Embed Size (px)
Citation preview

Aus der Abteilung für Biochemische EndokrinologieInstitut für Physiologische Chemie desUniversitätskrankenhauses Eppendorf in HamburgAbteilungsdirektor Professor Dr. med. Hans Joachim Seitz
SCHNELLE EFFEKTE DER SCHILDDRÜSENHORMONEUND SPEZIELL VON 3,5-L-DIJODTHYRONIN ( 3,5-L-T2 )
AUF DEN INTERMEDIÄRSTOFFWECHSELDER LEBER UND DER NIERE
ODER
DAS EINMALEINS DER LEBER- UND NIERENPERFUSION
D I S S E R T A T I O N
ZUR ERLANGUNG DES GRADES EINES DOKTORS DER MEDIZIN
dem Fachbereich Medizin der Universität Hamburg vorgelegt von
NIELS KÖSTER AUS HAMBURG
HAMBURG 1998

Yvette und Thiess-Richard,meiner Familie,meinen Großeltern,Jan Peter, Annemarie und Günther

ERKLÄRUNG
Ich versichere ausdrücklich, daß ich die Arbeit selbständig und ohne fremde Hilfe verfaßt,
andere als die von mir angegebenen Quellen und Hilfsmittel nicht benutzt und die aus den
benutzten Werken wörtlich oder inhaltlich entnommenen Stellen einzeln nach Ausgabe
( Auflage und Jahr des Erscheinens ), Band und Seite des benutzten Werkes kenntlich ge-
macht habe, und daß ich die Dissertation bisher nicht einem Fachvertreter an einer anderen
Hochschule zur Überprüfung vorgelegt oder mich anderweitig um Zulassung zur Promotion
beworben habe.
______________________
Niels Köster

Angenommen von dem Fachbereich Medizin
der Universität Hamburg am : 15. Oktober 1998
Gedruckt mit Genehmigung des Fachbereichs
Medizin der Universität Hamburg
Sprecher : Prof. Dr. med. H.-P. Leichtweiß
Referent : Prof. Dr. med. H.-J. Seitz
Korreferent : Prof. Dr. med. G. Mayr

Inhaltsverzeichnis4
I Einleitung 8
Physiologie und Pathologie der Schilddrüsenhormone 8
Hormonproduktion 10
Hormontransport, Metabolismus und Elimination 11
Hormonaktion 12
Fragestellungen 16
II Material und Methoden 17
1. Tierhaltung und Provokation der Hypothyreose 17
2. Rezyklierende Leberperfusion und ihre Standardisierung 18
3. Offene Leberperfusion und ihre Standardisierung 21
4. Isolierte Mitochondrien aus Rattenhepatozyten und ihre
Standardisierung 22
5. Isolierte Nierenperfusion und ihre Standardisierung 25
6. Bestimmung des Sauerstoffverbrauchs 27
7. Spektrophotometrische Bestimmungen 28
Protein-Bestimmung 28
Substratbestimmungen 28
Bestimmung von Enzymaktivitäten 30
8. Substanzen 31
9. Statistische Auswertung 32
III Ergebnisse 33
1. Rezyklierende Leberperfusion 33
1.1 Sauerstoffverbrauch der isoliert rezyklierend perfundierten Leber
hypothyreoter, männlicher und gehungerter Ratten 33
1.2 Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 35

Inhaltsverzeichnis5
1.3 Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei
perfundierten Lebern weiblicher Ratten 37
1.4 Vergleich der Stimulation des Sauerstoffverbrauchs bei männ-
lichen und weiblichen Versuchstieren 38
1.5 Steigerung der eingesetzten Hormonkonzentration 40
1.6 Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei isoliert
rezyklierend perfundierten Lebern männlicher, hypothyreoter
Ratten nach kohlenhydratreicher Diät 42
1.7 Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei isoliert
rezyklierend perfundierten Lebern weiblicher, hypothyreoter,
kohlenhydratreich ernährter Ratten 46
Zusammenfassung der Ergebnisse III.1 52
2 Kombiniert offene und rezyklierende Leberperfusion 54
2.1 Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei offen
und rezyklierend perfundierten Lebern männlicher, hypothyre-
oter und gehungerter Ratten 55
2.2 Steigerung der Glucoseproduktion bei offener und rezyklierender
Leberperfusion durch 3,5-L-T2 58
2.3 Steigerung der Harnstoffproduktion bei offener und rezyklieren-
der Leberperfusion durch 3,5-L-T2 59
2.4 Perfusionsdruck und Kaliumkonzentration im Perfusat 61
Zusammenfassung der Ergebnisse III.2 62
3 Einfluß von Ionenkanalblockern auf die Stimulation des Sauerstoff-
verbrauchs isoliert rezyklierend perfundierter Lebern hypothyre-
oter, gehungerter, männlicher Ratten durch 10-10 M 3,5-L-T2 63
3.1 Einfluß von Ouabain auf die Stimulation des Sauerstoffverbrauchs
durch 3,5-L-T2 63
3.2 Einfluß von Nifedipin auf die Stimulation des Sauerstoffverbrauchs
durch 3,5-L-T2 65

Inhaltsverzeichnis6
3.3 Einfluß von SITS auf die Stimulation des Sauerstoffverbrauchs
durch 3,5-L-T2 67
3.4 Einfluß von Amilorid auf die Stimulation des Sauerstoffverbrauchs
durch 3,5-L-T2 69
Zusammenfassung der Ergebnisse III.3 70
4 Einfluß von 3,5-L-T2 auf die oxydative Phosphorylierung bei
isolierten Mitochondrien der Rattenleber 72
Zusammenfassung der Ergebnisse III.4 75
5 Isolierte Nierenperfusion 76
5.1 Einfluß von 3,5-L-T2 bei isolierter Nierenperfusion
hypothyreoter Ratten 76
5.2 Einfluß von 3,5-L-T2 bei isolierter Nierenperfusion
euthyreoter Ratten 77
5.3 Einfluß von 3,5-L-T2 auf kultivierte Mesangiumzellen 78
Zusammenfassung der Ergebnisse III.5 79
IV Diskussion 81
1 Induzierte Hypothyreose durch Na131J i.p. 81
2 Die isolierte Leberperfusion als Modell der Wahl zur Untersu-
chung der schnellen Wirkungsweise von Schilddrüsenhormonen 82
3 Der Effekt von 3,5-L-T2 auf Gluconeogenese und O2-Verbrauch 83
4 Einfluß der Ernährung auf den Effekt von 3,5-L-T2 86
5 Einfluß des Geschlechts auf den Effekt von 3,5-L-T2 87
6 Untersuchungen zum Mechanismus der Wirkung von 3,5-L-T2
bei rezyklierender und offener Leberperfusion 89
7 Einfluß von Ionenkanalblockern auf den Effekt von 3,5-L-T2 92
8 Wirkung von 3,5-L-T2 auf isolierte Mitochondrien aus Hepatozyten
in vivo 97

Inhaltsverzeichnis7
9 Organspezifität der Wirkung von 3,5-L-T2 100
V Zusammenfassung 103
VI Literaturverzeichnis 104
Anhang
Abkürzungsverzeichnis 110
Danksagungen 112
Lebenslauf 113

Einleitung8
I EINLEITUNG
Physiologie und Pathophysiologie der Schilddrüsenhormone
Zu den physiologischen Funktionen der Schilddrüse gehört die Sekretion der Schilddrüsen-
hormone Thyroxin ( Tetrajodthyronin, T4 ) und Trijodthyronin ( T3 ). Sie beeinflussen eine
ganze Reihe von metabolischen Prozessen, wie den Stoffwechselgrundumsatz, kardiale und
neurologische Funktionen sowie das Wachstum u.a. mehr.
Die Einordnung der Schilddrüsenhormone in den hormonellen Regelkreis von Synthese und
Sekretion zeigt Abb. 1.
Hypothalamus
Hypophysenvorderlappen
Schilddrüse
T3 Albumin/Präalbumin/TBG T4
Ziel-Gewebe
Abb. 1Hypothalamus-Hypophysen-Schilddrüsen-Achse : Schematische Darstellung der hormonellen
Steuerung von Produktion, Ausschüttung und Transport der Schilddrüsenhormone.
Schilddrüsenerkrankungen äußern sich in quantitativen oder qualitativen Veränderungen
der Hormonsekretion bzw. in einer Größenzunahme der Schilddrüse oder in beidem.
TRH
TSH

Einleitung9
Die Hypothyreose teilt man in angeborene und erworbene Formen ein. Eine Schilddrüsen-
insuffizienz prä- oder perinatal kann als irreversible Folge das Krankheitsbild des Kretinis-mus
verursachen, das sich im Kindesalter durch Entwicklungsstörungen besonders der
Sinnesorgane, der Haut und des Skeletts sowie einer geistigen Retardierung auszeichnet.
Im Erwachsenenalter verursacht eine Unterfunktion der Schilddrüse das klinische Erschei-
nungsbild der Hypothyreose oder Myxödem mit reduziertem kalorischen Verbrauch
( Hypometabolismus ) als prinzipiellem Erscheinungsbild. Meistens handelt es sich bei
der Hypothyreose um eine ungenügende Produktion einer primär gestörten Schilddrüsen-
funktion ( primäre Hypothyreose ). Bei unzureichender Stimulation oder Ausfall der
Stimulation der Schilddrüse durch niedriges oder fehlendes TSH handelt es sich um eine
sekundäre Hypothyreose, und bei mangeldem TRH liegt eine tertiäre Hypothyreose vor.
Symptome einer Schilddrüsenunterfunktion leiten sich von einer Minderung des Stoff-
wechselgrundumsatzes ab und hierdurch bedingt eine erniedrigte Wärmeproduktion. Mit einer
die ( primäre ) Hypothyreose begleitenden Fettstoffwechselstörung gehen typischer-weise
erhöhte Cholesterinspiegel einher. Klinisch kann bei ausgeprägter Unterfunktion der
Schilddrüse im Erwachsenenalter eine Herzinsuffizienz, wie auch eine eingeschränkte phy-
sische sowie geistige Leistung imponieren.
Nicht zuletzt kommt es aufgrund einer unzureichenden Sekretion von Thyroxin ( T4 ), deren
Folge eine permanente Stimulation der Schilddrüse zur Hormonproduktion ist, zur Hyper-
plasie der Drüse und zur Ausbildung eines Kropfes.
Eine Überfunktion der Schilddrüse mit erhöhten Hormonspiegeln von T3 und/oder T4 hat das
Krankheitsbild der Hyperthyreose, bzw. bei exzessiver Hormonsekretion die Thyreo-toxikose
zur Folge. Als Symptome des Hypermetabolismus resultieren Gewichtsabnahme,
Wärmeintoleranz, Tachykardie, arterieller Hypertonus und Nervosität. Klinisch imponieren
Patienten in ausgeprägten Fällen durch eine physische und psychische Unruhe.
Auch bei der Hyperthyreose kann es zu einer Vergrößerung der Schilddrüse ( Struma )
kommen, bis hin zur Ausbildung eines Kropfes. Besonders ausgeprägt ist dies bei der von

Einleitung10
Basedow´schen Erkrankung der Fall mit dem zusätzlichen typischen Symptom des Exoph-
thalmus.
Die Hyperthyreose führt graduell zu einer Steigerung des Stoffwechselgrundumsatzes und in
ausgeprägten Fällen zur Einstellung einer katabolen Stoffwechsellage. Hierbei kommt es zu
vermehrtem Proteinabbau und zur Mobilisierung von Fettreserven. Die damit einher-gehende
Erhöhung des Sauerstoffverbrauchs und die gesteigerte Wärmeproduktion läßt sich durch
Messung des Sauerstoffverbrauchs im Stoffwechselgrundumsatz ( BMR ) nachweisen ( 8 ).
Hormonproduktion
Die Tagesproduktion der Hormone der Schilddrüse beträgt 83-93 µg T4 ( 11, 67, 84 ), 9 µg T3
und 0,9 µg rT3 ( 11, 64 ). Das im Blut zirkulierende T4 wird vollständig in der Schild-drüse
gebildet. Die tägliche Gesamtproduktion für T3 beträgt jedoch 30 µg ( 22,6 - 44,8 µg) ( 7, 11,
57, 84 ) und für rT3 beträgt sie 28 µg ( 97,5 % extrathyroidale Dejodierung von L-T4 ) ( 11 ).
Zustande kommt diese Menge durch Monodejodierung des von der Schilddrüse produzierten
T4, davon ca. 41 % zu T3 und ca. 38 % zu rT3, in peripheren Geweben. Beim T3 sind nur etwa
20 %( 46 ) direkt thyroidaler Herkunft . 80% des T3 sind Produkt aus den peripheren Geweben
durch das verantwortliche Enzym Monodejodinase Typ I, welches
L-T4 zunächst in 5´-Position am Außenring unter Beteiligung eines R-SH vorwiegend in Leber
und Niere dejodiert ( 17 ). Von der Tagesgesamtproduktion des T3 werden zwischen 29 und
56 %, vom rT3 ca. 28 % weiter dejodiert. Weitere Untersuchungen zu den einzel-nen
Dejodierungsschritten ergaben, daß praktisch das gesamte 3,5-L-T2 durch Dejodierung am
Phenolring des 3,5,3´-L-T3 entsteht, katalysiert durch die Enzyme Monodejodinase Typ I und
II und täglich 0,6 - 2,9 µg beträgt ( 17, 18, 21, 50 ). Durch weitere Dejodierungs-schritte
entsteht zuletzt das L-Thyronin ( s. Abb. 2 ).
Die bekannten therapeutisch verwendeten antithyroidalen Agenzien, wie Derivate des
Thioharnstoffs und Mercaptoimidazole entfalten komplexe Wirkung auf die Hormonbio-
synthese. Sie inhibieren die initiale Oxidation, dh. organische Bindung von Jod an das
Tyrosingrundgerüst, sodaß die Konzentration von Dijodtyrosin ( DIT ) in Relation zu
3,3´,5,5´-Tetraiodo-L-Thyronin ( T4 )

Einleitung11
Monodejodinase Typ I und II Monodejodinase Typ I und III
3,5,3´-Tiiodo-L-thyronin ( T3 ) 3,3´,5´-Triiodo-L-thyronin ( rT3 )
3,5-Diiodo-L-thyronin ( T2 ) 3,3´-Diiodo-L-thyronin 3´,5´-Diiodo-L-thyronin
3-Iodo-thyronin 3´-Iodo-thyronin
Thyronin
Abb. 2Dejodierungsschema der Schilddrüsenhormone, modifiziert nach J. Köhrle ( 38 ). Neben der
Monodejodinase Typ I werden zwei weitere Typen II und III beschrieben, wobei Typ II in
verschiedenen Geweben, wie Hypophyse, Gehirn und braunem Fettgewebe vorkommt und
Typ III als Isoenzym bei neonatalen Ratten, sowie in der Placenta von Mensch und Ratte vor-
kommt ( 17, 48 ). Wie Auf´mkolk et al., sowie Leonard et al. und andere Arbeitsgruppen
zeigen konnten, ist die Monodejodinase Typ I durch Propylthiouracil ( PTU ) hemmbar
( 3, 17, 48 ).
Monojodtyrosin ( MIT ) sinkt. Dadurch wird die Verbindung der Iodotyrosine zu den
hormonell aktiven Iodothyroninen blockiert.
Hormontransport, Metabolismus und Elimination
Im Blut sind T3 und T4 an Plasmaproteine gebunden ( Thyroxin bindendes Globulin TBG,
T4-bindendes Präalbumin TBPA, Albumin) . Zwischen T4 und den Proteinen stellt sich ein
Bindungsgleichgewicht ein, wobei nur ca. 0,03 % des Hormones frei vorliegen. Beim T3 liegen
ca. 0,3 % in der freien Form vor. Nur die freien, ungebundenen Hormonanteile ge-langen in die
Gewebe und können dann ihre Wirkungen entfalten.
Primäre Schwankungen der Schilddrüsenhormone bewirken persistierende Verschiebungen
der Konzentration von freiem Hormon im Verhältnis zum Gesamthormon, da die homöosta-

Einleitung12
tischen Mechanismen die Konzentration des freien Hormones nicht durch eine Veränderung
der TBG Konzentration normalisieren können. Die Konsequenz ist eine Veränderung des
metabolischen Status.
Schilddrüsenhormone unterliegen metabolisch hauptsächlich einer schrittweisen Dejodie-rung
einzelner Jodatome ( Monodejodierung ) bis hin zum Thyroningerüst ( s. Abb. 2 ). Im Falle der
Monodejodierung von T4 wird T3 erzeugt ( T3-Neogenese ) . Und da ca. 30 % des T4 zu T3
konvertiert wird und zusätzlich T3 eine höhere metabolische Potenz verglichen mit T4
entwickelt, könnte man T4 als Prohormon und T3 als das eigentlich aktive Hormon be-
zeichnen, das durch die Monodejodierung erzeugt wird. 80 % des im Blut zirkulierenden T3
wird durch Monodejodierung in verschiedenen Organen, insbesondere der Leber und der Niere
aus T4 erzeugt, der Rest direkt durch die Schilddrüse.
40 % des gesamten T4 wird ebenfalls durch Monodejodinase Typ I in 5-Position dejodiert
( Innenring ); es entsteht 3,3´,5´-Trijod-L-thyronin ( rT3 ), welches, wenn überhaupt nur
geringe metabolische Aktivität entfaltet. Ein weiterer Stoffwechselweg von T4 und T3 ist die
Konjugation in der Leber, prinzipiell mit aktivierter Glucuronsäure und Schwefelsäure, wo-ran
sich Dejodierung und biliäre Elimination anschließen ( 17 ).
20 % der gesamten T4 und T3 Menge werden an der Alaninseitenkette oxidativ desaminiert und
decarboxyliert, wodurch Tetrajod- und Trijodthyroacetat ( Tetrac bzw. Triac ) entste-hen, die
nur eine geringe biologische Aktivität aufweisen.
Hormonaktion
Schilddrüsenhormone beeinflussen das Wachstum und die Entwicklung von Geweben, ins-
besondere bei Gehirn, Knochen und Muskulatur. Ebenso wird der Gesamtenergieverbrauch
und der Umsatz von Substraten, wie Glucose, Fettsäuren und Aminosäuren von den Hor-
monen der Schilddrüse gesteuert, inklusive ihrer selbst. Die genauen Mechanismen und
zellulären Ebenen, auf denen die Schilddrüsenhormone eingreifen, sind Ziel vieler wissen-
schaftlicher Untersuchungen.

Einleitung13
Auf der Ebene des Zellkerns wird die Genexpression durch Schilddrüsenhormone beein-flußt,
auf mitochondrialer Ebene die oxydative Phosphorylierung und an der Plasmamem-bran wird
auf den Flux von Substraten und Kationen, wie Kalium und Calcium Einfluß genommen. Von
besonderem Interesse ist der zeitliche Ablauf verschiedener Effekte nach Hormonexposition,
da er Aufschluß über die physiologische Bedeutung und Wirkungsweise des eingesetzten
Hormones geben kann.
Das Tetrajodthyronin, T4 und seine einzelnen Dejodierungsprodukte, inklusive der Trijod-
thyronine T3, rT3 und Dijodthyronine T2 sind auf eigene hormonelle Aktivität in verschie-denen
Versuchsmodellen untersucht worden. T4 selbst weist eine geringe biologische Wirk-samkeit
auf, sodaß es auch als Prohormon des wirksamen T3 beschrieben werden kann. Tri-
jodthyronin, T3 bewirkt an seinem Zellkernrezeptor eine Stimulation oder Repression der
Transkription spezifischer Genabschnitte und damit die Bildung oder Unterdrückung der
Synthese spezifischer mRNAs und dadurch eine spezifische Induktion bzw. Repression be-
stimmter Proteine; vergleiche die Übersichten, dargestellt von Dillmann et al. sowie Marish und
Oppenheimer ( 16, 51 ).
Auch mitochondriale Proteine werden durch die Schilddrüsenhormone induziert, wie die
Enzyme Glycerin-3-phosphat-dehydrogenase ( 47 ), die Pyruvat-Carboxylase ( 5 ) und das
Cytochrom C ( 70, in einer Übersicht siehe 65 ). Der Wirkungseintritt beginnt jeweils nach
Stunden bzw. Tagen, und ob eine direkte oder bei Wirkungseintritt erst nach Tagen indi-rekte
Wirkung vorliegt, ist unklar.
Beispielhaft für einen schnellen Wirkungseintritt nach Schilddrüsenhormongabe ist die
kernvermittelte Bildung der mRNA „spot-14“, die 30-60 min nach T3-Gabe im Zytosol
nachgewiesen werden konnte, für deren biologische Funktion aber bisher noch keine Er-
kenntnisse vorliegen. Für weitere Enzyme, wie die hepatische Glucokinase oder die hepa-tische
Phosphoenol-pyruvat-Carboxykinase ( PEPck ) konnte ein ähnlich schneller Wir-
kungsmechanismus aufgezeigt werden.
Neben den kernvermittelten Wirkungen des T3 sind seit Jahren auch schnellere und sehr
schnelle Effekte des Hormones in der Diskussion. Von besonderem Interesse ist, ob den

Einleitung14
Effekten von L-T3 binnen Minuten eine Wirkung auf die Zellmembran zugrunde liegt oder auf
die Mitochondrien und so die mitochondriale Aktivität stimuliert wird.
Es gab Hinweise auf zeitlich sehr rasche Reaktionen nach dem Einsatz von Schilddrüsen-
hormonen. Bei diesen Wirkungen wurden aufgrund ihrer kurzen zeitlichen Abfolge mito-
chondriale Mechanismen angenommen. So schlossen Seitz et al. auf schnelle mitochondriale
Effekte in der Leber bei einer durch L-T3 bewirkten Senkung des mitochondrialen ATP/ ADP-
Quotienten ( 75 ). Müller et al. berichteten von einer Steigerung der hepatischen
Gluconeogenese und Glucoseproduktion aus Alanin bei gleichzeitig erhöhtem Sauerstoff-
verbrauch ( 56 ); in beiden Fällen am Modell der isoliert-perfundierten Rattenleber. Hier wurde
aufgrund des zeitlich raschen Ablaufs eine primär nukleäre Mitbeteiligung ausge-schlossen.
Horst et al., aus derselben Arbeitsgruppe, beschrieben ein neues wirksames Jodothyronin 3,5-
L-T2, welches Dejodierungsprodukt von L-T3 ist und zu einer raschen Steigerung des
Sauerstoffverbrauchs im beschriebenen Versuchsaufbau der isoliert-perfun-dierten Leber
hypothyreoter Ratten führte, während T3 und T4 durch Hemmung der Mono-dejodinase Typ I
durch PTU ihre ursprünglich steigernde Wirkung auf den Sauerstoffver-brauch verloren.
Hieraus abgeleitet wird ein getrennter Wirkungsmechanismus der Schilddrüsenhormone dis-
kutiert. Hiernach entfalten T3 und T4 eine primär nukleäre und T2 eine direkte mitochondri-ale
Wirkung ( 32 ).
Die Arbeitsgruppe von Hummerich et al. stieß bei der Untersuchung des Effektes von T3 an
der isoliert-perfundierten Rattenleber auf eine gesteigerte Calcium-Aufnahme und einer da-mit
verbundenen schnellen mitochondrialen Aktivierung. Im Vergleich zu anderen Hormo-nen wie
Glukagon, Vasopressin, sowie T2 und T4, bewirkte T3 im zeitlichen Verlauf eine vermehrte
Calcium-Aufnahme in die Zellen parallel zu gesteigerter Atmung und gesteiger-ter
Gluconeogenese. Die gleichsinnige Veränderung des Calcium-Einstroms nach Exposi-tion mit
den verglichenen Hormonen läßt die Autoren eine direkte calciumvermittelte Wir-kung auf den
mitochondrialen Stoffwechsel vermuten ( 34 ).
Versuche weiterer Arbeitsgruppen an Mitochondrien, die 15 min nach intraperitonealer
Injektion von L-T3 aus Lebern hypothyreoter Ratten isoliert worden waren, wiesen anhand

Einleitung15
mehrerer Parameter mitochondrialer Aktivität euthyreote Werte auf : Bei der Transportrate
von Adeninnucleotiden über die innere Membran ( 54 ), dem Protonengradienten ∆pH
( 14 ), dem ADP/O- Quotienten ( 14, 62 ), dem H+/O- Quotienten ( 14 ), wie auch bei der
Zykluszeit der Mitochondrien, um vom ADP-limitierten State 4 über den ADP-stimulierten
State 3 wieder zum State 4 zurückzugelangen ( 13 ). Unverändert blieben das Membran-
potential ∆Ψ, die proton-motive-force ∆p, wie auch der Sauerstoffverbrauch ( 14 ).
Darüber hinaus beschrieben Sterling et al. ( 81 ), daß bei 30 min nach ip. Injektion von T3
isolierten Mitochondrien eine signifikante Steigerung der ATP-Produktion und des Sauer-
stoffverbrauchs, wie auch bei direkter Gabe von L-T3 zu submitochondrialen Partikeln einen
gesteigerten Einbau von 32P in generiertes ATP. Hieraus wurde der Schluß gezogen, daß eine
gesteigerte oxydative Phosphorylierung zugrundeliegt ( 83 ). Einigen anderen Arbeitsgruppen
gelang es allerdings nicht, diesen Befund zu reproduzieren. Ferner haben eine Reihe von
Autoren eine Bindung von L-T3 an mitochondriale Proteine beschrieben; die physiologische
Bedeutung blieb kontrovers ( 26, 28, 80 ).
Die neuen Befunde über schnelle Wirkungen der Schilddrüsenhormone, insbesondere von 3,5-
L-T2 auf den Stoffwechsel der Leber geben Anlaß zur Diskussion über ein neues Prin-zip der
Wirkung dieser Hormone und sind in der Literatur unterschiedlich bewertet worden. Auch
wird die physiologische Bedeutung der Wirkung von 3,5-L-T2 kontrovers diskutiert, da einige
Laboratorien die Effekte nicht reproduzieren konnten ( mündliche Mitteilung ) und in anderen
Laboratorien die Effekte nur klein ( < 10 % ) waren.
So wurde in den vorliegenden Experimenten nicht nur die bewährte rezyklierende Perfu-sion,
wie in der Vergangenheit angewandt, sondern in Kooperation mit einem anderen La-bor das
aufwendige Modell einer nicht rezyklierenden, offenen Perfusion zusätzlich einge-setzt, um
jeden Zweifel zum Befund selbst auszuräumen.
Aufgrund der vorliegenden Befunde über eine schnelle Wirkung von 3,5-L-T2, soll auch der
Mechanismus und seine physiologische Bedeutung näher untersucht werden.
Fragestellungen :

Einleitung16
1. Welchen Einfluß hat L-T2 auf die Zellatmung und die Gluconeogenese der Leber und
wie schnell tritt die Wirkung ein ?
2. Gibt es geschlechtsspezifische Unterschiede bei der Wirkung von L-T2 auf die Leber
und welche Rolle spielt der Ernährungsstatus der Versuchstiere ?
3. Wie ist der Ablauf der Wirkung von L-T2 bei der isoliert perfundierten Leber ?
4. Welches sind die zugrundeliegenden subzellulären Mechanismen ? Läßt sich eine
Wirkung auf mitochondrialer Ebene nachweisen ?
5. Gibt es auch andere Organe, bei denen sich eine vergleichbare Wirkung von L-T2
nachweisen läßt ?
Für die einzelnen Fragestellungen wurden die Modelle der isoliert perfundierten Ratten-leber,
in den Variationen der offenen und geschlossenen Perfusion, sowie die Mitochon-
drienpräparation und die isolierte Nierenperfusion ausgewählt.
Die Ergebnisse zeigen, daß es sich bei 3,5-L-T2 in der Tat ( neben L-T3 ) um ein neues, seine
Wirkung schnell entfaltendes Schilddrüsenhormon handelt.
_______________

