Embed Size (px)
Citation preview

12 Endokrinologie
Klaus-Dieter Spindler
12.1 Hormone und ihreBildungsorte
12.1.1 Neurosekretorische Zellenund Zentren
Neurosekretorische Zellen (NSZ) wurden zunächst lichtmikroskopisch aufgrund ihrer vermeintlich spezifischen histologischen Anfärbbarkeit oder im Transmissions-Elektronenmikroskopaufgrund der zahlreich vorhandenen Grana miteinem Durchmesser von etwa 100 bis 400 nmDurchmesser, nachgewiesen. Charakteristisch sindauch der hohe Gehalt an rauhem endoplasmatischem Retikulum, sowie die zahlreichen aufgeweiteten axonalen Endbläschen, die Speicherortefür die neurosekretorischen Grana darstellen unddie der Freisetzung über eine Calcium-vermittelteExocytose dienen. Inzwischen können die Inhaltsstoffe neurosekretorischer Zellen eindeutiger charakterisiert werden: Neuropeptide durch immunhistochemische Methoden und Massenspektroskopie und biogene Amine durch fluoreszenzmikroskopisehe Verfahren. Außerdem können dieseZellen durch Füllung mit Kobalt und fluorogenenSubstanzen lokalisiert werden. Diese Verfahrenwurden auch eingesetzt um Aktivitätszyklen vonNSZ zu erfassen. Eine eindeutige Beziehung zwischen Anfärbbarkeit und der Aktivität der NSZmuss aber nicht immer bestehen.
NSZ wurden bei allen Metazoen nachgewiesen.Es handelt sich um Zellen, die typische neuronaleAktivitäten aufweisen und zudem noch sekretorisch tätig sind. Konventionelle Neurone sendenihre Axone direkt zu den Zielorganen . Das chemische Signal wird über die sehr kurze Distanz dessynaptischen Spaltes (10 bis 50 nm) abgegebenund seine physiologisch wirksame Konzentrationwird rasch auf und wieder abgebaut. Neurosekretorische Zellen dagegen geben ihr(e) Sekretionsprodukt(e) an das Blut oder die Hämolymphe ab,die Wirkorte können weit entfernt sein. Die physiologisch wirksame Konzentration wird wesentlich langsamer auf- und abgebaut.
NSZ kommen bei Insekten an sehr unterschiedlichen Stellen vor (Abb. 12-1):
• Im Cerebralganglion sind im Protocerebrumausser den pars intercerebralis noch die lateralenNSZ sowie die tritocerebralen NSZ nachgewiesen, sowie auch vereinzelte NSZ im Deutocerebrum.
• In den Ganglien des Bauchmarks,• Im anterioren sympathischen Nervensystem
sind sie ebenfalls vorhanden.• Periphere NSZ gibt es vor allem in der Nähe des
Herzens und des Verdauungstraktes.
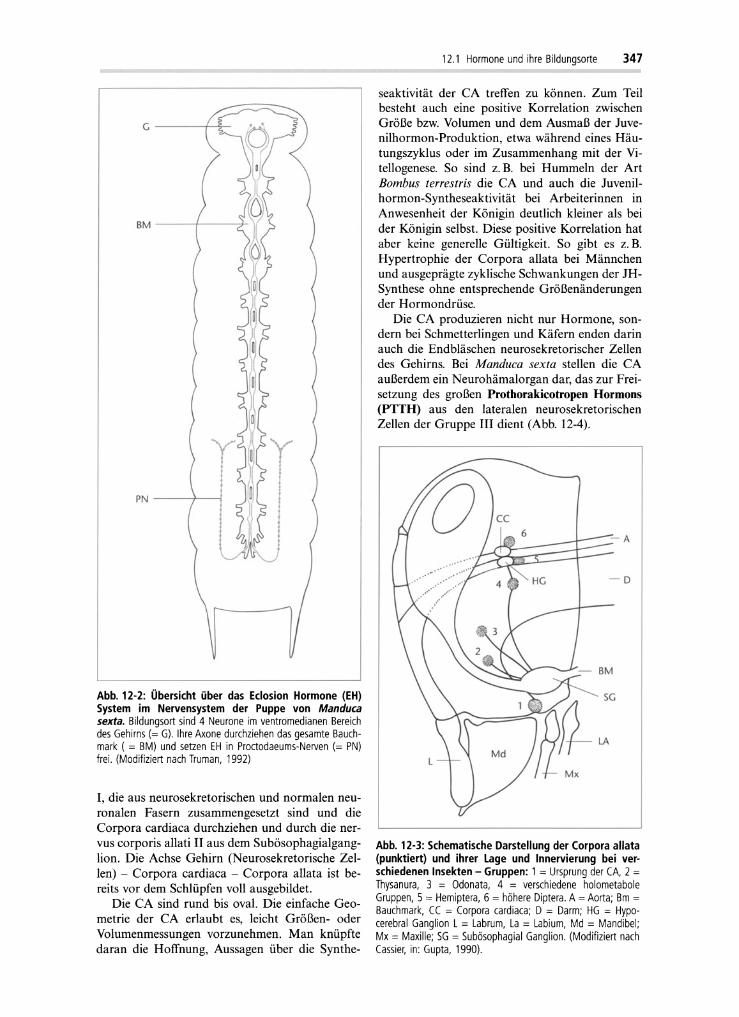
Produktionsort eines neurosekretorischen Hormons und die Stelle seiner Freisetzung aus demneurosekretorischen Endbläschen können weitauseinander liegen. Ein Beispiel aus der Gruppeder Insekten stellt das Eclosion Hormone (EH) beiManduca sexta dar (Abb. 12-2). Dieses Hormonwird in Larven in 2 Paar ventromedianen NSZ desGehirns gebildet, deren Axone ipsilateral durchdas gesamte ZNS ziehen und in den Proctodeumnerv projizieren und dort EH freisetzen. In adulten Tieren der gleichen Art ist diese Gehirn-Proc todeurn-Achse auch noch vorhanden. Bei denadulten Tieren wird jedoch EH zusätzlich noch inanderen NSZ im Gehirn gebildet und dann auchaus dem Corpora allata/Corpora cardiaca-Komplex freigesetzt. Nicht nur der Ort der Syntheseund der Freisetzung von EH ändern sich währendder Entwicklung, sondern auch die Regulation derHormonabgabe: In allen Entwicklungsstadien istfür die Freisetzung des Eclosion Hormons einabfallender Titer des Häutungshormones notwendig, bei den Adulten kommt noch die Regulationdurch eine circadiane Uhr hinzu.
