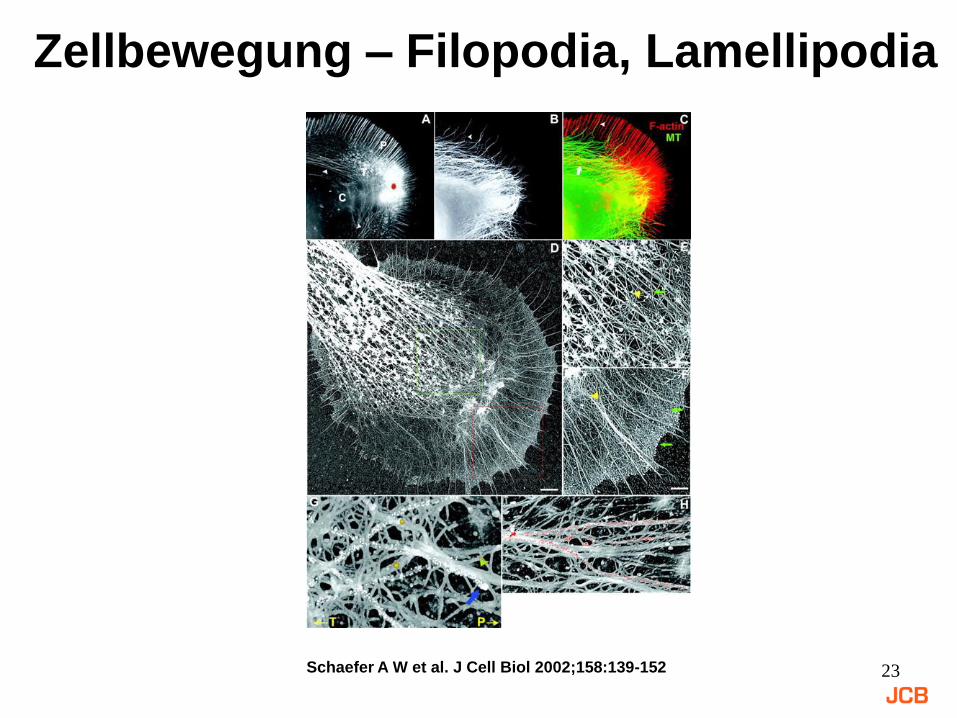
23Schaefer A W et al. J Cell Biol 2002;158:139-152
Zellbewegung – Filopodia, Lamellipodia
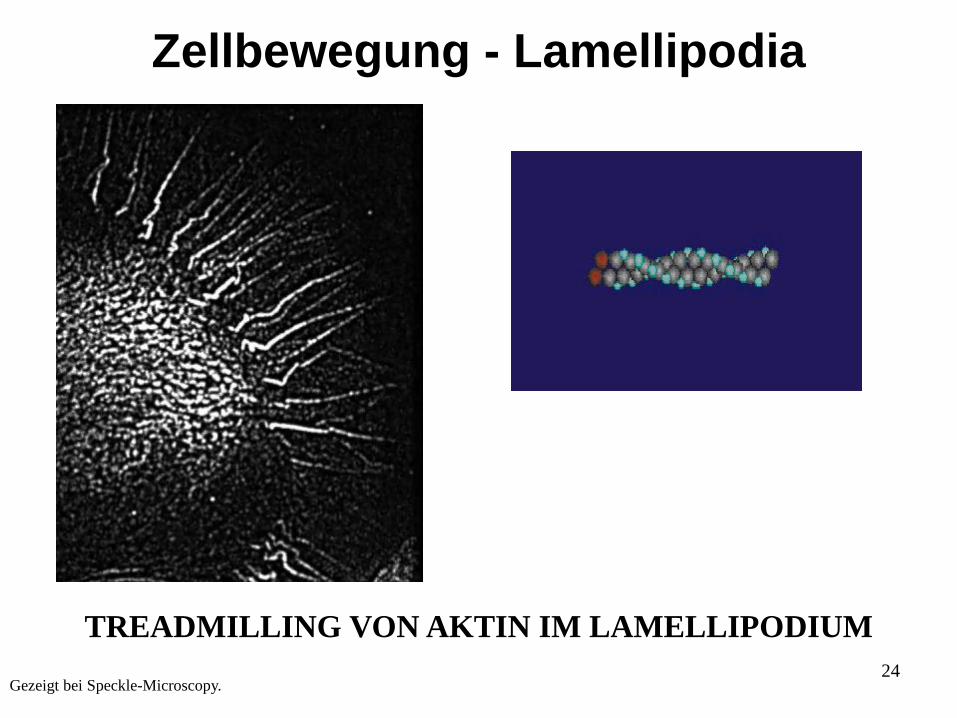
24Gezeigt bei Speckle-Microscopy.
TREADMILLING VON AKTIN IM LAMELLIPODIUM
Zellbewegung - Lamellipodia
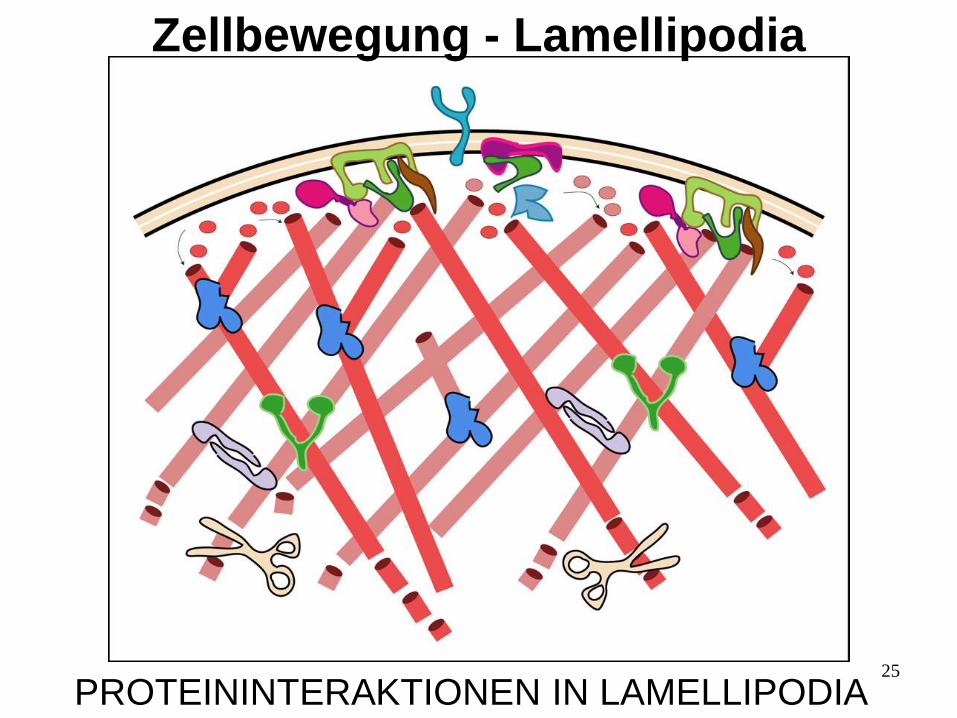
25
PROTEININTERAKTIONEN IN LAMELLIPODIA
Zellbewegung - Lamellipodia
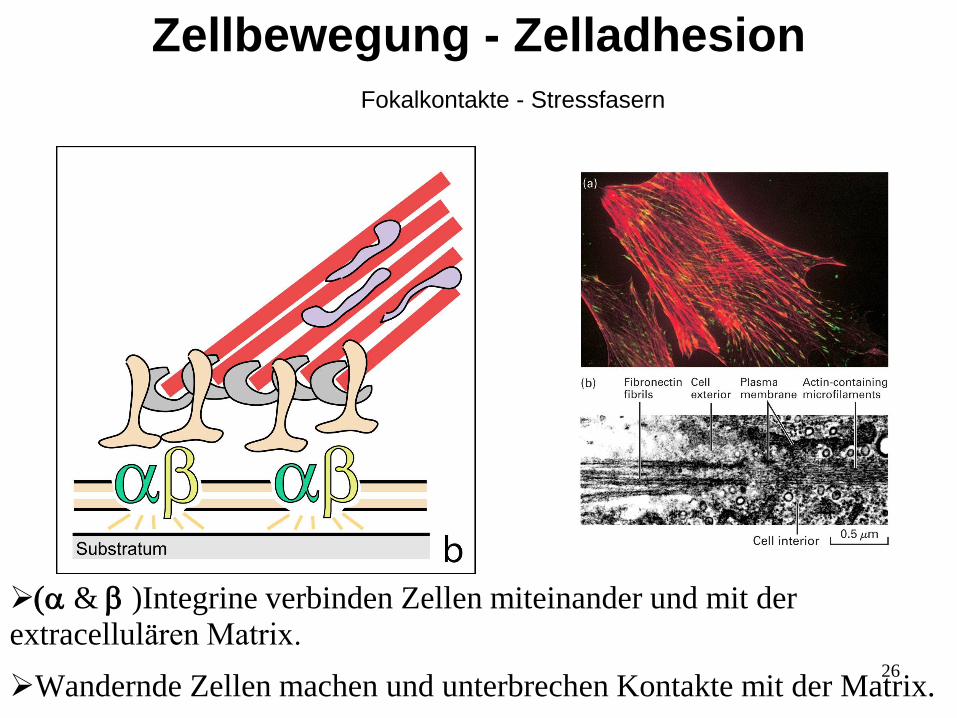
26
(a & b )Integrine verbinden Zellen miteinander und mit der extracellulären Matrix.
Wandernde Zellen machen und unterbrechen Kontakte mit der Matrix.
Zellbewegung - Zelladhesion
Fokalkontakte - Stressfasern
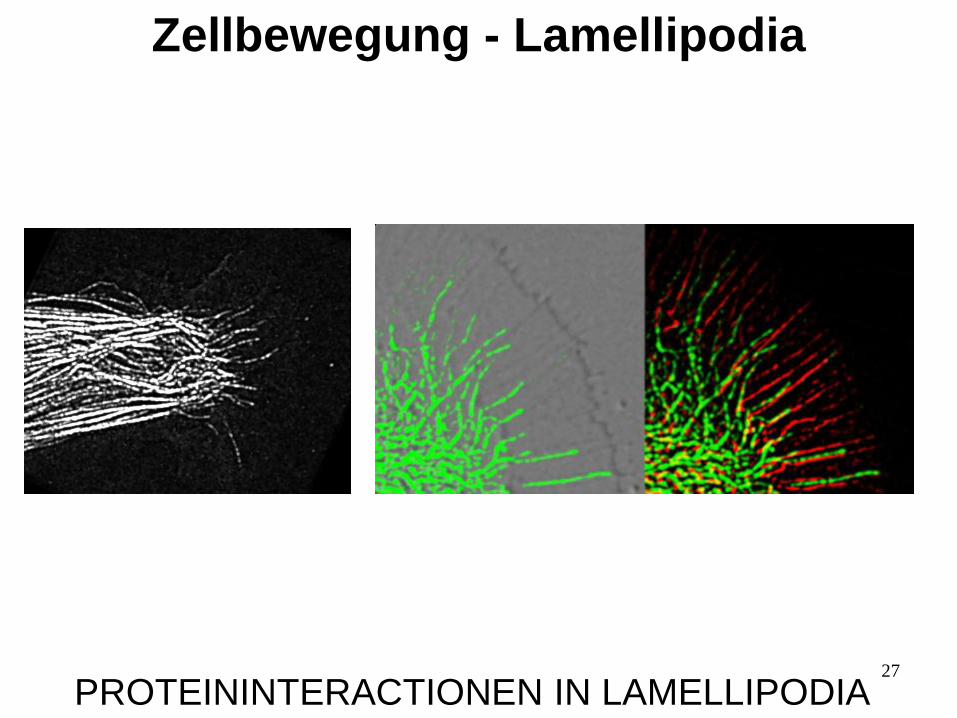
27
PROTEININTERACTIONEN IN LAMELLIPODIA
Zellbewegung - Lamellipodia

