Embed Size (px)
Citation preview

Aus der Klinik und Poliklinik für Mund-, Kiefer-, Gesichtschirurgie
der Friedrich-Alexander-Universität Erlangen-Nürnberg
Direktor: Prof. Dr. Dr. F.W. Neukam
Ossäre Regeneration eines experimentellen critical size-Defektes der Schweinekalotte mit einem biphasischen Knochenersatzmaterial und
biodegradierbarer Polyethylenglycol-Membran – Einfluss auf die Wnt-vermittelte, osteogene Rekrutierung und
Maturation mesenchymaler Osteoblast- und Osteoklastprogenitoren
Inaugural-Dissertation
zur Erlangung der Doktorwürde
an der Medizinischen Fakultät
der Friedrich-Alexander-Universität
Erlangen-Nürnberg
vorgelegt von
Monika Fruck aus Burglengenfeld
-2010-

Gedruckt mit Erlaubnis
der Medizinischen Fakultät der Friedrich-Alexander-Universität
Erlangen-Nürnberg
Dekan: Prof. Dr. med. Dr. h.c. Jürgen Schüttler
Referent: PD. Dr. med. Dr. med. dent Karl Andreas Schlegel
Koreferent: Prof. Dr .med. Dr. med. dent Friedrich Wilhelm Neukam
Tag der mündlichen Prüfung: 20.09.2010

3
Parentibus dedicatum

4
Abkürzungsverzeichnis
ABC-POX = Avidin-Biotin-Peroxidase
AD = Aqua Dest
APC = adenomatöse polyposis coli
BMP = bone morphogenetic protein
DKK = Dickkopf
Fzd = Frizzled
GC = Glucokortikoid
GSK-3β = Glycogensynthasekinase-3β
HA = Hydroxylapatit
HCl = Wasserstoffchlor
KEM = Knochenersatzmaterial
LEF = lymphoid enhancer factor
LiCl = Lithiumchlorid
Lrp5/6 = low-density lipoprotein receptor-related protein 5/6
LSAB = Labelled Streptavidin-Biotin
MEA = Methoxyethyl-acetat
MM = Multiples Myelom
OPG = Osteoprotegerin
PEG = Polyethylenglycol
RANK(L) = Receptor activator of nuclear factor-κB (ligand)
RT = Raumtemperatur
sFRP = secreted Frizzled-related protein
sRANKL = soluble RANKL
TCF = T-cell factor
TCP = Trikalziumphosphat
TGF-β = transforming growth factor beta
TNF = tumor necrosis factor
WIF-1 = Wnt inhibitory factor-1
Wnt = Wingless and Int-1

5
Inhaltsverzeichnis:
1. Zusammenfassung........................................................................................................... 7
1.1 Hintergrund und Ziele.................................................................................................. 7
1.2 Methoden...................................................................................................................... 7
1.3 Ergebnisse und Beobachtungen.................................................................................... 8
1.4 Praktische Schlussfolgerung......................................................................................... 8
2. Einleitung.......................................................................................................................... 11
2.1. Klinischer Hintergrund................................................................................................ 11
2.2. Stand der Forschung.................................................................................................... 13
2.2.1. Knochenersatzmaterialien.................................................................................. 13
2.2.1.1. Autografts................................................................................................. 13
2.2.1.2. Allografts.................................................................................................. 13
2.2.1.3. Alloplasts.................................................................................................. 14
a) Hydroxylapatit.......................................................................................... 15
b) Trikalziumphosphat.................................................................................. 15
c) BMP-2/4 und KEM.................................................................................. 16
2.2.2. Grenzen der KEMien.......................................................................................... 17
2.2.2.1. Verwendetes KEM................................................................................... 17
2.2.2.2. Verwendete Membranen.......................................................................... 18
2.2.3. GSK-3β als Schlüsselregulator der Knochendifferenzierung............................ 19
2.2.3.1. Interaktion mit pleiotropen Induktoren ossärer Differenzierung.............. 19
a) Wnt/β-catenin-Signalweg........................................................................ 19
b) Regulation des Wnt/β-catenin-Signalweges............................................ 20
c) Regulation der Knochenmasse über den Wnt-Weg................................. 21
d) Klinischer Bezug: Glucokortikoide und Lithium.................................... 23
2.2.3.2. Interaktion des Wnt-Weges mit dem TGF-β/BMP-Weg.......................... 24
2.2.3.3. HOX-Gene: Ossäre Regeneration im Kiefer und extrakranial................. 26
2.2.4. RANK(L)-OPG-System..................................................................................... 27
2.2.4.1. Relevanz des RANK(L)-OPG-Systems................................................... 27
a) RANKL................................................................................................... 28
b) RANK...................................................................................................... 28
c) OPG......................................................................................................... 29
2.2.4.2. Signalweg der Osteoklastogenese............................................................ 30
2.2.4.3. Interaktion des Wnt-Weges mit dem RANK(L)-OPG-System................ 30
2.2.4.4. Therapeutischer Ansatz am RANK(L)-OPG-System: Denosumab......... 31

6
3. Fragestellung und Ziele................................................................................................... 33
4. Material und Methoden.................................................................................................. 34
4.1. Versuchsablauf............................................................................................................ 34
4.2. Verwendete Materialien.............................................................................................. 34
4.3. Versuchsgruppen......................................................................................................... 35
4.4. Immunhistochemische Nachweisreaktionen............................................................... 35
4.4.1. Probenaufbereitung............................................................................................ 35
4.4.2. Methodenetablierung......................................................................................... 36
4.4.3. Angewendete immunhistochemische Methoden............................................... 36
4.4.4. Immunhistochemische Färbung nach der LSAB-Methode................................ 38
4.4.5. Immunhistochemie............................................................................................. 39
a) RANKL............................................................................................................. 39
b) DKK1................................................................................................................ 40
c) OPG................................................................................................................... 40
d) GSK-3β............................................................................................................. 40
4.4.6. Verwendete Reagenzien.................................................................................... 41
4.5. Qualitative und quantitative Analyseverfahren........................................................... 42
4.5.1. Qualitative und semiquantitative Expressionsbestimmung............................... 42
4.5.2. Statistische Analyse........................................................................................... 42
5. Ergebnisse........................................................................................................................ 43
5.1. Qualitative immunhistologische Ergebnisse............................................................... 43
5.2. Semiquantitative immunhistochemische Ergebnisse der Expression ......................... 45
5.2.1. RANKL............................................................................................................. 45
5.2.2. DKK1................................................................................................................. 48
5.2.3. OPG.................................................................................................................... 51
5.2.4. GSK-3β.............................................................................................................. 54
6. Diskussion......................................................................................................................... 58
7. Schlussfolgerung.............................................................................................................. 63
8. Literatur........................................................................................................................... 64
9. Anhang.............................................................................................................................. 72
9.1. Färbeprotokolle............................................................................................................ 72
9.2. Lebenslauf................................................................................................................... 76
9.3. Danksagung................................................................................................................. 77
9.4. Ehrenwörtliche Erklärung............................................................................................ 78

7
1. Zusammenfassung
1.1. Hintergrund und Ziele Knochenersatzmaterialien (KEMien) finden Verwendung, um das Problem der
limitierten Knochenverfügbarkeit zu umgehen und Donormorbiditäten in der
Entnahmeregion zu reduzieren. Biologisch degradierbare Matrizes können Haut- und
Mukosatransplantate ersetzen. Hierfür sind Vaskularisation, der zelluläre Einstrom
und die osteogene Differenzierung von Matrixstrukturen, die durch gesteuerte
Geweberegeneration erzeugt werden, entscheidende Faktoren. Dies rechtfertigt
Forschungen zur Verbesserung der zellulären Umgebung von critical size-Defekten.
Diese Studie zielte auf die Untersuchung der Einsetzbarkeit einer neuen
Polyethylenglycol(PEG)-Hydrogelmembran als temporäre Matrix und Abdeckung
während einer gesteuerten ossären Regeneration von critical size-Defekten der
Schweinekalotte ab, die mit einem biphasischen KEM aus Hydroxylapatit (HA) und
Trikalziumphosphat (TCP) gefüllt wurden. Es wurde untersucht, ob die PEG-
Membran als Träger von BMP-2 transfizierten Osteoblasten in der Lage ist, die
osteogene Stimulation kontrolliert an den Defekt abzugeben.
Im Vordergrund dieses Dissertationsprojektes stand die Untersuchung des Einflusses
dieser PEG-Membran auf die Wnt-vermittelte, osteogene Rekrutierung und
Maturation mesenchymaler Osteoblast- und Osteoklastprogenitoren und die
Evaluation der ossären Differenzierungs- und Proliferationsmarker OPG, RANKL,
GSK-3β und deren Antagonist DKK1.
Zusätzlich sollte die immunhistochemische Färbung von DKK1 und dessen
Nachweis am Tiermodell des Schweins und die immunhistochemische Färbung von
RANKL am Schwein etabliert werden.
1.2. Methodik Bei insgesamt 20 adulten Hausschweinen wurden je 9 critical size-Defekte (Defekte
A-I) gesetzt und mit biphasischer Straumann® Knochenkeramik (Defekte A-C und
E-H) und autologem Knochen (Defekt D) aufgefüllt. Der Defekt I ist als
Kontrolldefekt leer. Je 5 Tiere wurden zu 4 Observierungszeitpunkten 1, 2, 4 und 12
Wochen nach Operation geopfert. Es folgte die Probenentnahme, die Aufbereitung in
Paraformaldehyd und Technovit 9100® und die Erstellung der 3µm-Dünnschnitte.

8
Zur Beurteilung von Differenzierungs- und Proliferationsmarkern der
Osteoblastenrekrutierung und –maturation wie OPG, RANKL, GSK-3β und deren
Antagonist DKK1 wurden immunhistochemische Färbungen durchgeführt und deren
Expression semiquantitativ evaluiert.
1.3. Ergebnisse und Beobachtungen Die Wundheilung verlief ohne Anzeichen von Entzündungen und die Matrix wurde
im gewünschten Zeitraum integriert. Die Etablierung der immunhistochemischen
Färbung von DKK1 und dessen Nachweis am Tiermodell des Schweins ist gelungen.
Die immunhistochemische Färbung von RANKL am Schwein konnte etabliert
werden. Die PEG-Membran und die BMP-2 transfizierten Osteoblasten hatten zu
keinem der 4 Opferungszeitpunkte Einfluss auf das Expressionsverhalten von
RANKL. Die Expression von OPG stieg von der 1. – 4. Woche unabhängig von der
Defektbehandlung an. Nach anfänglicher Streuung stabilisierten sich die
Expressionswerte bis zur 12. Woche. In der 1. Woche zeigten sich hohe Werte von
OPG bei den mit BMP-2 behandelten Gruppen. Die Expression von GSK-3β ließ ein
Absinken von der 1. - 4. Woche erkennen, was auf den Übergang von anfänglicher
Fibrose zu Osteoneogenese hindeutet. In der 1. Woche bewirkte die
Periostabdeckung eine verminderte Expression von GSK-3β. GSK-3β zeigte zu
jedem Opferungszeitpunkt starke Streuungen der Expression.
1.4. Praktische Schlussfolgerung Aufgrund der gefundenen Ergebnisse kann der routinemäßige klinische Einsatz der
PEG-Membran zur positiven Beeinflussung der Wnt-vermittelten, osteogenen
Rekrutierung und Maturation mesenchymaler Osteoblast- und
Osteoklastprogenitoren nicht empfohlen werden. Die PEG-Membran eignet sich aber
als Carrier.

9
1. Abstract
1.1. Background and objectives Bone substitute materials are used to overcome the problem of limited bone
availability and to reduce donor site morbidity. Biologically degradable matrices can
replace dermal and mucosal transplants. Vascularization, cellular influx and
osteogenic differentiation of matrix structures, generated by guided tissue
regeneration have been shown to be crucial. Therefore clinical and experimental
approaches improving the cellular microenvironment of critical size defect
regeneration are justified.
The aim of this study was to evaluate the feasibility of a new PEG hydrogel
membrane as a temporary matrix and wound covering during guided osseous
regeneration of critical size defects in domestic pigs. The calvarial defects were filled
with a biphasic bone substitute material consisting of HA and TCP. It has been
addressed if the PEG membrane formulation is capable to serve as a carrier for bone
differentiation factors like BMP-2 and thus controlling the release of osteogenic
stimulation in the defect.
The main task of this dissertation was the examination of the influx of this PEG
membrane on the Wnt mediated osteogenic recruitment and maturation of
mesenchymal osteoblast and osteoclast progenitors and the evaluation of the
differentiation and proliferation markers OPG, RANKL, GSK-3β and their
antagonist DKK1.
Additionally the immunhistochemical stain of DKK1 should be established and the
objective evidence of DKK1 expression should be shown in the pig model. The
immunhistochemical stain of RANKL should be established for the pig model.
1.2. Study design 20 adult domestic pigs received 9 critical size defects in the os frontale and these
defects were filled with a biphasic Straumann® bone ceramic (defects A-C and E-H)
and autologous bone (defect D). Defect I was empty and served as control defect.
5 animals were sacrified 1, 2, 4 and 12 weeks after surgery. After harvesting the
samples they were prepared in an immersion fixation using 4% paraformaldehyd and
embedded in resin Technovit 9100®. The embedded samples were cut into 3µm

10
sections using a microtom. Immunhistochemistry was performed for the evaluation
of the differentiation and proliferation factors for osteoblast recruitment and
maturation like OPG, RANKL, GSK-3β and their antagonist DKK1. The
semiquantitative evaluation of the expression of these factors followed.
1.3. Results Wound healing passed off without any inflammatory processes and the matrix was
integrated within the desirable time period. The immunhistochemical stain of DKK1
could be established and the objective evidence of DKK1 expression was shown in
the pig model. The immunhistochemical stain of RANKL could be established for
the pig model. Neither the PEG membrane nor the BMP-2 osteoblasts had an
influence on the expression of RANKL at any timepoint. The expression of OPG
enhanced from timepoint 1 to timepoint 3 independent of the defect modulation.
After initial dispersal the expression stabilized at timepoint 4. The first week showed
high values for OPG in the BMP-2 treated defects. The expression of GSK-3β
decreased from timepoint 1 to timepoint 3 and that indicates the transition from
fibrosis to osteogenesis. The periost covering effected the decrease of GSK-3β in
week one. GSK-3β expression showed intense dispersal at each timepoint.
1.4. Conclusion Due to the shown results the routinely use of the PEG membrane can not be
recommended in order to influence Wnt mediated osteogenic recruitment and
maturation of mesenchymal osteoblast and osteoclast progenitors positively. But the
PEG membrane is suited as carrier.

11
2. Einleitung
2.1. Klinischer Hintergrund Ein ausreichendes periimplantäres Knochenangebot stellt eine Grundvoraussetzung
für den langfristig funktionellen Erhalt und ein befriedigendes ästhetisches Ergebnis
dentaler Implantate dar. Jedoch bedingen z.B. vorbestehender Zahnverlust mit
resultierender Inaktivitätsatrophie und ein ersatzschwaches Implantatlager nach
multimodaler Karzinomtherapie im Kieferbereich ein vermindertes Knochenangebot
[65, 86, 89]. Zur erfolgreichen Insertion dentaler Implantate ist jedoch ein
Mindestknochenangebot im Ober- und Unterkiefer von 6-8 mm in der Vertikalen und
5 mm in der Transversalen gefordert [64].
Durch augmentative Verfahren kann ein ungenügendes lokoregionäres
Knochenangebot verbessert werden [76]. Die Verwendung autologen Knochens zur
Augmentation gilt hierbei als „Gold-Standard“ [62, 76]. Nachteile stellen die
limitierte Verfügbarkeit [66], die Donormorbidität in der Entnahmeregion sowie die
Resorptionsneigung dar [18, 87]. Bei humanen, körperfremden Allograft-Materialien
hingegen sind die Infektionsgefahr für HIV, die Übertragungsmöglichkeit für
Prionen und die nicht vorhersagbare Osteoinduktivität negativ zu bewerten [10, 57,
58].
Durch die Entwicklung synthetischer, biokompatibler KEMien werden die eben
genannten Nachteile von humanem Knochenmaterial eliminiert [82, 83, 114]. Die
derzeit am häufigsten eingesetzten KEMien stellen Kalziumphosphat- und
Kalziumsulfatkeramiken dar [23-26, 61]. Die Substanzen HA und TCP haben sich
als klinisch langfristig erfolgreiche alloplastische Transplantate erwiesen [37, 73,
79].
Diese Studie zielte auf die Untersuchung der Einsetzbarkeit einer neuen PEG-
Hydrogelmembran als temporäre Matrix und Abdeckung während einer gesteuerten
ossären Regeneration von critical size-Defekten der Schweinekalotte ab, die mit
einem biphasischen KEM aus HA und TCP gefüllt wurden. Es wurde untersucht, ob
die PEG-Membran als Träger von BMP-2 transfizierten Osteoblasten in der Lage ist,
die osteogene Stimulation kontrolliert an den Defekt abzugeben. Im Vordergrund
dieses Dissertationsprojektes stand die Untersuchung des Einflusses dieser PEG-
Membran auf die Wnt-vermittelte, osteogene Rekrutierung und Maturation

12
mesenchymaler Osteoblast- und Osteoklastprogenitoren und die Evaluation der
ossären Differenzierungs- und Proliferationsmarker OPG, RANKL, GSK-3β und
deren Antagonist DKK1.
Da die Regeneration ossärer Defekte und die Knochenheilung an maturen
Knochenstrukturen als ein partieller Rückgriff auf embryonale
Differenzierungsmechanismen mit temporärer Reaktivierung der für die
Rekrutierung von Osteoblastprogenitoren und deren Maturation maßgeblichen
Signalwege aufgefasst wird, ist die Kenntnis der Interaktion von osteogener
Differenzierung und dem biokompatiblen KEM während der gesteuerten
Knochenregeneration zur Evaluation der osteokonduktiven und möglichen
osteoinduktiven Eigenschaften notwendig [35, 98]. Zudem bestehen Hinweise auf
eine enchondrale Ossifikation der Transitionszonen zwischen KEM und
ortsständigem Lagerknochen innerhalb des desmal ossifizierenden Kiefer- und
Schädelknochens [52, 54].
Um klinische und experimentelle Ansätze zur induzierten Knochenregeneration zu
evaluieren, ist die qualitative und quantitative Beschreibung der für die
Knochenneubildung und Regulation der Knochenmasse wichtigen Effektoren des
BMP- und Wnt-Weges sowie die Charakterisierung der Knochenmaturation über das
RANK(L)-OPG-System im Übergangsbereich zwischen KEM und Knochen
notwendig [35, 98].
Die Effektoren des Wnt-Weges sind GSK-3β und dessen Antagonist DKK1, die als
Bestandteile der Signalkaskade des Wnt-Weges die Osteoblastendifferenzierung
und -proliferation regulieren und zudem die Knochenresorption durch Interaktion mit
RANK(L) und OPG beeinflussen [30, 31, 74, 80].
Therapeutisch bietet die Suppression von GSK-3β einen experimentell
vielversprechenden Ansatz, da klinisch zugelassene GSK-3β-Hemmer wie Lithium
eine selektive Modulation von Osteoblasten realistisch erscheinen lassen [15, 35, 50,
95, 100]. Ein therapeutischer Eingriff in das RANK(L)-OPG-System zur
Bekämpfung von Knochenkrankheiten, wie der Osteoporose, ist durch Denosumab
gegeben, einem humanen, monoklonalen Antikörper, der die Rolle von OPG
funktionell imitiert, indem er an RANKL bindet und somit die Osteoklastogenese
nachweislich hemmt [71].

13
2.2. Stand der Forschung 2.2.1. Knochenersatzmaterialien
Die langfristig erfolgreiche Implantation im Kieferknochen mit einem
anspruchsvollen ästhetischen Ergebnis ist an eine ausreichende periimplantäre,
knöcherne Verankerung geknüpft [65]. Defizite des Hartgewebelagers betreffen
Knochenvolumen, Knochenstruktur, Knochenvitalität, Vaskularisation und Integrität
des bedeckenden Weichgewebes [65]. Zur erfolgreichen Insertion dentaler
Implantate ist ein Mindestknochenangebot von 6-8 mm in der Vertikalen und 5 mm
in der Transversalen im Ober- und Unterkiefer gefordert [64]. Allerdings bedingen
ein vorbestehender Zahnverlust, der durch das Ausbleiben von Krafteinwirkungen
auf den Alveolarfortsatz zur physiologischen Inaktivitätsatrophie führt, sowie ein
ersatzschwaches Implantatlager nach multimodaler Karzinomtherapie im
Kieferbereich insbesondere bei den Patienten, die von einer Implantatversorgung
profitieren, ein vermindertes Knochenangebot [65, 86, 89].
Ein lokoregionäres Knochendefizit lässt sich mithilfe augmentativer Verfahren
kompensieren [86]. Zur Augmentation stehen autologer Knochen (Autografts),
humane nicht körpereigene Transplantate (Allografts), bezüglich der Spezies
differente Transplantate (Xenografts) und bei geeigneter Indikation biokompatible
KEMien (Alloplasts) zur Auswahl [76].
2.2.1.1. Autografts
Autologer Knochen entstammt dem eigenen Körper und eignet sich wegen seiner
ihm charakteristischen Eigenschaften am besten als Augmentat [62, 76]. Jedoch steht
autologer Knochen nur in limitierter Menge zur Verfügung und dessen Gewinnung
bringt zusätzliche Operationsrisiken mit sich [66]. Negativ zu bewerten ist auch die
postoperativ zu erwartende Morbidität in der Transplantatentnahmeregion [18].
Zudem sind Unterschiede bezüglich der Mineralisation von autologen, ossären
Augmentaten in Abhängigkeit von der Donorregion, aus der sie gewonnen wurden,
festzustellen [87].
2.2.1.2. Allografts
Die Vorteile von genetisch differenten, humanen Allotransplantaten liegen in der
höheren quantitativen Verfügbarkeit, der Vermeidung der Donormorbidität in der

14
Transplantatentnahmeregion und der Vermeidung eines Zweiteingriffes bzw. der
Minimierung des Ausmaßes des Eingriffes und damit verbundenen Operationsrisiken
[57]. Allografts werden als Substitut für autogenen Knochen oder in Kombination
mit autologem Knochen verwendet [57]. Das Material wird vorzugsweise in
partikulierter Form eingesetzt [57]. Lyophylisierter Kadaverknochen wird bei -70°C
eingefroren und enthält biologisch aktive Wachstums- und Differenzierungsmarker
[92, 104]. Eine Entkalzifizierung und zusätzliche viruzide Behandlung des
lyophylisierten Knochens zeigt eine sichere Inaktivierung von HIV, die
Gefriertrocknung alleine nicht [10, 57, 58]. Allerdings weist der entkalkte
lyophylisierte Knochen eine geringere Formstabilität auf, so dass für die
Augmentation frischer Extraktionsalveolen, beim Sinus-Lift und zur Defektfüllung
nach Implantatverlust nicht entkalkter, formstabiler Knochen präferiert wird [56].
Die Nachteile der Allotransplantate zeigen sich demnach in der nicht sicher zu
eliminierenden Infektionsgefahr für HIV und der unkontrollierbaren
Übertragungsmöglichkeit von Prionen.
2.2.1.3. Alloplasts
Durch Fortschritte und Entwicklung im Bereich der Biomaterialien kommen
synthetische und kombiniert synthetisch-biomimetische Materialien als
Knochenersatz zum Einsatz, wodurch die Problematik der Limitation der autologen
Transplantate und die Nachteile der Allografts übergangen werden [82, 83, 114].
Synthetische Materialien sind osteokonduktiv ohne intrinsisches Potential für
Osteogenese oder Osteoinduktion [82]. Osteokonduktive Materialien bieten eine
Leitschiene für Endothelien, Kapillaren und nachfolgende Osteoprogenitorzellen zur
Knochenneubildung unter Degradation bzw. Persistenz des eingebrachten
alloplastischen Transplantates, abhängig von der Materialart [82]. Die unbegrenzte
quantitative Verfügbarkeit ist als größter Vorteil zu nennen [82]. Alloplastische
Materialien werden als poröse, synthetische Kalziumphosphat-Keramiken,
Kalziumkarbonat, Polymere, bioaktive Glaskeramiken sowie in Kombination mit
BMPs und Wachstumsfaktoren, wie auch in dieser Studie, klinisch eingesetzt [11,
34].
Kalziumphosphat- und Kalziumsulfat-Keramiken stellen die derzeit am häufigsten
eingesetzten KEMien dar [23-26, 61]. Gewonnen aus denaturiertem Knochen und
natürlicher, organischer Substanz aus Korallen stellen die Substanzen HA und TCP