Axonendigungen mehrerer NSZ können häufiggebündelt in enger räumlicher Nachbarschaft zurHämolymphe stehen und damit lokal begrenztNeurohormone abgeben. Solche Strukturen nenntman Neurohämalorgane. Das wohl bekanntesteNeurohämalorgan der Insekten stellen die Corpora cardiaca (s. 12.1.2) dar. Darüber hinaus gibtes noch mehrere andere Neurohämalorgane oder-Zonen, z. B. die perisympathetischen Organe,Aorta-Wände, bei manchen Arten auch die Umhüllung der Corpora allata (s. 12.1.2), sowie Neurohämalorgane im Gehirn , dem Suboesophagealganglion und in Abdomin alganglien (Abb. 12-1).Eine Reihe von NSZ endigen auch direkt an Effektorsystemen, z.B. an Muskeln, Darmzellen, Spei-

346 12 Endokrinologie
-........2--<:--- - nh
-t- - - ne
--,r::- np
Abb. 12-1: Schematische Darstellung von neurosekretori schen Zeilen und Zentren sowieOrten derFreisetzung neurosekretorischen Materials in Insekten . Punkte stellen neurosekretorische Zellen (NSZ) dar, die im Gehirn in der pars intercerebralis (pi), demlateralen (la)und dem tritocerebralen(tc) Bereich zu Komplexen zusammengefasst sind. CA = Corpora allata, CC =Corpora cardiaca, ne = NeuroeffektorEndigungen, nh= Neurohämal Zonen,np = Neuropil - neurosekretorische Endigungen, ps = perisympathische Organe. (Modifiziert nach Raabe, 1989).
cheldrüsen, Malpighischen Gefäßen, Spermatheken und dem Ovidukt. Aus elektronenmikroskopischen Untersuchungen weiß man, dass viele dieserNSZ peptiderge und aminerge Vesikel enthalten.NSZ von Insekten sind daher nicht nur ProduktionssteIlen "typi scher" peptiderger Neurohormone, sondern sie können auch peptiderge undaminerge Neurotransmitter und Neuromodulatoren sowie biogene Amine mit typischer Hormonfunktion produ zieren.
12.1.2 Corpora allata/Corporacardiaca-Komplex, Ringdrüse
Die Corpora allata (CA) sind Produktionsort derJ uvenilhormone. Sie entstehen als paarige Einstülpungen des Ektoderms im Bereich des Maxillarsegments oder zwischen Maxillar- und Mandibularsegment. Bei höher entwickelten Arten wandern sie aus ihrer ursprünglich mehr ventralenLage nach dorsal und treten dort in Kontakt mitden Anlagen der Corpora cardiaca oder mit Nerven, die aus den CC projizieren (Abb. 12-3). DieCA sind doppelt innerviert: Durch die aus demProtocerebrum stammenden nervi corporis allati
G ----+-7-
BM -----1'-----------,f-
PN - - -+- --1
Abb. 12·2: Übersicht über das Eclosion Hormone (EH)System im Nervensystem der Puppe von Manducasexta. Bildungsort sind 4 Neurone im ventromedianen Bereichdes Gehirns (= G). Ihre Axone durchziehen das gesamte Bauchmark ( = BM) und setzen EH in Proetodaeums-Nerven (= PN)frei. (Modifiziert nach Truman, 1992)
I, die aus neurosekretorischen und normalen neuronalen Fasern zusammengesetzt sind und dieCorpora cardiaca durchziehen und durch die nervus corporis allati II aus dem Subösophagi alganglion. Die Achse Gehirn (Neurosekretorische ZeIlen) - Corpora cardiaca - Corpora allata ist bereits vor dem Schlüpfen voll ausgebildet.
Die CA sind rund bis oval. Die einfache Geometrie der CA erlaubt es, leicht Größen- oderVolumenmessungen vorzunehmen . Man knüpftedaran die Hoffnung, Aussagen über die Synthe-
12.1 Hormone und ihre Bildungsorte 347
seaktivität der CA treffen zu können. Zum Teilbesteht auch eine positive Korrelation zwischenGröße bzw. Volumen und dem Ausmaß der Juvenilhormon-Produktion, etwa während eines Häutungszyklus oder im Zusammenhang mit der Vitellogenese. So sind z. B. bei Hummeln der ArtBambus terrestris die CA und auch die Juvenilhormon-Syntheseaktivität bei Arbeiterinnen inAnwesenheit der Königin deutlich kleiner als beider Königin selbst. Diese positive Korrelation hataber keine generelle Gültigkeit. So gibt es z. B.Hypertrophi e der Corpora allata bei Männchenund ausgeprägte zyklische Schwankungen der JHSynthese ohne entsprechende Größenänderungender Hormondrüse.
Die CA produzieren nicht nur Hormone, sondern bei Schmetterlingen und Käfern enden darinauch die Endbläschen neurosekretorischer Zellendes Gehirns. Bei Manduca sex ta stellen die CAaußerdem ein Neuroh ämalorgan dar, das zur Freisetzung des großen Prothorakicotropen Hormons(PTTH) aus den lateralen neurosekretorischenZellen der Gruppe III dient (Abb. 12-4).
Abb. 12-3: Schematische Darstellung der Corpora allata(punktiert) und ihrer lage und Innervierung bei verschiedenen Insekten - Gruppen: 1 = Ursprung der CA, 2 =Thysanura, 3 = Odonata, 4 = verschiedene holometaboleGruppen, 5 = Hemiptera, 6 = höhere Diptera. A = Aorta; Bm =Bauchmark, CC = Corpora cardiaca; D = Darm; HG = HypocerebraI Ganglion l = labrum, la = labium, Md = Mandibel;Mx= Maxille; SG = Subösophagial Ganglion. (Modifiziert nachCassier, in: Gupta, 1990).

348 12 Endokrinologie
111
CC
CA
~":::::'=--- TN
r=:::::;:::::::~~/,'I7~~- NCC-NR
NCC 5
Abb. 12-4: Schematische Darstellung des cerebralen neurosekretorischen Systems und der Innervie·rung der Corpora cardiaca (CCl in5. larvenstadien oder adultenSchmetterlingen. I bis IV: Gruppenneurosekretorischer Zellen (schraffiert).Die Gruppe I projiziert ipsilateral, dieGruppen 11 und 111 contralateral über dienervi corpori cardiaci 1+2 (NCC 1+2) inden Corpus cardiacum-Corpus allaturn(CA)-Komplex. Die tritocerebrale GruppeIV projiziert über die nervi corpori cardiaci 3 (NCC 3) zum Corpus cardiacum-Corpus allatus-Komplex. A= aorta; F=frontal ganglion; G=Gehirn; NCC 3, =nervus corporis cardiaci 3; NCC 5 =nervus corporis cardiaci ventralis; NCCNR = nervi corpori cardiaci - nervusrecurrens; NR = nervus recurrens; SG =Suboesophagial Ganglion; TN = Tegumental Nerv. (Modifiziert nach Ichikawa,J. exp. Biol. 161 ,217, 1991 und Copenhaver and Truman, J. Neurosci. 6,1738, 1986).
Die Corpora cardiaca (CC) (Abb. 12-4)sind einkomplexes Organ. Sie entstehen embryonal auspaarigen Zellgruppen , die aus dem dorsalen Dachdes Stomodaeums evaginieren und während desdorsalen Rückenschlusses ihre definitive Lage einnehmen. Schon vor dem Schlüpfen der Larve sindin Zellen der CC Zeichen neurosekretorischer Aktivität festzustellen. In Imagines bestehen die CCaus mindestens 3 verschiedenen Zelltypen: Gliazellen, neuronalen und glandulären Zellen. Dieneuronalen Zellen werden auch als extrinsischeZellen bezeichnet. Sie speichern und sezernierenNeurohormone, die in den Perikaryen typischerNSZ aus dem Gehirn oder aus Ganglien gebildetwerden. Axone von NSZ aus dem medianen Bereich des Gehirns (pars intercerebralis) sind gebündelt und stellen die beiden nervi corporis cardiaci I (NCC I) dar, solche aus lateralen Bereichendie nervi corporis cardiaci 11 (NCC 11) und fallsaus dem Tritocerebrum noch Axone neurosekretorischer Zellen in die CC ziehen, sind diese zu dennervi corporis cardiaci III gebündelt. NCC I und11 können auch verschmolzen sein. Die glandulären - auch intrinsisch genannten - Zellen habenihre Zellkörper in den CC selbst und produzierenauch ihre eigenen Sekretionsprodukte, die adipokinetischenHormone (s. 12.2.5). Diese neurosekretorischen Zellen sind innerhalb der CC dispers verteilt und nur bei Locusta bilden sie einen zusammengehörigen und vom Speicherteil der CC getrennten glandulären Lobus. Bei Schmetterlingenund Käfern ziehen die NCC durch die CC durch
und enden mit ihren neurosekretorischen Endbläschen entweder an der Aortawand oder in einerHülle um die CA.
