Embed Size (px)
Citation preview

NMR-Spektroskopische Untersuchungen
von
Körperflüssigkeiten
(Kinderurin)
DISSERTATION
Zur Erlangung des Grades eines Doktors
der Naturwissenschaften
-Dr. rer. nat.-
Dem Fachbereich Biologie/Chemie der
Universität Bremen
vorgelegt von
Simone Tamoschus-Witt
Bremen
September, 2007
1. Gutachter: Prof. Dr. Dieter Leibfritz
2. Gutachter: Prof. Dr. Wolf-Dieter Stohrer

Meinen Eltern
In der Wissenschaft gleichen wir alle nur den Kindern, die am Rande des Wissens hie und da
einen Kiesel aufheben, während sich der weite Ozean des Unbekannten vor unseren Augen
erstreckt.
- Isaac Newton (1643-1727) -

Danksagung
I
Danksagung
Mein besonderer Dank gilt Herrn Prof. Dr. D. Leibfritz für die Themenstellung und die
Unterstützung und Förderung, die es mir ermöglicht haben, diese Arbeit fertig zu stellen.
Herrn Prof. Dr. Stohrer danke ich für die Bereitschaft zur Begutachtung und Diskussion
dieser Arbeit.
Bei Herrn Dipl.-Ing. Johannes Stelten und Dr. Wieland Willker bedanke ich mich für die
Unterstützung bei technischen Problemen und NMR-spektroskopischen Fragestellungen.
Frau Dipl.-Ing. Dorit Kemken danke ich für die schnelle und erfolgreiche Einarbeitung in die
HPLC.
Vielen Dank an alle Mitarbeiter der Arbeitsgruppe Prof. Dr. D. Leibfritz für die Hilfe und die
Ratschläge im Rahmen der praktischen Arbeit, sowie die fördernden und unterhaltsamen
Gespräche.
Ich danke allen Mitarbeitern und Freunden, die mir bereitwillig Proben zur Verfügung gestellt
haben.
Ein spezieller Dank gilt meinen Freunden und meiner Schwester, die mich begleitet, meine
Launen ertragen und mich immer wieder motiviert haben.
Ganz besonders danke ich meinen Eltern für ihr Verständnis und die liebevolle Unterstützung
in jeder Hinsicht.
Danke Torben, für die unendliche Geduld und Liebe.

Inhalt
II
1. Einleitung.........................................................................................................5
2. Darstellung der Methoden
2.1. Kernmagnetische Resonanzspektroskopie......................................................................7
2.1.1. Einführung.............................................................................................................7
2.1.2. Physikalische Grundlagen.....................................................................................7
2.1.3. Allgemeine Experimente.......................................................................................8
2.1.4. Anwendungsgebiete..............................................................................................9
2.1.5. Aufarbeitung von Körperflüssigkeiten für die NMR-Spektroskopie..................10
2.1.5.1. Aufarbeitung von Urin.........................................................................10
2.2. Festphasenextraktion.......................................................................................................11
2.2.1. Einführung...........................................................................................................11
2.2.2. SPE-Phasen.........................................................................................................12
2.2.3. Methodenentwicklung.........................................................................................13
2.2.4. Durchführung der Trennung................................................................................14
2.3. Hochdruckflüssigchromatographie................................................................................16
2.3.1. Einführung...........................................................................................................16
2.3.2. Methodenentwicklung.........................................................................................17
2.3.3. Durchführung der Trennung................................................................................18
2.4. Ergebnisse der Trennungen............................................................................................18
2.4.1. Festphasenextraktion...........................................................................................19
2.4.1.1. Chromabond C18..................................................................................20
2.4.1.2. Chromabond EASY..............................................................................21
2.4.2. Hochdruckflüssigchromatographie.....................................................................22
2.4.3. Zusammenfassung...............................................................................................24
3. Identifizierung unbekannter Metabolite
3.1. Anatomie und Funktion der Niere.................................................................................25
3.2. Einführung........................................................................................................................27
3.2.1. Allgemeine Durchführung...................................................................................28
3.3. Vorstellung der Metabolite.............................................................................................29
3.3.1. Aromatische Verbindungen.................................................................................30
3.3.1.1. Aromaten..............................................................................................30

Inhalt
III
3.3.1.2. Heteroaromaten....................................................................................35
3.3.1.2.1. Nukleoside.........................................................................................35
3.3.1.2.2. Pyridine..............................................................................................39
3.3.1.2.3. Indole und Imidazole.........................................................................42
3.3.2. Proteinogene Aminosäuren.................................................................................44
3.3.2.1. Unpolare, aliphatischeAminosäuren.....................................................44
3.3.2.2. Aromatische Aminosäuren...................................................................46
3.3.2.3. Polare, ungeladene Aminosäuren.........................................................48
3.3.2.4. Positiv geladene Aminosäuren.............................................................50
3.3.3. Kohlenhydrate.....................................................................................................52
3.3.3.1. Monosaccharide....................................................................................53
3.3.3.2. Disaccharide.........................................................................................57
3.3.3.3. Zuckeralkohole.....................................................................................58
3.3.4. Biogene Amine....................................................................................................60
3.3.4.1. Quartäre Ammoniumverbindungen......................................................60
3.3.4.2. Methylamine.........................................................................................62
3.3.4.3. Guanidin-Verbindungen.......................................................................63
3.3.4.4. Primäre Amine......................................................................................65
3.3.5. Carbonsäuren.......................................................................................................67
3.3.5.1. Dicarbonsäuren.....................................................................................67
3.3.5.2. Tricarbonsäuren....................................................................................70
3.3.5.3. Aminocarbonsäuren..............................................................................72
3.3.5.4. Hydroxycarbonsäuren...........................................................................73
3.3.5.5. Oxocarbonsäuren..................................................................................75
3.3.5.6. Andere Verbindungen..........................................................................76
3.4. Zusammenfassung............................................................................................................78
4. Einfluss der Ernährung auf die Zusammensetzung des Urins
4.1. Einführung........................................................................................................................80
4.2. Allgemeine Durchführung...............................................................................................81
4.3. Untersuchte Nahrungsmittel...........................................................................................81
4.3.1. Bananen...............................................................................................................82
4.3.2. Pekanüsse............................................................................................................84
4.3.3. Orangen...............................................................................................................85

Inhalt
IV
4.4. Zusammenfassung............................................................................................................89
5. Anhang............................................................................................................90
5.1. Lagerung und Vorbereitung der Urinproben...............................................................90
5.2. Durchführung der NMR-Messungen.............................................................................90
5.3. NMR-Parameter von Urin-Metaboliten (pH 5,85).......................................................91
6. Abkürzungen................................................................................................107
7. Literatur.......................................................................................................109

Einleitung
5
1. Einleitung
Die Urinuntersuchung ist eine sehr alte Methode zur Diagnose von Krankheiten. Bereits im
alten Ägypten (etwa 2500 v. Chr.) waren Polyurie und Hämaturie bekannt. Hindu-Ärzte
untersuchten den Urin mit allen fünf Sinnen und die Griechen deuteten Veränderungen des
Urins als Symptom verschiedener Krankheiten. Im 16. Jahrhundert wurde die Uroskopie zu
einem diagnostischen Universalmittel. Sie galt als wichtigste Tätigkeit des Arztes. Das
kolbenförmige Harnglas wurde zum Standessymbol der Ärzteschaft. Ende des 18.
Jahrhunderts wurden mit der chemischen Analyse präzisere Methoden für die
Urinuntersuchung entwickelt. In den vierziger Jahren des 19. Jahrhunderts kam die
mikroskopische Betrachtung des Urins hinzu. Heute sind die Untersuchungen weitestgehend
automatisiert. Der Urin dient nicht nur zur Diagnose von Krankheiten, er wird auch für
Drogentests und zur Medikamentenforschung eingesetzt. Geringe Mengen reichen aus, um im
Labor eine Vielzahl von Krankheitsparametern zu bestimmen.
Die Probleme einer solchen Untersuchung liegen in dem hohen Zeit- und Kostenaufwand. Für
die Analyse ist eine aufwendige Probenvorbereitung notwendig, bis hin zur Isolierung
einzelner Metabolite. Dadurch ist die Wahrscheinlichkeit von Substanzverlusten sehr groß.
Verschiedene Untersuchungen können auch an nativen Urinproben erfolgen. Diese Methoden
sind jedoch zumeist nicht sehr spezifisch, so dass ähnliche Substanzen die Ergebnisse
verfälschen können. Zudem erfordert die Untersuchung von Einzelsubstanzen eine
Vorauswahl der zu untersuchenden Verbindungen. Dabei können unerwartete oder
unbekannte Metabolite nicht berücksichtigt werden.
Die NMR-Spektroskopie bietet hier verschiedene Vorteile. Das Protonenspektrum einer
Urinprobe stellt sämtliche niedermolekularen, protonenhaltigen Metabolite dar. Damit liefert
es einen nahezu vollständigen Überblick über die Zusammensetzung des Urins. Die
Messungen erfordern, je nach Fragestellung, nur wenig Probenvorbereitung und können in
kurzer Zeit durchgeführt werden. Mustererkennungsverfahren ermöglichen eine zügige
Auswertung der Spektren. Mit ihrer Hilfe können mögliche Abweichungen im
Metabolitenprofil des Urins ermittelt werden. Darüber hinaus ermöglicht diese Methode die
Aufklärung unbekannter Substanzen.
Im Rahmen dieser Arbeit wurden verschiedene Urinproben gesunder Probanden mit Hilfe der
NMR-Spektroskopie untersucht. Diese Untersuchungen sind nützlich, um neue Kenntnisse
über enthaltene Metabolite und deren Konzentrationen zu gewinnen. Mit diesem Wissen
können pathologische Veränderungen erkannt und Stoffwechselstörungen identifiziert
werden.

Einleitung
6
Zu Beginn werden die Grundlagen der NMR-Spektroskopie und zweier Trennmethoden,
Festphasenextraktion (SPE) und Hochdruckflüssigchromatographie (HPLC), beschrieben. Die
Festphasenextraktion wurde angewendet, um verschiedene Substanzklassen der komplexen
Urinproben zu separieren. Anschließend wurde mit Hilfe der HPLC eine Fraktion der SPE
weiter aufgetrennt. Auf diese Weise konnte die Anzahl der Signale in den NMR-Spektren
reduziert, und die Strukturaufklärung unbekannter Substanzen erleichtert werden.
In den folgenden Kapiteln werden verschiedene Metabolite vorgestellt, die in den Spektren
der Urinproben identifiziert werden konnten. Für diese Untersuchungen wurde Kinderurin
verwendet. Hier ist die Wahrscheinlichkeit, ungewöhnliche Substanzen (z. B. durch
Zigaretten- oder Kaffeekonsum hervorgerufen) zu finden, wesentlich geringer als bei
Erwachsenen. Die Identifizierung der Metabolite erfolgte über Literaturrecherche,
Datenbanken und Strukturaufklärung.
Ein weiterer Teil dieser Arbeit beschäftigt sich mit dem Einfluss der Ernährung auf die
Zusammensetzung von Urin. Hierfür wurden, nach dem Verzehr verschiedener
Nahrungsmittel, in bestimmten Zeitabständen Proben genommen und vermessen. Durch
Vergleich der Protonenspektren konnten Abweichungen in den Proben erkannt werden. Die
Änderungen der Konzentrationen wurden über die Intensitäten der Signale relativ zum
Creatininsignal bestimmt.

Darstellung der Methoden
7
2. Darstellung der Methoden
2.1. Kernmagnetische Resonanzspektroskopie
2.1.1. Einführung
Den Grundstein für die Kernmagnetische Resonanzspektroskopie (NMR) legte der
Amerikaner Otto Stern im Jahr 1933. Er fand heraus, dass Protonen, genau wie Elektronen,
magnetische Eigenschaften besitzen und wurde dafür 1943 mit dem Nobelpreis für Physik
ausgezeichnet. 1939 wies Isidor Isaac Rabi den Spin von Atomkernen nach und erhielt dafür
1944 den Nobelpreis für Physik. Felix Βloch und Edward Will Purcell gelang es 1945
unabhängig voneinander, erstmals NMR-Experimente in flüssiger und fester Phase
durchzuführen. Sie wurden dafür 1952 gemeinsam mit dem Nobelpreis für Physik
ausgezeichnet. Die Einführung der gepulsten Fourier-Spektroskopie durch den Schweizer
Richard Robert Ernst (Nobelpreis für Chemie 1991) und die Entwicklung supraleitender
Magnete, sowie zweidimensionaler Techniken, machen die NMR heute zu einem der
wichtigsten Analyseverfahren in der Physik, Chemie, Biochemie und Medizin (Schlemmer,
2005).
2.1.2. Physikalische Grundlagen
Nahezu alle Atomkerne haben einen Drehimpuls (Spin) I, der von der Anzahl der Protonen
und Neutronen abhängig ist. Dieser Spin ist durch eine Proportionalitätskonstante γ
(gyromagnetische Konstante) mit einem magnetischen Moment µ verknüpft. Wird nun ein
Kern mit einem Kernspin ≠ 0 in ein äußeres, angelegtes Magnetfeld B0 gebracht, richten sich
das magnetische Moment, und damit auch der Spin, im Magnetfeld aus und präzediert
kegelförmig um die Magnetfeldachse. Bei Kernen mit einem Spin = ½ kann der Spin sich
dabei entweder parallel (energiearm) oder antiparallel (energiereich) zur Magnetfeldachse
ausrichten. Die Geschwindigkeit der Präzession wird Lamor-Frequenz ωL genannt. Sie ist von
dem Produkt aus Magnetfeld und gyromagnetischem Verhältnis abhängig.
Die Summe der im Magnetfeld auftretenden magnetischen Momente wird als Magnetisierung
bezeichnet. Sie ist eine makroskopisch messbare Größe. Die magnetischen Momente verteilen
sich gleichmäßig über den Präzessions-Kegel, wobei die Anzahl der energieärmeren, parallel
zum Feld angeordneten Spins überwiegt. Durch diese Verteilung bildet sich eine
makroskopische Magnetisierung entlang der Magnetfeldrichtung (Z-Achse) aus. Wird
senkrecht zum Magnetfeld ein Hochfrequenz-Impuls eingestrahlt, kann die Magnetisierung
entsprechend der Pulsdauer aus seiner Gleichgewichtslage gebracht werden. Dies ist
allerdings nur möglich, wenn die Anregungsfrequenz des Pulses mit der Lamor-Frequenz

Darstellung der Methoden
8
übereinstimmt, da nur Strahlungsenergie dieser Frequenz absorbiert werden kann. Bei einem
90°-Puls beträgt die Auslenkung der Magnetisierung genau 90° zur Gleichgewichtslage. Ist
der Puls beendet, präzediert die Magnetisierung in der x-y-Ebene, also senkrecht zum
Magnetfeld. Diese Präzession erzeugt ein oszilllierendes Feld, welches in einer
Empfangsspule in y-Richtung wahrgenommen werden kann. Da jetzt nur noch das
Magnetfeld auf die Magnetisierung wirkt, kehren die Spins in den Ausgangszustand zurück
(Relaxation). Dabei wird die überschüssige Energie im gleichen Frequenzbereich wie die der
Anregung wieder abgegeben. Dieses abklingende Emissionssignal in y-Richtung wird als FID
(free induction decay) bezeichnet. Durch eine Fourier-Transformation wird der FID in ein
Spektrum der Frequenzdomäne umgewandelt.
Die Abhängigkeit der Larmorfrequenz eines Kernspins von seiner chemischen Umgebung
wird als chemische Verschiebung δ bezeichnet. Sie beschreibt die Änderung der
Resonanzfrequenz relativ zu einer Referenzsubstanz und wird unabhängig vom Magnetfeld in
ppm (parts per million) angegeben. Für 1H und 13C wird fast immer Tetramethylsilan (TMS)
als Referenz verwendet. Bei isolierten Atomkernen hängt die Resonanzfrequenz
ausschließlich vom äußeren Magnetfeld und ihrem gyromagnetischen Verhältnis ab. In realen
Molekülen wird der Kernspin durch Elektronen abgeschirmt. Die Elektronen besitzen ein
magnetisches Moment, welches dem äußeren Feld entgegengerichtet ist. Dadurch wird die
Resonanzfrequenz verkleinert. Ein Atomkern mit kleiner Verschiebung wird als abgeschirmt
(hohes Abschirmfeld) bezeichnet, ein Kern mit großer Verschiebung ist dagegen entschirmt
(tiefes Abschirmfeld).
In der hochauflösenden NMR-Spektroskopie kann eine Aufspaltung verschiedener Signale zu
Multipletts beobachtet werden. Diese Aufspaltung erfolgt durch indirekte oder skalare Spin-
Spin-Kopplung von Kernspins über kovalente Bindungen. Die Größe der Aufspaltung wird
als Kopplungskonstante J (Hz) bezeichnet und entspricht dem Frequenzabstand von zwei
Multiplettübergängen. Kopplungsmuster sind essentiell, um bestimmte Spinsysteme in einem
Molekül zu identifizieren und seine chemische Struktur aufzuklären.
2.1.3. Allgemeine Experimente
In der NMR-Spektroskopie werden hauptsächlich eindimensionale Experimente eingesetzt.
Dabei werden überwiegend die Kerne Wasserstoff (1H) und Kohlenstoff (13C), aber auch
Phosphor (31P), Stickstoff (15N) und Fluor (19F) gemessen. Um störende Signale zu
verhindern, werden deuterierte Lösemittel verwendet.

Darstellung der Methoden
9
Ein Protonenspektrum enthält diverse Informationen, die zur Strukturaufklärung
herangezogen werden können. Die chemische Verschiebung der Signale gibt Auskunft über
einzelne Strukturfragmente, die Integrale geben die Zahl der Protonen vor und die
Multiplizitäten der Signale ermöglichen Angaben über die Zahl der benachbarten Protonen.
Kohlenstoffspektren geben durch die chemischen Verschiebungen ebenfalls Auskunft über
Strukturfragmente. Über ein DEPT-Experiment (distortionless enhancement by polarization
transfer) kann die Anzahl der direkt gebundenen Wasserstoffe am Kohlenstoff bestimmt
werden. Quartäre Kohlenstoffe geben im DEPT-Spektrum kein Signal und können durch
Vergleich mit einem einfachen Kohlenstoffspektrum ebenfalls identifiziert werden.
Daneben kommen auch verschiedene zweidimensionale Verfahren zum Einsatz, die durch
Korrelation der einzelnen Signale zusätzliche Informationen liefern und
Resonanzüberlagerungen bei größeren Molekülen oder Substanzgemischen entzerren können.
Zum einen gibt es homonukleare Experimente, welche das skalare Wasserstoff-
Kopplungsnetzwerk darstellen. Dazu gehören das COSY-Experiment (correlation
spectroscopy), welches die Kopplung zweier Wasserstoffatome im Molekül über zwei
(geminal) oder drei (vicinal) Bindungen aufzeigt, und das TOCSY (total correlation
spectroscopy), mit dem sich alle Signale eines Spinsystems in Verbindung bringen lassen.
Daneben gibt es auch noch das NOESY (nuclear Overhauser enhancement spectroscopy),
welches räumlich benachbarte Wasserstoffatome erfasst.
Zu den heteronuklearen Experimenten zählen u. a. das HSQC-Experiment (heteronuclear
single quantum coherence), das HMBC (heteronuclear multiple bond correlation) und das
HSQC-TOCSY. Im ersten Experiment werden die Heteroatome und ihre direkt gebundenen
Wasserstoffatome korreliert, mit dem zweiten lässt sich die Kopplung eines Protons zu einem
Heteroatomen und die Kopplungen zu benachbarten Protonen aufzeigen. Dadurch ist es z. B.
möglich, quaternäre Kohlenstoffe zu identifizieren. Das dritte Experiment stellt sowohl die
direkt an das Heteroatom gebundenen Wasserstoffatome als auch Protonen, die zwei oder
mehr Bindungen entfernt sind, dar.
2.1.4. Anwendungsgebiete
Ein großer Vorteil der NMR liegt darin, dass die Proben während der Messung nicht zerstört
werden. So stehen sie hinterher für weitere Analysemethoden zur Verfügung. Deshalb findet
die NMR-Spektroskopie in vielen naturwissenschaftlichen Bereichen Anwendung. In der
organischen Synthese wird sie zur Reinheits- und Produktkontrolle genutzt. Mit ihrer Hilfe

Darstellung der Methoden
10
können Strukturen sowohl in Lösung als auch im Festkörper mit atomarer Auflösung
bestimmt werden. In der Biologie ermöglicht sie die dreidimensionale Aufklärung von
Proteinstrukturen und Enzym-Substratbindungen (Takeuchi und Wagner, 2006; Vaynberg und
Qin, 2006). Die MRT (Magnetresonanztomographie) basiert auf den Prinzipien der NMR und
ist heutzutage in der medizinischen Diagnostik unverzichtbar (Valencia und Castillo, 2006).
Ein weiteres, wichtiges Einsatzgebiet in der klinischen Chemie und Toxikologie ist die
Untersuchung von Körperflüssigkeiten (Blut, Urin, Zerebrospinalflüssigkeit, etc) auf
Abbauprodukte von Medikamenten und Drogen, Infektionen und Stoffwechselstörungen
(Hodavance et al, 2006; Coen et al, 2005). Diese Untersuchungen sind üblicherweise mit
einem geringen Aufwand an Vorbereitungs- und Messzeit verbunden und liefern einen
Überblick über alle enthaltenen Substanzen der Körperflüssigkeiten.
2.1.5. Aufarbeitung von Körperflüssigkeiten für die NMR-Spektroskopie
Körperflüssigkeiten können in verschiedene Klassen eingeteilt werden. Eine Möglichkeit ist
dabei die Einteilung in sekretorische Flüssigkeiten und Ultrafiltrate. Die sekretorischen
Flüssigkeiten enthalten Lipide und Proteine. Diese Substanzen erschweren die Auswertung
der NMR-Spektren, da sie breite Signale erzeugen, welche andere überlagern können. Es ist
deshalb sinnvoll, die Lipide vor der Messung zu extrahieren und getrennt zu vermessen.
Proteine können vor der Messung durch Ultrafiltration entfernt werden. Alternativ dazu ist es
aber auch möglich, ihre Signale mit einer geeigneten Pulssequenz wie CPMG- oder NOESY-
Pulsfolgen spektroskopisch zu unterdrücken.
Körperflüssigkeiten, die eine semipermeable Membran, wie zum Beispiel die Niere,
durchlaufen haben, werden als Ultrafiltrate bezeichnet. Sie besitzen einen sehr kleinen
Proteinanteil. Die Bestandteile sind niedermolekular und normalerweise gering konzentriert.
Durch Lyophilisation (Gefriertrocknung) ist es möglich, diese Flüssigkeiten um ein
Vielfaches aufzukonzentrieren. Die Probe wird vollständig getrocknet und der Rückstand
anschließend in kleinen Mengen deuterierten Wassers wieder aufgenommen. Auf diese Weise
wird auch das störende Wassersignal reduziert.
2.1.5.1. Aufarbeitung von Urin
Die Nieren halten das Milieu im Körper konstant, indem sie alle überflüssigen, organischen
und anorganischen Substanzen über den Urin ausscheiden. Eine Änderung im Körper,
beispielsweise durch einen Infekt oder eine Stoffwechselstörung, spiegelt sich direkt im
Ausscheidungsprofil wider. Das macht Urin zu einer sehr komplexen, diagnostisch wertvollen

Darstellung der Methoden
11
Flüssigkeit. Außerdem ist Urin leicht und nicht invasiv zugänglich und damit gerade für
Untersuchungen bei Kleinkindern geeignet.
Bei vielen Fragestellungen reicht es aus, die Urinproben nativ, ohne Vorbereitung zu
vermessen. Das resultierende Wassersignal kann mit Hilfe geeigneter Pulssequenzen
unterdrückt werden. Bei gering konzentrierten Proben ist diese Methode jedoch nur bedingt
zu empfehlen. Je größer der Wasseranteil im Verhältnis zu den Konzentrationen der
enthaltenen Substanzen, desto schwieriger und unvollständiger wird die spektroskopische
Wasserunterdrückung. Ein weiterer Nachteil der nativen Messung ist die Tatsache, dass viele
Metabolite aufgrund ihrer geringen Konzentration kaum oder gar nicht detektierbar sind.
Zur Aufkonzentrierung dieser Substanzen kann der Urin lyophilisiert werden. Auf diese
Weise wird auch das Wassersignal reduziert. Andererseits steigen die Intensitäten der Signale
im Hochfeldbereich. Die Anzahl der Signale ist in diesem Bereich so groß, dass es zu
Überlagerungen kommt, die eine Auflösung bis zur Basislinie verhindern.
Macht die Fragestellung den Vergleich mehrerer Spektren erforderlich, ist es notwendig, den
pH-Wert konstant zu halten. Die Resonanzfrequenzen verschiedener Substanzen sind stark
pH-abhängig, schon leichte Abweichungen führen zu teilweise großen Signalverschiebungen.
Dadurch werden die Identifizierung der Signale und die Auswertung der Spektren erschwert.
In der vorliegenden Arbeit wurde der Urin stets lyophilisiert und der pH-Wert mit dem
K2HPO4/KH2PO4-Puffer auf 5,75-5,95 eingestellt.
2.2. Festphasenextraktion
2.2.1. Einführung
Die Festphasenextraktion (SPE) ist eine schnelle und effiziente Methode zur Auftrennung von
Substanzgemischen. Sie wird hauptsächlich in der klinischen Analytik zur Vorbereitung von
Blut, Urin und anderen biologischen Proben eingesetzt (Jain et al, 2006; Persson et al, 2006;
Kato et al, 2005), aber auch in der Umweltanalytik (Botitsi et al, 2006; Zorita et al, 2006)
oder Lebensmittelchemie (Mortensen et al, 2005; Tribalat et al, 2006). Mit ihrer Hilfe können
störende Probenbestandteile, wie z. B. Proteine, entfernt werden und sie ermöglicht die
Aufkonzentrierung einzelner Substanzen bzw. Substanzklassen.
Die SPE basiert wie auch die HPLC (Hochdruckflüssigchromatographie) auf
Migrationsprozessen, bei denen die Komponenten adsorbiert und eluiert werden. Allerdings
kommen hier nur starke, reversible Wechselwirkungen zum Einsatz. Die
Extraktionskartuschen (Säulen) sind mit unterschiedlichen Materialien gepackt, die ionische,
hydrophile und lipophile Wechselwirkungen mit den Proben ausbilden können. Während die

Darstellung der Methoden
12
Probe durch die Kartusche hindurchgesaugt wird, binden die Substanzen an das Sorbens und
werden nach eventuell folgenden Waschschritten mit einem geeigneten Lösemittel wieder
eluiert. Dadurch ergeben sich zwei Möglichkeiten zur Trennung. Die gewünschte Substanz
kann entweder auf dem Sorbens zurückgehalten, und damit auch aufkonzentriert werden, oder
sie läuft durch, während störende Bestandteile adsorbieren. Gegenüber der Flüssig-Flüssig-
Extraktion (LLE) besitzt die SPE gleich mehrere Vorteile. Der Verbrauch an Lösemittel ist
gegenüber der LLE sehr niedrig. Damit sinken auch die Kosten und das Risiko von
Verunreinigungen. Gleichzeitig ist die Wiederfindungsrate größer und der zeitliche Aufwand
wesentlich geringer. Außerdem ist bei der SPE eine Automatisierung möglich, so kann sie z.
B. direkt vor eine HPLC geschaltet werden.
2.2.2. SPE-Phasen
Generell lassen sich die Materialien, mit denen die Extraktionskartuschen gepackt sind, in
verschiedene Klassen einteilen. In Umkehrphasen (RP), normale Phasen (NP),
Ionenaustauscher- und Größenausschlussphasen.
Umkehrphasen bestehen aus unpolaren Materialien und extrahieren organische Substanzen
aus polaren Lösemitteln. Die Substanzen werden über lipophile Wechselwirkungen (Van der
Waals-Kräfte) an das Sorbens gebunden und mit organischen Lösemitteln eluiert. Übliche
Materialien sind Kohlenwasserstoffketten oder andere lipophile Reste, die an Kieselgel
gebunden werden. Daneben sind auch freie Silanolgruppen vorhanden, welche bei hohen pH-
Werten dissoziieren und auf diese Weise zusätzliche ionische Wechselwirkungen einbringen.
Sie können aber auch mit Trimethylsilylgruppen enthaltenden Reagenzien, wie zum Beispiel
Trimethylchlorsilan geblockt werden (end-capping). Beispiele für RP-Materialen sind C18-
oder C8-Kartuschen, aber auch vernetzte Kopolymere wie Styrol-Divinylbenzol.
Normale Phasen isolieren polare Verbindungen über ein polares Sorbens aus unpolaren
Lösemitteln. Dabei werden hauptsächlich Dipol-Dipol-Wechselwirkungen und
Wasserstoffbrückenbindungen, aber auch π-π-Wechselwirkungen ausgebildet. Typische
Materialien sind Kieselgel-gebundene Aminopropyl- oder Cyanopropylgruppen.
Ionenaustauscher extrahieren polare Verbindungen aus polaren Lösemitteln über Coulomb-
Wechselwirkungen. Es gibt starke und schwache Ionenaustauscher, wobei die schwachen pH-
Wert abhängig sind. Typische Materialien für starke Anionenaustauscher sind quartäre
Amine, für schwache primäre oder sekundäre Amine. Sie werden eingesetzt, um azide
Verbindungen zu isolieren. Starke Kationenaustauscher enthalten Sulfonsäuregruppen,
schwache Carbonsäuregruppen. Der pH-Wert der Probe muss so eingestellt werden, dass die

Darstellung der Methoden
13
gewünschten Substanzen in ionisierter Form vorliegen. Sie werden dann an dem Sorbens
neutralisiert und anschließend mit einem niedrigeren bzw. höheren pH-Wert eluiert.
Größenausschlussphasen trennen Proben über die Porengröße auf. Große Moleküle, z. B.
Proteine, können diese nicht durchlaufen und werden zurückgehalten. In den Poren können
Materialien integriert sein, welche zusätzliche Wechselwirkungen mit der Probe eingehen und
eine weitere Trennung ermöglichen. Daneben gibt es Mischphasen, die zwei oder mehr dieser
Funktionen vereinen (Walker und Mills, 2002).
2.2.3. Methodenentwicklung
Die allgemeine Vorgehensweise bei der Festphasenextraktion kann in vier Hauptschritte
unterteilt werden: Konditionierung des Sorbens, Auftragen der Probe, Waschen des Sorbens
und Elution der Analyte. Die Konditionierung dient der Benetzung des Sorbens und schafft
damit eine Umgebung, die zur Adsorption der Analyten geeignet ist. Sie wird auch als
Solvatisierung bezeichnet. Unpolare Sorbentien werden üblicherweise mit einem
Lösungsmittel, welches mit Wasser mischbar ist, gefolgt von der reinen Matrix konditioniert.
Bei polaren Sorbentien werden unpolare Lösemittel zur Konditionierung eingesetzt. Im
Anschluss daran erfolgt die Probenaufgabe. Hierbei sollte die Menge so gewählt werden, dass
es nicht zu einer Volumen- oder Massenüberladung kommt. Der pH-Wert der Probe sollte
dem Sorbens entsprechend gewählt und eingestellt sein. Das Waschen des Sorbens wird mit
geeigneten Lösemitteln durchgeführt. Bei wässrigen Proben wird hierzu in der Regel Wasser
eingesetzt. Dieser Schritt dient dazu, störende Verunreinigungen und Nebenbestandteile der
Probe zu entfernen. Die Elution mit einem passenden Lösemittel sollte nicht zu schnell
durchgeführt werden, damit die Wiederfindungsrate möglichst hoch ist. Hier ist eine
Durchflussrate von etwa 3ml pro Minute geeignet.
Die Entwicklung einer Trennmethode mittels SPE setzt einige Überlegungen voraus. So
sollte bekannt sein, welche funktionellen Gruppen die Analyten tragen, ob sie ionisierbar
sind, in welcher Konzentration sie in etwa vorliegen und in welchem Lösemittel sie löslich
sind. Bei der Matrix ist es von Interesse, ob es sich um eine wässrige oder eine organische
Lösung handelt, welchen pH-Wert diese Lösung besitzt und ob störende Verunreinigungen
vorliegen. Wenn diese Informationen vorliegen, kann ein geeignetes Sorbens gewählt werden.
Die Wahl der Waschlösungen und Eluenten richtet sich nach dem Sorbens und den Analyten.
Sie lassen sich nur durch Versuche optimieren. Da die Entwicklung und Optimierung einer
Trennmethode kompliziert und zeitaufwändig ist, legen viele Hersteller ihren Kartuschen
generelle Applikationsvorschriften bei. Außerdem besteht bei einigen Firmen die

Darstellung der Methoden
14
Möglichkeit, im Internet oder in Papierform auf Applikations-Datenbanken zuzugreifen, um
geeignete Verfahren zu ermitteln.
2.2.4. Durchführung der Trennung
Die Festphasenextraktion wurde im Rahmen dieser Arbeit eingesetzt, um komplexe
Urinproben aufzutrennen und damit die Zahl der Signale in den NMR-Spektren zu reduzieren.
Zum einen sollten die Kohlenhydrate abgetrennt werden, da ihre Resonanzfrequenzen dicht
beieinander liegen und im Protonenspektrum zu Signalüberlagerungen führen. Zum anderen
sollten die aromatischen Verbindungen isoliert und aufkonzentriert werden. Sie sind nur
gering konzentriert und führen im Protonenspektrum zu Signalen mit zum Teil sehr
schwachen Intensitäten.
ppm (t1) 1.02.03.04.05.06.07.08.09.0
Abbildung 1: Protonenspektrum einer Urinprobe. Im aromatischen Bereich des Spektrums zwischen 6,5ppm und 9,5ppm sind die Signalintensitäten z. T. sehr schwach. Im Bereich der Kohlenhydrate zwischen 3,5ppm und 4,0ppm sind dagegen Überlagerungen vieler Signale zu beobachten.
Für die Trennung wurden verschiedene RP- und NP-Kartuschen von zwei Herstellern
getestet. Von Supelco kommen die Phasen DPA-6S und SC-Ph endcapped. DPA-6S ist eine
normale Phase, deren polares Material aus einem Polyamidharz besteht. Die SC-Ph-Phase ist
eine Umkehrphase, bestehend aus Kieselgel-gebundenen Phenylresten. Freie Silanolgruppen
wurden hier blockiert. Die übrigen Kartuschen, C8, C18, C18ec, C18hydra, CN, NH2 und
EASY kommen von Macherey-Nagel. Es handelt sich ausschließlich um Chromabond-Säulen
aus Polypropylen. C8, C18, C18ec und C18hydra sind Umkehrphasen, die sich in ihrer

Darstellung der Methoden
15
Kettenlänge und speziellen Modifikationen unterscheiden. C8 und C18 sind Silika-gebundene
Octyl- bzw. Octadecylreste. Die freien Silanolgruppen können sekundäre ionische
Wechselwirkungen einbringen, welche bei der C8-Phase aufgrund der kürzeren Kette stärker
zum Tragen kommen. C18ec kann diese ionischen Wechselwirkungen nicht einbringen, da
keine freien Silanolgruppen zur Verfügung stehen. Dadurch ist sie unpolarer als die C18-
Phase. C18hydra besitzt eine spezielle, vom Hersteller nicht näher benannte Modifikation der
Octadecylreste, die sie gegenüber den anderen Umkehrphasen hydrophiler macht.
Daneben wurden noch die normalen Phasen CN, NH2, und EASY eingesetzt. CN besteht aus
Kieselgel-gebundenen Cyanopropylresten. Die Phase ist polar bis mittelpolar und kann über
die hohe Elektronendichte an der CN-Gruppe sekundäre Wechselwirkungen ausbilden. Die
NH2-Phase ist polar und wirkt auch als schwacher Ionenaustauscher. Sie besteht aus
Kieselgel-gebundenen Aminopropylresten. EASY besteht aus einer polaren, bifunktionell
modifizierten Polystyrol-Divinylbenzol-Phase, welche einen schwachen Ionenaustauscher
enthält.
Die Kartuschen wurden alle auf die gleiche Weise konditioniert, indem zuerst Methanol und
dann Wasser aufgetragen wurde, um die Säulen auf die wässrige Probe vorzubereiten. Die
Mengen richteten sich dabei nach den Größen der Kartuschen. Die Probe wurde bei
verschiedenen pH-Werten aufgetragen, bei pH3, pH9 und nativ. Gewaschen wurden alle
Kartuschen mit Wasser, wobei die pH-Werte 3 und 9 für die Waschlösungen übernommen
wurden. Als Eluenten wurden verschiedene Lösemittel getestet. Zum einen wurden
verschiedene Wasser/Methanol Gemische in den Verhältnissen 85:15, 60:40 und 40:60 (%;
v/v) in gegebener Reihenfolge eingesetzt. Zum anderen erfolgte die Elution durch Ethylacetat
bzw. einer Tetrahydrofuran (THF) / Methanol Mischung im Verhältnis 1:1 (v/v). Die
Volumina der einzelnen Waschlösungen und Eluenten richteten sich nach den Größen der
Kartuschen. Sie wurden den Herstellerangaben entnommen. Von den Urinproben wurden
jeweils 3mL aufgetragen. Da ausreichend Probenmaterial zur Verfügung stand, wurde auf
Vorversuche mit weniger komplexen Gemischen verzichtet.
Im Allgemeinen erfolgt die Beurteilung einer Extraktion über die Wiederfindungsrate. Diese
Beurteilung ist aber nur sinnvoll, wenn einzelne, bekannte Verbindungen isoliert werden,
deren Konzentration in der Probe bekannt ist. In diesem Fall sollten keine einzelnen
Substanzen isoliert, sondern Stoffklassen möglichst genau voneinander getrennt werden. Die
Auswertung der Methoden erfolgte deshalb mittels NMR-Spektroskopie. Es wurde ein
Protonenspektrum der Urinprobe vor der Extraktion aufgenommen und anschließend mit den
Spektren der einzelnen Fraktionen verglichen. Diese Form der Auswertung lässt keine

Darstellung der Methoden
16
Aussagen über einzelne Substanzen zu. Im Hinblick auf die Zielsetzung ist sie jedoch
sinnvoll, da eine mögliche Reduzierung der Signalzahl sofort zu erkennen ist.
2.3. Hochdruckflüssigchromatographie
2.3.1. Einführung
Nachdem die HPLC in den siebziger Jahren entwickelt wurde, hat sie eine sprunghafte
Entwicklung durchgemacht. Aufgrund verschiedenartiger Säulenmaterialien und des
Einsatzes von Computern gehört sie heute zu den meistgenutzten Analysemethoden. Sie
findet in allen Bereichen Anwendung, in denen Proben mit geringen Konzentrationen
bearbeitet werden, wie der Biochemie, Pharmazie, Toxikologie oder der Umweltforschung.
Die HPLC kann sowohl zur qualitativen als auch zur quantitativen Analyse genutzt werden.
Mit ihrer Hilfe ist es möglich, chemisch ähnliche Verbindungen zu isolieren und
Zielverbindungen aus komplexen Gemischen zu extrahieren. Sie kann auch als
Reinigungsverfahren im präparativen Maßstab eingesetzt werden.
Der Begriff Chromatographie umfasst diverse physikalische Methoden, bei denen eine
Stofftrennung durch Verteilung zwischen einer ruhenden (stationären) Phase und einer
bewegten (mobilen) Phase erfolgt. Im Laufe der Jahre wurden verschiedene Methoden,
darunter die Gaschromatographie, die Dünnschichtchromatographie und die
Flüssigchromatographie, entwickelt. In der Hochdruckflüssigchromatographie wird die
Trennleistung durch den Einsatz von Partikeln mit sehr kleinen Korngrößen gesteigert. Die
mobile Phase wird mit Druck durch die stationäre Phase gepumpt, um den dadurch
entstehenden Strömungswiderstand zu überwinden.
Die HPLC arbeitet hauptsächlich mit Adsorptions- und Verteilungsgleichgewichten. Bei der
Adsorptionschromatographie kommt es zu reversiblen Bindungen der Analyten an die
stationäre Phase. Die Geschwindigkeit, mit der die Substanzen durch die Säule laufen, hängt
von zwei Faktoren ab. Zum einen von der Affinität der Analyten zur stationären Phase und
zum anderen von der Fähigkeit der mobilen Phase die Substanzen von den Bindungsstellen
des Sorbens zu verdrängen.
Die Verteilungschromatographie nutzt die unterschiedliche Löslichkeit der Analyten in der
mobilen und der stationären Phase. Durch die unterschiedliche Polarität der beiden Phasen
stellt sich ein Verteilungsgleichgewicht ein. Je stärker das Gleichgewicht auf der Seite des
Sorbens liegt, desto länger verbleiben die Moleküle auf der Säule.
Als Totzeit wird die Aufenthaltsdauer der Substanzen in der mobilen Phase bezeichnet. Die
Zeit, welche die Moleküle auf der stationären Phase verbleiben, nennt man

Darstellung der Methoden
17
Nettoretentionszeit. Die Gesamt- oder Bruttoretentionszeit gibt die Dauer vom Einspritzpunkt
bis zum Signalmaximum wieder. Retentionszeiten sind von den Eigenschaften der Analyten
abhängig und machen eine Aussage über die Identität der Stoffe. Die Zahl der theoretischen
Stufen wird über die Retentionszeit und die Signalbreite berechnet und ist ein Merkmal für
die Qualität einer Trennsäule. Je größer die Zahl der theoretischen Stufen, desto komplexere
Gemische können mit der Säule getrennt werden. Hier liegt, neben der Säulenlänge, der große
Unterschied zur Festphasenextraktion. Beide Methoden basieren auf den gleichen
Trennprinzipien, aber während in der SPE nur Trennstufenzahlen bis 50 erreicht werden, sind
in der HPLC Zahlen um die 10.000 gebräuchlich. Dadurch lässt sich die deutlich
empfindlichere und selektivere Trennung mittels HPLC erklären.
Die Identifizierung der Analyten erfolgt über geeignete Detektoren, zum Beispiel UV- oder
Massenspektrometer, mittels NMR oder elektrochemisch. Quantifiziert werden die
Substanzen mit Hilfe von Standardverbindungen bekannter Konzentration. Diese werden
entweder einzeln gemessen oder direkt zur Probe gegeben. Durch Integration der
Signalflächen können anschließend die Konzentrationen ermittelt werden.
2.3.2. Methodenentwicklung
Die Entwicklung einer Trennmethode beginnt mit der Auswahl einer geeigneten stationären
Phase. Dafür sind, wie in der SPE, einige Informationen über die Probe notwendig. In der
HPLC werden vorwiegend wässrige Proben analysiert, deshalb finden hauptsächlich RP-
Materialien Verwendung. Da diese Phasen polare Laufmittel erfordern, werden überwiegend
Gemische aus Wasser und Acetonitril oder Methanol eingesetzt. Die Zusammensetzung
richtet sich nach den Eigenschaften der Probe und muss experimentell ermittelt werden.
Zusätzliches Ansäuern der mobilen Phase mit Ameisensäure kann die Trennung weiter
verbessern. Die Elution mit einem Laufmittel konstanter Zusammensetzung wird auch als
isokratische Elution bezeichnet. Bei der Chromatographie von Substanzen mit stark
unterschiedlichen chromatographischen Eigenschaften kann die Zusammensetzung des
Laufmittels auch während der Trennung verändert werden. Die so genannte Gradientenelution
reduziert die Unterschiede in den Retentionszeiten der Analyte. Auch hier kann die
Optimierung nur experimentell erfolgen. Nachdem die Methodenentwicklung abgeschlossen
ist, ist es sinnvoll, die Reproduzierbarkeit der Trennung zu testen. Dazu wird die Probe
mehrmals vermessen und anschließend die Chromatogramme miteinander verglichen.
Alternativ besteht in der HPLC die Möglichkeit, genau wie in der SPE,
Applikationsvorschriften vom Hersteller oder aus Datenbanken zu erhalten. Allerdings