15
etablierte, klinisch langfristig erfolgreiche alloplastische Transplantate dar [37, 73,
79]. Diese Materialien zeichnen sich durch hohe Druck- und geringe Zugfestigkeit,
ähnlich natürlichem Knochen, aus [61]. Die Knochenneubildung entlang der
Oberfläche dieser osteokonduktiven Materialien wurde gezeigt [81].
a) Hydroxylapatit
Bei der Verwendung von HA bestimmen steuerbare physikalische und physiko-
chemische Parameter (pH-Wert, Porosität, Oberfläche, Dotierung mit Fremdionen)
des synthetisch, hydrothermal hergestellten kristallinen Materials die Resorption und
Degradationskinetik [61]. Poröses Material wird schneller von Knochenmatrix
erschlossen als kompakter HA [61]. Neben den bereits längerfristig etablierten
HA-basierten KEMien unter hauptsächlicher Verwendung von gesintertem HA, sind
aktuell Materialien, basierend auf ungesintertem HA in Pasten-Formulation
verfügbar [39]. Bei diesen nanopartikulären Materialien wird eine höhere
Löslichkeit, eine vollständige Resorbierbarkeit und eine initial vergrößerte, aktive
Oberfläche postuliert [39]. Die, bedingt durch die verkleinerte Partikelgröße von
ungesintertem HA, vergrößerte Bindungskapazität für freies Wasser in der
Inflammationsphase der Einheilung begünstigt den Influx zellulärer Elemente und
die frühzeitige Kapillarisierung [39]. Die ossäre Integration des Materials erfolgt
über primäre Kollagensynthese und Kalzifizierung, die vom bindegewebig
erschlossenen HA-Partikel ausgehen [99].
Ein Nachteil der porösen Keramiken ist die exponentielle Abnahme der
mechanischen Belastbarkeit mit der Porosität [26]. Die Sinusbodenaugmentation mit
HA zeigte in mehreren klinischen Studien eine anderen KEMien überlegene
Dimensionstreue [40, 42, 85, 88, 94].
b) Trikalziumphosphat
TCP, ein synthetisches KEM, liegt in den beiden Gitterformen α-TCP und β-TCP,
die sich hinsichtlich ihrer Degradationskinetik unterscheiden, vor. α-TCP wird
innerhalb von sieben Monaten zu 70% degradiert, β-TCP zu 80% [113]. Nach vier
Monaten sind sequestrierende TCP-Partikel mit umgebenden Makrophagen und
mehrkernigen Riesenzellen nachweisbar [49, 59]. Bei synthetischen
Kalziumphosphatkeramiken findet keine echte Resorption, sondern eine
Fragmentierung und Degradation in Abhängigkeit von den Materialeigenschaften

16
statt [113]. Bedingt durch seine Degradation im zeitlichen Verlauf, eignet sich der
Einsatz des osteokonduktiven TCPs im ersatzstarken Knochenlager, da die
entstehenden Resorptionsdefekte durch neugebildeten Knochen ersetzt werden [43,
61].
Sowohl HA als auch TCP zeichnen sich durch Biokompatibilität aus. Jedoch wird
keine mechanisch belastbare Verbindung zum Knochen erreicht, so dass eine
Augmentation großer Defekte mit HA und TCP problematisch ist [21]. Klinische
Langzeitbeobachtungen zeigten eine persistierende, unterschiedlich kristalline
Struktur natürlichen Knochens im Vergleich zu integrierten HA-Augmentaten [97].
Es bestehen Hinweise auf eine schnellere Knochenneubildung in HA-augmentierten
Defekten als in TCP-gefüllten Bereichen [63]. Die Aktivität der alkalischen
Phosphatase, sezerniert von mononukleären Zellen während des
Knochenremodellings, zeigte sich HA-assoziiert früher und erhöht als nach
vergleichbarer Zeitspanne in TCP-Augmentaten [63].
c) BMP-2/4 und KEM
Klinisch evaluierte Erkenntnisse zum Einsatz von Wachstumsfaktoren und
Zytokinen in Kombination mit KEMien liegen für die Morphogene der TGFβ-
Gruppe, insbesondere für BMP-2/4 vor [88, 108]. BMP-2/4, ein phylogenetisch hoch
konserviertes Signalprotein, ist neben seiner pleiotropen Beteiligung an multiplen
Induktionsprozessen während der embryonalen Entwicklung ein essentieller Faktor
der Osteogenese und der Knochenreparation [68, 107]. Die ektope Knochenbildung
im Muskelgewebe durch BMP-2/4-Expressionsinduktion wurde bereits gezeigt
[107]. Die für die Ossifikation maßgebliche Funktion von BMP-2/4 liegt in der
Rekrutierung mesenchymaler Stammzellen für eine osteogene Differenzierung sowie
in der Induktion der für die Proliferation und Maturation von Osteoblasten
notwendigen Transkriptionsfaktoren Msx-1, Runx-2 und der alkalischen Phosphatase
in Osteoblasten [68]. Der Einsatz von BMP-2/4 als Protein bzw. dessen stimulierte
Expression durch Gentransfer stellt eine Möglichkeit zur Osteoinduktion dar; die
Applikation von BMP-2/4 resultierte klinisch in einer induzierten
Knochenneubildung [14]. Derzeit ungeklärt ist jedoch die für klinische
Anwendungen optimale, lokale BMP-2/4-Konzentration, da endogenes BMP-2/4 in
kaum messbar niedriger Konzentration an der Regulation der physiologischen

17
Prozesse beteiligt ist, klinisch-therapeutisch und experimentell jedoch in einer
10.000-100.000-fach höheren Konzentration benötigt wird [69, 70, 87].
2.2.2. Grenzen der KEMien
Durch den Einsatz von KEMien wird das Problem der limitierten Verfügbarkeit und
der Donormorbidität in der Entnahmeregion umgangen [8, 20, 46, 85, 86]. Eine
Vielzahl von kommerziell verfügbaren KEMien findet derzeit ihren Einsatz, um
verschiedene Möglichkeiten einer gesteuerten Geweberegeneration auszutesten [85,
86, 101, 102]. Der Einfluss von Mikrostruktur, Material, Oberflächenarchitektur und
beschichteten Substanzen wurde in zahlreichen klinischen und experimentellen
Studien angesprochen [60, 111]. Dennoch gibt es einige ungelöste Probleme. Die
Integration von alloplastischen Materialien in den Knochen und ins Weichgewebe ist
nicht optimiert und besonders die Mikroumgebung von solchen Ersatzmaterialien
wird nicht moduliert, um Knochenformation, -differenzierung und –reifung zu
fördern [75, 77, 90, 93, 101]. Biodegradierbare in-vitro Matrizes oder gelartige
Lösungen könnten als Träger für Knochendifferenzierungsfaktoren wie BMP dienen,
um so die Knochenregeneration zu induzieren und die ossäre Integration von
KEMien zu ermöglichen, was die Defektregeneration verbessert und beschleunigt
[29, 103]. Eine Kombination aus KEMien von fester Beschaffenheit wie z.B. TCP-
basierten Materialien und Hydrogel-Polymer-Gemischen könnte die Möglichkeit
einer kontrollierten Abgabe von Differenzierungsfaktoren, Zytokinen und
Wachstumsfaktoren liefern [29].
2.2.2.1. Verwendetes KEM
Die biologischen Eigenschaften des biphasischen Kalziumphosphats, der
Straumann® Bone Ceramic, sind ein HA/TCP-Verhältnis von 60/40, eine
Kristallinisation von 100%, eine Porengröße von 100-500 µm, eine absolute
Porosität von 90% und eine Partikelgröße von 0,5-1 mm. Nach 24 Wochen heilte das
HA/TCP-Material mit reifem Lamellenknochen und direktem Kontakt zwischen
Knochen und verbleibendem Ersatzmaterial [43].

18
2.2.2.2. Verwendete Membranen
PEG-Hydrogele mit hydrolysierbaren Esterbindungen können durch das Reagieren
eines mehrarmigen PEG-Akrylats mit einem mehrarmigen PEG-Thiol erzeugt
werden. In schwach basischer Umgebung verbinden sich die Enden der PEG-
Moleküle durch eine höchst selektive Additionsreaktion, indem sie ein elastisches
Gel-Netzwerk formen [28].
Die Reaktion ist eine Additionsreaktion, d.h. bei der Vernetzung werden keine
Moleküle abgespalten [28].
Weil diese Vernetzungsreaktion schnell und nur bei physiologischem pH und
physiologischer Temperatur fortläuft (Thiole sind einige 100mal reaktiver gegenüber
Akrylaten als Aminen) [28], kann sie in der Gegenwart von Gewebe und Zellen
ablaufen. Triethanolamin/HCl dient als basischer Puffer. Die Gelierungszeit hängt
vom pH und von der Triethanolaminkonzentration ab [28]. Bei physiologischen
Bedingungen bildet sich das Hydrogel in einigen Minuten [28].
Die Esterbindungen der Akrylatgruppe werden bei Wasserkontakt hydrolysiert:
Die abschließenden Abbauprodukte sind mehrarmige PEG-OH und mehrarmige
PEG-Thioetherpropionsäure-Moleküle [28].
Die Hydrolyserate hängt, außer vom pH und der Temperatur, hauptsächlich von der
Vernetzungsdichte des Gels ab, z.B. von der Anzahl und Länge der Arme der PEG-
Moleküle. Zwei verschiedene Gelarten sind verfügbar, welche sich zu homogenen
klaren und farblosen Lösungen innerhalb von ca. 11 (MX10) und 120 Tagen
(Membragel) abbauen, wenn sie in PBS bei 37°C und pH 7,4±0,1 aufbewahrt
werden.

19
2.2.3. GSK-3β als Schlüsselregulator der Knochendifferenzierung
2.2.3.1. Interaktion mit pleiotropen Induktoren der ossären Differenzierung
a) Wnt/β-catenin-Signalweg
Die Glykogensynthasekinase-3β ist eine allgegenwärtig exprimierte Kinase, die
diverse zelluläre Prozesse reguliert, angefangen vom Stoffwechsel bis zur
Spezifizierung des Zellschicksals [44]. Obwohl sie zuerst als Kinase beschrieben
wurde, die die Glykogensynthese reguliert, hat man jetzt verstanden, dass GSK-3β
Signalwege der Entwicklung einschließlich des Wnt-Weges reguliert [44].
Wnts sind eine Familie von 19 abgesonderten Kohlenhydrat- und Lipid-modifizierten
Proteinen, die eine wichtige Rolle in der Entwicklung und Erhaltung von Organen
und Geweben, einschließlich Knochen, spielen [5]. Obwohl Wnt-Proteine in
mehreren Signalwegen wirken, um Zellwachstum, -differenzierung, -funktion
und -tod zu regulieren, scheint der Wnt/β-catenin-Signalweg besonders wichtig für
die Knochenbiologie zu sein [50]. Der Wnt/β-catenin-Signalweg wird auch canonical
Wnt-Weg genannt.
Eine Aktivierung des Wnt/β-catenin-Weges erfolgt durch Bindung von Wnt an den
aus 7-transmembranären Domänen bestehenden Rezeptor Frizzled (Fzd), welche eine
intrazelluläre Signalkaskade auslöst [12, 13, 47, 50]. Diese Bindung bewirkt die
Bildung des Transmembranrezeptorkomplexes aus Fzd und dem Korezeptor Low-
density lipoprotein receptor-related protein 5 oder 6 (Lrp 5 oder 6) [12, 13, 47, 50].
Der zytoplasmatische Teil von Fzd interagiert mit und aktiviert das Disheveled-
Protein, welches die Aktivität von GSK-3β hemmt [12, 13, 47, 50]. GSK-3β
verhindert in aktiver Form, welche bei Fehlen von Wnt besteht, die Dissoziation von
β-catenin vom Multiprotein-Komplex bestehend aus APC (adenomatöse polyposis
coli), Axin und GSK-3β [12, 13, 47, 50]. Dies bewirkt die Phosphorylierung,
Ubiquitinylierung und proteosomale Degradierung von β-catenin [12, 13, 47, 50]. Ist
die Aktivität von GSK-3β durch Disheveled gehemmt, kann β-catenin vom APC-
Komplex dissoziieren und in den Nukleus migrieren [12, 13, 47, 50]. Im Nukleus
angekommen assoziiert β-catenin mit den DNA-Bindungsproteinen T-cell
factor/lymphoid enhancer factor (TCF/LEF) und wird zu einem aktiven
Transkriptionsfaktor [12, 13, 47, 50]. Dieser Komplex aktiviert die Transkription
einer Vielzahl von Genen. Läuft der Wnt-Weg im Osteoblastprogenitor ab, ist das
Endprodukt die Proliferation und Differenzierung von knochenspezifischen Zellen
[12, 13, 47, 50].

20
GSK-3β ist ein Inhibitor des canonical Wnt-Weges, der selbst vom Wnt-Signal
unterdrückt wird (siehe Abb.1) [47].
b) Regulation des Wnt/β-catenin-Signalweges
Der Wnt-Weg kann extrazellulär, an der Zellmembran, im Zytosol und im Nukleus
reguliert werden [5, 30, 50, 72, 112]. Induziert wird der β-catenin-Signalweg durch
die Bindung eines Wnt-Moleküls an seinen aus Fzd und Lrp5 oder Lrp6 bestehenden
Rezeptors [5, 30, 50, 72, 112]. Ein wichtiger Regulationsmechanismus beinhaltet die
löslichen DKK-Proteine, welche an den Korezeptor Lrp5/6 binden und damit die
Formation des Fzd-Lrp5/6-Wnt-Komlexes verhindern, so dass die Induktion des
Wnt-Weges ausbleibt [5, 30, 50, 72, 112]. Desweiteren kann sich in Anwesenheit der
Transmembranproteine Kremen 1 oder Kremen 2 ein dritter Komplex zwischen
Kremen, DKK und Lrp5/6 bilden. Dies reduziert zudem die Verfügbarkeit des
Korezeptors Lrp5/6 für seinen Liganden Wnt (siehe Abb. 1) [5, 30, 50, 72, 112].
Alternative extrazelluläre Inhibitoren des Wnt-Weges sind die Mitglieder der
secreted frizzled-related proteins (sFRPs), die eine Domäne besitzen, die an Wnts
bindet. Somit kann Wnt nicht mehr an seinen Rezeptorkomplex binden [5, 30, 72].
In ähnlicher Weise agiert der Wnt inhibitory factor-1 (WIF-1), der eine
Unterdrückung des Wnt-Weges bewirkt [5, 30, 50, 72].
Zudem lässt sich SOST (Sclerostin) als extrazellulärer Wnt-Antagonist anführen.
SOST bindet an Lrp5/6 und verhindert so die Bildung des Komplexes aus Lrp5/6,
Fzd und Wnt [5, 30, 50, 72] .
Im Zytosol liegen Regulationen auf dem Level einer jeden Komponente des tertiären
GSK-3β-Axin-APC-Komplexes vor, die zur Phosphorylierung und Degradierung
von β-catenin führen [5, 30, 50, 72].
Schließlich lässt sich der Wnt-Weg auf nuklearer Ebene durch die Verfügbarkeit der
Trankskriptionsfaktoren TCF, LEF und Kofaktoren beeinflussen [5, 30, 50, 72].
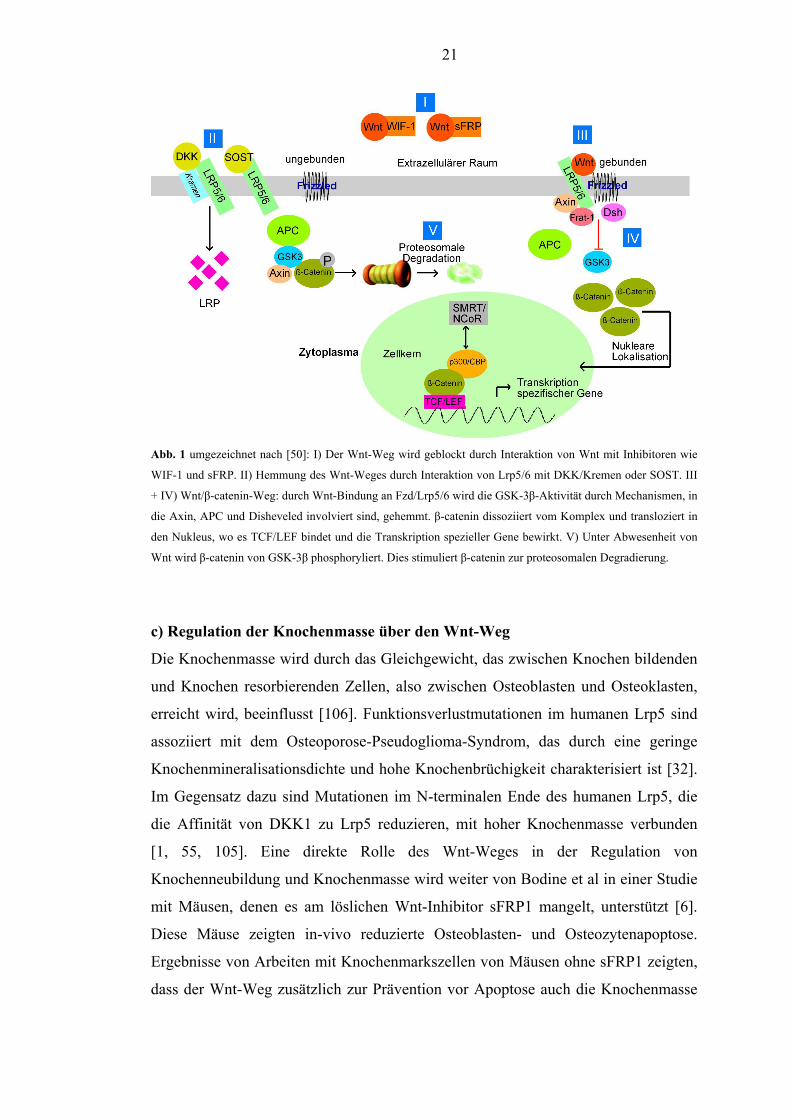
21
Abb. 1 umgezeichnet nach [50]: I) Der Wnt-Weg wird geblockt durch Interaktion von Wnt mit Inhibitoren wie
WIF-1 und sFRP. II) Hemmung des Wnt-Weges durch Interaktion von Lrp5/6 mit DKK/Kremen oder SOST. III
+ IV) Wnt/β-catenin-Weg: durch Wnt-Bindung an Fzd/Lrp5/6 wird die GSK-3β-Aktivität durch Mechanismen, in
die Axin, APC und Disheveled involviert sind, gehemmt. β-catenin dissoziiert vom Komplex und transloziert in
den Nukleus, wo es TCF/LEF bindet und die Transkription spezieller Gene bewirkt. V) Unter Abwesenheit von
Wnt wird β-catenin von GSK-3β phosphoryliert. Dies stimuliert β-catenin zur proteosomalen Degradierung.
c) Regulation der Knochenmasse über den Wnt-Weg
Die Knochenmasse wird durch das Gleichgewicht, das zwischen Knochen bildenden
und Knochen resorbierenden Zellen, also zwischen Osteoblasten und Osteoklasten,
erreicht wird, beeinflusst [106]. Funktionsverlustmutationen im humanen Lrp5 sind
assoziiert mit dem Osteoporose-Pseudoglioma-Syndrom, das durch eine geringe
Knochenmineralisationsdichte und hohe Knochenbrüchigkeit charakterisiert ist [32].
Im Gegensatz dazu sind Mutationen im N-terminalen Ende des humanen Lrp5, die
die Affinität von DKK1 zu Lrp5 reduzieren, mit hoher Knochenmasse verbunden
[1, 55, 105]. Eine direkte Rolle des Wnt-Weges in der Regulation von
Knochenneubildung und Knochenmasse wird weiter von Bodine et al in einer Studie
mit Mäusen, denen es am löslichen Wnt-Inhibitor sFRP1 mangelt, unterstützt [6].
Diese Mäuse zeigten in-vivo reduzierte Osteoblasten- und Osteozytenapoptose.
Ergebnisse von Arbeiten mit Knochenmarkszellen von Mäusen ohne sFRP1 zeigten,
dass der Wnt-Weg zusätzlich zur Prävention vor Apoptose auch die Knochenmasse

22
durch Stimulation der Osteoblastendifferenzierung und –replikation erhöhen kann
[6]. Eine Studie von Day et al zeigte, dass sich mesenchymale Progenitoren in der
Abwesenheit von β-catenin während der embryonalen Skelettogenese in
Chondrozyten anstatt Osteoblasten differenezieren [19]. Dies deutet darauf hin, dass
β-catenin erforderlich ist, um die Chondrogenese zu unterdrücken [19, 80].
Ein Mechanismus, durch den der Wnt-Weg die Knochenbildung steigert, ist die
Stimulation der Entwicklung von Osteoblasten [3, 50, 51]. Eine Rolle des Wnt/β-
catenin-Signalweges in diesem Prozess wird durch einen in-vitro Beweis unterstützt
[3, 50, 51]. Die Hemmung der enzymatischen Aktivität von GSK-3β mit
Lithiumchlorid (LiCl) stimuliert zum Beispiel mesenchymale Vorläufer, sich in
Osteoblasten zu differenzieren [3, 41, 50, 51]. Diese Auffassung wird durch
Beobachtungen mit Wnt3a, Wnt1, Wnt10b und dem wesentlich aktiveren β-catenin,
die alle den β-catenin-Signalweg aktivieren und die Osteoblastogenese stimulieren,
unterstützt, während DKK1, Inhibitor dieses Weges, die Osteoblastogenese
herabsenkt [3, 41, 50, 51]. Die Aktivierung des Wnt-Weges hemmt auch die
Adipogenese von mesenchymalen Vorläufern (siehe Abb. 2), was klinische
Wichtigkeit bei gegebener positiver Wechselbeziehung zwischen Knochenmark von
adipösem Gehalt und Knochenbrüchen hat [4, 50, 67, 78]. Aktives β-catenin wurde
von Glass et al in einer Studie mit Mäusen in Osteoblasten überexprimiert [31].
Diese Mäuse manifestierten einen Phänotyp von Osteopetrose; allerdings waren
keine Veränderungen in der Osteoblastenaktivität zu beobachten [31]. Anstatt dessen
waren Knochenresorption und Osteoklastogenese infolge von erhöhter Expression
von OPG, ein Köderrezeptor für RANKL, mangelhaft [31]. Der β-catenin-Weg
reguliert demnach die Osteoklastogenese durch seine Wirkung auf die Expression
von OPG und RANKL [31, 38, 50].

23
Abb. 2 umgezeichnet nach [50]: Der Wnt/β-catenin-Signalweg reguliert die Osteogenese durch mehrere
Mechanismen. Wnts unterdrücken alternative mesenchymale Differenzierungswege wie die Adipozyten- und
Chondrozytendifferenzierung und begünstigen die Osteoblastendifferenzierung, -proliferation
und -mineralisationsaktivität während sie die Osteoblastenapoptose blockieren. Durch den Anstieg des
Verhältnisses von OPG zu RANKL unterdrückt β-catenin die Osteoklastogenese.
Grüne Pluszeichen zeigen die positiven, rote Minuszeichen die hemmenden Effekte von Wnt (MSC =
mesenchymale Stammzelle).
d) Klinischer Bezug: Glucokortikoide und Lithium
Glucokortikoide (GC) werden ausgiebig für die Behandlung von Autoimmun- und
Entzündungskrankheiten genutzt [95, 96]. Außerdem finden GC in Kombination mit
anderen Medikamenten vielfachen Einsatz, um Entzündungen im Zusammenhang
mit Tumoren des Blutsystems und anderen Tumoren zu reduzieren und um
entzündlich bedingte Implantatabstoßungen zu vermeiden [95, 96]. Der größte
Nebeneffekt der GC-Behandlung ist ein schneller Verlust der Mineralisation und das
ansteigende Risiko für Knochenbrüche [95, 96]. Obwohl behauptet wurde, dass die
Knochenresorption zur GC-induzierten Osteoporose beiträgt, weiß man heute, dass
der Hauptmechanismus, der zum Knochenmineralisationsverlust, der durch die
Langzeitbehandlung mit GC induziert wird, führt, die Schwächung der
Osteoblastenfunktion und der Knochenneubildung ist [95, 96].
In einer Studie von Smith et al wurde gezeigt, dass die GC-vermittelte Aktivierung
von GSK-3β zur Hemmung des Wnt-Signalweges und der TCF/LEF-
Transkriptionsaktivität in differenzierenden Osteoblasten führt [96]. GC verursachen
also über eine Aktivierung von GSK-3β eine Hemmung der Knochenformation, was
langfristig zur Osteoporose führt.

