Embed Size (px)
Citation preview

Schwermetalle und Pflanzen - einekomplizierte Beziehung (II)
Schwermetall-Hyperakkumulation im Wilden Westenmodified from:

hem
men
d
Effe
kt
förd
erlic
h
Konzentration
nur Schadstoff (z.B. Quecksilber) Mikronährstoff (z.B. Kupfer)
Die Dosis macht das Gift - auch bei Schwermetallen!

Ein scheinbar intakter, natürlicherBachlauf - die Elodea canadensisdarin war jedoch an Zn-Vergiftungeingegangen, die ihr Chlorophyll inZn-Chlorophyll umgewandelt hatte.
Küpper H, Küpper F, Spiller M (1996) Journal of Experimental Botany 47 (295), 259-66
Umweltrelevanz von Schwermetall-Toxizität

Umweltrelevanz von Schwermetall-Toxizität (II)
Wo? Wodurch? Für Wen?
• Natürlicherweise auf schwermetallhaltigen Böden(Cu: z.B. in Zaire, Afrika; Zn/Cd: rel. häufig, auch inMitteleuropa; Nickel: rel. häufig, Serpentinböden z.B.in Afrika, Australien, Nord- und Mittelamerika): Schwermetallkonzentrationen hochgenug für Toxizität gegen die meisten Organismen.
• Natürlicherweise in kupferhaltigen Bereichen der Ozeane (z.B. Sargasso-See):Kupferkonzentrationen im nanomolaren Bereich hemmen bereits empfindlicheCyanobakterien.
• Anthropogen wegen Einsatzes von Schwermetallsalzen (z.B. CuSO4,z.B. Zn-Phosphid, Zn-Borat, Zn-Fluosilikat): Konzentrationen immikromolaren Bereich sind toxisch für viele Pflanzen, v.a. Wasserpflanzen inangrenzenden Gewässern
• Anthropogen durch Erz-Abbau und -Verhüttung, Konzentrationen in der Umgebungvon Minen, Hütten und Abraumhalden können extrem hoch und für alle Organismentoxisch sein.
• Anthropogen in geringerem Umfang auch durch andere Industrien. Der Rhein enthieltin den 70er Jahren bis zu 0.5 µM Kupfer, was für empfindliche Wasserpflanzen wieStratiotes oder Elodea bei längerer Einwirkung letal ist.

Wirkungsmechanismen von Schwermetall-Toxizitätund Resistenz-Mechanismen, die diese gezielt verhindern
--> direkte Hemmung der Photosynthese
--> oxidativer Stress, entweder direkt durch dieSchwermetall hervorgerufen oder durch dieFehlfunktion der Photosynthese induziert.
--> Hemmung der Wurzelfunktion
--> Hemmung der Atmung und anderer wenigersensitiver Prozesse
--> Substitution von aktiven Zentren in Enzymen

1. Hemmung der Wurzelfunktion
Warum Wurzeln?
• Bei terrestrischen Pflanzen ist die Wurzel generell das erste Organ der Pflanze, dasmit den Schwermetallen in Berührung kommt.
• Bei Schwermetallen mit relativ geringer Mobilität, z.B. Cu+, werden auch die höchstenKonzentrationen in der Wurzel erreicht
Mechanismen
• Kompetition in der Aufnahme mit weniger verfügbaren essentiellen Spurenelementen,da z.T. von denselben Proteinen transportiert: umstritten da widersprüchliche Daten
• Vermutlich Hemmung von Transportproteinen
• dazu diverse relativ unspezifische Inhibierungen von cytoplasmatischen Enzymen
• und Hemmung der Zellteilung (Wirkmechanismus unbekannt)
• Also Folge der Wurzeltoxizität sterben zunächst die Wurzelhaare und Wurzelspitzen ab

1. Hemmung der Wurzelfunktion:Resistenzmechanismen
Strategien
• Verringerung der unspezifischen Durchlässigkeit der Wurzel für unerwünschteSchwermetalle: Expression von Peroxidasen verstärkt Lignifizierung
• Aktive (ATP-abhängige) Ausschleusung durch Efflux-Pumpen: Wurde für Cu in Silenevulgaris und für diverse Metalle in Bakterien gefunden.