Darstellung der Methoden
18
müssen diese Vorschriften größtenteils noch optimiert werden, da die HPLC viel
empfindlicher arbeitet als die SPE.
2.3.3. Durchführung der Trennung
Im Rahmen dieser Arbeit wurde die HPLC eingesetzt, um damit eine Fraktion der
Festphasenextraktion aufzutrennen. Diese Auftrennung sollte eine weitere Reduzierung der
Signale in den NMR-Spektren zur Folge haben.
Als Säule wurde die Sphinx von Macherey/Nagel ausgewählt. Dabei handelt es sich um eine
RP-Phase, die bifunktionell belegt ist. Sie bringt sowohl hydrophobe Wechselwirkungen über
C18-Ketten als auch π-π-Wechselwirkungen über kurzkettige Phenylreste ein. Durch diese
Eigenschaften zeigt sie, besonders bei aromatischen Verbindungen mit polaren Eigenschaften,
eine gute Trennung. Die Methode wurde mit Hilfe einer analytischen Säule erarbeitet, um den
Proben- und Laufmittelverbrauch gering zu halten. Aufgetragen wurde die dritte Fraktion
(Elution) der SPE, um die darin enthaltenen aromatischen Verbindungen zu isolieren. Die
Probe wurde nach der Elution getrocknet und in 1ml Wasser wieder aufgenommen. Für die
HPLC wurde sie zentrifugiert, filtriert und mit Wasser im Verhältnis 1:1000 verdünnt. Es
wurden verschiedene Laufmittel getestet, um eine ausreichende und reproduzierbare
Trennung zu erreichen. Zum einen Gemische aus Wasser und Acetonitril bzw. Methanol mit
und ohne einem Zusatz von Ameisensäure, zum anderen verschiedene Gradienten des
Wasser/Methanol Gemisches. Da die Probe lediglich fraktioniert werden sollte, war es nicht
notwendig, die Signale bis zur Basislinie zu trennen. Aromatische Verbindungen absorbieren
UV-Frequenzen bei 254nm, deshalb erfolgte die Detektion über ein integriertes UV/VIS-
Spektrometer.
Die eigentliche Auftrennung wurde mit Hilfe einer semi-präparativen Säule mit einer
Durchflussrate von 4,8ml pro Minute durchgeführt. Dadurch musste die Probe nicht verdünnt
werden und es konnten jeweils 100µl aufgetragen werden. Anhand der UV-Spektren wurden
geeignete Zeitpunkte zum Schneiden der Fraktionen gewählt. Die Fraktionen wurden bei
jedem Durchlauf gesammelt, vereint und anschließend für die NMR getrocknet und in 0,5ml
D2O aufgenommen.
2.4. Ergebnisse der Trennungen
2.4.1. Festphasenextraktion
Eine vollständige Trennung der im Urin enthaltenen Stoffklassen ist nahezu unmöglich, da
sehr viele Verbindungen enthalten sind, deren chemische Eigenschaften sich überschneiden.
So sind zum Beispiel relativ polare (durch gebundene Hydroxygruppen) oder glukuronierte
Darstellung der Methoden
19
Aromaten vorhanden, Nukleoside tragen Ribose-Einheiten und langkettige organische Säuren
besitzen lipophile Eigenschaften. Deshalb wurde eine Trennung als ausreichend angesehen,
wenn eine deutliche Reduzierung der Signale in den NMR-Spektren vorlag. Die meisten
Phasen erreichten dieses Ziel unter den gewählten Bedingungen nicht. Möglicherweise
wurden nur Substanzen unterhalb der NMR-Nachweisgrenze (ca. 10µmol/L) adsorbiert oder
nur wenige Substanzen, die sich unter den gegebenen Konditionen nicht wieder eluieren
ließen. Nur zwei Phasen (EASY, C18) lieferten Spektren, welche sich deutlich von dem
unbehandelten Urins unterschieden.
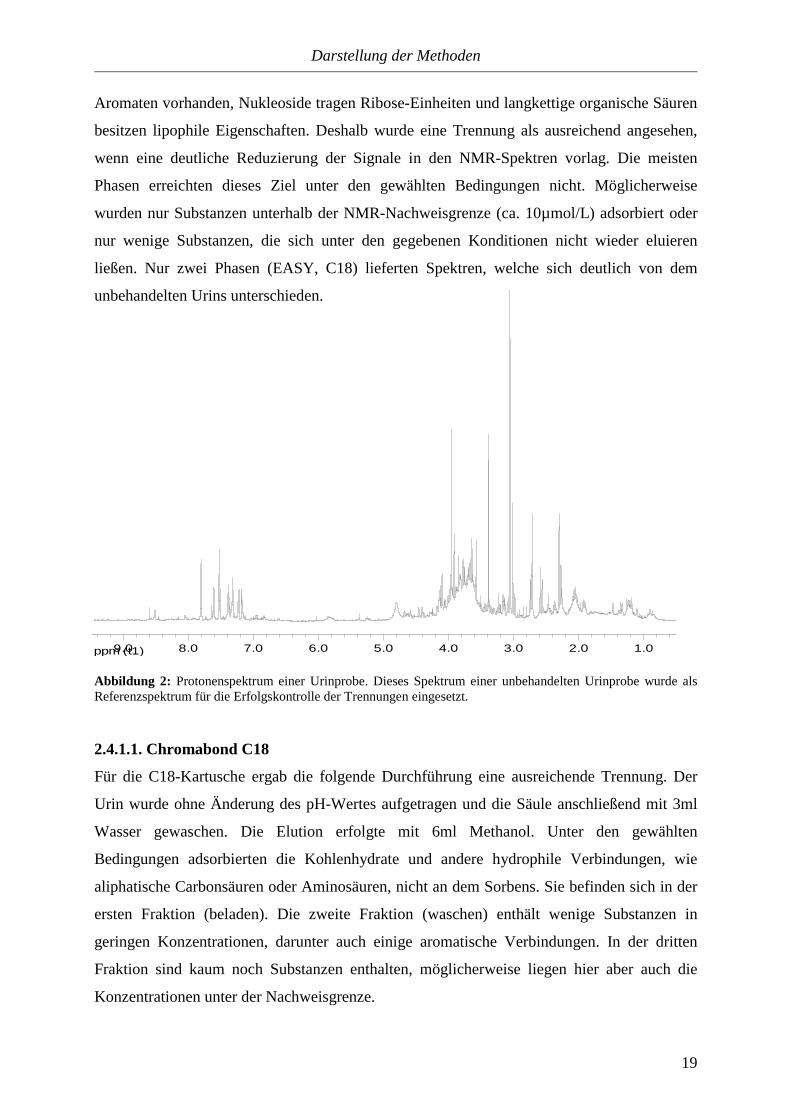
ppm (t1) 1.02.03.04.05.06.07.08.09.0
Abbildung 2: Protonenspektrum einer Urinprobe. Dieses Spektrum einer unbehandelten Urinprobe wurde als Referenzspektrum für die Erfolgskontrolle der Trennungen eingesetzt.
2.4.1.1. Chromabond C18
Für die C18-Kartusche ergab die folgende Durchführung eine ausreichende Trennung. Der
Urin wurde ohne Änderung des pH-Wertes aufgetragen und die Säule anschließend mit 3ml
Wasser gewaschen. Die Elution erfolgte mit 6ml Methanol. Unter den gewählten
Bedingungen adsorbierten die Kohlenhydrate und andere hydrophile Verbindungen, wie
aliphatische Carbonsäuren oder Aminosäuren, nicht an dem Sorbens. Sie befinden sich in der
ersten Fraktion (beladen). Die zweite Fraktion (waschen) enthält wenige Substanzen in
geringen Konzentrationen, darunter auch einige aromatische Verbindungen. In der dritten
Fraktion sind kaum noch Substanzen enthalten, möglicherweise liegen hier aber auch die
Konzentrationen unter der Nachweisgrenze.
Darstellung der Methoden
20
A
B
C
ppm (t1) 1.02.03.04.05.06.07.08.09.0
A
B
C
ppm (t1) 1.02.03.04.05.06.07.08.09.0
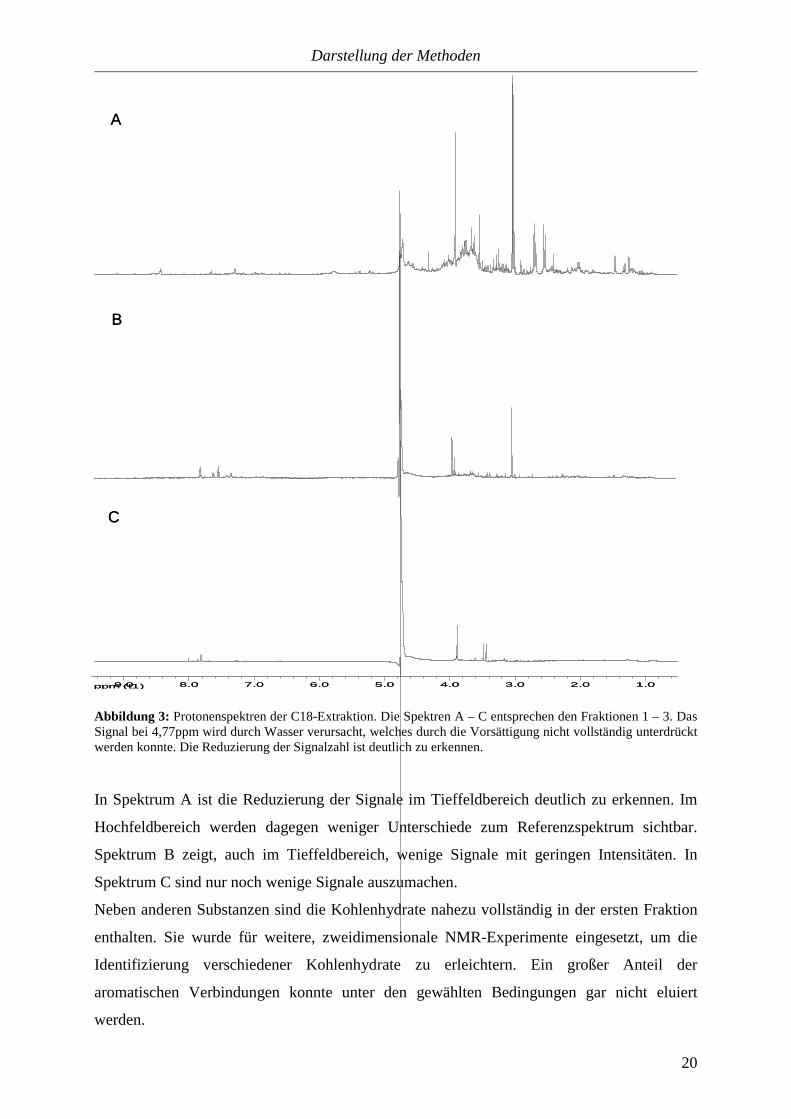
Abbildung 3: Protonenspektren der C18-Extraktion. Die Spektren A – C entsprechen den Fraktionen 1 – 3. Das Signal bei 4,77ppm wird durch Wasser verursacht, welches durch die Vorsättigung nicht vollständig unterdrückt werden konnte. Die Reduzierung der Signalzahl ist deutlich zu erkennen.
In Spektrum A ist die Reduzierung der Signale im Tieffeldbereich deutlich zu erkennen. Im
Hochfeldbereich werden dagegen weniger Unterschiede zum Referenzspektrum sichtbar.
Spektrum B zeigt, auch im Tieffeldbereich, wenige Signale mit geringen Intensitäten. In
Spektrum C sind nur noch wenige Signale auszumachen.
Neben anderen Substanzen sind die Kohlenhydrate nahezu vollständig in der ersten Fraktion
enthalten. Sie wurde für weitere, zweidimensionale NMR-Experimente eingesetzt, um die
Identifizierung verschiedener Kohlenhydrate zu erleichtern. Ein großer Anteil der
aromatischen Verbindungen konnte unter den gewählten Bedingungen gar nicht eluiert
werden.
Darstellung der Methoden
21
2.4.1.2. Chromabond EASY
Die Chromabond EASY erbrachte nach folgender Vorschrift ein ausreichendes
Trennergebnis. Der pH-Wert des Urins wurde auf 3 eingestellt und die Säule nach dem
Auftragen mit 3ml Wasser (pH3) gewaschen. Die Elution erfolgte mit 6ml THF/Methanol.
Die Kohlenhydrate adsorbieren hier ebenfalls kaum an dem Sorbens, sie sind nur in den
ersten beiden Fraktionen (Beladen und Waschen) zu finden. Aromatische Substanzen werden
relativ gut adsorbiert und erst mit dem Eluenten von dem Sorbens gelöst. Die Spektren der
Fraktionen sind in Abbildung 4 dargestellt.
A
B
C
ppm (t1) 1.02.03.04.05.06.07.08.09.0
A
B
C
ppm (t1) 1.02.03.04.05.06.07.08.09.0
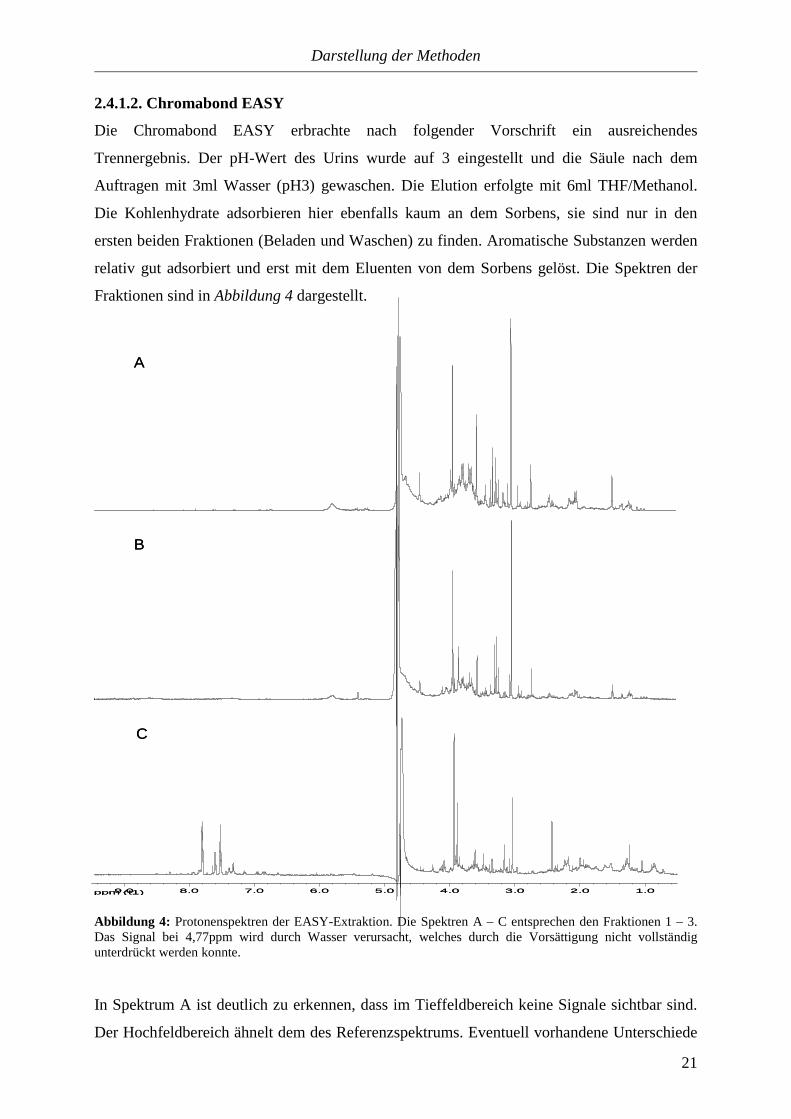
Abbildung 4: Protonenspektren der EASY-Extraktion. Die Spektren A – C entsprechen den Fraktionen 1 – 3. Das Signal bei 4,77ppm wird durch Wasser verursacht, welches durch die Vorsättigung nicht vollständig unterdrückt werden konnte.
In Spektrum A ist deutlich zu erkennen, dass im Tieffeldbereich keine Signale sichtbar sind.
Der Hochfeldbereich ähnelt dem des Referenzspektrums. Eventuell vorhandene Unterschiede

Darstellung der Methoden
22
zur ersten Fraktion der C18-Extraktion sind wegen den Signalüberlagerungen nicht zu
erfassen. Spektrum B weist ebenfalls keine Signale im Tieffeldbereich auf. Der
Hochfeldbereich ähnelt der ersten Fraktion, nur sind hier die Intensitäten der Signale geringer.
In Spektrum C sind Signale im Tieffeldbereich zu sehen. Die vielen Signale im
Hochfeldbereich lassen vermuten, dass auch noch andere Stoffklassen in dieser Fraktion
enthalten sind. Die dritte Fraktion wurde für weitere NMR-Experimente eingesetzt, mit ihrer
Hilfe sollten aromatische Verbindungen identifiziert werden. Bei der Auswertung dieser
Spektren wurde deutlich, dass auch hier Substanzen auf dem Sorbens haften geblieben sind.
So sind zum Beispiel in allen Fraktionen keine Imidazole zu finden, obwohl im Urin welche
vorkommen.
2.4.2. Hochdruckflüssigchromatographie
Die in der HPLC eingesetzte Fraktion der EASY-Extraktion ist weniger komplex als die
Urinprobe, enthält jedoch immer noch verschiedene Substanzklassen. Mit Hilfe der HPLC
sollten die NMR-Spektren weiter vereinfacht werden, das Hauptziel hierbei war eine weitere
Auftrennung der aromatischen Verbindungen. Die Optimierung des Laufmittels ergab eine
Zusammensetzung aus Wasser, Methanol und 0,1% Ameisensäure mit einem Gradienten von
5% Methanol pro Minute bei einer Anfangskonzentration von 5% Methanol und einer
Endkonzentration von 100% Methanol. Mit diesem Laufmittel gelang es, das UV-Spektrum
so weit zu entzerren, dass die Signale zumindest teilweise bis zur Basislinie getrennt waren
und geeignete Zeitpunkte zum Fraktionieren gewählt werden konnten. Ein Durchlauf der
HPLC dauerte 35 Minuten, es wurde aber nur in den ersten 15 Minuten fraktioniert, da
hinterher keine UV-Signale mehr detektiert wurden. Die Spektren der Fraktionen sind in
Abbildung 5 dargestellt.
Darstellung der Methoden
23
A
B
C
D
E
ppm (t1) 1.02.03.04.05.06.07.08.09.0
A
B
C
D
E
ppm (t1) 1.02.03.04.05.06.07.08.09.0
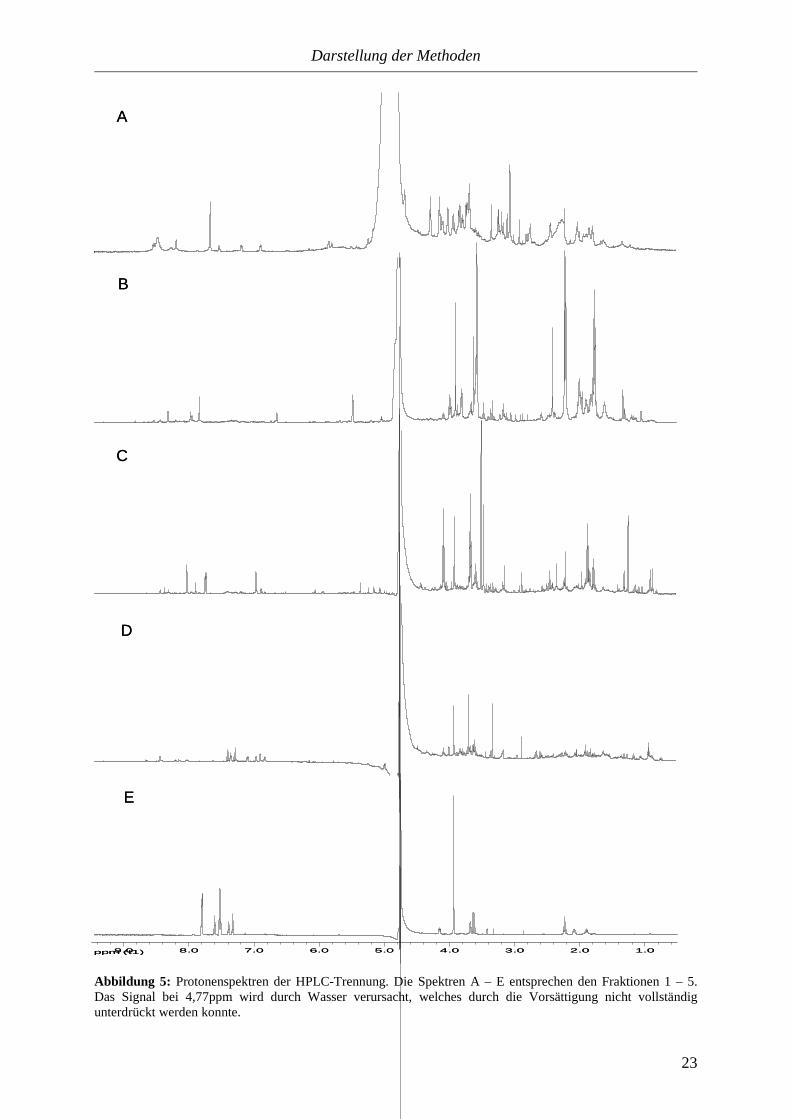
Abbildung 5: Protonenspektren der HPLC-Trennung. Die Spektren A – E entsprechen den Fraktionen 1 – 5. Das Signal bei 4,77ppm wird durch Wasser verursacht, welches durch die Vorsättigung nicht vollständig unterdrückt werden konnte.

Darstellung der Methoden
24
Die Spektren der einzelnen Fraktionen zeigen, dass die aromatischen Verbindungen
tatsächlich aufgetrennt wurden. Sie weisen charakteristische Unterschiede im Tieffeldbereich
auf. Die in den einzelnen Fraktionen enthaltenen Verbindungen besitzen auch strukturelle
Übereinstimmungen. So sind zum Beispiel in der dritten Fraktion drei verschiedene
Nikotinamid-Derivate enthalten. In allen Spektren sind auch noch Signale von aliphatischen
Carbonsäuren zu erkennen, obwohl erwartet wurde, dass diese vollständig in der ersten
Fraktion gesammelt werden. Möglicherweise ist die Ameisensäure Ursache für diese
Erscheinung.
Schon während der Festphasenextraktion sind Substanzverluste unvermeidbar, welche durch
die HPLC noch erhöht werden. Deshalb sind die Intensitäten der Signale sehr gering. Es ist
auch sehr wahrscheinlich, dass einige Substanzen nicht mehr detektiert werden, da sie
unterhalb der Nachweisgrenze liegen.
2.4.3. Zusammenfassung
Ob eine Auftrennung des Urins mit Hilfe der Festphasenextraktion erfolgreich ist oder nicht,
hängt sehr von der Fragestellung ab. Die Phasenvielfalt in Verbindung mit den vielen
Substanzklassen im Urin, deren Eigenschaften sich zum Teil überschneiden, macht eine
Methodenentwicklung nicht einfach. Für das hier gewählte Ziel der Signalreduzierung waren
die Extraktionen auf jeden Fall ausreichend, da zumindest einzelne Fraktionen für die
Strukturaufklärung herangezogen werden konnten. Die Spektren sind zwar immer noch
komplex, weisen aber weniger Signalüberlagerungen auf. Auch die Abtrennung einzelner
Substanzklassen war erfolgreich.
Die Durchführung einer HPLC mit Urinproben ist ohne vorhergehende SPE kaum möglich,
da es leicht zu einer Überladung der HPLC-Säule kommen kann. Die Chromatographie der
gewählten Fraktion hat gute Ergebnisse gezeigt. Mit ihrer Hilfe konnte die Strukturaufklärung
vereinfacht werden. Die Spektren der Fraktionen sind sehr hilfreich bei der Identifizierung
von unbekannten Substanzen. Es gibt kaum noch Überlagerungen und die grobe Einteilung
aufgrund möglicher struktureller Übereinstimmungen erleichtert die Zuordnung unbekannter
Signalgruppen. Es ist hier auch zum ersten Mal möglich, einige Kopplungskonstanten und
Multiplizitäten zu bestimmen. Sowohl in den Spektren der unbehandelten Urinprobe als auch
in den Spektren der Festphasenextraktionen kommen dafür zu viele Überlagerungen vor.

Identifizierung unbekannter Metabolite
25
3. Identifizierung unbekannter Metabolite
Urin ist für die Medizin eine sehr interessante Körperflüssigkeit, da ihre Zusammensetzung
nahezu alle Stoffwechselwege des menschlichen Körpers widerspiegelt. Schädliche
Substanzen und überschüssige Stoffwechselprodukte im Blut werden von den Nieren
abfiltriert und mit dem Urin ausgeschieden.
3.1. Anatomie und Funktion der Niere
Die Nieren sind rötliche, bohnenförmige Organe, die unterhalb des Zwerchfells zwischen dem
Peritoneum (Bauchfell) und der rückwärtigen Abdominalwand (Bauchwand) liegen. Sie
werden teilweise vom 11. und 12. Rippenbogen geschützt. Die rechte Niere liegt etwas tiefer
als die linke, da die Leber den Platz darüber beansprucht.
Jede Niere ist etwa 12cm lang, 7cm breit und 4cm dick. Ihr Gewicht beträgt 120 bis 200g. Sie
ist von drei Gewebeschichten umgeben. Die unterste (Capsula fibrosa) besteht aus dichtem
unregelmäßigem Bindegewebe, welches sich in der Außenhülle des Ureters fortsetzt. Sie
schützt vor Verletzungen und bewahrt die äußere Form der Niere. Die mittlere Schicht
(Capsula adiposa) wird aus einer Fettgewebemasse gebildet. Sie dient ebenfalls als Schutz vor
Verletzungen und hält die Niere an ihrem Platz in der Bauchhöhle. Die oberste Schicht
(Fascia renalis) besteht wiederum aus Bindegewebe, dass die Niere an den umgebenden
Strukturen und der hinteren Abdominalwand verankert. Ein Längsschnitt durch die Niere
zeigt zwei Schichten, außen die Nierenrinde und innen das Nierenmark. Das Nierenmark wird
durch Säulen der Rindensubstanz in etwa 12 Nierenpyramiden unterteilt. Die Spitzen dieser
Pyramiden werden als Nierenpapillen bezeichnet und sind von schlauchförmigen
Nierenkelchen überzogen. Sie fangen den fertigen Harn auf und leiten ihn in den
Sammelraum des Nierenbeckens.
Abbildung 6: Querschnitt einer Niere. Quelle: Roche Lexikon Medizin; 5. Auflage (www.gesundheit.de/roche/)
Identifizierung unbekannter Metabolite
26
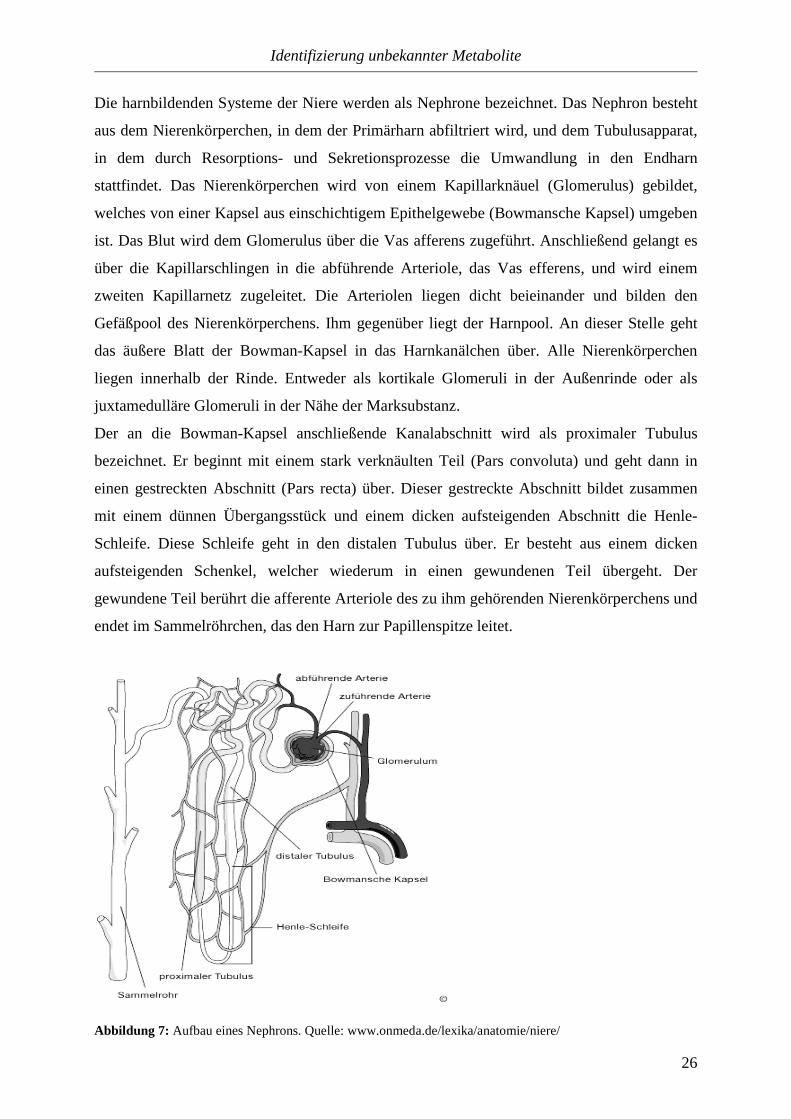
Die harnbildenden Systeme der Niere werden als Nephrone bezeichnet. Das Nephron besteht
aus dem Nierenkörperchen, in dem der Primärharn abfiltriert wird, und dem Tubulusapparat,
in dem durch Resorptions- und Sekretionsprozesse die Umwandlung in den Endharn
stattfindet. Das Nierenkörperchen wird von einem Kapillarknäuel (Glomerulus) gebildet,
welches von einer Kapsel aus einschichtigem Epithelgewebe (Bowmansche Kapsel) umgeben
ist. Das Blut wird dem Glomerulus über die Vas afferens zugeführt. Anschließend gelangt es
über die Kapillarschlingen in die abführende Arteriole, das Vas efferens, und wird einem
zweiten Kapillarnetz zugeleitet. Die Arteriolen liegen dicht beieinander und bilden den
Gefäßpool des Nierenkörperchens. Ihm gegenüber liegt der Harnpool. An dieser Stelle geht
das äußere Blatt der Bowman-Kapsel in das Harnkanälchen über. Alle Nierenkörperchen
liegen innerhalb der Rinde. Entweder als kortikale Glomeruli in der Außenrinde oder als
juxtamedulläre Glomeruli in der Nähe der Marksubstanz.
Der an die Bowman-Kapsel anschließende Kanalabschnitt wird als proximaler Tubulus
bezeichnet. Er beginnt mit einem stark verknäulten Teil (Pars convoluta) und geht dann in
einen gestreckten Abschnitt (Pars recta) über. Dieser gestreckte Abschnitt bildet zusammen
mit einem dünnen Übergangsstück und einem dicken aufsteigenden Abschnitt die Henle-
Schleife. Diese Schleife geht in den distalen Tubulus über. Er besteht aus einem dicken
aufsteigenden Schenkel, welcher wiederum in einen gewundenen Teil übergeht. Der
gewundene Teil berührt die afferente Arteriole des zu ihm gehörenden Nierenkörperchens und
endet im Sammelröhrchen, das den Harn zur Papillenspitze leitet.
Abbildung 7: Aufbau eines Nephrons. Quelle: www.onmeda.de/lexika/anatomie/niere/

Identifizierung unbekannter Metabolite
27
Jeder anatomische Abschnitt eines Nephrons hat eine bestimmte Funktion bei der
Harnbereitung. In den Nierenkörperchen wird durch Ultrafiltration des Blutplasmas der
Primärharn gebildet. Dieser Primärharn besteht aus einer proteinfreien, wässrigen Lösung
aller im Blutplasma enthaltenen Stoffe. Innerhalb von 24 Stunden werden ca. 150l
Primärharn filtriert. Durch Resorption, Sekretion und Exkretion wird der Primärharn stark
verändert während er die Tubuli durchläuft. In dem gewundenen Abschnitt des proximalen
Tubulus befindet sich eine Natriumpumpe. Die Entfernung des Natriums aus dem Primärharn
begründet ein osmotisches Druckgefälle, weswegen Wasser dem Natrium passiv folgt.
Dadurch werden etwa 80% des Wassers ins Blut rückresorbiert. Eine weitere
Harnkonzentrierung erfolgt in der Henle´schen Schleife. Hier werden noch einmal 18-19%
des Wassers resorbiert.
Harnpflichtige Substanzen gelangen durch die Primärharnbildung in den Urin,
Stoffwechselprodukte und körperfremde Stoffe werden dagegen von den Zellen der Tubuli
zur Ausscheidung an den Harn abgegeben. Umgekehrt werden einige Stoffe (Glucose,
Aminosäuren Chloride etc.) von den Tubulizellen ans Blut abgegeben. Bei diesen Stoffen
spricht man von Schwellenstoffen. Sie werden nur über den Urin ausgeschieden, wenn ihre
Konzentration im Blut einen bestimmten Schwellenwert überschreitet. Der Endharn gelangt
aus den Sammelrohren ins Nierenbecken und von dort über den Ureter in die Harnblase
(Leutert, 1994; Jecklin, 1986).
3.2. Einführung
Die Untersuchung von Urinproben ist heute ein gängiges Diagnoseverfahren, da sich
pathologische Veränderungen des Stoffwechsels unter anderem auf die Zusammensetzung des
Urins auswirken. Jede Stoffwechselstörung wirkt sich auf die Ausscheidung weniger
Substanzen aus und verursacht so charakteristische Änderungen der Zusammensetzung. Diese
Substanzen werden auch als Markermoleküle bezeichnet, da mit ihrer Hilfe eine Diagnose
gestellt werden kann. Im Allgemeinen erfolgen die Untersuchungen des Urins über
Einzelnachweise bestimmter Verbindungen, was mit einem hohen Zeit- und Kostenaufwand
verbunden ist. Zudem macht es eine Vorauswahl der zu untersuchenden Metabolite
erforderlich.
Die NMR-Spektroskopie bietet hier Vorteile. Neben einer schnellen und unkomplizierten
Probenvorbereitung ist die Messung, je nach Fragestellung, in kurzer Zeit durchführbar und
das resultierende Spektrum liefert einen metabolischen „Fingerabdruck“. Da sämtliche NMR-
aktiven Substanzen des Urins im Spektrum Signale liefern, kann auf diese Weise ein nahezu

Identifizierung unbekannter Metabolite
28
komplettes Stoffwechselprofil dargestellt werden. Der Vergleich verschiedener Spektren
macht eventuell vorhandene Auffälligkeiten sichtbar, die auf eine veränderte
Zusammensetzung des Urins schließen lassen.
Ein NMR-Spektrum enthält eine große Datenmenge. Um die Auswertung und den Vergleich
mehrerer Spektren zu erleichtern, können Mustererkennungsverfahren eingesetzt werden. Sie
bestehen im Grunde aus drei Schritten, der Datenreduktion, der Klassifizierung und der
Visualisierung. In der NMR-Spektroskopie werden Mustererkennungsverfahren eingesetzt um
in den Spektren Muster zu erkennen, die eine Einteilung in verschiedene Gruppen
ermöglichen und, die verantwortlichen Signale zu identifizieren. Sie finden in der klinischen
Diagnostik Anwendung, da sie nach einer Lernphase dazu in der Lage sind, die Spektren von
Urinproben verschiedenen Gruppen zuzuordnen. Je nach Vorgabe können auf diese Weise
kranke Probanden von gesunden unterschieden oder Patienten verschiedenen
Krankheitsstadien zugeordnet werden. Der Einsatz von Mustererkennungsverfahren ist jedoch
nur sinnvoll, wenn die auffälligen Signale auch einer Substanz zugeordnet werden können.
Deshalb ist es von Interesse, das NMR-Spektrum von Urin möglichst vollständig aufzuklären
und die Metabolite zu identifizieren.
3.3. Allgemeine Durchführung
Die Zuordnung der Signale in den NMR-Spektren erfolgte über Literaturrecherche,
Spektrendatenbanken und Referenzmessungen. Daneben konnten einige Metabolite mit Hilfe
der Strukturaufklärung identifiziert werden. Dazu wurden verschiedene 2D-Verfahren, i. e.
HSQC, COSY und TOCSY, eingesetzt.
Die Strukturaufklärung ist in komplexen Gemischen schwieriger als bei Einzelsubstanzen, da
verschiedene Methoden nicht verwendet werden können. So kommen in den
Protonenspektren von komplexen Gemischen so viele Signalüberlagerungen vor, dass
Integrale, Multiplizitäten und Kopplungskonstanten nur selten bestimmt werden können. 13C-
und DEPT-Spektren können wegen der hohen Signaldichte ebenfalls nicht vollständig
ausgewertet werden. Es ist in vielen Fällen aufgrund der Signalüberlagerungen auch nicht
möglich, die Wasserstoff-Kopplungsnetzwerke der einzelnen Verbindungen im COSY oder
TOCSY vollständig zu verfolgen. Selbst im HSQC erschweren die Überlagerungen in einigen
Bereichen eine eindeutige Zuordnung der Signale. Das HSQC mit TOCSY-Transfer und das
HMBC zeigen dieselben Probleme. Auch hier erschweren Signalüberlagerungen die
Auswertung. Außerdem sind bei den letzten zwei Methoden sehr lange Messzeiten
notwendig, um auswertbare Spektren zu erhalten, da sie relativ unempfindlich sind.

Identifizierung unbekannter Metabolite
29
Die Strukturaufklärung erfolgte hauptsächlich über die chemischen Verschiebungen der
Protonen und Kohlenstoffe. Diese geben zum Teil eindeutige Informationen über vorhandene
Strukturelemente. Mit ihrer Hilfe können, vor allem im aromatischen Bereich, zumindest die
Grundgerüste der Verbindungen ermittelt werden. Vorhandene Substituenten lassen sich zum
Teil ebenfalls über die chemischen Verschiebungen ermitteln und aliphatische Seitenketten
im COSY identifizieren.
Festphasenextraktion und HPLC verringern das Problem der Signalüberlagerungen,
verdünnen aber auch die Proben, so dass einige Substanzen nicht mehr detektiert werden
können. Die aus der HPLC resultierenden Proben waren so stark verdünnt, dass von den 2D-
Verfahren lediglich das COSY-Experiment auswertbare Spektren ergab. Dennoch
ermöglichte die Bestimmung der Kopplungskonstanten, Integrale und Multiplizitäten die
Identifizierung der Pyridin-Derivate und anderer Verbindungen. Die Korrelation der Protonen
zu den gebundenen Kohlenstoffen erfolgte in dem HSQC der unbehandelten Probe.
3.3. Vorstellung der Metabolite
In den folgenden Kapiteln werden die identifizierten Metabolite vorgestellt. Die gewählte
Einteilung der Substanzen ist variabel, da die meisten Substanzen mehrere Eigenschaften
vereinen (z. B. aromatische Carbonsäuren). Die mit den Metaboliten verbundenen
Stoffwechselstörungen sind beispielhaft und nicht vollständig. Viele Enzymdefekte sind bis
heute nicht aufgeklärt und die meisten Stoffwechselstörungen verursachen
Konzentrationsänderungen von mehreren Substanzen. So ist es zum Beispiel möglich, dass
die Konzentrationszunahme einer Verbindung mit der Konzentrationsabnahme einer anderen
Verbindung verknüpft ist. In einigen Fällen wird jedoch nur auf eine Substanz untersucht, so
dass die Zusammenhänge nicht zwingend erfasst werden.
In der Vorstellung der Verbindungen werden nicht alle Signale der einzelnen Metabolite
erwähnt. Die Beschreibung und die Darstellung der Spektren beschränken sich auf Signale,
die in den Spektren leicht und eindeutig identifiziert werden können und damit für analytische
Fragestellungen geeignet sind. Eine vollständige Aufzählung findet sich in Tabelle 1 im
Anhang 5.2..
Die Experimente wurden entsprechend Anhang 5.2. an einem 600MHz-Gerät durchgeführt.
Es wurden verschiedene, zu unterschiedlichen Zeiten genommene Urinproben eines Kindes
vermessen, um festzustellen, ob die identifizierten Metabolite ständig im Urin zu finden sind.

Identifizierung unbekannter Metabolite
30
3.3.1. Aromatische Verbindungen
Die meisten aromatischen Verbindungen im Urin sind nur sehr gering konzentriert, da sie
aufgrund ihrer strukturellen Eigenschaften nicht sehr hydrophil sind.
3.3.1.1. Aromaten
Benzol-Derivate machen einen Teil der aromatischen Verbindungen aus. Durch gebundene
Hydroxyl- und Carboxylgruppen wird die Hydrophilie dieser Verbindungen gesteigert.
Dadurch ist es möglich, sie mit dem Urin auszuscheiden. Zu den Aromaten, die im Urin
identifiziert werden konnten, gehören Vanillinsäure, Homovanillinsäure, 4-
Hydroxyphenylessigsäure, 3-Hydroxyphenylessigsäure, Phenylessigsäure, 4-Kresol, 4-
Hydroxybenzamid, 4-Hydroxybenzoesäure und Hippursäure.
Vanillinsäure ist ein Abbauprodukt von Tyrosin, Catecholaminen und von Vanillin, welches
mit der Nahrung aufgenommen wird. Bei Patienten mit Leberzirrhose sind erhöhte
Konzentrationen im Urin zu finden (Liebich und Pickert, 1985).
Homovanillinsäure ist ebenfalls ein Abbauprodukt im Catecholamin-Stoffwechsel. Vermehrte
Ausscheidung im Urin wird bei verschiedenen Tumoren, wie z. B. Phäochromozytomen,
Neuroblastomen und Ganglioneuromen beobachtet (Shi et al, 1998; Gerlo und Sevens, 1994).
4-Hydroxyphenylessigsäure ist ein deaminiertes Oxidationsprodukt von 4-Tyramin und
Tyrosin. Leberzirrhose führt unter anderem zu einer vermehrten Ausscheidung von 4-
Hydroxyphenylessigsäure in den Urin (Liebich und Pickert, 1985).
3-Hydroxyphenylessigsäure, ein Metabolit von Tyrosin und Rutin, wirkt als Antioxidans im
menschlichen Organismus. Stoffwechselstörungen oder andere Krankheiten, die zu erhöhten
Konzentrationen im Urin führen, sind bis heute nicht bekannt.
Phenylessigsäure ist ein Abbauprodukt von Phenylethylamin. Ihre Struktur ähnelt denen der
Neurotransmitter Dopamin und Adrenalin, weshalb sie ebenfalls anregende Wirkung besitzt.
Die Konzentration von Phenylessigsäure im Urin ist bei depressiven Menschen signifikant
erniedrigt (Szabo et al, 2001).
4-Kresol ist ein Endprodukt des Proteinstoffwechsels durch den Abbau von Tyrosin und
Phenylalanin. Die Ausscheidung in den Urin ist abhängig von der Proteinaufnahme. Benzol
oder Toluol werden unter anderem zu 4-Kresol abgebaut, welches dadurch vermehrt mit dem
Urin ausgeschieden wird.
Über 4-Hydroxybenzamid im Urin ist nichts bekannt, es könnte über die Nahrung in den
menschlichen Stoffwechsel gelangen.

Identifizierung unbekannter Metabolite
31
4-Hydroxybenzoesäure ist ein Stoffwechselprodukt von verschiedenen Substanzen, darunter
Catechine, Parabene und Polyphenole. Parabene sind in Kosmetika enthalten und werden
hauptsächlich über die Haut aufgenommen. Catechine und Polyphenole werden mit der
Nahrung aufgenommen. Als Biomarker im Urin ist 4-Hydroxybenzoesäure aufgrund der
vielen Quellen nur bedingt geeignet.
Hippursäure wird, in Folge der Entgiftungsfunktion von Leber und Niere, durch Konjugation
von Benzoesäure mit Glykokoll gebildet. Auch Toluol wird zum größten Teil zu Hippursäure
abgebaut. Niereninsuffizienz führt zu verringerter, Stoffwechselstörungen zu vermehrter
Ausscheidung mit dem Urin. Natriumbenzoat ist ein Konservierungsmittel in vielen
Lebensmitteln, welches ebenfalls zu Hippursäure abgebaut wird. Deshalb ist die
Konzentration im Urin zum Teil auch von der Ernährung abhängig (Zuppi et al, 1998).
Aufgrund ihrer geringen Konzentrationen geben Aromaten nur schwache Signale im
Protonenspektrum, die sich zum Teil nur wenig von der Basislinie abheben. Außerdem ist im
HSQC deutlich zu erkennen, dass viele Signale überlagert sind und im Protonenspektrum
deshalb nicht separiert dargestellt werden. Die chemische Verschiebung der aromatischen
Kohlenstoffe liegt, je nach Substituenten, zwischen 100ppm und 135ppm. Sie enthalten nur
wenig strukturelle Informationen. Hauptsächlich lassen sich die einzelnen Aromaten mit Hilfe
des COSY identifizieren. In vielen Fällen sind auch die Kopplungssignale der aromatischen
Protonen zu den Protonen alipahtischer Seitenketten zu beobachten, was für die
Strukturaufklärung sehr hilfreich ist. Die Resonanzfrequenzen der aliphatischen Seitenketten
liegen in dem überlagerten Bereich der Kohlenhydrate und sind für analytische Zwecke wenig
geeignet.
Identifizierung unbekannter Metabolite
32
ppm (t2) 7.007.508.00
110.0
115.0
120.0
125.0
130.0
135.0
ppm (t1)
ppm (t2) 7.007.508.00
110.0
115.0
120.0
125.0
130.0
135.0
ppm (t1)
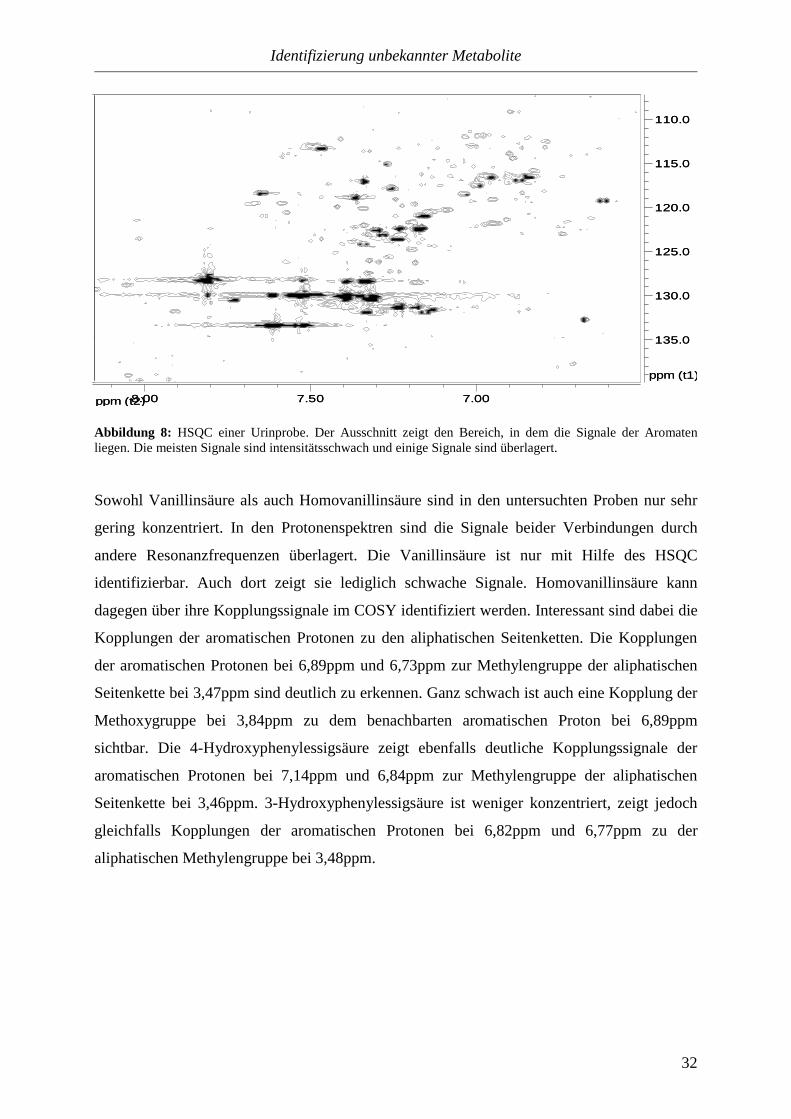
Abbildung 8: HSQC einer Urinprobe. Der Ausschnitt zeigt den Bereich, in dem die Signale der Aromaten liegen. Die meisten Signale sind intensitätsschwach und einige Signale sind überlagert.
Sowohl Vanillinsäure als auch Homovanillinsäure sind in den untersuchten Proben nur sehr
gering konzentriert. In den Protonenspektren sind die Signale beider Verbindungen durch
andere Resonanzfrequenzen überlagert. Die Vanillinsäure ist nur mit Hilfe des HSQC
identifizierbar. Auch dort zeigt sie lediglich schwache Signale. Homovanillinsäure kann
dagegen über ihre Kopplungssignale im COSY identifiziert werden. Interessant sind dabei die
Kopplungen der aromatischen Protonen zu den aliphatischen Seitenketten. Die Kopplungen
der aromatischen Protonen bei 6,89ppm und 6,73ppm zur Methylengruppe der aliphatischen
Seitenkette bei 3,47ppm sind deutlich zu erkennen. Ganz schwach ist auch eine Kopplung der
Methoxygruppe bei 3,84ppm zu dem benachbarten aromatischen Proton bei 6,89ppm
sichtbar. Die 4-Hydroxyphenylessigsäure zeigt ebenfalls deutliche Kopplungssignale der
aromatischen Protonen bei 7,14ppm und 6,84ppm zur Methylengruppe der aliphatischen
Seitenkette bei 3,46ppm. 3-Hydroxyphenylessigsäure ist weniger konzentriert, zeigt jedoch
gleichfalls Kopplungen der aromatischen Protonen bei 6,82ppm und 6,77ppm zu der
aliphatischen Methylengruppe bei 3,48ppm.
Identifizierung unbekannter Metabolite
33
ppm (t2) 7.00
3.00
3.50
4.00
ppm (t1)
Homovan
4-OH-PES 3-OH-PES
ppm (t2) 7.00
3.00
3.50
4.00
ppm (t1)
Homovan
4-OH-PES 3-OH-PES
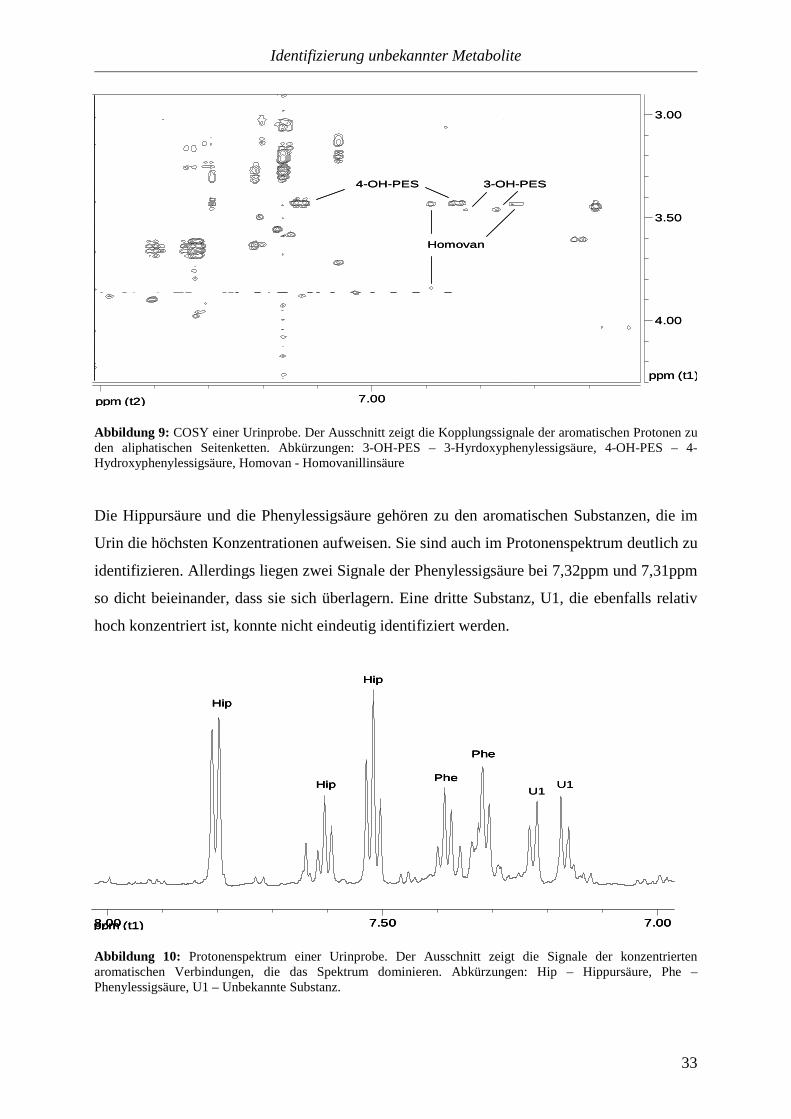
Abbildung 9: COSY einer Urinprobe. Der Ausschnitt zeigt die Kopplungssignale der aromatischen Protonen zu den aliphatischen Seitenketten. Abkürzungen: 3-OH-PES – 3-Hyrdoxyphenylessigsäure, 4-OH-PES – 4-Hydroxyphenylessigsäure, Homovan - Homovanillinsäure
Die Hippursäure und die Phenylessigsäure gehören zu den aromatischen Substanzen, die im
Urin die höchsten Konzentrationen aufweisen. Sie sind auch im Protonenspektrum deutlich zu
identifizieren. Allerdings liegen zwei Signale der Phenylessigsäure bei 7,32ppm und 7,31ppm
so dicht beieinander, dass sie sich überlagern. Eine dritte Substanz, U1, die ebenfalls relativ
hoch konzentriert ist, konnte nicht eindeutig identifiziert werden.
ppm (t1) 7.007.508.00
Hip
Hip
Hip
Phe
PheU1
U1
ppm (t1) 7.007.508.00
Hip
Hip
Hip
Phe
Phe
ppm (t1) 7.007.508.00
Hip
Hip
Hip
Phe
PheU1
U1
Abbildung 10: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale der konzentrierten aromatischen Verbindungen, die das Spektrum dominieren. Abkürzungen: Hip – Hippursäure, Phe – Phenylessigsäure, U1 – Unbekannte Substanz.