Material und Methoden17
II MATERIAL UND METHODEN
1. Tierhaltung und Provokation der Hypothyreose
Als Versuchstiere wurden Wistar-Ratten verwandt, spez. pathogen-frei, bezogen von der
Firma Wiga ( Charles River ), männlich und weiblich, denen, wenn nicht anders angege-ben,
am Vormittag des dem Versuch vorausgegangenen Tages das Futter entzogen wurde.
Als Futter wurde Altromin Standarddiät verwandt. Die Tiere hatten freien Zugang zum
Wasser. Die Raumtemperatur betrug konstant 22 ± 2°C, bei einem 12-stündigen Hell- Dunkel-
Wechsel.
Die Hypothyreose wurde erzeugt durch intraperitoneale Injektion von 0,25 mCi Na131J bei ca.
140 g schweren Tieren. Es war nach etwa 10 Tagen eine weitestgehende Wachstums-
stagnation zu verzeichnen ( zu diesem Zeitpunkt zwischen 200-240 g KG ). Das Leberge-
wicht ( LG ) der in den Versuch genommenen Tiere lag unter dem von euthyreoten Tieren
( <7 g ). Darüber hinaus machte sich der hypothyreote Status bemerkbar durch Hypother-
Tab. 1 Zusammenfassung der Befunde hypothyreoter im Vergleich zu euthyreoten und hyperthyreoten
Versuchstieren
Parameter Einheit Hypothyreose Euthyreose Hyperthyreose
Serum
L-T3 ng/ml 0,43 ± 0,04 0,65 ± 0,04 2,40 ± 0,20
L-T4 ng/ml 9,7 ± 1,0 23 ± 1 116 ± 14
Leber
Gewicht g/100 g KG 2,9 ± 0,2 4,1 ± 0,3 3,8 ± 0,1
Malic-Enzym U/g 0,12 ± 0,05 3,3 ± 0,1 9,6 ± 0,1
O2-Verbrauchin Perfusion, t = 0 min
µmol/g/min 1,71 ± 0,08 3,80 ± 0,21

Material und Methoden18
mie ( kalte Extremitäten ) und eine auffallende Hypomobilität der Tiere im Vergleich zu
euthyreoten. Die hypothyreoten Tiere wurden nach etwa vier Wochen in den Versuch ge-
nommen.
2. Rezyklierende Leberperfusion und ihre Standardisierung
Die anästhesierten Tiere ( Nembutal , Ceva, 60mg/ml, in einer Dosierung von 6mg/100g KG
) wurden laparotomiert, und nach Präparation und Darstellung der Vena cava caudalis wurde
diese zwischen linker und rechter Vena renalis mit einer Ligatur umschlungen, noch nicht
ligiert. Nach Ligieren der V. renalis dex. wurde rechts nephrektomiert. Nach Dar-stellung der
Vena portae wurde auch hier eine Ligatur gelegt und 2mm distal der Ligatur inzidiert und eine
Knopfkanüle eingebracht und mittels der liegenden Ligatur fixiert. Zu diesem Zeitpunkt wurde
die Leber mit vorher bereitetem und begastem Puffer perfundiert
( Fluß 20 ml/min, Krebs-Henseleit-Puffer ( 21 ), Carbogen-begast O2/CO2 95%/5%, 37°C,
pH=7,4, Heparin-Zusatz 1000 IE/1,5 l ) bei gleichzeitiger Eröffnung der V. cava caud. zur
Ausblutung und Freispülung der Leber, da die Lebern stets zellfrei perfundiert wurden. Jetzt
wurde das Diaphragma eröffnet und die V. cava proximal der Leber ligiert, was die
Flußrichtung von Knopfkanüle in V. portae via Leberbett in Richtung V. cava caud. be-
stimmte. Nach Kanülierung der eröffneten V. cava caud. mittels eines dünnen, kurzen Plas-
tikschlauches proximal der V. testicularis dex. und Fixierung durch die bereits liegende
Ligatur, konnte die Leber gänzlich freipräpariert und als intaktes Organ entnommen wer-den.
Abschließend wurde die so präparierte Leber in die vorbereitete und temperierte Per-
fusionskammer unter Konnektierung des hier bereits zirkulierenden Perfusionssystems ein-
gebracht.
Die isolierte Perfusion war stets zellfrei. Als Perfusionspuffer diente Krebs-Henseleit-Puffer (
21 ), der konstant temperiert war auf 37°C, dem als Substrat Alanin 10mM zugesetzt
wurde und nach einstündiger Perfusion, um eine Substratverarmung auszuschließen weitere
5mM Alanin. So war es möglich die Organfunktion, gemessen am Sauerstoffverbrauch und der
Gluconeogenese, sowie der Harnstoffsynthese für mindestens 3 Stunden stabil zu hal-ten.
37°C

Material und Methoden19
Abb. 3 Schematische Darstellung der isoliert rezyklierend perfundierten Rattenleber in der Perfu-
sionsapparatur nach Schimassek. Aufbau der Perfusionskammer 100x100x50 cm3 ( W.
Krannich KG, Göttingen ) s. Schema ; Temperatur 37°C, Verwendung von Glasröhren und
einem Magnetrührer im Reservoirgefäß.
Nach der Leberpassage wurde der Perfusionspuffer direkt an einer Clark-Elektrode vorbei-
geführt ( angeschlossen an einen Säure-Basen-Analysator, Typ PHM 71 b, Radiometer
Copenhagen; Nullpunkteinstellung durch 30 min N2-begastes H2O ), durch die kontinuier-lich
der Sauerstoffgehalt des Puffers gemessen und aufgezeichnet wurde. Die Elektrode wurde vor
jeder Perfusion neu auf Krebs-Henseleit Puffer, Carbogen-begast ( s.o. ), für 37°C geeicht (
830 µmol O2/Liter bei 95% O2/ 5% CO2 äquilibrierten Puffer ). Durch weitestgehende
Perfusionspufferführung durch Glasrohre wurde der Gasverlust minimiert und im
Perfusionssystem kontrolliert eingestellt auf 800 µmol O2/l. Die O2-Sättigung des Puffers
wurde zum Perfusionsbeginn und nach Perfusionsende kontrolliert.
Nach Konnektierung der Leber in der Perfusionskammer wurde nach 30 Minuten Vorper-
fusionszeit als Äquilibrierungsphase im Anschluß der Versuchsbeginn als Zeitpunkt

Material und Methoden20
t = 0 min festgelegt und dann, wenn nicht anders beschrieben, 90 Minuten perfundiert.
Während der Perfusion wurde zu im einzelnen angegebenen Zeitpunkten Perfusat zur Be-
stimmung verschiedener Parameter entnommen. Zum Zeitpunkt t = 0 min wurde, wenn nicht
anders beschrieben, das zu untersuchende Schilddrüsenhormon in der angegebenen
Konzentration zugesetzt.
Aus dem Perfusat wurden verschiedene Substrate und Enzyme bestimmt : Alaninverbrauch,
Glucose- und Harnstoffproduktion, GOT oder LDH u.a. zu den im einzelnen angegebenen
Tab. 2 Zusammenfassung der gemessenen Substrate als Stoffwechselparameter bei geschlossener
und offener Leberperfusion am Ende der Perfusionszeit; vergleiche hierzu auch die Kapitel
III.1.2 und III.2.2 und II.2.3
Parametergeschlossen-rezyklierende
Leberperfusionoffene
Leberperfusion
Glucose 28,6 ± 3,3 µmol/g 9,0 ± 0,7 µmol/ml/g
Harnstoff 74,6 ± 6,8 µmol/g 52,3 ± 13,4 µmol/ml/g
Alaninverbrauch 210 ± 30 µmol/g nicht gem.
Zeitpunkten. Mittels der gewählten Parameter ließ sich die Lebersyntheseleistung und die
Intaktheit der Zellen des Organs nachweisen. Die im Zytosol vorkommende GOT signali-
sierte bei erhöhtem Austritt ins Perfusat eine höhergradige Schädigung der Zytoplasma-
membran. Ein Anstieg der Produktion von Glucose und Harnstoff setzt eine abgelaufene
Gluconeogenese und Harnstoffsynthese, resp. intaktes Zytosol und intakte Mitochondrien
voraus.
Es fand sich bei intakten Lebern in den einzelnen Versuchsgruppen unter den dort angege-
benen Versuchsbedingungen nach 30minütiger Vorperfusion, zum Zeitpunkt t = 0 min, ein
einheitlicher Sauerstoffverbrauch ( s. auch Tab.1 und Kapitel Ergebnisse ).

Material und Methoden21
Abschließend wurde die Leber aus der Perfusionsapparatur entnommen, von Bindegewebs-
resten freipräpariert, abgetrocknet und das Feuchtgewicht ermittelt, um es in Bezug zu den
einzelnen Parametern zu setzen. Gleichzeitig erfolgte die Konnektierung der Perfusionsap-
paratur ohne die Leber, um abschließend die Linearität der Sauerstoffmessung zu überprü-fen,
als letzte Kontrolle im System.
3. Offene Leberperfusion und ihre Standardisierung
Die Präparation der Leber gestaltete sich w. o. beschrieben bis zur Entnahme des Organes. Die
Perfusionsapparatur ist in Abb. 4 schematisch dargestellt. Der Unterschied zur vorbe-
schriebenen Perfusion ist der offene Abfluß, sodaß das Perfusat nicht ein zweites mal zur
Abb. 4 Schema der offen isoliert perfundierten Rattenleber; gestrichelte Linie kennzeichnet Um-
wandlung zur geschlossenen Perfusion
Leber gelangt. Notwendigerweise wird das Pufferreservoir entsprechend groß gewählt
O2 Clark-Elektrode
pH-Elektrode
K+-Elektrode
∆p
Reservoir,Krebs-Henseleit-Puffer
Leber
Pumpe
37°C
Fritte,Carbogen-begast
Auffangbehälter
Perfusions-block

Material und Methoden22
( 5 Liter ), und in diesem Versuchsaufbau wird die Temperatur des Perfusionspuffers so
gewählt, daß sie bei Erreichen der Leber 37° C beträgt. Auch ist der Perfusionsblock ( s. Abb.
4 ) auf 37° C temperiert.
Vor der Passage des Perfusionspuffers durch die Leber wurde mittels eines Druckaufneh-mers
der Perfusionsdruck ∆p kontinuierlich aufgezeichnet. Direkt nach der Leberpassage wurde
mittels der oben beschriebenen Clark-Elektrode kontinuierlich der Sauerstoffgehalt des
Perfusates gemessen, sowie auch parallel dazu mithilfe einer pH-Elektrode ∆ pH und durch
eine weitere Elektrode der Kaliumausstrom bestimmt. Wie oben schon beschrieben wurden
auch hier als Stoffwechselparameter Glucose, Harnstoff und LDH aus dem Perfu-sionspuffer
bestimmt. Vergleiche dazu die Daten der Tabellen 1 und 2.
4. Isolierte Mitochondrien aus Rattenhepatozyten und ihre Standardisierung
Gehungerten, männlichen hypothyreoten Ratten wurde zu angegebenen Zeitpunkten vor
Dekapitierung intraperitoneal NaCl ( 0,9% bei Kontrollen ), bzw. als Schilddrüsenhormon
3,5-L-T2 ( in angegebener Konzentration ) injiziert.
Nach Dekapitierung der Tiere und anschließendem Ausbluten der Lebern wurden diese mit
Saccharose-Lösung ( 0,25 M Saccharose, pH 7,4 ) abgespült, von Bindegewebs- und Ge-
fäßresten befreit, gewogen und wie folgt beschrieben auf Eis oder bei 4°C weiterverarbei-tet.
Die Präparation der Mitochondrien wurde nach den für Zellfraktionierungen üblichen Ver-
fahren durchgeführt. Dazu wurde das Gewebe zunächst schonend homogenisiert mittels
Potter-Homogenisator und anschließend die Mitochondrien durch Differentialzentrifugation
isoliert ( s. schematische Darstellung in Abb. 5 ).
Homogenat
5min, 600 g ( 3000 rpm )

Material und Methoden23
Pellet Überstand ( Zelltrümmer, Zellkerne ) ( Mitochondrien, Lysosomen, Mikrosomen, Zytosol )
5min, 5100 g ( 9000 rpm )
Überstand Pellet ( Zytosol, Lysosomen, Mikrosomen ) ( Mitochondrien )
1,5 min, 12300 g ( 14000 rpm )
Überstand Pellet ( Mitochondrienbruchstücke ) ( Mitochondrien )
Aufnahme in 4 ml Saccharose-Lösung
Abb. 5 Schematische Darstellung der Mitochondrienpräparation
Messung mitochondrialer Atmung
Die Sauerstoffkonzentration im Inkubationsmedium wurde wiederum w. o. beschrieben mit
einer Clark-Elektrode bestimmt. Um einen definierten Sauerstoffgehalt im Meßmedium zu
erreichen wurde es zuvor 30 min im Thermostaten mit einer Pumpe belüftet. Die sich dabei
einstellende Sauerstoffkonzentration im Medium beträgt bei 30°C 195,5 µmol/l.
In den Versuchsansätzen wurde zunächst die Intaktheit der Präparation überprüft. Anschlie-
ßend nach Spülen des Reaktionsgefäßes mit destilliertem Wasser wurden dann 1,9 ml des
belüfteten, vortemperierten Inkubationsmediums ( 110mM Mannitol, 10mM KH2PO4, 60 mM
KCl, 60 mM Tris-HCl, 5 mM MgCl2, 0,5 mM EDTA, pH = 7,4 ) in das Reaktions-
gefäß pipettiert und die Rührung der Meßzelle eingeschaltet. Zuerst wurde der Meßwert für
100%ige Luftsättigung des Mediums über 60 Sekunden registriert.

Material und Methoden24
Nach Zusatz von 50 µl Mitochondriensuspension wurde das Reaktionsgefäß verschlossen.
Jetzt wurde die endogene Atmung für 60 sek. registriert und dann die Substrate ( 30 µl
Abb. 6 Schematische Darstellung der polarographischen Meßzelle
Succinat 1M bzw. 30 µl Glutamat 1M ) zugesetzt. Der ADP-Zusatz ( 15 µl ADP 0,05 M )
erfolgte dann erst nachdem die Atmungsgeschwindigkeit für mindestens 60 sek. konstant war.
Nach der Zugabe von ADP wurde abgewartet, bis die Mitochondrien vom aktiven
phosphorylierenden Zustand ( hohe Atmungsgeschwindigkeit ) in den kontrollierten Zustand
zurückgekehrt waren und eine konstante ( geringere ) Atmungsgeschwindigkeit erreicht hatten.
Bei der Durchführung der Atmungsversuche wurde jede Atmungskurve bis zum Erreichen des
anaeroben Zustandes registriert und vor jeder neuen Messung das Reaktionsgefäß gründlich
gereinigt.
Zur Berechnung des Verhältnisses der Atmungsgeschwindigkeit im aktiven phosphorylier-ten
Zustand ( nach ADP-Zusatz ) und dem im kontrollierten Zustand ( nach Phosphorylie-rung des
zugesetzten ADP ) wurde die Atmungsgeschwindigkeit aus dem Anstieg der Tan-genten
bezogen auf mg Protein / ml Mitochondriensuspension der Substrate wie folgt be-stimmt :
391 nmol O / ml ⋅ 30 mm / min ⋅ 2 ml Succinat V resp. = ⋅ tan α
Kanüle für Zusätze
Verschlußstempel
Temperiermantel
Inkubationsraum
Rührer Sauerstoffelektrode

Material und Methoden25
217 mm ⋅ mg Protein
= 108,1 nmol O / min ⋅ tan α / mg Protein
391 nmol O / ml ⋅ 10 mm / min ⋅ 2 ml Glutamat V resp. = ⋅ tan α 217 mm ⋅ mg Protein
= 36,0 nmol O / min ⋅ tan α / mg Protein
5. Isolierte Nierenperfusion und ihre Standardisierung
Zur isoliert rezyklierend perfundierten Nierenperfusion kamen Nieren von hypothyreoten, für
24 Stunden gehungerten, männlichen Ratten zum Einsatz, analog zu den für isolierte
Leberperfusion verwendeten Tiere.
Die Perfusion der isolierten Nieren erfolgte in einem Rezirkulationssystem mit Dialyse. Es
wurde mit dem gleichen Perfusionsmedium perfundiert, das auch schon bei der isolierten
Leberperfusion zum Einsatz kam ( Krebs-Henseleit-Puffer ( 21 ), Carbogen-begast O2/CO2
95%/5%, 37°C, pH 7,4, Heparin-Zusatz 1000 IE/5l ).
Apparativer Aufbau
Vom Vorratsgefäß wurde das Perfusionsmedium mit einer Rollenpumpe durch einen kom-
merziellen Kapillarplattendialysator gepumpt ( SECON, Göttingen, Typ 101 ) und in das
Vorratsgefäß zurückgeleitet. Das Perfusionsmedium wurde durch 5000 ml eines begasten
Dialysats kontinuierlich regeneriert ( Dialysatreservoir ). Eine Akkumulation von schädi-
genden Peroxiden konnte so durch die Dialyse vermieden werden. Damit war es möglich, die
Organfunktion für ca. 2 Stunden stabil zu halten.

Material und Methoden26
Der Dialysator diente auch als Oxygenator - „ dialung“ ; über Glasfritten wurde vorge-
wärmtes, wasserdampfgesättigtes Carbogen in das Dialysat geleitet. Die Rezirkulation des
Dialysats erfolgte mit 360 ml/min durch eine Zentrifugalpumpe.
Aus dem Perfusatkreislauf entnahm eine zweite Rollenpumpe unmittelbar hinter dem Dialy-
sator das für die Nierenperfusion benötigte dialysierte und aufoxygenierte Perfusat. Es ge-
langte nach Passieren eines 12 µm Zellulosenitrat-Filters und einer Wärmeschlange mit
Windkessel zu der am Präparationstisch fixierten doppelläufigen Perfusionskanüle. Wäh-rend
die Außenkanüle der Zuleitung des Perfusionsmediums diente, war der innere Teil der Kanüle
zur Registrierung des Perfusionsdruckes an einen Druckaufnehmer angeschlossen
( GOULD STATHAM INSTRUMENTS, USA, Typ P 50 ). So war es möglich, die Niere
fluß- und druckkonstant zu perfundieren.
Die isolierte Niere selbst lag in einer temperierten Metallschale, die ebenfalls am Präpara-
tionstisch fixiert war. Zum Auffangen des venösen Effluates wurde ein Doppeltrichter unter die
Nierenschale geschwenkt.
Die Zurückleitung des venösen Effluates in das Vorratsgefäß erfolgte luftblasenfrei über ein
geregeltes Quetschventil. Mit Hilfe einer weiteren Rollenpumpe wurde ein Aliquot des ve-
nösen Effluates ( 0,5 ml / min ) einer Tripel-Durchflußmeßeinheit zugeführt ( ESCHWEI-LER,
Kiel, Elektrodentyp MTD ) und anschließend in das Vorratsgefäß zurückgeleitet; pO2, pCO2
und pH wurden damit kontinuierlich gemessen.
Operationstechnik
Die Versuchstiere entsprachen den selben Anforderungen, wie sie bei der isolierten Leber-
perfusion zum Einsatz kamen. Zur Perfusion wurde jeweils die rechte Niere verwendet; das
Gewicht der dekapsulierten linken Niere diente als Referenzgewicht.
Nach intraperitonealer Anästhesie mit Nembutal wurde das Tier in Rückenlage auf einen
temperierten Präparationstisch aufgelegt. Nach medianer Laparotomie wurde die rechte Niere
zusammen mit dem Ureter von dem umgebenden Gewebe sowie der Nebenniere befreit. Für
den Gefäßanschluß der Niere an den Perfusionskreislauf wurden folgende Ge-fäße

Material und Methoden27
freipräpariert und legiert : Aorta abd., A. renalis sin. et dex., A. mesenterica sup., V. cava.
Nach dem Abbinden des Ureters wurde dieser ca. 1 cm distal des Nierenbeckens mit einem 1
cm langen Polypropylenschlauch, sehr sorgfältig unter Vermeidung von Blutungen
katheterisiert. Nach Einbettung der rechten Niere in eine temperierte Metallschale in situ folgte
das Abbinden der A. renalis sin., der A. mesenterica und der Aorta abd. ca. 1,5 cm kaudal der
A. renalis sin. und dann die Abklemmung der Aorta abd. kaudal der rechten Nierenarterie. Die
doppelläufige Perfusionskanüle wurde distal der Klemme in die Aorta abd. eingeführt. Die
Perfusion wurde in situ begonnen, sobald die Aorta abd. kranial der A. mesenterica
abgebunden und die Gefäßklemme entfernt war. Danach wurde die Perfu-sionskanüle fest
eingebunden, die Gefäßverbindungen zum Tier getrennt, das Tier entfernt und die Blutreste mit
warmen Perfusat abgespült. Ein temperierter Doppeltrichter wurde zum Auffangen des
venösen Effluates unter die Nierenschale geschwenkt. Die V. cava inf. wurde mit einem am
Trichterblock schwenkbar angeordneten V2A-Rohr kanüliert; das kau-dale Ende der V. cava
wurde abgebunden.
6. Bestimmung des Sauerstoffverbrauchs
Die Sauerstoffmessung während geschlossener, sowie während offener Leberperfusion wur-de
mit Hilfe eines Säure-Basen-Analysators Typ PHM 71 b Radiometer Copenhagen und einer
angeschlossenen Clark-Elektrode bestimmt und durch einen Schreiber aufgezeichnet. Vor den
Messungen wurde der Nullpunkt mittels N2-begasten Wassers vorgenommen. An-schließend
erfolgte die Einstellung auf 37°C temperierten, Carbogen-begasten ( 95% O2 / 5% CO2 )
Krebs-Henseleit-Puffer ( 21 ) : In einem Liter Puffer werden 830 µmol/ l Sauer-stoff gelöst (
35 ). Trotz weitestgehender Führung in Glasrohren waren geringfügige Gas-verluste zu
verzeichnen, sodaß sich ein kontrollierter Ausgangswert von 800 µmol/l O2 für den
Sauerstoffgehalt vor Leberperfusion ergab.
Am Anfang und am Ende jeden Experimentes wurde der Sauerstoffgehalt des Puffers bzw.
Inkubationsmediums überprüft, um erhöhten Gasverlust, Elektrodendriften o.a. auszu-
schließen.

Material und Methoden28
Bei den Versuchen mit isolierten Mitochondrien ( siehe dazu Kapitel III.4 ) wurde die Sau-
erstoffkonzentration im Inkubationsmedium wiederum mit einer Clark-Elektrode bestimmt.
Nach 30 minütiger Belüftung durch eine Pumpe betrug der Sauerstoffgehalt des Mediums bei
30 °C im Thermostaten 195,5 µmol/l als Referenzwert.
7. Spektrophotometrische Bestimmungen
Protein-Bestimmung
Zu 50 µl BSA-Standard bzw. Probe wurden 2 ml 1/5 verdünntes Bio-Rad-Reagenz pipettiert
und nach Ablauf von mindestens 15 min bei 595 nm die Proteinkonzentration ermittelt.
Substratbestimmungen
Alanin-Bestimmung ( 2 )
Nach dem Testprinzip :
ALANIN - DH
L - Alanin + NAD+ + H2O Pyruvat + NH4+ + NADH + H+
Testansatz : 980 µl Tris-Hydrazin-Puffer, pH 1020 µl 0,1 M NAD200 µl H2O300 µl Probeneinsatz
Die Bestimmung von E1 erfolgte nach 45 min bei 334 nm, die enzymatische Umsetzung mit 10
µl L-Alanin-Dehydrogenase ( Alanin-DH, 5 mg / ml ) gestartet und E2 nach weiteren
120 min bestimmt.
Glucose-Bestimmung ( 2 )
Nach dem Testprinzip :

Material und Methoden29
HEXOKINASE
Glucose + ATP Glucose - 6 - P + ADP
GLUCOSE - 6 - P - DH
Glucose - 6 - P + NADP Gluconat-6-Phosphat + NADPH + H+
Testansatz : 1340 µl 0,4 M TRA - Puffer, pH 7,210 µl 0,5 M MgSO4
10 µl 0,1 M ATP40 µl 0,02 M NADP100 µl Probe
Die Bestimmung von E1 erfolgte bei 334 nm. Anschließend wurde die enzymatische Umset-
zung mit 10 µl Hexokinase / Glucose - 6 - Phosphat - Dehydrogenase ( 3 mg / ml ) gestartet
und E2 nach 20 min bestimmt.
Harnstoff-Bestimmung ( 2 )
Nach dem Testprinzip :
UREASE
Harnstoff + H2O 2 NH4+ + CO2
GLU - DH
α - Ketoglutarat + NADH + NH4+ Glutamat + NAD+ + H2O
Testansatz : 500 µl 0,4 M TRA - Puffer, pH 8,010 µl 0,55 M α - Ketoglutarat70 µl 0,01 M NADH20 µl Glutamat - DH ( 10 mg / ml in Glycerin )1400 µl H2O100 µl Probe
Die Bestimmung von E1 erfolgte bei 334 nm. Anschließend wurde die enzymatische Umset-
zung mit 50 µl Urease ( 2mg / ml ) gestartet und E2 nach 60 min bestimmt.
Bestimmung von Enzymaktivitäten

Material und Methoden30
Bestimmungen von Enzymaktivitäten wurden bei 25 °C vorgenommen und kontinuierlich
aufgezeichnet. Die photometrische Auswertung erfolgte bei 334 nm.
Glutamat-Oxalacetat-Transaminase ( GOT ) - Bestimmung ( 2 )
Nach dem Testprinzip :
GOT
L - Aspartat + α - Ketoglutarat Oxalacetat + L - Glutamat
MDH
Oxalacetat + NADH + H+ Malat + NAD+
Testansatz : 750 µl 0,2 M KH2PO4, pH 7,450 µl 0,01M NADH10 µl Malat - Dehydrogenase ( 1 mg / ml )580 µl H2O200 µl Perfusat
Zunächst wurde die Reaktion mit 50 µl 0,55 M α - Ketoglutarat gestartet; nach 5 min er-folgte
der zweite Start mit 500 µl 0,25 M Aspartat.
Lactat-Dehydrogenase ( LDH ) - Bestimmung
Nach dem Testprinzip :
LDH
NADH + H+ + Pyruvat NAD + + Lactat
Testansatz : 1000 µl 0,4 M TRA - Puffer, pH 8,070 µl 0,01 M NADH20 µl 0,5 M Pyruvat500 µl H2O100 µl Probe
Die Reaktion wurde mit 70 µl 0,01 M NADH gestartet und die Messung der Extinktion bei
366 nm bei Raumtemperatur per Schreiber über mindestens 5 min registriert.
Malat-Enzym - Bestimmung ( 17, 27 )

Material und Methoden31
Nach dem Testprinzip :
MALAT - ENZYM
L - Malat + NADP Pyruvat + CO2 + NADPH + H+
Vorbereitet zur Messung wird 1 g Leber im siebenfachen Volumen eiskalter 0,154 M KCl
gepottert und bei 100 000 g für 30 min zentrifugiert.
Testansatz : 350 µl 0,4 M TRA - Puffer, pH 7,210 µl 0,5 M L - Malat670 µl 0,01 M MnCl2
870 µl H2O100 µl Überstand bei Lebern hypothyreoter Ratten
Gestartet wurde die Reaktion mit 20 µl 0,02 M NADP.
8. Substanzen
Es wurden nur die L - Formen der angegebenen Schilddrüsenhormone eingesetzt. Die
verwendeten Schilddrüsenhormone waren ein Geschenk der Firma Henning Berlin und wiesen
einen Reinheitsgrad von über 99,5 % auf.
Die Schilddrüsenhormone wurden nach dem Auswiegen in einem Tropfen 1 N NaOH
aufgenommen und anschließend mit H2O weiter auf angegebene Konzentrationen verdünnt.
Die verwendeten Enzyme wurden von der Firma Boehringer Mannheim bezogen.
Alle anderen verwendeten Substanzen wurden, wenn nicht anders angegeben, von den Firmen
Merck, Sigma Chemie oder Boehringer Mannheim bezogen.
9. Statistische Auswertung

Material und Methoden32
Die statistische Auswertung erfolgte nach dem Student´s t - Test für unabhängige bzw.
gepaarte Werte ( 33 ). Angegeben sind Mittelwerte ± Standardabweichung des Mittelwertes (
SEM ).