2 H2O O2 + 4 H+
P700*
P700
A0 A1
P680*
P680
PheoQA QB
WSC 4 e-
EChlChlPQ FeS/
Rieske
PC
electrontransport
AntennaChl-proteincomplexes,
main protein:LHCII4 h·ν
excitationenergytransfer
ChlChl
Chl
heavy metals
2.1. Angriffspunkte in den photosynthetischenLichtreaktionen
2. Direkte Hemmung der Photosynthese

“Sonnen-Reaktion”In hoher Lichtintensität ist dagegen nur einkleiner Anteil der Chlorophylle der Mg-Substitution zugänglich, statt dessenfindet die Hemmung direkt im PSIIRCstatt. Der größte Anteil der Chlorophyllebleicht im Zusammenhang mit derZerstörung des Photosyntheseapparatesaus.
“Schatten-Reaktion”Bei geringer Lichtintensität mit einerNachtphase ist der größte Teil derAntennenchlorophylle (v.a. LHCII) der Mg-Substitution zugänglich. Werden dabeistabile Schwermetallchlorophylle wie z.B.Kupferchlorophyll gebildet, bleiben diePflanzen auch nach ihrem Absterbengrün.
Küpper H, Küpper F, Spiller M (1998) Photosynthesis Research 58, 125-33
2.1. Der Mechanismus der Hemmung der photosynthetischenLichtreaktionen ist in Chlorophyta lichtabhängig

Warum sind Schwermetallchlorophylle untauglich zurPhotosynthese?
• Instabiler S1-Zustand: Aufgenommene Anregungsenergie wird vor allem thermischrelaxiert, so dass die anderen Prozesse (Fluoreszenz, Energie- undLadungsübertragung) nicht oder nur mit geringer Effizienz ablaufen
• Blauverschobene Absorptionsbande des S1-Zustandes: verringert die Überlappung derAbsorptions/Emissionsbanden in dem auf Mg-Chl abgestimmten System, wodurcherneut die Effizienz der Energieübertragung sinkt
• Wegen der vorgenannten Gründe wirken Schwermetallchlorophylle in einem Mg-Chl-basierten System als „schwarze Löcher“ für Anregungsenergie, so dass nur wenige (3-10) Prozent des Gesamtchlorophylls in Schwermetallchlorophyll umgewandelt werdenmüssen, um komplette Hemmung der Photosynthese zu erreichen.
• Schwermetall-Chlorophylle haben eine geringere Tendenz als Mg-Chl, eineLadungstrennung wie in den Reaktionszentren erforderlich durchzuführen.
• Schwermetall-Chlorophylle binden, im Gegensatz zu Mg-Chl, axiale Liganden nurschwach (z.B. Zn-Chl) oder gar nicht (Cu-Chl), was die Bindung im Protein und damitdie ganze Proteinstruktur (zer)stört.

Vergleich von Gausaustausch und steady state-Fluoreszenzin Elodea gestresst mit 0.2 µM Cu2+ im Schwachlicht
0 1 2 3 4 5 6 7 8 9 10 11 12
-20
0
20
40
60
80
100
photosynthesis
net p
hoto
synt
hetic
oxy
gen
rele
ase
days of heavy metal stress
0
20
40
60
80
100
µ y
fluor
esce
nce
fluorescence

UV/VIS -Spektroskopie: Sonnen vs. SchattenreaktionSpektren von E. canadensis Extrakten
Küpper H, Küpper F, Spiller M (1996) Journal ofExperimental Botany 47 (295), 259-66
In vivo Spektren von E. canadensis
Küpper H, Küpper F, Spiller M (1998) PhotosynthesisResearch 58, 125-33

BraunalgeEctocarpus siliculosus:Chl a/c-LHC immer derSubstitution zugänglich
--> immer Schattenreaktion
RotalgeAntithamnion plumula:
Phycobilis: kein Chl--> immer Sonnenreaktion
Küpper H, Šetlík I, Spiller M, Küpper FC, Prášil O (2002) Journal of Phycology 38(3), 429-441
UV/VIS -Spektroskopie: nicht-Chlorophyta

Hemmung des PSIIRC
Diskutierte Mechanismen
• Hemmung des wasserspaltendenKomplexes
• Hemmung der primärenLadungstrennung:- durch Bindung ans D1-Protein- durch Insertion ins Pheophytin
• Hemmung des Elektronentransports- durch Bindung an Qa-Domänedes D1-Proteins- durch Bindung an nicht-Häm-Fe-Dömäne des D1-Proteins