Identifizierung unbekannter Metabolite
34
Die Aufspaltung der Signale der unbekannten Substanz im Protonenspektrum weist darauf
hin, dass es sich um eine para-substituierte Verbindung handelt. Im COSY zeigt sie
Kopplungen der aromatischen Protonen bei 7,23ppm und 7,17ppm zu einer Methylgruppe bei
2,31ppm, die nahe bei denen des 4-Kresols liegen. Die Signale der aromatischen Protonen
sind gegenüber dem 4-Kresol tieffeldverschoben, was darauf hinweist, dass in der para-
Position keine freie Hydroxygruppe gebunden ist. Daneben ist im TOCSY eine schwache
Kopplung zu 4,62ppm zu erkennen. Das lässt darauf schließen, dass es sich hier um ein
Konjugat des 4-Kresols handelt, möglicherweise ein Glucuronid. Konjugierte Verbindungen
können allein mit Hilfe der NMR-Spektroskopie nicht identifiziert werden, da in der Regel
keine Kopplungen zu den gebundenen Kohlenhydraten oder Glucuroniden beobachtet werden
(siehe 3.1.2.1. Nukleoside). Für eine genaue Identifizierung sind deshalb weitere
Untersuchungen notwendig.
4-Kresol ist nicht sehr konzentriert, die Signale sind im Protonenspektrum überlagert. Im
COSY ist es dagegen eindeutig zu identifizieren. Die Kopplungen der Signale bei 7,16ppm
und 6,84ppm zu dem Signal der Methylgruppe bei 2,26ppm sind eindeutig zu erkennen.
Daneben sind auch die Kopplungssignale der unbekannten Substanz zu sehen.
ppm (t2) 7.0007.0507.1007.1507.2007.250
2.050
2.100
2.150
2.200
2.250
2.300
2.350
2.400
2.450ppm (t1)
U1 U1
4-Kresol 4-Kresol
ppm (t2) 7.0007.0507.1007.1507.2007.250
2.050
2.100
2.150
2.200
2.250
2.300
2.350
2.400
2.450ppm (t1)
ppm (t2) 7.0007.0507.1007.1507.2007.250
2.050
2.100
2.150
2.200
2.250
2.300
2.350
2.400
2.450ppm (t1)
U1 U1
4-Kresol 4-Kresol
Abbildung 11: COSY einer Urinprobe. Der Ausschnitt zeigt die Kopplungssignale der aromatischen Protonen zu den aliphatischen Seitenketten von 4-Kresol und der unbekannten Substanz U1.
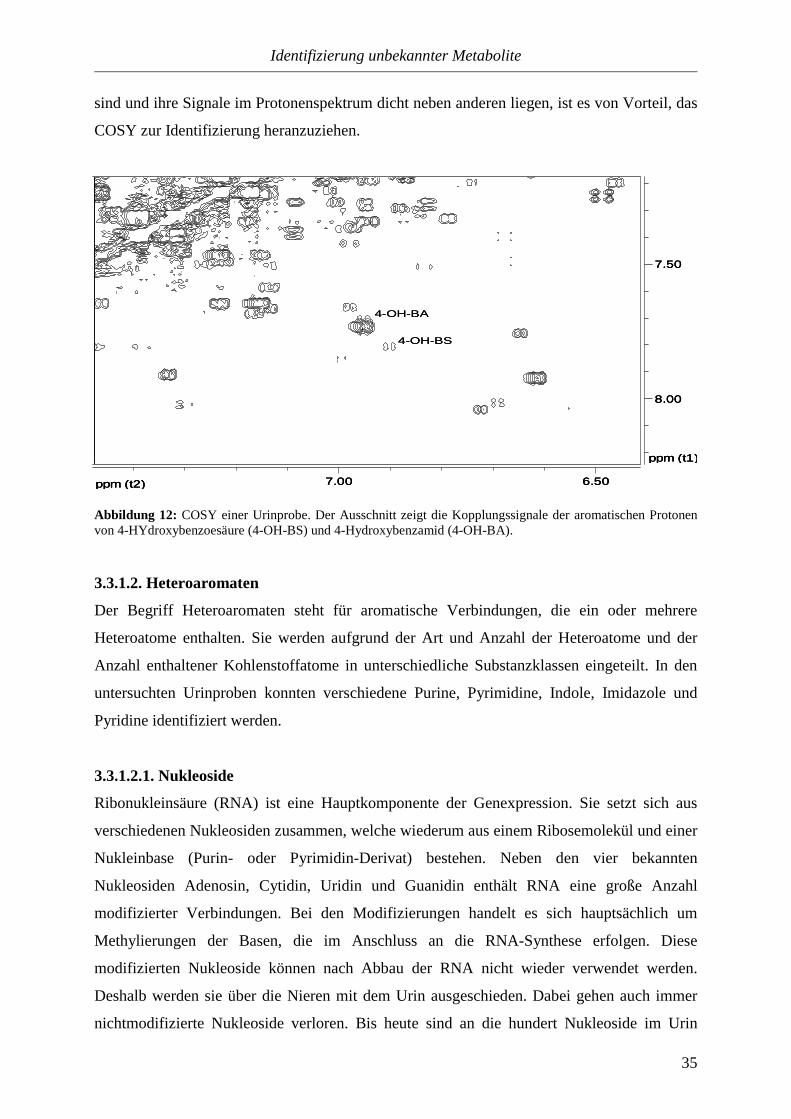
4-Hydroxybenzoesäure und 4-Hydroxybenzamid besitzen sehr ähnliche Strukturen. Deshalb
liegen ihre Signale auch dicht beieinander. Da beide Verbindungen relativ gering konzentriert
Identifizierung unbekannter Metabolite
35
sind und ihre Signale im Protonenspektrum dicht neben anderen liegen, ist es von Vorteil, das
COSY zur Identifizierung heranzuziehen.
ppm (t2) 6.507.00
7.50
8.00
ppm (t1)
4-OH-BA
4-OH-BS
ppm (t2) 6.507.00
7.50
8.00
ppm (t1)
ppm (t2) 6.507.00
7.50
8.00
ppm (t1)
4-OH-BA
4-OH-BS
Abbildung 12: COSY einer Urinprobe. Der Ausschnitt zeigt die Kopplungssignale der aromatischen Protonen von 4-HYdroxybenzoesäure (4-OH-BS) und 4-Hydroxybenzamid (4-OH-BA).
3.3.1.2. Heteroaromaten
Der Begriff Heteroaromaten steht für aromatische Verbindungen, die ein oder mehrere
Heteroatome enthalten. Sie werden aufgrund der Art und Anzahl der Heteroatome und der
Anzahl enthaltener Kohlenstoffatome in unterschiedliche Substanzklassen eingeteilt. In den
untersuchten Urinproben konnten verschiedene Purine, Pyrimidine, Indole, Imidazole und
Pyridine identifiziert werden.
3.3.1.2.1. Nukleoside
Ribonukleinsäure (RNA) ist eine Hauptkomponente der Genexpression. Sie setzt sich aus
verschiedenen Nukleosiden zusammen, welche wiederum aus einem Ribosemolekül und einer
Nukleinbase (Purin- oder Pyrimidin-Derivat) bestehen. Neben den vier bekannten
Nukleosiden Adenosin, Cytidin, Uridin und Guanidin enthält RNA eine große Anzahl
modifizierter Verbindungen. Bei den Modifizierungen handelt es sich hauptsächlich um
Methylierungen der Basen, die im Anschluss an die RNA-Synthese erfolgen. Diese
modifizierten Nukleoside können nach Abbau der RNA nicht wieder verwendet werden.
Deshalb werden sie über die Nieren mit dem Urin ausgeschieden. Dabei gehen auch immer
nichtmodifizierte Nukleoside verloren. Bis heute sind an die hundert Nukleoside im Urin

Identifizierung unbekannter Metabolite
36
bekannt (Schram, 1998; Seidel et al, 2006). Mit ihrer Hilfe können Aussagen über den RNA-
Stoffwechsel und über den Proteinstatus gemacht werden. Außerdem sind viele heute als
Tumormarker bekannt (Zheng et al, 2002; Kammerer et al, 2005; Seidel et al, 2006). Die
meisten Nukleoside und Nukleinbasen sind im Urin von gesunden Individuen jedoch so
gering konzentriert, dass sie mit Hilfe der NMR nicht detektiert werde können (Zheng et al,
2002; Hofmann et al, 2003). In den untersuchten Proben konnten Adenosin, Cytidin, Cytosin,
Dihydrouracil, Guanosin, Pseudouridine, Thymin, Xanthin, Uracil und Uridin identifiziert
werden.
Pseudouridine ist unter anderem ein Marker für das Wachstum von Lymphknotentumoren
und wird in Verbindung mit anderen Nukleosiden für die Anwesenheit verschiedener Tumore
als Marker verwendet (Kvist et al, 1990; Tu et al 1995).
Uracil und Dihydrouracil werden häufig als Indikator für die Enzymaktivität der
Dihydropyrimidin-Dehydrogenase eingesetzt. Wenn Tumore mit Uracilanaloga, wie zum
Beispiel 5-Fluoruracil, behandelt werden, müssen die Medikamente über die
Dihydropyrimidin-Dehydrogenase abgebaut werden. Ist die Enzymaktivität eingeschränkt,
werden die Medikamente angereichert und können zu Vergiftungen führen (Kuhara et al,
2001).
Nukleinbasen und Nukleoside sind im Urin nur gering konzentriert. Deshalb besitzen die
Signale in den Spektren auch nur schwache Intensitäten. Kopplungssignale der Nukleinbasen
zu den Ribosemolekülen werden weder im COSY noch im TOCSY beobachtet. Die
Ribosemoleküle der Nukleoside können jedoch mit Hilfe der α-Wasserstoffe identifiziert
werden. Diese zeigen charakteristische Resonanzfrequenzen zwischen 5,8ppm und 5,9ppm
und die dazugehörenden Kohlenstoffe zwischen 88ppm und 91ppm. Gegenüber den α-
Wasserstoffen anderer Kohlenhydrate sind diese Signale deutlich tieffeldverschoben.
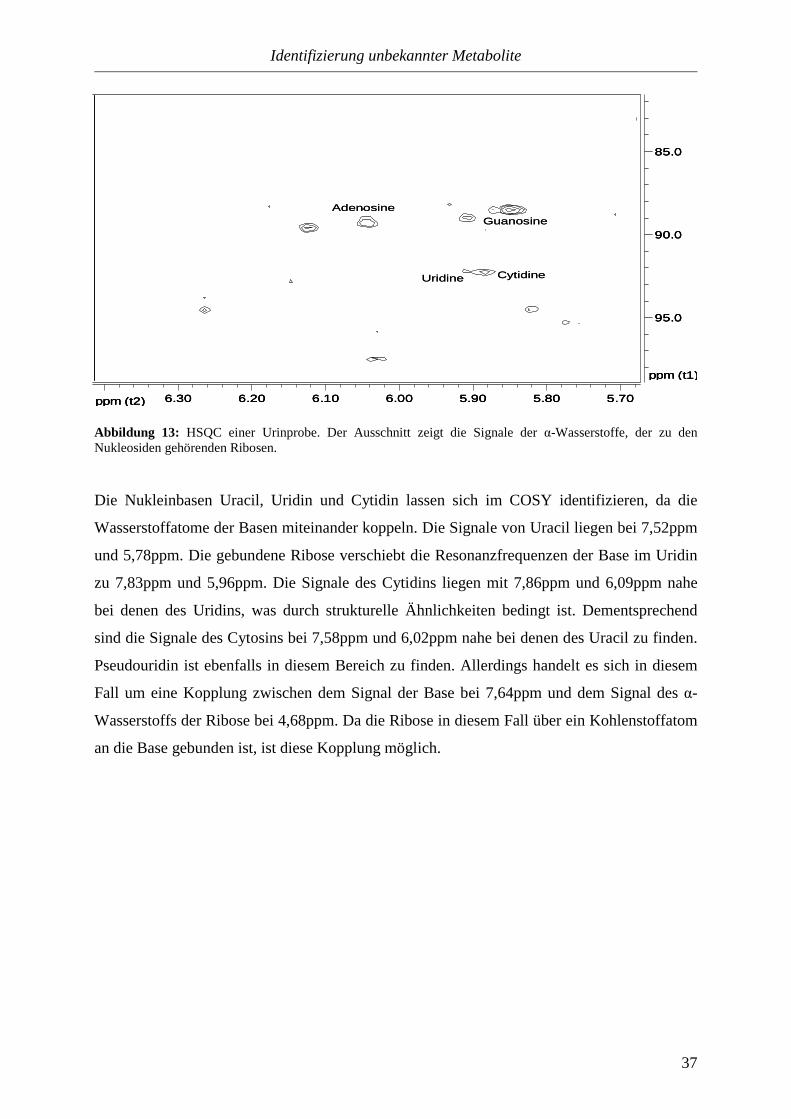
Identifizierung unbekannter Metabolite
37
ppm (t2) 5.705.805.906.006.106.206.30
85.0
90.0
95.0
ppm (t1)
Adenosine
Uridine
Guanosine
Cytidine
ppm (t2) 5.705.805.906.006.106.206.30
85.0
90.0
95.0
ppm (t1)
ppm (t2) 5.705.805.906.006.106.206.30
85.0
90.0
95.0
ppm (t1)
Adenosine
Uridine
Guanosine
Cytidine
Abbildung 13: HSQC einer Urinprobe. Der Ausschnitt zeigt die Signale der α-Wasserstoffe, der zu den Nukleosiden gehörenden Ribosen.
Die Nukleinbasen Uracil, Uridin und Cytidin lassen sich im COSY identifizieren, da die
Wasserstoffatome der Basen miteinander koppeln. Die Signale von Uracil liegen bei 7,52ppm
und 5,78ppm. Die gebundene Ribose verschiebt die Resonanzfrequenzen der Base im Uridin
zu 7,83ppm und 5,96ppm. Die Signale des Cytidins liegen mit 7,86ppm und 6,09ppm nahe
bei denen des Uridins, was durch strukturelle Ähnlichkeiten bedingt ist. Dementsprechend
sind die Signale des Cytosins bei 7,58ppm und 6,02ppm nahe bei denen des Uracil zu finden.
Pseudouridin ist ebenfalls in diesem Bereich zu finden. Allerdings handelt es sich in diesem
Fall um eine Kopplung zwischen dem Signal der Base bei 7,64ppm und dem Signal des α-
Wasserstoffs der Ribose bei 4,68ppm. Da die Ribose in diesem Fall über ein Kohlenstoffatom
an die Base gebunden ist, ist diese Kopplung möglich.
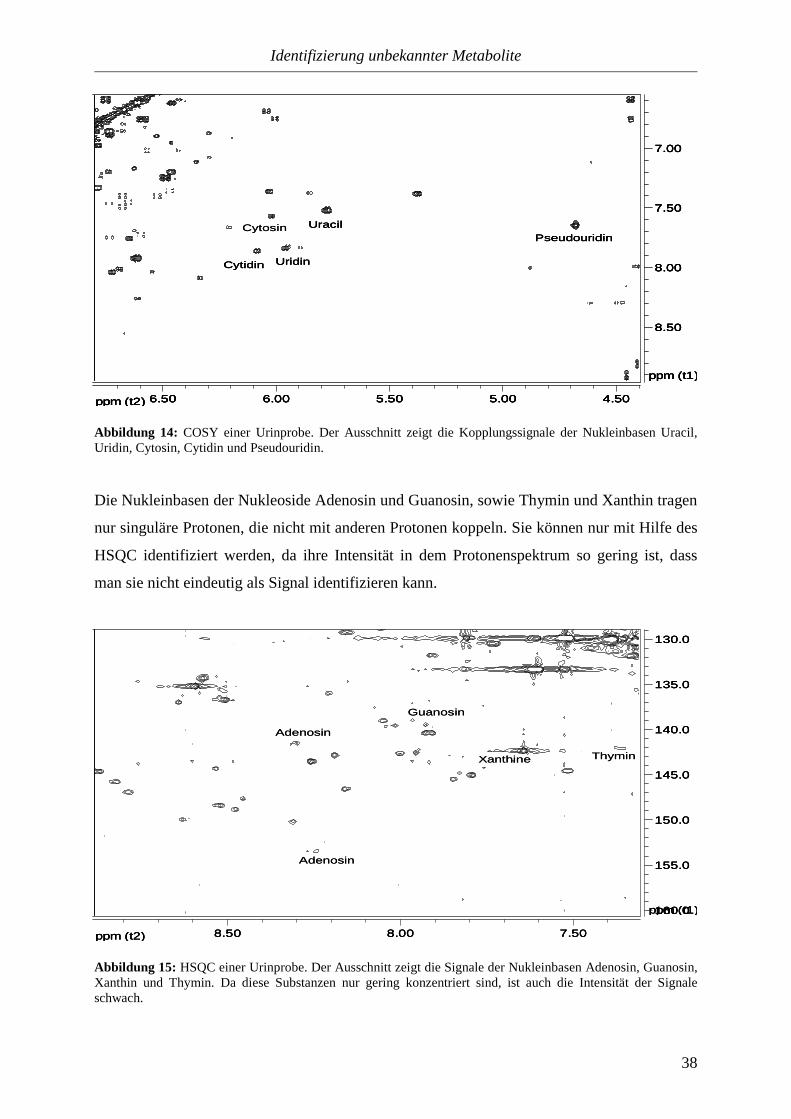
Identifizierung unbekannter Metabolite
38
ppm (t2) 4.505.005.506.006.50
7.00
7.50
8.00
8.50
ppm (t1)
Uridin
Uracil
Cytidin
PseudouridinCytosin
ppm (t2) 4.505.005.506.006.50
7.00
7.50
8.00
8.50
ppm (t1)
Uridin
Uracil
Cytidin
Pseudouridin
ppm (t2) 4.505.005.506.006.50
7.00
7.50
8.00
8.50
ppm (t1)
ppm (t2) 4.505.005.506.006.50
7.00
7.50
8.00
8.50
ppm (t1)
Uridin
Uracil
Cytidin
PseudouridinCytosin
Abbildung 14: COSY einer Urinprobe. Der Ausschnitt zeigt die Kopplungssignale der Nukleinbasen Uracil, Uridin, Cytosin, Cytidin und Pseudouridin.
Die Nukleinbasen der Nukleoside Adenosin und Guanosin, sowie Thymin und Xanthin tragen
nur singuläre Protonen, die nicht mit anderen Protonen koppeln. Sie können nur mit Hilfe des
HSQC identifiziert werden, da ihre Intensität in dem Protonenspektrum so gering ist, dass
man sie nicht eindeutig als Signal identifizieren kann.
ppm (t2) 7.508.008.50
130.0
135.0
140.0
145.0
150.0
155.0
160.0ppm (t1)
Adenosin
Adenosin
Guanosin
ThyminXanthine
ppm (t2) 7.508.008.50
130.0
135.0
140.0
145.0
150.0
155.0
160.0ppm (t1)
ppm (t2) 7.508.008.50
130.0
135.0
140.0
145.0
150.0
155.0
160.0ppm (t1)
Adenosin
Adenosin
Guanosin
ThyminXanthine
Abbildung 15: HSQC einer Urinprobe. Der Ausschnitt zeigt die Signale der Nukleinbasen Adenosin, Guanosin, Xanthin und Thymin. Da diese Substanzen nur gering konzentriert sind, ist auch die Intensität der Signale schwach.

Identifizierung unbekannter Metabolite
39
Die Resonanzfrequenzen von Dihydrouracil liegen nicht bei denen der anderen Nukleinbasen.
Da Dihydrouracil keine aromatische Struktur besitzt, liegen seine Signale in dem
aliphatischen Bereich bei 2,65ppm und 3,43ppm und sind durch viele Überlagerungen nicht
einfach zu identifizieren.
3.3.1.2.2. Pyridine
Neben den Purin- und Pyrimidin-Nukleosiden gibt es auch Pyridin-Nukleoside, welche
allerdings nicht in RNA oder DNA eingebaut werden. Eines davon konnte in den
untersuchten Urinproben identifiziert werden. Es handelt sich hierbei um 4-Pyridon-3-
Carboxamid-1-β-D-Ribonukleosid. Dieses Nukleosid wurde zuerst im Urin von Leukämie-
Patienten gefunden (Dutta et al, 1979). Die genaue Funktion im humanen Stoffwechsel ist bis
heute nicht bekannt. Die Konzentrationen im Blut und im Urin sind bei
Nierenfunktionsstörungen erhöht. Auch bei AIDS-Patienten wurden höhere Konzentrationen
festgestellt (Slominska et al, 2006).
Auch hier ist die Ribose zur Identifizierung nicht geeignet, da sie keine Kopplung zu dem
Pyridin-Derivat zeigt und sich ihre Signale in dem überlagerten Bereich befinden. Das 4-
Pyridon-3-Carboxamid-1-β-D-Ribonukleosid ist nur gering konzentriert. Sowohl im
Protonenspektrum als auch im HSQC sind die Intensitäten der Signale sehr schwach. Für eine
eindeutige Identifizierung ist deshalb das COSY hilfreich. Dort sind die Kopplungssignale
von 8,04ppm zu 6,72ppm und zu 8,76ppm zu erkennen.
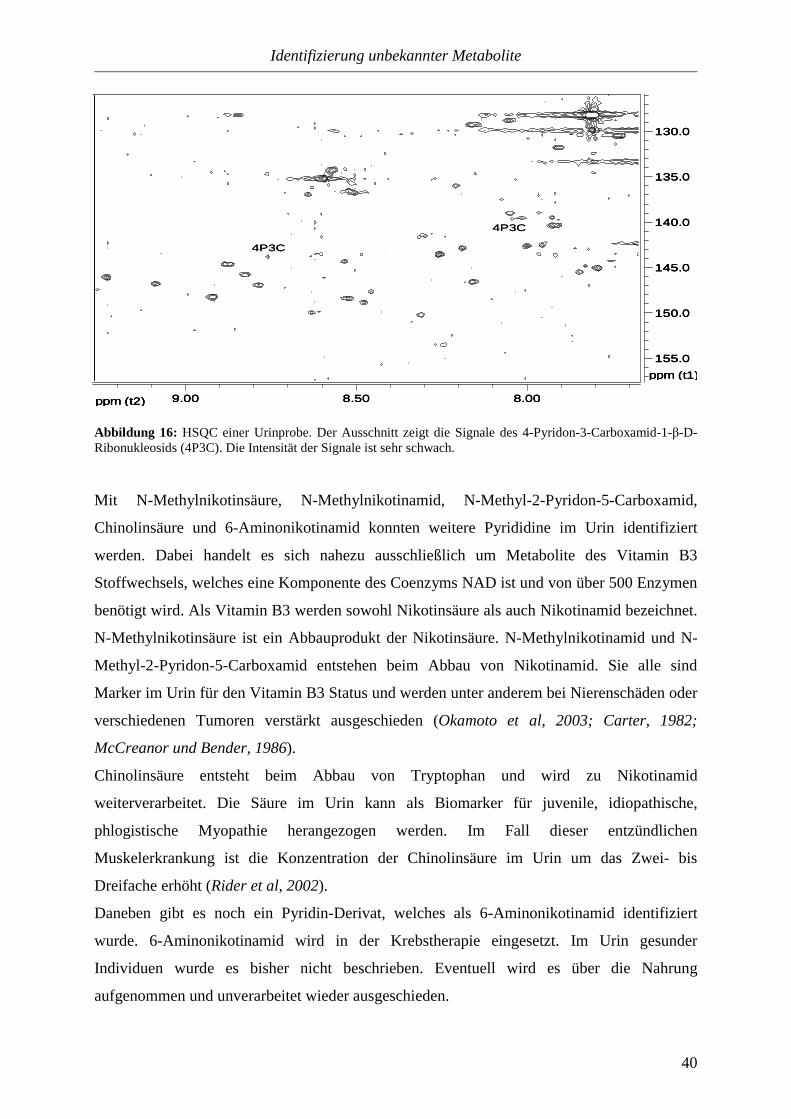
Identifizierung unbekannter Metabolite
40
ppm (t2) 8.008.509.00
130.0
135.0
140.0
145.0
150.0
155.0
ppm (t1)
4P3C
4P3C
ppm (t2) 8.008.509.00
130.0
135.0
140.0
145.0
150.0
155.0
ppm (t1)
ppm (t2) 8.008.509.00
130.0
135.0
140.0
145.0
150.0
155.0
ppm (t1)
4P3C
4P3C
Abbildung 16: HSQC einer Urinprobe. Der Ausschnitt zeigt die Signale des 4-Pyridon-3-Carboxamid-1-β-D-Ribonukleosids (4P3C). Die Intensität der Signale ist sehr schwach.
Mit N-Methylnikotinsäure, N-Methylnikotinamid, N-Methyl-2-Pyridon-5-Carboxamid,
Chinolinsäure und 6-Aminonikotinamid konnten weitere Pyrididine im Urin identifiziert
werden. Dabei handelt es sich nahezu ausschließlich um Metabolite des Vitamin B3
Stoffwechsels, welches eine Komponente des Coenzyms NAD ist und von über 500 Enzymen
benötigt wird. Als Vitamin B3 werden sowohl Nikotinsäure als auch Nikotinamid bezeichnet.
N-Methylnikotinsäure ist ein Abbauprodukt der Nikotinsäure. N-Methylnikotinamid und N-
Methyl-2-Pyridon-5-Carboxamid entstehen beim Abbau von Nikotinamid. Sie alle sind
Marker im Urin für den Vitamin B3 Status und werden unter anderem bei Nierenschäden oder
verschiedenen Tumoren verstärkt ausgeschieden (Okamoto et al, 2003; Carter, 1982;
McCreanor und Bender, 1986).
Chinolinsäure entsteht beim Abbau von Tryptophan und wird zu Nikotinamid
weiterverarbeitet. Die Säure im Urin kann als Biomarker für juvenile, idiopathische,
phlogistische Myopathie herangezogen werden. Im Fall dieser entzündlichen
Muskelerkrankung ist die Konzentration der Chinolinsäure im Urin um das Zwei- bis
Dreifache erhöht (Rider et al, 2002).
Daneben gibt es noch ein Pyridin-Derivat, welches als 6-Aminonikotinamid identifiziert
wurde. 6-Aminonikotinamid wird in der Krebstherapie eingesetzt. Im Urin gesunder
Individuen wurde es bisher nicht beschrieben. Eventuell wird es über die Nahrung
aufgenommen und unverarbeitet wieder ausgeschieden.
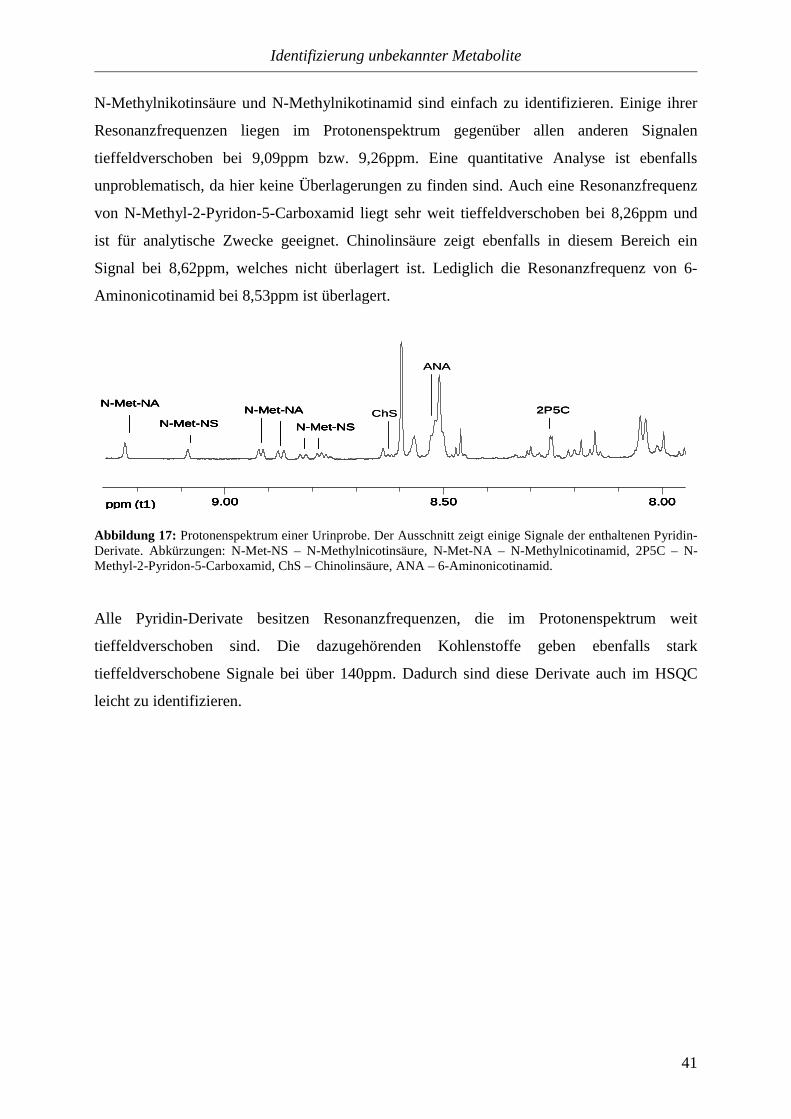
Identifizierung unbekannter Metabolite
41
N-Methylnikotinsäure und N-Methylnikotinamid sind einfach zu identifizieren. Einige ihrer
Resonanzfrequenzen liegen im Protonenspektrum gegenüber allen anderen Signalen
tieffeldverschoben bei 9,09ppm bzw. 9,26ppm. Eine quantitative Analyse ist ebenfalls
unproblematisch, da hier keine Überlagerungen zu finden sind. Auch eine Resonanzfrequenz
von N-Methyl-2-Pyridon-5-Carboxamid liegt sehr weit tieffeldverschoben bei 8,26ppm und
ist für analytische Zwecke geeignet. Chinolinsäure zeigt ebenfalls in diesem Bereich ein
Signal bei 8,62ppm, welches nicht überlagert ist. Lediglich die Resonanzfrequenz von 6-
Aminonicotinamid bei 8,53ppm ist überlagert.
ppm (t1) 8.008.509.00
N-Met-NA
N-Met-NSN-Met-NA
N-Met-NS
2P5CChS
ANA
ppm (t1) 8.008.509.00
N-Met-NA
N-Met-NSN-Met-NA
N-Met-NS
2P5C
ppm (t1) 8.008.509.00
N-Met-NA
N-Met-NSN-Met-NA
N-Met-NS
ppm (t1) 8.008.509.00
N-Met-NA
N-Met-NSN-Met-NA
N-Met-NS
2P5CChS
ANA
Abbildung 17: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt einige Signale der enthaltenen Pyridin-Derivate. Abkürzungen: N-Met-NS – N-Methylnicotinsäure, N-Met-NA – N-Methylnicotinamid, 2P5C – N-Methyl-2-Pyridon-5-Carboxamid, ChS – Chinolinsäure, ANA – 6-Aminonicotinamid.
Alle Pyridin-Derivate besitzen Resonanzfrequenzen, die im Protonenspektrum weit
tieffeldverschoben sind. Die dazugehörenden Kohlenstoffe geben ebenfalls stark
tieffeldverschobene Signale bei über 140ppm. Dadurch sind diese Derivate auch im HSQC
leicht zu identifizieren.
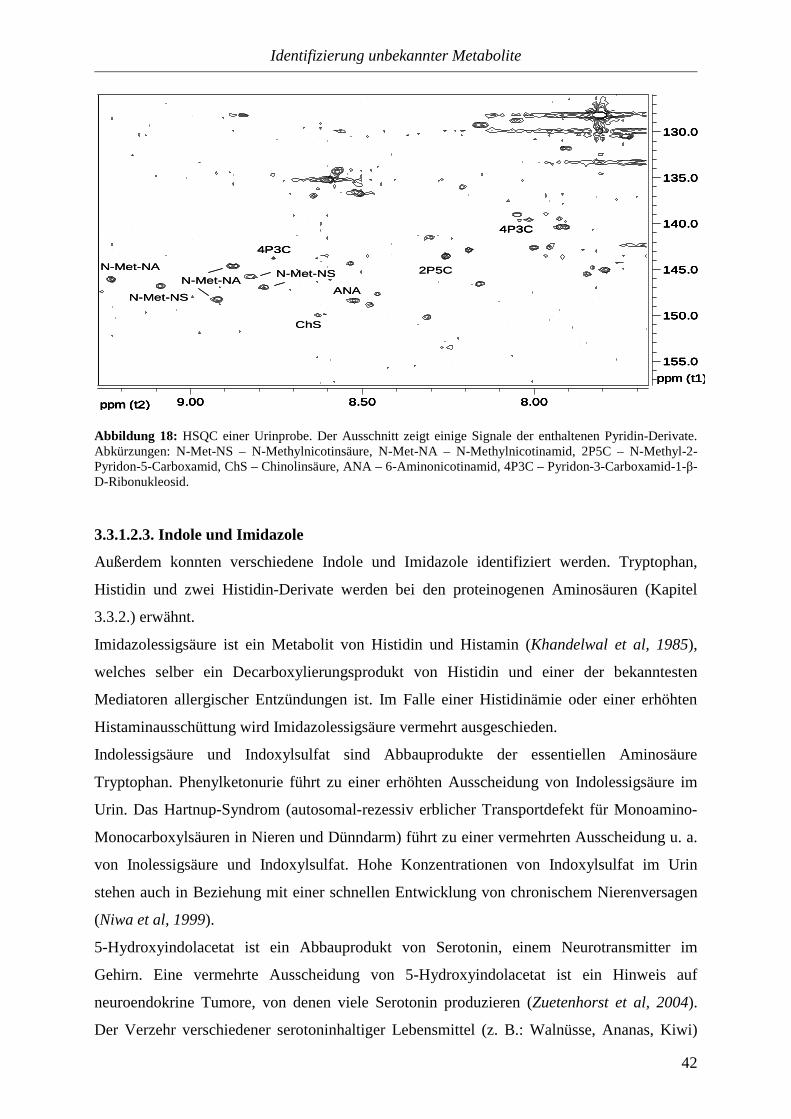
Identifizierung unbekannter Metabolite
42
ppm (t2) 8.008.509.00
130.0
135.0
140.0
145.0
150.0
155.0
ppm (t1)
4P3C
4P3C
N-Met-NS
N-Met-NA
ChS
ANA
2P5CN-Met-NA
N-Met-NS
ppm (t2) 8.008.509.00
130.0
135.0
140.0
145.0
150.0
155.0
ppm (t1)
4P3C
4P3C
ppm (t2) 8.008.509.00
130.0
135.0
140.0
145.0
150.0
155.0
ppm (t1)
ppm (t2) 8.008.509.00
130.0
135.0
140.0
145.0
150.0
155.0
ppm (t1)
4P3C
4P3C
N-Met-NS
N-Met-NA
ChS
ANA
2P5CN-Met-NA
N-Met-NS
Abbildung 18: HSQC einer Urinprobe. Der Ausschnitt zeigt einige Signale der enthaltenen Pyridin-Derivate. Abkürzungen: N-Met-NS – N-Methylnicotinsäure, N-Met-NA – N-Methylnicotinamid, 2P5C – N-Methyl-2-Pyridon-5-Carboxamid, ChS – Chinolinsäure, ANA – 6-Aminonicotinamid, 4P3C – Pyridon-3-Carboxamid-1-β-D-Ribonukleosid.
3.3.1.2.3. Indole und Imidazole
Außerdem konnten verschiedene Indole und Imidazole identifiziert werden. Tryptophan,
Histidin und zwei Histidin-Derivate werden bei den proteinogenen Aminosäuren (Kapitel
3.3.2.) erwähnt.
Imidazolessigsäure ist ein Metabolit von Histidin und Histamin (Khandelwal et al, 1985),
welches selber ein Decarboxylierungsprodukt von Histidin und einer der bekanntesten
Mediatoren allergischer Entzündungen ist. Im Falle einer Histidinämie oder einer erhöhten
Histaminausschüttung wird Imidazolessigsäure vermehrt ausgeschieden.
Indolessigsäure und Indoxylsulfat sind Abbauprodukte der essentiellen Aminosäure
Tryptophan. Phenylketonurie führt zu einer erhöhten Ausscheidung von Indolessigsäure im
Urin. Das Hartnup-Syndrom (autosomal-rezessiv erblicher Transportdefekt für Monoamino-
Monocarboxylsäuren in Nieren und Dünndarm) führt zu einer vermehrten Ausscheidung u. a.
von Inolessigsäure und Indoxylsulfat. Hohe Konzentrationen von Indoxylsulfat im Urin
stehen auch in Beziehung mit einer schnellen Entwicklung von chronischem Nierenversagen
(Niwa et al, 1999).
5-Hydroxyindolacetat ist ein Abbauprodukt von Serotonin, einem Neurotransmitter im
Gehirn. Eine vermehrte Ausscheidung von 5-Hydroxyindolacetat ist ein Hinweis auf
neuroendokrine Tumore, von denen viele Serotonin produzieren (Zuetenhorst et al, 2004).
Der Verzehr verschiedener serotoninhaltiger Lebensmittel (z. B.: Walnüsse, Ananas, Kiwi)
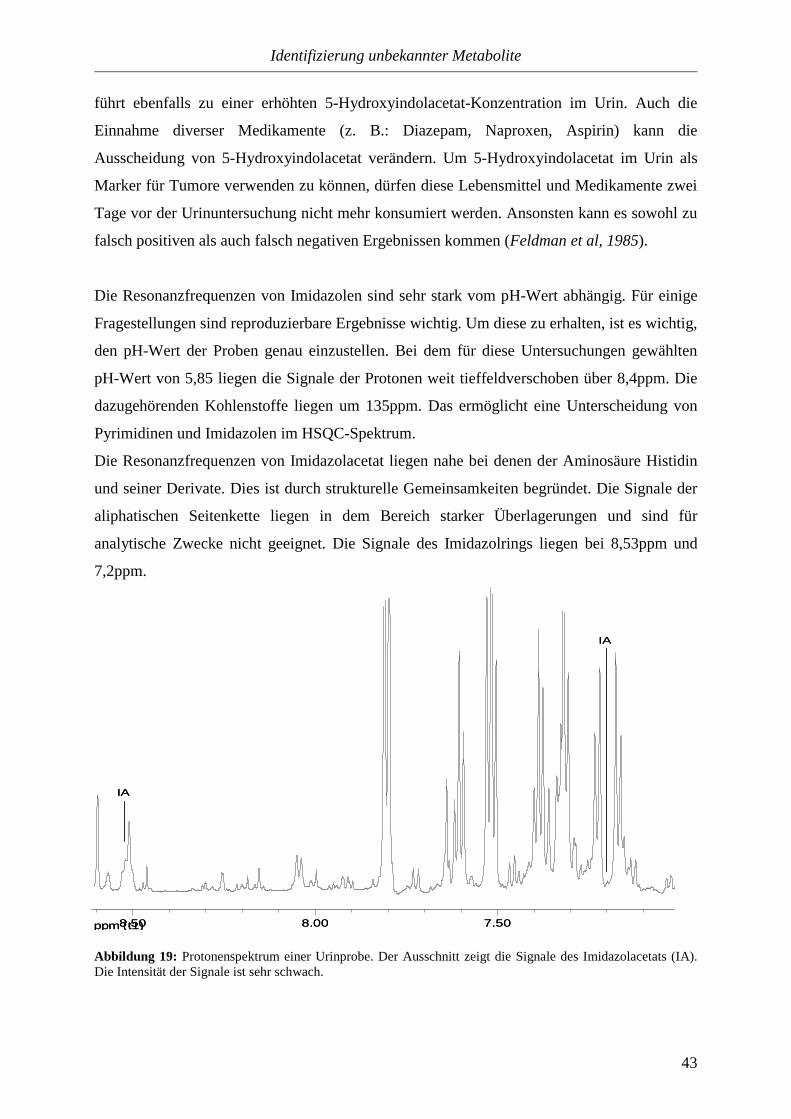
Identifizierung unbekannter Metabolite
43
führt ebenfalls zu einer erhöhten 5-Hydroxyindolacetat-Konzentration im Urin. Auch die
Einnahme diverser Medikamente (z. B.: Diazepam, Naproxen, Aspirin) kann die
Ausscheidung von 5-Hydroxyindolacetat verändern. Um 5-Hydroxyindolacetat im Urin als
Marker für Tumore verwenden zu können, dürfen diese Lebensmittel und Medikamente zwei
Tage vor der Urinuntersuchung nicht mehr konsumiert werden. Ansonsten kann es sowohl zu
falsch positiven als auch falsch negativen Ergebnissen kommen (Feldman et al, 1985).
Die Resonanzfrequenzen von Imidazolen sind sehr stark vom pH-Wert abhängig. Für einige
Fragestellungen sind reproduzierbare Ergebnisse wichtig. Um diese zu erhalten, ist es wichtig,
den pH-Wert der Proben genau einzustellen. Bei dem für diese Untersuchungen gewählten
pH-Wert von 5,85 liegen die Signale der Protonen weit tieffeldverschoben über 8,4ppm. Die
dazugehörenden Kohlenstoffe liegen um 135ppm. Das ermöglicht eine Unterscheidung von
Pyrimidinen und Imidazolen im HSQC-Spektrum.
Die Resonanzfrequenzen von Imidazolacetat liegen nahe bei denen der Aminosäure Histidin
und seiner Derivate. Dies ist durch strukturelle Gemeinsamkeiten begründet. Die Signale der
aliphatischen Seitenkette liegen in dem Bereich starker Überlagerungen und sind für
analytische Zwecke nicht geeignet. Die Signale des Imidazolrings liegen bei 8,53ppm und
7,2ppm.
ppm (t1) 7.508.008.50
IA
IA
ppm (t1) 7.508.008.50
IA
IA
Abbildung 19: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale des Imidazolacetats (IA). Die Intensität der Signale ist sehr schwach.
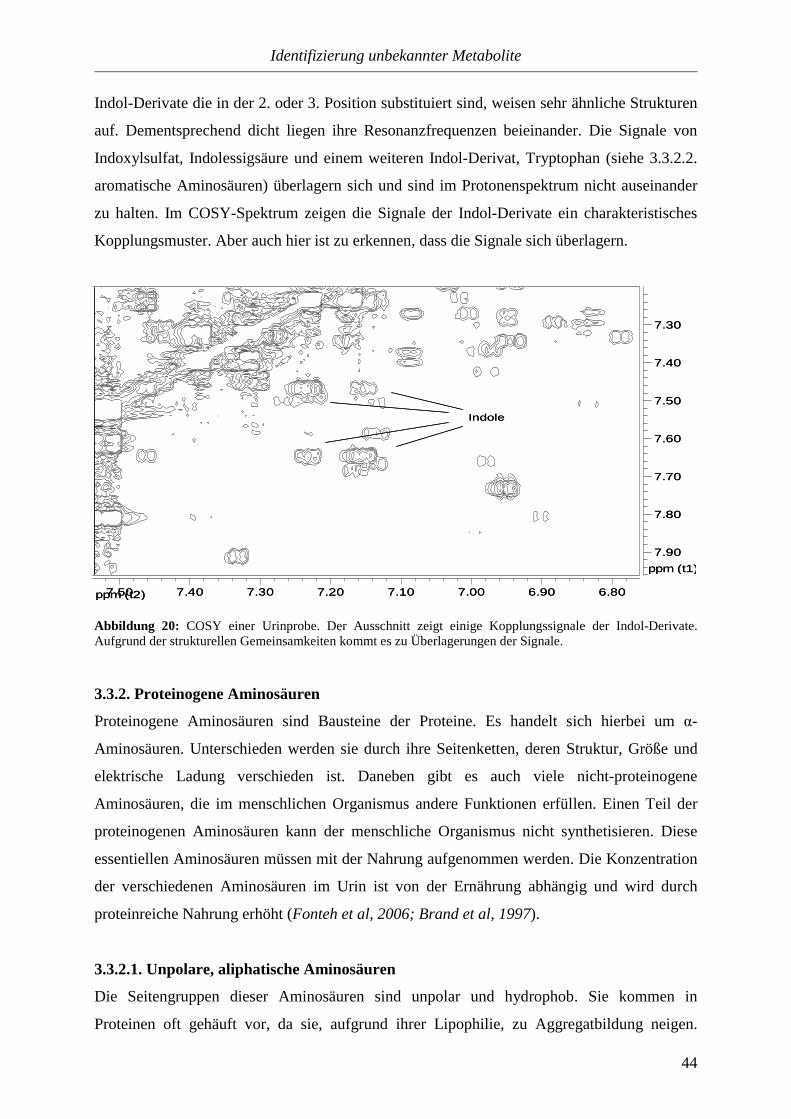
Identifizierung unbekannter Metabolite
44
Indol-Derivate die in der 2. oder 3. Position substituiert sind, weisen sehr ähnliche Strukturen
auf. Dementsprechend dicht liegen ihre Resonanzfrequenzen beieinander. Die Signale von
Indoxylsulfat, Indolessigsäure und einem weiteren Indol-Derivat, Tryptophan (siehe 3.3.2.2.
aromatische Aminosäuren) überlagern sich und sind im Protonenspektrum nicht auseinander
zu halten. Im COSY-Spektrum zeigen die Signale der Indol-Derivate ein charakteristisches
Kopplungsmuster. Aber auch hier ist zu erkennen, dass die Signale sich überlagern.
Indole
ppm (t2) 6.806.907.007.107.207.307.407.50
7.30
7.40
7.50
7.60
7.70
7.80
7.90
ppm (t1)
Indole
ppm (t2) 6.806.907.007.107.207.307.407.50
7.30
7.40
7.50
7.60
7.70
7.80
7.90
ppm (t1)
Abbildung 20: COSY einer Urinprobe. Der Ausschnitt zeigt einige Kopplungssignale der Indol-Derivate. Aufgrund der strukturellen Gemeinsamkeiten kommt es zu Überlagerungen der Signale.
3.3.2. Proteinogene Aminosäuren
Proteinogene Aminosäuren sind Bausteine der Proteine. Es handelt sich hierbei um α-
Aminosäuren. Unterschieden werden sie durch ihre Seitenketten, deren Struktur, Größe und
elektrische Ladung verschieden ist. Daneben gibt es auch viele nicht-proteinogene
Aminosäuren, die im menschlichen Organismus andere Funktionen erfüllen. Einen Teil der
proteinogenen Aminosäuren kann der menschliche Organismus nicht synthetisieren. Diese
essentiellen Aminosäuren müssen mit der Nahrung aufgenommen werden. Die Konzentration
der verschiedenen Aminosäuren im Urin ist von der Ernährung abhängig und wird durch
proteinreiche Nahrung erhöht (Fonteh et al, 2006; Brand et al, 1997).
3.3.2.1. Unpolare, aliphatische Aminosäuren
Die Seitengruppen dieser Aminosäuren sind unpolar und hydrophob. Sie kommen in
Proteinen oft gehäuft vor, da sie, aufgrund ihrer Lipophilie, zu Aggregatbildung neigen.

Identifizierung unbekannter Metabolite
45
Durch diese hydrophoben Wechselwirkungen wird die Proteinstruktur stabilisiert. In den
Urinproben konnten die unpolaren Aminosäuren Glycin, N, N-Dimethylglycin, Alanin, Valin,
Leucin und Isoleucin identifiziert werden.
Glycin ist die einfachste Aminosäure. Sie wird aus Serin, Threonin oder Glyoxylat gebildet
und ist hauptsächlich Bestandteil der Skleroproteine (z. B. Kollagen). Glycin ist ein wichtiges
Zwischenprodukt der Biosynthese von Porphyrinen, Purinen, Creatin und Glutathion. Es wird
über Glyoxylsäure zu Oxalsäure oder über Serin abgebaut. Vermehrte Glycinausscheidung in
den Urin kann unter anderem durch einen erblichen Defekt im Glycintransport im proximalen
Tubulus erfolgen. Nicht-ketotische Hyperglycinämie kann durch verschiedene Enzymdefekte
verursacht werden und führt ebenfalls zu erhöhten Konzentrationen im Urin (Tada und
Hayasaka, 1987).
N, N-Dimethylglycin ist ein Derivat der Aminosäure Glycin. Es entsteht beim Abbau von
Cholin zu Glycin und ist auch ein Nebenprodukt des Homocystein-Stoffwechsels.
Dimethylglycin Dehydrogenase Mangel führt zu einer vermehrten Urinausscheidung von N,
N-Dimethylglycin (Binzak et al, 2001; Moolenaar et al, 1999).
Alanin wird durch Pyruvattransaminierung oder Spaltung der Dipeptide Carnosine und
Anserin gebildet. Es ist vor allem im Muskelgewebe konzentriert und wird dort zur
Energiegewinnung freigesetzt. Das Hartnup-Syndrom ist ein erblicher Transportdefekt für
Monoaminocarboxyl-Säuren in Nieren und Dünndarm (Bröer et al, 2005). Alanin ist eine von
mehreren Aminosäuren, die bei diesem Syndrom vermehrt mit dem Urin ausgeschieden
werden. Daneben gibt es verschiedene Enzymdefekte, z. B. Pyruvat-Dehydrogenase-Mangel,
die ebenfalls zu erhöhten Konzentrationen im Urin führen.
Valin, Leucin und Isoleucin gehören zu den essentiellen Aminosäuren. Sie werden als
verzweigtkettige Aminosäuren bezeichnet und kommen in geringen Mengen in allen
Proteinen vor. Valin ist eine Biosynthesevorstufe für Panthothensäure und unentbehrlich für
Nerven- und Muskelfunktion. Leucin ist wichtig für den Aufbau und Erhalt von
Muskelgewebe. Es unterstützt die Proteinsynthese in Muskelgewebe und Leber und hemmt
den Abbau von Muskelprotein. Isoleucin ist für den Muskelaufbau von Bedeutung, da seine
hydrophoben Eigenschaften für die Ausbildung der Sekundärstruktur hilfreich sind. Rund ein
Drittel der Muskulatur besteht aus dieser Aminosäure. Es gibt viele Stoffwechselstörungen
dieser Aminosäuren, die bekannteste wird als Ahornsirupkrankheit bezeichnet (Chuang et al,
2006; Harris et al, 2005). Diese Krankheit führt unter anderem zu einer vermehrten
Ausscheidung von Valin, Leucin und Isoleucin mit dem Urin. Auch das Hartnup-Syndrom
verursacht erhöhte Urinkonzentrationen dieser Aminosäuren (Bröer et al, 2005).