28
Zellbewegung - Zelladhesion
Microtubuli sind notwendig für den Abbau von
Fokaladhesionen und Fokalkomplexen während der
Zellwanderung.
Vorderseite (Lamellipodium) Hinterseite

29
Schematic representation of the actin cytoskeleton in a polarised fibroblast. The different organisational forms
of actin filaments are depicted: diagonal actin filament meshwork in the lamellipodium (Lam.), with associated
radial bundles (filopodia, Fil); contractile bundles of actin (stress fibres, S.F.) in the cell body and at the cell
edge; and a loose actin network throughout the cell. Sites of adhesion of the cell with the substrate are also
indicated: focal complexes (Fx) associated with lamellipodia and filopodia and focal adhesions (F.A.) at the
termini of actin stress fibre bundles.
STRESS FIBERS werden
geformt, wenn die Zelle mit
dem Substratum eine stabile
Verbindung aufbaut.
Während der Kontraktion
der stress fibers Myosin trägt
zum Gleiten der
antiparallelen Aktinfilamente
bei.
Kontraktion der stress
fibers zieht das Zellende.
Zellbewegung

30
Microtubuli sind notwendig für den Abbau von
Fokaladhesionen und Fokalkomplexen während der
Zellmigration.
Zellbewegung – Zelladhesion und
Mikrotubuli

Listeria monocytogenes moving in PtK2 cells
These pathogenic bacteria grow directly in the host cell cytoplasm. The phase-dense streaks behind the bacteria are the actin-rich comet tails.
Actin-based motility is also used in cellular motility; this cell is using it's cytoskeleton to crawl toward the lower right-hand corner. Speeded up
150X over real time. Julie Theriot & Dan Portnoy
Viele Pathogene
benutzen das Aktin der
Wirtszelle, um sich in
der infizierten Zelle zu
bewegen und zu einer
anderen Zelle
übertragen zu werden.
Sie lassen Aktin-
Schwänze wachsen.
Pathogene und das Aktin-Cytoskelett
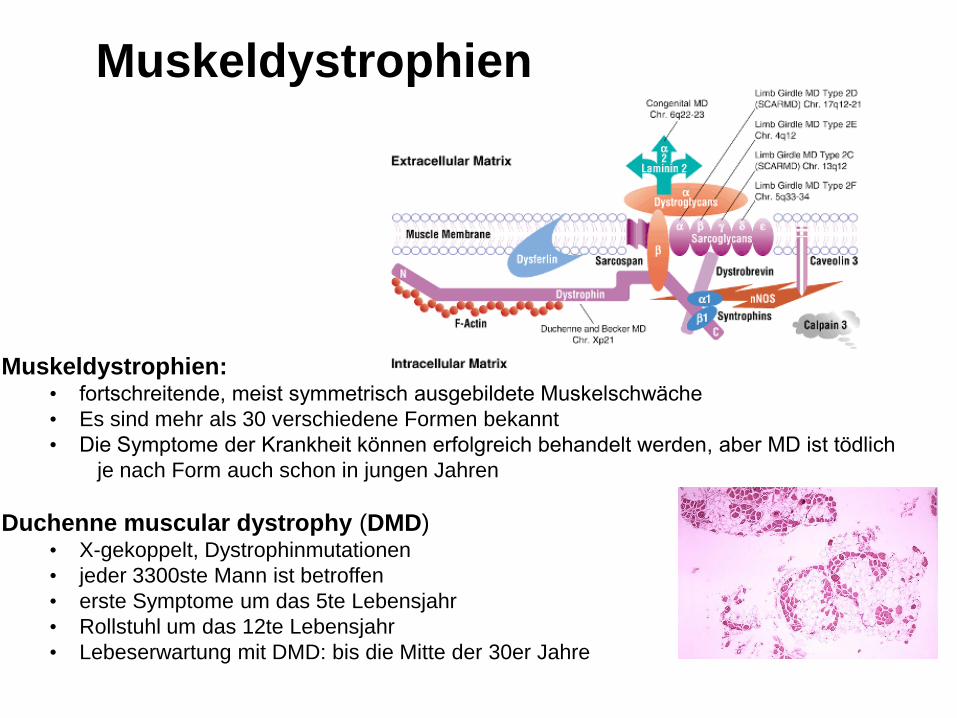
Muskeldystrophien:• fortschreitende, meist symmetrisch ausgebildete Muskelschwäche
• Es sind mehr als 30 verschiedene Formen bekannt
• Die Symptome der Krankheit können erfolgreich behandelt werden, aber MD ist tödlich
je nach Form auch schon in jungen Jahren
Duchenne muscular dystrophy (DMD)• X-gekoppelt, Dystrophinmutationen
• jeder 3300ste Mann ist betroffen
• erste Symptome um das 5te Lebensjahr
• Rollstuhl um das 12te Lebensjahr
• Lebeserwartung mit DMD: bis die Mitte der 30er Jahre
Muskeldystrophien

