
Aus der Medizinischen Klinik I
der Universität zu Lübeck
Direktor: Prof. Dr. med. H. Lehnert
Ghrelin moduliert die Baroreflex-vermittelte Regulation der
muskulären sympathischen Nervenaktivität (MSNA) beim Menschen
Inauguraldissertation
zur
Erlangung der Doktorwürde
der Universität zu Lübeck
- Aus der Sektion Medizin –
vorgelegt von
Alexander Friedrich Wilhelm Krapalis
aus Lübeck
Lübeck 2013

2
1. Berichterstatter: Prof. Dr. med. Dr. h.c. Hendrik Lehnert
2. Berichterstatter: Prof. Dr. med. Kerstin Oltmanns
Tag der mündlichen Prüfung: 13.9.2013
Zum Druck genehmigt. Lübeck, den 13.9.2013
- Promotionskommission der Sektion Medizin -

3
Inhaltsverzeichnis
Inhaltsverzeichnis
Inhaltsverzeichnis………………………………………………………………………… 3
Tabellenverzeichnis…………………………………………………………………..….. 6
Abbildungsverzeichnis…………………...……………………………………………... 7
Abkürzungsverzeichnis…………………………………………………………………. 8
1 Einleitung und Fragestellung…………………………..…......………………….. 10
1.1 Ghrelin…………………………......………………….………….……………..... 10
1.1.1 Struktur, Synthese und Metabolismus..................................................... 10
1.1.2 Vorkommen und Regulation.................................................................... 11
1.1.3 Rezeptor.................................................................................................. 11
1.1.4 Effekte auf die Inkretion anderer Hormone.............................................. 11
1.1.5 Orexigene, gastrointestinale und metabolische Wirkungen.................... 12
1.1.6 Kardiovaskuläre und weitere Effekte....................................................... 13
1.2 Sympathisches Nervensystem und Blutdruckregulation……..………… …... 13
1.2.1 Kontrolle des kardiovaskulären Systems durch das sympathische
Nervensystem.......................................................................................... 13
1.2.2 Arterieller Baroreflex................................................................................ 14
1.2.3 Mechanismen der langfristigen Blutdruckregulation................................ 16
1.2.4 Existenz und Regulation eines zentralen Blutdruck-Sollwertes............... 16
1.2.5 Wechselwirkungen des sympathischen Nervensystems mit dem
endokrinen System.................................................................................. 17
1.2.6 Messung der Aktivität des sympathischen Nervensystems..................... 18
1.2.7 Bestimmung von Setpoint und Sensitivität des arteriellen Baroreflex..... 22
1.2.8 Gesteigerte Aktivität des sympathischen Nervensystems als
pathogenetischer Faktor.......................................................................... 23
1.3 Fragestellung/Hypothesen…….....…………….............…….…………..…..... 23
1.3.1 Studienlage zur Bedeutung von Ghrelin für die Regulation des
sympathischen Nervensystems............................................................... 23
1.3.2 Formulierung der Hypothesen der vorliegenden Arbeit........................... 24
2 Methoden und Material………………..……………………………….....………... 25
2.1 Versuchspersonen……………………….………………….………………….... 25
2.2 Design und Ablauf der Studie………………....…………….………………...... 25
2.2.1 Design..................................................................................................... 25

4
Inhaltsverzeichnis
2.2.2 Instrumentierung...................................................................................... 26
2.2.3 Versuchsablauf........................................................................................ 26
2.3 Aufzeichnung hämodynamischer Parameter………..…...…………….……... 28
2.4 Mikroneurographie......................................................................................... 28
2.4.1 Plazierung der Elektrode......................................................................... 28
2.4.2 Signalprozessierung................................................................................ 29
2.4.3 Identifizierung der bursts......................................................................... 29
2.4.4 Burst-Rate............................................................................................... 30
2.5 Pharmakologischer Baroreflextest................................................................. 30
2.5.1 Modified Oxford protocol (MOP).............................................................. 31
2.5.2 Mathematische Analyse.......................................................................... 31
2.6 Analyse der Herzfrequenzvariabilität............................................................. 33
2.7 Laboranalytik................................................................................................. 33
2.8 Statistik.......................................................................................................... 33
3 Ergebnisse......................................................................................................... 35
3.1 Klinische Beobachtungen.............................................................................. 35
3.2 Hämodynamische Parameter........................................................................ 35
3.2.1 Mittlerer arterieller Blutdruck.................................................................... 38
3.2.2 Herzfrequenz........................................................................................... 38
3.2.3 Muskuläre sympathische Nervenaktivität (MSNA).................................. 39
3.3 Sensitivität des arteriellen Baroreflex............................................................ 39
3.3.1 Vaskulärer Baroreflex.............................................................................. 39
3.3.2 Kardialer Baroreflex................................................................................. 41
3.4 Herzfrequenzvariabilität................................................................................. 44
3.5 Endokrine Parameter..................................................................................... 47
4 Diskussion......................................................................................................... 50
4.1 Biphasisches Modell der sympathischen und kardiovaskulären
Ghrelinwirkung………………………………………………………………........ 50
4.2 Direkte Vasodilatation als dominierender Mechanismus der akuten Phase
der Ghrelinwirkung……………………………………………………................ 50
4.3 Zentrale Suppression der SNS-Aktivität in der verzögerten Phase der
Ghrelinwirkung……………………………………………………………………. 51

5
Inhaltsverzeichnis
4.3.1 Modulation des zentralen Setpoint der sympathischen
Blutdruckregulation…………………………………………………………... 51
4.3.2 Reduktion Stress-induzierter sympathischer feed-forward-Signale……. 52
4.4 Beeinflussung anderer Hormone und Transmitter……………………………. 53
4.5 Gesteigerte Sensitivität des MSNA-vermittelten arteriellen Baroreflex nach
Ghrelininjektion…………………………………………………………………… 54
4.6 Gegenüberstellung mit der aktuellen Studie von Lambert et al. zum Thema
Ghrelin und SNS............................................................................................ 55
4.7 Wirkung von zirkulatorischem Ghrelin auf die efferente kardiale
Sympathikus- und Vagusaktivität.................................................................. 56
4.7.1 Kardiale Vagusaktivität............................................................................ 56
4.7.2 Analyse der Herzfrequenzvariabilität....................................................... 57
4.8 Pathophysiologische und klinische Implikationen.......................................... 58
4.9 Limitationen................................................................................................... 58
4.10 Perspektive.................................................................................................. 59
5 Zusammenfassung............................................................................................ 61
6 Literaturverzeichnis.......................................................................................... 63
7 Ergänzende Materialien..................................................................................... 89
8 Lebenslauf.......................................................................................................... 95
9 Veröffentlichung................................................................................................ 97
10 Danksagungen................................................................................................. 98

6
Tabellenverzeichnis
Tabellenverzeichnis
Tab. 1 MSNA, Herzfrequenz und arterieller Blutdruck im Verlauf des Experiments
unter Ghrelin/Placebo.................................................................................. 36
Tab. 2 Vaskulärer Baroreflex. Mittelwerte der lineareren Faktoren (slopes) der
mithilfe des modified Oxford protocol (MOP) gewonnen den arteriellen
Baroreflex beschreibenden Regressionsgeraden vor und nach
Ghrelin/Placebo.......................................................................................... 41
Tab. 3 Kardialer Baroreflex. Mittelwerte der lineareren Faktoren (slopes) der
mithilfe des modified Oxford protocol (MOP) gewonnen den arteriellen
Baroreflex beschreibenden Regressionsgeraden vor und nach
Ghrelin/Placebo......................................................................................... 42
Tab. 4 Herzfrequenzvariabilität.............................................................................. 45
Tab. 5 Endokrinologische Laborparameter............................................................ 48

7
Abbildungsverzeichnis
Abbildungsverzeichnis
Abb. 1 Studienprotokoll.......................................................................................... 27
Abb. 2 Beispiel von MSNA und arteriellem Blutdruck bei experimenteller
Auslösung des arteriellen Baroreflex nach dem modified Oxford protocol
(MOP)......................................................................................................... 32
Abb. 3 MSNA, Herzfrequenz und arterieller Blutdruck vor und nach Applikation
von Ghrelin/Placebo................................................................................... 37
Abb. 4 Charakterisierung der Sensitivität des vaskulären arteriellen Baroreflex
vor und nach Applikation von Ghrelin/Placebo........................................... 43
Abb. 5 Herzfrequenzvariabilität - HF...................................................................... 46
Abb. 6 Herzfrequenzvariabilität - LF/HF................................................................. 47
Ergänzende Abb. I Struktur des Peptidhormons Ghrelin........................................ 89
Ergänzende Abb. II Neuronale Verschaltung des arteriellen Baroreflex in
Hirnstamm und Medulla spinalis.................................................................. 90
Ergänzende Abb. III Schematische Darstellung der mathematischen
Charakteristik des arteriellen Baroreflex....................................................... 91
Ergänzende Abb. IV MSNA der einzelnen Probanden........................................... 92
Ergänzende Abb. V Linearer Faktor (slope) des vaskulären Baroreflex
der einzelnen Probanden............................................................................. 93
8
Abkürzungsverzeichnis
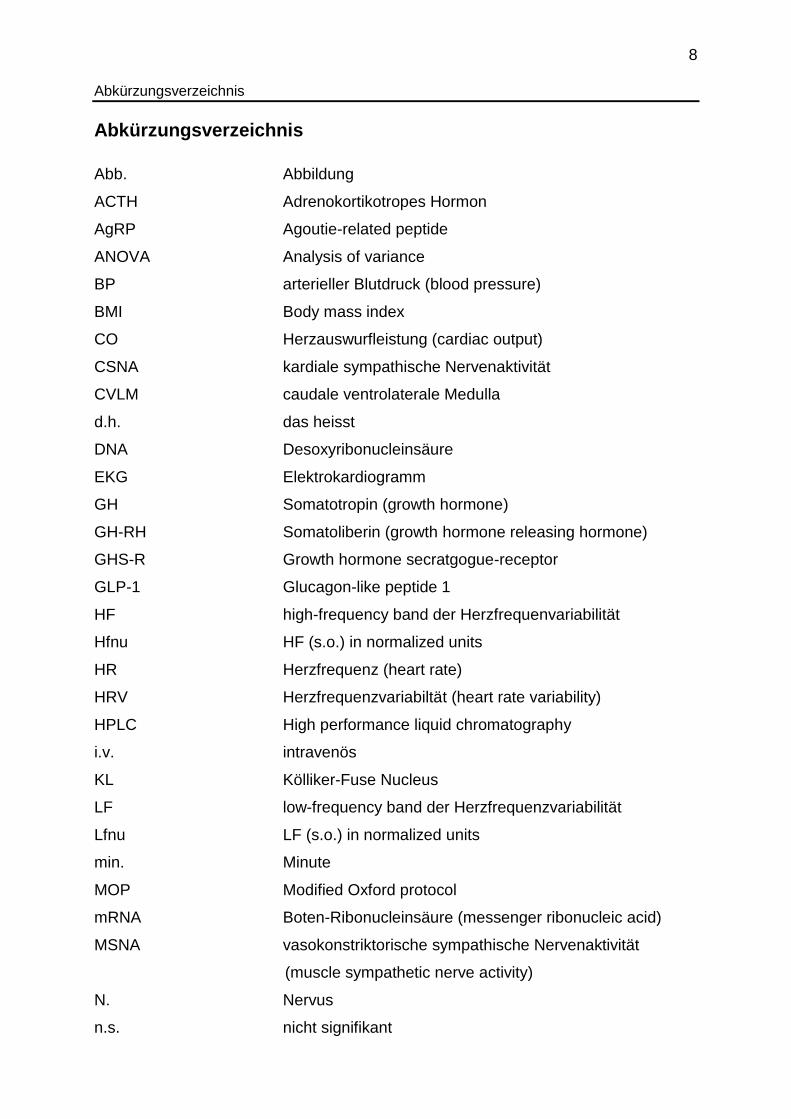
Abkürzungsverzeichnis
Abb. Abbildung
ACTH Adrenokortikotropes Hormon
AgRP Agoutie-related peptide
ANOVA Analysis of variance
BP arterieller Blutdruck (blood pressure)
BMI Body mass index
CO Herzauswurfleistung (cardiac output)
CSNA kardiale sympathische Nervenaktivität
CVLM caudale ventrolaterale Medulla
d.h. das heisst
DNA Desoxyribonucleinsäure
EKG Elektrokardiogramm
GH Somatotropin (growth hormone)
GH-RH Somatoliberin (growth hormone releasing hormone)
GHS-R Growth hormone secratgogue-receptor
GLP-1 Glucagon-like peptide 1
HF high-frequency band der Herzfrequenvariabilität
Hfnu HF (s.o.) in normalized units
HR Herzfrequenz (heart rate)
HRV Herzfrequenzvariabiltät (heart rate variability)
HPLC High performance liquid chromatography
i.v. intravenös
KL Kölliker-Fuse Nucleus
LF low-frequency band der Herzfrequenzvariabilität
Lfnu LF (s.o.) in normalized units
min. Minute
MOP Modified Oxford protocol
mRNA Boten-Ribonucleinsäure (messenger ribonucleic acid)
MSNA vasokonstriktorische sympathische Nervenaktivität
(muscle sympathetic nerve activity)
N. Nervus
n.s. nicht signifikant

9
Abkürzungsverzeichnis
NA Noradrenalin
NP Nitroprussid-Natrium
NO Stickstoffmonoxid
NPY Neuropeptid Y
NTS Nucleus tractus solitarii
PE Phenylephrin
POMC Proopiomelanocortin
PVH Nucleus paraventricularis hypothalami
s.o. siehe oben
s.u. siehe unten
RAS Renin-Angiotensin-System
RVLM Rostrale ventrolaterale Medulla
RSNA renale sympathische Nervenaktivität
SEM Standardfehler des arithmetischen Mittels
SNS Sympathisches Nervensystem
SSNA kutane sympathische Nervenaktivität (skin sympathetic
nerve activity)
SVRI Systemischer vaskulärer Widerstandsindex
t Zeitpunkt (time)
Tab. Tabelle
TP total power der Herzfrequenzvariabilität
vs. versus
ZNS Zentrales Nervensystem
Zeitangaben im Versuchsprotokoll
Zeitpunkte innerhalb des Versuchsprotokolls sind durch den Buchstaben t
gekennzeichnet. Ein negatives Vorzeichen kennzeichnet einen Zeitpunkt vor
Injektion der Versuchssubstanz (Ghrelin oder Placebo), ein positives Vorzeichen
einen Zeitpunkt danach. Die angegebene Zahl gibt den Abstand in Minuten vom
Zeitpunkt der Applikation der Versuchssubstanz an. Beispielsweise bedeutet die
Zeitangabe „t -25 min“ den Zeitpunkt 25 Minuten vor Injektion der Versuchssubstanz.

10
1 Einleitung und Fragestellung
1 Einleitung und Fragestellung
1.1 Ghrelin
Das Peptidhormon Ghrelin wurde erstmals 1999 von Kojima et al. beschrieben
(79). Zuvor war die Existenz des growth-hormone secretagogue Rezeptors (GHS-R)
(71; 96) bekannt geworden, der die Ausschüttung von GH (growth hormone) über
einen von GH-RH unabhängigen Mechanismus vermittelt. Für diesen Rezeptor
waren nur synthetische Liganden bekannt (3; 124). Kojima unternahm eine
systematischen Suche nach dem endogenen Liganden von GHS-R, indem er
Gewebeextrakte schrittweise mittels high performance liquid chromatography (HPLC)
fraktionierte. Mithilfe einer eigens konstruierten GHS-R exprimierenden Zelllinie
überprüfte er die isolierten Peptide auf ihre Eigenschaften als Ligand. Es gelang die
Charakterisierung der Aminosäure- und DNA-Sequenz sowie einer wesentlichen
posttranslationalen Modifikation (s.u.) eines bis dahin unbekannten Peptidhormons,
des Ghrelins.
1.1.1 Struktur, Synthese und Metabolismus
Ghrelin besteht aus 28 Aminosäuren. Die Aminosäure Serin in Position 3 ist n-
octanoyliert (siehe ergänzende Abb. I) Die bislang als einzigartig beschriebene
Acylierung eines Peptidhormons wird durch das Enzym Ghrelin-O-Acyl-Transferase
vermittelt (61; 150). Nach gegenwärtiger Kenntnis ist nur das acylierte Peptid
endokrin regulatorisch aktiv (80; 81). Anderweitige Wirkungen von Des-Acyl-Ghrelin
insbesondere auf Effektororgane des kardiovaskulären Systems wurden jedoch
beschrieben (7). In vitro wird Ghrelin im Plasma mit einer Halbwertszeit von ca. 45
Minuten deacyliert (15). Die Deacylierung wird durch das Enzym Acyl-Protein
Thioesterase 1 (APT1)/Lysophospholipase 1 katalysiert (111). In vivo beträgt die
Plasmakonzentration von n-octanoyl-Ghrelin mit 10-20 fmol/ml etwa ein Zehntel der
Gesamtkonzentration von acyliertem und deacyliertem Ghrelin (100-150 fmol/ml)
(80). Nach Injektion von endokrin aktivem, acyliertem Ghrelin beträgt die
Halbwertszeit des acylierten Ghrelins etwa 9-13 Minuten, die des entstehenden
deacylierten Ghrelins etwa 27-31 Minuten (2).

11
1 Einleitung und Fragestellung
1.1.2 Vorkommen und Regulation
Ghrelin wird überwiegend von der Mukosa des Magens sezerniert und gelangt
von dort in den Blutkreislauf (5). Eine Synthese des Peptidhormons kann jedoch
auch im zentralen Nervensystem, insbesondere im Nucleus arcuatus des
Hypothalamus (89), speziellen Neuronen in Nachbarschaft des dritten Ventrikels (23)
und in der Hypophyse (82) nachgewiesen werden.
Die Sekretion von Ghrelin in den Blutkreislauf unterliegt einer circadianen
Rhythmik mit nächtlichem Maximum und pulsatiler Ausschüttungscharakteristik
(153). Ihre Regulation wird vermutlich durch sympathische Efferenzen zur gastralen
Mukosa vermittelt (98). Nach Fasten steigt der Ghrelinspiegel im Plasma (24; 134).
Postprandial kommt es zu einem raschen Abfall des Spiegels (135). Aufnahme von
Glucose (95; 121) oder einer fettreichen Mahlzeit (59) reduziert die Inkretion von
Ghrelin. Schlanke Individuen weisen im Vergleich zu Adipösen einen höheren
Ghrelinspiegel auf (12; 109). Nach Gewichtsverlust kann ein Anstieg des
Ghrelinspiegels beobachtet werden (25; 66).
1.1.3 Rezeptor
Der Ghrelin-Rezeptor, GHS-R, ist ein G-Protein-gekoppelter Rezeptor. Über
eine Stimulation der Phosphokinase C vermittelt er einen Anstieg des intrazellulären
Inositoltriphosphats und Calciums (71). Eine ausgeprägte Expression von GHS-R-
mRNA findet sich im Nucleus arcuatus und den ventromedialen Nuclei des
Hypothalamus, im Hippocampus und der Hypophyse (60; 71). Ein Nachweis gelingt
auch in zahlreichen peripheren Geweben, insbesondere Myokard und Gefäßsystem
(55; 100).
1.1.4 Effekte auf die Inkretion anderer Hormone
Ghrelin bewirkt eine Ausschüttung von GH, die mit der durch GH-RH
vermittelten Sekretion quantitativ vergleichbar ist (6; 130). Da die Blut-Hirn-Schranke
für Ghrelin in gewissem Maße durchlässig ist (8; 9), kann das von der gastralen
Mukosa in den Blutkreislauf sezernierte Peptidhormon zentral wirksam werden.
Alternativ wird die Hypothese diskutiert, dass die zentrale Wirkung von
zirkulatorischem Ghrelin über vagale Afferenzen vermittelt wird (29; 81; 146).

12
1 Einleitung und Fragestellung
Nach i.v.-Applikation von Ghrelin ist ferner ein substantieller Anstieg der
Plasmaspiegel von Prolaktin, ACTH, Cortisol und Adrenalin beschrieben (100).
1.1.5 Orexigene, gastrointestinale und metabolischen Wirkungen
Das orexigene Peptidhormon Ghrelin hat eine Schlüsselstellung in der
Energiehomöostase des menschlichen Organismus inne. Es kann in gewisser Weise
als physiologischer Antagonist des Leptin aufgefasst werden (120; 123). Sowohl bei
intracerebroventrikulärer als auch intravenöser experimenteller Applikation wird das
Nahrungsaufnahmeverhalten erheblich gesteigert (103; 134; 147). Ghrelin stimuliert
in hypothalamischen Kerngebieten (Nucleus arcuatus und paraventricularis) die
Ausschüttung der orexigenen Neuropeptide NPY und AgRP und inhibiert die
Ausschüttung des anorexigenen Peptids POMC (80). Nucleus arcuatus und
paraventricularis erhalten Projektionen von neuronalen Populationen, die das Peptid
Ghrelin als Neurotransmitter ausschütten (50). Ob diesbezüglich das peripher-
zirkulatorische oder das zentral-neuronale Signal funktional überwiegt, ist nicht
abschließend geklärt. Kurz vor Beginn einer Mahlzeit kommt es zu einem scharfen
Anstieg der Plasma-Ghrelinkonzentration, gefolgt von einem Abfall innerhalb einer
Stunde nach Nahrungsaufnahme. Es wird angenommen, dass Ghrelin das
Nahrungsaufnahmeverhalten initiiert (24). Ferner stimuliert das Peptidhormon die
Sekretion von Magensäure und die gastrale Motilität (92). Diese Wirkung scheint
über den Nucleus tractus solitarii und vagale Efferenzen gesteuert zu werden (31).
Ghrelin hat differenzierte Wirkungen auf Glucosehomöostase und
Insulinsekretion, die noch nicht vollständig geklärt sind (80). Nach Bolusinjektion
bewirkt das Peptidhormon beim nüchternen Probanden eine Erhöhung des
Blutglucosespiegels bei gleichzeitiger Reduktion der Insulininkretion (16). Unter
kontinuierlicher Infusion von Ghrelin hingegen wurde ein erhöhter Blutglucosespiegel
bei zugleich erhöhtem Insulinspiegel und verminderter Insulinsensitivität beobachtet
(141). Im Experiment mit isolierten Langerhansinseln bewirkte Ghrelin beim
Vorliegen eines erhöhten Glucosespiegels in der Umgebungsflüssigkeit (8,3 mmol/l)
eine vermehrte Sekretion von Insulin, die bei niedrigerem Glucosespiegel (2,8
mmol/l) nicht nachweisbar war (30).

13
1 Einleitung und Fragestellung
Auch der Effekt von Ghrelin auf lipolytische Prozesse unterliegt einem
differenzierten Wirkmechanismus. Beim fastenden menschlichen Probanden bewirkt
die intravenöse Applikation von Ghrelin eine Zunahme der freien Fettsäuren im
Plasma (72; 140). Hingegen inhibiert Ghrelin die durch Betasympathomimetika
induzierte Lipolyse bei der Ratte (20).
1.1.6 Kardiovaskuläre und weitere Effekte
Das Peptidhormon Ghrelin weist ausgeprägte Effekte auf das kardiovaskuläre
System auf. Seine Wirkung auf arteriellen Blutdruck, Herzfrequenz und kardiale
Auswurfleistung werden gemeinsam mit den Effekten auf das sympathische
Nervensystem weiter unten ausführlich diskutiert (siehe 1.3.1). Darüber hinaus
inhibiert Ghrelin in der Zellkultur die Apoptose von Kardiomyozyten und
Gefäßendothelzellen (7) und stimuliert die Proliferation von Kardiomyozyten (108).
Nach experimenteller Induktion eines Myokardinfarktes im Tierexperiment vermag
die Applikation von Ghrelin das linksventrikuläre Remodelling zu reduzieren (125).
Die mit Ghrelin behandelten Tiere wiesen ein geringeres linksventrikuläres
enddiastolisches Volumen, einen niedrigeren enddiastolischen linksventrikulären
Druck und im weiteren Verlauf einen geringeren Kollagenanteil in den nicht
infarzierten Regionen des Myokards auf.
Ghrelin besitzt antiinflammatorische Eigenschaften. Insbesondere
antagonisiert das Peptidhormon im T-Zellmodell die durch Leptin induzierte
Ausschüttung proinflammatorischer Zytokine (37). Auch bei experimentell induzierter
Sepsis vermindert die Applikation von Ghrelin die Ausschüttung proinflamatorischer
Zytokine (148).
Neueren Studien zufolge ist das Neuropeptid Ghrelin an Prozessen der
Gedächtnisbildung beteiligt (18).
1.2 Sympathisches Nervensystem und Blutdruckregulation
1.2.1 Kontrolle des kardiovaskulären Systems durch das sympathische
Nervensystem
Das sympathische Nervensystem (SNS) ist von entscheidender Bedeutung für
die integrative Kontrolle des kardiovaskulären Systems. Um eine adäquate Perfusion

14
1 Einleitung und Fragestellung
aller Körpergewebe sicherzustellen ist ein entsprechender Perfusionsdruck
erforderlich. Auf Ebene des Gesamtorganismus entspricht dem der arteriellen
Blutdruck. Dieser wiederum ist ein Produkt der beiden Faktoren kardiale
Auswurfleistung (cardiac output, CO) und Gefäßwiderstand. Die kardiale
Auswurfleistung wird von den drei Variablen myokardiale Kontraktilität, Herzfrequenz
und enddiastolisches Volumen determiniert. Die ersteren beiden unterliegen der
Regulation durch kardiale sympathische Efferenzen (cardial sympathetic nerve
activity, CSNA); sie werden zusätzlich durch parasympathische (vagale)
Komponenten des vegetativen Nervensystems beeinflusst. Letztere ist abhängig vom
venösen Vasotonus, welcher von vaskulären sympathischen Efferenzen geregelt
wird. Der Gefäßwiderstand unterliegt neben der Regulation durch zirkulierende
vasoaktive Hormone und lokale Faktoren wie Metabolite und endotheliale Mediatoren
vor allem der Aktivität efferenter sympathischer vasokonstriktorischer Nerven, der
muskulären sympathischen Nervenaktivität (MSNA) (62; 143).
1.2.2 Arterieller Baroreflex
Kurzfristige Blutdruckschwankungen werden durch den arteriellen
Baroreflexbogen (siehe unten) korrigiert. Das Blutdruckniveau, auf dessen Erhaltung
diese reflektorische Korrektur innerhalb einer Feedback-Schleife abzielt, wird auch
als Setpoint bezeichnet (siehe ergänzende Abb. III). Es unterliegt einer ständigen
Anpassung an die jeweiligen physiologischen Erfordernisse, dem sogenannten
Resetting (27; 36; 143). Diese Anpassung wird von hypothalamischen, limbischen
und kortikalen Hirnregionen im Sinne eines feed-forward gesteuert. Diese wiederum
sind Teil eines komplexen integrierten neuroendokrinen Netzwerkes (siehe unten).
Der arterielle Baroreflexbogen läßt sich modellhaft als kybernetisches
Feedback-System beschreiben . Die Regelgröße (Ist-Wert) stellt der arterielle
Blutdruck dar. Er wird von Barorezeptoren im Sinus caroticus und Aortenbogen sowie
kardiopulmonalen Rezeptoren erfasst. Der mechanische Dehnungsreiz wird in ein
elektrisches neuronales Signal transduziert. Dieses erreicht über den N.
glossopharyngeus und den N. vagus den Hirnstamm. Es erfolgt der Abgleich mit
einer zentralnervös repräsentierten Führungsgröße (Soll-Wert). Dieser Wert wird
auch als Setpoint bezeichnet. Über sympathische Efferenzen werden die
Steuergrößen Vasokonstriktion und Herzminutenvolumen (cardiac output, CO)