Identifizierung unbekannter Metabolite
46
Diese unpolaren Aminosäuren sind in dem Protonenspektrum relativ einfach zu identifizieren.
Sie tragen aliphatische Seitenketten, deren Resonanzfrequenzen weit hochfeldverschoben
liegen. Diese Signale sind auch für analytische Zwecke geeignet, da dort nur wenige
Überlagerungen vorliegen. Eine Ausnahme bilden hier Glycin und N, N-Dimethylglycin, da
sie keine langen Seitenketten besitzen. Ihre Signale liegen bei 3,57ppm bzw. 3,72ppm und
2,91ppm. Alanin zeigt ein deutliches Dublett bei 1,47ppm und Valin zwei Dubletts bei
1,03ppm und 0,98ppm. Die beiden Dubletts von Leucin bei 0,93ppm und 0,95ppm liegen im
Protonenspektrum sehr dicht beieinander. Isoleucin ist nur sehr gering konzentriert, so dass
sein Signal bei 1,97ppm im Protonenspektrum kaum sichtbar ist.
ppm (t1) 1.001.502.00
Ala
ValLeu
Ile
ppm (t1) 1.001.502.00
Ala
ValLeu
Ile
Abbildung 21: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt einige Signale der unpolaren Aminosäuren Alanin (Ala), Valin (Val), Leucin (Leu) und Isoleucin (Ile).
3.3.2.2. Aromatische Aminosäuren
Die Seitengruppen dieser Aminosäuren sind relativ unpolar und können ebenfalls an
hydrophoben Wechselwirkungen beteiligt sein. In den Urinproben konnten die aromatischen
Aminosäuren Phenylalanin, Tyrosin und Tryptophan identifiziert werden.
Phenylalanin gehört zu den essentiellen Aminosäuren. Es ist an der Synthese von Adrenalin,
L-Dopa und anderen Hormonen beteiligt und dient als Edukt für weitere Substanzen, z. B. für
die Botenstoffe Dopamin, Serotonin oder Tyramin im Gehirn. Das Hartnup-Syndrom (Bröer
et al, 2005), Phenylketonurie und mildere Formen von Hyperphenylalaninämie verursachen
neben dem Erscheinen von Phenylbrenztraubensäure eine vermehrte Ausscheidung von
Phenylalanin im Urin (Kitagawa et al, 1975).
Tyrosin ist ein zentraler, intermediärer Metabolit für Schilddrüsenhormone, Catecholamine
und Melanine. Erhöhte Tyrosin Konzentrationen im Urin werden unter anderem durch das
Hartnup-Syndrom (Bröer et al, 2005) oder Hypertyrosinämie (Balato et al, 1986; Machino et
al, 1983) verursacht.
Tryptophan gehört zu den essentiellen Aminosäuren. Es ist Bestandteil vieler Proteine,
Vorläufer verschiedener biogener Amine wie Tryptamin, Melatonin und Serotonin und am
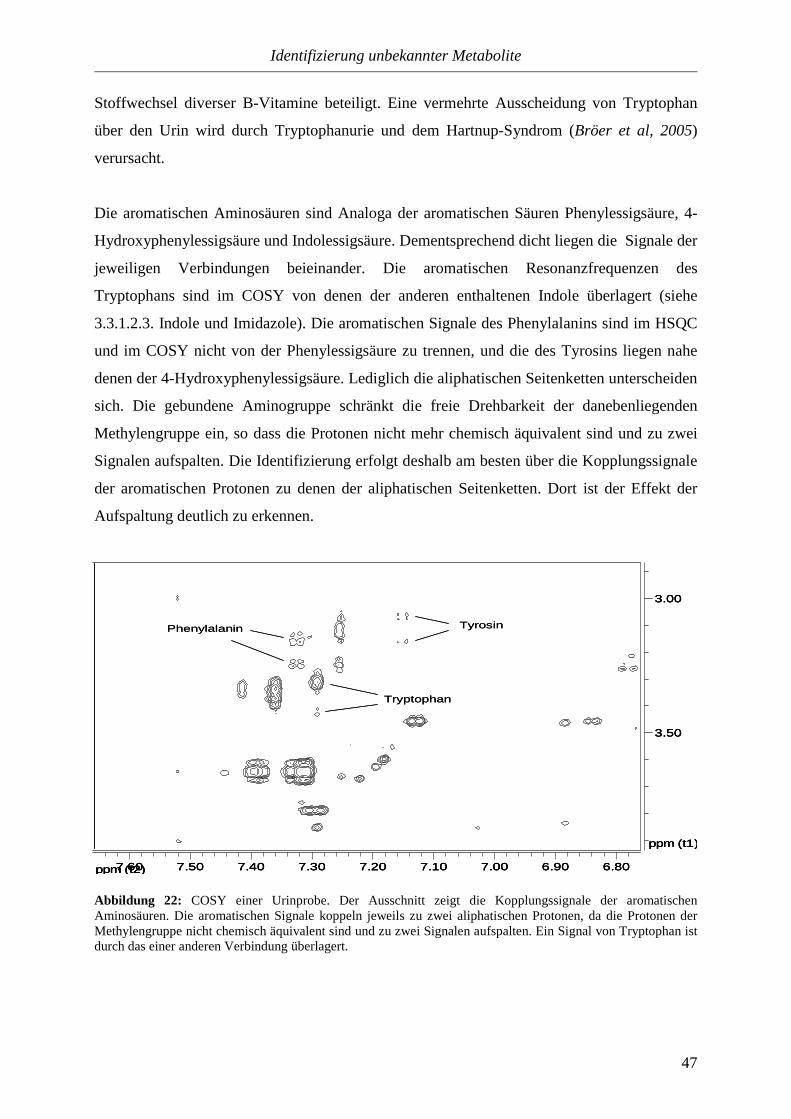
Identifizierung unbekannter Metabolite
47
Stoffwechsel diverser B-Vitamine beteiligt. Eine vermehrte Ausscheidung von Tryptophan
über den Urin wird durch Tryptophanurie und dem Hartnup-Syndrom (Bröer et al, 2005)
verursacht.
Die aromatischen Aminosäuren sind Analoga der aromatischen Säuren Phenylessigsäure, 4-
Hydroxyphenylessigsäure und Indolessigsäure. Dementsprechend dicht liegen die Signale der
jeweiligen Verbindungen beieinander. Die aromatischen Resonanzfrequenzen des
Tryptophans sind im COSY von denen der anderen enthaltenen Indole überlagert (siehe
3.3.1.2.3. Indole und Imidazole). Die aromatischen Signale des Phenylalanins sind im HSQC
und im COSY nicht von der Phenylessigsäure zu trennen, und die des Tyrosins liegen nahe
denen der 4-Hydroxyphenylessigsäure. Lediglich die aliphatischen Seitenketten unterscheiden
sich. Die gebundene Aminogruppe schränkt die freie Drehbarkeit der danebenliegenden
Methylengruppe ein, so dass die Protonen nicht mehr chemisch äquivalent sind und zu zwei
Signalen aufspalten. Die Identifizierung erfolgt deshalb am besten über die Kopplungssignale
der aromatischen Protonen zu denen der aliphatischen Seitenketten. Dort ist der Effekt der
Aufspaltung deutlich zu erkennen.
ppm (t2) 6.806.907.007.107.207.307.407.507.60
3.00
3.50
ppm (t1)
Phenylalanin Tyrosin
Tryptophan
ppm (t2) 6.806.907.007.107.207.307.407.507.60
3.00
3.50
ppm (t1)
ppm (t2) 6.806.907.007.107.207.307.407.507.60
3.00
3.50
ppm (t1)
Phenylalanin Tyrosin
Tryptophan
Abbildung 22: COSY einer Urinprobe. Der Ausschnitt zeigt die Kopplungssignale der aromatischen Aminosäuren. Die aromatischen Signale koppeln jeweils zu zwei aliphatischen Protonen, da die Protonen der Methylengruppe nicht chemisch äquivalent sind und zu zwei Signalen aufspalten. Ein Signal von Tryptophan ist durch das einer anderen Verbindung überlagert.

Identifizierung unbekannter Metabolite
48
3.3.2.3. Polare, ungeladene Aminosäuren
Die Seitengruppen dieser Aminosäuren sind hydrophil. Ihre funktionellen Gruppen können
Wasserstoffbrückenbindungen ausbilden und dadurch die Wasserlöslichkeit der Proteine
steigern. In den Urinproben konnten die Aminosäuren Serin, Threonin, Asparagin, Glutamin,
Cystin und Pyroglutamat identifiziert werden.
Serin kommt in Phospholipiden von Zellmembranen vor. Die Biosynthese von Serin erfolgt
ausgehend von 3-Phosphoglycerat oder Glycin. Es kann aber auch durch Protein- oder
Phospholipidabbau freigesetzt werden. Serin spielt eine zentrale Rolle beim Zellwachstum
und wird zum Aufbau von Cholin, Cystein und Purin-Nukleotiden benötigt. Außerdem ist es
wichtig für die Entwicklung und Funktion des Gehirns.
Threonin gehört zu den essentiellen Aminosäuren. Es wird für den Harnsäure-Stoffwechsel
und das Immunsystem benötigt, ist an der Biosynthese von Vitamin B12 beteiligt und spielt
eine Rolle im Stoffwechsel von Prophyrin. Erhöhte Konzentrationen im Urin werden unter
anderem durch das Hartnup-Syndrom verursacht (Bröer et al, 2005).
Asparagin wird aus Ammoniak und Asparaginsäure gebildet. Es wirkt wachstumsfördernd,
spielt eine Rolle in der Stoffwechselüberwachung in Hirn- und Nervengewebe und stellt eine
NH2-Quelle im Intermediärstoffwechsel für die Harnstoffsynthese dar. Stoffwechselstörungen
oder andere Krankheiten, die zu veränderten Urin Konzentrationen führen, sind bisher nicht
bekannt.
Glutamin wird aus Glutaminsäure gebildet. Es ist für den Hirnstoffwechsel von Bedeutung,
wird als NH2-Donor bei Transaminierungen eingesetzt und für NH3-Bindung und -Transport
benötigt. Erhöhte Konzentrationen im Urin werden unter anderem durch das Hartnup-
Syndrom (Bröer et al, 2005) oder dem LOWE-Syndrom verursacht. Ein Glutamin-Synthetase-
Mangel (Haberle et al, 2005) führt dagegen zu einer stark reduzierten Ausscheidung von
Glutamin.
Cystin ist eine schwefelhaltige Aminosäure. Es ist das Oxidationsprodukt von zwei
Cysteinmolekülen, welche über eine Disulfidbindung verknüpft sind. Cystein wird aus
Methionin gebildet und über Aminoacrylat zu Pyruvat abgebaut. Im extrazellulären Raum
liegt nur die oxidierte Form vor. Cystin ist hauptsächlich in Haaren und Haut zu finden und
spielt eine wichtige Rolle im Immunsystem. Erhöhte Konzentrationen im Urin treten bei
Cystinurie, einer Störung des renalen Transportsystems, auf (Font-Llitjos et al, 2005).
Die zyklisierte Glutaminsäure Pyroglutamat bzw. 5-Oxoprolin steht am Anfang von
Peptidketten und entsteht als Artefact bei der Lyophylisierung. Der Enzymdefekt Glutathion-
Synthetase-Mangel (Creer et al, 1989) führt zu erhöhten Konzentrationen im Urin.
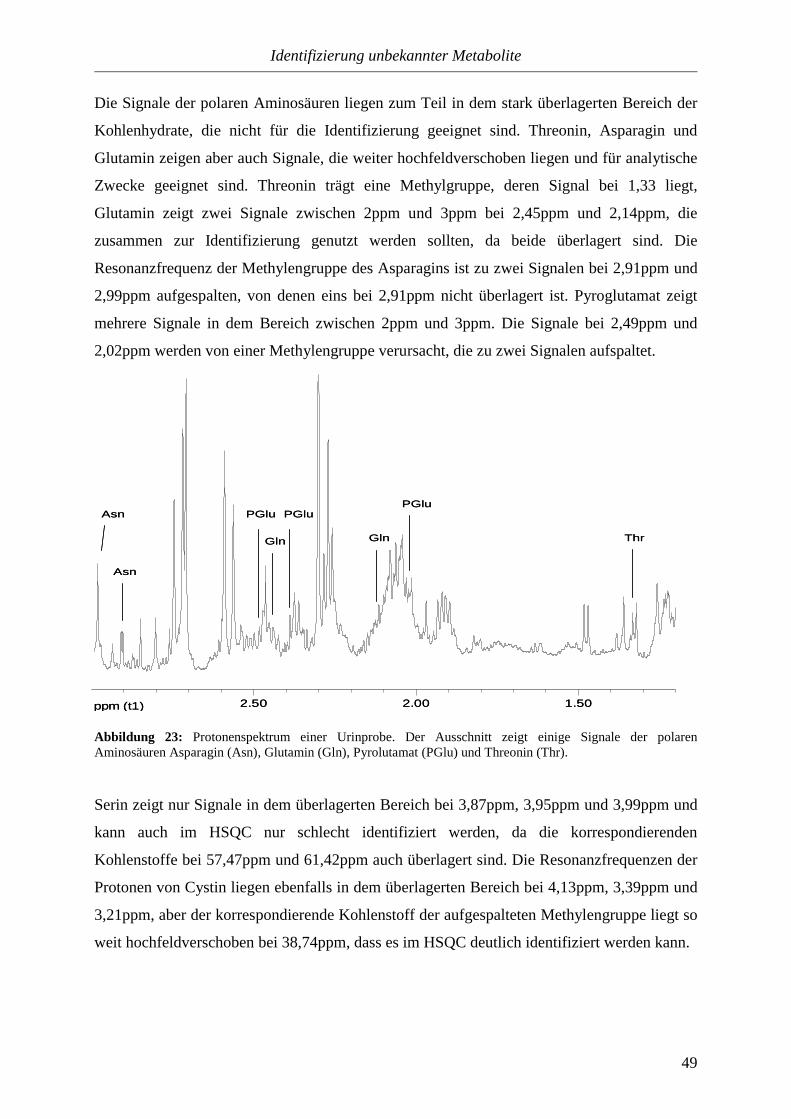
Identifizierung unbekannter Metabolite
49
Die Signale der polaren Aminosäuren liegen zum Teil in dem stark überlagerten Bereich der
Kohlenhydrate, die nicht für die Identifizierung geeignet sind. Threonin, Asparagin und
Glutamin zeigen aber auch Signale, die weiter hochfeldverschoben liegen und für analytische
Zwecke geeignet sind. Threonin trägt eine Methylgruppe, deren Signal bei 1,33 liegt,
Glutamin zeigt zwei Signale zwischen 2ppm und 3ppm bei 2,45ppm und 2,14ppm, die
zusammen zur Identifizierung genutzt werden sollten, da beide überlagert sind. Die
Resonanzfrequenz der Methylengruppe des Asparagins ist zu zwei Signalen bei 2,91ppm und
2,99ppm aufgespalten, von denen eins bei 2,91ppm nicht überlagert ist. Pyroglutamat zeigt
mehrere Signale in dem Bereich zwischen 2ppm und 3ppm. Die Signale bei 2,49ppm und
2,02ppm werden von einer Methylengruppe verursacht, die zu zwei Signalen aufspaltet.
ppm (t1) 1.502.002.50
Asn
Asn
PGlu
Gln
PGlu
Gln
PGlu
Thr
ppm (t1) 1.502.002.50
Asn
Asn
PGlu
Gln
PGlu
Gln
PGlu
Thr
Abbildung 23: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt einige Signale der polaren Aminosäuren Asparagin (Asn), Glutamin (Gln), Pyrolutamat (PGlu) und Threonin (Thr).
Serin zeigt nur Signale in dem überlagerten Bereich bei 3,87ppm, 3,95ppm und 3,99ppm und
kann auch im HSQC nur schlecht identifiziert werden, da die korrespondierenden
Kohlenstoffe bei 57,47ppm und 61,42ppm auch überlagert sind. Die Resonanzfrequenzen der
Protonen von Cystin liegen ebenfalls in dem überlagerten Bereich bei 4,13ppm, 3,39ppm und
3,21ppm, aber der korrespondierende Kohlenstoff der aufgespalteten Methylengruppe liegt so
weit hochfeldverschoben bei 38,74ppm, dass es im HSQC deutlich identifiziert werden kann.

Identifizierung unbekannter Metabolite
50
ppm (t2) 3.003.504.00
35.0
40.0
45.0
50.0
55.0
60.0
ppm (t1)
Cys2
Ser
ppm (t2) 3.003.504.00
35.0
40.0
45.0
50.0
55.0
60.0
ppm (t1)
Cys2
Ser
Abbildung 24: HSQC einer Urinprobe. Der Ausschnitt zeigt die aufgespaltenen Signale vom Cystin (Cys2). Das Signal von Serin (Ser) ist nicht so leicht zu identifizieren, da es überlagert ist.
3.3.2.4. Positiv geladene Aminosäuren
Geladene Seitengruppen besitzen die größte Hydrophilie. Die Ladungen ermöglichen auch
häufig enzymatisch katalysierte Reaktionen. Die positiv geladenen Aminosäuren Histidin, 3-
Methylhistidin, 1-Methylhistidin, Carnosin und Arginin konnten in den Urinproben
identifiziert werden.
Histidin ist eine halbessentielle Aminosäure. Sie kommt im Globin des Blutfarbstoffes vor
und ist in vielen Enzymen Bestandteil des aktiven Zentrums. Eine erhöhte Histidin
Konzentration im Urin kann verschiedene Ursachen haben. Dazu gehören die durch
Enzymdefekte verursachte Histidinämie, das Hartnup Syndrom (Bröer et al, 2005) und
Histidinurie, welche durch einen Defekt im tubulären Transportsystem hervorgerufen wird
(Nyhan und Hilton, 1992).
3-Methylhistidin ist Bestandteil von Actin und Myosin. Bei der Zersetzung dieser
Muskelproteine wird es freigesetzt und mit dem Urin ausgeschieden. Deshalb gilt es als
Marker für den Katabolismus von Muskelgewebe. 3-Methylhistidin wird aber auch über die
Nahrung aufgenommen. Um genaue Aussagen zu treffen, wie viel 3-Methylhistidin endogen
freigesetzt wurde, ist es hilfreich, zusätzlich die Konzentration von 1-Methylhistidin zu
ermitteln.
1-Methylhistidin bildet mit β-Alanin das Dipeptid Anserin. Es kommt im menschlichen
Gewebe kaum vor, wird aber ebenfalls mit der Nahrung aufgenommen. Seine Konzentration
korreliert gut mit der Konzentration von 3-Methylhistidin und ermöglicht dadurch eine
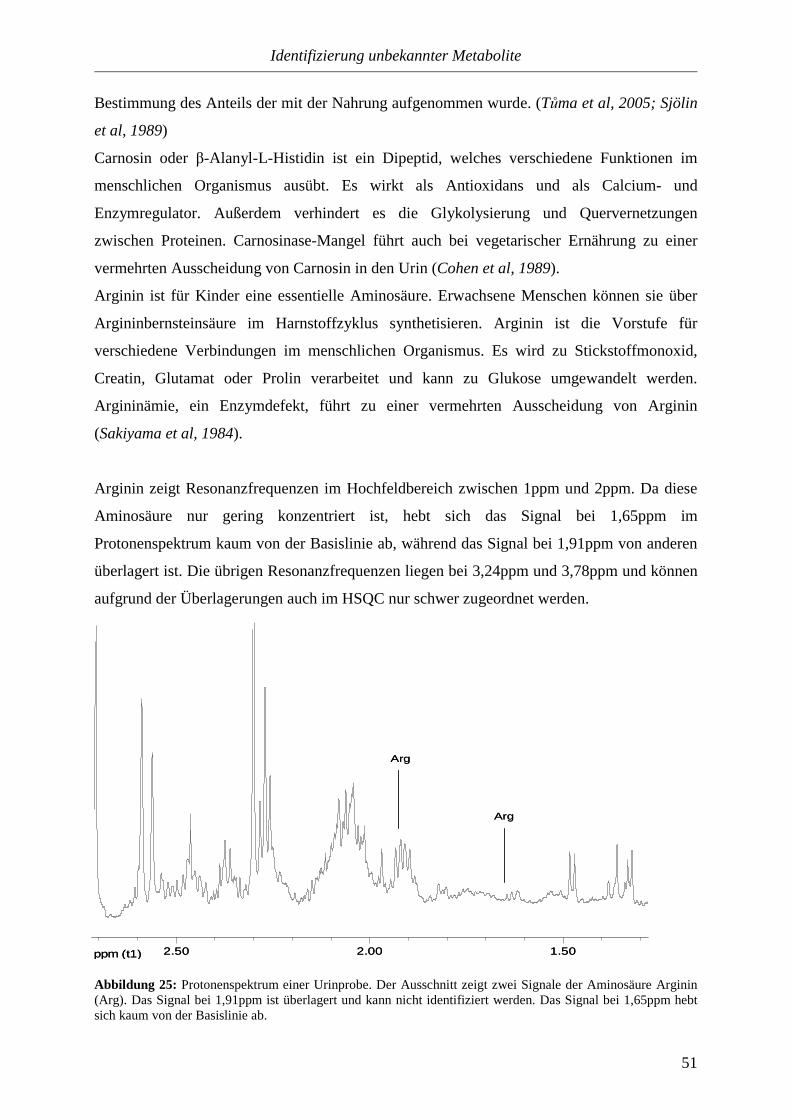
Identifizierung unbekannter Metabolite
51
Bestimmung des Anteils der mit der Nahrung aufgenommen wurde. (Tůma et al, 2005; Sjölin
et al, 1989)
Carnosin oder β-Alanyl-L-Histidin ist ein Dipeptid, welches verschiedene Funktionen im
menschlichen Organismus ausübt. Es wirkt als Antioxidans und als Calcium- und
Enzymregulator. Außerdem verhindert es die Glykolysierung und Quervernetzungen
zwischen Proteinen. Carnosinase-Mangel führt auch bei vegetarischer Ernährung zu einer
vermehrten Ausscheidung von Carnosin in den Urin (Cohen et al, 1989).
Arginin ist für Kinder eine essentielle Aminosäure. Erwachsene Menschen können sie über
Argininbernsteinsäure im Harnstoffzyklus synthetisieren. Arginin ist die Vorstufe für
verschiedene Verbindungen im menschlichen Organismus. Es wird zu Stickstoffmonoxid,
Creatin, Glutamat oder Prolin verarbeitet und kann zu Glukose umgewandelt werden.
Argininämie, ein Enzymdefekt, führt zu einer vermehrten Ausscheidung von Arginin
(Sakiyama et al, 1984).
Arginin zeigt Resonanzfrequenzen im Hochfeldbereich zwischen 1ppm und 2ppm. Da diese
Aminosäure nur gering konzentriert ist, hebt sich das Signal bei 1,65ppm im
Protonenspektrum kaum von der Basislinie ab, während das Signal bei 1,91ppm von anderen
überlagert ist. Die übrigen Resonanzfrequenzen liegen bei 3,24ppm und 3,78ppm und können
aufgrund der Überlagerungen auch im HSQC nur schwer zugeordnet werden.
ppm (t1) 1.502.002.50
Arg
Arg
ppm (t1) 1.502.002.50
Arg
Arg
Abbildung 25: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt zwei Signale der Aminosäure Arginin (Arg). Das Signal bei 1,91ppm ist überlagert und kann nicht identifiziert werden. Das Signal bei 1,65ppm hebt sich kaum von der Basislinie ab.

Identifizierung unbekannter Metabolite
52
Histidin, 3-Methylhistidin, 1-Methylhistidin, und Carnosin lassen sich am besten über die
Resonanzfrequenzen der Imidazolringe identifizieren. Im Protonenspektrum liegen die
Signale dicht beieinander, sind aber deutlich separiert. Lediglich das Signal von 1-
Methylhistidin bei 8,52ppm ist etwas von dem Signal einer anderen Verbindung überlagert.
Die Signale von Histidin bei 8,60ppm, 3-Methylhistidin bei 8,64ppm und Carnosin bei
8,58ppm werden nicht überlagert.
ppm (t1) 8.008.509.00
His
1-Met-His
3-Met-His
Car
ppm (t1) 8.008.509.00
His
1-Met-His
3-Met-His
Car
Abbildung 26: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale der Histidin-Derivate. Abkürzungen: Car – Carnosin, His – Histidin, 1-Met-His – 1-Methylhistidin, 3-Met-His – 3-Methylhistidin
3.3.3. Kohlenhydrate
Zucker und deren chemische Abkömmlinge werden als Kohlenhydrate bezeichnet. Sie sind
hochwertige Energielieferanten und Baustoffe, die im Körper schnell verwertet werden. Sie
können aber auch in polymerer Form (Glycogen) gespeichert werden. Es werden einfache
Kohlenhydrate (Monosaccharide) und zusammengesetzte Kohlenhydrate (Disaccharide
Oligosaccharide, Polysaccharide) unterschieden. Außerdem gibt es noch Zuckeralkohole und
Uronsäuren. Daneben kommen auch konjugierte Verbindungen wie Glykolipide und
proteingebundene Kohlenhydrate vor.
Das Protonenspektrum der Urinprobe weist im Bereich der Kohlenhydrate zwischen 3,5ppm
und 4,2ppm sehr viele Überlagerungen auf. Im HSQC sind die Signale ebenfalls nicht
separiert. Die korrespondierenden Kohlenstoffe geben ebenfalls dicht beieinander liegende
Signale zwischen 60ppm und 75ppm. Die Überlagerungen erschweren die Identifizierung der
Substanzen und machen eine quantitative Analyse nahezu unmöglich. Deshalb werden im
folgenden Abschnitt nur die Signale dargestellt, die für eine Analyse hilfreich sind.
Unbekannte konjugierte Verbindungen können alleine mit Hilfe der NMR nicht identifiziert
werden, da in der Regel im COSY keine Kopplung zwischen dem Kohlenhydrat und der
zugehörenden Substanz erkennbar ist (siehe 3.3.1.2.1. Nukleoside).

Identifizierung unbekannter Metabolite
53
3.3.3.1. Monosaccharide
Monosaccharide sind hydrolytisch nicht weiter aufspaltbare Zucker. Sie kommen in vielen
Lebensmitteln vor und werden durch die Nahrung dem humanen Stoffwechsel zugeführt. Der
Ringschluß dieser Verbindungen führt dazu, dass jeweils zwei Isomere (alpha und beta)
vorhanden sind. Zu den Monosacchariden, die im Urin identifiziert werden konnten, gehören
Glucose, Fructose, Fucose, Galactose, Xylose und N-Acetylneuraminsäure.
Glucose ist eine Schlüsselsubstanz im Kohlenhydratstoffwechsel. Sie wird als Glycogen in
Muskel und Leber gespeichert und steht in ihrer stoffwechselaktiven Form, Glukose-6-
phosphat, der Energiegewinnung durch Glycolyse, Citratzyklus und oxidativer
Phosphorylierung oder durch den Pentosephosphatzyklus zur Verfügung. Im Urin wird
Glucose hauptsächlich zur Diagnose von Diabetes mellitus und zur Kontrolle der
Insulintherapie (Kabadi, 1992; Koch, 2004) genutzt.
Fructose kommt in Früchten, Honig und gebunden mit Glucose als Saccharose vor, kann aber
auch im Körper synthetisiert werden. Sie wird zu Glycogen umgewandelt, oder nach
Phosphorylierung in der Leber abgebaut. Im Urin ist Fructose nach Verzehr nachweisbar
(Luceri et al, 1996). Fructokinase-Mangel bedingte Fructosämie oder Fructoseintoleranz
führen zu erhöhten Konzentrationen im Urin (Froesch, 1976).
Im menschlichen Organismus kommt Fucose unter anderem in Muco- und Glykoproteinen
vor (Kornfeld und Kornfeld, 1976). Verschiedene Krebsarten führen zu erhöhten Fucose-
Konzentrationen im Urin, darunter Leber-, Magen-, Lungen- und Pankreaskrebs (Suzuki et al,
1992, Sakai et al, 1990), außerdem auch Magengeschwüre und Leberzirrhose (Sakai et al,
1990; Yamauchi et al, 1993).
Galactose ist in Gangliosiden, Cerebrosiden, Glykolipiden und Glykosaminoglykanen zu
finden und wird hauptsächlich in der Leber abgebaut. Im Urin sind vor allem im Fall einer
Galactosämie erhöhte Galactosekonzentrationen nachweisbar (Easley et al, 2003).
Die Bestimmung von Xylose im Urin erfolgt hauptsächlich bei Absorptionstests. Mit der
Nahrung aufgenommene Xylose wird im oberen Dünndarm resorbiert und etwa 25% der
aufgenommenen Menge unverändert über den Urin ausgeschieden. Im Fall eines
Malabsorptionssyndroms ist Xylose in erniedrigten Konzentrationen im Urin zu finden (Rana
et al, 2007). Die Ursachen dafür sind unterschiedlich, so verursacht z. B. Zöliakie
(glutenbedingte Erkrankung der Darmschleimhaut), virale Gastroenteritis (Entzündung von
Magen- und Darmschleimhaut) oder Ankylostomiasis (Hakenwurmkrankheit) eine
Malabsorption.
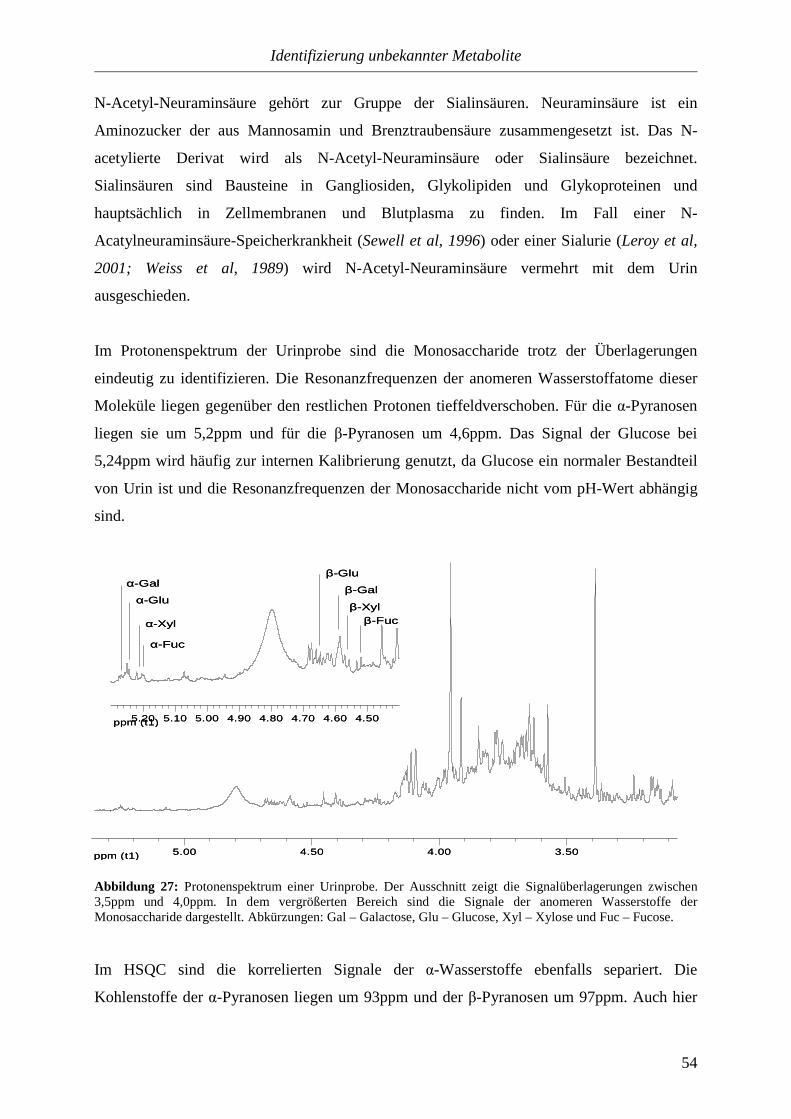
Identifizierung unbekannter Metabolite
54
N-Acetyl-Neuraminsäure gehört zur Gruppe der Sialinsäuren. Neuraminsäure ist ein
Aminozucker der aus Mannosamin und Brenztraubensäure zusammengesetzt ist. Das N-
acetylierte Derivat wird als N-Acetyl-Neuraminsäure oder Sialinsäure bezeichnet.
Sialinsäuren sind Bausteine in Gangliosiden, Glykolipiden und Glykoproteinen und
hauptsächlich in Zellmembranen und Blutplasma zu finden. Im Fall einer N-
Acatylneuraminsäure-Speicherkrankheit (Sewell et al, 1996) oder einer Sialurie (Leroy et al,
2001; Weiss et al, 1989) wird N-Acetyl-Neuraminsäure vermehrt mit dem Urin
ausgeschieden.
Im Protonenspektrum der Urinprobe sind die Monosaccharide trotz der Überlagerungen
eindeutig zu identifizieren. Die Resonanzfrequenzen der anomeren Wasserstoffatome dieser
Moleküle liegen gegenüber den restlichen Protonen tieffeldverschoben. Für die α-Pyranosen
liegen sie um 5,2ppm und für die β-Pyranosen um 4,6ppm. Das Signal der Glucose bei
5,24ppm wird häufig zur internen Kalibrierung genutzt, da Glucose ein normaler Bestandteil
von Urin ist und die Resonanzfrequenzen der Monosaccharide nicht vom pH-Wert abhängig
sind.
ppm (t1) 3.504.004.505.00
ppm (t1) 4.504.604.704.804.905.005.105.20
α-Gal
α-Glu
α-Xyl
α-Fuc
β-Glu
β-Gal
β-Xylβ-Fuc
ppm (t1) 3.504.004.505.00
ppm (t1) 4.504.604.704.804.905.005.105.20
α-Gal
α-Glu
α-Xyl
α-Fuc
β-Glu
β-Gal
β-Xylβ-Fuc
Abbildung 27: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signalüberlagerungen zwischen 3,5ppm und 4,0ppm. In dem vergrößerten Bereich sind die Signale der anomeren Wasserstoffe der Monosaccharide dargestellt. Abkürzungen: Gal – Galactose, Glu – Glucose, Xyl – Xylose und Fuc – Fucose.
Im HSQC sind die korrelierten Signale der α-Wasserstoffe ebenfalls separiert. Die
Kohlenstoffe der α-Pyranosen liegen um 93ppm und der β-Pyranosen um 97ppm. Auch hier
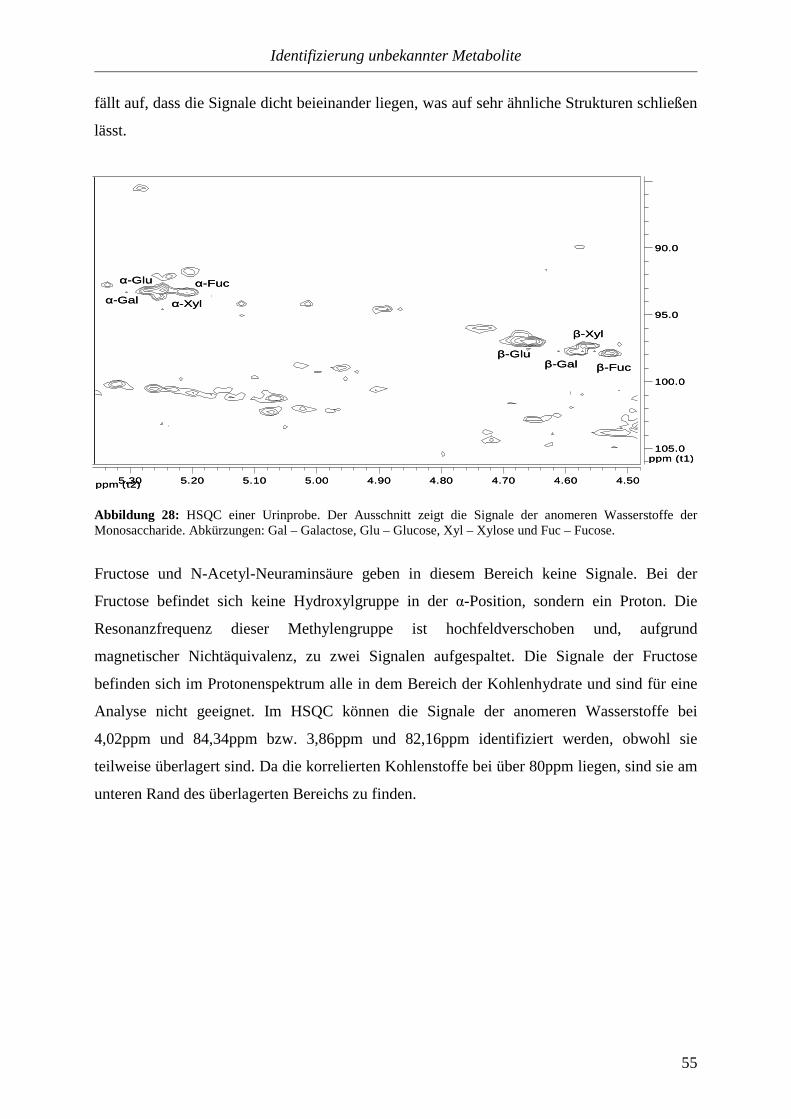
Identifizierung unbekannter Metabolite
55
fällt auf, dass die Signale dicht beieinander liegen, was auf sehr ähnliche Strukturen schließen
lässt.
ppm (t2) 4.504.604.704.804.905.005.105.205.30
90.0
95.0
100.0
105.0ppm (t1)
α-Glu
α-Gal α-Xyl
α-Fuc
β-Gluβ-Gal
β-Xyl
β-Fuc
ppm (t2) 4.504.604.704.804.905.005.105.205.30
90.0
95.0
100.0
105.0ppm (t1)
α-Glu
α-Gal α-Xyl
α-Fuc
β-Gluβ-Gal
β-Xyl
β-Fuc
Abbildung 28: HSQC einer Urinprobe. Der Ausschnitt zeigt die Signale der anomeren Wasserstoffe der Monosaccharide. Abkürzungen: Gal – Galactose, Glu – Glucose, Xyl – Xylose und Fuc – Fucose.
Fructose und N-Acetyl-Neuraminsäure geben in diesem Bereich keine Signale. Bei der
Fructose befindet sich keine Hydroxylgruppe in der α-Position, sondern ein Proton. Die
Resonanzfrequenz dieser Methylengruppe ist hochfeldverschoben und, aufgrund
magnetischer Nichtäquivalenz, zu zwei Signalen aufgespaltet. Die Signale der Fructose
befinden sich im Protonenspektrum alle in dem Bereich der Kohlenhydrate und sind für eine
Analyse nicht geeignet. Im HSQC können die Signale der anomeren Wasserstoffe bei
4,02ppm und 84,34ppm bzw. 3,86ppm und 82,16ppm identifiziert werden, obwohl sie
teilweise überlagert sind. Da die korrelierten Kohlenstoffe bei über 80ppm liegen, sind sie am
unteren Rand des überlagerten Bereichs zu finden.
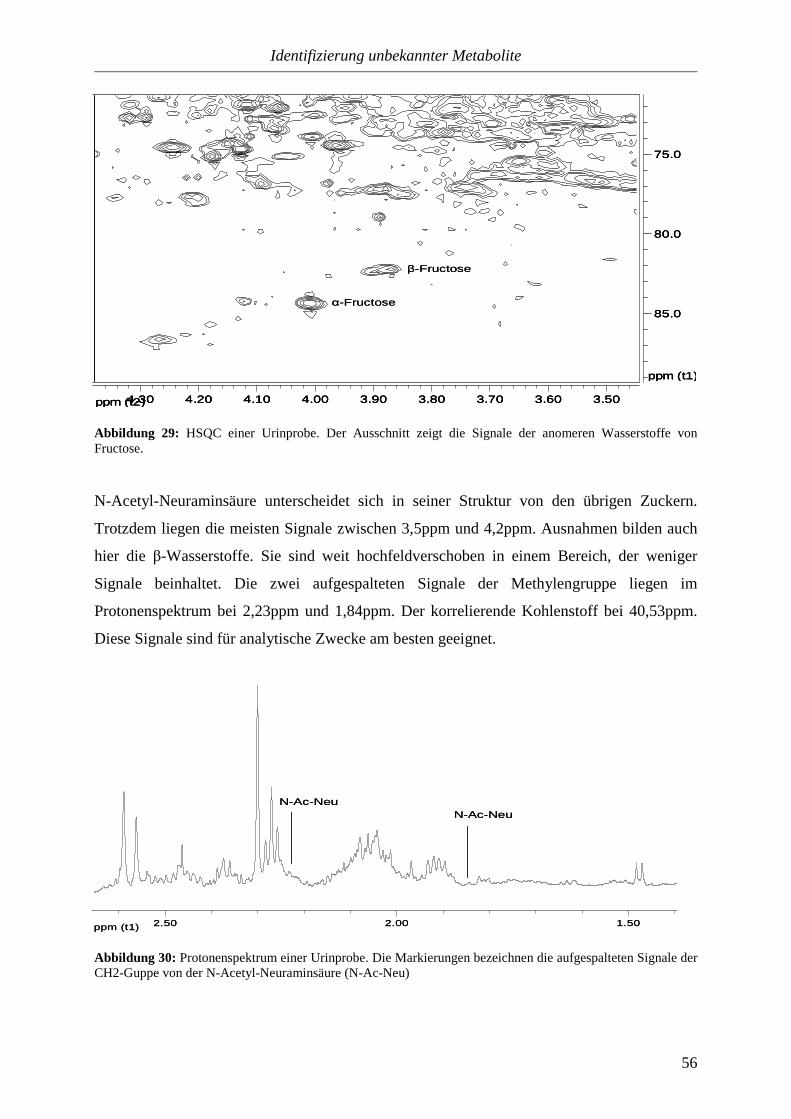
Identifizierung unbekannter Metabolite
56
ppm (t2) 3.503.603.703.803.904.004.104.204.30
75.0
80.0
85.0
ppm (t1)
α-Fructose
β-Fructose
ppm (t2) 3.503.603.703.803.904.004.104.204.30
75.0
80.0
85.0
ppm (t1)
ppm (t2) 3.503.603.703.803.904.004.104.204.30
75.0
80.0
85.0
ppm (t1)
α-Fructose
β-Fructose
Abbildung 29: HSQC einer Urinprobe. Der Ausschnitt zeigt die Signale der anomeren Wasserstoffe von Fructose.
N-Acetyl-Neuraminsäure unterscheidet sich in seiner Struktur von den übrigen Zuckern.
Trotzdem liegen die meisten Signale zwischen 3,5ppm und 4,2ppm. Ausnahmen bilden auch
hier die β-Wasserstoffe. Sie sind weit hochfeldverschoben in einem Bereich, der weniger
Signale beinhaltet. Die zwei aufgespalteten Signale der Methylengruppe liegen im
Protonenspektrum bei 2,23ppm und 1,84ppm. Der korrelierende Kohlenstoff bei 40,53ppm.
Diese Signale sind für analytische Zwecke am besten geeignet.
ppm (t1) 1.502.002.50
N-Ac-NeuN-Ac-Neu
ppm (t1) 1.502.002.50
N-Ac-NeuN-Ac-Neu
Abbildung 30: Protonenspektrum einer Urinprobe. Die Markierungen bezeichnen die aufgespalteten Signale der CH2-Guppe von der N-Acetyl-Neuraminsäure (N-Ac-Neu)

Identifizierung unbekannter Metabolite
57
3.3.3.2. Disaccharide
Disaccharide bestehen aus zwei Monosacchariden, die über eine glycosidische Bindung
verknüpft sind. Die Monosaccharide können an verschiedenen Stellen miteinander verbunden
sein und unterschiedliche Stereochemie aufweisen. Dadurch ergibt sich eine Vielzahl an
möglichen Disacchariden. In der Urinprobe konnten Saccharose und Laktose identifiziert
werden.
Saccharose ist ein Disaccharid, welches sich aus Glucose und Fructose zusammensetzt. Die
Konzentration an Saccharose im Urin hängt von der Ernährung ab (Tasevska et al, 2005;
Luceri et al, 1996).
Lactose besteht aus Glucose und Galactose. Lactase-Mangel führt zu einer vermehrten
Ausscheidung von Lactose mit dem Urin.
Die Resonanzfrequenzen der Saccharose unterscheiden sich von denen der jeweiligen
Monosaccharide, da die Isomerie durch die Verknüpfung aufgehoben wird. Sie lässt sich am
einfachsten im HSQC durch das α-Wasserstoffatom der Glucose bei 5,40ppm und 93,42ppm
identifizieren. Bei der Lactose ist dagegen die Stereochemie erhalten geblieben. Die
gebundene Glucose besitzt ihre Hydroxylgruppe in der α-Position noch und begründet eine
Auftrennung in die α- und β-Form der Lactose. Dabei sind die Resonanzfrequenzen der
Galactose bei beiden Isomeren identisch. Die Signale der α-Wasserstoffe der Glucose liegen
sehr dicht bei denen der freien Glucose. Zur Identifizierung sind sie deshalb nicht so gut
geeignet. Die Resonanzfrequenz des α-Wasserstoffes der Galactose, deren Isomerie
aufgehoben ist, liegt bei 4,45ppm und 104,03ppm. Dieses Signal ist für analytische Zwecke
besser geeignet, ermöglicht aber keine Unterscheidung der beiden Anomere.

Identifizierung unbekannter Metabolite
58
ppm (t2) 4.505.00
90.0
95.0
100.0
105.0
ppm (t1)
Saccharose
Lactose
ppm (t2) 4.505.00
90.0
95.0
100.0
105.0
ppm (t1)
ppm (t2) 4.505.00
90.0
95.0
100.0
105.0
ppm (t1)
Saccharose
Lactose
Abbildung 31: HSQC einer Urinprobe. Der Ausschnitt zeigt die Signale der Disaccharide Saccharose und Lactose, die sich nicht in dem überlagerten Bereich der Kohlenhydrate befinden.
3.3.3.3. Zuckeralkohole
Zuckeralkohole oder mehrwertige Alkohole werden durch Reduktion der Monosaccharid-
Aldosen bzw. -Pentosen gebildet und mittels der Anzahl der Kohlenstoffatome klassifiziert.
Über die Funktion und den Stoffwechsel der meisten Zuckeralkohole ist nur wenig bekannt,
ihre Ausscheidung ist alters- und geschlechtsabhängig (Lee und Chung, 2005). In den
untersuchten Urinproben konnten die Zuckeralkohole Arabinitol, Erythrol, Erythronsäure-1,4-
Lakton, Galactitol, Mannitol, Myo-Inositol, Threitol, Sorbitol, Ribitol, Fucitol und Xylitol
identifiziert werden.
Arabinitol bzw. Arabitol ist das Oxidationsprodukt von Xylulose und Bestandteil des Pentose-
Stoffwechsels. Im Fall einer systemischen Candidiasis (Pilzerkrankung) ist die Konzentration
an D-Arabinitol im Urin erhöht. Für eine eindeutige Diagnose muss das Verhältnis von D-
Arabinitol zu L-Arabinitol bestimmt werden (Sigmundsdóttir et al, 2000; Lehtonen et al,
1996). Störungen im Pentose-Stoffwechsel können ebenfalls zu einer vermehrten
Ausscheidung in den Urin führen (Verhoeven et al, 2001).
Erythritol ist als Süßstoff geeignet, da es nicht über den Darm ausgeschieden wird, sondern
über Magen und Zwölffingerdarm an die Niere, und damit an den Urin abgegeben wird
(Munro et al, 1998). Es gibt Beweise, dass Erythritol in humanen Geweben und
Körperflüssigkeiten existiert (Brusati et al, 2005), die Funktion ist aber bis heute unbekannt.
Störungen im Pentose-Stoffwechsel können zu erhöhten Konzentrationen an Erythritol im
Urin führen (Verhoeven et al, 2001).