Mikrovilli

34
MYOSINE

35
Skelettmuskulatur
36
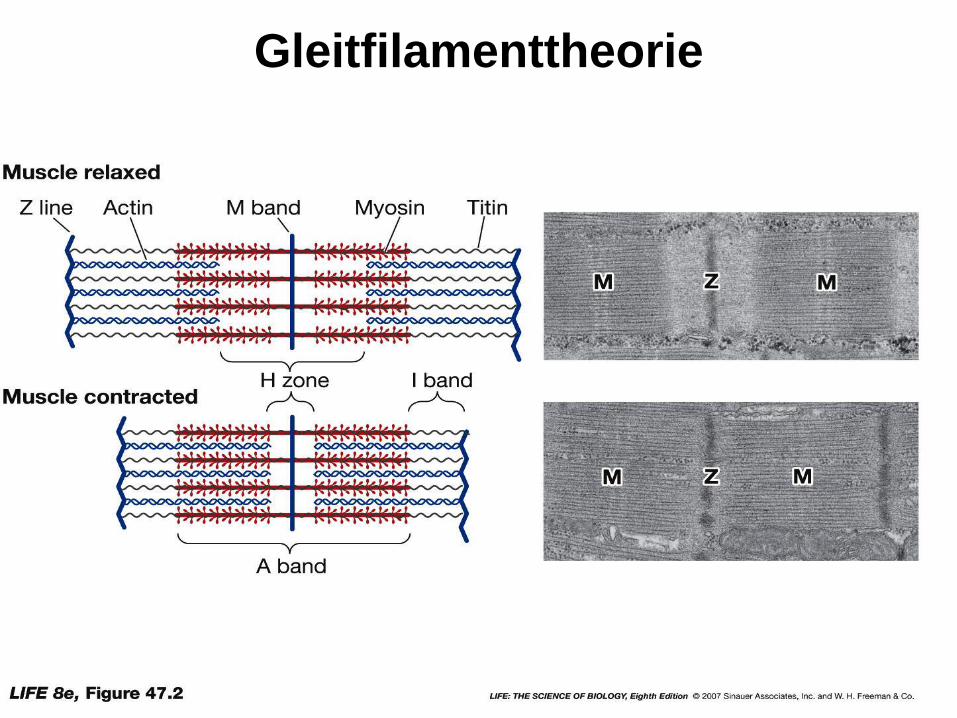
Gleitfilamenttheorie

37
MYOSINE
Aktin-bindende Motorproteine

38
Gleitfilamenttheorie
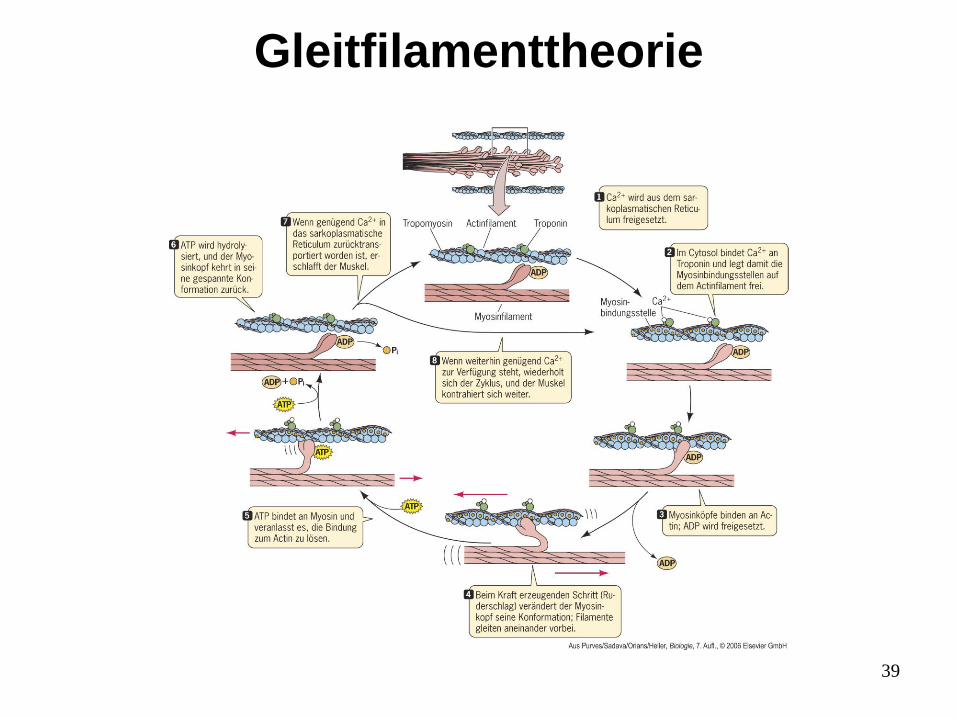
39
Gleitfilamenttheorie

40
RIGOR MORTIS
(Totenstarre)

41
MIKROTUBULI

42
• Mikrotubuli sind lange zylinderförmige Gebilde, die aus Tubulin gebildet werden. Tubulin besteht aus zwei Untereinheiten :
a-tubulin und b-tubulin
• Mikrotubuli verlängern oder verkürzen sich, indem Tubulin-Dimere zugefügt oder entfernt werden.
• Durch das Verkürzen von Mikrotubuli werden die Chromosomen
bei der Zellteilung bewegt.
• Wechselwirkungem zwischen Mikrotubuli ermöglichen Zellbewegungen.
• Mikrotubuli dienen als „Gleise” für die Beforderung von Vesikeln.
Tubulin Dimer
b-Tubulin
Monomer
a-Tubulin
Monomer
Mikrotubuli
(Makrofilamente)

43
MIKROTUBULI
(MT)
Dynamische Instabilität
MT-assiziierte Proteine
beeinflussen die
dynamische
Instabilität.
Zytostatika: Substanzen,
die die Zellteilung hemmen
Colchizin, Taxol,
Vinblastin-
Stabilisatoren, in
grosser Konzentration
depolymerisieren
- Ende + Ende
Aufbau
POLIMERISIERUNG
DEPOLIMERISIERUNG
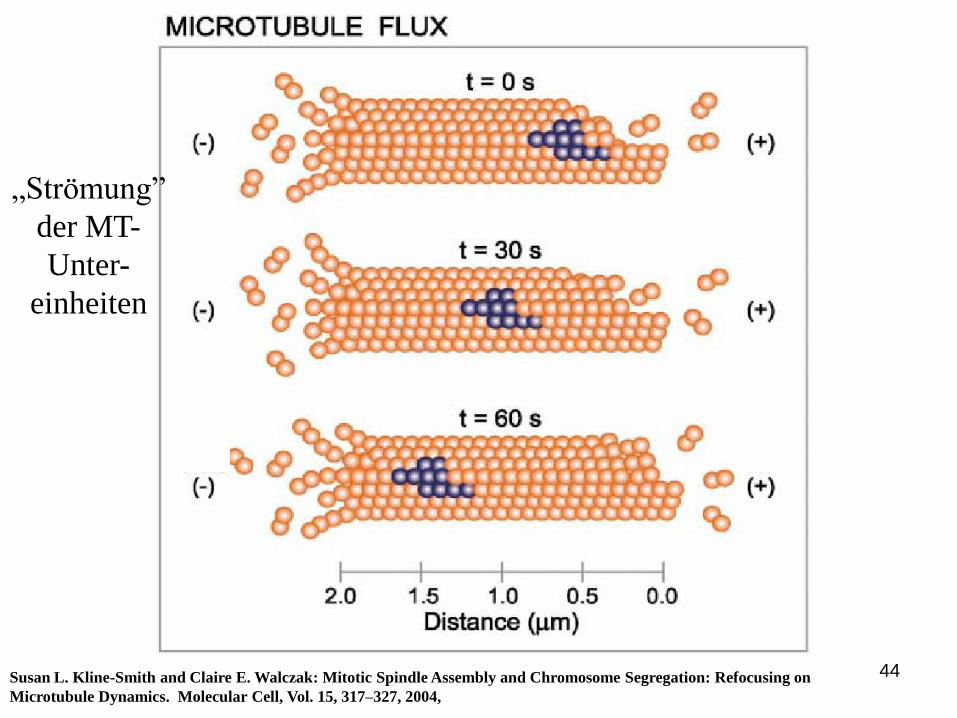
44Susan L. Kline-Smith and Claire E. Walczak: Mitotic Spindle Assembly and Chromosome Segregation: Refocusing on
Microtubule Dynamics. Molecular Cell, Vol. 15, 317–327, 2004,
„Strömung”
der MT-
Unter-
einheiten

45
Mikrotubuli sind dynamisch an
den + Enden
46
Motorproteine
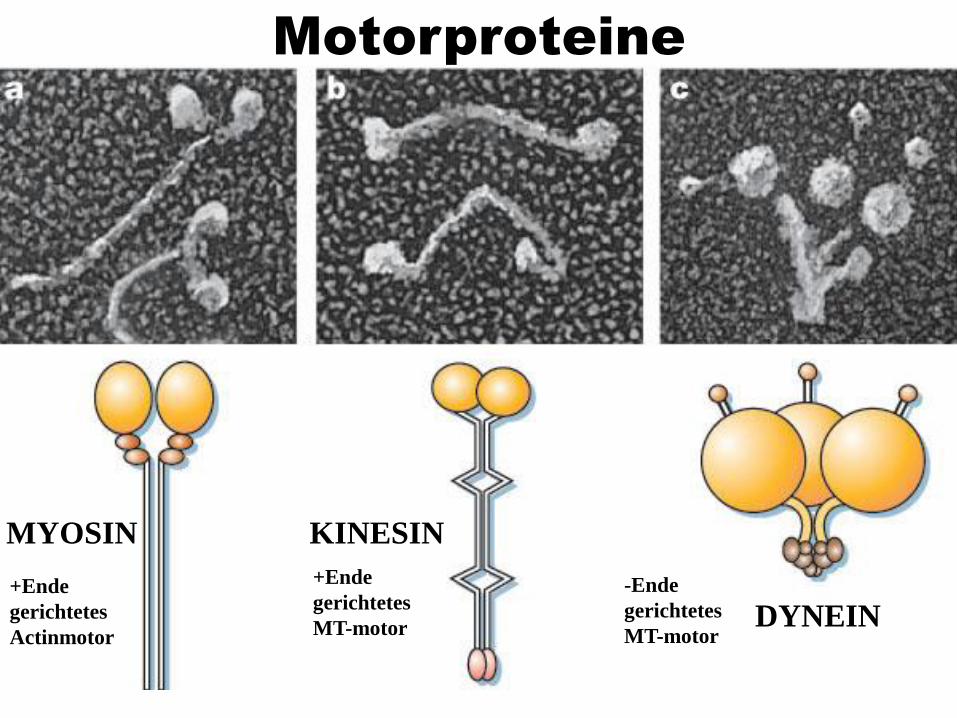
47
MYOSIN
DYNEIN
KINESIN
+Ende
gerichtetes
Actinmotor
Motorproteine
+Ende
gerichtetes
MT-motor
-Ende
gerichtetes
MT-motor
48
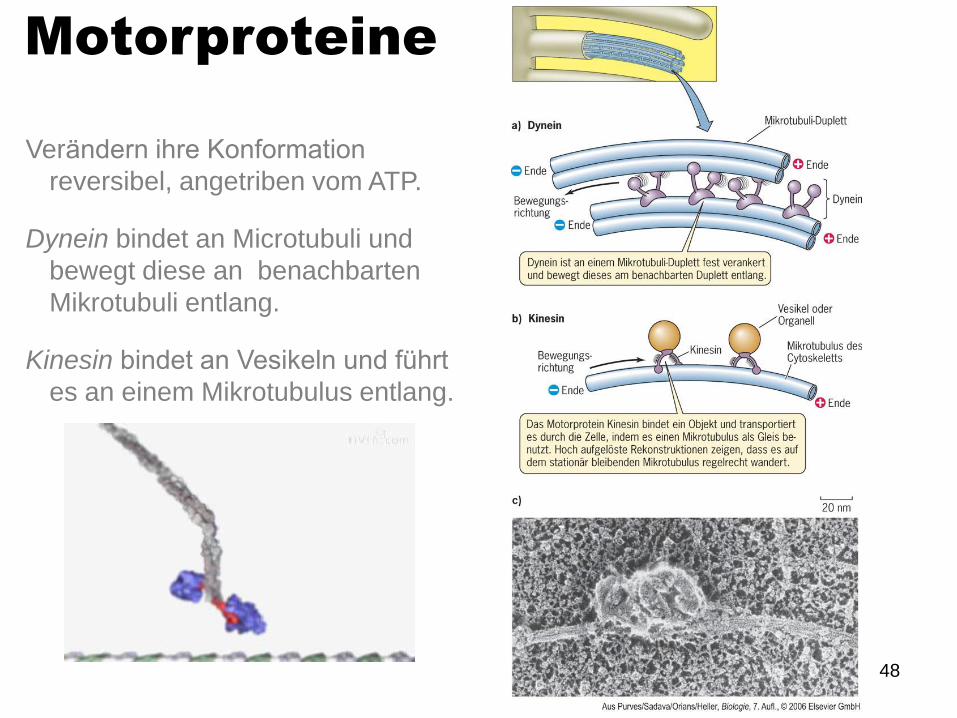
Motorproteine
Verändern ihre Konformation
reversibel, angetriben vom ATP.
Dynein bindet an Microtubuli und
bewegt diese an benachbarten
Mikrotubuli entlang.
Kinesin bindet an Vesikeln und führt
es an einem Mikrotubulus entlang.