2.2. Hemmung der Dunkelreaktionen der Photosynthese (Calvin-Cyclus)
• Indizien aus Messungen der Chlorophyll-Fluoreszenz-Kinetik
• Mechanismus und Bedeutung unklar

3. Oxidativer Stress
Mechanismen
• Direkt: Katalysiert von redox-aktiven Metallionen (Fe2+, Cu+) werden ausWasserstoffperoxid durch die Fenton-Reaktion reaktive Sauerstoff-Radikale:
• Indirekt: Durch Fehlfunktion der Photosynthese und Atmungskette können reaktiveSauerstoff-Spezies entstehen. Deswegen kann auch bei in vivo redox-inertenMetallionen wie Zn2+ und Cd2+ oxidativer Stress auftreten.
• Oxidativer Stress kann zur Oxidation von Lipiden in Membranen und somit zu derenUndichtigkeit führen. Populärer aber in seiner Bedeutung in vivo umstrittenerMechanismus.
Bedeutung
• Umstritten: Studien mit geringen aber noch toxischen Schwermetall-Konzentrationenzeigen oft keinen oxidativen Stress

Oxidativer Stress durch Chlorophylle: Transfer vonAnregungsenergie auf Singulett-Sauerstoff
chlorophyll oxygen
S0
S2
S1T1
T1
S1h·ν
h·ν
intersystem crossing
absorption
absorptionfluorescence
intersystem crossingintersystem crossing
intersystem crossing
phosphorescence
phosphorescence
intersystem crossing EET
photochemistry

NIR-Lumineszenz-Studie: Sind Schwermetallchlorophylle amschwermetallinduzierte oxidativen Stress beteiligt?
020406080
100120140160180200
Chl a derivatives Chl b derivativeseffic
ienc
y of
sin
glet
oxy
gen
prod
uctio
n/ %
of M
g-C
hl a
Mg2+ H+ (=pheophytin) Cu2+ Zn2+
0
20
40
60
80
100
120
140
lifet
ime
of C
hl tr
iple
t exc
ited
stat
e/ %
of M
g-C
hl a
0
20
40
60
80
100
120
140
Chl a derivatives Chl b derivatives
Mg2+ H+ (=pheophytin) Cu2+ Zn2+
lifet
ime
of s
ingl
et o
xyge
n/ %
of M
g-C
hl a
--> Schwermetall-Chlorophylle habenniedrigere oder gleiche Effizienz derBildung von Singulett-O2 (1O2 ) wie Mg-Chl, aber sie quenchen dieses weniger.Pheophytin erzeugt mehr und quenchtweniger 1O2 als Mg-Chl.
Küpper H, Dedic R, Svoboda A, Hála J, Kroneck PMH (2002) Biochim Biophys Act 1572, 107-113

3. Resistenzmechanismen gegen oxidativen Stress
• Verstärkte Expression von Enzymen, die reaktive Sauerstoffspezies entgiften, d.h.Superoxid-Dismutase und Katalase. Nicht immer möglich, da z.B. Cd-Stress die Zn-Aufnahme und damit die Expression von Zn-abhängigen Enzymen wie der SOD hemmt.
• Synthese von nicht-Enzym-Antioxidantien, z.B. Ascorbat und Glutathion
• Umbau der Zellmembranen, um sie resistenter gegen den Angriff von reaktivenSauerstoff-Spezies zu machen:- Lipide mit weniger ungesättigten Bindungen- Austausch von Phosphatidylcholin durch Phosphytidyl-Ethanolamin als Lipid-“Kopf“- Verringerter Anteil von Lipiden und erhöhter Anteil von stabilisierenden Proteinen inder Membran

4. Allgemeine Resistenz-Mechanismen4.1. Schwermetall-Entgiftung mit starken Liganden
Phytochelatine
• Binden mit sehr hoher Affinität Cd2+, andere Metallionen aber wenig (höchsten noch fürHg und As relevant).
• Speziell zur Cd2+-Bindung synthetisiert von γ-Glutamylcystein-Dipeptidyl-Transpeptidase = Phytochelatin-Synthase
• Sind der wichtigste Cd-Resistenzmechanismus in den meisten Pflanzen (außerHyperakkus...) und vielen Tieren
• Phytochelatin-Synthase wird von (z.B. durch Cd-Bindung) blockierten Thiolen vonGlutathion und ähnlichen Peptiden aktiviert
• Phytochelatine binden das Cd2+ im Cytoplasma, woraufhin der gesamte Komplex aktivin die Vakuole befördert wird.
• In der Vakuole bilden sich große Phytochelatin-Cd-Aggregate