Identifizierung unbekannter Metabolite
59
Erythronsäure-1,4-Lakton wurde als normaler Bestandteil im Urin gesunder Kinder und
Erwachsener identifiziert (Thompson et al, 1975). Über die Funktion und den Stoffwechsel ist
bis heute nichts bekannt.
Galactitol bzw. Dulcitol ist ein Produkt von Galactose, welches nicht weiter metabolisiert
wird. Es ist im Fall einer Galactosämie, aufgrund eines Mangels an Galactokinase oder
Galactose-1-Phosphat-Uridyl-Transferase, in erhöhten Konzentrationen zu finden (Schwarz et
al, 1985; Ridel et al, 2005).
Mannitol ist normaler Bestandteil von Urin. Es spielt eine Rolle bei der Osmoregulation des
menschlichen Organismus. In Verbindung mit Lactulose wird Mannitol häufig für
Permeabilitätsuntersuchungen der Darmwand eingesetzt (Wring et al, 1998; Bao et al, 1996).
Außerdem findet Mannitol als Abführmittel und als Osmotherapeutikum zur
Gewebeentwässerung Verwendung.
Myo-Inositol ist ein Vorläufer des Second-Messengers Phosphatidylinositol. Es wird in vielen
menschlichen Geweben synthetisiert und zusätzlich mit der Nahrung aufgenommen. Mitunter
wird es als Wuchsstoff zum Vitamin-B2-Komplex gerechnet. Im Urin kann Myo-Inositol
alleine (Sarashina et al, 2004) oder im Verhältnis zu Chiro-Inositol als Marker für eine
Glucoseintoleranz genutzt werden (Jung et al, 2005). Threitol ist als normaler Bestandteil von
Urin bekannt (Wamelink et al, 2005). Es ist das Hauptprodukt des Xylose-Stoffwechsels, der
in der Leber stattfindet. Bei Patienten mit Leberzirrhose ist die Konzentration an Threitol im
Urin signifikant verringert (Pitkanen, 1977).
Sorbitol bzw. Glucitol wird aus Glucose gebildet und kann zu Fructose weiter metabolisiert
werden. Frauen zeigen höhere Sorbitol Konzentrationen im Urin als Männer (Lee und Chung
2005). Bei Diabetes Mellitus kann es durch den Abbau von Glucose zu einer vermehrten
Ausscheidung in den Urin kommen (Nakano et al, 2003).
Ribitol ist das Oxidationsprodukt der Ribose und wird, genau wie Arabinitol und Erythritol,
bei Störungen des Phosphopentose-Stoffwechsels vermehrt mit dem Urin ausgeschieden
(Verhoeven et al, 2001).
Fucitol ist normaler Bestandteil von Urin (Haga und Nakajima, 1989), über den Stoffwechsel
und die Funktion von Fucitol ist bis heute nichts bekannt. Xylitol wird als Zwischenprodukt
bei der Umwandlung von L-Xylose zu D-Xylose gebildet. Mit der Nahrung aufgenommenes
Xylitol wird zum Teil über den Urin ausgeschieden. Der Rest wird in der Leber über Xylulose
als Zwischenprodukt zu Glucose umgewandelt. Leberzirrhose führt zu stark erhöhten,
Hepatitis zu leicht erhöhten Xylitol Konzentrationen im Urin (Oka et al, 1976)

Identifizierung unbekannter Metabolite
60
Sowohl im Protonenspektrum als auch im HSQC liegen die Signale der mehrwertigen
Alkohole alle sehr dicht beieinander in dem überlagerten Bereich. Eine quantitative Analyse
ist hier nicht möglich.
ppm (t2) 3.504.00
60.0
65.0
70.0
75.0
ppm (t1)
ppm (t2) 3.504.00
60.0
65.0
70.0
75.0
ppm (t1)
Abbildung 32: HSQC einer Urinprobe. Die Markierungen heben die Bereiche hervor, in denen die Signale der mehrwertigen Alkohole liegen. Einzelne Signale können nicht dargestellt werden. In der oberen Markierung liegen die Signale der endständigen Methylengruppen, in der unteren die Signale der Hydroxymethylengruppen.
3.3.4. Biogene Amine
Amine sind Derivate des Ammoniaks, bei dem die Wasserstoffe durch Alkyl- oder
Arylgruppen ersetzt wurden. Es gibt primäre, sekundäre und tertiäre Amine. Ein möglicher,
vierter Substituent verursacht am Stickstoff eine positive Ladung.
3.3.4.1. Quartäre Ammoniumverbindungen
Quartäre Ammoniumverbindungen sind meistens wichtige Träger von Methylgruppen, die für
Methylierungen verwendet werden. In den Urinproben konnten Betain, Carnitin und Cholin
identifiziert werden.
Betain ist ein Abbauprodukt von Cholin und ein wichtiger Methylgruppen-Donor für
Transmethylierungen. Bei Diabetikern wurde eine vermehrte Ausscheidung in den Urin
beobachtet (Lever et al, 1994). Die Glukose-Konzentration im Urin wird jedoch bevorzugt zur
Diagnose und Therapiekontrolle eingesetzt.
Carnitin wird aus Lysin gebildet. Es transportiert Fettsäuren in die Mitochondrien von
Muskelzellen, wo sie zur Energiegewinnung oxidiert werden. Mit der Nahrung
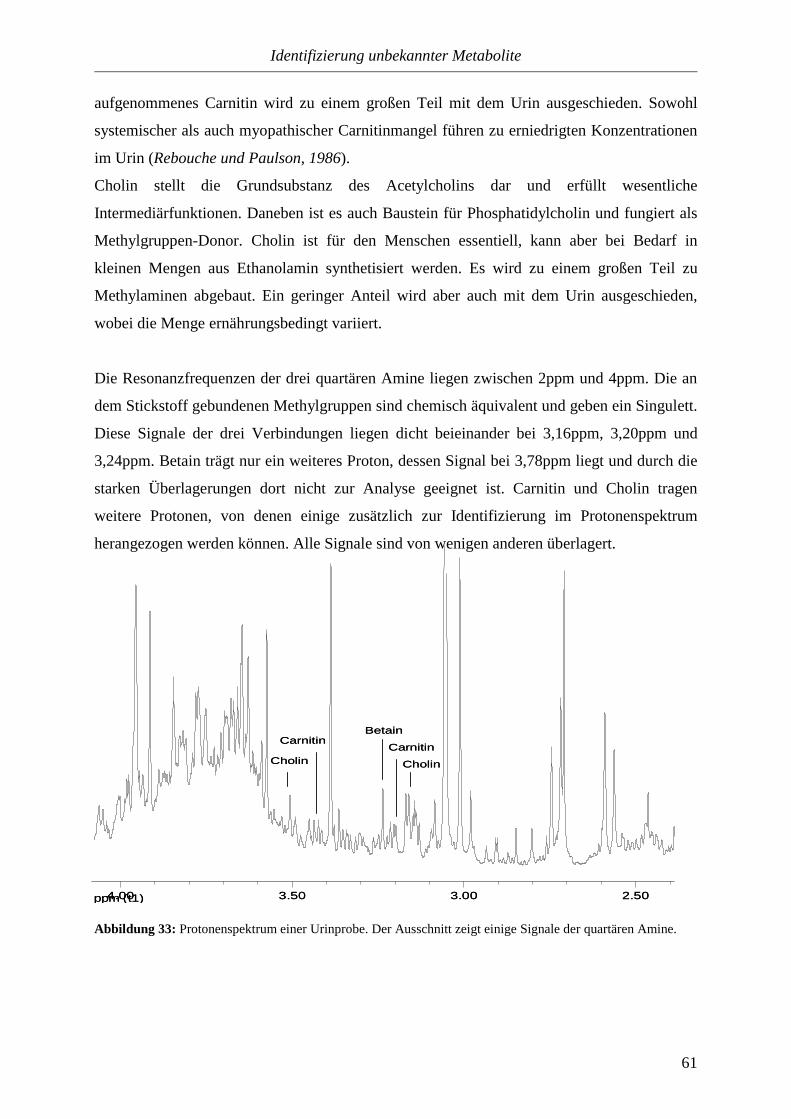
Identifizierung unbekannter Metabolite
61
aufgenommenes Carnitin wird zu einem großen Teil mit dem Urin ausgeschieden. Sowohl
systemischer als auch myopathischer Carnitinmangel führen zu erniedrigten Konzentrationen
im Urin (Rebouche und Paulson, 1986).
Cholin stellt die Grundsubstanz des Acetylcholins dar und erfüllt wesentliche
Intermediärfunktionen. Daneben ist es auch Baustein für Phosphatidylcholin und fungiert als
Methylgruppen-Donor. Cholin ist für den Menschen essentiell, kann aber bei Bedarf in
kleinen Mengen aus Ethanolamin synthetisiert werden. Es wird zu einem großen Teil zu
Methylaminen abgebaut. Ein geringer Anteil wird aber auch mit dem Urin ausgeschieden,
wobei die Menge ernährungsbedingt variiert.
Die Resonanzfrequenzen der drei quartären Amine liegen zwischen 2ppm und 4ppm. Die an
dem Stickstoff gebundenen Methylgruppen sind chemisch äquivalent und geben ein Singulett.
Diese Signale der drei Verbindungen liegen dicht beieinander bei 3,16ppm, 3,20ppm und
3,24ppm. Betain trägt nur ein weiteres Proton, dessen Signal bei 3,78ppm liegt und durch die
starken Überlagerungen dort nicht zur Analyse geeignet ist. Carnitin und Cholin tragen
weitere Protonen, von denen einige zusätzlich zur Identifizierung im Protonenspektrum
herangezogen werden können. Alle Signale sind von wenigen anderen überlagert.
ppm (t1) 2.503.003.504.00
Betain
Carnitin
Cholin
Carnitin
Cholin
ppm (t1) 2.503.003.504.00
Betain
Carnitin
Cholin
Carnitin
Cholin
Abbildung 33: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt einige Signale der quartären Amine.

Identifizierung unbekannter Metabolite
62
3.3.4.2. Methylamine
Unter Methylaminen werden Verbindungen zusammengefasst, bei denen, ausgehend vom
Ammoniak, Wasserstoffe durch Methylgruppen substituiert sind. In den Urinproben konnten
Methylamin, Dimethylamin, Trimethylamin und Trimethylaminoxid identifiziert werden.
Methylamin ist ein Abbauprodukt von Creatinin. Deshalb führen creatininhaltige
Lebensmittel zu einer vermehrten Ausscheidung von Methylamin in den Urin.
Stoffwechselstörungen und andere Krankheiten, die zu veränderten Konzentrationen im Urin
führen sind bis heute nicht bekannt.
Dimethylamin, eine Vorstufe des Carcinogens N-Nitrosodimethylamin, wird auf
verschiedenen Wegen synthetisiert. Zum einen über die N-methylierung von Methylamin
welches ein Abbauprodukt von Sarcosine und Glycin ist. Zum anderen ist Dimethylamin ein
Abbauprodukt von N, N-Dimethylarginin, ein endogener Inhibitor der
Stickstoffmonoxidsynthese aus Arginin (Tsikas et al, 2007). Außerdem wird Dimethylamin
auch mit der Nahrung aufgenommen. Als Biomarker im Urin ist Dimethylamin nicht
geeignet, da unterschiedliche Krankheiten, z. B. Erkrankungen der Herzkranzgefäße oder der
Leber, zu einer vermehrten Ausscheidung führen (Tsikas et al, 2007).
Trimethylamin ist ein Abbauprodukt von Cholin, Carnitin und Trimethylaminoxid.
Umgekehrt wird mit der Nahrung aufgenommenes Trimethylamin zu Trimethylaminoxid
abgebaut und hauptsächlich in dieser Form mit dem Urin ausgeschieden. Trimethylaminurie,
ein Enzymdefekt, ruft bei den Betroffenen einen charakteristischen Geruch nach Fisch hervor.
Dieser Geruch wird durch vermehrt mit dem Urin ausgeschiedenes Trimethylamin verursacht
(Bain et al, 2005; Mitchell und Smith, 2001). Trimethylaminoxid wird dementsprechend im
Fall einer Trimethylaminurie weniger ausgeschieden. Erhöhte Konzentrationen im Urin
können nach dem Verzehr von Fisch beobachtet werden (Zeisel und DaCosta, 1986).
Da die am Stickstoff gebundenen Methylruppen chemisch äquivalent sind, geben sie jeweils
nur ein Singulett. Trimethylaminoxid und Dimethylamin bei 3,39ppm bzw. 2,71ppm sind
aufgrund ihrer Konzentrationen leicht zu identifizieren. Trimethylamin und Methylamin bei
2,87ppm bzw. 2,61ppm sind dagegen sehr gering konzentriert.

Identifizierung unbekannter Metabolite
63
ppm (t1) 2.503.003.50
MA
DMA
TMA
TMAO
ppm (t1) 2.503.003.50
MA
DMA
TMA
TMAO
Abbildung 34: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale der Methylamine. Abkürzungen: TMAO – Trimethylaminoxid, TMA – Trimethylamin, DMA – Dimethylamin, MA – Methylamin.
3.3.4.3. Guanidin-Verbindungen
In den Urinproben konnten Creatin, Phosphocreatin, Creatinin und Guanidinoacetat
identifiziert werden.
Creatin, ein Zwischenprodukt des intermediären Stoffwechsels, wird in der Leber in
annähernd konstanter Menge aus Glycin, Arginin und Methionin gebildet. Creatin wird zum
großen Teil im Muskelgewebe gelagert, wo es in energiereiches Creatinphosphat
umgewandelt wird. Der Abbau erfolgt zu Creatinin. Auf einen Enzymdefekt basierende
Creatin-Mangel-Syndrome sind mit niedrigen Konzentrationen im Urin verbunden.
Mangelsyndrome, die auf eine Störung im tubulären Transport zurückzuführen sind,
verursachen dagegen erhöhte Creatin-Konzentrationen im Urin (Sykut-cegielska et al, 2004).
Phosphocreatin ist der aus Creatin gebildete Energiespeicher im Muskel.
Stoffwechselstörungen oder andere Krankheiten, die zu veränderten Konzentrationen im Urin
führen, sind bis heute nicht bekannt.
Guanidinoacetat ist ein Metabolit im Harnstoffzyklus und im Stoffwechsel der Aminosäuren
Glycin, Serin, Threonin, Arginin und Prolin. Es ist auch ein Vorläufer von Creatin und damit
essentiell für den Energie-Stoffwechsel der Muskeln. Guanidinoacetat-Methyltransferase-
Mangel bedingt eine erhöhte Konzentration im Urin (Sykut-cegielska et al, 2004),
Nierenversagen erniedrigte Konzentrationen (Shirokane et al, 1987).
Creatinin ist das Abauprodukt des Creatinphosphats und wird im Muskelgewebe gebildet. Die
Synthese und Ausscheidung des harnpflichtigen Anhydrids erfolgt relativ konstant und ist
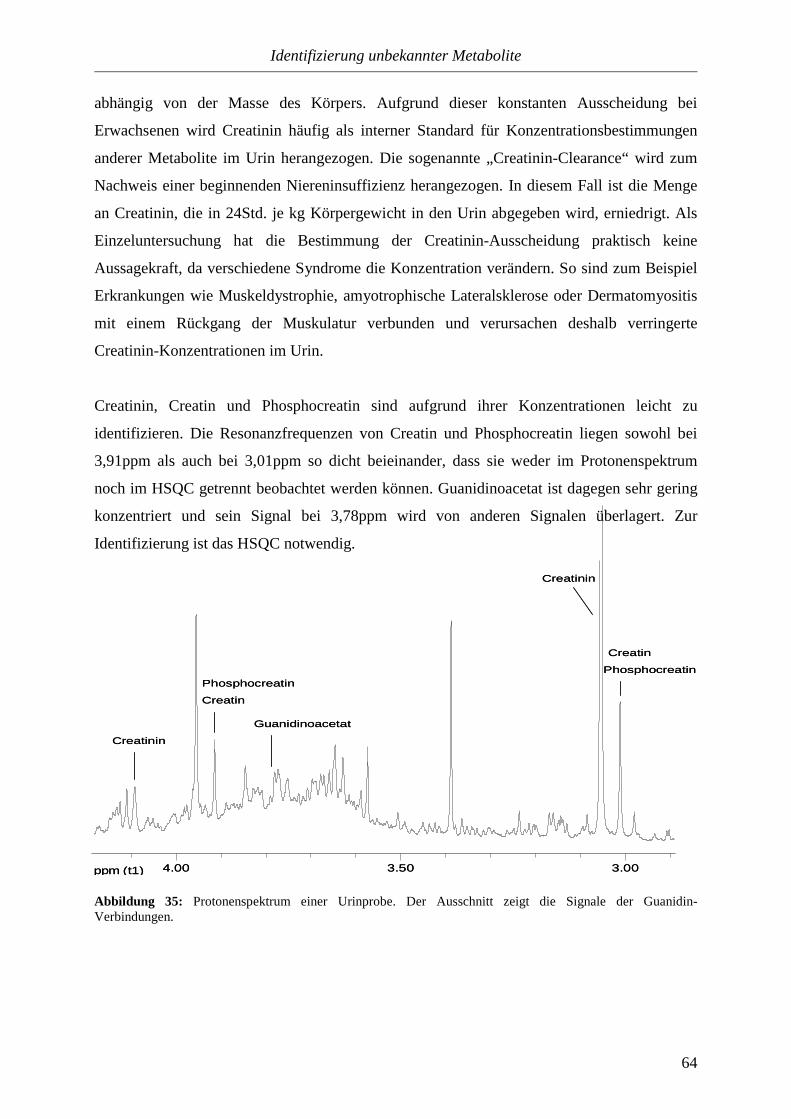
Identifizierung unbekannter Metabolite
64
abhängig von der Masse des Körpers. Aufgrund dieser konstanten Ausscheidung bei
Erwachsenen wird Creatinin häufig als interner Standard für Konzentrationsbestimmungen
anderer Metabolite im Urin herangezogen. Die sogenannte „Creatinin-Clearance“ wird zum
Nachweis einer beginnenden Niereninsuffizienz herangezogen. In diesem Fall ist die Menge
an Creatinin, die in 24Std. je kg Körpergewicht in den Urin abgegeben wird, erniedrigt. Als
Einzeluntersuchung hat die Bestimmung der Creatinin-Ausscheidung praktisch keine
Aussagekraft, da verschiedene Syndrome die Konzentration verändern. So sind zum Beispiel
Erkrankungen wie Muskeldystrophie, amyotrophische Lateralsklerose oder Dermatomyositis
mit einem Rückgang der Muskulatur verbunden und verursachen deshalb verringerte
Creatinin-Konzentrationen im Urin.
Creatinin, Creatin und Phosphocreatin sind aufgrund ihrer Konzentrationen leicht zu
identifizieren. Die Resonanzfrequenzen von Creatin und Phosphocreatin liegen sowohl bei
3,91ppm als auch bei 3,01ppm so dicht beieinander, dass sie weder im Protonenspektrum
noch im HSQC getrennt beobachtet werden können. Guanidinoacetat ist dagegen sehr gering
konzentriert und sein Signal bei 3,78ppm wird von anderen Signalen überlagert. Zur
Identifizierung ist das HSQC notwendig.
ppm (t1) 3.003.504.00
Creatin
Creatin
PhosphocreatinPhosphocreatin
Creatinin
Creatinin
Guanidinoacetat
ppm (t1) 3.003.504.00
Creatin
Creatin
PhosphocreatinPhosphocreatin
Creatinin
Creatinin
Guanidinoacetat
Abbildung 35: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale der Guanidin-Verbindungen.

Identifizierung unbekannter Metabolite
65
3.3.4.4. Primäre Amine
Bei primären Aminen ist nur ein Wasserstoff des Ammoniaks durch eine Alkylgruppe
substituiert. In den Urinproben konnten Ethanolamin, Phosphoethanolamin und Taurin
identifiziert werden.
Ethanolamin ist die unmethylierte Vorstufe des Cholins und ein Decarboxylierungsprodukt
von Serin. Es ist ein Baustein in Plasmalogenen und in Glycerophosphatiden.
Stoffwechselstörungen oder andere Krankheiten, die veränderte Konzentrationen im Urin
verursachen, sind bisher nicht bekannt.
Phosphoethanolamin, der Phosphorsäure-Ester des Ethanolamins, ist Bestandteil der
Phosphatidylamine und Abbauprodukt des Sphingosin-1-Phosphates. Hyperphosphatasie, ein
Enzymdefekt, bedingt eine vermehrte Ausscheidung an Phosphoethanolamin mit dem Urin
(Igbal et al, 2000). Taurin ist ein Abbauprodukt von Cystin und Methionin. Es fungiert als
Neurotransmitter im Gehirn, stabilisiert Zellmembranen und wird für den Ionentransport von
Natrium, Kalium, Magnesium oder Calcium genutzt. Sulfit-Oxydase-Mangel oder Hyper-β-
Alaninämie verursachen vermehrte Taurin Ausscheidung mit dem Urin.
Die primären Amine sind nur gering konzentriert und geben Signale mit schwacher Intensität.
Ihre Resonanzfrequenzen liegen zum Teil in dem stark überlagerten Bereich der
Kohlenhydrate. Aber jede Verbindung zeigt zumindest ein Signal, welches für analytische
Zwecke verwendet werden kann. Die Signale von Taurin bei 3,29ppm und 3,43ppm, und das
Signal von Ethanolamin bei 3,14ppm, sowie von Phosphoethanolamin bei 3,25ppm sind nur
wenig überlagert. Aufgrund der schwachen Signalintensitäten ist es sinnvoll, das HSQC zur
Identifizierung heranzuziehen.
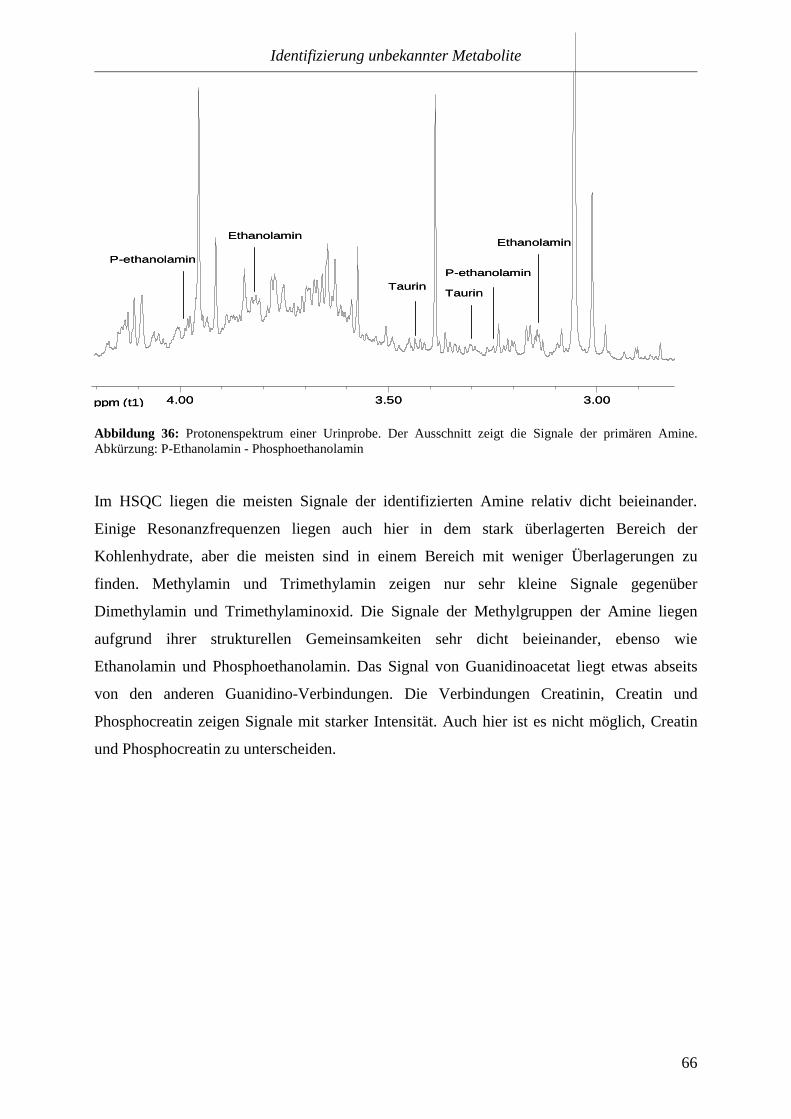
Identifizierung unbekannter Metabolite
66
ppm (t1) 3.003.504.00
Taurin
Ethanolamin
P-ethanolamin
Ethanolamin
Taurin
P-ethanolamin
ppm (t1) 3.003.504.00
Taurin
Ethanolamin
P-ethanolamin
Ethanolamin
Taurin
P-ethanolamin
Abbildung 36: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale der primären Amine. Abkürzung: P-Ethanolamin - Phosphoethanolamin
Im HSQC liegen die meisten Signale der identifizierten Amine relativ dicht beieinander.
Einige Resonanzfrequenzen liegen auch hier in dem stark überlagerten Bereich der
Kohlenhydrate, aber die meisten sind in einem Bereich mit weniger Überlagerungen zu
finden. Methylamin und Trimethylamin zeigen nur sehr kleine Signale gegenüber
Dimethylamin und Trimethylaminoxid. Die Signale der Methylgruppen der Amine liegen
aufgrund ihrer strukturellen Gemeinsamkeiten sehr dicht beieinander, ebenso wie
Ethanolamin und Phosphoethanolamin. Das Signal von Guanidinoacetat liegt etwas abseits
von den anderen Guanidino-Verbindungen. Die Verbindungen Creatinin, Creatin und
Phosphocreatin zeigen Signale mit starker Intensität. Auch hier ist es nicht möglich, Creatin
und Phosphocreatin zu unterscheiden.

Identifizierung unbekannter Metabolite
67
ppm (t2) 1.502.002.503.003.50
30
40
50
60
ppm (t1)
QA
MA
DMA
TMA
TMAO
Cn
Cr / PCrGA
Ta
EA
PEA
ppm (t2) 1.502.002.503.003.50
30
40
50
60
ppm (t1)
ppm (t2) 1.502.002.503.003.50
30
40
50
60
ppm (t1)
QA
MA
DMA
TMA
TMAO
Cn
Cr / PCrGA
Ta
EA
PEA
Abbildung 37: HSQC einer Urinprobe. Der Ausschnitt zeigt die Signale der identifizierten Amine. In diesem Bereich ist zumindest ein Signal von jeder Verbindung zu finden. Die Markierung hebt die Signale der Methylgruppen der vorhandenen quartären Amine hervor. Abkürzungen: GA – Guanidinoacetat, Cn – Creatinin, Cr – Creatin, PCr – Phosphocreatin, MA – Methylamin, DMA – Dimethylamin, TMA – Trimethylamin, TMAO – Trimethylaminoxid, PEA – Phosphoethanolamin, EA – Ethanolamin, QA – quartäre Amine.
3.3.5. Carbonsäuren
Carbonsäuren sind organische Verbindungen, die aus einer Kohlenwasserstoffkette und einer
funktionellen Gruppe, der Carboxyl-Gruppe, bestehen. Sie sind Intermediate oder
Endprodukte im Stoffwechsel der Aminosäuren, Kohlenhydrate, Fettsäuren und der biogenen
Amine.
3.3.5.1. Dicarbonsäuren
In den Urinproben konnten die Dicarbonsäuren Adipinsäure, Ameisensäure, Fumarsäure,
Glutarsäure, Suberinsäure, Maleinsäure, Malonsäure, Methylmalonsäure und
Carbamoylaspartat identifiziert werden.
Adipinsäure ist ein normaler Bestandteil von Urin. Sie entsteht bei der Fettsäureoxidation und
wird mit der Nahrung aufgenommen. Eine erhöhte Syntheserate von Glutaryl-CoA inhibiert
die Dehydrierung von Adipyl-CoA und führt zu einer vermehrten Ausscheidung von
Adipinsäure im Urin. Dieser Fall kann bei einer Glutarsäureurie Typ II auftreten.
Ameisensäure ist ein Metabolit im Intermediärstoffwechsel.
Fumarsäure entsteht im Citratzyklus durch Oxidation der Bernsteinsäure, im Harnstoffzyklus
durch Spaltung der Argininbernsteinsäure und beim Abbau von Phenylalanin, Tyrosin und

Identifizierung unbekannter Metabolite
68
Asparaginsäure. Fumarase-Mangel führt zu einer vermehrten Ausscheidung in den Urin
(Deschauer et al, 2006).
Glutarsäure ist ein Abbauprodukt des Lysins und ist als Glutaryl-CoA im Fettsäure-
Stoffwechsel zu finden. Verschiedene Enzymdefekte führen zu erhöhten Konzentrationen im
Urin, darunter Glutaryl-CoA-Dehydrogenase-Mangel (Zschocke et al, 2000).
Suberinsäure ist ein Metabolit aus dem Fettsäure-Stoffwechsel. Störungen in diesem
Stoffwechsel führen zu erhöhten Konzentrationen im Urin.
Methylmalonsäure ist ein Abbauprodukt der Aminosäuren Isoleucin und Valin und als
MethylmalonylCoA im Fettsäure-Stoffwechsel zu finden. Methylmalonyl-CoA-Isomerase-
Mangel (Drennan et al, 1996) oder Vitamin B12-Mangel (Bjorke Monsen und Ueland, 2003)
führen zu einer vermehrten Ausscheidung in den Urin.
Ureidobernsteinsäure ist ein Intermediärprodukt der Pyrimidin-Biosynthese. Ein Mangel an
Ornithin-Transcarbamylase oder Argininosuccinase führt zu einer erhöhten Konzentration an
Ureidobernsteinsäure im Urin (van Kuilenburg et al, 2004).
Glutarsäure, Adipinsäure und Suberinsäure sind Carbonsäuren mit CH2-Ketten. Die Signale
dieser CH2-Ketten liegen im Hochfeldbereich. Aufgrund der ähnlichen Strukturen liegen die
α-Wasserstoffe der drei Säuren dicht beieinander bei 2,21ppm, 2,22ppm und 2,20ppm. Da die
Wasserlöslichkeit durch die Lipophilie der CH2-Ketten eingeschränkt ist, sind die Säuren nur
gering konzentriert und geben Signale mit sehr schwacher Intensität.
ppm (t1) 1.502.00
Adipinsäure
Adipinsäure
Glutarsäure
GlutarsäureSuberinsäure
SuberinsäureSuberinsäure
ppm (t1) 1.502.00
Adipinsäure
Adipinsäure
Glutarsäure
GlutarsäureSuberinsäure
SuberinsäureSuberinsäure
Abbildung 38: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale der Dicarbonsäuren Adipinsäure, Glutarsäure und Suberinsäure.
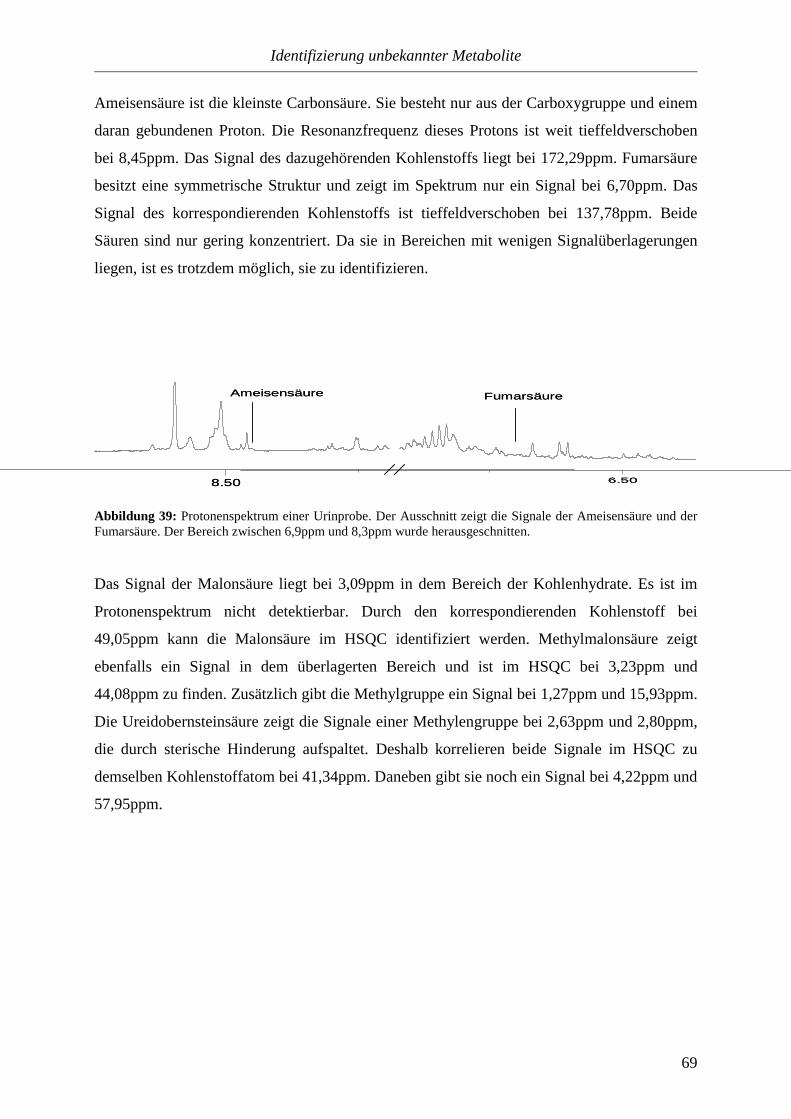
Identifizierung unbekannter Metabolite
69
Ameisensäure ist die kleinste Carbonsäure. Sie besteht nur aus der Carboxygruppe und einem
daran gebundenen Proton. Die Resonanzfrequenz dieses Protons ist weit tieffeldverschoben
bei 8,45ppm. Das Signal des dazugehörenden Kohlenstoffs liegt bei 172,29ppm. Fumarsäure
besitzt eine symmetrische Struktur und zeigt im Spektrum nur ein Signal bei 6,70ppm. Das
Signal des korrespondierenden Kohlenstoffs ist tieffeldverschoben bei 137,78ppm. Beide
Säuren sind nur gering konzentriert. Da sie in Bereichen mit wenigen Signalüberlagerungen
liegen, ist es trotzdem möglich, sie zu identifizieren.
8.50 6.50
Ameisensäure Fumarsäure
8.50 6.50
Ameisensäure Fumarsäure
Abbildung 39: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale der Ameisensäure und der Fumarsäure. Der Bereich zwischen 6,9ppm und 8,3ppm wurde herausgeschnitten.
Das Signal der Malonsäure liegt bei 3,09ppm in dem Bereich der Kohlenhydrate. Es ist im
Protonenspektrum nicht detektierbar. Durch den korrespondierenden Kohlenstoff bei
49,05ppm kann die Malonsäure im HSQC identifiziert werden. Methylmalonsäure zeigt
ebenfalls ein Signal in dem überlagerten Bereich und ist im HSQC bei 3,23ppm und
44,08ppm zu finden. Zusätzlich gibt die Methylgruppe ein Signal bei 1,27ppm und 15,93ppm.
Die Ureidobernsteinsäure zeigt die Signale einer Methylengruppe bei 2,63ppm und 2,80ppm,
die durch sterische Hinderung aufspaltet. Deshalb korrelieren beide Signale im HSQC zu
demselben Kohlenstoffatom bei 41,34ppm. Daneben gibt sie noch ein Signal bei 4,22ppm und
57,95ppm.

Identifizierung unbekannter Metabolite
70
ppm (t2) 2.503.003.50
35.0
40.0
45.0
50.0
ppm (t1)
MA
MMA
UBS
ppm (t2) 2.503.003.50
35.0
40.0
45.0
50.0
ppm (t1)
ppm (t2) 2.503.003.50
35.0
40.0
45.0
50.0
ppm (t1)
MA
MMA
UBS
Abbildung 40: HSQC einer Urinprobe. Der Ausschnitt zeigt die Signale der Malonsäure (MA), Methylmalonsäure (MMA) und der Ureidobernsteinsäure (UBS).
3.3.5.2. Tricarbonsäuren
In den Urinproben konnten die Tricarbonsäuren Citronensäure, cis-Aconitsäure und trans-
Aconitsäure identifiziert werden.
Citronensäure ist eine Tricarbonsäure aus dem Citratzyklus. Nierensteine und nephrogene
Azidose führen zu einer verringerten Ausscheidung in den Urin. Aconitsäure kommt in zwei
isomeren Formen vor. Die cis-Aconitsäure ist ein wichtiges Glied des Citratzyklus. Trans-
Aconitsäure gelangt wahrscheinlich über die Nahrung und die Darmflora in den Urin.
Die Citronensäure gehört zu den Verbindungen, die in größeren Mengen im Urin vorkommen.
Die Signale der Citronensäure bei 2,58ppm und 2,73ppm werden durch zwei
Methylengruppen verursacht, deren Protonen magnetisch nicht äquivalent sind. Diese Signale
sind aufgrund ihrer Intensität im Protonenspektrum gut zu erkennen.
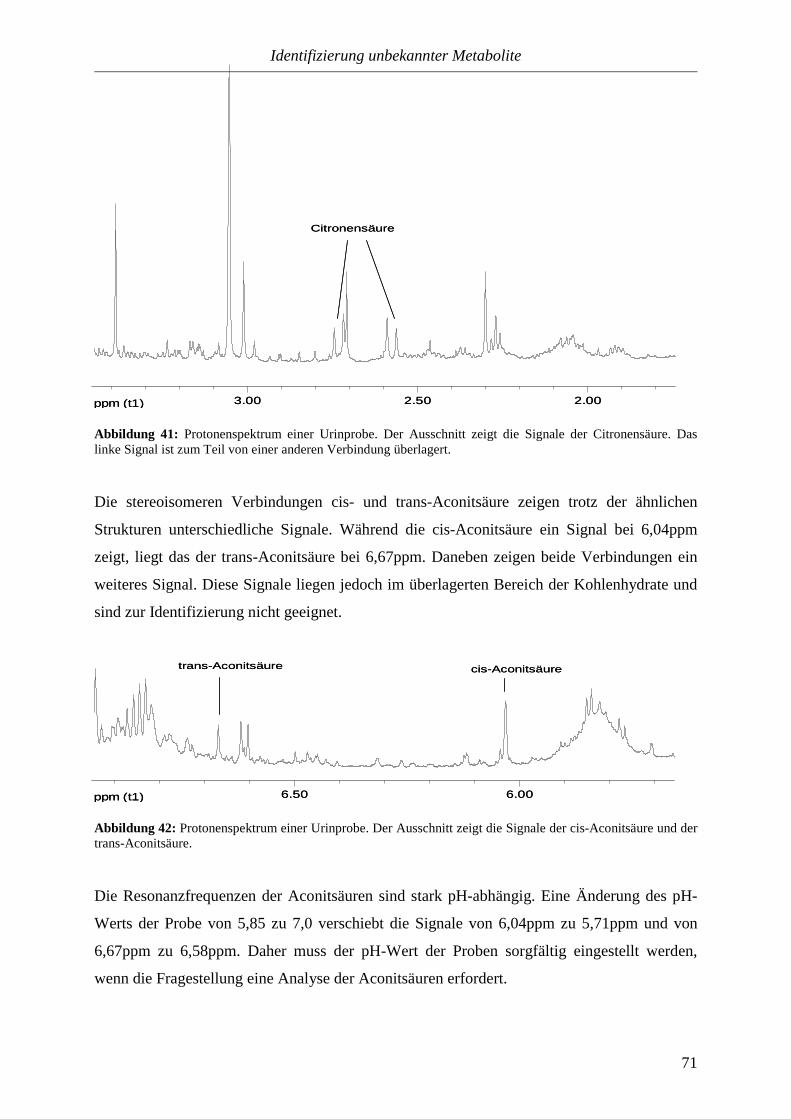
Identifizierung unbekannter Metabolite
71
ppm (t1) 2.002.503.00
Citronensäure
ppm (t1) 2.002.503.00
Citronensäure
Abbildung 41: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale der Citronensäure. Das linke Signal ist zum Teil von einer anderen Verbindung überlagert.
Die stereoisomeren Verbindungen cis- und trans-Aconitsäure zeigen trotz der ähnlichen
Strukturen unterschiedliche Signale. Während die cis-Aconitsäure ein Signal bei 6,04ppm
zeigt, liegt das der trans-Aconitsäure bei 6,67ppm. Daneben zeigen beide Verbindungen ein
weiteres Signal. Diese Signale liegen jedoch im überlagerten Bereich der Kohlenhydrate und
sind zur Identifizierung nicht geeignet.
ppm (t1) 6.006.50
trans-Aconitsäure cis-Aconitsäure
ppm (t1) 6.006.50
trans-Aconitsäure cis-Aconitsäure
Abbildung 42: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale der cis-Aconitsäure und der trans-Aconitsäure.
Die Resonanzfrequenzen der Aconitsäuren sind stark pH-abhängig. Eine Änderung des pH-
Werts der Probe von 5,85 zu 7,0 verschiebt die Signale von 6,04ppm zu 5,71ppm und von
6,67ppm zu 6,58ppm. Daher muss der pH-Wert der Proben sorgfältig eingestellt werden,
wenn die Fragestellung eine Analyse der Aconitsäuren erfordert.
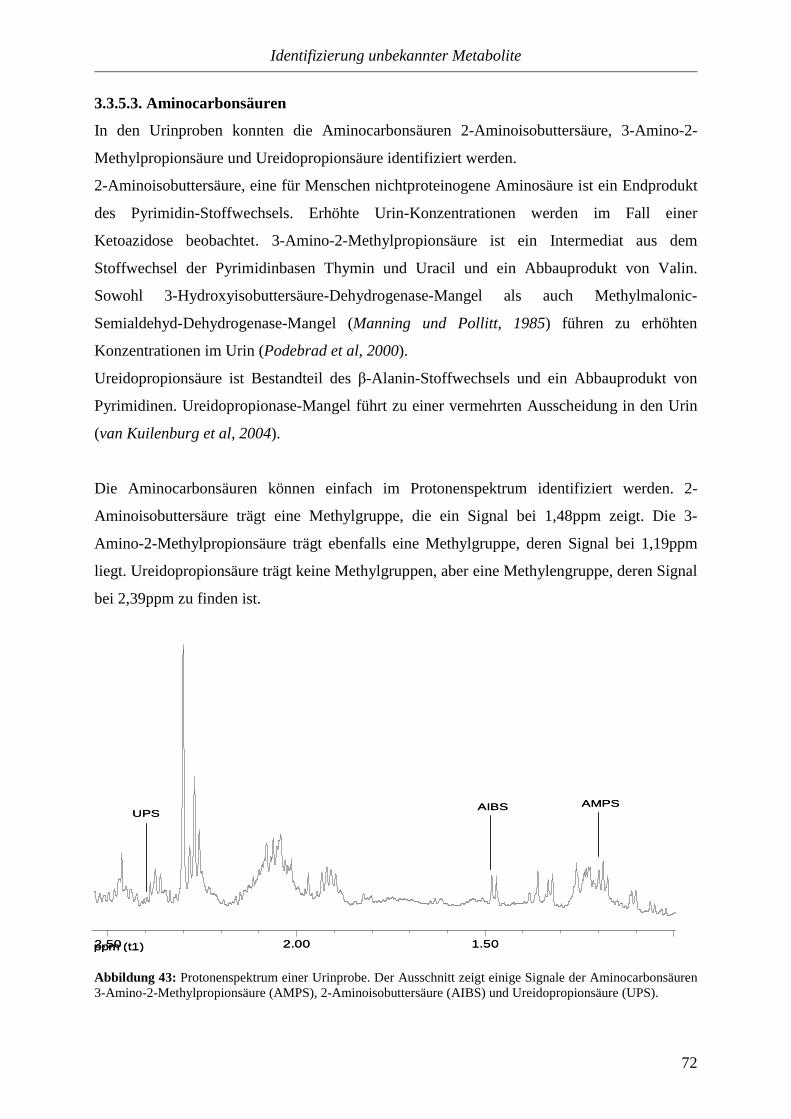
Identifizierung unbekannter Metabolite
72
3.3.5.3. Aminocarbonsäuren
In den Urinproben konnten die Aminocarbonsäuren 2-Aminoisobuttersäure, 3-Amino-2-
Methylpropionsäure und Ureidopropionsäure identifiziert werden.
2-Aminoisobuttersäure, eine für Menschen nichtproteinogene Aminosäure ist ein Endprodukt
des Pyrimidin-Stoffwechsels. Erhöhte Urin-Konzentrationen werden im Fall einer
Ketoazidose beobachtet. 3-Amino-2-Methylpropionsäure ist ein Intermediat aus dem
Stoffwechsel der Pyrimidinbasen Thymin und Uracil und ein Abbauprodukt von Valin.
Sowohl 3-Hydroxyisobuttersäure-Dehydrogenase-Mangel als auch Methylmalonic-
Semialdehyd-Dehydrogenase-Mangel (Manning und Pollitt, 1985) führen zu erhöhten
Konzentrationen im Urin (Podebrad et al, 2000).
Ureidopropionsäure ist Bestandteil des β-Alanin-Stoffwechsels und ein Abbauprodukt von
Pyrimidinen. Ureidopropionase-Mangel führt zu einer vermehrten Ausscheidung in den Urin
(van Kuilenburg et al, 2004).
Die Aminocarbonsäuren können einfach im Protonenspektrum identifiziert werden. 2-
Aminoisobuttersäure trägt eine Methylgruppe, die ein Signal bei 1,48ppm zeigt. Die 3-
Amino-2-Methylpropionsäure trägt ebenfalls eine Methylgruppe, deren Signal bei 1,19ppm
liegt. Ureidopropionsäure trägt keine Methylgruppen, aber eine Methylengruppe, deren Signal
bei 2,39ppm zu finden ist.
ppm (t1) 1.502.002.50
UPSAIBS AMPS
ppm (t1) 1.502.002.50
UPSAIBS AMPS
Abbildung 43: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt einige Signale der Aminocarbonsäuren 3-Amino-2-Methylpropionsäure (AMPS), 2-Aminoisobuttersäure (AIBS) und Ureidopropionsäure (UPS).

Identifizierung unbekannter Metabolite
73
3.3.5.4. Hydroxycarbonsäuren
In den Urinproben konnten die Hydroxycarbonsäuren 3-Hydroxy-2-Methylpropionsäure, 3-
Hydroxyisovaleriansäure, Milchsäure, Pantothensäure, Glycolsäure, Glycerinsäure,
Threonsäure, 2-Hydroxyisobuttersäure und Glucuronsäure identifiziert werden.
3-Hydroxy-2-Methylpropionsäure ist, wie 3-Amino-2-Methylpropionsäure, ein Abbauprodukt
von Valin, Thymin und Uracil. Beide Substanzen sind mit seltenen Enzymdefekten
verbunden. Sowohl 3-Hydroxyisobuttersäure-Dehydrogenase-Mangel als auch
Methylmalonic-Semialdehyd-Dehydrogenase-Mangel (Manning und Pollitt, 1985) führen zu
erhöhten Konzentrationen der beiden Verbindungen im Urin (Podebrad et al, 2000).
3-Hydroxyisovaleriansäure ist ein normaler Bestandteil von Urin. Verschiedene
Enzymdefekte, darunter Biotidinase-Mangel, 3-Methylcrotonyl-CoA-Carboxylase-Mangel
(Baumgartner et al, 2001) und 3-Methylglutaconsäure-Azidose, führen zu einer vermehrten
Ausscheidung in den Urin.
Milchsäure ist ein glykolytisches Hydrierungsprodukt der Brenztraubensäure im
Intermediärstoffwechsel. Laktoseintoleranz und Laktase-Mangel führen zu einer vermehrten
Ausscheidung in den Urin.
Pantothensäure bzw. Vitamin B3 ist ein Coenzym-A-Baustein. Die Konzentration im Urin
hängt zum Teil von der Ernährung ab.
Glykolsäure wird zu Glyoxylatsäure oxidiert und weiter zu Glycin umgesetzt. In einigen
Fällen der Hyperoxalurie Typ I wird sie vermehrt in den Urin ausgeschieden (Danpure,
2005; Dietzen et al, 1997).
Glycerinsäure ist das Oxidationsprodukt von Glycerin und spielt als Phosphoglycerat eine
wichtige Rolle im Kohlenhydratstoffwechsel. Zwei verschiedene Enzymdefekte führen zu
erhöhten Konzentrationen im Urin. Im Fall eines Glycerat-Kinase-Mangels wird
ausschließlich D-Glycerinsäure vermehrt ausgeschieden (Fontaine et al, 1989), im Fall eines
Glycerat-Dehydrogenase-Mangels (auch Hyperoxalurie Typ II) L-Glycerinsäure (Dietzen et
al, 1997).
Threonsäure ist ein Abbauprodukt der Ascorbinsäure. Ascorbinsäure kommt ebenfalls im
Urin vor, konnte jedoch mit Hilfe der NMR-Spektroskopie nicht nachgewiesen werden.
Stoffwechselstörungen oder andere Krankheiten, die zu veränderten Konzentrationen der
Threonsäure führen, sind bis heute nicht bekannt. Carbamoylaspartat ist ein
Intermediärprodukt der Pyrimidin-Biosynthese. Ein Mangel an Ornithin-Transcarbamylase
oder Argininosuccinase führt zu einer erhöhten Konzentration an Carbamoylaspartat im Urin
(van Kuilenburg et al, 2004).

Identifizierung unbekannter Metabolite
74
2-Hydroxyisobuttersäure ist ein normaler Bestandteil von Urin. Außerdem ist es ein Metabolit
von Methyl-tertiär-Butyl-Ether (Amberg et al, 2000). Eine vermehrte Ausscheidung in den
Urin ist bei Isovaleriatämie zu beobachten (Shigematsu et al, 1982).
Glucuronsäure ist Bestandteil von Glukosaminoglykanen und bildet, vor allem in der Leber,
mit vielen schädlichen Substanzen wasserlösliche Glucuronide, die dann über Niere und
Leber ausgeschieden werden können.
Die Signale der Hydroxycarbonsäuren Glykolsäure, Glycerinsäure und Threonsäure liegen bei
3,97ppm, 3,86ppm und 4,32ppm bzw. 3,69ppm, 3,62ppm, 3,98ppm und 4,07ppm im Bereich
der Kohlenhydrate und sind nicht einfach zu identifizieren.
3-Hydroxy-2-Methylpropionsäure, 3-Hydroxyisovaleriansäure, Milchsäure, Pantothensäure,
und 2-Hydroxyisobuttersäure tragen jeweils mindestens eine Methylgruppe. Die Signale
dieser Methylgruppen können zur Identifzierung der Hydroxycarbonsäuren herangezogen
werden. Das Signal der Methylgruppe von 3-Hydroxy-2-Methylpropionsäure liegt bei
1,06ppm, das der Milchsäure bei 1,34ppm und das der 2-Hydroxyisobuttersäure bei 1,37ppm.
Die Methylgruppen der 3-Hydroxyisovaleriansäure sind chemisch äquivalent und geben ein
Signal bei 1,32ppm. Die Methylgruppen der Pantothensäure sind nicht chemisch äquivalent.
Ihre Signale liegen bei 0,87ppm und 0,90ppm.
ppm (t1) 1.001.50
PSHMPS
HIVSMS
HIBS
ppm (t1) 1.001.50
PSHMPS
HIVSMS
HIBS
Abbildung 44: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt einige Signale der Carbonsäuren 2-Hydroxyisobuttersäure (HIBS), Milchsäure (MS), 3-Hydroxyisovaleriansäure (HIVS), 3-Hydroxy-2-Methylpropionsäure (HMPS) und Pantothensäure (PS)
Die Glucuronsäure ähnelt in ihrer Struktur der Glucose. Wie bei der Glucose existieren zwei
Diastereomere der Glucuronsäure, die ebenfalls anomere Wasserstoffe tragen. Die Signale
dieser α-Wasserstoffe liegen dicht neben denen der Glucose bei 5,26ppm und 4,66ppm und
können zur Identifizierung genutzt werden.

