Embed Size (px)
Citation preview

Cyr61: Ein durch Östrogen reguliertes Markergen der Endometriose
Inaugural-Dissertation
zur
Erlangung des Doktorgrades
Dr. rer. nat.
des Fachbereichs
Bio- und Geowissenschaften, Landschaftsarchitektur
an der
Universität Duisburg-Essen
vorgelegt von
Yvonne Absenger
aus Wuppertal
April 2003

Die der vorliegenden Arbeit zugrundeliegenden Experimente wurden am Institut für Anatomie
der Universität Duisburg-Essen durchgeführt.
1. Gutachter: Prof. Dr. E. Winterhager
2. Gutachter: Priv. Doz. Dr. L. Klein-Hitpaß
Vorsitzender des Prüfungsausschusses: Prof. Dr. H. Burda
Tag der mündlichen Prüfung: 17. September 2003
I

Danksagung
An dieser Stelle möchte ich allen danken, die mich bei der Durchführung dieser Doktorarbeit
unterstützt haben.
Ein besonderer Dank gilt Frau Prof. Dr. Elke Winterhager für die Überlassung des
interessanten Themas, die gute Betreuung dieser Arbeit sowie ihrer ständigen Bereitschaft
zur Diskussion.
Ebenso bedanke ich mich bei Herrn Dr. Holger Hess-Stumpp und Frau Dr. Anja Schmidt der
Schering AG für die Betreuung dieser Arbeit und die Überlassung von Testsubstanzen.
Der Abteilung für Gynäkologie des Universitätsklinikums Essen unter der Leitung von Herrn
Prof. Dr. Schindler bzw. Herrn Prof. Dr. Kimmig danke ich für die Bereitstellung der
Gewebeproben und der dazugehörigen Hormonwerte.
Bei Herrn Dr. Norbert Schütze der Orthopädischen Universitätsklinik in Würzburg möchte ich
mich für die Überlassung des Antiserums bedanken.
Ich bedanke mich bei Daniela Kottmann, Natalie Knipp und Georgia Rauter für die
Unterstützung bei der Durchführung vieler Versuche, sowie bei Dave Kittel für die Hilfe bei
der Bildbearbeitung.
Ich danke allen Mitarbeitern der Arbeitsgruppe Winterhager und dabei ganz besonders
Alexandra Gellhaus und Mark Kibschull, die mir jederzeit mit Hilfestellungen zur Verfügung
standen.
Der größte Dank gilt meinen Eltern und meinem Freund Marc Absenger, die mich mit viel
Geduld während der gesamten Zeit dieser Doktorarbeit unterstützt haben.
Die vorliegende Arbeit wurde durch die Schering AG gefördert.
II

Inhaltsverzeichnis
1 Einleitung............................................................................................................1
1.1 Endometriose................................................................................................1
1.1.1 Morphologie und Lokalisation der Endometrioseherde ..........................3
1.1.2 Klassifikation der Endometriose.............................................................4
1.1.3 Symptome der Endometriose und Therapieansätze ..............................4
1.1.4 Infertilität ................................................................................................7
1.1.5 Einfluß von Steroidhormonen auf die Endometriose..............................9
1.2 Angiogenese ...............................................................................................12
1.2.1 Angiogenetische Faktoren ...................................................................13
1.2.2 Angiogenese im Uterus........................................................................15
1.2.3 Angiogenese in der Endometriose .......................................................16
1.3 Cyr61, ein Protein der CCN-Familie............................................................18
1.4 Tiermodelle für die Untersuchung der Endometriose..................................20
2 Zielsetzung .......................................................................................................22
3 Material und Methoden ....................................................................................23
3.1 Feinchemikalien, Enzyme und Verbrauchsmaterialien................................23
3.2 Humane Gewebeproben .............................................................................25
3.3 Molekularbiologische Methoden..................................................................26
3.3.1 RNA-Isolierung.....................................................................................26
3.3.2 Konzentrationsbestimmung von Nukleinsäuren...................................26
3.3.3 DNase-Verdau .....................................................................................27
3.3.4 Reverse Transkription (RT)..................................................................27
3.4 Genexpressionsanalysen............................................................................28
3.4.1 Gene-Arrays.........................................................................................28
3.4.2 Polymerase-Kettenreaktion (PCR).......................................................28
3.4.2.1 RT-PCR ........................................................................................28
3.4.2.2 Real-time RT-PCR........................................................................30
3.5 Proteinbiochemische Methoden..................................................................32
3.5.1 Protein-Isolierung und Konzentrationsbestimmung..............................32
3.5.2 Fällung von Proteinen ..........................................................................32
3.5.3 SDS-Gelelektrophorese .......................................................................33
3.5.4 Western Blot ........................................................................................33
III

3.6 Untersuchungen im Nacktmausmodell........................................................35
3.6.1 Aufarbeitung des Gewebes..................................................................35
3.6.2 Versuchstiere .......................................................................................35
3.6.3 Transplantation der endometrialen Fragmente ....................................36
3.6.4 Behandlung der Nacktmäuse...............................................................36
3.6.5 Entnahme der transplantierten endometrialen Fragmente...................37
3.6.6 Histologische Aufarbeitung der Fragmente..........................................38
3.7 Immunhistochemische Untersuchungen .....................................................38
3.7.1 Immunhistochemischer Nachweis mit 3,3´-Diaminobenzidin (DAB) ....39
3.7.2 Immunhistochemischer Nachweis durch Fluoreszenz .........................39
3.7.3 Immunhistochemischer Nachweis von Ki67.........................................40
3.8 Statistische Auswertung..............................................................................40
4 Ergebnisse........................................................................................................41
4.1 Genexpressionsanalysen der Endometrien von Frauen mit und ohne
Endometriose..............................................................................................41
4.1.1 Validierung der Ergebnisse der „Gene-Array“ Analysen ......................42
4.1.1.1 Herunterregulierte Gene ...............................................................42
4.1.1.2 Hochregulierte Gene.....................................................................45
4.1.2 Cyr61-Expression in den endometriotischen Läsionen ........................51
4.1.3 Cyr61-Expression im Verlauf des Menstruationszyklus .......................52
4.2 Expression von Cyr61 auf Proteinebene.....................................................53
4.2.1 Western Blot ........................................................................................53
4.2.2 Immunhistochemie...............................................................................54
4.3 Untersuchungen zur hormonellen Regulation von Cyr61............................54
4.3.1 Validierung des Nacktmausmodells .....................................................55
4.3.1.1 Uterus- und Ovargewichte der Nacktmäuse .................................55
4.3.1.2 Zellhöhe und Proliferation des Uterusepithels der behandelten
Nacktmäuse..................................................................................56
4.3.2 Einfluß der hormonellen Behandlung auf die transplantierten
Fragmente ...........................................................................................58
4.3.2.1 Expression der Östrogenrezeptoren ERα und ERβ ......................59
4.3.2.2 Morphologie ..................................................................................61
4.3.2.3 Genexpression..............................................................................64
IV

4.4 Lokale Östrogensynthese in ektopen endometriotischen Läsionen? ..........69
4.4.1 Aromatase-Expression.........................................................................69
4.4.2 Expression von 17β-Hydroxysteroid Dehydrogenase Typ 1 ................70
4.4.3 Expression von 17β-Hydroxysteroid Dehydrogenase Typ 2 ................72
4.4.4 Expression von Cyr61 in GnRH behandelten endometriotischen
Läsionen ..............................................................................................72
4.5 Angiogenese ...............................................................................................74
4.5.1 Expression angiogenetischer Faktoren im eutopen Endometrium.......74
4.5.2 Untersuchungen zur Angiogenese im Nacktmausmodell.....................75
4.5.2.1 Zeitlicher Verlauf der Angiogenese...............................................76
4.5.2.2 Angiogenetische Faktoren im Nacktmausmodell ..........................78
4.5.2.3 Hormonelle Regulation von VEGF und bFGF...............................80
4.5.2.4 Behandlung der Nacktmäuse mit einem VEGF-Antagonisten ......82
5 Diskussion ........................................................................................................85
5.1 Veränderung des Genmusters der Endometrien der Sekretionsphase von
Endometriosepatientinnen ..........................................................................85
5.2 Regulation der Cyr61-Expression im Endometrium durch Östrogen...........87
5.3 Validierung des Nacktmausmodells für funktionelle Untersuchungen.........90
5.4 Validierung des Nacktmausmodells für die Angiogenese ...........................96
5.5 Mögliche Funktionen von Cyr61 in der Pathogenese der Endometriose...100
6 Ausblick ..........................................................................................................104
7 Zusammenfassung ........................................................................................105
8 Literatur...........................................................................................................107
9 Anhang............................................................................................................125
9.1 Primer-Sequenzen ....................................................................................125
9.2 Ergebnisse der „Gene-Array“ Analysen ....................................................126
V

Abkürzungen
17β-HSD 17β-Hydroxysteroid Dehydrogenase Abb. Abbildung aFGF „acidic fibroblast growth factor“ bFGF „basic fibroblast growth factor“ bGFG-R „basic fibroblast growth factor receptor“ β-ME β-Mercaptoethanol BSA Rinder- („bovine“-) Serumalbumin bp Basenpaare cAMP zyklisches Adenosinmonophosphat cDNA komplementäre („complementary“) DNA CT „Threshold-Cycle“ CTGF „connective tissue growth factor“ Cyr61 „cysteine rich protein 61“ Cx Connexin DAB 3,3´-Diaminobenzidin-Tetrahydrochlorid DMEM Dulbecco´s Modified Eagle Medium DNA Desoxyribonukleinsäure dNTP Desoxynukleosid-5´Triphosphat DTT Dithiothreitol EDTA Ethylendiamin-N,N,N´,N´-tetraessigsäure ER Östrogenrezeptor EtOH Ethanol FGF-R-1 „fibroblast growth factor receptor 1“ FITC Fluoreszeinisothiocyanat FSH Follikelstimulierendes Hormon, Follitropin GnRH „Gonadotropin Releasing Hormon“, Gonadoliberin HMVEC „human microvascular endothelial cells”, Endothelzellinie HUVEC „human umbilical vein endothelial cells”, Endothelzellinie IGFBP „insulin-like growth factor binding protein“ IgG Immunglobulin kDa Kilodalton LH Luteinisierendes Hormon, Lutropin LHRH Luteinisierendes Hormon Releasing Hormon min Minute mRNA Boten- („messenger“-) RNA Nov „nephroblastoma overexpressed“ OD optische Dichte PCR Polymerase-Kettenreaktion p.o. per oral PR Progesteronrezeptor RNA Ribonukleinsäure rpm Umdrehungen pro Minute RT Raumtemperatur sec Sekunde s.c. subkutan SCID „severe combined immunodeficient“ SDS Natrium- („sodium“-) dodecylsulfat SERM selektiver Östrogenrezeptor-Modulator Std. Stunde Tab. Tabelle Tris Tris-(hydroxymethyl)-aminomethan TSP-1 „thrombospondin type 1 repeat“ U Unit ü.N. über Nacht
VI

VEGF „vascular endothelial growth factor“ VEGF-R/KDR „vascular endothelial growth factor receptor“ VPF „vascular permeability factor“ vWC „von Willebrand factor type C repeat“ WISP „Wnt-1-induced secreted protein” w/v Gewichtsprozent
VII

Einleitung
1 Einleitung
1.1 Endometriose
Bei Endometriose handelt es sich um eine benigne Krankheit, die durch das Vorkommen von
endometrialem Gewebe außerhalb des Uterus charakterisiert ist. Sie ist eine der häufigsten
gynäkologischen Erkrankungen, von der fast 10 % aller Frauen im reproduktiven Alter
betroffen sind (Mahmood und Templeton, 1991). Endometriose wurde erstmals vor über 300
Jahren als „peritoneale Geschwüre” beschrieben, die auf der Oberfläche der Blase, des
Darms und des Uterus gefunden wurden (Shroen, 1690). Im 18. Jahrhundert schilderten
Ärzte dann zunächst einen Zusammenhang zwischen Vernarbungen, Gewebe-
Schädigungen und Unterbauchschmerzen, aber erst im 19. Jahrhundert konnte mit dem
Fortschritt der mikroskopischen Technik das Wachstum von ektopem endometrialem
Gewebe identifiziert werden (Witz, 2002). Heute werden drei verschiedene Formen der
Endometriose unterschieden (Donnez et al., 1992), die peritoneale Endometriose, die
ovarielle Endometriose und die rektovaginale Endometriose. Die Ätiologie und die
Pathogenese der Endometriose sind bis heute nicht geklärt.
Es werden unterschiedliche Theorien zur Entstehung vor allem der peritonealen
Endometriose in der Literatur diskutiert. Die älteste Vorstellung beruht auf der Annahme, daß
Endometriose in situ entweder aus Überresten des Müller-Gangs oder durch Metaplasie
entsteht (Ridley 1968; Lauchlan, 1972). Die Theorie der Metaplasie postuliert, daß
pluripotente Zellen des Coelomepithels durch Stimuli eine metaplastische Umwandlung zu
Endometrium vollziehen (Meyer 1919, Thomas 1992). Mit dieser Theorie kann die
Entstehung jeder Endometriose erklärt werden, die dort vorkommt, wo Mesothel vorhanden
ist (Suginami, 1991). In mehreren Studien wurde bereits das Auftreten von
Endometrioseherden außerhalb des Bauchraumes, z.B. in der Pleura beschrieben (Hobbs
und Bortnick, 1940; Cassina et al., 1997; Van Schil et al., 1996; Bhatia 35, 1998; Jubanyik
und Comite, 1997), was durch die Coelom-Metaplasie erklärt werden kann. Zum anderen
gibt es Beschreibungen von Männern, die wegen eines Prostatakarzinoms mit hohen
Östrogendosen therapiert wurden und während dieser Behandlung Endometrioseherde
entwickelten (Pinkert et al., 1979; Schrodt et al., 1980). Abgesehen von diesen Fallstudien
fehlt allerdings der experimentelle oder klinische Beweis für die Richtigkeit der Metaplasie-
Theorie (van der Linden, 1997). Endometrioseherde in der Pleura könnten auch dadurch
erklärt werden, daß endometriale Zellen und Fragmente über die Blut- oder Lymphbahn
transportiert worden sind (Javert, 1949; Leyendecker et al., 1995).
Die am häufigsten angenommene Implantationstheorie zur Entstehung von Endometriose,
auch als Sampson´s Theorie bezeichnet, postuliert, daß endometriale Fragmente während
der Menstruation durch die Eileiter in den Bauchraum gelangen, hier adhärieren und in
1

Einleitung
umgebendes Gewebe invadieren, so daß endometriotische Läsionen entstehen (Sampson,
1927) (Abb. 1A). Schon in den 1950er Jahren konnte gezeigt werden, daß das
Menstruationssekret vitale endometriale Zellen enthält, die zum einen kultiviert werden
können (Kettel und Stein, 1951), zum anderen in der Lage sind, an ektopischen Stellen zu
adhärieren und zu proliferieren (Ridley und Edwards, 1958). Auch durch Tierversuche an
Primaten konnte bestätigt werden, daß durch den Rückfluß von Menstruationssekret in den
Bauchraum Endometriose induziert werden kann (TeLinde und Scott, 1950; D´Hooghe et al.,
1995). Daß die retrograde Menstruation eine mögliche Ursache für die Entstehung der
Endometriose ist, wird auch durch die Beobachtung unterstützt, daß Frauen mit einer
abweichenden Anatomie, die den Abfluß des Menstruationssekrets über die Vagina hemmt,
ein erhöhtes Risiko für die Entwicklung einer Endometriose besitzen (Olive und Henderson,
1987; Sanflippo et al., 1986).
E
d
e
re
vo
E
in
T
Abb. 1A: Schematische Darstellung desUterus mit Tuben und Ovarien zurVerdeutlichung der retrogradenMenstruation. B: Die sich bildendenendometriotischen Läsionen verteilensich mit unterschiedlicher Häufigkeit imgesamten Bauchraum (Schweppe,1989).
s gibt allerdings auch einige Hinweise dafür, daß das Konzept der Endometriose
ifferenzierter gesehen werden muß. Möglicherweise liegen die Ursachen der Entstehung
ndometriotischer Läsionen in einem nicht adäquat differenzierten Endometrium. Denn
trograde Menstruation scheint ein physiologischer Vorgang zu sein, der in allen Frauen
rkommt (Liu und Hitchcock, 1986). Dennoch entstehen nicht bei jeder Frau
ndometrioseherde. Zudem ist die Rezidivrate der Endometriose sehr hoch, und es kommt
sehr vielen Fällen nach einer vermeintlich erfolgreichen hormonellen und/oder operativen
herapie innerhalb eines Jahres zum erneuten Auftreten endometriotischer Läsionen
2

Einleitung
(Wheeler und Malinak, 1983). Ein weiterer Hinweis dafür, daß im Endometrium selbst die
Ursachen der Krankheit liegen könnten, ist die Korrelation von Endometriose und Infertilität.
Bei fast 60 % der Patientinnen, die sich aufgrund von Fertilitätsstörungen operativ
untersuchen lassen, wird Endometriose diagnostiziert (Ledger, 1999). Ob und welche
molekularen Unterschiede zwischen den Endometrien von Frauen mit und ohne
Endometriose bestehen, ist bis heute nicht geklärt.
1.1.1 Morphologie und Lokalisation der Endometrioseherde
Das makroskopische Bild der Endometriose kann sehr unterschiedlich sein. Die Bandbreite
der beobachteten Veränderungen ist vielgestaltig und enthält in unterschiedlicher
Zusammensetzung Drüsen- und Stromaelemente. Die optisch erkennbaren Läsionen haben
eine durchschnittliche Größe von 1-4 mm und umfassen wasserklare blasige Auflagerungen,
rote Papeln, umgeben von einer mehr oder weniger ausgeprägten Vaskularisation, gelb-
braune, blaue, schwarze und weiße Areale (Abb. 2). Diesen farblich unterschiedlichen
Gewebeveränderungen kommt eine divergente biologische Aktivität zu. Aktive Läsionen
einer peritonealen Endometriose erscheinen rot, sind hypervaskularisiert und werden
begleitet von Ödemen und entzündlichen Zellinfiltraten (Schweppe, 2002). Sie zeigen in den
Drüsen und im Stroma proliferative Aktivität, was sich anhand der Mitoserate darlegen läßt.
Außerdem sind sie parakrin aktiv, sezernieren Adhäsionsmoleküle und Wachstumsfaktoren
und induzieren Neoangiogenese. Sind die Läsionen dagegen schwarz/braun und enthalten
wenige Blutgefäße, sind sie eher inaktiv (Schweppe, 2002). Neben den peritonealen
Läsionen sind erstmals im 19. Jahrhundert auch zystische Formationen im Bereich der
Ovarien mit hämorrhagischem Inhalt als eine Form der Endometriose beschrieben worden
(Russel, 1899). Diese Zysten enthalten häufig eine zähflüssige, dunkelbraune Masse und
werden aufgrund ihres Aussehens als Schokoladenzysten bezeichnet. Sie können eine
Größe von bis zu 10 cm erreichen und erfordern eine andere Behandlung als die peritoneale
Endometriose (Schweppe, 2002).
Abb. 2: Endoskopische Darstellung einer endometriotischen Läsion, lokalisiert auf der Harnblase. Das Bild wurde übernommen aus dem Internet: www.endogyn.at/endometriose.htm.
3

Einleitung
Die Läsionen der peritonealen Endometriose treten bei etwa 60 % der Patientinnen an den
Ligamenta sacrouterina auf, man findet sie aber auch häufig im Douglas-Raum (28 %), auf
der Harnblase (15 %) oder dem Rektum (12 %) (Abb. 1B) (Schweppe, 1989). Weniger häufig
sind Läsionen am Dick- und Dünndarm, am Ureter, an der Bauchdecke, am Nabel, am
Diaphragma, in der Pleura, an der Lunge, der Gallenblase, der Milz, am Magen oder an der
Niere (Luciano, 1982) zu finden.
Eine maligne Entartung der Endometriose wird selten beobachtet, nach Schätzungen ist es
etwa 1 % (Mostoufizadeh und Scully, 1980; Scully, 1966). Die Malignome sind überwiegend
im Bereich von Ovar oder Blase anzutreffen (Schreiner, 1985). Bei höchstens 5 % der Fälle
kann sich aus einem ovariellen Endometrioseherd ein endometrioides Adenokarzinom
entwickeln (Scully, 1978). Es besteht jedoch immer die Möglichkeit, daß überall dort, wo
auch endometriale Läsionen beobachtet werden, maligne Transformationen entstehen, z.B.
am Spatium rectovaginale, am Nabel oder der Pleura (Crum et al., 1981; Christmann und
Strobel, 1985).
1.1.2 Klassifikation der Endometriose
Die Endometriose wird eingeteilt in vier verschiedene Stadien, die heute weltweit
gebräuchlichste Klassifikation ist die „Revised American Society Classification 1985” (rAFS)
(American Fertility Society, 1985). Bei der Untersuchung des kleinen Beckens wird die
Anzahl, Größe und Lokalisation (Peritoneum, Ovarien, Eileiter) der ektopen endometrialen
Läsionen vermerkt und entsprechend dem Schweregrad eine festgelegte Anzahl von
Punkten zugeordnet (Hucke und Distler, 1989). Die ermittelte Punktzahl kennzeichnet das
entsprechende Stadium der Endometriose (I = gering, II = mäßig, III = schwer, IV =
ausgedehnt). In dieser Klassifikation nicht berücksichtigt sind die unterschiedlichen
biologischen Aktivitätsmerkmale der einzelnen Läsionen. Ebensowenig werden Läsionen im
Bereich von Vagina, Darm, Blase oder anderen Lokalisationen gewichtet, sie können
allenfalls unter Besonderheiten aufgeführt werden. Um diese Unzulänglichkeiten zu
beseitigen, wird derzeit eine intensive Diskussion um die Erneuerung des
Klassifikationssystems geführt. Für vergleichende internationale Studien zur Abschätzung
der Prognose und des einzuschlagenden therapeutischen Weges scheint allerdings eine
einheitliche Klassifizierung unerläßlich, damit unterschiedliche Ergebnisse vergleichbar
gemacht werden können.
1.1.3 Symptome der Endometriose und Therapieansätze
Endometriose ist assoziiert mit einer Vielzahl von Symptomen, viele Frauen leiden vor allem
unter chronischen Unterbauchschmerzen. Diese äußern sich als sekundäre Dysmenorrhoe,
Dysparunie oder nichtzyklische Rückenschmerzen. Andere Symptome wie zyklische rektale
4

Einleitung
Blutungen, Hämaturie oder Handrang sind häufig abhängig von der Lokalisation der
endometriotischen Läsionen oder der peritonealen Entzündungsreaktion. Untypische
Lokalisationen der Läsionen, wie z.B. an der Lunge, können ungewöhnliche Symptome wie
Brustschmerzen, Pneumothorax oder zyklisches Bluthusten hervorrufen (Foster et al., 1981).
Analog dazu können sehr selten auftretende Endometrioseherde im Gehirn zu typisch
rezidivierenden Kopfschmerzen und neurologischen Ausfallerscheinungen führen (Muse,
1988). Als typisch für eine Endometriose gilt, daß sich alle Beschwerden, gleichgültig wo sie
auftreten, zur Menstruationszeit verstärken. Die Symptome können sowohl vereinzelt als
auch in Kombination auftreten, ansonsten scheint es jedoch keinen Zusammenhang zu
geben, zwischen dem Umfang des Krankheitsbildes und der Intensität des empfundenen
Schmerzes (Fedele et al., 1990). Eine eindeutige Diagnose der Endometriose ist meist nur
durch direkte Betrachtung des Abdominalraumes möglich, bei der zusätzlich häufig eine
Gewebeprobe entnommen wird. Als beste Methode zur Diagnose der Endometriose gilt die
Laparoskopie, in ungünstigen Fällen kann auch eine Laparatomie notwendig werden (Hucke
und Distler, 1989).
Seitdem die Endometriose als eine hormonabhängige Krankheit identifiziert wurde (Dizerega
et al., 1980), wurde versucht, neben der operativen Sanierung eine möglichst effektive
hormonelle Therapie zu entwickeln. Daher werden am häufigsten orale Kontrazeptiva gegen
die Symptome der Endometriose eingesetzt, seit den 1960er Jahren sind einige
hochdosierte Empfängnis-Verhütungsmittel als Behandlung gegen Endometriose anerkannt.
Leider fehlen in der Literatur Ergebnisse von klinischen Studien, die die Wirksamkeit dieser
Therapie belegen. In einer Vergleichsstudie konnte lediglich gezeigt werden, daß die
Anwendung von oralen Kontrazeptiva im Vergleich zu dem „Gonadotropin-Releasing
Hormon”- (Gonadoliberin, GnRH-) Agonisten Goserelin gleiche Erfolge in der Linderung von
menstruationsunabhängigen Schmerzen erzielen konnte, in der Behandlung von Dysparunie
war jedoch das Goserelin wirksamer (Vercellini et al., 1993).
In den 1970er Jahren wurde Danazol, ein Derivat des 17-α Ethinyltestosteron, als Therapie
gegen die Endometriose eingeführt. Durch negative Rückkopplung wird durch dieses
Androgen-Analog die Produktion des Follikelstimulierenden Hormons (Follitropin, FSH) und
des Luteinisierenden Hormons (Lutropin, LH) reduziert und das Hypothalamus-Hypophysen-
Gonaden-System zum Teil herunterreguliert. Danazol beeinflußt weiterhin das Immunsystem
durch die Supprimierung von Antikörpern gegen autologes Endometrium (Dmowski, 1995).
Die Wirkung von Danazol in der Behandlung von Endometriose-Beschwerden konnte in
vielen internationalen klinischen Studien bewiesen werden (Wheeler et al., 1992, 1993). Eine
wirksame Dosis Danazol ruft jedoch ein erhebliches Spektrum an Nebenwirkungen hervor,
wie androgene Nebenwirkungen (z.B. Gewichtszunahme, fettige Haut), sowie geringe
hypoöstrogene Nebenwirkungen und meist erträgliche systemische Nebenwirkungen wie
5

Einleitung
Kopfschmerzen, Muskelkrämpfe und Übelkeit (Shaw, 1992; Wheeler et al., 1993). Trotz
dieser Nebenwirkungen gilt Danazol als Maßstab für die Behandlung von Endometriose, der
für alle nachfolgenden Therapien einen Bezugspunkt setzt (Miller und Anderson, 1997).
Erstmals in den 1980er Jahren wurden GnRH-Agonisten zur Endometriosebehandlung
eingesetzt. Nach einer kurzen Stimulationsphase bewirken GnRH-Agonisten eine
Desensitivierung und/oder Herunterregulation der Hypophyse durch Rezeptorblockade und
damit zum Wirkungsverlust des weiterhin pulsatil abgegebenen „Luteinisierendes Hormon
Releasing Hormon” (LHRH). Als Konsequenz dieser chemischen Kastration nimmt die FSH-
und LH- Freisetzung ab, die Ovarien werden nicht mehr stimuliert, und es resultiert eine
hypogonadotrope hypoöstrogene Amenorrhoe. Die gute Wirksamkeit in der
Schmerzlinderung von GnRH-Agonisten, vor allem des häufig angewendeten
Leuprorelinacetat, konnte in klinischen Studien bewiesen werden (Dlugi et al., 1990).
Verglichen mit Danazol wurde nach sechs Monaten durch beide Therapien gleich effektiv die
Anzahl der endometriotischen Läsionen sowie die Schmerzsymptomatik reduziert (Rivlin et
al., 1990; Wheeler et al., 1992). Hauptsächlich auftretende Nebenwirkungen während der
Behandlung mit GnRH-Agonisten sind vor allem abnehmende Knochendichte und
Hitzewallungen, die hervorgerufen werden durch den niedrigen Östrogenlevel der mit GnRH-
Agonisten behandelten Patientinnen. Eine längere Therapie mit GnRH-Agonisten ist daher
nur als sog. „Add-Back“-Therapie möglich, bei der den Patientinnen gerade soviel Östrogen
verabreicht wird, um die gefährlichen Nebenwirkungen zu unterdrücken. Laut einer Studie
kann es bis zu zwei Jahre dauern, bis die Dichte der Knochen nach 6monatiger Behandlung
mit GnRH-Agonisten wiederhergestellt ist (Paoletti et al., 1996). Trotz vieler Therapien
existiert bis heute keine Behandlungsmethode, die eine vollständige Heilung der
Endometriose bewirkt, die Rezidivrate dieser Krankheit ist sehr hoch. Nach Behandlung mit
Leuprorelinacetat kommt es innerhalb eines Jahres bei 60 % der Patientinnen zum erneuten
Auftreten von Beschwerden, 28 % müssen sich einer erneuten medikamentösen bzw.
operativen Endometriosebehandlung unterziehen (Regidor et al., 1996).
6

Einleitung
1.1.4 Infertilität
Eines der am häufigsten diskutierten Symptome der Endometriose ist die Infertilität. Der
Zusammenhang von Endometriose und Infertilität scheint jedoch durch eine Anzahl von
Studien zumindest bei den Patientinnen bestätigt zu werden, die keine morphologischen
Veränderungen des Reproduktionstraktes als Ursache für die Fertilitätsstörungen aufweisen
(Garrido et al., 2000). Es existieren zudem einige Hinweise darauf, daß selbst die mildeste
Form der Endometriose Infertilität induzieren kann. Bei Frauen, die unter Infertilität leiden,
wird zunächst sehr viel häufiger Endometriose diagnostiziert als im Durchschnitt. Bis zu 60 %
der Frauen, die sich aufgrund von Fertilitätsproblemen einer Laparoskopie unterziehen,
haben Endometriose (Ledger, 1999), während im Allgemeinen ca. 10 % der Frauen im
reproduktiven Alter betroffen sind (Mahmood und Templeton, 1991). Des weiteren wurde in
Tiermodellen demonstriert, daß durch eine induzierte Endometriose die Fruchtbarkeit
reduziert wird (Schenken und Asch, 1980; Vernon und Wilson, 1985; Barragan et al., 1992).
Zudem wurde in Mäusen eine geringere Implantationsrate beobachtet, nachdem den Tieren
Peritonealflüssigkeit von Endometriosepatientinnen injiziert wurde. Mäusen, denen
Peritonealflüssigkeit von fertilen Frauen injiziert wurde, zeigten keine reduzierte
Implantationsrate (Illera et al., 2000).
Der genaue Einfluß der Endometriose auf die reduzierte Fertilität der Patientinnen ist noch
ungeklärt. Durch schwere Formen der Krankheit können Läsionen am Eileiter oder am Ovar
entstehen, die mechanisch den Eisprung oder den Eitransport beeinträchtigen. Die Ursachen
für die Fertilität in Patientinnen mit milderen Formen der Endometriose könnten unter
anderem in Fehlfunktionen bei der Entwicklung der Oozyte liegen. Tummon et al. (1988)
detektierten Fehlfunktionen des Ovars in Endometriosepatientinnen, die sich unter anderem
in einer erhöhten Anzahl von dabei durchschnittlich kleineren Follikeln äußerten. Auch
wurden geringere Östrogenspiegel in der präovulatorischen Phase und nach dem LH-Peak
gemessen (Tummon et al., 1988). Fehlregulationen der Hypophysen-Gonaden-Achse in
Patientinnen mit Endometriose wurden ebenfalls von Cahill et al. (1995) diskutiert. Diese
äußerten sich in veränderten endokrinen Funktionen der Granulosazellen, sowohl die
Progesteron-Produktion als auch die Aromatase-Funktion war herunterreguliert. In einer
anderen Studie mit ähnlichem experimentellem Design dagegen wurde eine Überproduktion
von Progesteron in der Follikelflüssigkeit und in kultivierten Granulosazellen von Frauen mit
Endometriose detektiert (Pellicer et al., 1998). Auch Defekte in der Rezeptivität des
Endometriums von Endometriosepatientinnen, die die Implantation des Embryos verhindern
könnten, werden diskutiert (Garrido et al., 2002). Kontroverse Ansichten herrschen hier vor
allem über die Expression von Integrinen, die als sensitive Indikatoren des endometrialen
Status gelten, da sich ihr Expressionsmuster während der Implantation verändert (Lessey et
al., 1994a). Während Lessey et al. (1994b) eine Herunterregulation des Integrins αvβ3 im
7

Einleitung
eutopen Endometrium von Endometriosepatientinnen beschreiben, konnte diese
Beobachtung durch andere Studien nicht bestätigt werden (Bridges et al., 1994; Creus et al.,
1998; Hii und Rogers, 1998). Zudem werden Störungen des Immunsystems mit der
Ausbildung von Antikörpern gegen das Endometrium oder Ovarialgewebe diskutiert (Kreiner
et al., 1986; Gleicher et al., 1987). Allerdings konnte bis heute nicht vollständig geklärt
werden, ob tatsächlich Antikörper existieren, die auf endometriale Proteine reagieren (Wild
und Shivers, 1985; Gleicher et al., 1987; Kennedy et al., 1990; Switchenko et al., 1991;
Fernandez-Shaw et al., 1993). Odukoya et al. (1995) detektierten in der Hälfte der Seren von
Endometriosepatientinnen Antikörper der Klasse IgG, die gegen Antigene des eutopen
Endometriums gerichtet waren. Zusätzlich zeigte sich in dieser Studie bezüglich des
Vorkommens solcher Antikörper ein signifikanter Unterschied zwischen Frauen mit und ohne
Endometriose (Odukoya et al., 1995). Diese Resultate wurden zuletzt bestätigt, dennoch ist
nicht geklärt, welche Rolle die gegen das Endometrium gerichteten Antikörper bei der
Infertilität spielen könnten (Tanaka et al., 2000).
Obwohl die Ursachen der Fertilitätsstörungen von Patientinnen mit milder Form von
Endometriose wahrscheinlich in der Krankheit begründet liegen, ist es durch die Reduktion
der Größe oder der Aktivität der endometriotischen Läsionen nicht möglich, die Fertilität der
Frauen wiederherzustellen. In klinischen Studien wurde gezeigt, daß weder die Behandlung
mit GnRH-Agonisten noch mit Danazol die Fertilitätsrate in Endometriosepatientinnen
steigern kann (Thomas und Cooke, 1987). Studien, die die Schwangerschaftsraten nach
alleinigem operativen Entfernen der Läsionen durch Laparoskopie mit den Resultaten
verglichen, nach denen die Patientinnen zunächst supprimiert und dann operiert wurden,
demonstrierten, daß die Suppression der ovariellen Hormonproduktion keine geeignete
Therapie zur Behandlung der Endometriose-assoziierten Infertilität darstellt (Hughes et al.,
1993). Diese Ergebnisse wurden durch Adamson und Pasta (1994) bestätigt, die eine
Behandlung von Endometriose nicht für sinnvoll erachten, wenn außer Infertilität keine
weiteren Symptome der Krankheit auftreten. Allerdings bestreiten auch sie nicht die
Möglichkeit, daß eine medikamentöse Behandlung vor einem operativen Eingriff von Vorteil
sein kann (Adamson und Pasta, 1994). Denn es gibt zahlreiche Hinweise dafür, daß eine
länger andauernde Behandlung mit GnRH-Agonisten vor einer Sterilitätstherapie zu einer
deutlichen Verbesserung der Schwangerschafts- und Lebendgeburtrate führt (Marcus und
Edwards, 1994; Nakamura et al., 1992; Yovich, 1991). Diese Verbesserung wird dadurch
begründet, daß die uterine Rezeptivität möglicherweise nach einem Östrogenentzug deutlich
erhöht ist (Marcus und Edwards, 1994).
8

Einleitung
1.1.5 Einfluß von Steroidhormonen auf die Endometriose
Es gibt viele Hinweise darauf, daß die vom Ovar produzierten Steroidhormone eine große
Rolle bei der Entstehung von Endometriose spielen. Erste Symptome der Krankheit
entstehen normalerweise erst nach der ersten Menstruationsblutung und nur in seltenen
Fällen wurde Endometriose in postmenopausalen Frauen beobachtet (Thomas, 1995). Wie
bereits erwähnt, zielen auch die bis heute angewandten Therapien gegen die Endometriose,
wie die Behandlung mit GnRH-Agonisten, darauf ab, die ovarielle Hormonproduktion zu
unterdrücken und damit eine Linderung der Symptome zu erzielen.
Der Phänotyp des Endometriums unterliegt zyklischen Veränderungen, die hormonell durch
das Ovar gesteuert werden. Die Wirkung der vom Ovar gebildeten Hormone Östrogen und
Progesteron wird durch die Östrogenrezeptoren ERα und ERβ sowie durch die
Progesteronrezeptoren PR-A und PR-B vermittelt. Beide Östrogenrezeptor-Isoformen, ERα
und ERβ gehören zur Superfamilie der Steroidrezeptoren. Sie sind Transkriptionsfaktoren
(Parker et al., 1993) und spielen wahrscheinlich eine wichtige Rolle in der
Wachstumsregulation des Endometriums sowie der endometriotischen Läsionen. Die beiden
wichtigsten Domänen der Rezeptoren sind eine DNA-Bindungsdomäne und eine Liganden-
Bindungsdomäne, die in den verschiedenen Rezeptoren konserviert sind (Dhingra, 1999).
Die DNA-Bindungsdomäne enthält zwei Zinkfinger-Motive, die die Bindung der Rezeptoren
an spezifische DNA-Regionen, sog. „estrogen response elements”, vermitteln (Kumar et al.,
1987). Die wichtigste Funktion der Östrogenrezeptoren ist die Aktivierung der Transkription
bestimmter Gene nach Bindung von Östrogen an den Rezeptor (Dhingra, 1999). Die beiden
Progesteronrezeptoren PR-A und PR-B gehören ebenfalls zur Familie der
Hormonrezeptoren, die in den Nukleus übertreten können (Beato, 1989). Bei PR-A handelt
es sich um ein Protein von 94 kDa, während PR-B 164 zusätzliche Aminosäuren am N-
Terminus besitzt und ein Molekulargewicht von 114 kDa aufweist (Horwitz und Alexander,
1983; Alexander et al., 1989; Lessey et al., 1983). Ob beide Isoformen durch alternatives
Splicen einer mRNA (Conneely et al., 1987) oder durch verschiedene Promotoren (Kastner
et al., 1990) entstehen, ist bis heute ebenso ungeklärt wie die genaue Funktion der beiden
Isoformen. Während PR-B als Aktivator von progesteronregulierten Genen fungiert, scheint
PR-A ein Repressor von PR-B zu sein (Tung et al., 1993; Vegeto et al., 1993).
Wie das eutope Endometrium reagieren auch die ektopen endometriotischen Läsionen
sensitiv auf die Anwesenheit von Östrogenen, sie scheinen jedoch nicht auf die gleiche
Weise auf diese Hormone anzusprechen wie das eutope Endometrium. Durch
immunhistochemische Studien, in denen parallel ektope Läsionen und das
korrespondierende eutope Endometrium einer Patientin untersucht wurden, wurden sowohl
unterschiedliche Morphologien als auch verschiedene Rezeptorenausstattungen zwischen
9

Einleitung
den Geweben festgestellt (Lessey et al., 1989; Bergquist und Ferno, 1993a). Während im
eutopen Endometrium die Expression der Östrogenrezeptoren zyklischen Schwankungen
unterworfen ist (Koji und Brenner, 1993; Matsuzaki et al., 1999), konnte dies in
endometriotischen Läsionen nicht nachgewiesen werden (Lessey et al., 1989). Beide
Östrogenrezeptor-Isoformen wurden im prämenopausalen und fetalen Uterus detektiert
(Brandenberger et al., 1997), wobei im zyklischen Endometrium vorwiegend ERα exprimiert
wird (Enmark et al., 1997). In isolierten Stromazellen aus endometriotischen Läsionen
konnten ebenfalls beide Östrogenrezeptoren nachgewiesen werden, allerdings zeigte sich
hier eine andere Verteilung der Isoformen verglichen mit Stromazellen aus eutopen
Endometrien (Brandenberger et al., 1999). In den endometriotischen Läsionen wurde eine
geringere Expression von ERα beobachtet als im eutopen Endometrium, zudem war auch
das Verhältnis von ERα zu ERβ in den Läsionen reduziert (Brandenberger et al., 1999). Da
die beiden Rezeptor-Isoformen unterschiedliche Bindungsaffinitäten zu ihren Liganden
besitzen (Kuiper et al., 1997; Tong und Perkins, 1997), könnte das unterschiedliche
Expressionsverhältnis in den endometriotischen Läsionen verglichen mit dem eutopen
Endometrium funktionelle Folgen haben. Auch wäre es therapeutisch möglich, durch
Interaktion mit den Signaltransduktionswegen beider Isoformen das Wachstum und die
Persistenz der endometriotischen Läsionen zu beeinflussen (Brandenberger et al., 1999).
Die Verteilung der Progesteronrezeptoren in endometriotischen Läsionen scheint sich
ebenfalls von der Verteilung im eutopen Endometrium zu unterscheiden. Beide
Progesteronrezeptor-Isoformen konnten im eutopen Endometrium detektiert werden und
scheinen während des Zyklus unterschiedlich stark exprimiert zu werden (Mangal et al.,
1997; Mote et al., 1999). In endometriotischen Läsionen dagegen wurde ein geringerer
Expressionslevel der Progesteron-Rezeptoren beobachtet (Prentice et al., 1992; Bergqvist
und Ferno, 1993a, b). Attia et al. (2000) konnten sogar nur PR-A in endometriotischen
Läsionen nachweisen, PR-B dagegen wurde in dieser Studie nicht detektiert. Da beide
Progesteronrezeptor-Isoformen zum einen unterschiedlichen Einfluß auf die Genexpression
haben, zum anderen sowohl zelltypspezifisch als auch promotorspezifisch induziert werden,
könnte die abweichende Expression in den endometriotischen Läsionen die Wirkung von
Progesteron in den Läsionen verändern (Attia et al., 2000). Die in den endometriotischen
Läsionen detektierte und von eutopen Endometrien abweichende Expression von
Steroidhormonrezeptoren könnte zudem ein Grund für die hohe Rezidivrate von
Endometriose sein. Durch die geringeren Expressionslevel der Hormonrezeptoren reagieren
die endometriotischen Läsionen möglicherweise nicht sensitiv genug auf endokrine
Therapien (Revelli et al., 1995).
Neben der unterschiedlichen hormonellen Ansprechbarkeit könnte allerdings auch eine
lokale Östrogensynthese in den endometriotischen Läsionen ein Grund für die hohe
10

Einleitung
Rezidivrate der Endometriose sein (Zeitoun und Bulun, 1999; Bulun et al., 2001). Bei Frauen
im reproduktiven Alter sind die Ovarien der wichtigste Ort der Östrogensynthese, diese wird
katalysiert durch das Enzym Aromatase. Aromatase wird in verschiedenen Geweben des
menschlichen Körpers exprimiert, in den Granulosazellen des Ovars, dem
Synzytiotrophoblasten der Plazenta, im Fettgewebe und im Gehirn (Simpson et al., 1994).
Durch die pulsatorische Ausschüttung des im Hypothalamus gebildeten GnRH werden von
der Adenohypophyse LH und FSH freigesetzt. Nach der Bindung des FSH an seinen G-
Protein gekoppelten Rezeptor in der Zellmembran der Granulosazellen, steigt die
intrazelluläre cAMP-Konzentration an, wodurch die Bindung der zwei Transkriptionsfaktoren
SF-1 („steroidogenic factor-1“) und CREB („cAMP response element binding protein“) an den
proximalen Promotor II des Aromatase-Gens erhöht wird (Michael et al., 1995, 1997).
Dadurch wird die Expression der Aromatase aktiviert und Östrogen im präovulatorischen
Follikel synthetisiert (Simpson et al., 1994; Michael et al., 1995). Aromatase katalysiert die
Umwandlung von Androstendion, welches in der Nebenniere oder direkt im Ovar gebildet
wird, zu Östron. Östron hat keine ausgeprägte östrogene Wirkung und wird daher durch das
Enzym 17β-Hydroxysteroid Dehydrogenase Typ 1 (17β-HSD1) zu aktivem Östrogen, dem
17β-Östradiol, reduziert (Isomaa et al., 1993; Labrie et al., 1989). Ein weiteres Isoenzym,
17β-Hydroxysteroid Dehydrogenase Typ 2 (17β-HSD2) katalysiert die Umwandlung von 17β-
Östradiol zu Östron und Androstendion zu Testosteron in verschiedenen Geweben wie der
Plazenta und der Leber (Wu et al., 1993). Ein hoher mRNA-Level von 17β-HSD2 wurde auch
in den Drüsenepithelzellen des humanen Endometriums während der Sekretionsphase
nachgewiesen, weshalb angenommen wird, daß dieses Enzym durch Progesteron stimuliert
wird (Casey et al., 1994; Mustonen et al., 1998). Es scheint, als sei die Inaktivierung von
Östradiol durch 17β-HSD2 im Endometrium der Sekretionsphase ein wichtiger
Schutzmechanismus dieses durch Östrogen regulierten Gewebes.
Es wurde postuliert, daß auch in endometriotischen Läsionen lokal Östrogen synthetisiert
werden kann. Während im normalen eutopen Endometrium keine Aromatase-Expression
nachweisbar ist (Noble et al., 1997), konnte in endometriotischen Läsionen ein hoher mRNA
Expressionslevel von Aromatase gefunden werden (Kitawaki et al., 1997). Weiterhin wurde
in kultivierten Stromazellen aus endometriotischen Läsionen nach Inkubation mit einem
cAMP-Analog hohe Aromatase-Aktivität detektiert (Noble et al., 1997). Zusätzlich konnte
eine Unregelmäßigkeit in der Expression der Enzyme 17β-HSD1 und 17β-HSD2 in den
endometriotischen Läsionen beobachtet werden. Während 17β-HSD1 in den Läsionen
exprimiert wird (Zeitoun et al., 1998; Labrie et al., 1989), kann 17β-HSD2 weder auf mRNA-
Ebene noch auf Proteinebene detektiert werden (Zeitoun et al., 1998). Durch die
vorhandenen Enzyme Aromatase und 17β-HSD1 kann daher zirkulierendes Androstendion
in Östradiol umgewandelt werden, der Abbau des Östradiol in das weniger östrogene Östron
11

Einleitung
wäre durch die fehlende 17β-HSD2 jedoch kaum effizient. Dadurch kann es lokal in den
endometriotischen Läsionen zu einer Synthese von Östradiol kommen, die unabhängig von
der ovariellen Östrogensynthese ist. Damit wäre es auch nicht möglich, diese
Östrogensynthese durch die medikamentöse Behandlung mit GnRH-Agonisten zu
blockieren, was eine weitere Erklärung für die hohe Rezidivrate der Endometriose wäre. Eine
postmenopausale Patientin wurde daher mit Anastrozol, einem Aromatase-Inhibitor, gegen
wiederkehrende Beschwerden bei bekannter Endometriose behandelt (Takayama et al.,
1998; Zeitoun und Bulun, 1999).
1.2 Angiogenese
Neben der hormonellen Versorgung scheint die Angiogenese bei der Entstehung der
Endometriose ebenfalls eine große Rolle zu spielen. Als Angiogenese wird der Prozeß
bezeichnet, bei dem aus einem bereits bestehenden Kapillarnetzwerk neue Blutgefäße
entstehen. Angiogenese geschieht vor allem während der embryonalen Entwicklung, im
adulten Organismus findet dieser Vorgang physiologisch nur in wenigen Geweben statt,
hierzu gehört vor allem der weibliche Reproduktionstrakt. In ovariellen Follikeln, im Corpus
luteum sowie im Endometrium erfolgt Angiogenese (Findlay, 1986; Gordon et al., 1995).
Pathologisch spielt die Bildung neuer Blutgefäße aber auch eine große Rolle während des
Tumorwachstums (Goldberg und Rosen, 1997; Zetter, 1998) und in weiteren Krankheiten
(Folkman, 1995), zu denen auch die Endometriose zählt (Smith, 1997).
Abb. 3: Schematische Darstellung derBildung neuer Blutgefäße, übernommen ausFujimoto et al. (1999). Angiogenese ist einkomplexer Vorgang, der folgende Prozesseumfaßt: Auflösung der Basalmembran durchProteasen, Migration und Proliferation vonEndothelzellen und die Ausbildunglumenhaltiger Gefäße.
12