24
Diese hemmende Wirkung wird allerdings bei Lithiumgabe nicht mehr beobachtet
[96]. Lithiumionen werden in Form von LiCl oder Lithiumkarbonat abgegeben und
sind klinisch zugelassene GSK-3β-Inhibitoren [15, 100]. Lithium wurde ursprünglich
bei Problemen mit der Gehirnfunktion und bei periodischen
Stimmungsschwankungen wie Depressionen eingesetzt, bis man herausfand, dass es
die enzymatische Aktivität von GSK-3β hemmt, was seinen Einsatz bei
Osteoporosepatienten und begleitend zur GC-Gabe nachvollziehbar macht [45]. In
einer tierexperimentellen Studie von Clement-Lacroix et al wurde der
pharmakologische GSK-3β-Inhibitor LiCl verwendet, um zu zeigen, dass Wnt über
den Wnt/β-catenin-Signalweg die Knochenmasse vermehrt [15]. Das Auftragen von
LiCl für 4 Wochen steigerte die Knochenneubildungsrate und die Anzahl der
Osteoblasten stark [15]. Die Hemmung von GSK-3β reduzierte ebenfalls die Anzahl
von Adipozyten im Knochenmark über diesen Zeitraum [15]. LiCl beeinflusst auch
andere Signalwege neben dem Wnt-Weg und GSK-3β reguliert viele Proteine neben
β-catenin. Die Tatsache allerdings, dass LiCl β-catenin stabilisiert, die TCF/LEF
basierte Transkriptionsakivität und die Expression derer Zielgene steigert,
unterstützt nachhaltig einen Mechanismus, der durch den Wnt/β-catenin-Signalweg
vermittelt wird [15, 35, 50, 95, 100].
2.2.3.2. Interaktion des Wnt-Weges mit dem TGF-β/BMP-Weg
Die gegenseitige Beeinflussung des TGF-β/BMP- und Wnt-Weges ist seit langer Zeit
bekannt [13]. Die beiden Signalwege wirken das ganze Leben hindurch aufeinander
und auf molekularer Ebene interagieren sie auf mehreren Levels [13]. Erstens
regulieren TGF-β/BMP und Wnt ihre Ligandenproduktion, die kritisch für das
Aufbauen eines extrazellulären Gradienten dieser Morphogene während der
embryonalen Entwicklung ist, gegenseitig [13]. In einer Studie von Chen et al wurde
gezeigt, dass die Expression von Wnt7a, Wnt10b, Wnt11 und Wnt12 mRNA in einer
Dosis abhängigen Weise durch BMP-2 hochreguliert wird [13]. In ähnlicher Weise
stieg die Expression der Wnt-Rezeptoren Fzd1, Fzd3, Fzd10 und Lrp6 durch BMP-2-
Behandlung an [13]. Zweitens befindet sich die am besten bestimmte Lokalisation
der TGF-β/BMP/Wnt-Interaktion im Nukleus, wo der Smad/β-catenin/LEF-
Proteinkomplex eine Vielzahl von gemeinsam benutzten Zielgenen, oft in
synergistischer Art und Weise, reguliert [13]. Im Nukleus wird die Interaktion
zwischen Smad4 und β-catenin durch LEF1 vermittelt [36]. Ihre Kooperation ist

25
auch in Säugetierzellen bekräftigt [36]. Drittens haben jüngste Forschungen eine
zytoplasmatische Interaktion zwischen den Komponenten dieser Wege als neue
Mechanismen für die Feinabstimmung ihrer jeweiligen Signalwege identifiziert [13,
36]. Hierbei interagieren Smad7, Komponente des TGF-β/BMP-Weges und
β-catenin bzw. Axin, Komponenten des Wnt-Weges [36]. Smad7 kann β-catenin
direkt binden, dessen Degradierung fördern und dadurch die Wnt-Aktivität
reduzieren [36]. Ein jüngster Bericht beschrieb, dass Smad7 an Axin binden kann
[36]. Dies lässt β-catenin vom Axin-APC-GSK-3β-Komplex dissoziieren und
stabilisiert β-catenin. Zudem wurde von einer Smad7-Axin-Interaktion berichtet, die
das Ubiquitinylieren und die Degradierung von Smad7, induziert durch Axin,
bewirkt (siehe Abb.3) [36]. Die molekularen Details solcher Interaktionen sind oft
von Spezies zu Spezies erhalten [36]. Dies hebt die biologische Relevanz von
übergreifenden Signalwegen zudem hervor. Die beiderseitige Regulation der TGF-
β/BMP- und Wnt-Liganden sind wichtig und vorherrschend während der frühen
Entwicklung, beispielsweise bei der Festlegung der Rechts-Links-Körperachse, der
Formgebung des Mesoderms oder der neuralen Entwicklung, allerdings bestehen sie
auch im ausgewachsenen Gewebe [36, 91].
Abb.3 umgezeichnet nach [36]: TGF-β/BMP- und Wnt-Weg mit Interaktionen: TGF-β/BMP und Wnt können
über die Ligandenproduktion des anderen bestimmen. Proteininteraktionen im Zytoplasma, wie die Smad7-Axin-
Bindung, verbinden die beiden Signalwege ebenfalls. Die häufigste Wechselwirkung zwischen beiden Wegen
ereignet sich im Nukleus, wo Smad und LEF/β-catenin synergistisch eine Reihe von gemeinsamen Zielgenen
regulieren.

26
2.2.3.3. HOX-Gene: Ossäre Regeneration im Kiefer und extrakranial
Hox-Gene sind hoch konservierte Gene, deren Expression segmental gegliedert ist
(siehe Abb.4) [52]. Eine Ausnahme von der segmentalen Gliederung stellt das
Gehirn dar [52]. Dieser Bereich exprimiert keine Hox-Gene, wie auch der
1. Kiemenbogen und die daraus entstehende Mandibula [52]. Dies bestätigt eine
Studie von Creuzet et al, in der Hoxa2, Hoxa3 und Hoxb4 in den Hox-negativen
Bereich transpositioniert wurden, mit dem Ergebnis von schweren Defekten im
fazialen Skelett, wie das Fehlen von Ober- und Unterkiefer [17]. Der Hox-Status
bestimmt den Bodyplan. Die Hox-Gene geben vor, welches Gewebe sich in welchem
Bereich entwickelt und bestimmen die Wirkung der einzelnen Zytokine, z.B. wie
BMP, in den verschiedenen Segmenten [52]. So bewirkt Hox10 die Bildung von
lumbalen, Hox11 von sakralen Wirbeln [110]. Hoxa13 ist für die regelrechte Bildung
der distalen Extremitäten verantwortlich und reguliert die BMP-2 und -7-Expression
[48]. Jedes Hox-Gen trägt eine bestimmte Aufgabe in der Entwicklung des
menschlichen Körpers [53]. Kleinste Störungen während der Entwicklung führen zu
Abnormalitäten, die fast jeder Mensch unwissentlich in sich hat, oder gar
Missbildungen [53].
Abb.4 umgezeichnet nach [52]: Die segmentale Gliederung der Hox-Genexpression entspricht dem Bodyplan.
Die Hox-Gene spielen zudem bei der skelettalen Entwicklung und der
Knochenregeneration eine entscheidende Rolle [54]. Das fötale Skelett bildet sich
aus der Neuralleiste und dem Mesoderm [54]. Beide Abstammungsarten bringen eine
eigene Stammzellpopulation hervor, um verletztes Gewebe zu regenerieren [54].

27
Leucht et al konnten zeigen, dass die aus der kranialen Neuralleiste stammende
Mandibula mit Stammzellen aus der Neuralleiste heilt, wohingegen die aus dem
Mesoderm stammende Tibia, also extrakranialer Röhrenknochen, auf mesodermale
Stammzellen zurückgreift [54]. Der Austausch der skelettalen Stammzellen
gegeneinander zeigte, dass sich neurale Stammzellen bei Transplantation sowohl in
die Hox-positive Tibia als auch in die Hox-negative Mandibula zu Osteoblasten
differenzieren [54]. Mesodermale Stammzellen hingegen differenzieren sich nur bei
Transplantation in Röhrenknochen zu Osteoblasten, im Mandibuladefekt
differenzieren sie sich zu Chondrozyten [54]. Eine Ungleichheit des exprimierten
Hox-Status der Spender- und Empfängerregion ist für diese Abweichung der
Knochenreparatur verantwortlich [54]. Knochenvorläuferzellen aus der Mandibula
sind Hox-negativ, die allerdings bei Transplantation in einen Tibiadefekt aufgrund
ihrer hohen Plastizität und Verformbarkeit einen Hoxa11-positiven Status annehmen
[54]. Umgekehrt sind Progenitoren aus extrakranialem Röhrenknochen Hox-positiv
und behalten diesen Hox-Status bei Transplantation in einen Hox-negativen
Mandibuladefekt bei [54]. Osteoprogenitoren beider Herkunft unterscheiden sich,
zusätzlich zum Hox-Status und dem embryonalen Ursprung, in der osteogenen
Plastizität und der Proliferation [54]. Dies führt zu einer robusteren in-vivo
Knochenregeneration durch Stammzellen aus der Neuralleiste [54]. Somit sind
Kieferknochen und extrakranialer Röhrenknochen hinsichtlich der Anatomie,
Proliferation, Signaltransduktion und des Hox-Status verschieden [54]. Diese
Charakteristika beeinflussen den Vorgang der Knochenregeneration beim
Erwachsenen und bieten einen Ansatz zur Erklärung der immer stattfindenden
enchondralen Ossifikation auch desmalen Knochens [52, 54, 106].
2.2.4. RANK(L)-OPG-System
2.2.4.1. Relevanz des RANK(L)-OPG-Systems
Knochen wird ständig durch die Synthese einer Knochenmatrix durch Osteoblasten
und die Knochenresorption durch Osteoklasten umgebaut [106]. Störungen in
Entzündungszytokinen, Wachstumsfaktoren und Hormonen verursachen ein
Ungleichgewicht zwischen Osteoblasten- und Osteoklastenaktivitäten und können zu
skelettalen Abnormitäten, wie Osteoporose oder Osteopetrose, führen [106].
Osteoporose ist eine Krankheit, die durch geringe Knochendichte charakterisiert ist
und häufig bei älteren Menschen, besonders Frauen, und bei bettlägrigen Patienten

28
auftritt [71, 106]. Im Gegensatz dazu ist Osteopetrose von abnormal gesteigerter
Knochendichte gekennzeichnet und tritt hauptsächlich als ein Ergebnis von seltenen
erblichen Störungen auf [2, 9, 16, 71, 106]. Die Entdeckung der Einflussfaktoren auf
die Kontrolle der Osteoklasten, und damit auf die Osteoporose, haben die
Knochenforschung weit nach vorne gebracht. Die wichtigsten Faktoren sind der
Receptor activator of nuclear factor-κB (RANK), dessen Ligand RANKL und
Osteoprotegerin (OPG), der Köderrezeptor für RANKL [2, 9, 16, 106].
a) RANKL
RANKL ist ein homotrimeres, transmembranes Protein der tumor necrosis factor
(TNF)-Rezeptor-Superfamilie, das von Osteoblasten, Fibroblasten und T-Zellen
exprimiert wird [9, 71, 106]. RANKL existiert auch in löslicher, nicht
membrangebundener Form (sRANKL) [9, 71, 106]. Die lösliche Form von RANKL
besitzt eine stärkere Aktivität und spielt eine wichtige Rolle bei der durch Tumore
induzierten Osteolyse. sRANKL leitet sich von der membran-gebundenen Form als
Ergebnis entweder eines proteolytischen Spaltprodukts oder alternativ von Spleißen,
ab [106]. Die Expression von RANKL im Osteoblasten wird durch den Wnt- und
BMP-2-Weg reguliert. Eine Unterdrückung der RANKL-Expression geht mit einer
Erhöhung der OPG-Expression einher und umgekehrt [2, 9, 71, 106].
Osteoklastvorläufer interagieren mit Osteoblasten, damit sie sich zu reifen
Osteoklasten differenzieren können. RANKL kontrolliert durch Bindung an RANK
die Differenzierung, Rekrutierung, Aktivität und Lebensdauer der Osteoklasten und
auch vieler anderer Zellen (siehe Abb. 5) [71].
b) RANK
RANK wird neben Osteoklasten auch in der Brustdrüse, Prostata, in Immunzellen,
im skelettalen Muskel, Thymus, Leber, Darm, Prankreas, u.a. exprimiert [2, 9, 31,
72, 106]. Die Bindung von RANKL an RANK hat in den verschiedenen Zellen
unterschiedliche Folgen (siehe Abb.5). RANK ist ein homotrimeres transmembranes
Protein der Osteoklastprogenitormembran [106]. Die Bindung von RANKL an
RANK bewirkt die Aktivierung einer intrazellulären Signalkaskade im
Osteoklastprogenitor, die eine Differenzierung des Osteoklastvorläufers zum
maturen, mehrkernigen Osteoklasten zur Folge hat (siehe Abb.6) [2, 9, 31, 72, 106].
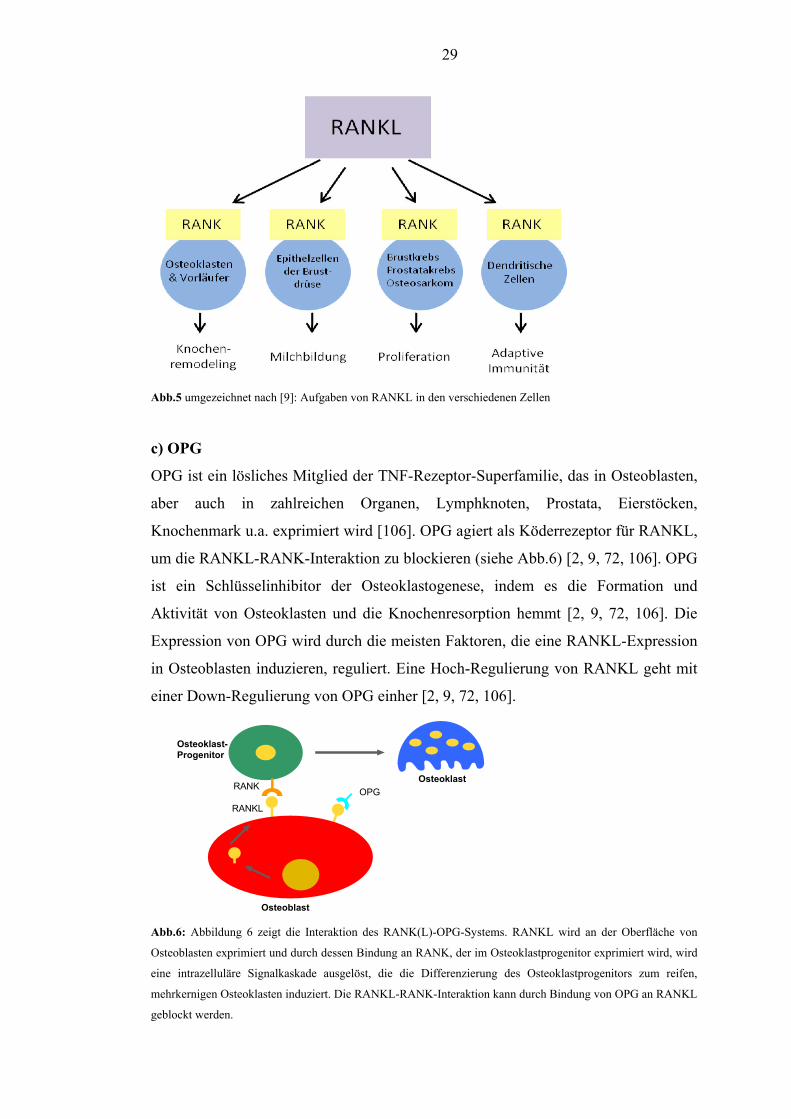
29
Abb.5 umgezeichnet nach [9]: Aufgaben von RANKL in den verschiedenen Zellen
c) OPG
OPG ist ein lösliches Mitglied der TNF-Rezeptor-Superfamilie, das in Osteoblasten,
aber auch in zahlreichen Organen, Lymphknoten, Prostata, Eierstöcken,
Knochenmark u.a. exprimiert wird [106]. OPG agiert als Köderrezeptor für RANKL,
um die RANKL-RANK-Interaktion zu blockieren (siehe Abb.6) [2, 9, 72, 106]. OPG
ist ein Schlüsselinhibitor der Osteoklastogenese, indem es die Formation und
Aktivität von Osteoklasten und die Knochenresorption hemmt [2, 9, 72, 106]. Die
Expression von OPG wird durch die meisten Faktoren, die eine RANKL-Expression
in Osteoblasten induzieren, reguliert. Eine Hoch-Regulierung von RANKL geht mit
einer Down-Regulierung von OPG einher [2, 9, 72, 106].
Abb.6: Abbildung 6 zeigt die Interaktion des RANK(L)-OPG-Systems. RANKL wird an der Oberfläche von
Osteoblasten exprimiert und durch dessen Bindung an RANK, der im Osteoklastprogenitor exprimiert wird, wird
eine intrazelluläre Signalkaskade ausgelöst, die die Differenzierung des Osteoklastprogenitors zum reifen,
mehrkernigen Osteoklasten induziert. Die RANKL-RANK-Interaktion kann durch Bindung von OPG an RANKL
geblockt werden.
Osteoklast- Progenitor
Osteoklast RANK
RANKL
Osteoblast
OPG

30
2.2.4.2. Signalweg der Osteoklastogenese
Unter physiologischen Bedingungen bindet RANKL, das von Osteoblasten
produziert wird, an RANK, das an der Oberfläche des Osteoklastvorläufers
exprimiert wird [9, 106]. RANK rekrutiert das Adapterprotein TRAF6, welches
NF-κB aktiviert [9]. NF-κB transloziert in den Nukleus und steigert dort die c-Fos-
Expression [9]. C-Fos interagiert mit dem Transkriptionsfaktor nuclear factor of
activated T cells c1 (NFATc1), um die Transkription von Osteoklasten spezifischen
Genen auszulösen [9]. OPG kann durch Bindung von RANKL dessen Interaktion mit
RANK blockieren, so dass im Osteoklastprogenitor die Aktivierung der
intrazellulären Signalkaskade über TRAF6 ausbleibt und somit die Differenzierung
zur maturen, mehrkernigen Riesenzelle, dem Osteoklasten (siehe Abb.7) [9, 22] .
Abb.7 umgezeichnet nach [9]: Signalweg der Osteoklastogenese stark vereinfacht dargestellt
2.2.4.3. Interaktion des Wnt-Weges mit dem RANK(L)-OPG-System
In einer Studie von Fujita et al wurde gezeigt, dass der Wnt-Weg mit dem RANK(L)-
OPG-System interagiert [30]. Wnt/β-catenin fördert die Knochenneubildung, indem
es als positiver Regulator von Osteoblasten und als negativer Regulator der
Osteoblasten abhängigen Osteoklastogenese durch das Hoch-Regulieren von OPG
und Down-Regulieren von RANKL in Knochenvorläuferzellen fungiert [30]. In
dieser Studie wurden mesenchymale Progenitoren, die eine Differenzierung zu

31
Osteoblasten in-vitro durchlaufen, verwendet, um zu zeigen, dass die Wnt-
Inhibitoren DKK1 und DKK2 die Osteoklastogenese von frühen Osteoprogenitoren
erhöhen. DKK1 und DKK2 sind befähigt, die Wnt/β-catenin induzierte Down-
Regulierung von RANKL und die gleichzeitige Hoch-Regulierung seines Inhibitors
OPG zu mindern [30, 74]. Die BMP-2 abhängige Unterdrückung von OPG wurde
durch DKK1 und DKK2 allerdings nicht beeinflusst [30, 74].
Eine Studie von Sato et al belegte die Auffassung, dass die OPG- und RANKL-
Expression durch den Wnt/β-catenin-Weg reguliert wird [80]. Es wurde gezeigt, dass
sowohl der intrazelluläre Signalweg von Wnt/β-catenin als auch von BMP-2 eine
wichtige Rolle bei der Regulation der Expression von OPG und RANKL spielen.
BMP-2, der Wnt/β-catenin-Weg, wie auch die Inaktivierung von GSK-3β erhöhen
die OPG-Expression und unterdrücken die RANKL-Expression [80]. Der
Wnt/β-catenin-Weg reguliert die OPG-Transkription über eine Promoter-Region, die
die Bindungsstellen 2 und 4 enthält, welche auf die zwei β-catenin spezifischen
TCF/LEF1-Bindungsstellen ansprechen [80]. BMP-2 steigert die Wnt/β-catenin
abhängige transkriptionale Aktivität des OPG-Promoters [80]. Diese BMP-2-
Stimulation bewirkt, dass Smad1 und Smad4, Mitglieder der BMP-Signalkaskade,
mit der Wnt/β-catenin spezifischen TCF/LEF1-Bindungsstelle interagieren [80].
OPG ist als Zielgen des Wnt/β-catenin- und BMP-Weges identifiziert [31]. Die
OPG-Transkription wird durch die TCF/LEF1-Bindungsstellen des OPG-Promoters
reguliert, da auch Smad4, Mitglied des BMP-Weges, und β-catenin, Mitglied des
Wnt-Weges, an TCF/LEF1 binden [31, 38, 80, 98].
2.2.4.4. Therapeutischer Ansatz am RANK(L)-OPG-System: Denosumab
Der postmenopausale Knochendichteverlust lässt sich hauptsächlich auf einen
Östrogenmangel zurückführen [71]. Osteoklasten, Osteoblasten und Osteozyten
tragen reichlich passende Hormonrezeptoren [71]. Prämenopausal wird der
Knochenstoffwechsel durch reichlich zirkulierendes Östrogen im Gleichgewicht
gehalten, mit zunehmendem Alter und damit verbundener Abnahme der
Ovarialfunktion sinkt der Östrogenspiegel [71]. Der Knochenumbau kippt in
Richtung Osteoporose, obwohl neben der Osteoklasten- auch die
Osteoblastenaktivität zunimmt [71]. Der OPG-Spiegel sinkt und gleichzeitig ist eine
zwei- bis dreifach erhöhte Menge von RANKL im Knochenmark zu finden [71].

32
Mit Entdeckung des RANK/RANKL/OPG-Systems vor rund 11 Jahren begann die
Suche nach einer therapeutischen Eingriffsmöglichkeit in dieses System, um die
pathologischen Anzeichen der Osteoporose zu stoppen, zu verringern oder gar zu
beheben [71]. Die Behandlung von humanen Osteoblasten in-vitro mit 17β-Östradiol
bewirkte einen Anstieg der OPG-Produktion, was die Osteoklastenaktivität durch
Blockierung von RANKL verminderte [71]. Nachdem bereits vor vielen Jahren
mittels verschiedener Tiermodelle das Potenzial eines pharmakologischen Eingriffs
in das RANKL/OPG-System gezeigt werden konnte, hatte die pharmazeutische
Industrie 2004 mit einer gezielten Produktentwicklung und –testung am Menschen
begonnen [71]. Der humane, monoklonale Antikörper Denosumab, der mit hoher
Affinität an RANKL bindet, die Rolle von OPG demnach funktionell imitiert, und
somit den Osteoklasten blockiert, hat die Phase-I-, Phase-II- und Phase-III-Studien
an postmenopausalen osteoporotischen Frauen erfolgreich durchlaufen [71]. Die
Phase-I-Studie zeigte Denosumab (vorgesehener Handelsname Prolia®) als effektive
und sichere osteoprotektive Substanz, die nach einmaliger Applikation einen lang
anhaltenden, jedoch reversiblen Effekt auf die Osteoklasten ausübt [71]. Nach diesen
überzeugenden Ergebnissen folgte 2006 die publizierte Dosisfindungsstudie, die eine
effektive Dosierung bei dreimonatlicher Gabe von 30 mg und halbjährlicher Gabe
von 60 mg an den Tag legte [71]. Erneut wurde der positive Effekt von Denosumab
auf den Knochenstoffwechsel gezeigt, der nun auch durch die Zunahme der
Knochenmineralisationsdichte bestätigt wurde. Eine Langzeitverträglichkeit der
Denosumab-Therapie über 48 Monate, die Reversibilität der Einflüsse auf die
Osteoklastogenese sowie das erneute Ansprechen nach Absetzen der Therapie wurde
gezeigt [71]. Bei der 2009 erfolgreich abgeschlossenen Phase-III-Studie wurde
postmenopausalen osteoporotischen Frauen zweimal jährlich 60 mg als subkutane
Injektion verabreicht [27]. Die Inzidenz von Frakturen wurde dadurch reduziert und
die Knochenmineralisationsdichte an allen untersuchten Skelettstellen wurde erhöht.
Die Marktzulassung von Denosumab (Prolia®) wird aufgrund dieser positiven
Ergebnisse noch im Jahr 2010 erwartet [27].
Denosumab stellt nach derzeitiger Datenlage eine ausgezeichnete, verträgliche
osteoprotektive Substanz dar, die Osteoporosepatienten hoffen lässt [71].

33
3. Fragestellung und Ziele
Das Ziel dieses Dissertationsprojektes war die Untersuchung der Einsetzbarkeit einer
neuen PEG-Hydrogelmembran als temporäre Matrix und Abdeckung während einer
gesteuerten ossären Regeneration von critical size-Defekten der Schweinekalotte, die
mit einem biphasischen KEM aus HA und TCP gefüllt wurden. Es wurde untersucht,
ob die PEG-Membran als Träger von BMP-2 transfizierten Osteoblasten in der Lage
ist, die osteogene Stimulation kontrolliert an den Defekt abzugeben.
Im Vordergrund stand hierbei die Untersuchung des Einflusses dieser PEG-Membran
auf die Wnt-vermittelte, osteogene Rekrutierung und Maturation mesenchymaler
Osteoblast- und Osteoklastprogenitoren und die Evaluation der ossären
Differenzierungs- und Proliferationsmarker OPG, RANKL, GSK-3β und deren
Antagonist DKK1.
Dazu wurden zunächst die immunhistochemischen Färbungen von DKK1, GSK-3β
und RANKL etabliert. Anschließend folgte die immunhistochemische Auswertung
der ossären Differenzierungs- und Proliferationsmarker OPG, RANKL, GSK-3β und
deren Antagonist DKK1. Ein weiteres Ziel stellte der Vergleich der verschiedenen
Defektbehandlungen mit dem Kontrolldefekt und dem autologen Knochen dar.
Die zu klärenden Fragen und Ziele lassen sich wie folgt zusammenfassen:
1. Etablierung der immunhistochemischen Färbung von DKK1 und dessen
Nachweis am Tiermodell des Schweins an formalinfixiertem, Technovit
9100®-präserviertem und entakryliertem Knochen
2. Etablierung der immunhistochemischen Färbung von RANKL am Schwein
3. Immunhistochemische, semiquantitative Evaluation der ossären
Differenzierungs- und Proliferationsmarker OPG, RANKL, GSK-3β und
deren Antagonist DKK1
4. Einfluss der biodegradierbaren PEG-Membran auf die Differenzierungs- und
Proliferationsmarker OPG, RANKL, GSK-3β und deren Antagonist DKK1
5. Einfluss von BMP-2 transfizierten Osteoblasten auf die Differenzierungs-
und Proliferationsmarker OPG, RANKL, GSK-3β und DKK1
6. Vergleich des Expressionsverhaltens der Differenzierungs- und
Proliferationsmarker nach Defektbehandlung mit BMP-2 und PEG-Membran
mit dem Kontrolldefekt und dem autologen Knochen.