15
1 Einleitung und Fragestellung
moduliert, um die Regelgröße dem Setpoint anzugleichen. Die efferenten Signale
projizieren von der Medulla oblongata über den spinalen Hinterseitenstrang in die
intermediäre Zone der grauen Substanz der Medulla spinalis. Dort erfolgt die
synaptische Umschaltung auf präganglionäre sympathische Neurone, welche ihre
Axone zu den Ganglien des paravertebralen Grenzstrangs entsenden. Dort steuern
sie die postganglionären Neurone, welche die Effektororgane innervieren. Das
basale sympathische Aktivitätsniveau des Baroreflexbogens wird von exzitatorischen
und inhibitorischen Neuronen im Nucleus tractus solitarii (NTS) sowie in der rostralen
und caudalen ventrolateralen Medulla oblongata erzeugt (27; 62) (siehe ergänzende
Abb. II). Hypothalamische Neurone projizieren in diese Strukturen und modulieren
das Aktivitätsniveau (154). Ein starkes afferentes Signal von den Barorezeptoren her
bewirkt eine Suppression der efferenten sympathischen Aktivität. Das Verhältnis der
Änderung der efferenten Sympathikusaktivität zur sie auslösenden Änderung des
arteriellen Blutdruckes beschreibt die Sensitivität des arteriellen Baroreflex (siehe
unten).
Der einfache Regelkreis des arteriellen Baroreflex unterliegt einer Reihe von
beeinflussenden Faktoren, so dass insgesamt ein in seinen Details noch
unverstandenes komplexes integrales kybernetisches System vorliegt. 1) Die
Regelgröße Blutdruck stellt keinen festen Wert dar, sondern folgt einer zyklisch-
pulsatilen Druckkurve mit Maxima und Minima (systolischer und diastolischer
Blutdruck). Es ist nicht abschließend geklärt, welcher Abschnitt der Druckkurve für
die Aktivierung der Barorezeptoren maßgeblich ist (143). 2) Das sympathische
Nervensystem regelt zwei Stellgrößen. Die muskuläre sympathische Nervenaktivität
(MSNA) moduliert den Gefäßwiderstand und ist damit maßgeblich für das
diastolische Blutdruckniveau. Die kardiale sympathische Nervenaktivität (CSNA)
moduliert gemeinsam mit vagalen Efferenzen die kardiale Auswurfleistung und damit
den systolischen Blutdruck (127). Die Sensitivität des Baroreflex hinsichtlich beider
Effektorstrecken des sympathischen Nervensystems scheint unterschiedlich reguliert
zu sein (40). Ferner bewirkt der Gefäßwiderstand im arteriolären Stromgebiet der
Extremitäten eine Pulswellenreflexion und –augmentation (11). Folglich sollte 3) der
an den Barorezeptoren gemessene Stimulus Gefäßdehnung zu unbekannten
Anteilen gemeinsam durch kardiale und vaskuläre sympathische Efferenzen
beeinflußt werden (143). 4) Die Hirnstammareale NTS und ventrolaterale Medulla, in

16
1 Einleitung und Fragestellung
denen die Regulation der efferenten Sympathikusaktivität lokalisiert sind, werden
durch andere afferente periphere Signale moduliert. So unterliegen sie der
Beeinflussung durch muskuläre Dehnungs- und Metaborezeptoren (74) und durch
Chemorezeptoren (65). Im NTS findet man eine hohe Dichte von Adrenorezeptoren
(151).
1.2.3 Mechanismen der langfristigen Blutdruckregulation
Die langfristige Blutdruckregulation wird vorrangig durch die renale Steuerung
des Wasser- und Salzhaushaltes vermittelt (63; 75). In der Vergangenheit war die
vorherrschende Überzeugung, dass das sympathische Nervensystem für die
langfristige Blutdruckregulation von untergeordneter Bedeutung sei (22; 63). Diese
Vorstellung wird zunehmend zugunsten des Konzeptes verlassen, dass die
langfristige Blutdruckkontrolle von einem übergeordneten zentralnervösen
sympathisch-neuroendokrinen Regelnetz aufrechterhalten wird, in dem das SNS eine
entscheidende integrierende Funktion innehat (76; 106; 154). Die Nieren stehen über
sympathische Afferenzen und Efferenzen in Interaktion mit zentralen
Blutdruckregulationsmechanismen (35; 83). Eine Verschiebung der renalen
Blutdruck-Natriurese-Beziehung bei arteriellem Hypertonus wird sowohl durch den
renalen Sympathikotonus als auch durch das Renin-Angiotensin-System (RAS), das
wiederum mit dem sympathischen Nervensystem wechselseitig interagiert, vermittelt
(10; 90). So führt eine Radiofrequenzablation des renalen sympathischen
Nervenbündels entlang des Nierenstiels bei Patienten mit therapieresistenter
arterieller Hypertonie zu einer nachhaltigen Blutdrucksenkung (83; 116).
Exemplarisch wurde gezeigt, dass diese Therapie eine Reduktion der muskulären
sympathischen Nervenaktivität (MSNA) bewirkt (117). Dieser Befund deutet auf die
Reduktion eines zentral vorgegebenen Blutdrucksollwertes durch besagte
Intervention hin.
1.2.4 Existenz und Regulation eines zentralen Blutdruck-Sollwertes
Kurzfristige Schwankungen des arteriellen Blutdrucks werden durch den
arteriellen Baroreflex mit Verschaltung auf Hirnstammebene ausgeglichen. Der
Blutdruck-Sollwert, auf deren Einstellung die Feedback-Korrektur hin erfolgt
(Baroreflex-Setpoint) und die Baroreflex-Sensitivität unterliegen einer ständigen

17
1 Einleitung und Fragestellung
Anpassung an die situativen Erfordernisse (27; 46; 62; 143). Signale aus
übergeordneten zerebralen Strukturen, sogenannte „feed-forward“-Signale, bewirken
eine veränderte efferente sympathische Aktivität, ohne dass dem eine Veränderung
des arteriellen Blutdruck ursächlich vorausgegangen sein muss. Eine feed-forward
Regulation findet statt im Zusammenhang mit orthostatischer Belastung (17),
körperlicher Aktivität (105), mentalem Stress und Schlaf (siehe unten). Ursprung der
feed-forward-Signale sind hypothalamische, limbische und kortikale Hirnregionen
(27; 62). Von herausgehobener Bedeutung sind die Nuclei paraventriculares
hypothalami. Ihre sympathoexzitatorische Funktion wird durch verhaltensabhängige
Stimuli wie (Nahrungsaufnahme, körperliche Aktivität und Schlaf), emotionale
Einflüsse und Hormone wie Angiotensin-II und Leptin moduliert (siehe unten). Trotz
aller Einflüsse bleibt der individuelle Setpoint von arteriellem Blutdruck und MSNA
des wachen, liegenden Probanden im physiologischen Fall über Jahre auf
normotensivem Niveau konstant. Dies weist auf die Existenz eines übergeordneten
zentralen Blutdrucksollwertes hin (76; 106; 154).
1.2.5 Wechselwirkungen des sympathischen Nervensystems mit dem endokrinen
System
Das sympathische Nervensystem unterliegt vielfachen Wechselwirkungen mit
dem endokrinen System. Beide können als Bestandteile eines integrierten
Netzwerkes aufgefasst werden. ACTH etwa stimuliert die MSNA (39), die Gabe von
Hydrokortison hingegen supprimiert sie über einen vermutlich zentralen
Mechanismus (38). Das SNS stimuliert über β1-Rezeptoren die Reninausschüttung
im juxtaglomerulären Apparat. Angiotensin-II wiederum erhöht die
Sympathikusaktivität sowohl auf peripherer als auch auf zentraler Ebene.
Angiotensin-II hemmt den Noradrenalin-Reuptake an der peripheren sympathischen
Synapse (34). Zirkulierendes Angiotensin-II kann über zirkumventrikuläre Organe die
Blut-Hirn-Schranke überwinden und zentral wirksam werden. In den Nuclei
paraventriculares des Hypothalamus sowie der Area postrema und den Nuclei
tractus solitarii des Hirnstamms aktiviert es sympathomimetische Mediatorkaskaden
(27; 62; 154). Zentrale baroreflexmodulierende Eigenschaften besitzen noch etliche
andere Hormone, etwa Aldosteron (69), Insulin (13), Glucagon (131), GLP-1 (14),
Melatonin (78) und Vasopressin (115).

18
1 Einleitung und Fragestellung
1.2.6 Messung der Aktivität des sympathischen Nervensystems
Zur Messung der Aktivität des sympathischen Nervensystems können direkte
und indirekte Verfahren dienen. Beim Menschen ist aufgrund anatomischer und
technischer Schwierigkeiten nur die muskuläre sympathische Nervenaktivität (MSNA)
einer direkten elektrophysiologischen Ableitung zugänglich. Im Gegensatz zum
Tierexperiment kann die Aktivität der kardialen und renalen sympathischen
Efferenzen nur anhand indirekter Parameter beurteilt werden (56; 57; 139; 143). Dies
gilt auch für die splanchnische und adrenale sympathische Innervation. Wegen des
hohen Grades an komplexer Integration der unterschiedlichen Faktoren
sympathischer Blutdruckregulation sind alle Meßgrößen nur im Kontext der übrigen
zu interpretieren.
Arterieller Blutdruck
Der nach Riva-Rochi am Oberarm gemessene Blutdruck kommt den
zentralen, an den Barorezeptoren wirksamen Drücken nah. Das Verfahren ist gut
reproduzierbar und gewährleistet eine gute interindividuelle Vergleichbarkeit. Die
Messung der Druckantwort auf einzelne Herzaktionen („beat-to-beat-Analyse“) ist
nicht möglich. Daher gestattet diese Methode nicht, gewisse Merkmale der
spontanen Baroreflexregulation wie zum Beispiel die sogenannten Mayer-Wellen zu
beobachten (21). Die Fingerplethysmographie ermöglicht eine „beat-to-beat“-
Messung des arteriellen Blutdrucks. Sie erlaubt nur eine unzuverlässige Messung der
absoluten Blutdruckhöhe, da inter- und intraindividuell variierende Faktoren wir
Gefäßelastizität und Vasotonus Einfluß nehmen. Die Gabe von vasoaktiven
Substanzen wie sie bei Verfahren zur Charakterisierung des Baroreflex verwendet
werden (siehe unten) kann die Messwerte beeinflussen. Die
fingerplethysmographische Messung hat ihren Wert für die Messung kurzfristiger
intraindividueller Blutdruckverläufe. Invasive Blutdruckmessungen mithilfe von
intraarteriellen Kathetern in unmittelbarer Nähe der Barorezeptoren (z.B. im
Aortenbogen), theoretisch die aussagekräftigste Messmethode, sind aufgrund der
inhärenten Risiken im Humanexperiment nur in Ausnahmefällen ethisch zu
rechtfertigen.

19
1 Einleitung und Fragestellung
Muskuläre sympathische Nervenaktivität (MSNA)
Die Mikroneurographie der muskulären sympathischen Nervenaktivität
(muscle sympathetic nerve activity, MSNA) ist zur Zeit die einzige zur Verfügung
stehende Methode zur Messung der efferenten postganglionären sympathischen
Nervenaktivität beim Menschen (139; 143). Die Spitze einer unipolaren
Wolframelektrode wird im sympathischen Faszikel des motorischen Anteils eines
gemischten Extremitätennervs platziert. Aufgrund ihrer subcutanen Verlaufsstrecke
kommen hierfür insbesondere N. peroneus und N. tibialis infrage. Nach Aufarbeitung
des Signals durch Verstärkung, Filterung und Integration über ein definiertes
Zeitintervall können sympathische Summenpotentiale in Form sogenannter „bursts“
dargestellt werden. Die Rate (Anzahl pro Zeiteinheit) und Amplitude dieser Bursts
stellen ein Maß für den zentralnervösen sympathischen Signalfluss zu den
arteriolären Widerstandsgefäßen der Skelettmuskulatur dar (muskuläre
sympathische Nervenaktivität, MSNA). Letzterer ist maßgebend für die Regulation
des Gesamtgefäßwiderstands, einer der beiden entscheidenden Stellgrößen des
Baroreflex-Regelkreises. Da eine hohe Konkordanz der efferenten muskulären
Sympathikusaktivität der unterschiedlichen Extremitätenregionen besteht, darf die
Ableitung an nur einem Extremitätennerv als repräsentativ gewertet werden (126;
138). Unter standardisierten Ruhebedingungen ist die MSNA-Burstrate
intraindividuell über lange Zeiträume gut reproduzierbar. Daher können Effekte
unterschiedlicher Interventionen an unterschiedlichen Tagen verglichen werden (49;
76; 127). Interindividuell ist die Burstrate sehr variabel. Sie korreliert bei jungen
gesunden Probanden nicht mit der Höhe des arteriellen Blutdrucks (104; 142).
Ursächlich könnten Unterschiede in der noradrenergen Gefäßreagibilität sein (19;
142), die durch eine unterschiedliche postsynaptische Adrenorezeptordichte oder
genetische Adrenozeptorpolymorphismen begründet sind. Die Qualität der
mikroneurographischen Ableitung wird von der intraneuralen Positionierung der
Elektrodenspitze beeinflußt. Burst-Amplitude und „total activity“, d.h. die Fläche unter
den Bursts pro Zeiteinheit, sind kritisch von der exakten Positionierung im efferenten
vasokonstriktorischen sympathischen Faszikel abhängig. Bereits eine geringfügige
Dislokation der Elektrode verändert beide Größen entscheidend. Daher setzt diese
Methode einen ruhig liegenden Probanden voraus. Eine exakt gleiche
Elektrodenposition ist in zwei unterschiedlichen Versuchssitzungen kaum zu

20
1 Einleitung und Fragestellung
erreichen, so dass Burst-Amplitude und „total activity“ in entsprechenden
Studiendesigns nur erschwert auszuwerten sind. Die Burst-Rate, d.h. die Anzahl der
bursts pro Zeiteinheit, stellt ein robusteres Maß für die MSNA dar, da sie nicht
entsprechend sensibel auf geringfügige Differenzen in der Elektrodenposition
reagiert.
Herzfrequenz und Herzfrequenzvariabilität
Primärer elektrischer Taktgeber der Herzaktion ist der Sinusknoten (Nodus
sinuatrialis). Die dort lokalisierten diastolisch spontan depolarisierenden Zellen des
Erregungsbildungs- und leitungssystems des Herzens generieren eine autonome
Sinusknotenfrequenz von 60-100/min. Sie unterliegen ständig modulierenden
Einflüssen durch das vegetative Nervensystem. Kardiale Efferenzen des
sympathischen Nervensystems („cardial sympathetic nerve activity“, CSNA) bewirken
über den Transmitter Noradrenalin einen frequenzsteigernden (positiv chronotropen)
Effekt. Ferner beschleunigen sie am AV-Knoten (Nodus atrioventricularis) die
Reizüberleitung (positive Dromotropie), steigern am Myokard die Kontraktilität
(positive Inotropie) und erhöhen in der Summe dieser Effekte das Herzzeitvolumen .
Vagale Efferenzen vermitteln über den Transmitter Acetylcholin eine Reduktion der
Herzfrequenz (negative Chronotropie) und verzögern am AV-Knoten die
Reizüberleitung (negative Dromotropie). In Ruhe sind letztere beim jungen gesunden
Probanden mit einer Herzfrequenz von 60-70/min vorherrschend (86). Die
Herzfrequenz dient als Summenmaß sympathischer und vagaler efferenter Aktivität
zum Herzen der Charakterisierung des arteriellen Baroreflex im Allgemeinen. Unter
Berücksichtigung der vagalen Einflüsse gestattet sie auch die Beurteilung des
sympathischen Schenkels des arteriellen Baroreflex und der CSNA im Besonderen.
In Bezug auf den Baroreflexbogen lassen sich die modulierenden Einflüsse des
vegetativen Nervensystems auf die Herzfrequenz anhand der respiratorischen
Oszillationen der Herzperiode (Schlag-zu-Schlag-Intervall) beobachten. Die durch
respirationsbedingt wechselnde kardiale Füllungsdrücke bedingten unterschiedlichen
Schlagvolumina werden über den Baroreflexbogen durch Modulation der
Schlagperiode beantwortet. Die Aktivität efferenter kardialer sympathischer und
parasympathischer Neurone unterliegt einer ständigen Fluktuation, die eine rasche
und eine langsame Oszillationen der Herzperiode bewirkt (42). Die Spektralanalyse

21
1 Einleitung und Fragestellung
der Herzfrequenzvariabilität (heart rate variability, HRV) erlaubt eine mathematische
Beschreibung dieser Oszillationen (133). Das HF-Band (high frequency; 0,15 – 0,4
Hz; Einheit ms2) reflektiert vor allem parasympathische Einflüsse auf das Herz. Das
LF-Band (low frequency; 0,04 – 0,15 Hz, Einheit ms2) reflektiert sympathische und
parasympathische Einflüsse (4; 133). Ihm scheinen sowohl Fluktuationen der
zentralen sympathischen Aktivität (107) als auch Resonanzeffekte des arteriellen
Baroreflex, die Fluktuationen der MSNA einbeziehen (4), zugrundezuliegen. HF- , LF-
und ein weiteres VLF (very low frequence; < 0,04 Hz)-Band ergeben in der Summe
die TP (total power). Die Übereinstimmung des LF-Bandes mit Herzfrequenz und
kardialer Noradrenalin-Kinetik ist unzureichend (42; 57), so dass spezifische
Aussagen über die efferente sympathische Aktivität zum Herzen anhand des LF-
Bandes allein nicht möglich sind. Um die Unzulänglichkeit der Parameter der HRV-
Analyse bei der Beschreibung der Aktivität vegetativer Efferenzen zum Herzen
zumindest teilweise auszugleichen, definiert man normalisierte Parameter (HFnu =
HF/(TP-VLF) und LFnu = LF/(TP-VLF) und den Quotienten LF/HF, der das Verhältnis
sympathischer und parasympathischer Nervenaktivität abbilden soll (97; 133).
Katecholamin-Plasmaspiegel
Das Signal der postganglionären efferenten sympathischen Neurone wird vom
Neurotransmitter Noradrenalin (NA) an das jeweilige Zielorgan übermittelt. Der
überwiegende Anteil des Noradrenalins unterliegt einer präsynaptischen
Wiederaufnahme („reuptake“). Nur ein geringerer Anteil von ca. 10-20% der
synaptisch ausgeschütteten NA-Menge tritt in die Zirkulation über („spillover“). Somit
ist der NA-Plasmaspiegel nicht nur von der adrenergen Nervenaktivität, sondern
auch von der synaptischen Clearance und den lokalen Zirkulationsbedingungen
abhängig (44). Die unterschiedlichen Äste des sympathischen Nervensystems,
insbesondere zur Niere („renal sympathetic nerve activity“, RSNA), zum Herzen
(CSNA), zu den Gefäßen der Skelettmuskulatur (MSNA) und zum
Splanchnikusgebiet, unterliegen einer differenzierten Regulation. Unter
physiologischen Bedingungen entstammen etwa 20% des venösen Plasma-NA den
sympathischen Efferenzen zu den Gefäßen der Skelettmuskulatur und korrellieren
mit der dort mikroneurographisch gemessenen MSNA (43; 144). Folglich ist die
Aussagekraft des venösen Plasma-NA hinsichtlich der sympathischen

22
1 Einleitung und Fragestellung
Kreislaufregulation eingeschränkt. Rückschlüsse sind nur unter differenzierter
Berücksichtigung aller genannten Faktoren zulässig. Prinzipiell besteht die
Möglichkeit, mithilfe selektiver Venenkatheterisierung den organbezogenen NA-
Spillover zu messen. In liegender Position korrelieren die kardialen und renalen
Spillover-Raten mit der MSNA (57). In Deutschland wird das invasive Verfahren zur
Grundlagenforschung aus ethischen Gründen am menschlichen Probanden im
Allgemeinen nicht durchgeführt. Die Bestimmung von Noradrenalin im Urin gestattet
eine globale Aussage über den systemischen NA-Spillover. Eine Organzuordnung ist
nicht möglich. Die Zeitliche Auflösung ist gering.
Die Ausschüttung von Adrenalin unterliegt der Regulation durch das
sympathische Nervensystem. Sie erfolgt fast ausschließlich im Nebennierenmark.
Das Hormon gelangt über die Zirkulation an seine Zielorgane. Die Messung der
Adrenalin-Plasmakonzentration bietet daher ein gutes Maß für die adrenomedulläre
Aktivität. Adrenalin weist wie Noradrenalin eine Plasmahalbwertszeit von wenigen
Minuten auf. Daher gestattet ihre Bestimmung eine gute zeitliche Auflösung.
1.2.7 Bestimmung von Setpoint und Sensitivität des arteriellen Baroreflex
Der Setpoint des arteriellen Baroreflex (siehe oben) wurde in der vorliegenden
Studie wie üblich definiert als Relation von Ruheblutdruckniveau und zugehöriger
Ruhe-MSNA der entspannt liegenden, wachen Versuchsperson (27; 36). Sie ist
intraindividuell gut reproduzierbar (siehe oben). Die Sensitivität des arteriellen
Baroreflexbogens kann durch pharmakologische Modulation des Blutdruckes bei
gleichzeitiger Messung der MSNA charakterisiert werden. Dazu werden direkt
vasodilatierende oder –konstringierende Pharmaka appliziert. Dies kann als Bolus
(„modified Oxford protocol“, siehe unten) oder unter kontinuierlicher Applikation als
„steady-state“-Infusionsprotokoll geschehen (143). Die Beziehung von arteriellem
Blutdruck und MSNA folgt einer sigmoiden Kurve (73; 132). Die Steigung des nahezu
linearen Kurvenabschnitts um den Setpoint herum beschreibt die Sensitivität des
arteriellen Baroreflex. Diese Technik erlaubt spezifische Aussagen über den die
MSNA als Regelgröße involvierenden Anteil des arteriellen Baroreflexbogens. Dieser
soll im Folgenden auch vereinfachend als „vaskulärer Baroreflex“ bezeichnet werden.
Korreliert man analog zum geschilderten Vorgehen die Herzfrequenz mit dem
arteriellen Blutdruck, erhält man eine Größe, die die Sensitivität des von kardialen

23
1 Einleitung und Fragestellung
sympathischen Efferenzen vermittelten Anteils des arteriellen Baroreflexes
beschreibt. Dieser kann vereinfachend auch als „kardialer Baroreflex“ benannt
werden.
1.2.8 Gesteigerte Aktivität des SNS als pathogenetischer Faktor
Die gesteigerte Aktivität des die Blutdruckregulation betreffenden
sympathischen Nervensystems ist pathogenetischer Faktor einer Reihe von
bedeutenden Erkrankungen. Viele Formen der arteriellen Hypertonie sind mit einer
erhöhten vasokonstriktorischen Sympathikusaktivität assoziiert. Ein kausaler
Zusammenhang wird vermutet („neuroadrenergic hypothesis in hypertension“) (56;
58). Schlafstörungen führen zu einem Ausbleiben der physiologischen nächtlichen
Blutdrucksenkung (non-dipping). Sie sind mit der Entstehung eines arteriellen
Hypertonus, inflammatorischen und metabolischen Veränderungen assoziiert (53; 70;
85). Die gesteigerte Aktivität des sympathischen Nervensystems scheint dabei ein
wesentlicher pathogenetischer Faktor zu sein. Nachgewiesen ist dies beim
obstruktiven Schlafapnoesyndrom (51; 52). Ein epidemiologischer Zusammenhang
zwischen nächtlichem non-dipping und kardiovaskulärer Morbidität und Mortalität ist
belegt (26; 67; 137).
1.3 Fragestellung/Hypothesen
1.3.1 Bedeutung von Ghrelin für die Regulation des sympathischen Nervensystems
Etliche Studien mit dem Ziel der Charakterisierung der Eigenschaften von
Ghrelin zeigten, dass das Peptidhormon eine ausgeprägte Wirksamkeit auf einige
zirkulatorische Parameter besitzt. Nach experimenteller intravenöser Applikation sinkt
der arterielle Blutdruck, ohne dass eine reflektorische Tachykardie auftritt, während
das Herzschlagvolumen zunimmt (100; 101). Die anhand der Parameter peak
systolic myocardial velocity und tissue tracking gemessene myokardiale Kontraktilität
nimmt signifikant zu (140). Diese Befunde könne nur teilweise durch einen lokalen
vasodilatorischen Effekt von Ghrelin auf die Blutgefäße (145; 149) oder einen lokalen
Effekt auf das Myokard (100) (siehe oben) erklärt werden. Da der arterielle Blutdruck
wesentlich durch die muskuläre (MSNA) und kardiale (CSNA) sympathische
Nervenaktivität geregelt wird, besteht ein möglicher Wirkmechanismus von Ghrelin in
einer Verminderung der efferenten Aktivität des sympathischen Nervensystems. Das

24
1 Einleitung und Fragestellung
Ausbleiben einer Zunahme der Herzfrequenz nach Abfall des arteriellen Blutdrucks
deutet auf eine Modulation des arteriellen Baroreflexes auf zentraler Ebene hin.
Tatsächlich bewirkt die intracerebroventrikuläre Gabe von Ghrelin im
Tierexperiment eine Reduktion der efferenten renalen sympathischen Aktivität
(RSNA) und eine vermehrte Sensitivität des arteriellen Baroreflex (94). Eine Injektion
des Neuropeptids in den Nucleus tractus solitarii senkt Herzfrequenz, arteriellen
Blutdruck und RSNA (88). Nach experimentell herbeigeführtem Myokardinfarkt
reduziert systemisch appliziertes Ghrelin die allgemeine (125) und kardiale (119)
efferente Aktivität des sympathischen Nervensystems.
1.3.2 Formulierung der Hypothesen der vorliegenden Arbeit
Aufgrund oben genannter Daten formulierten wir die Hypothesen, deren
Richtigkeit im Rahmen dieser Arbeit empirisch überprüft werden sollte.
1. Ghrelin bewirkt auf zentraler Ebene eine Reduktion der muskulären
sympathischen Nervanaktivität (MSNA) und damit des arteriellen Blutdrucks.
2. Ghrelin moduliert die Sensitivität des arteriellen Baroreflex.
Zum Zeitpunkt der Durchführung der Studie existierte in der Literatur keine
Publikation zur Wirkung von Ghrelin auf die MSNA und keine humanexperimentelle
Arbeit bezüglich der Beeinflussung der Sensitivität des arteriellen Baroreflex durch
das Peptidhormon.

