Embed Size (px)
Citation preview

Botanisches Institut der Tierarztlichen Hochschule Hannover, BRD
Der Einflu6 monovalenter Kationen auf die Nitratreduktion von Spinacia oleracea L.
The influence of monovalent cations on the reduction of nitrate in Spina cia oleracea 1.
REINHARD PFLUGER und REINHARD WIEDEMANN
Mit 4 Abbildungen
Eingegangen am 29. Marz 1977 . Angenommen am 14. Juni 1977
Summary
Experiments were carried out to determine the influence of different univalent cations on the activity of nitrate reductase from spinach in vitro and in vivo.
Nitrate reductase activity was maximal at pH 7.75 and 28°C. Under these conditions the alkali ions Li+, Na+, K+ and Rb+ had no different effect on the activity of the enzyme in vitro.
In further investigations it could be shown that monovalent cations had clearly different effects on the activity of nitrate reductase in vivo. In order to get better results of the cations plant material of a low salt status was used. Compared to the control K+ induced, after a lag phase of 3 h, a remarkable increase in the rates of nitrate reduction. Rb+ had a stimulating effect, too, but was less effective. Na+ and Li+ were without effect.
These differences are not attributable to different concentrations of cations within the cell. For example, under the conditions used for estimating the activity of the enzyme (50 mM K+, or Na+ respectively), the rate of uptake into the leaf tissue was similar for K+ and Na+.
The investigations show that the inductive formation of nitrate reductase caused by nitrate depends on the presence of certain cations, i.e. under natural conditions there is a strict requirement for K+.
Key words: nitrate reductase, inductive formation, monovalent cations.
Einleitung
Nitrat- und Ammoniumstickstoff sind fur die Pflanze die wichtigsten Stickstoffquellen. Zwar vermag die Pflanze auch organische Stickstoffverbindungen aufzunehmen, doch kommt diesen, mit Ausnahme des Harnstoffes, keine praktische Bedeutung zu. Wahrend der Einbau von NH/ in die organische Substanz einen verhaltnismamg einfachen Prozeg darstellt, mug N03- erst bis zur Stufe des NH3 reduziert werden,
z. Pflanzenphysiol. Bd. 85. S. 125-133. 1977.

126 REINHARD PFLUGER und REINHARD WIEDEMANN
bevor es tiber die reduktive Aminierung in den Stoffwechsel einbezogen werden kann. Somit stellt die assimilatorische Nitratreduktion fur die Pflanze einen Vorgang von zentraler Bedeutung dar.
Der erste Schritt in dieser Reaktionskette, die Reduktion von Nitrat zu Nitrit, wird durch das Enzym Nitratreduktase katalysiert (EVANS und NASON, 1953). Die Reduktion des Nitrits zum Ammonium findet ohne weitere fagbare Intermediate statt (SANDERSON und COCKING, 1964).
Es wird als gesichert angenommen, dag durch das Substrat Nitrat die Bildung von Nitratreduktase induziert wird (BEEVERS und HAGEMAN, 1969; KNYPL, 1973; SLUITER-SCHOL TEN, 1973). Dabei spielen als Cofaktoren einwertige Kationen eine Rolle. Nach Untersuchungen von NITSOS und EVANS (1966) an Neurospora crassa kommt dabei dem Kalium eine besondere Bedeutung zu.
In einer zusammenfassenden Darstellung geben EVANS und SORGER (1966) einen Oberblick uber die Rolle von einwertigen Kationen fur eine Reihe von Stoffwechselprozessen. Danach wird die Aktivitat von mehr als 40 Enzymen durch sie stimuliert.
Der Mechanismus der Aktivierung enzymatischer Reaktionen durch einwertige Kationen ist noch weitgehend ungeklart. WILSON und EVANS (1968) vermuten, dag die Wirkung mit der lonengroge zusammenhangen konnte und tiber Konformationsanderungen der Enzymstrukturen erfolgt. Auf diese Weise sollen dem Substrat durch die «kleinen» Alkaliionen K+ und Rb+ mehr reaktive Stellen am Enzymkomplex zuganglich gemacht werden als durch die «grogen» Alkaliionen Na+ und Li+. Am Beispiel der Induktion der Nitratreduktase diskutieren NITSOS und EVANS (1966) eine Wirkung von K+ auf die Synthese von Proteinen.