Einleitung
Die Neubildung von Kapillaren wird von verschiedenen angiogenetischen Faktoren initiiert
und beginnt mit einem Anstieg der Kapillardurchlässigkeit (Dvorak et al., 1995) (Abb. 3).
Fibrinogen und andere Serumproteine sind dadurch in der Lage, die Kapillaren zu verlassen
und es entsteht eine lockere perivaskuläre Fibringelmatrix. In diese Fibringelmatrix können
nach einer geordneten Fragmentierung der Basalmembran durch Metalloproteinasen
Endothelzellen mit Hilfe von Adhäsionsmolekülen und Plasminogenaktivatoren einwandern.
Beim Kontakt mit der extrazellulären Matrix rollen sich die Endothelzellen ein und bilden
lumenhaltige Kapillargefäße, in die allmählich Blut einzuströmen beginnt. Der Prozeß der
Angiogenese wird mit der Neusynthese der Basalmembran abgeschlossen. Damit
verschwindet auch die vorübergehend erhöhte Gefäßdurchlässigkeit wieder, die Fibrinmatrix
wird abgebaut und durch ein reifes kollagenhaltiges Granulationsgewebe ersetzt. Die
wichtigsten angiogenetischen Faktoren, die eine Schlüsselrolle bei der Entstehung neuer
Blutgefäße spielen, sind der „basic fibroblast growth factor” (bFGF) (Klein et al., 1997) und
der „vascular endothelial growth factor” (VEGF) (Ferrara und Davis-Smith, 1997; Ferrara,
1999).
1.2.1 Angiogenetische Faktoren
Die Familie der Fibroblasten-Wachstumsfaktoren (FGF) umfaßt bis zum jetzigen Zeitpunkt
neun Mitglieder, von denen die wichtigsten aFGF (acidic FGF) und bFGF sind. Die FGF-
Wachstumsfaktoren können mit mindestens vier unterschiedlichen Membranrezeptoren
interagieren, bei denen es sich um Tyrosinkinase-Rezeptoren handelt. FGF-Rezeptor 1
(FGF-R-1) ist der am häufigsten vorkommende Rezeptor, der in den unterschiedlichsten
Zelltypen exprimiert wird. Durch Bindung eines Wachstumsfaktors an den Rezeptor,
dimerisiert dieser und aktiviert eine Tyrosinkinase, die den Rezeptor autophosphoryliert. Der
aktivierte Rezeptor stimuliert eine Reihe von Signaltransduktionswegen in den
verschiedenen Zellen (Klint und Claesson-Welsh, 1999). bFGF war der erste aufgereinigte
Wachstumsfaktor, für den gezeigt werden konnte, daß er Angiogenese induziert (Shing et
al., 1984). bFGF stimuliert die Proliferation, Migration und Differenzierung von
Endothelzellen, glatten Muskelzellen und Fibroblasten. In diesen Zelltypen konnten auch
Rezeptoren für bFGF nachgewiesen werden (Klein et al., 1997). bFGF wurde im normalen,
nicht-schwangeren Uterus verschiedener Spezies nachgewiesen, wie im Menschen (Ferriani
et al., 1993; Sangha et al., 1997), im Affen (Samathanam et al., 1998), im Schaf (Reynolds et
al., 1998a), im Schwein (Gupta et al., 1997), in der Maus (Wordinger et al., 1994) und in der
Ratte (Rider et al., 1997). In den einzelnen Studien wurde bFGF immunhistochemisch in
unterschiedlichen Zelltypen lokalisiert, wie der Basallamina der uterinen Blutgefäße im
Endometrium und im Myometrium, in uterinen Stromazellen und der extrazellulären Matrix
sowie der Basallamina von Drüsen- und Oberflächenepithelzellen (Hyder und Stancel, 1999).
13

Einleitung
Auch der Rezeptor für bFGF, FGF-R-1, konnte in Drüsenepithelzellen, stromalen
Fibroblasten und Endothelzellen des humanen Endometriums detektiert werden (Sangha et
al., 1997). Es besteht daher die Möglichkeit, daß bFGF Einfluß auf verschiedene intrakrine,
autokrine und/oder parakrine Reaktionen ausübt. Während die Lokalisation von bFGF im
Uterus sehr genau untersucht wurde, ist die Regulation dieses Wachstumsfaktors durch
Steroidhormone bis heute ungeklärt. In kultivierten humanen Fibroblasten des Endometriums
(Fujimoto et al., 1997), verschiedenen endometrialen Krebs-Zellinien (Presta, 1988) und in
Kaninchen (Rider et al., 1997) wurde die Expression von bFGF durch Östrogene auf mRNA-
und Proteinebene gesteigert, während Progesteron in einigen Zellinien die
östrogeninduzierte Stimulation herunterregulierte (Fujimoto et al., 1997). Auf molekularer
Ebene konnte bisher nicht eindeutig geklärt werden, ob die Regulation von bFGF durch
Steroidhormone ein primärer oder ein sekundärer Effekt ist. Die Sequenz des humanen
bFGF-Gens enthält sowohl ein „progestrone response element” als auch ein „estrogen
response element” (Rider et al., 1997), funktionelle Studien sind bisher jedoch noch nicht
durchgeführt worden.
Die VEGF-Familie besteht bis zum heutigen Zeitpunkt aus fünf verschiedenen Mitgliedern
(Ferrara und Davis-Smyth, 1997; Achen et al., 1998; Carmeliet und Collen, 1998; Eriksson
und Alitalo, 1999), hierzu zählen VEGF-A (Leung et al., 1989), VEGF-B (Olson et al., 1994),
VEGF-C (Joukov et al., 1996; Lee et al., 1996) VEGF-D (Orlandini et al., 1996; Achen et al.,
1998) und PLGF („placental growth factors“) (Maglione et al., 1991). Die Bezeichnungen
VEGF-A und VEGF werden heute synonym verwendet (Breier, 2000). VEGF ist ein
multifunktionales Zytokin, daß ursprünglich in Tumorzellen identifiziert wurde, in denen es
eine Erhöhung der Kapillarpermeabilität induziert (Dvorak et al., 1995), daher wurde es
zunächst auch als VPF („vascular permeability factor“) bezeichnet. Später wurde entdeckt,
daß VEGF selektiv auf Endothelzellen wirkt und hier Angiogenese stimuliert (Ferrara und
Davis-Smyth, 1997; Ferrara, 1999). Sowohl das humane als auch das murine VEGF-Gen
besteht aus acht Exons mit sieben Introns, wobei unterschiedliche Splice-Varianten
existieren (Ferrara, 1999), die sich durch ihre Fähigkeit der Heparinbindung unterscheiden.
Anders als bFGF besitzt das VEGF-Gen eine Signalsequenz, so daß VEGF von
verschiedenen Zellen sezerniert werden kann und sowohl autokrine als auch parakrine
Funktionen bei der Angiogenese erfüllt (Hyder und Stancel, 1999). VEGF kann mit zwei
unterschiedlichen Rezeptoren interagieren, VEGF-R1 (auch als flt bezeichnet) und VEGF-R2
(auch als flk oder KDR bezeichnet). Es handelt sich hierbei um Tyrosinkinase-Rezeptoren,
die hauptsächlich aber nicht ausschließlich auf Endothelzellen exprimiert werden. Bindung
von VEGF an den Rezeptor führt zum einen zur Autophosphorylierung des Rezeptors, zum
anderen zur Phosphorylierung weiterer Faktoren, wie der Phospholipase Cγ oder dem
GTPase aktivierenden Ras-GAP. Die Aktivierung des VEGF-Rezeptors hat eine Vielzahl von
14

Einleitung
zellulären Antworten zur Folge, die eine Rolle bei der Angiogenese oder der Gewebe-
Umbildung spielen (Ferrara, 1999). Wichtigste Aufgabe des VEGF ist vor allem seine
Fähigkeit, Kapillarfenestration in vivo zu induzieren (Roberts und Palade, 1995). Während
bFGF verschiedene Zelltypen aktiviert, wirkt VEGF selektiv auf Endothelzellen. 1993 konnte
VEGF erstmals im Uterus der Maus (Shweiki et al., 1993), des Menschen (Charnock-Jones
et al., 1993) und der Ratte (Cullinan-Bove und Koos, 1993) nachgewiesen werden, seitdem
wurde es auch im Schaaf (Reynolds et al., 1998a), im Kaninchen (Das et al., 1997) und im
Affen (Greb et al., 1997) gefunden. VEGF und sein Rezeptor sind in diesen Studien im
Uterus hauptsächlich im Endometrium lokalisiert worden, seltener konnte der
Wachstumsfaktor im Myometrium identifiziert werden (Charnock-Jones et al., 1993;
Harrison-Woolrych et al., 1995; Brown et al., 1997). Auch die Expression von VEGF scheint
im weiblichen Reproduktionstrakt durch Östrogen reguliert zu werden, dies konnte für
endometriale Adenokarzinomzellen (Charnock-Jones et al., 1993), für primäre Kulturen
humaner uteriner Stromazellen (Shifren et al., 1996) sowie für Rattenuteri in vivo (Cullinan-
Bove und Koos, 1993; Hyder et al., 1996) nachgewiesen werden. Es scheint sich hier um
primäre Effekte des Östrogens zu handeln, die durch den Östrogen-Rezeptor induziert
werden, da die Induktion des VEGF sehr schnell erfolgt und durch reine Anti-Östrogene
blockiert werden kann (Hyder et al., 1997). Allerdings war es bisher nicht möglich, ein
„estrogen response element” im VEGF-Gen zu identifizieren (Hyder und Stancel, 1999).
Weniger bekannt ist bis heute die Wirkung von Progestinen auf die Expression von VEGF, in
einigen Studien wurde gezeigt, daß VEGF in humanen Stromazellen durch Progestine
induziert werden kann (Greb et al., 1997; Shifren et al., 1996).
1.2.2 Angiogenese im Uterus
Im humanen Uterus spielt Angiogenese eine sehr wichtige Rolle beim Wiederaufbau des
Stratum functionale des Endometriums nach der Menstruation und bei der Vorbereitung des
Endometriums auf die Implantation und Plazentation (Torry et al., 1996). Es ist bis heute
nicht geklärt, durch welche Mechanismen das Wachstum der Gefäße im Endometrium
kontrolliert wird oder in welcher Phase des Menstruationszyklus die Bildung neuer
Blutgefäße stattfindet (Rogers, 1996). Immunhistochemische Studien weisen darauf hin, daß
im Endometrium keine klassische Angiogenese stattfindet (Abb. 3), sondern durch
proliferierende Endothelzellen innerhalb eines Gefäßes neue Gefäße entstehen (Rogers und
Gargett, 1999). Die proliferierenden Endothelzellen wandern in das Lumen des Gefäßes ein
und teilen dieses. So entstehen aus einem Gefäß zwei Gefäße, die entweder parallel
verlaufen oder ein Netzwerk bilden können (Risau, 1997). Die wichtigsten Regulatoren der
uterinen Gefäßfunktionen und damit auch der endometrialen Angiogenese scheinen die
zirkulierenden Steroidhormone Östrogen und Progesteron zu sein (Magness, 1998; Hyder
15

Einleitung
und Stancel, 2000; Reynolds und Redmer, 2001). Die beiden wichtigsten angiogenetischen
Faktoren bFGF und VEGF werden im Uterus von ovariektomierten Schafen wenige Stunden
nach Applikation von Östrogen um das 3-10fache hochreguliert (Reynolds et al., 1998a, b).
Mit dieser Hochregulation auf mRNA-Ebene geht auch eine Steigerung der endometrialen
bFGF und VEGF Proteinexpression einher, ebenso wie eine Zunahme der uterinen
Angiogenese und der Durchblutung dieses Organs (Magness, 1998; Reynolds et al., 1998a,
b).
1.2.3 Angiogenese in der Endometriose
Endometriales Gewebe, das außerhalb des Uterus am Peritoneum adhäriert und
endometriotische Läsionen bildet, ist auf eine ausreichende Blutversorgung angewiesen,
daher erscheinen diese Läsionen unabhängig von der Morphologie und der Lokalisation in
endoskopischen Aufnahmen immer reich vaskularisiert (Abb. 2). Ähnlich wie Tumore, die
ohne eine ausreichende Blutversorgung nicht größer werden können als 3 mm3 (Folkman,
1995), scheinen auch die endometriotischen Läsionen, die außerhalb des Uterus wachsen,
eine angiogenetische Kaskade in dem Gewebe zu induzieren, an das sie adhärieren. In
laparoskopischen Untersuchungen konnte gezeigt werden, daß endometriotische Läsionen
mit Blut des umgebenden Peritoneums versorgt werden (Shaw, 1993). An diesen
angiogenetischen Prozessen, die zur Entstehung von endometriotischen Läsionen führen,
sind scheinbar sowohl bFGF als auch VEGF beteiligt, allerdings erfüllen sie unterschiedliche
Funktionen. bFGF konnte immunhistochemisch in Endometrien von Frauen mit und ohne
Endometriose sowie in endometriotischen Läsionen hauptsächlich in den
Drüsenepithelzellen nachgewiesen werden (Ferriani et al., 1993). Veränderungen in der
Expression von bFGF wurden weder während des Menstruationszyklus noch zwischen den
verschiedenen Gewebetypen detektiert (Ferriani et al., 1993). In isolierten Stromazellen
normaler Endometrien und endometriotischer Läsionen konnte bFGF ebenso nachgewiesen
werden wie sein Rezeptor, bFGF-R (Di Blasio et al., 1995). Die Expressionsstärke von bFGF
war im allgemeinen in den Zellen aus endometriotischen Läsionen geringer als im eutopen
Endometrium, allerdings zeigten Zellen aus größeren Läsionen eine stärkere bFGF-
Expression (Di Blasio et al., 1995). In der Peritonealflüssigkeit von Endometriosepatientinnen
dagegen war die Konzentration von bFGF so gering, daß eine Reaktion mit seinem Rezeptor
nicht möglich war (Huang et al., 1996). bFGF scheint daher die Entwicklung von
Endometriose über autokrine und parakrine Mechanismen zu beeinflussen, die unabhängig
sind von der hormonellen Situation (Fujimoto et al., 1999).
Die Peritonealflüssigkeit mit den enthaltenen Makrophagen, Wachstumsfaktoren und
angiogenetischen Faktoren könnte bei der Entstehung von Endometriose eine große Rolle
spielen (Ramey und Archer, 1993; Halme et al., 1987). Die Peritonealflüssigkeit füllt den
16

Einleitung
Bauchraum, wo die meisten endometriotischen Läsionen gefunden werden und könnte daher
direkten Einfluß haben auf die Adhäsion der Läsionen aber auch auf deren Persistenz und
Vaskularisierung sowie die Ausweitung der Krankheit (Ramey und Archer, 1993; Koninckx et
al., 1998). Höhere Konzentrationen von VEGF wurden in der Peritonealflüssigkeit von
Frauen mit Endometriose verglichen mit Frauen ohne Endometriose detektiert (McLaren et
al., 1996b).
Es stellt sich daher die Frage, ob durch anti-angiogenetische Therapien (Leenders, 1998) die
Entwicklung von endometriotischen Läsionen verhindert werden kann. Durch den komplexen
Prozeß der Angiogenese bieten sich unterschiedliche therapeutische Ansatzpunkte, die die
VEGF-Produktion, die Bindung von VEGF an seinen Rezeptor, die Auflösung der
extrazellulären Matrix, die Proliferation oder Migration der Endothelzellen und die
Kapillarneubildung einschließen (McLaren, 2000). Untersuchungen an transplantierten
Tumoren im Tiermodell zeigten, daß vor allem die Inhibition der Bildung von VEGF und
dessen Bindung an seinen Rezeptor effektive Reduktionen des Tumorwachstums und der
Vaskularisierung zur Folge haben (Saleh et al., 1996; Benjamin und Keshet, 1997; Millauer
et al., 1996). Auch Versuche, in denen die Verfügbarkeit von VEGF durch Transfektion von
Tumorzellen mit dem löslichen VEGF-Rezeptor inhibiert wurde, zeigten nach Transplantation
der transfizierten Zellen in Nacktmäuse eine signifikante Inhibition der Implantation und des
Wachstums der Tumore (Goldman et al., 1998). Ob die in der Zukunft für die Behandlung
von Tumoren entwickelten Therapien allerdings auch bei der Behandlung der Endometriose
eingesetzt werden können, müssen weitere Versuche zeigen.
17

Einleitung
1.3 Cyr61, ein Protein der CCN-Familie
Ein weiterer angiogenetischer Faktor, der Einfluß auf die Entstehung von endometriotischen
Läsionen haben könnte, ist Cyr61 („cysteine-rich protein”), ein Protein der CCN-Familie. Die
CCN-Familie besteht bis zum heutigen Tag aus sechs strukturell homologen Proteinen, die
verschiedenste Funktionen erfüllen (Babic et al., 1998, 1999) (Abb. 4).
Abb. 4: Schematische Darstellung der strukturellen Domänen der CCN-Proteine, übernommen aus Lau und Lam (1999). IGFBP: Domäne homolog zu „insulin-like growth factor binding“-Proteinen, vWC: Domäne homolog zum „von Willebrand factor type C repeat“, TSP-1: Domäne homolog zu „thrombospondin-1“, CT: C-terminale Domäne.
Cyr61, CTGF („connective tissue growth factor”) und Nov („nephroblastoma overexpressed”)
waren die ersten drei Proteine dieser Familie, die entdeckt wurden, daher entstand der
Name „CCN”-Familie (Bork, 1993). Proteine der CCN-Familie werden sekretiert, sind an die
extrazelluläre Matrix assoziiert und regulieren zelluläre Vorgänge, wie Adhäsion, Migration
und Differenzierung. Sie sind aber auch an komplexeren Vorgängen wie der Angiogenese
und Knorpelentwicklung beteiligt, außerdem spielen sie eine Rolle während der
Wundheilung, beim Tumorwachstum und in weiteren Krankheiten (Lau und Lam, 1999).
Cyr61 war das erste Mitglied der CCN-Familie, das kloniert wurde (Lau und Nathans, 1985).
Das humane Cyr61 Gen ist lokalisiert auf dem kurzen Arm des Chromosoms 1 (1p22-31)
(Jay et al., 1997) und kodiert für ein Protein von 381 Aminosäuren mit einem
Molekulargewicht von 42 kDa (Brigstock, 1999). Das Cyr61 Protein besteht aus strukturellen
Domänen, die jeweils von einem separaten Exon kodiert werden (Lau und Lam, 1999) (Abb.
4). Durch diese einzelnen Domänen ist Cyr61 in der Lage, verschiedene Funktionen zu
induzieren. Die erste Domäne besteht aus einem Signalpeptid, das bewirkt, daß Cyr61 von
der Zelle sezerniert wird. Die zweite Domäne ist homolog zu N-terminalen Regionen von
„insulin-like growth factor binding”-Proteinen (IGFBP). Sie fungiert wahrscheinlich als
Rezeptor für Wachstumsfaktoren, und induziert vor allem Migration (Grzeszkiewicz et al.,
2002). Das dritte Exon kodiert für eine Domäne, die Homologien zum von Willebrand Faktor
Typ C (vWC) beinhaltet, mit verschiedenen Integrinen interagieren kann und durch diese
Interaktionen Migration induziert. Vom gleichen Exon kodiert wird eine variable Region, die
18

Einleitung
wahrscheinlich als Verbindung dient zwischen dem N-Terminus und dem C-Terminus.
Darauf folgt eine Domäne, die homolog ist zum Thrombospondin Typ 1 (TSP-1). Hier bindet
das Integrin α6β1, durch das Cyr61 nach Interaktion Zell-Adhäsion vermittelt. Es folgt der C-
Terminus, der den C-Termini anderer extrazellulärer Proteine wie dem von Willebrand Faktor
oder Muzinen ähnelt. Das Integrin αMβ2 und Heparansulfat Proteoglykane interagieren mit
dem C-Terminus von Cyr61 und induzieren möglicherweise die Adhäsion von Monozyten an
Cyr61 (Schober et al., 2002).
Cyr61 ist ein sekretiertes, Heparin-bindendes Protein, das in Maus-Fibroblasten durch
verschiedene Stimuli wie Serum, Wachstumsfaktoren oder Zytokinen innerhalb von Minuten
induziert werden kann (Lau und Nathans, 1985, 1987; Brunner et al., 1991). Cyr61 reguliert
verschiedene Prozesse wie Proliferation, Migration, Differenzierung und Angiogenese, alles
Vorgänge, die beim Tumorwachstum und der Metastasierung eine Rolle spielen (Lau und
Lam, 1999). Cyr61 konnte bereits in vielen Tumorzellinien identifiziert werden und scheint
wichtig zu sein für die Tumor-Genese (Babic et al., 1998). Dem widerspricht allerdings die
Beobachtung, daß Cyr61 in Prostata-Tumoren herunterreguliert ist (Pilarsky et al., 1998).
Neben Wachstumsfaktoren und Zytokinen wird Cyr61 auch durch Hormone induziert, in Uteri
von ovariektomierten Ratten steigt die Cyr61-Expression nach Zugabe von 17β-Östradiol
innerhalb weniger Stunden um das 10fache (Rivera-Gonzales et al., 1998). Sampath et al.,
(2001a) konnten zeigen, daß auch in myometrialen Fragmenten ex vivo nach Zugabe von
17β-Östradiol Cyr61 innerhalb einer Stunde um das zweifache ansteigt. Diese Induktion wird
durch den Östrogenrezeptor ERα vermittelt, da sie durch das pure Anti-Östrogen ICI inhibiert
werden kann (Sampath et al., 2001a), welches die durch den Östrogenrezeptor ERα
induzierte Transkriptionsaktivierung blockiert (Hermenegildo und Cano, 2000). Auch in der
humanen Brustkrebszellinie MCF-7 wird die Cyr61-Expression durch Östradiol innerhalb
einer Stunde stark hochreguliert und ist hier wichtig für die hormonabhängige Zellproliferation
(Sampath et al., 2001b). Zusätzlich konnte beobachtet werden, daß Cyr61 zwar nicht im
gesunden Brustgewebe exprimiert wird, aber in Tumoren von Patientinnen mit invasiven
Karzinomen (Sampath et al., 2001b). Diese Studien belegen, daß es sich bei Cyr61 um ein
östrogenreguliertes Gen handelt, allerdings konnte bisher nicht geklärt werden, ob es sich
bei dieser Reaktion um einen primären Effekt des Östrogens handelt. Nur im murinen Cyr61-
Gen konnte bis heute ein „estrogen response element” detektiert werden (Sampath et al.,
2001a), da das humane Gen jedoch in 93 % mit der Maussequenz übereinstimmt (Jay et al.,
1997), kann die Existenz dieses Elementes nicht ausgeschlossen werden.
Laut Sampath et al. (2001a) wird Cyr61 im humanen Endometrium in den
Drüsenepithelzellen exprimiert. Nähere Untersuchungen zur Regulation von Cyr61 im
19

Einleitung
Endometrium und seiner möglichen Rolle bei der Entstehung von Endometriose sind bisher
jedoch noch nicht erfolgt.
1.4 Tiermodelle für die Untersuchung der Endometriose
Für die Entstehung einer spontanen Endometriose ist die Abstoßung des Endometriums
während der Menstruation Voraussetzung, daher findet man diese physiologisch nur beim
Menschen und bei Primaten. Die geringe Verbreitung von spontaner Endometriose in Affen
und die hohe Kostenintensität dieser Versuche, limitieren den Nutzen des Affen als
Tiermodell für die Erforschung der Endometriose (MacKenzie und Casey, 1975; Fanton und
Hubbard, 1983). Um andere Tiere als experimentelles Modell nutzen zu können, ist die
operative Verpflanzung des autologen Endometriums an ektopische Orte außerhalb des
Uterus erforderlich. Solche Versuche wurden bereits an Kaninchen, Ratten oder auch Affen
durchgeführt (Schenken und Asch, 1980; Vernon und Wilson, 1985; Dunselmann et al.,
1989; Schenken et al., 1991; Sharpe et al., 1991; D´Hooghe et al., 1995). Es bestehen
allerdings signifikante phylogenetische und biochemische Unterschiede zwischen humanem
und tierischem Endometrium, so daß Beobachtungen aus diesen Versuchen nicht
zwangsläufig auf den Menschen übertragbar sind (Awwad et al., 1999). Auch die hormonelle
Regulation unterscheidet sich zwischen Mensch und Tier. Proteine, die im Endometrium von
Nagetieren durch Östrogen induziert werden, können im humanen Endometrium durch
Progesteron beeinflußt werden (Awwad et al., 1999). Als sinnvoll erscheinen daher die
Modelle, bei denen humanes Endometrium in Tiere transplantiert werden kann. Für diese Art
von Versuchen eignen sich besonders immundefiziente Mäuse. Die SCID-Maus („severe
combined immunodeficient“) besitzt weder ein intaktes T-Zellsystem, noch verfügt sie über
ein intaktes B-Zellsystem. Es konnte bereits gezeigt werden, daß verschiedene
Transplantate in diesen Mäusen wachsen, ohne abgestoßen zu werden (Phillips et al.,
1989). Awwad et al. (1999) injizierten endometriale Fragmente in die Bauchhöhle von
weiblichen SCID-Mäusen und konnten nach 14 Tagen in 96,5 % der Fälle implantiertes
Gewebe beobachten. In diesen Implantaten konnten histologisch sowohl endometriale
Drüsen als auch stromales Gewebe nachgewiesen werden, und eine hormonelle
Beeinflussung war gegeben. Andere Studien zeigten, daß auch die Nacktmaus ein
geeignetes Modell zur Untersuchung von Endometriose darstellt (Zamah et al., 1984;
Bergqvist et al., 1985; Zaino et al., 1985; Bruner et al., 1997; Grümmer et al., 2001). Die
homozygote Nacktmausmutante (nu/nu) hat eine angezüchtete Thymusaplasie, was zur
Folge hat, daß diese Mäuse nicht über ein intaktes T-Zellsystem verfügen (Wortis, 1971). Sie
besitzen jedoch ein B-Zell-Abwehrsystem, auch die „Natural Killer”-Zellen (NK-Zellen) sind
aktiv (Hebermann, 1978). Trotz dieser restlichen Immunabwehr können endometriotische
Läsionen in der Bauchhöhle von Nacktmäusen nach Injektion von endometrialen Zellen
20

Einleitung
gefunden werden (Tabibzadeh et al., 1999). Die Adhäsion und Implantation der Zellen wurde
in diesen Versuchen durch die Zugabe von Peritonealflüssigkeit von
Endometriosepatientinnen zusätzlich erhöht. Nach der Transplantation von endometrialen
Fragmenten in die Bauchhöhle von Nacktmäusen, konnten Nisolle et al. (2000a) in 87 % der
untersuchten Fälle nach 21 Tagen endometriotische Läsionen aus Drüsen und Stromazellen
detektieren. Es zeigte sich, daß die transplantierten Fragmente in der Nacktmaus
Angiogenese induzieren und einen hohen Level an VEGF aufweisen (Nisolle et al., 2000a).
Die Entstehung der Blutgefäße und das Einwachsen in die humanen Fragmente wurden von
Grümmer et al. (2001) untersucht. Bereits nach zwei Tagen konnte die Adhäsion der
humanen Fragmente an verschiedenen Organen der Nacktmaus beobachtet werden. Das
Einwachsen von Maus-Blutgefäßen in die humanen Fragmente begann nach 4 Tagen,
unabhängig vom Ort der Adhäsion der Fragmente. Insgesamt zeigte es sich, daß das
Nacktmausmodell bis zu einem Zeitraum von 14 Tagen ein geeignetes Modell zur
Untersuchung von Endometriose ist, da die humanen Fragmente ihre morphologischen
Charakteristika sowie Differenzierungsparameter behalten (Grümmer et al., 2001). Bis zu
diesem Zeitpunkt ist es damit möglich, den Einfluß verschiedener Substanzen auf die
Entwicklung und die Persistenz der transplantierten endometrialen Fragmente zu
untersuchen.
21

Zielsetzung
2 Zielsetzung
Die molekularen Mechanismen, die die Pathogenese von Endometriose beeinflussen, sind
bis heute nicht vollständig geklärt. Es existieren viele verschiedene Hinweise dafür, daß die
Ursache für die Entstehung endometriotischer Läsionen bereits in unterschiedlichen
Genexpressionsprofilen der Endometrien von Endometriosepatientinnen im Vergleich zu den
Endometrien von Frauen, die keine Endometriose entwickeln, begründet liegt. Mit Hilfe von
„Gene-Array“ Analysen, die bei Beginn dieser Doktorarbeit vorlagen, sollten daher gezielt
Veränderungen im endometrialen Genexpressionsmuster von Frauen mit und ohne
Endometriose erfaßt und mögliche Dedifferenzierungen im Endometrium von
Endometriosepatientinnen detektiert werden. Unterschiedlich exprimierte Gene, die durch die
Array-Analysen identifiziert wurden, sollten zunächst anhand weiterer Gewebeproben
überprüft werden, um falsche Resultate auszuschließen. Mögliche Kandidatengene sollten
anschließend in zusätzlichen Versuchen lokalisiert und näher charakterisiert werden, um ihre
Rolle bei der Entstehung der Endometriose zu klären.
Ein etabliertes Nacktmausmodell sollte genutzt werden, um den Einfluß und die Regulation
dieser Gene experimentell zu überprüfen. Weiterhin sollte in diesem Modell zum einen die
Bedeutung der Angiogenese für die Entstehung der Endometriose untersucht und zum
anderen neue Therapieansätze getestet werden. Durch Behandlung der Nacktmäuse mit
unterschiedlichen Substanzen sollte der Einfluß von Hormonen und Anti-Hormonen auf die
Morphologie und die Genexpression der transplantierten endometrialen Fragmente getestet
werden. Zur Untersuchung der Funktion von Angiogenese für die Persistenz der Fragmente,
sollten zunächst verschiedene angiogenetische Faktoren identifiziert werden, die in den
transplantierten Fragmenten exprimiert werden und eine Rolle bei der Bildung neuer
Blutgefäße und dem Einwachsen dieser Gefäße in die Fragmente spielen könnten.
Anschließend sollte nach der Inhibition des angiogenetischen Faktors VEGF die Morphologie
und die Persistenz der transplantierten Fragmente untersucht werden.
22

Material und Methoden
3 Material und Methoden
3.1 Feinchemikalien, Enzyme und Verbrauchsmaterialien
Alle verwendeten Chemikalien entsprachen dem höchsten käuflichen Reinheitsgrad.
Neben den gebräuchlichen Laborchemikalien wurden verwendet:
Agarose peqLab (Erlangen) Albumin Bovine (BSA) Sigma (Deisenhofen) Antibody diluent with background reducing components DAKO (Hamburg)
Aprotinin Sigma (Taufkirchen) Arachisöl Henry Lamotte GmbH (Bremen) Micro BCA Protein Assay Reagent Kit Pierce (Rockford, Illinois, USA) Bromphenolblau Sigma (Taufkirchen) β-Mercaptoethanol Merck (Darmstadt) DAB + liquid chromogen solution (Diaminobenzidin) DAKO (Hamburg)
DMEM Gibco BRL (Karlsruhe) DMSO Merck (Darmstadt) 100 bp DNA Ladder Genecraft (Münster) 1Kb DNA-Ladder Genecraft (Münster) DNase I Invitrogen (Karlsruhe) dNTP-Mix Genecraft (Münster) DTT Invitrogen (Karlsruhe) Eosin (gelb) Merck (Darmstadt) 17β-Estradiol Sigma (Taufkirchen) Ethidiumbromid Serva (Heidelberg) Formaldehyd (37 %) Merck (Darmstadt) Glycerin Sigma (Taufkirchen) Ham’s F-12 Biochrom KG (Berlin) Histomount (Xylolersatz-Eindeckmedium) Shandon (Frankfurt) Instant Hämatoxylin Shandon (Frankfurt) Milchpulver Biorad (Hercules, Californien) 2-Methyl-2-Butanol (t-Amylalkohol) Sigma (Taufkirchen) M-MLV Reverse Transcriptase + Puffer Invitrogen (Karlsruhe) Nahtmaterial Ethicon (Norderstedt) Na-Deoxycholat Sigma (Taufkirchen)
Nitrocellulose-Membran Amersham Lifescience (Buckinghamshire, UK)
Nonidet P-40 Sigma (Taufkirchen) Oligo(dT)16 MWG (Ebersberg) PBS Dulbecco Seromed (Berlin) Penicillin Boehringer (Mannheim) Ponceau S-Lösung Sigma (Taufkirchen) Protease inhibitor cocktail tablets Boehringer (Mannheim) RNeasy Mini Kit QIAGEN (Hilden) RNeasy Midi Kit QIAGEN (Hilden) Streptomycin Boehringer (Mannheim) Suprapur Wasserstoffperoxid (30 %) Merck (Darmstadt) 2,2,2- Tribromethanol Aldrich (Taufkirchen) Taq Polymerase + Puffer Genecraft (Münster)
Tissue Tek Sakura O.C.T.TM Compound (Niederlande)
23

Material und Methoden
Toluidinblau Sigma (Taufkirchen) Tween 20 Sigma (Taufkirchen) Vecta shield Vector Laboratories (Peterbourgh, UK)Whatman-Papier Schleicher und Schuell (Kassel)
24

Material und Methoden
3.2 Humane Gewebeproben
Das für die Experimente verwendete Gewebematerial wurde von der gynäkologischen
Abteilung des Universitätsklinikums Essen (Prof. Schindler, Prof. Kimmig) zur Verfügung
gestellt. Mit dem Routinelabor wurden die Östrogen- und Progesteronserumwerte der
Patientinnen 24-48 Std. vor der Operation erfaßt, die zur Bestimmung der Zyklusphase
herangezogen wurden. Ein Progesteronserumwert von 1 ng/ml wurde als Grenzwert
festgelegt. Lag der gemessene Progesteronwert unter 1 ng/ml befanden sich die
Patientinnen definitionsgemäß in der Proliferationsphase, lag der Wert darüber, befanden sie
sich in der Sekretionsphase. Diese sowie weitere Informationen über vorherige
Behandlungen, die allgemeine Anamnese sowie über Medikamente, die vor der
Gewebeentnahme eingenommen wurden, wurden den Krankenakten entnommen. Die
Gewebeentnahme sowie die anschließende Bearbeitung wurden von der örtlich zuständigen
Ethikkomission genehmigt.
Insgesamt wurden 96 verschiedene Endometrien in den unterschiedlichen Versuchen
verwendet, 49 dieser Endometrien stammten von Frauen ohne Endometriose, 47
Endometrien stammten von Endometriosepatientinnen.
Die für die folgenden Versuche verwendeten Endometrien stammten aus Strichkürettagen
von prämenopausalen Frauen im Alter zwischen 19 und 48 Jahren aus der
Proliferationsphase und der Sekretionsphase, die sich aufgrund von Fertilitätsproblemen,
chronischen Unterbauchschmerzen oder unregelmäßigen Blutungen einer laparoskopischen
Behandlung unterziehen mußten. Auch von Patientinnen, bei denen aufgrund eines Uterus
myomatosus eine Hysterektomie durchgeführt wurde, wurde Gewebe verwendet. Hier wurde
das Endometrium aus dem Uterus nach der Operation entnommen.
Einige Patientinnen mußten sich wegen des Verdachts auf Endometriose einer Operation
unterziehen. Die 16 hierbei entfernten und für die Versuche verwendeten endometriotischen
Läsionen stammten vor allem von den Ligamenta sacrouterina, von der Blase oder aus dem
Douglas-Raum. Von diesen Patientinnen wurden sowohl die endometriotischen Läsionen als
auch, sofern möglich, das Endometrium untersucht. Zusätzlich wurden 3 endometriotische
Läsionen von Patientinnen verwendet, die zuvor 6x mit 3,57 mg Leuprorelin, einem GnRH-
Agonisten (Enantone-Gyn, Takeda) behandelt wurden.
Bei zwei Patientinnen wurden im Rahmen der operativen Behandlung die Ovarien bzw. ein
Teil des Brustdrüsengewebes entnommen. Bei einer weiteren Patientin konnte nach einer
Hysterektomie Myometrium entnommen werden. Diese Gewebe wurden für verschiedene
Versuche als Kontrollgewebe genutzt.
25

Material und Methoden
Die Gewebe wurden unmittelbar nach der Entnahme in kalte Moscona-Lösung (4°C)
überführt und für alle weiteren Versuche in das Labor des Instituts für Anatomie des
Universitätsklinikums Essen gebracht. Hier wurden die Proben unter sterilen Bedingungen
mit Moscona-Lösung gespült, um Verunreinigungen durch Blut zu vermeiden. Für
Untersuchungen der Genexpression wurde das Gewebe in flüssigem Stickstoff (-196°C)
gefroren und bis zur weiteren Verwendung bei -80°C gelagert. Um das Endometrium im
Tiermodell untersuchen zu können, wurde frisches Gewebe sofort nach der Entnahme mit
einem Skalpell in 1-2 mm große Fragmente zerschnitten und in Nährlösung bei 37°C und 5
% CO2 für kurze Zeit aufbewahrt (s. 3.6.1). Für immunhistochemische Färbungen wurde das
Gewebe für die Paraffineinbettung in 10 % Formalin fixiert bzw. frisch für Kryostatschnitte
(Leitz) in Tissue Tek eingefroren und bei -20°C gelagert (s. 3.6.6).
3.3 Molekularbiologische Methoden
3.3.1 RNA-Isolierung
Die Isolierung von Gesamt-RNA aus den Endometrien, die für die Genexpressionsanalysen
genutzt wurden sowie dem Ovar-Gewebe, das als Kontrolle diente, erfolgte mit dem
„RNeasy Midi Kit“ der Firma QIAGEN. Das bei -80°C gelagerte Gewebe wurde je nach
Menge in 2 oder 4 ml RLT-Lysepuffer mit Hilfe eines Homogenisators (Kinematika) bei einer
Geschwindigkeit von 19000 rpm homogenisiert. Die weitere Aufreinigung erfolgte nach dem
Protokoll „RNeasy Midi Protocol for Isolation of Total RNA from Animal Tissues“ (QIAGEN).
Die Gesamt-RNA wurde in 150 µl RNase freiem Wasser (Qiagen) eluiert.
Aus endometriotischen Läsionen sowie endometrialen Fragmenten, die in die Nacktmaus
transplantiert wurden und dem nicht-transplantierten Kontrollgewebe wurde die Gesamt-RNA
mit Hilfe des „RNeasy Mini Kit“ (Qiagen) isoliert. Die Gewebe wurden in 1 ml RLT-Lysepuffer
mit Hilfe eines Homogenisators (Kinematika) bei einer Geschwindigkeit von 19000 rpm
homogenisiert. Zur feineren Lyse wurden die Homogenate anschließend mehrfach durch
eine Kanüle (Terumo Neolus, 20Gx1 1/2’’, 0,9x40 mm) aufgezogen. Die weitere Aufreinigung
erfolgte nach dem Protokoll „RNeasy Mini Protocol for Isolation of Total RNA from Animal
Tissues“ (QIAGEN). Die Gesamt-RNA wurde in 30 µl RNase freiem Wasser (Qiagen) eluiert.
3.3.2 Konzentrationsbestimmung von Nukleinsäuren
Die Konzentrationsbestimmung von Nukleinsäuren erfolgte über die Messung der optischen
Dichte (OD) bei einer Wellenlänge von 260 nm. Bei dieser Wellenlänge haben Nukleinsäuren
in Lösung ihr Absorptionsmaximum. Die Konzentration der Lösung ist proportional zur
gemessenen Extinktion und kann unter Berücksichtigung der eingesetzten Verdünnung
berechnet werden. Eine Extinktion von 1,0 entspricht 40 µg/ml RNA bzw. 50 µg/ml
26

Material und Methoden
doppelsträngiger DNA. Die Bestimmung der Konzentration von RNA wurde in Quarzküvetten
mit Hilfe des RNA/DNA Calculator Gene Quant II (Pharmacia Biotech) durchgeführt.
3.3.3 DNase-Verdau
DNase I ist ein Enzym, das in der Lage ist, einzel- und doppelsträngige DNA abzubauen,
ohne die während der Reaktion vorhandene RNA zu schädigen. Zur Beseitigung von
möglichen DNA-Kontaminationen in der isolierten Gesamt-RNA wurde daher ein DNase-
Verdau durchgeführt. Nach dem Protokoll von Invitrogen wurden 2 µg Gesamt-RNA in 1x
DNase I-Puffer mit 2 U DNase I (Invitrogen) für 15 min bei RT inkubiert. Zur Inaktivierung des
Enzyms wurden anschließend 2 µl EDTA (25 mM) zugesetzt. Die RNA konnte dann direkt in
die Reverse Transkriptions- (RT-) Reaktion eingesetzt werden.
Nach der Isolierung von Gesamt-RNA aus endometriotischen Läsionen oder endometrialen
Fragmenten, die in die Nacktmaus transplantiert worden waren, war es teilweise wegen der
geringen Ausbeute nicht möglich, die Konzentration der Gesamt-RNA zu bestimmen. Dann
wurde die gesamte isolierte RNA in der SpeedVac (Savant) eingedampft und komplett in den
DNase-Verdau und die anschließende reverse Transkription eingesetzt.
3.3.4 Reverse Transkription (RT)
Bei der reversen Transkription wird aus RNA cDNA synthetisiert, die anschließend als
Template für PCRs dienen soll. Die 2 µg mit DNase I verdaute Gesamt-RNA wurde zur
Aufhebung von Sekundärstrukturen zunächst in einem Thermocycler für 10 min bei 65°C
denaturiert und anschließend für 2 min auf Eis gekühlt. Nach Zugabe des RT-Mixes erfolgte
bei 37°C im Thermocycler innerhalb einer Stunde die Synthese des komplementären DNA-
Stranges durch das Enzym „Moloney Murine Leukemia Virus Reverse Transcriptase“ (M-
MLV RT) (Invitrogen). Als Primer wurden Oligo-dT16-Primer (MWG) verwendet. Zur
Inaktivierung des Enzyms und zur Denaturierung des RNA/DNA-Doppelstranges wurde der
RT-Ansatz anschließend für 5 min bei 90°C erhitzt. Die erhaltene cDNA konnte dann sofort
für verschiedene PCR-Experimente eingesetzt werden.
Zusammensetzung des RT-Mixes: 2 µg RNA
1x First-Strand Buffer (5x) (Invitrogen) 10 mM dNTP-Mix (Genecraft) 0,5 µg Oligo dT16 (MWG) 0,1 M DTT (Invitrogen) 200 U M-MLV RT (Invitrogen)
ad 50 µl RNase freies Wasser
27

Material und Methoden
3.4 Genexpressionsanalysen
3.4.1 Gene-Arrays
In Kooperation mit der Schering AG (Berlin) wurden im dortigen „Labor für Fertilitätskontrolle
und Hormontherapie-Forschung“ zum Vergleich von Genexpressionsmustern der
Endometrien von Frauen mit und ohne Endometriose aus der Proliferationsphase und der
Sekretionsphase „Gene-Array“ Analysen mit den Arrays U95A der Firma Affymetrix mit mehr
als 12000 Genen durchgeführt. Die verwendeten Endometrien stammten aus der
gynäkologischen Abteilung des Universitätsklinikums Essen. Mit jeder Gewebeprobe wurde
eine Array-Analyse durchgeführt und zur Identifizierung unterschiedlich exprimierter Gene
wurde jedes normale Endometrium mit jedem Endometrium einer Endometriosepatientin der
gleichen Zyklusphase verglichen.
Die Validierung der Array-Ergebnisse wurde im Rahmen dieser Doktorarbeit am Institut für
Anatomie des Universitätsklinikums Essen mittels RT-PCR und real-time RT-PCR
durchgeführt.
3.4.2 Polymerase-Kettenreaktion (PCR)
3.4.2.1 RT-PCR
Zum Nachweis der Transkripte und zur relativen Quantifizierung der Expression
unterschiedlicher Gene wurde die PCR in einem Thermocycler mit beheizbarem Deckel
(Biometra) nach dem unten angegeben Programm angewendet. 4 µl des RT-Ansatzes (s.
3.3.4) wurden mit genspezifischen Primern (Sequenzen s. Anhang) in die PCR eingesetzt. In
einem Reaktionsansatz wurden gleichzeitig das gewünschte Zielgen und ein konstitutiv
exprimiertes Gen, hier β-Aktin, amplifiziert. Bedingt durch die unterschiedlichen
Fragmentgrößen der Zielgene wurden zwei verschiedene β-Aktin-Primer-Paare verwendet,
die Fragmente von 698 bp oder 214 bp amplifizierten (Tab. 1, Sequenzen s. Anhang). Um
sicherzustellen, daß die Amplifikation beider PCR-Produkte auch im letzten Zyklus noch
exponentiell verlief, wurden die Primerkonzentrationen sowie die Anzahl der Zyklen in den
verschiedenen Reaktionen variiert. Auch die Annealing-Temperatur wurde für jedes
zielgenspezifische Primerpaar individuell verändert (Tab. 1). Nach der Amplifikation wurden
die Transkripte mit DNA-Probenpuffer (5x) versetzt, in einem 2 %igen Agarose-Gel
elektrophoretisch aufgetrennt und durch Ethidiumbromid (Serva) unter UV-Licht sichtbar
gemacht (Gel Imager, Intas). Die Bestimmung der Bandenstärke nach Abzug des
Hintergrundes erfolgte densitometrisch mit Hilfe des Programms Gelscan Professional V4.0.
Der Expressionslevel des β-Aktins wurde als 1 gesetzt und der relative Expressionslevel des
Zielgens wurde gegen diesen Wert abgeglichen und so normalisiert. Diese Normalisierung
erfolgte in Excel.
28

Material und Methoden
Tab. 1: PCR-Bedingungen und Primerkombinationen Zielgen-Primer Konzentration β-Aktin-1/2 β-Aktin-F/R Annealing Zyklen
Nidogen-F/R je 25 pmol - je 2,5 pmol 62°C 27 p27-F/R je 25 pmol - je 2,5 pmol 62°C 27 Fibronektin-F/R je 25 pmol - je 2,5 pmol 62°C 27 Elastin-F/R je 25 pmol - je 2,5 pmol 62°C 27 KollIV-F/R je 25 pmol - je 2,5 pmol 62°C 27 PAC1-F/R je 25 pmol je 2 pmol - 60°C 28 CTGF-F/R je 25 pmol je 2 pmol - 57°C 30 Cyr61-up/dw2 je 25 pmol je 2 pmol - 59°C 30-34 hCx43-F/R je 25-50 pmol - je 2,5 pmol 63°C 30-34 hCx26-F/R je 25-50 pmol - je 2,5 pmol 68°C 30-34 hVEGF-d/k je 12,5-25 pmol - je 2,5 pmol 65°C 30-34 bFGF-F/R je 12,5-25 pmol - je 2,5 pmol 65°C 30-34 ERα-F/R je 50 pmol je 2 pmol - 59°C 34 ERβ-F/R je 50 pmol je 2 pmol - 56°C 34 Aromatase-F/R je 50 pmol je 2 pmol - 56°C 34 17β-HSD1-F/R je 50 pmol je 2 pmol - 58°C 34 17β-HSD2-F/R je 50 pmol je 2 pmol - 58°C 34 bFGFR-F/R je 12,5 pmol - je 2,5 pmol 65°C 30 KDR-F/R je 12,5 pmol - je 2,5 pmol 65°C 30
PCR-Ansatz: 4 µl RT-Ansatz (2 µg RNA in RT-Reaktion eingesetzt)
1x PCR-Puffer 10 mM dNTP-Mix
je x pmol antisense/sense genspezifische Primer je x pmol antisense/sense β-Aktin Primer
2,5 U Biotherm DNA-Polymerase (Genecraft) ad 50 µl H2O
PCR-Programm im Thermocycler: 94°C 10 min 94°C 1 min
Annealing 45 sec 72°C 90 sec 72°C 10 min
x Zyklen
DNA-Probenpuffer (5x): 25 mM Tris/HCl (pH 7,0)
150 mM EDTA 0,05 % Bromphenolblau
25 % Glycerin
29