25
2 Material und Methoden
2 Material und Methoden
2.1 Versuchspersonen
Die vorliegende Studie wurde von der Ethikkommission der Universität zu
Lübeck geprüft und für zulässig erachtet (Aktenzeichen 09-166, Datum des
Ethikvotums 19.11.2009). Die Versuche wurden im neurophysiologischen Labor der
Medizinischen Klinik I der Universität zu Lübeck durchgeführt. Alle Probanden
wurden ausführlich mündlich und schriftlich über die Untersuchungen und ihre
möglichen Risiken aufgeklärt und gaben ihr schriftliches Einverständnis zur
Teilnahme. Die Probanden waren überwiegend aus der Studierendenschaft über
Aushänge angeworben worden. An der Untersuchung nahmen 12 gesunde,
normalgewichtige (BMI 20-25 kg/m2) Männer im Alter von 22 bis 30 Jahren teil. Die
Probanden waren Nichtraucher und standen unter keiner regelmäßigen Medikation.
Da Nüchternheit und postprandiale Phase den Baroreflex modifizieren (47), wurden
die Versuchsteilnehmer aufgefordert, die letzte Mahlzeit am Abend vor dem
jeweiligen Versuchstermin einzunehmen. Ferner sollten sie mindestens 24 Stunden
vor dem Versuch auf Alkoholkonsum und koffeinhaltige Getränke verzichten.
2.2 Design und Ablauf der Studie
2.2.1 Design
Die vorliegende Untersuchung war als randomisierte, doppelblinde,
placebokontrollierte Studie im Rahmen eines Crossover-Designs angelegt. Die
Probanden erhielten an zwei Versuchstagen im Abstand von mindestens einer
Woche einen i.v.- Bolus Ghrelin oder Placebo. Das Bolus-Design orientierte sich an
dem physiologischen pulsatilen Sekretionsmodus von Ghrelin (24). Als primäre
Outcome-Parameter wurden MSNA, Blutdruck, Herzfrequenz und Sensitivität des
arteriellen Baroreflex, ausgedrückt als linearer Faktor einer linearen Modellfunktion
(s. unten), definiert. Als sekundäre Outcome-Parameter wurden mehrere endokrine
Laborparameter erfasst. Das Crossover-Design erlaubte insbesondere den
intraindividuellen Vergleich („within-subject“) interindividuell sehr variabler, jedoch
intraindividuell stabiler Parameter wie der MSNA unter beiden Versuchsbedingungen.

26
2 Material und Methoden
2.2.2 Instrumentierung
Die Versuche begannen um 7:30 morgens und endeten etwa um 12:30
mittags. Die Probanden wurden in bequemer Rückenlage gelagert. Elektroden zur
Ableitung eines 3-poligen Standard-EKG nach Eindhoven und eine Manschette zur
kontinuierlichen Blutdruckmessung mittels Fingerplethysmographie wurden angelegt.
Zusätzlich wurde eine Oberarmmanschette zur oszillometrischen Messung des
arteriellen Blutdrucks angelegt. Die kontinuierliche Blutdruckmessung wurde anhand
der oszillometrischen Messwerte kalibriert. Eine Venenverweilkanüle wurde zum
Zweck wiederholter Blutentnahmen in der Vena cubitalis des rechten Arms plaziert.
Eine zweite Venenverweilkanüle wurde in die Vena cubitalis des linken Arms gelegt.
Dort erfolgte die Applikation der Testsubstanz (Ghrelin oder Placebo). Die jeweilige
Substanz wurde von einem Mitarbeiter, der nicht an der weiteren
Versuchsdurchführung beteiligt war, in einem separaten Raum zubereitet und in
einem neutralen Behälter zur Verfügung gestellt. Die Kanüle des linken Arms wurde
zusätzlich zur Verabreichung vasoaktiver Substanzen im Rahmen der Untersuchung
des Baroreflex genutzt. Ferner wurde eine Mikroneurographienadel zur Messung der
muskulären sympathischen Aktivität (MSNA) plaziert (s. unten).
2.2.3 Versuchsablauf
Nach Instrumentierung und einer 10-minütigen Erholungsphase erfolgte
zunächst eine 10-minütige baseline-Aufzeichnung von MSNA, Blutdruck und
Herzfrequenz (t -25 min, „pre-injection baseline“). Anschließend wurde eine
Baroreflex-Testung nach dem modifizierten Oxford-Protokoll (s. unten) durchgeführt
(t -5 min, „MOP 1“). Nach kurzer Kreislaufrestitutionsphase wurde die Testsubstanz
(Ghrelin oder Placebo) intravenös appliziert. Weitere Messungen der
Baroreflexparameter unter Ruhebedingungen erfolgten unmittelbar (5-minütige
Intervalle, t +2 min und t +7 min, „immediate post-injection baseline 1+2“) und
deutlich verzögert (10-minütiges Intervall, t +85 min, „delayed post-injection
baseline“) nach Substanzgabe. Weitere pharmakologische Baroreflextestungen nach
dem modifizierten Oxford-Protokoll wurden jeweils 15, 35 und 65 Minuten nach
Substanzgabe durchgeführt („MOP 2-4“ zu t +15 min, t +35 min und t +65 min). Zu
Beginn der MOP-Testungen wurden alle Parameter unbeeinflusst aufgezeichnet
(siehe unten). Diese Messperioden wurden als zusätzliche baseline-Aufzeichungen

27
2 Material und Methoden
genutzt. Ferner wurden wiederholt Blutentnahmen zur Erfassung der
Katecholaminantwort und anderer endokriner Veränderungen durchgeführt (t -5 min,
t +15 min, t +30 min, t +60 min, t +110 min). Der Ablauf eines Experiments ist
schematisch in Abb. 1 dargestellt. Um eine konstante Qualität der MSNA-Ableitung
zu gewährleisten, verblieben die Mikroelektroden während des gesamten Versuches
in ihrer intraneuralen Position. Klinische Beobachtungen und spontane Äußerungen
der Probanden wurden notiert.
Abb. 1 Protokoll der vorliegenden Studie. Hämodynamische Parameter und MSNA
wurden vor (pre-injection), unmittelbar (immediate post-injection) und später (delayed
post-injection) nach intravenöser Injektion von Ghrelin oder Placebo gemessen. Die
Sensitivität des arteriellen Baroreflex wurde mittels Modulation des Blutdrucks durch
vasoaktive Substanzen charakterisiert (modified Oxford protocol, MOP 1-4).
Zusätzlich erfolgten Blutentnahmen zur endokrinologischen Analytik. (Aus eigener
Publikation, siehe unten).

28
2 Material und Methoden
2.3 Aufzeichnung hämodynamischer Parameter
Für die kombinierte digitalisierte Aufzeichnung der Daten der wurde ein
multifunktionales elektronisches Interface genutzt (PowerLab®, ADInstruments,
Heidelberg). Mit Hilfe einer speziellen Software (Chart® 5 Pro Version 5.5.6,
ADInstruments, Heidelberg) konnten Mikroneurographiesignal (s. unten), EKG (Bio
Amp, ADInstruments, Heidelberg), kontinuierlich fingerplethysmographisch
gemessene Blutdruckwerte (Finometer midi®, FMS, Amsterdam, Niederlande) und
thorakale Atemexkursion (Pulmobelt®, Ohmeda Medical, Ohio, USA) gemeinsam
dargestellt und in einem gemeinsamen Dokument für die nachfolgende Analyse
gespeichert werden. Die Aufzeichnung erfolgte jeweils kontinuierlich für einen
Versuchstag über alle später ausgewerteten Messintervalle hinweg ohne
Veränderung der Instrumentierung. Die oszillometrische Messung des arteriellen
Blutdrucks erfolgte mittels eines geeichten hochqualitativen Gerätes (Vital Signs
Monitor 300®, Series 53NTP-E1, Skaneateles Falls, New York, USA).
2.4 Mikroneurographie
2.4.1 Plazierung der Elektroden
Zur Ableitung der MSNA fanden Wolfram-Mikroelektroden Verwendung
(Göran Pegenius, Mölndal, Schweden). Während der 0,2 mm durchmessende
Elektrodenschaft elektrisch isoliert ist, fehlt diese Isolation im Bereich der wenige
Mikrometer feinen Elektrodenspitze. Dieser Aufbau ermöglichte die gezielte Ableitung
des Potentials efferenter muskulärer sympathischer Nerven (MSNA). Eine
Referenzelektrode wurde im nahen subcutanen Fettgewebe plaziert. Die Impedanz
der Ableitelektrode (20-100 kΩ) übertraf die Impedanz der Referenzelektrode. Beide
Elektroden wurden vor Verwendung unter dem Mikroskop auf Intaktheit der
Oberfläche untersucht und im Autoklav sterilisiert.
Die Ableitung der MSNA erfolgte entweder aus dem N. peroneus oder dem N.
tibialis in ihrem Verlauf dorsal des Fibulaköpfchens. Zur Lokalisierung des Nerven
wurde zunächst eine transkutane elektrische Stimulation (Stimulator S48, Grass
Instrument Company, Quincy, Massachusetts, USA) mit einer Spannung von 30-70
V, einer Reizdauer von 0,1 ms und einer Stimulationsfrequenz von 1 Hz
durchgeführt. Anhand der Beobachtung evozierter Muskelkontraktionen der

29
2 Material und Methoden
Zielmuskulatur des jeweiligen Nerven konnte dessen Verlauf bestimmt und markiert
werden. Eine Referenzelektrode wurde ca. 2 cm entfernt im subcutanen Fettgewebe
platziert. Anschließend wurde die Ableitelektrode perkutan in Richtung des
Zielnerven eingestochen. Bei Annäherung an den Nerven konnten mittels erneuter
elektrischer Stimulation (1-4 V) unwillkürliche Muskelkontraktionen im peronealen
oder tibialen Innervationsgebiet ausgelöst werden. Bei ausreichender Annäherung
reichte hierfür eine Spannung von 1 V aus. Die abschließende intraneurale
Feinadjustierung der Elektrodenspitze in unmittelbarer Nähe der efferenten
sympathischen Leitungsbahnen zur Gefäßmuskulatur erfolgte unter Beobachtung der
akustisch und visuell am Monitor dargestellten Signale, bis charakteristische
sympathische Entladungen (bursts) erkennbar waren. Eine hinreichende signal to
noise-ratio wurde als gegeben betrachtet, wenn die burst-Amplitude das
Grundrauschsignal um mindestens das 3-fache übertraf.
2.4.2 Signalprozessierung
Das mikroneurographische Rohsignal wurde einer Verstärkung unterzogen.
Das Verstärkersystem (T. Karlsson, Göteborg, Schweden) bestand aus einem
Vorverstärker (preamplifier, 1000-fache Verstärkung) und einem Hauptverstärker
(amplifier, 50-fache Verstärkung). Das verstärkte Signal wurde gefiltert (Bandfilter,
700-2000 Hz) und anschließend integriert (Zeitkonstante 0,1 sec.). Digitalisierung,
Speicherung und Visualisierung erfolgten mit der zuvor beschriebenen Hard- und
Software.
2.4.3 Identifizierung der bursts
Im aufgearbeiteten Signal konnten die efferenten muskulären sympathischen
Summenaktionspotentiale (bursts) anhand ihrer charakteristischen Morphologie (138)
visuell identifiziert werden. Alle mikroneurographischen Aufzeichnungen wurden vom
selben Beobachter, der bezüglich der Versuchsbedingung (Ghrelin/Placebo)
verblindet war, ausgewertet.
N. peroneus und N. tibialis sind gemischte Nerven. Bei den sympathischen
Efferenzen, welche die baroreflektorische Vasokonstriktion vermitteln, handelt es sich
um C-Fasern, die in motorischen Faszikeln verlaufen. Es wurde eine Reihe von
Prüfkriterien genutzt, die charakteristische Eigenschaften muskulärer

30
2 Material und Methoden
Sympathikusefferenzen nutzen und insbesondere eine Abgrenzung vom Signal
kutaner Sympathikusefferenzen gestatten (32; 33). Kennzeichen muskulärer
Sympathikusanteile sind sie Auslösbarkeit eines Signals bei Beklopfen des
innervierten Muskels durch Reizung von Dehnungsrezeptoren und Stimulierbarkeit
der MSNA durch inspiratorische Apnoe. Charakteristisch für kutane
Sympathikusanteile sind ein Signal beim Bestreichen der Haut und die Evozierbarkeit
von Potentialen durch externe Arousalsignale. Es wurden nur Ableitungen akzeptiert,
die alle Kriterien muskulärer Sympathikusefferenzen erfüllten und zugleich kein
kutanes sympathisches Signal enthielten. Kontinuität und Vergleichbarkeit der
Ableitungsqualität über den Messzeitraum wurden jeweils durch Apnoephasen zu
Beginn und Ende der mikroneurographischen Messungen überprüft. Die MSNA ist,
vermittelt über den Baroreflex, eng mit dem Herzzyklus verknüpft (64; 126). Die
Latenz eines Bursts zum zugehörigen Herzzyklus resultiert aus der Zeit bis zum
Eintreffen der Pulswelle an den Barorezeptoren sowie der Dauer der Reizleitung in
den afferenten und efferenten Sympathikusfasern. Bei normalwüchsigen
Erwachsenen beträgt diese Zeitspanne etwa 1,3 s bezogen auf die R-Zacke des
EKG (48). Bei der Auswertung der Mikroneurographie wurde auf eine eindeutige
Kopplung von MSNA und EKG geachtet.
2.4.4 Burst-Rate
Es stehen im Wesentlichen drei Parameter zur quantitativen Beschreibung der
MSNA zur Verfügung (143). Burst-Amplitude und Integral über der Zeit (area under
the curve) einer MSNA-Ableitung hängen jedoch kritisch von der Position der
Elektrode zum sympathischen Faszikel ab. Diese kann in wiederholten Versuchen
gegebenenfalls nicht exakt reproduziert werden. Daher eignen sich diese Parameter
weniger für das Design der vorliegenden Studie. Die burst-Rate, d.h. die Anzahl der
bursts pro Minute hingegen ist sehr gut reproduzierbar und wurde daher als für das
vorliegende Studiendesign geeignetster MSNA-Parameter ausgewählt.
2.5 Pharmakologischer Baroreflextest
Der Setpoint des Baroreflex wird definiert als dasjenige Niveau der Relation
von arteriellem Blutdruck und MSNA (vaskulärer Baroreflex) bzw. von arteriellem

31
2 Material und Methoden
Blutdruck und Herzfrequenz (kardialer Baroreflex), um das diese Parameter bei
entspannter Ruhe im Liegen in engen Grenzen pendeln.
2.5.1 Modified Oxford protocol (MOP)
Der pharmakologische Baroreflextest ist ein Provokationsmaneuver, durch das
Setpoint und Sensitivität des Baroreflex ermittelt werden können. Dafür wird der
arterielle Blutdruck zunächst mittels vasoaktiver Substanzen gesenkt und dann über
das Ruheniveau angehoben. Die gegenregulatorischen MSNA- und
Herzfrequenzänderungen werden in Bezug zur erzielten Änderung des arteriellen
Blutdrucks dokumentiert. In dieser Studie fand eine kombinierte Methode
Anwendung. Sie verwendete Elemente zweier zu diesem Zweck gut etablierter
Verfahren. Das eine ist das steady state-Protokoll unter Verwendung vasoaktiver
Substanzen, das von unserer Arbeitsgruppe bereits mehrfach zur Anwendung
gebracht wurde (112-114). Das andere ist das modified Oxford protocol (MOP) (41;
110). Das verwendete Protokoll zur Charakterisierung des Baroreflex bestand aus 3
Abschnitten und soll im Folgenden kurz als „MOP“ bezeichnet werden. Zunächst
erfolgte die Aufzeichnung von MSNA, Herzfrequenz und arteriellem Blutdruck in
einem unbeeinflussten Zeitintervall von einer Minute. Dann erfolgte die i.v. Injektion
eines Bolus von 150 µg Nitroprussid-Natrium, eines direkten NO-vermittelten
Vasodilatators. Nach einer weiteren Minute erfolgte die i.v. Injektion eines Bolus von
150 µg Phenylephrin, eines α1-Agonisten.
2.5.2 Mathematische Analyse
Zur Analyse wurden MSNA, arterieller Blutdruck und Herzfrequenz jeweils für
60-Sekunden-Intervalle bestimmt 1. vor Injektion der vasoaktiven Substanzen, 2. 30
bis 90 Sekunden nach Bolusinjektion von Nitroprussid-Natrium und 3. 90 bis 150
Sekunden nach Injektion von Phenylephrin. Für jeden Messzyklus (MOP) wurden
jeweils das arithmetische Mittel der burst-Rate der MSNA beziehungsweise der
Herzfrequenz in den genannten Zeitintervallen gegen den zugehörigen mittleren
arteriellen Blutdruck aufgetragen (vaskulärer und kardialer Baroreflex). Anschließend
wurde eine lineare Regressionsanalyse nach der Methode der kleinsten Quadrate
durchgeführt. Ein Messzyklus wurde akzeptiert, sofern der Determinationskoeffizient
R2 größer oder gleich 0,7 war. Der lineare Faktor (slope) der resultierenden linearen
32
2 Material und Methoden
Modellfunktion wurde genutzt, um die Sensitivität des Baroreflex zu charakterisieren
und der weiteren Statistik unterworfen. Ein Beispiel von MSNA und
hämodynamischen Parametern unter ungestörten (baseline) Bedingungen und nach
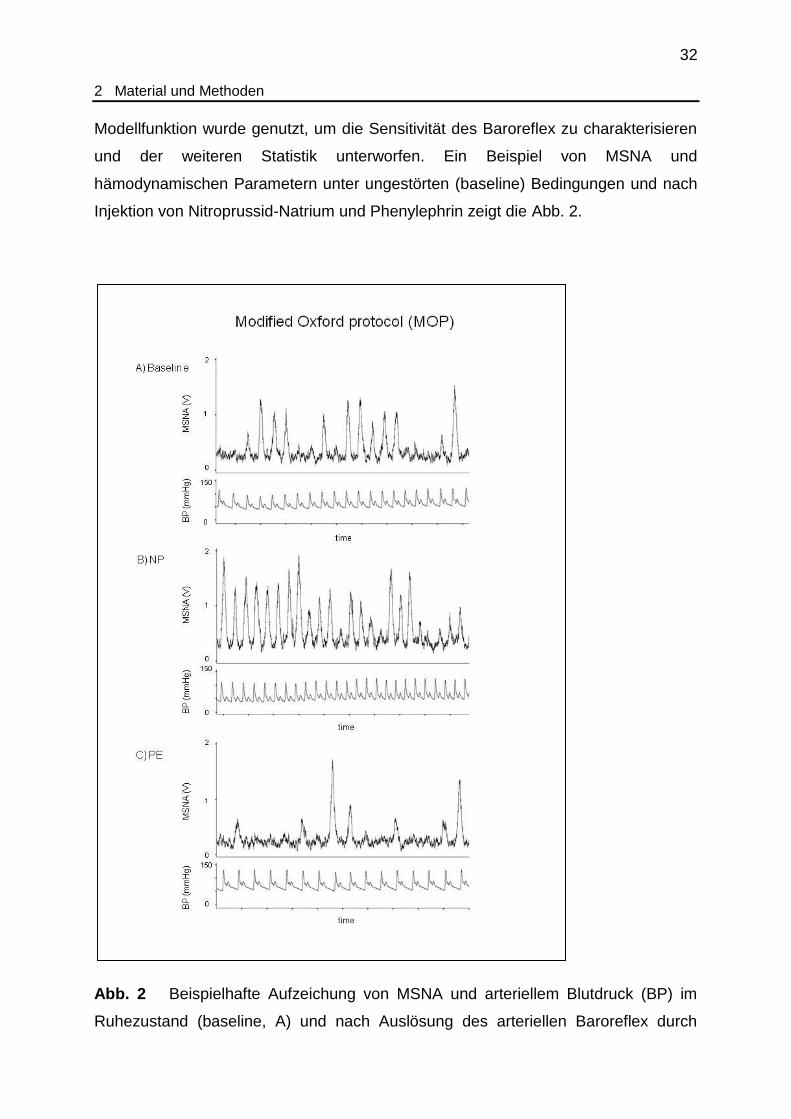
Injektion von Nitroprussid-Natrium und Phenylephrin zeigt die Abb. 2.
Abb. 2 Beispielhafte Aufzeichung von MSNA und arteriellem Blutdruck (BP) im
Ruhezustand (baseline, A) und nach Auslösung des arteriellen Baroreflex durch

33
2 Material und Methoden
Nitroprussid-Natrium (NP, B) beziehungsweise Phenylephrin (PE, C) nach dem
modified Oxford protocol (MOP). Man beachte die charakteristischen
Summenaktionspotentiale (bursts) der MSNA. Ein Abschnitt auf der Zeitachse
repräsentiert 2 Sekunden. (Aus eigener Publikation, siehe unten.)
2.6 Herzfrequenzvariabilität
Eine Auswertung der Herzfrequenzvariabilität erfolgte für drei Intervalle von
jeweils 5 Minuten vor (t -25 min), unmittelbar nach (t +2 min) und später nach (t +85
min) Injektion der Testsubstanz. Die Analyse wurde mithilfe des HRV-Moduls von
Chart® 5 Pro (Version 5.5.6, ADInstruments, Heidelberg) aus den digitalisierten
EKG-Daten (siehe oben) nach der frequency domain-Methode durchgeführt. Sie
folgte den Leitlinien der „Task Force of The European Society of Cardiology and The
North American Society of Pacing and Electrophysiology“(133).
2.7 Laboranalytik
Blutproben wurden zu den Zeitpunkten t -10 min, t +10 min, t+30 min, t +60
min und am Ende des Experimentes (t +110 min) über eine periphere
Venenverweilkanüle entnommen. Sie wurden umgehend in einer Kühlzentrifuge mit
4000 U/min über 10 Minuten zentrifugiert und abpipettiert. Anschließend wurden die
Proben umgehend gekühlt und bei -80°C gelagert. Die Laboranalytik erfolgte im
gemeinsamen Forschungslabor der Medizinischen Klinik I und des Instituts für
Neuroendokrinologie (medizinisch-technische Assistenz: Martina Grohs und Heidi
Ruf). Analysen wurden durchgeführt für Ghrelin (total ghrelin RIA Kit, Millipore,
Billerica, Massachusetts, USA), Renin (Renin III Generation, Cisbio, Codolet,
Frankreich), Katecholamine (HPLC, Chromsystems, München), GH, ACTH, Cortisol
(alle Immulite, Siemens, Llanberis, Vereinigtes Königreich) und Copeptin (Thermo
Fisher Scientific BRAHMS GmbH, Henningsdorf). In der Regel wurden alle Proben
doppelt mit demselben Verfahren bestimmt und validiert.
2.8 Statistik
Hämodynamische Variablen, MSNA und Laborwerte zeigen in aller Regel eine
parametrische Verteilung. Die Darstellung dieser stetigen, intervallskalierten

34
2 Material und Methoden
Merkmale erfolgte als arithmetischer Gruppenmittelwert +/- Standardfehler des
Mittelwertes (SEM). Es wurde das Signifikanzniveau α = 0,05 definiert. Als primäre
statistische Analyse wurde jeweils eine Varianzanalyse (ANOVA = analysis of
variance) mit dem Messwiederholungsfaktor „Messzeitpunkt“ sowie dem
Gruppenfaktor „Behandlung“ (Ghrelin vs. Placebo) durchgeführt. Es erfolgte eine
Korrektur der Freiheitsgrade nach Greenhouse-Geisser. Wurde in der ANOVA eine
Signifikanz für einen der beiden Faktoren oder ihre Interaktion festgestellt, erfolgte
eine post hoc-Analyse mittels paarweisen Vergleiches der Einzelwerte unter
Verwendung des T-Tests nach Student. Es wurde dann sowohl ein Vergleich beider
Versuchsbedingungen (Ghrelin/Placebo) zu jeweils einem Messzeitpunkt als auch
ein Vergleich der Parameter innerhalb jeweils einer Versuchsbedingung zu
verschiedenen Meßzeitpunkten, d.h. insbesondere vor und nach Injektion der
Testsubstanz durchgeführt. Die statistischen Analysen erfolgten unter Verwendung
der Software PASW Statistics 18. Die Kalkulation von Stichprobengröße, Power und
Effektgröße erfolgte mit Hilfe von G*Power 3.1.3 (Franz Faul, Universität Kiel). Der α-
Fehler wurde nicht an die Anzahl der Tests angepasst. Aus diesem Grund verstehen
sich alle inferenzstatistischen Auswertungen letztlich deskriptiv (1).