49
Kinezin Stammbaum
MT-MOTOREN: KINEZINE
Klassifizierung der Kinesine: (Position des
Motor-Domäns)
(N:N terminus, I:Innere Region, C: C-terminus):
Kin N (+ Ende-gerichtete Kinesine),
Kin I (MT-Depolymerasen)
Kin C (+ Ende-gerichtete Kinesine)
Monomer-, Dimer-, Tetramer-Formen

50
Kinezin családfa
MT MOTOROK: KINEZINEK
Funktionen der Kinesine:
Transport von Zellorganellen
• Vesikeln
• Pigmente
• ER und Golgi-Apparat
• Chromosomen
51
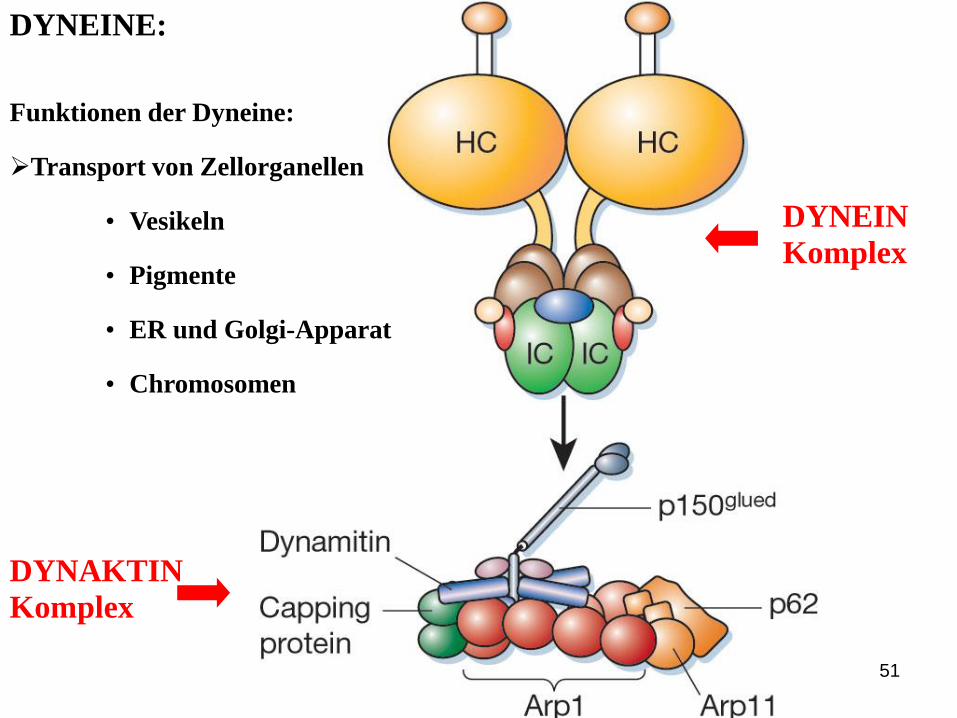
DYNEINE:
DYNAKTIN
Komplex
DYNEIN
Komplex
Funktionen der Dyneine:
Transport von Zellorganellen
• Vesikeln
• Pigmente
• ER und Golgi-Apparat
• Chromosomen

52
Figure 4.22 Sliding Microtubules Cause Cilia to Bend
Geisseln (Flagellen) und Cilien

53
Geisseln (Flagellen) und Cilien

54

55
56
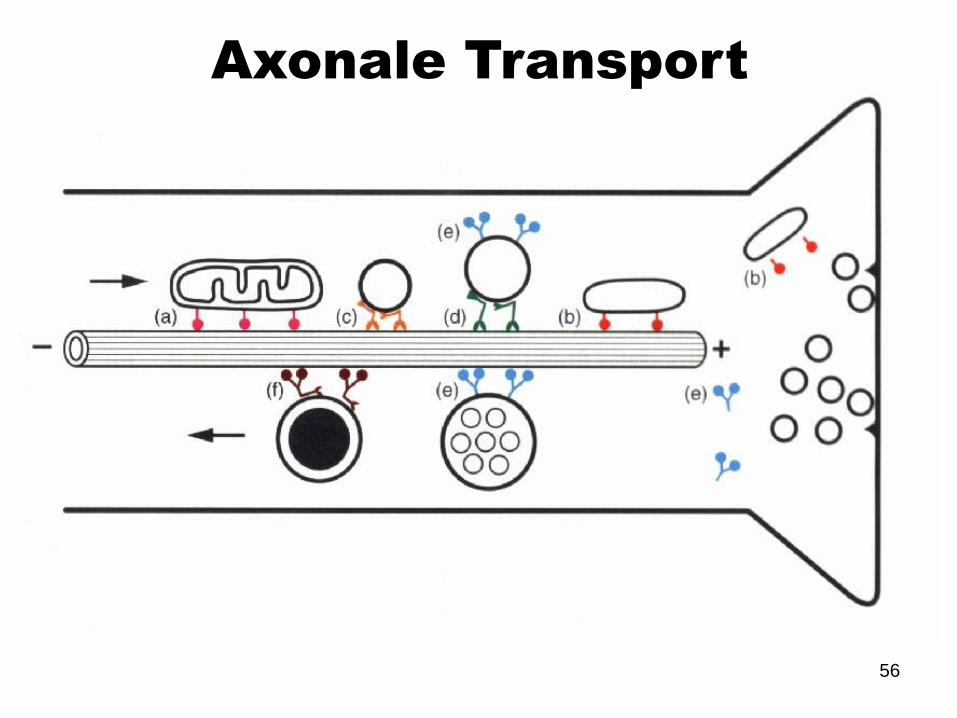
Axonale Transport
57
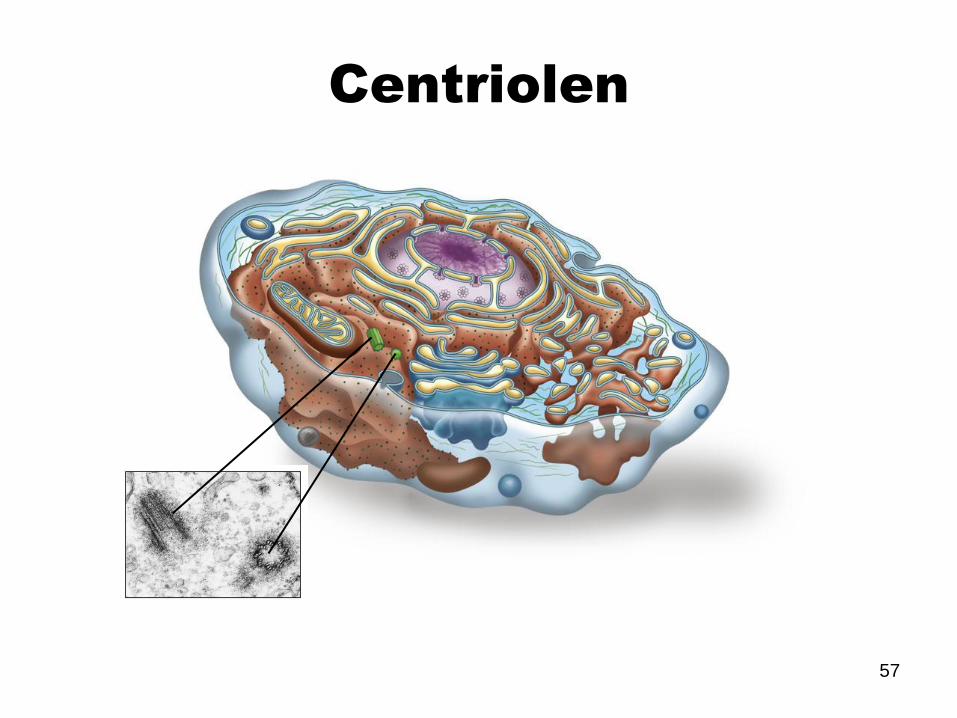
Centriolen
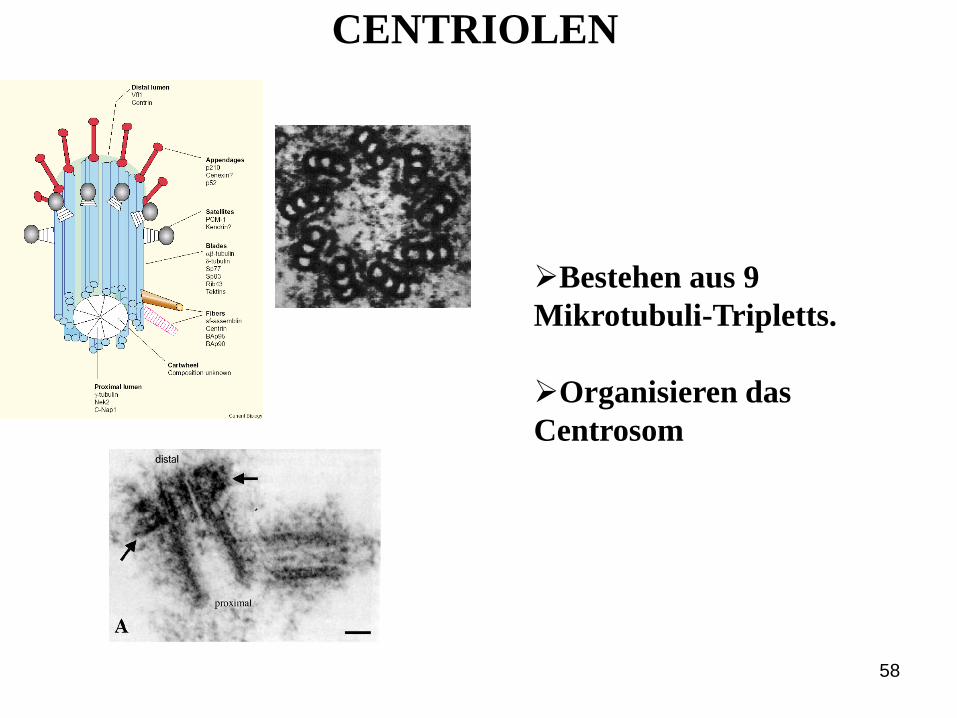
58
Bestehen aus 9
Mikrotubuli-Tripletts.
Organisieren das
Centrosom
CENTRIOLEN