In der vorliegenden Arbeit wurde untersucht, welch en Einflug die verschiedenen monovalenten Kationen auf die Aktivitat der Nitratreduktase in vitro und in vivo bei Spinat haben. Dieses Objekt ist insofern von besonderem Interesse, als Spinat zu den Pflanzen gehort, die fur ein optimales Wachstum Natrium benotigen. So sollte gleichzeitig der Frage nachgegangen werden, ob die positive Wirkung von Natrium in diesem Abschnitt des Stoffwechsels zu suchen ist.
Material und Methoden
Als Versuchspflanze diente Spinacia oleracea L. Die Anzucht der Pflanzen erfolgte in Klimakammern.
In vitro-Untersuchungen: Die Aktivitat der Nitratreduktase wurde aus dem Rohextrakt bestimmt. Aufarbeitung, Gewinnung des Extraktes und Bestimmung der Enzymaktivitat erfolgte kombiniert nach den Angaben von SANDERSON und COCKING (1964) und HAGEMAN und HUCKLESBY (1971). Urn niedermolekulare Bestandteile wie Nitrat und die zu testenden Salze weitgehend auszuschliegen, wurde eine Reinigung des Rohextraktes mittels einer Sephadex-G-25-Saule vorgenommen. Die Proteinbestimmung erfolgte nach LOWRY et a1. (1951). Ais Standard diente Serumalbumin.
In vivo-Untersuchungen: Fur diese Experimente wurden an Nahrstoffen verarmte Pflanzen verwendet, urn deutlichere Effekte nach Zusatz der zu testenden Kationen zu erzielen. Zur Verarmung wurden normal ernahrte Pflanzen fUr eine Woche in eine 5 mM CaC03-
Lasung iiberfiihrt. Flir die Versuche wurden die Blatter in ca. 4 mm2 kleine Stlickchen ge-
Z. PJlanzenphysiol. Bd. 85. S. 125-133. 1977.
Nitrate reduction stimulated by cations 127
schnitten und bei 28°C und 13 000 Lux inkubiert. Das Inkubationsmedium setzte sich aus 25 mM Ca(NOa)2, 50 mM TRIS-HCl 7,75 pH und den bei den einzelnen Experimenten angegebenen Kationenkonzentrationen zusammen. Nach entsprechender Inkubationszeit, die bei den Ergebnissen angefiihrt ist, wurde das Inkubationsmedium mit n-Propanol versetzt (Endkonzentration 4010, JAWORSKI, 1971) und weitere 30 Minuten stehengelassen. Das in dieser Zeit gebildete und in das Medium abgeschiedene Nitrit diente als MaB flir die Aktivitat der Nitratreduktase.
Die Bestimmung der K+- und Na+-Gehalte in der pflanzlichen Substanz erfolgte mittels atomarer Absorption (Pye Unicam SP 90). Die Nitratgehalte wurden nach der BrucinMethode bestimmt (Lit: Die Untersuchung von Wasser. - E. Merck, Darmstadt).
Die Bestimmung der Aufnahmeraten von K+ und Na+ in das Blattgewebe wurde unter den oben angegebenen Inkubationsbedingungen unter Zusatz von 50 mM 42KCl bzw. 24NaCl vorgenommen. Nach 4 h wurden die Blattscheibchen mit Wasser und inaktiver KCl- bzw. NaCl-Losung wiederholt gewaschen, urn die nicht aufgenommenen markierten Ionen gegen unmarkierte auszutauschen. Die Bestimmung der in das Gewebe aufgenommenen K+ - und Na+-Ionen erfolgte im Szintillationsziihler (FHT 770 D).