Material und Methoden
3.4.2.2 Real-time RT-PCR
Zur Untersuchung von Expressionsunterschieden uns interessierender Gene im
Endometrium und in endometriotischen Läsionen wurde die semiquantitative real-time RT-
PCR angewendet. In der Reaktion wurden 1 µl des RT-Ansatzes (s. 3.3.4) mit
genspezifischen Primern (s.u., Sequenzen s. Anhang) eingesetzt. Die Konzentrationen der
Primer wurde zuvor in einer sog. Primermatrix ausgetestet. Bei der real-time RT-PCR erfolgt
der Nachweis der PCR-Produkte über einen fluoreszierenden Farbstoff, SYBR Green I, der
in doppelsträngige DNA interkaliert, wodurch die Fluoreszenz meßbar erhöht wird. Da SYBR
Green I an jede doppelsträngige DNA bindet, konnten das Zielgen und β-Aktin, was auch
hier als „Housekeeping-Gen“ verwendet wurde, nur in getrennten Reaktionsansätzen
amplifiziert werden. Zusätzlich wurden die PCR-Produkte in einem 2 %igen Agarose-Gel
elektrophoretisch aufgetrennt und so kontrolliert, daß keine unspezifischen Produkte
amplifiziert wurden. Für jede Gewebeprobe wurden die Reaktionen für Cyr61 und β-Aktin
dreimal durchgeführt, jede dritte Probe wurde auf das Gel aufgetragen.
Der PCR-Lauf wurde in einem ABI Prism 7700 Sequence Detection System (Applied
Biosystems) in einer 96er well-Platte (96-well optical reaction plate, Applied Biosystems) mit
optischen Deckeln (MicroAmp optical caps, Applied Biosystems) durchgeführt. Bei diesem
Gerät handelt es sich um einen Thermocycler für die PCR-Reaktion, der zusätzlich mit einem
Laser ausgestattet ist, der die Fluoreszenz des Farbstoffes SYBR Green I anregt. Die
Messung der Fluoreszenzstärke erfolgt durch einen CCD-Detektor („charge-coupled
device“). Während des PCR-Laufs akkumuliert das PCR-Produkt und die Menge des
gebundenen SYBR Green I nimmt mit der Menge des PCR-Produktes zu. Bei dem
Threshold-Cycle (CT-Wert) handelt es sich um den PCR-Zyklus, bei dem sich das
Fluoreszenzsignal gerade deutlich vom Hintergrund abhebt. Zu diesem Zeitpunkt ist die
Vermehrung des PCR-Produktes noch exponentiell, daher wird der CT-Wert für die spätere
Auswertung herangezogen. Je kleiner der gemessene CT-Wert ist, desto größer muß die
Ausgangsmenge an cDNA gewesen sein.
Für die relative Quantifizierung konnte die vergleichende CT-Methode (∆∆CT-Methode)
herangezogen werden, da das Zielgen und das „Housekeeping-Gen“ gleich effizient
amplifiziert wurden. Bei dieser Methode wurde zur Relativierung der Expressionsstärke des
Zielgens der für β-Aktin gemessene CT-Wert jeder einzelnen Probe von dem für das Zielgen
bestimmte CT-Wert der gleichen Probe abgezogen und man erhielt den ∆CT-Wert. Bei der
vergleichenden CT-Methode wird die Expressionsstärke der Proben mit einem sog.
Calibrator verglichen, der eigenständig festgesetzt wird. Die ∆CT-Werte, d.h. die gegen β-
Aktin abgeglichenen Expressionslevel der einzelnen Gewebetypen, wurden für jede
Zyklusphase als Mittelwert zusammengefaßt und der ∆CT-Wert des festgesetzten Calibrators
30

Material und Methoden
abgezogen, man erhielt so den ∆∆CT-Wert. Wurde die Expression eines Gens in
Endometrien von Frauen mit und ohne Endometriose verglichen, wurde der ∆CT-Wert der
gesunden Frauen als Calibrator festgelegt und als 1 gesetzt. Beim Vergleich der
Endometrien von Frauen mit Endometriose mit der Expression in endometriotischen
Läsionen, wurde das Endometrium als Calibrator bestimmt. Für die Untersuchungen der
Genexpression während des Menstruationszyklus, fungierte das Endometrium aus der
Proliferationsphase als Calibrator. Mit der Formel 2-∆∆CT (berechnet wird ∆∆CT+s und ∆∆CT-s,
wobei s der Standardabweichung entspricht) wurde die xfache Genexpression eines
Gewebes verglichen mit dem Calibrator berechnet. Bei dieser Formel wurden die
Standardabweichungen einbezogen, dargestellt wurde der Mittelwert des berechneten
Bereiches ohne Standardabweichungen. Die Auswertung (ABI PRISM 7700 Sequence
Detection System User Bulletin #2: Relative Quantitation of Gene Expression, P/N 4303859)
wurde mit dem Programm Excel durchgeführt.
Zielgen-Primer Konzentration β-Aktin-F/R Annealing Zyklen Cyr61-F/R 900/300 nM je 50 nM 60°C 40
Real-time RT-PCR-Reaktionsansatz: 1 µl RT-Ansatz (2 µg RNA in RT-Reaktion eingesetzt)
x nM Forward-Primer x nM Reverse-Primer
12,5 µl SYBR Green Master Mix (2x, Applied Biosystems) ad 25 µl H2O
SYBR Green Master Mix: SYBR Green I dye AmpliTaq Gold DNA Polymerase dNTPs (mit dUTP) Referenz-Farbstoff Puffer
Real-time RT-PCR-Programm: 50°C 2 min 95°C 10 min 95°C 15 sec 60°C 1 min 40 Zyklen
31

Material und Methoden
3.5 Proteinbiochemische Methoden
3.5.1 Protein-Isolierung und Konzentrationsbestimmung
Myometrium, Brustdrüsengewebe sowie Endometrien von Frauen mit und ohne
Endometriose wurden in 2 ml modifiziertem RIPA-Puffer mit einem Homogenisator
(Kinematika) bei einer Geschwindigkeit von 19000 rpm homogenisiert. Nichtlösliche
Zellbestandteile wurden anschließend durch Zentrifugation bei 14000 rpm für 10 min bei 4°C
(Eppendorf-Tischzentrifuge) entfernt. Die Bestimmung der Proteinkonzentration in den
Zellysaten erfolgte mit Hilfe des „Micro BCA™ Protein Assay Reagent Kit“ (Pierce). Hierbei
wird BCA („bicinchoninic acid“) zur Detektion von Cu+1 genutzt, das entsteht, wenn Cu+2 in
Anwesenheit von alkalischen Reagenzien durch Proteine reduziert wird. Es entsteht ein
purpurroter, wasserlöslicher Komplex aus BCA und Cu+1, der eine hohe Absorption bei 562
nm aufweist, die linear mit steigender Proteinkonzentration zunimmt. Die Reaktionslösung
WR („working reagent“) wurde durch Mischung der Lösungen MA, MB und MC im Verhältnis
25:24:1 („Micro BCA™ Protein Assay Reagent Kit“, Pierce) hergestellt. Nach Verdünnung der
Proben mit Wasser (1:100, 1 ml) wurden diese mit 1 ml der Reaktionslösung WR versetzt,
für 1 Std. bei 60°C inkubiert und dann auf Raumtemperatur gebracht. Die Messung der
Absorption erfolgte bei 562 nm gegen einen Wasserabgleich im Spektrophotometer
(Beckman). Unter Verwendung von BSA als Eichprotein wurde eine Standardkurve erstellt
und die Proteinkonzentrationen bestimmt.
RIPA-Puffer: 50 mM Tris/HCl (pH 7,4)
150 mM NaCl 1 mM EDTA
frisch dazu: 0,5 % SDS
1 % Nonidet P-40 0,5 % Na-Deoxycholat
je 1/50 ml „Protease inhibitor cocktail tablet”
3.5.2 Fällung von Proteinen
Bei zu geringen Proteinkonzentrationen wurden 20 µg der Zellysate mit dem 3fachen
Volumen eiskaltes Aceton bei -20°C ü.N. gefällt. Die ausgefallenen Proteine wurden durch
Zentrifugation bei 14000 rpm für 30 min (4°C) pelletiert, mit 500 µl eiskaltem Aceton
gewaschen und erneut zentrifugiert (14000 rpm, 20 min, 4°C). In einer SpeedVac (Savant)
wurden die Pellets anschließend getrocknet.
32

Material und Methoden
3.5.3 SDS-Gelelektrophorese
Die analytische Auftrennung von Proteinen wurde unter denaturierenden Bedingungen in der
Gegenwart von Natriumdodecylsulfat (SDS) nach Laemmli (Laemmli, 1970) durchgeführt.
Der Gellauf erfolgte in einer Gelkammer der Firma Biorad bei einer maximalen Spannung
von 150 V und einer durchschnittlichen Stromstärke von 30-40 mA. Vor dem Auftragen auf
die 10 %igen Polyacrylamidgele (50 mm x 80 mm x 1 mm) wurden die Proteinproben mit 25
µl SDS-Probenpuffer versetzt und für 5-10 min bei 100°C im Heizblock denaturiert.
Sammelgelpuffer: 0,5 M Tris/HCl (pH 6,8) 0,4 % SDS
Trenngelpuffer:
1,5 M Tris/HCl (pH 8,8) 0,4 % SDS
SDS-Probenpuffer (2X): 62,5 mM Tris/HCl, pH 6,8
10 % Glycerin 2,3 % SDS
0,002 % Bromphenolblau 3 % β-ME (frisch dazu)
Laufpuffer:
25 mM Tris/HCl (pH 8,8) 95 mM Glycin 0,1 % SDS
3.5.4 Western Blot
Für den Nachweis von Proteinen durch Antikörper wurden die Proteine mit Hilfe der Blotting-
Technik von einem SDS-Gel auf eine Nitrocellulose-Membran (Amersham Lifescience)
übertragen. Western Blots wurden in einer Elektroblotkammer von Biorad durchgeführt.
20 µg Proteinlysat wurden auf ein 10 %iges SDS-Gel aufgetragen und elektrophoretisch
aufgetrennt (s. 3.5.3). Die in Blot-Puffer äquilibrierte Nitrocellulose-Membran wurde
luftblasenfrei auf das Gel aufgebracht, Membran und Gel mit je 3 Lagen Whatman-
Filterpapier (Schleicher und Schuell) und Vlies bedeckt und in die mit Eis gekühlter
Elektroblotkammer eingespannt. Der Blotvorgang erfolgte bei RT für 2 Std. bei 60 mA unter
ständigem Rühren. Anschließend wurde das Gel zur Kontrolle für 20 min in einer
Coomassie-Lösung (Lowry et al., 1951) gefärbt und ü.N. entfärbt. Die Membran wurde zur
Sichtbarkeit der Proteine für 5 min in einer 0,2 %igen Ponceau S-Lösung (Sigma) inkubiert,
kurz in H2O gewaschen und anschließend in 5 %igem Milchpulver (Biorad) ü.N. bei 4°C und
33

Material und Methoden
am nächsten Morgen für 1 Std. bei RT zur Absättigung unspezifischer Bindungsstellen
geblockt. Nach einem kurzen Waschschritt mit TBS-T erfolgte die Inkubation des
Primärantikörpers für 1 Std. bei RT. Vor und nach der anschließenden einstündigen
Inkubation des Sekundärantikörpers bei RT wurde die Membran mit TBS-T für 5 min, TBS-T-
S für 5 min und erneut mit TBS-T für 15 min gewaschen. Die Entwicklung der Blots erfolgte
mit Hilfe des ECL+Plus Kits von AmershamPharmacia unter Verwendung eines
Röntgenfilms.
Blot-Puffer (pH 8,2-8,8): 50 mM Tris 50 mM Borsäure
TBS-T:
20 mM Tris/HCl (pH 7,6) 137 mM NaCl 0,15 % Tween 20
TBS-T-S:
TBS-T mit 0,015 % SDS
Primärantikörper rabbit anti-Cyr61 antiserum (Labor für Molekulare Experimentelle Orthopädie, Orthopädische Universitätsklinik, Würzburg), 1:2000 Sekundärantikörper goat anti-rabbit, HRP-gekoppelt, sc-2030 (SantaCruz), 1:15000
34

Material und Methoden
3.6 Untersuchungen im Nacktmausmodell
3.6.1 Aufarbeitung des Gewebes
Insgesamt wurden 16 verschiedene endometriale Gewebe für Versuche im
Nacktmausmodell verwendet. Von diesen 16 Geweben stammten 10 von Frauen ohne
Endometriose und 6 von Frauen, bei denen Endometriose diagnostiziert wurde.
Das Endometrium wurde zunächst steril mit Moscona-Lösung gewaschen, um
Verunreinigungen durch Blut zu entfernen. Anschließend wurde es mit einem Skalpell in
Fragmente von 1-2 mm Durchmesser zerschnitten und bis zur Transplantation in die
Nacktmaus in Nährmedium bei 37°C und 5 % CO2 gelagert. Von jedem Gewebe wurde als
Kontrolle zur RNA-Isolierung ein Fragment in flüssigem Stickstoff (-196°C) schockgefroren
und bei -80°C bis zum Ende des Versuchs gelagert. Die Transplantation des übrigen
Gewebes in die Nacktmaus erfolgte meist sofort, spätestens jedoch nach drei Stunden.
Moscona-Lösung: 140 mM NaCl
4 mM KCl 12 mM NaHCO3
0,2 mM KH2PO4 0,4 mM NaH2PO4xH2O,
9 mM D-Glucose Monohydrat
Nährmedium: 500 ml Dulbecco´s modifiziertes Eagle Medium 500 ml Ham´s F12
100 IU/ml Penicillin 100 µg/ml Streptomycin
3.6.2 Versuchstiere
Für die Versuche wurden weibliche zyklische Nacktmäuse (Han: NMRI nu/nu) aus der Zucht
des zentralen Tierlaboratoriums der Universitätsklinik Essen unter der Leitung von Prof. Dr.
Militzer verwendet. Bei Versuchsbeginn hatten die Nacktmäuse ein Alter von mind. fünf
Wochen. Die Nacktmäuse wurden in einer keimfreien Umgebung unter kontrollierten
Umweltbedingungen und regulierten Tag- / Nachtzyklen (je 12 Stunden) gehalten. Futter,
Einstreu und alle in den Raum gebrachten und verwendeten Geräte wurden autoklaviert.
35

Material und Methoden
Es wurden insgesamt 57 Tiere für die folgenden Versuche verwendet. 12 Nacktmäusen
wurden endometriale Fragmente ohne weitere Behandlung implantiert, 4 Tieren wurde
zusätzlich ein VEGF-Antagonist (Schering, ZK 202650) appliziert. 10 Tiere wurden mit einem
reinen Anti-Östrogen (Schering, ZK 191703), 9 mit 17β-Östradiol (Sigma) behandelt.
Weiteren 9 Nacktmäusen wurde ein selektiver Östrogenrezeptor-Modulator (SERM;
Schering, ZK 186619) appliziert. Als Kontrolle wurde in jeder Versuchsreihe einer Maus die
entsprechende Lösungssubstanz, das Vehikel, verabreicht (13 Tiere).
3.6.3 Transplantation der endometrialen Fragmente
Das Endometrium wurde wie unter Punkt 3.6.1 beschrieben vorbereitet.
Die Nacktmäuse wurden mit einer intraperitonealen Injektion von 17 µl 2,5 % Avertin/g
Körpergewicht betäubt und unter sterilen Bedingungen laparatomiert. Ein endometriales
Fragment wurde in einer Schlinge aus Nahtmaterial (Ethicon, Vicryl, 6/0) befestigt und an der
inneren Seite der Bauchwand angenäht. Insgesamt wurden in jede Nacktmaus 3-6
Fragmente transplantiert. Es wurde darauf geachtet, keine unnötigen Reizungen und
Verletzungen zu verursachen. Anschließend wurde zunächst die Bauchwand, dann die Haut
mit Einzelknopfnähten (Ethicon, Vicryl, 5/0) verschlossen und die Wunde einer täglichen
Kontrolle unterzogen.
100 % Avertin: 10 g 2,2,2 tribromethylalkohol (Aldrich)
10 ml tert-amylalkohol (Sigma)
3.6.4 Behandlung der Nacktmäuse
In drei Versuchsreihen mit Geweben von verschiedenen Patientinnen wurden die
Nacktmäuse keiner weiteren Behandlung unterzogen. Die endometrialen Fragmente einer
Patientin wurden jeweils auf vier Mäuse verteilt und in einer Zeitreihe wurde jeweils nach 2,
4, 7 und 14 Tagen eine Maus getötet, die Fragmente entnommen und auf Angiogenese
untersucht.
In drei weiteren Versuchsreihen mit ebenfalls unterschiedlichen Geweben wurde das
endometriale Gewebe einer Patientin jeweils auf zwei bzw. in einer Versuchsreihe auf drei
Mäuse verteilt und den Nacktmäusen zusätzlich täglich 1,5 mg (50 mg/kg) eines VEGF-
Antagonisten (Schering, ZK 202650) verabreicht. Die Substanz wurde in einem Gemisch aus
DMSO und 100 % Ethanol (1:1) gelöst. Als Trägersubstanz wurde eine 0,9 %ige NaCl-
Lösung mit 0,085 % Myrj (Schering) versetzt, der angelöste VEGF-Antagonist zugegeben
und das Ganze mit Hilfe von einigen sterilen Glaskugeln im Reagenzglas für 15 min im
Ultraschallbad („Ultrasonic cleaner“, Sino) suspendiert. Diese Suspension wurde den
36
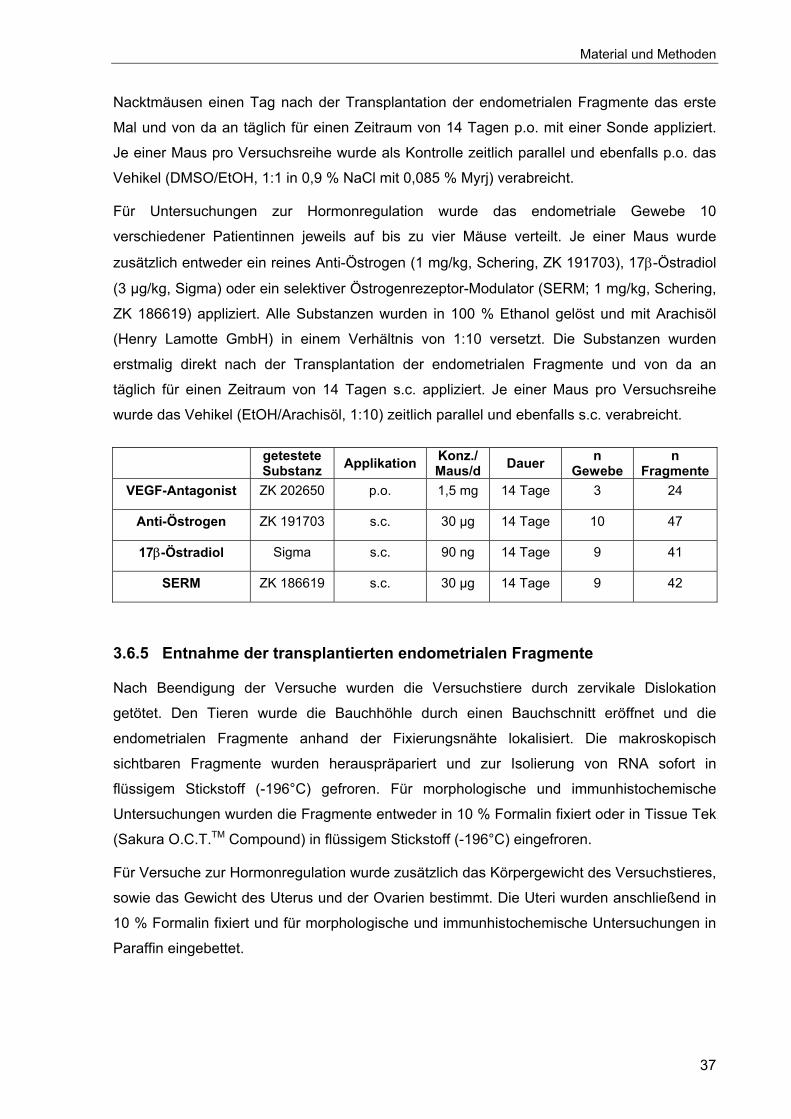
Material und Methoden
Nacktmäusen einen Tag nach der Transplantation der endometrialen Fragmente das erste
Mal und von da an täglich für einen Zeitraum von 14 Tagen p.o. mit einer Sonde appliziert.
Je einer Maus pro Versuchsreihe wurde als Kontrolle zeitlich parallel und ebenfalls p.o. das
Vehikel (DMSO/EtOH, 1:1 in 0,9 % NaCl mit 0,085 % Myrj) verabreicht.
Für Untersuchungen zur Hormonregulation wurde das endometriale Gewebe 10
verschiedener Patientinnen jeweils auf bis zu vier Mäuse verteilt. Je einer Maus wurde
zusätzlich entweder ein reines Anti-Östrogen (1 mg/kg, Schering, ZK 191703), 17β-Östradiol
(3 µg/kg, Sigma) oder ein selektiver Östrogenrezeptor-Modulator (SERM; 1 mg/kg, Schering,
ZK 186619) appliziert. Alle Substanzen wurden in 100 % Ethanol gelöst und mit Arachisöl
(Henry Lamotte GmbH) in einem Verhältnis von 1:10 versetzt. Die Substanzen wurden
erstmalig direkt nach der Transplantation der endometrialen Fragmente und von da an
täglich für einen Zeitraum von 14 Tagen s.c. appliziert. Je einer Maus pro Versuchsreihe
wurde das Vehikel (EtOH/Arachisöl, 1:10) zeitlich parallel und ebenfalls s.c. verabreicht.
getestete Substanz Applikation Konz./
Maus/d Dauer n Gewebe
n Fragmente
VEGF-Antagonist ZK 202650 p.o. 1,5 mg 14 Tage 3 24
Anti-Östrogen ZK 191703 s.c. 30 µg 14 Tage 10 47
17β-Östradiol Sigma s.c. 90 ng 14 Tage 9 41
SERM ZK 186619 s.c. 30 µg 14 Tage 9 42
3.6.5 Entnahme der transplantierten endometrialen Fragmente
Nach Beendigung der Versuche wurden die Versuchstiere durch zervikale Dislokation
getötet. Den Tieren wurde die Bauchhöhle durch einen Bauchschnitt eröffnet und die
endometrialen Fragmente anhand der Fixierungsnähte lokalisiert. Die makroskopisch
sichtbaren Fragmente wurden herauspräpariert und zur Isolierung von RNA sofort in
flüssigem Stickstoff (-196°C) gefroren. Für morphologische und immunhistochemische
Untersuchungen wurden die Fragmente entweder in 10 % Formalin fixiert oder in Tissue Tek
(Sakura O.C.T.TM Compound) in flüssigem Stickstoff (-196°C) eingefroren.
Für Versuche zur Hormonregulation wurde zusätzlich das Körpergewicht des Versuchstieres,
sowie das Gewicht des Uterus und der Ovarien bestimmt. Die Uteri wurden anschließend in
10 % Formalin fixiert und für morphologische und immunhistochemische Untersuchungen in
Paraffin eingebettet.
37

Material und Methoden
3.6.6 Histologische Aufarbeitung der Fragmente
Die in 10 % Formalin fixierten Fragmente wurden nach einer Fixierungszeit von 24 Std. (4°C)
mit Hilfe eines Einbettautomaten (Shandon) zunächst in einer aufsteigenden Alkoholreihe
entwässert und anschließend in Paraffin eingebettet. Die Herstellung der Paraffinblöckchen
erfolgte in einem „Histocentre“ (Shandon). Von den Paraffinblöckchen wurden dann mit
einem Mikrotom (Jung AG) 7 µm dicke Schnitte abgenommen und diese für morphologische
Untersuchungen mit Hämatoxylin und Eosin (HE) angefärbt.
Von den in Tissue Tek (Sakura O.C.T.TM Compound) eingefrorenen Fragmenten wurden in
einem Kryostaten (Leitz) bei -20°C ebenfalls 7 µm dicke Schnitte abgenommen. Diese
wurden entweder zur morphologischen Untersuchung mit Toluidin angefärbt oder für
immunhistochemische Untersuchungen genutzt.
In Kooperation mit der Schering AG (Berlin) wurden in den dortigen Laboratorien ebenfalls
HE-Färbungen durchgeführt. Paraffinblöckchen der humanen endometrialen Fragmente aus
den Nacktmaus-Versuchen wurden nach Beendigung der morphologischen Untersuchungen
im Institut für Anatomie der Schering AG übergeben. Dort wurden Schnitte abgenommen,
HE-Färbungen durchgeführt und in diesen Präparaten die Epithelhöhe des Drüsengewebes
bestimmt. Zusätzlich wurden Paraffinblöckchen der Nacktmaus-Uteri ebenfalls nach Berlin
geschickt, dort histologisch aufgearbeitet und die Uterusepithel-Höhe bestimmt.
3.7 Immunhistochemische Untersuchungen
Die Immunhistochemie wurde dazu genutzt, durch eine Antikörper-Reaktion spezifisch
Proteine in Gewebeschnitten nachzuweisen. Der Gewebeschnitt wurde hierbei zunächst mit
einem Primärantikörper gegen ein im Schnitt vorhandenes nachzuweisendes Protein
inkubiert. Der bei der Antigen-Antikörper-Reaktion im Gewebeschnitt entstandene Komplex
wurde durch indirekte Nachweismethoden sichtbar gemacht. Gegen den unmarkierten
Primärantikörper wurde ein entweder mit Biotin oder mit einem Fluoreszenzfarbstoff
markierter Sekundärantikörper eingesetzt, der spezifisch gegen den im Antigen-Antikörper-
Komplex im Gewebe gebunden Primärantikörper gerichtet ist. Nach Zugabe des
Fluoreszenzfarbstoff-gekoppelten Sekundärantikörpers konnte der markierte Komplex sofort
fluoreszenzmikroskopisch untersucht werden. Biotin-markierte Sekundärantikörper wurden
durch StreptAB-Komplexe, lösliche Komplexe aus Streptavidin und biotinylierter Meerrettich-
Peroxidase, nachgewiesen. Durch Zugabe des Substrats Diaminobenzidin (DAB) entstand
ein braunes Reaktionsprodukt, das lichtmikroskopisch nachgewiesen werden konnte.
38

Material und Methoden
3.7.1 Immunhistochemischer Nachweis mit 3,3´-Diaminobenzidin (DAB)
In einem Kryostaten wurden 7 µm dicke Gefrierschnitte abgenommen, auf Deckgläschen (15
mm) aufgezogen und auf einer Heizplatte (42°C) für 15-30 min getrocknet. Die Schnitte
wurden in eine 12er well-Platte (Beckton Dickinson) überführt und für 5 min in eiskaltem 96
%igem Ethanol fixiert. Nach einem Waschschritt mit PBS für 5 min wurden die Schnitte für
10 min in 3 %iger H2O2-Lösung inkubiert, um endogene Peroxidase-Aktivität zu blockieren.
Nach einem erneuten Waschen mit PBS (5 min) wurden für mind. 30 min (maximal ü.N.)
unspezifische Bindungsstellen durch Inkubation mit PBS/BSA (0,5 %) abgesättigt.
Anschließend wurden die Schnitte für 1 Std. in einer feuchten Kammer mit dem in
Antikörperverdünnung (Dako) angesetzten Primärantikörper inkubiert. Nach weiteren
Waschschritten erfolgte die Inkubation mit einem Biotin-gekoppelten Sekundärantikörper für
30 min, ebenfalls in einer feuchten Kammer. Unter Verwendung eines Detektionssystems
(StreptABComplex/HRP, Dako) wurden die Schnitte erst für 30 min mit einem Strept. A/B-
Komplex, dann für 5 min mit dem chromogenen Substrat für die Peroxidase (DAB) inkubiert,
in einer aufsteigenden Alkoholreihe entwässert und mit Histomount eingedeckt. Die
Auswertung erfolgte an einem Zeiss Axiophot.
Primärantikörper rat anti-mouse panendothelial Antigen (PAN), 09921D (Pharmingen), 1:100 Sekundärantikörper rabbit anti-rat, biotinyliert, E 0468 (Dako), 1:400 Detektion StreptABComplex/HRP (biotinylated horse radish peroxidase), Z 0113 (Dako)
3.7.2 Immunhistochemischer Nachweis durch Fluoreszenz
An humanen endometrialen Fragmenten aus den Nacktmausversuchen wurden
immunhistochemische Untersuchungen an Kryostatschnitten durchgeführt. In einem
Kryostaten wurden 7 µm dicke Schnitte abgenommen, auf Deckgläschen aufgezogen und
auf einer Heizplatte bei 42°C für 15-30 min getrocknet. Die Schnitte wurden in eine 12er well-
Platte (Beckton Dickinson) überführt, für 5 min mit eiskaltem Ethanol fixiert, 10 min in PBS
gewaschen und für mind. 30 min in PBS/BSA (0,5 %) zur Absättigung unspezifischer
Bindungsstellen inkubiert. Nach der Inkubation des Primärantikörpers für 1,5 Std. folgten drei
Waschschritte für je 10 min in PBS/BSA. Die Schnitte wurden anschließend für 45 min mit
einem Fluoreszenzfarbstoff- (FITC-) gekoppelten Sekundärantikörper inkubiert. Für diesen
und alle weiteren Inkubationsschritte wurde die well-Platte wegen der Lichtempfindlichkeit
des Antikörpers mit Alufolie bedeckt. Die Schnitte wurden anschließend 3x für 10 Minuten
mit PBS/BSA und 3x für 5 Minuten mit A.bidest gewaschen und anschließend mit dem
39

Material und Methoden
fluoreszenzverstärkenden Eindeckmittel Vectashield (Vector, Kat. No. H- 1000) eingedeckt.
Die Auswertung erfolgte an einem Zeiss Axiophot.
Primärantikörper goat (polyklonal) anti-Cyr61, sc-8561 (SantaCruz), 1:100 Sekundärantikörper swine anti-rabbit, FITC konjugated, F 0205 (Dako), 1:40
In Kooperation mit dem „Labor für Molekulare Experimentelle Orthopädie“ der
Orthopädischen Universitätsklinik in Würzburg wurden dort zusätzliche
Immunhistochemische Untersuchungen an 7 µm dicken Paraffinschnitten durchgeführt.
Eutope Endometrium sowie endometriotischen Läsionen wurden auf Super Frost
Objektträger (Menzel) aufgezogen und nach Würzburg geschickt. Dort wurde mit Hilfe der
APAAP-Färbung nach Verwendung des Primärantikörpers (anti mouse CYR antibody, Munin
Corp, Chicago, USA, 1:100) Cyr61 nachgewiesen.
3.7.3 Immunhistochemischer Nachweis von Ki67
In Kooperation mit der Schering AG (Berlin) wurde in den Uteri der hormonbehandelten
Nacktmäuse der Proliferationsmarker Ki67 immunhistochemisch nachgewiesen. Ki67 ist im
Zellkern lokalisiert und wird in der späten G1-, S-, M- und G2-Phase des Zellzyklus exprimiert.
In der G0-Phase ist dieser Marker hingegen nicht detektierbar (Schlüter et al., 1993).
3.8 Statistische Auswertung
Die Signifikanz der Expressionsunterschiede zwischen Endometrien von Frauen mit und
ohne Endometriose sowie zwischen den Endometrien aus den einzelnen Zyklusphasen, die
mittels real-time RT-PCR ermittelt wurden, wurde mit dem Einstichproben-Median-Test nach
Wilcoxon berechnet. Der Median-Test prüft die Nullhypothese, die davon ausgeht, daß zwei
zu vergleichende Stichproben aus Messungen mit identischem Median stammen. Wird der
Median-Test signifikant (p≤0,05), ist davon auszugehen, daß sich die Mediane der
zugrundeliegenden Messungen unterscheiden.
Die Signifikanz der Expressionsunterschiede zwischen Endometrien von Frauen mit und
ohne Endometriose, zwischen eutopen Endometrien und endometriotischen Läsionen oder
zwischen endometrialen Fragmenten aus den Nacktmaus-Versuchen, die mittels RT-PCR
ermittelt wurden, wurde mit dem (Studentischen) t-Test berechnet. Bei einem p-Wert kleiner
als 0,05 (p≤0,05) wurden statistisch signifikante Unterschiede angenommen.
40

Ergebnisse
4 Ergebnisse
4.1 Genexpressionsanalysen der Endometrien von Frauen mit und ohne Endometriose
Durch Genexpressionsanalysen mit Hilfe von „Gene-Arrays“ der Firma Affymetrix (U95A)
konnten insgesamt 95 Gene identifiziert werden, die in den Endometrien von
Endometriosepatientinnen einen signifikant verschiedenen Expressionslevel aufwiesen als in
den Endometrien von gesunden Frauen (s. Anhang). In die Auswertung mit einbezogen
wurden alle Gene, die sich in der Expression mindestens um den Faktor 1,5 unterschieden.
Von diesen 95 Genen konnte bei insgesamt 64 Genen eine reduzierte Expression gezeigt
werden, wobei 35 (55 %) dieser Gene in der Sekretionsphase herunterreguliert waren. Vor
allem extrazelluläre Matrixproteine wie Fibronektin, Nidogen oder Kollagen IV zeigten in den
Endometrien von Frauen mit Endometriose in der Sekretionsphase eine herunterregulierte
Expression (Tab. 2).
Die übrigen 31 der 95 unterschiedlich regulierten Gene wurden in den Endometrien von
Endometriosepatientinnen mindestens 1,5fach stärker exprimiert als in den Endometrien
gesunder Frauen (s. Anhang). Diese Expressionsunterschiede zeigten sich vor allem in der
Sekretionsphase. Unter den 23 (74 %) in der Sekretionsphase hochregulierten Genen
wurden insbesondere östrogenabhängig exprimierte Gene gefunden wie der TR3 Orphan-
Rezeptor, ein Mitglied der Hormonrezeptor-Superfamilie (Uemura et al., 1995) oder das
„early growth response protein“ EGR-1, das in der Literatur ebenso wie die
Transkriptionsfaktoren c-fos und jun-B als östrogenreguliertes Gen beschrieben wurde
(Thompson et al., 2002; Duan et al., 2002; Cicatiello et al., 1992) (Tab. 3). Die deutlichsten
Expressionsunterschiede zwischen den Endometrien von Frauen mit und ohne
Endometriose konnten für Cyr61 detektiert werden, ein Gen der CCN-Familie. Das Cyr61-
Gen war in 18 von 20 Vergleichen in Endometrien der Sekretionsphase von
Endometriosepatientinnen im Mittel 5fach stärker exprimiert als in Endometrien von
gesunden Frauen.
41

Ergebnisse
4.1.1 Validierung der Ergebnisse der „Gene-Array“ Analysen
4.1.1.1 Herunterregulierte Gene
Tab. 2: In den Endometrien von Endometriosepatientinnen aus der Sekretionsphase herunterregulierte Gene
42

Ergebnisse
An jeweils fünf Endometrien von Frauen mit und ohne Endometriose aus der
Sekretionsphase wurde überprüft, ob sich die mit Hilfe der Array-Analysen ermittelten
Veränderungen im Expressionsmuster der Gene Nidogen, p27, Fibronektin, Elastin und
Kollagen IV verifizieren lassen.
Für die semiquantitative RT-PCR wurde aus den Geweben die Gesamt-RNA isoliert und
mittels Reverser Transkription cDNA synthetisiert (s. 3.3.4). In der anschließenden PCR, die
für jedes Gen zweimal mit jeder Gewebeprobe im Thermocycler durchgeführt wurde, wurde
in einem Reaktionsansatz gleichzeitig das zu untersuchende Gen und β-Aktin amplifiziert.
Die hierfür verwendeten Primer sowie die PCR-Bedingungen für die einzelnen Gene sind in
Tab. 1 angegeben. Nach elektrophoretischer Auftrennung in einem 2 %igen Agarosegel
wurde die Bandenstärke der PCR-Produkte densitometrisch mit Hilfe des Programms
Gelscan Professional V4.0 bestimmt. Die Signalstärke des zu untersuchenden Gens konnte
nach Abzug des Hintergrunds in Relation zum β-Aktin-Signal derselben Probe gesetzt
werden. Die relativen Expressionswerte des zu untersuchenden Gens wurden jeweils für die
Endometrien von Frauen mit und ohne Endometriose zusammengefaßt. Der mittlere
Expressionslevel der normalen Endometrien wurde als Kontrollwert genommen und 1
gesetzt. Der Expressionslevel der Endometrien von Endometriosepatientinnen wurde
entsprechend berechnet und als xfache Expression bezogen auf das normale Endometrium
dargestellt (Abb. 5).
43

Ergebnisse
Abb. 5, A-F: Validierung der Ergebnisse der herunterregulierten Gene Nidogen (A und B), p27 (C), Fibronektin (D), Elastin (E) und Kollagen IV (F) aus den Array Analysen mittels RT-PCR. Der mittlere Expressionslevel der Endometrien von Endometriosepatientinnen wurde als xfache Expression bezogen auf das normale Endometrium plus Standardabweichung dargestellt. In den getesteten Gewebeproben aus der Sekretionsphase konnten anders als in den Array-Analysen keine Unterschiede in der Expression der Gene zwischen den Kontrollendometrien und den Endometrien von Endometriosepatientinnen detektiert werden. A: Agarosegel, Spur 1: 100 bp Marker, Spur 2, 4, 6, 8, 10: normale Endometrien, Spur 3, 5, 7, 9, 11: Endometrien von Endometriosepatientinnen. Die Positionen der Transkripte β-Aktin (214 bp) und Nidogen (534 bp) wurden gekennzeichnet. B-F: Excel-Diagramme.
Die Ergebnisse der Array-Analyse konnten für die Gene Nidogen, p27, Fibronektin, Elastin
und Kollagen IV nicht bestätigt werden. Nach Auswertung der „Gene-Arrays“ zeigte die
Expression der hier untersuchten Gene in den Endometrien von Endometriosepatientinnen in
der Sekretionsphase in 10-16 von 20 Fällen eine Herunterregulation (Tab. 2). In den von uns
mittels RT-PCR untersuchten Gewebeproben konnte kein signifikanter Unterschied in der
Expression der Gene zwischen den Kontrollendometrien (n=5) und den Endometrien von
Endometriosepatientinnen (n=5) detektiert werden (Abb. 5). Während in der Expression von
p27 und Fibronektin kein Unterschied zwischen den Geweben festgestellt werden konnte
(Abb. 5C und D), war der Expressionslevel von Kollagen IV in den Endometrien der
Endometriosepatientinnen leicht, aber nicht signifikant, reduziert (Abb. 5F). Die Nidogen- und
die Elastin-Expression dagegen waren in den Endometriosepatientinnen sogar leicht erhöht
44

Ergebnisse
(Abb. 5B und E). Weitere Untersuchungen zur Regulation von Nidogen, p27, Fibronektin,
Elastin und Kollagen IV wurden aufgrund dieser Ergebnisse nicht durchgeführt.
4.1.1.2 Hochregulierte Gene
Tab. 3: In den Endometrien von Endometriosepatientinnen aus der Sekretionsphase hochregulierte Gene
45
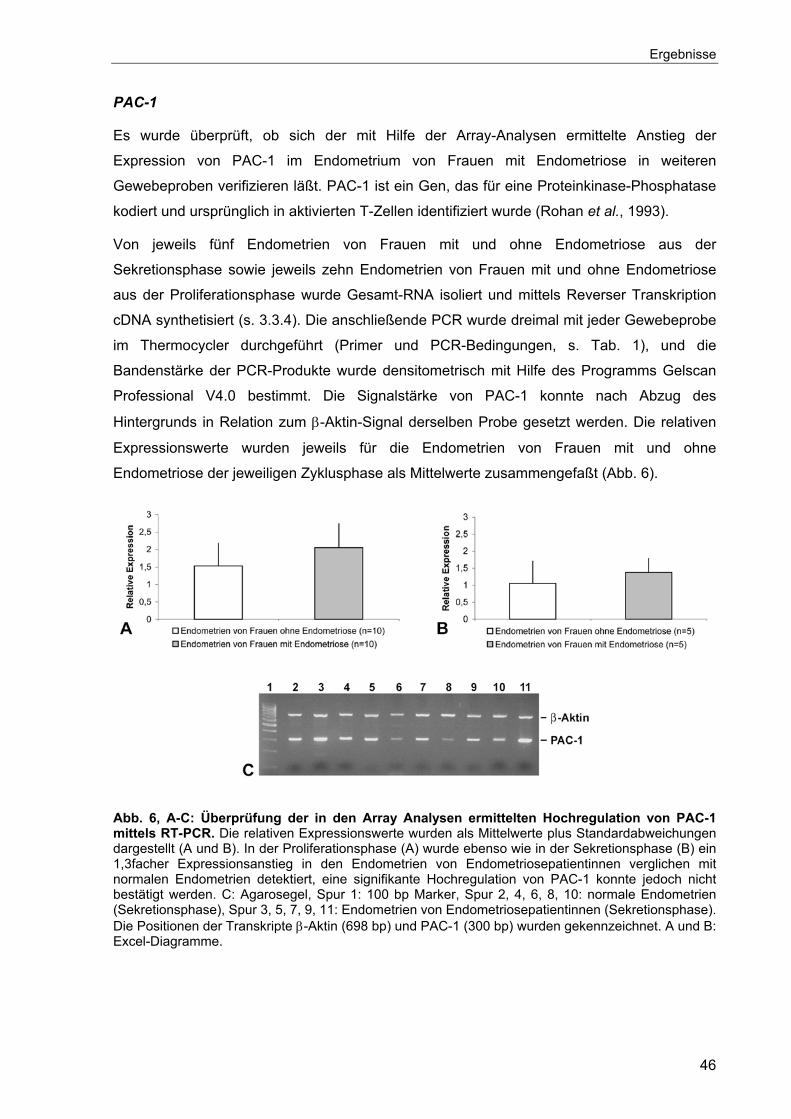
Ergebnisse
PAC-1
Es wurde überprüft, ob sich der mit Hilfe der Array-Analysen ermittelte Anstieg der
Expression von PAC-1 im Endometrium von Frauen mit Endometriose in weiteren
Gewebeproben verifizieren läßt. PAC-1 ist ein Gen, das für eine Proteinkinase-Phosphatase
kodiert und ursprünglich in aktivierten T-Zellen identifiziert wurde (Rohan et al., 1993).
Von jeweils fünf Endometrien von Frauen mit und ohne Endometriose aus der
Sekretionsphase sowie jeweils zehn Endometrien von Frauen mit und ohne Endometriose
aus der Proliferationsphase wurde Gesamt-RNA isoliert und mittels Reverser Transkription
cDNA synthetisiert (s. 3.3.4). Die anschließende PCR wurde dreimal mit jeder Gewebeprobe
im Thermocycler durchgeführt (Primer und PCR-Bedingungen, s. Tab. 1), und die
Bandenstärke der PCR-Produkte wurde densitometrisch mit Hilfe des Programms Gelscan
Professional V4.0 bestimmt. Die Signalstärke von PAC-1 konnte nach Abzug des
Hintergrunds in Relation zum β-Aktin-Signal derselben Probe gesetzt werden. Die relativen
Expressionswerte wurden jeweils für die Endometrien von Frauen mit und ohne
Endometriose der jeweiligen Zyklusphase als Mittelwerte zusammengefaßt (Abb. 6).
Abb. 6, A-C: Überprüfung der in den Array Analysen ermittelten Hochregulation von PAC-1 mittels RT-PCR. Die relativen Expressionswerte wurden als Mittelwerte plus Standardabweichungen dargestellt (A und B). In der Proliferationsphase (A) wurde ebenso wie in der Sekretionsphase (B) ein 1,3facher Expressionsanstieg in den Endometrien von Endometriosepatientinnen verglichen mit normalen Endometrien detektiert, eine signifikante Hochregulation von PAC-1 konnte jedoch nicht bestätigt werden. C: Agarosegel, Spur 1: 100 bp Marker, Spur 2, 4, 6, 8, 10: normale Endometrien (Sekretionsphase), Spur 3, 5, 7, 9, 11: Endometrien von Endometriosepatientinnen (Sekretionsphase). Die Positionen der Transkripte β-Aktin (698 bp) und PAC-1 (300 bp) wurden gekennzeichnet. A und B: Excel-Diagramme.
46

Ergebnisse
Aus den Array-Analysen ergab sich, daß die PAC-1-Expression in den Endometrien von
Endometriosepatientinnen in der Sekretionsphase in 13 von 20 Vergleichen höher war als in
den normalen Endometrien (Tab. 3). Auch in der Proliferationsphase zeigte sich eine
Regulation, in 10 von 30 Fällen war PAC-1 im Endometrium von Endometriosepatientinnen
stärker exprimiert (Daten nicht gezeigt). Diese Ergebnisse konnten mit Hilfe der RT-PCR nur
zum Teil bestätigt werden. Sowohl in der Proliferationsphase (Abb. 6A) als auch in der
Sekretionsphase (Abb. 6B) wurde in den Endometrien von Endometriosepatientinnen eine im
Mittel 1,3fach höhere Expression von PAC-1 beobachtet als in den normalen Endometrien.
Diese Hochregulation war allerdings statistisch nicht signifikant. In den untersuchten
Gewebeproben konnten zwischen den Endometrien von Frauen mit und ohne Endometriose
nur tendenzielle Expressionsunterschiede detektiert werden, so daß keine weiteren
Untersuchungen bezüglich der PAC-1-Expression durchgeführt wurden.
CTGF
In den Array-Analysen wurde ein Anstieg der Expression von CTGF, einem Mitglied der
CCN-Familie, in den Endometrien von Endometriosepatientinnen in der Sekretionsphase
gefunden. Diese Expressionsunterschiede zwischen den Endometrien von Frauen mit und
ohne Endometriose wurden in weiteren Gewebeproben mittels RT-PCR überprüft.
Aus Endometrien von Frauen mit und ohne Endometriose aus der Proliferationsphase und
der Sekretionsphase wurde Gesamt-RNA isoliert und mittels Reverser Transkription cDNA
synthetisiert (s. 3.3.4). Die anschließende PCR wurde dreimal mit jeder Gewebeprobe im
Thermocycler durchgeführt (Primer und PCR-Bedingungen, s. Tab. 1), und die Bandenstärke
der PCR-Produkte wurde densitometrisch mit Hilfe des Programms Gelscan Professional
V4.0 bestimmt. Die Signalstärke von CTGF konnte nach Abzug des Hintergrunds in Relation
zum β-Aktin-Signal derselben Probe gesetzt werden. Die relativen Expressionswerte wurden
jeweils für die Endometrien von Frauen mit und ohne Endometriose der jeweiligen
Zyklusphase als Mittelwerte zusammengefaßt (Abb. 7).
47