Ergebnisse33
III ERGEBNISSE
1. Rezyklierende Leberperfusion
Standardisierung des Modells
Die in die Versuche genommenen Tiere waren stets hypothyreot, männlich und hungerten 24
Stunden vor Versuchsbeginn, sodaß der Glykogengehalt der Lebern vernachlässigbar gering
war und bei Kontrollversuchen, wie auch den Versuchen mit Einsatz von Schild-
drüsenhormonen die Gluconeogenese anhand der Glucoseproduktion gemessen werden
konnte.
1.1 Sauerstoffverbrauch der isoliert rezyklierend perfundierten Leber hypothyreoter,
männlicher und gehungerter Ratten
Nach Installation der präparierten Lebern hypothyreoter, männlicher, gehungerter Ratten in die
Perfusionskammer wurde standardisiert für 30 Minuten vorperfundiert und dieser Zeit-punkt t
= 0 min gesetzt. Zu diesem Zeitpunkt erfolgte bei den Kontrollen kein weiterer Zu-satz, bei
weiteren Versuchen wurden Schilddrüsenhormone in angegebenen Konzentrationen
hinzugefügt und im weiteren der Sauerstoffverbrauch kontinuierlich gemessen, wie auch
mittels Perfusatentnahmen zu angegebenen Zeitpunkten die hierin enthaltenen Substrate bzw.
Enzyme bestimmt. Die Perfusionszeit betrug, wenn nicht anders angegeben, 90 Mi-nuten. Nach
Beendigung der Perfusion wurde das Feuchtgewicht der Lebern bestimmt, um den
Sauerstoffverbrauch über die Zeit ins Verhältnis zu setzen.
Das Perfusat enthielt bei allen Leberperfusionen, soweit nicht anders angegeben, Alanin in
einer Ausgangskonzentration von 10 mM, gelöst in Krebs-Henseleit-Puffer ( 21 ), tempe-riert
auf 37 °C, Carbogen - begast ( 95 % O2 / 5% CO2 ) bei konstantem pH 7,4. Nach 30-
minütiger Äquilibrierung, zum Zeitpunkt t = 0 min, wurde den Kontrollperfusionen kein

Ergebnisse34
weiteres Agens zugesetzt. Lediglich nach 60 minütiger Gesamtperfusionszeit ( zum Zeit-punkt
t = 30 min ) wurden weitere 5 mM Alanin, um einer Substratverarmung vorzubeu-gen, bei
allen Perfusionen in das Pufferreservoir nachgegeben. Die Ergebnisse der Perfusat-
bestimmungen zu den Zeitpunkten 0, 30, 60 und 90 Minuten ( Glucose, Harnstoff, GOT ) faßt
Tab. 2 auf Seite 20 zusammen.
Die Kontrollperfusionen von Lebern hypothyreoter, männlicher, gehungerter Ratten
( n = 11 ) sind in Abb. 7 graphisch zusammengefaßt. Der Ausgangs-Sauerstoffverbrauch
betrug zum Zeitpunkt t = 0 min 1,71 ± 0,08 µmol/min/g und stieg über 90 min Perfusions-
zeit um 0,53 ± 0,05 auf 2,24 ± 0,07 µmol/min/g signifikant, p < 0,01 an. Prozentual stieg
t in min0 20 40 60 80 100
∆ O 2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 20 40 60 80 100
O2-
Ve
rb
ra
uc
h i
n µ
mo
l/m
in/
g
1,5
2,0
2,5
3,0
Abb. 7Sauerstoffverbrauch isoliert rezyklierend perfundierter Lebern männlicher, hypothyreoter,
gehungerter Ratten
o Kontrollen ( MW ± SEM, n = 11 )
Dargestellt sind der relative Sauerstoffverbrauch ( gr. Graphik ), sowie der absolute
Sauerstoffverbrauch ( kl. Graphik ) in µmol/min/g.

Ergebnisse35
der Sauerstoffverbrauch hierbei im Vergleich zum Ausgangswert um 31 % nach 90 min.
Der Alaninverbrauch aus dem Perfusat, wie auch die abgelaufene Gluconeogenese mit Glu-
coseproduktion sowie die Harnstoffsynthese signalisieren funktionstüchtige Zellkomparti-
mente und eine GOT Aktivität von stets unter 1,5 U/l beweisen die Intaktheit der Zellen.
1.2 Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2
Der Effekt des Zusatzes von 3,5-L-Dijodthyronin ( 3,5-L-T2 ) in einer Konzentration von
10-12 M, entsprechend der von Pinna et al. ( 66 ) bei Probanden gefundenen physiologi-
t in min0 20 40 60 80 100
∆ O 2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 20 40 60 80 100
O2-
Ve
rb
ra
uc
h i
n µ
mo
l/m
in/
g
1,5
2,0
2,5
3,0
*
Abb. 8Sauerstoffverbrauch isoliert rezyklierend perfundierter Lebern männlicher, hypothyreoter,
gehungerter Ratten, ∗ = p < 0,01 .
o Kontrollen ( MW ± SEM, n = 11 )
n 10 -12 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 9 )
Dargestellt sind der relative Sauerstoffverbrauch ( gr. Graphik ), sowie der absolute
Sauerstoffverbrauch ( kl. Graphik ) in µmol/min/g.

Ergebnisse36
schen Konzentration, zum Perfusionspuffer zum Zeitpunkt t = 0 min im Modell der isoliert
rezyklierend perfundierten Leber hypothyreoter, männlicher, gehungerter Ratten steigerte den
Sauerstoffverbrauch innerhalb von 90 min Perfusion vom Ausgangswert 1,62 ± 0,12
µmol/min/g um 1,28 ± 0,12 µmol/min/g signifikant, p<0,01 auf 2,90 ± 0,18 µmol/min/g. Am
Ende der Perfusionszeit war der Sauerstoffverbrauch, verglichen mit den Kontrollen um 0,66
µmol/min/g oder 29,5 % über diese angestiegen, vergleiche dazu Abb. 8.
Wie in Abb. 8 ( kl. Graphik ) dargestellt entspricht dies im absoluten Vergleich gegenüber dem
Anstieg der Kontrollen von 31 %, nach beschriebener Schilddrüsenhormongabe einer
Stimulation des Sauerstoffverbrauchs um 79 % und damit mehr als dem zweieinhalbfachen.
Der Ausgangs-Sauerstoffverbrauch lag nach der Vorperfusionszeit zum Zeitpunkt t = 0 min
geringfügig unter dem der Kontrollen. Bis zur 30. Minute verliefen beide Kurven weitge-
hend parallel. Danach stieg der Sauerstoffverbrauch in der Versuchsgruppe im Vergleich zum
Kontrollkollektiv jedoch deutlich höher und steiler an. Die Kurvenverläufe im direkten
Vergleich unterscheiden sich bereits nach 45 min statistisch signifikant voneinander.
Tab. 3 Steigerung der Glucose- und Harnstoffproduktion rezyklierend perfundierter Rattenlebern durch
3,5-L-T2 10-12 M zum Zeitpunkt t = 0 min im Vergleich zu Kontrollen.
Angaben in µmol/g Leber Feuchtgewicht.
* p < 0,05, ** p < 0,01
Kontrollen 0 30 60 90 ( min ) n
Glucose 4,8 ± 1,1 8,9 ± 1,8 17 ± 3 29 ± 3 5
Harnstoff 8,1 ± 2,9 31 ± 9 58 ± 9 75 ± 7 5
3,5-L-T2 0 30 60 90 ( min ) n
Glucose 4,8 ± 1,1 10 ± 2 * 27 ± 2 ** 42 ± 7 ** 4
Harnstoff 8,1 ± 2,9 43 ± 6 * 84 ± 3 ** 136 ± 9 ** 4

Ergebnisse37
Wie der Sauerstoffverbrauch unterschieden sich auch die Stoffwechselparameter von denen des
Kontrollkollektivs. Die Harnstoffproduktion steigerte sich nach Hormonzusatz von 30,6 ± 8,8
auf 43,0 ± 5,9 µmol/g, wie auch die Glucoseproduktion nach Gabe von 10 -12 M 3,5-
L-T2 von 8,9 ± 1,8 auf 11,6 ± 1,6 µmol/g bereits nach 30 min an ( p < 0,05 ), um nach 60 min
ein Signifikanzniveau von p < 0,01 zu erreichen. Vergleiche dazu Tab. 3.
1.3 Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei perfundierten Lebern
weiblicher Ratten
Im folgenden wurde untersucht, ob es geschlechtsspezifische Unterschiede in der Wirkung von
3,5-L-T2 auf den Stoffwechsel der perfundierten Lebern gibt. Die Perfusion der Lebern
weiblicher Ratten vollzog sich analog denen der männlichen Tiere. Die Versuchsbedingun-gen
waren mit gehungertem Status, induzierter Hypothyreose sowie rezyklierender Perfu-sion
identisch.
Abb. 9 faßt die Versuchsergebnisse der weiblichen Kontrollperfusionen ohne Schilddrüsen-
hormonzusatz und die Versuchsgruppe mit Einsatz von 10-12 M 3,5-L-T2 zum Zeitpunkt
t = 0 min zusammen. Bei den Kontrollperfusionen betrug der Ausgangs-Sauerstoffver-brauch
2,08 ± 0,19 µmol/min/g und stieg im Verlauf der 90 minütigen Perfusion um 0,47 ± 0,03
µmol/min/g , entsprechend 22,6 % auf 2,55 ± 0,19 µmol/min/g an.
Nach Zusatz von 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min stieg der Sauerstoffverbrauch,
nach 15 Minuten gleichmäßigen Verlaufs mit den Kontrollen, dann steiler an als diese, um ein
signifikant höheres Niveau nach bereits 25 Minuten Perfusion zu erreichen mit p<0,01. Nach
einem zu Perfusionsbeginn niedrigeren Ausgangsverbrauch als die Kontrollgruppe von 1,97 ±
0,13 µmol/min/g und einer Steigerung um 1,03 µmol/min/g erreichte der Sauer-stoffverbrauch
nach 90minütiger Perfusion 3,00 ± 0,10 µmol/min/g, entsprechend 52,3 % über dem
Vergleichszeitpunkt der Kontrollen.

Ergebnisse38
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 20 40 60 80 100
O2-V
er
br
au
ch
in
µm
ol/
min
/g
2,0
2,5
3,0
*
Abb. 9Sauerstoffverbrauch isoliert rezyklierend perfundierter Lebern weiblicher, hypothyreoter,
gehungerter Ratten, ∗ = p < 0,01.
¡ Kontrollen ( MW ± SEM, n = 4 )
l 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 5 )
Dargestellt sind der relative Sauerstoffverbrauch ( gr. Graphik ), sowie der absolute
Sauerstoffverbrauch ( kl. Graphik ) in µmol/min/g.
1.4 Vergleich der Stimulation des Sauerstoffverbrauchs bei männlichen und
weiblichen Versuchstieren
Die Kontrollgruppen der männlichen und weiblichen Versuchstiere verlaufen in der Dar-
stellung des relativen Sauerstoffverbrauchs fast identisch; vergleiche dazu Abb. 10 a. Wie in
der männlichen Kontrollgruppe steigt der Sauerstoffverbrauch bei den weiblichen Tieren
geringfügig aber kontinuierlich an. Im Vergleich der absoluten Werte fällt jedoch trotz ins-
gesamt parallelen Verlaufs eine deutliche Betragsdifferenz auf; vergleiche dazu Abb. 10 b.

Ergebnisse39
Die Lebern weiblicher, hypothyreoter, gehungerter Ratten wiesen einen signifikant höheren
Sauerstoffverbrauch auf, als die männlichen Versuchstiere. Sowohl Ausgangs-Sauerstoffver-
brauch, wie auch der Wert nach 90 Minuten Perfusionszeit liegen bei den weiblichen Tieren
fast parallel verschoben um ca. 0,34 µmol/min/g höher als die männliche Kontrollgruppe mit
einem Signifikanzgrad von p<0,01.
Die prozentuale Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 beträgt in der Gruppe
der weiblichen Versuchstiere 17,7 % und fällt damit geringer aus als die Stimulation bei den
männlichen Versuchstieren ( 29,5 % ), wie anhand des Vergleichs der absoluten Werte (
dargestellt in Abb. 10 b ) sichtbar wird. Auch das Ausgangsniveau des Sauerstoffver-brauchs
der beiden Versuchsgruppen ist unterschiedlich und liegt bei den weiblichen Ratten signifikant
höher als bei den männlichen Versuchstieren, p<0,01 ( siehe Abb. 10 b ).
t in min0 20 40 60 80 100
∆ O 2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 20 40 60 80 100
O2-V
er
br
au
ch
in
µm
ol/
min
/g
1,5
2,0
2,5
3,0
Abb. 10 a Abb. 10 b
relativer Sauerstoffverbrauch in µmol/min/g absoluter Sauerstoffverbrauch in µmol/min/g
Stimulation des Sauerstoffverbrauchs isoliert rezyklierend perfundierter Lebern hypothyreoter, gehungerter
Ratten
o Kontrollen, männlich ( MW ± SEM, n = 11 )
n 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min, männlich ( MW ± SEM, n = 9 )
¡ Kontrollen, weiblich ( MW ± SEM, n = 4 )
l 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min, weiblich ( MW ± SEM, n = 5 )

Ergebnisse40
Zum Perfusionsende befinden sich dann aber beide Versuchsgruppen nach Stimulation des
Sauerstoffverbrauchs durch 10-12 M 3,5-L-T2 im Vergleich beim absoluten Verbrauch fast auf
demselben Endniveau nach 90 min; vergleiche hierzu Abb. 10 b.
Ein weiterer Unterschied liegt in der Dynamik der hormonstimulierten Verlaufskurven. Der
Zeitpunkt, zu dem nach Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 das Niveau der
Kontrollgruppe verlassen wird liegt im Falle der Versuchsgruppe der weiblichen Tiere deutlich
früher als bei den männlichen Tieren. So erreicht der Sauerstoffverbrauch der hor-
monstimulierten Versuchsgruppe bei den weiblichen Tieren früher ein signifikant erhöhtes
Niveau im Vergleich zu ihren Kontrollen, als es bei der Vergleichsgruppe der männlichen Tiere
erreicht wird. Auch erreicht der Sauerstoffverbrauch bei den perfundierten Lebern weiblicher
Ratten nach Stimulation durch 10-12 M 3,5-L T2 früher eine Einstellung auf ein Endniveau als
es bei den männlichen Versuchstieren erlangt wird.
1.5 Steigerung der eingesetzten Hormonkonzentration
Um eine Aussage darüber machen zu können, ob das geringere Ausmaß der Stimulation des
Sauerstoffverbrauchs der Ursache nach an der Erschöpfung der Atmungskapazität der Leb-ern
weiblicher, hypothyreoter, gehungerter Ratten lag, oder eine weitere Steigerbarkeit, auf ein ∆
O2, wie bei den männlichen Versuchstieren erreicht werden konnte, wurde die Kon-zentration
von 3,5-L-T2 auf 10-10 M erhöht und die Ergebnisse in Abb. 11 dargestellt.
Eine Steigerung der Konzentration von 3,5-L-T2 auf 10-10 M bewirkte im Endbetrag keine
weitere Zunahme des Sauerstoffverbrauchs im Vergleich zur Stimulation durch die niedri-gere
Konzentration von 10-12 M. Im Vergleich des relativen Sauerstoffverbrauchs der per-fundierten
Lebern lag nach 90 minütiger Perfusion der Verbrauch sogar geringfügig unter dem nach
Einsatz der geringeren Konzentration des Schilddrüsenhormones. Lediglich im Kurvenverlauf
liegt bei erhöhter Hormonkonzentration eine geringfügig früher signifikante Stimulation vor,
mit p<0,01 nach 20 Minuten; im Vergleich zum selben Signifikanzniveau nach Einsatz von 10-
12 M 3,5-L-T2 wurde dies nach 25 Minuten erreicht.

Ergebnisse41
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 20 40 60 80 100
O2-V
er
br
au
ch
in
µm
ol/
min
/g
2,0
2,5
3,0
*
Abb. 11 Sauerstoffverbrauch isoliert rezyklierend perfundierter Lebern weiblicher, hypothyreoter,
gehungerter Ratten, ∗ p < 0,01
¡ Kontrollen ( MW ± SEM, n = 4 )
l 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 5 )
t 10-10 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 4 )
Dargestellt sind der relative Sauerstoffverbrauch ( gr. Graphik ). sowie der absolute
Sauerstoffverbrauch ( kl. Graphik ) in µmol/min/g.
Der Ausgangssauerstoffverbrauch lag in der Versuchsgruppe mit 2,24 ± 0,16 µmol/min/g
geringfügig über dem Ausgangswert der Kontrollgruppe, wie auch über der durch 10-12 M T2
stimulierten Gruppe. Im weiteren Verlauf unterschieden sich die beiden hormonstimu-lierten
Versuchsgruppen jedoch kaum voneinander. Der Sauerstoffverbrauch der isoliert
rezyklierend perfundierten Lebern weiblicher, hypothyreoter, gehungerter Ratten scheint durch
Erhöhung der Hormonkonzentration von 3,5-L-T2 nicht weiter steigerbar ist.
Frühere Befunde unserer Arbeitsgruppe waren bei rezyklierender Perfusion männlicher

Ergebnisse42
Lebern identisch; auch hier konnte durch Steigerung der Hormonkonzentration keine wei-
tere Steigerung des Sauerstoffverbrauchs erzielt werden.
1.6 Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei isoliert rezyklierend
perfundierten Lebern männlicher, hypothyreoter Ratten nach kohlenhydratrei-
cher Diät
Es ist bekannt, daß der Serum-T3-Spiegel in Phasen des Hungers abfällt, um anschließend nach
Wiederfütterung mit kohlenhydratreicher Nahrung kurzfristig wieder anzusteigen. Daher stellt
sich die Frage, ob der Effekt von 3,5-L-T2 auch durch den Ernährungsstatus der Versuchstiere
modulierbar ist. Für 3,5-L-T2 sind keine Serum-Spiegel im Hunger bekannt.
Um den Einfluß des gehungerten Status der Versuchstiere auf die Steigerung des Stoffwech-
sels durch 3,5-L-T2 der perfundierten Lebern zu untersuchen, wurden die gehungerten Tie-re
mit einer Versuchsgruppe kohlenhydratreich ernährter Tiere verglichen.
Die Leberperfusion unterschied sich lediglich durch den nicht gehungerten Status der Tiere von
den unter III.1.1 und III.1.2 beschriebenen Versuchsanordnungen. Statt dessen wurden die
hypothyreoten, männlichen Tiere nach einem Tag Hungern die darauffolgenden zwei Tage mit
einem kohlenhydratreichen Futter versorgt. Der nicht gehungerte Status machte sich in Bezug
auf das Gewicht der Lebern kaum bemerkbar. Gehungerte Tiere wiesen ein Lebergewicht von
5,8 ± 0,01 g auf; die Lebern der Tiere dieser Versuchsanordnung wogen 5,8 ± 0,21 g.
Der Sauerstoffverbrauch der perfundierten Lebern hypothyreoter, männlicher, kohlenhy-
dratreich ernährter Ratten lag zum Zeitpunkt t = 0 min mit 1,38 ± 0,12 µmol/min/g O2 und
einem Anstieg über 90 Minuten um 0,47 ± 0,03 µmol/min/g auf 1,85 ± 0,10 µmol/min/g
signifikant und annähernd parallel verschoben unter den zu vergleichenden Kontrollen der
gehungerten Tiere aus dem Versuch III.1.1, p<0,01.
Die Stimulation des Sauerstoffverbrauchs wiederum durch 10-12 M 3,5-L-T2 bewirkte, bei
gleichem Ausgangsniveau der Versuchsgruppe mit 1,31 ± 0,07 µmol/min/g, eine schon nach 20

Ergebnisse43
Minuten signifikante Steigerung der Atmung, die nach 90 minütiger Perfusion, um 1,02 ± 0,14
µmol/min/g erhöht, auf 2,32 ± 0,15 µmol/min/g anstieg. Nach Hormonzugabe unterscheidet
sich diese Versuchsgruppe bei Aufzeichnung der Verlaufskurve des ∆ O2-Ver-brauchs durch
einen früheren Anstieg von den gehungerten Lebern. Der Anstieg des Sauer-stoffverbrauchs
beträgt bei der Kontrollguppe 34 %, verglichen mit 77 % nach Stimulation der Atmung durch
das eingesetzte Schilddrüsenhormon. Während die Kontrollgruppe ab der 60. Minute
Perfusionszeit ein Plateau zu erreichen scheint, deutet sich dieses bei hormonell stimulierter
Perfusion erst ab der 80. oder 90. Minute an; siehe dazu auch Abb. 12.
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 20 40 60 80 100
O2-V
er
br
au
ch
in
µm
ol/
min
/g
1,5
2,0
*
Abb. 12 Sauerstoffverbrauch isoliert rezyklierend perfundierter Lebern männlicher, hypothyreoter,
kohlenhydratreich ernährter Ratten, ∗ p < 0,01.
² Kontrollen ( MW ± SEM, n = 4 )
u 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 6 )
Dargestellt sind der relative Sauerstoffverbrauch ( gr. Graphik ), sowie der absolute
Sauerstoffverbrauch ( kl. Graphik ) in µmol/min/g.
Im Vergleich zu den Perfusionen gehungerter Ratten unterscheiden sich die Kontrollen beider
Versuchsgruppen im Verlauf kaum. Lediglich ein dezent späterer Anstieg der Kon-trollen der

Ergebnisse44
kohlenhydratreich ernährten Tiere, sowie der stets niedrigere relative Sauer-stoffverbrauch
machen die geringfügigen Unterschiede aus, wie in Abb. 13 a zum Ausdruck kommt.
Die Darstellung des absoluten Sauerstoffverbrauchs in der Abb. 13 b zeigt jedoch einen
Betragsunterschied von im Verlauf fast konstant 18 % niedrigerem, entsprechend 0,36
µmol/min/g O2-Verbrauch, gegenüber dem gehungerten Status; jeweils bezogen, wie an-
gegeben auf das Lebergewicht in Gramm.
Berücksichtigt man bei kohlenhydratreich ernährten Tieren die durch Glykogenspeicherung
bedingte geringere Konzentration atmungsaktiver Kompartimente, muß der Sauerstoffver-
brauch auf Milligramm Protein pro Gramm Leber bezogen werden. Die gehungerten Le-
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 20 40 60 80 100
O2-V
er
br
au
ch
in
µm
ol/
min
/g
1,5
2,0
2,5
3,0
Abb. 13 a Abb. 13 b
relativer Sauerstoffverbrauch in µmol/min/g absoluter Sauerstoffverbrauch in µmol/min/g
Stimulation des Sauerstoffverbrauchs isoliert rezyklierend perfundierter Lebern männlicher, hypothyreoter
Ratten
o Kontrollen, gehungert ( MW ± SEM, n = 11 )
n 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min, gehungert ( MW ± SEM, n = 9 )
² Kontrollen, kohlenhydratreiche Diät ( MW ± SEM, n = 4 )
u 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min, kohlenhydratreiche Diät ( MW ± SEM, n = 6 )
bern wiesen im Mittel 332 ± 10 mg Protein / g Leber ( n = 5 ) auf und die kohlenhydrat-reich
ernährten Tiere 269 ± 5 mg Protein / g Leber ( n = 10 ) auf; p<0,01. Bezogen auf das Protein
glichen sich die Sauerstoffverbrauchswerte fast an.

Ergebnisse45
Die Lebern gehungerter hypothyreoter Ratten wiesen zu Beginn der Perfusion einen Sauer-
stoffverbrauch von 5,15 ± 0,24 nmol/min/mg Protein und im Vergleich dazu, nach kohlen-
t in min0 20 40 60 80 100
∆ O 2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 20 40 60 80 100
O2-V
er
br
au
ch
in
µm
ol/
min
/g
1,5
2,0
Abb. 14 Sauerstoffverbrauch isoliert rezyklierend perfundierter Lebern männlicher, hypothyreoter,
kohlenhydratreich ernährter Ratten
² Kontrollen ( MW ± SEM, n = 4 )
u 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 6 )
t 10-10 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 2 )
Dargestellt sind der relative Sauerstoffverbrauch ( gr. Graphik ), sowie der absolute
Sauerstoffverbrauch ( kl. Graphik ) in µmol/min/g.
hydratreicher Diät einen Sauerstoffverbrauch von 5,12 ± 0,45 nmol/min/mg Protein auf. Nach
der abgelaufenen Stimulation erreichten die vorher gehungerten Lebern nach 90 Mi-nuten
Perfusionszeit einen O2-Verbrauch von 8,74 ± 0,54 nmol/min/mg Protein, vergleich-bar den
Lebern der zuvor kohlenhydratreich ernährten Tiere die am Ende der Perfusion bei 8,62 ± 0,56
nmol/min/mg Protein lagen.

Ergebnisse46
Die graphische Darstellung des absoluten Sauerstoffverbrauchs in Abb. 13 b veranschau-licht
noch einmal die Betragsdifferenzen der verglichenen Ernährungsstatus bei isoliert
rezyklierender Perfusion. Nach kohlenhydratreicher Kost kann das Atmungsniveau gehun-
gerter Lebern nach Stimulation nicht ganz erreicht werden. Die prozentuale Berechnung des
Sauerstoffverbrauchs nach Stimulation durch das Schilddrüsenhormon in den beiden Ver-
suchsgruppen beträgt bei gehungerten Versuchstieren 29,5 % und nach kohlenhydratreicher
Diät 25,4 % und fällt damit nahezu identisch aus.
In einem ausgewählten Experiment wurde die eingesetzte Konzentration von 3,5-L-T2 auf 10-10
M gesteigert, um auszuschließen, daß eine fehlende Steigerbarkeit des Sauerstoff-verbrauchs
auf einen Hormonmangel zurückzuführen ist. Wie in der Abb. 14 dargestellt, liegt der
Sauerstoffverbrauch nach erhöhter Hormonkonzentration bei hypothyreoten Ratten nach
kohlenhydratreicher Diät jedoch nicht über dem der Versuchsgruppe mit geringerem
Hormoneinsatz, sondern gerinfügig darunter. Auch ist die Stimulation des O2-Verbrauchs
gegenüber der Kontrollgruppe nicht früher signifikant gesteigert.
1.7 Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei isoliert rezyklierend
perfundierten Lebern weiblicher, hypothyreoter, kohlenhydratreich ernährter
Ratten
Um den Einfluß kohlenhydratreicher Diät auf den Sauerstoffverbrauch weiblicher, hypothy-
reoter Ratten zu untersuchen, wurde den Tieren nach Futterentzug für einen Tag, an den
darauffolgenden zwei Tagen kohlenhydratreiche Kost ad libitum angeboten. Am vierten Tag
wurden die Lebern in den Versuch genommen und perfundiert. Das Lebergewicht der ge-
hungerten Tiere von 4,17 ± 0,12 g unterschied sich nur gering von dem der nicht gehun-gerten
glykogenspeichernden Lebern, die im Mittel 4,56 ± 0,21 g Feuchtgewicht aufwiesen und damit
nur 9,4 % schwerer waren.
Der kontinuierlich aufgezeichnete Sauerstoffverbrauch der perfundierten weiblichen Lebern
hypothyreoter, kohlenhydratreich ernährten Ratten ohne Hormonzusatz betrug 1,50 ± 0,08
µmol/min/g zum Zeitpunkt t = 0 min. Verglichen mit den gehungerten Tieren aus Versuch

Ergebnisse47
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
t in min0 20 40 60 80 100
O2-V
er
br
au
ch
in
µm
ol/
min
/g
1,5
2,0
2,5
*
Abb. 15 Sauerstoffverbrauch isoliert rezyklierend perfundierter Lebern weiblicher, hypothyreoter,
kohlenhydratreich ernährter Ratten, ∗ = p < 0,01.
↵ Kontrollen ( MW ± SEM, n = 3 )¬ 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 3 )Dargestellt sind der relative Sauerstoffverbrauch ( gr. Graphik ), sowie der absolute
Sauerstoffverbrauch ( kl. Graphik ) in µmol/min/g.
III.1.3, bei deren Kontrollgruppe der Sauerstoffverbrauch 2,08 ± 0,19 µmol/min/g betrug,
entspricht dies einer Senkung um 27,9 %, die nicht allein durch das erhöhte Lebergewicht
erklärt ist, wie Abb. 16 b zeigt. Wie bei den Lebern der gehungerten weiblichen Ratten, stieg
bei den Kontrollen der gefütterten Tiere der Sauerstoffverbrauch im Verlauf der 90 minütigen
Perfusion signifikant um 0,55 ± 0,06 µmol/min/g O2 an, auf den Endbetrag von 2,05 ± 0,11
µmol/min/g, prozentual ausgedrückt um 36,7 %, p<0,01, vergleiche dazu Abb. 15.
Nach Zusatz von 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min stieg der Sauerstoffverbrauch von
1,52 ± 0,12 µmol/min/g binnen 90 minütiger Perfusion um 0,94 ± 0,09 µmol/min/g, bzw. 61,2

Ergebnisse48
% auf 2,45 ± 0,05 µmol/min/g signifikant und deutlich höher an, als das bei der Kontrollgruppe
zu verzeichnen war; siehe dazu Abb. 15.
Wie bei den gehungerten Tieren war der basale O2-Verbrauch durch 3,5-L-T2 zu steigern.
Bei Darstellung des relativen Sauerstoffverbrauchs als ∆-O2 scheint sich die Versuchsgrup-pe
bereits nach 20 min von der Kontrollgruppe zu unterscheiden. Die Auftragung des abso-luten
Sauerstoffverbrauchs jedoch macht deutlich, daß ein wirklicher Unterschied als signi-
t in min0 20 40 60 80 100
∆ O 2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 20 40 60 80 100
O2-V
er
br
au
ch
in
µm
ol/
min
/g
1,5
2,0
2,5
3,0
Abb. 16 a Abb. 16 b
relativer Sauerstoffverbrauch in µmol/min/g absoluter Sauerstoffverbrauch in µmol/min/g
Stimulation des Sauerstoffverbrauchs isoliert rezyklierend perfundierter Lebern weiblicher hypothyreoter
Ratten
¡ Kontrollen, gehungert ( MW ± SEM, n = 4 )
l 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min, gehungert ( MW ± SEM, n = 5 )
↵ Kontrollen, kohlenhydratreiche Diät ( MW ± SEM, n = 3 )¬ 10 -12 M 3,5-L-T2 zum Zeitpunkt t = 0 min, kohlenhydratreiche Diät ( MW ± SEM, n = 3 )
fikante Stimulation jedoch auch erst nach 40 min erreicht ist mit p<0,01, der sich bis zur 90.
Minute kontinuierlich weiter steigert.
Im direkten Vergleich zu den gehungerten weiblichen, hypothyreoten Versuchstieren aus dem
Kapitel III.1.3, dargestellt in Abb. 9 fallen mehrere Unterschiede auf. Zunächst scheint bereits
die hormonell nicht stimulierte Kontrollgruppe der kohlenhydratreich ernährten Tie-re im O2-
Verbrauch später anzusteigen, als die gehungerten Tiere. Nach 20 und 30 Minuten Perfusion