4. Allgemeine Resistenz-Mechanismen
4.1. Schwermetall-Entgiftung mit starken Liganden
Metallothionine
• MTs vom Typ I und II binden mit sehr hoherAffinität Cu2+ und scheinen an dessen Entgiftungbeteiligt zu sein.
• Die Hauptrolle der MTs in Pflanzen scheint abernicht Schwermetallresistenz, sondern Metall-Verteilung während des normalen (nicht-gestressten) Stoffwechsels zu sein.
Glutathion
• Auch Glutathion selbst, der Baustein der Phytochelatine, könnte Schwermetalle bindenund somit entgiften. Ob dies in vivo aber von Bedeutung ist, scheint eher fraglich.

4. Allgemeine Resistenz-Mechanismen
4.1. Schwermetall-Entgiftung mit starken LigandenFreie Aminosäuren
• Wie beim Metalltransport unter normalen physiologischen Bedingungen, scheinen freieAminosäuren wie Histidin, Nicotianamin und Prolin eine Rolle bei der Entgiftung vonSchwermetallen zu spielen.
• Die Bedeutung von Prolin als Schwermetall-Ligand ist aber umstritten; einige Autorenstellen es eher in einen Zusammenhang mit Trockenresistenz
Andere Liganden
• Diverse Proteine wurden als Schwermetall-Entgiftungs-Liganden vorgeschlagen, z.B.ATFP (A. thaliana farnesylated protein)
• Anthocyane: Scheinen in Brassicaceen Molybdän zu binden (zur Entgiftung? zurSpeicherung?)
• Zellwand
• Manche Algen scheiden unter Cu-Stress noch unidentifizierte Thiol-Liganden aus

4.2. Schwermetall-Entgiftung durch Kompartimentierung
Mechanismen
• Generell sind hier aktive Transportprozesse gegen denKonzentrationsgradienten, also Transportproteine beteiligt.- CDF‘s- Nramp‘s- CAX‘s
• Ausschluss aus der Zellen:- in Algen: bei Braunalgen beobachtet- in Wurzeln: siehe Wurzeln
• Einlagerung in die Vakuole:- Pflanzen-spezifischer Mechanismus (Tiere haben keine Vakuolen...)- Sehr wirksam, da die Vakuole keine zu hemmenden Enzyme enthält- Spart den Aufwand der Synthese von starken Liganden wie Phytochelatinen- wichtigster Mechanismus in Hyperakkus
• Einlagerung in wenig sensitive Gewebe, z.B. Epidermis ohne statt Mesophyll mitPhotosynthese

4.3. Weitere Resistenz-Mechanismen
• Reduktion durch Reduktasen, z.B. Hg2+ --> Hg0, Cu2+ --> Cu+
• Ausfällung unlöslicher Sulfide außerhalb der Zelle (auf der Zellwand)
• Methylierung, z.B. von Arsen

--> Biologische Funktion der Hyperakkumulation
--> Mechanismen der Hyperakkumulation
--> Anwendung von Hyperakkumulatoren zur Bodenreinigung und zur Erzgewinnung
Cadmium zu allen Mahlzeiten: Pflanzen mit einem ungewöhnlichen Appetit
Schwermetall-Hyperakkumulation

Akkumulation der Metalle in Hyperakkumulatoren
• Größter Bioakkumulationskoeffizient bei kleinen Schwermetall-Konzentrationen(--> Tafelbild)
• Aktiv, durch natürliche Überexpression von Metall-Transportproteinen
• Viele Transporter überexprimiert, aber bisher unklar, was primäre und was dadurchverursachte sekundäre Überexpression ist
• Regulation der Überexpression unbekannt