Ergebnisse
Abb. 7, A-C: Validierung der Resultate der Gene-Array Analysen für CTGF mittels RT-PCR. Die relativen Expressionswerte wurden als Mittelwerte plus Standardabweichungen dargestellt (A und B). In der Proliferationsphase (A) konnten ebenso wie in der Sekretionsphase (B) keine Expressionsunterschiede in den Endometrien von Endometriosepatientinnen verglichen mit normalen Endometrien nachgewiesen werden. Es zeigte sich allerdings in den endometriotischen Läsionen beider Zyklusphasen eine stärkere Expression verglichen mit den eutopen Endometrien. C: Agarosegel mit Geweben aus der Sekretionsphase, Spur 1: 100 bp Marker, Spur 2-5: normale Endometrien, Spur 6-10: Endometrien von Endometriosepatientinnen, Spur 11 und 12: endometriotische Läsionen. Die Positionen der Transkripte β-Aktin (698 bp) und CTGF (451 bp) wurden gekennzeichnet. A und B: Excel-Diagramme.
Nach Auswertung der Array-Analysen wurde CTGF in den Endometrien von
Endometriosepatientinnen in der Sekretionsphase in 13 von 20 Vergleichen stärker
exprimiert als in den normalen Endometrien der gleichen Zyklusphase (Tab. 3). In der
Proliferationsphase dagegen zeigte sich keine Regulation (Daten nicht gezeigt). Diese
Ergebnisse konnten mit Hilfe der RT-PCR nur zum Teil bestätigt werden. In den zusätzlich
untersuchten Gewebeproben sowohl aus der Proliferationsphase (Abb. 7A) als auch aus der
Sekretionsphase (Abb. 7B) konnten zwischen den Endometrien von Frauen mit
(Proliferationsphase: n=4, Sekretionsphase: n=5) und ohne Endometriose
(Proliferationsphase: n=4, Sekretionsphase: n=4) keine Expressionsunterschiede detektiert
werden. In beiden Zyklusphasen wurde in den Endometrien von Endometriosepatientinnen
die gleiche Expressionsstärke nachgewiesen wie in den normalen Endometrien (Abb. 7A und
B). In den zusätzlich untersuchten endometriotischen Läsionen (Proliferationsphase: n=3,
Sekretionsphase: n=3) dagegen wurde sowohl in der Proliferationsphase als auch in der
Sekretionsphase mehr CTGF exprimiert als in den eutopen Endometrien (Abb. 7A und B). Im
Mittel konnte in beiden Zyklusphasen ein 1,7facher Anstieg der CTGF-Expression in den
endometriotischen Läsionen im Vergleich zum eutopen Endometrium beobachtet werden.
Aufgrund größerer Schwankungen im Expressionsniveau der einzelnen Versuche ergaben
sich allerdings keine signifikanten Expressionsunterschiede.
48

Ergebnisse
Cyr61
Um die in den Array-Analysen ermittelten Expressionsunterschiede von Cyr61 in den
Endometrien von Frauen mit Endometriose verglichen mit normalen Endometrien zu
verifizieren, wurden zusätzliche Gewebeproben mittels real-time RT-PCR untersucht. Bei
dieser Methode erfolgte der Nachweis des PCR-Produktes durch einen fluoreszierenden
Farbstoff, SYBR Green I, der sich in die entstehende doppelsträngige DNA einlagert (Abb.
8A). Wegen der unspezifischen Bindung von SYBR Green I an jede doppelsträngige DNA
mußten Cyr61 und β-Aktin in getrennten Ansätzen amplifiziert werden. Zusätzlich wurden die
Proben nach dem PCR-Lauf in einem 2 %igen Agarosegel aufgetrennt und es konnte
gezeigt werden, daß keine unspezifischen Amplifikationen stattgefunden haben (Abb. 8B). In
allen PCR-Ansätzen wurde nur ein einziges Fragment amplifiziert, das der Größe nach
entweder Cyr61 oder β-Aktin zugeordnet werden konnte.
Abb. 8: A: Schematische Darstellung der Nachweismethode durch SYBR Green I - - bei der real-time RT-PCR, übernommen aus „Der Experimentator: Molekularbiologie/Genomics“ (Mülhardt, 2002). B: Agarosegel nach der real-time RT-PCR. Es konnte gezeigt werden, daß keine unspezifischen Amplifikationen stattgefunden haben. In den untereinander liegenden Spuren wurden jeweils die gleichen Proben aufgetragen. Spur 1: 100 bp-Marker, Spur 2, 4, 6, 8, 10: normale Endometrien (Proliferationsphase), Spur 3, 5, 7, 9, 11: Endometrien von Endometriosepatientinnen (Proliferationsphase), Spur 12, 14, 16: normale Endometrien (Sekretionsphase), Spur 13, 15, 17: Endometrien von Endometriosepatientinnen (Sekretionsphase). Die Positionen der Transkripte von β-Aktin (214 bp) und Cyr61 (223 bp) wurden gekennzeichnet.
Bei der real-time RT-PCR wurden die CT-Werte der einzelnen Proben ermittelt und für die
Auswertung herangezogen. Der CT-Wert ist der PCR-Zyklus, an dem sich das
Fluoreszenzsignal gerade vom Hintergrund abhebt, zu diesem Zeitpunkt ist die Vermehrung
des PCR-Produktes noch exponentiell. Von dem CT-Wert lassen sich Rückschlüsse ziehen
auf die ursprünglich vorhandene cDNA-Menge. Je geringer der CT-Wert ist, desto größer
muß die Menge an cDNA gewesen sein. Die relative Quantifizierung erfolgte wie unter
49

Ergebnisse
3.4.2.2 beschrieben nach der vergleichenden CT-Methode. Zunächst wurden durch
Subtraktion der CT-Werte die Expressionsstärken der einzelnen Proben für Cyr61 gegen β-
Aktin abgeglichen und auf diese Weise relativiert. Diese relativierten Werte wurden jeweils
für die normalen Endometrien und die Endometrien von Endometriosepatientinnen aus jeder
Zyklusphase als Mittelwerte zusammengefaßt. Die normalen Endometrien wurden als
Calibrator genutzt, daher wurde der ermittelte Expressionslevel als 1 gesetzt. Der xfache
Expressionslevel der Endometrien von Endometriosepatientinnen bezogen auf das normale
Endometrium wurde durch Subtraktion des Calibrators und Einsetzen des erhaltenen Wertes
in die Formel 2-∆∆CT berechnet.
In den Array-Analysen war Cyr61 in 18 von 20 Vergleichen in den Endometrien von
Endometriosepatientinnen in der Sekretionsphase im Mittel 5fach stärker exprimiert als in
den normalen Endometrien der gleichen Zyklusphase (Tab. 3). Diese Regulation konnte in
weiteren Gewebeproben mittels real-time RT-PCR bestätigt werden. In der Sekretionsphase
wurde hier eine im Mittel 3fach stärkere Expression (maximal 15fach) von Cyr61 in den
Endometrien von Frauen mit Endometriose (n=10) im Vergleich zu normalen Endometrien
(n=8) detektiert (Abb. 9B). Abweichend von den Array-Analysen konnte in den mit Hilfe der
real-time RT-PCR untersuchten Geweben auch in der Proliferationsphase eine signifikant
stärkere Cyr61-Expression nachgewiesen werden. In den Endometrien von
Endometriosepatientinnen aus der Proliferationsphase (n=15) war der Expressionslevel von
Cyr61 im Mittel 2fach höher (maximal 6fach) als in den Endometrien von Frauen ohne
Endometriose (n=16) (Abb. 9A). Wegen dieser signifikanten Hochregulation von Cyr61 in
beiden Zyklusphasen fokussierten sich die weiteren Versuche auf das Cyr61-Gen.
Abb. 9, A und B: Validierung der Resultate der Array Analysen für Cyr61 mittels real-time RT-PCR. Der mittlere Expressionslevel der Endometrien von Endometriosepatientinnen bezogen auf normale Endometrien wurde ohne Standardabweichung dargestellt. In der Proliferationsphase (A) wurde ebenso wie in der Sekretionsphase (B) ein signifikanter Expressionsanstieg von Cyr61 in den Endometrien von Endometriosepatientinnen verglichen mit normalen Endometrien detektiert. A und B: Excel-Diagramme, *p≤0,05.
50

Ergebnisse
4.1.2 Cyr61-Expression in den endometriotischen Läsionen
Die Expression von Cyr61 wurde zusätzlich in den endometriotischen peritonealen Läsionen
der Proliferationsphase und der Sekretionsphase im Vergleich zum eutopen Endometrium
von Endometriosepatientinnen untersucht. Diese Versuche wurden ebenfalls mit Hilfe der
real-time RT-PCR durchgeführt. Wie für die endometrialen Gewebe beschrieben (s. 3.4.2.2
und 4.1.1.2) wurde die relative Expressionsstärke von Cyr61 in den endometriotischen
Läsionen gegen β-Aktin der jeweils gleichen Probe abgeglichen und die relativierten CT-
Werte anschließend für die einzelnen Zyklusphasen als Mittelwerte zusammengefaßt. Als
Calibrator wurde die bereits ermittelte mittlere Expression der Endometrien von
Endometriosepatientinnen genutzt (s. 4.1.1.2) und hier als 1 gesetzt.
Obwohl Cyr61 im Endometrium von Endometriosepatientinnen im Vergleich zum normalen
Endometrium bereits sehr viel stärker exprimiert war, konnte in den endometriotischen
Läsionen ein zusätzlicher Anstieg in der Expression von Cyr61 gezeigt werden (Abb. 10). In
der Sekretionsphase wurde in den Läsionen (n=4) eine im Mittel 20fach (maximal 70fach)
stärkere Expression detektiert als in den eutopen Endometrien (n=10) (Abb. 10B). Auch in
der Proliferationsphase war Cyr61 in den endometriotischen Läsionen (n=5) zusätzlich
signifikant hochreguliert, hier wurde eine im Mittel 4fach (maximal 10fach) höhere Expression
beobachtet als in den eutopen Endometrien (n=15) (Abb. 10A).
Abb. 10, A und B: Expression von Cyr61 in endometriotischen Läsionen, ermittelt mit Hilfe von real-time RT-PCR. Der mittlere Expressionslevel der endometriotischen Läsionen bezogen auf das Endometrium von Frauen mit Endometriose wurde ohne Standardabweichung dargestellt. In der Sekretionsphase wurde in den ektopen Läsionen eine im Mittel 20fach stärkere Expression detektiert als in den eutopen Endometrien (B). In der Proliferationsphase wurde in den Läsionen eine im Mittel 4fach höhere Cyr61-Expression detektiert als in den eutopen Endometrien (A). A und B: Excel-Diagramme, *p≤0,05.
51

Ergebnisse
4.1.3 Cyr61-Expression im Verlauf des Menstruationszyklus
Neben den Expressionsunterschieden zwischen den Endometrien von gesunden Frauen und
Endometriosepatientinnen zeigte sich, daß Cyr61 in den einzelnen Phasen des
Menstruationszyklus im Endometrium unterschiedlich stark exprimiert wird.
Dies ergab sich aus den Untersuchungen mit Hilfe der real-time RT-PCR. Die unter 4.1.1.2
für Cyr61 erhaltenen Ergebnisse der normalen Endometrien und der Endometrien von
Endometriosepatientinnen wurden hier den Zyklusphasen entsprechend verglichen. So
konnte festgestellt werden, daß sich die Cyr61-Expression im Laufe des Menstruationszyklus
veränderte, sowohl in Endometrien von Frauen mit als auch ohne Endometriose. Für diese
Vergleiche wurden die Endometrien aus der Proliferationsphase jeweils von gesunden
Frauen und von Endometriosepatientinnen als Calibrator genutzt und der hierfür ermittelte
mittlere Expressionslevel als 1 gesetzt.
In der Proliferationsphase wurde sowohl im normalen Endometrium als auch im
Endometrium von Endometriosepatientinnen eine stärkere Expression detektiert als in der
Sekretionsphase (Abb. 11). In den Endometrien von gesunden Frauen wurde in der
Sekretionsphase (n=8) im Mittel 3fach weniger Cyr61 exprimiert als in der
Proliferationsphase (n=16) (Abb. 11A). In den Endometrien von Endometriosepatientinnen
aus der Sekretionsphase (n=10) war der Expressionslevel von Cyr61 im Mittel 2fach geringer
als in den Endometrien aus der Proliferationsphase (n=15) (Abb. 11B). Allerdings konnte nur
in den Endometrien von gesunden Frauen eine statistisch signifikante Reduktion detektiert
werden, in den Endometrien von Frauen mit Endometriose war die Herunterregulation des
Cyr61-Gens statistisch nicht signifikant.
Abb. 11, A und B: Veränderungen der Cyr61-Expression während des Menstruationszyklus. Die mittels real-time RT-PCR ermittelten relativierten Expressionsstärken für die normalen Endometrien und die Endometrien von Endometriosepatientinnen wurden für jede Zyklusphase zusammengefaßt. Der mittlere Expressionslevel der Endometrien aus der Sekretionsphase jeweils von Frauen mit und ohne Endometriose bezogen auf das jeweilige Endometrium aus der Proliferationsphase wurde ohne Standardabweichung dargestellt. Sowohl im normalen Endometrium (A) als auch im Endometrium von Frauen mit Endometriose (B) wurde Cyr61 in der Proliferationsphase stärker exprimiert als in der Sekretionsphase. Dieser Expressionsunterschied war jedoch nur in den normalen Endometrien signifikant. A und B: Excel-Diagramme, *p≤0,05.
52

Ergebnisse
4.2 Expression von Cyr61 auf Proteinebene
4.2.1 Western Blot
Das für den Western Blot verwendete Antiserum wurde freundlicherweise vom „Labor für
Molekulare Experimentelle Orthopädie“ der orthopädischen Universitätsklinik in Würzburg
zur Verfügung gestellt. Mit Hilfe dieses Western Blots konnte Cyr61 auch auf Proteinebene
im eutopen Endometrium von Frauen mit und ohne Endometriose detektiert werden (Abb.
12). Als Kontrolle wurde humanes Myometrium und Brustdrüsengewebe verwendet, in denen
Cyr61 mittels Western Blot bereits nachgewiesen werden konnte (Sampath et al., 2001a, b)
(Abb. 12, Spur 2 und 3). Der verwendete Sekundärantikörper reagierte mit den einzelnen
Banden des Markers, so daß es möglich war, die Größe der erhaltenen Banden direkt am
Blot zu bestimmen. In Übereinstimmung mit der Studie von Sampath et al. (2001a) wurde
nach der Antikörperreaktion eine einzelne Bande auf der Höhe von 42 kDa detektiert, was
dem Molekulargewicht von Cyr61 entspricht (Abb. 12). Das verwendete Antiserum scheint
zusätzlich eine unspezifische Reaktion einzugehen, da eine weitere schwächere Bande auf
der Höhe von 110 kDa in einigen Spuren nachgewiesen wurde. Über posttranslationale
Modifikationen des Cyr61-Proteins ist bis heute nichts bekannt.
Abb. 12: Western Blot zum Nachweis von Cyr61 in eutopen Endometrien von Frauen mit und ohne Endometriose aus der Proliferationsphase. Spur 1: Marker, Spur 2: humanes Myometrium, Spur 3: humanes Brustdrüsengewebe, Spur 4+5: normale Endometrien, Spur 6-8: Endometrien von Endometriosepatientinnen. Die Position des Cyr61-Proteins (42 kDa) wurde gekennzeichnet.
Es war nicht möglich, einen Abgleich des nachgewiesenen Proteins gegen β-Aktin
durchzuführen, um eine gleichmäßige Auftragung der Proteine in den einzelnen Spuren
nachzuweisen. Dadurch war es auch nicht möglich, eine Aussage über die tatsächlichen
Bandenstärken und damit über Expressionsunterschiede zwischen normalen Endometrien
(Proliferationsphase: n=2) und Endometrien von Endometriosepatientinnen
(Proliferationsphase: n=3) zu treffen. Unter der Annahme, daß die gleichen Proteinmengen in
jeder Spur aufgetragen wurden, war jedoch erkennbar, daß in den Endometrien gesunder
Frauen weniger Cyr61 exprimiert wurde als in den Endometrien von
Endometriosepatientinnen (Abb. 12, Spur 4-8). Es zeigte sich dann allerdings auch eine
53
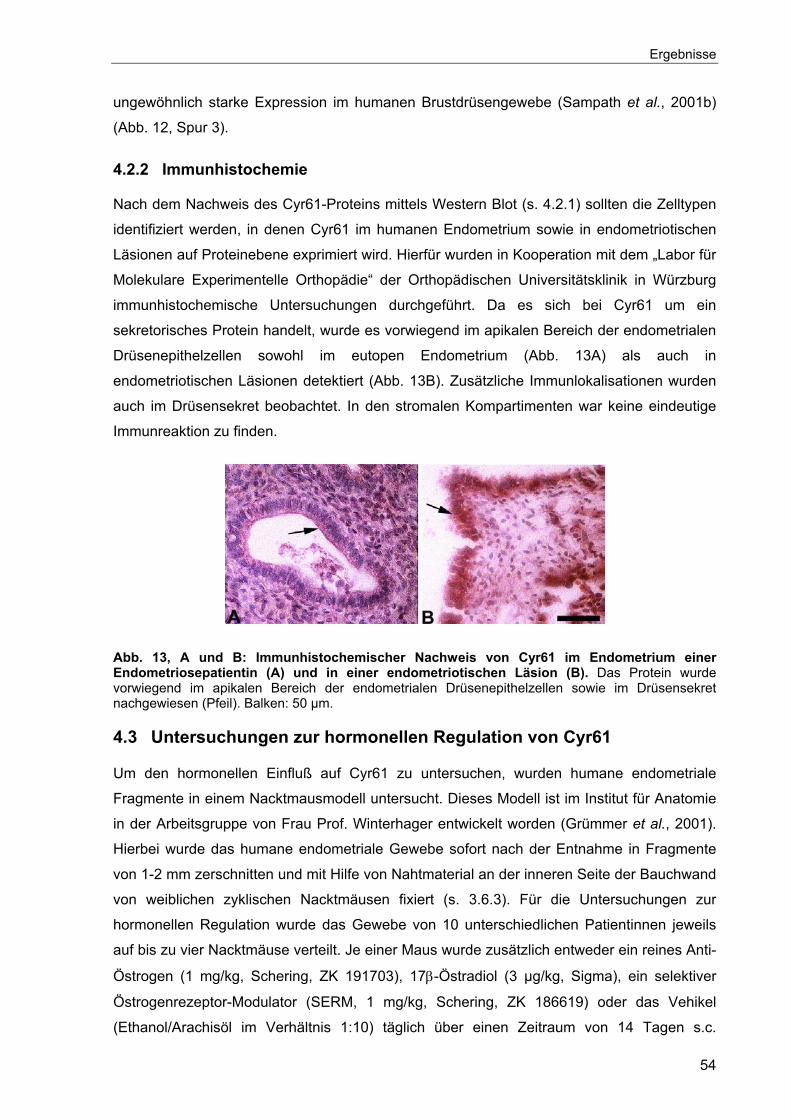
Ergebnisse
ungewöhnlich starke Expression im humanen Brustdrüsengewebe (Sampath et al., 2001b)
(Abb. 12, Spur 3).
4.2.2 Immunhistochemie
Nach dem Nachweis des Cyr61-Proteins mittels Western Blot (s. 4.2.1) sollten die Zelltypen
identifiziert werden, in denen Cyr61 im humanen Endometrium sowie in endometriotischen
Läsionen auf Proteinebene exprimiert wird. Hierfür wurden in Kooperation mit dem „Labor für
Molekulare Experimentelle Orthopädie“ der Orthopädischen Universitätsklinik in Würzburg
immunhistochemische Untersuchungen durchgeführt. Da es sich bei Cyr61 um ein
sekretorisches Protein handelt, wurde es vorwiegend im apikalen Bereich der endometrialen
Drüsenepithelzellen sowohl im eutopen Endometrium (Abb. 13A) als auch in
endometriotischen Läsionen detektiert (Abb. 13B). Zusätzliche Immunlokalisationen wurden
auch im Drüsensekret beobachtet. In den stromalen Kompartimenten war keine eindeutige
Immunreaktion zu finden.
Abb. 13, A und B: Immunhistochemischer Nachweis von Cyr61 im Endometrium einer Endometriosepatientin (A) und in einer endometriotischen Läsion (B). Das Protein wurde vorwiegend im apikalen Bereich der endometrialen Drüsenepithelzellen sowie im Drüsensekret nachgewiesen (Pfeil). Balken: 50 µm.
4.3 Untersuchungen zur hormonellen Regulation von Cyr61
Um den hormonellen Einfluß auf Cyr61 zu untersuchen, wurden humane endometriale
Fragmente in einem Nacktmausmodell untersucht. Dieses Modell ist im Institut für Anatomie
in der Arbeitsgruppe von Frau Prof. Winterhager entwickelt worden (Grümmer et al., 2001).
Hierbei wurde das humane endometriale Gewebe sofort nach der Entnahme in Fragmente
von 1-2 mm zerschnitten und mit Hilfe von Nahtmaterial an der inneren Seite der Bauchwand
von weiblichen zyklischen Nacktmäusen fixiert (s. 3.6.3). Für die Untersuchungen zur
hormonellen Regulation wurde das Gewebe von 10 unterschiedlichen Patientinnen jeweils
auf bis zu vier Nacktmäuse verteilt. Je einer Maus wurde zusätzlich entweder ein reines Anti-
Östrogen (1 mg/kg, Schering, ZK 191703), 17β-Östradiol (3 µg/kg, Sigma), ein selektiver
Östrogenrezeptor-Modulator (SERM, 1 mg/kg, Schering, ZK 186619) oder das Vehikel
(Ethanol/Arachisöl im Verhältnis 1:10) täglich über einen Zeitraum von 14 Tagen s.c.
54

Ergebnisse
appliziert. Insgesamt wurden 7 normale Endometrien und 3 Endometrien von
Endometriosepatientinnen transplantiert. Nach Beendigung der Versuche stellte sich heraus,
daß ein normales Endometrium einer postmenopausalen Frau verwendet wurden, weitere
zwei normale Endometrien stammten von Frauen, die zur Zeit der OP die Pille einnahmen.
Diese Gewebe wurden dennoch in die Auswertung mit einbezogen, da sich keine
Abweichungen zeigten.
4.3.1 Validierung des Nacktmausmodells
4.3.1.1 Uterus- und Ovargewichte der Nacktmäuse
Zur Kontrolle, ob die s.c. verabreichten Hormone und Anti-Hormone ihre Zielorgane
erreichten, wurden der Uterus und die Ovarien der Nacktmäuse entnommen und das
Gewicht der Organe bestimmt (Lan und Katzenellenbogen, 1976). Die Organgewichte
wurden jeweils auf das Körpergewicht der Maus bezogen und anschließend für die einzelnen
Behandlungen als Mittelwerte zusammengefaßt (Abb. 14).
Abb. 14, A und B: Ovar- und Uterusgewichte der hormonbehandelten Nacktmäuse. Die mittleren Organgewichte plus Standardabweichungen wurden dargestellt. Die Gewichte der Ovarien wurden durch die verabreichten Substanzen nicht beeinflußt (A). Die Uterusgewichte dagegen nahmen nach Applikation des Anti-Östrogens signifikant ab und nach Behandlung mit Östradiol signifikant zu (B). C: Die Effekte der applizierten Hormone war auch makroskopisch erkennbar, die Uteri der mit Anti-Östrogen behandelten Tiere waren sehr viel dünner als die Uteri der Tiere, denen Östradiol verabreicht wurde. A und B: Excel-Diagramme, *p≤0,05, C: Photo.
Es zeigte sich, daß das Gewicht der Ovarien in den unterschiedlich behandelten Tieren nicht
variierte (Abb. 14A), die verabreichten Hormone zeigten also keine Nebenwirkungen. Das
Uterusgewicht dagegen wurde entsprechend von den verabreichten Hormonen beeinflußt.
Mäuse, denen ein Anti-Östrogen appliziert wurde, hatten ein signifikant geringeres
Uterusgewicht als die Kontrolltiere, das Organgewicht wurde fast um die Hälfte reduziert
(Abb. 14B). Durch die Applikation von Östradiol wurde das Uterusgewicht im Vergleich zum
55

Ergebnisse
Vehikel signifikant gesteigert (Abb. 14B). Die Uterusgewichte der Tiere, denen das SERM
verabreicht wurde, zeigten eine 20 %ige Reduktion gegenüber den Kontrolltieren, diese
Reduktion war jedoch nicht signifikant (Abb. 14B). Die Effekte der applizierten Hormone war
auch makroskopisch erkennbar, die Uteri der mit Anti-Östrogen behandelten Tiere waren
sehr viel dünner als die Uteri der Tiere, denen Östradiol verabreicht wurde (Abb. 14C). Nach
Applikation des SERM zeigten sich makroskopisch dickere Uteri als in den mit Anti-Östrogen
behandelten Tieren aber dünnere Uteri als in den der mit Östradiol behandelten Tieren (Abb.
14C). Die applizierten Hormone hatten damit das Zielorgan, den Uterus, entsprechend
beeinflußt und so die lokale Wirkung bestätigt.
4.3.1.2 Zellhöhe und Proliferation des Uterusepithels der behandelten Nacktmäuse
Epithelhöhen
In Kooperation mit der Schering AG (Berlin) wurden die Epithelhöhen der Maus-Uteri
bestimmt, denn ein weiterer Parameter für östrogene Wirkung ist die Höhe des
Uterusepithels. An Paraffin-Schnitten der Maus-Uteri 6 verschiedener Versuchsreihen
wurden die Höhen der Uterusepithelien ermittelt und für die einzelnen Behandlungen als
Mittelwerte zusammengefaßt. Durch die Bestimmung der Uterusgewichte konnte bereits
demonstriert werden, daß die den Nacktmäusen applizierten Hormone das Zielorgan, den
Uterus, erreichen und beeinflussen (s. 4.3.1.1).
Diese Ergebnisse bestätigten sich nur teilweise in den gemessenen Epithelhöhen. Durch die
Applikation des Anti-Östrogens wurden die Uterusepithel-Höhen in diesen Tieren signifikant
um durchschnittlich 35 % reduziert (Abb. 15). Der Entzug von Östrogen wirkte sich also auch
auf die Höhe des Uterusepithels aus. Allerdings konnte durch die Behandlung der zyklischen
Tiere mit zusätzlichen Mengen Östradiol kein entsprechender Effekt erzeugt werden. In den
Tieren, denen Östradiol appliziert wurde, zeigte sich ebenfalls eine leichte aber nicht
signifikante Reduktion der Epithelhöhen gegenüber dem Vehikel (Abb. 15). Im Durchschnitt
waren die Uterusepithelien der mit SERM behandelten Tiere knapp 30 % niedriger
verglichen mit den Kontrolltieren, bedingt durch hohe Schwankungen in den einzelnen
Versuchsreihen war diese Reduktion jedoch nicht signifikant (Abb. 15).
Die durch die Gewichtsbestimmung der Uteri ermittelten Einflüsse der verabreichten
Hormone ließen sich für die Epithelhöhen nur tendenziell bestätigen. Es konnte zwar durch
die Applikation des Anti-Östrogens die erwartete signifikante Reduktion erzielt werden, die
Behandlung mit Östradiol hatte jedoch auf das Uterusepithel keinen Effekt. Auffällig waren
die starken Schwankungen zwischen den einzelnen Versuchsreihen.
56

Ergebnisse
Abb. 15: Epithelhöhen der Maus-Uteri nach hormoneller Behandlung. Die Epithelhöhen aus 6 verschiedenen Versuchsreihen wurden als Mittelwerte plus Standardabweichungen dargestellt. Die Uterusepithel-Höhen der mit Anti-Östrogen behandelten Tiere wurden signifikant reduziert. In den Tieren, die mit Östradiol oder SERM behandelt wurden, konnten keine signifikanten Änderungen der Epithelhöhen verglichen mit dem Vehikel detektiert werden. Excel-Diagramm, *p≤0,05.
Proliferation
Zusätzlich wurde in den Uteri der hormonbehandelten Mäuse 5 verschiedener
Versuchsreihen mittels immunhistochemischer Färbung die Expression des
Proliferationsmarkers Ki67 untersucht, der in allen Phasen der Mitose exprimiert wird, mit
Ausnahme der G0-Phase (Schlüter et al., 1993). In allen Fällen wurde Ki67 hauptsächlich im
Uterusepithel nachgewiesen, Proliferation im Stroma konnte nur sehr selten beobachtet
werden (Abb. 16). Eindeutige Effekte der applizierten Hormone konnten nicht detektiert
werden, da sich die Ergebnisse der einzelnen Versuchsreihen sehr stark voneinander
unterschieden. In der ersten Versuchsreihe zeigte das Uterusepithel der mit Vehikel
behandelten Maus etwa eine gleich starke Proliferation wie das Uterusepithel der Maus,
denen Östradiol verabreicht wurde (Abb. 16A). In der mit Anti-Östrogen behandelten Maus
konnte dagegen keine Ki67-Färbung nachgewiesen werden (Abb. 16A). In dem Uterus der
Maus, der das SERM appliziert wurde, wurde nur geringe Proliferationsaktivität detektiert
(Abb. 16A). Die Ki67-Färbung war hier deutlicher als in dem mit Anti-Östrogen behandelten
Tier, jedoch nicht so stark wie in den Mäusen, denen das Vehikel bzw. Östradiol verabreicht
wurde.
In einer weiteren Versuchsreihe war die Proliferationsaktivität genau umgekehrt verteilt. In
diesem Fall wurde die stärkste Ki67-Färbung im Uterus der mit Anti-Östrogen behandelten
Maus detektiert (Abb. 16B). In den mit Vehikel oder SERM behandelten Tieren zeigte sich
eine schwächere Proliferationsaktivität, während im Uterus der mit Östradiol behandelten
Maus kein Ki67 nachgewiesen werden konnte (Abb. 16B). Auch in den drei übrigen
Versuchsreihen konnte die Proliferationsaktivität im Uterusepithel nicht den applizierten
Substanzen zugeordnet werden (Daten nicht gezeigt). Obwohl nach der Messung der
Uterusepithel-Höhen eine statistisch signifikante Reduktion in den mit Anti-Östrogen
behandelten Tieren im Vergleich zum Vehikel detektiert wurde (Abb. 15), wurde die
57

Ergebnisse
Proliferationsrate im Uterusepithel der Mäuse durch die applizierten Substanzen nicht
beeinflußt (Abb. 16) und stellte damit keinen Marker für östrogene Wirkung dar.
Abb. 16, A und B: Immunhistochemischer Nachweis von Ki67 in den Nacktmaus-Uteri nach hormoneller Behandlung zur Darstellung der Proliferationsaktivität. A: In der dargestellten
len Behandlung auf die transplantierten Fragmente
bb. 17A: Photo einer weiblichen Nacktmaus, der humane endometriale Fragmente an der
inneren Bauchwandseite fixiert wurden. B: Die Fragmente konnten anhand der Fixierungsnähte am
Versuchsreihe wurde die deutlichste Proliferationsaktivität in den Uterusepithelien der mit Vehikel und Östradiol behandelten Tiere nachgewiesen. B: In dieser Versuchsreihe zeigte sich eine umgekehrte Verteilung. Die deutlichste Proliferationsaktivität wurde hier in den mit Anti-Östrogen behandelten Tieren detektiert. Balken: 300 µm.
4.3.2 Einfluß der hormonel
Die an die Bauchwand angenähten humanen endometrialen Fragmente wurden anhand der
Fixierungsnähte lokalisiert (Abb. 17B). Für morphologische Untersuchungen wurden die
Fragmente zusammen mit einem Teil der Bauchwand entnommen, in Formalin fixiert und in
Paraffin eingebettet (s. 3.6.6).
A
Ende des Versuchs lokalisiert und entnommen werden. Die Pfeile weisen auf die vier transplantierten Fragmente.
58

Ergebnisse
Zur Untersuchung der Genexpression wurden die Fragmente direkt in flüssigem Stickstoff
gefroren. Bei der Entnahme der Fragmente wurde darauf geachtet, daß kein
Bauchwandgewebe der Nacktmaus mit entfernt wurde (3.6.5). Aus den Fragmenten wurde
Gesamt-RNA isoliert, der komplette Ansatz einem DNase-Verdau unterzogen und in die RT-
Reaktion eingesetzt. Die anschließende PCR wurde wegen der geringen Ausbeute an RNA
nur einmal mit jedem Fragment im Thermocycler durchgeführt (Primer und PCR-
Bedingungen, s. Tab. 1), und die Bandenstärke der PCR-Produkte wurde densitometrisch
mit Hilfe des Programms Gelscan Professional V4.0 bestimmt. Die Signalstärke des Zielgens
konnte nach Abzug des Hintergrunds in Relation zum β-Aktin-Signal derselben Probe
gesetzt werden. Die relativen Expressionswerte der Fragmente aus den Nacktmäusen,
denen das Vehikel appliziert wurde, wurden für jede Versuchsreihe als Kontrolle genutzt und
1 gesetzt. Die Expressionslevel in den Fragmenten der mit Östradiol, Anti-Östrogen und
SERM behandelten Tiere wurden in jeder Versuchsreihe gegen das jeweilige Vehikel der
gleichen Versuchsreiche abgeglichen, anschließend für jede Substanz als Mittelwert
zusammengefaßt und als xfache Expression bezogen auf das Vehikel dargestellt.
In einigen Versuchsreihen konnte wegen zu geringer Ausbeute an RNA aus den Fragmenten
des mit Vehikel behandelten Tieres keine PCR durchgeführt werden. Dann wurde der
Expressionslevel in den Fragmenten der mit Östradiol, Anti-Östrogen oder SERM
behandelten Tiere der entsprechenden Versuchsreihe gegen den Mittelwert der relativen
Expressionswerte aus den mit Vehikel behandelten Tiere der übrigen Versuchsreihen
abgeglichen und in die weiteren Berechnungen mit einbezogen. Es war aufgrund der
geringen Ausbeute an RNA auch nicht möglich, die Expression jedes Gens in jeder der 10
Versuchsreihen zu untersuchen.
4.3.2.1 Expression der Östrogenrezeptoren ERα und ERβ
ERα
Die Expression eines Östrogenrezeptors ist Voraussetzung für die Wirkung der applizierten
Substanzen, daher wurde untersucht, ob in den humanen endometrialen Fragmenten nach
Transplantation in die Nacktmaus die Östrogenrezeptoren ERα und ERβ exprimiert werden.
Der Östrogenrezeptor ERα konnte in allen untersuchten humanen endometrialen
Fragmenten, die in die Nacktmaus transplantiert wurden, nachgewiesen werden (Abb. 18A).
Durch die applizierten Substanzen wurde die Menge des exprimierten Gens allerdings nicht
beeinflußt (Abb. 18B). In den Fragmenten der Tiere, die mit Östradiol behandelt wurden,
zeigte sich im Mittel eine gleich starke Expression wie in den Fragmenten der Tiere, denen
das Vehikel verabreicht wurde (Abb. 18B). Durch die Applikation des Anti-Östrogens wurde
die Expression von ERα verglichen mit den Fragmenten der Kontrolltiere reduziert, allerdings
59

Ergebnisse
nicht signifikant (Abb. 18B). Nach Behandlung der Tiere mit dem SERM, zeigte sich eine im
Durchschnitt um den Faktor 1,3 erhöhte Expression verglichen mit dem Vehikel. In den
einzelnen Versuchsreihen wurde ERα in den mit SERM behandelten Tieren sehr
unterschiedlich stark exprimiert, woraus eine hohe Standardabweichung resultierte (Abb.
18B). Eine signifikante Änderung der Expression von ERα konnte daher auch in diesen
Fragmenten nicht nachgewiesen werden.
Der Östrogenrezeptor ERα wurde in allen transplantierten Fragmenten nachgewiesen, eine
Wirkung der östrogenwirksamen Substanzen auf die transplantierten Fragmente war daher
möglich. Signifikante Änderungen des Rezeptors ERα durch die verabreichten Hormone
konnten nicht beobachtet werden.
Abb. 18, A und B: Expression des Östrogenrezeptors ERα in humanen endometrialen Fragmenten nach Kultivierung in der Nacktmaus. Die mit Hilfe von RT-PCR ermittelten mittleren Expressionswerte der hormonell behandelten Tiere wurden als xfache Expression bezogen auf das Vehikel plus Standardabweichungen dargestellt (B). Der Östrogenrezeptor ERα konnte in allen untersuchten Fragmenten nachgewiesen werden (A), eine signifikante Beeinflussung des Expressionslevels durch die applizierten Substanzen wurde allerdings nicht beobachtet (B). A: Agarosegel, Spur 1: 100 bp-Marker, Spur 2: nicht transplantiertes Kontrollendometrium, Spur 3: Fragment aus einem mit Vehikel behandelten Tier, Spur 4: Fragment aus einem mit Anti-Östrogen behandelten Tier, Spur 5: Fragment aus einem mit Östradiol behandelten Tier, Spur 6: Fragment aus einem mit SERM behandelten Tier. Die Positionen der Transkripte von β-Aktin (698 bp) und ERα (227 bp) wurden gekennzeichnet. B: Excel-Diagramm.
60

Ergebnisse
ERβ
Der Östrogenrezeptor ERβ konnte unabhängig von den applizierten Substanzen (Abb. 19,
Spur 3-6) nicht in den transplantierten endometrialen Fragmenten nachgewiesen werden.
Nur in den Kontrollendometrien, die nicht transplantiert wurden, wurde ERβ schwach
exprimiert (Abb. 19, Spur 2).
Abb. 19: Expression des Östrogenrezeptors ERβ in humanen endometrialen Fragmenten nach Kultivierung in der Nacktmaus. Nur im Kontrollendometrium, das nicht transplantiert wurde, konnte eine schwache ERβ-Expression nachgewiesen werden (Spur 2). In den Fragmenten aus den Tieren, die mit Vehikel (Spur 3), Anti-Östrogen (Spur 4), Östradiol (Spur 5) oder SERM (Spur 6) behandelt wurden, konnte ERβ nicht detektiert werden. Spur 1: 100 bp-Marker. Die Positionen der Transkripte von β-Aktin (698 bp) und ERβ (279 bp) wurden gekennzeichnet.
4.3.2.2 Morphologie
Trotz der Behandlung der Nacktmäuse mit Östradiol, einem Anti-Östrogen oder einem
SERM zeigten sich keine Veränderungen in der Morphologie der transplantierten humanen
endometrialen Fragmente.
In den meisten der untersuchten Fragmente wurden endometriale Drüsen und stromales
Gewebe identifiziert (Abb. 20). Weiterhin adhärierte in allen untersuchten Fragmenten das
stromale Gewebe an die anliegende Bauchwand der Nacktmaus, nie das Drüsenepithel
(Abb. 20B). In den Fragmenten der Kontrolltiere wurden sowohl englumige Drüsen mit
zylindrischem bis hohem Epithel gefunden als auch weitlumige zystenartige Drüsen mit
flachem Epithel (Abb. 20A, Ausschnittsvergrößerung), das Stroma war meist zellreich und
von Lymphozyten durchsetzt. Die Fragmente aus den mit Östradiol behandelten Tieren
zeigten ein ähnliches Bild (Abb. 20E, Ausschnittsvergrößerung).
Andererseits zeigte sich eine sehr hohe Varianz zwischen den untersuchten Fragmenten, die
jedoch in keinem Zusammenhang stand mit der verabreichten Substanz. Wurden mehrere
Fragmente aus einer Maus histologisch aufgearbeitet, konnten zwischen diesen
morphologische Unterschiede detektiert werden. Die zwei in Abbildung 20C und 20D
gezeigten Fragmente stammten beide aus der gleichen mit Anti-Östrogen behandelten
Maus. Während das eine Fragment sehr klein war und hier nur eine weitlumige dilatierte
61

Ergebnisse
Drüse vorhanden war (Abb. 20C, Ausschnittsvergrößerung), war das andere Fragment sehr
viel größer, und es konnten mehrere auch englumige Drüsen mit zylindrischem Epithel
beobachtet werden (Abb. 20D, Ausschnittsvergrößerung). Charakteristisch für die mit dem
SERM behandelten Fragmente waren langgezogene englumige Drüsen mit einem hohen
Epithel (Abb. 20F, Ausschnittsvergrößerung). Diese Drüsen, die dem Querschnitt eines
Maus-Uterus glichen, wurden nur in den mit SERM behandelten Fragmenten beobachtet,
allerdings nicht in allen.
Abb. 20, A-F: Morphologie der humanen endometrialen Fragmente nach Kultivierung in der Nacktmaus und unterschiedlicher hormoneller Behandlung. Die Fragmente aus den Tieren, denen das Vehikel (A und B), Anti-Östrogen (C und D), Östradiol (E) oder SERM (F) appliziert wurde, wiesen keine morphologischen Unterschiede auf. B: Invasion der Stromazellen (S) in die anliegende Bauchwand der Nacktmaus. BW: Bauchwand der Nacktmaus, F: transplantiertes humanes Fragment, N: Fixierungsnaht. Balken: in A und C-F: 160 µm, in B: 25 µm, in den Ausschnittsvergrößerungen A-E: 100 µm, in der Ausschnittsvergrößerung F: 80 µm.
62
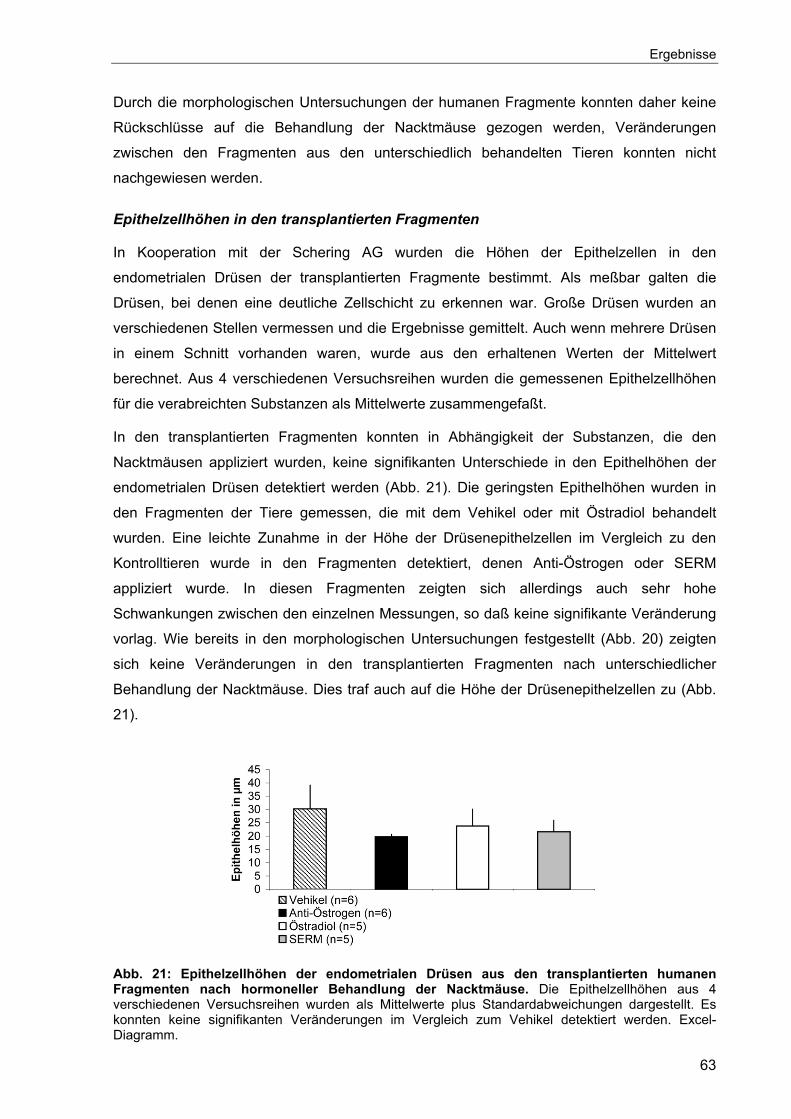
Ergebnisse
Durch die morphologischen Untersuchungen der humanen Fragmente konnten daher keine
Rückschlüsse auf die Behandlung der Nacktmäuse gezogen werden, Veränderungen
zwischen den Fragmenten aus den unterschiedlich behandelten Tieren konnten nicht
nachgewiesen werden.
Epithelzellhöhen in den transplantierten Fragmenten
In Kooperation mit der Schering AG wurden die Höhen der Epithelzellen in den
endometrialen Drüsen der transplantierten Fragmente bestimmt. Als meßbar galten die
Drüsen, bei denen eine deutliche Zellschicht zu erkennen war. Große Drüsen wurden an
verschiedenen Stellen vermessen und die Ergebnisse gemittelt. Auch wenn mehrere Drüsen
in einem Schnitt vorhanden waren, wurde aus den erhaltenen Werten der Mittelwert
berechnet. Aus 4 verschiedenen Versuchsreihen wurden die gemessenen Epithelzellhöhen
für die verabreichten Substanzen als Mittelwerte zusammengefaßt.
In den transplantierten Fragmenten konnten in Abhängigkeit der Substanzen, die den
Nacktmäusen appliziert wurden, keine signifikanten Unterschiede in den Epithelhöhen der
endometrialen Drüsen detektiert werden (Abb. 21). Die geringsten Epithelhöhen wurden in
den Fragmenten der Tiere gemessen, die mit dem Vehikel oder mit Östradiol behandelt
wurden. Eine leichte Zunahme in der Höhe der Drüsenepithelzellen im Vergleich zu den
Kontrolltieren wurde in den Fragmenten detektiert, denen Anti-Östrogen oder SERM
appliziert wurde. In diesen Fragmenten zeigten sich allerdings auch sehr hohe
Schwankungen zwischen den einzelnen Messungen, so daß keine signifikante Veränderung
vorlag. Wie bereits in den morphologischen Untersuchungen festgestellt (Abb. 20) zeigten
sich keine Veränderungen in den transplantierten Fragmenten nach unterschiedlicher
Behandlung der Nacktmäuse. Dies traf auch auf die Höhe der Drüsenepithelzellen zu (Abb.
21).
Abb. 21: Epithelzellhöhen der endometrialen Drüsen aus den transplantierten humanen Fragmenten nach hormoneller Behandlung der Nacktmäuse. Die Epithelzellhöhen aus 4 verschiedenen Versuchsreihen wurden als Mittelwerte plus Standardabweichungen dargestellt. Es konnten keine signifikanten Veränderungen im Vergleich zum Vehikel detektiert werden. Excel-Diagramm.
63

Ergebnisse
4.3.2.3 Genexpression
Das Modell wurde zusätzlich genutzt, um eine mögliche Östrogenregulation der Gene Cyr61,
CTGF, Cx43, Cx26, bFGF und VEGF zu untersuchen (s. 4.3.2).
Cyr61
Es zeigte sich, daß die Behandlung der Nacktmäuse mit unterschiedlichen Hormonen
starken Einfluß hatte auf die Cyr61-Expression in den kultivierten humanen endometrialen
Fragmenten. Durch die Verabreichung des reinen Anti-Östrogens, das eine ähnliche Wirkung
hat wie ICI und die durch den Östrogenrezeptor induzierte Transkriptionsaktivität blockiert
(Hermenegildo und Cano, 2000), konnte die Expression des Cyr61 signifikant inhibiert
werden (Abb. 22A, Spur 4 und Abb. 22B). In sieben von zehn Versuchsreihen wurde in den
Fragmenten aus den mit Anti-Östrogen behandelten Tieren eine 2-6fach reduzierte
Expression im Vergleich zum Vehikel detektiert. In zwei Versuchsreihen dagegen war die
Expression in den mit Anti-Östrogen behandelten Fragmenten etwa vergleichbar mit der
Expression des Vehikels, in einer weiteren Versuchsreihe zeigte sich sogar eine 2fach
erhöhte Expression des Cyr61 im Anti-Östrogen behandelten Tier (Abb. 22C und D).
Durch die Applikation des Anti-Östrogens war es allerdings nicht möglich, die Expression von
Cyr61 vollständig zu inhibieren, eine schwache Genexpression war in den Fragmenten
immer nachweisbar (Abb. 22A, Spur 4). Auch konnte die den zyklischen Nacktmäusen
zusätzlich applizierte Menge an Östradiol die Expression von Cyr61 in den humanen
Fragmenten im Vergleich zum Vehikel nur in einem Fall um den Faktor 2,5 steigern, in den
übrigen Versuchsreihen entsprach sie in etwa dem Vehikel. Die Schwankungen der Cyr61-
Expression in den mit Östradiol behandelten Fragmenten waren vergleichsweise hoch (Abb.
22B).
Durch die Applikation des SERM wurde die Expression von Cyr61 in den humanen
Fragmenten ebenfalls signifikant reduziert. In den einzelnen Versuchen zeigte sich eine bis
zu 4fach geringere Expression in den mit SERM behandelten Fragmenten verglichen mit den
Kontrolltieren. Der Expressionslevel von Cyr61 war daher in den mit SERM behandelten
Fragmenten erniedrigt gegenüber dem Vehikel, jedoch leicht erhöht gegenüber den mit Anti-
Östrogen behandelten Fragmenten (Abb. 22B).
64