Ergebnisse49
liegen die kohlenhydratreich gefütterten Tiere im Sauerstoffverbrauch noch unter-
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
t in min0 20 40 60 80 100
O2-V
er
br
au
ch
in
µm
ol/
min
/g
1,5
2,0
2,5
Abb. 17 Sauerstoffverbrauch isoliert rezyklierend perfundierter Lebern weiblicher, hypothyreoter,
kohlenhydratreich ernährter Ratten
↵ Kontrollen ( MW ± SEM, n = 3 )¬ 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 3 )t 10-10 M 3,5-L- T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 2 )
Dargestellt sind der relative Sauerstoffverbrauch ( gr. Graphik ), sowie der absolute
Sauerstoffverbrauch ( kl. Graphik ) in µmol/min/g.
halb der gehungerten Ratten, erreichen aber nach 90 minütiger Perfusion dasselbe Niveau des
O2-Verbrauchs. In der Darstellung des absoluten Sauerstoffverbrauchs unterscheiden sich die
beiden Versuchsgruppen, jedoch aufgrund des schon unterschiedlichen Ausgangs-niveaus, wie
oben beschrieben, gravierend voneinander.
Nach hormoneller Stimulation stiegen die gehungerten Lebern jedoch signifikant früher im
Sauerstoffverbrauch an als die Vergleichsgruppe der stimulierten Lebern nach kohlenhy-
dratreicher Diät. Nachdem die gehungerten Lebern bereits nach 25 Minuten signifikant mehr
Sauerstoff verbrauchten, ist dies bei den gefütterten Tieren erst nach 40 Minuten der Fall.
Auch betrug das ∆ des O2-Verbrauchs bei den gehungerten Tieren im Mittel 0,56 µmol mehr

Ergebnisse50
Sauerstoff nach Gabe von 10-12 M 3,5-L-T2, im Vergleich zu einer Steigerung um 0,39 µmol
O2 als Mittelwert der nicht gehungerten Versuchstiere. Prozentual fällt der Vergleich jedoch
nicht so gravierend aus. Aufgrund des geringeren basalen Sauerstoffver-brauchs beträgt die
Stimulation nach kohlenhydratreicher Wiederfütterung 61,2 %. Vergli-chen mit der bei
gehungerten Tieren fällt diese hier mit 52,3% geringfügig niedriger aus.
In einem weiteren Experiment wurde die Konzentration des eingesetzten Schilddrüsen-
hormones 3,5-L-T2 auf 10-10 M erhöht, um zu prüfen, ob die Stimulation des Sauerstoff-
verbrauchs in diesem Versuchsaufbau zu steigern, bzw. zu beschleunigen war. Abb. 17 stellt
das Versuchsergebnis vom Einfluß einer Erhöhung des Hormonzusatzes bei isoliert
rezyklierend perfundierter Lebern weiblicher, kohlenhydratreich ernährter Ratten dar. Aus der
Darstellung des ∆ O2-Verbrauchs wird ersichtlich, daß praktisch im gesamten Verlauf der
Perfusion keine weitere Steigerung des Sauerstoffverbrauchs möglich war. Erst nach 80- bis
90-minütigem nahezu identischem Anstieg, als bei dem geringeren Hormonzusatz die
Verlaufskurve in ein Plateau zu münden scheint, ist dies bei Einsatz von 10-10 M 3,5-L-T2 noch
nicht zu registrieren. Bei Auftragung der absoluten Werte des Sauerstoffverbrauchs ( siehe
Abb. 17, kl. Graphik ) wird es noch deutlicher, daß zu diesem Zeitpunkt noch kein Plateau
erreicht zu sein scheint. Im Vergleich zu dem vorherigen Experiment betrug der Ausgangs-
Sauerstoffverbrauch zum Zeitpunkt t = 0 min 1,58 ± 0,03 µmol/min/g und wurde binnen 90
Minuten Perfusion um 1,15 ± 0,13 und damit um 81,0 % signifikant auf 2,86 ± 0,09
µmol/min/g O2 gesteigert, p<0,01.
Der abschließende Vergleich männlicher und weiblicher hypothyreoter Ratten nach kohlen-
hydratreicher Diät ist in den Abb. 18 a und b zusammengefaßt. Hier ist der basale Sauer-
stoffverbrauch, wie auch die Stimulation des Sauerstoffverbrauchs durch 10-12 M 3,5-L-T2
nebeneinander dargestellt. Wie schon im Kapitel III.1.4 beschrieben und in den Abb. 10 a und
b dargestellt, liegt der basale Sauerstoffverbrauch bei weiblichen, gehungerten, hypo-thyreoten
Ratten höher als bei männlichen. Genauso fällt diese Differenz zugunsten eines höheren
Atmungsniveaus bei weiblichen, kohlenhydratreich ernährten Versuchstieren im Vergleich zu
den männlichen aus.

Ergebnisse51
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
t in min0 20 40 60 80 100
O2-V
er
br
au
ch
in
µm
ol/
min
/g
1,5
2,0
2,5
Abb. 18 a Abb. 18 b
relativer Sauerstoffverbrauch in µmol/min/g absoluter Sauerstoffverbrauch in µmol/min/g
Stimulation des Sauerstoffverbrauchs isoliert rezyklierend perfundierter Lebern kohlenhydratreich ernährter
Ratten
² Kontrollen, männlich ( MW ± SEM, n = 4 )u 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min, männlich ( MW ± SEM, n = 6 )↵ Kontrollen, weiblich ( MW ± SEM, n = 3 )¬ 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min, weiblich ( MW ± SEM, n = 3 )
Bei den weiblichen Tieren liegt der Sauerstoffverbrauch mit 1,52 ± 0,12 µmol/min/g auch in
dieser Versuchsanordnung wiederum höher als bei den männlichen Ratten mit 1,31 ± 0,07
µmol/min/g; vergleiche dazu die Abb. 18 a und b. Durch die Darstellung des ∆ O2-Verbrauchs
wird anschaulich gemacht, daß sich sowohl männliche als auch weibliche Kon-trollen im
Verlauf gleichen. Auch die Steigerung des Sauerstoffverbrauchs während 90 mi-nütiger
Perfusion auf 0,55 ± 0,06 µmol/min/g ohne Zusatz von 3,5-L-T2 bei den weiblich-en, sowie auf
0,47 ± 0,03 µmol/min/g bei den männlichen Versuchstieren unterscheidet sich kaum
voneinander.
Nach Einsatz von 10-12 M 3,5-L-T2 ist der Sauerstoffverbrauch männlicher Lebern nach 25-30
Minuten etwas früher signifikant erhöht, als der der weiblichen Versuchstiere, die sich erst
nach 40 Minuten von ihrer Kontrollgruppe unterscheiden. Im weiteren Verlauf der 90
minütigen Perfusion sind die Unterschiede der verglichenen Versuchsgruppen gering.

Ergebnisse52
_______________
Zusammenfassung der Ergebnisse III.1
3,5-L-T2 stimuliert signifikant den Sauerstoffverbrauch isoliert rezyklierend perfundierter
Lebern hypothyreoter männlicher Ratten. Korrespondierend zu diesem Befund ist die
Gluconeogeneserate, wie auch die Harnstoffproduktion der Lebern im Vergleich zu den
Kontrollperfusionen gesteigert.
Im Vergleich zu weiblichen Versuchstieren fällt die Stimulation des Sauerstoffverbrauchs der
männlichen Tiere höher aus, wobei jedoch das Ausgangsniveau des O2-Verbrauchs der
männlichen Tiere bei Perfusionsbeginn niedriger liegt als bei den weiblichen. So ist der Betrag
der Atmungsstimulation ∆ O2 nach Zusatz von 3,5-L-T2 bei männlichen gehungerten Ratten
höher als bei weiblichen.
Der gefütterte Status der männlichen und weiblichen Tiere hat keinen Einfluß auf die Höhe des
Sauerstoffverbrauchs im Ausgangsniveau, wie auch nicht nach Stimulation der Atmung durch
3,5-L-T2, wenn die Werte auf das hepatische Protein bezogen werden, daß bei Le-bern nach
kohlenhydratreicher Diät geringer ausfällt. Die Steigerung des Sauerstoffver-brauchs ist aber
früher signifikant erhöht, als das bei den gehungerten Tieren zu verzeich-nen war.
Eine Steigerung der zugesetzten Hormonkonzentration hat, unabhängig von den sonstigen
Versuchsbedingungen, nie eine weitere Steigerung oder einen wesentlich früheren Anstieg des
Sauerstoffverbrauchs der perfundierten Lebern zur Folge gehabt.

Ergebnisse53
2. Kombiniert offene und rezyklierende Leberperfusion
Für ausgewählte Experimente wurde der apparative Perfusionsaufbau verändert, um den
Mechanismus des Effektes von 3,5-L-T2 näher zu untersuchen. Im Gegensatz zur geschlos-sen
rezyklierenden Perfusion, die in einem Kreissystem den Perfusionspuffer immer wieder zur
Leber geleitet, wird er bei der offenen Perfusion lediglich einmal zur Leber geleitet und direkt
dahinter an verschiedenen Elektroden vorbeigeführt, mittels derer der Sauerstoffver-brauch,
wie auch die Ionenströme gemessen werden können. Anschließend wird der Perfu-sionspuffer

Ergebnisse54
abgeleitet, sodaß Perfusatproben entnommen werden können, und der Über-schuß in einem
Sammelbehälter aufgefangen.
Diese Perfusionsapparatur eignet sich besonders zur Untersuchung des Stoffwechsels der
perfundierten Organe, da die Stoffwechselprodukte zu jedem Zeitpunkt bestimmt werden
können, ohne daß sie in einem rezyklierenden System akkumulieren. Eine weitere Beson-
derheit dieses Systems liegt in der Möglichkeit, nach entsprechender Vorperfusion zur
Äquilibrierung zunächst, wie oben beschrieben, eine Zeit offen zu perfundieren und dann das
System umzustellen, um anschließend für den gleichen Zeitraum geschlossen rezyklie-rend
weiterperfundieren zu können. Technisch bedarf es hierzu lediglich eines Kurzschlie-ßens des
Systems, indem der Puffer nach Oxygenierung wieder dem Perfusionsorgan zuge-führt wird;
siehe dazu die schematische Darstellung der Abb. 4 in Kapitel II. 3. Freigesetz-te Mediatoren,
die den Stoffwechsel beeinflussen gelangen bei offener Perfusion nach Durchströmen des
Perfusionsorganes nicht wieder zu diesem zurück, sodaß bei einem nach-zuweisenden Einfluß,
dieser durch das Umschalten auf rezyklierende Perfusion aufgedeckt werden kann.
Die Vorperfusionszeit betrug 20 Minuten und analog zur vorher beschriebenen rezyklieren-den
Perfusion wurde nach deren Ablauf dieser Zeitpunkt t = 0 min gesetzt und bei Kon-
trollperfusionen kein Zusatz, bzw. bei den Versuchen mit Hormoneinsatz zu diesem Zeit-punkt
3,5-L-T2 zum Perfusionspuffer zugegeben wurde. Aus den Daten der vorangegan-genen
Experimente geht hervor, daß eine Konzentration von 10-9 M 3,5-L-T2 sicher im maximal
stimulierenden Bereich liegt, sodaß sie hier für diese ausgewählten Experimente eingesetzt
wurde.
Das Versuchsprotokoll sah die ersten 60 Minuten offene Perfusion vor und im direkten
Anschluß mit der selben Leber, durch Umschalten des Systems für weitere 60 Minuten
geschlossen rezyklierende Perfusion. Während der gesamten Perfusionszeit wurde der
Sauerstoffverbrauch kontinuierlich durch eine Clark-Elektrode aufgezeichnet. Durch eine
Kalium-Elektrode wurde der Kaliumausstrom der Leberzellen gemessen und durch einen
Druckaufnehmer der Perfusionsdruck ∆p kontrolliert. In 10 minütigem Abstand gewonnene
Perfusatproben dienten zur Bestimmung von Glucose, Harnstoff und der LDH, als Parame-ter
des Zellschadens des perfundierten Organs. Das Umschalten der Perfusionapparatur ge-

Ergebnisse55
währleistete, daß weder Sauerstoffmessung, noch die Bestimmung der Substrate bzw. der
LDH unterbrochen werden mußten.
Um die Intaktheit der perfundierten Lebern zu kontrollieren, wurde abschließend nach 140
Minuten Perfusion Adrenalin in einer Konzentration von 10-6 M dem Perfusionmedium zu-
gesetzt. In allen Versuchen fiel binnen Sekunden der Sauerstoffverbrauch der perfundierten
Lebern dramatisch ab, um anschließend ebenso schnell und steil deutlich über das vorherbe-
stehende Atmungsniveau hinaus wieder anzusteigen. Der initiale Abfall erklärt sich durch die
α1-Rezeptor vermittelte Vasokonstriktion, während nach erneuter Öffnung der Gefäße der
rapide Anstieg des Sauerstoffverbrauchs die Intaktheit der perfundierten Organe, auch nach
140 minütiger Perfusion bewies.
2.1 Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei offen und rezyklierend
perfundierten Lebern männlicher, hypothyreoter und gehungerter Ratten
Der Sauerstoffverbrauch der Kontrollperfusionen betrug zum Zeitpunkt t = 0 min 2,69 ± 0,14
µmol/min/g und war damit fast identisch mit dem Ausgangswert der Versuchsgruppe zum
selben Zeitpunkt, der 2,72 ± 0,12 µmol/min/g betrug als 3,5-L-T2 dem Perfusions-puffer
zugesetzt wurde.
Bei offener Perfusion verliefen die Kontrollen fast linear auf dem Ausgangsniveau und be-
trugen am Ende der 60 minütigen Perfusion 2,71 ± 0,20 µmol/min/g ( Abb. 19 ). Zu die-sem
Zeitpunkt begann die rezyklierende Perfusion und der Sauerstoffverbrauch stieg bei
den Kontrollen, identisch zu den vorherigen Versuchen zur rezyklierenden Perfusion, auch hier
um 0,55 µmol/min/g nach dann insgesamt 120 Min Perfusion auf 3,26 ± 0,22 µmol/ min/g an,
entsprechend 20,3 % ( vergleiche dazu die Befunde aus III.1.1 ).

Ergebnisse56
t in min0 20 40 60 80 100 120 140 160 180
O2-Verbrauch in µmol/min/g
2,0
2,5
3,0
3,5
4,0
*
** *
of fen of fenrezykl ierend
Abb. 19 Sauerstoffverbrauch isoliert offen perfundierter ( t = 0 bis 60 min ) und isoliert rezyklierend
perfundierter ( t = 60 bis 120 min ) Lebern männlicher, hypothyreoter, gehungerter Ratten.
o Kontrollen ( MW ± SEM, n = 4 )
n 10-9 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 4 )
Zur Kontrolle nach 15 min offener Perfusion zum Zeitpunkt t = 135 min Zusatz von Adrenalin
in einer Konzentration von 10-6 M; ∗ p < 0,01 ∗∗ p < 0,05.
Die weitere offene Perfusion ließ den Sauerstoffverbrauch auf 2,79 ± 0,14 µmol/min/g im
Verlauf von weiteren 15 min abfallen. Damit pendelte sich der Sauerstoffverbrauch nach
60 Minuten rezyklierender Perfusion und daran anschließender offener Perfusion nach kur-zer
Zeit wieder auf dem Niveau der offenen Perfusionsperiode ein.
Abschließend wurde die Kontrolle der Intaktheit der perfundierten Lebern, wie oben be-
schrieben durch den Zusatz von 10-6 M Adrenalin überprüft. Der Sauerstoffverbrauch fiel bei
den Kontrollen binnen Sekunden um 0,69 µmol/min/g auf 2,10 ± 0,19 µmol/min/g ab, um im
weiteren Verlauf genauso schnell wieder anzusteigen. Nach 10 Minuten, zum Zeit-punkt t =

Ergebnisse57
145 min, betrug der O2-Verbrauch dann 3,11 ± 0,31 µmol/min/g und fiel in der
darauffolgenden Zeit allmählich auf das Niveau bei offener Perfusion zurück.
Während der 60 minütigen offenen Perfusion bewirkte 3,5-L-T2 einen langsameren allmäh-
lichen Anstieg des Sauerstoffverbrauchs von 2,72 ± 0,12 µmol/min/g um 0,22 µmol/min/g auf
2,94 ± 0,02 µmol/min/g, entsprechend 8,1 % nach 60 min; p<0,05.
Anschließend folgte die rezyklierende Perfusion, die im Unterschied zu den vorherigen
Versuchen, dargestellt in III.1.2, einen sofortigen Anstieg des Sauerstoffverbrauchs zeigte.
Dieser war bereits nach 10 Minuten signifikant erhöht mit 3,34 ± 0,06 µmol/min/g, gegen-über
2,89 ± 0,24 µmol/ min/g O2-Verbrauch der Kontrollen; p<0,01; vergleiche dazu Abb. 19.
Im weiteren Verlauf der 60 min geschlossen rezyklierender Perfusion ist der Sauerstoffver-
brauch nach Stimulation durch das Schilddrüsenhormon 3,5-L-T2 stets signifikant höher als
derjenige des Kontrollkollektivs mit p<0,01 und scheint, ab der 40. Minute ( entspricht der
100. Minute im Versuchsprotokoll ) ein Plateau zu erreichen. Nach 120 min Perfusion ( 60 min
offen und 60 min rezyklierend ) beträgt der Sauerstoffverbrauch der 3,5-L-T2-stimu-lierten
Versuchsgruppe 3,86 ± 0,02 µmol/min/g, gegenüber einem Sauerstoffverbrauch von 3,26 ±
0,22 µmol/min/g der Kontrollen. Die Stimulation durch 3,5-L-T2 bewirkte damit nach weiteren
60 Minuten Perfusion einen um 18,4 % höheren O2-Verbrauch. Vergleiche hierzu auch die
vorherigen Befunde in III.1.2 und Abb. 8, bei denen die Steigerung des O2-Verbrauchs erst
nach 45 Minuten statistisch signifikant war, die Zunahme des Sauerstoff-verbrauchs jedoch
29,5 % über den Kontollen lag.
Nach Ablauf von 120 min Perfusion folgten erneut 15 min offene Perfusion, in der der Sau-
erstoffverbrauch nach dann 135 min Gesamtperfusionszeit um 0,64 µmol/min/g auf 3,22 ± 0,09
µmol/min/g abfiel. Im Gegensatz zu den Kontrollen, die fast auf das Ausgangsniveau abfielen,
lag er damit immer noch 15,4 % höher als der Endwert der offenen Perfusions-periode.
Der Zusatz von 10-6 M Adrenalin, wie bei den Kontrollen hatte wiederum einen sofortigen
Abfall des Sauerstoffverbrauchs zur Folge. Nach bereits 2 Minuten sank der O2-Verbrauch auf
2,31 ± 0,04 µmol/min/g, um daran anschließend ebenso schnell wieder anzusteigen und mit
dem Kontrollkollektiv verglichen, zum Zeitpunkt t = 145 min ein wesentlich höheres Niveau
von 3,42 ± 0,19 µmol/min/g O2 zu erreichen. Zum Zeitpunkt des Abfalls des Sauer-

Ergebnisse58
stoffverbrauchs war auch der durch den Druckaufnehmer gemessene Perfusionsdruck er-höht,
was auf den durch α1-Rezeptoren vermittelten vasokonstringierenden Effekt des Adre-nalins
zurückzuführen ist; siehe auch hierzu Abb. 19.
2.2 Steigerung der Glucoseproduktion bei offener und rezyklierender Leberperfusion
durch 3,5-L-T2
Als weiteren Stoffwechselparameter der perfundierten Leber nach Stimulation durch das
Schilddrüsenhormon 3,5-L-T2 wurde aus beschriebenen Perfusatproben die Glucoseproduk-
t in min0 10 20 30 40 50 60 70 80 90 100 110 120
0
5
10
15
20
Glucose-Produktion in µmol/g
* *
a b
Abb. 20 a offene Perfusion Abb. 20 b geschlossene Perfusion
o Kontrollen ( MW ± SEM, n = 4 )n 10-9 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 4 )
Glucoseproduktion isoliert perfundierter Lebern männlicher, hypothyreoter und gehungerter Ratten. OffenePerfusion ( t = 0 bis 60 min ) und geschlossen rezyklierende Perfusion ( t = 60 bis 120 min ); ∗ p < 0,01tion bestimmt. Den Glucosespiegel des Perfusates nach Leberpassage während offener Per-
fusion stellt die Abb. 20 a dar. Wie hier zu sehen, war die Steigerung der Gluconeogenese in
den ersten 60 min offener Perfusion aus dem angebotenen Alanin durch 3,5-L-T2 nach 40 min
signifikant höher als der Glucosespiegel der Kontrollgruppe; p<0,01. In der graphi-schen
Darstellung lag die Glucoseproduktion zwar stets über derjenigen der Kontrollgruppe, der

Ergebnisse59
Befund ist aber weniger ausgeprägt, als in der anschließenden rezyklierenden Leberper-fusion
von der 60. bis zur 120. Minute; vergleiche dazu Abb. 20 b.
Nach Beginn der rezyklierenden Perfusion vom Zeitpunkt t = 60 min bis 120 min war die
Steigerung der Glucoseproduktion bereits nach 10 min ( in der 70. Minute im Versuchspro-
tokoll ) in der durch 3,5-L-T2 stimulierten Versuchsgruppe signifikant gegenüber den Kon-
trollen erhöht; p < 0,01. Die Glucoseproduktion stieg steil an, bis sie nach 40 Minuten ( in der
100. Minute im Versuchsprotokoll ) ein Plateau zu erreichen schien; vergleiche dazu Abb. 20
b. Dieser Befund korrespondierte genau mit der Kinetik des Sauerstoffverbrauchs, der sich
zum selben Zeitpunkt einem Plateau näherte. Der Vergleich der Glucoseproduktion bei
rezyklierender und offener Leberperfusion ist in Tab. 2 auf Seite 20 zusammengestellt. Für den
direkten Vergleich geht in die Berechnung bei rezyklierender Perfusion die Zeit mit ein, bei
offener Perfusion zusätzlich die Flußgeschwindigkeit.
Wie Abb. 20 a veranschaulicht, lag die 3,5-L-T2 stimulierte Glucoseproduktion stets über der
Kontrollgruppe, parallel zum Sauerstoffverbrauch. Wiederum wurde nach rezyklieren-der
Perfusion ab dem Zeitpunkt t = 120 min weiter offen perfundiert und nach 15 min zum
Zeitpunkt t = 135 min Adrenalin in einer Konzentration von 10-6 M zugesetzt, um die
Intaktheit der perfundierten Lebern zu kontrollieren ( siehe oben ).
2.3 Steigerung der Harnstoffproduktion bei offener und rezyklierender
Leberperfusion durch 3,5-L-T2
Die Harnstoffsynthese ist, genauso wie die Glucoseproduktion, ein wichtiger Stoffwechsel-
parameter für die perfundierte Leber. Wie oben beschrieben wurde aus den gewonnenen
Perfusatproben auch der Harnstoff bestimmt. Die Ergebnisse sind in den Abb. 21 a für den
Zeitraum der offenen ( t = 0 bis 60 min ) und 21 b für den Zeitraum der rezyklierenden
( t = 60 bis 120 min ) Perfusion dargestellt. Wie hier zu sehen verlief die Harnstoffproduk-tion
der Kontrollen sowohl bei offener, wie auch bei rezyklierender Perfusion nahezu auf

Ergebnisse60
t in min
0 10 20 30 40 50 60 70 80 90 100 110 120
Ha
rn
st
of
f-P
ro
du
kt
ion
in
µm
ol/
g
25
50
75
100
125
150 ba
*
*Abb. 21 a offene Perfusion Abb. 21 b geschlossene Perfusion
o Kontrollen ( MW ± SEM, n = 4 )n 10-9 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 4 )
Harnstoffproduktion isoliert perfundierter Lebern männlicher, hypothyreoter und gehungerter Ratten. Offene
Perfusion ( t = 0 bis 60 min ) und geschlossen rezyklierende Perfusion ( t = 60 bis 120 min ); ∗ p < 0,01
einem gleichbleibenden Niveau. Nach Zugabe des Schilddrüsenhormones 3,5-L-T2 stieg die
Harnstoffsynthese schon bei offener Leberperfusion an und unterschied sich signifikant nach 40
Minuten von den Kontrollen; p < 0,01. Auch im weiteren Verlauf lag sie zu je-dem Zeitpunkt
über dem Niveau der Kontrollen.
Nach Beginn der rezyklierenden Perfusion vom Zeitpunkt t = 60 bis 120 min war die Stei-
gerung der Harnstoffproduktion bereits nach 20 Minuten ( in der 80. Minute im Versuchs-
protokoll ), in der durch 3,5-L-T2 stimulierten Versuchsgruppe signifikant gegenüber den
Kontrollen erhöht; p < 0,01. Wie bei der Glucoseproduktion und dem Sauerstoffverbrauch
stieg auch die Harnstoffproduktion steil an und schien nach 30 Minuten ( 90. Minute im
Versuchsprotokoll ) ein Plateau zu erreichen; vergleiche dazu Abb. 19 und 20 b, sowie

Ergebnisse61
21 b. Der Vergleich der Harnstoffproduktion bei rezyklierender und offener Leberperfusion ist
in Tab. 2 auf Seite 20 zusammengestellt. Für den direkten Vergleich geht in die Berech-nung
bei rezyklierender Perfusion die Zeit mit ein, bei offener Perfusion zusätzlich die
Flußgeschwindigkeit.
Wiederum wurde nach rezyklierender Perfusion ab dem Zeitpunkt t = 120 min weiter offen
perfundiert und nach 15 min zum Zeitpunkt t = 135 min Adrenalin in einer Konzentration von
10-6 M zugesetzt, um die Intaktheit der perfundierten Lebern zu kontrollieren ( siehe oben ).
2.4 Perfusionsdruck und Kaliumausstrom in das Perfusat
Die kontinuierliche Registrierung des Perfusionsdruckes ∆p wies zu keinem Zeitpunkt der
Perfusionen weder des Kontrollkollektivs, noch der durch 3,5-L-T2 stimulierten Versuchs-
gruppe je einen Unterschied auf. Lediglich nach beschriebenem Zusatz von 10-6 M Adre-nalin
stieg der Perfusionsdruck kurzfristig an, aufgrund der durch α1-Rezeptoren vermittel-ten
Vasokonstriktion ( Werte nicht dargestelt ).
Auch bei der Messung des Kalium-Ausstromes der Leberzellen in das Perfusionsmedium war
keine Änderung festzustellen. Zu diesem Parameter ergab sich bei den Kontrollen, wie auch bei
der Versuchsgruppe nach Zusatz von 10-9 M 3,5-L-T2 kein Hinweis auf einen Ef-fekt ( Werte
nicht dargestellt ).
_______________
Zusammenfassung der Ergebnisse III.2
Bei der offenen Leberperfusion stieg der Sauerstoffverbrauch nach Stimulation durch 3,5-L-T2
langsamer an, als dies bei von Beginn an rezyklierender Perfusion der Fall war. Korres-

Ergebnisse62
pondierend zu diesem Befund verhielt es sich mit der Glucose- und Harnstoffproduktion, die
beide erst nach 40 minütiger Perfusion signifikant über den Kontrollen lagen.
Als jedoch nach 60 minütiger offener Perfusion das System umgestellt wurde auf rezyklie-
rende Perfusion, stieg der Sauerstoffverbrauch ohne erneute Hormonzugabe im Vergleich zu
den Kontrollen sofort steil an. Nach weiterer Perfusion unterschieden sich die hormon-
stimulierten Lebern bereits nach 10 min signifikant von den Kontrollen und erreichten nach 40
bis 60 minütiger rezyklierender Leberperfusion ein Plateau, welches sofort nach Rück-stellung
auf offene Perfusion wieder abfiel.
Auch Glucose- und Harnstoffproduktion beschrieben den selben Verlauf. Sie stiegen signi-
fikant bereits nach 10 bzw. 20-minütiger rezyklierender Perfusion an und erreichten, wie der
Sauerstoffverbrauch nach 40 bzw. 30 Minuten ein Plateau.
Bei den anderen gemessenen Parameter, wie dem Perfusionsdruck und insbesondere dem
Kaliumausstrom lies sich ein Einfluß von 3,5-L-T2 nicht nachweisen.
3. Einfluß von Ionenkanalblockern auf die Stimulation des Sauerstoffverbrauchs
isoliert rezyklierend perfundierter Lebern hypothyreoter, gehungerter,
männlicher Ratten durch 10-10 M 3,5-L-T2