59
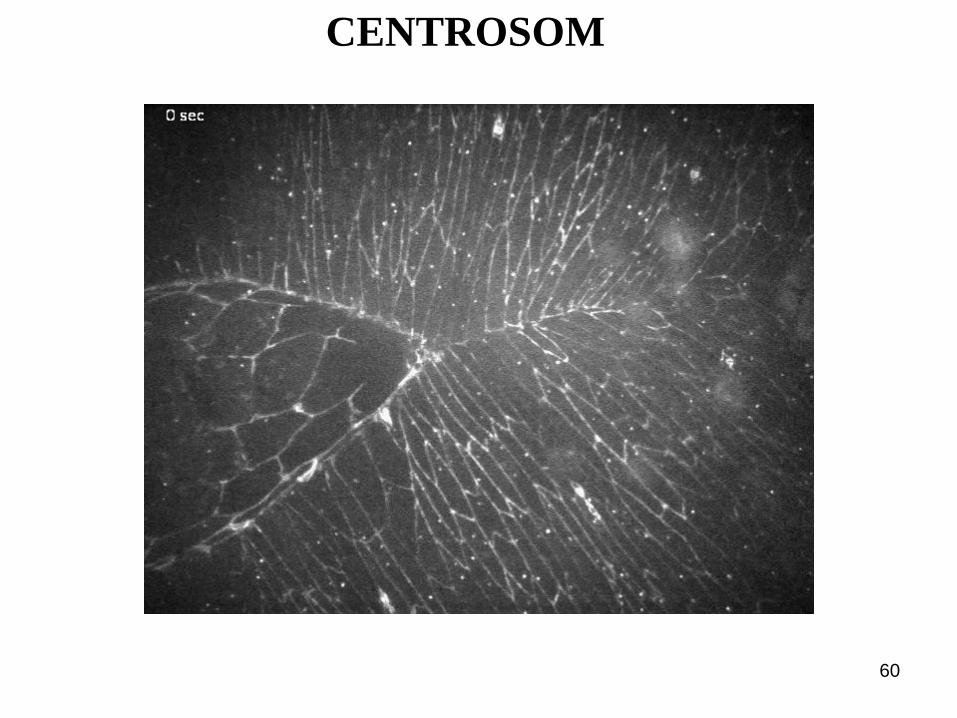
CENTROSOM
Marie Delattre and Pierre Gönczy: The arithmetic of centrosome biogenesis. Journal of Cell Science 117, 1619-1629 2004
CENTROSOM:
CENTRIOLEN
und PCM
g-tubulin ring
complex :
MT-Nucleation
Microtubuli-
Organisations-
centrum (MTOC)
60
CENTROSOM

61
Zellteilung
MITOSE & MEIOSE

62
Mitose und Meiose

63
Mitose und Meiose

64
Jonathan M. Scholey, Ingrid
Brust-Mascher & Alex
Mogilner: Cell division
NATURE VOL 422, 2003
Anaphase A
Anaphase B
TelophaseMetaphase
Prometaphase
ProphaseMitose

65
Der Struktur der Spindel

66
Chromosomen
Gene
Centromer
Tochterchromatiden
Dupliziertes
Chromosom
Telomer
Aufbau der Chromosomen
Chromatin:
Nicht kondensierte DNA mit Proteinen (in der
Interphase des Zellzyklus
Chromosom: kondensierte DNA mit Proteinen
verpackt (in der M-Phase des Zellzyklus)Karyogram
(human)
„a” Allel„A” Allel
Homologe Chromosomen
1.♀ 1.♂2.♀ 2.♂

67
KINETOCHOR – „Search and capture” Modell
C
E
N
T
R
O
M
E
R
DYNAKTIN Complex DYNEIN Complex Kinetochorproteine und Adaptorproteine
Kinetochor: Struktur aus Proteinen und DNA-Abschnitten, die dem Zentromer
aufsitzt und bei Kernteilungsvorgängen als Ansatzstelle für die Fasern des
Spindelapparates dient.

68
Rebecca W. Heald: Burning the spindle at both ends NATURE VOL 427 2004
Bewegung der Chromosomen
metafázis
anafázis
kinetokór MT
kinetokór
centroszóma
kromoszóma
Aufrollen
Pac-Man

69Rogers et. al.: Two mitotic kinesins cooperate to drive sister chromatid separation during anaphase NATURE | VOL 427 | 22 J 2004
MT –
depolymerisierendes
KinesinDynein
Kinetochor
Centrosom
Dynaktin
MT-Flux
Microtubuli und die Segregation der Chromosomen
MT –
depolymerisierendes
Kinesin

70
Dynein
Kinesin
Microtubuli und die Entstehung der Spindel

71
Spindel ohne Centrosom
Pflanzenzellen Weibliche MeioseMeiste Tierzellen
Patricia Wadsworth and Alexey Khodjakov: E pluribus unum:
towards a universal mechanism for spindle assembly. TRENDS in
Cell Biology Vol.14 No.8 2004

72Oliver J. Gruss and Isabelle Vernos: The mechanism of spindle assembly: functions of Ran and its target TPX2. The Journal of Cell
Biology, Volume 166, Number 7, 2004 949–955
Wie entstehen Spindel ohne Centrosom?
NUKLEATION ORGANISATION AUSBILDUNG DER POLEN

73Andreas Merdes and Don W. Cleveland: Pathways of Spindle Pole Formation: Different Mechanisms; Conserved Components
The Journal of Cell Biology 138, Number 5, September 8, 1997, 953–956
Mit Centrosom Ohne Centrosom
Microtubuli und die Entstehung der Spindel
Chromosomengeheftetes
Kinesin Tetramer Kinesin
Dynein Complex

74
Meiose I
Meiose II
oder
2 Ry : :
Heterozygous diploid
cell (YyRr) to
undergo meiosis
yy
y
y y
y
yy
y
Y
YY
Y
YY
Y
Y Y
R
R
R r
r
y
r
r R
R
Y
R
RR
R
y
R
r
rr
r
Y
r
Yy
Rr
R
rr
2 rY 2 ry 2 RY
Trennung der Chromosomen in der Meiose
Yy, rR