35
3 Ergebnisse
3. Ergebnisse
3.1 Klinische Beobachtungen
Die intravenöse Verabreichung sowohl des Ghrelin- als auch des
Placebobolus wurde im Allgemeinen gut toleriert. Nach Gabe von Ghrelin berichteten
die Probanden von Hunger (N=6), Wärmegefühl (N=3), Schwindel (N=2), Müdigkeit
(N=3), Magenknurren (N=4) und Schwitzen (N=5). Während das Hungergefühl nach
Ghrelingabe bis zum Ende des Versuches persistierte, verflüchtigten sich alle
übrigen mit Ghrelin assoziierten Symptome innerhalb der ersten 30 Minuten nach
Applikation. Unter Placebobedingung berichtete lediglich ein Proband über
Müdigkeit.
3.2 Hämodynamische Parameter
Arterieller Blutdruck und Herzfrequenz konnten für alle 12 Probanden
analysiert werden. Bei zwei Probanden ging im Verlauf des Experimentes die
Position der Mikroneurographieelektrode im sympathischen Faszikel verloren, so
dass eine valide Ableitung nicht mehr möglich war. Daher gingen in die Analyse der
MSNA nur die Daten von 10 Probanden ein. Alle Ergebnisse sind in Tab. 1 und Abb.
3 angegeben.
Vor Injektion der Testsubstanz (t -25 min, t -5 min) befanden sich arterieller
Blutdruck, Herzfrequenz und MSNA unter beiden Versuchsbedingungen auf
gleichem Niveau.
Die ANOVA erbrachte eine signifikante Interaktion des
Messwiederholungsfaktors „Messzeitpunkt“ und des Gruppenfaktors „Behandlung“
(Ghrelin vs. Placebo) für die Parameter MSNA (P=0,037) und Herzfrequenz
(P<0,001). Hinsichtlich des Parameters arterieller Blutdruck zeigte sich ein starker
Trend (P=0,057).
36
3 Ergebnisse
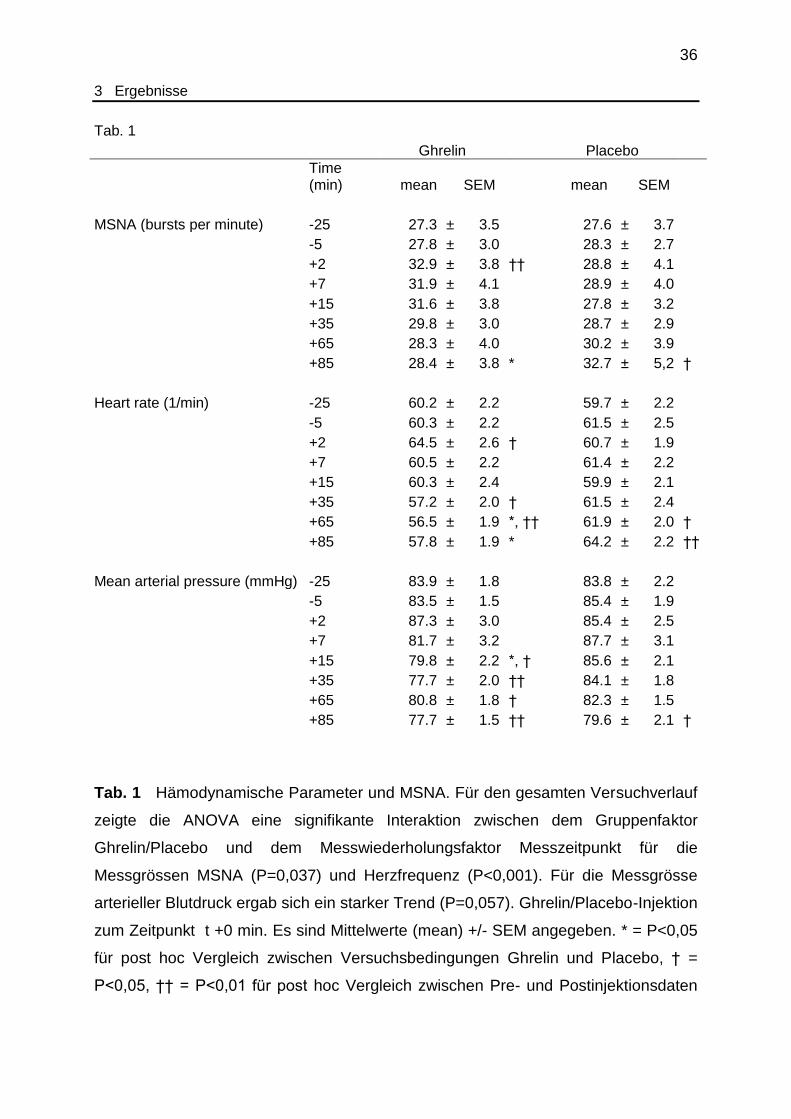
Tab. 1
Ghrelin Placebo
Time (min) mean SEM mean SEM
MSNA (bursts per minute) -25 27.3 ± 3.5 27.6 ± 3.7
-5 27.8 ± 3.0 28.3 ± 2.7
+2 32.9 ± 3.8 †† 28.8 ± 4.1
+7 31.9 ± 4.1 28.9 ± 4.0
+15 31.6 ± 3.8 27.8 ± 3.2
+35 29.8 ± 3.0 28.7 ± 2.9
+65 28.3 ± 4.0 30.2 ± 3.9
+85 28.4 ± 3.8 * 32.7 ± 5,2 †
Heart rate (1/min) -25 60.2 ± 2.2 59.7 ± 2.2
-5 60.3 ± 2.2 61.5 ± 2.5
+2 64.5 ± 2.6 † 60.7 ± 1.9
+7 60.5 ± 2.2 61.4 ± 2.2
+15 60.3 ± 2.4 59.9 ± 2.1
+35 57.2 ± 2.0 † 61.5 ± 2.4
+65 56.5 ± 1.9 *, †† 61.9 ± 2.0 †
+85 57.8 ± 1.9 * 64.2 ± 2.2 ††
Mean arterial pressure (mmHg) -25 83.9 ± 1.8 83.8 ± 2.2
-5 83.5 ± 1.5 85.4 ± 1.9
+2 87.3 ± 3.0 85.4 ± 2.5
+7 81.7 ± 3.2 87.7 ± 3.1
+15 79.8 ± 2.2 *, † 85.6 ± 2.1
+35 77.7 ± 2.0 †† 84.1 ± 1.8
+65 80.8 ± 1.8 † 82.3 ± 1.5
+85 77.7 ± 1.5 †† 79.6 ± 2.1 †
Tab. 1 Hämodynamische Parameter und MSNA. Für den gesamten Versuchverlauf
zeigte die ANOVA eine signifikante Interaktion zwischen dem Gruppenfaktor
Ghrelin/Placebo und dem Messwiederholungsfaktor Messzeitpunkt für die
Messgrössen MSNA (P=0,037) und Herzfrequenz (P<0,001). Für die Messgrösse
arterieller Blutdruck ergab sich ein starker Trend (P=0,057). Ghrelin/Placebo-Injektion
zum Zeitpunkt t +0 min. Es sind Mittelwerte (mean) +/- SEM angegeben. * = P<0,05
für post hoc Vergleich zwischen Versuchsbedingungen Ghrelin und Placebo, † =
P<0,05, †† = P<0,01 für post hoc Vergleich zwischen Pre- und Postinjektionsdaten
37
3 Ergebnisse
innerhalb der Versuchsbedingung Ghrelin oder Placebo (T-Test). (Aus eigener
Publikation, siehe unten.)
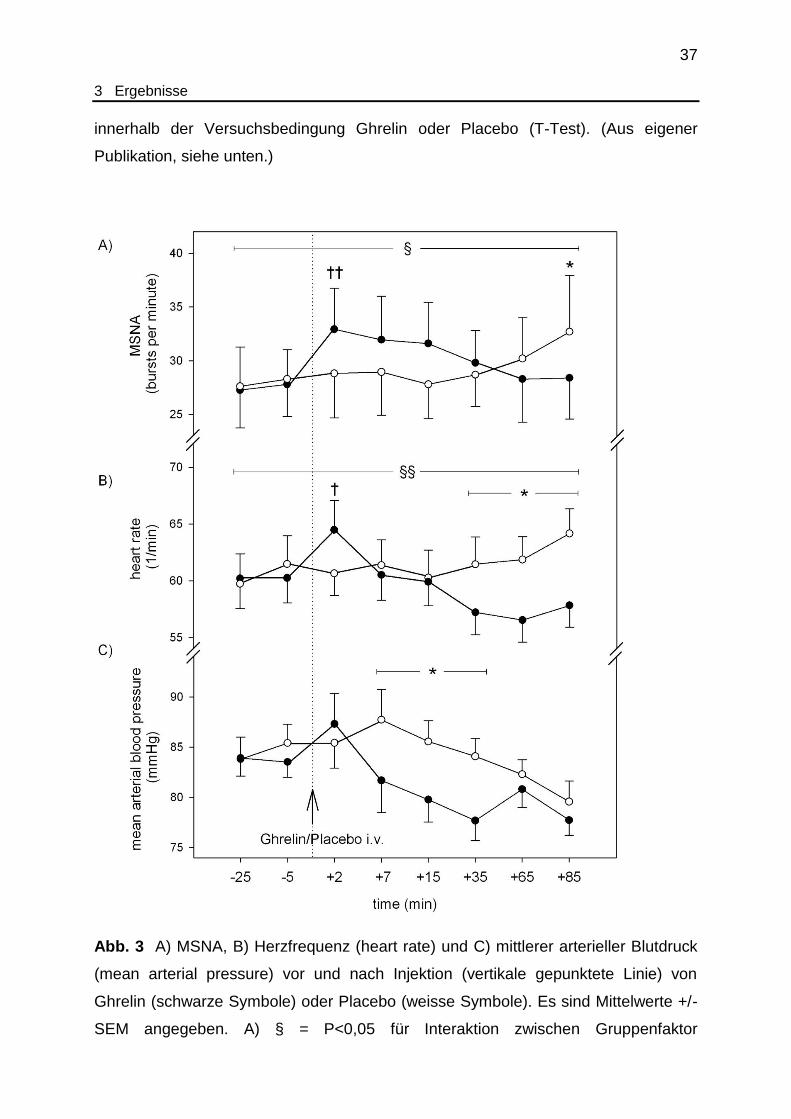
Abb. 3 A) MSNA, B) Herzfrequenz (heart rate) und C) mittlerer arterieller Blutdruck
(mean arterial pressure) vor und nach Injektion (vertikale gepunktete Linie) von
Ghrelin (schwarze Symbole) oder Placebo (weisse Symbole). Es sind Mittelwerte +/-
SEM angegeben. A) § = P<0,05 für Interaktion zwischen Gruppenfaktor

38
3 Ergebnisse
Ghrelin/Placebo und Messwiederholungsfaktor Messzeitpunkt (ANOVA, N=10);
* = P<0,05 für Vergleich zwischen Ghrelin und Plazebo (T-Test); †† = P<0,01 für
Vergleich zwischen Pre- und Postinjektionsdaten innerhalb der Ghrelin-
Versuchsbedingung (T-Test). B) §§ = P<0,001 für Interaktion zwischen
Gruppenfaktor Ghrelin/Plazebo und Messwiederholungsfaktor Messzeitpunkt
(ANOVA, N=12); * = P<0,05 für Faktor Ghrelin/Plazebo, t +35 min bis t + 85 min
(ANOVA); † = P<0,05 für Vergleich zwischen Pre- und Postinjektionsdaten innerhalb
der Ghrelin-Versuchsbedingung (T-Test). C) P=0,057 für Interaktion zwischen
Gruppenfaktor Ghrelin/Plazebo und Messwiederholungsfaktor Messzeitpunkt
(ANOVA, N=12); * = P<0,05 für Faktor Ghrelin/Plazebo, t +7 min bis t +35 min
(ANOVA). (Aus eigener Publikation.)
3.2.1 Mittlerer arterieller Blutdruck
Nach Injektion von Ghrelin (t +7 min und folgende Werte) zeigte sich ein im
Vergleich zur Vorinjektionsperiode (pre-injection baseline) verminderter mittlerer
arterieller Blutdruck. Im Vergleich zu den jeweils zeitlich korrespondierenden
Placebodaten war der mittlere arterielle Blutdruck frühzeitig nach Injektion von
Ghrelin signifikant vermindert (t +7 min bis t +35 min P=0,014, ANOVA für
Gruppenfaktor Ghrelin/Placebo). Später näherte sich der mittlere arterielle Blutdruck
nach Ghrelinapplikation wieder dem Placebo-Niveau an (t +65 min und folgende
Werte).
3.2.2 Herzfrequenz
Die Herzfrequenz stieg unmittelbar nach Injektion von Ghrelin vorübergehend
an (t +2 min: P=0,018 im Vergleich zum Wert vor Injektion bei t – 25 min; P=0,117 im
Vergleich zur Placebobedingung; jeweils T-Test). Später (t +35 min und folgende
Werte) war die Herzfrequenz nach Ghrelininjektion im Vergleich mit den
Vorinjektionsdaten signifikant vermindert. Unter Placebobedingung hingegen kam es
während der späteren Messzeitpunkte zu einem Anstieg der Herzfrequenz. Der
Vergleich zwischen Ghrelin- und Placebobedingung in der späteren
Postinjektionsphase zeigte eine nach Ghrelin signifikant reduzierte Herzfrequenz

39
3 Ergebnisse
(t + 35 min bis t +85 min: P=0,043, ANOVA für den Faktor Ghrelin/Placebo, siehe
Abb. 3).
3.2.3 Muskuläre sympathische Nervenaktivität (MSNA)
Unmittelbar nach Injektion von Ghrelin kam es zu einem im Vergleich zum
Vorinjektionswert signifikanten Anstieg der MSNA (t +2 min versus t -25 min;
P=0,005, T-Test; siehe ergänzende Abb. IV). Nach Placeboinjektion blieb ein
entsprechender Effekt aus. Obwohl dieser Anstieg der MSNA für mehr als 15
Minuten persistierte, zeigte sich im statistischen Vergleich der Ghrelin- mit den
korrespondierenden Placebodaten lediglich ein Trend (t +2 min: P=0,088, T-Test). Zu
den späteren Messzeitpunkten (ab t + 35 min) nach Ghrelinapplikation kehrte die
MSNA auf das Niveau vor Injektion zurück und verharrte dort bis zum Ende des
Versuches (t +85 min). Im Gegensatz dazu beobachteten wir unter
Placebobedingung unmittelbar nach Injektion der Testsubstanz eine unveränderte
MSNA, während in der späteren Phase (t +85 min) ein Anstieg der MSNA mit
statistischer Signifikanz sowohl im Vergleich zur Ghrelinbedingung (28,4 versus 32,7
bursts pro Minute, P=0,047, T-Test) als auch zum Vorinjektionswert (P=0,016, T-
Test) zu beobachten war.
3.3 Sensitivität des arteriellen Baroreflex
Um differenzierte Effekte von Ghrelin auf den Baroreflex näher zu
charakterisieren, wurden wie oben beschrieben vasoaktive Substanzen verabreicht
(MOP-Zyklen) und der lineare Faktor (slope) einer linearen Modellfunktion berechnet.
Die den Baroreflex beschreibenden linearen Faktoren waren vor Injektion der
Testsubstanz (MOP 1, t -5 min) für beide Bedingungen (Ghrelin/Placebo) sowohl für
den MSNA-vermittelten vaskulären als auch für den herzfrequenzvermittelten
kardialen Baroreflex vergleichbar.
3.3.1 Vaskulärer Baroreflex
Unmittelbar nach Injektion der Testsubstanz (MOP 2, t +15 min) genügten nur
die Daten von N=5 Probanden dem geforderten Kriterium eines
Determinationskoeffizienten R2 grösser oder gleich 0,7. Ein signifikanter Unterschied

40
3 Ergebnisse
des linearen Faktors zwischen den Versuchsbedingungen Ghrelin und Placebo war
bei wesentlich zu geringer Teststärke der ANOVA nicht nachzuweisen (siehe Tab. 2).
Für die späteren Messzyklen (MOP 3, t +35 min und MOP 4, t +65 Minuten)
deuteten die Daten einen Unterschied des linearen Faktors zwischen den
Versuchsbedingungen Ghrelin und Placebo an, ohne dass die ANOVA bei geringer
Anzahl oben genanntem Kriterium genügender Datensätze signifikant wurde (N=6 für
MOP 3, N=7 für MOP 4). Die Daten der hämodynamischen Parameter rechtfertigen
die Annahme, dass die Messzyklen MOP 3 und 4 gemeinsamen physiologischen
Bedingungen hinsichtlich der Ghrelinwirkung unterliegen (verzögerte Phase der
Ghrelinwirkung oder delayed ghrelin phase, siehe unten). Daher führten wir die
Daten aus MOP 3 und 4 zusammen, indem wir einen gepoolten „delayed post-
injection“- Faktor einführten. Er ist definiert als arithmetisches Mittel der vorhandenen
gültigen linearen Faktoren aus MOP 3 und 4 jeweils einer Versuchsbedingung eines
individuellen Probanden.
Unter Verwendung dieses gepoolten Faktors für MOP 3 und 4 (t +35 min und
t +65 min) wird ein signifikanter Anstieg des Betrags der linearen Faktoren und somit
eine signifikante Zunahme der Sensitivität des vaskulären Baroreflex in der späten
Phase nach Ghrelininjektion sichtbar (siehe Abb. 4, Tab. 2; Mittelwert des linearen
Faktors -2,25 versus -1,66 nach Ghrelin bzw. Placebo; ANOVA: P=0,036 für die
Interaktion zwischen dem Gruppenfaktor Ghrelin/Placebo und dem
Messwiederholungsfaktor „Messzeitpunkt“; N=10; siehe ergänzende Abb. V).

41
3 Ergebnisse
Tab. 2 vascular baroreflex
A)
MOP 1 MOP 2
mean SEM mean SEM
slope of baroreflex ghrelin -1.79 0.29 -1.36 0.20
placebo -1.70 0.11 -1.62 0.40
B)
MOP 1 MOP 3/4
mean SEM mean SEM
slope of baroreflex ghrelin -1.78 0.22 -2.25 0.41
placebo -1.70 0.16 -1.66 0.27
Tab. 2 Linearer Faktor (slope) der Regressionsgeraden des vaskulären, MSNA-
vermittelten arteriellen Baroreflex. A) Lineare Faktoren von MOP 1 (t -5 min) vs. MOP
2 (t +15 min); N=5, n.s. B) Lineare Faktoren von MOP 1 vs. MOP 3/4 (gepoolter
Faktor von MOP 3 und 4, siehe Text); ANOVA: P=0,036 für die Interaktion zwischen
dem Gruppenfaktor Ghrelin/Placebo und dem Messwiederholungsfaktor
Messzeitpunkt; N=10. Angegeben sind Mittelwerte +/- SEM. i.v.-Applikation von
Ghrelin bzw. Placebo nach MOP 1 zum Zeitpunkt t +0 min.
3.3.2 Kardialer Baroreflex
Ähnlich der Situation beim vaskulären Baroreflex genügten unmittelbar nach
Injektion der Testsubstanz (MOP 2, t +15 min) nur die Daten von N=2 Probanden
dem geforderten Kriterium eines Determinationskoeffizienten R2 grösser oder gleich
0,7. Wiederum war ein signifikanter Unterschied der linearen Faktoren zwischen den
Versuchsbedingungen Ghrelin und Placebo bei wesentlich zu geringer Teststärke
der ANOVA nicht nachzuweisen (siehe Tab. 3).
Bezüglich der späteren Phase nach Injektion der Testsubstanz (MOP 3 und 4,
t +35 min und t +65 min) wurde wie oben für den vaskulären Baroreflex geschildert
ein gepoolter Faktor für MOP 3 und 4 eingeführt. Die statistische Analyse erbrachte
keinen signifikanten Unterschied zwischen der Ghrelin- und der Placebobedingung
(ANOVA: n.s.; N=8, siehe Tab. 3). Eine post hoc-Analyse zeigte jedoch, dass die
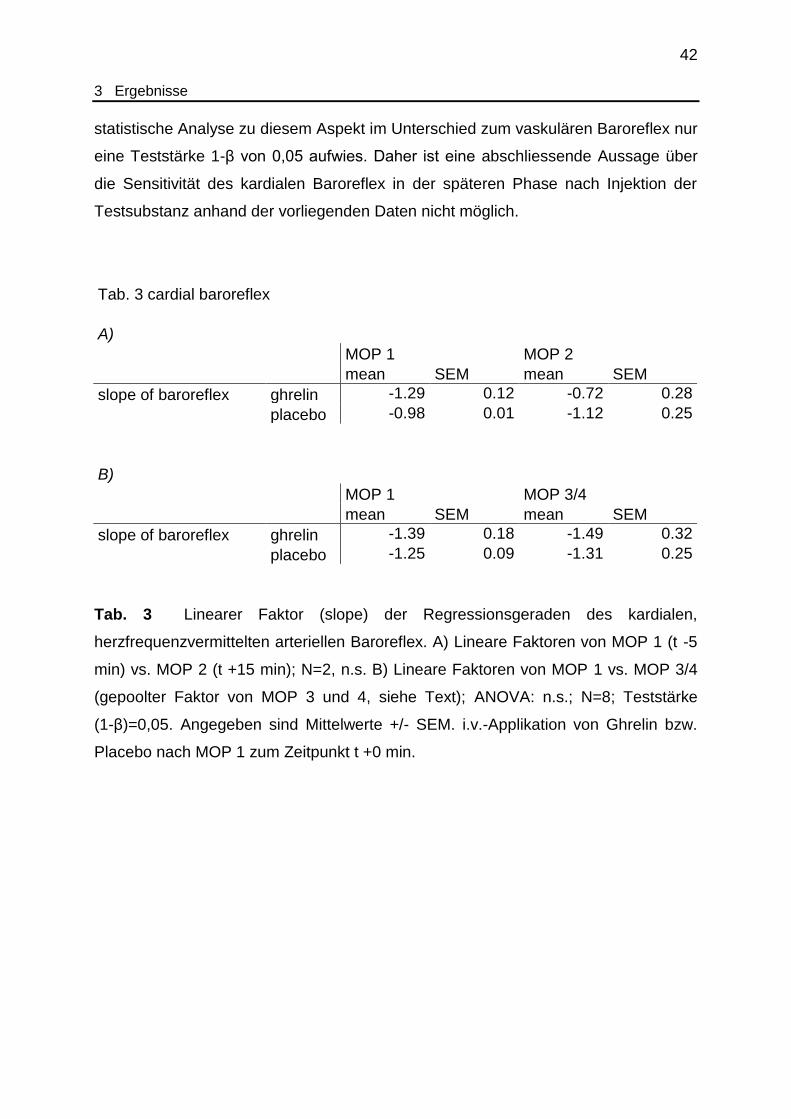
42
3 Ergebnisse
statistische Analyse zu diesem Aspekt im Unterschied zum vaskulären Baroreflex nur
eine Teststärke 1-β von 0,05 aufwies. Daher ist eine abschliessende Aussage über
die Sensitivität des kardialen Baroreflex in der späteren Phase nach Injektion der
Testsubstanz anhand der vorliegenden Daten nicht möglich.
Tab. 3 cardial baroreflex
A)
MOP 1
MOP 2
mean SEM mean SEM
slope of baroreflex ghrelin -1.29 0.12 -0.72 0.28
placebo -0.98 0.01 -1.12 0.25
B)
MOP 1
MOP 3/4
mean SEM mean SEM
slope of baroreflex ghrelin -1.39 0.18 -1.49 0.32
placebo -1.25 0.09 -1.31 0.25
Tab. 3 Linearer Faktor (slope) der Regressionsgeraden des kardialen,
herzfrequenzvermittelten arteriellen Baroreflex. A) Lineare Faktoren von MOP 1 (t -5
min) vs. MOP 2 (t +15 min); N=2, n.s. B) Lineare Faktoren von MOP 1 vs. MOP 3/4
(gepoolter Faktor von MOP 3 und 4, siehe Text); ANOVA: n.s.; N=8; Teststärke
(1-β)=0,05. Angegeben sind Mittelwerte +/- SEM. i.v.-Applikation von Ghrelin bzw.
Placebo nach MOP 1 zum Zeitpunkt t +0 min.
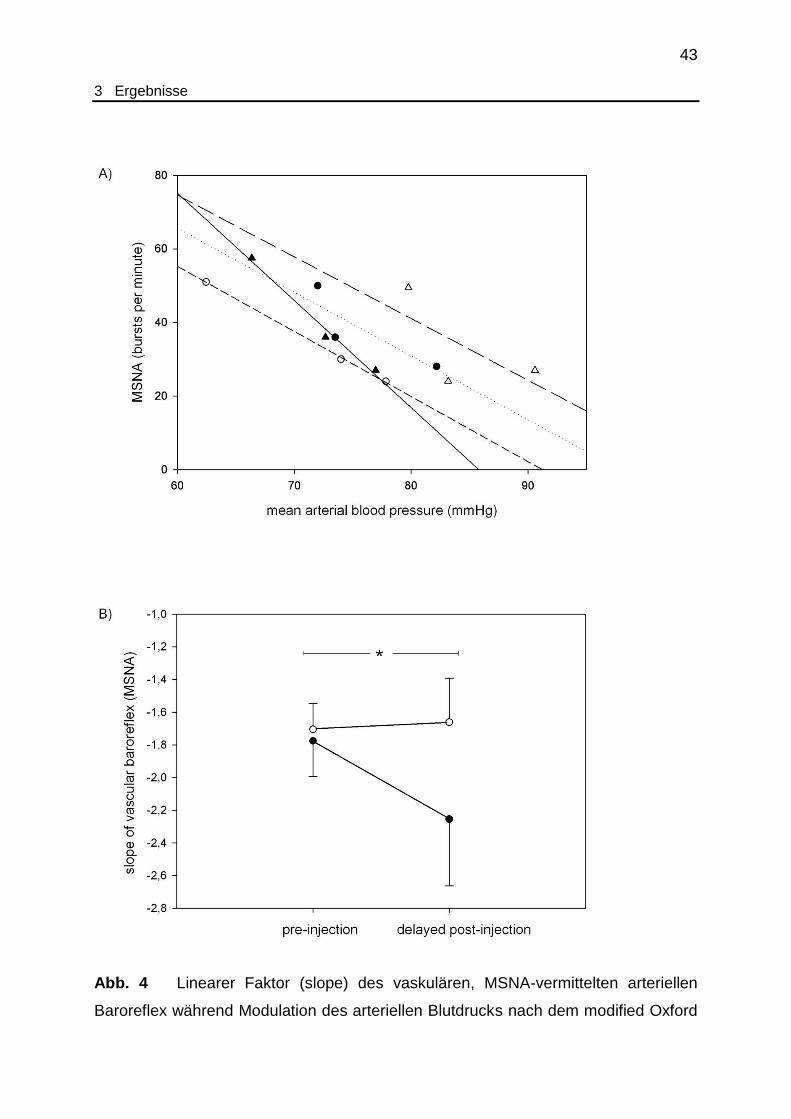
43
3 Ergebnisse
Abb. 4 Linearer Faktor (slope) des vaskulären, MSNA-vermittelten arteriellen
Baroreflex während Modulation des arteriellen Blutdrucks nach dem modified Oxford

44
3 Ergebnisse
protocol (MOP), jeweils vor (pre-injection) und in der Spätphase nach Injektion
(delayed post-injection) von Ghrelin/Placebo. Für jeden individuellen Messzyklus
wurde eine lineare Regression durchgeführt. Ein Messzyklus wurde akzeptiert, wenn
der Determinationskoeffizient R2 ≥ 0.7. Die Daten von MOP 3 und 4 wurden gepoolt
(delayed post-injection, verzögerte Phase der Ghrelinwirkung, siehe Text).
A) Beispielhafte Darstellung der Daten eines individuellen Probanden.
Schwarze/weisse Symbole: Ghrelin/Placebo-Bedingung. Kreise/Dreiecke: Vor/nach
Injektion von Ghrelin oder Placebo. Regressionsgeraden für Ghrelin vor/nach
Injektion (gepunktet/solid), Placebo vor/nach Injektion (kurz/lang gestrichelt).
B) Statistik für linearen Faktor (slope) des Baroreflex . Angegeben sind Mittelwerte
+/- SEM. Schwarze/weisse Kreise: Ghrelin/Placebo-Bedingung. * = P<0,05 für
Interaktion zwischen Gruppenfaktor Ghrelin/Placebo und Messwiederholungsfaktor
Messzeitpunkt (ANOVA; N=10). Der grössere Betrag des linearen Faktors nach
Ghrelininjektion zeigt eine gesteigerte Sensitivität des arteriellen Baroreflex an. (Aus
eigener Publikation).
3.4 Herzfrequenzvariabilität
Die Herzfrequenzvariabilität (heart rate variability, HRV) wurde für jeweils 5-
minütige Intervalle vor (t -25 min), unmittelbar (t +2 min) und später (t +85 min) nach
Injektion der Testsubstanz bestimmt. Alle Probanden gingen in die Analyse ein
(N=12). Die ANOVA wies in der post hoc-Analyse lediglich für den Parameter HF
eine hinreichende Teststärke (1-β) > 0,9 auf. Für alle übrigen Parameter war die
Teststärke der ANOVA zu gering (TP 0,81; LF/HF 0,61; HFnu und LFnu 0,42; LF
0,19).
Hinsichtlich der HF-Domäne, Mass für die Aktivität des efferenten kardialen
Vagusnerven, zeigte die ANOVA eine signifikante Interaktion zwischen dem
Gruppenfaktor Ghrelin/Placebo und dem Messwiederholungsfaktor Messzeitpunkt
(Tab. 4, Abb. 5). Nach Injektion von Ghrelin war der HF-Wert vermindert (Vergleich
Ghrelinbedingung t -25 min, t +2 min: T-Test P=0,092). Anschliessend stieg HF
wieder an (Vergleich Ghrelinbedingung t +2 min, t +85 min: T-Test P=0,118). Zu t
+85 min zeigte sich ein starker Trend zu einem höheren HF-Wert unter