34
4. Material und Methoden
4.1. Versuchsablauf In diese Studie wurden nach Genehmigung durch die Regierung von Mittelfranken
20 Hausschweine eingeschlossen. Pro Tier wurden 9 Defekte gesetzt, in die das
KEM eingebracht wurde. Die 9 Defekte entsprechen den Versuchsgruppen A-I (siehe
4.3.).
Es wurden je 5 Tiere nach 1, 2, 4 und 12 Wochen Einheilzeit geopfert und
untersucht.
Abb. 8: Versuchsablaufschema
4.2. Verwendete Materialien Verwendet wurden zwei verschiedene Gels, die sich innerhalb von 11 (MX10) oder
120 Tagen (MembraGel) in homogene klare und farblose Lösungen abbauen. MX10
wurde auch in Kombination mit Wachstumsfaktoren verwendet.
Folgende Materialkombinationen kamen zum Einsatz:
a) Straumann® Bone Ceramic: Biphasische Kalzium-Phosphat Keramik (60%
Hydroxylapatit, 40% Trikalziumphosphat, Partikelgröße 0,5 - 1 mm)
b) PEG Hydrogel Membran MX10 (vollständiger Abbau in ca. 11 Tagen)
c) PEG Hydrogel Membran MX10 mit Wachstumsfaktor
d) PEG Hydrogel Membran Membragel (vollständiger Abbau in ca. 120 Tagen)

35
4.3. Versuchsgruppen Im Rahmen dieses Dissertationsprojektes wurden die fett markierten Gruppen
verwendet. Die Gruppen F und H kamen im Rahmen weiterer Dissertationsprojekte
zum Einsatz.
A: Straumann® Bone Ceramic mit PEG MX10
B: Straumann® Bone Ceramic mit PEG MX10 und Membragel
C: Straumann® Bone Ceramic mit Membragel
D: Autologer Knochen mit Membragel
E : Straumann® Bone Ceramic mit PEG MX10 mit BMP-2-transfizierten
Zellen versetzt und Membragel
F : Straumann® Bone Ceramic mit PEG MX10 mit PDGF versetzt und
Membragel
G: Straumann® Bone Ceramic mit PEG MX10 mit BMP-2-transfizierten
Zellen versetzt
H: Straumann® Bone Ceramic mit PEG MX10 mit PDGF versetzt
I: Kontrolldefekt (leer)
Abb. 9: Verteilung der Defekte an der Schädelkalotte
4.4. Immunhistochemische Nachweisreaktionen 4.4.1. Probenaufbereitung Die gewonnenen Proben wurden unverzüglich in 4%igem Paraformaldehyd fixiert,
um so die biologische Aktivität der in den Proben befindlichen Proteine zu erhalten,
und in Technovit 9000® eingebettet. Seit gezeigt wurde, dass die PEG-Membran
löslich in Xylen ist, wurde Aceton anstelle von Xylen für die Rehydratation
verwendet. Darauf folgte eine aufsteigende Alkoholreihe im Entwässerungsbad
(Exakt Apparatebau GmbH, Norderstedt, Deutschland). Eine Pilotstudie sollte die

36
Möglichkeit des Einbettens der Präparate in Paraffin nach Demineralisation zeigen.
Hierbei blieb unglücklicherweise die Morphologie der Defekte und des
Knochengewebes zu den frühen Regenerationszeitpunkten nicht erhalten. Deshalb
wurden alle weiteren Proben ausschließlich in Technovit 9100®, ein
Kaltpolymerisationssystem auf Methylmethacrylatbasis, eingebettet und wie vorher
beschrieben weiterbehandelt. Von den eingebetteten Proben wurden 3µm-
Dünnschnitte am Mikrotom (Leica Mikrosysteme, Heidelberg, Germany) angefertigt.
Für alle weiteren Schritte wurden die Probenschnitte auf Glasobjektträgern, deren
Oberfläche speziell behandelt ist, fixiert.
4.4.2. Methodenetablierung
Zur Detektion der OPG-, GSK-3β-, RANKL- und DKK1 - Expression im
entsprechenden Probenmaterial wurde die Avidin-Biotin-Peroxidase-Methode (ABC-
POX-Methode) und die Labelled Streptavidin-Biotin-Methode (LSAB-Methode)
vergleichend erprobt. Das verwendete Färbekit (Dako REAL Detektion System)
basierend auf der LSAB-Methode zeigte für OPG, GSK-3β, RANKL und DKK1 ein
sensitiveres Färbeergebnis, so dass dieses Verfahren verwendet wurde. Die
immunhistochemische Färbung von DKK1 wurde an formalinfixiertem, Technovit
9100®–präserviertem und entakryliertem Knochen des Hausschweins etabliert. Die
immunhistochemische Färbung für RANKL wurde am Schweinegewebe
selbstständig erarbeitet.
4.4.3. Angewendete immunhistochemische Methoden
ABC-POX-Methode
Die ABC-POX-Methode als indirektes Detektionsverfahren nutzt die hohe Affinität
von Avidin zu Biotin [7]. Der mehrstufige Ablauf ist in Abbildung 10 schematisch
dargestellt. Nach Bindung des Primärantikörpers an das entsprechende Epitop erfolgt
die Inkubation mit einem biotinylierten Sekundärantikörper [7]. Biotin, ein
wasserlösliches Vitamin, eignet sich optimal zur Kopplung an den
Brückenantikörper [7]. Als dritter Schritt wird der Avidin-Biotin-Enzymkomplex
zugesetzt. Im verwendeten Komplex ist die Bindung von drei Biotinmolekülen pro
Molekül Avidin möglich [7]. Somit können sowohl der biotinmarkierte
Sekundärantikörper als auch die biotinmarkierten Meerrettichperoxidasemoleküle

37
binden [7]. Abschließend erfolgt die chromogene Nachweisreaktion durch
enzymatische Umsetzung eines Farbsubstrates [7].
Abb.10 umgezeichnet nach [7]: Die ABC-POX-Methode erfolgt in 3 Schritten: Primärantikörper (Schritt 1), Sekundärantikörper (Schritt 2), Avidin-Biotin-Enzymkomlpex (Schritt 3)
LSAB - Methode
Die LSAB-Methode, ebenfalls ein indirektes Färbeverfahren, verläuft in den ersten
beiden Reaktionsschritten analog zur ABC-POX-Methode [7]. Nach Auftragen des
Primärantikörpers erfolgt in einem zweiten Schritt die Inkubation mit einem
biotinylierten Sekundärantikörper, wobei im verwendeten Kit der
Sekundärantikörper mittels Spacerarm aus sieben Atomen mit Biotin markiert wurde
[7]. So kann jedes biotinylierte Antikörpermolekül mit mehreren
Streptavidinmolekülen reagieren und damit die Empfindlichkeit des Systems steigern
[7].
Das verwendete Streptavidin wird aus dem Bakterium Streptomyces avidinii isoliert
und zeigt gegenüber Avidin weniger unspezifische Reaktionen [7].
Bei der LSAB-Methode liegt das Streptavidin direkt mit einem Enzym verbunden als
Streptavidin-Enzymkonjugat vor [7]. Im vorliegenden Protokoll wird das Enzym
alkalische Phosphatase eingesetzt, wobei auch Peroxidase verwendet werden kann
[7]. Abbildung 11 zeigt schematisch das immunhistochemische Verfahren in drei
Schritten [7].
Die Farbreaktion erfolgt anschließend mit einem geeigneten Substrat-Chromogen.
Schritt 1 Schritt 2 Schritt 3

38
Abb. 11 umgezeichnet nach [7]: LSAB-Methode in 3 Schritten: Primärantikörper (Schritt 1), biotinylierter Brückenantikörper (Schritt 2), Enzymmarkiertes Streptavidin (Schritt 3)
4.4.4. Immunhistochemische Färbung nach der LSAB-Methode
Zur Vorbereitung für die immunhistochemische Färbung im Autostainer durchliefen
die Proben eine aufwendige Vorbehandlung. Zu Beginn erfolgte die Entakrylierung
(4 x 15 Minuten) mittels frischem Methoxyethyl-acetat (MEA) und darauf die
absteigende Alkoholreihe (Ethanol 100% 2 x 2 min, 96% 2 x 2 min, 90% 1 x 2 min,
70% 2 x 2 min). Nach kurzem Spülen in AD (3 min) folgte die Entkalkung in
10%igem EDTA für 20 Minuten. Das anschließende Spülen der Gewebeschnitte
unter fließendem Leitungswasser (10 min), AD (3 x 2 min) und DAKO Waschpuffer
pH 7,4 (3 x 2 min) dient der Rehydrierung. Die Antigendemaskierung erfolgte bei
GSK-3β, OPG und DKK1 als HIER-hitzeinduziertes Epitop Retrieval durch
Citratpuffer (pH 6,0, DAKO S 2369), bei RANKL mittels Target Retrieval Puffer
(pH 9,0, DAKO S 2367). Diese Antigendemaskierung ist nötig, um während der
Fixierung entstehende Aldehydvernetzungen aufzuheben. Diese Vernetzungen,
meistens mit Formalin, können die dreidimensionale Struktur des Antigens
verändern.
Bereits zu Beginn wurde der Puffer in einer Glasküvette im Wasserbad je nach
Färbeprotokoll auf 95° C oder 100° C erhitzt.
Die Proben wurden für 15 bzw. 30 Minuten in den entsprechenden Puffer gegeben.
Nach der Entnahme der Küvette aus dem Wasserbad kühlten die Objektträger vor der
Weiterbehandlung für 15 Minuten bei Raumtemperatur (RT) ab. Kurzes Spülen im
Waschpuffer beendete die Vorbehandlungen (3 x 2 min).
Schritt 1 Schritt 2 Schritt 3

39
Im Autostainer wirkte zunächst der serumfreie Proteinblock (DAKO, Hamburg,
Germany) für 20 Minuten, um unspezifische Proteinbindungen und eine damit
verbundene Hintergrundfärbung zu vermeiden. Der mit „Antibody diluent“ (DAKO
S 2022) verdünnte Primärantikörper reagierte nun 1 Stunde bei RT mit dem Gewebe.
„Antibody diluent“ diente auch zum Befeuchten der für jede Probe mitgeführten
Negativkontrolle, die von hier ab identisch weiterbehandelt wurden. Alle Schnitte
wurden daraufhin mit dem biotinylierten Sekundärantikörper (DAKO, Hamburg,
Germany) und anschließend mit einem Streptavidin-HRP Komplex (Streptavidin
Alkaline Phosphatase) für je 15 Minuten inkubiert. Zwischen den Schritten wurde
mit Waschpuffer gespült. Die Gewebeschnitte reagierten nun zunächst für 8 Minuten
mit dem Chromogen (DAKO, Hamburg, Germany) und nach dem Spülen mit
Waschpuffer erneut für 4 Minuten. Das Chromogen darf maximal 10 Minuten vor
Gebrauch aus einer bestimmten Menge Substratpuffer, den einzelnen Chromogenen
E-F in gleicher Menge und dem Levamisol angemischt werden. Levamisol dient der
Blockierung der endogenen alkalischen Phosphatase. Zwischen den einzelnen
Schritten muss das Gemisch mittels Vortexer vermischt werden. Der Autostainer
beendete sein Programm mit einer AD-Spülung.
Für die Gegenfärbung wurden die Objektträger für 5 Minuten in Hämalaunlösung
getaucht, 5 Minuten mit Leitungswasser gebläut und weitere 5 Minuten in AD
gewässert. Hauptanliegen der Gegenfärbung ist die Kontrasterhöhung, um eine
Verbesserung der Beurteilung des Färbeergebnisses zu erzielen. Abschließend
wurden die Schnitte mit einem Fixierungsmittel auf wässriger Basis eingedeckt
(Aquatex).
4.4.5. Immunhistochemie
Alle folgenden Färbeprotokolle sind im Anhang unter 9.1. hinterlegt.
a) RANKL
RANKL wurde mithilfe eines Primärantikörpers der Firma Abcam (sRANKL ab
62516-100 abcam, Lot:496856) nachgewiesen. Der polyklonale Antikörper,
gewonnen aus dem Kaninchen, war bei einer Verdünnung mit „Antibody diluent“
von 1:150 ideal auszuwerten. Der spezifische Schritt dieser Färbung ist die
Vorbehandlung der Gewebeschnitte im Target Retrieval Puffer pH=9,0 im 95°C
heißen Wasser für 15 Minuten.

40
b) DKK1
Zur Lokalisation einer Expression von DKK1 wurde ein polyklonaler
Primärantikörper der Firma Abcam verwendet (DKK1 ab 61034-50 abcam,
Lot:576372), der aus dem Kaninchen gewonnen wird. Eine Verdünnung mit
„Antibody diluent“ von 1:100 lieferte für die Expressionsbestimmung gute
Ergebnisse. Eine Expression von DKK1 konnte am Defektmodell nachgewiesen
werden. Die spezifischen Schritte dieser Färbung sind die Vorbehandlung im
kochenden Wasserbad für 30 Minuten in Citratpuffer pH=6,0 und im Autostainer die
Behandlung der Schnitte mit Avidin und Biotin je für 15 Minuten.
c) OPG
Dem Expressionsnachweis von OPG diente ein Primärantikörper der Firma Santa
Cruz Biotechnology (OPG (H-249):sc-11383, Lot # C2305). Es handelt sich um
einen polyklonalen Antikörper, der aus dem Kaninchen gewonnen wird. Eine
Verdünnung mit „Antibody diluent“ (DAKO S 2022) von 1:50 erwies sich für die
Expressionsbestimmung als ideal. Der für diese Färbung spezifische Schritt ist die
Vorbehandlung der Schnitte im kochenden Wasserbad für 15 Minuten in Citratpuffer
pH=6,0.
d) GSK-3β
Zur Lokalisation einer Expression von GSK-3β wurde ein Primärantikörper der
Firma Santa Cruz Biotechnology verwendet (p-GSK-3β (Ser 9):sc-11757-R, Lot
# E2508). Der polyklonale Antikörper wird aus dem Kaninchen gewonnen. Eine
Verdünnung mit „Antibody diluent“ von 1:50 brachte gute qualitative Ergebnisse.
Der für diese Färbung spezifische Schritt ist die Vorbehandlung der Schnitte im
kochenden Wasserbad im Citratpuffer pH=6,0 für 15 Minuten.

41
4.4.6. Verwendete Reagenzien Verwendete Primärantikörper
Gesuchtes Protein Antikörper (polyklonal) Verdünnung Hersteller
OPG Kaninchen antihuman/
Schwein OPG-IgG
1:50 Santa Cruz
Biotechnology, USA
GSK-3β Kaninchen antihuman/
Schwein GSK-3β-IgG
1:50 Santa Cruz
Biotechnology, USA
sRANKL Kaninchen antihuman/
Schwein RANKL-IgG
1:150 Abcam,
Cambridge, UK
DKK1 Kaninchen antihuman/
Schwein DKK1-IgG
1:100 Abcam,
Cambridge, UK
Tab. 1: Verwendete Primärantikörper. Für die immunhistochemischen Nachweisreaktionen wurden die
angegebenen polyklonalen, affinitätsgereinigten Primärantikörper in den angegebenen Konzentrationen und
Spezifitäten eingesetzt.
Alle weiteren Reagenzien sind der Tabelle 2 zu entnehmen. Citratpuffer, Target
Retrieval Puffer und Waschpuffer wurden vor Gebrauch mit AD im Verhältnis 1:9
mit anschließender pH-Einstellung verdünnt. Hämalaun wurde vor Verwendung
filtriert, um kristallinisierte Partikel zu eliminieren.
Reagenz Hersteller
Dako REAL Biotinylated Secondary Antibodies
Dako REAL Streptavidin Alkaline Phosphatase
Dako REAL Chromogen
Dako REAL AP Substrate Buffer
Dako REAL Levamisole
Dako REAL Detection System Alkaline,
K 5005, Phosphatase/RED,Rabbit/Mouse
Dako Cytomation, Carpinteria, USA
Antibody Diluent with backround reducing
components, S 2022
Dako Corporation, Carpinteria, USA
Serumfreier Proteinblock DAKO, Hamburg, Germany
Hämalaun, S 3301 Dako Corporation, Carpinteria, USA
Target Retrieval Solution Citrate pH 6,0 (x10),
S 2369
Dako Corporation , Carpinteria, USA
Target Retrieval Puffer pH 9,0 (x10), S 2367 DAKO, Glostrup, Dänemark
Avidin, X 0590 DAKO, Hamburg, Germany
Biotin, X 0590 DAKO, Hamburg, Germany
Ethanol Carl Roth GmbH, Karlsruhe, Germany
MEA Merck Schuchardt OHG, Hohenbrunn, Germany
Waschpuffer pH 7,4 (x10) Dako Corporation, Carpinteria, USA
Tab. 2: Verwendete Materialien für die Immunhistochemie

42
4.5. Qualitative und quantitative Analyseverfahren 4.5.1. Qualitative und semiquantitative Expressionsbestimmung
Bei 400-facher Vergrößerung wurde im Hellfeldmikroskop (Axioskop, Zeiss, Jena,
Deutschland) die Verteilung und Expression von OPG, GSK-3β, RANKL und DKK1
im Übergangsbereich vom ortsständigen Knochen zum KEM beurteilt. Zur
Quantifizierung des Expressionsgrades wurden je Probe zwei Gesichtsfelder (Boden
+ Rand rechts oder links) bei 400-facher Vergrößerung betrachtet (siehe Abb.12).
Mittels der AxioCam MRc 5 und dem Softwareprogramm Carl Zeiss AxioVision
Release 4.5 wurden zwei repräsentative Gesichtsfelder je Gewebeschnitt digitalisiert.
Zur Ermittlung des Labeling Index erfolgte die Auszählung der Zellzahl pro
Gesichtsfeld nach der Methode des randomisierten, systematischen subsamplings
[109]. Hierbei wurde das Verhältnis der positiven Zellen zur Gesamtzellzahl je
Bildausschnitt ermittelt. Als positive Zellen gelten die Antikörper exprimierenden
Zellen, die an einem sie umgebenden roten Hof zu erkennen sind. Zur Zählung
wurde ein Gitternetz mit cm-Einteilung verwendet. Die Zellzahlen lagen pro
Bildausschnitt im Median bei 300 ± 50. Um eine objektive Zählung zu
gewährleisten, erfolgte diese ohne Beachtung von Lokalisation, Opferungszeitpunkt,
Defektbehandlung und Antikörperapplikation.
Abb. 12: Gesichtsfelder am Übergangsbereich vom Knochenlager zum KEM
4.5.2. Statistische Analyse
Für die qualitative Analyse wurde der Labeling Index je Gesichtsfeld, Probe, Tier
und Tag für den Übergangsbereich zwischen ortsständigem Knochenlager und KEM
errechnet. Die Daten wurden nach Lokalisation, Defektbehandlung,
Opferungszeitpunkt und Antikörperapplikation zusammengefasst. Ohne Annahme
einer Normalverteilung wurden Median und Standardabweichung bestimmt. Die
graphische Darstellung erfolgte mittels Boxplots. Zur Prüfung der Unterschiede
zwischen den Gruppen wurde der ANOVA-Test auf dem Signifikanzniveau p < 0,05
durchgeführt. Alle Berechnungen erfolgten mit dem Programm SPSS 12.0 für
Windows (SPSS Inc., Chicago, USA).
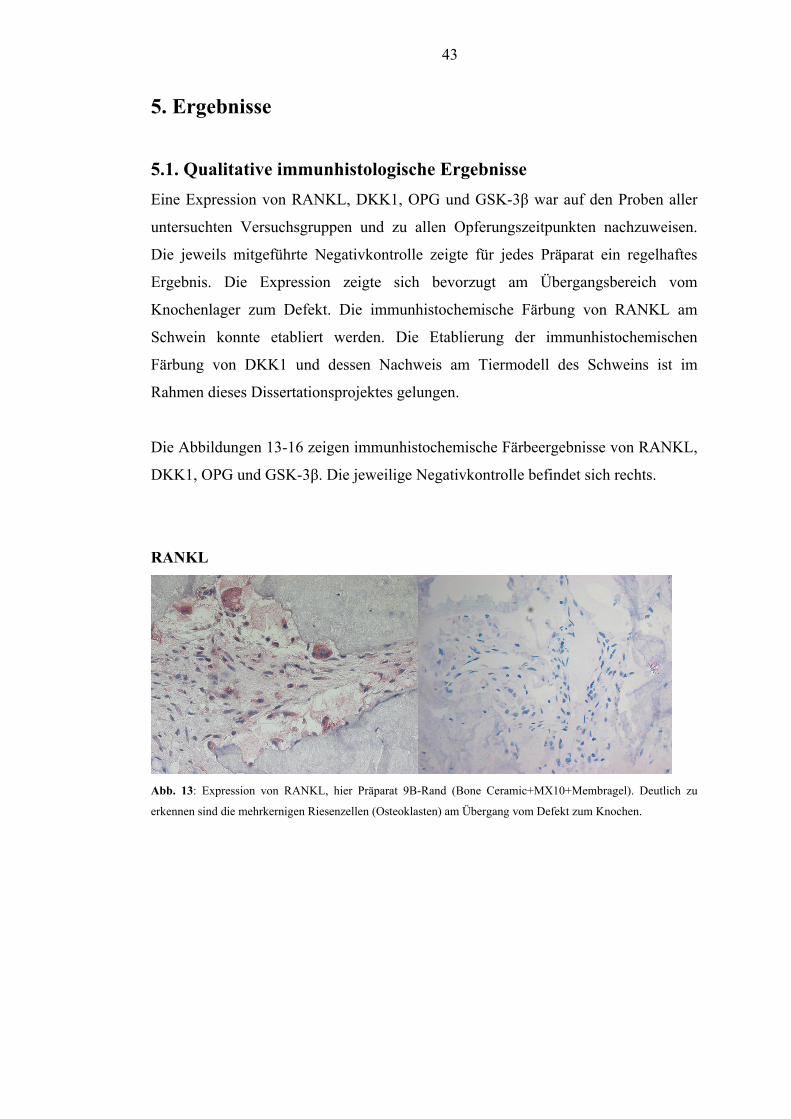
43
5. Ergebnisse
5.1. Qualitative immunhistologische Ergebnisse Eine Expression von RANKL, DKK1, OPG und GSK-3β war auf den Proben aller
untersuchten Versuchsgruppen und zu allen Opferungszeitpunkten nachzuweisen.
Die jeweils mitgeführte Negativkontrolle zeigte für jedes Präparat ein regelhaftes
Ergebnis. Die Expression zeigte sich bevorzugt am Übergangsbereich vom
Knochenlager zum Defekt. Die immunhistochemische Färbung von RANKL am
Schwein konnte etabliert werden. Die Etablierung der immunhistochemischen
Färbung von DKK1 und dessen Nachweis am Tiermodell des Schweins ist im
Rahmen dieses Dissertationsprojektes gelungen.
Die Abbildungen 13-16 zeigen immunhistochemische Färbeergebnisse von RANKL,
DKK1, OPG und GSK-3β. Die jeweilige Negativkontrolle befindet sich rechts.
RANKL
Abb. 13: Expression von RANKL, hier Präparat 9B-Rand (Bone Ceramic+MX10+Membragel). Deutlich zu
erkennen sind die mehrkernigen Riesenzellen (Osteoklasten) am Übergang vom Defekt zum Knochen.
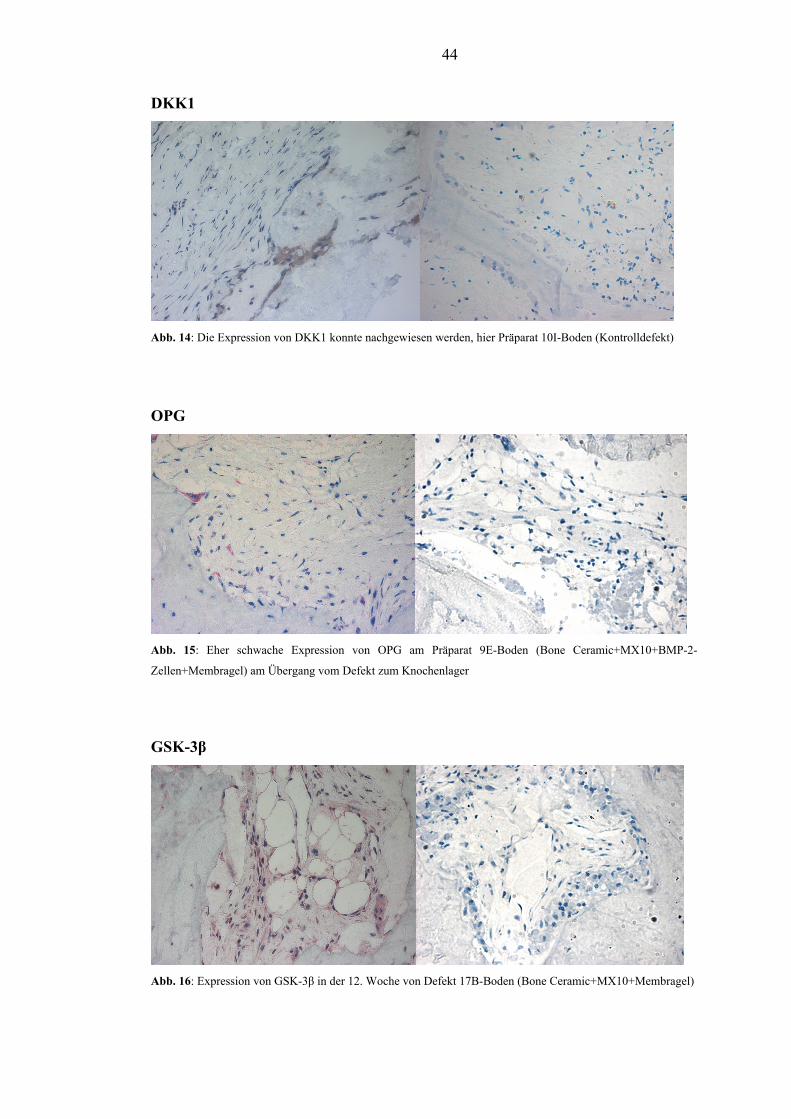
44
DKK1
Abb. 14: Die Expression von DKK1 konnte nachgewiesen werden, hier Präparat 10I-Boden (Kontrolldefekt)
OPG
Abb. 15: Eher schwache Expression von OPG am Präparat 9E-Boden (Bone Ceramic+MX10+BMP-2-
Zellen+Membragel) am Übergang vom Defekt zum Knochenlager
GSK-3β
Abb. 16: Expression von GSK-3β in der 12. Woche von Defekt 17B-Boden (Bone Ceramic+MX10+Membragel)