Identifizierung unbekannter Metabolite
75
ppm (t1) 4.505.00
α-Glucuronsäure
β-Glucuronsäure
ppm (t1) 4.505.00
α-Glucuronsäure
β-Glucuronsäure
Abbildung 45: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale der anomeren Wasserstoffe der Glucuronsäure.
3.3.5.5. Oxocarbonsäuren
In den Urinproben konnten die Oxocarbonsäuren 2-Ketoglutarsäure, Brenztraubensäure,
Oxalacetat und Acetoacetat identifiziert werden.
2-Ketoglutarsäure, eine Dicarbonsäure, ist eine Schlüsselsubstanz des Citratcyklus und eine
Vorstufe von Glutaminsäure und Glutamin. Enzymdefekte, wie zum Beispiel 2-Ketoglutarat-
Dehydrogenase-Mangel (Dunckelmann et al, 2000) oder Glykogenose, führen zu einer
vermehrten Ausscheidung von 2-Ketoglutarsäure in den Urin.
Brenztraubensäure ist die einfachste Ketosäure und ein wichtiger Metabolit im
Intermediärstoffwechsel der Kohlenhydrate (Glykolyse, Citratzyklus). Durch
Sauerstoffmangel, Vitaminmangel oder Gifte verursachte Zellschädigungen führen ebenso zu
einer Pyruvat-Anreicherung im Urin, wie Pyruvat-Dehydrogenase-Mangel und andere
enzymatische Defekte.
Oxalacetat ist, wie 2-Ketoglutarat, ein zentraler Metabolit im Citratcyclus.
Stoffwechselstörungen oder andere Krankheiten, die zu veränderten Konzentrationen im Urin
führen, sind bis heute nicht bekannt. Pantothensäure bzw. Vitamin B3 ist ein Baustein des
Coenzym-A. Die Konzentration im Urin hängt zum Teil von der Ernährung ab
Acetoacetat entsteht im Intermediärstoffwechsel aus ketoplastischen Aminosäuren und im
Fettstoffwechsel. Eine Stoffwechselentgleisung bei Diabetes mellitus führt zu erhöhten
Acetoacetat-Konzentrationen im Urin (Bales et al, 1984).
Die Signale von 2-Ketoglutarsäure, Brenztraubensäure, Oxalacetat und Acetoacetat liegen
alle im Hochfeldbereich. 2-Ketoglutarsäure zeigt zwei Signale bei 2,48ppm und 3,02ppm.
Das Signal der Brenztraubensäure ist bei 2,28ppm zu finden, das des Oxalacetats bei
2,38ppm. Die beiden Signale vom Acetoacetat liegen bei 2,27ppm und 3,44ppm.
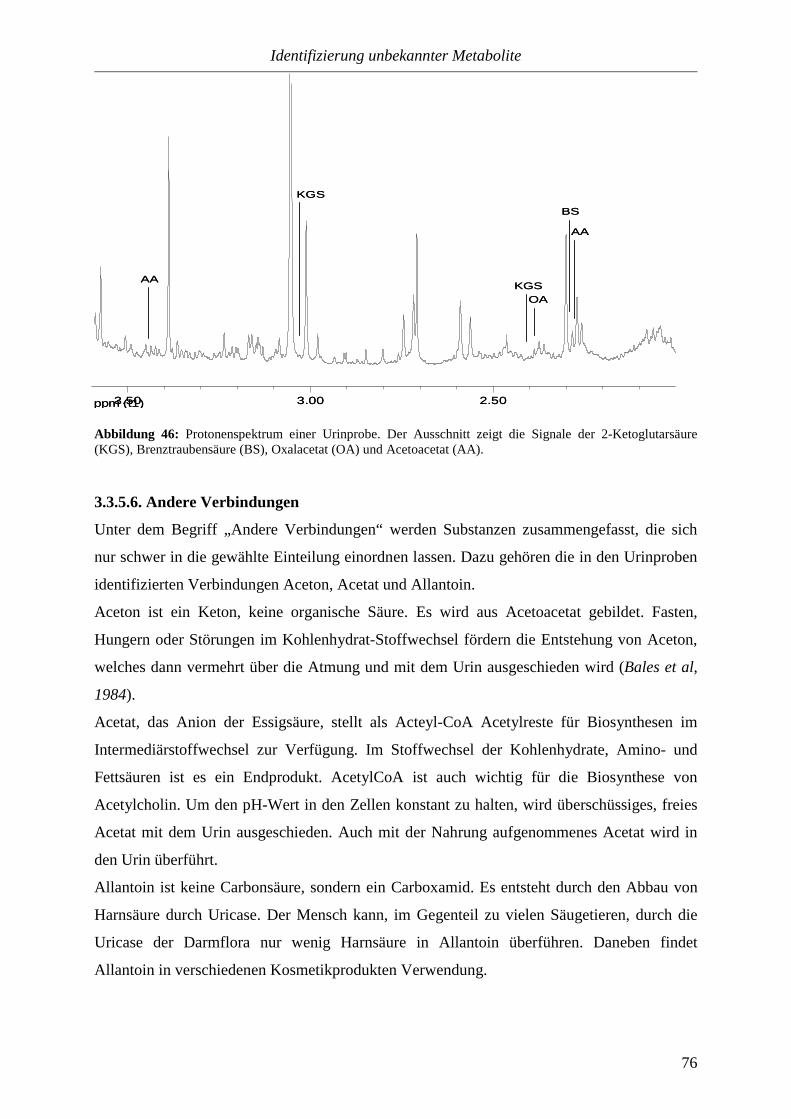
Identifizierung unbekannter Metabolite
76
ppm (t1) 2.503.003.50
KGS
KGS
BS
OA
AA
AA
ppm (t1) 2.503.003.50
KGS
KGS
BS
OA
AA
AA
Abbildung 46: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale der 2-Ketoglutarsäure (KGS), Brenztraubensäure (BS), Oxalacetat (OA) und Acetoacetat (AA).
3.3.5.6. Andere Verbindungen
Unter dem Begriff „Andere Verbindungen“ werden Substanzen zusammengefasst, die sich
nur schwer in die gewählte Einteilung einordnen lassen. Dazu gehören die in den Urinproben
identifizierten Verbindungen Aceton, Acetat und Allantoin.
Aceton ist ein Keton, keine organische Säure. Es wird aus Acetoacetat gebildet. Fasten,
Hungern oder Störungen im Kohlenhydrat-Stoffwechsel fördern die Entstehung von Aceton,
welches dann vermehrt über die Atmung und mit dem Urin ausgeschieden wird (Bales et al,
1984).
Acetat, das Anion der Essigsäure, stellt als Acteyl-CoA Acetylreste für Biosynthesen im
Intermediärstoffwechsel zur Verfügung. Im Stoffwechsel der Kohlenhydrate, Amino- und
Fettsäuren ist es ein Endprodukt. AcetylCoA ist auch wichtig für die Biosynthese von
Acetylcholin. Um den pH-Wert in den Zellen konstant zu halten, wird überschüssiges, freies
Acetat mit dem Urin ausgeschieden. Auch mit der Nahrung aufgenommenes Acetat wird in
den Urin überführt.
Allantoin ist keine Carbonsäure, sondern ein Carboxamid. Es entsteht durch den Abbau von
Harnsäure durch Uricase. Der Mensch kann, im Gegenteil zu vielen Säugetieren, durch die
Uricase der Darmflora nur wenig Harnsäure in Allantoin überführen. Daneben findet
Allantoin in verschiedenen Kosmetikprodukten Verwendung.
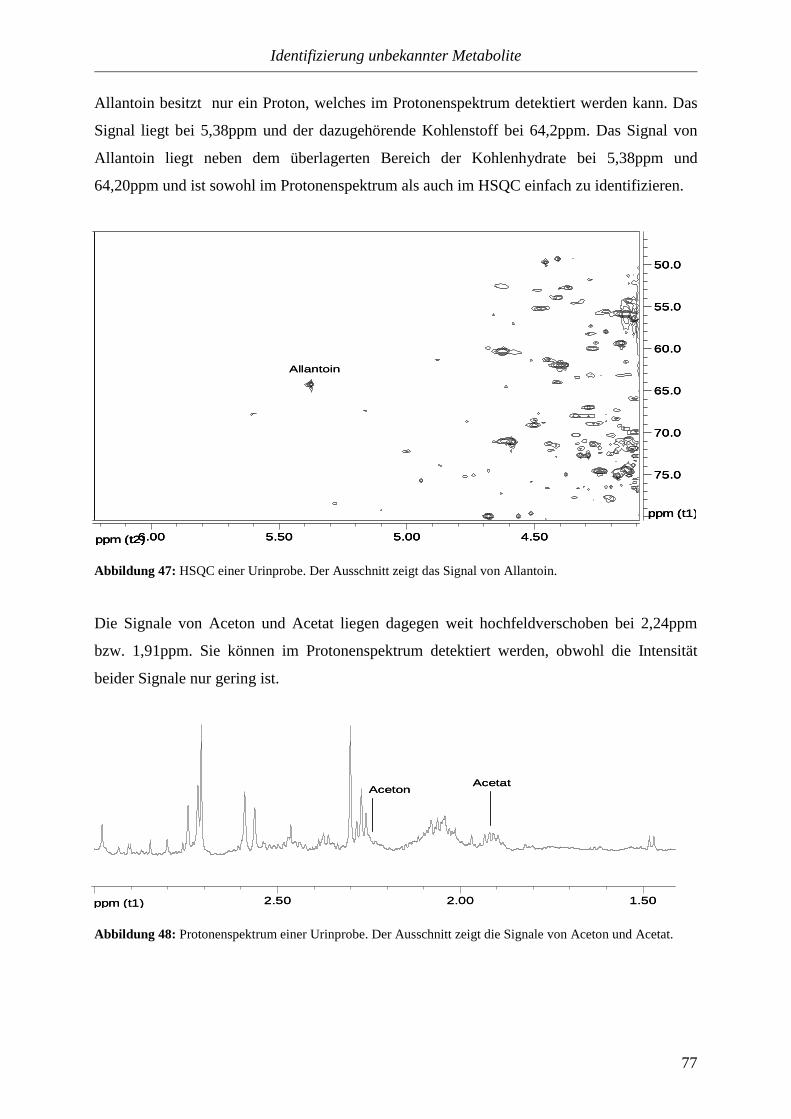
Identifizierung unbekannter Metabolite
77
Allantoin besitzt nur ein Proton, welches im Protonenspektrum detektiert werden kann. Das
Signal liegt bei 5,38ppm und der dazugehörende Kohlenstoff bei 64,2ppm. Das Signal von
Allantoin liegt neben dem überlagerten Bereich der Kohlenhydrate bei 5,38ppm und
64,20ppm und ist sowohl im Protonenspektrum als auch im HSQC einfach zu identifizieren.
ppm (t2) 4.505.005.506.00
50.0
55.0
60.0
65.0
70.0
75.0
ppm (t1)
Allantoin
ppm (t2) 4.505.005.506.00
50.0
55.0
60.0
65.0
70.0
75.0
ppm (t1)
ppm (t2) 4.505.005.506.00
50.0
55.0
60.0
65.0
70.0
75.0
ppm (t1)
Allantoin
Abbildung 47: HSQC einer Urinprobe. Der Ausschnitt zeigt das Signal von Allantoin.
Die Signale von Aceton und Acetat liegen dagegen weit hochfeldverschoben bei 2,24ppm
bzw. 1,91ppm. Sie können im Protonenspektrum detektiert werden, obwohl die Intensität
beider Signale nur gering ist.
ppm (t1) 1.502.002.50
AcetonAcetat
ppm (t1) 1.502.002.50
AcetonAcetat
Abbildung 48: Protonenspektrum einer Urinprobe. Der Ausschnitt zeigt die Signale von Aceton und Acetat.

Identifizierung unbekannter Metabolite
78
3.4. Zusammenfassung
Die hier vorgestellten Metabolite wurden auf verschiedenen Wegen identifiziert. Ein Teil der
Signalzuordnung erfolgte über Literaturrecherche. So konnte ein Großteil der proteinogenen
Aminosäuren, diverse Monosaccharide, Indole, Amine und ein Teil der organischen Säuren
mit Hilfe verschiedener Artikel identifiziert werden (Fan, 1996; Nicholson und Foxall, 1995;
Holmes et al, 1997). Die Signalzuordnung der Zuckeralkohole erfolgte ebenfalls über
Literaturrecherche (Hawkes und Lewis, 1984; Angyal und Wheen, 1980; Angyal und Le Fur,
1980). Außerdem konnten mittels Literatur die Substanzen N-Acetyl-Neuraminsäure (Gervay
und Batta, 1994), Hippursäure (Fan, 1996), Pseudouridin (Grey et al, 1971) identifiziert
werden.
Andere Signale konnten mit Hilfe der Datenbank SDBS zugeordnet werden. Über diese
Datenbank konnte ein weiterer Teil der organischen Säuren und der Amine identifiziert
werden. Die Signalzuordnung der meisten Nukleoside erfolgte ebenfalls über die Datenbank.
Hier waren jedoch zusätzliche Referenzmessungen notwendig, da die Signale dieser
Verbindungen sehr dicht beieinander liegen. Außerdem wurden die Metabolite 1-Methyl-
Nicotinamid, 1-Methylnicotinsäure, 4-Hydroxybenzamid, Fucose, Xylose, Lactose,
Saccharose, Guanidinoacetat und Allantoin mit Hilfe der Datenbank identifiziert.
Ein Großteil der aromatischen und heteroaromatischen Verbindungen und einige organische
Säuren wurden durch Strukturaufklärung identifiziert. So konnten Histidin, 1-Methylhistidin,
3-Methylhistidin und Carnosine nur mit Hilfe der Strukturaufklärung identifiziert werden, da
bei gegebenen pH-Wert keine Signalzuordnung vorliegt. Im Fall des 4-Pyridon-3-
Carboxyamid-1-beta-D-Ribonucleosid wurde die Strukturaufklärung durch anschließende
Literaturrecherche (Slominska et al, 2006) unterstützt.
Die meisten Metabolite, die in den Urinproben identifiziert wurden, können als
Markermolekül mit Stoffwechselstörungen in Verbindung gebracht werden. Viele von ihnen
können über ihre Signale in einem Protonenspektrum zugeordnet werden, andere müssen mit
Hilfe des HSQC oder COSY identifiziert werden. Mit Hilfe von NMR und
Mustererkennungsverfahren ist es möglich, die hier genannten Substanzen zur Diagnose
pathologischer Stoffwechseländerungen einzusetzen. Einige von ihnen sind nur gering
konzentriert, so dass sie unter normalen Bedingungen durch die Datenreduzierung der
Mustererkennungsverfahren nicht mehr erfasst werden. Erhöhte Konzentrationen können
jedoch detektiert werden.
Einzelne Substanzen, wie zum Beispiel Harnsäure tragen keine Protonen und geben im NMR
keine Signale. Deshalb sind sie nicht detektierbar. Andere Metabolite sind an Glucuronsäuren,

Identifizierung unbekannter Metabolite
79
Ribosen oder andere Moleküle gebunden, die die Hydrophilie steigern. Diese gebundenen
Moleküle können ebenfalls nicht identifiziert werden, da eine Kopplung der Protonen über
Heteroatome nur sehr selten beobachtet wird.
Es gibt noch eine Vielzahl an Signalen, die aufgrund der Überlagerungen bisher noch nicht
identifiziert werden konnten. Hier wäre eine höhere Magnetfeldstärke oder ein
Kryoprobenkopf hilfreich. Beide Möglichkeiten verbessern die Auflösung der Spektren und
können dadurch die Überlagerungen entzerren. Weitere Festphasenextraktionen und HPLC-
Methoden können durch Reduzierung der Signalzahl ebenfalls die Identifizierung zusätzlicher
Substanzen erlauben.

Einfluss der Ernährung auf die Zusammensetzung des Urins
80
4. Einfluss der Ernährung auf die Zusammensetzung des Urins
4.1. Einführung
Die Zusammensetzung biologischer Flüssigkeiten unterliegt immer natürlichen
Schwankungen. So hängen auch die Konzentrationen der im Urin enthaltenen Metabolite von
verschiedenen physischen und psychischen Faktoren ab. Neben Stress, Bewegung, Hormonen
und anderen Einflüssen wirkt sich auch die Ernährung auf die Zusammensetzung des Urins
aus. Zum Beispiel ist Hippursäure ein Abbauprodukt der Benzoesäure (Kubota et al, 1991).
Diese findet, in Form des Natriumbenzoats, als Konservierungsmittel in Lebensmitteln
Verwendung. Deshalb wird die Hippursäure nach dem Verzehr konservierter Lebensmittel
vermehrt ausgeschieden. Für die klinische Diagnostik ist es wichtig, die Ursachen möglicher
Konzentrationsänderungen bestimmter Metabolite zu kennen. Werden andere, nicht
pathologische Einflüsse auf die Zusammensetzung des Urins vernachlässigt, kann es leicht zu
Fehldiagnosen kommen.
Der Einfluss diverser Nahrungsmittel auf die Zusammensetzung von Urin ist Inhalt
verschiedener Untersuchungen (Ito et al, 2005; Brevik et al, 2004). Hierbei werden in den
meisten Fällen HPLC und Massenspektrometrie zur Analyse verwendet. So wurden zum
Beispiel der Konsum von Schokolade und mögliche Auswirkungen auf die Zusammensetzung
des Urins untersucht (Rios et al, 2003). Dabei zeigte sich, dass die in der Schokolade
enthaltenen Polyphenole durch die Darmflora zu aromatischen Carbonsäuren abgebaut
werden. Nach dem Verzehr konnten erhöhte Konzentrationen von verschiedenen
Benzoesäure- und Phenylessigsäure-Derivaten im Urin detektiert werden. Andere
beschäftigten sich mit dem Einfluss von Weizenkleie auf die Konzentration bestimmter
Metabolite im Urin (Kern et al, 2003). Dabei zeigte sich, dass dem Konsum eine erhöhte
Ausscheidung der Ferulasäure (3-Methoxy-4-Hydroxy-Zimtsäure) und des Feroylglycins
folgte. Eine weitere Studie untersuchte die Effekte von grünem Tee auf den menschlichen
Metabolismus (Lee et al, 1995). Grüner Tee enthält verschiedene Catechine, die als
Glucuronat oder Sulfat in den menschlichen Organismus gelangen. Wenige Stunden nach
dem Konsum konnten konjugierte Formen von Epigallocatechin und Epicatechin im Urin
detektiert werden.
Die NMR-Spektroskopie wird hier überwiegend eingesetzt, um Stoffwechselprofile von
Probanden zu erstellen. Die Untersuchungen beschäftigen sich unter anderem mit ethnischen
Metabolitenprofilen, die durch unterschiedliche Ernährungsgewohnheiten verursacht werden.
Eine Studie verglich britische und schwedische Einwohner (Lenz et al, 2004) und konnte die
beiden Völker anhand der Zusammensetzung der Urinproben gut voneinander unterscheiden.

Einfluss der Ernährung auf die Zusammensetzung des Urins
81
Die Abweichungen werden hauptsächlich durch unterschiedliche Ernährungsgewohnheiten
verursacht. Eine weitere Untersuchung beschäftigt sich mit dem Vergleich arktischer
Einwohner mit denen aus Rom (Zuppi et al, 1998). Auch hier verursachen die
verschiedenartigen Ernährungsgewohnheiten unterschiedliche Metabolitenprofile. Da
Bewohner der Arktis generell mehr Fisch und konservierte Lebensmittel zu sich nehmen, sind
bei ihnen erhöhte Konzentrationen von Trimethylaminoxid und Hippursäure im Urin zu
finden. Römer ernähren sich dagegen eher von kohlenhydratreichen Lebensmitteln. Dadurch
werden vermehrt Metabolite des Citrat-Cyclus ausgeschieden.
Für klinische Studien ist es hilfreich, die Variabilität von Urinproben möglichst gering zu
halten. Auf diese Weise wird ein Vergleich der Proben erleichtert. Das kann durch eine
Standard-Diät ermöglicht werden. Sofern alle Probanden einer Studie die gleiche Diät
erhalten, sind die Unterschiede in den Urinproben deutlich reduziert (Walsh et al, 2006; Lenz
et al, 2003).
4.2. Allgemeine Durchführung
Für die Untersuchungen wurden die Urinproben von zwei Probanden eingesetzt. Die erste
Probennahme erfolgte unmittelbar vor dem Verzehr der gewählten Nahrungsmittel.
Anschließend wurden vier weitere Proben nach 6, 12, 18 und 24 Stunden genommen. Die
Proben wurden lyophillisiert und der pH-Wert auf 5,75 – 5,95 eingestellt. Anschließend
wurden Protonenspektren der Urinproben aufgenommen und die Spektren auf das Signal der
Hippursäure bei 7,81ppm kalibriert.
Zur Bestimmung möglicher Konzentrationsänderungen wurden relative Veränderungen der
Integrale herangezogen. Creatinin diente hierbei als interner Standard. Die Bildung dieses
Metaboliten ist unmittelbar mit der vorhandenen Muskelmasse verknüpft. Deshalb kann die
Konzentration von Creatinin im Urin unter Normalbedingungen als konstant betrachtet
werden. Der Integralwert des Creatinins wurde auf 1000 gesetzt und so alle anderen Integrale
auf diesen Wert normiert. Es wurde ein Integral von 1000 gewählt, damit auch die
intensitätsschwachen Signale im Tieffeldbereich erfasst werden konnten.
4.3. Untersuchte Nahrungsmittel
Für die Untersuchungen wurden verschiedene Obstsorten ausgesucht, da sie relativ schnell
verdaut werden. Mögliche Änderungen in der Zusammensetzung des Urins sind auf diese
Weise schon nach kurzer Zeit zu erkennen. Nach dem Verzehr der eher schwer verdaulichen
Pekanüsse zeigen sich die Abweichungen deutlich später.

Einfluss der Ernährung auf die Zusammensetzung des Urins
82
4.3.1. Bananen
Bananen sind serotoninhaltige Nahrungsmittel. Untersuchungen mittels HPLC und MS
zeigten, dass der Verzehr zu einer vermehrten Ausscheidung des Serotoninabbauprodukts 5-
Hydroxyindolessigsäure führt (Feldman et al, 1989). Die NMR-Spektren der Urinproben
konnten dieses Ergebnis bestätigen.
Serotonin (5-Hydroxytryptamin) ist ein Indolamin und fungiert im Organismus als
Neurotransmitter. Es ist auch in verschiedenen Nahrungsmitteln enthalten. Ein Verzehr dieser
Lebensmittel führt zu einer vermehrten Ausscheidung von 5-Hydroxyindolacetat, wobei der
Abbau enzymatisch erfolgt. Das Enzym Monoaminoxygenase baut Serotonin zu 5-
Hydroxyindolacetaldehyd ab. Dieser Aldehyd wird anschließend von dem Enzym
Aldehyddehydrogenase-2 weiter abgebaut zur 5-Hydroxyindolacetat.
NH
OH
NH2
NH
OH
O
NH
OH
O
OH
Monoaminoxygenase
Aldehyddehydrogenase
5-Hydroxyindolessigsäure
5-Hydroxyindolacetaldehyd
Serotonin
NH
OH
NH2
NH
OH
O
NH
OH
O
OH
Monoaminoxygenase
Aldehyddehydrogenase
5-Hydroxyindolessigsäure
5-Hydroxyindolacetaldehyd
Serotonin
Abbildung 49: Enzymatischer Abbaumechanismus von Serotonin über 5-Hydroxyindolacetaldehyd zur 5-Hydroxyindolessigsäure.
Der Abbau von Serotonin und die Ausscheidung des Abbauproduktes sind von Interesse, da
Neuroendokrine Tumore Serotonin produzieren und über die vermehrte Ausscheidung des
Abbauprodukts 5-Hydroxyindolessigsäure diagnostiziert werden. Wird die Konzentration der
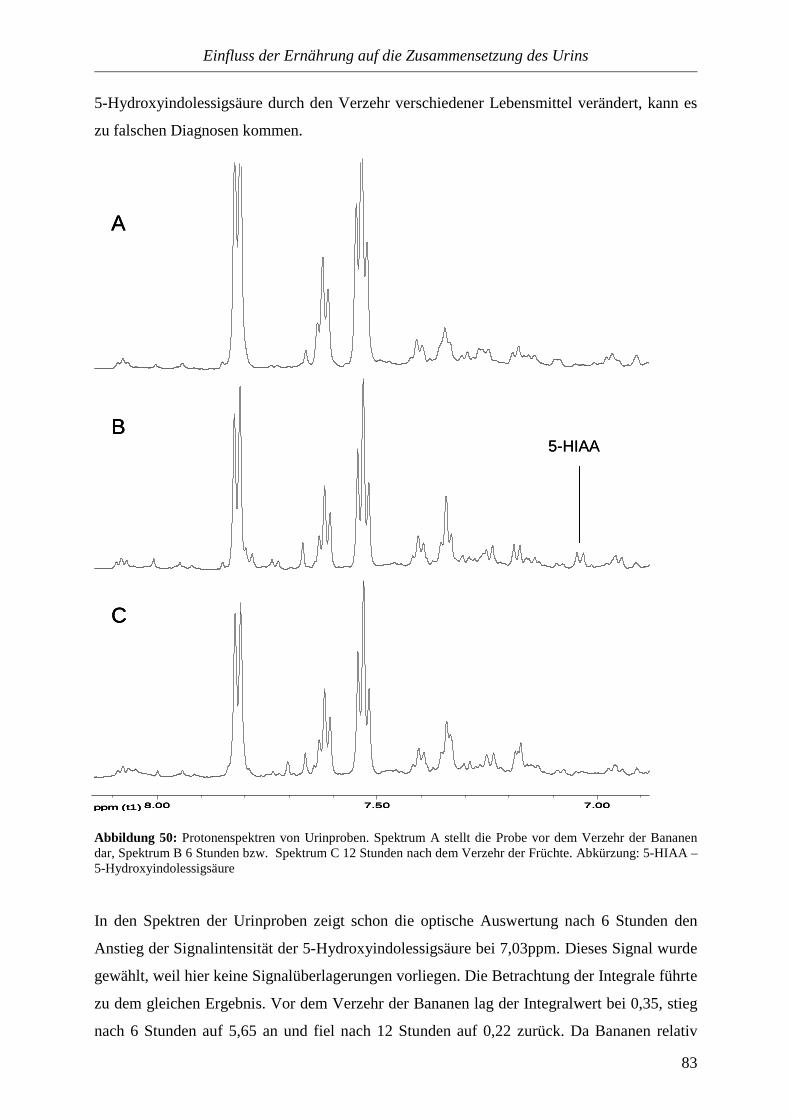
Einfluss der Ernährung auf die Zusammensetzung des Urins
83
5-Hydroxyindolessigsäure durch den Verzehr verschiedener Lebensmittel verändert, kann es
zu falschen Diagnosen kommen.
ppm (t1) 7.007.508.00
A
B
C
5-HIAA
ppm (t1) 7.007.508.00
A
B
C
ppm (t1) 7.007.508.00
A
B
C
5-HIAA
Abbildung 50: Protonenspektren von Urinproben. Spektrum A stellt die Probe vor dem Verzehr der Bananen dar, Spektrum B 6 Stunden bzw. Spektrum C 12 Stunden nach dem Verzehr der Früchte. Abkürzung: 5-HIAA – 5-Hydroxyindolessigsäure
In den Spektren der Urinproben zeigt schon die optische Auswertung nach 6 Stunden den
Anstieg der Signalintensität der 5-Hydroxyindolessigsäure bei 7,03ppm. Dieses Signal wurde
gewählt, weil hier keine Signalüberlagerungen vorliegen. Die Betrachtung der Integrale führte
zu dem gleichen Ergebnis. Vor dem Verzehr der Bananen lag der Integralwert bei 0,35, stieg
nach 6 Stunden auf 5,65 an und fiel nach 12 Stunden auf 0,22 zurück. Da Bananen relativ
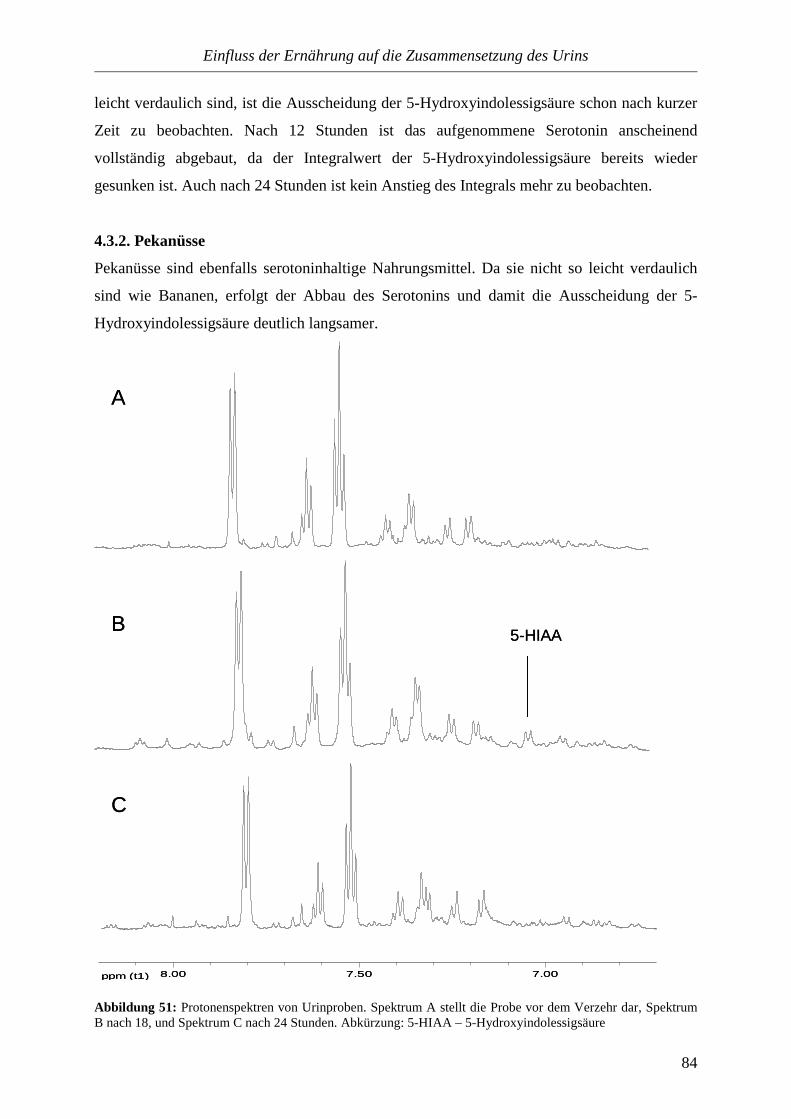
Einfluss der Ernährung auf die Zusammensetzung des Urins
84
leicht verdaulich sind, ist die Ausscheidung der 5-Hydroxyindolessigsäure schon nach kurzer
Zeit zu beobachten. Nach 12 Stunden ist das aufgenommene Serotonin anscheinend
vollständig abgebaut, da der Integralwert der 5-Hydroxyindolessigsäure bereits wieder
gesunken ist. Auch nach 24 Stunden ist kein Anstieg des Integrals mehr zu beobachten.
4.3.2. Pekanüsse
Pekanüsse sind ebenfalls serotoninhaltige Nahrungsmittel. Da sie nicht so leicht verdaulich
sind wie Bananen, erfolgt der Abbau des Serotonins und damit die Ausscheidung der 5-
Hydroxyindolessigsäure deutlich langsamer.
ppm (t1) 7.007.508.00
5-HIAA
A
B
C
ppm (t1) 7.007.508.00
5-HIAA
A
B
C
Abbildung 51: Protonenspektren von Urinproben. Spektrum A stellt die Probe vor dem Verzehr dar, Spektrum B nach 18, und Spektrum C nach 24 Stunden. Abkürzung: 5-HIAA – 5-Hydroxyindolessigsäure

Einfluss der Ernährung auf die Zusammensetzung des Urins
85
In den Spektren der Urinproben steigt der Integralwert des Signals bei 7,03ppm von 2,56 auf
13,09 an. Allerdings erst nach 18 Stunden. Nach 24 Stunden ist die Intensität mit 13,96 immer
noch erhöht. Die Verdauung der Nüsse nimmt also mehr Zeit in Anspruch.
4.3.3. Orangen
Orangen gehören zu den polyphenolreichen Nahrungsmitteln. Sie enthalten vor allem große
Mengen an Hesperitin und Naringenin. Nach dem Verzehr von Orangen konnten mittels
HPLC und MS erhöhte Konzentrationen dieser Verbindungen im Urin detektiert werden (Ito
et al, 2005).
Hesperitin und Naringenin gehören zu den Flavanonen, die wiederum den Flavonoiden
zugeordnet werden. Flavonoide bestehen aus drei Kohlenstoffringen mit zwei aromatischen
(A und B) und einem Sauerstoff-heterocyclischen Ring (C). Anhand struktureller
Unterschiede am C-Ring werden die Flavonoide in verschiedene Gruppen (z. B. Flavanone)
eingeteilt.
O
OH
O
OH
OH
OH
O
OH
OOH
OH
A C C
BB
A
Hesperitin Naringenin
O
OH
O
OH
OH
OH
O
OH
OOH
OH
A C C
BB
A
O
OH
O
OH
OH
OH
O
OH
OOH
OHO
OH
O
OH
OH
OH
O
OH
OOH
OH
A C C
BB
A
Hesperitin Naringenin
Abbildung 52: Strukturen der Flavanone Hesperitin und Naringenin. Die Kohlenstoffringe sind mit den Buchstaben A, B und C beschriftet.
Aufgrund der drei Kohlenstoffringe besitzen Flavanone, wie alle Flavonoide, eine relativ
unpolare Struktur. Deshalb werden sie nur in sehr geringen Mengen mit dem Urin
ausgeschieden. Auch nach dem Verzehr von Orangen liegen die Konzentrationen von
Hesperitin und Naringenin im Urin unterhalb der Nachweisgrenze für die NMR-
Spektroskopie. Daher konnten sie auch in den hier vermessenen Proben nicht identifiziert
werden.
Ein Teil der aufgenommenen Flavonone wird durch die Darmflora abgebaut. Dabei wird der
C-Ring im Flavonoidgerüst gespalten (Rechner et al, 2004). Anschließend werden die
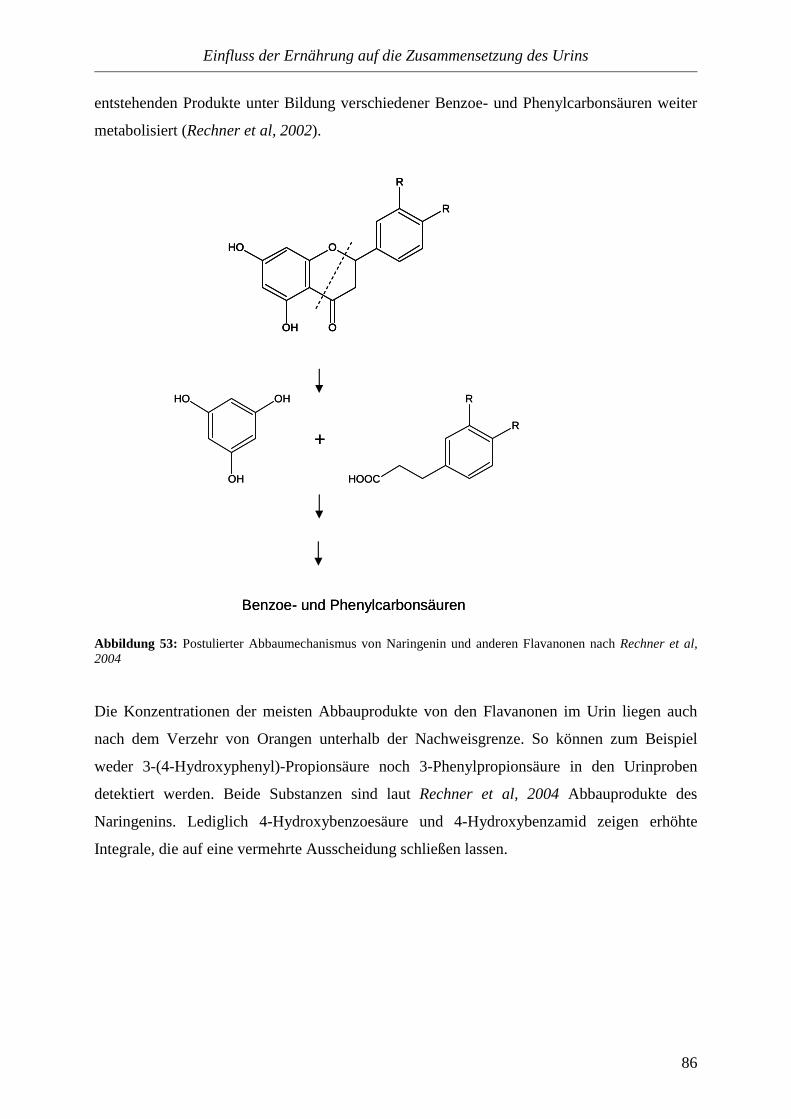
Einfluss der Ernährung auf die Zusammensetzung des Urins
86
entstehenden Produkte unter Bildung verschiedener Benzoe- und Phenylcarbonsäuren weiter
metabolisiert (Rechner et al, 2002).
O
R
R
O
OH
OH
OHOH
OH
R
R
HOOC
+
Benzoe- und Phenylcarbonsäuren
O
R
R
O
OH
OH
O
R
R
O
OH
OH
OHOH
OH
R
R
HOOC
+
Benzoe- und Phenylcarbonsäuren
Abbildung 53: Postulierter Abbaumechanismus von Naringenin und anderen Flavanonen nach Rechner et al, 2004
Die Konzentrationen der meisten Abbauprodukte von den Flavanonen im Urin liegen auch
nach dem Verzehr von Orangen unterhalb der Nachweisgrenze. So können zum Beispiel
weder 3-(4-Hydroxyphenyl)-Propionsäure noch 3-Phenylpropionsäure in den Urinproben
detektiert werden. Beide Substanzen sind laut Rechner et al, 2004 Abbauprodukte des
Naringenins. Lediglich 4-Hydroxybenzoesäure und 4-Hydroxybenzamid zeigen erhöhte
Integrale, die auf eine vermehrte Ausscheidung schließen lassen.
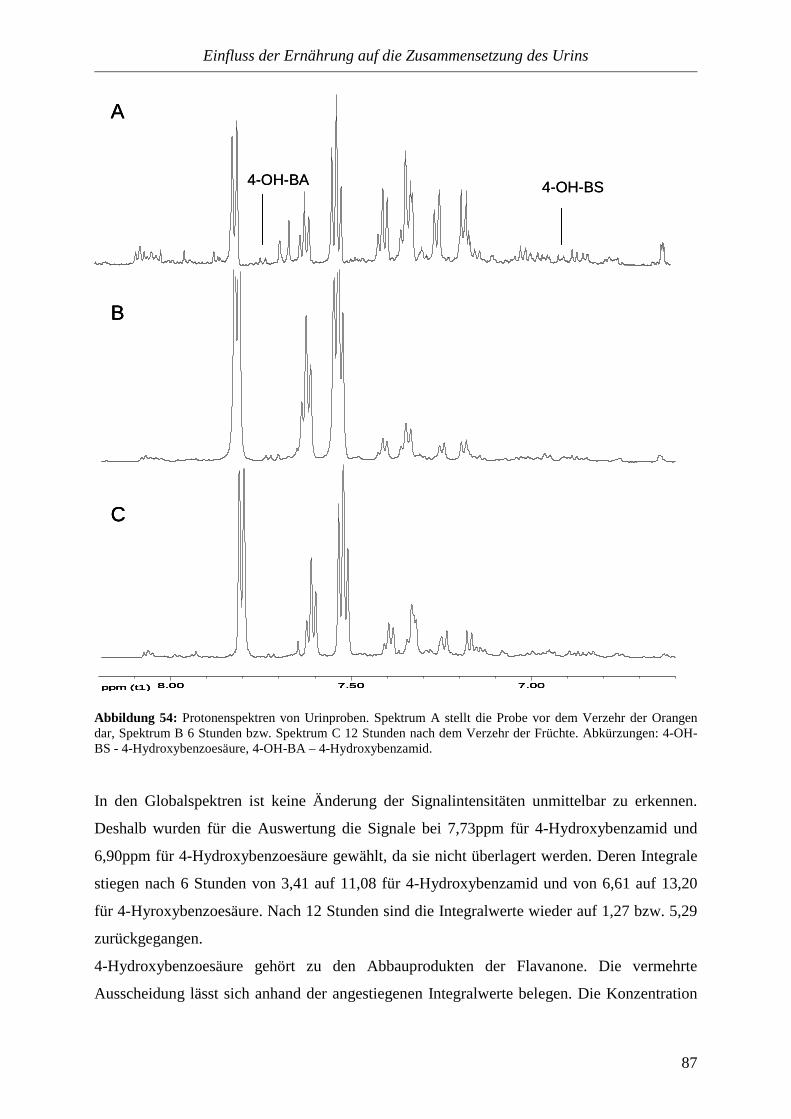
Einfluss der Ernährung auf die Zusammensetzung des Urins
87
ppm (t1) 7.007.508.00
A
B
C
4-OH-BA 4-OH-BS
ppm (t1) 7.007.508.00
A
B
C
ppm (t1) 7.007.508.00
A
B
C
4-OH-BA 4-OH-BS
Abbildung 54: Protonenspektren von Urinproben. Spektrum A stellt die Probe vor dem Verzehr der Orangen dar, Spektrum B 6 Stunden bzw. Spektrum C 12 Stunden nach dem Verzehr der Früchte. Abkürzungen: 4-OH-BS - 4-Hydroxybenzoesäure, 4-OH-BA – 4-Hydroxybenzamid.
In den Globalspektren ist keine Änderung der Signalintensitäten unmittelbar zu erkennen.
Deshalb wurden für die Auswertung die Signale bei 7,73ppm für 4-Hydroxybenzamid und
6,90ppm für 4-Hydroxybenzoesäure gewählt, da sie nicht überlagert werden. Deren Integrale
stiegen nach 6 Stunden von 3,41 auf 11,08 für 4-Hydroxybenzamid und von 6,61 auf 13,20
für 4-Hyroxybenzoesäure. Nach 12 Stunden sind die Integralwerte wieder auf 1,27 bzw. 5,29
zurückgegangen.
4-Hydroxybenzoesäure gehört zu den Abbauprodukten der Flavanone. Die vermehrte
Ausscheidung lässt sich anhand der angestiegenen Integralwerte belegen. Die Konzentration

Einfluss der Ernährung auf die Zusammensetzung des Urins
88
im Urin ist allerdings nur wenig erhöht. Eventuell liegen die Konzentrationen der anderen
Abbauprodukte deshalb weiterhin unterhalb der Nachweisgrenze.
Die vermehrte Ausscheidung des 4-Hydroxybenzamids kann nicht durch die Metabolisierung
der Flavanone erklärt werden. Möglicherweise ist diese Substanz selber ein Bestandteil von
Orangen oder ein Abbauprodukt einer anderen Verbindung. Der Konzentrationsanstieg findet
parallel zu dem der 4-Hydroxybenzoesäure statt, was vermuten lässt, dass die beiden
Verbindungen den gleichen Ursprung haben.
4.4. Zusammenfassung
Die Messungen zeigen, dass der Einfluss von Nahrungsmitteln auf die Zusammensetzung des
Urins auch mit Hilfe der NMR aufgezeigt werden kann. Die Konzentrationsänderungen
aromatischer Substanzen sind jedoch relativ gering und nur über die Integralwerte
nachzuweisen. Verschiedene Substanzen sind gar nicht detektierbar, da ihre Konzentrationen
unterhalb der Nachweisgrenze liegen.
Die Einflüsse der Nahrung auf die Zusammensetzung des Urins sind deutlich zu erkennen.
Deshalb müssen sie bei klinischen Studien bedacht werden. Eine Standardisierung der
Ernährung bei den Probanden ist hier hilfreich. Auf diese Weise kann die Variabilität der
Urinproben reduziert und die Auswertung der Messungen erleichtert werden. Die Diagnose
von Stoffwechselstörungen wird durch den Einfluss der Ernährung ebenfalls erschwert. Auch
hier kann, je nach Fragestellung, eine Diät vor der Probennahme helfen.