45
3 Ergebnisse
Ghrelinbedingung (T-Test P=0,061), dessen Aussagekraft allerdings dadurch
eingeschränkt wurde, dass bereits vor Injektion der Testsubstanz (t -25 min) unter
Ghrelin ein höherer HF-Wert vorlag als unter Placebobedingung.
Für den Quotienten LF/HF, Mass für die sympathovagale Balance, erbrachte
die ANOVA einen schwachen Trend für die Interaktion zwischen dem Gruppenfaktor
Ghrelin/Placebo und dem Messwiederholungsfaktor Messzeitpunkt (P=0,115). LF/HF
zeigte im zeitlichen Verlauf unter Placebobedingung einen Anstieg, der unter
Ghrelinbedingung ausblieb (P=0,085 für T-Test, Vergleich t -25 min zu t +85 min
unter Placebobedingung, siehe Abbildung 6).
TP, die Summe der zeitlichen Domänen der Herzratenvariabilität, zeigte in der
ANOVA einen Trend für die Interaktion zwischen dem Gruppenfaktor Ghrelin/Placebo
und dem Messwiederholungsfaktor Messzeitpunkt (P=0,07). TP wies unter
Ghrelinbedingung im Vergleich zur Placebobedingung sowohl vor Injektion der
Testsubstanz (T-Test P=0,054) als auch danach (T-Test P=0,057) einen geringeren
Wert auf. Die Ursache dieser Differenz bleibt unklar, kann aber nicht der jeweiligen
Testsubstanz zugeschrieben werden.
Tab. 4 heart rate variability
ghrelin
placebo
time (min) mean SEM mean SEM
TP -25 11757.06 2785.83 7731.17 1393.04 a), b)
+2 10945.75 2697.93 9216.86 1383.07
+85 14604.81 3127.93 7994.76 625.60 c)
HF -25 3179.28 1204.24 e) 1738.30 365.39 d)
+2 2362.47 968.91 f) 1817.49 417.65
+85 3012.02 901.96 1265.78 168.84 g)
HFnu -25 0.41 0.05
0.43 0.05
+2 0.38 0.04
0.40 0.05
+85 0.42 0.03
0.34 0.05
LF -25 3716.20 1084.50 2301.94 386.67
+2 3237.00 827.05 2609.87 391.48
+85 4339.80 1303.68 2698.48 449.24
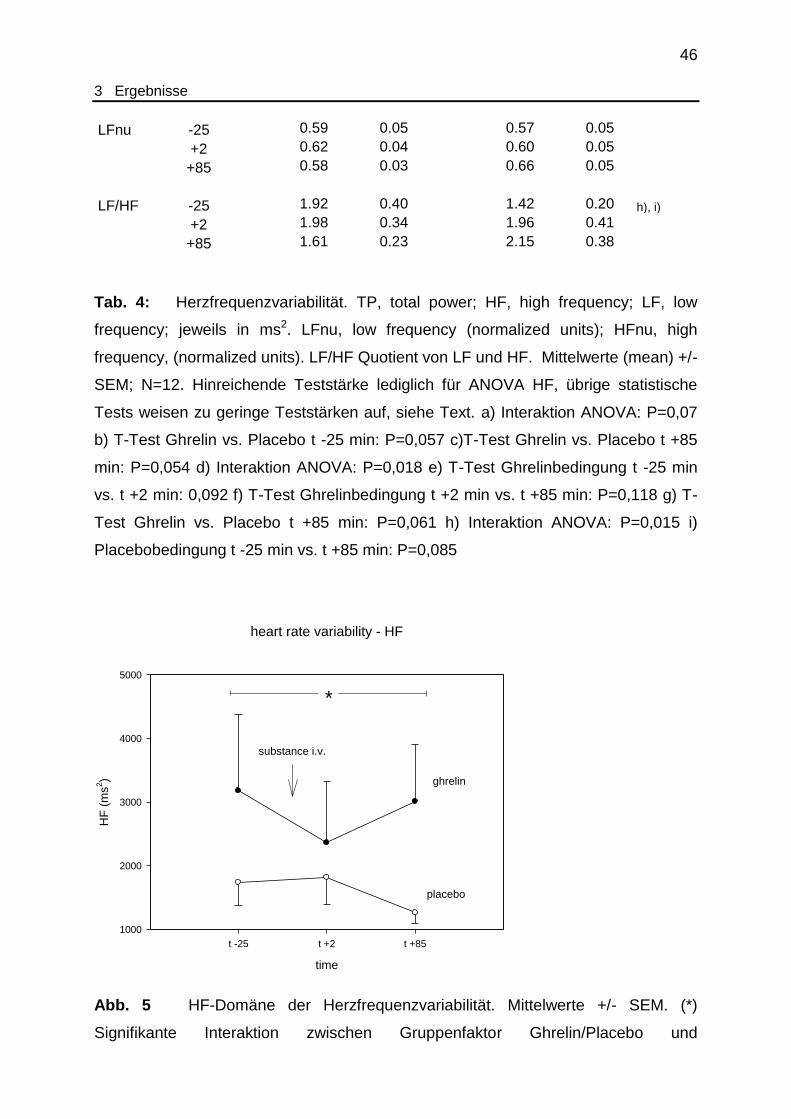
46
3 Ergebnisse
LFnu -25 0.59 0.05
0.57 0.05
+2 0.62 0.04
0.60 0.05
+85 0.58 0.03
0.66 0.05
LF/HF -25 1.92 0.40
1.42 0.20 h), i)
+2 1.98 0.34
1.96 0.41
+85 1.61 0.23
2.15 0.38
Tab. 4: Herzfrequenzvariabilität. TP, total power; HF, high frequency; LF, low
frequency; jeweils in ms2. LFnu, low frequency (normalized units); HFnu, high
frequency, (normalized units). LF/HF Quotient von LF und HF. Mittelwerte (mean) +/-
SEM; N=12. Hinreichende Teststärke lediglich für ANOVA HF, übrige statistische
Tests weisen zu geringe Teststärken auf, siehe Text. a) Interaktion ANOVA: P=0,07
b) T-Test Ghrelin vs. Placebo t -25 min: P=0,057 c)T-Test Ghrelin vs. Placebo t +85
min: P=0,054 d) Interaktion ANOVA: P=0,018 e) T-Test Ghrelinbedingung t -25 min
vs. t +2 min: 0,092 f) T-Test Ghrelinbedingung t +2 min vs. t +85 min: P=0,118 g) T-
Test Ghrelin vs. Placebo t +85 min: P=0,061 h) Interaktion ANOVA: P=0,015 i)
Placebobedingung t -25 min vs. t +85 min: P=0,085
*
heart rate variability - HF
time
t -25 t +2 t +85
HF
(m
s2)
1000
2000
3000
4000
5000
substance i.v.
ghrelin
placebo
Abb. 5 HF-Domäne der Herzfrequenzvariabilität. Mittelwerte +/- SEM. (*)
Signifikante Interaktion zwischen Gruppenfaktor Ghrelin/Placebo und
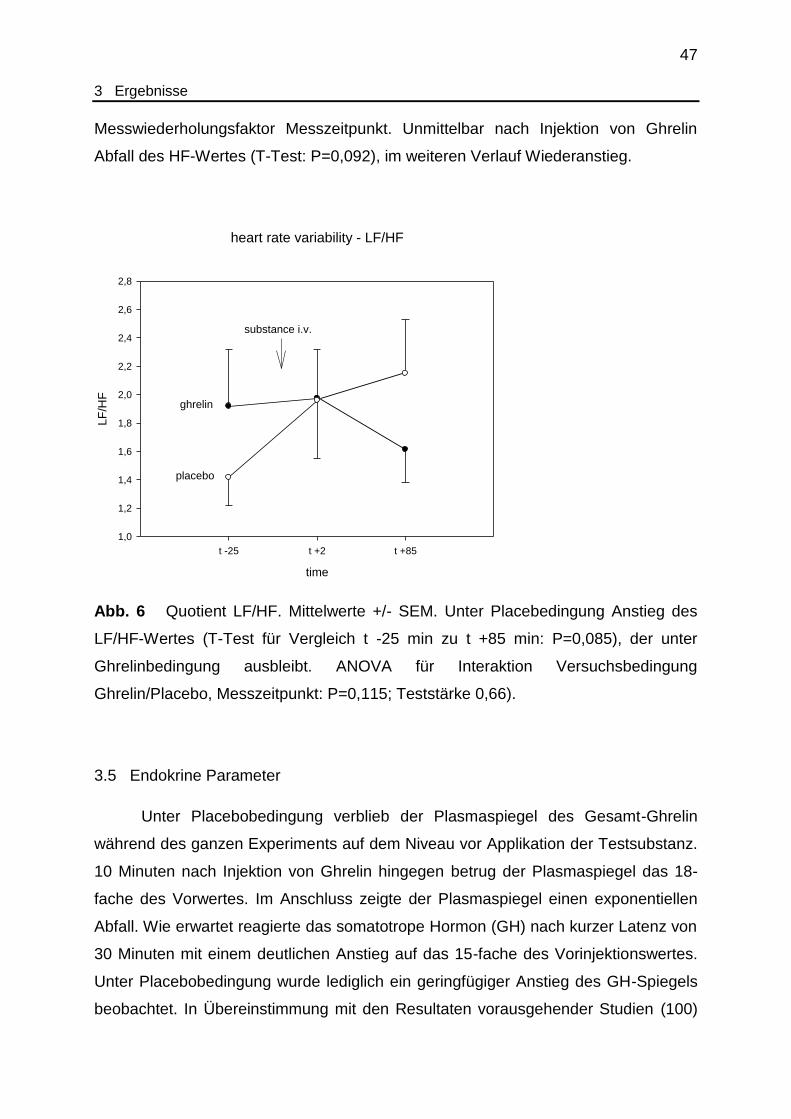
47
3 Ergebnisse
Messwiederholungsfaktor Messzeitpunkt. Unmittelbar nach Injektion von Ghrelin
Abfall des HF-Wertes (T-Test: P=0,092), im weiteren Verlauf Wiederanstieg.
heart rate variability - LF/HF
time
t -25 t +2 t +85
LF
/HF
1,0
1,2
1,4
1,6
1,8
2,0
2,2
2,4
2,6
2,8
substance i.v.
ghrelin
placebo
Abb. 6 Quotient LF/HF. Mittelwerte +/- SEM. Unter Placebedingung Anstieg des
LF/HF-Wertes (T-Test für Vergleich t -25 min zu t +85 min: P=0,085), der unter
Ghrelinbedingung ausbleibt. ANOVA für Interaktion Versuchsbedingung
Ghrelin/Placebo, Messzeitpunkt: P=0,115; Teststärke 0,66).
3.5 Endokrine Parameter
Unter Placebobedingung verblieb der Plasmaspiegel des Gesamt-Ghrelin
während des ganzen Experiments auf dem Niveau vor Applikation der Testsubstanz.
10 Minuten nach Injektion von Ghrelin hingegen betrug der Plasmaspiegel das 18-
fache des Vorwertes. Im Anschluss zeigte der Plasmaspiegel einen exponentiellen
Abfall. Wie erwartet reagierte das somatotrope Hormon (GH) nach kurzer Latenz von
30 Minuten mit einem deutlichen Anstieg auf das 15-fache des Vorinjektionswertes.
Unter Placebobedingung wurde lediglich ein geringfügiger Anstieg des GH-Spiegels
beobachtet. In Übereinstimmung mit den Resultaten vorausgehender Studien (100)
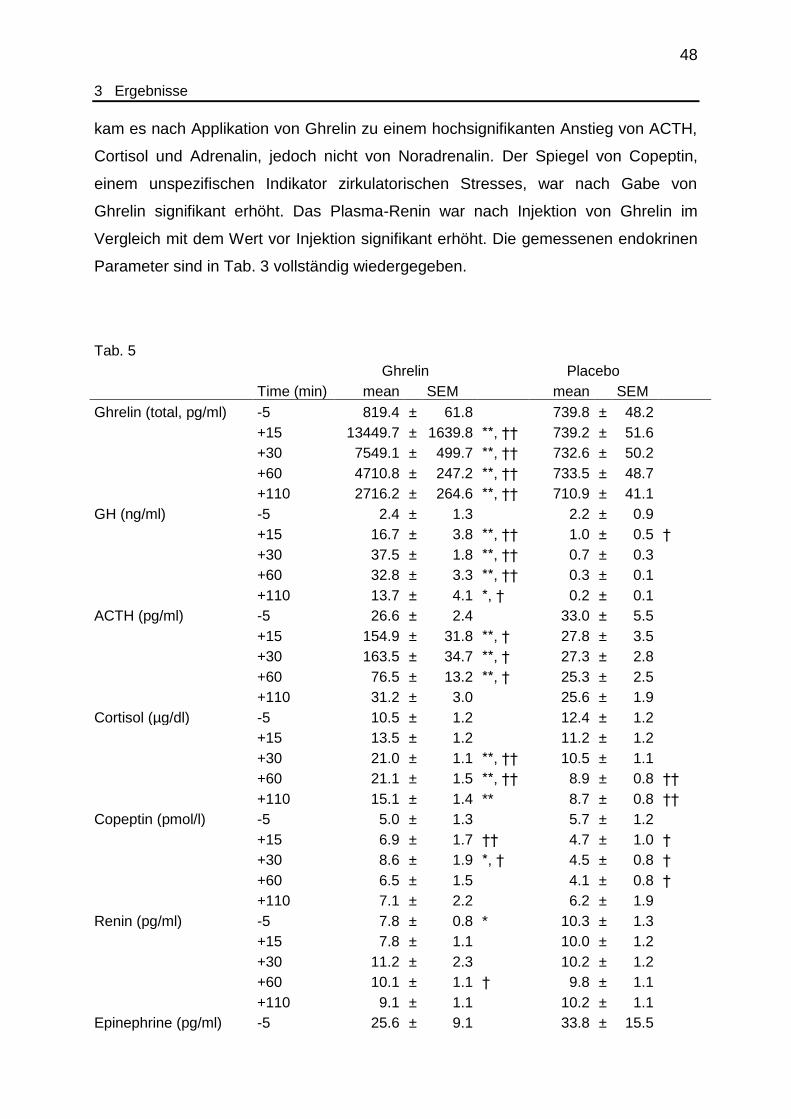
48
3 Ergebnisse
kam es nach Applikation von Ghrelin zu einem hochsignifikanten Anstieg von ACTH,
Cortisol und Adrenalin, jedoch nicht von Noradrenalin. Der Spiegel von Copeptin,
einem unspezifischen Indikator zirkulatorischen Stresses, war nach Gabe von
Ghrelin signifikant erhöht. Das Plasma-Renin war nach Injektion von Ghrelin im
Vergleich mit dem Wert vor Injektion signifikant erhöht. Die gemessenen endokrinen
Parameter sind in Tab. 3 vollständig wiedergegeben.
Tab. 5
Ghrelin Placebo
Time (min) mean SEM mean SEM
Ghrelin (total, pg/ml) -5 819.4 ± 61.8 739.8 ± 48.2
+15 13449.7 ± 1639.8 **, †† 739.2 ± 51.6
+30 7549.1 ± 499.7 **, †† 732.6 ± 50.2
+60 4710.8 ± 247.2 **, †† 733.5 ± 48.7
+110 2716.2 ± 264.6 **, †† 710.9 ± 41.1
GH (ng/ml) -5 2.4 ± 1.3 2.2 ± 0.9
+15 16.7 ± 3.8 **, †† 1.0 ± 0.5 †
+30 37.5 ± 1.8 **, †† 0.7 ± 0.3
+60 32.8 ± 3.3 **, †† 0.3 ± 0.1
+110 13.7 ± 4.1 *, † 0.2 ± 0.1
ACTH (pg/ml) -5 26.6 ± 2.4 33.0 ± 5.5
+15 154.9 ± 31.8 **, † 27.8 ± 3.5
+30 163.5 ± 34.7 **, † 27.3 ± 2.8
+60 76.5 ± 13.2 **, † 25.3 ± 2.5
+110 31.2 ± 3.0 25.6 ± 1.9
Cortisol (µg/dl) -5 10.5 ± 1.2 12.4 ± 1.2
+15 13.5 ± 1.2 11.2 ± 1.2
+30 21.0 ± 1.1 **, †† 10.5 ± 1.1
+60 21.1 ± 1.5 **, †† 8.9 ± 0.8 ††
+110 15.1 ± 1.4 ** 8.7 ± 0.8 ††
Copeptin (pmol/l) -5 5.0 ± 1.3 5.7 ± 1.2
+15 6.9 ± 1.7 †† 4.7 ± 1.0 †
+30 8.6 ± 1.9 *, † 4.5 ± 0.8 †
+60 6.5 ± 1.5 4.1 ± 0.8 †
+110 7.1 ± 2.2 6.2 ± 1.9
Renin (pg/ml) -5 7.8 ± 0.8 * 10.3 ± 1.3
+15 7.8 ± 1.1 10.0 ± 1.2
+30 11.2 ± 2.3 10.2 ± 1.2
+60 10.1 ± 1.1 † 9.8 ± 1.1
+110 9.1 ± 1.1 10.2 ± 1.1
Epinephrine (pg/ml) -5 25.6 ± 9.1 33.8 ± 15.5
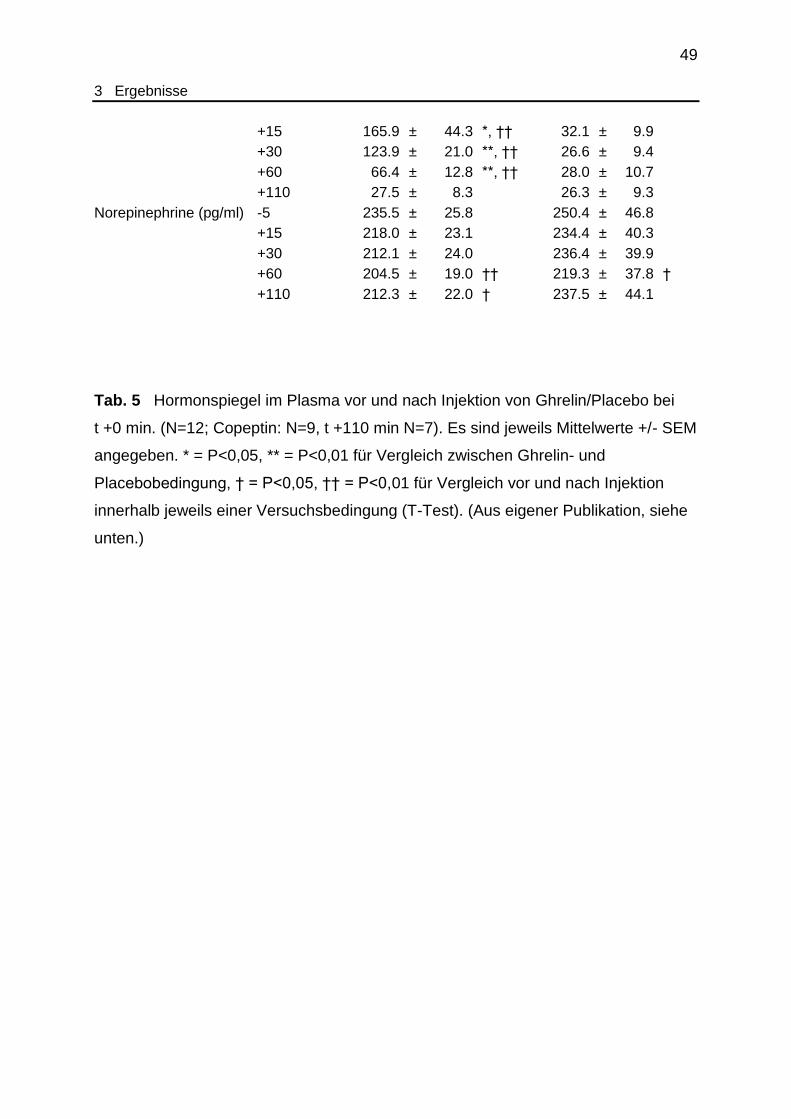
49
3 Ergebnisse
+15 165.9 ± 44.3 *, †† 32.1 ± 9.9
+30 123.9 ± 21.0 **, †† 26.6 ± 9.4
+60 66.4 ± 12.8 **, †† 28.0 ± 10.7
+110 27.5 ± 8.3 26.3 ± 9.3
Norepinephrine (pg/ml) -5 235.5 ± 25.8 250.4 ± 46.8
+15 218.0 ± 23.1 234.4 ± 40.3
+30 212.1 ± 24.0 236.4 ± 39.9
+60 204.5 ± 19.0 †† 219.3 ± 37.8 †
+110 212.3 ± 22.0 † 237.5 ± 44.1
Tab. 5 Hormonspiegel im Plasma vor und nach Injektion von Ghrelin/Placebo bei
t +0 min. (N=12; Copeptin: N=9, t +110 min N=7). Es sind jeweils Mittelwerte +/- SEM
angegeben. * = P<0,05, ** = P<0,01 für Vergleich zwischen Ghrelin- und
Placebobedingung, † = P<0,05, †† = P<0,01 für Vergleich vor und nach Injektion
innerhalb jeweils einer Versuchsbedingung (T-Test). (Aus eigener Publikation, siehe
unten.)

50
4 Diskussion
4. Diskussion
4.1 Biphasisches Modell der sympathischen und kardiovaskulären Ghrelinwirkung
Die Frage nach der Richtigkeit der dieser Studie zugrundeliegenden
Hypothesen erfordert eine differenzierte Beantwortung. Nach Injektion von Ghrelin
konnten im zeitlichen Verlauf zwei unterschiedliche Konstellationen der
hämodynamischen Parameter beobachtet werden. Wir interpretierten diese Daten im
Sinne zweier von jeweils unterschiedlichen physiologischen Mechanismen
dominierten Phasen der Wirkung von Ghrelin und definierten eine akute sowie eine
verzögerte Phase der Ghrelinwirkung (“immediate ghrelin phase” und “delayed
ghrelin phase”).
Kurze Zeit nach Injektion von Ghrelin wurde ein Abfall des arteriellen
Blutdrucks beobachtet. Zugleich zeigte sich ein Anstieg der muskulären
sympathischen Aktivität (MSNA) und ein vorübergehender Anstieg der Herzfrequenz
(akute Phase der Ghrelinwirkung, “immediate ghrelin phase”). 35 Minuten nach
Injektion war die MSNA auf ihr Ausgangsniveau zurückgekehrt und die Herzfrequenz
supprimiert, während der arterielle Blutdruck weiterhin vermindert war. Zum Ende
des Versuches befand sich der arterielle Blutdruck nach Ghrelingabe auf
Placeboniveau, während MSNA und Herzfrequenz im Vergleich mit der
Placebobedingung signifikant supprimiert waren (verzögerte Phase der
Ghrelinwirkung, “delayed ghrelin phase”).
4.2 Direkte Vasodilatation als dominierender Mechanismus der akuten Phase der
Ghrelinwirkung
Dem weithin akzeptierten kybernetischen Modell des arteriellen Baroreflex
zufolge (siehe oben) sollte ein verminderter arterieller Blutdruck eine gesteigerte
efferente sympathische Aktivität zum Herzen (CSNA) und den Gefäßen (MSNA) hin
bewirken, um den arteriellen Blutdruck wieder auf dem Niveau der Zielgröße
(Setpoint) einzustellen. Umgekehrt sollte ein arterieller Blutdruck oberhalb des
Setpoint von einer supprimierten MSNA und reduzierten Herzfrequenz beantwortet
werden, es sei denn, der Setpoint selbst ist verschoben worden (27; 62). Die
Konstellation der Parameter arterieller Blutdruck, MSNA und Herzfrequenz in der
akuten Phase der Ghrelinwirkung kann durch eine direkte Vasodilatation der

51
4 Diskussion
Widerstandsgefäße erklärt werden. Der resultierende Abfall des arteriellen Blutdrucks
bewirkt über den arteriellen Baroreflex einen Anstieg der MSNA und eine kurzfristige
Steigerung der Herzfrequenz. Es ist bekannt, dass Ghrelin direkt den systemischen
arteriellen Widerstand reduziert (101). Dieser Effekt könnte von einer durch Ghrelin
vermittelten Aktivierung der endothelialen NO-Synthetase hervorgerufen werden
(149). Auch ein NO-unabhängiger Mechanismus wird diskutiert, der auf einer
Endothelin-1-antagonistischen Wirkung von Ghrelin beruht (145). Interessant ist,
dass der reflektorische Anstieg der MSNA in Bezug auf den Abfall des arteriellen
Blutdrucks bereits in dieser Phase relativ gering ausfällt. In früheren Studien
resultierte ein NO-vermittelter Blutdruckabfall ähnlicher Größenordnung in einer
deutlich stärkeren Aktivierung des sympathischen Nervensystems (114; 132). Diese
Beobachtung könnte auf einen schnell eintretenden zentral sympatholytischen Effekt
bereits in der akuten Phase der Ghrelinwirkung hinweisen.
4.3 Zentrale Suppression der SNS-Aktivität in der verzögerten Phase der
Ghrelinwirkung
In der verzögerten Phase der Ghrelinwirkung konnte zunächst ein
verminderter arterieller Blutdruck beobachtet werden, der mit einer MSNA auf
Placeboniveau und einer reduzierten Herzfrequenz einherging (t +35 min). In der
Folge kehrte der arterielle Blutdruck auf Placeboniveau zurück, während MSNA und
Herzfrequenz nach Ghrelin im Vergleich zur Placebobedingung signifikant reduziert
waren (t +65 und +85 min).
4.3.1 Modulation des zentralen Setpoint der sympathischen Blutdruckregulation
Eine schlüssige Interpretation letztgenannter Konstellation besteht in der
Verschiebung des zentralen Setpoints der sympathischen Blutdruckregulation zu
einem niedrigeren Wert hin (eine schematische Darstellung zeigt die ergänzende
Abb. III). Im Unterschied zu den tatsächlich gemessenen Parametern müsste bei
unverändertem Setpoint ein erniedrigter arterieller Blutdruck vom Baroreflexbogen
stets mit einem Anstieg von MSNA und Herzfrequenz (CSNA) beantwortet werden (t
+ 35 min). Bei unverändertem Setpoint und arteriellem Blutdruck auf Placeboniveau
(t +85 min) sollte der Baroreflexbogen auch eine MSNA und Herzfrequenz auf
Placeboniveau bewirken, nicht aber eine signifikant reduzierte MSNA und

52
4 Diskussion
Herzfrequenz, wie es die Daten zeigen. Eine Reihe früherer Studien, die sich mit der
Interaktion zwischen Ghrelin und dem sympathischen Nervensystem befassen,
erbrachten Ergebnisse, die mit dem vorliegenden Interpretationsvorschlag gut
vereinbar sind. So bewirkte die Applikation von Ghrelin bei gesunden jungen
Probanden einen Abfall des arteriellen Blutdrucks ohne signifikannte Änderung der
Herzfrequenz, allerdings begleitet von einem erhöhten Herzindex (100). In einer
anderen Studie erhielten Patienten mit chronischer Herzinsuffizienz zweimal täglich
für drei Wochen eine intravenöse Ghrelingabe. Dies führte zu einer Reduktion des
Noradrenalin im Plasma, eines Markers der globalen Sympathikusaktivierung, bei
zugleich numerisch vermindertem arteriellen Blutdruck (102). Im Tierversuch an
wachen Kaninchen bewirkte die intracerebroventrikuläre Applikation von Ghrelin eine
Reduktion von arteriellem Blutdruck, Herzfrequenz und mikroneurographisch
gemessener renaler sympathischer Aktivität (RSNA). In letzterer Studie wurde eine
Zunahme der Sensitivität des arteriellen Baroreflex durch Ghrelin beobachtet (94).
Nach experimentell herbeigeführtem Myokardinfarkt an der Ratte bewirkte die
intravenöse Applikation von Ghrelin eine Reduktion der efferenten sympathischen
Aktivität zum Herzen (CSNA) (118; 119; 125) und vermag die Inzidenz maligner
Arrhythmien zu vermindern (91).
Die Konstellation zum Ende des Versuchsablaufs (t +85 min, arterieller
Blutdruck auf Placeboniveau und im Vergleich zu Placebobedingung signifikant
supprimierte MSNA) ist vereinbar mit der Annahme eines zu niedrigerem arteriellem
Blutdruck und verminderter MSNA hin verschobenen Setpoint des Baroreflex, sofern
zugleich ein vom Baroreflex unabhängiger den Blutdruck erhöhender Faktor vorliegt.
Dabei könnte es sich um auf Injektion von Ghrelin hin ausgeschüttete vasoaktive
Hormone handeln (siehe unten). Der arterielle Blutdruck oberhalb des neuen
Setpoint wird dann durch den Einfluss des vasopressorischen Hormons bewirkt, der
arterielle Baroreflex reagiert darauf mit einer Suppression der MSNA.
4.3.2 Reduktion Stress-induzierter sympathischer feed-forward-Signale
Eine alternative Interpretationsmöglichkeit der Kreislauf- und MSNA-Daten in
der verzögerten Phase der Ghrelinwirkung besteht darin, dass die Applikation von
Ghrelin stressinduzierte feed-forward-Signale der sympathischen Blutdruckregulation
abgeschwächt haben könnte. In der vorliegenden Studie zeigte sich unter