45
5.2. Semiquantitative immunhistochemische Ergebnisse der Expression 5.2.1. RANKL
Die immunhistochemische Färbung von RANKL am Schwein konnte selbstständig
erarbeitet werden. RANKL lieferte im Vergleich zu OPG, DKK1 und GSK-3β die
gleichmäßigsten Werte und zeigte sich unbeeinflusst, sowohl von der hier
verwendeten Defektbehandlung, als auch von der Zeit. Knochenregeneration kommt
durch das Gleichgewicht von Knochenneubildung und Knochenresorption zustande
[106]. Osteoblasten sind für die Knochenregeneration ebenso wichtig wie
Osteoklasten, solange keine pathologische Überexpression vorliegt [106]. In dieser
Arbeit konnte gezeigt werden, dass die Expression von RANKL weder durch
osteogene Stimulation über BMP-2 noch durch Periostabdeckung oder den zeitlichen
Verlauf beeinflusst wird. Autologer Knochen und Kontrolldefekt zeigten in der
1. Opferungswoche mit 18,3% (±3,1) und 19,9% (±2,1) erniedrigte und zur
4. Opferungswoche mit 20,0% (±2,8) und 17,8% (±5,4) leicht erhöhte
Expressionswerte gegenüber den übrigen Gruppen. Signifikant zeigten sich zum
2. Opferungszeitpunkt die Gruppen D gegen E (p=0,006), E gegen I (p=0,007),
E gegen C (p=0,008) und E gegen A (p=0,006). Zum 4. Opferungszeitpunkt zeigten
die Gruppen D gegen C (p=0,012), D gegen E (p=0,004) und D gegen G (p=0,006)
signifikante Unterschiede. Tabelle 3 zeigt die Ergebnistabelle der einzelnen Gruppen
zu den 4 Opferungszeitpunkten, Abbildung 17 die Labeling Indizes der sRANKL
exprimierenden Zellen mit Signifikanzen und Abbildung 18 immunhistochemische
Färbeergebnisse von RANKL.
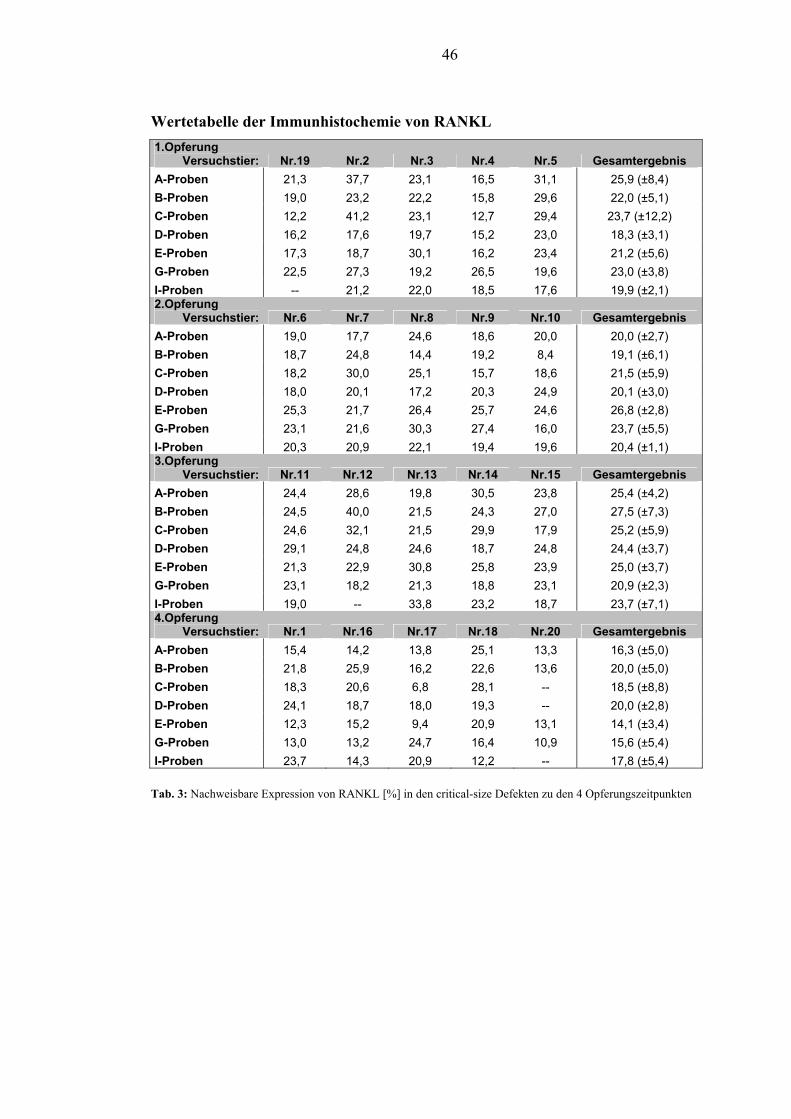
46
Wertetabelle der Immunhistochemie von RANKL 1.Opferung
Versuchstier: Nr.19 Nr.2 Nr.3 Nr.4 Nr.5 Gesamtergebnis A-Proben 21,3 37,7 23,1 16,5 31,1 25,9 (±8,4) B-Proben 19,0 23,2 22,2 15,8 29,6 22,0 (±5,1) C-Proben 12,2 41,2 23,1 12,7 29,4 23,7 (±12,2) D-Proben 16,2 17,6 19,7 15,2 23,0 18,3 (±3,1) E-Proben 17,3 18,7 30,1 16,2 23,4 21,2 (±5,6) G-Proben 22,5 27,3 19,2 26,5 19,6 23,0 (±3,8) I-Proben -- 21,2 22,0 18,5 17,6 19,9 (±2,1) 2.Opferung
Versuchstier: Nr.6 Nr.7 Nr.8 Nr.9 Nr.10 Gesamtergebnis A-Proben 19,0 17,7 24,6 18,6 20,0 20,0 (±2,7) B-Proben 18,7 24,8 14,4 19,2 8,4 19,1 (±6,1) C-Proben 18,2 30,0 25,1 15,7 18,6 21,5 (±5,9) D-Proben 18,0 20,1 17,2 20,3 24,9 20,1 (±3,0) E-Proben 25,3 21,7 26,4 25,7 24,6 26,8 (±2,8) G-Proben 23,1 21,6 30,3 27,4 16,0 23,7 (±5,5) I-Proben 20,3 20,9 22,1 19,4 19,6 20,4 (±1,1) 3.Opferung
Versuchstier: Nr.11 Nr.12 Nr.13 Nr.14 Nr.15 Gesamtergebnis A-Proben 24,4 28,6 19,8 30,5 23,8 25,4 (±4,2) B-Proben 24,5 40,0 21,5 24,3 27,0 27,5 (±7,3) C-Proben 24,6 32,1 21,5 29,9 17,9 25,2 (±5,9) D-Proben 29,1 24,8 24,6 18,7 24,8 24,4 (±3,7) E-Proben 21,3 22,9 30,8 25,8 23,9 25,0 (±3,7) G-Proben 23,1 18,2 21,3 18,8 23,1 20,9 (±2,3) I-Proben 19,0 -- 33,8 23,2 18,7 23,7 (±7,1) 4.Opferung
Versuchstier: Nr.1 Nr.16 Nr.17 Nr.18 Nr.20 Gesamtergebnis A-Proben 15,4 14,2 13,8 25,1 13,3 16,3 (±5,0) B-Proben 21,8 25,9 16,2 22,6 13,6 20,0 (±5,0) C-Proben 18,3 20,6 6,8 28,1 -- 18,5 (±8,8) D-Proben 24,1 18,7 18,0 19,3 -- 20,0 (±2,8) E-Proben 12,3 15,2 9,4 20,9 13,1 14,1 (±3,4) G-Proben 13,0 13,2 24,7 16,4 10,9 15,6 (±5,4) I-Proben 23,7 14,3 20,9 12,2 -- 17,8 (±5,4) Tab. 3: Nachweisbare Expression von RANKL [%] in den critical-size Defekten zu den 4 Opferungszeitpunkten
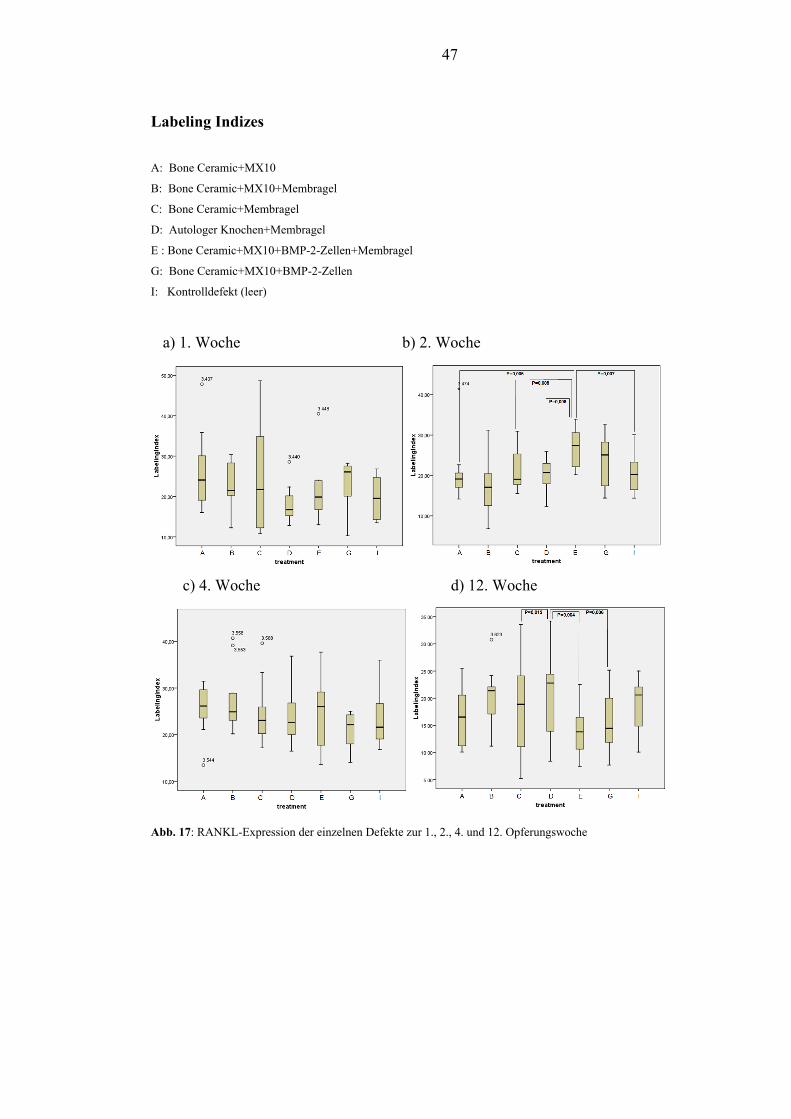
47
Labeling Indizes A: Bone Ceramic+MX10
B: Bone Ceramic+MX10+Membragel
C: Bone Ceramic+Membragel
D: Autologer Knochen+Membragel
E : Bone Ceramic+MX10+BMP-2-Zellen+Membragel
G: Bone Ceramic+MX10+BMP-2-Zellen
I: Kontrolldefekt (leer)
a) 1. Woche b) 2. Woche
c) 4. Woche d) 12. Woche
Abb. 17: RANKL-Expression der einzelnen Defekte zur 1., 2., 4. und 12. Opferungswoche
48
3E-Boden (x=102)
16A-Boden (x=67) Abb. 18: konstante Expression von RANKL, oben zur 1. Woche von Defekt 3E-Boden (Bone Ceramic+MX10+BMP-2-Zellen+Membragel), unten zur 4. Woche von Defekt 16A-Boden (Bone Ceramic+MX10), rechts entsprechende Negativkontrollen (x=Anzahl positiver Zellen)
5.2.2. DKK1
Die Hauptaufgabe bei DKK1 lag in der Etablierung der Färbung und in dessen
Nachweis am Tiermodell des Schweins. Hierfür wurden die Gruppen B (Bone
Ceramic+MX10+Membragel), G (Bone Ceramic+MX10+BMP-2-Zellen) und
I (Kontrolldefekt) der 2. Opferungswoche immunhistochemisch behandelt. Der
2. Opferungszeitpunkt wurde gewählt, da DKK1 durch dessen Rolle als natürlicher
Antagonist des Wnt-Weges zur hierfür spezifischen Knochenproliferation beiträgt
[30, 72, 112]. Die Expression von DKK1 zeigte sich unabhängig von der
Defektbehandlung zwischen 16,4% (±3,1) und 17,6% (±1,4). Es wurden keine
Signifikanzen festgestellt. Anschließend sind die Ergebnistabelle (Tab.4), die
Labeling Indizes (Abb.19) und immunhistochemische Färbeergebnisse von DKK1
(Abb. 20) dargestellt.
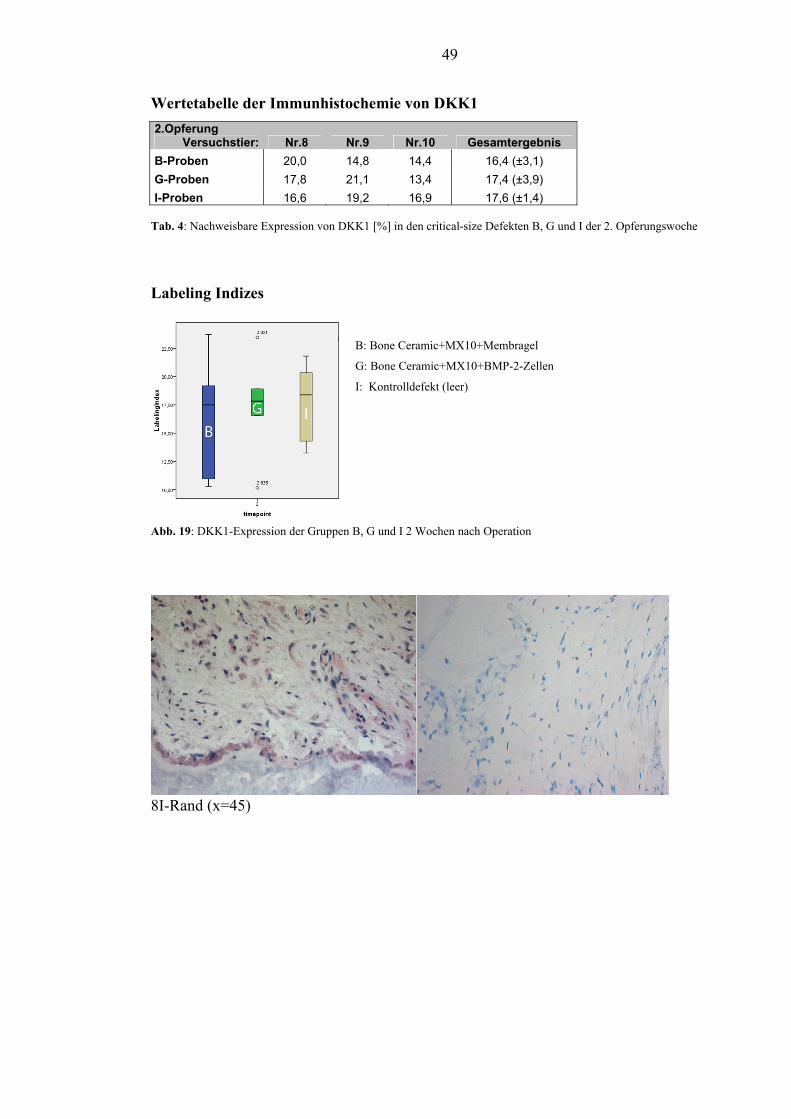
49
Wertetabelle der Immunhistochemie von DKK1 2.Opferung
Versuchstier: Nr.8 Nr.9 Nr.10 Gesamtergebnis B-Proben 20,0 14,8 14,4 16,4 (±3,1) G-Proben 17,8 21,1 13,4 17,4 (±3,9) I-Proben 16,6 19,2 16,9 17,6 (±1,4)
Tab. 4: Nachweisbare Expression von DKK1 [%] in den critical-size Defekten B, G und I der 2. Opferungswoche
Labeling Indizes
B: Bone Ceramic+MX10+Membragel
G: Bone Ceramic+MX10+BMP-2-Zellen
I: Kontrolldefekt (leer)
Abb. 19: DKK1-Expression der Gruppen B, G und I 2 Wochen nach Operation
8I-Rand (x=45)
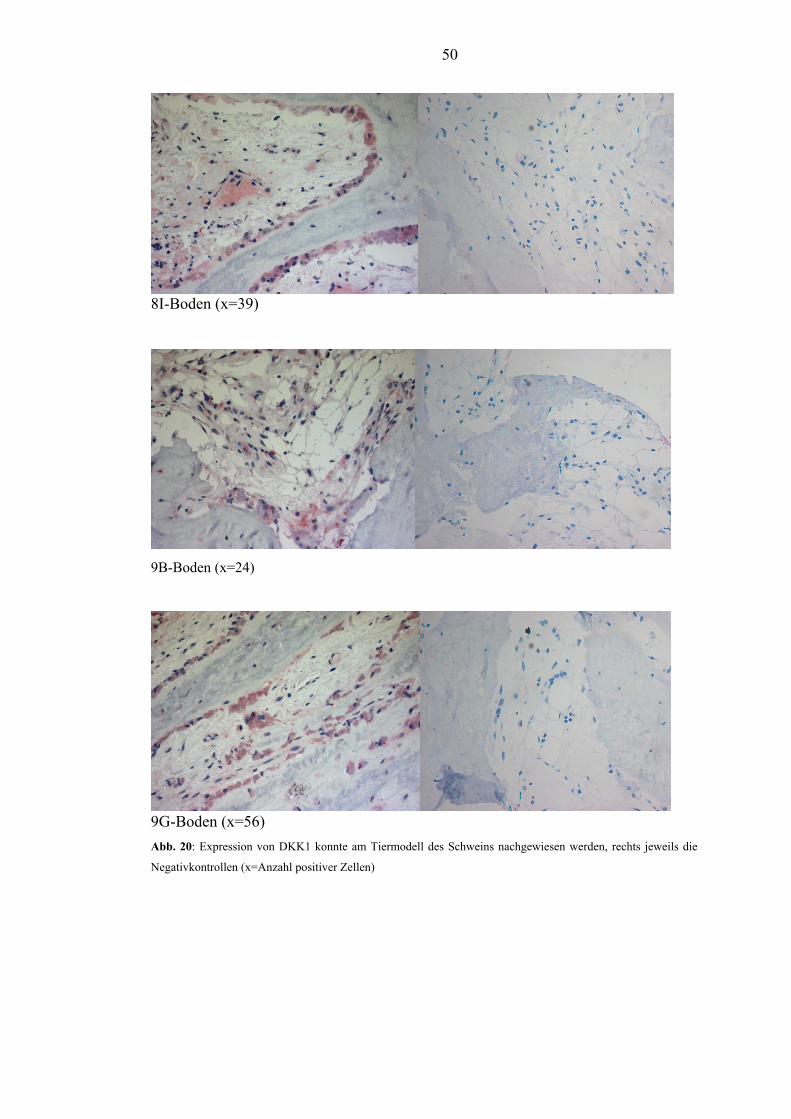
50
8I-Boden (x=39)
9B-Boden (x=24)
9G-Boden (x=56) Abb. 20: Expression von DKK1 konnte am Tiermodell des Schweins nachgewiesen werden, rechts jeweils die
Negativkontrollen (x=Anzahl positiver Zellen)