Anhang
89
5. Anhang
5.1. Lagerung und Vorbereitung der Urinproben
Alle für diese Arbeit verwendeten Urinproben wurden bis zur Aufarbeitung bei -80°C
gelagert. Nach dem Auftauen wurde jeweils ein Aliquot von 10ml entnommen und
lyophillisiert. Eventuell vorhandene, unlösliche Bestandteile wurden vorher durch
zentrifugieren der Proben (3000U/min, 20°C, 5min) entfernt. Anschließend wurde die
getrocknete Probe in 1ml D2O aufgenommen und mit 80µl eines 1,5M KH2PO4 / K2HPO4-
Puffers (7:1) in D2O versetzt. Nachdem der pH-Wert der Proben mit DCl bzw. NaOD auf
5,85-5,95 eingestellt wurde, konnten die Proben gemessen werden.
5.2. Durchführung der Messungen
Die NMR-Spektroskopische Messung der Urinproben erfolgte mit der zgpr-Pulssequenz
(Bruker, 600MHz AVANCE-Gerät). Einer eindimensionalen Methode mit Vorsättigung des
Wassersignals. Die Parameter der Messungen blieben nahezu konstant. Lediglich der Offset
des Wassersignals und der 90°-Puls wurden für jede Probe optimiert.
O1: Frequenz des Wassersignals
AQ: 1,827s
D1: 3s
DE: 6µs
DS: 8
NS: 64
TD: 32k
TE: 300K
p1: 90°
pl9: 65dB
RG: 256
SWH: 10964,912Hz
Die zweidimensionalen Messungen erfolgten mit entsprechenden Pulssequenzen von Bruker.
Auch hier wurde der 90°-Puls für Protonen optimiert. Für 13C wurde ein Geräte-spezifischer
Wert übernommen.
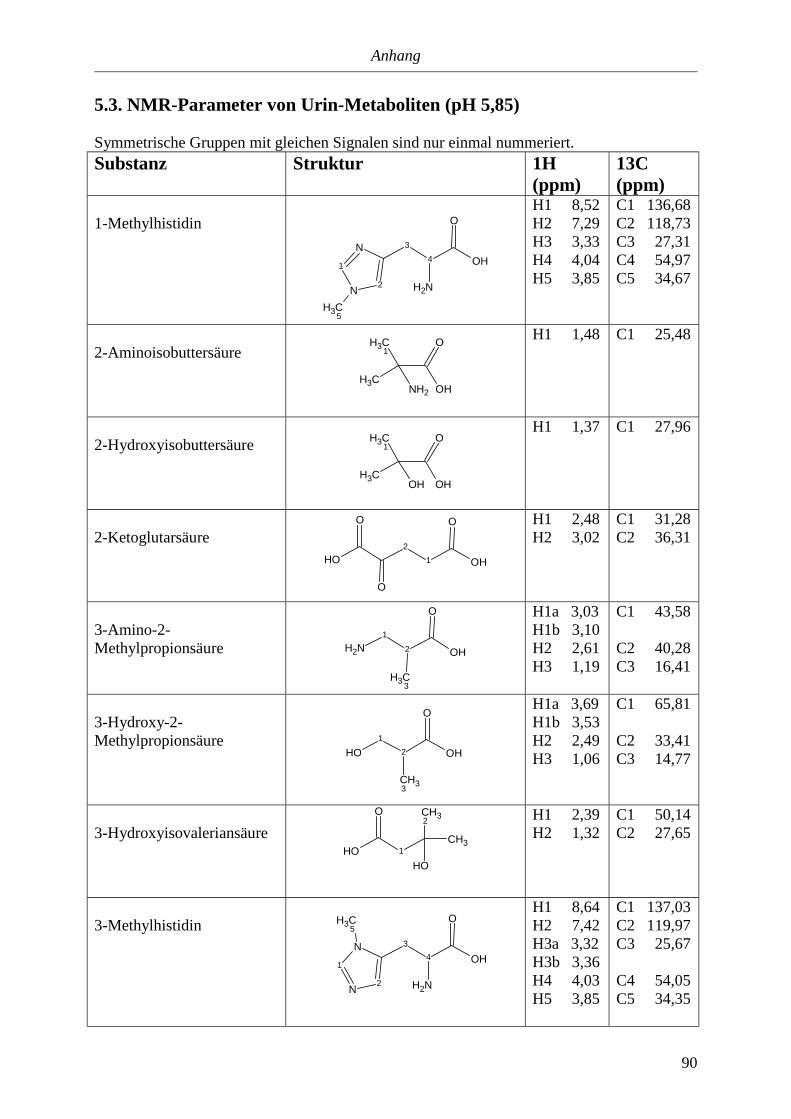
Anhang
90
5.3. NMR-Parameter von Urin-Metaboliten (pH 5,85)
Symmetrische Gruppen mit gleichen Signalen sind nur einmal nummeriert. Substanz Struktur 1H
(ppm) 13C (ppm)
1-Methylhistidin H1 8,52
H2 7,29 H3 3,33 H4 4,04 H5 3,85
C1 136,68 C2 118,73 C3 27,31 C4 54,97 C5 34,67
2-Aminoisobuttersäure H1 1,48 C1 25,48
2-Hydroxyisobuttersäure H1 1,37 C1 27,96
2-Ketoglutarsäure H1 2,48
H2 3,02 C1 31,28 C2 36,31
3-Amino-2-Methylpropionsäure
H1a 3,03 H1b 3,10 H2 2,61 H3 1,19
C1 43,58
C2 40,28 C3 16,41
3-Hydroxy-2-Methylpropionsäure
H1a 3,69 H1b 3,53 H2 2,49 H3 1,06
C1 65,81
C2 33,41 C3 14,77
3-Hydroxyisovaleriansäure H1 2,39
H2 1,32 C1 50,14 C2 27,65
3-Methylhistidin H1 8,64
H2 7,42 H3a 3,32 H3b 3,36 H4 4,03 H5 3,85
C1 137,03 C2 119,97 C3 25,67
C4 54,05 C5 34,35
1
N
N2
CH35
3
4
NH2
OH
O
OH2
1 OH
O
O
O
NH2
1
2 OH
CH33
O
OH1
2 OH
CH33
O
1
N
N2
CH35
3
4
NH2
OH
O
OH 1
O CH32
OH
CH3
OH
OCH31
NH2
CH3
OH
OCH31
OHCH3
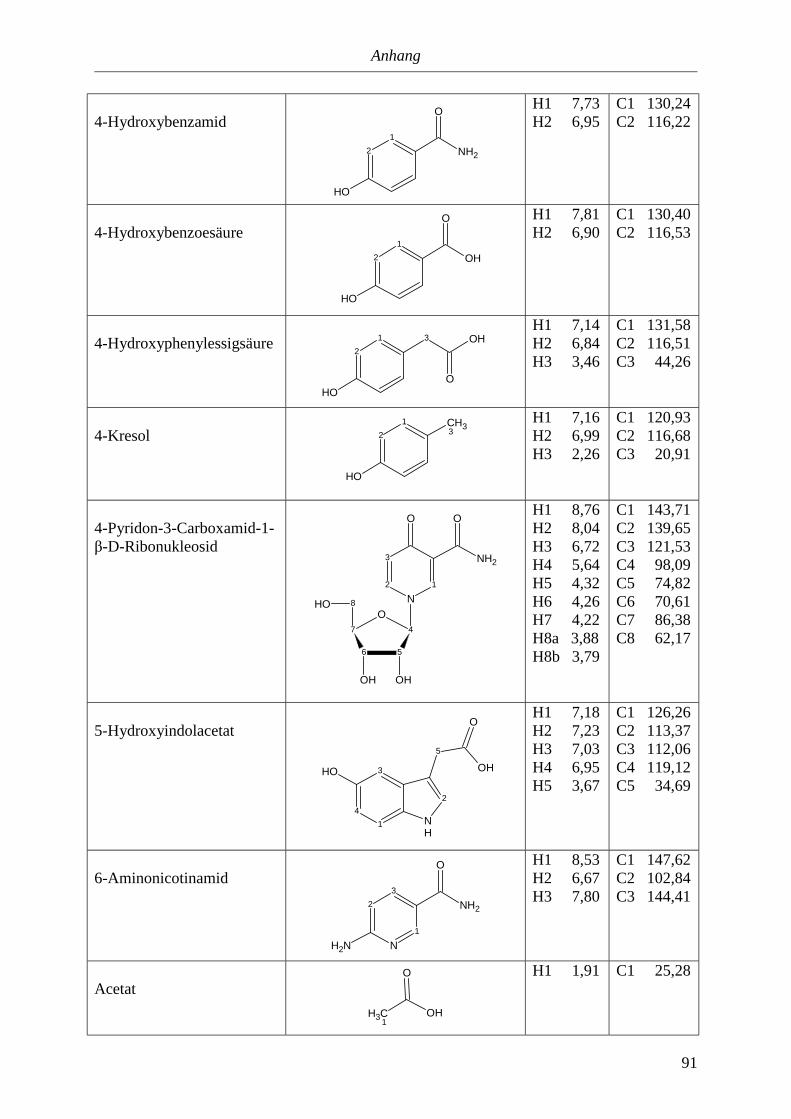
Anhang
91
4-Hydroxybenzamid H1 7,73
H2 6,95 C1 130,24 C2 116,22
4-Hydroxybenzoesäure H1 7,81
H2 6,90 C1 130,40 C2 116,53
4-Hydroxyphenylessigsäure H1 7,14
H2 6,84 H3 3,46
C1 131,58 C2 116,51 C3 44,26
4-Kresol H1 7,16
H2 6,99 H3 2,26
C1 120,93 C2 116,68 C3 20,91
4-Pyridon-3-Carboxamid-1-β-D-Ribonukleosid
H1 8,76 H2 8,04 H3 6,72 H4 5,64 H5 4,32 H6 4,26 H7 4,22 H8a 3,88 H8b 3,79
C1 143,71 C2 139,65 C3 121,53 C4 98,09 C5 74,82 C6 70,61 C7 86,38 C8 62,17
5-Hydroxyindolacetat H1 7,18
H2 7,23 H3 7,03 H4 6,95 H5 3,67
C1 126,26 C2 113,37 C3 112,06 C4 119,12 C5 34,69
6-Aminonicotinamid H1 8,53
H2 6,67 H3 7,80
C1 147,62 C2 102,84 C3 144,41
Acetat H1 1,91 C1 25,28
1
2
OH
NH2
O
1
2
OH
OH
O
1
2
OH
3 OH
O
1
2
OH
CH33
1
N
3
2
O
NH2
O
4
5
O
6
7
8OH
OHOH
3
1
4
2
NH
5
O
OHOH
1
3
N
2
NH2
NH2
O
CH31
OH
O
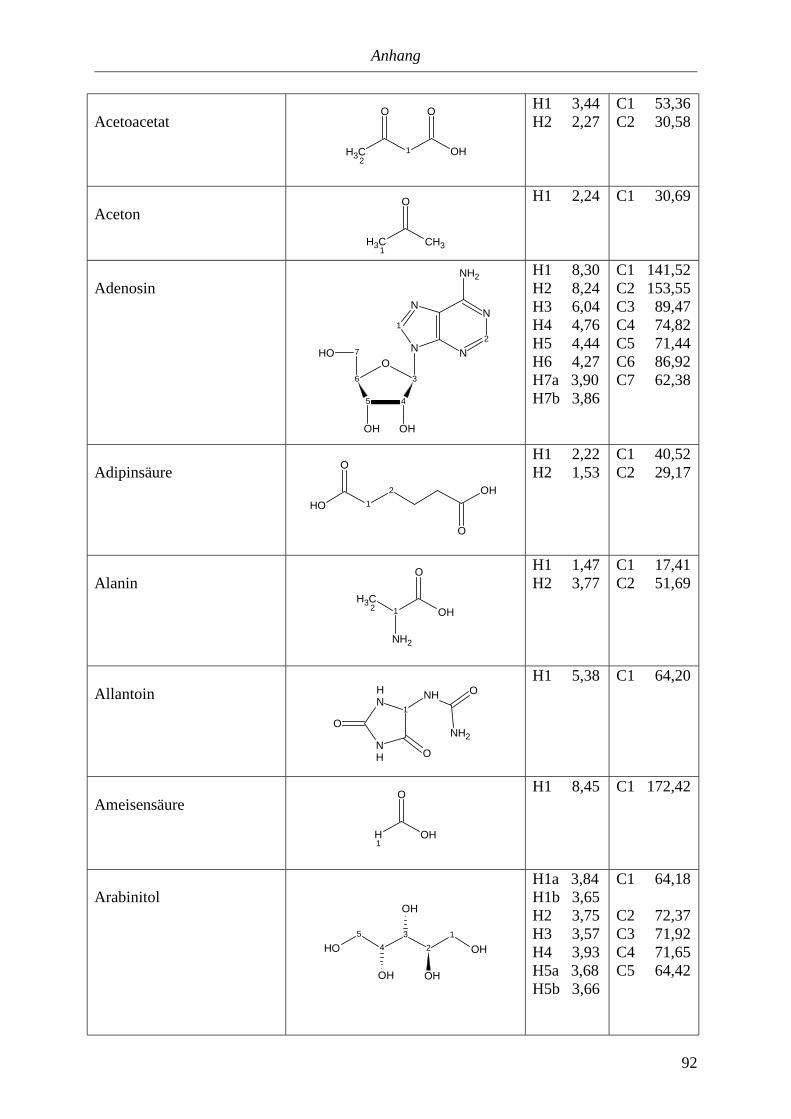
Anhang
92
Acetoacetat H1 3,44
H2 2,27 C1 53,36 C2 30,58
Aceton H1 2,24 C1 30,69
Adenosin H1 8,30
H2 8,24 H3 6,04 H4 4,76 H5 4,44 H6 4,27 H7a 3,90 H7b 3,86
C1 141,52 C2 153,55 C3 89,47 C4 74,82 C5 71,44 C6 86,92 C7 62,38
Adipinsäure H1 2,22
H2 1,53 C1 40,52 C2 29,17
Alanin H1 1,47
H2 3,77 C1 17,41 C2 51,69
Allantoin H1 5,38 C1 64,20
Ameisensäure H1 8,45 C1 172,42
Arabinitol H1a 3,84 H1b 3,65 H2 3,75 H3 3,57 H4 3,93 H5a 3,68 H5b 3,66
C1 64,18
C2 72,37 C3 71,92 C4 71,65 C5 64,42
CH32
1 OH
OO
CH31
CH3
O
N
N
2N
1
N
NH2
3
4
O
5
6
7OH
OHOH
OH 1
2 OH
O
O
O
1
NH2
CH32 OH
NH
NH
1
O
O
NH O
NH2
H1
OH
O
OH5
4
3
2
1
OH
OH
OH
OH
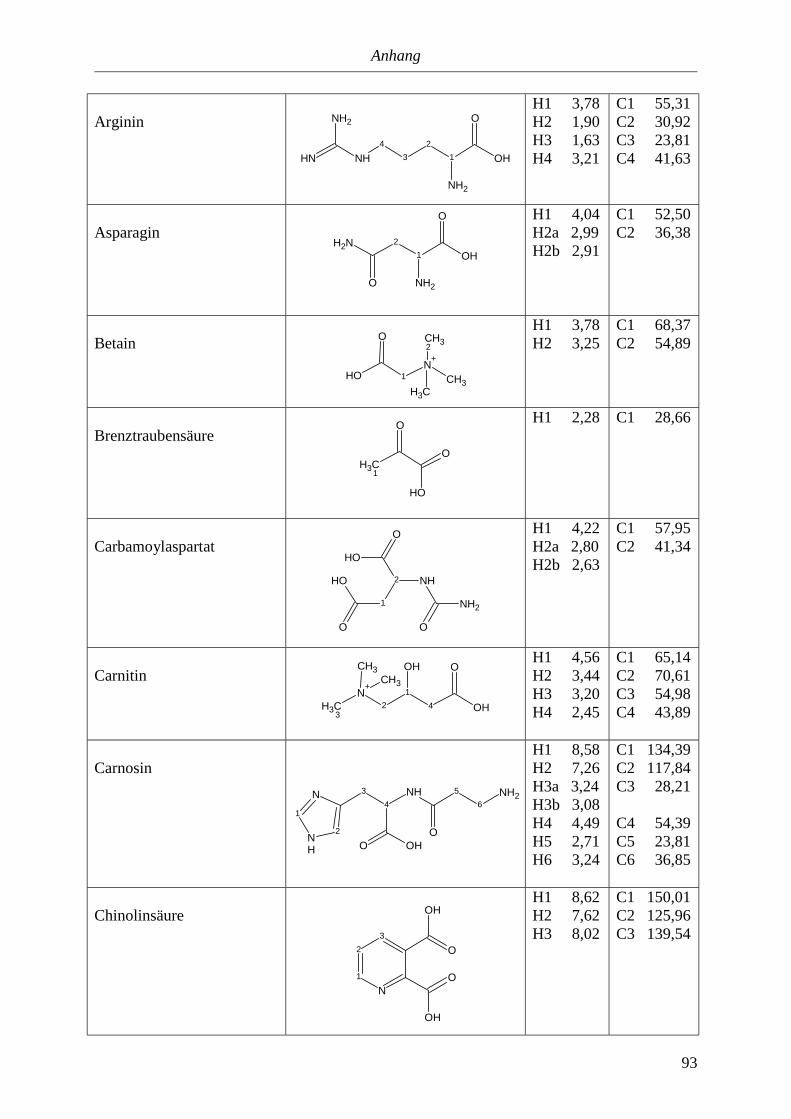
Anhang
93
Arginin H1 3,78
H2 1,90 H3 1,63 H4 3,21
C1 55,31 C2 30,92 C3 23,81 C4 41,63
Asparagin H1 4,04
H2a 2,99 H2b 2,91
C1 52,50 C2 36,38
Betain H1 3,78
H2 3,25 C1 68,37 C2 54,89
Brenztraubensäure H1 2,28 C1 28,66
Carbamoylaspartat H1 4,22
H2a 2,80 H2b 2,63
C1 57,95 C2 41,34
Carnitin H1 4,56
H2 3,44 H3 3,20 H4 2,45
C1 65,14 C2 70,61 C3 54,98 C4 43,89
Carnosin H1 8,58 H2 7,26 H3a 3,24 H3b 3,08 H4 4,49 H5 2,71 H6 3,24
C1 134,39 C2 117,84 C3 28,21
C4 54,39 C5 23,81 C6 36,85
Chinolinsäure H1 8,62 H2 7,62 H3 8,02
C1 150,01 C2 125,96 C3 139,54
NH
O
NH 1
NH2
NH2
4 2
3 OH
2
1
O
O
NH2
NH2
OH
OH 1N
+
CH3
O CH32
CH3
CH31
O
O
OH
OH
2 NH
NH2
O
1
OO
OH
CH33
N+
2
1
4 OH
OOHCH3CH3
1
N
NH
2
3
4NH
O OH
5
O
6NH2
3
N
2
1
OH
O
O
OH
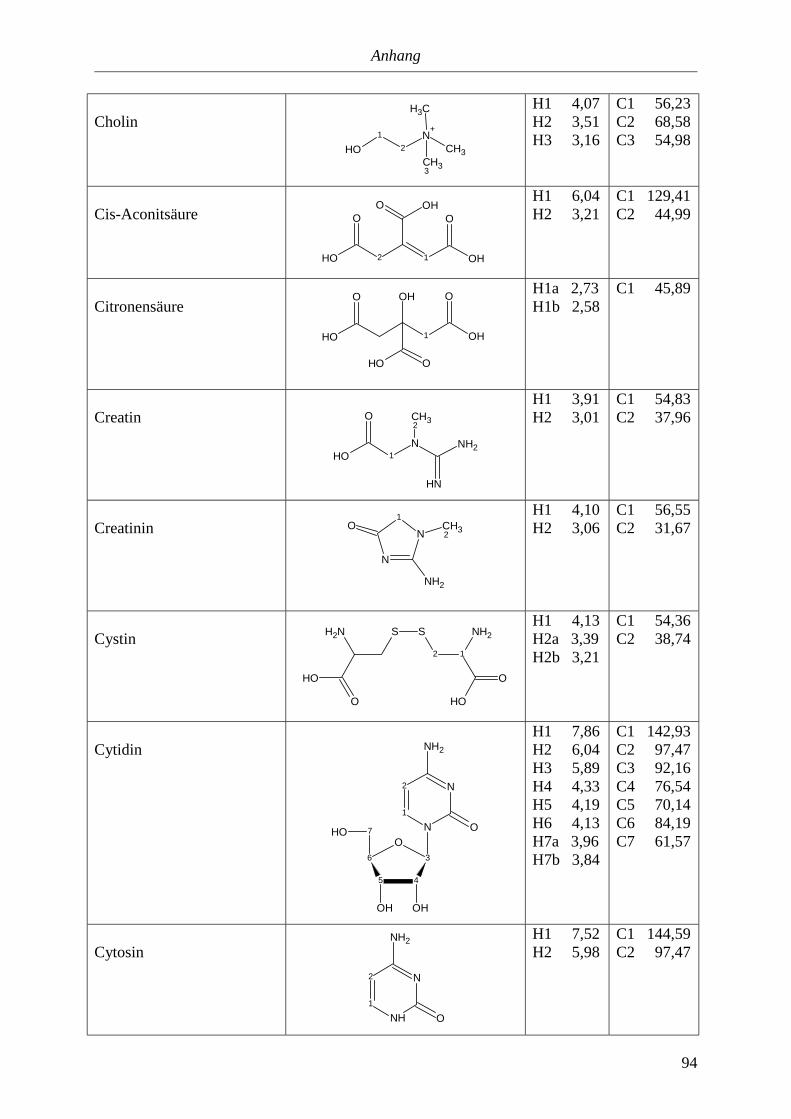
Anhang
94
Cholin H1 4,07
H2 3,51 H3 3,16
C1 56,23 C2 68,58 C3 54,98
Cis-Aconitsäure H1 6,04
H2 3,21 C1 129,41 C2 44,99
Citronensäure H1a 2,73
H1b 2,58 C1 45,89
Creatin H1 3,91
H2 3,01 C1 54,83 C2 37,96
Creatinin H1 4,10
H2 3,06 C1 56,55 C2 31,67
Cystin H1 4,13
H2a 3,39 H2b 3,21
C1 54,36 C2 38,74
Cytidin H1 7,86 H2 6,04 H3 5,89 H4 4,33 H5 4,19 H6 4,13 H7a 3,96 H7b 3,84
C1 142,93 C2 97,47 C3 92,16 C4 76,54 C5 70,14 C6 84,19 C7 61,57
Cytosin H1 7,52
H2 5,98 C1 144,59 C2 97,47
OH
1
2N
+
CH3
CH3
CH33
OH 2 1 OH
O OOHO
OH 1N NH2
O
NH
CH32
OH 1 OH
O OOH
OH O
1
N
NO
NH2
CH32
OH
O
NH2 S S
2 1
NH2
OH
O
N
N
2
1
NH2
O
3
4
O
5
6
7OH
OHOH
N
NH
2
1
NH2
O
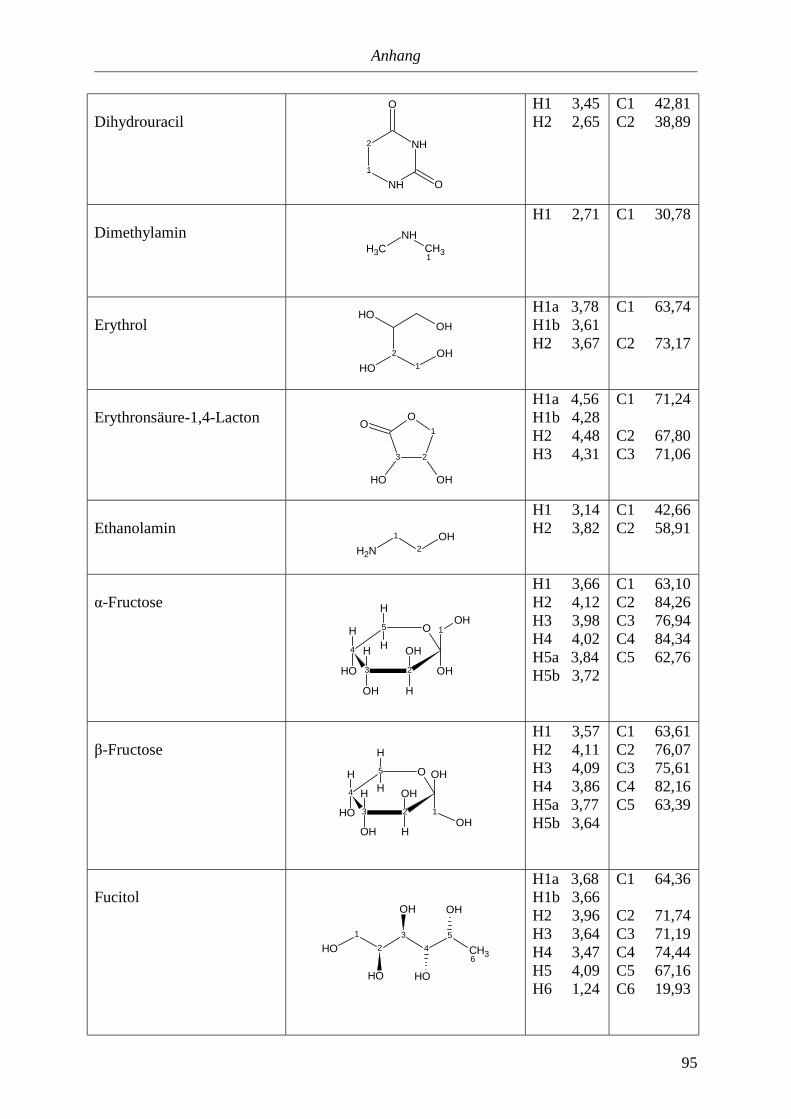
Anhang
95
Dihydrouracil H1 3,45
H2 2,65 C1 42,81 C2 38,89
Dimethylamin H1 2,71 C1 30,78
Erythrol H1a 3,78
H1b 3,61 H2 3,67
C1 63,74
C2 73,17
Erythronsäure-1,4-Lacton H1a 4,56 H1b 4,28 H2 4,48 H3 4,31
C1 71,24
C2 67,80 C3 71,06
Ethanolamin H1 3,14
H2 3,82 C1 42,66 C2 58,91
α-Fructose H1 3,66 H2 4,12 H3 3,98 H4 4,02 H5a 3,84 H5b 3,72
C1 63,10 C2 84,26 C3 76,94 C4 84,34 C5 62,76
β-Fructose H1 3,57 H2 4,11 H3 4,09 H4 3,86 H5a 3,77 H5b 3,64
C1 63,61 C2 76,07 C3 75,61 C4 82,16 C5 63,39
Fucitol H1a 3,68 H1b 3,66 H2 3,96 H3 3,64 H4 3,47 H5 4,09 H6 1,24
C1 64,36
C2 71,74 C3 71,19 C4 74,44 C5 67,16 C6 19,93
2
1
NH
NH
O
O
NHCH3 CH3
1
OH
2
OH 1OH
OH
O
3
1
2
O
OH OH
NH2
1
2OH
5 O
23
4
OH
OHHH
OH
H
OH H
1
HOH
5 O
23
4
1
OHHH
OH
H
OH H
OH
H
OH
OH1
2
3
4
5
CH36
OH
OH
OH
OH
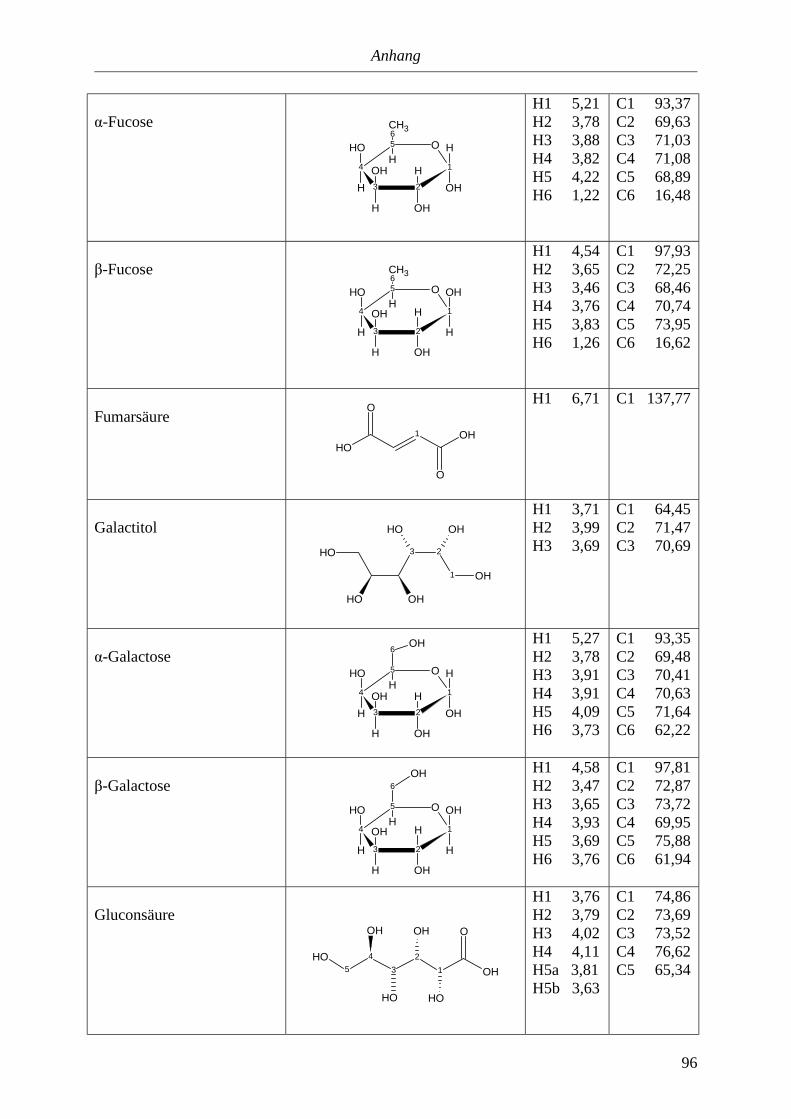
Anhang
96
α-Fucose H1 5,21 H2 3,78 H3 3,88 H4 3,82 H5 4,22 H6 1,22
C1 93,37 C2 69,63 C3 71,03 C4 71,08 C5 68,89 C6 16,48
β-Fucose H1 4,54 H2 3,65 H3 3,46 H4 3,76 H5 3,83 H6 1,26
C1 97,93 C2 72,25 C3 68,46 C4 70,74 C5 73,95 C6 16,62
Fumarsäure H1 6,71 C1 137,77
Galactitol H1 3,71
H2 3,99 H3 3,69
C1 64,45 C2 71,47 C3 70,69
α-Galactose H1 5,27
H2 3,78 H3 3,91 H4 3,91 H5 4,09 H6 3,73
C1 93,35 C2 69,48 C3 70,41 C4 70,63 C5 71,64 C6 62,22
β-Galactose H1 4,58
H2 3,47 H3 3,65 H4 3,93 H5 3,69 H6 3,76
C1 97,81 C2 72,87 C3 73,72 C4 69,95 C5 75,88 C6 61,94
Gluconsäure H1 3,76 H2 3,79 H3 4,02 H4 4,11 H5a 3,81 H5b 3,63
C1 74,86 C2 73,69 C3 73,52 C4 76,62 C5 65,34
5 O
1
23
4
OH
HH
OH
H
OH
H OH
H
CH36
5 O
1
23
4
H
HH
OH
H
OH
H OH
OH
CH36
OH
O
1 OH
O
5 O
1
23
4
OH
HH
OH
H
OH
H OH
H
6OH
OH
OH
3 2
OH
1
OH
OH
OH
5 O
1
23
4
H
HH
OH
H
OH
H OH
OH
6OH
OH5
4
3
2
1 OH
O
OH
OH
OH
OH
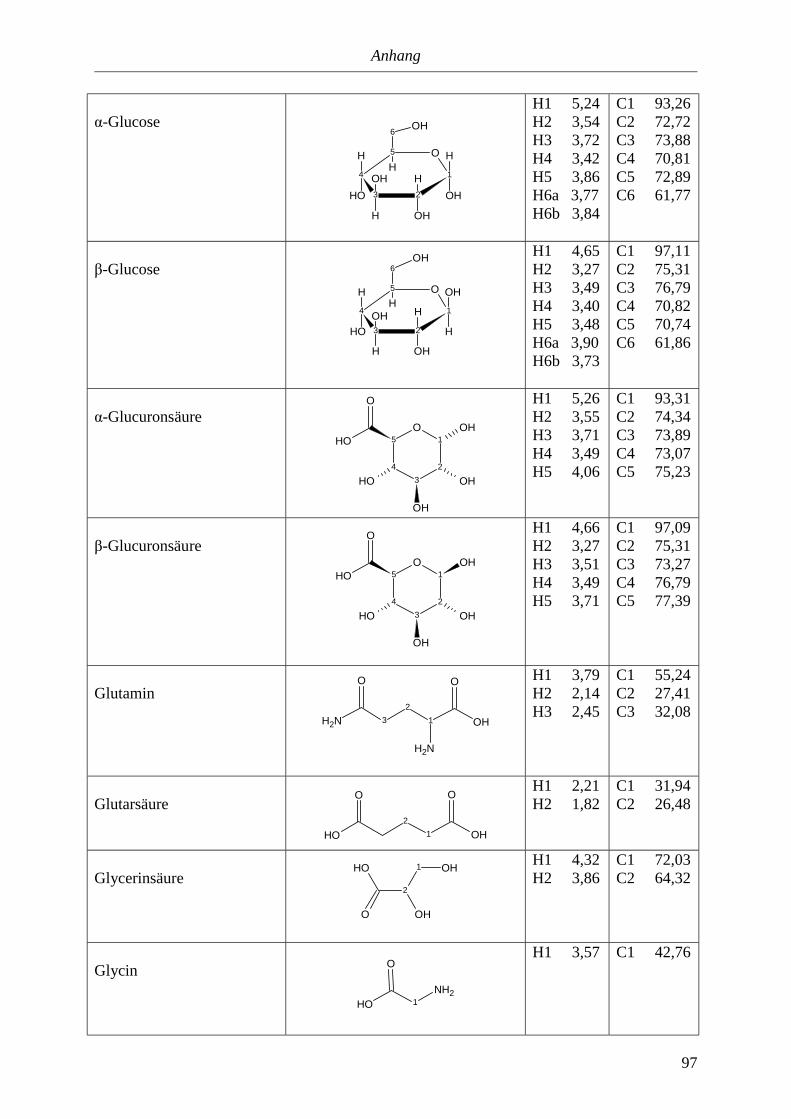
Anhang
97
α-Glucose H1 5,24 H2 3,54 H3 3,72 H4 3,42 H5 3,86 H6a 3,77 H6b 3,84
C1 93,26 C2 72,72 C3 73,88 C4 70,81 C5 72,89 C6 61,77
β-Glucose H1 4,65
H2 3,27 H3 3,49 H4 3,40 H5 3,48 H6a 3,90 H6b 3,73
C1 97,11 C2 75,31 C3 76,79 C4 70,82 C5 70,74 C6 61,86
α-Glucuronsäure H1 5,26
H2 3,55 H3 3,71 H4 3,49 H5 4,06
C1 93,31 C2 74,34 C3 73,89 C4 73,07 C5 75,23
β-Glucuronsäure H1 4,66
H2 3,27 H3 3,51 H4 3,49 H5 3,71
C1 97,09 C2 75,31 C3 73,27 C4 76,79 C5 77,39
Glutamin H1 3,79
H2 2,14 H3 2,45
C1 55,24 C2 27,41 C3 32,08
Glutarsäure H1 2,21
H2 1,82 C1 31,94 C2 26,48
Glycerinsäure H1 4,32
H2 3,86 C1 72,03 C2 64,32
Glycin H1 3,57 C1 42,76
5 O
1
23
4
OH
HH
H
OH
OH
H OH
H
6OH
1
2
O
3
5
4
OH
OH
OH
OH
O
OH
OH2
1 OH
O O
2
OH
O OH
1 OH
NH21OH
O
5 O
1
23
4
H
HH
H
OH
OH
H OH
OH
6OH
1
2
O
3
5
4
OH
OH
OH
OH
O
OH
NH2 3
2
1 OH
O O
NH2
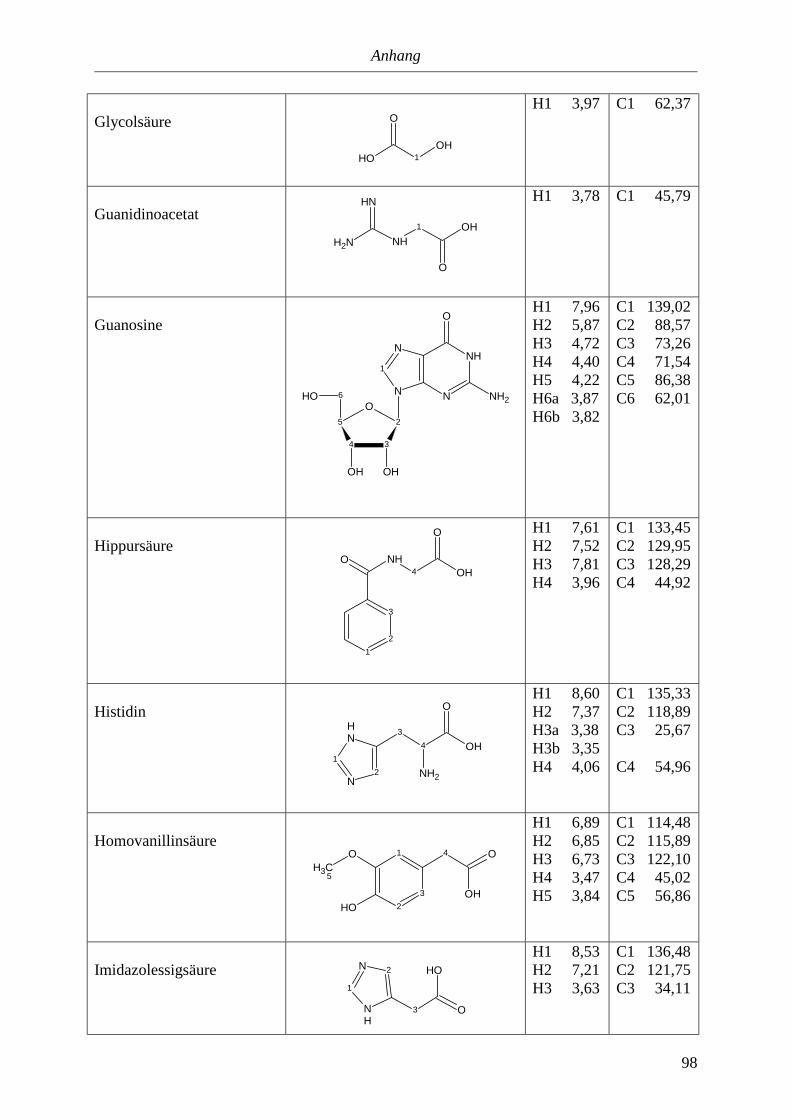
Anhang
98
Glycolsäure H1 3,97 C1 62,37
Guanidinoacetat H1 3,78 C1 45,79
Guanosine H1 7,96
H2 5,87 H3 4,72 H4 4,40 H5 4,22 H6a 3,87 H6b 3,82
C1 139,02 C2 88,57 C3 73,26 C4 71,54 C5 86,38 C6 62,01
Hippursäure H1 7,61
H2 7,52 H3 7,81 H4 3,96
C1 133,45 C2 129,95 C3 128,29 C4 44,92
Histidin H1 8,60
H2 7,37 H3a 3,38 H3b 3,35 H4 4,06
C1 135,33 C2 118,89 C3 25,67
C4 54,96
Homovanillinsäure H1 6,89 H2 6,85 H3 6,73 H4 3,47 H5 3,84
C1 114,48 C2 115,89 C3 122,10 C4 45,02 C5 56,86
Imidazolessigsäure H1 8,53
H2 7,21 H3 3,63
C1 136,48 C2 121,75 C3 34,11
OH 1OH
O
NH2 NH
1 OH
O
NH
N
NH
N
1
N
NH2
O
2
3
O
4
5
6OH
OHOH
3
2
1
NHO4 OH
O
O
4
3
NH2
NH
1
N2
OH
3
1
2OH
OCH35
4 O
OH
1
N
NH
2
3 O
OH
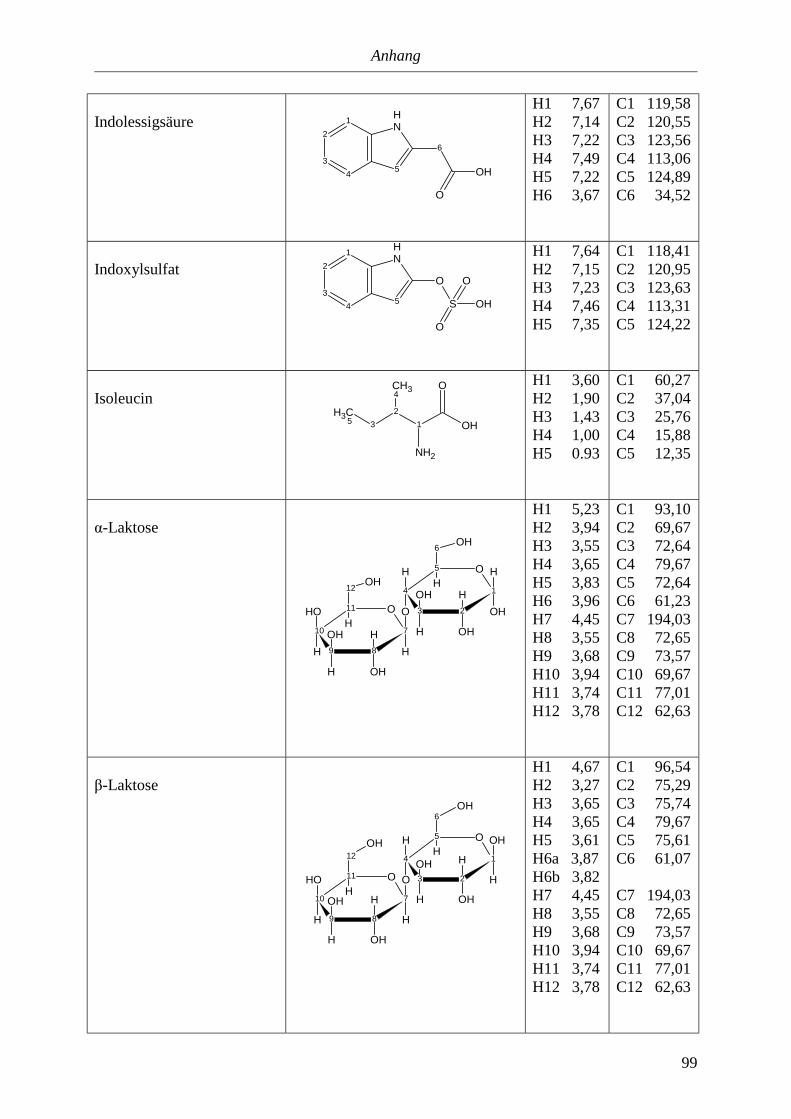
Anhang
99
Indolessigsäure H1 7,67
H2 7,14 H3 7,22 H4 7,49 H5 7,22 H6 3,67
C1 119,58 C2 120,55 C3 123,56 C4 113,06 C5 124,89 C6 34,52
Indoxylsulfat H1 7,64
H2 7,15 H3 7,23 H4 7,46 H5 7,35
C1 118,41 C2 120,95 C3 123,63 C4 113,31 C5 124,22
Isoleucin H1 3,60
H2 1,90 H3 1,43 H4 1,00 H5 0.93
C1 60,27 C2 37,04 C3 25,76 C4 15,88 C5 12,35
α-Laktose H1 5,23 H2 3,94 H3 3,55 H4 3,65 H5 3,83 H6 3,96 H7 4,45 H8 3,55 H9 3,68 H10 3,94 H11 3,74 H12 3,78
C1 93,10 C2 69,67 C3 72,64 C4 79,67 C5 72,64 C6 61,23 C7 194,03 C8 72,65 C9 73,57 C10 69,67 C11 77,01 C12 62,63
β-Laktose H1 4,67 H2 3,27 H3 3,65 H4 3,65 H5 3,61 H6a 3,87 H6b 3,82 H7 4,45 H8 3,55 H9 3,68 H10 3,94 H11 3,74 H12 3,78
C1 96,54 C2 75,29 C3 75,74 C4 79,67 C5 75,61 C6 61,07
C7 194,03 C8 72,65 C9 73,57 C10 69,67 C11 77,01 C12 62,63
1
2
3
45
NH
6
OH
O
1
2
3
45
NH
O
S OH
O
O
1
O
NH2
2
3
CH34
CH35 OH
5 O
1
23
4
OH
HH
H
O
OH
H OH
H
6OH
11 O
7
89
10
H
HH
OH
H
OH
H OH
12OH
5 O
1
23
4
H
HH
H
O
OH
H OH
OH
6OH
11 O
7
89
10
H
HH
OH
H
OH
H OH
12OH
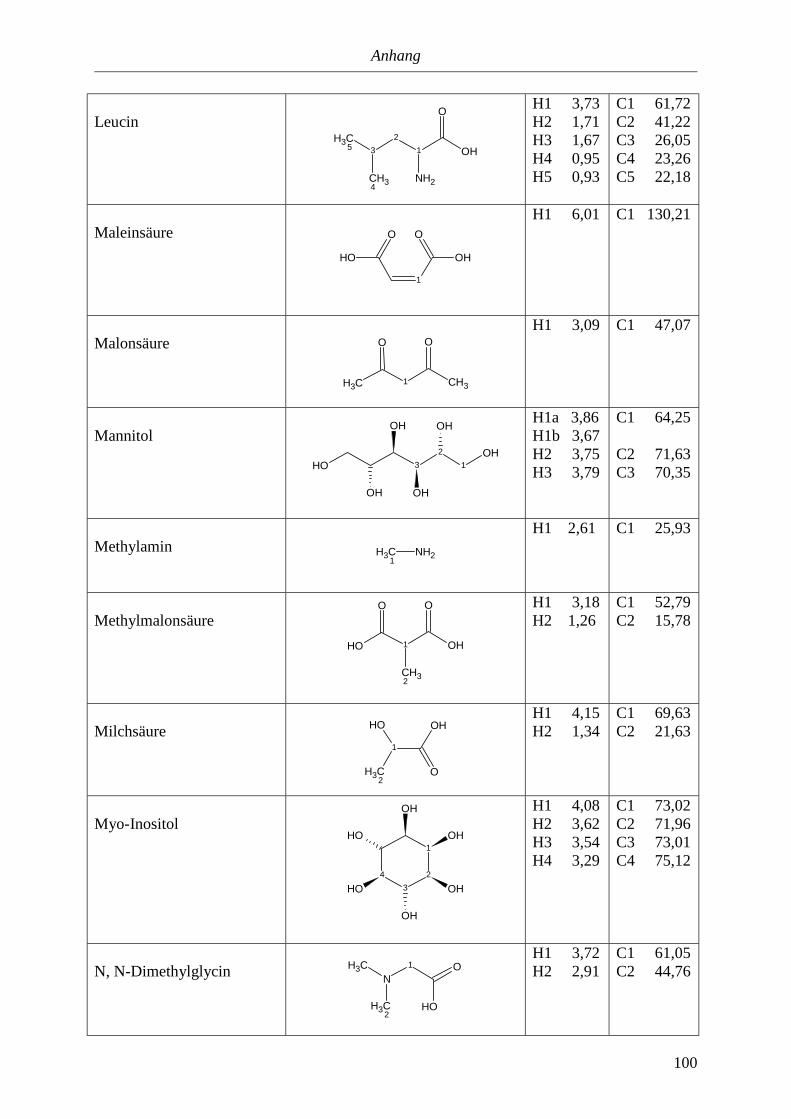
Anhang
100
Leucin H1 3,73
H2 1,71 H3 1,67 H4 0,95 H5 0,93
C1 61,72 C2 41,22 C3 26,05 C4 23,26 C5 22,18
Maleinsäure H1 6,01 C1 130,21
Malonsäure H1 3,09 C1 47,07
Mannitol H1a 3,86
H1b 3,67 H2 3,75 H3 3,79
C1 64,25
C2 71,63 C3 70,35
Methylamin H1 2,61 C1 25,93
Methylmalonsäure H1 3,18
H2 1,26 C1 52,79 C2 15,78
Milchsäure H1 4,15
H2 1,34 C1 69,63 C2 21,63
Myo-Inositol H1 4,08
H2 3,62 H3 3,54 H4 3,29
C1 73,02 C2 71,96 C3 73,01 C4 75,12
N, N-Dimethylglycin H1 3,72
H2 2,91 C1 61,05 C2 44,76
1
O
2
NH2
3CH35
CH34
OH
1
OH
O
OH
O
CH31 CH3
O O
OH 3
2
1OH
OH
OH
OH
OH
CH31
NH2
OH 1 OH
OO
CH32
CH32
1
OH
O
OH
1
2
3
4
OH
OH
OHOH
OH
OH
N1CH3 O
OHCH32
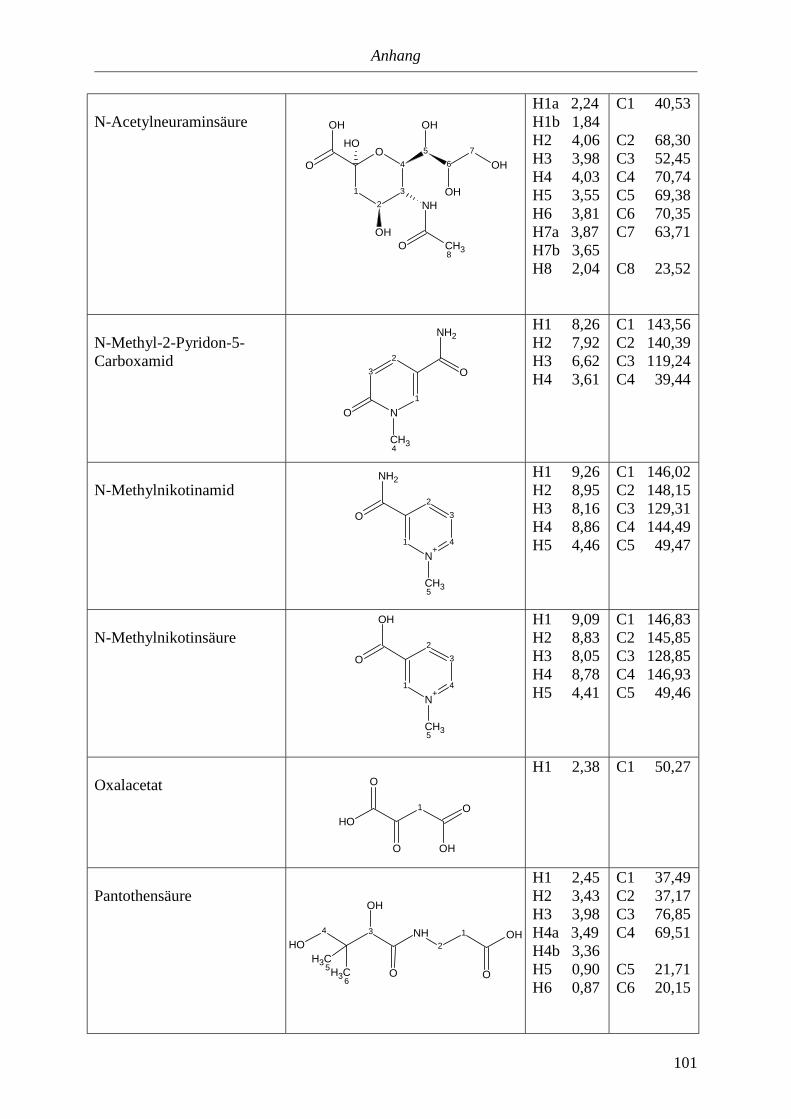
Anhang
101
N-Acetylneuraminsäure H1a 2,24 H1b 1,84 H2 4,06 H3 3,98 H4 4,03 H5 3,55 H6 3,81 H7a 3,87 H7b 3,65 H8 2,04
C1 40,53
C2 68,30 C3 52,45 C4 70,74 C5 69,38 C6 70,35 C7 63,71
C8 23,52
N-Methyl-2-Pyridon-5-Carboxamid
H1 8,26 H2 7,92 H3 6,62 H4 3,61
C1 143,56 C2 140,39 C3 119,24 C4 39,44
N-Methylnikotinamid H1 9,26
H2 8,95 H3 8,16 H4 8,86 H5 4,46
C1 146,02 C2 148,15 C3 129,31 C4 144,49 C5 49,47
N-Methylnikotinsäure H1 9,09
H2 8,83 H3 8,05 H4 8,78 H5 4,41
C1 146,83 C2 145,85 C3 128,85 C4 146,93 C5 49,46
Oxalacetat H1 2,38 C1 50,27
Pantothensäure H1 2,45
H2 3,43 H3 3,98 H4a 3,49 H4b 3,36 H5 0,90 H6 0,87
C1 37,49 C2 37,17 C3 76,85 C4 69,51
C5 21,71 C6 20,15
4
3
O
2
1
OH
OH
O
OH
NH
CH38
O
5
6
OH
OH
7
OH
1
2
N
3
CH34
O
O
NH2
3
4
2
N+
1
CH35
NH2
O
3
4
2
N+
1
CH35
OH
O
OH1 O
OHO
O
OH4 3 NH
2
1 OH
CH36
CH35
OH
O O
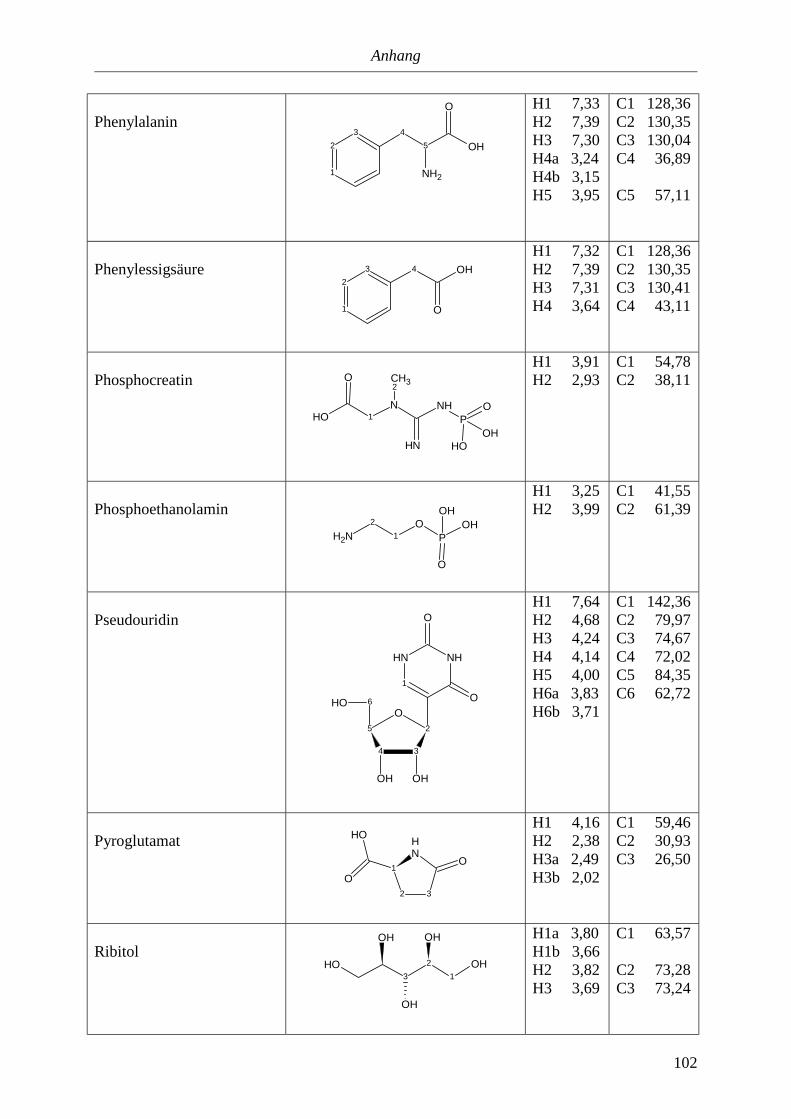
Anhang
102
Phenylalanin H1 7,33
H2 7,39 H3 7,30 H4a 3,24 H4b 3,15 H5 3,95
C1 128,36 C2 130,35 C3 130,04 C4 36,89
C5 57,11
Phenylessigsäure H1 7,32
H2 7,39 H3 7,31 H4 3,64
C1 128,36 C2 130,35 C3 130,41 C4 43,11
Phosphocreatin H1 3,91
H2 2,93 C1 54,78 C2 38,11
Phosphoethanolamin H1 3,25
H2 3,99 C1 41,55 C2 61,39
Pseudouridin H1 7,64
H2 4,68 H3 4,24 H4 4,14 H5 4,00 H6a 3,83 H6b 3,71
C1 142,36 C2 79,97 C3 74,67 C4 72,02 C5 84,35 C6 62,72
Pyroglutamat H1 4,16
H2 2,38 H3a 2,49 H3b 2,02
C1 59,46 C2 30,93 C3 26,50
Ribitol H1a 3,80
H1b 3,66 H2 3,82 H3 3,69
C1 63,57
C2 73,28 C3 73,24
5
O
4
NH2
3
2
1
OH
43
2
1
OH
O
OH 1N NH
P
O CH32
NH
O
OHOH
NH2
2
1O
POH
O
OH
NH
1
NH
O
O
2
3
O
4
5
6OH
OHOH
32
1
NH
O
OH
O
OH1
2
3OH
OH
OH
OH
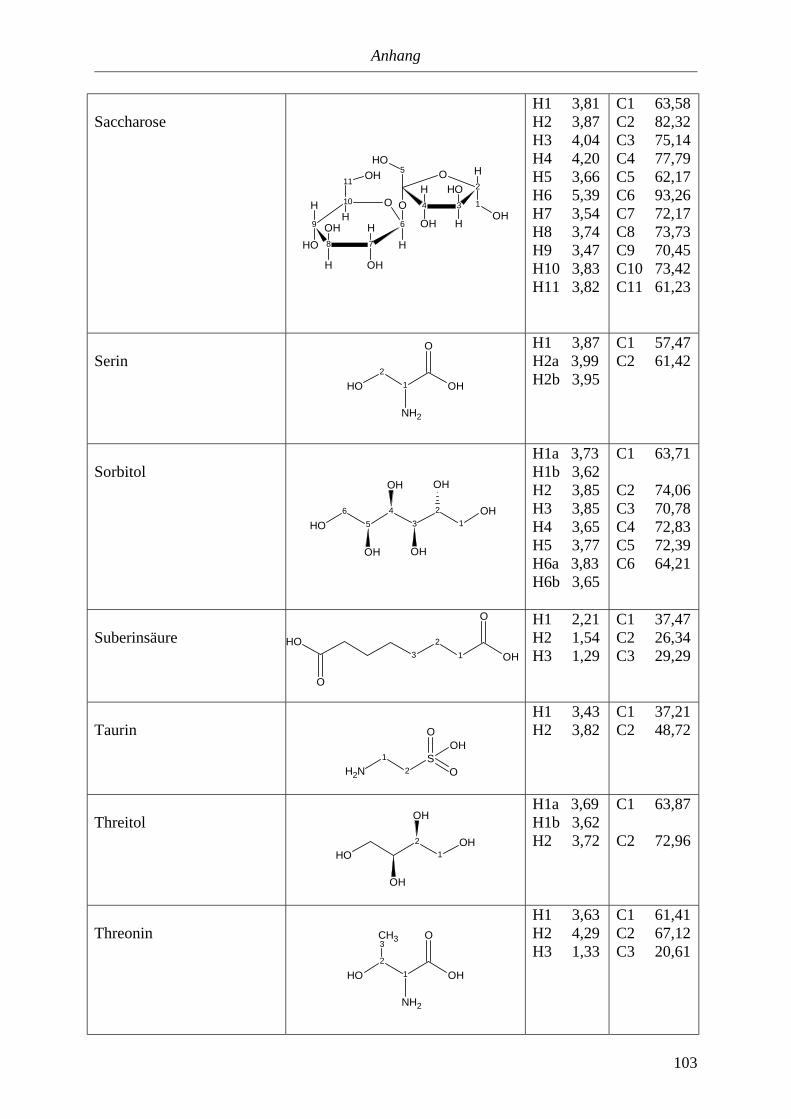
Anhang
103
Saccharose H1 3,81
H2 3,87 H3 4,04 H4 4,20 H5 3,66 H6 5,39 H7 3,54 H8 3,74 H9 3,47 H10 3,83 H11 3,82
C1 63,58 C2 82,32 C3 75,14 C4 77,79 C5 62,17 C6 93,26 C7 72,17 C8 73,73 C9 70,45 C10 73,42 C11 61,23
Serin H1 3,87
H2a 3,99 H2b 3,95
C1 57,47 C2 61,42
Sorbitol H1a 3,73
H1b 3,62 H2 3,85 H3 3,85 H4 3,65 H5 3,77 H6a 3,83 H6b 3,65
C1 63,71
C2 74,06 C3 70,78 C4 72,83 C5 72,39 C6 64,21
Suberinsäure H1 2,21
H2 1,54 H3 1,29
C1 37,47 C2 26,34 C3 29,29
Taurin H1 3,43
H2 3,82 C1 37,21 C2 48,72
Threitol H1a 3,69
H1b 3,62 H2 3,72
C1 63,87
C2 72,96
Threonin H1 3,63
H2 4,29 H3 1,33
C1 61,41 C2 67,12 C3 20,61
4 3
2O H
1
H
OH
OH
H
5
O
OH
OH10 O
6
78
9
H
HH
H
OH
OH
H OH
11OH
1
O
NH2
2
OH OH
OH
6
5
4
3
2
1OH
OH
OH
OH
OH
OH3
2
1 OH
O
O
NH2
1
2S
O
OOH
OH2
1OH
OH
OH
1
O
2
NH2
OH
CH33
OH
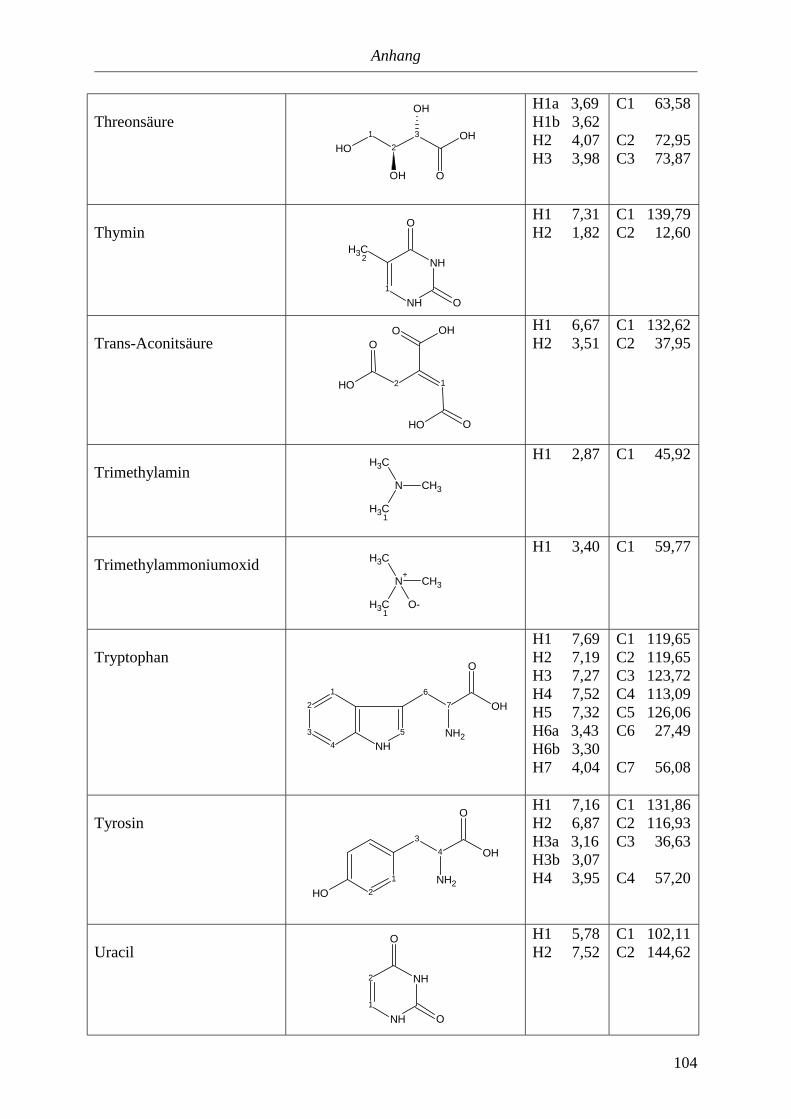
Anhang
104
Threonsäure H1a 3,69
H1b 3,62 H2 4,07 H3 3,98
C1 63,58
C2 72,95 C3 73,87
Thymin H1 7,31
H2 1,82 C1 139,79 C2 12,60
Trans-Aconitsäure H1 6,67
H2 3,51 C1 132,62 C2 37,95
Trimethylamin H1 2,87 C1 45,92
Trimethylammoniumoxid H1 3,40 C1 59,77
Tryptophan H1 7,69
H2 7,19 H3 7,27 H4 7,52 H5 7,32 H6a 3,43 H6b 3,30 H7 4,04
C1 119,65 C2 119,65 C3 123,72 C4 113,09 C5 126,06 C6 27,49
C7 56,08
Tyrosin H1 7,16
H2 6,87 H3a 3,16 H3b 3,07 H4 3,95
C1 131,86 C2 116,93 C3 36,63
C4 57,20
Uracil H1 5,78
H2 7,52 C1 102,11 C2 144,62
OH1
2
3 OH
O
OH
OH
1
NH
NH
O
O
CH32
OH 2 1
O
OH O
OHO
N CH3
CH3
CH31
N+
CH3
CH3
CH31
O-
O
7
6
NH25
NH
1
4
3
2 OH
O
4
3
NH21
2OH
OH
2
1
NH
NH
O
O
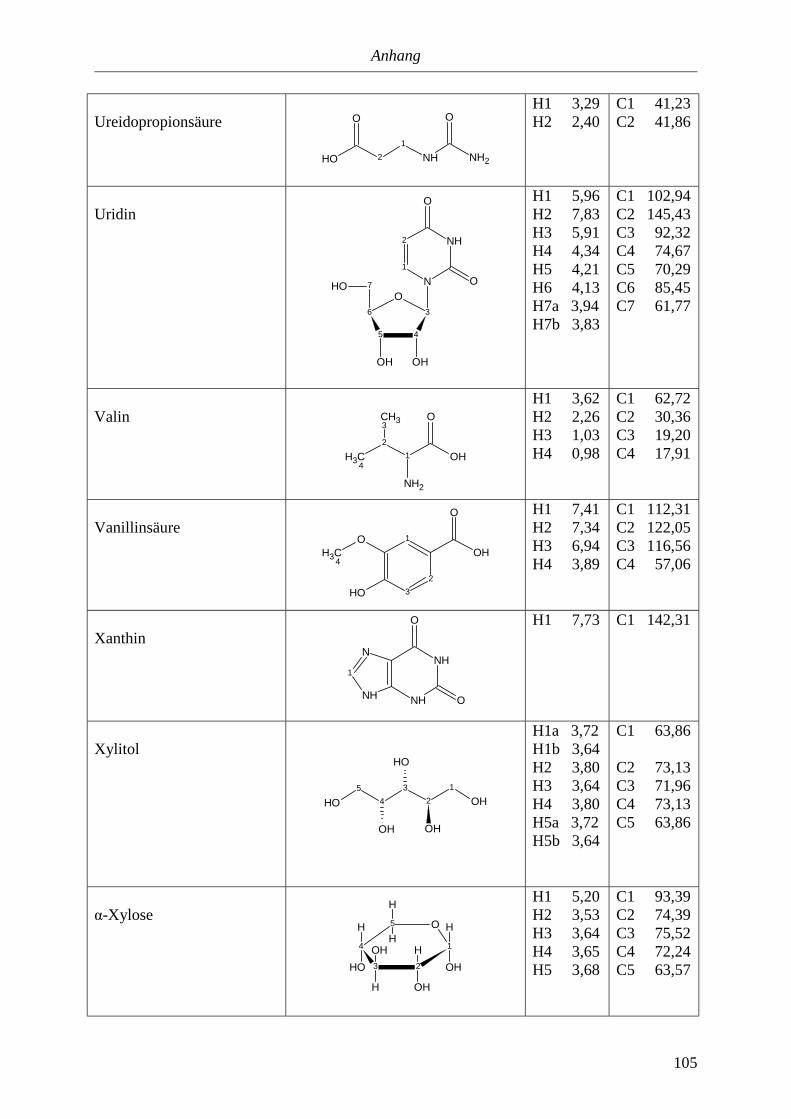
Anhang
105
Ureidopropionsäure H1 3,29
H2 2,40 C1 41,23 C2 41,86
Uridin H1 5,96
H2 7,83 H3 5,91 H4 4,34 H5 4,21 H6 4,13 H7a 3,94 H7b 3,83
C1 102,94 C2 145,43 C3 92,32 C4 74,67 C5 70,29 C6 85,45 C7 61,77
Valin H1 3,62
H2 2,26 H3 1,03 H4 0,98
C1 62,72 C2 30,36 C3 19,20 C4 17,91
Vanillinsäure H1 7,41
H2 7,34 H3 6,94 H4 3,89
C1 112,31 C2 122,05 C3 116,56 C4 57,06
Xanthin H1 7,73 C1 142,31
Xylitol H1a 3,72
H1b 3,64 H2 3,80 H3 3,64 H4 3,80 H5a 3,72 H5b 3,64
C1 63,86
C2 73,13 C3 71,96 C4 73,13 C5 63,86
α-Xylose H1 5,20
H2 3,53 H3 3,64 H4 3,65 H5 3,68
C1 93,39 C2 74,39 C3 75,52 C4 72,24 C5 63,57
OH 2
1
NH NH2
OO
2
1
N
NH
O
O
3
4
O
5
6
7OH
OHOH
1
O
NH2
2
CH33
CH34
OH
2
1
3OH
OCH34
OH
O
NH
NH
NH
1
N
O
O
OH5
4
3
2
1
OH
OH
OH
OH
5 O
1
23
4
OH
HH
H
OH
OH
H OH
H
H