53
4 Diskussion
Placebobedingung im Zeitverlauf ein Anstieg der MSNA. Im Rahmen
mikroneurographischer MSNA-Aufzeichung längerer Dauer wird dieses Phänomen
häufiger beobachtet. Es wird üblicherweise als Auswirkung zunehmender
Harnblasenfüllung und körperlicher Missempfindungen infolge des langen Liegens in
unveränderter Körperposition aufgefasst (114). In der vorliegenden Studie äusserten
die Probanden allerdings unter Placebobedingung kein vermehrtes subjektives
Stressempfinden. Im Gegensatz zum MSNA-Anstieg nach Placebo zeigte sich zum
Ende des Experiments (t +85min) nach Ghrelingabe eine im Vergleich zu Placebo
signifikant reduzierte MSNA und Herzfrequenz bei unverändertem Blutdruck. Die
MSNA stellt einen direkten Messwert der zentralen efferenten sympathischen
Aktivität dar. Daher könnten die vorliegenden Daten darauf hindeuten, dass Ghrelin
eine stressinduzierte Aktivierung des sympathischen Nervensystems unterdrückt.
Diese Interpretation ist vereinbar mit den Ergebnissen eines kürzlich publizierten
Humanexperiments, in dem unter Ghrelingabe eine verminderte Aktivierung des
sympathischen Nervensystems auf mentalen Stress hin nachgewiesen werden
konnte (84).
4.4 Beeinflussung anderer Hormone und Transmitter
Mikroneurographische Ansätze zur Messung der Sensitivität des Baroreflex
nutzen ein vereinfachtes kybernetisches Modell der Regulation von arteriellem
Blutdruck und MSNA. Ghrelin könnte jedoch beide Parameter auch unabhängig von
einer direkten Beeinflussung des Baroreflex-Regelkreises beeinflussen. So wurde im
Rahmen dieser Arbeit in Übereinstimmung mit früheren Studien (100) nach
Ghrelingabe ein erheblicher transienter Anstieg mehrerer vasoaktiver Hormone wie
Adrenalin und Cortisol gemessen. Auch der Plasmaspiegel von Copeptin, einem
Marker der Vasopressin-Ausschüttung, zeigte im Vergleich zur Placebobedingung
nach Ghrelingabe einen signifikanten Anstieg. Diese hormonellen Effekte können
durch die Existenz von Ghrelin-Rezeptoren (GHS-R) in Hypothalamus, Hypophyse
und Nebennierenmark erklärt werden (54; 84). Im Gegensatz zum Plasmaspiegel
von Adrenalin war der von Noradrenalin nach Ghrelingabe nicht erhöht. Dieses
Muster der Katecholaminantwort zeigte sich auch schon in früheren Studien, in
denen zum Teil deutlich höhere Dosen von bis zu 10 µg/kg KG Ghrelin intravenös
verabreicht wurden (100). Nach wiederholter Injektion von Ghrelin über einen

54
4 Diskussion
Zeitraum von 3 Wochen in einer Dosis, die mit der in der vorliegenden Studie
verwendeten vergleichbar ist, kam es sogar zu einem Abfall des Noradrenalin-
Plasmaspiegels (99). Das an den Synapsen des sympathischen Nervensystems als
Transmitter sezernierte Noradrenalin gelangt nur in variablen und geringen Mengen
in die Zirkulation (“spill over”, siehe oben). Ferner entstammt das Noradrenalin im
Plasma sympathischen Ästen zu unterschiedlichen Endorganen. Das Ausbleiben
einer Erhöhung des Plasmaspiegels dieses groben Markes der sympathischen
Gesamtaktivität stimmt mit den MSNA-Daten überein und stützt somit die zuvor
abgeleitete Argumentation.
Ghrelin bewirkt in der Hypophyse eine Ausschüttung von GH, die mit der
durch GH-RH vermittelten Sekretion quantitativ vergleichbar ist (siehe oben). Bei
Krankheitsbildern, die durch einen Mangel an GH gekennzeichnet sind, kann eine
erhöhte Aktivität des sympathischen Nervensystems festgestellt werden (129). Unter
therapeutischer Substitution von GH kommt es zu einem Rückgang der MSNA (128).
Daher erscheint die Vermutung gerechtfertigt, dass GH eine Reduktion der Aktivität
des sympathischen Nervensystems bewirkt. Auch Glucocorticoide, deren
Plasmaspiegel nach Injektion von Ghrelin deutlich ansteigen, bewirken eine akute
Verminderung der MSNA (38). Daher könnte alternativ zu den oben geschilderten
Mechanismen auch der Anstieg der Plasmaspiegel von GH oder Cortisol die in der
vorliegenden Studie beobachtete Reduktion der sympathischen Aktivität nach
Injektion von Ghrelin bewirkt haben.
4.5 Gesteigerte Sensitivität des MSNA-vermittelten arteriellen Baroreflex nach
Ghrelininjektion
Die vorliegende Studie zeigte bei Testung des arteriellen Baroreflex während
der verzögerten Phase der Ghrelinwirkung (35 und 65 Minuten nach Injektion) eine
signifikante Zunahme der Steilheit der linearen Regressionsfunktion von MSNA und
arteriellem Blutdruck im Vergleich zu Placebo. Dies bedeutet, dass zirkulatorisches
Ghrelin die Sensitivität des vaskulären arteriellen Baroreflex steigert (eine
schematische Darstellung zeigt die ergänzende Abb. III). In Übereinstimmung damit
wurde im Tiermodell ein ähnlicher Befund für die renale sympathische Aktivität
(RSNA) erhoben (93). Diese Ergebnisse unterstützen die These, dass Ghrelin die
Blutdruckkontrolle auf zentralnervöser Ebene einschließlich hypothalamischer und

55
4 Diskussion
Hirnstammstrukturen moduliert(88). Da Ghrelin die Blut-Hirn-Schranke passieren
kann (8; 9) und eine intracerebroventrikuläre Applikation sehr geringer Mengen des
Peptids qualitativ entsprechend wirksam ist (94; 152), kann davon ausgegangen
werden, dass zirkulatorisches Ghrelin nach Übertritt in zerebrale Strukturen dort
direkt wirksam wird. Möglicherweise wird die zentrale Wirkung von zirkulatorischem
Ghrelin jedoch auch über vagale Afferenzen vermittelt (28; 29; 91). Die
Fragestellung, auf welchem Weg zirkulatorisches Ghrelin zentral wirksam wird,
erfordert weitere Untersuchungen. Auch eine weitere Differenzierung der Funktion
zerebral als Neurotransmitter sezernierten Ghrelins in Abgrenzung zur Wirkung des
hormonellen, in der Zirkulation befindlichen Ghrelins ist wünschenswert (50).
4.6 Gegenüberstellung mit der aktuellen Studie von Lambert et al. zum Thema
Ghrelin und SNS
In einer kürzlich erschienen Publikation beschrieben Lambert et al., dass die
kontinuierliche intravenöse Infusion von Ghrelin über eine Stunde einen signifikanten
Abfall des arteriellen Blutdrucks in Vergleich zum Zeitpunkt vor Infusion und zur
Placebobedingung bewirkte (84). Der Blutdruckabfall wurde von einem signifikanten
Anstieg der MSNA begleitet, was auf das Wirksamwerden einer durch den arteriellen
Baroreflex gesteuerten Gegenregulation des sympathischen Nervensystems
hinweist. Das Design der vorliegenden Studie unterscheidet sich substantiell von
dem von Lambert et al. gewählten, da im Gegensatz zu einer kontinuierlichen
Infusion des Neuropeptids die Applikation als Bolus erfolgte. Unter physiologischen
Bedingungen folgt die Sekretion von Ghrelin einem pulsatilen Muster (24). Das
Peptidhormon unterliegt einer Elimination mit sehr kurzer Halbwertszeit (siehe oben).
Daher erscheint das in dieser Studie implementierte Bolusdesign angemessen, um
die physiologischen Bedingungen abzubilden. Darüber hinaus wurden die
Experimente unter Ghrelin- und Placebobedingung in der vorliegenden Studie an
unterschiedlichen Tagen im Abstand von mindestens einer Woche durchgeführt. In
der Arbeit von Lambert et al. wurden Ghrelin- und Placebobedingung im Abstand von
nur einer Stunde untersucht. Dieses Vorgehen birgt die vermehrte Gefahr einer
Beeinflussung der Daten durch einen Carryover-Effekt. Die vordergründig
widersprüchlich erscheinenden Daten von Lambert et al. und dieser Studie lassen
eine einheitliche Interpretation zu. Vermutlich sind die in der Studie von Lambert et

56
4 Diskussion
al. unter kontinuierlicher Infusion von Ghrelin wirksamen physiologischen
Mechanismen vergleichbar mit denen, die in der akuten Phase der Ghrelinwirkung
nach Bolusapplikation wirksam sind. Lambert et al. beschreiben, dass unter
kontinuierlicher Infusion von Ghrelin ein signifikanter Anstieg der Herzfrequenz
ausbleibt. Ferner fanden die Autoren, dass unter Ghrelin die sympathische
Aktivierung auf mentalen Stress hin vermindert war. Beides deutet darauf hin, dass
auch in dieser Studie ein zentralnervöser, das sympathische Nervensystem
dämpfender Effekt von Ghrelin zu beobachten war, wie die vorliegende Arbeit ihn für
die verzögerte Phase der Ghrelinwirkung beschreibt.
4.7 Wirkung von zirkulatorischem Ghrelin auf die kardiale Sympathikus- und
Vagusaktivität
Neben der kardialen sympathischen Aktivität unterliegt das Herz efferenten
vagalen Einflüssen, die an der Frequenzregulation auch im Rahmen des arteriellen
Baroreflex beteiligt sind (62). Bei der Interpretation der Reaktion der Herzfrequenz
auf die Gabe der Testsubstanz sind daher sowohl sympathische als auch vagale
Einflüsse zu berücksichtigen.
4.7.1 Kardiale Vagusaktivität
Intravenös appliziertes Ghrelin stimuliert die efferente Aktivität des N. vagus
und vermehrt auf diesem Wege zum Beispiel die Pankreassekretion(87). Eine neuere
Studie belegt, dass zentral appliziertes Ghrelin im Tierexperiment die efferente
kardiale Vagusaktivität stimuliert (122). Im Rahmen der vorliegenden Arbeit konnte
nach Ghrelininjektion eine signifikante Reduktion der Herzfrequenz beobachtet
werden (t +35 min bis t +85 min), teilweise bei zugleich vermindertem arteriellem
Blutdruck (t + 35 min). Zuvor (t + 7 min, t + 15 min) zeigte sich unter
Ghrelinbedingung bereits eine Herzfrequenz auf Placeboniveau trotz signifikant
verminderten arteriellen Blutdrucks. Die Suppression der Herzfrequenz erscheint
quantitativ ausgeprägter als die der MSNA und geht ihr zeitlich voraus (Suppression
der Herzfrequenz ab t +7 min, der MSNA ab t +35 min). Diese ausgeprägte und frühe
Reduktion der Herzfrequenz lässt vermuten, dass über eine Suppression der CSNA
hinaus zusätzlich eine durch Ghrelin bewirkte Steigerung der efferenten kardialen
Vagusaktivität vorlag.

57
4 Diskussion
4.7.2 Analyse der Herzfrequenzvariabilität
Um die Wirkung von Ghrelin auf die Aktivität der efferenten sympathischen
und vagalen Innervation des Herzens zu differenzieren, erfolgte in der vorliegenden
Studie eine Analyse der Herzfrequenzvariabilität. Die mithilfe dieser Methode
gewonnenen Daten weisen ein hohes Mass an Streuung auf. Im Design dieser Arbeit
stand methodisch die sehr aufwendige mikroneurographische Messung der MSNA im
Vordergrund. Daher konnte nur eine begrenzte Zahl von Probanden untersucht
werden. Aus diesen Gründen wurde nur für die statistische Analyse der HF-Domäne
der Herzfrequenzvariabilität eine hinreichende Teststärke (power) erzielt. Hinsichtlich
aller anderen HRV-Parameter ergab die post hoc-Analyse eine zu geringe
Teststärke. Daher sind Aussagen über diese Parameter auf Grundlage des
vorhandenen Datenmaterials nur mit grossem Vorbehalt möglich.
Die HF-Domäne der Herzfrequenzvariabilität ist ein Mass für die efferente
Aktivität des kardialen N. vagus. Die diesbezügliche ANOVA erbrachte eine
signifikante Interaktion zwischen der Versuchsbedingung (Ghrelin oder Placebo) und
dem Faktor Zeitpunkt der Messung (Abb. 5). Unmittelbar nach Injektion von Ghrelin
(t +2 min) zeigte sich ein Abfall von HF (Trend), gefolgt von einem Wiederanstieg auf
Placeboniveau (schwacher Trend). Vermutlich bewirkt das Peptidhormon also in der
akuten Phase der Ghrelinwirkung (siehe oben) ausser einer vermehrten Aktivität des
sympathischen Nervensystems eine Suppression der kardialen vagalen Aktivität. Es
erscheint plausibel, dass dieser Frühwirkung wie zuvor für das sympathische
Nervensystem beschrieben eine direkte vasodilatatorische Wirkung von Ghrelin mit
kompensatorischer Aktivierung des arteriellen Baroreflex zugrundeliegt.
Der Quotient LF/HF ist ein Mass für die sympathovagale Balance der
vegetativen kardialen Innervation. Trotz zu geringer Teststärke zeigte die ANOVA
bezüglich LF/HF einen schwachen Trend für die Interaktion zwischen der
Versuchsbedingung (Ghrelin oder Placebo) und dem Faktor „Messzeitpunkt“. Unter
Placebobedingung wurde ein Trend zu einer Zunahme von LF/HF zum Ende des
Versuches hin beobachtet, der unter Ghrelinbedingung ausblieb. Dies deutet darauf
hin, dass sich die sympathovagale Balance unter Placebobedingung zunehmend in

58
4 Diskussion
Richtung einer vermehrten Aktivität der sympathischen kardialen Innervation und
einer verminderten vagalen kardialen Innervation verschob. Unter Ghrelinbedingung
hingegen schien diese Verschiebung auszubleiben. Dieser Befund stützt die zuvor
getroffene Aussage, dass Ghrelin im Vergleich zu Placebo in der verzögerten Phase
der Ghrelinwirkung die efferente Aktivität des sympathischen Nervensystems
supprimiert.
4.8 Pathophysiologische und klinische Implikationen
Die empirischen Daten der vorliegenden Studie stützen die Hypothese, dass
hohe zirkulatorische Spiegel des hauptsächlich von der Magenmukosa sezernierten
Peptidhormons Ghrelin beim Menschen sowohl die Aktivität des sympathischen
Nervensystems als auch den arteriellen Blutdruck senken und Sensitivität des
arteriellen Baroreflex steigern. Erniedrigte Ghrelin-Spiegel, wie sie bei adipösen
Menschen beschrieben werden (136), könnten daher zur bekannten vermehrten
Aktivität des sympathischen Nervensystems bei Adipositas (45), einem Kennzeichen
der Adipositas-assoziierten arteriellen Hypertonie, beitragen. Andererseits könnten
die hohen physiologischen Ghrelinspiegel im Tiefschlaf (slow wave sleep) an der
nächtlichen Absenkung des arteriellen Blutdrucks (dipping) und dem Resetting des
arteriellen Baroreflex während des Nachtschlafes beteiligt sein (112). Ferner
verbesserte die therapeutische Gabe von Ghrelin klinische Parameter bei Patienten
mit chronischer Herzinsuffizienz (102) und Kachexie infolge chronisch obstruktiver
Lungenerkrankung (99), Krankheitsbildern, die mit einer chronisch vermehrten
Aktivierung des sympathischen Nervensystems einhergehen. In den zitierten Studien
wurde Ghrelin für drei Wochen zwei mal täglich intravenös appliziert. Die Dosis von 2
µg/kg Körpergewicht entsprach der auch in dieser Studie gewählten. Der
therapeutische Nutzen von Ghrelin in diesen beiden Studien könnte zumindest
teilweise von einer auf zentralnervöser Ebene lokalisierten Inhibition des
sympathischen Nervensystems durch Ghrelin bewirkt worden sein.
4.9 Limitationen
Diese Studie zielte darauf ab, die akute Wirkung von Ghrelin auf die
Baroreflex-vermittelte sympathische Blutdruckregulation zu charakterisieren. Die
MSNA ist der physiologisch bedeutendste vasokonstriktive Stimulus. Ihre

59
4 Diskussion
Aufzeichung gestattet eine direkte Messung der efferenten sympathischen Aktivität
zu den Widerstandsgefäßen. Daher konnten im Rahmen dieser Arbeit sehr
spezifische Schlüsse auf das Verhalten dieses Aspekts des arteriellen Baroreflex
gezogen werden. Andere Aspekte des Regelkreises, insbesondere die Funktion des
kardialen Baroreflexes, konnten in dieser Studie hingegen nicht mit entsprechender
Spezifität charakterisiert werden. Aufgrund der mit hohem Aufwand verbundenen
Methodik konnte nur eine geringe Anzahl von Probanden untersucht werden.
Dennoch erlaubte das randomisierte, placebokontrollierte Crossover-Design valide
Aussagen bezüglich der formulierten Fragestellungen. Die statistischen Tests wiesen
eine hinreichende Teststärke (Power) auf, um Veränderungen des vaskulären
(MSNA-vermittelten) arteriellen Baroreflex nachzuweisen. Die statistische Analyse
bezüglich Unterschieden in der kardialen Baroreflexschleife gestattete aufgrund einer
zu geringen Teststärke keine entsprechende Aussage. Der in dieser Studie
verwendete Baroreflex-Test stellt einen vereinfachten Ansatz dar. Technisch
komplexere dynamische Methoden zur Charakterisierung des Baroreflex könnten
noch detailliertere Informationen erbringen (68; 77), die die Grundaussage dieser
Arbeit jedoch vermutlich nicht verändern werden.
4.10 Perspektive
Diese Arbeit schlägt ein zweiphasiges Modell des Effekts von Ghrelin auf die
sympathische Kreislaufregulation vor. In einer akuten Phase der Ghrelinwirkung
bewirkt das Peptidhormon eine direkte periphere Vasodilatation mit konsekutivem
Abfall des arteriellen Blutdrucks. Dies wird über eine Aktivierung des arteriellen
Baroreflex mit einer Steigerung der efferenten Aktivität des sympathischen
Nervensystems beantwortet. Später folgt eine neurohumorale verzögerte Phase der
Ghrelinwirkung, die durch eine im Vergleich zur Placebobedingung verminderte
vasokonstriktorische sympathischen Aktivität (MSNA) und eine gesteigerte
Sensitivität des vaskulären arteriellen Baroreflex charakterisiert ist. Dieses
zweiphasige Modell gewährleistet eine einheitliche Interpretation sich scheinbar
widersprechender Daten vorhergehender Studien zu diesem Thema. Um ein
besseres Verständnis der Interaktion zwischen Ghrelin und arteriellem Baroreflex
über längere Zeiträume zu erlangen, ist die Durchführung einer Studie
wünschenswert, innerhalb derer die in dieser Arbeit untersuchten Parameter vor und

60
4 Diskussion
nach chronischer Applikation von Ghrelin gemessen werden. Es ist zu vermuten,
dass die längerfristige Gabe von Ghrelin eine noch ausgeprägtere Inhibition der
muskulären sympathischen Nervenaktivität bewirkt als für den in der vorliegenden
Arbeit kurzen Zeitraum gezeigt wurde. Ferner ist es von höchstem Interesse, die
potentiell positiven Effekte von Ghrelin auf die MSNA bei Krankheiten mit permanent
erhöhter Aktivität des sympathischen Nervensystems wie chronischer
Herzinsuffizienz, Leberzirrhose, morbider Adipositas oder obstruktivem
Schlafapnoesyndrom experimentell zu überprüfen.

61
5 Zusammenfassung
5. Zusammenfassung
Ghrelin ist ein Peptidhormon, das überwiegend von Zellen der Magenmukosa
sezerniert wird. Es stimuliert in der Hypophyse die Ausschüttung von
Wachstumshormon und hat orexigene, d.h. die Nahrungsaufnahme stimulierende
Eigenschaften. Darüber hinaus sind in jüngerer Zeit bedeutende pleiotrope Effekte
auf das kardiovaskuläre System bekannt geworden. Eine zunehmende Anzahl von
empirischen Studien deutet darauf hin, dass diese Effekte zu einem erheblichen
Anteil vom sympathischen Nervensystem vermittelt werden. Die vorliegende Arbeit
zielt darauf ab, die akute Wirkung von Ghrelin auf die efferente Aktivität des
sympathischen Nervensystems zu den Blutgefäßen der Skelettmuskulatur (muscle
sympathetic nerve activity, MSNA) und die Baroreflex-vermittelte Regulation des
arteriellen Blutdrucks bei gesunden menschlichen Probanden aufzuklären. Im
Rahmen eines randomisierten, doppelblinden Crossover-Designs erhielten 12
normalgewichtige junge Männer eine einzelne Dosis von 2 µg/kg Ghrelin oder
Placebo. MSNA, Herzfrequenz und arterieller Blutdruck wurden kontinuierlich von 30
Minuten vor bis 90 Minuten nach Verabreichung der Testsubstanz aufgezeichnet. Die
Sensitivität des arteriellen Baroreflex wurde wiederholt mithilfe der Injektion
vasoaktiver Substanzen nach dem modified Oxford protocol (MOP) gemessen.
Innerhalb der ersten 30 Minuten nach Injektion wurde ein signifikanter Abfall des
arteriellen Blutdrucks beobachtet, welcher von einer vorübergehenden
reflektorischen Zunahme von MSNA und Herzfrequenz begleitet wurde. Im weiteren
Verlauf des Experiments (>30 bis 85 Minuten) näherte sich der arterielle Blutdruck
wieder dem Placeboniveau an, während MSNA und Herzfrequenz im Vergleich zur
Placebobedingung signifikant vermindert waren. Die Sensitivität des vaskulären,
MSNA-vermittelten arteriellen Baroreflex war 35-65 Minuten nach Injektion von
Ghrelin im Vergleich zu Placebo signifikant erhöht. Eine Interpretation dieser Daten
besteht in der Definition zweier unterschiedlicher zeitlicher Phasen der Wirkung von
Ghrelin, in denen unterschiedliche physiologische Mechanismen dominieren. In der
akuten Phase der Ghrelinwirkung (immediate ghrelin phase) ist der arterielle
Blutdruck vermutlich aufgrund direkter vasodilatierender Eigenschaften des
Peptidhormons vermindert. Daraufhin bewirkt der arterielle Baroreflexbogen einen
gegenregulatorischen Anstieg von MSNA und Herzfrequenz. In der verzögerten
Phase der Ghrelinwirkung ist die Aktivität des sympathischen Nervensystems

62
5 Zusammenfassung
vermindert und die Sensitivität des arteriellen Baroreflex erhöht. Beides wird
vermutlich über einen zentralnervösen Wirkmechanismus des Peptidhormons
vermittelt.

63 6 Literaturverzeichnis
6. Literaturverzeichnis
Reference List
1. Abt K. Descriptive data analysis: a concept between confirmatory and
exploratory data analysis. Methods Inf Med 26: 77-88, 1987.
2. Akamizu T, Takaya K, Irako T, Hosoda H, Teramukai S, Matsuyama A,
Tada H, Miura K, Shimizu A, Fukushima M, Yokode M, Tanaka K and
Kangawa K. Pharmacokinetics, safety, and endocrine and appetite effects of
ghrelin administration in young healthy subjects. Eur J Endocrinol 150: 447-
455, 2004.
3. Akman MS, Girard M, O'Brien LF, Ho AK and Chik CL. Mechanisms of
action of a second generation growth hormone-releasing peptide (Ala-His-D-
beta Nal-Ala-Trp-D-Phe-Lys-NH2) in rat anterior pituitary cells. Endocrinology
132: 1286-1291, 1993.
4. Akselrod S, Gordon D, Madwed JB, Snidman NC, Shannon DC and
Cohen RJ. Hemodynamic regulation: investigation by spectral analysis. Am J
Physiol 249: H867-H875, 1985.
5. Ariyasu H, Takaya K, Tagami T, Ogawa Y, Hosoda K, Akamizu T, Suda M,
Koh T, Natsui K, Toyooka S, Shirakami G, Usui T, Shimatsu A, Doi K,
Hosoda H, Kojima M, Kangawa K and Nakao K. Stomach is a major source
of circulating ghrelin, and feeding state determines plasma ghrelin-like

64 6 Literaturverzeichnis
immunoreactivity levels in humans. J Clin Endocrinol Metab 86: 4753-4758,
2001.
6. Arvat E, Maccario M, Di VL, Broglio F, Benso A, Gottero C, Papotti M,
Muccioli G, Dieguez C, Casanueva FF, Deghenghi R, Camanni F and
Ghigo E. Endocrine activities of ghrelin, a natural growth hormone
secretagogue (GHS), in humans: comparison and interactions with hexarelin,
a nonnatural peptidyl GHS, and GH-releasing hormone. J Clin Endocrinol
Metab 86: 1169-1174, 2001.
7. Baldanzi G, Filigheddu N, Cutrupi S, Catapano F, Bonissoni S, Fubini A,
Malan D, Baj G, Granata R, Broglio F, Papotti M, Surico N, Bussolino F,
Isgaard J, Deghenghi R, Sinigaglia F, Prat M, Muccioli G, Ghigo E and
Graziani A. Ghrelin and des-acyl ghrelin inhibit cell death in cardiomyocytes
and endothelial cells through ERK1/2 and PI 3-kinase/AKT. J Cell Biol 159:
1029-1037, 2002.
8. Banks WA, Burney BO and Robinson SM. Effects of triglycerides, obesity,
and starvation on ghrelin transport across the blood-brain barrier. Peptides 29:
2061-2065, 2008.
9. Banks WA, Tschop M, Robinson SM and Heiman ML. Extent and direction
of ghrelin transport across the blood-brain barrier is determined by its unique
primary structure. J Pharmacol Exp Ther 302: 822-827, 2002.