51
5.2.3. OPG
Die Expression von OPG bewegte sich in der 1. Woche zwischen 4,7% (±3,5) und
11,1% (±3,2), stieg innerhalb der 2. Woche auf Werte zwischen 10,9% (±2,6) und
18,3% (±0,7). Zur 4. Woche hin stabilisierte sich die Expression aller Gruppen bei
rund 20%. In der 12. Woche lag sie bei ca. 12,5%.
Diese Stabilisation aller Gruppen auf einem Niveau zeigt die Unabhängigkeit einer
OPG-Über- oder Unterexpression von der Defektmodalität und deren Nicht-Eignung
als therapeutischen Ansatz. In der 1.Woche zeigten sich hohe Werte für die mit
BMP-2 behandelten Gruppen E und G. Dies deutet auf die indirekte Stimulation der
OPG-Expression durch BMP-2 hin [81], die ab der 2. Woche ausblieb. Autologer
Knochen (Defekt D) und der Kontrolldefekt (Defekt I) zeigten keine Besonderheiten
bezüglich der Expression. Signifikant zeigten sich zum 1. Opferungszeitpunkt die
Gruppen E gegen B (p=0,006), E gegen D (p=0,003) und E gegen I (p=0,007) und
zum 2. Opferungszeitpunkt die Gruppen A gegen E (p=0,003). Zum
3. Opferungszeitpunkt waren signifikante Unterschiede der Gruppen E gegen B
(p=0,022), E gegen D (p=0,009) und E gegen I (p=0,009) und zum
4. Opferungszeitpunkt der Gruppen E gegen B (p=0,004), E gegen C (p=0,011),
E gegen D (p=0,009) und E gegen I (p=0,006) zu erkennen. Nachfolgend sind die
Ergebnistabelle (Tab.5), die Labeling Indizes mit Signifikanzen (Abb.21) und
immunhistochemische Färbeergebnisse von OPG (Abb.22) dargestellt.
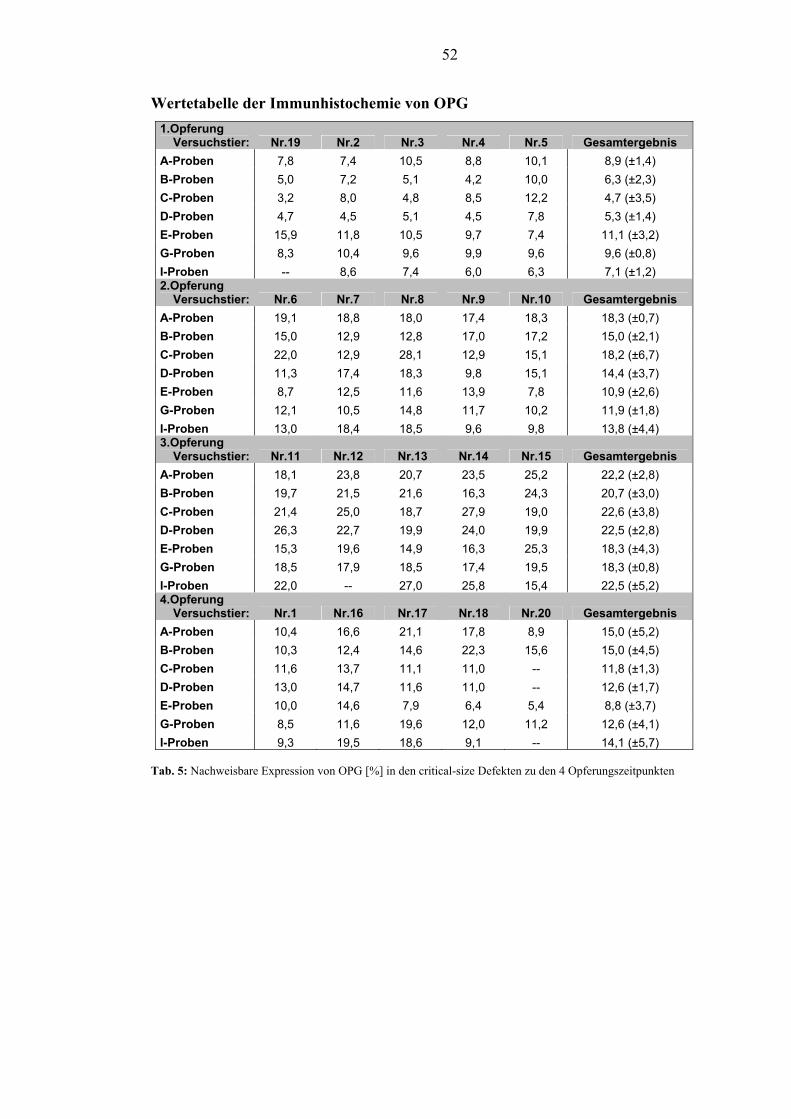
52
Wertetabelle der Immunhistochemie von OPG 1.Opferung
Versuchstier: Nr.19 Nr.2 Nr.3 Nr.4 Nr.5 Gesamtergebnis A-Proben 7,8 7,4 10,5 8,8 10,1 8,9 (±1,4) B-Proben 5,0 7,2 5,1 4,2 10,0 6,3 (±2,3) C-Proben 3,2 8,0 4,8 8,5 12,2 4,7 (±3,5) D-Proben 4,7 4,5 5,1 4,5 7,8 5,3 (±1,4) E-Proben 15,9 11,8 10,5 9,7 7,4 11,1 (±3,2) G-Proben 8,3 10,4 9,6 9,9 9,6 9,6 (±0,8) I-Proben -- 8,6 7,4 6,0 6,3 7,1 (±1,2) 2.Opferung
Versuchstier: Nr.6 Nr.7 Nr.8 Nr.9 Nr.10 Gesamtergebnis A-Proben 19,1 18,8 18,0 17,4 18,3 18,3 (±0,7) B-Proben 15,0 12,9 12,8 17,0 17,2 15,0 (±2,1) C-Proben 22,0 12,9 28,1 12,9 15,1 18,2 (±6,7) D-Proben 11,3 17,4 18,3 9,8 15,1 14,4 (±3,7) E-Proben 8,7 12,5 11,6 13,9 7,8 10,9 (±2,6) G-Proben 12,1 10,5 14,8 11,7 10,2 11,9 (±1,8) I-Proben 13,0 18,4 18,5 9,6 9,8 13,8 (±4,4) 3.Opferung
Versuchstier: Nr.11 Nr.12 Nr.13 Nr.14 Nr.15 Gesamtergebnis A-Proben 18,1 23,8 20,7 23,5 25,2 22,2 (±2,8) B-Proben 19,7 21,5 21,6 16,3 24,3 20,7 (±3,0) C-Proben 21,4 25,0 18,7 27,9 19,0 22,6 (±3,8) D-Proben 26,3 22,7 19,9 24,0 19,9 22,5 (±2,8) E-Proben 15,3 19,6 14,9 16,3 25,3 18,3 (±4,3) G-Proben 18,5 17,9 18,5 17,4 19,5 18,3 (±0,8) I-Proben 22,0 -- 27,0 25,8 15,4 22,5 (±5,2) 4.Opferung
Versuchstier: Nr.1 Nr.16 Nr.17 Nr.18 Nr.20 Gesamtergebnis A-Proben 10,4 16,6 21,1 17,8 8,9 15,0 (±5,2) B-Proben 10,3 12,4 14,6 22,3 15,6 15,0 (±4,5) C-Proben 11,6 13,7 11,1 11,0 -- 11,8 (±1,3) D-Proben 13,0 14,7 11,6 11,0 -- 12,6 (±1,7) E-Proben 10,0 14,6 7,9 6,4 5,4 8,8 (±3,7) G-Proben 8,5 11,6 19,6 12,0 11,2 12,6 (±4,1) I-Proben 9,3 19,5 18,6 9,1 -- 14,1 (±5,7)
Tab. 5: Nachweisbare Expression von OPG [%] in den critical-size Defekten zu den 4 Opferungszeitpunkten
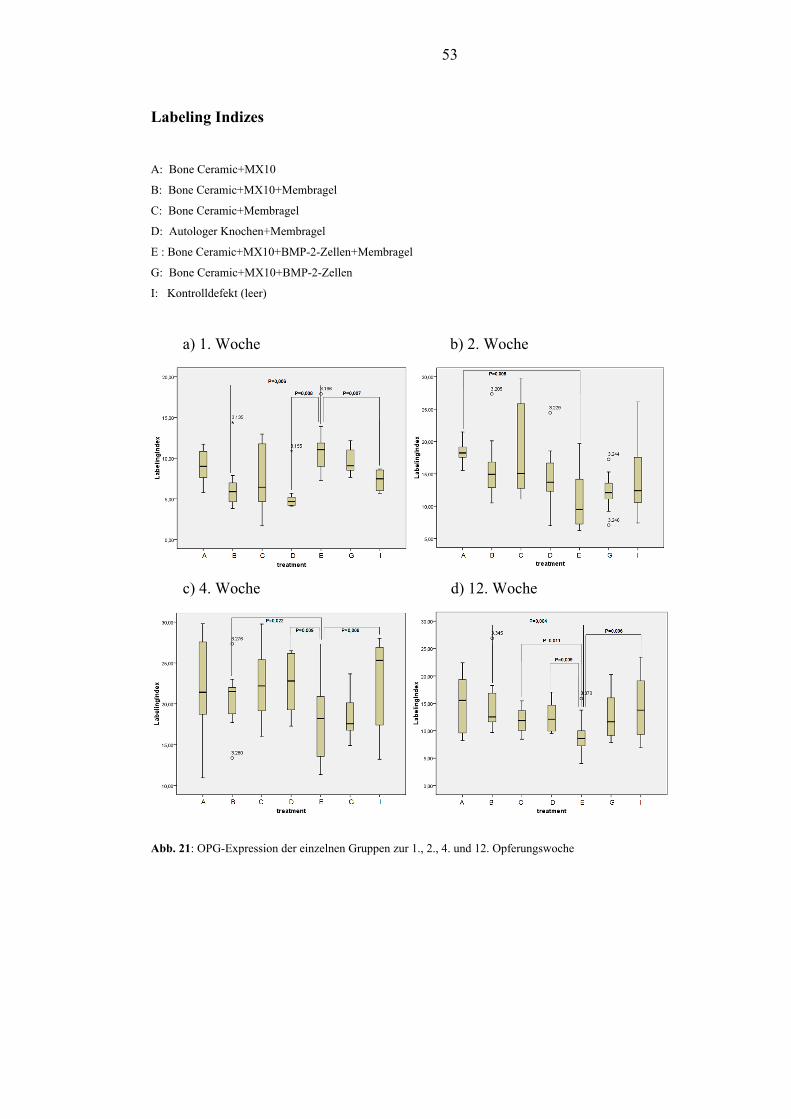
53
Labeling Indizes
A: Bone Ceramic+MX10
B: Bone Ceramic+MX10+Membragel
C: Bone Ceramic+Membragel
D: Autologer Knochen+Membragel
E : Bone Ceramic+MX10+BMP-2-Zellen+Membragel
G: Bone Ceramic+MX10+BMP-2-Zellen
I: Kontrolldefekt (leer)
a) 1. Woche b) 2. Woche
c) 4. Woche d) 12. Woche
Abb. 21: OPG-Expression der einzelnen Gruppen zur 1., 2., 4. und 12. Opferungswoche
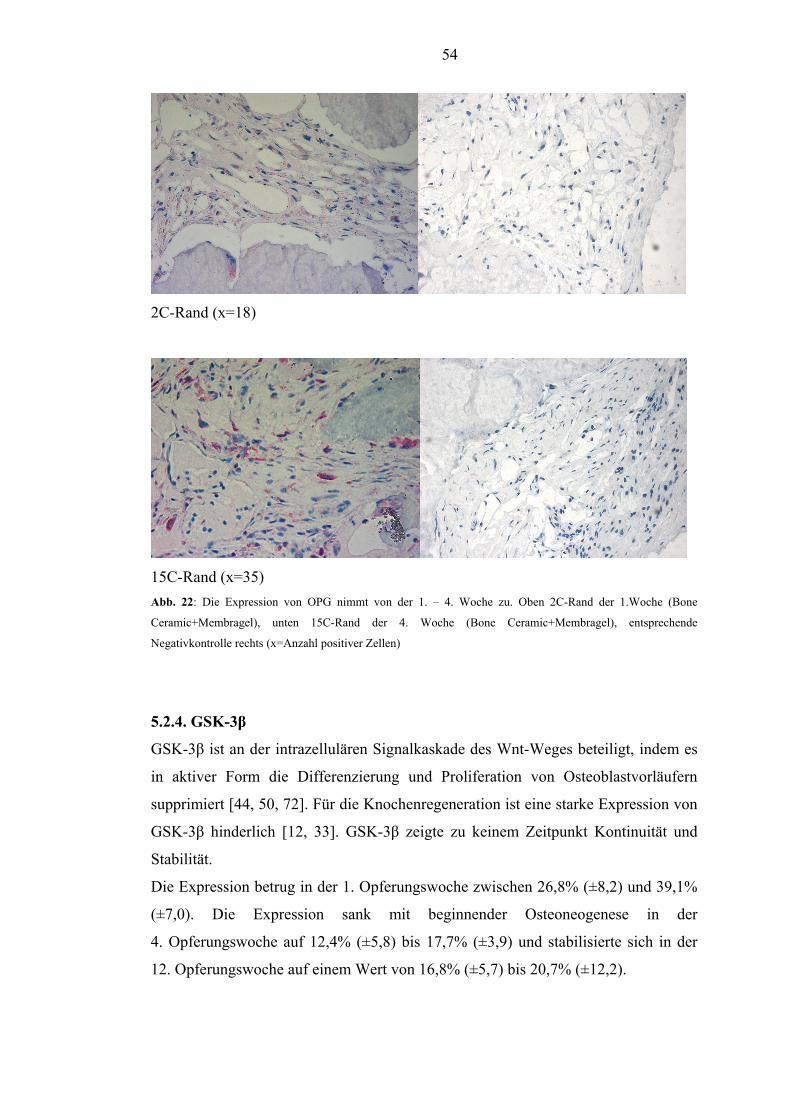
54
2C-Rand (x=18)
15C-Rand (x=35) Abb. 22: Die Expression von OPG nimmt von der 1. – 4. Woche zu. Oben 2C-Rand der 1.Woche (Bone
Ceramic+Membragel), unten 15C-Rand der 4. Woche (Bone Ceramic+Membragel), entsprechende
Negativkontrolle rechts (x=Anzahl positiver Zellen)
5.2.4. GSK-3β
GSK-3β ist an der intrazellulären Signalkaskade des Wnt-Weges beteiligt, indem es
in aktiver Form die Differenzierung und Proliferation von Osteoblastvorläufern
supprimiert [44, 50, 72]. Für die Knochenregeneration ist eine starke Expression von
GSK-3β hinderlich [12, 33]. GSK-3β zeigte zu keinem Zeitpunkt Kontinuität und
Stabilität.
Die Expression betrug in der 1. Opferungswoche zwischen 26,8% (±8,2) und 39,1%
(±7,0). Die Expression sank mit beginnender Osteoneogenese in der
4. Opferungswoche auf 12,4% (±5,8) bis 17,7% (±3,9) und stabilisierte sich in der
12. Opferungswoche auf einem Wert von 16,8% (±5,7) bis 20,7% (±12,2).
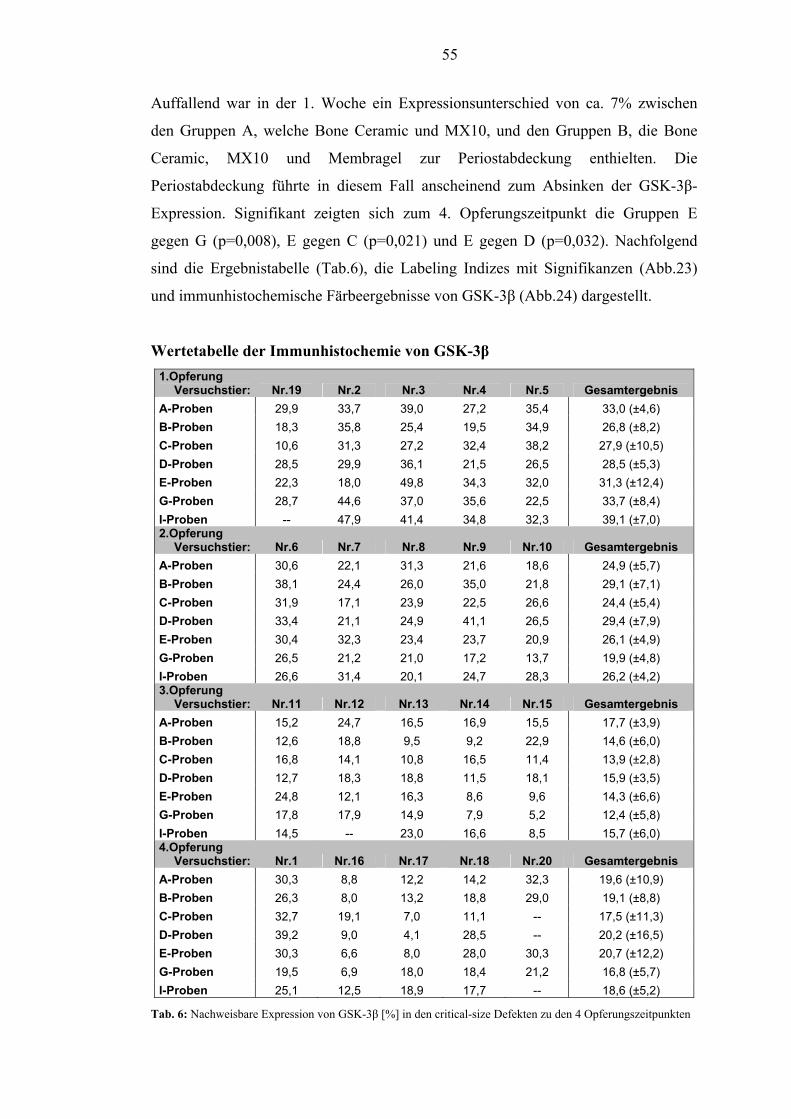
55
Auffallend war in der 1. Woche ein Expressionsunterschied von ca. 7% zwischen
den Gruppen A, welche Bone Ceramic und MX10, und den Gruppen B, die Bone
Ceramic, MX10 und Membragel zur Periostabdeckung enthielten. Die
Periostabdeckung führte in diesem Fall anscheinend zum Absinken der GSK-3β-
Expression. Signifikant zeigten sich zum 4. Opferungszeitpunkt die Gruppen E
gegen G (p=0,008), E gegen C (p=0,021) und E gegen D (p=0,032). Nachfolgend
sind die Ergebnistabelle (Tab.6), die Labeling Indizes mit Signifikanzen (Abb.23)
und immunhistochemische Färbeergebnisse von GSK-3β (Abb.24) dargestellt.
Wertetabelle der Immunhistochemie von GSK-3β 1.Opferung
Versuchstier: Nr.19 Nr.2 Nr.3 Nr.4 Nr.5 Gesamtergebnis A-Proben 29,9 33,7 39,0 27,2 35,4 33,0 (±4,6) B-Proben 18,3 35,8 25,4 19,5 34,9 26,8 (±8,2) C-Proben 10,6 31,3 27,2 32,4 38,2 27,9 (±10,5) D-Proben 28,5 29,9 36,1 21,5 26,5 28,5 (±5,3) E-Proben 22,3 18,0 49,8 34,3 32,0 31,3 (±12,4) G-Proben 28,7 44,6 37,0 35,6 22,5 33,7 (±8,4) I-Proben -- 47,9 41,4 34,8 32,3 39,1 (±7,0) 2.Opferung
Versuchstier: Nr.6 Nr.7 Nr.8 Nr.9 Nr.10 Gesamtergebnis A-Proben 30,6 22,1 31,3 21,6 18,6 24,9 (±5,7) B-Proben 38,1 24,4 26,0 35,0 21,8 29,1 (±7,1) C-Proben 31,9 17,1 23,9 22,5 26,6 24,4 (±5,4) D-Proben 33,4 21,1 24,9 41,1 26,5 29,4 (±7,9) E-Proben 30,4 32,3 23,4 23,7 20,9 26,1 (±4,9) G-Proben 26,5 21,2 21,0 17,2 13,7 19,9 (±4,8) I-Proben 26,6 31,4 20,1 24,7 28,3 26,2 (±4,2) 3.Opferung
Versuchstier: Nr.11 Nr.12 Nr.13 Nr.14 Nr.15 Gesamtergebnis A-Proben 15,2 24,7 16,5 16,9 15,5 17,7 (±3,9) B-Proben 12,6 18,8 9,5 9,2 22,9 14,6 (±6,0) C-Proben 16,8 14,1 10,8 16,5 11,4 13,9 (±2,8) D-Proben 12,7 18,3 18,8 11,5 18,1 15,9 (±3,5) E-Proben 24,8 12,1 16,3 8,6 9,6 14,3 (±6,6) G-Proben 17,8 17,9 14,9 7,9 5,2 12,4 (±5,8) I-Proben 14,5 -- 23,0 16,6 8,5 15,7 (±6,0) 4.Opferung
Versuchstier: Nr.1 Nr.16 Nr.17 Nr.18 Nr.20 Gesamtergebnis A-Proben 30,3 8,8 12,2 14,2 32,3 19,6 (±10,9) B-Proben 26,3 8,0 13,2 18,8 29,0 19,1 (±8,8) C-Proben 32,7 19,1 7,0 11,1 -- 17,5 (±11,3) D-Proben 39,2 9,0 4,1 28,5 -- 20,2 (±16,5) E-Proben 30,3 6,6 8,0 28,0 30,3 20,7 (±12,2) G-Proben 19,5 6,9 18,0 18,4 21,2 16,8 (±5,7) I-Proben 25,1 12,5 18,9 17,7 -- 18,6 (±5,2)
Tab. 6: Nachweisbare Expression von GSK-3β [%] in den critical-size Defekten zu den 4 Opferungszeitpunkten
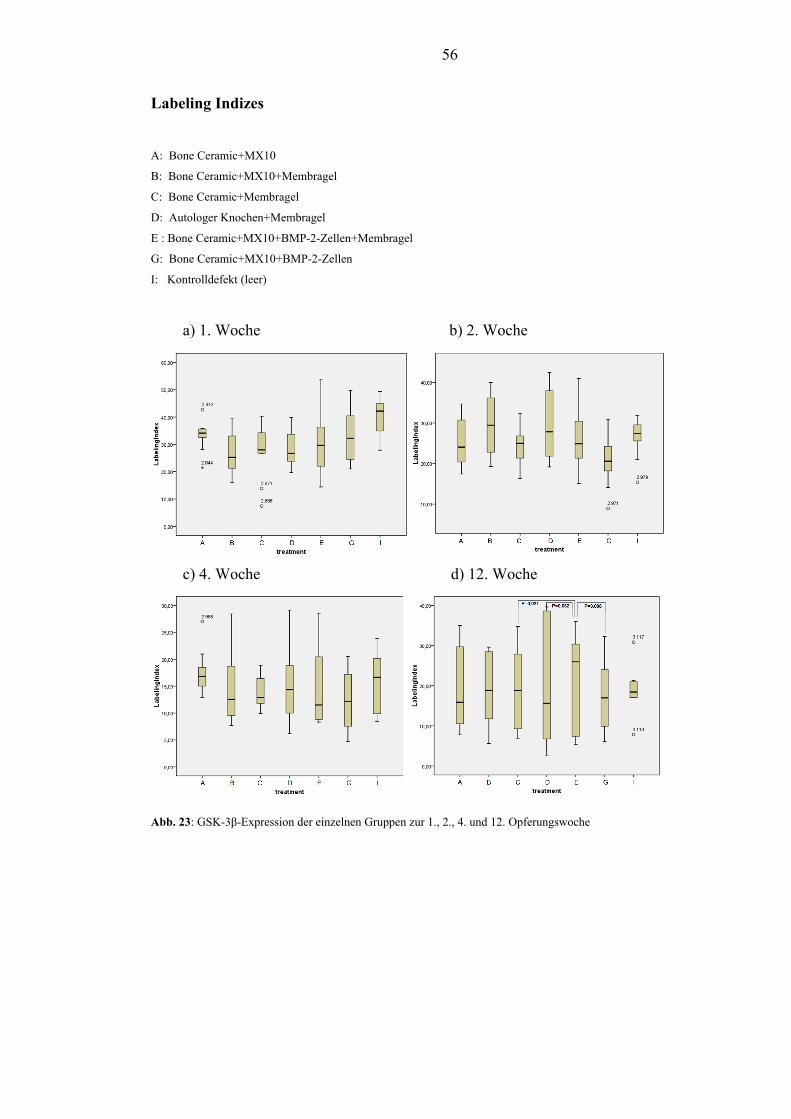
56
Labeling Indizes
A: Bone Ceramic+MX10
B: Bone Ceramic+MX10+Membragel
C: Bone Ceramic+Membragel
D: Autologer Knochen+Membragel
E : Bone Ceramic+MX10+BMP-2-Zellen+Membragel
G: Bone Ceramic+MX10+BMP-2-Zellen
I: Kontrolldefekt (leer)
a) 1. Woche b) 2. Woche
c) 4. Woche d) 12. Woche
Abb. 23: GSK-3β-Expression der einzelnen Gruppen zur 1., 2., 4. und 12. Opferungswoche
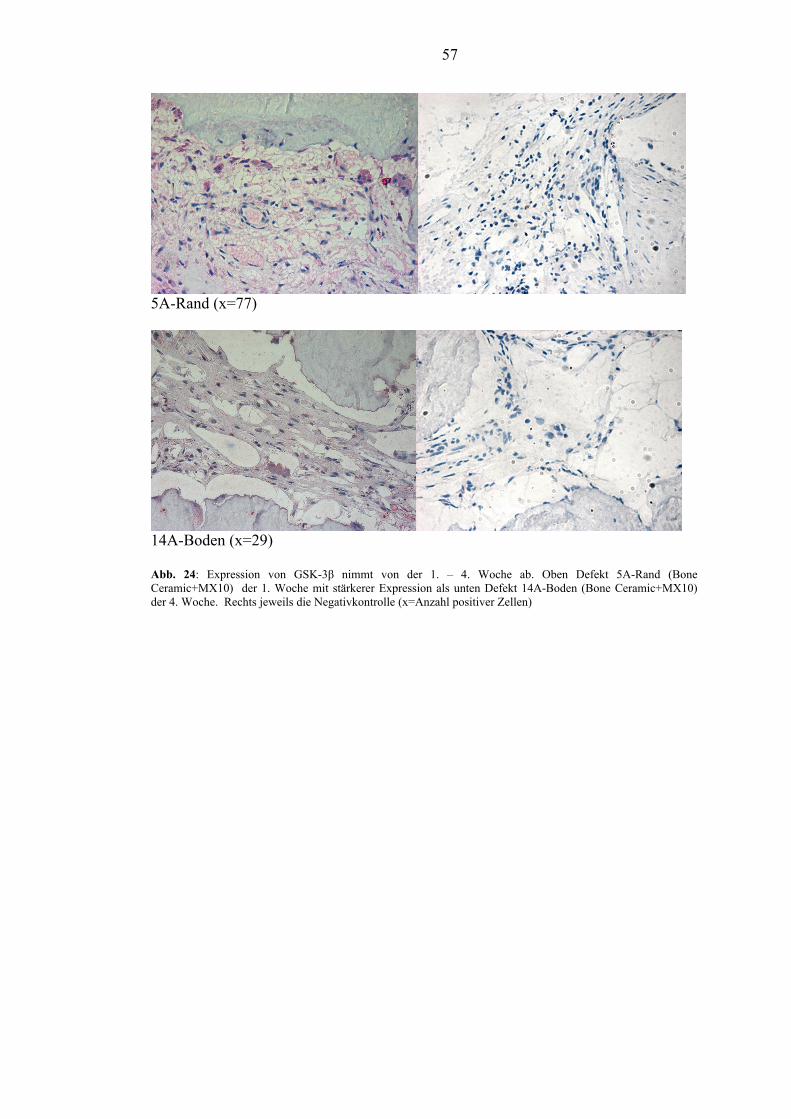
57
5A-Rand (x=77)
14A-Boden (x=29) Abb. 24: Expression von GSK-3β nimmt von der 1. – 4. Woche ab. Oben Defekt 5A-Rand (Bone Ceramic+MX10) der 1. Woche mit stärkerer Expression als unten Defekt 14A-Boden (Bone Ceramic+MX10) der 4. Woche. Rechts jeweils die Negativkontrolle (x=Anzahl positiver Zellen)

58
6. Diskussion
Das Ziel dieses Dissertationsprojektes war die Evaluation der ossären
Differenzierungs- und Proliferationsmarker OPG, RANKL, GSK-3β und deren
Antagonist DKK1. Da GSK-3β als Schlüsselregulator des Wnt-Signalweges die
ossäre Regeneration steuert, wurde somit auch die Wnt-vermittelte, osteogene
Rekrutierung und Maturation mesenchymaler Osteoblast- und
Osteoklastprogenitoren untersucht [5, 12, 13, 31, 44, 50, 112].
Es wurde der Einfluss einer biodegradierbaren PEG-Membran zur temporären
Abdeckung von critical size-Defekten der Schweinekalotte während gesteuerter
Defektregeneration mit einem biphasischen KEM unter zusätzlichem Einsatz von
transient liposomal BMP-2 transfizierten Osteoblasten auf die oben genannten
ossären Differenzierungs- und Proliferationsmarker evaluiert.
Demnach wurde die Genetik der intrazellulären Signaltransduktion bei
Osseointegration eines KEMs unter Anwendung einer PEG-Membran mit der
Fragestellung, ob diese Signalwege im Vergleich zur Einheilung eines
Knochentransplantates alternieren, untersucht.
RANKL
Knochenregeneration kommt durch das Gleichgewicht von Knochenneubildung über
Osteoblasten und Knochenresorption über Osteoklasten zustande [106]. Probleme
treten bei pathologischer Überexpression auf, was zu Knochenkrankheiten wie
Osteoporose, Osteopetrose oder Knochenmetastasen bei Tumorerkrankung führt
[106]. Um das Gleichgewicht von Knochenneubildung und Knochenresorption in
Richtung Knochenneubildung beeinflussen zu können, war die Entdeckung von
RANKL/RANK und deren Inhibitor OPG vor rund 10 Jahren ausschlaggebend [71].
Dies öffnete den Weg für weitere Knochenforschungen und die Entwicklung von
therapeutischen Mitteln [71]. Die Kontrolle von RANKL und RANK auf
Rezeptorebene und des RANKL/RANK-Signalweges ist ein möglicher Ansatzpunkt
zur Behandlung von systemischem oder lokalem Knochenmineralisationsverlust
[71]. Die Hemmung von RANKL als therapeutische Strategie, um neue
Medikamente gegen pathologisch erhöhte Knochenresorption zu entwickeln, gelang
mit der Entwicklung des humanen, monoklonalen anti-RANKL-Antikörpers