Ergebnisse63
Die vorangegangenen Experimente zeigten einen deutlichen stimulierenden Effekt von 3,5-L-
T2 auf den Sauerstoffverbrauch, Glucose- und Harnstoffproduktion. Über den Mechanis-mus
dieser Wirkung ist noch wenig bekannt. Um zu prüfen, ob der Effekt durch Elektrolyt-ströme (
z.B. Kalium- oder Calciumeinstrom, Natriumausstrom ) zu erklären ist, wurden in den
folgenden Experimenten spezifische Hemmer der Ionenkanäle in das Perfusat gegeben und der
T2-Effekt auf den Sauerstoffverbrauch gemessen.
Die folgenden Versuche beschreiben den Einfluß unterschiedlicher Ionenkanäle auf die Sti-
mulation des Sauerstoffverbrauchs der isoliert rezyklierend perfundierten Lebern hypothy-
reoter, gehungerter, männlicher Ratten durch 3,5-L-T2 . Die eingesetzte Konzentration des
Hormons wurde im Vergleich zu den vorher beschriebenen Experimenten im Kapitel III.1 auf
10-10 M erhöht und liegt im sicher maximal stimulierenden Bereich. Die Versuchsanord-nungen
sind die selben, wie in den vorbeschriebenen Experimenten des Kapitels III.1 zur
rezyklierenden Leberperfusion.
3.1 Einfluß von Ouabain auf die Stimulation des Sauerstoffverbrauchs durch
3,5-L-T2
Die Na+-K+-Pumpe reguliert die Konzentration der Na+- und K+-Ionen in der Zelle. Das
Transportsystem ist in der Plasmamembran so ausgerichtet, daß sie Na+ aus der Zelle hin-aus-
und K+ in die Zelle hineinpumpt. Um die Pumpe zu betreiben muß im Inneren der Zelle ATP
vorhanden sein. Ouabain ist ein spezifischer Inhibitor der Pumpe, der nur dann wirksam ist,
wenn es sich außerhalb der Zelle befindet.
Vor Installation der freipräparierten Lebern in die Perfusionsapparatur wurde dem Perfu-
sionspuffer Ouabain in einer Konzentration von 10-6 M zugesetzt. Nach anschließender
Äquilibrierungszeit von 30 min wurde den Kontrollen kein weiteres Agenz zugesetzt; der
Versuchsgruppe wurde 3,5-L-T2 in der Konzentration von 10-10 M zugesetzt.
Die Abb. 22 b faßt die Ergebnisse des Versuches unter Blockade des Na+/K+-Kanals durch
Ouabain ohne und mit Stimulation durch 3,5-L-T2 zusammen und im Vergleich dazu ist in

Ergebnisse64
Abb. 22 a dieselbe Versuchsanordnung ohne Ionenkanalblockade aus III.1.2 noch einmal
dargestellt.
Der basale Sauerstoffverbrauch der perfundierten Lebern zum Zeitpunkt t = 0 min wurde durch
den Zusatz von 10-6 M Ouabain zum Perfusat geringfügig gesenkt. Dies traf auf die Kontroll-,
wie auch auf die Versuchsgruppe zu. Im Verlauf der Perfusion stieg dann weder,
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 30 60 90
O2 µ
mo
l/m
in/
g
1,5
2,0
2,5
3,0
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 30 60 90
O2 µ
mo
l/m
in/
g
1,5
2,0
2,5
3,0*
Abb. 22 a, ohne Zusatz zum Perfusat Abb. 22 b, Zusatz von 10-6 M Ouabain
o Kontrollen ( MW ± SEM, n = 11 ) o Kontrollen ( MW ± SEM, n = 2 )n 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min n 10-10 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 9 ), ∗ p < 0,01 ( MW ± SEM, n = 5 )
Wirkung von Ouabain auf die Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei isoliert perfundierten
Lebern männlicher, hypothyreoter, gehungerter Ratten.
Dargestellt sind der relative Sauerstoffverbrauch ( gr. Graphik ), sowie der absolute Sauerstoffverbrauch
( kl. Graphik ) in µmol/min/g.
wie in den Vorexperimenten, der Sauerstoffverbrauch der Kontrollen, noch derjenige der durch
3,5-L-T2 stimulierten Versuchsgruppe an. Sowohl bei der Darstellung des absoluten
( kl. Graphik ), wie auch des relativen Sauerstoffverbrauchs ( gr. Graphik ) in der Abb.
22 b wurde die inhibierende Wirkung auf eine Steigerung des Sauerstoffverbrauchs durch 3,5-
L-T2 deutlich. Die Kontrollen zum Zeitpunkt t = 0 min verbrauchten 1,59 ± 0,16 µmol/min/g
und nach 90 Minuten Perfusion dann 1,75 ± 0,09 µmol/ min/g O2, und damit lediglich 0,16
µmol/min/g mehr; vergleiche dazu auch Abb. 22 a ohne Ouabain. Nach Zu-

Ergebnisse65
satz von 3,5-L-T2 zum Ouabain-haltigen Perfusionspuffer unterschied sich die Steigerung des
Sauerstoffverbrauchs von 1,37 ± 0,09 µmol/min/g um 0,21 µmol/min/g auf 1,57 ± 0,11
µmol/min/g nicht von den Kontrollen.
Nach Blockade der Na+/K+-Pumpe durch Ouabain in einer Konzentration von 10-6 M war also
durch 3,5-L-T2 bei isoliert rezyklierender Leberperfusion der Sauerstoffverbrauch hy-
pothyreoter, gehungerter, männlicher Ratten nicht zu steigern.
Um zu prüfen, ob die Lebern bezüglich des Sauerstoffverbrauchs überhaupt noch stimulier-bar
waren, wurde am Ende der Perfusionen jeweils Adrenalin in einer Konzentration von 10-6 M
dem Perfusat zugesetzt, was stets den vorbeschriebenen rapiden Anstieg des Sauer-
stoffverbrauchs zur Folge hatte und die Intaktheit der perfundierten Lebern auch zum Ver-
suchsende bewies; siehe dazu auch Abb. 19 im Kapitel III.2.1.
3.2 Einfluß von Nifedipin auf die Stimulation des Sauerstoffverbrauchs durch
3,5-L-T2
Bei Nifedipin handelt es sich um einen selektiven Blocker des Calciumkanals, der dem Per-
fusionsmedium, analog dem oben beschriebenen Versuch aus III.3.1, vor Installation der
Lebern zugegeben wurde. Nifedipin, in der Konzentration von 10-6 M, blockiert den Calci-
umeinstrom in die Zelle.
Nach der Äquilibrierungszeit verbrauchten die Kontrollen zum Zeitpunkt t = 0 min 2,17
± 0,08 µmol/min/g O2, dargestellt in Abb. 23 b. Der Ausgangs-Sauerstoffverbrauch lag mit
Nifedipin-Zusatz im Medium signifikant höher als bei den Kontrollen ohne Nifedipin-Zu-satz;
vergleiche dazu die Kontrollgruppe in Abb. 23 a. Die Zunahme des O2-Verbrauchs war mit
0,65 µmol/min/g den Kontrollen ohne Calciumkanalblocker vergleichbar und stieg nach 90 min
Perfusion auf 2,82 ± 0,04 µmol/min/g an.

Ergebnisse66
t in min0 20 40 60 80 100
∆ O 2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 20 40 60 80 100
∆ O 2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 30 60 90
O2 µ
mo
l/m
in/
g
1,52,02,53,03,5
t in min0 30 60 90
O2
µmo
l/m
in/
g
1,52,02,53,03,5
* *
Abb. 23 a, ohne Zusatz zum Perfusat Abb. 23 b, Zusatz von 10-6 M Nifedipin
o Kontrollen ( MW ± SEM, n = 11 ) o Kontrollen ( MW ± SEM, n = 3 )n 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min n 10-10 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 9 ) ( MW ± SEM, n = 2 )
Wirkung von Nifedipin auf die Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei isoliert perfundierten
Lebern männlicher, hypothyreoter, gehungerter Ratten.
Dargestellt sind der relative Sauerstoffverbrauch ( gr. Graphik ), sowie der absolute Sauerstoffverbrauch
( kl. Graphik ) in µmol/min/g; ∗ p < 0,01.
Die Versuchsgruppe mit 3,5-L-T2-Zusatz zum Zeitpunkt t = 0 min wies unter Einfluß von
Nifedipin einen niedrigeren Ausgangs-Sauerstoffverbrauch auf, stieg aber in noch größerem
Ausmaß um 1,62 ± 0,17 µmol/min/g von 1,92 ± 0,04 µmol/min/g auf 3,54 ± 0,13 µmol/ min/g
an. Die Steigerung des Sauerstoffverbrauchs lag somit noch über der vergleichbaren durch 3,5-
L-T2 stimulierten Gruppe ohne Nifedipin-Zusatz; vergleiche dazu die Abb. 23 a und b.
Der Zusatz des Calciumkanalblockers Nifedipin zum Perfusionsmedium konnte die Stimula-
tion des Sauerstoffverbrauchs durch 3,5-L-T2 nicht inhibieren. Eher schien ein zusätzlich
potenzierender Effekt vorzuliegen, da bereits nach 40 min Perfusion, verglichen mit den
Kontrollen, ein signifikant höherer O2-Verbrauch vorlag. Die Steigerung des Sauerstoffver-
brauchs gegenüber den Kontrollen betrug am Perfusionsende 25,5 %.
Um zu prüfen, ob die Lebern bezüglich des Sauerstoffverbrauchs überhaupt noch stimulier-bar
waren, wurde am Ende der Perfusionen jeweils Adrenalin in einer Konzentration von 10-6 M

Ergebnisse67
zum Perfusat gegeben, was stets den vorbeschriebenen rapiden Anstieg des Sauer-
stoffverbrauchs zur Folge hatte und die Intaktheit der perfundierten Lebern auch zum Ver-
suchsende bewies; siehe dazu auch Kapitel III.2.1.
3.3 Einfluß von SITS auf die Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2
Bei SITS handelt es sich um einen selektiven K+/H+-Kanalblocker, der dem Perfusions-
medium in einer Konzentration von 10-6 M zugesetzt wurde. Die Versuchsergebnisse sind
in Abb. 24 b zusammengefaßt.
Nach Zusatz von SITS zum Perfusionspuffer betrug der Ausgangs-Sauerstoffverbrauch 1,93 ±
0,03 µmol/min/g in der Kontrollgruppe und liegt damit nur geringfügig oberhalb der
Kontrollgruppe ohne diesen K+/H+-Blocker; vergleiche dazu Abb. 24 a und b. Und auch im
weiteren Verlauf der 90 minütigen Perfusion wiesen die Kontrollen einen nahezu iden-
tischen Verlauf auf. Die Zunahme des Sauerstoffverbrauchs betrug 0,56 µmol/min/g und
erreichte 2,48 ± 0,03 µmol/min/g.
Nach Zusatz von 10-10 M 3,5-L-T2 zum Zeitpunkt t = 0 min stieg der Sauerstoffverbrauch erst
spät an und unterschied sich erst in der 60. Minute signifikant von den Kontrollen. Der
Ausgangs-Sauerstoffverbrauch lag mit 1,79 ± 0,05 µmol /min/g zwar unterhalb dem der
Kontrollgruppe, wie aber die Darstellung des relativen Sauerstoffverbrauchs in Abb. 24 b
verdeutlicht unterschieden sich Versuchs- und Kontrollgruppe bezogen auf den Ausgangs-
verbrauch zum Zeitpunkt t = 0 min innerhalb der ersten 60 min jedoch kaum voneinander.
Dennoch erreichte der stimulierte O2-Verbrauch 2,82 ± 0,07 µmol/min/g und und lag damit
1,03 ± 0,11 µmol/min/g, bzw. 13,7 %, signifikant über den Kontrollen; p<0,01. Im Ver-
gleich zur Perfusion ohne Ionenkanalblocker fiel die Stimulation des Sauerstoffverbrauchs
durch 3,5-L-T2 jedoch geringer aus; vergleiche dazu die Abb. 24 a und b.

Ergebnisse68
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 30 60 90
O2
µmo
l/m
in/
g
1,5
2,0
2,5
3,0
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 30 60 90
O2 µ
mo
l/m
in/
g
1,5
2,0
2,5
3,0
*
*
Abb. 24 a, ohne Zusatz zum Perfusat Abb. 24 b, Zusatz von 10-6 M SITS
o Kontrollen ( MW ± SEM, n = 11 ) o Kontrollen ( MW ± SEM, n = 3 )n 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min n 10-10 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 9 ) ( MW ± SEM, n = 2 )
Wirkung von SITS auf die Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei isoliert perfundierten
Lebern männlicher, hypothyreoter, gehungerter Ratten.
Dargestellt sind der relative Sauerstoffverbrauch ( gr. Graphik ), sowie der absolute Sauerstoffverbrauch
( kl. Graphik ) in µmol/min/g; ∗ = p < 0,01.
Um zu prüfen, ob die Lebern bezüglich des Sauerstoffverbrauchs überhaupt noch stimulier-bar
waren, wurde am Ende der Perfusionen jeweils Adrenalin in einer Konzentration von 10-6 M
zum Perfusat gegeben, was stets den vorbeschriebenen rapiden Anstieg des Sauer-
stoffverbrauchs zur Folge hatte und die Intaktheit der perfundierten Lebern auch zum Ver-
suchsende bewies; siehe dazu auch III.2.1.
3.4 Einfluß von Amilorid auf die Stimulation des Sauerstoffverbrauchs durch
3,5-L-T2
Amilorid blockiert selektiv den Kanal, über den Na+- und H+-Ionen ausgetauscht werden.

Ergebnisse69
Es wurde dem Perfusionsmedium in einer Konzentration von 10-3 M zugesetzt. Hiernach
betrug bei der Kontrollgruppe ohne Zugabe von 3,5-L-T2 der Sauerstoffverbrauch zum
Zeitpunkt t = 0 min 1,48 ± 0,03 µmol/min/g. Er fiel damit niedriger aus als bei der Kon-
trollgruppe ohne Amiloridzusatz, wie auch schon im Ouabain-Experiment beschrieben. Im
weiteren Verlauf unterschied sich die Amilorid-Gruppe von allen anderen bisherigen Versu-
chen durch einen allmählichen Abfall des Sauerstoffverbrauchs auf 1,28 ± 0,04 µmol/min/g
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 30 60 90
O2
µmo
l/m
in/
g
1,5
2,0
2,5
3,0
t in min0 20 40 60 80 100
∆ O2-Verbrauch in µmol/min/g
0,0
0,5
1,0
1,5
t in min0 30 60 90
O2 µ
mo
l/m
in/
g
1,5
2,0
2,5
3,0
*
*
Abb. 25 a, ohne Zusatz zum Perfusat Abb. 25 b, Zusatz von 10-3 M Amilorid
o Kontrollen ( MW ± SEM, n = 11 ) o Kontrollen ( MW ± SEM, n = 2 )n 10-12 M 3,5-L-T2 zum Zeitpunkt t = 0 min n 10-10 M 3,5-L-T2 zum Zeitpunkt t = 0 min ( MW ± SEM, n = 9 ) ( MW ± SEM, n = 4 )
Wirkung von Amilorid auf die Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei isoliert perfundierten
Lebern männlicher, hypothyreoter, gehungerter Ratten.
Dargestellt sind der relative Sauerstoffverbrauch ( gr. Graphik ), sowie der absolute Sauerstoffverbrauch
( kl. Graphik ) in µmol/min/g, ∗ = p < 0,01.
nach 90 Minuten Perfusion, p<0,01; vergleiche dazu Abb. 25 a und b.
Nach Zusatz von 10-10 M 3,5-L-T2 zum Zeitpunkt t = 0 min, bei einem O2-Verbrauch von 1,38
± 0,07 µmol/min/g war dieser nach 10 min sehr schnell signifikant erhöht, p<0,01; siehe Abb.
25 b. Die Steigerung des Sauerstoffverbrauchs in der Versuchsgruppe erreichte schon nach 20
min ihr Maximum mit 1,85 ± 0,10 µmol/min/g, und fiel anschließend nach der 20. Minute
langsam bis zum Perfusionsende, parallel zum Verlauf der Kontrollen, kon-

Ergebnisse70
tinuierlich ab.
Der O2-Verbrauch nach 90 Minuten rezyklierender Perfusion betrug noch 1,53 ± 0,14 µmol
/min/g und war bei Auftragung der absoluten Werte, verglichen mit dem Ausgangswert nicht
mehr signifikant erhöht. Wie bei den Leberperfusionen mit Zusatz von Ouabain ( sie-he III. 3.1
und Abb. 22 a und b ), wurde auch hier durch Blockade eines Ionenkanals der am Na+-
Transport beteiligt ist, nicht nur der basale Sauerstoffverbrauch gesenkt, sondern auch seine
Steigerung durch 3,5-L-T2 verhindert.
_______________
Zusammenfassung der Ergebnisse III.3
Um zu prüfen, ob der Wirkungsmechanismus der Sauerstoffstimulation durch 3,5-L-T2 von
Ionenströmen über die Zellmembran abhängig ist wurden dem Perfusionspuffer verschie-
dene selektive Blocker der Ionenkanäle zugesetzt.
Während die Zusätze von Nifedipin und SITS kaum einen Einfluß auf die Hormonwirkung
hatten, war eine Stimulation des Sauerstoffverbrauchs isoliert rezyklierend perfundierter
Lebern nach Ouabain-Zusatz zum Perfusat nicht mehr nachweisbar. Ouabain ist ein selek-tiver
Hemmer der Na+-K+-Pumpe, sodaß der stimulierende Effekt durch 3,5-L-T2 auf den O2-
Verbrauch von diesem Ionenkanal abhängig war.
Amilorid, als ein Hemmer des Na+/H+-Kanals, hat auch einen, im Vergleich zu Ouabain
geringeren, inhibierenden Einfluß auf die Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2.
Da weder Nifedipin noch SITS einen Einfluß auf die Stimulation des O2-Verbrauchs hatten ist
ein Calcium- bzw. Kalium-vermittelter Effekt ausgeschlossen, wie das auch für das Ka-lium in

Ergebnisse71
den Experimenten der offenen Leberperfusion gezeigt werden konnte; vergleiche dazu Kapitel
III.2.
4. Einfluß von 3,5-L-T2 auf die oxidative Phosphorylierung bei isolierten
Mitochondrien der Rattenleber
Die vorangegangenen Experimente zeigten überzeugende Befunde zur schnellen Wirkung von
3,5-L-T2 an der isoliert perfundierten Rattenleber im Sinne einer Steigerung des Sauer-
stoffverbrauchs und damit korrespondierend einer Stimulation der Gluconeogenese aus dem

Ergebnisse72
angebotenen Alanin sowie der Harnstoffsynthese. Diese Effekte waren bei offener Leber-
perfusion weniger ausgeprägt, als nach der anschließenden rezyklierenden Perfusion, was
darauf hindeutete, daß die Wirkung von 3,5-L-T2 auf die Lebern keine direkte ist. Um auf-
zudecken, welche Mechanismen in diese Wirkung involviert sind wurden, unter der Vor-
stellung, daß Ionenströme über die Zellmembran den Effekt vermitteln könnten, die Expe-
rimente mit dem Einsatz selektiver Ionenkanalblocker durchgeführt. Die Untersuchungen
ergaben, daß die Wirkung von 3,5-L-T2 auf den Sauerstoffverbrauch mit dem transmembra-
nen Flux der Na+-Ionen und der Na+/K+-ATPase korrespondieren.
Zur weiteren Abklärung des Mechanismus einer stimulierenden Wirkung von 3,5-L-T2 auf die
Atmung der perfundierten Lebern wurden Experimente an isolierten Mitochondrien
durchgeführt. Vorversuche unserer Arbeitsgruppe zeigten nach direkter Zugabe von 3,5-L-T2
in vitro zu isolierten Mitochondrien aus Lebern männlicher, hypothyreoter, gehungerter Ratten
keinen Einfluß auf deren Atmungsaktivität ( Werte nicht dargestellt ). Deshalb wurde, unter
der Vorstellung, daß die Wirkung auf die Mitochondrien analog der oben beschriebenen
Befunde eine indirekte sein könnte, das Hormon intraperitoneal gespritzt,
in vivo, und die Mitochondrien anschließend isoliert, um ihre Atmungsaktivität zu messen.
Untersucht wurde hierzu der stimulierende Einfluß von 3,5-L-T2 auf die oxidative Phospho-
rylierung unter Einsatz der Substrate Succinat und Glutamat bei isolierten Mitochondrien aus
Rattenlebern. Männlichen, hypothyreoten, gehungerten Wistar-Ratten wurde intraperi-toneal
250 µg 3,5-L-T2/100 g KG und den Kontrollen NaCl 0,9% gleichen Volumens inji-ziert, um
nach 30, 60 und 90 Minuten die Mitochondrien zu isolieren und im Anschluß de-ren
Atmungsaktivität zu messen. Nach Inkubation der Mitochondrien bei 30° C in 110mM
Mannitol mit den Substraten Succinat ( 10 mM ) bzw. Glutamat ( 5 mM ) wurde nach Zu-satz
von 0,05 M ADP die mitochondriale Atmung gemessen. Das Ergebnis zeigen die Abb. 26 und
27.
Die Kontrollen, denen intraperitoneal NaCl 0,9% injiziert wurde wiesen nach Succinat-Zusatz
einen Sauerstoffverbrauch von 154 ± 8 nmol/min/mg Protein auf; vergleiche dazu Abb. 26,
zum Zeitpunkt t = 30 min. Nach Zusatz von Glutamat betrug der Sauerstoffver-brauch der
Kontrollen 104 ± 4 nmol/min/mg Protein; vergleiche dazu Abb. 27, zum Zeit-punkt t = 30 min.

Ergebnisse73
30 Minuten nach intraperitonealer Injektion von 250 µg 3,5-L-T2/100 g KG und Zusatz von
Succinat stieg der Sauerstoffverbrauch auf 173 ± 4 nmol/min/mg Protein signifikant an;
p<0,01, vergleiche hierzu Abb. 26.
nm
ol
O2/
min
/m
g P
ro
te
in
Kontrollen
nach 30 min
0
120
140
160
180Oxidative Phosphorylierung, Succinat
*
3,5-L-T2
nach 30 min
Abb. 26 Sauerstoffverbrauch isolierter Mitochondrien aus Lebern männlicher, hypothyreoter, gehun-
gerter Ratten. Den Kontrollen wurde 30 min vor der Mitochondrienpräparation NaCl 0,9%
und der Versuchsgruppe 250 µg 3,5-L-T2 / 100g KG in vivo i. p. injiziert.
Substrat zur oxidative Phosphorylierung war jeweils Succinat 10 mM.
Beide Versuchsgruppen n = 3; ∗ p<0,01
Ebenso stieg der Sauerstoffverbrauch 30 Minuten nach intraperitonealer Injektion von 250 µg
3,5-L-T2/100 g KG und Zusatz von Glutamat auf 118 ± 6 nmol/min/mg Protein auch
signifikant an; p<0,0125, vergleiche dazu Abb. 27.
60 Minuten nach intraperitonealer Injektion der gleichen Menge von 3,5-L-T2 lag der Sau-
erstoffverbrauch nach Zusatz von Succinat mit 162 ± 3 nmol/min/mg Protein im Bereich
der Kontrollen. Bei Zusatz von Glutamat als Substrat der oxidativen Phosphorylierung be-

Ergebnisse74
0
90
100
110
120
Kontrollen
nach 30 min
nm
ol
O2/
min
/m
g P
ro
te
in
Oxidative Phosphorylierung, Glutamat *
3,5-L-T2
nach 30 min
Abb. 27 Sauerstoffverbrauch isolierter Mitochondrien aus Lebern männlicher, hypothyreoter, gehun-
gerter Ratten. Den Kontrollen wurde 30 min vor der Mitochondrienpräparation NaCl 0,9%
und der Versuchsgruppe 250 µg 3,5-L-T2 / 100g KG in vivo i. p. injiziert.
Substrat zur oxidative Phosphorylierung war jeweils Glutamat 5 mM.
Beide Versuchsgruppen n = 3; ∗ p<0,0125
trug der O2-Verbrauch nach 3,5-L-T2-Applikation 113 ± 1 nmol/min/mg Protein und lag
damit wiederum signifikant höher als die Kontrollen; p<0,01, Werte nicht dargestellt.
Und auch 90 Minuten nach intraperitonealer Injektion von 250 µg 3,5-L-T2 je 100 g KG lag
der Sauerstoffverbrauch bei Zusatz beider verwendeten Substrate signifikant über den Kon-
trollen. Nach Succinat-Zusatz betrug der O2-Verbrauch 181 ± 14 nmol/min/mg Protein;
p<0,0125. Nach Glutamat-Zusatz stieg der Sauerstoffverbrauch auf 127 ± 3 nmol/min/mg
Protein und lag erneut signifikant über dem der Kontrollen; p<0,01, Werte nicht darge-stellt.
Der vereinheitlichte Versuchsansatz lies mittels beschriebener Methodik der Messung der
Atmung isolierter Mitochondrien aus Lebern männlicher, hypothyreoter gehungerter Ratten
sehr gleichmäßige Einzelergebnisse erzielen. Bereits 30 min nach intraperitonealer Injektion
von 250 µg 3,5-L-T2 je 100g KG stieg der Sauerstoffverbrauch der Mitochondrien sowohl bei
Succinat als auch bei Glutamat als Substrat der oxidativen Phosphorylierung signifikant im
Vergleich zu den Kontrollen an. Nach 60 min war der Sauerstoffverbrauch bei Succinat als

Ergebnisse75
Substrat in seinem Anstieg weniger ausgeprägt, als bei dem Substrat Glutamat ( keine Abb.,
vergleiche die Werte oben ). Nach 90 min lag der O2-Verbrauch wieder signifikant bei beiden
verwendeten Substraten hoch über den Kontrollen.
_______________
Zusammenfassung der Ergebnisse III.4
In vivo durch 3,5-L-T2 stimulierte isolierte Mitochondrien aus Lebern männlicher, hypothy-
reoter, gehungerter Ratten wiesen nach 30 Minuten eine signifikante Steigerung des Sauer-
stoffverbrauchs und damit eine gesteigerte oxydative Phosphorylierung bei Einsatz der Sub-
strate Succinat, wie auch Glutamat auf.
Die Wirkung von 3,5-L-T2 auf den Anstieg der mitochondrialen Atmung hatte nach 30 Mi-
nuten ein Niveau erreicht, daß zu späteren Zeitpunkten der Messung ( nach 60 und 90 min )
nicht mehr wesentlich steigerbar war.
5. Isolierte Nierenperfusion

Ergebnisse76
Nach den bisherigen Experimenten lies sich ein schneller Effekt von 3,5-L-T2 auf die Sti-
mulation des Sauerstoffverbrauchs, sowie auf die Glucose- und Harnstoffproduktion am
Versuchsmodell der isoliert perfundierten Leber nachweisen. In weiteren Versuchen wurde
dann der Mechanismus dieses Effektes an verschiedenen Versuchsmodellen näher unter-sucht.
Neben der Leber stellt die Niere ein im Stoffwechsel der Schilddrüsenhormone wichtiges
Organ dar; siehe dazu auch S. 10 in der Einleitung. Die folgenden Experimente sollen die
Frage beantworten, ob die Effekte von 3,5-L-T2 auf den Stoffwechsel der Leber begrenzt sind
oder sich auch eine Wirkung im Sinne einer Stimulation des Sauerstoffverbrauchs an der Niere
nachweisen läßt. Als Versuchsmodell wurde die isoliert perfundierte Niere von hypothyreoten
Ratten gewählt.
Die in den Versuch genommenen Tiere waren stets männlich, hypothyreot und hungerten 24
Stunden vor dem Versuchsbeginn. Dem Perfusionspuffer wurde 3,5-L-T2 in einer Kon-
zentration von 10-9 M zugesetzt, die sicher im maximal stimulierenden Bereich liegt, wie aus
vorangegangenen Experimenten hervorging.
5.1 Einfluß von 3,5-L-T2 bei isolierter Nierenperfusion hypothyreoter Ratten
Im beschriebenen Versuchsmodell der isolierten Nierenperfusion ( siehe Material und Me-
thoden S. 25 ff. ) wurde zum Referenzgewicht der Nieren der Sauerstoffverbrauch ( O2 in
µmol/min/g ) gemessen, sowie glomeruläre Filtrationsrate ( GFR in µl/min/g ), Natrium-
Filtrationsrate ( Na FR in µmol/min/g ), Natrium-Reabsorptionsrate ( Na RR in µmol/min /g ),
Kalium-Filtrationsrate ( K FR in µmol/min/g ), Kalium-Reabsorptionsrate ( K RR in
µmol/min/g ), Glucose-Reabsorptionsrate ( Gl RR in µmol/min/g ) und der Quotient aus
Natriumreabsorption / O2-Verbrauch. Die Ergebnisse sind in Tabelle 4 zusammengefaßt.
Sie zeigten, daß durch den Zusatz von 3,5-L-T2 nach 20 minütiger Äquilibrierungszeit in
einer Konzentration von 10-9 M zum Perfusionspuffer keine Steigerung des Sauerstoffver-
Tab. 4 Zusammenfassung der Befunde zum Einfluß von 3,5-L-T2 auf die isolierte Nierenperfusion;
angegebene Werte als MW ± SEM; Kontrollen n = 19; 10-9 M 3,5-L-T2; n = 10

Ergebnisse77
Parameter Einheit Kontrollen 10-9 M 3,5-L-T2
O2-Verbrauch µmol/min/g 8,3 ± 0,5 7,8 ± 0,8
GFR µl/min/g 761 ± 50 693 ± 58
Na FR µmol/min/g 108 ± 7 100 ± 9
Na RR µmol/min/g 96 ± 6 88 ± 8
K FR µmol/min/g 3,7 ± 0,2 3,4 ± 0,3
K RR µmol/min/g -0,2 ± 0,3 0,7 ± 0,3
GI RR µmol/min/g 5,1 ± 0,4 4,7 ± 0,4
Na RR/O2-Verbrauch --- 11,8 ± 0,7 11,6 ± 0,6
brauchs der isoliert perfundierten Nieren zu verzeichnen war. Auch die übrigen gemesse-nen
Funktionsparameter der Nieren waren im Verlauf nicht signifikant verändert; verglei-che hierzu
Tab 4.
5.2 Einfluß von 3,5-L-T2 bei isolierter Nierenperfusion euthyreoter Ratten
Bei Voruntersuchungen an isoliert perfundierten Nieren von euthyreoter Ratten war nach
Zusatz von 10-9 M 3,5-L-T2 eine Reduktion der Natrium-Rückresorption ( Na RR ), sowie des
Quotienten aus Natrium-Rückresortion/O2-Verbrauch zu verzeichnen. Im einzelnen betrug die
Na RR bei Kontrollen 87,2 ± 10,3 µmol/min/g ( n = 3 ) und nach Zusatz von
10-9 M 3,5-L-T2 nur noch 77,6 ± 2,1 µmol/min/g ( n = 8 ); p<0,05. Der Quotient aus
Na RR/O2-Verbrauch betrug bei Kontrollen 11,2 ± 1,0 ( n = 3 ) und nach Hormonzusatz
wie oben nur noch 9,9 ± 0,3 ( n = 8 ); p<0,01 ( Werte nicht dargestellt ). Die übrigen
gemessenen Parameter unterschieden sich auch in diesem Versuchsansatz nicht signifikant
voneinander. Eine Steigerung des Sauerstoffverbrauchs durch 3,5-L-T2, war auch in dieser
Versuchsanordnung nicht nachzuweisen.