75
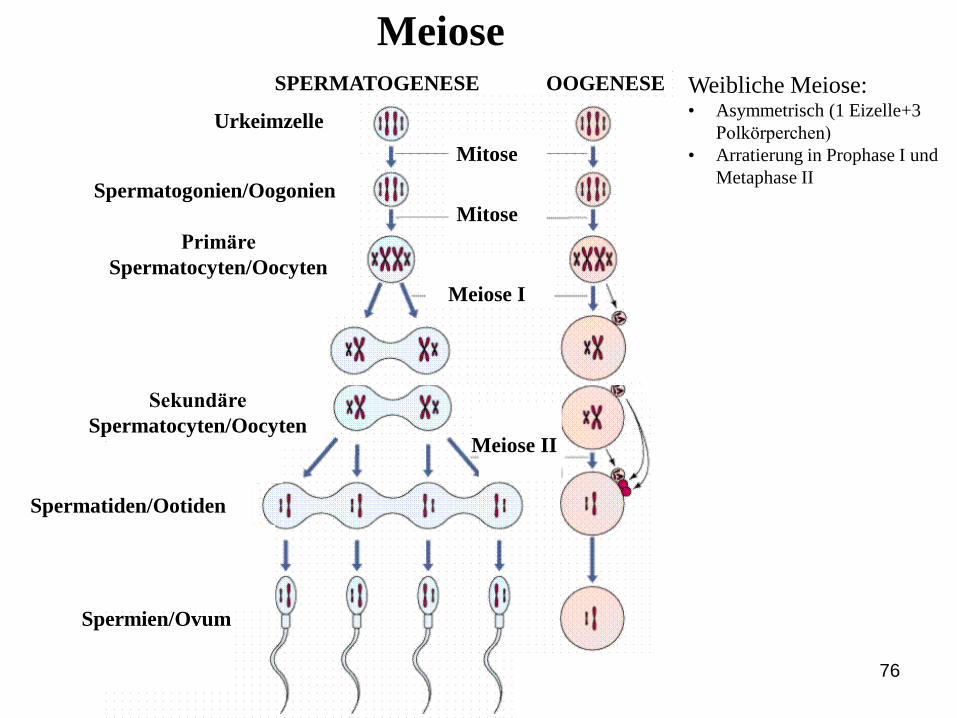
MEIOSE
76
MeioseSPERMATOGENESE OOGENESE
Urkeimzelle
Spermatogonien/Oogonien
Primäre
Spermatocyten/Oocyten
Sekundäre
Spermatocyten/Oocyten
Spermatiden/Ootiden
Spermien/Ovum
Meiose I
Meiose II
Mitose
Mitose
Weibliche Meiose:• Asymmetrisch (1 Eizelle+3
Polkörperchen)
• Arratierung in Prophase I und
Metaphase II

77
Fehler bei der Meiose
Nondisjunktion:
Ein homologes Chromosomenpaar wird
bei der Meiose I nicht getrennt.
Die Chromatiden werden bie der
Meiose II nicht getrennt.
Die Folge ist Aneuploidie: Es fehlen ein oder mehrere Chromosomen, oder sind
überzählig vorhanden (Nullisomie, Monosomie, Trisomie).

78
Michael Glotzer :ANIMAL CELL CYTOKINESIS Annu. Rev. Cell Dev. Biol. 2001. 17:351–86
CYTOKINESE – Der kontraktile Ring
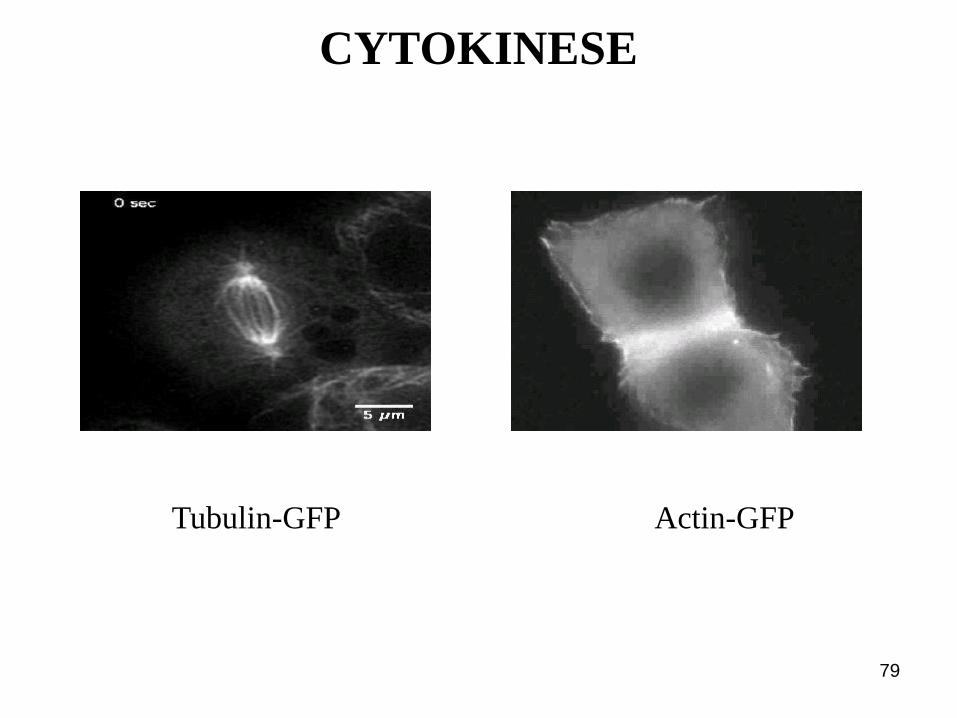
79
Tubulin-GFP Actin-GFP
CYTOKINESE

80
Regulierung des Zellzyklus

8181

82
Der Zellzyklus bei Eukaryoten
Die meisten Zellen teilen sich
nie:differenzierte Zellen
(Nervenzellen, Muskelzellen)
Manche Zellen teilen sich oft
und schnell
(embrionale Zellen, Stammzellen)
Sogar diese Zellen verbringen die
meiste Zeit in der Interphase

83
Verlassen des Zellzyklus – G0
G0
Interphase
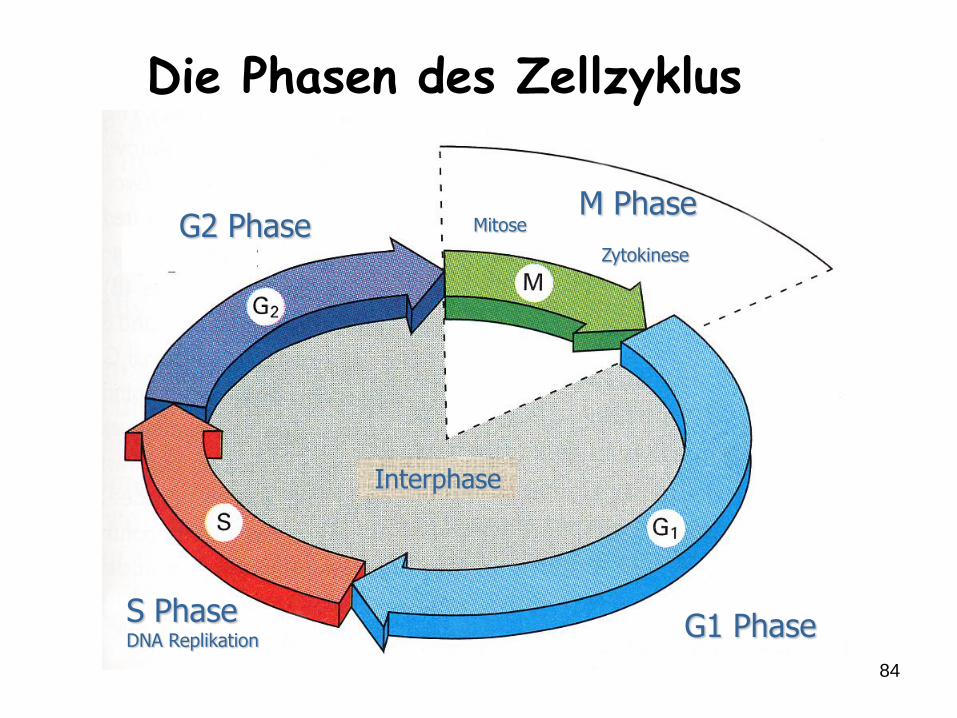
84
G1 Phase
G2 Phase
S PhaseDNA Replikation
Interphase
M PhaseMitose
Zytokinese
Die Phasen des Zellzyklus
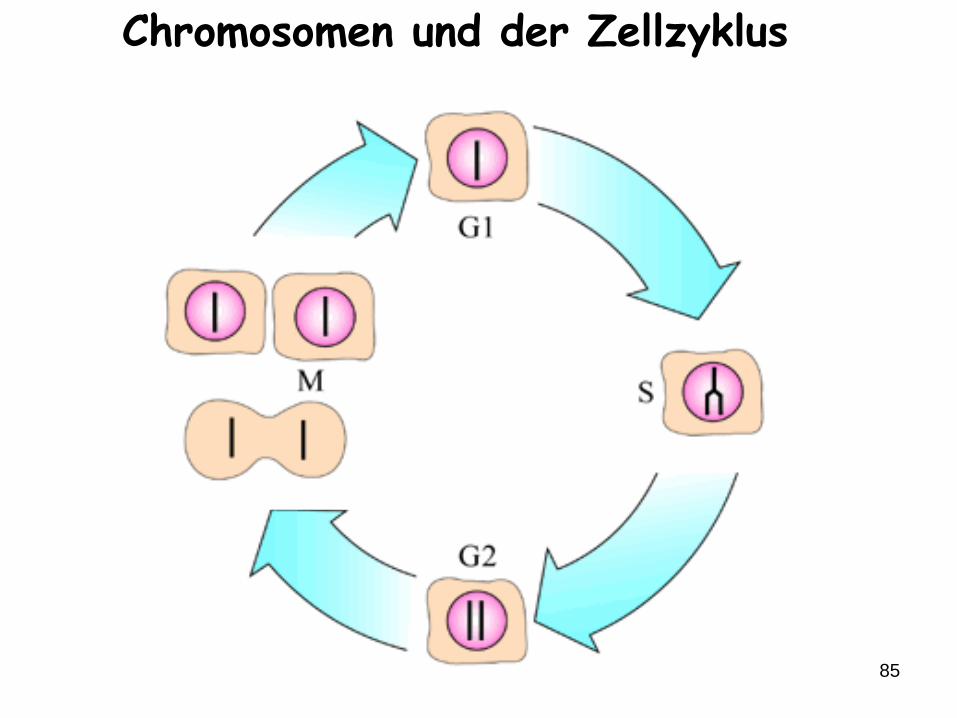
85
Chromosomen und der Zellzyklus