Ergebnisse
Abb. 22, A-F: Beeinflussung der Expression der CCN-Proteine Cyr61 (A-D) und CTGF (E und F)
von RT-PCR ermittelten mittleren Expressionswerte der hormonell behandelten Tiere wurden als
E und F: Die Expression von CTGF wurde in den transplantierten endometrialen Fragmenten durch
nt aus einem mit SERM behandelten Tier. Die Positionen der Transkripte von β-Aktin (698 bp), 197 bp) und CTGF (451 bp) wurden gekennzeichnet. B, D, F: Excel-Diagramme, *p≤0,05.
ehikel behandelten Fragmente ein ähnliches
Expressionsmuster von Cyr61 wie die eutopen Endometrien (Abb. 23A). Cyr61 wurde auch
hier im apikalen Bereich der endometrialen Drüsenepithelien detektiert, zusätzlich zeigte sich
allerdings auch eine schwache Expression im Stroma. Nach Behandlung der Nacktmäuse
mit einem Anti-Östrogen konnte Cyr61 dagegen kaum nachgewiesen werden (Abb. 23B).
in den transplantierten Fragmenten durch die Applikation verschiedener Hormone. Die mit Hilfe
xfache Expression bezogen auf das Vehikel plus Standardabweichungen dargestellt. A-D: Die Cyr61-Expression in den transplantierten Fragmenten wurde nach Applikation des Anti-Östrogens signifikant reduziert (A und B). In drei weiteren Versuchsreihen konnten diese Resultate nicht bestätigt werden (C und D).
die hormonelle Behandlung nicht verändert. A, C, E: Agarosegele, Spur 1: 100 bp-Marker, Spur 2: nicht transplantiertes Kontrollendometrium, Spur 3: Fragment aus einem mit Vehikel behandelten Tier, Spur 4: Fragment aus einem mit Anti-Östrogen behandelten Tier, Spur 5: Fragment aus einem mit Östradiol behandelten Tier, Spur 6: FragmeCyr61 (
Cyr61 schien in den transplantierten endometrialen Fragmenten trotz zwei Ausreißern
östrogenabhängig exprimiert zu werden, durch Behandlung der Nacktmäuse mit einem Anti-
Östrogen oder SERM wurde die Expression signifikant reduziert. Diese Reduktion konnte
auch immunhistochemisch nachgewiesen werden. Nach Behandlung der Fragmente mit
einem gegen Cyr61 gerichteten Antikörper und Nachweis durch einen Fluoreszenzfarbstoff-
markierten Sekundärantikörper zeigten die mit V
65

Ergebnisse
66
Abb. 23, A und B: Immunhistochemischer Nachweis von Cyr61 in transplantierten humanen Fragmenten nach hormoneller Behandlung der Nacktmäuse. In den mit Vehikel behandelten Tieren zeigte Cyr61 in den transplantierten Fragmenten ein ähnliches Expressionsmuster wie im eutopen Endometrium (A). In den Tieren, denen das Anti-Östrogen appliziert wurde, konnte Cyr61 kaum detektiert werden (B). A und B: Fluoreszenz. Balken: 50 µm.
CTGF
In den humanen endometrialen Fragmenten, die für 14 Tage in der Nacktmaus kultiviert
wurden, konnte CTGF nachgewiesen werden (Abb. 22E), eine Beeinflussung der Expression
durch die verabreichten Hormone war jedoch nicht gegeben. Keine der den Nacktmäusen
applizierten Substanzen konnte die Expression von CTGF in den endometrialen Fragmenten
verändern (Abb. 22F). Im Mittel entsprachen die Expressionswerte in den Fragmenten der
Transplantation in die Nacktmaus durch hormonelle Behandlung der Tiere beeinflußt
er
weiteren Versuchsreihe war Cx43 in den mit Östradiol behandelten Tieren stark
herunterreguliert (Abb. 24C und D). In diesen Fragmenten der beiden Versuchsreihen wurde
mit Anti-Östrogen, Östradiol oder SERM behandelten Tiere denen der Kontrolltiere. In den
einzelnen Versuchsreihen wurden bezüglich der CTGF-Expression kaum Schwankungen
beobachtet. Eine Regulation von CTGF im Endometrium durch Östrogen konnte nicht
nachgewiesen werden.
Cx43
Die Expression des östrogensensitiven Gap Junction-Proteins Connexin 43 (Cx43)
(Grümmer et al., 1999) konnte in den humanen endometrialen Fragmenten nach
werden. Es zeigte sich ein ähnliches Expressionsmuster wie bei Cyr61. In den Fragmenten
der mit Anti-Östrogen behandelten Nacktmäuse war die Expression des Cx43 in 7 von 9
Versuchsreihen bezogen auf die Expression in den Fragmenten der mit Vehikel behandelten
Tiere reduziert (Abb. 24A und B). In zwei dieser Versuchsreihen war Cx43 in den mit Anti-
Östrogen behandelten Tieren kaum nachweisbar, hier zeigte sich gegenüber dem Vehikel
eine bis zu 20fache Reduktion. Bedingt durch größere Schwankungen war die Reduktion
durch das Anti-Östrogen jedoch nicht signifikant (Abb. 24B).
In einem Fall wurde eine 2fach höhere Cx43-Expression in den mit Anti-Östrogen
behandelten Fragmenten detektiert als in den mit Vehikel behandelten Fragmenten, in ein

Ergebnisse
auch Cyr61 anders exprimiert als erwartet (Abb. 22D), d.h. es bleibt zu klären, wie diese
paradoxe Genexpression in zwei Versuchsreihen zustande kommen konnte.
Die den zyklischen Nacktmäusen zusätzlich applizierte Menge an Östradiol konnte auch die
Expression von Cx43 in den humanen Fragmenten im Vergleich zum Vehikel nicht signifikant
steigern (Abb. 24B). Ähnlich der Expression des Cyr61 wurde auch das Cx43 in den
Fragmenten der mit Östradiol behandelten Tiere in den einzelnen Versuchsreihen
zeigten sich unterschiedlich hohe Cx43-Expressionswerte in den mit SERM behandelten
nten, auch hier ergab sich dadurch eine sehr hohe Standardabweichung. Im Mittel
unterschiedlich stark exprimiert.
Die Behandlung der Nacktmäuse mit dem SERM hatte auf die Cx43-Expression in den
humanen endometrialen Fragmenten keinen Einfluß (Abb. 24B). In den einzelnen Versuchen
Fragme
entsprach die Expression von Cx43 in den mit SERM behandelten Fragmenten in etwa dem
Vehikel (Abb. 24B).
67

Ergebnisse
Abb. 24, A-F: Beeinflussung der Expression der Gap Junction-Proteine Cx43 (A-D) und Cx26 (E und F) in den transplantierten humanen Fragmenten durch die Applikation verschiedener Hormone. Die mit Hilfe von RT-PCR ermittelten mittleren Expressionswerte der hormonell behandelten Tiere wurden als xfache Expression bezogen auf das Vehikel plus Standardabweichungen dargestellt. A-D: Die Expression von Cx43 war in den Fragmenten der mit Anti-Östrogen behandelten Tiere am geringsten (A und B). In zwei weiteren Versuchsreihen konnten diese Resultate nicht bestätigt werden (C und D). E und F: Die Expression von Cx26 wurde in den transplantierten Fragmenten nicht durch die verabreichten Hormone beeinflußt. A, C, E: Agarosegele, Spur 1: 100 bp-Marker, Spur 2: nicht transplantiertes Kontrollendometrium, Spur 3: Fragment aus einem mit Vehikel behandelten Tier, Spur 4: Fragment aus einem mit Anti-Östrogen behandelten Tier, Spur 5: Fragment aus einem mit Östradiol behandelten Tier, Spur 6: Fragment aus einem mit SERM behandelten Tier. Die Positionen der Transkripte von β-Aktin (214 bp), Cx43 (811 bp) und Cx26 (556 bp) wurden gekennzeichnet. B, D, F: Excel-Diagramme.
Cx26
Obwohl bekannt ist, daß das Gap Junction-Protein Connexin 26 (Cx26) durch Östrogene
reguliert wird (Grümmer et al., 1999), konnten die den Nacktmäusen verabreichten Hormone
die Expression von Cx26 in den humanen endometrialen Fragmenten nicht beeinflussen
(Abb. 24E und F). Cx26 wurde in den Fragmenten der einzelnen Versuchsreihen unabhängig
von der verabreichten Substanz sehr unterschiedlich stark exprimiert. Dadurch entstanden
hohe Standardabweichungen, eine durch die applizierten Hormone verursachte Regulation
des Cx26 konnte nicht nachgewiesen werden (Abb. 24F). Im Mittel entsprachen die
Expressionslevel in den Fragmenten der mit Östradiol und Anti-Östrogen behandelten Tiere
dem der Kontrollfragmente. In den Fragmenten der Tiere, denen SERM appliziert wurde,
konnte eine 1,7fache Reduktion der Cx26-Expression detektiert werden, die statistisch
jedoch nicht signifikant war (Abb. 24F).
68

Ergebnisse
4.4 Lokale Östrogensynthese in ektopen endometriotischen Läsionen?
Durch real-time RT-PCR Analysen konnte gezeigt werden, daß Cyr61 im Laufe des
Menstruationszyklus in Abhängigkeit der auf das Endometrium einwirkenden Hormone
Östrogen und Progesteron unterschiedlich stark exprimiert wird (Abb. 11). Auch mit Hilfe des
Nacktmausmodells konnten Expressionsunterschiede nach Applikation eines reinen Anti-
Östrogens nachgewiesen werden (Abb. 22B). Zusätzlich wurde festgestellt, daß Cyr61 in
endometriotischen Läsionen sehr viel stärker exprimiert wird als im eutopen Endometrium
(Abb. 10). Diese Regulation war möglicherweise unabhängig von den im Ovar synthetisierten
Steroidhormonen. Zudem wird von Zeitoun und Bulun (1999) eine lokale Östrogensynthese
in den endometriotischen Läsionen durch eine abweichende Expression von Aromatase und
ein Ungleichgewicht der ebenfalls an der Östrogensynthese beteiligten Enzyme 17β-HSD1
und 17β-HSD2 postuliert. Zur Überprüfung dieser Theorie wurde die Expression von
Aromatase sowie von 17β-HSD1 und 17β-HSD2 in den Endometrien von Frauen mit und
ohne Endometriose und in endometriotischen Läsionen untersucht.
Aus Endometrien von Frauen mit und ohne Endometriose sowie aus endometriotischen
isten (Enantone-Gyn, Takeda) behandelt wurden. Die
gungen im Thermocycler durchgeführt, und die Bandenstärke der PCR-Produkte
(Gelscan Professional V4.0). Die Signalstärke der Zielgene
jeweiligen
Zyklusphase als Mittelwerte zusammengefaßt.
Läsionen wurde Gesamt-RNA isoliert und mittels reverser Transkription cDNA synthetisiert
(s. 3.3.4). Zusätzlich wurden 3 endometriotische Läsionen von Frauen untersucht, die über 6
Monate mit einem GnRH-Agon
anschließende PCR wurde dreimal mit jeder Gewebeprobe nach den in Tab. 1 angegebenen
PCR-Bedin
wurde densitometrisch bestimmt
konnte nach Abzug des Hintergrunds in Relation zum β-Aktin-Signal derselben Probe
gesetzt werden. Die relativen Expressionswerte wurden jeweils für die Endometrien von
Frauen mit und ohne Endometriose sowie für die endometriotischen Läsionen der
4.4.1 Aromatase-Expression
Transkripte der Aromatase konnten in keinem der untersuchten Gewebe gezeigt werden
(Abb. 25). Insgesamt wurden jeweils 4 Endometrien von Frauen mit und ohne Endometriose
sowie 3 endometriotische Läsionen aus der Proliferationsphase und jeweils 4 Endometrien
von Frauen mit und ohne Endometriose sowie 4 endometriotische Läsionen aus der
Sekretionsphase untersucht. Nur im Ovar, das als Kontrollgewebe genutzt wurde, wurde
Aromatase exprimiert, in den untersuchten Endometrien und endometriotischen Läsionen
konnte keine Aromatase-Expression detektiert werden (Abb. 25). Die Abb. 25 zeigt als
Beispiel die Resultate einiger getesteter Gewebe.
69

Ergebnisse
Abb. 25: Durch RT-PCR ermittelte Expression von Aromatase, dargestellt als Agarosegel. Nur im Ovar-Gewebe (Spur 2) konnte Aromatase nachgewiesen werden. In normalen Endometrien (Spur
endometriotischen Läsionen (Spur 11: Sekretionsphase, Spur 12-14: Proliferationsphase) konnte keine Aromatase-Expression detektiert werden. Spur 1: 100 bp-Marker. Die Positionen der Transkripte von β-Aktin (698 bp) und Aromatase (376 bp) wurden gekennzeichnet.
4.4.2 Expression von 17β-Hydroxysteroid Dehydrogenase Typ 1
17β-HSD1 wird in der Proliferationsphase in den Endometrien von normalen Frauen (n=4) in
etwa gleich stark exprimiert wie in den Endometrien von Endometriosepatientinnen der
gleichen Zyklusphase (n=4) (Abb. 26A). Es konnten also keine
3-6, Sekretionsphase), Endometrien von Endometriosepatientinnen (Spur 7-10, Sekretionsphase) und
endometriosebedingten
SD1 in den endometriotischen Läsionen der
Sekretionsphase (n=3) allerdings im Mittel um den Faktor 2 stärker exprimiert als in den
eutopen Endometrien von Endometriosepatientinnen (Abb. 26B). Diese Hochregulation war
Die Expression von 17β-HSD1 war also in den Endometrien sowohl von gesunden Frauen
als auch von Frauen mit Endometriose zyklusabhängig exprimiert. Die im allgemeinen
geringe Expression wurde in den Endometrien der Sekretionsphase zusätzlich
herunterreguliert. In den endometriotischen Läsionen dagegen war 17β-HSD1 in der
Sekretionsphase sehr viel stärker exprimiert als in den endometriotischen Läsionen aus der
Proliferationsphase. Die zusätzlich untersuchten endometriotischen Läsionen der
Patientinnen, deren ovarielle Östrogensynthese durch einen GnRH-Agonisten für 6 Monate
blockiert wurde, zeigten ein ähnliches Expressionsmuster wie die endometriotischen
Unterschiede in der Expression von 17β-HSD1 detektiert werden. In den endometriotischen
Läsionen der Proliferationsphase (n=5) dagegen war das Enzym sehr schwach exprimiert,
verglichen mit dem eutopen Endometrium von Endometriosepatientinnen war die Expression
signifikant um den Faktor 3,5 reduziert (Abb. 26A).
Obwohl in allen untersuchten Geweben die Expression von 17β-HSD1 nur sehr gering war,
war die Expression des Enzyms in den Endometrien aus der Sekretionsphase im Mittel
weiter um den Faktor 2 reduziert, verglichen mit den Endometrien aus der
Proliferationsphase (Abb. 26A und B). Auch in den Geweben aus der Sekretionsphase
konnte kein Unterschied in der 17β-HSD1-Expression zwischen normalen Endometrien (n=6)
und den Endometrien von Frauen mit Endometriose (n=6) detektiert werden (Abb. 26B).
Anders als in der Proliferationsphase war 17β-H
jedoch statistisch nicht signifikant.
70

Ergebnisse
Läsionen aus der Proliferationsphase. Auch hier wurde das Enzym 17β-HSD1 nur sehr
schwach exprimiert (Abb. 26C).
und endometriotischen Läsionen aus der Proliferationsphase (A und D) und der
Läsionen von Patientinnen, die über 6 Monate mit einem GnRH-Agonisten behandelt wurden, konnten
Abb. 26, A-F: Expression von 17β-HSD1 (A-C) und 17β-HSD2 (D-F) in endometrialen Geweben
Sekretionsphase (B und E). Mittels RT-PCR konnten keine Expressionsunterschiede zwischen normalen Endometrien und Endometrien von Endometriosepatientinnen detektiert werden, weder für 17β-HSD1 noch für 17β-HSD2. In den endometriotischen Läsionen der Sekretionsphase zeigte sich eine Unregelmäßigkeit in der Expression der beiden Enzyme (B und E). In endometriotischen
beide Enzyme kaum detektiert werden (C und F). A-F: Die mittleren Expressionslevel plus Standardabweichungen wurden als Excel-Diagramme dargestellt, *p≤0,05.
71

Ergebnisse
4.4.3 Expression von 17β-Hydroxysteroid Dehydrogenase Typ 2
17β-HSD2 ist ein weiteres Enzym, das an der Synthese von Östradiol beteiligt ist. Auch
bezogen auf die Expression von 17β-HSD2 konnten keine signifikanten Unterschiede
zwischen den Endometrien von Frauen mit und ohne Endometriose detektiert werden, weder
in der Proliferationsphase (jeweils n=4) (Abb. 26D) noch in der Sekretionsphase (jeweils
n=6) (Abb. 26E). Allerdings wurde das Enzym erwartungsgemäß im Laufe des
Menstruationszyklus unterschiedlich stark exprimiert. In der Sekretionsphase war der
Expressionslevel im Mittel um den Faktor 1,5 höher als in der Proliferationsphase.
In den endometriotischen Läsionen beider Zyklusphasen (Proliferationsphase: n=7,
Sekretionsphase: n=3) konnte 17β-HSD2 kaum nachgewiesen werden und war bezogen auf
das eutope Endometrium von Frauen mit Endometriose signifikant herunterreguliert (26D
und E). Vergleiche der Expressionsstärke von 17β-HSD2 in endometriotischen Läsionen der
Proliferationsphase und der Sekretionsphase unbehandelter Frauen mit der Expression in
den endometriotischen Läsionen von Frauen, die mit einem GnRH-Agonisten behandelt
wurden, ergaben, daß 17β-HSD2 in den Läsionen der GnRH behandelten Frauen fast nicht
detektierbar war (Abb. 26F).
Durch diese Untersuchungen wurde ein Ungleichgewicht der beiden Enzyme 17β-HSD1 und
17β-HSD2 in den endometriotischen Läsionen der Sekretionsphase detektiert. Während 17β-
HSD1 in den Läsionen exprimiert wurde, war 17β-HSD2 kaum detektierbar (Abb. 26B und
D). Das Enzym Aromatase konnte in keinem der untersuchten Gewebe nachgewiesen
werden (Abb. 25).
4.4.4 Expression von Cyr61 in GnRH behandelten endometriotischen Läsionen
Es sollte zudem überprüft werden, ob sich die Expression von Cyr61 in endometriotischen
Läsionen verändert, wenn die Frauen über einen Zeitraum von 6 Monaten mit einem GnRH-
Agonisten behandelt wurden, der die ovarielle Östrogensynthese inhibiert. Es wurde daher
die Expression von Cyr61 in endometriotischen Läsionen aus GnRH behandelten Frauen mit
er Expression in hormonell unbehandelten endometriotischen Läsionen aus der
Proliferationsphase und der Sekretionsphase verglichen. Diese Versuche wurden mit Hilfe
für die behandelten
Läsionen (n=3) als Mittelwerte zusammengefaßt. Als Calibrator wurde der Expressionslevel
d
der real-time RT-PCR durchgeführt. Wie für die endometrialen Gewebe beschrieben (s.
3.4.2.2 und 4.1.1.2) wurde die relative Expressionsstärke von Cyr61 in den
endometriotischen Läsionen gegen β-Aktin der jeweils gleichen Probe abgeglichen und die
relativierten CT-Werte anschließend für die unbehandelten Läsionen aus der
Proliferationsphase (n=5) und der Sekretionsphase (n=4) sowie
72

Ergebnisse
der hormonell unbehandelten endometriotischen Läsionen aus der Proliferationsphase
genutzt und hier als 1 gesetzt. Der xfache Expressionslevel der unbehandelten Läsionen aus
der Sekretionsphase und der GnRH behandelten endometriotischen Läsionen bezogen auf
die unbehandelten Läsionen aus der Proliferationsphase wurde durch Subtraktion des
Calibrators und Einsetzen des erhaltenen Wertes in die Formel 2 T berechnet.
Läsionen
beobachtet werden (Abb. 27). Auch nach der Blockierung der ovariellen Östrogensynthese
war das östrogenabhängige Gen Cyr61 in diesen Läsionen genauso stark exprimiert wie in
den unbehandelten Läsionen aus der Proliferationsphase. Nur in den unbehandelten
Abb. 27: Expression von Cyr61 in endometriotischen Läsionen von Patientinnen, deren
Unterschied in der Expression von Cyr61 verglichen mit hormonell unbehandelten endometriotischen
-∆∆C
Obwohl Cyr61 in den unbehandelten endometriotischen Läsionen sehr viel stärker exprimiert
war als in den eutopen Endometrien (Abb. 10), konnte in den GnRH behandelten
endometriotischen Läsionen kein signifikanter Unterschied zu den unbehandelten
endometriotischen Läsionen aus der Sekretionsphase wurde Cyr61 um den Faktor 2,5
stärker exprimiert (Abb. 27), hier wurde aber auch ein sehr viel stärkerer Anstieg in der
Expression verglichen mit den eutopen Endometrien detektiert als in der Proliferationsphase
(Abb. 10). Obwohl kein Östrogen durch die Ovarien produziert wurde, zeigte sich auch in den
GnRH behandelten endometriotischen Läsionen eine hohe Cyr61-Expression (Abb. 27).
ovarielle Östrogensynthese für 6 Monate durch einen GnRH-Agonisten blockiert wurde. Der mittlere Expressionslevel der unbehandelten Läsionen aus der Sekretionsphase sowie der GnRH-behandelten Läsionen bezogen auf die unbehandelten Läsionen aus der Proliferationsphase wurde ohne Standardabweichung dargestellt. In den GnRH-behandelten Läsionen zeigte sich kein
Läsionen aus der Proliferationsphase. In den unbehandelten Läsionen aus der Sekretionsphase wurde Cyr61 dagegen sehr viel stärker exprimiert. Excel-Diagramm.
73

Ergebnisse
4.5 Angiogenese
In „Gene-Array“ Analysen wurde VEGF im Endometrium von Endometriosepatientinnen der
Sekretionsphase als hochreguliertes Gen identifiziert (Tab. 3). Daher wurde die
4.5.1 Expression angiogenetischer Faktoren im eutopen Endometrium
Expression
der angiogenetischen Faktoren VEGF und bFGF im humanen eutopen Endometrium von
Frauen mit und ohne Endometriose untersucht.
die
Bandenstärke der PCR-Produkte wurde densitometrisch bestimmt (Gelscan Professional
V4.0). Die Signalstärke von bFGF und VEGF konnte nach Abzug des Hintergrunds in
Relation zum β-Aktin-Signal derselben Probe gesetzt werden. Die relativen
Expressionswerte wurden jeweils für die Endometrien von Frauen mit und ohne
Endometriose der jeweiligen Zyklusphase als Mittelwerte zusammengefaßt.
Aus den „Gene-Array“ Analysen ergab sich, daß VEGF in 10 von 20 Fällen in den
Endometrien von Endometriosepatientinnen in der Sekretionsphase hochreguliert war. Diese
Ergebnisse konnten nicht bestätigt werden. In den von uns mittels RT-PCR untersuchten
ewebeproben konnte kein signifikanter Unterschied in der Expression von VEGF zwischen
wurde in den Endometrien von Endometriosepatientinnen aus der Sekretionsphase (n=5)
ebenfalls nicht stärker exprimiert als in den Endometrien von Frauen ohne Endometriose der
gleichen Zyklusphase (n=5) (Abb. 28D). In der Proliferationsphase konnten ebenfalls keine
Unterschiede in der bFGF-Expression zwischen den verschiedenen Geweben (jeweils 9
Endometrien von Frauen mit und ohne Endometriose) festgestellt werden (Abb. 28C).
Allerdings wurden im Verlauf des Menstruationszyklus Expressionsunterschiede der beiden
Angiogenese-Faktoren im Endometrium detektiert. Sowohl VEGF (Abb. 28A, B) als auch
Jeweils 5 Endometrien von normalen Frauen und Endometriosepatientinnen aus der
Sekretionsphase wurden auf die Expression von VEGF und bFGF mittels RT-PCR getestet.
Aus der Proliferationsphase wurde der Expressionslevel von VEGF in jeweils 10
unterschiedlichen Endometrien von Frauen mit und ohne Endometriose untersucht, die
Expression von bFGF wurde in jeweils 9 verschiedenen Endometrien von gesunden Frauen
und Endometriosepatientinnen aus der Proliferationsphase getestet. Aus den einzelnen
Geweben wurde Gesamt-RNA isoliert und mittels Reverser Transkription in cDNA
umgeschrieben (s. 3.3.4). Die anschließenden PCR wurde dreimal mit jeder Gewebeprobe
im Thermocycler durchgeführt (Primer und PCR-Bedingungen, s. Tab. 1), und
G
den Kontrollendometrien (n=5) und den Endometrien von Endometriosepatientinnen (n=5) in
der Sekretionsphase detektiert werden (Abb. 28B). Auch in Geweben aus der
Proliferationsphase entsprach die VEGF-Expression in den Endometrien von
Endometriosepatientinnen (n=10) der Expression in den Kontrollendometrien (n=10) (Abb.
28A). Ähnliche Resultate wurden auch in bezug auf die bFGF-Expression erzielt. bFGF
74
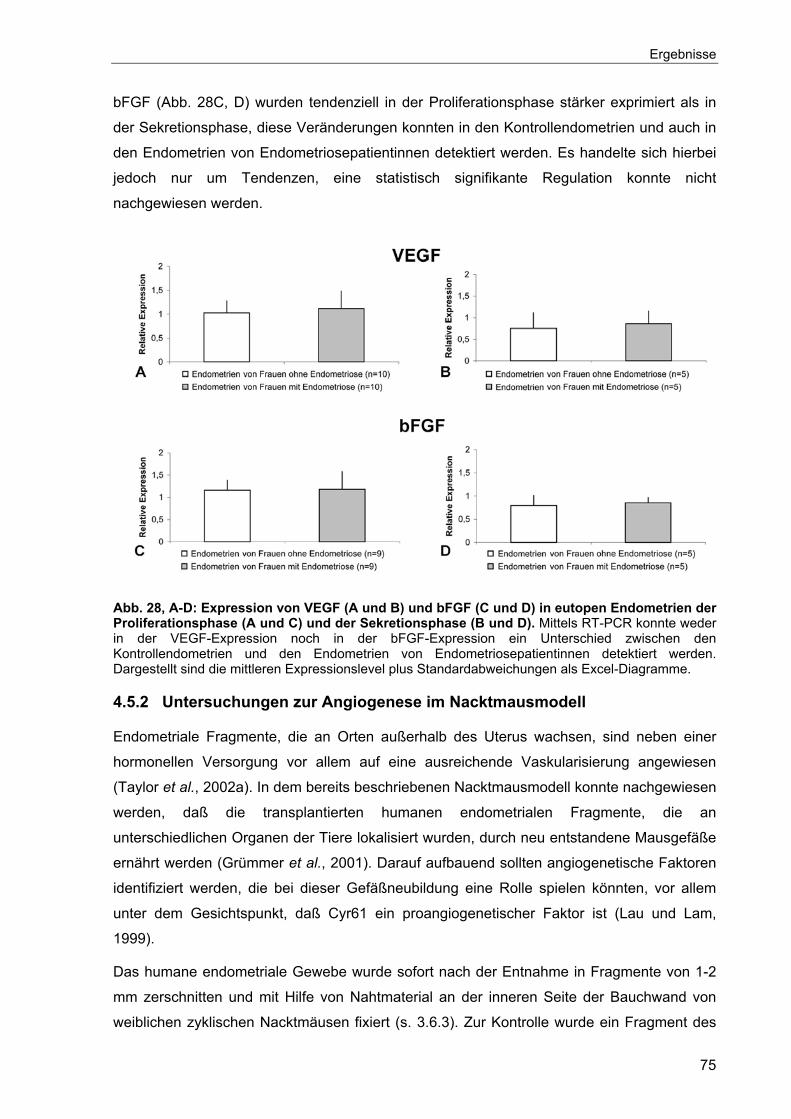
Ergebnisse
bFGF (Abb. 28C, D) wurden tendenziell in der Proliferationsphase stärker exprimiert als in
der Sekretionsphase, diese Veränderungen konnten in den Kontrollendometrien und auch in
den Endometrien von Endometriosepatientinnen detektiert werden. Es handelte sich hierbei
Endometriale Fragmente, die an Orten außerhalb des Uterus wachsen, sind neben einer
hormonellen Versorgung vor allem auf eine ausreichende Vaskularisierung angewiesen
(Taylor et al., 2002a). In dem bereits beschriebenen Nacktmausmodell konnte nachgewiesen
werden, daß die transplantierten humanen endometrialen Fragmente, die an
unterschiedlichen Organen der Tiere lokalisiert wurden, durch neu entstandene Mausgefäße
ernährt werden (Grümmer et al., 2001). Darauf aufbauend sollten angiogenetische Faktoren
identifiziert werden, die bei dieser Gefäßneubildung eine Rolle spielen könnten, vor allem
unter dem Gesichtspunkt, daß Cyr61 ein proangiogenetischer Faktor ist (Lau und Lam,
1999).
on 1-2
mm zerschnitten und mit Hilfe von Nahtmaterial an der inneren Seite der Bauchwand von
weiblichen zyklischen Nacktmäusen fixiert (s. 3.6.3). Zur Kontrolle wurde ein Fragment des
jedoch nur um Tendenzen, eine statistisch signifikante Regulation konnte nicht
nachgewiesen werden.
Abb. 28, A-D: Expression von VEGF (A und B) und bFGF (C und D) in eutopen Proliferationsphase (A und C) und der Sekretionsphase (B und D). Mittels RT-P
Endometrien der CR konnte weder
in der VEGF-Expression noch in der bFGF-Expression ein Unterschied zwischen den Kontrollendometrien und den Endometrien von Endometriosepatientinnen detektiert werden. Dargestellt sind die mittleren Expressionslevel plus Standardabweichungen als Excel-Diagramme.
4.5.2 Untersuchungen zur Angiogenese im Nacktmausmodell
Das humane endometriale Gewebe wurde sofort nach der Entnahme in Fragmente v
75

Ergebnisse
Ausgangsgewebes direkt in flüssigem Stickstoff gefroren und bis zum Ende des Versuchs
bei -80°C gelagert. Für die Untersuchungen zur Angiogenese wurde das Gewebe von 3
unterschiedlichen Endometriosepatientinnen jeweils auf 4 Nacktmäuse verteilt, pro Tier
wurden 4-6 endometriale Fragmente transplantiert. Die Mäuse blieben während des
Versuchszeitraums unbehandelt. In einer Zeitreihe wurde jeweils 2, 4, 7 und 14 Tage nach
Transplantation der humanen endometrialen Fragmente eine Maus getötet. Die Fragmente
wurden entnommen und entweder zur Isolierung von Gesamt-RNA direkt in flüssigem
Stickstoff gefroren oder für immunhistochemische Untersuchungen in Tissue Tek (OCT
Compound Sakura) in flüssigem Stickstoff gefroren.
4.5.2.1 Zeitlicher Verlauf der Angiogenese
Zur Untersuchung, nach welcher Zeit das Einwachsen neu gebildeter Mausgefäße von der
Bauchwand in die hier fixierten humanen endometrialen Fragmente beginnt, wurden Maus-
Endothelzellen immunhistochemisch nachgewiesen (s. 3.7.1).
Zwei Tage nach der Transplantation konnten noch keine von der Bauchwand der Nacktmaus
ausgehenden Blutgefäße in den humanen Fragmenten nachgewiesen werden (Abb. 29B,
Ausschnittsvergrößerung). Im Wirtsgewebe allerdings, das dem transplantierten Fragment
anliegt, wurden zahlreiche Gefäße detektiert. Bereits nach 4 Tagen konnten dann auch im
Transplantat Maus-Endothelzellen in einwachsenden Gefäßen beobachtet werden (Abb.
9D, Ausschnittsvergrößerung). Abhängig von der Größe des transplantierten Fragments
Anteil eingewachsene Mausgefäße detektiert werden (Abb. 29H, Ausschnittsvergrößerung).
2
invadierten die Gefäße 7 Tage nach Versuchsbeginn zunächst den Transplantatanteil, der
der Bauchwand anlag, oder sie durchsetzten bereits das ganze Fragment (Abb. 29F,
Ausschnittsvergrößerung). Nach 14 Tagen konnten in allen Fragmenten auch im innersten
76

Ergebnisse
Abb. 29: Immunhistochemischer Nachweis von Maus-Endothelzellen in transplantierten humanen endometrialen Fragmenten nach 2 Tagen (A und B), 4 Tagen (C und D), 7 Tagen (E und F) und 14 Tagen (G und H). Erst nach 4 Tagen konnten Mausgefäße in den humanen Fragmenten nachgewiesen werden (D, Ausschnitt), nach 7 Tagen waren die Fragmente bereits vollständig mit Gefäßen durchsetzt (F, Ausschnitt). A, C, E, G: Toluidinblau-Färbung; B, D, F, H: immunhistochemische Färbung. BW: Bauchwand der Nacktmaus, F: transplantiertes humanes Fragment, N: Fixierungsnaht. Balken: in A-H: 160 µm, in den Ausschnittsvergrößerungen von B, D und H: 200 µm, in der Ausschnittsvergrößerung von F: 50 µm.
77

Ergebnisse
4.5.2.2 Angiogenetische Faktoren im Nacktmausmodell
Zusätzlich sollten in diesen Versuchsansätzen die wichtigsten angiogenetischen Faktoren
untersucht werden, die bei der Gefäßneubildung in den transplantierten endometrialen
Fragmenten eine Rolle spielen könnten. Mittels RT-PCR wurde die Expression von bFGF,
VEGF, Cyr61 und die Expression je einer der Rezeptoren für bFGF und VEGF (bFGF-R-1
und KDR) in den Fragmenten in einer Zeitreihe untersucht. Nach Beendigung der Versuche
wurden die Fragmente entnommen und sofort in flüssigem Stickstoff gefroren. Dabei wurde
darauf geachtet, daß kein Bauchwandgewebe der Nacktmaus mit entfernt wurde.
Aus den transplantierten Fragmenten und dem Ausgangsgewebe, das nicht transplantiert
worden ist, wurde Gesamt-RNA isoliert, der komplette Ansatz einem DNase-Verdau
unterzogen und in die RT-Reaktion eingesetzt. Die anschließende PCR wurde wegen der
geringen Ausbeute an RNA nur einmal mit jedem Fragment im Thermocycler durchgeführt
(Primer und PCR-Bedingungen, s. Tab. 1), und die Bandenstärke der PCR-Produkte wurde
densitometrisch mit Hilfe des Programms Gelscan Professional V4.0 bestimmt. Die
Signalstärke des Zielgens konnte nach Abzug des Hintergrunds in Relation zum β-Aktin-
Signal derselben Probe gesetzt werden. Die relativen Expressionswerte des
Ausgangsgewebes wurden für jede der drei Versuchsreihen als Kontrolle genutzt und als 1
gesetzt (Tag 0). Die relativen Expressionswerte in den Fragmenten der Tiere, die nach 2, 4,
7 oder 14 Tagen getötet wurden, wurden in jeder Versuchsreihe gegen das jeweilige
Ausgangsgewebe der gleichen Versuchsreiche abgeglichen. Da in der ersten Versuchsreihe
die Expression aller Gene im Ausgangsgewebe sehr gering war, erfolgte der Abgleich hier
gegen den Mittelwert aller drei Ausgangsgewebe. Die Expressionslevel der Gene wurden
nach Abgleich gegen das Ausgangsgewebe für die drei Versuchsreihen als Mittelwerte
zusammengefaßt und als xfache Expression bezogen auf das Ausgangsgewebe dargestellt.
Cyr61/bFGF/VEGF
Alle drei angiogenetischen Faktoren, Cyr61, bFGF und VEGF, konnten sowohl im humanen
Endometrium, das als Ausgangsgewebe für die Versuche im Nacktmausmodell diente (Tag
0), als auch in den transplantierten endometrialen Fragmenten nachgewiesen werden (Abb.
30A). Die Neubildung der Blutgefäße in der Bauchwand der Nacktmaus beginnt vermutlich
am zweiten Tag nach der Transplantation der endometrialen Fragmente (Abb. 29B und D).
Zur gleichen Zeit veränderte sich auch die Expression der getesteten Angiogenese-Faktoren
). Cyr61 wurde im Mittel 1,6fach
in den endometrialen Fragmenten. Bei einsetzender Angiogenese in den humanen
Fragmenten in der Nacktmaus wurden vermehrt die angiogenetischen Faktoren Cyr61 und
bFGF exprimiert. Ein deutlicher Anstieg der Expression beider Faktoren konnte am Tag 2
nach der Transplantation in den endometrialen Fragmenten verglichen mit dem
Ausgangsgewebe (Tag 0) detektiert werden (Abb. 30A
78

Ergebnisse
stärker exprimiert verglichen mit dem Ausgangsgewebe, die Expression von bFGF stieg im
geringsten
auch die der Rezeptoren. Die Expression der Gene nach 2, 4, 7 und 14 Tagen wurden als xfache
Mittel um den Faktor 1,5 an. Auch im weiteren Versuchsverlauf blieb Cyr61 von den drei
getesteten angiogenetischen Faktoren der am stärksten exprimierte. Die Expression von
Cyr61 nahm im weiteren Versuchsverlauf ab und war an Tag 14 auf den Wert des
Ausgangsgewebes abgefallen (Abb. 30A). Im Vergleich dazu hatte die bFGF-Expression
bereits nach 7 Tagen den gleichen Level erreicht wie das Ausgangsgewebe, an Tag 14
wurde eine 1,4fach reduzierte Expression verglichen mit Tag 0 nachgewiesen (Abb. 30A).
Die Expression von VEGF zeigte dagegen in den endometrialen Fragmenten die
Veränderungen der getesteten Faktoren. Eine stärkere Expression von VEGF verglichen mit
dem Ausgangsgewebe konnte weder zu Beginn noch während des weiteren
Versuchsverlaufs beobachtet werden. Am Tag 2 wurde die gleiche Expressionsstärke
detektiert wie an Tag 0, im weiteren Verlauf nahm die Expression ab, bis an Tag 14 1,5fach
weniger VEGF exprimiert wurde als im Ausgangsgewebe (Abb. 30A). Wegen hoher
Schwankungen in den einzelnen Versuchsreihen konnten keine statistisch signifikanten
Expressionsänderungen nachgewiesen werden.
Während der Neubildung von Maus-Blutgefäßen und dem Einwachsen dieser Gefäße in die
transplantierten humanen endometrialen Fragmente wurde in diesen Fragmenten eine
Zunahme der angiogenetischen Faktoren Cyr61 und bFGF detektiert. Im weiteren
Kultivierungsverlauf zeigte Cyr61 eine verstärkte Expression bis zu 14 Tagen.
Abb. 30: Expression der angiogenetischen Faktoren bFGF, VEGF und Cyr61 (A) sowie der Rezeptoren bFGF-R-1 und KDR (B) in transplantierten humanen endometrialen Fragmenten nach 2, 4, 7 und 14 Tagen Kultivierung in der Nacktmaus. Zu Beginn der Angiogenese an Tag 2 nach der Transplantation veränderte sich sowohl die Expression der angiogenetischen Faktoren als
Expression bezogen auf das Ausgangsgewebe (Tag 0) plus Standardabweichungen dargestellt. A und B: Excel-Diagramme.
bFGF-R/VEGF-R
Die Expression der Rezeptoren bFGF-R-1 für bFGF und KDR für VEGF wurde in diesen
Versuchsansätzen ebenfalls untersucht. Auch bFGF-R-1 und KDR konnten sowohl im
humanen endometrialen Ausgangsgewebe (Tag 0) als auch in den transplantierten
endometrialen Fragmenten nachgewiesen werden (Abb. 30B). Am Tag 2 veränderten sich
79

Ergebnisse
neben den Expressionswerten der getesteten Angiogenese-Faktoren auch die
Expressionswerte der entsprechenden Rezeptoren. Bei beginnender Angiogenese in den
humanen Fragmenten in der Nacktmaus wurde vermehrt der untersuchte Rezeptor für VEGF
exprimiert. Ein deutlicher Anstieg der Expression von KDR konnte am Tag 2 nach der
Transplantation verglichen mit dem Ausgangsgewebe (Tag 0) nachgewiesen werden (Abb.
30B). KDR wurde in den Fragmenten im Mittel 2fach stärker exprimiert als im
Ausgangsgewebe. Im Versuchsverlauf reduzierte sich zwar die Expression des Rezeptors,
4.5.2.3 Hormonelle Regulation von VEGF und bFGF
Da Cyr61 als proangiogenetischer Faktor bekannt ist und durch bFGF induziert wird, wurde
in diesem Modell auch die hormonelle Regulation von VEGF und bFGF in den kultivierten
endometrialen Fragmenten untersucht (s. 4.3.2).
Die Expression von VEGF in den humanen endometrialen Fragmenten konnte durch die
Applikation des Östradiols im Mittel um den Faktor 1,5 gegenüber den Kontrollfragmenten
duziert werden (Abb. 31B), diese Reduktion war statistisch jedoch nicht signifikant. Die
s VEGF-Expressionslevels war wegen hoher Schwankungen in den
sreihen jedoch statistisch nicht signifikant. Zusammengefaßt bewirkten die
den Nacktmäusen applizierten östrogenen und anti-östrogenen Hormone nur tendenziell
eine Veränderung der VEGF-Expression in den humanen endometrialen Fragmenten.
in drei Versuchsreihen eine bis zu 2fache Erhöhung der bFGF-Expression in den
an Tag 14 konnte jedoch noch die gleiche Expressionsmenge nachgewiesen werden wie an
Tag 0 (Abb. 30B). Die Expression des untersuchten Rezeptors für bFGF wurde dagegen in
den endometrialen Fragmenten nach der Transplantation kaum verändert. An Tag 2 konnte
keine Steigerung der Expression von bFGF-R-1 gefunden werden, im Mittel entsprach dieser
dem Ausgangsgewebe. Auch im weiteren Versuchsverlauf zeigte sich keine wesentliche
Veränderung im Vergleich zum Ausgangsgewebe (Abb. 30B). Wegen hoher Schwankungen
in den einzelnen Versuchsreihen konnten keine statistisch signifikanten
Expressionsänderungen detektiert werden.
Wie für die angiogenetischen Faktoren bFGF und VEGF konnten bei einsetzender
Angiogenese auch Veränderungen in der Expression der Rezeptoren bFGF-R-1 und KDR
detektiert werden. Es zeigte sich, daß im weiteren Versuchsverlauf vor allem der untersuchte
Rezeptor für VEGF stark exprimiert wurde.
re
Verabreichung von Anti-Östrogen führte zu keiner Beeinflussung der VEGF-Expression im
Vergleich zum Vehikel (Abb. 31B). Die Behandlung der Nacktmäuse mit dem SERM bewirkte
dagegen in den vier einzelnen Versuchsreihen eine bis zu 4fache Herunterregulation der
VEGF-Expression in den endometrialen Fragmenten verglichen mit dem Vehikel (Abb. 31B).
Diese Reduktion de
einzelnen Versuch
Die Behandlung der zyklischen Nacktmäuse mit einer zusätzlichen Menge Östradiol bewirkte
80

Ergebnisse
endometrialen Fragmenten verglichen mit dem Vehikel, in den übrigen vier Fällen konnte
keine Beeinflussung nachgewiesen werden, die Änderung des Expressionslevels war
statistisch nicht signifikant. In den Fragmenten der mit Anti-Östrogen behandelten Tiere war
die Expression von bFGF im Mittel knapp 1,5fach reduziert verglichen mit den
Kontrollfragmenten (Abb. 31D). Auch diese Expressionsänderung war nicht signifikant, es
konnte jedoch in fünf von sechs Versuchsreihen eine Reduktion der bFGF-Expression
nachgewiesen werden. Durch die Applikation des SERM wurde die Expression von bFGF in
den endometrialen Fragmenten nicht beeinflußt, im Mittel entsprach sie der Expression in
den mit Vehikel behandelten Fragmenten (Abb. 31D). Zusammenfassend läßt sich sagen,
daß die Expression von bFGF in den humanen endometrialen Fragmenten durch die
Behandlung der Nacktmäuse mit Hormonen nicht signifikant verändert wurde. Eine
Regulation durch Östrogen war zu erkennen, da in fast allen Versuchsreihen nach
Applikation des Anti-Östrogens die bFGF-Expression leicht reduziert war. Die zusätzliche
Östradiol-Gabe konnte die bFGF-Expression in den Fragmenten verglichen mit dem Vehikel
dagegen leicht steigern. Diese Resultate konnten statistisch nicht abgesichert werden, da
hohe Schwankungen in den einzelnen Versuchsreihen vorhanden waren.
und bFGF (C und D) in den transplantierten Fragmenten durch Applikation verschiedener
signifikant verändert.
Östrogen behandelten Tier, Spur 5: Fragment aus einem mit Östradiol behandelten Tier, Spur 6:
Abb. 31, A-D: Beeinflussung der Expression der angiogenetischen Faktoren VEGF (A und B)
Hormone. Die mit Hilfe von RT-PCR ermittelten mittleren Expressionswerte der hormonell behandelten Tiere wurden als xfache Expression bezogen auf das Vehikel plus Standardabweichungen dargestellt. A und B: Die VEGF-Expression wurde in den Fragmenten durch die Behandlung der Tiere nicht
C und D: In den mit Anti-Östrogen behandelten Tieren zeigte sich eine Reduktion der bFGF-Expression gegenüber dem Vehikel. A und C: Agarosegele, Spur 1: 100 bp-Marker, Spur 2: nicht transplantiertes Kontrollendometrium, Spur 3: Fragment aus einem mit Vehikel behandelten Tier, Spur 4: Fragment aus einem mit Anti-
Fragment aus einem mit SERM behandelten Tier. Die Positionen der Transkripte von β-Aktin (698 bp), VEGF (341 bp) und bFGF (313 bp) wurden gekennzeichnet. B und D: Excel-Diagramme.
81

Ergebnisse
4.5.2.4 Behandlung der Nacktmäuse mit einem VEGF-Antagonisten
Da die Vaskularisierung für die Persistenz der transplantierten humanen endometrialen
Fragmente eine große Rolle zu spielen scheint, sollten nach Inhibition der Angiogenese die
Auswirkungen auf die Adhäsion und die Morphologie der Fragmente untersucht werden. Bei
dem den Nacktmäusen applizierten VEGF-Antagonisten handelte es sich um einen Inhibitor
der Tyrosinkinase-Aktivität der beiden VEGF-Rezeptoren flt und KDR. Durch diese Inhibition
sollte exprimiertes VEGF keine Rolle bei der Bildung neuer Blutgefäße spielen können.
Das endometriale Gewebe wurde sofort nach der Entnahme in Fragmente von 1-2 mm
zerschnitten und mit Hilfe von Nahtmaterial an der inneren Seite der Bauchwand von
weiblichen zyklischen Nacktmäusen fixiert (s. 3.6.3). Zur Inhibition der Angiogenese wurde
das Gewebe von 3 unterschiedlichen Patientinnen ohne Endometriose in zwei
Versuchsreihen auf 2 Nacktmäuse, in einem Fall auf drei Nacktmäuse, verteilt, pro Tier
wurden 6 endometriale Fragmente transplantiert. Über einen Zeitraum von 14 Tagen wurde
den Mäusen täglich 1,5 mg eines VEGF-Antagonisten (Schering, ZK 202650) p.o.
verabreicht. In jeder Versuchsreihe wurde einer Maus zeitlich parallel das Vehikel (s. 3.6.4)
appliziert. Die Fragmente wurden nach Beendigung des Versuchs entnommen und für
immunhistochemische Färbungen in Tissue Tek (OCT Compound Sakura) in flüssigem
Stickstoff gefroren oder für morphologische Untersuchungen in 10 % Formalin fixiert und in
Paraffin eingebettet.
Morphologie
Durch diese Untersuchungen sollten mögliche morphologische Veränderungen in den
Fragmenten der mit dem VEGF-Antagonisten behandelten Tiere detektiert werden. Durch
die Behandlung der Nacktmäuse mit dem VEGF-Antagonisten wurden in diesen Fragmenten
allerdings in keiner der drei Versuchsreihen derartige morphologische Veränderungen im
Vergleich zu den Kontrolltieren beobachtet. Die Abbildung 32 zeigt als Beispiel jeweils ein
Fragment aus den mit dem VEGF-Antagonisten behandelten Tieren (C) und ein Fragment
aus den Kontrolltieren (A). Das Drüsenepithel des behandelten Fragments war abgeflacht,
ie Drüsen insgesamt waren zystisch vergrößert (Abb. 32C). Ein ähnliches Bild zeigte sich
wurden.
d
auch in den Kontrollfragmenten (Abb. 32A), zusätzlich konnten zahlreiche Blutgefäße
nachgewiesen werden (Abb. 32A, Ausschnittsvergrößerung, Pfeile). Sowohl in den
behandelten Fragmenten als auch in den Fragmenten aus den Kontrolltieren adhärierte das
Stroma an die Bauchwand der Nacktmaus, nicht das Drüsenepithel. Es konnten keine
morphologischen Veränderungen beobachtet werden, die durch die Behandlung mit dem
VEGF-Antagonisten hervorgerufen
82