Ergebnisse78
5.3 Einfluß von 3,5-L-T2 auf kultivierte Mesangiumzellen
Nachdem es nicht gelungen war an der Niere als ganzem Organ in der isolierten Perfusion
einen Einfluß von 3,5-L-T2 nachzuweisen stellte sich die Frage, ob dies auch auf zellulärer
Ebene zutrifft. Mesangiumzellen sind spezialisierte glatte Muskelzellen der Niere und regu-
lieren funktionell durch ihren Kontraktionszustand den Ultrafiltrationskoeffizienten und da-mit
die glomeruläre Filtrationsrate ( GFR ). Mesangiumzellen von Ratten können kultiviert werden
und kontrahieren sich nach Zusatz vasoaktiver Hormone.
Der Kontraktionszustand der Zellen in Kultur wurde mikroskopisch beobachtet und jeweils vor
Zusatz von 3,5-L-T2 in angegebener Konzentration und 5 min bzw. 8 min nach Zusatz von 3,5-
L-T2 photographiert. Die Ergebnisse sind in der Tab. 5 zusammengefaßt. Es wur-den pro
Versuch 20 Zellen ausgewählt und in ihrem Kontraktionszustand beurteilt und aus-gemessen.
Die Ergebnisse sind angegeben als Prozentsatz der kontrahierten Mesangium-zellen.
Tab. 5 Ausmaß der Kontraktion kultivierter Mesangiumzellen nach Zusatz von Schilddrüsenhormonen
in angegebenen Konzentrationen; Angaben jeweils in %
SD-HormoneKonz.
L-T4 L-T3 L-T2
10-13 M <15 <15 44
10-9 M <15 <15 64
Die Kontraktion der Zellen trat bereits 4 Minuten nach Zusatz von 3,5-L-T2 ein und ist bereits
nach 8 Minuten abgeschlossen. L-T4 und L-T3 in angegebenen Konzentrationen bewirkten nur
einen geringen Anteil kontrahierter Zellen.
Nach Zusatz von 3,5-L-T2 zu kultivierten Mesangiumzellen kontrahierten sich die Zellen in
starkem Ausmaß. Dieser Effekt war durch Zusatz verschiedener Analoga zu L-T2 nicht wei-ter
steigerbar ( Werte nicht dargestellt ). Besonders eine Dosisreduktion bis auf 10-13 M 3,5-L-T2

Ergebnisse79
mit einer 44%igen Kontraktionsrate ist in diesem Ausmaß nur vergleichbar der be-kannten
kontrahierenden Wirkung von Angiotensin II in einer Konzentration von 10-8 M auf
Mesangiumzellen.
Diese Befunde würden einen Einfluß von 3,5-L-T2 auf die glomeruläre Filtrationsrate
( GFR ) erwarten lassen. Im Versuchsmodell der isoliert perfundierten Niere war dies bei
hyothyreoten Ratten nicht nachweisbar und bei euthyreoten Ratten gerade nicht mehr signi-
fikant ( Werte nicht dargestellt ).
_______________
Zusammenfassung der Ergebnisse III.5
Bei isolierter Nierenperfusion männlicher, hypothyreoter, gehungerter Ratten waren nach
Zusatz von 3,5-L-T2 in einer Konzentration von 10 -9 M zum Perfusionsmedium die gemes-
senen Parameter im Vergleich zu den Kontrollen nahezu unverändert, sodaß eine Wirkung auf
den Stoffwechsel, vergleichbar derjenigen bei der isolierten Leberperfusion, nicht nach-
zuweisen ist.
Lediglich einen Nebenbefund stellte die Induktion einer ausgeprägten Kontraktion von kulti-
vierten Mesangiumzellen dar, der sogar noch bei einer Konzentration von 10-13 M des ein-
gesetzten Schilddrüsenhormones 3,5-L-T2 nachzuweisen war. Der analoge Befund eines
Einflusses auf die glomeruläre Filtrationsrate bei der isolierten Perfusion des ganzen Or-gans
fehlte, so daß sich hier keine physiologische Bedeutung ableiten lies.

Diskussion81
IV DISKUSSION
1. Induzierte Hypothyreose durch Na131J i.p.
In den beschriebenen Versuchen wurden vornehmlich hypothyreote Ratten verwendet, um die
Versuchsansätze weitest möglich zu standardisieren und damit vergleichbar zu machen.
Die durch intraperitoneale Injektion von Na131J induzierte Hypothyreose ist ein gängiges
Verfahren, wobei die Dosierung des Isotops mit 0,25 mCi im Vergleich zu anderen Arbeits-
gruppen z.T. niedriger ausfällt. Müller et. al. ( 55 und 56 ) verwendeten dieselbe Dosie-rung;
Menahan et al. ( 53 ) setzten zur Induktion der Hypothyreose 0,5 mCi ein und Sestoft et al. (
77 ) dosierten zur Induktion der Hypothyreose bei Ratten 2 mCi und nahmen die Tiere nach 3
Wochen in den Versuch. Die nach Ablauf von 4 Wochen zum Versuchsbeginn gemessenen
Schilddrüsenhormonspiegel, betrugen 0,43 ± 0,04 ng/ml für L-T3 bzw. 9,7 ± 1,0 ng/ml für L-
T4 und bestätigten die hypothyreote Stoffwechsellage. Im Vergleich bestimmten Müller et al. (
56 ) ebenfalls nach i.p. verabreichten Na131J für L-T4 < 8 ng/ml und Cooper et al. ( 12 ) bei
MMI-hypothyreoten Ratten für L-T3 0,2 ng/ml und L-T4 4,8 ng/ml. Bei euthyreoten Tieren
betrugen die Hormonspiegel bei L-T3 0,65 ± 0,04 und bei L-T4 23 ± 1 ng/ml.
Weitere gemessene Parameter signalisierten eine Schilddrüsenunterfunktion der Versuchs-tiere,
wie das im Verhältnis zum Körpergewicht reduzierte Lebergewicht im Vergleich zu
euthyreoten Tieren. Es betrug bei hypothyreoten Ratten 2,9 ± 0,2%, identisch bei Menahan et
al.( 53 ) und bei Sestoft et al. 3,2 % ( 76 ). Bei euthyreoten Ratten ergab sich ein Ver-hältnis
von 4,1 ± 0,3 % ( siehe Tab. 1 ).
Es gab noch weitere Hinweise auf den hypothyreoten Status im Vergleich zu euthyreoten
Tieren : Na131J-gespritzte Ratten boten neben stets kalten Extremitäten auch eine ausgepräg-te
Hypomobilität, ein reduziertes Abwehrverhalten, ein struppigeres Fell, sowie insgesamt auch
eine Stagnation des Wachstums 10 bis 14 Tage nach der Na131J-Behandlung. Die Malat-
Enzym-Aktivität in der Leber, als ein Parameter des thyreoten Status ( 51 ), war bei
hypothyreoten Tieren deutlich niedriger als bei euthyreoten. Und vor allem der niedrige
Sauerstoffverbrauch der perfundierten Lebern sprach für eine deutliche Reduzierung der
Schilddrüsenhormon-Spiegel ( vergleiche auch Tab. 1 auf Seite 17 ).

Diskussion82
2. Die isolierte Leberperfusion als Modell der Wahl zur Untersuchung der schnellen
Wirkungsweise von Schilddrüsenhormonen
Das Modell der isolierten Leberperfusion mit Lebern hypothyreoter Ratten ist besonders gut
geeignet zur Untersuchung des Einflusses von Schilddrüsenhormonen auf den Lebermetabo-
lismus. Nach Präparation der Lebern ( vergleiche dazu Kapitel II.2, Seite 18 ff. ) und In-
stallation in die Perfusionsapparatur wurde sowohl beim Versuchsaufbau der rezyklieren-den,
wie auch der offenen Perfusion eine konstante Perfusionstemperatur durch die beheizte
Perfusionskammer nach Schimassek ( siehe Abb. 3, Seite 19 ), bzw. durch den beheizten
Perfusionsblock bei der offenen Perfusion ( siehe Abb. 4, Seite 21 ) und in beiden Ansätzen
durch den auf 37°C temperierten Krebs-Henseleit-Puffer gewährleistet.
Die kontinuierliche Begasung des Krebs-Henseleit-Puffers mit Carbogen ( 95% O2/ 5%
CO2 ) stellte den pH konstant auf 7,4 ein und gewährleistete einen Sauerstoffgehalt von
über 0,8 µmol/ml. Die weitestgehende Führung des Perfusionspuffers durch Glasrohre ver-
hinderte einen nennenswerten Gasverlust. Bei Probeläufen, ohne dazwischengeschaltete Le-
bern, konnte ein konstant hoher Sauerstoffgehalt des Puffers aufgezeichnet werden. Bei einem
Perfusionsfluß von ca. 5 ml/min/g bedeutete das ein Sauerstoffangebot von über 4 µmol/min/g,
sodaß ein Sauerstoffmangel der perfundierten Lebern ausgeschlossen werden konnte. Eine
weitere Steigerung des Perfusionsflusses hätte einen höheren Perfusionsdruck zur Folge, was
dann einerseits zu einer Schädigung der Leberzellen und einem Anstieg der gemessenen GOT-
bzw. LDH-Aktivität geführt hätte und andererseits zu einer Shunt-Perfu-sion im Sinne einer
Mehrperfusion der Hauptgefäße unter Umgehung der Leberkapillaren.
Direkt nach der Leberpassage wurde der Sauerstoffgehalt des Perfusionspuffers durch eine
Clark-Elektrode mit Säure-Basen-Analysator aufgezeichnet und daraus der Sauerstoffver-
brauch der perfundierten Lebern berechnet. Die Gruppe der Kontrollperfusionen wies bei
männlichen, hypothyreoten, gehungerten Ratten bei rezyklierender Perfusion einen Sauer-
stoffverbrauch von 1,71 µmol/min/g auf ( siehe dazu Abb. 7, Seite 34 ) und entsprach den
Werten von Horst et al. ( 32 ) mit 1,9 µmol/min/g im gleichen Versuchsmodell, wie auch den

Diskussion83
gemessenen Werten von Müller et al. ( 56 ) mit 1,62 µmol/min/g, ebenfalls bei isoliert
perfundierten Lebern hypothyreoter, gehungerter männlicher Ratten unter Verwendung ei-nes
Fluorocarbonmediums zur Perfusion. Für Lebern euthyreoter Ratten geben Sestoft et al. ( 77 )
3,8 µmol/min/g O2-Verbrauch an, wobei es sich hier um kleine Ratten mit einem
Körpergewicht von 170 g handelte.
Auch die gemessenen Substrate gaben Aufschluß über den intakten Stoffwechsel der perfun-
dierten Lebern ( vergleiche dazu Tab. 2, Seite 20 ). Das mit dem Perfusionspuffer angebo-tene
Alanin lag mit 10 mM über der von Exton et al. ( 19 ) beschriebenen, für die Gluco-neogenese
limitierenden Konzentration. Die Glucoseproduktion der Kontrollen lag mit 29 µmol/g bei
rezyklierender und 22 µmol/g bei offener Leberperfusion etwas über den gemes-senen Werten
von Müller et al. ( 55 ); wie auch die Harnstoffsynthese mit 75 µmol/g bzw. 98 µmol/g im
Vergleich zur selben Arbeitsgruppe etwas höher ausfiel. Der Unterschied könnte an einer
niedrigeren Gluconeogeneserate und Harnstoffproduktion aufgrund des ver-wendeten
Perfusionsmediums ( Fluorocarbon bei Müller et al. ) liegen.
3. Der Effekt von 3,5-L-T2 auf Gluconeogenese und O2-Verbrauch
Im Laufe der isoliert rezyklierenden Leberperfusion bei männlichen, hypothyreoten, gehun-
gerten Kontrollen stieg der Sauerstoffverbrauch ohne Hormonzusatz über den Zeitraum von 90
Minuten langsam von 1,71 auf 2,24 µmol/min/g an ( Abb. 7, Seite 34 ). Durch die ab-gelaufene
Gluconeogenese, aus dem im Perfusionspuffer in einer Konzentration von 10 mM gelösten
Alanin, stieg die Glucoseproduktion der nach 24 Stunden Hunger glykogenleeren Lebern, wie
auch die Harnstoffproduktion parallel an ( Tab. 3, Seite 36 ). Nach Ablauf der
Äquilibrierungszeit, zum Zeitpunkt t = 0 min betrug die Glucoseproduktion der Kontrollen 4,8
µmol/g und stieg über die 90-minütige Perfusion kontinuierlich auf 29 µmol/g. Gleich-zeitig
stieg die Harnstoffproduktion von 8,1 auf 75 µmol/g in der selben Perfusionszeit.
Nach Zusatz von 3,5-L-T2 in einer Konzentration von 10-12 M, die der von Pinna et al.
( 66 ) im Serum von gesunden Probanden gefundenen entspricht, stieg der Sauerstoffver-
brauch auf 2,90 µmol/min/g an und lag im Verlauf nach 50 Minuten bereits signifikant über den

Diskussion84
Kontrollen. Parallel dazu war auch die Gluconeogeneserate und die Harnstoffproduk-tion nach
Hormonzusatz erhöht, jedoch bereits nach 30 Minuten signifikant über den Kon-trollen und
stieg im weiteren Verlauf der 90 Minuten Perfusion auf 42 µmol/g bzw. 136 µmol/g an.
Somit konnte der Sauerstoffverbrauch der isoliert rezyklierend perfundierten Lebern männ-
licher, hypothyreoter, gehungerter Ratten um 30 % gesteigt werden. Die Steigerung der
Gluconeogenese betrug 47 % und bei der gemessenen Harnstoffsynthese sogar 82 %. In der
Bilanz wird also mehr Harnstoff produziert als Glucose, da ein Teil der Glucose in Pyruvat
umgewandelt und zu CO2 und H2O abgebaut wird.
Für 3,5-L-T2, welches durch Monodejodierung aus L-T3 entsteht, beschrieben Horst et al.
( 32 ) einen schnellen Effekt auf den Sauerstoffverbrauch von perfundierten Lebern männli-
cher, hypothyreoter, gehungerter Ratten 30 Minuten nach Hormonzusatz; einen Effekt der
durch L-T3 und L-T4 bei gleichzeitiger Inhibierung ihrer Dejodierung durch Propylthioura-cil (
PTU ) in diesem Ausmaß nicht hervorzurufen war. Hieraus schlossen die Autoren auf einen für
L-T2 spezifischen schnellen Effekt, der über eine direkte mitochondriale Wirkung vermittelt
wird. Kvetny ( 41 ) beschreibt eine Stimulation des Sauerstoffverbrauchs bei hu-manen
Monozyten ebenfalls durch 3,5-L-T2, ebenfalls schnell innerhalb von 30 Minuten. Eine
Steigerung der Glucoseaufnahmen in die Zellen konnte er jedoch nicht nachweisen.
Im zeitlichen Ablauf unterscheidet sich die 3,5-L-T2-Wirkung von anderen bekannten Hor-
monen, wie Glukagon oder Adrenalin erheblich. Diese bewirken an der Plasmamembran durch
die Stimulierung der Adenylatcyclase die Bildung von cAMP als zweiten Boten im Cytosol,
der verschiedene Reaktionskaskaden und zelluläre Prozesse in Gang setzt, u. a. steigt auch der
Sauerstoffverbrauch ( vergleiche hierzu auch Abb. 19, Seite 56 ). Im Ver-gleich laufen diese
Prozesse sofort, binnen Sekunden bis wenigen Minuten ab. Da die Stei-gerung des
Stoffwechsels durch 3,5-L-T2 langsamer abläuft ( ab 30 min ), scheint sie nicht einem
unmittelbar vergleichbaren Reaktionsprinzip zu unterliegen, welches wie ein Schalter entweder
an- oder ausgeschaltet werden kann.
Es ist bekannt, daß unter Einfluß des Schilddrüsenhormones L-T3 eine Steigerung des Inter-
mediärstoffwechsels über eine vermehrte Synthese spezifischer Proteine über den nukleären
Stoffwechsel gesteuert wird. Den Einfluß der Proteinbiosynthese auf die schnellen Effekte von

Diskussion85
Schilddrüsenhormonen untersuchten Horst et al. ( 32 ) am Modell der isolierten Leber-
perfusion durch Zusatz von Cycloheximid in einer Konzentration von 20 µg/ml zum Perfu-
sionsmedium. Die schnelle Steigerung des Sauerstoffverbrauchs durch L-T3 konnte durch den
Cycloheximidzusatz nicht verringert werden, woraus die Autoren den Schluß zogen, daß ein
durch Proteinsynthese vermittelten Effekt in diesem Versuchsansatz nicht vorlag.
In der selben Arbeit wurde untersucht, ob durch eine Steigerung der Hormonkonzentration der
Effekt auf den Sauerstoffverbrauch noch früher zu erzielen bzw. weiter steigerbar ist. Das
Ergebnis zeigte, daß oberhalb einer Konzentration von 10-12 M 3,5-L-T2 keine wesent-liche
Steigerung des Sauerstoffverbrauchs gemessen werden konnte. Dieser Befund ent-spricht auch
den Ergebnissen dieser Arbeit, als bei weiblichen Ratten und kohlenhydrat-reich gefütterten
männlichen Tieren eine weitere Steigerung des Sauerstoffverbrauchs der perfundierten Lebern
durch eine Erhöhung der eingesetzten Hormonkonzentration nicht zu erzielen war.
Die Untersuchungen einzelner Dejodierungsschritte durch Engler et al. ( 17 ) und Faber et al. (
20 ) ergaben, daß etwa 5 % des geamten 3,5,3´-L-T3 durch 3´-Monodejodierung zu 3,5-L-T2
reduziert werden und entsprechend eine Serumkonzentration von 0,4 - 5,3 ng/dl anzunehmen
ist. Für die tatsächliche physiologische Konzentration von 3,5-L-T2 im menschlichen Serum
gesunder Probanden bestimmten Pinna et al. ( 66 ) 16,2 ± 6,4 ∗ 10-12 M/l ( einen
Normalbereich von 3,4 - 29 ∗ 10-12 M/l, entsprechend dem MW ± 2 Standard-abweichungen ).
Bei hypothyreoten Patienten betrug der Wert für 3,5-L-T2 0,75 ± 1,6 ∗
10-12 M/l. Somit entsprach das in den Versuchen eingesetzte Hormon der physiologischen
Konzentration im unteren Normalbereich. Inwieweit sich möglicherweise das Bild durch die
dabei noch nicht berücksichtigte Proteinbindung an Albumin ändert müssen weitere Unter-
suchungen zeigen.
4. Einfluß der Ernährung auf den Effekt von 3,5-L-T2

Diskussion86
In den vorangegangenen Experimenten hatten die hypothyreoten Ratten vor Versuchsbeginn
stets 24 Stunden gehungert, da die Glucoseproduktion der perfundierten Lebern einen sensi-
tiven Parameter für ihren Stoffwechsel darstellte.
Es ist aber bekannt, daß es eine Interaktion zwischen der Wirkung von Schilddrüsenhormo-nen
und kohlenhydratreicher Ernährung gibt, da die Enzymausstattung der Zellen durch den
Ernährungszustand bestimmt wird. Mariash et al. ( 52 ) zeigten eine unterschiedliche Induk-
tion von hepatischen Enzymen der Lipogenese ( ME, FAS, G6PD, 6PGD ) nach kohlenhy-
dratreicher Diät bei hypo- und euthyreoten Ratten. Die Aktivität dieser Enzyme war bei hy-
pothyreoten Ratten nach kohlenhydratreicher Diät gegenüber normaler Diät bereits erhöht. Für
eine weitere Stimulation durch L-T3 waren nach kohlenhydratreicher Diät nur geringe und
weitaus niedrigere Konzentrationen des Hormones notwendig als nach normaler Diät. Die
Autoren interpretierten die Ergebnisse als eine synergistische Wirkung bei der Induk-tion
hepatischer Enzyme der Lipogenese durch Schilddrüsenhormone und nach kohlenhy-
dratreicher Diät. Als mögliche Ursachen wurden sowohl eine erhöhte metabolische Clea-rance
von L-T3, als auch eine durch die kohlenhydratreiche Diät bedingte gesteigerte relati-ve
Affinität zum nukleären Rezeptor ausgeschlossen.
Bei isoliert rezyklierender Leberperfusion hypothyreoter Ratten nach kohlenhydratreicher Diät
fiel bei der Auftragung der auf das Lebergewicht bezogenen Werte für den Sauerstoff-
verbrauch auf, daß die gehungerten Tieren sowohl beim Vergleich der Kontrollgruppen, als
auch nach Stimulation durch 3,5-L-T2 einen höheren O2-Verbrauch aufwiesen ( vergleiche
Abb. 13, Seite 44 ). Berücksichtigt man bei kohlenhydratreich ernährten Tieren die durch
Glykogenspeicherung bedingte geringere Dichte atmungsaktiver Kompartimenten, so muß man
den Sauerstoffverbrauch auf Milligramm Protein pro Gramm Leber beziehen. So um-gerechnet
ergaben sich für den Sauerstoffverbrauch der Lebern gehungerter Tiere mit 5,15
nmol/min/mgProtein und nach kohlenhydratreicher Diät 5,12 nmol/min/mgProtein gleiche Aus-
gangswerte.
Nach Zugabe von 3,5-L-T2 war der Sauerstoffverbrauch der Lebern nach kohlenhydratrei-cher
Diät bereits nach 30 Minuten signifikant im Vergleich zur Kontrollgruppe gesteigert. Durch
den gefütterten Zustand der Tiere lief die Stimulation des Sauerstoffverbrauchs of-fensichtlich
rascher ab als bei gehungerten Tieren. Auch hier könnte die Erklärung dafür eine bereits nach

Diskussion87
kohlenhydratreicher Diät vorbestehende Enzyminduktion sein, die nach Hormonexposition den
Sauerstoffverbrauch früher ansteigen läßt.
Vergleicht man nun das Ausmaß der Steigerung des Sauerstoffverbrauchs durch 3,5-L-T2,
ergaben sich bei gehungerten und kohlenhydratreich ernährten hypothyreoten Versuchstie-ren
keine Unterschiede. Bezogen auf den Ausgangswert betrug die Zunahme der Atmung 30%,
bzw. 25 %.
Bei Untersuchungen zur Monodejodierung von T3 und rT3 fanden Burger et al. ( 10 ) nach
hochkalorischer Diät bei sonst gesunden Probanden eine 4- bis 5-%ige Dejodierung von L-T3
an 3´-Position ( und dadurch Bildung von 3,5-L-T2; vergleiche Abb. 2, Seite 11 ) bei ei-nem
insgesamt erhöhten L-T3-Plasmaspiegel. Darüber hinaus stellten die Autoren bei dieser
Ernährung eine deutlich gesteigerte Plasma-Clearance-Rate von 3,5-L-T2 fest. Dies inter-
pretierten sie als eine Alteration des peripheren Stoffwechsels der Schilddrüsenhormone durch
die hochkalorische Ernährung im Sinne einer Zunahme der Dejodierung am Innen-ring. Sollte
diese Hypothese zutreffen, wäre sie eine Erklärung für die fehlende Zunahme der Stimulation
des Sauerstoffverbrauchs nach kohlenhydratreicher Ernährung. Nach Zusatz einer höheren
Konzentration von 3,5-L-T2 müßte dann jedoch der vollständige vorbeschrie-bene Effekt zu
beobachten sein.
Aber auch durch eine Erhöhung der eingesetzten Hormonkonzentration auf 10-10 M konnte
keine weitere Steigerung des Sauerstoffverbrauchs, sowie auch keine schneller ablaufende
Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 erzielt werden ( vergleiche dazu Abb. 14,
Seite 45 ).
5. Einfluß des Geschlechts auf den Effekt von 3,5-L-T2
In den vorangegangenen Experimenten kamen ausschließlich männliche hypothyreote Ver-
suchstiere zum Einsatz. Der Stoffwechsel der Schilddrüsenhormone ist bei den Geschlech-tern
jedoch nicht einheitlich. Faber et al. ( 21 ) fanden heraus, daß die physiologisch vor-
kommenden Serumspiegel von 3,5-L-T2 bei männlichen Ratten höher sind als bei weibli-chen.