Effekte von Ni2+ auf das Wachstumund Ni2+ -Konzentrationen im Sproß
(a)
Ni added to the substrate (mg kg-1)
0 1000 2000 3000 4000
Shoo
t dry
wei
ght (
g)
0
2
4
6
8
10
12
14 Thlaspi goesingenseAlyssum bertoloniiAlyssum lesbiacum
Alyssum lesbiacum
(b)
Ni added to the substrate (mg kg-1)
0 1000 2000 3000 4000
Ni c
once
ntra
tion
(µg
g-1)
0
5000
10000
15000
20000
25000
30000Thlaspi goesingenseAlyssum bertoloniiAlyssum lesbiacum
Küpper H, Lombi E, Zhao FJ, Wieshammer G, McGrath SP(2001) J Exp Bot 52 (365), 2291-2300
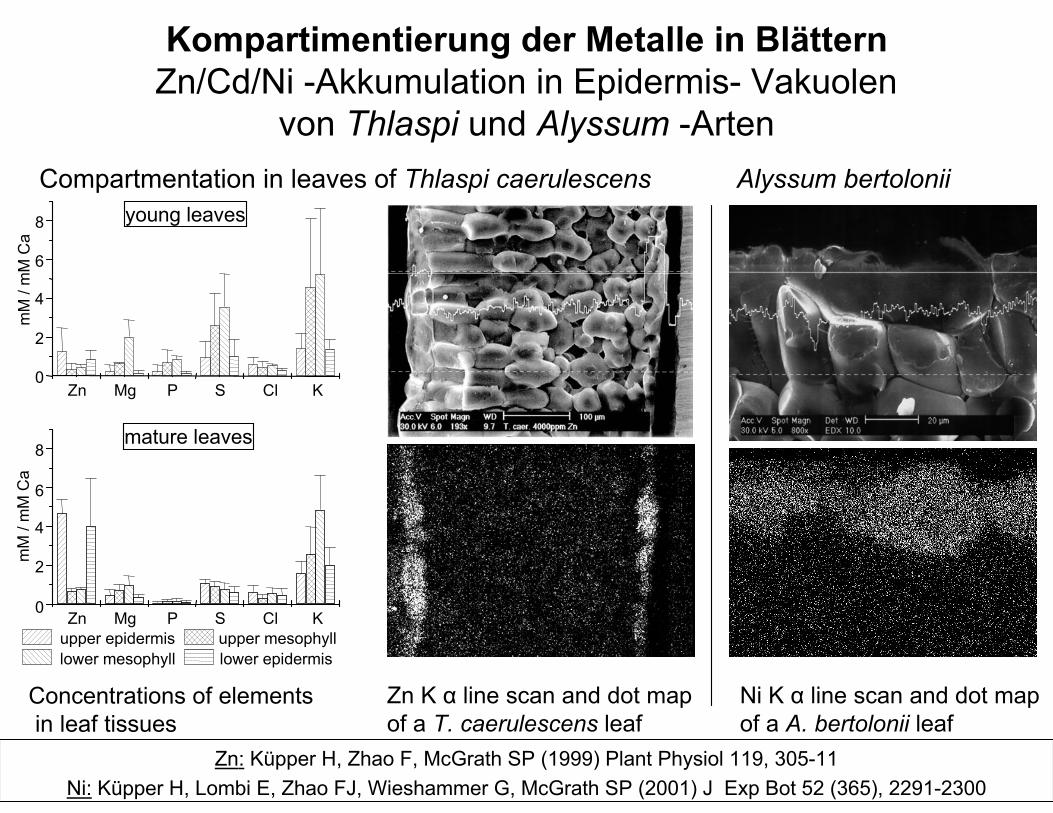
Kompartimentierung der Metalle in BlätternZn/Cd/Ni -Akkumulation in Epidermis- Vakuolen
von Thlaspi und Alyssum -ArtenCompartmentation in leaves of Thlaspi caerulescens Alyssum bertolonii
Zn Mg P S Cl K0
2
4
6
8mature leaves
mM
/ m
M C
a
Zn Mg P S Cl K0
2
4
6
8 young leaves
mM
/ m
M C
a
upper epidermis upper mesophyll lower mesophyll lower epidermis
Concentrations of elements in leaf tissues
Zn K α line scan and dot mapof a T. caerulescens leaf
Ni K α line scan and dot mapof a A. bertolonii leaf
Zn: Küpper H, Zhao F, McGrath SP (1999) Plant Physiol 119, 305-11Ni: Küpper H, Lombi E, Zhao FJ, Wieshammer G, McGrath SP (2001) J Exp Bot 52 (365), 2291-2300