12.1.3 Häutungsdrüsen
Die Häutungsdrüsen der Insekten (Abb. 12-5)werden in der Embryogenese noch vor dem dorsalen Rückenschluss angelegt. Ektodermale Zellenaus dem Labialsegment invaginieren und bildenzunächst eine schlauchförmige Struktur aus. DieseAnlagen lagern sich im weiteren Entwicklungsverlauf den ebenfalls im Labialsegment gebildetenSpeicheldrüsen an. Sie wachsen aus, werden kompakt und haben, je nach Insektenart, schon amEnde der Blastokinese die endgültige Zellzahl erreicht. Die Häutungsdrüsen können an der Kopfkapsel als Ventraldrüsen angeheftet im Kopf verbleiben. Sie können aber auch caudad auswachsen,mit der Kopfkapsel verwachsen oder frei sein. Indiesen Fällen bezeichnet man sie dann als Prothoraxdrüsen. Ursprünglichere Taxa weisen Ventraldrüsen auf, während Blattoidea, Hemipteroidea und alle holometabolen Insekten Prothoraxdrüsen besitzen. Prothoraxdrüsen legen sich häufig eng an Tracheen oder auch Muskelfasern anund umwachsen diese zum Teil. Sie sind innerviert, meist vom Prothorakal- und Suboesophagialganglion, bei Lepidopteren noch zusätzlichvom Mesothorakalganglion. Bei Dipteren schlie-

12.1 Hormone und ihre Bildungsorte 349
A
c
B
o
rI'\, ~••.. ' , . . ....
......- "~'
1·····,
.J T
r 'c-: 'l..
F
SG
MG-~~J--4-
G
PG
Abb.12·5: Häutungsdrüsen und assoziierte Organe in Larvenstadien verschiedener Insekten - Gruppen: A =Ephemeroptera, B= Isoptera, C=Lepidoptera (Bombyx moii), D=Lepidoptera (Hyalophora), E=Orthoptera (Gryllus bimaculatus),F= höhere Diptera, G= Dictyoptera (Periplaneta americana). CA =Corpora allata, CC = Corpora card iaca, MG = MesothorakaiGanglion, PD = Prothorax Drüse, PG = Prothorakal Ganglion, SG = Subösophagial Ganglion, T = Tracheen. (Modifiziert nachBeaulaton, in: Gupta, 1990).
Ben sich die Prothoraxdrüsen zu einem Corporaallata/Corpora cardiaca -Komplex zur so genannten Ringdrüse zusammen. Lage und Aussehen derHäutungsdrüsen sind daher sehr vielgestaltig(Abb. 12-5).
Es ist bis jetzt nicht eindeutig untersucht worden, zuwelchem Zeitpunkt die Häutungsdrüsen erstmals Hormone synthetisieren. Licht- und elektronenmikroskopisch kann man schon während der Embr yogenese 2
Aktivitätszyklen der Prothoraxdrüsen feststellen, diezeitlich mit der Ablagerung der 2. und 3. (larvalen)embryonalen Kutikula korrelieren. Diese zyklischen Änderungen spiegeln sich auch in Titeränderungen an Ecdysteroiden wider, wobei nicht nur die Synthese, sondernauch die Freisetzung von Ecdysteroiden aus Konjugatenentscheidend ist. Während der postembryonalen Entwicklung nimmt die Größ e und bei einigen Gruppenauch noch die Zellzahl der Häutungsdrüsen zu. In derPuppe oder häufiger in den Imagines kommt es aber

350 12 Endokrinologie
meist zu einer Degeneration der Häutungsdrüsen. Hierbei wirken die Häutungshormone selbst als Auslöser fürApoptose und Autophagie.
Die Feinstruktur der Häutungsdrüsen wurde beimehreren Arten intensiv untersucht. Charakteristische Bestandteile sind: ein hoher Anteil glattenendoplasmatischen Reticulums (sER) und vielefreie Ribosomen, wie sie für Steroide produzierende Zellen typisch sind . Es kommt aber auchraues endoplasmatisches Reticulum (rER) vor, dasal~ Produktionsort für Ecdysteroid-Transportproteme angesehen wird . Die Plasmamembran der~äutungsdrüsen weist starke Einfaltungen auf, dieem ausgeprägtes Lakunensystem bilden können.Bei mehreren Arten (Tenebrio molitor, Aeschnacyanea, Galleria mellonella) wurden auch Lipidvakuolen beschrieben, die als Speicher für Cholesterin angesehen werden, das als Vorstufe für dieEcdys~eroidsynthese benötigt und von Arthropoden nicht selbst synthetisiert werden kann, in derNahrung aber reichlich vorhanden ist. Im Verlaufeines Häutungszyklus kommt es zu ausgeprägtenVeränderungen der Feinstruktur. Die Membraneinfaltungen nehmen an Größe zu, wenn die Drüsen Ecdysteroide synthetisieren und sie werdenreduziert kurz vor einer Häutung, wenn der Ecdysteroidspiegel bereits abgefallen ist. Ein gleichartiger Verlauf ist auch für das sER beobachtbar.Im Verlauf der Ecdysteroidsynthese nehmen dieLipidvakuolen und auch die Glykogengrana ab.Wie für alle Drüsen, die Steroidhormone produzieren, gilt auch für die Häutungsdr üsen, dass sieEcdysteroide nach Bedarf synthetisieren und keineSpeicherorte für diese Hormone darstellen.