Diskussion88
Bei höherer Syntheserate der weiblichen Tiere findet sich bei diesen aber gleichzeitig auch eine
höhere Hormon-Clearance, woraus ein insgesamt niedrigerer Spiegel resultiert.
Die Leberperfusion hypothyreoter, gehungerter weiblicher Ratten wies nur wenige Unter-
schiede im Vergleich zu der Perfusionen von Lebern männlicher Versuchstiere auf. Der
Ausgangs-Sauerstoffverbrauch weiblicher Rattenlebern lag aber mit 1,97 µmol/min/g
signifikant höher als bei den männlichen Tieren ( vergleiche dazu Abb. 10, Seite 39 ). Und auch
die Steigerung des Sauerstoffverbrauchs unterschied sich nach Zusatz von 3,5-L-T2 bereits
nach 40 Minuten signifikant von den Kontrollen. Sie fiel jedoch mit 18 % geringer aus als bei
der männlichen Vergleichsgruppe, bei der die Steigerung 30 % betrug.
Ursache für den zu Perfusionsbeginn höheren Sauerstoffverbrauch der weiblichen hypothy-
reoten Ratten kann eine im Vergleich zu den männlichen Tieren veränderte Rezeptorausstat-
tung für Schilddrüsenhormone der einzelnen Zellkompartimente der Lebern und/oder auch eine
unterschiedliche Enzymausstattung der Zellen sein, die durch die Steroidhormone der
Geschlechter ( Androgene und Östrogene ) gesteuert wird. Ebenso wäre dies eine Erklärung
für die geringfügig früher signifikante Stimulation des Sauerstoffverbrauchs bei den weib-lichen
Ratten. Die von Faber et al. ( 21 ) beschriebene höhere Hormon-Clearance für 3,5-L-T2 ist
möglicherweise der Grund dafür, daß der Sauerstoffverbrauch der weiblichen hy-pothyreoten
Tiere nicht um den selben Prozentsatz steigerbar war, wie bei den männlichen Tieren. Denn
auch eine Steigerung der Hormonkonzentration auf 10-10 M hatte weder einen höheren Anstieg
des Sauerstoffverbrauchs zur Folge, noch fiel dieser früher signifikant aus, als bei den
Kontrollen ( vergleiche dazu Abb. 11, Seite 41 ). Insgesamt scheint bei der Betrachtung der
Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 der Geschlechter in der absoluten
Darstellung die höhere Steigerbarkeit bei den männlichen Tiere das höhere Ausgangsniveau der
weiblichen Tiere auszugleichen.
Parallel zu den Experimenten kohlenhydratreich ernährter männlicher Versuchstiere, wur-den
analog zur Beantwortung der Frage, ob die Zeit bis zu einer signifikanten Steigerung des
Sauerstoffverbrauchs bei weiblichen Tieren durch ihren Ernährungszustand noch weiter
verkürzt werden kann, Experimente mit weiblichen hypothyreoten kohlenhydratreich er-
nährten Ratten durchgeführt ( vergleiche dazu Abb. 18, Seite 51 ).

Diskussion89
Die Ergebnisse zeigten, daß der Sauerstoffverbrauch auch in dieser Versuchsgruppe nach 40
Minuten signifikant anstieg, was im Vergleich zu den gehungerten weiblichen Tieren keine
zusätzliche Beschleunigung der Wirkung von 3,5-L-T2 darstellte.
Auch bei diesem Experiment fiel der berechnete Sauerstoffverbrauch bezogen auf das Le-
bergewicht im Vergleich zum gehungerten Status geringer aus, was durch die Glykogenver-
dünnung der atmungsaktiven Zellkompartimente zu erklären ist. Im Vergleich lag sogar das
Lebergewicht der gefütterten Tiere mit 4,56 g um 9,4 % meßbar höher, als das der gehun-
gerten mit 4,17 g.
6. Untersuchungen zum Mechanismus der Wirkung von 3,5-L-T2 bei rezyklierender
und offener Leberperfusion
Für ausgewählte Experimente an männlichen, hypothyreoten, gehungerten Ratten wurde der
Versuchsaufbau der Leberperfusion verändert, um den Mechanismus des Effektes durch 3,5-L-
T2 näher untersuchen zu können.
Das Modell der kombiniert offenen und rezyklierenden Leberperfusion ermöglichte die pa-
rallele Messung von Sauerstoffverbrauch, Kaliumausstrom aus den Leberzellen, Perfusions-
druck sowie zur Kontrolle der Intaktheit des Stoffwechsels der perfundierten Lebern die Pa-
rameter Harnstoffsynthese und Glucoseproduktion. In dieser Versuchsanordnung folgte der
zunächst offenen, anschließend eine Phase rezyklierender Perfusion.
Im Vergleich zu den in Kapitel IV.3 diskutierten Experimenten lag der Ausgangswert des
Sauerstoffverbrauchs mit 2,69 µmol/min/g ( vergleiche Abb. 19, Seite 56 ) über den gemes-
senen Werten des Ausgangsniveaus bei rezyklierender Perfusion. Verglichen mit den Wer-ten
bei euthyreoten Versuchstieren ( vergleiche Tab. 1, Seite 17 ) spricht auch dieser Wert für eine
deutliche Hypothyreose, wie auch die zur Kontrolle gemessenen Serumkonzentra-tionen der
Schilddrüsenhormone der in den Versuch genommenen Tiere. Auch bei diesem
Ausgangsniveau war, wie in den Versuchen zu sehen und im Folgenden diskutiert, durch 3,5-
L-T2 noch eine deutliche Stimulation des Sauerstoffverbrauchs zu erzielen.

Diskussion90
Bei der zunächst 60-minütigen offenen Perfusion lief die Kurve des O2-Verbrauchs bei den
Kontrollen annähernd auf dem Ausgangsniveau geradeaus. Nach Stimulation durch 10-9 M 3,5-
L-T2, einer sicher im maximal stimulierenden Bereich liegenden Konzentration, stieg der
Sauerstoffverbrauch im Verlauf weitaus weniger steil an, als bei den vorangegangenen
Experimenten ( vergleiche Abb. 8, Seite 35 und Abb. 19, Seite 56 ) und erreichte nach 60
Minuten Perfusion ein gerade signifikant höheren Wert als die Kontrollen.
Nach Umschalten auf rezyklierende Perfusion stieg der Sauerstoffverbrauch jedoch bereits
nach abgelaufenen 10 Minuten steil an, und erreichte im weiteren Verlauf ab der 40. Minu-te (
entsprechend der 100. Minute Gesamtperfusionzeit ) ein Plateau ( vergleiche Abb. 19, Seite 56
). Im Vergleich zu den Kontrollen war der O2-Verbrauch bei rezyklierender Perfu-sion schon
nach 10 Minuten und anschließend bis zum Perfusionsende signifikant erhöht.
Nach abschließend erneut offener Perfusion fiel der Sauerstoffverbrauch der hormonstimu-
lierten Versuchsgruppe ab, verblieb aber immer noch auf einem deutlich höheren Niveau als die
Kontrollen, die sich wieder in der Nähe des Ausgangsverbrauchs einpendelten
( Abb. 19 ).
Der Verlauf der Stimulation des Sauerstoffverbrauchs durch 3,5-L-T2 bei im Wechsel offe-ner,
rezyklierender und wieder offener Perfusion legte den Schluß nahe, daß nicht das Hor-mon
selbst die Wirkung vermittelt, da sonst schon bei zuerst offener Perfusion eine deutli-chere
Stimulation zu erwarten gewesen wäre ( vergleiche auch die Adrenalin-Wirkung nach
abschließend offener Perfusion, Abb. 19 ). Anscheinend führte 3,5-L-T2 an den Zellen eine
Veränderung herbei oder setzte eine Substanz X frei, die erst bei Rezirkulation des Medi-ums
wirksam wird. Ferner ist diese entfaltete Wirkung nach Unterbrechung der Rezirkula-tion (
durch abschließende offene Perfusion ), möglicherweise durch Auswaschen der Sub-stanz X,
weitgehend reversibel. Die vorangegangenen Experimente, mit ausschließlich re-zyklierender
Perfusion ( vergleiche dazu Kapitel IV.3, Seite 83 ff. ), machten des weiteren deutlich, daß das
Prinzip dieses Effektes nicht dem eines Schalters unterliegt, der entweder an- oder
ausgeschaltet wird, sondern offenbar einer kontollierten Modulation.

Diskussion91
Die gemessene Glucoseproduktion verlief gleichsinnig wie der Sauerstoffverbrauch: wäh-rend
der zunächst offenen Leberperfusion stieg nach Stimulation durch 3,5-L-T2 die Gluco-
seproduktion nach 40 Minuten auf ein signifikant höheres Niveau als die Kontrollen. Nach
Umschalten auf rezyklierende Perfusion war der Anstieg wiederum sofort nach 10 Minuten
signifikant höher als bei Kontrollperfusionen und erreichte, korrespondierend zum Sauer-
stoffverbrauch, ab der 40. Minute ( 100. Minute Gesamtperfusionszeit ) ein Plateau ( ver-
gleiche dazu Abb. 20 a und b, Seite 58 ).
Ebenfalls verlief die Harnstoffproduktion der perfundierten Lebern gleichsinnig zum O2-
Verbrauch. Bei zunächst offener Perfusion war der Anstieg 30 Minuten nach Zusatz von 3,5-
L-T2 verglichen mit den Kontrollen zwar schon signifikant; aber weitaus deutlicher fiel auch
hier die Steigerung der Harnstoffsynthese nach Umschalten auf rezyklierende Perfu-sion nach
Ablauf von 60 Minuten aus. Erneut stieg die Harnstoffproduktion bereits nach 10 Minuten steil
an, unterschied sich im Verlauf immer deutlicher von den Kontrollen und erreichte bereits nach
30 Minuten ( 90. Minute Gesamtperfusionszeit ) ein Plateau ( verglei-che dazu Abb. 21 a und
b, Seite 60 ), wie zuvor schon beim Sauerstoffverbrauch und der Glucoseproduktion
beschrieben.
Die kontinuierliche Registrierung des Perfusionsdruckes ∆p wies zu keinem Zeitpunkt der
Perfusionen, weder des Kontrollkollektivs, noch der hormonstimulierten Versuchsgruppe einen
Unterschied auf. Ein vasoaktiver Effekt des eingesetzten Hormones war daher auszu-schließen.
Gleiches gilt für die Messung des Kaliumausstromes der Leberzellen in das Perfusionsmedi-um.
Auch bei diesem Parameter ergaben sich bei den Kontrollen, wie auch der Versuchs-gruppe
nach Zusatz von 10-9 M 3,5-L-T2 keine Hinweise auf eine Änderung. Somit scheint ein Kalium-
Signal als Vermittler des stimulierenden Effektes von 3,5-L-T2 auf den Sauer-stoffverbrauch,
die Steigerung der Harnstoffproduktion sowie der Gluconeogenese auszu-scheiden.
Bei der Untersuchung des Wirkungsmechanismus von 3,5-L-T2 auf die isoliert perfundierte
Leber ist zu berücksichtigen, daß die Leber kein einheitliches Organ darstellt. Sie ist die größte
„Drüse“ des menschlichen Organismus und ihr Funktionsspektrum außerordentlich groß. So
konnten Hummerich et al. ( 34 ) bei isolierten Hepatozyten nach Zusatz von 3,5-L-T2 keinen
Effekt feststellen. Der strukturelle Aufbau umfaßt aber neben den Hepatozyten und den

Diskussion92
interlobulären Gallengängen auch Epithelzellen und die Kupffer-Zellen ( organspe-zifische
Makrophagen ), die ebenfalls als mögliche Wirkorte des Hormones zu diskutieren sind und die
gemessenen Effekte vermittelt haben könnten.
7. Einfluß von Ionenkanalblockern auf den Effekt von 3,5-L-T2
Zur Abgrenzung der Vermittlung des auslösenden Effektes von 3,5-L-T2 durch Ionenströme
über die Zellmembran bei rezyklierend perfundierter Lebern hypothyreoter Ratten, wurden dem
Perfusionsmedium verschiedene, im Folgenden diskutierte selektive ionenkanalblockie-rende
Agenzien zugesetzt.
Für die Experimente mit Einsatz selektiver Ionenkanalblocker kamen wiederum männliche,
hypothyreote, gehungerte Ratten zum Einsatz, und für 3,5-L-T2 wurde eine maximal stimu-
lierende Konzentration von 10-10 M gewählt.
Na+-K+-ATPase-Aktivität (Na+/K+-Kanal ) und Ouabain
Die durch Schilddrüsenhormone induzierte Thermogenese wird vermittelt durch die Stimu-
lation des aktiven Na+-Transportes der plasmamembranständigen Na+-K+-ATPase. Wie Asano
und Edelman ( 2 ) beschrieben, sind Na+-K+-ATPase-Aktivität und Sauerstoffver-brauch nach
Stimulation durch L-T3 bei Skelettmuskelpräparaten direkt proportional. Ferner beschrieben
Kalderon et al. ( 37 ) eine erhöhte Dichte der Na+-K+-ATPase auf der hepati-schen
Plasmamembran hyperthyreoter Ratten.
Ismail-Beigi und Edelman ( 36 ) führten die Stimulation der Na+-Pumpe durch die
Schilddrüsenhormone auf entweder eine Aktivierung der mitochondrialen ATP-Synthese und
einem daraus resultierenden lokalen Anstieg der ATP-Konzentration zurück oder auf eine
direkte Aktivierung des Enzymes. Die Autoren leiten daraus auch einen möglichen
Regulationsmechanismus für den Stoffwechsel durch Schilddrüsenhormone in verschiedenen
physiologischen Status ( Kälte, Hunger, Fieber, Katecholaminausschüttung wie bei Sepsis etc.

Diskussion93
) ab. Wenn es einen Zusammenhang dieses Regulationsprinzips auch mit 3,5-L-T2 gibt, müßte
die Stimulation der Atmung durch die Blockade des Na+/K+-Kanals unterdrückt werden
können.
Krenning et al. ( 40 ) beschrieben bei isolierten Hepatozyten für den aktiven Transport von
Schilddrüsenhormonen eine Abhängigkeit vom Na+-Gradienten an der Plasmamembran und
einen reduzierten Transport bei Inhibierung der Aktivität der Na+-K+-ATPase durch Oua-bain.
Die Autoren leiteten daraus, neben der Dejodierung der Schilddrüsenhormone, ein mögliches
weiteres Regulationsprinzip von Wirkung und Metabolismus der Hormone ab.
Bei Ouabain handelt es sich um einen selektiven Blocker des Na+/K+-Kanals, der in einer
Konzentration von 10-6 M vor Einsatz des Hormons dem Perfusionsmedium zugesetzt wur-de
( vergleiche dazu Abb. 22 b, Seite 64 ). Der Ausgangs-Sauerstoffverbrauch wurde so-wohl bei
der Kontrollgruppe, als auch bei der Versuchsgruppe durch Ouabain geringfügig gesenkt.
Im weiteren Verlauf konnte dann jedoch durch 3,5-L-T2, auch in der eingesetzten sicher
maximal stimulierenden Konzentration von 10-10 M über die gesamte Perfusionszeit keine
Steigerung des Sauerstoffverbrauchs verzeichnet werden. Nach abschließender Überprüfung
der Intaktheit der perfundierten Lebern durch Zusatz von Adrenalin und einer darunter
sofortigen Steigerung des Sauerstoffverbrauchs war eine toxische Ursache als Erklärung für
den vorher fehlenden Anstieg des Sauerstoffverbrauchs ausgeschlossen.
Somit scheint die Na+-K+-ATPase in die Vermittlung der Steigerung des Sauerstoffver-brauchs
durch 3,5-L-T2, wie auch schon für die anderen Schilddrüsenhormone gezeigt wur-de ( 9, 37 ),
bei der isoliert rezyklierend perfundierten Leber hypothyreoter Ratten invol-viert zu sein.
Hierbei müßte dann 3,5-L-T2 selbst, oder eine durch 3,5-L-T2 freigesetzte Substanz X an die
Zellmembran binden, um die beobachteten Effekte hervorzurufen wo-durch die Steigerung der
Gluconeogeneserate und des Alaninverbrauchs durch 3,5-L-T2 noch nicht erklärt ist.
In weiteren Experimenten kamen Nifedipin als selektiver Blocker des Ca++-Kanals, SITS als
selektiver Blocker des K+/H+-Kanals und Amilorid als selektiver Blocker des Na+/H+-Kanals
zum Einsatz, bei ansonsten identischen Versuchsanordnungen zu dem oben beschrie-benen
Einsatz von Ouabain.

Diskussion94
Ca++-Kanal und Nifedipin
Als einen schnellen Effekt von Schilddrüsenhormonen an der Plasmamembran von Ratten-
thymozyten, beschrieben Segal et al. ( 72, 73, 74 ), die Aufnahme von Glucose und Ca++ in die
Zelle und charakterisierten das Ca++ als „ersten Boten“ für die prompte Wirkung von
Schilddrüsenhormonen. Müller et al. ( 55 ) und Hummerich et al. ( 34 ) stellten nach
Schilddrüsenhormongabe eine Stimulation der Aufnahme von Aminosäuren und Ca++ in
Leberzellen fest. Die Autoren korrelierten diesen Effekt bei gehungerten Ratten mit einer
Steigerung des Sauerstoffverbrauchs und der Gluconeogenese und bei gefütterten Ratten mit
der Stimulation von Atmung und Glykolyse ( 34 ).
In der selben Arbeit wird auch von einer gleichgerichteten Wirkung von 3,5-L-T2 bei ge-
hungerten Versuchstieren auf den Ca++-Einstrom, sowie O2-Verbrauch und Gluconeoge-nese
berichtet und die Stimulation durch die Hormone auf eine Änderung der mitochondri-alen,
sowie der cytosolischen Ca++-Konzentrationen zurückgeführt. An isolierten Erythro-
cytenmembranen stellten Davis et al. ( 15 ) eine durch Schilddrüsenhormone und einige
-analoga stimulierte Ca++-ATPase-Aktivität fest.
Um eine mögliche Beteiligung der Vermittlung des Effektes von 3,5-L-T2 durch einen Ca++-
Einstrom in die Leberzellen zu überprüfen, wurde dem Perfusionspuffer Nifedipin zugesetzt
und der Sauerstoffverbrauch der Lebern hypothyreoter, gehungerter Ratten ge-messen (
vergleiche dazu Abb. 23 a und b, Seite 66 ). Der Einsatz von Nifedipin, einem wirksamen
selektiven Blocker des Ca++-Kanals, in einer Konzentration von 10-6 M hatte einen steigernden
Effekt auf das Ausgangsniveau des Sauerstoffverbrauchs der beiden Ver-suchsgruppen.
Nach Hormonzusatz stieg der Sauerstoffverbrauch früher auf signifikant höhere Werte als bei
der Kontrollgruppe, und sogar früher als bei den Perfusionen ohne Nifedipin-Zusatz. Auch das
Ausmaß der Stimulation erreichte einen signifikant höheren Sauerstoffverbrauch als ohne
Nifedipinzusatz, trotz eines höheren Ausgangsniveaus. Hieraus folgt, daß der Zu-satz von
Nifedipin zum Perfusionsmedium dem stimulierenden Effekt von 3,5-L-T2 bei re-zyklierender
Leberperfusion eher Vorschub leistet, auf keinen Fall jedoch inhibiert.

Diskussion95
Somit scheint Ca++ für den Hormoneffekt keine wesentliche Rolle zu spielen. Auch die zeitliche
Lücke zwischen dem Beginn des beschriebenen Ca++-Einstromes [ nach 2 Minu-ten ( 34 )] und
dem Anstieg des Sauerstoffverbrauchs [ nach 20 Minuten ] ist nicht plausi-bel.
K+/H+-Kanal und SITS
Zur Überprüfung eines Einflusses des K+/H+-Kanals auf die Stimulation des Sauerstoffver-
brauchs durch 3,5-L-T2 wurde dem Perfusionsmedium der selektive Blocker SITS zugesetzt (
vergleiche dazu Abb. 24 a und b, Seite 68 ). Der Einsatz von SITS in einer Konzentration von
10-6 M zum Perfusionsmedium hatte nur eine geringe Erhöhung des Ausgangsniveaus des
Sauerstoffverbrauchs zur Folge. Nach Zusatz von 3,5-L-T2 stieg der Sauerstoffver-brauch,
vergleichbar den vorherigen Experimenten ohne SITS-Zusatz zum Perfusionsmedi-um, an. Für
einen inhibierenden Effekt von SITS und damit ein Einfluß des K+/H+-Kanals auf den 3,5-L-T2
Effekt gibt es somit keinen Hinweis.
Auch die vorangegangenen Experimente der kombiniert offenen und rezyklierenden isolier-ten
Leberperfusion, bei denen eine Kaliumelektrode zur Messung des K+-Ausstromes aus den
Leberzellen zum Einsatz kam ( vergleiche Kapitel IV.6, Seite 89 ), hatten bereits erge-ben, daß
ein Kaliumsignal als Vermittler des 3,5-L-T2 Effektes nicht wahrscheinlich ist.
Na+/H+-Kanal und Amilorid
Zur Überprüfung eines Einflusses des Na+/H+-Kanals auf die Stimulation des Sauerstoff-
verbrauchs durch 3,5-L-T2 wurde dem Perfusionsmedium der selektive Blocker Amilorid
zugesetzt ( vergleiche dazu Abb. 25 a und b, Seite 69 ). Der Einsatz von Amilorid in einer
Konzentration von 10-3 M zum Perfusionsmedium hatte eine Reduktion des Sauerstoffaus-
gangsniveaus zur Folge. Der O2-Verbrauch der Kontrollgruppe fiel im weiteren Verlauf der
Perfusion stetig leicht ab im Unterschied zu den Kontrollen ohne Amiloridzusatz, bei denen der
Sauerstoffverbrauch leicht anstieg.

Diskussion96
Nach Zugabe von 3,5-L-T2 stieg der Sauerstoffverbrauch bereits nach 10 Minuten auf ein
gegenüber Kontrollen signifikantes Niveau und noch für die darauf folgenden 10 Minuten
weiter an. Nach insgesamt 20 Minuten Perfusionszeit fiel die Kurve stetig, den Kontrollen von
Perfusionsbeginn an vergleichbar, bis zum Perfusionsende ab. Nach Amiloridzusatz zum
Perfusionsmedium schien aufgrund einer den Na+-Strom über die Zellmembran blok-kierenden
Wirkung erneut eine hormoninduzierte Steigerung des Sauerstoffverbrauchs wirk-sam
verhindert worden zu sein. Die anfängliche Steigerung des O2-Verbrauchs könnte durch den
nicht-blockierten Na+/K+-Kanal ermöglicht worden sein, wenn durch diesen Kanal initial ein (
reflektorisch ) erhöhter Na+-Einstrom stattgefunden hat, der im weiteren Ver-lauf der
Perfusion langsam abklang.
So zeigten die Experimente der isolierten rezyklierenden Leberperfusion mit Einsatz ver-
schiedener selektiver ionenkanalblockierender Substanzen, daß die Natrium-Kanäle wahr-
scheinlich einen Einfluß auf den durch 3,5-L-T2 vermittelten Effekt haben.
Jedoch weder die Calcium- noch die Kalium-Kanäle, korrespondierend zu den vorherigen
Experimenten bei kombiniert offener und rezyklierender Leberperfusion ( vergleiche Kapi-tel
IV.6, Seite 89 ), scheinen die hormonelle Wirkung zu vermitteln.
8. Wirkung von 3,5-L-T2 auf isolierte Mitochondrien aus Hepatozyten in vivo
Die Regulierung der Atmung durch Schilddrüsenhormone betrifft die Zellkompartimente Kern
und Mitochondrien. Schilddrüsenhormone bewirken eine Steigerung der Expression
bestimmter Genabschnitte, die für Proteine der Atmungskette kodieren; möglicherweise
werden auch Membranlipide verändert. Sterling et al. ( 79, 82 ) fanden eine Proteolipid-
fraktion der inneren Mitochondrienmembran isoliert aus Rattenhepatozyten, die eine spe-
zifische feste Bindung mit T3 eingeht. Biologische Effekte konnten mit der Bindung nicht
korreliert werden.

Diskussion97
Vom selben Autor sind auch schnelle Effekte der Schilddrüsenhormone auf die oxidative
Phosphorylierung, wie bei submitochondrialen Vesikeln durch direkten nanomolaren Zusatz
von T4 beschrieben worden ( 83 ); wie auch eine alternative Interpretation der von ihm
gefundenen schnellen Effekte durch eine Bindung von L-T3 an die Adeninnukleotidtrans-lokase
( ANT ) der inneren Mitochondrienmembran.
Oppenheimer und Silva ( 60 ) stellten fest, daß Schilddrüsenhormone Schlüsselenzyme kon-
trollieren, die letztlich die Bereitstellung von Reduktionsäquivalenten für die mitochondriale
Matrix erhöhen. In der Folge kommt es zu einer Steigerung der Atmungskapazität und ATP-
Synthese.
Soboll beschrieb in einer Übersichtsarbeit ( 78 ), daß Schilddrüsenhormone die für die Zell-
atmung zuständigen Gene beeinflussen. Hierbei findet die Modulation hauptsächlich auf der
Ebene der Transkription statt.
Darüber hinaus bewirken Schilddrüsenhormone aber auch schnelle Effekte auf die mito-
chondriale Aktivität, die binnen Minuten auftreten und von der Genexpression, der für die
Zellatmung verantwortlichen Abschnitte unabhängig sind. Die schnellen Effekte durch
Schilddrüsenhormone auf die Atmung resultieren aus der direkten Interaktion mit Kompo-
nenten des mitochondrialen Atmungsapparates ( 45 ), bzw. seinen inneren Membranen
( 37 ). Permeabilitätsänderungen der inneren Mitochondrienmembran können die Atmungs-
aktivität und damit die ATP-Produktion der Mitochondrien dem Energiebedarf der Zelle
entsprechend beeinflussen und stellen somit einen möglichen Regulationsmechanismus des
Stoffwechsels durch die Schilddrüsenhormone dar ( 25 ).
Bei der Adeninnukleotidtranslokase ( ANT ) handelt es sich um ein in die innere Mitochon-
drienmembran integriertes Protein, daß den Austausch von Adeninnukleotiden zwischen dem
Cytosol und der mitochondrialen Matrix kontrolliert. Durch mitochondriale oxydative
Phosphorylierung generiertes ATP ( 6 ) wird durch die ANT ins Cytosol transferiert, um so für
energieverbrauchende Reaktionen ( Muskelkontraktion, Proteinbiosynthese, Gluconeo-genese
u.a. ) zur Verfügung zu stehen. Damit ist die ANT ein wichtiges Verbindungsglied zwischen
ATP-Synthese und Energieverbrauch und stellt mit ihrer postulierten Beeinfluß-barkeit durch
die Schilddrüsenhormone ein weiteres mögliches Regulationsprinzip dar.

Diskussion98
Die Stimulation mitochondrialer Aktivität und die Steigerung der Aktivität der ANT durch
Schilddrüsenhormone, speziell L-T3, haben Höppner et al. in einer Arbeit ( 29 ) zusammen-
gefaßt. Hiernach ist offen, ob eine direkte Bindung von L-T3 an mitochondriale Proteine oder
eine Bindung an spezifische Proteine der Zellmembran mit konsekutiver Alteration des
Ionentransportes zugrundeliegt. Ausgeschlossen wurde von ihnen jedoch ursächlich die ANT
Genexpression und eine direkte Bindung der Schilddrüsenhormone an das ANT-Mole-kül.
Auch Mowbray et al. ( 54 ) konnten zeigen, daß die bei thyroidektomierten Ratten reduzier-te
ANT-Aktivität 15 Minuten nach einer einzelnen intravenösen Gabe von L-T3 in physiolo-
gischer Konzentration auf normale Werte angehoben werden konnte. Hieraus schlossen die
Autoren, daß die Kontrolle über die Adeninnucleotidtranslokase eine Komponente des Me-
chanismus sei, über den die schnelle und direkte Wirkung der Schilddrüsenhormone auf die
oxydative Phosphorylierung vermittelt wird.
Nach den Untersuchungen von Horst et al. ( 32 ) an isoliert perfundierten Lebern hypothy-
reoter Ratten war die Stimulation der Atmung durch L-T3 und L-T4 nach Hemmung ihrer
Dejodierung durch PTU nicht mehr nachweisbar. Die Stimulation der Atmung durch 3,5- L-T2
war dagegen unbeeinträchtigt. Hieraus leiteten die Autoren ab, daß die Wirkung der
Schilddrüsenhormone auf eine Steigerung der Atmung durch 3,5-L-T2 vermittelt wird.
Um einen Einfluß von 3,5-L-T2 auf die oxydative Phosphorylierung und mitochondriale At-
mung zu untersuchen, wurden die Experimente an isolierten hepatischen Mitochondrien hy-
pothyreoter, gehungerter Ratten durchgeführt. Nachdem Halangk et al. aus unserer Arbeits-
gruppe zunächst nach Gabe von 3,5-L-T2 in vitro zu isolierten Mitochondrien keinen Effekt
nachweisen konnten ( Werte nicht dargestellt ), wurden die beschriebenen Versuche in vivo
durchgeführt.
In vivo war 30 Minuten nach intraperitonealer Injektion von 250 µg 3,5-L-T2 /100 g KG bei im
Anschluß isolierten Mitochondrien von gehungerten, hypothyreoten, männlichen Ratten eine
signifikante Steigerung des Sauerstoffverbrauchs und damit der oxydativen Phosphory-lierung
bei Zusatz der Substrate Succinat, wie auch Glutamat zu beobachten ( vergleiche dazu die
Abb. 26 und 27 auf den Seiten 73 und 74 ). Zu späteren Zeitpunkten ( 60 und 90 Minuten ) ist

Diskussion99
keine wesentliche Zunahme des Sauerstoffverbrauchs aufgetreten, sodaß der vollständige
Effekt von 3,5-L-T2 nach 30 Minuten erreicht ist.
Nachdem in vitro der Zusatz von 3,5-L-T2 zu isolierten Mitochondrien keinen Einfluß auf den
Sauerstoffverbrauch hatte, in vivo jedoch binnen 30 Minuten eine signifikante Steige-rung des
Sauerstoffverbrauchs und damit der oxydativen Phosphorylierung nachzuweisen war, kann der
Effekt kein direkter sondern muß ein vermittelter sein. Wie oben schon dis-kutiert kommt
dafür eine durch 3,5-L-T2 bewirkte Veränderung an den Zellen oder die Freisetzung einer
Substanz X in Frage, die zu einer Steigerung der mitochondrialen Atmung und damit des
Sauerstoffverbrauchs führt ( vergleiche dazu Kapitel IV.6, Seite 89 ff. ).
Der zeitliche Ablauf der Stimulation des Sauerstoffverbrauchs isolierter Mitochondrien kor-
respondiert wiederum mit den Befunden des stimulierten Sauerstoffverbrauchs und der ge-
steigerten Gluconeogenese bei isolierter Leberperfusion ( vergleiche dazu Kapitel IV.3, Sei-te
83 ff. ).
O´Reilly und Murphy ( 61 ) beschrieben im selben Modell, bei in vivo isolierten Mitochon-
drien aus Lebern männlicher, hypothyreoter Ratten ebenfalls eine Stimulation der mitochon-
drialen Atmung durch 3,5-L-T2 nach 60 Minuten. Nach Inhibierung der Dejodinase mit PTU
konnte durch L-T3 kein vergleichbarer Effekt erzielt werden, woraus die Autoren den Schluß
zogen, daß die stimulierende Wirkung von L-T3 auf die mitochondriale Atmung erst nach
seiner Dejodierung zu 3,5-L-T2 vermittelt wird.
Ebenfalls in einem Versuchsaufbau mit isolierten Mitochondrien von Hepatozyten euthyre-oter
Ratten beschrieben Lombardi et al. ( 49 ) 60 Minuten nach Einmalinjektion von 3,5-L-T2 eine
30%-ige Steigerung der mitochondrialen Atmung. Hierbei waren weder der Proto-nenfluß über
die innere Mitochondrienmembran, noch das phosphorylierende System von 3,5-L-T2
beeinflußt. Die Autoren diskutierten, daß die Steigerung der mitochondrialen At-mung durch
eine Steigerung der Substratoxydation hervorgerufen wird. Sie postulierten ei-nen prinzipiell
unterschiedlichen Wirkmechanismus von 3,5-L-T2 und L-T3 auf die Stimula-tion der
mitochondrialen Atmung. In der Interpretation der Befunde stellten sie die Ge-schwindigkeit
der Atmungssteigerung nach 3,5-L-T2 heraus, die physiologische Bedeutung haben könnte,
wenn schnelle Energiebereitstellung für die Zelle notwendig ist.