Kompartimentierung von Zn und Ni in BlätternFunktionelle Differenzierung von Epidermis-Zellen in Thlaspi
15 20 25 30 35 40 45 500
2
4
6
8
regression line confidence limits
data points
Linear Regression: Y = A + B * X
Parameter Value Error-----------------------------------------------A -0.057 1.027B 0.153 0.034-----------------------------------------------
R SD N P-----------------------------------------------0.816 1.079 12 0.00118-----------------------------------------------
mM
Zn
/ mM
Ca
cell width
Upper leaf surface of T. goesingense
Dot map of the Ni K alpha lineCorrelation between cell size and zinc concentration
Zn: Küpper H, Zhao F, McGrath SP (1999) Plant Physiol 119, 305-11Ni: Küpper H, Lombi E, Zhao FJ, Wieshammer G, McGrath SP (2001) J Exp Bot 52 (365), 2291-2300

Kompartimentierung von Metallen in BlätternZn/Cd-Akkumulation in Blatthaaren von Arabidopsis halleri
Küpper H, Lombi E, Zhao FJ, McGrath SP (2000) Planta 212, 75-84

Kompartimentierung von Metallen in BlätternKorrelation zwischen Metallkonzentrationen
im Mesophyll von Arabidospis halleri
0 2 4 6 8 100
5
10
15
20
25
data pointsregression line (R = 0.79; P < 0.0001) 95% confidence limits
Mg
/ mM
Cd / mM0 20 40 60
0
2
4
6
8
10
data points regression line (R = 0.94; P < 0.0001) 95% confidence limits
Cd
/ mM
Zn / mM
Küpper H, Lombi E, Zhao FJ, McGrath SP (2000) Planta 212, 75-84
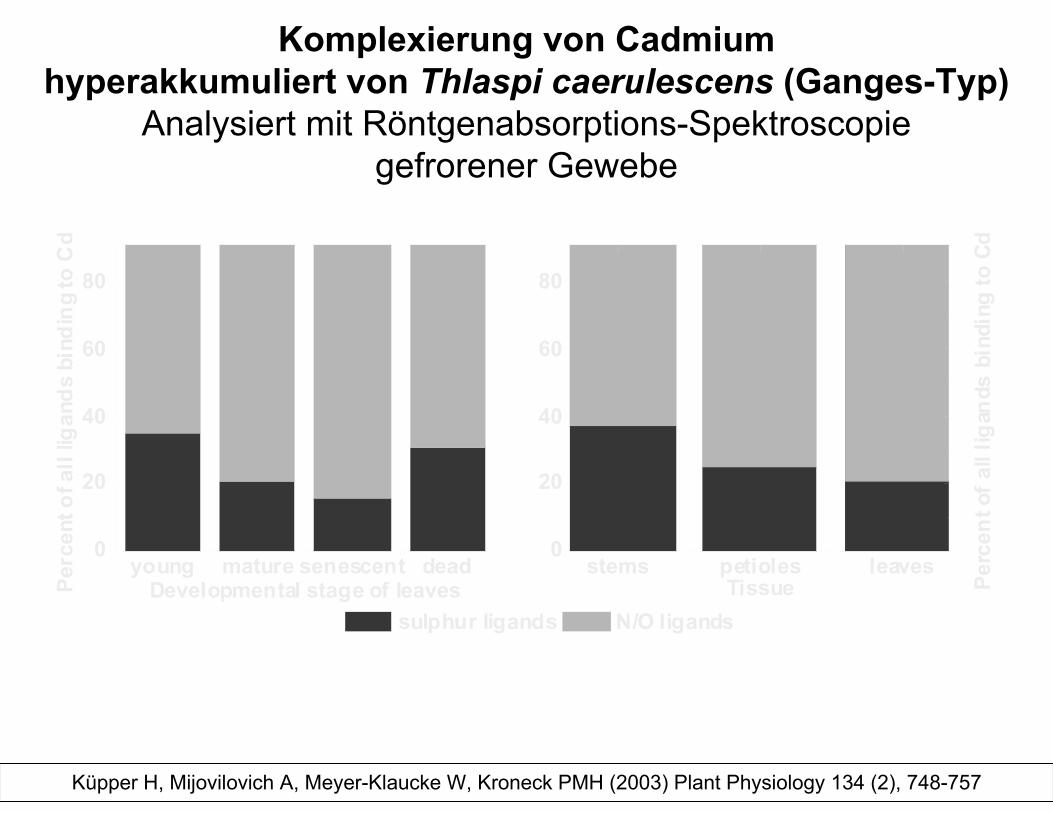
Komplexierung von Cadmiumhyperakkumuliert von Thlaspi caerulescens (Ganges-Typ)
Analysiert mit Röntgenabsorptions-Spektroscopiegefrorener Gewebe
young mature senescent dead0
20
40
60
80
Perc
ent o
f all
ligan
ds b
indi
ng to
Cd
Developmental stage of leaves sulphur ligands N/O ligands
stems petioles leaves0
20
40
60
80
Per
cent
of a
ll lig
ands
bin
ding
to C
d
Tissue
Küpper H, Mijovilovich A, Meyer-Klaucke W, Kroneck PMH (2003) Plant Physiology 134 (2), 748-757