12.1.4 Reproduktionsorgane
Im Gegensatz zu Vertebraten ist die Synthese vonHormonen in den Fortpflanzungsorganen der Insekten bisher nur bei wenigen Arten nachgewiesenworden, obwohl bei vielen Insekten - aber auchanderen Arthropoden - in Gonaden Ecdysteroideund Juvenilhormone gefunden wurden. Die Synthese von Ecdysteroiden ist in Hoden einigerSchmetterlinge und Orthopteren und in den FollikelzelIen der panoistischen Ovarien von Locustamigratoria, Gryllus bimaculatus und Nauphoeta cinerea gezeigt worden. In diesen Arten wird in denvitelIogenen Oocyten der Block der Meiose während der Prophase durch Ecdyson aufgehoben.O~arien produzieren zudem noch Peptidhormone,WIe z.B. Neb~TMOF (s. 12. 2. I), die u.a. anhormonellen Regulationen, etwa der Eireifungbeteiligt sind. '
12.1.5 Andere Gewebe alsHormonproduzenten
Am B:ispiel der Reproduktionsorgane (12.1.4)und bel der Besprechung pleiotroper Effekte vonHäutungshormonen (12.2.1) wird offensichtlichdass außer Häutungsdrüsen auch andere Geweb~Ecdysteroide zu synthetisieren vermögen. AusserOvar und Testis sind noch Oenocyten und dasIntegument in der Lage, Ecdysteroide in geringenMengen zu synthetisieren. Die physiologische Bedeutung dieser zusätzlichen Ecdysteroidsyntheseist nicht in allen Fällen klar. Zwar gibt es Befundedass im frühen letzten Larvenstadium isoliert~Abdomina ohne Prothoraxdrüse eine puppale undsogar adulte Häutung durchlaufen können; die~cdyster~idspiegel sind aber deutlich niedriger alsIm GanztIer. Ecdysteroide aus der Epidermis spiele~ vermutlich eher eine Rolle als auto- oder para~nne. Media~oren, die in der Epidermis über gapjunctions weitergeleitet werden.
Zu den Hormondrüsen und neurosekretorischen Zellen kommt noch das sogenannte "diffuseendokrine System" hinzu . Es handelt sich hierbeium Zellen, die zwischen die VerdauungszelIen desMitteldarmes eingestreut sind und deren Feinstruktur den endokrinen Zellen des Darmes vonVertebraten entspricht. Außerdem wurden in diesen Insektenzellen mit immuncytochemischen Methoden bei Vertebraten vorkommende und metabolisch wirksame Peptidhormone wie Gastrin<?holcystokinin, Pankreatisches Polypeptid, Insu~Im, Glucagon, Vasopressin, aber auch Opioide wieß-Endorphin .und Enkephaline, sowie peptidergeNeurotransmitter der FMRFamid-Familie nachgewiesen.
Bereits vor der Ausbildung der Hormondrüsensind die entsprechenden Hormone schon währendder Embryogenese nachzuweisen. Die Herkunftdieser Hormone wie Ecdysteroide, Juvenilhormonund Neuropeptidhormone (PTTH in Manducasexta und Bombyx mori, sowie Diapausehormoni~ B. ~ori) in Embryonen vor der Ausbildung dereigentlichen Hormondrüsen ist unterschiedlich.H.äufig wurde ein maternaler Ursprung nachgeWIesen. Darüber hinaus besteht die Möglichkeitder Synthese durch andere Gewebe. So kann JHIII ~~ der Serosa von Locusta migratoria synthetisiert werden , bevor die Corpora allata angelegt sind. Erst in späteren Stadien der Embryogenese kommt die Hormonsynthese der dann bereits ausgebildeten endokrinen Drüsen hinzu.

12.1.6 Hormonrezeptoren
Zu einem Hormonsystem gehören definitionsgemäß außer der Hormondrüse als Bildungsort unddem Transportweg über die Hämolymphe nochder oder die Wirkorte, die durch den Besitz spezifischer Erkennungsstrukturen oder Rezeptoren fürein entsprechendes Hormon ausgezeichnet sind .Für Insekten liegen umfassendere Kenntnisse überdie Struktur, Funktion und Verteilung von Rezeptoren nur für den Ecdysteroidre~ept~r vor. Derfunktionelle Ecdysteroidrezeptor Ist ein Heterodimer aus dem Ecdysteroidrezeptor (EcR) undeinem zweiten Transkriptionsfaktor, ultraspiracle(USP) . Beide Proteine zählen zur großen .Familieder nukleären Rezeptoren. Der Ecdysteroidrezeptor konnte autoradiographisch mithilfe des radioaktiv markierten Ecdysteroidagonisten Ponasteron A oder immuncytochemisch in vielen Geweben, z. B. in spezifischen Zielorganen wie derEpidermis und dem Nervensystem und in einemausgeprägt entwicklungsabhängigen Muster nachgewiesen werden. Ecdysteroidreze~toren findetman u. a. auch in den neurosekretonsehen Zellen,die das prothoracicotrope Hormon (PTTH) ausschütten, in den Häutungsdrüsen seIbst, aber auchin Geweben, die Ecdysteroide metabolisieren wiedem Fettkörper und den Malpighischen Gefäßen.Diese Befunde machen es wahrscheinlich, dassEcdysteroide ihre eigene Synthese und ihren ~eta
bolismus über Ecdysteroidrezeptor-vermittelteProzesse regulieren . Der Ecdysteroidrezeptorkann bei einer Art in 2 bis 3 Isoformen vorkommen . Für Drosophila melanogaster wurde gezeigt, dass die Expression einer spezifischen Is~
form charakteristisch ist für Zellen im ZNS, diebeim Übergang von der Puppe zum adulten Tiereinem programmierten Zelltod unterliegen. ~cdy~teroidrezeptoren wurden neuerdings auch In ~l
nem charakteristischen Verteilungsmuster an Riesenchromosomen von Chironomus tentans detektiert: sie sind auch notwendig für die Oogenese.Die' Induktion von Puffs durch Ecdysteroideführte vor über 30 Jahren zu dem allgemeingültigen Konzept der Steroidhormonwirkung in Formvon Regulation der Transkription.
Für einige wenige Peptidhormone wurden neuerdings membranständige, mit G-Proteinen gekoppelte Rezeptoren nachgewiesen , die. über second messenger wie cAMP, cGMP sowie Hydrolyseprodukte aus dem Signaltransduktionswegüber Phosphatidylinositol wirken .
12.2 Funktionen einiger ausgewählter Hormone 351
12.2 Funktionen einigerausgewählter Hormone
12.2.1 Häutungshormone und dieRegulation ihrer Synthese
Häutungshormone oder Ecdysteroide (Abb. 12-6)spielen eine zentrale Rolle beim Häutungsg.esc~e
hen aller Arthropoden, sind aber weder dIe einzigen an diesem Prozess beteiligten Hormone,noch ist dies ihre einzige Aufgabe. Neben denmorphogenetischen Effekten in Form von Hä~
tung, Metamorphose und Evagination von Im~g~
nalscheiben können sie noch Wirkungen auf Eireifung und Embryogenese, Vitellogenese, Ovulation,Spermiogenese, Pheromon-Biosynthese, Syntheseund Aufnahme von Arylphorinen in den Fettkörper, Synthese unterschiedlichster Proteine undEnzyme, Farbwechsel, und das Verhalten ausüben,sowie einige nicht-genomische Effekte verursachen . Nicht alle diese Wirkungen treten aber ineiner Art auf.