Diskussion100
Befunde von Lanni et al. ( 42 ) zeigten, daß es einen unterschiedlichen Mechanismus der
Wirkung von L-T3 und L-T2 auf die Mitochondrien von Rattenhepatozyten gibt. Während L-T3
auf die mitochondriale Masse und Aktivität Einfluß hat, entfaltet L-T2 seine Wirkung direkt auf
den Cytochrom-Oxydase-Komplex. Der selbe Autor beschrieb aber in weiteren
Untersuchungen ( 43, 44 ), im Gegensatz zu den Befunden oben, eine spezifische Bindungs-
stelle für 3,3´-L-T2 an isolierten Mitochondrien, an die auch weniger spezifisch 3,5-L-T2
bindet. Eine physiologische Bedeutung für 3,5-L-T2 wurde nicht diskutiert.
Ob für die beschriebene Wirkung von 3,5-L-T2 eine spezifische Bindung an Proteine der
Zellmembran verantwortlich ist oder analog den Befunden zu L-T3 auch ein Einfluß von 3,5-L-
T2 auf ANT vorliegt, muß in weiteren Untersuchungen gezeigt werden. Nach fehlen-der
direkter Wirkung von 3,5-L-T2 in vitro ist das Vorhandensein eines spezifischen intra-
mitochondrialen Rezeptors, den Wrutniak et al. ( 85 ) für L-T3 an der mitochondrialen Ma-trix
beschrieben, nicht wahrscheinlich.
9. Organspezifität der Wirkung von 3,5-L-T2
Nachdem die bisher beschriebenen Versuche zur Wirkung von 3,5-L-T2 ausschließlich auf
das Organ, bzw. subzelluläre Organellen der Leber beschränkt waren, sollte im weiteren
untersucht werden, ob auch an anderen, für den Stoffwechsel der Schilddrüsenhormone
wichtigen Organen, wie der Niere eine Wirkung des Hormones nachweisbar ist.
Isolierte Nierenperfusion
Die Niere, als ein wichtiges Organ im Stoffwechsel der Schilddrüsenhormone, stellt neben
der Leber den hauptsächlichen Ort ihrer peripheren Dejodierung dar, was auch in den zahl-
reichen Untersuchungen zu diesem Thema zum Ausdruck kommt.
Ferguson et al. ( 22 ) zeigten, daß die isoliert perfundierte Niere eine hohe Kapazität zur
Dejodierung von L-T4 zu L-T3 hat, wobei die L-T3-Produktion proportional zur L-T4-Dejo-

Diskussion101
dinase-Aktivität und L-T4-Aufnahme ist. Daher konnte die Seite der 5´-Dejodinase auf kon-
traluminaler Oberfläche des renalen Tubulus lokalisiert werden. Bei diesen Untersuchungen
fand er keine Abhängigkeit der glomerulären Filtrationsrate ( GFR ) oder Na+-Rückresorp-tion
( Na RR ) von der Konzentration der eingesetzten Schilddrüsenhormone.
Adlkofer und Schurek ( 1 ) stellten fest, daß nach tubulärer Filtration die Reabsorptionska-
pazität für L-T3 und L-T4, auch bei einer Konzentration der eingesetzten Hormone, die die
physiologische um das 100-fache übersteigt, nicht sättigbar war. Der Sauerstoffverbrauch der
perfundierten Nieren lag trotz euthyreoter Versuchstiere im gleichen Bereich, wie bei den
durchgeführten Versuchen mit hypothyreoten, gehungerten Tieren ( vergleiche dazu Tab. 4,
Seite 77 ).
Bei hypothyreoten Versuchstieren fanden Holmes et al. ( 30 ) einen erhöhten Na+-Verlust der
perfundierten Nieren. Diesen führten sie auf eine geringere Atmung und Glykolyse zu-rück und
eine dadurch verringerte Energiebereitstellung für die Na+-Rückresorption der Nieren. Die
Autoren leiteten daraus eine wichtige Bedeutung der Schilddrüsenhormone für den
Nierenstoffwechsel ab, im Sinne einer Regulierung der Energiebereitstellung für die
Nierenrinde, wie auch für das Nierenmark durch anaerobe Glykolyse.
Die Versuche an der isoliert perfundierten Niere hypothyreoter, männlicher Ratten zeigten, daß
nach dem Zusatz von 3,5-L-T2 zum Perfusionsmedium in beschriebener Versuchsan-ordnung
keine signifikante Stimulation des Sauerstoffverbrauchs zur Folge hatte ( verglei-che dazu Tab.
4, Seite 77 ).
Auch eine ganze Reihe funktioneller Parameter der isolierten Nierenperfusion, wie der
glomerulären Filtrationsrate ( GFR ), der Natriumfiltration ( Na FR ) und -reabsorption
( Na RR ), wie auch der Kaliumfiltration ( K FR ) und -reabsorption ( K RR ) blieben un-
beeinflußt, oder der Betrag der entfalteten Wirkung ist so gering, daß er durch den be-
schriebenen Versuchsansatz nicht erfaßt wird.
Lediglich der singuläre Befund an kultivierten Mesangiumzellen, bei denen sogar nach Zu-satz
von 3,5-L-T2 in einer Konzentration von nur 10-13 M Kontraktionen nachgewiesen wer-den

Diskussion102
konnten, die in diesem Ausmaß sonst nur nach Zusatz von Angiotensin II vorzufinden sind,
bleibt zu diskutieren.
Funktionell regulieren Mesangiumzellen als Abkömmlinge der glatten Muskulatur durch ih-ren
Kontraktionszustand die glomeruläre Filtrationsrate durch Beeinflussung des Ultrafiltra-
tionskoeffizienten. Wenn bei isolierter Nierenperfusion keine Änderung der glomerulären
Filtrationsrate nach Zusatz von 3,5-L-T2 nachweisbar war, so ist dies möglicherweise durch
den Versuchsaufbau erklärbar, der bei dieser sensiblen Funktionsgröße durch den Perfu-
sionsdruck nicht differenziert genug beurteilt werden kann. Weitere Versuche in vivo und in
vitro müssen zeigen, wie schnell der Effekt von 3,5-L-T2 zu beobachten und worauf er
zurückzuführen ist.
Zusammengefaßt unterscheiden sich die Befunde zum Sauerstoffverbrauch der isoliert per-
fundierten Niere nach Zusatz von 3,5-L-T2 nicht von denen der anderen Schilddrüsenhor-mone
L-T3 und L-T4.
Somit ist eine Wirkung von 3,5-L-T2 auf den Sauerstoffverbrauch, oxydative Phosphorylie-
rung und Gluconeogenese, vergleichbar derjenigen auf die Leber, an der Niere nicht nach-
weisbar und weitere Versuche an spezifischeren Modellen müssen zeigen, ob die Wirkun-gen
von 3,5-L-T2 einzig auf die Leber beschränkt sind.

Zusammenfassung103
V Zusammenfassung
Die vorliegenden Experimente an der offen und rezyklierend perfundierten Leber zeigeneindeutig, daß es neben den schon lange bekannten Hormonen T3 und T4 ein weiteresSchilddrüsenhormon gibt, nämlich 3,5-L-T2, welches auf eine bisher nicht vollständig geklärteWeise binnen 30 Minuten mitochondriale Atmung, Glucose- und Harnstoff-Produktion in derLeber stimuliert. Mögliche Wirkungsmechanismen werden diskutiert.
Beteiligt sind die Zellkompartimente Zellmembran und Mitochondrien. An der Zellmem-bransteht der Wirkungsmechanismus in einem direkten Zusammenhang mit der plasma-membranständigen Na+/K+-ATPase, denn deren Hemmung verhinderte eine Stimulation desSauerstoffverbrauchs durch 3,5-L-T2. Aufgrund der Geschwindigkeit der eintretendenWirkung von 3,5-L-T2 kann diese nicht kernvermittelt und muß unabhängig von der Pro-teinbiosynthese sein.
Der durch das Hormon vermittelte Effekt trat bei rezyklierender Perfusion weitaus schnellerein und erreichte höhere Werte als bei offener Perfusion. Anscheinend wird das Hormon nachdem erstem Kontakt mit der Leber modifiziert oder es vermittelt seine Wirkung durchFreisetzung einer Substanz X, die dann ihrerseits die beschriebene Steigerung des Stoff-wechsels auslöst. Diese Wirkung ist reversibel, wie die Versuche bei offener Leberperfu-sionzeigten, als am Versuchsende wieder hormonfrei perfundiert wurde.
Darüberhinaus entfaltet das Hormon seine atmungssteigernde Wirkung über die Mitochon-drien. Auch hier wurde der Effekt bei isolierten Mitochondrien nicht direkt durch 3,5-L-T2
erzielt, wie die Experimente in vitro zeigten, sondern nach in vivo Applikation des Hormo-neswar die Atmung wiederum bereits nach 30 Minuten signifikant gesteigert.
Dem Prinzip dieses Effektes von 3,5-L-T2 liegt offenbar kein Schalter zugrunde, der entwe-deran- oder ausgeschaltet wird, sondern eher eine kontrollierte Regulation, an der auch einspezifischer Rezeptor beteiligt sein könnte. Eine Reihe von schnellen Effekten, die früherL-T3 zugeschrieben worden sind, müssen, wie die Experimente mit Einsatz von PTU zei-gen,als Wirkung von 3,5-L-T2 aufgefaßt werden.
Durch die veränderte Enzymausstattung der Zellen, wie nach kohlenhydratreicher Ernäh-rungoder unter Einfluß der unterschiedlichen Steroidhormone der Geschlechter, ist die Ge-schwindigkeit des Effektes nur geringfügig steigerbar unabhängig von der eingesetzten Hor-monkonzentration.
Bei Mesangiumzellen, spezialisierten glatten Muskelzellen, die für die Regulation der GFR derNiere verantwortlich sind, wurde durch 3,5-L-T2 in einer Konzentration von lediglich 10-13 Meine starke Kontraktion ausgelöst, vergleichbar mit Angiotensin II. Wurden die Experimenteauf das Gesamtmodell der isoliert perfundierten Niere ausgedehnt konnte nur eine geringeWirkung festgestellt werden.
3,5-L-T2 ist das Produkt aus der 3´-Dejodierung von 3,5,3´-L-T3 durch das Enzym Mono-dejodinase Typ I. Die tägliche Produktion beträgt beim Menschen 0,6 - 2,9 µg und die phy-siologische Serumkonzentration 16,2 ± 6,4 ∗ 10-12 Mol/l bzw. 0,4 - 5,3 ng/dl. Die Serum-spiegel von 3,5-L-T2 unterliegen einer hohen Hormon-Clearance.

Literaturverzeichnis104
VI LITERATURVERZEICHNIS
1. Adlkofer F, Schurek HJ, Sörje H, 1980, Horm Metab Res 12 : 400
2. Asano Y, Liberman UA, Edelman IS, 1976, J Clin Invest 57 : 368
3. Auf´mkolk M, Koehrle J, Hesch RD, Cody V, 1986, J Biol Chem 261 : 11623
4. Bergmeyer HU, 1974, Methoden der Enzymatischen Analyse, 3. Auflage,
Verlag Chemie
5. Böttger I, Kriegel H, Wieland O, 1970, Eur J Biochem 13 : 253
6. Brand MD, Murphy MP, 1987, Biol Rev 62 : 141
7. Braverman LE, Vagenakis A, Downs P, Foster AE, Sterling K, Ingbar SH, 1973,
J Clin Invest 52 : 1010
8. Burger AG, 1981, Int J Obesity 5 Suppl 1 : 69
9. Burger AG, Berger M, Wimpfheimer K, Danforth E, 1980, Acta Endocr 93 : 322
10. Burger AG, O´Connell M, Scheidegger K, WooR, Danforth jr. E, 1987,
J Clin Endocrin Metab 65 : 829
11. Chopra IJ, 1976, J Clin Invest 58
12. Cooper DS, Kieffer JD, Saxe V, Mover H, Maloof F, Rigdway EC, 1983,
Endocr 114 : 786
13. Corrigall J, Tselentis BS, Mowbray J, 1984, Eur J Biochem 141 : 435

Literaturverzeichnis105
14. Crespo-Armas A, Mowbray J, 1987, Biochem J 241 : 657
15. Davis FB, Cody V, Davis PJ, Borzynski LJ, Blas SD, 1983,
J Biol Chem 258 : 12373
16. Dillmann WH, Barrieux A, Neeley WE, Contreras P, 1983, J Biol Chem 258 : 7738
17. Engler D, Burger AG, 1984, Endocr Rev 5 : 151
18. Engler D, Merkelbach U, Steiger G, Burger AG, 1984,
J Clin Endocrin Metab 58 : 49
19. Exton JH, Park CR, 1967, J Biol Chem 242 : 2622
20. Faber J, Kirkegaard C, Lumholtz IB, Siersbaek-Nielsen K, Friis T, 1979,
J Clin Endocrin Metab 48 : 611
21. Faber J, Kirkegaard C, Lumholtz IB, Siersbaek-Nielsen K, Friis T, 1982,
J Clin Endocrin Metab 55 : 8
22. Ferguson DC, Jennings AS, 1983, Am J Physiol 245(Endocr Met 8) : E220
23. Guernsey DL, Edelman IS, 1983, in „ Molecular basis of thyroid hormone action,
Acad. Press : 293
24. Haber RS, Ismail-Beigi F, Loeb JN, 1988, Endocr 123 : 238
25. Harper ME, Ballantyne JS, Leach M, Brand MD, 1993,
Biochem Soc Trans 21 : 784
26. Hashizume K, Ichikawa K, 1982, Biochem Biophys Res Comm 106 : 920
27. Hashizume K, Ichikawa K, Kobayashi M, 1984, Endocr. Jpn. 31 : 311

Literaturverzeichnis106
28. Hashizume K, Kobayashi M, Miyamoto T, Yamauchi K, 1986, Endocr. 119 : 1063
29. Höppner W, Horst C, Rasmussen UB, Seitz HJ, June 1987, Int Symp Reisensburg
30. Holmes jr. EW, Discala VA, 1971, Am J Physiol 221 : 839
31. Horst C, Hummerich H, Soboll S, Seitz HJ, 1989, „ Rapid effect of thyroid
hormones on hepatic energy metabolism“ in : „Hormones, Thermogenesis and
Obesity“, Els. Sci. Publ. Co.; Inc. : 311
32. Horst C, Rokos H, Seitz HJ, 1989, Biochem J 261 : 945
33. Hsu RY, Lardy HA, 1969, Methods Enzymol 8 : 230
34. Hummerich H, Soboll S, 1989, Biochem J 258 : 363
35. Ichikawa K, Hashizume K, 1991, Life Sci 49 : 1513
36. Ismail-Beigi F, Edelman IS, 1970, Proc. Nat. Acad. Sci. USA 67 : 1071
37. Kalderon B, Hermesh O, Bartana J, 1995, Endocr 136 : 3552
38. Köhrle J, in : „The low T3 syndrome“, 1981, Edit. Hesch RD, Academic Press : 27
39. Krebs HA, Henseleit K, 1932, Hoppe-Seyler Z 210 : 33
40. Krenning E, Docter R, Bernard B, Visser T, Hennemann G, 1981, Biochim Biophys
Acta 676 : 314
41. Kvetny J, 1992, Horm Metab Res 24 : 322
42. Lanni A, Moreno M, Cioffi M, Goglia F, 1992, Mol Cell Endocr 86 : 143

Literaturverzeichnis107
43. Lanni A, Moreno M, Cioffi M, Goglia F, 1993, J Endocr 136 : 59
44. Lanni A, Moreno M, Horst C, Lombardi A, Goglia F, 1994, FEBS Lett. 351 : 237
45. Lanni A, Moreno M, Lombardi A, Goglia F, 1994, Mol Cell Endocr 99 : 89
46. Larsen PR, 1975, J Clin Endocrinol Metab 41 : 1098
47. Lee YP, Takemor AE, Lardy H, 1959, J Biol Chem 234 : 3051
48. Leonard JL, Visser TJ, „Biochemistry of deiodination“ in : „Thyroid Hormone
Metabolism“, 1986, Edit. Hennemann G, Marcel Dekker Inc., : 189
49. Lombardi A, Lanni A, Moreno M, Brand MD, Goglia F, 1997, 24th ETA München
Poster Nr. 226
50. Maciel RMB, Chopra IJ, Ozawa Y, Geola F, Solomon DH, 1979,
J Clin Endocrin Metab 49 : 399
51. Marish CN, Oppenheimer JH, General review in : „Molecular basis of thyroid
hormone action“, 1983, Academic Press, : 266
52. Mariash CN, Kaiser FE, Schwartz HL, Towle HC, Oppenheimer JH, 1980,
J Clin Invest 65 : 1126
53. Menahan LA, Wieland O, 1969, Eur J Biochem 10 : 188
54. Mowbray J, Corrigall J, 1984, Eur J Biochem 139 : 95
55. Müller MJ, Seitz HJ, 1980, Life Sci 27 : 827
56. Müller MJ, Seitz HJ, 1981, Life Sci 28 : 2243

Literaturverzeichnis108
57. Nicoloff JT, Low JC, Dussault JH, Fisher DA, 1972, J Clin Invest 51 : 473
58. Ochoa S, 1955, Methods Enzymol 1 : 739
59. Oppenheimer JH, 1979, Science 203 : 971
60. Oppenheimer JH, Silva E, Schwartz HC, Surks MI, 1977, J Clin Invest 59 : 517
61. O´Reilly I, Murphy MP, 1992, Biochem Soc Trans 20 : 59 S
62. Palacios-Romero R, Mowbray J, 1979, Biochem J 184 : 527
63. Pennock GD, Raya TE, Bahl JJ, Goldman S, Morkin E, 1992,
J Pharma Exp Ther 263 : 163
64. Pfannstiel P, in : „Schilddrüsenkrankheiten - Diagnose und Therapie“, 1985,
Grosse Verlag Berlin, S. 10
65. Pillar TM, Seitz HJ, 1997, Eur J Endocrinol, 1997, 136 : 231
66. Pinna G, Hiedra L, Höll T, Stoltenburg G, Eravci M, Finke R, Meinhold H,
Baumgartner A, 1997, 24th ETA München, Poster Nr. 227
67. Pittman CS, Chambers Jr JB, Read VH, 1971, J Clin Invest 50 : 1187
68. Rasmussen H, Köhrle J, Rokos H, Hesch RD, 1989, FEBS Let 255 : 385
69. Sachs L, Statistische Methoden, 1982, Springer Verlag
70. Scarpulla CR, Kilar MC, Scarpulla KM, 1986, J Biol Chem 261 : 4660
71. Scholz R, in : „Stoffwechsel der isoliert perfundierten Leber“, 1968, Hrsg. Staib und
Scholz, Springer Verlag, S. 25

Literaturverzeichnis109
72. Segal J, 1988, Biochemistry 27 : 2586
73. Segal J, 1988, Endocr 122 : 2240
74. Segal J, Ingbar SH, 1985, J Clin Invest 76 : 1575
75. Seitz HJ, Müller MJ, Soboll S, 1985, Biochem J 227 : 149
76. Sestoft L, 1974, Biochim Biophys Acta 343 : 1
77. Sestoft L, Bartels PD, Fleron P, Folke M, Gammeltoft S, Kristensen LÖ, 1977,
Biochim Biophys Acta 499 : 119
78. Soboll S, 1993, Biochim Biophys Acta 1144 : 1
79. Sterling K, 1979, N Engl J Med 300 : 117 und 173
80. Sterling K, 1986, Endocr 119 : 292
81. Sterling K, Brenner MA, Sakurada T, 1980, Science 210 : 340
82. Sterling K, Lazarus JH, Milch PO, Sakwada T, Brenner MA, 1978, Science
( Wash. DC ) 201 : 1126
83. Sterling K, Milch PO, Brenner MA, Lazarus JH, 1977, Science 197 : 996
84. Surks MI, Schadlow AR, Stock JM, Oppenheimer JH, 1973, J Clin Invest 52 : 805
85. Wrutniak C, Cassar-Malek I, Marchal S, Rascle A, Heusser S, Keller JM, Fléchon
J, Dauca M, Samarut J, Ghysdael J, Cabello G, 1995, J Biol Chem 270 : 16347

Anhang110
ABKÜRZUNGEN
ADP Adenosindiphosphat
ANT Adeninnukleotidtranslokase
ATP Adenosintriphosphat
BMR Basal Metabolic Rate, Stoffwechselgrundumsatz
cAMP zyklisches Adenosinmonophosphat
Chol Cholesterin
DIT Dijodtyrosin
FAS Fettsäuresynthetase
FR Filtrationsrate
g Gramm
GFR Glomeruläre Filtrationsrate
GOT Glutamat-Oxalacetat-Transaminase
G6PD Glucose-6-phosphat-Dehydrogenase
i.p. intra peritoneal
KG Körpergewicht
l Liter
LDH Lactat-Dehydrogenase
LG Lebergewicht
M Mol
ME Malic Enzyme
min Minute

Anhang111
MIT Monojodtyrosin
MMI 2-Mercapto-1-Methylimidazol
MW Mittelwert
p Druck
PEPck Phosphoenolpyruvat-Carboxykinase
6PGD 6-Phosphogluconat-Dehydrogenase
PTU 6-n-Propyl-2-Thiouracil
eingetragener Warenname
RNA Ribonukleinsäure
RR Rückresorptionsrate
R-SH Sulfidseitengruppen-tragender Cofaktor
SEM Standard Error of the Mean
SITS 4-Acetamido-4´-isothiocyanatostilbene-2,2´-disulfon-säure
T1 3-Monojodthyronin, L-T1
T2 3,5-Dijod-L-thyronin, L-T2
T3 3,5,3´-Trijod-L-thyronin, L-T3
rT3 reverses L-T3
T4 3,5,3´,5´-Tetra-L-thyronin, L-Thyroxin
TBG Thyroxin bindendes Globulin
TBPH T4 bindendes Präalbumin
Tetrac Tetrajodthyroacetat
TRH TSH Releasing Hormone
TSH Thyroidea stimulierendes Hormon

Anhang112
DANKSAGUNGEN
Ich danke Herrn Professor Dr. med. Hans Joachim Seitz, Institut für Physiologische Chemie,
Abteilung für Biochemische Endokrinologie, für die Überlassung des Themas und die
Betreuung der Arbeit.
Meine weiterer Dank gilt
Herrn Prof. Dr. med. Christoph Weiss (em.), Institut für Physiologie der Universität Lübeck,
und Herrn Dr. Horst Pagel für die Gastfreundschaft und Unterstützung bei der Durchführung
der Experimente an der isoliert perfundierten Niere;
Herrn Prof. Dr. med. Dieter Häussinger, Klinik für Innere Medizin der Universität Düsseldorf,
vormals Universität Freiburg, für Gastfreundschaft und Unterstützung bei der Durchführung
der Experimente an der offenen Leberperfusion;
Herrn Priv.-Doz. Dr. rer. nat. Peter Schönfeld, Institut für Physiologische Chemie der
Universität Magdeburg für die Unterstützung bei der Durchführung der Experimente an den
isolierten Mitochondrien;
der Firma Henning Berlin, die die verwendeten Schilddrüsenhormone dem Institut für
Physiologische Chemie der Universität Hamburg als Geschenk überließen.

Anhang113
LEBENSLAUF
Niels Köster
geb. am 24. März 1965 in Hamburg
1971 bis 1975 Grundschule Hasselbrook, Hamburg
1975 bis 1984 Gymnasium Borgfelde, Hamburg, Abitur
1985 bis 1992 Studium der Medizin an der Universität Hamburg,
Staatsexamen
1988 bis 1994 Experimentelle Arbeit zur Erstellung der Promotion, zunächst in der
Abt. Biochemische Endokrinologie, am Institut für Physiologische
Chemie an der Universität Hamburg; später Zusammenarbeit mit
Arbeitsgruppen aus Laboratorien der Universitäten Lübeck,
Magdeburg und Freiburg
1992 bis 1993 wissenschaftlicher Mitarbeiter am Institut für Physiologische Chemie,
Abt. Biochemische Endokrinologie bei
Herrn Prof. Dr. H.-J. Seitz, an der Universität Hamburg, Fachbereich
Medizin
1993 bis 1994 Arzt im Praktikum in der Abt. für Anästhesiologie und operative
Intensivmedizin bei Herrn Dr. med. P. Hoffmann,
Allgemeines Krankenhaus Barmbek in Hamburg
seit 1994 Assistenzarzt in der Abt. Anästhesiologie, operative Intensivmedi-
zin bei Herrn Dr. med. P. Hoffmann, Allgemeines Krankenhaus
Barmbek in Hamburg;
seit 1998 Leitender Arzt Herr Dr. med. Hp. Moecke
Hamburg, den 27. Februar 1998






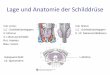











![53. Rundversuch „TUMOR - qmimed.oequasta.atqmimed.oequasta.at/GAWOUT/11/Soll_11_053.pdf · Rundversuch „TUMOR – MARKER“ Probe 1 = A Probe 2 = B CEA [µg/l] AFP [µg/l]](https://img.pdfslide.org/doc/110x75/5d65e24c88c993bc348bb0a1/53-rundversuch-tumor-rundversuch-tumor-marker-probe-1-a-probe.jpg)




![i 39 Labor information · Transferrinsättigung = [Serumeisen (µg/dl):TEBK (µg/dl)] x 100]; soll unter EPO > 20 % sein Triglyceride halbjährlich Kardiovaskuläre Risikoabschätzung,](https://img.pdfslide.org/doc/110x75/5d47081188c993c55b8ba3f7/i-39-labor-information-transferrinsaettigung-serumeisen-gdltebk-gdl.jpg)
![Bericht zu PM10-Tagesmittelwerten und Überschreitungen …...28.04.2011 PM10 [µg/m³] 1 58 05.11.2011 PM10 [µg/m³] 5 62 12.11.2011 PM10 [µg/m³] 3 102 23.11.2011 PM10 [µg/m³]](https://img.pdfslide.org/doc/110x75/5feb2fd0c3ceb232dc68d90f/bericht-zu-pm10-tagesmittelwerten-und-oeberschreitungen-28042011-pm10-gm.jpg)