Ergebnisse
Abb. 32, A-D: Morphologie (A und C) und immunhistochemischer Nachweis (B und D) von Maus-Endothelzellen in transplantierten humanen endometrialen Fragmenten nach Behandlung mit dem Vehikel (A und B) oder einem VEGF-Antagonisten (C und D). Trotz der Inhibition von VEGF konnten keine morphologischen Unterschiede in den behandelten Fragmenten (C) im Vergleich zur Kontrolle (A) detektiert werden. Auch auf die Menge der eingewachsenen Maus-Blutgefäße (Pfeil in Ausschnittsvergrößerung A) hatte die Inhibition von VEGF keinen Einfluß (B und
-Färbung, B und D: immunhistochemische Färbung. Balken: in A, C, D: 160 µm, in B: 50 µm, in der Ausschnittsvergrößerung von A: 100 µm. D). A und C: HE
Angiogenese in den transplantierten Fragmenten
Die Wirkung des VEGF-Antagonisten wurde auch durch die Menge der neu gebildeten und
in die Fragmente eingewachsenen Maus-Blutgefäße durch immunhistochemischen
Nachweis von Maus-Endothelzellen untersucht (s. 3.7.1).
Durch den immunhistochemischen Nachweis von Maus-Endothelzellen konnte in den
Kontrollfragmenten gezeigt werden, daß die transplantierten humanen endometrialen
Fragmente von Blutgefäßen der Maus durchsetzt sind (Abb. 32B). Auch in den Fragmenten
der mit dem VEGF-Antagonisten behandelten Tiere wurden Mausgefäße detektiert (Abb.
32D). Trotz Inhibition von VEGF konnte Angiogenese stattfinden und die behandelten
Fragmente waren ebenso durchdrungen mit eingewachsenen Blutgefäßen wie die
Fragmente aus den Vehikel-Tieren. In keinem der untersuchten Fragmente aus den drei
Versuchsreihen konnte nach Applikation des VEGF-Antagonisten eine Reduktion der
83

Ergebnisse
Angiogenese detektiert werden. Auffällige Unterschiede zu den Fragmenten der mit Vehikel
behandelten Tiere wurden nicht beobachtet.
84

Diskussion
5 Diskussion
5.1 Veränderung des Genmusters der Endometrien der Sekretionsphase von Endometriosepatientinnen
Die molekularen Mechanismen der Pathogenese von Endometriose sind bis heute ungeklärt.
Viele Hinweise wie die hohe Rezidivrate der Krankheit (Wheeler und Malinak, 1983) oder die
Korrelation zwischen Endometriose und Infertilität (Ledger, 1999) sprechen allerdings dafür,
daß die Ursachen der Entstehung endometriotischer Läsionen in einem nicht adäquat
differenzierten Endometrium liegen. Daher wurden Genexpressionsprofile der Endometrien
von Frauen mit und ohne Endometriose erstellt, um Gene zu identifizieren, die im
Endometrium von Endometriosepatientinnen abweichend vom normalen Endometrium
exprimiert werden. Mit Hilfe dieser Gene könnte es möglich sein, Einblicke in den
Entstehungsmechanismus der Krankheit zu bekommen und mögliche Kandidatengene für
therapeutische Ansätze zu identifizieren. Für diese Versuche wurden „Gene-Arrays“ der
Firma Affymetrix benutzt, mit denen es möglich ist, gleichzeitig über 12000 Gene zu
untersuchen. Es zeigte sich, daß einige Gene, von denen bekannt ist, daß ihre Transkription
durch Östrogen reguliert wird, im Endometrium von Endometriosepatientinnen aus der
Sekretionsphase stärker exprimiert waren als im Endometrium von gesunden Frauen der
gleichen Zyklusphase. Zu den in der Sekretionsphase hochregulierten östrogenabhängigen
Genen gehörten der TR3 Orphan-Rezeptor (Uemura et al., 1995), das „early growth
response protein“ EGR-1 (Thompson et al., 2002), sowie die Transkriptionsfaktoren c-fos
(Duan et al., 2002) und jun-B (Cicatiello et al., 1992). Ähnliche Ergebnisse wurden auch in
der Studie von Lundeen et al. (2002, eingereicht) erzielt. Mittels „Differential Display“-PCR
wurde auch hier das „early growth response-1 gene“ (EGR-1) im Endometrium von
Endometriosepatientinnen als hochreguliertes Gen identifiziert, diese
Expressionsunterschiede wurden allerdings für die Proliferationsphase nachgewiesen.
Zudem konnte in ektopen endometriotischen Läsionen ein zusätzlicher Expressionsanstieg
des untersuchten Gens verglichen mit dem eutopen Endometrium detektiert werden
(Lundeen et al., 2002, eingereicht).
In den Studien von Taylor et al. (2002b) zeigten wie in unserer Studie hormonregulierte
Gene ein abweichendes Expressionsmuster in den Endometrien von
Endometriosepatientinnen. Mittels „subtractive cDNA hybridization“ wurde Glycodelin-A als
ein spezifisches Genprodukt des sekretorischen Endometriums identifiziert (Vaisse et al.,
1990), was durch spätere „Gene-Array“ Analysen bestätigt werden konnte (Giudice et al.,
2001; Kao et al., 2002). Es zeigte sich, daß Glycodelin-A im gesunden Endometrium in der
Sekretionsphase am stärksten exprimiert wird, im Endometrium von
Endometriosepatientinnen dagegen war die Expression von Glycodelin-A im sekretorischen
Endometrium verglichen mit normalen Endometrien reduziert (Giudice et al., 2001; Kao et
85

Diskussion
al., 2002). Die genaue Funktion des Glycodelin-A ist nicht bekannt, es wird jedoch postuliert,
daß dieses Protein für die Rezeptivität des Endometriums bei der Implantation eine wichtige
Genexpression bei der Entstehung von endometrialen Karzinomen wurde beobachtet, daß
liferationsphase nachgewiesen. Die Ursache dafür liegt
Rolle spielt (Mueller et al., 2000). Auch in Untersuchungen zur Veränderung der
Gene, die im normalen Endometrium hormonabhängig exprimiert werden, in endometrialen
Adenokarzinomen ein abweichendes Expressionsprofil aufwiesen (Mutter et al., 2001). Die
Genmuster der Tumore glichen eher dem des normalen Endometriums aus der
Proliferationsphase als dem Endometrium aus der Sekretionsphase (Mutter et al., 2001).
Das normale Endometrium ist ein sehr dynamisches Gewebe, dessen Phänotyp sich
während des Menstruationszyklus in Abhängigkeit der zirkulierenden Hormone Östrogen und
Progesteron ständig ändert. Diese zyklischen Veränderungen unterliegen der direkten
hormonellen Steuerung durch das Ovar. Da sich die Serumhormonwerte von Frauen mit und
ohne Endometriose nicht unterscheiden, scheint es, als ob das Endometrium von
Endometriosepatientinnen nicht adäquat auf vorhandenes Östrogen reagieren würde.
Dadurch wäre es auch nicht möglich, im Endometrium die Voraussetzungen für die
Implantation eines Keims zu schaffen, wodurch sowohl die verminderte Fertilität von
Endometriosepatientinnen (Garrido et al., 2002) als auch die geringe Implantationsrate von
Endometriosepatientinnen nach in vitro Fertilization (Barnhart et al., 2002) erklärt werden
könnte. Des weiteren könnte das Menstruationssekret durch den Einfluß von Östrogen in der
Sekretionsphase vitale Zellen enthalten, die nach retrograder Menstruation an das
umgebende Peritoneum adhärieren und Läsionen bilden könnten (Ridley und Edwards,
1958).
Das in den Array-Analysen am stärksten regulierte Gen war Cyr61, das in den Endometrien
von Endometriosepatientinnen der Sekretionsphase in 90 % der Fälle im Mittel 5fach
hochreguliert war. Durch Validierung der Array-Analysen mittels real-time RT-PCR konnten
diese Ergebnisse bestätigt werden. Abweichend von den Array-Analysen wurde in diesen
Versuchen aber auch eine signifikant höhere Expression von Cyr61 in den Endometrien von
Endometriosepatientinnen in der Pro
möglicherweise in der Auswertung der verwendeten Arrays. Um die Anzahl falsch-positiver
und falsch-negativer Resultate möglichst gering zu halten, wurde jede einzelne
Gewebeprobe auf einem separaten Array untersucht. Dadurch sollte sichergestellt werden,
daß gemessene Unterschiede zwischen den einzelnen Arrays tatsächlich durch
Unterschiede in den Genexpressionsmustern hervorgerufen werden, nicht durch
verschiedene Hintergrundsignale. Dennoch können falsche Ergebnisse nicht vollständig
ausgeschlossen werden, so daß es immer nötig ist, die identifizierten Regulationen in
weiteren Proben zu verifizieren (Cook und Rosenzweig, 2002). Zudem wurde bekannt, daß
die spezifischen Sequenzen der untersuchten Gene auf den Arrays teilweise fehlerhaft sind.
86

Diskussion
In einer Charge von Arrays zur Untersuchung von mausspezifischen Genen waren bis zu 30
% der Sequenzen falsch, durchschnittlich sind auf einem Array zwischen 1 % und 5 % der
verwendeten Sequenzen nicht korrekt (Knight, 2001). Auch aus diesen Gründen ist es
notwendig, die durch die Array-Analysen erhaltenen Resultate zu überprüfen. Dies zeigte
sich auch bei einigen Genen, die nach den Array-Analysen im Endometrium von
Endometriosepatientinnen aus der Sekretionsphase herunterreguliert waren. Mittels RT-PCR
konnten die Ergebnisse für die Gene Nidogen, p27, Fibronektin, Elastin und Kollagen IV in
zusätzlichen Geweben nicht verifiziert werden, abweichend von den Array-Analysen konnten
Expressionsstärke von Cyr61 im Laufe des Menstruationszyklus verändert. Während
der Proliferationsphase, wenn vorwiegend Östrogen im Endometrium vorhanden ist, wurden
höhere Expressionswerte von Cyr61 detektiert als in der Sekretionsphase, wenn vor allem
Progesteron auf das Endometrium wirkt. Nur in normalen Endometrien war diese Regulation
signifikant, in Endometrien von Endometriosepatientinnen dagegen konnte nur eine Tendenz
beobachtet werden, keine signifikante Reduktion. Dies ist ein weiterer Hinweis darauf, daß
das Endometrium von Endometriosepatientinnen möglicherweise nicht adäquat auf
zirkulierendes Östrogen reagiert. Derartige Expressionsunterschiede während des Zyklus
konnten in Untersuchungen des Myometriums dagegen nicht beobachtet werden (Sampath
et al., 2001a). Zwar wurde in myometrialen Fragmenten, die ex vivo mit Östrogen behandelt
wurden, bereits innerhalb einer Stunde die Cyr61-Expression als „immediate early gene“ um
den Faktor 2 gesteigert, Unterschiede zwischen Geweben aus der Proliferationsphase und
der Sekretionsphase wurden dagegen nicht nachgewiesen (Sampath et al., 2001a). Im
Endometrium wurde im Rahmen dieser Studie eine umgekehrte Reaktion beobachtet. In
endometrialen Fragmenten ex vivo konnte innerhalb einer Stunde keine Steigerung der
Cyr61-Expression durch Zugabe von Östrogen induziert werden (Daten nicht gezeigt),
keine Expressionsunterschiede zwischen normalen Endometrien und Endometrien von
Endometriosepatientinnen nachgewiesen werden. Ähnliche Beobachtungen wurden auch in
anderen Studien gemacht, nicht immer ließen sich alle durch Array-Analysen identifizierten
Genregulationen mit Hilfe anderer Methoden verifizieren (Cook et al., 2002).
5.2 Regulation der Cyr61-Expression im Endometrium durch Östrogen
In früheren Studien wurde bereits gezeigt, daß es sich bei Cyr61 um ein östrogenreguliertes
Gen handelt. In den Uteri von ovariektomierten Ratten wurde Cyr61 innerhalb kürzester Zeit
nach Zugabe von 17β-Östradiol bis zu 10fach stärker exprimiert (Rivera-Gonzales et al.,
1989). Auch im Myometrium sowie in humanen Brustkrebszellen wurde Cyr61 als ein Gen
identifiziert, das östrogenabhängig exprimiert wird (Sampath et al., 2001a, b). Im Rahmen
dieser Doktorarbeit konnte gezeigt werden, daß die Expression von Cyr61 auch im humanen
Endometrium durch Östrogen reguliert wird. Erste Hinweise lieferte die Beobachtung, daß
sich die
87

Diskussion
während sich im Laufe des Menstruationszyklus in Abhängigkeit der zirkulierenden Hormone
die Expression änderte. Möglicherweise wird Cyr61 daher im Endometrium anders reguliert
als im Myometrium, die genauen Regulationsmechanismen von Cyr61 im Endometrium oder
im Myometrium sind nicht bekannt.
Cyr61 scheint wegen des erhöhten Expressionslevels in den Endometrien von
Endometriosepatientinnen zum einen ein Markergen für Endometriose zu sein, zum anderen
könnte es auch eine Rolle bei der Pathogenese spielen. Darauf deutet die zusätzliche
signifikante Hochregulation der Expression von Cyr61 die in den endometriotischen Läsionen
beider Zyklusphasen hin. In der Sekretionsphase war der Expressionslevel in den
endometriotischen Läsionen im Mittel 20fach erhöht gegenüber den eutopen Endometrien, in
der Proliferationsphase wurde im Mittel ein 4facher Anstieg detektiert. Da es sich bei Cyr61,
wie bereits erwähnt, um ein östrogenreguliertes Gen handelt, könnte dieser zusätzliche
e
in diesen Geweben zusammenhängen. In früheren Studien wurde beschrieben, daß in den
Drüsenepithelzellen von endometriotischen Läsionen dagegen konnte 17 -HSD2 nicht
nachgewiesen werden (Zeitoun et al., 1998). Diese Fehlregulation in den endometriotischen
Läsionen soll zusammen mit der sonst im Endometrium nicht vorhandenen Aromatase lokal
zur Synthese von Östrogen führen (Zeitoun und Bulun, 1999).
daß ein methodischer Fehler ausgeschlossen werden konnte. Eine lokale Synthese von
Expressionsanstieg in den endometriotischen Läsionen mit einer lokalen Östrogensynthes
endometriotischen Läsionen aufgrund der abweichenden Expression von Aromatase lokal
Östrogen synthetisiert wird (Zeitoun und Bulun, 1999). Das Enzym Aromatase, das die
Umwandlung von C19-Steroiden zu Östrogen katalysiert, wird in verschiedenen Geweben
exprimiert wie im Ovar, in der Plazenta, im Fettgewebe, in der Haut oder auch im Gehirn
(Simpson et al., 1994). Im eutopen Endometrium dagegen ist Aromatase nur sehr schwach
exprimiert oder in den meisten Fällen nicht detektierbar (Noble et al., 1996, 1997). 17β-HSD
1 und 2 sind zwei weitere Enzyme, die an der Synthese von Östrogen beteiligt sind. 17β-
HSD1 katalysiert die Umwandlung des nur sehr schwach östrogen wirkenden Östron zum
wirksamen Östradiol in den Granulosazellen des Ovars. Die Expression von 17β-HSD1
konnte sowohl im eutopen Endometrium als auch in endometriotischen Läsionen während
der Proliferationsphase und der Sekretionsphase nachgewiesen werden (Zeitoun et al.,
1998). 17β-HSD2 wurde in den Drüsenepithelzellen des eutopen Endometriums verstärkt
während der Sekretionsphase detektiert und spielt vermutlich eine Rolle bei der Inaktivierung
des Östradiols in dieser Phase (Casey et al., 1994, Mustonen et al., 1998). In den
β
Diese Resultate konnten im Rahmen dieser Studie nur teilweise bestätigt werden. Die
Expression von Aromatase wurde weder in eutopen Endometrien von Frauen mit und ohne
Endometriose noch in ektopen endometriotischen Läsionen nachgewiesen. Nur im humanen
Ovar-Gewebe, das als Kontrolle verwendet wurde, konnte Aromatase detektiert werden, so
88

Diskussion
Östrogen wäre daher in den endometriotischen Läsionen nicht möglich, da aufgrund der
fehlenden Aromatase-Expression zirkulierendes Androstendion nicht zu Östron umgewandelt
werden könnte. Allerdings wurde in unserer Studie wie bei Zeitoun et al. (1998) eine
unausgewogene Expression der Enzyme 17β-HSD 1 und 2 in den endometriotischen
utopen Endometrien von
Läsionen der Sekretionsphase beobachtet. Während 17β-HSD1 in den endometriotischen
Läsionen nachgewiesen werden konnte, war es kaum möglich, das Enzym 17β-HSD2 zu
detektieren. Zirkulierendes Östron könnte also in den Läsionen durch 17β-HSD1 zu
wirksamem Östradiol umgewandelt werden, der Abbau des Östradiol durch 17β-HSD2
dagegen wäre kaum effektiv. Diese abweichende Enzym-Expression könnte die Synthese
von Östradiol lokal in den endometriotischen Läsionen der Sekretionsphase begünstigen und
damit eine Ursache für die extrem starke Expression des östrogenregulierten Gens Cyr61
sein. In den endometriotischen Läsionen aus der Proliferationsphase dagegen konnte eine
derart abweichende Expression von Enzymen, die an der Östrogensynthese beteiligt sind,
nicht beobachtet werden. Zwar wurde auch in den endometriotischen Läsionen aus der
Proliferationsphase 17β-HSD2 nicht exprimiert, allerdings konnte auch nur eine sehr
schwache Expression von 17β-HSD1 nachgewiesen werden. Zirkulierendes Östron könnte
also in den endometriotischen Läsionen der Proliferationsphase kaum zu wirksamem
Östradiol umgewandelt werden, so daß das Fehlen der 17β-HSD2 hier kaum eine Rolle
spielt. Dennoch ist auch in endometriotischen Läsionen der Proliferationsphase Cyr61
stärker, wenn auch schwächer als in der Sekretionsphase, exprimiert als in den eutopen
Endometrien von Endometriosepatientinnen der gleichen Zyklusphase. Möglicherweise sind
neben der hormonellen Regulation weitere Faktoren wie die Wachstumsfaktoren bFGF oder
TGF-β ausschlaggebend für die höhere Expression des Gens in diesem Gewebe (Lau und
Lam, 1999) (s. 5.5). Dafür würde auch das Expressionsmuster sprechen in
endometriotischen Läsionen aus Patientinnen, deren ovarielle Östrogensynthese für 6
Monate durch Behandlung mit einem GnRH-Agonisten blockiert wurde. Vergleicht man die
Expression von Cyr61 in diesen Läsionen mit der Expression in hormonell unbehandelten
endometriotischen Läsionen, zeigt sich, daß die ektopen Läsionen der Frauen, die mit dem
GnRH-Agonisten behandelt wurden, den gleichen Expressionslevel von Cyr61 aufweisen wie
unbehandelte ektope Läsionen aus der Proliferationsphase. Unabhängig davon, daß die
ovarielle Östrogensynthese blockiert wurde, wurde in diesen Läsionen dennoch eine starke
Cyr61-Expression beobachtet, stärker als in den e
Endometriosepatientinnen. Diese Expressionsrate wurde vermutlich jedoch sowohl in den
unbehandelten Läsionen der Proliferationsphase als auch in den mit GnRH behandelten
Läsionen nicht durch eine lokale Östrogensynthese hervorgerufen. Denn die
Enzymausstattung der mit GnRH behandelten Läsionen war ähnlich wie die der Läsionen
aus der Proliferationsphase. Sowohl 17β-HSD1 als auch 17β-HSD2 konnten in den
89

Diskussion
behandelten Läsionen kaum nachgewiesen werden. Daher wäre es auch in den mit GnRH
behandelten Läsionen nicht möglich, zirkulierendes Östron in wirksames Östradiol
umzuwandeln. Auch hier könnte eine Regulation durch Wachstumsfaktoren für die starke
Expression von Cyr61 verantwortlich sein (s. 5.5).
Mit Hilfe eines etablierten Nacktmausmodells (Grümmer et al., 2001) sollte der Einfluß von
Östrogen auf die Expression des Cyr61 und die Entwicklung endometriotischer Läsionen
näher untersucht werden. Für einen Zeitraum von 14 Tagen bietet dieses Modell die
Möglichkeit verschiedene Aspekte der Endometriose zu untersuchen, da die transplantierten
humanen endometrialen Fragmente ihre morphologischen Charakteristika und
Differenzierungsparameter beibehalten (Grümmer et al., 2001). Weiterhin ist es in diesem
Modell möglich, den Einfluß von Hormonen auf die transplantierten Fragmente und deren
Genexpressionsmuster zu untersuchen. Daher wurde den Nacktmäusen nach der
Transplantation der Fragmente entweder ein reines Anti-Östrogen, Östradiol oder ein
selektiver Östrogenrezeptor-Modulator (SERM) appliziert und die Effekte der Substanzen auf
die Morphologie der Fragmente und die Genexpression, vor allem der Expression des Cyr61,
ermittelt.
Anti-Östrogene sind Substanzen, die die Funktion des Östrogens blockieren können. Nach
MacGregor und Jordan (1998) werden zwei verschiedene Typen von Anti-Östrogenen
unterschieden, Typ I und Typ II. Das in dieser Studie verabreichte SERM gehört zu den Anti-
Östrogenen des Typ I, die gewebespezifisch und speziesspezifisch sowohl östrogene als
auch anti-östrogene Wirkung haben können. Typ I Anti-Östrogene sind kompetetive
Inhibitoren der Östrogenbindung an seinen Rezeptor. Für Raloxifen konnte bereits gezeigt
werden, daß diese Verbindungen scheinbar einen Komplex mit dem Östrogenrezeptor
eingehen, wodurch dieser durch unvollendete Konformationsänderungen des Komplexes
teilweise aktiviert bleibt (Brzozowski et al., 1997). Die biologischen Funktionen dieser
Substanzen sind abhängig von der Spezies, dem Gewebe oder den untersuchten Zielgenen
(Bryant und Dere, 1998). Die zweite Gruppe der Anti-Östrogene, zu denen auch das in
dieser Studie verabreichte Anti-Östrogen zählt, sind die reinen Anti-Östrogene (Typ II). Reine
Anti-Östrogene wurden bereits Mitte der 1980er Jahre entdeckt (Wakeling und Bowler,
1987), sie besitzen keinerlei östrogene Eigenschaften. Auch die Typ II Anti-Östrogene
agieren als kompetetive Inhibitoren des Östrogen/Rezeptor-Komplexes, und es konnte
gezeigt werden, daß beide Östrogenrezeptoren ERα und ERβ inhibiert werden (Barkhem et
al., 1998; Tremblay et al., 1998). Weiterhin wurde für das reine Anti-Östrogen ICI 182780
nachgewiesen, daß es in der Lage ist, neu synthetisierten Östrogenrezeptor im Zytoplasma
zu binden und damit dessen Transport in den Nukleus zu verhindern. Der unwirksame
5.3 Validierung des Nacktmausmodells für funktionelle Untersuchungen
90

Diskussion
Rezeptor wird dann wahrscheinlich durch Lysosomen zerstört (Davois et al., 1993). Der
Unterschied zwischen reinen Anti-Östrogenen und SERMs liegt also darin, daß SERMs die
Menge an Östrogenrezeptor im Nukleus anhäufen, während reine Anti-Östrogene sie
verringern (Devin-Leclerc et al., 1998).
Voraussetzung für den Einfluß der applizierten Hormone auf die transplantierten humanen
endometrialen Fragmente war die Expression von Östrogenrezeptoren auch nach der
In allen endometrialen Fragmenten konnte auch nach 14 Tagen Kultivierung in der
Nacktmaus der Östrogenrezeptor ERα nachgewiesen werden, ERβ dagegen wurde nur in
den nicht-transplantierten Ausgangsgeweben detektiert. Es zeigte sich allerdings keine
Regulation des Rezeptors in Abhängigkeit der verabreichten Hormone. Grümmer et al.
gegen konnten den Östrogenrezeptor ERα immunhistochemisch nur in zyklischen
Nacktmäusen nachweisen. Auch nach 14 Tagen wurde ERα sowohl in Drüsenepithelzellen
Expression und der Regulation der Östrogenrezeptoren ER und ER zwischen eutopem
Endometrium und ektopischen Läsionen wurden schon in anderen Studien beschrieben.
Matsuzaki et al. (2000a) konnten mittels in situ Hybridisierung und RT-PCR in allen
untersuchten endometriotischen Läsionen des Ovars ERα detektieren, ERβ dagegen nur in
63 % bzw. in Abhängigkeit der Methode nur in 47 % der untersuchten Läsionen. In allen
eutopen Endometrien dagegen wurden beide Östrogenrezeptoren nachgewiesen, wobei ein
höherer Expressionslevel von ERα detektiert wurde als von ERβ (Matsuzaki et al., 2000a, b).
et al.
Sekretionsphase. Eine solche Veränderung in Abhängigkeit des hormonellen Status konnte
in den endometriotischen Läsionen des Ovars dagegen nicht beobachtet werden (Matsuzaki
et al., 2000b). Die genaue Funktion beider Östrogenrezeptoren ist bis heute nicht vollständig
geklärt, es konnte allerdings gezeigt werden, daß beide Isoformen verschiedene
Transplantation in die Nacktmaus. Die Hauptaufgabe der Östrogenrezeptoren ist die
Aktivierung der Expression bestimmter Gene (Dhingra 1999). Nach der Bindung von
Östrogen an den Rezeptor kommt es zur Phosphorylierung und Dimerisierung des
Rezeptors (Lieberman, 1997; Fawell et al., 1990). Dieses Rezeptor-Dimer bindet an
„estrogen response elements“, eine Palindrome Sequenz aus 13 bp, die sich upstream des
Transkriptionsstarts von östrogenregulierten Genen befindet (Klein-Hitpass et al., 1986).
(2001) da
als auch in Stromazellen exprimiert. In ovariektomierten Mäusen konnten dagegen keine
Östrogenrezeptoren nachgewiesen werden (Grümmer et al., 2001). Unterschiede in der
α β
Diese Ergebnisse konnten in einer weiteren Studie, in der die Expression der
Östrogenrezeptoren mittels real-time RT-PCR untersucht wurden, bestätigt werden
(Matsuzaki , 2000b). Zudem wurde nachgewiesen, daß sich nur im eutopen
Endometrium die Expression beider Östrogenrezeptoren während des Menstruationszyklus
änderte, in der Proliferationsphase wurde eine stärkere Expression nachgewiesen als in der
91

Diskussion
Expressionsmuster in fetalen Organen, einschließlich des Uterus und der Ovarien, aufweisen
und möglicherweise unterschiedliche, organspezifische Funktionen erfüllen (Brandenberger
et al., 1998). Zudem wurde nachgewiesen, daß ERα und ERβ in den einzelnen Organen
sowohl zusammen als auch gegeneinander wirken können, abhängig von der Bildung von
rus
Homodimeren und/oder Heterodimeren zwischen den Rezeptor-Isoformen (Paech et al.,
1997; Ogawa et al., 1998; Watanabe et al., 1998). Die genaue Rolle von ERβ oder einer
Fehlregulation des Rezeptors bei der Entstehung endometriotischer Läsionen ist bis heute
nicht geklärt (Matsuzaki et al., 2000a).
Durch den Nachweis des Östrogenrezeptors ERα konnte gewährleistet werden, daß die
verschiedenen Hormone und Anti-Hormone auf die transplantierten Fragmente einwirken
können. Zusätzlich sollte in verschiedenen Vorversuchen sichergestellt werden, daß die den
Nacktmäusen s.c. verabreichten Substanzen tatsächlich das Zielorgan, nämlich den Ute
erreichen und entsprechend beeinflussen. Dafür wurden zum einen die Ovar- und
Uterusgewichte der Nacktmäuse bestimmt (Lan und Katzenellenbogen, 1976), zum anderen
wurden die Zellhöhe und die Proliferationsaktivität des Uterusepithels der Mäuse untersucht.
Es konnte gezeigt werden, daß das Uterusgewicht durch das applizierte Östrogen signifikant
gesteigert und durch das Anti-Östrogen signifikant reduziert wurde. Nebenwirkungen wurden
nicht beobachtet, da das Gewicht der Ovarien durch die Hormone nicht verändert wurde. Die
den Tieren verabreichten Substanzen gelangten also ins System der Nacktmaus und zeigten
die erwarteten Effekte am Zielorgan, dem Uterus.
Diese Resultate konnten durch die Messungen der Uterusepithel-Höhen allerdings nur
teilweise bestätigt werden. Nach Applikation des Anti-Östrogens zeigte sich eine signifikante
Reduktion der Zellhöhe des Uterusepithels im Vergleich zu den mit Vehikel behandelten
Tieren. Allerdings konnte in den Uteri der zyklischen Tiere durch die Verabreichung
zusätzlicher Mengen Östradiol keine Steigerung der Zellhöhe erreicht werden. Statt dessen
zeigte sich auch in den mit Östradiol behandelten Tieren eine leichte, nicht signifikante
Reduktion der Uterusepithel-Höhen. Ähnliche Beobachtungen wurden auch bei der
Bestimmung der Proliferationsraten nach immunhistochemischem Nachweis von Ki67 in den
Uterusepithelien der hormonell behandelten Nacktmäuse gemacht, hier zeigten sich
ebenfalls keine einheitlichen Ergebnisse. In einer Versuchsreihe wurde erwartungsgemäß
die stärkste Proliferation im Uterus der mit Östradiol behandelten Nacktmaus und keine Ki67-
Färbung im Uterus des mit Anti-Östrogen behandelten Tieres detektiert. In einer weiteren
Versuchsreihe allerdings zeigte sich keine Proliferationsaktivität im Uterusepithel der mit
Östradiol behandelten Nacktmaus, die stärkste Ki67-Färbung wurde in diesem Fall im Uterus
der mit Anti-Östrogen behandelten Maus beobachtet. In diesem Modell war daher weder die
Höhe noch die Proliferationsaktivität der Uterusepithelien ein zuverlässiger östrogener
92

Diskussion
Marker. Wie diese entgegengesetzten Ergebnisse zustande kamen, konnte nicht geklärt
werden. Möglicherweise war die Verwendung von zyklischen Nacktmäusen verantwortlich für
diese Resultate, mögliche Ursachen werden im folgenden näher diskutiert.
In diesem Nacktmausmodell wurde auch die Genexpression in den transplantierten
humanen endometrialen Fragmenten bezüglich der Östrogenregulation untersucht. Zur
Kontrolle wurde die Expression der beiden Gap Junction-Proteine Cx43 und Cx26 in
Abhängigkeit der applizierten Hormone getestet, die als östrogenregulierte Markergene des
humanen Endometriums bekannt sind (Jahn et al., 1995). In den meisten Geweben zeigen
Beeinflussung der Cx43-Expression durch SERMs konnte zudem nicht nachgewiesen
werden (Heikaus et al., 2002). In unserer Studie konnten diese Beobachtungen nur teilweise
bestätigt werden. Die Expression von Cx43 wurde in 7 von 9 Versuchsreihen in den
transplantierten humanen endometrialen Fragmenten durch das den Nacktmäusen
verabreichte Anti-Östrogen im Mittel um den Faktor 2 im Vergleich zum Vehikel
herunterreguliert. Allerdings wurden in den einzelnen Versuchsreihen starke Schwankungen
Im Gegensatz zu anderen Studien war es zudem nicht möglich, morphologische
Unterschiede in den kultivierten Fragmenten nachzuweisen, die auf die applizierten Hormone
zurückzuführen waren. Bereits 1985 untersuchten Bergquist et al. den Einfluß von
Steroidhormonen auf die Morphologie subkutan transplantierter humaner endometrialer
Fragmente in der Nacktmaus und wiesen nach, daß keines der Fragmente histologisch mit
dem Ausgangsgewebe übereinstimmte. Veränderungen des Stromas und des
Drüsenepithels in den Fragmenten ließen darauf schließen, daß diese auf eine hormonelle
Behandlung ähnlich wie eutopes Endometrium reagierten (Bergquist et al., 1985). Auch
Nisolle et al (2000a) konnten histologische Veränderungen der endometrialen Fragmente
nach Transplantation in die Nacktmaus beobachten. In allen Fragmenten zeigten sich nach
der Transplantation morphologische Charakteristika der Proliferationsphase, unabhängig
Cx-Gene eine wenig regulierte Expression, im Uterus allerdings, insbesondere im
Myometrium (Petrocelli und Lye, 1993; Lye et al., 1993) aber auch im Endometrium
(Grümmer und Winterhager, 1998), werden sie durch Steroidhormone reguliert. In früheren
Studien konnte bereits für das Ratten-Endometrium gezeigt werden, daß die Expression von
Cx26 sehr sensitiv durch Östrogen beeinflußt wird (Grümmer et al., 1999). Auch nach
Zugabe von SERMs veränderte sich der Expressionslevel von Cx26 (Heikaus et al., 2002).
Die Regulation von Cx43 durch Östrogen ist weniger sensitiv (Grümmer et al., 1999), eine
in der Cx43-Expression beobachtet, so daß keine signifikante Regulation vorhanden war.
Cx43 konnte durch das applizierte SERM nicht beeinflußt werden. Die Expression des Cx26
wurde in den transplantierten endometrialen Fragmenten durch keine der applizierten
Substanzen beeinflußt. Es war daher nicht möglich, eine eindeutige Östrogenregulation der
Gene Cx43 und Cx26 in den kultivierten endometrialen Fragmenten nachzuweisen.
93

Diskussion
davon, aus welcher Zyklusphase das verwendete Ausgangsmaterial stammte. Diese
Ergebnisse konnten in unserer Studie nicht bestätigt werden. Es wurden weder einheitliche
morphologische Charakteristika beobachtet, die einer bestimmten Zyklusphase zugeordnet
d in das
Gewebe invadierten. Zellen des Drüsenepithels konnten dagegen nie in direktem Kontakt mit
werden konnten, noch wurden Unterschiede zwischen transplantierten Geweben aus der
Proliferationsphase und der Sekretionsphase nachgewiesen. Die transplantierten Fragmente
waren unabhängig von den applizierten Substanzen morphologisch gut erhalten, fast immer
konnten endometriale Drüsen detektiert werden, die von zellreichem Stroma umgeben
waren. Allerdings wurden in den meisten Fragmenten auch Drüsen gesehen, die zystisch
vergrößert waren und ein sehr flaches Epithel aufwiesen. Das Vorkommen solcher Drüsen
korrelierte jedoch nicht mit der Behandlung der Nacktmäuse. Es war daher auch in den
transplantierten Fragmenten wie im Uterusepithel der Nacktmäuse nicht möglich, die Höhe
der Drüsenepithelzellen als diagnostischen Marker zu nutzen, im Mittel unterschieden sich
die Epithelzellhöhen der Drüsen aus den Fragmente der behandelten Tiere nicht von
Fragmenten aus den Kontrolltieren. In anderen Studien wurde ebenfalls das Vorkommen
zystisch vergrößerter Drüsen in transplantierten endometrialen Fragmenten beschrieben.
Nisolle et al. (2000b) beobachteten bereits 5 Tage nach der Transplantation von
Endometrium, das während der Menstruation entnommen wurde, daß das Epithel der
vorhandenen Drüsen abflachte und die Drüsen ein dilatiertes Aussehen annahmen. Auch
Grümmer et al. (2001) beschrieben eine derartige Veränderung der endometrialen Drüsen
nach Transplantation der Fragmente in die Nacktmaus unabhängig von der hormonellen
Behandlung der Tiere oder der Lokalisation der Fragmente. Für die zystische Vergrößerung
der Drüsen mit zunehmender Versuchsdauer könnte die fehlende Abflußmöglichkeit der
weiterhin sekretorisch aktiven endometrialen Drüsen verantwortlich sein (Awaad et al.,
1999). In Übereinstimmung mit der Studie von Nisolle et al. (2000b) konnte im Rahmen
dieser Studie gezeigt werden, daß einheitlich in allen untersuchten Fragmenten die
Stromazellen an das anliegende Bauchwandgewebe der Nacktmaus adhärierten un
dem tierischen Gewebe beobachtet werden. Das endometriale Stroma scheint bei der
Adhäsion des endometrialen Gewebes in diesem Modell und möglicherweise auch bei der
Entstehung der Endometriose im Peritoneum eine große Rolle zu spielen (Nisolle et al.,
2000b, Witz et al., 1999).
Obwohl morphologisch keine Veränderungen in den transplantierten Fragmenten beobachtet
werden konnten und keine signifikante Regulation der östrogensensitiven Gene Cx43 und
Cx26 nachgewiesen werden konnte, war es dennoch möglich, Unterschiede in der
Genexpression von Cyr61 in den transplantierten humanen endometrialen Fragmenten nach
Behandlung der Tiere mit verschiedenen Hormonen zu beobachten. Die Expression des
Gens Cyr61 wurde in den Fragmenten der Tiere, die mit Anti-Östrogen behandelt wurden,
94

Diskussion
signifikant reduziert. In 7 von 10 Versuchsreihen mit verschiedenen Geweben wurde die
Cyr61-Expression im Mittel 3fach herunterreguliert. Auch nach Applikation des SERM wurde
die Expression von Cyr61 signifikant herunterreguliert, im Mittel konnte eine 2fach geringere
Expression detektiert werden. Es war allerdings nicht möglich, die Transkription des Gens
vollständig zu inhibieren, eine schwache Expression war in allen Versuchsreihen
detektierbar. Des weiteren zeigte sich nur in einem Fall eine Steigerung des
Expressionslevels nach der Applikation von zusätzlichen Mengen Östradiol. Im Mittel
entsprach die Expression von Cyr61 in den Fragmenten der mit Östradiol behandelten Tiere
der Expression in den Kontrollfragmenten, in den einzelnen Versuchsreihen wurden jedoch
hohe Schwankungen beobachtet. Diese Ergebnisse konnten allerdings in drei
Versuchsreihen nicht bestätigt werden. In zwei Fällen entsprach die Expressionsstärke des
Cyr61 in den Fragmenten der mit Anti-Östrogen und SERM behandelten Tiere dem der
Kontrolltiere, in einem Fall zeigte sich nach Applikation des Anti-Östrogens sogar ein 2facher
Anstieg der Expression. Dieser Effekt kann nicht durch das Fehlen des Östrogenrezeptors
erklärt werden, da ERα in allen untersuchten Fragmenten exprimiert wurde. Auch in den für
diese Versuchsreihen verwendeten unbehandelten Ausgangsgeweben von Frauen ohne
Endometriose konnte auf mRNA-Ebene sowohl eine starke Expression des
Östrogenrezeptors ERα als auch des Cyr61 nachgewiesen werden.
Für zukünftige Hormon-Versuche sollte dieses Nacktmausmodell weiter entwickelt und unter
anderem die Serumwerte der Nacktmäuse bestimmt werden. Zur Untersuchung der
Hormonregulation bestim
Die Ursache für die starken Schwankungen in den einzelnen Versuchsreihen scheint die
Verwendung zyklischer Nacktmäuse gewesen zu sein. Der Zyklus einer Nacktmaus dauert
im Durchschnitt 4-5 Tage, so daß die Tiere während der Versuchsdauer von 14 Tagen bis zu
drei Zyklen durchlaufen. Es wurden keine Hormonwerte im Serum der Nacktmäuse
bestimmt, so daß nicht geklärt werden konnte, in welchem Zyklusstadium sich die einzelnen
Mäuse zu Versuchsbeginn befanden. Die möglicherweise unterschiedlichen
Serumhormonwerte der Tiere könnten die Ursache für die starken Schwankungen in den
einzelnen Versuchsreihen sein, denn die endogen produzierten Hormone schienen die
Wirkung der exogen zugeführten Hormone zu beeinflussen.
mter Gene könnte es von Vorteil sein, die Nacktmäuse zu
ovariektomieren, um den endogenen Zyklus auszuschalten. Gleichzeitig wäre es jedoch
nötig, die Tiere hormonell zu substituieren, zumindest dann, wenn die Effekte eines Anti-
Östrogens untersucht werden sollen. Damit könnten die Schwankungen der endogenen
Zyklen, die bereits hormonelle Unterschiede zwischen den einzelnen Mäusen hervorrufen,
vermieden werden.
95

Diskussion
Dennoch konnte mit Hilfe des Nacktmausmodells demonstriert werden, daß Cyr61 auch im
Endometrium östrogenabhängig exprimiert wird. Diese Regulation zeigte sich nicht nur auf
mRNA-Ebene, sondern auch auf Proteinebene. Ebenso wie im eutopen Endometrium und in
endometriotischen Läsionen war Cyr61 in Fragmenten aus Nacktmäusen, die mit dem
Vehikel behandelt wurden, vor allem apikal in den Drüsenepithelzellen exprimiert. Diese
Lokalisation läßt sich vor allem dadurch erklären, daß es sich bei Cyr61 um ein sekretiertes
Protein handelt, das mit der extrazellulären Matrix assoziiert ist (Yang und Lau, 1991). Es
handelt sich hierbei um eine nicht-kovalente Bindung, die wahrscheinlich durch Interaktionen
mit Heparan-Proteoglykanen zustande kommt (Lau und Lam, 1999). In transplantierten
humanen endometrialen Fragmenten nach Behandlung mit Anti-Östrogen ist es dagegen
kaum möglich, Cyr61 zu detektieren. Die auf mRNA-Ebene detektierte Regulation von Cyr61
durch zirkulierendes Östrogen konnte damit auch auf Proteinebene bestätigt werden.
5.4 Validierung des Nacktmausmodells für die Angiogenese
Neben dem hormonellen Status ist auch eine ausreichende Versorgung mit Blutgefäßen
Voraussetzung für das Anwachsen von endometrialem Gewebe außerhalb des Uterus
(Taylor et al., 2002a). Bei der durch die endometrialen Fragmente induzierte Angiogenese
(Shaw, 1983) scheinen die angiogenetischen Faktoren bFGF und VEGF eine große Rolle zu
weist darauf hin, daß VEGF und
bFGF eine Rolle bei der Angiogenese während der Entstehung und der Persistenz
von Endometriosepatientinnen gefunden werden (Halme et al., 1983). In
neuesten Studien konnte zudem gezeigt werden, daß lösliches VEGF auch im Serum von
spielen. Beide Faktoren konnten sowohl im eutopen Endometrium als auch in
endometriotischen Läsionen nachgewiesen werden (Donnez et al., 1998; Ferriani et al.,
1993). In dieser Studie wurde mittels „Gene-Array“ Analysen VEGF in den Endometrien von
Endometriosepatientinnen aus der Sekretionsphase in 10 von 20 Fällen stärker exprimiert
als in den normalen Endometrien. Zwar war es nicht möglich, diese Ergebnisse mittels RT-
PCR in weiteren Geweben zu verifizieren, dennoch wurden sowohl für VEGF als auch für
bFGF in den Endometrien von Endometriosepatientinnen hohe Expressionswerte
beobachtet. Zudem konnte die Expression beider Faktoren in ektopen endometriotischen
Läsionen nachgewiesen werden (Daten nicht gezeigt). Dies
endometriotischer Läsionen spielen könnten. Des weiteren wurde VEGF in der
Peritonealflüssigkeit von Frauen mit Endometriose zyklusabhängig exprimiert, die höchste
Konzentration wurde während der Proliferationsphase gemessen, wenn das Peritoneum
endometrialen Fragmenten durch retrograde Menstruation ausgesetzt ist (McLaren et al.,
1996a). Das in der Peritonealflüssigkeit detektierte VEGF wird zusammen mit anderen
Wachstumsfaktoren vor allem von aktivierten Makrophagen produziert, (Olive et al., 1991;
Chao et al., 1995; McLaren et al., 1996a), von denen bekannt ist, daß sie vermehrt in der
Peritonealflüssigkeit
96

Diskussion
Endometriosepatientinnen im Vergleich zu gesunden Frauen signifikant erhöht ist
(Metalliotakis et al., 2003). Nach 6 Monaten Behandlung mit Danazol konnte der Level von
VEGF signifikant reduziert werden (Metalliotakis et al., 2003). Das von aktivierten
Makrophagen in die Peritonealflüssigkeit sekretierte VEGF könnte daher direkten Einfluß
haben auf die Angiogenese in entstehenden endometriotischen Läsionen (Fujimoto et al.,
1999).
Es ist schon länger bekannt, daß das Wachstum von Tumoren und die Bildung von
Metastasen abhängig sind von Angiogenese (Folkman, 1990; Fidler und Ellis, 1994). Schon
früh wurde zudem die Möglichkeit erkannt, durch Inhibition von Angiogenese das
Tumorwachstum zu stoppen (Folkman, 1971). Eine solche anti-angiogenetische Therapie
könnte einige Vorteile gegenüber konventionellen Therapien haben. Zum einen herrscht in
vielen Tumoren ein starker Innendruck, der das Eindringen chemotherapeutischer
gegen wirken
hauptsächlich auf die Endothelzellen, die die Gefäße auskleiden, so daß der Tumor-
Substanzen in den Tumor behindert. Anti-angiogenetische Substanzen da
Innendruck hier keine Barriere darstellt (Jain, 1987). Zum anderen ist es unwahrscheinlich,
daß sich eine genetisch bedingte Resistenz gegen diese Art der Therapie entwickelt (Kerbel,
1997). Weiterhin sollten bei der entsprechenden Spezifikation die Nebenwirkungen einer
anti-angiogenetischen Therapie gering sein, da im adulten Körper Angiogenese
physiologisch nur bei der Wundheilung und während des menstruellen Zyklus im Uterus und
im Ovar stattfindet. Während der letzten Jahre wurden unterschiedliche anti-angiogenetische
Therapien entwickelt, die entweder die vorhandene Gefäßversorgung des Tumors durch
Toxine attackieren oder verschiedene Prozesse der Angiogenese inhibieren, um so die
Entstehung neuer Gefäße zu verhindern (Westphal et al., 2000). Zur Inhibition der
Entstehung neuer Gefäße wurden vor allem Therapien entwickelt, die die Synthese, die
Verfügbarkeit oder die Rezeptorbindung von VEGF verhindern (McLaren, 2000), da VEGF
wahrscheinlich eine wichtige Funktion bei der Tumorangiogenese hat. In vielen Studien
konnte bereits die Wirksamkeit von Antikörpern gegen VEGF bewiesen werden, die das
Tumorwachstum reduzieren (Kim et al., 1993; Asano et al., 1995; Brogstrom et al., 1996;
Leenders, 1998), so daß einige dieser Substanzen in klinischen Studien untersucht werden
(Kim et al., 1993). Eine weitere Möglichkeit besteht darin, die Tyrosinkinase-Rezeptoren zu
blockieren und damit die Signalkaskade von VEGF zu unterbrechen. Es konnte bereits
gezeigt werden, daß durch Inhibition der VEGF-Rezeptoren flt und KDR in einem
Mausmodell das Tumorwachstum reduziert wurde (Millauer et al., 1996), so daß Rezeptor-
Antagonisten effektive anti-angiogenetische Substanzen darstellen könnten. Die Substanz
SU5416 blockiert beispielsweise die durch VEGF induzierte Phosphorylierung des
Rezeptors. Hierbei handelt es sich um einen selektiven Inhibitor, dessen Wirksamkeit bereits
97