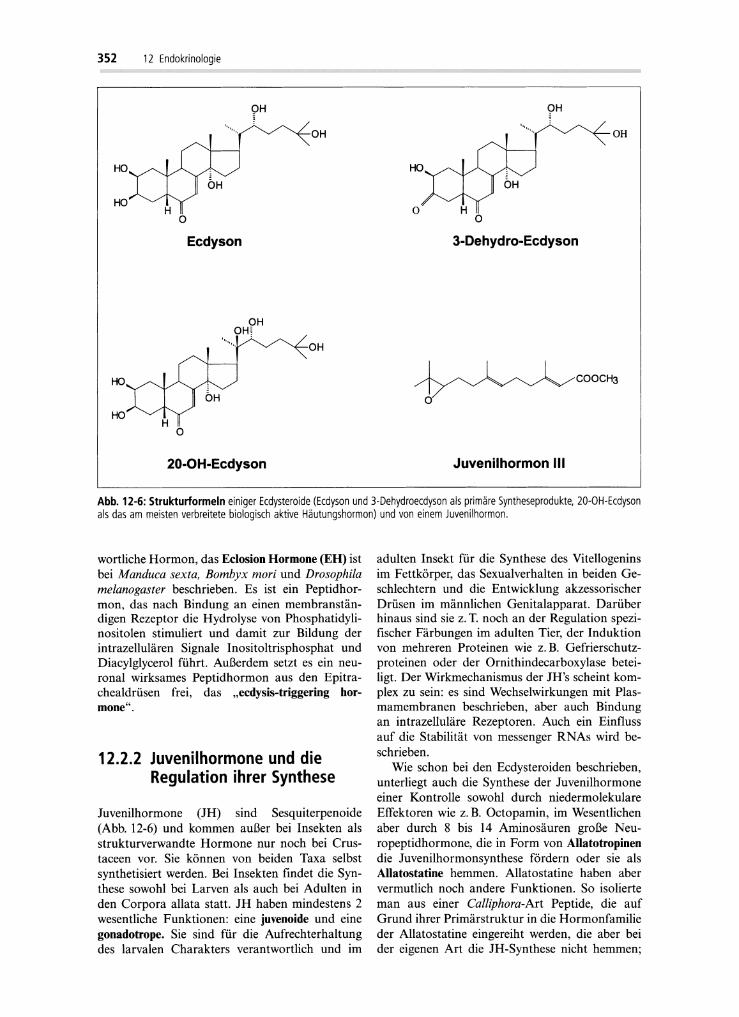
Das in den Häutungsdrüsen oder anderen Ecdysteroid-synthetisierenden Organen aus Cholesterin gebildete Ecdysteroid ist meist Ecdyson undhäufig zusätzlich 3-Dehydroecdyson (Abb. 12-6),die in den Zielorganen zu biologisch aktiverenEcdysteroiden, z. B. dem am weitesten verbreiteten20-0H-Ecdyson (Abb. 12-6) umgewandelt werden. Die Synthese der Ecdysteroide wird durchPeptidhormone aus dem Gehirn, die,soge.nan~ten
prothorakicotropen Hormone (PTTH s) stIm~hert.
In den meisten Arten fand man sowohl meder(4-7 kDa; PTTH-II) als auch hochmolekulare(22-30 kDa; PTTH-I) PTTH's; beide PTTH'skommen in einer Art in verschiedenen molekularen Varianten vor. PTTH's wirken über membranständige Rezeptoren; cAMP ist second messenger für beide prothoracicotrope Ho~mone'.Dieniedermolekulare Form fördert aber mcht bei allen Arten die Ecdysteroidsynthese. Dies trifft z. B.für den Seidenspinner, Bombyx mori, zu, dessenPTTH-Il eine erstaunlich hohe Sequenz- undStrukturähnlichkeit zum Insulin aufweist . Nebenden PTTH's sind auch noch andere Hormone ander Regulation der Ecdysteroidsynthese beteiligt:zum einen Ecdysteroide selbst, zudem noch Juvenilhormone und - ganz neu entdeckt das Hexapeptid Neb-TMOF (Neobellieria (=Sarcophaga)bullata Trypsin Modulating Oostatic Factor), dasu. U. die Ecdysteroidsynthese hemmt. Außer derEinleitung von Häutungsprozessen durch Häutungshormone ist auch der letzte Schritt, das Abwerfen der Exuvie, der Reste des alten Exoskeletts,bei der Ecdysis hormonell reguliert . Das verant-
352 12 Endokrinologie
HO
HO
~H
OH
o
'. ~H· · · · ·.~OH
o
Ecdyson
OHOHj
.......,
HO
HO
20-0H-Ecdyson
OH
3-0ehydro-Ecdyson
COOCH3
Juvenilhormon 111
Abb. 12-6:Strukturformeln einiger Ecdysteroide(Ecdyson und 3-Dehydroecdyson alsprimäre Syntheseprodukte, 20-0H-Ecdysonalsdas am meisten verbreitete biologisch aktive Häutungshormon) und von einem Juvenilhormon.
wortliehe Hormon, das Eclosion Hormone (EH) istbei Manduca sex ta, Bombyx mori und Drosophilamelanogaster beschrieben. Es ist ein Peptidhormon, das nach Bindung an einen membranständigen Rezeptor die Hydrolyse von Phosphatidylinositolen stimuliert und damit zur Bildung derintrazellulären Signale Inositoltrisphosphat undDiacylglycerol führt. Außerdem setzt es ein neuronal wirksames Peptidhormon aus den Epit raehealdrüsen frei, das "ecdysis-triggering hormone" ,
12.2.2 Juvenilhormone und dieRegulation ihrer Synthese
Juvenilhormone (JH ) sind Sesquiterpenoide(Abb. 12-6) und kommen außer bei Insekten alsstrukturverwandte Hormone nur noch bei Crustaceen vor. Sie können von beiden Taxa selbstsynthetisiert werden. Bei Insekten findet die Synthese sowohl bei Larven als auch bei Adulten inden Corpora allata statt. JH haben mindestens 2wesentliche Funktionen: eine juvenoide und einegonadotrope. Sie sind für die Aufrechterhaltungdes larvalen Charakters verantwortlich und im
adulten Insekt für die Synthese des Vitellogeninsim Fettkörper, das Sexualverhalten in beiden Geschlechtern und die Ent wicklun g akzessor ischerDrü sen im männlichen Gen italapparat. Darüberhinaus sind sie z.T. noch an der Regulation spezifischer Färbungen im adulten Tier, der Induktionvon mehreren Proteinen wie z.B. Gefrier schutzproteinen oder der Orn ith indecarboxyl ase beteiligt. Der Wirkmechanismus der JH 's scheint komplex zu sein: es sind Wechselwirkungen mit Plasmamembranen beschrieben , aber auch Bindungan intrazelluläre Rezeptoren. Auch ein Einflussauf die Stabilität von messenger RNAs wird beschrieben.