59
Denosumab [71]. Dieser Antikörper imitiert die Rolle von OPG funktionell, indem
Denosumab mit hoher Affinität an RANKL bindet [71]. Der positive Effekt von
Denosumab (Prolia®) auf den Knochenstoffwechsel von postmenopausalen,
osteoporotischen Frauen mit Zunahme der Knochenmineralisationsdichte, die
Langzeitverträglichkeit der Denosumab-Therapie über 48 Monate, die Reversibilität
der Einflüsse auf die Osteoklastogenese sowie das erneute Ansprechen nach
Absetzen der Therapie wurden in den erfolgreichen Phase-I- und Phase-II-Studien
gezeigt [71]. Die 2009 erfolgreich abgeschlossene Phase-III-Studie zeigte, dass durch
die halbjährliche subkutane Injektion von 60 mg Prolia® die Inzidenz von Frakturen
reduziert und die Knochenmineralisationsdichte erhöht wurden [27].
Neben den tragenden Rollen von RANKL und RANK bei der Osteoporose, könnten
RANKL und RANK auch Ziele einer Behandlung von Knochenverlust verursacht
durch Krebs, Knochenmetastasen, Sepsis, rheumatoide Arthritis oder Parodontitis
sein [106]. RANKL ist zum Beispiel in malignen Tumorzellen hoch exprimiert und
ist an der Knochendestruktion beteiligt [106]. Das RANKL-OPG-Verhältnis ist bei
maligner osteolytischer Pathogenese oder Knochenmetastasen bei Prostata- und
Brustkrebs verändert [9, 106]. Bei humaner Parodontitis wurde eine durch
Mikroorganismen ausgelöste Induktion der RANKL-Expression in CD4+ T-Zellen
und die konsequente Aktivierung von Osteoklasten beobachtet [2, 9, 106].
Tatsächlich wurde in einem Mausmodell zum Thema Parodontitis der
Alveolarknochenverlust durch die Hemmung von RANKL mittels OPG reduziert
[2, 9, 106]. Demzufolge ist die Inhibition von RANKL eine mögliche viel
versprechende, neue Therapie zur Kontrolle von Knochendichteverlusten [2, 9, 106].
In unserer Untersuchung lieferte die Expressionsbestimmung von RANKL
gleichbleibend stabile, konstante Werte. Die Expression von RANKL zeigte sich
weder für die osteogene Stimulation über BMP-2 noch die Periostabdeckung oder
den zeitlichen Verlauf sensitiv. Dies wurde in einer Studie von Qiang et al bestätigt,
die zeigte, dass das durch multiples Myelom induzierte DKK1 die Wnt regulierte
OPG- und RANKL-Produktion in Osteoblasten unterbricht [74]. Wnt3a reguliert
sowohl die OPG- als auch die RANKL-Expression und DKK1 unterbricht diesen
Prozess [74]. Die Wnt3a-Stimulation hatte hierbei einen stärkeren Effekt auf die
OPG- als auf die RANKL-Expression, wobei OPG erhöht und RANKL gehemmt
wurde [74]. Ein Anti-DKK1-Antikörper stellte die durch DKK1 unterdrückte

60
OPG-Expression in Osteoblasten, die mit primären MM-Zellen kokultiviert wurden,
wieder her, wohingegen der Effekt auf den durch DKK1 vermittelten Anstieg von
RANKL ausblieb. Dies zeigt, dass OPG sensitiver auf den Wnt-Signalweg reagiert
als RANKL [74].
OPG
Die Expression von OPG bewegte sich in der 1. Opferungswoche zwischen 4,7%
(±3,5) und 11,1% (±3,2), stieg innerhalb der 2. Opferungswoche auf Werte zwischen
10,9% (±2,6) und 18,3% (±0,7) an. Zur 4. Opferungswoche hin stabilisierte sich die
Expression aller Gruppen bei rund 20% und in der 12. Woche bei ca. 12,5%. Diese
Stabilisation aller Gruppen auf einem Niveau zeigt die Unabhängigkeit einer OPG-
Über- oder Unterexpression von der Defektmodalität und deren Nicht-Eignung als
therapeutischen Ansatz.
In der 1. Woche zeigten sich für die mit BMP-2 behandelten Gruppen höhere Werte
im Vergleich zu den übrigen Defekten, was auf die indirekte Stimulation der OPG-
Expression durch BMP-2 hindeutet [80], die ab der 2. Woche ausblieb. Eine Studie
von Sato et al belegte die Auffassung, dass die OPG- und RANKL-Expression durch
den BMP-2-Signalweg und den Wnt/β-catenin-Weg reguliert werden [80]. BMP-2,
der Wnt/β-catenin-Weg, wie auch die Inaktivierung von GSK-3β erhöhen die OPG-
Expression und unterdrücken die RANKL-Expression [80]. Im Speziellen bedeutet
dies, dass der Wnt/β-catenin-Weg die OPG-Transkription über eine Promoter-Region
reguliert, die die Bindungsstellen 2 und 4 enthält, welche auf die zwei β-catenin
spezifischen TCF/LEF1-Bindungsstellen ansprechen [80]. BMP-2 steigert die Wnt/β-
catenin abhängige transkriptionale Aktivität des OPG-Promoters [80]. Diese BMP-2-
Stimulation bewirkt, dass Smad1 und Smad4, Mitglieder der BMP-Signalkaskade,
mit der Wnt/β-catenin spezifischen TCF/LEF1-Bindungsstelle interagieren [80].
OPG ist als Zielgen des Wnt/β-catenin- und BMP-Weges identifiziert [31]. Die
OPG-Transkription wird durch die TCF/LEF1-Bindungsstellen des OPG-Promoters
reguliert, da auch Smad4, Mitglied des BMP-Weges, und β-catenin, Mitglied des
Wnt-Weges, an TCF/LEF1 binden [31, 38, 80, 98].

61
DKK1
Die Expression von DKK1 konnte am Tiermodell des Schweins nachgewiesen
werden. DKK1 kann als löslicher Inhibitor des canonical Wnt-Weges sowohl die
Osteoblastendifferenzierung und –proliferation als auch die über Wnt regulierte
OPG- und RANKL-Expression in Osteoblasten beeinflussen. Dies wurde in einer
Studie von Qiang et al belegt, die zeigte, dass das durch MM induzierte DKK1 die
Wnt regulierte OPG- und RANKL-Produktion in Osteoblasten unterbricht [74]. Das
Krankheitsbild des MMs ist charakterisiert durch osteolytische Knochenläsionen, die
als Folge von Osteoblasteninaktivierung und Osteoklastenaktivierung bei dem
Knochen entstehen, der an die Tumorherde angrenzt [74]. Der Wnt-Signalweg in
Osteoblasten reguliert die Osteoklastogenese durch die unterschiedliche Aktivierung
und Inaktivierung von RANKL als positiver und OPG als negativer Regulator der
Osteoklastogenese [30, 74]. Die Wnt3a induzierte OPG-Expression wurde in
Osteoblasten verringert, die mit einer MM-Zelllinie, die DKK1 exprimiert,
kokultiviert wurden [74]. Zudem wurde gezeigt, dass Knochenmarksseren von 21
MM-Patienten die Wnt3a induzierte OPG-Expression in Osteoblasten in einer DKK1
abhängigen Weise unterdrückten und die RANKL-Expression erhöhten [74]. Diese
Ergebnisse deuten darauf hin, dass DKK1 eine Schlüsselrolle in der Entwicklung von
osteolytischen Knochenläsionen, die mit MM assoziiert sind, spielt - durch direktes
Unterbrechen der Wnt regulierten Osteoblastendifferenzierung und indirektes
Erhöhen der Osteoklastogenese durch einen DKK1-vermittelten Anstieg des
RANKL-OPG-Verhältnisses [74]. In dieser Studie wurde auch gezeigt, dass ein
Medium, das mit mesenchymalen Stammzellen konditioniert wurde, MM-Zelllinien
induzieren kann, DKK1 zu produzieren und dass diese mesenchymalen Stammzellen
auch hohe Levels von Interleukin-6, einem potenten Wachstumsfaktor des MMs,
produzieren [74]. Interleukin-6 abhängige MM-Zelllinien breiteten sich in einem
Medium, dass mit mesenchymalen Stammzellen konditioniert wurde, stark aus und
dieses Wachstum konnte gehemmt werden, indem ein neutralisierender Antikörper
gegen Interleukin-6 der Kultur hinzugefügt wurde [74]. Dies deutet darauf hin, dass
das von MM induzierte DKK1 auch zum Tumorüberleben durch Regulierung der
Interleukin-6-Produktion durch mesenchymale Stammzellen beiträgt [74]. All diese
Daten suggerieren, dass DKK1 ein wichtiges therapeutisch lenkbares Ziel darstellt,
wie es in vorklinischen Studien zu MM und rheumatoider Arthritis behauptet wurde
[30, 74].

62
GSK-3β
GSK-3β ist an der intrazellulären Signalkaskade des Wnt-Weges beteiligt, indem es
in aktiver Form die Differenzierung und Proliferation von Osteoblastvorläufern
supprimiert [44, 50, 72]. Für die Knochenregeneration ist eine starke Expression von
GSK-3β hinderlich [12, 33]. Die GSK-3β-Expression zeigte zu keinem Zeitpunkt
Kontinuität oder Stabilität. Die Expression war in der 1. Woche am höchsten. Die
Expression sank mit beginnender Osteoneogenese bis zur 4. Woche und stabilisierte
sich in der 12. Woche. Auffallend war in der 1. Woche ein Expressionsunterschied
von ca. 7% zwischen den Gruppen A, welche Bone Ceramic und MX10, und den
Gruppen B, die Bone Ceramic, MX10 und Membragel zur Periostabdeckung
enthielten. Die Periostabdeckung führte in diesem Fall anscheinend zum
erwünschten Absinken der GSK-3β-Expression. Dies wurde durch die Ergebnisse
begleitender Dissertationsprojekte bestätigt [84]. Eine Periostabdeckung ist
erforderlich, wenn keine exogene ossäre Stimulation durch beispielsweise BMP-2
transfizierte Zellen vorliegt [84]. BMP-2 induziert die ossäre Differenzierung, aber
reduziert die Proliferation von Knochenprogenitorzellen [84]. Daher ist eine
Applikation von Zellen, die BMP-2 in hoher Dosis exprimieren, nur in Defekten, die
eine Quelle von proliferierenden Zellen enthalten oder Defekten, die nicht vom
Periost getrennt sind, sinnvoll, was Ergebnisse aus begleitenden
Dissertationsprojekten ergeben haben [84].
Die GSK-3β-Expression kann nicht nur experimentell vermindert werden. Die GSK-
3β-Aktivität kann therapeutisch durch LiCl moduliert werden, um einen positiven
Einfluss auf die Proliferation von knochenbildenden Osteoblasten zu erreichen
[36, 100]. GC werden ausgiebig für die Behandlung von Autoimmun- und
Entzündungskrankheiten genutzt [96]. GC finden in Kombination mit anderen
Medikamenten, wie LiCl, vielfachen Einsatz [36, 46]. Der größte Nebeneffekt der
GC-Behandlung ist ein schneller Knochenmineralisationsverlust und ein
ansteigendes Risiko für Knochenbrüche [95, 96]. GC verursachen über eine
Aktivierung von GSK-3β eine Hemmung des Wnt-Signalweges, der TCF/LEF-
Transkriptionsaktivität in differenzierenden Osteoblasten und somit der
Knochenformation, was langfristig zur Osteoporose führt [96]. Diese hemmende
Wirkung wird allerdings bei Lithiumgabe nicht mehr beobachtet [96]. LiCl steigert
die Knochenneubildungsrate und die Anzahl der Osteoblasten in großer Menge [96].
Lithiumionen werden in Form von LiCl oder Lithiumkarbonat abgegeben und sind

63
klinisch zugelassene GSK-3β-Inhibitoren [15, 100]. Lithium findet seinen Einsatz
bei Osteoporosepatienten und begleitend zur GC-gabe [36, 46, 100].
7. Schlussfolgerung
Die vorliegende experimentelle Studie zeigt, dass aufgrund der gefundenen
Ergebnisse der routinemäßige klinische Einsatz der PEG-Membran zur positiven
Beeinflussung der Wnt-vermittelten, osteogenen Rekrutierung und Maturation
mesenchymaler Osteoblast- und Osteoklastprogenitoren nicht empfohlen werden
kann.
Die PEG-Membran trägt nicht zur Knochenregeneration bei, eignet sich aber als
Carrier.
Die Etablierung der immunhistochemischen Färbung von DKK1 und dessen
Nachweis am Tiermodell des Schweins ist gelungen.
Die immunhistochemische Färbung von RANKL am Schwein konnte selbstständig
erarbeitet werden.
Die immunhistochemische, semiquantitative Evaluation der ossären
Differenzierungs- und Proliferationsmarker OPG, RANKL, GSK-3β und deren
Antagonist DKK1 gelang.
Die Expression von RANKL und DKK1 konnte weder durch die BMP-2-Stimulation
noch die Periostabdeckung beeinflusst werden. Die Expression von GSK-3β ließ sich
beim 1. Opferungszeitpunkt durch die Periostabdeckung verringern, wodurch die
Osteoblastendifferenzierung und –proliferation gefördert wurde. Die OPG-
Expression in der 1. Woche zeigte aufgrund der indirekten Stimulation durch BMP-2
hohe Werte in den mit BMP-2 behandelten Defekten.
Die Proliferationsmarker bieten großes Potential als therapeutischen Ansatzpunkt.
Ein Eingriff in das RANK(L)-OPG-System kann durch den humanen monoklonalen
Antikörper Denosumab erfolgen. GSK-3β und dessen Antagonist DKK1 können
durch Lithium moduliert werden, v.a. während der Einnahme von GC.

64
8. Literatur
[1] Ai M, Holmen SL, Van Hul W, Williams BO, Warman ML. (2005) Reduced affinity to and inhibition by DKK1 form a common mechanism by which high bone mass-associated missense mutations in LRP5 affect canonical Wnt signaling. Mol Cell Biol. 25(12):4946-4955.
[2] Asagiri M, Takayanagi H. (2007) The molecular understanding of osteoclast differentiation. Bone. 40(2):251-264.
[3] Bennett CN, Longo KA, Wright WS, Suva LJ, Lane TF, Hankenson KD, MacDougald OA. (2005) Regulation of osteoblastogenesis and bone mass by Wnt10b. Proc Natl Acad Sci U S A. 102(9):3324-3329.
[4] Bennett CN, Ross SE, Longo KA, Bajnok L, Hemati N, Johnson KW, Harrison SD, MacDougald OA. (2002) Regulation of Wnt signaling during adipogenesis. J Biol Chem. 277(34):30998-31004.
[5] Bodine PV. (2008) Wnt signaling control of bone cell apoptosis. Cell Res. 18(2):248-253.
[6] Bodine PV, Zhao W, Kharode YP, Bex FJ, Lambert AJ, Goad MB, Gaur T, Stein GS, Lian JB, Komm BS. (2004) The Wnt antagonist secreted frizzled-related protein-1 is a negative regulator of trabecular bone formation in adult mice. Mol Endocrinol. 18(5):1222-1237.
[7] Boenisch T, Key, M., Welcher, R., Harvey, R., Atwood, K.N., Stead, A.J.F., Stead, R.H., ed. Immunhistochemische Färbemethoden. Dako Cytomation ed 2003.
[8] Boivin G, Meunier PJ. (2002) Changes in bone remodeling rate influence the degree of mineralization of bone. Connect Tissue Res. 43(2-3):535-537.
[9] Boyce BF, Xing L. (2007) Biology of RANK, RANKL, and osteoprotegerin. Arthritis Res Ther. 9 Suppl 1:S1.
[10] Buck BE, Resnick L, Shah SM, Malinin TI. (1990) Human immunodeficiency virus cultured from bone. Implications for transplantation. Clin Orthop Relat Res. (251):249-253.
[11] Burchardt H. (1983) The biology of bone graft repair. Clin Orthop Relat Res. (174):28-42.
[12] Cadigan KM, Liu YI. (2006) Wnt signaling: complexity at the surface. J Cell Sci. 119(Pt 3):395-402.
[13] Chen Y, Whetstone HC, Youn A, Nadesan P, Chow EC, Lin AC, Alman BA. (2007) Beta-catenin signaling pathway is crucial for bone morphogenetic protein 2 to induce new bone formation. J Biol Chem. 282(1):526-533.
[14] Chung EH, Gilbert M, Virdi AS, Sena K, Sumner DR, Healy KE. (2006) Biomimetic artificial ECMs stimulate bone regeneration. J Biomed Mater Res A. 79(4):815-826.
[15] Clement-Lacroix P, Ai M, Morvan F, Roman-Roman S, Vayssiere B, Belleville C, Estrera K, Warman ML, Baron R, Rawadi G. (2005) Lrp5-independent activation of Wnt signaling by lithium chloride increases bone formation and bone mass in mice. Proc Natl Acad Sci U S A. 102(48):17406-17411.
[16] Cohen MM, Jr. (2006) The new bone biology: pathologic, molecular, and clinical correlates. Am J Med Genet A. 140(23):2646-2706.

65
[17] Creuzet S, Couly G, Vincent C, Le Douarin NM. (2002) Negative effect of Hox gene expression on the development of the neural crest-derived facial skeleton. Development. 129(18):4301-4313.
[18] Cricchio G, Lundgren S. (2003) Donor site morbidity in two different approaches to anterior iliac crest bone harvesting. Clin Implant Dent Relat Res. 5(3):161-169.
[19] Day TY, BS and Yingzi; PhD. (2008) Wnt and Hedgehog Signaling Pathways in Bone Development. The Journal of Bone and Joint Surgery (America).90:19-24.
[20] Ersanli S, Olgac V, Leblebicioglu B. (2004) Histologic analysis of alveolar bone following guided bone regeneration. J Periodontol. 75(5):750-756.
[21] Fetner AE, Hartigan MS, Low SB. (1994) Periodontal repair using PerioGlas in nonhuman primates: clinical and histologic observations. Compendium. 15(7):932, 935-938; quiz 939.
[22] Filvaroff E, Derynck R. (1998) Bone remodelling: a signalling system for osteoclast regulation. Curr Biol. 8(19):R679-682.
[23] Frame JW. (1987) Hydroxyapatite as a biomaterial for alveolar ridge augmentation. Int J Oral Maxillofac Surg. 16(6):642-655.
[24] Frame JW, Brady CL. (1987) The versatility of hydroxyapatite blocks in maxillofacial surgery. Br J Oral Maxillofac Surg. 25(6):452-464.
[25] Frame JW, Edmondson HD, Furniss A. (1987) Mandibular reconstruction using split autogenous bone grafts. Br J Oral Maxillofac Surg. 25(1):1-8.
[26] Frame JW, Laird WR, McLaughlin EA. (1987) Augmentation of localised alveolar bone loss using porous hydroxyapatite particles. Restorative Dent. 3(4):80, 82, 84-85.
[27] Freischem J. (2009) "Positive Opinion" für Denosumab. Osteoporose News, wwwosteoporosede.
[28] Friedman M, Cavins, J., Wall, J. (1965) Relative nucleophilic reactivities of amino groups and mercaptide ions in addition reactions with alpha-,beta-unsaturated compounds. J Am Chem Soc.87: 3672-3682.
[29] Friess W, Uludag H, Foskett S, Biron R. (1999) Bone regeneration with recombinant human bone morphogenetic protein-2 (rhBMP-2) using absorbable collagen sponges (ACS): influence of processing on ACS characteristics and formulation. Pharm Dev Technol. 4(3):387-396.
[30] Fujita K, Janz S. (2007) Attenuation of WNT signaling by DKK-1 and -2 regulates BMP2-induced osteoblast differentiation and expression of OPG, RANKL and M-CSF. Mol Cancer. 6:71.
[31] Glass DA, 2nd, Bialek P, Ahn JD, Starbuck M, Patel MS, Clevers H, Taketo MM, Long F, McMahon AP, Lang RA, Karsenty G. (2005) Canonical Wnt signaling in differentiated osteoblasts controls osteoclast differentiation. Dev Cell. 8(5):751-764.
[32] Gong Y, Slee RB, Fukai N, Rawadi G, Roman-Roman S, Reginato AM, Wang H, Cundy T, Glorieux FH, Lev D, Zacharin M, Oexle K, Marcelino J, Suwairi W, Heeger S, Sabatakos G, Apte S, Adkins WN, Allgrove J, Arslan-Kirchner M, Batch JA, Beighton P, Black GC, Boles RG, Boon LM, Borrone C, Brunner HG, Carle GF, Dallapiccola B, De Paepe A, Floege B, Halfhide ML, Hall B, Hennekam RC, Hirose T, Jans A, Juppner H, Kim CA, Keppler-Noreuil K, Kohlschuetter A, LaCombe D, Lambert M, Lemyre E, Letteboer T, Peltonen L, Ramesar RS, Romanengo M, Somer H, Steichen-Gersdorf E, Steinmann B, Sullivan B, Superti-Furga A, Swoboda W, van den Boogaard

66
MJ, Van Hul W, Vikkula M, Votruba M, Zabel B, Garcia T, Baron R, Olsen BR, Warman ML. (2001) LDL receptor-related protein 5 (LRP5) affects bone accrual and eye development. Cell. 107(4):513-523.
[33] Gordon MD, Nusse R. (2006) Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors. J Biol Chem. 281(32):22429-22433.
[34] Greenspan DS, Northrup H, Au KS, McAllister KA, Francomano CA, Wenstrup RJ, Marchuk DA, Kwiatkowski DJ. (1995) COL5A1: fine genetic mapping and exclusion as candidate gene in families with nail-patella syndrome, tuberous sclerosis 1, hereditary hemorrhagic telangiectasia, and Ehlers-Danlos Syndrome type II. Genomics. 25(3):737-739.
[35] Gregory CA, Perry AS, Reyes E, Conley A, Gunn WG, Prockop DJ. (2005) Dkk-1-derived synthetic peptides and lithium chloride for the control and recovery of adult stem cells from bone marrow. J Biol Chem. 280(3):2309-2323.
[36] Guo X, Wang XF. (2009) Signaling cross-talk between TGF-beta/BMP and other pathways. Cell Res. 19(1):71-88.
[37] Hansen RC. (1984) Blindness, anonychia, and oral mucosal scarring as sequelae of the Stevens-Johnson syndrome. Pediatr Dermatol. 1(4):298-300.
[38] Holmen SL, Zylstra CR, Mukherjee A, Sigler RE, Faugere MC, Bouxsein ML, Deng L, Clemens TL, Williams BO. (2005) Essential role of beta-catenin in postnatal bone acquisition. J Biol Chem. 280(22):21162-21168.
[39] Huber FX, Berger I, McArthur N, Huber C, Kock HP, Hillmeier J, Meeder PJ. (2008) Evaluation of a novel nanocrystalline hydroxyapatite paste and a solid hydroxyapatite ceramic for the treatment of critical size bone defects (CSD) in rabbits. J Mater Sci Mater Med. 19(1):33-38.
[40] Hurzeler MB, Kohal RJ, Naghshbandi J, Mota LF, Conradt J, Hutmacher D, Caffesse RG. (1998) Evaluation of a new bioresorbable barrier to facilitate guided bone regeneration around exposed implant threads. An experimental study in the monkey. Int J Oral Maxillofac Surg. 27(4):315-320.
[41] Jackson A, Vayssiere B, Garcia T, Newell W, Baron R, Roman-Roman S, Rawadi G. (2005) Gene array analysis of Wnt-regulated genes in C3H10T1/2 cells. Bone. 36(4):585-598.
[42] Jensen OT, Sennerby L. (1998) Histologic analysis of clinically retrieved titanium microimplants placed in conjunction with maxillary sinus floor augmentation. Int J Oral Maxillofac Implants. 13(4):513-521.
[43] Jensen SS, Broggini N, Hjorting-Hansen E, Schenk R, Buser D. (2006) Bone healing and graft resorption of autograft, anorganic bovine bone and beta-tricalcium phosphate. A histologic and histomorphometric study in the mandibles of minipigs. Clin Oral Implants Res. 17(3):237-243.
[44] Kapadia RM, Guntur AR, Reinhold MI, Naski MC. (2005) Glycogen synthase kinase 3 controls endochondral bone development: contribution of fibroblast growth factor 18. Dev Biol. 285(2):496-507.
[45] Kast RE. (2008) How lithium treatment generates neutrophilia by enhancing phosphorylation of GSK-3, increasing HIF-1 levels and how this path is important during engraftment. Bone Marrow Transplant. 41(1):23-26.
[46] Kessler P, Thorwarth M, Bloch-Birkholz A, Nkenke E, Neukam FW. (2005) Harvesting of bone from the iliac crest--comparison of the anterior and posterior sites. Br J Oral Maxillofac Surg. 43(1):51-56.