Ergebnisse
I. Untersuchungen zur Charakterisierung der Nitratreduktase in vitro
Die Versuche hierzu wurden mit vorgereinigtem Rohextrakt durchgefiihrt.
1. Abhangigkeit der Nitratreduktaseaktivitat vom pH-Wert, der Temperatur und der NOa--Konzentration des Inkubationsmediums
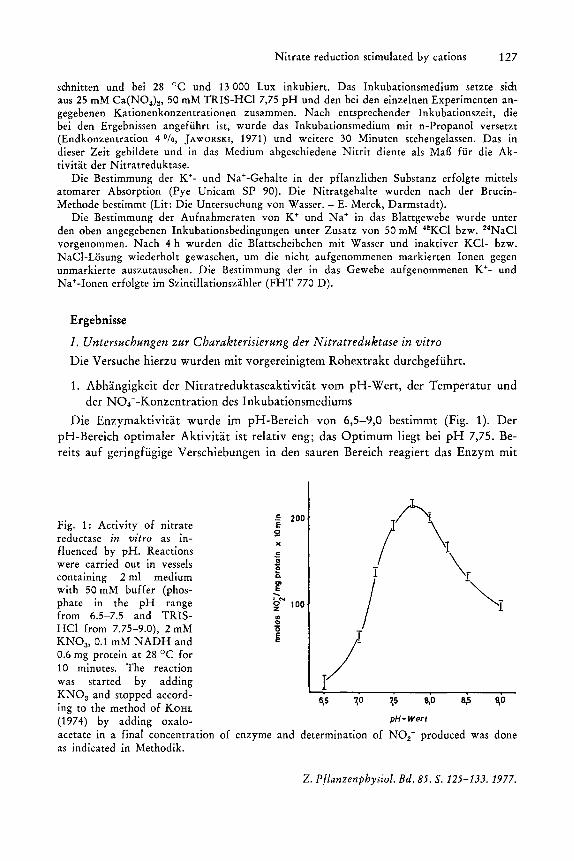
Die Enzymaktivitat wurde im pH-Bereich von 6,5-9,0 bestimmt (Fig. 1). Der pH-Bereich optimaler Aktivitat ist relativ eng; das Optimum liegt bei pH 7,75. Bereits auf geringfiigige Verschiebungen in den sauren Bereich reagiert das Enzym mit
Fig. 1: Activity of nitrate reductase in vitro as influenced by pH. Reactions were carried out in vessels contammg 2 ml medium with 50 mM buffer (phosphate in the pH range from 6.5-7.5 and TRISHCl from 7.75-9.0), 2 mM KNOa, 0.1 mM NADH and 0.6 mg protein at 28°C for 10 minutes. The reaction was started by adding KNOa and stopped accord-ing to the method of KOHL (1974) by adding oxaloacetate in a final concentration as indicated in Methodik.
c 200 /1" E / \ ~
c '0
I ~r ~ Ii '"
I ~ E .... 'N 0 100 z UI <> '0 ~ J I
5,5 ~o ~5 8,0 8,5 9,0
pH-WE'rt
of enzyme and determination of N02- produced was done
Z. Pjlanzenphysiol. Bd. 85. S. 125-133. 1977.
128 REINHARD PFLUGER und REINHARD WIEDEMANN
einer steil en Aktivitatsabnahme. Basischere Bedingungen fuhren gleichfalls zu einem Verlust an Aktivitat, doch ist hier der Abfall weniger steil.