Wie schon bei den Ecdysteroiden beschrieben,unterliegt auch die Synthese der Juvenilhormoneeiner Kontrolle sowohl durch niedermolekulareEffektoren wie z. B. Octopamin, im Wesentlichenaber durch 8 bis 14 Aminosäuren große Neuropeptidhormone, die in Form von Allatotropinendie Juvenilhormonsynthese fördern oder sie alsAllatostatine hemmen . Allatostatine haben abervermutlich noch andere Funktionen. So isolierteman aus einer Calliphora-Art Peptide, die aufGrund ihrer Primärstruktur in die Hormonfamilieder Allatostatine eingereiht werden, die aber beider eigenen Art die JH-Synthese nicht hemmen;
12.2 Funktionen einiger ausgewählter Hormone 353
LIV LV ppp
I
III
ECDYSIS
III,.-......I ............_.-......III
o 2
IIII
ECDYSIS WANDERPHASE
~
,,I
ECDYSIS, ECDYSISIII,II
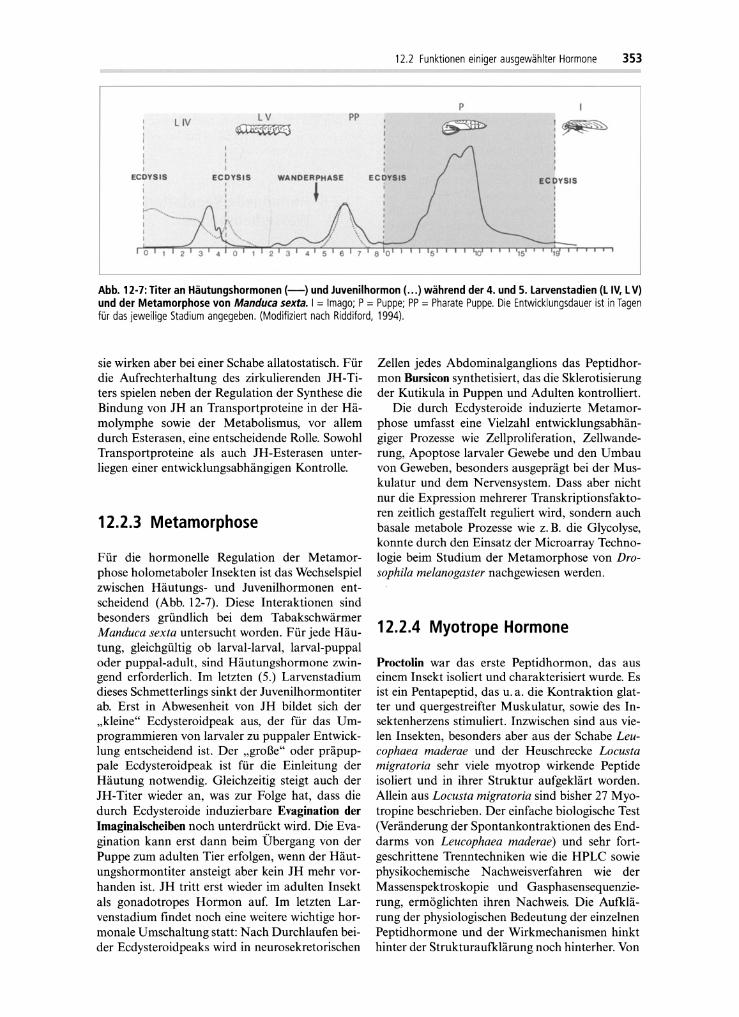
Abb. 12·7:Titer an Häutungshormonen (-) und Juvenilhormon (... ) während der 4. und 5. Larvenstadien (L IV, LV)und der Metamorphose von Manduca sexta. I = Imago; P= Puppe; PP = Pharate Puppe. Die Entwicklungsdauer ist inTagenfür das jeweilige Stadium angegeben. (Modifiziert nach Riddiford, 1994).
sie wirken aber bei einer Schabe allatostatisch. Fürdie Aufrechterhaltung des zirkulierenden JH-Titers spielen neben der Regulation der Synthese dieBindung von JH an Transportproteine in der Hämolymphe sowie der Metabolismus, vor allemdurch Esterasen , eine entscheidende Rolle. SowohlTransportproteine als auch JH-Esterasen unterliegen einer entwicklungsabhängigen Kontrolle.
12.2.3 Metamorphose
Für die hormonelle Regulation der Metamorphose holometaboler Insekten ist das Wechselspielzwischen Häutungs- und Juvenilhormonen entscheidend (Abb. 12-7). Diese Interaktionen sindbesonders gründlich bei dem TabakschwärmerManduca sexta untersucht worden. Für jede Häutung, gleichgültig ob larval-larval, larval-puppaloder puppal-adult, sind Häutungshormone zwingend erforderlich. Im letzten (5.) Larvenstadiumdieses Schmetterlings sinkt der Juvenilhormontiterab. Erst in Abwesenheit von JH bildet sich der"kleine" Ecdysteroidpeak aus, der für das Umprogrammieren von larvaler zu puppaler Entwicklung entscheidend ist. Der "große" oder präpuppale Ecdysteroidpeak ist für die Einleitung derHäutung notwendig . Gleichzeitig steigt auch derJH-Titer wieder an, was zur Folge hat, dass diedurch Ecdysteroide induzierbare Evagination derImaginalscheiben noch unterdrückt wird. Die Evagination kann erst dann beim Übergang von derPuppe zum adulten Tier erfolgen, wenn der Häutungshormontiter ansteigt aber kein JH mehr vorhanden ist. JH tritt erst wieder im adulten Insektals gonadotropes Hormon auf. Im letzten Larvenstadium findet noch eine weitere wichtige hormonale Umschaltung statt: Nach Durchlaufen beider Ecdysteroidpeaks wird in neurosekretorischen
Zellen jedes Abdominalganglions das Peptidhormon Bursicon synthetisiert , das die Sklerotisierungder Kutikula in Puppen und Adulten kontrolliert.
Die durch Ecdysteroide induzierte Metamorphose umfasst eine Vielzahl entwicklungsabhängiger Prozesse wie Zellproliferation, Zellwanderung, Apoptose larvaler Gewebe und den Umbauvon Geweben, besonders ausgeprägt bei der Muskulatur und dem Nervensystem . Dass aber nichtnur die Expression mehrerer Transkriptionsfaktoren zeitlich gestaffelt reguliert wird, sondern auchbasale metabole Prozesse wie z.B. die Glycolyse,konnte durch den Einsatz der Microarray Technologie beim Studium der Metamorphose von Drosophila melanogaster nachgewiesen werden.
12.2.4 Myotrope Hormone
Proctolin war das erste Peptidhormon, das auseinem Insekt isoliert und charakterisiert wurde. Esist ein Pentapeptid, das u.a. die Kontraktion glatter und quergestreifter Muskulatur, sowie des Insektenherzens stimuliert. Inzwischen sind aus vielen Insekten , besonders aber aus der Schabe Leucophaea maderae und der Heuschrecke Locustamigratoria sehr viele myotrop wirkende Peptideisoliert und in ihrer Struktur aufgeklärt worden.Allein aus Locusta migratoria sind bisher 27 Myotropine beschrieben. Der einfache biologische Test(Veränderung der Spontankontraktionen des Enddarms von Leucophaea maderae) und sehr fortgeschrittene Trenntechniken wie die HPLC sowiephysikochemische Nachweisverfahren wie derMassenspektroskopie und Gasphasensequenzierung, ermöglichten ihren Nachweis. Die Aufklärung der physiologischen Bedeutung der einzelnenPeptidhormone und der Wirkmechanismen hinkthinter der Strukturaufklärung noch hinterher. Von

354 12 Endokrinologie
der Primärstruktur aus lassen sich einige Gruppenan myotropen Hormonen zusammenfassen: Tachykinine, die Verwandtschaft zu den Tachykininen von Vertebraten aufweisen und die auch beiInsekten über G-Protein-gekoppelte Rezeptorenwirken. Außerdem findet man Kinine, Sulfakinine,die ein sulfatiertes Tyrosin aufweisen und einenhohen Grad an Sequenzähnlichkeit zu den gastrointestinalen Hormonen Gastrin II und Cholecystokinin der Vertebraten aufweisen, sowie diekurzkettigen FMRF-Amid-ähnlichen Peptide, diein allen Tiergruppen vorkommen und häufig neuromodulatorisch wirksam sind.