65 6 Literaturverzeichnis
10. Barrett CJ, Ramchandra R, Guild SJ, Lala A, Budgett DM and Malpas SC.
What sets the long-term level of renal sympathetic nerve activity: a role for
angiotensin II and baroreflexes? Circ Research 92: 1330-1336, 2003.
11. Baulmann J, Nürnberger J, Slany J, Schmieder R, Schmidt-Trucksäss A,
Baumgart D, Cremerius P, Hess O, Mortensen K and Weber T. Arterial
stiffness and pulse wave analysis. Dtsch Med Wochenschr 135: S4-S14,
2010.
12. Bellone S, Rapa A, Vivenza D, Castellino N, Petri A, Bellone J, Me E,
Broglio F, Prodam F, Ghigo E and Bona G. Circulating ghrelin levels as
function of gender, pubertal status and adiposity in childhood. J Endocrinol
Invest 25: RC13-RC15, 2002.
13. Benedict C, Dodt C, Hallschmid M, Lepiorz M, Fehm HL, Born J and Kern
W. Immediate but not long-term intranasal administration of insulin raises
blood pressure in human beings. Metabolism 54: 1356-1361, 2005.
14. Bharucha AE, Charkoudian N, Andrews CN, Camilleri M, Sletten D,
Zinsmeister AR and Low PA. Effects of glucagon-like peptide-1, yohimbine,
and nitrergic modulation on sympathetic and parasympathetic activity in
humans. Am J Physiol (Regul Integr Comp Physiol) 295: R874-R880, 2008.
15. Blatnik M and Soderstrom CI. A practical guide for the stabilization of
acylghrelin in human blood collections. Clin Endocrinol (Oxf) 74: 325-331,
2011.

66 6 Literaturverzeichnis
16. Broglio F, Arvat E, Benso A, Gottero C, Muccioli G, Papotti M, van der
Lely AJ, Deghenghi R and Ghigo E. Ghrelin, a natural GH secretagogue
produced by the stomach, induces hyperglycemia and reduces insulin
secretion in humans. J Clin Endocrinol Metab 86: 5083-5086, 2001.
17. Burke D, Sundlöf G and Wallin BG. Postural effects on muscle nerve
sympathetic activity in man. J Physiol 272: 399-414, 1977.
18. Carlini VP, Ghersi M, Schioth HB and de Barioglio SR. Ghrelin and
memory: differential effects on acquisition and retrieval. Peptides 31: 1190-
1193, 2010.
19. Charkoudian N, Joyner MJ, Sokolnicki LA, Johnson CP, Eisenach JH,
Dietz NM, Curry T and Wallin BG. Vascular adrenergic responsiveness is
inversely related to tonic activity of sympathetic vasoconstrictor nerves in
humans. J Physiol 2006.
20. Choi K, Roh SG, Hong YH, Shrestha YB, Hishikawa D, Chen C, Kojima M,
Kangawa K and Sasaki S. The role of ghrelin and growth hormone
secretagogues receptor on rat adipogenesis. Endocrinology 144: 754-759,
2003.
21. Cohen MA and Taylor JA. Short-term cardiovascular oscillations in man:
measuring and modelling the physiologies. J Physiol 542: 669-683, 2002.

67 6 Literaturverzeichnis
22. Cowley AW, Liard JF and Guyton AC. Role of baroreceptor reflex in daily
control of arterial blood pressure and other variables in dogs. Circ Research
32: 564-576, 1973.
23. Cowley MA, Smith RG, Diano S, Tschop M, Pronchuk N, Grove KL,
Strasburger CJ, Bidlingmaier M, Esterman M, Heiman ML, Garcia-Segura
LM, Nillni EA, Mendez P, Low MJ, Sotonyi P, Friedman JM, Liu H, Pinto S,
Colmers WF, Cone RD and Horvath TL. The distribution and mechanism of
action of ghrelin in the CNS demonstrates a novel hypothalamic circuit
regulating energy homeostasis. Neuron 37: 649-661, 2003.
24. Cummings DE, Purnell JQ, Frayo RS, Schmidova K, Wisse BE and
Weigle DS. A preprandial rise in plasma ghrelin levels suggests a role in meal
initiation in humans. Diabetes 50: 1714-1719, 2001.
25. Cummings DE, Weigle DS, Frayo RS, Breen PA, Ma MK, Dellinger EP and
Purnell JQ. Plasma ghrelin levels after diet-induced weight loss or gastric
bypass surgery. N Engl J Med 346: 1623-1630, 2002.
26. Cuspidi C, Sala C, Valerio C, Negri F and Mancia G. Nocturnal blood
pressure in untreated essential hypertensives. Blood Press 20: 335-341, 2011.
27. Dampney RA, Coleman MJ, Fontes MA, Hirooka Y, Horiuchi J, Li YW,
Polson JW, Potts PD and Tagawa T. Central mechanisms underlying short-
and long-term regulation of the cardiovascular system. Clin Exp Pharmacol
Physiol 29: 261-268, 2002.

68 6 Literaturverzeichnis
28. Date Y. Ghrelin and the vagus nerve. Methods Enzymol 514: 261-269, 2012.
29. Date Y, Murakami N, Toshinai K, Matsukura S, Niijima A, Matsuo H,
Kangawa K and Nakazato M. The role of the gastric afferent vagal nerve in
ghrelin-induced feeding and growth hormone secretion in rats.
Gastroenterology 123: 1120-1128, 2002.
30. Date Y, Nakazato M, Hashiguchi S, Dezaki K, Mondal MS, Hosoda H,
Kojima M, Kangawa K, Arima T, Matsuo H, Yada T and Matsukura S.
Ghrelin is present in pancreatic alpha-cells of humans and rats and stimulates
insulin secretion. Diabetes 51: 124-129, 2002.
31. Date Y, Nakazato M, Murakami N, Kojima M, Kangawa K and Matsukura
S. Ghrelin acts in the central nervous system to stimulate gastric acid
secretion. Biochem Biophys Res Commun 280: 904-907, 2001.
32. Delius W, Hagbarth K-E, Hongell A and Wallin BG. General characteristics
of sympathetic activity in human muscle nerves. Acta Physiol Scand 84: 65-
81, 1972.
33. Delius W, Hagbarth K-E, Hongell A and Wallin BG. Manoeuvres affecting
sympathetic outflow in human skin nerves. Acta Physiol Scand 84: 177-186,
1972.

69 6 Literaturverzeichnis
34. Dendorfer A, Raasch W, Tempel K and Dominiak P. Interactions between
the renin-angiotensin system (RAS) and the sympathetic system. Basic Res
Cardiol 93: 24-29, 1998.
35. DiBona GF. Neural control of the kidney: past, present, and future.
Hypertension 41: 621-624, 2003.
36. DiCarlo SE and Bishop VS. Central baroreflex resetting as a means of
increasing and decreasing sympathetic outflow and arterial pressure. Ann NY
Acad Sci 940: 337, 2001.
37. Dixit VD, Schaffer EM, Pyle RS, Collins GD, Sakthivel SK, Palaniappan R,
Lillard JW, Jr. and Taub DD. Ghrelin inhibits leptin- and activation-induced
proinflammatory cytokine expression by human monocytes and T cells. J Clin
Invest 114: 57-66, 2004.
38. Dodt C, Keyser B, Molle M, Fehm HL and Elam M. Acute suppression of
muscle sympathetic nerve activity by hydrocortisone in humans. Hypertension
35: 758-763, 2000.
39. Dodt C, Wallin BG, Fehm HL and Elam M. The stress hormone
adrenocorticotropin enhances sympathetic outflow to the muscle vascular bed
in humans. J Hypertens 16: 195-201, 1998.

70 6 Literaturverzeichnis
40. Dutoit AP, Hart EC, Charkoudian N, Wallin BG, Curry TB and Joyner MJ.
Cardiac baroreflex sensitivity is not correlated to sympathetic baroreflex
sensitivity within healthy, young humans. Hypertension 56: 1118-1123, 2010.
41. Ebert TJ and Cowley AW, Jr. Baroreflex modulation of sympathetic outflow
during physiological increases of vasopressin in humans. Am J Physiol 262:
H1372-H1378, 1992.
42. Eckberg DL. Physiological basis for human autonomic rhythms. Ann Med 32:
341-349, 2000.
43. Esler M. The sympathetic nervous system and catecholamine release and
plasma clearance in
normal blood pressure control, in aging, and in hypertension. In: Hypertension:
Pathopysiology, Diagnosis, and Management, edited by Laragh JH and
Brenner BM. New York: Raven Press, Ltd., 1995.
44. Esler MD, Jennings G, Lambert E, Metz CN, Hornyak M and Eisenhofer G.
Overflow of catecholamine neurotransmitters to the circulation: source, fate,
and functions. Physiol Rev 70: 963-985, 1990.
45. Esler MD, Straznicky N, Eikelis N, Masuo K, Lambert G and Lambert E.
Mechanisms of sympathetic activation in obesity-related hypertension.
Hypertension 48: 787-796, 2006.

71 6 Literaturverzeichnis
46. Fadel PJ and Raven PB. Human investigations into the arterial and
cardiopulmonary baroreflexes during exercise. Exp Physiol 97: 39-50, 2012.
47. Fagius J. Sympathetic nerve activity in metabolic control--some basic
concepts. Acta Physiol Scand 177: 337-343, 2003.
48. Fagius J, Sundlof G and Wallin BG. Variation of sympathetic reflex latency
in man. J Auton Nerv Syst 21: 157-165, 1987.
49. Fagius J and Wallin BG. Long-term variability and reproducibility of resting
human muscle nerve sympathetic activity at rest, as reassessed after a
decade. Clin Auton Res 3: 201-205, 1993.
50. Ferrini F, Salio C, Lossi L and Merighi A. Ghrelin in central neurons. Curr
Neuropharmacol 7: 37-49, 2009.
51. Fletcher EC. Sympathetic over activity in the etiology of hypertension of
obstructive sleep apnea. SLEEP 26: 15-19, 2003.
52. Friedman O and Logan AG. Sympathoadrenal mechanisms in the
pathogenesis of sleep apnea-related hypertension. Curr Hypertens Rep 11:
212-216, 2009.
53. Fung MM, Peters K, Redline S, Ziegler MG, Ancoli-Israel S, Barrett-
Connor E, Stone KL and Osteoporotic Fractures in Men Research Group.

72 6 Literaturverzeichnis
Decreased slow wave sleep increases risk of developing hypertension in
elderly men. Hypertension 58: 596-603, 2011.
54. Ghigo E, Arvat E, Broglio F, Giordano R, Gianotti L, Muccioli G, Papotti
M, Graziani A, Bisi G, Deghenghi R and Camanni F. Endocrine and non-
endocrine activities of growth hormone secretagogues in humans. Horm Res
51 Suppl 3: 9-15, 1999.
55. Gnanapavan S, Kola B, Bustin SA, Morris DG, McGee P, Fairclough P,
Bhattacharya S, Carpenter R, Grossman AB and Korbonits M. The tissue
distribution of the mRNA of ghrelin and subtypes of its receptor, GHS-R, in
humans. J Clin Endocrinol Metab 87: 2988, 2002.
56. Grassi G. Assessment of Sympathetic Cardiovascular Drive in Human
Hypertension: Achievements and Perspectives. Hypertension 54: 690-697,
2009.
57. Grassi G and Esler MD. How to assess sympathetic activity in humans. J
Hypertens 17: 734, 1999.
58. Grassi G, Seravalle G and Quarti-Trevano F. The 'neuroadrenergic
hypothesis' in hypertension: current evidence. Exp Physiol 95: 581-586, 2009.
59. Greenman Y, Golani N, Gilad S, Yaron M, Limor R and Stern N. Ghrelin
secretion is modulated in a nutrient- and gender-specific manner. Clin
Endocrinol (Oxf) 60: 382-388, 2004.

73 6 Literaturverzeichnis
60. Guan XM, Yu H, Palyha OC, McKee KK, Feighner SD, Sirinathsinghji DJ,
Smith RG, Van der Ploeg LH and Howard AD. Distribution of mRNA
encoding the growth hormone secretagogue receptor in brain and peripheral
tissues. Brain Res Mol Brain Res 48: 23-29, 1997.
61. Gutierrez JA, Solenberg PJ, Perkins DR, Willency JA, Knierman MD, Jin
Z, Witcher DR, Luo S, Onyia JE and Hale JE. Ghrelin octanoylation
mediated by an orphan lipid transferase. Proc Natl Acad Sci U S A 105: 6320-
6325, 2008.
62. Guyenet PG. The sympathetic control of blood pressure. Nat Rev Neurosci 7:
335-346, 2006.
63. Guyton AC. Blood pressure control--special role of the kidneys and body
fluids. Science 252: 1813-1816, 1991.
64. Hagbarth K-E and Vallbo AB. Pulse and respiratory grouping of sympathetic
impulses in human muscle-nerves. Acta Physiol Scand 74: 96-108, 1968.
65. Halliwill, J. R., Morgan, B. J., and Charkoudian, N. Peripheral chemoreflex and
baroreflex interactions in cardiovascular regulation in humans. J Physiol 552,
295-302. 2003.
Ref Type: Journal (Full)

74 6 Literaturverzeichnis
66. Hansen TK, Dall R, Hosoda H, Kojima M, Kangawa K, Christiansen JS
and Jorgensen JO. Weight loss increases circulating levels of ghrelin in
human obesity. Clin Endocrinol (Oxf) 56: 203-206, 2002.
67. Hansen TW, Li Y, Boggia J, Thijs L, Richart T and Staessen JA. Predictive
role of the nighttime blood pressure. Hypertension 57: 3-10, 2011.
68. Hart EC, Joyner MJ, Wallin BG, Karlsson T, Curry TB and Charkoudian N.
Baroreflex control of muscle sympathetic nerve activity: a nonpharmacological
measure of baroreflex sensitivity. Am J Physiol (Heart Circ Physiol ) 298:
H816-H822, 2010.
69. Heindl S, Holzschneider J, Hinz A, Sayk F, Fehm HL and Dodt C. Acute
Effects of Aldosterone on the Autonomic nervous Sytem and the Baroreflex
Function in Healthy Humans. J Neuroendocrin 18: 115-121, 2006.
70. Higashi Y, Nakagawa K, Kimura M, Noma K, Hara K, Sasaki S, Goto C,
Oshima T, Chayama K and Yoshizumi M. Circadian variation of blood
pressure and endothelial function in patients with essential hypertension:a
comparison of dippers and non-dippers. J Am Coll Cardiol 40: 2039-2043,
2002.
71. Howard AD, Feighner SD, Cully DF, Arena JP, Liberator PA, Rosenblum
CI, Hamelin M, Hreniuk DL, Palyha OC, Anderson J, Paress PS, Diaz C,
Chou M, Liu KK, McKee KK, Pong SS, Chaung LY, Elbrecht A,
Dashkevicz M, Heavens R, Rigby M, Sirinathsinghji DJ, Dean DC, Melillo

75 6 Literaturverzeichnis
DG, Patchett AA, Nargund R, Griffin PR, DeMartino JA, Gupta SK,
Schaeffer JM, Smith RG and Van der Ploeg LH. A receptor in pituitary and
hypothalamus that functions in growth hormone release. Science 273: 974-
977, 1996.
72. Huda MS, Dovey TM, Wong SP, English PJ, Halford JC, McCulloch P,
Cleator J, Martin B, Cashen J, Hayden K, Ghatei MA, Bloom SR, Wilding
JP and Pinkney JH. Ghrelin does not orchestrate the metabolic changes
seen in fasting but has significant effects on lipid mobilisation and substrate
utilisation. Eur J Endocrinol 165: 45-55, 2011.
73. Hunt BE and Farquhar WB. Nonlinearities and asymmetries of the human
cardiovagal baroreflex. Am J Physiol (Regul Integr Comp Physiol) 288: R1339-
R1346, 2005.
74. Ichinose M, Saito M, Kondo N and Nishiyasu T. Baroreflex and muscle
metaboreflex: control of muscle sympathetic nerve activity. Med Sci Sports
Exerc 40: 2037-2045, 2008.
75. Joyner MJ, Charkoudian N and Wallin BG. A sympathetic view of the
sympathetic nervous system and human blood pressure regulation. Exp
Physiol 93: 715-724, 2008.
76. Joyner MJ, Charkoudian N and Wallin BG. Sympathetic nervous system
and blood pressure in humans: individualized patterns of regulation and their
implications. Hypertension 56: 10-16, 2012.

76 6 Literaturverzeichnis
77. Kienbaum P, Karlsson T, Sverrisdottir YB, Elam M and Wallin BG. Two
sites for modulation of human sympathetic activity by arterial baroreceptors? J
Physiol 531: 861-869, 2001.
78. Kitajima T, Kanbayashi T, Saitoh Y, Ogawa Y, Sugiyama T, Kaneko Y,
Sasaki Y, Aizawa R and Shimisu T. The effects of oral melatonin on the
autonomic function in healthy subjects. Psychiatry and Clinical Neurosciences
55: 299-300, 2001.
79. Kojima M, Hosoda H, Date Y, Nakazato M, Matsuo H and Kangawa K.
Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature
402: 656-660, 1999.
80. Kojima M and Kangawa K. Ghrelin: structure and function. Physiol Rev 85:
495-522, 2005.
81. Kojima M and Kangawa K. Ghrelin: more than endogenous growth hormone
secretagogue. Ann N Y Acad Sci 1200: 140-148, 2010.
82. Korbonits M, Kojima M, Kangawa K and Grossman AB. Presence of
ghrelin in normal and adenomatous human pituitary. Endocrine 14: 101-104,
2001.
83. Krum H, Schlaich M, Whitbourn R, Sobotka PA, Sadowski J, Bartus K,
Kapelak B, Walton A, Sievert H, Thambar S, Abraham WT and Esler M.
Catheter-based renal sympathetic denervation for resistant hypertension: a

77 6 Literaturverzeichnis
multicentre safety and proof-of-principle cohort study. Lancet 373: 1275-1281,
2009.
84. Lambert E, Lambert G, Ika-Sari C, Dawood T, Lee K, Chopra R,
Straznicky N, Eikelis N, Drew S, Tilbrook A, Dixon J, Esler M and
Schlaich MP. Ghrelin Modulates Sympathetic Nervous System Activity and
Stress Response in Lean and Overweight Men. Hypertension 2011.
85. Leproult R and Van Cauter E. Role of sleep and sleep loss in hormonal
release and metabolism. Endocr Dev 17: 21, 2010.
86. Levy MN. Sympathetic-parasympathetic interactions in the heart. Circ
Research 29: 437-445, 1971.
87. Li Y, Wu X, Zhao Y, Chen S and Owyang C. Ghrelin acts on the dorsal vagal
complex to stimulate pancreatic protein secretion. Am J Physiol Gastrointest
Liver Physiol 290: G1350-G1358, 2006.
88. Lin Y, Matsumura K, Fukuhara M, Kagiyama S, Fujii K and Iida M. Ghrelin
acts at the nucleus of the solitary tract to decrease arterial pressure in rats.
Hypertension 43: 977-982, 2004.
89. Lu S, Guan JL, Wang QP, Uehara K, Yamada S, Goto N, Date Y, Nakazato
M, Kojima M, Kangawa K and Shioda S. Immunocytochemical observation
of ghrelin-containing neurons in the rat arcuate nucleus. Neurosci Lett 321:
157-160, 2002.

78 6 Literaturverzeichnis
90. Mancia G, Dell'Oro R, Quarti-Trevano F, Scopelliti F and Grassi G.
Angiotensin - sympathetic system interactions in cardiovascular and metabolic
disease. J Hypertens 24: S51-S56, 2006.
91. Mao Y, Tokudome T, Otani K, Kishimoto I, Nakanishi M, Hosoda H,
Miyazato M and Kangawa K. Ghrelin prevents incidence of malignant
arrhythmia after acute myocardial infarction through vagal afferent nerves.
Endocrinology 153: 3426-3434, 2012.
92. Masuda Y, Tanaka T, Inomata N, Ohnuma N, Tanaka S, Itoh Z, Hosoda H,
Kojima M and Kangawa K. Ghrelin stimulates gastric acid secretion and
motility in rats. Biochem Biophys Res Commun 276: 905-908, 2000.
93. Matsumura K, Tsuchihashi T and Abe I. Central orexin-A augments
sympathoadrenal outflow in conscious rabbits. Hypertension 37: 1382-1387,
2001.
94. Matsumura K, Tsuchihashi T, Fujii K, Abe I and Iida M. Central ghrelin
modulates sympathetic activity in conscious rabbits. Hypertension 40: 694-
699, 2002.
95. McCowen KC, Maykel JA, Bistrian BR and Ling PR. Circulating ghrelin
concentrations are lowered by intravenous glucose or hyperinsulinemic
euglycemic conditions in rodents. J Endocrinol 175: R7-11, 2002.

79 6 Literaturverzeichnis
96. McKee KK, Palyha OC, Feighner SD, Hreniuk DL, Tan CP, Phillips MS,
Smith RG, Van der Ploeg LH and Howard AD. Molecular analysis of rat
pituitary and hypothalamic growth hormone secretagogue receptors. Mol
Endocrinol 11: 415-423, 1997.
97. Montano N, Ruscone TG, Porta A, Lombardi F, Pagani M and Malliani A.
Power spectrum analysis of heart rate variability to assess the changes in
sympathovagal balance during graded orthostatic tilt. Circulation 90: 1826-
1831, 1994.
98. Mundinger TO, Cummings DE and Taborsky GJ, Jr. Direct stimulation of
ghrelin secretion by sympathetic nerves. Endocrinology 147: 2893-2901, 2006.
99. Nagaya N, Itoh T, Murakami S, Oya H, Uematsu M, Miyatake K and
Kangawa K. Treatment of cachexia with ghrelin in patients with COPD. Chest
128: 1187-1193, 2005.
100. Nagaya N, Kojima M, Uematsu M, Yamagishi M, Hosoda H, Oya H,
Hayashi Y and Kangawa K. Hemodynamic and hormonal effects of human
ghrelin in healthy volunteers. Am J Physiol Regul Integr Comp Physiol 280:
R1483-R1487, 2001.
101. Nagaya N, Miyatake K, Uematsu M, Oya H, Shimizu W, Hosoda H, Kojima
M, Nakanishi N, Mori H and Kangawa K. Hemodynamic, renal, and
hormonal effects of ghrelin infusion in patients with chronic heart failure. J Clin
Endocrinol Metab 86: 5854-5859, 2001.

80 6 Literaturverzeichnis
102. Nagaya N, Moriya J, Yasumura Y, Uematsu M, Ono F, Shimizu W, Ueno K,
Kitakaze M, Miyatake K and Kangawa K. Effects of ghrelin administration on
left ventricular function, exercise capacity, and muscle wasting in patients with
chronic heart failure. Circulation 110: 3674-3679, 2004.
103. Nakazato M, Murakami N, Date Y, Kojima M, Matsuo H, Kangawa K and
Matsukura S. A role for ghrelin in the central regulation of feeding. Nature
409: 194-198, 2001.
104. Narkiewicz K, Phillips BG, Kato M, Hering D, Bieniaszewski L and
Somers VK. Gender-selective interaction between aging, blood pressure, and
sympathetic nerve activity. Hypertension 45: 522-525, 2005.
105. Ogoh S, Fisher JP, Raven PB and Fadel PJ. Arterial baroreflex control of
muscle sympathetic nerve activity in the transition from rest to steady-state
dynamic exercise in humans. Am J Physiol (Heart Circ Physiol ) 293: H2202-
H2209, 2007.
106. Osborn JW, Jacob F and Gunzman P. A neural set point for the long-term
control of arterial pressure: beyond the arterial baroreceptor reflex. Am J
Physiol (Regul Integr Comp Physiol) 288: R846-R855, 2005.
107. Pagani M and Malliani A. Interpreting oscillations of muscle sympathetic
nerve activity and heart rate variability. J Hypertens 18: 1709-1719, 2000.

81 6 Literaturverzeichnis
108. Pettersson I, Muccioli G, Granata R, Deghenghi R, Ghigo E, Ohlsson C
and Isgaard J. Natural (ghrelin) and synthetic (hexarelin) GH secretagogues
stimulate H9c2 cardiomyocyte cell proliferation. J Endocrinol 175: 201-209,
2002.
109. Rosicka M, Krsek M, Matoulek M, Jarkovska Z, Marek J, Justova V and
Lacinova Z. Serum ghrelin levels in obese patients: the relationship to serum
leptin levels and soluble leptin receptors levels. Physiol Res 52: 61-66, 2003.
110. Rudas L, Crossman AA, Morillo CA, Halliwill JR, Tahvanainen KU,
Kuusela TA and Eckberg DL. Human sympathetic and vagal baroreflex
responses to sequential nitroprusside and phenylephrine. Am J Physiol 276:
H1691-H1698, 1999.
111. Satou M, Nishi Y, Yoh J, Hattori Y and Sugimoto H. Identification and
characterization of acyl-protein thioesterase 1/lysophospholipase I as a ghrelin
deacylation/lysophospholipid hydrolyzing enzyme in fetal bovine serum and
conditioned medium. Endocrinology 151: 4765-4775, 2010.
112. Sayk F, Becker C, Teckentrup C, Fehm HL, Struck J, Wellhoener JP and
Dodt C. To dip or not to dip: on the physiology of blood pressure decrease
during nocturnal sleep in healthy humans. Hypertension 49: 1070-1076, 2007.
113. Sayk F, Teckentrup C, Becker C, Heutling D, Wellhoner P, Lehnert H and
Dodt C. Effects of selective slow-wave sleep deprivation on nocturnal blood

82 6 Literaturverzeichnis
pressure dipping and daytime blood pressure regulation. Am J Physiol Regul
Integr Comp Physiol 298: R191-R197, 2010.
114. Sayk F, Vietheer A, Schaaf B, Wellhoener P, Weitz G, Lehnert H and Dodt
C. Endotoxemia causes central downregulation of sympathetic vasomotor tone
in healthy humans. Am J Physiol Regul Integr Comp Physiol 295: R891-R898,
2008.
115. Scheuer DA and Bishop VS. Effect of vasopressin on baroreflex control of
lumbar sympathetic nerve activity and hindquarter resistance. Am J Physiol
270: H1963-H1971, 1996.
116. Schlaich MP, Krum H and Sobotka PA. Renal sympathetic nerve ablation:
the new frontier in the treatment of hypertension. Curr Hypertens Rep 12: 39-
46, 2010.
117. Schlaich MP, Sobotka PA, Krum H, Lambert E and Esler MD. Renal
sympathetic-nerve ablation for uncontrolled hypertension. N Engl J Med 361:
932-934, 2009.
118. Schwenke DO, Tokudome T, Kishimoto I, Horio T, Cragg PA, Shirai M
and Kangawa K. One dose of ghrelin prevents the acute and sustained
increase in cardiac sympathetic tone after myocardial infarction. Endocrinology
153: 2436-2443, 2012.

83 6 Literaturverzeichnis
119. Schwenke DO, Tokudome T, Kishimoto I, Horio T, Shirai M, Cragg PA
and Kangawa K. Early ghrelin treatment after myocardial infarction prevents
an increase in cardiac sympathetic tone and reduces mortality. Endocrinology
149: 5172-5176, 2008.
120. Sharma V and McNeill JH. The emerging roles of leptin and ghrelin in
cardiovascular physiology and pathophysiology. Curr Vasc Pharmacol 3: 169-
180, 2005.
121. Shiiya T, Nakazato M, Mizuta M, Date Y, Mondal MS, Tanaka M, Nozoe S,
Hosoda H, Kangawa K and Matsukura S. Plasma ghrelin levels in lean and
obese humans and the effect of glucose on ghrelin secretion. J Clin Endocrinol
Metab 87: 240-244, 2002.
122. Shimizu S, Akiyama T, Kawada T, Sonobe T, Kamiya A, Shishido T,
Tokudome T, Hosoda H, Shirai M, Kangawa K and Sugimachi M. Centrally
administered ghrelin activates cardiac vagal nerve in anesthetized rabbits.
Auton Neurosci 162: 60-65, 2011.
123. Shintani M, Ogawa Y, Ebihara K, Aizawa-Abe M, Miyanaga F, Takaya K,
Hayahi T, Inoue G, Hosoda K, Kojima M, Kangawa K and Nakao K.
Ghrelin, an endogenous growth hormone secretagogue, is a novel orexigenic
peptide that antagonizes leptin action through the activation of hypothalamic
neuropeptide Y/Y1 receptor pathway. Diabetes 50: 227-232, 2001.