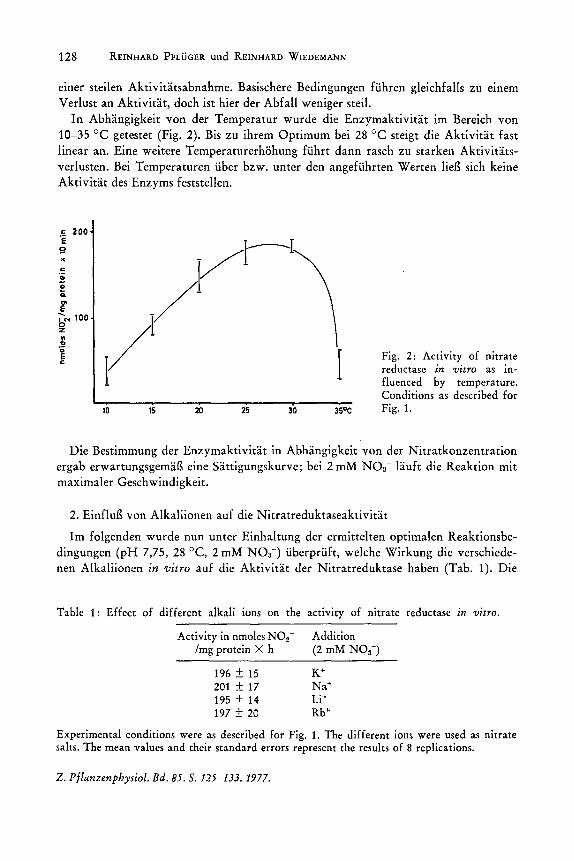
In Abhangigkeit von der Temperatur wurde die Enzymaktivitat im Bereich von 10-35 °C getestet (Fig. 2). Bis zu ihrem Optimum bei 28°C steigt die Aktivitat fast linear an. Eine weitere Temperaturerhohung fuhrt dann rasch zu starken Aktivitatsverlusten. Bei Temperaturen uber bzw. unter den angefuhrten Werten lieg sich keine Aktivitat des Enzyms feststellen.
c 200 E
" c '0 f Q.
r i{< 100 z ~ '0 E c
10 15 20 25
I 30
Fig. 2: Activity of nitrate reductase in vitro as influenced by temperature. Conditions as described for Fig. 1.
Die Bestimmung der Enzymaktivitat in Abhangigkeit von der Nitratkonzentration ergab erwartungsgemag eine Sattigungskurve; bei 2 mM N03- lauft die Reaktion mit maximaler Geschwindigkeit.
2. Einflug von Alkaliionen auf die Nitratreduktaseaktivitat
1m folgenden wurde nun unter Einhaltung der ermittelten optimal en Reaktionsbedingungen (pH 7,75, 28°C, 2 mM N03-) uberpruft, welche Wirkung die verschiedenen Alkaliionen in vitro auf die Aktivitat der Nitratreduktase haben (Tab. 1). Die
Table 1: Effect of different alkali ions on the activity of nitrate reductase in vitro.
Activity in nmoles N02-
Img protein X h
196 ± 15 201 ± 17 195 ± 14 197 ± 20
Addition (2 mM NOn
Experimental conditions were as described for Fig. 1. The different ions were used as nitrate salts. The mean values and their standard errors represent the results of 8 replications.
Z. Pjlanzenphysiol. Bd. 85. S. 125-133. 1977.

Nitrate reduction stimulated by cations 129
Resultate zeigen, daB Li+, Na+, K+ und Rb+ sich in ihrem EinfluB auf die Raten der Nitratreduktion nicht unterscheiden.
II. Untersuchungen zum Einfluft verschiedener Kationen auf die Aktivitat der Nitratreduktase in vivo
Pflanzen, die an Nahrstoffen verarmt sind, zeigen nach Zusatz der zu untersuchenden lonen klarere Resultate. Daher wurde zunachst die Abnahme der lonengehalte im Blattgewebe nach dem Nahrstoffentzug ermittelt.
1. Verarmung der Versuchspflanzen an Nahrstoffen
Am 1., 3. und 7. Tage nach Beginn des Nahrstoffentzuges wurden die Kalium-, Natrium- und Nitratgehalte der Pflanzen bestimmt (Tab. 2), in dieser Zeit sinkt der Kaliumgehalt um fast die Halfte, der von Natrium um 2/a und der von Nitrat auf 1/4 der Anfangsgehalte. Die Abnahme von K+ und Na+ im Gewebe ist nicht allein auf eine Verdiinnung durch Wachstum zuriickzufiihren; ein betrachtlicher Teil diirfte auch iiber die Wurzeln in das AuBenmedium effluiert sein. Die reduzierten Nitratgehalte beruhen auf Metabolisierung. Fiir die nachstehend geschilderten Experimente wurden Pflanzen verwendet, die 7 Tage ohne Nahrstoffe gehalten worden waren.