12.2.5 Adipokinetische Hormone
Bis heute sind 28 Peptide bekannt, die als adipokinetischeHormone (AKH) beschrieben werden. DieNamensgebung erfolgte nach dem ersten beschriebenen Hormon aus Schistocerca gregaria, das imFettkörper Fette abbaut. Alle beschriebenen AKHweisen einen hohen Grad an struktureller Ähnlichkeit auf, sie können aber sehr unterschiedlicheFunktionen ausüben . AKH bestehen aus 8, 9 oder10Aminosäuren . Sowohl die N-termin ale als auchdie C-terminale Aminosäure sind blockiert inForm von pyro-Glutamyl, bzw. als Amid.
Namensgebung und Hauptfunktion müssennicht übereinstimmen, wie dies im Falle von Allatostatinen (12.2.2) und Neb-TMOF (12.2.1) bereits beschrieben wurde. In diese Hormonfamilie(AKH-RPCH-Familie) gehört auch noch das ersteaus einem Wirbellosen isolierte und charakterisierte Peptidhormon, das Red Pigment Concentrating Hormone (RPCH) aus Crustaceen . In vielen bisher untersuchten Insektenarten kommengleichzeitig 2 verschiedene AKH vor, wobei diephysiologische Notwendigkeit unbekannt ist. Inden meisten Arten haben die beiden Hormonekeine qualitativ, sondern allenfalls quantitativ unterschiedlichen Effekte. Im wesentlichen beruhendie Wirkungen der AKH auf einer Mobilisierungvon Energie. Dabei können im gleichen Tier sowohl Lipid- als auch Glykogenspeicher betroffensein, zusätzlich wird der Proteinstoffwechse1 beeinflusst. Die mengenmäßigen Anteile dieser Energieträger sind artspezifisch. Auslöser für die Aktivierung des Energiestoffwechsels ist z. B. der Beginn des Fluges, der zu einer Ausschüttung vonAKH führt. Da aber auch nichtfliegende Insektenwie etwa die Stabheuschrecke Carausius morosusoder Larvenstadien von Heuschrecken, sowie einige sehr langsam bewegliche Käferarten überAKH verfügen, kann die Bereitstellung von Energie für den Flug durch AKH nicht die alleinigeAufgabe dieser Hormone sein. Eine Bedeutung für
den Lipidtransport, sowie den Metabolismus unddie Aufnahme von Lipiden in den Flugmuskelwurden für Heuschrecken nachgewiesen. EinigeMitglieder der AKH/RPCH-Familie weisen auchmyotrope Aktivität auf.
12.2.6 Hormonelle Regulation desWasserhaushaltes
Die Regulation des Wasser- und Mineralhaushaltes ist für landlebende Insekten von enormer Bedeutung. Neben Baumerkmalen, die Wasserverlust verhindern helfen wie beispielsweise Cuticulaund Tracheenatmung, ist aber umgekehrt beiPflanzensaft- oder Blutsaugenden Insekten einewirksame Abgabe überschüssiger Flüssigkeit notwendig (s. 4.5.4). So kann z. B. eine weiblicheAnopheles bei einer Blutmahlzeit mehr als dasZweifache des eigenen Körpergewichtes an Flüssigkeit aufnehmen und mit der Exkretion vonWasser schon während der Mahlzeit beginnen. Beiallen diesen Regulationen spielen Hormone einegroße Rolle. Sie können dabei im wesentlichen an 2Stellen angriffen: a) diuretische Hormone (DH)stimulieren die Produktion des Primärharnes inden Malpighischen Gefäßen und b) antidiuretischeHormone (ADH), fördern die selektive Reabsorption durch den Enddarm. Als Ausnahme ist beider Grille Acheta domesticus ein ADH bekannt,das antagonistisch zum DH direkt an den Malpighischen Gefäßen angreift.
Die Aufklärung der funktionellen Bedeutungund der Wirkmechanismen der DH und ADHwird u.a. dadurch erschwert, dass z.B. die diuretische Wirkung in ein und derselben Art durchmehrere Hormone verursacht sein kann . So sindin Locusta migratoria 3 verschiedene diuretischePeptidhormone beschrieben: eines, das Ähnlichkeit mit dem Corticotropin Re1easing Faktor(CRF) hat, ein zweites, das eine hohe Sequenzähnlickeit mit dem ebenfalls aus Vertebraten beschriebenen Hypophysen - Hinterlappen HormonArginin-Vasopressin (bzw. Arginin-Vasotocin;AVP, bzw. AVT) hat und ein drittes, das aus 46Aminosäuren besteht (Lom-DH = Locusta migratoria Diuretisches Hormon). Allen gemeinsamist, dass sie an membranständige Rezeptoren binden und daraufhin in den Malpighischen Gefäßenvermehrt cAMP als second messenger gebildetwird und als Folge die Sekretionsrate erhöht wird.Das AVP-ähnliche diuretische Hormon wird inden Thorakal- und Subösophagealganglien gebildetes. Es ist als Monomeres biologisch unwirksamund wirkt nur als Dimeres; die beiden Monomeresind antiparallel kovalent miteinander verknüpft .Lom-DH kommt im Gehirn, im wesentlichen aber

in den Speicherloben der Corpora cardiaca vorund erreicht eine maximale Wirkung bereits beieiner Konzentration von 5 x 10-12 M.
12.3 Eingriffe in das Hormonsystem als Mittel zurSchädlingsbekämpfung
Das Vorkommen Arthropoden-spezifischer Hormone, wie Juvenilhormone und Ecdysteroide bietet einen Ansatzpunkt für die Entwicklung vonInsektiziden mit geringer Wirbeltier-Toxizität.Aufgrund der einfacheren chemischen Struktur imVergleich zu Ecdysteroiden werden Juvenilhormonanaloge schon seit über 30 Jahren synthetisiert und erfolgreich zur Schädlingsbekämpfungeingesetzt. Durch chemische Modifikationen derJH's konnten Agonisten gewonnen werden , die imVergleich zu den endogenen JH 's sowohl photochemisch stabiler als auch schlechter metabolischabbaubar sind . Aufgrund der Metamorphosehemmenden Wirkung der JH kann allerdings dasProblem auftreten, dass unter dem Einfluss dernoch vorhandenen Ecdysteroide zusätzliche undgrößere Larvenstadien induziert werden, was imFalle von Arten, bei denen die Larven Fraßschädlinge sind , unerwünscht ist. Zudem sind auchschon die ersten Resistenzen gegen Juvenilhormon-Analoge nachgewiesen worden. InsektizideWirkung von mehreren Substanzen, die entwederzu einem Verlust oder einer Hemmung der JHSynthese führen, wurde ebenfalls nachgewiesen.Da für eine wirkungsvolle Bekämpfung aber höhere Dosen notwendig sind und da zudem einigeder Substanzen z. B. bei den meisten holometabolen Insekten nicht wirken , haben sich diese Substanzen zwar als wertvolle Hilfsmittel für das Labor erwiesen, aber nicht als Insektizide für dasFreiland.