84 6 Literaturverzeichnis
124. Smith RG, Cheng K, Schoen WR, Pong SS, Hickey G, Jacks T, Butler B,
Chan WW, Chaung LY, Judith F and . A nonpeptidyl growth hormone
secretagogue. Science 260: 1640-1643, 1993.
125. Soeki T, Kishimoto I, Schwenke DO, Tokudome T, Horio T, Yoshida M,
Hosoda H and Kangawa K. Ghrelin suppresses cardiac sympathetic activity
and prevents early left ventricular remodeling in rats with myocardial infarction.
Am J Physiol Heart Circ Physiol 294: H426-H432, 2008.
126. Sundlöf G and Wallin BG. The variability of muscle nerve sympathetic
activity in resting recumbent man. J Physiol 272: 383-397, 1977.
127. Sundlöf G and Wallin BG. Human muscle nerve sympathetic activity at rest.
Relationship to blood pressure and age. J Physiol 274: 621-637, 1978.
128. Sverrisdottir YB, Elam M, Caidahl K, Soderling AS, Herlitz H and
Johannsson G. The effect of growth hormone (GH) replacement therapy on
sympathetic nerve hyperactivity in hypopituitary adults: a double-blind,
placebo-controlled, crossover, short-term trial followed by long-term open GH
replacement in hypopituitary adults. J Hypertens 21: 1905-1914, 2003.
129. Sverrisdottir YB, Elam M, Herlitz H, Bengtsson BA and Johannsson G.
Intense sympathetic nerve activity in adults with hypopituitarism and untreated
growth hormone deficiency. J Clin Endocrinol Metab 83: 1881-1885, 1998.

85 6 Literaturverzeichnis
130. Takaya K, Ariyasu H, Kanamoto N, Iwakura H, Yoshimoto A, Harada M,
Mori K, Komatsu Y, Usui T, Shimatsu A, Ogawa Y, Hosoda K, Akamizu T,
Kojima M, Kangawa K and Nakao K. Ghrelin strongly stimulates growth
hormone release in humans. J Clin Endocrinol Metab 85: 4908-4911, 2000.
131. Takayama S, Nakajima Y, Toma S and Sakamoto T. Increased muscle
sympathetic nerve activity after glucagon administration in man. J Auton Nerv
Syst 54: 171-175, 1995.
132. Tank J, Diederich A, Szchech E, Luft FC and Jordan J. Baroreflex
Regulation of Heart Rate and Sympathetic Vasomotor Tone in Women and
Men. Hypertension 45: 1159-1164, 2005.
133. Task Force of The European Society of Cardiology. Heart rate variability.
Standards of measurement, physiological interpretation, and clinical use. Task
Force of the European Society of Cardiology and the North American Society
of Pacing and Electrophysiology. Eur Heart J 17: 354-381, 1996.
134. Tschop M, Smiley DL and Heiman ML. Ghrelin induces adiposity in rodents.
Nature 407: 908-913, 2000.
135. Tschop M, Wawarta R, Riepl RL, Friedrich S, Bidlingmaier M, Landgraf R
and Folwaczny C. Post-prandial decrease of circulating human ghrelin levels.
J Endocrinol Invest 24: RC19-RC21, 2001.

86 6 Literaturverzeichnis
136. Tschop M, Weyer C, Tataranni PA, Devanarayan V, Ravussin E and
Heiman ML. Circulating ghrelin levels are decreased in human obesity.
Diabetes 50: 707-709, 2001.
137. Tsioufis C, Andrikou I, Thomopoulos C, Petras D, Manolis A and
Stefanadis C. Comparative prognostic role of nighttime blood pressure and
nondipping profile on renal outcomes. Am J Nephrol 33: 277-288, 2011.
138. Vallbo AB, Hagbarth KE, Torebjork HE and Wallin BG. Somatosensory,
proprioceptive, and sympathetic activity in human peripheral nerves. Physiol
Rev 59: 919-957, 1979.
139. Vallbo AB, Hagbarth K-E and Wallin BG. Microneurography: how the
technique developed and its role in the investigation of the sympathetic
nervous system. J Appl Physiol 96: 1262-1269, 2004.
140. Vestergaard ET, Andersen NH, Hansen TK, Rasmussen LM, Moller N,
Sorensen KE, Sloth E and Jorgensen JO. Cardiovascular effects of
intravenous ghrelin infusion in healthy young men. Am J Physiol Heart Circ
Physiol 293: H3020-H3026, 2007.
141. Vestergaard ET, Hansen TK, Gormsen LC, Jakobsen P, Moller N,
Christiansen JS and Jorgensen JO. Constant intravenous ghrelin infusion in
healthy young men: clinical pharmacokinetics and metabolic effects. Am J
Physiol Endocrinol Metab 292: E1829-E1836, 2007.

87 6 Literaturverzeichnis
142. Wallin BG. Regulation of sympathetic nerve traffic to skeletal muscle in
resting humans. Clin Auton Res 16: 262-269, 2006.
143. Wallin BG and Charkoudian N. Sympathetic neural control of integrated
cardiovascular function: insights from measurement of human sympathetic
nerve activity. Muscle Nerve 36: 595-614, 2007.
144. Wallin BG, Sundlof G, Eriksson BM, Dominiak P, Grobecker H and
Lindblad LE. Plasma noradrenaline correlates to sympathetic muscle nerve
activity in normotensive man. Acta Physiol Scand 111: 69-73, 1981.
145. Wiley KE and Davenport AP. Comparison of vasodilators in human internal
mammary artery: ghrelin is a potent physiological antagonist of endothelin-1.
Br J Pharmacol 136: 1146-1152, 2002.
146. Williams DL, Grill HJ, Cummings DE and Kaplan JM. Vagotomy dissociates
short- and long-term controls of circulating ghrelin. Endocrinology 144: 5184-
5187, 2003.
147. Wren AM, Seal LJ, Cohen MA, Brynes AE, Frost GS, Murphy KG, Dhillo
WS, Ghatei MA and Bloom SR. Ghrelin enhances appetite and increases
food intake in humans. J Clin Endocrinol Metab 86: 5992, 2001.
148. Wu R, Dong W, Cui X, Zhou M, Simms HH, Ravikumar TS and Wang P.
Ghrelin down-regulates proinflammatory cytokines in sepsis through activation
of the vagus nerve. Ann Surg 245: 480-486, 2007.

88 6 Literaturverzeichnis
149. Xu X, Jhun BS, Ha CH and Jin ZG. Molecular mechanisms of ghrelin-
mediated endothelial nitric oxide synthase activation. Endocrinology 149:
4183-4192, 2008.
150. Yang J, Brown MS, Liang G, Grishin NV and Goldstein JL. Identification of
the acyltransferase that octanoylates ghrelin, an appetite-stimulating peptide
hormone. Cell 132: 387-396, 2008.
151. Yao ST. a-adrenergic receptors in the nucleus tractus solitarii: fitting a new
piece to a complex puzzle. Exp Physiol 94: 771-772, 2009.
152. Yasuda T, Masaki T, Kakuma T and Yoshimatsu H. Centrally administered
ghrelin suppresses sympathetic nerve activity in brown adipose tissue of rats.
Neurosci Lett 349: 75-78, 2003.
153. Yildiz BO, Suchard MA, Wong ML, McCann SM and Licinio J. Alterations in
the dynamics of circulating ghrelin, adiponectin, and leptin in human obesity.
Proc Natl Acad Sci U S A 101: 10434-10439, 2004.
154. Zanutto BS, Valentinuzzi ME and Segura ET. Neural set point for the control
of arterial pressure: role of the nucleus tractus solitarius. BioMedical
Engineering OnLine 9: 2010.

89 7 Ergänzende Materialien
7 Ergänzende Materialien
Ergänzende Abb. I: Struktur des Peptidhormons Ghrelin. Angegeben ist die
Aminlsäuresequenz jeweils für Mensch und Ratte. Die n-Octanoylgruppe an Ser3 ist
entscheidend für die biologische Wirksamkeit. Nach Kojima und Kangawa (80).
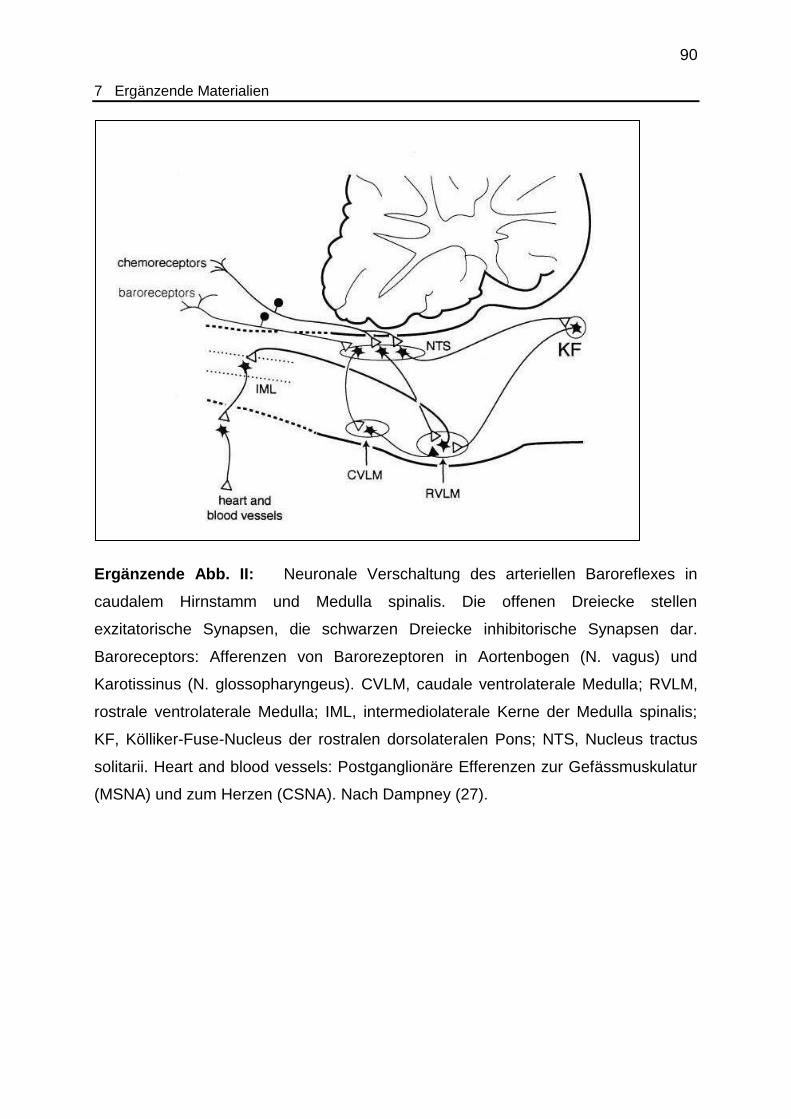
90 7 Ergänzende Materialien
Ergänzende Abb. II: Neuronale Verschaltung des arteriellen Baroreflexes in
caudalem Hirnstamm und Medulla spinalis. Die offenen Dreiecke stellen
exzitatorische Synapsen, die schwarzen Dreiecke inhibitorische Synapsen dar.
Baroreceptors: Afferenzen von Barorezeptoren in Aortenbogen (N. vagus) und
Karotissinus (N. glossopharyngeus). CVLM, caudale ventrolaterale Medulla; RVLM,
rostrale ventrolaterale Medulla; IML, intermediolaterale Kerne der Medulla spinalis;
KF, Kölliker-Fuse-Nucleus der rostralen dorsolateralen Pons; NTS, Nucleus tractus
solitarii. Heart and blood vessels: Postganglionäre Efferenzen zur Gefässmuskulatur
(MSNA) und zum Herzen (CSNA). Nach Dampney (27).
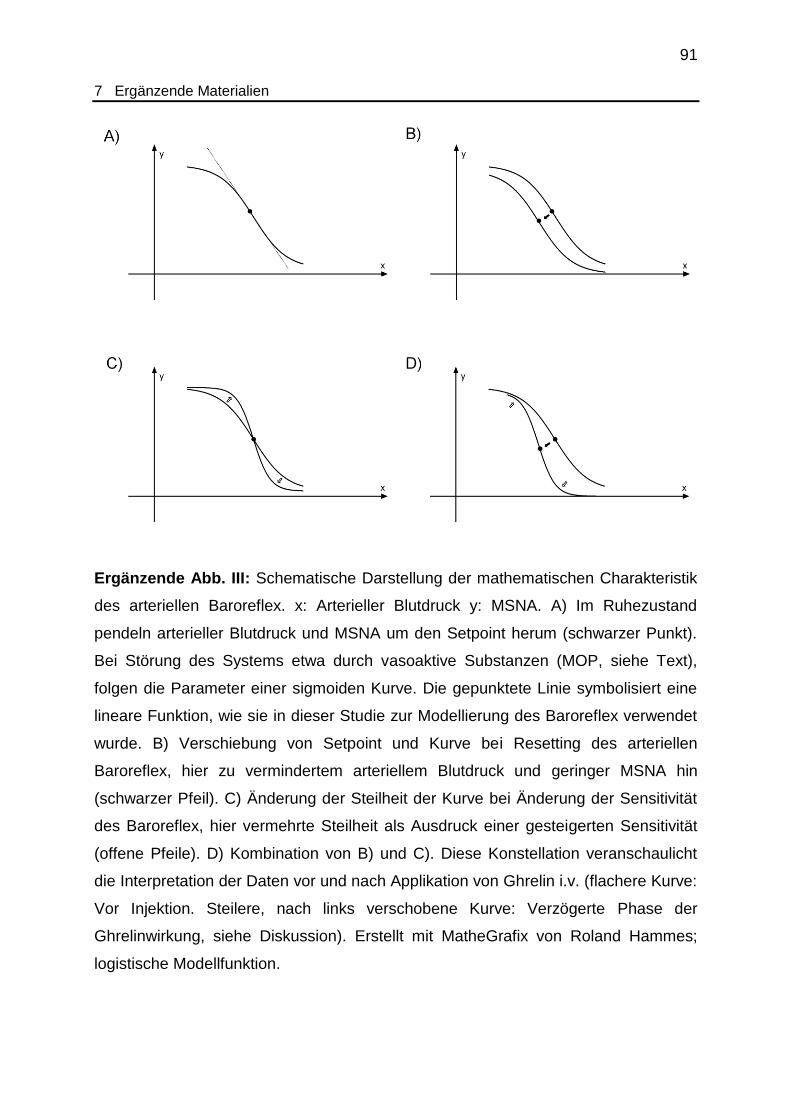
91 7 Ergänzende Materialien
Ergänzende Abb. III: Schematische Darstellung der mathematischen Charakteristik
des arteriellen Baroreflex. x: Arterieller Blutdruck y: MSNA. A) Im Ruhezustand
pendeln arterieller Blutdruck und MSNA um den Setpoint herum (schwarzer Punkt).
Bei Störung des Systems etwa durch vasoaktive Substanzen (MOP, siehe Text),
folgen die Parameter einer sigmoiden Kurve. Die gepunktete Linie symbolisiert eine
lineare Funktion, wie sie in dieser Studie zur Modellierung des Baroreflex verwendet
wurde. B) Verschiebung von Setpoint und Kurve bei Resetting des arteriellen
Baroreflex, hier zu vermindertem arteriellem Blutdruck und geringer MSNA hin
(schwarzer Pfeil). C) Änderung der Steilheit der Kurve bei Änderung der Sensitivität
des Baroreflex, hier vermehrte Steilheit als Ausdruck einer gesteigerten Sensitivität
(offene Pfeile). D) Kombination von B) und C). Diese Konstellation veranschaulicht
die Interpretation der Daten vor und nach Applikation von Ghrelin i.v. (flachere Kurve:
Vor Injektion. Steilere, nach links verschobene Kurve: Verzögerte Phase der
Ghrelinwirkung, siehe Diskussion). Erstellt mit MatheGrafix von Roland Hammes;
logistische Modellfunktion.
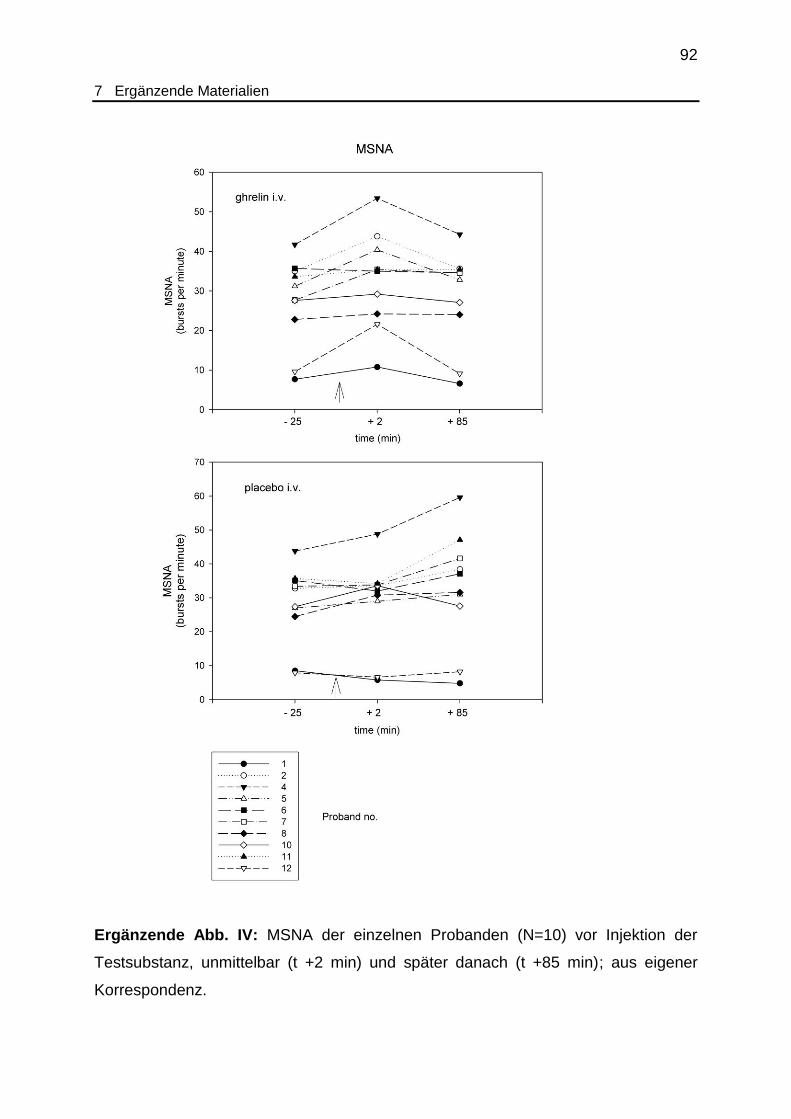
92 7 Ergänzende Materialien
Ergänzende Abb. IV: MSNA der einzelnen Probanden (N=10) vor Injektion der
Testsubstanz, unmittelbar (t +2 min) und später danach (t +85 min); aus eigener
Korrespondenz.
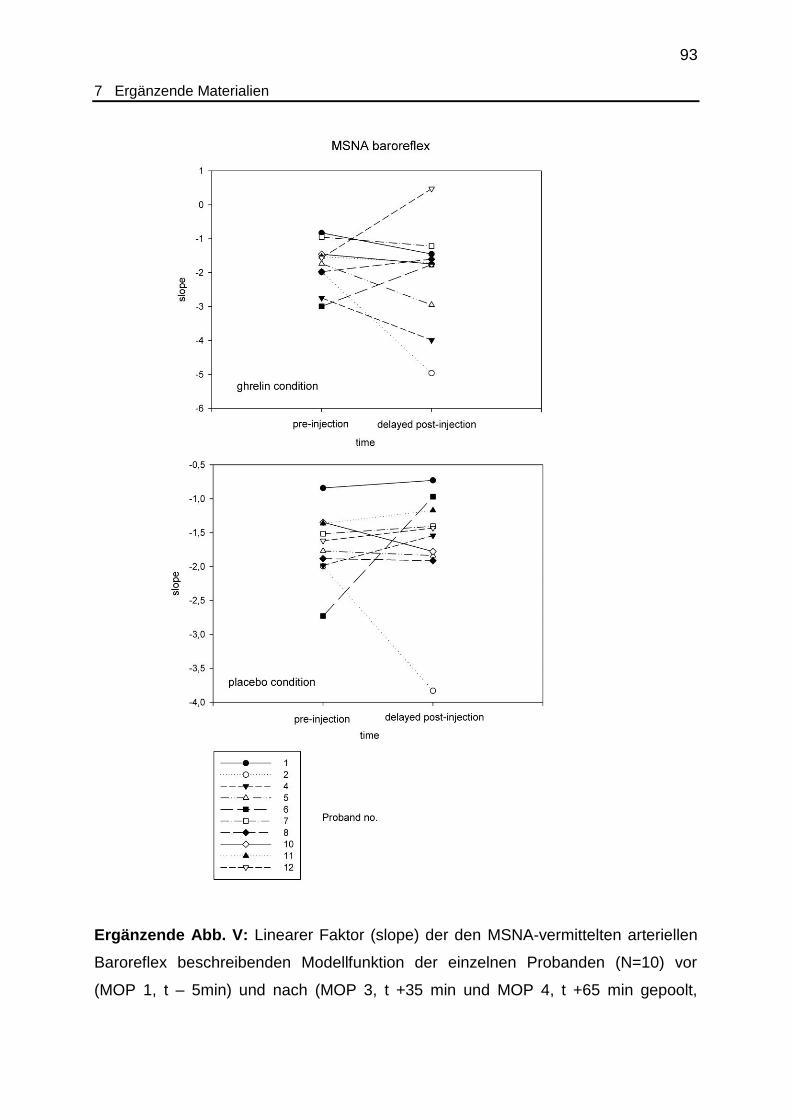
93 7 Ergänzende Materialien
Ergänzende Abb. V: Linearer Faktor (slope) der den MSNA-vermittelten arteriellen
Baroreflex beschreibenden Modellfunktion der einzelnen Probanden (N=10) vor
(MOP 1, t – 5min) und nach (MOP 3, t +35 min und MOP 4, t +65 min gepoolt,

94 7 Ergänzende Materialien
delayed post-injection, siehe Text) Injektion der Testsubstanz; aus eigener
Korrespondenz.

95
8 Lebenslauf
8 Lebenslauf
Angaben zur Person
Name Alexander Friedrich Wilhelm Krapalis
Eltern Margund Krapalis geb. Kroll, Lehrerin
Wilfried Krapalis, Lehrer
Geboren 08.09.1970 in Lübeck
Nationalität Deutsch
Kinder Friedrich Johannes, geboren 11/9/2007
Richard Alexander, geboren 25/7/2010
Facharzt und Zusatzbezeichnung
12/2008 Zusatzbezeichnung Notfallmedizin
05/2010 Facharzt für Allgemeinmedizin
Schulbildung
1976 - 1980 Luisenhof-Grundschule in Lübeck
1980 - 1990 Trave-Gymnasium in Lübeck, Abitur
Zivildienst
1990 - 1991 Rettungssanitäter bei der Johanniter-
Unfallhilfe
Studium
1991 - 1993 Mathematik und Physik an der Christian-
Albrechts-Universität zu Kiel
1993 - 1995 vorklinisches Studium an der medizinischen
Universität zu Lübeck
1995 - 1999 klinisches Studium an der medizinischen

96
8 Lebenslauf
Universität zu Lübeck
1999 - 2000 Praktisches Jahr an der medizinischen
Universität zu Lübeck, Wahlfach
Anästhesiologie
klinische Tätigkeit
02/2001 - 04/2001 AIP in der medizinischen Klinik I der
medizinischen Universität zu Lübeck,
Hämatologie und Onkologie
09/2002 - 05/2005 AIP und Assistenzarzt in der Praxis Dres.
Lübbert, Kube und Heinelt, Internisten in
Mölln
02/2006 - 06/2008 Assistenzarzt im Zentrum für Innere Medizin
und Intensivmedizin der Sana Kliniken
Ostholstein, Klinik Eutin, Chefarzt
Dr.Gützkow
10/2008 - 05/2009 Assistenzarzt in der Klinik für Allgemein- und
Visceralchirurgie des UKSH, Campus
Lübeck, Klinikdirektor Prof. Bruch
10/2009 - 10/2010 Assistenzarzt in der Klinik für Kinder- und
Jugendmedizin des UKSH, Campus Lübeck,
Klinikdirektor Prof. Herting
09/2009 - 9/2012 Mitarbeiter der Medizinischen Klinik I,
Campus Lübeck des UKSH, Klinikdirektor
Prof. Lehnert. Durchführung der dieser Arbeit
zugrundeliegenden experimentellen Studie.
Klinische Tätigkeit in der Notaufnahme.
ab 10/2012 Freiberufliche Tätigkeit (Praxisvertretungen
und NEF-Dienst). Niederlassung als
Hausarzt angestrebt.
Persönliches
Christliches Glaubensbekenntnis; Interesse für klassische Musik, englische und
amerikanische Literatur; Ausdauersport (Schwimmen, Laufen, Radfahren).

97
9 Veröffentlichung
9 Veröffentlichung
Ghrelin modulates baroreflex-regulation of sympathetic vasomotor tone in
healthy humans; Alexander F. Krapalis, Jacqueline Reiter, Felix Machleidt, K.
Alexander Iwen, Christoph Dodt, Hendrik Lehnert and Friedhelm Sayk; Am J Physiol
Regul Integr Comp Physiol 302:R1305-R1312, 2012. First published 4 April 2012;
doi: 10.1152/ajpregu.00663.2011

98
10 Danksagungen
10 Danksagungen
Herrn Dr. med. Friedhelm Sayk gilt mein besonderer Dank. Auf ihn geht die
wissenschaftliche Initiative und Konzeption dieser Arbeit zurück. Ferner unterstützte
er mich unermüdlich in allen praktischen und theoretischen Belangen. Herrn Prof. Dr.
med. Hendrik Lehnert danke ich sowohl für die Bereitstellung des institutionellen
Rahmens als auch für die Unterstützung bei Publikation unserer Studie. Herrn Prof.
Dr. med. Christoph Dodt danke ich für die fachliche Beratung in allen Abschnitten der
wissenschaftlichen Arbeit. Frau Jacqueline Reiter danke ich herzlich für ihre
engagierte und zuverlässige Mitarbeit bei den umfangreichen Versuchen. Ebenso
danke ich Herrn Felix Machleidt für seine freundliche Hilfe. Herrn Dr. med. Alexander
Iwen danke ich für die geleistete wissenschaftliche Beratung in endokrinologischen
Fragestellungen. Den medizinisch-technischen Assistentinnen Frau Heidi Ruf und
Frau Martina Grohs danke ich für die professionelle Durchführung der
Laboruntersuchungen und die immer freundliche Zusammenarbeit.
Mein Dank und meine Anerkennung gilt allen studentischen Probanden, die
sich für die die manchmal anstrengenden Versuche zur Verfügung gestellt haben.
Nicht zuletzt danke ich meiner Familie und besonders meinen Söhnen
Friedrich und Richard, die mir in den Arbeitspausen Freude und neue Kraft
geschenkt haben.
Bei der Finanzierung dieser Studie war die Deutsche Forschungsgemeinschaft
im Rahmen des SFB 654/B4 behilflich.
Recommended