Table 2: Decrease of potassium, sodium and nitrate content in dry matter of leaves after removing nutrients.
Days after removing nutrients
3 7
,umoles/mg dry matter K+ Na+
2.70 2.29 1.51
0.58 0.45 0.18
nmoles NOa-/mg dry matter
0.59 0.18 0.15
2. EinfluB von Alkaliionen auf die Aktivitat der Nitratreduktase in vivo
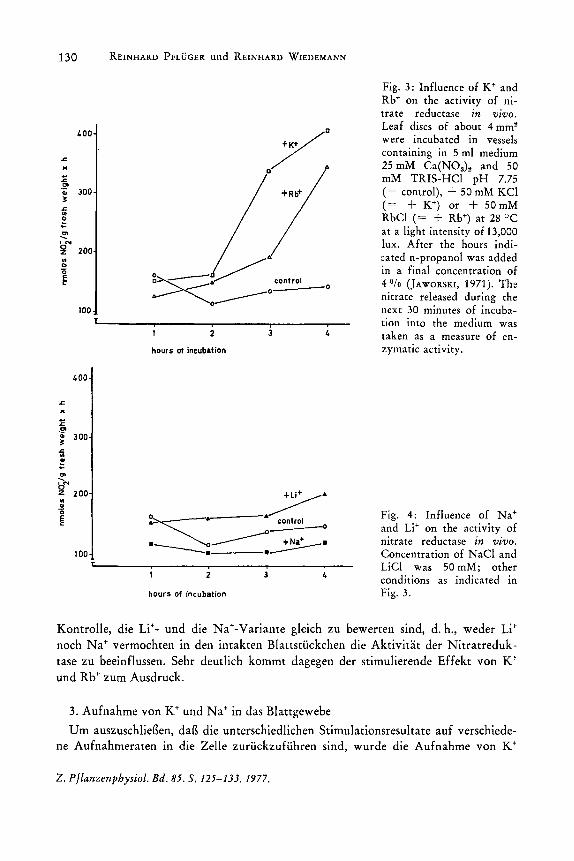
In Vorversuchen war ermittelt worden, daB bei Arbeiten mit Blattstiickchen die Reaktionsgeschwindigkeit bei 50 mM NOa- ihren maximalen Wert erreicht; diese Konzentration wurde durch 25 mM Ca(NOah vorgelegt (= Kontrolle) und die Alkaliionen als Chloridsalze zugesetzt. Um die Kinetik der Wirkung der Kationen auf die Aktivitat des Enzyms zu verfolgen, wurde das Blattgewebe 4 h inkubiert. Nach jeder Stunde erfolgte eine Aktivitatsbestimmung. Gegeniiber der Kontrolle bewirkte K+ nach einer Induktionszeit von 3 heine beachtliche Erhohung der Aktivitat der Nitratreduktase (Fig. 3). Gleiches gilt fiir Rb+, das ebenfalls eine merkliche Stimulierung bewirkte. Durch Li+ und Na+ erfolgte weder eine deutliche Erhohung noch eine Erniedrigung der Enzymaktivitat (Fig. 4). Die dargestellten MeBpunkte sind Mittelwerte aus 8 Wiederholungen. In Tabelle 3 sind die Einzelwerte der Aktivitatsmessungen nach 4stiindiger Inkubation zusammengestellt; die Angaben zeigen die Schwankungsbreite. Trotz der groBen Varianz der Werte wird ersichtlich, daB die
z. Pjlanzenphysiol. Bd. 85. S. 125-133. 1977.