Diskussion
in Tiermodellen bewiesen wurde und der nun in klinischen Studien getestet wird (Fong et al.,
1999).
Wie bereits erwähnt spielt Angiogenese auch beim Wachstum endometrialer Fragmente
außerhalb des Uterus eine große Rolle. Daher sollte im Rahmen dieser Studie mit Hilfe des
Nacktmausmodells untersucht werden, ob es durch die Inhibition von VEGF auch möglich
ist, die Angiogenese in den transplantierten humanen endometrialen Fragmenten zu
außerhalb des Uterus wachsen, wurden ebenfalls mit Hilfe des Nacktmausmodells
inhibieren und damit die Morphologie und die Persistenz der Fragmente zu beeinflussen.
Nach der Transplantation der Fragmente wurden dafür die weiblichen zyklischen
Nacktmäuse mit einem Inhibitor der Tyrosinkinase-Aktivität der beiden humanen VEGF-
Rezeptoren flt und KDR behandelt. Dieser VEGF-Inhibitor der Firma Schering wurde dort
bereits in einem Mausmodell getestet und war in der Lage, die ovarielle Angiogenese zu
unterdrücken. Auch im Rahmen dieser Studie sollte die Signalkaskade des VEGF blockiert
und Angiogenese verhindert werden. Nach einer Versuchsdauer von 14 Tagen konnten
jedoch keine Unterschiede in den Fragmenten detektiert werden, unabhängig davon, ob die
Tiere mit dem VEGF-Antagonisten oder dem Vehikel behandelt wurden. Alle untersuchten
Fragmente wiesen neu gebildete Mausgefäße auf. Anders als beim Tumorwachstum
scheinen bei der Entstehung endometriotischer Läsionen auch andere Faktoren als VEGF
eine große Rolle zu spielen. Anti-angiogenetische Therapien bergen daher die Gefahr, daß
nach der Inhibition eines einzelnen angiogenetischen Faktors das Gewebe alternative
Faktoren synthetisiert, die dessen Funktionen erfüllen (Giavazzi und Taraboletti, 1999). Dies
sollte bei der Entwicklung einer möglichen anti-angiogenetischen Therapie gegen die
Endometriose ebenso berücksichtigt werden wie mögliche Nebenwirkungen. Das
ursprünglich gegen Übelkeit und Kopfschmerzen während der Schwangerschaft entwickelte
Medikament Thalidomid hat ebenfalls anti-angiogenetische Eigenschaften. Es inhibiert die
Proliferation von Endothelzellen und Tumorzellen in vitro und besitzt damit reduzierende
Wirkungen auf das Tumorwachstum und die Metastasenbildung (Moreira et al., 1999;
Minchinton et al., 1996). In klinischen Studien konnte die Wirksamkeit von Thalidomid in der
Therapie gegen multiple Myelome bestätigt werden (Juliusson et al., 2000; Zomas et al.,
2000). Dennoch muß berücksichtigt werden, daß diese anti-angiogenetischen Eigenschaften
nach der Einnahme von Schwangeren zu Fehlbildungen des Embryos führten, die durch die
Inhibition der embryonalen Gefäßentwicklung erklärt werden können (D´Amato et al., 1994).
Es muß daher überprüft werden, ob die für die Tumortherapie entwickelten anti-
angiogenetischen Substanzen auch in der Behandlung der Endometriose eingesetzt werden
können, da viele Endometriosepatientinnen unter Infertilität leiden und eine Therapie gegen
die Endometriose mit einer möglichen Schwangerschaft vereinbar sein muß.
Weitere Untersuchungen zur Angiogenese in endometrialen Fragmenten, die an Stellen
98

Diskussion
durchgeführt. Humane endometriale Fragmente wurden in weibliche zyklische Nacktmäuse
transplantiert und nach 2, 4, 7 und 14 Tagen wurde je eines der unbehandelt belassenen
lauf der VEGF-
Tiere getötet. Grümmer et al. (2001) hatten bereits den zeitlichen Verlauf der Entstehung
neuer Blutgefäße in der Nacktmaus und das Einwachsen dieser Gefäße in die
transplantierten humanen Fragmente an verschiedenen Organen untersucht. Am Tag 4 nach
der Transplantation konnten neu gebildete Blutgefäße der Maus in den humanen
endometrialen Fragmenten detektiert werden, die an der Leber, am Fettgewebe oder am
Darm der Nacktmaus angewachsen waren. Eine Korrelation zwischen der Anzahl der
detektierten Blutgefäße und der Lokalisation der Fragmente bestand dabei nicht. Durch
immunhistochemische Färbung des von-Willebrand Faktors konnte zusätzlich bestätigt
werden, daß die Anzahl der Gefäße humanen Ursprungs mit zunehmender
Kultivierungsdauer abnahm, so daß nach 12 Tagen keine humanen Gefäße mehr
nachgewiesen werden konnten (Grümmer et al., 2001). Im Rahmen dieser Doktorarbeit
konnten diese Ergebnisse bestätigt werden. Auch in humanen endometrialen Fragmenten,
die an der Bauchwand weiblicher zyklischer Nacktmäuse fixiert wurden, konnten die ersten
einwachsenden Blutgefäße der Maus nach 4 Tagen in den Fragmenten detektiert werden.
Bereits nach 7 Tagen waren kleinere Fragmente vollständig mit neu entstandenen
Mausgefäßen durchsetzt. Um nachzuweisen, welche Faktoren hierbei eine Rolle spielen
könnten, wurde in dieser Studie zusätzlich die Expression der bekannten angiogenetischen
Faktoren VEGF und bFGF sowie von Cyr61, das als proangiogenetischer Faktor
beschrieben wurde (Lau und Lam, 1999), untersucht. Alle drei angiogenetischen Faktoren
wurden sowohl im endometrialen Ausgangsgewebe exprimiert, das nicht transplantiert
wurde, als auch in den kultivierten endometrialen Fragmenten. In Übereinstimmung mit dem
zeitlichen Verlauf der Angiogenese wurde ein Anstieg der Expressionslevel der
angiogenetischen Faktoren bFGF und Cyr61 ebenfalls an Tag 2 beobachtet. Vor allem
Cyr61 wurde auch im weiteren Versuchsverlauf stark exprimiert. Die Expression von VEGF
dagegen zeigte keinen Anstieg in den transplantierten endometrialen Fragmenten im
Vergleich zum Ausgangsgewebe, statt dessen sank der Expressionslevel im Versuchsverlauf
um den Faktor 1,5.
Nisolle et al. (2000a) dagegen konnten zeigen, daß nach immunhistochemischer Färbung
große Mengen an VEGF sowohl in den Drüsenepithelzellen als auch im Stroma humaner
endometrialer Fragmente nach 3 Wochen Kultivierung in der Nacktmaus exprimiert wurden.
Im Drüsenepithel zeigte sich dabei eine stärkere Expression als im Stroma. Dennoch
müssen diese Ergebnisse nicht zwangsläufig denen unserer Studie widersprechen. Es muß
berücksichtigt werden, daß von Nisolle et al. (2000a) nicht der zeitliche Ver
Expression untersucht wurde und zudem die exprimierte VEGF-Menge nicht quantifiziert
oder mit der Expressionsmenge des Ausgangsgewebes verglichen wurde. Des weiteren
99

Diskussion
wurde VEGF in unserer Studie auch an Tag 14 noch in den transplantierten endometrialen
Fragmenten exprimiert, unklar bleibt, ob der Expressionslevel der von Nisolle et al. (2000a)
detektierten Menge entspricht.
Diese Ergebnisse machen deutlich, daß bei der Angiogenese der endometrialen Fragmente
im Nacktmausmodell und möglicherweise auch bei der Angiogenese der endometriotischen
Läsionen im humanen Peritoneum neben VEGF weitere angiogenetische Faktoren wie bFGF
und Cyr61 eine Rolle spielen könnten. Die in dieser Studie nicht untersuchten
angiogenetischen Faktoren Angiopoietin 1 und 2 (Ang1 und Ang2) könnten ebenfalls beteiligt
sein. Bei Ang1 und Ang2 handelt es sich um sekretierte Wachstumsfaktoren, die an den
Tyrosinkinase-Rezeptor Tie2 binden, der auf Endothelzellen sowohl im embryonalen als
auch im adulten Gefäßsystem exprimiert wird (Breier, 2000). Bei der möglichen Entwicklung
anti-angiogenetischer Therapien gegen die Endometriose sollte die Vielfalt der
angiogenetischen Faktoren berücksichtigt werden. Möglicherweise wäre eine Kombination
mehrerer Strategien zur Inhibition verschiedener Faktoren eine Alternative zur alleinigen
Inhibition von VEGF (Giavazzi und Taraboletti, 1999).
5.5 Mögliche Funktionen von Cyr61 in der Pathogenese der Endometriose
Diese Studie ergab, daß Cyr61 in endometriotischen Läsionen sowohl aus der
Proliferationsphase als auch aus der Sekretionsphase sehr stark exprimiert wird und zudem
in diesen Geweben einen höheren Expressionslevel aufweist als in den eutopen
Endometrien. Des weiteren konnte gezeigt werden, daß Cyr61 in der induzierten
Endometriose im Tiermodell bei der Angiogenese eine wichtige Rolle spielen könnte. Daher
könnte Cyr61 auch eine wichtige Funktion bei der Angiogenese in den endometriotischen
Läsionen im humanen Peritoneum haben. Daß Cyr61 ein wichtiger angiogenetischer Faktor
ist, konnten zuletzt Mo et al., (2002) zeigen, die das Cyr61-Gen in der Maus ausknockten.
Alle Cyr61 KO-Mäuse starben, 70 % von ihnen, weil durch das Fehlen von Cyr61 die
Gefäßverzweigung in der sich entwickelnden Plazenta gestört war (Mo et al., 2002). Cyr61
induziert zudem Adhäsion, Chemotaxis und DNA-Synthese in Endothelzellen (Kireeva et al.,
6 1
Adhäsion, Migration, das Überleben der Zellen und die Gefäßbildung durch das Integrin v 3
induziert (Leu et al., 2002). Einige Mitglieder der Integrin-Familie wie die Integrine α1, α4 und
β3 werden im Laufe des Menstruationszyklus im Drüsenepithel des Endometriums exprimiert
1996, 1997). Es konnte gezeigt werden, daß diese angiogenetischen Funktionen durch
verschiedene Integrine vermittelt werden. Cyr61 induziert Zell-Adhäsion und die Bildung
lumenhaltiger Gefäße durch das Integrin α β in humanen Endothelzellen aus der
Umbilicalvene (HUVEC) aus frühen Passagen. In Endothelzellen, in denen die Integrine
durch Phorbolester oder VEGF aktiviert wurden, werden die durch Cyr61 geförderte
α β
100

Diskussion
(Lessey et al., 1992; Tabibzadeh, 1992). Die Integrin-Untereinheit β3, die mit αv und gpIIIa
interagiert (Fitzgerald et al., 1987), konnte beispielsweise ab dem Tag 20 und während der
gesamten Sekretionsphase im humanen Endometrium nachgewiesen werden. Über die
eine Rolle bei der Entstehung von Endometriose. CTGF besitzt den gleichen strukturellen
Aufba Mitglieder der CCN-Familie, das humane Gen besteht aus 349
weitere Expression der Integrine herrschen kontroverse Ansichten. Während Lessey et al.
(1994b) eine Herunterregulation des Integrins αvβ3 beschrieben, detektierten Hii und Rogers
(1998) in den endometrialen Gefäßen von Endometriosepatientinnen signifikant höhere
Expressionsmengen des Integrins αvβ3 als in den Kontrollendometrien. Bisher war es nicht
möglich, einen spezifischen Rezeptor für Cyr61 zu identifizieren, dennoch kann nicht
ausgeschlossen werden, daß ein solcher Rezeptor existiert. Alle zellulären und
physiologischen Funktionen von Cyr61 könnten jedoch auch durch die Interaktion mit
Integrinen vermittelt werden (Lau und Lam, 1999, Leu et al., 2002). Dies würde bedeuten,
daß durch die stärkere Expression des Cyr61 in den Endometrien von
Endometriosepatientinnen verglichen mit normalen Endometrien die Entwicklung
endometriotischer Läsionen durch vermehrte Zelladhäsion und Angiogenese, vermittelt
durch das Integrin αvβ3, begünstigt werden könnte.
Möglicherweise spielen neben Cyr61 auch weitere Mitglieder der CCN-Familie wie CTGF
u wie die übrigen
Aminosäuren und besitzt alle 38 konservierten Cysteine (Brunner et al., 1991; Ryseck et al.,
1991; Brigstock et al., 1997). CTGF wurde bisher auf mRNA-Ebene unter anderem in
Fibroblasten, Endothelzellen, Gefäßmuskelzellen und Epithelzellen detektiert (zur Übersicht
s. Brigstock, 1999). Wie Cyr61 wird auch CTGF von einem „immediate early gene“ kodiert. In
ruhenden Fibroblastenzellen wird CTGF nur sehr schwach exprimiert (Lin et al., 1998;
Steffen et al., 1998), ein Anstieg der CTGF-Transkription kann jedoch innerhalb kurzer Zeit
nach Induktion der Zellproliferation durch Zugabe von Serum oder TGF-β beobachtet werden
(Ryseck et al., 1991; Igarashi et al., 1993). In Untersuchungen zur Mausentwicklung wurde
zudem nachgewiesen, daß CTGF und Cyr61 in verschiedenen Organen wie dem
kardiovaskulären System, in der Haut und in der Plazenta zusammen exprimiert werden
(Kireeva et al., 1997).
Aus den Array-Analysen im Rahmen dieser Studie ergab sich, daß neben Cyr61 auch CTGF
in den Endometrien von Endometriosepatientinnen der Sekretionsphase stärker exprimiert
wird als in normalen Endometrien, während in der Proliferationsphase keine Veränderungen
im Expressionslevel von CTGF zwischen Endometrien von Frauen mit und ohne
Endometriose detektiert werden konnten. Obwohl es nur teilweise möglich war, diese
Resultate in RT-PCR Experimenten zu verifizieren, zeigte sich hier eine tendenziell stärkere
Expression von CTGF in den endometriotischen Läsionen beider Zyklusphasen im Vergleich
zum eutopen Endometrien. Eine signifikante Hochregulation von CTGF konnte anders als
101

Diskussion
bei Cyr61 in den Läsionen allerdings nicht gezeigt werden, ein Grund hierfür könnte die
geringere Anzahl getesteter Gewebe oder die weniger sensitive Methode der RT-PCR
gegenüber der real-time RT-PCR sein. Dennoch könnte CTGF in den endometriotischen
Läsionen eine Rolle bei der Entstehung und der Persistenz dieser Läsionen spielen. In
früheren Studien konnte CTGF bereits im Endometrium der Maus, des Schweins und des
Menschen nachgewiesen werden (zur Übersicht s. Brigstock, 1999). Möglicherweise spielt
CTGF eine Rolle bei der Dezidualisierung und der Implantation, denn zu diesem Zeitpunkt
zeigten sich in der Maus Veränderungen in der Expression von CTGF. Zunächst wurde
CTGF im Uterusepithel stark reduziert, während sich in den nächsten zwei Tagen nach der
Implantation dann aber eine deutliche CTGF-Expression in den dezidualisierenden
Stromazellen zeigte (Surveyor et al., 1998). Immunhistochemisch wurde CTGF im humanen
Endometrium der Proliferationsphase im Drüsenepithel und in Endothelzellen lokalisiert, im
Stroma dagegen konnte CTGF kaum nachgewiesen werden (Uzumcu et al., 2000). Während
des Zyklus und zu Beginn der Schwangerschaft allerdings zeigten sich auch hier
Veränderungen sowohl in der Expressionsstärke als auch in der Lokalisation. CTGF konnte
auch im Endometrium der Sekretionsphase in Drüsenepithelzellen und Endothelzellen
splantierten humanen endometrialen Fragmenten im Vergleich zum
v 3
deren Transkription durch Wachstumsfaktoren wie bFGF oder TGF induziert wird (zur
nachgewiesen werden, in der späten Sekretionsphase und während der Schwangerschaft
wurde CTGF aber zusätzlich in dezidualisierten Stromazellen detektiert (Uzumcu et al.,
2000). Dies ist ein Hinweis dafür, daß CTGF auch bei Veränderungen des humanen
Endometriums eine Rolle spielen könnte. Fehlregulation von CTGF im Endometrium von
Endometriosepatientinnen könnte auch hier die Dezidualisierung des Endometriums und
damit auch die Implantation beeinflussen. Zur genauen Expression von CTGF im
Endometrium und seiner möglichen Funktion in der Endometriose müssen weitere
Untersuchungen erfolgen. Auch der genaue Regulationsmechanismus der CTGF-Expression
ist bis heute nicht geklärt und müßte näher untersucht werden, möglicherweise spielen
Glukokortikoide bei der Induktion des CTGF eine Rolle (Dammeier et al., 1998). Eine
Regulation durch Östrogen konnte im Rahmen dieser Studie durch Untersuchungen mit Hilfe
des Nacktmausmodells ausgeschlossen werden. Hier zeigte sich nach Behandlung der Tiere
mit einem reinen Anti-Östrogen, Östradiol oder einem SERM keine Veränderung der CTGF-
Expression in den tran
Vehikel.
Möglicherweise erfüllt CTGF zusammen mit Cyr61 eine Funktion in der Angiogenese
endometriotischer Läsionen. In früheren Studien konnte gezeigt werden, daß neben Cyr61
auch CTGF Zelladhäsion und Migration in „human microvascular endothelial cells“ (HMVEC)
induziert und auf diese Zellen ebenfalls durch das Integrin α β wirkt (Babic et al., 1998,
1999). Zudem wurden sowohl Cyr61 als auch CTGF als „immediate early genes“ identifiziert,
β
102

Diskussion
Übersicht, s. Lau und Lam, 1999), von denen bekannt ist, daß sie Angiogenese in vivo
induzieren (Klein et al., 1997; Roberts und Sporn, 1989). Ein hypothetisches Modell
postuliert daher, daß Wachstumsfaktoren ihre angiogenetische Wirkung zumindest zum Teil
durch die Induktion von Cyr61 und CTGF entfalten (Lau und Lam, 1999). bFGF und TGFβ
könnten demnach entweder eine direkte Wirkung auf die Endothelzellen ausüben oder in
Fibroblasten die Expression von Cyr61 und CTGF stimulieren. Die freigesetzten Proteine
Cyr61 und CTGF könnten dann durch das Integrin αvβ3 verschiedene Funktionen in den
Endothelzellen induzieren (Lau und Lam, 1999).
Dadurch wird deutlich, daß im humanen Endometrium und in endometriotischen Läsionen
neben Östrogen möglicherweise auch verschiedene Wachstumsfaktoren an der Regulation
der Cyr61- und CTGF-Expression beteiligt sind. Weitere Untersuchungen müssen erfolgen,
um die genauen Regulationsmechanismen dieser Proteine im Endometrium und ihrer
möglichen Funktion bei der Pathogenese der Endometriose zu klären.
103

Ausblick
6 Ausblick
In zukünftigen Experimenten soll die Funktion von Cyr61 in der Entstehung der
Endometriose geklärt werden. Dazu steht zum einen die endometriale Ishikawa-Zellinie zur
Verfügung, zum anderen die Kultivierung endometrialer Fragmente in der Nacktmaus. In
angiogenetischen Assays wird der mögliche Einfluß von Cyr61 auf die Bildung endometrialer
Blutgefäße näher untersucht. Nach Induktion des Cyr61 durch z.B. bFGF oder nach
Repression des Cyr61 durch z.B. Anti-Östrogene werden die Auswirkungen auf kultivierte
endometriale Fragmente in der Nacktmaus getestet. Eventuell nachgewiesene
Allerdings muß das Nacktmausmodell dahingehend validiert werden, daß sich die
hormonellen Schwankungen der Maus nicht auf die humanen endometrialen Fragmente
auswirken. Weitere Substanzen sollen zudem untersucht werden, die die Entstehung und die
Persistenz endometriotischer Läsionen beeinflussen könnten.
Die Expression weiterer Gene, die nach den Array-Analysen in Endometrien von
Endometriosepatientinnen unterschiedlich reguliert wurden, werden in zusätzlichen
Gewebeproben mit anderen Methoden untersucht. Vor allem soll geklärt werden, welche
Rolle CTGF oder bFGF zusammen mit Cyr61 in der Entstehung von Endometriose spielen.
angiogenetische Wirkungen des Cyr61 sollen zur möglichen Entwicklung anti-
angiogenetischer Strategien gegen die Endometriose genutzt werden.
104

Zusammenfassung
7 Zusammenfassung
sion im Vergleich zu
. In diesen Versuchen wurde
allerdings nicht nur in den Endometrien von Endometriosepatientinnen der Sekretionsphase
exprimiert wird. In Endometrien von gesunden Frauen wurde Cyr61 in der
roliferationsphase signifikant stärker exprimiert als in der Sekretionsphase. In Endometrien
von Endometriosepatientinnen zeigte sich ebenfalls diese Tendenz, hier konnte jedoch keine
signifikante Regulation nachgewiesen werden. Weitere Hinweise ergaben Untersuchungen,
die mit Hilfe eines etablierten Nacktmausmodells durchgeführt wurden. Auch in humanen
endometrialen Fragmenten, die an die innere Bauchwand weiblicher zyklischer Nacktmäuse
transplantiert wurden, wurde die Cyr61-Expression nach Behandlung der Mäuse mit einem
reinen Anti-Östrogen oder einem SERM signifikant reduziert. Allerdings war es nicht möglich,
einen Anstieg des Expressionslevels in den Fragmenten der mit zusätzlichen Mengen
Östradiol behandelten Tiere gegenüber den mit Vehikel behandelten Tieren zu erzielen.
Damit scheint Cyr61 nicht nur ein Markergen für Endometriose zu sein, möglicherweise spielt
es auch eine Rolle bei der Entstehung der endometriotischen Läsionen. Denn in den ektopen
Läsionen beider Zyklusphasen wurde ein signifikant höherer Expressionslevel von Cyr61
detektiert als in den eutopen Endometrien von Endometriosepatientinnen. Diese
Hochregulation in den endometriotischen Läsionen der Sekretionsphase könnte durch eine
lokale Östrogensynthese in diesem Gewebe begünstigt werden, da eine Fehlregulation der
an der Östrogensynthese beteiligten Enzyme 17β-Hydroxysteroid Dehydrogenase Typ1 und
2 detektiert wurde. In den endometriotischen Läsionen der Proliferationsphase und in
Obwohl die molekularen Mechanismen, die zur Entstehung endometriotischer Läsionen
führen, bis heute nicht vollständig geklärt sind, existieren mehrere Hinweise darauf, daß
bereits das Endometrium von Endometriosepatientinnen Dedifferenzierungen aufweist. Ein
Ziel dieser Arbeit war es daher, mit Hilfe der Array-Technologie Gene zu identifizieren, die im
Endometrium von Endometriosepatientinnen abweichend vom normalen Endometrium
exprimiert werden. Aus diesen Analysen wurde ersichtlich, daß die hormonelle
Ansprechbarkeit des Endometriums von Frauen mit Endometriose gegenüber der von
gesunden Frauen verschoben ist. Östrogenregulierte Gene zeigten in den Endometrien von
Endometriosepatientinnen der Sekretionsphase eine stärkere Expres
normalen Endometrien der gleichen Zyklusphase. Eines dieser Gene war Cyr61, ein
sekretiertes Protein der CCN-Familie, das bei der Zelladhäsion, der Zellmigration aber auch
bei der Angiogenese eine Rolle spielt. Die Ergebnisse der Array-Analysen konnten in
weiteren Geweben mittels real-time RT-PCR bestätigt werden
eine signifikante Hochregulation der Expression von Cyr61 detektiert, sondern zusätzlich
auch in den Endometrien von Endometriosepatientinnen der Proliferationsphase.
Zudem konnte nachgewiesen werden, daß Cyr61 im Endometrium östrogenabhängig
P
105

Zusammenfassung
endometriotischen Läsionen von Frauen, deren ovarielle Östrogensynthese über einen
Zeitraum von sechs Monaten blockiert wurde, konnten ebenfalls hohe Expressionslevel von
Cyr61 nachgewiesen werden. Hier scheinen allerdings andere Regulationsmechanismen als
Östrogen eine Rolle zu spielen, denn die Enzyme 17β-Hydroxysteroid Dehydrogenase Typ1
und 2 konnten in beiden Gewebetypen kaum nachgewiesen werden.
tehung und
der Persistenz endometriotischer Läsionen spielen. Die genauen zellbiologischen Funktionen
dieser Faktoren für die Pathogenese der Endometriose müssen an endometrialen Zellinien
und mit Hilfe der Kultivierung endometrialer Fragmente in der Nacktmaus weiter geklärt
werden.
Möglicherweise spielt Cyr61 eine Rolle bei der Angiogenese der endometriotischen
Läsionen, da für deren Persistenz die Bildung neuer Blutgefäße von großer Bedeutung ist.
Mit Hilfe des Nacktmausmodells konnte gezeigt werden, daß neben VEGF und bFGF auch
Cyr61 in den transplantierten endometrialen Fragmenten mit einsetzender Angiogenese
induziert wird. Zudem war es nicht möglich, nach der Inhibition von VEGF die Angiogenese
in diesem Modell zu blockieren und damit die Morphologie oder die Persistenz der
transplantierten Fragmente zu beeinflussen. Dies könnte ein weiterer Hinweis dafür sein,
daß nicht nur VEGF in der Angiogenese der endometriotischen Läsionen eine Rolle spielt,
sondern auch bFGF und Cyr61. Möglicherweise sind weitere Proteine der CCN-Familie wie
CTGF an der Entstehung von Endometriose beteiligt. Es wurden höhere
Expressionsmengen von CTGF in den endometriotischen Läsionen beider Zyklusphasen
nachgewiesen. CTGF könnte daher zusammen mit Cyr61 eine Rolle bei der Ents
106

Literatur
8 Literatur
(1998): Vascular endothelial growth factor D (VEGF-D) is a ligand for the tyrosine kinase VEGF receptor 2 (Flk-1) and VEGF receptor 3 (Flt-4). Proc. Natl. Acad. Sci. USA 95:548-
Achen MG, Jeltsch M, Kukk K, Makinen T, Vitali A, Wilks AF, Alitalo K, Stacker SA
553
Adamson GD, Pasta DJ (1994): Surgical treatment of endometriosis-associated infertility:
progesterone receptor gene expression in human breast cancer cells. Mol. Endocrinol.
and metastasis by an immunoneutralizing monoclonal antibody to human vascular
85:2897-2902
Awwad JT, Sayegh RA, Tao XJ, Hassan T, Awwad ST, Isaacson K (1999): The SCID
Natl. Acad. Sci. USA 95:6355-6360
Babic AM, Chen CC, Lau LF (1999):
elial cell survival, and induces angiogenesis in vivo. Mol. Cell. Biol. 19:2958-2966
Barkhem T, Carlsson B, Nilsson Y, Enmark G, Gustafsson J, Nilsson S (1998): Differential response of estrogen receptor alpha and estrogen receptor beta to partial
estrogen agonists/antagonists. Mol. Pharmacol. 54:105-112
Barnhart K, Dunsmoor-Su R, Coutifaris C (2002): Effect of endometriosis on in vitro fertilization. Fertil. Steril. 77:1148-1155
Barragan JC, Brotons J, Ruiz JA, Acien P (1992): Experimentally induced endometriosis in rats: effects on fertility and the effects of pregnancy and lactation on the ectopic endometrial tissue. Fertil. Steril. 58:1215-1219
Beato M (1989): Gene regulation by steroid hormones. Cell 56:335-344
Benjamin LE, Keshet C (1997): Conditional switching of vascular endothelial growth factor (VEGF) expression in tumours: induction of endothelial cell shedding and regression of hemangioblastoma-like vessels by VEGF withdrawal. Proc. Natl. Acad. Sci. USA 94:8761- 8766
Bergquist A, Jeppson S, Kullander S (1985): Human endometrium transplanted into nude mice. Histologic effects of various steroid hormones. Am. J. Pathol. 119:336-344
Bergquist A, Ferno M (1993a): Oestrogen and progesterone receptors in endometriotic tissue and endometrium: comparison of different cycle phases and ages. Hum. Reprod. 8:2211-2217
Bergquist A, Ferno M (1993b): Estrogen and progesterone receptors in endometriotic tissue and endometrium: comparison according to localization and recurrence. Fertil. Steril. 60:63-68
meta-analysis compared with survival analysis. Am. J. Obstet. Gynecol. 171:1488-1505
Alexander IE, Clarke CL, Shine J, Sutherland RL (1989): Progestin inhibition of
3:1377-1386
American Fertility Society (1985): Revised American Fertility Society classification of endometriosis:1985. Fertil. Steril. 43:351-352
Asano A, Yukita A, Matsumoto T, Kondo S, Suzuki H (1995): Inhibition of tumor growth
endothelial growth factor/vascular permeability factor 121. Cancer Res. 55:5296-5301
Attia GR, Zeitoun K, Edwards D, Johns A, Carr BR, Bulun SE (2000): Progesterone receptor isoform A but not B is expressed in endometriosis. J. Clin. Endocrinol. Metab.
mouse: an experimental model for endometriosis. Hum. Reprod. 14:3107-3111
Babic AM, Kireeva ML, Kolesnikova TV, Lau LF (1998): CYR61, product of a growth factor-inducible immediate-early gene, promotes angiogenesis and tumor growth. Proc.
Fisp12/mouse connective tissue growth factor mediates endothelial cell adhesion and migration through integrin αvβ3, promotes endoth
107

Literatur
Bhatia DS, McFad South. Med. J. 91
den PM, Kline RC (1998): Recurrent catamenial hemopneumothorax. :398-401
estrogen receptor alpha (ER-α) and beta (ER-β) mRNA in the midgestational human
Brandenberger AW, Tee MK, Jaffe RB (1998): Estrogen receptor alpha (ER- ) and beta
R-beta isoforms in normal endometrial and
in uterine
tissue growth factor/cysteine-rich 61/nephroblastoma
r
-Arispe ML (1997): Uterine
. Lab. Invest. 76:245-255
n of a gene family regulated by transforming growth factor-β. DNA Cell Biol. 10:293-300
i AM, Pike AC, Dauter Z, Hubbard RE, Bonn T, Engstrom O, Ohman L,
Maile L, Harlow C, Hull M (1995): Pituitary-ovarian dysfunction as a
Bork P (1993): The modular architecture of a new family of growth regulators related to connective tissue growth factor. FEBS Lett. 327:125-130
Brandenberger AW, Tee MK, Lee JY, Chao V, Jaffe RB (1997): Tissue distribution of
fetus. J. Clin. Endocrinol. Metab. 82:3509-3512
α (ER-β) mRNAs in normal ovary, ovarian serous cystadenocarcinoma and ovarian cancer cell lines: down-regulation of ER-β in neoplastic tissues. J. Clin. Endocrinol. Metab. 83:1025-1028
Brandenberger AW, Lebovic DI, Tee MK, Ryan IP, Tseng JF, Jaffe RB, Taylor RN (1999): Oestrogen receptor (ER)-alpha and E endometriosis-derived stromal cells. Mol. Hum. Reprod. 5:651-655
Breier G (2000): Angiogenesis in embryonic development – A review. Placenta 21 Suppl. A:S11-S15
Bridges J, Prentice A, Roche W, Englefield P, Thomas EJ (1994): Expression of integrin adhesion molecules in endometrium and endometriosis. Br. J. Obstet. Gynaecol. 101:696- 700
Brigstock DR, Steffen DL, Kim GY, Vegunta RK, Diehl JR, Harding PA (1997): Purification and characterization of novel heparin-binding growth factors secretory fluids: identification as heparin-regulated Mr 10000 forms of connective tissue growth factor. J. Biol. Chem. 272:20275-20282
Brigstock DR (1999): The connective overexpressed (CCN) family. Endocr. Rev. 20:189-206
Brogstrom P, Hillan KJ, Sriramarao P, Ferrara N (1996): Complete inhibition of angiogenesis and growth of microtumors by anti-vascular endothelial growth facto neutralizing antibodies. Novel concepts of angiostatic therapy from intravital videomicroscopy. Cancer Res. 56:4032-4039
Brown LF, Detmar M, Tognazzi K, Abu-Jawdeh G, Iruela smooth muscle cells express functional receptors (flt-1 and KDR) for vascular permeability factor/vascular endothelial growth factor
Bruner KL, Matrisian LM, Rodgers WH (1997): Suppression of matrix metalloproteinases inhibits establishment of ectopic lesions by human endometrium in nude mice. J. Clin. Invest. 99:2851-2857
Brunner A, Chinn J, Neubeuer M, Purchio AF (1991): Identificatio
Bryant HU, Dere WH (1998): Selective estrogen receptor modulators: an alternative to hormone replacement therapy. Proc. Soc. Exp. Biol. Med. 217:45-52
Brzozowsk Greene GL, Gustafsson JA, Carlquist M (1997): Molecular basis of agonism and antagonism in the oestrogen receptor. Nature 389:753-758
Bulun SE, Yang S, Fang Z, Gurates B, Tamura M, Zhou J, Sebastian S (2001): Role of aromatase in endometrial disease. J. Steroid. Biochem. Mol. Biol. 79:19-25
Cahill D, Wardle P, cause for endometriosis-associated and unexplained infertility. Hum. Reprod. 10:3142- 3146
108

Literatur
Carmeliet P, Collen D (1998): Role of vascular endothelial growth factor and vascular endothelial growth factor receptors in vascular development. Curr. Top. Microbiol.
roxysteroid dehydrogenase type
hroder S, Weder W (1997): Catamenial
S, Sharkey AM, Rajput-Williams J, Burch D, Schofield JP, Fountain
cur. Clinical follow-up over 17 years. Geburtshilfe Frauenheilkd. 45:792-796
terine cells. J. Steroid. Biochem. Mol. Biol. 41:523-528
2): Transcriptional effects of chronic Akt
Implications for cardiovascular
uterine capillary permeability and growth. Endocrinology 133:829-
e is an inhibitor of
ne is a novel potent
997): Expression of vascular
Immunol. 237:133-157
Casey ML, MacDonald PC, Andersson S (1994): 17β-Hyd 2: chromosomal assignment and progestin regulation of gene expression in human endometrium. J. Clin. Invest. 94:2135-2141
Cassina PC, Hauser M, Kacl G, Imthurn B, Sc hemoptysis. Diagnosis with MRI. Chest 111:1447-1450
Chao TC, Van Alten PJ, Greager JA, Walter RJ (1995): Steroid sex hormones regulate the release of tumor necrosis factor by macrophages. Cell. Immunol. 160:43-49
Charnock-Jones D SA, Boocock CA, Smith SK (1993): Identification and localization of alternately spliced mRNAs for vascular endothelial growth factor in human uterus and estrogen regulation in endometrial carcinoma cell lines. Biol. Reprod. 48:1120-1128
Christmann D, Strobel E (1985): Stromal endometriosis. A rare semimalignant tumor with a tendency to re
Cicatiello L, Ambrosino C, Coletta B, Scalona M, Sica V, Bresciani F, Weisz A (1992): Transcriptional activation of jun and actin genes by estrogen during mitogenic stimulation of rat u
Conneely OM, Maxwell BL, Toft DO, Scharder WT, O´Malley BW (1987): The A and B forms of a chicken progesterone receptor arise by alternate initiation of translation of the unique mRNA. Biochem. Biophys. Res. Commun. 149:493-501
Cook SA, Matsui T, Li L, Rosenzweig A (200 activation in the heart. J. Biol. Chem. 277:22528-22533
Cook SA, Rosenzweig A (2002): DNA microarrays: medicine. Circ. Res. 91:559-564
Creus M, Balasch J, Ordi J, Fabregues F, Casamitjana R, Qunito L, Coutifaris C, Vanrrell JA (1998): Integrin expression in normal and out-of-phase endometria. Hum. Reprod. 13:3460-3468
Crum CP, Wible J, Frick HC, Fenoglio CM, Richart RM, Williamson S (1981): A case of extensive pelvic endometriosis terminating in endometrial sarcoma. Am. J. Obstet. Gynecol. 140:718-719
Cullinan-Bove K, Koos RD (1993): Vascular endothelial growth factor/vascular permeability factor expression in the rat uterus: rapid stimulation by estrogen correlates with estrogen- induced increases in 837
D´Amato RJ, Loughnan MS, Flynn E, Folkman J (1994): Thalidomid angiogenesis. Proc. Natl. Acad. Sci. USA 91:4082-4085
Dammeier J, Beer H-D, Brauchle M, Werner S (1998): Dexamethaso inducer of connective tissue growth factor expression. Implications for glucocorticoid therapy. J. Biol. Chem. 273:18185-18190
Das SK, Chakraborty I, Wang J, Dey SK, Hoffman LH (1 endothelial growth factor (VEGF) and VEGF-receptor messenger ribonucleic acids in the peri-implantation rabbit uterus. Biol. Reprod. 56:1390-1399
Davois S, White R, Parker MG (1993): The antiestrogen ICI 182780 disrupts estrogen receptor nucleocytoplasmic shuttling. J. Cell Sci. 106:1377-1388
109

Literatur
Devin-Leclerc J, Meng X, Delahaye F, Leclerc P, Baulieu EE, Catelli MG (1998): Interaction and dissociation by ligands of estrogen receptor and Hsp90: the antiestrogen RU 58668 induces a protein synthesis-dependent clustering of the receptor in the
causes endometriosis in baboons
i (1995): Basic
Role of ovarian steroids in
98): Vascular endothelial
strogen regulation of c-fos gene
ls. Biochem. Biophys. Res. Commun. 294:384-394
tmar M, Dvorak AM (1995): Vascular permeability factor/vascular
M, Grandien K, Lagercrantz J, Fried G, Nordenskjold M,
r endothelial growth factors. Curr. Top. Microbiol. Immunol. 237:41-57
cytoplasm. Mol. Endocrinol. 12:842-854
Dhingra K (1999): Antiestrogens – tamoxifen, SERMs and beyond. Invest. New Drugs 17:285-311
D´Hooghe TM, Bambra CS, Raeymaekers BM, De Jonge I, Lauweryns JM, Koninckx PR (1995): Intrapelvic injection of menstrual endometrium (Papio cynocephalus und Papio anubis). Am. J. Obstet. Gynecol. 173:125-134
D Blasio AM, Centinaio G, Carniti C, Somigliana E, Vigano P, Vignali M fibroblast growth factor messenger ribonucleic acid levels in eutopic human endometrial stromal cells as assessed by competetive polymerase chain reaction amplification. Mol. Cell. Endocrinol. 115:169-175
Dizerega GS, Barber DL, Hodgen GD (1980): Endometriosis: initiation, maintenance and suppression. Fertil. Steril. 33:649-653
Dlugi AM, Miller JD, Knittle J (1990): Lupron depot in the treatment of endometriosis: a randomized, placebo-controlled, double blind study. Fertil. Steril. 54:419-427
Dmowski WP (1995): Immunological aspects of endometriosis. Int. J. Gynaecol. Obstet. 50:S3-10
Donnez J, Nisolle M, Casanas-Roux F (1992): Three-dimensional architectures of peritoneal endometriosis. Fertil. Steril. 57:980-983
Donnez J, Smoes P, Gillerot S, Casanas-Roux F, Nisolle M (19 growth factor (VEGF) in endometriosis. Hum. Reprod. 13:1686-1690
Duan R, Xie W, Li X, McDougal A, Save S (2002): E expression through phosphatidylinositol-3-kinase-dependent activation of serum response factor in MCF-7 breast cancer cel
Dunselman GA, Willebrand D, Land JA, Bouckaert PX, Evers JL (1989): A rabbit model of endometriosis. Gynecol. Obstet. Invest. 27:29-33
Dvorak HF, Brown LF, De endothelial growth factor, microvascular hyperpermeability, and angiogenesis. Am. J. Pathol. 146:1029-1039
Enmark E, Pelto-Huikko Gustafsson JA (1997): Human estrogen receptor β-gene structure, chromosomal localization, and expression pattern. J. Clin. Endocrinol. Metab. 82:4258-4265
Eriksson U, Alitalo K (1999): Structure, expression and receptor-binding properties of novel vascula
Fanton JW, Hubbard GB (1983): Spontaneous endometriosis in a cynomolgus monkey (Macaca fascicularis). Lab. Anim. Sci. 33:57-59
Fawell SE, Lees JA, White R, Parker MG (1990): Characterization and colocalization of steroid binding and dimerization activities in the mouse estrogen receptor. Cell 60:953- 962
Fedele L, Parazzini F, Bianchi S, Arcaini L, Candiani GB (1990): Stage and localization of pelvic endometriosis and pain. Fertil. Steril. 53:155-158
Fernandez-Shaw S, Hicks BR, Yudkin PL, Kennedy S, Barlow DH, Starkey PM (1993): Antiendometrial and antiendothelial autoantibodies in women with endometriosis. Hum. Reprod. 8:310-315
110

Literatur
Ferrara N, Davis-Smith T (1997): The biology of vascular endothelial growth factor. Endocr. Rev. 18:4-25
Ferrara N (1999): Vascular endothelial growth factor: molecular and biological aspects. Curr. Top. Microbiol. Immunol. 237:1-30
Ferriani RA, Charnock-Jones DS, Prentice A, Thomas EJ, Smith SK (1993): Immunohistochemical localization of acidic and basic fibroblast growth factors in normal
therapy of
SC Jr, Lo SS, Phillips DR (1987): Protein sequence of
1/KDR) that inhibits
ymal
an steroids regulate the expression of
H, Hirose R, Wen H, Tamaya T (1999): Angiogenesis in
od. Update
2002): The
N, el-Roeiy A, Confino E, Friberg J (1987): Is endometriosis an autoimmune
genesis. EXS. Birkhäuser Verlag,
human endometrium and endometriosis and the detection of their mRNA by polymerase chain reaction. Hum. Reprod. 8:11-16
Fidler IJ, Ellis LM (1994): The implications of angiogenesis for the biology and cancer metastasis. Cell 79:185-188
Findlay JK (1986): Angiogenesis in reproductive tissues. J. Endocrinol. 111:357-366
Fitzgerald LA, Steiner B, Rall endothelial glycoprotein IIIa derived from a cDNA clone. Identity with platelet glycoprotein IIIa and similarity to ”integrin”. Biol. Chem. 262:3936-3939
Folkman J (1971): Tumor angiogenesis: Therapeutic implications. N. Engl. J. Med. 285:1182-1186
Folkman J (1990): What is the evidence that tumors are angiogenesis dependent? J. Nat. Cancer Inst. 82:1-3
Folkman J (1995): Angiogenesis in cancer, vascular, rheumatoid and other diseases. Nat. Med. 1:27-31
Fong TA, Shawver LK, Sun L, Tang C, App H, Powell TJ, Kim YH, Schreck R, Wang X, Risau W, Ullrich A, Hirth KP, McMahon G (1999): SU5416 is a potent and selective inhibitor of the vascular endothelial growth factor receptor (FLK- tyrosine kinase catalysis, tumor vascularisation, and growth of multiple tumor types. Cancer Res. 59:99-106
Foster DC, Stern JL, Buscema J, Rock JA, Woodruff JD (1981): Pleural and parench pulmonary endometriosis. Obstet. Gynecol. 58:552-556
Fujimoto J, Hori M, Ichigo S, Tamaya T (1997): Ovari basic fibroblast growth factor and its mRNA in fibroblasts derived from uterine endometrium. Ann. Clin. Biochem. 34:91-96
Fujimoto J, Sakaguchi endometriosis and angiogenic factors. Gynecol. Obstet. Invest. 48:14-20
Garrido N, Navarro J, Remohí J, Simón C, Pellicer A (2000): Follicular hormonal environment and embryo quality in women with endometriosis. Hum. Repr 6:67-74
Garrido N, Navarro J, García-Velasco J, Remohí J, Pellicer A, Simón C ( endometrium versus embryonic quality in endometriosis-related infertility. Hum. Reprod. Update 8:95-103
Giavazzi R, Taraboletti G (1999): Angiogenesis and angiogenesis inhibitors in cancer. Forum (Genova) 9:261-272
Gleicher disease? Obstet. Gynecol. 70:115-122
Goldberg ID, Rosen EM (eds) (1997): Regulation of angio Basel, 79
111

Literatur
Goldman CK, Kendall RL, Cabrera G, Soroceanu L, Heike Y, Gillespie GY, Siegal GP, Mao X, Bett AJ, Huckle WR, Thomas KA, Curiel DT (1998): Paracrine expression of a
dometrium is regulated by oestrogen-receptor and
): Regulation of gap junction connexins in the
743
nctions of CYR61: A structural analysis. Abstracts from the Second
LC, Yang J, Taylor RN, Osteen K, Lessey BD (2001): Global gene
uterine and conceptus tissues. Biol. Reprod. 56:1527-1536
iosis. Am. J. Obstet. Gynecol.
of endometriosis. Am. J. Obstet. Gynecol.
95): Localization
RB (1978): Natural cell-mediated cytotoxicity in nude mice. In: Fogh J.,
ager E, Traub O, Grümmer R (2002): Responsiveness of endometrial
nd xenoestrogens. J. Mol. Endocrinol. 29:239-249
ular and glandular expression of integrins αvβ3 in
tnick AR (1940): Endometriosis of the lungs: experimental and clinical study. Am. J. Obstet. Gynecol. 40:832-833
native soluble vascular endothelial growth factor receptor inhibits tumor growth, metastasis, and mortality rate. Proc. Natl. Acad. Sci. USA 95:8795-8800
Gordon JD, Shifren JL, Foulk RA Taylor RN, Jaffe RB (1995): Angiogenesis in the human female reproductive tract. Obstet. Gynecol. Surv. 50:688-697
Greb RR, Heikinheimo O, Williams RF, Hodgen GD, Goodman AL (1997): Vascular endothelial growth factor in primate en progesterone-receptor ligands in vivo. Hum. Reprod. 12:1280-1292
Grümmer R, Winterhager E (1998 endometrium during early pregnancy. Cell Tissue Res. 293:189-194
Grümmer R, Traub O, Winterhager E (1999): Gap junction connexin genes cx26 and cx43 are differentially regulated by ovarian steroid hormones in rat endometrium. Endocrinology 140:2509-2516
Grümmer R, Schwarzer F, Bainczyk K, Hess-Stumpp H, Regidor PA, Schindler AE, Winterhager E (2001): Peritoneal endometriosis: validation of an in-vivo model. Hum. Reprod. 16:1736-1
Grzeszkiewicz TM, Leu S-J, Chen N, Chen C-C, Todorovic V, Lam SC-T, Lau LF (2002): Integrin-mediated fu International Workshop on the CCN Family of Genes, eingereicht.
Guidice LC, Kao expression profiling of human endometrium from women with endometriosis using genechip microarray technology (abstract). In: NICHD Conference on endometriosis: Emerging research and intervention strategies. Bethesda MD:62-63
Gupta A, Bazer FW, Jaeger LA (1997): Immunolocalization of acidic and basic fibroblast growth factors in porcine
Halme J, Becker S, Hammond MG, Raj MH, Raj S (1983): Increased activation of pelvic macrophages in infertile women with mild endometr 145:333-337
Halme J, Becker S, Haskill S (1987): Altered maturation and function of peritoneal macrophages: possible role in pathogenesis 156:783-789
Harrison-Woolrych ML, Sharkey AM, Charnock-Jones DS, Smith SK (19 and quantification of vascular endothelial growth factor messenger ribonucleic acid in human myometrium and leiomyomata. J. Clin. Endocrinol. Metab. 80:1853-1858
Hebermann Giovanella B.C. eds.: The nude mouse in experimental and clinical research. New York: Academic Press, 135-166
Heikaus S, Winterh genes Connexin26, Connexin43, C3 and clusterin to primary estrogen, selective estrogen receptor modulators, phyto- a
Hermenegildo C, Cano A (2000): Pure anti-oestrogens. Hum. Reprod. Update 6:237-243
Hii L, Rogers P (1998): Endometrial vasc women with and without endometriosis. Hum. Reprod. 13:1030-1035
Hobbs JE, Bor
112