Ecdysteroide können insektizide Wirkung haben, wenn sie in hoh en Dosen oder zu "falschen"Zeiten im Häutungszyklus gegeben werden . Einechemische Synthese von Ecdysteroiden ist zwarmöglich , aber wirtschaftlich unrentabel; gleichesgilt auch für die Isolierung aus pflanzlichem Material. Erst durch die Entwicklung einfacher gebauter, nicht-steroidaler Häutungshormon-Agonisten,den Benzoylhydrazin-Derivaten, ist dieser Angriffspunkt attraktiv geworden . Diese Substanzen binden mit hoher Affinität an den Ecdysteroidrezeptor und lösen damit ecdysteroid-vermittelte Reaktionen aus, z.B. zu früh induzierte Häutungen.Eine Wechselwirkung mit Steroidhormonrezeptoren aus Wirbeltieren konnte ausgeschlossen werden. Die Toxizität gegenüber Säugern ist außer-
Verzeichnis weiterführender Literatur 355
ordentlich gering . Zumindest im Labormaßstabsind aber Resistenzen gegenüber diesen Hormonagonisten beobachtet worden , die auf verändertenEcdysteroidrezeptoren beruhen. Eine Hemmungder Synthese oder Veränderungen des Metabolismus der Ecdysteroide, z. B. durch Extrakte desNeem Baumes in denen als wirksame Komponenten Azadirachtine vorhanden sind, wird neuerdings als Schädlingsbekämpfungsmaßnahme inAfrika und Indien angewandt.
Attraktiv erscheint auch ein Eingriff in dasSystem der Peptidhormone, da hier nicht nur Arthropoden-, sondern sogar Artspezifität vorliegt.Rezeptorblocker oder Mimetika für Neuropeptidhormone, Antagonisten und Superagonisten, sowie Enzyminhibitoren, sowohl für die Synthese alsauch den Abbau werden als mögliche Insektizidediskutiert. Hohe Gestehungskosten für Peptide,proteolytischer Abbau und die Verfügbarkeit derentsprechenden Substanzen am Wirkort stellennoch Probleme für eine Anwendung dar. Ein Einbau von Genen, die für Peptidhormone aus Insekten kodieren, entweder in Pflanzen, die vonInsekten befallen werden, oder in Baculoviren, diez. T. art- oder zumindest gruppenspezifisch Insekten befallen , ist zumindest im Labormaßstabbereits erfolgreich durchgeführt worden. Versuche,virale Insektizide dadurch noch zu verbessern,dass in das Genom des Baculovirus zusätzlich dasGen für die JH-Esterase integriert wurde, erbrachten bisher aber nicht den gewünschten Effekt .
Verzeichnis weiterführender Literatur(Bücher und Übersichtsartikel)
Birkenbeil, H. (1990) Die ultrastrukturellen Grundlagender Ecdysonbildung in den Häutungsdrüsen vonCrust aceen und Insekten . Zool. Jb. Physiol. 94:409-444
Coast, G. M. and Webster, S.G. (1998) Recent advancesin arthropod endocrinology. Cambridge UniversityPress, Cambridge
Dhadialla, T.S., Carlson, G. R., and Le, D. P., (1998)New insecticides with ecdysteroidal and juvenile hormone activity. Annu . Rev. Entomol. , 43: 545-569
Gäde, G., Hoffmann, K.-H., and Spring, 1.H. (1997)Hormonal regulation in insects: facts, gaps, and future directions. Physiol. reviews, 77: 963-1031
Gäde, G. (1996) The revolution in insect neuropeptidesilIustrated by the adipokinetic hormone/red pigmentconcentrating hormone family of peptides. Z. Naturforsch. 51c: 607-617
Gupta, A. P. (1990/91) Morphogenetic hormones of arthropods, 3 Bände, Rutgers University Press, NewBrunswick
Kerkut, G.A. and Gilbert, L. 1. (1985) Comprehensiveinsect physiology, biochemistry, and parmacology.Vol 7 und 8 (Endocrinology I und II). PergamonPress, Oxford

356 12 Endokrinologie
Henrich , V.C; Rybczynski , R., and Gilbert L. I. (1999).Peptide Hormones, Steroid Hormones and Puffs: Mechanisms and Model s in Insect Development. Vitamins and Hormones, 55: 73-125
Koolman, 1. (1989) Ecdysone - from chemistry to modeof action . Georg Thieme Verlag, Stuttgart
Laufer, H. and Downer, R. G. H. (1983) Invertebrateendocrinology I - Endocrinology of insects. Alan R.Liss., New York
Masler, E. P., Kelly, T.1. and Menn , 1.1. (1993) Insectneuropeptides: Discovery and application in insectmanagement. Arch. Insect Biochem. PhysioI. 22:87-111
Nijhout, H. F. (1994) Insect Hormones. Princeton Uni versity Press, Princeton
Ohni shi, E. and Ishizaki , H. (1990). Molting and metamorphosis. Springer Verlag, Heidelberg
Raabe, M. (1989) Recent developments in insect neurohormones. Plenum Press, New York
Riddiford, L. (1994) Cellular and molecular actions ofjuvenile hormone I. General considerations and premetamorphic actions. Adv. Insect PhysioI. 24:213-274
Schoofs, L., Vanden Broeck, 1. and de Loof, A. (1993)The myotropic peptides of Locusta migratoria: Structures, distribution, functions and receptors. InsectBiochem. Molec . BioI. 23: 859-881
Spindler-Barth, M. (1992). Endocrine strategies for insect vector and pest controI. ParasitoI. Res. 78:89-95
Spindler-Barth, M. and Spindler, K.-D. (1998) Morphogenetic actions and mode of action of ecdysteroids ina dipteran cell line. Current Trends Steroid Res. I:73- 81
Spindler-Barth, M. and Spindler, K.- D. (1987) Antiecdystero ids and receptors. In : Receptor mediated antisteroid action (ed.: M. K. Agarwal) W. de Gruyter,Berlin,497-511
Spindler-Barth, M. and Spindler, K.-D. (2000) Hormonal regulation of larval moulting and metamorphosis- Molecular aspects. In: Progress in developmentalendocrinology. (Dorn, A., ed.) Wiley-Liss, New York,117-144
Spindler, K.-D. (1997) Vergleichende Endokrinologie Regulation und Mechanismen. G. Thieme Verlag,Stuttgart
Spindler, K.-D., Quack, S. and Spindler-Barth, M. (1993)Insect cell Iines as tools for insecticide screening .Trends in Comparat. Biochem. PhysioI. I:1045-1056
Spindler, K.-D, Przibilla , S., and Spindler-B arth, M.(2001) Moulting hormones of arthropods: molecularmechanisms. Zoology 103: 189-201
Truman, 1.W. (1992) The eclosion hormone system ofinsects. Progr. in Brain Res. 92: 361-374
White, K. P., Rifkin, S.A., Hurban, P. and Hogness, D. S.(1999) Microarray analy sis of Drosophila development during metamorphosis. Science 286:2179-2184
![Lehrbuch Persönlichkeitspsychologie · : [Lehrbuch]__Schuetz__Persoenlichkeitspsychologie__[Druck-PDF]/04.05.2016/Seite 2 Lehrbuch Persönlichkeitspsychologie Astrid Schütz, Martin](https://img.pdfslide.org/doc/110x75/5e1ef9515ee7da6b5b6d41fc/lehrbuch-persnlic-lehrbuchschuetzpersoenlichkeitspsychologiedruck-pdf04052016seite.jpg)




