Anhang
106
β-Xylose H1 4,56
H2 3,24 H3 3,42 H4 3,60 H5a 3,93 H5b 3,31
C1 97,37 C2 75,26 C3 77,01 C4 72,44 C5 66,36
5 O
1
23
4
H
HH
H
OH
OH
H OH
OH
H

Abkürzungen
107
6. Abkürzungen
γ gyromagnetische Konstante SPE Festphasenextraktion
δ chemische Verschiebung RNA Ribonukleinsäure
µ magnetischer Moment THF Tetrahydrofuran
π π-Elektronen TMS Tetramethylsilan
ωL Lamor-Frequenz TOCSY total correlation spectrosc.
B0 Magnetfeld UV Ultraviolett
CoA Coenzym A v/v Volumenprozent
COSY correlation spectroscopy VIS Visible
CPMG Carr-Pucell-Meiboom-Gill
Experiment
DEPT Distortionless Enhancement by
Polarisation Transfer
DNA Desoxyribonukleinsäure
FID freier Induktionsabfall (free
induction decay)
HMBC Heteronuclear Multiple Bond
Correlation
HPLC Hochdruckflüssigchromatographie
HSQC Heteronuclear Single Quantum
Coherence
Hz Hertz
I Drehimpuls
J Kopplungskonstante
LLE Flüssig-Flüssig-Extraktion
MHz Megahertz
MRT Magnetresonanztomographie
nm Nanometer
NMR Kernmagnetische Resonanz
NOESY nuclear Overhauser enhancement
Spectroscopy
NP Normale Phase
ppm parts per million
RP Umkehrphase (reversed phase)

Literatur
108
7. Literatur
Amberg, A.; Rosner, E.; Dekant, W. Biotransformation and kinetics of excretion of ethyl
tert-butyl ether in rats and human. Toxicol. Sci. (2000); 53: 194-201
Angyal, S. J.; Le Fur, R.The 13C-NMR spectra of alditols. Carbohydrate research (1980);
84: 201-209
Angyal, S. J.; Wheen, R. G.The Composition of reducing sugars in aqueos solution:
Glyceraldehyde, Erythrose, Threose. Aust. J. Chem. (1980); 33: 1001-1011
Bain, M. A.; Fornasine, G.; Evans, A. M. Trimethylamine: Metabolic, pharmacokinetic and
safety aspects. Curr. Drug Metab. (2005); 6: 227-240
Balato, N.; Cusano, F.; Lembo, G.; Santoianni, P. Tyrosinemia type II in two cases
previously reported as Richner-Hanhart syndrome. Dermatologica (1986); 173: 66-74
Bales, J. R.; Higham, D. P.; Howe, I.; Nicholson, J. K.; Sadler, P. J. Use of high-
resolution proton nuclear magnetic resonance spectroscopy for rapid multi-component
analysis of urine. Clin. Chem. (1984); 30: 426-432
Bao, Y.; Silva, T. M. J.; Guerrant, R. L.; Lima, A. A. M.; Fox, J. W. Direct analysis of
mannitol, lactulose and glucose in urine samples by high performance anion-exchange
chromatography with pulse amperometric detection. Clinical evaluation of intestinal
permeability in human immunodeficiency virus infection. J. Chromatogr. B (1996);
685: 105-112
Baumgartner, M. R.; Almashanu, S. Suormala, T.; Obie, C.; Cole, R. N.; Packman, S.;
Baumgartner, E. R.; Valle, D. The molecular basis of human 3-methylcrotonyl-CoA
carboxylase deficiency. J. Clin. Invest. (2001); 107: 495-504
Binzak, B. A.; Wevers, R. A.; Moolenaar, S. H.; Lee, Y. M.; Hwu, W. L.; Poggi-Bach, J.;
Engelke, U. F.; Hoard, H. M.; Vockley, J. G.; Vockley, J. Clonig of
dimethylglycine dehydrogenase and a new human inborn error of metabolism,
dimethylglycine dehydrogenase deficiency. Am. J. Hum. Genet. (2001); 68: 839-847

Literatur
109
Bjorke Monsen, A. L.; Ueland, P. M. Homocysteine and methylmalonic acid in diagnosis
and risk assessment from infancy to adolescence. Am. J. Clin. Nutr. (2003); 78: 7-21
Botitsi, E.; Frosyni, C.; Tsipi, D. Determination of pharmaceuticals from different
therapeutic classes in wastewaters by liquid chromatography-electrospray ionization-
tandem mass spectrometry. Anal. Bioanal. Chem. (2007); 387: 1317-1327
Brand, H. S.; Jörning, G. G. A.; Chamuleau, R. A. F. M.; Abraham-Inpijn, L. Effect of a
protein-rich meal on urinary and salivary free amino acid concentrations in human
subjects. Clin. Chim. Acta (1997); 264: 37-47
Brevik, A.; Rasmussen, S. E.; Drevon, C. A.; Andersen, L. F. Urinary excretion of
flavonoids reflects even small changes in the dietary intake of fruits and vegetables.
Cancer Epidemiol. Biomarkers Prev. (2004); 13: 843-849
Bröer, S.; Cavanaugh, J. A.; Rasko, J. E. Neutral amino acid transport in epithelial cells
and its malfunction in Hartnup disorders. Biochem. Soc. Trans. (2005); 33: 233-236
Brusati, V.; Jozwik, M.; Jozwik, M.; Teng, C.; Paolini, C.; Marconi, A. M.; Battaglia, F.
C. Fetal and maternal non-glucose carbohydrates and polyols concentrations in normal
human pregnancies at term. Pediatr. Res. (2005); 58: 700-704
Carter, E. G. A. Quantitation of urinary niacin metabolites by reversed-phase liquid
chromatography. Am. J. Clin. Nutr. (1982); 36: 926-930
Chuang, D. T.; Chuang, J. L.; Wynn, R. M. Lessons from genetic disorders of branched-
chain amino acid metabolism. J. Nutr. (2006); 136: 243S-249S
Coen, M.; O´Sullivan, M.; Bubb, W. A.; Kuchel, P. W.; Sorrel, T. Proton nuclear
magnetic resonance-based metabonomics for rapid diagnosis of meningitis and
ventriculitis. Clin. Infect. Dis. (2005); 41: 1582-1590

Literatur
110
Cohen, M.; Hartlage, P. L.; Krawiecki, N.; Roesel, R. A.; Carter, A. L.; Hommes, F. A.
Serum carnosinase deficiency: A non-disabling phenotype. J. Ment. Defic. Res.
(1985); 29: 383-389
Creer, M. H.; Lau, B. W.; Jones, J. D.; Chan, K. M. Pyroglutamic academia in an adult
patient. Clin. Chem. (1989); 35: 684-686
Danpure, C. J. Molecular etiology of primary hyperoxaluria type I: new directions for
treatment. Am. J. Nephrol. (2005); 25: 303-310
Deschauer, M.; Gizatullina, Z.; Schulze, A.; Pritsch, M.; Knoppel, C.; Knape, M.; Zierz,
S.; Gellerich, F. N. Molecular and biochemical investigations in fumarase deficiency.
Mol. Genet. Metab. (2006); 88: 146-152
Dietzen, D. J.; Wilhite, T. R.; Kenagy, D. N.; Milliner, D. S.; Smith, C. H.; Landt, M.
Extraction of glyceric and glycolic acids from urine with tetrahydrofuran: utility in
detection of primary hyperoxaluria. Clin. Chem. (1997); 43: 1315-1320
Drennan, C. L.; Matthews, R. G.; Rosenblatt, D. S.; ledley, F. D.; Fenton, F. D.; Ludwig,
M. L. Molecular basis for dysfunction of some mutant forms of methylmalonyl-CoA
mutase: Deductions from the structure of methionine synthase. Proc. Natl. Acad. Sci.
U S A (1996); 93: 5550-5555
Dunckelmann, R. J.; Ebinger, F.; Schulze, A.; Wanders, R. J.; Rating, D.; Mayatepek,
E. 2-ketoglutarate dehydrogenase deficiency with intermittent 2-ketoglutaric aciduria.
Neuropediatrics (2000); 31: 35-38
Dutta, S. P.; Crain, P. F.; McCloskey, J. A.; Chheda, G. B. Isolation and characterization
of 1-beta-D-ribofuranosylpyridin-4-one-3-carboxamide from human urine. Life Sci.
(1979); 24: 1381-1388
Easley, C. J.; Jin, L. J.; Presto Elqstoen, K. B.; Jellum, E.; Landers, J. P.; Ferrance, J. P.
Capillary electrophoresis with laser-induced fluorescence detection for laboratory
diagnosis of galactosemia. J. Chromatogr. A (2003); 1004: 29-37

Literatur
111
Fan, T. W.-M. Metabolite profiling by one- and two-dimensional NMR analysis of complex
mixtures. Prog. Nuc. Magn. Res. Spec. (1996); 28: 161-219
Feldman, J. M.; Lee, M. D.; Lee, E. Serotonin content of foods: effect on urinary excretion
of 5-hydroxyindoleacetic acid Am. J. Clin. Nutr. (1985); 42: 639-643
Fontaine, M.; Porchet, N.; Largilliere, C.; Marrakchi, S.; Lhermitte, M.; Aubert, J.-P.;
Degand, P. Biochemical contribution to diagnosis and study of a new case of D-
glyceric academia/aciduria. Clin. Chem. (1989); 35: 2148-2151
Fonteh, A. N.; Harrington, R. J.; Harrington, M. G. Quantification of free amino acids and
dipeptides using isotope dilution liquid chromatography and electrospray ionization
tandem mass spectrometry. Amino Acids (2007); 32: 203-212
Font-Llitjos, M.; Jimenez-Vidal, M.; Bisceglia, L.; Di Perna, M.; de Sanctis, L.;
Rousaud, F.; Zelante, L.; Palacin, M.; Nunes, V. New insights into cystinuria: 40
new mutations, genotype-phenotype correlation, and digenic inheritance causing
partial phenotype. J. Med. Genet. (2005); 42: 58-68
Fribolin, H. Ein- und Zweidimensionale NMR-Spektroskopie: Eine Einführung. 3. Auflage,
Weinheim: Wiley-VCH (1999)
Froesch, E. R. Disorders of fructose metabolism. Clin. Endocrinol. Metab. (1976); 5: 599-
611
Gerlo, E. A.; Sevens, C. Urinary and plasma catecholamines and urinary catecholamine
metabolites in pheochromocytoma: Diagnostic value in 19 cases. Clin. Chem. (1994);
40: 250-256
Gervay, J.; Batta, G. pH dependent C6 and C8 13C chemical shift assignment in N-Acetyl
Neuraminic acid. Tetrahedron Letters (1994); 35: 3009-3012

Literatur
112
Grey, A. A.; Smith, I. C. P.; Hruska, F. E. A model for the molecular conformation of α-
Pseudouridine from nuclear magnetic resonance data. J. Am. Chem. Soc. (1971); 93:
1765-1769
Haberle, J.; Gorg, B.; Rutsch, F.; Schmidt, E.; Toutain, A.; Benoist, J. F.; Gelot, A.; Suc,
A. L.; Hohne, W.; Schliess, F.; Haussinger, D.; Koch, H. G. Congenital glutamine
deficiency with glutamine synthetase mutations. N. Engl. J. Med. (2005); 353: 1926-
1933
Harris, R. A.; Joshi, M.; Jeoung, N. H.; Obayashi, M. Overview of the molecular and
biochemical basis of branched-chain amino acid catabolism. J. Nutr. (2005); 135:
1527S-1530S
Hawkes, G. E.; Lewis, D. 1H nuclear magnetic resonance spectra and conformations of
alsitols in deuterium oxide. J. Chem. Soc. Perkin Trnas. II (1984); 395: 2073-2078
Hodavance, M. S.; Ralston, S. L.; Pelcer, I. Beyond blood sugar: the potential of NMR-
based metabonomics for type 2 human diabetes, and the horse as a possible model.
Anal. Bioanal. Chem. (2006); 387: 533-537
Hofmann, U.; Schwab, M.; Seefried, S.; Marx, C.; Zanger, U. M.; Eichelbaum, M.;
Mürdter, T. E. Sensitive method fort the quantification of urinary pyrimidine
metabolites in healthy adults by gas chromatography-tandem mass spectrometry. J.
Chroamtogr. B (2003); 791: 371-380
Holmes, E.; Foxall, P. J. D.; Spraul, M.; Farrant, R. D.; Nicholson, J. K.; Lindon, J. C.
750MHz 1H NMR spectroscopy characterisation of the complex metabolic pattern of
urine from patients with inborn errors of metabolism: 2-Hydroxyglutaric aciduria and
maple syrup urine disease. J. Pharm. Biomed. Anal. (1997); 15: 1647-1659
Igbal, S. J.; Davies, T.; Cole, R.; Whitaker, P.; Chapman, C. Neutrophil alkaline
phosphatase (NAP) score in the diagnosis of hyperphosphatasia. Clin. Chem. (2000);
302: 49-57

Literatur
113
Ito, H.; Gonthier, M.-P.; Manach, C.; Morand, C.; Mennen, L.; Rémésy, C.; Scalbert, A.
Polyphenol levels in human urine after intake of six different polyphenol-rich
beverages. Brit. J. Nutr. (2005); 94: 500-509
Jain, D. S.; Subbaiah, G.; Sanyal, M.; Jain, V. K.; Shrivastav, P. A rapid and specific
approach for direct measurement of Pravastatin concentration in plasma by LC-MS /
MS employing solid-phase extraction. Biomed. Chromatogr. (2007); 21: 67-78
Jecklin, E. Arbeitsbuch Anatomie und Physiologie: für Krankensschwestern, Krankenpfleger
und andere Medizinalfachberufe. 4. Auflage, Stuttgart: Fischer (1986)
Jung, T. S.; Hahm, J. R.; Kim, J. J.; Kang, M. Y.; Moon, S. W.; Lee, K. W.; Kim, H. C.;
Lee, J. D.; Kim, J. H.; Kim, D. R.; Chung, S. I. Determination of urinary Myo-
/chiro-inositol ratios from Korean diabetes patients. Yonsei Med. J. (2005); 46: 532-
538
Kabadi, U. M. Urine glucose testing: Reliable backup for whole blood glucose monitoring. J.
Fam. Pract. (1992); 34: 495-497
Kammerer, B.; Frickenschmidt, A.; Müller, C. E.; Laufer, S.; Gleiter, C. H.; Liebich, H.
Mass spectrometric identification of modified urinary nucleosides used as potential
biomedical markers by LC-ITMS coupling. Anal. Bioanal. Chem. (2005); 382: 1017-
1026
Kato, K.; Silva, M. S.; Needham, L. L.; Calafat, A. M. Determination of 16 phthalate
metabolites in Urine using automated sample preparation and on-line preconcentration
/ high-performance liquid chromatography / tandem mass spectrometry. Anal. Chem.
(2005); 77: 2985-2991
Kern, S. M.; Bennet, R. N.; Mellon, F. A.; Kroon, P. A.; Garcia-Conesa, M.-T.
Absorption of hydroxycinnamates in humans after high-bran cereal consumption. J.
Agric. Food Chem. (2003); 51: 6050-6055

Literatur
114
Khandelwal, J. K.; Kline, T.; Green, J. P. Measurement of imidazoleacetic acid in urine by
gas chromatography-mass spectrometry. J. Chromatogr. (1985); 343: 249-257
Kitagawa, T.; Smith, B. A.; Brown, E. S. Gas-liquid chromatography of phenylalanine and
its metabolites in serum and urine of various hyperphenylalaninemic subjects, their
relatives, and controls. Clin. Chem. (1975); 21: 735-740
Koch, B. The role of urine glucose testing in the management of diabetes mellitus. Can. J.
Diabetes (2004); 28: 238-245
Koelbing, H. M. Die antiken Grundlagen der Harnschau. Der Urin im medizinischen Denken.
Basel: Geigy (1967)
Kornfeld, R.; Kornfeld, S. Comparative aspects of glycoprotein structure. Annu. Rev.
Biochem. (1976); 45: 217-237
Kubota, K.; Ishizaki, T. Dose-dependent pharmacokinetics of benzoic acid following oral
administration of sodium benzoate to humans. Er. J. Clin. Pharmcol. (1991); 41: 363-
368
Kuhara, T.; Phdoi, C.; Ohse, M. Simple gas chromatographic-mass spectrometric procedure
for diagnosing pyrimidine degradation defects for prevention of severe anticancer side
effects. J. Chromatogr. B (2001); 758: 61-74
Kvist, E.; Sjolin, K. E.; Iversen, J. Excretion patterns of pseudouridine and beta-
aminoisobutyric acid in patients with tumours of the upper urinary tract. Scand. J.
Urol. Nephrol. (1990); 24: 287-292
Lee, J.; Chung, B. C. Simultaneous measurement of urinary polyols using gas
chromatography/mass spectrometry. J. Chromatogr. B (2005); 831: 126-131
Lee, M.-J.; Wang, Z.-Y.; Li, H.; Chen, L.; Sun, Y.; Gobbo, S.; Balentine, D. A.; Yang, C.
S. Analysis of plasma and urinary tea polyphenols in human subjects. Cancer
Epidemiol. Biomarkers Prev. (1995); 4: 393-399

Literatur
115
Lehtonen, L.; Anttila, V.-J.; Ruutu, T.; Salonen, J.; Nikoskelainen, J.; Eerola, E.;
Ruutu, P. Diagnosis of disseminated candidiasis by measurement of urine D-
arabinitol/L-arabinitol ratio. J. Clin. Micobiol. (1996); 34: 2175-2179
Lenz, E. M.; Bright, J.; Wilson, I. D.; Hughes, A.; Morrisson, J.; Lindberg, H.; Lockton,
A. Metabonomics, dietary influences and cultural differences: A 1H NMR-based study
of urine samples obtained from healthy british and Swedish subjects. J. Pharm.
Biomed. Anal. (2004); 36: 841-849
Lenz, E. M.; Bright, J.; Wilson, I. D.; Morgan, S. R.; Nash, A. F. P. A 1H NMR-based
metabonomic study of urine and plasma samples obtained from healthy human
subjects. J. Pharm. Biomed. Anal. (2003); 33: 1103-1115
Leroy, J. G.; Seppala, R.; Huizing, M.; Dacremont, G.; De Simpel, H.; Van Coster, R.
N.; Orvisky, E.; Krasnewich, D. M.; Gahl, W. A. Dominant inheritance of sialuria,
an inborn error of feedback inhibition. Am. J. Hum. Genet. (2001); 68: 1419-1427
Leutert, G. Systematische und funktionelle Anatomie des Menschen: für medizinische
Assistenzberufe. 2. Auflage, Berlin: Ullstein-Mosby (1994)
Liebich, H. M.; Pickert, A. Gas chromatographic profiling of phenolic acids in urine of
patients with cirrhosis of the liver. J. Chromatogr. (1985); 338: 25-32
Luceri, C.; Caderni, G.; Lodovici, M.; Spagnesi, M. T.; Monserrat, C.; Lanvioni, L.;
Dolara, P. Urinary excretion of sucrose and fructose as a predictor for sucrose intake
in dietary intervention studies. Cancer Epidemiol. Biomarkers Prev. (1996); 5: 167-
171
Machino, H.; Miki, Y.; Kawatsu, T.; Kida, K.; Matsuda, H. Successful dietary control of
tyrosinemia II. J. Am. Acad. Dermatol. (1983); 9: 533-539
Manning, N. J.; Pollitt, R. J.; Tracer studies of the interconversion of R- and S-
methylmalonic semialdehydes in man. Biochem. J. (1985); 231: 481-484

Literatur
116
McCreanor, G. M.; Bender, D. A. The metabolism of high intakes of tryptophan,
nicotinamide and nicotinic acid in the rat. Brit. J. Nutr. (1986); 56: 577-586
Mitchell, S. C.; Smith, R. L. Trimethylaminuria: The fish malodour syndrome. Drug Metab.
Dispos. (2001); 29: 517-521
Moolenaar, S. H.; Poggi-Bach, J.; Engelke, U. F.; Corstiaensen, J. M.; Heershap, A.; de
Jong, J. G.; Binzak, B. A.; Vockley, J.; Wevers, R. A. Defect in dimethylglycine
dehydrogenase, a new inborn error of metabolism: NMR spectroscopy study. Clin.
Chem. (1999); 45: 459-464
Mortensen, G. K.; Main, K. M.; Andersson, A.-M.; Leffers, H.; Skakkebæk, N. E.
Determination of phthalate monoesters in human milk, consumer milk, and infant
formula by tandem mass spectrometry (LC-MS-MS). Anal. Bioanal. Chem. (2005);
382: 1084-1092
Munro, I. C.; Bernt, W. O.; Borzelleca, J. F.; Flamm, G.; Lynch, B. S.; Kennepohl, E.;
Bär, E. A.; Modderman, J. Erythritol: An interpretive summary of biochemical,
metabolic, toxicological and clinical data. Food and Chemical Toxicology (1998); 36:
1139-1174
Nakano, I.; Tsugawa, T.; Shinohara, R.; Watanabe, F.; Fujita, T.; Nagata, M.; Kato, T.;
Himeno, Y.; Kobayashi, T.; Fujiwara, K.; Nagata, M.; Itoh, M.; Nagasaka, A.
Urinary sorbitol measurement and the effect of an aldose reductase inhibitor on its
concentration in the diabetic state. J. Diabetes Complications (2003); 17: 337-342
Nelson, D. L.; Cox, M. M. Lehninger Biochemie. 3. Auflage, Berlin Heidelberg: Springer
(2001)
Nicholson, J. K.; Foxall, P. J. D. 750MHz 1H and 1H- 13C NMR spectroscopy of human
blood plasma. Anal. Chem. (1995); 67: 793-811

Literatur
117
Niwa, T.; Aoyama, I.; Takayama, F.; Tsukushi, S.; Miyazaki, T.; Owada, A.; Shiidai, T.
Urinary indoxyl sulphate is a clinical factor that affects the progression of renal
failure. Miner. Electrolyte Metab. (1999); 25: 118-122
Nyhan, W. L.; Hilton, S. Histidinuria: defective transport of histidine. Am. J. Med. Genet.
(1992); 44: 558-561
Oka, H.; Suzuki, S.; Suzuki, H.; Oda, T. Increased urinary excretion of L-xylulose in
patients with liver cirrhosis. Clin. Chim. Acta (1976); 67: 131-136
Okamoto, H.; Ishikawa, A.; Yoshitake, Y.; Kodama, N.; Nishimuta, M.; Fukuwatari, T.;
Shibata, K. Diurnal variations in human urinary excretion of nicotinamide catabolites:
effects of stress on the metabolism of nicotinamide. Am. J. Clin. Nutr. (2003); 77: 406-
410
Oyanagi, K.; Tsuchiyama, A.; Itakura, Y.; Tamura, Y.; Nakao, T.; Fujita, S.; Shiono, H.
Clinical, biochemical and enzymatic studies in type I hyperprolinemia associated with
chromosomal abnormality. Tohoku J. Exp. Med. (1987); 151: 465-475
Persson, E.; Löfgren, L.; Hansson, G.; Abrahamsson, B.; Lennernäs, H.; Nilsson, R.
Simultaneous assessment of lipid classes and bile acids in human intestinal fluid by
solid phase extraction and HPLC methods. J.Lipid Res. (2007); 48: 242-251
Pitkanen, E. The conbersion of D-xylose into D-threitol in patients without liver disease and
in patients with portal liver cirrhosis. Clin. Chim.Acta (1977); 80: 49-54
Podebrad, F.; Heil, M.; Beck, T.; Mosandl, A.; Sewell, A. C.; Bohles, H.
Stereodifferentiation of 3-hydroxyisobutyric- and 3-aminoisobutyric acid in human
urine by enantioselective multidimensional capillary gas chromatography-mass
spectrometry. Clin. Chim. Acta. (2000); 292: 93-105
Rana, S. V.; Thapa, B. R.; Pal, R. Comparison of D-xylose hydrogen breath test with
urinary D-xylose test in indian children with celiac disease. Dig. Dis. Sci. (2007);
ahead of print.

Literatur
118
Rebouche, C. J.; Paulson, D. J. Carnitine metabolism and function in humans. Ann. Rev.
Nutr. (1986); 6: 41-66
Rechner, A. R.; Smith, M. A.; Kuhnle, G.; Gibson, G. R.; Debnam, E. S.; Srai, S. K.;
Moore, K. P.; Rice-Evans, C. A. Colonic metabolism of dietary polyphenols:
influence of structure on microbial fermentation products. Free Radic. Biol. Med.
(2004); 36: 212-225
Rechner, A. R.; Kuhnle, G.; Bremner, P.; Hubbard, G. P.; Moore, K. P.; Rice-Evans, C.
A. The metabolic fate of dietary polyphenols in humans. Free Radic. Biol. Med.
(2002); 33: 220-235
Ridel, K. R.; Leslie, N. D.; Gilbert, D. L. An update of the long-term neurological effects of
galactosemia. Pediatr. Neurol. (2005); 33: 153-161
Rider, L. G.; Schiffenbauer, A. S.; Zito, M.; Lim, K. L.; Ahmed, A.; Zemel, L. S.;
Rennebohm, R. M.; Passo, M. H.; Summers, R. M.; Hicks, J. E.; Lachenbruch, P.
A.; Heyes, M. P., Miller, F. W. Neopterin and quinolinic acid are surrogate measures
of disease activity in the juvenile idiopathic inflammatory myopathies. Clin. Chem.
(2002); 48: 1681-1688
Rios, L. Y.; Gonthier, M.-P.; Rémésy, C.; Mila, I.; Lapierre, C.; Lazarus, S. A.;
Williamson, G.; Scalbert, A. Chocolate intake increases urinary excretion of
polyphenol-derived phenolic acids in healthy human subjects. Am. J. Clin. Nutr.
(2003); 77: 912-918
Sakai, T.; Yamamoto, K.; Yokota, H.; Hakozaki-Usui, K.; Hino, F.; Kato, I. Rapid,
simple enzymatic assay of free L-fucose in serum and urine, and its use as a marker
for cancer, cirrhosis and gastric ulcers. Clin. Chem. (1990); 36; 474-476
Sakiyama, T.; Nakabayashi, H.; Shimizu, H.; Kondo, W.; Kodama, S.; Kitagawa, T. A
successful trial of enzyme replacement therapy in a case of argininemia. Tohoku J.
Exp. Med. (1984); 142: 239-248

Literatur
119
Sarashina, G.; Yamakoshi, M.; Noritake, M.; Takahashi, M.; Kure, M.; Katsura, Y.;
Shiomi, H.; Tsuboi, I.; Kawazu, S.; Yamagata, F.; Tominaga, M.; Matsuoka, T.A
study of urinary myo-inositol as a sensitive marker of glucose intolerance. Clin. Chim.
Acta (2004); 344: 181-188
Schlemmer, H.-P. Vom Magneteisenstein zum Magnetresonanztomographen. Radiologe
(2005); 45: 356-362
Schram, K. H. Urinary nucleosides. Mass spectrum. Rev. (1998); 17: 131-251
Schwarz, H. P.; Schaefer, T.; Bachmann, C. Galactose and galactitol in the urine of
children with compound heterozygosity for duarte variant and classical galactosemia
(GtD/gt) after an oral galactose load. Clin. Chem. (1985); 31: 420-422
Seidel, A.; Brunner, S.; Seidel, P.; Fritz, G. I.; Herbarth, O. Modified nucleosides: An
accurate tumour marker for clinical diagnosis of cancer, early detection and therapy
control. Br. J. Cancer (2006); 94: 1726-1733
Sewell, A. C.; Poets, C. F.; Degen, I.; Stoss, H.; Pontz, B. F. The spectrum of free
neuraminic acid storage disease in childhood: Clinical, morphological and biochemical
observations in three non-Finnish patients. Am. J. Med. Genet. (1996); 63: 203-208
Shi, R. Z.; Ho, Y. P.; Yeung, J. H.; Or, P. M.; To, K. K.; Lau, M. W.; Arumanayagam,
M. Development of an enzyme-linked immunosorbent assay with monoclonal
antibody for quantification of homovanillic in human urine samples. Clin. Chem.
(1998); 44: 1674-1679
Shigematsu, Y.; Sudo, M.; Momoi, T.; Inoue, Y.; Suzuki, Y.; Kameyama, J. Changing
plasma and urinary organic acid levels in a patient with isovaleric acidemia during an
attack. Pediatr. Res. (1982); 16: 771-775
Shirokane, Y.; Utsushikawa, M.; Nakajima, M. A new enzymic determination of
guanidinoacetic acid in urine. Clin. Chem. (1987); 33: 394-397

Literatur
120
Sigmundsdóttir, G.; Christensson, B.; Björklund, L. J.; Håkansson, K.; Pehrson, C.;
Larrson, L. Urine D-arabinitol/L-arabinitol ratio in diagnosis of invasive candidiasis
in newborn infants. J. Clin. Microbiol. (2000); 38: 3039-3042
Sjölin, J.; Stjernström, H.; Henneberg, S.; Hambraeus, L.; Friman, G. Evaluation of
urinary 3-methylhistidine excretion in infection by measurements of 1-methylhistidine
and the creatinine ratios. Am. J. Clin. Nutr. (1989); 49: 62-70
Slominska, E. M.; Carrey, E. A.; Foks, H.; Orlewska, C.; Wieczerzak, E.; Sowinski, P.;
Yacoub, M. H.; Marinaki, A. M. Simmonds, H. A.; Smolenski, R. T. A novel
nucleotide found in human erythrocytes, 4-pyridone-3-carboxyamide-1-beta-D-
ribonucleoside triphosphate. J. Biol. Chem. (2006); 281: 32057-32064
Suzuki, J.; Akihiro, K.; Ikunoshin, K.; Sumihiro, H.; Tokuji, I. Assay of urinary free
fucose by fluorescence labelling and high-performance liquid chromatography. Clin.
Chem. (1992); 38: 752-755
Sykut-Cegielska, J.; Gradowska, W.; Mercimek-Mahmutoglu, S.; Stockler-Ipsiroglu, S.
Biochemical and clinical characteristics of creatine deficiency syndromes. Acta
Biochim. Pol. (2004); 51: 875-882
Szabo, A.; Billett, E.; Turner, J. Phenylethylamine, a possible link to the antidepressant
effects of exercise? Br. J. Sports Med. (2001); 35: 342-343
Tada, K.; Hayasaka, K. Non-ketotic hyperglycinaemia: Clinical and biochemical aspects.
Eur. J. Pediatr. (1987); 146: 221-227
Takeuchi, K.; Wagner, G. NMR studies of protein interactions. Curr. Opin. Struct. Biol.
(2006); 16: 109-117
Tasevska, N.; Runswick, S. A.; McTaggart, A.; Bingham, S. A. Urinary sucrose and
fructose as biomarkers for sugar consumption. Cancer Epidemiol. Biomarkers Prev.
(2005); 14: 1287-1294

Literatur
121
Thompson, J. A.; Markey, S. P.; Fennessey, P. V. Gas-chromatographic/mass-
spectrometric identification and quantitation of tetronic and deoxytetronic acids in
urine from normal adults and neonates. Clin. Chem. (1975); 21: 1892-1898
Tribalat, L.; Paisse, O.; Dessalces, G.; Grenier-Loustalot, M.-F. Advantages of LC-MS-
MS compared to LC-MS for the determination of nitrofuran residues in honey. Anal.
Bioanal. Chem. (2006); 386: 2161-2168
Tsikas, D.; Thum, T.; Becker, T.; Pham, V. V.; Chobanyan, K.; Mitschke, A.;
Beckmann, B.; Gutzki, F. M.; Bauersachs, J.; Stichtenoth, D. O. Accurate
quantification of dimethylamine (DMA) in human urine by gas chromatography-mass
spectrometry as pentafluorbenzamide derivative: Evaluation of the relationship
between DMA and ist precursor asymmetric dimethylarginine (ADMA) in health and
disease. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. (2007); epub ahead of
Tu, Z.; Xu, S.; Wu, M. Clinical value of urinary and serum pseudouridine in diagnosis and
monitoring of primary liver cancer. Chin. Med. J. (Engl.) (1995); 108: 204-208
Tůma, P.; Samcová, E.; Balínová, P. Dtermination of 3-methylhistidine and 1-
methylhistidine in untreated erine samples by capillary electrophoresis. J.
Chromatogr. B (2005); 821: 53-59
Valencia, M. P.; Castillo, M. MRI findings in posttraumatic spinal cord wallerian
degeneration. Clin. Imaging (2006); 30: 431-433
Van Kuilenburg, A. B. P.; Meinsma, R.; Beke, E.; Assmann, B.; Ribes, A.; Lorente, I.;
Busch, R.; Mayatepek, E.; Abeling, N. G.; van Cruchten, A.; Stroomer, A. E.;
van Lenthe, H.; Zoetekouw, L.; Kulik, W.; Hoffmann, G. F.; Voit, T.; Wevers, R.
A.; Rutsch, F.; van Gennip, A. H. Beta-ureidopropionase deficiency: An inborn error
of pyrimidine degradation associated with neurological abnormalities. Hum. Mol.
Genet. (2004); 13: 2793-2801

Literatur
122
Van Kuilenburg, A. B. P.; van Lenthe, HH.; Löffler, M.; van Gennip, A. H. Analysis of
pyrimidine synthesis „de novo“ intermediates in urine and dried urine filter-paper
strips with HPLC-electrospray tandem mass spectrometry. Clin. Chem. (2004); 50:
2117-2124
Vaynberg, J.; Qin, J. Weak protein-protein interactions as proped by NMR spectroscopy.
Trends Biotechnol. (2006); 24: 22-27
Verhoeven, N. M.; Huck, J. H. J.; Roos, B.; Struys, E. A.; Salomons, G. S.; Douwes, A.
C.; van der Knaap, M. S.; Jakobs, C. Transaldolase deficiency: Liver cirhosis
associated with a new inborn error in the pentose phosphate pathway. Am. J. Hum.
Genet. (2001); 68: 1086-1092
Walker, V.; Mills, G. A. Solid-phase extraction in clinical biochemistry. Ann. Clin. Biochem.
(2002); 39: 464-477
Walsh, M. C.; Brennan, L.; Malthouse, J. P. G.; Roche, H. M.; Gibney, M. J. Effect of
acute dietary standardization on the urinary, plasma, and salivary metabolomic profiles
of healthy humans. Am. J. Clin. Nutr. (2006); 84: 531-539
Wamelink, M. M. C.; Smith, D. E. C.; Jakobs, C.; Verhoeven, N. M. Analysis of polyols
in urine by liquid chromatography-tandem mass spectrometry: A usefull tool for
recognition of inborn errors affecting polyol metabolism. J. Inherit. Metab. Dis.
(2005); 28: 951-963
Weiss, P.; Tietze, F.; Gahl, W. A.; Seppala, R.; Ashwell, G. Identification of the metabolic
defect in sialuria. J. Biol. Chem. (1989); 264: 17635-17636
White, W. A new look at the role of Urinalysis in the history of diagnostic medicine. Clin.
Chem. (1991); 37: 119-125
Wring, S. A.; Terry, A.; Causon, R.; Jenner, W. N. The electroanalysis of mannitol, xylose
and lactulose at copper electrodes: coltammetric studies and bioanalysis in human

Literatur
123
urine by means of HPLC with electrochemical detection. J. Pharm: Biomed. Anal.
(1998); 16: 1213-1224
Yamauchi, M.; Kimura, K.; Maezawa, Y. Ohata, M.; Mizuhara, Y.; Hirakawa, J.;
Nakajima, H.; Toda, G. Urinary level of L-fucose as a marker of alcoholic liver
disease. Alcohol. Clin. Exp. Res. (1993); 17: 268-271
Zeisel, S. H.; DaCosta, K.-A. Increase in human exposure to methylamine precursors of N-
Nitrosamines after eating fish. Cancer Res. (1986); 46: 6136-6138
Zheng, Y.-F.; Xu, G.-W.; Liu, D.-Y.; Xiong, J.-H.; Zhang, P.-D.; Zhang, C.; Yang, Q.;
Lv, S. Study of nucleosides as biological marker in cancer patients analyzed by
micellar electrokinetic capillary chromatography. Electrophoresis (2002); 23: 4104-
4109
Zorita, S.; Westbom, R.; Thörneby, L.; Björklund, E.; Mathiasson, L. Development of a
combined solid-phase extraction – supercritical fluid extraction procedure for the
determination of polychlorinated biphenyls in wastewater. Anal. Sci. (2006); 22: 1455-
1459
Zschocke, J.; Quak, E.; uldber, P.; Hoffmann, G. F. Mutation analysis in glutaric aciduria
type I. J. Med. Genet. (2000); 37: 177-181
Zuetenhorst, J. M.; Korse, C. M.; Bonfrer, J. M.; Peter, E.; Lamers, C. B.; Taal, B. G.
Daily cyclic changes in the urinary excretion of 5-hydroxyindoleacetic acid in patients
with carcinoid tumors. Clin. Chem. (2004); 50: 1634-1639
Zuppi, C.; Messana, I.; Forni, F.; Ferrari, F.; Rossi, C.; Giardina, B. Influence of feeding
on metabolite excretion evidenced by urine 1H NMR spectral profiles: A comparison
between subjects living in Rome and subjects living at arctic latitudes (Svaldbard).
Clin. Chim. Acta (1998); 278: 75-79
Zuppi, C.; Messana, I.; Forni, F.; Ferrari, F.; Rossi, C.; Giardina, B. Influence of feeding
on metabolite excretion evidenced by urine 1H NMR spectral profiles: A comparison

Literatur
124
between subjects living in Rome and subjects living at arctic latitudes (Svaldbard).
Clin. Chim. Acta (1998); 278: 75-79
SDBS http://riodb01.ibase.aist.go.jp/sdbs/cgi-bin/direct_frame_top.cgi


















![Chromatographische, NMR-spektroskopische und ...€¦ · 2+ und cis-[PtCl 2(NH3)2] Dissertation zur Erlangung des Grades eines Doktors der Naturwissenschaften der Fakultät für Chemie](https://img.pdfslide.org/doc/110x75/5aff25557f8b9a444f8fc9c8/chromatographische-nmr-spektroskopische-und-2-und-cis-ptcl-2nh32-dissertation.jpg)