130 REINHARD PFLUGER und REINHARD WIEDEMANN
'00
.c
1: .S!' .. 300 ~ .c on
~ '" ;;.-. z 200 on C> "0 E c:
100
yo. ~,,,,,,----o. conlrol
.~~------:~ T
i i
2 3 , hours of Incubation
Fig. 3: Influence of K+ and Rb+ on the activity of nitrate reductase in VIVO.
Leaf discs of about 4 mm2
were incubated in vessels containing in 5 ml medium 25 mM Ca(NOah and 50 mM TRIS-HCl pH 7.75 (= control), + 50 mM KCl (= + K+) or + 50mM RbCl (= + Rb+) at 28°C at a light intensity of 13,000 lux. After the hours indicated n-propanol was added in a final concentration of 4 Ofo (JAWORSKI, 1971). The nitrate released during the next 30 minutes of incubation into the medium was taken as a measure of enzymatic activity.
Fig. 4: Influence of Na+ and Li+ on the activity of nitrate reductase in vivo. Concentration of NaCl and LiCl was 50 mM; other conditions as indicated in Fi~. 3.
Kontrolle, die Li+- und die Na+-Variante gleich zu bewerten sind, d. h., weder Li+
noch Na+ verrnochten in den intakten Blattstlickchen die Aktivitat der Nitratreduk-· tase zu beeinflussen. Sehr deutlich kornrnt dagegen der stirnulierende Effekt von K· und Rb+ zurn Ausdruck.
3. Aufnahrne von K+ und Na+ in das Blattgewebe
Urn auszuschliegen, dag die unterschiedlichen Stirnulationsresultate auf verschiedene Aufnahrneraten in die Zelle zuriickzuflihren sind, wurde die Aufnahrne von K+
z. Pjlanzenphysiol. Bd. 85. S. 125-133. 1977.

Nitrate reduction stimulated by cations 131
Table 3: Effect of different alkali ions on the activity of nitrate reductase in vivo.
Addition (50 mM)
none (control) K+ Rb+ Li+ Na+
Activity in nmoles N02 -/g fresh weight X h
120-164 310-500 266-424 133-220 95-137
In this table the data of Fig. 3 and 4 after an incubation time of 4 h are summarized. The given values are the limits of variability.
und Na+ untersucht. Das Experiment erfolgte unter den gleichen lnkubationsbedingungen wie die Untersuchungen iiber den Einflug der Alkaliionen auf die Aktividit der Nitratreduktase. Mit 66,8,uMol Na+/g FS X 4 h und 68,4,uMol K+/g FS X 4 h wurden beide lonen etwa gleich rasch aufgenommen. Daher ist ein auf unterschiedlichen Aufnahmeraten der lonen beruhender Sekundareffekt auf die Enzymaktivitat auszuschliegen.
Diskussion
Die Untersuchungen zur Charakteristik der Nitratreduktase aus Spin at ergaben ein pH-Optimum von 7,75 und ein Temperaturoptimum von 28 DC. Diese Werre stehen in guter Dbereinstimmung mit Befunden anderer Autoren (DALLING et aI., 1972; KOHL et aI., 1974).
Als mogliche Wirkungsweise einwertiger Kationen auf enzymatisch katalysierte Reaktionen wird die unterschiedliche Beeinflussung der sterischen Konformation eines Enzymkomplexes diskutiert, wobei die verschiedenen lonenradien eine entscheidende Rolle spielen sollen. Es soli bei diesem Vorgang weniger die Michaeliskonstante fiir das Substrat verandert werden; dagegen soli die maxima Ie Reaktionsgeschwindigkeit erhoht werden (HIATT, 1964; WILSON und EVANS, 1968). Fiir das untersuchte Enzym Nitratreduktase ist das nicht der Fall. Aile vier in ihrer Wirkung auf die Aktivitat des Enzyms getesteten Alkaliionen (Li+, Na+, K+, Rb+) zeigten bei den Untersuchungen mit dem isolierten Enzym den gleichen Effekt. Dieser Befund stimmt mit Ergebnissen von AUSTENFELD (1974) iiberein, der ahnliche Untersuchungen mit dem Halophyten Salicornia europaea durchfiihrte.