67
[47] Kim JB, Leucht P, Lam K, Luppen C, Ten Berge D, Nusse R, Helms JA. (2007) Bone regeneration is regulated by wnt signaling. J Bone Miner Res. 22(12):1913-1923.
[48] Knosp WM, Scott V, Bachinger HP, Stadler HS. (2004) HOXA13 regulates the expression of bone morphogenetic proteins 2 and 7 to control distal limb morphogenesis. Development. 131(18):4581-4592.
[49] Kondo N, Ogose A, Tokunaga K, Umezu H, Arai K, Kudo N, Hoshino M, Inoue H, Irie H, Kuroda K, Mera H, Endo N. (2006) Osteoinduction with highly purified beta-tricalcium phosphate in dog dorsal muscles and the proliferation of osteoclasts before heterotopic bone formation. Biomaterials. 27(25):4419-4427.
[50] Krishnan V, Bryant HU, Macdougald OA. (2006) Regulation of bone mass by Wnt signaling. J Clin Invest. 116(5):1202-1209.
[51] Kulkarni NH, Onyia JE, Zeng Q, Tian X, Liu M, Halladay DL, Frolik CA, Engler T, Wei T, Kriauciunas A, Martin TJ, Sato M, Bryant HU, Ma YL. (2006) Orally bioavailable GSK-3alpha/beta dual inhibitor increases markers of cellular differentiation in vitro and bone mass in vivo. J Bone Miner Res. 21(6):910-920.
[52] Lappin TRJ, Grier, D.G., Thompson, A., Halliday, H.L. (2006) Hox genes: Seductive science, mysterious mechanisms. Ulster Med J.75:23-31.
[53] Leroi AM, ed. Tanz der Gene München: Elsevier, Spektrum akademischer Verlag 2004.
[54] Leucht P, Kim JB, Amasha R, James AW, Girod S, Helms JA. (2008) Embryonic origin and Hox status determine progenitor cell fate during adult bone regeneration. Development. 135(17):2845-2854.
[55] Little RD, Recker RR, Johnson ML. (2002) High bone density due to a mutation in LDL-receptor-related protein 5. N Engl J Med. 347(12):943-944; author reply 943-944.
[56] Meffert RM. (1998) Current usage of bone fill as an adjunct in implant dentistry. Dent Implantol Update. 9(2):9-12.
[57] Mellonig JT. (1992) Autogenous and allogeneic bone grafts in periodontal therapy. Crit Rev Oral Biol Med. 3(4):333-352.
[58] Mellonig JT, Prewett AB, Moyer MP. (1992) HIV inactivation in a bone allograft. J Periodontol. 63(12):979-983.
[59] Merten HA, Wiltfang J, Honig JF, Funke M, Luhr HG. (2000) [Intra-individual comparison of alpha- and beta-TCP ceramics in an animal experiment]. Mund Kiefer Gesichtschir. 4 Suppl 2:S509-515.
[60] Mikos AG, Bao Y, Cima LG, Ingber DE, Vacanti JP, Langer R. (1993) Preparation of poly(glycolic acid) bonded fiber structures for cell attachment and transplantation. J Biomed Mater Res. 27(2):183-189.
[61] Misch CE, Dietsh F. (1993) Bone-grafting materials in implant dentistry. Implant Dent. 2(3):158-167.
[62] Moore WR, Graves SE, Bain GI. (2001) Synthetic bone graft substitutes. ANZ J Surg. 71(6):354-361.
[63] Nagahara K, Isogai M, Shibata K, Meenaghan MA. (1992) Osteogenesis of hydroxyapatite and tricalcium phosphate used as a bone substitute. Int J Oral Maxillofac Implants. 7(1):72-79.
[64] Neukam FW, Hausamen, J.E. and Scheller, H. (2004) Implantology. Mund-Kiefer-Gesichtschirurgie.4 Suppl 1: S249 - S256.

68
[65] Neukam FW, Hausamen, J.E. and Scheller, H. (1989) Possibilities and limitations of implantology in older patients. Dtsch Zahnärztl Z.44: S490 - S492.
[66] Nkenke E, Radespiel-Troger M, Wiltfang J, Schultze-Mosgau S, Winkler G, Neukam FW. (2002) Morbidity of harvesting of retromolar bone grafts: a prospective study. Clin Oral Implants Res. 13(5):514-521.
[67] Nuttall ME, Gimble JM. (2000) Is there a therapeutic opportunity to either prevent or treat osteopenic disorders by inhibiting marrow adipogenesis? Bone. 27(2):177-184.
[68] Oka K, Oka S, Sasaki T, Ito Y, Bringas P, Jr., Nonaka K, Chai Y. (2007) The role of TGF-beta signaling in regulating chondrogenesis and osteogenesis during mandibular development. Dev Biol. 303(1):391-404.
[69] Park J, Gelse K, Frank S, von der Mark K, Aigner T, Schneider H. (2006) Transgene-activated mesenchymal cells for articular cartilage repair: a comparison of primary bone marrow-, perichondrium/periosteum- and fat-derived cells. J Gene Med. 8(1):112-125.
[70] Park J, Lutz R, Felszeghy E, Wiltfang J, Nkenke E, Neukam FW, Schlegel KA. (2007) The effect on bone regeneration of a liposomal vector to deliver BMP-2 gene to bone grafts in peri-implant bone defects. Biomaterials. 28(17):2772-2782.
[71] Pietschmann P, Diel, I.J., Fahrleitner-Pammer, A., Griesmacher, A., Gruber, R., Obermayer-Pietsch, B., Patsch, J., Schett, G., Sipos, W., Woloszczuk, W., ed. Osteoimmunologie - Zytokine und Biologika in der Osteologie. Bremen: Uni-Med 2008.
[72] Piters E, Boudin E, Van Hul W. (2008) Wnt signaling: a win for bone. Arch Biochem Biophys. 473(2):112-116.
[73] Precheur HV. (2007) Bone graft materials. Dent Clin North Am. 51(3):729-746, viii.
[74] Qiang YW, Chen Y, Stephens O, Brown N, Chen B, Epstein J, Barlogie B, Shaughnessy JD, Jr. (2008) Myeloma-derived Dickkopf-1 disrupts Wnt-regulated osteoprotegerin and RANKL production by osteoblasts: a potential mechanism underlying osteolytic bone lesions in multiple myeloma. Blood. 112(1):196-207.
[75] Radu FA, Bause M, Knabner P, Lee GW, Friess WC. (2002) Modeling of drug release from collagen matrices. J Pharm Sci. 91(4):964-972.
[76] Rissolo AR, Bennett J. (1998) Bone grafting and its essential role in implant dentistry. Dent Clin North Am. 42(1):91-116.
[77] Robson MC, Krizek TJ, Koss N, Samburg JL. (1973) Amniotic membranes as a temporary wound dressing. Surg Gynecol Obstet. 136(6):904-906.
[78] Ross SE, Hemati N, Longo KA, Bennett CN, Lucas PC, Erickson RL, MacDougald OA. (2000) Inhibition of adipogenesis by Wnt signaling. Science. 289(5481):950-953.
[79] Roy DM, Linnehan SK. (1974) Hydroxyapatite formed from coral skeletal carbonate by hydrothermal exchange. Nature. 247(438):220-222.
[80] Sato MM, Nakashima A, Nashimoto M, Yawaka Y, Tamura M. (2009) Bone morphogenetic protein-2 enhances Wnt/beta-catenin signaling-induced osteoprotegerin expression. Genes Cells. 14(2):141-153.
[81] Schepers EJ, Ducheyne P, Barbier L, Schepers S. (1993) Bioactive glass particles of narrow size range: a new material for the repair of bone defects. Implant Dent. 2(3):151-156.

69
[82] Schlegel AK, Donath K. (1998) BIO-OSS--a resorbable bone substitute? J Long Term Eff Med Implants. 8(3-4):201-209.
[83] Schlegel AK, Donath K, Weida S. (1998) Histological findings in guided bone regeneration (GBR) around titanium dental implants with autogenous bone chips using a new resorbable membrane. J Long Term Eff Med Implants. 8(3-4):211-224.
[84] Schlegel AK, Neukam, F.W., Nkenke, E., Wehrhan, F., Lutz, R., Srour, S. Feasibility study to evaluate the efficacy of a hydrogel-biphasic calcium phosphate system for bone regeneration in a pig calvaria model. Report on experimental study; 2008, 1-26.
[85] Schlegel KA, Fichtner G, Schultze-Mosgau S, Wiltfang J. (2003) Histologic findings in sinus augmentation with autogenous bone chips versus a bovine bone substitute. Int J Oral Maxillofac Implants. 18(1):53-58.
[86] Schlegel KA, Kloss FR, Kessler P, Schultze-Mosgau S, Nkenke E, Wiltfang J. (2003) Bone conditioning to enhance implant osseointegration: an experimental study in pigs. Int J Oral Maxillofac Implants. 18(4):505-511.
[87] Schlegel KA, Schultze-Mosgau S, Wiltfang J, Neukam FW, Rupprecht S, Thorwarth M. (2006) Changes of mineralization of free autogenous bone grafts used for sinus floor elevation. Clin Oral Implants Res. 17(6):673-678.
[88] Schlegel KA, Zimmermann R, Thorwarth M, Neukam FW, Klongnoi B, Nkenke E, Felszeghy E. (2007) Sinus floor elevation using autogenous bone or bone substitute combined with platelet-rich plasma. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 104(3):e15-25.
[89] Schliephake H, Neukam FW, Wichmann M. (1997) Survival analysis of endosseous implants in bone grafts used for the treatment of severe alveolar ridge atrophy. J Oral Maxillofac Surg. 55(11):1227-1233; discussion 1233-1224.
[90] Schultze-Mosgau S, Blatz MB, Wehrhan F, Schlegel KA, Thorwart M, Holst S. (2005) Principles and mechanisms of peri-implant soft tissue healing. Quintessence Int. 36(10):759-769.
[91] Shao JS, Aly ZA, Lai CF, Cheng SL, Cai J, Huang E, Behrmann A, Towler DA. (2007) Vascular Bmp Msx2 Wnt signaling and oxidative stress in arterial calcification. Ann N Y Acad Sci. 1117:40-50.
[92] Shigeyama Y, D'Errico JA, Stone R, Somerman MJ. (1995) Commercially-prepared allograft material has biological activity in vitro. J Periodontol. 66(6):478-487.
[93] Sittinger M, Bujia J, Rotter N, Reitzel D, Minuth WW, Burmester GR. (1996) Tissue engineering and autologous transplant formation: practical approaches with resorbable biomaterials and new cell culture techniques. Biomaterials. 17(3):237-242.
[94] Small S, Arnosti DN, Levine M. (1993) Spacing ensures autonomous expression of different stripe enhancers in the even-skipped promoter. Development. 119(3):762-772.
[95] Smith E, Coetzee GA, Frenkel B. (2002) Glucocorticoids inhibit cell cycle progression in differentiating osteoblasts via glycogen synthase kinase-3beta. J Biol Chem. 277(20):18191-18197.
[96] Smith E, Frenkel B. (2005) Glucocorticoids inhibit the transcriptional activity of LEF/TCF in differentiating osteoblasts in a glycogen synthase kinase-3beta-dependent and -independent manner. J Biol Chem. 280(3):2388-2394.

70
[97] Spector M. (1994) Anorganic bovine bone and ceramic analogs of bone mineral as implants to facilitate bone regeneration. Clin Plast Surg. 21(3):437-444.
[98] Spencer GJ, Utting JC, Etheridge SL, Arnett TR, Genever PG. (2006) Wnt signalling in osteoblasts regulates expression of the receptor activator of NFkappaB ligand and inhibits osteoclastogenesis in vitro. J Cell Sci. 119(Pt 7):1283-1296.
[99] Springer IN, Nocini PF, Schlegel KA, De Santis D, Park J, Warnke PH, Terheyden H, Zimmermann R, Chiarini L, Gardner K, Ferrari F, Wiltfang J. (2006) Two techniques for the preparation of cell-scaffold constructs suitable for sinus augmentation: steps into clinical application. Tissue Eng. 12(9):2649-2656.
[100] Stambolic V, Ruel L, Woodgett JR. (1996) Lithium inhibits glycogen synthase kinase-3 activity and mimics wingless signalling in intact cells. Curr Biol. 6(12):1664-1668.
[101] Thorwarth M, Schultze-Mosgau S, Kessler P, Wiltfang J, Schlegel KA. (2005) Bone regeneration in osseous defects using a resorbable nanoparticular hydroxyapatite. J Oral Maxillofac Surg. 63(11):1626-1633.
[102] Thorwarth M, Schultze-Mosgau S, Wehrhan F, Srour S, Wiltfang J, Neukam FW, Schlegel KA. (2005) Enhanced bone regeneration with a synthetic cell-binding peptide--in vivo results. Biochem Biophys Res Commun. 329(2):789-795.
[103] Uludag H, Gao T, Porter TJ, Friess W, Wozney JM. (2001) Delivery systems for BMPs: factors contributing to protein retention at an application site. J Bone Joint Surg Am. 83-A Suppl 1(Pt 2):S128-135.
[104] Urist MR, Huo YK, Brownell AG, Hohl WM, Buyske J, Lietze A, Tempst P, Hunkapiller M, DeLange RJ. (1984) Purification of bovine bone morphogenetic protein by hydroxyapatite chromatography. Proc Natl Acad Sci U S A. 81(2):371-375.
[105] Van Wesenbeeck L, Cleiren E, Gram J, Beals RK, Benichou O, Scopelliti D, Key L, Renton T, Bartels C, Gong Y, Warman ML, De Vernejoul MC, Bollerslev J, Van Hul W. (2003) Six novel missense mutations in the LDL receptor-related protein 5 (LRP5) gene in different conditions with an increased bone density. Am J Hum Genet. 72(3):763-771.
[106] Wada T, Nakashima T, Hiroshi N, Penninger JM. (2006) RANKL-RANK signaling in osteoclastogenesis and bone disease. Trends Mol Med. 12(1):17-25.
[107] Warnke PH, Wiltfang J, Springer I, Acil Y, Bolte H, Kosmahl M, Russo PA, Sherry E, Lutzen U, Wolfart S, Terheyden H. (2006) Man as living bioreactor: fate of an exogenously prepared customized tissue-engineered mandible. Biomaterials. 27(17):3163-3167.
[108] Wehrhan F, Grabenbauer GG, Rodel F, Amann K, Schultze-Mosgau S. (2004) Exogenous modulation of TGF-beta(1) influences TGF-betaR-III-associated vascularization during wound healing in irradiated tissue. Strahlenther Onkol. 180(8):526-533.
[109] Weibel ER. (1989) Measuring through the microscope: development and evolution of stereological methods. J Microsc. 155(Pt 3):393-403.
[110] Wellik DM, Capecchi MR. (2003) Hox10 and Hox11 genes are required to globally pattern the mammalian skeleton. Science. 301(5631):363-367.

71
[111] Werner J, Linner-Krcmar B, Friess W, Greil P. (2002) Mechanical properties and in vitro cell compatibility of hydroxyapatite ceramics with graded pore structure. Biomaterials. 23(21):4285-4294.
[112] Williams BO, Insogna KL. (2009) Where Wnts went: the exploding field of Lrp5 and Lrp6 signaling in bone. J Bone Miner Res. 24(2):171-178.
[113] Wiltfang J, Merten HA, Schlegel KA, Schultze-Mosgau S, Kloss FR, Rupprecht S, Kessler P. (2002) Degradation characteristics of alpha and beta tri-calcium-phosphate (TCP) in minipigs. J Biomed Mater Res. 63(2):115-121.
[114] Wiltfang J, Schlegel KA, Schultze-Mosgau S, Nkenke E, Zimmermann R, Kessler P. (2003) Sinus floor augmentation with beta-tricalciumphosphate (beta-TCP): does platelet-rich plasma promote its osseous integration and degradation? Clin Oral Implants Res. 14(2):213-218.

72
9. Anhang
9.1. Färbeprotokolle
RANKL
Vorbehandlungen 4x15’ MEA zum Entakrylieren
Je 2’
Absteigende Ethanolreihe (verdünnt mit AD):
100% - 100% - 96% - 96% - 90% - 70% - 70%
5’ AD
20’ EDTA 10% zum Entkalken
10’ Fließendes Leitungswasser
3x2’ AD
3x2’ DAKO Waschpuffer
15’ 95°C Target Retrieval Puffer pH 9,0 im Wasserbad
15’ RT Abkühlen im Target Retrieval Puffer
3x2’ DAKO Waschpuffer
Im Autostainer
20’ Proteinblock, anschließend abblasen
1h RT Primärantikörper: sRANKL 1:150 verdünnt mit „Antibody diluent“
Spülen mit Waschpuffer
15’ RT Biotinylierter Sekundärantikörper
Spülen mit Waschpuffer
15’ RT Tertiärantikörper: Streptavidin Alkaline Phosphatase
Spülen mit Waschpuffer
8’ Chromogen
Spülen mit Waschpuffer
8’ bis 22°C
4’ > 22°C
Chromogen
Spülen mit AD
10’ AD
Spülen mit AD
Von Hand
5’ Hämalaun (Kernfärbung)
5’ Fließendes Leitungswasser
5’ AD
Eindecken mit Aquatex

73
DKK1 Vorbehandlungen
4x15’ MEA zum Entakrylieren
Je 2’
Absteigende Ethanolreihe (verdünnt mit AD):
100% - 100% - 96% - 96% - 90% - 70% - 70%
5’ AD
20’ EDTA 10% zum Entkalken
10’ Fließendes Leitungswasser
3x2’ AD
3x2’ DAKO Waschpuffer
30’ kochen Citratpuffer pH 6.0 im Wasserbad
15’ RT Abkühlen im Citratpuffer
3x2’ DAKO Waschpuffer
Im Autostainer
15’ Avidin
Spülen mit Waschpuffer
15’ Biotin
Spülen mit Waschpuffer
20’ Proteinblock, anschließend abblasen
1h RT Primärantikörper: DKK 1:100 verdünnt mit „Antibody diluent“
Spülen mit Waschpuffer
15’ RT Biotinylierter Sekundärantikörper
Spülen mit Waschpuffer
15’ RT Tertiärantikörper: Streptavidin Alkaline Phosphatase
Spülen mit Waschpuffer
8’ Chromogen
Spülen mit Waschpuffer
8’ bis 22°C
4’ > 22°C
Chromogen
Spülen mit AD
10’ AD
Spülen mit AD
Von Hand
5’ Hämalaun (Kernfärbung)
5’ Fließendes Leitungswasser
5’ AD
Eindecken mit Aquatex

74
OPG Vorbehandlungen
4x15’ MEA zum Entakrylieren
Je 2’
Absteigende Ethanolreihe (verdünnt mit AD):
100% - 100% - 96% - 96% - 90% - 70% - 70%
5’ AD
20’ EDTA 10% zum Entkalken
10’ Fließendes Leitungswasser
3x2’ AD
3x2’ DAKO Waschpuffer
15’ kochen Citratpuffer pH 6.0 im Wasserbad
15’ RT Abkühlen im Citratpuffer
3x2’ DAKO Waschpuffer
Im Autostainer
20’ Proteinblock, anschließend abblasen
1h RT Primärantikörper: OPG 1:50 verdünnt mit „Antibody diluent“
Spülen mit Waschpuffer
15’ RT Biotinylierter Sekundärantikörper
Spülen mit Waschpuffer
15’ RT Tertiärantikörper: Streptavidin Alkaline Phosphatase
Spülen mit Waschpuffer
8’ Chromogen
Spülen mit Waschpuffer
8’ bis 22°C
4’ > 22°C Chromogen
Spülen mit AD
10’ AD
Spülen mit AD
Von Hand
5’ Hämalaun (Kernfärbung)
5’ Fließendes Leitungswasser
5’ AD
Eindecken mit Aquatex

75
GSK-3β Vorbehandlungen
4x15’ MEA zum Entakrylieren
Je 2’
Absteigende Ethanolreihe (verdünnt mit AD):
100% - 100% - 96% - 96% - 90% - 70% - 70%
5’ AD
20’ EDTA 10% zum Entkalken
10’ Fließendes Leitungswasser
3x2’ AD
3x2’ DAKO Waschpuffer
15’ kochen Citratpuffer pH 6.0 im Wasserbad
15’ RT Abkühlen im Citratpuffer
3x2’ DAKO Waschpuffer
Im Autostainer
20’ Proteinblock, anschließend abblasen
1h RT Primärantikörper: GSK-3β 1:50 verdünnt mit „Antibody diluent“
Spülen mit Waschpuffer
15’ RT Biotinylierter Sekundärantikörper
Spülen mit Waschpuffer
15’ RT Tertiärantikörper: Streptavidin Alkaline Phosphatase
Spülen mit Waschpuffer
8’ Chromogen
Spülen mit Waschpuffer
8’ bis 22°C
4’ > 22°C
Chromogen
Spülen mit AD
10’ AD
Spülen mit AD
Von Hand
5’ Hämalaun (Kernfärbung)
5’ Fließendes Leitungswasser
5’ AD
Eindecken mit Aquatex

76
9.2. Lebenslauf
Angaben zur Person:
Name: Monika Christine Fruck
Anschrift: Saaß 7,
93133 Burglengenfeld
Geburtstag: 15.07.1983
Geburtsort: Burglengenfeld
Vater: Harald Fruck, Industriemeister
Mutter: Renate Fruck
Geschwister: Matthias Fruck, Inspektor im Notardienst
Alexander Fruck, Schüler
Familienstand: ledig
Nationalität: deutsch
Schulabschluss: Abitur
Lebensdaten:
1990 – 1994 Geschwister Scholl Grundschule, Burglengenfeld
1994 – 2003 Johann-Michael-Fischer Gymnasium, Burglengenfeld
10/03-03/04 Studium der Wirtschaftsmathematik an der FAU Erlangen
04/04-07/09 Studium der Zahnheilkunde an der FAU Erlangen-Nürnberg
08/09-12/09 Staatsexamen Zahnmedizin
01/10 Approbation als Zahnarzt
03/10 Antritt der Assistentenstelle in Schwandorf, Praxis Dr. Saur
Burglengenfeld, den 01.06.2010

77
9.3. Danksagung
Mein besonderer Dank gilt meinem Doktorvater Herrn Priv.-Doz. Dr. med. Dr. med.
dent. K. A. Schlegel für die Planung des Versuchsvorhabens und die Überlassung des
interessanten Themas sowie für seine Unterstützung bei der Bearbeitung des
Dissertationsprojektes.
Bei Herrn Prof. Dr. med. Dr. med. dent. F. W. Neukam möchte ich mich für die
Schaffung der finanziellen Möglichkeiten bedanken, insbesondere in der
Etablierungsphase der immunhistochemischen Nachweismethoden.
Meinem wissenschaftlichen Lehrer und Betreuer, Herrn Dr. med. Dr. med. dent. F.
Wehrhan danke ich für die freundschaftliche Betreuung und die jederzeit
wohlwollende Unterstützung bei der Bearbeitung des Dissertationsprojektes. Bei der
Erlernung eines effizienten wissenschaftlichen Arbeitsstiles, der Planung und
Durchführung der Experimente und der Schaffung eines wissenschaftlich
leistungsfähigen Umfeldes von Kooperationspartnern war er mir ein Vorbild.
Für die liebevolle und geduldige Unterstützung bei der Erlernung der Abläufe in der
Immunhistochemie danke ich in besonderer Weise der medizinisch-technischen
Assistentin Frau H. Heider.
Für die gegenseitige fachliche und emotionale Untertstützung während der
praktischen Arbeit im Labor, den intellektuellen Austausch und die Anregungen zur
Ergebnisinterpretation danke ich besonders meinen Freunden Nadine Knoll und Felix
Kölpin.
Für die Hilfe bei der Anfertigung der schriftlichen Arbeit und deren Korrektur, das
Verständnis für die Einschränkung der gemeinsamen Freizeit und die emotionale
Unterstützung in schwierigen Situationen danke ich in besonderem Maße meinem
Lebenspartner Oliver Bressler.
Meinen Eltern gilt mein besonderer Dank für die in ihrem Rahmen mögliche
finanzielle Unterstützung während des Studiums und die Motivation zur
Fertigstellung des gewählten Dissertationsprojektes.

78
9.4. Ehrenwörtliche Erklärung
Hiermit erkläre ich,
dass mir die Promotionsordnung der Medizinischen Fakultät der Friedrich-
Alexander-Universität bekannt ist,
dass ich die Dissertation selbst angefertigt habe und alle von mir benutzten
Hilfsmittel, persönlichen Mitteilungen und Quellen in meiner Arbeit angegeben sind,
dass mich folgende Person bei der Auswahl und Auswertung des Materials sowie bei
der Herstellung des Manuskripts unterstützt hat: Dr. Dr.med.dent. F. Wehrhan,
dass die Hilfe eines Promotionsberaters nicht in Anspruch genommen wurde und
dass Dritte weder unmittelbar noch mittelbar geldwerte Leistungen von mir für
Arbeiten erhalten haben, die im Zusammenhang mit dem Inhalt der vorgelegten
Dissertation stehen,
dass ich die Dissertation noch nicht als Prüfungsarbeit für eine staatliche oder andere
wissenschaftliche Prüfung eingereicht habe und
dass ich die gleiche, eine in wesentlichen Teilen ähnliche oder eine andere
Abhandlung nicht bei einer anderen Hochschule als Dissertation eingereicht habe.
Burglengenfeld, den 01.06.2010





