Normal: Sequestration inepidermal storage cells
Acclimated: Enhancedsequestration in epidermalstorage cells
Stressed: additional sequestrationin selected mesophyll cells
vvv
Küpper H, Šetlík I (2004) in preparation
1. Heterogenität als Resistenzmechanismus?

Übersicht Schwermetall-Toxizität und -Resistenz
Heavy metals in soil or water
root uptake exclusionmechanisms
damage to roots
root
± root to shoottranslocation
stemepidermis
regulation byboundary cells?
stemxylem
stemphloem
leaf xylemleaf phloem
photosynthetic cells(e.g. mesophyll cells,stomatal guard cells)
epidermis cells(not present e.g. in Elodea)
vacuole
cell walls
cell walls vacuole
in the case ofsubmerged
aquatic plants
metal-induced damage tophotosynthesis
(e.g. Mg-substitution)
cyto-plasm
vacuole cytoplasm
cell walls
oxidative stressand other damagecomplexation by strong
ligands (e.g.phytochelatins)
complexation by strongligands (e.g.
phytochelatins)
Küpper H, Kroneck PMH (2004) MIBS 44 (Sigel et al., eds), chapter 5

Anwendung von Hyperakkumulatoren zur Bodenreinigung:Phytoremediation
Abraumhalde: zu giftig für normalePflanzen
Aufnahme des Metalls inHyperakkumulatoren. Nach deisemProzess ist das Metall durch den hohenBioakkumulations-Koeffizienten derHyperakkus in einem viel kleinerenVolumen konzentriert. Die geerntetenPflanzen werden verbrannt; die Aschewird entweder deponiert oder erneut zurMetallgewinnung eingesetzt.
Die gleiche Halde nach der grünenBodenentgiftung: normalesPflanzenwachstum ist wieder möglich.

Vegetation auf natürlich nickelreichemBoden (Serpentin). Solcher Boden istweder für die Landwirtschaft (Ni-Konz.viel zu hoch) noch für diekonventionelle Erzgewinnung tauglich(Ni-Konz zu niedrig).
Werden Nickel-Hyperakkumulatorenauf solchen Böden angebaut, reichernsie mehrere Prozent ihrerTrockenmasse an Nickel an. NachVerbrennung enthält die Asche 10 bis20% Nickel, so dass sie als “Bio-Erz”genutzt werden kann.
So eine “Pflanzenmine” kann,Feldstudien unter kommerziellanwendbaren Bedingungen zufolge,etwas 170 kg Nickel pro Hektar undJahr produzieren. Beim derzeitigenNickelpreis von gut 11 € pro KiloRohnickel sind das etwa 2000 €.ha-1.
Anwendung von Hyperakkumulatoren zur Metallgewinnung:Phytomining

Alle Folien meiner Vorlesungen im Internet unter
http://www.uni-konstanz.de/FuF/Bio/kuepper/Homepage/AG_Kuepper_education.html
weiterführende Literatur als pdf auf Anfrage












![Bakterielle Resistenzmechanismen – Spezielle Formen der Resistenz · 2019. 8. 20. · Carbapeneme, Monobaktame], Glykopeptide wie Vancomycin) • Inhibierung der bakteriellen Proteinsynthese](https://img.pdfslide.org/doc/110x75/60c9a1343afb1115981d336b/bakterielle-resistenzmechanismen-a-spezielle-formen-der-resistenz-2019-8-20.jpg)