Die in vivo-Experimente mit Blattstiickchen brachten dagegen ganz andere Resultate. Hier konnten nach einer <<lag phase» durch K+, aber auch durch Rb+, die Raten der Nitratreduktion beachtlich erhoht werden; dagegen konnte weder durch Na+ noch durch Li+ gegeniiber der Kontrolle eine Steigerung der Enzymaktivitat beobachtet werden. Die unterschiedlichen lnduktionen sind nicht darauf zuriickzufiihren, dag die einzelnen Kationen verschieden schnell von den Blattstiickchen aufge-
z. Pflanzenphysiol. Bd. 85. S. 125-133. 1977.

132 REINHARD PFLUGER und REINHARD WIEDEMANN
nommen wurden und SO in der Zelle in ungleichen Konzentrationen vorlagen. Aus den Versuchsergebnissen geht deutlich hervor, dag unter den gewahlten Inkubationsbedingungen (50 mM Kation) K+ und Na+ in etwa gleicher Rate aufgenommen werden.
Erstaunlich ist die Tatsache, dag in der Kontrolle, in der NOs- als Ca-Salz vorgelegt wurde und in der bis auf die verarmten endogenen Gehalte keine monovalenten Kationen vorhanden waren, wahrend der Induktionszeit von 4 h keine Knderung in der Aktivitat der Nitratreduktase zu beobachten war; gleiches gilt auch fur die Na+- und Li+-Variante. NITSOS und EVANS (1966) berichten von ahnlichen Ergehnissen mit Neurospora crassa, wo nach einer langeren K+-freien Inkubationszeit schlieg}ich doch noch eine leichte Aktivitatssteigerung zu beobachten war; sie fuhren dies auf eine in der Z wischenzeit erfolgte K + -Anreicherung zuruck, die auf K + -V erunreinigungen der verwendeten Chemikalien beruhte. Sie diskutieren ihre Befunde als eine Wirkung von Kationen auf die Synthese des Enzymproteins Nitratreduktase.
Eingangs wurde festgestellt, dag durch Nitrat die Bildung von Nitratreduktase induziert wird (z. B. BEEVERS und HAGEMA.N, 1969). Diese Vorstellung ist dahingehend zu erweitern, dag der Induktionsprozeg nur dann optimal ablaufen kann, wenn entsprechende einwertige Kationen in ausreichender Konzentration als Cofaktoren zur Verfugung stehen.
Es ist jedoch unwahrscheinlich, dag speziell fur die Biosynthese der Nitratreduktase monovalente Kationen erforderlich sind. Nach LUBIN (1964) hat das K+-Ion einen Einflug auf die Synthese von Proteinen. Die Bindung der t-RNS an das Ribosom und die Enzymreaktion der Peptidyltransferase scheinen hier die Kalium bedurftigen Prozesse darzustellen. Der genaue Wirkungsmechanismus einwertiger Kationen bleibt nach SUELTER (1970) jedoch immer noch weitgehend ungeklart.
Von Rb+ ist bekannt, dag bei Kurzzeitversuchen K+ zum Teil durch dieses Ion ersetzt werden kann; seine ahnliche Wirkung hangt mit der chemischen Verwandtschaft beider Kationen zusammen. In der Pflanze kommt Rb+ jedoch nur in kaum megbaren Mengen vor (MENGEL, 1969), so dag unter naturlichen Bedingungen nur dem K+-Ion entscheidende Bedeutung fur die induktive Bildung der Nitratreduktase zukommt.
Literatur
AUSTENFELD, F.-A.: Der EinfluE des NaCI und anderer Alkalisalze auf die Nitratreduktaseaktivitat von Salicornia europaea L. Z. Pflanzenphys. 71, 288-296 (1974).