86
Regulierung der Zellzyklus
szabályozószabályozó
Entstehung der Spindel
Abschluss der Zellteilung
S Anfang
M Anfang M Ende
DNS replikációDNA-Replikation

87
M Anfang M Ende
S Anfang
G1/S Kontrollpunkt
G2/M Kontrollpunkt Metaphasen- Kontrollpunkt
Ist die DNA-Replikation abgeschlossen?
Ist die replizierte DNA beschädigt?
Wurde die mitotische Spindel aufgebaut?
Sind alle Kinetochore mit Transportfasern des
Spindelapparates verbunden?
Sind die Umweltbedingungen günstig?
Regulierung der Zellzyklus

88
Cdk 1
Cyklin B
M-Phase promoting ComplexCyklin-abhängige Kinase (CDK)+Cyklin
Aktiviert/inaktiviert viele Targetproteine durch Phosphorylierung, wodurch
die M- Phase eingeleitet wird.

89MPF ist nur bei hocher Cyklinkonzentration aktiv
Mitotisches
Cyklin
MPF-Aktivität
idő
Relative
Konzentration
M-Phase promoting ComplexCyklin-abhängige Kinase (CDK)+Cyklin

90
CDK-Cyklin Complexe der höheren Eukaryoten
M G1 G2S M G1
Cdk-s
D E A B(A)Cykline
Cdk4 Cdk2 Cdk1

91
Regulationsmechanismen der Cyklin-abhängigen Kinasen
1. CYKLIN-Bindung
4. CDK-INHIBITOR 2. AKTIVIERENDE PHOSPHORYLIERUNG
3. HEMMENDE PHOSPHORYLIERUNG

92
Pozitive
Rückkopplung
Aktivierende
Phosphirilierung
Inaktíve
M-Cdk
Inaktíve
M-Cdk
aktíve M-
Cdk
M-Cyklin
Cdk1
inaktive
Phosphatase
PHOSPHORYLIERUNG - DEPHOSPHORYLIERUNG
Cdk-
Aktivator
Kinase
Cdk-Inhibitor
Kinase
Regulationsmechanismen der Cyklin-abhängigen Kinasen
Hemmende
Phosphirilierung

93
Inaktives
Cyklin-Cdk-p27
Complex
Aktíves
Cyklin-Cdk
Complexp27
CyklinCdk
CDK- INHIBITOREN (CDKI)
Regulationsmechanismen der Cyklin-abhängigen Kinasen

94
APC-abhängige Proteolyse
aktives
APC
Aktivierende Untereinhet
(Cdc20)
inaktives
APC
Ubiquitinierungsenzyme
Polyubiquitinkette
Cdk inhibítor fehérje
(CDKI)
M-Cyiklin
Abbau
UBIQUITIN-abhängiger Proteinabbau
Regulationsmechanismen der Cyklin-abhängigen Kinasen
Proteasom

95
Protoonkogene sind Gene die den normalen
Ablauf des Zellzyklus fördern. Durch
dominante Mutationen werden sie zu
Onkogene.
Dominante (hypermorphe) Mutationen von
CDKs und Cykline können zur Tumorenbildung
führen.