Literatur
Horwitz KB, Alexander PS (1983): In situ photo linked nuclear progesterone receptors of human breast cancer cells: subunit molecular weights after transformation and translocation. Endocrinology 113:2195-2201
Huang JC, Papasakelariou C, Dawood MY (1996): Epidermal growth factor and basic
w of controlled trial in
y L, Makela S (1996):
ta C, Murthy L, Stancel GM (1997): Selective inhibition of estrogen-
(1999): Regulation of angiogenic growth factors in the female
y
Hum. Reprod. 10:2666-2670
n t4
ynecol. Clin. North.
u enhoff S, Adriansson M, Malm C (2000): Frequent good partial remissions from thalidomide including best response ever in patients with advanced refractory and relapsed myeloma. Br. J. Haematol. 109:89-96
fibroblast growth factor in peritoneal fluid of women with endometriosis. Fertil. Steril. 65:931-934
Hucke J, Distler W (1989): Klinische Manifestation und Klassifizierung der Endometriose. Gynäkologe 22:289-293
Hughes EG, Fedorkow DM, Collins JA (1993): A quantitative overvie endometriosis-associated infertility. Fertil. Steril. 59:963-970
H der SM, Stancel GM, Chiappetta C, Murthy L, Boettger-Tong H Uterine expression of vascular endothelial growth factor is increased by estradiol and tamoxifen. Cancer Res. 56:3954-3960
Hyder SM, Chiappet regulated gene expression in vivo by the pure antiestrogen ICI 182,780. Cancer Res. 57:2547-2549
Hyder SM, Stancel GM reproductive tract by estrogens and progestins. Mol. Endocrinol. 13:806-811
H der SM, Stancel GM (2000): Regulation of VEGF in the reproductive tract by sex-steroid hormones. Histol. Histopathol. 15:325-334
Igarashi A, Okochi H, Bradham DM, Grotendorst GR (1993): Regulation of connective tissue growth factor gene expression in human skin fibroblasts and during wound repair. Mol. Biol. Cell 4:637-645
Illera MJ, Juan L, Stewart CL, Cullinan E, Ruman J, Lessey BA (2000): Effect of peritoneal fluid from women with endometriosis on implantation in the mouse model. Fertil. Steril. 74:41-48
Isomaa VV, Ghersevich SA, Maentausta OK, Peltoketo EH, Poutanen MH, Vihko RK (1993): Steroid biosynthetic enzymes: 17 beta-hydroxysteroid dehydrogenase. Ann. Med. 25:91-97
Jahn E, Classen-Linke I, Kusche M, Beier HM, Traub O, Grümmer R, Winterhager E (1995): Expression of gap junction connexins in the human endometrium throughout the menstrual cycle.
Jain RK (1987): Transport of molecules in the tumor interstitium: A review. Cancer Res. 47:3039-3051
Javert CT (1949): Pathogenesis of endometriosis based on endometrial homeoplasia, direct extension, exfoliation and implantation, lymphatic and hematogenous metastasis. Including five case reports of endometrial tissue in pelvic lymph nodes. Cancer 2:399-410
Jay P, Berge-Lefranc JL, Marsollier C, Mejean C, Taviaux S, Berta P (1997): The human growth factor-inducible immediate early gene, CYR61, maps to chromosome 1p. Oncogene 14:1753-1757
Joukov V, Pajusola K, Kaipainen A, Chilov D, Lahtinen I, Kukk E, Saksela O, Kalkkine N, Alitalo K (1996): A novel endothelial growth factor, VEGF-C, is a ligand for the Fl (VEGFR-3) and KDR (VEGFR-2) receptor tyrosine kinases. EMBO J. 15:290-298
Jubanyik KJ, Comite F (1997): Extrapelvic endometriosis. Obstet. G Am. 24:411-440
J liusson G, Celsing F, Turesson I, L
113

Literatur
Kao LC, Tulac S, Lobo S, Imani B, Yang J, Germeyer A, Osteen K, Taylor RN, Lessey BA, Giudice LC (2002): Global gene profiling in human endometrium during the window of implantation. Endocrinology 143:2119-2138
ative translational initiation from a single mRNA as the mechanism
, Sargent IL, Hicks BR, Barlow DH (1990): Antiendometrial
44
ol. Cell. Biol.
hen CC, Yang GP, Abler AS, Lau LF (1997):
atsu T, Maeda K, Tsukamoto K, Yamamoto T, Fushiki S,
, Schorpp M, Wagner U, Ryffel CU (1986): An estrogen response element
uction by fibroblast growth factor receptors.
en the chips are down. Nature 410:860-861
FB, Richardson DA, Kenigsberg D (1986): Endometrial
tafsson
s α and β. Endocrinology 138:863-870
Kastner P, Bocquel MT, Turcotte B, Garnier JM, Horwitz KB, Chambon P, Gronemeyer H (1990): Transient expression of human and chicken progesterone receptors does not support altern generating two receptor isoforms. J. Biol. Chem. 265:12163-12167
Kennedy SH, Starkey PM antibodies in endometriosis measured by enzyme linked immunosorbent assay before and after treatment with danazol and nafarelin acetate. Obstet. Gynecol. 75:914-918
Kerbel RS (1997): A cancer therapy resistant to resistance. Nature (Lond.) 390:335-335
Kettel WC, Stein RJ (1951): The viability of the cast-off menstrual endometrium. Am. J. Obstet. Gynecol. 61:440-442
Kim KJ, Li B, Winer J, Armanini M, Gillet N, Phillips HS, Ferrara N (1993): Inhibition of vascular endothelial growth factor-induced angiogenesis suppresses tumor growth in vivo. Nature 362:841-8
Kireeva ML, Mo FE, Yang GP, Lau LF (1996): Cyr61, product of a growth factor-inducible immediate-early gene, promotes cell proliferation, migration, and adhesion. M 16:1326-1334
Kireeva ML, Latinkic BV, Kolesnikova TV, C Cyr61 and Fisp12 are both ECM-associated signaling molecules: activities, metabolism, and localization during development. Exp. Cell Res. 233:63-77
Kitawaki J, Noguchi T, Am Osawa Y, Honjo H (1997): Expression of aromatase cytochrome P450 protein and messenger ribonucleic acid in human endometriotic and adenomyotic tissues but not in normal endometrium. Biol. Reprod. 57:514-519
Klein S, Roghani M, Rifkin DB (1997): Fibroblast growth factors as angiogenesis factors: new insights into their mechanism of action. EXS. 79:159-192
Klein-Hitpass L derived from the 5´ flanking region of the Xenopus vitellogenin A2 gene functions in transfected human cells. Cell 46:1053-1061
Klint P, Claesson-Welsh L (1999): Signal transd Front. Biosci. 4:D165-177
Knight J (2001): Wh
Koji T, Brenner RM (1993): Localization of estrogen receptor messenger ribonucleic acid in rhesus monkey uterus by nonradioactive in situ hybridization with digoxigenin-labelled oligodeoxynucleotides. Endocrinology 132:382-392
Koninckx PR, Kennedy SH, Barlow DH (1998): Endometriotic disease: the role of peritoneal fluid. Hum. Reprod. Update 4:741-751
Kreiner D, Frommowitz immunofluorescence associated with endometriosis and inflammatory disease. Fertil. Steril. 46:243-246
Kuiper GGJM, Carlsson B, Grandien K, Enmark E, Haggblad J, Nilsson S, Gus JA (1997): Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptor
Kumar V, Green S, Stack G, Berry M, Jin JR, Chambon P (1987): Functional domains of the human estrogen receptor. Cell 51:941-951
114

Literatur
Labrie F, Luu-The V, Labrie C, Berube D, Couet J, Zhao HF, Gagne R, Simard J (1989): Characterization of two mRNA species encoding human estradiol 17β-dehydrogenase and assignment of the gene to chromosome 17. J. Steroid Biochem. 34:189-197
hort- and long-acting
s in
e secondary Müllerian system. Obstet. Gynecol. Survey 27:133-146
n by chromatography and photoaffinity
89): Immunohistochemical
J. Clin. Invest. 90:188-195
, Sun J (1994a): Further
, Schinnar R, Bilker W, Strom BL
6248-46255
etriose - ein neues Modell ihrer
Germany, pp. 8-13
Laemmli UK (1970): Cleavage of structural proteins during the assembly of the head of bacteriophage A. Nature 227:680-685
Lan NC, Katzenellenbogen BS (1976): Temporal relationships between hormone receptor binding and biological responses in the uterus: studies with s derivates of estriol. Endocrinology 98:220-227
Lau LF, Nathans D (1985): Identification of a set of genes expressed during G0/G1 transition of cultured mouse cells. EMBO J. 4:3145-3151
Lau LF, Nathans D (1987): Expression of a set of growth-related immediate early gene BALBc3T3 cells coordinate regulation with c-fos or c-myc. Proc. Natl. Acad. Sci. USA 84:1182-1186
Lau LF, Lam SC-T (1999): The CCN family of angiogenic regulators: the integrin connection. Exp. Cell Res. 248:44-57
Lauchlan SC (1972): Th
Ledger WL (1999): Endometriosis and infertility: an integrated approach. Int. J. Gynaecol. Obstet. 64:S33-40
Lee J, Gray A, Yuan J, Luoh SM, Avraham H, Wood WI (1996): Vascular endothelial growth factor-related protein: a ligand and specific activator of the tyrosine kinase receptor Flt 4. Proc. Natl. Acad. Sci. USA 93:1988-1992
Leenders WPJ (1998): Targeting VEGF in anti-angiogenic and anti-tumour therapy. Int. J. Exp. Pathol. 79:339-346
Lessey BA, Alexander PS, Horwitz KB (1983): The subunit structure of human breast cancer progesterone receptors: characterizatio labeling. Endocrinology 112:1267-1274
Lessey BA, Metzger DA, Haney AF, McCartney KS Jr (19 analysis of estrogen and progesterone receptors in endometriosis: comparison with normal endometrium during the menstrual cycle and the effect of medical therapy. Fertil. Steril. 51:409-415
Lessey BA, Damjanovich L, Coutifaris C, Castelbaum A, Albelda SM, Buck CA (1992): Integrin adhesion molecules in the human endometrium. Correlation with the normal and abnormal menstrual cycle.
Lessey BA, Castelbaum AJ, Buck CA, Lei Y, Yowell CW characterization of endometrial integrins during the menstrual cycle and in pregnancy. Fertil. Steril. 62:497-506
Lessey BA, Castelbaum AJ, Sawin SW, Buck CA (1994b): Aberrant integrin expression in the endometrium of women with endometriosis. J. Clin. Endocrinol. Metab. 79:643-649
Leu SJ, Lam SC, Lau LF (2002): Pro-angiogenic activities of CYR61 (CCN1) mediated through integrins alphavbeta3 and alpha6beta1 in human umbilical vein endothelial cells. J. Biol. Chem. 48:4
Leung DW, Cachianes G, Kuang WJ, Goeddel DV, Ferrara N (1989): Vascular endothelial growth factor is a secreted angiogenic mitogen. Science 246:1306-1309
Leyendecker G, Wildt L, Plath T, Kunz G (1995): Endom Entstehung. In: Der Frauenarzt, Endometriose. Schweppe KW, Schindler AE, Semm K, Runnebaum B (eds) Demeter Verlag, Balingen,
115

Literatur
Lieberman BA (1997): The estrogen receptor activity cycle: dependence on multiple protein- protein interactions. Crit. Rev. Eucaryotic Gene Expression 7:43-59
Lin J, Liliensiek B, Kanitz M, Schimanski U, Böhrer H, Waldherr R, Martin E, Kauffmann
, fibroblasts and smooth muscle cells. Cardiovasc.
tet. Gynaecol. 93:859-862
n-progesterone ratio. Endocrinology 132:2380-2386
ey HW (1975): Animal model of human disease: endometriosis in
the vascular permeability factor.
rnal cardiovascular and other physiological responses to the
evalence and genesis of endometriosis. Hum.
63:195-202
eaction assay. Fertil. Steril. 74:753-759
rkey AM, Smith peritoneal fluid
G, Ziegler R, Nawroth PP (1998): Molecular cloning of genes differentially regulated by TNF-α in bovine aortic endothelial cells Res. 38:802-803
Liu DT, Hitchcock A (1986): Endometriosis: its association with retrograde menstruation, dysmenorrhoea and tubal pathology. Br. J. Obs
Lowry OH, Rosebrough NJ, Farr AL Randall RJ (1951): Protein measurement with the folin phenol reagent. J. Biol. Chem. 193:265-275
Luciano AA (1982): The treatment of endometriosis. Contemp. Obstet. Gynecol. 19:211
Lundeen et al. (2002), eingereicht
Lye SJ, Nicholson BJ, Mascarenhas M, MacKenzie L, Petrocelli T (1993): Increased expression of connexin-43 in the rat myometrium during labour is associated with an increase in plasma estroge
MacGregor JI, Jordan VC (1998): Basic guide to the mechanisms of antiestrogen action. Pharmacol. Rev. 50:151-196
MacKenzie WF, Cas rhesus monkeys. Am. J. Pathol. 80:341-344
Maglione D, Guerriero V, Viglietto G, Delli-Bovi P, Persico MG (1991): Isolation of a human placenta cDNA encoding for a protein related to Proc. Natl. Acad. Sci. USA 88:9267-9271
Magness RR (1998): Mate endocrinology of pregnancy. In: The endocrinology of pregnancy., Ed: FW Bazer. Totowa, NJ: Humana Press; pp. 507-539
Mahmood TA, Templeton A (1991): Pr Reprod. 6:544-549
Mangal RK, Wiehle RD, Poindexter III AN, Weigel NL (1997): Differential expression of uterine progesterone receptor forms A and B during the menstrual cycle. J. Steroid Biochem. Mol. Biol.
Marcus SF, Edwards RG (1994): High rates of pregnancy after long-term down-regulation of women with severe endometriosis. Am. J. Obstet. Gynecol. 171:812-717
Matsuzaki S, Fukaya T, Suzuki T, Murakami T, Sasano H, Yajima A (1999): Oestrogen receptor alpha (ERα) and beta (ERβ) mRNA expression in human endometrium throughout the menstrual cycle. Mol. Hum. Reprod. 5:559-564
Matsuzaki S, Fukaya T, Uehara S, Murakami T, Sasano H, Yajima A (2000a): Characterization of messenger RNA expression of estrogen receptor-α and -β in patients with ovarian endometriosis. Fertil. Steril. 73:1219-1225
Matsuzaki S, Uehara S, Murakami T, Fujiwara J, Funato T, Okamura K (2000b): Quantitative analysis of estrogen receptor alpha and beta messenger ribonucleic acid levels in normal endometrium and ovarian endometriotic cysts using a real-time reverse transcription-polymerase chain r
McLaren J, Prentice A, Charnock-Jones DS, Milican SA, Muller KH, Sha SK (1996a): Vascular endothelial growth factor is produced by macrophages in endometriosis and is regulated by ovarian steroids. J. Clin. Invest. 98:482-489
116

Literatur
McLaren J, Prentice A, Charnock-Jones DS, Smith SK (1996b): Vascular endothelial growth factor (VEGF) concentrations are elevated in peritoneal fluid of women with
oumenou AG, Koumantakis GE, Neonaki MA, Koumantakis EE,
i-endometriosis
Adenomyositis und Adenomatome im und
ashi Ki, Simpson ER (1995): Ad4 BP/SF-1 regulates
REB and
e. Mol. Cell. Endocrinol. 134:147-156
er Res. 56:1615-1620
low KA, Hayes MM (1996): The effect of
nt and vascular integrity. Mol. Cell. Biol. 22:8709-8720
e inhibit endothelial cell proliferation in vitro. J. Neurooncol. 43:109-
endometrium
Muse K (1988): Clinical manifestations and classification of endometriosis. Clin. Obstet.
ogenase type 2
ilcox M (2001): Global expression changes of constitutive and hormonally regulated genes during endometrial neoplastic transformation. Gynecol. Oncol. 83:175- 176
endometriosis. Hum. Reprod. 11:220-223
McLaren J (2000): Vascular endothelial growth factor and endometriotic angiogenesis. Hum. Reprod. Update 6:45-55
Metalliotakis IM, G Dionyssopoulou E, Athanassakis I, Vassiliadis S (2003): Serum concentrations of growth factors in women with and without endometriosis: the action of ant medicines. Int. Immunopharmacol. 3:81-89
Meyer R (1919): Über den Stand der Frage allgemeinen und insbesondere über Adenomyositis serosoepithelialis Adenomyometritis sarcomatosa. Zbl. f. Gynäk. 43:745-750
Michael MD, Kilgore MW, Moroh cyclic AMP-induced transcription from the proximal promotor (PII) of the human aromatase P450 (CYP19) gene in the ovary. J. Biol. Chem. 270:13561-13566
Michael MD, Michael LF, Simpson ER (1997): A CRE-like sequence that binds C contributes to cAMP-dependent regulation of the proximal promotor of the human aromatase P450 (CYP19) gen
Millauer B, Lunghi MP, Plate KH (1996): Dominant-negative inhibition of Flk-1 suppresses the growth of many tumor types in vivo. Canc
Miller JD, Anderson MG (1997): Therapeutic effects of leuprorelin microspheres on endometriosis and uterine leiomyomata. Adv. Drug Deliv. Rev. 28:139-155
Minchinton AI, Fryer KH, Wendt KR, C thalidomide on experimental tumors and metastases. Anticancer Drugs 7:339-343
Mo FE, Muntean AG, Chen CC, Stolz DB, Watkins SC, Lau LF (2002): CYR61 (CCN1) is essential for placental developme
Moreira AL, Friedlander DR, Shif B, Kaplan G, Zagzag D (1999): Thalidomide and a thalidomide analogu 114
Mostoufizadeh M, Scully RE (1980): Malignant tumors arising in endometriosis. Clin. Obstet. Gynecol. 23:951-963
Mote PA, Balleine RL, McGowan EM, Clarke CL (1999): Colocalization of progesterone receptors A and B by dual immunofluorescent histochemistry in human during the menstrual cycle. J. Clin. Endocrinol. Metab. 84:2963-2971
Mueller MD, Vigne J-L, Vaisse C, Taylor RN (2000): Glycodelin: a pane in the implantation window. Semin. Reprod. Med. 18:289-298
Mülhardt C (2002): Der Experimentator: Molekularbiologie/Genomics. 3. Aufl. – Heidelberg; Berlin: Spektrum, Akad. Verl. 2002
Gynecol. 31:813-822
Mustonen MVJ, Isomaa VV, Vaskivou T, Tapanainen J, Poutanen MH, Stenback F, Vihko RK, Vihko PT (1998): Human 17β-hydroxysteroid dehydr messenger ribonucleic acid expression and localization in term plazenta and in endometrium during the menstrual cycle. J. Clin. Endocrinol. Metab. 83:1319-1324
Mutter GL, Baak JP, Fitzgerald JT, Gray R, Neuberg D, Kust GA, Gentleman R, Gullans SR, Wei LJ, W
117

Literatur
Nakamura H, Oosawa M, Kondou J, Inagaki S, Shibata H, Narita O, Suganuma N, Tomoda Y (1992): Menotropin stimulation after prolonged Gonadotropin releasing hormone agonist pretreatment for in vitro fertilization in patients with endometriosis. J.
, Marbaix E, Jadoul P, Donnez J (2000a): Transplantation of
un SE (1996): Aromatase expression in
tes aromatase expression in
m. Reprod.
uman. Nucleic Acids
lies. Obstet. Gynecol.
itioned media
Proc. Natl. Acad. Sci. USA 93:11675-11680
vation of estrogen receptors ER α and ER β at AP1 sites.
-releasing hormone
n P, White R (1993): Structure and function of
C, Albert C, Bonilla-Musoles F, Remohí J, Simón C
Assit. Reprod. Genet. 9:113-117
Nisolle M, Casanas-Roux F cultured explants of human endometrium into nude mice. Hum. Reprod. 15:572-577
Nisolle M, Casanas-Roux F, Donnez J (2000b): Early-stage endometriosis: adhesion and growth of human menstrual endometrium in nude mice. Fertil. Steril. 74:306-312
Noble LS, Simpson ER, Johns A, Bul endometriosis. J. Clin. Endocrinol. Metab. 81:174-179
Noble LS, Takayama K, Putman JM, Johns DA, Hinshelwood MM, Agarwal VR, Zhao Y, Carr BR, Bulun SE (1997): Prostaglandin E2 stimula endometriosis-derived stromal cells. J. Clin. Endocrinol. Metab. 82:600-606
Odukoya OA, Wheatcroft N, Weetman AP, Cooke ID (1995): The prevalence of endometrial immunoglobulin G antibodies in patients with endometriosis. Hu 10:1214-1219
Ogawa S, Inoue S, Watanabe T (1998): Molecular cloning and characterization of human estrogen receptor betacx: a potential inhibitor of estrogen action in h Res. 26:3505-3512
Olive DL, Henderson DY (1987): Endometriosis and mullerian anoma 69:412-415
Olive DL, Montoya I, Riehl RM, Schenken RS (1991): Macrophage cond enhance endometrial stromal cell proliferation in vitro. Am. J. Obstet. Gynecol. 164:953- 958
Olson TA, Mohanraj D, Carson LF, Ramakrishnan S (1994): Vascular permeability factor gene expression in normal and neoplastic human ovaries. Cancer Res. 54:276-280
Orlandini M, Marconcini L, Feruzzi R, Oliviero S (1996): Identification of a c-fos-induced gene that is related to the platelet-derived growth factor/vascular endothelial growth factor family.
Paech K, Webb P, Kuiper GG, Nilsson S, Gustafsson J, Kushner PJ, Scanlan TS (1997): Differential ligand acti Science 277:1508-1510
Paoletti AM, Serra GG, Cagnacci A, Vacca AM, Guerriero S, Solla E, Melis GB (1996): Spontaneous reversibility of bone loss induced by gonadotropin analog treatment. Fertil. Steril. 65:707-710
Parker MG, Arbuckle N, Dauvois S, Danielia the estrogen receptor. Ann. N.Y. Acad. Sci. 684:119-126
Pellicer A, Valbuena D, Bauset (1998): The follicular endocrine environment in stimulated cycles of women with endometriosis: steroid levels and embryo quality. Fertil. Steril. 69:1135-1141
Petrocelli T, Lye S (1993): Regulation of transcripts encoding the myometrial gap junction protein, connexin-43, by estrogen and progesterone. Endocrinology 133:284-290
Phillips RA, Jeweit MA, Gallie BL (1989): Growth of human tumors in immunodeficient SCID mice and nude mice. Curr. Top. Microbiol. Immunol. 152:259-266
Pilarsky CP, Schmidt U, Eissrich C, Stade J, Froschermaier SE, Haase M, Faller G, Kirchner TW, Wirth MP (1998): Expression of the extracellular matrix signaling molecule Cyr61 is downregulated in prostate cancer. Prostate 36:85-91
118

Literatur
Pinkert TC, Catlow CE, Straus R (1979): Endometriosis of the urinary bladder in a man with prostatic carcinoma. Cancer 43:1562-1567
Prentice A, Randall BJ, Weddell A, McGill A, Henry L, Horne CH, Thomas EJ (1992): Ovarian steroid receptor expression in endometriosis and in two potential parent epithelia:
tor in
antone®-Gyn
linical aspects. Obstet. Gynecol. Surv. 50:747-754
-620
the uterine response to estradiol-17beta in ovariectomized ewes: Uterine
A review of facts and fancies. Obstet.
e. Science 259:1763-1766
endometrium and peritoneal mesothelium. Hum. Reprod. 7:1318-1325
Presta M (1988): Sex hormones modulate the synthesis of basic fibroblast growth fac human endometrial adenocarcinoma cells: implications for the neovascularization of normal and neoplastic endometrium. J. Cell. Physiol. 137:593-597
Ramey JW, Archer DF (1993): Peritoneal fluid: its relevance to the development of endometriosis. Fertil. Steril. 60:1-14
Regidor PA, Schindler AE, Bühler K, Gerhard I, Kimmig R, Meinen K, Hoffmann G, Lübben G, Kienle E (1996): Ergebnisse eines Langzeit Follow-up bei der Behandlung der Endometriose mit dem GnRH-Agonisten Leuprorelinacetat-Depot (En Monats-Depot). Zentralbl. Gynakol. 118:283-290
Revelli A, Modotti M, Ansaldi C, Massobrio M (1995): Recurrent endometriosis: A review of biological and c
Reynolds LP, Kirsch JD, Kraft KC, Redmer DA (1998a): Time-course of the uterine response to estradiol-17beta in ovariectomized ewes: expression of angiogenic factors. Biol. Reprod. 59:613
Reynolds LP, Kirsch JD, Kraft KC, Knutson DL, McClaflin WJ, Redmer DA (1998b): Time-course of growth and microvascular development. Biol. Reprod. 59:606-612
Reynolds LP, Redmer DA (2001): Angiogenesis in the placenta. Biol. Reprod. 64:1033- 1040
Rider V, Carlone DL, Foster RT (1997): Oestrogen and progesterone control basic fibroblast growth factor mRNA in the rat uterus. J. Endocrinol. 154:75-84
Ridley JH, Edwards IK (1958): Experimental endometriosis in the human. Am. J. Obstet. Gynecol. 76:783-790
Ridley JH (1968): The histogenesis of endometriosis. Gynecol. Survey 23:1-23
Risau W (1997): Mechanisms of angiogenesis. Nature 386:671-674
Rivera-Gonzalez R, Petersen DN, Tkalcevic G, Thompson DD, Brown TA (1998): Estrogen-induced genes in the uterus of ovariectomized rats and their regulation by droloxifene and tamoxifene. J. Steroid Biochem. Mol. Biol. 64:13-24
Rivlin ME, Miller JD, Krueger RP, Patel RB, Bower JD (1990): Leuprolide acetate in the management of ureteral obstruction caused by endometriosis. Obstet. Gynecol. 75:532- 536
Roberts AB, Sporn MB (1989): Regulation of endothelial cell growth, architecture, and matrix synthesis by TGF-β. Am. Rev. Respir. Dis. 140:1126-1128
Roberts WG, Palade GE (1995): Increased microvascular permeability and endothelial fenestration induced by vascular endothelial growth factor. J. Cell Sci. 108:2369-2379
Rogers PAW (1996): Structure and function of endometrial blood vessels. Hum. Reprod. Update 2:57-62
Rogers PAW, Gargett CE (1999): Endometrial angiogenesis. Angiogenesis 2:287-294
Rohan PJ, Davis P, Mokaluk CA, Kearns M, Krutzsch H, Siebenlist U, Kelly K (1993): PAC-1: A mitogen-induced nuclear protein tyrosine phosphatas
119

Literatur
Russel WW (1899): Aberrant portions of the Müllerian duct found in an ovary. Ovarian cysts of Müllerian origin. Bull. John Hopkins Hosp. 10:8-10
Biol. Reprod. 59:491-496
d overexpression in
peritoneal cavity. Am. J. Obstet. Gynecol. 14:422-469
factor receptor-1 is a
t growth factor and FGF-R1 in human endometrium during the menstrual cycle
e rabbit: effects
RF, Hodgen GD (1991): Experimental endometriosis in monkeys.
hlenberg C, Becker MHG, Key G, Flad HD, Gerdes J (1993): very large, ubiquitous
nuclear protein with numerous repeated elements, representing a new kind of cell
Ugarova TP, Ye RD, Lau LF, Lam SC-T (2002): Identification of integrin alpha(M)beta(2) as an adhesion
-4465
eue Therapiemöglichkeiten durch Buserelin. Schindler AE, Schweppe
Ryseck RP, Macdonald-Bravo H, Mattéi MG, Bravo R (1991): Structure, mapping and expression of fisp-12, a growth-factor-inducible gene encoding a secreted cysteine-rich protein. Cell Growth Differ. 2:225-233
Saleh M, Stacker SA, Wilks AF (1996): Inhibition of growth of C6 glioma cells in vivo by expression of antisense vascular endothelial growth factor sequence. Cancer Res. 56:393-401
Samathanam CA, Adesanya OO, Zhou J, Wang J, Bondy CA (1998): Fibroblast growth factors 1 and 2 in the primate uterus.
Sampath D, Zhu Y, Winneker RC, Zhang Z (2001a): Aberrant expression of Cyr61, a member of the CCN (CTGF/Cyr61/NOVH) family, and dysregulation by 17β-estradiol and basic fibroblast growth factor in human uterine leiomyomas. J. Clin. Endocrinol. Metab. 86:1707-1715
Sampath D, Winneker RC, Zhang Z (2001b): Cyr61, a member of the CCN family, is required for MCF-7 cell proliferation: regulation by 17β-estradiol an human breast cancer. Endocrinology 142:2540-2548
Sampson J (1927): Pertioneal endometriosis due to the menstrual dissemination of endometrial tissue into the
Sanflippo JS, Wakim NG, Schikler KN, Yussman MA (1986): Endometriosis in association with uterine anomaly. Am. J. Obstet. Gynecol. 154:39-43
Sangha RK, Li XF, Shams M, Ahmed A (1997): Fibroblast growth critical component for endometrial remodeling: localization and expression of basic fibroblas and decreased FGF-R1 expression in menorrhagia. Lab. Invest. 77:389-402
Schenken RS, Asch RH (1980): Surgically induction of endometriosis in th on fertility and concentration of peritoneal fluid prostaglandins. Fertil. Steril. 34:581-587
Schenken RS, William Ann. N.Y. Acad. Sci. 622:242-255
Schlüter C, Duchrow M, Wo The cell proliferation-associated antigen of antibody Ki-67: A
cycle-maintaining proteins. J. Cell. Biol. 123:513-522
Schober JM, Chen N, Grzeszkiewicz TM, Jovanovic I, Emeson EE,
receptor on peripheral blood monocytes for Cyr61 (CCN1) and connective tissue growth factor (CCN2): immediate-early gene products expressed in atherosclerotic lesions. Blood 99:4457
Schreiner WE (1985): Gefahren und Komplikationen der Endometriose. In: Litschgi M (Hrsg), Genitalendometriose. Karger, Basel, pp. 87
Schrodt GR, Alcorn MO, Ibanez J (1980): Endometriosis of the male urinary system: a case report. J. Urol. 124:722-723
Schweppe KW (1989): Ätiologie, Histologie und Pathophysiologie der Endometriose. In: Endometriose- n KW (eds), de Gruyter, Berlin, pp.3-22
Schweppe KW (2002): Therapie der Endometriose unter Berücksichtigung der Aktivitätsgrade. Gynäkologe 35:255-261
120

Literatur
Scully RE (1966): The development of malignancy in endometriosis. Clin. Obstet. Gynecol. 9:384-411
Scully RE (1978): Tumors of the ovary and maldeveloped gonads. In: Hartmann (ed) Atlas of tumorpathology. Washington
Sharpe KL, Bertera MC, Muse KN, Vernon MW (1991): Spontanous and steroid-induced
: An open randomized comparative study of the effect of goserelin depot
Group, Carnforth, UK
ndometriosis. J. Clin. Endocrinol. Metab. 81:3112-3118
Shweiki D, Itin A, Neufeld G, Gitay-Goren H, Keshet E (1993): Patterns of expression of
5-2243
Steffen CL, Ball-Mirth DK, Harding PA, Bhattacharyya N, Pillai S, Brigstock DR (1998):
in vitro. Growth Factors 15:199-213
Surveyor GA, Wilson AK, Brigstock DR (1998): Localization of connective tissue growth
w
2): Patterns of expression of integrin molecules in human endometrium
p PG (1999): An experimental model
, Sasano H, Carr BR, Bulun SE (1998): Treatment of
a ujino Y, Ogita S (2000): Antiendometrial IgM
recurrence of endometriosis after suppression by a gonadotropin-releasing hormone agonist in the rat. Am. J. Obstet. Gynecol. 164:187-194
Shaw RW (1992) and danazol in the treatment of endometriosis. Fertil. Steril. 58:265-272
Shaw RW (1993): An atlas of endometriosis. Parthenon Publishing
Shifren JL, Tseng JF, Zaloudek CJ, Ryan IP, Meng YG, Ferrara N, Jaffe RB, Taylor RN (1996): Ovarian steroid regulation of vascular endothelial growth factor in the human endometrium: implications for angiogenesis during the menstrual cycle and in the pathogenesis of e
Shing Y, Folkman J, Sullivan R, Butterfield C, Murray J, Klagsbrun M (1984): Heparin affinity: purification of a tumor-derived capillary endothelial cell growth factor. Science 223:1296-1299
Shroen D (1690): Disputatio inauguralis medica de ulceribus uteri. Jena: Krebs:6-17
vascular endothelial growth factor (VEGF) and VEGF receptors in mice suggest a role in hormonally regulated angiogenesis. J. Clin. Invest. 91:223
Simpson ER, Mahendroo MS, Means GD, Kilgore MW, Hinshelwood MM, Graham- Lorence S, Amarneh B, Ito Y, Fisher CR, Michael MD, Mendelson CR, Bulun SE (1994): Aromatase cytochrome P450, the enzyme responsible for estrogen biosynthesis. Endocr. Rev. 15:342-355
Smith SK (1997): Angiogenesis. Semin. Reprod. Endocrinol. 15:221-227
Characterization of cell-associated and soluble forms of connective tissue growth factor (CTGF) produced by fibroblast cells
Suginami H (1991): A reappraisal of the coelomic metaplasia theory by reviewing endometriosis occuring in unusual sites and instances. Am. J. Obstet. Gynecol. 165:214- 218
factor during the period of embryo implantation in the mouse. Biol. Reprod. 59:1207-1213
S itchenko AC, Kauffman RS, Becker M (1991): Are there antiendometrial antibodies in sera of women with endometriosis? Fertil. Steril. 56:235-241
Tabibzadeh S (199 throughout the menstrual cycle. Hum. Reprod. 7:876-882
Tabibzadeh S, Miller S, Dodson WC, Satyaswaroo for the endometriosis in athymic mice. Front. Biosci. 4:C4-9
Takayama K, Zeitoun K, Gunby RT severe postmenopausal endometriosis with an aromatase inhibitor. Fertil. Steril. 69:709- 713
T naka T, Umesaki N, Mizuno K, F autoantibodies in endometriotic patients: a preliminary study. Clin. Exp. Obstet. Gynecol. 27:133-137
121

Literatur
Taylor RN, Lebovic DI, Mueller MD (2002a): Angiogenic factors in endometriosis. Ann. N.Y. Acad. Sci. 955:89-100
8
mptomatic endometriosis: does it
triosis: the best of
ts and response
btypes. Endocrinology 138:4022-4025
ascular
98): Em-800, a novel
women with minimal endometriosis or unexplained infertility.
-receptors. Mol.
of human TR3 orphan receptor gene. A member of steroidreceptor superfamily. J.
z
Taylor RN, Lundeen SG, Giudice LC (2002b): Emerging role of genomics in endometriosis research. Fertil. Steril. 78:694-69
TeLinde RW, Scott RB (1950): Experimental endometriosis. Am. J. Obstet. Gynecol. 60:1147-1173
Thomas EJ, Cooke ID (1987): Successful treatment of asy benefit infertile women? Br. Med. J. (Clin. Res. Ed.) 294:1117-1119
Thomas EJ (1992): Combining medical and surgical treatment for endome both worlds? Br. J. Obstet. Gynaecol. 99:5-8
Thomas EJ (1995): Endometriosis, 1995 – confusion or sense? Int. J. Gynaecol. Obstet. 48:149-155
Thompson CJ, Tam NN, Joyce JM, Leav I, Ho SM (2002): Gene expression profiling of testosterone and estradiol-17 beta-induced prostatic dysplasia in Noble ra to the antiestrogen ICI 182,780. Endocrinology 143:2093-2105
Tong W, Perkins R (1997): QSAR models for binding of estrogenic compounds to estrogen receptor α and β su
Torry DS, Holt VJ, Keenan JA, Harris G, Caudle MR, Torry RJ (1996): V endothelial growth factor expression in cycling human endometrium. Fertil. Steril. 66:72- 80
Tremblay A, Tremblay GB, Labrie C, Labrie F, Giguere V (19 antiestrogen, acts as a pure antagonist of the transcriptional functions of estrogen receptors alpha and beta. Endocrinology 139:111-118
Tummon IS, Maclin VM, Radwanska E, Binor Z and Dmowski WP (1988): Occult ovulatory dysfunction in Fertil. Steril. 50:716-720
Tung L, Mohamed MK, Hoeffler JP, Takimoto GS, Horowitz KB (1993): Antagonist- occupied human progesterone B-receptors activate transcription without binding to progestrone response elements and are dominantly inhibited by A Endocrinol. 7:1256-1265
Uemura H, Mizukami A, Chang C (1995): Identification of a new enhancer in the promotor region Biol. Chem. 270:5427-5433
U umcu M, Homsi MF, Ball DK, Coskun S, Jaroudi K, Hollanders JM, Brigstock DR (2000): Localization of connective tissue growth factor in human uterine tissues. Mol. Hum. Reprod. 6:1093-1098
Vaisse C, Atger M, Portier B, Milgrom E (1990): Human placental protein 14 gene: sequence and characterization of a short dublication. DNA Cell Biol. 9:401-413
Van der Linden PJQ (1997): Endometriosis: A review of its pathogenesis. Front. Biosci. 2:48-52
Van Schil PE, Vercauteren SR, Vermeire PA, Nackaerts YH, Van Marck EA (1996): Catamenial pneumothorax caused by thoracic endometriosis. Ann. of Thorac. Surg. 62:585-586
Vegeto E, Shahbaz MM, Wen DX, Goldman ME, O´Malley BW, McDonnell DP (1993): Human progesterone receptor A form is a cell- and promotor-specific repressor of human progesterone receptor B function. Mol. Endocrinol. 7:1244-1255
122

Literatur
Vercellini P, Trespidi L, Colombo A, Vendola N, Marchini M, Crosignani MG (1993): A gonadotropin-releasing hormone agonist versus a low dose oral contraceptive for pelvic
dies on the surgical induction of endometriosis in the
er J (1987): Steroidal pure antioestrogens. J. Endocrinol. 112:R7-10
romotor contex, and
5
us danazol in treatment of
9): Whole explants of peritoneum
Bell C, Chang IF (1994): The immunolocalization of basic
ssing 20 -hydroxysteroid dehydrogenase activity.
ingapur
pain associated with endometriosis. Fertil. Steril. 60:75-79
Vernon MW, Wilson EA (1985): Stu rat. Fertil. Steril. 44:684-694
Wakeling AE, BowlWatanabe T, Inoue S, Ogawa S, Ishi Y, Hiroi H, Ikeda K, Orimo A, Muramatsu M (1998): Agonistic effect of tamoxifen is dependent on cell type, ERE-p estrogen receptor subtype: functional difference between estrogen receptors α and β. Biochem. Biophys. Res. Commun. 236:140-14
Westphal JR, Ruiter DJ, De Waal RMW (2000): Anti-angiogenic treatment of human cancer: pitfalls and promises. Int. J. Cancer 86:870-873
Wheeler JM, Malinak LR (1983): Recurrent endometriosis: incidence, management, and prognosis. Am. J. Obstet. Gynecol. 146:247-253
Wheeler JM, Knittle JD, Miller JD (1992): Depot leuprolide vers women with symptomatic endometriosis. I. efficiacy results. Am. J. Obstet. Gynecol. 167:1367-1371
Wheeler JM, Knittle JD, Miller JD (1993): Depot leuprolide acetate versus danazol in the treatment of women with symptomatic endometriosis: a multi-centre, double-blind randomized clinical trial. Am. J. Obstet. Gynecol. 169:26-33
Wild RA, Shivers CA (1985): Endometrial antibodies in patients with endometriosis. Am. J. Reprod. Immunol. Microbiol. 8:84-87
Witz CA, Monotoya-Rodriguez IA, Schenken RS (199 and endometrium: a novel model of the early endometriosis lesion. Fertil. Steril. 71:56-60
Witz CA (2002): Pathogenesis of endometriosis. Gynecol. Obstet. Invest. 53:52-62
Wordinger RJ, Smith KJ, fibroblast growth factor in the mouse uterus during the initial stages of embryo implantation. Growth Factors 11:175-186
Wortis HH (1971): Immunological response of ”nude” mice. Clin. Exp. Immunol. 8:305-317
Wu L, Einstein M, Geissler WM, Chan HK, Elliston KO, Andersson S (1993): Expression, cloning and characterization of human 17β-hydroxysteroid dehydrogenase type 2, a microsomal enzyme posse α J. Biol. Chem. 268:12964-12969
Yang GP, Lau LF (1991): Cyr61, product of a growth factor-inducible immediate early gene, is associated with the extracellular matrix and the cell surface. Cell Growth & Differ. 2:351- 357
Yovich J (1991): Endometriosis and assisted reproduction. Abstract 0369, FIGO, S
Zaino RJ, Satyaswaroop PG, Mortel R (1985): Histologic response of normal human endometrium to steroid hormones in athymic mice. Hum. Pathol. 16:867-872
Zamah NM, Dodson MG, Stephens LC, Buttram VC Jr, Besch BK, Kaufman RH (1984): Transplantation of normal and ectopic human endometrial tissue into athymic nude mice. Am. J. Obstet. Gynecol. 149:591-597
123

Literatur
Zeitoun K, Takayama K, Sasano H, Suzuki T, Moghrabi N, Andersson S, Johns A, Meng L, Putman M, Carr B, Bulun SE (1998): Deficient 17β-hydroxysteroid dehydrogenase type 2 expression in endometriosis: failure to metabolize 17β-estradiol. J. Clin. Endocrinol.
: Aromatase: a key molecule in the pathophysiology of
splant. 25:1319-1320
Metab. 83:4474-4480
Zeitoun K, Bulun SE (1999) endometriosis and a therapeutic target. Fertil. Steril. 72:961-969
Zetter RR (1998): Angiogenesis and tumor metastasis. Annu. Rev. Med. 49:407-424
Zomas A, Anagnostopoulus N, Dimopoulus MA (2000): Successful treatment of multiple myeloma relapsing after high-dose therapy and autologous transplantation with thalidomide as a single agent. Bone Marrow Tran
124
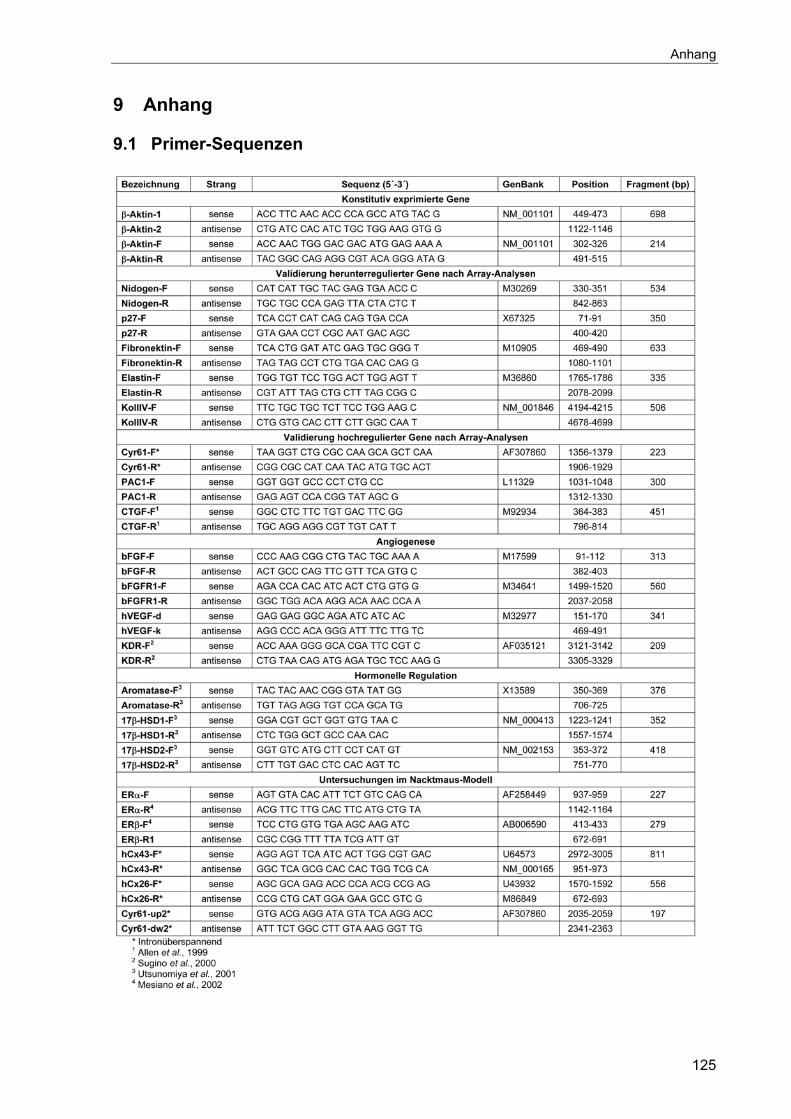
Anhang
9 Anhang
9.1 Primer-Sequenzen
125

Anhang
9.2 Ergebnisse der „Gene-Array“ Analysen
126
Anhang
127

Anhang
128

Lebenslauf
Name: Yvonne Janßen
Anschrift: Löhrerlen 28 42279 Wuppertal Geburtsdatum: 15.03.1976
Geburtsort: Wuppertal
Staatsangehörigkeit: deutsch
Familienstand: ledig
Konfession: evangelisch
Schulbildung: 1982-1986 Grundschule Kratzkopfstraße, Wuppertal
1986-1995 Städt. Gymnasium an der Siegesstraße, Wuppertal
Schulabschluß: Abitur
Studium: 1995-2000 Ruhr-Universität Bochum Diplomstudiengang Biologie
Thema der Diplomarbeit:
„Strukturelle und funktionelle Charakterisierung von Konformationsänderungen der cAMP- abhängigen Proteinkinase“
Studienabschluß: Diplom-Biologin
Doktorarbeit: seit Mai 2000 Biologie-Doktorandin am Institut für Anatomie, Universitätsklinikum Essen bei Frau Prof. Dr. Elke Winterhager
Titel der Dissertationsschrift: „Cyr61: Ein durch Östrogen reguliertes Markergen der Endometriose“ Wuppertal, 08. April 2003 Yvonne Janßen

Erklärung:
Hiermit erkläre ich, gem. §6 Abs. 2, Nr. 7 der Promotionsordnung der Fachbereiche 6 bis 9 Thema „Cyr61: Ein durch
Östrogen reguliertes Markergen der Endometriose“ zuzuordnen ist, in Forschung und Lehre
Prof. Dr. Elke Winterhager
mich keiner anderen als der angegebenen Hilfsmittel bedient habe.
Yvonne Janßen
is 9
e Arbeit von keiner anderen Fakultät abgelehnt worden ist.
en
zur Erlangung des Dr. rer. nat., daß ich das Arbeitsgebiet, dem das
vertrete und den Antrag von Frau Yvonne Janßen befürworte.
Essen, 08. April 2003
Erklärung:
Hiermit erkläre ich, gem. §6 Abs. 2, Nr. 6 der Promotionsordnung der Fachbereiche 6 bis 9 zur Erlangung des Dr. rer. nat., daß ich die vorliegende Dissertation selbständig verfaßt und
Essen, 08. April 2003
Erklärung:
Hiermit erkläre ich, gem. §6 Abs. 2, Nr. 8 de Promotionsordnung der Fachbereiche 6 btionen bzw.
r zur Erlangung des Dr. rer. nat., daß ich keine anderen PromoPromotionsversuche in der Vergangenheit durchgeführt habe und daß dies
Essen, 08. April 2003 Yvonne Janß