BEEVERS, L. and R. H. HAGEMAN: Nitrate reduction in higher plants. Ann. Rev. Plant Physio!. 20, 495-522 (1969).
DALLING, M. ]., N. E. TOLBERT, and R. H. HAGEMAN: Intracellular location of nitrate reductase and nitrite reductase. 1. Spinach and tobacco leaves. Biochim. Biophys. Acta 283, 505-512 (1972).
EVANS, H. ]. and A. NASON: Pyridine nucleotid-nitrate reductase from extracts of higher plants. Plant Physio!. 28, 233-254 (1953).
z. Pflanzenphysiol. Bd. 85. S. 125-133. 1977.

Nitrate reduction stimulated by cations 133
EVANS, H. and G. J. SORGER: Role of mineral elements with emphasis on the univalent cations. Ann. Rev. Plant PhysioI. 17,47-76 (1966).
HAGEMAN, R. H. and D. FLESHER: Nitrate reductase activity in corn seedlings as affected by light and nitrate content of nutrient media. Plant PhysioI. 35, 700-708 (1960).
HAGEMAN, R. H. and D. P. HUCKLESBY: Nitrate reductase from higher plants. Methods EnzymoI. 23 A, 491-503 (1971).
HIATT, A. J.: Further studies on the activation of acetic thiokinase by magnesium and univalent cations. Plant PhysioI. 39, 475-479 (1964).
JAWORSKI, E. G.: Nitrate reductase assay in intact plant tissue. Biochem. Biophys. Res. Comm. 43, 1274-1279 (1971).
KNYPL, J. S.: Synergistic induction of nitrate reductase activity by nitrate and benzylaminopurine in detached cucumber cotyledons. Z. PflanzenphysioI. 70, 1-11 (1973).
KOHL, J.-G., E. SOMMERFELD, G. DUDEL und R. IRMLER: Der EinfluB der exogenen Nitratkonzentration auf die Nitratreduktaseaktivitat in Wurzel und Blatt von Zea mays L. Biochem. PhysioI. Pflanzen 165, 123-136 (1974).
LOWRY, O. H., N. J. ROSEBROUGH, A. C. FARR, and R. J. RANDALL: Protein measurement with the Folin phenol reagent. J. BioI. Chern. 193,265-275 (1951).
LUBIN, M. and H. L. ENNIS: On the role of intracellular potassium in protein synthesis. Biochem. Biophys. Acta 80, 614-663 (1964).
MENGEL, K.: Funktionen anorganischer Verbindungen in der lebenden Zelle. Vitalstoffe 5 (1969).
NITsos, R. E. and H. J. EVANS: Effect of univalent cations on the inductive formation of nitrate reductase. Plant PhysioI. 41,1499-1504 (1966).
SANDERSON, G. W. and E. C. COCKING: Enzymic assimilation of nitrate in tomato plants. I. Reduction of nitrate to nitrite. Plant Physiol. 39, 416-422 (1964).
- - Enzymic assimilation of nitrate in tomato plants. II. Reduction of nitrite to ammonia. Plant PhysioI. 39, 423-431 (1964).
SLUITERS-SCHOLTEN, C. M. TH.: Effect of chloramphenicol and cycloheximide on the induction of nitrate reductase and nitrite reductase in bean leaves. Planta 113, 229-240 (1973 ).
SUELTER, C. H.: Enzymes activated by univalent cations. Science 168,789-795 (1970). WILSON, R. H. and H. J. EVANS: The effect of potassium and other univalent cations on the
conformation of enzymes. In: KILMER, V. J. (ed): The role of potassium in agriculture, 189-202, Madison, 1968.
Dr. R. PFLUGER, Botanisches Institut der Tierarztlichen Hochschule Hannover, Biinteweg 17 d, D-3000 Hannover-Kirchrode, Bundesrepublik Deutschland.
Z. Pflanzenphysiol. Ed. 85. S. 125-133. 1977.





