96
G2/M
Kontrollpunkt
Metaphasen-Kontrollpunkt
G1/S
Kontrollpunkt
Zellgrösse
Nährstoffe
Wachstumsfaktoren
DNA-Schäden
Zellgrösse
DNA-Schäden
Chromosomen-Spindel-Bindung
G0
Kontrollenpunkte des Zellzyklus
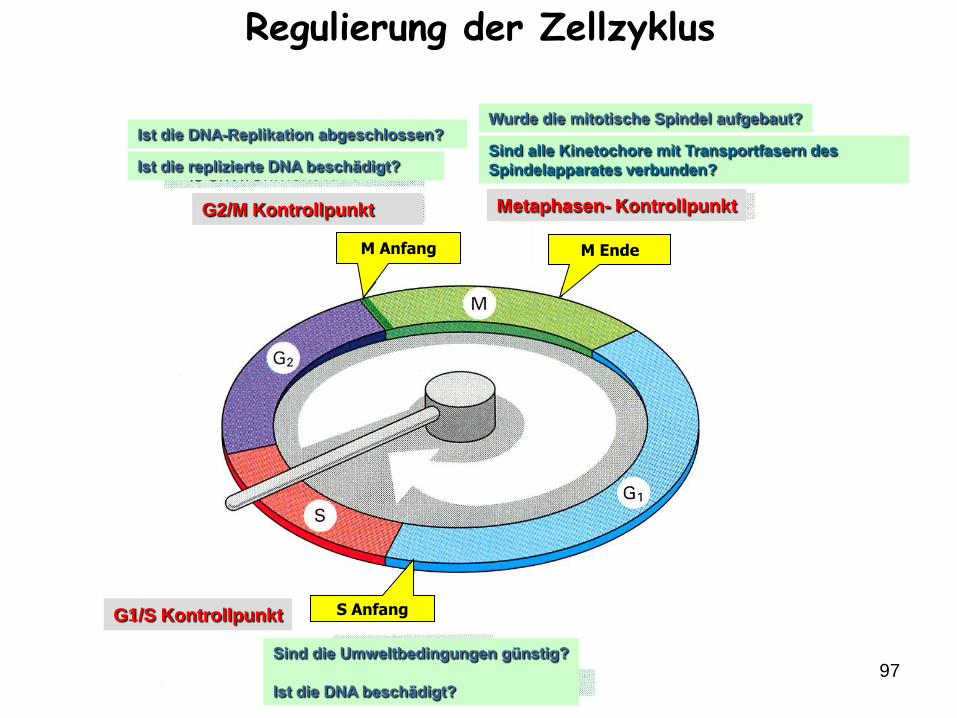
97
M Anfang M Ende
S AnfangG1/S Kontrollpunkt
G2/M Kontrollpunkt Metaphasen- Kontrollpunkt
Ist die DNA-Replikation abgeschlossen?
Ist die replizierte DNA beschädigt?
Wurde die mitotische Spindel aufgebaut?
Sind alle Kinetochore mit Transportfasern des
Spindelapparates verbunden?
Sind die Umweltbedingungen günstig?
Ist die DNA beschädigt?
Regulierung der Zellzyklus
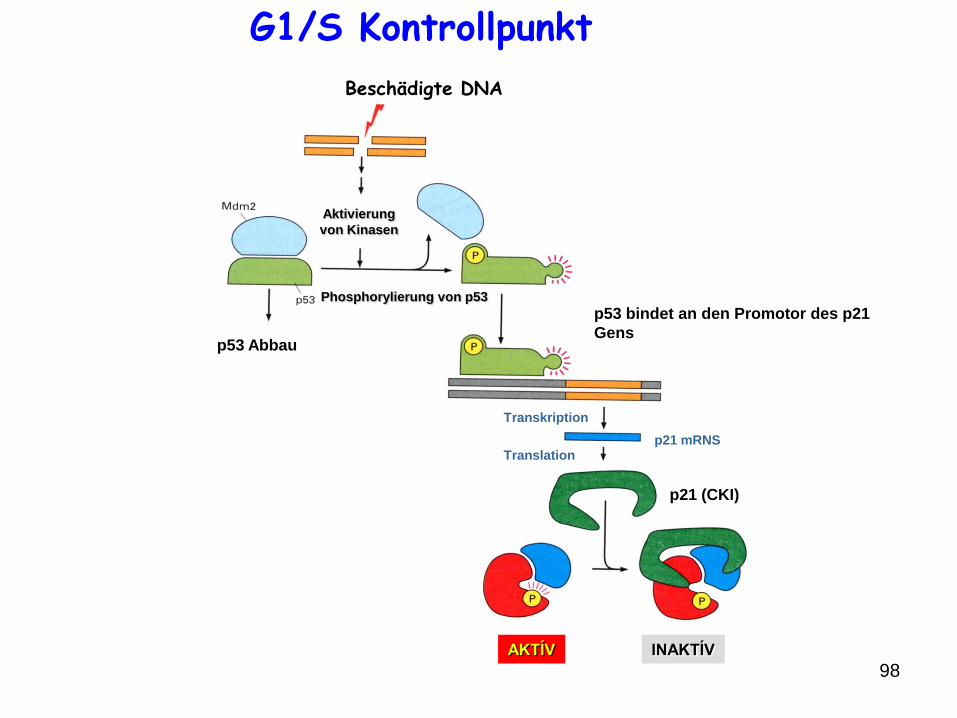
98
DNS
Aktivierung
von Kinasen
Phosphorylierung von p53
p53 Abbau
Stabil,
Aktív p53
p53 bindet an den Promotor des p21
Gens
Transkription
Translationp21 mRNS
p21 (CKI)
AKTÍV INAKTÍV
G1/S Kontrollpunkt
Beschädigte DNA
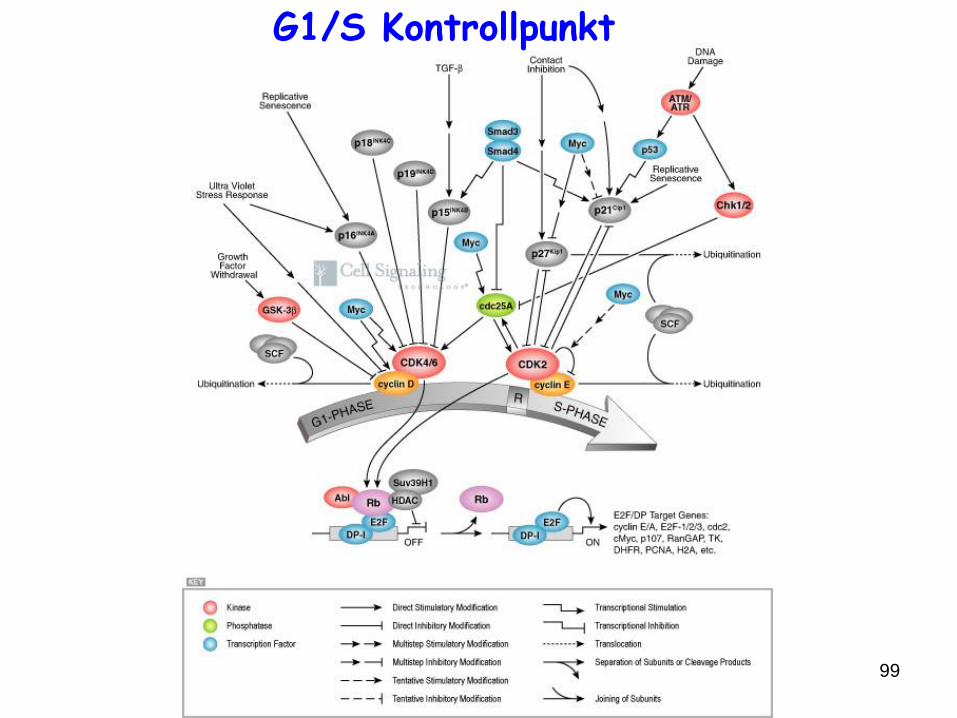
99
G1/S Kontrollpunkt
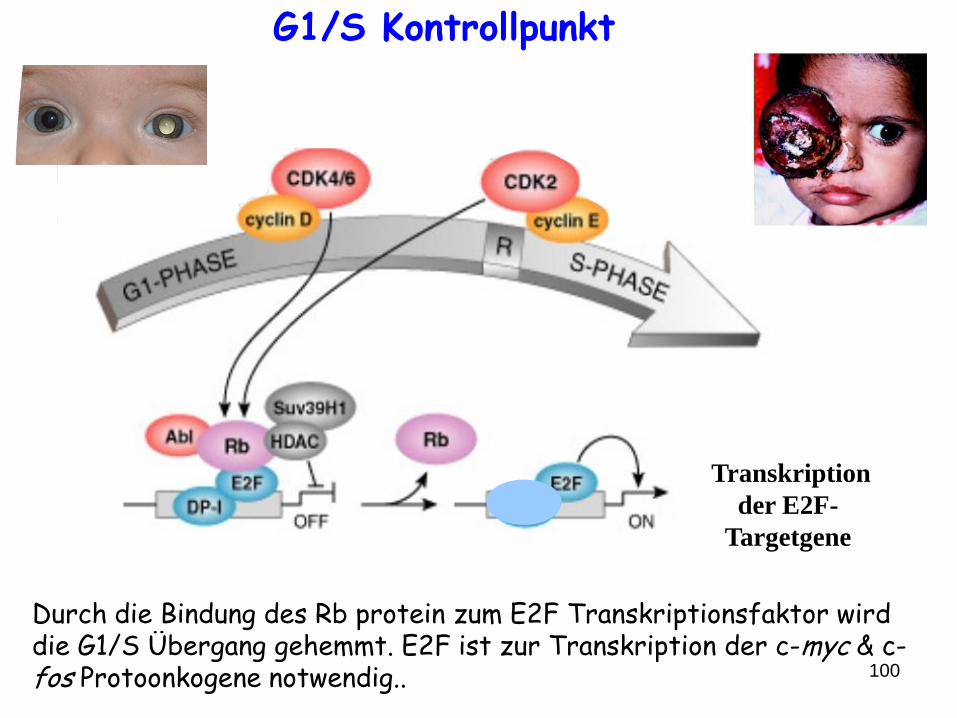
100
Durch die Bindung des Rb protein zum E2F Transkriptionsfaktor wird die G1/S Übergang gehemmt. E2F ist zur Transkription der c-myc & c-fos Protoonkogene notwendig..
Transkription
der E2F-
Targetgene
G1/S Kontrollpunkt

101Symptome vom ataxia telangiektázia. Das ATM-Protein spielt eine
wichtige Rolle in der Detektierung der DNA-Schäden.)
ATM (ataxia telangiectasia mutated)
Sterilität
Strahlunds-
empfindlichkeit
Neurodegeneration
telangiektasia
Immunprobleme
Krebs-
bildung

102
G2/M
Kontrollpunkt
Metaphasen-Kontrollpunkt
G1/S Kontrollpunkt
Zellgrösse
Nährstoffe
Wachstumsfaktoren
DNA-Schäden
Zellgrösse
DNA-Schäden
Chromosomen-Spindel-Bindung
G0
Kontrollenpunkte des Zellzyklus

103
DNA-Replikation ist abgeschlossen
Kontrollpunkt-
proteine
MitoseArretierung
in G2
Kontrollpunkt-
proteine
G2/M KontrollpunktDNA-Replikation ist nicht abgeschlossen / DNA-Schäden

104
cdk1
G2/M Kontrollpunkt

105
G2/M
Kontrollpunkt
Metaphasen-Kontrollpunkt
G1/S Kontrollpunkt
Zellgrösse
Nährstoffe
Wachstumsfaktoren
DNA-Schäden
Zellgrösse
DNA-Schäden
Chromosomen-Spindel-Bindung
G0
Kontrollenpunkte des Zellzyklus

106
Schwester-chromatiden Cohesin
anafázisMetaphase
Anaphasen
promoting
Complex
(APC)
MPF
(cdk-Cyclin-Complex)
Abgebautes
mitotisches Cyclin
Abgebautes
Securin
Separin
Separase
Securin
Anaphase
Metaphasen-Kontrollpunkt (Metaphase-Anaphase-Übergang)

107Anaphase wird nicht eingeleitet, wenn nicht alle Chromosmen an
Spindelfasern gebunden sind.
AnaphaseMetaphase
Metaphasen-Kontrollpunkt (Metaphase-Anaphase-Übergang)

108
Was wurde ohne die
Metaphasen-Kontrollpunkt
passieren?
Aneuploide Zellen
Tumorzellen sind alle aneuploid!
Abnormale
Spindel
Anaphase
Cytokinese
Extra
Chromosom
Chromosomen-
verlust
Metaphasen-Kontrollpunkt (Metaphase-Anaphase-Übergang)

109
APC wird durch den Metaphasen-Kontrollpunkt inaktiviert
HTLV-1 vírus Tax Protein des HTLV- Virus
bindet und inaktiviert Mad, und verursacht
dadurch T-Zellen Leukaemie.
Kontrollpunkt inaktiv
Kontrollpunkt aktiv
Kinetochor ist nicht an
beiden Seiten an
Spindelfasern gebunden
Kinetochor ist an
beiden Seiten an
Spindelfasern
gebunden
Anaphase
wird
gehemmt
Anaphase beginnt
Metaphasen-Kontrollpunkt (Metaphase-Anaphase-Übergang)

110
Tumorsupressor Gene hemmen den normalen Zellzyklus.
Recessive Mutationen der Tumorsupressor Gene führen zur Krebsbildung.
![01.Die Materie [Kompatibilit si m d]web.med.u-szeged.hu/mdbio/ger/material/...l/1/1/01.Die_Materie.ppt.pdf · Die Materie Die Eigenschaften der Elementarteilchen erklären es wie](https://img.pdfslide.org/doc/110x75/5d668aac88c993ec358b48c2/01die-materie-kompatibilit-si-m-dwebmedu-die-materie-die-eigenschaften.jpg)